








































































Author: Мори Ф.Р.
Tags: лесное хозяйство лесоводство биология ботаника дендрология рост деревьев ксилология общая ботаника эволюция древесных форм
Year: 1980




Филип Р Мори Как
растут
Филип Р Мори
Как
растут
ДЕРЕВЬЯ
Перевод с английского
А. Ю. Клячко
Москва
Издательство «Лесная промышленность»
1980
УДК 630*161.4 (03.20)
The Institute of Biology’s
Studies in Biology no. 39
How Trees Grow
by Philip R. Morey Ph. D.
Associate Professor of Biology
Texas Tech. University
London, Edward Arnold
Мори Ф. P. Как растут деревья.— Пер. с англ. Л. Ю. Кляч-
ко.—М.: Лесная пром-сть, 1980.—80 с.
В этой небольшой книге суммированы основные данные
о росте дерева. На современном научном уровне известный аме-
риканский физиолог Ф. Р. Мори излагает анатомию и морфоло-
гию ствола, корня и побегов, подробно рассматривает процесс об-
разования и химический состав древесины и коры. Автор обоб-
щил работы, посвященные роли регуляторов роста в иницииро-
вании камбиальной активности и дифференциации различных
тканей у древесных растений; он отметил основополагающее зна-
чение ауксина в образовании креневой древесины у голосеменных
растений и тяговой у покрытосеменных. Представляют интерес
палеоботанические данные о происхождении и эволюции древес-
ных форм.
Книга представляет большой интерес для инженерно-техниче-
ских работников лесоводственного профиля, студентов и препо-
давателей, а также для озеленителей и садоводов, желающих оз-
накомиться с биологией древесных растений.
Табл. 2, ил. 29, библиогр.— 68 назв.
40502—139
М 039(01)—80 БЗ—71—14—80 3903000000
© Philip R. Morey, 1973, reprinted 1976, 1978
© Перевод на русский язык. Издательство «Лесная промышлен-
ность», 1980
Предисловие
к русскому
изданию
Основная задача лесного хозяйства — ускорение роста
и повышение продуктивности лесов. Для управления про-
цессами роста и развития деревьев нужно хорошо знать
их морфологию, анатомию и физиологию. Предлагаемая
книга Филипа Ричарда Мори — хорошее пособие по ос-
новам роста дерева, впервые изданное в серии «Изучение
биологии» в 1973 г.
Процессы роста и развития растений издавна были
в центре внимания многих ученых мира. Большой вклад
внесли в науку Ю. Сакс, К. А. Тимирязев, В. В. Гарнер,
Г. А. Аллард, Н. Г. Холодный, Ф. В. Вент, Д. А. Сабинин
и др. Интенсивному расширению знаний о предмете спо-
собствовала научно-техническая революция, вызвавшая
к жизни высокочувствительные физико-химические, ки-
бернетические и прочие методы исследований.
Ф. Мори — один из ведущих американских исследова-
телей гормональной регуляции роста деревьев. Эта тема
проходит красной нитью через всю книгу при рассмотре-
нии таких вопросов, как рост дерева в высоту, иницииро-
вание камбиальной активности и вторичного роста кор-
ней, регуляция образования ксилемы и флоэмы, годич-
ного кольца, установление причин образования реактив-
ной древесины, определение перспективных направлений
в исследованиях коры. Даже в заключительном палеобо-
таническом экскурсе Ф. Мори показывает значение аук-
сина для эволюции побегов и образования годичных ко-
лец. При изложении материала о происхождении и эво-
люции древесных форм автор использует данные о со-
держании лигнина у разных растительных групп из
монографии С. М. Манской и Т. В. Дроздовой «Геохимия
органического вещества», переведенной в США в 1968 г.
Клячко Л. Ю.
3
Общее
предисловие
к серии
В настоящее время невозможно в одном руководстве
охватить целую область биологии, поэтому Институт био-
логии начал издание этой серии, чтобы дать представле-
ние преподавателям и студентам об основных направле-
ниях работ. Восторженный прием опубликованных вы-
пусков серии «Изучение биологии» показывает, что
такие книги обеспечивают достаточно полное рассмотре-
ние биологических тем. Основная задача серии — дать
рекомендации по применению тех или иных методов
и выбору литературы для дальнейшего чтения, а также
информацию для практической работы.
Замечания читателей будут с благодарностью приняты
учебной комиссией института.
1978 Институт биологии, Лондон
Предисловие
Литература по росту деревьев так разнообразна, что
неспециалисту просто невозможно составить общее пред-
ставление об этом предмете. Настоящая книга объясняет
основные процессы развития, лежащие в основе роста
деревьев, в том числе рост в длину и радиальный прирост
стебля и корня, образование коры. Кроме того, здесь рас-
сматриваются структура древесины и последовательность
ее развития, включая дифференциацию древесных кле-
точных оболочек, а также эволюция форм роста деревьев,
что прослеживается по палеонтологической летописи.
Задача этой книги — ознакомление учащихся, исследо-
вателей нелесных специальностей и любителей с состоя-
нием современных научных представлений о росте дре-
весных растений. Надеемся, что вопросы, обсуждаемые
в нашем коротком обзоре, заинтересуют читателя, и он
пожелает ознакомиться с литературой, предложенной для
дальнейшего чтения.
Лаббок, шт. Техас, 1973 г. Филип Ричард Мори
4
1. Рост в высоту
1.1. МОРФОЛОГИЯ РОСТА В ВЫСОТУ
Все многообразие форм роста побегов у деревьев уме-
ренных широт обычно сводят в четыре типа (Zimmer-
mann and Brown, 1971). Для первого типа характерен
рост в высоту за один цикл, что свойственно клену, ясе-
ню, каштану конскому, пихте и некоторым видам сосны
(например, Pinus lambertiana) У зимующей верхушеч-
ной почки зачатки всех органов находятся в эмбриональ-
ном состоянии. Весной почки распускаются и побеги на-
чинают удлиняться и развиваться. Рост в высоту у де-
ревьев этого типа происходит в течение короткого и рано
заканчивающегося периода роста. У других сосен (напри-
мер, у Pinus taeda) рост повторный с двумя или боль-
шим числом отчетливых циклов удлинения побегов,
разделенных периодами покоя. Третий тип роста побегов,
называемый непрерывным, свойствен деревьям (на-
пример, Ginkgo), развивающим заранее сформированный
побег сразу после завершения роста в длину, включая
заложение и морфогенез новых листьев. У деревьев, отно-
сящихся к четвертому типу, после распускания почки
и окончания периода роста в длину верхняя часть кончи-
ка побега обычно опадает (сирень обыкновенная, акация
белая, вяз и др.). Опавшую главную ветвь замещает
побег, образованный из верхней пазушной почки, в ре-
зультате его ось приобретает типичную у-форму; это—
симподиальный характер роста.
Ход развития побегов из почек у карии Бу клея (центр
США), пихты одноцветной и сосны Ламберта (виды
пихты и сосны с запада США), у которых бывает обыч-
но только один годичный цикл роста в высоту, рассмот-
рим поподробнее.
1 Сосна обыкновенная и дуб черешчатый также относятся к это-
му типу роста (Прим, перев.)
5
У карии Буклея (Сагуа buckleyi var. arkansana) мо-
ноподиальная система побегов характеризуется тем, что
укороченные боковые ветви вырастают из пазушных по-
чек, расположенных немного ниже кончика побега на
выделяющемся главном стволе. Доминирующая ось на-
зывается удлиненным побегом, а боковые — уко-
роченными. Верхушечные почки удлиненных побе-
гов, покрытые сверху 9—11 почечными чешуями, состоят
из 5—11 зачатков листьев и 2 дополнительных маленьких
зачатков почечных чешуй, непосредственно граничащих
с апикальной меристемой (Foster, 1931).
Почечные чешуи, или катафиллы (нем. Nie-
derblatt — нижние листья; Romberger, 1963),—это видо-
измененные листья, основная функция которых заключа-
ется в защите от пересыхания зимующей почки, так как
в результате замерзания почвенная влага становится
недоступной для корневой системы. Годичный прирост
в высоту определяют по системно расположенным на
стволе зонам сближенных рубцов от почечных чешуй
(рис. 1). Большие расстояния между рубцами от после-
довательно образующихся листьев характеризуют энер-
гичный рост удлиненных побегов. Зимующая верхушечная
почка укороченных побегов карии Буклея обычно состоит
из 9 наружных почечных чешуй, 4 зачатков листьев
и 2 зачатков почечных чешуй; этот орган легко поддается
экспериментальному исследованию (Foster, 1932).
Верхушечные почки на укороченных побегах с 22 мар-
та начинают набухать, а наружные почечные чешуи
с этого же времени и до 22 апреля постепенно отгибают-
ся кнаружи, из-за чего молодые листья, еще не начавшие
рост в длину, остаются незащищенными. В середине ап-
реля междоузлия заметно удлиняются и листья разви-
ваются, а к концу месяца почечные чешуи опадают.
В апикальной меристеме в период роста побега быстро
сменяются органообразовательные процессы. В конце
марта из 2 зимующих зачатков почечных чешуй апикаль-
ной меристемы развиваются почечные чешуи, располага-
ющиеся в верхнем наружном слое верхушечной почки,
распускающейся в следующем году. К середине апреля
образуется еще 7 почечных чешуй, затем развиваются
2 маленьких зачатка их и 4 зачатка листьев. Новая вер-
хушечная почка, теперь хорошо освещаемая в результате
распускания и роста в длину побега текущего года, сло-
жена из ожидаемых 9 наружных почечных чешуй, 4 за-
6
Почечная
чешуя
Пазушная
почка
Постовой
рубец
Рубец
от почечной
чешуи
РИС. 1. Морфология побе-
гов карии Бук лея (Foster.
1931):
а' — удлиненного, •— XI, 0,5;
б — укороченного — XI, 8 с
2-летним приростом (междоуз-
лия короче, чем у удлиненных
побегов)
За чатни чешуи
верхушечной
почни
боновая
почна
Унороченный
побег
Стерильные
почечные
чешуи
X
Чешуи я,
верхушечной
почни
РИС. 2. Схема продольного среза верхушечной почки пихты од-
ноцветной с зачатками хвои и почечными чешуями
РИС. 3. Строение нераспустившейся верхушечной почки Pinus.
Самые верхние стерильные (без пазушных образований) почеч-
ные чешуи называют чешуями верхушечной почки. Почечные
чешуи, постепенно приближаясь к вершине побега, защищают
сначала укороченные побеги, а затем и боковые почки. Зачаточ-
ные чешуи верхушечной почки появляются по соседству с апи-
кальной меристемой. Чешуи верхушечной почки закладываются
за 2 года до распускания (Sacher, 1954)
чатков листьев и 2 недоразвитых внутренних почечных
чешуй. После этого органогенез в апикальной меристеме
прекращается до следующей весны. Итак, у кари1и Бук-
лея верхушечная почка образуется с середины марта до
середины мая, захватывая только небольшую часть пери-
ода роста в Оклахоме.
У пихты одноцветной (Abies concolor) зимующая
верхушечная почка состоит из 20—30 почечных чешуй,
окружающих заложившийся побег с 50—60 зачатками
хвои (рис. 2, Parke, 1959). Распускание почек начинает-
ся в апреле с удлинения междоузлий с хвоей. Почечные
чешуи, плотно покрывающие быстро удлиняющуюся вер-
хушечную почку, отходят от узлов и со временем опада-
ют. В апикальной меристеме органогенез начинается
с образования зачатков почечных чешуй, только когда
новый побег достигает нескольких сантиметров. Органо-
генез зачатков чешуй продолжается до середины июня,
после чего закладываются зачатки хвои. Переход от об-
разования зачатков чешуй к образованию зачатков хвои
коррелирует с увеличением самой апикальной меристемы
и уменьшением прироста в длину у побега последнего
года. К сентябрю в новую верхушечную почку входит за-
ложившийся побег с 50—60 зачатками хвои, окруженны-
ми оболочкой |из 20—30 почечных чешуй. В отличие от
карии Буклея у пихты одноцветной органообразователь-
ная стадия развития длится большую часть периода роста
и не коррелирует тесно с ростом в длину побега послед-
него года.
Зимующая почка у сосны Ламберта (Pinus lamberti-
апа), как и у пихты одноцветной, состоит из предвари-
тельно сформированного заложившегося побега. Но отме-
чаются и некоторые важные конструктивные различия.
Хвоя образуется на оси главного побега не сразу. После-
довательная органообразовательная активность апикаль-
ной меристемы направлена на формирование почечных
чешуй, которые располагаются в определенном порядке
от нижней до верхней части нераспустившейся почки
(Sacher, 1954). Зимующая почка обернута многочислен-
ными расположенными по спирали стерильными че-
шуями (без пазушной почки), из которых самые верх-
ние известны как верхушечные почечные че-
шуи (рис. 3). Следующие серии почечных чешуй содер-
жат в своих пазухах укороченные побеги, на которых
в свою очередь образуются зачатки хвои. Система боковых
8
ветвей у нераспустившейся верхушечной почки образует-
ся из пазушных почек почечных чешуй, занимающих
еще более высокое положение на оои. И наконец, боль-
шинство соседних с апикальной меристемой зачатков
стерильных почечных чешуй зимующей почки следующе-
го года становятся верхушечными почечными чешуямш
У сосны Ламберта рост верхушечной почки начинает-
ся в начале апреля с роста в длину междоузлий главной
оси, удлинения и быстрого развития зачатков хвои на
укороченных Побегах. Позднее в этом же месяце апикаль-
ная меристема начинает закладывать новые стерильные
почечные чешуи. Вместе с зачатками верхушечных по-
чечных чешуй зимующей почки они развиваются за пе-
риод роста и становятся кроющими защитными чешуями
зимующей верхушечной почки следующего года. Вслед
за образованием стерильных почечных чешуй иницииру-
ется ряд почечных чешуй, каждая из которых покрывает
укороченный побег, а затем и боковые почки. Наконец
поздней осенью, когда активность роста апикальной ме-
ристемы ослабевает, развиваются маленькие зачатки сте-
рильных почечных чешуй. Через год они становятся
верхушечными почечными чешуями зимующей почки
следующего года.
О механизмах, ответственных за рост в длину междо-
узлий, и сезонной динамике образования стерильных по-
чечных чешуй, почечных чешуй, покрывающих укорочен-
ные побеги и боковые почки, и чешуй без пазушных по-
чек известно немного; наши знания носят в основном
описательный характер. Для дальнейшего углубленного
ознакомления с морфогенезом побегов отсылаем читателя
к подробным работам Romberger (1963) и Zimmermann
and Brown (1971).
Для некоторых пород деревьев умеренных широт,
например у Ginkgo, Populus, характерно запоздалое об-
разование верхушечных почек, рост побегов у них про-
должается до конца сезона. Листья на удлиненных побе-
гах Ginkgo biloba, распускающиеся из верхушечных по-
чек, называют ранними; по морфологии они значительно
отличаются от листьев, развивающихся позже и называ-
емых поздними (Critchfield, 1970). Это явление отно-
сят к гетероф и ллии (разнолистности). Ранние ли-
стья у Ginkgo — типично цельные или двулопастные, от-
личающиеся по форме от многолопастных поздних листь-
ев (рис. 4). У сеянцев этого вида листья морфологически
2 2640
9
очень похожи на поздние листья удлиненных побегов
взрослых деревьев.
Все заранее заложенные зачатки листьев в зимующей
почке Ginkgo на удлиненных и укороченных побегах
развиваются в ранние л!истья. Удлиненные побеги стано-
вятся отличимыми от укороченных только после их рас-
пускания, когда закладываются новые зачатки листьев
и удлиняются междоузлия. Усиленный прирост удлинен-
ных побегов проходит летом до формирования новой вер-
хушечной зимующей почки. Многолопастные поздние
а б
РИС. 4. Листья Gin-
kgo biloba:
а — ранние; б — позд-
ние (у них пластинка
сильно рассечена)
листья закладываются апикальной меристемой и разви-
ваются за один сезон роста. Critchfield (1970) считает,
что гетерофиллию лучше всего различать по срокам сле-
дования онтогенеза листьев. Для поздних листьев Ginkgo,
как и для листьев сеянцев, характерно непрерывное раз-
витие от заложения до состояния зрелости. С другой сто-
роны, развитие ранних листьев прерывается периодом
зимнего покоя, необходимого для их заложения и дости-
жения зрелости.
Гетерофиллия отмечается также у пород с одним
циклом роста. Особые условия (например, сильный
дождь) могут стимулировать повторный цикл роста. По-
бег, появившийся в результате повторного распускания
почки, называют ламмасовым, так как его возникновение
часто приурочено к дню Ламмаса (1 августа). Листья
ламмасовых побегов по форме отличаются от листьев
обычного цикла роста. Гетерофиллия здесь, по-видимому,
обусловлена различиями онтогенеза у развивающихся
листьев.
1.2. РОСТ В ВЫСОТУ ТРОПИЧЕСКИХ ДЕРЕВЬЕВ
В некоторых тропических регионах для большинства
пород характерен прерывистый рост побегов даже в по-
стоянно благоприятных условиях. Так, дерево какао
10
(Theobroma cacao) растет ритмично: периоды роста
в длину его побегов, включающие всю активность орга-
ногенеза, чередуются с периодами покоя (Greathouse, La-
etsch and Phinney, 1971). Ритмичный рост побегов у ка-
каового дерева продолжается даже в камере при регули-
руемых условиях среды: 12-часовом фотопериоде и по-
стоянном температурном режиме, обусловливающих про-
текание внутренних процессов. Однако система побегов
всего дерева растет асинхронно: одни из них растут,
другие — находятся в состоянии покоя. Поэтому даже при
ритмичном росте побегов регуляция внутренних процес-
сов у отдельных ветвей отличается от таковой у деревьев
умеренных широт.
Borchert (1969) описывает интересный ритмичный
характер роста побегов у Огеорапах, вечнозеленого дерева
тропиков, встречающегося в горах Колумбии; почечные
чешуи у него — составная часть нормальной динамики
роста. В период активного роста развиваются листья
больших размеров и побеги сильно удлиняются. Размеры
позже появляющихся листьев постепенно уменьшаются,
и также постепенно уменьшается прирост по длине несу-
щих их междоузлий. Наконец при наступлении состоя-
ния покоя на вершинке побега закладывается розетка из
листьев самых маленьких размеров с сильно утолщенным
основанием черешков. Почечные чешуи в этой стадии по-
являются только вблизи апикальной меристемы в виде
очень маленьких зачатков. Отличие развития Огеорапах
состоит в том, что у него вершинка побега вступает
в состояние покоя, когда почечные чешуи еще недораз-
виты. Верхушечная почка, покрытая зрелыми чешуями,
при возобновлении роста появляется из розетки. При
этом растут и другие почечные чешуи и связанные с ни-
ми междоузлия. От появления верхушечной почки до ее
распускания обычно проходит 6 недель. Все это время
при быстром усилении активности органогенеза вершйн-
ка побега покрыта защитными почечными чешуйками,
которые опадают после распускания верхушечной почки
и роста нового облиственного побега. Огеорапах с нахо-
дящейся в состоянии покоя верхушкой побега отличается
от древесных растений умеренной зоны тем, что у него
нет заранее заложенных зародышевых плоских листьев.
В связи с этим ему свойствен более длительный период
развития новых зародышевых листьев побега после вы-
хода его из состояния покоя.
11
Как и у пород умеренных широт, побег Огеорапах
растет ритмично и формирует почечные чешуи. Однако
период покоя у него начинается до развития верхушеч-
ной почки. Borchert предполагает, что покоящиеся почки
могли возникнуть у растений в областях, пограничных
с тропиками, и что их переселен'ие в экваториальные зо-
ны с относительно постоянными условиями роста могло
обусловить сложные процессы выпадения стадии образо-
вания типичных покоящихся верхушечных почек.
1.3. РОЛЬ АУКСИНА
Давно известно, что ауксин участвует в регуляции
роста побегов в длину. Так, улавливаемый ауксин, на-
пример у Ginkgo, получают из растущих побегов и осо-
бенно из растущих в длину междоузлий, в то время как
в зимующих почках его нет.
Система побегов у Ginkgo biloba состоит из укорочен-
ных (примерно 10 мм) и удлиненных (от 2 до 75 см)
побегов (Critchfield, 1970). На ранних этапах развития
заложившиеся удлиненные и укороченные побеги очень
похожи; они мало различаются по размерам или внешне-
му виду во время распускания почек в начале апреля
(Массачусетс, США). После раскрывания и опадания
в мае почечных чешуй быстро вырастают ранние листья
при незначительном (или отсутствии) росте в длипу
междоузлий. Теперь становится очевидным путь разви-
тия укороченных и удлиненных побегов, так как рост
первых заканчивается формированием 8—14 почечных
чешуй. В это же время междоузлия удлиненных побегов
начинают быстро расти, активная апикальная меристема
инициирует появление новых листьев — поздних. Быст-
рый рост в длину продолжается и в июне с образованием
новых листьев и междоузлий.
По мере распускания почек из кончиков за ложивших-
ся удлиненных и укороченных побегов можно получить
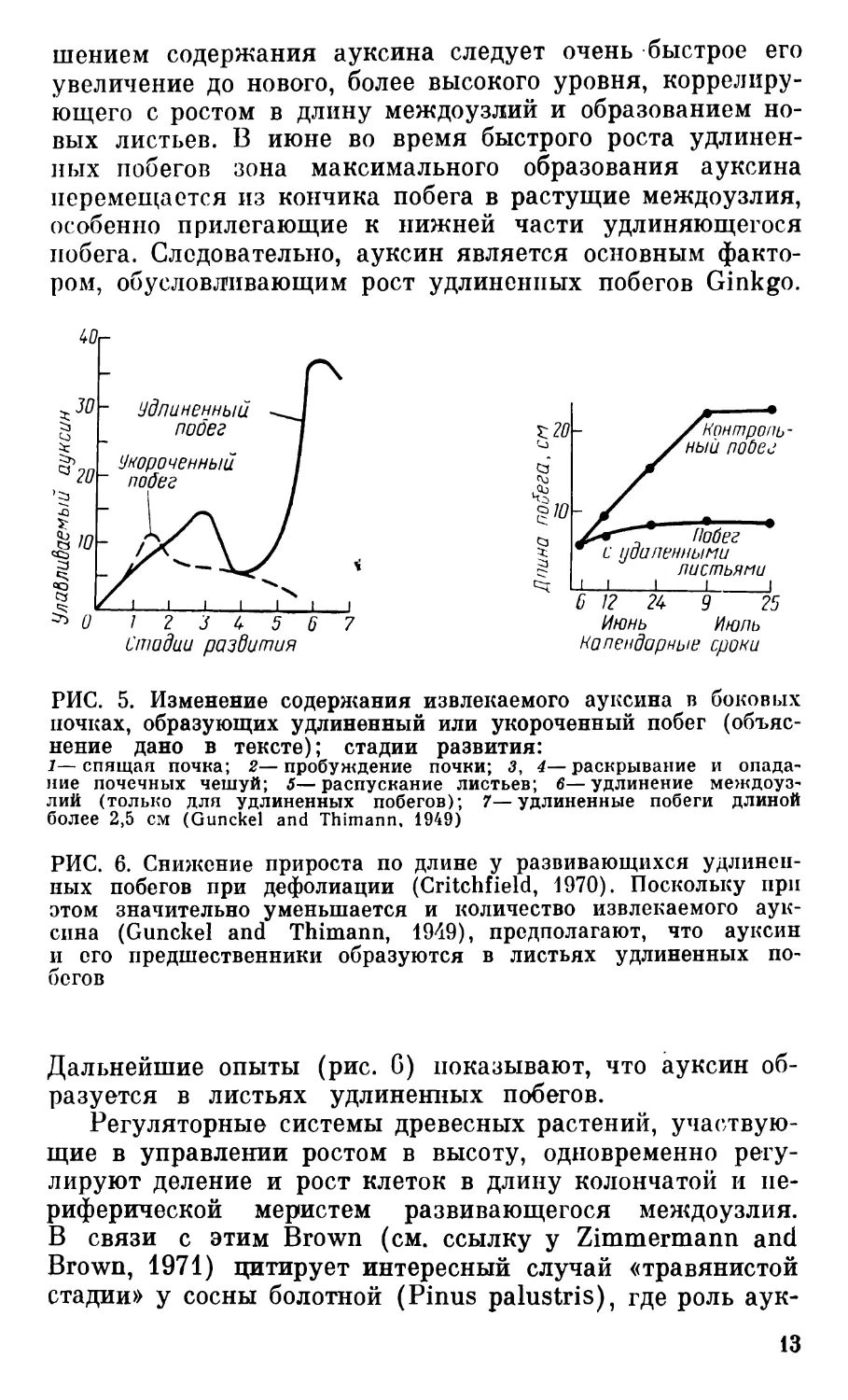
улавливаемый ауксин (рис. 5). Содержание его увеличи-
вается и достигает максимума в конце апреля; в это же
время дифференцируется ксилема жилок зародышевых
листьев, еще находящихся в почках (Critchfield, 1970).
Впоследствии, при распускании почек уровень улавли-
ваемого ауксина в укороченных побегах, определяемых
по прекратившим рост в длину междоузлиям, постепенно
снижается; в удлиненных побегах за временным умень-
1?
шением содержания ауксина следует очень быстрое его
увеличение до нового, более высокого уровня, коррелиру-
ющего с ростом в длину междоузлий и образованием но-
вых листьев. В июне во время быстрого роста удлинен-
ных побегов зона максимального образования ауксина
перемещается из кончика побега в растущие междоузлия,
особенно прилегающие к нижней части удлиняющегося
побега. Следовательно, ауксин является основным факто-
ром, обусловливающим рост удлиненных побегов Ginkgo.
Стадии разбития
РИС. 5. Изменение содержания извлекаемого ауксина в боковых
почках, образующих удлиненный или укороченный побег (объяс-
нение дано в тексте); стадии развития:
1—спящая почка; 2—пробуждение почки; 3, 4—раскрывание и опада-
ние почечных чешуй; 5—распускание листьев; б—удлинение междоуз-
лий (только для удлиненных побегов); 7—удлиненные побеги длиной
более 2,5 см (Gunckel and Thimann, 1949)
Июнь Июль
Календарные срони
РИС. 6. Снижение прироста по длине у развивающихся удлинен-
ных побегов при дефолиации (Critchfield, 1970). Поскольку при
этом значительно уменьшается и количество извлекаемого аук-
сина (Gunckel and Thimann, 1949), предполагают, что ауксин
и его предшественники образуются в листьях удлиненных по-
бегов
Дальнейшие опыты (рис. G) показывают, что ауксин об-
разуется в листьях удлиненных побегов.
Регуляторные системы древесных растений, участвую-
щие в управлении ростом в высоту, одновременно регу-
лируют деление и рост клеток в длину колончатой и пе-
риферической меристем развивающегося междоузлия.
В связи с этим Brown (см. ссылку у Zimmermann and
Brown, 1971) цитирует интересный случай «травянистой
стадии» у сосны болотной (Pinus palustris), где роль аук-
13
сина в регуляции роста в длину установлена более точно.
До 15 лет главный побег этой сосны ежегодно растет как
укороченный, в это время сильно ингибировано удлинение
междоузлий. Brown отмечает, что ауксин можно выде-
лить из главного укороченного побега, но для этого нель-
зя использовать диффузионный способ. Иначе говоря,
в травянистой стадии отсутствует полярное перемещение
ауксина: его невозможно диффундировать в агаровый
блок.
Нанесение индолилуксусной кислоты (ИУК) па апекс
побега вызывает отмирание меристематической ткани;
этим в основном объясняется отсутствие перемещения
гормона в укороченных побегах, так как он накапливает-
ся в токсичных для роста количествах. В травянистой
стадии укороченный побег, конечно, не полярен; во время
превращения его в удлиненный побег убедительно дока-
зано наличие ауксина в главном побеге сосны болотной.
В связи с этим основные черты укороченного побега свя-
зывают с отсутствием полярного перемещения ауксина.
Механизм стимуляции ауксином роста в длину с поляр-
ным его перемещением через эмбриональную ткань меж-
доузлий — это задача современных физиологических ис-
следований, которая выходит за пределы компетенции
нашей книги (см. Wareing and Phillips, 1970).
1.4. СОСТОЯНИЕ ПОКОЯ
Для большинства древесных растений характерны пе-
риоды приостановки деятельности точек роста или состоя-
ние покоя. У пород средних широт состояние покоя имеет
три последовательно сменяющиеся фазы: коррелятивно-
го ингибирования, глубокого и затем вынужденного покоя
(Romberger, 1963; Wareing and Phillips, 1970).
Верхушечная почка летом не растет в длину, но это
не истинный покой, а скорее всего ее рост ингибирован
деятельностью других органов, например полностью рас-
пустившихся листьев. Но в случаях сильного поврежде-
ния и дефолиации побега насекомыми у верхушечной
почки отмечается хороший рост. Другой пример потен-
циальной возможности роста у кажущихся бездеятельны-
ми верхушечных почек — развитие ламмасовых побегов
после сильного дождя или удобрения. Осенью у верху-
шечных почек наступает состояние глубокого покоя,
в это время их рост в дллшу нельзя вызвать даже опыт-
14
ной обработкой наподобие дефолиации. Это обусловлено
состоянием покоя самой почки, и у большинства древесных
растений рост побегов начинается только по окончании
периода зимнего охлаждения. После выхода побегов из
состояния глубокого покоя их рост затормаживают небла-
гоприятные условия среды. Обусловленный внешними
причинами покой известен как вынужденный.
Отличным примером, показывающим различия между
глубоким и вынужденным покоем, служит сбор по не-
мецкому обычаю ветвей вишн!и в день св. Барбары
(4 декабря), а начинают они цвести к Новогоднему
празднику (Romberger, 1963). Если же ветви собраны
в более ранние сроки, даже в ноябре, то в помещении они
не зацветают. В первом случае ветви находятся в состоя-
нии вынужденного покоя и поэтому реагируют на благо-
приятные для роста условия; во втором они не были
выведены из состояния глубокого покоя, так как не про-
шли период с пониженной температурой.
У большинства пород рост побегов зависит от фото-
периода: длинный день стимулирует рост, короткий —
ингибирует (Wareing, 1969; Romberger, 1963). Например,
рост сеянцев Robinia в режиме короткого дня приоста-
навливается, кончики их побегов затем опадают. Если
же кончики побегов держать в режиме длинного дня,
а листья — короткого, экстенсивный рост все равно пре-
кращается. Но при создании противоположных условий
опыта рост побега продолжается, поэтому предполагают,
что развившиеся листья являются местом восприятия
растением фотопериодической реакции. Wareing с сот-
рудниками показал, что прекращение роста побега в ре-
жиме короткого дня связано с образованием в развивших-
ся листьях ингибиторов.
Это причинное соотношение подтверждено опытами:
изменение режима длинного дня на режим короткого дня
вызвало значительное увеличение уровня ингибиторов
в кончиках побега незадолго до закладки верхушечной
почки (Wareing, 1969). Например, у сеянцев черной смо-
родины (Ribes nigrum), выдержанных в режиме длинного
дня, а затем переведенных в режим короткого дня, быст-
ро возрастала концентрация ингибиторов в кончиках
побега (Wareing and Phillips, 1970). Кроме того, в этом
случае, как предполагают, уменьшается содержание гиб-
береллина, поскольку наступление состояния покоя регу-
лируется увеличением соотношения и системе ин^иби-
13
тор — гиббереллин. Если нераспустившиеся почки, содер-
жащие ингибиторы и Гиббереллины, проходят период
зимнего охлаждения, то, как обнаружили, концентрация
гиббереллинов постепенно возрастает, а ингибиторов —
снижается. Следовательно, выход зимующей почки из
состояния глубокого покоя связан с низким соотношением
в системе ингибитор — гиббереллин.
Природный ингибитор, образующийся в листьях в ре-
жиме короткого дня, определили как сесквитсрпепоид-
абсцизовую кислоту (АБК). АБК, нанесенная па листья
сеянцев черной смородины, вызывала прекращение роста
побегов и заложение верхушечной ночки с почечными че-
птуями (Wareing and Phillips, 1970). При обработке рас-
тения АБК в течение нескольких педель его верхушечная
почка вступает в состояние глубокого покоя; прекраще-
ние обработки пе способствует возобновлению ее роста.
Недавние работы показали, что АБК участвует в регули-
ровании вступления в состояние покоя у многих покры-
то- и голосеменных растений. АБК, без сомнения, один из
факторов, регулирующих состояние покоя у многих по-
род, особенно отзывчивых па изменение фотопериода.
С другой стороны, у некоторых пород периодичность рос-
та побегов регулируется всецело отличающимися меха-
низмами. Например, у Огеорапах (см. 1.2) Borchert сом-
невается в значении АБК для периодичности роста, так
как экстрагируемые ингибиторы отсутствовали в течение
всего цикла роста побегов.
2. Древесина
2.1. СТРУКТУРА ДРЕВЕСИНЫ
Еще во времена неандертальцев древесину использо-
вали в качестве топлива и, вероятно, для изготовления
инструментов и оружия. В настоящее время ее широко
применяют в строительных конструкциях, при произ-
водстве бумаги и пр. Древесина образуется в стволах,
корнях и ветвях голосеменных и древесных покрытосе-
менных растений. Для производства бумаги и строитель-
ных конструкций особенно ценятся породы, относящиеся
к хвойным голосеменном (с мягкой древесиной): для
красивой мебели и панельной обшивки — породы, относя-
щиеся к двудольным покрытосеменным (с твердой дре-
весиной). Прочно утвердившиеся в американской литера-
16
type названия мягко- и твердодревесные породы не могут
быть строгим критерием их физической твердости; на-
пример, у некоторых твердодревесных (бальза заячья)
в действительности древесина гораздо мягче, чем у боль-
шинства мягкодревесных пород.
Вторичное тело растения, древесина которого являет-
ся главной составной частью, образуется в результате
деятельности двух латеральных меристем — васкулярного
камбия 1 и феллогена, или пробкового камбия. Для обще-
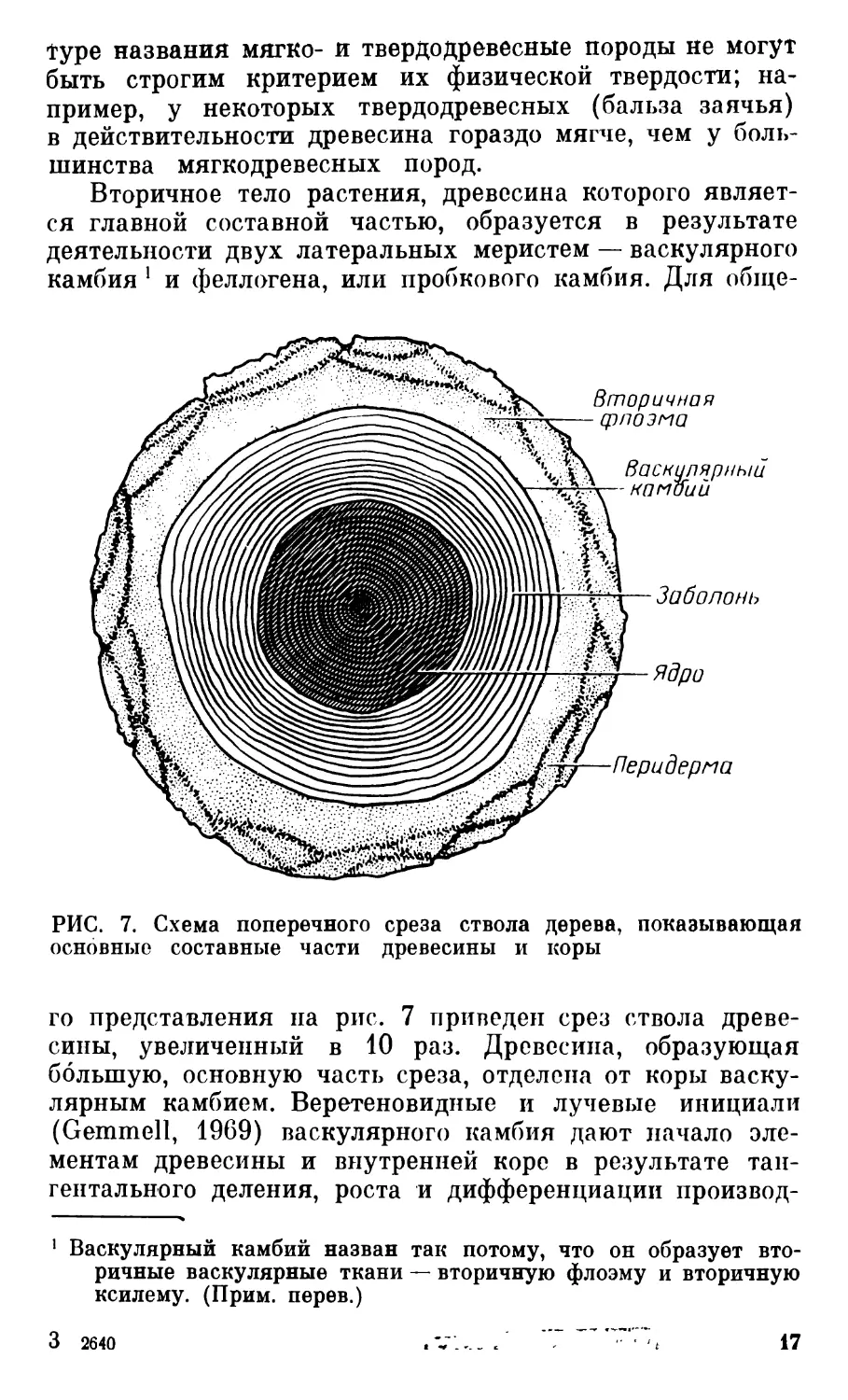
РИС. 7. Схема поперечного среза ствола дерева, показывающая
основные составные части древесины и коры
го представления на рис. 7 приведен срез ствола древе-
сины, увеличенный в 10 раз. Древесина, образующая
большую, основную часть среза, отделена от коры васку-
лярным камбием. Веретеновидные и лучевые инициали
(Gemmell, 1969) васкулярного камбия дают начало эле-
ментам древесины и внутренней коре в результате тан-
гентального деления, роста и дифференциации производ-
1 Васкулярный камбий назван так потому, что он образует вто-
ричные васкулярные ткани — вторичную флоэму и вторичную
ксилему. (Прим, перев.)
3 2640 .VV„ / ”, ч 17
пых клеток1. Кора — не специальный термин, относя-
щийся ко всем тканям, расположенным кнаружи от
камбия сосудов, и обозначающий как вторичную флоэму,
так и перидерму или ритидом, если охватывается не
только перидерма. Органические ассимиляты проводит
только тонкий слой флоэмы, большей частью заново диф-
ференцировавшийся из производных васкулярного кам-
бия. Перидерма образуется пробковым камбием и выпол-
няет защитную функцию (см. разд. 5). Некоторые породы
можно различать по окраске частей древесины: перифе-
рической заболони и центрального ядра. Заболонь физио-
логически активна в накоплении и передвижении пита-
тельных и минеральных веществ и воды из корней
в крону. В ядре нет живых клеток, оно выполняет только
механическую функцию придания прочности.
Древесина состоит из двух взаимопроникающих сис-
тем клеток, одна из которых ориентирована продольно,
другая — радиально. Элементы осевой, или продольной,
системы клеток, состоящие из члеников сосудов2, трахе-
ид, волокон и паренхимы, возникли из веретеновидных
ннициалей. Радиальная система клеток, состоящая из
лучей, образована лучевыми инициалями васкулярного
камбия, в основном она паренхиматозная; исключение
составляют некоторые хвойные, например сосна, у кото-
рой встречаются лучевые трахеиды.
Древесина хвойных пород — структурно однородна,
так как у них трахеиды составляют почти 90% элемен-
тов оси, а у лиственных более разнородна и состоит из
члеников сосудов, волокон и иногда (например, у дуба)
трахеид. Помимо несходства в клеточном составе у хвой-
ных и лиственных пород имеются различия в развитии
некоторых осевых клеток. У хвойных пород длина трахе-
ид примерно 3—4 мм, тогда как у лиственных длина
волокон 1 — 1,5 мм. Далее, если трахемды слабо увеличи-
ваются и по длине превосходят исходные инициали толь-
ко на 20—30%, то зрелые.волокна в 5 раз длиннее вере-
теновидных ипициалей, из которых они произошли. Во-
1 Производная клетка образуется делением меристематической
клетки таким образом, что она вступает на путь дифферен-
циации в протопласт; ее сестринская клетка может оставать-
ся в меристеме. (Прим, перев.)
2 Членик сосуда — один из компонентов клеток сосуда, называе-
мый также элементом сосуда и сегментом полного сосуда.
(Прим, перев.)
18
заикает вопрос, почему при дифференциации волокна
так сильно удлиняются. Чтобы дать окончательный ответ,
нужно изучить, как поблизости от васкулярного камбия
взаимодействуют генетическая потенциальная способность
дифференцирующейся клетки и в основном физиологиче-
ская последовательность смены среды.
Немного увеличив продолжительность обычного цикла
роста деревьев, т. е. развития волокон и трахеид, выясни-
ли, что генетический контроль не такой уж жесткий,
чтобы предотвратить явление изменчивости. Много лет
тому назад Bailey (Esau, 1965) наблюдал, что структур-
ная вариабельность элементов древесины осуществляется
не только на уровне рода и вида, но даже в пределах
одного организма. Так, у хвойных пород в одном и том
же годичном кольце длина трахеид зависит от места их на-
хождения — в древесине ствола или ветви. В месте при-
крепления ветвей к стволу трахеиды короче расположен-
ных выше или ниже. Даже на одной высоте ствола длина
трахеид постепенно увеличивается от первого к почти
60-летнему годичному кольцу. После этого возраста их
размеры остаются более или менее постоянными.
2.2. ХИМИЧЕСКИЙ СОСТАВ
Структура и многие свойства древесины обусловлены
спецификой устройства составных частей ее клеток. Да-
же износ мебели из клена зависит от относительно одно-
родного распределения члеников сосудов и волокон в дре-
весине каждого годичного кольца. С другой стороны,
нарядные узоры у мебели из дуба также обусловлены
местным расположением очень широких члеников сосу-
дов в самой ранней части каждого годичного кольца.
Структуру древесины можно понять по устройству ее
клеток, а еще лучше по организации и химии веществ
их оболочек.
Клеточные оболочки волокон и трахеид разнородны
и по структуре и по химическому составу. Первичной
оболочкой клетки называют первоначальную оболочку
камбия. Первичная оболочка клетки растет в поверхност-
ной зоне, когда, например, членик сосуда увеличивается по
диаметру. Смежные производные камбия в ранней стадии
дифференциации разделены только двумя тонкими пер-
вичными оболочками, соединяемыми межклетным веще-
ством, называемым срединной и л а с т и и к о й. У зре-
19
°\ /ои
хс Фенилаланин-
I аммиак-
о он
Феруловая
кислота
Коричная
Фенилаланин кислота
Восстановление
ОН
I
н-с-н
I
н-с
II
н-с
ОН
I
н-с—н
I
н-с
ОСН3
ОН
Окисление g
--------Со
Кониферилодый спирт
О
он
Синаповый спирт
б
лых трахеид и волокон первичная клеточная ооолочка —
самая верхняя часть клеточной оболочки (рис. 8), состав-
ляющая незначительную долю процентного отношения
к веществу всей оболочки. Толстая вторичная о б о-
л очка клетки откладывается на внутренней стороне
вскоре после прекращения поверхностного роста первич-
ной оболочки. Вторичная оболочка, несомненно, содержит
большую часть веществ клеточной оболочки.
Первичную и вторичную оболочки клетки составляют
3 главных компонента: целлюлоза и гемицеллюлоза, отно-
сящиеся к полисахаридам, и лигнин — ароматический
полимер, образованный из строительных блоков фенил-
проиапа (Сб — Сз-ряда, рис. 9). Целлюлозу можно срав-
нить с каркасным материалом, определяющим форму
и структуру клеточной оболочки (Wardrop, 1964а). Цел-
люлоза сначала внедряется в пористый аморфный мат-
рикс \ представленный гем'ицеллюлозой, затем эти поли-
сахариды инкрустируются и вследствие этого цементиру-
ются друг с другом лигнином, очень устойчивым трех-
мерным . полимером.
Целлюлоза. Давно известно, что целлюлоза не
только наиболее распространенное, естественно образую-
щееся органическое вещество, но и главная составная
часть оболочки растительной клетки1 2, что и определило
ее название. Физическое строение клеточной оболочки
зависит от целлюлозы. Ее длинные и прямые молекулы
состоят из нескольких тысяч глюкозных остатков, соеди-
ненных «голова с хвостом». Свыше 2000 молекул целлю-
лозы, сгруппированных параллельными рядами в тяжи,
называют микрофибриллами (см. рис. 8). Несколь-
ко сотен последних могут быть вытянутыми вместе в раз-
дельных образованиях, называемых м а к р о ф и б р и л-
л а м и, которые видны в световом микроскопе как тонкие
полосы в клеточных оболочках. Макрофибриллы, соеди-
РИС. 8. Схема строения клеточной оболочки у волокна древеси-
ны; слои оболочек в поляризованном свете и электронном микро-
скопе различают по расположению их микрофибрилл
РИС. 9. Схема образования предшественников лигнина:
а— кониферилового спирта из фенилаланина у голосеменных; б— конифе-
рилового и синапового спиртов
1 Матрикс — основное вещество. (Прим, перев.)
2 Клетка — cellula — лат. (Прим, перев.)
21
ненные бок о бок, называют ламеллам и, совокупность
которых составляет слой оболочки клетки. У одревеснев-
ших волокон вторичные клеточные оболочки состоят из
трех различающихся между собой слоев, обозначаемых
от внешней стороны но направлению к клеточной полости
как Si, S2 и S3 соответственно (рис. 10). Поэтому моле-
кулы целлюлозы по расположению представляют собой
один возрастающий по сложности ряд — от микрофиб-
рилл, видимых только па электронных микрофотографиях,
через макрофибрпллы и ламеллы до слоев, образующих
вторичную клеточную оболочку, размеры которой разли-
чимы невооруженным глазом.
Среди основных компонентов клеточной оболочки
только целлюлоза частично кристаллическая. Молекулы
целлюлозы в зоне микрофибрилл, называемые мп цел ла-
мп, вытянуты рядами в исключительно регулярном по-
рядке и образуют кристаллическую решетку. Молекулы,
составляющие решетку, вдоль разных осей располагаются
в разных направлениях: целлюлоза — анизотропна
(Esau, 1965). В связи с этим свойством целлюлозу легко
обнаруживают с помощью поляризационного микроскопа,
в который вставляют поляризационные фильтры по опти-
ческой оси: один под конденсор, другой в корпус тубуса.
Когда оба фильтра одинаково ориентированы, нлоскоио-
ляризованный свет1 выходит из первого фильтра, назы-
ваемого поляризатором, и проходит пеизменяющимся
через второй фильтр — анализатор. Если анализатор по-
вернут под углом 90° к поляризатору, свет не проходит,
поскольку плоскополяризованный свет гасится, и поле
зрения, видимое в окуляре,— темное2.
У целлюлозы две плоскости поляризации, т. е. она
оптически анизотропна. Поляризованный свет, падающий
на нее под любым угом, расщепляется на два компонента
колебаний в этих плоскостях. Компоненты колебаний вы-
ходят из фазы и не смешиваются, следовательно, первона-
чальный пучок не образуется. Компоненты проходят че-
рез анализатор, и образец выглядит светлым в поле зре-
ния, темным в других случаях. Оптическая анизотропия
целлюлозы наиболее очевидна, когда поляризованный
1 Поляризованный свет образуется световым пучком с колебани-
ями волн только в одной плоскости. (Прим, перев.)
2 При таком расположении поляроиды (т. е. анализатор и поля-
ризатор) называют скрещенными. (Прим, перев.)
22
РИС. 10. Поперечные срезы древесины:
а — клена красного в поляризованном микроскопе; в каждом волокне F
наблюдали две зоны оптической анизотропии, из них верхняя, светлая,
соответствовала первичной оболочке плюс Si слой, а тонкая внутренняя
— слою S3 (при такой ориентации слой S2 темный); б — микрофотография
среза клена красного с флуоресцирующим лигнином; распределение лиг-
нина в клеточной оболочке можно измерять по характерной синей или
сине-зеленой флуоресценции, излучающейся при поглощении УФ-света,
высокое содержание его в срединной пластинке и первичной оболочке
волокон древесины (Morey and Cronshaw, 1968); в—электронная микро-
фотография поперечного среза трахеид сосны колючей (Pinus pungens),
изображающая слой S2 и S3 вторичной оболочки (Cote and Day, 19G9)
свет падает под прямым углом к длинной оси целлюлоз-
ной цепочки, и не очевидна, если плоскость поляризации
падающего пучка параллельна одной из двух плоскостей
поляризации целлюлозы. Например, часть клеточной обо-
лочки волокна может оставаться темной при скрещенных
поляроидах, если целлюлозные цепочки, а значит, и одна
из плоскостей полимеризации ориентированы параллель-
но оптической оси микроскопа, которая в свою очередь
параллельна плоскости колебания падающего поляризо-
ванного света.
Поляризационные микроскопы можно использовать
для обнаружения и определения направленности целлю-
лозы в волокнах и трахеидах, так как только эта часть
клеточной оболочки оптически анизотропна. В связи
с этим на поперечном срезе между скрещенными поля-
роидами у слоя S2 выявлена лишь незначительная ани-
зотропия (см. рис. 10, а). Однако на продольном разрезе,
когда этот же слой расположен плоско на предметном
столике микроскопа, анизотропия выражена полностью;
слой S2 выглядит светлым, и цепочки целлюлозы в нем
ориентированы главным образом в осевом направлении
под незначительным углом к длинным осям клеток. Из-
мерения с помощью поляризационных и электронных
микроскопов показали, что эти углы у волокон и трахеид
составляют 10—30° (см. Roelofsen, 1959). Слои Si п S3
обычно светлые на поперечных срезах при использовании
скрещенных поляроидов (см. рис. 10, а). Целлюлоза этих
слоев построена в ряды под углами 60—90° к осям воло-
кон и трахеид (см. рис. 8, б).
Толстые вторичные оболочки клеток древесных расте-
ний гетерогенны в понятиях строения их основных ве-
ществ. Прочность древесины частично обусловлена ее
слоистой структурой, которую можно сравнить с клееной
фанерой, отдельные слои которой непрочные, по при сое-
динении образуют очень прочный комплекс.
Гемицеллюлоза. В оболочках клеток древесных расте-
ний на целлюлозу приходится немногим более половины их
полисахаридов. Гемицеллюлоза — другая важная
группа полисахаридов клеточной оболочки; она представ-
лена очень разнообразной, неопределенной группой поли-
меров с низкой молекулярной массой. Как и целлюлоза,
гемицеллюлоза нерастворима в воде, но она растворяется
в водных растворах щелочи. Хотя молекулы у гемицел-
люлозы в основном линейные, они могут разными спосо-
24
бами разветвляться, обеспечивая матрикс порами, обна-
руживаемыми вокруг микрофибрилл целлюлозы.
У покрыто- и голосеменных древесных растений ос-
новной химический состав древесины существенно разли-
чается. У лиственных пород обычно больше гемицеллю-
лозы, а у хвойных — лигнина. Интересно отметить, что
из основных составных частей древесины только целлю-
лоза относительно постоянное вещество; ее содержание
у лиственных и хвойных пород достигает 40—45%.
Некоторые различия в свойствах хвойных и лиственных
пород были выявлены при изучении химического состава
древесины. В одинаковых внешних условиях влажность
древесины у покрытосеменных растений выше, чем у го-
лосеменных, причем из основных частей клеточной обо-
лочки наиболее гигроскопична гемицеллюлоза (Roelofsen,
1959). Поскольку в древесине лиственных пород геми-
целлюлозы содержится относительно больше, этим и объ-
ясняется ее высокая влажность.
Лигнин. В заключительной фазе дифференциации тра-
хеид или волокон лигнин инкрустирует и делает жестки-
ми клеточные оболочки. Это — высокоароматическое ве-
щество и единственный главный неуглевод клеточной обо-
лочки. Основные строительные блоки лигнина — произ-
водные фенилпропана, связанные различным образом
вместе (например, углерод — углеродной и эфирной свя-
зями). У хвойных преобладающий строительный блок
лигнина — конифериловый спирт, тогда как у лиственных
(см. рис. 9) производные синапового и кониферилового
спиртов используются в равных количествах. Эти произ-
водные дегидрируются и впоследствии строятся в слож-
ные трехмерные полимеры, которые полностью инкрус-
тируют составные части оболочки из полисахаридов.
Лигнин устойчивый, жесткий полимер. По одной тех-
нологии для выделения твердого лигнина древесину об-
рабатывают сильной минеральной кислотой, например
72%-ной (в. ч.) серной кислотой, реагентом, легко гид-
ролизующим полисахариды клеточной оболочки. В серной
кислоте трахеиды хвойных хорошо сохраняют свою пер-
воначальную форму, что указывает на стойкое действие
лигнина в клеточной оболочке. На торфяных болотах
процент лигнина в растительных остатках обычно возра-
стает с увеличением глубины залегания, что иллюстриру-
ет относительную нейтральность лигнина среди основных
составных частей клеточной оболочки.
4 2640
25
Специалисты по химии древесины давно интересуются
содержанием лигнина в слоях клеточных оболочек тра-
хеид и волокон. Данные о распределении лигнина важны
для химического дефибрирования древесины, когда лиг-
пин экстрагируют и древесина разрушается на клеточные
составные части.
В УФ-спектре лигнин очень сильно поглощает свет
при характерном максимуме 212 и 280 нм (см. рис. 10, б).
Концентрацию лигнина в любой точке клеточной оболоч-
РИС. 11. Использование Уф-света для определения структуры
клеток:
а — распределение лигнина в клеточных оболочках трахеид, регистрируе-
мое микрофотографическим способом по поглощению УФ-света; б — мпк-
рофотодепситометрический спектр, показывающий высокое поглощение
срединной пластинкой (СП) и первичной оболочкой (ПО) и меньшее по-
глощение вторичной оболочкой (ВО) (Scott et al., 19(59)
ки специально можно определять по УФ-п о г л о щ е п и ю.
Используя срезы древесины ели черной толщиной 0,5 мк
и монохроматический источник излучения (280 нм), Scott
et al. (1969) с помощью УФ-микроскопа микрофотографи-
чески записал УФ-поглощение лигнина (рис. И, а). УФ-
аппаратура в основном схожа с обычным микроскопом
за исключением того, что оптика из стекла заменена
кварцевой, передающей УФ-спектр света. Микроденсито-
метр регистрирует УФ-поглощение по ходу прямой ли-
нии, пересекающей оболочку трахеиды (см. рис. И, б).
Обусловленное лигнином относительное УФ-поглоще-
нив первичной оболочкой и срединной пластинкой значи-
тельно выше, чем вторичной оболочкой. Первичная обо-
26
лочйа и срединная пластинка содержат в трахеидах ран-
ней древесины соответственно 0,5 и 0,85 г/г лигнина,
а вторичная оболочка — 0,23 г/г. Из этого можно сделать
вывод, что наиболее заметный слой оболочки волокон
и трахеид — вторичная клеточная оболочка (см. рис. 8).
У черной ели она занимает 87% объема оболочек трахеид
ранней древесины (Fergus et al., 1969). Если же теперь
вычислить распределение лигнина в процентном отноше-
нии к общему его количеству, то вторичная оболочка
клетки содержит 72%, а остальная часть его приходится
на первичную оболочку и срединную пластинку. Поэтому,
несмотря на относительно низкую концентрацию лигнина
во вторичной клеточной оболочке, в ней содержится боль-
шая его часть.
2.3. ОБРАЗОВАНИЕ КЛЕТОЧНОЙ ОБОЛОЧКИ
Трахеиды, членики сосудов, волокна, осевые и луче-
вые паренхимные клетки — все они дифференцируются
из инициалей камбия, проходя во время своего развития
четыре частично перекрывающихся этапа: 1) деление
клетки; 2) поверхностный рост; 3) утолщение оболочек;
4) лигнификацию (Wardrop, 1964а).
Делением веретеновидных и лучевых инициалей
в тангентальной плоскости создаются вторичная ксилема
и вторичная флоэма и этим же объясняется основное
расположение дифференцированных клеток в радиальные
ряды. С увеличением размеров древесного цилиндра воз-
растает и окружность васкулярного камбия. Это проис-
ходит в результате делений в радиальной плоскости
и увеличения числа инициалей камбия.
У первичной клеточной оболочки при дифференциа-
ции производных камбия начинается поверхностный
рост. Волокна у лиственных пород растут преимущест-
венно в продольном направлении; их длина в 5 раз больше
длины исходных инициалей. При утолщении кле-
точной оболочки образуется трехслой пая вторичная
оболочка клетки. Следует помнить, что поверхностный
рост и утолщение клеточной оболочки не только взаимно
не исключаются, но происходят одновременно. Например,
в период поверхностного роста кончиков волокон у кле-
ток, расположенных в более проксимальной части, откла-
дываются вторичные клеточные оболочки.
27
Лигнификация —заключительныйэтап дифферен-
циации клеточных оболочек у волокон и трахеид. В не-
давней работе по биохимии показано, что предшественник
лигнина возникает в самой лигнифицирующей клетке.
Решающий этап в биосинтезе лигнина — дезаминирова-
ние L-фенилаланина фенилаланин-аммиак-лиазой (см.
рис. 9). Коричная кислота преобразуется в конечном сче-
те на пути метаболизма в конифериловый спирт. Послед-
ний и родственные ему спирты дегидрогенизируются
окислительными ферментами, например пероксидазой,
и превращаются в лигнин. Применив простую анатомиче-
скую технику, Rubery and Northcote (1968) отделили
дифференцирующуюся ткань ксилемы с большим синте-
зом лигнина от камбиальной зоны, где лигнина образуется
мало. Дифференцирующаяся ксилема обладает высокой
активностью фенилаланин-аммиак-лиазы, тогда как в кам-
биальной зоне этого фермента мало. Более того, отделен-
ные дифференцирующиеся клетки легко присоединяют
к лигнину насыщенную тритием коричную кислоту,
а в камбиальной зоне ее включений мало. Дифференциру-
ющиеся производные камбия образуют собственных пред-
шественников лигнина, которые непосредственно включа-
ются в лигнин клеточных оболочек. После этого лигнифи-
кация протекает независимо от предшественников, по-
ступающих из камбиальной зоны.
3. Радиальный рост
3.1. ВАСКУЛЯРНЫЙ КАМБИЙ
Если рост в высоту связан с органообразовательной
деятельностью апикальной меристемы и удлинением
междоузлий (см. разд. 1), то радиальный рост зависит от
активности клеток васкулярного камбия. Самый харак-
терный, наверное, этап в радиальном росте — п ере кли-
па л ьное деление (в тангентальной плоскости) ини-
циальных клеток, которое обусловливает начало роста
примыкающих радиально расположенных элементов дре-
весины и флоэмы. Это приводит к постепенному переме-
щению и самой меристемы, так как производные клетки
дифференцируются внутрь и происходит приращение
цилиндра древесины.
При изучении поперечного среза древесины ствола
весной выявляют несколько слоев активно делящихся
28
камбиальных клеток, а также постоянство в дифферен-
циации клеток внутрь и кнаружи у элементов ксилемы
и флоэмы соответственно. Эти клетки васкулярного кам-
бия, подвергающиеся делению, образуют к а м б и а л ь-
п у ю з о и у. При таком размещении внутренние элементы
ксилемы дифференцируются и проходят в системном по-
рядке путь обмена веществ через поверхностный рост,
утолщение оболочек и лигнификацию. На этом основании
структурные особенности некоторых клеток камбиальной
зоны можно объяснить за счет камбиальных и п и-
циалей (Philipson et al., 1971). При периклинальном
делении клетки образуется одна дочерняя инициаль, под-
держивающая постоянство меристемы, второй отрез клет-
ки внутрь или кнаружи обычно создает соответственно
ксилему или флоэму. Другие клетки камбиальной зоны
внутрь и кнаружи от камбиальных инициален называют
материнскими клетками соответственно к с и л е-
мы и флоэмы. При делении материнской клетки в пе-
риклинальной плоскости образуются две производные,
которые позднее развиваются в элементы ксилемы и фло-
эмы.
Периклинальное деление удлиняющихся клеток кам-
биальной зоны неизбежно влечет за собой разъединение
частей оболочки на значительное расстояние; этот про-
цесс давно привлекал внимание ботаников. Деление кле-
ток можно условно разделить на две фазы: 1) митоз —
деление ядерного материала, 2) цитокинез — разделе-
ние цитоплазмы на 2 дочерние клетки (Esau, 1965).
У большинства растительных клеток полный процесс
клеточного деления завершается быстро, но у перикли-
нально делящихся удлиненных клеток камбия цитокинез
значительно растянут во времени и пространстве. В ста-
дии анафазы при митозе 2 группы дочерних хромосом
расходятся от так называемого фрагмопласта (веретено-
видного волокна) в экваториальной плоскости. В телофа-
зе образуется разделительная стенка — клеточная
пластинка между делящимися протопластами цент-
ральной зоны экваториальной плоскости.
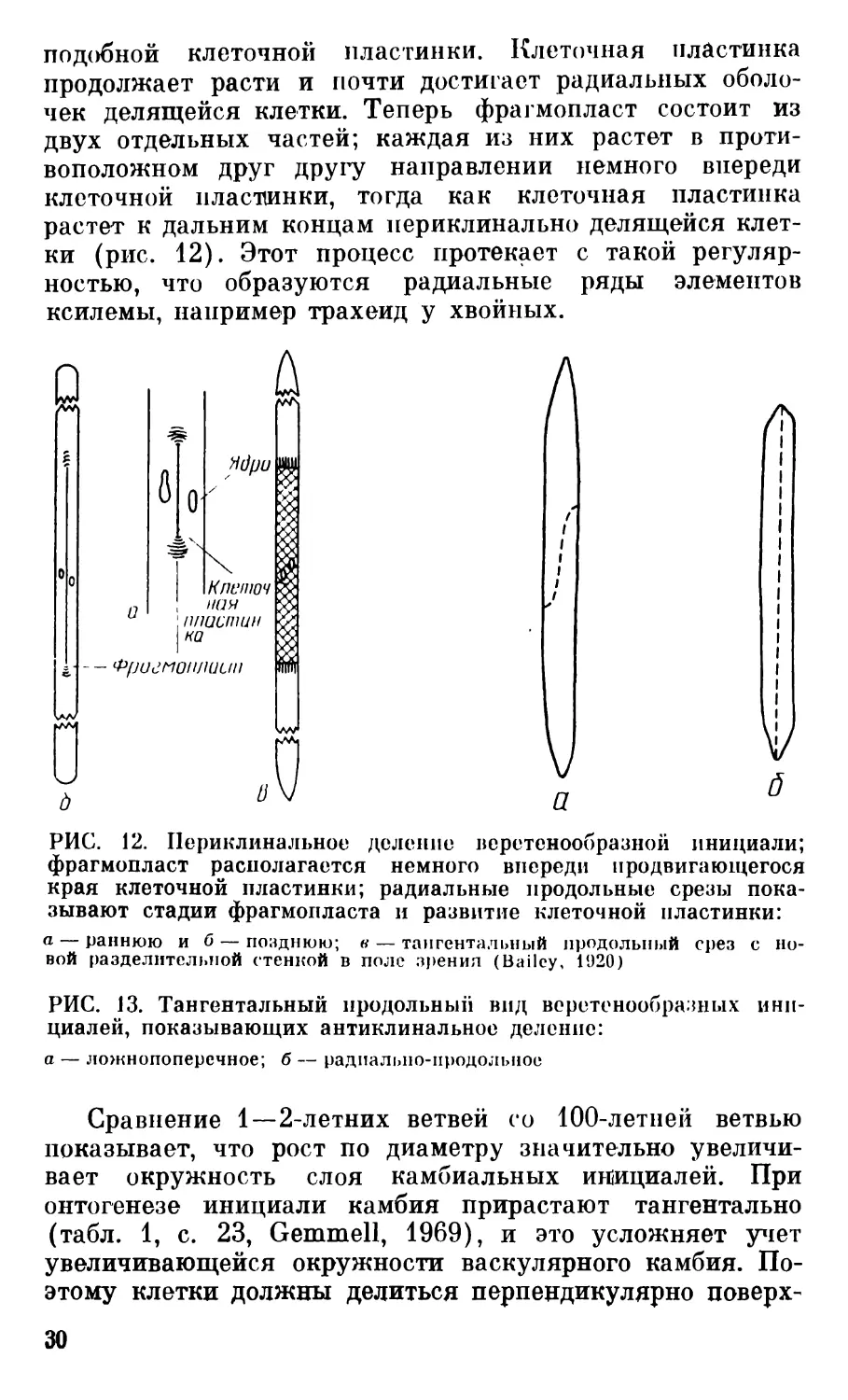
Вначале клеточная пластинка состоит из ограничива-
ющих клеточные мембраны дочерних протопластов вместе
с общей срединной пластинкой. Позднее у каждой дочер-
ней клетки выделяется целлюлозная первичная оболочка.
К концу телофазы фрагмопласт отходит к периферии
боковой стенЮи сосуда вблизи и немного впереди диско-
29
подобной клеточной пластинки. Клеточная пластинка
продолжает расти и почти достигает радиальных оболо-
чек делящейся клетки. Теперь фрагмопласт состоит из
двух отдельных частей; каждая из них растет в проти-
воположном друг другу направлении немного впереди
клеточной пластинки, тогда как клеточная пластинка
растет к дальним концам периклинально делящейся клет-
ки (рис. 12). Этот процесс протекает с такой регуляр-
ностью, что образуются радиальные ряды элементов
ксилемы, например трахеид у хвойных.
V
б
РИС. 12. Периклинальное деление веретенообразной инициали;
фрагмопласт располагается немного впереди продвигающегося
края клеточной пластинки; радиальные продольные срезы пока-
зывают стадии фрагмопласта и развитие клеточной пластинки:
а — раннюю и б — позднюю; в — тангентальный продольный срез с но-
вой разделительной стенкой в поле зрения (Bailey, 1920)
РИС. 13. Тангентальный продольный вид веретенообразных ини-
циалей, показывающих антиклинальное деление:
а — ложнопоперечное; б — радпалыю-продолыюе
Сравнение 1—2-летних ветвей со 100-летней ветвью
показывает, что рост по диаметру значительно увеличи-
вает окружность слоя камбиальных инициалей. При
онтогенезе инициали камбия прирастают тангентально
(табл. 1, с. 23, Gemmell, 1969), и это усложняет учет
увеличивающейся окружности васкулярного камбия. По-
этому клетки должны делиться перпендикулярно поверх-
30
пости ствола и ветвей; такие деления, называемые анти-
клинальными, обусловливают сохранение целостности
латеральной меристемы при радиальном росте васкуляр-
ного камбия. Антиклинальное деление обычно ограничи-
вается камбиальными инициалями.
В зависимости от угла наклона разделительной стенки,
образующейся при цитокинезе, антиклинальное деление
подразделяют па два типа. Для голосеменных и некото-
рых покрытосеменных (например, орехов) характерно
ложнопоперечное антиклинальное деление, когда
разделительная стенка расположена наклонно к длинной
оси клетки (рис. 13, а). Разделенные таким образом всрете-
повидпые инициали образуют 2 дочерние клетки, которые
вначале короче родительской клетки. Прирост по окруж-
ности осуществляется двумя путями: во-первых, вследст-
вие апикального роста в длину каждой дочерней клетки
возрастает общее число инициалей в любой поперечной
плоскости; во-вторых, при следующем делении увеличи-
вается латеральная меристема. Для других покрытосе-
менных (Robinia) характерно радиально-продоль-
ное антиклинальное деление, при котором веретеновид-
ная ипициаль сверху донизу пересекается в радиальной
плоскости разделительной стенкой (см. рис. 13, б). По
высоте дочерние инициали одинаковы с родительской
клеткой, и в дальнейшем у них обычно очень незначи-
тельная или совсем нет тенденции к апикальному росту
в длину. Таким образом, деревья с радиально-продольным
делением увеличиваются в диаметре просто тангенталь-
ным приращением дочерних инициалей.
3.2. СЕЗОННОЕ РАЗВИТИЕ
У пород, произрастающих в умеренных широтах, де-
тально изучена динамика развития васкулярного камбия.
Далее рассмотрим сезонную динамику периклинальных
делений инициалей, в результате чего образуются новые
ксилема и флоэма.
В провинции Онтарио (Канада) у туи западной (Thu-
ja occidentalis) период дифференциации ксилемы начи-
нается раньше, чем флоэмы. Перезимовавшая камбиаль-
ная зона состоит только из трех слоев клеток с более
верхним единственным слоем камбиальных инициалей
и двумя слоями материнских клеток ксилемы (рис. 14, а;
Ваппап, 1962). В середине апреля в камбиальной зоне
31
материнских клеток ксилемы начинаются первые пери-
клинальные деления (см. рис. 14,6). В мае деления силь-
но учащаются, к концу месяца камбиальная зона стано-
вится широкой и лежащие глубоко внутри производные
быстро дифференцируются в трахеиды (см. рис. 14, в).
В это же время камбиальные инициали, делясь произ-
вольно, образуют новые материнские клетки ксилемы
и флоэмы. При более позднем делении производные диф-
ференцируются в элементы флоэмы; к концу мая образу-
ются в среднем только 2 ее элемента, тогда как на дру-
пкк
<—
РИС. 14. Схема обычной последовательности при инициировании
активности камбия у туи западной (объяснения даны в тексте):
НИ — пницпаль камбии; МНН — материнская клетка ксилемы; МКФ — ма-
теринская клетка флоэмы; К—дифференцирующаяся клетка ксилемы
(Ваппап, 1962)
гой стороне камбиальные инициали образуют до 60 кле-
ток, находящихся в разных стадиях развития.
Последние изучения показывают, что по особенностям
сезонной активности сосны сильно отличаются от туи
западной. В шт. Висконсин (США) у сосны веймутовой
(Finns strobus) дифференциация клеток флоэмы начина-
ется на 6 недель раньше, чем ксилемы (Alfieri and Evert,
1968). В конце марта начинается образование флоэмы
с дифференциации ситовидных клеток в самой верхней
части камбиальной зоны без предшествующего деления
инициалей. В апреле происходит незначительное пери-
клинальное деление в камбиальной зоне, но до наступле-
ния мая материнские клетки ксилемы не дифференциру-
ются в трахеиды. Повышенная активность развития кси-
лемы продолжается с середины мая до середины июня;
она происходит в результате периклинального деления ее
32
Материнских клеток и быстрого созревания производных
клеток I Alfieri and Evert дополнительно сообщают, что
производные образуют осенью ситовидные зимующие
клетки, и функционирование задерживается до начала
следующего сезона роста>
В сезонном цикле камбиальной активности можно ус-
ловно выделить две особенности с учетом размеров и рас-
пределения сосудов в ксилеме годичного кольца. У неко-
торых пород с рассеяннососудистой древесиной1 2
(Acer), где сосуды одинаковых размеров однообразно
размещены в годичном кольце, дифференциация флоэмы
происходит на 4—6 недель раньше, чем дифференциация
ксилемы. У пород с кольцесосудистой древесиной
(Robinia), где самые крупные сосуды находятся исклю-
чительно в наиболее ранней части годичного прироста,
ксилема и флоэма развиваются более или менее одновре-
менно.
У рассеяннососудистого клена ясенелистного (Acer
negundo) членики ситовидных трубок дифференцируются
предыдущей осенью, перезимовывают и задерживают
функционирование до следующей весны (Tucker and
Evert, 1969). Камбиальная активность начинается в конце
марта (шт. Висконсин) с дифференциации клеток
с внешней поверхности камбиальной зоны непосредст-
венно в элементы флоэмы. В это же время у оставшихся
слоев камбиальной зоны, особенно с флоэмной стороны,
начинается периклинальное деление. Первые элементы
ксилемы созревают значительно позднее, только в середи-
не мая. Дальнейший цикл камбиальной активности у Acer
во многих отношениях схож с циклом у сосны. Например,
им свойственны круглогодичное присутствие функциони-
рующих ситовидных элементов, образование новой флоэ-
мы на месяц раньше кцилемы, начало роста новых фло-
эмных элементов ранней весной непосредственно из са-
мых верхних недифференцированных клеток камбиальной
зоны.
В середине апреля (шт. Висконсин) у кольцесосуди-
стой акации белой (Robinia pseudoacacia) происходят
1 Созревание производных клеток — достижение производными
клетками своего окончательного (дефинитивного) состояния.
(Прим, перев.)
2 Рассеяннососудистая древесина в отечественных работах прош-
лых лет называлась рассеяннопорой древесиной. (Прим, пе-
рев.)
2640
33
периклинальное деление верхних клеток камбиальной зонь!
и быстрая дифференциация элементов флоэмы (Derr and
Evert, 1967). В это же время начинается периклиналь-
ное деление у остальной части камбия. Однако созревают
ксилемные элементы медленно, до появления первых
члеников сосудов образуется несколько рядов члеников
ситовидных трубок. Такую беспорядочную изменчивость
во времени образования флоэмы и ксилемы у покрыто-
и голосеменных можно понять в категориях объединенной
гормональной теории.
3.3. ИНИЦИИРОВАНИЕ КАМБИАЛЬНОЙ
АКТИВНОСТИ
Кольце- и рассеяннососудистые породы имеют разли-
чия не только в начале образования ксилемы и флоэмы,
но и в последовательности инициирования камбиальной
активности в разных частях ствола и ветвей. Рассмотрим
результаты исследования Hanson и Brenke (1926) коль-
цесосудистого Fraxinus campestris и рассеяннососудистого
Acer saccharinum, растущих недалеко от Линкольна (шт.
Небраска, США). У ясеня активизация камбиальной зо-
ны начинается с середины апреля, а к 27 апреля нижняя
часть ствола и 1-летние ветви имеют одинаковое количе-
ство новой древесины. Это подтверждает положение, что
активность васкулярного камбия наступает одновременно
по всей высоте дерева. Отмечено также, что периклиналь-
ное деление в камбиальной зоне начинается незадолго до
распускания верхушечных почек. По всей вероятности,
у ясеня растущие побеги не стимулируют активность
камбиальной зоны.
В отличие от ясеня у клена верхушечные почки рас-
пускаются в середине апреля, но ксилема образуется
в нижней части ствола с задержкой на 1 — 2 недели.
Камбиальная активность на 1-футовой высоте ствола на-
ступает только 27 апреля, когда хорошо растут побеги
и размеры листьев составляют половину полностью раз-
вившихся. У вышерасположенных (па 2,4 и 5,4 м) час-
стей ствола в это же время активизируется васкулярный
камбий, о чем свидетельствует присутствие члеников со-
судов и волокон в стадии поверхностного роста и утол-
щения оболочек. Новая ксилема 1-летних ветвей клена
состоит из члеников частично лигнифицированных тол-
стостенных сосудов. У пород с рассеяннососудистой дре-
весиной, например у клена, волны камбиальной активно-
34
ст*и медленно опускаются йз верхних ветвей к основанию
ствола.
Более новые работы с рассеяннососудистыми порода-
ми указывают на тесную связь в период роста побегов
между инициированием камбиальной активности и обра-
зованием ауксина. Ее хорошо иллюстрирует сравнение
камбиальной активности у клена ложноплатанового (Acer
pseudoplatanus) с полностью удаленными почками и ин-
тактных растений. Предполагается, что камбиальная ак-
тивность у интактных пород инициируется в ветвях
и медленно продвигается вниз, а к концу мая (Лондон,
Англия) радиальный рост быстро прогрессирует по всей
длине ствола (Wareing, 1951). Однако при удалении
сучьев, ветвей и придаточных почек васкулярный камбий
остается неактивным и не образует новой ксилемы. Ис-
ключение составляет рост на стволе придаточных почек,
тогда камбиальная активность строго ограничивается
участком ниже места прикрепления почки. При сильном
росте побеги рассеяннососудистых пород контролируют
инициирование камбиальной активности, и это подтверж-
дает опосредствующую роль ауксина.
Digby и Wareing (1966b) изучили уровень ауксина,
экстрагируемого из камбиальных зон на разной высоте
ствола у Populus trichocarpa (рассеяннососудистая поро-
да) до и после окончания распускания почек. До распус-
кания почек ауксин из камбиальной зоны по всей высоте
дерева не экстрагируется; его содержание значительно
увеличивается в период распускания почек в верхней
части ствола рядом с удлиняющимися побегами, тогда как
в нижней части его мало иди совсем нет. Через 3 недели
после распускания почек, когда камбиальная активность
достаточна во всем стволе, уровень ауксина одинаково
велик по всей высоте дерева. Итак, медленное продвиже-
ние вниз камбиальной активности весной у рассеянносо-
судистых деревьев связано с соответствующим перемеще-
нием ауксина, образующегося в удлиняющихся междо-
узлиях и листьях.
Камбиальная активность у кольцесосудистых пород
контролируется более неуловимо. Например, Wareing
(1951) установил, что при удалении почек и у интактных
растений Fraxinus excelsior камбиальная активность
в стволах индуцируется в одно время; более того, новый
прирост ксилемы в обоих случаях состоит из широких
сосудов. Это подтверждает, что камбиальная активность
35
РИС. 15. Диаграм-
ма распростране-
ния новых сосу-
дов у черенков
ясеня; сосуды
диффере н ц и р у-
ются у маленьких
придаточных по-
чек, образующих-
ся на старом ли-
стовом рубце
(Wareing, 1951)
у кольцесосудистых пород стимулирует-
ся при очень низком содержании аук-
сина (при удалении почек он может
образовываться в крошечных почках,
связанных с листовыми рубцами) или
что его предшественник, уже находя-
щийся в васкулярном камбии, активи-
руется одновременно в стволе и ветвях.
У кольцесосудистых пород камби-
альную активность контролируют даже
совсем крошечные почки. Если черенки
ясеня, находящегося в состоянии по-
коя, внести в теплую лабораторию, ак-
тивность васкулярного камбия, особен-
но в основании черенков, пробуждается
(Wareing, 1951). При сдирании коры
новые широкие сосуды всегда проходят
кверху, но не выше прежнего местона-
хождения маленькой почки (рис. 15).
Это способствует большему радиально-
му приросту у основания черенка.
Роль почек не выяснена до конца, но
наблюдения показывают, что даже при
очень незначительных размерах они
включаются в контроль зарождения
камбиальной активности у кольцесосудистых пород.
Результаты исследований показывают, что одновре-
менное наступление камбиальной активности у кольце-
сосудистого Ulmus glabra зависит от активации предше-
ственника ауксина (Digby and Wareing, 1966b). В кам-
биальной зоне на любой высоте ствола в феврале отме-
чается высокий уровень регулятора роста, по всей веро-
ятности, триптофана; при распускании почек он превра-
щается в индолил-3-уксусную кислоту (ИУК), одновре-
менно в стволе инициируется камбиальная активность.
Необходимы дальнейшие исследования по выявлению
роли почек, а может быть и других факторов, в регуля-
ции превращения триптофана в ИУК.
3.4. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ
ОБРАЗОВАНИЯ КСИЛЕМЫ И ФЛОЭМЫ
Помимо инициирования камбиальной активности аук-
син включается в дифференциацию производных камбия.
Это показано в опытах при раздельном и совместном
нанесении ИУК и гибберелловой кислоты (ГК) в лано-
лине на стволики Acer platanus с удалением спящих
почек (Wareing, 1958). Изучение срезов через 1—3 не-
дели показало, что один ланолин не оказывает никакого
действия, васкулярный камбий остается неактивным.
При воздействии ИУК у ксилемы образуются тонкие зо-
ны из лигпифицированных члеников сосудов, но без зре-
лых волокон. Одна ГК стимулирует деление некоторых
клеток, но, в отличие от ИУК, дальнейшей дифференциа-
ции производных камбия не происходит. При совместном
применении обоих гормонов у ксилемы образуется широ-
кий прирост, похожий на обычную древесину; он состоит
из лигпифицированных волокон и члеников сосудов.
Затем в нормальное развитие вторичной ксилемы вклю-
чаются ауксин и гиббереллин. Гормоны различаются по
своему назначению: ауксин первично включается в диф-
ференциацию производных камбия, а гиббереллин стиму-
лирует деление клеток, обусловливающее образование
производных камбия.
Если ауксин стимулирует дифференциацию ксилемы,
то высокий уровень гиббереллина вызывает образование
флоэмы (Digby and Wareing, 1966а). Нанесение ГК на
стволики вышедшего из состояния покоя и с удаленными
почками Populus robusta вызвало образование утолщен-
ного прироста у флоэмы, при этом ксилема не дифферен-
цировалась. Применение только ИУК стимулировало
рост ксилемы, флоэма не закладывалась. Эти опыты под-
тверждают, что образованию ксилемы благоприятствует
высокое содержание ауксина в комплексе ауксин — гиб-
береллин, тогда как последний стимулирует дифференци-
ацию флоэмы.
Сезонная динамика образования ксилемы и флоэмы
зависит от изменения баланса ауксина и гиббереллина
у васкулярного камбия. В связи с этим Digby и Wareing
(1966b) измерили содержание ауксина и гиббереллина
в стволах и листьях у кольцесосудистого Ailanthus altis-
sima, выросшего в режиме длинного дня и затем пере-
веденного в режим короткого дня. Образование ксилемы
37
затормаживается через 3 недели после перевода дерева
в режим короткого дня, но новая флоэма формируется
еще 2 недели, в связи с чем продолжается прирост ствола
по диаметру. В режиме длинного дня ствол и листья
имеют высокий уровень ауксина и гиббереллмна. После
перехода на короткий фотопериод содержание ауксина
снижается и через 4 недели его уже очень мало, ксилема
больше не дифференцируется; уровень гиббереллина ос-
тается высоким, образуется одна флоэма. Эти опыты под-
тверждают участие в регуляции образования ксилемы
и флоэмы эндогенного уровня ауксина и гибберел-
лина. В дальнейших исследованиях можно показать, что
динамика образования ксилемы и флоэмы у покрыто-
и голосеменных (см. 3.2) зависит от сезонного изменения
баланса ростовых веществ в камбиальной зопе.
Исследования Wareing (1958) Acer pseudoplatanus
показывают, что совместное действие ауксина и гиббе-
реллина стимулирует нормальное развитие ксилемы.
Кроме того, очевидно, что процесс развития васкулярного
камбия связан с взаимодействием других факторов. На-
пример, у черепков ивы ломкой (Salix fragilis Robards
et al., 1969) обнаружили, что многие клетки в дифферен-
цирующейся ксилеме, сильно растущей в ширину (пред-
полагаемые членики сосудов), увеличиваются при раз-
дельном применении ряда веществ: ауксина, ГК, 6-фур-
фуриламинопурина (цитокинина), сахарозы и мио-ино-
зита (табл. 1). При использовании одновременно двух
веществ число члеников сосудов значительно увеличи-
лось дополнительно к ужо образовавшимся. Как указыва-
1. ДЕЙСТВИЕ НЕКОТОРЫХ ВЕЩЕСТВ НА ДИФФЕРЕНЦИАЦИЮ
КСИЛЕМЫ У ЧЕРЕНКОВ ИВЫ (ФАП — ФУРФУРИЛАМИНОПУРИН;
САХ — САХАРОЗА: ИНО — МИО-ИТТОЗИТ
(A. W. Robards, Е. Davidson and Р. Kidwai, 1969)
Вещество 1 Процент, члени- ков сосудов от дифференци- рующихся кле- ток ксилемы Вещество Процент члени- ков сосудов от дифференци- рующихся кле- ток ксилемы
ИУК 9,6±0,5 ИНО 13,1±1,7
ГК 9,2±1,1 ИУК + ГК + ФАП +
ФАП 7,2±0,5 + САХ 31,5±4,7
САХ 12,6±0,8 Контроль 2,8±0,6
38
ют Robards et al. (1969), не следует делать преждевремен-
ных выводов о зависимости дифференциации от какого-
либо одного вещества, поскольку она может зависеть от
эндогенного взаимодействия целого комплекса их.
3.5. ФИЗИОЛОГИЯ ОБРАЗОВАНИЯ
ГОДИЧНОГО КОЛЬЦА
При внимательном рассмотрении тонкого поперечного
среза древесины у деревьев в умеренных широтах можно
выявить явное структурное несоответствие в ксилеме,
сформированной в разное время вегетационного периода.
У трахеид ранней древесины голосеменных пород диа-
метр больше и вторичная клеточная оболочка толще, чем
у трахеид, дифференцировавшихся в этом же периоде
роста, но позднее (рис. 16, Gemmell, 1969). Аналогично
и у кольцесосудистых покрытосеменных диаметр сосудов
уменьшается от начала к концу годичного прироста кси-
лемы.
Названия «ранняя» и «п о з д н я я» древесина упот-
ребляют для различия соответственно первоначальной
менее плотной и последующей более плотной древесины,
образованной в динамике годичного прироста. Структур-
ные отличия поздней древесины одного сезона от ранней
древесины следующего года объясняются периодичностью
образования древесины, что ясно видно на примере го-
дичного кольца. Годичное кольцо — это прирост
ксилемы, включающий раннюю и позднюю древесину
и образовавшийся за одногодичный Цикл роста. У неко-
торых пород годичный прирост ксилемы у ветвей и ство-
лов может состоять из двух колец прироста, особенно
если радиальный рост временно прерывается сильной
летней засухой. Второе кольцо роста называют л о ж п ы м
годичнымкольцом (Esau, 1965).
Переход от ранней древесины к поздней связан с дву-
мя экспериментально разделимыми, но обычно совпада-
ющими явлениями дифференциации клеток ксилемы —
уменьшением размера клетки и возрастанием толщины
ее оболочки. Формирование поздней древесины начина-
ется в основании ствола и постепенно продвигается вверх
к ветвям. В связи с этим одно годичное кольцо вблизи
основания ствола содержит поздней древесины больше,
чем в выше расположенных его частях.
39
РИС. 16. Поперечный срез ствола Sphenophyllum. Вторичная кси-
лема сначала располагается треугольником в соответствии со
строением первичной флоэмы, позднее развивается цилиндриче-
ская древесная сердцевина (Morey, 19G8)
Ауксин, поступающий в ствол из ветвей, как считают,
регулирует в период образования древесины рост по диа-
метру дифференцирующихся клеток ксилемы. Известно,
что такие факторы среды, как короткий фотопериод и за-
суха, тормозящие экстенсивный рост побега, ускоряют
образование поздней древесины в нижней части ствола.
Развивающиеся листья и междоузлия — богатый источник
ауксина (см. 1.3). В связи с этим предполагают, что об-
разованный в побеге ауксин способствует росту по диа-
метру дифференцирующихся клеток ксилемы в располо-
женной под побегом части ствола. Отличным доказатель-
ством этому положению служит опыт по воздействию
фотопериода на образование древесины у молодых дере-
вьев сосны смолистой (Pinus resinosa). Образование ран-
ней древесины у деревьев, выросших в режиме длинного
дня (18-часовой фотопериод), коррелировало с высоким
содержанием ауксина у быстро растущего побега (Larson,
1962). Как рост побега, так и содержание ауксина умень-
шались при переводе растения на 2 недели в режим
короткого дня (8-часовой фотопериод); в этих условиях
образовалась поздняя древесина. Повторный перевод сос-
40
ны смолистой с режима длинного дня в режим короткого
дня вызвал вторичный рост побегов, увеличение содержа-
ния ауксина и отложение нового прироста ранней древе-
сины. Так была создана экспериментальная обстановка
для получения ложного кольца прироста.
Прямое подтверждение регуляции размера клетки
ауксином вытекает из опыта применения ИУК на дека-
питированные стволики сосны смолистой при их росте
в режиме короткого дня. Воздействие ИУК стимулирует
образование широких трахеид вместо узких, формирую-
щихся до воздействия (Larson, 1962). Таким образом,
обычное уменьшение содержания поздней древесины по
мере продвижения к вершине объясняется тем, что в ниж-
ней части ствола очень мало ауксина вследствие меньшего
числа ростовых побегов в нижней части кроны. Ствол и вет-
ви используют ауксин высокой концентрации, причем
продолжительное время и в значительном количестве.
Следовательно, динамике образования поздней древесины
свойствен акропетальный 1 характер, что следует хотя бы
из размера клеток.
Аналогичный опыт по регуляции диаметра члеников
сосудов проводили с кольцесосудистой Robinia pseudoaca-
cia (Digby and Wareing, 1966a). Растения в 2-летнем воз-
расте перевели на 3 недели в режим короткого дня; в ре-
зультате этого прекратились рост побегов, камбиальная
активность и образование ксилемы. Затем побег декапи-
тировали и наносили на него ИУК разной концентрации;
в этом случае при постоянно низком уровне ГК камби-
альная активность возобновлялась и образовывалась кси-
лема. У экспериментально стимулированной ксилемы
отмечена отчетливая взаимосвязь между диаметром чле-
ников сосудов и концентрацией ауксина. Далее можно
предполагать, что увеличение дифференцирующихся чле-
ников сосудов регулируется посезонно меняющимися
концентрациями эндогенного ауксина, передвигающегося
из кроны.
Утолщение клеточных оболочек в поздней древесине
связано с увеличенным поступлением продуктов фотосин-
теза по мере развития хвои побега. Эта связь отлично
видна у сосны смолистой, выращиваемой па протяжении
декабря в теплице в режиме 18-часового фотопериода.
Измерения показали участие 1-летней и вновь появив-
1 Акропетальный — направленный к вершине. (Прим, перев.)
41
шейся хвои у этой сосны в росте побегов и образовании
древесины. Для упрощения схемы опыта осенью пред-
шествующего года удалили всю хвою старше 1 года,
боковые ночки и ветви. В интервале 3—7 недель после
распускания почек новая хвоя растет очень быстро, затем
ее рост в длину замедляется. В этот период ассимиляты
поступают из 1-летней хвои в верхушечный побег и по-
глощаются, например, структурными компонентами новой
ткани хвои; одновременно они транспортируются в другие
части дерева, в том числе в основание ствола, где обра-
зуются тонкостенные трахеиды ранней древесины. Новая
хвоя в это время очень мало отдает углеводов другим
частям дерева. Толщина клеточных оболочек у диффе-
ренцирующихся трахеид в основании ствола увеличивает-
ся через 7—9 недель после распускания почек. Это свя-
зано с созреванием новой хвои, из которой теперь транс-
портируются ассимиляты также и в нижележащие ткани
ствола. Ассимиляты, образованные старой хвоей, не
нуждающейся больше в росте расположенных над ней
побегов, поглощаются нижележащими тканями ствола.
Итак, у сосны смолистой переход от ранней древеси-
ны к поздней причинно связан с двумя явлениями: быст-
рым включением в камбиальную зону продуктов фото-
синтеза при созревании хвои побегов текущего года
и уменьшением уровня содержания ауксина при прекра-
щении экстенсивного роста кроны. Однако общее пред-
ставление о том, что переход ранней древесины к поздней
регулируется исключительно наличием ауксина и ассими-
лятов, возможно, слишком упрощенное. Например, разви-
тие дифференцирующихся клеток ксилемы обусловливают
и многие иные вещества (см. 3.4). В конечном счете
в регуляцию образования годичного кольца могут вклю-
чаться помимо ауксина и ассимилятов более широкие
взаимосвязи гиббереллинов, цитокининов и ингибиторов
роста. Для всестороннего изучения образования колец
прироста можно рекомендовать книгу Zimmermann
и Brown (1971).
3.6. ГОДИЧНЫЕ КОЛЬЦА И КЛИМАТ
Годичные кольца образуются при воздействии ряда
переменных факторов среды, поэтому для кроны харак-
терна изменчивость роста. Например, при переводе сосны
смолистой с длинного на короткий фотопериод уменыпа-
42
ется рост побегов, снижается уровень ауксина и форми-
руются узкие клетки, свойственные поздней древесине.
Кроме того, на образование древесины влияют интенсив-
ность света, температура и дефолиация (насекомыми или
градом). В аридных районах, где бывают сильные засухи,
почвенная влага может ограничивать рост побегов и вы-
зывать образование очень узкого годичного кольца или
его отсутствие.
Влияние количества осадков на динамику образования
годичного кольца иллюстрируется сравнением роста де-
ревьев на побережье Калифорнии и в западном Техасе.
В окрестностях Сан-Франциско (шт. Калифорния) дожди
выпадают в основном зимой, летом их почти не бывает.
Самая высокая влажность почвы отмечается ранней вес-
ной, когда и начинают расти деревья, но затем из-за
отсутствия дополнительных осадков их рост прекращает-
ся. Годичные кольца здесь образуются регулярно и харак-
теризуются только одним циклом радиального роста при-
мерно одинаковой каждый год интенсивности и продол-
жительности. (Glock and Agerter, 1962). С другой сторо-
ны, в Лаббоке (шт. Техас) сезон роста начинается ранней
весной при низкой влажности почвы. В большинстве
своем осадки выпадают в конце веспы и летом, споради-
чески, периоды влажной погоды сменяются жарой и су-
хостью разной продолжительности. В таких условиях
динамика образования годичных колец наиболее вариа-
бельна. Годичный прирост в случае образования за год
нескольких колец роста в действительности может быть
множественным годичным кольцом. У других
деревьев развиваются неполные кольца прироста, так на-
зываемые линзы, так как камбиальная зона у деревьев
недостаточна, чтобы стать активной по всей окружности
ствола. В условиях продолжительной засухи, которая
в отдельные годы бывает на юго-западе Америки, почвен-
ной влаги для поддержания роста деревьев не хватает
и годичное кольцо отсутствует. По этой причине в запад-
ном Техасе очень трудно определить возраст старых де-
ревьев подсчетом годичных колец, несколько легче в Ка-
лифорнии.
При подсчете динамики годичных колец недавно уста-
новлено, что деревья сосны остистой (Pinus aristata) 1
1 Сейчас за этим видом закрепилось основное название — Pinus
longaeva — сосна долговечная. (Прим, перев.)
43
в Калифорнии самые старые из всех живущих на земле.
Они растут очень медленно на высоте 3000—3300 м над
ур. м. в Уайт-Маунтинсе (Калифорния) с сухим клима-
том. Необычный пример уменьшенного радиального при-
роста представляет кусочек древесины 12,7 см толщины,
содержащий 1100 годичных колец (Ferguson, 1968).
Хронология годичных колец, например у сосны остис-
той, основана на определении у относительно молодого
дерева точного календарного возраста, которому соответ-
ствует каждое годичное кольцо. Было замечено, что ра-
диальная толщина годичных колец ежегодно варьирует.
Более того, одинаковая динамика их прироста никогда
не повторяется. Два образца при тщательном изучении
динамики годичных колец можно кросс-датировать, на-
пример, путем сопоставления у них участков, выделяе-
мых па наиболее узких кольцах. Для продления хроно-
логии древесину с установленным возрастом кросс-дати-
руют с древесиной немного старшего возраста, у которой
частично совпадает динамика годичных колец. Самой
старой сосне остистой в Уайт-Маунтинсе около 4600 лет,
но при тщательном кросс-датировании по древесине давно
отмерших деревьев возраст сосны недавно продлили до
7100 лет (Ferguson, 1968). По всей вероятности, хроно-
логию можно дополнительно продлить па 2 тыс. лет, так
как с помощью радиоуглеродного анализа древесины
возраст сосны остистой установлен примерно 9000 лет.
Одно из самых важных положений дендрохронологии,
возможно, заключается, в регистрации местного климата
прошлых лет. Она получается в связи с тем, что годичные
кольца — хороший измеритель количества осадков, выпа-
дающих за вегетационный период. Таким образом, струк-
тура годичных колец сохраняет историческую запись
климата прошлых лет. Кроме того, древесину старейших
деревьев используют теперь в качестве стандарта при
определении возраста радиоуглеродным методом, что дает
возможность более точно устанавливать время археологи-
ческих артефактов в глубоких слоях.
4. Реактивная Древесина
4.1. СТРУКТУРА И РАСПРЕДЕЛЕНИЕ
РЕАКТИВНОЙ ДРЕВЕСИНЫ
При отклонении дерева от вертикального положения
у него постепенно изменяется направление роста; в ре-
зультате образуется древесина нетипичная по структуре
и химическому составу, так называемая реактивная1.
Она образуется по всей той или иной стороне наклонен-
ного ствола и, сжимаясь или растягиваясь, вызывает из-
РИС. 17. Изменение направле-
ния роста у горизонтально
размещенного саженца:
а — быстрое восстановление вер-
шинки побега (с участием в росте
только первичных тканей) путем
дифференцированного продольного
роста противоположных сторон
молодого стволика; б — медленное
изменение направления роста
стволика при изгибе
менение направления его роста. Обычно дерево снова
принимает вертикальное положение, но если саженцам
для этого достаточно нескольких месяцев, то зрелым де-
ревьям в лесу требуется много лет (рис. 17).
Потребителям хорошо знаком такой порок, как реак-
тивная древесина, образующийся при продольном сжатии
в процессе сезонного роста. Доски с этим пороком быстро
растрескиваются и коробятся. Такая древесина, образуе-
мая васкулярным камбием, лигнифицируется, становится
особо твердой и теряет способность к растяжению. В про-
цессе изгибания изменяется направление роста ствола
или ветви. По своей природе этот процесс полностью
отличается от движения органов растений с участием
только первичных тканей (например, у колеоптиля Ave-
na), когда изменение направления движения вызвано
дифференцированным продольным ростом.
У покрытосеменных древесных растений реактивная
древесина, называемая тяговой, образуется на верхней
1 Реактивная древесина (англ, reaction wood) — общее название
креневой (англ, compression wood), или древесины сжатия,
и тяговой (англ, tension wood), или древесины растяжения.
(Прим, перев.)
45
стороне стволов и ветвей в результате изменения направ-
ления роста при продольном сжатии в месте нахождения.
Шероховатость досок, полученных из свежесрубленного
строевого лиственного леса, объясняется наличием тяго-
вой древесины. Волокна ее, выделяясь в отдельные уча-
стки, препятствуют распиловке, строганию и пр. Другой
серьезный недостаток, возможный из-за наличия тяговой
древесины,— продольное расщепление конца бревна в мо-
мент валки в результате высвобождения сил внутреннего
напряжения, создаваемого этой тканью.
По сравнению с нормальной древесиной тяговая со-
держит больше целлюлозы и меньше лигнина. В ней не-
много члеников сосудов, причем небольших в диаметре
(рис. 18, а). Волокна тяговой древесины, называемые
иногда желатинозными, состоят из утолщенного, гелеоб-
разного внутреннего слоя S (G) вторичной клеточной
оболочки (см. рис. 18, б). Этот слой может быть допол-
нительным у обычной трехслойной вторичной оболочки
или может замещать слои S3 или S2 и S3 (Wardrop,
1964b). Целлюлоза слоя S (G) ориентирована параллельно
длинной оси волокна (см. рис. 18, в, г); лигнина в ней
мало или совсем пет. Высокое содержание целлюлозы
и низкое лигнина в тяговой древесине объясняется осо-
бенностями химического состава слоя S (G).
У голосеменных древесных растений реактивная дре-
весина, называемая креневой, образуется на нижней
стороне наклонного дерева. Она находится в стволах
и ветвях в состоянии продольного сжатия, и, как предпо-
лагается, вызывает изменение направления роста путем
растяжения места нахождения. У лиственных пород кре-
невая древесина серьезный порок из-за ее большей твер-
дости и плотности по сравнению с нормальной; она обла-
дает слабой гвоздеудерживающей способностью (Westing,
1965).
В целом реактивная древесина считается ненормаль-
ной, возникшей в результате необычного роста дерева. Но
существуют и явные исключения. Например, на востоке
Северной Америки у тсуги канадской (Tsuga canadensis)
верхушечный побег в начале сезона роста характерно
поникает и раскачивается от ветра, позднее он постепен-
но изгибается кверху, что связано с развитием креневой
древесины (Mergen, 1958). Сезонная особенность роста
лидера типичная для этого вида, она генетически пред-
46
РИС. 18. Срез креневой древесины клена красного:
а, б — поперечный, стрелкой обозначен толстый S (я) слой волокон;
в, г — тот же срез, что и на б, но под флуоресцентным и поляриза-
ционным микроскопом соответственно (в —отсутствие флуоресценции
у слоя S (£') указывает на низкий уровень содержания лигнина; г —
этот же слой, показывающий незначительное двойное преломление
или его отсутствие в зависимости от расположения осей целлюлозы)
47
определена, поэтому образование у него крейевой древе-
сины принимается за нормальный процесс роста дерева.
Трахеиды креневой и нормальной древесины значи-
тельно различаются по своей структуре. У первых Попе-
речный срез имеет круглое сечение, и их легко обнару-
жить по большим межклетным пространствам; их вторич-
ная клеточная оболочка состоит из верхнего слоя Si
и утолщенного внутреннего слоя S2. В отличие от трахеид
нормальной древесины (см. разд. 2) целлюлоза слоя S2
РИС. 19. Схема распределе-
ния микрофибрилл на по-
перечном срезе полностью
делигнифицированной тра-
хеиды креневой древесины
(Cote et al., 1968)
ориентирована под углом почти 45° к длинной оси кле-
ток; слой S3 у оболочки отсутствует.
По сравнению с нормальной древесиной креневая со-
держит лигнина больше, а целлюлозы меньше. Это обус-
ловлено строением ее трахеид. В самой верхней (от
полости клетки) части слоя S2 очень мало целлюлозы
и значительное количество лигнина. Такую лигнинообо-
гащенную зону называют S2 (L) (рис. 19). Cote et al.
(1968) изучали ультраструктуру и химический состав
слоя S2 (L) у Larix. Маленькие блоки древесины делиг-
нифицировали подкисленным хлоритом натрия, который
окисляет лигнин; полисахариды клеточной оболочки на
эту обработку почти не реагировали. Из других блоков
древесины целлюлозу удаляли при контролируемой
экспозиции плавиковой и соляной кислотами. Лигнин
к кислотам устойчив, а полисахариды клеточной оболоч-
ки, в том числе целлюлоза, легко гидролизуются. С по-
мощью электронного микроскопа исследовали древесину
с удаленной в результате гидролиза целлюлозой и уста-
новили, что слой S2 у оболочки структурно связан, осо-
бенно слой S2 (L) представляет собой очень плотный
остаток лигнина. В образцах с экстрагированным лигни-
ном в слое S2 (L) выявлена целлюлоза, правда, в незна-
чительных количествах; остаток ее в этом слое оболочки
очень пористый (см. рис. 19). Это исследование убеди-
48
тельно доказывает, что слой S2 (L) у трахеид креневой
древесины содержит исключительно свободно построен-
ный скелет из целлюлозы внутри очень плотного лигни-
фицированного матрикса.
4.2. ПРИЧИНЫ ОБРАЗОВАНИЯ РЕАКТИВНОЙ
ДРЕВЕСИНЫ
В начале XX в. анатомы растений установили, что
образование реактивной древесины — это реакция на
асимметрическое продольное напряжение. Предполагают,
что сопротивление на растяжение верхней стороны у на-
клоненного ствола покрытосеменных растений обуслов-
лено дифференциацией тяговой древесины. Однако у го-
лосеменных креневая древесина образуется па нижней
стороне изогнутых стволов, где максимальное напряжение
от сжатия. Концепция образования реактивной древесины
как реакции на напряжение получила частичную под-
держку у исследователей, изучающих воздействие физи-
ческих факторов на дифференциацию растительных тка-
ней. Jaccard (Sinnott, 1960) изучил интересное явление
у маленького корня, росшего над большим одревеснев-
шим корнем и, следовательно, находившегося при росте
по диаметру под давлением последнего. У элементов кси-
лемы маленького корня оболочки клеток были толще, чем
оболочки соответствующих клеток контрольного образца,
не испытывавшего давления.
Дальнейшие опыты показали, что образование реак-
тивной древесины — это реакция па гравитационный
стимул Например, если ствол молодого лиственного
деревца согнуть по вертикали в петлю и удерживать
в этом положении, тяговая древесина образуется на верх-
них сторонах ствола в верхней и нижней частях петли
(Wardrop, 1964b). Поэтому реакция древесины на верх-
ней стороне не зависит от воздействия на петлю про-
дольного растяжения или напряжения сжатия. Непосред-
ственное влияние гравитации на образование реактивной
древесины также видно из опытов со стволами, выведен-
ными из свойственного им вертикального положения.
Реактивная древесина обычно- отсутствует у стволов при
сохранении ими вертикального положения, но легко
1 Гравитация — сила земного тяготения, сила тяжести. (Прим,
перев.)
49
образуется, если ось отклонена от вертикали на 1—2°.
Количество реактивной древесины коррелирует со сте-
пенью отклонения ствола от вертикального положения.
Образование реактивной древесины в стволе объясня-
ют гравитационным стимулом, для ветвей же объяснение
этого образования более сложное. Sinnott (1960) устано-
вил, что креневая древесина образуется на верхней или
па нижней стороне ветвей Pinus strobus, отогнутых соот-
ветственно вниз или вверх (рис. 20), причем ее положе-
РПС. 20. Креневая древесина Я, образующаяся на нижней или
верхней стороне ветвей, изогнутых соответственно книзу или
кверху; в аналогичных опытах с покрытосеменными растениями
расположение реактивной древесины противоположное (Sinnott,
1960)
РИС. 21. Плагиотропные перемещения самых верхних ветвей Li-
quidambar styraciflua (окончательное положение ветви показано
сплошной линией):
а — у вертикально расположенного интактного контрольного растения от-
мечено незначительное перемещение петви в направления к главной оси;
б — у декапитированного контрольного растения (устранена ьпинастия)
ветвь сильно отклонилась кверху в результате образования тяговой дре-
весины (ТД) на верхней стороне ветви; в — у декапитированного растения
при вращении его в горизонтальной плоскости на клиностате (устройство
для устранения постоянно поддерживаемого направления гравитацион-
ной силы) ветвь незначительно отклоняется книзу без образования тяго-
вой древесины (Wardrop, 1964b)
ние не постоянно по отношению к гравитационной силе.
Креневая древесина появляется либо на верхней, либо
на нижней части ветви, так что восстанавливается преж-
няя (ненарушенная) ориентация роста. Такие опыты
показывают, что гравитационный стимул только один из
факторов, индуцирующих в ветвях реактивную древеси-
ну; могут также действовать и некоторые внутренние
факторы.
Образование реактивной древесины в ветвях можно
объяснить в зависимости от плагиотропизма. В соответст-
50
вии с плагиотропизмом направление ветви точно асим-
метрично образованию реактивной древесины и определя-
ется взаимодействием двух противоположных сил: внут-
ренней (эпинастии), от которой ветвь изгибается кверху,
и отрицательного геотропизма, от которого она стремится
занять более вертикальное положение. Эпинастия у боль-
шинства растений, как предполагают, связана с физиоло-
гической активностью верхушечной почки ствола, от
которого отходит ветвь.
Изменение направления движений самой верхней вет-
ви ликвидамбара смолоносного (Liquidambar styraciflua)
иллюстрирует плагиотропизм, отмечаемый при росте де-
рева (рис. 21). Двухлетние саженцы декапитировали
и выращивали 3 месяца в вертикальном положении или
в горизонтальном — на клиностате1 (Wardrop, 1964b).
В первом случае у самой верхушечной ветви проявлялся
сильный отрицательный геотропизм, и на ее верхней сто-
роне образовывалась тяговая древесина (см. рис. 21, б);
во втором направление роста изменялось совсем незна-
чительно — верхушечная ветвь отклонялась книзу, при-
чем тяговая древесина отсутствовала (см. рис. 21, в). По-
следнее объясняется тем, что были устранены гравита-
ционный и эпинастический стимулы в результате обра-
ботки растений соответственно па клиностате и декапита-
цией. У вертикальнорастущего Liquidambar при декапи-
тации был устранен только эпинастический фактор,
и ветвь реагировала на гравитационный стимул образова-
нием тяговой древесины на верхней стороне вследствие
сильного отрицательного геотропического изгиба. Поэтому
противоположно действующие эпинастическая и геотро-
пическая силы обусловливают расположение реактивной
древесины на той или другой стороне ветви. Thimann
(Wardrop, 1964b) обсуждает эту концепцию в связи
с общей гормональной теорией.
В последнее время отмечается значительный интерес
к объяснению образования реактивной древесины па гор-
мональной основе. Necesany (Wardrop, 1964b) сообщал
о более высоком уровне ауксина на нижней стороне гори-
зонтально расположенных ветвей у Populus и Pinus. При
горизонтальном положении стволов Populus копцеитра-
1 Клиностат — прибор, позволяющий выращивать растение, избе-
жав одностороннего влияния на него гравитации. (Прим,
перев.)
51
ция ауксина в верхней и нижней сторонах составляла
отношение 40:60 (Leach and Wareing, 1967). Поэтому
у покрыто- и голосеменных ауксин обнаружен в большем
количестве на нижней стороне наклоненных стволов
и ветвей. Однако реактивная древесина у голосеменных
образовывалась на стороне с высокой концентрацией
ауксина, а у покрытосеменных — с низкой.
Получено прямое экспериментальное доказательство,
что развитие креневой древесины в стволах опосредовано
реакцией ауксина. Нанесением а-нафтилуксусной кисло-
ты (НУК) в ланолине на прямоствольные сосны вызыва-
ли образование креневой древесины на участках с нор-
мальной древесиной (Wardrop, 1965). Креневая древеси-
на образовывалась выше и ниже участка, обработанного
ауксином, если же последний наносили па одну из сторон,
то через несколько недель ствол изгибался в направлении
противоположно м участку обработки.
Тяговая древесина, как показали опыты, образуется
при недостатке ауксина. Necesany (Wardrop, 1964b)
наносил ИУК в ланолине на верхнюю или нижнюю сто-
рону ствола Populus, согнутого в горизонтальное положе-
нно. Нанесение па нижнюю сторону ИУК мало влияло
па образование тяговой древесины на противоположной
стороне, тогда как нанесение ауксина той же концентра-
ци|и па верхнюю сторону оказывало на образование тяго-
вой древесины сильное ингибирующее действие. Недавно
было установлено, что тяговую древесину можно экспери-
ментально индуцировать у вертикальнорастущих стволов
Acer при обработке их 2, 3, 5-трийодбензойной кислотой
(ТИБК) — ингибитором полярного передвижения аукси-
на. Ниже обработанного места у вертикально растущих
стволов локально образовывалось сплошное кольцо тяго-
вой древесины (рис. 22). Если же наносили одновремен-
но ауксин и ТИБК, тяговой древесины было мало или она
совсем отсутствовала; из этого следует, что она образу-
ется при недостатке ауксина.
Таким образом, появление в стволах реактивной дре-
весины можно объяснить механизмом соотношения уров-
ней ауксина: у голосеменных она формируется при высо-
ком его содержании, у покрытосеменных связь обратная.
В наклоненных стволах деревьев ауксин накапливается
на нижней стороне; реактивная древесина формируется
на нижней стороне у голосеменных и на верхней у по-
крытосеменных. Присутствие реактивной древесины в вет-
52
РИС. 22. Поперечный срез ствола 8-недельного сеянца клена, об-
работанного за 2 недели ТИБК (в ланолине), нанесенной кольцом
вокруг стволика посредине первого междоузлия. Срез сделан на
вертикальнорастущем стволе примерно на 1 см ниже места обра-
ботки. За время обработки дифференцировалось сплошное кольцо
тяговой древесины. Стрелкой показаны волокна тяговой древеси-
ны (Morey and Cronshaw, 1968)
вях объясняют на основе двух противоположно действую-
щих сил — отрицательного геотропизма и эпинастии,
которые регулируются ауксином соответственно в ниж-
ней или верхней части. Взаимодействие эгих сил детер-
минирует распределение ауксина в ветви, место образо-
вания реактивной древесины и изменение направления
роста. Например, у ветви, изогнутой кверху, ауксин на-
капливается в верхней стороне, так как гравитационная
53
сила действует менее асимметрично и поэтому менее
эффективно регулирует поступление ауксина на нижнюю
сторону. В то же время эпинастия продолжает регулиро-
вать поступление ауксина в верхнюю часть, так что здесь
накапливается ауксин. В соответствии с этим, в верти-
кально изогнутых ветвях Pinus ауксин накапливается
в верхней части, вызывая образование креневой древеси-
ны и изменение направления роста ветви книзу в ее
прежнее положение (см. рис. 20). В будущих исследова-
ниях предстоит проверить эту концепцию на эксперимен-
тальной основе.
Значительные разработки требуются по механизму
передвижения ауксина с той или иной стороны в толстых
стволах и ветвях деревьев. Кроме того, нет достаточно
удовлетворительного объяснения ни на структурной, ни
на физиологической основе, почему покрыто- и голосемен-
ные обладают такими несхожими механизмами передви-
жения ауксина. У Drimys aromatica, примитивного покры-
тосеменного растения с мелкососудистой древесиной, ре-
активная древесина похожа на таковую у голосеменных,
по в отличие от нее формируется в верхней стороне от-
клоненного ствола (Westing, 1965). Изучение этого вида
представляет значительный интерес в эволюционном
отношении, так как регулирующий механизм у него,
способствующий развитию реактивной древесины, объ-
единяет четкие составные части покрыто- и голосеменных
растений.
5. Кора
5.1. СТРУКТУРА КОРЫ
Кору используют в производстве на протяжении мно-
гих веков. Римские мореплаватели изготовляли плоты из
коры пробкового дуба (Quercus suber) (Schery, 1952).
13 Китае из волокон коры получали бумагу задолго до
разработки современной технологии ио дефибрированию
древесины.
Формирование коры начинается только после оконча-
ния экстенсивного прироста ствола. Полностью удлинив-
шаяся ветвь поверх паренхиматической первичной коры 1
1 Первичная кора (англ, cortex) — первичная основная зона тка-
ни между васкулярной (проводящей) системой и эпидерми-
сом у стебля и корня. (Прим, перев.)
54
Покрывается эпидермисом, затем оба они оказываются
под тангентальным напряжением, вызванным иницииро-
ванием васкулярного камбия и заложением радиального
прироста. Вскоре эпидермальные и коровые ткани могут
подвергаться радиальному приросту при одновременно
происходящих антиклинальном делении и тапгентальном
приросте клеток.
Новая система защитных тканей, называемая пери-
дермой, возникает обычно в наружной первичной коре.
РИС. 23. Изображение на продольном срезе феллогена с радиаль-
но сгруппированными его производными
Например, внешний слой коровой паренхимы может
стать меристематическим по всей окружности. Клетки
ограничиваются в основном делением в периклинальной
плоскости, и закладывается новая латеральная меристема,
называемая феллогеном. Инициали феллогена дают
начало большому множеству производных кнаружи, на-
зываемых феллемой (пробка), а внутрь — меньшему
числу клеток, называемых феллодермой. Ткани фел-
логена и двух производных феллодермы составляют пери-
дерму (рис. 23). Производные феллогена подобно произ-
водным васкулярного камбия смыкаются в радиальные
ряды. Однако в отличие от васкулярного камбия у фел-
логена больше производных образуется кнаружи, чем
внутрь. Кроме того, клетки феллогена сами все одинако-
вые, у клеток перидермы нет ни осевых, ни радиальных
систем.
Пробка — отмершая ткань, защищающая лежащие
под ней живые части ствола и ветвей. Пробковые клетки
наполнены воздухом и плотно сгруппированы, поэтому
пробка широко применяется в производстве изоляцион-
ных плит. Помимо обычных компонентов клеточной
55
оболочки — целлюлозы и лигнина она содержит большое
количество воска и суберина. Суберин — гетерогенный
полиэфир, состоящий из множества жирных оксикислот,
таких, как феллоновая НО—СН2—(СН2)20—СООН ифлой-
оновая НООС-(СН2)7-(СНОН)2-(СН2)7-СООН. Вмес-
те с воском он делает феллему непроницаемой для боль-
шинства жидкостей (пригоден для бутылочной пробки)
и предопределяет защитную функцию перидермы.
Первую перидерму обычно замещают новые, развива-
ющиеся в нижерасположенных тканях коры. Существует
несколько исключений, например бук, у которого перво-
начальная перидерма может оставаться в течение столе-
тия. Посредством множественных антиклинальных деле-
ний феллоген бука отчетливо сохраняет свой прирост
наравне с радиальным приростом древесины. Но у боль-
шинства пород первоначальную перидерму вскоре вытес-
няют образующиеся из паренхимы в последовательных
слоях коры: вначале — в первичной коре, затем в первич-
ной флоэме и, наконец, во вторичной. По форме перидер-
ма в такой же последовательности обычно походит на
изогнутые раковины с обращенными кверху концами
(см. рис. 7). По мере созревания пробки в каждой новой
перидерме первичная кора и флоэма, изолированные об-
разованиями пробки, отрезаются от расположенных под
ними живых тканей и вскоре отмирают. Кора этого тина,
называемая чешуйчатой, позднее ошелушивается не-
большими частями в соответствии с динамикой строения
перидермы. Внешняя кора, или ритидом \ состоит из
ряда сменяющихся перидерм вместе с тканями (первич-
ной коры и поздней флоэмы), изолированными их обра-
зованиями. У секвойи вечнозеленой (Sequoia sempervi-
rens) па западе США при сменяющихся перидермах ри-
тидом ствола может достигать толщины 30 см.
Внутренняя кора — это все ткани внешней части
от камбиальной зоны, охватывающие наиболее внутрен-
нюю по расположению в стебле перидерму. Внутреняя
кора состоит из функционирующей и нефункциопирую-
щей флоэмы. Первая активна в передвижении сахаров,
вторая активности не проявляет, но содержит живые
осевую и лучевую паренхимы. Нефупкционирующая
флоэма находится под тангентальным напряжением в ре-
1 В отечественной литературе эту часть коры обычно называют
коркой. (Прим, перев.)
56
зультате дифференциации расположенным под пей вас-
кулярным камбием новой ксилемы и флоэмы. Танген-
тальное напряжение ослабляется, во-первых, перемещени-
ем самой внешней нефункционирующей флоэмы к обра-
зованию перидермы и последующим отслаиванием, во-вто-
рых, ростом расширения осевой и лучевой паренхимы1.
Например, у кленов и вязов осевая паренхима пефункци-
онирующей флоэмы делится и прирастает тангентально,
вызывая расширение ткани и обеспечивая тем самым
увеличение окружности у внешней поверхности внутрен-
ней коры (Esau, 1969).
По текстуре поверхности коры деревья даже одного
и того же семейства значительно различаются. Сравнение
бука (Fagus sylvatica) и дуба (Quercus robur), предста-
вителей семейства Fagaceae, показывает, что у первого
кора гладкая (феллема ошелушивается очень тонкими
полосками), тогда как у второго она трещиноватая и че-
шуйчатая. У бука поверхностная гладкая перидерма со-
храняется в течение многих лет в результате того, что
тангентальное напряжение в первичной коре и вторичной
флоэме ослабляется за счет значительного прироста рас-
ширением; у дуба же меньший рост расширением, а на-
ружная часть внутренней коры постоянно удаляется
сменяющимся перидермным образованием.
Whitmore (1963) предполагает, что та или иная по-
верхность коры зависит от скорости образования флоэмы.
Если скорость велика и тангентальное напряжение у внут-
ренней коры не ослабляется достаточным ростом расши-
рения, большая часть ткани удаляется при сменяющихся
образованиях перидермы, что приводит к развитию че-
шуйчатой коры. У пород с гладкой корой отторгается
мало тканей, так как у них малая скорость образования
флоэмы и сильный рост расширением. Ряд исследовате-
лей (Esau, 1969) указывают, что хотя предположения
Whitmore интересны, но его измерения скорости образо-
вания флоэмы основываются не на прямом наблюдении,
а скорее на допущениях сомнительной достоверности.
1 Рост расширением (англ, dilation, dilatation, growth) — рост па-
ренхимы за счет деления клеток у сердцевинных лучей или
осевой системы у васкулярных тканей, вызывающий увели-
чение окружности коры стебля и корня. (Прим, перев.)
57
5.2. НОВОЕ НАПРАВЛЕНИЕ ИССЛЕДОВАНИЙ
Васкулярный камбий и феллоген несхожи не только
по производным тканям, по также по времени их заложе-
ния и сезонной активности. Например, в ветвях Sassafras
васкулярный камбий закладывается в период первого се-
зона роста, но первая перидерма образуется через не-
сколько лет и притом только прерывисто вокруг ствола
и ветвей (W eiss, 1906). У деревьев, выросших в открытом
грунте вблизи Нью-Хейвена (шт. Коннектикут, США),
феллоген образуется первоначально па верхней стороне
ветвей и на южной стороне прямостоящих стволов. Толь-
ко через 3—4 года первая перидерме! формируется вокруг
всей окружности ствола или ветви. Итак, перидерма за-
кладывается значительно позднее васкулярного камбия,
и в ее индукцию включаются факторы внешней среды,
подобные инсоляции.
У большинства ветвей акации белой (Израиль) фелло-
ген активен на протяжении апреля и повторно в июле;
остальное время года оп остается в состоянии покоя
(Waisel et al., 1967). Васкулярный камбий активизиру-
ется на этих же ветвях в начале марта и действует бес-
прерывно до конца августа. В опытах с 3-летними сажен-
цами акации белой, выросшей при высокой температуре
в режиме длинного дня, феллоген не активизируется да-
же в случае инициирования васкулярного камбия и роста
дерева в высоту. Это подтверждает положение, что фел-
логен не активизируется даже при активном росте и пред-
положительно высоком уровне ауксина и гиббереллина
(см. 1.3).
Дальнейшие опыты с акацией белой показали, что при
опрыскивании ее ауксином или гиббереллином феллоген
закладывается в междоузлиях, причем дальше от вер-
шинки побега, чем у контрольных растений (Arzee et al.,
1968). Возможно, гормоны, образующиеся в период экс-
тенсивного роста побегов, действуют в качестве ингиби-
тора на формирование перидермы (Cutter, 1969). В то
же время развитие последней могут специфично стиму-
лировать и другие гормоны. Последние опыты показывают,
что цитокинин (бензиладенин) стимулирует у корней
Ophioglossum petiolatum формирование перидермопо-
добной ткани, тогда как у папоротника перидерма никог-
да не развивается (Peterson, 1971). Обработка бензил-
аденином наружных коровых кл еток индуцирует перикли-
58
нальное деление, приводящее к образованию отчетливых
радиальных рядов из производных. Гистохимический
анализ показал, что клеточные оболочки иеридсрмоподоб-
ной ткани суберинизовапы. Хотя бензиладепин и ио ин-
дуцирует феллоген, этот интересный опыт говорит о на-
правлениях будущей работы с растениями, у которых,
как правило, перидерма формируется.
5.3. ДУБ ПРОБКОВЫЙ
Пробку бутылочную, а также используемую в произ-
водстве верхних прокладок у бутылей, изоляционных
плит, основания при покрытии линолеумом и черепи-
цей — весь этот материал получают из отчетливой коль-
цеподобной перидермы дуба пробкового (Quercus suber).
Слой пробки в несколько дюймов толщины образуется
у него из феллогена только через 8 или 10 лет; у боль-
шинства деревьев этот слой тонкий, ритидом состоит
в основном из изолированной вторичной флоэмы.
При удалении пробкового слоя со ствола и самых
больших ветвей нужно быть очень внимательным, чтобы
не повредить живую ткань под феллогеном. Кору разре-
зают вертикально по всей ее длине специально загнутым
топором или пилой. Применяя для отслаивания вагу,
пробку вместе с расположенной под ней внутренней
корой снимают в форме цилиндра. Первое время незащи-
щенная поверхность ствола или ветвей имеет розово-ры-
жевато-коричневый цвет, но при подсыхании приобретает
почти черный (Metcalf, 1947). Новый феллоген образу-
ется глубоко во внутренней коре вскоре после снятия
пробки. Подтверждение деятельности феллогена можно
вскоре обнаружить по появлению вертикальных трещин
светло-рыжевато-коричневого цвета у нового пробкового
слоя, расположенного сразу под черной поверхностью
ствола. Второй слой лучшего качества, чем первый, для
снятия он готов примерно через 10 лет. Пробку самого
высокого качества (однородной структуры) получают
при третьем—шестом отслаивании; ее применяют для
изготовления бутылочных пробок.
Производство пробок для вин высшего качества заслу-
живает специального разъяснения. Пробка Quercus suber,
как и у других деревьев, не полностью водонепроницаема
во внешней среде. Точнее, у дышащих тканей внутренней
коры и наружного воздуха должен происходить газооб-
59
Рискцпярныц камбии
Дридесина //
РИС. 24. Строение некоторых структурных элементов ствола:
а — схема свободной структуры (комплементарные клетки) из чечевичек
у феллемы; б — поперечный срез коры Quercus sober с пробкой, феллоге-
ном, внутренней корой и древесиной, чечевички появляются как отчетли-
во выраженные линии, пересекающие пробку; « — пробка для закупорки,
вырезаемая из заготовки так. чтобы поверхности были под прямым углом
и чечевички пересекали пробку горизонтально ее поверхности
мен СОг и О2. Он осуществляется за счет чечевичек,
маленьких локальных участков перидермы, где клетки
пробки довольно свободно сцеплены и имеют большие меж-
клеточные пространства (рис. 24).У дуба пробкового чече-
вички длиной несколько дюймов (1 дюйм = 2,54 см) про-
ходят от феллогена до поверхности наружной коры, по-
этому пробку следует вырезать не с поверхностного слоя,
а под прямым углом к этой поверхности (Schery, 1952).
Таким образом, чечевички в пробке должны располагать-
ся горизонтально, а не вертикально, ибо в последнем
случае она может протекать.
6. Корень
6.1. МОРФОЛОГИЯ КОРНЯ
В условиях леса корневая система дерева должна
проникать на большую глубину. Помимо основной под-
держивающей функции корни должны ранней весной об-
разовывать корневые волоски, посредством которых вода
и минеральные вещества сразу же обеспечивают быстрый
рост побегов. Регуляция роста корней — сложный про-
цесс, зависящий не только от побегов (например, сезон-
ного налитая ассимилятов), но и от внешних факторов,
в том числе от температуры почвы, распределения пита-
тельных веществ и влаги в ней.
У пород с крупными семенами первый корень сеянца,
называемый первичным, быстро растет на большую
глубину и древеснеет. Первичные корни мимозки (Proso-
pis) и акации белой (Robinia) иногда залегают на глу-
бине соответственно 15 и 20 м (Lyr and Hoffman, 1967).
В периоды водного стресса первичные корни имеют боль-
шое значение для роста деревьев, так как могут исполь-
зовать влагу грунтовых вод даже при крайне низком ее
уровне в почве.
Не у всех пород корни глубоко проникают в почву.
На дне почвенных прикопок в сомкнутом древостое мно-
жество канатоподобных одревесневших корней, называе-
мых горизонтальными, и растущих в верхнем
1-футовом слое (1 фут=30,48 см). Эти корни простира-
ются кнаружи от ствола иногда на расстояние значитель-
но большее радиуса кроны дерева. У ели поверхностные
корпи горизонтального типа.
Многочисленные интенсивно ветвящиеся неодревес-
певшие тонкие корни закладываются на большом
первичном или на горизонтальных корнях; их значение
состоит в большой общей площади поверхности. У 1-лет-
пей акации белой корни диаметром менее 1 мм составля-
ют только 16% массы корневой системы, но 93% общей
ее длины, а по площади поверхности соотношение еще
большее (Lyr and Hoffman, 1967). Благодаря большой
площади поверхности тонкие корни играют исключитель-
но важную роль — всасывают воду и минеральные веще-
ства из почвы.
С учетом характерных особенностей диаметра, скорос-
ти роста и долговечности мелкие корни подразделяют на
61
два класса — на длинные и короткие (Wilcox, 1964).
Каркас корневой системы создается у лесных деревьев
из относительно толстых (до 2 мм) быстрорастущих
длинных корней. Позднее, по мере развития васку-
лярного камбия и вторичного тела растения, эти корни
могут стать долголетними. Они дают начало топким бо-
ковым корням, называемым короткими, с продолжи-
тельностью 1—2 сезона и часто имеющим микоризу.
Диаметр коротких корней сосны смолистой около 0,4 мм
(Wilcox, 1964).
Длинные корпи подразделяют па пионерские
и материнские. К первым относят более толстые
быстрорастущие корни, которые пронизывают большой
объем почвы и вскоре создают каркас корневой системы;
ответвления на них образуются очень редко. В противо-
положность пионерским материнские корни немного
тоньше, растут медленнее и дают начало множеству ко-
ротких корней.
Для роста деревьев разнородность в системе топких
корней представляет очевидное преимущество. На бедных
почвах, например на истощенных песчаных, адаптивная
способность дерева значительно лучше в случае образо-
вания длинных корней пионерского типа. Такая система
в поисках воды и питательных веществ способна быстро
пронизывать большой объем почвы; при достаточно бо-
гатом почвенном горизонте развиваются материнские
корни с короткими боковыми корнями, интенсивно ис-
пользующими доступную влагу и минеральные вещества.
6.2. СЕЗОННЫЙ РОСТ
В сезонной динамике роста побегов и корней у рас-
тений в умеренных широтах отмечают ряд интересных
различий. Например, у отдельных пород экстенсивный
рост тонких корней весной начинается раньше и осенью
заканчивается позже, чем рост побегов (Lyr and Hoffman,
1967). Доля тонких корней, активно растущих в какой-то
произвольно выбранный отрезок времени, также значи-
тельно различается в зависимости от сезона.
Методы изучения роста корней лесных деревьев про-
сты, но крайне трудоемки. Рост горизонтальных корней
можно наблюдать сверху, от места прикрепления их
к стволу до отдельных боковых корней. Для этого длин-
ные корни, находящиеся в нескольких дюймах от поверх**
62
ности почвы, обнажают по всей длине; положение кой-
чиков отмечают поставленными сбоку колышками (Ste-
vens, 1931), направление роста — впереди. На одну или
две недели корень присыпают почвой, затем кончик рас-
крывают, измеряют прирост в длину по расстоянию от
крайнего колышка до нового положения кончика корня.
Такие обмеры можно в вегетационный период повторять
много раз, не повреждая корень. Ростовые корни легко
отличить от неростовых по соответственно белой и ко-
ричневой окраске их кончиков.
Апрель Мий Июнь Июль Абг. Сет. Опт Нояб
РИС. 25. Сезонная динамика прироста корней сосны веймутовой;
высокий процент активно удлиняющихся корней приходится на
середину весны и начало осени, в середине лета большинство из
них не активны (Stevens, 1931)
У 4—5-летней сосны веймутовой в Нью-Гэмпшире
(на востоке США) длинные корни активны с середины
апреля до начала ноября (Stevens, 1931), но соотношение
активно удлиняющихся корней в течение сезона изменя-
ется (рис. 25). Отмечают два максимума в росте корней:
в первом (середина мая) почти все они активны, во вто-
ром (начало осени) больше половины уже заканчивают
рост в длину. В начале августа 60—80% выкопанных
корней оказываются неростовыми, что заметно по корне-
вым кончикам. Отсутствие в середине лета активности
у корней, видимо, связано с высокими температурами
и пересыханием верхнего горизонта почвы, в котором они
растут.
63
Аналогичную сезонную характеристику получили при
измерении ежедневного прироста только у активно удли-
няющихся корней (рис. 26). Максимальный показатель
получен в конце весны и затем снова в начале осени,
в середине лета он понижается. На рис. 26 показано, что
первая волна прироста корней совпадает с быстрым подъ-
емом прироста но высоте побегов в конце весны. Далее
ход прироста корней и побегов у сосны веймутовой раз-
личен. Только в конце лета отмечен незначительный при-
рост побегов, связанный, по-видимому, с созреванием
РИС. 26. Схема среднего ежедневного прироста по длине только
у растущих корней; скорость прироста активно растущих корней
в середине лета снижается, прирост корней и побегов имеет раз-
ные сезонные особенности (Stevens, 1931)
верхушечной почки (Stevens, 1931); корпи же в это вре-
мя и в конце осени сильно удлиняются, тем самым под-
тверждая автономность роста корня и побега. С другой
стороны, опыты по затенению и обрезке указывают, что
наличие ассимилятов, образовавшихся в побегах, сильно
влияет на рост корней (Lyr and Hoffman, 1967).
Изучению сезонной динамики роста корней у лесных
пород лабораторными методами посвящено меньше работ.
Один из лучших методов основан на обрезке горизонталь-
ных корней (Lyford and Wilson, 1966). Подрезанный кон-
чик корня, отходящего от ствола, обертывают влажной
почвой, и через 2—3 недели на одревесневшем корне ре-
генерирует большое число тонких корней. В ряде случаев
подрезанный горизонтальный корень прочно закрепляют
64
на дне длинного узкого деревянного лотка и присыпают
тонким слоем влажной почвы; вновь развивающиеся кор-
пи принимают форму лотка. Этот метод позволяет легко
изучать влияние на рост корней состава почвы, темпера-
туры и других факторов среды.
В развитие указанного метода создано специальное
сооружение — ризотрон (Lyford and Wilson, 1966). Тран-
шею небольшой глубины устраивают возле крупного де-
рева и накрывают недорогим покрытием. Тонкие корпи,
образовавшиеся на подрезанных горизонтальных корнях
у стенки траншей, выращивают в лотках в контролируе-
мых лабораторных условиях. По данным Lyford и Wilson
в Питерсгеме (шт. Массачусетс, США), тонкие корни
клена красного в необогреваемых лотках ризотрона не
активизируются в холодные зимние месяцы, но при 20°С
они непрерывно прирастают на 5—10 мм в день несмотря
на пребывание надземной части растения в состоянии
покоя.
Аналогичные результаты получены в опыте с сосной
веймутовой, помещенной в обогреваемую теплицу в кон-
це октября, когда обычно кончики корней неактивны
и принимают коричневую окраску (Stevens, 1931). У не-
которых корней за 4 месяца в подходящих для роста ус-
ловиях теплицы прирост возобновлялся (один корень
удлинился более чем на 22,5 см). В то же время побег
остается неактивным, пока покой не прерывается зимним
охлаждением. Такие опыты показывают, что прирост кор-
ней у деревьев в некоторые периоды не зависит от регу-
ляции их роста побегами. Более того, рост корней зимой
в обогреваемых лотках ризотрона или в обогреваемой
теплице показывает, что в отличие от побегов у них сос-
тояние покоя эндогенно не обусловлено. Рост корней про-
должается, пока его не ограничат температура, влаж-
ность и питание.
6.3. ВТОРИЧНЫЙ РОСТ
Длинные корни становятся постоянным компонентом
корневой системы посредством развития васкулярного
камбия и образования вторичных проводящих тканей.
Васкулярный камбий и его производные ткани у корней
и стебля по структуре в основном схожи, но у этих совер-
шенно разных органов отмечаются интересные отличия
и в характере инициации латеральных меристем. Изве-
65
стно, что в стеблях двудольных покрытосеменных расте-
ний васкулярный камбий инициируется в прокамбии пер-
вичного проводящего пучка, располагающегося отчасти
в периферической зоне стебля. В противоположность
этому в корнях, где проводящая ткань существует как
центральный цилиндр, васкулярный камбий залегает глу-
боко внутри.
По расположению первичной ксилемы в зависимости
от числа радиальных полюсов корни относят к д и а р х-
ным, триархным, тетрархным и т. д. (рис. 27,
РИС. 27. Реконструкция ствола
Lepidodendron:
а — ромбовидные листовые подушки
в верхней части ствола плотно сгруп-
пированы, у основания ствола ошелу-
шиваются; б — ствол состоит из круп-
ной перидермы и относительно не-
большой вторичной ксилемы (Eggert,
1961)
Gemmell, 1969). Ксилема в полюсах созревает первой
и называется протоксилемой, дифференциация более
центрально расположенной метаксилемы завершается
немного позднее. Первичная флоэма располагается в раз-
дельных тяжах у периметра ксилемного цилиндра между
соседними протоксилемными полюсами. При образовании
васкулярного камбия в корнях слой прокамбия остается
между флоэмными тяжами и ксилемой. Прокамбий во-
круг ксилемы прерывистый до тех пор, пока каждый про-
токсилемный полюс непосредственно примыкает к пери-
циклу. Этот слой окружает весь цилиндр первичной
ксилемы и флоэмы и в свою очередь окружен первичной
корой.
Васкулярный камбий происходит из прокамбия моло-
дого корня в виде ряда прерывистых слоев у внутренней
стороны каждого тяжа первичной флоэмы (Esau, 1965).
Сменяющиеся периклинальные деления в каждом камби-
альном слое вскоре образуют радиально сгруппированные
ряды производных. По мере того как вогнутые полости
между соседними протоксилемными полюсами заполня-
66
ются вторичной ксилемой, ксилема цилиндра постепенно
принимает цилиндрическую конфигурацию. Вскоре пери-
клинальные деления начинаются у перицикла: вначале
локально напротив протоксилемных полюсов, позднее по
всему слою. В результате образуется утолщенный пери-
цикл с радиально сгруппированными клетками. Лежащие
глубоко внутри периклинально делящиеся клетки пери-
цикла, граничащие с каждым протоксилемным полюсом,
соединяются камбиальными клетками с обеих сторон каж-
дого тяжа первичной флоэмы, давая начало васкулярно-
му камбию, который теперь замыкается поверх всего ци-
линдра ксилемы. Васкулярный камбий в течение многих
лет образует толстые сезонные приросты ксилемы и фло-
эмы, особенно вблизи ствола, где корни сильносбежистые.
Древесина в корнях обычно менее плотная, чем в стволах,
так как у нее волокна и трахеиды большого диаметра.
Утолщенный перицикл в корнях груши недолго сохра-
няет за собой площадь из-за расширения нижележащего
цилиндра, что обусловливается образованием вторичных
проводящих тканей (Esau, 1943). После тангентальпого
расширения самые дальние от центра клетки перицикла
суберинизируются, чем обеспечивают защитную функцию.
К этому времени лежащие сверху первичная кора и эпи-
дермис разрушаются и слущиваются. Позднее из внут-
реннего слоя делящегося перицикла развивается перидер-
ма L
Инициирование вторичного роста корней регулируется
взаимодействующими гормональными и метаболитиче-
скими факторами. У подрезанных корней редьки равно-
весная смесь ауксина (ИУК), цитокинина (кинетина),
циклита (мио-инозита) и сахарозы приводит в действие
процессы, вызывающие индукцию и максимальную
деятельность васкулярного камбия (Torrey and Loomis,
1967). Для большей эффективности химическую смесь
применяют, используя узкую стеклянную трубку, надева-
емую на нижний конец корня. Это доказывает, что ини-
циирование вторичного роста корней обычно регулирует-
ся стимулом, перемещающимся из побега вниз.
1 У большинства растений перидерма развивается из внешнего
слоя перицикла. (Прим, перев.)
67
1._ Палеоботаника роста деревьев
7.1' ПРОИСХОЖДЕНИЕ ДРЕВОВИДНЫХ ФОРМ
Возникновение древовидных форм стало возможным
только благодаря заложению камбиальных меристем
и формированию ригидифицированных, особенно лигни-
фицированных, вторичных тканей. Вначале рассмотрим
ряд растений девонского периода, не обязательно связан-
ных филогенетически, но тем не менее доказывающих
РИС. 28. Строение ксилемы на
поперечных срезах стеблей де-
вонских растении в период пе-
рехода от травянистой к древо-
видной форме (сплошь окрашена
первичная ксилема, штрихов-
кой — вторичная):
а — Rhynia — сплошной цилиндр пер-
вичной ксилемы; б — Asteroxylon —
первичная ксилема частично рассе-
чена; в — Shizopodium — трахеиды в
радиальных рядах расположены у
концов дольчатой ксилемы; г — Cal-
lixylon — крупная вторичная ксилема;
первичная ксилема представлена
кольцом из раздельных пучков, ок-
ружающих центральную сердцевину
эволюционное развитие систем тканей в филогенезе от
травянистых до древовидных форм. У самых простых ра-
стений палеозоя, как это установлено у представителя
псилофитовых Rhynia, рост происходил только за счет
верхушечной меристемы, а проводящая система была
примитивной организации (рис. 28, а). На поперечном
срезе безлиственного стебля можно обнаружить неболь-
шой сплошной цилиндр проводящей ткани (ксилема
и флоэма), называемый протостелью. Ствол был тон-
ким, например у R. Gwynne —vaughani, высота его была
примерно 0,2 м. У Asteroxylon, псилофпта или, возможно,
примитивного плауна, стебель моноподиальный, с малень-
кими листьями, а ксилема с заметными дольками и не-
много покрупнее, чем у Rhynia (рис. 28, б). Самые круп-
ные стволы у Asteroxylon достигали высоты 0,5 м и тол-
щины 1 см. Schizopodium, безлистный представитель пси-
лофитовых среднего девона, дает первое представление
о развитии вторичной ткани. Ксилема протостели дольча-
68
тая и не похожа на ксилему у Asteroxylon, трахеиды
у края долек располагались в радиальных рядах, под-
тверждая наличие латеральной меристемы (см. рис. 28, в).
По-видимому, у Schizopodium еще не было настоящего
васкулярного камбия, так как крайние трахеиды непо-
стоянны по размеру и форме. Вероятно, меристема, ко-
торая давала у этого растения начало радиально сгруппи-
рованным трахеидам, была переходной между прокамби-
ем и васкулярным камбием.
Настоящий васкулярный камбий появился вначале
у Tetraxylopteris, праголосемепного среднего девона
(Beck, 1970). Праголосеменные были группой бессемей-
ных сосудистых растений, имевших некоторые структур-
ные признаки, такие же, как у голосеменных, и, как
предполагается, дали начало семенным растениям. Листья
Tetraxylopteris вайяподобные \ ствол около 6,5 см толщи-
ны и с древесиной, похожей на древесину голосеменных.
Крупная древовидная форма развилась позднее у Archae-
opteris, праголосемепного верхнего девона с большими
составными вайями — до 1 м длиной. Одревесневший
ствол Archaeopteris (см. рис. 28, г), относившийся многие
годы к роду Callixylon, был огромных размеров (у неко-
торых экземпляров превышал в диаметре 1,5 м) и отли-
чался по деятельности васкулярного камбия. Древесина
у Archaeopteris (Callixylon) была несложной по структуре
и состояла только из трахеид и паренхимы, поэтому име-
ла большое сходство с современными голосеменными,
особенно с Araucaria. В радиальных стенках осевых тра-
хеид находили многочисленные группы круглых окайм-
ленных пор, характеризующих уровень эффективности
проводящей системы, образовавшейся в заболони этого
вида во времена девона. Лучи, шириной в 1—4 клет-
ки, состояли уже не только из паренхимы, но и из тра-
хеид (Beck, 1970).
Кажется удивительным, почему так быстро в девоне
возникла древовидная форма. В лучшем случае мы мо-
жем только предполагать, что прямостоящий одревеснев-
ший ствол эволюционировал соответственно двум древним
явлениям: 1) сплющивание и соединение дихотомически
сильно ветвящихся ветвей привело к образованию листьев
или вай; 2) развитие биохимических механизмов синтеза
1 Вайя (англ, frond) — перистый лист со спорофиллом у папорот-
ника. (Прим, перев.)
69
фенилпропаноидных молекул и их полимеризация в rtpit-
митивные лигниноподобные вещества.
Листья древовидных растений — главный источник
гормонального стимула, включающегося в удлинение меж-
доузлий, например у Ginkgo, и во многие звенья процесса
радиального прироста (см. разд. 3). Далее можно пред-
положить, что физиологические системы, необходимые
для развития вторичных тканей, возникают при эволю-
ции примитивного листа. Недавно было показано, что
инициирование васкулярного камбия регулируется воз-
действием тока из побега. Это отчетливо выяснено в опы-
те с прививкой укороченного мутанта и нормальной
формы томата настоящего (Caruso and Cutter, 1970). При
выращивании мутанта в стерильной культуре у него не
образовывались ни верхушечный побег, ни листья, ни
семядоли, как и васкулярный камбий в гипокотиле. Но
когда мутанту прививали кончик нормального побега,
в гипокотиле инициировался васкулярный камбий и фор-
мировались вторичные ксилема и флоэма. Кончик побега
обеспечивал гормоны и метаболиты, необходимые для
закладки у гипокотиля латеральной меристемы. Корням
редьки для инициирования васкулярного камбия совер-
шенно необходимы ауксин и цитокинин (Torrey and Loo-
mis, 1967). Поэтому очевидно, что древовидная форма
возникла в девоне при эволюции аналогичного биохими-
ческого механизма, регулирующего инициирование вас-
кулярного камбия у иных неодревесневших наземных
растений.
Первостепенное значение у древовидных форм имел
лигнин, так как он создает стойкость и жесткость других
компонентов клеточной оболочки. Следовательно, сохране-
ние большого прямостоячего древовидного тела у растения
зависело исключительно от прочности систем лигнифици-
рованных вторичных тканей. Для попытки разгадать древ-
ние явления, включаемые в эволюцию лигнина, Manskaya
и Drozdova (1968) проанализировали состав клеточных
оболочек у современных представителей нескольких
главных групп растений. Лигнин отсутствовал у водорос-
лей и несосудистых наземных растений, например у мхов;
в то же время у плаунов, папоротников, покрыто- и го-
лосеменных лигниновые строительные блоки присутство-
вали в разных количествах (табл. 2). Таким образом,
только сосудистые наземные растения получили возмож-
ность биосинтезировать лигнин, и он присутствует (или
70
2, ЛИГНИН У СОВРЕМЕННЫХ РАСТИТЕЛЬНЫХ ГРУПП. ОСТАТКИ
ЛИГНИНА (ВАНИЛИНА И СИРЕНЕВОГО АЛЬДЕГИДА) ПОЛУЧЕНЫ
ПУТЕМ ОКИСЛЕНИЯ ЩЕЛОЧНЫМ НИТРОБЕНЗОЛОМ
(Manskaya a. Drozdova,1968)
Материал Ванилин, %
Fucus serratus (водоросль) Следы
Polytrichum commu- ne (мох)
Lycopodium (плаун) 4
Alsophila australis (папоротник) 4
Araucaria bidwillii (голосеменное растение) 6
Casuarina stricta (покрытосемен- ное растение) 2 (+1 % сиреневого альдегида)
присутствовал раньше) лишь у групп с древовидными
формами. Более того, несмотря на присутствие уже
в среднем докембрии других главных компонентов обо-
лочки растительной клетки — целлюлозы и гемицеллюло-
зы (Barghoorn, 1964), крупные прямостоячие формы
растений появились значительно позднее, в девоне,
что совпало с развитием механизмов, необходимых для
синтеза лигнина.
7.2. ЭВОЛЮЦИЯ РАДИАЛЬНОГО ПРИРОСТА
К началу каменноугольного периода древовидные
формы получили большое распространение у многих
групп сосудистых растений, включая классы Lycopsida
и Sphenopsida. Одпако у некоторых из них системы вто-
ричных тканей имели необычную структуру, не свойст-
венную существующим в настоящее время породам. У ис-
копаемых Lycopsida вторичный рост основывался не на
преобладании производных васкулярного камбия, а, ско-
рее, на экстенсивном развитии перидермоподобной ткани.
Lepidodendron — крупные древовидные растения, до
30 м высоты, в верхней части которых на дихотомически
ветвящихся стеблях образовывались густособранные ли-
стья, расположенные в виде спирали. Анатомическая ре-
конструкция показала, что у Lepidodendron был настоя-
щий васкулярный камбий с ограниченной активностью
71
й соответственно только с небольшим колйчестйом сфор-
мировавшейся вторичной ксилемы (см. рис. 27). При
этом отмечен не только относительно низкий процент
древесины в стволе, но и то, что сами трахеиды были до-
вольно тонкостенными и имели меньшее механическое
значение, чем аналогичные клетки праголосеменнУх.
В крупной первичной коре закладывалась рассеянная
меристема, которая в процессе последовательных пери-
клинальных делений давала начало радиальному ряду
клеток, составлявших вторичную кору, или перидерму.
В отличие от перидермы у современных покрыто- и голо-
семенных (см. разд. 5) у Lepidodendron не доказано на-
личие самостоятельного феллогена (Eggert, 1961), но,
вероятнее всего, меристема была рассеянной, и ее произ-
водные становились иногда волокнами, может быть живы-
ми, и имели функции упрочения и поддержания. Широко
распространившиеся в каменноугольном периоде древо-
видные формы Lepidodendron в стрессовых условиях сре-
ды вымерли, по всей очевидности, из-за ограниченной
способности передвижения воды недостаточно развитой
вторичной ксилемой.
Как было точно установлено, к концу палеозоя у дре-
весных растений развились механизмы, участвующие
в регуляции радиального прироста. В качестве примера
рассмотрим Sphenophyllum из рода Sphenopsida — ма-
ленькое ветвящееся, одревесневшее, стелющееся наподо-
бие виноградной лозы растение с четко выраженными
узлами и междоузлиями, достигавшее в каменноугольный
период иногда 10 м длины. Протостель была треугольной
формы с тремя протоксилемпыми полюсами и размещен-
ной в центре метаксилемой (Darrah, 1968). Васкулярный
камбий закладывался непосредственно с внешней сторо-
ны первичной ксилемы и вначале при образовании одре-
весневших тканей принимал треугольную форму (см.
рис. 16); затем в процессе радиального прироста преобла-
дающей становилась цилиндрическая конфигурация. Пе-
реход от треугольной к цилиндрической вторичной ксиле-
ме осуществлялся увеличенным числом периклинальных
делений и (или) большим расширением камбиальных
производных в зонах напротив метаксилемы. Аналогич-
ным способом развивается цилиндр древесины поверх
вначале дольчатого цилиндра первичной ксилемы у кор-
ней некоторых современных древовидных растений.
72
7.3. ВЛИЯНИЕ УДЛИНЕННЫХ ПОБЕГОВ
Как уже упоминалось ранее, праголосеменному
Archaeopteris свойственна крупная вторичная ксилема.
Настоящие голосеменные (образующие голые семена)
возникли в каменноугольном периоде, когда они были
представлены родом Cordaites порядка Gordaitales. Это
было дерево 30-метровой высоты, с крупной, но простой
по строению древесиной. Клетки осевой системы состояли
только из трахеид, а паренхима радиальной системы —
из однорядных (шириной в одну клетку) лучей. Поэтому
древесина Cordaites имеет большое сходство с древесиной
современных хвойных. У Cordaites вырастали длинные
ветви и спирально расположенные листья с характерным
параллельным жилкованием (рис. 29). Их растущие вет-
ви вполне можно характеризовать, как удлиненные побе-
ги, означающие, что ось побега можно разделить на меж-
доузлия (на которые прирастал ранее стебель) и уз-
лы (точки прикрепления листьев). Определенное под-
тверждение удлинения междоузлия у стебля Cordaites
было получено по присутствию в отложениях сердцевины
ряда полостей, соответствующих междоузловым частям
стебля (Arnold, 1947). Подобным образом верхушечные
ветви Archaeopteris были удлиненными побегами (Beck,
1970), поэтому их оси также можно описать на основе
узлов и междоузлий. •
Ауксин включается в инициирование васкулярного
камбия и в различные звенья процесса радиального при-
роста. Похоже, что влияние верхушечного побега на ра-
диальный прирост опосредуется по крайней мере частич-
но улавливаемым ауксином, который образуется в рас-
пускающихся листьях и междоузлиях (Gunckel and
Thimann, 1949). Далее можно предположить, что мощное
развитие вторичной ксилемы у палеозойных Archaeopte-
ris и Cordaites связано с формой удлиненного побега, т. е.
со способом роста побегов, который остается у генеало-
гических линий голосеменных и в настоящее время. По-
явление удлиненного побега смогло обеспечить гормональ-
ный источник, способствовавший мощному развитию дре-
весины у этих архаических растений. В противополож-
ность им у Lepidodendron, которому свойственны неиз-
менные формы укороченного побега (см. рис. 27), удли-
нения оси между последовательно сменяющимися листо-
73
выми подушками здесь не
отмечаются, вторичная
ксилема развита незначи-
тельно.
7.4. ПРОИСХОЖДЕНИЕ
КОЛЕЦ ПРИРОСТА
У современных пород
из умеренных широт и у
некоторых из тропиков
проявляется ритмичность
в последовательности за-
кладывания васкулярным
камбием древесины с об-
разованием кольца приро-
ста. Наличие ритмичного
вторичного прироста и сла-
бого прироста колец про-
слеживается у Callixylon
erianum из верхнего дево-
на (Нью-Йорк) (Arnold,
1947). По данным Jeffrey
(1917),в кордаитовой дре-
весине с о-ва Принца Эду-
арда (46° с. ш.) вторич-
ный прирост однорядный,
хотя годичные кольца на-
ходили в аналогичной
широт северной Англии
РИС. 29. Параллельное жилко-
вание у листьев ростового по-
бега Cordaites, размещенных
на стволе с регулярными ин-
тервалами; появление побегов
с удлиненными междоузлия-
ми (побегами) вероятно кор-
релировало с образованием
крупной вторичной ксилемы *
(Darrah, 1960)
древесине из более высоких
(54° с. ш., Ланкашир). Ритмичный вторичный прирост
установлен в древесине из Ланкашира по уменьшенным
размерам трахеид вблизи грайицы каждого прироста.
Правда, нельзя быть уверенным в том, какие факторы
предопределяют эти древние ритмичные свойства, но об-
разование кольца прироста в кордаитовой древесине
(Ланкашир) может отражать сезонные изменения клима-
та, что, вероятно, происходило в более высоких широтах.
Однако кольца прироста были обнаружены в кордаитовой
древесине и из других отдаленных мест, например у Da-
doxylon indicum, палеозойского кордаитового вида из Ин-
дии. В перми (последний период палеозоя) и мезозое
кольца прироста с хорошо обозначенными границами ран-
ней и поздней древесины распространились очень щиро
74
ко, причем это совпадало с сезонными ритмами окружаю-
щей среды, включая изменения температуры и почвенной
влаги. Легко заметить, что различия во внешней среде
влияют на образование древесины посредством утолщения
оболочек клеток притоком ассимилятов и инициирования
ауксином клеточного расширения. Оба эти процесса под-
вержены изменениям факторами внешней среды, дейст-
вующими на прирост побегов.
В связи с этим кольцо прироста, первоначально обра-
зовавшееся в палеозое и мезозое, явилось прямым ре-
зультатом деятельности внешних факторов на физиоло-
гические системы растущего побега. В то же время на-
личие в настоящее время колец прироста у многочислен-
ных тропических пород подтверждает, что физиологиче-
ские механизмы, регулирующие периодичность радиаль-
ного прироста, позднее стали неотъемлемой частью роста
древесных растений, по крайней мере у некоторых де-
ревьев (Barghoorn, 1964). Аналогичным образом (см.
разд. 1) явление состояния покоя у тропической породы
Огеорапах могло зародиться ранее, когда ее род подвер-
гался сезонным ритмам внешней среды, произрастая,
по-видимому, в более высоких широтах.
Будущая работа, основанная на сравнении структур-
ных признаков ископаемых растений с механизмами рос-
та, действующими у современных представителей, позво-
лит разгадать способы и происхождение роста деревьев
в прошлом.
Список литературы
Alfieri, F. J. and Evert, R. F. (1968). Am. J. Bot, 55, 518—28.
Arnold, C. A. (1947). An Introduction to Paleobotany. New York,
McGraw-Hill.
Arzee, T., Liphschitz, N. and Waisel, Y. (1968). New Phytol. 67,87—93.
Bailey, I. W. (1920). J. Gen. Physiol, 2, 519-33.
Bannan, M. W. (1962). The Vascular Cambium and Tree-Ring Deve-
lopment. In Tree Growth. Edited by T. T. Kozlowski. New York,
Ronald Press.
Barghoorn, E. S. (1964). Evolution of Cambium in Geologic Time.
In The Formation of Wood in the Forest Trees. Edited by M. H.
Zimmermann. New York, Academic Press.
Beck, С. B. (1970). Biol. Rev, 45, 379-400.
Borchert, R. (1969). Am. J. Bot, 56, 1033-41.
Caruso, J. L. and Gutter, E. G. (1970). Am. J. Bot, 57, 420—29.
Cote, W. A. and Day, A. C. (1969). Technical Publication, NO 95.
State University College of Forestry at Syracuse University,
New York.
Cote, W. A, Jr, Day, A. C. and Timell, T. E. (1968). Wood Sci. Tech-
nol, 2, 13—37.
Critchfield, W. B. (1970). Bot. Gaz, 131, 150-62.
Cutter, E. G. (1969). Plant Anatomy. Part. I. London, Edward Arnold.
Darrah, E. L. (1968). Phalaentographica, B. 121, 87—101.
Darrah, W. C. (1960). Principles of Paleobotany. New York, Ronald
Press.
Derr, W. F. and Evert, R. F. (1967). Am. J. Bot, 54, 147—53.
Digby, J. and Wareing, P. F. (1966a). Ann. Bot. N. S, 30, 539—48.
Digby, J. and Wareing, P. F. (1966b) . Ann. Bot. N. S, 30, 607—22.
Eggert, D. A. (1961). Palaeontographica, B. 108, 43—92.
Esau, K. (1943). Hilgardia, 15, 299-324.
Esau, K. (1965). Plant Anatomy. New York, John Wiley.
Esau, K. (1969). The Phloem. Berlin, Gebruder Borntraeger.
Fergus, B. J., Procter, A. R., Scott, J. A. N. and Goring, D. A. I.
(1969). Wood Sci. Technol, 3, 117-38.
Ferguson, C. W. (1968). Science, 159, 839—46.
Foster, A. S. (1931). Am. J. Bot, 18, 864-87.
Foster, A. S. (1932). Am. J. Bot, 19, 75-99.
Gemmel, A. R. (1969). Developmental Plant Anatomy. London,
Edward Arnold.
Glock, W. S. and Agerter, S. R. (1962). Rainfall and Tree Growth.
In Tree Growth. Edited by T. T. Kozlowski. New York, Ronald
Press.
76
Gordon, J. C. and Larson, P. ft. (1968). PL Physiol., Lancaster, 43
1617—24.
Greathouse, D. C., Laetsch, W. M. and Phinney, B. 0. (1971). Am.
J. Bot., 58, 281-86.
Gunckei, J. E. and Thimann, К. V. (1949). Am. J. Bot., 36, 145—51.
Hanson, H. C. and Brenke, B, (1926). Bot. Gaz., 82, 286—305.
Jeffrey, E. C. (1917). The Anatomy of Woody Plants. Chicago, Uni-
versity Press.
Larson, P. R. (1962). Auxin Gradients and the Regulation of Cam-
bial Activity. In Tree Growth. Edited by T. T. Kozlowski. New
York, Ronald Press.
Leach, R. W. A. and Wareing, P. F. (1967). Nature, Lond., 214,
1025-27.
Lyford, W. H. and Wilson, B. F. (1966). Harvard Forest Paper, No
16. Petersham, Massachusetts.
Lyr, H. and Hoffmann, G. (1967). Growth Rates and Growth Periodi-
city of Tree Roots. In Int. Rev. For. Res., 2, Edited by J. A. Rom-
berger and P. Mikola. New York, Academic Press.
Manskaya, S. M. and Drozdova, T. V. (1968). Geochemistry of Organic
Substances. Translated, Edited by L. Shipiro and I. A. Breger.
Oxford, Pergamon Press.
Mergen, F. (1958). For. Sci., Washington, D. C., 98—109.
Metcalf. W. (1947). Econ Bot., I, 26-46.
Morey, P. R. and Gronshaw, J. (1968). Protoplasma. 65, 287—313.
Parke, R. V. (1959). Am. J. Bot., 46, 110-18.
Peterson, R. L. (1971). Ann. Bot. N. S., 35, 165—67.
Philipson, W. R., Ward, J. M. and Butterfield, B. G. (1971). The Vas-
cular Cambium. London, Chapman and Hall.
Robards, A. W., Davidson, E., and Kidwai, P. (1969). J. exp. Bot.,
20. 912-20.
Roelofsen, P. A. (1959). The Plant Cell Wall. Berlin, Gebruder Born-
traeger.
Romberger, J. A. (1963). Meristems, Growth and Development in
Woody Plants. Washington, D. C., U. S. Dept. Agric.
Rubery, P. H. and Northcote, D. H. (1968), Nature, Lond., 219,
1230-34.
Sacher, J. A. (1954). Am. J. Bot., 41, 749—59.
Schery, R. W. (1952). Plants for Man. Englewood Cliffs, Prentice
Hall.
Scott, J. A. N., Procter, A. R., Fergus, B. J. and Goring, D. A. I.
(1969). Wood Sci. Technol, 3, 73-92.
Sinnott, E. W. (1960). Plant Morphogenesis, New York, McGraw-
Hiell.
Stevens, C. L. (1931). Yale Univ. School Forestry Bull. NO 32. New
Haven, Connecticut.
Torrey, J. G. and Loomis, R. S. (1967). Am. J. Bot., 54, 1098—1106.
Tucker, С. M. and Evert, R. F. (1969). A. J. Bot., 56, 275-84.
Waisel, Y., Liphschitz, N. and Arzee, T. (1967). New Phytol., 66,
331-35.
Wardrop, A. B. (1964a). The Structure and Formation of the Cell
Wall in Xylem. In The Formation of Wood in Forest Trees.
Edited by M. H. Zimmermann. New York, Academic Press.
Wardrop, A. B. (1964b). The Reaction Anatomy of Arboscent An-
giosperms. In The Formation of Wood in Forest Trees. Edited
by M. H. Zimmermann. New York, Academic Press.
77
Wardrop, A. В. (1965). The Formation and Function of Reaction
Wood. In Cellular Ultrastructure of Woody Plants. Edited by
W. A. Cote Syracuse, University Press.
Wareing, P. F. (1951). Physiologia Pl., 4, 546—62.
Wareing, P. F. (1958). Nature, Lond., 181, 1744-45.
Wareing, P. F. (1969). Germination and Dormancy. In The Physi-
ology of Plant Growth and Development. Edited by M. B. Wil-
kins. London, McGraw-Hill.
Wareing, P. F. and Phillips, I. D. J. (1970). The Control of Growth
and Differentiation in Plants. Oxford, Pergamon Press.
Weiss, H. F. (1906). Bot, Gaz., 41, 434-44.
Westing, A. H. (1965). Bot. Rev., 31, 381—480.
Whitmore, T. C. (1963). New Phytol., 62, 161-69.
Wilcox, H. (1964). Xylem in Roots of Pinus resinosa Ait. in Rela-
tion to Heterorhizy and Growth Activity. In The Formation of
Wood in Forest Trees. Edited by M. H. Zimmermann. New
York, Academic Press.
Zimmermann, M. H. and Brown, C. L. (1971). Trees: Structure and
Function. New York, Springer — Verlag.
Содержание
ПРЕДИСЛОВИЕ К РУССКОМУ ИЗДАНИЮ......................... 3
ОБЩЕЕ ПРЕДИСЛОВИЕ К СЕРИИ............................. 4
ПРЕДИСЛОВИЕ.............................................. 4
1. РОСТ В ВЫСОТУ......................................... 5
1.1. Морфология роста в высоту...................... 5
1.2. Рост в высоту тропических деревьев............ 10
1.3. Роль ауксина.................................. 12
1.4. Состояние покоя............................... 14
2. ДРЕВЕСИНА........................................ 16
2.1. Структура древесины........................... 16
2.2. Химический состав ...................... 19
2.3. Образование клеточной оболочки........... 27
3. РАДИАЛЬНЫЙ РОСТ ... 28
3.1. Васкулярный камбий....................... 28
3.2. Сезонное развитие........................ 31
3.3. Инициирование камбиальной активности ... 34
3.4. Гормональная регуляция образования ксилемы
и флоэмы................................... 37
3.5. Физиология образования годичного кольца ... 39
3.6. Годичные кольца и климат................. 42
4. РЕАКТИВНАЯ ДРЕВЕСИНА............................ 45
4.1. Структура и распределение реактивной древесины 45
4.2. Причины образования реактивной древесины . . 49
5. КОРА ................................................ 54
5.1. Структура коры........................... 54
5.2. Новое направление исследований........... 58
5.3. Дуб пробковый ................................ 59
6. КОРЕНЬ.......................................... 61
6.1. Морфология корня............................. 61
6.2. Сезонный рост ................................ 62
6.3. Вторичный рост ............................... 65
7. ПАЛЕОБОТАНИКА РОСТА ДЕРЕВЬЕВ.................... 68
7.1. Происхождение древовидных форм................ 68
7.2. Эволюция радиального прироста................. 71
7.3. Влияние удлиненных побегов.................... 73
7.4. Происхождение колец прироста.................. 74
СПИСОК ЛИТЕРАТУРЫ....................................... 76
Филип Р. Мори
КАК РАСТУТ ДЕРЕВЬЯ
Перевод с английского А. Ю. Клячко
Редактор издательства Э. И, Снегирева
Обложка художника Б. К. Шаповалова
Художественный редактор В. Н. Журавский
Технический редактор Е. Б. Капралова
Корректор И. Б. Шеманская
ИБ № 1457
Сдано в набор 03.07.80. Подписано в печать 20.10.80. Формат
84ХЮ8/з2. Бумага типографская для множительных аппара-
тов. Гарнитура обыкновенная новая. Печать высокая. Усл.
печ. л. 4,20. Уч.-изд. л. 4,28. Тираж 1000 экз. Заказ 2640.
Цена 30 коп.
Издательство «Лесная промышленность», 101000, Москва,
ул. Кирова, 40а.
Типография им. Анохина Управления по делам издательств,
полиграфии и книжной торговли Совета Министров Карельской
АССР, Петрозаводск, ул. Правды, 4.