/














































































































































































































































Author: Чернов Ю.И.
Tags: тропический лес тундра издательство мысль главная редакция географической литературы полярный климат тундровый рельеф
Year: 1980




Text
1 Ю.И.Чернов ЖИЗНЬ
ТУНДРЫ
Э Р Am
\ W
ЮИЧернов ЖИЗНЬ
ТУНДРЫ
Москва
«Мысль»
1980
57.026
4-49
РЕДАКЦИИ
ГЕОГРАФИЧЕСКОЙ
ЛИТЕРАТУРЫ
© Издательство «Мысль». 1980.
Ттут 132-79- 1905000000
ПРЕДИСЛОВИЕ
Эту книгу автор посвящает столетию со дня выхода
в свет «Тропической природы» Альфреда Уоллеса,
ярко и самобытно описавшего жизнь тропического
леса.
Пышная природа тропиков и Арктика — царство
холода и снега. Казалось бы, что может быть более
несхожего? Но именно сравнение
противоположностей часто дает ключ к пониманию сложных явлений.
А. Уоллес в первых же строках своей книги
сопоставляет устойчивость тропического климата с
непостоянством полярного. Контрасты помогают лучше понять
суть. Поэтому изучение тундр, полярных пустынь и
ледников важно не только для познания природы
самой Арктики, но и для того, чтобы лучше понять
жизнь биосферы в целом.
В ландшафтах Заполярья особенно отчетливо
проявляется взаимообусловленность сил природы.
Впервые посетившего эти края поражают удивительные
формы тундрового рельефа — полигоны, сети, бугры
пучения — результат взаимодействия почвы,
растительности и воды. Чрезвычайно своеобразны
взаимоотношения организмов тундры, например зависимость
всех видов пернатых и четвероногих хищников от
леммингов. В суровых арктических условиях особенно
выпуклы закономерности приспособления организмов
к среде. Все это затрагивает многие общие принципы
естествознания. Именно поэтому изучающих
природу Арктики всегда тянуло к широкому охвату явлений
и их многоплановому анализу. Уже первые
исследователи Заполярья —А. Ф. Миддендорф, А. Г. Шренк,
Ф. Р. Чельман, А. Норденшельд — дали образцы
анализа общих особенностей жизни полярных областей
Земли.
Традиции многопланового, синтетического
изучения природы очень сильны в отечественном тундрове-
3
дении. Выдающийся тундровед Б. Н. Городков был
одновременно геоботаником, флористом, почвоведом.
Его работы содержат глубокий анализ флоры,
растительного покрова и почв. Многие из них,
выполненные несколько десятилетий назад, до сих пор
сохраняют научную ценность. Другой известный
исследователь Севера, Б. А. Тихомиров, в своих трудах освещал
такие вопросы, как роль растений в питании
животных и влияние животных на растительность,
зависимость растений и животных от структуры почвы и
влияние разных групп организмов на почву, рельеф
и т. д. Одно из первых в нашей стране биоценологи-
ческих исследований было выполнено в
субарктическом районе на Кольском полуострове В. Ю. Фридо-
линым более сорока лет назад (1936). Пример
истинно комплексного эколого-географического анализа
природы — широко известная «Субарктика» А. А.
Григорьева.
На Крайнем Севере особенно сильно ощущается
необходимость комплексного подхода к освоению
ландшафта и использованию природных ресурсов.
Жизнь человека в Арктике, как нигде более, зависит
от окружающей природы, противостоять которой
неизмеримо труднее, чем в других зонах. Изменять
природу Севера надо с большой осторожностью, ибо она
очень ранима, после нарушения восстанавливается
чрезвычайно медленно и часто оборачивается против
человека непредвиденными последствиями
неразумного обращения с нею. Освоение арктических районов
ставит множество специфических проблем, которым
нет аналогов в других ландшафтах. Специалисты,
разрабатывающие принципы и методы рационального
использования и охраны природных ресурсов,
должны учитывать множество фактов из самых различных
сфер изучения природы Арктики — климатологии,
почвоведения, мерзлотоведения, ботаники, зоологии
и т. д. Для этого нужны общие сводки и популярные
книги, освещающие разные стороны жизни
ландшафтов Заполярья.
Настоящую книгу автор рассматривает как
предварительный этап создания общедоступного
руководства по основам современного тундроведения. Она на-
4
писана в виде кратких очерков по некоторым
наиболее интересным эколого-географическим вопросам.
В процессе работы над книгой ему оказали большую
помощь ботаники Н. В. Матвеева, Б. А. Юрцев и
зоолог А. В. Кречмар, любезно предоставивший
фотографии птиц и зверей. В. Г. Ковалев и В С. Шишкин
сделали для книги несколько рисунков. Автор выра^-
жает всем глубокую благодарность.
ЧТО ТАКОЕ ТУНДРА?
Тундрой обычно называют безлесные ландшафты,
расположенные к северу от тайги — вдоль побережья
Северного Ледовитого океана и на его островах.
Слово «тундра» происходит от финского tunturi (плоская
безлесная возвышенность). Именно в таком смысле
оно употребляется на Кольском полуострове. Это
слово встречается в языке многих народов, живущих и
в лесных районах. На юге Сибири, Камчатке и
Сахалине тундрой называют различные болота.
Существует физико-географическое понятие
«горная тундра» — безлесные ландшафты в
высокогорьях. Горные тундры особенно широко распространены
в Сибири. Для гор, расположенных в более низких
широтах, например в субтропических, тундровый тип
растительности менее характерен. Выше уровня леса
там развиты так называемые альпийские луга.
Необходимо иметь в виду, что слово «тундра»
(так же как «тайга» и «степь») имеет двойной смысл.
С одной стороны, так для краткости называют всю
тундровую зону (аналогичным образом говорят
«тайга», подразумевая таежную зону), с другой стороны,
им обозначают совершенно определенные
растительные сообщества, которые надо отличать, например, от
пойменной растительности, болот или массивов леса,
расположенных среди тундр. Аналогичным образом в
таежной зоне далеко не все тайга. Там есть и луга, и
болота, и поймы рек. Нет основания относиться негативно
к двойственности этих понятий. Ниже слово «тундра»
будет использоваться в том и другом смысле.
В этой книге речь пойдет только о тех
территориях, которые относятся к природной зоне тундр в
физико-географическом понимании. Это территории, где
на равнинах развиты торфянисто-глеевые почвы и
растительность, в которой главную роль играют мхи,
образующие сплошную или разбитую на отдельности
дернину. Кроме мхов для растительного покрова
очень характерны лищайники, а из цветковых — в
«
первую очередь осоковые (осоки, пушицы), а также
злаки, кустарники (ивы, березки, ольховник),
кустарнички и полукустарнички (карликовые ивы, дриады,
вересковые, брусничные и др.), некоторые
двудольные травы. Но мхи—это основные «строители»
сообществ, или, как говорят экологи, эдификаторы
(так же как, например, ель в еловом лесу). Они
определяют в тундре и летние температуры почвы, и
глубину оттаивания мерзлоты, а также служат
местом обитания многих животных, травянистых
растений и кустарничков. Тундроведы часто приводят в
связи с этим яркие слова М. В. Ломоносова из его
сочинения «О слоях земных»: «...тундрами
называются места мохом зарослые».
Этим тундра сходна с тайгой, где мхи также
образуют сплошной покров. В литературе можно
даже встретить выражение, что тундра — это тайга
без деревьев. Причина этого—некоторое сходство
климатических условий (например, сильное
увлажнение, длительная холодная зима и др.), а также
единство истории этих территорий. Об этом
свидетельствует множество видов растений и животных,
общих для тайги и тундры.
Тундровая зона тянется вдоль побережья
Северного Ледовитого океана (см. рис. 1). Это, конечно, не
значит, что тундра — приморский ландшафт. Дело в
том, что побережье океана более или менее
соответствует широтам субарктического пояса, в котором
преимущественно развиты тундры. Вместе с тем
океан сильно влияет па тундровый ландшафт, смещая
его границы и изменяя растительность, почву,
животный мир.
Общая площадь равнинных тундр относительно
невелика, всего около 4 млн. км2, что составляет
небольшую часть суши — примерно 3%. Это результат
того, что большая часть тундр «срезана» океаном. Но
дело не только в этом. Сама протяженность высоких
широт относительно мала: расстояние по экватору—
40 076 км, а по параллели 70°—13 752 км. Более
половины площади тундровой зоны приходится на
территорию СССР.
Ширина полосы тундр в разных частях различна.
Так, на Таймыре она простирается с юга на север на
7
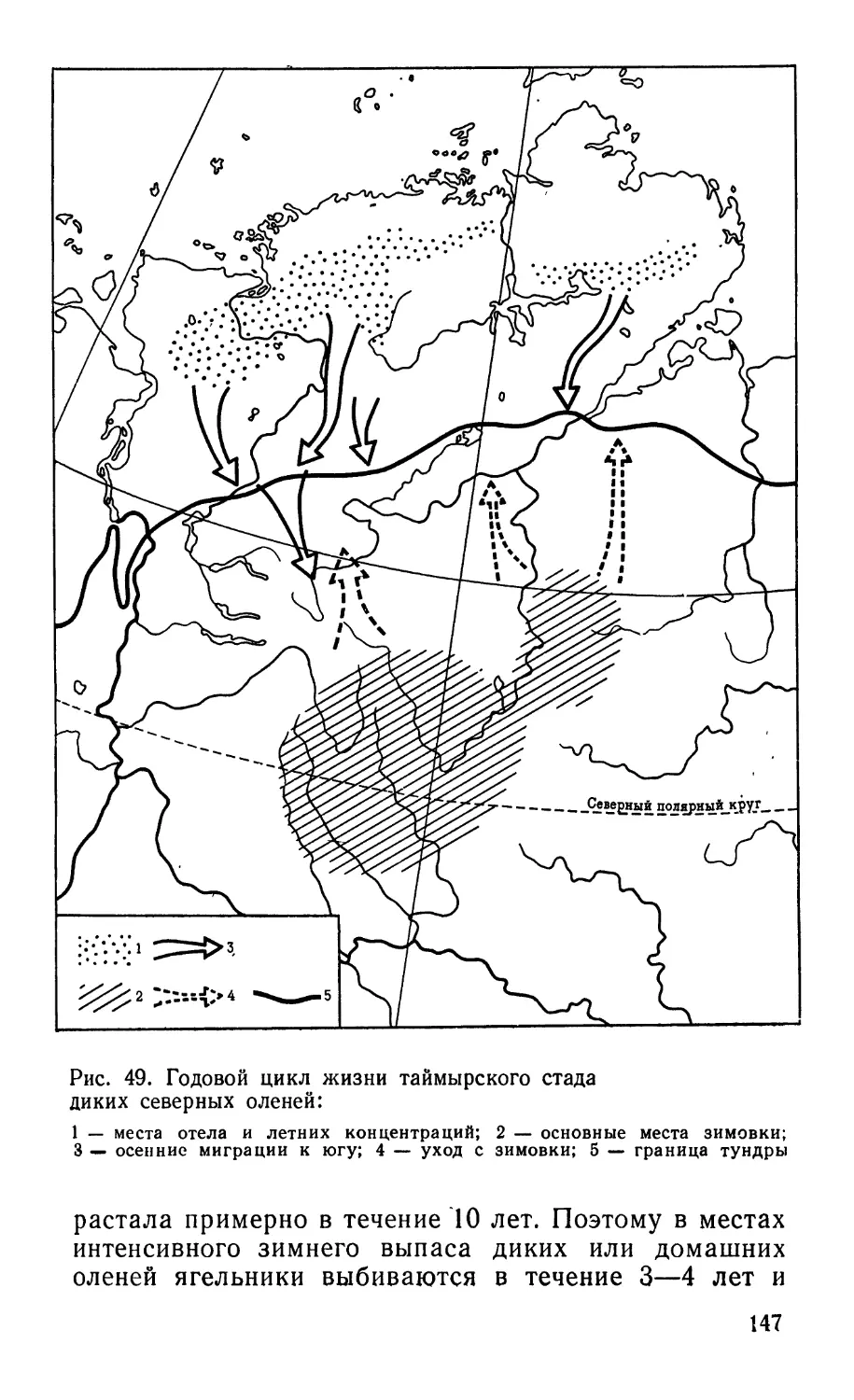
Рис. 1. Основные зональные ландшафты Арктики в Евразии:
1 -— полярные пустыни; 2 -— арктические, 3 — типичные, 4 — южные
600—700 км, контактирует на юге с лесотундрой, а
на севере — с полярными пустыэдми. Однако в
других местах материковые субарктические ландшафты
сильно «обрезаны» береговой линией. Так, к западу
от Югорского полуострова развита узкая лолоса
южных тундр. На Кольском полуострове она уже, а в
Скандинавии срезана полностью. Здесь имеются лишь
горные тундры. К востоку от Таймыра равнинная
тундра тянется вдоль побережья узкой полосой. Ее
самые северные варианты здесь представлены в
основном на островах (Новосибирские, Врангеля).
Еще меньшую площадь занимают полярные
пустыни (см. книгу В. Д. Александровой, 1977). Лишь ffa
Таймыре (мыс Челюскин) они контактируют с
тундрами. В других районах располагаются только на
островах (Земля Франца-Иосифа, Северный остров
8
(кустарниковые и кочкарные) тундры; 5 — южная граница лесотундры
Новой Земли, Северная Земля, острова Де-Лонга,
Шпицберген, часть Канадского Арктического
архипелага). Иногда к ним относят также и все
Новосибирские острова и остров Врангеля (см.
Геоботаническую карту СССР, 1956 г.), что явно
ошибочно. Их растительность соответствует всем
характерным признакам арктических тундр.
Полярные пустыни бесспорно должны выделяться
в качестве особой природной зоны. Они
характеризуются не только бедностью жизни. Много
своеобразного в структуре их ландшафта, видовом составе,
распределении и пищевых отношениях растений и
животных. Лишайники, мхи, цветковые там образуют не
сплошной покров, а лишь изолированные куртины.
Главные «продуценты» органического вещества в
полярных пустынях — одноклеточные водоросли, обиль-
9
ные на поверхности кажущихся безжизненными
оголенных грунтов. Они дают пищу также мелким
животным — ногохвосткам, личинкам комаров, червям —
энхитреидам и нематодам.
К югу от тундр располагается лесотундра. Многие
авторы называют ее зоной. Но едва ли это
правомочно. Настоящие, общепринятые зоны, например
таежная или тундровая, четко делятся на три подзоны —
две окраинные и одну срединную. Лесотундру нельзя
разделить на достаточно четкие широтные части, а
по занимаемой площади в равнинных районах и по
однообразию ландшафта она примерно равноценна
любой из тундровых или лесных подзон. Вероятно, ее
правильнее называть переходной полосой (между тундрой
и тайгой) или зоной второго порядка (Чернов, 1975).
Итак, две лесные зоны (тайга и лиственные леса)
на севере отделены лесотундрой от другой пары зон —
тундровой и полярнопустынной. Аналогично между
лесными и аридными территориями имеется
переходная полоса — лесостепь. Такие ландшафты в силу
своего промежуточного положения играют огромную
роль в формировании флоры и фауны, служат
поставщиками видов в сообществе соседних зон. Это
проявление так называемого пограничного эффекта —
повышения разнообразия жизни и интенсивности
процессов на границе между разными типами среды (так
же как, например, между водной и сухопутной).
Показано большое значение лесотундровых территорий
в формировании арктической флоры (Юрцев, 1966).
Все заполярные безлесные территории часто
называют Арктикой. В более узком (специальном)
понимании Арктика — бассейн Северного Ледовитого
океана с островами, покрытыми ледниками,
полярными пустынями и тундрами наиболее высокоширотных
вариантов. Территорию, лежащую южнее, где развиты
преимущественно тундровые ландшафты, называют
Субарктикой. А. А. Григорьев (1956) использовал
эти названия для обозначения типов
физико-географической среды, т. е. вкладывал в них
преимущественно геофизическое содержание.
Территориально Субарктика не соответствует полностью тундровой
зоне. А. А. Григорьев включал в ее состав также и
лесотундру. В Энциклопедическом словаре географи-
10
ческих терминов под редакцией С. В. Калесника
северная граница Субарктического пояса проходит по
среднеиюльской изотерме +5°, а южная — по
изотерме + 12°. На северной границе тундровой зоны сред-
неиюльская температура —1,5—3°, а на южной —
несколько ниже 12° (местами 10°). Таким образом, по
отношению к тундровой зоне границы Субарктики
несколько смещены к югу. На севере она не достигает
границ тундры, а на юге захватывает лесотундру.
В таком контексте Субарктика звучит как «нижняя»
Арктика, т. е. как часть Арктики. То, что находится
севернее Субарктики,— это собственно, или
«высокая», Арктика. Ниже название «Арктика» будет
употребляться и в широком, и в узком смысле, т. е. для
обозначения как цсей безлесной заполярной
территории, так и бассейна Северного Ледовитого океана с
островами, на которых развиты арктические пустыни
и ледники.
Зарубежные авторы под Субарктикой часто
понимают то, что находится южнее Арктики в широком
смысле этого слова. Так, некоторые американские
исследователи называют Субарктикой территорию,
лежащую к югу от северной границы леса, и включают
в нее лесотундровые, северо- и среднетаежные
районы. Едва ли это целесообразно. Использование
слова «арктика» (в любом сочетании) для наименования
настоящих лесных территорий вносит излишнюю
терминологическую путаницу. Что касается приоритета,
то он, бесспорно, за нашими отечественными
учеными, внесшими наибольший вклад в
физико-географическое изучение арктических районов. Наиболее
рационально употреблять название «Субарктика» в том
смысле, в котором его использовал А. А. Григорьев.
В качестве произвольных наименований арктических
и приарктических районов весьма удобны выражения
«Север», «Крайний Север», «Заполярье», не имеющие
строго физико-географического содержания.
ТЕПЛО И ВЛАГА В ТУНДРЕ
Природная зональность — это прежде всего
результат неравномерного распределения солнечного тепла.
Различают несколько форм солнечной радиации:
прямую, рассеянную, суммарную и др. Прямая
солнечная радиация — это тепло, доходящее до места
наблюдения непосредственно от солнца. Ее величина
может меняться в зависимости от прозрачности
атмосферы, высоты над уровнем моря, но главным
образом от широты местности. В высоких широтах она
минимальна. Достигая атмосферы, большая или
меньшая часть прямой радиации рассеивается (в
зависимости от облачности и влажности), а затем уже
поступает от небесного свода на земную поверхность.
Это рассеянная солнечная радиация. В полярных
районах вследствие высокой облачности рассеянная
радиация особенно велика.
Для характеристики тепловых условий важны
величины суммарной солнечной радиации и
радиационного баланса (см. табл. 1). Суммарная радиация —
это совокупность прямой и рассеянной. Она может
рассчитываться за сутки, месяц, сезон, год. Ее
величина колеблется от 60 ккал/см2/год в Арктике до
200 ккал/см2/год в тропиках. На равнинных
территориях ее изолинии в общем соответствуют широтному
направлению.
Главный показатель тепловых условий —
радиационный баланс земной поверхности, или остаточная
радиация,— разность между суммарной радиацией,
достигшей земной поверхности, и той ее частью,
которая излучается в мировое пространство. Иными
словами, это то количество тепла, которое может быть
использовано в различных процессах, проходящих в
ландшафтной оболочке. Годовые величины
радиационного баланса изменяются от 100 ккал/см2/год в
тропиках до нескольких единиц и даже
отрицательных величин у полюсов. Радиационный баланс
зависит от отражательной способности земной поверхнос-
12
ти — так называемого альбедо. Естественно, что оно
особенно велико в полярных районах, большую часть
года находящихся под снежным покровом, альбедо
которого — до 95%.
Таблица 1
Среднегодовые радиационные показатели
в разных зонах по линии Диксон—Кызыл, ккал/см2
Место измерения
Тундра
Северная тайга
Южная тайга
Степь
Суммарная
радиация
67
79
87
119
Радиационный баланс
16
25
32
49
Эффективное
излучение
18
24
25
34
Альбедо,
%
50
38
35
30
Источ ник. «Справочник по климату СССР». Л., 1967.
Суммарная радиация в тундровых районах около
70 ккал/см2/год. Изолиния этой величины в Евразии
проходит в основном по середине тундровой зоны,
опускаясь южнее на западе, в приатлантической
части, и поднимаясь к северу на востоке. У северных
границ тундры величина суммарной радиации около
63 ккал/см2/год. Изолинии радиационного баланса
более соответствуют направлениям границ зон и
подзон, чем широт. На территории тундровой зоны он
чаще всего колеблется от 10 до 20 ккал/см2/год. Если
принять его среднюю величину за 15 ккал, то это
составит примерно 21% от суммарной радиации (в
полярных пустынях около 16%). В более южных
широтах, например в лесной полосе, в степной и
пустынных зонах радиационный баланс составляет 30—
35% суммарной радиации. Это обусловлено в первую
очередь очень большой величиной альбедо полярных
ландшафтов, находящихся большую часть года под
снежным покровом.
Много своеобразия в сезонности теплового режима
тундровой зоны (см. рис. 2). Именно в этом и
заключаются главные характерные черты экологических
условий Субарктики. С ноября по февраль, во время
полярной ночи, среднемесячная суммарная радиация
13
Рис. 2. Сезонные
изменения интенсивности
солнечной радиации
в Болынеземельской
тундре (по А. А.
Григорьеву, 1956):
1 — суммарная радиация;
2 — радиационный баланс
подстилающей поверхности
в тундре около нуля. Общая доля зимней радиации
ничтожна (см. табл. 2). В марте она повышается до
3—4 ккал/см2 и почти вся расходуется на
эффективное излучение и альбедо. С наступлением полярного
дня, в апреле, суммарная радиация возрастает до
8—9, а в мае до 12 ккал. Но в это время еще лежит
сплошной снег, отражающий основную часть
радиации, поэтому тепловой баланс подстилающей
поверхности еще очень мал (в апреле — 0, а в мае—1—
3 ккал).
Погода в апреле на большей части Субарктики
вполне зимняя (сильные морозы и метели). В мае
также бывают сильные морозы, пурги, но
температуры сильно колеблются, иногда наступают оттепели.
Лишь с середины и с конца мая на юге тундровой
зоны появляются признаки весны: проталины, ясные,
теплые или пасмурные, с мокрыми снегопадами дни.
Однако еще в середине июня может быть длительная
14
пурга с низкими температурами. В июне суммарная
радиация возрастает всего до 14 ккал. Но в это
время резко снижается альбедо — следствие обнажения
почвы и уменьшения отражательной способности
тающего снега. Июньский радиационный баланс в
тундре — 5—6 ккал. Полностью стаивает снег в конце
июня — начале июля (не считая, конечно, длительно
тающих снежников).
С конца июня начинает уменьшаться солнечная
радиация в результате понижения высоты стояния
солнца. Среднеиюльская суммарная радиация около
13 ккал. Но радиационный баланс в июле еще
растет. Его максимум — в середине этого месяца. В
августе и сентябре он резко снижается и достигает
минимума в октябре.
Таблица 2
Соотношение величин годовой и летней суммарной
радиации в восточноевропейской Субарктике
(по А. А. Григорьеву, 1956)
Место измерения
Северная часть
тундровой зоны
Южная часть
тундровой зоны
Годовая
суммарная
радиация, ккал/см2/
год
64
70
Суммарная
радиация за апрель-
сентябрь
ккал/см2/
58
61
%
91
87
Суммарная
радиация за июнь-
август
ккал/см2/
35
36
%
55
51
Каковы же главные особенности теплового
режима Субарктики? Во-первых, кривая радиационного
баланса одновершинная. Круто поднимаясь с мая по
июль, она резко падает в августе и сентябре. Период
наиболее благоприятного теплового режима в тундре
очень короток. Этим определяется чрезвычайная
динамичность сезонного развития природы,
сближенность фенологических периодов. Во-вторых,
наблюдается резкое расхождение во времени максимумов
суммарной радиации и радиационного баланса. В апреле
и мае количество солнечной радиации в тундровой
зоне равно ее среднемесячной величине в субтропи-
15
ках, однако это огромное количество энергии не
может расходоваться на биологические процессы,
поскольку радиационный баланс еще очень мал. В то
время когда создаются условия для более
эффективного использования ландшафтом солнечной радиации,
она начинает снижаться.
В представлениях большинства людей Заполярье,
Арктика, тундра — царство холода. Но не холод сам
по себе (в смысле низких температур) определяет
главные особенности среды жизни в заполярных
районах. Наиболее холодные районы в северном
полушарии не тундровые, а таежные. Например, всем
известен так называемый «полюс холода» — Верхоянск,
расположенный южнее тундровых районов. Конечно,
лето там гораздо теплее, чем в Арктике. Вместе с
тем в Восточной Сибири, за полярным кругом, у
южных границ тундры, а местами и в ее пределах,
встречаются настоящие степные сообщества с
характерными растениями и животными, для которых нужно
жаркое сухое лето (Юрцев, 1974). И действительно,
лето в этих районах, например на севере Якутии,
бывает очень жарким: в июле температура поднимается
до 30°.
Что же в первую очередь определяет развитие
ландшафтов тундровой зоны, особенности ее флоры
и фауны?
Общее годовое количество тепла, приходящее с
солнечными лучами, обусловливает радиационный
баланс земной поверхности. Но не все его части
равнозначны для жизни. Наибольшую^ роль играют
летние тепловые условия. Именно в летних температурах
в первую очередь отражается главный фактор
зональности— величина радиационного баланса; зимние —
сильно зависят от степени континентальности —
удаленности от теплых, незамерзающих морей. Основная
часть тундровой зоны — приморские прибрежные
районы. Северный Ледовитый океан большую часть года
покрыт льдом и поэтому климатически составляет
единое целое с материком. Здесь формируются
воздушные массы с очень низкими температурами. На
западе — мощный климатический фактор —
незамерзающая Атлантика. Ее влияние в холодное время
перекрывает действие радиационных факторов.
16
Максимальные температуры в тундрах могут быть
весьма высокими, но непродолжительное время.
Например, на севере Таймыра в июле температура
воздуха часто бывает около 20°, а абсолютный максимум
на широте 73° — 29° С. В южных частях Субарктики
температура воздуха может в течение нескольких
дней держаться около 25°.
Но уровни максимальных температур — это еще
не решающий фактор развития органического мира
тундр. Главное, пожалуй, продолжительность теплого
периода. Отдельные виды животных, в основном птиц
и млекопитающих, могут быть активными в условиях
Арктики в течение всего года. Таковы песец, белый
медведь, тундряная куропатка, северный олень.
Некоторые даже могут размножаться зимой в тундре,
как это делают лемминги. Но основная часть
тундровых сообществ деятельна лишь в летнее время
(растительность, микроорганизмы, беспозвоночные
животные). Летом протекают и все основные
абиотические процессы в ландшафте: выветривание, эрозия,
оттаивание мерзлоты и т. д. Поэтому первостепенное
значение в жизни тундры имеет продолжительность
безморозного периода, который и определяет главные
особенности тундрового ландшафта, его
органического мира.
От длительности достаточно теплого периода
зависит возможность завершения жизненных циклов
животных и растений. Ниже будет показано, что именно
в жизненных циклах проявляются важнейшие
приспособительные особенности животных и растений
тундры.
Важная деталь теплового режима Арктики —
очень резкие широтные смены температур. Это легко
увидеть, если сравнить ход изолиний среднемесячных
температур теплого периода в тундровой зоне и в
более южных широтах (см. рис. 3). Так, на
территории Болынеземельской тундры на протяжении
300 км среднеиюльская температура увеличивается
с 6 до 12°. Примерно на такой же порядок (с 13 до
20°) изменяется июльская температура на всей
территории Русской равнины, на протяжении 2000 км от
полярного круга до пятидесятой параллели, где
сменяются четыре природные зоны. Резкая климатиче-
17
Рис. 3. Среднеиюльские изотермы (°С) на территории Русской
равнины (по Ф. Н. Милькову и Н. А. Гвоздецкому, 1976)
екая смена характерна не только для Заполярья. Это
характерная черта ландшафтов, развивающихся при
недостатке тепла или влаги. В пустынях смены
климата так же резки, как и в тундрах.
Резкие широтные изменения климата в Заполярье
отражаются в структуре ландшафта, в органическом
18
мире тундры и определяют сильные различия между
подзонами.
Известно, что в тундре много воды, высокая
влажность почвы и воздуха. Большие территории этой
зоны заняты болотами. Иногда ее называют даже тунд-
рово-болотной полосой. Но это не потому, что здесь
выпадает много осадков. Если бы увлажнение
определялось лишь количеством осадков, то тундру
следовало бы отнести к одному из самых сухих
ландшафтов (см. табл. 3). В разных районах Субарктики
выпадает от 150 до 350 мм/год осадков, т. е. столько,
сколько в полупустынях, сухих степях и саваннах. По
этому признаку расположенные к северу от тундр
полярные пустыни вполне оправдывают свое
название, так как там осадков выпадает столько же,
сколько в настоящих жарких пустынях,— менее 200 мм.
Тем не менее в тундре, да и в полярной пустыне,
влаги всегда много.
Обилие здесь влаги — прямое следствие малой
испаряемости, что определяется недостатком тепла.
Кроме осадков в виде дождя и снега в Субарктике
есть еще один мощный источник увлажнения —
оседание изморози из насыщенного влагой воздуха на
охлажденную поверхность снега. Это особенно часто
случается при вторжении более теплых воздушных
масс (Григорьев, 1956).
Таблица 3
Увлажнение в условиях различных широт
(по С. В. Калеснику, 1970 и А. М. Рябчикову, 1972)
Зональные типы ландшафтов
Полярные пустыни
Арктические тундры
Типичные тундры
Тайга
Смешанные леса
Лесостепь
Пустыни
Тропические леса
Количество
осадков в год, мм
ПО
200
250
370
450
380
100
1000
Радиационный
индекс сухости
0,1
0,2
0,3
0,6
0,8
1,0
3,0
0,9
19
Тундра увлажнена сильнее прочих ландшафтов
Земли. Только некоторые пространства заболоченных
таежных районов, например в Западной Сибири,
могут соперничать с ней по обилию воды. Нигде ланд-
шафтообразующая роль воды не выступает так
отчетливо, как в тундре. С самолета она часто имеет
вид сплошного кружева из-за бесчисленных озер,
которые образуются в результате протаивания мерзлого
грунта и подземных льдов. В тундрах много болот.
Их своеобразная структура (полигоны, мочажины,
торфяные бугры) также образуется в результате
криогенных процессов. Подземные льды, снег, талые
воды, туманы и длительные моросящие дожди — все
это мощнейшие экологические и ландшафтообразую-
щие факторы в тундре.
Тепло и влага в ландшафте тесно взаимосвязаны.
Чем больше тепла, тем больше может испариться
воды. На испарение расходуется тепло. Следовательно,
чем больше воды, тем сильнее охлаждение.
Влажность — результат не только количества осадков, но
и количества поступающего солнечного тепла. Она
может быть высокой и при большом количестве
осадков, и при очень малом. Поэтому для характеристики
климата используют не только величины тепла и
влаги, но и их соотношения — так называемые
гидротермические индексы. Широко известен «радиационный
индекс сухости» М. И. Будыко (1948, 1950)
—отношение радиационного баланса к годовой сумме осадков,
выраженных через скрытую теплоту испарения.
Величина индекса, равная единице, указывает на
соразмерность тепла и влаги, при которой
возможность испарения примерно соответствует количеству
выпавшей влаги. Территории с таким соотношением
тепла и влаги наиболее благоприятны для развития
органического мира. На них не бывает избыточного
увлажнения, сильных засух, как правило, достаточно
тепло (например, районы широколиственных лесов и
лесостепи).
В пустынях умеренного пояса этот индекс
возрастает до 3—5, а в тропических аридных
ландшафтах— до 10. К северу от лесостепи и
широколиственных лесов индекс меньше единицы: в тайге — около
0,6; в тундре — 0,3; в полярной пустыне — 0,2 и ни-
20
же. В этих зонах создаются условия для
накопления избытка влаги: тепла не хватает для испарения
годовых осадков. Территорию, куда входят зоны
смешанных лесов, тайги, тундр и полярных пустынь,
называют гумидным поясом (от латинского слова
humidus — влажный).
Величина индекса около 1 свойственна не только
ландшафтам умеренного пояса, но и дождевым
субтропическим и экваториальным лесам; величина
0,3 характерна как для тундр, так и для некоторых
субтропических и тропических территорий с
преобладанием заболоченных лесов. Эта повторяемость в
соотношении тепла и влаги в разных климатических
поясах описана в работах М. И. Будыко и А. А.
Григорьева под названием периодического закона
географической зональности. По этому закону тундра имеет
некоторые черты сходства с сильно
переувлажненными ландшафтами субтропиков и тропиков (например,
в некоторых элементах почвообразовательного
процесса). Одинаковое соотношение тепла и влаги в
этих, казалось бы, совсем непохожих ландшафтах,
бесспорно, должно определять сходство некоторых
особенностей их жизни. Это подчеркивает единство
ландшафтной оболочки Земли, взаимосвязь и
подчинение общим законам ее частей, находящихся даже
в диаметрально противоположных климатических
условиях.
Недостаток тепла определяет экстремность
условий жизни в Арктике. Но она иного типа, чем в
пустынях, где много тепла и мало влаги. Дело в том,
что вода в пустыне все-таки есть (и местами в
неограниченном количестве), хотя и распределена
локально. Чтобы жить в пустыне, нужно уметь добывать
воду, запасать ее впрок и сохранять длительное время.
Поэтому в процессе эволюции у организмов пустыни
вырабатываются бесконечно разнообразные, порой
удивительно тонкие приспособления для обеспечения
водой (длинные корневые системы, глубокие норы,
запасающие ткани, минимум испарения и т. д.).
Жизнь в пустыне не может существовать повсюду, она
локальна, но зато очень разнообразна.
Недостаток тепла в тундре нельзя существенно
восполнить никакими приспособлениями. Здесь могут
21
иметь значение лишь самые общие особенности
крупных групп организмов, например теплокровность (го-
мойотермность) или холоднокровность (пойкилотерм-
ность). Частные адаптации, направленные против
холода, заведомо ограничены и не могут быть столь
разнообразными, как приспособления к сухости
(знаменательно, что и в тундре многие яркие адаптивные
признаки растений связаны с условиями
водоснабжения, о чем речь пойдет ниже). Такие
морфологические особенности животных, как увеличение размеров
тела, усиление теплоизолирующих и теплозащищаю-
щих свойств покровов или биотопическое
перераспределение (обитание теплолюбивых форм на южном
склоне и т. п.), дают сравнительно небольшой эффект.
Тепло — главное условие протекания всех
биологических процессов, недостаток его тормозит жизнь
во всех ее проявлениях. Поэтому органический мир
тундровой зоны беден качественно (число видов,
приспособительных типов), но зато более равномерно
насыщает ландшафт.
РАЗНООБРАЗИЕ ЛАНДШАФТОВ
Тундры в разных частях зоны неодинаковы. Это
обусловлено широтно-зональными изменениями климата
(деление на подзоны), степенью его континентальнос-
ти, неоднородностью рельефа. Рассмотрим влияние
этих факторов на тундровые ландшафты с общих
географических позиций.
Подзоны
Выше говорилось о том, что наиболее важные для
органического мира температуры теплого периода в
Субарктике меняются с юга на север гораздо резче,
чем в умеренном поясе. Между тем при общем
недостатке тепла всякое повышение температуры сильно
сказывается на жизнедеятельности организмов и на
структуре ландшафта. Это выражается в резких
широтных различиях субарктических ландшафтов, в
четком делении тундровой зоны на подзоны. Обычно
выделяют три подзоны: южные (или кустарниковые),
типичные и арктические тундры (см. рис. 1).
В подзоне южных тундр на основных
площадях водоразделов развит кустарниковый ярус
высотой около полуметра, иногда до двух метров
(см. рис. 4). Этот ярус в разных местах образован
березками, ивами, ольховником. Под пологом
кустарников обильны травянистые растения (осоки, пушицы,
злаки), кустарнички и полукустарнички (карликовые
ивы, голубика, брусника, багульник). Еще ниже
располагается сплошной моховой покров, который
сформирован многими видами. Самые массовые из них —
Hylocomium splendens alaskanum, Aulacomnium tur-
gidum, Tomenthypnum nitens, Rhacomitrium lanugino-
sum, Ptilidium ciliare, Dicranum, Polytrichum
и др.
В подзоне южных тундр встречаются единичные
древесные растения, чаще всего лиственницы. Они низ-
23
Рис. 4. Ландшафт подзоны южных тундр на Таймыре
корослы, имеют искривленные тонкие стволы или
особую, стланиковую форму. Нередки толстые
полусгнившие пни, что наводит на мысль о похолодании
климата и отступании леса к югу или об интенсивном
вырубании деревьев человеком. Высота кустарников в
тундре определяется мощностью снежного покрова:
выступающие над снегом тонкие веточки зимой
подвергаются так называемой снежной корразии
(обтачиванию веток снегом, гонимым ветром). Но это
бывает не везде и не со всеми кустарниками.
Ольховник, например, на увалах достигает 2—2,5 м высоты,
что значительно превышает мощность снежного
покрова в этих местах.
Главное проявление суровости полярного климата
в этой подзоне — отсутствие здесь древесной
растительности. В остальном южные тундры —
относительно богатые сообщества. Здесь весьма разнообразны
флора и фауна. Кроме типичных тундровых видов
много обитателей средних широт. Например, в
европейских и сибирских южных тундрах из растений
повсюду можно встретить обычные в средней полосе —
сабельник болотный, селезеночник обыкновенный,
калужницу болотную и даже теплолюбивый тимьян
обыкновенный; из птиц — пеночку-весничку, дрозда-
белобровика, обыкновенного бекаса и болотную
сову. На озерах здесь гнездится шилохвость, а вместе с
типичными тундровыми грызунами обитает широко
распространенная полевка-экономка,
24
В южных тундрах очень разнообразен
растительный покров. На водоразделах перемежаются заросли
ив, березки (ерники), ольховника и тундры без
кустарников со сплошным моховым покровом или с
пятнами голого грунта. В понижениях развиты различные
болота — гипновые, сфагновые, плоские и с
торфяными буграми. На южных склонах, где скапливается
много снега, растительный покров из злаков, бобовых,
разнообразного разнотравья. На приподнятых
бровках— заросли ягодных кустарничков и
полукустарничков: брусники, голубики, водяники (Empetrum
nigrum), арктоуса (Arctous alpina) и т. д. Вблизи
воды, у озер и по берегам ручьев развиты различные
околоводные растительные группировки из осок,
хвощей, злаков (особенно характерна арктофила — Агс-
tophila fulva).
Такая пестрота растительности еще более
характерна для лесотундры. Сочетание на плакоре
разнотипных сообществ — общая черта переходных
зональных категорий. Вспомним лесостепь и полупустыню.
В лесостепи на водоразделах чередуются степные и
лесные сообщества, а в полупустыне — степные и
пустынные ценозы, кустарники и даже древесные
насаждения. Ландшафт таймырской южной тундры с
кустами ольховника поразительно напоминает
прикаспийскую полупустыню, где среди полынной
растительности в западинках развиты куртины спиреи.
На востоке субарктической части Евразии,
особенно на Чукотском полуострове, кустарниковые
сообщества в южной подзоне замещаются так
называемыми кочкарными тундрами, в них пушица влагалищная
(Eriophorum vaginatum) образует мощные
многолетние кочки, среди которых разреженно растут березки
и ивы.
Южные тундры во многом сходны с лесотундрой.
У них много общих видов растений и животных.
Выделяют специальную группу «гипоарктов», которые
особенно характерны для южнотундровых и
лесотундровых районов. Таковы полевка Миддендорфа (Mic-
rotus middendorffi), белая куропатка (Lagopus lago-
pus), кулики малый веретенник (Limosa lapponica) и
щеголь (Tringa erythropus), жужелица Carabus odo-
ratus, самый крупный из тундровых пауков — Lycosa
25
hirta, карликовая березка (Betula nana). Сходны у
них и основные эдификаторы сообществ. В южной
тундре, так же как и в лесотундре, различные
кустарники (березка, ольховник, ивы) образуют сомкнутые
заросли, их корни пронизывают торфянистую почву,
а листья осенью опадают сплошным слоем, что имеет
большое значение в жизни травянистых растений и
животных. Например, в южной тундре еще обычны
типичные обитатели лесной подстилки —
колониальные личинки комаров-бибионид, которые исчезают в
типичных тундрах, где листового опада очень мало.
Но едва ли целесообразно при ландшафтном
районировании отделять южную тундру с лесотундрой от
остальной части Субарктики, как это предлагают
некоторые авторы. Бесспорно, по многим важнейшим
особенностям структуры ландшафта, растительного
покрова и животного населения вся территория к
северу от лесотундры характеризуется многими
общими особенностями, что позволяет ее считать единой
зоной.
К северу от южных тундр располагается
подзона типичных тундр. Ее границы примерно
соответствуют изотермам июля: 8—11° — на юге и 4—
—5° — на севере. Потенциальная площадь этой
подзоны больше, чем площадь других подзон. На
Таймыре ширина ее 300—400 км, тогда как ширина подзоны
арктических тундр — 200 км, а южных— 150—200 км.
Однако линия берега Северного Ледовитого океана
отрезает большие куски суши, где должна была бы
располагаться эта подзона. В Евразии она хорошо
представлена на Таймыре, Ямале, Гыданском и
Югорском полуостровах, между Яной и Колымой, а на
остальной ее части — только небольшими, в основном
южными, фрагментами. Полностью она отсутствует
на материке к западу от Югорского полуострова.
Эта подзона — воплощение того типа ландшафта,
который называют тундрой. Здесь нет не только
деревьев, но и достаточно высоких кустарников на
водоразделах. Высота растительности полностью
определяется мощностью снежного покрова. Вследствие
снежной корразии зиму могут пережить только те
растения, которые спрятаны под снегом. Между тем
его толщина невелика, чаще всего 20—40 см. Кустар-
26
никовые заросли высотой до 1 м развиты в низинах,
в долинах ручьев и по берегам озер, где
скапливается много снега.
Типичные тундры — это царство мхов. Мощная
моховая дернина, сплошным слоем покрывающая
почву,— самая важная часть растительного покрова. Ее
толщина обычно 5—7 см, местами — до 12 см. Она
состоит из многих видов мха, среди которых около
десятка бесспорных лидеров. Особенно выделяются
три: Tomenthypnum nitens, Hylocomium splendens
•oar. alaskanum, Aulacomnium turgidum. Обильны
Rhacomitrium lanuginosum и печеночник Ptilidium
ciliare, а также различные дикрановые и политрихо-
вые (Dicranum, Polytrichum), образующие плотные
кочки и подушки. Почти все эти мхи широко
распространены и в более южных зонах, особенно в тайге.
Моховой покров играет огромную и
противоречивую роль в жизни тундры. Именно мхи
обеспечивают полную сомкнутость растительности на
водораздельных пространствах этой подзоны. Большое
влияние они оказывают на температуру почвы, влажность,
динамику сезонного протаивания грунтов. С одной
стороны, моховой покров задерживает оттаивание
мерзлоты, препятствует прогреванию почвы и тем
самым отрицательно действует на развитие организмов.
Чем он толще, плотнее и суше, тем холоднее почва и
выше уровень мерзлоты. В трещинах и западинках,
где нарастает особенно мощная дернина, лежащий
под ней грунт не оттаивает иногда до конца лета.
С другой стороны, моховой покров препятствует
возникновению термокарста (глубокому протаиванию
грунтов и подземных льдов) и тем самым оказывает
стабилизирующее действие на растительность.
Хорошо известны губительные последствия сдирания
моховой дернины в результате, например, движения
гусеничного транспорта.
В летнее время моховая дернина препятствует
иссушению верхних горизонтов почвы, что крайне важно
в связи с малым количеством осадков. В то же
время весной и осенью она как губка впитывает воду и
создает условия, близкие к болотным. В моховую
дернину погружены стебли, корневища, а часто и корни
цветковых. Благодаря ей в тундрах благоденствуют
27
некоторые длиннокорневищные виды. Для растений
моховых тундр характерно сильное ветвление в
горизонтальной плоскости в толще дернины. В
осенне-зимний период моховая дернина — хранилище для
семян, но летом проростки часто не успевают
укорениться и гибнут в результате иссушения и
недостатка питательных веществ, как бы «повисая» в толще
мха.
Моховая дернина служит средой обитания
богатого комплекса беспозвоночных, называемого геми-
эдафоном (полупочвенным). В него входит большое
число видов ногохвосток, клещей, пауков, жуков-ста-
филинов. Вместе с тем в моховом ярусе живут и
типичные почвенные формы, например дождевые
черви, энхитреиды, личинки комаров-долгоножек,
жужелиц и др. От мхов зависит жизнь леммингов —
массовых зверьков тундры. Они прокладывают в дернине
сложные лабиринты ходов, зимой питаются
мясистыми частями цветковых растений, скрытыми в ее
толще. Чтобы добраться до них, зверьки вынуждены
выгрызать огромное количество мхов.
Травяной ярус образуют в основном осоки, среди
которых выделяется Carex ensifolia — самый
массовый вид цветковых растений в тундрах Евразии, где
он представлен особым подвидом — arctisibirica.
Осоки— второй по значению (по эдификаторной роли)
компонент растительности типичных тундр. Поэтому
зональные сообщества этой подзоны следует
именовать осоково-моховыми тундрами (на последнем
месте в геоботанических названиях стоит главный эди-
фикатор). Кроме того, в типичных тундрах много
стелющихся кустарников (ивы), кустарничков и
полукустарничков (карликовые ивы, куропаточья трава,
Кассиопея, брусника и др.)- Иногда обильны пушицы,
злаки и двудольные травянистые растения:
камнеломки, грушанки, сложноцветные и др. Местами в
моховой дернине много лишайников самых различных
видов. В зависимости от обилия этих растений тундры
могут именоваться ивняково-осоково-моховыми, дриа-
дово-осоково-моховыми, лишайниково-моховыми и т. д.
В научной литературе сообщества этой подзоны
часто называют мохово-лишайниковыми тундрами.
Это название, несмотря на широкое его использова-
28
ние, следует считать неправильным. Дело в том, что,
хотя лишайники в тундрах весьма многочисленны,
они нигде не выступают в качестве эдификаторов.
Чаще всего ойи образуют заметную примесь в
моховой дернине. В подзоне типичных тундр
лишайники очень разнообразны. В каждом районе их
насчитывается до 200 видов самых различных жизненных
форм (листоватые, трубчатые, кустистые, накипные
и т. д.). Сообщества, где лишайники выступают в
качестве первостепенных доминантов, встречаются лишь
местами на песчаных и щебнистых грунтах.
Ландшафт подзоны типичных тундр весьма
однообразен (см. рис. 5): сглаженный полого-волнистый
рельеф, стабильный растительный покров на
водоразделах. Эта монотонность нарушается долинами рек,
крутыми берегами, заболоченными низинами,
озерными понижениями.
Кроме основных зональных сообществ со
сплошным моховым покровом в этой подзоне весьма
распространены так называемые пятнистые тундры (см.
рис. 6). Пятна оголенной почвы (диаметром обычно
40—50 см) окружены несколько приподнятым
валиком из сплошной моховой дернины. Валики
соседних пятен разделены понижениями — ложбинками,
заполненными торфом и рыхлой моховой дерниной.
Обычно пятнистые тундры приурочены к высоким
террасам. Их образование связано с процессами
растрескивания грунта, разрыва моховой дернины, морозного
«кипения» оголенной почвы, выдавливания на
поверхность переувлажненного грунта и т. д.
Оголенные грунты в пятнистых тундрах
постепенно зарастают. На одном участке можно встретить
пятна, совершенно оголенные и почти полностью
заросшие мхами и цветковыми растениями. Все это создает
большую пестроту экологических условий, благодаря
чему в пятнистых тундрах разнообразны
растительность и животный мир.
В подзоне типичных тундр еще можно встретить
виды животных и растений внеарктического
происхождения. Но относительно немногие из пришельцев
с юга могут здесь существовать нормально. Как
правило, это виды вообще широко распространенные,
обитающие в нескольких зонах. Одни из них как бы без-
29
.. .-^-.^,
Р&\0ЩЩ'
Рис. 5. Ландшафт подзоны типичных тундр на Западном Таймыре
А — полого-увалистый рельеф; Б — участок кочковатой осоково-моховой
тундры на месте заросшей пятнистой; В — поверхность ивняково-ocokobq-
моховой тундры; Г — зарастающее пятно голого грунта
Рис. 6. Общий вид осоково-моховой пятнистой тундры
в подзоне типичных тундр:
1 — пятно голого грунта; 2 — валик; 3 — ложбинка
различны к ландшафтным условиям, так называемые
эврибионты: горностай, ворон, семиточечная божья
коровка, различные виды мхов. Другие связаны со
специфическими биотопами, например с прибрежными
местообитаниями: калужница болотная, трясогузка
белая. Вместе с тем многие виды, обычные в южной
подзоне и лесотундре, в типичных тундрах
отсутствуют или редки. Например, здесь не встречаются такие
характерные обитатели лесотундровых и
южнотундровых ландшафтов, как гусь пискулька (Anser erythro-
pus), малый веретенник (Limosa lapponica) и
азиатский бекас (Gallinago stenura). Из мелких зверьков
остаются в основном лемминги, а многочисленные в
южных тундрах полевки — узкочерепная (Microtus
gregalis), Миддендорфа (М. middendorffi), экономка
(М. oeconomus), а также арктическая бурозубка (So-
rex arcticus)—отсутствуют или редки. Массовые в
южных тундрах ягодные кустарнички и
полукустарнички— морошка, брусника, голубика — здесь вотре-
31
чаются, но мало плодоносят. В самых северных
частях подзоны они, как правило, отсутствуют.
Кроме охарактеризованных выше основных пла-
корных осоково-моховых тундр в локальных
биотопах этой подзоны развиты весьма разнообразные
сообщества: множество вариантов болот, луга и заросли
кустарников по берегам рек и т. д. В целом
климатические условия в типичных тундрах можно считать
умеренно экстремными, поэтому и обеднение
животного и растительного мира здесь не так велико, как
это наблюдается еще дальше к северу.
Подзона арктических тундр развита на
Ямале, Гыдане, Таймыре, местами небольшими
участками на побережье от Анабара до Колымы, а также
на Вайгаче, Новой Земле, острове Белом,
Новосибирских островах, острове Врангеля. Название этой
подзоны, быть может, не совсем удачно, так как его
иногда используют для наименования всех тундр
северного полушария. Однако оно общепринято, и нет
основания от него отказываться. Данная подзона
располагается в области настоящего арктического
климата. У ее южной границы среднеиюльские
температуры составляют 4—5°, у северной — около 1,5°. В
течение всего лета здесь возможны температуры ниже 0°
и снегопады. Мощность снежного покрова
незначительна, поэтому зимние условия для животных и
растений особенно суровы.
Главная черта ландшафта арктических тундр —
повсеместное распространение оголенных грунтов. На
водоразделах развиты различные варианты
сообществ, в которых пятна голого грунта окружены
растительной дерниной. Их называют пятнистыми, ме-
дальонными, полигонально-пятнистыми и т. д. (см.
рис. 7). Оголенные грунты занимают в них примерно
50% площади. Моховая дернина с вкрапленными в
нее веточками карликовых ив, каменоломками,
злаками располагается по морозобоиной трещине
вокруг голого грунта. Внешне эти тундры напоминают
пятнистые тундры южных подзон. Но там голые
грунты вторичны, они образуются путем разрыва моховой
дернины. В арктических тундрах голые грунты
первичны, а моховой покров преимущественно вторичен.
Мхи поселяются в морозобоиной трещине, которая
32
Рис. 7. Арктическая полигонально-пятнистая тундра на
Восточном Таймыре (фото Н. В. Матвеевой)
постепенно расширяется в результате осыпания и
проседания краев. Арктические тундры весьма
разнообразны: каменистые, щебнистые, глинистые, с
правильной медальонной структурой, с растительным
покровом в виде куртин, полос, сетей и т. д.
Мерзлотные явления в подзоне арктических тундр очень
разнообразны и заметны повсеместно.
Ослабленное выветривание и интенсивные
криогенные (мерзлотные) процессы создают в арктических
тундрах весьма разнообразный, резко пересеченный
микро- и нанорельеф. Повсюду много скальных
обломков, щебня. Поверхность грунта покрыта
трещинами, ложбинками, бугорками. Хотя здесь климат
гораздо суровее, чем в типичных тундрах, но
ландшафт местами выглядит более пестрым и красочным
2 Ю. И, Чернов
33
из-за обилия цветущих растений. Условия обитания
в этих тундрах для некоторых групп организмов
более разнообразны и благоприятны, чем в южных
тундрах, где сплошной и однообразный моховой покров
создает монотонность среды. Не защищенные
моховым покровом грунты прогреваются относительно
хорошо, несмотря на низкие температуры. На
Таймыре в подзоне типичных тундр при среднеиюльской
температуре 10,5° глубина оттаивания мерзлоты на
водоразделе равна 48 см, а в подзоне
арктических тундр при среднеиюльской температуре 4° —
50 см.
Поэтому в арктических тундрах почвы хотя и
маломощные, но заселены животными не хуже, чем в
южных подзонах. Видов почвенных животных здесь
меньше, но численность их местами выше.
Оголенные грунты арктических тундр на первый взгляд
кажутся безжизненными, но на них развивается богатый
мир организмов. В верхнем слое почвы масса
одноклеточных водорослей и питающихся ими нематод,
кольчатых червей энхитреид, коллембол и более
крупных животных — дождевых червей, личинок комаров-
долгоножек. На поверхности много накипных
лишайников, имеющих вид плесени. Среди щебня
разбросаны цветковые растения — злаки, ожики, маки,
сиверсия, дриада, мытники, камнеломки, крупки,
незабудки и др. (см. рис. 8). Часть этих растений
образует плотные подушки или кочки, которые
служат местом накопления органического вещества,
почвообразовательных процессов.
У растений оголенных грунтов есть интересные
морфологические приспособления. Так, камнеломка
усатая (Saxifraga platysepala) размножается с
помощью отводков, «шагающих» по поверхности почвы.
Многие беспозвоночные арктической тундры имеют
приспособления для обитания на поверхности голого
грунта при сильных ветрах: у некоторых мух и
комаров отсутствуют или уменьшены крылья. Они быстро
ползают, но плохо летают, иногда используют крылья
как парус, с помощью которого, подгоняемые ветром,
быстро бегают по поверхности грунта. Мелкие
почвенные животные — клещи и коллемболы
представлены в основном относительно крупными видами с
34
Рис. 8. Арктическая разнотравно-злаковая трещиноватая тундра
на Восточном Таймыре
плотными темноокрашенными покровами, что
характерно для обитателей поверхности почвы.
В арктических тундрах совсем нет кустарников и
очень слабо развит травянистый ярус. Трав
сравнительно много, но они не формируют сомкнутого
покрова. Часто цветковые растут куртинками. Высота
большинства растений — 3—10 см, лишь единичные
особи злаков достигают 20 см. В травянистом ярусе
мало осок и пушиц, но зато обильны злаки. Например,
на Таймыре в арктических тундрах повсеместно
растет альпийский лисохвост (Alopccurus alpinus),
образующий местами сплошные поросли, похожие на
посевы. В этом главное отличие арктических тундр от
типичных, где, наоборот, много осок и пушиц, но мало
злаков. Другие важные отличия — общее
разнообразие видов двудольных трав, большое значение в
растительном покрове таких групп, как камнеломки,
крупки (из крестоцветных), маки, ожики (см.табл.4).
В типичных тундрах эти растения приурочены к
различным локальным биотопам — склонам, долинам
ручьев, каменистым россыпям, а водораздельных
моховых участков избегают.
2*
35
В арктических тундрах резко сокращается число
видов кустарничков, которые представлены здесь
дриадами и карликовыми ивами. Но все же эта
жизненная форма играет существенную роль в
растительном покрове. Дриада, преимущественно точечная
(Dryas punctata), сплошным ковром или плотными
подушками покрывает каменистые грунты. Карликовые
ивы, из которых особенно характерна ива полярная
(Salix polaris), в большом количестве растут в
моховой дернине вокруг пятен голого грунта. При этом
основная часть кустарничка скрыта в толще мха, а
на поверхности видны лишь верхушечные листочки
и соцветия.
Самые массовые мхи здесь те же, что и в
типичных тундрах. В арктических тундрах они не
образуют сплошного покрова, но все же сохраняют роль эди-
фикаторов. При растрескивании грунта мхи
поселяются по краям трещины. По мере ее расширения,
проседания и осыпания краев они заполняют ложбинку,
образовавшуюся на месте трещины и окружающую
Таблица 4
Роль различных групп цветковых растений
в сообществах на Таймыре
Группы растений
Злаки
Осоки
Ожики
Ивы кустарниковые
Ивы кустарничковые
Березы
Гвоздичные
Маки
Крестоцветные
Камнеломки
Розоцветные
Бобовые
Эрикоидные
Норичниковые
Сложноцветные
Типичные
тундры
+*
+++ +
+
++++
++
++
+•
+
+
+
++++
++
+++
++
++
Арктические
тундры
+++
+
++++
—
Н-+++
—
+
+++
+++
++++
++
+
++
+
Полярные
пустыни
+++
—
—
—
—
—
++ +
+
+
+ + +
—
—
—
* Глазомерная оценка роли в зональных сообществах по четырехбалльной
шкале.
36
пятно голого грунта. В толще мха поселяются
различные цветковые растения, в особенности
карликовая полярная ива, переплетающая дернину
стеблями и корнями. Нижняя часть дернины отмирает и
превращается в торфянистую массу. Под моховой
дерниной торфянистый слой и минеральный грунт
прогреваются плохо, здесь часто все лето сохраняются
линзы льда.
Главная особенность видового состава флоры и
фауны арктических тундр — очень высокий процент
видов, характерных именно для этой подзоны. Как и
в южных подзонах, здесь обитают растения и
животные, вообще широко распространенные, не
тундровые. Таковы многие мхи, лишайники, водоросли,
почвенные животные, для которых вообще характерны
очень обширные ареалы. Встречаются здесь и
широко распространенные животные, например белая
трясогузка, каменка, горностай. Но количество таких
видов в арктических тундрах во много раз меньше, чем
в типичных и южных. На водоразделах
подавляющее большинство цветковых растений, насекомых,
птиц — типичные арктические виды, характерные для
этой подзоны и именно здесь достигающие наибольшей
численности. Примерами могут служить тундряная
куропатка (Lagopus mutus), кулики — краснозобик
(Calidris testacea) и исландский песочник (С. са-
nutus), белая сова (Nyctea scandiaca),
комар-долгоножка Tipula carinifrons, ива полярная (Salix pola-
ris), камнеломка Saxifraga platysepala, крупка Draba
subcapitata. Даже в группах мелких почвенных
животных, для которых вообще характерно широкое
распространение, в этой подзоне преобладают настоящие
арктические виды. Так, на севере Таймыра среди
коллембол и панцирных клещей примерно половина
видов — типичные арктические. Особенно важно то,
что эти виды многочисленны и интенсивно заселяют
разнообразные биотопы.
В арктические тундры не заходят ни таежные, ни
лесотундровые, ни южнотундровые виды. Например,
здесь совсем нет таких видов, как карликовая
березка, водяника, арктоус альпийский, брусника,
голубика, морошка, белая куропатка, кулики —
щеголь и малый зеретенник, полевка Миддендорфа.
37
Более того, здесь малочисленны или отсутствуют
и многие характерные массовые обитатели подзоны
типичных тундр, такие, как кулик-воробей (Calidris
minuta), чернозобик (С. alpina), осока Сагех
ensifolia.
Все это подчеркивает чрезвычайную
специфичность и своеобразие климатического режима этой
подзоны. Для обитания здесь необходимы специальные
приспособления, позволяющие существовать в этих
суровых условиях.
Широтные изменения климата определяют наиболее
сильные преобразования ландшафтов Арктики, что
воплощается в существовании резко очерченных
подзон. Большую роль играет также степень континен-
тальности, которая проявляется прежде всего в
годовой амплитуде температуры воздуха. Вся территория
тундровой зоны расположена вблизи океана. Но в
разных частях побережья эта роЛь принципиально
различна. Восточные и западные окраины Евразии и
Америки находятся под влиянием водных масс
Атлантического и Тихого океанов. Особенно сильно
действие теплой Атлантики. В средней части береговой
линии севера Евразии материк контактирует с
холодным Северным Ледовитым океаном, который
большую часть года находится подо льдом и по существу
подобен суше. Поэтому здесь климат
континентальный.
Казалось бы, теплое море должно смягчать
климат и отодвигать границу тундры к северу. На деле
явление обратное. Именно на окраинах материков,
подверженных влиянию более теплых морей,
тундровый безлесный ландшафт простирается далеко к югу.
Так, на побережье Тихого океана тундры развиты
вплоть до 60° с. ш. На этой широте в глубине
материка располагаются среднетаежные леса. Даже на
Командорских островах, лежащих на широте 55° с. ш.,
ландшафт тундрового типа. В Европе южная граница
тундр опускается до 66,5° с. ш., тогда как на Таймыре
поднимается до 72° с. ш. Это определяется прежде
всего особенностями сезонного хода тепловых
процессов в условиях континентального и морского
климатов. В районах с высокой степенью континенталь-
38
ности меньше облачность, поэтому максимумы
температуры выше и меньше запаздывают относительно
времени высшего стояния солнца.
Очевидно, районы, сходные по зональному
положению, но лежащие в разных широтах, могут сильно
различаться по продолжительности светлого
времени, фенологических периодов, по уровню
максимальных температур и др. Все это определяет многие
особенности ландшафта разных районов тундровой
зоны, играет большую роль в распространении
животных и растений.
Рельеф и мерзлота
Различают несколько форм рельефа: макрорельеф
(горные хребты, низменности, равнины), мезорельеф
(холмы, увалы, террасы, долины, котловины),
микрорельеф (бугры, дюны, овраги), нанорельеф (мелкие
неровности земной поверхности — кочки, ложбинки,
промоины).
Наиболее мощный фактор преобразования
зональных ландшафтов — макрорельеф. В
высокогорьях в любой зоне могут предваряться ландшафты,
аналогичные тем, которые на равнинах развиваются в
более холодном климате. Растительность, близкая к
тундровому типу, в принципе может повторяться
вследствие высотной поясности на территории любой
зоны, кроме полярно-пустынной.
В Евразии наиболее крупные горные массивы на
территории тундровой зоны — горы Бырранга и
Чукотское нагорье. Кроме того, в пределы зоны
вклиниваются северные отроги Хибин, Урала,
Среднесибирского плоскогорья, хребтов Верхоянского и Черского,
Колымского нагорья. Горные системы сильно
нарушают распределение ландшафтов в соответствии с
природной зональностью. На хребтах, расположенных в
тундровой зоне, развиты аналоги полярных пустынь —
оголенные горные, породы, покрытые скудной
растительностью. По хребтам, выходящим за пределы Суб-
арктики, тундры распределяются далеко к югу.
От Белого моря до Оленека преобладают
равнинные ландшафты с весьма правильной широтной
39
структурой: четкое деление на подзоны, меньшая (чем
в других районах) изломанность их границ. Сдвиги
зональных границ к югу или северу здесь определяются
в первую очередь степенью континентальности.
Восточнее Оленека многочисленные горные
хребты буквально «подпирают» тундровую зону и местами
полностью ее вытесняют. Особенно сильно влияние
горных хребтов к востоку от Колымы. Здесь
тундровые зональные ландшафты ютятся лишь на узкой
прибрежной равнинной полосе. На некоторых картах
равнинные участки материкового побережья Восточно-
Сибирского и Чукотского морей отнесены к подзоне
арктических тундр. Едва ли это правомочно.
Береговая линия проходит здесь на уровне подзон типичных
или южных тундр. Арктические же в этом районе
развиты на острове Врангеля и на Новосибирских
островах.
Рассмотрим роль мезорельефа и
микрорельефа. Говоря о зональных особенностях какой-
либо местности, прежде всего имеют в виду
относительно выровненные горизонтальные части
водораздельного ландшафта, на которых нет ни подтока
грунтовых вод, ни повышенного сноса частиц. Такие
участки называют плакором. Именно на плакорах
развиты коренные зональные типы растительности:
хвойные или широколиственные леса, степи и т. д.
На элементах мезорельефа или микрорельефа, где
.имеется подток грунтовых вод, повышенный прогрев
вследствие южной экспозиции склонов или усиленный
дренаж, развиваются разнообразные интразональные
сообщества: луга, болота и т. д.
Два мощных дополнительных фактора определяют
специфику и особое разнообразие тундрового мезо- и
микрорельефа — криогенные процессы, связанные с
таянием мерзлых грунтов и ископаемых льдов, а
также ослабление выветривания, благодря чему на
поверхности сохраняется множество обломочного
материала, скального субстрата, каменистых обрывов
и т. д. Характер мезо- и микрорельефа в разных
подзонах сильно меняется. В подзоне южных тундр и
отчасти в типичных рельеф более выровненный,
пологий, в очертаниях водоразделов преобладают
слабоволнистые линии. В северной подзоне линии рельефа
40
ломаные, больше неровностей, резких перепадов,
особенно много выходов коренных пород и обломочного
материала.
Для тундровой зоны характерны следующие
формы мезорельефа: 1) водораздельные увалы с
плоскими или слегка выпуклыми вершинами и очень пологими
склонами (этот элемент ландшафта следует считать
плакором; настоящих плакоров, аналогичных
обширным плоским территориям более южных широт, в Суб-
арктике нет); 2) разнообразные заболоченные
понижения среди водораздельных увалов; 3) низкие
заболоченные побережья рек, морей и крупных озер (лайды);
4) берега рек с крутыми высокими склонами (яры) и
обширными песчаными и галечными косами; 5)
овраги (распадки) и долины временных ручьев; 6)
высокие приречные террасы и останцы; 7) гряды
обломочного скального субстрата.
Вершины и склоны плоских увалов занимают
характерные для подзон типы растительного покрова,
о которых речь шла выше. Кроме них развито
множество вариантов сообществ, которые нельзя назвать
тундрами — это различные интразональные
включения. Главные из них — болота. Они очень
разнообразны, часто их трудно отграничить от собственно
тундры. Для Субарктики в высшей степени
характерны полигональные болота (см. рис. 9). Они
приурочены к низким террасам, приозерным понижениям,
сточным котловинам.
Поверхность болотного массива разбита морозо
бойными трещинами на полигоны (часто тетрагоны)
7—15 м в диаметре. Центральная часть полигона
плоская или немного вогнутая. Полигон окаймляет
повышение в виде валика, приподнимающегося над
поверхностью днища на 0,7—1,0 м. Один полигон от
другого отделяется глубокими обводненными
ложбинами, которые располагаются вдоль трещин. В одном
массиве может быть от десятка до сотен таких
полигонов. Полигональные болота хорошо узнаются с
самолета: их клетчатый рисунок заметно выделяется на
фоне мелкоячеистой структуры плакорных тундр.
Своеобразны бугристые болота: на заболоченных
низинах рядами или группами развиты
плосковершинные торфяные бугры диаметром от 1 до 10 м, высотой
41
Б
• 23 м-
1ED2&Q4|2I|5E36EE|7
Рис. 9. Схема
строения полигонального
болота (А — план,
Б — вертикальный
профиль):
1 — пол и гоны-мочажины;
2, 3 — вершины и
склоны валиков,
разделяющих мочажины; 4 —
трещины, разделяющие
соседние полигоны; 5 —
озерко на месте
мочажины; б — уровень воды;
7 — уровень мерзлоты
от 0,5 до 1,5 м. Они состоят из торфа, образованного
разрастающимися на их поверхности мхами. Гряды
бугров отделены друг от друга мочажинами —
топкими обводненными участками. Эти болота наиболее
характерны для подзон южных и типичных тундр
западного сектора Субарктики Евразии. К северу, и
особенно в арктических тундрах, их становится все
меньше. Плоскобугристые комплексы не следует
путать с крупнобугристыми торфяниками лесотундры и
северной тайги, в которых высота бугров достигает
5—8 м при диаметре 25—100 м.
Другая своеобразная форма криогенного
микрорельефа в тундре — бугры-байджарахи (см. рис. 10).
Байджарахи — якутское название останцовых бугров,
возникающих при вытаивании ископаемых
полигонально-жильных льдов. Байджарахи встречаются
группами, располагаясь рядами. Поэтому их называют
сундучными рядами или могильными буграми.
Диаметр байджарахов — 3—10 м, высота — 0,5—5 м. Они
отделены друг от друга ложбинами. На пологих
склонах долин рек и ручьев можно видеть только слабо
намечающиеся ряды бугров высотой не более 0,5 м.
На крутых склонах они обычно имеют конусовидную
42
Рис. 10. Бугры-байджарахи на острове Котельном
(фото А. В. Кречмара)
форму и высоту от 1,5 до 3 м. Своеобразны так
называемые «байджараховые цирки» на крутых склонах
округлых котловин. По дну такой котловины течет
ручеек без явного русла: вода сочится между
побегами злаков и пушиц. Прорезав стенку котловины,
ручеек впадает в более крупный ручей. Если соседние
котловины соединяются, то массив байджарахов
занимает огромную площадь.
Географическое распространение байджарахов
тесно связано с развитием полигонально-жильных
льдов, которые распространены в Северо-Восточной
Азии и на Аляске, где подземное оледенение в
прошлом преобладало над надземным. Область развития
классических вариантов жильных льдов и
термокарста — Новосибирские острова.
Специфический элемент рельефа заполярных
ландшафтов — бугры-гидролакколиты. В Якутии их
43
называют булгунняхами, а в канадской Арктике — пин-
го. В отличие от байджарахов они встречаются
одиночно. Их высота меняется от 1 м (диаметром 2—5 м)
до 70 м (диаметром 150—200 м). Возникновение бул-
гунняхов объясняют пучением грунта в результате
образования подземной ледяной линзы. Снаружи бул-
гуннях покрыт торфяным пластом толщиной около
1 м. Под ним промерзший минеральный грунт,
состоящий из слоистых озерных отложений, мощностью от
одного до нескольких метров. Минеральный грунт
подстилается куполообразной массой льда. Это и
определяет форму булгунняха.
Что находится подо льдом — неизвестно,
поскольку пока нет ни одного полного разреза, доходящего
до основания булгунняха. Местные жители сообщали,
что иногда зимой со стороны бугра раздавался
оглушительный треск («взрыв»), после чего появлялась
надземная наледь. Вероятно, подо льдом находится
полость, наполненная водой, а может быть, отчасти и
газом, которые во время глубокого растрескивания
бугров выходят наружу. Хорошо развитые бугры
обычно имеют на вершине глубокую трещину. Ее
образование— начало разрушения булгунняха: вдоль
трещины отмирает растительная дернина, затем
разрушаются торф и почвенный слой, лед обнажается и
тает. Разрушающиеся бугры издалека кажутся
двухвершинными. Булгунняхи приурочены к почти
ровным, сильно заболоченным территориям. Чаще всего
они встречаются в котловинах на месте бывших озер.
Булгуннях, как правило, занимает наиболее
пониженную часть озерной котловины.
О скорости образования булгунняхов имеются
очень противоречивые суждения. Так, В. Н. Андреев
считает, что «процесс выпирания гидролакколита-бул-
гунняха растянут на сотни, а может, и тысячи лет».
Американские исследователи при описании бугров
пинго пишут, что они появляются внезапно зимой,
существуют несколько лет и затем в результате
таяния ледяного ядра разрушаются. В современных
климатических условиях крупные булгунняхи
образуются только в тайге, лесотундре и южных частях
тундровой зоны; в типичных тундрах это, как правило,
мертвые образования, следы того времени, когда кли-
44
^-«r-flfi
Рис. 11. Типичное тундровое озерко с зарослью пушицы
матические зоны были смещены к северу по крайней
мере на 2—3°.
Наиболее типичные формы тундрового
микрорельефа так или иначе связаны с криогенными
процессами, которые вызывают или пучение грунта, или его
растрескивание. При формировании более крупных
форм рельефа главную роль играют процессы
термокарста и эрозии. Типичный тундровый ландшафт —
это слабовсхолмленная равнина, прорезанная
многочисленными ручьями и распадками, образующими
густую сеть, со множеством небольших озер,
располагающихся группами в межувальных котловинах.
Озера возникают в результате местного протаива-
ния мерзлых грунтов на плоских поверхностях (см.
рис. И). Обычно мерзлые тонкозернистые отложения
содержат чистый лед в виде линз. После таяния льда
грунт оседает и образуется бассейн, в котором может
возникнуть озеро. Озера протаивания обычно
невелики, их длина редко больше 300 м, а глубина — 3 м.
Очертания озер округлые, овальные или грубопрямо-
угольные; группы озер обычно соединяются друг с
другом извилистыми водотоками. По мере
расширения речных долин и развития сети ручьев такие озера
45
получают связь с долиной основной реки,
оказываются спущенными и превращаются в болота.
Распадки—это короткие неглубокие овраги с
изрезанными склонами. Они выходят в долину большой
реки и не имеют постоянного водотока. Зимой
распадки забиты снегом до уровня водораздельной
поверхности; ручей, образующийся весной, питается талыми
водами, а затем пересыхает. Под действием талых
вод распадок расширяется, образуются боковые
отроги. В его верховье нередки обвалы грунта. Там
имеется уступ, по которому весной текут бурные
потоки воды. Тундровые распадки напоминают степные
балки.
С распадками, берегами рек и озер, точнее, с их
склонами связана растительность, совсем непохожая
на тундровую. Часто пишут и говорят о красочности
летней тундры, о травяных коврах с множеством
цветков. Это звучит некоторым парадоксом. Столь
суровый край, однообразные бедные тундры и вдруг
ковры буйной зелени и цветов. Типичная плакорная
тундра действительно однообразна и бедна красками, в
ней мало ярких и крупных цветков. Но стоит подойти
к крутому склону берега реки или оврага, как
взгляду открывается пестрый и богатый растительный
покров, который подчас напоминает наши обычные
лесные лужайки или сенокосные обочины дорог:
сплошная поросль злаков, масса буйно цветущих ромашек,
гвоздик, бобовых, незабудок, раковых шеек и еще
каких-то уже незнакомых жителям средней полосы
растений со своеобразными яркими цветками. Над ними
летают пестрые мухи-журчалки, гудят шмели. Все
это очень похоже на то, что у нас принято называть
лугом. И это действительно не тундра, а луг.
Ботаники до сих пор спорят, можно ли называть
эти сообщества тундровой зоны лугами. Часто их
называют луговинной тундрой. Это все равно, если бы
наши суходольные луга назвали луговым лесом.
Конечно, дело не в словах, а в сути явления. А она
такова: эта растительность ничего общего не имеет с
собственно тундрой. В этих сообществах хорошо гуму-
сированные почвы со сплошным злаковым покровом
(корни злаков образуют мощную дерновину), масса
разнотравья и мало или совсем нет мхов. Конечно,
46
часто можно видеть смешанные варианты, в которых
есть и тундровые черты (много мхов, оторфованная
почва и т. д.). Но и в средних широтах луга часто
зарастают лесом. Луга — это типичные интразональные
сообщества. Они могут быть в любой зоне — от
Арктики до тропиков и чаще всего развиты в каких-либо
локальных биотопах — на склонах, в поймах, на
берегах озер, в горах, на месте уничтоженного
зонального растительного покрова и т. д. Их
общая черта—доминирование мезофильных
(требующих умеренного увлажнения) злаков и
двудольных трав.
Есть на Земле места, где луговая растительность
выходит из локальных биотопов на плакор и занимает
большие площади водоразделов. Так, в европейской
черноземной лесостепи на плакорах развиты два
типа растительности — дубравы и травянистые
сообщества, которые называют то луговыми степями, то
остепненными лугами. Они действительно очень
напоминают настоящие луга — сплошной злаковый
покров, очень много разнотравья. В отличие от
настоящих степей они не выгорают летом, а буйно вегетиру-
ют до осени. Растительность лугового типа
формируется там, где достаточно тепла и нет избытка влаги.
Именно такое соотношение тепла и влаги характерно
для климата лесостепи: здесь радиационный индекс
сухости равен примерно единице, то есть осадков
выпадает столько, сколько их может испариться за счет
приходящего солнечного тепла.
В тундре радиационный индекс около 0,3; здесь
постоянно ощущается избыток влаги, но в отдельных
локальных биотопах могут быть такие условия, при
которых тепла несколько больше, а влаги меньше. Это
наблюдается, например, на крутых склонах с легкими
почвами. Южные склоны в тундровой зоне
прогреваются в 1,5—2 раза сильнее, чем горизонтальные
участки. Но этого еще недостаточно для того, чтобы
развились столь богатые и непохожие на тундры
сообщества. Для них неблагоприятно сильное промерзание
зимой, от которого может защитить только
достаточно мощный снежный покров. Там, где снега мало,
например на вершинах склонов, на бровках, откуда он
сдувается ветром, луга не образуются. Они развива-
47
ются на тех склонах, где зимой много снега. Однако
слишком большие, долго не стаивающие залежи
снега для них неблагоприятны.
Достаточно мощный слой снега, защищающий
почву от сильного промерзания зимой,—необходимое
условие формирования лугов в тундровой зоне, поэтому
такие луга называют нивальными. Чаще всего они
развиты по берегам рек, на крутых склонах. В их
формировании большую роль играют животные, особенно
лемминги, перекапывающие и удобряющие почву, а
также дождевые черви и другие почвенные
беспозвоночные, более обильные здесь, чем в прочих
местообитаниях тундры.
В подзоне арктических тундр тепла уже настолько
мало, что даже при самом благоприятном
микроклимате его недостаточно для формирования настоящей
луговой растительности. Здесь встречаются
фрагментарные разнотравно-злаковые группировки, которые с
некоторыми оговорками можно отнести к луговому
типу (см. рис. 12, 13). В полярных пустынях
луговины вообще отсутствуют.
Условия для нивальных лугов ухудшаются и на
юге Субарктики, хотя климат там гораздо
благоприятнее. В подзо«е южных тундр повсюду видны
разнотравные и злаковые группировки. Но они реже
бывают «чистыми», чем в подзоне типичных тундр.
Обычно в них много кустарников (ивы, березки) —
наиболее сильного здесь элемента растительности.
Кустарники на юге тундровой зоны вытесняют
луговую растительность из благоприятных для нее
местообитаний.
Тундровые луга представляют особенно большой
интерес. Именно по ним можно судить, на что
способна арктическая природа. Это самые богатые и
самые продуктивные в данных условиях сообщества.
В них наибольший для Субарктики годовой прирост
растительной массы. На Таймыре луга дают до
200 г/м2 сухой надземной массы в год, а осоково-мо-
ховые тундры — всего 25 г/м2.
Кроме нивальных в Субарктике есть и другие
варианты лугов. Например, прибрежные заросли злака
влаголюбивого — арктофилы рыжеватой (Arctophila
fulva) также можно отнести к категории лугов, в
48
Рис. 12. Луговое разнотравье на южном склоне увала
среди арктической щебнистой куртинной тундры на Вайгаче
Рис. 13. Вертикальные профили растительности в подзоне
арктических тундр на Таймыре:
А — луговина на южном склоне; Б — злаково-разнотравная тундра
данном случае — пойменных, или заливных. Особые
варианты луговой растительности формируются в
процессе деятельности позвоночных животных — на
песцовых норах, на кормовых «столиках» птиц, а
также под влиянием человека — около поселков, на
старых стойбищах и т. д.
Если на хорошо дренированном склоне с легкой
супесчаной или каменистой почвой снега мало, то на
нем развивается не луг, а сообщество из
кустарничков и полукустарничков, в самых малоснежных
местах— чистые дриадовые группировки. Дриада —
одно из самых характерных арктических растений. Это
полукустарничек с листьями ксероморфной
структуры, т. е. с признаками приспособления к недостатку
влаги. Листья узкие, с плотным кожистым
эпидермисом и подвернутыми краями; стебли стелющиеся,
деревянистые. В тундрах Евразии встречается в
основном два вида дриады: точечная {Dryas punctata)
и восьмилепестная (D. octopetala). В северной полосе
Субарктики в наиболее холодных и сухих местах —
на бровках высоких холмов и берегов, на каменистых
грядах — дриада образует сплошной, плотный,
прижатый к земле покров.
На юге зоны в таких биотопах формируются
более богатые группировки из разнообразных ягодных
кустарничков ксероморфного облика — брусники,
голубики, арктоуса, вороники. Для таких мест
характерны своеобразное субарктическое растение,
напоминающее по морфологии суккулент,— Кассиопея
четырехгранная (Cassiope tetragona), а также
оригинальная карликовая ива монетовидная (Salix nummu-
laria) с очень мелкими кожистыми листочками. Если
на склоне снега несколько больше, то кустарничко-
вые сообщества имеют примесь разнотравья и
злаков, т. е. луговых элементов.
Некоторые травянистые растения очень
характерны для сухих возвышенных местообитаний. Они
имеют ксероморфную структуру — жесткие узкие листья,
иногда сильно опушенные. Из злаков это тонконог
азиатский (Koeleria asiatica), зубровка альпийская
(Hierochloe alpina) и др. Некоторые характерные для
этих мест растения образуют плотные куртинки и
кочки. Таковы мытник Pedicularis dasyantha, остроло-
50
дочник Миддендорфа (Oxytropis middendorffii) и
растущий на щебнистых грунтах остролодочник
чернеющий (О. nigrescens).
Рассмотренные интразональные включения в
основной зональный фон тундрового ландшафта
объединяются в три основных типа: болота, луга и кустар-
ничковые группировки. Их географическое
распределение различно. Болота и луга в принципе могут быть
развиты в любой зоне земного шара. Поэтому их
иногда называют азональными или интраполизональными.
Аналогичный пример — растительность песчаных кос
и дюн, для которых характерен, например, колосняк
Leymus mollis, распространенный по морским
побережьям от Субарктики до тропиков. Описанная
растительность из ксероморфных кустарничков и
полукустарничков встречается преимущественно в
субарктических районах. Подобные элементы ландшафта
называют собственно интразональными или интра-
стенозональными (Чернов, 1975). Пример
аналогичных образований — солончаки в аридных районах.
Рассмотрим еще один вариант интразональных
включений в ландшафт. В балке среди степи может
быть дубовый лес, а на высоком холме среди
северного редколесья — участок тундры. В данных случаях
дубрава и тундра находятся за пределами «своей»
зоны (тундровой или лиственных лесов). Такие
элементы ландшафта именуют экстразональными.
Обычно они связаны со склонами, понижениями или
вершинами холмов. Механизм формирования
экстразональной растительности известен под названием
«правила предварения»: растения, обитающие на плакоре
и образующие зональные сообщества, на юге
переходят на северные склоны и в низины, а на севере — на
южные склоны. В пределах тундровой зоны могут
быть экстразональные участки как южных зональных
типов растительности (например, лесных), так и
более северных (полярных пустынь).
В южные районы тундровой зоны повсеместно
проникают островки леса, располагающиеся на речных
террасах, южных склонах водораздельных увалов. На
Таймыре и в бассейне Оленека они доходят до
72° с. ш. В подзоне типичных тундр часто
встречаются заросли ив, березки, ольховника. Это также
51
экстразональные элементы. Обычно они формируются
по склонам холмов, в приозерных низинах, т. е. там, где
скапливается много снега. Кустарниковая ольха
характерна для южных тундр, но проникает и в
типичные. На Таймыре, в бассейне Агапы, ее плотные
заросли опоясывают склоны высоких холмов. Очевидно,
это результат микроклиматических условий: ниже
скапливается холодный воздух и много снега, а выше
сильнее ветер и мало снега. В южных частях подзоны
типичных тундр кустарники заполняют любые
понижения, склоны и все долины. Севернее они становятся
более разреженными и спорадичными, в подзоне
арктических тундр отсутствуют.
Экстразональные полярно-пустынные участки в
тундровой зоне развиты на щебнистых вершинах
высоких холмов, каменистых грядах. На горных
массивах в Субаркгике также распространены аналоги
полярных пустынь — как проявление вертикальной
поясности. Но ее подчас трудно отделить от инверсий
растительности и почв под влиянием мезорельефа.
Дело в том, что в высоких широтах резкие смены
климата совершаются на гораздо меньшем пространстве,
чем в средних. Вместе с тем климатические сдвиги в
экстремных условиях имеют большее значение для
растительности и почв. Поэтому незначительные
поднятия или опускания рельефа, равно как и смещения
к югу или северу, сильно сказываются на облике
ландшафта.
В тундровой зоне бесконечно разнообразны формы
нанорельефа, обусловленного криогенными
процессами, идущими в слое грунта, оттаивающего
летом. Его мощность колеблется от 30 до 200 см и
зависит от экспозиции и крутизны склона, влажности
и механического состава грунта, мощности и
длительности стаивания снежного покрова, характера
растительности. На юге Субарктики глубина сезонного
протаивания на плакорах — 80—90 см, в типичных и
арктических тундрах — 40—60, в полярных
пустынях —- 20—40 см.
В зависимости от погоды разница в глубине
оттаивания в разные годы бывает 10—20 см, поэтому
можно различать три слоя: верхний, оттаивающий
ежегодно, периодически активный, оттаивающий только в
52
годы с наиболее теплым летом, и вечномерзлый в
данном климате. Поэтому летом в мерзлой почве можно
встретить корни растений, которые проникают туда
в более теплые годы. Минеральные и дренированные
почвы оттаивают быстрее и глубже, чем торфянистые
и переувлажненные. Оттаивание длится все лето, и
часто даже в конце июля уровень мерзлоты находится
на глубине 20—30 см. Мощность активного слоя
зависит от экспозиции и крутизны поверхности. В
подзоне типичных тундр, например на Таймыре, глубина
сезонного оттаивания грунта на плакоре примерно
50 см, а на южных склонах— 150 см. К северу
различия в прогреве плоских поверхностей и южных
склонов сглаживаются. В арктических тундрах и на
водоразделах, и на склонах активный слой может
быть 40—60 см.
В активном слое почвы весной, летом и осенью
идут разнообразные криогенные процессы: пучение,
растрескивание и сортировка грунта, выпирание
камней из мелкозема, солифлюкция (медленное течение
насыщенных водой почвенных масс), экспульсии
(выливание жидкого грунта на поверхность) и др. Все
это приводит к формированию различных форм
тундрового нанорельефа, которые в специальной
литературе называют структурными грунтами. Это
собирательное понятие объединяет в основном симметричные
формы полярного нанорельефа: круги, полигоны,
медальоны, сети, полосы и т. д.
Американский исследователь А. Л. Уошберн
(1968) в основу классификации нанорельефа
положил форму элементов и наличие или отсутствие
сортировки грунта. Например, круги — это структурные
грунты в виде разнообразных округлых ячеек. Они
делятся на отсортированные и неотсортированные в
зависимости от того, имеются ли вокруг них бордюры
из камней (см. рис. 14). К отсортированным кругам
относятся каменные круги и каменные кольца. Они
могут встречаться одиночно или группами, диаметр
их колеблется от 0,8 до 3,0 м и выше. Центральные
участки большинства отсортированных кругов
сложены мелкоземом, но некоторые — более крупным
гравием. Отсортированные круги характерны для полярных
пустынь, изредка встречаются в субарктических и
53
Рис. 14. Распределение камней и щебня в отсортированном круге
альпийских районах. Обычно на них совсем нет
растительного покрова или растут единичные
экземпляры растений.
К неотсортированным кругам американские
авторы относят образования без бордюра из камней. Они
также могут быть одиночными или встречаться
группами. В отечественной литературе такие участки
называют пятнистыми тундрами (см. рис. 6, 7, 15).
Пятна голого грунта со скудной растительностью, часто
покрытые корочками накипных лишайников,
окружены валиком хорошо развитой моховой дернины,
который приподнимается над поверхностью грунта на
5—10 см. Между валиками двух соседних пятен
имеется понижение в виде неглубокой ложбинки,
заполненной торфом и мхами с примесью цветковых
растений.
Полигоны — это структурные грунты, в которых
ячейки имеют форму правильных многоугольников.
Они не встречаются одиночно. Размеры их от
нескольких сантиметров до десятка метров. В
отсортированных полигонах крупные обломки образуют бордюр
вокруг мелкозема. Отечественные тундроведы
называли такие структуры каменными полигонами,
каменными сетями, полигональными почвами.
Распространены отсортированные полигоны преимущественно в
полярных пустынях или в горных тундрах. В
неотсортированных полигонах нет каменных бордюров.
Таковы полигональные тундры и полигональные
болота, о которых говорилось выше (см. рис. 9).
Сети — это форма структурных грунтов,
переходная между кругами и полигонами. Они также могут
54
I I J
ПТПП1 ШИ2 EE33 Е34
Рис. 15. Профиль пятна голого грунта в подзоне типичных
тундр:
1 — моховая дернина; 2 — торфянистая масса; 3 — поверхность голого
грунта; 4 — уровень мерзлоты
быть отсортированными или неотсортированными,
т. е. с каменными бордюрами или без них.
Растительность приурочена к трещинам между ячейками и
образует сеть. Особый тип неотсортированной сети —
плакорные мелкобугорковые тундры, в которых на-
норельеф представлен небольшими бугорками
высотой 10—30 см, разделенными извилистыми
ложбинками. Обычно в этих тундрах развита сплошная
мощная моховая дернина. Такой тип структурных грунтов
характерен для южных и типичных тундр и почти не
встречается в арктических.
Пятнистые и полигональные комплексы
поражают не только новичков, но и тех, кто бывал в
Арктике не один раз. Действительно, это поразительная
картина, когда среди сплошного покрова на равном
расстоянии друг от друга располагаются пятна
голого грунта, как «окна» или «глаза»; недаром даже в
научной литературе такие пятна называли
«устьицами земли». Размеры пятен, их форма, число на
единицу площади зависят от механического состава
грунта (на песках — мельче, на суглинках—крупнее), ог
положения в рельефе (на наклонных поверхностях
65
Рис. 16. Схема
распределения пятен голого
грунта в подзонах
типичных (А) и арктических (Б)
тундр (сторона
квадрата — 5 м)
они имеют вытянутую форму, на горизонтальных —
более округлую). К северу они становятся мельче, но
располагаются чаще (см. рис. 16). На Таймыре в
южных тундрах на площадке 10X10 м около 15
пятен, в типичных тундрах на такой же площадке их
около 30, в арктических — более 100. Уменьшаются
не только размеры пятен, но и ширина моховых
перемычек между ними: от 1—2 м на юге тундры до 20—
30 см на севере.
Причины образования пятен голого грунта
многообразны. Общее для южных районов —
появление их в результате разрыва некогда сплошной
растительной дернины. Образовавшиеся пятна
постепенно зарастают, в результате чего целостность
покрова восстанавливается. Правда, этот процесс очень
длителен, вероятно десятки и сотни лет. На севере
тундровой зоны и в полярных пустынях схема
образования пятнистых тундр иная; растения
поселяются на оголенных грунтах вдоль трещин, где зимой
задерживается снег. В результате образуется
растительный покров в виде сети с ячеями голого грунта
(см. рис. 17,18). В условиях постоянно сурового
климата растительность в таком виде существует
неопределенно долго.
Уже на ранних этапах изучения тундровой зоны
вопрос о происхождении структурных грунтов
интересовал многих. Большое внимание этой проблеме
уделяли русские ботаники, начиная с В. Н. Сукачева и
В. Н. Городкова. Было выдвинуто множество гипотез
образования пятнистых и полигональных тундр.
Причем если вначале эти явления стремились объяснить
действием какого-либо одного фактора, то в
настоящее время все чаще склоняются к полигенетической
66
A ;:V/:}V;'J!v'//.;.
Рис. 17. Схема
образования
полигонально-пятнистой
арктической тундры:
А — морозобойные
трещины на оголенном
грунте; Б — моховые
валики по оседающим краям
полигона; В — старое
пятно; на месте
трещины — широкая
ложбинка, заполненная
торфянистой массой и моховой
дерниной; 1 —
минеральный грунт; 2 — моховая
дернина; 3 — торф; 4 —
мерзлый грунт; 5 —
морозобоина я трещина
4 Ш»
Рис. 18. Три варианта полигональных грунтов с разной степенью
развития моховой дернины вдоль морозобойных трещин:
А — щебнистый участок с минимальным покрытием растительностью; Б —
сетчатая структура растительности, сформировавшейся в расширившихся
трещинах; В — еще более мощная дернина, заполняющая просевшие и
расширившиеся трещины
57
Рис. 19. Криогенные явления в пятнистых тундрах:
А — морозобойные трещины, щебенка и корешки на
поверхности голого грунта (подзона типичных тундр);
Б — корни ив, выдавленные на поверхность пятна голого
грунта (южная тундра)
концепции и происхождение структурных грунтов
связывают с разнообразными сторонами криогенеза.
Ряд гипотез основывается на явлении расширения
грунта в результате его промерзания. Этим,
например, объясняют выпирание камней из мелкозема. При
промерзании грунта максимальное расширение его
объема происходит в местах с наибольшей концен-
58
трацией мелкозема. Оно ведет к радиальному распи-
ранию породы. Во время протаивания и сжатия
мелкозем вследствие сцепления частиц оттягивается
назад, оставляя камни на том месте, где они
находились во время замерзания. Многократно повторяясь,
эти процессы приводят к тому, что камни
оказываются «вымороженными» на поверхность грунта (см.
рис. 19). Так объясняется отсортированность
материала в структурных грунтах.
На явлениях расширения и пучения грунтов
основываются представления о механизмах
образования некоторых вариантов пятнистых, или медальон-
ных, тундр. В местах, где дернина тонка или где
меньше снега, грунт промерзает быстрее, и там
может образоваться его местное пучение. В результате
поверхность выгибается. Под действием постоянного
давления дернина разрывается. В последующем
снежная, водная и ветровая эрозии довершают
уничтожение растений и приводят к появлению пятен голого
грунта. Пятнистые тундры в южных районах
тундровой зоны, вероятно, обязаны своим образованием
именно этому процессу.
В результате промерзания грунта в направлении
от дневной поверхности к уровню вечной мерзлоты
создается высокое гидростатическое давление на еще
не замерзший переувлажненный грязевидный грунт.
В результате он может выдавливаться на
поверхность через трещины. Именно на этом явлении
В. Н. Сукачев (1911) основал гипотезу образования
пятен голого грунта. Она неоднократно оспаривалась
в большинстве случаев из-за того, что
подразумевалось катастрофическое явление в виде быстрого
изливания жидкого грунта. Но совсем не обязательно
предполагать внезапное выливание полужидкого
материала на поверхность. Эта гипотеза хорошо
увязывается с явлением тиксотропности тундровых почв.
Тиксотропность — способность относительно
плотного тонкодисперсного грунта под действием
дополнительной нагрузки переходить в разжиженное,
плывунное состояние. Работающие в тундре знают, что,
равномерно нажимая на землю ногой, можно
«раскачать» участок на площади в несколько квадратных
метров. При снятии нагрузки грунт снова становится
59
плотным. Подобное явление может происходить и при
криостатическом давлении. Бесспорно, что некоторые
варианты пятнистых тундр образуются именно таким
путем (Говорухин, 1960).
Несколько гипотез образования структурных
грунтов основано на явлении сжатия вследствие
высыхания и влияния низких температур. Летом при
высыхании грунта на нем появляются небольшие
трещины, которые заполняются водой и в дальнейшем
расширяются при ее замерзании. Там, где возникла
система трещин, они обычно распространяются все
шире и шире. Так объясняют развитие мелких
полигональных структур.
Согласно другой гипотезе, полигоны образуются
вследствие сжатия мерзлой.почвы при очень низкой
температуре. Б. Н. Городков считал морозное
растрескивание грунтов основной причиной
формирования полигональных арктических тундр. Если в
мерзлой почве много льда, то в результате его сжатия при
температуре значительно ниже точки замерзания
могут появиться морозобойные трещины, а затем целая
полигональная система. Так объясняют
возникновение полигональных болот с диаметром полигонов до
10 м.
Таким образом, генезис тех или иных видов
структурных грунтов обусловлен различными процессами.
Полигональные структуры небольшого размера в
высокоширотных районах развиваются под влиянием
сжатия при высыхании и низких температурах. В
образовании округлых форм, свойственных средней
полосе тундровой зоны, большое значение приобретает
дифференциальное пучение. Сортировка грунтов
происходит путем выпирания обломков вследствие
многократных промерзаний. Возможно, что процессы
сжатия, пучения, экспульсии грунта могут сочетаться.
Чтобы полностью объяснить процессы формирования
современных и ископаемых форм структурных
грунтов, нужно больше детальных и точных
сведений. Очень желательны эксперименты в
холодильных камерах и полевые наблюдения в различные
времена года. Необходимы совместные исследования
геологов, физиков, мерзлотоведов, почвоведов,
ботаников.
60
Облик тундрового ландшафта так или иначе
связан с мерзлотой, с процессами ее сезонной и
многолетней динамики. Влияние ее на растительность в
целом можно оценить как отрицательное:
пониженная температура почвы, переувлажненность грунта,
механическое препятствие развитию корневых систем,
повреждение растительной дернины при различного
рода криогенных процессах. Все это нарушает
нормальную деятельность растений, ограничивает
возможность существования многих из них. Чем меньше
глубина сезонного протаивания, тем скуднее
растительность и больше площадь оголенного грунта (см.
рис. 16).
Но и растительный покров влияет на динамику
и глубину оттаивания грунтов. Между мерзлотой и
растительностью тундры существуют весьма сложные,
противоречивые взаимоотношения. Чем мощнее
растительная дернина, особенно сложенная мхами, тем
сильнее ее теплоизолирующее действие, тем ниже
температуры в почве и выше уровнь мерзлоты, а в
конечном счете хуже условия для развития цветковых
растений, имеющих корни. Оголенные почвы,
напротив, прогреваются лучше. Это хорошо проявляется в
пятнистых тундрах. Весной уровень мерзлоты быстрее
опускается под пятном и медленнее — под мощным
моховым покровом в ложбинках. Лишь в конце лета
поверхность мерзлоты под всеми элементами
пятнистой тундры выравнивается. Хорошо прогреваемые
оголенные грунты интенсивно заселяются
различными растениями, но формирующаяся сплошная
дернина постепенно ухудшает прогрев почвы, способствует
поднятию уровня мерзлого слоя. Одна из причин
красочности арктических тундр, обилия цветковых
растений в том и состоит, что в этих климатических
условиях не может формироваться сплошная и
достаточно мощная моховая дернина и поэтому почва
летом прогревается и аэрируется сравнительно хорошо.
При нарушении растительной дернины часто
начинается термокарст, т. е. протаивание льда и
оседание поверхности. В зависимости от массы льда,
степени льдистости грунта термокарст может принимать
различные размеры. Если это происходит на
небольшой площади, появляется углубление, заполненное
61
водой,— бочаг, в котором поселяются водяные мхи
(в особенности Calliergon), постепенно заполняющие
его и превращающие в блюдцеобразное болотце.
Разрастающиеся влаголюбивые мхи образуют мощную
дернину, которая все более изолирует грунт от
наружного тепла, и уровень мерзлоты поднимается. На
месте болотца может вновь возникнуть типичная
тундровая растительность.
Термокарст часто вызывается уничтожением
растительного покрова при строительных работах.
Губительное действие на растительный покров оказывает
гусеничный транспорт, столь широко используемый
на Севере. Гусеницы вездеходов сдирают
растительную дернину, при этом обнажается минеральный
грунт, начинается термокарст, в результате которого
образуются оползни, овраги.
Растительный покров и вечная мерзлота —
компоненты единой системы, которая в каждом конкретном
месте в условиях данной климатической обстановки
находится в состоянии некоторого равновесия.
Стабильность этого равновесия — залог устойчивого
состояния тундрового ландшафта.
РОЛЬ СНЕГА В ЖИЗНИ ТУНДРЫ
Снежный покров — очень сложное
физико-географическое явление. Его характеризуют: толщина,
плотность, теплопроводность, отражательная способность,
продолжительность залегания, структура
составляющих частиц и т. д. Наиболее важны толщина (или
мощность) и продолжительность залегания, между
которыми нет прямой зависимости. Часто большая
продолжительность залегания снежного покрова бывает
там, где его толщина относительно невелика. В
тундровой зоне снег лежит от 220 до 280 дней — гораздо
дольше, чем в других зонах (в средней полосе
Европейской части СССР—120—160 дней, в степях —
60—80 дней).
Толщина снежного покрова в тундровой зоне
колеблется от 10 до 50 см, на большей ее части—20—
30 см. Примерно столько же снега в лесостепи.
Мощность снежного покрова меняется отнюдь не в
соответствии с широтами и зональными границами. В
Сибири, в том числе и на субарктической территории,
более существенны изменения с запада на восток, чем
с севера на юг. Это связано со степенью
континентальное™, с общим количеством осадков и с
распределением их в течение года. В приатлантической Субарк-
тике, на полуострове Канин, в Малоземельской и
Большеземельской тундрах высота снега около 50 см,
а на востоке, от северного Ямала до Чукотки,— чаще
всего 20 — 30 см. Продолжительность залегания
снежного покрова, напротив, максимальна в
центральных и восточных частях тундровой зоны и
минимальна в приатлантической: на Таймыре снег лежит
от 260 дней на юге до 280 на севере, в
Большеземельской тундре и на юге Новой Земли — 240 дней, а
на севере Кольского полуострова — всего 200, т. е.
почти столько же, сколько в степных районах
Восточной Сибири. Таким образом, часто трудно
отделить то, что характерно для зоны, от того, что
обусловлено континентальностью или близостью к океану.
63
В западной, приатлантической, части Субарктики,
смягченной Гольфстримом, продолжительность
снежного периода относительно невелика. Но вследствие
обильных зимних осадков на склонах холмов, среди
скал, в узких долинах могут образовываться не
стаивающие за один сезон снежники. Накапливаясь в течение
нескольких лет, снег уплотняется, превращается в
фирн, а затем и в лед. Эти процессы в настоящее
время интенсивно идут в областях современного
оледенения, например в Гренландии.
Для сибирских тундр характерны следующие
величины мощности снежного покрова: в южных —
около 50 см, в типичных — около 25 см, в арктических —
10—20. При толщине слоя, характерной для
арктических тундр, любые, даже самые незначительные,
поднятия микрорельефа зимой и ранней весной
оказываются обнаженными и подвергаются продолжительному
действию низких температур. Таким образом, в
тундрах снега сравнительно мало, но лежит он очень
долго. То и другое — отрицательные факторы в
развитии растительного и животного мира.
Очень важная особенность снежного покрова
Арктики — его неравномерность. Снег в тундре очень
«сухой» и сыпучий. Постоянные сильные ветры
сдувают его с вершин холмов и увалов, и там остается
слой толщиной 2—5 см, не прикрывающий даже
низкорослые травы. Зато в низинах, распадках, на
склонах образуются мощные снежные толщи. Под
действием ветра снег сильно уплотняется. Уже в декабре
он держит человека без лыж. В дальнейшем
становится более плотным, спрессовывается настолько, что
с трудом поддается железной лопате. Часто
образуются особенно плотные корки (наст),
перемежающиеся с более рыхлыми слоями. Даже весной, в период
интенсивного таяния, по снежникам можно ходить
не проваливаясь.
Высокая плотность и хорошие
теплоизолирующие свойства снега в Арктике позволяют
использовать его для постройки жилищ. Это наиболее развито
на американском Севере и в Гренландии. В
эскимосских иглу, сложенных из снежных кирпичей, при
наружной температуре около —40° можно
поддерживать положительную температуру с помощью костра
64
или печки. Эскимосы делают это очень искусно, не
допуская таяния снежных стен. Вследствие большого
запаса холода в толстой снежной стене иглу при
отапливании медленно обтаивает ее внутренний слой.
Капли воды сразу замерзают, покрывая изнутри стены
ледяной коркой, что .увеличивает прочность снежной
хижины.
Роль снега в тундрах многообразна: участие в
формировании теплового режима, в частности
отражение солнечной радиации (альбедо) и поглощение
тепла на таяние; уменьшение процессов
выветривания и денудации; предохранение растений и животных
от зимних холодов; снежная корразия; ограничение
сроков активной жизни и т. д. Широко известна
роль снега как теплоизолятора, защищающего
почву, растительность и животных от низких
зимних температур. Теплоизолирующая роль снега
зависит от его теплопроводности, а последняя — от
плотности, т. е. от количества содержащегося в нем
воздуха. Минимальная теплопроводность — у свежего
снега, выпавшего крупными хлопьями в тихую и
относительно теплую погоду; максимальная — у сильно
уплотненного и фирнизированного. Плотность свеже-
выпавшего и сильно уплотненного снега отличается
раз в 10, а теплопроводность — примерно в 100.
Талый снег в 2—3 раза теплопроводнее сухого при
равной их плотности (Рихтер, 1945).
Теплообмен между покрытой снегом почвой и
атмосферой почти отсутствует. Суточные колебания
температуры наблюдаются в снежной толще лишь до
глубины 25 см. Кратковременные изменения
температуры воздуха четко отражаются в слое 0—5 см.
В средних широтах при мощности снежного покрова
50—70 см почва может оставаться незамерзшей в
течение всей зимы. Даже под слоем 20 см могут
сохраняться положительные температуры. Однако в
тундрах зимой почва, по-видимому, не может оставаться
незамерзшей. Даже если снег быстро покроет талую
землю, она промерзнет, поскольку холод будет
распространяться снизу (в мерзлоте на глубине 1,5—
2 м температура около 1—3° ниже нуля). Все же
зимой под снегом условия вполне благоприятные не
только для сохранения животных и растений в покоя-
3 Ю. И. Чернов
65
щемся состоянии, но и для активной жизни
теплокровных— леммингов, других полевок, землероек,
горностая, ласки. Более того, лемминги и тундровые
полевки в это время успешно размножаются. Зимний
период жизни тундровых грызунов детально изучила
на Ямале Т. Н. Дунаева (1948).
Полевки Миддендорфа (Microtus middendorffi)
и узкочерепная (Microtus gregalis) распространены
преимущественно в южных тундрах, где снежный
покров весьма мощный и рыхлый. Обский, или
сибирский (Lemmus sibiricus), и копытный (Dicrostonyx
torquatus) лемминги обитают по всей территории
зоны и в полярных пустынях. В районах, где снега
немного, они переселяются на зиму в места с более
мощными снежными скоплениями — на склоны, в
долины ручьев, в западины между каменными грядами.
Копытный лемминг, вероятно, лучше приспособлен
к жизни в плотном снегу. У него развиваются
своеобразные вильчатые когти («копытца»), которые, как
считают, облегчают прокладывание ходов. В тихую
погоду можно слышать, как копытный лемминг
скребется под снегом.
В течение долгой зимы лемминги прокладывают в
моховой дернине многочисленные ходы по радиусам
на десять и более метров от гнезда. Они питаются в
основном мясистыми прикорневыми частями осок и
пушиц, скрытыми в толще дернины. Чтобы добраться
до них, зверьки «срезают» Ъгромное количество мхов,
раскапывают и выгрызают их плотные куртины и
кочки. Сухие части осок и пушиц не едят, но строят
из них в толще снега шаровидные гнезда. Диаметр
гнезда — 20—30 см. По распределению вытаявших
гнезд можно многое узнать о зимней жизни
леммингов. Гнезда располагаются на значительном
расстоянии друг от друга: даже в годы высокой численности
этих зверьков — примерно на 50 м, а местами — на
100—150 м. Это, вероятно, определяется размером
кормового участка семей. Иногда можно найти 2—3
гнезда на расстоянии 2—5 м друг от друга. Очевидно,
они принадлежат одной семье. Гнезда располагаются
только там, где рано устанавливается постоянный и
достаточно глубокий снежный покров (не меньше
50 см),— в понижениях между холмами, по склонам
66
долин и распадков, на болотах между торфяными
буграми, берегах озер и т. д.
Сроки наиболее интенсивного размножения
леммингов под снегом точно не выяснены. Считается, что
оно проходит в основном с апреля по июнь. Имеются
данные о рождении потомства в январе и феврале.
Об этом, в частности, свидетельствуют находки
молодых зверьков в желудках песцов, добытых зимой.
Размножение глубокой зимой более характерно для
обского лемминга, что, вероятно, связано с лучшей
терморегуляцией у этого вида.
Под снежным покровом тундры живут горностай
и ласка. Они охотятся за полевками и леммингами,
используя их же норы, ходы и гнезда, в которых
можно встретить обильные остатки трапезы этих
хищников. Они избегают водораздельных тундр, а чаще
поселяются в оврагах, на склонах холмов, где много
снега и он более рыхлый.
Снег — важнейший фактор зимней жизни крупных
растительноядных млекопитающих и птиц —
северного оленя, овцебыка, зайца-беляка, белой и тундряной
куропаток (см. рис. 20, 21). Все они должны каким-то
образом добираться до скрытой под снегом
растительности. В южной половине тундровой зоны заяц-
беляк зимой поедает торчащие из-под снега ветки
кустарников. Зайцев в тундре немного, и им вполне
хватает этой скудной и грубой пищи. Но для оленя и
куропатки здесь корма недостаточно. Они не могут
разрыть мощный слой очень плотного снега и
откочевывают осенью на юг, в лесотундру и тайгу, где
снег рыхлый и где больше кормов.
Однако местами в самых северных частях
тундровой зоны, например на островах Белом, Новай Земле,
Новосибирских и Врангеля, олени живут круглый год.
Конечно, условия в столь высоких широтах
чрезвычайно суровы, пищи очень мало. Но зато в
арктической тундре мало снега, есть сильно обдуваемые
бесснежные места, где олени могут добывать корм и
зимой. Их пищей в это время года служат веточки ив,
прикорневые сочные части и спящие почки пушицы,
камнеломок, злаков и т. д. (Кищинский, 1971).
Можно только поражаться тому, как им удается
насыщаться суровой зимой этой малорослой скудной рас-
67
Рис. 20. Тундряная куропатка — типичный обитатель тундровых
и высокогорных нивальных ландшафтов (фото А. В. Кречмара)
Рис. 21. Поведение
тундряной куропатки
при закапывании в снег
(по А. В. Андрееву,
1975)
1 — выбор места; 2 — начало
закапывания; 3 — рытье
туннеля; 4 — птица
осматривается, высунув голову из-под
снега; 5 — положение
куропатки под снегом во время
отдыха
тительностью. Но ясно одно, что в арктической тундре
олени могут прожить зимой более успешно, чем в
типичной или южной.
Белая куропатка обитает в основном в южной
половине Субарктики. Зимой она откочевывает в
лесотундру и тайгу. Однако имеются указания на то, что
она иногда зимует в арктической тундре, например на
68
Новосибирских островах, хотя в этой подзоне ее
гнездование нехарактерно и плотность невелика.
Только один вид крупных растительноядных
млекопитающих сумел хорошо приспособиться к жизни
в условиях арктических тундр и даже полярных
пустынь. Это овцебык, который некогда был широко
распространен по всей Арктике, а теперь сохранился
лишь в американском секторе. Именно благодаря ма-
лоснежности арктических тундр это крупное
млекопитающее может жить здесь оседло.
Какова роль снега в жизни растений тундры,
хорошо видно на примере распределения кустарников
на склоне оврага. Самые высокие кусты растут в
нижней или средней части склона. У вершины они как бы
прижаты к земле, словно подстрижены. Если зимой
или ранней весной промерить здесь глубину снега, то
окажется, что ей точно соответствует высота кустов.
Причина этого не только утепляющее влияние снега.
Часто можно встретить кусты березки, ивы и ольхи
с сухими вершинками, на которых кора содрана в
результате снежной корразии. Эти веточки
постепенно иссушаются и отмирают. Уровень снега
ограничивает рост кустарников, у которых повреждаются
верхушечные почки. Это в свою очередь усиливает
ветвление верхних побегов. Поэтому тундровые
кустарники имеют вид постоянно подстриженных.
В южных тундрах можно встретить лиственницу,
имеющую вид сильно разветвленного, прижатого к
земле куста, возвышающегося всего на 30—50 см. Это
так называемая стланиковая форма, которую в Суб-
арктике образуют многие древесные породы. Иногда
они формируют густые, труднопроходимые заросли.
Стланики особенно характерны для горных районов и
дальневосточного Севера, где тундровый ландшафт
опускается в очень низкие широты и захватывает
ареалы многих древесных пород.. Там повсюду широко
распространен кедровый стланик, который считают
то разновидностью кедровой сосны, то особым видом.
В зарослях стлаников создаются благоприятные
условия для зимующих зверьков: под лежащим поверх
густых кустов снегом множество пустот, местами
открыта поверхность подстилки или почвы. Это
облегчает передвижение и добывание пищи,
09
Рис. 22. Флагообразная
крона кустарниковой
ольхи в результате
снежной корразии
(подзона южных тундр
на Таймыре)
Не всегда высота кустарников ограничена
толщиной снежного покрова. Например, ольховник в южных
тундрах достигает высоты 3 м, и его вершины зимой
торчат из-под снега. Но по расположению ветвей
хорошо заметен его уровень. Над густым
разветвленным прикомлевым кустом (спрятанным зимой под
снегом) поднимаются прямые стволы, лишенные
боковых веток. У вершины они разветвляются (см.
рис. 22). Подобную двукроновую форму часто имеют
и лиственницы. Их называют деревьями в «юбках».
Такая форма кроны обусловлена тем, что снежная
корразия особенно сильна у поверхности снега. На
некоторой высоте, где она ослабевает, тонкие веточки
сохраняются и формируют вторую крону,
называемую флагообразной. Часто лиственницы, ели, березы
имеют форму «ходылей»—низкорослых деревьев с
многочисленными искривленными оголенными
тонкими стволами.
Снежный покров исключает зимой процессы
выветривания и денудации, переноса минеральных
частиц. Но в тундрах, особенно в северных подзонах,
много мест, где снег полностью сдувается и
обнажается почва (обычно на вершинах бугров и бровках
береговых яров). Эти участки подвергаются
интенсивной снежной корразии Поселяющиеся здесь
кустарнички имеют сильно искривленные стелющиеся
побеги, с которых часто бывает содрана кора. Если почва
на таких буграх легкая —- песчаная или супесчаная,
то местами она выдувается, развевается, при этом
ГО
обнажаются корни растений, которые отмирают. На
таких местах образуются так называемые ярей —
котловины глубиной до 1 м. Участки с глинистой,
суглинистой или каменистой почвой более устойчивы, на
них часто образуется плотный покров из вечнозеленой
дриады, которая противостоит зимней денудации
благодаря упругой и плотной листве.
Малоснежные, рано обнажающиеся весной
участки подвергаются еще одному своеобразному
воздействию суровой арктической природы, так называемому
«морозному кипению» (frost boils — американских
авторов). В первые солнечные весенние дни такие
участки периодически оттаивают и вновь
промерзают при понижениях температуры,
растрескиваются, в трещинах и полостях почвы образуются
ледяные клинья, кристаллы и «стебельки». Одновременно
наблюдаются вертикальные перемещения почвенного
раствора, периодическое замерзание талой почвы и
ее пучение. В результате всех этих процессов
поверхность почвы приобретает ноздреватую и
слоисто-пористую структуру, имеет вид кипящей жидкости.
Главная причина экстремности среды тундровой
зоны для животно-растительных сообществ —
краткость вегетационного периода. Она определяется не
только радиационными условиями, недостатком
солнечного тепла, но и задержкой снежного покрова,
поглощением и отражением им солнечных лучей.
Таяние снега в тундре начинается еще при
отрицательных температурах воздуха. Местами уже в середине
мая он сходит на вершинах бугров и бровках
береговых яров. В южных тундрах интенсивное стаивание
снега начинается в конце мая, а в типичных — в
начале или даже в середине июня. Скорость схода
снега зависит от микро- и нанорельефа (см. рис. 23).
Обычно в конце мая — начале июня стоит ясная
погода с небольшими морозами. Затем задувает
сильный ветер, приходит пурга, после которой наступает
оттепель и начинается бурное таяние. Иногда снег
сходит за три-четыре дня. Так, в 1967 г. на Таймыре
5 июня под снегом было 95% площади тундры, 7-го
наступила оттепель и к концу этого же дня снег
стаял на 50%, 8 июня он покрывал всего 10% тундры,
9 июня — 2%. Но не каждый год и не везде в тундре
71
Рис. 23. Весной в пятнистой тундре снег стаивает раньше
на валиках и пятнах, задерживаясь в ложбинках
снег сходит так быстро. Часто весна бывает
затяжной. В низинах, на склонах, в долинах ручьев снег
лежит до июля, а самые мощные снежники — до августа.
Обитатели тундр стремятся как можно
эффективнее использовать для активной жизни период ранней
весны, когда высока солнечная радиация, но еще
низки температуры воздуха и много снега. Весной жизнь
в тундре развивается очень бурно и зависит от
интенсивности таяния снега, от распределения проталин и
снежников, от потоков талых вод, повторных
снегопадов, различных физических свойств снега. В общем
снег в это время играет огромную роль в жизни всех
групп организмов тундры. В сезонном развитии
растительности и животного населения особенно важно, с
какой скоростью и в какой последовательности
сходит снег с разных частей рельефа.
В описаниях весеннего периода жизни тундры
часто упоминают так называемые снежные парнички и
отмечают их большую роль в развитии
растительности (Тихомиров, 1963). Снежные парнички — это
небольшие полости между почвой и поверхностью сне-
72
га, образующиеся в результате нагревания
поверхности почвы проникающими через слой снега
солнечными лучами. Нижние слои снега вытаивают,
возникают небольшие полости. Снег над полостью перекри-
сталлизовывается и превращается в ледяную
пластину, покрывающую, как стеклом, этот миниатюрный
парник. В нем происходит то, что в физике
называется тепличным эффектом: попадая в полость через
прозрачную крышу, солнечные лучи нагревают
темные предметы, а возникающее при этом тепловое
излучение многократно отражается от стенок, повышая
температуру, которая может быть больше наружной
на 10° (чаще—на 2—5°). Парнички очень
разнообразны, одни из них покрыты тонкой прозрачной
оплавленной пластинкой льда, другие — толстым
полупрозрачным слоем смерзшегося снега. Некоторые
образуются вокруг растений, побеги или листья которых
пронизывают ледяную крышку.
Парнички образуются ранней весной, когда
температура воздуха держится ниже 0°. Они характерны
для'подзоны арктических тундр, где снежный покров
маломощен и при стаивании его образуется
множество чередующихся друг с другом проталин и
небольших снежников. Вероятно, в этой подзоне они имеют
определенное значение в развитии растений,
способствуя вегетации в период отрицательных температур
воздуха. В. Д. Александрова (1961) на острове Бол.
Ляховском наблюдала вегетацию в парничках яско-
лок, лапчаток, камнеломок, маков в первых числах
июня при температуре воздуха ниже 0°. У растений,
зимующих зелеными, в парничках появляется анто-
циановая окраска, начинается рост побегов и
распускание почек.
Таким образом, благодаря тепличному эффекту
растения, вероятно, могут использовать солнечное
тепло для вегетации при отрицательных температурах
воздуха. В период интенсивного таяния снега и
размораживания почвы парнички теряют свое значение.
Растения начинают интенсивно вегетировать на
проталинах. Но если последние образуются на местах,
где были парнички, вегетация, возможно, ускоряется.
К югу от арктических тундр с увеличением
мощности снежного покрова его таяние приобретает па-
73
водковый характер, с верховодкой и обнажением
одновременно больших площадей. В этих условиях пар-
нички, очевидно, не образуются вовсе или не имеют
реального значения для растений. Да и вообще роль
тепличного эффекта в сезонном развитии тундровых
сообществ пока не ясна. Эти вопросы заслуживают
более детального изучения с постановкой
специальных экспериментов. Не раз высказывавшееся мнение
о большой роли парничков в жизни тундровых
растений подкреплялось в основном краткими
натуралистическими наблюдениями. Думается, что
привлекательность этого своеобразного явления сыграла
определенную роль в позитивной оценке его значения.
Проникающие сквозь снег тепловые лучи играют
существенную роль в жизни тундровых макрофитов —
кустарников. В подзоне типичных тундр
кустарниковые ивы растут на склонах и в долинах, где
образуются мощные залежи снега, которые долго не
стаивают. Ветви, находящиеся в толще снега, нагреваются
весенним солнцем и быстро вытаивают. И хотя корни
еще погружены в мерзлую почву, начинают набухать
и распускаться цветочные почки, появляются
пушистые сережки.
С давних пор в качестве одного из интереснейших
явлений жизни Арктики описываются массовые
скопления на снегу и льдах различных насекомых — в
первую очередь «снежных блох» — мелких бескрылых
насекомых, называемых ногохвостками, или подурами,
или коллемболами. Насекомые встречаются на снегу
во всех широтах, но особенно это заметно в
Заполярье и высокогорьях. На весеннем снегу в тундре
можно встретить десятки видов ногохвосток, жуков-
стафилинид, различных двукрылых — зеленушек,
береговушек, снежных комаров и др.
Очевидно, зимующие насекомые весной
согреваются под снегом солнечными лучами и выходят на
поверхность. Не совсем ясно, как они преодолевают
снежную толщу. Часто их можно встретить на
поверхности сплошного снежного покрова большой
мощности. Скорее всего они пробираются вверх по
обтаивающим растениям, приподнимающимся над землей.
Если весной выкопать из-под снега ветки ивы с
засохшими листьями и сережками или высокие побеги
74
Рис. 24.
Ногохвостки, наиболее часто
встречающиеся
на снегу в тундре:
1 — Ballistura; 2 — Ро
dura aquatica; 3 — Нуро-
gastrura; 4 — /sotomurus;
5 — I so to та
травянистых растении, например полыни, и положить
их в тепле на лист бумаги, то можно убедиться, как
много мелких насекомых зимует в толще снега. На
бумагу начнут выпрыгивать ногохвостки, мухи, на
ездники и др. Создается впечатление, что они
специально располагаются ближе к поверхности снега, к
весенним лучам солнца.
Ногохвостки, встречающиеся на снегу, интенсивно
окрашены в темно-синий или фиолетово-черный цвет.
Тело их покрыто густыми волосками, чешуйками и не
смачивается водой. Поэтому они могут находиться на
талом снегу, на льду или на поверхности воды, по
которой даже передвигаются, отталкиваясь прыга-
тельной вилкой (см. рис. 24). По-видимому, главный
смысл выхода на снег — более полное использование
солнечного весеннего тепла. Благодаря этому
удлиняется период активной жизни, увеличивается
возможность завершения жизненного цикла.
Коллемболы, в особенности их наиболее древние
и примитивные группы, вообще очень характерны для
полярных и высокогорных ландшафтов. Здесь они
достигают максимальной численности. Возможно, чго
снеголюбивость, или, как принято говорить, хионо-
фильность,— первоначальная, исходная черта
биологии данного отряда насекомых. Эта адаптация,
бесспорно полезная в различных ландшафтах, возможно,
75
сформировалась именно в полярных и высокогорных
нивальных условиях или связана с ледниковыми
периодами нашей планеты.
Скорость схода снега и его распределение в
весенней тундре очень много значат для гнездования
птиц. У крупных хищников, например соколов,
мохноногого канюка и белой совы, а также у гусей
период от постройки гнезда до подъема молодняка на
крыло длится до 3,5 месяца. При такой его
продолжительности птицы должны начать гнездиться где-то
в конце мая — начале июня, поскольку в сентябре
наступают холода, выпадает снег, и они должны
улететь на юг.
В конце мая — начале июня в тундре лежит
сплошной снег и еще мало подходящих для
гнездования мест. Этим некоторые авторы объясняют
известное сожительство хищных и «мирных» птиц
(рядом с гнездами канюка, сапсана, белой совы часто
находятся гнезда гусей, казарок). Считается, что
весной на первых подходящих проталинах
вынуждены селиться вместе птицы нескольких видов. При этом
как вторичное явление возможна и защитная роль
пернатого хищника. Вблизи своего гнезда у него не
появляется охотничьего инстинкта, и он не трогает
сидящих рядом птиц. Вместе с тем он защищает от
врагов, например от песца, свое гнездо, а заодно и
расположенные рядом гусиные.
Медленное освобождение тундры от снега имеет
особенно большое значение в жизни белого гуся
(Anser caerulescens), гнездящегося на острове
Врангеля (см. рис. 25). Колонии его занимают строго
определенные территории, но вместе с тем пары не
могут гнездиться слишком плотно, каждая из них
должна иметь свой собственный участок. Чтобы успешно
вырастить потомство, белые гуси должны приступить
к гнездованию здесь в конце мая — начале июня.
В отдельные годы к этому времени большие площади
тундры освобождаются от снега, и гуси равномерно
распределяются на гнездовой территории. Но чаще
к началу июня появляются лишь небольшие
проталины. В таких случаях возникает жестокая
конкуренция за гнездовые участки. Гуси постоянно дерутся
друг с другом. Построив гнездо, пара ревностно охра-
76
Рис. 25. Белые гуси отыскивают освободившиеся от снега
пригодные участки и сразу приступают к постройке гнезд.
Остров Врангеля (фото А. В. Кречмара)
няет его. Не всем хватает места на территории,
занятой колонией. Некоторые пары остаются без гнезд.
«Бездомные» гусыни зорко следят за более
счастливыми парами, и, если заметят гнездо, оставленное без
присмотра, занимают и начинают охранять как свое
собственное или быстро откладывают в него яйцо и
улетают. В результате в некоторых гнездах
накапливается до 20 яиц (при нормальной кладке из трех —
шести яиц). Но не все созревшие яйца «бездомные»
гусыни успевают подбросить в чужие гнезда. Часть
из них они вынуждены отложить где попало. Обычно
на территории разбросано множество таких яиц.
Период таяния снега — катастрофа в жизни
мелких зверьков — леммингов, полевок, землероек,
отчасти горностая и ласки. В это время гибнет много
молодняка, родившегося зимой, затопляется талой
водой потомство, находящееся еще в гнездах.
Подснежные гнезда вытаивают и разрушаются паводком.
77
Летние норы и ходы в толще моховой дернины
бывают залиты водой или еще с зимы забиты ледяными
пробками. Главная забота леммингов во время
паводка — переселение из зимних «квартир» в летние,
т. е. с низких мест и со склонов с мощным снеговым
покровом на увалы и водоразделы — в моховую
тундру. Здесь по мере оттаивания дернины лемминги
прокладывают новые ходы и роют норы или подновляют
старые. В наилучшем положении оказываются
зверьки, которые занимают норы на высоких южных
склонах с легкими почвами. Эти участки раньше
освобождаются от снега и быстро просыхают.
Тяжкое событие в жизни растений и животных
тундры — возврат холодов и поздние летние
снегопады. В южной тундре это наблюдается реже, а в
арктической снег может выпадать в течение всего
лета. Весенние и летние снегопады вредят цветкам,
гнездящимся птицам и некоторым насекомым, в
особенности шмелям, начавшим выкармливать
потомство.
Вот как описывает В. Д. Александрова (1961)
влияние летних снегопадов на растительность
арктической тундры острова Бол. Ляховского. В 1956 г.
летние снегопады наблюдались там неоднократно.
Четыре из них сопровождались отрицательными
температурами воздуха. Первый летний снегопад был
12 июля при средней температуре воздуха —0,4°. Снег
покрыл всю тундру, затем быстро растаял и не оказал
видимого влияния на растительность.
Вторая атака мороза — 20 июля. К вечеру этого
дня пошел снег. К утру он покрыл тундру слоем до
10 см, местами намело сугробы. Из-под снега
виднелись побеги лисохвоста, цветки мака, лютиков.
Вокруг их венчиков образовалась ледяная корочка. 21 и
22 июля снег шел с перерывами. Лишь 23 июля он
прекратился. К вечеру этого дня снег сошел.
Температура воздуха в эти дни снижалась до —1,5°.
Большинство растений не пострадало. Когда с цветков
лютиков стаяла ледяная корочка, они продолжали
цвести. Но цветки маков съежились, стали черными,
приобрели изумрудный оттенок. 29 и 30 июля была
настоящая пурга с температурой воздуха около —0,7°,
Вся тундра оделась снегом, ручьи замерзли.
78
Когда 31 июля снег растаял, вновь расцветшие
цветки маков оказались погибшими. Но остальные
растения продолжали жить без заметных изменений.
Четвертая атака холода и снега наблюдалась 5
августа. К этому времени появилось много новых цветков
маков, которые в третий раз за лето были
повреждены морозом. Снег шел 6 августа, а 7_го температура
опустилась до —2°. 8 августа снег растаял, но
заморозок окончательно погубил цветки маков.
Распускание их прекратилось, хотя еще были бутоны. На
несколько дней наступила устойчивая теплая погода.
Но после 10 августа появилась осенняя окраска
листьев, быстро отцветали уцелевшие цветки,
растения стали готовиться к зиме.
Это описание дает представление о том,
насколько суровы условия в арктических тундрах. По
существу все лето в столь высоких широтах — это
затянувшаяся весна. Вместе с тем растения поразительно
устойчивы к резким переменам погоды. Лишь
отдельные из них, например маки, не способны переносить
сильный холод во время цветения. Интересно, что
полярные маки — группа наиболее типичных
арктических растений, в целом хорошо приспособленных к
суровым условиям Заполярья.
Снег в тундре очень плотный, поэтому талые воды
мощными потоками текут по его поверхности. Эта так
называемая верховодка несет различные
растительные продукты подснежной деятельности леммингов:
остатки пищи, материал гнезд. Талые воды все это
перемещают на большие расстояния и отлагают на
склонах холмов и оврагов в виде валов,
напоминающих скошенное косой сено. Помет леммингов
намывается по берегам ручьев кучами по нескольку
килограммов. В большом количестве воды несут
растительный опад, особенно листья березки и ив, которые
также откладываются по берегам ручьев и на
склонах оврагов. Это одна из причин того, что в тундрах
не образуется подстилка. Накопление растительных
остатков на поверхности мохового покрова могло бы
существенно менять характер ценотических
отношений и направление почвообразовательного процесса.
Особую роль играют воды, вытекающие из
медленно тающих снежников, которые иногда сохраняются
79
до осени. В таких местах развиваются своеобразные
приснежниковые ценозы. Для них особенно
характерны влаголюбивые и холодолюбивые мхи, например
Drepanocladus revolvens, Dicranum spadiceum, из
кустарничков— полярная ива (Salix polaris), из
травянистых— напоминающая нашу мать-и-мачеху нар-
досмия холодная (Nardosmia frigida) и несколько
своеобразных видов лютиков, например очень мелкие
снежный (Ranunculus nivalis), крошечный (/?. pyg-
maeus) и более крупный с мясистыми листьями
сернистый (R. sulphureus), и др.
На месте долго лежащих снежников формируется
низкорослая и разреженная растительность. Моховой
покров и кустарники отсутствуют, преобладают
мелкие травы и кустарнички — лютики, камнеломки,
кисличник, полярная ива. Они очень медленно растут,
многие из них не цветут или не плодоносят.
Вегетативное размножение у них также подавлено.
Растения выглядят угнетенными, недоразвитыми.
От снега зависит тепловой режим тундры. Весной
почти вся солнечная радиация отражается в
пространство, расходуется на тепловое излучение и
таяние. Отражательная способность снега зависит от
его загрязнения, насыщенности водой и т. д. С этим
связано резкое увеличение радиационного баланса в
мае. В это время снег подтаивает, фирнизируется,
обнажается скопившийся на нем мусор. В результате
резко уменьшается его альбедо. Вокруг темных
частиц снег протаивает быстрее, образуются глубокие
дыры. Поверхность снега увеличивается, так как
становится ноздреватой. Это еще более ускоряет его
таяние. К моменту, когда снег полностью стаивает,
солнечная радиация достигает своего максимума, а
затем начинает снижаться.
Итак, различные свойства снежного покрова —
глубина, продолжительность залегания, плотность,
теплопроводность и многие другие — оказывают
крайне разнообразные и противоречивые влияния на
почву, растительность и животный мир тундры. Чем
тоньше слой снега и суровее зимние условия, тем
беднее видовой состав растительности и животного
населения. Однако отдельные, хорошо приспособленные к
суровому арктическому режиму виды могут давать
80
при этом максимальную численность. Чем больше
снега, тем плодороднее почва, богаче растительный
покров (примером могут служить нивальные луговые
сообщества). Однако в местах особенно мощных
снежных наносов, например в нижних частях
северных склонов, где таяние слишком затягивается,
экологические условия ухудшаются, растительность
обедняется. Здесь опять преимущество получают
единичные виды, хорошо приспособленные к данным
своеобразным условиям.
Арктика — это нивальные ландшафты, мир снега
и льда. Продолжительность залегания снежного
покрова — главный отрицательный фактор в жизни
большинства животных и растений. В то же время
снег играет огромную положительную роль,
определяет возможность существования многих видов,
предохраняя их от зимних холодов. Защищая биотопы
от зимней стужи, снег способствует обитанию в
тундровой зоне видов более южного происхождения. В тех
районах, где снега мало, жизнь беднее, но
усиливается процесс формирования холодостойких форм,
хорошо приспособленных к арктическим условиям. Все
это увеличивает разнообразие флоры и фауны
Севера. А в этом залог процветания и устойчивости
тундровых сообществ.
ПРИСПОСОБЛЕНИЯ ОРГАНИЗМОВ
К УСЛОВИЯМ ТУНДРОВОЙ ЗОНЫ
Примеры удивительной приспособленности к среде
млекопитающих, птиц и некоторых растений Арктики
известны нам со школьной скамьи. Этому вопросу
посвящена обширная научная литература. Вместе с тем
пока еще очень слабо изучены общие закономерности
адаптации организмов к суровым условиям
Заполярья. Например, не ясно, есть ли адаптивные
особенности, присущие всем арктическим животным или
растениям, отдельным их крупным группам и
отличающие их от обитателей других зон. В данном
разделе, не останавливаясь на частностях, сосредоточим
внимание на главных путях освоения организмами
тундровых ландшафтов.
Уникальны ли условия жизни
в тундре?
Природная среда тундровой зоны в высшей степени
своеобразна. Поэтому заманчиво выявить ее главные
особенности и наметить единый путь освоения
организмами этого ландшафта. И биологи пытались найти
у арктических растений или животных
приспособительные формы, отличающие их от обитателей
других зон. Но это не так просто, как кажется на первый
взгляд. Несмотря на общее своеобразие климата в
тундровой зоне, едва ли существуют какие-либо
специфические факторы ее среды, уникальные по
отношению ко всем группам организмов.
Недостаток тепла, низкие температуры — это
характерные черты условий жизни в ландшафтах
Заполярья. Вместе с тем для подавляющего большинства
животных и растений наиболее низкие температуры
в тундре не имеют значения, поскольку здесь почти
нет организмов, которые подвергались бы
непосредственному влиянию зимних холодов. Зимой они либо
82
недеятельны, либо живут под снегом, либо
откочевывают к югу. Да и сами морозы в тундре не
сильнее, чем в более южных континентальных районах
(см. табл. 5), где зимуют многие птицы и
млекопитающие, в том числе откочевывающие из тундры,—
северный олень, белая сова, пуночка, белая и
тундряная куропатки. Вместе с типичными таежниками —
кукшей, кедровкой, гаичкой, белкой, росомахой,
соболем — они подвергаются действию сильнейших
морозов. Таким образом, наиболее холодостойкими
должны быть обитатели тайги, а не тундры. Растения
тундры зимой укрыты снегом, а таежные деревья и
кустарники переносят жестокие морозы. Вероятно, не
различаются существенно по уровню холодостойкости
также и беспозвоночные Арктики и более южных
ландшафтов (Лозина-Лозинский, 1972).
Чуть ли не главной особенностью среды
Заполярья считают круглосуточный летний день. Однако
в жизни большинства организмов он имеет
наибольшее значение, пожалуй, не в тундре, а в лесотундре и
северной тайге, особенно в районах с
континентальным климатом, например в низовьях рек Хатанги,
Анабара, Лены, где лесные ландшафты поднимаются
до 71—72° с. ш. Там разгар вегетации растений и
активной жизни животных (в особенности процессов,
связанных с размножением) приходится на самое
светлое время года — на июнь и начало июля. На
большей части тундровой зоны настоящее лето
приходит в начале или середине июля, когда ночная
освещенность уже падает.
В таймырских типичных тундрах (73° с. ш.) в
середине июля, т. е. в разгар вегетации и цветения,
роста и размножения животных, при еще не заходящем
солнце освещенность в полночь в 10 раз меньше, чем
в полдень. В соответствии с этим в поведении
животных проявляется четкий суточный ритм (см. рис. 26).
С 8 августа на этой широте солнце заходит за
горизонт. На широте 68°, на которой в западном секторе
Субарктики развиты южные тундры, полярный день
кончается 19 июля.
Пожалуй, главный отрицательный фактор жизни
в полярных районах — краткость теплого периода,
когда возможны вегетация растений, активная жизнь
83
50
40
30
20|
10
г
L
Ь
iiii
**ы
i
A
/\
/ \
/\
/A\
С ' \ \
J 1 I 1 ] i—¦Г
6 9 12 15
18 21 24
часы суток
Рис. 26. Суточный ход
освещенности (1) и
интенсивности лёта
шмелей (2) на Таймыре
(27 июля 1969 г.,
широта 73°)
Таблица 5
Температуры воздуха в тундровом и таежных районах Сибири
Многолетние температуры
Среднегодовая
Среднеянварская
Среднеиюльская
Самая низкая
Самая высокая
Типичные тундры в
низовьях Пясины на
Таймыре
— 14,0
—35,0
+ 10,5
—50,0
+29,0
Верхоянск
-15,7
-48,6
+ 15,2
—67,8
+34,6
Якутск
— 10,3
—43,2
+ 18,7
-64,0
+ 38,3
Источник. «Справочник по климату СССР». Л., 1967.
пойкилотермных животных, процессы размножения,
роста и развития большинства теплокровных. Но этот
фактор также относителен и для разных групп
организмов имеет неодинаковое значение. Выше уже
отмечалось, что некоторые животные и растения
способны вести активную жизнь и использовать
солнечную радиацию еще при сплошном снежном покрове и
отрицательных температурах воздуха. Это, вероятно,
направлено на своевременное завершение
жизненного цикла и процессов размножения в условиях
короткого лета. Для леммингов лучшая пора размноже-
84
ния — зима. Под снегом они приносят несколько
пометов. Именно зимнее размножение определяет
уровень их численности. Напротив, весной и в начале
лета, когда вода затопляет ходы и норы леммингов,
наступает самая тяжелая пора для этих зверьков.
Краткость полярного лета создает большие
трудности для размножения многих птиц, но не для всех.
Крупные птицы, особенно хищные (кречет, сапсан,
зимняк, белая сова) и водоплавающие (гуси,
казарки, лебеди), действительно едва успевают за
короткое полярное лето завершить цикл размножения.
Поэтому они должны как можно раньше отложить яйца.
Их гнездовая жизнь начинается при почти сплошном
снежном покрове. И часто кладки гибнут при
возврате холодов и снегопадов. Но для мелких воробьиных
и куликов краткость полярного лета сама по себе не
имеет большого значения. Так, у воробьиных
гнездовой период, от постройки гнезда до вылета
молодняка, длится около 30 дней. Если подорожник отложит
яйца в начале июля, птенцы полностью вырастут и
будут способны к самостоятельной жизни к середине
августа, когда еще тепло и много корма. Вот вторая
кладка, наблюдаемая у многих птиц в средних
широтах, в условиях тундры невозможна.
Тундровые растения не только успевают
завершить жизненный цикл, но и дают несколько сезонных
аспектов. Так, в подзоне типичных тундр по времени
зацветания выделяются ранневесенние растения
(кустарниковые ивы, супротиволистная камнеломка),
весенние (чернеющий остролодочник, мытник Эдера),
летние (подавляющее большинство тундрового
разнотравья), осенние или позднелетние (копеечник
арктический— Hedysarum arcticum, живокость Мидден-
дорфа — Delphinium middendorffii, соссурея Тилезиу-
са — Saussurea tilesii). Ко времени зацветания,
например, живокости у чернеющего остролодочника уже
созревают плоды.
Таким образом, условия тундрового ландшафта,
несмотря на их общую суровость, обеспечивают
возможность обитания видов с различным образом жизни
и несовпадающей фенологией, ибо организмы живут
не в ландшафте вообще, а в каких-либо конкретных
местах обитания: на склонах или на плакоре, среди
85
камней или в моховой дернине и в почве. В
разнообразных местах обитания общие климатические
условия претерпевают большую или меньшую
трансформацию. На южных склонах подходящие условия
могут найти весьма теплолюбивые виды, а вблизи
снежников — холодолюбивые.
Несмотря на общее обеднение органического мира
тундры, в нем есть виды с разным строением тела,
физиологией, образом жизни и поведения. И едва ли
имеются какие-либо уникальные и специфичные
приспособительные формы, которые резко отличали бы
животных и растения тундры от обитателей других
зон.
Психрофиты и криофиты
Многие арктические растения имеют признаки,
характерные для обитателей местностей с жарким и
сухим климатом: жесткие с плотной кожицей,
мясистые или иглообразные листья, плотно прижатые к
стеблю (см. рис. 27). Такие особенности строения
называются ксероморфизмом или ксероморфностью.
Их экологический смысл — уменьшить испарение
воды. Ботаники выделяют две группы тундровых
растений с такими признаками: психрофиты и криофиты.
Психрофиты — это растения, физиологически цриспо-
собленные к обитанию в холодных и влажных
условиях. Криофиты — растения холодных и сухих
местообитаний. Эти две группы разделяются недостаточно
четко. Часто растения одного вида с ксероморфными
признаками растут и в сухих и в более влажных
местах. Возможно, все тундровые растения с такой
структурой целесообразно называть психрофитами, так
как они очень редко оказываются в условиях
истинной физической сухости.
Характерные ксероморфные психрофиты и
криофиты — многолетники, кустарнички и
полукустарнички, такие, как дриады, Кассиопея, супротиволистная
и усатая камнеломки, водяника (шикша), ива моне-
товидная, различные злаки — луговик сизый (Descha-
mpsia glauca), тонконог азиатский (Koeleria asia-
tica), овсяница коротколистная (Festuca brachyphy-
lla) и др. Большинство из них наиболее широко
86
1 2 3
Рис. 27. Тундровые растения с типичной ксероморфной
структурой листьев:
1 — Кассиопея четырехгранная; 2 — камнеломка супротиволистная; 3 —
дриада точечная; 4 — камнеломка усатая
распространено в подзонах южных и типичных тундр,
лишь единичные, такие, как супротиволистная и
усатая камнеломки, характерны для арктических.
Формирование ксероморфизма в местообитаниях
с высокой влажностью объясняется так называемой
физиологической сухостью — затрудненностью
поступления воды в растения в условиях низких
температур, при которых замедляются физиологические
процессы, уменьшается сосущая сила. На первый
взгляд кажется, что психрофиты— характерные
растения тундр, так как почвы там всегда влажные и
холодные. Однако самые типичные психрофиты,
имеющие явную ксероморфную структуру органов,
больше свойственны гипоарктической полосе (южной
тундре, лесотундре, северной тайге, северным горным
районам), чем настоящей тундре. Именно там наиболее
обильны такие растения, как вороника, подбел,
багульник, брусника.
Многие типичные психрофиты — настоящие
болотные растения: подбел, клюква, багульник. Причины
ксероморфизма болотных растений ясны. В болоте
много влаги, но в то же время это холодный биотоп.
Мощная дернина и торф препятствуют прогреву.
Условия в равнинной тундре близки к болотным.
Поэтому многие болотные растения проникают в
тундру. Однако в арктических тундрах эти растения
отсутствуют. Нет там и таких характерных для более
87
сухих биотопов, как Кассиопея, брусника, ива монето-
видная, обычных в южной половине Субарктики.
Хвойные — основные эдификаторы тайги — это
тоже своего рода психрофиты. Черты ксероморфной
структуры листьев у них налицо. Происхождение этих
признаков у хвойных не связано с условиями
увлажнения. Они обусловлены особенностями проводящей
системы голосеменных, отсутствием у них сосудов, что
замедляет передвижение воды по стволу к листьям.
Однако у северных хвойных ксероморфизм сыграл
роль важнейшего приспособления к суровым зимним
условиям, когда особенно опасно иссушение,
поскольку в это время вода не поступает к листьям. Имеются
данные о том, что ксероморфизм может
формироваться в условиях азотной недостаточности.
В тундровой зоне ксероморфизм в большей мере
свойствен растениям интразональных биотопов, в
особенности сухих или каменистых эродированных
склонов, вершин бугров и др. Именно в таких местах
обильны супротиволистная камнеломка, дриада,
Кассиопея, водяника. В плакорных тундрах их гораздо
меньше.
Среди типичных тундровых растений есть
множество видов, у которых отсутствуют признаки
ксероморфизма. Таковы почти все бобовые, кустарниковые
ивы, многие мытники, камнеломки, лютики и т. д.
Рядом с дриадой и Кассиопеей растут типичные
мезофиты и гигрофиты — селезеночник, морошка,
одуванчики, кисличник, астрагалы. Лишь на песчаных
или каменистых вершинах бугров доминируют крио-
фиты с ярко выраженной ксероморфной структурой —
дриада, мятлик сизый (Роа glauca), зубровка
альпийская (Hierochloe alpina). В таких местах
физиологическая сухость почвы может усугубляться и ее
периодической физической сухостью. То же можно
сказать об оголенных щебнистых грунтах арктических
тундр, для которых характерны, например, типичные
криофиты — усатая и супротиволистная
камнеломки.
Как считает известный специалист по арктической
флоре Б. А. Юрцев, у многих тундровых растений,
имеющих родичей среди степняков, ксероморфизм
даже ослабевает. Так, тундровые виды полыней, осок и
88
овсяниц менее ксероморфны, чем близкие к ним
степные формы.
Таким образом, ксероморфизм арктических
растений — явление в большей мере биотопическое, чем
зональное. В данном случае налицо общая
закономерность: проблема приспособления к любому
экологическому фактору разрешается разными группами
по-разному. Во всяком случае физиологическая
сухость тундровых почв крайне неодинаково
сказывается на строении растений. В одних случаях в их
облике отчетливо отражается именно эта особенность
среды, в других ведущую роль играют иные факторы.
Существует ли общий путь
адаптации животных
к условиям тундры?
Зоологи неоднократно пытались представить дело так,
что животным тундры, их большим группам,
например млекопитающим, птицам, свойственны строго
определенные общие черты строения и физиологии,
которые достаточно резко отличают их от животных
южных ландшафтов. К таким признакам относили
повышенную холодостойкость, ускоренное развитие,
быстрый рост, общую высокую интенсивность
жизненных процессов, иными словами, все те особенности
образа жизни, которые повышают автономность
организма, делают его менее зависимым от капризов
полярной природы. Были даже попытки отнести всех
позвоночных тундры к единой жизненной форме
арктического животного, понимая под этим некий тип
строения и физиологии, находящийся в полном
соответствии с условиями существования в тундровом
ландшафте. Иными словами, зоологи пытались найти
единый путь адаптации организмов к условиям
Заполярья.
Но эти попытки казались убедительными лишь на
уровне общих рассуждений и умозрительных
заключений. Как только были сделаны конкретные
исследования, выявилось множество фактов,
противоречащих таким взглядам. Так, изучение адаптивных
особенностей арктических и высокогорных позвоночных
89
животных (см., например, работы С. С. Шварца,
Н. Н. Данилова, В. Н. Большакова, М. Н. Денисовой
и 3. В. Артамоновой) показали, что у тундровых птиц
и млекопитающих часто наблюдается пониженный
обмен веществ, замедленные рост и половое
созревание, небольшая плодовитость. Причем все это
свойственно древним и типичным обитателям тундрового
ландшафта. Напротив, у представителей южных фаун,
проникающих в тундры, часто можно обнаружить
черты более высокой жизнедеятельности —
повышение интенсивности роста, скорости развития,
созревания. Благодаря этому они осваивают южные
районы тундровой зоны, но оказываются неспособными к
обитанию в более суровых условиях ее северных
частей.
Приспособительные особенности тундровых
животных зависят не только от климатических,
ландшафтных и биотопических условий, но и от особенностей
биологии, размеров организма, истории данного вида,
уровня филогенетического развития, разнообразия в
данной группе и т. д. Каждый вид по-своему
разрешает проблему приспособления к суровым арктическим
условиям, и часто у форм со сходным образом жизни
бывают прямо противоположные типы адаптации.
Некоторые группы тундровой фауны своими
приспособительными особенностями как бы опровергают
кажущиеся безусловными общие принципы
адаптивной эволюции. Так, бесспорно, что разнообразие и
яркость нарядов птиц увеличиваются от высоких
широт к тропическим. Одна из причин этого —
нарастание богатства фауны. При этом повышается значение
распознавательных признаков.
Но вот кулики дают прямо противоположный
пример. Большинство из них в средних и низких широтах
окрашены однообразно и скромно, с преобладанием
серых, коричневых и пепельных тонов. В тундровой
зоне, напротив, окраска куликов весьма разнообразна
и ярка. Таковы окрашенные в ржаво-красные тона с
разнообразным сложным рисунком краснозобик,
исландский песочник, плосконосый плавунчик, хрустан,
пестрые камнешарка, тулес, золотистая и бурокрылая
ржанки, разноцветный турухтан и др. Это следствие
высокого разнообразия и обилия куликов на единице
90
площади тундрового ландшафта. Здесь повышается
роль распознавательных признаков, точно так же как
у многих других птиц в тропиках.
Хороший пример разнообразия адаптации к
условиям тундры и видовой специфики дают шмели. Как
показал в своих классических исследованиях
И. Д. Стрельников (1940), шмели — это своего рода
«теплокровные» насекомые. Благодаря крупному и
густоопушенному телу они способны накапливать
теплоту, вырабатываемую в полете. Поэтому они
бывают активными при очень низких температурах.
Естественно предположить, что крупные размеры
шмелей, способствующие лучшему сохранению тепла
(вследствие меньшей относительной поверхности
тела),— полезный признак в арктических условиях.
Действительно, в тундровой зоне весьма много
крупных видов, таких, как Bombus hyperboreus, В. pyrrho-
pygus.
Неизбежное следствие больших размеров —
увеличение длительности развития. С этим связана важная
особенность полярных шмелей — сокращение
числа рабочих особей или полная их утрата. При этом
матка сама выкармливает других самок и самцов.
Однако в тундровой зоне широко распространен и
многочислен один из самых мелких видов шмелей —
В. lapponicus. У него имеются нормальные рабочие
особи. Очевидно, развитие этого мелкого шмеля
проходит значительно быстрее, чем у других видов.
Поэтому он успевает выращивать рабочие особи в
условиях непродолжительного полярного лета. Таким
образом, мелкие размеры, неблагоприятные с точки
зрения адаптации к холоду, позволяют сохранить
полиморфизм шмелиной семьи в суровых условиях
Заполярья. Крупные виды, имея преимущество в
терморегуляционном отношении, вынуждены поступиться
своими рабочими особями.
Этот пример еще раз свидетельствует о том, что
пути приспособления к условиям тундровой зоны
часто диаметрально противоположны. Мелкие и крупные
размеры, замедленное и ускоренное развитие в
зависимости от других особенностей могут играть
положительную роль в освоении суровых субарктических
условий.
м
В тундровой зоне успешно реализуются самые
разнообразные приспособления организмов, подчас
противоположные. Субарктические условия экстрем-
ны, однако они достаточно разнообразны для
обитания различных жизненных форм, образующих весьма
насыщенные и сложные сообщества. В
приспособлениях тундровых организмов отражаются и прошлое
ландшафта, и история видов, и пути заселения ими
Субарктики, и дифференцированность арктической
среды, и специфика самих видов — их физиология,
образ жизни, а также взаимоотношения с другими
организмами.
Роль истории видов
Часто признаки, сформировавшиеся не в
субарктических, а в иных условиях, оказываются особенно
полезными в тундровой зоне. Хороший пример —
подснежное размножение тундровых грызунов —
леммингов и узкочерепной полевки. С. С. Шварц (1963)
считал, что подснежное размножение — одно из тех
приспособлений, которое позволило этим грызунам
успешно освоить тундру, давать полноценное и
обильное потомство независимо от капризов полярного лета.
Но эта черта биологии возникла явно не как
результат приспособления к тундровым условиям.
Родина узкочерепной полевки — степная зона. Там
живет особый подвид, который также может
размножаться зимой под снегом. Это свойственно и другим
степным грызунам. Заселив тундру, узкочерепные
полевки образовали другой подвид, у которого
подснежное размножение сохранилось как очень полезная в
данных условиях адаптация. Из степи происходят и
предки тундровых леммингов. Ближайший предок
копытного лемминга (см. рис. 28) в период последнего
оледенения был обычным обитателем так называемых
перигляциальных степей на территории от
Центральной Европы до Восточной Сибири.
Вообще во флоре и фауне Субарктики весьма
много таких случаев, когда вид не приспосабливался к
полярным условиям, а оказался к ним
приспособленным в силу совпадения уже имевшихся у него свойств
92
Рис. 28. Копытный лемминг — потомок степных грызунов
(фото А. В. Кречмара)
с требованиями данного типа среды. Такое явление
называется преадаптацией. Попросту говоря, это
предрасположенность организма к обитанию в тех или
иных условиях. В равной мере это относится и к
отдельным видам, и к целым группам животных и
растений. Так, общеизвестна удивительная
приспособленность лишайников и мхов к суровым арктическим
условиям. Но нет никаких оснований считать, что
чрезвычайная холодостойкость лишайников и мхов —
результат непосредственной адаптации к суровому
холодному климату. Главная причина процветания в
тундрах мхов и лишайников — их общая
пониженная жизнедеятельность, способность прекращать рост
в любой момент и продолжать его без всяких
изменений, когда позволят условия. Эти качества могли
возникнуть как приспособления и к сухому жаркому
климату. Возможно, что определенную роль здесь
играет в общем невысокий филогенетический уровень
этих групп, их относительная примитивность,
способность к пониженной интенсивности жизненных
процессов.
В. М. Швецова и В. Л. Вознесенский (1970) не
нашли каких-либо специфических особенностей
фотосинтеза тундровых растений. Среди них, так же как и
93
¦1 2
18 21 24
часы суток
Рис. 29. Изменение
интенсивности пения
обыкновенной каменки
на Западном
Таймыре (1) и под
Воронежем (2) (по данным
В. И. Подаруевой)
среди лесных, есть виды с низкой и с высокой
интенсивностью фотосинтеза. Более того, у некоторых
растений тундры верхняя температурная граница
фотосинтеза— около 50°. Таких температур не бывает в
Субарктике. Возможно, что это тот случай, когда
растение сохраняет приспособление, возникшее в
условиях других, гораздо более теплых ландшафтов. В
данном случае это свойство не имеет значения для жизни
в тундре и может рассматриваться как атавизм.
В этом плане интересны особенности суточной
активности животных. В Заполярье у некоторых широко
распространенных видов сохраняется суточный ритм
поведения, свойственный более южным популяциям,
обитающим в иных условиях освещенности (см.
рис. 29).
У видов, обитающих в Субарктике, могут
реализоваться такие свойства, которые, казалось бы, не
соответствуют тундровым условиям. Так, спячка, летняя
или зимняя, характерна для исконных обитателей
аридных ландшафтов — сусликов, сурков,
тушканчиков. Обитателями высоких широт этот тип адаптации
широко использоваться не может, так как очень
трудно накопить столько жира, чтобы его хватило на
длительную зиму. Но один впадающий в спячку грызун
все же освоил приполярные ландшафты и обитает в
тундрах Чукотки, Аляски и Канады. Это
американский длиннохвостый суслик (Citellus parryi) (см.
рис 30), близкий другому суслику — азиатскому
94
Рис. 30. Американский длиннохвостый суслик в чукотской тундре
(фото А. В. Кречмара)
длиннохвостому (С. undulatus). Эти зверьки —
исконные обитатели степей и высокогорных лугов.
Ландшафт Чукотки, где обитает американский
суслик, представляет собой вариант южной тундры.
Там он выбирает разнотравные и луговые участки по
берегам рек, по склонам грив, холмов, среди скальных
обнажений. Этот крупный грызун нуждается в
обильном сочном зеленом корме. Именно способность
прекращать активную жизнь в зимнее время позволила
ему заселить тундру. Однако накопленного летом
жира едва хватает на время спячки в течение
длительной полярной зимы. В связи с этим осенью суслики
засыпают очень поздно. Они активны еще при сильных
морозах вплоть до выпадения глубокого снега.
Весной выходят на поверхность при сплошном снежном
покрове и в поисках пищи протаптывают в нем
глубокие тропы.
Итак, многие обитающие в тундровой зоне виды
фактически преадаптированы к ее суровым условиям.
Естественно, что та или иная черта их биологии,
оказавшаяся полезной при освоении тундр, в дальней-
95
шем могла изменяться и усиливаться по мере
специализации видов уже как типичных обитателей
Заполярья.
Роль размеров тела
Чем мельче организм, тем легче ему найти
благоприятные условия. Одноклеточная водоросль может
обитать в морях, лужах и пленках воды, окружающих
почвенные частицы. Для крупных водных животных
нужны определенные типы постоянных водоемов.
Среди микроскопических организмов тундры много видов,
распространенных в лесах, степях и даже пустынях.
Таковы различные почвенные клещи, коллемболы,
нематоды. Мелкие организмы часто не имеют
каких-либо специальных приспособлений для обитания в
тундре. Даже если эти виды характерны для тундровой
зоны и встречаются только на ее территории, они
очень похожи на своих собратьев из южных
ландшафтов.
Мелкий зверек, например лемминг или горностай,
проводит зиму под снегом и не чувствует сильных
морозов. Но северный олень и песец не могут
спрятаться от зимней стужи и должны переносить полярные
холода. Выше говорилось о влиянии снежной
корразии на деревья и кустарники. Однако это явление не
имеет значения для подавляющего большинства
травянистых растений и кустарничков, скрытых зимой под
снегом. Таким образом, чем крупнее организм, тем
больше он подвергается непосредственному
воздействию климатических условий тундры, тем большими
способностями он должен обладать, чтобы
противостоять им.
Низкий рост растений тундры, их «карликовость»,
вероятно, один из самых заметных внешних
признаков. Единичные деревья, заходящие в южные районы,
зоны, низкорослы и часто приобретают стланиковую
форму с пригнутыми к земле искривленными
стволами. Высота тундровых кустарников обычно 0,5—1 м.
Таковы карликовая березка, ивы — ползучая и
мохнатая. Большинство видов ив имеют стелющуюся
форму. Их стебли скрыты в моховом покрове или
приподнимаются над ним на 10—15 см, как, например, у
96
арктической (Salix arctica), ивы красивой (S. pul-
shra).
Еще меньшие размеры у тундровых
кустарничков — настоящих хозяев тундры. Это и знакомые
жителям умеренного климата брусника, голубика, и
настоящие «тундровики»— Кассиопея, куропаточья
трава, и многие другие. Обычно их высота около
10 см, а часто 3—5 см. Крошечные размеры имеет
один из самых маленьких в мире кустарничков — ива
полярная (Salix polaris). Тонкие стебли этого
растения скрыты в моховой дернине, и лишь по два-три
небольших листочка, величиной не более копеечной
монеты, приподнимаются над мхами. Высота трав,
злаков, осок, разнотравья в среднем 10—15 см, и
только генеративные побеги к моменту плодоношения
обычно вытягиваются до 20 см.
Одна из самых характерных особенностей
развития тундровых растений — цветение при
минимальных размерах цветоносного стебля. Многие растения
начинают цвести, как только земля освободится от
снега. В это время рост еще не начинается, и
цветоносы очень короткие. Это явление, бесспорно, имеет
приспособительное значение. Температура воздуха
выше всего в приземном слое, что благоприятствует
цветению в прохладную весеннюю погоду. Для
распространения семян, которые у большинства
растений разносятся ветром, более целесообразны длинные
цветоносы. У весенних видов они удлиняются (в 4—
5 раз) после цветения, а у летних — в период
бутонизации. Полярные маки имеют довольно длинные
цветоносы, около 8—12 см. Однако в арктических
тундрах они часто лежат, слегка извиваясь, на более
теплой по сравнению с воздухом поверхности почвы,
и лишь сами цветки немного приподняты.
С малыми размерами тундровых растений
связаны кажущиеся большими величина и яркость их
цветков. Сами по себе цветки арктических растений
не крупнее лесных, но по отношению к размерам
стеблей и листьев воспринимаются как большие.
Именно поэтому тундровые растения — благодатный
объект для фотографирования. Крупным планом
можно заснять все растение. Многие из них имеют
действительно довольно крупные цветки — диаметром
97
около 3 см, например новосиверсия ледяная (Novosie-
versia glacialis)', мак Papaver pulvinatum, яскрлка
крупная (Cerastium maximum). Часто мелкие цветки
собраны в крупные соцветия, от чего они становятся
более заметными (астрагалы, мытники).
Низкий рост арктических растений связан с общей
суровостью климата, дефицитом тепла, более
благоприятными температурными условиями в приземном
слое воздуха, небольшой глубиной снежного покрова,
с недостатком азота и т. д. С высоким ростом
сопряжен большой расход питательных веществ; низкий,
напротив, способствует их экономии, столь важной
для вызревания семян и завершения жизненного
цикла.
Карликовость тундровых популяций многих видов
закреплена генетически. Это было показано
американскими исследователями в опыте с маленьким
растением из семейства крестоцветных (Cardamine belli-
difolia). Семена этого растения были собраны в
альпийском поясе гор Аляски и на островах Канадского
Арктического архипелага между 80—82° с. ш. При
выращивании в одинаковых умеренных
температурных условиях альпийские растения вырастали
высокими, а арктические оставались такими же, как в
тундре,— маленькими, с большим количеством мелких
листьев.
Небольшие размеры тела выгодны и арктическим
беспозвоночным. Чем мельче животное, тем оно легче
находит подходящие условия, тем скорее
завершаются его рост и развитие. В связи с этим среди
тундровых насекомых и других беспозвоночных
относительное количество мелких видов значительно
больше, чем в фаунах южных зон. Особенно отчетливо
прослеживается это в полярных пустынях, где нет
сколько-нибудь крупных насекомых, таких, как
дневные бабочки, жужелицы, шмели, жуки-плавунцы
и т. д. Там остаются лишь самые мелкие их
представители — коллемболы, комары-звонцы, снежные и
грибные комары, мелкие ночные бабочки, наездники.
Но в общем процесс «измельчания» в животном мире
высоких широт менее выражен, чем в растительном.
В подзоне арктических тундр еще достаточно много
крупных насекомых, таких, как шмели, комары-дол«
98
гоножки, некоторые мухи, например оводы, журчалки.
Причем долгоножки, или типулиды,— самые крупные
среди комаров, относятся к одной из процветающих
групп фауны этой подзоны.
Очень сложны в высоких широтах закономерности
изменения размеров позвоночных теплокровных
животных. Согласно известному правилу К. Бергмана,
установленному еще в прошлом веке, величина тела
во многих группах родственных видов или подвидов
томойотермных увеличивается от экватора к
полярным районам (см. табл. 6). Чем крупнее тело, тем
меньше относительная поверхность и, следовательно,
меньше теплоотдача. Иными словами, чем крупнее
животное, тем легче ему сохранять тепло. Свое
правило К. Бергман формулировал очень осторожно, как
общую тенденцию, имеющую множество исключений.
Таблица 6
Северные пределы распространения
и величина тела медведей (Чернов, 1975)
Вид
Белый медведь
Бурый медведь
Черный медведь
Малайский медведь
Примерная
северная
граница
ареала, °с. ш.
85°
72°
53°
20°
Максимальная длина
тела, м
3,0
2,5
1,7
1,4
Максимальный вес, кг
1000
750
150
65
Он понимал всю сложность затрагиваемых им
вопросов и подчеркивал, что величина тела — лишь один
из множества факторов приспособления к тому или
иному типу климата, в том числе и к арктическому.
На ряде примеров он показал, что проблема
терморегуляции у каждого вида разрешается по-своему.
В частности, кроме величины тела огромную роль
играет шерстный и перьевой покровы, убежища,
спячка и т. д.
В дальнейшем некоторые авторы трактовали это
правило в более категоричной форме. Так,
американский ученый Д. Аллен писал: «Как правило, размеры
99
млекопитающих и птиц Северной Америки возрастают
с юга на север. Это справедливо не только в
отношении индивидуумов данного вида, но и вообще самые
крупные виды каждого рода или семейства являются
более северными». В литературе неоднократно
приводились примеры более крупных арктических видов
или подвидов. Вместе с тем имеется множество
случаев с прямо противоположной закономерностью.
Преимущество крупного тела у теплокровного
животного в холодном климате бесспорно. Однако
зональные изменения размеров птиц и зверей
чрезвычайно сложны и противоречивы. При сопоставлении
размеров птиц, обитающих в высоких и более низких
широтах, пожалуй, отчетливее прослеживается
тенденция уменьшения тела полярных видов по
сравнению с их южными родичами. Тундряная куропатка,
обитающая севернее, меньше белой. Южный
(лесостепной) подвид белой куропатки — наиболее
крупный. Тундровый лебедь (Cygnus bewickii)
—наименьший представитель рода; самые мелкие виды среди
гусиных — арктические, например черная казарка
(Branta bernicla), краснозобая казарка (В. ruficollis),
пискулька (Anser erythropus см. табл. 7). Самые
крупные птицы в тундровой зоне населяют в основном
ее южную полосу (см. рис. 31, 32).
В книге С. С. Шварца (1963) приведены
следующие сведения о размерах тундровых зверей. Из 20
видов млекопитающих, распространенных как в Суб-
арктике, так и за ее пределами, большие размеры
особей заполярных популяций наблюдаются у девяти
видов, меньшие (по сравнению с лесными
популяциями) — у пяти, у трех видов нет различий по размерам,
по остальным данные отсутствуют. У некоторых видов
млекопитающих размеры тела уменьшаются или
увеличиваются независимо от зональных изменений
климата во всех направлениях от определенного
района. Так, общая длина черепа у зайца-беляка меньше
всего в Восточной Европе и Западной Сибири. Она
увеличивается не только к северу, но и к востоку,
западу, югу от этих районов. Самый мелкий подвид
зайца-беляка обитает в наиболее суровых
климатических условиях — на севере Восточной Сибири.
Показательны и такие примеры: размеры особей север-
100
Рис. 31. Лебедь-кликун (А), белоклювая гагара (Б), канадский
журавль (В) — самые крупные птицы в тундре (фото А. В. Креч-
мара)
Рис. 32. Черная (А) и краснозобая (Б) казарки —
самые мелкие виды гусиных (фото А В. Кречмара)
ных популяций горностая больше, а ласки — меньше.
Ее тундровый подвид так и назван малой лаской
(см. рис. 33). Тундровый волк относительно крупный,
но все же он меньше лесного.
Таблица 7
Максимальные показатели веса и продолжительности
развития некоторых видов гусиных птиц
(по Е. В. Сыроечковскому, 1978)
Вид
Лебедь-шипун
Лебедь-кликун
Тундровый лебедь
Гуменник
Белый гусь
Пискулька
Краснозобая
казарка
Черная казарка
Вес,
кг
16,0
13,0
6,0
3,7
2,9
2,5
1,7
1,4
Продолжительность
инкубации яиц,
сут.
36
40
30
29
26
28
26
26
Длительность
роста
птенцов, сут
150
120
45
45
49
40
40
40
Общий период
развития, сут.
186
160
75
74
75
68
66
66
Почему же вопреки явной выгоде крупного тела
размеры многих теплокровных животных в высоких
широтах меньше? Скорее всего основная причина
этого — необходимость завершения цикла
размножения, воспитания потомства в течение одного летнего
сезона. Наиболее отчетливо это проявляется именно
у птиц, которые обязательно должны закончить рост
и улететь к югу. Чем мельче размеры тела, тем короче
периоды роста и развития (см. табл. 7). У крупных
видов птиц период гнездовой жизни и роста птенцов
соизмерим с продолжительностью теплого времени
в тундре (бывает и длиннее, и короче). Лебедь-
кликун не может жить в типичных, а тем более в
арктических тундрах уже потому, что ему просто
недостаточно безморозного периода для воспитания
потомства независимо от прочих условий. Но и гуси
едва успевают вырастить птенцов и поэтому
начинают гнездиться очень рано, при крайне
неблагоприятной погоде. Бесспорно, в отдельных случаях опреде-
103
Рис. 33. Самый мелкий хищный зверь — ласка проникает
далеко в тундровую зону (фото А. В. Кречмара)
ленное значение имеют и запасы пищи, возможность
насыщения, которая более сложна для крупного
организма.
Краткость полярного лета, вероятно, служит
причиной преобладания мелких форм среди насекомых и
других беспозвоночных высоких широт. Правда, здесь
картина несколько иная. Один из главных путей их
приспособления к суровым условиям тундры —
способность к замедленному, полностью подчиненному
погодным условиям развитию. Многие насекомые
тундры развиваются два-три, а то и пять сезонов.
Таким образом, в некоторой степени они не зависят от
краткости полярного лета и от капризов погоды. Но
и в этом случае мелкие размеры более выгодны,
поскольку развитие крупных беспозвоночных в крайне
суровых условиях арктических тундр и полярных
пустынь может растягиваться на неопределенно долгое
время.
104
Принцип экономии
поверхности тела
Широко известна еще одна общая закономерность
географического изменения признаков теплокровных
животных — уменьшение относительной величины
выступающих частей тела с похолоданием климата.
Эта закономерность именуется правилом Аллена. Она
связана с проблемой терморегуляции. Чем больше у
животного выступающих частей и чем они длиннее
(уши, хвост, лапы), тем больше теплоотдача и
сильнее охлаждение тела. Меньшие размеры их выгодны
в холодных условиях, сильное развитие — в жарких.
Обычно это правило иллюстрируется примером
разных видов группы лисиц. Самые короткие уши у
песца, несколько длиннее — у лисицы, еще длиннее — у
степного корсака и самые длинные — у пустынного
фенека.
Это правило — частный случай более общего
явления, которое можно назвать принципом экономии
поверхности тела. В холодных условиях, вероятно,
наиболее выгодная форма тела — шар, у которого
минимальная относительная поверхность и,
следовательно, наименьшая теплоотдача. В связи с этим для
теплокровных обитателей Заполярья наряду с
различными утепляющими приспособлениями в виде
меха, пера характерна более компактная округлая
форма тела с минимумом выступающих частей (см.
рис. 34).
Явление экономии поверхности в определенной
степени свойственно и тундровым растениям. У
многих цветковых основная масса листьев собрана в
прикорневые розетки или подушки. Мхи и лишайники
часто разрастаются в виде округлых плотных куртин.
При продвижении на север зоны это проявляется
особенно заметно.
Плотные розетки листьев образуются у маков,
крупок, камнеломок, новосиверсии и многих других.
Чем суровее условия, тем заметнее эти особенности
роста растений. Например, у экземпляров дриады,
растущих в моховой дернине, более длинные стебли
и более рыхло расположенные листья, чем у особей
этого же вида на щебнистых участках, бесснежных в
105
Рис. 34. Обский, или сибирский, лемминг — характернейший
представитель тундровых млекопитающих (фото А. В. Кречмара)
зимнее время, где они формируют плотно прижатую
шпалеру с густо облиственными стеблями. Незабу-
дочник (Eritrichium villosum) на юге тундровой зоны
образует очень рыхлые, распадающиеся на отдельные
стебли дернинки. На севере они почти шаровидные.
Многие виды камнеломок, например поникающая
(Saxifraga сетиа), болотная (S. hirculus), дернистая
{S. caespitosa), в южных и типичных тундрах мало-
стебельны, с небольшой розеткой прикорневых
листьев, в арктических тундрах они плотные
многостебельные, дернистые (см. рис. 35, 36). Самые крупные и
плотные подушки у новосиверсии ледяной (см.
рис. 37). Отдельные их экземпляры достигают
60—80 см в диаметре и 15 см высоты.
Некоторые виды образуют плотно прижатую к
земле, распластанную дернину. Такие шпалеры
образует, например, в арктических тундрах минуарция
(Minuartia macrocarpa). Однако на юге тундровой
зоны это растение имеет немногочисленные рыхлооб-
лиственные стебли. Сильно меняется форма одного из
самых раннецветущих растений —¦ камнеломки супро-
тиволистной. В южных районах тундры это
травянистое растение с мягкими стеблями, на которых на
расстоянии примерно 1 см сидят листья. На севере
или в горах развивается одревесневший главный сте-
100
Рис. 35. Плотная куртинка камнеломки дернистой
(фото Н. Р. Матвеевой)
Рис. 36. Куртинки и подушки камнеломок, маков,
незабудочника в арктической тундре
бель с жесткими укороченными побегами,
образующими плотную подушку. Такие примеры можно
продолжить.
В чем же экологический смысл таких форм? По-
видимому, наиболее существенна защита почек
возобновления в осенне-зимнее время как от снеговой
абразии, так и от иссушения. Можно было бы
ожидать, что почки арктических растений хорошо защи-
107
Рис. 37. Многолетние кочки, образованные побегами
новосиверсии ледяной
щены твердыми чешуями, предохраняющими от
механического повреждения и от иссушения. Однако они
чаще всего покрыты лишь старыми листьями или же
вообще не защищены. По-видимому, твердые
почечные чешуи препятствуют быстрому весеннему
развитию и, следовательно, не выгодны растению в
условиях краткого арктического вегетационного периода.
В плотных подушках или розетках точки роста
надежно укрыты и старыми отмершими, и
многочисленными живыми зимующими листьями. В таких
подушках лучше тепловой режим в весеннее время,
когда снег только начинает сходить.
Вот как об этом пишет известный исследователь
Севера Б. А. Тихомиров (1963): «...отмершие листья
Novosieversia glacialis, долгое время оставаясь на
растении, при неблагоприятных условиях служат
защитой для корневища и почек возобновления...
Возвышаясь над поверхностью почвы на 8—10 см, кусты
108
новосиверсии, благодаря обильным остаткам старых
побуревших листьев, прогреваются солнечными
лучами еще при наличии снежного покрова». Образование
розеточных и подушковидных форм у арктических
растений, так же как их малый рост, позволяет им
функционировать в наиболее благоприятном для
жизни приземном слое воздуха летом.
Пути формирования
флоры и фауны
Тундра и полярные пустыни — самые молодые из
зональных ландшафтов Земли. В течение большей
части третичного периода на территории современной
Субарктики были развиты лиственные и хвойные
леса. Тундровая растительность начала
формироваться на северных оконечностях Евразии и Америки
в связи с похолоданием климата во время
четвертичных оледенений. При этом данные палеоботаники и
палеозоологии свидетельствуют о том, что уже в
самом начале четвертичного периода существовал
типичный тундровый ландшафте характерными фито-
ценозами и набором приспособленных к этим
условиям видов животных и растений. Это заставляет
предположить, что ядро арктической флоры и фауны, во
всяком случае их ближайшие предшественники,
сформировалось до оледенений, что быстро помогло
освоить новый тип ландшафта. Большинство
ботаников считает, что основными предшественниками
тундровой флоры были растения, обитавшие под пологом
леса в таежной зоне, а также в альпийском поясе
высокогорий. Они и составили базу тундровых флор в
северных частях Евразии и Америки.
Во время даже максимальных оледенений
обширные пространства северных частей Азии и Америки
оставались свободными от ледника. Это прежде
всего районы Восточной Сибири и Аляски, которые были
своего рода флоро- и фауногенетическими
лабораториями, поставлявшими виды, способные к
формированию сообществ тундрового типа. Они и сыграли
основную роль в происхождении современного
органического мира Субарктики. Древние флора и фауна
этих территорий видоизменялись, заселяя освобож-
109
давшиеся от ледников территории, осваивая новые
типы ландшафтов. Наряду с этим большую роль
играло и по сей день играет вселение в субарктические
ландшафты более южных видов, а также
космополитов. Процессы расселения по тундровой зоне и
формирования видового состава ее животного населения и
растительности не закончились и в настоящее время,
и часто можно быть свидетелем расширения ареалов,
вхождения новых форм в состав сообществ Заполярья.
Экспансия в пределы тундровой зоны в первую
очередь свойственна видам, вообще широко
распространенным, обитающим во многих зонах. Примерами
могут служить обычные в тундровой зоне горностай,
белая куропатка, сокол-сапсан, семиточечная божья
коровка и др. Подобные виды называют
полизональными. Напротив, типичные обитатели северных
лесных ландшафтов менее интенсивно заселяют Субарк-
тику. Более того, даже характерные представители
южнотундровой фауны, так называемые гипоарктиче-
ские виды, незначительно проникают в более высокие
широты. Часто их распространение резко обрывается
у границ подзоны типичных тундр. Так,
полизональные обыкновенный бекас и горностай очень широко
распространены по Субарктике до ее северных
границ, тогда как характерные для лесотундры
азиатский бекас и полевка Миддендорфа обычны в южных
тундрах, но исчезают в типичных. Получается
парадоксальная ситуация: типичные обитатели
заполярных ландшафтов заселяют тундру менее интенсивно,
чем многие южные формы — явно вторичные вселен-
цы в Субарктику.
Это можно объяснить следующим образом.
Адаптация к зональным условиям — всегда определенная
специализация, ограничивающая возможность
обитания в других зонах. Она тем глубже, чем суровее
климат. Можно считать, что лесостепные или лесные
виды менее специализированы, чем пустынные,
тундровые, а тем более полярно-пустынные. Поэтому
виды, сформировавшиеся в умеренном климате,
заведомо будут иметь больше возможности к вселению в
тундровую зону, чем тундровые виды — в лесную.
Еще меньше способность к экспансии у типичных
обитателей полярных пустынь.
110
Все это справедливо не только по отношению к
разным зонам, но и к частям одной. Многие виды
умеренного пояса гораздо шире распространены по
лесной полосе, чем арктические по тундровой.
Значительное число видов хорошо приспособлено к обитанию
только в одной или двух смежных подзонах тундры.
Причина этого — резкие широтные смены условий на
территории Субарктики. Южные, типичные и
арктические тундры резко отличаются друг от друга
особенностями климата и ландшафта, что хорошо видно на
примере изменения среднеиюльских температур (см.
рис. 3).
Тундра — молодой ландшафт, но многие
заселяющие ее виды обладают чрезвычайно глубокими адап-
тациями к суровым заполярным условиям и
выделяются в самостоятельные роды, не имеющие близких
родичей. Таковы лапландский подорожник, полярная
сова, северный олень, новосиверсия ледяная. Это
свидетельствует о том, что они прошли длительный путь
развития в условиях тундровых или сходных с ними
ландшафтов. Современные представления о скорости
эволюции не позволяют предположить, что эти виды
сформировались после возникновения тундры как
природной зоны. Скорее всего они появились гораздо
раньше. Вновь парадокс: типичный тундровый вид
оказывается более древним, чем тундровый
ландшафт. Очевидно, вселенцы в тундровую зону где-то
преадаптировались к ее условиям, иными словами,
они должны были обитать в биотопах, сходных с
тундрой. В условиях более мягкого климата,
господствовавшего на Земле в дочетвертичное время, в
плиоцене, это могло быть только в высокогорьях. И
действительно, среди характерных обитателей тундры
много потомков горных животных и растений. При
этом тундровые популяции чаще всего имеют лишь
подвидовой ранг. Примерами могут служить кулик
глупая сивка (хрустан), тундряная куропатка,
лилейное растение ллойдия поздняя.
Здесь вновь наблюдается строгая специфичность
процессов эволюции, приспособления к среде у
разных групп организмов. Арктоальпийских видов
больше всего в типе цветковых растений и в классе
насекомых. Среди птиц их около шести видов, а среди
111
млекопитающих нет совсем. По этому поводу
С. С. Шварц (1963) писал: «...один из важнейших
путей формирования фауны тундры (за счет
альпийских элементов) для млекопитающих по каким-то
причинам оказался недоступным». Никто из горных
видов млекопитающих, распространенных на
территории Субарктики, не спустился с гор на равнины, не
стал типичным тундровым животным, не вошел в
настоящие тундровые зюнальные биоценозы. Все они
остаются горными. Речь идет о таких зверях, как
горные бараны, сурки, пищухи, красно-серая и
северосибирская полевки. Возможно, что здесь
определенную роль сыграло то, что тундровый ландшафт
освоило вообще весьма ограниченное число видов
млекопитающих. При этом существенное значение
имели общий уровень развития и особенности
поведения организмов. Для млекопитающих с их сложным
образом жизни и поведения высокогорья и тундра,
несмотря на определенное сходство, все же в своей
основе разные биотопы. Поэтому виды, глубоко
адаптированные к условиям альпийских поясов, не смогли
перейти в тундру.
В тундре обитают многие представители фауны
как будто совершенно не схожего с ней ландшафта —
степного. В одних случаях это те же виды, что и в
степи, например клоп Chiloxanthus pilosus, рогатый
жаворонок, узкочерепная полевка. Естественно, что
в высоких широтах они приобрели какие-то
особенности в биологии, морфологии, окраске, размерах.
Обычно их выделяют в особые подвиды. В других
случаях это самостоятельные виды, явные потомки
«степняков». Так, ближайшие родичи копытного
лемминга — типичные степные виды пеструшек (род
Lagurus). Выходцы из степей приспособились к
условиям тундры благодаря разным чертам биологии.
У полевок это подснежный образ жизни, у рогатого
жаворонка — комплекс приспособлений к обитанию в
открытом травянистом ландшафте, политрофность,
способность переходить с одного корма на другой.
Современные климаты тундры и степи резко
различаются, и родство видов, представляющих их
фауны, на первый взгляд кажется странным. Этот вопрос
имеет давнюю историю. Еще в прошлом веке ученые,
т
изучающие ископаемые остатки животных
плейстоцена — новейшей геологической эпохи Земли,
обнаружили своеобразное явление. В различных районах
Северной Евразии вместе находились остатки
животных, ареалы которых в настоящее время разобщены.
Одни из них обитают на территории современной
тундры, другие — в степных или полупустынных
районах. Встречались среди них и лесные виды. Эта
своеобразная ископаемая фауна была названа
«смешанной». Она включала таких, как сайгак, овцебык,
лемминг, степные пеструшки, северный олень, дикая
лошадь, песец и др. 1
Для объяснения ее происхождения высказывались
различные точки зрения. Одни ученые считали
причиной этого сезонные миграции млекопитающих,
другие— выровненный климат тех времен, в результате
чего животные имели вообще более обширные
ареалы, а с появлением резко выраженной зональности
современного типа «разошлись» по разным зонам.
В настоящее время наиболее обоснованной
признается точка зрения, высказанная еще в 1928 г.
известным орнитологом А. Я. Тугариновым, который
объяснял смешение арктических и степных видов
следующим образом: «...климатические и экологические
условия были таковы, что они в равной мере
удовлетворяли жизненным потребностям как северных, так
и южных обитателей».
Действительно, по данным палеоклиматологии, в
ледниковые периоды на больших пространствах
Северной Евразии господствовали сухие и холодные
воздушные массы. Под их влиянием формировались так
называемые перигляциальные (приледниковые)
ландшафты, в частности тундростепи, совмещающие
черты тундр и степей. Их климат был в целом холодным
(но с жарким летом). Недостаток влаги для
растений усиливался физиологической сухостью. В этих
своеобразных ландшафтах и формировались виды
одновременно холодостойкие и суховыносливые или
с разным сочетанием тех и других качеств (в
зависимости от конкретных условий биотопов, пищевых
1 Подробнее об этой проблеме см. в книге Э, А. Вангенгейм
(1977)
ИЗ
отношений). По мере эволюции послеледниковых
ландшафтов отдельные виды заселили настоящие
степи, некоторые — тундры или одновременно степи и
тундры и в дальнейшем преобразовались в
самостоятельные родственные степные и тундровые формы
(подвиды и виды).
Реальность этой картины подтверждается не
только палеонтологическими материалами. Ландшафты,
аналогичные тем, о которых говорилось, известны и в
настоящее время. На севере Якутии и крайнем
Северо-Востоке СССР, за полярным кругом у самых
границ тундры, а местами и в ее пределах встречаются
сообщества степного типа или такие, которые можно
назвать тундростепными. Они не занимают больших
площадей, а распределены на небольших участках,
где не могут жить крупные степные животные, но где
во множестве обитают типичные степные растения и
насекомые. Так, в растительном покрове встречаются
ковыль изменчивый (Stipa decipiens), овсяница
ленская (Festuca lenensis), пырей гребенчатый (Agropy-
ron cristatum), а в почве живут личинки типичных
степных жуков рода Eodorcadion из семейства усачей.
Не вполне ясно, в какой мере эти сообщества
можно считать реликтовыми, а в какой — современными.
Во всяком случае их существование поддерживается
высокой континентальностью климата, которая
усиливается местным климатом межгорных впадин и
долин. В этих районах, особенно на севере Якутии,
лето очень жаркое, температуры часто бывают около
30°, повышена сухость. Интересно, что небольшие
участки тундростепного типа встречаются даже на
острове Врангеля (Юрцев, 1974).
Приспособленными к тундровым условиям
оказались и некоторые исконно лесные виды. Естественно,
что все дендробионты, т. е. связанные тесно с
деревьями, в тундре отсутствуют. Некоторые типичные
лесные виды, такие, как росомаха, лось, эпизодически
заходят в тундру с юга. Но вот северный олень освоил
тундру гораздо шире. В настоящее время в диком
состоянии сохранились в основном его тундровые
популяции Между тем это безусловно исконно лесное
животное. Об этом свидетельствует и большая
площадь лесной части его ареала, и миграции на вре-
U4
IfSJ
^i
"4
li
Рис. 38. Самка комара-долгоножки
Tipula glaucocinerea, откладывающая
яйиа под лишайниково-моховую дернину
Рис. 39. Характерный представитель
насекомых арктических тундр —
комар-долгоножка Tipula carinifrons
(короткокрылая самка)
мя зимы из тундры в тайгу и лесотундру, и более
крупные размеры лесных форм, и место
происхождения.
Северные олени (род Rangifer) произошли в
начале четвертичного периода в лесных районах Северной
Америки, а затем широко расселились по бореаль-
ным ландшафтам. В Евразию они проникли,
вероятно, через Берингию, а также через Гренландию и
Исландию по льдам. Олень не связан
непосредственно с древесной растительностью. Он питается
преимущественно травами, лишайниками, кустарниками
и кустарничками. С точки зрения кормности летняя
тундра даже благоприятнее для оленя, чем тайга.
В освоении животными и растениями тундрового
ландшафта огромную роль играют свойства биотопа.
Так, почвенные животные менее зависимы от
климата, чем поверхностные формы. Климатические
условия в почве выравниваются, сглаживаются. Ее
обитатели могут совсем не подвергаться воздействиям
максимальных или минимальных температур воздуха.
Это одна из причин того, что почвенные животные
распространены очень широко. Например, среди кол-
лембол и клещей тундровой зоны большинство —
полизональные виды. Поверхностно обитающие
беспозвоночные, напротив, должны вырабатывать
различные специальные приспособления для обитания в
суровых условиях тундр (см. рис. 38, 39).
Условия обитания в болотах разных зон также
весьма стабильны, поэтому многие болотные
животные и растения беспрепятственно проникают в
тундру. Примером могут служить хорошо известные
жителям средней полосы калужница болотная,
селезеночник, сабельник болотный, обыкновенный бекас,
желтая трясогузка. Много ярких примеров
вторжения в пределы тундровой зоны среди обитателей пес-
чано-галечных побережий рек и морей.
Южные виды проникают в Субарктику по
хорошо прогреваемым склонам, особенно песчаным, на
которых развиты разнотравные сообщества.
Например, до подзоны типичных тундр встречается
теплолюбивый тимьян обыкновенный (Thymus serpyllutn).
Благодаря таким местообитаниям в тундровой зоне
процветают некоторые группы организмов, в целом
116
термофильные и солнцелюбивые. Примером может
служить семейство мух-журчалок, представленное в
субарктической фауне многими яркоокрашенными
видами, питающимися на цветках нектаром и пыльцой.
Таким образом, используя разнообразные интра-
зональные биотопы, животные и растения южного
происхождения, не имеющие специальных адаптации
к обитанию в арктических условиях, преодолевают
климатические рубежи и вселяются в тундровый
ландшафт. Это экологический путь приспособления к
среде, который заключается в выборе подходящих
биотопов и избегании неблагоприятных воздействий.
Более важны физиологические механизмы
адаптации. Очевидно, у организма есть два способа
приспособиться к суровым условиям: противостоять им
или подчиниться. Первый выражается, например, в
теплокровности птиц и млекопитающих, которые
могут сохранять внутреннюю постоянную температуру
(но для этого им необходим постоянный приток
энергии в виде пищи); второй — в холоднокровности
беспозвоночных, которые могут быть деятельными лишь
при достаточно высокой температуре (но зато в
покоящемся состоянии переносят любые
неблагоприятные условия). Первый способ можно назвать
активным путем приспособления к среде, второй
пассивным. Элементы того и другого свойственны самым
различным группам организмов, и часто у
беспозвоночных животных обнаруживаются признаки
активного типа адаптации, а у млекопитающих — пассивного.
Морфофизиологические адаптации активного
типа — это усиление общей жизнедеятельности и регу-
ляторных процессов, ускорение темпов роста и
развития. Такие адаптации в целом увеличивают
сопротивляемость неблагоприятным влияниям среды, но в
то же время сужают возможность существования в
резко пессимальных условиях, так как ставят вид в
зависимость от критического порогового значения
определенных факторов среды.
Пассивные эколого-физиологические
приспособления подчиняют жизнедеятельность организмов
окружающей среде, но в то же время повышают
резистентность их к неблагоприятным воздействиям и
способствуют более экономному использованию энер-
117
гии. Для многих животных, как теплокровных, так и
холоднокровных, это наиболее приемлемый путь
приспособления к суровым условиям Заполярья.
Работами С. С. Шварца и его сотрудников было
выявлено, что элементы активного пути адаптации
(более интенсивная работа сердца, повышенная пло-.
довитость, ускоренные рост и развитие) в условиях
Субарктики свойственны в основном широко
распространенным видам, пришельцам с юга. Но все они,
как правило, не доходят до северных границ тундры,
а осваивают лишь ее южные районы. Напротив,
типичным арктическим (так же как и
специализированным высокогорным) видам млекопитающих и птиц
свойственны пониженный метаболизм, невысокая
плодовитость, замедленные рост и развитие. Эти факты
опровергли давно уже высказываемые представления
о том, что для арктических позвоночных особенно
характерны черты биологии, обусловливающие
большую автономность организма (повышенный
метаболизм, ускоренное развитие и т. д.).
Сходные закономерности были выявлены и у
беспозвоночных (Чернов, 1975). Оказалось, что для
самых типичных высокоарктических видов характерно
замедленное, часто растянутое на несколько лет (до
5 лет) развитие, пониженная интенсивность питания
и роста, невысокая плодовитость, отсутствие четкой
сезонной локализации жизненного цикла (развитие
может завершиться за один сезон, но при
неблагоприятных условиях может растянуться на несколько
сезонов). Все эти черты способствуют увеличению
стойкости организмов к неблагоприятным погодным
условиям, большей энергетической экономичности
и т. д. Среди беспозвоночных тундры активный путь
освоения среды (повышенная плодовитость,
ускоренное развитие, быстрый рост) также свойствен в
основном южным видам или представителям в целом
южных групп. Таковы многие виды насекомых —
листоедов, долгоносиков, двукрылых и т. д.
Вследствие замедленных темпов развития многих
эварктов годовой прирост их популяций невелик.
В результате и продуктивность тундровых сообществ
в целом относительно низка. Этому на первый взгляд
противоречит довольно высокий уровень накопляемой
118
в них биомассы. Например, общий вес почвенных
беспозвоночных в арктических тундрах может быть в
2—3 раза выше, чем в южных, где условия гораздо
благоприятнее (Чернов, 1978). Однако противоречие
это кажущееся, поскольку к накоплению биомассы
почвенных обитателей приводят растянутость их
жизненных циклов и замедленный оборот поколений.
Происходит как бы концентрация биомассы при общей
низкой продуктивности, т. е. при очень небольшом
ежегодном приросте популяций.
С активными и пассивными путями
приспособления к арктическим условиям можно связать
особенности систематического состава животного мира.
Большинство благоденствующих в тундровой зоне
крупных групп животных находится на весьма низком
филогенетическом уровне и характеризуется
определенной примитивностью. Так, среди отрядов насекомых
самые процветающие в арктических ландшафтах — но-
гохвостки, или коллемболы (один из наиболее
примитивных форм класса). Из отряда двукрылых
особенно успешно освоили тундру древние примитивные
комары-долгоножки и комары-звонцы; из отряда
жуков — жужелицы и стафилины; из
перепончатокрылых—пилильщики. Из класса птиц в Арктике наиболее
разнообразны виды также весьма примитивных и
древних отрядов — гагар, гусеобразных, ржанкооб-
разных (кулики, чайки, чистиковые). Именно
представители таких групп осваивают самые
высокоширотные районы. Более прогрессивные отряды
представлены в арктической фауне, как правило,
единичными родами или видами, иногда очень хорошо
приспособленными к суровым условиям, но подчас не
достигающими самых высоких широт. Таковы,
например, среди насекомых различные высшие двукрылые,
единичные листоеды из жуков, шмели из
перепончатокрылых, несколько видов воробьиных птиц.
Эти особенности состава фауны Арктики,
вероятно, обусловлены прежде всего приспособительной
ценностью пассивного пути адаптации в суровых
условиях. Низкий уровень организации, примитивность и
связанная с ней способность вполне подчиняться
среде в пессимальных условиях оказываются наиболее
выгодными,
РАСПРОСТРАНЕНИЕ ЖИВОТНЫХ
И РАСТЕНИЙ
Область распространения вида, или его ареал,
формируется под влиянием климатических,
ландшафтных и биотопических условий, взаимоотношений с
ближайшими родичами, врагами, конкурентами, в
результате изменений среды в прошлом и т. д. В общем
ареал отражает историю вида (место возникновения,
пути расселения) и современные условия.
Исторические факторы часто проявляются в меридиональных
границах, а современные — преимущественно в
широтных, т. е. в характере заселения видами зон, подзон
и т. д. Естественно, что отделить исторические
факторы от современных часто невозможно. Так, обский
лемминг распространен от арктических районов
Северной Америки и Чукотского полуострова до
побережья Белого моря, а далее к западу, на Кольском
полуострове и в Скандинавии, замещается близким
видом — норвежским леммингом (Lemmus lemmus).
В данном случае граница между двумя видами
одного рода может быть обусловлена и местом
возникновения, и путями расселения, и антагонизмом по
отношению друг к другу, что в равной степени играло
роль в прошлом и играет в настоящем. Обский
лемминг — типичный тундровый вид, а на Кольском
полуострове и в Скандинавии развиты лишь лесотундра и
горные тундры, которые для него менее благоприятны
и к которым, очевидно, более приспособлен
норвежский лемминг.
Роль истории вида, места его возникновения и
путей расселения особенно хорошо видна в тех
случаях, когда ареал простирается на территории
нескольких зон и в то же время имеет отчетливые
меридиональные границы. Крупная нырковая утка черный
турпан — обычная птица в европейских тундрах.
В Скандинавии, европейской части СССР и в
Западной Сибири турпан заселяет тундровые,
лесотундровые, таежные, а местами и лесостепные районы,
120
Рис. 40. Ареал черного турпана
(«Птицы Советского Союза», т. 4)
но на востоке его ареал обрывается примерно по
линии от низовий Енисея до верховий Оби (см.
рис. 40).
В сибирских тундрах, от Югорского полуострова
до Чукотки, живет единственный вид дождевых
червей— Eisenia nordenskioldi. На юг он распространен
до Приморья, Монголии и, вероятно, до Китая,
заходит в Восточную Европу и на Кавказ. Однако на
территории Центральной и Западной Европы он
отсутствует, нет его и в тундрах Кольского полуострова и
в Скандинавии. Очевидно, этот вид расселялся на
запад широким фронтом, от тундр до степей. На
большей территории Сибири конкурентов у него не было,
и здесь он до сих пор единственный вид семейства.
Но в Европе обитает весьма богатая и разнообразная
121
Рис. 41. Ареал бесстебельной смолевки
(«Арктическая флора СССР», вып. VI)
фауна дождевых червей, что, возможно, и
затормозило расширение его ареала.
Хороший пример значения места возникновения и
путей расселения вида в формировании ареала дает
бесстебельная смолевка (Silene acaulis) —
представитель семейства гвоздичных. Это мелкое растение с
очень коротким стеблем в тундре образует дернинки,
которые, уплотняясь и нарастая в течение многих лет,
превращаются в подушки, сплошь покрывающиеся
ковриком ярких цветков (см. рис. 41). Бесстебельная
смолевка широко распространена в тундрах и
высокогорьях Америки и Европы, а в Азии — лишь на
северо-востоке (Чукотский полуостров, Камчатка,
Командорские острова). Ареал вида образует
незамкнутое кольцо с разрывом от Ямала до Колымы.
Ботаники считают, что это древний высокогорный вид,
возникший, вероятно, в горах Европы и расселившийся
в тундры: через Атлантику — в Америку, через
Америку — на северо-восток Азии. -
Виды, распространенные в арктических районах и
на высокогорьях, расположенных южнее, называют
арктоальпийскими (см. рис. 42). От настоящих аркто-
альпийских ареалов надо отличать случаи
распространения, при которых арктические виды проникают в
соседние с тундровой зоной высокогорья в силу
сходства современных условий. Многие типичные
тундровые виды заходят на сибирские хребты —
Верхоянский, Черского, Колымское нагорье, плато Путорана
122
Рис. 42. Типичное арктоальпийское распространение лилейного
растения (Lloydia serotina) (по А. И. Толмачеву, 1974)
и др. Так, обычнейшая во всех тундрах ива ползучая
(Salix reptans) встречается повсюду и в высокогорьях
северной Сибири. Типичный тундровый кулик — буро-
крылая ржанка расселена по сибирским хребтам до
Байкала и Амура. В горах, непосредственно
примыкающих к тундровой зоне, например в северных
отрогах Путорана, можно встретить практически любое
животное или растение тундры. В отличие от
типичного арктоальпийского такое распространение
обусловлено сходством современных условий в тундровых
и высокогорных ландшафтах. Близость этих
ландшафтов способствует постоянному взаимному обмену
видами.
В тундровой зоне Евразии много видов, ареалы
которых простираются от Забайкалья, Монголии и
Алтая до арктического побережья и часто идут на
запад до Урала или Печоры. Виды животных с
такими ареалами относят к так называемой ангарской,
или восточносибирской, фауне. Ее формирование
связано с древней Ангарской сушей, слабо
подвергавшейся оледенению (современные территории Алтая, Саян,
Забайкалья и др.). Эти территории в прошлом —
важнейшие центры образования видов многих групп
животных и, очевидно, растений. В частности, ангар-
123
екая фауна была основным источником современной
таежной фауны.
Многие виды, не связанные тесно с лесом,
проникли и в тундры Сибири. Лишь единичные из них
расселились за Урал. На Таймыре встречаются виды,
распространенные до Забайкалья и Монголии, но
отсутствующие в Европе. Таковы листовой долгоносик
Phytonomus ornatus, личинки которого развиваются в
плодах и цветках бобовых; единственный в высоких
тундрах вид многоножек — костянка Monotarsobius
alticus, пилильщик Pontania crassipes, образующий
на листьях ив крупные галлы, и др. Эти примеры
показывают, что животный и растительный мир
Арктики формировался в тесной связи с процессами
генезиса фаун и флор на огромных территориях,
охватывающих несколько современных климатических
поясов.
В тундровой зоне много видов, ареалы которых
сравнительно невелики, но ориентированы в
меридиональном направлении и пересекают зональные
границы. Иными словами, они в первую очередь
свойственны какому-либо району, но не типу ландшафта.
Видов с такими ареалами особенно много на северо-
востоке Азии, где сильно пересеченная местность,
наличие горных хребтов и крупных рек способствуют
изоляции видов. Примерами могут служить песчанка
чукотская (Arenaria tschuktschorum), распространенная
от Чаунской губы до верховьев Колымы и Анадыря;
даурский прострел (Pulsatilla dahurica),
встречающийся от тундр Восточной Сибири между Леной и
Анадырем до Амура; кулик американский бекасовид-
ный веретенник (Limnidromus griseus), обитающий от
Чукотки и тундр Аляски до юга таежной зоны в
Америке. Это примеры типичного эндемизма, т. е.
обитания в каком-либо строго определенном районе. Такие
ареалы могут отражать место возникновения вида
или представлять собой остаток некогда более
обширного распространения.
Несколько иные случаи эндемизма — когда вид
заселяет какой-либо район, целиком помещающийся в
пределах тундровой зоны. Здесь совмещаются два
явления — приуроченность к району и связь с
определенным типом ландшафта, в данном случае тундро-
124
Рис. 43. Ареал одного из самых типичных тундровых растений —
Draba glacialis («Арктическая флора СССР», вып. VII)
вым. В пределах тундровой зоны встречаются виды с
очень небольшими ареалами. Представитель
крестоцветных— крупка Чельмана (Draba kjellmanii)
известна только на Вайгаче и Новой Земле. Вид этого же
семейства — сердечник Cardamine hyperborea
встречается в тундрах на побережье между Чаунской
губой и Чукотским полуостровом и на острове
Врангеля. Мак Ушакова (Papaver uschakovi) найден на
острове Врангеля и Чукотском полуострове. Кулик-лопа-
тень (Eurynorhynchus pygmeus) обитает на
северо-восточном побережье Чукотского полуострова, а желто-
зобик (Tryngites subruficollis) также и в тундрах
Аляски.
Однако в тундровой зоне таких видов
относительно немного. Чаще ареалы охватывают более или
менее значительные отрезки зоны. Один из самых
характерных видов арктической флоры — крупка ледяная
распространена от полуострова Канина до
Новосибирских островов и отсутствует во всех прочих
районах тундровой зоны (см. рис. 43). Очковая гага
(Somateria fischeri) гнездится на побережье
Северного Ледовитого океана, от Оленека до Аляски.
Многие виды животных и растений
распространены по всей тундровой зоне Евразии и Америки. Такое
125
Рис. 44. Ареал крупки Draba ^ubcapitata — кругополярного
высокоарктического вида (по А. И. Толмачеву, 1974)
распространение называют циркумполярным или кру-
гополярным (см. рис. 44, 45). Типичное
циркумполярное распространение у песца, белой совы,
пуночки, гаги-гребенушки, черной казарки, длиннохвостого
поморника и т. д. Около 40% видов арктических
цветковых имеют циркумполярные ареалы Виды с
подобными (круговыми) ареалами есть и в тайге
(росомаха, белка, лось), но там их гораздо меньше, чем в
тундре. Среди типичных степняков лишь единицы
обитают в обоих полушариях. В тропиках мало даже
родов, общих разным материкам. Одна из причин
этого — история формирования современных материков.
По широко известной теории А. Вегенера, они образо-
126
Рис. 45. Ареал кулика тулеса — характерного обитателя
зональных арктических и типичных тундр
(«Птицы Советского Союза», т. 3)
вались в результате расхождения единого древнего
материка. Оно началось с южных частей (Австралия,
Южная Америка, Южная Африка). Азия и Америка
были соединены еще совсем недавно в районе
Берингова пролива. В кайнозое они соединялись и
разъединялись несколько раз, и через Берингию происходил
обмен между фаунами и флорами обоих материков.
Но тут дело не только в возможностях обмена
видами. В тундровой зоне вообще относительно высок
процент видов с широкими зональными ареалами
независимо от того, переходят они с одного материка на
другой или нет. Причины этого еще недостаточно вы-
127
яснены. Расселению по территории тундровой зоны,
формированию циркумполярных ареалов, бесспорно,
способствует сравнительно небольшая протяженность,
т. е. компактность, сближенность (вследствие
шарообразности Земли) арктических районов. Так,
протяженность параллели 70° почти в 3 раза меньше
протяженности экватора. Это важное обстоятельство
часто забывается при биогеографических построениях.
Большое значение для зонального
распространения имеет бедность, невысокое видовое разнообразие
фауны и флоры заполярных районов. Это ослабляет
конкуренцию между разными видами и тем самым
способствует расселению в пределах зоны. Оно может
быть тем успешнее и быстрее, чем меньше видов со
сходной биологией, занимающих одни и те же места
обитания (экологические ниши). Действительно,
наиболее типичное циркумполярное распространение
свойственно видам, не имеющим близких
родственников и образующим отдельные самостоятельные
монотипические (включающие единственный вид) роды.
Таковы песец, белая сова, лапландский подорожник,
пуночка, морянка.
Естественно, что хорошо обособленный вид,
отличающийся определенными приспособительными
особенностями, не будет испытывать конкуренции со
стороны других видов в процессе расселения по
тундровой зоне. Но широкое зональное распространение
таких видов можно объяснить и тем, что это, бесспорно,
древние обитатели тундрового ландшафта,
прошедшие вместе с ним длительную историю
Некоторые роды, включающие вообще несколько
видов, в тундровой зоне представлены единичными
видами, которые также распространены весьма
широко, иногда циркумполярно (полярный лебедь,
мохноногий канюк, грушанка крупноцветная — Pyrola gran-
di flora).
Чем больше родственных видов обитает в
тундровой зоне, тем разнообразнее и дробнее их ареалы.
В этих случаях часто наблюдается замещение одного
вида другим. Это явление называют викариировани-
ем. В одних случаях виды замещаются в разных
подзонах, в других — в разных меридиональных отрезках
зоны. Часто викариирование бывает неполным, т. е.
128
Рис. 46. Распространение четырех видов куликов-песочников
в Евразии:
1 __ песчанка; 2 — кулик-воробей; 3 — острохвостый песочник; 4 — бер-
дов песочник
ареал одного вида перекрывает часть ареала другого.
Но в целом бесспорна общая закономерность: чем
больше видов рода, семейства обитает в одних и тех
же ландшафтах, тем разнообразнее ареалы.
Яркий пример: один из самых характерных родов
арктической фауны — кулики-песочники (Calidris).
К этому роду относится, по данным разных авторов,
14—17 видов. И все они распространены
преимущественно в заполярных районах: в тундровой зоне,
частично в лесотундре, на севере тайги и в приарктиче-
ских высокогорьях. Из них только один вид —
чернозобик (Calidris alpina)—имеет циркумполярный
ареал. У остальных ареалы крайне разнообразны, от
занимающих значительные отрезки тундровой зоны до
узкоэндемичных (см. рис. 46). Это отражает сложную
историю рода — процессы видообразования в нем,
дивергенцию признаков, захват различных
экологических ниш, расселение в разных направлениях и т. д.
Большинство видов этого рода обитает по обе
стороны Берингова пролива — на Аляске, на
северо-востоке Азии (см. табл. 8). На относительно небольшой
территории дальневосточной Субарктики (включая
остров Врангеля, Чукотский подуостров и тундры к
б Ю. И. Чернов
129
западу до Колымы) встречается 13—14 видов этого
рода, на Таймыре — 9, а в западносибирских и во*
сточноевропейских тундрах (Ямал, Югорский
полуостров, Новая Земля) —только 4 вида. Бесспорно, что
Северо-Восток Азии входил в область наиболее
интенсивного видообразования, а возможно, и
формирования этого рода. Отсюда шло расселение: на
восток— по Американскому континенту, на запад — по
Евразии. Там, где достаточно много видов песочников,
Таблица в
Распределение куликов-песочников в Евразии
Вид
Краснозобик
Чернозобик
Кулик-воробей .
Белохвостый песочник
Песочник-красношейка
Длиннопалый песочник
Бердов песочник
Кулик-дутыш
Острохвостый песочник
Исландский песочник
Большой песочник
Морской песочник
Берингийский песочник
Перепончатопалый песочник
Песчанка
Всего видов
Новая Земля
и Югорский
п-ов
+
+
+
—
—
—
—
—
—
+
—
—
4
Таймыр
+
+
+
+
+
—
—
+
—
+
+
—
—
+
9
Чукотский
п-ов
-г-
+¦
-г-
+
-t-
4-
+¦
-f-
¦f-
-h
-f-
—
+
+
13
П римечание. Плюс означает, что вид гнездится в данном
районе; минус—не гнездится.
они живут в разных биотопах или в разных подзонах.
На Таймыре всего девять видов, но в каждой подзоне
лишь три — шесть. В распространении видов этого
рода отчетливо проявляются все формы
территориального разграничения (меридионального, широтного,
биотопического). При этом особенно отчетлива роль
исторического фактора — места формирования и
направления расселения видов.
130
Меньше закономерности и больше пестроты в
распространении и соотношении видовых ареалов
наблюдается в том случае, если взять не один род, а
группу высшего таксономического ранга. Так, из
подсемейства гусиных в тундровой зоне Евразии обитает
десять видов, относящихся к пяти родам: три вида
рода гусей — белолобый гусь (Anser albifrons),
гуменник (A fabalis) и пискулька (A. erythropus)\ три
вида рода казарок — черная (Branta bernicla),
белощекая (В. leucopsis) и канадская (В. canadensis);
два монотипичных рода — белый гусь (Chen caerules-
cens) и белошей (Philacte canadica). Картина их
распространения в тундровой зоне чрезвычайно пестра.
Три вида Anser обитают на всем протяжении
тундровой зоны Евразии, и их ареалы в общем
перекрываются. Но у каждого из них имеются особенности в
зональном распределении. Белолобый гусь встречается
от южных границ тундры до северных. Гуменник
населяет не только всю тундровую зону, но и
лесотундровые и таежные районы. Пискулька распространена
в основном в южных тундрах и лесотундре. Черная
казарка — птица высокоарктических побережий.
У прочих видов ареалы строго региональные.
Белощекая казарка населяет арктическое побережье
Атлантики, канадская — заполярные и отчасти
умеренные широты Северной Америки. Краснозобая
казарка — эндемик Таймыра и Гыдана, белошей — эндемик
Чукотки и западной Аляски, белый гусь
распространен в прибрежных тундрах от острова Врангеля до
Баффиновой земли. В этой группе гораздо меньше
взаимной зависимости распространения видов.
Особенно отчетливо викариируют виды со сходной
биологией. Яркие примеры — виды купальниц (см.
рис. 47) и два вида куликов-ржанок — бурокрылая и
золотистая — обитателей сухих плакорных тундр.
Одно из интереснейших биогеографических
явлений в фауне и флоре Арктики — разрывы в ареалах
или дробление их на отдельные участки. Последнее
очень характерно для обитателей наиболее высоких
широт—полярных пустынь и арктических тундр.
В этих случаях разобщенность частей ареала
объясняется чаще всего тем, что полярные пустыни и
арктические тундры представлены фрагментами на островах
б*
131
Рис. 47. Распространение видов купальниц на севере Евразии
(«Арктическая флора СССР», вып. VI):
1 — Trollius europaeus; 2 — Т. apertus; 3 — Т. asiattcus; 4 — Т boreosibi-
ricus; 5—7. membranostylis
и полуостровах арктического бассейна. Таково
распространение исландского песочника, песчанки,
морского песочника. Аналогична фрагментарность
распространения новосиверсии ледяной — одного из
характерных высокоширотных растений. В данном
случае причина прерывистого распространения —
прерывистость самого ландшафта, в котором обитает вид.
Как уже не раз говорилось, характерный элемент
фауны и флоры Заполярья — прибрежные приморские
виды. Это различные чайки, чистиковые, гаги,
казарки, разнообразные насекомые и растения. Некоторые
из них распространены очень широко в
субарктических и собственно арктических ландшафтах, но имеют
разрыв на побережье морей Карского и Лаптевых,
чаще всего на Таймыре. Такой разрыв в ареале имеют,
например, обыкновенная гага, несколько видов
прибрежных мух, развивающихся в выброшенных на
берег водорослях. Нет оснований считать, что это
обусловлено историческими причинами,
незаконченностью расселения. В этих районах береговая линия
материка заходит в очень высокие широты, на
Таймыре— до 77° с. ш. Омывающие его моря очень
холодные и ледовитые. Их литораль практически
безжизненна. Нет бурых водорослей, столь обильных в дру-
132
гих частях арктического побережья, на мелководьях
отсутствуют массовые поселения мидий и других
беспозвоночных. Иными словами, нет пищи ни для гаги,
питающейся беспозвоночными в прибрежных водах, ни
для мух, личинки которых развиваются в гниющих
водорослях.
Один из итогов изучения ареалов (их величины,
формы, взаиморасположения, истории
формирования)— зоо- и фитогеографическое районирование
Земли. Его основная задача — отразить процессы
формирования животного и растительного мира, т. е.
выделить такие территории, на которых
видообразование происходило более или менее изолированно.
Самые крупные категории такого районирования —
области, например, Австралийская, Неотропическая
(Южная Америка), Эфиопская (Африка без северной
части). Зоогеографическое и фитогеографическое
районирование в значительной степени совпадают,
поэтому часто говорят о едином биогеографическом
районировании.
Арктические территории зоологи и ботаники
относят к самой крупной области — Голарктике,
включающей Северную Америку, Европу, северную часть
Африки и Азию, кроме ее южных районов (Индии,
Индокитая). На всей этой огромной территории
животный и растительный мир развивался в течение
длительных геологических периодов более или менее
самостоятельно и изолированно от других областей.
Между Северной Америкой и Азией проходили обмены
видами через древнюю Берингию, а частично и
другими путями, например через Атлантику. В зоогеогра-
фических схемах районирования Голарктика часто
делится на две самостоятельные области — Палеарк-
тику (Евразия) и Неарктику (Сев. Америка). Но и в
этом случае понятие Голарктика сохраняется. Иногда
ее называют царством и делят на области,
подобласти, провинции.
Большинство зоологов и ботаников относят все
арктические территории к единой Арктической
подобласти (или области) Голарктики. Для этого как будто
есть все основания. Действительно, множество видов
и родов животных и растений распространено на
территории тундровой зоны в обоих полушариях, что под-
ш
черкивает флористическое и фаунистическое единство
заполярных районов. Однако при более внимательном
рассмотрении этот вопрос оказывается не таким
простым. Выше были приведены примеры, когда виды
обитают в относительно небольших отрезках
тундровой и более южных зон. Таких видов особенно много
на северо-востоке Азии. Их ареалы часто
захватывают и северозападную оконечность Америки. Данные
территории были центром формирования и
расселения видов ряда групп растений и животных. В связи
с этим выделяют самостоятельный биогеографический
регион — Берингию, объединяющую северо-восток
Азии, Аляску и прилежащие районы, включая как
тундровые, так и более южные таежные районы (Юр-
цев, 1974).
Среди животных и растений тундр многие виды
имеют западные границы на Таймыре, в низовьях
Енисея, который считается одной из важнейших
биогеографических границ. Некоторые из таких видов —
типичные тундровые, иные обитают также и в более
южных таежных и горных районах Сибири. Выше
приведено несколько примеров такого распространения.
В связи с этим выделяют самостоятельную Восточно-
Сибирскую (Ангарскую) подобласть Палеарктики,
объединяющую как тундровые, так и таежные
районы. Такого мнения придерживался, например,
известный зоогеограф И. И. Пузанов.
Выделение таких категорий, как Берингия и
Ангарская подобласть, осложняет представление о
флористическом и фаунистическом единстве всех
арктических территорий. В настоящее время еще
недостаточно разработан вопрос о критериях установления
ранга биогеографических категорий. Не ясно, каков
должен быть процент эндемиков, характерных видов
или иных таксонов для того, чтобы придать той или
иной территории ранг области, подобласти или
провинции. Так, А. Л. Тахтаджян (1978) в отличие от
большинства ботаников относит арктические районы
всего лишь к провинции Голарктики. Столь
противоречивые мнения обусловлены не недостатком
фактических данных о распространении растений и
животных, а нечеткостью основных принципов
районирования. В частности, пока еще не ясно, как соотносятся
134
регионы, выделенные на основании структуры
растительного покрова, сообществ в целом и генезиса
фауны и флоры. Изучение в этом плане тундровой зоны
бесспорно даст много ценного для разработки общих
принципов биогеографического районирования.
Выше было рассмотрено распространение
животных и растений Заполярья в основном с точки зрения
целостных ареалов. При этом основной упор сделан
на те случаи, когда распространение вида отражает
его историю, исконные географические связи. Другой
аспект биогеографии — анализ распределения
организмов того или иного вида в пределах ареала в
зависимости от современных условий, которые
выступают в виде климатических и высотных поясов,
природных зон, ландшафтов, биотопов, разных форм
рельефа.
Арктическими называют виды с крайне
различными ареалами и зональным распределением. Так,
обский лемминг заселяет всю тундровую зону и
большую часть полярных пустынь, а полевка Мидден-
дорфа обитает в основном в южных тундрах и
лесотундре. Маленький злак фиппсия холодная (Phippsia
algida) весьма широко распространен в арктических
районах, а местами уходит за их пределы, проникая
в лежащие южнее высокогорья. Он играет большую
роль в растительном покрове полярных пустынь,
обилен в арктических тундрах. В типичных тундрах
растет в самых холодных местах, например у снежников.
Этот вид — характерный обитатель высокой Арктики.
Другой злак — альпийский лисохвост (Alopecurus
alpinus) в южных районах тундровой зоны растет на
северных склонах, по участкам с подтоком холодных
вод, в ложбинах с залежами снега. В арктической
тундре обилен повсеместно, образует сплошные
заросли, напоминающие посевы. Кулик-воробей —
характернейшая массовая птица в типичных тундрах,
исчезает или малочислен в арктических и южных.
Итак, многие животные и растения приурочены
к отдельным широтным полосам тундры — подзонам
или даже частям их. Поэтому общее понятие
«арктический вид» следует конкретизировать, разделить на
несколько более частных. Можно выделить по край-
135
ней мере четыре варианта зонального распределения,
используя приставки из греческого языка: гипер
(сверх), эу (настоящий), геми (полу), гипо (под),—
гипераркты, эуаркты, гемиаркты, гипоаркты. Первый
включает характерных обитателей полярных пустынь,
три других отражают приуроченность к подзонам
тундр. Эти категории зонального распределения
наиболее четко проявляются среди млекопитающих, птиц,
высших насекомых, в меньшей мере среди цветковых
растений, лишь на единичных видах прослеживаются
у низших растений, совсем не выражены в ряде групп
микроскопических беспозвоночных.
Гипераркты, или высокоарктические виды,
распространены преимущественно на островах полярного
бассейна в зоне полярных пустынь и в подзоне
арктических тундр. Высокоарктические позвоночные
животные в большинстве случаев тесно связаны с морем:
белый медведь, обычнейшие птицы арктических
птичьих базаров — люрик (Alle alle), белая чайка (Pago-
phila eburnea). К этой группе можно отнести и
черную казарку, куликов — исландского и морского
песочников, хотя местами их ареал опускается в более
низкие широты. Среди растений можно назвать
несколько видов, явно тяготеющих к самым высоким
широтам, однако типичных представителей гиперарк-
тов, аналогичных, например, белой чайке, среди них,
вероятно, нет. Примером растений, наиболее
тяготеющих к полярным пустыням, могут быть злак фиппсия
холодная, крупка головчатая (см. рис. 44).
Эуаркты (эваркты), или собственно арктические
виды, обитают в северной полосе тундровой зоны, в
подзоне арктических и частично типичных тундр.
Некоторые из них распространены по всей тундровой
зоне до ее южных границ, но их экологический
оптимум явно в арктических и в северных типичных
тундрах. Характерные представители — пуночка, красно-
зобик, белая сова, копытный и обский лемминги,
комар-долгоножка Tipula carinifrons (см. рис. 39). Из
растений — лютик Сабина (Ranunculus sabinii),
альпийская крупка (Draba alpina) и звездчатка Эдвардса
(Stellaria edwardsii), несколько видов камнеломок,
например дернистая (Saxifraga caespitosa). Эти виды
воплощают наиболее характерные черты арктической
136
фауны и флоры. Обычно у них имеются
определенные адаптации к жизни в столь суровых условиях
(например, уменьшенные размеры тела, многолетнее
развитие, приспособление к обитанию на обнаженных
каменистых грунтах, бескрылость и т. д.).
Гемиаркты, или типичные тундровые виды,—
характерные обитатели подзоны типичных тундр. Они
обычно не доходят до северных границ зоны и
малочисленны в южных. По высокогорьям часто
проникают далеко на юг. К этой группе относятся многие
широко известные обитатели тундр, например
чернозобик, кулик-воробей, шмель Bombus lapponicus, ива
ползучая, дриада точечная, Кассиопея четырехгранная.
Гипоаркты — обитатели южнотундровых,
лесотундровых и отчасти северотаежных ландшафтов. Их
ареалы часто тянутся узкой полосой по югу Субарк-
тики, иногда простираются далеко на юг в таежную
зону и особенно в горные районы. Типичные
представители— полевка Миддендорфа, белая куропатка,
кулики — малый веретеник и щеголь, гусь пискулька,
овсянка-крошка, самый крупный тундровый паук —
полярный тарантул (Lycosa hirta), самый крупный1
тундровый жук — жужелица Carabus odoratus;
карликовая березка (ерник), водяника, морошка. В
лесной полосе обычно обитают в интразональных
биотопах (на болотах, хребтах, гривах, холмах). К северу
от южной тундры численность гипоарктов резко
снижается. В средней части подзоны типичных тундр их
очень мало, а в арктических нет вообще.
Не все виды могут быть отнесены к этим группам.
Так, у большинства низших растений, особенно
микроскопических, у многих видов цветковых, почвенных
животных, насекомых и даже у некоторых птиц и
млекопитающих нет сколько-нибудь заметных
зональных привязанностей. Часто виды распространены во
многих зонах, например на территории всей Голарк-
тики или Палеарктики, а иногда населяют почти весь
земной шар. Такие виды называют транспалеарктами,
трансголарктами, космополитами, а когда хотят
подчеркнуть отношение к зонам — полизональными,
реже—мультизональными. Типичные примеры
полизональных видов в тундровой зоне — горностай,
семиточечная божья коровка, болотная калужница, селезе-
137
«очник. В тундре встречаются и типичные
космополиты, такие, как сокол-сапсан, различные мелкие
почвенные членистоногие, например панцирный клещ
Liochthonius sellnicki, ногохвостка Ceratophysella
armata и др.
Обитание на территориях нескольких зон должно
свидетельствовать о безразличии к смене условий, т. е.
о широком экологическом диапазоне вида. Это в
полной мере относится к таким видам, как горностай и
семиточечная божья коровка. Они распространены на
огромной территории почти всего северного
полушария и населяют самые разнообразные биотопы —
пустыни, степи, леса, тундры, альпийские луга и т. д.
Хотя они и предпочитают некоторые биотопы
(например, горностай чаще селится по берегам водоемов),
но могут жить в самых разнообразных местах.
Однако многие полизональные виды на
протяжении своего ареала выбирают строго определенные
биотопы. Таковы различные прибрежные виды, селящиеся
в тундровой и в иных зонах по берегам рек, озер, по
окраинам болот, на галечных косах, на песчаных
дюнах и т. д. Калужница и селезеночник встречаются на
территории почти всей нашей страны по берегам
ручьев, на болотах, по сырым лугам и т. д.
Жужелица тинник Elaphrus riparius обитает на песчаных
и галечных косах рек от юга лесной полосы до тундр.
Огромна меридиональная протяженность ареалов
нескольких видов мух, личинки которых развиваются в
выброшенных на морской берег водорослях. Такова
муха Fucomyia frigida, распространенная от
тропических широт до севера Гренландии, Новой Земли и
Чукотки. Крупный злак колосняк мягкий (Leytnus
mollis)—типичный обитатель приморских песчаных
дюн, по которым распространен от тропиков до
Чукотского полуострова.
Обыкновенная каменка — птица семейства
дроздовых распространена от Африки до подзоны
арктических тундр, но нигде на этом огромном пространстве
не гнездится в настоящих зональных сообществах.
Гнездо она устраивает под обрывами, среди скал,
под камнями, в норах, по берегам рек, на свалках,
в поселках и т. д. В южной половине тундровой зоны
обычна желтая трясогузка, которая распространена
138
на всей территории Азии и повсеместно связана с
болотами, заболоченными лугами и кустарниковыми
зарослями.
Таким образом, на протяжении огромного ареала
все эти виды связаны с интразональными частями
ландшафта. Вместе со своими биотопами они
распространены очень неравномерно, иногда отсутствуют на
больших территориях, а затем вновь появляются, но
в целом очень широко, независимо от зональных
границ.
Условия на приморских песчаных дюнах, на
болотах, в прибрежной полосе у уреза воды или на
скальных субстратах весьма специфичны и требуют
совершенно определенных и глубоких адаптации. И если
вид приспособился к этим условиям, то прочие
факторы для него играют меньшую роль. Кроме того,
в таких биотопах как бы сглаживаются
климатические колебания. Например, независимо от того, где
находится побережье реки — в тундре или пустыне, на
нем всегда достаточно влажно, и в то же время
близость воды в сильную жару способствует охлаждению,
а в холод — согреванию, поскольку водные массы
медленно остывают. Вследствие однообразия
растительности и животного населения весьма стабильны и
биотические отношения.
Это, конечно, не значит, что все обитатели таких
биотопов распространены независимо от зональных
границ. Речь идет лишь о том, что они меньше
зависят от смены климатических факторов, чем животные
и растения плакоров, зональных сообществ.
Множество характерных обитателей болот, побережий,
скальных субстратов живет только в одной зоне. Часто
среди форм, связанных с интразональными
биотопами, встречаются типичные эндемики,
распространенные в небольшом районе одной зоны. Но каждый
такой, случай можно объяснить какими-либо
особенностями истории вида, биологии, взаимоотношений с
другими видами и т. д.
Большую роль в распространении животных и
растений играют экстразональные сообщества, т. е.
находящиеся за пределами «своей» зоны или подзоны:
островки леса в южной тундре, кустарники в
типичной и т. д. В южной подзоне тундр обычны многие
139
характерные обитатели лесотундровых, таежных или
вообще лесных ландшафтов. Здесь они, как правило,
связаны с зарослями кустарников или
изолированными участками леса по склонам холмов, в долинах рек,
ручьев. Таковы два вида купальниц — европейская и
азиатская, овсянка-крошка, пеночка-таловка, пеночка-
весничка, различные виды насекомых. Местами
заросли высоких кустарников и даже леса проникают
очень далеко на север. Так, в Большеземельской
тундре встречаются острова ивовых лесов, а в
центральной части Таймыра, вблизи озера Таймыр,—
ивняки высотой в рост человека. В таких биотопах
обитает много южных животных и растений, не
свойственных зональным тундровым сообществам.
Описанные здесь явления — это по существу
разные пути обогащения бедного тундрового
органического мира южными формами. Бесспорно, что те виды,
которые широко распространены не только в тундрах,
но и в более южных зонах,— это, как правило,
вторичные вселенцы в молодые субарктические
ландшафты.
ВЗАИМООТНОШЕНИЯ ОРГАНИЗМОВ
Бесконечно разнообразные отношения между
организмами обычно объединяют в несколько основных
типов: конкуренцию, симбиоз, комменсализм, или на-
хлебничество, поедание одного другим, паразитизм
и др. Главную роль, бесспорно, играют пищевые
связи, в процессе которых организм получает
необходимую энергию для поддержания жизни. Зеленые
растения (автотрофы) синтезируют основную массу
органического вещества. В тундре это водоросли (в том
числе и в лишайниках), мхи и цветковые растения.
Органическое вещество, образующееся в растительном
покрове, называется первичной продукцией. Обычно
она рассчитывается в единицах веса за год.
Первичная продукция используется как самим растением, так
и различными гетеротрофными организмами, которые
образуют вторичную продукцию. Гетеротрофы делятся
на сапрофитных (грибы, некоторые бактерии),
использующих продукты разложения сложных органических
веществ, и голозойных (животные), нуждающихся в
наборе сложных органических веществ. Голозойные
в свою очередь делятся на сапрофагов (потребителей
отмерших растительных частей), фитофагов
(растительноядных), хищников, некрофагов (поедающих
трупы других животных). Все это составляет предмет
продукционной, или энергетической, экологии,
принципы которой еще очень мало использовались в
исследованиях тундровых сообществ.
Животные и микроскопические
растения
Животных, питающихся микроскопическими
растениями, называют микрофитофагами, или микробофагами.
Пищевые связи их, особенно в заполярных условиях,
изучены очень слабо. Самое трудное и важное для
экологов — отделить потребителей автотрофных
141
микроскопических растений (водорослей) от
потребителей гетеротрофных организмов (бактерий, грибов,
простейших).
Микроскопические водоросли занимают видное
место в сообществах тундры. Вследствие постоянно
высокой влажности почвы они находят здесь весьма
благоприятные условия. Будучи характерными для
первичных примитивных почв, водоросли развиваются
в огромном количестве на оголенных грунтах
пятнистых и арктических тундр, а также полярных пустынь.
В этих биотопах они образуют основную часть
первичной продукции. Почба от их обилия часто
прокрашивается в зеленый цвет на несколько сантиметров.
В этих же грунтах в большом количестве живут
различные животные, питающиеся водорослями (альго-
фаги): нематоды, энхитреиды, коллемболы, личинки
комаров-звонцов.
Личинки звонцов — это хорошо известный
рыболовам и любителям аквариумных рыб «мотыль».
У большинства видов звонцов личинки живут в воде
и имеют ярко-красную окраску. Но есть и сухопутные
виды, не столь ярко окрашенные, чаще зеленоватые
или светло-серые. Они особенно многочисленны в
арктических почвах (до нескольких сот на квадратном
метре), где питаются микроскопическими
водорослями, так же как и водные виды.
Почвенные нематоды — это огромный
разнообразный мир. Кроме типичных микрофитофагов,
питающихся водорослями и, вероятно, бактериями, среди
них есть хищники, паразиты животных и растений,
виды, поедающие грибной мицелий (микофаги), а
также разнообразную пищу (политрофы). Специалисты
различают характер питания того или иного вида
нематод по строению ротового аппарата, органов
пищеварения, форме тела и содержимому кишечника.
В тундрах преобладают микрофитофаги. На их долю
приходится около половины, часто до 70%, видов и
особей. Много также политрофов. Лишь в тундровых
лугах микрофитофагов сравнительно мало. Там
преобладают паразиты высших растений.
Численность нематод в тундрах от 1 до
5 млн. экз/м2, а в субарктических нивальных лугах —
около 10 млн. На возможных потребителей микроор-
142
ганизмов — водорослей и бактерий в тундрах и лугах:
приходится примерно 2 млн. экз/м2, что составляет 1,5 г
сырой массы, или 0,5 г сухой. Можно рассчитать
примерные величины выедания нематодами
микроскопических растений. Для этого надо знать не только
величину массы этих организмов на данный момент, но
и их продукцию, г. е. превышение массы в единицу
времени. По данным О. М. Паринкиной (1973),
продукцию бактерий в пятнистых тундрах можно оценить
2 кг/м2 в год (сырая масса). Для нематод
величина годовой продукции порядка 45 г/м2. Допустим,
что, питаясь бактериями, нематоды усваивают 70%
съеденной массы, 90% усвоенной пищи тратится на
поддержание их жизни и лишь 10% идет на прирост.
Из этого следует, что нематоды выедают примерно
32% годовой продукции бактериальной массы. Пока
нет данных о запасах и приросте массы водорослей
в тундрах. Неизвестно также, в какой мере они
уничтожаются нематодами. В тех случаях, когда
нематоды питаются в основном водорослями, они выедают,
вероятно, не менее половины продукции этих
микроскопических растений.
Кроме нематод водорослями питаются и более
крупные животные, в особенности кольчатые черви эн-
хитреиды, размер которых 0,5—3 см. Скорее всего,
они заглатывают водорослевые клетки вместе с
детритом, остатками растений. Теснее связаны с
водорослями водные энхитреиды, живущие в болотах. Их
кишечники часто бывают сплошь набиты зелеными и
сине-зелеными водорослями. Водорослями питаются
также панцирные клещи, коллемболы,
разнообразные личинки двукрылых — мух и комаров, из которых
выше уже упоминались личинки звонцов.
Еще одна группа микрофитофагов, совершенно не
исследованная в тундровой зоне,— это простейшие.
Известно лишь, что они в большом количестве живут
в тундровых почвах. Их можно увидеть в микробных
«обрастаниях» на специальных микробиологических
стеклянных пластинках. Но каков их образ жизни в
этих условиях и чем они питаются — неизвестно.
Можно предполагать, что комплекс водорослей с их
потребителями играет огромную роль в жизни тундры.
Животных, которые питаются водорослями, поедают
143.
хищные насекомые, пауки, клещи, а также птицы, в
особенности кулики, а последних — хищные птицы и
млекопитающие, например песец, горностай. Так
выглядит пищевая цепь, которая начинается с
микроскопических автотрофных организмов. Особенно велико
их значение в самых северных вариантах полярных
сообществ — в подзоне арктических тундр и в
полярных пустынях. Здесь микрофитофаги дают основную
-часть продукции животной массы.
Животные и лишайники
Связи животных с лишайниками — интереснейшая и
<слабо изученная экологическая проблема. Она
особенно важна при изучении экологии тундры, где лишай-
лики играют наибольшую, чем где бы то ни было,
роль в растительном покрове. Они представлены там
множеством видов и жизненных форм в виде кустиков
<с плоскими (Cetraria) или трубчатыми (Cladonia)
слоевищами, волосовидных пучков (Alectoria), полых
пальцевидных или червеобразных трубок (Dactylina,
Thamnolia), листоватых пластин (Peltigera), плесне-
бидных налетов и т. д. Они встречаются разбросанно
в моховой дернине, образуют пучки и подушки,
поселяются на голых грунтах и на каменистых субстратах.
Взаимоотношения животных с лишайниками в
тундрах весьма разнообразны. Для многих мелких
беспозвоночных, например клещей, ногохвосток, их кур-
тинки и подушки служат местом обитания. Под
слоевищами лишайников, поселяющихся на оголенных
грунтах или на скалах и камнях, концентрируются
мелкие почвенные животные, которые способствуют
образованию первичных почвенных пленок.
Отмирающие части лишайников, очевидно, служат пищей жи-
вотным-сапрофагам. Многие птицы, например
кулики-ржанки, куропатки, выстилают лишайниками
гнезда. Особенно часто используют мягкие белые трубочки
тамнолии червеобразной (Thamnolia vermicularis) и
полые пальцевидные талломы дактилины арктической
(Dactylina arctica). Лемминги иногда делают из
лишайников зимние гнезда. В щебнистых арктических
тундрах, и особенно в полярных пустынях, где травя*
J44
нистые растения растут разреженно и их часто не
хватает для гнезд, зверьки используют для этой цели
крупные подушки цетрарий Делиса (Cetraria delisei).
Найдя под снегом подушку диаметром 15—30 см,
лемминг изгрызает ее и из образовавшейся трухи
устраивает шарообразное гнездо.
Лишайники весьма питательны. Грибные гифы,
образующие слоевище, состоят в основном из особого
полисахарида, называемого лишайниковым
крахмалом. Встречаются и другие полисахариды, а также
дисахариды. Из азотсодержащих веществ обнаружены
многие аминокислоты. Имеются в них и различные
витамины, например аскорбиновая кислота, кобал-
амин (Bi2), никотиновая кислота и др. Очень
разнообразны минеральные вещества, входящие в состав тела
лишайников. Часто состав и количество минеральных
веществ зависят от того субстрата, на котором
поселяются лишайники. В них много так называемых
лишайниковых веществ, обладающих разными формами
биологического действия (например, они могут
подавлять рост грибов и мхов, всхожесть некоторых
цветковых растений). Вероятно, они оказывают
действие и на животных, но этот вопрос пока не изучен.
Пищевые связи животных с лишайниками изучены
плохо. Очень мало известно видов беспозвоночных,
для которых они служат основным кормом. Например,
среди бабочек с их бесконечно разнообразными
трофическими связями с растениями описано всего
несколько видов, гусеницы которых развиваются на
лишайниках. Почти ничего не известно о питании
беспозвоночных тундры этой группой растений.
Все растительноядные птицы, живущие в тундре,—
гуси, куропатки — лишайников, по-видимому, не едят,
если не считать незначительного их количества,
попадающего в желудок как примесь при заглатывании
основного корма.
Известно несколько видов зверей, для которых
лишайники играют роль важных кормов в зимнее время.
Обитатель восточносибирской тайги кабарга и
отчасти изюбр поедают эпифитные лишайники, растущие
на ветвях хвойных деревьев. Северный олень —
единственный вид позвоночных, который может нормально
жить в течение длительного времени, питаясь только
6 Ю. И. Чернов
145
Рис. 48. Дикий северный олень (Таймыр, фото А. В. Кречмара)
этой группой растений. Он поедает различные их
виды, но главным образом ягели (род Cladonia).
Однако едва ли можно сказать, что олень
специализировался на питании лишайниками. Он может жить и
размножаться, питаясь одними лишайниками, но
легко обходится и без них. Более того, многие
травянистые корма он ест с большей охотой.
Ягель — это в основном зимний корм в
заполярных горнотундровых, лесотундровых и северотаежных
районах. Именно там эти лишайники образуют
сплошные покровы на больших площадях. Очень богатые
ягельники известны на Кольском полуострове и в
Эвенкии, где проводят зиму стада северных оленей
(см. рис. 48, 49). Олень выкапывает ягель из-под
снега, роя глубокие «копанки». Благодаря этой
способности олени смогли освоить заполярные ландшафты.
Лишайники зимой не отмирают, не увядают и не
теряют своих питательных качеств. Во влажной среде
их ткани мягки и сочны, легко скусываются и
размельчаются. Они образуют сплошные ковры, что
позволяет животным насыщаться с небольшой площади,
а это особенно важно при зимнем подснежном
питании.
Лишайники растут очень медленно. Ягель
кладония приальпийская {Cladonia alpestris) за год
отрастает всего на 4,8 мм. Продуктивность ягельников
мала —около 7 ц/га в год. О. И. Семенов-Тян-Шан-
ский (1977) считает, что, скусывая ягель, олень
уничтожает за один раз ту часть лишайника, которая
пане
Рис. 49. Годовой цикл жизни таймырского стада
диких северных оленей:
1 — места отела и летних концентраций; 2 — основные места зимовки;
3 — осенние миграции к югу; 4 — уход с зимовки; 5 — граница тундры
растала примерно в течение 10 лет. Поэтому в местах
интенсивного зимнего выпаса диких или домашних
оленей ягельники выбиваются в течение 3—4 лет и
147
начинают угнетаться и деградировать; на их месте
возникают другие варианты растительного покрова,
например злаковые. Лишь на шестой год передышки
ягели на потравленном пастбище начинают заметно
отрастать. Для полного восстановления сильно
выбитого зимней пастьбой ягельника нужно 15—
20 лет. Летом на ягельниках часто бывают пожары,
после чего они могут восстановиться лишь через
70 лет.
В образе жизни и поведении северных оленей
имеются особенности, предотвращающие быстрое
стравливание зимних пастбищ. Дикие олени летом
покидают зимние пастбища и перекочевывают в другие
места. Так, таймырское стадо уходит для отела на
север Таймыра. При оседлом образе жизни в южной
Субарктике, например на Кольском полуострове, они
распределяются на большей площади.
Лишайники — вполне полноценный, но все же
далеко не самый излюбленный корм северного оленя.
Летом олени охотнее едят разнотравье, особенно
бобовые, злаки, осоки, пушицу, а также листья
кустарников и кустарничков. Поедают они и грибы, и даже
различных животных — леммингов, птенцов. Роль
лишайников в питании оленей уменьшается от южных
районов Субарктики к северным. В наиболее
северных частях ареала, в подзонах типичных и
арктических тундр она незначительна. Здесь олени питаются
в основном травянистыми растениями и
кустарничками.
Летом в тундрах лишайники лишь примесь в
корме оленей. Это обусловлено несколькими причинами.
Во-первых, в высокоширотных тундрах нет настоящих
ягелей. Здесь преобладают иные виды лишайников,
чаще всего с грубыми, жесткими телами, которые
менее охотно поедаются оленями: цетрарии, другие
виды кладоний, стереокаулоны, разнообразные
накипные и т. д. Во-вторых, в тундрах лишайники не
образуют сплошных порослей, ковров, что особенно важно
для зимнего питания. Они или вкраплены в моховой
покров, или растут в виде изолированных куртин.
Лишайники летом обычно сильно высыхают, становятся
ломкими, жесткими и колючими. Поэтому олени едят
их в основном в пасмурную погоду, когда их таллом
148
набухает и вновь становится мягким. То же
наблюдается в лесотундре и тайге.
Таким образом, питание лишайниками — это
своего рода приспособление северного оленя к зимним
условиям. Именно благодаря питанию лишайниками
олени могут зимовать в лесотундровых и
северотаежных районах. Южные и типичные тундры, где снега
много и он очень плотный, а лишайники не образуют
сплошных покровов, они вынуждены на зиму
покидать.
Не ясен вопрос о питании лишайниками
леммингов. Как и северные олени, они охотно едят
шляпочные грибы. Поэтому можно предположить, что
лемминги питаются и^лишайниками, значительную часть
тела которых образует грибной мицелий. Но пока нет
надежных сведений о том, какое место занимают
лишайники в их питании. Т. Н. Дунаева (1948) на
основании длительных наблюдений на Ямале и
экспериментального скармливания разных кормов пришла к
выводу, что оба вида леммингов лишайников не едят.
Иные данные приводят Б. С. Юдин, В. Г. Кривошеев,
В. Г. Беляев (1976) по леммингам дальневосточных
тундр. На острове Врангеля лишайники составляют
до 60% веса кормов, запасаемых копытным
леммингом. Но авторы не установили, что именно ест
лемминг из этих запасов.
Таким образом, лишайники в тундре, как и в
других зонах, очень мало используются
растительноядными животными. Причины этого пока не ясны. Для
фитофагов неблагоприятна очень сильная зависимость
содержания воды в лишайниках от влажности
окружающей среды, быстрое высыхание их в сухой
атмосфере, когда они становятся ломкими и очень
жесткими. Возможно, что определенную роль играет и
своеобразие химизма лишайников, в частности
наличие у них большого количества специфических
биологически активных веществ.
Кормовые отношения насекомых с лишайниками,
вероятно, в значительной мере предопределены
эволюционными факторами. Эволюция насекомых
теснейшим образом связана с высшими растениями, в
особенности с покрытосеменными. Лишайники остались
в стороне от основных направлений кормовых
149
специализаций насекомых-фитофагов. При этом в
тундре отсутствуют те небольшие группы насекомых,
у которых сформировались пищевые связи с
лишайниками.
Животные и мхи
Мхи в тундре играют огромную средообразующую
роль. Выше неоднократно упоминалось о значении
моховой дернины как фактора температурного режима
почвы, как места обитания беспозвоночных и
концентрации проростков и почек возобновления цветковых.
Однако до сих пор очень мало надежных сведений
об их роли в питании животных. Бесспорно лишь то,
что они поедаются плохо.
На территории северных тундровых подзон автор
отметил единственный вид насекомых, трофически
связанный с мохообразными растениями. Это личинки
мухи Omphalophora grandis из семейства бекасниц,
питающиеся слоевищами печеночного мха
маршанции. Эти личинки ярко-зеленые — под цвет слоевищ.
Они встречались в нескольких местах на севере
Таймыра. Но настоящие мхи насекомыми, вероятно, не
поедаются. При бесконечном разнообразии пищевых
связей насекомых с растениями самого различного
состава тканей, в том числе и ядовитыми, трудно
предположить, что тут дело в химизме.
Как неизбежная примесь, мхи, конечно,
заглатываются питающимися в толще дернины или в почве
личинками насекомых, дождевыми червями. Однако
пока очень мало достоверных фактов, которые
подтверждали бы питание беспозвоночных именно этой
группой растений. Этот вопрос требует специального
изучения в условиях тундры. Во всяком случае
выпадение из пищевых связей столь важного компонента
растительного покрова тундры составляет одну из
экологических загадок.
Живущие в тундре растительноядные птицы —
гуси, куропатки — также не едят мхов. Причем они их
определенно избегают. Выщипывая из моховой
дернины сочные части пушицы, осок и злаков, гуси
ухитряются почти совсем не проглатывать мхов. Как де-
150
значительная примесь они встречаются в желудках
белой и тундряной куропаток.
В литературе постоянно отмечается поедание мхов
оленями. Некоторые авторы даже относят мхи к
важным составным частям их питания. Это мнение
основано преимущественно на том, что в желудках
убитых оленей всегда можно обнаружить остатки мхов.
Но какова их пищевая ценность для оленей, не ясно.
Не ясно также и то, едят ли олени мхи специально,
или это примесь, попавшая в желудок вместе с
кормовыми растениями. Различные травы и кустарнички
своими наиболее сочными и питательными частями
погружены в мох или прижаты к поверхности
дернины, поэтому олени, выщипывая их, неизбежно
должны захватывать и мох. О. И. Семенов-Тян-Шанский
(1977), длительное время изучавший диких оленей на
Кольском полуострове, пб этому вопросу ограничился
следующими фразами: «Известно два случая
поедания мхов... Мхи, вероятно, часто захватываются
вместе с другим кормом».
В литературе неоднократно отмечалось, что мхи
поедают лемминги. Более того, существует мнение,
что мхи необходимы для их нормальной жизни. Его
придерживался, например, С. С. Шварц (1963).
Однако Т. Н. Дунаева (1948), исследовавшая экологию
леммингов на Ямале, высказывала противоположное
мнение: «Лишайники и мхи они совершенно не
употребляли в пищу, а пользовались ими только для
устройства гнезда».
В качестве доказательства поедания мхов
леммингами упоминают «плешины» — участки, с которых эти
зверьки зимой полностью «выстригают» моховой
покров. Этот довод приводят, например, Б. С. Юдин,
В. Г. Кривошеее, В. Г. Беляев (1976). «Плешины»
действительно обычны в тундрах, но они
свидетельствуют о противоположном. Если внимательно
приглядеться к местам зимнего питания леммингов, то
можно заметить, что масса срезанных мхов
остается неиспользованной. Совершенно очевидно, что
для питания леммингам не нужно такого количества
мхов.
Мхи срезаны сплошь, буквально «под корень», на
большой площади. Если бы лемминги питались
151
мхами, они срывали бы их живые зеленые верхушки и
не трогали основную грубую и частично
омертвевшую нижнюю часть. Между тем в остатках леммин-
говых погрызов легко рассмотреть массу мхов с
целыми верхушками. Для чего же лемминги
выгрызают такое огромное количество мха? В толще
моховой дернины, прикрытой сверху плотным снегом,
скрыты излюбленные корма леммингов —
прикорневые мясистые части осок и пушиц, молодые побеги,
листья и почки возобновления кустарничков и трав.
Чтобы добраться до них зимой под снегом, лемминги
вынуждены буквально вгрызаться в дернину.
Проделывая огромную работу, зверьки оказывают
колоссальное влияние на растительность тундры,
превращая моховой покров на огромных территориях в
труху. Этим они повышают динамичность
растительности, способствуют ее обновлению и повышению
продуктивности. Известно, что моховой покров при
чрезмерном развитии играет отрицательную роль (как
«войлок» в степи): ухудшает прогрев почвы,
заглушает проростки и т. д.
Резюмируя все известные факты, можно сделать
заключение, что типичные тундровые лемминги мхи
специально не едят, а лишь заглатывают их как
неизбежную примесь. Правда, возможна их балластная
роль в желудке. Этот вопрос требует более
детального исследования. Многое о значении мхов для
леммингов может проясниться при изучении особенностей
питания на разных этапах многолетней динамики
численности тундровых грызунов.
Птицы, звери и цветковые
растения
Взаимоотношения животных с цветковыми
растениями в тундровой зоне, как и повсюду, гораздо
разнообразнее, чем со споровыми. Здесь можно встретить
различные формы кормовой специализации, сводящиеся
к поеданию определенных органов растений. Это
достаточно отчетливо проявляется среди насекомых.
О многих птицах и млекопитающих тундры можно
сказать лишь то, что это растительноядные виды, так
152
Рис. 50. Гусь-гуменник — типичная растительноядная
птица тундры (фото А. В. Кречмара)
как они чаще всего поедают травянистое растение
целиком или в различные периоды года разные его
части. Но и среди них есть примеры определенной
специализации (некоторые зерноядные воробьиные
или зеленоядные гуси).
Общее число зеленоядных позвоночных в
тундровой зоне Евразии относительно велико, около 25
видов. Главные из них, достаточно широко
распространенные и многочисленные,— северный олень, обский
и копытный лемминги, гуменник (см. рис. 50),
белолобый гусь, белая и тундряная куропатки. В отдельных
районах высокой плотности достигают популяции
таких видов, как полевки Миддендорфа, узкочерепная,
красно-серая, экономка, норвежский лемминг,
длиннохвостый суслик, белый гусь, черная, краснозобая и
153
белощекая казарки, пискулька, тундровый лебедь и
лебедь-кликун и др.
Сравнительно недавно здесь обитал и овцебык —
один из самых специализированных видов зверей
Арктики. Вероятно, он был истреблен древним
человеком (на Таймыре можно встретить хорошо
сохранившиеся черепа). Сейчас его вновь завезли из
Америки и поселили на Таймыре и на острове Врангеля.
Полностью исчез еще один крупный фитофаг
тундры— мамонт, который жил менее 10 тыс. лет назад.
В тундровой зоне отсутствуют настоящие семено-
ядные и плодоядные млекопитающие, такие, как
лесные мыши, белка, бурундук. Птицы, питающиеся
семенами и плодами, представлены сравнительно
хорошо. Правда, наиболее типичные их формы обитают
в основном в южных районах зоны. Самые
специализированные семенояды — чечетки — массовые
обитатели лесотундры и кустарниковых тундр. Чечеток
обычно делят на два трудноразличимых вида —
обыкновенную (Acanthis flammea) №*тундряную (A. home-
manni), хотя веских оснований для этого нет. Летом
они поедают и насекомых, но большую часть года
питаются семенами различных кустарников и трав.
Даже птенцов выкармливают частично семенами. Менее
специализированные семенояды — обитатели южных
тундр Евразии — овсянка-крошка и полярная овсянка
(Emberiza pallasi).
Наиболее массовые воробьиные птицы — виды со
смешанным питанием, поедающие как семена, так и
животную пищу — насекомых. Это три самых
характерных представителя воробьиных птиц в
арктической фауне — подорожник, рогатый жаворонок и
пуночка (см. ниже). Они образуют самостоятельные
монотипические роды. Подорожник и пуночка не
выходят за пределы тундр и не имеют близких родичей.
Это говорит об их длительной связи с тундровым
ландшафтом и о высокой степени адаптации к его
условиям. Летом эти виды питаются в основном
насекомыми, которыми выкармливают и птенцов, осенью,
а иногда и летом в пасмурную погоду переходят на
питание семенами осок, ситников, камнеломок и др.
Семена, сочные ягоды, «луковички» живородящих
растений (горца, некоторых камнеломок) служат
154
важным дополнительным кормом куликам —
турухтану, ржанкам (золотистой, бурокрылой, тулесу).
Нырковые утки в большом количестве поедают крупные
семена водного лютика Палласа. В южных тундрах
осенью многие птицы едят голубику, морошку,
бруснику, и особенно охотно — куропатки, поморники,
речные утки. В типичных тундрах роль ягод ничтожна,
а в арктических они отсутствуют. ~ <
В пищевых отношениях позвоночных с
растительностью тундры много своеобразного и даже
парадоксального. Все типичные растительноядные птицы и
млекопитающие Субарктики— в принципе типичные
политрофы. Это значит, что они могут поедать самые
разнообразные корма, и не только растительные. Так,
олени едят грибы, леммингов, птичьи яйца и др.
Лемминги могут есть самые различные травы,
кустарнички, кустарники, листья, корневища, ягоды, цветки;
куропатки— листья, ягоды, цветки, почки и т. д. Ни у
одного из исконных обитателей тундры нет
морфологических или физиологических признаков, которые
можно было бы считать показателем их узкой
кормовой специализации (например, приспособления для
добывания какого-либо строго определенного корма
или для его переваривания). Северный олень,
лемминги, гуси, куропатки — это растительноядные
животные в широком смысле. Более
специализированные фитофаги, например потребители веточных
кормов (заяц-беляк) или типичные семенояды
(чечетки),— это представители по существу не
тундровых фаун, они заселяют преимущественно южные
части Субарктики.
Однако в определенном районе или в определенное
время года в питании каждого из типичных
обитателей Субарктики, как правило, выделяется небольшое
число кормов, которые играют главную роль. Так,
гуси— гуменники и белолобые — могут поедать самые
разнообразные растительные корма, в основном
сочные зеленые части. Но чаще всего их желудки бывают
набиты одним-двумя видами злаков, например
прибрежной арктофилой рыжеватой (Arctophila fulva),
пушицей (узколистной — Eriophorum angustifolium,
влагалищной — Е. vaginatum), осокой прямостоячей
(Carex stans) и др. Самый излюбленный корм гусей—
155
сочные прикорневые части пушиц и осок, которые они
искусно выдергивают из моховой дернины. Обычно
следы питания гусей мало заметны. Однако в
отдельных местах, где пушицы и злаки особенно крупны и
сочны, они бывают сильно потравлены: поверхность
дернины устлана остатками трапезы гусей —
листьями, которые они бросают, откусывая прикорневые
части.
Белая куропатка на Таймыре зимой питается почти
исключительно побегами и почками ив, а тундряная —
почками и сережками ольхи. Весной оба вида поедают
в основном листья дриады.
Таким образом, круг основных кормов типичных
фитофагов действительно весьма ограничен. Многие
исследователи, изучавшие пищевые связи позвоночных
в Субарктике, из этих факторов делали вывод о том,
что тундровым позвоночным свойственна более узкая
кормовая специализация, чем обитателям
ландшафтов, расположенных южнее. Например, леммингов
многие авторы относят к стенофагам, т. е. к
животным, приспособленным к питанию строго
определенным, ограниченным набором кормов. Такое мнение
высказано во всех сводках по экологии тундровых
позвоночных (например, в работах Т. Н. Дунаевой
(1948), С. С. Шварца (1963), Н. Н. Данилова (1966),
Б. С. Юдина и др. (1976) и т. д.).
Но это, конечно, не специализация питания, не сте-
нофагия. Употребляя эти термины, обычно имеют в
виду адаптацию к питанию определенной пищей.
Здесь же по существу следствие полифагии. Это не
что иное, как способность питаться теми кормами,
которые в данный момент наиболее питательны и
доступны. Зимой единственно возможная пища
куропаток— веточки кустарников, торчащие из-под снега.
Весной во время прилета их в тундры первыми
обнажаются из-под снега вершины бугров, поросшие
дриадой, и куропатки едят ее. Холостые гуси могут
выбирать для кормежки участки тундры с обилием пушицы,
но сидящие на яйцах питаются только тем, что растет
вблизи от гнезда.
Северный олень с зеленого летнего корма переходит
на ягель и веточный корм в местах зимовок. Обский
лемминг может есть самые разнообразные растения,
156
но основное поголовье этого вида живет за счет все
тех же мясистых частей осок и пушиц — самых
массовых цветковых растений моховой тундры, в
которой других видов трав просто не хватает этим
зверькам. В полярной пустыне, где нет осок и пушиц,
лемминги едят злаки, камнеломки, ясколки, маки,
крупки.
Самые массовые воробьиные тундры —
подорожник, пуночка, рогатый жаворонок в течение сезона
могут питаться в основном семенами, насекомыми, а то
и тем и другим. Весной они кормятся в поселках
отбросами. Пуночка на арктических островах питается
тем, что выбрасывает море. Для птиц характерно
переключение на питание кормами, появляющимися в
массе. Так, в южной тундре в годы урожая морошки
или голубики их в массе поедают речные утки, куро*
патки, поморники, гуси и др.
Конечно, велико значение и видовых особенностей
фитофагов. Это хорошо видно при сравнении
родственных видов. Так, обский лемминг более постоянен
в выборе кормов, теснее связан с типичными
массовыми растениями тундры — пушицами и осоками.
Питание копытного разнообразнее. Он больше ест листьев
кустарничков и разнообразных трав (ив, березки,
голубики, дриады, морошки, брусники и т. д.). При этом
у копытного лемминга отмечена способность запасать
на зиму корм, так же как это делают, например,
пищухи. Это особенно резко выражено у зверьков,
живущих на острове Врангеля, где в его «кладовых»
отмечено более 50 видов растений (Юдин и др.,
1976).
Другой пример существенных различий в питании
близких видов — белая и тундряная куропатки (см.
табл. 9). В каждый сезон питание этих видов
существенно различается: зимой белая куропатка
предпочитает веточки ив, а тундряная — сережки и почки
ольхи.
Итак, однообразие кормов тундровых фитофагов
определяется бедностью арктической флоры.
Растительноядные звери и птицы едят то, за счет чего в
данный момент мджио успешнее просуществовать
Это не стенофагия и не специализация питания, а
одно из проявлений потенциальной полифагии, спо-
157
собности использовать самую разнообразную пищу
в зависимости от сезона, погоды, фазы жизненного
цикла состояния кормовой базы.
Таблица 9
Соотношение кормов белой и тундряной куропаток на Таймыре
(по данным Б. М. Павлова, 1974), %
Корм
Л истья дриады
Почки и листья березки
Листья, почки, веточки ив
Почки и сережки ольхи
Семена и ягоды
Белая
куропатка
12
23
44
1
10
Тундряная
куропатка
10
7
2
77
2
Растительноядные насекомые
Растительноядные насекомые очень разнообразны в
пустынях, степях, лесах. Многие из них питаются
какими-либо определенными частями растений: корнями
(ризофаги), древесиной (ксилофаги), листьями (фил-
лофаги, или зеленояды), семенами и плодами (карпо-
фаги), цветками (антофаги) или выделяемым ими
нектаром (нектарофаги, антофилы) и т. д. В тундровой
зоне нет ксилофагов — короедов, усачей, рогохвостов,
хотя в принципе некоторые из них могли бы селиться
на ветвях кустарников. Массовые в южных широтах
потребители травянистых растений — саранчовые
доходят лишь до лесотундры. Очень мало здесь сосущих
насекомых, образующих колонии (тли, червецы,
щитовки).
В связи с суровыми условиями жизни в почве
численность ризофагов невысока. Во всяком случае они
не имеют здесь того огромного значения, как в других
зональных сообществах. Например, личинки жуков-
щелкунов, так называемые проволочники, встречаются
лишь до южных границ подзоны типичных тундр.
Личинки жукоз-пилюльщиков (Byrrhus), которые также
поедают корни, обитают почти на всей территории
158
Рис. 51. Представители ризофагов тундровой зоны:
1 — личинка долгоносика рода Lepyrus; 2 — гусеница листовертки Oleth-
reutes; 3 — гусеница листовертки ЕрСЫета
южных и типичных тундр, но повсюду малочисленны.
Самые обычные в Субарктике корнееды — личинки
долгоносиков рода Lepyrus, которые питаются
корнями ив, а возможно, и других растений (см. рис. 51). На
разнотравных и кустарничковых склонах их
численность достигает нескольких десятков на 1 м2. В южной
половине зоны на корнях бобовых изредка
встречаются личинки клубеньковых долгоносиков рода Sltona.
Важная группа субарктических ризофагов —
гусеницы бабочек-листоверток. В мясистых корнях
мытников (Pedicularis oederi, P. dasyantha) часто
встречаются гусеницы рода Olethreutes. Развитие этих
гусениц прерывается длительной диапаузой, поэтому из
них очень трудно вывести имаго, а это затрудняет
159
установление видовой принадлежности. На корнях
полыней встречаются гусеницы другого рода бабочек-
листоверток— Epiblema. На Таймыре они
представлены Е. guentheri.
На корнях мытников часто встречаются оранжевые
личинки комариков рода Bradysia (семейство Sciari-
dae). К этому роду относятся известные
сельскохозяйственные вредители, например огуречный комарик,
который, кстати, широко расселился в арктических
поселках с комнатной культурой огурцов.
В водораздельных моховых тундрах, особенно
заболоченных, ризофаги крайне малочисленны. Здесь
можно встретить на корнях различных растений
личинок мелких двукрылых — галлиц, комариков, а также
ногохвосток. Но все они поселяются, вероятно, на
отмерших или отмирающих корнях и корневищах,
возможно уже подвергнувшихся нападению нематод,
грибов.
Как это ни парадоксально, но обилие корнеедов
несколько увеличивается в подзоне арктических тундр,
где холоднее, но нет сплошного мохового покрова,
затрудняющего прогрев почвы и заглушающего
проростки цветковых растений. Эти тундры очень
красочны, в них много разнотравья, в том числе видов
с разнообразными типами корней и корневищ
(камнеломки, крупки, мытники, злаки, паррия, ллойдия
и т. д.). В почве почти совсем нет
специализированных корнеедов, таких, как личинки долгоносиков,
корневые листовертки, но многочисленны животные,
относящиеся к сборной группе сапрофагов и
употребляющие в пищу различные растительные остатки,
мертвые и живые корни. Таковы личинки комаров-
долгоножек, из них особенно характерны Tipula cari-
nifrons, Т. glaucocinerea, T. lionota. Их численность
достигает сотен экземпляров на 1 м2.
Как и в других зонах, в тундре среди
растительноядных насекомых по числу видов преобладают зе-
ленояды. Но здесь эта группа сильно обеднена, а в
арктических тундрах вообще утратила роль важного
компонента биоценозов. Наиболее характерная черта
тундрового комплекса зеленоядов — очень малая роль
сосущих насекомых, питающихся соками растений
160
с помощью колющего хоботка (тли, червецы,
цикадовые, листоблошки, клопы). По 2—3 вида клопов, ли-
стоблошек, тлей и цикадок обитают в подзоне
типичных тундр; в арктических тундрах они отсутствуют.
Больше этих насекомых в южных тундрах, но и там
они не играют существенной роли, свойственной им
в лесах, степях, пустынях.
Лишь на отдельных видах растений они могут
размножаться в больших количествах. Так, на нолынях
в массе встречаются клопы-слепняки рода Orthotylus.
На кустарниковых видах ив, особенно на сережках ив
ползучей и мохнатой, в изобилии поселяются
листоблошки рода Psylla, в особенности P. palmeni и P. zai-
cevi. Местами на злаковых участках многочисленны
мелкие цикадки. На Таймыре они представлены Strep-
tanus arctous, Hardya youngi, H. taimyrica. На
прикорневых частях некоторых растений встречаются
колонии тлей, а в толще моховой дернины — широко
распространенный по всему Северу крупный
пластинчатый червец Arctorthezia cataphracta.
Угнетенность сосущих растительноядных
насекомых обусловлена здесь отнюдь не характером связей
с кормовыми растениями. Скорее, наоборот, их способ
питания и образ жизни (малая подвижность, колони-
альность, способность многих видов к партеногенети-
ческому размножению) выгодны в суровых
заполярных условиях. Тут дело в характере их развития. Эти
насекомые развиваются без полного метаморфоза,
т. е. у них нет стадий настоящей личинки и куколки,
как у жуков, бабочек, мух. Из яйца выходит
маленькое насекомое, похожее на взрослое и обычно
питающееся той же пищей. Таким образом, уже в яйце у них
формируются основные органы взрослого организма.
Этим они отличаются от насекомых с полным
превращением, личинки которых совсем непохожи на
взрослых. Они должны долго расти, линять, чтобы
приобрести черты имаго.
У насекомых с неполным превращением яйца
снабжены большим количеством питательного материала,
за счет которого зародыш и проходит все
необходимые стадии. Для развития таких яиц нужно много
тепла, чего как раз и не хватает в тундре. Поэтому
161
большинство представителей отрядов насекомых с
неполным превращением (тараканы, прямокрылые,
кожистокрылые, сеноеды, равнокрылые,
полужесткокрылые) не находят для себя подходящих условий.
У насекомых с полным превращением яйца бедны
питательным веществом. Для их развития требуется
меньше тепла. Дальнейшее развитие вышедших из
яйца личинок, ведущих принципиально иной, чем
имаго, образ жизни, может происходить ускоренно или
замедленно в зависимости от погоды, свойств биотопа,
обилия пищи и т. д. Полный метаморфоз более
подвижен, лабилен и поэтому выгоден в суровых условиях
Заполярья. Основную часть энтомофауны тундровой
зоны составляют насекомые, развивающиеся с
полным превращением,— представители отрядов
двукрылых, перепончатокрылых, чешуекрылых, ручейников,
жесткокрылых.
Весьма богата тундровая фауна жуков-листоедов.
Большинство из них относится к роду Chrysolina (до
недавнего времени он именовался Chrysomela). Виды
этого рода заходят далеко на север, в том числе и в
арктические тундры. Самый обычный листоед в
тундрах Евразии — Ch. septentrionalis. Жуки и личинки
этого вида поедают листья лютиков, живокости,
арники, незабудок, мытников и других, а также бутоны
и завязи. Личинки растут очень медленно, в течение
2—4 лет, и зимуют в разных возрастах. В Евразии
встречается еще несколько арктических видов,
относящихся к этому роду,— Ch. subsulcata, Ch. tolli, Ch. bun-
gei. Эти листоеды на юге зоны обитают в локальных
биотопах, в основном на разнотравных склонах, а в
арктических тундрах выходят на водоразделы и заселяют
зональные сообщества, что опять-таки связано с
обилием и разнообразием в этом ландшафте цветковых
травянистых растений.
С юга в тундры проникает несколько видов
листоедов, питающихся листьями кустарниковых ив. Самые
обычные из них — Phratora polaris и Phytodecta
pallida. Оба этих вида в массе встречаются в лесотундре
и южной тундре.
В лесных районах хорошо известны крупные яркие
листоеды, относимые до недавнего времени к роду
Melasoma, теперь переименованные в Chrysomela
162
(ольховый, осиновый, тополевый листоеды и др.)-
Долго виды этого рода не были известны в тундрах.
Лишь в самое последнее время на Таймыре, крайнем
Северо-Востоке СССР и Аляске было описано сразу
несколько видов. На Таймыре, в полосе типичных
тундр, обитает СА. taimyrensis. Личинки этого вида
развиваются в основном на иве арктической, имеющей
бордюрную форму. При воспитании в лабораторных
условиях личинки поедали и другие местные ивы:
ползучую, красивую, мохнатую, полярную. На всех этих
видах развивались вполне жизнеспособные жуки.
Лишь на одной из них — монетовидной, с грубыми
мелкими листьями — личинки гибнут. Таким образом,
этот листоед — факультативный монофаг,
предпочитающий один вид кормового растения, но способный
развиваться на нескольких. Численность его местами
высока, личинки выедают до 50% массы листьев.
В состоянии беспокойства личинки выделяют по
бокам тела капельки беловатой жидкости со
специфическим резким запахом камфоры. В тихую погоду он
чувствуется на значительном расстоянии, и по нему
можно узнать о наличии личинок.
В отличие от типичных тундровых листоедов рода
Chrysolina этот вид развивается очень быстро, в
течение одного лета. Личинки интенсивно питаются и
заканчивают развитие к середине августа. Зимуют
только взрослые жуки. Жизненный цикл этого вида
должен обязательно завершиться за одно лето. Такой
тип развития отражает южное происхождение вида.
В суровых условиях Заполярья он невыгоден.
Вероятно, поэтому эти листоеды не проникают в тундры так
далеко, как виды рода Chrysolina.
Важнейшие филлофаги — личинки пилильщиков.
В моховых тундрах на их долю приходится половина
общей массы обитателей травостоя, на разнотравных
луговых участках — около четверти. Большинство их
относится к подсемейству Nematinae и связано с
ивами. Почти все тундровые ивы поражаются галловыми
пилильщиками (см. рис. 52, 1—3). Галлы
(болезненные мясистые наросты на листьях, в которых живут
личинки) особенно обильны на кустарниковых ивах
(ползучей, мохнатой), меньше их на стелющихся
арктической, красивой и полярной. На Таймыре галлы на
163
всех ивах вызывает один вид пилильщика — Pontania
crassipes. Он широко распространен по территории
Северной Европы и Сибири. В тундрах его развитие
завершается за один сезон. Зимует на стадии пред-
куколки в галле, который вместе с листом падает на
поверхность почвы. Многие виды тундровых
пилильщиков живут на листьях открыто.
Личинок пилильщиков часто называют ложногусе-
ницами. Они действительно похожи на гусениц
бабочек, ведут такой же образ жизни, питаясь зелеными
частями растений. Между экологически сходными
группами организмов часто наблюдаются обратные
соотношения: там, где много видов одной группы,
мало другой. В степях преобладают гусеницы, в
широколиственных лесах их больше, но много и
пилильщиков, а в тундрах преобладают пилильщики.
Особенно заметно это в типичных тундрах, где
численность пилильщиков высока, а гусениц—мала. В
арктических тундрах, где численность и видовое
разнообразие пилильщиков еще достаточно высоки, из
бабочек обитают единичные малочисленные виды.
Сведений о пищевых связах гусениц бабочек в
тундровой зоне очень мало. Например, почти ничего
не известно о питании самых обычных арктических
видов дневных чешуекрылых — перламутровок рода
Brenthis. Единственный в северной половине Субарк-
тики вид сатиров — Erebia fasciata, вероятно, связан
со злаками. Предположительно на злаках развивается
и редкий в сибирских и европейских тундрах Oeneis
аттоп. На различных бобовых встречаются гусеницы
желтушек (Colias nastes, С. hecla).
В тундрах довольно много так называемых ночных
бабочек — совок, пядениц, огневок, медведиц,
волнянок и т. д. В целом их видовой состав выяснен
достаточно хорошо, но экология почти не изучена. Нет
сведений о том, какими растениями питаются гусеницы
тех или иных видов, какой вред они наносят, каково
их распределение. Подавляющее большинство ночных
бабочек — мелкие невзрачные формы, летающие мало
и в течение короткого периода.
Гусеницы ведут скрытый образ жизни. Многие
питаются корнями, плодами, проделывают ходы внутри
стеблей и т. д. Но есть среди них и крупные, яркоок-
164
Рис. 52. Характерные повреждения растений насекомыми
на Таймыре:
1 — 3 — галлы на листьях ив, вызываемые личинками пилильщика Pontania
crassipes; 4 — 5 — сережки арктической и ползучей ив, пораженные
личинками пилильщиков рода Pontopristia; 6 — лист арктической ивы,
объеденный личинками листоеда Chrysomela taimyrensis; 7 — повреждение
корневой шейки мытника Эдера личинками мух семейства Muscidae; 8 — корень
мытника Pedicularis dasyantha, поврежденный гусеницами
бабочки-листовертки Olsthreutes; 9 — метелка злака арктического мятлика, объеденная
жужелицей А тага alpina
рашенные и хорошо заметные виды. Самая крупная
бабочка в сибирских тундрах — волнянка Gynaephora
lugens. Ее волосатая гусеница обитает на поверхности
моховой дернины, питаясь различными цветковыми
растениями — березкой, ивой, пушицей, зимует в
плотном шерстистом коконе и окукливается весной.
Бабочки выводятся в конце июля. Самки массивные,
почти не летают. Иногда они остаются рядом с
коконом, куда прилетают самцы. После спаривания самка
откладывает яйца здесь же, на коконе. Самцы очень
активны и слетаются во множестве к самкам. Это
можно наблюдать, если содержать самок в садках.
Встречаются в тундре крупные яркоокрашенные
бабочки-медведицы с мохнатыми гусеницами,
например Hyphoraia subnebulosa, обитающая на болотах и
питающаяся мясистыми, скрытыми во мху частями
осок и пушиц.
Сравнительно много в тундрах бабочек-совок.
Гусеницы ярко-пестрой Polia richardsoni питаются,
вероятно, брусникой и голубикой. На возвышенных
сухих местах встречается сильно опушенная серо-рыжая
Graphiphora liquidaria. Обычны также весьма крупные
пяденицы Cidaria, Aspilates, огневки Scoparia и др.
О характере их пищевых связей можно только
догадываться на основании того, что известно о биологии
близких им видов в южных районах.
От южных тундр к арктическим зеленоядных
насекомых становится все меньше. В арктических
сохраняются лишь единичные виды пилильщиков, бабочек и
листоедов. Однако они шире распределены в
водораздельных зональных сообществах. Это обусловлено большим
разнообразием цветковых травянистых растений.
Еще одна важная группа растительноядных
насекомых— антофаги, поедающие цветки. Наиболее
типичные представители этой группы — листоеды рода
Hydrothassa, из которых дальше всех на север
заходит Н. hannoverana. Этот жук в средних широтах
живет на калужнице, а в тундрах также и на лютиках,
например на северном (Ranunculus borealis),
сернистом (R. sulphureus). Жук и его личинки поедают
лепестки, пыльники, завязи (рис. 53, 4).
К антофагам относится распространенный от
Монголии до сибирских тундр листовой долгоносик Phyto-
166
Рис. 53. Представители антофилов и антофагов типичной тундры:
1 — муха из семейства мусцид на цветке дриады; 2 — муха из семейства
толкунчиков на цветке дриады; 3 — самец комара Aedes, сосущий нектар
на цветке лютика; 4 — листоед Hydrothassa hannoverana, поедающий
венчик лютика; 5—6 — жужелица А тага alpina на цветках незабудки и
лапчатки
nomus ornatus. Его своеобразные личинки, похожие
больше на гусениц бабочек, чем на личинок других
долгоносиков, развиваются на астрагалах, остролодоч-
167
никах, поедая цветки еще на стадии бутона и переходя
по мере отцветания на питание плодами. Местами
они повреждают до 20% цветков остролодочников
и астрагалов. Окукливаются личинки в конце
лета, в период созревания плодов, в прочном
полупрозрачном плетеном коконе. С цветками бобовых
связаны также мелкие оранжевые личинки комариков —
галлиц. Особенно в большом количестве они заселяют
венчики зонтичного астрагала. В некоторые годы
бывают поражены все цветки этого растения.
Массовый антофаг во многих районах — жужелица
Amara alpina. В ночные часы, при снижении
освещенности и достаточно высокой температуре (не ниже
8°), взрослые жуки поднимаются на различные
растения и объедают верхушечные цветоносные побеги,
бутоны, цветки, молодые плоды (см. рис. 53). Особенно
интенсивно они объедают метелки арктического
мятлика (Роа arctica). Часто на одной метелке можно
увидеть одновременно несколько жуков. Охотно
поедают они пыльники и другого арктического злака —
альпийского лисохвоста (Alopecurus alpinus),
встречаются на соцветиях единственного в высоких
широтах зонтичного — Pachypleurum alpinum. На Таймыре
удалось обнаружить более 20 видов растений,
соцветия которых поедают жуки Amara alpina.
К антофагам примыкают карпофаги. Упомянутые
выше представители антофагов — долгоносик Ph.orna-
tus и жужелица A. alpina — поедают также завязи и
созревающие семена. Поэтому они в такой же мере
могут быть отнесены и к группе карпофагов. Много
карпофагов среди мелких комариков — галлиц. Их
личинки развиваются в плодах десятков видов
тундровых растений. Очень сильно поражаются плоды
мытников, особенно двух видов, растущих в подзоне
типичных тундр на болотах,— судетского (Pedicularis
sudetica) и волосистого (P. hirsuta). Плоды одного из
характернейших арктических бобовых —
остролодочника чернеющего (Oxytropis nigrescens), растущего
куртинами на щебнистых участках, поедают гусеницы
мелких ночных бабочек, близких к молям (их
определение требует специального изучения этой группы).
Плоды различных ив, бобовых, мытников и
сложноцветных поедают личинки настоящих мух (Muscidae).
168
Особенно сильно повреждаются корзинки Saussurea
iilesii. К сожалению, несмотря на многократные
попытки, автору так и не удалось вывести имаго из этих
личинок.
Плоды тундровых ив сильно поражают
пилильщики рода Pontopristla. Пораженные сережки легко
узнать по плотному тампону пуха, образующемуся
в месте нахождения личинки (см. рис. 52, 4,5). На
Таймыре удалось вывести имаго из сережек четырех
видов ив. На каждом из них отмечен определенный
вид пилильщика. Это свидетельствует о большой
специализации карпофагов по сравнению, например, с
филлофагами. Так, упоминавшийся выше галловый
пилильщик развивается на многих видах ив.
В лесной полосе особенно много карпофагов среди
долгоносиков. В тундровой зоне встречаются
единичные виды этих жуков. Кроме Phytonomus ornatus с
семенами бобовых связаны недавно описанные виды
семяедов: Apion tschernovi и A. wrangelianum. В
южных частях тундровой зоны на ивах встречаются се-
режковые долгоносики Dorytomus, например D. flavi-
pes, D. nordenskioldi, D. salicinus в европейских
тундрах.
Все трофические группы растительноядных
насекомых подвержены прогрессирующему обеднению от
южных границ Субарктики к северным. В ряде
случаев резко снижается число видов группы независимо
от наличия кормовых растений. Примером могут
служить сосущие насекомые, саранчовые, многие
чешуекрылые. Причинами угнетения группы могут быть
недостаток тепла, краткость летнего периода,
отсутствие необходимой для фотопериодизма смены дня и
ночи и т. д. Распространение фитофагов лимитируется
также кормовыми растениями (ксилофильные
насекомые, зеленоядные виды, связанные с кустарниковыми
ивами, и т. д.).
Вместе с тем в тундре прослеживается широкое
расселение фитофагов с их кормовыми растениями.
При этом часто сталкиваешься с распространением,
почти не зависящим от зональных границ. Это
особенно характерно для тех случаев, когда растение и
связанное с ним насекомое обитают в интразональных
биотопах, например на болотах, по берегам ручьев.
169
Наиболее яркий случай — листоед Hydrothassa han-
noverana. Он распространен на огромной территории
от лесостепи до тундр и живет на калужнице
болотной. Ботаники выделяют в тундровой зоне два вида
калужниц: кроме обычной в ее южных районах
болотной еще арктическую (Caltha arctica). Листоед не
только сохраняется на этих растениях в тундрах, но
и становится более многочисленным, чем в лесной
полосе, а кроме того, переходит на питание различными
лютиками. Этот пример относится к тем случаям,
когда вид на пределе распространения реализует все
свои экологические возможности.
Несколько иной случай — распространение и
пищевые связи окаймленного листоеда (Chrysolina margi-
nata). Этот вид обитает от тундр до степей и
полупустынь и связан с различными сложноцветными —
тысячелистником, ромашкой, поповником и т. д. В
тундровой зоне его кормовые связи сужаются.
Например, на Таймыре он встречается в основном на полы-
нях. В арктических тундрах, где полыни крайне
малочисленны, этот листоед отсутствует.
Окаймленный листоед — пример олигофага, т. е.
вида, связанного с растениями одной группы (в
данном случае со сложноцветными). Другие типичные
олигофаги — листовой долгоносик, развивающийся на
бобовых, ивовый галловый пилильщик, листоед
таймырский и т. д. Все это внеарктические по своему
происхождению виды. Именно олигофаги наиболее
интенсивно проникают в тундровую зону с юга. Это,
конечно, возможно лишь в том случае, если они
находят здесь подходящие кормовые растения.
Узкоспециализированные монофаги, связанные с
одним видом растений, равно как и полифаги,
живущие на различных растениях, в тундры проникают
гораздо слабее. Самые типичные полифаги, широко
распространенные в лесной полосе (например,
саранчовые, различные многоядные клопы, чешуекрылые,
листоеды), ограничиваются в распространении на север
лесотундрой или северной тайгой.
Среди типичных тундровых насекомых есть и
полифаги. Таковы все листоеды рода Chrysolina,
описанная выше волнянка и некоторые другие чешуекрылые.
Есть и монофаги (возможно, все виды сережковых пи-
170
лилыдиков Pontopristia). Кроме того, наблюдается
факультативная монофагия, когда насекомое, в
принципе способное питаться многими растениями, живет
в основном на одном виде. Примером может служить
листоед Chrysomela taimyrensis. Вопрос о степени
специализации растительноядных насекомых тундры
очень интересен и важен, например, в связи с
возможным формированием вредителей по мере развития
полярного земледелия.
Большинство олигофагов связано с важнейшими
группами тундровых растений — ивами, бобовыми,
норичниковыми, сложноцветными, лютиковыми.
Относительное количество видов насекомых, питающихся
этими растениями, не меньше, а в ряде случаев
больше, чем в южных широтах. Например, очень богат
комплекс потребителей ив. Это разнообразные
пилильщики, сережковые и корневые долгоносики,
листоеды, гусеницы бабочек, двукрылые и шмели,
питающиеся пыльцой и нектаром, и т. д. Много насекомых
живет за счет бобовых (семеноядные и корневые
клубеньковые долгоносики, гусеницы бабочек и личинки
мух, развивающиеся в плодах, опылители — шмели
и др.).
В то же время очень бедна энтомофауна таких
важнейших групп растений, как крестоцветные и
злаки. В более южных широтах с крестоцветными
связано огромное число насекомых (бабочки-белянки,
огневки, совки, различные пилильщики, листоеды, в
особенности разнообразные земляные блошки и др.).
В тундрах растения этого семейства обильны и весьма
разнообразны (крупки, паррия, сердечник, брайя, ло-
жечница, ярутка и др.). Между тем насекомых на них
мало. Цветки их нерегулярно посещают антофилы.
В южных и отчасти типичных тундрах встречается
несколько видов бабочек-белянок, развивающихся на
крестоцветных, например брюквенница (Pieris napi),
альпийская белянка (Synchloe callidice). Но они
малочисленны и нехарактерны для тундры. Отсутствие
специализированной энтомофауны крестоцветных
особенно заметно в арктических тундрах, где эти
растения чрезвычайно обильны.
Из богатейшей фауны злаковых растений в
северной полосе Субарктики сохраняются лишь единичные
171
виды, обитающие преимущественно в ее южных
районах. Это злаковые мухи, личинки которых живут в
стеблях, а также несколько видов гусениц,
питающихся листьями. Почти нет сведений о фауне
насекомых, связанных с другой важнейшей группой
тундровых однодольных растений — осоками. Можно
предполагать, что она чрезвычайно бедна.
Интересен вопрос о масштабах деятельности
растительноядных насекомых в условиях Заполярья, о
степени повреждения ими растений. Сведения об этом
ограниченны. Надо заметить, что влияние
растительноядных насекомых на тундровые сообщества
перекрывается во много раз более интенсивной
деятельностью леммингов, которые являются главными
потребителями растительности и оказывают на нее столь
большое воздействие, что все прочие кажутся
незначительными. Лишь в отдельных биотопах насекомые
могут соперничать с леммингами по количеству
потребляемой растительной массы. Например, в южной
части зоны в зарослях ив листоеды и личинки
пилильщиков объедают около 12% массы листвы. Очень
сильно повреждаются бобовые растения, в
особенности их цветки и плоды. Личинки долгоносиков
иногда выедают до 70% плодов остролодочников.
Эти вопросы нуждаются в более глубоком
изучении. Расширяется хозяйственное освоение Арктики,
все дальше на север распространяется земледелие.
В ближайшее время перед учеными и практическими
работниками встанет проблема регуляции численности
растительноядных насекомых. В этом деле особенно
важны сведения о том, как ведут себя насекомые,
каковы их взаимоотношения с растениями в условиях
не тронутой человеком субарктической природы.
Насекомые-опылители и цветки
От описанных выше антофагов надо отличать антофи-
лов — насекомых, которые прилетают на цветки за
нектаром или пыльцой и участвуют в перекрестном
опылении. Растения, опыляемые насекомыми,
называют энтомофилами (слова «антофилы» и «энтомофи-
лы» образованы от греческих «антос» — цветок, «энто-
172
мон»—насекомое, «филео»—люблю).
Взаимоотношения антофильных насекомых и энтомофильных
растений— классический пример симбиоза, оказавшего
огромное влияние на эволюцию обеих групп
организмов.
Опыление растений в арктических условиях
издавна привлекало внимание исследователей. Еще в
конце XIX в. крупные ботаники того времени Е. Вар-
минг, О. Экстам, Ф. Чельман опубликовали
результаты наблюдений над опылением тундровых растений
(обзор этих работ дан в статьях В. Ф. Шамурина,
1966 и Ю. И. Чернова, 1966). Уже тогда были
выявлены своеобразные особенности энтомофильных
растений в Арктике. Отмечалось, что многие из них, хотя
и имеют крупные яркие цветки, могут обходиться без
перекрестного опыления, ибо они способны к
автогамии (самоопылению). Переход к самоопылению
обычно связывается с малым количеством опылителей и
неблагоприятными условиями для их активности. Авто-
гамность должна отражаться на строении цветка,
способствовать его упрощению, утрате признаков энтомо-
фильности (уменьшению размеров венчика, яркости
окраски, количества нектара и т. д.). Однако многие
растения, у которых наблюдалась или предполагалась
автогамия, сохраняют цветки типичного энтомофиль-
ного облика. Это в первую очередь относится к
мытникам— одной из наиболее важных групп арктической
флоры. Их крупные яркие цветки собраны в плотные
соцветия, еще больше усиливающие
привлекательность для антофилов. Тем не менее именно у этих
растений неоднократно наблюдалось самоопыление.
Считается, что это результат молодости арктической
флоры: перейдя к автогамии, растения не успели
утратить внешние признаки энтомофильносги.
Среди энтомофилов наиболее интересны растения
с цветками, устроенными так, что они могут
посещаться и опыляться лишь строго определенными
насекомыми, имеющими для этого приспособления.
Типичными примерами таких растений во флоре средней
полосы могут служить шалфей, льнянка, чина. У них
нектар помещается в особых углублениях, карманах,
шпорцах; расположение пыльников и рылец
рассчитано на опыление насекомыми определенных размеров
173
и формы. Обычно это шмели, пчелы, крупные мухи
или бабочки с их длинными хоботками.
Такие энтомофилы представлены в арктической
флоре в основном видами трех родов семейства
бобовых (астрагалы — Astragalus, остролодочники — Оху-
tropis, копеечники — Hedysarum) и родом мытников
{Pedicularis) семейства норичниковых. Всего в
тундровой зоне известно примерно 15 видов бобовых и 11
видов мытников. К специализированным энтомофи-
лам относится и представитель семейства
лютиковых— живокость (Delphinium). Этот род во флоре
тундр представлен несколькими видами и формами,
из которых наиболее обычна живокость Миддендорфа.
К этой же группе энтомофилов можно отнести и
обладающие длиннотрубчатыми цветками горечавки (Gen-
tiana), наиболее обычные в восточном секторе тундр
Евразии.
Большинство энтомофильных растений имеет
цветки, которые могут опыляться самыми различными
насекомыми. У таких цветков неглубокий, обычно
раздельнолепестный венчик, открыто расположенные
пыльники, рыльца и нектарники (типичный пример —
цветок лютика). Нектар и пыльцу на них могут
поедать любые насекомые, как типичные антофилы,
например шмели, так и различные жуки, комары, мухи,
наездники. Условимся называть такие растения
неспециализированными энтомофилами. Среди них в
тундрах преобладают розоцветные, лютиковые,
сложноцветные, гвоздичные, камнеломковые, крестоцветные.
Такие семейства, как маковые, зонтичные,
валериановые, толстянковые, бурачниковые, синюховые,
лилейные, представлены единичными, но массовыми видами.
Для того чтобы выяснить особенности
взаимоотношений насекомых с цветками, важно знать количество
энтомофилов во флоре и число видов их возможных
опылителей. Рассмотрим в этом плане состав флоры
одного из тундровых районов.
На Таймыре в подзоне типичных тундр в
окрестностях поселка Тарея отмечено 236 видов цветковых
растений (Полозова и Тихомиров, 1971). Из них 60
видов— злаки, осоки, ситниковые, для которых
опыление насекомыми не играет никакой роли. Среди
оставшихся 176 видов 19 могут быть отнесены кслучай-
174
ным элементам флоры, 11—очень редки, 9 —
представители групп, для которых нет основания
предполагать энтомофилию (березка, полыни, щавели). За
вычетом всех этих видов, остается 137 видов местной
потенциально энтомофильной флоры. Из них 86 видов
достаточно многочисленны и играют большую роль в
растительном покрове. Из специализированных энто-
мофилов в этом районе встречается менее 20: 8 видов
бобовых, 8 мытников, один вид живокости, один вид
горечавки. Все остальные — представители энтомо-
фильного разнотравья с неспециализированными
цветками.
В этом же районе изучен и состав опылителей
(Чернов, 1978). В окрестностях поселка Тарея на
цветках растений встречается 140—150 видов
насекомых. Из них примерно 90—100 видов обладают
характерными признаками потенциальных опылителей
(питание пыльцой или нектаром, достаточно
активный полет, относительно крупное и опушенное тело).
Реальное значение для опыления наиболее типичных
энтомофилов местной флоры имеет не более двух
десятков видов: 3 вида шмелей (Bombus hyperboreus,
В. pyrrhopygus, В. pleuralis), 5—6 видов журчалок
(Conosyrphus tolli, Helophilus groenlandicus, Syrphus
tarsatus, S. dryadis, Platychirus hirtipes), вид синих
мясных мух (Boreellus atriceps), 5—6 видов из
семейств настоящих мух, цветочниц и толкунчиков,
2—3 вида ночных бабочек и 3—4 вида пилильщиков
(из них в первую очередь Tenthredo).
Как видно, общее число энтомофильных растений
в типичной тундре примерно равно числу видов анто-
фильных насекомых. В арктических тундрах это
соотношение смещается в сторону преобладания
энтомофильных растений. Так, в окрестностях Диксона
встречается всего один вид шмелей, два вида крупных
журчалок и около десятка видов растений, которые
могут опыляться только этими насекомыми. В более
южных зонах антофильная фауна бесспорно богаче
энтомофильной флоры. В ельниках на цветках одной
кислицы можно встретить несколько десятков видов
мух-журчалок. В луговой степи видов шмелей и пчел
примерно столько, сколько растений-энтомофилов.
Этими соотношениями определяются многие
175
особенности энтомофилии в тундровой зоне, в
частности, создаются предпосылки для конкуренции
растений из-за опылителей. Вероятно, результатом этой
конкуренции следует считать такие особенности
биологии многих субарктических растений, как
способность к куртинному разрастанию, образованию
пышных подушек или ковров с концентрацией большого
количества цветков. Немаловажное значение имеют
и сроки цветения, некоторые привлекающие
насекомых детали цветка, запахи и т. д. Все эти вопросы
требуют дополнительного изучения.
Привлекательность энтомофильных растений для
насекомых крайне различна. Это хорошо заметно при
наблюдении за посещением шмелями цветков разных
растений на одном участке (см. табл. 10). В данном
опыте наиболее привлекательными оказались
остролодочники Миддендорфа и Адамса. У этих растений
крупные яркие цветки собраны в головчатые соцветия.
Они выделяют много нектара. За один-два дня хорошей
погоды шмели посещают большинство цветков этих
видов. Это, естественно, способствует успешному
опылению. Совершенно очевидно, что именно такие виды
привлекают к себе большинство шмелей и тем самым
уменьшают возможность опыления других растений.
Плотное произрастание, концентрация цветков
способствуют привлечению опылителей. Однако очень
высокая численность растения на больших площадях
может иметь значение и «утомляющего» фактора,
провоцировать переключение шмелей на другие
растения. Так, в приведенном опыте астрагал полярный
(Astragalus subpolaris)—самый массовый вид среди
бобовых, а мытник мутовчатый (Pedicularis verticilla-
ta)—среди норичниковых. Оба этих растения —
типичные энтомофилы. Однако именно их шмели
посещали меньше, чем другие.
Во взаимоотношениях антофильных насекомых с
растениями велика роль их биотопического
распределения. Основная масса антофилов и энтомофилов
приурочена к локальным биотопам, занимающим
небольшие площади (склоны берегов рек, озер, оврагов
и т. д.). Все это повышает эффективность опыления
растений. На больших пространствах водораздельных
тундр мало опылителей. Здесь обитают в основном не-
176
Таблица 10
Интенсивность посещаемости шмелями цветков
на участке разнотравно-лугового сообщества
(Таймыр, 27 июля 1969 г.)
Вид растения
Астрагал полярный
Остролодочник Адамса
Остролодочник Мидден-
дорфа
Мытник мутовчатый
Мытник Эдера
Живокость Миддендорфа
Число
цветков под
наблюдением
35 475
1593
770
6 601
640
35
Посещаемость
шмелями,
%
10
24
96
6
14
34
Минимальное
число дней,
необходимое для
посещения всех
цветков на участ^
ке
10
4
2
17
7
3
крупные мухи, иногда совсем нет шмелей. Недостаток
насекомых — одно из препятствий проникновения эн-
томофильного разнотравья в плакорные моховые
тундры, в чем можно убедиться, проделав такой опыт.
Типичные энтомофилы, например бобовые,
опыляющиеся шмелями и крупными мухами-журчалками,
еще до раскрытия бутонов выкапываются вместе с
почвой и переносятся с берега реки или склона оврага
в водораздельную моховую тундру на расстояние 2—
3 км. Подсчет количества образовавшихся плодов
после отцветания показывает, что многие цветки не
опылились. Во всяком случае образовывалось
значительно меньше плодов, чем в том биотопе, откуда
было взято растение.
Семенное возобновление бобовых в тундре
невозможно без участия насекомых. Основные их
опылители — шмели и крупные мухи-журчалки. В типичных
тундрах обитает всего три вида мух-журчалок,
которые могут питаться пыльцой и нектаром на цветках
бобовых. Это крупные и яркоокрашенные Conosyrphus
iolli, Helophilus borealis и Н. groenlandicus.
О значении насекомых в опылении бобовых
свидетельствуют результаты изоляции их соцветий от ан-
7 Ю. И. Чернов
177
тофилов (см. табл. 11). Бобовым свойственна
энтомофилия. При изоляции от насекомых у них почти не
образовывалось плодов. Формирование единичных
плодов у остролодочника Миддендорфа, вероятно,
случайность. Можно предположить и другое, что этот
вид, как самый специализированный,
приспособленный к опылению почти исключительно крупными
шмелями, сохранил (или выработал) резервные способы
опыления, которые используются при отсутствии
энтомофилии.
Как уже говорилось, у полярных мытников
неоднократно отмечалась автогамия. В том, что она
действительно свойственна видам этой группы, убеждает даже
внешний вид некоторых из них. Так, у мытника
волосистого (Pedicularis hirsuta) цветки мелкие, с плохо
раскрывающимся венчиком, сильно опушенные,
прижатые к стеблю. При изоляции его цветков от
насекомых плоды образовывались в таком же количестве,
как и без изолятора. Совершенно очевидно, что это
типичный автогам.
Таблица 11
Результаты изоляции соцветий. Западный Таймыр (1969 г.)
Вид растения
Остролодочник чернеющий
Остролодочник Адамса
Остролодочник Миддендорфа
Астрагал полярный
Копеечник арктический
Мытник Эдера
Мытник судетский
Мытник головчатый
Мытник мутовчатый
Живокость Миддендорфа
Число
цветков
под
изоляторами
219
119
18
220
112
120
127
83
73
29
Число
развившихся плодов
под
изоляторами
0
0
1
0
2
2
15
12
4
0
Процент
завязывания
плодов в
контроле
93
—
71
68
67
64
83
—
79
92
У большинства видов мытников процессы
опыления гораздо сложнее. При изоляции некоторое
количество плодов образовывалось (см. табл. 11), однако
их было меньше, чем в контроле (без изоляции). Кро-
178
ме того, плоды под изолятором, т. е.
предположительно возникающие в результате автогамии, отличались
меньшими размерами или были явно
недоброкачественными. Так, у мытника судетского средний вес
плодов под изолятором — 29 мг, а в контроле — 62. Этот
вид растет на полигональных болотах, вдали от
разнотравных склонов, концентрирующих основную
массу опылителей, и автогамия может иметь
существенное значение для сохранения семенной продукции
в годы, неблагоприятные для насекомоопыления.
Основной его опылитель — журчалка Conosyrphus tolli,
личинки которой развиваются здесь же, в мочажинах
болот.
Своеобразно опыление у мытника головчатого
(P. capitata). Этот вид растет небольшими группами,
что, вероятно, связано с паразитизмом, характерным
для норичниковых. Его цветки типичного энтомофиль-
ного типа, крупные. Но насекомые их посещают
сравнительно редко. Все же изоляция цветков показала,
что перекрестное опыление для этого вида имеет
существенное значение. Так, под изоляторами
завязалось всего 14% плодов. Большинство цветков к
моменту распускания имеет расположение рыльца и
пыльников, вполне соответствующее энтомофильности.
Однако у некоторых цветков этого мытника еще до
распускания из венчика высовывается длинный
столбик с рыльцем (см. рис. 54, 2), и цветок производит
впечатление типичного анемофила (опыляемого
ветром). У отдельных цветков после некоторого
периода цветения сильно отрастают все четыре тычинки,
которые колеблются на длинных нитях и сильно
пылят. В таком случае рыльце, если оно было вытянуто
раньше, к этому времени засыхает, что можно
рассматривать как приспособление, препятствующее
самоопылению. Все это говорит о том, что у
головчатого мытника существенную роль играет ветроопы-
ление, выполняющее функцию резервного способа
переноса пыльцы.
Сходные явления (отрастание рыльца и тычинок,
пыление) наблюдаются и у цветков мытника Эдера.
Признаки ветроопыляемых цветков обычно
появляются не сразу с их распусканием, а спустя некоторое
время, по прошествии которого не совершилось пере-
7*
179
Рис. 54. Цветки
мытника Pedicular is capi-
tata:
1 — распускающийся цветок
типичного энтомофильного
облика; 2 — цветок с сильно
отросшим и выставленным
наружу столбиком пестика;
3 — цветок с завядшим
рыльцем, вытянувшимися
тычиночными нитями и пылящими
пыльниками
носа пыльцы насекомыми. Вероятно, анемофилия —
резервный способ опыления, страхующий основной —
энтомофилию. Кроме того, есть все основания считать,
что этот вид способен и к самоопылению.
Таким образом, главной особенностью биологии
цветения мытников можно считать совмещение у них
нескольких способов опыления, что, вероятно, и
обусловило процветание этой группы растений в суровых
условиях тундровой зоны. Вообще совмещение разных,
страхующих друг друга способов опыления
характерно для многих растений различных зон. На это
указывал еще А. Кернер (1903): «...здесь на случай порчи
одной машины в запасе держат еще другую».
Опыление насекомыми у тундровых видов этого рода
безусловно имеет большое значение. Так, из шести
наиболее обычных видов мытников в подзоне типичных
тундр Таймыра у четырех отчетливо преобладает
энтомофилия. Автогамное опыление дает лишь
небольшое число семян.
Все это заставляет признать, что тенденция
полярных мытников к автогамии в литературе явно
преувеличена. Более того, ряд данных свидетельствует об
обратном, а именно что в Субарктике по сравнению с
более южными широтами сильнее проявляется энто-
180
мофильность этой группы. Наряду с автогамными
видами здесь имеются энтомофилы, такие, как мытники
Эдера, Адамса, судетский, мутовчатый. Яркие
крупные цветки этих видов весьма привлекательны для
насекомых, в том числе для шмелей. Особого внимания
заслуживает тесная связь мытника судетского с жур-
чалкой Conosyrphus tolli — одним из характернейших
эндемиков тундровой зоны. Можно предполагать
сопряженную эволюцию этой журчалки и болотных
мытников. Автогамия отмечалась и у многих видов
энтомофильного тундрового разнотравья с
неспециализированными цветками. Типичные автогамы —
многочисленные виды камнеломок. Среди них лишь
единичные виды, например супротиволистная
камнеломка,— энтомофилы. Бесспорна склонность к
самоопылению у крупок, сложноцветных и др. Но все же и
среди неспециализированного разнотравья есть
настоящие энтомофильные растения, опыляемые
главным образом мухами — журчалками, цветочницами,
толкунчиками, синими мясными и т. д. Нет оснований
сомневаться в наличии энтомофилии у многих
розоцветных, у некоторых лютиковых, гвоздичных,
крестоцветных и др.
Опыты по изоляции различных растений в период
их цветения от насекомых с помощью сетчатых
колпаков подтвердили предположение, что многие виды
тундрового разнотравья действительно могут
обходиться без опылителей. Так, под изоляторами
завязывались вполне полноценные, жизнеспособные плоды
у камнеломок — точечной (Saxifraga punctata),
снежной (S. nivalis), ястребинколистной (5. hieracifolia),
у арники Ильина (Arnica iljinii), мелколепестника
одноглавого (Erigeron eriocephalus), пиретрума дважды-
перистого (Pyrethrum bipinnatum), у армерии
арктической (Armeria arctica), лапчатки Potentilla stipula-
ris, ясколки крупной (Cerastium maximum), пролом-
ника северного (Androsace septentrionalis) и др.
Однако у некоторых представителей разнотравья
без доступа опылителей плоды не завязывались
совсем или были недоразвиты. Это наблюдалось у
незабудки азиатской, лютика северного, дриады
точечной, ллойдии поздней и др. В ряде случаев плоды под
изолятором образовывались, но в значительно мень-
181
шем количестве, чем в контроле. Это наблюдалось у
синюх, некоторых лютиков, камнеломок. Вообще
камнеломки очень разнообразны по своей биологии и
способам опыления. Среди них есть и типичные авто-
гамы, и ветроопыляемые формы, и настоящие энтомо-
филы.
Знакомство со специальной литературой и личные
исследования не убедили автора в том, что автогамия
среди тундровых растений распространена в большей
степени, чем в сообществах других зон. Недавно были
получены данные о значительном распространении
самоопыления среди растений, характерных для
таежных лесов. Даже в лесостепи процент типичных энто-
мофилов не очень высок. Между тем там численность
и видовое разнообразие опылителей максимальны для
умеренного пояса. Очевидно, наличие резервных
способов опыления (автогамии, анемофилии и др.) —
общая черта большинства энтомофилов. Лишь
отдельные виды и небольшие группы выходят победителями
в конкуренции за опылителей и имеют возможность
сохранять и совершенствовать облигатную
энтомофилию.
Птицы и беспозвоночные
животные
Многих птиц называют насекомоядными, или энтомо-
фагами. Это не совсем точно, ибо любая птица,
питающаяся насекомыми, может поедать и других
беспозвоночных, например пауков, моллюсков,
многоножек и т. д. И все же этими привычными терминами
удобно пользоваться (понимая их условность). В
тундровой зоне насекомых и других беспозвоночных
поедают все кулики, воробьиные, нырковые утки,
некоторые чайки, в отдельных случаях хищные птицы
(подробнее см. Чернов, 1967).
Кулики составляют примерно треть арктической
орнитофауны. Так, в тундрах Евразии встречается
около 30 видов куликов, а всего птиц — около 100
видов. Практически все тундровые кулики — типичные
энтомофаги. Лишь отдельные поедают и растения.
Например, турухтан в большом количестве ест плоды
tt*2
лютика Палласа, «луковички» горца живородящего
и т. д. Желудок турухтана сравнительно велик и
мускулист, в нем постоянно встречается галька (признаки
растительноядной птицы). Ржанки иногда поедают
зеленые листья, плоды и цветки.
Среди куликов есть виды, связанные с озерами,
болотами, и «сухопутные», населяющие высокие
моховые тундры. По составу пищи и способам ее
добывания куликов тундровой зоны можно разделить на
несколько групп.
В качестве первой группы рассмотрим
интереснейших птиц тундры — плавунчиков, приспособленных к
питанию водными организмами, которых они
добывают преимущественно с поверхности воды. В тундрах
Евразии их два вида — круглоносый (Phalaropus loba-
tus) и плосконосый (Ph. fulicarius). Плосконосый
плавунчик отсутствует в материковых тундрах к
западу от Енисея. Эти изящные кулички прекрасно
плавают и, передвигаясь по воде короткими толчками,
склевывают с ее поверхности мелких беспозвоночных.
Иногда они кружатся на одном месте, продолжая
быстро склевывать вокруг себя попадающих в
водоворот животных.
Обычно эти кулички охотятся на небольших
озерках, у самого берега, ловко лавируя среди водных
растений. К ним в пищу попадают не только водные
беспозвоночные, такие, как дафнии, жуки-плавунцы,
гаммарусы, личинки кровососущих комаров, но и
сухопутные насекомые — мухи, комары, жужелицы,
листоеды, бабочки и т. д., которые сносятся в водоемы
ветром, вымываются из прибрежной моховой дернины
волнами и скапливаются вместе с мусором в
прибойной полосе. В таких местах любят кормиться эти
кулики.
Плавунчики не ограничиваются той пищей,
которую они склевывают с поверхности воды, а добывают
ее и из залитой водой моховой дернины. Они поедают
много личинок комаров-долгоножек (типулид) рода
Prionocera. В одном желудке кулика можно встретить
около 20 этих личинок (рис. 55). Типулиды живут в
погруженной или сильно переувлажненной моховой
дернине тундровых болот и прибрежной части
озерков. На заднем конце у них имеется венчик снабжен-
Ш
Рис. 55. Личинки комаров-долгоножек — основной корм
многих птиц-энтомофагов:
1 — общий вид личинки рода Tipula; 2 — куколка комара-долгоножки;
3 — задний конец почвенной личинки Tipula; 4 — задний конец водной
личинки Prionocera
ных волосками плоских лопастей, окружающих два
дыхательных отверстия,(стигмы). С помощью этих
лопастей личинки подвешиваются к поверхностной
пленке воды, а остальная часть их тела остается в
моховой дернине. Очевидно, плавунчики хорошо
различают личинок в таком состоянии, чему, вероятно,
помогают темные крупные стигмы.
В тундровых водоемах в массе обитают личинки
комаров-звонцов. Плавунчики добывают их в
небольшом количестве на мелководьях. Перед выплодом
взрослых комаров куколки звонцов поднимаются кпо-
184
верхности воды, где и становятся добычей
плавунчиков.
Плавунчики поедают в огромном количестве
микроскопических беспозвоночных, например ногохво-
сток (коллембол), обитающих на поверхностной
пленке воды в бочагах, озерках (это всем известная
водная подура — Podura aquatica, а также еще несколько
других видов). Они едят пресноводных рачков —
дафний и циклопов, а также морских — каланусов.
Желудки их бывают сплошь набиты этими мелкими
животными без заметных примесей. Очевидно, у
плавунчиков чрезвычайно острое зрение и удивительно
точная координация движения. Можно только
поражаться тому, как они безошибочно различают и
схватывают этих мельчайших подвижных животных на
колеблющейся водной поверхности, где много мусора.
Биология плавунчиков тесно связана с тундровыми
ландшафтами, с обилием мелких водоемов,
насыщенных жизнью и дающих достаточно пищи этим
удивительным птицам.
Вторая группа — болотные виды: кулик-воробей
(Calidris minuta), чернозобик (С. alpina), дутыш
(С. melanotos), острохвостый песочник (С.
acuminata), кулик-красношейка (С. ruficollis), турухтан
(Philomachus pugnax) и некоторые другие,
проникающие с юга. Эту группу представляют самые массовые
в тундре кулики, особенно многочисленны
кулик-воробей, чернозобик, турухтан.
Виды этой экологической группы наиболее
характерны для двух южных подзон. В арктических
тундрах одни из них отсутствуют, другие малочисленны.
Гнездятся эти кулики на болотах, лайдах, в
приозерных низинах, иногда и в водораздельных сухих
тундрах. Они отыскивают добычу во влажной моховой
дернине и на ее поверхности. При этом им помогает
зрение, осязание, возможно, обоняние. Погружая в
мох клюв, кулики, вероятно, ориентируются по
колебаниям, создаваемым движением беспозвоночных.
Они питаются и на мелководьях озерков и речек, а
иногда и моря. Это особенно часто наблюдается
в конце лета и осенью на пролете.
В пище этих куликов в гнездовой период
преобладают личинки комаров-долгоножек рода Prionocera.
185
Эти личинки составляют не менее половины массы
кормов. Кулики поедают и куколок Prionocera, а в
дни массового выплода их желудки бывают набиты
взрослыми комарами.
Личинки типулид развиваются весьма медленно:
в типичных тундрах, вероятно, не менее трех лет. При
этом в отдельные годы наблюдается особенно
интенсивный выплод этих комаров. На следующий год
массового лёта не бывает. Это, вероятно, то самое
явление, которое известно и у других насекомых с
многолетним развитием, например у майских жуков.
Основная часть особей развивается синхронно, и за
массовыми вылетами следуют годы, в течение которых
подрастает новое поколение. В годы с низкой
численностью личинок Prionocera резко снижается и
количество гнездящихся куликов. Из других кормов в
питании этих куликов наибольшее значение имеют
личинки комаров-звонцов, ручейников, комаров-лимо-
ниид (ближайших родичей типулид), жуки, водные
малощетинковые черви, а также растительные корма,
особенно плоды водного лютика Палласа.
К этой группе куликов, вероятно, можно отнести
белохвостого песочника, грязовика, обыкновенного и
азиатского бекасов, гаршнепа, малого веретеника,
щеголя и некоторых других, проникающих в тундры с
юга. У каждого из них характерные особенности
биологии, способа питания и биотопа. Данные по их
питанию очень скудны. Можно лишь привести
некоторые интересные факты. Так, бекасы, известные своей
способностью добывать пищу из влажной почвы,
остаются верными себе и в тундре: именно в желудках
обыкновенного и азиатского бекасов чаще всего
встречался дождевой червь Норденшельда, которого
плохо поедают тундровые птицы.
Третья группа —«сухопутные» кулики, заселяющие
наиболее высокие и сухие участки. Наиболее
типичные представители — ржанки. Самые сухолюбивые —
тулес (Charadrius squatarola) и бурокрылая ржанка
{Ch. dominicus). Тулес — кругополярный вид,
характерная птица северной половины зоны. Бурокрылая
ржанка распространена в типичных тундрах Сибири и
Америки. В европейских тундрах она замещается
золотистой ржанкой {Ch. apricaria), которая, однако
186
менее «арктична». Она не доходит до севера
тундровой зоны, но обычна в лесотундре. Четвертый вид
ржанок — хрустан (Charadrius morinellus) —
обитатель горных хребтов и арктических тундр.
К этой же группе относятся и типичные обитатели
подзоны арктических тундр и отчасти полярных
пустынь— краснозобик (Calidris testacea), исландский
песочник (С. canutus), песчанка (С. alba) и морской
песочник (С. maritima). В гнездовое время эти
кулики — наиболее типичные обитатели зональных
ландшафтов. В отличие от болотных видов в этой группе
нет четкой картины пищевых отношений. Состав их
пищи сильно варьирует в разные сезоны и в разных
подзонах.
В типичных тундрах, где фауна насекомых
достаточно богата, пища сухолюбивых ржанок очень
разнообразна. В основном это взрослые насекомые, в
особенности жуки, крупные двукрылые,
комары-долгоножки, пауки и др. В желудках тулеса, бурокрылой
ржанки, хрустана можно обнаружить набор
различных жуков, подчас весьма малочисленных, которых с
трудом можно найти при длительных экскурсиях
(например, жужелиц рода Carabus, крупных видов Pte-
rostichus, листоедов, долгоносиков Lepyrus и т. д.).
Хрустан часто питается и на болотцах, где интенсивно
поедает личинок Prionocera. Все виды этой группы
в том или ином количестве поедают дождевых червей.
Тулес и бурокрылая ржанка также ловят много
личинок типулид, но не Prionocera, а сухолюбивых,
типичных почвенных, относящихся к роду Tipula.
С этими личинками особенно тесно связан
краснозобик— один из характернейших видов орнитофауны
арктических тундр. На Таймыре основную часть его
пищи составляют личинки столь же характерного
обитателя этого ландшафта — комара-долгоножки Tipula
carinifrons. Они живут в поверхностном слое слабо
заросших и оголенных грунтов и становятся легкой
добычей красиозобика с его длинным и чувствительным
клювом. Вероятно, личинок этого же комара поедают
исландский песочник, песчанка и тулес. Морской
песочник обитает в самых северных частях тундровой
зоны и в полярных пустынях, где местами остается
единственным представителем куликов. Там он пита-
J87
ется главным образом наземными
комарами-звонцами и их личинками, живущими в грунтах полярных
пустынь.
Таким образом, состав пищи сухолюбивых
тундровых куликов меняется от очень разнообразного на юге
зоны до крайне бедного на севере, четко отражает
степень разнообразия фауны беспозвоночных.
Отметим еще одну особенность питания этих куликов: у них
чаще, чем у болотных видов, желудки бывают пусты
или содержат много непищевых остатков в виде
детрита, мертвых веточек, песка и т. д. Это наводит на
мысль, что сухолюбивые кулики хуже обеспечены
пищей и им чаще приходится голодать.
Четвертая группа — зуйки, обитатели песчано-га-
лечного побережья рек и морей. В тундровой зоне
повсеместно обычен галстучник (Charadrius hiaticula).
Его можно встретить на берегах рек, моря, а иногда
и в водораздельных щебнистых тундрах. Он
распространен далеко к югу, обитает в субтропических и
тропических широтах. На Чукотском полуострове
встречается еще один вид зуйков — монгольский (С/г.
mongolus). Первичными стациями предков всех
ржанок были берега морей или внутренних водоемов с
твердым грунтом, а исходным способом добычи
пищи—'отыскивание ее с помощью зрения, охота за
ползающими и взлетающими насекомыми «в угон».
Именно так ведет себя галстучник. Он питается
разнообразными насекомыми, обитающими на поверхности
галечных и песчаных побережий: мухами,
развивающимися в выброшенных на берег водорослях,
комарами-долгоножками, жужелицами-тинниками,
прибрежными клопами из семейства Saldidae и т. д.
В большом количестве он поедает и водных
беспозвоночных—жуков и личинок плавунцов, личинок
комаров-звонцов, а также личинок мух, которые
живут в толще моховой дернины, где они не могут быть
найдены с помощью только зрения. Вероятно, эти
кулики обнаруживают пищу и «вслепую», с помощью
осязания. В орнитологической литературе описан
особый способ отыскивания пищи зуйками путем
«зондирования» грунта ногой. Потревоженные личинки
начинают двигаться и, создавая при этом колебания в
почве, песке, обнаруживают себя.
188
В более южных ландшафтах подавляющее
большинство пернатых относится к отряду воробьиных.
В тундровой зоне воробьиные обеднены гораздо
сильнее, чем другие отряды птиц. Так, тундровые кулики
составляют около 40% всего видового состава этой
группы на территории СССР, а воробьиные — всего
лишь 6% (22 вида). Столь сильное обеднение этого
отряда в тундрах в первую очередь связано с
условиями питания. Воробьиные менее других птиц
приспособлены к отыскиванию пищи в почве, под мхом, они
питаются главным образом тем, что находится
открыто, на поверхности. Но главное, они не способны
добывать пищу из водоемов — самых «кормных» в
тундровой зоне биотопов, с которыми связаны массовые
тундровые животноядные птицы — кулики и нырковые
утки. Нет среди воробьиных и видов, способных
поедать грубые растительные корма, которыми питаются,
например, куропатки. Лишь единичные виды
воробьиных освоили тундровые зональные ландшафты в
такой же мере, как многие кулики. Таких видов всего
около пяти; если подойти более строго, то это число
можно сократить до трех видов.
Большинство воробьиных в той или иной мере
поедают насекомых и других беспозвоночных.
Некоторые только птенцов выкармливают животной пищей.
Можно выделить три весьма условные группы: виды
преимущественно растительноядные (семеноядные
или зерноядные), виды со смешанным питанием и
типичные энтомофаги. Абсолютно растительноядных
воробьиных птиц, таких, как клесты, в тундровой
зоне нет. Преимущественно семеноядными птицами
являются чечетки, которые обычны в южных
кустарниковых тундрах, а местами и в типичных.
Из группы со смешанным питанием в северных
подзонах широко распространены три вида:
лапландский подорожник (Calcarius lapponicus), рогатый
жаворонок, или рюм (Eremophila alpestre), и пуночка
(Plectrophenax nivalis) (см. рис. 56). Их можно
считать наиболее характерными представителями
тундровых воробьиных. Этим видам свойствен четко
выраженный смешанный характер питания. Основной
пищей взрослых служат семена различных растений,
насекомые — существенная добавка в их питании. Во
189
Рис. 56. Типичные тундровые воробьиные птицы —
виды со смешанным питанием:
1 -— пуночка; 2 — лапландский подорожник; 3 — рогатый жаворонок
время высокой активности насекомых они могут
питаться как настоящие энтомофаги. В годы с затяжной
весной и холодным летом взрослые птицы
употребляют в пищу преимущественно семена, но птенцов во
всех случаях выкармливают насекомыми. Наиболее
растительнояден — подорожник. Эти птицы не могут ^
выбирать насекомых из моховой дернины, а тем
более из почвы; их добычей в основном становятся
малоподвижные или плохо летающие формы: комары-
долгоножки, жужелицы, в меньшей мере ручейники,
пилильщики, различные мухи и др.
К группе видов со смешанным питанием
относятся также овсянка-крошка (Emberiza pusilla),
полярная овсянка (Е. pallasi), дрозд Науыанна (Turdus
naumanni) и др., проникающие в южные районы Суб-
арктики.
Из типичных насекомоядных воробьиных в
тундровой зоне встречаются краснозобый конек (Anthus
cervinus), белая (Motacilla alba), желтая (М. flava) и
желтоголовая (М. citreola) трясогузки, каменка
(Oenanthe oenanthe), варакушка (Luscinia svecica),
весничка (Phylloscopus trochilus), теньковка {Ph. col-
lybita) и некоторые другие.
Среди них резко выделяется краснозобый конек —
единственный из этой группы массовый обитатель
водораздельных ландшафтов. Но он явно тяготеет к
южной полосе Субарктики. Самая высокая его
численность в южных тундрах и лесотундре. Пища крас-
нозобого конька очень разнообразна. В основном это
190
насекомые, которых он выбирает из моховой дернины,
листового опада под кустарниками и злаковой
дерновины. В большом количестве поедает гусениц,
личинок пилильщиков, листоедов, обитающих на листьях
ив и на травянистых растениях, а также очень мелких
животных, например клещей.
Варакушку можно встретить повсюду, где есть хотя
бы незначительные по площади участки ивняков;
местами заходит на север дальше границы
кустарниковых ив и гнездится на кочковатых или каменистых
склонах. Иногда ведет себя как настоящий синантроп
и строит гнезда среди досок, стружек, мотков провода
и т. д. Все это характеризует ее как чрезвычайно
пластичный в экологическом отношении вид. Там, где
имеются кустарники, варакушка питается главным
образом различными обитателями ивняков:
подстилочными формами (в основном жужелицами),
насекомыми верхнего яруса. Особенно часто в ее желудках
встречаются гусеницы бабочек-листоверток, сереж-
ковые долгоносики, пилильщики, листоблошки и др.
Молодые особи в конце лета посещают поселки, где
поедают расползающихся из мясных отбросов для
окукливания личинок и только что вылупившихся
синих мясных мух.
По всей зоне распространена каменка. Ее
гнездовые стации — обрывы и каменистые россыпи,
галечные косы с плавником, крутые берега и осыпающиеся
склоны. Пища каменки ограничивается
поверхностными малоподвижными формами. Как и все тундровые
энтомофаги, она в большом количестве поедает
долгоножек в период их массового лёта. В ее питании
большую роль играют жуки, в особенности крупные
виды: жужелицы (Pterostichus vermiculosus),
мертвоеды (Thanatophilus lapponicus), долгоносики (Lepyrus
arcticus), водные жуки (Colymbetes и Graphoderes).
Характерная черта пищевых отношений каменки —
потребление в большом количестве шмелей, которых
редко едят другие птицы. В желудках каменки
встречены Bo'mbus hyperboreus, В. lapponicus, В. balteatus.
Ее гнездовые стации совпадают с основными
местообитаниями шмелей (прибрежные разнотравные
группировки, крутые склоны, поросшие энтомофильным
разнотравьем). В целом у каменки наблюдается тен-
191
денция к поеданию наиболее крупных насекомых.
Известная склонность этого вида к синантропизму в
высоких широтах проявляется в еще большей
степени. Если в южных широтах это обусловлено,
по-видимому, гнездовыми условиями, то в тундровой зоне не
меньшее значение имеет и питание. Гнездящиеся в
поселках каменки поедают личинок и имаго мясных
мух, мертвоедов.
Широко распространена в тундровой зоне и белая
трясогузка. Как и на всем протяжении своего
огромного ареала, она связана преимущественно с
побережьями и поселениями человека.
Другие типичные насекомоядные воробьиные
обитают лишь в южных частях тундровой зоны, где
условия для энтомофагов весьма благоприятны.
Высокой численности достигает здесь как болотная желтая
трясогузка, так и связанная с кустарниками пеночкам
весничка.
Таким образом, в северных подзонах среди
воробьиных доминируют виды со смешанным питанием,
способные в любой период года переходить на
растительные корма. К ним относятся: пуночка,
подорожник, рогатый жаворонок. Видов с преобладающим
животным питанием в Субарктике зарегистрировано
более десяти. Большинство из них в своем
распространении на север ограничивается южными
тундрами. Такие виды, как варакушка, каменка, белая
трясогузка, занимают экологические ниши в интразо-
нальных частях ландшафта. Краснозобый конек —
единственный вид из этой группы, широко
распространенный в зональных сообществах, но все же он
явно тяготеет к южной полосе Субарктики.
Кустарниковый ярус южных тундр создает
качественно иные условия питания, наиболее
благоприятные для энтомофагов: с одной стороны, подстилочный
комплекс беспозвоночных, а с другой — обилие откры-
тообитающих форм в кустарниковом ярусе (тли, ли-
стоблошки, личинки пилильщиков,
бабочки-листовертки, масса ранневесенних форм, связанных с
сережками ив,— журчалки, шмели и др.).
Из отряда чаек в тундровой зоне гнездится девять
видов. Всю ее заселяют три вида поморников
(средний, короткохвостый и длиннохвостый), серебристая
192
чайка, полярная крачка. Полярная чайка
(бургомистр) тяготеет к приарктическим районам. Сизая
чайка в тундровую зону заходит на Канине. В
отдельных районах заполярной Сибири распространены
розовая и вилохвостая чайки.
Поморники — преимущественно хищники,
питающиеся грызунами, птицами, рыбой. Насекомые в их
питании играют второстепенную роль. Меньше всего
их поедает самый крупный из них — средний
поморник. Напротив, в питании мелкого длиннохвостого
значение насекомых иногда весьма велико. В
большом количестве он поедает взрослых типулид в
период их массового выплода, а также различных
хорошо заметных малоподвижных жуков — жужелиц,
листоедов, мертвоедов. Энтомофагия у этого вида
особенно сильно проявляется в годы с низкой
численностью леммингов. В жизни крупных чаек —
серебристой и бургомистра —- энтомофагия не имеет
значения, хотя в их желудках можно встретить в
большом количестве личинок мясных мух, которых они
проглатывают вместе с трупами леммингов.
Плохо изучены пищевые связи, как и вообще
экология, двух интереснейших видов чаек — вилохвостой
(Хеша sabini) и розовой (Rhodostethia rosea). Эти
изящные небольшие птицы с очень красивым
оперением гнездятся в тундрах, а зимуют севернее, кочуя в
полярную ночь по разводьям Северного Ледовитого
океана. В гнездовое время это типичные энтомофаги.
Их пища — как сухопутные насекомые, в особенности
взрослые комары-долгоножки, которых они в массе
поедают в период выплода имаго, так и различные
болотные и водные беспозвоночные — щитни,
моллюски, бокоплавы, личинки комаров-звонцов,
ручейников.
Полярная крачка (Sterna paradisaea) —*
узкоспециализированный вид. Ее пища ограничивается
специфическим кормовым биотопом, которым служит
водная поверхность. Кроме настоящих водных форм
существенную роль в ее питании играют наземные
насекомые, которые в большом количестве сносятся
ветром в водоемы. Большую роль играют куколки хи-
рономид, которых она поедает в период их массового
подъема к поверхности воды перед выплодом имаго.
193
Около половины потребляемых крачками кормов
приходится на рыбу.
При сравнении пищевых отношений различных
видов тундровых чаек наблюдается обратная
зависимость между размерам птиц и степенью их энтомо-
фагии. Так, среди поморников существенна роль
насекомых в питании наименьшего из них —
длиннохвостого. Самые мелкие чайки — розовая, вилохвостая,
а также полярная крачка — в гнездовое время
преимущественно энтомофаги.
Видное место среди тундровых энтомофагов
занимают нырковые утки: морянка, морская чернеть,
сибирская гага, гага-гребенушка, очковая гага и др.
Их прилет с моря на места гнездовий и переход на
питание пресноводными беспозвоночными начинается
по мере протаивания до дна мелких тундровых
водоемов. На Югорском полуострове личинки комаров-
звонцов встречались в желудках морянок уже 25 мая,
а в восточных секторах Субарктики, как правило,
позднее.
Летом основу содержимого желудков нырковых
уток повсеместно составляют личинки звонцов. В
желудках гаг их бывает до 2000 экз. при весе до 22 г.
Существенную роль в питании нырковых уток играют
также водные личинки комаров-долгоножек. В
желудках морянок их бывает до 30 штук. В большом
количестве они поедают личинок ручейников (у гаги-гре-
бенушки встречается до 100 экз. в одном желудке).
Прочие группы кормов в летний период —
жуки-плавунцы, личинки мух, бокоплавы. Из растительных
кормов нырковые утки едят семена лютиков Палласа.
Вегетативные части растений — это обычно примесь,
заглатываемая при добывании бентоса.
Можно выделить несколько групп насекомых,
имеющих особенно большое значение в питании
насекомоядных птиц. Прежде всего это комары-звонцы и
комары-долгоножки. Первые составляют основу
кормовой базы всех позвоночных, связанных с
тундровыми озерами. По-видимому, гнездовая плотность
нырковых уток (как и запасы озерных сиговых рыб) в
первую очередь определяется обилием личинок
звонцов. Как дополнительные корма они используются
194
куликами, в фазе куколки их в массе поедает
полярная крачка. Взрослых комаров-звонцов потребляют
все наземные птицы.
Большую роль в трофических связях тундровых
птиц играют долгоножки. Многие птицы в равной
мере питаются и личинками долгоножек, и взрослыми
комарами. Их полуводные личинки составляют
основу кормовой базы многих куликов. В некоторых
районах обилие этих личинок, вероятно, определяет
гнездовую плотность болотных куликов. Взрослых
долгоножек во время их массового лёта, который проходит
в сжатые сроки и очень интенсивно, поедают все
кулики, воробьиные, чайки. Эти комары обладают
целым рядом особенностей, обусловливающих их
большое трофическое значение для насекомоядных птиц.
Прежде всего следует отметить крупные размеры,
высокую численность, обитание на поверхности тундры,
малую подвижность. Последнее особенно характерно
для долгоножек, которые обычно ползают по земле
или медленно перелетают на небольшие расстояния.
Для некоторых типичных арктических массовых
видов характерна редукция крыльев у самок.
В условиях тундровой зоны, несмотря на крайне
напряженные трофические отношения, большая часть
важных групп животных не включается в пищевые
связи птиц или используется далеко не
пропорционально своей численности. Так, мало поедаются
почвенные беспозвоночные. Лишь в желудках красно-
зобика, тулеса, бурокрылой ржанки, а также
проникающих с юга бекасов регулярно встречаются
почвенные личинки долгоножек и дождевые черви.
Болотные птицы мало едят крупных водных мало-
щетинковых червей, обитающих в залитой водой
моховой дернине. Эти подвижные черви прячутся среди
переплетенных мхов и легко обрываются при попытке
их вытащить. Вероятно, поэтому они не попадают в
желудки кормящихся на этих участках куликов.
Не занята также экологическая ниша птиц,
охотящихся в воздухе. Гнездовой ареал этой группы птиц
на Севере ограничен южной лесотундрой. Под
Норильском еще обычны городская ласточка и
береговушка. Залетные экземпляры деревенских ласточек
и черных стрижей автор встречал на Югорском полу-
195
острове. Проникновению этих энтомофагов дальше на
север, очевидно, препятствуют сильные колебания
активности насекомых и очень короткий период их
массового лёта.
Примеры наибольшей пищевой специализации
наблюдаются среди птиц, в той или иной мере
связанных с водной средой, приспособленных к питанию
водными организмами. Так, в группе чаек наиболее
узкоспециализированный вид—полярная крачка. Среди
куликов наиболее специализированными следует
признать плавунчиков. Для сухолюбивых куликов,
например ржанок, характерна высокая политрофность.
Самые многочисленные воробьиные (пуночка,
лапландский подорожник, рогатый жаворонок) — это
типичные эврифаги. У других видов воробьиных
специализация не идет дальше общей энтомофагии.
Водные беспозвоночные включены в пищевые
отношения птиц гораздо полнее, чем наземные. Это
обусловлено их доступностью в мелких тундровых
водоемах. Существенная для птиц особенность водных
беспозвоночных — сильное смещение максимума их
сезонного развития по сравнению с наземными. Это
связано с продолжительным таянием льда и более
длительным остыванием водных масс по сравнению
с поверхностным слоем почвы. В конце полярного
лета, в августе, наземные насекомые заканчивают
развитие, погибают, уходят на зимовку или скрытно
живут в почве и моховой дернине. В это время в
водоемах разгар жизни: здесь в огромном количестве
размножаются мелкие рачки, личинки звонцов. На
конец лета приходится и массовое развитие морского
планктона, в особенности рачков каланусов.
С подъемом молодняка на крыло большинство
куликов переходит на питание водными
беспозвоночными. Птицы, не способные добывать корм из воды, в
конце гнездового периода начинают поедать
растительную пищу. Примечательно, что только три-четыре
вида птиц в течение всего пребывания в тундрах
питаются наземными насекомыми. Из них только крас-
нозобый конек сравнительно широко распространен
по тундровым ландшафтам, а прочие распределены
локально, в основном в интразональных частях
ландшафта.
196
Хищные птицы
Сведениями о пищевых связях хищных птиц тундр
мы обязаны прежде всего В. И. Осмоловской,
опубликовавшей в 1948 г. работу по хищникам Ямала.
Автор настоящей книги во время полевых работ смог
убедиться в правильности основных положений,
высказанных В. И. Осмоловской. Они нашли
подтверждение и при анализе пищевых отношений птиц с
насекомыми (Чернов, 1967).
В тундрах Евразии встречается до десятка видов
птиц, которых можно отнести к хищникам. Это шесть
видов дневных хищников, в том числе: из
ястребиных — мохноногий канюк, или зимняк (Buteo lago-
pus), орлан-белохвост (Haliaeetus albicilla) и полевой
лунь (Circus cyaneus)\ из соколиных — сапсан (Falco
peregrinus), кречет (F. gyrfalco) и дербник (F. colum-
barius)\ два вида сов — белая (Nyctea scandiaca) и
болотная (Asio flammeus); три вида поморников —
длиннохвостый (Stercorarius longicaudus),
короткохвостый (S. parasiticus) и средний (S. pomarinus).
Ястребиные, соколиные и совы тундры питаются
почти исключительно теплокровными животными —
птицами и грызунами. Поморники, близкие по
систематическому положению к чайковым птицам,
питаются более разнообразной пищей, в том числе и
растительной. Строго говоря, это эврифаги, или поли-
трофы, т. е. способные к поеданию самых
разнообразных кормов. Но все же хищные наклонности у них
превалируют.
Кроме перечисленных видов здесь встречается
ворон, который также известен своим хищничеством.
Понятие «хищник» применимо и к рыбоядным
птицам — чайкам, гагарам, но это уже относится к
жизни водоемов, которые не рассматриваются в данной
книге.
Из названных выше видов далеко не все играют
существенную роль в здешних биоценозах. Кречета
часто ошибочно относят к типичным обитателям
тундры. Этот крупный сокол распространен
преимущественно в лесотундровых, северотаежных, а также в
приморских и горных субарктических районах. В
настоящее время кречет сильно истреблен человеком и
197
повсюду очень редок. Широко распространенные во
многих зонах орлан-белохвост, полевой лунь, дерб-
ник и болотная сова регулярно встречаются в южных
частях Субарктики. Орлан часто кочует по тундрам,
но редко гнездится. Полевой лунь обычен в
лесотундре и лишь изредка залетает севернее. Дербник — это
типично лесная птица и в Субарктике гнездится
только там, где есть хотя бы одиночные деревья или
высокие кустарники. Болотная сова в южной подзоне
гнездится регулярно, в годы, изобилующие
грызунами,— в гораздо большем количестве; в типичных
тундрах редка.
Только шесть видов хищных птиц — непременные
и обычные обитатели тундры, имеющие
существенное значение в жизни ее биоценозов. Это белая сова,
зимняк, сапсан и три вида поморников. Из них белая
сова — самый типичный обитатель высоких широт.
Основная область ее гнездования — арктические
тундры. Зимняк — вид с несколько большей
экологической амплитудой, гнездится по всей Субарктике при
наибольшей плотности в типичных тундрах.
Сапсан — космополит. Но нигде его численность
не достигает столь высокого уровня, как в
Субарктике. Совершенно очевидно, что тундра — наиболее
благоприятный для этого вида ландшафт: открытые
пространства, обилие птиц, отсутствие конкурентов,
множество подходящих мест для гнездования (обрывов,
уступов, скал). Однако нет основания связывать
происхождение этого вида с ландшафтами Заполярья.
Все три поморника — типичные эваркты,
распространенные по всей территории зоны. Из них самый
массовый — длиннохвостый. Средний более других
тяготеет к высоким широтам. Наибольшая его
гнездовая плотность в арктической тундре.
Добыча пернатых хищников — птицы и мелкие
зверьки, в основном лемминги, а в южных районах
зоны и другие полевки. Здесь нет лягушек, ящериц,
змей и крупных насекомых — этой существенной
прибавки к основной пище ястребиных и соколиных птиц в
более южных широтах. Перечисленные хищники
охотятся разными способами, что зависит от их
специализации, поведения и численности жертв. В. И.
Осмоловская выделила четыре основных способа:
198
1. С налета или в угон — свойственный в основном
соколам. Они настигают птиц в воздухе
стремительным броском под углом сверху и бьют лапами.
Именно так охотится сапсан. Этот способ следует
рассматривать как наиболее совершенный тип охоты
пернатых. Но он ставит хищника в зависимость от
поведения жертвы, ибо может быть применен только на
открытой местности и только тогда, когда жертва
находится в воздухе.
2. Подкарауливание — наиболее характерно для
полярной совы, которая подолгу сидит на
возвышении, высматривая леммингов. Этим способом часто
пользуются и поморники, а иногда и зимняк.
3. Высматривание с воздуха—обычно для зимня-
ка. Он медленно летает над землей, изредка
останавливаясь в воздухе, трепеща крыльями (как это
делает пустельга). Также ведут себя и поморники.
4. Скрадывание — низкий полет на высоте 3—5 м
и внезапное нападение на застигнутую врасплох
жертву— характерно для болотной совы. Также
охотятся поморники, а иногда зимняк и полярная сова.
У каждого вида есть свой излюбленный способ
охоты, в котором он достиг совершенства. Однако для
тундровых хищников очень характерно совмещение
разных способов, что позволяет полнее использовать
кормовую базу, которую им предоставляет тундра.
В этом особенно преуспевают поморники. Они очень
подвижны, постоянно «шныряют» по тундре,
выискивая добычу (см. рис. 57). Их излюбленная пища —
лемминги, они часто нападают и на птиц, разоряют
их гнезда, поедая яйца и птенцов. Воздушными
пиратами назвал их известный исследователь животных
тундры В. М. Сдобников (1953). Весьма разнообразно
охотится и зимняк. Наряду с основной пищей —
леммингами он ловит горностая, молодняк зайца-беляка,
самых различных птиц от куропаток до мелких
воробьиных. Более консервативна белая сова, которая
в гнездовой период питается почти исключительно
леммингами и лишь в очень небольшом количестве
ловит птиц (см. рис. 58).
В рацион сапсана (см. рис. 59) входят белая и
тундряная куропатки, нырковые и речные утки,
кулики и воробьиные. Как правило, преобладают птицы
199
Рис. 57. Поморники — настоящие разбойники тундры
(фото А. В. Кречмара):
А — короткохвостый; Б — длиннохвостый
средних размеров, в особенности кулики (турухтан,
золотистая и бурокрылая ржанки, бекасы, щеголь,
малый веретеник). Но и сапсан не ограничивается
своим главным способом охоты и часто берет пищу
с земли, о чем свидетельствуют многочисленные
остатки леммингов у его гнезд.
Менее характерный для того или иного хищника
способ охоты чаще используется в случае нехватки
основных кормов. Так, в годы низкой численности
леммингов зимняк больше охотится на птиц, а
сапсан в период затяжного ненастья или сильных ветров
чаще ловит леммингов. В зависимости от состояния
кормовой базы меняется и размер охотничьего участ-
200
Рис. 58. Белая сова питается в тундре почти исключительно
леммингами (фото А. В. Кречмара)
ка. При обилии леммингов полярные совы и зимняки
охотятся поблизости от гнезда, иногда в радиусе
2—3 км; при низкой численности грызунов они
вынуждены облетать большие пространства. Размеры
охотничьего участка сапсана зависят от состава
птичьего населения. В районах, изобилующих
мелкими птицами, охотничий участок меньше. Иногда сокол
возвращается в гнездо с добычей через 3—5 минут
после вылета. Там, где сокол больше ловит куропаток
и уток, он может улетать на расстояние десятков
километров.
Пищевой рацион хищников меняется в разных
подзонах. Это в первую очередь связано с видовым
составом их возможных жертв. В южных тундрах
кроме леммингов обитает еще несколько видов
полевок — узкочерепная, Миддендорфа, местами
экономка, водяная и др. Эти грызуны в том или ином коли-
201
Рис. 59. Сокол-сапсан — главный враг птиц средних размеров
(фото А. В. Кречмара)
честве поедаются зимняком, болотной совой,
поморниками. Однако в пище этих птиц преобладают самые
массовые зверьки — лемминги. В северной половине
тундровой зоны из грызунов обитают только
лемминги. Для сапсана наиболее благоприятны кормовые
условия в южной половине тундровой зоны, где много
белой куропатки и куликов средних размеров.
Севернее он чаще добывает нырковых уток, особенно
морянку, а также мелких воробьиных, и в первую
очередь лапландского подорожника.
Обычно в рационе хищников преобладают
массовые виды птиц или грызунов. Однако в ряде случаев
наблюдается явная избирательность, т. е. добывание
видов жертв не в той пропорции, в которой они
встречаются в природе. Различают активную и пассивную
202
избирательности. Пассивная определяется поведением
самой жертвы. Просматривая различные
литературные данные по питанию тундровых хищников, легко
заметить, что из мелких птиц к ним в пищу чаще
всего попадает лапландский подорожник. Например,
по данным В. И. Осмоловской, дербник ловит
подорожников почти в 2 раза больше, чем прочих
воробьиных, вместе взятых. В основном этот вид воробьиных
птиц встречается в пище зимняка и белой совы.
Часто его ловит и сапсан.
Лапландский подорожник — массовый вид. Однако
дело тут не только в численности. Это — обитатель
открытых мест, птица с относительно слабым
полетом. Самец в гнездовой период много токует,
поднимаясь с песней в воздух и медленно опускаясь.
Именно в этот момент он и становится добычей'
сокола. В конце гнездового периода взрослые
подорожники интенсивно линяют и очень плохо летают. В это
время их ловят поморники. Другие воробьиные —
краснозобый конек, рогатый жаворонок и пуночка,
несмотря на их высокую численность, значительно
реже становятся добычей хищников. Конек и
жаворонок весьма подвижны, а пуночка гнездится в местах,
где много укрытий (каменистые россыпи, скалы,
береговые обрывы) и в любой момент можно
спрятаться.
Пассивная выборочность наблюдается в питании
зимняка. В южной тундре полевки Миддендорфа и
узкочерепная местами более многочисленны, чем
лемминги. Между тем в рационе мохноногого канюка
всегда преобладают лемминги. По сравнению с ними
полевки ведут более скрытный образ жизни и
подвижны. В меньшем количестве поедается узкочерепная
полевка, которая живет колониями и строит систему
взаимосвязанных нор, в которые при необходимости
быстро прячется.
Лемминги также различаются по степени
истребления хищниками. Копытный менее подвижен,
поэтому уничтожается интенсивнее обского и преобладает
в пище зимняка и полярной совы. Обский лемминг
подвижен, в местах поселений он прокладывает
множество ходов, помогающих ему быстро скрываться
от врагов. По наблюдениям В. И. Осмоловской, совы
203
и зимняки чаще ловят обских леммингов во время
кормежки, когда их легче застать врасплох, а
копытного — во время перебежек по открытой тундре.
В пищевых отношениях животных велика роль
покровительственной окраски й формы тела жертвы, ее
умения затаиваться. Эти свойства формируются в
процессе естественного отбора под влиянием
хищника. Они отчетливее у копытного лемминга,
подверженного более интенсивному истреблению. Летом
благодаря пестрой желто-буровато-серой окраске он
скрывается на фоне лишайниково-моховой тундры.
Зимой у него мех белый, маскирующий на снегу.
Покровительственная окраска, как правило, не
может быть абсолютной защитой от врагов,
поскольку в процессе эволюции совершенствуется и
способность хищника обнаруживать жертву. Однако
имеются примеры очень тонких и действенных защитных
приспособлений. В. И. Осмоловская отметила
интересную деталь в питании хищников. В течение по
крайней мере двух недель в тундре очень много
пуховых нелетающих птенцов куликов, которые по
своему поведению и размерам, казалось бы, должны
успешно ловиться птицами, добывающими леммингов.
Между тем они практически не попадают в число
жертв пернатых хищников. Это, вероятно, результат
их совершенной покровительственной окраски,
полностью сливающейся с расцветкой моховой тундры,
а также способности быстро затаиваться при
опасности.
В процессе естественного отбора у птенцов
куликов выработались защитные приспособления,
исключившие их из сферы пищевых отношений хищных
птиц. У последних не возникает встречных
приспособлений потому, что главные их охотничьи адаптации
направлены на добычу мышевидных грызунов. Если
бы у пуховичков не было таких действенных способов
защиты, то массовые хищники тундры, используя их
даже в качестве второстепенного корма, могли бы
уничтожать большую часть популяций куликов еще
в раннем возрасте, поскольку гнездовая плотность
этих птиц чрезвычайно мала по сравнению с тем
огромным количеством леммингов, которое съедают
тундровые хищники.
204
Эти примеры иллюстрируют пассивную
избирательность, которая наиболее широко распространена
в тундровой зоне. Активная избирательность — это
способность хищника выбирать среди доступных
кормов наиболее полноценный. Например, многие
насекомоядные птицы активно отыскивают личинок с
мягкими покровами, в частности гусениц, и не трогают
жуков, муравьев. Проявление такой способности
возможно в тех случаях, когда вообще много доступных
кормов. В Субарктике она ограничена малым
разнообразием кормовых объектов. Обычно в питании
тундровых плотоядных животных отчетливо
прослеживается преобладание наиболее доступной добычи.
Отчасти это свойственно и такому
специализированному хищнику, как сокол-сапсан. Он ловит птиц в
тундре главным образом по принципу «кто первый
попался». Выше было показано, что так же ведут себя
и насекомоядные птицы тундры.
Безусловно, у хищников тундры в той или иной
степени проявляется и активная избирательность, но
точных данных пока нет. Вероятно, именно активной
избирательностью обусловлено преобладание в пище
сапсана птиц средних размеров. Их добыча
наиболее выгодна с точки зрения рациональной траты
энергии. Крупную птицу, например утку, сокол не может
целиком принести в гнездо или съесть сразу, а
мелкие птицы дают слишком малое количество пищи.
Между тем на их поимку и перенос в гнездо
тратится не меньше времени и энергии.
По составу основной пищи тундровые хищники
делятся на две группы — орнитофагов (потребителей
птиц) и миофагов (потребителей грызунов). К
первой относятся соколы, ко второй — совы, зимняк,
поморники. Эти группы отличаются многими
особенностями экологии, динамики популяций. Гнездовая
плотность растительноядных и насекомоядных птиц,
которыми питаются соколы, мало меняется по годам
(что отражает стабильность их кормовой базы).
Колебания численности отдельных видов не могут
сильно отразиться на обеспеченности соколов кормами,
поскольку их добыча весьма разнообразна. В связи
с этим и численность орнитофагов в тундре постоянна
в течение многих лет.
205
Наблюдаемое в последнее время сокращение
численности сапсана и кречета обусловлено
антропогенными причинами — прямым уничтожением этих птиц
или подрывом их кормовой базы, например
истреблением белой куропатки — главной пищи крупных
соколов в южных тундрах. При отсутствии влияния
человека плотность сапсанов в Субарктике отличается
исключительным постоянством. Из года в год они
занимают одни и те же гнездовые участки, часто одни
и те же гнезда, расположенные на определенном
расстоянии друг от друга. Это отмечали Б. М. Житкова
(1913) и В. И. Осмоловская (1948) на Ямале,
А. В. Кречмар (1966)—на Таймыре. При наличии
подходящих мест (обрывистые берега, скалы) гнезда
сапсанов в южной подзоне располагаются на
расстоянии примерно 8 км друг от друга, а в типичных
тундрах— на расстоянии 15—20 км. Однако на огромных
пространствах полого-увалистых водораздельных
тундр эти птицы часто не гнездятся совсем.
Простейшие расчеты показывают, что при такой
плотности гнездования соколы могут изъять лишь
небольшую часть популяции тундровых птиц (не
более 5%). При этом особенно важно то, что набор
добываемых видов птиц весьма велик и меняется в
зависимости от местности, состава птичьего
населения, сезона и т. д. Совершенно очевидно, что
стабильный и в общем невысокий уровень гнездовой
плотности соколов в тундре закреплен отбором, как
оптимальный вариант системы «хищник — жертва» в
данных условиях. Бесспорно, соколы могут иметь только
положительное значение в жизни видов своих жертв,
так как в первую очередь уничтожают больных и
ослабленных особей.
Иная картина во взаимоотношениях хищных птиц
с грызунами. Копытный и обский
лемминги—главный корм большинства хищников тундры (пернатых
и четвероногих). Полярная сова, зимняк, поморники,
равно как горностай и песец, живут в основном за
счет этих грызунов. Особенно велика роль леммингов
в период выкармливания птенцов. Взрослые птицы
еще могут разнообразить свой корм, поедая птиц,
насекомых, а в южной подзоне — других грызунов и
ягоды (длиннохвостый поморник). Но птенцов они
206
выкармливают в основном леммингами, что особенно
характерно для полярной совы и зимняка.
Однако численность леммингов неустойчива,
подвержена резким годовым колебаниям. Вопрос этот
еще недостаточно выяснен, хотя ему и посвящена
большая литература. Одни авторы отмечают
цикличность в изменении численности леммингов и
устанавливают ее 3—4-летнюю периодичность. Другие
приводят данные, опровергающие строгую цикличность.
Несомнецно лишь одно, что численность этих
грызунов сильно колеблется. В год ее максимума лемминги
встречаются в тундре в огромном количестве
(основное их размножение происходит зимой под снегом)
и чрезвычайно сильно выедают растительность. В это
время меняется их поведение. Они становятся менее
осторожными, иногда даже агрессивными, не
прячутся, а нападают с криком на проходящего человека.
Иногда бывают вялыми и малоподвижными.
Вероятно, это признаки каких-то эпизоотии,
распространяющихся в переуплотненной популяции. В этот период
леммингов в массе едят все хищники тундры и даже
северный олень. Весной много леммингов попадает в
озера, реки, где их поедают нырковые утки, например
гаги, а также хищные рыбы — голец, нельма. Автору
удавалось ловить крупных гольцов, у которых
желудки были буквально набиты леммингами (до десятка
в одном желудке).
После достижения максимума численность
леммингов начинает резко снижаться. Причины этого
весьма различны. С одной стороны, падает их
плодовитость, с другой — среди зверьков
распространяются болезни. Недавно у леммингов и других полевок
на Ямале и Таймыре были обнаружены туляремия и
лептоспирозы. Разумеется, эпизоотии не могут
оказать решающего воздействия на численность
зверьков. Так, по данным Т. Н. Дунаевой (1978), среди
мертвых леммингов погибшие от туляремии составили
всего 7%. Очевидно, помимо болезней действуют
многие другие факторы: хищники, нехватка кормов,
снижение плодовитости самок и количества
размножающихся зверьков (как следствие переуплотнения
популяции), гибель от намокания и переохлаждения во
время интенсивных перемещений по тундре и т. д.
207
После массового размножения леммингов в тундре
остается множество трупов зверьков. Их съедают
личинки мух, из которых особенно характерен массовый
арктический вид синих мясных мух — Boreellus atri-
ceps. На оставшихся костях поселяется мох Tetraplo-
don mnioides. Его округлые ярко-зеленые куртинки
во множестве вкраплены в тундровой дернине.
Имеется несколько специализированных трупных видов
клещей, нематод.
Итак, кормовая база хищников-миофагов очень
неустойчива, что определяет сильные колебания
гнездовой плотности и плодовитости белой совы, зимняка
и поморников в разные годы. Меняется и характер
их пищевых связей. В годы максимальной
численности леммингов гнезда зимняков располагаются на
расстоянии всего 2—3 км друг от друга при охотничьей
территории в среднем около 7 км2. Часто поблизости,
на той же каменной гряде, можно найти гнездо
белой совы, а по соседству в ровной тундре —
поморников. По расчетам В. И. Осмоловской, одной семье
зимняка за летний сезон необходимо около 2000
леммингов. При плотности 20—40 зверьков на 1 га это
составляет около 10% популяции грызунов. Все виды
хищников вместе, вероятно, могут уничтожать до
30% грызунов. Это весьма ощутимая цифра. При
такой доле изъятия поголовья леммингов птицы могут
играть роль существенного фактора снижения их
численности.
В годы высокой численности грызунов в тундре
гнездится подавляющее большинство половозрелых
хищных птиц. При этом они откладывают много яиц,
гораздо больше, чем их южные сородичи. Так, у
зимняка в годы обилия леммингов бывает обычно 5 яиц
в кладке, а иногда и 7; у полярной совы — чаще 5—9,
а иногда до 11. У гнездящегося в средней полосе
обыкновенного канюка кладка, как правило, состоит
из двух— четырех яиц. Большинство лесных сов
откладывают около пяти яиц. В годы низкой
численности леммингов снижается плотность гнездования
хищников. Многие взрослые птицы не гнездятся.
Уменьшается количество яиц в кладке: у зимняка —
до двух-трех, у полярной совы — до трех. У
поморников в кладке почти всегда два яйца, и они реаги-
208
руют на снижение численности леммингов в основном
уменьшением количества гнездящихся пар.
Часто в литературе высказывается мнение о
высокой плодовитости тундровых птиц, в том числе
хищных. Действительно, тундровые хищники в годы
обилия леммингов откладывают сравнительно много яиц
и выращивают большое потомство. Это позволяет
полнее использовать богатые кормовые возможности,
которые дают им грызуны в годы массового
размножения. Но в периоды низкой численности зверьков
кладки уменьшаются или размножение полностью
прекращается. Высокая плодовитость лишь
восполняет ущерб, наносимый нехваткой корма в годы
депрессий. Таким образом, главная особенность
плодовитости тундровых хищных птиц заключается не
в ее высоком уровне, а в сильных колебаниях в
зависимости от состояния кормовой базы. Это
отражает приспособление к непостоянству запасов
пищи.
Еще один механизм регуляции плотности
популяции хищных птиц в зависимости от количества
пищи— растянутая кладка и насиживание сразу после
откладывания первого яйца. В результате птенцы
вылупляются неодновременно (разница во времени
между появлением первого и последнего может
достигать двух недель). В гнезде совы можно увидеть
одновременно крупных птенцов с распустившимися
маховыми перьями и только что вылупившихся
пуховиков. При нехватке корма он достается в основном
более активным старшим птенцам, а молодые
гибнут. Это облегчает выкармливание оставшегося
потомства. Иногда из большой кладки у сов или
канюков вырастает всего один-два птенца. Бывают случаи
каннибализма: при голодовках крупные птенцы
съедают маленьких.
В год самой низкой численности (депрессии)
леммингов зимняк и полярная сова совсем не
гнездятся. На огромных пространствах тундр можно
встретить только одиночных особей, которые питаются ос*
тавшимися леммингами и птицами. Большинство
хищников в такие годы откочевывает в другие райо*
ны. Например, зимняк концентрируется в южных рай*
онах Субарктики, где кроме леммингов обитает еще
209
несколько видов полевок. Поморники переходят на
питание птицами, на морском побережье едят рыбу,
отходы промысла. Длиннохвостый поморник в южных
тундрах поедает много ягод (голубики, морошки,
брусники), насекомых.
Гнездовая плотность миофагов определяется
обилием леммингов, которые относятся ко второму
трофическому уровню тундровой экосистемы, т. е. к
непосредственным потребителям зеленых автотрофных
растений (первый трофический уровень — сами
зеленые растения). Таким образом, хищники-миофаги
обычно занимают третий трофический уровень. На
нем же находится и подавляющее большинство
прочих птиц (питающиеся беспозвоночными — кулики,
воробьиные, нырковые утки), которые могут стать
добычей хищника. Лишь куропатки и гуси (как и
грызуны)—типичные фитофаги. Но их сравнительно
редко добывают миофаги.
Переключаясь в годы депрессий численности
леммингов на питание птицами, тундровые хищники
переходят с третьего уровня на четвертый.
Продукционные возможности, т. е. способность увеличивать
поголовье, наращивать биомассу (в расчете на единицу
площади сообщества) определяется положением в
системе трофических уровней. Обычно на каждом
верхнем трофическом уровне накапливается биомассы в
10 раз меньше, чем на нижнем. Хищники-миофаги,
питаясь леммингами, достигают такой плотности,
которая не может поддерживаться за счет птиц. Тем
более они не могут сохранять свойственные им
плодовитость и темпы прироста популяции. Это и
обусловливает неизбежное ограничение или прекращение
размножения, а также гибель потомства, миграции в
другие районы и как результат снижение плотности
хищников на данной территории.
Кого больше в тундре —
хищников или фитофагов?
Орнитологи неоднократно отмечали, что среди
тундровых птиц по сравнению с фауной более южных
широт резко преобладают формы, питающиеся животной
210
пищей (хищники, рыбоядные, насекомоядные). Об
этом, например, писал А. А. Бируля в 1907 г. Это
мнение весьма логично, поскольку птицы, будучи
высокоорганизованными животными, с очень большой
интенсивностью физиологических процессов,
успешнее могут осваивать Заполярье, питаясь более
калорийной пищей. Это положение в общем правильно,
но в то же время нуждается в существенных
оговорках, если рассматривать птиц тундры не только в
целом, но и по группам и по подзонам.
Плотоядность свойственна всем наиболее
процветающим в высоких широтах группам птиц. Таковы
гагары, чистики, кулики, чайки. Показателен такой
пример. Растительноядные речные, или настоящие,
утки весьма многочисленны в лесотундре и южной
тундре (чирки, шилохвость, свиязь). Но среди них
нет ни гемиарктических, ни гипоарктических видов.
В более высоких широтах они исчезают, замещаясь
животноядными нырковыми утками (гаги, турпаны,
морянка), которые остаются единственными
представителями подсемейства утиных в типичных и
арктических тундрах. Среди нырковых есть ряд
характерных арктических видов, в том числе настоящие
эваркты.
Интересны данные об относительном числе видов
того или иного отряда. В тундровой зоне Евразии
обитает около 40% видов куликов фауны СССР,
около 30 — чаек, 100 — гагар и только 9 — куриных и
6%—воробьиных. Куриные — это типичные
фитофаги, а среди воробьиных растительноядные
составляют значительную часть (имеются в виду и виды со
смешанным питанием).
Со сходными соотношениями приходится
сталкиваться и при рассмотрении других групп
организмов. Например, все характерные для высоких широт
рыбы: хариусы, гольцы, ряпушки, пыжьян, муксун,
чир, омуль, нельма, сибирский осетр и др., а также
проникающие с юга окунь, щука — типичные
плотоядные формы. Растительноядные рыбы в тундровой
зоне отсутствуют. Даже виды со смешанным питанием,
такие, как плотва, елец, язь, доходят лишь до
лесотундры,
8*
211
Аналогичные примеры дают некоторые группы
насекомых и паукообразных. Среди жуков в тундре
наиболее широко распространены хищные жужелицы и
стафилины, из клопов далее всего на север
проникают прибрежные хищные виды из семейства сальдид.
В Субарктике резко сокращается разнообразие
растительноядных клещей, зато хорошо представлены
пауки и хищные клещи.
Как ни кажутся убедительными приведенные
выше примеры и цифры, положение о преобладании в
животном мире тундры животноядных форм
нуждается в существенных оговорках. Многое здесь еще не
ясно. Хищникам надо чем-то питаться. Если в тундре
хорошо представлена группа плотоядных, то еще
более многочисленными должны быть
растительноядные. Действительно, в тундре разнообразны и
обильны многие типичные растительноядные насекомые,
прежде всего пилильщики, жуки-листоеды,
потребители почвенных водорослей нематоды и личинки
двукрылых.
Но главную массу беспозвоночных, как правило,
составляют не фитофаги, а сапрофаги, питающиеся
мертвыми растительными остатками — опадом. Это
различные кольчатые черви (дождевые, энхитреиды),
клещи, личинки двукрылых. Однако среди огромного
мира животных, относимых обычно к сапрофагам,
многие поедают не только растительные остатки, но и
бактерий и грибы, которые в свою очередь также
живут за счет мертвых растительных тканей. Получены
данные о том, что в тундрах преобладают группы
почвенных животных именно с такими пищевыми
связями, а истинных сапрофагов мало. Так, питание
грибами характерно для коллембол. Бактериями
питаются многие нематоды и простейшие. Вместе с детритом
их поедают кольчатые черви. Таким образом,
большинство беспозвоночных тундры, составляющих
основную часть животной массы, находится не на втором,
а на третьем трофическом уровне (бактерии и
грибы— это типичные гетеротрофы). Итак, в этом случае
также наблюдается своего рода смещение на более
высокие гетеротрофные уровни. Но здесь еще много
неясного, и мы рискуем ступить на путь догадок и
необоснованных суждений. Для глубокого и объектив-
212
ного анализа этого вопроса нужны более надежные
данные о питании беспозвоночных, относимых к
комплексу сапрофагов.
Вернемся к птицам. Перечисляя их группы,
наиболее процветающие в тундровой зоне (гагары, чайки,
кулики, нырковые утки), легко заметить, что все это
водные или околоводные формы или сухопутные лишь
в определенное время года, но способные питаться и
на воде (поморники, большинство куликов, чайки).
Таким образом, преобладание плотоядных видов — это
черта не всей орнитофауны тундры, а в первую
очередь птичьего населения водоемов. Все отряды,
включающие настоящих наземных птиц, плотоядных и
растительноядных (куриные, совы, дневные хищники,
воробьиные), представлены в тундре примерно в
одинаковой степени (6—10% видов отряда на
территории СССР). Каждый из этих отрядов в Субарктике
сильно обеднен, но представлен характернейшими
гемиарктами или эварктами: куриные — белой и
тундряной куропаткой; совы — полярной совой; дневные
хищники — зимняком; воробьиные — подорожником,
рюмом, пуночкой.
Среди воробьиных тундры по числу видов
преобладают энтомофаги, однако, как было сказано выше,
это или заходящие на юг Субарктики или связанные
с интразональными биотопами. У самых ярких
тундровиков— рюма, подорожника, пуночки — смешанное
питание.
К отряду пластинчатоклювых, из которого выше
упоминались нырковые и речные утки, относятся еще
и гусиные — преимущественно сухопутные,
кормящиеся наземными растениями. Это бесспорно одна из
процветающих групп тундровой орнитофауны (10—11
видов). Таким образом, среди пластинчатоклювых в
тундровой зоне лучше представлены
растительноядные наземные гусиные и животноядные водные
нырковые, хуже — растительноядные водные настоящие
утки.
Среди зверей, встречающихся на территории
Субарктики, весьма много хищных и вообще плотоядных:
волк, лисица, росомаха, бурый медведь, ласка,
горностай, несколько видов землероек. Это не раз
отмечалось как одна из характерных черт фауны млекогт-
21$
Рис. 60. Песец — единственный настоящий тундровый вид
хищных млекопитающих (фото А, В. Кречмара)
тающих тундры. Однако все перечисленные виды —
пришельцы из других зон, заселяющие южные районы
Субарктики или распространенные по всей ее
территории весьма спорадично. Среди хищных
млекопитающих всего два представителя истинно арктической
фауны — эваркт песец и гипераркт белый медведь.
Песец —единственный исконно тундровый вид
хищных зверей, имеющий существенное значение в
биоценозах Заполярья (см. рис. 60). Зато среди
растительноядных грызунов и копытных встречаем
наибольшее число характернейших тундровых эндемиков. Это
эваркты — копытный и обский лемминги, овцебык и
гипоаркты — северный олень, узкочерепная полевка
и полевка Миддендорфа.
Затронутые здесь вопросы относятся к очень
сложным и мало разработанным сферам современной
экологии — к проблемам трофоэнергетических связей и
соотношения структуры и функций в сообществах.
Здесь нет возможности углубляться в эту тему. От*
И«
метим лишь, что приведенные факты ЛодчеркйвакУГ
необходимость строго конкретного анализа
закономерностей жизни тундровых сообществ. Экосистемы
тундры, несмотря на их обедненность и, казалось бы,
простую структуру, полны противоречий и своеобразия
в каждой своей части и в каждом звене. Познание
закономерностей строения сообществ тундры,
взаимоотношений их частей — насущная задача
современности. Без этого невозможно рациональное
использование ее ресурсов, равно как и охрана природной
среды.
ТУНДРА И ЧЕЛОВЕК
Заселение человеком послеледниковых ландшафтов
тундровой зоны этнографы относят ко временам
позднего палеолита — раннего неолита. Это примерно 7—
8 тыс. лет назад. Вся ранняя история человека на
Крайнем Севере Евразии была теснейшим образом
связана с охотой на северного оленя. Ловля рыбы,
использование в пищу съедобных растений, охота на
пернатую дичь и морского зверя на первых этапах
заселения тундры имели явно второстепенное
значение. Только северный олень мог дать достаточно пищи
и все необходимое для жизни. Используя
экологическую терминологию, можно сказать, что древнейший
человек тундры был монофагом.
Со временем совершенствовались орудия и способы
охоты на оленя. Особенно важный этап развития
культуры древних охотников — возникновение способа
добычи оленя на воде во время его переправ через реки
с применением лодок. Появление лодки — само по себе
очень важное событие и в истории жителей Заполярья.
Основная масса оленей стала добываться осенью, во
время миграций их на зимние пастбища в лесотундру
и тайгу, меньшее количество — весной при движении
к местам отела и летнего пребывания. Добыча
большого количества оленей за короткий срок требовала
участия множества людей, выполняющих самые
разнообразные функции. Этот способ охоты при изобилии
оленей позволял добывать практически
неограниченное количество мяса, что обусловило быстрый рост
населения тундровой зоны и привлечение различных
южных племен, которые, смешиваясь, формировали
этнические группы.
Численность оленя длительное время определяла
уровень плотности населения древних охотников. По
подсчетам Ю. Б. Симченко (1976), предельная
численность тундровых популяций диких северных оленей
Евразии была около 3 млн. голов при годовом
приросте стад около 7%. В связи с этим население древ-
216
них охотников могло достигать не более 11 тыс.
человек. Дальнейший рост коренного населения тундры
был связан с более полным использованием других
ресурсов (рыбы, морского зверя), с развитием
оленеводства и товарообменом с южными племенами.
У населения тундры архаичный уклад жизни
сохранялся дольше, чем у народностей средней полосы.
Этнографы объясняют это однообразием и
устойчивостью производственной базы, в первую очередь
популяций северного оленя. Способы добычи оленей
были весьма совершенными уже на раннем этапе
развития культуры древних охотников. С этим
связывается длительная неизменность материальной
культуры после оформления наиболее рационального
способа охоты — поколки оленей на воде. Но тут дело
не столько в стабильности производственной базы,
сколько в ее неустойчивости по отношению к
интенсивной эксплуатации.
При высокой плотности оленей в тундре годовой
прирост их стад все же невелик, гораздо ниже, чем
у копытных в южных широтах. Кроме того, надо
учесть, что олень был основным или даже
единственным источником жизни древних охотников. Оленье
мясо нечем было заменить. Поэтому нормальное
существование было возможным лишь при
гарантированной добыче достаточного количества оленей, т. е.
при их избыточной по сравнению с потребностями
человека плотности. Таким образом, длительное
сохранение низкой интенсивности производства и
сравнительно небольшой плотности населения — единственно
возможный в тех условиях путь поддержания
равновесия между человеческим обществом и окружающей
природой. Древнее население не могло
эксплуатировать производственную базу «на грани возможного»,
т. е. изымать предельно допустимую часть поголовья
оленей.
С охотой на оленей связан кочевой образ жизни
населения Крайнего Севера. Для него характерен
особый тип миграций, который можно назвать
«челночным»: из года в год летом охотники приходили за
оленями в одни и те же места у переправ, а осенью
возвращались в районы зимовок оленей в лесотундру и
северную тайгу. Совершенно очевидно, что древние
217
люди могли жить в материковой тундре только летом.
Едва ли можно было запастись мясом на всю зиму.
Кроме того, в тундре невозможно найти достаточного
количества дров. Подледный лов рыбы был еще
недоступен первобытным охотникам.
Важный этап развития населения Субарктики —
приручение оленя и возникновение оленеводства.
Сначала олени использовались во время охоты на диких,
а затем в целях массового разведения. С развитием
оленеводства в укладе жителей тундр появляется
множество новых черт, но в центре их хозяйственной
деятельности оставался олень. Наряду с этим у разных
племен совершенствовались охота на других
животных, рыбная ловля. Этому способствовало
лавинообразное сокращение численности дикого оленя,
которое началось, вероятно, еще в неолите.
Очевидно, уже в далекие времена стоял вопрос
о том, что выгоднее — охота на оленя или его
разведение? Вопрос о рациональном соотношении поголовья
диких и домашних популяций не потерял
актуальности и в настоящее время. От
практиков-оленеводов и ученых иногда можно услышать
отрицательные отзывы о «дикаре». Обычно ему приписывается
два «греха»: стравливание пастбищ и увод домашних
животных во время контактов стад. Так, известный
тундровед В. Н. Андреев на страницах журнала
«Экология» (1977) пишет о диком олене как о вредителе
оленеводства, считая, что он якобы наносит вред
пастбищам, осложняет содержание стад домашних,
вызывает потери их поголовья. В связи с этим он
предлагает ограничить численность дикого оленя и
переселить его на пастбища, непригодные для
выпаса домашнего (арктические и высокогорные
тундры).
Тем, кто бывал в тундре, где есть дикие олени,
наблюдал их пастьбу и миграции, эта проблема
кажется надуманной. По данным В. Н. Андреева, в
настоящее время в нашей стране имеется около 2,5 млн.
голов домашнего оленя (при потенциальной емкости
пастбищ 3,4 млн.) и около 800 тыс. диких (по данным
других авторов — около 600 тыс.). Дикий олень
сохранился в основном в виде трех стад: на Кольском
полуострове в условиях заповедного режима, на Тай-
21§
мыре и на севере Якутии. Территория, занимаемая
этими стадами, по отношению к общей площади зоны
оленеводства невелика.
Самое большое стадо таймырское (см. рис. 49).
Места его основных летних миграций и отела
(северная половина полуострова Таймыр) лежат за
пределами оленеводческих районов, т. е. там, где выпас
домашних явно нерентабелен. Только дикая форма
способна успешно использовать обширные
малопродуктивные пастбища этих суровых высокоширотных
ландшафтов, не вызывая существенных нарушений
растительного покрова. Горные районы Путорана, где
дикий олень концентрируется на зимовку, также мало
пригодны для использования оленеводческими
хозяйствами. Контакты диких и домашних оленей в этих
районах возможны лишь в относительно
непродолжительные периоды.
Таким образом, регулирование взаимоотношений
домашних оленей с таймырским стадом может быть
разрешено самым простейшим способом:
незначительным разграничением зон их миграций. Таймырское
стадо, насчитывающее 400 тыс. голов,— наша
национальная гордость, пример успешного сохранения
интенсивно истреблявшегося вида. Теперь оно
эксплуатируется охотничьими хозяйствами, дает доход
государству и бесспорно заслуживает того, чтобы уступить
ему небольшую часть территории, которая,
возможно, и представляет интерес для оленеводства.
Противоречит духу времени постановка вопроса В.
Н.Андреевым о выселении дикого оленя в места,
«непригодные для стадного выпаса домашних оленей». При
этом имеются в виду арктические и высокогорные
тундры, в которых олень может влачить лишь жалкое
существование.
Охота на морского зверя — тюленей, моржей,
китов—могла развиться на более позднем этапе, чем
охота на оленя. Для нее нужны были большие,
прочные лодки, мощные гарпуны; сам процесс охоты был
весьма сложен и требовал многих сложных навыков.
Морской зверь не мог обеспечить древнего человека
всем необходимым, в частности легкой и теплой
одеждой. Кроме того, в морях Карском, Лаптевых,
Восточно-Сибирском зверя или очень мало, или его трудно
219
добыть, особенно зимой. Лишь в более теплых
атлантических и тихоокеанских водах морской зверь мог
составить достаточно надежную базу для жизни
человека. Именно на Чукотке и в Гренландии и
обосновались первые приморские поселенцы.
С охотой на морского зверя связано и потребление
на Крайнем Севере растительной пищи. Мясо моржей
и тюленей очень жирное, его химический состав
сравнительно беден. Между тем для нормального
переваривания и усвоения жирной пищи необходимы
углеводы и разнообразные неорганические соли. Поэтому
при питании жирным мясом морского зверя человек
испытывает потребность в растительной пище как в
дополнительном источнике различных веществ и в
ферментативной приправе. В связи с этим
использование в пищу растений широко практикуется именно
у прибрежных жителей, например у береговых чукчей
и эскимосов. Употребление в пищу трав и традиции их
сбора складывались на протяжении веков. В женских
эскимосских погребениях, относящихся ко второму
тысячелетию до н. э., находят костяные мотыги, сходные
с теми, которыми пользуются для выкапывания
растений и в настоящее время.
Чукчи используют в пищу около 20 видов
растений. Они кладут их как приправу в суп, употребляют
в свежем виде, иногда заливая жиром,
заготавливают впрок в сушеном или квашеном виде (Соколова,
1961). Их наиболее излюбленные растения —
камнеломка Нельсона (Saxifraga nelsoniana), кисличник
(Oxyria digyna), арктический щавель (Rumex arcti-
cus), горец (Polygonum ellipticum), родиола (Rhodiola
atropurpurea), листья некоторых ив (Salix pulchra,
S. arctica), цветки зонтичного астрагала (Astragalus
umbellatus). Собирают они и морскую капусту —
бурые морские водоросли (ламинарию). Чукотские
женщины хорошо различают все эти растения, знают
время, когда лучше всего собирать тот или иной
вид.
Однако многие народности тундровой зоны
растительную пищу почти не едят и, очевидно, не
испытывают потребности в ней. Это прежде всего относится
к питающимся оленьим мясом. Оно не слишком
жирное и очень богато различными химическими соединен
220
ниями. Кто впервые пробует суп из свежей летней
оленины, поражается его запаху. Он напоминает то
запах грибов, то душистого травяного отвара.
Непосредственные взаимоотношения человека с
растительным миром на Крайнем Севере весьма
ограниченны. Здесь нет тех форм деятельности, которые
приводят к направленным и часто необратимым
изменениям растительного покрова (как, например, при
распашке, вырубании лесов, сенокошении). Однако
воздействия человека на растительный мир тундры
весьма многообразны. Известный исследователь
растительности Арктики Ф. Чельман писал в 1883 г.:
«Против своей воли и, по-видимому, совершенно
бессознательно чукча становится растениеводом. Почти
повсюду вокруг его чума густыми сомкнутыми
группами произрастают виды растений, из которых одни
совершенно отсутствуют в окрестностях, другие
встречаются поблизости, но в очень небольшом количестве
или произрастают рассеянно. Не подлежит сомнению,
что часть их проникла сюда без участия чукчей и,
найдя благоприятную для произрастания среду на
кучах отбросов, которые накопляются с течением
времени вокруг жилых мест, сохранилась здесь и
размножилась. Другие, несомненно, обязаны своим
присутствием чукчам, которые собрали их где-то далеко
и частично разбросали в виде отбросов здесь, где они
вслед за тем укоренились и размножились» (цит. по
Е. В. Дорогостайской, 1972).
Изменения растительного покрова в местах
поселения человека в тундре и занос им более южных
видов— интереснейшие экологические проблемы.
Особенно важен вопрос о формировании комплекса
сорняков на северных форпостах земледелия. Эти вопросы
в течение нескольких лет изучала Е. В. Дорого-
стайская, опубликовавшая в итоге специальную
монографию. Она разработала классификацию тундровой
антропофильной (связанной с человеком) флоры.
Приведем лишь две самые крупные группы: апофиты —
местные виды, переходящие в населенные пункты и
нарушенные биотопы, успешно там приживающиеся,
размножающиеся в большем количестве, чем в
природных местообитаниях, и антропохоры, занесенные в
тундровую зону человеком.
221
Какие же виды становятся апофитами в тундровой
зоне? Существует мнение, что растительность
антропогенных местообитаний, в том числе так называемая
рудеральная, свойственная замусоренным местам
(например, хорошо известные лопух, крапива),
сформировалась первоначально на зоогенных местообитаниях.
Это понятно, так как основные формы воздействия
на среду со стороны животных и человека в
принципе сходны: уничтожение растительной дернины,
рыхление почвы, обогащение ее питательными
веществами, в особенности азотом, и т. д. В связи с этим
состав растительности на территориях поселков весьма
близок к тому, что можно видеть на песцовых норах,
птичьих базарах, в местах интенсивной деятельности
грызунов.
Апофитную флору можно разделить на нитрофилов
(любящих избыток азота) и эрозиофилов
(приуроченных к местам с нарушенной разрыхленной почвой,
с содранной растительной дерниной).
Типичные нитрофилы в тундровой зоне — лютик
гиперборейский, альпийский лисохвост, крестовник
арктический, арктофила рыжеватая. Они особенно
обильны в загрязненных местах, часто образуют
заросли на помойках, свалках. Эрозиофилы в
природных условиях связаны с рыхлыми супесчаными и
песчаными эродируемыми грунтами. В таких местах они
закрепляются благодаря способности быстро
разрастаться с помощью корневищ и мощных корней,
противостоять механическому воздействию подвижного
грунта. Типичный апофит-эрозиофил — полынь Тиле-
зиуса, обильная на различных участках с
осыпающимся грунтом.
Большинство эрозиофилов в то же время относится
и к нитрофилам. Именно такие виды самые
многочисленные в антропогенных биотопах. Яркий их
представитель— ромашка крупноцветная (ранее широко
известная как Matricaria grandiflora, теперь—Tripleu-
rospermum phaeocephalum). Она образует сплошные
заросли в арктических поселках, обильна на месте
разрушенных строений, на зарастающих рытвинах,
у помоек и т. д. Отличается буйным цветением.
Представителями нитрофилов-эрозиофилов можно считать
также крестовник арктический, мятлики арктический
222
и высокогорный, арктический хрен (Cochlearla arctica),
хвощ полевой, дескуранию Софии, пиретрум дважды-
перистый и др.
Тундровые апофиты — преимущественно виды ин-
тразональных природных местообитаний
(эродированных склонов, побережий, нивальных луговин и т. д.).
Среди них почти нет растений, которые были бы
характерными для плакорных зональных сообществ.
Склонность к заселению антропогенных биотопов
проявляют некоторые влаголюбивые растения, например
нардосмия холодная — типичный обитатель торфяных
заболоченных участков, побережий ручьев
(аналогичные примеры есть и среди насекомых).
На вытаптываемых и удобряемых отходами
участках, на которых уничтожается или угнетается
типичная тундровая дернина, развиваются особые варианты
растительности, напоминающие обычные рудеральные
заросли в средней полосе. Местами наблюдается то,
что принято называть залужением тундры, т. е.
образование растительного покрова, аналогичного
луговому. Такую растительность можно встретить в
небольших поселках, на местах давнишних стойбищ,
временных поселений охотников и рыбаков. Злаки,
формирующие луговую дерновину, обильны там, где
мало мхов, где они угнетены вытаптыванием или
уничтожены полностью.
Известный энтузиаст освоения Севера А. В. Жу-
равский видел в залужении путь повышения
биологической продуктивности тундры и широко
пропагандировал его. В статье под красноречивым названием
«Полярные окраины в новом освещении» (1915 г.) он
писал: «Тундра, следственно, есть обыкновенная
русская «пустошь», вовсе не сопровождающаяся
специфическою «скудностью почв», ибо флора и фауна
моховых и ягельных формаций в коренной лесной зоне,
в главной массе, те же, обусловливаясь мшаным
покровом, причем мхи, не использывая почвенных
продуктов окисления и разложения, не определяют и не
могут определять своей наличностью состава почвы,
лишь «консервируя» последнюю... Так ли, иначе ли, но
все, без изъятий факты, фактические материалы по
всем группам фауны, по флоре, геологии, этнографии,
и все мои наблюдения вынуждают меня считать луго-
223
водство, скотоводство и маслоделие — географически
специфичными отраслями хозяйства Севера, и севера
«полярного»—преимущественно...»
А. В. Журавский явно увлекался и переоценивал
возможность этим путем добиться резкого повышения
продуктивности тундры. Пока практически
обнадеживающие результаты экспериментов по залужению
коренных сообществ получены лишь в лесотундровых
районах, например в районе Салехарда. Здесь на
месте кустарниково-мохового сообщества сформированы
сеяные луга, дающие ежегодный урожай сена по 30—
40 ц/га (Филиппова, 1963). Доказана перспективность
создания в этом районе культурных пастбищ. В
типичных и даже южных тундрах это дело будущего.
Залужению здесь поддаются лишь высокие
дренированные участки, на которых и естественная
растительность отличается обилием разнотравья, злаков.
Злаки-апофиты очень чувствительны к степени
нарушения биотопа. Они поселяются лишь в том случае,
если уничтожаются мхи, но сохраняется торфяной
горизонт. На вспаханных и длительно
культивируемых участках тундровые апофиты, как правило, не
приживаются. Е. В. Дорогостайская показала, что на
северных форпостах земледелия практически ни один
вид местной флоры не вселяется на распаханные
участки, на которых приживаются лишь привезенные
с юга культурные или сорные растения. Этот факт
косвенно свидетельствует о том, что тундровые
апофиты могут иметь лишь ограниченное значение в
процессах залужения и рекультивации. Надо также иметь
в виду, что мхи — более сильные конкуренты злаков
в Заполярье.
С поселениями людей связаны различные антропо-
хоры, среди которых можно выделить несколько групп
по степени приживаемости в антропогенных биотопах:
случайные, встречающиеся единично вдалеке от
основного ареала и не способные к самостоятельному
существованию в тундровой зоне; временные,
заселяющие успешно лишь строго определенные специфичные
биотопы, например теплицы, парники, и
натурализовавшиеся, давно занесенные на Север и обильно
произрастающие в различных антропогенных биотопах.
Пути проникновения антропохоров в тундровую зону
224
различны: преднамеренный завоз культивируемых
видов и непреднамеренный (с фуражом, посевным
материалом, пищевыми продуктами).
Е. В. Дорогостайская описывает случай,
показывающий, в каких количествах заносятся человеком
южные растения в Заполярье. В Норильске на месте,
где разгружалось сено, она насчитала 19 видов веге-
тировавших растений, находящихся за сотни и тысячи
километров от их ареала. Однако лишь очень
немногие антропохоры способны прижиться в Арктике на
значительный срок. Чаще всего они существуют
некоторое время в отдельных биотопах, не давая
полноценных семян, а затем вымирают. Лишь в южных
районах Субарктики отмечается успешная
натурализация многих антропохоров. Так, в лесотундровых и
некоторых южнотундровых районах на территориях
поселков можно найти такие растения, как крапива
двудомная, полынь обыкновенная, клевер ползучий,
мокрица, подорожник большой, спорыш и т. д. Однако
суровые условия собственно тундр, особенно
типичных, создают для этих видов непреодолимые
препятствия.
Единичные виды проявляют большую активность
в расселении по антропогенным биотопам Субарктики.
Можно привести пример американской ромашки
Matricaria matricaroides, которая, попав в Воркуту, в
течение нескольких лет размножилась там в огромных
количествах, образовав густые заросли. Другой вид —
крестовник обыкновенный, который также быстро
расселился в тундровых поселках.
Некоторые из антропохорных растений переходят
в различные интразональные биотопы — на береговые
отмели, обрывы, осыпи. На таких местах они не
испытывают конкуренции со стороны исконйо тундровых
растений и часто успешно приживаются. Таковы
пижма обыкновенная, костер безостый, полынь
обыкновенная, ромашка непахучая и др. Однако в зональные
тундровые биотопы антропохорные виды не
внедряются.
Сходные закономерности выявляются и при
анализе формирования животного населения
антропогенных биотопов. В этом плане очень показателен
комплекс так называемых синантропных (живущих с
225
человеком) мух. Их субарктическую фауну можно
разбить на несколько групп, отражающих основные пути
формирования: типичные синантропы, проникающие
в тундровую зону непосредственно с человеком
(комнатная муха, малая комнатная муха, зубоножка,
рыжая навозница и др.); широко распространенные, в
том числе и в Субарктике, мясные мухи — Protophor-
mia terraenovae, Cynomyia mortuorum; арктические
виды, в той или иной степени склонные к обитанию
в поселениях человека, преимущественно падальные
формы, связанные с отходами промыслов.
Специфика природных условий Субарктики столь
велика, что ни один из типичных синантропов (первая
группа) не может размножаться в значительных
количествах в поселках Заполярья. Самые
многочисленные в тундровой зоне синантропные мухи —
представители второй группы, а также единичные виды из
типичных эварктов. Это прежде всего синяя мясная
муха Boreellus atriceps и мелкая, развивающаяся в
различных отбросах Neoleria prominens.
Такие же соотношения наблюдаются и среди птиц.
В населенных пунктах на всей территории тундровой
зоны обычны широко распространенные обитатели ин-
тразональных биотопов — белая трясогузка и каменка.
Типичные синантропные птицы, проникающие в
высокие широты непосредственно с человеком (полевой
воробей, деревенская ласточка, скворец и др.),
ограничиваются в расселении на север южными районами
Субарктики. Значительное число видов аборигенной
фауны в той или иной мере проявляет склонность
к синантропизму, но из них только пуночка —
постоянный и массовый обитатель полярных поселков, в
которых она заменяет воробьев.
Приведенные выше примеры свидетельствуют о
том, что влияния человека на самые различные
группы организмов подчиняются сходным
закономерностям. В этом можно видеть предпосылки
регулирования его отношений с животным и растительным
миром тундры.
Биологические ресурсы Крайнего Севера при
общем пониженном разнообразии жизни весьма
обильны. Исходная (до использования человеком)
численность здесь может быть выше, чем в ландшафтах,
226
расположенных южнее. Однако это не результат
высокой продуктивности, т. е. годового прироста, а
следствие своего рода накопления популяций при низкой
интенсивности биологических процессов, а также
концентрации основной части продукции того или иного
трофического уровня небольшим числом видов.
Именно поэтому внешне богатые ресурсы Севера так
быстро подрываются и медленно восстанавливаются.
В Арктике наблюдается, казалось бы, парадоксальная
ситуация: при очень низкой плотности людей многие
виды животных истребляются ими быстрее, чем где бы
то ни было. Достаточно назвать таких животных, как
бескрылая гагарка, морж, белый медведь, овцебык,
еще совсем недавно населявший тундру Евразии,
а также мамонт, скорее всего истребленный древним
человеком.
Именно поэтому освоение Севера человеком с
самого начала носило пассивный характер, скорее
экстенсивный, чем интенсивный. Люди стремились
перемещаться, осваивать новые районы, но не промышлять
долго на одном месте, не создавать большие и
длительные нагрузки на используемые ресурсы. Уклад
жизни многих северных народностей был таков, что
о нем можно сказать: люди в здешних условиях не
эксплуатируют запасы животных, а подлаживаются
под их образ жизни. Так, у нганасан отношения
между членами племени и семьи, выбор места поселения,
распределение труда определялись целиком жизнен-
ным циклом, сезонными миграциями стад северного
оленя.
Отмеченные выше особенности популяций
тундровых животных обусловливают необходимость
специфических мер их использования и охраны, строго
дифференцированного подхода, более осторожного изъятия
прироста. Один из путей рациональной
эксплуатации тундрового животного мира — экстенсивный:
равномерное распределение промысловых нагрузок по
территории, строгое чередование изъятия приплода и
«отдыха» популяции, переложенный тип
использования угодий и т. д.
Важнейшая черта сообществ Арктики, имеющая
прямое отношение к практике рационального
использования и охране ее ресурсов,— очень резкая широт-
227
ная дифференциация ландшафтов. Выше было
показано, что существенные градиенты климата здесь
проявляются на гораздо меньшей площади, чем в южных
широтах. Соответственно этому состав флоры и
фауны, облик сообществ на протяжении тундровой зоны,
от ее северных границ к южным, изменяются
чрезвычайно резко. Три подзоны тундр (арктические,
типичные и южные) отличаются друг от друга важнейшими
особенностями. Например, многие доминирующие в
арктических тундрах животные совсем не встречаются
в южных и наоборот. Все это также требует строго
дифференцированного подхода как к сообществам,
так и к ландшафтам в целом. Кроме широтной
зональности в Арктике очень сильна секторная
дифференциация ландшафтов. Атлантика, Тихоокеанское
побережье, сибирские тундры — все эти районы резко
отличаются друг от друга по структуре сообществ, по
составу флоры и фауны.
При использовании ресурсов Арктики надо
учитывать еще одну очень важную особенность. Ее
природа— это своеобразный резонатор (Крючков, 1976).
Часто небольшие нарушения здесь резко усиливаются
и приводят к необратимым коренным изменениям
сообществ, ландшафтов. Так, небольшая колдобина и
выбоина в вечной мерзлоте может вызвать
оползневые и термокарстовые явления. Небольшие понижения
температуры приводят к местным оледенениям.
С аналогичными явлениями приходится
сталкиваться и при воздействии на тундровые сообщества.
В их структуре особенно важно явление
супердоминантности, когда один вид (или группа животных или
растений) играет очень большую роль в жизни многих
других компонентов сообществ. Таковы лемминги —
основная пища всех хищников тундры; комары-
долгоножки (всего 2—4 вида) —кормовая база
большинства наземных энтомофагов, а личинки хироно-
мид — база водных позвоночных. Подрыв популяций
таких видов и групп неизбежно вызовет необратимые
отрицательные изменения в популяциях одновременно
многих видов.
Относительно простая структура сообществ
Арктики, сильная зависимость большого количества
компонентов от немногих эдификаторов приводят к быст-
228
рому распространению в сообществах ядовитых
веществ, радионуклидов. Так, исследования последствий
применения инсектицидов для борьбы с гнусом
показали, что они очень быстро распределяются по
основным членам сообществ тундры, мигрируют по цепям
питания и в течение короткого времени оказываются
в теле различных беспозвоночных, леммингов, оленей,
птиц, в растениях и т. д. (Шилова и др., 1973). В то
же время выведение ядовитых веществ из экосистем,
процессы их самоочищения в условиях тундр сильно
понижены. Естественно, что применение ядовитых
веществ в экстремных условиях Севера требует особой
осторожности.
Уязвимость природы Севера, ее способность чутко
реагировать на все воздействия и усиливать их
вынуждает к необходимости незамедлительной
разработки системы мероприятий, строго специфичных и
дифференцированных, направленных на поддержание
равновесия в экосистемах Арктики и Субарктики.
Развитие народного хозяйства в СССР все в
большей степени опирается на ресурсы Сибири и Севера.
Значение этих районов в экономике нашей страны
будет неуклонно возрастать. В связи с этим необходимо
широко развернуть глубокое и всестороннее изучение
влияния производственной деятельности человека на
природные процессы Заполярья.
В решениях XXV съезда КПСС мероприятия по
охране окружающей среды, рациональному
использованию и воспроизводству природных ресурсов
названы одними из основных задач развития народного
хозяйства на 1976—1980 гг. В условиях Севера
особенно важна детальная разработка принципов освоения
ресурсов с учетом всех возможных последствий,
поскольку его природа крайне чувствительна к
деятельности человека.
Вопросам освоения Севера была посвящена
выездная сессия научного совета Академии наук СССР,
состоявшаяся в июле 1978 г. в Якутске. Видные
советские ученые, исследователи Севера и руководители
научных учреждений выработали ряд рекомендаций,
направленных на охрану окружающей среды, наметили
планы дальнейших исследований этих проблем,
обратились к соответствующим ведомствам с просьбой
229
организовать на севере Сибири ряд заповедников,
заказников, обеспечить очистку сточных вод в городах,
взять под контроль применение ядохимикатов,
создать новые виды транспортных средств, снижающих
нарушение почвенного покрова, разработать
комплексную медико-биологическую программу по
проблемам адаптации человека на Севере.
Непосредственное отношение к проблемам
рационального использования и охраны природных
ресурсов Севера имеет и долгосрочная программа
«Сибирь», разработанная Сибирским отделением
Академии наук СССР. В нее включены узловые проблемы
комплексного освоения минерально-сырьевых,
земельных, лесных и водных ресурсов, и специальное
внимание уделяется вопросам охраны окружающей среды.
Значительная часть этой программы посвящена
разработке научно обоснованных мероприятий по охране
биосферы в районах интенсивного развития
добывающей промышленности и крупного строительства.
Предусматривается разработка новых средств защиты
биосферы от загрязнений и техногенных нарушений,
методов рационального использования, реконструкции
и охраны растительного и животного мира Сибири, и
Севера в частности.
К дальнейшему освоению Севера непосредственно
относится и важнейшая общая проблема
использования биологических ресурсов. Издавна деятельность
человека в Заполярье базировалась на промысле диких
животных. Между тем сейчас становится все более
очевидным, что эра простого изымания нужных
продуктов из биосферы безвозвратно уходит. Получая
что-либо от природы, необходимо позаботиться о
восстановлении данного ресурса. Это можно выразить
формулой: переход от промысла к хозяйству. Иными
словами, настало время, когда человек вынужден
переходить от принципов охоты к принципам
культурного хозяйствования. Под этим понимается
полувольное и вольерное разведение промысловых животных и
полное приручение их, введение аквакультур,
создание мощных рыборазводных комплексов, питомников
для выращивания молоди различных видов,
селекционно-генетических станций и т. д. Для решения этой
проблемы потребуется комплексное обширное иссле-
230
дование процессов размножения, роста, развития и
трофических отношений организмов в- своеобразных
арктических условиях. Необходима разработка
акклиматизационных и реакклиматизационных
мероприятий в широких масштабах.
Все большее значение приобретает проблема
рекультивации земель на Крайнем Севере, например
восстановление сильно потравленных и выбитых
оленьих пастбищ, почвенного покрова на местах
строек, промышленных комплексов, добывающих
предприятий. Для этого потребуется детальное изучение
закономерностей экологических сукцессии в
экосистемах Субарктики. Только глубокое знание законов
протекания сукцессии в экстремных условиях
позволит правильно ориентировать восстановительные
процессы.
Освоение Севера ставит множество сложнейших
медико-биологических проблем. Жизнь человека в
экстремных условиях Заполярья сопровождается
напряженностью адаптационных процессов в
организме. В связи с этим особенно большое значение
приобретают методы профессионального отбора для работы
на Крайнем Севере, ранняя, а затем систематическая'
диагностика состояния человека в новых условиях и
разработка оптимальных профилактических
мероприятий. Необходимо дальнейшее всестороннее развитие
исследований экологии человека в условиях Арктики
и Субарктики. На территории Заполярья, в том числе
и в тундровых районах, выявлены предпосылки
возникновения природно-очаговых болезней (туляремии,
лептоспироза). В связи с этим нужно изучить
возможности их развития при дальнейшем росте плотности
населения, а также разработать методы
профилактики. Все более очевидной становится необходимость
всемерно учитывать гигиенические и экологические
критерии при проектировании
территориально-производственных комплексов в Заполярье.
Наконец, при использовании ресурсов Заполярья
и воздействии на его природу следует иметь в виду,
что Арктика оказывает огромное влияние на климат
и погоду всей Земли. От состояния полярных морских
льдов и ледников зависит уровень Мирового океана.
Изменения температурного режима арктических тер-
231
риторий резко отражаются на циркуляции атмосферы
и влагообороте в более южных районах и могут
вызвать засухи или периоды повышенного увлажнения
в умеренных широтах. Поэтому неблагоприятные
влияния человека на заполярные ландшафты, на воды,
почвы, мерзлые грунты, атмосферу чреваты
серьезными последствиями для всей нашей планеты. В связи
с этим перед советскими учеными стоит задача
дальнейшего совершенствования единой комплексной
системы освоения природных ресурсов страны.
ЛИТЕРАТУРА
Александрова В. Д. Влияние снежного покрова на растительность
в арктической тундре Якутии. — В сб.: Материалы по
растительности Якутии. Л., 1961.
Александрова В. Д. Геоботаническое районирование Арктики
и Антарктики. Л., 1977.
Андреев А. В. Зимняя жизнь и питание тундряной куропатки
на крайнем Северо-Востоке СССР. — «Зоол. журн.», 1975, т. L1V,
вып. 5.
Большаков В. Н. Пути приспособления мелких
млекопитающих к горным условиям. М., 1972.
Бируля А. А. Очерки из жизни птиц полярного побережья
Сибири. —«Зап. Акад. наук, сер. 8», 1907, т. XVIII, № 2.
Будыко М. И. О закономерностях поверхностного физико-
географического процесса. —«Метеорология и гидрология», 1948,
№4.
Вангенгейм Э. А. Палеонтологическое обоснование
стратиграфии антропогена Северной Азии. М., 1977.
Говорухин В. С. Пятнистые тундры и пликативные почвы
Севера. —«Землеведение», 1960, вып. 5.
Городков Б. Н. Вечная мерзлота и растительность. — В сб.:
Вечная мерзлота. М. — Л., 1930.
Городков Б. Н. Растительность Арктики и горных тундр
СССР. —В кн.: Растительность СССР, т. 1. М. — Л., 1938.
Городков Б. Н. Морозная трещиноватость грунтов на
Севере. —«Изв. Всес. геогр. об-ва», 1950, т. 82, вып. 5.
Григорьев А. Л. Субарктика. М., 1956.
Данилов Н. Н. Пути приспособления наземных позвоночных
животных к условиям существования в Субарктике. Птицы.
Свердловск, 1966.
Дорогостайская Е. В. Сорные растения Крайнего Севера
СССР. Л., 1972.
Дунаева Т. Н. Сравнительный обзор экологии тундровых
полевок полуострова Ямал. —«Труды Ин-та географии АН СССР»,
1948, вып. XLI.
Дунаева Т. Н. К экологии леммингов в северо-западной
Якутии. —«Бюлл. Моск. об-ва испыт. прир., отд. биол.», 1978,
т. 83, вып. 2.
Житков Б. М. Полуостров Ямал. —«Зап. Русск. геогр. об-ва»,
1913.
Кищинский А. А. Современное состояние популяции дикого
северного оленя на Новосибирских островах. —«Зоол. журн.»,
1971, т. L, вып. 1.
Калесник С. В. Общие географические закономерности Земли.
М., 1970.
Караваев М. Н. Конспект флоры Якутии. М. — Л., 1958.
Кернер А, Жизнь растений. Спб., 1903.
233
Кренмар А. В. Птицы Западного Таймыра. —«Труды Зоолог.
йн-ta АН СССР», 1966, т. XXXIX.
Крючков В В. Чуткая Субарктика М, 1976.
Лозина-Лозинский Л. /С Очерки по криобиологии. Л., 1972.
Матвеева Н. В., Чернов Ю. И. Арктические тундры на
северо-востоке Таймыра.—«Бот. журн.», 1977, т 62, № 7.
Миддендорф А. Ф. Путешествие на север и восток Сибири.
Спб., 1860—1867.
Мильков Ф. Н., Гвоздецкий Н. А. Физическая география
СССР. М., 1976.
Осмоловская В. И. Экология хищных птиц полуострова
Ямал.— «Труды Ин-та географии АН СССР», 1948, вып. XL1.
Павлов Б. М. Белая и тундряная куропатки Таймыра. Авто-
реф. канд. дис. М., 1974.
Паринкина О. М Биологическая продуктивность
бактериальных сообществ тундровых почв. — В сб.: Биогеоценозы
таймырской тундры и их продуктивность, 2. Л., 1973.
Полозова Т. Г., Тихомиров Б. А. Сосудистые растения
района таймырского стационара. — В сб.: Биогеоценозы таймырской
тундры и их продуктивность. Л., 1971.
Рихтер Г. Д. Снежный покров, его формирование и свойства.
М — Л., 1945.
Рябчиков А. М. Структура и динамика геосферы и ее
естественное развитие и изменение человеком. М., 1972.
Сдобников В. М. По арктической тундре. М., 1953.
Семенов-Тян-Шанский О. И. Северный олень. М, 1977.
Симченко Ю. Б. Культура охотников на оленей северной Ев*
разии. М., 1976.
Соколова Т. Г. К вопросу об использовании чукотским
населением дикой флоры в районе мыса Дежнева. —«Зап.Чукот. крае*
вед. музея». Магадан, 1961.
Стрельников И. Д. Значение солнечной радиации в экологии
высокогорных насекомых. —«Зоол. журн.», 1940, т. XIX, вып. 2.
Сыроечковский Е. В. Размеры лебедей, гусей и казарок в
связи с адаптацией к полярным условиям.—«Зоол. журн.», 1978,
т. LVII, вып. 5.
Тахтаджян А. П. Флористические области Земли. Л., 1978.
Тихомиров Б. А. Взаимосвязи животного мира и
растительного покрова тундры. М. — Л., 1959.
Тихомиров Б. А, Очерки по биологии растений Арктики.
М.— Л., 1963.
Толмачев А. И. Введение в географию растений. Л., 1974.
Тугаринов А. #. О происхождении арктической фауны. —
«Природа», 1928, № 7—8.
Уошберн А. П. Классификация структурных грунтов и обзор
теорий их происхождения. — В кн.: Мерзлые горные породы
Аляски и Канады. М., 1958.
Филиппова Л. Н. Проблемы залужения тундры в
Ямало-Ненецком национальном округе. —«Труды Науч.-исслед. ин-та сельск.
хоз-ва Крайнего Севера», т. XI. Норильск, 1963.
Фридолин В. Ю. Животно-растительное сообщество горной
страны Хибин. — «Труды Кольской базы АН СССР», 1936, 3.
Чернов Ю. И. Комплекс антофильных насекомых в тундровой
зоне. — «Вопросы географии», 1966, вып. 69.
234
Чернов Ю. И. Трофические связи птиц с насекомыми в
тундровой зоне. —«Орнитология», 1967, вып. 8.
Чернов Ю. И. Природная зональность и животный мир суши.
М., 1975.
Чернов Ю. И. Структура животного населения Субарктики.
М., 1978.
Чернов Ю. И. Антофильные насекомые в подзоне типичных
тундр Западного. Таймыра и их роль в опылении растений.—
В сб.: Структура и функции биогеоценозов таймырской тундры.
Л., 1978.
Шамурин В. Ф. Сезонный ритм и экология цветения растений
тундровых сообществ на севере Якутии. — В сб.: Приспособление
растений Арктики к условиям среды. М. — Л., 1966.
Шварц С. С. Пути приспособления наземных позвоночных
животных к условиям существования в Субарктике, т. 1.
Млекопитающие. Свердловск, 1963.
Швецова В. М., Вознесенский В. Л. Суточные и сезонные
изменения интенсивности фотосинтеза у некоторых растений
Западного Таймыра. — «Бот. журн.», 1970, т. 55, № 1.
Шилова С. А., Денисова А. В. и др. О последействии
применения инсектицидов в Субарктике. — «Зоол. журн.», 1973, т. LII,
вып. 7.
Юдин Б. С, Кривошеее В. Г., Беляев В. Г. Мелкие
млекопитающие севера Дальнего Востока. Новосибирск, 1976.
Юрцев Б. А. Гипоарктический ботанико-географический пояс
и происхождение его флоры. —«Комаровские чтения», 1966, XIX.
Юрцев Б. А. Проблемы ботанической географии
Северо-Восточной Азии. Л., 1974.
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ 3
ЧТО ТАКОЕ ТУНДРА? 6
ТЕПЛО И ВЛАГА В ТУНДРЕ 12
РАЗНООБРАЗИЕ ЛАНДШАФТОВ 23
Подзоны —
Рельеф и мерзлота 39
РОЛЬ СНЕГА В ЖИЗНИ ТУНДРЫ 63
ПРИСПОСОБЛЕНИЯ ОРГАНИЗМОВ К УСЛОВИЯМ
ТУНДРОВОЙ ЗОНЫ 82
Уникальны ли условия жизни в тундре? ... —
Психрофиты и криофиты 86
Существует ли единый путь адаптации животных к
условиям тундры? 89
Роль истории видов 92
Роль размеров тела 96
Принцип экономии поверхности тела 105
Пути формирования флоры и фауны 109
РАСПРОСТРАНЕНИЕ ЖИВОТНЫХ И РАСТЕНИЙ . . 120
ВЗАИМООТНОШЕНИЯ ОРГАНИЗМОВ 141
Животные и микроскопические растения —
Животные и лишайники 144
Животные и мхи 150
Птицы, звери и цветковые растения 152
Растительноядные насекомые 158
Насекомые-опылители и цветки 172
Птицы и беспозвоночные животные 182
Хищные птицы 197
Кого больше в тундре — хищников или фитофагов? 210
ТУНДРА И ЧЕЛОВЕК 216
ЛИТЕРАТУРА
233
Чернов Ю. И,
4-49 Жизнь тундры, — М.: Мысль, 1980. —236 с,
ил., карт., схем.
80 к.
Автор, известный биогеограф и эколог, обобщил результаты
своей многолетней работы в различных районах Субарктики, а
также проанализировал классическую и новейшую литературу по
проблемам Севера. На оригинальном материале впервые
анализируются многие вопросы приспособления организмов к условиям
Субарктики, структуры фауны и флоры, взаимоотношения
различных групп организмов в этой специфической природной среде.
Рассматриваются проблемы биологической продуктивности тундр
и подходы к рациональному использованию природных
ресурсов.
57.026
Ч Ш"М_ 132-79
004(01)-80
ИБ №> 760
Чернов Юрий Иванович
ЖИЗНЬ ТУНДРЫ
Заведующий редакцией А. П. Воронин
Редактор Ю. П. Митяева
Редактор карт Д. Г. Фаттахова
Младший редактор С. И. Ларичева
Оформление художника Т. К. Самигулина
Художественный редактор С. М. Полесицкая
Технический редактор О. А. Барабанова
Корректор Н. С. Приставко
Сдано в набор 29.12.78. Подписано в печать 31.07.79. А08177. Формат
84х1087з2. Бумага типограф. № 2. Литерат. гарнит. Высокая печать. Усл.
печатных листов 12,6. Учетно-издательских листов 12,1. Тираж 30 000 экз.
Заказ JSfe 384. Цена 80к.
Издательство «Мысль». 117071. Москва, В-71, Ленинский проспект, 15.
Ордена Октябрьской Революции, ордена Трудового Красного Знамени
Ленинградское производственно-техническое объединение «Печатный Двор»
имени А. М Горького «Союзполиграфпрома» при Государственном
комитете СССР по делам издательств, полиграфии и книжной торговли.
197136, Ленинград, П-136. Чкаловский пр., 15.