/






































































































































































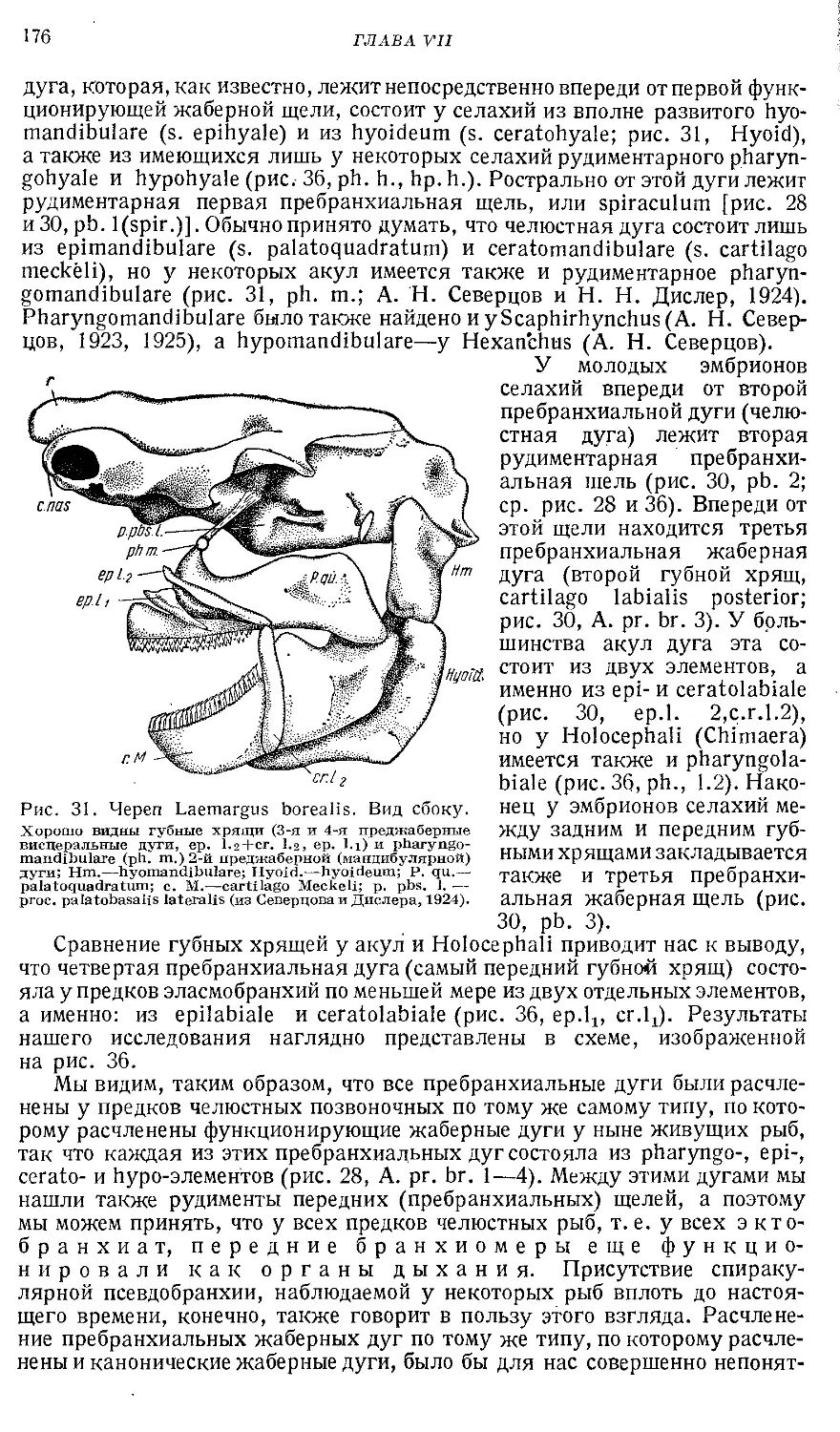






























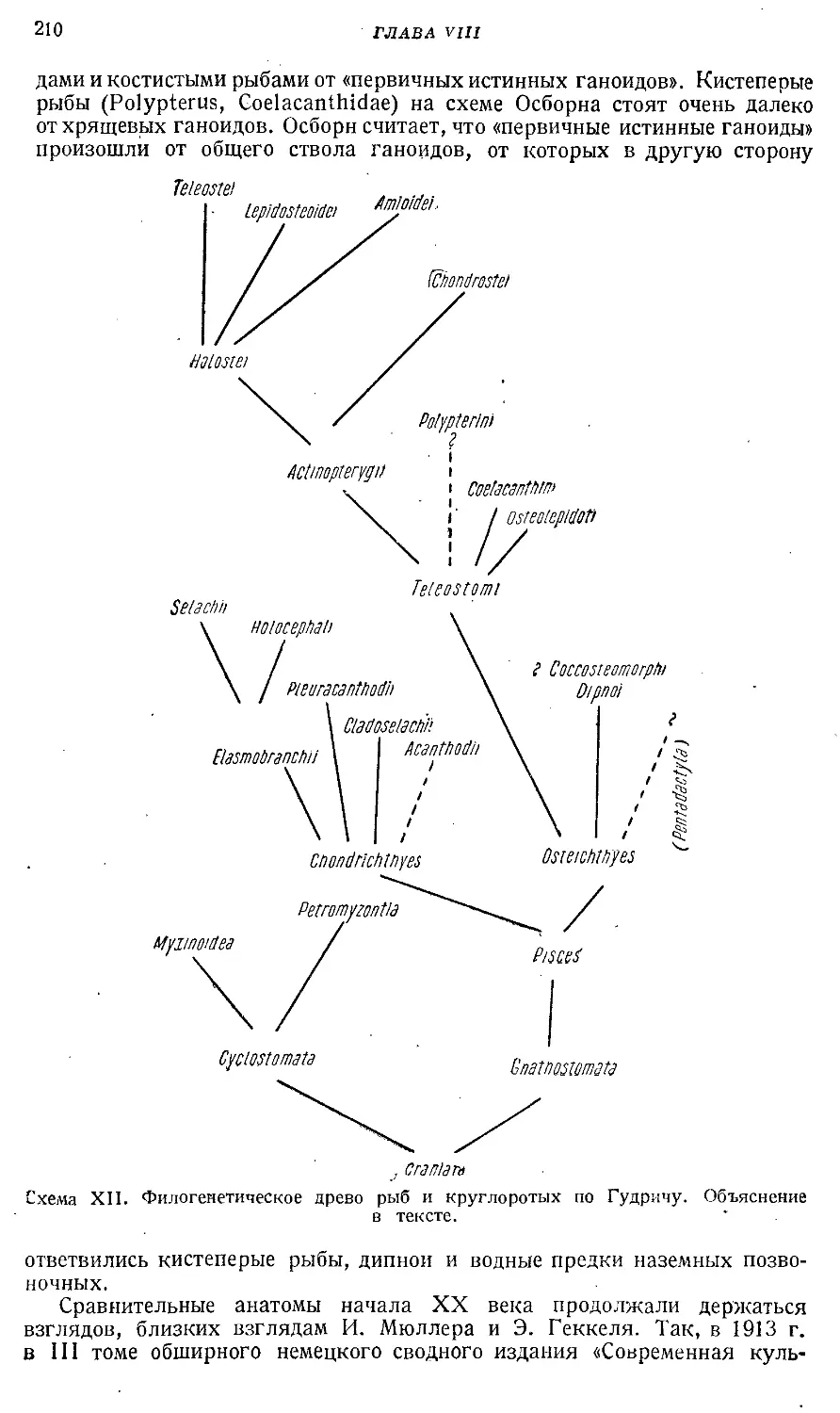























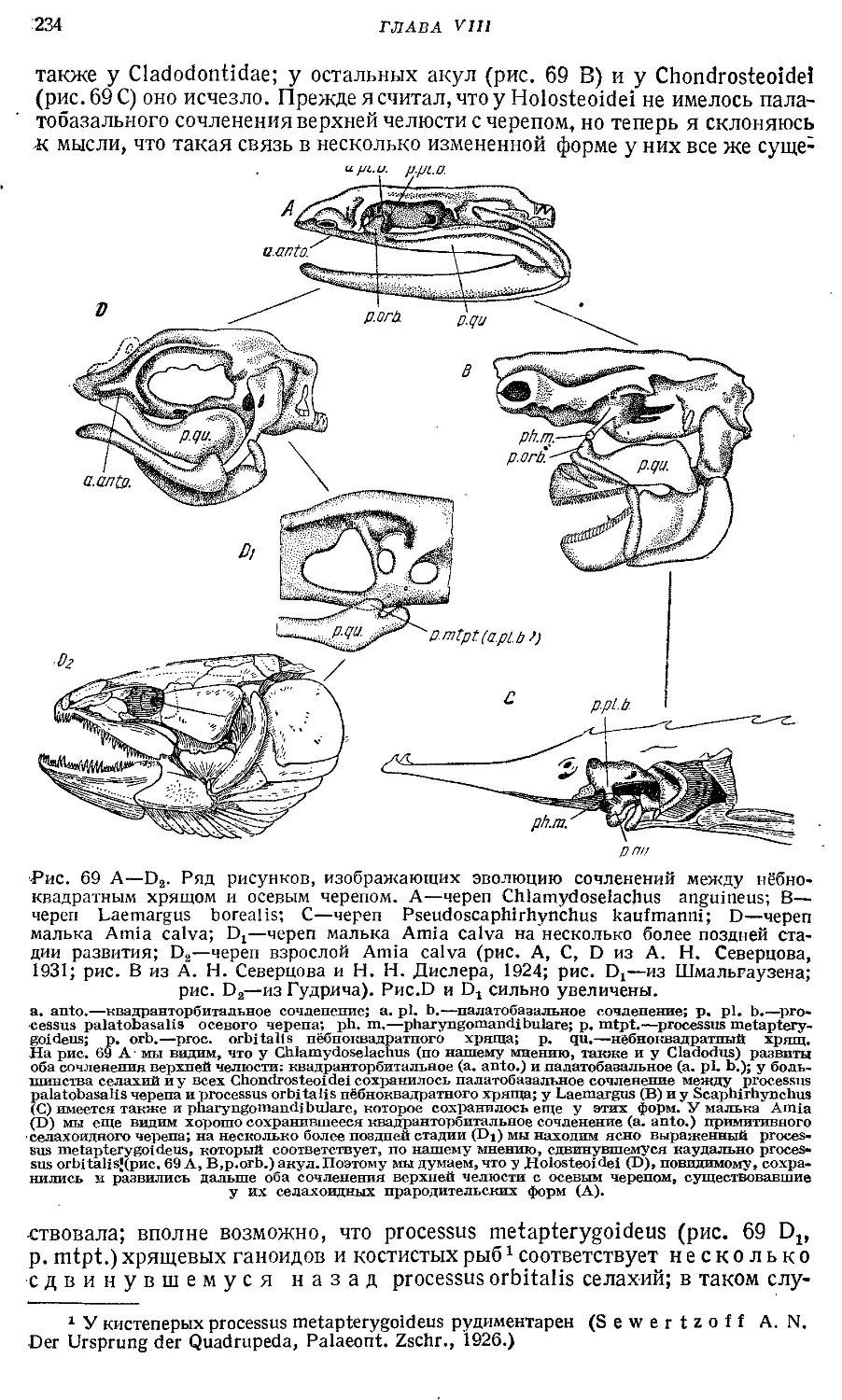





































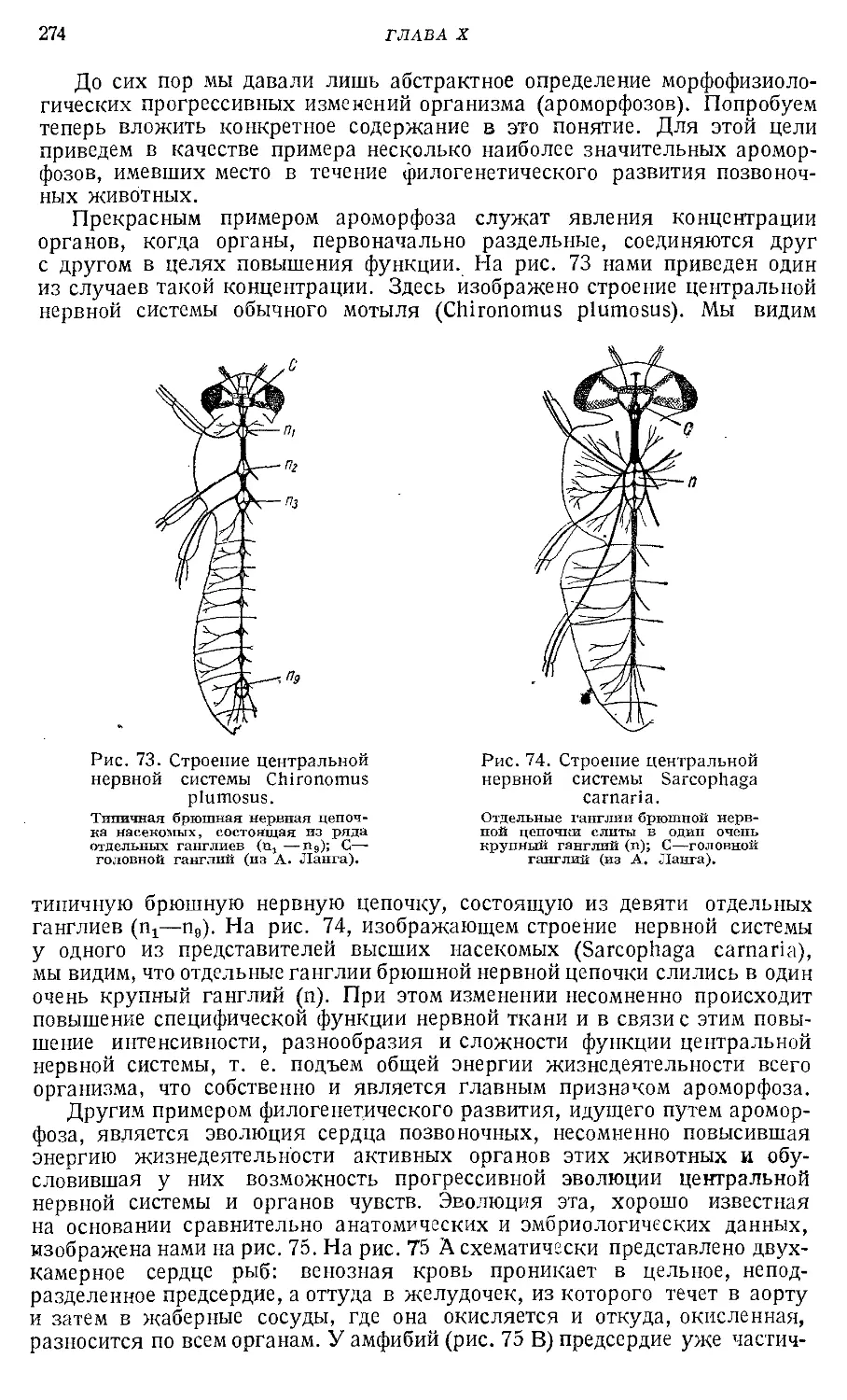






































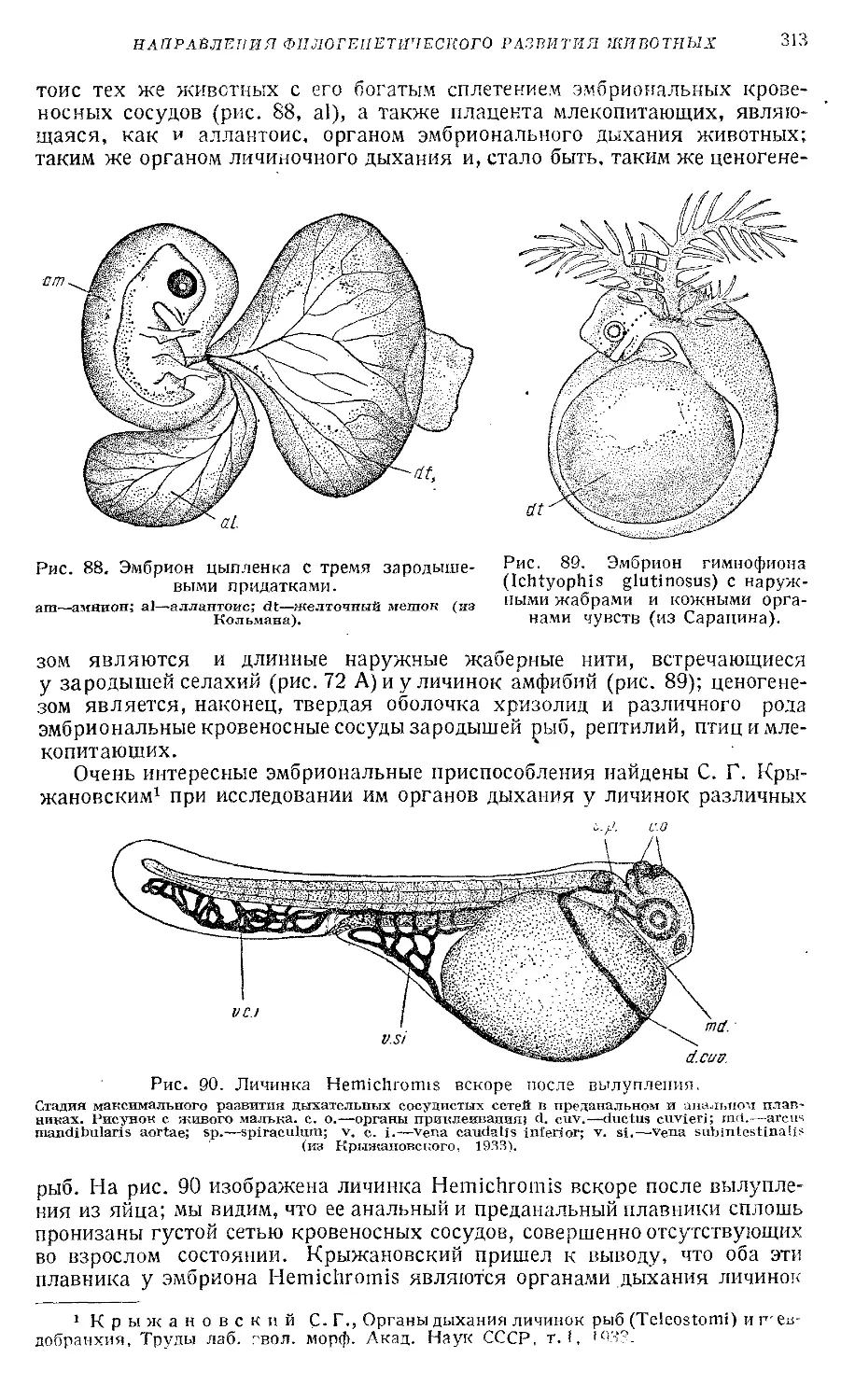



































































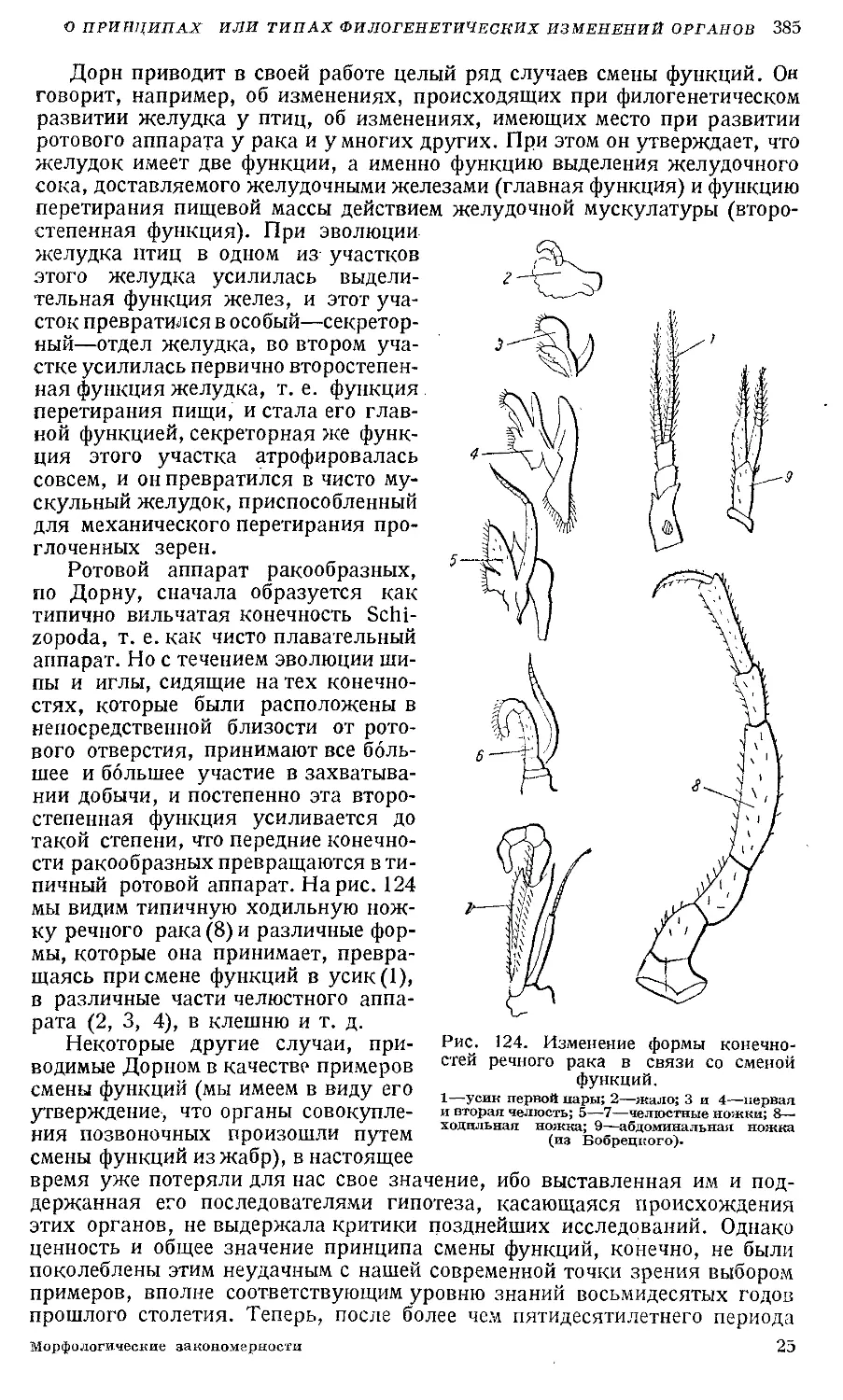




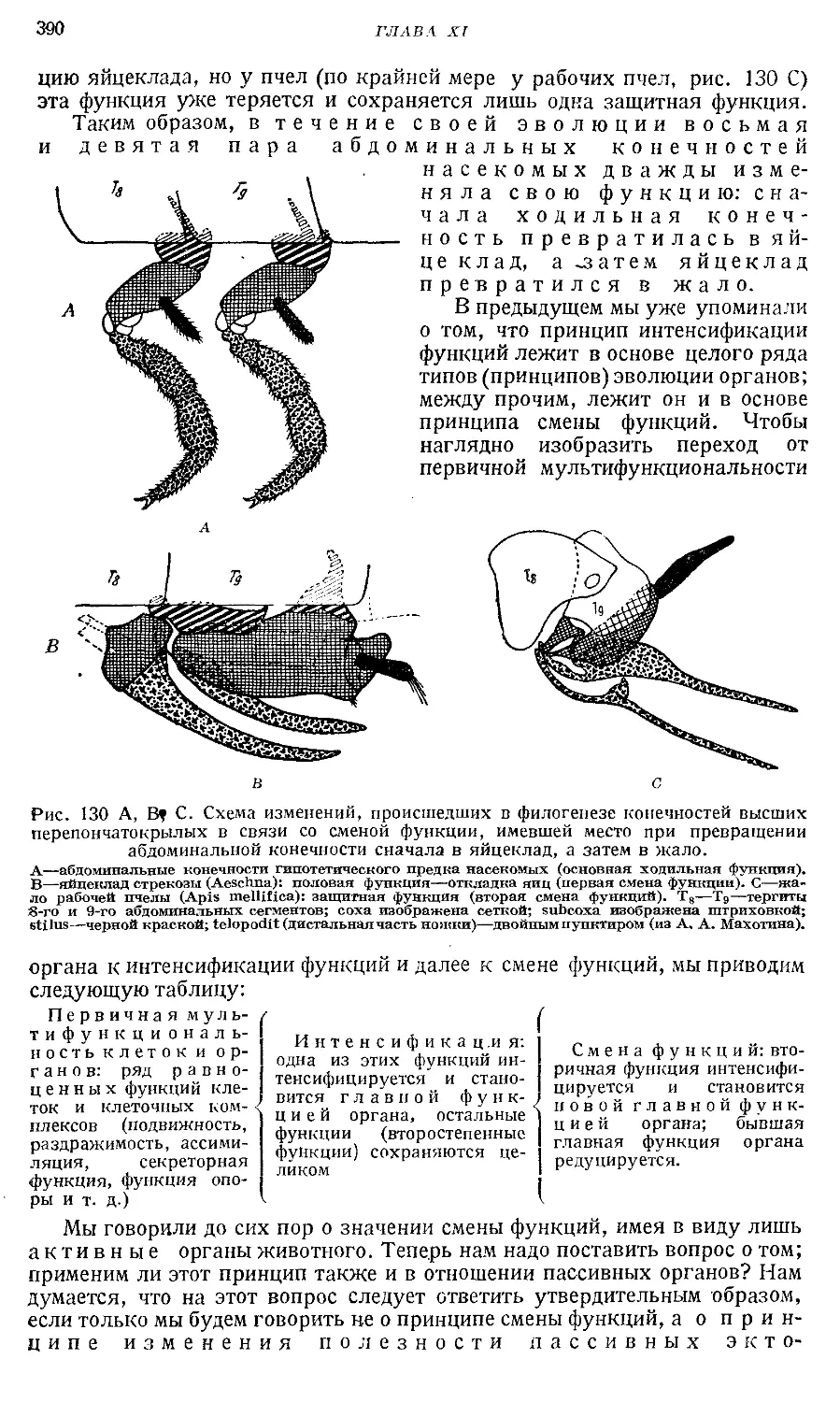


















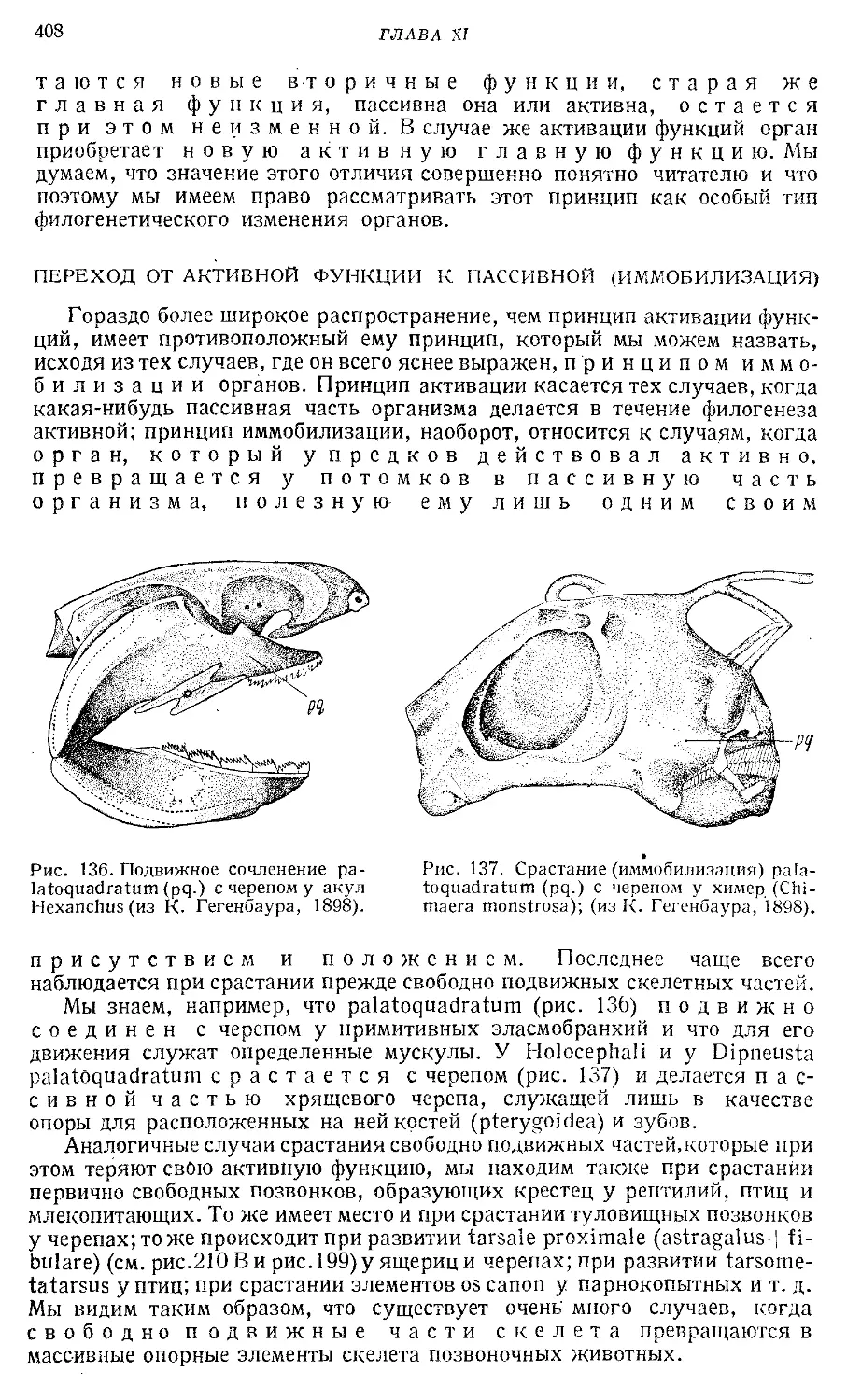




























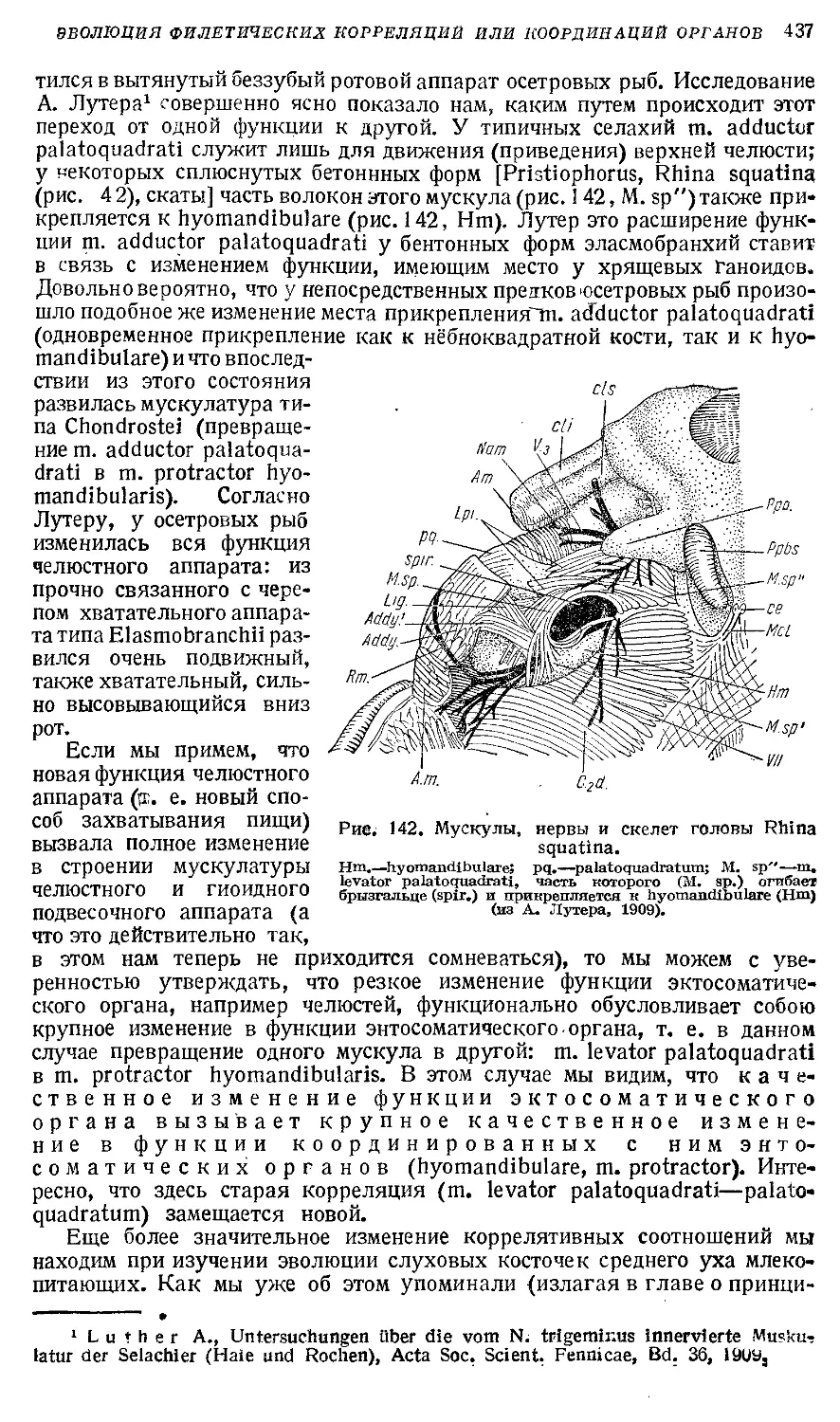

































































































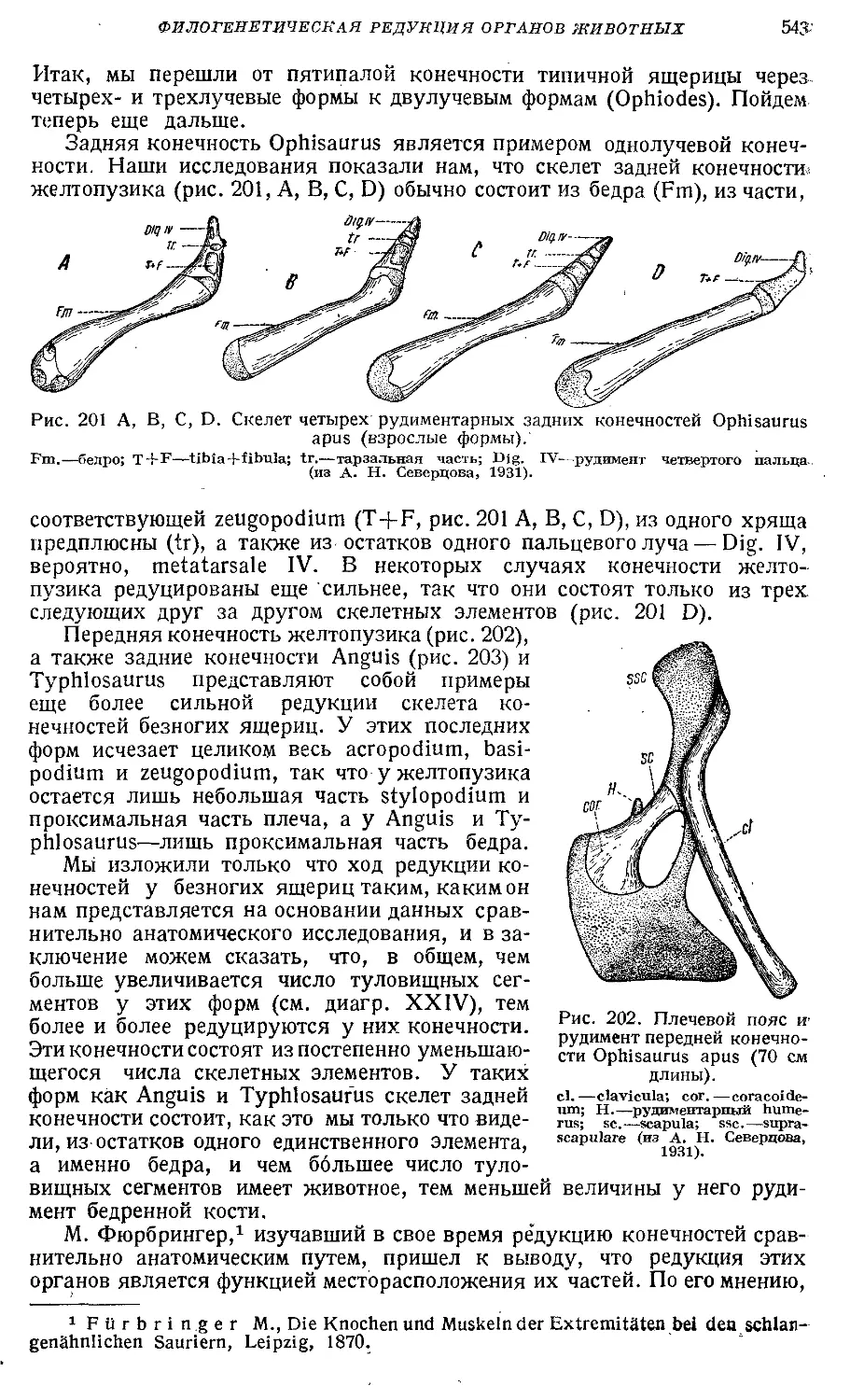
























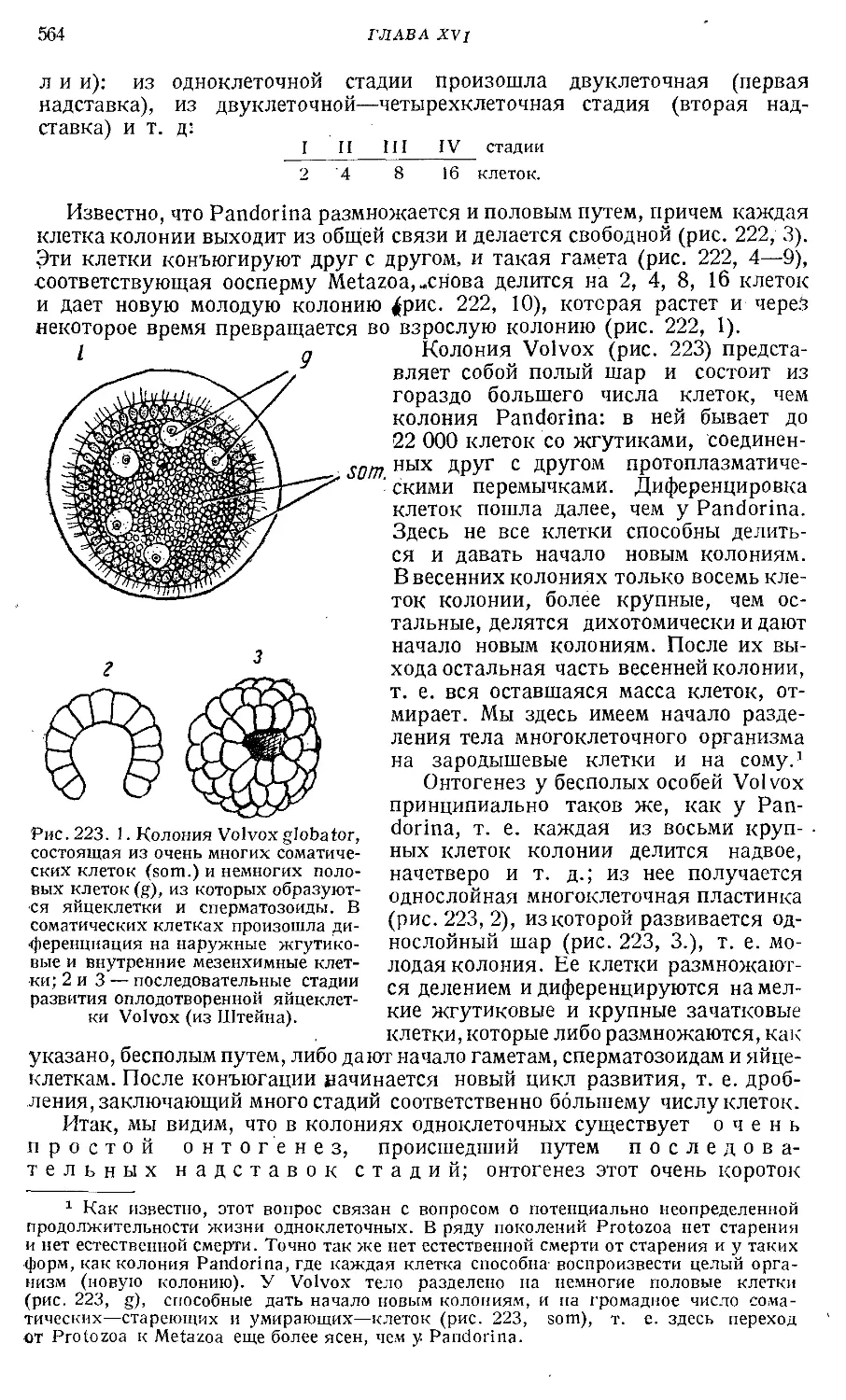





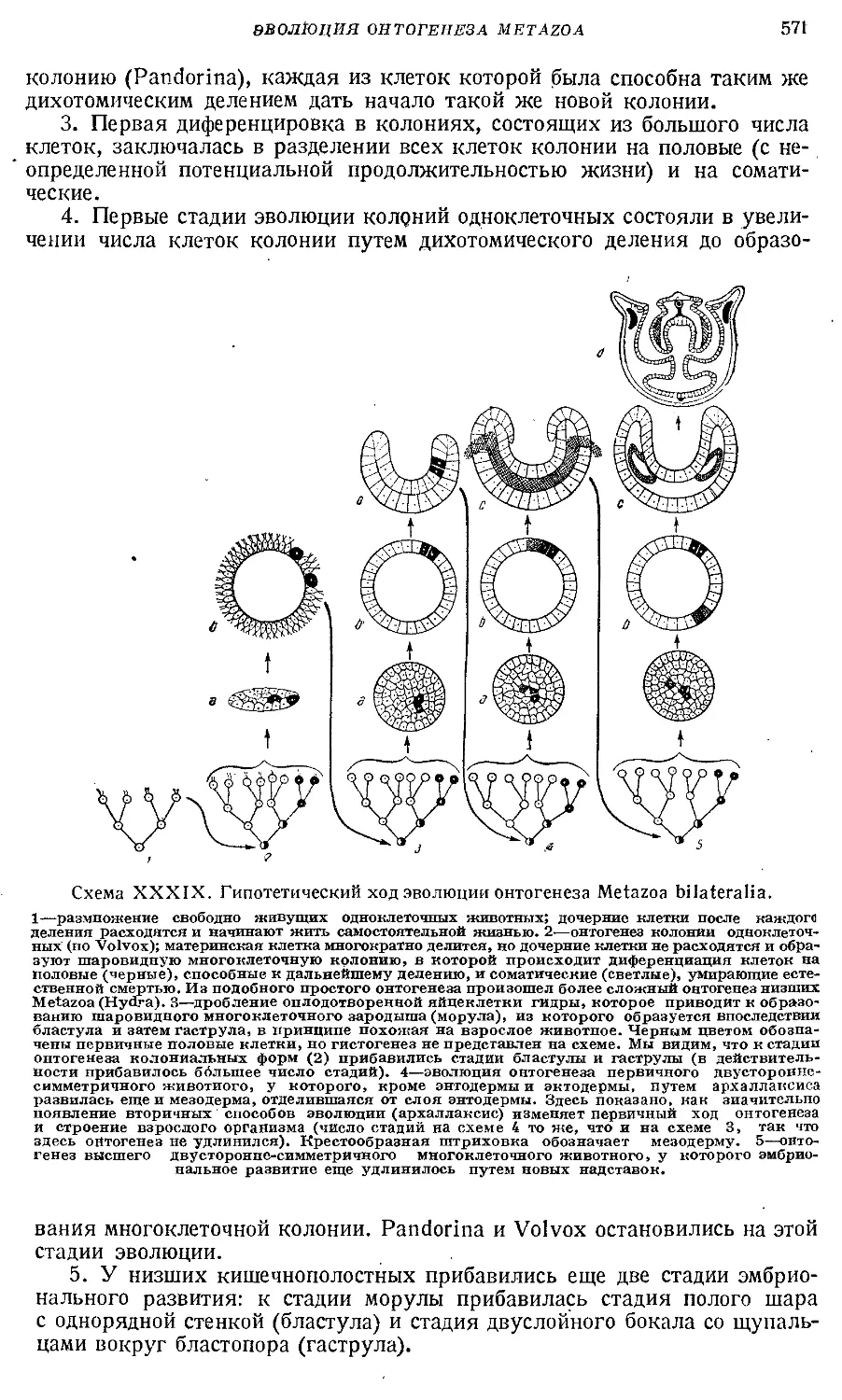

















































Text
A. H. СЕВЕРЦОВ
ДЕЙСТВИТЕЛЬНЫЙ ЧЛЕН АКАДЕМИИ НАУК СССР
МОРФОЛОГИЧЕСКИЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА — 1939 — ЛЕНИНГРАД
Ответственные редакторы проф. Б. С. Матвеев и С. В. Емельянов.
Технический редактор И. П. П о тп е ш у л и н. Корректор А. И. Архангельский.
Сдано в набор 20/VI 1938 г. Подписано к печати 13/IX 1939 г. Формат 70X108*/iS. Объем 38*/4 п. л. (20 бум. л.). Вкл. 9. В 1 п. л. 58 000 печ. зн., 51 уч.-авт. л. Тираж 3000 акз. Уполн. Главлита № А-767. АНИ 1212, РИСО 922. Заказ № 3921. Цена 18 руб., перепл. 3 руб.
Набрано и сматрицировано в 16-й тип. треста «Полиграфкнига». Москва, Трехпрудный, 9.
Отпечатано в 1-й Образцовой тип. Огиза РСФСР треста «Полиграфкнига». Москва, Валовая, 28.
ОТ РЕДАКЦИИ
Книга «Морфологические закономерности эволюции» является послед-нимэтапом 45-летней научной деятельности акад. А. Н. Северцова. В этой книге уже на закате своей жизни он попытался в окончательной форме сформулировать все свои основные теоретические выводы по различным вопросам эволюционного учения и обосновать их конкретными данными из всех своих специальных исследований и работ своих учеников.
Несмотря на то, что разработка теоретических основ дарвинизма имеет громадное значение не только для правильного построения теоретической биологии, но и для развития социалистического строительства, среди советских биологов не так много можно назвать ученых, которые своими трудами сделали существенный вклад в материалистическое развитие теории дарвинизма. Еще меньше сделано в этом направлении в Западной Европе, где широко развиты различные идеалистические направления в биологии, причем большинство западноевропейских «теоретиков-биологов» свое мировоззрение строит на базе фашистской идеологии. Свои фашистские выводы они стараются обосновывать закономерностями эволюционного процесса, взятыми из биологии. В морфологии на западе культивируется идеалистическая морфология виталистического направления (А. Нэфа, Е. Якобсгагена и др.). Среди биологов развивается «теория холизма» Дж. Смётса (1927) и Ад. Майера (1926, 1935), объясняющая целостность организма как особое свойство, находящееся вне организма, т. е. в понимание целостности вносится идеалистическое толкование. Среди генетиков и эволюционистов проповедуется «теория преадаптации» Л. Кено (1932), А. Парра (1926), Дэвенпорта и др., считающих, что «приспособление» в организме предшествует изменению среды и возникает независимо от среды. По воззрениям сторонников теории преадаптации приспособление возникает не в результате отбора изменения, полезного виду для его существования в данных условиях среды, а в результате выбора подходящей среды для уже возникшей в организме структуры, иначе говоря, по этим взглядам «форма возникает независимо от среды». Идеалистический характер носят также теоретические выводы «органи-цистов» (Рессель, 1930; Берталамфи, 1932); наконец, палеонтологи Бейр-лен (1932 и 1936), Дакке (1935) приходят к полному отрицанию дарвиновского понимания эволюции и возвращаются к настоящей средневековой мистике. Отсюда становится ясным, насколько важной задачей является материалистическая разработка дарвинизма в СССР.
На фоне широко распространенных идеалистических толкований эволюционного процесса на Западе особенно четко вырисовывается значение теоретических взглядов акад. А. Н. Северцова как крупнейшего теоретика эволюционного учения в СССР, развивающего материалистические основы учения Дарвина. Для теоретических взглядов акад. А. Н. Северцова чрезвычайно характерно, что каждое теоретическое суждение является обобщением, вытекающим из ряда конкретных фактических исследований автора или его учеников. Соответственно этому построено и изложение его книги.
1*
4
ОТ РЕДАКЦИИ
Другой характерной чертой акад. А. Н. Северцова, отразившейся в данной Книге, является то, что он очень мало времени и места уделяет полемике со своими противниками. Из каждого разбираемого автора А. Н. Северцов берет лишь то, что по его мнению представляет объективную ценность, не тратя сил на доказательство неправильности неверных положений. Эти две черты настоящей книги резко отличают ее от большинства книг, посвященных общим вопросам эволюции. Большинство авторов пользуется другим методом изложения; они для доказательства своих взглядов подбирают из разных книг подходящие примеры, оперируют главным образом с чужим фактическим материалом и, с другой стороны, много места уделяют полемике со взглядами различных авторов и доказательству несостоятельности их воззрений.
Основной задачей своих исследований А. Н. Северцов ставит разрешение вопроса, как происходит эволюция животных и каким законам она подчиняется, а не вопроса, почему происходит эволюция животных. Однако первая часть книги посвящена изложению теоретических взглядов автора по вопросу о причинах эволюции. С первых же страниц А. Н. Северцов занимает четкую позицию сторонника дарвиновского понимания эволюционного процесса и подчеркивает, что «причины филетических изменений в организме животных следует искать в изменениях внешней среды», при этом «влияние косвенного (не прямого) воздействия среды на организм (дарвиновский фактор эволюции) имеет неизмеримо большее значение, чем все еще недоказанное влияние непосредственного воздействия условий внешней среды». Отсюда следуют выводы об эктогенетическом характере эволюции и о несостоятельности аутогенеза, критика ламаркизма и неоламаркизма, критика принципа целесообразной реакции.
Не менее четко А. Н. Северцов занимает позицию сторонника монофилетической эволюции и сразу же ставит вопрос о приспособительном характере эволюционных изменений и о значении коррелятивных изменений как вторичных соотносительных изменений, функционально связанных с первичными адаптативными изменениями.
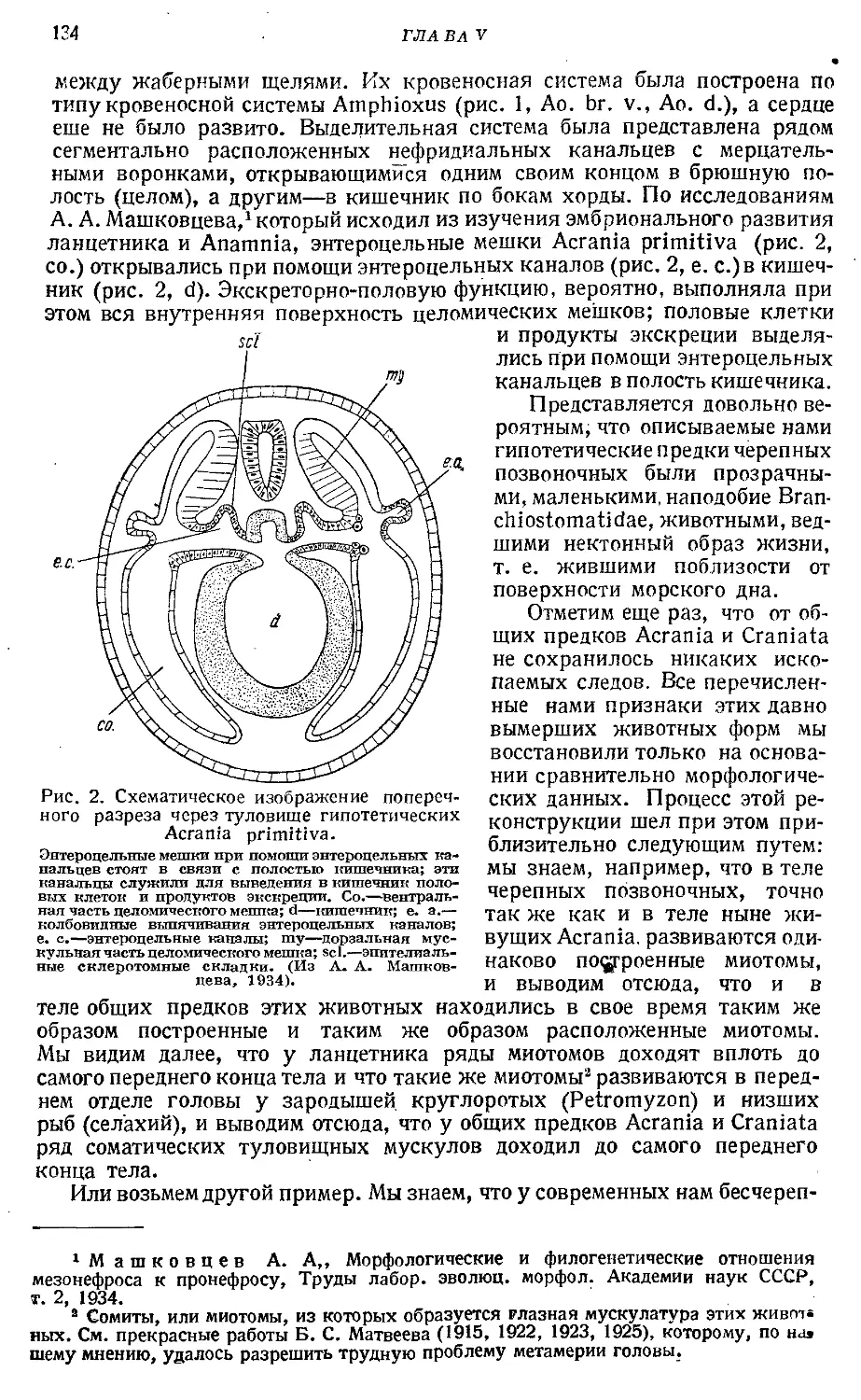
Вторая часть книги посвящена обзору эволюции низших позвоночных, начиная от древнейших предков хордовых животных и кончая обособлением от рыбообразных предков первичных наземных четвероногих позвоночных. В этой части А. Н. Северцов подводит итоги многочисленным сравнительно анатомическим и эмбриологическим исследованиям советских морфологов и восстанавливает последовательную картину филогенеза низших хордовых животных. Филогенетические построения А. Н. Северцова характеризуются новым подходом эволюционного морфолога, а не филогенетика геккелевского направления. Во-первых, низко организованные группы животных не являются в глазах А. Н. Северцова формами статическими, остановившимися в своем эволюционном развитии. Отсюда следует анализ первичных анцестральных признаков и вторичных новых приспособительных изменений к современным условиям существования. Во-вторых, ставится задача изучить эволюцию не отдельных органов или систем органов, но составить представление о ходе эволюции целых организмов, живущих в определенных условиях существования, органы которых несут определенную функцию. В-третьих, на основании изложенных принципов построения, между современными формами восстанавливаются промежуточные анцестральные предки(Асгаша primitiva, Рго-tocraniata, Entobrachiata, Ectobranchiata и т. д.), от которых отделились боковые ветви потомков.
Исследование эволюции низших позвоночных, сопоставленное с данными об эволюции высших групп, хорошо известных по новейшим данным палеонтологии, дает возможность А. Н. Северцову поставить
ОТ РЕДАКЦИИ
5
вопрос об общих путях и направлениях эволюционного процесса. Таким образом он переходит к рассмотрению морфологических закономерностей филогенетического развития животных, что и должно являться основной задачей современного морфолога—сравнительного анатома или эволюционного морфолога.
А. Н. Северцов при анализе морфологических закономерностей эволюции выделяет две теории: морфобиологическую теорию путей эволюции и теорию филэмбриогенеза.
Разбирая общее направление эволюционного процесса, А. Н. Северцов отмечает, что филогенетическое развитие в общем и целом представляет собой приспособительный процесс к изменяющимся условиям существования. При этом биологический прогресс определяется в конечном итоге филогенетическим развитием отдельных приспособлений, связанных с отдельными факторами внешней среды, а не с внешней средою, взятой в Целом. Однако, принимая во внимание, что части организма связаны друг с другом в одно целое,—в результате отдельного приспособительного изменения получается целая цепь коррелятивных вторичных изменений в других частях организма.
Весьма большое методологическое значение имеет дальнейший разбор путей и направлений биологического прогресса и регресса (теория ароморфозов, идиоадаптаций, дегенерации и ценогенезов), где дается дальнейший анализ эволюции как приспособительного процесса. Здесь с большой ясностью определяется взаимная роль прогрессивных и регрессивных изменений, направляющих, благодаря естественному отбору, эволюцию в сторону дальнейшего филогенетического развития вида. Особенно важно подчеркнуть этот разбор главных направлений эволюционного процесса как разработку дарвиновского понимания эволюционного развития в противовес западноевропейским идеалистическим теориям: теории направленности эволюции по пути постепенного совершенствования^. Плате, В. Франц и др.), теории внезапных взрывов творческого начала эволюции (Шиндевольф, Бейрлен, Дакке) и другим анти-дарвинистическим теориям, главенствующим в фашистских странах.
Не меньшим сдвигом в биологических науках различных специальностей является раздел книги о типах (принципах) филогенетических изменений органов. Здесь А. Н. Северцов с большим размахом теоретика эволюционного учения ставит на очередь крупнейшую проблему биологии о соотношении формы и функции в процессе эволюции. Основной предпосылкой учения о типах филогенетических изменений является функциональная множественность каждого органа—его мультифункциональность. Соответственно изменениям во внешней среде меняются взаимоотношения этих функций, и в связи с этим в филогенетическом развитии, благодаря естественному отбору, отбираются соответственные филогенетические преобразования органов как при прогрессивной, так и при регрессивной эволюции. Эта общая предпосылка позволяет провести анализ различных типов этих преобразований: усиления, ослабления, расширения, сужения, смены функций, различных видов субституции и т. д. В этой области открывается широкое поле для дальнейших исследований. Морфологи призываются к изучению строения органов в связи с функцией у организма, живущего в определенных условиях среды, изменяющихся в процессе эволюции; физиологи призываются к изучению не только отправлений органов отдельного животного, но к изучению эволюции их функций опять же во взаимоотношении с окружающей средою; экологи призываются к учету тех морфологических и функциональных преобразований, которые происходят при изменении экологических условий существования.
Ламаркисты пытались связать воззрения А. Н. Северцова о форме и функции с защитой им якобы идей ламаркизма, т. е. прямого воздействия
Б ОТ РЕДАКЦИИ
среды на эволюцию организма, но это неправильно. А. Н. Северцов настолько четко проводит дарвинистическое понимание эволюционного процесса, что этот вопрос не’ требует разъяснения. Последним разделом морфобиологической теории является глава о филетических корреляциях, которые А. Н. Северцов выделяет от индивидуальных или физиологических корреляций в особую группу взаимосвязей, названных им координациями. Это деление корреляций на две группы вносит ясность в сложную систему корреляций и определяет их роль в эволюционном развитии. Однако, как подчеркивает и сам А. Н. Северцов, этот важный отдел морфологических закономерностей в понимании организма как целостной системы еще далек от своего полного разрешения.
Второй теорией, выдвинутой А. Н. Северцовым, является теория филэмбриогенеза, поставившая на новые рельсы проблему взаимоотношений онтогенеза и филогенеза. По существу эта теория была им установлена еще в 1910—1912 гг., и, уже пользуясь ею, А. Н. Северцов разобрал другие морфологические закономерности. Теория филэмбриогенеза восстановила значение эмбриологических данных в теории эволюции, поколебленное односторонностью и механистичностью формулировок биогенетического закона, данных Э. Геккелем.
А. Н. Северцов ставит вопрос о роли изменений хода онтогенеза при возникновении новых признаков, а часто и новых органов для процесса видообразования. Он заново восстанавливает учение Ф. Мюллера, впервые определившего, что наследственные изменения хода эмбрионального развития органов могут затрагивать любые стадии онтогенеза и в различной степени влияют на строение потомков. «Филогенез является, таким образом, функцией онтогенеза». Разрабатывая значение роли эмбриональных изменений в процессе эволюции в течение 26 лет (1910—1936), А. Н. Северцов создал стройную теорию филэмбриогенеза, в которой он объединил высказывания других авторов (Ф. Мюллер, Седжвик, А. Нэф, Р. Гарстанг, В. Франц, Де Беер, А. Наук и др.). В ней он разработал различные способы филогенетических преобразований органов в онтогенезе при прогрессивной и регрессивной эволюции и их влияние на рекапитуляцию признаков предков в онтогенезе. Сюда относятся как общие процессы (частный и общий рост, гистогенез, странствование клеток и др.), так и отдельные способы или модусы (анаболии, девиации и архаллаксисы), а также сдвиги во времени (гетерохронии) и т. д. Длительность разработки данной проблемы нашла некоторое отражение в этой книге. В последних работах А. Н. Северцов стал значительно шире понимать теорию филэмбриогенеза, чем это было сформулировано им раньше. Поэтому в некоторых местах возможны несоответствия. .
В заключение нельзя не отметить также особого раздела теоретических выводов акад. А. Н. Северцова, посвященного процессу редукции органов, разработанного им с точки зрения типов филогенетических изменений органов и теории филэмбриогенеза.
Из всего вышеизложенного становится вполне ясным то громадное значение, какое представляет для советских биологов выход в свет настоящей книги. Здесь читатель впервые может в целостной связной форме представить роль, которую сыграл в советской науке покойный А. Н. Северцов. Мы знаем, что идеи А. Н. Северцова нашли среди советских биологов широкий отклик. Специалисты самых различных отраслей биологии в своей творческой исследовательской работе развивают дальше теоретическое наследие А. Н. Северцова. Можно надеяться, что эта книга явится призывом к еще более широкому развитию материалистических основ дарвинизма и исканию путей, связывающих закономерности эволюционного процесса с нуждами практической жизни.
Проф. Б. Матвеев.
«Морфологические закономерности эволюции» выходят в свет через два года после смерти автора. Так как Алексей Николаевич Северцов, изнуренный тяжелой болезнью, продолжавшейся в течение двух последних лет его жизни, не мог принимать непосредственного участия в окончательной редакции своей книги, то я считаю нужным сказать несколько слов о том", как подготовлялась к печати настоящая книга.
В основу «Морфологических закономерностей эволюции» положен полный перевод книги А. Н. Северцова «Morphologische Gesetzmassigkeiten der Evolution», изданной в Иене в 1931 г. Текст этого перевода, сделанного мной, был полностью проредактирован А. Н. Северцовым. В тексте перевода, по сравнению с изданием 1931 г., им было сделано мало изменений, но в самую книгу внесено очень много добавлений.
В первую часть, по указанию А. Н. Северцова, были введены две добавочные главы: глава первая—«Эволюционное учение после Дарвина и задачи филогенетического исследования», ранее напечатанная в несколько ином виде в Зоологическом журнале (1932), и глава вторая—«О неоламаркизме», нигде не опубликованная. В четвертую главу он внес наново написанный раздел «Полифилетическая теория».
Значительные видоизменения и дополнения были введены в восьмую главу филогенетической части, для которой А. Н. Северцовым был наново написан «Очерк вопроса о классификации рыб». Здесь же, в качестве приложения, дана, согласно его желанию, его последняя морфологическая работа «О покровных костях черепа». Эта работа, носящая характер предварительного сообщения, представляет собой одно из тех исследова-ваний, которые были им задуманы (и частично выполнены) для опровержения некоторых гипотез Стеншиб, с которыми он не был согласен. С этой же целью А. Н. Северцов сделал и ряд вставок в раздел «Эволюция Holosteoidei actinopterygii и Holosteoidei crossopterygii» в той же восьмой главе, в филогенетической части книги.
Коренной переработке подверглась, по указаниям А. Н. Северцова, глава десятая третьей части книги, излагающая учение о различных направлениях эволюции животных. Объем этой главы, по сравнению с книгой «Morphologische Gesetzmassigkeiten der Evolution», увеличился почти вдвое. В главу одиннадцатую .(О принципах, или типах филогенетического изменения органов) введен совершенно новый раздел: «Принцип выпадения промежуточной функции», разработанный А. Н. Северцовым в его работе «Эволюция брюшных плавников и принцип выпадения промежуточных функций» (1933). Сюда же, в ряд глав, посвященных морфобиологической теории эволюции, отнесена в настоящем издании глава об «Эволюции филетических корреляций, или координаций органов», стоявшая несколько особняком в «Morphologische Gesetzmassigkeiten der Evolution».
Значительно переработана теория филэмбриогенеза. Здесь, на основании своих позднейших работ, а также работ своих учеников, А. Н. Северцовым внесено очень много вставок и дополнений почти во все разделы этой части. В частности, совершенно новым является ранее не опубликованный раздел «Периодическое изменение хода развития органов»
8
ОТ РЕДАКЦИИ
в главе четырнадцатой. Наново введены и очерк «О гистологической рекапитуляции», помещенный в разделе «Рекапитуляция признаков органов и тканей» той же главы, а также вся шестнадцатая глава—«Эволюция онтогенеза Metazoa», ранее опубликованная в виде отдельной статьи (1934). Наконец, совершенно особое место занимает в книге глава семнадцатая—«О соотношении между принципами эволюции и модусами филэмбриогенеза». Задуманная в самые последние годы его жизни, эта глава не могла быть разработана А. Н. Северцовым вследствие болезни: он успел набросать ее лишь в виде конспекта. Сделать эту работу, а тем более литературно ее оформить, ему уже не пришлось. Я все же решилась поместить эти наброски, несмотря на их схематичный характер, потому что выраженной в них мысли, представляющей собой попытку объединить в одно целое морфобиологическую теорию эволюции с теорией филэмбриогенеза, А. Н. Северцов придавал большое значение. Кроме того, в этой главе поставлен ряд интереснейших вопросов и намечены пути для целой серии новых исследований, которые еще могут быть сделаны в этом направлении.
По предложению редакции, этой книге предпослана биография А. Н. Северцова, написанная мною.
Мне осталось сказать несколько слов о той большой помощи, которую оказал мне Институт эволюционной морфологии имени А. Н. Северцова в деле окончательного оформления его книги. Особенно обязана я трем старшим ученикам А. Н. Северцова: акад. И. И. Шмальгаузену, проф. Б. С. Матвееву и проф. М. М. Воскобойникову, к которым я всегда могла свободно обращаться за разрешением своих сомнений. Ряд ценных указаний сделал мне С. Г. Крыжановский. С большой тщательностью* книга была проредактирована Б. С. Матвеевым и С. В. Емельяновым. Много материала для вставок, иллюстрирующих новыми примерами ряд основных положений А. Н. Северцова или представляющих собою дальнейшее развитие этих положений, было предоставлено мне после его смерти В. В. Васнецовым, А. А. Машковцевым, Б. С. Матвеевым, С. Н. Боголюбским, Д. Ф. Федотовым, А. Н. Дружининым, С. В. Емельяновым, А. А. Махотиным и А. Г. Рындзюнским. Ряд новых препаратов и рисунков был сделан Н. О. Ланге.
Всем перечисленным мною лицам выражаю свою глубокую и искреннюю благодарность.
Л. Северцова
АЛЕКСЕИ НИКОЛАЕВИЧ
СЕВЕРЦОВ
БИОГРАФИЧЕСКИЙ ОЧЕРК
АЛЕКСЕЙ НИКОЛАЕВИЧ СЕВЕРЦОВ
Биографический очерк* 1
I
Алексей Николаевич Северцов родился в Москве 11 сентября 1866 г., но раннее детство его протекало в деревне, в Воронежской губ., в с. Петровском. Он жил там у деда, подполковника Алексея Петровича Северцова, участника Бородина, потерявшего в сражении руку. У деда жил он с отцом Николаем Алексеевичем Северцовым, знаменитым орнитологом, путешественником и исследователем Туркестана, и матерью Софьей Александровной; иногда,—во время длительных среднеазиатских экспедиций отца,— с одной только матерью.
В детстве Алексей Николаевич был слабым и нервным ребенком. Часто хворал. И только после зимы, проведенной с родителями по совету врачей в Швейцарии, он окреп, развился и стал подвижным и ловким мальчиком. Единственный сын у родителей, он рос одиноким ребенком, шаловливым и беспечным, страстным фантазером и мечтателем. Жадно увлекался чтением. Он часто забирался на высокие дубы, окружающие усадьбу деда, и долгие часы проводил там с книжкою в руках. Особенно любил он детские исторические романы и рассказы о путешествиях. Вероятно, здесь сказалось влияние отца, рассказывавшего мальчику о своих путешествиях к границам далекого Памира, по киргизским степям, тогда еще совсем диким, и о своем пребывании в плену у кокандцев, сильно изранивших его пиками во время пленения. Рассказы эти, ведшиеся в спокойном и простом тоне, повествующие о действительно героическом, как о чем-то очень будничном и обыкновенном, производили на мальчика огромное, на всю жизнь сохранившееся впечатление.
Вместе с отцом, коллекционировавшим в то время птиц Воронежской губернии, он часто ходил на охоту и уже тогда научился хорошо стрелять. Тогда же выучился он верховой езде, плаванью и гребле. Ко всему этому мальчик относился со страстным увлечением: эхо было началом его будущих спортсменских успехов.
В 1876 г- родители Алексея Николаевича окончательно переселились в Москву и отдали его в частную Поливановскую гимназию. Л. И. Поливанов, талантливейший педагог и глубокий знаток русской литературы, пушкинист и шекспирист, имел на Алексея Николаевича сильное влияние, и, вероятно, именно им была привита Алексею Николаевичу та особенная его любовь к Пушкину, которая у него сохранилась на всю жизнь.
1 В качестве материалов при составлении биографии А. Н. Северцова служили мне:
1) его «Воспоминания», доведенные им до 1909—1910 гг., 2) его многочисленные ко мне письма, 3) личные мои воспоминания о нем.
12
БИОГРАФИЧЕСКИЙ ОЧЕРК
В гимназии Алексей Николаевич сначала учился плохо. Ленился и нервничал в классах, часто терялся, когда его вызывали на уроках. Несколько лучше пошли занятия в средних классах, когда из экспедиции вернулся отец; совсем хорошо он учился лишь в последнем классе гимназии. Вообще период после возвращения Николая Алексеевича из последней экспедиции вплоть до его смерти был временем особенно тесного сближения между отцом и сыном. Под непосредственным воздействием Николая Алексеевича складывался в те годы характер Алексея Николаевича и создавалось его юношеское мировоззрение. С огромным уважением и любовью говорил он всегда о своем отце,—«большом ученом и мыслителе»,—и охотно и много рассказывал о нем.
Алексей Николаевич был в VIII классе гимназии, когда внезапно скончался Николай Алексеевич. 27 января 1885 года, переправляясь в санях через Дон, он попал-в полынью и, плотно закутанный в тяжелую доху, стал тонуть. Из воды кучер его вытащил, но Николай Алексеевич внезапно сильно ослабел и на берегу упал. Кучер бросился в соседнее село за помощью, когда же он вернулся, Николай Алексеевич был уже мертв: смерть, повидимому, последовала от апоплексического удара.
Смерть отца тяжело поразила Алексея Николаевича, но пример матери, с большим мужеством и достоинством переносившей свое горе, заставил его взять себя в руки и помог ему выправиться. С тех пор он особенно сблизился с матерью: они стали большими друзьями и дружба эта продолжалась всю ее долгую жизнь, центром которой являлся единственный сын.
Со смертью отца меняется весь прежний уклад жизни. Были тяжелые долги, и Алексей Николаевич вместе с матерью оказываются в очень стесненном материальном положении. Пришлось оставить московскую квартиру и переселиться в меблированные комнаты, пришлось высчитывать буквально каждую копейку. В этот тяжелый год стал определяться характер Алексея Николаевича. Он весь подобрался, возмужал, стал вдумчив, серьезен и сдержан. Много перечитал и еще больше передумал. Мужественный по природе, он поставил перед собою целью закалить свою волю, упражняя ее, научиться скрывать боль, не признавать ее,—ни физическую, ни нравственную. Твердо и непреклонно занялся он самовоспитанием и, как показало будущее, преуспел в этом.
Кроме Л. И. Поливанова, в гимназический период его жизни на Алексея Николаевича имел большое влияние С. А. Усов, ближайший друг его покойного отца. Профессор зоологии московского университета С. А. Усов был в то же время большим знатоком искусства и в последние годы пребывания Алексея Николаевича в гимназии читал на квартире у J1. И. Поливанова лекции по истории искусства для некоторых поли-вановцев. Эти лекции, первые лекции в его жизни, Алексей Николаевич слушал с захватывающим интересом: можно смело сказать, что в отношении восприятия произведений искусства они оставили на нем след на всю жизнь. Своим глубоким знанием истории и философии искусства, своим тонким пониманием живописи он, несомненно, был обязан этим лекциям, прослушанньш им в тот период жизни, когда «все впечатления бытия» воспринимаются с особенной остротой.
Окончив гимназию, Алексей Николаевич осенью 1885 г. поступил в московский университет на физико-математический факультет. Это был тяжелый год в жизни московского университета, время очередного усиления реакции, время введения пресловутого устава 1884 г. Но вместе с тем это было и время, когда в университет, впервые в его истории, свежей струей ворвались революционизирующие идеи дарвинизма и материализма, идеи, гонимые духовенством и официальными представителями власти, но горячо проповедуемые по всей России крупнейшими биологами
БИОГРАФИЧЕСКИЙ ОЧЕРК
13
конца прошлого столетия: братьями В, О. и А. О. Ковалевскими, И. И. Мечниковым, Н. М. Сеченовым и К. А. Тимирязевым.
В момент поступления Алексея Николаевича в университет состав профессуры на физико-математическом факультете был очень хорош. Зоологию позвоночных он слушал сначала у С. А.Усова, затем у М. А. Мензбира, зоологию беспозвоночных—у А. П. Богданова, ботанику—у И. Н. Горожанкина и К. А. Тимирязева, анатомию—у Д. П. -Зернова, геологию—у молодого тогда А. П. Павлова, физику—у А. Г. Столетова, химию — у В. В. Марковникова. Таким образом, Алексей Николаевич с первого же курса учился у целого ряда выдающихся русских ученых. Вкусы его определились очень скоро, и он со второго курса стал увлекаться биологическими науками, в частности—зоологией. Этому много способствовали превосходные, насквозь пропитанные духом эволюционизма лекции М. А. Мензбира по сравнительной анатомии и зоологии позвоночных.
М. А. Мензбира Алексей Николаевич знал с детства: он был другом и учеником его отца и ко времени поступления Алексея Николаевича в университет обрабатывал орнитологические коллекции Николая Алексеевича, что, естественно, еще больше сближало с ним Алексея Николаевича. Однако, несмотря на влияние отца и Мензбира, Алексей Николаевич не пошел по их пути: он не стал ни орнитологом, ни зоогеографом, но, избрав однажды предметом своего изучения сравнительную морфологию, остался верен своему выбору всю жизнь. В этом выборе, вероятно, сказался природный склад его ума, уже тогда пытливого и смелого, стремящегося к широким обобщениям и далеко идущим выводам, сравнительная морфология,—эта наиболее философская область биологии, ставящая своей конечной целью установление общих закономерностей эволюции,— давала ему в этом отношении очень много.
В кабинете сравнительной анатомии работами Алексея Николаевича и его друзей: П. П. Сушкина.и Н. А. Иванцова, а позже М. М. Хомякова и Н. К. Кольцова, руководил талантливый, рано умерший эмбриолог В. Н. Львов; он обучал их гистологической технике и тонкой анатомической пр'епаровке. Вместе с Сушкиным Алексей Николаевич изготовлял ценные препараты для музея сравнительной анатомии и вместе с ним работал над предложенной факультетом темой для медального сочинения: «Свод сведений по организации и систематике гимнофион». Оба друга занимались усердно и получили за эту работу по золотой медали каждый.
В жизни Алексея Николаевича эта первая научная студенческая работа имела решающее значение. Принимаясь за нее (это был конец 1887 г.), он переживал трудное время. Не внешними обстоятельствами трудное, но тем переломом, которым у него сопровождался переход от юности к возмужалости. У него были тяжелые сомнения относительно того, можно ли в его время заниматься теоретической наукой, действительно ли это теперь «нужно людям»? Его мучил и вопрос о том, способен ли он сам заниматься этой наукой, хватит ли у него упорства и выдержки, чтобы довести раз начатое дело до конца? В этом отношении эта первая работа была для него пробным камнем, сдав этот внутренний экзамен успешно, он определил, что ему стоит «заниматься и дальше». К этому времени у него был решен и другой, гораздо более важный вопрос, вопрос о том, имеет ли он право заниматься той наукой, к которой его тянуло неудержимо, но которая в данный момент «не приносит непосредственной пользы». Решение это ему далось не легко: лишь медленно и постепенно зрела в нем уверенность, что только благодаря успехам теоретической науки, являющейся предпосылкой практического, прикладного знания, «людям легче жить станет, т. е. люди станут и сытее, и здоровее, и сильнее». Другой путь, ведущий к той же цели,—путь социальной революции
БИОГРАФИЧЕСКИЙ ОЧЕРК
П4
ему тогда еще не представлялся ясным; поэтому, несмотря на кажущуюся незатейливость этой житейской философии, она была дорога Алексею Николаевичу, ибо оправдывала в его глазах непобедимое стремление заниматься теоретической наукой, ту жажду «дознаться до пределов человеческого знания», которою он был обуреваем в двадцать лет.
В кабинете сравнительной анатомии работали усердно, не считаясь со временем. Приходили рано утром и занимались до 12, когда к кабинетскому чаю сходились все работавшие в лаборатории. Чай этот был интимным центром лабораторной жизни: после напряженных утренних занятий сходились товарищи, чтобы за стаканом крепкого чая с неизменными цукатными булками поделиться очередными результатами текущей работы,—потолковать о только что законченной книге, обсудить последнее заседание Общества испытателей природы или Психологического общества, усердными посетителями которого они являлись, а главное, чтобы поболтать и поспорить, посмеяться и поостро-словить. После чая занимались снова до вечера, пока позволял свет.
Работая у Мензбира, Алексей Николаевич стал впервые увлекаться рисованием. Вместе со своими приятелями П. П. Сушкиным, Н. А. Иванцовым и П. С. Усовым он, по совету М. А. Мензбира, настаивавшего на умении хорошо срисовывать препараты и чучела, начал работать в студии художника Н. А. Мартынова. У Алексея Николаевича оказались большие способности к живописи: он быстро овладел техникой рисования карандашом и сепией; легко далась ему также и акварель. Каждый, кто впоследствии когда-либо просматривал рисунки Алексея Николаевича (а знали об их существовании очень немногие), неизменно выражал сожаление, что этому искусству ему пришлось уделить так мало времени. В живописи он был, конечно, дилетант, но его легкие, изящные, полные тончайшего юмора рисунки пером и тушью, всегда фантастические и всегда оригинальные, надолго пленяли воображение зрителя.
В свои студенческие годы Алексей Николаевич не только учился и работал. Он был здоров, общителен и жизнерадостен и полон жадного интереса ко всему окружающему. Вращался он в самых разнообразных кругах тогдашнего московского общества. Больше всего бывал среди товарищей студентов, участвовал во всех их собраниях (а иногда и в кутежах в татьянинские дни), участвовал и в. студенческих волнениях, хотя особенно активно в них себя не проявлял. Однако от товарищей отнюдь не отставал, а в знаменитой «Брызгаловской истории»,1 когда у манежа ему пришлось резко осадить зарвавшегося казачьего офицера и, схватив его лошадь под уздцы, посадить ее на задние ноги (чем воспользовался ряд студентов, чтобы добраться до безопасного места), он даже оказался неожиданно для себе героем дня.
Часто бывал Алексей Николаевич и в профессорской среде, особенно у Усовых и Лопатиных. Посещал «Поливановские субботы» и во всех этих местах встречал много крупных и интересных людей, считавшихся «цветом» тогдашней Москвы: В. О. Ключевского, С. А. Юрьева, А. Н. Веселовского, Н. В. Бугаева, Н. И. Стороженко, В. Я. Цингера, М. Н. и Л. М. Лопатиных, Я. А. Борзенкова, М. А. Мензбира и многих других. Посещал он и чинное светское общество, где, правда, ему бывало очень скучно, но где зато можно было встретить иногда Л. Н. Толстого, а упускать эти случаи Алексею Николаевичу не хотелось, ибо Толстым он тогда увлекался страстно и влияние на него Толстого в ту пору его жизни было огромно.
1 Брызгалов был весьма нелюбимый студентами инспектор Московского университета, которому один из студентов публично на концерте дал пощечину.
БИОГРАФИЧЕСКИЙ ОЧЕРК
15-
Конец гимназических лет и студенческие годы Алексея Николаевича были временем его сильнейшего увлечения спортом и гимнастикой. Именно тогда он развил в себе ту огромную физическую силу, которой впоследствии славился много лет. Зимою процветало фехтование, гимнастика, катание с ледяных гор на коньках, летом верховая езда, гребля, а главное, охота, которая в те годы была особенно хороша в целинных Воронежских степях.
Алексей Николаевич окончил университет в 1889 г., но государственные экзамены, которые были тогда только что введены, они с Сушкиным решили отложить на год, чтобы получше к ним подготовиться. Экзамены были благополучно сданы в 1890 г., после чего оба друга были оставлены при университете по кафедре сравнительной анатомии для «подготовки к профессорскому званию».
Вскоре после окончания государственных экзаменов, т. е. в 1890 г., Алексей Николаевич женился на дочери С. А. -«Усова, М. С. Усовой, которую знал с детства. В следующем году появилась в печати его первая самостоятельная работа.1
Собрав необходимый материал в Саратовской и Воронежской губ., Алексей Николаевич стал исследовать чисто анатомическим путем последовательные стадии постэмбрионального развития черепа головастика; при этом ему удалось открыть существование самостоятельного supraoc-cipitale, сливающегося на более поздних стадиях с черепом (как известно, отличительным признаком черепа Anura считалось именно отсутствие supraoccipitale). Но для него лично имело значение другое: здесь он впервые подошел к мысли о том, что у высоко стоящих специализованных форм сохраняются многие признаки примитивной организации предков. Об этой работе он прочел свой первый доклад в Обществе испытателей природы.
Приблизительно в это же время Алексей Николаевич начинает вместе с Сушкиным готовиться к магистерским экзаменам. Мензбировская программа, мастерски составленная, была очень трудна, и на подготовку к экзаменам у обоих друзей ушло свыше двух лет.Об этом времени им, однако, не пришлось сожалеть, так как в результате этих занятий фундамент специальных естественно-исторических знаний у них был заложен основательно и прочно.
После экзамена Алексей Николаевич прочитал, как это было тогда принято, две пробных лекции, был избран приват-доцентом и стал читать в университете специальный курс сравнительной анатомии для небольшой группы студентов. Вместе с тем он с жаром принимается за исследовательскую работу, прерванную подготовкой к магистерским экзаменам. Он снова набирает материал по Pelobates, но на этот раз исследует лишь самые ранние стадии его развития, и для сравнения исследует те же стадии у Bombinator igneus и Siredon. Задачей этого второго исследования было изучение сегментации головной мезодермы у Anura и Urodela. Чтобы не пропадало летнее время, Алексей Николаевич устроил у себя в деревне маленькую лабораторию. Выписал из Москвы микротом, при помощи деревенского слесаря соорудил термостат и принялся весьма энергично фиксировать, заливать и резать собранный материал. В результате у него получилась интересная, хотя и небольшая по объему работа, вышедшая из печати в 1892 г.1 2
Занимаясь исследованиями над Pelobates, Алексей Николаевич пришел к мысли, что все основные работы по метамерии головы позвоночных, центральной проблемы морфологии того времени, выполненные до сих
1 Ueber einige Eigentiimlichkeiten in der Entwicklung und im Bau des Schadels vom Pelobates fuscus. Бюлл. Моск. общ. испыт. прир., 1891.
2 Zur Frage fiber die Segmentierung des Kopfsmesoderms bei Pelobates fuscus. Бюлл. Моск. общ. испыт. прир., 1892.
16
БИОГРАФИЧЕСКИЙ ОЧЕРК
пор лишь на одних акуловых рыбах, являются недостаточными для решения этого вопроса и что всю эту проблему надо переисследовать на другом материале. В качестве такового он выбрал хрящевых ганоидов и решил отправиться вместе с М. М. Хомяковым на Волгу, чтобы собрать эмбриологический материал по стерлядям. Денег на эту поездку у него не было, и ему пришлось для нее заложить единственную имевшуюся в его распоряжении ценность—свою золотую университетскую медаль.
Эмбриологический материал в то время добывался самым примитивным образом, но Алексею Николаевичу удалось хорошо провести искусственное оплодотворение на плотах, после чего они с Хомяковым выращивали мальков в гостинице, остроумно приспособив для этого резиновые трубки и деревянные кадушки. В результате они получили огромный, превосходно зафиксированный материал, который Алексей Николаевич перевез к себе в деревню и стал обрабатывать с большим увлечением. Закончил он эту работу в Москве, где она была напечатана в Ученых записках Московского университета в 1895 г. под заглавием: «Развитие затылочной области низших позвоночных». В следующем году она появилась в немецком переводе в Бюллютенях Общества испытателей природы.
Это была первая работа Алексея Николаевича, завоевавшая себе прочное место в литературе и сразу обратившая на себя внимание специалистов: М. Фюрбрингер, один из крупнейших анатомов того времени, писал о ней, что «одно это исследование А. Н. Северцова дало для решения вопроса о Метамерии головы позвоночных больше, чем все исследования, произведенные до сих пор». Нечего и говорить о том, какое значение имела эта работа для самого Алексея Николаевича, она вплотную ввела его в методику детального филогенетического исследования и поставила его впервые перед лицом тех важнейших вопросов эволюционной теории, о которых до этого времени он знал только по книгам и над решением которых впоследствии работал всю жизнь.
Исследование метамерных образований затылочной области низших позвоночных, т. е. метамерных мускулов, нервов и скелета, очень много дало ему в смысле понимания явлений рекапитуляции. Так как в этом отделе головы наиболее рудиментарные сегменты передней части затылочной области соединены рядом последовательных переходов с вполне сформированными позвонками туловищного характера ее задней части, то перед Алексеем Николаевичем здесь как бы воочию восстанавливалось само течение эволюционного процесса. Для иллюстрации своих выводов он составил особые диаграммы, наглядно представляющие происходящие здесь изменения, и этот способ графического изображения теперь прочно вошел в литературу. Свою работу «О развитии затылочной области низших позвоночных» Алексей Николаевич представил в факультет в качестве магистерской диссертации. Его оппонентами на диспуте были М. А. Мензбир и А. А. Тихомиров. Диспут сошел хорошо, Алексей Николаевич получил искомую степень и вскоре после этого уехал в двухгодичную заграничную командировку, которую для него выхлопотал в министерстве народного просвещения М. А. Мензбир.
II
Защитой магистерской диссертации заканчиваются «ученические годы» Алексея Николаевича. Он едет за границу уже вполне сложившимся человеком, с установившейся, несмотря на его молодость (ему было только 30 лет), репутацией исследователя, с которым уже приходится серьезно считаться, с более или менее определившимся мировоззрением.
Каково же было это мировоззрение? Положительная наука, «Наука» с большой буквы, стояла в центре его миропонимания. Он считал, что наука не только определяет собою прогресс мировой техники, но коренным
БИОГРАФИЧЕСКИЙ ОЧЕРК
17
образом меняет и философское мировоззрение современного человека, внося постоянные поправки во все его старые взгляды, во все догматические утверждения философии и этики, причем поправки эти являются общеобязательными, как всякая научная истина. Такое отношение к науке у Алексея Николаевича отнюдь не было теоретическим. Уже в ту пору своей жизни он действительно жил ею, жил в самом простом и буквальном смысле этого слова. Его страстно интересовало и радовало всякое научное открытие, всякий успех положительного знания, и мысль его была постоянно настороже, отыскивая, что могло дать ему лично, для его текущей работы, то или иное открытие в области эволюционной биологии, ибо вопросы эволюции постоянно стояли в центре его внимания. Это особое, страстное отношение к науке вообще и к проблемам эволюции в частности характеризовало Алексея Николаевича не только в период его молодости. С годами оно утверждалось и крепло все более и более и достигло своего высшего расцвета в период его полной научной и философской зрелости.
За границу Алексей Николаевич стремился, чтобы познакомиться с жизнью западноевропейских лабораторий, окунуться в атмосферу новых научных школ, очутиться в центре столкновений новых научных гипотез. Морфология в то время переживала период бурного расцвета: Гегенбаур, Гексли и Геккель заканчивали свои классические работы; Ковалевский и Гатчек своими исследованиями над ланцетником, Ван-Бенеден и Купфер—над асцидиями, Семпер и Дорн—над аннелидами дали громадный материал для работ над вопросом о происхождении позвоночных. Проблемы метамерии головы, происхождения конечностей, происхождения органов чувств глубоко интересовали молодых ученых и оживленно дебатировались в лабораториях. Это было время расцвета неодарвинизма, когда Вейсман был в апогее своей славы, а работы Гельмгольца, М. Фер-ворна и Дюбуа Реймона властвовали над умами. Возникал неовитализм, появились первые труды Дриша и Франсе, и Алексей Николаевич сразу почувствовал себя их яростным противником.
Алексей Николаевич уехал за границу ранней весной 1896 г. Своим первым местопребыванием в Европе он избрал маленькую зоологическую станцию на границе Франции и Испании—Баньюль. Здесь он собирал материал по акулам, экскурсировал в море, знакомился с прибрежной фауной. Исследовательскую работу на станции ему вести не пришлось. Лишь однажды рыбак принес ему большого одноглазого ската. Проанато-мировав его, он написал о нем заметку, которую вставил в свой отчет о командировке.1 Случай этот интересен, конечно, не заметкой, но тем, что изучение строения мускулов и нервов на стороне, лишенной глаза, впервые навело его на мысль о громадном значении корреляции в эволюции, мысль, развить которую ему удалось лишь много лет спустя в целом ряде работ.
Из Баньюля Алексей Николаевич переехал на зоологическую станцию в Виллафранке. Здесь он продолжал собирать эмбрионов акул и скатов, но его главным приобретением был превосходный материал по гекконам, которых он собирал вместе сМ. М. Давыдовым, причем на долю каждого из них досталась полная серия в 400 эмбрионов.
По пучив телеграмму, извещавшую его о смерти маленькой дочери, Алексей Николаевич бросает работу и возвращается в Россию, ио через месяц уже снова едет в Мюнхен работать в лаборатории известного морфолога Купфера. Казалось бы, что для Алексея Николаевича самым естественным было поехать не к Купферу, но к знаменитому Гегенбауру Но этого именно он не сделал. Здесь сказалось его всегдашнее стремление
1 Отчет о заграничной кома-ндяровке магистра зоологии Алексея Северцова за I полугодие 1896 г. Уч. зап. оск. универ., вым. 14. 1898.
18
БИОГРАФИЧЕСКИЙ ОЧЕРК
к самостоятельности в работе. У Гегенбаура и в смысле выбора темы и в смысле ее обработки существовало определенное давление, т е. именно то, чего ему хотелось избежать. У Купфера этого не было, и, кроме того, у Купфера работал в качестве ассистента близкий друг Давыдова, А. А. Бём, у которого Алексею Николаевичу хотелось пройти специальную гистологическую технику. Вспоминая свою молодость и годы своего странствования по чужим землям, Алексей Николаевич часто упоминал о Бёме, к которому питал большое расположение, и в его рассказах, сквозь тонкий юмор веселых анекдотов (о Бёме их можно было рассказывать без конца), всегда просвечивало чувство глубокого уважения к Бёму, этому «страшно талантливому и страшно милому человеку».
Из приезжих в лаборатории Купфера, кроме Алексея Николаевича, работали А. Г. Гурвич и двое американцев: проф. Г. Нил и мисс Платт; последние, как и он, занимались исследованием метамерии головы. Купфер знал Алексея Николаевича, и ему хотелось, чтобы он взял у него работу на заданную им тему. Но Алексей Николаевич на это не согласился, отговариваясь тем, что ему хотелось бы предварительно пройти специальный курс гистологии у Бема. Это он и сделал, но параллельно и, так сказать, неофициально, предпринял самостоятельную работу по материалу, набранному в Виллафранке исследуя развитие хрящевого черепа акул.
В этой работе Алексей Николаевич детально сравнил развитие черепа у акул и у-осетровых, причем ему удалось установить наличие трабекул у селахий и существование эмбриональной коррелятивной связи, объясняемой гетерохронией, между закладкой головного мозга и закладкой черепа, развивающегося позднее мозга. При этом оказалось, что положение trabeculae cranii зависит от степени изгиба мозга. В то время это было настоящим открытием, вторым научным открытием Алексея Николаевича, прочно вошедшим в литературу.
Закончив работу, Алексей Николаевич написал о ней короткое предварительное сообщение1 и понес его к Купферу. Купфер, несмотря на то, что работа шла в разрез с теоретическими установками его школы, пришел от нее в восторг, но заявил, что необходимо исправить стиль немецкого текста и что за это он возьмется сам, несмотря на свою обремененность административной работой (он был тогда ректором университета). В благодарность за это Алексей Николаевич, уже в бытность свою профессором в Юрьеве, напечатал эту работу in extenso в юбилейном сборнике ко дню 70-летия проф. Купфера.
В Мюнхене Алексей Николаевич много работал, много читал, пользуясь его великолепными библиотеками, и много веселился. По вечерам бывал в театре и страстно увлекался Вагнером; по воскресным дням, в бесплатные часы, ходил в пинакотеки, в глиптотеку и в маленькую Шаховскую галлерею, где впервые познакомился с немецкой романтической живописью начала XIX в: со Швиндтом и Рётелем, с модными тогда Бёклином, Штуком и Клингером, и это его увлечение гармонировало с давнившим его увлечением немецкой романтической литературой того же времени: Гофманом, Тиком, Фуке, Брентано и Уландом.
Занимался он также и спортом: боксировал в лаборатории с Нилом, фехтовал со студентами. Смотрел вместе с Нилом студенческие мензуры, которые на них обоих произвели очень неприятное впечатление. Ходил в погребок пить пиво с ассистентами Купфера: Неймайером и Бёмом.
Весною 1897 г. Алексей Николаевич отправился вместе с Нилом в Неаполь. Здесь он собрал большой эмбриологический материал по акулам и электрическим скатам, которых тут же начал обрабатывать, а также и по миногам (в речке Сарно, впадающей недалеко от Неаполя в залив).
* Beitragzur Entwicklungsgeschichtedes Wirbeltierschadels. Anat. Anz. Bd. 13, 1897.
БИОГРАФИЧЕСКИЙ ОЧЕРК
19
На зоологической станции он встретился со многими интересными для него людьми: с некоторыми из них, например, с известным протистологом Дженингсом и с директором станции знаменитым А. Доном, он довольно близко сошелся; иные, например, известный неовиталист Дриш и его ярый последователь Гёрбст, произвели на него резко отрицательное впечатление. Познакомился он также с В. Геккером и П. Мейером; у последнего, как у большого специалиста этого дела, Алексей Николаевич учился консервированию материала и технике эмбриологического исследования.
Закончив рабочий сезон на станции, Алексей Николаевич направился в Киль, где встретился с Н. К. Кольцовым, приехавшим работать у Флемминга. У Флемминга Алексей Николаевич работал по цитологии, принявшись за изучение сперматогенеза Blatta germanica, а у Мёвеса, ассистента Флемминга, с которым он вскоре сдружился,—по гистологии органов. Несмотря на то, что занятия у Флемминга и Мёвеса далеко выходили за пределы его специальности, он очень ценил свое пребывание в Киле, ибо и его собственная работа и частые разговоры с Флеммингом и Мёвесом позволили ему непосредственно ознакомиться с тем материалом, который' кладется в основу гипотез о наследственности. Постоянный же просмотр их препаратов дал ему возможность судить об этих гипотезах из первых рук, а не только по литературным данным, чего он всегда старался избежать.
В Киле, как и в Мюнхене, Алексей Николаевич, параллельно с официальной работой в лаборатории, продолжал вести и самостоятельное, начатое им в Неаполе исследование по метамерии головы электрического ската. В этой работе он пытался разрешить одно из основных разногласий теории метамерии головы, а именно вопрос о первичном числе головных сомитов. Известно, что Бальфур и фан Вай признавали первичным лишь небольшое число головных сомитов, между тем как Киллиан и Дорн считали первичным очень большое число сомитов. Однако Алексей Николаевич смог лишь частично разобраться в этих противоречиях, ибо в то время он еще не пришел к мысли о существовании диплоспондильных сегментов в голове (самостоятельно развить и обработать эту мысль удалось лишь много лет спустя его ученику Б. С. Матвееву). Ошибкой этой работы, исправленной впоследствии Кольцовым в его монографии о метамерии головы миног, по признанию Алексея Николаевича, является то, что он отнес два передних сомита головы акул к висцеральной мускулатуре головы; ее большим «козырем» то, что ему удалось установить наличие замечательных корреляций между мозгом, глазом и глазными мускулами в голове у скатов.
Написав эту работу в Киле, Алексей Николаевич послал ее в Москву для напечатания в Ученых записках Московского университета.1 Когда печатание закончилось, он вернулся в Россию, чтобы защитить ее в качестве докторской диссертации. Защита, состоявшаяся в Москве в 1898 г., сошла прекрасно, причем оппонентами на диспуте снова были М. А. Мензбир и А. А. Тихомиров.
III
Алексей Николаевич вернулся в Москву с твердым намерением тотчас же приняться за обработку собранного им за границей материала. Но намерению этому не суждено было исполниться. В университете в это время началась постройка нового здания института сравнительной анатомии, сам же институт временно перебрался на небольшую квартиру, где зани
1 Очерки по развитию головы позвоночных. I. Метамерия головы электрического ската. Уч, зап. Моск, универе., вып. 15, 1898.
2*
БИОГРАФИЧЕСКИЙ ОЧЕРК
маться было и неудобно и очень тесно: Алексею Николаевичу фактически негде было работать. В это время М. А. Мензбир сообщил ему, что в Юрьеве учреждается параллельная кафедра по зоологии для русских студентов и предложил ему выставить свою кандидатуру. Отсутствие лаборатории и материальные затруднения заставили его согласиться на это предложение, хотя ему и очень не хотелось снова уезжать из Москвы так скоро после возвращения из-за границы.
В Юрьеве Алексей Николаевич был выбран на кафедру сплошь белыми шарами и в конце 1899 г. приступил к работе в качестве сверхштатного экстраординарного профессора зоологии.
Неприятной стороной пребывания в Юрьеве была проводившаяся там грубыми административными мерами обрусительная политика, вызвавшая со стороны местного населения глубокую ненависть к царскому правительству. В университете, где преподавание велось на немецком языке, эта политика выразилась в том, что туда стали принимать в неогра-ченном количестве и без всякого экзамена русских семинаристов, для которых и были приглашены русские профессора, читавшие им параллельные курсы на русском языке. Многие из этих профессоров были преподавателями-чиновниками, некоторые были ярыми консерваторами, но была среди них и прогрессивно-либеральная группа молодых ученых, в которую входили М. А. Дьяконов, Е. Г. Пассек, Ф. Ю. Левинсон-Лессинг, Н. А. Андрусов, Г. В. Хлопин, В. Э. Грабарь .К этой группе, естественно, примкнул и Алексей Николаевич, став довольно скоро одним из ее наиболее уважаемых и наиболее ценимых сочленов.
Положение Алексея Николаевича в Юрьевском университете на первых порах было не легкое. У него не было помещения для лаборатории, и ему пришлось ютиться в трех маленьких комнатах, предоставленных ему проф. Чермаком в институте гистологии; приходилось пользоваться его микроскопами для занятий со студентами и т. д. Первое время пребывания в Юрьеве у него и у его молодого ассистента М. М. Воскобойникова сплошь ушло на организацию лаборатории, на оборудование ее всем необходимым, на чтение самых разнообразных курсов. Алексею Николаевичу сразу же пришлось читать семь различных курсов, в том числе также зоологию, сравнительную анатомию и эмбриологию беспозвоночных. Не мудрено поэтому, что для исследовательской работы у него вначале не оставалось времени. Зато преподавание было поставлено прекрасно, студенты охотно посещали лекции и с увлечением работали на практических занятиях. В лаборатории появились первые ученики-специалисты.
Когда все было налажено, Алексей Николаевич с жаром принялся за научные исследования. Прежде всего он дополнил новыми наблюдениями сделанную им в Мюнхене работу над черепом селахий и отослал ее в сборник, посвященный Купферу.1 Не замеченная в качестве предварительного сообщения, здесь эта работа обратила на себя большое внимание и с тех пор постоянно цитируется в специальной литературе.
Затем Алексей Николаевич предпринял исследование над хрящевым черепом гекконов по материалу, набранному в Виллафранкс, но об этой работе им было написано лишь короткое предварительное сообщение,1 2 так как, получив от акад. В. В. Заленского редкий материал по австралийскому цератоду, он увлекся исследованием его черепа, зубов и мускулатуры.3 Эта работа много ему дала для понимания строения черепа низших телеостом и оказалась весьма ценной для его последующих обобщений по филогенезу рыб. Не успел он ее закончить, как принялся за новое
1 Die Entwicklung des Selachierschadels. Festschr. f. C. v. Kupffer. Jena, 1899.
2 Zur Entwicklungsgeschichte von Ascalabotes fascicularis. Anat. Anz., Bd. 18, 1900.
» Zur Entwicklungsgeschichte des Ceratodus forsteri. Anat. Anz., Bd. 21, 1902.
БИОГРАФИЧЕСКИЙ ОЧЕРК
21
большое исследование, посвященное развитию скелета, мускулов и нервов конечностей рептилий, но эту работу ему удалось завершить лишь несколько лет спустя, уже в Киеве.
Одновременно стала налаживаться и исследовательская работа в лаборатории Алексея Николаевича. Стал самостоятельно работать М. М. Воскобойников, которому Алексей Николаевич посоветовал заняться вопросом о развитии черепа и висцеральной мускулатуры у костистых рыб; стал работать над развитием присосок на лапках геккона и другой его ученик, Куницкий. Его работа интересна тем, что является первой работой, вышедшей из лаборатории Алексея Николаевича, сделанной в духе его будущих исследований над развитием приспособлений у специализован-ных форм.
В Юрьеве Алексей Николаевич занимался не только наукой и преподаванием: его заинтересовала и общественная деятельность. Он стал впервые читать публичные лекции: объявил курс по дарвинизму, к которому усердно готовился и который имел большой успех, если судить по тем «овациям», которые ему устраивали по старому дерптскому обычаю. «Овации» эти заключались в том, что вдоль всей лестницы, ведущей от актового зала, где происходили лекции, к выходу выстраивалась публика и неистово аплодировала проходившему сквозь ее ряды лектору.
В Юрьеве Алексей Николаевич прожил около четырех лет. Вероятно-это были лучшие годы его жизни: он был еще молод, был вполне здоров и очень жизнерадостен, и в этом маленьком иноземном городке сумел устроить для себя живую, полную разнообразных интересов жизнь. Его научно-исследовательская работа,—суть и смысл этой жизни,—шла хорошо; появились первые ученики; радостно возбуждали первые успехи как преподавателя и лектора. Число друзей его, из которых многие впоследствии стали выдающимися русскими учеными, возрастало с каждым годом; семейная жизнь шла счастливо; чувство бодрости и ощущение физической силы, развиваемое постоянными физическими упражнениями, все крепло.
IV
В 1902 г. Алексей Николаевич переходит в Киевский университет в качестве заместителя проф. Н. В. Бобрецкого, вышедшего в отставку. С отъездом из Юрьева кончается молодость Алексея Николаевича. Киевский период и первые годы московского периода его жизни уже являются годами полной его зрелости: физической, научной и философской.
Киевский университет в те годы был типичным провинциальным университетом большого губернского города. Среди его профессуры были крупные научные силы, особенно на физико-математическом факультете: тот же Бобрецкий, С. Г. Навашин, уже тогда европейски известный ученый, А. А. Коротнев, основатель Виллафранкской зоологической станции. Прекрасными преподавателями были А. Н. Реформатский и И. И. Ко-соногов, авторы известных учебников по химии и физике. Не менее крупные силы подвизались и на историко-филологическом факультете. Очень многие профессора были ярыми консерваторами, но существовала и небольшая группа «либералов», куда вошел Алексей Николаевич. Между обеими группами профессуры шла открытая борьба, обычно заканчивавшаяся победой консерваторов, бойких карьеристов, задававших общий тон университетской жизни.
В Киеве Алексей Николаевич прежде всего занялся устройством лаборатории, которая была в большом забросе. Научных работ в ней не велось, а делами лаборатории и ее хозяйством ведали по преимуществу служителя. С помощью М. М. Воскобойникова, которого он пригласил к себе в качестве старшего ассистента из Юрьева, Алексей Николае
БИОГРАФИЧЕСКИЙ ОЧЕРК
вич быстро навел порядок, реорганизовал лабораторию, наладил преподавание и научно-исследовательскую работу. Вскоре у него появились многочисленные ученики, быстро заполнившие свободные места не только в предназначенных для них комнатах, но и в лабораторных залах, где велись практические занятия со студентами, и даже в музее, расположенном рядом с лабораторией. Таким образом, если в Юрьеве были лишь единичные ученики, то в Киеве уже образовалась настоящая школа работавших в определенном направлении специалистов.
Сам Алексей Николаевич при первой возможности возобновляет свою прерванную переездом и организацией лаборатории научно-исследовательскую работу. Прежде всего он заканчивает начатую им в Юрьеве большую монографию (430 стр.) по развитию скелета, мускулов и нервов пятипалой конечности рептилий. Материалом для нее послужили набранные им в Виллафранке зародыши гекконов и других ящериц, зародыши сепса и черепахи, а также некоторые современные и ископаемые амфибии. Работа эта, восстанавливающая ход филогенеза пятипалой конечности, ценна не только своим фактическим исследованием, тщательно и детально продуманным и выполненным, но также и своими выводами: Алексей Николаевич создает здесь свою теорию происхождения пятипалой конечности наземных позвоночных из семи-десятилучевой конечности предков, образовавшейся в свою очередь из многолучевого плавника рыбообразного существа. В настоящее время она прочно вошла в научную литературу и считается общепринятой теорией происхождения пятипалой конечности.
Монография эта, предварительное сообщение о которой появилось еще в 1904 г.,1 вышла из печати только в 1908 г.:1 2 это случилось потому, что работа над нею была прервана тяжелою болезнью Алексея Николаевича, уложившею его на долгое время в постель. Плохо поддававшаяся лечению ангина Ludovisii осложнилась острым нефритом, последствием же незалеченного нефрита был склероз почек—источник тяжелых страданий в последние годы его жизни, приведший Алексея Николаевича к смерти.
Еще не вполне закончив писание монографии, Алексей Николаевич взялся за новое, еще более широкое исследование, а именно за изучение развития головы круглоротых. Эта работа, сделанная вчерне в Киеве, была закончена и напечатана гораздо позднее, уже после переезда в Москву. В Киеве же он сделал еще одну специальную работу, а именно исследование о нервах, иннервирующих жаберные дуги рыб.3 * s
Все эти исследования, точно так же как и объединенные одним общим планом исследования его тогдашних учеников, подвели Алексея Николаевича вплотную к проблеме эволюции низших позвоночных, а также к вопросу о методике филогенетического исследования, в частности— к вопросу о биогенетическом законе.
Что заставило его остановиться на этих именно проблемах? По его .словам—его всегдашняя заинтересованность общими законами эволюционного процесса животного мира и уверенность, что эти законы могут быть найдены только двумя путями: путем экспериментальных исследований и путем исследования эволюции больших групп животного царства. Алексей Николаевич останавливается на этом последнем способе и принимается вместе со своими учениками за детальную филогенетическую разработку наименее изученной группы среди позвоночных, большой
1 Die Entwicklung der pentadaktylen Extremitat der Wirbeitiere. Anat. Anz. fid. 25, 1904.
2 Studien fiber die Entwicklung der Muskeln, Ncrvcn und des Skeletts der Extremi-
taten der niederen Tetrapoda. Beitrage zu einer Theorie der pentadaktylen Extremitaten
•der Wirbeitiere. Бюлл. Моск. общ. испыт. прир., 1908.
s Die Kiemenbogennerven der Fische. Anat. Anz. Bd. 38, 1911,
БИОГРАФИЧЕСКИЙ ОЧЕРК
23
группы рыб и амфибий, где так мало было сделано и так много оставалось еще темного и непонятного. Ему казалось, что если бы удалось свести результаты предпринятых им исследований над низшими позвоночными с результатами новейших американских и европейских исследований над рептилиями, птицами и млекопитающими, то это дало бы возможность в будущем сделать ряд выводов относительно общих законов эволюционного процесса, тех выводов, к которым пока, за неимением достаточно обширного материала, обработанного с филогенетической точки зрения, он подойти еще не решался.
Но здесь перед ним с особенной остротой встал вопро^. о методике филогенетического исследования. Если при изучении эволюции групп животных, имеющих хорошо развитый костный скелет, всегда можно было пользоваться палеонтологическими данными, то при изучении эволюции предков низших позвоночных с недостаточно развитым костным скелетом для Алексея Николаевича отпадал, или почти отпадал, этот наиболее достоверный метод филогенетического исследования, ибо от эпохи, когда происходила эволюция этих низших форм, почти не осталось палеонтологических следов. Таким образом, в своем исследовании ему пришлось довольствоваться комбинацией двух оставшихся методов: сравнительно анатомического и сравнительно эмбриологического, и было совершенно естественно проверить их методологическую ценность. Это было тем более уместно, что «биогенетический закон», основа сравнительно эмбриологического метода, в то время подвергся резким нападкам со стороны О. Гертвига, Кейбеля, Оппеля, Гарбовского и др., указывавших, что при применении его исследователь наталкивается на слишком большие трудности и что, опираясь на него, можно в лучшем случае притти к построению лишь весьма гипотетических схем.
Алексей Николаевич прерывает поэтому свои исследования над эволюцией низших позвоночных и принимается за самую тщательную проверку теории рекапитуляции. Это привело его к изменению биогенетического закона и к установлению нового понятия «филэмбриогенеза». Результаты этого своего методологического исследования он доложил Киевскому обществу естествоиспытателей еще в 1907 г., а затем, более подробно, изложил их в блестящей речи, произнесенной на общем собрании XII съезда естествоиспытателей и врачей в Москве в 1910 г.1 Этой речью Алексей Николаевич кладет основы одной из своих самых важных работ,—теории филэмбриогенеза.
С теорией филэмбриогенеза произошла своеобразная вещь: ее основное положение, гласящее, что «филогенетические изменения строения органов взрослых животных обусловлены изменением хода их онтогенеза», почти одновременно с Алексеем Николаевичем было опубликовано Седж-виком, но оба они тогда не подозревали о том, что эта мысль уже была высказана за сорок слишком лет до них Ф. Мюллером в его небольшой книжке «Fiir Darwin» (1866). Таким образом, с Ф. Мюллером случилось то же, что и с Менделем, известные законы которого были вновь пере-открыты Корренсом и де Фризом: Алексей Николаевич и Седжвик сыграли ту же роль по отношению к Мюллеру. Но Седжвик, однажды высказав эту мысль, к ней больше не возвращался, Алексей же Николаевич дальнейшей ее разработкой был занят всю жизнь: более или менее завершенной мы находим ее только в книге, ныне предлагаемой вниманию читателей.
Основные мысли, высказанные в этих двух докладах, Алексей Николаевич подробно развил в книге «Этюды по теории эволюции. Индивидуальное развитие и эволюция», вышедшей в Киеве в 1912 г. и переизданной в Бер
1 Эволюция и эмбриология. Москва, 1910.
24
БИОГРАФИЧЕСКИЙ ОЧЕРК
лине Госиздатом в 1922 г. Здесь, кроме детальной критики биогенетического закона, мы уже находим, как об этом будет более подробно сказано ниже, и первые наброски морфобиологической теории эволюции, и достаточно разработанную теорию корреляции, и детально разобранные основные положения теории филэмбриогенеза. Здесь же Алексей Николаевич подвергает глубокому анализу вопрос о ценогенезах и о рекапитуляции ценогенетических признаков, классифицируя последние по различным категориям.
Время пребывания Алексея Николаевича в Киеве было временем полного рассвета его лекторского таланта и сильнейшего увлечения педагогической деятельностью. Лекции свои в университете и на высших женских курсах он читал превосходно: умно и строго, и подкупающе искренне. Слова его были вески, полны убеждения, образы—незабываемо ярки. Особенно любил он читать о Дарвине. Он широко использовал его биографию и «Переписку с друзьями», и образ Дарвина живо вставал перед слушателями—образ большого ученого, простого, мудрого, неотразимо обаятельного человека. Еще лучше удавались ему публичные лекции, которые собирали такое количество народу, что администрация университета (эти лекции происходили в актовом зале) обычно вызывала полицию для «наведения порядка». В такой обстановке было трудно читать, но она не могла и не влиять на лектора особенным образом: и действительно, возбужденный жадным вниманием слушателей, ожидая каждую минуту, что лекцию придется прервать, Алексей Николаевич читал особенно хорошо: с подъемом и подлинным вдохновением. И необыкновенно художественно, просто. Как стеклянные бусы, нанизывались один за другим интересные, яркие, неожиданно новые факты, и блестящею нитью тянулась сквозь них руководящая мысль, сквозила так ясно, так неопровержимо убедительно, что необходимый вывод естественно возникал в уме слушателя еще задолго до того, как к нему подходил сам лектор.
На свою преподавательскую деятельность и особенно на свои публичные выступления, являвшиеся горячей пропагандой материалистического миропонимания и дарвинизма, Алексей Николаевич всегда смотрел как на свою прямую общественную обязанность. Что это было действительно так, видно из появившейся уже после отъезда Алексея Николаевича из Киева брошюры-памфлета, написанной по поводу его публичных лекций профессором богословия Киевского университета,1 в которой автор горько сетует и негодует на те большие «огорчения и хлопоты», которые доставлял ему в свое время своими «шарлатанскими лекциями» Алексей Николаевич. Яростно боролся он против него, и имел успех; когда в 1909 г. всем ученым миром праздновалось в Кембридже 50-летие со дня выхода в свет «Происхождения видов» Дарвина, Киевский университет, следуя его воздействию, «по мотивам научным (!), религиозным и общественным (!)» отказался от участия в праздновании Дарвиновского юбилея и отменил командировку Алексея Николаевича, как руководящего дарвиниста и представителя этого университета, в Англию.
Самый популярный из киевских профессоров, Алексей Николаевич на экзаменах был неумолимо строг и грозен, и до такой степени «страшен», что многие студенты, даже не плохо знавшие предмет, иногда совсем отказывались от экзамена. Большинство стремилось попасть к ассистентам. Зато какая марка была получить у Северцова «весьма»! До некоторой степени это предопределяло научную карьеру экзаменующегося.
1 П. Я. Светлов. Перед разбитым кумиром (по поводу просветительной деятельности «седящих во тьме» дарвинистов). С. Петербург, 1912.
БИОГРАФИЧЕСКИЙ ОЧЕРК
Еще большего спрашивал Алексей Николаевич со своих непосредственных учеников. Беспощадно строгий по отношению к самому себе, требуя от себя во всяком деле достижения максимально возможного для него совершенства, Алексей Николаевич был неумолимо требователен и по отношению к этим ученикам-специалистам, постоянно добиваясь от них «настоящей работы». И несмотря на это, как любила его всегда молодежь! Молодежь чутка и прекрасно разглядела, что перед нею друг, что за его небрежной манерой говорить, за беспощадно острой, уничтожающей шуткой, за угрюмо-недоступным видом сурового профессора кроется такое понимание, такое к ней доверие и уважение, такое вдумчивое, чуткое отношение к человеку, какое не часто встречается в жизни.
Несчастная русско-японская война, жестокое подавление революции 1905 г. царским правительством и наступившая после этого тяжелая реакция, конечно, не могли не отразиться на Алексее Николаевиче, болезненно и напряженно переживавшем вместе со всей передовой интеллигенцией то, что творилось вокруг. Лето 1905 г. он, как всегда, проводил у себя в Воронежской губ. Освободительное крестьянское движение, разразившееся со стихийной силой, выразилось там главным образом в разгроме и поджоге помещичьих усадеб. Но пожары, свирепствующие в округе, не коснулись усадьбы Алексея Николаевича, и это лучше всяких слов говорило об отношении к нему местных крестьян. Когда же наступила реакция и повсюду свирепствовали карательные экспедиции, Алексей Николаевич с ужасом и отвращением отшатнулся от них и, созвав крестьян, объявил им, чтобы они были спокойны, что этих экспедиций он к себе никакой ценою не допустит. И нечего говорить о том, как крепко и честно сдержал он свое слово. Более того, угнетенный впечатлением, карательных экспедиций и не желая более владеть землей, он тут же на очень льготных условиях продал свое имение крестьянам, оставив за собой только дом, построенный его отцом, и сад—дачу, где он с семьей ежегодно проводил летние каникулы.
В 1910 г. М. А. Мензбир предложил Алексею Николаевичу перейти в Московский университет. Как ни хорошо жилось ему в Киеве, но перспектива вернуться на кафедру в свой родной университет так улыбалась. Алексею Николаевичу, что он сразу дал свое согласие на баллотировку Выборы в совете университета и в факультете прошли блестяще, но он не мог среди года бросить свою лабораторию и оставить в университете и на курсах свои кафедры незамещенными, поэтому переехать в Москву ему удалось только через год после избрания.
Тем временем произошел известный разгром Московского университета министром Кассо, вызвавший уход в отставку ряда лучших профессоров. Когда Алексей Николаевич переехал, наконец, в Москву, последствия этого разгрома уже вполне сказались: научная жизнь лабораторий замерла, лекции читались с перебоями, практические занятия со студентами шли кое-как.
Из Киева Алексей Николаевич пригласил к себе в качестве ассистентов И. И. Шмальгаузена и М. Е. Макушка. В Москве к ним присоединились. С. Н. Боголюбский и Д. П. Филатов. Очень скоро появились и первые ученики, составившие впоследствии ядро «московской школы Северцова»: Б. С. Матвеев, В. В. Васнецов, Е. В. Рылкова, С. Г. Крыжановский, А. И. Кривецкий, А. А. Машковцев, С. А. Северцов; через некоторое время, по мере окончания университета, к ним присоединяются С. В. Емельянов, А. Н. Дружинин, Н. Н. и О. Н. Дислер, А. Г. Рындзюнский, Е. Ф. Еремеева и целый ряд лиц, имена которых перечислены ниже, в списке работ-учеников Алексея Николаевича.
БИОГРАФИЧЕСКИЙ ОЧЕРК
Все они, точно так же, как и сам Алексей Николаевич, по определенному, заранее намеченному им плану принялись за систематическое ’изучение большой группы низших позвоночных, детально исследуя историю развития различных систем органов у систематически близких форм. Работы эти можно рассматривать как подготовительные исследования, чтобы иметь возможность перейти от воссоздания филогенеза отдельных органов и систем органов к воссозданию филогенеза целых организмов, видов и родов, а затем и всей группы низших позвоночных. Сам Алексей Николаевич прежде всего возобновляет начатое им в Киеве обширное исследование о развитии головы круглоротых, разрабатывая в первую очередь вопрос о развитии черепа, висцеральной мускулатуры, висцерального скелета, жаберных нервов и жаберных мешков у миног и- отчасти у миксин.1 Эту работу он лично считал одной из своих лучших работ, и ее теоретическим выводам придавал (для себя) большое значение. Впоследствии он положил эти выводы в основу своей монографии об организации предков современных позвоночных,1 2 в которой ему удалось детально реконструировать не только строение целого ряда систем органов гипотетических предков круглоротых и акуловых рыб, но и определить их функциональное значение, а также восстановить внешнюю форму и образ жизни этих примордиальных гипотетических форм.
Работа эта страстно увлекает его, но дается ему не легко. Занятый педагогическими и общественными делами и другими очередными работами, Алексей Николаевич со свойственным ему упорством снова и снова возвращается к ней и постоянно переделывает ее. Интересно отметить, что некоторые из гипотетических предположений, сделанные им здесь на основании сравнительно эмбриологических и сравнительно анатомических исследований, много лет спустя были подтверждены новейшими палеонтологическими находками. Большую радость однажды доставил Алексею Николаевичу Стеншиб, когда в 1927 г. показал ему в Стокгольме на одном из экземпляров своих шпицбергенских остракодерм те самые пребранхиаль-ные щели, расположенные на том самом месте, где это было предопределено Алексеем Николаевичем на основании его работ 1915—1917 гг. О подобных же палеонтологических подтверждениях теоретически восстановленных •им признаков говорили ему на международных конгрессах в Веймаре .(1925) и Падуе (1930) палеонтологи Иекель и Паттен.
Занимаясь все это время специальными исследованиями, Алексей Николаевич все острее и острее чувствовал потребность в более широких эволюционных обобщениях; однако, он не давал себе воли в этом отношении: он сознавал, что для этого у него все еще не имеется достаточно обильного и достаточно проверенного фактического материала.- И' вот он затевает (и выполняет) все новые и новые специальные исследования, кропотливо пополняя пробелы; засаживает своих учеников за новые и новые темы, дающие ему материал для разрешения того или иного, попутно возникшего в его исследовании вопроса. Так, он исследует конечности хамелеона, и, хотя эта работа и носит, по его словам, эпизодический характер, он находит в ней много новых данных, поясняющих пути своеобразной адаптации этих конечностей.3 Он работает над жабрами и жаберными сосудами рыб4 и много времени и труда уделяет вопросу о развитии и преобразовании висцерального аппарата у элас-мобранхий.
1 Исследование об эволюции низших позвоночных. I. Морфология скелета и мус-’кулатуры головы циклостом. Русск. Арх. анат., гист. и эмбр., т. I, 1916.
2 Исследование об эволюции низших позвоночных. II. Организация ближайших предков современных позвоночных. Русск. арх. анат., гист. и эмбр., т. 2, 1917.
а Развитие конечностей Chamaeleo bilineatus. Зоол. вести., т. 1. 1916.
4 Развитие жабр и жаберных сосудов рыб. Русск. зоол. журн., т. 2, 1917.
БИОГРАФИЧЕСКИЙ ОЧЕРК
27
Но стремление к обобщениям становится все сильнее и сильнее, и Алексей Николаевич, наконец, разрешает себе свободно работать над общими проблемами филогенеза. Что же он делает в этом направлении эти годы? Прежде всего он много работает над вопросом о происхождении позвоночных, много пишет по этому поводу, но не опубликовывает написанного. Единственный след, который мы находим в печати об этой большой работе—это доклад, прочитанный им на эту тему на общем собрании XIII Съезда естествоиспытателей и врачей в Тифлисе в 1913 г. Зато другим вопросам эволюционной теории., а именно вопросам о корреляции, о параллелизме, о дивергентной и конвергентной эволюции, о прогрессе и регрессе, о монофилии и полифилии он посвящает целую книжку.1 Наконец, он снова возвращается к вопросу о факторах продолжительности жизни, разработку которого он начал еще в Киеве, и об этой работе печатает предварительное сообщение 1 2 в незадолго до этого основанном им совместно с В. С. Елпатьевским «Русском зоологическом, журнале».
Наступившая империалистическая война застала Алексея Николаевича в Швейцарии. Больной, с большими трудностями, на палубе 3-го класса возвращался он через Дарданеллы на родину. Война наложила длинную черную тень на жизнь Алексея Николаевича. Мучительно переживал он развертывавшиеся события. К тому же младшему сыну его пришлось итти на войну, и с тех пор каждый день его жизни был отравлен тревогой за жизнь любимого сына. Стареет и быстро слабеет его мать, с каждым месяцем катастрофически ухудшается здоровье жены. Его личная жизнь становится невыносимо тяжела. В довершение всего в самом начале 1917 г. три смерти близких людей одна за другой поражают Алексея Николаевича. умирает друг его детства, проф. П. С. Усов, вслед за ним умирает жена, и, наконец, любимый сын.
VI
Алексею Николаевичу шел 52-й год, когда наступила великая Октябрьская социалистическая революция. В таком возрасте люди его склада редко внезапно меняют привычный строй мысли. Долго и напряженно, тщательно и честно продумывал Алексей Николаевич совершавшиеся события, и прошло не мало времени прежде, чем он по-настоящему осмыслил и воспринял Октябрьскую революцию, но, однажды приняв, он уже не изменил своей точки зрения до конца жизни.
Что более всего повлияло на него в этом отношении? Влияло многое, но больше всего единственный в истории беспримерно быстрый расцвет науки у нас в Союзе. С каким удовлетворением отмечал он, бывало, вновь воздвигающиеся научные институты, неудержимо растущие исследовательские кадры, нарождающиеся научные школы. С какой гордостью следил за успехами своих учеников, создающих свои собственные научные школы в различных уголках нашей обширной родины. Но главным толчком в этом направлении была поразившая его на первых же его публичных выступлениях, на первых же популярных лекциях, прочтенных им красногвардейцам и железнодорожным рабочим, та небывалая, та страстная жажда знания, что пробудились в народе. Он не мог не видеть и не учесть того, какие огромные усилия, несмотря на разруху, на гражданскую войну, на холод и на голод, клались у нас на то, чтобы эту жажду удовлетворить как можно лучше и как можно полнее, а что больше этого говорит уму и сердцу настоящего ученого?
1 Современные задачи эволюционной теории. Москва, 1914.
2 О факторах, определяющих продолжительность жизни многоклеточных живот-: ных. Русск. зоол. жури., т. 2, 1917,
БИОГРАФИЧЕСКИЙ очерк
В начале Октябрьской революции, в годы гражданской войны, разрухи и голода, Алексею Николаевичу материально жилось очень плохо, хуже, чем многим его друзьями знакомым, ибо он был житейски абсолютно непрактичным человеком, хотя и искренне обижался, когда кто-либо позволял себе усомниться в его практичности. Поэтому голодал он честно и порою сильно страдал от холода. Иногда от холода у него болели и опухали пальцы, и это мешало микроскопированию и зарисовке препаратов. Он никогда не терял спокойствия, никогда не жаловался на это. И ни на один день не прекращал работы. Когда пересматриваешь список его трудов и останавливаешься на исследованиях, сделанных за эти годы, невольно поражаешься и тому, как мог один человек сделать так много за такое относительно короткое время, и тому, как цельна и целеустремленна была его научная работа.
Прежде всего он взялся за продолжение своих специальных исследований над развитием висцерального скелета акуловых рыб, причем в качестве объекта для сравнения взял скафиринха и химеру. Открытие у селахий рудиментов двух преджаберных щелей, которым он придавал большое теоретическое значение,—так как благодаря этому для него осветился вопрос о нервах и мускулах передней части головы,—и детальное изучение развития жаберных дуг навели его на мысль, что жаберный аппарат, состоявший у предков гнатостом из еще нерасчлененных дужек, доходил у них до самого рта; на следующей стадии филогенеза шло расчленение этих дужек, одинаковое как в бранхиальной, так и в пребран-хиальной части жаберного аппарата, еще функционировавшей в качестве органа дыхания; в течение последующей эволюции произошло преобразование передних висцеральных дуг в ротовой аппарат хватательного типа. Работу эту, сделанную в течение 1917—1919 гг., Алексей Николаевич заканчивает печатанием лишь в 1925 г.1 (в 1927 г. она выходит в переводе на французский язык в Неаполе) и этим завершает свою трехтом-иую монографию об «Эволюции низших позвоночных».
Это исследование показало Алексею Николаевичу, какое огромное значение с точки зрения развития функций дыхания имеет висцеральный аппарат в эволюции позвоночных. Поэтому в своей следующей работе он принимается за детальное исследование дыхательного аппарата, т. е.жабр и жаберных сосудов, и попутно блестяще разрешает знаменитый парадокс А. Гетте. В результате ему удалось восстановить исходную форму жабр челюстных позвоночных. Эта работа, сделанная позднее предыдущей, была, однако, напечатана раньше ее, а именно в 1922 г.1 2'
Следующая работа Алексея Николаевича посвящена исследованию развития осевого черепа, висцерального скелета и мышц Acipenser ru-thenus и Amia calva; в качестве материала для сравнения он исследовал развитие черепа у Lepidosteus и Polypterus. Здесь ему удалось показать, что по характеру своего эмбрионального развития и по строению черепа хрящевые ганоиды стоят очень близко к низшим акулам и что Holostei и Crossopterygii, будучи в этом отношении весьма близки друг к другу, сильно отличаются от Chondrostei. Мысль эта легла в основу его позднейшей работы относительно положения хрящевых ганиодов в системе, вышедшей сначала на русском, а затем в Америке на английском языке.3 Впоследствии Алексей Николаевич еще раз проверил это положение на более
1 Исследование об эволюции низших позвоночных. III. Первичное строение висцерального скелета эласмобранхий. Русск. арх. анат., гист. и эмбр., т. 3, 1925.
2 Die Entwicklung der Kiemen und Kiemenbogengefasse der Fische. Zschr. f. wiss. Zool. Bd. 121, 1922.
3 Положение хрящевых гаиоидов в системе. Русск. зоол. журп., т. 3, 1922. The place of the cartilaginous Ganoids in the system and the Evolution of the Osteichthyes. Journ. of Morph., v. 38. 1923.
БИОГРАФИЧЕСКИЙ ОЧЕРК
обширном материале. Он переисследовал хрящевой череп, висцеральный скелет и мускулы стерляди и, сравнив их развитие с развитием тех же органов у Scymnus, Heptanchus и Chlamydoselachus, с одной стороны, и Amia, Lepidosteus, Polypterus и Palaeoniscidae—с другой, целиком подтвердил свои прежние выводы. Эта последняя работа напечатана в Швеции в 1928 г.1
Все эти исследования, равно как и последующие работы над костным черепом, позволили Алексею Николаевичу построить свое первое родословное древо низших позвоночных. Своей неполнотой оно отличается от того древа, которое мы находим в настоящей книге, но вместе с тем в нем заметен шаг вперед по сравнению с тем, что мы видим у других авторов: здесь между низшими группами позвоночных уже восстановлен ряд промежуточных звеньев, детально охарактеризованных с морфологической и биологической сторон.1 2
Далее следует цикл работ, посвященных вопросу о покровном скелете, которому Алексей Николаевич придавал большое теоретическое значение. Начав со сравнительно морфологического исследования костных чешуй осетровых и установив сходство в развитии покровных костей черепа и туловищных чешуй стерляди, Алексей Николаевич определил исходный тип анцестральных чешуй как чешуй, имевших ромбическую форму и медианный гребень на поверхности. Эта работа много значила для Алексея Николаевича, она оказалась отправной точкой, исходя из которой ему впоследствии действительно удалось перестроить всю теорию костного черепа. Начатая еще в 1918—1919 гг., она увидела свет лишь в 1926 г.3
Расширяя это исследование, Алексей Николаевич в своей следующей работе приходит к выводу, что в эволюции предков хрящевых ганоидов была стадия, когда их голова и туловище были сплошь покрыты такими ромбическими чешуями с гребнем. В течение дальнейшей эволюции произошла дифференциация этих чешуй, различная в туловище и в голове, причем в голове предков хрящевых ганоидов число покровных костей было значительно больше, чем у современных форм (срастание). Закончил и напечатал эту работу он в том же 1926 г.4
Через некоторое время, получив материал по. кистеперым, которого он тщетно добивался несколько лет, Алексей Николаевич исследует сначала чешуи и своеобразный спинной плавник,5 а затем череп6 Polypterus и приходит к выводу, что данные, полученные им относительно костного черепа селахий, распространяются и на кистеперых. Наконец он принимает решение еще раз проверить свои выводы на костных ганои-дах и детально исследует в этом отношении Amia calva7; результаты этой работы еще раз подтверждают правильность его предположений. И только после всего этого, сопоставляя все полученные им результаты, Алексей Николаевич в последней из работ этого цикла8 окончательно завершает построение теории костного черепа и приходит к выводу, что у отдаленных предков Chondrostei, Holostei, Teleostei и Crossopterygii, т. е. у Osteichthyes
1 The head skeleton and muscles of Acipenser ruthenus. Acta zool., 1928.
2 См. родословное древо в «Главных направлениях эволюционного процесса». Москва, 1925.
3 The development of the scales of Acipenser ruthenus. Journ. of Morph., v. 42, 1926.
4 Studies on the bony skull of fishes. I. Structure and development of the bony skull of Acipenser ruthenus. II. Primary structure of the bony skull of fishes. Quart. Journ. of Mier Scien., v. 70, 1926.
5 The development of the dorsal fin of Polypterus delhesi. Journ. of Morph., v. 38, 1924.
6 Zur Morphologic des Schadels von Polypterus delhesi. Anat. Anz., Bd 59, 1924/1925,
’ Развитие покровных костей черепа Amia calva. Бюлл. испыт. прир. 1925.
8 Beitrage zu einer Theorie des knochernen Schadels der Wirbeltiere. Palaeont. Ztschr., Bd. 8, 1926.
30
БИОГРАФИЧЕСКИЙ ОЧЕРК
primitivi, череп был построен по одному общему типу: он весь состоял из большого числа ромбических чешуи с продольными гребнями на поверхности.
Едва закончив этот ряд работ, Алексей Николаевич приступает к исследованию нового вопроса,—вопроса, которым он интересовался уже давно, а именно проблемы происхождения наземных позвоночных. Это исследование он, естественно, начинает с изучения эволюции парных плавников: сначала он исследует развитие брюшных плавников стерляди,1 а затем—развитие грудных плавников различных рыб.1 2 Первая из этих работ показала ему, что развитие плавников идет не в направлении распадения, как это принято было думать, но в направлении срастания первоначально раздельных элементов скелета и что в исходной своей форме плавник представляет собой образование метамерное. Во второй работе Алексей Николаевич установил, что такой метамерный плавник переходит в течение эволюции в унисериальный, а не бисериальный архиптеригий, и остается таким у селахий и у хрящевых и костных ганоидов. При переходе предков кистеперых и двоякодышащих от нектонного к бентонному образу жизни, сопровождавшемуся прогрессивным развитием метаитери-гоидной впадины, развился бисериальный тип плавника этих рыб, а из этого последнего в течение дальнейшей эволюции, а именно при переходе низших позвоночных из воды на сушу, образовались конечности наземных позвоночных. Конечные выводы из этих исследований, бросающих свет на происхождение наземных позвоночных, изложены им в небольшой статье, появившейся в печати тоже в 1926 г.3
Накопив путем собственных исследований и исследований своих учеников достаточный фактический материал, Алексей Николаевич почувствовал, что теперь он снова имеет право перейти к той работе обобщения, в которой полагал весь смысл своей деятельности как ученого и от которой сознательно воздерживался столько лет, хотя постоянное обуздывание себя в этом отношении ему не легко давалось. За этот период он печатает три теоретических работы,4 явившиеся в известной мере подготовительными работами к книге «Морфологические закономерности эволюции». Так как я имею в виду ниже подробно рассмотреть историю возникновения и создания этой книги, то здесь я на них останавливаться не стану. Отмечу только, что в «Эволюции и психике» Алексей Николаевич впервые говорит о «полиморфизме функций» (впоследствии—это «мультифункциональность органов»), которому он придавал большое теоретическое значение, полагая, что всякий орган имеет не одну, но много функций и что от развития той или иной из этих функций зависит направление его последующей эволюции. Здесь же он рассматривает и вопрос о роли ненаследственной деятельности «разумного типа» в прогрессивной эволюции высших млекопитающих и человека, подчеркивая, что роль этого фактора биологически до сих пор еще не была достаточно учтена. В то время как рефлекс и инстинкт являются приспособлением к медленным изменениям внешней среды, психическая деятельность «разумного типа» является ответом организма на быстрые, но незначительные изменения внешней среды. В этих случаях высоко организованные животные отвечают на изменения среды не медленным изменением своей организации, но быстрым изменением своего поведе
1 Development of the pelvic fins of Acipenser ruthenus. New data for the theory of the paired fins of fishes. Jottrn. of Morph., v. 41, 1926.
2 Die Morphologie der Brustlfossen der Fische. Jen. Ztschr., 1926.
3 Der Ursprung der Quadrupeda. Paleont. Ztschr Bd. 8, 1926.
4 Эволюция и психика. Москва, 1922.
Факторы прогрессивного развития низших позвоночных. Русск. зоол. жури., 1924.
Главные направления эволюционного процесса. Прогресс, регресс и адаптация. Москва, 1925.
БИОГРАФИЧЕСКИЙ ОЧЕРК
ния. В эволюции человека этот фактор имел особое значение, ибо здесь-над его наследственной приспособляемостью появилась надстройка индивидуальной приспособляемости поведения.
Исследовательской работой, конечно, не ограничивалась деятельность-Алексея Николаевича в Москве и в Московском университете. Идо и, особенно, после революции он горячо отдавался педагогической и общественной деятельности. Много времени уходило у него на заседания в совете и факультете, на студенческие и магистерские экзамены, на диспуты. Много сил положил он на чтение публичных лекций, на организацию всероссийских, а потом всесоюзных съездов зоологов, анатомов и гистологов,, на издание «Русского зоологического журнала», а после революции— на работу в различных комиссиях, предметных и квалификационных, и различных научных обществах (президентом одного из них, а именно-Общества любителей естествознания, антропологии и этнографии, он состоял несколько лет), на организацию Научно-исследовательского института зоологии при Московском университете, директором которого он также был несколько лет, и позже на организацию Лаборатории эволюционной морфологии при Академии Наук.
Преобразованная впоследствии в Институт эволюционной морфологии и названная постановлением ВЦИК ко дню 70-летия Алексея Николаевича его именем, она была его любимым детищем. По мысли Алексея Николаевича, лаборатория эта, основанная им еще в 1930 г и первоначально образовавшаяся из основного ядра его московских учеников, должна была разрабатывать общие закономерности эволюционного процесса, иначе говоря, изучать эволюционную теорию, ее пути и ее законы, т. е. продолжать его работу, начатую им за много лет до этого времени.
Другим начинанием Алексея Николаевича, которым он очень дорожил, был основанный им в самом начале революции семинар по эволюционной морфологии. На каждом заседании этого семинара делалось лишь по одному докладу (исключительно о собственных исследованиях докладчика), который всеми участниками разбирался весьма детально. Критика, доброжелательная по характеру, по существу всегда бывала необычайно жестка. Немудрено поэтому, что доклады эти имели большое научно-воспитательное значение для молодых ученых и что они ценили свой семинарий. Сам Алексей Николаевич, лишь только, бывало, закончит новое исследование, шел докладывать о нем на семинар своим ученикам и сотрудникам, и они разбирали и критиковали его работы, может быть, несколько более почтительно, чем работы своих товарищей, но столь же жестко, столь же обстоятельно и детально.
У Алексея Николаевича вообще были особые, не совсем обычные отношения со своими учениками. Внешне он никогда не возился с ними, никогда не обучал их технике (это делали его ассистенты и старшие специалисты среди студентов), никогда не «няньчил» их, как он имел обыкновение выражаться, но, учитывая способности, характер и наклонности ученика, всегда долго и тщательно продумывал тему работы, которую ему давал и с постоянным, глубоким интересом следил за ее выполнением. А главное, он работал в лаборатории сам, работал открыто и на виду у всех, не отгораживаясь и не запираясь у себя в кабинете; он любил делиться ходом своих работ с близкими ему учениками, рассказывал им о них, обсуждал с ними вместе различные затруднения, встречавшиеся ему на пути исследования. И на этом они больше всего и учились,— учились тому, как ученому надо «работать над своей работой». Открытый нрав А. Н., общность работы делали то, что ни в одной из его лабораторий—ни в юрьевской, ни в киевской, ни в московской, университетской и академической,—никогда не бывало интриг, ссор и столкновений, никакой «склоки» и зависти друг к другу: был крепкий, здоровый, рабочий, товарищеский дух.
БИОГРАФИЧЕСКИЙ ОЧЕРК
В 1919 г умер академик В. В. Заленский, и на его кафедру в Академию Наук в 1920 г. был избран почти единогласно (один черный шар на Общем 'Собрании) Алексей Николаевич. Но избрание в Академию не вызвало, как это было тогда принято, его переезда в Петербург. Привязанный к своей лаборатории и к своим ученикам, Алексей Николаевич не покинул Москвы и стал лишь ездить туда на заседания Отделения и Общего собрания.
В 1925 г. он был избран действительным членом Украинской Академии Наук.
В 1925 г. исполнилось 35-летие научно-педагогической деятельности Алексея Николаевича. В это время в Москве происходил II Съезд зоологов, анатомов и гистологов, и чествование Алексея Николаевича решено было приурочить ко дням созыва этого Съезда, председателем которого он был избран. От официального празднования своего юбилея он уклонился, но- неофициальная его часть, оживленная и сердечная, доставила ему много удовольствия: в тот вечер, тряхнув стариной, он был особенно блестящ и остроумен.
Ко времени юбилея Алексею Николаевичу шел 60-й год. Кончался период зрелости и подходила старость. Каким был, каким видим мы Алексея Николаевича в это время?
И в зрелом возрасте и в старости он отличался необыкновенной ровностью характера, простотой и сдержанностью манер. Он никогда не торопился, никогда не суетился, почти никогда не сердился. Однако в нем было что-то, что заставляло и слушаться и бояться его. Нервности, в смысле •слаббнервности, в нем не было и следа, но огромное нервное напряжение чувствовалось во всем, и этот внутренний огонь, сдержанный и сосредоточенный, в сочетании с глубоким спокойствием, с удивительной простотой движения и речи, создавал впечатление несокрушимой нравственной силы. Впечатление это усугублялось его внешностью. Не слишком высокий, но казавшийся громадным, с очень широкими плечами и длинными руками, внешне неуклюжий (когда он двигался, всегда казалось, что он непременно что-нибудь опрокинет, разобьет или смахнет), но внутренне весь подобранный, с точными, ловкими, сильными движениями, с четким, скупым и поэтому, быть может, особенно выразительным жестом длинных, тонких пальцев—физически он казался таким же несокрушимо крепким и устойчивым, как и духовно. Особенно бросалось в глаза его лицо,—худое, желтовато бледное, скуластое, монгольского типа, некрасивое в общепринятом смысле слова, но значительное, всегда поражавшее художников лицо— с великолепным, громадным лбом, горячим взглядом затененных очками на редкость умных и красивых глаз, с пленительной своей необычайной искренностью улыбкой.
Алексея Николаевича как-то не видишь перед собой иначе, как с книгою в руках. Интересуясь всеми отраслями знания, он с глубоким вниманием просматривал .каждую книгу, попадавшую ему под руку (с величайшим увлечением читал он даже детские книги своих внучат), и оторвать его от такого чтения (оно происходило обычно за утренним кофе или за вечерним чаем) могла лишь какая-нибудь особенно спешная работа, но отнюдь не присутствие его домашних или приход обычных посетителей. Чтение было его главным отдыхом после работы, особенно с тех пор как спорт стал ему недоступен. Больше всего интересовал его фольклор: народные сказания, героический эпос и сказки, сказки всех времен и народов были его любимым чтением. Очень любил исторические романы и романы приключений (последние—обязательно с благополучным концом), любил старых русских классиков, особенно Толстого и Тургенева, и постоянно перечитывал их. Часто перечитывал Плутарха и Геродота. Очень любил поэзию, и эпос—больше, чем лирику. О Пушкине говорить не приходится:
БИОГРАФИЧЕСКИЙ ОЧЕРК
33
его он перечитывал каждый день и действительно почти всего знал наизусть.
Свои работы Алексей Николаевич обдумывал непрерывно—в самом простом и самом прямом смысле этого слова: это была сущность, основной тон его жизни. Что бы он ни делал, чем бы ни был занят, как бы ни был увлечен разговором с интересным собеседником—за всем этим и на ряду со всем этим шла привычная, напряженная, упорная работа мысли. Отражение этой работы запечатлелось в его многочисленных тетрадях и записных книжках, куда он заносил приходившие ему в голову мысли, поразившие его наблюдения. Все это был обычный, повседневный фон его жизни. Но бывали и частые периоды особенно напряженной работы мысли. Случалось это всегда, когда Алексей Николаевич задумывал новое исследование и приступал к работе. В такие дни (и в такие ночи) он отмеривал медленными, большими, тяжелыми шагами длинные версты по своему кабинету, от окна до библиотеки и от библиотеки снова к окну, был суров и сосредоточен и не терпел, чтобы ему мешали. Утомившись от ходьбы, он усаживался на диване и раскладывал на преддиванном столике бесконечные пасьянсы. «Пасьянсы мои почти никогда не выходят», говорил он, бывало, с усмешкой, «но мысли надумываются, и работа начинает выходить»...
VII
После своего юбилея Алексей Николаевич получил годичную командировку для лечения и для научной работы и осенью 1925 г уехал за границу.
Здесь он прежде всего направился в Киссинген, но, не успев как следует закончить курс лечения водами и едва окрепнув, поспешил на палеонтологический конгресс в Веймар. В Веймаре на общих собраниях съезда Алексей Николаевич прочел два доклада. «О теории костного черепа» и «О происхождении наземных позвоночных». Доклады эти возбудили большой интерес и, несмотря на то, что на съезде присутствовали (и в немалом количестве) виднейшие палеонтологи Европы, были единодушно признаны «гвоздем» конгресса. Ряд ученых подходил к Алексею Николаевичу знакомиться, и по окончании докладов он был так окружен, что к нему было трудно пробраться.
Успех на конгрессе радовал Алексея Николаевича, ибо он сознавал всю ответственность этого первого своего выступления за границей, чувствовал настороженное внимание к каждому своему слову и понимал, что его личный успех является отраженным успехом советской науки вообще и, в частности, успехом близкой его сердцу, им взращенной и взлелеянной эволюционной морфологии, родившейся и выросшей в нашей стране.
Из Веймара, побывав проездом в отстоящей от него в нескольких минутах езды Иене,'—куда его привлекал знаменитый филетический музей,—Алексей Николаевич отправился в Мюнхен. Здесь он пробыл больше двух месяцев. Основной задачей его пребывания там было знакомство с новейшей палеонтологической техникой, зародившейся на Западе в последние годы мировой войны и блестяще усовершенствованной Стен-шиб'и Киэром. Овладев этой техникой, он предпринимает самостоятельную пропедевтическую работу по палеонтологии. Ио так как палеонтологический материал мюнхенского музея был не тот, который был нужен Алексею Николаевичу, а необходимые ему ископаемые (Holostei и Dipnoi) находились в Вене, то из Мюнхена он переезжает в Вену Здесь Алексей Николаевич исследует Phaneropleuron и делает небольшую работу по ребрам этой и некоторых других ископаемых рыб. Об этой работе он докладывает в Венском университете, но напечатана она никогда не была. Морфологические закономерности 3
34
БИОГРАФИЧЕСКИЙ ОЧЕРК
И в Мюнхене и в Вене Алексей Николаевич несколько раз выступал с докладами о своих работах. Так, в Мюнхене в Анатомическом обществе он докладывает о своей работе над костным черепом хрящевых ганоидов, а в Биологическом обществе—«О полифилетической теории». Доклады эти были интересны, и, вопреки немецкому обычаю, прения по ним затянулись до глубокой ночи. Но особенно большой успех имел он в Вене, где его доклад «О различных направлениях эволюционного процесса», на котором он впервые за границей изложил основы своей морфобиологической теории эволюции, обратил на себя большое внимание и оживленно дебатировался на заседании, объединившем три научных общества.
Считая в то время свою морфобиологическую теорию в общих чертах законченной и сознавая, что в теории филэмбриогенеза имеются еще пробелы, Алексей Николаевич решил отправиться на зоологическую станцию в Неаполь, чтобы там набрать недостававший ему материал для дальнейшей разработки этого вопроса. Но, переутомленный работою в Вене, он по дороге снова заболевает, и притом настолько серьезно, что два раза, в Венеции и во Флоренции, прерывает свой путь и отлеживается в гостиницах.
Это не помешало ему, однако, по приезде в Неаполь сразу же приступить к работе. Весенний сезон на станции еще не наступил, работающих было мало, и станционные рыбаки, более свободные, в изобилии доставляли материал. Главною целью работы Алексея Николаевича на станции была дальнейшая разработка теории филэмбриогенеза, основные положения которой он решил еще раз проверить на новом сравнительно эмбриологическом материале. Кроме того, его глубоко интересовал известный закон К. Э. Бэра, и у него явилась мысль о возможности связать этот закон с некоторыми проблемами филэмбриогенеза. Исследование это, о котором подробно будет сказано ниже, далось Алексею Николаевичу не легко: только в одном Неаполе работал он над ним около девяти месяцев, работал с величайшим увлечением, не покладая рук, отказывая себе в отдыхе, часто уклоняясь от участия в интереснейших экскурсиях, устраиваемых станцией. Не отказывался он только от поездок в Помпею. В том году здесь шли большие раскопки и было отрыто несколько новых вилл с прекрасно сохранившимися фресками.
Крайне неприятной стороной его пребывания в Неаполе была постоянная, проводившаяся совершенно открыто слежка со стороны фашистской охранки. Она сильно раздражала Алексея Николаевича и порою очень мешала ему сосредоточиться на работе.
На станции работало немного народу. Интереснее других вначале казался Алексею Николаевичу А. Нэф, с которым он много разговаривал на различные морфологические темы. Разговоры эти, однако, почти всегда переходили в более или менее ожесточенные споры, ибо Алексей Николаевич и Нэф стояли на диаметрально противоположных точках зрения. Идеалистическая морфология Нэфа была абсолютно неприемлема для него, стоявшего на точке зрения естественнонаучного материализма, и в спорах Нэфу не везло. Его нападки на Алексея Николаевича встречали несокрушимый и жесткий отпор. Он уходил, посрамленный, но через несколько дней, обдумав, возвращался снова, нападал вдвое напористее, снова получал «должное», как говорил Алексей Николаевич, а - затем вся история повторялась сначала. •
Тем временем разгоралось и кончалось лето. В Неаполе настала жестокая жара. Станция опустела. Все, кто мог, уехали в горы, и только Алексей Николаевич писал, не отрываясь, работая из последних сил. Наконец, и его силы истощились, и к концу пребывания в Неаполе он совсем свалился. Ему пришлось снова поехать в Киссинген. Поправившись, он на пути в Москву еще заехал в Гёттинген, чтобы принять участие
БИОГР АФИЧЕСКИИ ОЧЕРК
35
в состоявшемся там в это время втором палеонтологическом съезде. •Познакомился здесь с рядом выдающихся ученых, но докладов на этот раз не читал.
Результатами своей первой после войны и революции командировки Алексей Николаевич остался очень доволен: он' овладел техникой палеонтологического исследования, сделал в Неаполе большую работу по фил-эмбриогенезу, которую справедливо считал своей основной работой в этом вопросе, и вывез оттуда громадный и весьма ценный эмбриологический материал, собранный на станции; кроме того, благодаря своим с-пешным выступлениям на конгрессе и в различных научных обществах, он ознакомил европейских ученых с результатами своих последних филогенетических исследований и с основными установками своей морфобиологической теории эволюции; этими выступлениями он показал, что в «советской России» научное творчество и развитие материалистических идей дарвинизма не только не ослабело, но, наоборот, нашло наиболее подходящую обстановку; наконец, ознакомившись, путем усиленного чтения и личного общения с наиболее выдающимися учеными Европы, с последними новинками в своей дисциплине, Алексей Николаевич снова вошел в курс современной мировой науки.
Вернувшись в Москву, он с удвоенным жаром принимается за научную работу, за педагогическую и общественную деятельность. Он продолжает читать лекции в университете, состоит директором двух институтов: сравнительно-анатомического и научно-исследовательского зоологического, ездит в Ленинград, на заседание Академии Наук, работает в квалификационной комиссии ЦЕКУБУ, председательствует в заседаниях научных обществ, редактирует «Русский зоологический журнал» и «Большую советскую энциклопедию», впоследствии принимает участие в планировании науки на первую пятилетку.
Научная деятельность Алексея Николаевича шла в эти годы в трех направлениях: в направлении чисто филогенетическом, в направлении развития морфологической теории эволюции и в направлении разработки теории филэмбриогенеза. Но так как о филогенетических работах этого периода уже было сказано, а о работах, посвященных мэрфобиологической теории и теории филэмбриогенеза будет подробно сказано ниже, то здесь мы на них останавливаться не будем. Отметим, однако, что именно к этому времени относится разработка теории принципов эволюции и проблемы редукции органов и что именно этот период 1926—1931 гг., т. е. время, протекшее с его возвращения из первой заграничной командировки до напечатания «Morphologische uesetzmSssigkeiten der Evolution», -нам, несо-шенно, приходится рассматривать как кульминационный период в жизни Алексея Николаевича. О степени его работоспособности в эти годы дает нам представление количество ежегодно печатаемых им и приготовленных к печати работ
Может быть, Алексею Николаевичу и не удалось бы сделать всего того, что он сделал для своей науки в эти последние десять лет жизни,—когда здоровье его так сильно расшаталось,—если бы бытовые условия не сложились бы для него так благоприятно. Не говоря уже о i ом, что Комиссия содействия ученым при Совнаркоме СССР специально приставила к нему, как и ко всем крупным ученым Союза, врача, наблюдавшего за его здоровьем, постоянно давала ему возможность отдыхать в «Узком» и лечиться за границей,—много помощи, реальной практической помощи, оказали ему его ученики и сотрудники. Пока Алексей Николаевич был здоров, он обходился без личного ассистента и все мелочи технической работы проделывал сам, на что у него, конечно, уходило много времени. Он, правда, не тяготился этой работой, ибо любил исследовательскую технику, и труд этот доставлял ему наслаждение. Когда же он заболел, ему стали з*
БИОГРАФИЧЕСКИЙ ОЧЕРК
помогать в технической работе его более молодые ученики. В старости личное обаяние Алексея Николаевича было так велико, а отношение к нему учеников так исключительно, что вряд ли среди них можно было найти человека, который не постарался бы помочь ему в работе чем и как только мог.
Осенью 1927 г. Алексей Николаевич снова едет в Киссинген. С этих пор ему уже вообще приходится почти каждый год пользоваться водами то в Киссингене, то в Карлсбаде, то в Мариенбаде.
Закончив курс лечения в Киссингене, Алексей Николаевич через Берлин, где в Зоологическом обществе уже был объявлен его доклад «О различных направлениях филогенетического развития», направляется в Швецию, куда его уже давно усиленно приглашали Н. Гольмгрен и Э. Стеншиб.
Пребывание в Стокгольме и Упсале было для Алексея Николаевича очень продуктивно и интересно. Здесь, пользуясь богатым материалом лаборатории Гольмгрена, он проверил еще раз свои выводы относительно покровных костей черепа рыб с костным скелетом; кроме того, что было для него особенно интересно, просмотрел знаменитые препараты Стеншиб, из-за которых, в сущности, и приехал в Швецию. Стеншиб сам показывал их Алексею Николаевичу, подробно рассказывая о методике своих палеонтологических исследований и детально знакомя его с результатами своих работ по эволюции Ostracodermi. Эти частые и долгие беседы со Стеншиб были, по словам Алексея Николаевича, одними из самых интересных, которые ему пришлось вести в жизни. Иногда соглашаясь, а чаще споря, они со страстным увлечением обсуждали эмбриологическую и палеонтологическую методику, вопрос о происхождении костного вещества и черепа, вопрос о систематическом положении круглоротых и остракодерм, и надо было видеть, как бывали довольны оба, когда результаты, полученные ими столь различными путями, совпадали. Тем не менее, в некоторых отношениях взгляды их расходились, особенно в вопросе о происхождении осетровых: здесь Алексей Николаевич, чувствуя твердую почву под ногами, стойко держался своей точки зрения.
В научных кругах Стокгольма имя Алексея Николаевича было, невидимому, хорошо известно, и приездом его были заинтересованы, особенно после его доклада («О прогрессивной и регресивной эволюции»), прочитанного им в Стокгольмском биологическом обществе. Но, как всегда, больше всего льнула к нему молодежь. Так же, как в Мюнхене и в Вене, так же, как впоследствии в Упсале, он был постоянно окружен молодыми биологами, вводившими его в круг своих работ и наперебой приглашавшими его к своим столам, на которых уже заранее были выставлены особенно интересные препараты. Ни одному из них Алексей Николаевич не отказал в совете или в указаниях: совсем как в Москве, переходил он от стола к столу, знакомясь с работами, разглядывая рисунки, обсуждая сомнения и решая научные споры.
После возвращения в Москву Алексей Николаевич с удвоенным усердием принимается за работу. Три последующих года, вплоть до напечатания его книги «Morphologische Gesetzmassigkeiten der Evolution», он работал непрерывно в самом буквальном смысле этого слова. Его частые наезды в подмосковный дом отдыха для ученых—«Узкое»—отнюдь не прерывали работы: менялась обстановка, но чемоданы с книгами и рукописями и я со своей пишущей машинкой сопровождали его повсюду.
Именно в «Узком» Алексей Николаевич больше всего работал над своею книгой о морфологических закономерностях эволюции. Писал он ее в течение трех лет, писал с подъемом и увлечением, постоянно изменяя и переделывая отдельные части, многое выкидывая и многое дополняя. В 1928 г. он приводит в порядок филогенетическую часть. В 1929 г. переходит к
БПОГРЛФ11ЧЕСЮ1Й ОЧЕРК
разработке морфобиологической теории эволюции, сосредоточив свое внимание на проблеме принципов эволюции. В 1930г. пишет последнюю часть книги: теорию филэмбриогенеза, которую заканчивает уже в Бордигере
В Бордигеру Алексей Николаевич попал из Падуи, куда был командирован Академией Наук для участия в XI Международном конгрессе зоологов. Он прочел на этом конгрессе два больших доклада, один на секции («Рудиментация и полная редукция органов»), другой на общем собрании съезда («О значении принципа субституции и некоторых других принципов в филогенезе»). Кроме того, ему пришлось в течение всего конгресса дважды в день председательствовать на секции анатомии и эмбриологии и резюмировать доклады, читавшиеся на английском, французском, немецком и итальянском языках, что требовало большого напряжения внимания. Все это его чрезвычайно утомило, и это было тем более досадно, что сам конгресс для Алексея Николаевича был мало интересен: научно-деловая его сторона была организована слабо, в парадной же, казовой его стороне, которая, к слову сказать, была поставлена весьма торжественно, он участия почти не принимал.
Между прочим, на этом конгрессе Алексей Николаевич снова столкнулся с Нэфом, в то время профессором Каирского университета. На одном из заседаний между ними произошел интереснейший диспут, во время которого Алексей Николаевич, защищая свои методологические позиции и материалистическое мировоззрение дарвинизма, со свойственным ему остроумием и четкостью блестяще доказал несостоятельность идеалистического миропонимания своего противника.
После конгресса Алексей Николаевич прожил некоторое время в Сиене, а затем переехал в Бордигеру. Здесь он продолжал писание своей книги и стал готовить ее к печати. Продолжалась эта работа всю зиму 1930/1931 г. Все это время он вел очень тихий и размеренный образ жизни: рано вставал, рано ложился, ни с кем не встречался, не предпринимал никаких экскурсий или поездок. Только ежедневно выходил на час—полторт для прогулки и, не переставая, усиленно работал и писал. Когда эта работа была закончена, Алексей Николаевич направился через Фрейбург (здесь он познакомился со Шпеманом и по приглашению местного Общества естествоиспытателей, прочел у них доклад: «Об отношении филогенеза к онтогенезу») в Иену, чтобы печатать там свою книгу у Фишера.
Пребывание в Иене было одним из самых трудных, самых напряженных периодов в жизни Алексея Николаевича. Для того, чтобы книга была издана поскорее и получше, он оставался во все время печатания в Иене, тут же, на месте правя корректуры и давая необходимые указания. В это же время, как на беду, на него лавиной посыпались корректуры «Этюдов по редукции органов»,1 поэтому он работал дни и ночи в самом прямом и простом смысле этого слова, и к концу пребывания в Иене так устал, так нервно был напряжен, так ослабел от переутомления и, вероятно, от кровопускания, которое делали ему йенские врачи, что порою как-то не верилось, что он живым выберется из Иены. Наконец, закончились и последние корректуры, и книга была готова. По своим размерам она была почти в два раза меньше настоящей книги и состояла из двух частей: специальной (филогенетической) и общей, заключавшей в себе морфобиологическую тёорию эволюции и теорию филэмбриогенеза.
Когда печатание совершенно закончилось, Алексей Николаевич был так рад, что, наконец, разделался с книгой, что после очередного лечения в Карлсбаде на радостях решил отдохнуть. Отдых заключался в поездке на генетический конгресс в Мюнхен, где ему не пришлось ни председательствовать, ни читать докладов, но лишь присутствовать на заседаниях
1 Studien uber die Reduktion der Organe der Wirbeitiere. Zool. Jahrb., 1931.
38
БИОГРАФИЧЕСКИЙ ОЧЕРК
в качестве свободного гостя. Это действительно составляло для него приятный отдых, ибо конгресс, на котором собрались виднейшие генетики Европы, был очень интересен в научном отношении.
Из Мюнхена он заехал в Дрезден, где остановился в Зоологическом саду в доме директора сада и каждое утро отправлялся на весь день к зверям наблюдать их повадки и нравы и изучать движения брадипуса. Оттуда Алексей Николаевич направился домой в Москву. Это была его последняя поездка в Германию.
В 1931 г. общее внутреннее положение в Германии при поверхностном взгляде оставалось как будто таким же, каким оно было в первое десятилетие после немецкой революции, но политическая атмосфера начинала сгущаться, и в ней появилось что-то («наци»!), тогда еще едва уловимое, что, однако, сильно не понравилось Алексею Николаевичу.
«Думается мне, что я уже больше в Германию никогда не поеду,—говорил в вагоне Алексей Николаевич, задумчиво глядя на удалявшиеся огоньки Берлина,—нехорошо здесь станет». «Жаль немцев, очень, очень жаль...» прибавил он через некоторое время.
VIII
Уже на следующий день по приезде в Москву началась обычная трудовая жизнь. Пошли заседания Академии Наук, на которые все еще приходилось ездить в Ленинград (лекции в университете после 1931 г. Алексей Николаевич уже не возобновлял, передав их целиком проф. Б. С. Матвееву), начались работы по планированию биологических наук на вторую пятилетку для Госплана, начались доклады на семинаре и в научных обществах, и, главное, началась спокойная, планомерная исследовательская работа в лаборатории, на которую, стосковавшись после годичного перерыва, ушедшего на подготовку к печати и на издание книги, Алексей Николаевич с жадностью набросился.
Работу эта была непосредственно связана с подготовкою нового, советского издания его книги, которое, по его словам, должно было быть «вдвое больше и вдвое лучше немецкого», вмещая в себе все то новое в смысле развития его идей и доказательства его положений, что сложилось у него в голове во время печатания книги, когда он весь был полон мыслями о ней. Так как почти все крупные теоретические работы Алексея Николаевича («Этюды по теории эволюции», «Современные задачи эволюционной теории», «Эволюция и психика») вышли только на русском языке, то он решил выпустить в свет свою новую книгу одновременно на русском и на немецком языках. Поэтому еще в Бордигере он засаживает меня за перевод основного текста на русский язык (начав писание своей книги по-русски с тем, чтобы я переводила ее на немецкий язык, он впоследствие изменил свой план и стал писать ее на немецком языке, которым владел в совершенстве, потому что переводить мне ее с немецкого языка на русский было несравненно легче, чем с русского на немецкий). По приезде в Москву перевод этот продолжался, сам же Алексей Николаевич вновь принимается за переработку, проверку и дальнейшее развитие своих выводов.
Для этого он предпринимает две новых работы филогенетического характера: первая—исследование чешуй у Polypterus delhesi,1—представляя собой непосредственное продолжение его работ 1924, 1925 и 1926 гг., относя
1 Die Entwicklung der Knochenschuppen von Polypterus delhesi. Jen. Ztschr., Bd. 67, 1932.
БИОГРАФИЧЕСКИЙ ОЧЕРК
щихся к вопросу об эволюции костного скелета, не выходит из ряда его обычных филогенетических работ, вторая—об эволюции брюшных плавников рыб1—носит исключительный характер и занимает особое положение среди филогенетических работ Алексея Николаевича.
Проследив на громадном сравнительно анатомическом материале строение и развитие брюшных плавников у ископаемых и современных акул, у хрящевых и костных ганоидов, у палеонисцид, кистеперых и костистых рыб (последнее—по материалу, собранному в Неаполе в 1926 г.), Алексей Николаевич строит схему происхождения и развития плавников, начиная от гипотетической исходной формы и кончая плавниками высших рыб. Он устанавливает, что эволюция этого важнейшего органа все время шла в прогрессивном направлении, в результате чего плавники потомков начинали функционировать раньше, чем плавники предков, и лучше, чем они. Новое для него самого в этой работе заключалось в том, что при этом он проводил глубокий и тонкий анализ процесса эволюции изучаемого органа, разбирая различные этапы прогрессивного хода его развития и устанавливая, путем каких принципов и каких модусов филэмбриогенеза оно осуществлялось. Шаг за шагом прослеживает он, как в течение эволюции развивались, следуя тому или иному модусу филэмбриогенеза, различные части плавников, как в различные эпохи, на разных стадиях филогенеза один модус сменялся другим и возвращался снова, как сменялись и сочетались друг с другом различные принципы эволюции.
В течение двух следующих лет,т. е. в 1933 и 1934, Алексей Николаевич продолжает разработку «Морфологических закономерностей эволюции», расширяя и уточняя отдельные положения, заполняя пробелы, которые он находит в своей книге. Отражение этой работы в части, касающейся морфобиологической теории эволюции, выразилось в появлении второго, значительно переработанного и вдвое расширенного издания его книги «Главные направления эволюционного процесса», вышедшей в свет в Москве в 1934 г. Что же касается до последующей разработки теории филэмбриогенеза, о которой подробно будет сказано ниже, то она шла в различных направлениях. Алексей Николаевич прежде всего пытается классифицировать модусы филэмбриогенеза,1 2 затем он пробует применить методы исследования филогенеза органов к исследованию филогенеза тканей, т. е. к гистологии, причем ему удалось установить наличие рекапитуляций в эволюции различных тканей,3 и, наконец, ставит себе целью разрешить вопрос о том, как возник и как развивался онтогенез в течение эволюции Metazoa.4 Этот последний вопрос чрезвычайно увлекает Алексея Николаевича, и он читает на эту тему два общих доклада: один у себя на семинаре, другой в Московском Доме ученых. Кроме всего этого, он принимается за разработку проблемы соотношения между принципами эволюции и модусами филэмбриогенеза и пишет подробный конспект своих предполагаемых исследований в этом направлении. Судя по ширине охвата затронутых в конспекте тем, эта работа,—если бы Алексею Николаевичу удалось ее осуществить,—вероятно, была бы наиболее значительной из его работ общего характера.
После этого периода теоретических обобщений Алексея Николаевича, как это всегда бывало в таких случаях, снова неудержимо потянуло к специальной, чисто исследовательской работе, той работе, которая давала в его руки новые факты. В 1934 г. он опять принимается за осет
1 Эволюция брюшных плавников рыб'И принцип выпадения промежуточных функций. «Природа», 1933. Evolution der Bauchflossen det Fisc lie. Zool. Jahrb., Bd 58, 1934.
2 Модусы филэмбриогенеза. Зоол. журн., т. 14, 1935.
8 Морфологическая и гистологическая рекапитуляция. Доклады Акад. Наук 1935.
4 Морфологические закономерности эволюционного процесса. Социал, реконстр. и наука (Сорена), 1934.
40
БИОГРАФИЧЕСКИЙ ОЧЕРК
ровых рыб и исследует, или вернее, переисследует их осевой череп и костный скелет головы и туловища (чешуи и жучки) (Acipenser ruthenus, Acipen-ser stellatus, Acipenser gOldenstadtii, Huso huso). Работа эта была им предпринята со специальной целью опровергнуть некоторые положения Стен-шиб, относительно которых у него с ним было резкое расхождение во взглядах. Результаты этого исследования Алексей Николаевич еще успел доложить на семинаре своим ученикам и сотрудникам. Это была его последняя работа и его последний доклад. Она вошла в «Морфологические закономерности эволюции» в качестве отдельного раздела в главе восьмой.
Следующую свою филогенетическую работу, для которой ему удалось получить редчайший эмбриологический материал по Polyodon, добытый американской экспедицией на Миссисипи, Алексею Николаевичу закончить уже не пришлось. Этот материал, так же как и весь огромный эмбриологический материал, собранный им в течение многих лет на различных биологических станциях, так же как и все препараты и оттиски отдельных работ, присылавшихся ему в течение всей его жизни различными авторами со всех концов света, он завещал Институту своего имени в Академии Наук.
В течение двух слишком лет после выхода в свет «Morphologische Gesetzmassigkeiten der Evolution» в советской и заграничной печати появлялись многочисленные на нее рецензии. Рецензии эти, написанные на всех европейских языках, включая шведский, голландский и мадьярский, не могли не доставить Алексею Николаевичу глубокого удовлетворения: среди специалистов книга имела несомненный успех. Мнения рецензентов разделились, главным образом, в том отношении, что одним казалась наиболее ценной филогенетическая часть книги, тогда как другие полагали центр тяжести в морфобиологической теории эволюции и особенно в принципах (наибольший успех имел «принцип фиксации фаз»), а третьи (и таких было большинство)—в теории филэмбриогенеза. Все, однако, сходились в одном, считая особенно ценным, что обе эти теории были построены автором на основании личных исследований и исследований его учеников, и авторитет «школы Северцова в новой созданной им лаборатории эволюционной морфологии Академии Наук СССР, где автор и его ученики продолжают вести свои дальнейшие исследования», прочно укрепился в научном мире.
В отзывах говорилось о том, что «книга Северцова должна быть горячо рекомендована как творение, будящее мысль»,* 1 что она «открывает новое поле исследований не только для морфолога, но и для генетика, антрополога, эколога, анатома и зоолога-экспериментатора»; что «изучение его труда не только знакомит читателя с мыслями гениального исследователя, но представляет в его распоряжение неистощимый запас новых рабочих гипотез»;2 что благодаря его трудам «вся старая, частью уже оставленная область исследования (оставленная не потому, что она оказалась бесплодной, но потому, что никто не взял на себя труд отыскать в ней новые пути), снова оказалась новой...», ибо «то, что излагает Северцов в своей книге, является фундаментом новой, молодой науки, разработку которой должны взять на себя морфологи, так как морфология здесь показала всем, кто сомневается в ее ценности, что она представляет собою нескошенную ниву, что она работает не над мертвым, но над живым материалом, что она раскрывает законы жизни и только жизни»?
Не менее горячо приветствуют книгу и представители других биологических дисциплин. Так, известный энтомолог Г. Гаас пишет, что
1 De Beer, Nature, v. 129, 1932.
1 L. Neumayer Neues Jahrb. f. Miner 1932.
® E. Th. Nauck. Morphol. Jahrb-, 1932.
биографически!! очерк 41
«это произведение, охватывающее собой результаты многочисленных, проведенных со строгой закономерностью трудов как самого автора, так и его учеников, имеет исключительное значение». Особенно важна и значительна, по его мнению, теория филэмбриогенеза. Вообще, «книга эта во всех своих важнейших чисто морфологических выводах имеет огромное значение и играет выдающуюся роль благодаря логичности своих построений, а также тому обстоятельству, что вся она базируется на собственных и с с л е д о в а и и я х1 автора. Ею должны будут руководствоваться все те генетики и все те исследователи механики развития, которые будут работать над построением типов филогенетических изменений».1 2
Эти последние отзывы были важны и ценны для Алексея Николаевича, ибо он болезненно реагировал на то пренебрежение, с которым с начала нынешнего столетия многие относились к морфологии, которую было принято считать наукой отсталой, наукой без будущности и без перспектив. Всеобщее увлечение экспериментальным направлением в различных его видах затерло историческое направление в биологии и заставило морфологические науки отступить на задний план. Поэтому Алексея Николаевича, глубоко убежденного в большом теоретическом значении морфологии, радовало, что его книга произвела известный поворот в этом отношении в научнообщественном мнении и привела к признанию того, что морфология, став на новый путь, им указанный и разработанный, еще многое может дать в будущем в смысле установления общих закономерностей эволюции.
IX
Быстро надвигающаяся старость, постоянное недомогание и физическая слабость заставили Алексея Николаевича вести все эти годы очень тихую и замкнутую жизнь. Он редко выходил из дома, стал меньше бывать в лаборатории. На его связи с лабораторией это, впрочем, не отразилось, так как теперь его ученики и сотрудники стали сами приходить в его домашний кабинет для обсуждения своих работ, для получения его советов и указаний.
Но когда, вследствие всё усиливавшейся болезни Алексея Николаевича эта связь действительно грозила порваться (а это должно было случиться при переезде Академии Наук из Ленинграда в Москву, когда он мог получить для своей академической лаборатории, ютившейся до тех пор в непосредственной близости от его квартиры в трех комнатах Института сравнительной анатомии Московского университета, прекрасное новое помещение в здании Академии Наук), то эту возможность разрыва Алексей Николаевич переживал очень остро и тяжело. Нельзя без волнения читать бумаг, адресованных им в различные инстанции Академии Наук, где он просит Академию отвести ему хотя бы самую маленькую квартиру при новом помещении лаборатории, и не лишать его таким образом возможности принимать участие в ее работах: «Если при переезде лаборатории,—пишет он в одной из этих бумаг,-—мне придется остаться на своей старой квартире или даже переселиться в один из академических домов, то, по состоянию моего здоровья, мне придется порвать всякую связь с лабораторией: это значит для меня в значительной степени сократить свою исследовательскую работу, а для лаборатории в значительной степени потерять мое непосредственное руководство. Даже если бы мне дали в мое личное пользование автомобиль, то и это мало поправило бы дело, так как ездить каждый ден&, в особенности зимою, опять-таки по состоянию моего здоровья, мне было бы совершенно невозможно. Таким образом»
1 Подчеркнуто рецензентом.
2 G. Haas. Die Naturwiss., Bd. 47, 1932.
42
БИОГРАФИЧЕСКИЙ ОЧЕРК
связь моя с основанной мною лабораторией грозит сделаться чисто номинальной, что было бы чрезвычайно вредно для дела».
Возможно, что Академия Наук и согласилась бы на удовлетворение просьбы Алексея Николаевича, но когда произошел фактический переезд Академии в Москву и переезд лаборатории Алексея Николаевича в новое помещение,—переезд самого Алексея Николаевича на новую квартиру уже не мог осуществиться: к этому времени, вследствие неизлечимой его болезни (быстро прогрессирующего склероза почек), уже не могло быть и речи о его работе в лаборатории.
Алексей Николаевич слег в конце февраля 1935 г. в «Узком». Почу-ствовав опасность своего положения, он потребовал к себе в постель рукопись, над которой в это время работал. Удостоверившись, что писать он уже не может, он пытался диктовать мне дальнейший текст: он не мог допустить мысли о том, чтобы оставить незаконченной очередную работу. Но и диктовка вскоре стала невозможной: ему становилось все хуже и хуже. Так продолжалось недели две. Наконец, консилиум врачей решил перевезти его в Кремлевскую больницу, предупредив, однако, что 95% за то, что этого переезда он не перенесет. «Но так как конец,—говорили они,— может наступить и здесь каждую минуту», а 5% оставалось за то, что переезд он все же выдержит и, может быть, даже поправится, то было решено вернуться в Москву. Действительно, после этого Алексей Николаевич, хотя и неизлечимо больной, прожил еще около двух лет.
В больнице бывали дни, когда жизнь его висела на волоске, бывали дни, жогда его физические страдания становились невыносимыми. Он никогда не жаловался на них и поражал окружающих, врачей и сестер, своей выдержкой, своим спокойствием, своим терпением. Едва он пришел в себя и окреп настолько, что мог сидеть в постели, как снова потребовал свою рукопись. Так как ему абсолютно было запрещено работать, а мысль о ненапечатанной рукописи просто мучила его, то он потребовал, чтобы я написала за него предварительное сообщение, использовав его черновики и рисунки. Это было сделано там же, в больнице, в часы, когда он спал. Ясность, точность, определенность его отдельных указаний были при этом поразительны. Это была последняя из напечатанных работ Алексея Николаевича, называлась она «Морфологическая и гистологическая рекапитуляция»; ниже о ней будет сказано более подробно.
После выхода из больницы Алексей Николаевич зимой жил в Москве, летом в «Узком». Здесь я не могу не вспомнить с огромной благодарностью деятельность Комиссии содействия ученым при Совнаркоме по отношению к Алексею Николаевичу: благодаря ее попечению для него было установлено дежурство сестер, был приставлен специальный врач, все время живший с нами в «Узком».
Медленно и постепенно угасал Алексей Николаевич. Иногда, когда ему бывало лучше, он еще пытался работать: ему хотелось завершить прерванное болезнью исследование над чешуями осетровых и в особенности—над страстно интересовавшим его Polyodon. Каждый день к нему приходила его ассистентка Н. О. Ланге все с новыми и новыми рисунками, и каждый день он проверял ее работу и давал ей новые задания. Таким образом, хотя и медленно, работа все же двигалась вперед.
Весною 1936 г. в Академии Наук состоялась конференция по эволюции. Вместе с тем это был и общественный просмотр Института эволюционной морфологии. Алексею Николаевичу хотелось присутствовать на этой конференции и лично сделать доклад об общем направлении работ своего института. Зимою он тщательно продумывал речь, которую собирался произнести, писал подробный ее конспект, но быть на конференции ему уже не пришлось. Однако он так интересовался ею, что, несмотря «а до, что об этой конференции ему подробно докладывали его ученики,
♦
БИОГР АФИЧЕСКИИ ОЧЕРК
несмотря на то, что обычно он в это время уже не отпускал меня от себя ни на шаг,—он все же потребовал, чтобы я ездила на все заседания конференции и детально рассказывала ему обо всем, что на ней происходило.
Вскоре Алексею Николаевичу пришлось совсем отказаться от работы; тогда он стал еще больше читать. Когда же и чтение начало его утомлять, ему много читали вслух. Оставаясь часто один (себя и дежурную сестру я в расчет не принимаю), он очень ушел в себя: стал еще более покоен, еще более молчалив, еще более сдержан и сосредоточен. Все мелкое, повседневное, будничное отошло в сторону; все стало на место. И выступило наружу то, что он, всю жизнь замкнутый и застенчивый, скрывал за семью замками: большая мудрость большого ученого, большая доброта большого человека.
24 сентября 1936 г. Алексею Николаевичу минуло 70 лет. Он жил в это время в «Узком». Из Москвы приехали ученики, привезли цветы, адреса, подарки. Они собрались в соседней комнате, чтобы не беспокоить больного, но Алексею Николаевичу захотелось их видеть, и маленькими группами, одни вслед за другими, входили они к нему в комнату. Внимательно выслушав приветственную речь официального представителя Академии Наук, Алексей Николаевич еще нашел в себе силы, чтобы ответить: «Благодарю вас за приветствие», сказал он ему, «я очень болен теперь, и мне трудно говорить. Поэтому скажу вам одно: я всю жизнь работал и сделал все, что мог».
19 декабря в 9 ч. 46 м. вечера Алексей Николаевич скончался. Еще накануне вечером он просил меня читать ему и долго слушал мое чтение, пока не уснул.
В ночь с 19 на 20 декабря, около 4 часов утра мы вчетвером: я, дежурная сестра, дочь Алексея Николаевича и Б. С. Матвеев, приехавшие в этот день из Москвы, перевезли его из «Узкого» домой. Несмотря иа поздний час, на квартире уже собрался народ: пришли его ученики, все приготовившие для последней встречи. Трудно без волнения вспоминать о том, что они сделали для Алексея Николаевича в эти дни: как распределили между собою ночные и денные дежурства у гроба, как хоронили его, как и что говорили о нем на траурном митинге в Академии Наук и на гражданской панихиде в университете.
Иногда человеку выпадает на долю счастье умереть героической смертью. Такая смерть легка, проста и ясна, как ясен светлый день. Иное дело умирать долго и медленно, болея, страдая и слабея с каждым днем, с каждым днем уходя от жизни все дальше и дальше, каждый-день ожидая неизбежного конца. В том, как переживал Алексей Николаевич все это— свой закат, свою болезнь, свои страдания, свои последние предсмертные дни,—сказался он весь, во весь свой рост. Слабело тело, но дух крепнул, и с каждым днем становился он спокойнее и умиротвореннее, тверже, проще и мудрее.
Прекрасна была жизнь Алексея Николаевича, простая человеческая жизнь, с ее горями и радостями, с ее вдохновенным трудом, длинная трудовая жизнь большого человека и большого ученого, и под стать этой жизни была его смерть.
X
Нам остается выполнить еще одну задачу проследить историю возникновения и оформления книги, ныне предлагаемой вниманию читателя. Об ее основной—филогенетической—части много говорить не приходится, в сущности, к построению ее, за исключением нескольких единичных работ, вели все изложенные нами выше специальные работы Алексея Николаевича, предпринятые им на протяжении своей более чем сорока-
БИОГРАФИЧЕСКИЙ ОЧЕРК
летней научной деятельности. В особенности это касается его трехтомной монографии о «Происхождении низших позвоночных», серии работ по покровному скелету и серии работ по происхождению и эволюции плавников и пятипалой конечности. В «Морфологических закономерностях эволюции» изложен их конечный результат- воссоздан конкретный ход эволюции низших позвоночных, начиная от гипотетических Acrania primitiva и кончая высшими рыбами и предками наземных позвоночных. При этом, в результате комплексного изучения, восстановлен не только филогенез отдельных органов со всеми их функциями, но и эволюция цельных организмов со свойственным им образом жизни. Только читатель, хорошо знакомый со специальными работами Алексея Николаевича, может увидеть, как глубоко обосновано здесь каждое его филогенетическое построение: иногда лишь к двум-трем абзацам сведены итоги многолетних исследований. Так обстоит дело с теорией конечностей, с теорией костного черепа, с вопросом об эволюции висцерального скелета, с вопросом о происхождении наземных позвоночных.
Что касается остальных частей его книги, построенных на основании всех этих работ и трактующих об общих эмпирических закономерностях эволюции, то здесь нам придется рассмотреть два отдельных вопроса: вопрос о том, как сложилась теория филэмбриогенеза, и вопрос о том, как сложилась морфобиологическая теория эволюции.
Как это уже было отмечено выше, первым толчком, заставившим Алексея Николаевича обратить внимание на соотношение между онтогенезом и филогенезом, было его желание проверить в методологических целях правильность биогенетического закона Геккеля. Еще в первой работе, сделанной в этом направлении, он приходит к противопоставлению понятию ценогенеза своего понятия о филэмбриогенезах как о «филетичес-ких изменениях эмбриональных органов, определяющих изменения строения взрослых органов». Основные мысли этой работы он излагает в докладе, сделанном в Киевском обществе естествоиспытателей в 1907 г., а затем в речи, произнесенной на общем собрании Съезда естествоиспытателей и врачей в Москве в 1910 г. В этой речи мы находим первые зачатки будущей теории филэмбриогенеза. Уже здесь Алексей Николаевич устанавливает, что филетические изменения органов образуются в онтогенезе двумя способами: способом эмбриональных изменений первичных зачатков (в будущем архаллаксисы) й способом надставки стадий (в будущем анаболии), причем рекапитуляция в смысле Геккеля имеет место лишь при этом последнем способе развития.
Основные мысли, выраженные в этих двух докладах, Алексей Николаевич подробно развил в своей книге «Этюды по теории эволюции. Индивидуальное развитие и эволюция» (1912, 1922). Здесь, в отличие от того, что мы видим в «Морфологических закономерностях эволюции», он еще считает, что путем изменений зачатков органов у взрослых животных достигаются одинаково направленные изменения общего характера, путем изменения конечных стадий морфогенеза—ч а с т н ы е и ограниченные изменения. Здесь же Алексей Николаевич дает и свою первую классификацию филэмбриогенезов, также сильно отличающуюся от той классификации модусов филэмбриогенеза, которую он приводит в «Морфологических закономерностях эволюции». Он подразделяет филэмбрио-генезы на первичные (в будущем те же архаллаксисы) и на вторичные (в будущем те же анаболии), причем первичные филэмбриогенезы подразделяет на соответственные (когда изменения зачатков носят тот же характер, что и изменения взрослых органов) и несоответственные (когда этого не наблюдается). Вместе с тем он снова подтверждает, что рекапитуляции в своем полном виде наблюдаются лишь при развитии, идущем путем вторичных филэмбриогенезов, т. е. путем надставки стадий; при
развитии же, идущем путем первичных филэмбриогенезов, т. е. путем эмбриональных изменений, они или не существуют вовсе, или существуют как повторение очень ранних эмбриональных стадий.
Так обстояло дело в 1912 г. После этого в разработке теории филэмбриогенеза наступает большой перерыв. Алексей Николаевич возобновляет работу над этим вопросом только в 1926 г., во время своего пребывания на зоологической станции в Неаполе, куда он направился специально с этой целью. Решив еще раз проверить теорию филэмбриогенеза, сложившуюся у него на основании исследования эволюции конечностей рептилий и амфибий, на совершенно новом материале, он берет в качестве такового костистых рыб с эксцессивно развитыми органами (Belone, Sargus, Lo-phius, Gobius, Centriscus, Syngnathus, Hippocampus, Trigla) и тщательно исследует сравнительно эмбриологическим путем их внешнюю форму (внешняя форма выбрана для большей наглядности). Объектом для сравнения служили Salmo и Atherina как формы с нормально развитыми органами. ,’В результате Алексей Николаевич полностью подтвердил основные положения теории филэмбриогенеза, устанавливая, что прогрессивная эволюция органов взрослых животных происходит либо путем надставки конечных стадий морфогенеза, или анаболии, либо путем изменения первых стадий морфогенеза, или архаллаксиса. Термины анаболия и архал-лаксис здесь упоминаются впервые, о модусе девиации еще не говорится ни слова. Центральным пунктом этой работы является та тесная связь, которую удалось установить Алексею Николаевичу между известным законом К. Бэра о порядке появления признаков различных систематических категорий в онтогенезе и анаболией. С предельной ясностью и исчерпывающей полнотой изъясняет он здесь, каким образом чисто эмпирический, до тех пор еще никак не объясненный закон К. Бэра с естественной и неизбежной очевидностью вытекает из обычного хода монофилетической дивергентной эволюции, идущей путем анаболии. Другим результатом эволюции, идущей путем анаболии, является факт рекапитуляции исчезнувших анцестральны.х признаков у потомков.
Исследование это, которое Алексею Николаевичу далось не легко (больше года работал он над ним с огромным напряжением сил), появилось в печати в 1927 г.1
С этих пор Алексей Николаевич уже не оставляет разработку теории филэмбриогенеза. В 1929 и 1930 гг. он совместно с Е. Ф. Еремеевой изучает развитие органов обоняния у целого ряда форм (Acipenser ruthenus, Onchorhynchus keta, Sargus vulgaris, Belone acus) и приходит в выводу, что прогрессивная эволюция животных может совершаться также и третьим путем, а именно, как это предполагал уже Ф. Мюллер, по способу девиации, или отклонения (изменения) хода эмбрионального развития на средних стадиях морфогенеза. Результаты этого исследования он опубликовывает в книге «Morphologische Gesetzmassigkeiten der Evolution» (1931), где приводит ряд примеров эволюции, идущей этим путем.
Следующим этапом в развитии теории филэмбриогенеза было исследование, посвященное проблеме редукции органов. Считая, что прогрессивное развитие органов происходит тремя способами положительного филэмбриогенеза—анаболией, архаллаксисом и девиацией—, Алексей Николаевич теперь ставит вопрос, применимы ли основные положения его теории и для понимания явлений регрессивной эволюции органов? Это было пробным камнем для проверки ценности всей теории. В качестве материала для этого обширного исследования он берет ряд ящериц, начиная от форм с хорошо развитыми конечностями и кончая безногими
1 Ueber die Beziehungen zwischen der Ontogenese und der Phylogenese der Tiere. Jen. Ztschr., Bd. 63, 1927.
ВИОГРАФИЧЕСКИЛ ОЧЕРК
формами: Scincus officinalis, Eurneces schneideri, Lygosoma punctulatum, Chalcides ocellatus, Chalcides sepoides, Typhlosaurus, Ophiomorus tridactylus, Seps chalcides, Ophiodes striatus, Ophisaurus apus, Anguis fragi-lis. Материал этот он получает частично в Академии Наук, частично его собирает в Кизлярских степях его ученик А. Г. Рындзюнский. Кроме того, Алексей Николаевич изучает явление редукции спинной мускулатуры черепах и поручает исследование дегенерации зубов у стерляди одной из своих учениц—О. Н. Дислер.
Исследование всего этого материала показало Алексею Николаевичу, что филетическая редукция органов также происходит путем изменения хода эмбрионального развития предков, а не путем суммирования вариаций взрослых форм, как это принято было думать. Установив два типа редукции органов: рудиментацию органов и полную их редукцию (аннигиляцию, или афанизию, как он назвал ее впоследствии), Алексей Николаевич нашел, что и рудиментация и афанизия осуществляются теми же модусами филэмбриогенеза, что и прогрессивное изменение органов, но со знаком минус, т. е. что рудиментация происходит путем отрицательного архаллаксиса, а афанизия—посредством отрицательной анаболии. Таким образом, изучение дегенерации органов показало Алексею Николаевичу, что теория филэмбриогенеза вполне применима и для объяснения регрессивной эволюции животных, и это обстоятельство, конечно, послужило веским подтверждением всей теории.
Об этой работе Алексей Николаевич прочел доклад на общем собрании Международного съезда зоологов в Падуе в 1930 г.,1 а затем напечатал ее in extenso в 1931 г.1 2 * «Этюды по редукции органов» имели большой успех, и в сущности только после появления их в печати теория филэмбриогенеза получила надлежащее признание.
В «Morphologische.Gesetzmassigkeiten der Evolution» (1931) мы находим теорию филэмбриогенеза уже во вполне разработанном виде. Однако Алексей Николаевич не останавливается и на этом. Детально разбирая (в последующей своей работе об эволюции брюшных плавников),8 какими модусами филэмбриогенеза осуществлялась в различных частях органа и на разных стадиях филогенеза эта эволюция, он показывает, что она отнюдь не идет, все время каким-нибудь одним определенным путем, что обычно один модус филэмбриогенеза исторически сменяется другим, снова возвращается (вторичные анаболии и вторичные архаллаксисы) и снова сменяется. Он показывает, как бесконечно сложен процесс эволюции: эволюция не течет покойно и ровно; про какой-нибудь орган нельзя сказать, что он всегда и вообще развивался, следуя тому или иному принципу,4 * * * осуществленному тем или иным модусом филэмбриогенеза. Наоборот, на разных этапах эволюции филогенетический процесс шел разными путями: различными направлениями, различными принципами, различными модусами. Задача филогенетического исследования, по мысли •Алексея Николаевича, заключается в том, чтобы, детально изучая историю развития организмов и органов, установить, каково было направле
1 Rudimentation und vollstandige Reduction der Organe der Tiere. Archivio zool. ital., v. 16, 1931.
2 Studien tiber die Reduction der Organe der Wirbeitiere. Zol. Jahrb., Bd. 53, 1931
8 Evolution der Bauchflossen der Fische. Zool. Jahrb., Bd. 58, 1934.
4 Во избежание недоразумения, могущего возникнуть в связи со своеобразным
применением Алексеем Николаевичем термина «принцип эволюции», считаю необ-
ходимым отметить, что здесь понятие «принцип» не имеет ничего общего с тем по-
нятием, которое обычно вкладывается в это слово. Следуя авторам, впервые применившем его в этом смысле (Дорн, Клейненберг, Плате), Алексей Николаевич под
«принципами эволюции» подразумевает частные типы филогенетических изменений строения и функций органов, о которых более подробно будет сказано ниже.
БИОГРАФИЧЕСКИЙ ОЧЕРК
ние этого развития на каждом этапе эволюции и каким путем и способом, т. е. какими принципами и какими модусами филэмбриогенеза оно осуществлялось.
Мысль о чрезвычайной сложности эволюционного процесса, о невозможности построения упрощенных схем и проведения резких граней между отдельными принципами и модусами, переходящими в течение эволюции один в другой и сменяющими друг друга, красной нитью проходит через все работы Алексея Николаевича последнего периода его жизни (1931—-1936).
В своей работе «О модусах филэмбриогенеза», написанной в это время,1 излагая последний из открытых им модусов—способ периодического изменения хода развития органов—Алексей Николаевич пытается построить первую, еще грубую, классификацию модусов филэмбриогенеза, включая в их число, кроме модусов, указаных выше (анаболия, девиация, архаллаксис—положительные и отрицательные) также переселение клеток, равномерный и неравномерный рост, гистогенез и гетерохронию, т. е. более ранние и более поздние закладки органов. Говоря о том, что им сделано для теории филэмбриогенеза и что еще можно и должно для нее сделать, он пишет: «Идя по этому пути, морфолог может в будущем открыть еще несколько модусов филэмбриогенеза и дать им рациональную классификацию, но, конечно, он должен сознавать, что его теория есть только первое приближение к полной теории способов эволюции. Этой полной теории у нас в настоящее время еще нет, и мы можем только догадываться о том, как она будет создана. Нам думается, что в ближайшее время, на основании наших исследований, к созданию такой теории должны будут подойти, базируясь на своих исследованиях, экологи, генетики и биологи-экспериментаторы. Созданная ими теория, вероятно, будет вторым приближением к полной теории эволюции».
Но не только на экологов, генетиков и биологов-экспериментаторов рассчитывал Алексей Николаевич для создания этого «второго приближения к полной теории эволюции». Еще в большей мере возлагал он надежды на физиологов и на гистологов. В своих беседах с физиологами он неоднократно убеждал их в необходимости ввести исторический метод в обычные их способы исследования, в желательности поставить одною из целей этого исследования изучение эволюции функций, эволюции физиологических признаков, подобно тому как морфологи ставят своей целью изучение эволюции формы, эволюции морфологических признаков. Считая, что это является «труднейшей задачей», Алексей Николаевич тем более был доволен, что именно у нас в Союзе эта его пропаганда нашла наиболее живой отклик: в настоящее время уже две молодых школы физиологов (проф. Коштоянц и проф. Анохин) сознательно и планомерно идут по этому пути. Недавно вышедшая книга акад. И. И. Шмальгаузена1 2 показывает, что и механика развития в свою очередь начинает перебрасывать мост к основным проблемам эволюционной морфологии, пытается произвести широкий синтез между работами этих двух различных дисциплин.
Сам Алексей Николаевич в своей статье «Морфологическая и гистологическая рекапитуляция»3 затронул новую и интересную область исследования, а именно изучение эволюционных закономерностей гистогенеза. Ему удалось показать, что эволюция некоторых клеток (красных кровяных клеток и униполярных нервных клеток) и тканей (эпителиальной ткани, ткани электрических органов ската) шла по модусу анаболии, и что река-
1 Модусы филэмбриогенеза. Зоол. жури., т. 14, 1935.
s И. И. Шмальгаузен. Организм как целое в индивидуальном и историческом развитии. Изд. Акад. Наук СССР Москва, 1938.
3 Морфологическая и гистологии, рекапитуляция.Доклады Акад. Наук СССР,1935,.
БИОГРАФИЧЕСКИЙ ОЧЕРК
пигуляции в такой же мере существуют при гистогенетическом развитии клеток и тканей, как и при онтогенетическом развитии органов.
Наконец, Алексей Николаевич ставит коренной вопрос о том, как возник в течение эволюции онтогенез, как возникли и вариировали различные модусы филэмбриогенеза? При этом он приходит к выводу, что примитивный онтогенез впервые появился у первичных колониальных форм и удлинялся путем анаболии по мере развития и усложнения организации высших Metazoa. Примитивные архаллаксисы и девиации исторически появились позднее. Развитие онтогенеза Алексей Николаевич рассматривает как единственное средство, при помощи которого было достигнуто появление высоких форм жизни на земле.1
Что касается морфобиологической теории эволю-ц и и, то первые ее зачатки мы находим уже в «Этюдах по теории эволюции» (1912). Здесь Алексей Николаевич впервые устанавливает свои критерии прогрессивной и регрессивной эволюции животных, полное определение которых он дает лишь в «Главных направлениях эволюционного процесса» (1925).1 2 Определенно оставаясь на позиции эволюциониста-дарвиниста, сторонника эктогенетического толкования эволюции, Алексей Николаевич резко отмежевывается от идеалистического аутогенетического объяснения причин эволюционного прогресса.
Отправной точкой, исходя из которой Алексей Николаевич построил •свою морфобиологическую теорию эволюции, объединяющую проблему направлений эволюции, проблему принципов эволюции и проблему корреляций, был простой и логический для него вопрос, вопрос о том, почему в современной фауне существует такое громадное количество примитивных, специализованных и регрессивных форм, почему они не вымерли в течение эволюции, когда их близкие родичи развивались в прогрессивном направлении, почему не были вытеснены этими более высокими конкурентами? Ответом явилась разработка проблемы биологического прогресса и тех путей (направлений), которыми он осуществляется. Примитивные, специализованные и регрессивные формы потому не вымирают и не вытесняются прогрессивными формами, что все они в одинаковой мере идут по пути биологического прогресса, который, однако, далеко не всегда совпадает с прогрессом морфологическим и которому Алексей Николаевич дает совершенно четкое определение: «если какая-нибудь форма животных находит обильный корм, быстро размножается, мало истребляется врагами и, благодаря этому, сильно увеличивается численно, то она начинает расселяться и постепенно захватывать все новые и новые ареалы распространения; если она при этом распадается на разновидности, превращающиеся в новые виды, то данная группа изменяется прогрессивно, и этот прогресс носит биологический характер». При биологическом прогрессе организация животных может не только усложняться, но и упрощаться: явления увеличения численности, прогрессивного расселения и распадения на разновидности наблюдаются также и при превращении свободно живущих животных в паразитов, причем организация их сильно упрощается.
Какими общими путями достигаются эволюция приспособлений и биологический прогресс? В 1925 г. в первом издании «Главных направлений эволюционного процесса», Алексей Николаевич считал, что в природе существуют только три основных способа эволюции, три направления, следуя которым животные достигают биологического прогресса, или победы
1 Морфологические закономерности эволюционного процесса. Социал, рекопстр. и наука (Сорена), 1934.
2 В книге «Современные проблемы эволюции» (1914) Алексей Николаевич также подробно развивает эти мысли, но здесь его внимание сосредоточено главным образом на проблеме корреляций.
БИОГРАФИЧЕСКИЙ ОЧЕРК
49
в борьбе за существование. Это—ароморфоз, идиоадаптация и общая дегенерация. Только гораздо позже, в «Morphologische Gesetzmassigkeiten der Evolution», т. e. в 1931 г., он к этим трем направлениям прибавляет четвертое направление—ценогенезы.
Следуя по пути ароморфоза, животные приобретают такие морфофизиологические изменения организации, которые подымают их на более высокую ступень, повышая общую энергию их жизнедеятельности. Следуя по пути идиоадаптаций, животные приобретают такие приспособительные признаки, при которых организация их меняется, и иногда довольно резко, но общая энергия жизнедеятельности не повышается. Следуя по пути эмбриональных приспособлений (ценогенезов), зародыши приобретают такие приспособления, при которых общая энергия жизнедеятельности взрослых особей не повышается, но увеличивается их число (в связи с лучшим выживанием зародышей), что также ведет к биологическому прогрессу.. Следуя по пути общей дегенерации, организация животных упрощается, энергия их жизнедеятельности понижается, но способы защиты от врагов и способность размножения сильно повышаются, что обусловливает их победу в борьбе за существование.
В течение эволюции животные иногда переходят от одного направления к другому, от ароморфоза к идиоадаптации или к специализации, от ароморфоза к ценогенезу, от идиоадаптации к ароморфозу или к дегенерации и т. д. Однако случаев перехода от дегенерации к ароморфозу Алексей Николаевич не отмечает. Переход этот, т. е. изменение направления эволюционного процесса, зависит от характера и высоты организации изменяющегося животного в период, когда наступает изменение внешней среды, вызывающее соответственное изменение животного, от интенсивности и скорости изменения этой внешней среды и от количественного соотношения между изменениями среды и организацией и функциями животного. Этой работой, которая страшно его интересовала, Алексей Николаевич был занят (с перерывами) около семи-восьми лет.
Следующим этапом в развитии морфобиологической теории эволюции была разработка проблемы принципов эволюции. Этими принципами, или типами филогенетических изменений органов, Алексей Николаевич начинает заниматься примерно с 1926 года.
Если ароморфозы, идиоадаптации, ценогенезы и дегенерации представляют собой лишь очень общие пути биологического прогресса и определяют собою эволюцию организма, взятого как целое, то теория принципов эволюции говорит нам о том, какими частными филогенетическими изменениями строения и функций органов эти общие изменения направлений эволюции достигаются. Анализируя эти частные типы, или, следуя исторически сложившемуся названию, принципы филогенетических изменений, Алексей Николаевич в дополнение к трем известным принципам Дорна, Клейненберга и Плате, устанавливает в это время еще два новых принципа преобразования органов, именно принцип субституции функций и принцип фиксации фаз.
Проблема соотношения формы и функции до чрезвычайности занимает Алексея Николаевича, он весь отдается работе над принципами филогенетических изменений органов. Не счесть верст, отмеренных в большом его кабинете, не счесть пасьянсов, начатых и брошенных на преддиван-ном столике, когда с огромным напряжением, с упорным, «скрученным» вниманием обдумывал он эти принципы. Он поглощен ими. Он докладывает о них повсюду. На семинарии—своим ученикам и сотрудникам, в Обществе любителей естествознания, антропологии и этнографии, в Научно-исследовательском институте зоологии. В конце 1927 г. он едет в Ленинград на Съезд и докладывает о них ленинградскому Обществу Морфологические закономерности 4
БИОГРАФИЧЕСКИЙ ОЧЕРК
естествоиспытателей. В том же году, во время своей заграничной командировки, он снова делает о них доклад берлинскому Зоологическому обществу и Биологическому обществу в Стокгольме. Пишет о них предварительное сообщение1 и, связывая принцип субституции функций с разрабатываемой им в это время теорией редукции органов, докладывает о них на общем собрании Академии Наук,1 2 а затем на общем собрании Международного съезда зоологов в Падуе.3 Наконец, в целях пропаганды своих идей также и на Западе, дополняет и печатает в Швеции перевод своих «Главных направлений эволюционного процесса».4
С той поры Алексей Николаевич уже не оставляет работу над дальнейшим развитием проблемы принципов. В «Morphologische Gesetzmas-sigkeiten der Evolution» (1931) он делает огромный шаг вперед в. этом направлении, устанавливая еще четыре новых принципа эволюции: принцип активации функций, принцип иммобилизации частей, принцип симиляции и принцип разделения частей. При этом он подразделяет все известные в настоящее время типы (принципы) филетических изменений органов (теперь их 12) на две большие группы: типы (принципы) филетических изменений органов, которые сопровождаются лишь количественным изменением функции,—ее усилением или ослаблением, и типы (принципы) филетических изменений органов, которые сопровождаются качественным изменением функции.
В период 1932—1934 гг., при работе над эволюцией брюшных плавников, Алексею Николаевичу удалось установить еще один последний принцип эволюции, который он назвал «принципом выпадения промежуточной функции». Имея своим результатом усиление главной функции органа, он относится к первой группе принципов.
Что касается проблемы корреляций, то, хотя этот вопрос всегда горячо интересовал Алексея Николаевича,5 однако, предметом специального исследования, как это было, например, с вопросом о направлениях и принципах эволюции, или особенно с вопросом о филэмбриогенезе, он для него не служил никогда.
Первые обобщения, касающиеся корреляций, мы встречаем в «Этюдах по теории эволюции» (1912), где Алексей Николаевич, устанавливая понятие об экто- и энтосоматических органах, подразделяет изменения, образующиеся в них под воздействием внешней среды, на первичные (проталлаксисы) и на коррелятивно связанные с ними вторичные изменения (дейталлаксисы). Образующиеся при этом корреляции он расчленяет на прогрессивные и регрессивные, на соответственные и обратные.
В 1914 г. в книге «Современные проблемы эволюции» Алексей Николаевич снова затрагивает вопрос о корреляциях. Он подробно развивает «гипотезу корреляции», согласно которой эволюция идет путем подбора, соответственно меняющейся среде, лишь немногих наследственных изменений, непосредственно связанных с данной стороной среды. С такого рода изменениями связан ряд коррелятивных изменений в органах, функционально связанных с органами, испытавшими первичное изменение. Здесь Алексею Николаевичу удалось наметить правильное разрешение проблемы коадаптации органов, в свое время доставившей
1 О принципах филогенеза. Русск. зоол. жури., т. 8, 1928.
2 Дегенерация органов и субституция. Изв. Акад. Наук СССР, 1930.
3 Ueber die Bedeutungdes Prinzips der Substitution und einiger anderer Prinzipien in der Phylogenese. Arch. zool. ital., v. 16, Padova, 1931
4 Directions of Evolution. Acta zool. Bd. 10, 1929.
5 Мы упоминали уже о том, что, начиная с заметки об одноглазом скате (1898) и особенно с работы об эволюции черепа (1899), Алексей Николаевич постоянно изучал встречающиеся ему на пути исследования корреляции и обращал внимание своих учеников на громадное значение коррелятивных взаимоотношений органов в процессе эволюции.
БИОГРАФИЧЕСКИЙ ОЧЕРЕ
51
много неприятных затруднений дарвинистам. С той поры он надолго оставляет разработку этого вопроса. Только в «Morphologische Gesetz-massigkeiten der Evolution» (1931) он снова возвращается к проблеме корреляций и дает более или менее полную сводку этого вопроса, впервые вводя понятие о филетических координациях, которые он противопоставляет индивидуальным или физиологическим корреляциям. Филе-тические координации Алексей Николаевич подразделяет на морфофизиологические (цепи функционально связанных между собою органов и соотношений) и топографические, когда органы связаны между собою только по их взаимоположению. Подробно разбирая и те и другие, он устанавливает при этом различные типы филогенетических изменений корреляционных цепей.
Наконец, в самые последние годы своей жизни Алексей Николаевич , пытается объединить в одно целое морфобиологическую теорию эволюции с теорией филэмбриогенеза. Это удалось ему частично выполнить лишь в отделе, излагающем взаимоотношения частных типов (принципов) филогенетических изменений органов и модусов филэмбриогенеза. Он пытается разобраться в вопросе о том, в каких соотношениях стоят различные способы филэмбриогенеза к известным ему типам филогенетических изменений органов, иначе говоря, пытается определить, какими модусами филэмбриогенеза осуществляется тот или иной принцип эволюции. Здесь мы имеем перед собою не готовое исследование, но конспект будущих работ, рассчитанных на много лет-вперед, программу того, что он предполагал сделать (и что ему удалось фактически сделать только для одного принципа выпадения промежуточной функции1) в смысле дальнейшей разработки своих теорий.
Но это дело будущего, дело молодой смены, тех учеников и сотрудников, которым Алексей Николаевич посвятил труд своей жизни, свою книгу о «Морфологических закономерностях эволюции». Посмотрим, что дают нам в настоящем теория корреляций, установление общих направлений эволюции и разработка проблемы принципов, или частных изменений строения и функций органов? Дают уже теперь не мало. Как это полагал Алексей Николаевич, морфобиологическая теория эволюции должна внести распорядок в наше, пока еще довольно хаотическое, представление об общем ходе эволюционного процесса. Она позволяет нам для каждого частного случая эволюции установить, в каком направлении шли общие изменения организации той или иной животной формы при ее филогенетическом развитии; она помогает нам лучше, чем прежде, представать себе закономерную последовательность в развитии каждой конкретной формы.
Морфобиологическая' теория и теория филэмбриогенеза тесно связаны между собой, дополняя'друг друга. Первая раскрывает общие и частные закономерности прогрессивной и регрессивной эволюции, вторая разъясняет нам тот путь, которым возникают изменения в органах животного в течение его зародышевой жизни.
Морфобиологическая теория эволюции ставит исследование филогенеза на новые рельсы, делая упор не на форму, но на функцию исследуемого органа. Иначе говоря, она ставит на иную почву проблему соотношения между формой и функцией, перенося центр тяжести с изучения анатомических изменений органа на их функциональные изменения, вызванные адаптацией органов к изменяющимся в процессе эволюции условиям внешней среды. Изучая эволюцию какой-либо животной формы, или, более частно, какого-либо органа, мы можем теперь с достаточной подробностью наметить общий путь этой эволюции, определив, шла ли она
1 Evolution der Bauchflossen der Fische. Zool. Jahrb., Bd. 58, 1934.
БИОГРАФИЧЕСКИЙ ОЧЕРК
—и когда—путем ароморфоза, или идиоадаптации, или каким-либо иным из четырех известных нам путей; и если шла, то шла ли она этим путем всегда, или ее общее направление менялось, и если менялось, то когда и в какую сторону,—в сторону ли ценогенеза или общей дегенерации. Мы можем определить и причину, почему это произошло. Мы можем определить, путем каких частных изменений строения органов и функций, т. е. путем каких принципов эволюции, осуществилось то или иное изменение в ее общем направлении: принципом ли субституции функций, или смены их, принципом ли активации, симиляции или фиксации фаз, или, может быть, еще каким-либо новым, доселе еще не открытым принципом эволюции? Наконец,—и это, вероятно, самое главное,—руководствуясь теорией филэмбриогенеза как методической основой исследования, мы можем конкретно определить и то, как все это произошло, т. е. путем каких изменений в индивидуальном развитии органов предков, определяющих, по Алексею Николаевичу, филогенетическое развитие потомков, путем ли архаллаксиса, анаболии или девиации, некогда осуществилось у анцестральных зародышей то или иное частное изменение органов, .эволюирующих по тому или иному принципу эволюции.
В этом синтетическом сочетании теории направления эволюции, теории принципов эволюции, теории корреляций и теории филэмбриогенеза, в сущности, и кроется основное ядро того нового, оригинального и углубленного понимания процесса эволюции, раскрытие которого составляло жизненную задачу Алексея Николаевича Северцова.
Л. Северцова.
МОИМ УЧЕНИКАМ И СОТРУДНИКАМ
ПРЕДИСЛОВИЕ А. Н СЕВЕРЦОВА1
Задача и общий характер этой книги наиболее ясно вырисуются перед читателем, если он ознакомится с историей ее возникновения. Поэтому я позволю себе рассказать в нескольких словах о том, как я пришел к мысли о необходимости написать эту книгу.
Мои первые сравнительно анатомические работы (А. Н. Северцов, 1891—1908), так же как и работы большинства морфологов последарвинов-ского периода, имели целью исследовать ход филогенеза различных органов позвоночных животных. Около 1908 г., когда я закончил свое большое исследование по филогенезу конечностей наземных позвоночных, я поставил перед собой вопрос: не представляется ли возможным, на основании имеющегося в настоящее время сравнительно морфологического материала, подойти несколько ближе к проблеме исследования морфологических закономерностей эволюции и попытаться привести в некоторую систему огромное разнообразие филогенетических процессов? На первый взгляд эта задача казалась мне так трудно достижимой, что я долгое время оставался в нерешительности. Однако мысль, что некоторым морфологам XIX и XX столетия удавалось в своих специальных работах—стало быть, совершенно попутно—открыть некоторые морфологические закономерности эволюционного процесса, давала мне надежду, что, может быть, подобное мое начинание не окажется вполне бесплодным. К. Бэр открыл закон соотношения между временем появления признаков в эмбриональном развитии животных и степенью общности этих признаков, Ф. Мюллер открыл закон рекапитуляций, А. Дорн и Н. Клейненберг установили принципы смены функций и субституции органов, Л. Долло открыл принцип необратимости эволюции, Г. Осборн—«адаптивную радиацию» и т. д. Все эти полученные ими обобщения позволяли надеяться на благоприятный результат в том случае, если сознательно поставить перед собою целью исследование морфологических закономерностей эволюции.
Мне казалось наиболее правильным исходить из того, что уже является известным, поэтому я начал с переисследования закона рекапитуляций Мюллера-Геккеля. Результатом этой работы явилась книга «Этюды по теории эволюции. Индивидуальное развитие и эволюция», вышедшая в свет в 1912 г.
Работа над этой книгой показала мне, что наилучшим способом изучения интересующей меня проблемы является путь личного исследования детально обработанного возможно более обширного сравнительно морфологического материала по филогенезу органов какой-либо крупной группы Metazoa. Этого можно было достигнуть, во-первых, путем ряда личных планомерно проведенных исследований, во-вторых, путем использования результатов детально известных мне исследований моих сотрудников и учеников. Этот метод работы имел для меня то преимущество, что собран
1 Так как А. Н. Северцов не успел написать предисловия к настоящей книге, то мы берем в качестве такового предисловие, написанное им для «Morphologische Gesetzmassigkeiten der Evolution» (1931). (Прим, ped.)
56
ПРЕДИСЛОВИЕ
ный таким путем материал был мне известен вплоть до мельчайших подробностей, известен гораздо более, чем материал, почерпнутый из литературы. Этот метод работы давал мне также и то преимущество, что позволял расширить и углубить исследование в том направлении, какое представляло для меня в данный момент особый теоретический интерес. Результаты всех этих исследований (см. список литературы на стр. 59—67) послужили мне отправной точкой для моих дальнейших теоретических построений, касающихся морфологических закономерностей филогенетических изменений органов. Само собою разумеется, что при этом принимались во внимание также факты и выводы, взятые из литературы. Недостатком изложенного метода является его длительность. Предпринятые мною и моими учениками работы продолжались с 1908 по 1930 г. За это время было предпринято несколько экспедиций для сбора необходимого материала в различные области СССР: на Мурман, на Кавказ, в Туркестан, в Сибирь и на Дальний Восток. Я собирал материал также и на различных европейских зоологических станциях: в Неаполе, Виллафранке и т. д.
В качестве материала нам послужило большое число низших позвоночных: круглоротые, эласмобранхии (акулы, скаты, Holocephali), хрящевые ганоиды (Acipenseridae, Pseudoscaphirhynchus), костные ганоиды (Amia, Lepidosteus), кистеперые (Polypterus), очень многие костистые рыбы, бесхвостные и хвостатые амфибии и некоторые рептилии (многие ящерицы и некоторые черепахи).
Изобилие материала позволило произвести не только сравнительно анатомические, но и детальные сравнительно эмбриологические исследования. Результаты этих специальных исследований были опубликованы в период с 1908 по 1931 г. Из списка литературы читатель увидит, что очень многие работы касаются скелета позвоночных животных. Это произошло потому, что задачей наших работ было исследование филетических соотношений между различными формами позвоночных; для этого являлось весьма желательным иметь возможность сопоставить результаты сравнительно анатомических и сравнительно эмбриологических исследований с результатами палеонтологических работ, которые главным образом касаются скелетной системы животных. Вместе с тем мне представлялось важным понять функции реконструируемых нами анцестраль-ных органов; поэтому многие из упомянутых нами работ посвящены изучению органов движения (скелет и мускулатура), т. е. той системы органов, функция которой является для нас наиболее понятной.
В своих чисто филогенетических исследованиях я пришел к результатам, отличающимся от обычных взглядов. Эти результаты изложены в III—VI главах этой книги.1
Все эти исследования производились мною и моими учениками сначала в Зоотомической лаборатории Киевского университета, затем в Институте сравнительной анатомии Московского университета. Я должен упомянуть при этом имена И. И. Шмальгаузена, М. М. Воскобойникова, Д. Е. Беллинга, Б. С. Матвеева, С. А. Северцова, В. В. Васнецова, Е. В. Рылковой, С. Г. Крыжановского, М. Е. Макушка, А. А. Машковцева, С. Н. Боголюбского, А. Н. Дружинина, Н. Н. Дислера, А. Г. Рын-дзюнского, О. Н. Дислер, С. В. Емельянова, Е. Ф. Еремеевой, В. Ф. Еремеева, О. В. Белобородовой, А. И. Кривецкого, И. И. Бугаева, Т. С. Расса, В. А. Ракова. Эта исследовательская работа продолжалась, как это уже было сказано, более 20 лет (1908—1931) и продолжается и сейчас в том же направлении в руководимой мною лаборатории эволюционной морфологии Академии Наук СССР.
1 Главы V—VIII настоящей книги. (Прим, ред.)
ПРЕДИСЛОВИЕ
Как уже было указано, основная цель настоящего исследования состояла в установлении морфологических закономерностей филогенеза. Но можно поставить вопрос, возможно ли здесь вообще говорить о каких-либо закономерностях? Я подчеркиваю, что употребляю термин «закономерность» в том самом смысле, в каком он был употреблен моими предшественниками. Закон К. Бэра, принцип смены функций А. Дорна и т. п. вносят определенный распорядок в наши представления о том, как протекал процесс филогенетического изменения многих органов; обычно принимается, что этот процесс является результатом некоторых закономерных явлений. По моему мнению, несущественно,—каким термином мы обозначим эти или подобные этим обобщения: назовем ли мы их правилом, принципом или законом. Значение подобных обобщений заключается в том, что они позволяют нам лучше классифицировать и лучше понять ход филогенетического изменения органов животных, чем это можно было сделать раньше.
Основной план изложения результатов моих исследований был продиктован мне тем общим соображением., что надо исходить от известного к неизвестному. В первой части этой книги, после некоторых вводных замечаний, касающихся морфологической методики филогенетического исследования, я изложил сначала монофилетическую теорию в той форме, в какой я ее принимаю (гл. I, II),1 а зател; пришел к описанию результатов моих исследований по эволюции низших позвоночных (Acrania primitiva— Quadrupeda).1 2 Эти исследования могут служить читателю в качестве примера того материала, из которого я исходил в своих теоретических построениях, касающихся закономерностей филогенеза.
Здесь же я изложил некоторые сравнительно морфологические теории, к построению которых я пришел на основании результатов моих специальных исследований: теорию пребранхиальных висцеральных сегментов (бранхиомерии) головы, теорию происхождения покровных костей черепа, теорию сочленения челюстной дуги с осевым черепом и т. д. На основании результатов тех же специальных исследований я внес также и некоторые изменения в классификацию низших позвоночных, в особенности в классификацию Osteichthyes.
В главе VII3 я даю общую схему хода адаптивной эволюции, такой, какою она мне представляется на основании моих филогенетических исследо-ний. В VIII главе4 излагается учение об общих направлениях эволюционного процесса. В главе IX5 я пытаюсь дать общий предварительный очерк частных типов филогенетических изменений строения и функций органов животных. Глава Xе примыкает по своему содержанию к предшествующей главе: здесь я пытался дать морфобиологическую теорию редук
1 Глава IV настоящей книги (Прим. ред.).
2 В этой части нашей книги приведено несколько схем, иллюстрирующих предполагаемое строение гипотетических предков современных животных (схемы протокраниат, энтобранхиат, эктобранхиат и т. д.). На основании сравнительно морфологического исследования можно было восстановить многие признаки строения этих гипотетических форм. Излагая результаты своих исследований, я мог бы вместо схем просто привести самые наблюдения (эмбриологические, сравнительно анатомические и палеонтологические) и предоставить читателю самому сопоставить полученные результаты. Но это значительно увеличило бы объем филогенетической части книги и затруднило бы чтение. Я предпочел поэтому проделать эту работу сам и представить ее результаты в виде простых схем, дающих в наглядной и упрощенной форме краткий вывод из длительной работы научного исследования. Мне не нужно, я думаю, особенно подчеркивать то, что приведенные мною схемы являются именно схемами, а не реконструкциями.
3 Глава IX настоящей книги. (Прим, ред.)
4 Глава X настоящей книги. (Прим, ред.)
5 Глава XI настоящей книги. (Прим, ред.)
8 Глава XII настоящей книги. (Прим, ред.)
ПРЕДИСЛОВИЕ
ции органов. В двух следующих главах изложена проблема соотношений между филогенезом и онтогенезом как при прогрессивной (глава XI1), так и при регрессивной (глава XII1 2) эволюции органов многоклеточных животных. В главе XIII3 изложена трудная и наименее мною разработанная проблема филетических корреляций. Последняя глава (гл. XIV4) заключает в себе краткий обзор общих выводов, полученных в результате всей работы.
В этой книге нами изложены две тесно связанные между собой общие теории: морфобиологическая теория хода эволюции, раскрывающая общие и частные закономерности хода прогрессивной эволюции животных, и теория филэмбриогене-з а. Мне кажется, что нам не приходится особенно подчеркивать того, что обе эти теории являются морфологическими теориями эволюции, не занимающимися вопросами причинной связи между различными эволюционными процессами, но имеющими целью привести в порядок и объяснить эти многочисленные и разнообразные процессы. С моей точки зрения обе эти теории представляют собою необходимые предпосылки для каузального исследования филогенеза.
В особенности я подчеркиваю то, что обобщения, которые читатель найдет в этой книге, я не рассматриваю как законченные теории. Больше чем кто-либо я сознаю, как много в этих обобщениях гипотетичного и как много еще не достаточно проработано. Возможно, что некоторые мои выводы при дальнейшем исследовании будут изменены, но так как я всегда старался быть объективным и не связывать себя общепринятыми взглядами, то мне кажется, что моя попытка упорядочить наше представление о ходе эволюции не окажется бесплодной и что, исходя из него, можно будет и дальше разрабатывать намеченные здесь проблемы. Я убежден, что для морфологов, работающих над вопросами филогении, здесь открывается широкая и мало изученная область иследования, где еще можно найти много нового и много интересного.
Мне хочется закончить свое предисловие выражением моей искренней и глубокой благодарности всем моим ученикам и сотрудникам, которые оказали мне помощь в этой далеко не легкой работе. В особенности это касается моей жены, Л. Б. Северцовой, которая была мне верной помощницей как при сборе необходимого материала, так и при его обработке. Посвящение этой книги моим ученикам и сотрудникам пусть явится выражением этого чувства признательности.
Я очень благодарен также и тем моим коллегам, кто либо прислал мне материал для исследования, либо дал нужные литературные указания, либо помогал советом при работе над этой книгой, в особенности же проф. Н. Гольмгрену (Стокгольм), проф. Е. Стеншиб (Стокгольм), проф. Л. Ней-майеру (Мюнхен), проф. Г. Нилу (США).
1 Глава XIV настоящей книги (Прим, ред.)
2 Глава XV настоящей книги. (Прим, .ред.)
3 Глава XIII настоящей книги. (Прим, ред.)
4 Глава XVIII настоящей книги. (Прим, ред.)
СПИСОК ТРУДОВ А. Н. СЕВЕРЦОВА И ЕГО УЧЕНИКОВ,1 ИСПОЛЬЗОВАННЫХ ДЛЯ НАСТОЯЩЕЙ КНИГИ
Андреева Е. Г. О строении костей некоторых диких и домашних животных. Тр. Лаб. генет. Акад. Наук СССР, 1933.
— Морфологический анализ черепов взрослых гибридов и исходных форм Bovinae. Тр. Инет, гибрид, и акклим. животных (ВАСХНИЛ), т. 2, 1935.
— О пропорциях костей конечностей некоторых Bovinae. Докл. Акад. Наук СССР, т. 4, 1936.
— Окостенение скелета конечностей эмбрионов курдючной овцы. Докл. Акад. Наук СССР, т. 16, 1937.
— Палеофауна домашних и промысловых животных Кондраковского городища. Тр. Комисе, по происх., эвол. и породообр. дом. жив. Акад. Наук СССР, 1939.
— Анализ структуры костной ткани в поляризованном свете на примере изучения пястья лошади Пржевальского. Тр. Инет, эволюц. морф, им. А. Н. Северцова Акад. Наук СССР, т. II, вып. 4, 1939.
Беллинг Д. Е. Der Bau der vorderen paarigen Extremitaten u. des Schultergiirtels der Trigla im Zusammenhang mit dem Bau entspre-chender Organe beianderen Teleostei. Бюлл. Моск. общ. испыт. прир., 1911.
Белобородова О. В. К морфологии нервов-коллекторов Elasmo-branchii. Русск. зоол. журн., т. 8, 1928.
Боголюбский С. Н. Brustbein- u. Schultergiirtelentwicklung bei einigen Lacertilien. Zeitschr. f. wiss. Zool., Bd. 110, 1914.
— Регенерация подъязычного аппарата и нижней челюсти у тритона. Русск. зоол. журн., т. 4, 1924.
— К сравнительной морфологии подъязычного аппарата амфибий, ч. I—IV. Русск. зоол. журн., тт. 4 и 5, 1924 и 1925.
— Материалы по сравнительной краниологии северных собак (лаек). Бюлл. моек. общ. испыт. прир., 1925.
— Об аналогичных признаках в черепе собак. Русск. зоол. журн., т. 8, 1928.
— Происхождение верблюдов. Тр. общ. изучения Казахстана. Алма-Ата, 1929.
— Материалы по морфогенезу черепа Bovinae. Русск. зоол. журн., т. 10, 1930.
— Проблема доместикации в морфологии. Тр. IV Всесоюзн. съезда зоол., анат. и гистол. Киев, 1930.
— Проблема происхождения домашних животных. Тр. Лаб. генет. Акад. Наук СССР, 1933.
1 В список трудов учеников А. Н. Северцова внесены также работы, напечатанные после смерти А. Н. Северцова. Это сделано потому, что работы эти были ему известны с момента их возникновения, за много времени до того, как они появились в печати, а также и потому, что он учитывал их в своих выводах и обобщениях. (Прим, ред.)
60
СПИСОК ТРУДОВ А. И. СЕВЕРЦОВА И ЕГО УЧЕНИКОВ
Боголюбский С. Н. Эволюционная морфология и генетика. Природа, 1933.
— О типах хвостов овец и их развитии. Тр. Лаб. эволюц. морф. Акад. Наук, т. 2, 1934.
— Доместикационные изменения грудного аппарата баранов. Тр. Лаб. эволюц. морф. Акад. Наук, т. 2, 1934.
— Материалы по морфогенезу черепов гибридов Bovinae. Тр. Инет, гибрид, и акклим. животных ВАСХНИЛ, т. 2, 1935.
— Проблемы эволюционной морфологии домашних животных. Изв. Акад.Наук СССР, сер. биол., 1936.
— Опыт анализа комплекции разводимых Canidae. Тр. Инет, эволюц. морф. им. А. Н. Северцова Акад. Наук. СССР т. II, вып. 4, 1939.
— О путях овладения эволюцией домашних животных. Тр. Комисе, по происх., эвол. и породообразовании дом. жив. Акад. Наук СССР, 1939.
Бугаев. И. И. Uber das Pharyngmandibulare der Knorpelganoiden. Anat. Anz. Bd. 67, 1929.
— Ober den Ban des Oberkieferapparates bei den Acipenseroidei und niederen Haifischen. Anat. Anz., Bd. 68, 1930.
Васнецов В. В. О функции mesocoracoideum у костистых рыб. Русск. зоол. жури., т. 3, 1922—1923.
— Экспериментальные исследования функции мышц грудного плавника Teleostei, обладающих mesocoracoideum. Русск. зоол. жури., т. 5, 1925.
— Мускулатура грудного плавника Lophius piscatorius. Бюлл. Моск, общ. испыт природы, 1925.
— Развитие первичного плечевого пояса и скелета грудного плавника Lepidosteus osseus. Русск. зоол. жури., т. 7, 1927.
— Развитие плечевого пояса Amia calva. Русск. зоол. жури., т. 8, 1928.
— Zur Frage fiber die Evolution der Brustflosse bei Knochenfischen. Anat. Anz., Bd. 66, 1928.
— Zur Frage uber die Morphologic der Schwimmblase. Anat. Anz., Bd. 66, 1928.
— Uber die Morphologic der Schwimmblase. Zool. Jahrb., Bd. 56, 1932.
— Эволюция окраски костистых рыб. Тр. Лаб. эволюц. морфол. Акад. Наук СССР, т. 1, 1934.
— Опыт сравнительного анализа роста карповых рыб. Зоол. жури., т. 13, 1934.
— К морфологии жирового плавника костистых рыб. Тр. Лаб. эволюц. морфол. Акад. Наук СССР, т. 2, 1935.
— Пути филогенетических исследований костистых рыб. Извест. Акад. Наук СССР, сер. биол., № 5, 1936.
— Эволюция глоточных зубов карповых рыб. Сб. памяти А. Н. Северцова, т. 1, 1939.
Воскобойников М. М. Развитие висцерального скелета костистых рыб. Зап. Киевск. общ. естествоиспыт., т. 21, 1909.
— Этюды по бранхиомерии позвоночных. Ч. I, II, III Записки киевск. общ. естествоиспытат., т. 24, 1914.
— Висцеральный скелет Gnathostomata. Тр. I Всеросс. съезда зоол., анат. и гистол. Петроград, 1923.
— Эволюция функции жаберных лепестков рыб. Тр. I Всеросс. съезда зоол., анат и гистол. Петроград, 1923.
— О механизме кровообращения в жаберных лепестках Teleostei. Тр. II Всеросс. съезда зоол., анат и гистол. Москва, 1927.
— Эволюция функции жаберных лепестков рыб. Тр. II Всеросс. съезда зоол.. анат и гистол. Москва, 1927.
СПИСОК ТРУДОВ А. И. СЕВЕРЦОВА И ЕГО УЧЕНИКОВ
01
Войскобойников М. м. Аппарат дыхания у рыб. Тр. III Всеросс. съезда зоол., анат. и гистол. Ленинград, 1928.
— Закономерности в эволюции висцерального аппарата позвоночных. Тр. III Всеросс. съезда зоол., анат. и гистол. Ленинград, 1928.
— Der Apparat der Kiemenatmung bei den Fischen. Zool. Jahrb., Bd. 55, 1932.
Геро нимус E. C. Die Enwicklung des Brustflossenskeletts von Amia calva. Anat. Anz., Bd. 39, 1911.
Дислер H. H. и Северцов A. H. Das Pharyngomandibulare der Selachier Anat. Anz.. Bd. 58, 1924.
Дислер H. H. Строение кожных органов чувств головы акул. Сб. памяти А. Н. Северцова, т. 2 (в печати).
Дислер О. Н. Morphologic und Entwicklung der Augenmuskeln bei Knorpelganoiden. Anat. Anz., Bd. 70, 1930.
Домбровский Б. А. Сравнительно анатомический очерк проводящего звук аппарата земноводных.Зап. Киевск. общ. естествоиспытат., 1917.
— Барабанная полость Anura и брызгальце рыб. Русск. зоол. жури., т. 2, 1918.
— Строение слухового аппарата у ящериц. Русск. зоол. журн., т. 4, 1.924.
— Uber einige Gesetzmassigkeiten im Aufbau des schalleitenden u. Kiefferapparates der Tetrapoda. Zeitschr. f. Anat. u. Entwick-lungsgesch., Bd. 75, 1924.
— Слуховой аппарат у змей, черепах и крокодилов. Русск. зоол. журн., т. 4, 1924.
— Очерк среднего уха птиц. Русск. зоол. журн., т. 5, 1925.
— До бюморфологи слошв. Зап. юевск. вет. зоот. hict., 1925.
— Ein Versuch der Analyse einiger Korrelationen. Zeitschr. f. Anat? u. Entwicklungsgesch., Bd. 79, 1926.
— Der Suspensorialapparat der Ichthyopsiden. Ibid., Bd. 82, 1927.
— Zur Phylotektonik der Respirationsmuskulatur der Reptilien u. Saugetiere. Ibid., Bd. 93, 1930.
— Попытка классификации брюшной мускулатуры амниот. Тр. IV Всесоюзн. съезда зоол., анат. и гистол. Киев, 1930.
Дружинин А. Н. К вопросу о строении, функции и генезисе пояса конечностей у Delphinus del phis. Русск. зоол. журн,, т. 4, 1924. __=; | Новые данные об исходном положении конечностей Tetrapoda.
'Тр. II Всеросс. съезда зоол., анат. игистол. Москва, 1927.
— ' Общее в строении конечностей Dipnoi и Quadrupeda. Тр. Лаб. эволюц. морфол. Акад. Наук СССР, т. 1, 1932.
— К вопросу о строении и эволюции локтевого сустава млекопитающих. Докл. Акад. Наук СССР, т. 3 (12), 1936.
—К познанию анатомии яка (Poephagus grunniens L.). Докл. Акад. Наук СССР, т. 4(13), 1936.
— Сравнительно анатомическое описание яка и его гибридов. Извест. Акад. Наук СССР, 1937.
Емельянов С. В. К вопросу о развитии позвоночника у амфибий. Русск. зоол. журн., т. 5, 1925.
— Развитие ребер и соотношения их с позвоночником: ч. I. Развитие ребер амфибий. Русск. зоол. журн., т. 5, 1925; ч. II. Развитие ребер Acipenser, Amia, Lepidosteus и Polypterus. Ibid., т. 6, 1926; ч. III. Развитие ребер акуловых рыб. Ibid., т. 6, 1926; ч. IV. Развитие ребер и мускульных косточек у костистых рыб. Ibid. т. 8, 1928.
— Развитие ребер наземных позвоночных. Тр. Лаб. эволюц. морфол. Акад. Наук. СССР, т. 1, 1933.
СПИСОК ТРУДОВ А. н. северцова и его учеников
Емельянов С. В. Die Morphologie der Fischrippen. Zool. Jahrb., Bd. 61, 1935.
— Die Morphologie der Tetrapodenrippen. Zool. Jahrb., Bd. 62, 1936.
— Развитие и строение некоторых мало изученных элементов позво-М ночника рыб, их связь с непарными плавниками и некоторые вопросы теории конечностей. Сборник памяти акад. А. Н. Северцова т. 1, 1939.
Еремеева Е. Ф. Морфология и развитие глазных мускулов и межглазничной области костистых рыб. Зоол. журн., т. 11, 1932.
— Строение скелета парных плавников Palaeoniscidae Кара-Тау. Сб. памяти А. Н. Северцова, т. 2 (в печати).
Кривецкий А. И. К вопросу о морфологии элементов гиоидной дуги селахий. Русск. зоол. журн., т. 2, 1917.
— Мускулатура подъязычной дуги скатов. Русск. зоол. журн , т 5 1925.
К р ы ж а н о в с к и й С. Г. Развитие скелета парных плавников Pristiu-rus melanostomus. Тр. Инет, сравн. анат. Моск, унив., вып. 12, 1917.
— Развитие парных плавников Acipenser, Amia и Lepidosteus и вопросы теории конечностей. Тр. Научно-исслед. инет. зоол. Моск. унив. т. 1, 1925.
— Развитие артерий передних висцеральных сегментов Teleostomi. Тр. Ill Всеросс. съезда зоол., анат. и гистол. Ленинград, 1928.
— О биологическом значении псевдобранхии. Тр. Гидробиол. станции на Глубоком Озере, 1932.
— Органы дыхания личинок рыб и псевдобранхия. Тр. Лаб. эволюц. морф. Акад. Наук СССР, т. 1, 1933.
— Die Atmungsorgane der Fischlarven. Zool. Jahrb., Bd. 58, 1934.
— О зависимости миграции рыбца и шемаи от особенностей их развития. Тр. Новорос. биолог, станции, т. 2, 1936.
— Некоторые результаты гетерогенной гибридизации рыб. Тр. Новорос. биол. станции, т. 1 (н. с.). 1937.
— Принцип рекапитуляции и условия исторического понимания развития. Сб. памяти А. Н. Северцова, т. 1, 1939.
Курепина М. М. Развитие первичных хоан у амфибий: ч. I. Anura. Русск. зоол. журн., т. 6, 1926; ч. II. Urodela. Ibid., т. 7, 1927.
— К вопросу о сосудистом сплетении мозговых желудочков. Тр. III Всеросс. съезда зоол., анат. и гистол., Ленинград, 1928.
Ма кушо к М. Е. Zur Frage liber die phylogenetische Entwicklung der Lungen bei den Wirbeltieren. T. I—IV. Anat. Anz. 1911—1914.
— Ueber genetische Beziehungen zwischen Schwimmblase und Lungen. Anat. Anz., Bd. 44, 1913.
Матвеев Б. С. Метамерия головы селахий. Тр. Инет, сравн. анат. Моск, унив., т. 9, 1915.
— Сегментация головной мезодермы у амфибий. Русск. зоол. журн., т. 3, 1922—1923.
— Первичная сегментация головной мезодермы позвоночных животных. Тр. I Всерос. съезда зоол., анат. и гистол. Петроград, 1923.
— Развитие хрящевого черепа рыб. Бюлл. Моск. общ. испыт. прир., 1923.
— Строение эмбрионального осевого черепа низших рыб. Бюлл. Моск, общ. испыт. прир., т. 34,1925.
— Отношение сегментальных органов в голове позвоночных. Тр. II Всерос. съезда зоол., анат и гистол. Москва, 1927.
— Некоторые данные по развитию черепа черепахи и других позвоночных. Тр. II Всеросс. съезда зоол., анат. и гистол., Москва, 1927
— К вопросу о границе затылочной области позвоночных. Тр. 111 Всеросс. съезда зоол., анат. и гистол. Ленинград, 1928.
С ПИСОН ТРУДОВ А. Н. СЕВЕРЦОВА И ЕГО УЧЕНИКОВ
63
Матвеев Б. С. О явлении диплоспондилии в позвоночнике Cyprinidae при развитии. Тр. III Всерос. съезда зоол., анат. и гистол., Ленинград, 1928.
— Die Entwicklung der vorderen Wirbel u. des Weber’schen Appara-tes bei Cypriniden. Zool. Jahrb., Bd. 51, 1929.
— Zur Theorie der Rekapitulation (Ueber den Ursprung der Schuppen, Federn u. Haare auf dem Wege embryonal er Veranderungen). Zool. Jahrb., Bd. 54, 1932.
— Об-эволюции кожных покровов позвоночных путем эмбриональных изменений (К теории рекапитуляции). Зоол. журн., т. 12, 1933.
— Достижения эволюционной морфологии за 15 лет. Природа, 1933.
— О заимствовании материала при развитии органов. Бюлл. Научно-исслед. инет. зоол. Москов. унив., 1935.
— К вопросу о расхождении признаков в онтогенезе. Тр. Новорос. биол. станц., т 2, 1936.
— Современные задачи эволюционной морфологии. Извест. Акад. Наук СССР, сер. биол., № 5, 1936. л
— Задачи проблемы о соотношении онтогенеза и филогенеза. Извест. Акад. Наук СССР, сер. биол., № 1, 1937
Махотин А. А. Морфологическое значение придатков девятого абдоминального сегмента самцов Odonata. Зоол. журн., т. 13, 1934.
— К явлениям редукции в морфологии imagines зимней пяденицы (Operophtera brumata L.). Докл. Акад. Наук СССР, т. 2 (11), 1936. Машковцев A. A. Entwicklung des Urogenital systems bei den Knorpelganoiden. Anat. Anz., Bd. 59, 1925.
— Zur Phylogenie des Urogenital systems der Wirbeltiere auf Grund der Entwicklung des Mesoderms, des Pronephros, der Analbffnung und der Abdominalporen bei Acipenser stellatus. Zool. Jahrb., Bd. 48, 1926.
— Entfernung, Transplantation u. Entwicklung des Pronephros in vitro bei Siredon pisciformis u. Rana temporaria. Zool. Jahrb., Bd. 54, 1934.
— Der Genitalapparat der Acipenseridae, Zool. Jahrb., Bd. 56, 1934.
— - Zur Phylogenese der Geschlechtsdriisen u. der Geschlechtsausfuhr-gange bei Vertebratae. Zool. Jahrb., Bd. 56, 1934.
— Морфологические и филогенетические отношения мезонефроса к пронефросу. Тр. Лаб. эволюц. морф. Акад. Наук СССР, т. 2, 1934.
— Влияние горного климата на конституцию млекопитающих. Тр. Лаб. эволюц. морф. Акад. Наук СССР, т. 2, 1935.
— Роль функции при развитии органов позвоночных животных. Тр. Лаб. эволюц. морф. Акад. Наук СССР, т. 2, 1935.
— - Смена эндогенных и экзогенных факторов эмбрионального развития в онтогенезе и филогенезе (Эволюция онтогенеза). Извест. Акад. Наук СССР, Сер. биол., № 5, 1936.
— Морфофизиологические закономерности в развитии позвоночных животных. Роль внутренних и внешних морфогенетических факторов в развитии наружных жабр амфибий. Сб. памяти А. Н. Северцова, т. 1, 1939.
» Паншин Б. A. Die peripheren Nerven des Hechtes. Anat. Anz., Bd. 35, 1910.
Раков В. A. Zur Frage der Verknocherungen des Knorpelschadels der Acipeneriden. Zool. Jahrb., Bd. 57, '1933.
Расс T. С. Морфология крыши промежуточного мозга костистых рыб. Русск. зоол. журн., т. 8, 1928.
— Zur Morphologie des Gehirns der Knochenfische. Anat. Anz., Bd. 68, 1929.
64
СПИСОК ТРУДОВ А. И. СЕВЕРЦОВА И ЕГО УЧЕНИКОВ
Xj Р ы л к о в а Е. В. Некоторые данные по вопросу о трансплантации конечностей Amphibia anura. Тр. I Всеросс. съезда зоол., анат, и гистол., Петроград, 1923.
— К вопросу о происхождении и развитии полимелии у амфибий. Русск. зоол. журн., т. 4, 1924.
— Die Entwicklung der Schultermuskeln bei urodelen Amphibien, Zeitschr. f. wiss. Zool., Bd. 22, 1925.
Рындзюнский А. Г. Развитие воротной системы почек Trygon. Русск. зоол. журн., т. 9, 1929.
— К морфологии миотома рыб. Тр.Инст. эволюц. морфол. им. А. Н. Северцова, Акад. Наук СССР, т. II, вып. 4, 1939.
— Типы связей между преобразованиями органов в филогенезе. Сб. памяти А. Н. Северцова, т. 1, 1939.
Северцов А. Н. Uber einige Eigentumlichkeiten in der Entwicklung u. im Bau des Schadels von Pelobates fuscus. Бюлл. Моск. общ. испыт. прир., 1891.
Zur Frage fiber die Segmentierung des Kopfmesoderms bei Pelobates * fuscus. Бюлл. Моск. общ. испыт. прир., 1892.
— Die Entwicklung der Occipital region der niederen Vertebraten. Бюлл. Моск. общ. испыт. прир., 1895.
— Развитие затылочной области низших позвоночных в связи с вопросом о метамерии головы. Уч. зап. Моск, унив., вып. 11, 1895.
— Beitrag zur Entwicklungsgeschichte des Wirbeltierschadels. Anat. Anz., Bd., 13, 1897.
— Die Metamerie des Kofpes von Torpedo. Anat. Anz., Bd. 14, 1898.
— Очерки по истории развития головы позвоночных. I. Метамерия головы электрического ската. Уч. зап. Моск, унив., вып. 15, 1898.
— Studien zur Entwicklungsgeschichte des Wirbeltierkojjfes. I. Die Metamerie des Kopfes des Elektrischen Rochens. Бюлл. Моск. общ. испыт. прир., 1899.
— Die Entwicklung des Selachierschadels. Festschrift fiir C. v. Kupffer, Jena, 1899.
— Zur Entwicklungsgeschichte von Ascalabotes fascicularis. Anat. Anz., Bd. 18, 1900.
x, — Развитие пятипалой конечности позвоночных. Дневник XI Съезда 44 русских естествоиспытат. и врачей, 1901.
— Zur Entwicklungsgeshichte des Ceratodus Forsteri. Anat. Anz., Bd. 21, 1902.
4< — Die Entwicklung der pentadaktylen Extremitat der Wirbeltiere. Anat. Anz., Bd. 25, 1904.
— Studien fiber die Entwicklung der Muskeln, Nerven u. des Skeletts der Extremitaten der niederen Tetrapoda. Beitrage zu einer Theorie der pentadactylen extremitaten der Wirbeltiere. Бюлл. Моск. общ. испыт. прир., Москва, 1907 (отд. изд. 1908).
— Эволюция и эмбриология. Речь, произнесенная на соединенном заседании XII Съезда русских естествоиспыт. и врачей и Общ. испытат. природы, 1910.
— Die Kiemenbogennerven der Fische. Anat. Anz., Bd. 38., 1911.
— Висцеральный скелет циклостом. Протоколы Киевск. общ. естествоиспытат. Москва, 1911.
— Этюды по теории эволюции. Индивидуальное развитие и эволюция. Киев, 1912.
— Das Visceralskelett der Cyclostomen. Anat. Anz., Bd. 45, 1913.
— Современные задачи эволюционной теории. Изд. Bios. Москва, 1914.
— Развитие конечностей Chamaeleo bilineatus. Зоол. вестник, т. I, 1916.
СПИСОК ТРУДОВ А. II. северцова и его учеников
Северцов А. Н. Исследования об эволюции низших позвоночных. I. Морфология скелета и мускулатуры головы циклостом. Русск. арх. анат., гистол. и эмбр., т. 1, 1916.
— Исследования об эволюции низших позвоночных. И. Организация ближайших предков современных позвоночных. Русск. арх. анат., гистол. и эмбр., т. 2, 1917.
— О факторах, определяющих продолжительность жизни многоклеточных животных. Русск. зоол. журн., т. 2, 1917.
— Развитие жабр и жаберных сосудов рыб. Русск. зоол. журн., т. 2, 1917.
— Этюды по теории эволюции. 2-е изд. Госиздат, Берлин, 1922.
— Die Entwicklung der Kiemen u. Kiemenbogengefasse der Fiscbe. Zeitschr. ftir wissensch. Zool., Bd. 121, 1922.
— Эволюция и психика, изд. Сабашниковых, Москва, 1922.
— Положение хрящевых ганоидов в системе. Русск. зоол. журн., т 3, 1922.
— Die Morphologic des Visceralapparates der Elasmobranchier, Anat. Anz., Bd. 56, 1923.
— On the place of the cartilaginous Ganoids in the System andon the Evolution of the Osteichthyes. Journ. of Morph., v. 38, 1923.
— Das Pharyngomandibulare der Selachier (совм. с H. H. Дислером). Anat. Anz., Bd. 58, 1924.
— The development of the dorsal fin of Polypterus delhesi. Journ. of Morph., v. 38, 1924.
— Факторы прогрессивной эволюции у низших позвоночных. Русск. зоол. журн., т. 4, 1924.
— Zur Morphologic des Schadels von Polypterus delhesi. Anat. Anz., Bd. 59, 1924/1925.
— Uber die verschiedenen Richtungen des Evolutionsprocesses. Verh. der Zool.-Bot. Ges. Wien, Bd. 74—75, 1924/1925.
— Главные направления эволюционного процесса. Прогресс, регресс и адаптация. Москва, изд. Думнова, 1925.
— Исследования об эволюции низших позвоночных. III. Первичное строение висцерального скелета эласмобранхий. Русск. арх. анат., гистол. и эмбриол., т. 3, 1925.
— Обзор исследований по сравнительной морфологии позвоночных. Тр. Научно-иссл. инет. зоол. Моск, унив., т. 1, 1925.
— Развитие покровных частей черепа Amia calva. Бюлл. Моск. общ. испыт. прир., т. 34, 1925.
— Beitrage zu einer Theorie des Knochenschadels der Wirbeitiere. Anat. Anz., Bd. 60, 1925/1926.
— Development of the pelvic fins of Acipenser ruthenus. New data for the theory of the paired fins of fishes. Journ. of Morph.'and Physiol., v. 41, 1926.
— Die Morhologie der Brustflossen der Fische. Jen. Zeitschr., 1926.
— Studies on the bony skull of fishes. I Structure and development of the bony skull of Acipenser ruthenus. II. Primary structure of the bony scull of fishes. Quarterly Journ. of Microscop. Science, v 70, 1926.
— The development of the scales of Acipenser ruthenus. Journ. of Morph , v. 42, 1926.
— Der Ursprung der Quadrupeda. Palaontolog. Zeitschr., Bd. 8, 1926.
— Uber den Bau u. die Funktion der Rippen von Polypterus. Anat. Anz., Bd. 61, 1926.
— Beitrage zu einer Theorie des knochernen Schadels der Wirbeitiere. Palaont. Zeitschr., Bd. 8, 1926.
Морфологические закономерности
66 СПИСОК ТРУДОВ Л. И. СЕВЕРЦОВА И ЕРО УЧЕНИКОВ
Северцев А. Н. Uber die Beziehungen zwischen der Ontogenese u. der Phylogenese der Tiere. Jen. Zeitschr., Bd. 63, 1927.
— Etudes sur devolution des Vertebres inferieurs. III. Structure primitive de 1’appareil visceral des Elasmobranches. Publicazioni della Staz. Zool. di Napoli, v 8, 1927.
— Die Richtungen der phylogenetischen Entwicklung. Sitzungsber. der Ges. naturf. Freunde vom 11. Oktober. Jena. 1927.
— The head skeleton and muscles of Acipenser ruthenus. Acta zool., Bd. 9, 1928.
, — Einige Bemerkungen fiber die systematischen Beziehungen von Ana-spida zu den Cyklostomen u. Fischen. Palaont. Zeitschr., Bd. 10, 1928.
— О принципах филогенеза. Русск. зоол. журн., т. 8, 1928.
— Историческое направление в зоологии. Наука и техника СССР, т. 2, 1928.
— Directions of evolution. Acta zool., Bd. 10, 1929.
— Дегенерация органов и субституция. Извест. Акад. Наук СССР, 1.930.
— Uber die Bedeutung des Prinzips der Substitution u. einiger anderer Prinzipien in der Phylogenese. Arch. zool. ital., v. 16, 1931.
— Morphologische Gesetzmassigkeiten der Evolution. G. Fischer. Jena, 1931. ‘
— Studien uber die Reduktion der Organe der Wirbeltiere. Zool. Jahrb., Bd. 53, 1931.
— Rudimentation u. vollstandige Reduktion der Organe der Tiere. Arch. zool. ital., v 16. 1931.
— Die Entwicklung der Knochenschuppen von Polypterus delhesi. Jen. Ztschr., Bd. 67, 1932.
— Эволюционное учение после Дарвина (1859—1932). Зоол. журн., т. 11, 1932.
— - Естественный подбор и филэмбриогенез. Природа, 1932.
— Основные направления работ Лаборатории эволюционной морфологии Академии Наук СССР в области изучения морфологических закономерностей эволюционного процесса (за время 1931—1932 г ) Доклады Акад. Наук СССР, 1932.
— Эволюция брюшных плавников рыб и принцип выпадения промежуточных функций. Природа, 19.33.
— Главные направления эволюционного процесса. Биомедгиз, Москва, 1934.
— Evolution der Bauchflossen der Fische. Zool. Jahrb., Bd. 58, 1934.
— Морфологические закономерности эволюционного процесса. Социал, реконстр. и наука, 1934.
— Модусы филэмбриогенеза. Зоол. журн., т. 14, 1935.
— Морфологическая и гистологическая рекапитуляция. Докл. Акад. Наук. СССР, 1935.
Северцов С. А. История развития языка Chamaeleo bilineatus. Русск. зоол. журн., т. 3, 1922/1923.
— Биологический викариат и морфологические параллелизмы у млекопитающих. Тр. II Всерос. съезда зоол., анат. и гистол., Москва, 1927.
— Порядок появления прогрессивных признаков у млекопитающих. Тр. II Всерос. съезда зоол., анат. и гистол. Москва, 1927.
— О соотношении между плодовитостью и продолжительностью жизни млекопитающих. Извест. Акад. Наук. СССР, 1930.
— Материалы по биологии размножения тетеревиных; ч. I. Тр. Лаб. прикл. зоол. Акад. Наук СССР, 1932.
— Vom Massenwechsel bei den Wirbeltieren. Biol. Centralbl.,Bd.54,1934.
— Динамика стада у позвоночных. Извест. Акад. Наук. СССР, 1932.
СПИСОК ТРУДОВ А. Н. СЕВЕРЦОВА И ЕГО УЧЕНИКОВ
67
С верцов С. А. Материалы по биологии размножения тетеревиных, ч. II. Зоол. журн., т. 13, 1934.
— Морфологический прогресс и борьба за существование. Извест. Акад. Наук СССР, Сер. биол., № 5, 1936.
— Эволюционное учение и некоторые проблемы народного хозяйства. Извест. Акад. Наук СССР, Сер. биол., № 5, 1936.
Шахунянц Р. М. Морфологический анализ подъязычных аппаратов, у Amphibia anura. Тр. II Всеросс. съезда зоол., анат. и гистол. Москва, 1927.
— Развитие висцерального скелета у Amphibia anura в момент метаморфоза. (К вопросу о происхождении подъязычного аппарата,)* Русс. зоол. журн., т. 6, 1926.
Шмальгаузен И. И. Die Entwicklung der Lungen bei Tropidonotus natrix. Anat. Anz., Bd. 31, 1905.
\— Die Entwicklung des Skeletts der vorderen Extremitat der anuren Amphibien. Anat. Anz., Bd. 31, 1907.
— Die Entwicklung des Skeletts der hinteren Extremitat der anuren Amphibien. Anat. Anz., Bd. 33, 1908.
— Zur Morphologic des Saugetierfusses. Anat. Anz., Bd. 33, 1908.
— Die Entwicklung des Extremitatenskeletts von Salamandrella Kays. Anat. Anz., Bd. 37, 1910.
— Непарные плавники рыб и их филогенетическое развитие. Зап. , Киевск. общ. естествоиспыт., т. 23, 1913.
ч) — Развитие конечностей амфибий и их значение в вопросе о происхождении конечностей наземных позвоночных. Учен. зап. Моск, унив., 1915.
— On the median fin of the Dipnoi. Русск. зоол. журн., т. 1, 1916. О функциональном значении плавников рыб. Русск. зоол. журн., т. 1, 1916.
— О покровных костях плечевого пояса амфибий. Русск. зоол. журн., т. 2, 1917.
— О конечностях Ranidens sibiricus Kessl. Русск. зоол. журн., т. 2, 1917.
— Ober die Autostylie der Dipnoi u. Tetrapoda. Anat. Anz., Bd. 56, 1923.
— Der Suspensorialapparat der Fische u. das Problem der Gehorknochel-chen. Anat. Anz., Bd. 56, 1923.
— Значение аутостилии у Dipnoi и Tetrapoda. Русск. зоол. журн т. 3, 1923.
— Подвесочный аппарат рыб. Тр. I Всеросс. съезда зоол., анат. игистол. Петроград. 1923.
— Преобразование подвесочного аппарата у наземных позвоночных. Тр. 1 Всеросс. съезда зоол., анат. и гист. Петроград, 1923.
— Ober die Beeinflussung der Morphogenese der Extremitaten vom Axolotl durch verschiedene Factoren. Arch.f.Entw.Meeh., Bd. 105, 1925.
— К феногенетике некоторых морфологических признаков у домашних кур. Докл. Акад. Наук СССР, т 1, 1934.
— Коррелящя в розвитку деяких расових ознак у курей. Тр. 1нст„ зоол. та бюл. Всеукр. Акад. Наук, 1935.
— Основы сравнительной анатомии, 3-е изд. Москва, 1938.
— Значение корреляций в эволюции организмов. Сб. памяти А. I!.. Северцова, т. 1, 1939.
— Организм как целое в индивидуальном и историческом развитии.. Изд. Акад. Наук СССР, 1938.
ЧАСТЬ ПЕРВАЯ
ВВЕДЕНИЕ
In nova tert animus mutatas dicere formas Corpora.
P; Ovidius Naso, Metamorphoses, L. I.
ГЛАВА 1
ЭВОЛЮЦИОННОЕ УЧЕНИЕ ПОСЛЕ ДАРВИНА И ЗАДАЧИ ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИИ
С начала прошлого века цели и задачи морфологической науки претерпели многие и важные перемены. Если мы возьмем сравнительную анатомию Р. Оуэна или Ж. Кювье, то увидим, что задача тогдашней морфологической науки заключалась главным образом в раскрытии плана творения животного мира и в реконструкции архетипов (Р. Оуэн) больших групп животных. Сравнительная анатомия в этом раннем периоде своего развития открыла целый ряд гомологий в органах у систематически далеко отстоящих друг от друга животных, гомологий, которые при последующих проверках оказались правильными. В этом заключается и поныне ее огромное значение. Можно было бы сказать, что это направление сравнительной анатомии носило геометрический характер, иными словами, что закономерности гомологии, постигнуть которые стремились первые сравнительные анатомы, носили тот же характер сходства по местоположению, как и закономерности геометрии Эвклида (сходство в фигурах, формах и т. д.).
В последарвиновское время, т. е. примерно с шестидесятых годов прошлого столетия, морфология животных превратилась в историческую науку.
В «Происхождении видов» Дарвин сделал то, чего не удалось сделать Ламарку: он самым убедительным для современников образом доказал существование процесса эволюции в природе. На основании громадного числа систематически собранных палеонтологических, сравнительно анатомических, эмбриологических и зоогеографических данных ему удалось показать, что единственно вероятным объяснением всего характера современной фауны и флоры земного шара является гипотеза постепенного и очень медленного прогрессивного изменения строения живших в прежние геологические эпохи и в общем более'просто построенных предков современных животных и растений, т. е. сделать убедительной теорию постепенной прогрессивной эволюции.
Не останавливаясь на голом доказательстве факта эволюции, он дал кроме того в своей теории естественного подбора и наиболее вероятное, сохранившее свое научное значение и поныне, объяснение эволюционного процесса.
Сущность этого объяснения, как известно, заключается в следующем. У всех современных животных и растений существуют индивидуальные наследственные вариации, которые биологически являются полезными, индиферентными или вредными для их обладателей. При переменах условий существования, т. е. при усилении интенсивности борьбы за существование, животные, обладающие полезными вариациями, выживают в измененных условиях, т. е. организация потомков новой формы путем
72
накопления этих полезных вариаций изменяется в направлении большей приспособленности к новым условиям. Если условия среды в течение данного времени меняются все в том же направлении, то суммирование мелких наследственных вариаций приводит к крупным изменениям всего строения потомков, т. е. к образованию новых видов, родов, семейств ит. д., другими словами—к постепенной прогрессивной эволюции. В этом и заключается простая и гениальная основная мысль теории переживания наиболее приспособленных в борьбе за существование, или теории естественного подбора Ч. Дарвина, оказавшая столь значительное влияние на современников.
Действительно, мы видим, что противниками эволюционной теории и дарвинизма явились люди, a priori не допускавшие самой мысли о возможности какой-либо эволюции, т. е. главным образом представители протестантского и католического духовенства и их сторонники. Для характеристики этой оппозиции напомним знаменитый спор епископа Виль-берфорса с Т. Г. Гексли на съезде Британской ассоциации в Оксфорде, спор, окончившийся полным торжеством сторонников эволюции. Эта первая борьба за эволюционную идею длилась недолго: знаменитый геолог Ляйель, наиболее крупные биологи Англии (Гексли, Гукер, Ромене, Карпентер и др.), континента (Геккель, Фр. Мюллер, К. Фохт) и Америки (Аза-Грей и др.) приняли новую теорию, и к ним примкнуло большинство рядовых исследователей: зоологов, ботаников и палеонтологов.
Первая задача, которая встала перед сторонниками эволюционизма, состояла в восстановлении истории животного и растительного мира. Было ясно, что говорить об эволюции, не зная истории изменений, происшедших в строении организмов в течение прошлых геологических эпох, невозможно. В «Происхождении видов» Дарвин привел доказательства факта эволюции на ряде отдельных примеров, но филогенеза современных животных он не воссоздал, да одному человеку это было и не под силу. Однако без знания филогенеза невозможно было разрабатывать его теорию далее.
Задача эта, т. е. построение родословного древа» животного царства, представляет громадные трудности, и даже теперь, после 75 лет усиленной работы наиболее талантливых морфологов и палеонтологов Европы и Америки, мы хорошо знаем ход эволюции лишь крупных групп животного царства; эволюция мелких групп, т. е. происхождение видов в собственном смысле этого слова, изучена еще совершенно недостаточно.
Главной заслугой Э. Геккеля является то, что он вскоре после появления «Происхождения видов» осознал неотложность этой задачи и построил на основании всего бывшего в его распоряжении материала первое родословное древо животного царства. Его построения вызывали немало возражений. Были сделаны многочисленные попытки дискредитировать его выводы, но в настоящее время мы можем только удивляться морфологическому таланту Геккеля, так как длительная работа последующих поколений морфологов и палеонтологов в общем подтвердила результаты, полученные им еще в 1866 г.
Морфологические науки—сравнительная анатомия и эмбриология— прониклись эволюционными идеями и стали в центре интересов большинства зоологов 60—90-х годов прошлого века. Это время мы можем обозначить как филогенетический период зоологической науки.
Из представителей этого направления упомянем, кроме Геккеля, Т. Г. Гексли и Р. Ланкастера в Англии, К. Гегенбаура, М. Фюрбрингера, братьев О. и Р. Гертвигов, А. Дорна, К. Рабля в Германии, Годри во Франции, Долло и фан Вайя в Голландии, братьев А. О. и В. О. Ковалевских, И. И. Мечникова, В. В. Заленского в России, Копа, Марша и Осборна в Америке; к ним можно было бы прибавить много других имен ученых,
ЭВОЛЮЦИОННОЕ УЧЕНИЕ ПОСЛЕ ДАРВИНА
13
исследовательскими работами которых создано современное родословное древо животных.
Мы можем сказать, что относительно филогенетических соотношений большинства крупных систематических групп, т. е. классов, отрядов, в некоторых случаях семейств, входящих в главные типы животного царства, достигнуто соглашение между крупнейшими специалистами в этой области, так что теперь уже вполне возможно установить основы филогенетической классификации крупных групп животного царства.
Громадное значение для эволюционной теории имела упорная работа сравнительных анатомов и сравнительных эмбриологов этой эпохи над. филогенезом отдельных систем органов беспозвоночных и особенно позвоночных животных: на основании этих исследований удалось установить подробные филогенезы многих систем органов позвоночных и построить ряд важных морфологических теорий, дающйх ясное и детальное представление о ходе эволюции этих органов. Упомянем, например, теорию метамерии головы позвоночных, теорию позвоночника, теорию эволюции парных плавников и пятипалой конечности, теорию происхождения черепа и висцерального скелета, теорию эволюции мозга, нервов и органов чувств, теорию происхождения жабр и легких, выделительной системы и т. д. Значение всех этих разработанных сотнями исследователей морфологических теорий заключается не только в том, что они позволили проследить ход сложных эволюционных преобразований отдельных частей организмов и связать изменения формы и функции эволюировавших в течение прошлых геологических эпох органов, но и в том, что они дали возможность открыть ряд общих морфологических законов эволюционного процесса.
Так, Фр. Мюллером еще в 1864 г. был, во-первых, открыт закон рекапитуляции признаков предков в онтогенезе потомков, закон, обобщенный и разработанный Э. Геккелем и его учениками под именем «основного' биогенетического закона», а во-вторых—установлена связь между эмбриональным развитием и эволюцией взрослых животных. Фр. Мюллер высказал важную мысль, что ход эволюции органов взрослых потомков обусловлен изменениями хода эмбрионального и постэмбрионального развития органов их предков. Мысль эта, не оцененная и забытая его современниками, в настоящее время становится руководящей теорией эволюционноморфологического исследования. Затем А. Дорном и Н. Клейненбергом' были открыты два чрезвычайно важных принципа (типа) филогенетических изменений: принцип смены функций и принцип субституции (замещения) органов. Л. Долло открыл принцип необратимости эволюционного процесса, Осборн—принцип адаптивной радиации и т. д. Эти попутно сделанные открытия положили основание изучению морфологических закономерностей эволюционного процесса.
Несколько особое положение среди исследователей второй половины XIX века занял А. Вейсман. Дарвин признавал, что основным фактором, эволюции является естественный подбор мелких наследственных вариаций животных и растений, но допускал, что в некоторых случаях и изменения, происходящие в организмах вследствие непосредственного воздействия изменившихся внешних условий существования (факторы Ламарка и Э. Жоффруа Сент-Илера), могут сделаться наследственными и обусловить эволюционные изменения животных. Геккель, Спенсер и особенно американские зоологи и палеонтологи придали этим факторам первостепенное значение при объяснении причин эволюции (неоламаркизм). Вейсман же, частью на основании опытов, частью на основании теоретических соображений, пришел к отрицанию значения наследования таких «благоприобретенных изменений» и, вместе со вторым творцом теории естественного подбора—А.Р. Уоллесом, сделался основателем так называемого неодарви-
74
ГЛАВА I
низма, т. е. теории, признающей естественный подбор выгодных наследственных вариаций не только главным, но и единственным фактором эволюции. В связи со взглядом, что только наследственные изменения лежат в основе эволюционного процесса, Вейсман подробно разработал гипотезу наследственности; всем известна его теория строения зародышевой плазмы (биофоры, детерминанты, иды и т. д.), оказавшая большое влияние на теоретические взгляды современных генетиков (Морган и его последователи). К направлению Вейсмана примкнули многие зоологи в Англии и на континенте.
Эволюционное направление давало тон не только в области морфологических наук и учения о наследственности, но и в работах зоогеографов и зоологов-систематиков. Первые подошли в своих исследованиях к вопросам о первоначальных центрах видообразования и путях расселения совре-.менных животных форм с исторической, т . е. эволюционной точки зрения; вторые стали задаваться вопросами построения филогенетической систематики.
Меньшее влияние эволюционное учение оказало на работы палеонтологов 60—90-х годов. Хотя мы и можем указать на ряд блестящих филогенетических исследований в этой области,—назовем сделавшиеся классическими работы В . О. Ковалевского, Долло, Годри, Копа, Марша и др.,— однако очень большое число, палеонтологов сознательно отстранялось от филогенетических выводов, либо продолжая придерживаться традиций времен Кювье, либо, подобно К. Циттелю и его школе, признавая, что имеющиеся в распоряжении палеонтологов материалы недостаточны для филогенетических обобщений и выводов.
Сводя все сказанное об исследовательской работе зоологов в течение второй половины XIX века, мы должны признать, что эволюционное учение давало тон научной мысли и стояло в центре научно-теоретических обобщений биологов этого времени. В конце XIX и в начале XX века положение изменилось: на первый план биологических наук выступили новые научные дисциплины и—что, может быть, важнее,—новые методы исследования.
Не вдаваясь в подробности, отметим расцвет экспериментальной эмбриологии или механики развития, поставившей своей задачей экспериментальное изучение физиологии эмбриональных процессов развивающихся многоклеточных организмов (В. Ру, Вильсон, Дриш, Гербст, Бовери, Шпеман и др.). Перед зоологами и анатомами открылось громадное, до тех пор неведомое поле исследований, в котором каждая новая работа давала новые факты и ставила новые и интересные задачи. Вполне естественно, что масса молодых исследователей набросилась на изучение и разработку этой новой многообещающей дисциплины.
В 1900 г. Корренсом, де Фризом и Чермаком были вновь открыты законы Менделя и обоснована другая, не менее важная дисциплина— экспериментальное изучение явлений наследственности, или генетика. Мы упоминали, что уже Вейсман пытался в своей теории зародышевой плазмы связать явления наследственности со строением зародышевых клеток. Блестящие успехи цитологии позволили связать открытия генетиков с наблюдениями над структурой ядра половых клеток и создать цитогенетическую теорию наследственности (Бовери, Вильсон, Морган и др.).
Эти новые дисциплины отвлекли большое число исследователей в Европе и Америке от эволюционных проблем, а усиленный интерес к экспериментальным работам изменил их взгляды на значение фиологенетических исследований. Среди зоологов-экспериментаторов распространилось мнение, что ценных и, главное, точных результатов в биологических науках можно достигнуть только применением экспериментального метода исследования и что результаты, полученные описательными науками—сравни
ЭВОЛЮЦИОННОЕ УЧЕНИЕ ПОСЛЕ ДАРВИНА 75
тельной анатомией и эмбриологией, палеонтологией и зоогеографией— не точны, не научны, многие говорили—фантастичны. Эта оценка касалась главным образом филогенетических теорий. Даже очень крупные представители экспериментального направления, с мнением которых рядовым ученым приходилось считаться, придерживались этих взглядов. Так, наиболее выдающийся генетик Англии—Бэтсон высказался в том смысле, что эволюционная теория есть предмет научно недоказанной веры; аналогичное мнение было высказано и знаменитым исследователем чистых линий Иогансеном. Это, конечно, немало способствовало дискредитированию эволюционных и в частности филогенетических исследований, и новое поколение зоологов стало гораздо меньше интересоваться ими, чем их предшественники.
Аналогичное влияние имело увлечение зоологов конца XIX века возродившейся в это время виталистической натурфилософией (Дриш, Радль, отчасти Паули, Франсе, Дакке и др.), которая оказалась в общем враждебной эволюционным исследованиям.
Усиленная работа исследователей над вопросами эволюции, ознаменовавшая 60—90-е годы, ослабела; конец XIX и начало XX века являются периодом застоя эволюционных исследований зоологов.
Но постепенное углубление исследований экспериментального направления, главным образом генетиков, привело исследователей к тем же проблемам, которые стояли перед зоологами и ботаниками в 50-х годах прошлого века, т. е. к проблеме эволюции и ее причинного объяснения.
Именно усиленная исследовательская работа генетиков и громадные успехи, достигнутые ими, и привели их к постановке эволюционной проблемы. В начале своих работ они, как и сравнительные анатомы и зоологи-систематики времен Кювье, могли оставаться на статической точке зрения и изучать вопросы наследственности только у современных животных, не задумываясь над историческими проблемами. Однако, когда законы эти были более или менее изучены, возник вопрос о том, какую роль они играли в прошлом, т. е. в процессе эволюции. Можно было либо сознательно отказаться от решения этих вопросов, либо попытаться применить законы генетики к эволюционному процессу. Изучение чистых линий (Иогансен) сначала привело генетиков к воззрению, что наследственные факторы не менялись во времени, т. е. к гипотезе постоянства видов. Однако изучение внезапно возникающих мутаций и открытие зависимости наследственных мутаций от изменений внешних условий (температура, влажность, рентгеновские лучи и т. д.) привели к возможности снова подойти к эволюционным вопросам. Поэтому в настоящее время мы отнюдь не должны забывать громадного положительного значения исследований генетиков для объяснения эволюционного процесса, т. е. для дарвиновской теории естественного подбора. Учение Дарвина принимает, что в борьбе за существование переживают и дают потомство особи, обладающие полезными для них вариациями, что эти вариации передаются по наследству их потомкам, суммируются ит. д. Дарвин и его ближайшие последователи знали, конечно, факт наследования признаков, но законы наследования, которыми им приходилось оперировать, были им мало известны. Менделем же и его преемниками эти законы (законы преобладания, расщепления и самостоятельности признаков) были исследованы очень подробно. Были открыты спонтанные изменения наследственных факторов, новые мутации, и эти открытия дали новый материал для теоретиков наследственности и для морфологов и палеонтологов, которые не могли отказаться от результатов всей предшествующей работы.
В настоящее время началась усиленная разработка учения о доминантных и рецессивных мутациях, о частоте их и о причинах их возникновения. В этих работах, повидимому, начинает оживать дарвинистическое напра
76
ГЛАВА г
вление, к которому палеонтологи, склонные к неоламаркистским взглядам, относились очень отрицательно. Основные теории хода эволюционного процесса, т. е. теории Ламарка и Дарвина, были построены в эпоху, когда явления наследования индивидуальных вариаций, положенные Дарвином в основу теории естественного подбора, были мало известны. Мы имеем полное основание думать, что обширный и ценный материал, уже полученный генетиками, и результаты дальнейших исследований относительно мутаций будут использованы сторонниками естественного подбора.
В период, когда сравнительные анатомы и эмбриологи усиленно работали над филогенетическими проблемами, очень многие палеонтологи отстранились от этих вопросов, занимаясь вопросами палеонтологической систематики и выдвигая на первый план своих исследований проблемы исторической геологии (стратиграфии), для которой остатки ископаемых животных служили лишь подсобным материалом. Но в самом конце прошлого века и в начале текущего в руках палеонтологов накопилось громадное количество хорошо изученных ископаемых остатков прошлых фаун, и эволюционные вопросы сами собой выдвинулись и для них на первый план. Большое значение при этом имели исследования американской школы (Коп, Осборн, Грегори и др.) и новое палеобиологическое направление, основанное Долло и усиленно разрабатываемое Абелем и его школой.
В конце XIX и в начале XX века палеонтологическое исследование достигло громадных успехов как в направлении описания новых форм вымерших животных, так и в направлении изучения их биологии и родственных связей. Оставляя в стороне ценные результаты подробного изучения некоторых богато представленных палеонтологически типов беспозвоночных—кораллов, моллюсков, иглокожих и членистоногих,—остановимся очень коротко лишь на позвоночных животных. Обширные работы Р. Трэквера, А. С. Вудварда и Л. Долло обогатили наши сведения по систематике и анатомии ископаемых рыб громадным числом новых форм, строение и филогенетические соотношения которых были изучены с большой подробностью. Блестящие исследования шведских и норвежских палеонтологов, особенно Э. Стеншиб (1927) и И. Киэра, с неожиданной полнотой выяснили не только строение скелета, но и целого ряда внутренних органов—нервной системы, органов чувств, кровеносной системы, органов дыхания ит. д.—древнейших водных позвоночных—Osteostraci. Оказалось, что фауне рыб, от которых произошли все высшие наземные позвоночные, предшествовала в силуре и девоне большая, очень богатая представителями и крайне разнообразная фауна водных позвоночных, происшедших от общих с рыбами предков; единственными остатками этой богатой фауны в настоящее время являются круглоротые наших пресных и морских вод— миноги и миксины. Весьма интересно, что результаты последних палеонтологических исследований вполне подтверждают результаты новейших сравнительно морфологических работ (А. Н. Северцов, 1910—1917). Исследования того же Стеншиб и некоторых немецких палеонтологов восстанавливают с большой подробностью анатомию и филогенетические соотношения древнейших родичей настоящих рыб—Arthrodira и Antiarchi.
Вопрос о происхождении низших наземных позвоночных от рыб разработан главным образом благодаря исследованиям американских палеонтологов (Грегори и его сотрудников), которые показали, что низшие амфибии (стегоцефалы) произошли, по всей вероятности, от форм, близких к палеозойским кистеперым рыбам (Sauripterus), причем отделение их от ствола рыб имело место еще в верхнем силуре. Изучение строения стегоцефалов было продвинуто работами Ватсона, Кэза и Виллистона.
Массу нового дала усиленная разработка палеонтологами филогене
ЭВОЛЮЦИОННОЕ УЧЕНИЕ ПОСЛЕ ДАРВИНА Я
тических соотношений рептилий. В настоящее время установлено, например, что еще очень рано (в начале каменноугольного периода) от примитивных панцырных амфибий (стегоцефалов) отделились предки рептилий, которые в течение пермского периода и триаса распались на большое число приспособленных к различным условиям обитания групп. Некоторые формы приспособились к различным родам наземного образа жизни, другие (мозозавры, ихтиозавры, плезиозавры) снова перешли к жизни в водной среде, из которой вышли их амфибийные предки, наконец третьи перешли к воздушному образу жизни и сделались летающими животными (рамфоринхи, птеродактили и т. д.). Филогенез этих весьма разнообразных форм теперь, благодаря интенсивной исследовательской работе палеонтологов и богатству и полноте найденных ископаемых остатков, известен с большой подробностью.
Меньше успехов, вследствие бедности сохранившихся остатков, достигнуто в отношении филогенеза птиц, ответвившихся от своих рептилийных предков еще в юре. Зато очень интересные и ценные результаты получены относительно эволюции главных групп млекопитающих и, в частности, человека. Благодаря громадному числу новых находок ископаемых остатков, филогенетические соотношения многочисленных групп современных и вымерших млекопитающих в настоящее время выяснились в значительной степени,и палеонтологи уже могут построить родословное древо не только отрядов, но и большинства семейств этого класса.
Трудный вопрос о происхождении млекопитающих от рептилий теперь также выяснен: предками первичных млекопитающих являются пермские териодонты, у которых были развиты многие характерные для млекопитающих признаки. Эти рептилийные предки млекопитающих в позднепермские времена или в начале триаса превратились в настоящих, по всей вероятности, еще яйцекладных млекопитающих.
Боковыми ветвями этого раннего ствола мезозойских млекопитающих являются ископаемые Multituberculata и современные Prototheria (австралийские утконос и ехидна) и большая группа сумчатых млекопитающих. Весьма вероятно, что еще в мезозое развились типичные плацентарные млекопитающие, представленные большим числом мелких лазающих форм, близких к насекомоядным.
Знаменитые центральноазиатские экспедиции, организованные нью-иоркским естественно-историческим музеем (1916—1927), в числе многих ценных находок привезли также найденные в меловых отложениях пустыни Гоби черепа двух мелких, близких к насекомоядным млекопитающих (Zalambdalestes, Deltatherium), бывших современниками гигантских динозавров мелового периода. Эти формы принадлежат к предкам плацентарных млекопитающих, вытеснивших в течение кайнозоя всю богатую реп-тилийную фауну мезозойской эры.
Не вдаваясь в подробности долгой эволюции отдельных отрядов млекопитающих, остановимся очень коротко на новых данных относительно эволюции человека, т. е. на вопросе, который особенно интересовал Дарвина. В настоящее время, благодаря усиленной исследовательской работе палеонтологов и анатомов и новым находкам, вопрос о родословной человека может считаться решенным.
Можно признать, что отдаленными предками Hominidae были эоценовые приматы, близкие к Notharctus, от которых в олигоцене произошли антропоидные обезьяны (Propliopitheci). Непосредственными предшественниками человекоподобных форм была группа Dryopitheci. Потомками живших на деревьях Dryopitheci являются питекантропы, которые в строении зубов и черепа еще сохранили обезьяноподобный характер, но перешли уже к двуногому способу передвижения, т. е. к хождению и беганию по земле.
78
ГЛАВА I
Сомнения относительно строения и систематического положения яванского Pithecanthropus (Вирхов и др.) теперь, благодаря новым находкам, разрешены в пользу взглядов Е. Дюбуа. На материке Азии, близ Пекина, при раскопках холма Шу-Кути-эн было найдено несколько хорошо сохранившихся черепов формы если не тождественной, то чрезвычайно близкой к питекантропу—Sinanthropus pekinensis. Тщательное изучение этих черепов позволило восстановить строение и установить систематическое положение этой группы антропоидов.
Теперь мы можем с уверенностью сказать, что Дюбуа действительно нашел переходную форму между человеком и антропоидными обезьянами, форму, более примитивную, чем все известные нам формы человека, но более высокую, чем все известные нам современные и ископаемые формы обезьян. Очень важно и то, что теперь мы знаем, что питекантропоидные предки человека были очень широко распространенными животными. Географическая область их распространения простиралась от центрального Китая до Зондского архипелага, т. е. до самого южного края громадного азиатского материка.
Таким образом, вопрос о происхождении человека теперь разрешен в том смысле, как это предполагал Дарвин еще в 1871 г. в своем «Происхождении человека».
Мы до сих пор говорили только о филогенетических результатах современной палеонтологии. Необходимо отметить и крупные достижения наиболее новой ветви этой науки—палеобиологии.
Это направление идет от работ В. О. Ковалевского. Л. Долло, продолжая работу Ковалевского, поставил себе задачу ’связать фиологенетиче-ские исследования с изменениями той биологической и органической среды, в которой обитали эволюировавшие формы животных, т. е., другими словами, проследить филогенез образа жизни вымерших форм. Начав, с палеобиологии водных животных, Долло в ряде работ наметил главные пути этой новой дисциплины. Продолжателями этого направления являются О. Абель и его многочисленные ученики. На основании обстоятельного изучения, ими восстановлены строение и функции органов весьма многих вымерших животных, причем им удалось (на основании сравнения с современными формами) установить также способы их передвижения, роды пищи и т. д. Критическое изучение слоев, где были найдены ископаемые остатки, позволило выяснить биологическую обстановку, в которой жили эти формы, а также все изменения строения и обстановки, происшедшие в течение филогенеза. Не останавливаясь на громадном значении этой дисциплины, напомним читателю о последнем исследовании О. Абеля в этой области, а именно об исследовании по эволюции пещерных медведей (Ursus spelaeus) Миксницкой пещеры, где впервые жизнь вида, условия его расцвета, упадка и вымирания были прослежены шаг за шагом за очень долгий период времени—приблизительно за 170 000 лет.1
Некоторые результаты исследований палеонтологов начала XX века были подкреплены и дополнены исследованиями сравнительных анатомов и эмбриологов, которые не прекратили своей исследовательской работы во время упомянутого периода застоя на границе XIX и XX столетий и прибавили много новых данных по филогенезу животных.
Упомянем ряд крупных имен морфологов, работавших в начале нашего века. Отметим работы Э. Гудрича (Оксфорд) о выделительной системе и кожном скелете низших позвоночных, к которым примыкают работы его ученика де Беера; весьма интересны ценные эмбриологические исследования Г. Кэрра по низшим рыбам (Crossopterygii, Dipnoi) и Эджеворта по мускулатуре.
1 A b е 1 О. u. Kyrle I., Die Drachenhoie bei Mixnitz, Wien, 1931.
ЭВОЛЮЦИОННОЕ УЧЕНИЕ ПОСЛЕ ДАРВИНА
В Австрии надо отметить филогенетические исследования И. Вершлейса (I. Versluys) по морфологии черепа рептилий, в Германии—Л. Неймайера по центральной и периферической нервной системе, Г. Маркуса по развитию* органов гимнофион, В. Франца по строению и развитию ланцетника, О. Вейта.по развитию скелета низших Teleostomi и т. д. В Швеции многочисленны и весьма ценны работы Н. Гольмгрена и его учеников по морфологии центральной нервной системы. В Голландии интересны неврологические работы палеонтолога Е. Дюбуа. Из американских морфологов, отметим исследования Г. В. Нила по метамерии головы, Джонстона по морфологии центральной нервной системы и Элиса—по скелету и мускулатуре рыб.
Исследования только что названных авторов (мы упомянули лишь, наиболее известных) показывают, что эволюционное направление в работах морфологов в занимающую нас эпоху, хотя и ослабело под влиянием, роста экспериментальных дисциплин, но далеко не прекратилось.
- Е нашей стране общий интерес к вопросам эволюции и, в частности, филогенеза в начале века сильно понизился, но ряд морфологов продолжал упорно заниматься филогенетическими исследованиями, причем ими поставлены- перед собой и некоторые новые цели исследования. Изложению этих целей, методов их выполнения и достигнутых в этом отношении результатов и посвящена книга, ныне предлагаемая вниманию читателя.
Для нашего времени весьма примечательно то сближение, которое наблюдается между тремя морфологическими науками: сравнительной анатомией, палеонтологией и эмбриологией. Исследователю, работающему в настоящее время над палеонтологическим материалом, зачастую приходится прибегать к результатам, добытым эмбриологическим и сравнительно анатомическим исследованием, и, наоборот, сравнительный анатом и эмбриолог, лишь только он коснется в своих работах филогенетических проблем,, не может не считаться с результатами палеонтологического исследования. Повидимому, уже приближается то время, когда эти три в прежнее время столь отдаленные друг от друга морфологические дисциплины составят' одну науку—ф илогенетическую морфологию. Главнейшей целью этой науки будет начатое еще со времен Геккеля построение общего родословного древа животного мира со всеми его важнейшими ответвлениями, иными словами—установление филогенетических связей между всеми известными нам современными и вымершими животными. Результаты этого направления исследования выразятся в появлении полной филогенетической систематики животного царства.
Совершенно ясно, что подобная цель является вполне законной для всякого исследователя, занимающегося эволюционной проблемой, и что филогенез животных должен быть исследован со всей возможной точностью и подробностью всеми способами, имеющимися в нашем распоряжении; иначе всякая теория эволюции будет представлять собою лишь абстрактную схему, лишенную конкретного содержания.
Мы знаем, что многие биологи преимущественно заняты теориями, дающими каузальное объяснение происхождению животного мира (дарвинизм и неодарвинизм, ламаркизм и неоламаркизм, различные ортогенетические воззрения и т. д.); ясно, однако, что все эти теории при отсутствии филогенетической базы могут оперировать лишь одними абстрактными понятиями. Поэтому мы считаем, что поставленная Геккелем задача построения филогенетического родословного древа и филогенетической систематики и в настоящее время остается правильной. Но мы оставляем за собой право поставить вопрос о том, исчерпываются ли одним этим все цели и все проблемы филогенетического исследования?
Уже довольно давно явилась у нас мысль, что настало время, когда.
«о
ГЛАВА I
на основании филогенетических исследований можно было бы вплотную подойти к изучению морфологических закономерностей филогенеза. В связи с этим изменилась цель наших исследований. Реконструирование хода филогенеза и последовательности в наступлении изменений ванцестральных органах сменилось в наших исследованиях целеустремленным исканием обобщений, вносящих известный закономерный распорядок в великое разнообразие филогенетического процесса. Детальное изложение этих обобщений читатель найдет в третьей и четвертой частях нашей книги, посвященных обоснованию морфобиологической теории хода эволюции и теории филэмбриогенеза.
В настоящее время перед эволюционным учением стоят две основные проблемы, а именно: 1) изучение хода эволюции и ее закономерностей и 2) изучение причин филогенетических изменений организмов. Наш очень краткий обзор показал, что в течение 75 лет, протекших со времени выхода в свет «Происхождения видов» Дарвина, палеонтологами и морфологами немало сделано для выяснения первой из этих задач, т. е. построения родословного древа животного царства; ими же положено начало систематическому изучению закономерностей филогенеза. Если их исследовательская работа пойдет дальше в том же направлении, то можно надеяться, что ее очередная задача, т. е. раскрытие филогенеза малых систематических групп, скоро приблизится к разрешению.
Хуже обстоит дело со второй задачей дарвинизма, а именно с причинным объяснением эволюции. Выше мы отметили, что, несмотря на блестящие успехи теории наследственности, результаты генетических исследований мало применялись для разрешения эволюционных вопросов. Еще меньше сделано в этом направлении эмбриологами-экспериментаторами.
И в настоящее время, как во времена, непосредственно следовавшие за выходом в свет основных работ Дарвина, происходят споры о двух основных теориях причинного объяснения эволюции—дарвинизме и ламаркизме, мало переменивших свое содержание при их превращении в неодарвинизм и неоламаркизм. К сожалению, мы видим, что разработка этих теорий мало продвинута зоологами-экспериментаторами; то же мы можем сказать и о разработке биологической стороны эволюционного учения, т. е. о дисциплине, занимающейся изучением биологических соотношений между современными животными и той средой, в которой они живут. Несмотря на значение таких исследований, как работы Р. Гессе, для причинного объяснения эволюционного процесса здесь сделано еще очень мало.
О причинном объяснении эволюции нам предстоит высказаться в следующей главе, разбирая вопрос о несостоятельности неоламаркизма.
ГЛАВА II
НЕОЛАМАРКИЗМ
АУТОГЕНЕЗ И ЭКТОГЕНЕЗ
Хотя непосредственной задачей наших личных исследований является вопрос не о том, почему происходит эволюция животных, а о том, как она происходит и каким законам она подчиняется, однако, прежде чем перейти к обсуждению проблемы морфологических закономерностей эволюционного процесса, являющейся предметом настоящей книги, нам представляется необходимым изложить нашу точку зрения по вопросу о причинах эволюции. Для этого мы считаем нужным, коснувшись вкратце вопроса об аутогенезе, который в настоящее время уже не имеет актуального значения, подробно остановиться на вопросах об ортогенезе и особенно о неоламаркизме, ибо в последние годы некоторая часть западноевропейских ученых снова пытается выдвинуть неоламаркистскую теорию.
Мы не будем останавливаться на теории естественного подбора, во-первых, потому, что мы упоминали о ней в предыдущей главе, а во-вторых, потому, что наша критика аргументации неоламаркизма, мне кажется, с достаточной ясностью выразит дарвинистическую точку зрения автора.
Как известно, некоторыми исследователями и натурфилософами считается принятым, что эволюция приспособлений происходит в силу действия известного внутреннего, так сказать, имманентного эволюционного принципа, теоретически стоящего вне всякой зависимости от изменений, имеющих место во внешней среде (аутогенез, теория К. Бэра, батмизм Копа, воззрения многих представителей неовитализма и т. д.). Другие исследователи считают, что главным стимулом филогенетических изменений являются изменения, происходящие в окружающей среде (эктогенетические теории Бюффона, Ламарка, отчасти Дарвина и его новейших последователей).
Мне лично существование имманентного аутогенетического принципа в эволюции всегда казалось невероятным именно потому, что, как это мы прекрасно знаем и как об этом будем еще говорить подробно ниже, все организмы вплоть до мельчайших деталей приспособлены к окружающей среде. Если мы примем наличие аутогенетического принципа в эволюции, то нам придется вместе с тем принять, что в живой и неживой природе существует некоторая предопределенная гармония изменений в смысле Лейбница, т. е. мы должны будем предположить, что каждому изменению среды (а они бесчисленны) соответствует заранее предустановленное, вполне определенное и целесообразное изменение организации животного. При этом (по посылке) эти изменения должны быть независимы друг от друга, т. е., употребляя известный пример, мы имели бы здесь нечто аналогичное двум часовым механизмам, которые, не будучи связаны друг с другом, так изначально регулированы искусным часовщиком, что всегда идут согласно, не отставая и не опережая друг друга.
Морфологические закономерности
б
82
ГЛАВА II
Но в живой природе мы должны были бы предположить существование невероятно большого числа самых разнообразных сортов часовых механизмов (живые организмы), из которых каждый урегулирован по целому ряду других механизмов иного устройства (изменяющиеся внешние условия) и притом так, что все эти часы не только идут верно, но и запаздывают и спешат гармонично друг с другом. Такое истолкование эволюционного процесса является, по-моему, по меньшей мере невероятным, и если мы в него вдумаемся, то увидим, что в смысле «объяснения» оно нам ничего не дает, так как в сущности мы в нем находим только констатирование факта приспособленности, но не объяснение его.
Мне кажется поэтому, что многое говорит гораздо более в пользу экто-генетической гипотезы эволюции, чем в пользу аутогенеза. По всей видимости, причины филетических изменений в организации животных следует искать в изменениях внешней среды. Если мы этого принципа не примем, то нам останется совершенно непонятным тот общий факт, что организация и функции всех известных нам животных приспособлены до мельчайших подробностей к бесконечно разнообразным условиям среды, окружающей этих животных. Мы считаем при этом, что влияние косвенного (не прямого) воздействия среды на организм (дарвиновский фактор эволюции) имеет неизмеримо большее значение, чем все еще недоказанное, как мы это увидим ниже, влияние непосредственного воздействия условий внешней среды и чем, в особенности, еще более голословное утверждение наследования результатов упражнения и неупражнения органов (факторыЖ. Сент-Илера, Бюффона и Ламарка).
ЛАМАРКИЗМ И НЕОЛАМАРКИЗМ
Изучая эволюционную литературу, мы можем сказать, что к дарвинизму в настоящее время склоняются ученые, занимающиеся преимущественно филогенетическими и генетическими исследованиями, теории же Ламарка (точнее неоламаркизма) придерживаются некоторые систематики, физиологи и палеонтологи.
Ознакомление с литературой показывает, что эти теории являются далеко не равноценными по своей разработанности, т. е. в смысле приложения их к объяснению различных случаев приспособительных филогенетических изменений организации животных и растений. Более детально и широко разработана теория Дарвина. Самим Дарвином, затем Уоллесом и целым рядом выдающихся дарвинистов1 теория естественного подбора, как принципа, объясняющего развитие приспособительных изменений животных и растений, разобрана на громадном числе примеров, а многочисленные возражения на нее критически обсуждены.
Теория Ламарка находится в гораздо менее благоприятном положении в смысле разработанности. Первое ее изложение, сделанное Ламарком в его «Философии зоологии» (1809), было дано в очень общей и мало убедительной форме как для современников, так и для последующих поколений ученых. И позже, когда по почину Э. Геккеля интерес эволюционистов обратился к принципам Ламарка и в биологии возникло неоламаркистское движение, сторонники неоламаркизма обратили свое внимание на вопросы о том, наследуются ли результаты упражнения и неупражнения органов и последствия изменений, вызванных непосредственным влиянием внешней среды на организмы, но сравнительно мало занимались разработ
1 Отметим изложение дарвинизма в лекциях Вейсмана (Vortrage Uber Descen-denztheorie, О. Fischer, Jena, 1902) и в сводке Л. Плате (Plate L., Selectionprin-cip„ 1913).
НЕОЛАМАРКИЗМ
83
кой самой теории, как системы, объясняющей эволюционный процесс: предполагалось, что если будут опровергнуты возражения, сделанные против неоламаркизма неодарвинистами, то получится теория, объясняющая все явления эволюции без остатка, т. е. подтвердится целиком первоначальное воззрение Ламарка. И в настоящее время мы не имеем в достаточной мере проработанного и подробного изложения ламаркизма. Ученый, интересующийся этой теорией и относящийся к ней объективно, мог бы не без основания сказать, что ламаркизм слаб не только потому, что плохо доказаны такие положения, как принцип наследования результатов упражнения органов, но в значительной мере и потому, что сама теория недостаточно обстоятельно разработана ее сторонниками и недостаточно показано, как различные виды приспособлений, происходивших в течение филогенеза, объясняются на основании принципов Ламарка.
Можно также сказать, что натурфилософский характер сочинений некоторых сторонников этой системы, например психоламаркистов Паули, Франсе и др., отталкивал от неоламаркизма исследователей, для которых прежде всего убедительны фактические доказательства, а не априорные рассуждения.
Приступая .к разбору теории неоламаркизма, мы отмечаем, что, по нашему мнению, доказать или опровергнуть ее можно только путем опытного исследования, ставящего своей прямой целью решение эволюционных вопросов и проводимого в течение достаточно долгого времени.1 Мы лично подходим к данной проблеме не с этой экспериментальной точки зрения. Мы ставим перед собой другой вопрос, а именно вопрос о том, что может дать эта теория, в современной ее постановке и при принятии ее предпосылок, для объяснения процессов филогенеза, т. е. в какой мере она может служить рабочей теорией для эволюциониста-филогенетика.
Мы уже отметили, дто подробной сводки теории неоламаркизма мы не имеем. Сделать такую, правда, слишком короткую и неполную сводку попытался анатом Ф. Вейденрейх.2 В виду того, что его книга встретила сочувственный прием со стороны некоторых крупных палеонтологов, являющихся сторонниками неоламаркизма (назовем, например, О. Абеля3), мы можем думать, что она достаточно верно передает взгляды более позитивно мыслящих неоламаркистов. Поэтому мы можем положить ее в основу нашего изложения современного состояния этой теории.
Старый ламаркизм, т. е. ламаркизм самого Ламарка, принимал, что эволюция организмов определяется двумя основными принципами, а именно: 1) принципом упражнения и неупражнения органов, вызывающим изменения организации животных и 2) принципом изменения условий существования, благодаря которому изменяется организация растений. Эти положения остаются в силе и для современных неоламаркистов, но доказательства для них берутся уже из современных экспериментально-зоологических и ботанических исследований.
Передавая содержание книги Вейденрейха, мы остановимся главным образом на ее положительной стороне, т. е. на доказательствах теории неоламаркизма, а не на многочисленных полемических выпадах против сторонников теории естественного подбора и современных теоретиков наследственности, часто затемняющих у него, как и у многих его единомышленников, положительную сторону работы.
1 Мы подчеркиваем, что случайные наблюдения, сделанные попутно при опытах, преследующих другие цели, до сих пор мало дали для решения вопроса о причинах эволюции. Именно поэтому многие опыты, приводимые для доказательства наследования благоприобретенных признаков, имеют мало значения для теории Ламарка.
“WeidenreichF., Das Evolutionsproblem und der individuelle Gestaltungs-anteil am Entwicklungsgescheen, Berlin, V. F. Springer, 1921.
8 A b e 1 O., Das biologische Tragheitgesetz, Biol. Oener., Bd. 4, 1928.
84
. ГЛАВА II
В начале своей книги Вейденрейх останавливается на вопросе об индивидуальности животных организмов и пытается показать, что реакции различных особей одного и того же вида на одно и то же раздражение во многих случаях различны и что различные изменения, вызванные этими раздражениями, должны, по его мнению, какими-либо способами передаваться по наследству потомкам, т. е., другими словами, так называемые благоприобретенные признаки должны передаваться по наследству следующим поколениям.
Этот взгляд нашего автора несовместим с распространенной среди современных теоретиков наследственности теорией непрерывности зародышевой плазмы (Вейсман), по которой половые клетки являются неизмененными потомками оплодотворенной яйцеклетки. Вейденрейх на основании явлений регенерации отвергает эту теорию, так как у некоторых ленточных червей (Moniezia) половые органы регенерируют не только из паренхимы червя, но и из диференцированных мускульных клеток. С другой стороны, исследования Дриша показали, что весь организм, а следовательно, и его половые клетки, регенерируют у асцидий из выделенного из организма жаберного аппарата. Таким образом, по мнению Вейденрейха, половые клетки с наследственной плазмой, от строения которой зависит передача наследственных особенностей, образуются в каждом поколении заново из диференцированных клеток и передают изменения, вызванные в организме влиянием внешних условий.1
Высказавши свой взгляд на наследственность, наш автор обращается к основным положениям неоламаркизма и указывает, что изменения условий внешней среды, в которой живут данные организмы, вызывают изменения в потребностях животных; каждое изменение в потребностях вызывает изменение в действиях и ведет к новым привычкам; далее, всякая новая потребность, ведущая к новым действиям,.направленным к удовлетворению этой потребности, вызывает учащенное употребление соответствующих частей организма или ведет к изменению характера деятельности этих частей: это последнее изменение вызывается в организме незаметно вследствие деятельности внутреннего чувства3 (eines inneren Gefiihls). Другими словами, в основу неоламаркизма кладется старый ламарковский принцип усиленного упражнения и неупраж-нения органов, которое здесь сближается с теорией функционального приспособления Вильгельма Ру.
Непосредственных доказательств того, что .вследствие упражнения органы животных разиваются прогрессивно и что эти изменения передаются по наследству, наш автор не приводит и ограничивается общими соображениями. Обыкновенно неоламаркисты ссылаются на некоторые примеры, которые, по их мнению, косвенно доказывают наследование результатов упражнения органов, в частности на исследования Семона над толщиной эпидермиса на подошве ноги человека. Известно, что у человека на подошвенной стороне ноги эпидермис толще, чем в других местах ноги: принимают, что он сделался таким вследствие давления веса тела на подошвенную поверхность ноги при стоянии и ходьбе. Так как уже у эмбрионов человека эпидермис на подошве утолщен, то предполагают, что здесь имеет место результат упражнения, т. е., что утолщения эпидермиса при ходьбе сделались наследственными и перешли из взрослого состояния в эмбриональное. В качестве аналогичного подтверждения теории о наследовании результатов упражнения приводится также факт образования мозолей на коленных суставах у бородавчатой свиньи (Phacochoerus), 1 2
1 Механизм этой передачи автором совершенно не выясняется; не разбирается и отношение его взглядов к теории хромосом как носителей наследственных свойств.
2 Подчеркнуто мною. А. Н. С.
НЕОЛАМАРКИЗМ
85
которая, кормясь, ползает на коленях; у этого животного, по исследованиям Лехе, мозоли образуются уже в течение эмбриональной жизни.
Едва ли, однако, эти примеры могут считаться вполне убедительными, так как помимо ламаркистского толкования, что мозолистые утолщения развились у взрослых животных от трения о землю и, сделавшись наследственными, сдвинулись на эмбриональный период жизни, возможно и другое толкование, а именно, предположение, что такие привычки могли развиться только у животных, у которых кожа на коленях была утолщена от природы и у которых на ногах могли образоваться мозоли, а не ссадины и раны. По моим исследованиям1 сдвигание признаков, развившихся у взрослых животных, на ранние стадии эмбрионального развития мало вероятно и во всяком случае не доказано.
Ход филогенетических изменений по Вейденрейху представляется таким образом: изменение условий существования вызывает изменение потребностей животных и усиление, ослабление или изменение функций,1 2 а следовательно и количественное и (при изменении функций) качественное изменение строения соответствующих органов, причем результаты этих изменений строения и функций органов передаются потомкам по наследству.
В настоящее время слабым местом неоламаркизма является вопрос о наследовании результатов упражнения и неупражнения органов: вопрос этот, во времена Ламарка казавшийся таким простым, что о нем не стоило даже рассуждать, теперь, после исследований Менделя и его последователей, крайне осложнился: многочисленные экспериментальные исследования, по мнению генетиков, говорят против наследования благоприобретенных признаков и, в частности, против наследования результатов упражнения и неупражнения органов. Вейденрейх пытается обойти это затруднение ссылками на факты длительных модификаций, вызванных влияниями изменения условий среды: если альпийский эдельвейс (Leontopodium alpinum) пересадить в низменную местность, то он, как говорят, вырождается: уменьшается волосатость растения, цветки делаются меньше и т. д. Многие горные растения, пересаженные в низменные места, теряют свой альпийский характер, но при обратной пересадке в горы утраченные признаки альпийских растений снова развиваются. Capsella bursa pastoris, пересаженная в горную местность, приобретает признаки горного растения, т. е. длинные, глубоко врастающие в землю корни, ксерофитные листья, короткий стебель и т. п. При обратной пересадке в равнинную местность эти признаки исчезают не сразу: некоторые из них, например короткость стебля, удерживаются в течение четырех поколений и только затем заменяются признаками равнинной формы. В этой задержке горных признаков растений неоламаркисты видят начало закрепления новоприобретенных под влиянием горных условий признаков; предполагается, что если измененные условия будут действовать более долгое время, то новые признаки сделаются вполне стойкими.3
ПРИНЦИП ЦЕЛЕСООБРАЗНОЙ РЕАКЦИИ
В тех случаях, когда изменения строения организмов не могут быть объяснены упражнением и неупражнением органов, неоламаркизм обращается к гипотезе или принципу так называемой целесообразной реакции.
1 Sewertzoff A. N., Ueberdie Beziehungen zwischen der Ontogenese und der Phylogenese der Tiere, Jen. Zschr., Bd. 63, 1927.
2 Как и почему функции при упражнении не только усиливаются, но и изменяются, автор не объясняет, и это, конечно, является слабым пунктом его гипотезы: трудно понять, почему от упражнения, например от упражнения жевательной мускулатуры, изменяется строение ее, изменяется функция ротового аппарата.
8 Эти соображения не объясняют нам, однако, почему «альпийские» признаки таких растений, как эдельвейсы, которые, несомненно, очень давно подвергаются влиянию горной среды, не закрепились и не сделались наследственными.
86
ГЛАВА II
Сущность этой гипотезы заключается в предположении, что и у растений и у животных существует основная способность целесообразно реагировать на изменения условий среды, т. е. спонтанно вырабатывать нужные при каждой новой обстановке приспособления к измененным условиям существования. Для доказательства этого положения Вейденрейх ссылается на только что приведенные ботанические примеры пересадок горных растений в низменные места и равнинных растений в горные местности. При этих пересадках сразу под влиянием измененных условий климата и почвы вырабатываются приспособительные изменения, например ксерофитные листья, глубоко погружающиеся в землю корни и т. д. у растений, пересаженных из долин в альпийскую обстановку, и соответственные изменения у растений горных мест при обратных пересадках. Расширяя это положение, Вейденрейх предполагает, что этим принципом целесообразной или приспособительной реакции объясняются все приспособительные изменения, которые нельзя объяснить влиянием упражнения и неупражнения органов. При этом само собдй предполагается, что измененное строение, вызванное измененными условиями среды, со временем закрепляется в организмах и делается наследственным.
Та же способность целесообразной реакции обусловила, по мнению Вейденрейха, прогрессивную эволюцию органов животных. Правда, для доказательства этого положения он приводит только косвенные аргументы, что, конечно, еще менее убедительно, чем эксперименты ботаников, о которых мы только что говорили. Так, предки хвостатых амфибий первоначально жили в воде, дышали наружными жабрами и иногда плавательным пузырем и имели приспособленный для плавания хвост с вертикально поставленным плавником; потомки этих водных амфибий перешли к наземному образу жизни, причем у них развилось легочное дыхание и изменилось строение хвоста, который сделался круглым. Мы знаем, что личинки наземных хвостатых амфибий живут в воде и дышат жабрами, так же как и их водные взрослые предки. По мнению Вейденрейха, у современных амфибий (точнее у их личинок) сохранилась двойная способность реагировать на раздражения, т. е. пребывание личинок в водной среде обусловливает у них развитие жабр, а переход на сушу, точнее—пребывание в мелкой воде и возможность выползать на берег, вызывает редукцию жабр и развитие легочного дыхания; эти изменения в строении органов дыхания сопровождаются соответственными изменениями в строении хвоста. Доказательства этой сложной приспособительной реакцицна раздражения, исходящие из внешней среды, автор видит, например, в том, что дышащий жабрами и нормально размножающийся в водной среде мексиканский аксолотль (Siredon mexicanus aut.) при некоторых изменениях условий среды превращается в безжаберную, дышащую легкими саламандру—амблистому (Amblystoma sp.). Наоборот, у личинок некоторых нормально дышащих легкими наземных амфибий удается путем изменения условий, в которых они живут в течение личиночного периода, подавить тенденцию к превращению в наземную форму: так, Каммереру удалось заставить личинку жабы-повитухи (Alytes obstetricans) достигнуть половой зрелости при сохранении жаберного дыхания и других личиночных признаков.
Далее Вейденрейх поясняет действие способности к целесообразной реакции при измененных условиях существования животных на некоторых других примерах, например на восстановлении глаз протея под влиянием света и на развитии асимметрии тела у камбаловых рыб.
У протея (Proteus anguineus), этой слепой хвостатой амфибии, живущей в водоемах глубоких пещер Каринтии и Крайны, очень маленькие глаза, настолько редуцированные, что у взрослого животного они носят совершенно эмбриональный характер. Каммерер воспитывал личинок
НЕОЛАМАРКИЗМ
87
протеев на свету,1 причем оказалось, что глаза не остановились в своем развитии, но развились как у типичных наземных амфибий, размножающихся на свету, до нормальных размеров. Из этого факта Каммерер (а за ним и Вейденрейх) делают вывод, что нормальное развитие и дегенерация глаз амфибий стоят в прямой зависимости от влияний света и темноты, т. е. представляют собой целесообразные реакции на раздражения светом и темнотой.
Камбаловые рыбы, как известно, во взрослом состоянии являются животными асимметричными, ведущими донный образ жизни: рыбы в покое лежат на дне моря, на боку. Это положение отразилось у них весьма сильно на форме и строении тела и вызвало асимметрию целого ряда органов. Глаз той стороны тела, которая обращена ко дну моря, передвинулся на ту сторону, которая обращена кверху, и вследствие этого череп сделался асимметричным; точно так же ряд органов боковой линии сдвинулся на обращенную кверху сторону головы; плавник стороны, обращенной ко дну, атрофировался и произошел ряд других изменений симметрии, связанных с изменением положения, т. е. с тем, что камбалы нормально лежат на боку и плавают, обращаясь к поверхности воды не спинной стороной, а боком. Обыкновенно все рыбы данного вида камбал ложатся на дно одной и той же стороной, преимущественно левой (Pleuronectes platessa), но у некоторых видов, например у Pleuronectes flesus еще симметричные личинки ложатся на дно то правой, то левой стороной, причем процент ложащихся на правую сторону довольно велик (от 5.36 до 36.4%). От того, на какую сторону ложится личинка камбалы, зависит развитие асимметрии, т. е. то, что у одних правый глаз сдвигается на левую сторону, у других левый—на правую.
По мнению Вейденрейха, у камбал наследственностью предопределена наклонность (инстинкт) вести бентонный образ жизни и ложиться при этом на бок, но крупные морфологические изменения, происходящие при этом, т. е. сдвигание глаза, асимметрия черепа, покровительственная окраска обращенной к свету стороны тела ит. д., не наследственный представляют повторяющуюся в каждом поколении заново реакцию на факт лежания рыбы на одном боку. Эти процессы еще не зафиксированы наследственностью у современных камбаловых рыб.
Как совершилось изменение строения этих форм в течение филогенеза, т. е. произошло ли оно внезапно, как реакция на изменение положения тела, или путем постепенных изменений в течение целого ряда поколений, Вейденрейхом не разобрано. Но для пояснения того, как происходили вообще филогенетические изменения организации, он ссылается на выдвинутый В. Ру принцип функционального раздражения.
Согласно В. Ру, жизнь каждого животного можно разделить на четыре периода: первые стадии индивидуального развития определяются только наследственными факторами; за ними следует промежуточный период, в течение которого развитие обусловливается частью наследственными факторами, частью раздражениями, идущими из внешней среды; в течение третьего периода характер развития определяется только внешними раздражениями, т. е. гипертрофией одних частей вследствие деятельности и атрофией других от бездействия; наконец, четвертый период Ру является периодом старости или увядания, который для эволюции органов не имеет значения. Для неоламаркизма важен главным образом третий период, так как предполагается, что изменения в условиях среды именно в течение этого периода вызывают изменения в строении развивающихся органов эволюирующих животных и обусловливают их иное, сравнительно с предками, строение.
’ 1 В естественной обстановке молодь протея развивается в полной темноте пещер-
ных водоемов.
88 ГЛАВА II
Указанными двумя принципами, т. е. старым ламарковским принципом упражнения и неупражнения органов и принципом целесообразной реакции на раздражения, исходящие от измененных условий среды, неоламаркизм находит возможным объяснить эволюционный процесс в целом: по мнению сторонников неоламаркизма, эти принципы не нуждаются в других дополнительных объяснениях (например в теории естественного подбора, аутогенетического ортогенеза и т. д.) и объясняют все филогенетические изменения организма без остатка.
Попробуем теперь на основании данных сравнительно-анатомического и филогенетического исследования разобрать то, что эта теория дает положительного для объяснения известных нам фактов эволюции, чтобы установить, в какой мере мы можем пользоваться ею как общей теорией, объясняющей филогенез.
НЕСОСТОЯТЕЛЬНОСТЬ ПРИНЦИПА УПРАЖНЕНИЯ ДЛЯ ОБЪЯСНЕНИЯ ПРОГРЕССИВНОЙ эволюции
Остановимся сначала на вопросе об упражнении и неупражнении органов и на изменениях, вызываемых в организмах этими процессами. В статьях по неоламаркизму этот вопрос трактуется обыкновенно в очень общей и довольно неопределенной форме. Принято говорить, что органы от упражнения усиливаются и развиваются прогрессивно, а от неупражнения дегенерируют, но примеров таких изменений приводят мало. Какого-либо систематического обзора изменений морфологических и гистологических, вызываемых упражнением, а тем более классификации этих изменений, мы в неоламаркистской литературе на нашли. Самые изменения описываются очень коротко, как вещи всем известные, и с изменениями, которые происходили в течение эволюции различных групп животных, обычно не сопоставляются. Обыкновенно при упоминании о прогрессивных изменениях, вызываемых упражнением, ссылаются на усиление мускулов при гимнастических упражнениях или при физической работе, и, невидимому, считается, что этих общеизвестных фактов достаточно для объяснения прогрессивной эволюции мускулатуры, происходившей в течение филогенеза хотя бы позвоночных животных. С таким заключением едва ли можно согласиться.
Мы знаем, что при усиленном упражнении мускулы делаются толще, а при напряжении тверже и в общем сильнее, и мы можем констатировать, что самые волокна поперечнополосатых мышц от упражнения делаются толще, что сопровождается увеличением в них числа ядер; в некоторых случаях удалось также установить, что в гладких мускульных волокнах появляется поперечная полосатость.
Допуская вместе с неоламаркистами, что результаты упражнения мышц со временем делаются наследственными (что до сих пор экспериментально не доказано), мы, может быть, могли бы допустить, что усиление поперечнополосатых мышц и в некоторых случаях превращение гладкой мускулатуры в поперечнополосатую могло бы быть объяснено усиленным упражнением данных групп мускулов. Это, например, применимо к объяснению хода изменений при превращении гладкой висцеральной мускулатуры кишечного канала в поперечнополосатые мышцы жаберного аппарата.
Но мы знаем, что в большинстве случаев филогенетическое усиление мускулатуры состояло не только в утолщении мускульных волокон, но и в увеличении числа этих волокон: так, грудные мускулы птиц (mm. pectora-les у Carinatae) усилились вследствие громадного увеличения числа мускульных волокон, на что указывает сравнение этих мускулов птиц с соответствующими мускулами рептилий. Зная, что число волокон при упражнении не увеличивается, мы едва ли имеем право ссылаться на принцип упражнения для объяснения тех многочисленных случаев, когда при ко
НЕОЛАМАРКИЗМ
89
личественном изменении объема мышц мы имеем дело с увеличением числа волокон.
И многие другие прогрессивные изменения в строении мускулов позвоночных животных нам трудно объяснить принципом упражнения. Так, мы знаем, что во многих случаях прогрессивное развитие мускулатуры •выразилось в увеличении числа мускульных пучков данного отдела мускулатуры. Например, при превращении акулоподобных предков скатов в типичных скатов их грудные* плавники сильно увеличились, и это было обусловлено сильным увеличением числа сегментальных мускульных пучков в каждом плавнике. Зная, что даже от усиленного упражнения число сегментальных мускулов не увеличивается, мы не имеем никакого основания предполагать, что у скатов увеличение числа сегментальных мускулов и лучей грудных плавников было обусловлено упражнением: эта гипотеза невероятна. Таких случаев увеличения числа сегментальных мускулов в течение эволюции мы знаем очень много и объяснить их принципом упражнения мы не можем.
Точно так же трудно объяснить упражнением факт разделения первоначально однородных сплошных мускулов на отдельные мускульные индивидуумы, растущие в различных направлениях и прикрепляющиеся к различным, часто удаленным друг от друга частям скелета, что мы, например, находим при исследовании эволюции проксимальных мускулов пятипалой конечности рептилий.1 Мы видим, таким образом, что принцип упражнения объясняет сравнительно весьма немногие случаи количественного изменения мускулов позвоночных животных.
Сторонники разбираемой нами теории для подтверждения своих взглядов часто ссылаются на изменения, происходящие при усиленной функции в эпителиальных органах, в частности в железах. Так, при удалении одной почки другая начинает усиленно функционировать и значительно увеличивается в размерах, что зависит от усиленного размножения клеток почечного эпителия и. увеличения числа мальпигиевых телец. По аналогии мы имеем право предполагать, что и в других железах при усиленном функционировании происходят прогрессивные количественные изменения, но объяснять (при недоказанном предположении, что вызванные упражнением изменения желез передаются по наследству) филогенетическое увеличение размеров желез принципом упражнения мы не можем. Изменения строения желез объяснять этим принципом мы, при теперешнем состоянии наших сведений, тоже не имеем права.
Под понятие морфологических изменений, вызванных упражнением, обыкновенно подводят некоторые изменения в строении костного скелета, происходящие после его повреждения. Уже давно было обращено внимание на то, что костные перекладины в губчатых частях костей расположены целесообразно по отношению к действующей на них силе тяжести; например, в головке бедра человека ряды костных перекладин губчатого вещества лежат так, что с механической точки зрения они представляют наиболее выгодное расположение для сопротивления главным направлениям давления, которым подвергается данная кость. Кроме того, при переломах костей новые перекладины костного вещества в длинных костях человека и млекопитающих располагаются при срастании по тому же механическому закону, т. е. представляют собой силовые линии, наиболее выгодно расположенные в соответствии с новыми направлениями давления, образовавшимися после срастания частей переломленной кости.
На основании этих наблюдений В. Ру пришел к выводу, что в этих случаях функция, т. е. сопротивление давлению и растяжению срастаю
1 Sewertzoff, A. N., Studien iiber die Entwicklung der Muskeln, Nerven und des Skeletts der Extremitaten der niederen Tetrapoda, Bull, de la Soc. Imp. des Nat. de Moscou, 1908.
so
ГЛАВА II
щихся костей, определяет целесообразное расположение костных перекладин губчатого вещества костей. Некоторые другие наблюдения и опыты подтверждают это заключение: форма и величина костей позвоночных, повидимому, определяются их функцией, т. е. давлением и натяжением, испытываемыми ими; так, перерезка нервов лицевых мускулов молодых млекопитающих, вызывавшая атрофию соответствующих мускулов, обусловила асимметрию при развитии лицевых костей.
Э. Фульд ампутировал передние лапы трехмесячных щенят. Последствием этого было изменение способа передвижения: оперированные молодые собаки приучались передвигаться, прыгая как кенгуру на задних лапах, кости голени у них удлинялись, мускулы-оттягиватели тазобедренного сочленения усиливались и кожа голени делалась мозолистой. Мы видим, таким образом, что усиленная функция до известной степени вызывает изменение строения и, что особенно интересно, изменение функций некоторых частей костного скелета. Относительно того, наследуются ли эти изменения, мы ничего не знаем. По аналогии с изменениями, вызываемыми травматическими повреждениями, этого едва ли можно ожидать.
Резюмируя все приведенные нами до сих пор факты, мы можем сказать, что упражнение некоторых органов, а именно усиленное упражнение мускулов, кожи, желез и костного скелета, вызывает довольно значительные, по большей части количественные изменения упражняемых частей. Некоторое изменение функции наблюдается и при экспериментах над изменением строения костей. Но ничто не указывает на то, что эти изменения являются наследственными.
Еще более трудно применить принцип усиленного упражнения для объяснения прогрессивной эволюции некоторых других систем органов, например центральной нервной системы. Мы знаем, что центральная нервная система высших позвоночных при упражнении испытывает прогрессивные функциональные изменения. Так, у человека при упражнении в заучивании усиливается память; сообразительность, при упражнении ее в определенном направлении (например, в решении математических задач), усиливается именно в этом направлении; напомним также об усилении, вследствие упражнения, прирожденных способностей художников, музыкантов ит. д. Можем ли мы воспользоваться этими общеизвестными фактами (при допущении их наследственной передачи потомкам) для объяснения прогрессивной эволюции центральной нервной системы?
Мы знаем, что при прогрессивной эволюции головного мозга увеличились определенные отделы головного мозга. При превращении предков рептилий в птиц увеличились и изменили свое строение большие полушария мозга и мозжечок. При превращении головного мозга рептилийных предков млекопитающих в мозг типичных млекопитающих и при прогрессивных изменениях мозга в ряду млекопитающих (до человека включительно) некоторые отделы, в первую очередь большие полушария и, в особенности, крыша полушарий и мозжечок, также сильно увеличились, и мы можем с полной определенностью сказать, что это увеличение зависело как от увеличения числа ганглионарных клеток этих отделов мозга, так и отувеличения числа и сложности нервных путей, связующих отдельные центры мозга друг с другом и с органами высших чувств. Но мы знаем, что в головном мозгу нервные клетки взросл ыхживот-ных не размножаются делением. На этом основании мы имеем полное право сказать, что наблюдаемое при прогрессивной эволюции головного мозга увеличение числа ганглиозных клеток (невро-нов) не может быть объяснено упражнением, так как, насколько мы знаем, упражнение не вызывает деления ганглионарных клеток, т. е. образования новых нервных центров в мозгу у взрослых, т. е. в тот период, когда это упражнение происходит.
неоламаркизм
91
Точно так же мы не можем сказать, что прогрессивное развитие глаз может быть объяснено принципом упражнения, ибо клетки ретины взрослых животных и человека также н е размножаются. Мы не имеем положительных наблюдений, показывающих, что чувствующие клетки органа слуха размножаются у взрослых позвоночных животных, а строение органа слуха у человека и млекопитающих делает такое предположение весьма мало вероятным. То же мы можем сказать и относительно органа обоняния.
Во всяком случае сторонникамнеоламаркистской гипотезы надо былобы путем наблюдения показать, что клетки чувствующего эпителия органов зрения, слуха и обоняния размножаются во взрослом состоянии, и что усиленная функция вызывает размножение этих клеток. Пока это не доказано, мы не имеем основания объяснять эволюцию органов высших чувств принципами неоламаркизма.
Едва ли возможно применить принцип усиленного упражнения органов к объяснению прогрессивной эволюции нервов позвоночных, так как число нервных волокон находится в определенных соотношениях с числом нервных клеток в спинном и головном мозгу и в ганглиях, с которыми данные нервы связаны морфологически и функционально. Едва ли также возможно объяснить этим принципом и случаи, когда нервы меняют свой ход, дают новые ветви и т. д. Во всяком случае, нам с применением этого принципа к объяснению эволюции центральной и периферической нервной системы прйдется обождать до тех пор, пока неоламаркисты не приведут непосредственных доказательств того, что строение и функции нервной системы изменяются под влиянием упражнения в тех направлениях, в которых они изменялись в течение филогенеза позвоночных.
Большое затруднение для неоламаркистской гипотезы представляют так называемые пассивные органы животных, т. е. такие органы, которые полезны для животных не своей активной деятельностью, а самим фактом своего присутствия: сюда относятся очень многие органы защиты, как, например, костные чешуи и панцыри рыб (Chondrostei, Holostei, Cros-sopterygii), скорлупы и раковины моллюсков и ракообразных, хитиновые скелеты многих насекомых и т. д. Представить себе, что все эти органы изменяют свое строение вследствие упражнения, мы никак не можем.
Точно так же трудно представить себе, что прогрессивно развивались вследствие упражнения многие органы нападения животных, как, например, клыки, бивни, шипы, пила пилы-рыбы, зуб единорога (Monodpn) и рога полорогих копытных; мы знаем, что эти органы пускаются в дело животными сравнительно редко, и уподобить употребление этих органов гимнастике, от которой они развивались бы, мы никоим образом не можем. То же самое нам пришлось бы сказать о сложно построенных, развившихся из частей брюшных плавников, мужских совокупительных органах позвоночных, например акуловых рыб, или насекомых, например жуков. Трудно представить себе, чтобы эти органы, функционирующие в течение очень короткого периода один раз в год и остальное время года остающиеся без функции, образовались на основании принципа усиленного упражнения.
Резюмируя все сказанное нами относительно значения упражнения органов как принципа, объясняющего прогрессивную эволюцию животных, нам приходится признать, что лишь сравнительно небольшое число филогенетических изменений в строении мускулатуры, кожи, желез и костного скелета может быть объяснено, и то с большой натяжкой, на основании этого принципа. При этом функции органов при усиленном упражнении качественно остаются теми же, так что многочисленные случаи изменений строения органов на основании этого принципа объяснены быть не могут. Совершенно то же можно сказать и относительно много
92
ГЛАВА It
численных случаев субституции органов и субституции функций: почему вследствие упражнения спинная струна низших рыб была вытеснена костными позвонками; или хрящевой череп заместился эндохондральными костями, или меккелев хрящ высших позвоночных был вытеснен покровными костями и т. д., для нас (с точки зрения гипотезы упражнения) остается непонятным. Мы видели далее, что не можем пользоваться этим принципом и для объяснении эволюции тех органов, клетки которых не размножаются у взрослых животных, т. е. не размножаются в то время, когда органы функционируют по преимуществу (центральная нервная система и органы чувств). Так же мало допустимо применение этого принципа для объяснения эволюции пассивных органов и органов биологически важных, но сравнительно редко функционирующих.
В общем мы видим, что объяснить эволюцию сложно построенных органов путем усиленного упражнения едва ли возможно.
НЕСОСТОЯТЕЛЬНОСТЬ ПРИНЦИПА НЕУПРАЖНЕНИЯ ДЛЯ ОБЪЯСНЕНИЯ РЕГРЕССИВНОЙ ЭВОЛЮЦИИ
Мы до сих пор говорили только о прогрессивных изменениях органов при усиленном упражнении и не касались вопроса о редукции органов вследствие неупражнения, которой неоламаркисты придают большое значение. Предположение, что многочисленные случаи редукции органов объясняются неупражнением этих органов, было бы очень удобно для объяснений явлений атрофии органов, и, на первый взгляд, многие факты говорят в пользу такого объяснения. Ежедневный опыт показывает, что некоторая редукция мускулов происходит при неупражнении их (мускулы ног людей, ведущих сидячий образ жизни и т. д.). Обыкновенно предполагают, что неупражнение данного органа в течение целого ряда поколений может привести к полной атрофии его. Некоторые опыты также, невидимому, подтверждают это положение: так, по Каптереву,1 у дафний, воспитывавшихся в темноте, атрофировались глаза, и, сопоставляя этот факт с атрофией глаз пещерных животных (пещерные амфибии, ракообразные, рыбы), можно предположить, что у дафний глаза атрофировались от неупотребления при пребывании их в постоянной темноте. Правда, некоторые другие опыты и наблюдения показывают, что не всегда пребывание в полной темноте ведет к атрофии глаз: так у Drosophila, размножавшихся в течение 69 поколений в темноте, глаза не атрофировались (Пайн). Наряду со слепыми пещерными амфибиями в тех же пещерах живут близкие к ним амфибии с еще развитыми глазами. Эти факты как будто говорят о том, что темнота не всегда одинаково влияет на- разных животных. Мы знаем, что среди Drosophila, живущих и развивающихся на свету, появляются слепые особи с редуцированными глазами, и это, повидимому, указывает на то, что редукция глаз может происходить и независимо от влияния темноты, т. е. без всякого влияния неупражнения.
Другие опыты, однако, как будто несколько изменяют значение только что упомянутых опытов Каптерева и наводят на мысль, что редукция глаз у Cladocera зависит не от темноты, а от недостаточного питания. Это, повидимому, подтверждается также и опытами Н. В. Ермакова,хпри которых велся точный учет количества пищи и которые показали, что дегенерация глаз стоит в прямой связи не с неупраж-
1 Кар ter е w Р., Experimentale Untersuchungen fiber die Frage vom Einfiusse der Dunkelheit auf die Geftihlsorgane der Daphnien, Biol. Zbl., Bd. 30, 1910.
Idem. Ober den Einflussder Dunkelheit auf das Daphnien Auge, Biol. Zbl., 32,1912.
“Ермаков H. В., К вопросу об изменении глаза у Simocephalus octulus под влиянием темноты и голода. Работы Волжской биологической станции, 1924.
НЕОЛАМАРКИЗМ
93
нением при пребывании животных в темноте, а с голоданием: глаза атрофируются у голодающих животных и на свету и в темноте.
Этот результат весьма интересен, так как он ясно показывает, что определенное влияние изменения условий среды, в которой живут животные, существует, но что это влияние не то, которого можно было бы ожидать на основании неоламаркистской гипотезы: на глаза влияет не отсутствие света, а понижение питания.
Сравнительно морфологическое исследование редуцированных органов не говорит в пользу гипотезы неупражнения как причины редукции. Мы находим, что атрофируются многие пассивные органы, которые вообще не упражняются, как, например, части твердого наружного скелета. Так, мы знаем, что у акуловых рыб все тело (включая и брюшную поверхность) покрыто плакоидными чешуями. У многих скатов, которые несомненно произошли от акул, чешуи вполне атрофировались на брюшной поверхности—той, которой скат, лежа на дне моря, прикасается к песку и камням и которая раздражается механически несомненно более, чем спинная поверхность животного, обращенная к поверхности воды.
У симметричных предков камбаловых рыб костные чешуи были развиты равномерно на обеих сторонах тела: у Pleuronectus flesus мы видим, что на правой (верхней) поверхности чешуи хорошо развиты, а на левой, обращенной ко дну моря, на которой рыба нормально лежит, в области головы чешуи вполне исчезли; в туловище же на этой стороне тела их редукция началась: многие чешуи утратили характерные для них шипы и скрылись под кожей.
Мы видим, таким образом, что у рыб чешуи редуцируются именно на той стороне, которая наиболее подвергается механическим раздражениям и сохраняются там, где этих раздражений нет (на верхней стороне тела). Эти факты говорят не в пользу гипотезы неупражнения, как причины редукции органов.
То же мы можем сказать и относительно тех случаев, когда у животных атрофируются зубы, как это имело место, например, у предков черепах или у предков птиц. Очень вероятно, что у этих животных изменилась пища, но совершенно несомненно, что они все же продолжали есть, т. е. что они двигали челюстями и надавливали на редуцирующиеся зубы, так что эти последние, точнее клетки зубной пульпы, раздражались постоянно при каждой еде, и, несмотря на это, зубы атрофировались. Это показывает, что постоянное функциональное раздражение не препятствует редукции раздражаемых органов.
Едва ли существует много органов, так активно функционирующих, как части кровеносной системы, т. е. сердце и сосуды. Изучая развитие сосудов, мы часто видим, что функционирующий сосуд начинает утончаться, просвет его сужается, и через некоторое время данный участок сосуда атрофируется и исчезает совершенно. Так, при исследовании развития жаберных сосудов акуловых рыб1 я нашел, что дорзальный участок артерии жаберной дуги срастается с дорзальным концом задней жаберной вены и что затем часть артерии, лежащая вентрально от новообразо-вавшейся анастомозы сосудов, делается тоньше и совсем атрофируется. Совершенно такая же редукция происходит при развитии дорзальных комиссур между передними и задними жаберными венами (v. communi-cantes dorsales arcuum branchialium) каждых двух соседних бранхио-меров личинки акулы. Само собой разумеется, что в течение этого процесса жабер«ые сосуды не перестают функционировать и ток крови в них не прекращается. Напомню также о редукции дистальных концов длин-
1 Sewer tzoff A. N., Die Entwicklung der Kiemen und Kiemenbogengefas-se, Zschr. f. wiss. Zool., Bd. 71, 1923.
94
ГЛАВА II
них жаберных нитей акул и скатов, которые продолжают функционировать и в которых ток крови вообще не прерывается и в течение процесса редукции и после его окончания. Во всех этих случаях атрофия органов происходит при сохранении их функции: мы ни в коем случае не можем думать, что причиной редукции было неупражнение атрофирующихся органов (InactivitStsatrophie). Как это будет подробно разобрано ниже, изучение явлений редукции органов (в частности «пассивных» органов) приводил нас к выводу, что органы атрофируются не тогда, когда они перестают функционировать, а тогда, когда они становятся биологически бесполезными для их обладателей.
Мы видим, таким образом, что сравнительно очень немногие случаи эволюции органов, как прогрессивной, так и регрессивной, могут быть объяснены принципами упражнения и неупражнения органов. Во времена Ламарка, когда вопрос о приспособительных изменениях был почти сосовершенно не затронут исследованием, казалось, что эти принципы объясняют очень многие (если не все) явления прогрессивной и регрессивной эволюции. В настоящее же время, когда, благодаря шестидесятилетним упорным исследованиям зоологов, сравнительных анатомов и палеонтологов, нам стал известен ход эволюции громадного числа приспособлений, не известных Ламарку, нам становится совершенно ясным, что даже при допущении столь оспариваемого принципа наследования последствий упражнения и неупражнения органов мы можем объяснить принципами неоламаркизма сравнительно лишь очень небольшое число известных нам приспособительных изменений; громадное же большинство этих изменений объяснению посредством этих принципов не поддается.
КРИТИКА ПРИНЦИПА ЦЕЛЕСООБРАЗНОЙ РЕАКЦИИ
Те приспособительные изменения, которые не могут быть объяснены старыми ламарковскими принципами упражнения и неупражнения органов, неоламаркистам приходится объяснять изложенным нами принципом целесообразной (приспособительной) реакции организмов на изменения внешней среды. Как мы уже отметили, по этому принципу организм, попадая в измененные условия существования, реагирует целесообразным изменением своего строения, т. е. спонтанно меняет строение тех органов, которые имеют отношение к измененньш условиям, так что эти органы оказываются приспособленными к новым условиям среды.
Вейденрейх в качестве доказательства этого положения приводит упомянутые нами выше примеры горных растений, приспособляющихся при пересадке в долины и низменные местности к условиям этих мест, и растений низменных мест, приспособляющихся при пересадке на горы к горной среде и соответственно этому меняющих свое строение. В некоторых случаях новые признаки, приобретенные растением после пересадки, удерживаются его потомками в течение нескольких поколений; в этом неоламаркисты видят начало закрепления наследственностью новоприоб.ретенных приспособительных изменений.
Становясь на точку зрения неоламаркистского учения, мы должны принять, что все приспособительные изменения организации растений произошли на основании деятельности только этого принципа, так как, очевидно, принцип упражнения и неупражнения органов к растениям не применим. В таком случае этот единственный принцип объяснения эволюции всего растительного царства заслуживает гораздо более подробного разбора и подтверждения примерами и, если возможно, опытами, чем это делалось до сих пор. Очень интересно было бы, например, чтобы неоламаркисты указали, какие именно изменения среды вызвали в каче
НЕОЛАМАРКИЗМ
95
стве непосредственной реакции развитие шипов и колючек, защищающих растения от топтания и поедания растительноядными животными? Какие изменения среды вызвали развитие плодов с крючочками, прицепляющимися к шерсти млекопитающих, благодаря которым многие растения способны распространять свои семена на далекое расстояние от мест, где они растут, т. е. расселяться (напомним наши репейники)? Какие изменения среды вызвали развитие летучек на семенах ковыля и других подобных форм? Какие изменения среды обусловили развитие вкусной мякоти у покрытых твердой оболочкой зерен (вишен и слив), которые проглатываются и непереваренные извергаются птицами, благодаря чему эти растения расселяются на далекие расстояния, что несомненно является признаком весьма полезным для данных видов? Должны ли мы себе представить эволюцию колючек акации, алоэ и наших колючих растений таким образом, что травоядные млекопитающие ели и топтали предков этих растений и что в качестве реакции на это изменение среды у них развились острые колючки и шипы?
• Мы можем свободно себе представить, что на общее изменение климатических условий в сторону сухости растения и животные могут ответить целесообразной реакцией своих наружных покровов: растения—ксе-рофитным изменением своих поверхностных слоев, животные—ороговением эпидермиса,1 но нам очень трудно представить себе, какими изменениями среды вызваны те изменения, которые имеют непосредственное отношение к изменениям в жизни других- растительных и животных организмов.
Принцип целесообразной реакции принимает два положения: 1) изменение условий среды производит заметные изменения в организации растений и животных, подвергающихся этим измененным условиям, и 2) эти изменения являются целесообразными, т. е. полезными относительно тех именно неблагоприятных сторон данного изменения среды, которые их вызывают. Таким образом, данный принцип принимает, что при наступлении холодного климата в организмах развиваются такие черты строения, которые защищают организм от холода, а при изменении условий освещения развиваются в органах зрения приспособления к изменившимся условиям света, например к видению при слабом свете (телескопические глаза и т. п.). При переселении наземного животного в водную среду реакция (по предпосылке) должна состоять в спонтанном развитии органов дыхания, приспособленных к дыханию в водной среде, органов движения, приспособленных к плаванию, и т. д.
Посмотрим, насколько изменения среды действительно вызывают изменения именно такого рода. Экологические исследования последнего времени показали, что некоторые изменения условий существования действительно вызывают ряд изменений в строении организмов. Много интересных примеров таких изменений приводит в своей работе «Роль климата в эволюции птиц»П. В. Серебровский.1 2 По его исследованиям большое влияние на окраску птиц оказывает влажность и в несколько меньшей степени температура тех местностей, в которых живут данные птицы. При этом автор отмечает, что бурые и черные тона окраски зависят главным образом от влажности, красные и желтые—чувствительнее к температуре. Как правило, северные подвиды крупнее южных; континентальный климат содействует увеличению роста, умеренный—уменьшению. Из слагаемых климата наибольшее значение имеет температура и степень континентальности, наименьшее—влажность. До известной степени изменения климата повлияли и на другие признаки строения птиц, например
1 Мы здесь совершенно оставляем в стороне вопрос о том. действительно ли эти изменения среды вызывают такие реакции, и, если они действительно их вызывают то всегда ли и у всех ли форм данной группы происходят подобные изменения.
2 Бюллетень Московского общества испытателей природы, т. 34, 1925.
96
ГЛАВА II
на форму клюва и на изменение формы крыльев, на увеличение перьев всего туловища и отдельных частей тела, на удлинение хохлов, рожков и украшающих выростов и т. д.
Много примеров непосредственного влияния внешних условий на признаки животных приводит Р. Гессе в своей «Зоогеографии».1 Так, многие бабочки, живущие в сыром климате, отличаются темной окраской (мела-низм). Многие костистые рыбы, живущие в южных морях, отличаются от своих сородичей, обитающих в северных морях, меньшим числом позвонков: южноатлантические камбалы (Ancylopsetta quadrocel-lata, Xistreuris liolepis, Paralichthys californicus и др.) имеют 35—37 позвонков, между тем как северноатлантические формы (H'ppoglossoides platessoides, Asteresthes stomias, Hippoglossus hippoglossus) имеют 45, 49 и 50 позвонков. Большая часть тропических Acanthopterygii имеет 24 поз-цонка, а соответственные формы из более холодных областей—большее число позвонков (от 27 до 39).
Величина многих беспозвоночных в холодных морях больше, чем в теплых (фораминиферы, кольчатые черви, иглокожие, моллюски и ракообразные).
Известно, что существует определенное соотношение между объемом тела и его поверхностью у теплокровных животных: отдача тепла происходит через поверхность тела, и чем больше поверхность, тем больше потеря тепла. У небольших животных (при той же форме тела) отдача тепла больше, чем у крупных, так как отношение поверхности тела животного к объему у мелких животных больше, чем у крупных. Чтобы понять это положение, представим себе, что мы имеем три куба с величиной ребра в 1, 2 и 3 см; поверхности этих кубов будут 6, 24 и 54 см1 2, а объемы 1, 8 и 27 см3. Мы видим, что поверхности увеличиваются пропорционально квадратам, а объемы—пропорционально кубам; таким образом, у крупных форм поверхность относительно меньше, чем у мелких, т. е. мелкие животные охлаждаются при холодной температуре сильнее, чем крупные. Это правило называется по имени открывшего его ученого правилом Бергмана. Гессе установил, что у животных различных климатов соотношение между поверхностью тела и объемом регулируется при переходе из одного климата в другой изменением величины кожных придатков, увеличивающих величину поверхности тела (ушей, хвостов). Так, у полярной лисицы (Canis lagopus) очень маленькие уши, у нашей лисицы (Canis vulpes), живущей в умеренных широтах, уши больше, а у лисиц жаркого климата (Canis cerdo) уши очень велики: другими словами, у форм, обитающих в жарком климате, отдающая тепло поверхность сильно увеличена, благодаря чему животные легко переносят жару. Повидимому, эта особенность развивается непосредственно под влиянием изменения температуры: мыши и крысы, разводимые при высокой температуре, делаются меньше ростом, но уши их увеличиваются и хвосты удлиняются, т. е. у них под влиянием жары происходит приспособительная реакция. Точно так же у кроликов высокая температура способствует развитию больших висячих ушей..
Весьма интересны опыты над влиянием изменения в пище животных на размеры и строение их кишечного канала. Старые авторы (Семпер,3 Гюнтер, Эдмондстон) принимали, что строение желудка птиц очень сильно меняется под влиянием изменения корма: при кормлении хищной птицы твердой растительной пищей (зернами) тонкие мускульные стенки желудка, по их мнению, делались очень толстыми, а внутри желудка развивался роговой эпителиальный слой, приспособленный к перетиранию зерен; другими словами, путем перемены пищи можно заставить желудок хйщ-
1 Hesse R., Tiergeographie auf oekologischer Grundlage, Jena, 1924.
2 Cp. Semper G., Die natiirlichen Existenzbedingungen der Tiere, 1880.
НЕОЛАМАРКИЗМ
97
ной птицы превратиться в желудок зерноядной. Обратное изменение получалось, согласно взглядам этих авторов, при кормлении зерноядных птиц мясом: мускульные стенки желудка делались тонкими, роговой слой атрофировался и весь желудок принимал характер желудка хищной птицы. •
Экспериментальные исследования Брандеса1 заставили усомниться в этих результатах и вызвали новые исследования в этом направлении. Работы В. Руа и его ученика Шепельмана1 * 3 4 5 показали, что при перемене пищи изменения в строении желудка действительно происходят, но что они далеко не так значительны, как предполагали прежние авторы. Оказывается, что при кормлении гусей мягкой и очень питательной пищей (манной кашей, клецками) происходит некоторая редукция стенок мускульного желудка, но толщина рогового слоя желудка не уменьшается, хотя величина' самой роговой пластинки становится несколько меньше и по консистенции пластинка делается менее твердой. По Шепельману, у гусей при кормлении их мясом объем мышц мускульного желудка не уменьшается, а делается больше: таким образом, мнение о превращении желудка типа зерноядных птиц в тонкостенный желудок типа хищников под влиянием мясной пищи не подтвердилось.1
Вообще мы видим, что этими исследованиями предположение о приспособительном изменении стенок желудка птиц под влиянием изменения состава пищи не подтверждается.
Больше дают для неоламаркизма весьма тщательно поставленные опыты Бабака.6 Этот исследователь кормил головастиков бесхвостых амфибий (Rana fusca, Rana arvalis, Pelobates fuscus) мясной (мясо лягушек, раков, лошадиное мясо и т. д.) и растительной (Stellaria media) пищей. В результате оказалось, что кишечный канал головастиков, питавшихся растительной пищей, удлинился почти вдвое сравнительно с кишечным*каналом головастиков, питавшихся животной пищей. Мы видим, что здесь род пищи несомненно влияет на относительные размеры кишечника, так как эпителиальная стенка кишечника под влиянием раздражения, исходящего от менее питательной пищи, сильно растет в длину и просвет ее становится несколько уже. Эти изменения, невидимому, носят приспособительный характер, так как при менее питательной растительной пище усваивающая поверхность канала увеличивается, а пища, вследствие удлинения и сужения пищеварительного тракта, проходит медленнее и, благодаря этому, лучше переваривается и усваивается.
Как и в предыдущем случае, мы имеем здесь дело с количественным изменением органа, т. е. с простым увеличением его размеров. Происходит ли здесь изменение функции, т. е. изменение в химическом составе выделения кишечных желез, мы не можем сказать.
В этих опытах весьма интересна одна сторона, а именно то, что изменение строения кишечного канала имеет до известной степени приспособительный характер: мало питательная растительная пища в длинном и узком пищеварительном канале полнее усваивается, чем в более коротком и широком, так что мы можем сказать, что при измене
1 Brandes G., Ueber den vermeintlicben Einfluss veranderter Ernahrung auf die Structur des Vogelmagens, Leopoldina, Bd. 32, 1896.
! Roux W., Ueber die functionelle Anpassung des Muskelmagens der Gans, Arch, f. entw. Meeh., 21, 1906.
dSchepelmann E., Ueber die gestaltende Wirkung verschiedener Ernahrung auf die Gans etc., Arch. f. entw. Meeh., Bd. 21 u. 23, 1906 и 1907.
4 Шепельман высказывает гипотезу, что при многолетнем кормлении гусей мясной пищей это превращение может совершиться.
5 Babak, Experi men telle Untersuchungen uber die Variabilitat der Verdaungs-rohre, Arch. f. entw. Meeh., 1909.
Морфологические закономерности 7
S8
ГЛАВА II
нии рода пищи у головастиков происходит ауторегуляция строения кишечного канала соответственно роду пищи. Выработалась ли эта ауторегуляция в течение эволюции личинок бесхвостых амфибий или она представляет собой свойство, характерное для пищеварительного тракта всех позвоночных, мы без дальнейших опытов, касающихся других животных, в первую очередь личинок хвостатых амфибий, сказать не можем. Изложенные исследования над изменением пищеварительного канала птиц в связи с изменением пищи, невидимому, говорят против предположения, что изменения строения кишечного канала всегда имеют приспособительный характер.
Мы рассмотрели сравнительно большое число изменений признаков животных под влиянием измененных условий внешней среды, и они показали нам, что в природных условиях изменение среды при достаточно длительном воздействии оказывает во многих случаях изменяющее влияние на организацию животных. Отметим, что в тех случаях, которые нами приведены, и в весьма многих аналогичных случаях такое изменяющее влияние имеют лишь очень общие изменения условий неорганической среды, главным образом изменения климата и до известной степени того, что географы называют ландшафтом.
Мы, в общем, можем согласиться с первой частью приведенного выше тезиса неоламаркизма и принять, что некоторая часть внешней среды непосредственно влияет изменяющим образом на строение организмов. Но нерешенными остаются следующие важные для данной проблемы вопросы:
1. Наследственны ли изменения, вызванные непосредственным влиянием среды?
2. В какой мере изменения, вызванные в организме изменениями среды, являются приспособительными?
3. Все ли изменения среды, и неорганической и биологической, непосредственно влияют изменяющим образом на организмы?
Относительно наследственности изменений, вызванных непосредственным влиянием измененных условий среды, вопрос в настоящее время не может считаться решенным, ибо неоламаркистам до сих пор не удалось убедить своих оппонентов в том, что наследование так называемых благоприобретенных признаков является правилом, а не исключением.
Что касается вопроса о приспособительном характере изменений, вызванных общими изменениями неорганической среды, то этот важный для неоламаркизма тезис едва ли может считаться доказанным. В случаях, когда для нас ясно, что данные признаки животных развились в зависимости от непосредственного влияния внешних условий, приспособительный характер этих признаков далеко не является правилом. Большая часть изменений окраски птиц и насекомых, развивающихся под влиянием климата или при экспериментах, насколько мы можем судить о них с биологической точки зрения, являются признаками индиферентными, т. е. если не вредными, то и не полезными при данных условиях существования.1 Только относительно очень немногих изменений этого рода мы можем сказать, что они носят приспособительный характер: сюда относятся только, что разобранные изменения длины кишечного канала у личинок бесхвостых амфиий (Бабак) и изменения величины тела и размеров кожных придатков (Гессе).
Таким образом, нам очень трудно принять положение, что все приспособительные изменения, эволюцию которых трудно объяснить на основа
1 Мы могли бы сказать, что изменения окраски животных, а также изменения формы являются признаками приспособительными, если бы мы видели, что таким образом развиваются покровительственные окраски и формы (как, например, форма гусениц пядениц, прямокрылых насекомых, похожих на сухие ветки, и т.. д.);-одиако этого в изменениях окраски и формы животных под влиянием климата мы не находим.
НЕОЛА МАРКИЗМ
99
нии принципа упражнения и неупражнения органов, развились на основании принципа целесообразной реакции организма на изменение среды, ибо наблюдение показывает нам, что целесообразная реакция является не общим правилом, а скорее исключением.
Так, мы не можем указать, какая причина обусловливает уменьшение числа позвонков у высших костистых рыб при их распределении по направлению с севера на юг; мы не можем также сказать, какое общебиологическое (приспособительное) значение имеет это крупное морфологическое изменение строения скелета данных форм. Но в рассмотренных нами случаях мы видим, что общие и сильные изменения условий неорганической среды оказывают на организм животных изменяющее влияние и что некоторые из таких изменений имеют приспособительный характер; это позволяет заключить на основании аналогии, что некоторые приспособления к изменениям климата выработались таким путем.
Мы можем себе представить, что некоторые очень общие и простые приспособления количественного характера развились как непосредственные реакции на воздействия изменении неорганической среды. Для более сложных изменений такое объяснение, если не вкладывать в него, некоторого мистического значения, применить трудно. Мы вполне можем, например, понять биологическое значение третьего века птиц, т. е. той боковой прозрачной складки кожи, которая при помощи особого мускула надвигается на роговицу глаза и, не препятствуя зрению, ослабляет действие слишком ослепительного освещения. Для неоламаркиста казалось естественным признать, что это изменение произошло при переходе предков птиц к жизни на ярком солнечном свету, т. е. при развитии способности летать. Нам такая «целесообразная реакция» была бы понятна, если бы изменение в освещении вызвало изменение в пигментных клетках и клетках ретины, подвергающихся действию света и реагирующих на него, но почему изменение в интенсивности света вызвало ряд сложных изменений в эпителии, в соединительной ткани кожи и в мускулах, нормально не реагирующих на свет, для нас непонятно. Функция век, защищающих роговицу глаз от повреждений и раздражений, вполне понятна; но как эти раздражения роговицы вызвали развитие век, ресниц и мускулатуры век, т. е. частей, которых раздражения роговицы не касаются, нам , опять-таки непонятно. Далее, мы знаем, что глазная мускулатура по-’ звоночных, благодаря которой животные по своей воле могут направлять глаза на привлекающие их внимание предметы, развилась из передних мускульных сегментов тела, двигавших у их предков еще гибкий передний конец тела вправо и влево; эти передние (I, II и III, по фан Вайю) миотомы головы изменили свою функцию, прикрепились своими дистальными концами к глазному яблоку и сделались глазными мускулами. Нам вполне понятно, что подвижность глаз с образным зрением была крайне полезна для предков черепных позвоночных, но какое изменение условий среды вызвало изменение строения и функций передних глазных миотомов,1 мы совершенно не можем себе представить.
Мы знаем также, что у предков млекопитающих квадратная кость верхней челюсти и сочленовная кость нижней челюсти (quadratum и arti-culare) превратились в слуховые косточки среднего уха (молоточек и нако- вальню). Мы в праве поставить вопрос о том, какой переменой среды неоламаркизм объясняет смену функций и строения этих элементов?
Мы видели, что неоламаркисты объясняют посредством принципа приспособительной реакции, вызванной изменениями условий среды, приспособления животных и растений к изменениям этой среды (кли-
1 Об упражнении' глазных передних миотомов головы как о принципе, объясняющем превращение передних миотомов в глазные мускулы, говорить довольно трудно.
7*
100
ГЛАВА II
мат и т. д.). Но мы знаем, что у животных, точно так же как у растений, существует очень много приспособлений к измененным условиям биологической среды, т. е. к изменениям в отношениях данного животного или растения к растениям и животным, с которыми ему в течение своей жизни приходится вступать в те или иные биологические (т. е. неин-диферентные) отношения. Мы имеем полное право требовать у неоламаркистов, чтобы они показали на примерах, когда и как такие изменения отношений между данной формой и другими животными и растениями являлись стимулами, вызвавшими определенные «приспособительные реакции».
Мы видели, что эволюцию целого ряда пассивных органов как позвоночных, так и беспозвоночных животных нельзя объяснить принципом усиленного упражнения, и поэтому нам, оставаясь на почве неоламаркизма, приходится принять, что прогрессивная эволюция таких органов объясняется принципом «целесообразной реакции». При этом мы должны поставить вопрос о том, в чем состояла та перемена условий среды, которая вызвала, например, развитие панциря, состоящего из костных чешуи и покрывающего тело и голову кистеперых рыб? Какие внешние условия обусловили эволюцию сложных отростков и связок, которыми эти чешуи сочленяются друг с другом, т. е. замков, которыми каждая нижняя чешуя сочленяется с непосредственно над ней лежащей верхней чешуей?1
Такие костные панцыри очень полезны для рыб (кистеперые, палеонис-циды, костные ганоиды) тем, что они защищают их от укусов хищных животных, главным образом других рыб, нападающих на них. Мы с полной вероятностью можем предположить, что появление таких хищников и было стимулом, вызвавшим развитие сложно построенных костных чешуйчатых панцырей. С точки зрения неоламаркизма мы, повидимому, должны были бы признать, что поражения кожи, наносимые хищниками, оказались раздражениями, на которые предки кистеперых и па-леонисцид реагировали развитием сложно построенных чешуй. Прежде чем'принять такое толкование эволюционного процесса, желательно было бы, чтобы нам привели примеры того, что последствием поранений у рыб является развитие на всем теле (а не только в местах поранений) костного вещества. Конечно, такого доказательста неоламаркисты привести • не могут.
Мы привели примеры приспособительных изменений организации животных, которые нам трудно объяснить «принципом приспособительной реакции» и где нелегко определить, в чем состояло изменение среды, вызвавшее развитие сложно построенных органов. Число этих примеров нетрудно увеличить. Так, мы можем спросить у сторонников данного принципа, в чем состояли те изменения среды, которые в качестве раздражения вызвали развитие ядовитых зубов у змей с их сложно построенными ядовитыми железами, с каналом, пронизывающим зуб по длине, и с приспособлениями, благодаря которым эти зубы двигаются? Интересно было бы получить ответ и на вопрос о том, какие изменения среды вызвали в качестве целесообразной реакции образование крючков у головок репейника и ему подобных растений? Интересно было бы узнать у неоламаркистов, какое раздражение вызвало в качестве целесообразной реакции развитие колючек кактусов и алое, акаций и терновника, защищающих эти растения от поедания растительноядными животными и от топтания и ломания со стороны крупных млекопитающих? Таких затруднений, т. е. случаев эволюции, которые трудно объяснить на основании принци.
1 Каждая чешуя сочленяется с чешуей, лежащей над ней, посредством костного шипа, входящего в углубление на нижнем крае лежащей над ней чешуи.
НЕОЛАМАРКИЗМ
101
пов неоламаркизма, в частности принципа приспособительной (целесообразной) реакции, мы находим очень много.
Вдумываясь в значение этого принципа, мы видим, что положение о целесообразной реакции, в сущности, не дает нам никакого объяснения того, почему и как происходят приспособительные изменения в организации животных. Нам говорят, что у животных есть такое «свойство», в силу которого под влиянием раздражений у них развиваются «целесообразные изменения организации», т. е., другими словами, такие изменения, которые полезны именно при данных условиях среды. Почему организм реагирует именно «целесообразно», неоламаркисты не объясняют. Эта целесообразная реакция является некоторой «способностью», очень напоминающей нам «virtus sanativa» или «facultas occulta» медицины мольеровских времен.
Почему у животных под влиянием изменения внешней среды появляются особенности строения, полезные именно при этих измененных условиях, нам также не объясняют, но говорят, что это их «первичное свойство», и по аналогии в тех случаях, когда мы не наблюдаем перемены условий, но видим совершившееся приспособление, мы должны принять, что это приспособление развилось вследствие основной способности животных изменять свою организацию в приспособительном направлении вследствие прирожденной им способности целесообразной реакции.
Но с принципом аналогии, как известно, надо обращаться очень осторожно. Приведенное умозаключение, может быть, было бы правильно, если бы мы могли сказать, что все животные (и растительные) организмы всегда отвечают на изменение условий среды целесообразной реакцией: мы знаем, однако, что такие реакции отнюдь не являются правилом, а скорее могут быть названы счастливыми исключениями. Очень многие изменения организации, вызванные изменениями среды, является индиферентными и только сравнительно очень немногие—полезными для их обладателей. Мы видели, что эти приспособительные изменения (изменения длины кишечника головастиков бесхвостых амфибий, изменения величины поверхности тела млекопитающих) являются реакциями на очень общие изменения среды, т. е. на резкие изменения характера пищи, климата и т. д.; к тому же это очень простые, главным образом количественные изменения строения (удлинение кишечного канала, увеличение поверхности тела и т. д.). Едва ли мы имеем право заключать на основании этих сравнительно редких полезных реакций животных организмов, что и такие сложные приспособительные изменения организации, как изменения строения костных чешуй или развитие рогов оленей, появились как целесообразные реакции организмов на изменения среды.
Когда мы переходим к изменениям организации, которые являются приспособлениями к изменениям других животных или растений (к изменениям врагов, добычи, конкурентов или биологической обстановки, например леса, болота и т. д.), то, прежде чем признать, что эти приспособления развились как целесообразные реакции, мы имеем право требовать, чтобы нам показали действительные, а не гипотетические примеры таких реакций, т. е. показали, что при появлении опасного врага действительно развиваются чешуи или покровительственная оКраска, или же новые органы обороны или нападения и т. д. Но этих примеров нам не приводят, и поэтому мы едва ли имеем право предположить, что такие целесообразные реакции организмов при изменениях биологической среды действительно происходят. Это тем более невероятно, что обычно такие изменения биологической среды на животных непосредственно не влияют и влиять не могут: новый враг, например, влияет на добычу только тогда когда он ее убивает, т. е. когда для нее уже поздно изменяться.
В виду этих соображений мы едва ли можем пользоватьсяпринципом
102
ГЛАВА II
целесообразной реакции как общим принципом, объясняющим развитие всех тех признаков, эволюция которых не объясняется путем принципа усиленного упражнения органов. Предполагать, что этим принципом объясняется эволюция сложных приспособлений животных к изменениям их биологической среды, при настоящем состоянии наших знаний—значит признавать за животными некоторую facultas occulta, вызывающую развитие сложных приспособлений, т. е. лишать принцип целесообразной реакции всякого научного значения. Сказать, что глаз или ухо позвоночного животного развились потому, что у этих животных была способность под влиянием раздражения развить глаз или ухо, не значит дать объяснение, а значит отказаться от всякого объяснения.
Мы на этом можем покончить с нашим разбором принципа целесообразной реакции. Мы видим, что аиют принцип, как и принцип упражнения и неупражнения органов, очень мало дает для объяснения явлений эволюции. Только очень немногие и сравнительно очень простые факты филогенеза могут быть объяснены принципами неоламаркизма в том виде, как их теперь предлагают сторонники этой теории.
Возвращаясь к вопросу о наследственности благоприобретенных признаков, замечу, что для эволюциониста далеко не все аргументы, приводимые для доказательства и опровержения этого положения, являются одинаково убедительными и равноценными. Мы могли бы, например, признать доказанным, что явления меланизма, полученные Штандфусом или Фишером под влиянием изменения температуры, наследственны, но имеем ли мы право экстраполировать это положение настолько, чтобы предположить, что мы имеем здесь доказательство наследственности результатов упражнения органов? Делает ли оно вероятным факт, что дегенерация II и IV пальцев у предков лошади, происшедшая от того, что один , из предков лошади на них опирался слабее, сделалась наследственной? Как отразилось это в сущности ничтожное изменение величины запястных и предплюсневых косточек и фаланг на яйцеклетках и сперматозоидах потомков этих лошадей? Каким образом эти маленькие количественные изменения костей II и IV пальцев изменили строение половых клеток так, что через очень большое число клеточных делений это изменение отразилось на костях metacarpus, metatarsus и фаланг тех же пальцев потомков этих лошадей в том же направлении, т. е. в направлении уменьшения?
Сделать эти выводы из факта меланизма вследствие холода едва ли возможно, и поэтому прежде чем утверждать, что результаты упражнения и неупражнения органов наследственны, естественно было бы спросить, почему неоламаркистами не сделано прямых опытов относительно наследственности признаков, развившихся при упражнении и неупражнении органов? Нам думается, что без таких опытов этот очень важный для неоламаркизма принцип будет для беспристрастных исследователей казаться неубедительным. Здесь нам еще раз приходится повторить сказанное нами в начале этой главы относительно недостаточной разработанности неола-маркистской теории со стороны опыта и наблюдения.
При том состоянии, в котором эта теория находится в настоящее время, надо признать, что она очень, мало дает для объяснения эволюционного процесса и во всяком случае не может претендовать на то, чтобы быть единственной и общей теорией, объясняющей эволюцию животных. В лучшем случае, на основании неоламаркизма мы могли бы объяснить только некоторые очень простые примеры эволюционных изменений органов, которые к тому же вполне возможно объяснить и на основании других принципов, например на основании теории естественного подбора.
ГЛАВА III
МЕТОДЫ ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
Прежде чем перейти к изложению того фактического материала, из которого мы исходили при установлении морфологических закономерностей эволюции, нам придется сначала остановиться на рассмотрении тех методов реконструкции анцестральных признаков, которыми мы пользовались в наших филогенетических исследованиях.
Как известно, ученый, занимающийся проблемами морфологической эволюции животных, может работать тремя методам: палеонтологическим, сравнительно анатомическим и сравнительно эмбриологическим. Разберем в кратких чертах каждый из этих методов отдельно и остановимся прежде всего на методе палеонтологическом.
ПАЛЕОНТОЛОГИЧЕСКИЙ МЕТОД ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
Обычно принято думать, что палеонтологический метод филогенетического исследования является тем единственным прямым методом исследования, при помощи которого удается непосредственно установить наличность филогенетических изменений в организации данных групп животных: многие ученые, палеонтологи в особенности, считают, что только этим путем мы и можем притти к безусловно верным результатам относительно происхождения животных.
Однако мы не можем согласиться с этой точкой зрения и полагаем, что, независимо от его огромного научного значения, палеонтологический метод совсем не является способом непосредственного исследования филогенетических изменений организации животных. Это зависит прежде всего от того, что палеонтологии доступно лишь изучение окаменелостей и что поэтому ей приходится преимущественно заниматься только одной системой органов, а именно скелетом, и реконструировать другие системы органов животных уже только на основании сравнительно анатомических законов, т. е. исходя из принципа корреляции частей. Для этого палеонтологии приходится прибегать к построению целого ряда специальных гипотез, как, например, гипотезы об отношении мускулов к скелету, гипотезы об отношении зубов к органам пищеварения, ребер к органам дыхания и т. д. Мы отнюдь не склонны отрицать вероятность и научное значение подобного рода выводов, но мы все же должны согласиться с тем, что гипотетический элемент здесь играет очень большую роль и чтсг поэтому нам никак не приходится говорить о непосредственном наблюдении эволюционного процесса.
Надо сказать, что ценность палеонтологических гипотез, конечно, в значительной мере зависит от того состояния, в котором сохранились остатки скелета ископаемых животных, а также и от самого количества остатков, дошедших до нас. В известной степени она зависит также и от сходства между ископаемыми остатками и скелетом современных
104
ГЛАВА III
животных, а также и от их строения. Так как остатки вымерших животных очень часто находят разбитыми, неполными и разрозненными, то нельзя не признать, что реконструирование их представляет собою большие трудности и что многое в этой работе является весьма гипотетичным.
Когда эта предварительная реконструктивная работа может считаться удачно завершенной, палеонтолог приступает ко второй части своего исследования, именно к установлению генетических отношений между реконструированной животной формой, ее предками и ее более поздними потомками.
Палеонтолог строит свои филогенетические ряды на основании тех же законов, что и сравнительный анатом, т. е. на основании тех признаков сходства и различия, которые он находит в организации исследуемых форм. При этом он обычно принимает, что формы животных данного типа, встречающиеся в более молодых (верхних) слоях, являются потом-д ками тех форм, лежащих в более древних (ниж-
1 них) слоях, которые по общей сумме своих морфо-
I логических признаков наиболее на них похожи.
Однако, если в ряду предков Aj—>(А2)—>А9 (схема I) форма (А2) не найдена вовсе или пред-г д' ставлена лишь в виде разрозненных остатков, но
1 зато имеется в наличии боковая ветвь А', то ис-
I s' следователю всегда грозит опасность установить
s' вместо истинного филогенетического ряда предков
s' Aj—>(А2)—>А3 ложный филогенетический ряд, а
Схема I. Объяснение в именно ряд Aj—»А'—>А3, представив формы А' иА3, тексте. не связанные между собой непосредственным род-
ством, как близкие члены одного ряда Aj—>А'—>А3.
Трудность построения лишенного таких пробелов филогенетического ряда из имеющихся в наличии остатков вымерших животных, реконструированных на основании только что изложенных принципов, не была^ конечено, недооценена и палеонтологами, и они обычно резко отличают подобные филогенетические ряды предков «Ahnenreihen» (О. Абель), от так называемых «Formenreihen», или сравнительно анатомических рядов. Вникнув в сущность построения филогенетических рядов чисто палеон4 тологическим путем, мы, конечно, увидим, что «филогенетические ряды» животных построены не на основании прямого наблюдения, как это мно4 гим кажется, но, наоборот, на основании целого ряда критически провес ренных логических умозаключений и по существу являются результатом трудной теоретической работы. Во всяком случае, мы имеем дело не с непосредственным наблюдением эволюционного процесса, а лишь с теоретическими и отчасти гипотетическими выводами. В случаях^ когда эти логические выводы прошли через достаточно строгую само-i критику, они имеют очень большую научную ценность, и мы можем^ опираясь на них, смело итти вперед.
СРАВНИТЕЛЬНО АНАТОМИЧЕСКИЙ МЕТОД ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
Сравнительно анатомический метод филогенетического исследования,! совершенно так же, как и палеонтологический метод, отнюдь не является способом непосредственного познания филогенетического развития животных. Сравнительно анатомическое исследование заключается в том, что обычно стараются найти ряд переходных форм между гомологичными («гомогенетичными», по Гудричу) органами современных животных. Так, например, при исследовании скелета конечностей непарнокопытных; млекопитающих в начале сравнительно анатомического ряда ставят
МЕТОДЫ ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ 105-
четырехпалую конечность дамана (Нугах, схема И, А), а в конце— однопалую конечность современной лошади ( D), и эти крайние группы соединяются между собой промежуточными формами, представленными четырехпалой конечностью тапира (В) и трехпалою конечностью носорога (С). Мы знаем, что в конечности современной лошади имеются следы трехпалого строения (грифельные косточки соответствуют рудиментарным metacarpalia II и IV); поэтому мы имеем основание считать, что однопалая конечность лошади произошла от трехпалой конечности типа носорога (Rhinoceros), которая сама произошла от четырехпалой конечности типа тапира, происшедшей в свою очередь от типичной пятипалой конечности. Вместе с тем было impiicite принято, что эволюция всех этих форм все время шла в одном и том же направлении, т. е. в направлении редукции краевых пальцев (I и V, потом II и IV) и прогрессивного развития среднего пальца, причем эволюция носорога и тапира остановилась на определенной стадии их филогенетического развития и не пошла дальше.
Схема II показывает нам, что все перечисленные нами животные А, В, С и D произошли от одной примитивной формы млекопитающих—X определенном, указанном стрелкою,
и что все они развивались в одной направлении. У дамана (Нугах, А) строение конечностей изменилось относительно мало (степень изменчивости строения конечности символизирована здесь отклонением линий X—А, X—В, X—С и X—D), у лошади (D) оно изменилось больше всего. Принято думать, что формы конечностей А, В, С, т. е. конечности дамана, носорога и тапира, приблизительно соответствуют по своему строению филогенетическим стадиям Ар В1, Сх, в свое время пройденным предками лошади. Только что приведенный сравнительно анатомический ряд А. В. С,
Схема II. Объяснение в тексте.
D имеет значение только для одной какой-нибудь системы органов, в данном случае—для скелета конечностей. Если мы захотели бы изучить ход филогенетического развития черепа у тех же непарнокопытных животных, то нам пришлось бы построить другой ряд, ибо череп носорога не является промежуточной ступенью между черепом тапира и черепом лошади. Иными словами, определенное направление в эволюции одного органа или одной системы органов отнюдь не предопределяет собою идентичного направления в эволюции другого органа или другой системы органов.
Сравнительный анатом, строящий свой ряд современных животных форм (например, только что приведенный нами ряд промежуточных форм между однопалой и пятипалой конечностью млекопитающих), вполне сознает, конечно, что он имеет здесь дело не с настоящим филогенетическим рядом, т. е. не с рядом предков—потомков, но лишь с рядом родственных современных форм, иллюстрирующим путь и направление эволюции данных органов.
. Главная ценность сравнительно анатомического метода исследования заключается в том, что если подобного рода «сравнительно анатомические ряды» дают нам вполне отчетливую картину эволюции отдельных систем органов (достоверность этой картины может быть проверена палеонтологическим или эмбриологическим путем), то это всегда позволяет нам понять и физиологическое и биологическое значение происшедших в этих орга
106
ГЛАВА lit
нах морфологических изменений. Так, возвращаясь к приведенному нам|1 выше примеру, мы можем, пользуясь этим методом, непосредственно изучать функцию конечностей, а также строение мускулов, нервов и сосудов конечностей, не сохранившихся палеонтологически.
Здесь нам необходимо подчеркнуть то особенное значение, которое имеют для изучения филогенеза рудиментарные органы современных животных. Когда сравнительному анатому приходится исследовать рудиментарный орган какой-либо ныне живущей формы, например рудиментарные задние конечности кита или змеи, то он может смело принятй, что именно у прямых предков этой формы эти конечности находились во вполне развитом и функционировавшем состоянии.
Не входя в дальнейшее обсуждение метода сравнительно анатомического исследования, мы хотели бы только еще раз обратить внимание читателя на то сходство, которое существует между сравнительно анатомическим и палеонтологическим способами исследования. Сравнительный анатом, на основании сходства в признаках ныне живущих взрослых животных,1 устанавливает между этими последними отношения родства: он приходит к выводу, что животные, органы которых Можно уложить в стройные, связанные между собой сравнительно анатомические ряды происходят от общих предков (схема II, X); палеонтолог сравнивает между собой различные формы вымерших животных, лежащие друг подле друга в различных геологических формациях, и, совершенно так же, как и сравнительный анатом, устанавливает на основании признаке^ сходства родственные соотношения между ископаемыми животными определенной геологической формации и их непосредственными предками. Ценность палеонтологических родственных линий, таким образом, в знай чительной степени зависит от числа промежуточных животных форм, найденных в различных формациях, а также от степени сходства между этими формами.
СРАВНИТЕЛЬНО ЭМБРИОЛОГИЧЕСКИЙ МЕТОД ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
Мы перейдем теперь к обсуждению сравнительно эмбриологического метода филогенетического исследования, вызвавшего, как известно, наиболее резкие возражения со стороны различных авторов. В четвертой части этой книги, в главах, посвященных проблеме филэмбриогенеза, читатель найдет детальный анализ явлений рекапитуляции признаков; поэтому сейчас мы не будем на них останавливаться подробно и отметим лишь методологическое значение закона Мюллера—Геккеля.
Наши исследования показали нам, что закон рекапитуляции признаков носит иной, более узкий характер, чем тот, который приписывал ему Геккель и его последователи. Прежде было принято думать, что повторение признаков взрослых предков в онтогенезе потомков представляет собой общее правило и что исключения из этого правила, все так называемые «искажения» биогенетического закона, вызываются исключительно явлениями ценогенеза, т. е. разнообразными приспособлениями зародышей к различным условиям эмбриональной жизни.
Что касается меня лично, я пришел к другому заключению. Я считаю, что существуют различные виды или типы филогенетических изменений признаков предков и что только при одном из них, именно при ’ эволюции, идущей путем надставок, конечных стадий в онтогенезе:
1 В настоящее время сравнительно анатомический метод в его чистом виде, т. е. без одновременного применения сравнительно эмбриологического исследования, употребляется довольно редко.
МЕТОДЫ ФИЛОГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
107
(анаболии), имеет место повторение анцестральных признаков взрослых предков1 в онтогенезе потомков. При всех других типах филогенетических видоизменений такой рекапитуляции признаков взрослых рредков не происходит.
В тех случаях, когда рекапитуляция действительно имеет мы не можем, однако, говорить о повторении целых стадий развития, т. е. о повторении всей суммы признаков анцестрального организма, а лишь о повторении некоторых отдельных признаков. При таком толковании изменение у потомков анцестрального хода онтогенезу вследствие ценогенезов имеет лишь второстепенное значение, отчего; естественно, отпадают всякие основания говорить о «фальсификациях» 0 смысле Мюллера и Геккеля.
Так как эволюция посредством надставки конечных стадий онтогенеза чрезвычайно распространена, то мы находим при наших сравн эмбриологических исследованиях очень большое числ питуляций; поэтому этот метод реконструкции признаков исчезнувших предков современных животных несомненно имеет большое практическое значение: в этом заключается его преимущество перед двумя другими методами филогенетического исследования: сравнительно анатомическим и палеонтологическим.
Мы уже упоминали о том, что при исследовании с помощью палеонтологического метода всегда существует некоторая опасность принять более или менее отдаленную родственную форму предков данного жил т. е. боковую линию предков, за его прямых предков и приписать этим последним признаки, свойственные формам, принадлежащим к упомянутой боковой линии. Но если мы находим какой-нибудь признак данной формы в эмбриональном развитии этой формы, то мы може ждать с полной определенностью, что признак этот принадлежал прямым предкам данной формы и отнюдь не относился к одной из ее боковых прародительских ветвей. Так, например, если мы видим, что tarsale proximale современных рептилий состоит у зародышей ящерицы из двух отдельных tarsalia: из astragalus и fibulare, которые в бофее поздних стадиях развития срастаются в один цельный элемент, то мы можем с уверенностью сказать, что в tarsus прямых предков рептилий были развиты и astragalus и fibulare. Если же мы будем постоянно наблюдать в течение онтогенеза некоторую определенную последовательность в развитии изменений признаков какого-нибудь органа жи то мы можем с определенностью сказать, что данная эмбриональная последовательность соответствует последовательности в филогенетическом развитии этих признаков упрямых предков давно" животных, что с филогенетической точки зрения имеет очень значение.
Резюмируя вкратце то, что мы сказали относительно общей методики филогенетического исследования, мы видим, что все три метода, имеющиеся в нашем распоряжении, не являются прямыми мет (одами исследования и что ни один из них не ведет нас к непосредственному познанию эволюционного процесса. В своих исследованиях нам постоянно приходится прибегать к критическому сопоставлении) результатов нашего непосредственного наблюдения, добытого всеми тремя способами, для того чтобы иметь возможность судить о ходе филогенетического
место,
ительно о река-
ротного,
предков м утвер-
вотных,2
й формы большое
1 Точнее говоря, повторение поздних эмбриональных признаков, сходных с признаками взрослых предков.
2 Это относится, само собою разумеется, только к последовательности Признаков (рекапитуляции) в одном каком-нибудь органе животного, например в передних конечностях, и отнюдь не касается случайного совпадения в развитии признаков в различных органах.
в различ-
108 ГЛАВА III
процесса, т. е. постоянно иметь дело с тщательной логической работой филогенетической реконструкции.
Каким именно путем может выполняться эта работа по реконструкции процесса филогенетического развития, нам было указано уже давно' еще Э. Геккелем. Это старый путь так называемого тройного п а р а л л е л и з м а: он заключается в том, что результаты палеонтологического, сравнительно анатомического и сравнительно эмбриологического исследований критически сопоставляются для каждого конкретного случая; и только тогда, когда данные, добытые этими |ремя методами исследования, вполне согласуются между собою, мы д^ожем говорить о достижении верного результата в нашей филогенетической работе.
ГЛАВА IV
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ
Прежде чем обратиться к вопросу о происхождении низших позвоночных, мы должны сказать несколько слов о той точке зрения, с которой мы подходим к разрешению этой проблемы. Наша цель в настоящий момент заключается в том, чтобы составить себе по возможности отчетливое представление о том, полифилетическим или монофилетическим (моногенети-ческим) путем шла эволюция крупных групп животного мира, в частности эволюция той крупной группы, представителями которой являются низшие позвоночные животные.
МОНОФИЛЕТИЧЕСКАЯ ТЕОРИЯ
Исследователи второй половины девятнадцатого столетия Придерживались преимущественно монофилетической теории, принимая, что каждая крупная группа многоклеточных животных (вплоть до типов)! происходит от единого корня, т. - - - -
Схематически такое воззрение на происхождение животных групп можно было бы представить себе следующим образом. Вообразим себе, что вид А живет в определенном месте, имеет определенное географическое распространение и окружен определенной средой М [схема Ш, А (М)] и что этот вид, вследствие каких-либо изменений в окружающей среде (М', М"), — изменений, на которых мы сейчас останавливаться не будем,—распался на два или более подвида (а', а"), приспособившихся в свою очередь к новым, несколько отличным друг от друга условиям среды (М', М"). Если окружающая среда будет изменяться и дальше в том же направлении, то, согласно монофилетической гипотезе, образовавшиеся подвиды а' и а",'изменяясь далее, постепенно превратятся в виды А' и А", уже не способные к спариванию между собою или к скрещиванию с родоначальным видом А и не способные давать с ними плодучее потомство.
В случаях, когда родоначальный вид А оказывается, по этой теории, менее приспособленным к новой среде (М', М"), чем происходящие от него дочерние виды (А', А"), и когда он, вследствие этого обстоятельства, вымирает, систематики, имеющие непосредственную возможность [наблюдать
е.
от одного единственного
6^)4' А2(М’)
2
(М')А'
(М')а'
вида.
А"<м2") а’(м2)
А’(М’)
а"(М")
3----;------‘.............//W ;
Схема III. Монофилетическая эволюция групп животных А;—As' Для упрощения схемы нами! взят лишь один случай дихотомического деления (см.; объяснение в тексте).
А—исходный вид предков; a', a", a'j, a'g, a*j, a'S,—дочерние и внучатые подвиды; A', A", A'i, А'2, A"i, А"2—дочерние и внучатые виды; М, М', М", М'1, М'2, M"j, М"2—изменяющиеся условия окружающей среды.
но
ГЛАВА IV
лишь эти дочерние виды (схема III, 2), скажут нам, что виды А' и А* принадлежат к одному роду (genus) и назовут общие унаследованные ими от вида А признаки родовыми признаками, а признаки, которыми виды А' и А" отличаются между собой,—видовыми признаками.
Если условия окружающей среды будут продолжать изменяться и дальше (М'1; М'2, М"1; М"2), то каждый из дочерних видов А', А" снова распадается на подвиды (а'х а'2, а"2), которые при дальней-
ших изменениях среды в том же направлении в свою очередь превратятся во внучатые виды А'1; А'2, А\, А"2 (схема III, 3), и т. д. Систематики же нам скажут тогда, что эти последние виды (А'1( А'2, А\, А"2) принадлежат уже к двум различным родам, характеризующимся признаками А'и А"(2)и принадлежащим к семейству А (1). Если мы продолжим этот ход мыслей далее, то нам легко будет представить себе, как могли произойти высшие систематические группы животного царства монофилетическим путем.
полифилетическая теория
Полифилетическая теория не имела первоначально среди.зоологов и палеонтологов того значения, которое придается ей в настоящее время ее наиболее рьяными поборниками. Во многих зоологических и палеонто-
логических трудах можно найти утверждение, что та или иная группа животных имеет полифилетическое происхождение. Однако, если более глубоко вдуматься в каждый отдельный случай, то нельзя не увидеть, что иногда вместо предполагаемой полифилии исследователь имеет перед собой явления совершенно другого порядка.
Так, например, М. Фюрбрингер (1888)1 на основании своих сравнительно анатомических исследований над мускулами птиц приходит к выводу, что подкласс бескилевых птиц (Ratitae), к которому принадлежат современные страусы, нанду, казуары и киви, а также вымершие Aepiornls, DinornisH Hesperornis, филогенетически развился полифилети-ческим путем. С другой стороны,глубокий знаток анатомии птиц Гадов (1893)1 2 принимает, что современные и некоторые из вымерших бескилевых птиц отделились от общего корня с С гур turi formes, Galliformes и Gruifor-mes и превратились в бегающих птиц (см. схему IV); при этом у них редуцировались крылья и киль, а задние конечности прогрессивно развились.
1 Fiirbri tiger М., Untersucliung zur Mopliologie und Systematik der Vogel zugleich ein Beitrag zur Anatomie der Stutz- und Bewegungsorgane, Jena, 1888.
2 G a d о w H., Bronn’s Klassen und Ordnungen des Tierreichs, VI Bd., IV Abt., Vogel, Leipzig, 1893.
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ • Ш
Предки же гесперорнитид ответвились от общего корня с Colymbiformis, Ardeiformes и Anseriformes (В), причем у них также редуцировались крылья и киль грудины и они также превратились в бескилевых птиц.
Однако Гадов и Фюрбрингер в сущности оба принимают, что птицы филогенетически развились монофилетическим путем, и считают, что от общего родоначальника птиц X (схема IV) ответвились две дивергентных ветви А и В, от которых впоследствии, в свою очередь, ответвились группы Aj и Bj; эти последние, попав в аналогичные условия существования, эволюировали в конвергентных направлениях и приобрели поэтому целый ряд сходных признаков (редукция Киля, редукция крыльев, аналогичные изменения в оперении, прогрессивное развитие ног и таза ит. д.). Вследствие этого эти две группы nTmx(Ratitae и Hesperornitidae) сделались так похожи друг на друга, что систематики сочли возможным соединить их в одну группу, обозначив ее как Ratitae auctorum. Лишь сравнительной анатомии удалось в свое время раскрыть сомнительное происхождение этой своеобразной группы, так как, иссследуя различные формы этих птиц, удалось обнаружить среди них ряд признаков, сближающих некоторых птиц из рода Ratitae auctorum с птицами из рода Gruiformes (корень А), и ряд других признаков, сближающих их с родом Anseriformes (корень В). Таким образом, мы видим, что имеем здесь дело отнюдь не с типичной полифилией, не с происхождением данной группы от многих корней, но лишь со своеобразным случаем ^энвергентноого монофилетического развития.
В настоящее время теория полифилии в собственном смысле этого слова поддерживается среди палеонтологов главным образом Штейнма-ном1 и Дакке,1 2 а среди ботаников—Лотси.3 Среди приверженцев этой теории в СССР мы можем назвать систематика и географа Л. С. Берга4 и палеонтолога Д. Н. Соболева.5
Теория эта в ее чистом виде, какой ее выдвигают и поддерживают Штейнман и Лотси, целиком отвергает принцип дивергентного развития и принимает, что все количество известных нам форм животных и растений произошло от точно такого же количества неизвестных нам простейших форм («плазм», по терминологии Лотси), которые развивались параллельно друг другу. При этом оба автора также принимают, что различные виды анцестральных «плазм» первично столь же сильно отличались между собою, как и их нынешние потомки.
Надо сказать, что логически можно защищать лишь эту последнюю-крайнюю форму полифилетической теории, так как ее, если так можно выразиться, компромиссная форма переходит незаметными чертами, как мы это сейчас увидим, к монофилетическому толкованию явлений филогенеза. В самом деле, в тех случаях, когда сторонники полифилетической теории, как это иногда с ними бывает, принимают для толкования некоторых фактов принцип дивергенции, они вместе с этим принимают и то, что потомки исходной группы А, эволтоируя в течение долгого периода без распадения на подчиненные систематические группы, все же, в конце концов, в какой-то определенный момент их эволюции начинают диверги-ровать и распадаются на два или более отдельных видов, характеризующихся неспособностью скрещиваться между собою и давать плодучее
1 S tei nmann G., Die geologische Grundlagen der Abstammungslehre, 1908. Idem., Einfiihrung in die Palaeontologie. Leipzig, 1907.
2 Dacque E., Der Dessendenzgedanke und seine Geschichte, Miinchen, 1913.
8 L о t s у , J. P., Vorlesungen liber Deszendenztheorien, Jena, 1906, I, 1908, II.
4 Берг JI. С., Номогенез или эволюция на основе закономерностей, 1922.
‘Соболев Д. Н., Наброски по филогении гониотид, Известия Варшавского политехнического института, вып. I, 1913
Его же, Начала исторической биогенетики, 1924.
112
ГЛАВА IV
потомство. При этом у них нет оснований утверждать, что процесс .дивергенции на этом должен непременно остановиться и что он не может иметь места также и у потомков вышеупомянутых групп. Однако, если это так, то имеется возможность и дальнейшей дивергенции т. е дальнейшего распадения на внучатые группы дочерних групп. Так как время, в течение которого совершается эволюция, на самом деле чрезвычайно велико, то сторонникам этого «компромиссного» взгляда поневоле приходится итти далее и допустить, что фазы дивергентной эволюции многократно повторялись на протяжении эволюционного процесса и что таким именно путем образовались такие крупные систематические группы, как семейства и отряды. Но тогда надо установить, почему дивергенция не может итти еще далее, итти до конца, и почему еще более высокие систематические группы (классы и типы) не могут также произойти этим путем?
Принятием такой смешанной, частично параллельной, частично дивергентной эволюции стройное здание полифилетической теории лишается своего основного фундамента, и поэтому с логической точки зрения Штейнман вполне прав в том, что он принимает и проводит эту гипотезу в ее чистой и крайней форме. В последующем мы и будем говорить именно о чистой полифилетической теории, в той ее форме, в какой она представлена у Лотси и Штейнмана.
Итак; если мы примем полифилетическую теорию в ее чистом виде и предположим, что ^се современные и вымершие виды животных произошли независимо друг от друга от такого же числа первичных протозоообразных форм (Штейнман), то мы неизбежно натолкнемся на весьма значительные затруднения. При этом нам прежде всего придется принять два следующих основных положения: 1) каждый вид развивается параллельно всякому другому, виду, причем его представители диференцируются гистологически и морфологически с различною степенью интенсивности, и 2) существует значительная разница в темпе филогенетического развития, так что определенная стадия эволюции одним родом животных достигается быстрее, чем другим.
Далее мы знаем, что все животные, принадлежащие к одной и той же крупной систематической группе, характеризуются рядом общих, свойственных всем им признаков. Если это так, то как объяснить, что совершенно одинаковые признаки развиваются иногда и у животных, генети-тически вовсе не связанных между собою? Ведь в самом деле, следуя полифилетической гипотезе, мы должны были бы признать, например, что млечные железы плацентарных млекопитающих со всеми деталями их гистологического и морфологического строения развились совершенно независимо друг от друга у различных видов этого подкласса или что волосы, которые у самых разнообразных млекопитающих имеют абсолютно аналогичную гистологическую структуру, образовались также совершенно независимо друг от друга в тех различных группах рептилий, от которых, согласно этой гипотезе, произошли эти млекопитающие. Мы должны были бы также принять, что одинаковое строение перьев у различных птиц также образовалось совершенно независимо в течение эволюции птиц из их рептилийных предков; и совершенно то же нам пришлось бы допустить относительно строения скелета и мускулатуры крыльев и задних конечностей этих птиц. Из этого следует, что мы должны допустить, что все то огромное количество сходных и одинаковых признаков, которые мы наблюдаем у особей, принадлежащих к любой крупной систематической группе животных, образовалось и развилось параллельно и независимо друг от друга. Однако нам очень трудно себе представить, как это могло реально произойти. Для объяснения этого явления авторы, защищающие полифилетическую точку зрения, либо принимают, что при этом действовал некий внутренний эволюционный принцип, который позво
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ
ИЗ
лил одинаковым признакам развиться параллельно у несвязанных между собою кровным родством животных индивидуумов (аутогенез), либо утверждают, что сходные признаки возникают здесь вследствие сходства в условиях той внешней среды, среди которых растут и развиваются упомянутые животные.
Чтобы пояснить эту мысль примером, укажем, что в первом случае (аутогенез) авторам полифилетической гипотезы пришлось бы принять, что у всех рыбообразных предков четвероногих животных (число видов которых, согласно этой гипотезе, должно было бы соответствовать числу всех вымерших и всех современных видов амфибий, рептилий, птиц и млекопитающих) должна была существовать некоторая внутренняя тенденция к видоизменению их грудных плавников в пятипалую (и отнюдь не в трехпалую и не в шестипалую!) конечность; такая же внутренняя тенденция должна была привести к образованию одинаково построенных тазовых и плечевых поясов, одинаково построенных скелетов плеча, предплечья и кисти, бедра, голени и плюсны, одинаково построенных мускулов и нервов.
Позднее эти неисчислимые амфибиообразные предки1 высших Quadru-peda перешли в рептилийную стадию их филогенетического развития; при этом у всех у них от неизвестных внутренних причин увеличилось в яйце количество желтка, образовались одинаково построенные амнион и аллантоис, одинаково ороговела кожа и т. д. Если мы примем в соображение все то неизмеримое количество сходных признаков, которое, по этой гипотезе, развилось параллельно и независимо друг от друга у позвоночных животных, то мы увидим, что к аутогенетическому принципу предъявляются слишком большие требования и что он действительно превращается в чисто абстрактный, далеко отстоящий от научного исследования принцип. При всем этом весьма интересно отметить, что сторонниками полифилетической гипотезы никогда не приводилось доказательств в пользу реального существования аутогенетического принципа в эволюции: он всегда носил чисто спекулятивный характер.
В одной из моих прежних работ я указал еще на одну трудность, с которой приходится считаться при допущении «внутреннего» принципа в истолковании эволюционного процесса. Я имею в виду невозможность для нас, при допущении этого принципа, понять, каким образом может случиться то, что одна какая-нибудь группа животных, эволюируя в течение определенного геологического периода в прогрессивном направлении, вдруг, как это часто случается, меняет свое первоначальное направление и начинает развиваться в регрессивном порядке. При этом нам пришлось бы допустить, что один и тот же внутренний эволютивный стимул может действовать сначала в одном направлении, а затем в обратном, что было бы очевидной бессмыслицей.
Другие последователи полифилетической теории, как, например, Эймер,2 объясняют параллелизм в развитии сходных признаков тем, что считают животных чрезвычайно подверженными непосредственному воздействию внешней среды; они считают, что сходные условия внешнего воздействия, непосредственно влияющие на сходные организмы, вызывают исходные изменения в строении этих организмов. Но и эта гипотеза наталкивается на резкий отпор со стороны генетиков, которые, как известно, пришли к выводу, что все изменения, вызванные непосредственным действием окружающей среды, точно так.же, как и изменения, вызванные упражнением и неупражнением органов, не передаются по наследству.
Если мы примем это возражение в расчет, то нам придется согласиться,
1 Т. е. все те, кто не вымерли, и, остановившись в своем развитии, не превратились в современных амфибий.
!Eimer Th., Die Entstehung der Arten 1888. Id., Orthogenesis der Schmet-terlinge, Leipzig, 1897.
Морфологические закономерности. 8
114
ГЛАВА IV
что в сущности мы имеем право требовать от полифилетиков более веских доказательств своих воззрений, чем обычно приводимые ими заключения по аналогии. Поэтому мы будем продолжать держаться нашего отрицательного отношения к этой гипотезе до тех пор, пока доказательства в пользу этой теории не примут несколько иного характера.
Принятие принципа наследования приобретенных признаков (Эймер) мало помогает делу, так как принцип этот не в состоянии объяснить многих хорошо известных сравнительным анатомам фактов. Становясь на эту точку зрения, мы не можем, например, понять, вследствие каких внешних влияний среды или вследствие какого сильного, упражнения могли возникнуть перегородки в сердцах птиц и млекопитающих? Или, что еще более примечательно, вследствие каких внешних условий могла возникнуть в течение филогенеза редукция у птиц левой, а у млекопитающих правой дуги аорты? Или, наконец, каковы были те изменения среды, которые могли вызвать у многочисленных видов амфибий образование амниона и аллантоиса типа рептилий? С другой стороны, едва, ли мыслимо предположить, что тысячи и тысячи видов рептилий, являющихся, по разбираемой нами гипотезе, предками всех птиц и млекопитающих и населяющих огромные пространства земли, жили бы все в одинаковой обстановке и в совершенно одинаковых условиях внешней среды, вызывающих совершенно одинаковые изменения в их организации. То, что мы знаем о географической обстановке, в которой жили эти ископаемые животные, отнюдь не поддерживает этого представления. Мы видим, таким образом, что всякая попытка систематически провести полифилетическую точку зрения на каком-нибудь конкретном примере приводит к совершенно несообразным заключениям, и нам становится поэтому вполне понятным, почему авторы, отстаивающие эти гипотезы, говорят о них лишь в самых общих чертах: со своей точки зрения они поступают вполне резонно, ибо всякая попытка к детальной разработке этих теорий лишь делает их совершенно неприемлемыми.
Не меньшие трудности встречает на своем пути полифилетическая теория и тогда, когда ее начинают критиковать с палеонтолого-геологической точки зрения. Принимая ее в ее чистой, так сказать, штейнмановской форме (так как лишь в этой ее форме она не наталкивается на внутренние противоречия), нам придется допустить, что в определенный геологический период существовало огромное число различных рептилий, готовых превратиться в такое же огромное число различного рода птиц; совершенно то же мы должны были бы сказать и относительно общего состояния предков млекопитающих в течение мезозоя. Однако, хотя нам знакомо очень большое число видов мезозойных рептилий, тем не менее среди этой многочисленной фауны непосредственных предков млекопитающих и птиц мы пока не находим. Штейнман, повидимому, учел это затруднение и попытался поэтому построить филогенез высших позвоночных со своей полифилетической точки зрения. Однако лучше бы ему не делать этой попытки, так как при этом он приходит к выводам, являющимся абсолютно неприемлемыми для большинства морфологов. Дело в том, что все филогенетические построения Штейнмана базируются на сходстве между некоторыми отдельными признаками, находимыми им у далеко отстоящих друг от друга видов животных; общий же комплекс всех остальных признаков данной животной формы им совершенно не принимается в расчет. Следуя этому методу, Штейнман принимает на основании сходства в строении костного панцыря, что броненосцы (Loricata) непосредственно происходят от панцырных мезозойных рептилий(Polacanthus), совершенно упуская при этом из виду все чрезвычайно важные различия, которые имеются в других частях костного скелета этих животных, как то: различия в строении слуховых косточек, различия в строении нижней челюсти
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ
115
различия в строении костей черепа и т. д. Бездоказательные и невероятные вспомогательные гипотезы, к которым он при этом прибегает (Штейнман допускает, например, что у всех этих вымерших рептилий, Polacanthus и др., существовали развившиеся совершенно самостоятельно и независимо друг от друга млечные железы типа млекопитающих, того же типа волосы, слуховые косточки и т. д.), настолько мало помогают делу, что он сам упоминает о них лишь вскользь. То же самое в сущности приходится сказать о предполагаемом Штейнманом происхождении мегатериев от игуанодонов, дельфинов от ихтиозавров, кашалотов от талат-тозавров; не более обосновано и воззрение Штейнмана на происхождение летучих мышей от летучих рептилий.
Я не вдаюсь здесь в более детальную критику филогенетических построений Штейнмана, ибо мои морфологические возражения будут мало понятны читателю, не знакомому со сравнительной анатомией; читателю же, которому хорошо известны сравнительно анатомические данные, они окажутся излишними, так как вся беспочвенность штейнмановских филогенезов ему и без того будет ясна.
Некоторые ученые, как, например, Соболев и Дакке, основываются в своих рассуждениях относительно полифилетической теории на фактах, почерпнутых ими из исследований, произведенных над моллюсками, в частности над головоногими. Но для сравнительного морфолога метод этих доказательств совершенно недопустим: существует одно хорошо обоснованное правило филогенетического и морфологического исследования, а именно, что никакой филогенез и никакая классификация не могут быть построены на основании изучения структуры только одного какого-нибудь органа или только одного какого-нибудь признака, например на основании строения кожи и ее придатков (чешуй, перьев) или на основании окраски животных и т. д. (По филогенезу, построенному на основании такого принципа, может, например, оказаться, что тигр и зебра являются родственными формами!). Тем не менее филогенез моллюсков, который, по Соболеву и Дакке, должен был служить решительным доказательством в пользу полифилетической гипотезы, построен именно на основании изучения одного единственного органа, т. е. раковины, выделяемой мантией этих животных. Между тем если мы сопоставим сравнительно простое строение мантии с весьма сложной структурой остальных частей тела этих моллюсков, например со строением их органов движения, органов выделения, органов чувств, со строением их нервов и кровеносной системы, то мы увидим, что имеем полное право усомниться в том, можно ли действительно всю сложную филогению моллюсков строить т о л ь к о на основании данных конхиологии, т. е. на основании изучения одних раковин. Если бы мы филогенез рыб, о котором речь будет впереди, стали строить только на основании кожного скелета этих животных, который можно рассматривать как аналог раковин моллюсков, то можно было бы справедливо усомниться в правильности наших выводов, и поэтому и по отношению к филогенезу моллюсков, нам думается, требуется не меньшая осторожность.1
Резюмируя все то, что нами было сказано относительно полифилетической теории, мы видим, что принятие ее необходимо требует от нас
1 Мы вполне признаем, что палеонтологи могут судить о филогенезе ископаемых моллюсков только на основании строения их раковин, так как от остальных органо» этих животных в ископаемом состоянии не сохранилось ничего, и мы отнюдь не сомневаемся в значении моллюсков как руководящих ископаемых; мы сомневаемся лишь в том, что на основании исследования одного органа можно воссоздать филогенез всего животного; поэтому все то, что нами сказано об этом вопросе, относится, сам» собой разумеется, к опенке исключительно палеонтологического метода исследования и нисколько не касается теории филогенеза моллюсков, построенной на основании сравнительно морфологических и эмбриологических исследований,
8»
116
ГЛАВА IV
в дальнейшем допущения существования двух весьма богатых фаун, состоящих в одном случае из форм, переходных между рептилиями и птицами, а в другом—из форм, переходных между рептилиями и млекопитающими, причем обе фауны эти вымерли, не оставив после себя никаких палеонтологических следов. Это последнее обстоятельство кажется нам весьма странным, особенно если мы вспомним о том, какое огромное число ископаемых остатков найдено нами от тех рептилий, которые вымерли, не оставив после себя потомков!
Для монофилетической теории этот пункт не представляет собою особых затруднений, так как совершенно ясно, что трудно ожидать найти палеонтологические остатки там, где предполагается существование малочисленной и географически ограниченной группы, от которой произошли рептилии, между тем как сильно размножившиеся потомки этой группы имеют много шансов на то, чтобы сохраниться в окаменелом состоянии. Для полифилетической же теории, предполагающей, что предки птиц, млекопитающих и рептилий были исключительно многочисленны, здесь имеется серьезный камень преткновения. Таким образом, перечисленные трудности (а также ряд других, о которых мы здесь еще не упомянули), стоящие на пути полифилетической гипотезы, не дают нам возможности примкнуть к ней.1
ЗНАЧЕНИЕ МОНОФИЛЕТИЧЕСКОЙ ТЕОРИИ
Чтобы быть правильно понятыми, мы должны определенно высказаться относительно того, в какой форме мы лично принимаем монофилетическую теорию. Некоторые авторы, в особенности уже упомянутые сторонники полифилетической точки зрения, когда они хотят доказать неприемлемость монофилии, утверждают будто все новые формы (подвиды и виды) являются, согласно монофилетической теории, потомками одной •единственной пары животных^ т. е., иными словами, что представители подвидов а' и а” (потомки вида А, схема III) представляют собою родных братьев и сестер. Вместе с большинством сторонников монофилетической теории я не принимаю такого ультрамонофилетического воззрения и признаю, что мы имеем право говорить о монофилии также и в тех случаях, когда подвиды а' и а" появляются в отличных от вида А условиях обитания (М' М"), как потомки индивидуумов, которые могут свободно спариваться между собою и производить плоду ч ее потомство. При этом для нашего воззрения не имеет значения, появляются ли признаки, характерные для подвидов а' и а", вначале у единичных особей или у большого числа их.
Первый признак появления дивергентной эволюции заключается в том, что определенный вид (фенотип) распадается на две (или более) группы особей, которые уже не способны спариваться между собой и производить плодучее потомство. При этом совершенно безразлично с биологической точки зрения, обусловлена ли эта неспособность к взаимному скрещиванию внутренними (строение зародышевых клеток) или внешними (различные периоды размножения) причинами. Именно с того времени, когда прекращается спаривание представителей различных подвидов (а' и а"), начинается их самостоятельная эволюция и их превращение в отдельные виды А' и А".
Нам не представляется возможным дать общее и непосредственное доказательство монофилетической теории в ее только что приведенной форме,
1 Можно было бы думать, что так как теория эта весьма слабо обоснована, тЪ подробное обсуждение возражений, которые -можно было бы выставить против нее, совершенно излишне; однако в виду того, что среди молодых зоологов, не занимавшихся специально морфологией, она встречает многих приверженцев, по возможности беспристрастная критика здесь является далеко не бесполезной.
ТЕОРИЯ монофилетической эволюции
117
но мы можем считать доказанным, что в большинстве случаев (правильнее сказать—во всех случаях), когда мы имеем достаточно полную палеонтологическую запись, эволюция протекает в дивергентных направлениях, так что линии, которыми мы символически обозначаем анцестральные ряды, почти соприкасаются своими корнями; однако сближаются ли они между собою настолько, что мы действительно можем говорить об одном первичном прародительском вид е,—об этом в точности мы еще не знаем ничего.
Отметим, что не один только палеонтологический материал указывает нам на наличность дивергентной эволюции; к совершенно аналогичным результатам приходим мы и тогда, когда исследуем эволюцию при помощи сравнительно анатомического или сравнительно эмбриологического метода.
Схема V. Объяснение в тексте. Схема VI. Объяснение в тексте.
Наша точка зрения сделается еще более понятной, если мы иллюстрируем ее при помощи двух прилагаемых схем (V и VI).
Рассматривая эти схемы, мы можем сказать, что эволюция в период, у—2 во всяком случае шла дивергентным путем, но обычно мы не знаем, как протекала она в предшествующее время, т. е. в период х—у. Можно предположить, что дивергенция появилась лишь в периоде у—z и что раньше, т. е. в период х—у, ряды предков (А,—Ai, А2—Аг, А3—Аз) шли параллельными линиями, не сближаясь своими корнями (схема V), или можно предположить, что эти ряды шли в период х—у в том же .направлении, что и позже, т. е. в период у—z, так что в период х—у они сближались между собою в одном пункте А (схема VI); это будет признаком того, что дивергентная эволюция началась с вида А. В этом последнем случае мы имеем дело с примером такой формы монофилетической эволюции, когда из одного вида А развиваются три более высоко стоящих систематических группы АГ, АГ и АГ (ср. схему III).
Что же касается первого нашего предположения, а именно, что ряды предков Аг—А1, А2—Аг, А3—АГ разделенные в течение бесконечно длинного периода х—у, изменялись все это время параллельно друг другу, то оно нам кажется, как мы это уже отметили выше, весьма невероятным; поэтому обычно принято думать, что и в предшествующую эпоху эволюция шла в том же дивергентном направлении, в каком она шла и в более поздний период у—z (схема VI). Надо сказать, что палеонтологические и эмбриологические факты во всяком случае решительно говорят в пользу этого последнего (монофилетического) толкования эволюционного процесса, хотя мы можем привести лишь очень ограниченное число прямых и безупречных примеров такой эволюции. К таким примерам относится, между прочим, цитируемый Плате (1925)1 ряд предков Planorbis multi for-
1 Р 1 a t е L., Allgemeine Zoology, 1925.
11'8
ГЛАВА IV
mis, всем известный анцестральный ряд Equidae (схема VIII) и, может быть, ряд титанотериев, приводимый Осборном, а также некоторые немногочисленные ряды других млекопитающих.
Итак, палеонтологические и сравнительно анатомические факты, подтверждающие дивергентную эволюцию, логически приводят нас к выводу, что монофилетическая теория является правильной теорией и что дивергентное развитие начинается в пределах какого-нибудь одного прародительского вида А. Противоположная точка зрения, а именно взгляд, что анцестральные ряды —А',, А2—А'а, А3—А'3 развивались не дивер-гентно, но параллельно в течение периода х—у (схема V), является также не чем иным, как логическим умозаключением, но умозаключением чрезвычайно невероятным.
АДАПТИВНЫЕ ПРИЗНАКИ КРУПНЫХ СИСТЕМАТИЧЕСКИХ ГРУПП
Если мы обратим свое внимание на разницу, лежащую между различными систематическими группами, то мы увидим, что у высших систематических групп—у семейств, отрядов и классов—э та разница относится главным образом к физиологически и биологически важным приспособительным признакам. Я должен напомнить при этом читателю об осборнов-ских примерах «адаптивной радиации».1 Он приходит, например, к выводу, что конечности всех млекопитающих развивались из короткой, стопоходящей, снабженной когтями пятипалой конечности; эта примитивная форма конечностей, непрерывно изменяясь в течение эволюции, перешла в целый ряд адаптивных конечностей, характерных для различных крупных групп млекопитающих. Так, мы имеем: 1) роюший тип конечностей у кротов, слепышей (Spalacidae), Chrysochloris и др.; 2) плавающий тип конечностей у тюленей, моржей, морских коров, китов и др.; 3) лазающий тип конечностей у обезьян, ленивцев, лемуров; 4) летательный тип конечностей (у летучих мышей, летучих собак); 5) бегающий тип конечностей, из которого развилась однопалая и двупалая конечность копытных. Во всех этих характерных для отдельных отрядов млекопитающих изменениях конечностей мы имеем дело с адаптивными изменениями, т. е. с приспособлениями скелета конечностей к различным условиям окружающей среды, той среды, в которой приходится жить и двигаться всем этим животным.
Совершенно тот же характер изменений встречаем мы и у зубов различного рода млекопитающих; развившись из зубов насекомоядного типа (Осборн), они превратились в зубы хищного, всеядного, травоядного и мура-вьеядного типа (myrmephagous, тип с редуцированными зубами). Наконец, точно такую же адаптивную эволюцию встречаем мы и при исследовании дивергентного развития формы тела и формы конечностей у крупных групп амфибий (Anuга, Urodela, Apoda) и рептилий. Напомним также о разнице в форме тела и в скелете между веретенообразными акулами и сплюснутыми в дорзовентральном направлении скатами, которая обусловлена разницей между нектонным и бентонным образом жизни этих рыб.
Всякий сравнительный анатом легко может привести огромное число примеров такой адаптивной дивергентной эволюции крупных животных групп, но здесь' мы на них останавливаться не будем.1 2 Как мы это только что отметили, все эти адаптивные признаки характеризуют собою именно крупные систематические группы животных —семейства, отряды и классы.
1 О s b о г n Н. F., The age of Mammals in Europe, Asia and North America, New York, 1910.
2 Cai. Plate, Allgemeine Zoologie, Bd. 1 и 2, где приведено большое количество такого рода примеров.
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ
119
АДАПТИВНЫЕ ПРИЗНАКИ МЕЛКИХ СИСТЕМАТИЧЕСКИХ ГРУПП 4
Если мы теперь обратимся к признакам, которыми характеризуются систематические группы низшего порядка (роды, виды и подвиды), то мы увидим, что приспособительный характер признаков, которыми они друг от друга отличаются, проявляется далеко не так отчетливо, как он должен был бы проявляться у признаков,/ развившихся вследствие дивергентной эволюции. Правда, есть случаи, когда мы можем сказать, что признаки различия между видами одного рода и между родами одного семейства имеют биологическое значение, как, например, случай, где однотонная окраска, которая отличает льва и пуму от их ближайших сородичей, является приспособлением к жизни в пустыне! здесь мы знаем, что эта окраска развилась из полосатого и чернопятнистого рисунка шкуры предков этих животных, некогда обитавших в джунглях и лесах. Но во многих случаях трудно говорить о видовых признаках как о приспособлениях. Сопоставляя признаки, на основании которых систематик различает роды, принадлежащие к одному семейству, или виды, принадлежащие к одному роду, мы находим, что эти группы животных часто характеризуются такими отличиями, которые очень трудно рассматривать как приспособления и которые скорее всего приходится отнести к категории «индиферентных» признаков. Таким образом, мелкие систематические группы чаще всего отличаются друг от друга именно «индиферентными» признаками. Но здесь обычно нельзя установить соотношения между родовыми и видовыми признаками, на основании которых мы строим наши систематические группы, и особенностями жизненных условий животных. Поэтому нам приходится спросить себя, не в результате ли приобретения именно этих мелких «индиферентных» признаков животные эволюировали в дивергентном направлении и постепенно приспособились к различным условиям окружающей среды?
Как известно, проблема «индиферентных» признаков рассматривается в биологической литературе главным образом в связи с дарвиновской теорией, причем обычно говорится, что теория естественного подбора не в состоянии объяснить происхождение этих признаков. Мы хотели бы поставить этот вопрос несколько иначе и спросить, действительно ли направление дивергентной эволюции в мелких систематических группах обусловлено теми биологически «индиферентными» признаками, которые мы употребляем для нашей классификации, т. е. действительно ли дивергентная эволюция следовала по пути наших систематических признаков, а не каких-то других признаков, не принятых во внимание нашими систематиками? Мы должны были бы ответить на этот вопрос утвердительно, если бы животные, принадлежащие к мелким систематическим группам— к подвидам, видам, родам,—отличались между собою только теми «индиферентными» признаками, которыми пользуются наши систематики. Но мы знаем, что это не так: в организации животных, принадлежащих к различным видам одного и того же рода, имеются вполне определенные и постоянные отличия, которые систематик, однако, оставляет в стороне, так как эти отличия являются признаками, не удобными для классификационных целей. Систематик, разумеется, выбирает признаки, удобные в диагностическом отношении, и поэтому обращает главное внимание на те наружные черты отличия, которые позволяют ему легко определить вид, не прибегая при этом к анатомическому или экологическому исследованию.
Однако систему животных можно было бы. построить и по другому принципу, основываясь преимущественно на внутренних признаках животного, и пример такой классификации мы находим у палеонтологов, построивших свою систему ископаемых форм на основании признаков строения
120
ГЛАВА IV
внутреннего скелета животных. В сущности выбор систематического признака, по которому классифицируются в системе мелкие систематические группы низшего порядка, часто является по преимуществу вопросом удобства.1
Мы говорим здесь о признаках, по которым виды отличаются один от другого, так как само собою разумеется, что мы объединяем сходных животных в один вид на основании биологического признака, т. е. исходя из их способности спариваться между собою и давать плодовитое потомство.
Итак, мы видим, что животные виды отличаются друг от друга не только теми признаками, которые служат для наших классификационных целей, но и целым рядом других морфологических и физиологических признаков, которыми наша классификация и наши определители не пользуются вовсе. Мы видели также, что многие классификационные- признаки могут быть охарактеризованы как физиологически и биологически индифе-рентные, и поставили вопрос о том, что когда произошло разделение (дивергенция) предков современных форм на виды, с которыми приходится иметь ныне дело зоологу, то не произошло ли это разделение по признакам, оставленным систематиками всех классификаций без внимания, но важным в физиологическом или биологическом отношении?
Чтобы ответить на этот последний вопрос, мы должны установить прежде всего, действительно ли виды, встречающиеся в природе, отличаются между собою биологически важными (адаптивными) признаками или нет.1 2
ПРИСПОСОБИТЕЛЬНО-ДИВЕРГЕНТНАЯ ЭВОЛЮЦИЯ
Все зоологи, занимающиеся биологией, знают, что виды отличаются друг от друга, помимо прочих признаков, также и своими инстинктами и привычками, местом своего обитания, своим образом жизни и т. д. Всякий опытный охотник сообщит нам о биологической разнице между беляком (Lepus timidus) и русаком (Lepus europeus); между гаршнепом (Gallinago gallinula), дуппелем (Gallinago major), бекасом (Gallinago gallinago) и вальдшнепом (Scolopax rusticola). Эти птицы, привычки и образ жизни которых нам хорошо знакомы, отличаются друг от друга своими инстинктами, местом своего обитания, своим гнездованием, своим более быстрым или более медленным полетом и т. д. Мы находим здесь много мелких, но’ биологически важных отличий, как, например, отличия в способах, к которым эти птицы прибегают для того, чтобы укрыться от врагов, отличия в способах, которыми они защищаются от их нападения, отличия во времени размножения, отличия в поисках и добывании пищи и т. п. Т^к, синица (Parus ater) живет в хвойных лесах, а весьма близкая к ней кавказская Parus ater prageri—в лиственных лесах. В биологическом отношении отличаются между собою также и близко родственные виды пищух (Certhia familiaris и Certhia brachydactyla), а также другие синицы: Parus communis и Parus atrecapyllus; отличаются также и гори-хвостка-лысушка (Phoenicurus phoenicurus) от горихвостки-чернушки (Phoenicurus ochrurus); сюда же относятся и биологические отличия между каменной и благородной куницей.
1 Если мы будем классифицировать животных только по внешним признакам сходства и отличия, то нам придется отличающихся резким диморфизмом самцов и самок некоторых животных, например Boneilia и др., поместить в различные виды и роды.
2 Мы говорим здесь о линнеевских видах и отнюдь не имеем ввиду тех чистых линий, которые выводятся зоологами экспериментаторами и ботаниками и не встречаются в природе.
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ
121
Особенно интересна для нас в этом отношении водяная крыса (Arvicola amphibius),—вид, обнаруживающий совершенно различные привычки в зависмости от того места, где он обитает. Те Arvicolae, которые живут поблизости от водных бассейнов, ведут себя как типичные водные животные: плавают в воде и роют свои норки и ходы на берегу; те же, что живут в сухой местности, роют, подобно кротам, длинные подземные ходы. Можно было бы сказать, что здесь мы имеем перед собою пример н а-чала дивергентной эволюции (схема III, а', а")-
Эти наблюдения, сделанные над хорошо знакомыми нам дикими животными, прекрасно подтверждаются наблюдениями над еще более знакомыми нам домашними животными. Здесь мы имеем перед собою целый ряд весьма характерных отличий, из которых многие имеют большое биологическое значение. Среди этих отличий отметим, например, различия в мускульной силе, в выносливости, в быстроте и величине, в злобности, в обонянии у различных рас наших собак. То же самое можем мы, конечно, сказать и о различных расах лошадей, овец, рогатого скота и т. д.
Мы знаем, что, помимо всех этих физиологических и морфологических признаков, различные расы домашних животных, иногда настолько близко стоящие друг к другу, что они могут даже скрещиваться между собою, различаются еще и своими наружными признаками, своим так называемым «экстерьером», и что этот «экстерьер» наследственно связан с внутренними признаками, о которых только что шла речь. Знатоки-животноводы на основании одного только внешнего вида животного могут безошибочно определить, много или мало молока даст корова и быстро или нет будет бежать лошадь или борзая собака. Селекционеры будут подбирать производителей по их общему габитусу, т. е. по их чисто внешним признакам, но цель, которую они при этом преследуют, будет, конечно, заключаться в том, чтобы получить быстрых рысаков, быструю гончую собаку, корову с большим удоем молока, сильное грузовое животное и т. п., т. е. в том, чтобы получить животных с физиологически и морфо.т’о-гически важными особенностями внутренних органов (костей, мускулов, легких, желез и т. д.). «Экстерьер» при этом служит лишь в качестве наружного указателя филетически связанных с ним коррелятивных изменений признаков внутренней организации.
Основываясь на всем известных фактах, касающихся только что упомянутых диких животных, мы можем принять, что все эти животные, которых мы различаем’и классифицируем по их таксономическим признакам, на самом деле отличаются между собою по их биологическим и физиологическим особенностям, т. е. по тем их внутренним морфологическим признакам, с которыми наружные признаки связаны лишь коррелятивно. Так, мы знаем, что различия в мускульной силе, в быстроте, в выносливости, в инстинктах и привычках зависят от совершенно определенных, хотя, и мало заметных отличий в строении мускулов, сердца, легких, нервной системы и органов чувств. Нам кажется поэтому, что при несколько лучшем знании биологии и сравнительной физиологии животных мы могли бы построить чисто биологическую систематику животных, которая базировалась бы на биологически и морфологически важных признаках, а также на коррелятивно связаных с ними особенностях структуры органов. Для музейных целей подобная систематика была бы, возможно, мало пригодна, но для нашего понимания эволюции животных она была бы, по всей вероятности, чрезвычайно полезна.
Если мы согласимся с тем, что живущие на свободе дикие виды действительно отличаются между собою вышеупомянутыми биологическими и физиологическими признаками, то мы имеем полное право заключить, что видообразование обусловлено большей частью не «индиферентными»-признаками, на которых м’ы основываем наши систематические категории,
122
ГЛАВА IV
но теми биологически важными отличиями, связанными с условиями существования, о которых мы говорили выше.
Если какой-нибудь вид разделяется сначала на два подвида (схема III а' и а"), а затем на два вида (А'и А"), то мы знаем, что это расчленение вызвано начинающимся приспособлением подчиненных систематических групп (подвидов или дочерних видов) к различным условиям существования, т. е. к несколько отличной от прежнего травоядной или мясной пище, к другим врагам, к другой структуре почвы, вообще говоря, к несколько иным условиям окружающей среды. Мы не говорим здесь о географической изоляции в собственном смысле этого слова, но единственно о приспособлении к несколько отличным от прежних
(1,.1г.13,а,,аг,з3) А
Схема VII
А—прародительский вид; Als Аз, A'j, А'2—дочерние и внучатые виды; а1; а», а3, а',, а'«, а'.
TV ~ * *
...а 3 —полезные, т. е. приспособительные признаки видов A, Aj, А2 , и т. Дii, i2, i3, i'j,
—индиферентные признаки, коррелятивно связанные с приспособительными признаками, условиям внешней среды, т. е. о приспособлении к жизни в пустыне, в болоте, в степи, в лесу с характерной для них флорой и фауной. При подобного рода приспособлении дочерние виды отличаются друг от друга именно теми своими биологическими особенностями, при помощи которых они приспособились к различным условиям существования, т. е. своей величиной, силой своих мускулов, своей способностью укрыться от врагов и нападать на них, своей выносливостью и т. п., а также соответствующими морфологическими отличиями в строении отдельных органов: в расцветке кожи и ее придатков, в строении мускулов и скелета, в строении легких и сердца, в строении органов защиты и нападения.
Именно эти-то отличия, которые, как мы это только что показали, всегда имеются налицо, и являются причиной дивергентной эволюции подчиненных систематических групп и их дальнейшего распадения на еще •более мелкие систематические группы.
Приводимая выше схема (VII) нагляднее пояснит нашу мысль.
В этой схеме мы принимаем, что в течение филогенеза у потомков вида А, т. е. у форм Аг и А2, существовали двоякого рода признаки, а именно: 1) адаптивные признаки, т. е. такие, которые при данных условиях существования были им полезны (у формы Ах—это признаки а',’а', а'; у формы А2—это признаки а", а", а" ; у формы А'—это признаки а"', а а у формы А'—это признаки а 7, а1^, а1^), и индиферентные признаки,
т. е. такие, которые с адаптивными признаками были связаны коррелятивно (у формы' Аг—это признаки i', i', i' ; у формы А2—это признаки i”, i2, i'3',;y формы А'—это признаки Г”, 1'Г> *7, и У формы А'—это признаки iry, iT2v, 1ГУ).
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ 123
Чем лучше и чем в большей степени такие дочерние виды приспособляются к новым условиям существования, тем лучше и тем в большей степени будут выражены в них и биологические различия (адаптации ах, а2, а3) и коррелятивно связанные с этими последними индиферентные признаки (iv i2, i3, позднее i'”, Г", i"', и i1/, i1^, iV), так что, наконец, развиваются и те резкие отличия в биологически важных признаках (адаптации а'^', а"'» а'з, и a\v, а1^, а1^ ), которыми отличаются между собой выше стоящие группы: роды, семейства, отряды и т. д.
Только что изложенное нами представление о видообразовании как о процессе приспособительной эволюции позволяет нам понять, каким образом могли развиться приспособительные признаки, которыми отличаются между собою крупные систематические группы.
Предпосылки такой дивергентной эволюции даны нам в постоянно нарастающих отличиях в местах обитания животных1 (схема III, М, М', М" и т. д.) и в связанных с ними условиях окружающей среды, а также в наличности физиологических и морфологических признаков организации данных форм.
ПЕРВИЧНЫЕ ПРИСПОСОБЛЕНИЯ И КОРРЕЛЯЦИИ
Монофилетическая теория принимает, таким образом, что признаки крупных систематических групп развиваются из приспособительных признаков прародительского вида. При этом надо заметить, что далеко не всегда легко сказать, в чем именно заключается первичное приспособление в каждом отдельном случае, так как, вообще говоря, приспособления сами по себе носят весьма разнообразный характер: существуют, например, приспособления наружных, т. е. эктосоматических, органов к определенным условиям существования, а также многочисленные приспособления внутренних органов к переменам, происшедшим в органах, имеющих непосредственное отношение к внешней среде, т. е. в только что указанных эктосоматических органах. Мы можем обозначить эти двоякого рода приспособительные изменения как изменения первичные и изменения координированные. Если мы возьмем в качестве примера скатов, тонам придется принять, что здесь первичное изменение будет заключаться в дорзовентраль-ном сплющивании тела, стоящем в связи с переходом этих животных к бен-тонному образу жизни. У предков скатов увеличиваются при этом грудные плавники (срастание pro- и metapterygium, увеличение числа первичных плавниковых лучей и их сочленений). Направленное вперед разрастание грудных плавников вызывает сдвигание жаберных отверстий на вентральную, а брызгальца(зр!гаси1ит)—на дорзальную сторону тела. Эти вторичные изменения являются в свою очередь причиной целого ряда третичных, координированных с ними, изменений, происходящих в форме и положении hyomandibula re, в челюстях, в жаберных лучах и т. д.
В данном случае сравнительно легко установить, в чем именно заключается первичное изменение у предков скатов (оно состоит в расширении и сплющивании тела) и как развиваются здесь филетические корреляции. В других случаях это представляет собой гораздо большие трудности.
Так, например, очень трудно сказать, в чем именно заключалось первичное изменение, вызвавшее определенное строение тела многих групп костистых рыб, так как совершенно несомненно, что весьма значительные перемены, происшедшие в их организации, носят отнюдь не первичный характер, но представляют собою лишь вторичные филетические корреляции, вызванные неизвестными первичными изменениями в организации предков современных форм.
1 Мы знаем из геоморфологии, что подобного рода различия всегда имеют место в природе и что с течением времени и по мере географического распространения животных, они делаются все более и более значительными.
124
ГЛАВА IV
Рассматривая характерные признаки крупных систематических групп, мы находим, что классы, отряды, а нередко и семейства отличаются друг от друга целым комплексом адаптивных признаков и что именно эти признаки встречаются в самых различных органах животных, даже если они функционально и слабо связаны между собою. Монофилетическая теория принимает, что признаки, являющиеся общими для какой-нибудь группы животных, наследуются от ее общего предка, из чего следует, что этот родоначальный вид некогда уже обладал всей суммой признаков или по крайней мере всей суммой ориментов признаков,1 характеризующих происшедшие от него группы животных (роды, семейства, отряды). Так как современные виды отличаются друг от друга главным образом лишь незначительными признаками, то возможность присутствия в одном прародительском виде целого ряда характерных признаков кажется мало понятной; поэтому я позволю себе разобрать этот вопрос на одном хорошо всем знакомом палеонтологическом примере, а именно на эволюции лошади.
Подсемейство Equidae характеризуется следующими признаками: глазные впадины сзади целиком отграничены костями; коренные зубы (molares) гипселодонтного типа; их жевательная поверхность покрыта складчатою эмалью (селенодонтный тип); ulna и radius сраслись друг с другом, причем нижний конец ulna чрезвычайно утончен; конечности— монодактильного типа, боковые же метаподиальные косточки (metacarpalia s. metatarsalia II, IV) превращены в тонкие грифелеобразные косточки; fibula редуцирована.
Предком Equidae1 2 и близко стоящих к ним Hyracotheria< был, по всей вероятности, принадлежащий к Condylarthra Tetraclaenodon (из сем. Phaenacodontidae, палеоцен). Тетракленодоны были сравнительно мелкими животными с длинным хвостом, с бунодонтными зубами и пятипалыми конечностями, заканчивающимися маленькими копытцами; они были полупальцеходными формами, т. е. касались земли при ходьбе нижней поверхностью своих фаланг, и не были поэтому такими хорошими бегунами, как копытные животные. Существование бунодонтных коренных зубов, т. е. зубов с тупыми бугорками, указывает на то, что эти животные были всеядными формами, близко стоящими к хищникам.
Эволюция потомков тетракленодонов шла совершенно определенным путем, а именно в направлении приспособления организации этих животных к быстрому передвижению по плоской поверхности степей и к переходу их от всеядного питания к чисто травоядному.
У потомка этих еще очень примитивных форм, у небольшого Eohip-pus3 ulna и radius были еще раздельны, но редукция краевых лучей конечностей уже наступила: передние конечности имели уже четыре пальца, а задние даже три; в коренных зубах наметился переход от бунодонтного типа к лофодонтному,4 что указывает на преимущественное употребление растительной пищи; metacarpalia и metatarsalia были сильно удлинены, и это дает нам повод предполагать, что Eohippus был уже довольно быстро бегающим животным.
От Eohippus произошел весьма близко стоящий к нему Orohippus, отличающийся от Eohippus лишь некоторыми незначительными изменениями, происшедшими в строении зубов.
1 Ср. относящуюся к этому вопросу теорию гомойологии Плате (Plate, Abstam-
mungslehre, S. 73, Jena, 1925).
3 Ср. очерк филогении лошади в работе О. Абеля, Das biologische TfSgheits-gesetz, Biol. Gen., Bd. 4, 1928.
3 Размер этих животных средний между кошкою и фокстеррьеро.м.
4 С V-образными складками эмали.
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ ЭВОЛЮЦИИ
125
В том же направлении изменялась и организация более поздних потомков Eohippus, а именно Epihippus, остатки которого найдены уже в верхнем эоцене, и Mesohippus, относящегося к нижнему и среднему олигоцену. У последнего глазница сзади еще не отграничена костями; все его четыре конечности были уже тридактильны, т. е. Mesohippus при передвижении касался земли всеми тремя пальцами; но его краевые пальцы сделались уже значительно тоньше, чем средний; травоядный тип зубов у него был более резко выражен, чем у предыдущих форм.
Прямым потомком Mesohippus был живший в верхнем олигоцене Miohip-pus, от которого ответвился в одну сторону Anchitherium, вымерший цели- ; ком в среднем миоцене, не оставив после себя потомства, в другую— Parahippus, живший в нижнем миоцене (его глазная впадина уже была целиком отграничена сзади костями, а боковые пальцы всех четырех конечностей сильно редуцированы), а в третью—Kalohippus, Archaeohippus и Hypohippus, вымершие также в нижнем миоцене, не оставив после себя потомства.
По прямой линии от Parahippus происходят непосредственные предки ] лошади, Meryhippus и Hipparion, у которых коренные зубы имели уже ясно выраженный гипселодонтный характер; ulna и fibula, еще довольно сильно развитые у Mesohippus и его предков, здесь стали резко утонченными в своих дистальных частях костями, причем ulna срастается с radius в одно образование, наподобие того, как это имеет место у современной лошади. Краевые пальцы еще у Meryhippus стали уже настолько короткими, что животное употребляло при быстром беге лишь один сильно развитый сред- I ний палец.
Все только что перечисленные нами ископаемые формы жили в свое время в Северной Америке, и лишь непосредственный предок’нашей лошади—трехпалый, величиною с осла гиппарион, переселился в Европу.
От гиппариона происходит настоящая лошадь (Equus), у которой боковые пальцы уже окончательно редуцируются и превращаются в маленькие грифельные косточки. Поселившись в одной из степных местностей Евразии, эти древние лошади распространились по всей Евразии и Африке, распавшись при этом на большое количество видов (зебры, квагги, джигетаи, тарпаны, лошади Пржевальского, дикие ослы и т. д.). Кроме европейской ветви потомков Meryhippus, от него произошло еще некоторое число плиоценовых и плейстоценовых американских форм: Neohipparion и Neohippus, Protohippus, Pliohippus и, наконец, Plesiohippus, от которого происходит южноамериканский однопалый Hippidium. Все эти американские формы вымерли еще в доисторическое время. Ныне живущие американские дикие лошади происходят, как известно, от одичавших европейских лошадей, в свое время вывезенных из Испании в Америку. Родословное древо лошади поедставлено в нижеследующей таблице (схема VIII),; заимствованной у Абеля.
Рассматривая историю эволюции вида Equus, мы видим прежде всего, что в общем в большей или меньшей степени изменились именно адаптивные признаки вида Equus и что процесс филогенетического развития лошади протекал весьма медленно (с начала эоцена вплоть до плейстоцена) и весьма постепенно.
Мы видели, что исходные предки лошади (Tetraclaenodontidae) были; мелкими пятипалыми всеядными животными. Эволюция потомков тетра-1 кленодонов заключалась в чрезвычайно медленном изменении главным] образом двух систем органов, именно черепа с его жевательным аппаратом] (челюсти, зубы и т. п.) и конечностей. Оба эти ряда изменений шли парал-i дельно друг другу и находились в совершенно определенных соотношениях с переменами, имевшими место в образе жизни этих животных. Основываясь на строении зубов тетракленодонов, можно думать, что эти
126
ГЛАВА IV.
медленно передвигающиеся животные обитали в местности, покрытой деревьями и кустарниками, и что они питались смешанной, частью растительной, частью животной (мясной) пищей.
Строение и общая организация их ближайших потомков, только что
Европа Сев Америка Южн. Америка
Голоцен Современный Ецийз Equus Equus
Плейстоцен Ледниковый период 1 Equus — -^Equus Neoiiippus— Neohippus Hippidium 1
Плиоцен Верю Neohip)vs 1
Средн. 1 Ptesihippus I Neohippprion ।
Нижи. 1 Hipparion — | 1 Protohippus Hipparion Neohipp^ion МЖррие^курцМрриз
Миоцен. Верлн Средн. An chitheruuxi Meryivppus L_ 1—J 1
Нт. — j Arcnaeotiippus Parahippus Kaiohippus | Hypohippus 4 111
Олигоцен Верхе
Средн. 1 Mesohippus
Нижн. 1 Mesohtpp'js
Эоцен Верю. Epihippus ♦'
Средн . 1
1 Orohippus
Нижн. i Eohippus I
Hyracotheriurn 1 Eohippus
Палеоцен Общий предок Hyracothenum и Eohippus ыГгруппы' Hyopsodus MarKmani Abel и Сион из нижнего эоцена Северной Америки Исходный тип. Tetraclaenodon (Новая Мексика)
Схема VIII. Филогенетическое древо лошадей по О. Абелю. Объяснение в тексте.
перечисленных нами предков лошади (Eohippus, Orohippus, Epihippus и т. д.), заставляют нас предполагать что формы эти уже в ту эпоху переселились в открытую местность, покрытую густой травой и мелким кустарником, и приспособились к условиям этой новой для них среды. Медленное приспособление заключалось в видоизменении ротовых органов (зубы, челюсти, жевательная мускулатура) и органов пищеварения этих животных, перешедших к питанию травою, а также в видоизменении строения
ТЕОРИЯ МОНОФИЛЕТИЧЕСКОЙ эволюции 127
конечностей1 в связи с необходимостью спасаться бегством от быстроногих степных хищников, от которых им негде было укрыться в открытой местности (изменения в строении пальцев). Эти чрезвычайно медленные и постепенные видоизменения организации, соответствующие переменам, происшедшим в окружающей среде (лес—степь), происходили все время в одном и том же определенном направлении, так что, в конце концов, вся организация Equidae сильно изменилась.
Общая величина изменений, имевших место в этих двух системах органов, представляет собою результат суммирования крайне незначительных и мелких изменений организации, происшедших в течение чрезвычайно длинного промежутка времени (с эоцена до конца плиоцена).
Факт, что различные системы органов (череп, зубы и конечности) изменялись одновременно и параллельно, объясняется тем обстоятельством, что упомянутые животные одновременно приспособились к различного рода переменам, параллельно происходившим в окружающей с р е д е, а именно к перемене в пище (переход от всеядного способа питания к травоядному) и к новым врагам (переход от преследования со стороны лесных хищников, от которых им приходилось скрываться в лесу, к преследованию со стороны быстро бегающих степных хищников). Таким образом, здесь изменения начались в различных, не связанных между собою системах. органов одновременно с началом изменений в окружающей среде, и совершались в дальнейшем параллельно во всех этих органах. Ь приведенном нами примере (схема VIII) морфологические признаки родоначального вида Equus развивались в происшедших от него дочерних видах путем постепенного накопления (суммирования) мелких, почти незаметных на первый взгляд видоизменений.
В других случаях изменения в различных функционально не связанных между собою системах органов развиваются не одновременно (как у Equidae), но в известной последовательности, одно после другого, причем они бывают отделены друг от друга более или менее значительными промежутками времени. Так, мы находим, например, что у некоторых ископаемых рептилий (Triceratops, Iguanodon), передние конечности были пятипалы, а задние трехпалы, и можем поэтому сказать, что трехпалость представляет собою новый признак, развившийся у рептилий гораздо позднее, чем пятипалость. У современных птиц мы находим, что их передние конечности превратились в крылья и что их челюсть стала беззубой. Так как существовали птицы, у которых зубы были выражены еще вполне ясно, то мы можем сказать с полной определенностью, что редукция зубов наступила в эволюции птиц гораздо позднее, чем развитие крыльев.
Мы имеем все основания признать, что и у млекопитающих характерные признаки развивались не одновременно, но одни после других, в различные геологические эпохи. Мы знаем, например, что превращение articulare и quadratum в слуховые косточки (malleus и incus) началось уже у Therocephalia, т. е. еще у типичных рептилий, так что у них нижняя челюсть (dentale) сочленялась непосредственно с височною областью, и можем поэтому считать, что остальные характерные черты млекопитающих (волосы, млечные железы, четырехкамерное сердце, редукция правой аортальной дуг и и т.д.) образовались уже позднее. К сожалению, последовательность в образовании только что перечисленных признаков еще не может считаться установленной специальными исследованиями.
С подобными и аналогичными примерами мы встретимся еще не раз в дальнейшем; поэтому я не буду сейчас на них задерживаться и лишь
» Вероятно, также легких и сердца.
128
ГЛАВА IV
отмечу, что различные комплексы приспособительных признаков, которыми животные группы характеризуются на определенной ступени их филогенетического развития, развиваются либо одновременно как приспособления одновременно наступающим переменам в окружающей среде, либо в известной последовательности как приспособления к по- ♦ следовательным переменам в окружающей среде, в которой эволюиру-ющие животные формы жили в различные периоды своего развития.
ВЫВОДЫ
На этом мы можем закончить изложение теоретических взглядов относительно монофилетической эволюции, резюмируя наши основные результаты в следующей форме:
1. Известные нам палеонтологические, сравнительно анатомические и эмбриологические факты целиком подтверждают монофилетическую теорию, т. е. тот взгляд, по которому все виды одного рода, все роды одного семейства и т. д. происходят из одного прародительского вида.
2. Полифилетическая теория, т. е. тот взгляд, по которому все современные и вымершие виды животных произошли независимо друг от .друга от такого- же количества первичных протозоообразных форм, не подтверждается фактами и поэтому не может быть признана правильной.
3. Мелкие систематические группы (подвиды, позднее виды), с которых начинается эволюция, отличаются между собою не только биологически индиферентными признаками, но также и весьма многочисленными приспособительными признаками, имеющими для животных большое биологическое и физиологическое значение. Образование дивергентных видов происходит именно по этим адаптивным признакам, ибо дочерние подвиды (а позднее—-дочерние виды), по всей вероятности, приспособляются к новым условиям существования при изменении окружающей среды именно при помощи этих приспособительных (мутации и геновариации) признаков (адаптивная монофилетическая эволюция).
4. При дальнейших изменениях в окружающей среде, изменениях, идущих в одном и том же направлении, суммируются мелкие наследственные приспособительные вариации, постепенно превращаясь в крупные адаптивные признаки,—такие признаки, которыми отличаются друг от друга более высокие систематические группы (семейства, отряды и т. д.).
5. Параллельно с эволюцией первичных адаптивных изменений идет эволюция функционально с ними связанных коррелятивных признаков. Некоторые из этих коррелятивных признаков имеют биологическое или физиологическое значение (полезные признаки), другие, наоборот, являются индиферентными. Направление эволюции каждой данной группы животных определяется эволюцией его первичных адаптивных признаков.
6. Параллелизм в эволюции различных функционально связанных между собою частей какой-либо системы органов (например, когда удлиняются скелетные элементы конечности и параллельно этому усиливаются соответствующие им мускулы) большей частью обусловливается корреляцией. В тех случаях, когда органы, мало связанные между собою функционально или даже вовсе не связанные, развиваются параллельно во время филогенеза (зубы и конечности у различных Equidae), этот параллелизм в эволюционном развитии объясняется одинаково направленными изме-нениями среды, в которой обитали животные.
7. Признаки, находящиеся у современных и вымерших форм в непосредственной близости, часто развиваются один после другого; при этом древние признаки характеризуют собою крупные систематические группы, более новые признаки—мелкие систематические группы.
ГЛАВА V
ЭВОЛЮЦИЯ ACRANIA И PROTOCRANIATA
В настоящей книге мы поставили перед собою задачу на основании имеющихся в нашем распоряжении результатов филогенетических исследований подойти несколько более детально, чем это было сделано до сих пор, к обсуждению общих закономерностей филогенеза животных. Чтобы дать читателю представление о том материале, который послужил фактической базой для наших обобщений, мы изложим в самой сжатой форме в пяти последующих главах изученную нами эволюцию низших позвоночных животных.
Выбор именно этого объекта исследования диктуется нам отнюдь не каким-либо особенным, специфическим характером изученного нами материала; наоборот, мы думаем, что пришли бы к тем же, изложенным нами в третьей и четвертой частях этой книги, общим выводам и в том случае, если бы стали изучать какую-либо иную крупную прогрессивную группу животных (например, высших позвоночных, ракообразных или насекомых). Наш выбор обусловлен другим соображением: многие затрагиваемые нами вопросы филогенеза низших позвоночных были в различное время изучены и так или иначе разрешены либо мною лично, либо моими ближайшими сотрудниками и учениками. Это именно обстоятельство позволило мне во многих случаях принять во внимание и тщательно проанализировать такие детали этой проблемы (детали, оказавшиеся впоследствии чрезвычайно существенными для построения общих выводов), которые прошли бы незамеченными для автора, строящего свои филогенетические схемы преимущественно на основании литературных данных.
Так как мы поставили себе целью изложить чисто филогенетический отдел настоящей книги в очень краткой форме, то мы принуждены воздержаться от критической оценки некоторых отдельных—часто очень интересных самих по себе—проблем филогенеза; такой критический разбор интересующийся этими вопросами читатель может найти в наших специальных трудах, перечень которых помещен в приложенном списке литературы. Здесь же в краткой и поэтому несколько догматической форме мы приводам лишь наиболее вероятные, по нашему мнению, филогенетические теории и выводы, полученные в результате чужих и личных исследований и наблюдений. Во всех тех случаях, когда наши личные выводы расходятся с общепризнанными воззрениями,1 мы приводим (насколько возможно, объективно) мнения авторов, не согласных с нами.
1 Такими отклоняющимися от общепринятых воззрениями являются, например, наши взгляды на филогенетические соотношения между хрящевыми ганоидами и другими Osteichthyes, на филогенетические соотношения между Crossopterygii и остальными рыбами и др.
9®
132
ГЛАВА V
Не вдаваясь в критику запутанной и пока еше темной для нас проблемы о происхождении Chordata,1 мы начинаем наш очерк филогенеза низших позвоночных с описания эволюции группы Branchiostomatidae.
ACRANIA PRIMITIVA
В настоящее время можно принять с большой долей вероятности, что все Branchiostomatidae (Amphioxus lanceolatus и родственные ему формы), а также все черепные позвоночные (Cyclostomata, Osteostraci, рыбы и Quadrupeda) произошли от общих предков, т. е. от некой, палеонтологически еще неизвестной формы Chordata, которую мы, однако, на основании детально изученной истории развития и анатомии Branchiostomatidae, Cyclostomata и низших рыб, можем охарактеризовать довольно подробно. Факт, что до сих пор не найдено никаких следов ископаемых предков бесчерепных и черепных позвоночных, является, само собою разумеется, обстоятельством, чрезвычайно невыгодным для всякого исследователя этой проблемы, так как в своих построениях ему приходится опираться в данном случае исключительно на данные сравнительно анатомического и сравнительно эмбриологического исследования. Поэтому
Рис. 1. Схема вероятного строения первичных бесчерепных позвоночных (Acrania primitiva), т. е. гипотетических предков современных бесчерепных (Branchiostomatidae) и черепных позвоночных (круглоротых, Ostracodermi и рыб).
ап—анальное отверстие; Ao. d—aorta branchialfs dorsalis; ao. br. v.—aorta branchialis ven trails; br.l— br. 17—жаберные Щели 1—17 (число жаберных щелей может быть дано'лишь приблизительно); Cart, ann—околоротовое кольцо; ch—хорда; Сг—оримепты головного мозга; err—околоротовое кольцо со щупальцами; Dr.—дорзальная ресничатая борозда; endost.—эндостиль; Нер.—печеночный мепюк; int.—кишечный тракт; М—ротовое отверстие; Myot.—мускульные сегменты (миотомы); Nc.—нервная трубка. На эту схему не нанесены жаберные дуги, которые, вероятно, уже существовали (ср. рис. 4); не изображены также в нервной трубке и глазки Гессе.
При построении этой схемы мы приняли, что мозг Acrania primitiva достиг приблизительно той стадии развития, на которой находится в настоящее время мозг ланцетника и что первая жаберная щель (первый бранхиомер) Acrania primitiva расположена непосредственно позади ротового отверстия (из А. Н. Северцова, 1929).
мы с особенной настойчивостью подчеркиваем, что иллюстрирующие наши выводы рисунки являются только простыми схемами, наглядно изображаю- j щими наши представления о соотношении различных органов у гипотети- < ческих предков,—схемами, отнюдь не претендующими на то, чтобы вое- ’ произвести истинную форму тела и подробное строение органов этих животных.
Что касается изложенных здесь воззрений относительно строения предков черепных позвоночных—Acrania primitiva и Protocraniata, то они представляют собою, само собой разумеется, лишь провизорные гипо-
1 Новейшее и, по нашему мнению, чрезвычайно интересное обсуждение проблемы происхождения хордат мы находим в статье Д. Третьякова (Tretyakoti D. Ursprungder Chordatae, Zschr. f. wiss. Zool., Bd. 134, 1929), где имеется также и сводка новейшей литературы, касающейся этого вопроса. Должен сказать, однако, что я сот ггласен далеко не со всеми выводами автора.
ЭВОЛЮЦИЯ ACRANIA И PROTOCRANIATA
133
*тезы. Однако для того, чтобы их построить, мы использовали все известные нам палеонтологические, сравнительно анатомические и сравнительно эмбриологические факты. Поэтому мы имеем основание твердо рассчитывать на то, что приводимые нами гипотезы заключают в себе некоторое зерно истины.
Гипотетические предки Branchiostomatidae и Craniata представляют собою свободно живущих, билатерально симметричных хордат с вполне развитой метамерией мезодермы и ее дериватов; так, мускульные сегменты тела (рис. 1, Myot.) доходили у них, так же как и у ланцетника и у зародышей Craniata, до самого переднего конца тела. Под расположенной на этом переднем конце тела центральной нервной системой (neurocoel, рис. 1, Nc.) лежала хорда (ch.), которая, по всей вероятности, не доходила до головного конца тела и имела такое же расположение, как и хорда ланцетника. На переднем конце нервной трубки, повидимому, образовалось уже два Мозговых желудочка .(рис. 1, Сг.). Сегментация спинномозговых нервов в общем соответствовала сегментации миотомов, и чувствующие, собственно смешанные (соматически-чувствующиеН-висцерально-двигательные) спинномозговые нервы были уже отделены от соматических двигательных нервов наподобие того, как это имеет место у ланцетника (фан Вай) и у круглоротых. Кожные органы чувств были представлены чувствующими клетками эпидермиса. В высокой степени вероятно, что чувствующие клетки передней части головы, иннервируемые разветвлениями самых передних чувствующих нервов, функционировали как органы обоняния.1 Внутри переднего отдела нейроцеля были расположены светочувствительные пигментированные клетки (глазки Р. Гессе),* 2 из которых у черепных потомков примитивных бесчерепных позвоночных развились клетки ретины.3 4
Что касается пищеварительного тракта, то в нем большой передний отдел кишечника функционировал как глоточная полость (рис. 1, Br. 1 — Вг. 17), а задний его отдел (int.), на вентральной стороне которого был развит слепой печеночный мешок (hep.), доходил непосредственно до анального отверстия (ап.). Ротовое отверстие (М) было окружено околоротовым скелетным кольцом (Cart, апп.), усаженным ресничками (err.). Жабры этих животных были построены по типу жабр ланцетника, и первая жаберная щель лежала, так же как и у последнего, непосредственно позади ротового отверстия; число жаберных щелей на каждой стороне тела было, однако, меньше, чем у ланцетника (вероятно, их было около 17). Эти жаберные щели открывались непосредственно наружу, и особой атриальной полости, столь характерной для ланцетника (рис. 4, atr.), у них не существовало вовсе. Ток воды, служащий им для дыхания, вероятно, направлялся, как и у ланцетника, движением ресничек жаберной полости и выходил через жаберные отверстия наружу. Питание этих гипотетических форм было, по всей вероятности, таким же пассивным, как и питание ныне живущих Branchiostomatidae: плавающие в воде пищевые частицы приносились вместе с током воды в глоточную полость, где они обволакивались выделяемой эндостальными железками (рис. 1, endost.)слизью и, склеенные друг с другом, проталкивались мерцательным движением ресничек (рис. 1, Dr.) в заднюю, пищеварительную часть кишечника.
Скелет Acrania primitiva состоял из хорды (ch.), из околоротового, снабженного ресничками кольца (Cart, апп.), состоящего из хрящеподоб-.ной ткани, и из некоторого числа нерасчлененных жаберных дуг,1 лежащих
‘Goldschmidt R., Amphioxides, Wiss. Erg. d. Tiefsee-Exp. «Valdivia», Bd. 12, 1905.
2 Hesse R., Sehorgane des Amphioxus, Zschr. f. wiss. Zool., Bd. 63, 1898.
8 В о v er i Th., Ober die phylogenetische Bedeutung der Sehorgane des Amphioxus, Festschr. f. A. Weismann, 1904.
4 He представленных на схематизированном рис. 1.
134
ГЛ A BA V
sct
Рис. 2. Схематическое изображение поперечного разреза через туловище гипотетических Acrania primitiva.
Энтероцельные мешки при помощи энтероцельных канальцев стоят в связи с полостью кишечника; эти канальцы служили для выведения в кишечник половых клеток и продуктов экскреции. Со.—вентральная часть целомического мешка; d—кишечник; е. а.— колбовидные выпячивания энтероцельных каналов; е. с.—энтероцельные каналы; ту—дорзальная мускульная часть целомического мешка; scl.—эпителиальные склеротомные складки. (Из А. А. Машковцева. 1934).
между жаберными щелями. Их кровеносная система была построена по типу кровеносной системы Ampbioxus (рис. 1, Ao. br. v., Ao. d.), а сердце еше не было развито. Выделительная система была представлена рядом сегментально расположенных нефридиальных канальцев с мерцательными воронками, открывающимися одним своим концом в брюшную полость (целом), а другим—в кишечник по бокам хорды. По исследованиям А. А. Машковцева,1 который исходил из изучения эмбрионального развития ланцетника и Anamnia, энтероцельные мешки Acrania primitiva (рис. 2, со.) открывались при помощи энтероцельных каналов (рис. 2, е. с.)в кишечник (рис. 2, d). Экскреторно-половую функцию, вероятно, выполняла при этом вся внутренняя поверхность целомических мешков; половые клетки и продукты экскреции выделялись при помощи энтероцельных канальцев в полость кишечника.
Представляется довольно вероятным, что описываемые нами гипотетические предки черепных позвоночных были прозрачными, маленькими, наподобие Вгап-chiostomatidae, животными, ведшими нектонный образ жизни, т. е. жившими поблизости от поверхности морского дна.
Отметим еще раз, что от общих предков Acrania и Craniata не сохранилось никаких ископаемых следов. Все перечисленные нами признаки этих давно вымерших животных форм мы восстановили только на основании сравнительно морфологических данных. Процесс этой реконструкции шел при этом приблизительно следующим путем: мы знаем, например, что в теле черепных позвоночных, точно так же как и в теле ныне живущих Acrania. развиваются одинаково построенные миотомы, и выводим отсюда, что и в
теле общих предков этих животных находились в свое время таким же образом построенные и таким же образом расположенные миотомы. Мы видим далее, что у ланцетника ряды миотомов доходят вплоть до самого переднего конца тела и что такие же миотомы* 2 развиваются в переднем отделе головы у зародышей круглоротых (Petromyzon) и низших рыб (селахий), и выводим отсюда, что у общих предков Acrania и Craniata ряд соматических туловищных мускулов доходил до самого переднего конца тела.
Или возьмем другой пример. Мы знаем, что у современных нам бесчереп
‘Машковцев А. А,, Морфологические и филогенетические отношения мезонефроса к пронефросу, Труды лабор. эволюц. морфол. Академии наук СССР, т. 2, 1934.
2 Сомиты, или миотомы, из которых образуется глазная мускулатура этих живот» ных. См. прекрасные работы Б. С. Матвеева (1915, 1922, 1923, 1925), которому, по на» шему мнению, удалось разрешить трудную проблему метамерии головы.
8В0Л10ЦИЯ ACRANIA И PROTOCRANIATA 135
ных позвоночных (ланцетник) спинные и брюшные нервы каждого сегмента тела проходят раздельно, и точно такое же положение мы находим у зародышей круглоротых и рыб. У циклостом спинные и брюшные корешки закладываются в непосредственной близости друг от друга, а у рыб нервные волокна спинных и брюшных корешков являются смешанными, так что каждый периферический нерв заключает в себе как двигательные, так и чувствующие волокна, которые лишь позднее отделяются друг от друга. Состояние, которое мы наблюдаем у взрослых бесчерепных и у зародышей циклостом и рыб, дает нам право заключить, что у общих предков бесчерепных, циклостом и гнатостом—с пинные и брюшные нервы проходили раздельно и что то положение, которое мы находим у циклостом и рыб, является новоприобрете-нием этих форм.
Р. Гессе обнаружил во внутренней стенке нейроцеля ланцетника присутствие особых светочувствительных пигментированных клеток, получивших название „глазки Гессе“. Животные получают при помощи их световые раздражения, причем световые волны на своем пути к светочувствительным клеткам проникают сквозь прозрачную ткань тела. Как известно, ретина глаза позвоночных представляет собой часть зародышевого промежуточного мозга, и ее светочувствительные клетки развиваются из тех внутренних эпителиальных клеток нейроцеля, которые по своему положению соответствуют глазкам Гессе. Бовери принял поэтому, что предки черепных позвоночных, т. е. общие предки круглоротых и рыб, прошли в своем филогенетическом развитии через стадию, соответствующую стадии бесчерепных позвоночных, когда зрительные органы этих последних были представлены лишь интрамедулярными светочувствительными клетками Гессе, из которых уже впоследствии развилась ретина круглоротых и гнатостом.
Все только что перечисленные нами признаки строения гипотетических родоначальников Branchiostomatidae и Craniata реконструированы нами на основании подобного рода весьма детальных сравнительно морфологических исследований, и они гипотетичны лишь в том смысле, что эти прародительские формы палеонтологически еще неизвестны. Однако мы уверены в том, что подобного рода прародительские формы с описанными нами признаками несомненно жили в какую-то весьма отдаленную от нас геологическую эпоху (кембрий или еще более ранний период). Факт их реального существования для нас столь же несомненен, как, скажем, несомненно существование водорода в тех созвездиях, спектр которых обнаруживает водородную линию. Его присутствие в созвездиях для нас не подлежит сомнению, хотя никто из астрономов на этих созвездиях не бывал и никто не подвергал исследованию их химический состав обычными методами химического анализа. Так и наша реконструкция, добытая методами сравнительно морфологического анализа, конечно, может быть, несовершенна в своих деталях, но в основном, в своих главных чертах, она безусловно верна.
Итак, гипотетические предки Acrania и Craniata многими чертами своего утроения похожи на современных Acrania. От этих примитивных предков произошли два ряда потомков:1 с одной стороны, предки ныне живущих Branchiostomatidae, с другой—предки всех черепных позвоночных.
1 Очень возможно, скажем более, очень вероятно, что от Acrania primitiva произошли не два, но гораздо более рядов потомков, которые вымерли целиком, не оставив после себя никаких палеонтологических следов. Однако методы сравнительно морфологического исследования позволяют нам говорить лишь о тех предках животных, потомки которых дожили до нашего времени.
136
IWABA V
ЭВОЛЮЦИЯ BRANCHIOSTOMATIDAE
Та линия потомков, которая привела в конечном итоге к образованию современных Branchiostomatidae1 (родословное дерево, схема XV) не пошла по пути прогрессивного развития, но приспособилась к совершенно особому образу жизни, позволившему представителям этой группы сохранить вплоть до наших дней многие примитивные признаки своих предков в почти неизменившемся состоянии (специализация). Предки ланцетников приобрели ряд особенностей, сделавших возможным столь характерный для них подводно-подземный образ жизни: они жили на дне моря, зарывшись белее или менее глубоко в морской песок.
Мне кажется, что теория Мак Брайда1 2 объясняет нам наилучшим образом, каким путем мог произойти переход от нектонного образа жизни предков Amphioxus и его сородичей к подводно-подземному образу жизни самого ланцетника. Этот автор предполагает, что предки бесчерепных позвоночных были первоначально нектонными животными, плавающими
Рис. 3. Схематическое изображение бептонной камбалообразной формы, переходной между Acrania primitiva и Branchiostomatidae.
Эта гипотетическая переходная форма изображена лежащей на левой стороне. Окаймленный щупальцами рот обращен ко дну. Семь пар жаберных щелей (правой и левой стороны), а также анальное отверстие, открываются сверху. На этом рисунке мы в точности воспроизводим схему Мак Брайда; по нашему мнению, предки Branchiostomatidae (см. рис. 1) имели большее число жаберных щелей (10—20), чем это принимал Мак Брайд (из Мак Брайда, 1909).
в глубине моря, поблизости от морского дна, и что они перешли постепенно к бентонному образу жизни; при этом животные эти приобрели сначала привычку лежать на дне, прижавшись к нему одной стороной своего тела. Это первичное изменение в нормальном положении тела обусловило, по мнению Мак Брайда (как это случилось позднее у камбал), смещение у этих анцестральных форм первично симметрично построенных органов, как, например, рта, жаберных щелей и т. п. При этом рот их переместился на левую сторону тела и сюда же сдвинулись и жаберные щели, первоначально расположенные на обеих сторонах туловища (ср. рис. 3, изображающий схему Мак Брайда). Следы этой асимметрической структуры тела предков Branchiostomatidae сохранились самым ясным образом в истории развития ныне живущих Branchiostomatidae (на стадиях личинок).
Позже эти донные животные перешли к подземному или, если можно так выразиться, к подземно-донному образу жизни и стали зарываться в рыхлое морское дно. Этот новый образ жизни вызвал в них ряд новых изменений: несимметрические органы становятся вторично вновь симметричными
1 Ср. Franz V., Morphologieder Akranier, Erg. anat. Entwicklungsgesch., Bd. 27, 1927.
2 Ma c Bride E. W., The formation of the layers in Amphioxus and its bearing on the interpretation of the early ontogenetic processus in the other Vertebrates, Quart. Journ. of Mier. Science, v. 54, 1909.
ЭВОЛЮЦИЯ ACRANIA И PROTOCRANIATA
137
у взрослых форм;1 передний конец спинной струны (рис. 4, Ch.) вырастает в длину и образует собою твердый опорный орган для рыла (рис. 4, R.), при помощи которого животные зарываются в песок. Так как при зарывании жаберные реснички животных, как и сами жабры, расположенные поперечно по отношению к продольной оси тела, легко повреждались, то у Branchiostomatidae постепенно развился аппарат, защищающий жабры, а, именно симметрично расположенные боковые атриальные складки, от сращения которых впоследствии произошла большая атриальная полость ланцетника (рис. 4, atr.).
Так как процесс дыхания у зарывшихся в морское дно животных оказался, вследствие недостатка там кислорода, несколько затрудненным, то у них постепенно произошло сильное увеличение поверхности; выразившееся в резком повышении количества жаберных щелей (рис. 4, Вг)— до 90 пар у ланцетника. По всей вероятности и образование довольноплотной жаберной решетки (рис. 4, Kgt.) также было обусловлено этим особенным, специфическим образом жизни современных Acrania.
Рис. 4. Схематическое изображение организации ланцетника ап.—анальное отверстие; at. р.—atrioporus; atr.—атриальная полость; Вт.—жаберные щели; Вт. а.— жаберные дуги; Вт. С.—жаберный отдел кишечника; Cart. ann.—околоротовое кольцо; Сг—мозг; Ch.—хорда; сгг.— околоротовые щупальцы (cirri); Hep.—печеночный мешок; int—кишечный тракт; Kgt.—жаберная решетка; М—ротовое отверстие; М. sp.—спинной мозг; Myot.—мускульные сег-менты{ миотомы); Nph.—сегментально расположенные выделительные канальцы; R—rostrum. Число жаберных щелей и жаберных дуг на этой схеме значительно преуменьшено (из Г. Паркера).
Экскреторная система при этом также претерпела некоторые изменения: кишечные отверстия нефридиальных канальцев постепенно передвинулись сначала в жаберные щели, а затем и в перибранхиальную (атриальную) полость. Чрезвычайно интересно при этом, что сходство в строении нефридиальных канальцев у кольчатых червей и Branchiostomatidae не обусловлено генетическим родством этих двух групп, а является параллелизмом; нефридии ланцетника не гомологичны нефридиям кольчатых червей, а развились самостоятельно из энтероцельных канальцев.
Branchiostomatidae представляют собою типичнейший пример специализации: животные перешли в такую среду и в такие условия существования, при которых они оказались вполне защищенными от врагов и опасностей, угрожающих их сородичам в открытом море. Среда эта была к тому же в известном смысле постоянна, ибо условия ее (как неорганические, так и биологические) оказались неизменными, или во всяком случае очень мало изменившимися в течение целых геологическх эпох. Таким образом, специализировавшиеся формы, однажды приспособившись к ее условиям, уже не имели причин для того, чтобы видоизменяться дальше морфологически или физиологически: они оставались неизменными, находясь, так сказать, в стороне от борьбы за существование. Поэтому-то мы и видим, что современные нам бесчерепные позвоночные сохранили в своем взрослом состоянии, наряду ,с различными чисто приспособительными признаками (атриальная полость—рис. 4, atr., атриальные поры—
. 1 Анцестральная асимметрия ближайших предков сохранилась у плавающих личинок Branchiostomatidae.
138
ГЛАВА V
рис. 4, at. р., большое число жаберных щелей—рис. 4, Ьг., рыло—рис. 4, R, и т. д.), еще целый ряд вполне примитивных признаков, в свое время характерных для их давным давно вымерших предков, т. е. для наших гипотетических Acrania primitiva. Такими признаками являются присутствие у них хорды, сегментация тела, примитивное строение нервной системы и органов чувств и т. д. Отметим, что некоторые из этих анцестраль-ных признаков, как, например, остатки передних миотомов в голове, закладка передних жаберных щелей и дуг, строение центральной нервной системы и спинномозговых нервов, сохранились также и у современных черепных позвоночных (у круглоротых, у низших рыб и т. д.), но не во взрослом состоянии, а лишь на ранних стадиях их эмбрионального, или личиночного развития. Это обстоятельство свидетельствует о том, что и эти формы происходят, также как и ланцетник, от Acrania primitiva. Подчеркнем, что для филогенетического исследования имеет громадное значение факт существования такой примитивной формы, как ланцетник, удержавшей вплоть до настоящего времени, несмотря на свою специализацию, ряд примитивнейших признаков своих вымерших предков, ибо это позволяет нам, на основании сравнения между собою ныне существующих форм, восстановить первичную организацию давным давно вымерших Acrania primitiva.
Другая линия потомков, происшедших от первичных бесчерепных позвоночных, именно Protocraniata, разветвилась в свою очередь в дивергентном направлении на две линии, из которых одна привела в конечном итоге к образованию группы современных круглоротых, а также к вымершим Osteostraci, а другая—к образованию группы низших рыб. К несчастью, от представителей этой линии не сохранилось, в виде живых свидетелей прошлого, таких примитивных, потомков, какими являются для Acrania только что рассмотренные нами Branchiostomatidae. Поэтому нам приходится, восстанавливая структуру общих предков этих животных, которых мы только что обозначили именем Protocraniata (см. родословное древо, схема XV), удовольствоваться главным образом результатами сравнительно анатомического и особенно сравнительно эмбриологического исследования.
ЭВОЛЮЦИЯ PROTOCRANIATA
Если мы сравним наиболее примитивных из черепных позвоночных, т. е. циклостом и низших эласмобранхий, со ставшими нам хорошо известными, благодаря превосходным исследованиям Стеншиб1 Osteostraci, то мы увидим, что все эти три группы животных сходны между собою в очень большом количестве признаков: отсюда мы заключаем, что сходные признаки этих форм унаследованы каждой из этих групп животных от каких-то общих всем им предков.
Такими сходными признаками будут прежде всего положение главных систем органов в теле животного, т. е. взаимное расположение хорды, нервной системы, кишечника и т. п.; затем метамерия миотомов и спинномозговых нервов, структура хорды и ее оболочек, присутствие осевого черепа и хрящевого висцерального скелета, общее строение головного и спинного мозга, структура и расположение парных глаз и слуховых пузырьков, положение и отхождение главных ветвей головных нервов, положение жаберных щелей и их отношение к головным нервам, образование двухкамерного сердца и главных артерий и вен тела, образование массивной печени и т. д. Но сравнительно анатомическое изучение рудиментарных органов и, особенно,’ сравнительно эмбриологическое
1 S t е n s i б Е., The Downtonian and Devonian Vertebrates of Spitzbergen, p. I, Family Cephalaspidae, Norsk. Vidensk. Akad., Oslo, 1927.
8В0ЛЮ11ИЯ acrania и protocraniata
139
их исследование открыло нам еще целый ряд общих признаков, хорошо развитых у предков этих трех групп животных, но находящихся в редуцированном состоянии или вовсе исчезнувших у их взрослых потомков. И наша задача заключается в том, чтобы, на основании всей суммы имеющихся в настоящее время в нашем распоряжении сведений, реконструировать строение гипотетических форм, названных нами Protocraniata—общих предков примитивных позвоночных животных.
Мы начинаем нашу реконструкцию с кожных покровов этих животных. Нам представляется чрезвычайно вероятным, что кожа Protocraniata состояла из многослойного эпидермиса с одноклеточными железами и органами боковой линии и из тонкого слоя cutis; по всей вероятности, у этих животных, так же как и у ныне живущих циклостом, не имелось никакого кожного скелета.
Факт, что палеозойские Osteostraci имели, как это утверждает Э. Стен-шиб, хорошо развитый кожный скелет и что они были родственны предкам Cyclostomata,1 наводит его на мысль, что и у предков Cyclostomata некогда также имелся твердый., содержащий известь кожный скелет, впоследствии исчезнувший вследствие редукции. Мы, однако, думаем, что, напротив, Cyclostomata, так же как.и их предки Protocraniata, были первоначально голыми животными. Это предположение имеет следующие основания: у Elasmobranchii зубы, лежащие в полости рта, развиваются из плакоидных чешуй, лежащих на поверхности тела и в полости рта, выстланной эктодермой, у ископаемых же Osteostraci существовал, по мнению Киэра,1 2 вполне развитый ротовой скелет (рис. 22), образованный из элементов кожного скелета, т. е. из видоизмененных чешуй. Мы можем поэтому думать, что если у предков Cyclostomata действительно существовал когда-либо кожный скелет, то и у них также зубы должны были сформироваться из костных, resp. плакоидных чешуй. Однако мы видим, что у миног, так же как и у миксин (ныне живущих потомков первичных Cyclostomata), имеются лишь чисто эпидермальные роговые зубы. Нам представляется поэтому в высокой степени невероятным, чтобы ротовой скелет общих предков Cyclostomata и родственных им Osteostraci, развившийся из кожных чешуй и необходимо являющийся постоянно функционирующим органом, мог бы редуцироваться целиком и замениться функционально подобным, но филогенетически совершенно отличным от него образованием, а именно роговыми зубами. Гораздо более правдоподобным является, мне кажется, предположение, что роговые зубы Cyclostomata представляют собою новообразование и что отдаленные предки круглоротых и остальных к р а-н и ат не имели в свое время ни зубов, ни содержащих известь кожных чешуй.
Вполне достоверно, что у Protocraniata имелась везикулярная хорда (рис. 5, Ch.) с двумя оболочками, и довольно вероятно, что у них были развиты характерные для наиболее низко организованных черепных позвоночных, т. е. для круглоротых и низших рыб, диплоспондильные верхние дуги позвонков (рис. 5, A. vrt.).
Исследование развития черепа круглоротых и рыб® приводит нас ^заключению, что осевой череп Protocraniata был построен чрезвычайно примитивно и что головной мозг обхватывался им снизу и сбоку; были развиты, по всей вероятности, parachordalia (рис. 5, Pch.), образовавшиеся из сросшихся между собою нейральных дуг, и, возможно, также trabeculae
1 S t е n s I б Е., 1. с.
2 К 1 а е г J., The structure of the Mouth of the oldest known Vertebrates. Pteras-pids and Cephalaspids, Palaeobiologica, vol. I, 1928.
’Северцов, A. H., Исследования об эволюции низших позвоночных, ч. 1, П, III, Русск. архив анат., гист. и эмбр., 1916, 1917, 1925.
140
ГЛАВА V
cranii (рис. 5, Тг. сг.), которые к черепу, однако, еще не приросли. Затылочная часть черепа также еще не образовалась, так что п. vagus лежал на задней границе черепа (рис. б, X,). Слуховые капсулы (рис. 5, С. aud.) уже существовали и были, повидимому, сращены с парахордалиями; наоборот, хрящевые носовые капсулы (рис. 5, С. nas.) едва развитые, еще не срослись с осевым черепом (А. Н. Северцов, 1916, 1917).
Aprbr1(=Hyoid)
Рис. 5. Схематическое изображение скелета, центральной нервной системы и органов высших чувств гипотетических Protocraniata.
А. Ьг. 1 — А. Ьг. 13 — жаберные дуги 1 — 13; А. рг. br. 1 — А. рг. Ьг. 4.— преджаберные дуги 1 —4; A. vrt.—диштоспондильные верхние дуги позвоночника; Ьг. 1 — Ьг. 13—жаберные щели 1—13 (число жаберных сегментов может быть здесь определено лишь приблизительно); Cart. arm.—около-ротовое кольцо; С. aud.—хрящевая слуховая капсула; Ch.—хорда; С. nas.—парные носовые капсулы; Dien.—промежуточный мозг; Н—сердце; Нур—гипофиз в виде еше трубчатой, открывающейся наружу железы; М.—рот; Ms. еп.—средний мозг; Mt. ев.—продолговатый мозг; М. sp.—спинной мозг; Mst.—миосепты между туловищными (головными) миотомами; Nas.—носовые ямки; od.—парные глаза; pb. 1 (spir)—pb. 3 преджаберные щели 1—3; Pch—парахордалии черепа; Т1. еп.—передний мозг; Тг. сг.—парные трабекулы, еще не сросшиеся с парахордалиями; I — и. olfactorius; II — п. opticus.
Настоящая схема построена на основании детального изучения строения и истории развития Branchio-stomatidae (ланцетника), круглоротых (миноги) и низших рыб (селахий и хрящевых ганоидов). Основные черты строения предков всех низших позвоночных животных: 1) диференциация главных отделов головного мозга и образование органов высших чувств (у протокраниат, вероятно, были развиты также функционирующие пинеальные глаза, не изображенные па нашей схеме); 2) отсутствие срастания трабекул с парахордалиями, т. е. отсутствие сплошного дна черепа; 3) образование дипло-спондильных верхних дуг позвоночника; 4) присутствие околоротового кольца со щупальцами; 5) у протокраниат ряд бранхиомеров, г. е. ряд жаберных щелей, жаберных дуг и т. д., начинался непосредственно позади рта, причем эти органы в передней части головы также функционируют еще в качестве органов дыхания (жабры); 6) вероятное образование мешковидного сердца (из А. Н. Северцова, 1917.; несколько изменено).
Число жаберных щелей было приблизительно то же, что и у Acrania primitiva (т. е. около 17, рис. 5, pb. 1—pb. 3, br. I—Ьг. 13),1 причем между каждою парою жаберных щелей лежала одна хрящевая нерасчлененная жаберная дужка (рис. 5, A. br. 1—А. Ьг. 13, А. рг. br. 1—4). Ряд жаберных дуг (и щелей) начинался непосредственно позади ротового отверстия,
1 В этом пункте мои взгляды резко расходятся с воззрениями Д. Третьякова (1929, I. с.). Последний придерживается мнения, что число жаберных щелей у первичных предков челюстных позвоночных, т. е. у предков Branchiostomatidae, было сравнительно не велико и равнялось, приблизительно, семи, причем шели эти принадлежали главным образом передней части головы: сюда относились четыре преджаберные висцеральные шели, а также первая (glossopharyngeus). вторая и третья (vagus 1) истинные жаберные щели типа минор и рыб. Задние жаберные щели миног и Heptanchus (а также задние жаберные мешки и щели Cephalaspida) произошли, по мнению Третьякова, путем отпочкования(?) от жабр, иннервируемых vagus. В отношении самой передней из пребранхиальных жаберных щелей я, разумеется, вполне с ним согласен (А. Щ Северцов, 1916, 1917), но считаю, что Третьяковым не достаточно учтено то обстоятельство, что у Bdellostomatidae имеется по 10—14 жаберных щелей и что у личинок Bdellostoma, по данным Г. Прайса, их закладывается целых 30. Вместе с тем факт, что у личинок Amphioxus, у Enteropneusta и у Bdellostoma stouti имеются также многочисленные жаберные щели, приводит меня к выводу, что протокраниаты имели во всяком случае гораздо ббльшее число жаберных щелей, чем это наблюдается у современных миног и нотиданид.
8В0ЛЮЦИЯ ACRANIA И PROTOCRANIATA
141
окруженного хрящевым ротовым кольцом (Cart. апп.)и,по всей вероятности, усаженного ресничками (cirri). Примитивному строению скелета Protocraniata соответствовало такое же примитивное строение их мускульной системы. Только немногие из дорзальных мускулов были у них специали-зованы, вся же остальная мускулатура туловища и головы состояла из примитивных и равномерно сегментированных миотомов. Из этих миотомов пока диференцировались лишь мускулы глаз и, может быть, гипо-бранхиальная мускулатура.1
Висцеральная мускулатура, происходящая из боковых пластинок, была построена также очень просто и состояла в каждом жаберном сегменте (бранхиомере) из одного констриктора и одного аддуктора соответствующей жаберной дуги (рис. 8 и 9). Сравнительно эмбриологическое исследование показало, что у общих предков круглоротых и челюстных рыб ряд жаберных дуг также доходил вплотную до самого рта (А. Н. Северцов, 1916, 1917, 1925); чрезвычайно интересно, что у личинок циклостом (Ammocoetes) ряд висцеральных мускульных сегментов (mm. constrictors et adductores) и лежащих в передней части головы жаберных дуг* 2 также достигает рта.
Как было указано выше, у протокраниат были развиты настоящие подвижные парные глаза типа циклостом, т. е. глаза с инвертированном ретиной и хрусталиком, уже способные к образному зрению, а также типичные глазные мускулы, происшедшие из миотомов передней части головы, и вполне развитые глазные нервы (nn. oculomotorius, trochlearis и abducens).
Мы здесь не будем подробно разбирать вопроса об эволюции парных глаз, а также не будем касаться вопроса о теменных глазах протбкраниат. Факт, что все круглоротые и челюстные рыбы обладают одинаково построенными глазами, т. е. глазами, снабженными эпидермальным хрусталиком, и одинаково расположенными и одинаково иннервированными мускулами (rm. 1, 11, III), заставляет нас признать, что и у общих предков циклостом и гнатостом, т. е. у протокраниат, были в свое время развиты диференцированные и инвертированные глаза с хрусталиком и типичными глазными мускулами.
Что касается до эволюции собственно парных глаз, то в этом вопросе я придерживаюсь, наряду с большинством сравнительных анатомов, теории, ведущей свое начало от Бовери и полагающей, что инвертированные глаза произошли от светочувствительных клеток (глазки Р. Гессе), лежавших первоначально внутри мозга первичных бесчерепных позвоночных. Не вдаваясь в подробности, отсылаю читателя, интересующегося этим специальным вопросом, к сводке этих проблем, сделанной в одной из работ Л. Плате.3 Весьма вероятно, что у протокраниат функционировали вполне развитые париетальные глаза (ср. Cyclostomata, Osteostraci). У них безусловно были развиты слуховые пузырьки, и вполне возможно, что в этих последних уже имелись два полукружных канальца. В голове и туловище их были развиты чувствующие почки органов боковой линии. Органы обоняния, в виде парных носовых ямок (рис. 5 и 6, Nas.), еще слабо диференцированных, были также развиты на переднем конце тела.
Мы имеем все основания думать, что головной мозг Protocraniata представлял собою уже довольно сложно диференцированное образование
• i L1 а т в е е в Б; С;, К вопросу о мезомерии головы селахий, Дневн. зоол. отд. О-ва Любителей Естеств., Антроп. и Этногр., 1915.
Матвеев Б. С. Сегментация головной мезодермы у амфибий. Русск. зоол. жур< т. 3, 1923.
Neal Н. V;, The development of the Hypoglossus musculature in Petromyzon and Squalus, Anat. Anz., Bd. 13, 1897.
2 По моей номенклатуре—пребранхиальные висцеральные дуги (1916, 1917),
’Plate L., Allgemeine Zoologie, Bd. 2. Jena, 1925.
142
ГЛАВА V
(рис. 5 и б). Прекрасные исследования И. Джонстона1 над круглоротыми и низшими рыбами указывают нам на то, что головной мозг общих предков этих форм (т. е., стало быть, головной мозг наших протокраниат) состоял из следующих частей; из telencephalon impar с ориментами парных передних полушарий, из mesencephalon и из metencephalon.
Можно думать, что для животных, занимавших в системе промежуточное положение между гипотетическими Protocraniata и гипотетическими Acrania primitiva, было характерно еще более примитивное строение их сегментального мозга, метамерия которого (нейромерия Г. Нила1 2)
Nnsczity ns.en Men\
Ep..
aera.
Hen.-.
С.паз.:\-
N3S
NSfi
; NtHSC n(Xst
Hrm bitty
: /ivacb(vn)
1 ; №.5m Nnsc6()(i}
-сп
'Ьгл 'Mbri 'Abd.
МЬГ12 : A bra bra
Сагшп: /-
Aprbft. ; pb-3 Д1 :Upb.t.
P Ьргйг,
Рис. 6. Схематическое изображение головных нервов, висцерального аппарата и висцеральной мускулатуры гипотетических Protocraniata. Этот рисунок дополняет собою рис. 5.
А.Ьг 1 — А. Ьг. 13—жаберные дуги 1—13; А. рг. br. 1 (A. vise. 4-Hyoid)—первая преджаберная дуга (соответствует гиоидной дуге челюстных и круглоротых); А. рг. Ьг. 4—четвертая преджаберная дуга; ЬгЛ — Ьг, 13 — жаберные щели 1—13; С. nas. —парные носовые капсулы; Ch.—хорда; Dien.—diencephalon; Ер. —эпифиз; Нур.—гипофиз; М—рот; М. Ьг. 1—М. Ьг. 12—жаберные мускулы 1—12; М. pb. 1—М. pb. 4—преджаберные мускулы 1—4 протокраниат (на этой схеме строение жаберных мускулов сильно упрощено; сегментация висцеральных мускульных сегментов едва намечена); М. sp.—спинной мозг; Ms. еп.—mesencephalon; Mt. en.—metencephalon; Nas.—носовые ямки; N. opht. (Vj)—n. ophtalmicus profundus trigemini l(Vt); N. vise. 1(VT)—висцеральная ветвь n. ophtalmicus, иннервирующая первый бранхиомер; N. vise. 2 (V8) —висцеральный нерв второго брахиомера (n. Maxillarius trigemini); N. vise. 3(V''a)r—висцеральный нерв третьего бранхиомера (n. mandibularis trigemini); n. vise. 4 (VII)—висцеральный нерв четвертого брапхио-мера (n. hyomandibularis facialis). [Мы не можем с уверенностью сказать, образовали ли нервы боковой линии головы (органы боковой линии, по всей вероятности, уже существовали упротокра-наит) единый комплекс с n. facialis, поэтому мы не нанесли их на нашу схему; отметим, что то же относится и к латеральным ветвям n. glossopharyngeus и n. vagus]; n. vise. 5(1Х>— висцеральный нерв пятого бранхиомера (n. glossopharyngeus современных черепных); n. vise. 6(Хх)—висцеральный нерв шестого бранхиомера (первая жаберная ветвь комплекса блуждающего нерва); n. vise. ll(Xg)—висцеральный иерв одиннадцатого бранхиомера (шестая жаберная ветвь блуждающего нерва современных семижаберных краниат); pb. I (spir)—первая преджаберная щель, соответствующая брызгальцу низших рыб; pb. 3—третья преджаберная щель; г. int— г. intestinalis vagi; Tl. en. —Telencephalon (из A. H. Северцова, 1917, несколько видоизменено).
соответствовала метамерии мезодермических сегментов головы. Из I нейромера образовался telencephalon impar, из II и III—diencephalon с париетальными и парными глазами, из IV и V нейромеров—mesencephalon, а из VI—X нейромеров—metencephalon.
Спинномозговые нервы Protocraniata были еще очень примитивны и состояли из чувствующих (вернее, смешанных) и чисто двигательных нервов, которые в туловище проходили раздельно друг от друга, наподобие того, что мы имеем у современных бесчерепных позвоночных, у круглоро
1 Johns ton I. В., The Brain of Petromyzon, Journ. Comp. Neur., v. 12, 1902; The Morphology of Vertebrate Head etc., ibid., v. 15, 1905; The Nervous System of Vertebrates, London, 1908.
2 N e a 1 H. V., Neuromeres and metameres, Journ. Morph., vol. 31, 1918. The segmentation of the head nervous system in Squalus Acanthias, Bull. Mus. Comp. Zool,, 1898.
ЭВОЛЮЦИЯ ACRANIA И PROTOCRANIATA
143
тых и у эмбрионов рыб. Что же касается головы этих гипотетических форм то в задней ее части, а также в слуховой области, соматически двигательные нервы были построены по тому же принципу, что и в туловище, но в глазничной области были уже диференцированы настоящие глазные нервы (III, IV, VI).
Очень вероятно, что сегментация жаберных нервов (рис. 6) соответствовала сегментации жаберных щелей и жаберных мускулов и что нервы органов боковой линии были уже вполне развиты; однако очень трудно сказать с уверенностью, были ли корешки этих последних нервов связаны с жаберными нервами (facialis, giossopharyngeus, vagus), или они образовали собою независимую систему нервов боковой линии. Эти последние нервы не обозначены на нашей схеме, изображенной на рис. 6.
N. vagus был, по всей вероятности, построен по типу нервов круглоротых и иннервировал все задние бранхиомеры [рис. б, n. vise. 6 (Хх), n. vise. 11 (Х6) ит. д.]. N. giossopharyngeus [рис. б, n. vise. 5 (IX)] иннервировал пятый бранхиомер (первую функционирующую жаберную щель современных рыб и куглоротых, а также вымерших Osteostraci); n. facialis [рис. 6. N. vise. 4 (VII)] иннервировал четвертый бранхиомер (первую преджаберную дугу, т. е. гиоидную дугу рыб); n. trigeminus, вероятно состоявший у этих форм из трех раздельных бранхиальных ветвей (N. vise. 1—3), иннервировал три передних бранхиомера.1
У Protocraniata несомненно уже имелось мешковидное сердце с предсердием, желудочком и сердечным клапаном (рис. 5, Н), а также главные кровеносные сосуды, построенные по общему типу кровеносной системы черепных позвоночных. Число жаберных щелей у Protocraniata, как это уже отмечено выше, было относительно велико (около 17). Мы приходим к этому положению на основании следующих соображений. У большинства круглоротых мы находим семь функционирующих жаберных щелей и мешков, иннервируемых n. giossopharyngeus, а также ветвями n. vagus; кроме того, мы находим, что у пескоройки впереди первой ее жаберной щели имеются по крайней мере два рудиментарных жаберных мешка и четыре типично построенных висцеральных мускульных сегмента (преджаберные бранхиомеры 1—4). Мы имеем все основания принять, что самому переднему из этих висцеральных мускульных сегментов соответствовал у предков циклостом свой жаберный мешок и своя жаберная щель: поэтому мы насчитываем у пескоройки четыре преджаберных (рис. 8, 9, A. pb. 1—4, С. pb. 1—4) плюс семь бранхиальных сегментов (рис. 7, Ы—Ь7), т. е. в общей сложности 11 бранхиальных сегментов. Но у Bdellostomatidae (Bdellostoma stouti, Bdellostoma polytrema) мы находим от 10 до 14 функционирующих жаберных щелей (и мешков); самые задние из них иннервируются жаберными ветвями n. vagus. Так как у нас нет никаких
1 R. maxillaris и г. mandibularis trigemini соответствовали, по всей вероятности, второму и третьему преджаберному бранхиомеру [рис. 6, N. vise. 2 (V'2), N. vise. 3 (V"2), а от r. ophtalmicus profundus trigemini ответвлялась ветвь [рис. 6, n. vise. 1 (Vx)], которая иннервировала первый бранхиомер. В моих работах 1916 и 1917 гг. я принимал, что у протокраниат (и у взрослых миног) существовала также и пятая пребран-хиальная висцеральная дуга, представленная у взрослых миног (Petromyzon) cartilage dorsalis anterior и соответствующая самому переднему бранхиомеру в теле протокраниат. Однако я не мог найти соответствующего этому бранхиомеру мускула, а так как его иннервация при помощи n. thalamicus возбуждала сомнение (см. Д. Третьяков, 1. с., 1929) и так как, с другой стороны, у гнатостом от этого бранхиомера не сохранилось никаких следов, то я думаю, что будет более осторожным пока не настаивать на существовании этого гипотетического бранхиомера (arcus praebranchialis 5) и начинать счет жаберных сегментов в теле протокраниат с четвертого пребранхналь-ного сегмента, т. е. считать за первый бранхиомер тот сегмент, который прежде (А. Н. Северцов,!. с., 1916, 1917) я обозначал как второй висцеральный сегмент или второй бранхиомер. Поэтому я изменяю здесь мои схемы (рис. 5 и 6), по сравнению с тем, что имело место в моих работах 1916 и 1917 гг.
144
ГЛАВА V
Рис. 7, 8 и 9. Реконструкции (по сагиттальным срезам) трех последующих стадий развития висцерального аппарата и головных нервов Ammocoetes.
№S АЬг5 АЬГ4. АОгЗ. А0Г.2. Abet Cpbt рб.1 М-2- .
Рис. 7. Относительно ранняя стадия развития Ammocoetes, когда происходит первая закладка сегментов висцеральной мускулатуры (A.pb.2, A.pb.3, C.pb.i—С.рЬ. 3, С.Ь.1—С.Ь.4) и еще не прорвавшихся наружу энтодермальных жаберных мешков (pb.l, pb.2, b.l-b.7).
Рис. 8. Более поздняя стадия развития Ammocoetes, где развитие висцеральной мускулатуры несколько продвинулось вперед.
С.Ь.З A.b.3 A.br.2 Cb.l A.bJ A pBr.l
Рис. 9. Еще более поздняя стадия развития Ammocoetes, на которой сегментация висцеральной мускулатуры в передней части головы уже вполне ясно выражена.
А.Ь.1—А.Ь.З'-тт. adductores branchiales функционирующих жаберных мешков. А.Ьг.1—A.br. 7—зачатки жаберных дуг Ammocoetes (Petromyzon); A. pbr. 1.—A. pbr. 3—преджаберные висцеральные дуги 1—3,A.pb. 1—А.рЬ. 4 — mm. adductores praebranchiales четырех передних (предслуховых) бранхиомеров головы; Au.*—слуховой пузырек; Ь.1—Ь-7—жаберные мешки 1—7.
8В0ЛЮЦИЯ ACRANIA И PROTOCRANIATA
145
оснований считать, что здесь произошло вторичное увеличение числа жаберных мешков, то нам остается лишь факультативно принять, что у общих предков ныне живущих круглоротых имелось в свое время приблизительно на шесть бранхиомеров больше, чем у их современных потомков, т. е. у миксин и миног. Таким образом, в общем у них имелось 17 жаберных сегментов. Задние из этих бранхиомеров редуцировались у большинства Cyclostomata, точно так же как они атрофировались у Osteostraci, имевших, по Стеншиб, только 10 жаберных мешков.
У нас имеются основания предполагать, что и у предков челюстных позвоночных имелись также четыре преджаберных бранхиомера1 и что у них задние бранхиомеры, т. е. бранхиомеры, лежащие каудально от седьмой функционирующей жаберной щели нотиданид, также редуцировались. Поэтому возможно, что и у общих предков круглоротых и рыб число функционирующих жаберных щелей, дуг, висцеральных мускулов и нервов(т. е. число бранхиомеров) было также довольно значительно (между 12 и 17).2 Приведенная ниже таблица (табл. 1) дает нам представление о бранхиоме-рии примитивных позвоночных животных (протокраниат, энтобранхиат и эктобранхиат) такой, какой она представляется на основании наших исследований (ср. рис. 5, 6, 11 и 13). О строении висцеральной мускулатуры мы судим по строению эмбриональной мускулатуры у Ammocoetes; но у протокраниат висцеральная мускулатура была, конечно, менее диферен-цирована. Нам проставляется невероятным, чтобы у протокраниат уже
1 Т. е. бранхиомеры, лежащие впереди от первого функционирующего жаберного сегмента (впереди от бранхиомера, иннервируемого п. giossopharyngeus).
2 Мы должны, однако, заметить, что изложенные нами выводы, вероятно, еще нуждаются в дальнейшей переработке. Так, например, Прайс [G. С. Price, Zur Ontogenie der Myxinoiden (Bdellostoma stouti Lockington), Sitzber. math.-phys. KL Bayer. Akad. d. Wiss., Bd. 26, 1906] утверждает, что у эмбрионов Bdellostoma имеется гораздо большее число жаберных щелей (до 30). Дальнейшее детальное изучение этой .проблемы, предпринятое над теми же Bdellostoma, было бы во всяком случае крайне желательно, так как очень коротко изложенные результаты Прайса не позволяют нам свести его выводы с тем, что нам известно относительно бранхиомерии у всех других циклостом. С другой стороны, Гольмгрен, которому удалось наблюдать стадии метаморфоза у пескоройки и который отрицает установление бранхиомерии по мускульным сегментам, считая здесь мускульную сегментацию за вторичное явление, утверждает, что впереди от первой жаберной дуги могло быть самое большое только три преджаберных бранхиомера (Holmgren u. Stensio, Kranium und Visceralskelett der Akranier, Cyclostomen und Fischen, Handb. der vergl. Anatomie, 1936). К такого же рода выводам приходит, невидимому, и Балабай.
С.аи—слуховая капсула; с.Ь.1—с. Ь. 6—mm. constrictores branchiales функционирующих жаберных мешков пескоройки (миноги); C.pb.l—С.рЬ. 4—mm. constrictores praebranchiales четырех передних бранхиомеров головы; Ch.—хорда; М.—зачаток рта, на более поздних стадиях— рот; Nh.—назогипофизарное отверстие; Ос.—глаз; Oph.—n. ophthalmicus profundus Trigemiin iCVj);*—рудиментарный висцеральный нерв четвертого преджаберного нейромера; pb.l—pb.2— первый и второй жаберные мешки; Pch.—парахордалии; Pch. а—передние парахордалии; r.md. V,—г. mandiblaris n. tregimini 2; r.mx. V2—г. maxillaris n. trigemini 2; VII—n. facialis; IX—n. giossopharyngeus; X. 1.—n. lateralis vagi; Xt—Xe—бранхиальные ганглии блуждающего нерва (из А,- Н. Северцова, 1916).
Если мы сравним между собою реконструкции 7, 8 и 9, то увидим, что у круглоротых (пескоройки) на относительно ранних стадиях развития имеется два преджаберных мешка: они лежат впереди от первой функционирующей жаберной щели (глоссофарингеальная щель; жаберные мешки pb. 1, pb. 2, рис. 7); здесь имеется также четыре вполне развитых преджаберных висцеральных мускульных сегмента, из которых каждый состоит из одного m. constrictor (С. pb. 1—С. pb. 4) и из одпого т. adductor (A. pb.—1 A. pb. 4; ср. схему, изображенную на рис. 6). Мы находим, кроме того, что трем задним мускульным сегментам соответствуют прохопдраль-ные преджаберные висцеральные дуги (A. pbr. 1, A. pbr. 2, A. pbr. 3, рис. 9). Из совокупности всех этих факторов мы заключаем, что у предков циклостом имелось во всяком случае четыре преджаберных бранхиомера, расположенных в переднем отделе головы, вполне гомодинамных бранхиомерам, расположенным в задней части головы. Каждый из этих бранхиомеров состоял из хрящевой висцеральной дуги, из сегмента висцеральной мускулатуры (mm. adductor и constrictor), из жаберного мешка и жаберной щели и из сегментального висцерального нерва. Этот вывод находится в полном согласии с результатами наших исследований над развитием бранхиомеров передней части головы у примитивных челюстных позвоночных (эласмобранхий). К только, что изложенному взгляду на бранхиомерию общих предков круглоротых мы пришли впервые еще в 1916—1917 гг., исследуя историю развития циклостом. В этой работе я еще принимал, что преджаберная дуга представлена у взрослой миноги губным хрящом (Cart. lateralis). Но так как для этого бранхиомера я не нашел ни соответственного висцерального мускула, ни не вызывающего сомнений рудимента соответственного висцерального нерва, то я думаю, что будет более осторожным считать, что у предков круглоротых было не более четырех преджаберных бранхиомеров.
Морфологические закономерниости. 10
146
ГЛАВА V
Таблица 1
Висцеральный скелет Висцеральная мускулатура Висцеральные нервы
Arcus visceralis 1 Arcus praebran- M. constrictor praebranchia- Рудиментарная
chialis 4? lis 4 висцеральная
M. adductor praebranchia- ветви n. ophtal-
lis 4 mici
Arcus visceralis 2 Arcus praebran- M. constrictor praebranchia-
chialis 3 lis 3 R.. maxillaris п.
M. adductor praebranchia- trigemini 2
Arcus visceralis 3 Arcus praebran-chialis 2 M. constrictor praebranchia-lis 2 R. mandibularis n. trigemini 2
M. adductor praebranchia-
lis 2
Arcus visceralis 4 Arcus praebran- M. constrictor praebranchia- R. hyomandibu-
chialis 1 lis 1 M. adductor praebranchia- laris n. facialis
lis 1
Arcus visceralis 5 Arcus branchia- M. constrictor branchialis 1 N. glossopharyn-
lis 1 M. adductor branchialis 1 geus
Arcus visceralis 6 Arcus branchia- M. constrictor branchialis 2 R. branchialis 1
lis 2 M. adductor branchialis 2 n. vagi
Arcus visceralis 7 Arcus branchia- M. constrictor branchialis 3 R. branchialis 2
lis 3 M. adductor branchiali^*3 П. vagi
Arcus visceralis 8 Arcus branchia- M. constrictor branchialis 4 R. branchialis 3
lis 4 M. adductor branchialis 4 n. vagi
Arcus visceralis 9 Arcus branchia- M. constrictor branchialis 5 R. branchialis 4
lis 5 M. adductor branchialis 5 n. vagi
Arcus visceralis 10 Arcus branchia- M. constrictor branchialis 6 R. branchialis 5
lis 6 M. adductor branchialis 6 n. vagi
Arcus visceralis 11 Arcus branchia- M. constrictor branchialis 7 R. branchialis 6
lis 7 M. adductor branchialis 7 n. vagi
После этого следуют бранхиомеры 12, 13, 14 и т. д., которые развиты у Bdellostoma stout! и иннервируются задними ветвями n. vagi.
существовали эктодермальные или энтодермальные кожные складки (жабры), увеличивающие дыхательную поверхность. Во всяком случае ни эмбриологических, ни сравнительно анатомических следов подобных образований мы найти не можем.
В глоточной полости Protocraniata находился эндостиль (thyroidea зародышей круглоротых и рыб), а также ресничатые борозды. Это обстоятельство, точно так же как и строение ротового кольца и как положение передних жаберных щелей, приводит нас к выводу, что питание Protocraniata, подобно питанию вымерших Acrania primitiva и питанию ныне живущих ланцетника и пескоройки, происходило пассивным путем.1 Несомненное присутствие массивной печени у Protocraniata наводит нас на мысль, что эти гипотетические животные формы должны были хорошо питаться; можно думать, что присутствие, у них органов высших чувств (глаз и органов обоняния) позволяло им отыскивать такие места в воде, где находилось наиболее обильное питание.
Мочеполовая система протокраниат, как это показывают результаты последних работ А. А. Машковцева1, эволюировала прогрессивно в совер-
1 Мы знаем, что у современных Cyclostomata и Gnathostomata из передних предслуховых жаберных дуг развились особые скелетные элементы, служащие им для активного питания (подвижные части сосущего ротового аппарата, челюсти гнатостом и т. д.), но у нас нет никакого основания предполагать, что у предков круглоротых и рыб когда-либо существовали также и иные скелетные образования, приспособленные для активного захватывания пищи и расположенные рострально от самых передних жаберных дуг, еще функционирующих как органы дыхания. Судя по расположению этих элементов, такое допущение является весьма невероятным (А. Н. Северцов, 1916, 1917).
ЭВОЛЮЦИИ ACRANIA И PROTOCRANIATA
147
«у.
Рис, 10. Схема строения мочеполовой системы гипотетических Protocraniatata, у которых из краниальной части шейки сомита развивались пронефрические канадцы, а из каудальной части шейки сомита—половые мешки.
Ь — пронефрическая камера (Боумевовекая капсула); ту—миотомы; р. с.—пронефрические канальцы; t—каудальный отдел целомического мешка; и. д.—пронефрический выводной проток (из А. А. Машковцева, 1934).
двух преанальных парах целомических
шенно ином направлении, чем это имело место у Branchiostomatidae. В течение эволюции протокраниат произошла постепенная редукция энтеро-цельных каналов и сложная морфологическая диференцировка целомических мешков. Изменения, которым подвергались целомические мешки, в основном свелись к отделению дорзальной части, давшей мышечные сегменты, от вентральной части, превратившейся собственно в целом. Для развития экскреторно-половой системы протокраниат особенно важное значение имел промежуточный участок между миотомом и целомом, который, потеряв связь с миотомом, распался на два участка: на каудальный, принявший на себя половую функцию (рис. 10, t) и на краниальный, из которого развились пронефрические канальцы (рис. 10, р. с.) и пронефрический проток (рис. 10, u. g.), открывающийся в задний конец кишечника.
В течение дальнейшей эволюции энтероцельные каналы сохраняются только
мешков, из которых первая пара с ее энтероцельными каналами осуществляет связь первичного мочеточника с кишечником, а из второй пары образуются абдоминальные поры. В результате описанных выше процессов возникла экскреторнополовая система протокраниат, схематически изображенная на рис. 10 и состоящая из сегментальных экскреторных канальцев (р. с.) и сегментальных мешков (t). Как продукты экскреции, так и половые клетки через первичный мочеточник выделялись в задний отдел кишечника.
Таким образом, мы видим, что сравнительно морфологическое исследование действительно позволяет нам восстановить довольно значительное количество признаков наших гипотетических Protocraniata, совершенно так же, как оно позволяет реконструировать и некоторые черты их образа жизни.2
От охарактеризованных нами Protocraniata произошли две крупные группы потомков, прогрессивно развившихся в дивергентных направлениях (см. родословное дерево, схема XV): одну из них, развитие которой привело в конечном итоге к образованию современных миног и миксин, мы обозначаем именем Entobranchiata, другую, современными потомками которой являются низшие рыбы,—именем Ectobranchiata. Разберем сначала эволюцию Entobranchiata—предков всех круглоротых и их сородичей.
1 А. М a s c h к о w z е f f, Zur Phylogenie des Urogenitalsystems der Wirbeitiere. p. 1 u. II, Zool. Jahrb., Bd. 48, 1926; Bd. 59, 1934. А. А. Машковцев. Морфологические и филогенетические отношения мезонефроса к пронефросу. Тр. Лаб. эв. морф. Ак. Наук СССР, т. 2, 1934.
1 Ясно, что между Acrania primitiva и Protocraftiata некогда существовал еще целый ряд промежуточных форм, и сравнительно морфологическое исследование раскрывает нам уже теперь многие черты их примитивной организации: нам известны, например, многие нейромеры их головного мозга, строение их примитивных глазных пузырьков и т. д. Однако, при современном состоянии наших знаний нам еще не представляется возможным дать полную и связную картину организации какой-либо из этих промежуточных форм.
1С*
ГЛАВА VI
ЭВОЛЮЦИЯ ENTOBRANCHIATA
СТРОЕНИЕ ENTOBRANCHIATA
От Entobranchiata произошли ныне живущие Cyclostomata (миноги и миксины), вымершие девонские Osteostraci и родственные им формы.
Мы имеем основание думать, что гипотетические Entobranchiata стояли во многих отношениях значительно выше, чем только что описанные нами Protocraniata. Это относится в первую голову к системе их органов дыхания: у общих предков Cyclostomata и Ostracodermi развились медиально от хрящевых жаберных дужек крупные энтодермические жаберные мешки (рис. 11, br. 1—Ьг. 11, pb. 1—pb. 3), выстланные изнутри склад-
Рис. 11. Схематическое изображение передней части тела с жаберной решеткой и эктодермальными Жаберными мешками гипотетических Entobranchiata.
A.br. 1—A.br. 7—жаберные Дуги 1—7 и следующие за ними; А.pr.br.1—А.рг. Ьг.4—пред-жаберные дуги 1—4; br. 1—Ьг. 11—жаберные щели (жаберные мешки) 1—11; Cart. апп.—хрящевое околоротовое кольцо; С. ап.—слуховая капсула. Ch.—хорда; C.d.a.—cartilago dorsalis anterior; C.ept.—эпитрематическая комиссура жаберной решетки; C.hpt.—гипотремагическая комиссура жаберной решетки; C.sch.—субхордальная комиссура жаберной решетки; C.v.—вентральная комиссура жаберной решетки; С. nas.—носовая капсула; Dien.—промежуточный мозг; Ер.—эпифиз с париетальными глазами; Нур.—гипофиз; М.—рот; Ms. еп.—средний мозг. Mt. еп.—продолговатый мозг: М. sp.—спинной мозг; Nas.—носовые ямки; Ос.—глаз; pb. 1—pb. 3—пребранхиальные жаберные щели (жаберные мешки); Pch.—parachordalia; Tl. еп.—передний мозг; Тг. сг.—trabeculae cranii. На этой схеме изображено первичное строение жаберной решетки, которая у предков круглоротых простиралась вплоть до самого рта (ср. рис. 13 и 14) и функционировала в передней части головы в качестве частей жаберного скелета (из А. Н. Северцова, 1917; несколько изменено).
ками кожи (эктодермальные жаберные лепестки, entobranchiae; рис. 12, Br. 1.), вследствие чего дыхательная поверхность у этих форм в значительной степени увеличилась, а функция дыхания повысилась. Ряд жаберных мешков начинался, как и у Protocraniata, непосредственно позади ротового отверстия (рис. 11, pb. 3), и число их, вероятно, соответствовало их числу у протокраниат. В связи с этим изменением структуры органов дыхания изменилось и строение жаберного скелета этих гипотетических форм: между концами жаберных дужек, а также под и над наруж
ЭВОЛЮЦИЯ ENTOBRANCHIATA
149
ными жаберными отверстиями развились продольные хрящевые перемычки или комиссуры (субхордальные—рис. 11, С. sch., эпитрематические— С. ept., гипотрематические—С. hpt. и вентральные—С. v.), и таким образом сформировалась довольно сложно построенная жаберная решетка типа Cyclostomata.
Строение висцерального скелета в передней части головы у личинок круглоротых (рис. 9) и у взрослых миног (рис. 13 и 14) совершенно ясно указывает нам на то, что жаберная решетка эта находилась у предков Cyclostomata и в голове, где она доходила до самого рта. Эмбриологическое исследование говорит нам также о том, что функционирующие жаберные мешки были развитый в передней части головы (рис. 7, pb. 1,рЬ.2).То, что мы находим в переднем отделе головы пескоройки и миног остатки (рис. 8, 9, 13, 14), наводит нас на мысль, лежащий непосредственно й отдел кишечника функцио-
жаберной решетки что у Entobranchiata позади рта передни нировал еще как глоточная полость, служа в то же время для целей дыхания; отсюда можно заключить, что питание этих гипотетических форм было еще пассивным. Присутствие у Ammocoetes хрящевого околоротового кольца (рис. 13, Cart, ann.), строение его эндостиля и ре-сничатых борозд целиком под-
Рис. 12. Энтодермальные жабры пескоройки (Ammocoetes). Рисунок несколько схематизирован.
А. Ьг.—жаберная дуга; af. — arteria afferens; Ao.d. — aorta dorsalis; Ao. v.—aorta ventralis; Atr.—предсердие; Br.c.—жаберный отдел кишечника; Br.l.—эндодермальные жаберные лепестки; Ch—хорда; end.— эпдостиль; ef.—vena efferens жабр; М. sp.—спинной мозг; vt.—желудочек сердца. На этом рисунке видно образование крупных эндодермальных жаберных мешков, стенки которых покрыты эктодермальными жаберными складками (Вг.1.); жаберные мешки расположены медиально от жаберных дуг.
тверждает этот вывод.
История развития Myxi-noidea пока еще мало исследована сравнительно эмбриологическим методом; поэтому мы затрудняемся сказать теперь что-либо определенное относительно структуры других систем органов энтобран-
хиат. Весьма вероятно, однако, что скелет этих гипотетических форм находился еще на очень низкой стадии своего развития, близко напоминающей то, что мы видим у протокраниат: их осевой череп (рис. 11, Pch.) и слуховые капсулы (рис. 11, С. аи.) были, по всей вероятности, еще такими же, как и у протокраниат, т. е. у них, повидимому, имелся лишь один парахордальный череп, a trabeculae cranii еще не принимали участия в образовании neurocranium (рис. 11, Тг. сг.). По исследованиям В. Па-тэна, подтвержденным и расширенным исследованиями Н. Гольмгрена (цитирую по работе Стеншиб 1927 г.), у личинок циклостом развит поверхностный, состоящий из слизистого хряща головной щит: можно думать поэтому, что подобного же рода слизисто-хрящевое образование было развито и у энтобранхиат.
В позвоночнике этих гипотетических форм несомненно существовали диплоспондильно расположенные верхние дуги.
Относительно центральной и периферической нервных систем Ento-branchiataMbi можем прибавить лишь очень мало к тому, что было сказано о строении этих органов в связи с вопросом о протокраниатах. Что же касается их органов чувств, то ухо Entobranchiata было, повидимому, уже
150
ГЛАВА VI
построено по типу слуховых органов Myxinoidea, т. е. в нем уже имелось два полукружных канала; парные глаза были вполне развиты, причем весьма вероятно, что и париетальные глаза также еще функционировали.
Рис. 13. Схематическое изображение головы и висцерального скелета миноги. На схеме гомодинампые части висцерального скелета графически обозначены одинаково (горизонтальной и вертикальной штриховкой, пунктиром и т. д.).
А.Ьг.1—А.Ьг.7— жаберные дуги 1—7; А.рг.Ьг.1—А.pr.br.л—пребранхиальные жаберные дуги 1—4;br. 1—Ьг. 7—жаберные щели 1—7; Cart. arm.—околоротвое кольцо; С. ап.—слуховая капсула; C.d.a.—cartilage dorsalis anterior; C.d.p.—cartilago dorsalb posterior; C. ept.—эпитрема-тическая комиссура; C. epti—эпитрематическая комиссура между первой и второй преджаберными дугами; C.epta, C.eptg—эпитрематические комиссуры между 2-й и 3-й и 3-й и 4-й преджаберными дугами; C.hpt.—гипогрематическая комиссура; C.hpt.'—гинотреманпеокая комиссура между 2-й и 3-й преджаберными дугами; C.nas.—хрящевая носовая капсула! С.sch.— субхордальная комиссура; С. v.—вентральная комиссура жаберной решетки; C.v/—вентральная комиссура между 1-й и 3-й преджаберными дугами, существующая только у пескоройки; VII—n. facialis ;IX— и. glos-sopharyngeus;X. 1.—n. lateralis vagi; X.br.b—X.br.6 — жаберные ветви блуждающего нерва. На этой схеме изображены (черным цветом) семь функционирующих жаберных дуг миноги (A.br.l—А.Ьг.7) и четыре хрящевых комиссуры, соединяющих каждую жаберную дугу с впереди от нее лежащей дугой и образующих все вместе жаберную решетку; эпитрематические комиссуры изображены черными точками на белом поле, субхордальные—черной краской с белыми точками; гипотрематические комиссуры изображены вертикальной штриховкой, а вентральные комиссуры— горизонтальной штриховкой. A.pr.br.l—A.pr.br.4—представляют собой части четырех пред-жаберных дуг (ср. рис. 9, A.pbr.l—3), функционирующие у миноги как части черепа кротового скелета; у взрослой миноги сохранились три элитрематических комиссуры: C.eptj ,C.ept2 (arcus subocularis), C.eptg! C. hpt' представляет собой остаток гипотрематической комиссуры между 2-Й и 3-й преджаберными дугами; несколько измененные вентральные комиссуры (С. v.) между первой и второй и второй и третьей преджаберными дугами развиваются лишь у поздних личинок миноги (Ammocoetes). Мы принимаем, таким образом, что: 1) в переднем отделе головы энтобран-хпат была развита жаберная решетка, состоящая из четырех пребранхиалъпых жаберных дуг и соответствующих им типичных продольных комиссур, и 2) некоторые части этой жаберной решетки (преджаберные дуги, хрящевые продольные комиссуры) сохранились в переднем отделе головы миног, а вероятно также и миксин1 (по А. Н. Севердову, 1916).
Зато весьма характерные изменения произошли в носовой и гипофизарных областях этих животных. Весьма вероятно, что гипофиз, представляющий собою еще простую мешковидную железу, наподобие той, которую мы находим теперь у молодых личинок Petromyzon и у эмбрионов селахий, открывался у протокраниат наружу либо непосредственно впереди рта, либо в области переднего неба (рис. 15, hyp.). Промежуточную область, лежащую между ротовым отверстием и отверстием гипофиза (рис. 15, С), мы можем обозначить как постгипофизарную область. У предков всех Cyclo
1 Как известно, было сделано немало попыток истолковать морфологическое значение скелетных частей, расположенных в передней части головы миАги. Считается более или менее общепризнаиым, что большинство этих толкований не выдерживает критики с точки зрения сравнительной анатомии. Причина этого лежит, по-моему, в том, что при истолковании строения скелета головы миноги обычно исходили из строения сравнительно анатомически более изученного краниального и висцерального скелета рыб. В нашей интерпретации частей головы миноги мы исходим не из принципа гомологии, но из принципа гомодинамии. Мы приняли, что скелетная решетка функционирующего жаберного отдела миноги (A. br. 1—А. Ьг. 7) со всеми своими типичными продольными комиссурами была развита также и в переднем отделе головы циклостом. На основании этого гипотетического, но весьма вероятного предположения (ср. рис. 7, 8, 9) мы сделали попытку истолковать строение предслуховых элементов висцерального скелета миноги, исходя из их сравнения с заслуховыми элементами голов,-кого скелета. Это позволило нам дать, невидимому, довольно правильную интерпретацию таких элементов, как, например, подглазничная дуга (С. ept2), processus cornualis, как элементы С. ept! С. ept3 и т. д. (рнс.13).К сожалению, однако, нам не удалось наблюдать процесса превращения пескоройки в миногу, поэтому я не могу сказать с определенностью, насколько далеко вперед доходила жаберная решетка в передней части головы первичных круглоротых. Мне не ясно также значение таких частей ротового скелета, как Cartilago lateralis anterior, Cartilago styloideum а особенно непарных частей ротового скелета (см. ниже).
ЭВОЛЮЦИЯ ENTOBRANCHIATA
151
stomata и Osteostraci, т. e. у разбираемых нами энтобранхиат, эта передняя часть головы стала усиленно расти (рис. 15—17, С.), причем в ней образовались широкие плоские хрящи [рис. 17, tr. cr. (С. dors, post.)],1 из которых впоследствии развилась так называемая верхняя губа Ammo-coetes. Вследствие разрастания.' постгипофизарной области выводное отверстие гипофиза приблизилось к носовым ямкам (рис. 15—17, hyp., f. nas.), а эти последние сблизились между собой настолько, что между ними осталась лишь тонкая перегородка (рис. 18, s. nas.). Раньше эти ямки открывались непосредственно наружу, теперь же, вследствие только что упомянутого разрастания постгипофизарной области, вторично образовалась особая назогипофизарная полость (рис. 17 и 18, С. nas. hyp.), в которую открывались обе эти ямки (рис. 17, f. nas.) и сам гипофиз (hyp.). Полость эта сообщалась с наружным миром при помощи особого медианного канальца, так называемого назогипофизарного прохода (рис. 18, Tr. nas. hyp.) или назогипофизарного отверстия (рис. 17 и 18, F. nas. hyp.).
Рис. 14. Скелет головы миноги (Petromyzon planeri). Не схематизирован.
Интересно сравнить этот рисунок со схемой, изображенной на рис. 13.Это позволит читателю убедиться в гомодинамии, существующей меясду элементами висцерального скелета передней части головы в элементами жаберной решетки. Обозначения те же, что и иа рис. 13 (из А. Н. Северцова, 1916).
Таким образом, вследствие этого филогенетического изменения, перестроилась вся структура передней части головы Entobranchiata: носовые ямки, которые первично (рис. 15, f. nas.) лежали, как они лежат сейчас у селахий, на вентральной поверхности головы, передвинулись на ее дорзальную сторону (рис. 17) и стали открываться уже не непосредственно наружу, но в особую, только что упомянутую нами назогипофизарную полость (рис. 17, 18, С. nas. hyp.), которая со своей стороны открывалась наружу посредством своего особого канала (рис. 17 и 18, F. nas. hyp.); в эту же полость открывался и гипофиз (hyp.).
Из постгипофизарного отдела (соответствующего части неба рыб) образовался новый, значительный по своим размерам, отдел передней части головы (так называемая верхняя губа Cyclostomata1 2), который можно обозначить как вторичный преназальный отдел головы; в нем лежат особые скелетные образования, а именно—cartilago dorsalis posterior et cartilage anterior миноги [рис. 17, tr. cr. (C. dors. post)].
Подводя итоги всему сказанному, мы видим, что в эволюции Entobranchiata произошли весьма существенные изменения: прежде всего у них появились крупные энтодермические жаберные мешки и, в соответствии
1 Хрящи эти—cartilago dorsalis posterior миноги—мы гомологизируем с trabeculae cranii гнатостом.
2 Так называемая верхняя губа циклостом, соответствует, следовательно, части нёба рыб
«
152
ГЛАВА VI
Рис. 15, 16, 17. Три медиальных разреза через голову личинки миноги на трех последовательных стадиях развития.
На всех этих трех рисунках участок кожи, лежащий первоначально впереди от носовой ямки (f. nas.), изображен черными точками на белом поле (А—преназальный отдел); участок кожи, первоначально лежащий между носовой ямкой (f. nas.) и гипофизом (hyp.), изображен сплошными белыми линиями на черном фоне (В—прегипофизарный отдел) и участок, первично лежавший каудально от гипофиза (С—постгипофизарный отдел),' графически изображен маленькими овалами. Ch.—хорда; С. nas. hyp.—назогипофизарная полость; ер.—эпифиз; f. nas.hyp.—назогипо-физарное отверстие» f.nas.—носовая ямка; hyp.—гипофиз; М — рот; tr.cr.(c.dors.post.)— cartilago dorsalis posterior (ср. рис. 14), который по своему положению, вероятно, гомологичен trabeculae cranii.
Рис. 15. Ранняя стадия развития личинки миноги. Здесь взаимоотношения между носовой ямкой, гипофизом и ротовым отверстием такие же, как и у зародышей челюстных позвоночных (Gnathostomata; ср. рис. 32).
Рис. 16. Несколько более поздняя стадия развития личинки миноги, когда начинает расти постгипофизарный отдел, так называемая верхняя губа пескоройки (С); этот отдел соответствует постгипофизарной части нёба зародышей челюстных позвоночных (Gnathostomata; ср. рис. 32).
Рис. 17. Значительно более поздняя стадия развития переднего отдела головы пескоройки На рисунке видно разрастание постгипофизарного отдела нёба (ср. рис. 15), образующего так называемую верхнюю губу («вторичная преназальная часть»). Вследствие этого рострального разрастания образовалась новая полость, назогипофизарная полость (С.nas.hyp.), в которую открывается гипофиз (hyp.) и обе носовые ямки (f.nas.). Назогипофизарная полость сама открывается наружу при помощи непарного назогипофизарного отверстия (f.nas.hyp.), которое прежними авторами обозначалось как носовое отверстие; истинное носовое отверстие находится у f. nas. (из А. Н. Северцова, 1917).
ЭВОЛЮЦИЯ ENTOBRANCHIATA
153
с ними, путем образования продольных хрящевых перемычек, сложно
построенная жаберная решетка сти головы, где также находились функционирующие жаберные мешки. В передней части головы имели место еще и другие крупные перемены: здесь колоссально разросся постгипофизарный отдел, причем из него образовалась преназальная часть головы, вследствие чего сформировалась назогипофизар-ная полость со своим протоком и наружным отверстием (обозначавшимся как непарное носовое отверстие); на дне этой полости открывались сближенные между собою носовые ямки и мешок гипофиза. Наряду с только что перечисленными переменами в организации Entobranchiata произошли и другие структур-
Решетка эта достигла передней ча-
Рис. 18. Поперечный срез через назогипэфи-зарный проток (Tr. nas. hyp.) и носовые ямки (F. nas.) еще более крупной пескоройки (ср. рис. 17).
С. п.—хрящевая носовая капсула; C.nas.hyp.—иа.зо-гипофизарпая полость; F.nas. hyp.—пазогипофизарное отверстие; F. nas. — носовая ямка; N. oil.—п. olfactorius; S. nas.—septum nasale.
На этом рисунке мы видим, что носовая ямка (F. nas.), так же как и у гнатостом, является парным образованием, разделенным носовой перегородкой (S. nas.) на две части. На рисунке видно также, что обе носовые ямки (F. nas.) открываются в назогипофизарную полость C.nas.hyp. (из А. И. Северцова, 1917).
ные изменения, о которых мы еще не можем сказать ничего конкретного, так как нам пока мало известна история развития семейства Myxinoidea.
От Entobranchiata произошли, по крайней мере, две линии потомков, а именно: 1) Protocyclostomata и 2) Osteostraci с их ближайшими сородичами.
ЭВОЛЮЦИЯ PROTOCYCLOSTOMATA
Мы объединяем под именем Protocyclostomata, или первичных циклостом, тех потомков Entobranchiata, от которых произошли ныне живущие миноги и миксины. Между гипотетическими протоциклостомами и непосредственными предками Ostracodermi, ответвившимися от энто-бранхиат в другую сторону, имеется ряд существенных различий, из которых отметим следующие.
Хотя предки современных круглоротых и перешли, подобно предкам вымерших Ostracodermi, от пассивного питания к активному, однако переход этот осуществился совершенно различным образом в двух названных выше группах: у предков Ostracodermi редуцировалось ротовое кольцо, а жевательный аппарат образовался за счет костного чешуйчатого панцыря, некогда покрывавшего у них с вентральной стороны переднюю часть головы (И. Киэр); у предков круглоротых ротовое кольцо сохранилось, но позади него из видоизменившихся передних висцеральных элементов развились парные и непарные хрящи сосущего рта. У протоциклостом все пребран-хиальные жаберные мешки редуцировались, а соответствующие им щели закрылись.1 У Ostracodermi, наоборот, спиракулярная и премандибуляр-
1 Включая ^пиракулярную, т. е. иннервируемую лицевым нервом, щель, которая сохранилась вплоть до настоящего времени у гнатостом. В этом отношении низшие эласмобранхии оказались более примитивно построенными, чем миноги и миксины.
154
ГЛАВА VI
ная жаберные щели, т. е. первая и вторая пребранхиальные щели, сохранились и функционировали как типичные жаберные щели.
Хрящевое кольцо протокраниат и энтобранхиат сохранилось в качестве опоры для края рта у протоцйклостом и имеется в наличности также у их отдаленных потомков, ныне живущих миксин и миног. У первых оно состоит из единого кольцеобразного хряща, на котором совершенно отсутствуют реснички (cirri), у последних оно образовалось срастанием отдельных хрящей, несущих на себе реснички (cirri). Можно думать, что то первичное состояние, которое мы находим у гипотетических энтобранхиат, сохранилось наиболее полным образом именно у миксин и что миноги являются в этом отношении вторично измененными формами.
Как у миксин, так и у миног имеются сходно построенные эпидермальные роговые зубы. Отсюда мы можем заключить, что и у их общих предков, протоцйклостом, в свое время существовали подобного же рода роговые зубы. Вполне возможно, что именно появление этих зубов и явилось стимулом к переходу этих форм от пассивного питания к активному.
В настоящее время представляется довольно затруднительным определить, каким путем могли возникнуть те непарные и парные хрящи, из которых образовался ротовой скелет миног (рис. 14, A.pr.br4.,C.lat. ant., Styl. и т. д.) и миксин, ибо сравнительно эмбриологическое изучение истории развития этих элементов пока еще слишком недостаточно и неполно. В своих работах об эволюции низших позвоночных 1916 и 1&17 гг. я пришел к выводу, что передние парные ротовые хрящи миног представляют собою видоизмененные элементы передней части жаберной решетки энтобранхиат (рис. 11). Я должен согласиться, однако, с тем, что взгляд этот является лишь гипотезой. Детальное развитие губных ротовых хрящей на стадиях метаморфоза личинок при переходе их во взрослое состояние как у миног, так, особенно, у миксин, по развитию которых у меня не было достаточного материала, является поэтому крайне желательным.1
Повидимому, и миноги и миксины (в особенности Bdellostomatidae) сохранили весьма многие примитивные черты строения своих отдаленных предков, так что представляется довольно затруднительным сказать с определенностью, которая из этих групп является наиболее примитивной. Можно думать, что жаберная решетка в бранхиальном отделе туловища передней части головы (рис. 11, 13 и 14) сохранилась у миног лучше, чем у миксин. Строение переднего отдела висцерального скелета миксин указывает на то, что у предков этих животных была развита жаберная решетка типа миног. В заслуховой области (отдел, иннервируемый блуждающим нервом) жаберный, скелет у миксин и у Bdellostomatidae сильно редуцировался; он представлен у этих форм лишь совершенно рудиментарными жаберным дугами. Поэтому мы имеем право предположить, что в жаберном скелете миксин произошел в свое время ряд вторичных дегенеративных < процессов.
Весьма вероятно, что у большинства круглоротых число жаберных мешков резко уменьшилось в течение филогенеза. У энтобранхиат и у ближайших предков современных круглоротых (Protocyclostomata) число это было еще довольно значительным (около 17): у них были развиты,
1 Во время подготовки к печатанию этой книги появилась статья Е. Stensio и N. Holmgren: «Kranium und Visceralskelett der Akranier, Cyclostomjn und Fische» (1936); Гольмгрен, на основании наблюдений над метаморфозом личинок миноги, отрицает эту гипотезу, утверждая, что указанные образования появились у миноги лишь после метаморфоза и поэтому не могут иметь филетического значения. Вопрос этот нуждается, однако, в дальнейшей проверке.
ЭВОЛЮЦИЯ ENTOBRANCHIATA
155
невидимому, четыре пребранхиальных бранхиомера.1 Жаберные мешки и щели этих бранхиальных сегментов атрофировались при переходе энто-бранхиат от пассивного к активному способу питания. С другой стороны, у миног и у миксин редуцировались также и задние жаберные сегменты, так что анцестральное состояние, характеризуемое относительно большим числом жаберных щелей (10—14), сохранилось лишь у некоторых бделло-стоматид (Bdellostoma stouti).1 2 * * 5
Сравнивая организацию миног со строением миксин, мы можем с к;#' зать, что миксины—и среди них в особенности Bdellostomatidae—в целом ряде признаков остановились в своей эволюции на гораздо более низкой ступени развития, чем миноги: вспомним, в особенности, строение их ротового кольца, снабженного ресничатыми выростами (cirri), строение их мозга и слухового органа, число их жаберных щелей и т. д. С другой стороны, в некоторых иных отношениях (присутствие особых выводных протоков в жаберных мешках у миксин, редукция жаберной решетки и т. д.) животные эти представляют собою специализованные формы.
Возвращаясь к организации протоциклостом, мы можем еще сказать, что их сравнительно просто построенный рот сосущего типа был снабжен роговыми зубами, при помощи которых они»прокусывали кожу животных, служивших для них добычей. Их пребранхиальные щели уже закрылись. В передней части головы имелась назогипофизарная полость, в которую открывались как гипофиз, так и обе носовые ямки. В бранхиальном отделе протоциклостом уже образовалась жаберная решетка, построенная по только что описанному нами типу (рис. 11).
Весьма вероятно, что главным изменением, имевшим место при переходе энтобранхиат в протоциклостомы, было именно изменение их ротового аппарата, который из органа, приспособленного к пассивому принятию пищи, превратился в орган активного питания особого рода, в бесчелюст-ный сосущий рот, снабженный крепкими и острыми роговыми зубами. Позади ротового скелета лежала жаберная решетка, состоящая из многочисленных дужек, соединенных между собою продольными перемычками. Число функционирующих жаберных мешков было довольно значительным. Остальные органы были построены приблизительно так же, как и у энтобранхиат, т. е. были гораздо более примитивны, чем соответствующие им органы у миксин и у миног.
1 Как уже было сказано выше, Гольмгреи в только что названной сволочной работе признает вместе со Стеншиб существование только трех преджаберных дуг.
2 Д. Третьяковым был высказан взгляд, что и у циклостом и у гнатостом число функционирующих жаберных щелей в течение филогенеза вторично возросло. Я придерживаюсь противоположной точки зрения и думаю, что значительное число истинных жаберных щелей у Bdellostoma stouti является первичным признаком, последним остатком состояния, некогда типичного для общих предков всех энто-и эктобран-хиат. Меня наводит на эту мысль несомненная и резко выраженная у всех позвоночных тенденция к сокращению числа их жаберных щелей. В течение эволюции как у кругло-потых, так и у челюстных рыб постепенно закрылись пребранхиальные жаберные щели; процесс редукции последней из них, а именно, спиракулярной щели, мы имеем возможность непосредственно наблюдать и сейчас. Редукция эта, вообще говоря, происходит в ростро-каудальном направлении. Но наряду с этим как у циклостом, так и у гнатостом жаберные щели редуцируются также и начиная с заднего конца ряда, т. е. редукция идет и в обратном, каудо-ростральном направлении. Весьма примитивные Bdellostomatidae имеют в общем от 10 до 14 жаберных щелей, остальные кругло-
„отые_7 Osteostraci—10; из акул: Heptanchus имеет семь жаберных щелей и руди-
мент восьмой, Hexanchus и Chlamydoselachus по б, остальные акулы по 5, Osteichthyes—
5_4 Quadrupeda—4—0. Можно думать поэтому, что значительное число жаберных
щелей (к мешков) у Bdellostoma stouti является не вторичным, но первичным, т. е.-аниестгальным, признаком. В пользу моего взгляда говорят и данные, приводимые Ппайсом (1. с., 1906), который утверждает, что у личинок Bdellostoma закладывается около 30 жаберных щелей, а также и тот факт, что задние жаберные щели Bdellostoma stouti так же построены и так же иннервируются, как и ее передние щели (па неопубликованным данным моей ученицы О. Белобородовой).
156
ГЛАВА VI
Все эти перемены сделали возможным для предков современных круглоротых, т. е. для предков современных миног и миксин, переход от пассивного способа питания к активному. Отметим сейчас же, что этот новый способ питания оказался в известном смысле специализованным, так как эти животные могли питаться лишь кровью других животных, у которых кожа была настолько слаба, что ее легко можно было прогрызть роговыми зубами. Совершенно ясно, что их пища при этом оказалась несравненно более питательной, чем пища, которой в свое время питались их предки при пассивном способе питания.
От этих гипотетических, по всей вероятности нектонных, форм произошли, как это уже было указано, две линии потомков (см. родословное древо, схема XV), эволюировавших дальше в несколько отличных друг от друга направлениях, а именно: Petromyzontes и Myxinoidea. У миног, сохранивших некоторые примитивные черты строения более отдаленных предков, т. е. энтобранхиат: жаберную решетку, диплоспондильные верхние дуги позвонков ит. д., развились в прогрессивном направлении мозг и орган слуха, у миксин, специально приспособившихся к питанию путем высасывания крови крупных рыб, редуцировалась жаберная решетка и дуги позвонков (специализация); зато у них ряд других признаков остановился в своем развитии на гораздо более примитивной ступени, чем это имеет место у миног.
ЭВОЛЮЦИЯ OSTRACODERMI
Мы рассматривали до сих пор эволюцию энтобранхиат такой, какой она нам представлялась на основании данных, полученных путем изучения сравнительной анатомии и истории развития этих животных; теперь нам предстоит рассмотреть результаты палеонтологических исследований, бросающих совершенно новый свет на интересующие нас проблемы. Мы имеем в виду новейшие труды двух скандинавских ученых, Е. А. Стен-шиб1 и И. Киэра, над так называемыми«панцырными рыбами» (Ostraco-dermi),—труды, внесшие в изучаемый нами вопрос так много нового, что нам пришлось основательно пересмотреть наши взгляды на систематическое положение и филогенетические соотношения этих загадочных, животных.
Здесь мы приведем в очень сжатой форме результаты блестящих исследований Стеншиб над Cephalaspida, остатки которых были недавно найдены на Шпицбергене, а затем сравним выводы, к которым он приходит на основании своих палеонтологических изысканий, с теми результатами, к которым пришли мы на основании наших сравнительно морфологических исследований.
Стеншиб детально обработал богатейший материал Ostracodermi из даун-тоновских слоев острова Шпицберген. Однако, прежде чем приводить фактические результаты его исследований, нам надо сказать несколько слов относительно метода его работы,—метода, который, по нашему мнению, составляет целую эпоху в истории палеонтологии.
Стеншиб удалось, благодаря исключительно тщательному отпрепари-рованию внутренних органов с помощью полной серии шлифов, изучить внутреннюю организацию Ostracodermi вплоть до мельчайших подробностей; ему удалось, например, детально реконструировать положение и прохождение таких органов, которые до негр оставались совершенно недоступными палеонтологам, т. е. нервов и кровеносных сосудов: он смог, таким образом, сравнить внутренние органы Ostracodermi с соответствующими внутренними органами современных круглоротых и рыб, и сравнение это было им выполнено мастерски.
1 S t е п s i б Е., Downtonian and Devonian Vertebrates, p. I, Cephalaspidae, Norsk. Vidensk. Akad. Oslo, 1927
ЭВОЛЮЦИЯ ENTOBRANCHIATA
157
Как известно, Cephalaspidae,Tai< же как и огромное большинство остальных Ostracodermi, были бентонными формами, т. е. животными,обитавшими на дне моря. Соответственно этому обстоятельству они обладали широким, сплюснутым в дорзовентральном направлении телом (рис. 19); их широкая, спереди закругленная голова была покрыта цельным костным щитом с парой крупных шипов назади. Неподвижная передняя часть туловища была защищена тем же щитом; задняя же часть туловища и гетероцеркальный хвост были покрыты костными чешуйками.
Передний головной щит не состоял, как это думали прежде, из отдель- ' ных пластинок, но представлял собою единый цельный панцырь, наружный слой которого состоял из дентина, а внутренний—из перихондральной костной ткани. Под этим щитом лежал вполне развитый плоский хрящевой череп, сквозь который проходили каналы нервов и кровеносных сосу-
Рис. 19. Cephalaspis murchisoni. Вид сверху (из Циттеля, 1923).
дов.1 На наружной поверхности головного мозга мы видим отверстие для глаз (рис. 20, orb.); между ними расположена непарная ямка пинеального органа (рис. 20, pin.), а несколько впереди от них—так называемая непарная йосовая ямка (рис. 20, f. сп.). Кроме того, на наружной поверхности головы обрисовываются три довольно крупных поля, которые Стеншиб истолковывает как электрические органы этих животных (рис. 20, 1. sf., d. sf.). Его толкование нам кажется вполне обоснованным и правдоподобным. Покрытый костным панцырем хрящевой череп Osteostraci, вероятно, соответствовал хордальному черепу энтобранхиат, сросшемуся с упомянутым нами выше головным щитом из слизистого хряща (В. Паттен, Н. Гольмгрен), гомологичным cartilago dorsalis posterior миног.
На нижней поверхности головы у Cephalaspidae находились расположенные по кругу узкие жаберные отверстия;1 2 круглое пространство между ними (т. е. нижняя поверхность жаберной полости) было покрыто мелкими чешуйками (рис. 21). Ротовое отверстие имело вентральное положение. По исследованиям И. Киэра,3 у описываемых животных имелся вполне развитый, состоящий из небольших покровных пластинок или костей ротовой скелет
1 Эти каналы, точно так же как и внутренняя полость осевого черепа, как слуховые капсулы и носовая ямка, были окружены тонким слоем'костной ткани; именно это обстоятельство и позволило Сеншиб восстановить положение и распределение всех этих образований.
2 Р a*t t е n W., On the Structure and Classification of the Tremataspidae, Mem. de 1’Acad. de St. Petersburg, v. 13. 1903.
8 К i a e r J., The Structure of the mouth of Pleraspids and Cephalaspids, Palae-biologica, Bd. 1, 1928.
158
ГЛАВА VI
rb г tel
ni.2
\pm
. l.sr
acpl
bpl
20. Передняя часть тела Kiae-
Рис.
raspis auchenaspidoides. Вид сверху.
d. sp.—спинной гребень панциря; d. st.— дорзальные электрические .органы; 1. sf.— боковые электрические органы; 1. сп.— I'ossa circumnasalis; nai—отверстие гипофиза; наг—носовое отверстие; orb.—орбитальное отверстие; pin.—пинеальное отверстие; р. s.—пекторальный синус (из Е.
Стеншиб, 1927).
Рис. 21. Передняя часть тела Tremataspis. Вид снизу.
Видны кругообразно расположенные жаберные отверстия. Между жаберными отверстиями правой и левой стороны лежат мелкие подвижные чешуи вентральной стороны тела (по В. Паттену, 1903; из Киэра, 1928).
Рис. 22. Передняя часть головы Pteraspis vogti sp. nov. Вид снизу. .
Виден ротовой скелет, состоящий из относительно малых чешуй. Ьр1.—жаберная пластинка; 1р1.— боковая пластинка, mtpl.-—максилярная, или челюстная пластинка, несущая на себе зубы; rb.—ростральновентральный отросток; opl. — ротовые пластинки; olpl.—боковые ротовые пластинки; ос. pl.—глазничные пластинки; vs.—вентральный щит (из Киэра, 192b).
Рис. 23. Горизонтальный разрез через брюшную часть головы Kiaeraspis, показывающий вероятное расположение жаберных мешков.
e.br.Ci — е.Ьг.Сю—наружные жаберные отверстия 1—10; ibs.i—ibs.9—скелетные перегородки между жаберными мешками; К. ebr.—третий жаберный мешок; Oes.—пищевод; psg. — преспиракулярный жаберный мешок; sg.—спиракулярный жаберный мешок; г.рт.—пребранхиальная борозда; vm„ vm.i, vm.a— висцеральные мускулы соответственных жаберных мешков; Vi, V2, VII, IX—выходы соответствующих нервов. Читатель обратит, внимание на тоьс какой подробностью Стеншиб восстанавливает детали строения головы этой ископаемой формы (из Стеншиб, 1927).
ЭВОЛЮЦИЯ ENTOBRANCHIATA 15»
(рис. 22, opl). Поэтому приходится думать, что вымершие Osteostraci некогда перешли к активному способу питания, точно так же как это сделали и ныне живущие Cyclostomata; при этом, однако, их ротовой скелет образовался совершенно иным путем, чем у круглоротых и у рыб: он развился не из передних жаберных дуг, как у этих последних форм, но из пластинок (чешуй) кожного скелета. Откуда могла произойти мускулатура такого ротового скелета—мы сказать не можем, но по ее положению можно-предположить, что она развилась из мускулов самых передних бранхиоме-ров, первого и второго. Если точка зрения И. Киэра правильна, а это-весьма вероятно, то у нас нет никаких оснований предполагать, что Cyclostoma ta произошли от Osteostraci, ибо в таком случае осталось бы совершенно непонятным развитие их ротового аппарата и элементов скелета сосущего рта.
Огромное теоретическое значение имеет открытое Стеншиб строение жаберного аппарата у Kiaeraspis: на рис. 23 мы видим положение десяти жаберных мешков (е. Ьг. Сг.—е. Ьг. С1о), из которых самый передний лежит непосредственно позади ротового отверстия. Каждый жаберный мешок отделен от соседних мешков, двумя скелетными перегородками (рис. 23, ibSj, ibs2 и т. д.) и чрезвычайно напоминает собою жаберные мешки типа круглоротых, т. е. миног и миксин. На основании их формы и положения Стеншиб принимает, что у Cephalaspidae были развиты именно э нт о д ер-ми ч е с к и е жаберные мешки, и, по моему мнению, трудно дать этим образованиям другое толкование: Cephalaspidae были несомненно э н то-бра нхиальны ми позвоночными и принадлежали поэтому к той же сам^й группе животных, что и круглоротые, т. е. к группе Entobranchiata. Реставрация Стеншиб, приведенная нами на рис. 23, показывает, как, по его представлению, были распределены на перегородках жаберные лепестки.
Мы уже упоминали, что у Cephalaspidae имелось значительное число жаберных щелей (10) и что Стеншиб удалось установить их гомологию с жаберными щелями круглоротых и рыб: на основании прохождения жаберных нервов он приходит к выводу, что третий жаберный мешок и третье жаберное отверстие (рис. 23, К. ebr.) Cephalaspidae соответствуют первому функционирующему жаберному мешку и первому наружному жаберному отверстию Cyclostomata, так что у Cephalaspidae имелось два ф у н к-ционирующих преджаберных мешка (жаберные щели). По мнению Стецшиб, дорзальные отрезки жаберных дуг срослись, у этих животных с вентральной поверхностью плоского примордиального черепа; отрезки эти представляют собою упомянутые выше межбранхиальные перегородки (рис. 23, ibs1? ibs2). Как была построена вентральная часть жаберного аппарата—мы не знаем; наш автор предполагает, что она состояла из подвижных частей, и с функциональной точки зрения его предположение кажется нам вполне вероятным.
На рисунках, которые приводит в своей работе Стеншиб, изображено положение и строение головного мозга и органов чувств, а также прохождение кровеносных сосудов и нервов головы Cephalaspidae: здесь мы воочию видим, в каких мельчайших подробностях удалось ему восстановить все эти образования; его искусство, техника и его талант наблюдателя вызывают чувство глубокого восхищения у тех, кому пришлось, как мне, видеть эти замечательные препараты.
На рис. 24 мы видим, например, реставрацию головного мозга Kiaeraspis,'сделанную с восковой модели, сконструированной в свою очередь, по серии шлифов. Мы видим, что здесь головной мозг решительно напоминает по типу головной мозг круглоротых и отличается от мозга миног только относительной величиной своего мозжечка (рис. 24, met.). У миксин, однако, имеется хорошо развитый мозжечек.
160
ГЛАВА VI
Орган слуха этих животных, как видно из рис. 25, был также построен 1 по типу слуховых органов циклостом: так же как и у них, в нем имелись только два полукружных канала, а именно, canali semicirculares posterior , et anterior (рис. 25, c.sem.post. ,c.sem.ant.). Canalis semicircularis exter- ' nus не был развит у Cephalaspidae, совершенно так же как он не. развит и у циклостом. Глаза этих ископаемых форм (рис. 19 и 20) были относительно велики.
Гораздо интереснее, однако, для нас строение их обонятельных органов. Стеншиб удалось показать, что у них имелась непарная продольная носовая ямка, лежащая несколько впереди от глаз (рис. 20, f. сп.), и что в эту носовую ямку (согласно нашему взгляду—в назогипофизарную полость, в которой лежат обе сильно сближенные между собою носовые ямки) открывается и назогипофизарный проток. Передняя часть этой продольной так назыв аемой «носовой ямки» представляет собою отверстие соб-
Рис. 24. Реставрация головного мозга Kiaeraspis. Вид сбоку.
die—diencephalon; ер.—эпфиз; f.rh.—fossa rhomboidalis; hab.—ganglion habenulae; hy. s.— гипофизарный мешок; ]at.—корешок проотического n. lateralis; mec.—mesencephalon; med.— medulla oblongata; mbr.—соединительнотканная перегородка между cavum cerebrale и обонятельным мешком; met—cerebellum; olfc.—обонятельный мешок; Spi.i d.—спинной корешок n. spinooccipitalis I; spi. sv.—брюшной корешок того же нерва; spi.2 d.,spi.2V.—спинной и брюшной корешки n. spinooccipitalis 2; tel.—telencephalon; I—n. olfactorius; II—n. opticus; III—n. oculomotorius; IV—n. trochiearis; V.i m—висцерально двигательный корешок n. profundi; Vg— кожная ветвь n. profundi; V.2 m—висцерально двигательный корешок n. trigemini; V.2g—кожная ветвь n. trigemini; VIII — корешки n. facialis, иннервирующие электрические органы; VIII p— задние корешки n. acusticus; IX + X—корешки n. giossopharyngeus и n. vagus (из Стеншиб, 1927).
ственно гипофиза (рис. 20, nSi), задняя ее часть—отверстие носовой ямки в собственном смысле этого слова (рис. 20, па2). Иначе говоря, у Cephalaspidae, точно так же как и у Cyclostomata, уже сформировался настоящий назогипофизарный проток (назогипофизарное отверстие).
Чрезвычайно подробно были изучены Стеншиб нервы головы у Cephalaspidae. Оказывается, что от ганглия n. facialis отходят к упомянутым выше боковым электрическигл органам шесть очень толстых нервных стволов, сильно разветвляющихся на своих концах. Отсюда можно заключить, что электрические органы эти развились из частей мускулатуры, иннервируемой п. facialis. Что касается остальных нервов головы, то они интересуют нас лишь в том отношении, что здесь первый жаберный мешок (преманди-булярный, или, по нашей терминологии, второй пребранхиальный) иннервируется первой ветвью тройничного нерва (n. profundus), которая заключает в себе висцерально двигательные волокна и, таким образом, имеет строение типичного бранхиального нерва. Второй жаберный мешок иннервируется собственно тройничным нервом, а третий—лицевым. Таким образом, расположение и иннервация головных нервов показывает нам весьма примитивный характер этих образований у Cephalaspidae; все эти нервы, даже самые передние из них, были еще простыми жаберными нервами. Такими же примитивными были и спинномозговые нервы этих животных. Их дорзально-чувствующие ветви еще не сплетались в один ствол с вент-
ЭВОЛЮЦИЯ ENTOBRANCHIATA
161
рально-двигательными, но проходили раздельно, наподобие того, что мы наблюдаем и сейчас у современных нам бесчерепных позвоночных и циклостом.
Мы не будем задерживаться на описании кровеносных сосудов головы Cephalaspidae, исследованных Стеншиб с не меньшей подробностью, чем
головные нервы, отметим только, что кровеносные сосуды этих ископаемых форм носили столь же примитивный характер, как и их нервы, и что они чрезвычайно напоминали собою кровеносные сосуды головы ныне живущих миксин и миног.
Как известно, у круглоротых не имеется ни грудных, ни брюшных плавников. У Cephalaspidae же позади головы, повидимому, имелся плавниковоподобный придаток. По крайней мере, Стеншиб находит костный отросток, отходящий от задней части примордиального черепа, который он рассматривает как приросший к черепу плечевой пояс. Он нашел здесь также два отверстия, которые он принимает за foramina arteriae et venae branchiales. От внутреннего скелета грудных плавников не сохранилось ни малейших следов; возможно, что скелет этот (если он вообще когда-либо существовал у этих форм) был хрящевым.
Итак, из этого краткого обзора результатов, достигнутых многолетней работою Стеншиб, мы видим, что взятая им на себя задача реконструировать внутреннюю ор-
Рис. 25. Слуховой лабиринт Kiaeraspis auchenaspidoides.
Фотография с восковой модели, сделанной по се* рии шлифов, a. snii и a. sma — каналы второй и третьей дорз о-латеральных поверхностных артерий; a. sp.—кан&л четвертой дорзо-латеральной поверхностной артерии; am. ant.—ampulla anterior; am. post.—ampulla posterior; c. sem. ant.— canalis semicircularis anterior слухового лабиринта; c. sem. post.—canalis semicircularis posterior слухового лабиринта; des.—канал, по которому проходит нерв и вена дорзального электрического органа; sel.2—sei.в—каналы, по которым проходят нервы электрических органов; v.dso. — канал слуховой вены; VII+Villa—канал, по которому проходят n. facialis и n. acusticus; VII+a.sa.— канал для выхода n. facialis и дорзо-латеральной поверхностной артерии; IX — канал для выхода n. glossopharyngeus; vest.—vestibulum слухового лабиринта (из Е. Стеншиб, 1927).
ганизацию Cephalaspidae выполнена им с изумительною для палеонтолога подробностью и точностью. Резюмируя его данные, мы можем утверждать с полною определенностью, что Cephalaspidae представляли собою весьма примитивно построенных и вместе с тем сильно специ-ализованных бентонных энтобранхиальных позвоночных, близко стоящих по многим признакам своего строения к современным нам Cyclostomata.
ВЗАИМООТНОШЕНИЯ МЕЖДУ CYCLOSTOMATA И OSTRACODERMI
Родственные цефаласпидам формы, принадлежащие к Osteostraci (Benneciaspis, Hoelaspis, Boreaspis, Kiaeraspis, а также семейство Trema-taspidae), имели, по Стеншиб, в общих чертах то же строение, что и сами Cephalaspidae.
Наиболее близко стоящие к ним Anaspida имели такое же назогипофи-зарное отверстие на дорзальной поверхности головы, как и Cephalaspidae, а также сходно расположенные многочисленные наружные жаберные отверстия и туловище, покрытое крепким костным панцырем. Общая форма их тела, указывающая на нектонный образ жизни, представляла собою вторичное приобретение этих форм. К этой же группе причисляет Стен-Морфологические закономерности 11
162
ГЛАВА VI
шиб и современных миног. Он считает при этом, что миноги непосредственно произошли от Anaspida. Описанный Гольмгреном у пескоройки головной щит, состоящий из слизистого хряща, он принимает за последний рудимент костного головного панцыря, имевшегося у их анаспидных предков.
Перечисленные выше формы, т. е. Cephalaspidae и их ближайшие сородичи—Osteostraci, Anaspida и ныне живущие Petromyzontia—образуют, по Стеншиб, все вместе один общий подкласс Cephalaspidomorphi.
Несколько менее подробно изучены Стеншиб представители другого, им же установленного подкласса, а именно подкласса Pteraspidomorphi (отряды Heterostraci, Palaeospondyloidea, Myxinoidea). Он считает, что Heterostraci в общем построены по тому же типу, что и Osteostraci, и что, подобно этим последним, у них имелись два полукружных канала в слуховом органе, непарное назогипофизарное отверстие и вполне развитый костный, покрытый дентином скелет головы. Так же как и у Osteostraci, у Heterostraci были развиты эктодермальные жаберные мешки с отверстиями, расположенными на вентральной стороне тела. По мнению Стеншиб, от Heterostraci произошли крайне загадочные формы, а именно Palaeospondyloidea, а от этих последних—ныне живущие Myxinoidea. Стеншиб обращает при этом особое внимание на то, что миксины стоят ближе к Osteostraci, чем к миногам, так как у миксин имеется этмоидальный ро-струм, вполне соответствующий роструму ископаемых Heterostraci.
Свои общие систематические и филогенетические выводы Стеншиб формулирует в следующих положениях:
1. Ostracodermi образуют особую группу
весьма примитивных бесчелюстных черепных позвоночных, которые в общем и целом обладали очень своеобразной организацией, не имеющей ничего общего с организацией Artrodira и Antiarchi, представлявших собою настоящих рыб, близко стоящих к эласмобранхиям.
2. Группа Ostracodermi primitivi заключает в себе Osteostraci, Anaspida, Heterostraci, Palaeospondylus и, что всего примечательнее, ныне живущих Petromyzontia и Myxinoidea.
3. Согласно этому Ostracodermi подразделяются на две большие группы, из которых одна заключает в себе Osteostraci, Anaspida и миног, а другая— Heterostraci, Palaeospondylus и миксин.
4. Таким образом, устанавливается не только факт родственности отношений между миногами, миксинами и Ostracodermi вообще, но определяется и их принадлежность к двум различным систематическим группам в пределах общего класса Ostracodermi. При этом ясно, что миноги произошли либо непосредственно от Anaspida, либо от какого-то предка, общего для Anaspida и Osteostraci, точнотак же как миксины произошли либо непосредственно от Palaeospondyloidea, либо от предка, общего для этих последних и Heterostraci,
5. Взаимоотношение между отдельными группами Ostracodermi, каким оно представляется нам на основании современного уровня наших знаний, может быть выражено приведенной выше схемой (схема IX). Классификация же Ostracodermi, согласно общей сумме наших о них знаний, укладывается в следующую таблицу:
Cephalaspidmrphi Pteraspidomorphi
Схема IX. Объяснение в тексте.
ЭВОЛЮЦИЯ ENTOBRANCHtATA
163
Vertebrata craniata Ветвь I. Agnathi
Класс Ostracodermi (Cyclostomata)
Подкласс A. Pteraspidomorphi
Отряд 1. Heterostraci
Отряд 2. Palaeospondyloidea
Отряд 3. Myxinoidea
Подкласс В. Cephalaspidomorphi
Отряд 1. Osteostraci
Отряд 2. Anaspida
•Отряд 3. Petromyzontia
Ветвь II. Gnathostomi.
Факт, что у наиболее древних Ostracodermi встречается вполне выраженная костная ткань не только в кожном, но и во внутреннем скелете, заставляет Стеншиб предположить, что и предки гнатостом обладали вполне развитым костным скелетом, впоследствии заменившимся вторично развившимся хрящевым скелетом.
Мы изложили результаты большой монографии Стеншиб настолько объективно и точно, насколько это оказалсь возможным сделать в столь краткой форме для такого объемистого и богатого фактическим и идейным материалом труда, каким является его книга. Мы видим, что Стеншиб рисует нам следующую картину происхождения и эволюции Agnathi, этой крупной и чрезвычайно древней группы позвоночных животных. Еще в досилурийское время появляются первичные Ostracodermi, которые, поскольку мы можем представить себе их строение, должны были быть бесчелюстными позвоночными, питающимися при помощи сосущего ротового аппарата, животными с относительно большим числом энтодермальных жаберных мешков (от 10 до 15) и с вполне развитым внутренним и наружным костным скелетом. Для нас наибольший интерес представляет то обстоятельство, что у всех этих форм, т. е. у всех Cephalaspidomorphi и Pteraspidomorphi, составляющих вместе группу Ostracodermi, или, как их еще иначе называют, «панцырных рыб», имелось, по крайней мере, две пары вполне развитых и, стало быть, функционирующих пребранхиаль-ных жаберных щелей (по терминологии Стеншиб, спиракулярная и преман-дибулярная жаберная щель). Таким'образом, наше утверждение, принятое нами на основании сравнительно эмбриологических исследований, что предки современных позвоночных некогда имели функционирующие пребранхиальные жаберные щели, дуги, мускулы и т. п., всецело подтверждается позднейшими палеонтологическими наблюдениями (ср. Стеншиб, 1927, А. Н. Северцов, 1916, 1917, 1925).
Стеншиб не дает нам точных указаний относительно образа жизни описываемых им ископаемых форм, но по некоторым данным можно думать, что это были бентонные животные с плоским телом, у которых дорзальная часть жаберных дуг была сращена с черепом.
Теперь я должен обратиться к тем сомнениям, которые возникли у меня при чтении филогенетических выводов автора, сомнениям, которые, однако, оговорюсь тут же, отнюдь не умаляют ценности его фактических исследований. Я рассмотрю сначала вопрос, имеющий непосредственное отношение к моим личным исследованиям.
Стеншиб считает, что у палеаспид не имелось жаберных щелей, расположенных впереди от премандибулярной (второй пребранхиальной) жаберной щели, и в этом пункте, мне кажется, он совершенно прав. Но он счи
11*
164
ГЛАВА VI
тает, что и у предков круглоротых тоже не имелось этих передних жаберных мешков, и в этом своем утверждении он ссылается на наблюдение Н. Гольмгрена,1 что премандибулярное хрящевое образование у пескоройки (по моей терминологии—третья пребранхиальная жаберная дуга) превращается при метаморфозе пескоройки в миногу не в передний хрящевой бугор подглазничного хряща («anterior pillar» подглазничной дуги Вудланда)2, но в cartilago lateralis anterior, лежащий непосредственно позади рта у взрослой миноги. Из этого наблюдения Гольмгрена Стеншиб выводит, что у предков циклостом впереди от пре-мандибулярной дуги вообще не существовало никаких скелетных элементов, которые можно было бы истолковать как пребранхиальные жаберные дуги. При этом он, однако, не принимает во внимание того, что моя теория пребранхиальных бранхиомеров круглоротых содержит собственно два, и притом далеко не равноценных, положения: во-первых, утверждение, что у общих предков круглоротых (энтобранхиат) в свое время существовало по меньшей мере четыре функционирующих пребранхиальных бранхиомера, и во-вторых, предположение, что у непосредственных предков циклостом, в частности у предков миног, образованная из пребранхиальных дуг жаберная решетка простиралась далеко вперед, достигая области рта. Первое из этих положений является непосредственным выводом, вытекающим из моих эмбриологических наблюдений над развитием висцеральной мускулатуры пескоройки (ср. рис. 7, 8, 9); второе представляет собою лишь гипотезу, построенную на основании сравнения строения ротового скелета взрослой миноги с таковым же у пескоройки и на основании сравнения передних частей висцерального скелета с его задними (бранхиальными) частями у той же миноги.
Если правильны наблюдения Н. Гольмгрена, что третья премандибу-лярная дуга пескоройки смещается во время метаморфоза в ростральном направлении и превращается в передний латеральный губной хрящ взрослой миноги (а в правильности наблюдения такого точного исследователя, каким является Гольмгрен, у нас вряд ли имеются основания сомневаться), то мое отождествление этой дуги с переднею частью подглазничного хряща (с «anterior pillar» подглазничной дуги Вудланда) неверно; при этом теряет под собою почву и мое толкование «anterior pillar» и двух наиболее передних губных хрящей взрослой миноги. Вполне возможно, что жаберная решетка у предков миног, т. е. у протоциклостом, не простиралась так далеко вперед, как я это принимал в моих работах 1916 и 1917 гг.
Наблюдения Гольмгрена не меняют, однако, ни в какой мере тех фактов, на основании которых я построил свою теорию передних бранхиомеров. Факты эти заключаются в том, что, как это уже отмечено выше, в передней части головы пескоройки наблюдается на ранних стадиях развития закладка сегментов висцеральной мускулатуры, которые при этом имеют совершенного же строение, что и лежащие позади них настоящие жаберные сегменты. Каждый из этих сегментов состоит из одного констриктора и одного аддуктора, причем передние сегменты висцеральной мускулатуры вполне гомодинамны задним, т. е. истинным жаберным сегментам и образуют с ними одно непрерывное целое. Таким образом, мы находим, что в передней части головы молодой пескоройки висцеральная жаберная мускулатура простирается вплоть до самого рта (ср. рис. 7, 8 и 9, A. pb. 1—4; С. pb. 1—4). Задним мускулам соответствуют зачатки двух жаберных мешков (рис. 7, pb. 2) и третьей более или менее рудиментарной жаберной дуги (рис. 9); для четвертого пребранхиального бранхиомера мы
1 Это наблюдение опубликовано Гольмгреном лишь в 1936 г. (1. с.)
’Woodland W. N. Е., On the supposed Gnathostome ancestry of the Marsi-pobranchii etc., Anat. Anz., Bd. 53, 1913.
ЭВОЛЮЦИЯ EHT0BBANCH1ATA
165
не находим соответствующей хрящевой висцеральной дуги, и позади него не имеется зачатка жаберного мешка. Я считаю, однако, что у предков циклостом это образование должно было быть развито, хотя у современных нам форм оно онтогенетически уже не закладывается.
В своих работах 1916 и 1917 гг. я еще принимал, что самая передняя из этих дужек представлена у взрослой миноги первым латеральным губным хрящем; теперь же, после наблюдений Гольмгрена, которому удалось исследовать стадии метаморфоза пескоройки при ее превращении в миногу, т. е. стадии, которые я тогда не имел возможности получить, я этого уже не могу утверждать, во всяком случае не могу До тех пор, пока мне не
удастся самому точно проследить развитие мускульных и скелетных образований этой области при превращении пескоройки в миногу.
Так как на ранних стадиях развития третий пребран-! хиальный хрящ (рис. 9, A. pbr. 3) соответствует у пескоройки третьему пре-. бранхиальному мускульному сегменту (рис. 9, A. pb. 3, С. pb. 3), то мы должны признать, что он представляет собою не что иное, как третью пре-бранхиальную жаберную дугу. Его перемещение в ростральном направлении на более поздних стадиях развития, т. е. во время метаморфоза, не имеет никакого отношения к его гомологизации, ибо в противном случае, т. е. если бы такое перемещение имело морфологическое значение, нам пришлось бы считать, что и югулярный брюшной плавник акантоптеригий, резко сдвигаясь во время своего онтогенетического развития вперед, этим самым уже становится не гомологичным брюшному плавнику примитивных костистых рыб, например сельдей.
Резюмируя все сказанное, мы можем изобразить бранхиомерию предков миног в следующий таблице (табл. 2):
Т аб лица 2
Arcus visceralis 1 ? М. constrictor praebranchialis 4=М. buccalis anterior ex-ternus пескоройки M. adductor praebranchialis 4=M. buccalis anterior profundus пескоройки
Arcus visceralis 2 ! Arcus praebranchialis 3 пескоройки, которая у взрослого животного становится, по Гольмгрену, cartilago 1а-bialis anterior M. constrictor praebranchialis 3 = M. buccalis posterior ex-ternus пескоройки M. adductor praebranchialis 3 = M. buccalis posterior profundus пескоройки
Arcus visceralis 3 Arcus.praebranchialis 2 (processus styloideus у миноги) M. constrictor praebranchialis 2 = M. constrictor velaris M. adductor praebranchialis 2 = M. velaris profundi
Arcus visceralis 4 Arcus praebranchialis 1 (ex-trahyale Паркера, передняя дуга жаберной решетки миноги) M. constrictor praebranchialis I M. adductor praebranchialis 1
Arcus visceralis 5 Arcus branchialis 1 M. constrictor branchialis 1
М. adductor branchialis 1
; Таким образом, эмбриологические наблюдения приводят нас к выводу, что у предков круглоротых имелось по крайней мере четыре функцио-; нирующих пребранхиальных бранхиомера. Если мы примем, что Palaea-: spida (и, по всей вероятности, все Ostracodermi) и круглоротые происходят общих энтобранхиальных предков, а это предположение после открытий Стеншиб является, по моему мнению, единственно верным, то нам при-s детсятакже принять,что Cephalaspidae, у которых, согласно Стеншиб, были развиты только два пребранхиальных бранхиомера, утратили в течение
/66
ГЛАВА VI
Рис. 7, 8 и 9. Реконструкции (по сагиттальным срезам) трех последующих стадий развития висцерального аппарата и головных нервов Ammocoetes.
Рис. 7. Относительно ранняя стадия развития Ammocoetes, когда происходит первая закладка сегментов висцеральной мускулатуры (A.pb.2, А.рЬ. 3, С. pb. 1—С. pb. 3, С. b. 1—СЬ. 4) и еще не прорвавшихся наружу эктодермальных жаберных мешков (pb.l, pb.2, Ь.1—Ь.7).
Рис. 8. Более поздняя стадия развития Ammocoetes, где развитие висцеральной мускулатуры несколько продвинулось вперед.
Рис. 9. Еще более поздняя стадия развития Ammocoetes, на которой сегментация висцеральной мускулатуры в передней части головы уже вполне ясно выражена.
А.Ь.1—АЬ.З—mm. adductores branchiales функционирующих жаберных мешков. A.br. 1— А.Ьг.7—зачатки м>аберных дуг Ammocoetes (Petromyzon); A.pbr.l.—A.pbr. 3—преджаберные висцеральные дуги 1—3; A. pb. 1—A. pb. 4—mm. adductores praebranchiales четырех передних (предслуховых) бранхиомеров головы; Au.—слуховой пузырек; b. 1—Ь.7—жаберные мешки 1—7; С. аи.—слуховая капсула; c.b. 1 — с.Ь.6 — mm. constrictores branchiales функционирующих жаберных мешков пескоройки (миноги); С.pb.l — C.pb.4 — mm. constrictores praebrgnchiales
ЭВОЛЮЦИЯ ENTOBRANCHIATA
167
филогенеза два самых передних бранхиомера. Другими словами, мы считаем, что у гипотетических форм, от которых некогда ответвились в одну сторону Cephalaspidae, а в другую—Cyclostomata, имелось, подобно тому, что мы видим у молодых личинок миног (пескороек), четыре пре-бранхиальных бранхиомера (т. е. четыре жаберных мешка, четыре жаберные щели, четыре жаберных мускульных сегмента и четыре жаберные дуги). Во время дальнейшей эволюции Cephalaspidae два самых передних из этих бранхиомеров редуцировались.
Мы упомянули при описании организации Cephalaspidae о том, что у этих ископаемых форм уже образовались грудные плавники. Стеншиб утверждает, что их костный плечевой пояс был сращен с затылочной областью черепа. У Cephalaspis lyelli эти плавники, напоминающие по своему строению плавники кита, были сплошь покрыты чешуями; плавниковые чешуи при этом, вероятно, не были развиты. Повидимому, плавники эти представляли собою весьма своеобразные и резко специализованные образования. От всяких попыток гомологизировать их с грудными плавниками рыб нам, конечно, приходится отказаться в настоящее время, так как пока нам еще не известно строение их скелета и еще не найдены переходные формы. При современном уровне наших знаний мы не можем также решить, развились ли грудные плавники Palaeaspidae и грудйые плавники рыб из каких-либо ориментов, существовавших у их общих предков, или нет. Факт, что ни Cyclostomata, ни Palaeospondyloidea, ни Anaspida1 не имеют вообще плавников, говорит, как будто, против такого предположения.
Стеншиб считает, что предки циклостом имели наружный и внутренний костный скелет, образованный типичным головным щитом с приросшими к нему жаберными дугами (интербранхиальные перегородки) и что поэтому современные круглоротые, имеющие, как известно, лишь один хрящевой скелет, должны представлять собой регрессивную линию филогенетического развития, идущую от остракодерм. Нам кажется, однако, мало вероятным, чтобы нектонные циклостомы (миноги и миксины) могли непосредственно произойти от таки^ резко специали-зованных форм, какими являются непосредственные предки Cephala-spidomorphi, к которым принадлежат Osteostraci, и Pteraspidomorphi,
1 Шипы, которые встречаются у Anaspida и которые Киэр истолковывает как плавники, отнюдь не являются таковыми (см. Sewertzoff A. N., Einige Berner-kungen uber die systematischen Beziehungen der Anaspida usw., Palaeont. Zschr., 1928).
четырех передних бранхиомеров головы; Ch.—хорда; М.—зачаток рта, на более поздних стадиях рот. Nh.—назогипофизарное отверстие; Ос.—глаз; Opii.—n. ophthalmicus profundus trigemini; рудиментарный висцеральный нерв четвертого преджаберного нейромера; pb.l, pb. 2—первый и второй жаберные мешки; Pch.—парахордалии; Pch. -а.—передние парахордалии; г. md. V2—г. mandibularis n. trigemini 2; r.max.Vs—г. maxillaris n.trigemini 2; VII—n. facialis; IX—n. glossopharyngeus; X.l.—n. lateralis vagi; Xi—Xe—бранхиальные ганглии блуждающего нерва (из А. Н. Северцова, 1916).
Если мы сравним между собою реконструкции 7, 8 и 9, то увидим, что у круглоротых (пескоройки) на относительно ранних стадиях развития имеется два преджаберных мешка; они лежат впереди от первой функционирующей жаберной щели (глоссофарингеальная щель; жаберные мешки pb 1, pb. 2, рис. 7); здесь имеется также четыре вполне развитых преджаберных висцеральных мускульных сегмента, из которых каждый состоит из одного т. constrictor (C.pb. 1 — С. pb. 4) и из одного т. adductor (A.pb. 1 — A. pb. 4) (ср. схему, изображенную на рис. 6). Мы находим, кроме того, что трем задним мускульным сегментам соответствуют прохондральные преджаберные висцеральные дуги (A. pbr. 1, A. pbr. 2, A.pbr.3, рис. 9). Из совокупности всех этих факторов мы заключаем, что у предков циклостом имелось во всяком случае четыре преджаберных бранхиомера, расположенных в переднем отделе головы, вполне гомодинамных бранхиомерам, расположенным в задней части головы. Каждый из этих бранхиомеров состоял из хрящевой висцеральной дуги, из сегмента висцеральной мускулатуры (mm. adductor и constrictor), из жаберного мешка и жаберной щели и из сегментального висцерального нерва. Этот вывод находится в полном согласии с результатами наших исследований над развитием бранхиомеров передней части головы у примитивных челюстных позвоночных (эласмобранхий). К только что изложенном взгляду на бранхиомерию общих предков круглоротых мы пришли впервые еще в 1916—1917 гг., исследуя историю развития циклостом. В этой работе я еще принимал, что преджаберная дуга представлена у взрослой миноги губным хрящом (Cart, lateralis). Но так как для этого бранхиомера я не йашел ни соответствующего висцерального мускула, ни не вызывающего сомнений рудимента соответственного висцерального нерва, то я думаю, что будет более осторожным считать, что у предков круглоротых было не более четырех преджаберных бранхиомеров. *
168
ГЛАВА VI
к которым принадлежат Heterostraci. Нам кажется также, что различия между миногами и миксинами далекр не так значительны, как это думает Стеншиб. Он считает, например, что задний дорзальный губной хрящ миноги является висцеральным образованием, тогда как ростральный хрящ миксин представляет собою «этмоидальное» образование. Нам, однако, это положение далеко не представляется доказанным: нельзя и неправильно противопоставлять cartilage dorsalis posterior миноги, как висцеральное образование, ростральному хрящу (palatine-(-«cornual cartilage» Bdellostoma, Гудрич, 1909) миксин. Эмбриональное развитие этого последнего хряща нам неизвестно, но его положение у Bdellostoma таково же, как и положение задних дорзальных губных хрящей у миноги. Различие между этими двумя образованиями заключается лишь в том, что ростальный хрящ сросся с черепной коробкой (parachordalia anteriora mihi) и что назоги-пофизарный проток у Bdellostoma гораздо длиннее, чем у миног. Оба эти различия между миногами и миксинами представляются мне явлениями вторичного характера; мне кажется, что до тех пор, пока нам точно неизвестна история развития головного скелета миксин, мы не можем делать каких-либо определенных выводов из только что приведенных фактов, которые, как мы видим, допускают различное толкование.
Некоторые анатомические факты говорят, повидимому, в пользу того мнения, что миноги и миксины ближе стоят по отношению друг к другу, чем по отношению к Osteostraci и Heterostraci. Едва ли можно принять, например, что одинаково и весьма своеобразно построенные роговые зубы миног и миксин могли развиться независимо друг от друга, а не быть унаследованными от какого-либо миногоподобного или бделлостомоподобного общего предка.1 Равным образом и общий тип строения ротового скелета у этих форм и в особенности наличие у них ротового кольца и непарных губных хрящей, а также особенности механизма сосательного аппарата— все это скорее указывает на происхождение миног и миксин от какого-то общего предка, отнюдь не похожего по своим признакам на описанных выше Ostracodermi.
Особенно трудно объяснимым, с точки зрения гипотезы Стеншиб, является факт, что современные нам круглоротые обладают жаберной решеткой, состоящей из тонких свободно лежащих хрящевых перемычек; очень мудрено себе представить, чтобы из слабо выраженных жаберных перегородок типа Cephalaspidae могло развиться такое гибкое образование, каким является скелет жаберной решетки миног и миксин.
Стеншиб принимает также, что костный скелет позвоночных филогенетически возник весьма рано: он, вероятно, образовался уже у общих предков гнатостом и циклостом, и его отсутствие у современных круглоротых и хрящевых рыб объясняется, по Стеншиб, его вторичной редукцией. Но эта гипотеза была бы приемлема только в том случае, если бы на основании общей суммы признаков организации этих форм было бы бесспорно доказано, что Cyclostomata действительно происходят от Osteostraci и Heterostraci; однако это предположение не является достаточно обоснованным, и мы, наоборот, думаем, что много фактов говорит против него. •
По нашему мнению, можно было бы скорее выставить следующую гипотезу о происхождении и филогенетическом соотношении этих спорных форм: можно, например, думать, что общие предки всех энтобран-хиальных нектонных позвоночных (я назвал бы их
1 Среди круглоротых Bdellostomatidae мне кажутся более примитивными формами, чем миксины; поэтому я не придавал бы такого большого значения сходству, которое существует, по мнению Стеншиб, между жаберным аппаратом Drepanaspidae и миксин.
ЭВОЛЮЦИЯ ENTOBRANCHIATA
169
скорее индиферентным именем «энтобранхиат», чем, как это делает Стеншиб, «панцирными рыбами»—Ostracodermi), были не к тонны ми животными с внутренним хрящевым скелетом без костей, с бесчелюстным ртом и пассивным образом питания^ с относительно большим числом жаберных мешков и щелей, из которых самые передние были расположены непосредственно позади ротового отверстия (пребранхиальные жаберные мешки или щели 1—4). Представляется также весьма вероятным, что эти гипотетические формы обладали также и поверхностным головным скелетом, состоящим из слизистого хряща, наподобие того, который можно наблюдать у пескоройки (Паттен, Гольмгрен).
Одни из потомков этих нектонных предков энтобранхиат перешли к бенто иному образу жизни и приобрели плоскую форму тела, у них редуцировались при этом два самых передних жаберных мешка и щели (первый и второй бранхиомер), а состоящий из слизистого хряща скелет головы развился прогрессивно и превратился в твердый, состоящий из дентина и кости панцырь; кроме того, у них образовались окостенения внутреннего скелета. Так, вероятно, произошла группа бентонных предков Osteostraci и Heterostraci (Ostracodermi primitivi Стеншиб).
Мне кажется, что к этой же группе вымерших бентонных панцырных энтобранхиат принадлежат и все Coelolepidae. Мнение Стеншиб, который считает, что Coelolepidae стояли. в близких родстренных отношениях к группе Drepanaspidae и Pteraspidae, я считаю совершенно правильным, но его гипотеза, что кожгйяй скелет Coelolepidae, состоящий из плакоидных чешуи, мог образоваться путем дегенерации за счет распада сплошного, состоящего из Дентина и кости панцыря типа Cephalaspidae и Pteraspidae, кажется мне мало вероятной. Невольно возникает вопрос, почему костный панцырь, покрывающий собой хрящевой череп, раз он становится ненужным, не просто дегенерирует, но превращается в новое образование, а именно, в плакоидные чешуи? Мне кажется, что мнение Трэквера, рассматривающего кожный скелет Coelolepidae (Thelodus) как образование примитивное, покоится на более веских основанях, чем эта гипотеза, и более вероятным является обратное предположение, что панцырь остальных Ostracodermi образовался путем срастания чешуи.
Если мы в своих выводах будем исходить, как это делает Стеншиб, из строения скелета головы Ostracodermi, то можно легко притти к мысли, что головной скелет образовался не путем срастания многочисленных дентинных или костных чешуи типа Thelodus или эласмобранхий, но что он развился путем распада из сплошного костного панцыря. Если же вместо этого исходить из фактов, которые наблюдаются при исследовании строения кожного скелета туловища водных челюстных позвоночных, у которых подвижность отдельных скелетных элементов является необходимой предпосылкой двигательной функции тела, то гораздо более вероятным становится взгляд Гудрича, по которому весь кожный скелет головы и туловища первоначально состоял из подвижных кожных чешуи.
После всего только что сказанного мы можем, мне кажется, принять, что от некоторых примитивных нектонных энтобранхиат (которые нам палеонтологически, к сожалению, еще не известны) ответвились две линии потомков: во-первых, предки всех Ostracodermi (как Osteostraci, так и Heterostraci), которые перешли к бентонному образу жизни, причем у них развился прочный головной панцырь, дорзовентрально сплющенная форма тела и другие признаки; во-вторых, Protocyclostomata, которые, по нашему мнению, сохранили нектонный образ жизни примитивных энтобранхиат, но развились прогрессивно в другом направлении, а именно, изменили свой способ питания: животные перешли от пассивного питания к активному,
170
ГЛАВА VI
причем у них из передних пребранхиальных частей висцерального скелета и из соответствующих этим частям висцеральных мускулов образовался сложно построенный скелет и мускулатура бесчелюстного сосущего рта типа круглоротых.
Большим затруднением для гипотезы, предполагающей, что круглоротые произошли от Osteostraci и Heterostraci, является то, что для нас остается непонятным, каким образом и из каких именно скелетных частей Ostracodermi мог развиться сложно построенный ротовой скелет миног и миксин. Нам приходится либо принять, что многочисленные мускулы и хрящи этого ротового скелета являются новообразованием, либо предположить, что у Ostracodermi впереди первой интербранхиальной перегородки некогда существовал сложно построенный и, вероятно, еще хрящевой ротовой скелет, что, по исследованиям того же Стеншиб, является довольно невероятным. При сильном окостенении, имевшем место в черепе Palaeaspidae, представляется совершенно невероятным, чтобы такой скелет со всеми своими нервами, сосудами и т. п. не оставил после себя никаких следов или чтобы эти следы ускользнули от острого взгляда Стеншиб.
В каких соотношениях с только что перечисленными нами группами стоят Anaspida и Palaeospondyloidea и действительно ли Anaspida принадлежат к Cyclostomata, как это думают Стеншиб и Киэр, решить с достоверностью мы еще, мне кажется, не можем. Я лично думаю, что^далеко не исключена возможность, что Anaspida не принадлежат к ряду круглоротых, а скорее относятся к челюстным рыбам. Это последнее наше предположение стоит в резком противоречии со взглядами Стеншиб и Киэра, и поэтому мы высказываем его с большой осторожностью, ибо мнение таких крупных исследователей, какими являются Стеншиб и Киэр, работам которых мы в значительной степени обязаны расширением и уточнением наших сведений относительно строения наиболее древних из известных нам позвоночных, мы, конечно, не можем не придавать большого значения. Тем не менее отметим, что наш взгляд опирается на довольно обстоятельное и притом добытое личными исследованиями знание истории развития циклостом; только поэтому мы позволяем себе его здесь высказать.
Только что изложенные исследования обоих скандинавских ученых говорят нам о том, что в силурийские и в до-силурийские времена существовала богатая фауна весьма примитивных энтобранхиальных позвоночных, остатки которых (циклостомы) сохранились вплоть до нашего времени. Главное отличие нашей точки зрения от точки зрения Стеншиб заключается в том, что он считает Osteostraci (и Heterostraci) самыми примитивными формами из всех форм этой «энтобранхиальной эпохи» и переносит их характерные признаки на группу гипотетических первичных Ostracodermi, в то время как мы принимаем, что в эту «энтобранхиальную эпоху», наряду с вышеописанными специализованными, приспособленными кбентонному образу жизни панцырными формами,1 вероятно, существовали также и другие, менее специализованные формы, характеризующиеся не костным, но хрящевым скелетом и поэтому не сохранившиеся в ископаемом состоянии.
Мы думаем также, что именно такие специализованные формы, какими были Ostracodermi, и должны были непременно вымереть с течением времени; наоборот, должны были выжить и сохраниться до нашей эпохи потомки менее специализованных нектонных энтобранхиат— современные нам круглоротые.
1 Которые именно вследствие своих приспособлений к жизни на дне (плоская форма тела, электрические органы, строение головного щита и т. п.) сохранились в ископаемом состоянии.
ЭВОЛЮЦИЯ ENTOBRANCHIATA
171
ВЫВОДЫ
Резюмируя все сказанное, мы представляем себе следующим образом филогенез только что рассмотренных нами форм (см. родословное древо, схема XV на стр. 213 и 247.
1. От протокраниат произошла группа не кт о н-н ы х, пассивно питающихся энтобранхиат с о т ночи с л о м эктодермальных поверхностно расположен-хрящевой жаберной решеткой.
— этих энтобранхиат эволюировали
сите л ь но большим жаберных мешков и с н
о й
2. Потомки
двух различных направлениях: у одной линии о т о м к о ем раз
в п н сосущий рот—это были форм чаем именем протоциклос от них
сосущие кровь миноги и м нымхрящевым
3. У второй линии потом вился костный панцырь;
вался за счет покров таким образом
в в
в
и
ротовое л и с ь
к о л ь ц роговые
свою
очередь
с к е л е т о
п
м.
о сохранилось, зубы ы, к
о м о и к с
т Р и
о в
И X
3 о п а н ц ы р я; Ostracodermi primitivi способу питания другим м е произошли
сто у
о т (Р
3 о и н
а
н
и образовался орые мы об rotocyclosto шли
ы с и
Н О
некто х а’р а
х
о з н а-m a t а); иные, к т е р-
э н т о б р
к ротовой с
ЦЫ X
т р а з-о б р а-
циклостом. От э все известные
а : к । костей первичные перешли к путем, чем тих первичных форм нам Ostracodermi.
н х и а е л е т
i кожного бентонные активно му это имело
ГЛАВА VII
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ ПОЗВОНОЧНЫХ
ЭВОЛЮЦИЯ ХАРАКТЕРНЫХ ПРИЗНАКОВ EGTOBRANCHIATA
Исследуя эволюцию предков круглоротых, мы видели, что направление их филогенетического развития определяется двумя чрезвычайно важными в проспективном отношении моментами, а именно: 1) повышением дыхательной функции этих животных и 2) их переходом от пассивного питания к активному. Прогрессивное развитие дыхательной функции заключалось в образовании эктодермальных жаберных мешков, жаберных лепестков и их сосудов и в развитии поверхностно расположенного жаберного скелета (жаберной решетки). Прогрессивное развитие функции питания заключалось в закрытии передних жаберных щелей, превращении передних элементов висцерального скелета в части сосущего ротового скелета, развитии роговых зубов (циклостомы) или покровных зубных пластинок («панцырные рыбы»). Все эти изменения оказали решительное влияние на последующую эволюцию энтобранхиат и их потомков,iCyclo-stomata и Ostracodermi. Чрезвычайно интересно, что при эволюции другой линии потомков протокраниат, а именно энтобранхиат, повышение функции дыхания и переход от пассивного питания к ‘активному также явились двумя определяющими моментами в их прогрессивном филогенетическом развитии.
Эмбриологические исследования (А. Гетте,1 Т. Моров,* 2 А. Н. Северцов3) показали, что жабры всех низших рыб (селахий, хрящевых ганоидов, кистеперых и дипной) закладываются в виде эктодермальных жаберных нитей. Нити эти (А. Н. Северцов, 1923) сидят в два ряда на наружной поверхности каждой жаберной дуги (рис. 26 и 27, Аг. Ь.). Жаберные сосуды здесь распределяются по иному типу, чем у циклостом: вдоль каждой жаберной, дуги проходит жаберная артерия (рис. 26 и 27, а. Ь.), несущая венозную кровь от сердца, и две жаберные вены(рис. 26, v. b. a., v. b. р.), несущие артериальную кровь, окислившуюся в капиллярных сосудах жаберных нитей. Основываясь на наших сравнительно морфологических исследованиях, мы можем принять, что здесь мы имеем перед собой рекапитуляцию филогенетического развития жаберного аппарата предков гнатостом и что сложно построенные жаберные нити и сосудистая система развились из относительно просто построенного жаберного аппарата протокраниат. Физиологическое значение этих изменений нам совершенно понятно: дыха
8 Goette A.; Uber die Kiemen der Fische, Zschr; f. wiss. Zool.^ Bd. 69, 1911.
2 Mo г о И Th., Uber die Entwicklung der Kiemen bei Fischen, Arch. f. mikr. Anat., Bd. 64, 1904.
8 S e w e r t z о f f A. N;-, Die Entwicklung der Kiemen und Kiemenbogengefasse der Fische, Zschr. f. wiss. Zool,, 1923.
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
173
тельная поверхность вследствие образования многочисленных эктодермальных жаберных нитей (лепестков, рис. 26) сильно увеличилась, вследствие чего резко интенсифицировалось окисление крови, иными словами, функция дыхания стала совершеннее. Изменения
Рис. 26. Графическая реконструкция (по фронтальным срезам) средней части первой жаберной дуги с двумя жаберными лепестками (Ь. а. и Ь. р.) довольно позднего зародыша Pristiurus melanostomus.
a. an. а.—артерия сосудистой петли (ansae anterioris) переднего жаберного лепестка; а. ап. р.—артерия сосудистой петли (ansae posterioris) заднего жаберного лепестка; a. b.—arteria arcus branchia-11 s; ar. b.—хрящевая жаберная дуга; b. a.—жаберный лепесток (бранхион) переднего ряда жаберной дуги; Ь. р.—жаберный лепесток (бранхион) заднего ряда жаберной дуги; са.—ср.—поперечные капиллярные сосуды жаберного лепестка переднего и заднего ряда; f. b. р.—зачаток задней жаберной нити; ЬГ, М"—жаберные мускулы; N—нерв жаберной дуги; г. Ь.—жаберный луч; s—septum branchiale; V. an. p.—вена ansae posterioris; v. b. a.—vena branchialis anterior; V. b. p.—vena branchialis posterior (из A. H. Северцова, 1923).
Рис. 27. Графическая реконструкция пары жаберных лепестков (бранхионов) из средней части второй жаберной дуги молодого экземпляра стерляди.
Обозначения те же, что и на рис. 26. Этот рисунок дополняет собою рис. 26 в том отношении, что здесь оба бранхиона (х—z и х'—-z') одинаково развиты, точно так же» как и оба жаберных луча (г. Ь. а. и г. Ь. р) (первичное состояние). Наличие только одной vena branchialis communis (v. b. с.) является вторичным приобретением сем. Chondrosteoidei (из А. Н. Северпова, 1923).
эти физиологически подобны изменениям, которые имели место у предков Cyclostomata и Ostracodermi, но происходят они иным путем, чем у этих последних: у энтобранхиат образовались эктодермальные мешки и щели, у челюстных позвон очны х— эктодермальные жаберные нити и лепестки. Так
174
ГЛАВА VII
как именно изменения в органах дыхания являются наиболее характер, ними для предков первичных челюстных, то мы и обозначили эту группу животных общим именем эктобранхиат.
A.prbrt A.prbr.2 А.рг.ЬгР
Рис. 28. Схематическая реконструкция строения передней части головы гипотетических Ectobranchi ata.
A.br.l—A.br.13 — жаберные Луги 1—13; A.pr.br. 1—A.pr.br. 4 — прелжаберные луги 1—4; Als— алисфеноидный хрящ; Аг. ос.—затылочные луги задней части головы; Ьг. 1—Ьг. 13—жаберные щели 1—13; Ch.—хорда; С. ап—хрящевая слуховая капсула; Cart. апп.—хрящевое около-ротовое кольцо со щупальцами; С. nas.—хрящевая носовая капсула; ср.—copulae жаберного скелета; сг. Ьг. — ceratobranchialia висцеральных дуг; Dien—промежуточный мозг; ер. Ьг.— epibranchialia висцеральных дуг; Ьр. Ьг.—hypobranchialia висцеральных луг; Нур.—гипофиз; М.—рот; Msen.—средний мозг; М. sp.—спинной мозг; Mten.—продолговатый мозг; Nas.— носовая ямка; Ос.—глаз; pb. 1—pb. 3—пребранхиальные жаберные щели 1 — 3; Pch.—parachor-dalia; ph. br.—pharyngobranchialia висцеральных луг; Tien.—передний мозг; Tr. сг.—трабекулы черепа. На этой схеме обратить внимание на число пребранхиальных жаберных щелей (3) и пребранхиальных жаберных дут (4), а также на расчленение как истинных, так и пребранхиальных жаберных дуг. Число истинных жаберных дуг гипотетично; мы только зиаем, что их было во всяком случае больше восьми; на нашу схему (рис. 5) мы нанесли 13 жаберных дуг. Гипотетичным является и присутствие хрящевого околоротового кольца. Только cirri сохранились у некоторых Gnathostomata. В черепе обратить внимание на срастание трабекул с парахордалиями, а также на.образование алис-феноидного хряща и затылочных дуг. В целях упрощения схемы мы не нанесли на нее диплоспон-дильных дуг позвоночника (из А. Н. Северцова, 1917).
п interarc MMecarc. dors, lot
Рис. 29. А. В. С. Развитие жаберной дуги и ее мускулатуры у селахий (сильно схематизировано).
Arc.br.—зачаток жаберной дуги на ранней сталии развитая; Ceratobr.—ceratobranchjale; Epibr.—epi branch] ale; Hypobr.—bypobranchiale; MJam.br.—еще не диференпированный зачаток висцеральной мускулатуры жаберной дуги; М. adduct.br. — m. adductor branchialis; М.const, dors., М. const, vent., M. const, med.—m. m. constrictores dorsales, ventrales и mediales; M. interarc.— m. m. interarcuales; M. interbr.—m. interbranchialis; M. interarc.dors.med.—m. interarcualis dorsalis medialis; M. interarc. dors. lat.—m. interarcualis dorsalis lateralis; Phar. br. — pharyngobranchiale (из A. H. Северпова, 1923).
У эктобранхиат, так же как и у круглоротых, изменения дыхательных органов сопровождались важными изменениями в строении жаберного скелета: первоначально сплошные жаберные дуги протокраниат распались у эктобранхиат на отдельные палочковидные отрезки (рис. 28 и 29).
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
175
r.lab.V.3 г.1аЬ.2.г L. lab.sup
A.pr.br. 4-) ep.l.l
pb.3
A.pr.br.3
c.r.L.2
6
ер.1.2
pb.1 (spir)
pb.2 r.md.'V.j
ep.md.
(Pqu.)
части
Рис. 30. Графическая реконструкция передней висцерального аппарата позднего зародыша Acanthi as vulgaris, на которой видны рудиментарные пребран-хиальные жаберные щели (pb. 2 и pb. 3) и дуги (А.рг.
Ьг.З и А.pr.br.4).
Обозначения такие же, как и на рис. 28. Кроме того: ст. md.— ceratomandibulare; ер. md. (pqu)—epimandlbulare; L. lab. sup.— m. levator lab. super!oris; r.lab. Va—r.lab.V3—rami labiales trigemini V, и Vs; r. md.' Vs, r. md.' V3—rami mandibulares trigemini (V3) (из A. H. Северцова, 1925).
В предыдущем мы пришли к выводу, что жаберные дуги протокраниат имели вид тонких, гибких хрящевых палочек. Эту первичную структуру они сохранили и у круглоротых (пескоройка, минога) и у родственных этим последним форм. При изменении жабр у общих предков челюстных каждая жаберная дуга расчленилась на четыре части, подвижно сочлененных между собой связками. Части эти можно обозначить как pharyngo-branchiale, epibranchiale, ceratobranchiale и hypobranchiale (рис. 28 и 29, ph. br., ep.br.,cr. br., hp.br.). Вследствие этого расчленения подвижность каждого отрезка дуги сильно возросла,
В связи с изменениями, происшедшими в строении жаберного скелета, стоят и изменения в структуре жаберных мускулов. Мы знаем, что жаберные мускулы у эмбрионов круглоротых построены так же просто, как и у эмбрионов рыб, и имеют форму сплошных мускульных пластинок, лежащих рядом с жаберными дугами (рис. 29 А, М. lam. br.). Отсюда мы заключаем, что жаберные мускулы общих предков гнатостом и циклостом, т. е. протокраниат, имели форму поверхностно расположенных сплошных мускуль-
ных тяжей, сгибавших хрящевые жаберные дужки. У предков циклостом, т. е. у протоциклостом, из этих мускульных лент образовались поверхностные m. т. constrictores и глубоко лежащие т. т. adductores. У первичных челюстных диференцировка висцеральной мускулатуры пошла гораздо дальше: для каждой жаберной дуги образовалась уже не одна пара, но целый ряд отдельных мускулов: m. т. constrictores, т. т. leva-tores, т. т. adductores, in. т. interarcuales и т. д. (рис. 29, С).
Наш дальнейший вывод заключался в том, что у протокраниат имелось, кроме типичных жаберных щелей, функционирующих и у ныне живущих рыб, также инекоторое число пребранхиальных, т. е. впереди от них лежащих щелей и дуг (бранхиомеров), которые были расположены между первой жаберной дугой современных рыб (так называемой глоссофарингеальной дугой) и ротовым отверстием. Естественно возникает вопрос: были ли эти передние (пребранхиаль-ные) щели развиты также и у непосредственных предков гнатостом и были ли расчленены лежащие между ними жаберные дуги по тому же типу, как и ныне функционирующие жаберные дуги современных рыб?
Эмбриологические исследования дают нам положительный ответ на оба эти вопроса (А. Н. Северцов, 1923, 1926; А. Н. Северцов и Н. Н. Дислер, 1924). У эмбрионов селахий мы находим рудименты трех пребранхиальных жаберных щелей (рис. 30, pb. 1—3, ср. рис. 28, pb. 1—3), а также несомненные доказательства того, что жаберные дуги, лежавшие между этими щелями, уже были сегментированы и состояли из четырех частей. Гиоидная
176
ГЛАВА VII
Рис. 31. Череп Laemargus borealis. Вид сбоку. Хорошо видны губные хрящи (3-я и 4-я преджаберные висцеральные дуги, ер. 1.г+сг. 1.2, ер. l.i) и pharyngomandibulare (ph. т.)2-й преджаберной (мандибулярной) дуги; Нт.—hyomandibulare; Hyoid.—hyoideum; Р. qu.— palatoquadratum; с. M.—-cartilago Meckeli; p. pbs. 1. — proc, palatobasaiis lateralis (из Северцова и Дислера, 1924).
дуга, которая, как известно, лежит непосредственно впереди от первой функционирующей жаберной щели, состоит у селахий из вполне развитого hyo-mandibulare (s. epihyale) и из hyoideum (s. ceratohyale; рис. 31, Hyoid), а также из имеющихся лишь у некоторых селахий рудиментарного pharyn-gohyale и hypohyale (рис.- 36, ph. h., hp. h.). Рострально от этой дуги лежит рудиментарная первая пребранхиальная щель, или spiraculum [рис. 28 и 30, pb. l(spir.)]. Обычно принято думать, что челюстная дуга состоит лишь из epimandibulare (s. palatoquadratum) и ceratomandibulare (s. cartilago meckeli), но у некоторых акул имеется также и рудиментарное pharyn-gomandibulare (рис. 31, ph. m.; А. Н. Северцов и Н. Н. Дислер, 1924). Pharyngomandibulare было также найденоиу8сарЫгЬупсЬиз(А. Н. Северцов, 1923, 1925), a hypomandibulare—у Hexanchus (А. Н. Северцов).
У молодых эмбрионов селахий впереди от второй пребранхиальной дуги (челюстная дуга) лежит вторая рудиментарная пребранхиальная шель (рис. 30, pb. 2; ср. рис. 28 и 36). Впереди от этой щели находится третья пребранхиальная жаберная дуга (второй губной хрящ, cartilago labialis posterior; рис. 30, А. рг. Ьг. 3). У большинства акул дуга эта со-, стоит из двух элементов, а именно из epi- и ceratolabiale (рис. 30, ер.1. 2,с.г.1.2), но у Holocephali (Chimaera) имеется также и pharyngola-biale (рис. 36, ph., 1.2). Нако-
нец у эмбрионов селахий между задним и передним губными хрящами закладывается также и третья пребранхиальная жаберная щель (рис.
30, pb. 3).
Сравнение губных хрящей у акул и Holocephali приводит нас к выводу, что четвертая пребранхиальная дуга (самый передний губной хрящ) состояла у предков эласмобранхий по меньшей мере из двух отдельных элементов, а именно: из epilabiale и ceratolabiale (рис. 36, ep.l,, cr.lj. Результаты нашего исследования наглядно представлены в схеме, изображенной на рис. 36.
Мы видим, таким образом, что все пребранхиальные дуги были расчле-
нены у предков челюстных позвоночных по тому же самому типу, по которому расчленены функционирующие жаберные дуги у ныне живущих рыб, так что каждая из этих пребранхиальных дуг состояла из pharyngo-, epi-, cerato- и hypo-элементов (рис. 28, А. рг. br. 1—4). Между этими дугами мы нашли также рудименты передних (пребранхиальных) щелей, а поэтому мы можем принять, что у всех предков челюстных рыб, т. е. у всех э кто-бранхиат, передние бранхиомеры еще функционировали как органы дыхания. Присутствие спираку-лярной псевдобранхии, наблюдаемой у некоторых рыб вплоть до настоящего времени, конечно, также говорит в пользу этого взгляда. Расчленение пребранхиальных жаберных дуг по тому же типу, по которому расчленены и канонические жаберные дуги, было бы для нас совершенно непонят-
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ ПОЗВОНОЧНЫХ П7
ним, если бы мы не считали, что и эти дуги некогда функционировали в качестве органов дыхания, подобно типичным жаберным дугам.
Мы приходим, таким образом, к выводу, что у эктобранхиат функционирующий жаберный аппарат, состоявший из типично расчлененных бранхиомеров, доходил до самого рта. Это ставит перед нами вопрос о строении рта у эктобранхиат.
К сожалению, мы не можем ответить на этот вопрос исчерпывающим образом, ибо эмбриологические исследования низших рыб не открыли нам до сих пор никаких следов существования здесь примитивного ротового скелета, который, по всей вероятности, некогда лежал рострально от самой передней из пребранхиальных висцеральных дуг.
В предыдущем мы пришли к выводу, что ротовой скелет протокраниат и ближайших предков круглоротых (энтобранхиат), у которых ряд бранхиомеров также начинался непосредственно позади рта, состоял лишь из хрящевого ротового кольца (рис. 11, Cart, ann.) типа современных кругло-ротых или челюстных и что эти гипотетические протокраниаты и энтобран-хиаты были животными с пассивным питанием. Так как у эктобранхиат ряд жаберных щелей (и жаберных дуг) также начинался непосредственно позади ротового отверстия, то нам очень трудно себе представить, чтобы у них уже мог образоваться приспособленный к активному питанию и более или менее сложно построенный ротовой скелет с диференцировавшейся мускулатурой: гораздо вероятнее предположение, что эти животные все еще питались пассивно.
Мы знаем, что ротовое отверстие ланцетника и миксин усажено рядом хрящевых выростов и что эти выросты (cirri) у миног иннервируются ветвями тройничного и лицевого нерва. Такие же cirri, расположенные впереди рта и иннервируемые теми же нервами, что и у миног, существуют также у низших хрящевых ганоидов, и это также несомненно говорит в пользу только что высказанного нами взгляда относительно способа питания примитивных эктобранхиат. Поэтому мы придерживаемся гипотезы, что древнейшие эктобранхиаты имели усаженное cirri ротовое кольцо (схема на рис. 28, Cart, ann.) и ч т о питание.их, подобно питанию современных бесчерепных, все еще носило пассивный характер.
Затем мы пришли к выводу, что между протокраниатами и челюстными позвоночными, т.е. гнатостомами в собственном смысле слова, некогда существовала промежуточная группа рыб (эктобранхиаты), у которой функция дыхания развилась прогрессивно (увеличение дыхательной поверхности путем образования эктодермальных жаберных нитей, многочисленные жаберные дуги, состоящие каждая из четырех элементов: pharyngo-, epi-, cerato- и hypo-branchialia). Эктобранхиаты отличались от гнатостом в собственном смысле слова тем, что у них п ре б ран хиа льные бранхиомеры еще .функционировали как органы дыхания и что жаберные щели этих передних сегментов еще оставались открытыми.
Не вдаваясь в подробную реконструкцию всех признаков строения i эктобранхиат, отметим еще некоторые наиболее существенные особенности их организации. В предыдущем мы уже указали на то, что у энтобранхиат и у их потомков, циклостом (а также, по всей вероятности, и у Osteostraci) существовал лишь парахордальный череп.1 Часть скелета, соответствующая по своему первоначальному положению trabeculae
1 Т. е. только передние и задние парахордальные хрящи и cornua parachordalia А. Н. Северцов, 1916).
Морфологические закономерности
12
178
ГЛАВА VII
cranii гнатостом, лежала у энтобранхиат впереди мозга вследствие разра^ станин преназального (постгипофизарного) отдела головы и не принимала участия в образовании neurocranium. Наоборот, у эктобранхиат, у которых постгипофизарная область не разрастается в преназальную часть головы, здесь, т. е. по обеим сторонам гипофиза, вентрально от п.
Рис. 32. Голова зародыша Pristiurus rnelanostomus. Медиальный разрез;
Преназальный отдел А, прегипофизарный отдел В и постгипофизарный отдел С обозначены так же» как и на рис. 15, 16, 17. Положение носовой ямки (f. nas.) проецировано на медианную плоскость» Ch.—хорда., Dien.—diencephalon; ер.—эпифиз; hyp.—гипофиз; М.—рот; Msen.—mesencephalon, Mten.—metencephalon; p. ch.-—парахордалии; Pr. e.—prosencephalon; tr. cr.—trabeculae cranii, или трабекулярный отдел черепа (из А. Н. Северцова, 1917).
opticus, развиваются парные trabeculae cranii (рис. 32, tr. сг.), морфологически соответствующие cartilago dorsalis posterior миног [рис. 17, tr. cr. (с. dors. post.)]. Трабекулы эти срастались с передними краями парахордалии, и из них была развита вся передняя часть черепной коробки челюстных позвоночных (рис. 33, tr. сг.). Здесь, как мы видим, имеется значитель-
Рис. 33. Медиальный разрез головы взрослого экземпляра Pristiurus rnelanostomus.
Обозначения те же, что и на рис. 32. Кроме того: Ер. еп.—epencephalon; R.—rostrum. Сравнение рис. 32 и 33 с рис. 15, 16 и 17 показывает, что у гнатостом прегипофизарный (В) и постгипофпзар-ный (С) отделы черепа занимают у взрослых животных приблизительно то же положение, что и у зародышей, но что преназальный отдел (А) сильно вырастает и образует ростральный отдел головы взрослой рыбы (из А. Н. Северцова, 1917).
ное отличие между энтобранхиатами и эктобранхиат а м и: у первых развит только парахордальный (рис. 8 и 9, Р. ch. a., Pch.), у последних пар а-х о р д а л ь н ы й-f-T рабекулярный (рис. 33, tr. сг.) нейро-к р а н и у м.
У потомков энтобранхиат боковая стенка глазницы образуется за счет cornua parachordalia, у гнатостом она состоит из самостоятельной хряще
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ ПОЗВОНОЧНЫХ
179
Рис. 34. Схема строения мочеполовой системы эктобранхиат.
Половые мешки срослись с мочеточниками, превратившись в мезонефрические канальцы. Ь—пронефрическая камера, k. f.— половые продукты; ту—миотомы; р. с.—пронефрические канальцы; u.g.—мезонефрический выводной проток (из Машковцева,
«
мешков (рис. 34). В течение филогенеза у экто-
вой пластинки, а именно, из алисфеноидного xpgnja1 (рис. 28, Als), который, возможно, начинает закладываться уже у энтобранхиат.
У энтобранхиат, как известно, носовые ямки первично открываются наружу в вентральном направлении (рис. 15—18) и лишь позднее, сблизившись, принимают дорзальное положение. У энтобранхиат, наоборот, они сохраняют свое вентральное положение и ле-жат далеко друг от друга. В слуховом органе энтобранхиат развиваются лишь два полукружных канала, тогда как у эктобранхиат к этим двум каналам прибавляется еще и третий (рис. 37). У энтобранхиат блуждающий нерв лежит как раз на границе между черепом и первой дугой позвоночника, у эктобранхиат несколько верхних дуг позвонков, лежащих позади блуждающего нерва, слились между собою и приросли вплотную к черепу (см. схему на рис. 28, Аг. Ос.), образуя новую часть черепа, а именно, затылочную его часть (pars occipitalis cranii).
Нам осталось теперь рассмотреть лишь те изменения, которые произошли в мочеполовой системе эктобранхиат. По данным моего ученика А. А. Машковцева1 2 3, переработавшего заново вопрос о филогенезе этой системы у низших позвоночных, у первичных эктобранхиат имелся экскреторный орган, пронефрос, состоявший из нескольких десятков сегментальных канальцев и сегментальных половых бранхиат произошла частичная редукция большинства каудальных пронефрических канальцев и сохранились лишь 9—10 самых передних канальцев. Взамен редуцировавшихся пронефрических канальцев из сегментальных половых мешков протокраниат путем их срастания с мочеточником (рис. 34, u.g.) филогенетически развилось второе поколение почечных канальцев, а именно мезонефрос.
Половые продукты, которые у протокраниат, как мы это видели выше, развивались в стенках половых сегментальных мешков, сдвинулись у эктобранхиат ближе к полости целома (рис. 34, k.f.), к целомическим отверстиям мезонефрических воронок. У эктобранхиат половые клетки падали в брюшную полость, откуда захватывались как воронками пронефроса, так и воронками мезонефроса и выводились в задний отдел кишечника через мочеточник.
Мы видим, таким образом, что эктобранхиаты (включая гнатостом) отличались от энтобранхиат не только строением своего жаберного аппарата, но и целым рядом других морфологически важных признаков.
Итак, резюмируя в нескольких словах сказанное выше, мы приходим к выводу, что между протокраниатами и первичными гнатостомами суще
1 Sewer zo ft A. N. Die Entwicklung des Selachierschadels, Festschrift usw. von K. v. Kupffer, 1899.
2 Машковцев А. А., Морфологические и филогенетические отношения мезоне-
фроса к пронефросу. Труды лаборатории эволюционной морфологии Академии наук СССР, т. 2, 1934.
12*
180
ГЛАВА VII
ствовала группа переходных форм, у которых еще не был развит челюстным аппарат. Эктобранхиальный жаберный скелет этих форм, поддерживавший органы дыхания, состоял из жаберных дуг, распавшихся на четыре сегмента й доходивших до рта, служившего для пассивного питания этих животных. Эти не имеющие челюстей гипотетические переходные формы мы обозначили именем эктобранхиат.
Перейдем теперь к потомкам эктобранхиат, к примитивным гнатостомам, или первичным челюстным рыбам.
ЭВОЛЮЦИЯ PROTOGNATHOSTOMATA
Превращение бесчелюстных эктобранхиат в гнатостом, т. е. в позвоночных животных, питающихся при помощи челюстей хватательного типа, сопровождалось важными изменениями, имевшими место в наружном скелете, в ротовом аппарате и в органах движения. Мы разберем поэтому в отдельности все изменения, происшедшие в трех главных системах органов
Рис. 35. А—плакоидная чешуя в медианном разрезе, В—плакоидные чешуи Scyllium canicula, вид сверху.
Ы.—кровеносный сосуд; Ьр!.—базальная пластинка; dent.—зубной каналец и верхняя часть полости пульпы; epid.—эпидермис; ob. сог.—поверхностный слой corium; pulp.—пульпа; pig.—пигмент scler.bl.—склеробласты; schm.—эмаль; tief.cor.—глубокие слои corium; z—зуб (по Клаатчу из
* Плате).
этих животных, а именно: 1) в кожном скелете и, в частности, в зубах, 2) в челюстном скелете, т. е. в скелете челюстной и гиоидной дуг, в их мускулах и т. д. и 3) в плавниках. Рассмотрим эволюцию каждой из этих систем органов в отдельности.
Мы имеем основание думать, что развитие этих трех систем органов не было синхронным. По всей вероятности, у непосредственных предков челюстных позвоночных первым образовался наружный скелет в форме маленьких, состоящих из дентина зубов (рис. 35 А и В, dent) с покрытыми эмалью и загнутыми назад остриями (рис. 35 A, schm.). Эти плакоидные зубы имели, вероятно, более простое строение, чем зубы примитивнейших эласмобранхий.
В настоящее время трудно с уверенностью ответить на вопрос о том, когда именно образовался этот наружный скелет плакоидного типа, и развился ли он самостоятельно у предков гнатостом или был ими унаследован от общих предков Gnathostomata, Cyclostomata и Osteostraci. Сходство в строении плакоидного скелета у таких форм, как Thelodus и Elasmobranchii, казалось бы, должно было нас привести к мысли, что эта система органов имеет общее происхождение и унаследована от протокраниат. Но против этой гипотезы говорит, однако, тот факт, что у всех циклостом при переходе их к активной форме питания развились не плакоидные, а роговые зубы; если же допустить, что у предков циклостом существовал плакоидный скелет, то можно было бы ожидать, что и у миксин и у миног разовьются также не роговые, а плакоидные зубы. Поэтому возможно, что плакоидные чешуи первичных Ostracodermi и Gnathostomata развились независимо и парад-
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
181
лельно друг другу, совершенно так же, как независимо и параллельно развилась, например, хрящевая ткань у моллюсков (radula) и у позвоночных животных.1 Однако, пока у нас нет новых фактов для суждения об этой трудной проблеме, нам приходится оставить открытым вопросом о происхождении плакоидных чешуи.
Одно мы можем принять с достоверностью, а именно то, что эволюция этих нлакоидных чешуи или зубов происходила на очень ранней стадии развития предков челюстных позвоночных.
Рис. 36. Схематическая реконструкция висцерального аппарата первичных челюстных предков эласмобранхий (Protognathostomata), на которой видно положение преджаберных висцеральных дуг и щелей.
Эта схема сконструирована на основании сопоставления результатов сравнительно анатомических и эмбриологических исследований над различными эласмобранхиями (ср. рис. 30 и 31). А.рг. Ьг.1—A.pr.br.4—пребранхиальныеЬ висцеральные дуги 1—4;Ъг. 1—первая истинная жаберная щель bs.br.l--basibranchiafe 1; bs.h.—basihyaie; bs.md.—basimandjbulare; cr.br. 1—ceratobranchiale 1; cr.li.—ceratohyale; cr.lj, cr.l2—ceratolabiale 1 и 2; cr.md. (c. Meeh)—ceratomandibulare, или меккелев хрящ; ep.br. 1—epibranchiale 1; ep. h.—epihyale; ep.li, ep.l2—epilabiale 1 и 2: ep. md. (P. qu.)—epimandibulare (palatoquadratum); hp. br. 1—hypobranchjale 1; hp.h.—hypohyale; hp. md,—hypomandibulare; M.—m. adductor челюстной дуги; M'—m. levator labiae superioris; Orb.— глазница; ph. br.1—pharyngobranchiale 1; ph.h.—pharyngohyale; ph.lg—pharyngolabjale 2; ph. md.—pharyngomandibulare; Pb. 1 (spir)—первая преджаберная щель, или дыхальце; pb.8> pb.8— вторая и третья преджаберныя щели; p.pbs.l.—processus palatobasalis lateralis.
На челюстной дуге(А.рг. br. 2) и на второй губной дуге (A.pr.br. 3) сидели зубы. Впереди первой жаберной щели (br. 1) лежит типично расчлененная висцеральная дута—подъязычная дуга, со своей мускулатурой (М) и с наполовину закрывшимся дыхальцем pb.l (spir.), phaiyngo- и hypo- элементы этой дуги стали рудиментарны; вся совокупность этих элементов составляет первый пребранхиаль-ный бранхиомер. Второй пребранхиальный бранхиомер состоит из четырехчленной челюстной дуги; на схеме pharyngomandibulare (ph. md.) и hypomandibulare рудиментарны или отсутствуют; вторая пребранхиальная щель (pb.a) не открывается наружу вовсе, точно так же как и третья (pb.8). Третья и четвертая преджаберные дуги (A.pr.br.3, A.pr. Ьг. 4) представлены расчлененными губными хрящами (ph. 1а, ер. 18 сга и ep.lj, cr.lj). Соотношения между basibranchialia преджаберных дуг и бранхио-мерами A.pr.br. 3—A.pr.br. 3 изображены на схеме рис. 28 (из А. Н. Северцова, 1931).
Изучение истории развития дает нам вполне ясное представление о том, каким путем шло образование ротового скелета у челюстных позвоночных. Прилагаемая здесь схема (рис. 36) иллюстрирует строение передней части головы эктобранхиат, которую мы реконструировали на основании наших сравнительно эмбриологических исследований. Превращение эктобранхиат в гнатостом, т. е. развитие челюстей и гиоидного аппарата, осуществляется прежде всего замыканием передних жаберных щелей (рис. 36, pb. 2, pb. 3), а затем превращением передних пребранхиальных жаберных дуг в части
1 См. теорию гистологического параллелизма у А. А. Заварзина. A. Sawarsin.
Zeitschr f. Wiss. Zool., Bd., 113, 1924; Bd. 124, 1925. ' '
182
ГЛАВА VII
ротового скелета, а передних висцеральных и спинальных мускулов, приводивших в движение эти дуги (т. е. подчелюстных мускулов), в челюстные и гиоидные мускулы.
Закрытие жаберных щелей шло в рострокаудальном направлении. Это видно из того, что самые передние жаберные щели не прорываются у современных форм, а задняя недоразвивается и остается в виде spiraculum [рис. 36, Pb.l.(spir.)j у многих селахий, хрящевых ганоидов и кистеперых рыб.
Что касается судьбы висцеральных дуг этого отдела, то две передние дуги редуцировались и сохраняются лишь у селахий в виде рудиментарных губных хрящей (рис. 31, ер. lj„2 , сг. 12); средние элементы двух следующих дуг, т. е. epi- и ceratomandibulare, а также epi- и ceratohyale, развились в прогрессивном направлении и стали больше и массивнее (рис. 36, А. рг. br. 1, А. рг. Ьг. 2), чем соответствующие части остальных жаберных дуг; дистальные же элементы этих двух дуг (pharyngomandibulare и pha-ryngohyale, hypomandibulare nhypohyale), напротив, у акул атрофировались.1
Благодаря всем этим изменениям два средних элемента второй пребран-хиальной дуги превратились в верхнюю и нижнюю челюсти [ср. рис. 36, ер. md. (Р. qu.) ,cr. md. (с. Meek.), с рис. 37, Р. qu., С. М.], т. е. развился новый ротовой скелет хватательного типа. Таким образом осуществился чрезвычайно важный в биологическом отношении переход от пассивной формы питания позвоночных животных к активной.
При дыхательных движениях рыб, в особенности эласмобранхий, благодаря действию жаберной мускулатуры (т. т. constrictores и adductores) epibranch ialia сближаются с ceratobranchialia. Мы знаем, что ранний зачаток челюстной мускулатуры чрезвычайно схож с зачатком жаберных мускулов, но на более поздних стадиях развития этот зачаток диференцируется в несколько ином направлении, чем зачаток жаберной мускулатуры, а именно из него образуются следующие четыре мускула: m. adductor mandibulae (рис. 37, add. m.), m. levator maxillae s. palatoquadrati (рис. 37, 1. p. qu), m. levator labiae superioris (рис. 37, l.l.s.)2 и M. constrictor ventralis (рис. 37, cn. v.).
Таким образом, способность средних скелетных элементов жаберных дуг (верхней и нижней челюсти, т. е. epi- и ceratomandibulare) к смыканию (хватательная функция) имела место еще на той стадии эволюции, когда челюстная дуга была жаберной дугой и служила лишь для дыхания.
Мы не знаем, почему именно вторая пребранхиальная дуга, а не одна из лежащих впереди нее, превратилась в челюстную дугу, но можно думать, что челюстная дуга развилась из той пребранхиальной дуги, которая находилась в области черепа, представляющей собой прочную опору для массивной верхней челюсти. Вероятно, передняя часть осевого черепа на стадии, когда происходило развитие челюстей, была недостаточно прочна для того, чтобы служить опорой для больших хрящевых челюстей.
В одной из моих предшествующих работ (А. Н. Северцов, 1917) я рассмотрел вопрос о развитии плакоидных чешуй на поверхности кожи у предков гнатостом и отметил, что эти образования генетически тесно связаны с эктодермальными, т. е. эпителиальными, слоями кожи: эмалевый
1 Рудиментарное pharyngomandibulare сохранилось еще у некоторых пятижаберных акул (Laemargi's, Mustelus), hypomandibulare сохранилось у Hexanchus.
8 Мы имеем некоторые основания предполагать, что m. levator labiae superioris (рис. 37, 1. 1. s.) первично принадлежал не челюстной (второй пребранхиальной) дуге, а заднему губному хрящу (третьей пребранхиальной дуге).
8В0ЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
183
слой кожных зубов развивается за счет эпителия кожи. Плакоидные зубы развиваются не только на поверхности кожных покровов, но и на тех частях рта, которые выстланы эктодермальным эпителием, причем здесь они постепенно превращаются уже в настоящие, приспособленные для кусания зубы.
Факт, что ротовая полость у всех Deuterostomia выстлана эктодермальным эпителием, указывает нам на чрезвычайную древность этого признака, характеризующего предков позвоночных животных, и мы можем поэтому думать, что только что упомянутое образование плакоидных зубов внутри полости рта у непосредственных предков гнатостом и было тем именно стимулом, который вызвал преобразование двух пребранхиальных дуг (второй и третьей) в части ротового скелета (в челюстную и гиоид-.
Рис. 37. Графическая реконструкция (по сагиттальным разрезам) челюстной и гиоидной мускулатуры позднего эмбриона Pristiurus melanostomus.
add.m.—зачаток m, adductor mandjbulae; Als.—алисфеноидвый хрящ; aud.—ухо; buc.VII—r.buc-calis липового нерва; C. aud.—зачаток слуховой капсулы; С. М.—меккелев хрящ; cn. v. —челюстной отдел m. constrictor ventralis; С. sc. а.—передний полукружный канал; С. sc. h.~горизонтальный полукружный; канал; С. sc. р.-—задний полукружный канал; d.end.1—ductus endolymphaticus; Hm.—hyomandybulare; hm.VH—r. hyomandibularis n. facialis; l,l,s,—зачаток m. levator labiae superioris; 1. p. qu.—зачаток m. levator palatoquadrati; M.—рот; M. by.—зачаток гиоидной мускулатуры; mi, md. V—r. maxillomandibularis n. trigemini; nas.—носовая ямка; oph. p V—r. ophtalmicus profundus n. trigemini; oph, s. VII—r. ophtalmicus superficialis n. facialis; Pqu,—-небноквадратный хрящ; Pr. en.—prosencephalon; sp.—брызгальце; Tr. cr.—трабекулы; II—n. opticus; V.—n. trigeminus; VII—n. facialis (из A. H. Северцова, 1925).
ную дуги). Диференцировка же самих зубов, т. е. увеличение их размеров и различного рода изменения, происшедшие в их форме, представляются, разумеется, уже явлениями вторичного характера.
Что касается предков гнатостом, то и у них также сначала развились плакоидные зубы и на поверхности кожи и в ротовой полости, и лищь после этого передние висцеральные (пребранхиаль-ные) дуги превратились в соответствующие части ротового скелета. С полной определенностью можем мы утверждать, что в челюстную и гиоидную дугу у предков гнатостом превратились две пре бранхиальные висцеральные дуги, и поэтому уже a priori является возможным, что висцеральная мускулатура этих дуг превратилась в челюстную и гиоидную мускулатуру.
Эта точка зрения подтверждается данными сравнительно анатомического наблюдения, а также историей развития этих животных. На рис. 29 (А—С) представлено развитие челюстной мускулатуры акул, и если мы сравним
184
ГЛАВА VII
этот рисунок с рис. 37, изображающим развитие жаберной мускулатуры у Pristiurus melanostomus, то мы тотчас же заметим, что зачатки челюстной и гиоидной мускулатуры, несмотря на общее их сходство с зачатками типичной жаберной мускулатуры, занимают, однако, несколько иное, чем эти последние, положение. Зачатки челюстной и гиоидной мускулатуры располагаются при закладке латерально от зачатков висцеральных дуг. При закладке же мускулов типичных жаберных дуг средний отрезок каждой мускульной ленты проходит сквозь среднюю часть прохондральной жаберной дуги там, где образуется продольная щель (нижний конец epi-и верхний конец ceratobranchiale) и переходит на медиальную сторону дуги (рис. 29 С). Из этого медиального отрезка мускульной ленты, который впоследствии отделяется от зачатка m. constrictor, образуется m. adductor arcus branchialis соответствующей дуги (рис. 29 С).
Следя за развитием челюстной и гиоидной мускулатуры, мы не находим ни на одной из стадий их развития такого расщепления прохондральной, или уже охрящевевшей, челюстной или гиоидной дуги (рис. 37, Pqu., С. М.); поэтому здесь мускулы лежат всегда латерально от дуги (рис. 37, add. m., l.l.s., l.p.qu.). Что касается челюстного аппарата эласмобранхий, то здесь из средней части мускульной ленты второго пре-бранхиального бранхиомера образуется m. adductor mandibulae (рис. 37, add. т.), из ее верхней части—т. levator palatoquadrati s. maxillae superioris (рис. 37, l.p.qu.), из ее переднего выроста—т. levator labiae superioris (рис. 37, l.l.s) и, наконец, из ее нижнего отрезка—т. constrictor ventralis (рис. 37, cn. v.).
Сравнительные анатомы долго спорили о гомологии m. adductor mandibulae: одни утверждали, что этот мускул гомодинамен m. т. adductores типичных жаберных дуг; другие считали, что он соответствует части m. constrictor arcuum. Мускул этот действительно развивается из той части мускульной ленты, которая лежит снаружи от висцеральной (челюстной) дуги, но различие заключается здесь в том, что он не расщепляет челюстную дугу по длине, подобно тому как это имеет место в жаберных дугах, а лежит от нее снаружи, подобно мускулу констриктору. Однако, так как он развивается из той части мускульной ленты, которая, в случае если бы произошло расщепление, соответствовала бы m. adductor, то я думаю, что будет более правильным считать его гомодинамным m. т. adductores arcuum истинных жаберных дуг. Диференциация же здесь, повидимому, произошла на ранней стадии эволюции предков гнатостом, именно тогда, когда расщепление жаберной дуги еще вообще не имело места и когда висцеральный мускул лежал еще латерально от висцеральной дуги. Таким образом, в челюстной и в гиоидной дугах сохранилось первичное соотношение между жаберной мускулатурой и жаберными дугами.
Резюмируя все сказанное, мы видим, что при переходе предков современных рыб от пассивного питания к активному оба средних скелетных элемента (epi- и ceratomandibulare) второй преджаберной дуги превратились в несущие зубы челюсти (palatoquadratum и меккелев хрящ). При этом весьма вероятно, что дорзальный конец жаберной дуги передвинулся в ростральном направлении и причленился к отростку орбитальной части черепа (processus palatobasalis lateralis, рис. 36, р. pbs. 1.) при помощи proc, orbitalis palatoquadrati (место причле-нения редуцированного pharyngomandibulare).1 Вместе с тем жаберная мускулатура челюстной дуги превратилась в челюстную мускулатуру.
Изменения, происшедшие в первой пребранхиальной (гиоидной) дуге,
1 Северцов А. Н., Этюды по эволюции низших позвоночных, ч. III, Русский арх. анат., гист. и эмбр. 1925.
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ ПОЗВОНОЧНЫХ 185
которая превратилась в подвесочный аппарат, стоят в тесной коррелятивной зависимости от только что описанных изменений, имевших место во второй преджаберной (челюстной) дуге: pharyngohyale атрофировалось, epi-hyale, наоборот, сильно увеличилось в размерах и превратилось в hyoman-dibulare, причем его вентральный конец сочленился с челюстной дугой. Повидимому, это сочленение обусловило собою частичную редукцию вентральной части спиракулярной (первой пребранхиальной) щели [рис. 28, pb. 1; ср. рис. 36 Pb.l(Spir)]; дорзальная часть, этой щели сохранилась как spiraculum.
Ниже мы увидим, что у первичных предков рыб, вероятно, также и третья пребранхиальная дуга, т. е. задняя пара губных хрящей, была усажена зубами.1 Возможно, что некоторые несущие на себе зубы кости челюстного аппарата Osteichthyes (praemaxilla, dentale) принадлежали не челюстной, но третьей преджаберной, или иначе—первой премандибулярной дуге.1 2 Поэтому, как мы это только что отметили, мы и можем предположить, что у примитивных гнатостом имелись две пары функционирующих, т. е. несущих зубы, челюстных дуг, расположенных одна позади другой, а именно, palatoquadratum-J-cartilago meckeli и cartilago labialis posterior superior-[-cartilago labialis posterior inferior (рис. 36, A pr. br. 2, A. pr. br.3).
Мы потому остановились несколько подробнее на изменениях, имевших место в первом и втором преджаберных бранхиомерах (в гиоидной и мандибулярной дугах), что именно эти морфологические изменения имели огромное биологическое значение для рассматриваемых нами форм. Это явствует хотя бы из того, что новообразовавшиеся челюсти оказались настолько стойкими, что, несмотря на все другие крупные морфологические изменения, происшедшие в передней части головы потомков первичных гнатостом, они остались неизменными в своих основных чертах даже у наиболее поздних потомков этих простейших форм.Действительно, ротовой аппарат хватательного типа, состоящий из верхней и нижней челюсти, сохранился у всех потомков первичных гнатостом, т. е. у всех рыб, амфибий, рептилий, птиц и млекопитающих. Зубы этих животных оказались признаком не менее постоянным по своей структуре и форме. Вместе с тем этот тип хватательного челюстного скелета оказался также и чрезвычайно пластичным образованием, приспособляющимся к самым разнообразным изменениям пищи.
Большое биологическое значение только что описанного хватательного или кусательного ротового аппарата понятно само собою; оно заключается по преимуществу в том, что этот новый ротовой скелет давал гнато-стомам возможность выбирать наиболее подходящую для них пищу, а также сравнительно легко приспособляться к ней. Выбор же наиболее подходящей пищи естественно приводит клучшему питанию и, стало быть,, к повышению энергии и общей жизнедеятельности организма животных. Значение кусательного ротового аппарата, как органа нападения и защиты, понятно само собой и в дальнейших пояснениях, конечно, не нуждается. Факт, что пла-коидные зубы и хватательные челюсти объединились в одной общей функции в ротовом скелете гнатостом, оказался особенно выгодным: весьма вероятно, что именно это обстоятельство, т. е. сочетание функции элементов кожного скелета с функциями частей жаберных дуг, и обусловило победу этих животных в борьбе за существование.
1 Воскобойников М. М., Развитие висцерального скелета костистых рыб, Киев, 1909.
2 Se w er t z о f f A. N., Head Skeleton and Muscles of Acipenser ruthenus, Acta Zoologies, Bd. 9, 1928.
•486
ГЛАВА VII
Чрезвычайно правдоподобным является предположение, что изменения, происшедшие в способе принятия пищи животными, вызвали соответствующие изменения и в их органах движения и органах чувств, так как поиски и выбор пищи, а также ловля соответствующей добычи потребовали иного рода движений и лучшей способности к ориентировке в пространстве, чем это требовалось раньше для пассивного питания. Это объясняет нам прогрессивное развитие непарных плавников и факт образования парных плавников у рыб. Превосходные исследования И. И. Шмальгау-зена1 разъяснили нам ход развития непарных плавников рыб, и мы знаем теперь, что различные формы хвостового плавника, т. е. главного органа движения этих животных при плавании, развились из простого симметрично построенного протоцеркального плавника. От него произошел, благодаря изгибанию хорды в дорзальном направлении и вследствие прогрессивного развития вентральной плавниковой лопасти, эпицеркальный тип плавников, характерный для многих примитивных рыб (Elasmobran-chii, Chondrostei). Эта специфическая форма хвостовых плавников наводит нас на мысль, что вымершие примитивные рыбы, повидимому, вели -бентонный образ жизни.
Рис. 38. Belone acus. Личинка длиной в 15 мм.
На рисунке виден преанальный плавник (ра), расположенный впереди от анального отверстия, а— анальный плавник; ап—анальное отверстие; с—хвостовой плавник; ds—задний спинной плавник; о—► верхняя челюсть; os—рот; pct.—грудные плавники; и—нижняя челюсть (из А. Н. Северцова, 1927).
Из дорзальной и вентральной непарной плавниковой складки развились непарный спинной и анальный плавники. Весьма возможно, что у предков гнатостом также имелся функционирующий преанальный плавник (рис. 38, ра); по крайней мере С. Г. Крыжановский2 нашел в личиночном преанальном плавнике у мальков Osteichthyes метамерно расположенные мускульные почки, что указывает на возможность существования преанального плавника, как функционирующего органа, также и у взрослых первичных гнатостом.
Трудная проблема эволюции парных плавников челюстных рыб в последнее время в значительной степени разъяснилась, и мы можем сообщить читателю, конечно в очень краткой форме, результаты новейших исследований по этому вопросу. Ставшие известными в последнее время эмбриологические и палеонтологические факты столь решительно говорят в пользу теории развития конечностей из боковых складок кожи, что теперь мы можем уже вполне уверенно сказать, что гипотеза двустороннего архиптеригия, выставленная в свое время Гегенбауром, имеет лишь историческое значение.
У всех рыб, даже у тех, которые характеризуются типично построенным архиптеригием (двоякодышащие), парные плавники закладываются в виде горизонтально расположенных боковых складок с широким основанием (эврибазальные плавники). На вентральных краях миотомов, лежащих против этих боковых складок, образуется ряд мускульных отрост
1Шмальгаузен И. И., Непарные плавники рыб и их филогенетическое развитие, Записки Киевского О-ва Естествоиспытателей, 1913.
’Крыжановский С. Г., Развитие парных плавников у Acipenser, Amia « Lepidosteus. Труды научно-иссл. инет, зоолоогии Моск. унив.,т, 1, 1925.
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ ПОЗВОНОЧНЫХ
187
ков (рис. 39 А), на которых впоследствии развиваются сегментально расположенные мускульные почки;1 эти почки отделяются от отростков
Рис. 39 А—Е. Эмбриональная закладка плавников у Scyllium (А) по Гудричу; у Aci* penser (В) по Купферу, у Amia (С) по Рейгарту, у Polypterus (D) по Грахам Кэрру и у Ceratodus (Е) по Pi Семону.
а—ростральный край зачаточного грудного плавника; р—его каудальный край. Сравнивая между собою эти рисунки, на которых изображены очень ранние стадии развития плавников, мы видим, что зачатки грудных плавников имеют всегда широкое основание (а—р, эврибазальные плавники) и приблизительно горизонтальное положение. Мы знаем, однако, что во взрослом состоянии плавники некоторых изображенных здесь рыб (например, D и Е) имеют узкое основание (стенобазальный тип плавника); из этого можно заключить, что плавники селахий, Chondrostei, Holostei (также Teleostei), Crossopterygii и Dipueusta развились в течение своего филогенеза из горизонтальной эври-базальпой формы плавника (А).
1 Прежде обычно считали, что из каждого мускульного отростка образуются две дорзальные и две вентральные почки. Исследования С; Г*. Крыжановского показали, однако; что первично из каждого мускульного отростка парных плавников образуется, вероятно, только одна дорзальная и одна вентральная почка, совершенно так же. как это имеет место в непарных плавниках.
188
ГЛАВА VII
и, проникая в упомянутые выше боковые складки, образуют в них пучку плавниковых мускулов (рис. 40, sec. et р. vm.). Из этих-то мускульных
отростков и развиваются те первично сегментированные мускульные почки, из которых состоит дорзальная и
sec.et.p.vm.
unf
Рис. 40. Графическая реконструкция вентральных мускульных почек (sec. et р. vm) из плавника зародыша Amia calva.
uni — отростки миотомов (из Крыжановского, 1927).
вентральная мускулатура плавника.
При этом возникает важный вопрос: как же был построен первичный скелет плавников у примитивных гнатостом? Структура сегментально расположенной мускулатуры и строение скелета плавников у наиболее примитивных из всех известных нам эла-смобранхий [грудные и брюшные плавники у Cladodus (рис. 46), брюшные плавники у Pleuracant-hus (рис. 41 С)] наводят нас на мысль, что первично скелет свободных плавников был так же ясно сегментирован, как и сама мускулатура: он состоял из раз-деленного на членики метаптеригия, лежащего
в широком основании плавника, и из метамерно расчлененных лучей, сидевших на члениках этого метаптеригия. Особенно ясно выражена сегментация метаптеригия в брюшном плавнике ископаемого Coccolepis sp. п. (рис. 42, Mtptv— Mtpt17), подробно исследованном моей ученицей Е. Ф. Еремеевой.
Рис. 41 А—D. А—скелет брюшного плавника малька Acipenser ruthenus; В—скелет брюшного плавника взрослого Acipenser ruthenus; С—скелет брюшного плавника Pleuracanthus; D—брюшной плавник Heptanchus (рис. С и D из Видерсгейма).
bd,—bd,—basalia distalia 1—7; bp,—bpe—basalia proximalia 1—6; Be.—basale commune, или тазовый пояс; mt.pt.—metapterygium; ppt.—propterygium; r,—r9 и г,—rlt—radialia proximalia 1—11. Строение плавников у малька стерляди является рекапитуляцией строения первичной правильно сегментированной исходной формы парных плавников. Метаптеригий прилегает к брюшной стенке тела рыбы (из А. Н. Северцова, 1926).
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
1S9
Рис. 42. Скелет брюшного плавника Соссо-lepis sp.n. из юрских отложений Кара-тау.
bspt—basipterygium; dlt—dl15—длинные трехугольные покровные косточки защитного аппарата брюшного плйвника; Iptr — лепидотрихии, Idk—боковая покровная кость; mdk—медианная покровная кость; Mtptt—Mtpt17—свободные базальные членики метаптеригия; т\—г21—плавниковые лучи 1—21.
Эмбриологические исследования над развитием скелета плавников у акуловых и осетровых рыб вполне подтверждают эту точку зрения.1 Следя за развитием брюшных плавников • у осетровых рыб, мы видим совершенно отчетливо, что метаптеригий образуется здесь благодаря частичному срастанию базальных члеников плавниковых лучей (рис. 41 А и В).
Так как гегенбауровская гипотеза двустороннего архиптеригия имеет большое историческое значение (Гегенбаур, как известно, считал, что плечевой и тазовый пояс представляли собою видоизмененные жаберные дуги), то история развития только что описанных образований приобретает особенный интерес именно с теоретической точки зрения.
Мне удалось показать (А. Н. Северцов, 1926), что у осетровых рыб как тазовый пояс, так и метаптеригий вначале состоят из самостоятельных, метамерно расположенных основных члеников (basalia) плавниковых лучей (рис. 44, Ьр! — bp3); эти basalia впоследствии срастаются между собой и образуют большой хрящ тазового пояса (рис. 41, 43 и 44 Ьс). Так как плечевой пояс гомодинамен тазовому поясу и так как все остальные части (т. е. лучи, сегментальные мускулы и нервы) обоих плавников, грудного и брюшного, закладываются совершенно одинаково, то мы имеем полное право предположить, что и у первичных гнатостом плечевой пояс конечно
стей также образовался путем срастания базальных члеников метамерно расположенных хрящевых лучей. При этом мы принимаем, что у названных гнатостом обе пары плавников были построены совершенно одинаково, а именно в виде горизонтальных широких боковых
складок кожи, внутри которых лежали сегментально расположенные му-скульные пучки, нервы и членистые хрящевые лучи (рис. 39 А—Е).
У этих животных мускулатура каждого плавника состояла из двух слоев метамерно расположенных мускульных пучков, дорзального и вентрального, иннервируемых метамерно расположенными ветвями спинальных нервов. Между обоими мускульными пучками находился скелет плавника, состоящий из метамерно расположенных лучей (рис. 41); каждый из этих членистых хрящевых лучей состоял из медиальных, прилежащих к мускульной стенке тела basalia и из членистого стержня свободно лежащего плавника (рис. 41). Первичное строение парных плавников, таким обазом, чрезвычайно близко напоминало собой первичное строение непарных плавников: как те, так и другие были построены по одному и тому же плану.
1 S е we г t zo f f A. N., Development of the pelvic fin of Acipenser ruthenus etc., Journ. of Morphol., vol. 41, 1926.
190
ГЛАВА VII
Мы только что вкратце очертили ход развития и изменения, имевшие место в последних двух весьма важных в биологическом отношении системах органов, а именно: в ротовом аппарате и в органах движения предков современных рыб. Теперь мы должны поставить очень трудный вопрос о том, происходили ли только что описанные изменения одновременно
Ьс
Рис. 43 и 44. Горизонтальные срезы через основание брюшного плавника малька Acipenser ruthenus на двух следующих друг за другом стадиях.
Ь.,— b.e—basalia distalia 1 — 6; be—basale commune; bpM—bp.e—basalia proximalia 1—6; rt — rft—radialia proximalia 1—6. На рис. 43 скелетные элементы плавника видны на той стадии их развития, когда они еще отделены друг от друга и расположены строго сегментально. На рис. 44 мы видим, что basalia плавников (basalia distalia et proximalia, b.t—b.4, bp.x—bp.8) срослись друг с другом и образовали брюшной пояс или basale commune (из А. Н. Северцова, 1926).
(в геологическом смысле этого слова) или же они совершались одно вслед за другим?
Мы легко можем представить себе, что превращение передних жаберных дуг в части ротового скелета имело место уже после того, как развились эктодермальные жабры, и после того как жаберные дуги расчленились на pharyngo-, epi-, cerato- и hypobranchialia, так как образование этих последних частей является необходимой предпосылкой для развития подвижной челюсти. Можно думать, с другой стороны, что у предков рыб сначала образовались челюсти хватательного типа и лишь после этого начали развиваться парные плавники. Эту мысль с известными оговорками я уже высказал однажды в моей работе, посвященной
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
19г
вопросу о главных направлениях эволюционного процесса (1925, 1934), хотя я уже тогда сознавал, что доказать ее не легко. Если бы мы могли с определенностью утверждать, что лишенные плавников Anaspida представляют собою видоизмененных пресноводных гнатостом, то мы могли бы с большой вероятностью отнести этих животных к бесплавниковым Gnathostomata apterygiata. Мне лично кажется, судя по строению их ротового скелета, что Anaspida обладали хватательными челюстями и что вследствие этого они были родственны примитивным гнатостомам. Присутствие у Anaspida анального плавника также говорит в пользу этого предположения. Против выставленной нами точки зрения говорит структура и положение носового отверстия, которое соответствует у этих животных дорзальному назогипофизарному отверстию циклостом, а также форма их жаберных щелей.
Мне кажется, что до тех пор, пока вопрос о принадлежности Anaspida к гнатостомам или к круглоротым в широком смысле этого слова не будет решен окончательно, более осторожно было бы не делать преждевременных выводов относительно строения примитивных гнатостом, исходя из структуры Anaspida, и поэтому я здесь отказался от введения в наше родословное древо (схема XV) гипотетических промежуточны животных форм—Proto-gnathostomata apterygiata.
Итак,"мы не можем с уверенностью сказать, обладали ли примитивные гнатостомы парными плавниками или нет; мы оставляем поэтому этот вопрос открытым. Если бы у Protognathostomata, о которых только что шла речь, действительно имелись парные плавники, то мы должны были бы признать, что они находились у них еще на очень низкой ступени филогенетического развития1 и, вероятно, были представлены лишь-двумя парами боковых складок с метамерно в них расположенными мускульными почками и в крайнем случае—с зачатками сегментально расположенных лучей.
Резюмируя все изложенное выше, мы можем охарактеризовать прото-гнатостом как еще чрезвычайно примитивных эктобранхиат, перешедших от пассивного способа питания к активному. У них имелся ротовой скелет хватательного типа, состоящий из челюстных дуг, еще весьма близко напоминающих жаберные дуги, и, вероятно, из третьей пребранхиальной дуги. У них имелся также протоцеркальный хвостовой плавник и, может быть, уже были намечены орименты парных плавников. От этих форм произошли непосредственные предки всех известных нам современных и ископаемых эласмобранхий и всех Osteichthyes, т. е. все те формы, которые мы обозначили именем Chondrichthyes selachoidei.
ЭВОЛЮЦИЯ CHONDRICHTHYES SELACHOIDEI И ELASMOBRANCH!!
С сравнительно морфологической точки зрения представляется довольно вероятным, что только что упомянутые предки эласмобранхий были схожи многими чертами своего строения с наиболее примитивными из неспециализованных акуловых рыб, например с Pleuropterygii s. Cla-dodontidae; при этом они во многих отношениях были, конечно, еще более примитивно построены, чем известные нам Pleuropterygii и примитивнейшие из современных Elasmobranchii—Notidanidae и Chlamydoselachii.
Организация известных нам эласмобранхий, как, например, Ichthyo-tomi (Pleuracanthodei), Holocephali и Plagiostomi (акулы и скаты), дает
1 Мы знаем, что у гораздо более высоко стоящих Pleuropterygii парные плавники были построены еще очень примитивно. То же Самое можно сказать и относительно-брюшных плавников таких сравнительно высоко стоящих форм, какими являлись Palaeoniscoidei и Chondrostei. Поэтому мы можем принять, что парные плавники у прародительских первичных гнатостом (если такие плавники вообще уже существовали) имели еще чрезвычайно примитивное строение.
192
ГЛАВА VII
нам возможность судить об организации и строении их вымерших плевро-птеригиообразных предков, т. е. Chondrichthyes selachoidei. Однако мы не можем с уверенностью утверждать то же и относительно таких форм, как Acanthodii или Arthrodira, которые, по мнению Стеншиб, также принадлежат к группе эласмобранхий. Не лишено вероятия предположение, что эти две последние группы, т. е. Acanthodii и Arthrodira, отделились от еще более ранних предков эласмобранхий, чем те плевро-птеригиальные формы, о которых мы только что говорили, и что они пошли по пути специализации.
На основании некоторых сравнительно морфологических исследований, к сожалению, пока страдающих еще многими пробелами, мы можем попытаться восстановить некоторые черты строения отдаленных предков, пт которых произошли все современные Elasmobranchii (т. е. все Holoce-phali, акулы и скаты), Pleuracanthodein, по всей вероятности, примитивные Osteichthyes. Форма тела этих гипотетических Chondrichthyes selachoidei, вероятно, напоминала собою форму тела современных акул; их непарные плавники были построены по типу непарных плавников акул, их хвостовой плавник еще был построен по протоцеркальному типу, а их парные плавники имели сильно выраженный эврибазальный характер. Мы можем также сказать, что число жаберных щелей у них значительно редуцировалось и что их было не более восьми; spiraculum существовал и, возможно, функционировал еще как жабра, т. е. служил для целей дыхания. Носовые ямки имели, по всей вероятности, индиферентное положение, и лежали по бокам на конце рыла. Все тело было покрыто маленькими плакоидными чешуями, и зубы во рту были уже вполне диференци-рованы.
Внутренний скелет этих животных был еще целиком хрящевым; в позвоночнике тела позвонков еще не диференцировались (существовала еще хорда с двумя оболочками), но верхние и нижние диплоспондильные дуги позвонков уже были развиты; возможно, и нижние ребра1 также существовали.
Что касается до осевого черепа, то он был представлен хрящевой коробкой с лежащей впереди дорзальной фонтанелью. Слуховые капсулы представляли одно целое с собственно осевым черепом. Существовал rostrum и целый ряд характерных выростов черепа. Processus occipitales (medialis et laterales), processus postorbitales, praeorbitales и antorbitales были уже типично выражены. Базальный угол черепа (processus palatobasalis medialis и processus palatobasales laterales) был, невидимому, также развит.
Жаберные дуги были построены по типу дуг эласмобранхий, так что каждая дуга и здесь состояла из четырех элементов: pharyngo-, epi-, cerato- и hypobranchialia;1 2 на каждой жаберной дуге сидели в два ряда тонкие хрящевые жаберные лучи. Между вентральными концами жаберных дуг, т. е. между концами hypobranchialia, помещались непарные basibranchialia s. copulae, которые представляли собою ряд хрящевых палочек, одинаковых по форме и по величине.3
Гиоидная дуга также состояла из четырех элементов,4 и hypohyale было хорошо развито.
1 Исследования С. В. Емельянова (Die Morphologic der Fischrippen, Zool. Jahrb., Bd. 60, 1935) показали, что акулы, точно так же как и двоякодышащие и ганоиды, обладают нижними, прилегающими к соматоплевре ребрами, которые вторично оказываются лежащими в туловищной мускулатуре.
2 Вероятно, supra- и infrapharyngobranchialia (И. И. Шмальгаузен, 1923).
3 Как известно, у эласмобранхий редуцируются передние copulae, у Osteichthyes-задние; поэтому вполне можно себе представить исходное состояние этого аппарата, как полный ряд одинаковых copulae.
4 Именно из pharyngo, epi-, cerato-и hypohyale. Hyomandibulare (epihyale), по всей вероятности, еще не было столь массивным образованием, каким оно является у акул типа нотиданид.
8В0ЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
(93
Вероятно, у Chondrichthyes selachoidei имелось несколько большее число жаберных щелей, чем у совремецных и ископаемых большей частью пятижаберных эласмобранхий. Изучение развития плавательного пузыря и легких, предпринятое Васнецовым и Неймаейром,1 показало, что у Chondrichthyes selachoidei имелось не менее шести, а может быть и семи жаберных щелей, расположенных позади spiraculum. Из одной из задних пар этих жаберных щелей развился у их потомков, а именно у Osteichthyes, дорзальный (или вентральный) плавательный пузырь.
Palatoquadratum этих животных прикреплялся к черепу в двух местах, а именно: в области глазницы к processus palatobasalis lateralis [art. paiatobasalis (рис. 45, p. ob., p. b. 1.)] ив анторбитальной области черепа [art. quadrantorbital is (рис. 45, a. ant. о.), А. И. Северцов, 1928]. Symphysis palatoquadrati также существовал; и pharyngomandibulare был еще
Рис. 45. Скелет головы Chlamydoselachus anguineus.
a. ant. о.—квадранторбитальное сочленение верхней челюсти с осевым черепом; с. М.—меккелев хрящ; сп—хрящевая носовая капсула; hd.—hyoideum; hm.—hyomandibulare; n—носовое отверстие» p.ant.o.—proc, antorbitalis; p.b.l.—processus palatobasalis lateralis; p. ос. 1.—processus occipi-tails lateralis; p.ocm.—processus occipitalis medialis; p.ob.—processus orbitalis; p. pro—processus praeorbitalis; pq.—нёбноквадратный хрящ; R—rostrum; symph.—symphisis palatoquadrati. Ha этом рисунке видны оба сочленения нёбноквадратного хряща (верхней челюсти) с черепом, из которых впоследствии образовались соответствующие сочленения современных эласмобранхий и современных Osteichthyes, а именно квадранторбитальное (a. ant. о.) и платобазальное (р. ob., p.b.l.) сочленения. Первое из этих сочленений сохранилось у Osteichthyes и у происшедших от них наземных четвероногих животных (Quadrupeda); второе сохранилось у современных селахий. Первичное состояние, из которого впоследствии развились оба типа сочленений, сохранилось только у Chlamydoselachus (из А. Н. Северцова, 1928).
развит. Губные хрящи, вероятно, не были столь рудиментарными образованиями, какими они являются у современных селахий.
Парные плавники были построены по типу плавников Cladodus (рис. 46); грудные плавники состояли из дуги плечевого пояса, с которой сочленялся длинный сегментированный metapterygium; на члениках (сегментах) metapterygium сидели также сегментированные хрящевые лучи. Propterygium и mesopterygium еще не образовались, так что передние плавниковые лучи причленялись непосредственно к дуге плечевого пояса. Потому же плану были построены и брюшные плавники: весьма вероятно, что образовавшийся из передних basalia тазовый пояс был еще слабо развит. Metapterygium как брюшного, так и грудного плавника прилегал всей своей длиной к брюшной туловищной муску-
1 Wasnetzoff W., Zur Frage uber die Morphologie der Schwimmblase, Anat. Anz., Bd. 66, 1928; Uber die Morphologie der Schwimmblase, Zool. Jahrb. (Anat.), Bd. 56, 1932;
NeumayerL., Die Entwicklung des Darmes von Acipenser, Acta Zool., Bd. 11, 1930.
Морфологические закономерности 13
194
ГЛАВА VII
Рис. 46. Грудной ппавник Cladodus Neilsoni.
с—s—грудной пояс; I—проксимальные членики передних лучей грудного плавника; ргг.—рг.—дистальные членики плавниковых лучей; mt. — ta.—метаптеригий грудного плавника. Обратить внимание на то, что базальные членики передних плавниковых лучей являются непосредственным продолжением сегментированного метаптеригия. Ср. рис. 41 А, В и рис. 43 и 44 (из Гудрича, 1909).
строение их плавников позволяет нам
латуре (рис. 47, mt. pt^-mt. pt2); incisura metapterygoidea еще не образовалась (А. Н. Северцов, 1926), и концентрация плавниковых лучей и мускулов еще не началась.
Центральная нервная система, повидимому, была более примитивна, чем центральная нервная система эласмобранхий. Сердце было построено, как у современных акуловых рыб, и conus arteriosus уже был развит. В кишечном канале спиральный клапан также уже образовался.
Как известно, жаберные сосуды ныне живущих эласмобранхий сегментированы по жаберным щелям, т. е. каждая пара вен, впадающая в соответствующий arcus aortae dorsalis, обхватывает с двух сторон, спереди и сзади, упомянутую щель. Однако чрезвычайно вероятно, что у примитивных Chondrichthyes selachoidei жаберные.сосуды были сегментированы не по щелям, но по жаберным дугам, как это имеет место у эмбрионов эласмобранхий и рыб с костным скелетом, где каждая пара впадающих в arcus aortae dorsalis жаберных вен проходит, подобно жаберным артериям, внутри соответствующих дуг.1
Исходя из сравнительно морфологического анализа современных и вымерших эласмобранхий, а также примитивных Os-teichthyes, мы могли бы восстановить еще много других признаков строения общих предков этих животных, но мне кажется, что и сказанного вполне достаточно для того, чтобы дать читателю представление о структуре этих форм’. В сущности, наиболее близко они стояли к ископаемым Pleuropterygii (С1а-dodus); форма и положение их челюстного аппарата, а также думать, что они вели нектон-
ный образ жизни.
От этих гипотетических плевроптеригиальных примитивных Chondrichthyes произошли все известные нам группы типичных эласмобранхий, а именно: Pleuropterygii, Pleuracanthodei, Holocephali и Plagiostomi (Squaloidei и Batoidei). Отделение предков этих групп от прародительского ствола Chondrichthyes произошло очень давно, еще в досилурий-скую эпоху.
Нам представляется вероятным, что группа ископаемых Pleuracanthodei перешла в течение своей эволюции к ползающему, донному образу жизни; уэтихживотных развилась в передних конечностях метаптеригиаль-
‘Северцов А. Н., 1923.
ЭВОЛЮЦИЯ ПРЕДКОВ ЧЕЛЮСТНЫХ позвоночных
195
ная выемка (Incisura metapterygoidea),1 так что плавники их стали свободно подвижными и превратились в своей дистальной части в бисериаль-ный архитптеригий; точно так же как и плавники современных двоякодышащих рыб (рис. 70 L), плавники этой формы (рис. 70 Е) служили уже не только для плавания, но й для того, чтобы животное могло опираться на них всей тяжестью своего тела при ползании по дну водного бассейна. У самцов, кроме того, из дистальных отрезков брюшного плавника образовался копулятивный аппарат, из чего мы можем заключить, что у них, так же как и у современных Plagiostomi, существовало внутреннее оплодотворение, что, конечно, было весьма выгодным в биологическом отношении.
Предки Holocephali приспособились к иной форме бентонного образа жизни. Правда, их плавники, также как и у акул, представляют собою типично плавательные конечности, но их ротовой аппарат приспособился
Рис. 47. Грудной плавник Scymnus lichia in situ. Вид снизу.
ms. pt.—mesopterygium; mt. ptt—2—metapterygium 1—2; n. sp.—спинномозговые нервы конечности, лежащие перпендикулярно к оси метаптеригия; pr.pt.—propterygium; —r15 — лучи 1—15. На этом рисунке видно, что метаптеригий плавника прилегаеФ всей своей длиной к мускульной стенке тела; inci sura metapterygoidea (рис. 70 К, L, М,оооо), находящаяся у высших рыб между брюшной стенкой тела и метаптеригиальной осью плавника, здесь еще не образовалась (из А. Н. Северцова,
к питанию моллюсками и ракообразными, живущими на дне моря, поэтому верхние челюсти (palatoquadratum) Holocephali прочно срослись с осевым черепом, а отдельные мелкие зубы слились и образовали крупные зубы, предназначенные для раздавливания раковин. Невидимому, все эти (и многие другие) изменения произошли на ранних стадиях филогенетического развития, так как многие черты организации Holocephali, например строение их позвоночника и т. д., остались весьма примитивными до сих пор. Среди Plagiostomi акулы (Squaloidei) удержали свой первичный нек-тонный образ жизни, причем некоторые из современных представителей этой группы (Hexanchus, Heptanchus, Chlamydoselachus) сохранили и многие первичные признаки организации своих отдаленных предков. Невидимому, это случилось'главным образом благодаря прогрессивному развитию некоторых важных органов, например центральной нервной системы и ряда эмбриональных приспособлений. Среди акуловых рыб к бентон-ному образу жизни перешли ответвившиеся от них скаты, причем у них изменился характер движения: колоссально разросшиеся грудные плавники взяли на себя локомоторную функцию, свойственную хвостовому плавнику
1 Этим названием мы обозначаем щель, отделяющую ось метаптеригия от боковой стенки тела и развившуюся у двоякодышащих и кистеперых рыб, а также у всех наземных позвоночных.
13*
ГЛАВА VII
их акулообразных предков. Ряд органов у них развился прогрессивно, например органы боковой линии, центральная нервная система, мочеполовая система и т. д.; другие признаки1 еще сохранили свой первичный характер.
Мы видим, таким образом, что многие представители эласмобранхий развивались в направлении специализации (бентонно-ползающие Pleura-canthodei, бентонно-плавающие Holocephali и скаты), и нам представляется чрезвычайно вероятным, что именно это обстоятельство и послужило причиною того, что примитивные признаки их отдаленных предков сохранились у этих животных так полно вплоть до наших дней.
1 Присутствие плакоидных чешуй, строение черепа и висцерального скелета^ присутствие spiraculum и т. д.
ГЛАВА VIII
ЭВОЛЮЦИЯ OSTEICHTHYES
ОБЩАЯ ХАРАКТЕРИСТИКА OSTEICHTHYES
Весьма вероятно, что общие предки всех рыб с костным скелетом— Osteichthyes primitivi, т. е. общие предки всех дипной и кистеперых, всех костных и хрящевых ганоидов, а также всех костистых рыб, произошли от только что разобранной нами группы Chondrichthyes selachoidei (см. родословное древо; схема XV). В другую сторону от Chondrichthyes selachoidei ответвились, как мы это только что разобрали подробно, элас-мобранхии.
Существуют некоторые признаки, которые являются характерными для всех Osteichthyes, вследствие чего их нужно рассматривать как признаки, унаследованные от общих предков всей этой группы рыб. Такими признаками являются:
1. Присутствие покровных костей, т. е. костных чешуй, имеющих весьма определенное, характерное для этих животных положение, формуираспределение.
2. Образование характерного operculum, защищающего жаберный аппарат.
3. Положение и форма носовых ямок.
4. Способ развития жаберных кровеносных с о с у д о в.
Так как эти признаки являются общими для всей этой группы рыб, то нам приходится принять, что все они характеризовали собою общих предков Osteichthyes, т. е. наших Osteichthyes primitivi, и, стало быть, образовались уже у селахоидных предков этих последних.
Конечно, не присутствие костной ткани в организме, но образование характерно расположенных и характерно оформленных костных чешуй и покровных костей является типичным для общих предков Osteichthyes. Костная ткань развивалась в свое время и у далеко отстоящих в систематическом и генеалогическом отношениях от рассматриваемой .нами группы Osteostraci; поэтому ее присутствие не является само по себе филогенетически важным признаком.1
Сравнительно анатомическое изучение костных чешуй ископаемых и современных рыб приводит нас к выводу, что все эти чешуи имели первично ромбическую форму и несли на своей поверхности продольный гребень. Эта форма сохранилась всего полнее в хвостовом отделе хрящевых и костных ганоидов, палеонисцид и кистеперых рыб (рис. 48, а, а').
1 В сущности, гистологические параллелизмы так распространены в мире позвоночных и беспозвоночных, что на основании их трудно делать какие бы то ни было филогенетические заключения. В самом деле, мы не можем, например, строить выводов о родстве позвоночных с насекомыми на основании сходства в гистологической структуре их нервной ткани.
198
ГЛАВА VIU
Вообще говоря, три признака, а именно: 1) метамерное расположение костных чешуй, 2) их ромбическая форма и 3) образование продольного гребня на их поверхности являются признаками, характеризующими собой чешуи
Рис. 48. Преобразование туловищных чешуй Acipenser ruthenus.
а—ромбическая чешуя с продольным гребнем из хвостового отдела стерляди, а'—поперечный разрез через эту же чешую; Ь, Ь', Ъ"—крупные чешуи дорзального ряда (под ними—их поперечные срезы); с, с'—чешуи боковой линии; d—чешуи нижнего бокового ряда; е, ё', е” ,е'"—маленькие .чешуи, лежащие между большими чешуями; f—лепидотрихии плавников; g—fulcra, состоящая из двух сросшихся собою чешуй; h.—шип грудного плавника, состоящий из целого ряда сросшихся между собою чешуй, несущих гребень на своей поверхности.
Типичную ромбическую чешую с продольным гребнем на поверхности (а, а') из хвостового •отдела туловища стерляди можно рассматривать как исходную форму для всех чешуй хрящевых ганбидов. Чешуи b, Ь', Ь", с, с', е, е', е", е"' представляют собой ряд переходных форм между этой первичной чешуей и различными крайними формами чешуй в теле стерляди. Изображенные рядом с чешуями их поперечные срезы показывают, что у чешуй, даже крайне уклоняющихся в своей форме от первичной формы (b", с', d, е'" f), сохраняется типичная структура с продольным гребнем на поверхности (из А. Н. Северцова, 1926).
и покровные кости примитивных Osteichthyes. Разберем несколько подробнее эти три признака.
Вопрос о метамерном расположении чешуй вкратце сводится к следую-ющему. Исследования Газе1 показали нам, Что чешуи костистых рыб распо-
1 Hase A., Uber das Schuppenkleid der Teleostier, Jen. Zschr., Bd. 35, 1907.
ЭВОЛЮЦИЯ OSTEICHTHYES
199
ложеньг сегментально, т. е. что каждому миосепту туловища соответствует определенный поперечный ряд чешуй. Изучая развитие чешуй у Acipen-seridae, я нашел,1 что крупные чешуи стерлядей, образующие на туловище пять продольных рядов, имеют совершенно то же сегментальное расположение, и, наконец, точно такое же расположение костных чешуй у Polypterus и у Lepidosteus, так что мы имеем полное что одинаковое, типично сегментальное расположение туловищных чешуй действительно имеет место у современных хрящевых и костных ганоидов, у кистеперых и у костистых рыб. Что же касается до ископаемых палеонисцид, двоякодышащих и кистеперых рыб, то мы не знаем, каково было у них соотношение между миосептами и чешуями, но чрезвычайное сходство в форме и расположении этих чешуй у различных представителей этих групп с формой и расположением чешуй у Lepidosteus и у Polypterus заставляет нас предполагать, что чешуи примитивно построенных представителей этих вымерших групп были также расположены метамерно по миосептам.
Вопрос о первичной форме чешуй также разрешается сравнительно просто. Как мы уже отметили выше, чешуи хвостовой области современных хрящевых ганоидов, Lepidosteus, Polypterus и Calamoichthys имеют типично ромбическую форму, и то же может быть сказано относительно чешуй хвостовой области многих ископаемых палеонисцид и кистеперых рыб. Сравнительное изучение туловищных чешуй хрящевых ганоидов (Aci-penseridae) показало мне, что различные, часто весьма своеобразные формы, которые принимают эти чешуи, развились все из основной типич-
я нашел’ сказать,
основание
so
К
bs.n
Рис. 49. Поперечный разрез через туловищную чешую Acipenser ruthenus.
cut.—cutjs; D.—дорзальная складка эпидермиса: sc., sc.—костная пластинка чешуи; sp. m. медианный продольный гребень чешуи (из А. Н. Северцова, 1926).
50. Basirostrale me-
Рис.
diale Acipenser stellatus.
На рисунке видно, что эта кость (bs. г. т.) состоит из нескольких продолговатых ромбических мелких косточек, несущих на своей поверхности более или менее ясно выраженные продольные гребни (К); sc.—пластинки мелких костей или чешуй. Позади отдельные маленькие косточки срастаются между собой, образуя сплошную пластинку (pt.) basirostrale mediate (из А. Н.
Северцова, 1926).
ной формы, т. е. из ромбической чешуи с продольным гребнем на ее наружной поверхности (рис. 49, sc., sp. m.). Исследование туловищных чешуй Lepidosteus и Polypterus в свою очередь указывает нам на то, что
1 Se wertzof f A. N., The development of the Scales of Acipenser ruthenus, Journ. of Morph., v. 42, 1926.
200
ГЛАВА VIII
чешуи эти являются видоизмененными ромбическими чешуями; наконец, к такому же выводу приводит нас и изучение туловищных чешуй ископаемых Crossopterygii и Palaeoniscidae. Поэтому мы можем вполне определенно принять, что все формы костных чешуй рыб сводятся, в конце концов, к основной ромбической чешуе (рис. 48, а), встречающейся, как это было только что указано, в хвостовом отделе очень многих ископаемых и современных Osteichthyes.
Вероятно, и покровные кости головы также имели первоначально ромбическую форму, так что у первичных Osteichthyes и туловище и голова были .покрыты ромбическими костными чешуями. Изучая морфологию
Рис. 51 А, В, С. Три поперечных разреза через чешуи боковой линии туловища Acipenser ruthenus на трех последовательных стадиях развития.
cl.—канал боковой линии на той стадии развития, когда он еще представлен открытой бороздой: cl.'—тот же канал боковой линии в форме трубки; ер.—эпидермис; N. 1.—nervus lateralis; sc.—пластинка чешуи; sc.i—часть пластинки чешуи, лежащая под каналом боковой линии; sp. m.—медианный костный гребень чешуи.
На ранней стадии развития (А) мы видим еще индиферентную чешую sc., sc.x с гребнем sp. m. (ср. рис. 49), лежащую под еще открытым каналом боковой линии (с!.); на несколько более поздней стадии развития (В) костный гребень sp. m. и часть чешуи sc.x загибаются вокруг еще открытого канала боковой линии с!.; на еще более поздней стадии развития (С) медианный гребень чешуи sp. m. и изогнутая часть пластинки чешуи (sc.x) срастаются между собой (образование вторичного гребня чешуи sp. m.'), образуя замкнутую костную трубку вокруг эпителиальной трубки канала боковой линии; из сравнения рисунков А, В и С видно, что костная трубка образуется из медианного гребня sp. m. и половины пластинки чешуи sc.i (из А. Н. Северцова, 1926).
чешуй и покровных костей головы осетровых рыб (А. Н. Северцов, 1926), я нашел у них в голове много костей ромбической формы с продольным гребнем на поверхности. В качестве примера подобного рода костей можно привести suprarostralia у Acipenser ruthenus и медианные basirostralia у Acipenser stellatus (рис. 50, bs. г.). Многие другие покровные кости в черепе осетровых рыб безусловно представляют собой видоизменившиеся ромбические кости или чешуи, большинство которых, в особенности кости молодых животных, несет продольные гребни на своей поверхности.
Весьма существенные изменения претерпели те чешуи, которые лежали на поверхности парных и непарных плавников. У селахоидных предков Osteichthyes, так же как и у современных акул, свободные края плавников поддерживались гибкими роговыми лучами (цератотрихиями); у самих же Osteichthyes эти роговые лучи заместились такими же гибкими, но более крепкими костными лучами (лепидотрихиями), состоящими из преобразованных и сериально расположенных костных чешуй. Интересно, чтомалень-
ЭВОЛЮЦИЯ OSTEICHTHYES
201
кие костные членики, из которых состоят лепидотрихии, несут на своей поверхности типичные продольные гребни (рис. 48, f).
Несколько более сложным является вопрос о том, всегда ли имелся продольный костный гребень на наружной поверхности чешуй тех примитивных Osteichthyes, от которых, по нашему мнению, произошли все рыбы с костным скелетом.
Чтобы решить зтот вопрос, нам приходится принять во внимание отно-
шение чешуй к каналам боковой линии. У личинок стерл'яди эпидер-
мальный канал боковой линии,
представленный на ранних стадиях продольной бороздой в эпидермисе (рис.
51 A, cl.) проходит латерально от костной пластинки чешуи (рис. 51 А, sc, sc.J так, что гребень чешуи (рис.
51 A, sp. m.) лежит рядом с эпидермальным каналом. На несколько более поздних стадиях развития та часть пластинки (рис. 51 В, sc.J, на которой
Рис. 52 А, В, С. Три ^перечных среза через хрящевой череп малька Acipenser ruthenus.
А—разрез через затылочную область, В—разрез через орбитальную область, С—разрез через ростральную область черепа ;bs.г. I.—basirostrale laterale; bs.r.m.—basirostrale mediale; chcr.—хондрокра-ниум; cl. ос.—затылочный канал боковой линии; c.l.sp.o.—супраорбитальный канал боковой линии; d.s.oc.—dermosupraoccipitale; Fr.—frontale mediale; Fr.t—frontale laterale; 1.—изогнутая часть пластинки, соответствующая части scM, рис. 51; pt.ос.—postoccipitale; R.—rostrum; Sp. r.—o. o. suprarostralia; sp. m.—медианный костный гребень; sp. m.'—медианный гребень кости, образующей костный канал боковой линии; sp. m."—медианный гребень frontale laterale, принимающей участие в образовании канала боковой линии; sp. m#1—медианный гребень frontale mediale. Из сравнения рисунков 52 и 51 видно, что образование костного канала боковой линии всегда происходит за счет медианных гребней, находящихся на костях или на чешуях (из А. Н. Северцова, 1926).
лежит канал боковой линии (рис. 51 В, с._ 1.), загибается вокруг этого канала и обхватывает его с одной стороны; таким образом, боковые стенки костного канала, в котором теперь лежит канал боковой линии, образованы, с одной стороны, продольным гребнем (рис. 51 В, sp. m.), а сдругой—изогнутой частью пластинки чешуи sc.v На еще более поздней стадии развития (рис. 51 С) продольный гребень (sp. m.) и изогнутая часть костной пластинки sc.-t срастаются вместе, образуя костный канал с находящимся внутри него эпителиальным каналом боковой линии (cl') и его чувствующими почками. Таким образом, здесь чешуя и ее продольный гребень (sp. m.) находятся в тесной связи с органом боковой линии.
' Как мы это только что отметили, многие покровные кости в голове Acipenseridae снабжены продольным костным гребнем на поверхности. И мы можем сказать, что целый ряд черепных костей этих рыб, как, например, postoccipitalia (рис. 52 A, pt. ос.), parietalia, supraor-
'202
ГЛАВА VIII
bitalia, frontalia (рис.- 52 В, Fr., Fr.n), praefronfalia, все suprarostralia (рис. 52 C, sp. r.), basirostralia (рис. 52 C, bs. г. 1., bs. r.’m.) и еще некоторые другие кости представляют по своей форме и структуре несколько измененные чешуи с гребнем (sp. m.) на поверхности.
Вполне вероятно, однако, что и остальные покровные кости черепа осетровых рыб образовались из таких же точно чешуй. Если мы проследим развитие тех костей головы, сквозь которые проходят каналы органов боковой линии, то мы увидим, что на ранних стадиях развития кости эти состоят из пластинки с продольным костным гребнем (рис. 52 А, В); эпидермальный канал боковой линии проходит рядом (рис. 52 A, cl. ос.; рис. 52 В, с. 1. sp. о.) с этим гребнем (sp. m.', sp. m."), совершенно таким же образом, как это имеет место в чешуях боковой линии туловища осетровых рыб (рис. 51 А, В, sp. m.). В течение дальнейшего развития часть костной пластинки (рис. 52 А, В, 1.), лежащая непосредственно под каналом боковой линии, сгибается и охватывает с одной стороны этот эпидермальный канал; еще позже края продольного гребня и изогнутой части пластинки срастаются между собой и образуют костную трубочку, внутри которой лежит канал боковой линии с чувствующими почками.
Этот процесс был мной прослежен на целом ряде черепных костей, и я могу сказать с полной определенностью, что posttemporalia, supratem-poralia, dermosupraoccipitale,1 squamosa, frontalia lateralia и nasalia являются на ранних стадиях развития Acipenser типичными костями с гребнями; позже, при дальнейшем развитии, эти кости превращаются в широкие кости без гребней.1 2
Кроме только что упомянутых костей, мы находим у осетровых рыб еще ряд других мелких косточек, сквозь которые также проходят каналы боковой линии и которые в боковом разрезе имеют полулунную (серповидную) форму: это—ossa postorbitalia и suborbitalia, а также многочисленные трубчатые basirostralia lateralia (рис. 52 С, bs. г. 1.). Мне удалось показать, что многие кости, через которые проходит канал боковой линии и которые закладываются у низших рыб (Acipenseridae) как кости с гребнями, у высших форм закладываются как типично полулунные ^серповидные) кости. Это дает нам право предполагать, что и полулуннь!?(серповидные) кости Acipenseridae закладывались первоначально как типичные кости с гребнями и что они лишь впоследствии вторично приобрели свою полулунную форму.
Только что вкратце изложенное исследование о происхождении костного черепа привело нас к выводу, что большинство покровных костей Acipenseridae 3 представляет собою видоизменение костных чешуй, которые первоначально имели ромбическую форму и несли на своей наружной поверхности продольный костный гребень.
Теперь возникает вопрос: имеем ли мы право расширить этот вывод и считать, что чешуи и покровные кости общих предков всех Osteichthyes были также построены по этому типу?
Мы знаем, что туловищные чешуи у ныне живущих костных ганоидов
1 История развития показывает, что dermosupraoccipitale осетровых рыб образуется путем срастания двух самостоятельных dermosupraoccipitalia. То же относится и к кости frontale auctorum, которая также образуется из двух костей с гребнями: frontale mediate и frontale iaterale.
2 Вообще говоря, типичные продольные гребни встречаются в покровных костях только на ранних стадиях развития; на более поздних стадиях их уже невозможно различить, вследствие развития скульптуры на поверхности костей.
3 Исключением здесь являются лишь кости, находящиеся в ротовой полости, т. е. несущие зубы maxillae, praemaxillae, pterygoidea и parasphenoideum, а также трубчатые кости, образующиеся из элементов жаберного скелета.
ЭВОЛЮЦИЯ OSTEICHTHYES
203
Рис. 53. Поперечный разрез через затылочную часть головы малька Amia calva.
c.l.—канал боковой линии; с.ос.—затылочная часть черепа; 1m.—костная пластинка posttemporale; 1m'.— часть этой пластинки, загибающаяся вокруг эпителиального канала боковой линии; т. — затылочная мускулатура; med. оЫ.—продолговатый мозг; р. tm.— posttemporale; sp. m.—медианный продольный гребень, находящийся на posttemporale. На этом рисунке видно, как из медианного костного гребня и изогнутой части костной пластинки posttemporale (ср. рис. 51 и 52) начинает образовываться костный канал боковой линии (из А. Н. Северцова, 1925).
(Lepidosteus), у кистеперых (Polypterus, Calamoichthys) и костистых рыб, а также у многих ископаемых рыб имеют либо ромбическую форму, либо форму, которая по всей видимости развилась из ромбической формы; у остальных рыб чешуи имеют циклоидную или ктеноидную форму. Большею частью поверхность этих чешуй бывает гладкой, иногда она покрыта скульптурой, причем в некоторых случаях покрыта ею настолько, что мы совершенно не имеем возможности судить о присутствии или отсутствии на чешуях продольного гребня, столь характерного для хрящевых ганоидов. То же самое приходится нам сказать и относительно покровных костей черепа у костных ганоидов, кистеперых рыб, палеони-сцид, дипной и т. д. Возникает вопрос: имели ли при этом чешуи и покровные кости черепа предков всех Osteichthyes такие же характерные костные гребни, как те, что мы находим у хрящевых ганоидов?
Сравнительно эмбриологические исследования дают нам безусловно утвердительный ответ на этот вопрос.
Нам известно, что покровные кости черепа взрослой Amia бывают либо гладкими, либо покрытыми беспорядочной скульптурой—бугорками и бороздками. Однако, если мы проследим развитие этих костей, то увидим, что у тех покровных костей черепа, которые прилегают к каналам органов боковой линии, продольные гребни закладываются совершенно таким же образом, как и у мальков Acipenser. Чтобы убедиться в этом, обратимся к рис. 53, представляющему поперечный разрез через posttemporale личинки Amia. Здесь мы увидим, что костный гребень (sp. m.) posttemporale (р. tm.) лежит рядом с каналом боковой линии (с. 1.), а пластинка самой кости изогнулась вокруг противоположной стороны эпидермального канала боковой линии (1m.'). Дальнейшее развитие этих образований протекает так же, как и у осетровых рыб. Аналогичным образом развиваются у Amia костные каналы, внутри которых проходят каналы органа боко
вой линии, и на других покровных костях черепа, например на squamosum, nasale, frontale и т. д.
Что касается других костей черепа Amia calva, как, например, postor-bitalia, suborbitalia, praeoperculum и supratemporale, то они развиваются из костей, которые мы обозначили у Acipenser как полулунные или серповидные кости, и мы имеем полное основание считать, что в этом случае перед нами—вторичное изменение только что описанного типа, т. е. костей с продольным гребнем.
Из всего этого следует, что у предков Amia несомненно были развиты в голове типичные покровные кости с гребнями; затем, в течение дальнейшей истории их развития, те из них, которые не имел и никакого отношения к каналам боковой линии, потеряли свои гребни
204
ГЛАВА VIII
(подобно тому, как это имело место у осетровых рыб), те же покровные кости с гребнями, которые послужили для образования костного канала боковой линии, сохранили у мальков типичные продольные гребни. Исследования истории развития туловищных чешуй у Amia показало мне, что здесь имеет место совершенно тот же процесс. Таким образом, мы имеем полное право принять, что все покровные кости предков Amia несли в свое время гребни на своей наружной поверхности.
Тот же тип развития покровных костей черепа мне удалось наблюдать и у Lepidosteus, что указывает нам на то, что и покровные кости предков Lepidosteus, в свою очередь, также развились из костей с продольными гребнями, т. е. из чешуй; тот же тип развития покровных костей черепа встречается и у Teleostei, где он наблюдался уже давно,1 но где на него до сих пор не было обращено достаточного внимания, и, наконец, тот же тип образования этих костей я нашел у личинок Polypterus,1 2 так что, пови-димому, и кистеперые рыбы также происходят от предков, характеризовавшихся покровными костями с продольным гребнем на своей наружной поверхности.
Мы потому остановились с такой подробностью на происхождении костных чешуй и покровных костей черепа рыб, что вопрос этот (А. Н. Северцов, 1924, 1925, 1926) приводит нас к весьма важным филогенетическим выводам. Нам представляется весьма вероятным ,что все Osteichthyes произошли от некоторых примитивных хрящевых рыб, у которых все тело было покрыто метамерно расположенными ромбическими костными чешуями, несущими продольные гребни на своей наружной поверхности. Такие же точно чешуи покрывали и всю поверхность головы э т и х р ы б.
Снаружи от этого костного скелета у предков Osteichthyes лежали древние, чрезвычайно многочисленные, плакоидные чешуи их акулоподобных предков. В тех местах, где костные чешуи лежали на хряще, т. е. в черепе, в поясе передних конечностей, в нижней челюсти, они—либо вследствие своего разрастания, либо в результате срастания мелких чешуй между собой—стали больше: таким образом, уже на ранних стадиях филогенеза развились такие крупные покровные кости черепа, как dermo-suproccipitalia (dextrum+sinistrum), squamosa, parietalia, frontalia (frontale mediale+frontale laterale) и т. д.
Эмбриологическое и сравнительно анатомическое исследование черепа современных осетровых рыб (А. Н. Северцов, 1926) выявило нам, таким образом, два важных факта: во-первых, что у этих рыб многие кости черепа представляют собой еще до сих пор чешуеподобные образования с продольным гребнем на поверхности, во-вторых, что целый ряд черепных костей, которые у других рыб являются едиными образованиями, здесь представляют собою результат сращения в онтогенезе первично самостоятельных кожных окостенений. Оба эти факта, с одной стороны, указывают нам на то, что костный череп осетровых рыб остановился в своем развитии на очень низкой ступени эволюции, с другой стороны, они дают нам право, на основании изложенных выше результатов, составить общую схему черепа гипотетических Osteichthyes primitivi, еще состоявшего из чешуй. Схема эта представлена на рис. 54.
1 См., например, рисунки в Handb. der vergl. Entwicklungsgesch. d. Wirbelt. von O. Herwig, Bd. 3, 1906.
2 Нужная стадия развития Polypterus была мне любезно представлена для исследования проф. Гольмгреном, которому я приношу здесь свою искреннюю благодарность.
ЭВОЛЮЦИЯ OSTEICHTHYES
205
Мы говорили до сих пор только о тех костях головы Osteichthyes, которые произошли из чешуй, теперь же нам предстоит разобрать, правда лишь в самых кратких чертах, вопрос о происхождении остальных покровных костей примитивных Osteichthyes. Мы знаем, что в ротовой полости примитивных Osteichthyes, кроме упомянутых нами покровных костей, существовали ещедругие кости, а именно кости, несущие зубы. Я думаю, что в отношении этих последних образований наиболее правильным является взгляд Гауппа,1 который полагает, что кости эти развились как резистент-
ная подстилка, на тех местах, где у предков Osteichthyes сидели зубы, и что они не имеют ничего общего с чешуями, развившимися на поверхности тела. Эти несущие зубы кости (maxillae, praemaxillae, pterygoidea, dentalia и т. д.) расположены у современных форм* главным образом на хряще челюстной дуги.
М. М. Воскобойников,2 на основании своих исследований над костистыми рыбами, пришел к выводу, что praemaxilla и dentale первоначально принадлежали не челюстной дуге, а премандибулярному бранхиомеру. Мои личные исследования над хрящевыми ганоидами и селахиями (А. Н. Северцов, 1928)3 побуждают меня присоединиться к этой точке зрения. Однако, если эта гипотеза верна, то мы должны принять, что у предков Osteichthyes не только челюстная, но и третья пре-мандибулярная дута (задняя губная дуга или задний губной хрящ) некогда несла на себе зубы и принадлежала челюстному аппарату. Трубчатые же кости, образующиеся вокруг хрящей и принадлежащие к гиоидной и челюстной дугам Osteichthyes, по всей вероятности произошли как самостоятельные кости, т. е. как новообразования.
Мы отметили выше, что вторым признаком, характеризующим всех Osteichthyes, является присутствие у них жаберной крышки (operculum), и что этот' признак, точно так же как и предыдущий, т. е. присутствие покровных костей, был унаследован ими от их общих предков—Osteichthyes primitivi. Признак этот важен в том отношении, что его возникновение прямо указывает на изменение у предков рыб с костным скелетом спо-
рно. 54. Схема гипотетического чешуйчатого черепа примитивных Osteichthyes на той стадии развития, когда он еще состоит из ромбических, несущих продолный гребень чешуй. f, Г.—frontalia (mediale и laterale)» m. d., m. d.—mediodorsalfa; m. r., m. r.—mediorostralia; n., n.—nasalia; pf., pf,—postfrontalia; pr. f«—praefr-ontalia; p., p.—parietalia; r., r.— rostralia; so., so. —supraorbitalia; s. o.lts. o.j—dermosupraoccipii dia; s. q., s. q.—squamosa; s. t., s. t.— supratemporalia. Эта схема построена главным образом на основании результатов эмбриологического исследования черепа осетровых рыб
(из А. Н. Северцова, 1925).
соба дыхания, что является фактом большого биологического значения.
В этом отношении мы совершенно согласны с исследованиями М. М. Воскобойникова,4 которые указывают нам на то, что образование подвижной
1 G au р р Е., Die Entwicklung des Kopfskeletts, Handb. der vergl. Entwicklungs-gesch. d. Wirbelt. von O. Hertwig. Bd. 3, 1906.
’Воскобойников M. M., Развитие «висцерального скелета костистых рыб, Киев, 1909.
3 Sew er tzo f f A. N., The Head Skeleton and Muscles of Acipenser ruthenus, Acta zool., Bd. 9, 1928.
4 Воскобойников M. M., Аппарат дыхания у рыб, Труды III. Всеросс. съезда зоологов и т. д., 1928.
Woskoboinikoff М. М., Der Apparat der Kiemenatmung bei den Fischen, Zool. Jahrb., (Anat.), Bd. 55, 1932.
206
ГЛАВА VIII
жаберной крышки, функционирующей наподобие насасывающего аппарата, ; явилось большим шагом вперед в развитии дыхательного аппарата рыб. '
Отметим, что у предков Osteichthyes, вероятно, уже существовал зачаток ' жаберной крышки. На эту мысль наводит нас образование оперкулярной складки у Holocephali и особенно у Chlamydoselachus, череп которого, как известно, чрезвычайно похож на череп Cladodus. Однако полного развит.и я operculum достигает, повидимому, лишь тогда, когда в оперкулярной складке начинают образовываться костные чешуи. К этому выводу, по крайней мере, приводит нас изучение развития типичных оперкулярных костей Osteichthyes, т. е. operculum, interoperculum 'j и suboperculum, очень похожих на чешуи осетровых рыб. 1
Образование оперкулярной складки вместе с поддерживающими ее j костями, иными словами, развитие нового способа жаберного дыхания, 5 привело со своей стороны к важным морфологическим изменениям во всех \ близлежащих органах: во-первых, во всех жаберных дугах, которые про- -двинулись вперед под осевой череп, а затем в дуге плечевого пояса, которая > плотно соединилась с черепом (связь posttemporalia с cleitrum).1 Мускулы ; жаберного аппарата также резко изменились по сравнению с тем, что имело место у эласмобранхий: на месте длинных и широких пластинкр- I образных жаберных мускулов (constrictores) селахий развились массивные mm. levatores arcuum и mm. interarcuales ventrales, а также многочисленные мелкие мускульные ленты mm. interbranchiales. Часть мускулатуры челюстной дуги, будучи связана с operculum, приобрела новую функцию; при этом она претерпела значительные перемены, а именно—дорзальная часть m. constrictor mandibuiaris, т. е. m. adductor palatoquadrati, изменила место своего прикрепления и превратилась в m. protractor hyoman-dibularis.1 2 3
Что касается гиоидной дуги, то и в ней произошли перемены: от hyoman-dibulare отчленилась вентральная часть и превратилась в symplecticum; то же самое произошло и с дорзальной частью ceratohyale, от которой отчленилось stylohyale.
Таковы те крупные коррелятивные изменения, которые произошли в соседних с operculum органах в связи с изменившимся способом дыхания этих форм.
Носовые ямки имеюту всех Osteichthyes дорзо-латеральное положение. Вполне возможно, однако, что у селахоидных предков Osteichthyes и у самых примитивных Osteichthyes эти носовые ямки лежали, так же как и у ныне живущих акул, на вентральной поверхности передней части головы» и что их дорзо-латеральное положение явилось результатом вторичного сдвигания.
Наружные носовые складки, характеризующие селахий, срастаются у Osteichthyes друг с другом и образуют перемычку, разделяющую носовую ямку на две части: верхние и нижние наружные ноздри.
Наконец, строение плавников у современных осетровых рыб и Palaeoni-scoidei говорит в пользу того, что у их гипотетических предков, т. е. у Osteichthyes primitivi, в грудных плавниках не имелось еще ни мезо- ни проптеригия, точно так же как не было их и в брюшных плавниках этих форм, причем весьма вероятно, что метаптеригий гипотетических Osteichthyes primitivi в то время состоял еще из некоторого числа несросшихся между собой basalia и был, таким образом, еще более или менее отчетливо
1 Достоверно известно, что эти кости, так же как и другие покровные кости плечевого пояса, представляют собою изменившиеся и сильно разросшиеся костные чешуи.
2 Северцов А. Н., 1927 и 1928.
3 Положение носовых ямок у эмбрионов Ceratodus и Quadrupeda говорит в пользу высказываемого нами взгляда. См. Sewertzoff A. N., Der Ursprung der Quadrupeda, Palaeont, Zschr., Bd.8 , 1926.
эволюция osteichthyes
207
сегментирован.1 Плавники у этих форм были несомненно построены, по эврибазальному типу, т. е. имели широкое основание.
Позвоночник этих животных состоял из хорды с оболочками и из верхних и нижних дуг позвонков, на которых сидели нижние ребра (Chondro-stei, Palaeoniscoidei и Dipneusta).
Хрящевой череп этих животных был построен по типу черепа Cladodon-tidae. и Chlamidoselachus (см. ниже), что также указывает нам на то, что примитивные Osteichthyes произошли от очень примитивных эласмобран-хообразных форм, во многих отношениях близких к Cladodontidae, но, может быть, еще более примитивных, чем они. В этом отношении моя точка зрения почти полностью совпадает со взглядами Гудрича1 2 с той, однако, разницей, что я принимаю за предков рыб (мои Chondrichthyes selachoidei, его Pisces) формы, по многим признакам похожие на селахий, чего он, повидимому, не делает.
Однако, что касается последующего хода эволюции различных потомков Osteichthyes, то здесь мои взгляды во многих отношениях резко расходятся с общепринятой точкой зрения на этот вопрос.
ЭВОЛЮЦИЯ CHONDROSTEOIDEI И ВОПРОС О КЛАССИФИКАЦИИ РЫВ
Прежде чем перейти к дальнейшему изложению эволюции рыб с костным скелетом, в частности эволюции Chondrosteoidei, нам необходимо остановиться на важном вопросе об общей классификации рыб.
Начало современной классификации рыб было дано еще И. Мюллером (1846), который уже тогда в основание своей системы поставил акуловых рыб, или эласмобранхий. Это свое примитивное положение эласмобранхий сохранили во всех последующих классификациях, начиная со знаменитой классификации Э. Геккеля (1866), и в настоящее время можно считать, общепринятым, что предками всех рыб были акуловые рыбы и их сородичи.
Таким образом, положение эласмобранхий в родословных древах не вызывает никаких сомнений; сомнения и споры возникают главным образом в связи с положением хрящевых ганоидов (Chondrosteoidei) в системе, и на этот именно вопрос мы и обратим в дальнейшем наше преимущественное внимание.
Зоологи первой половины XIX века вслед за И. Мюллером, который подразделил подкласс ганоидов на три отряда: Chondrostei (Acipenseridae,. Spatulariae), Holostei (Lepidosteini: Lepidosteus, Amia) и Polypterini (Polypterus), считали хрящевых ганоидов очень примитивной группой рыб с хрящевым скелетом, близкой по своему строению к акуловым рыбам.
В дарвиновское и последарвиновское время классификация Мюллера была положена в основу всех родословных древ, изображавших ход эволюции рыб. Первое такое древо было построено Э. Геккелем в 1868 г. в его «Generelle Morphologie», но здесь мы приводим (схема X) родословное древо, построенное им гораздо позднее и помещенное в его «Systema-tische Phyl< genie der Wirbeitiere» в 1895 г.3 В нем осетровые рыбы (Sturiones) помещены очень низко; вместе с так называемыми «панцирными рыбами» (Placodermi, Cephalaspides, Pteraspides) они, совершенно независимо от кистеперых рыб (Crossopterygii), ответвились от общих предков рыб с костным скелетом, а именно от Proganoides.
1 Об этом мы заключаем на основании существования сегментации метаптеригия в брюшном плавнике у взрослых Acipenseridae, Polyodontidae (A. N. Sewerzoff, 1926-и 1934) и Palaeoniscidae (Smith Woodward, 1889—1891).
.‘Goodrich E. S., Cyclostomes and Fisches, Treatise of Zoology, 1909.
3 Это родословное древо Геккель повторил в несколько сокращенном виде в своей «Antropogenia», изданной уже в 1903 г.
208
ГЛАВА VIII
Рассматривая филогенетическую таблицу Геккеля с нашей современной точки зрения, мы видим, что некоторые ее части несомненно неверны. На основании замечательных работ Киэра, Стеншиб и других современных палеонтологов оказалось, что так называемые «панцырные рыбы» (по совре-
менной классификации Ostracodermi) стоят очень близко к циклостомам и должны быть поэтому отнесены не к рыбам, но к бесчелюстным круглоротым, т. е. к совершенно другому классу позвоночных.
Однако, принимая во внимание, что этих палеонтологических открытий XX века Геккель в конце XIX века знать не мог, нам приходится считаться
ЭВОЛЮЦИЯ OSTEICHTHYES
209
с его взглядами, выражающими общее мнение морфологов эпохи расцвета эволюционной морфологии после Дарвина.
Я не привожу целого ряда родословных древ этого периода, т. е. схем, построенных сравнительными анатомами на основании изучения только одной или во всяком случае немногих систем органов, и отсылаю читателя к известной книге Бешфорда Дина,1 где все эти гипотезы и соответствующие им схемы приведены in extenso. Из этих схем рассмотрим только классификацию и схему Т. Бриджа,1 2 * * * которого английские авторы считают моим предшественником в этом направлении.
Бридж подразделяет ганоидных рыб на следующие группы: a) Selachoidei (Polyodon, Acipenser, Scaphirhynchus, Chondrosteus), b) Teleosteoidei. Эту классификацию он выразил также в виде родословного древа, представленного нами на схеме XI.
Teteostoitiei.
Selachoidei
Dipnoi Amphibia
Pneumatocoela
Chondrostew Acipenser Scaphirhynchus Polyodon
Elasmobranch/!
Apneumatocoeia '
Схема XI. Филогенетическое древо ганоидных рыб по Бриджу. Объяснение в тексте
К сходным результатам с Бриджем пришел на основании своих исследований над органами кожных чувств и Коллиндж.®
Э. Гудрич,1 на основании сопоставления обширного сравнительно анатомического и палеонтологического материала, дает следующую ниже схему филогенетических соотношений различных групп рыб (схема XII).
Мы видим, что он не производит высших рыб, в частности Chondro-stei, от кроссоптеригий, но отводит и тех и других от примитивных Teleo-stomi, причем непосредственными предками Chondrostei и Holostei считает гипотетических Actinopterygii.
Аналогичную схему соотношений между хрящевыми ганоидами и остальными рыбами мы находим также и у Осборна в его книге о происхождении и эволюции жизни (1918).6 На его родословном древе (схема XIII) видно, что Осборн производит хрящевых ганоидов вместе с костными ганои-
1 BashfordDean, Fishes living a. fossil, New York a. London, 1895.
2 В r i d g e T. W., On the Osteology of Polyodon folium, Phylos. Trans, of the Royal Society of London, 1879.
8 С о 1 11 n g e W. E., The Sensory Canal System of Fishes, Quart. Journ. of Mier, Scien., vol. 36.
‘Goodrich E. S., A Treatise on Zoologie, London, 1909.
‘Osborn H. F., The origin and evolution of life, London, 1918.
Морфологические закономерности 14
210
ГЛАВА VIII
дами и костистыми рыбами от «первичных истинных ганоидов». Кистеперые рыбы (Polypterus, Coelacanthidae) на схеме Осборна стоят очень далеко от хрящевых ганоидов. Осборн считает, что «первичные истинные ганоиды» произошли от общего ствола ганоидов, от которых в другую сторону
Схема XII. Филогенетическое древо рыб и круглоротых по Гудричу. Объяснение в тексте.
ответвились кистеперые рыбы, дипнои и водные предки наземных позвоночных.
Сравнительные анатомы начала XX века продолжали держаться взглядов, близких взглядам И. Мюллера и Э. Геккеля. Так, в 1913 г. в III томе обширного немецкого сводного издания «Современная куль
ЭВОЛЮЦИЯ OSTEICHTHYES
211
тура» помещена сводка по филогении позвоночных, написанная Боасом,1 в которой, как это видно из приводимой ниже схемы XIV, хрящевые га-ноиды производятся от селахоидных форм—предков химеровых рыб.
Отметим, что эта родословная на первый взгляд несколько отличается от родословного древа, построенного Геккелем в 1895 г. Так, Геккель вел линию хрящевых ганоидов (Sturiones) через гипотетических Proganoides к гипотетическим же Proselachii; Боас же ведет эту линию от примитивных селахий не прямо к предкам ганоидов, но через предков химер, т. е. одной из групп селахоидных рыб, от которых, по его мнению, ответвились химеры, к предкам двоякодышащих рыб (z) и через них к хрящевым ганоидам. Таким образом, Боас пытается построить филогенез хрящевых ганоидов несколько более подробно, чем это сделано у Геккеля. Основная же мысль обоих построений одна и та же.
§
£
Уетвертичная Третичная
Асци- Дшрлм рругло- Акулы Пм„ЛЯП11в дни orbs ротые а скаты Осетровые у-г VI А/ Н Hp/dosteus
Amia Гнос/лис-
-j тые
Двояко- п . . дышащие P°typt&us
Верхний мел
1-М \\Coelacan-wn/dae
Наземные\ оозвоноча : ные
Юра
Юоимигивные 1 осетровые
Первичные
Himhuu мел
Триас
костистые
Пермская
Карбон
.1
'Примитивные рептилии
Девон
Arthrodires С
Силур
~ "Различные X Ostracodermi
Примитивные ~ "Перво чньТе истинные ' первойн. двор ко- ^Кистелерые' акулы гзиоиды^^^ дышащие галоиды —
отвод ганеидов
Сроович
первичные, \ ostracodermi
"ствол cenaiul)
челюстные рыбы
Первичные * наземные
U03B0HQ4HV
КзрВин
ч
Кембрии
? Голыелорда ты
Схема Х1И. Происхождение и адаптивная радиация рыб по Осборну. Объяснение в тексте.
Интересно, что Боас, на основании своих исследований, обратил особенное внимание на признаки, обыкновенно не сохраняющиеся у ископаемых форм, а именно на присутствие и отсутствие клоаки, на способы дробления желтка у зародыша и т. д. Поэтому его филогенетическое древо страдает обычным недостатком филогенезов, построенных палеонтологами, и теми сравнительными анатомами, которые делают свои выводы на основании очень небольшого числа верных, но односторонне подобранных признаков: по мере изучения все большего и большего числа признаков эти построения приходится дополнять и исправлять. Идеал и тех и других (в настоящее время еще недостижимый) состоит в построении филогенетического древа на основании всей суммы признаков (анатомических, эмбриологических, палеонтологических и экологических) данной группы животных.
1 Boas I. V., Phylogenie der Wirbeitiere, в книге «Die Kultur des Gegenwart, ihre EntwicKlung und ihre Ziele», Bd. 3, 1913.
1'4*
212
ГЛАВА VIII
В 1923 г. я выступил со своей первой работой по филогенезу современных хрящевых ганоидов, явившейся началом длинного ряда исследований по этому вопросу. Эти исследования привели меня к выводам, в некоторых отношениях приближающимся к изложенным выше взглядам Бриджа, Гудрича и Осборна и сильно отличающимся от гипотез Копа, Трэквера, Смите Вудварда и некоторых других палеонтологов (см. ниже).
Отличия эти зависят от того, что я в своих выводах не только учитывал сравнительно анатомические данные, но и обращал особое внимание на эмбриологические факты, которых палеонтологи в соображение не принимают, а также и от того, что в своих построениях я исходил из теории
Схема XIV. Филогенетическое древо рыб по Боасу. Объяснение в тексте.
филэмбриогенеза, мало знакомой как палеонтологам, так и анатомам. В результате этих работ я пришел к следующей схеме филогенеза рыб (схема XV), подробно разбираемой мною ниже.
Сейчас нам следует обратить внимание лишь на положение хрящевых ганоидов в этой системе. Как это нами будет подробно рассмотрено ниже, хрящевые ганоиды сходны с низшими .эласмобранхиями в целом ряде существенных морфологческих признаков, и это заставляет признать их примитивность и близость к низшим селахиям. Кистеперые же рыбы (Crossopterygii), двоякодышащие (Dipneusta) и предки Quadrupeda, с одной стороны, и костные ганоиды (Holosteoidei) и примитивные костистые рыбы (Teleostei)—с другой, также сходны между собой в целом ряде признаков; вследствие этого мы объединяем всех первых в группу Holosteoidei crossopterygii, а всех вторыхв группу Holosteoidei actinopferygii и производими тех и других от гипотетических Holosteoidei primitivi. Гораздо более примитивные хрящевые ганоиды (Chondrosteoidei) ответвились очень рано от только что разобранных нами древних и примитивных «первичных кост-
ЭВОЛЮЦИЯ OSTEICHTHYES
213
ных рыб» (Osteichthyes primitivi), сохранивших еще довольно много признаков своих селахоидных предков.
На этом мы закончим разбор схем, построенных сравнительными анатомами, схем по основной своей мысли сходных друг с другом. Но параллельно с этим
рядом исследований шел другой
ряд филогенетических
(Hi
(S-)
Teleostei(H)
Dipneusta (S-)
Mynnoidei
Protoqnathostomata
Entobranchiata
Entobranchiata
Branchiostomatidae
Protocraniata
Ecrania primitiva
A mi a des (<d>
lepidosteidei
Ostracodermi
Ooadrupeda
PmtocycMmta
Holosteoidei actinopterygn
Holosteoidei erossopterygit
(Sl)Elasmobranchii
Holosteoidei primitivi
^Anaspida
Chrondrosleoidei
(S)
Osteichthyes primitivi
Chondrichthyes selachoidei
Схема XV. Схематическое изображение родословного древа низших позвоночных.
На этой диаграмме в виде обычной схемы родословного древа представлены вероятные, по нашему мнению, филогенетические соотношения между различными родоначальными группами низших позвоночных. Описанные нами в тексте промежуточные гипотетические формы (Protocraniata, Entobranchiata, Ectobranchiata и т. д.) также вошли в это древо. Черными кружками обозначены вымершие формы животных, светлыми кружками обозначены группы, дожившие до нашего времени. Одним восклицательным знаком (!) обозначены группы, не обнаруживающие признаков вымирания, знаком минус (—) обозначены группы, близкие к вымиранию. Буквой S изображены идиоадаптивные или специализованные группы, двумя восклицательными знаками (!!)—прогрессивные современные группы. Связь между Anaspida и Ectobranchiata я лично считаю возможней, но спорной; поэтому на линии, идущей от Ectobranchiata к Anaspida имеется вопросительный знак (?). Ра laeoniscidae совсем не вошли в наше родословное древо; их надо было бы поместить поблизости от Holosteoidei actinopterygii.
214
ГЛАВА VIII
построений, ряд родословных древ низших водных позвоночных (циклостом и рыб), созданный палеонтологами на основании своего материала и в некоторых отношениях отличный от только что разобранных нами рядов. Начало этих палеонтологических родословных древ положено было американским палеонтологом й зоологом Копом.
Родословное древо, изображающее филогенез рыб по Копу, представлено нами на схеме XVI. Рассматривая эту схему (взятую из упомянутой выше книги Башфорда Дина), мы видим, что Коп поместил хрящевых ганоидов (Chondrostei) на самом верху своего родословного древа, принимая, что они ответвились от потомков кроссоптеригий очень поздно, причем одна ветвь потомков кроссоптеригий дала хрящевых ганоидов, а другая—общих предков костистых рыб (Teleostei).
Вполне понятно, почему Коп считал кроссоптеригий за прародительские формы, от которых произошли все остальные Osteichthyes. Во-первых, потому, что они, действительно, являются очень древними (еще со времен девона) и очень примитивными формами, во-вторых, потому, что в то время, когда Коп и его последователи создавали свою теорию, гипотеза Гегенбаура относительно происхождения всех известных нам форм плавников рыб из бисериал ь-ного архиптеригия, весьма характерного для всех кроссоптеригий, была еще в полной силе.
Теперь же, когда мы имеем полное основание отказаться от гегенбауровской гипотезы архиптеригия, заменив ее теорией боковых складок, опирающейся на большое число эмбриологических и палеонтологических фактов, мне кажется, следовало бы признать по меньшей мере спорным положение относительно происхождения актиноптеригий
от кроссоптеригий, или, что то же самое, утверждение, что все известные нам формы плавников рыб развилисьиз бисериального архиптеригия. Во всяком случае, следовало бы основательно пересмотреть тот фактический материал, на котором основываются эти положения.1
К гипотезе Копа примкнул крупный английский палеонтолог, известный своими исследованиями над палеозойскими палеонисцидами, Р. Трэк-вер; его многочисленные работы относятся к периоду от 1875 до 1912 г. Именно Трэквером была впервые высказана распространенная в настоящее время среди палеонтологов гипотеза дегенерации хрящевых ганоидов в течение филогенеза. Он принял, что осетровые рыбы и лопатоносы произошли от палеонисцид путем вторичного упрощения их организации, т. е. путем дегенерации, которая выразилась во вторичном развитии хрящевого черепа, благодаря чему он снова сделался похож на череп акуловых рыб, во вторичном распадении крупных костей черепа
1 Для этой цели частью мною лично, частью моими учениками и сотрудниками был предпринят целый ряд специальных исследований над развитием конечностей и многих других важных в этом отношении органов. См. Sewertzoff A. N., Journ. of Morphol., 1926, Jen. Zschr., 1926, (К г у z а и о w s к у, 1. с., 1927), Васнецов В. В. (Развитие первичного плечевого пояса и грудных плавников у Lepidosteus, Русск. зоол. журн., т. 7, 1927).
ЭВОЛЮЦИЯ OSTEICHTHYES
215
на мелкие покровные кости, в развитии хрящевого рыла, в изменении положения ротового отверстия и в удлинении hyomandibulare. Надо сказать, что никаких доказательств того, что такая дегенерация действительно имела место, ни у Трэквера, ни у разделяющих его взгляды палеонтологов мы не находим.
Того же взгляда придерживается и А. Смите Вудвард,1 положивший его в основу установленной им классификации Teleostomi. Мы приводим здесь эту классификацию целиком, так как она служит примером ходячих классификаций этого рода.
Подкласс Teleostomi *
Отряд I. Crossopterygii
Подотряд 1. Haplistia
Подотряд 2. Rhipidistia
Подотряд 3. Actinistia
Подотряд 4. Cladistia
Отряд II. Actinopterygii
Подотряд I. Chondrostei
А. Прогрессивный ряд
Семейство 1. Palaeoniscidae
Семейство 2. Platysomatidae
Семейство 3. Catopteridae
В. Регрессивный’ ряд*
Семейство 1. Chondrosteidae
Семейство 2. Belonorhynchidae
Семейство 3. Acipenseridae
Семейство 4. Polyodontidae
Как мы видим, здесь implicite принимается, что наиболее примитивными Teleostomi были кроссоптеригий и что от них произошли два ряда потомков: прогрессивные Palaeoniscidae, Platisomatidae и Catopteridae и дегенеративные Chondrosteidae, Belonorhynchidae, Acipenseridae и Polyodontidae.
В последнее время Стеншиб (1932)1 2 также подверг этот вопрос подробному пересмотру и на основании своего палеонтологического материала пришел к выводам, согласным со взглядами Копа, Трэквера и Смите Вудварда. Так, он считает, что предки древних Actinopterygii произошли от древнейших девонских или верхнесилурских кроссоптеригий, и дает следующую классификацию этих первичных актиноптеригий.
Chond rostei
Группа А. Семейство Palaeoniscidae
Группа В. 1. Семейство Platysomidae
2. Семейство Dorypteridae
3. Семейство Bobasatraniidae
Группа С. Семейство Pholidopleuridae
Группа D. 1. Семейство Catopteridae
2. Семейство Perleididae
3. Семейство Ospiidae
4. Семейство Parasemionotidae
Группа Е. 1. Семейство Phanerorhynchidae
2. Семейство Saurichthyidae
Группа F. 1. Семейство Chondrosteidae
2. Семейство Acipenseridae
3. Семейство Polyodontidae
1 S m i t h W о d ward A., Catalogue of the fossil Fishes in the British Museum, vol. 1—4 London, 1891—1901.
2 S t e n s i 5 E. A., Triassic Fisches from East Greenland. Kobenhavn, 1932.
216
ГЛАВА VIII
Ближайшим родичем группы хрящевых ганоидов по восходящей линии Стеншиб считает совершенно определенную форму рыб, подробно им исследованную, а именно, нижнетриассовую Saurichthys. Схемы родослов-
Схема XVII. Родословное древо рыб, построенное Саве-Сбдербергом на основании данных Стеншиб . Объяснение в тексте.
ного древа сам Стеншиб не дает, но его ученик Саве-Сбдерберг,1 на основании его описания, построил родословно древо примитивных рыб с костным скелетом, изображенние на схеме XVII.
Схема XVIII. Родословное древо рыб по Грегори. Объяснение в тексте.
В последнее время ко взглядам Копа относительно происхождения и эволюции хрящевых ганоидов примкнул также и крупный американский
1 Save - Soderbergh, Some points of view concerning the evolution of the Vertebrates and the classification of this group, Arckiv for Zool., Bd. 26,Stockholm, 1934.
ЭВОЛЮЦИЯ OSTEICHTHYES
217
палеонтолог В. К. Грегори.1 На схеме XVIII приведено построенное им родословное древо.
Рассматривая эту схему, мы видим, что при построении своего родословного древа Грегори учел результаты также и новейших палеонтологических исследований над Marsipobranchii, обнимающих всех современных круглоротых (миног и миксин) и ископаемых Osteostraci, а также их девонских сородичей. Marsipobranchii представляют собою большую группу палеозойских (силурских и девонских) черепных, но еще бесчелюстных позвоночных, рано специализировавшихся и не являющихся предками низших челюстных позвоночных, т. е. рыб. Рыбы же ответвились от палеонтологически неизвестного общего корня всех черепных позвоночных
Схема XIX. Родословное древо позвоночных по Саве-Сбдербергу. Объяснение в тексте.
(диагр. XVIII) ив раннем палеозое были представлены специализованными панцирными формами (Antiarchi, Arthrodira, Anarthrodira) и предками акуловых рыб (Elasmobranchii), от которых, по мнению Грегори, очень рано ответвились предки наземных позвоночных (Tetrapoda), а затем и предки двоякодышащих, кистеперых рыб и Polypterini. От общего корня этой группы первичных кистеперых, т. е. от общих предков наземных позвоночных, двоякодышащих, палеозойских кистеперых и Polypterini, произошли палеонисциды и предки костистых рыб, т. е. Acti-nopterygii. От палеонисцид ответвились хрящевые ганоиды.
Обсуждая вопрос о положении хрящевых ганоидов в системе, Грегори примыкает к старому взгляду Трэквера, утверждавшего, что хрящевые ганоиды являются дегенерировавшими потомками палеонисцид, утратившими целый ряд признаков своей примитивной организации. К этим взглядам Грегори, так же как и ко взглядам Стеншиб, нам придется еще вернуться впоследствии при обсуждении вопроса об эволюции Chondrosteoidei и Holosteoidei и их ближайших потомков, поэтому здесь мы на них останавливаться более подробно не будем.
Новая филогенетическая схема (схема XIX) принадлежит ученику Стеншиб Саве-Сбдербергу (1934),1 2 который в общем придерживается
1Gregory W. К., Fish skulls: A study on the evolution of Natural Mechanisms^ Trans, of the Amer. Philos. Soc., 1933.
2Save-Soderberg, 1. c., 1934.
218
ГЛАВА VIII
взглядов своего учителя, но дополняет их собственными исследованиями над девонскими стегоцефалами, так называемыми ихтиостегидами. По мнению Саве-Сбдерберга, эласмобранхий, к которым он относит Arthrodira и Antiarchi, представляют собой боковую ветвь общего ствола гнатостом и не имеют отношений к остальным рыбам. От общего ствола челюстных позвоночных (Gnathostomata) ответвились, с одной стороны, все Actinopterygii, а с другой .стороны, рыбы с наружными и внутренними ноздрями (хоанами) —Choanata. От этих Choanata ответвились в одну сторону двоякодышащие рыбы и хвостатые амфибии, а в другую—кистеперые рыбы и предки остальных наземных позвоночных. Первичные четвероногие (Eutetrapoda) дали две ветви, первую из которых наш автор обозначает термином «батрахо-морфов» (лягвообразных), а вторую—«рептилиоморфов» (ящерообразных). От Batrachomorpha произошли девонские предки стегоцефалов—Ichthyo-stegalia, давшие начало различным группам типичных стегоцефалов и, наконец, бесхвостым амфибиям, а от Reptiliomorpha произошли все многочисленные группы вымерших и современных рептилий и, кроме того, все Anthracosauria, которых Саве-Сбдерберг относит к амфибиям. От типичных рептилий ответвились предки птиц и млекопитающих.
Филогенетическая схема Саве-Сбдерберга отличается от обычно принимаемых филогенетиками родословных схем тем, что этот автор, так же как и Н. Гольмгрен,1 принимает дифилетическое происхождение наземных позвоночных, производя хвостатых и бесхвостых амфибий от различных корней: хвостатых—от двоякодышащих, а бесхвостых—от кистеперых рыб.
Итак, мы сделали краткую сводку филогенетических построений второй половины XIX и начала XX века. Из этой сводки мы видим, что родословные схемы довольно правильно распадаются на две группы: на построения сравнительных морфологов и на построения палеонтологов, причем оба эти типа схем расходятся в целом ряде признаков. Конечно, мы еще не достигли общего идеала филогенетического построения, каким явилось бы построение филогенетического древа, основанное на всей сумме существующих в природе признаков, и, конечно, мы еще очень далеки от него. Тем не менее, мы несомненно приближаемся к нему: сравнивая хотя бы наше родословное древо (схема XV) с родословным древом Грегори (схема XVIII), мы видим, что эти две филогенетические схемы безусловно стоят ближе друг к другу, чем родственные им схемы Копа и Геккеля.
Теперь, разобрав в общих чертах вопрос о положении хрящевых ганоидов в классификационных системах различных авторов, обратимся к вопросу об эволюции Chondrosteoidei.
Отметим прежде всего, что как мои собственные исследования, так и исследования моих учеников и сотрудников, приводят меня к трем основным выводам: s
1. Хрящевые ганоиды— Chondrosteoidei (Chondrosteidae, Acipenseridae и Polyodontidae) являются не дегенерировавшими, но примитивными формами, сохранившими многие черты своего анцестральнрго (с е-лахоидного) строения; они не происходят от пале о н и с ц и д.
2. Остальные актиноптеригии и кроссопт е-ригии (а также двоякодышащие) произошли от примитивной группы Osteichthyes, которую мы можем обозначить как Holosteoidei primi-tivi. Эти формы ближе стоят друг к другу, чем к Chondrosteoidei. Holosteoidei primitivi
1 Н о 1 m gr е n N. On the origin of the tetrapod Limb, Akta zoologica.Bd. 14, 1933.
ЭВОЛЮЦИЯ OSTEICHTHYES
219
имели сегментально построенные горизонтальные эврибазальные грудные плавники.
3. Группы Chondrosteoidei nHolosteoidei primitivi равноценны с систематической точки зрен.ия и обе происходят от только что описанных гипотетических Osteichthyes primitivi.
Так как мы во всех этих пунктах значительно отклоняемся от широко распространенного представления относительно филогенеза рыб, то мне хочется привести в самых кратких чертах те соображения, на основании которых я пришел к этой, отличной от общепринятой, точке зрения.
Выше нами уже было указано, что покровный костный скелет Chondro-stei является в некоторых отношениях более примитивным, чем покровный скелет остальных Osteichthyes и, в особенности, чем скелет Crossopterygii. У них сохранились, например, в своей примитивной ромбической форме многие чешуи и покровные кости черепа и удержались продольные гребни на наружной поверхности некоторых костей. У большинства Ostei-chtyes, т. е. у костных ганоидов, у Crossopterygii и, по всей вероятности, у Palaeoniscidae1 гребни эти сохранились лишь в эмбриональном (личиночном) состоянии, и притом лишь на тех костях, которые имеют отношение к органам боковой линии. По числу, форме и положению костей головы, а также по форме костей плечевого пояса, покровный костный скелет современных Chondrostei является весьма примитивным образованием, более примитивным, чем покровный костный скелет всех остальных Osteichthyes. На это можно было бы возразить, что у других Osteichthyes, например у современных костных ганоидов (Lepidosteus) и у Crossopterygii, сохранились мелкие рудиментарные зубы и космин, т. е. ткань, которая, быть может, развилась из плакоидных чешуй. Это, конечно, примитивный признак, и приходится признать, что у Acipenseridae плакоидные чешуи редуцировались целиком.
Таким образом, мы видим, что хрящевые ганоиды (Acipenseridae) по строению своего кожного скелета, т. е. чешуй и покровных костей головы, являются формами, гораздо более примитивными, чем костные ганоиды, Crossopterygii, Teleostei и, по всей вероятности, Palaeoniscidae, ичто их примитивные признаки нельзя объяснить путем редукции.
Если мы перейдем теперь к строению хрящевого черепа и сравним череп Acipenseridae с черепом только что упомянутых групп рыб и с черепом эласмобранхий (см. А. Н. Северцов, 1928), то мы увидим, что хрящевой череп Acipenseridae многими чертами своего строения схож с черепом примитивных селахий и что именно эти признаки отсутствуют у Hoiostei, Palaeoniscoidei и Crossopterygii.
1. У. примитивных акул и. у хрящевых ганоидов имеется ясно выраженное хрящевое рыло (rostrum); у высших рыб (Hoiostei, Palaeoniscoidei, Crossopterygii, Dipneusta) оно отсутствует или рудиментарно.
2. У хрящевых ганоидов и у акул хрящевые носовые капсулы широко раздвинуты и носовая перемычка (septum nasale) отсутствует; у остальных Osteichthyes (Holosteoidei) носовые капсулы тесно прилегают друг к другу и разделяются типичной тонкой носовой перемычкой.
3. У Acipenseridae имеется базальный угол (processus palatobasalis
1 Насколько мне известно, у современных Dipneusta не сохранилось гребней на покровных костях даже у личинок, что вполне понятно, если мы примем в соображение, что у них не образуется во взрослом состоянии трубчатого костного скелета вокруг органов боковой линии. Наоборот, у стегоцефалов в черепе были развиты костные каналы боковой линии, и этот факт может служить аргументом-в пользу того, что стегоцефалы произошли от Crossopterygii.
220
ГЛАВА VIII
medialis) и ясно выраженные processus palatobasales laterales, служащие для подвешивания palatoquadratum; мы знаем, что эти признаки, весьма характерные для примитивных эласмобранхий (Notidanidae, Chlamydose-lachus), отсутствуют у всех Holosteoidei.
4. У хрящевых ганоидов, так же как и у всех эласмобранхий, образуется характерно выраженный symphysis palatoquadrati; это является важным признаком различия между эласмобранхиями и хрящевыми ганои-дами с одной стороны, и всеми высшими Osteichthyes (остальные Actino-pterygii, Crossopterygii и Dipnoi)—с другой.
5. Palatoquadratum хрящевых ганоидов по своей форме похоже на palatoquadratum акул.
б. У хрящевых ганоидов, так же как и у примитивных акул (у взрослых Laemargus и у эмбрионов Mustelus nScyllium) имеется рудиментарное pharyngomandibulare;1 у Holosteoidei pharyngomandibulare не найдено.
Плавники современных осетровых сохранили примитивный характер; boz многих отношениях они более примитивны, чем плавники современных селахий. Так же как и у этих последних, они еще имеют горизонтальное расположение и сидят на широком основании(эврибазальный тип плавника); incisura metapterygoidea, характерная для высших рыб, здесь только слегка обозначается. В грудных плавниках осетровых рыб базальные хрящи pro- и mesopterygium, которые у селахий образуются срастанием проксимальных члеников плавниковых лучей, здесь еще не образовались, так что все хрящевые лучи еще являются свободными, т. е. такими же, какими они, например, были у Cladodus. Брюшные плавники построены еще более примитивно, чем грудные: метаптеригий Acipenseridae (Acipenser ruthenus juv., Acipenser nudiventris, Acipenser giildenstadti, Huso huso, Pseu-doscaphirhynchus kaufmanni), так же как у Cladodus и Pleuracanthus, состоит из еще не сросшихся между собой хрящевых basalia плавниковых лучей. Это наводит на мысль, что Acipenseridae ответвились не от высших, а, наоборот, от низших, очень примитивных эласмобранхий типа Cladodus. Непарные плавники и гетероцеркальный хвостовой плавник также построены у осетровых по типу примитивных плавников первичных акул.
Позвоночник хрящевых ганоидов построен по очень примитивному типу. Здесь, помимо общеизвестных примитивных особенностей позвоночника (пожизненно сохраняющаяся хорда, наличие всех четырех пар дуг, отсутствие слияния отдельных элементов друг с другом), С. В. Емельянов2 отмечает еще ряд признаков, указывающих на примитивность строения Acipenseridae. У них, например, гемальные дуги в хвостовом отделе позвоночника не только не дают гемальных отростков, но даже не замыкаются совсем в большинстве сегментов; по этому признаку Acipenseridae стоят ниже современных эласмобранхий. Вместе с тем у Acipenseridae развиты отсутствующие у Holosteoidei, но имеющиеся у эласмобранхий непарные supradorsalia и парные supradorsalia lateralia.
По данным А. Г. Рындзюнского,3 хвостовые и туловищные миотомы хрящевых ганоидов, так же как и у акул, асимметричны по отношению к горизонтальной септе, совпадающей с их первичной плоскостью симметрии и имеют в качестве дефинитивной вторичную плоскость симметрии,
1 Palatoquadratum хрящевых ганоидов при помощи соединительнотканной связки, в которой лежит pharyngomandibulare, связано с proc, palatobasalis lateralis черепа. Судя по положению этой связки, можно предположить, что у Acipenseridae, так же как и у акул (Notidanidae, Laemargus, Spinaceidae), имелось несколько видоизмененное палатобазальное сочленение верхней челюсти с черепом.
’Емельянов С. В., Развитие и строение некоторых мало изученных элементов позвоночника рыб, их связь с непарными плавниками и некоторые вопросы теории конечностей. Сборник памяти акад. А. Н. Северцова, т. 1, 1939.
•Рындзюнский А. Г., Развитие формы миотома рыб, Труды Ин-та эволюц. морф. им. ак. Северцова Академии наук СССР, том II, вып 4. 1938.
8В0ЛЮЦИЯ OSTEICHTHYES
221
что отличает хрящевых ганоидов от всей группы Holosteoidei. При этом по ряду признаков миотомы имеют у осетровых более примитивное строение, чем даже у современных акул. Такого рода примитивными признаками являются: 1) меньшая мощность горизонтальной септы, 2) меньшая обособленность друг от друга их конусов, 3) отсутствие срединной складки, 4) меньшее приближение их ко вторичной симметрии.
Мы могли бы присовокупить к этому еще целый ряд эмбриональных признаков, свойственных как эласмобранхиям, так и хрящевым ганоидам и отсутствующих у высших Osteichthyes; сюда относится, например, характер срастания трабекул с парахордалиями,1 образование трабекулярного дна черепа и т. д. Мы думаем, однако, что и приведенных нами признаков достаточно для того, чтобы показать, что хрящевые ганоиды (Chondrostei) являются весьма примитивными Osteichthyes, довольно близко стоящими к низшим эласмобранхиям.
Мы считаем также, что гипотеза о дегенеративной эволюции Chondrostei1 2 не может объяснить селахоидного характера указанных выше признаков хрящевых ганоидов. Поэтому мы должны принять, что Chondrostei являются относительно мало измененными потомками примитивных Osteichthyes, обладающими многими селахоидными признаками. Г ипо-теза о происхождении хрящевых ганоидов от известных нам девонских палеонисцид является, следовательно, невероятной. В моей работе о развитии черепа Acipenseridae я подробно разбираю этот вопрос.3 4
О ПОКРОВНЫХ КОСТЯХ ЧЕРЕПА CHONDROSTEOIDEI1
Относительно систематического положения и филогенеза ганоидов были высказаны две гипотезы: 1) гипотеза, которой держатся преимуще
1 У хрящевых ганоидов и у селахий трабекулы прирастают к вентральной поверхности парахордалий, у остальных Osteichthyes они прирастают к их передним концам (А. Н. Северцов, 1928).
2 Исследователи, принимающие эту гипотезу, говорят лишь в очень общей форме о дегенеративных изменениях у Acipenseridae. Они не разбирают подробно отдельные дегенеративные признаки. Мы, конечно, не отрицаем, что у Acipenseridae (как и у всякой другой группы) имеются и дегенеративные признаки, к которым можно отнести следующие: 1) редукция зубов у взрослых Acipenseridae (у личинок они хорошо развиты); 2) возможное уменьшение величины большинства туловищных чешуй [мы говорим «возможное», ибо не ясно, имеем ли мы здесь редукционный процесс, поскольку мелкие чешуи Acipenseridae изменили свою форму вполне закономерно (А. Н. Северцов, 1926) и их число в течение филогенеза несомненно возросло]; 3) уменьшение величины глаз у Pseudoscaphirhynchus kaufmanni; 4) редукция второго дорзального конуса миотомов и некоторые другие признаки.
3 S е we г t z о f f A. N., Head Skeleton and muscles of Acipenser ruthenus, Acta Zool., Bd. 9, 1928.
4 В 1932 г. Стеншиб опубликовал работу о триасовых рыбах Гренландии (Triassic Fishes from East Greenland, Kovenhavn, 1932), в которой он возражает против взглядов А. Н. Северцова на положение в системе хрящевых ганоидов. Стеншиб принимает, что то большое число костей, которое развито в .черепе рыб (в частности, хрящевых ганоидов) произошло путем распадения крупных окостенений головы первичных форм. Эта гипотеза Стеншиб стоит в связи с его взглядом на хрящевых ганоидов как на дегенерировавшие формы. В ответ на работу Стеншиб А. Н. Северцов предпринял большое исследование по покровным костям Chondrostei, в котором он показывает, что большое число костей в черепе этих рыб является примитивным признаком, и что эволюция черепа рыб идет в сторону уменьшения, а не увеличения числа костей. Это исследование осталось не вполне законченным и не было опубликовано. Этой своей работе А. Н. Северцов придавал большое значение и считал необходимым включить ее результаты в настоящую книгу; Исполняя желание А. Н. Северцова, в книгу включены в качестве особого раздела этой главы оставшиеся после его смерти материалы по морфологии покровных костей Chondrostei.
(Прим, ред-)
222
ГЛАВА VIU
ственно сравнительные анатомы (И. Мюллер, Геккель и другие, в том числе и я) и которая принимает, что хрящевые ганоиды являются примитивными формами, близкими к селахиям, и 2) гипотеза, которой придерживаются палеонтологи (Коп, Трэквер, Ватсон и др.) и которая утверждает, что хрящевые ганоиды являются дегенерировавшими формами, происшедшими от более высоких Chondrosteoidei. Эта последняя гипотеза с некоторыми добавлениями была принята Стеншиб (1932),1 который выставляет против моих взглядов ряд возражений.
В основу своего взгляда Стеншиб положил свою гипотезу о происхождении и эволюции костного черепа, сильно отличающуюся от тех взглядов, к которым привели меня мои сравнительно анатомические и эмбриологические исследования. Стеншиб принимает, что то большое число костей, которое развито в черепе рыб, произошло путем распадения крупных окостенений головы первичных форм. Эта гипотеза Стеншиб стоит в полном согласии с его взглядами на происхождение хрящевых ганоидов путем дегенерации от примитивных кроссоптеригий. Он принимает, что Acipenseridae являются непосредственными потомками Saurichthyidae и что череп их развился из черепа этих последних. Черепные кости Acipenseridae произошли путем распадения костей черепа Saurichthyidae. Это приводит его к гипотезам, конечно, возможным, но мало вероятным, несогласным с сравнительно эмбриологическими фактами и выводами. Мы не будем подвергать его взгляды подробной критике, но изложим новые фактические данные, которые привели нас к выводам, отличным от палеонтологических построений Стеншиб. Наши исследования привели к выводу, который можно назвать теорией срастания костных элементов, основанную главным образом на анатомических и эмбриологических данных по костному черепу осетровых рыб.
Если мы посмотрим, как развиваются кости черепа Acipenser ruthenus, то найдем, что у него закладывается ряд продолговатых пластинок, костных чешуй, из которых каждая несет продольный гребень на поверхности. Эти кости в некоторых случаях срастаются друг с другом, образуя более крупные костные пластинки (frontale l-f-frontale 2). Мы нашли, что все они закладываются по одному определенному типу развития, подробно разобранному нами при описании покровных костей у Osteichthyes pri-mitivi .Именно в костях, лежащих рядом с каналами боковой линии,продольный костный гребень и боковая часть костной пластинки срастаются во время развития и образуют костную трубку, внутри которой проходит канал боковой линий (рис. 51 и 52). Этот же тип развития мы нашли в черепе у костистых рыб и костных ганоидов (Amia, Lepidosteus). Тип этот настолько своеобразен, что мы едва ли можем истолковать его иначе, как признав, что современные костные рыбы произошли от таких рыб, у которых череп состоял из многочисленных костей с продольными гребнями.
При исследовании других хрящевых ганоидов: Acipenser giildenstadti, Acipenser stellatus, Huso huso, Pseudoscaphirhynchus kaufmanni (й, вероятно, Polyodon folium) мы нашли тот же тип строения, что и у Acipenser ruthenus, те же чешуи с продольными гребнями на наружной поверхности, то же срастание и разрастание зачатков крупных костей и тот же ряд непарных медианных костей, в котором кости, лежащие в теменной области, рудиментарны. Мы нашли также, что и у этих форм кости с гребнями, лежащие около эпителиальных каналов боковой линии, участвуют в образовании костных каналов боковой линии: продольный гребень и часть пластинки, лежащая под каналом, срастаются и образуют костную трубку канала боковой линии.
Изучение костей черепа современных осетровых рыб показывает нам,
J S t е ns i б Е., Triassic Fishes from East Greenland, Kovenhavn, 1932.
ЭВОЛЮЦИЯ OCTEICHTHY'ES
223
что покровные кости черепа у них гораздо более многочисленны, чем покровные кости черепа костных ганоидов и костистых рыб.
Эти многочисленные покровные кости расположены в трех областях черепа: 1) в дермосупратемпоральной, 2) в супраростральной и 3) в суб-ростральной. В дермосупратемпоральной области по бокам от непарной dermosupraoccipitale постоянно лежат следующие кости: большая supra-temporale (рис. 55, stm-t), впереди которой помещается вариирующее число более мелких supratemporalia. У одного экземпляра Acipenser guldenstadti мы находим три небольших supratemporalia впереди от большего заднего
Рис. 55 А. Покровные кости черепа Acipenser guldenstadti (длина головы 10,2 см).
ds. ое.—dermosupraoccipitale; fr.—frontale; n.—nasale; par.—parietale; pr. fr. — prae-frontale; pt. fr. — postfrontale; pt. oc.— postoccipltale; sq.—squamosum; srl.—sup-rarostralia lateralia; srm.—suprarostralla medialia; stm.,—stm.5—supratemporalia 1-5
Рис. 55 В. Покровные кости черепа Acipenser guldenstadti (длина головы 12,8 см).
ds. ос.—dermosupraoccipitale; fr.—frontale; m.i, m.a> m,3—medialia; par. — parietale; pt. oc. — postocclpitale; sq. — squamosum; srl.—suprarostralla lateralia; srm.—supra-rostralia medialia; stm.,—stm.4—supratemporalia 1—4.
supratemporale 1 (рис. 55 A, stm4), t. e. supratemporalia 2, 3, 4 с левой стороны и четыре supratemporalia (рис. 55 A, stm2, stm3, stm4, stm5) с правой стороны. У другого экземпляра Acipenser guldenstadti (рис. 55 В) мы видим с левой стороны в супратемпоральной области черепа четыре маленьких supratemporalia (рис. 55 В, stm2, stm3, stm4, stm5), а с правой—три (рис. 55 В, stm2—stm4). Таким образом, мы видим, что у Acipenser guldenstadti число передних surpatemporalia вариирует.
Подобное же явление мы наблюдаем и у других осетровых рыб. Так, у молодого Acipenser stellatus (рис. 56 А) с левой стороны расположены четыре supratemporalia, с правой семь (рис. 56 A, stm4—stm7). У взрослого Acipenser stellatus (рис. 56 В) с левой стороны—семь, с правой—шесть supratemporalia. У Acipenser ruthenus(попрепаратами. О. Ланге, рис. 57)
224
ГЛАВА VIII
по бокам от dermosupraoccipitale лежит большое заднее supratemporale 1 (stmj и одно маленькое supratemporale 2 (stm2) с левой и два с правой стороны, до сих пор еще никем не описанные.
В супратемпоральной области черепа белуги (Huso huso) число этих костей также .сильно вариирует. Так, у одной молодой белуги (рис. 58 А) в супратемпоральной области с левой стороны мы видим шесть supratemporalia, с правой—четыре. У другой небольшой белуги (рис. 58 В) их имеется с левой и с правой сторон по четыре. У крупной белуги
(длина головы 37 см; рис. 58 С) мы находим с обеих сторон по пяти supratemporalia: большие задние supratemporalia (smtt), типичные для всех осетровых, и по четыре боковых supratemporalia (по две крупных и две мелких: stm2—stm5). Сравнивая количество и расположение supratemporalia крупной белуги (рис. 58 С) и маленьких молодых белуг (рис. 58 А и В), можно думать, что в течение роста рыбы некоторые из supratemporalia молодых белуг отстают в росте и редуцируются.
Морфологические закономерности
ЭВОЛЮЦИЯ OCTEICHTHYES
Рис. 58. А. Покровные кости черепа очень молодого экземпляра Huso huso (длина головы 8 см).
ds.ос.—dermosupraoccipitale; fr.—frontale; m.— mediale; par.—parietale; pt. oc.—post-occipitale; sq. — squamosum; srm.—supra-rostralia medialia; stm. x—stm. 6—supratem-poralia 1—6.
Рис. 58 В. Покровные кости черепа несколько более крупной белуги Huso huso (длина головы 8,7 см).
ds.ос.—dermosupraoccipitale; fr.—frontale; par.— parietale; pt.oc.—postoccipitale; sq.—squamosum; srm.—suprarostralia medialia; stm.x—stm.a — supratemporalia 1—4.
Рис. 58 С. Покровные кости черепа взрослой белуги Huso huso (длина головы 37 см.). ds.ос.—dermosupraoccipitale; fr.—frontale; m. — mediale; par.—parietale; pt. oc.—postoccipitale; sq. — squamosum; srm. — suprarostralia medialia; stm.; —stm. 5—supratemporalia 1—5.
to to Ut
226
ГЛАВА VIII
Рис. 59. Покровные кости черепа Pseu-doscaphirhynchus kaufmanni. ds.ос. —dermosupraoccipitale; fr.— frontale; m.—mediale; par.—parietale; pt. oc. — post-oecipitale; srl.—suprarostralia lateralia; srm.— suprarostralia medialia; sq. — squamosum;
stm.x—stm.B— supratemporalia 1—5.
У Pseudoscaphyrhinchus kaufmanni мы находим с каждой стороны одну большую supratemporale (рис. 59 stmj, три маленькие supratemporalia (stm2—stm4) с правой и четыре с левой стороны.
Новые и интересные данные мы получили при исследвании ростральных покровных окостенений в дорзальной и вентральной стороне рыла, т. е. в супраростральной и в субростральной областях черепа осетровых рыб. У Acipenseridae по моим исследованиям число suprarostra-П а и basirostralia сильно отличается от числаэтих костей, находимого нами у костных ганоидов и костистых рыб, а также у двоякодышащих икисте-п е р ы х рыб. На тупом рыле у Acipenser giildenstadti расположено большое число мелких suprarostralia. У экземпляра, изображенного на рис. 55 А, таких мелких suprarostralia имеется 54, а у изображенного на рис. 55 В — 56. У молодой севрюги (рис. 56 А) с длиной головы в 14,3 см. мы нашли 40 suprarostralia. Интересно и само строение этих костей у молодого экземпляра севрюги: на каждой из них имеется продольный гребень с направленным назад острием. В черепе более крупной севрюги с длиною головы в 30 см (рис. 56 В) имеется 54 suprarostralia, и на них видна типичная скульптура. У взрослой стерляди (рис. 57) имеется всего 34 suprarostralia.
Особенно интересно для нас число suprarostralia у белуги (Huso huso). У молодой белуги (рис. 58 А и В) мы находим большое число suprarostralia
с типичными гребнями на дорзальной поверхности, а именно: у одной 45, а у другой—54. У крупной белуги (рис. 58 С) мы видим гораздо меньшее число suprarostralia, а именно 22, причем здесь suprarostralia не соприкасаются между собой, как у молодых особей, а отделены друг от друга свободными промежутками. Таким образом, число их у взрослого экземпляра уменьшилось весьма значительно.
Изучение других взрослых белуг показывает нам, что уменьшение числа и относительной величины suprarostralia в течение индивидуального развития животного—о бщее правило.
Результаты, полученные нами при исследовании suprarostralia Acipenseridae подтверждаются результатами изучения basirostralia этих рыб. Оказывается, что число этих элементов в действительности также гораздо больше, чем это думали до сих пор.
У одного экземпляра Acipenser giildenstadti на нижней поверхности головы (рис. 60) имеется пять basirostralia (bs. г.); у других экземпляров число этих костей доходит до 10—12, причем расположение их остается тем же: эти косточки лежат впереди основной basirostrale mediale (рис. 60 bsr. m.).
эволюция, osteichthyes
227
У Acipenser stellatus имеется еще большее число basirostralia. Так, у экземпляра, изображенного на рис. 61, мы нашли 17 basirostralia, представляющих собою длинные пластинки с продольными гребнями, занимающие типичное положение между костными каналами боковой линии.
У изображенного-на рис. 62 Acipenser ruthenus мы находим 15 basiro-stralia, типичных по форме и по положению. /йВйк
• Рис. 60. Нижняя поверхность головы Acipenser guldenstadti (длина головы 4,9 см).
bsr. m.—basirostrale modi ale; bsr.y—bsr.5—baslrost-• ralia 1—5.
Рис. 61. Нижняя поверхность головы Acipenser stellatus (длина головы 30 см).
£sr.—basirostralia; bsr. m.—basirostrale-mediale; c. 1. 1.—костный канал боковой линии.
ности рыла имеются 32 basirostralia с гребнями, тесно прилегающими друг к другу. У крупной белуги (рис. 64, длина головы 46 см) мы находим всего 17 basirostralia, далеко отстоящих друг от друга. Сопоставление возрастных изменений supra- и basirostralia ясно показывает, что у белуги число этих элементов' в течение индивидуальной жизни уменьшается, что стоит в явном противоречии с гипотезой происхождения этих костей путем разделения покровных элементов черепа. Вернее говоря, мы здесь видим, что в течение индивидуальной жизни число supra- и basirostralia сначала увеличивается, так же как и у остальных хрящевых ганоидов (Acipenser guldenstadti, Acipenser stellatus, Acipenser ruthenus), а затем убывает, причем сами кости отодвигаются друг от друга.
15*
Рис. 64. Нижняя поверхность рыла Huso huso (длина головы 46 см).
bsr.—basirostralia; с. 1.1,—костный канал боковой линии.
ГЛАВА VIII
Рис. 62. Нижняя поверхность рыла Acipenser ruthenus (длина головы 10 см)-
bsr—basirostralia; bsr. m.—basirostrale mediale; e.1.1.—костный канал боковой
линии.
Рис. 63. Нижняя поверхность рыла Huso huso (длина головы 8 см), bsr.—basirostralia; bsr. m.—basirostrale mediate.
5 ВО ЛЮ НИ H OSTEICHTHYES
229
На рис. 59 представлена сверху голова Pseudoscaphirhynchus kauf-manni. Подсчет ростральных элементов показывает нам, что здесь имеется много suprarostralia (84 косточки), в то время как на нижней поверхности головы имеется только пять basirostralia (рис. 65), причем на всех basirostralia, не исключая и длинного заднего элемента, наблюдается типич
ная скульптура.
Polyodon на ранних стадиях развития (рис. 66 А и В) напоминает Pseudoscaphirhynchus. Его расширенное и удлиненное рыло (рис. 67 А и В) образуется позднее в течение индвидуального развития. Мои наблюдения
над костями черепа Polyodon1 несколько отличаются от наблюдений Брид-
жа (1879). Мы находим у Polyodon парные ра-rietalia (par.), в то время как по Бриджу имеется только одно непарное parietale. Позади от parietalia лежат две der-mosupraoccipitalia (ds. ос.). Впрочем, без эмбриологических наблюдений нельзяточно решить вопроса, имеются ли здесь только parietalia или же parietalia + der-mopostoccipitalia. Между ними и parietalia проходит поперечный затылочный канал боковой линии. Впереди от parietalia лежат парные fron-talia (рис. 67 A, fr.), между которыми расположено длинное mediale (ш.). Кнаружи от fron-talia лежат две небольшие квадратные кости, вероятно соответствующие костям х Huso huso (рис. 58 В и С). Кнаружи от кости х лежит большая squamosum
Рис. 65. Нижняя поверхность головы Pseudoscaphirhynchus kaufmanni.
bsr.—basirostralia; bsr. m—basirostrale mediale; c.I.l. костный канал боковой линии.
(sq), к которой прилегает глаз. Впереди от squamosum лежит nasale (п). Впереди от mediale расположены длинные заостренные средние suprarostralia medialia (srm); по бокам от этих средних rostralia лежат многочисленные звездчатые suprarostralia lateralia. Этих последних костей имеется
очень много—всего 98 suprarostralia lateralia.
На рис. 67 В изображен череп Polyodon folium с нижней стороны. Посредине видны вытянутые средние basirostralia medialia, числом 10; по бокам от них лежат звездчатые basirostralia lateralia, которых также имеется очень много: мы насчитали 137 косточек.
Мы видим, таким образом, что в супраростральном и субростральном отделе черепа Polyodon кости в общем расположены по тому же типу, как и у осетровых и у скафиринхов; но мы полагаем, что число и расположение
1 Я был тяжело болен, и препарат костей черепа Polyodon был для меня сделан моей ассистенткой Н. О. Ланге.
230
ГЛАВА VIII
этих костей изменилось вторично и что число их увеличилось сравнительно с тем, что мы наблюдаем у Acipenseridae.
На основании изложенных выше исследований можно притти к следующим выводам.
Предки рыб с костным скелетом, т. е. предки Acipenseridae, Holostei и всех Teleostei, а именно Osteichthyes primitivi, имели скелет головы, состоявший из большого числа сравнительно небольших ромбических чешуй, из которых каждая на своей поверхности несла продольный костный гребень. Чешуи, лежавшие вдоль каналов боковой линии, образованы из срастающихся гребней и частей пластинок, лежавших под эпителиальными каналами боковой линии. У современных хрящевых ганоидов некоторые из этих чешуй (костей) черепа разрослись, другие слились и т. д.; в общем они сделались типичными черепными костями.
Рис. 66 А. Малек Polyodon folium (20 мм). Вид сверху.
Рис. 66 В. Тот же малек Polyodon folium. Вид снизу.
Костные гребни, характерные для туловищных и черепных костей (чешуй) осетровых и костных ганоидов (Amia, Lepidosteus), у костистых рыб (все Teleostei) исчезли и только в течение эмбрионального развития сохраняются на чешуях, связанных с каналами боковой линии этих форм.
Таким образом, костный череп позвоночных животных представляет собой результат разрастания и слияния многочисленных ромбических костных чешуй с гребнями (расположенных на голове и в передней части туловища животного) и их последующего видоизменения. Этот вывод является результатом непосредственного наблюдения развития костей черепа, а не предвзятой палеонтологической гипотезы.
Сравнивая покровные кости головы осетровых с костями костных ганоидов (Amia, Lepidosteus) и костистых рыб, мы видим, что у этих последних череп построен совершенно иначе и что целый ряд покровных окостенений,
ЗВ0Л10ЦИЯ OSTEICHTHYES
231
л
Рис. 67 А. Верхняя поверхность головы Polyodon folium (длина тела 19,5 см), ds. ос.—dermosupraoccipitalla; fr.—frontale; m.— mediale; n.—nasale; par.—parletalia; sc.—squamosum; srl.—suprarostralia lateralia; srm.—suprarostralia medialia; stm.i—supratemporale 1.
Рис. 67 В. Нижняя поверхность головы того же экземпляра Polyodon folium.
bsr. 1.—basirostralia lateralia; bsr. m.—basiro-
например многочисленные supra- и basirostralia, типичные для хрящевых ганоидов, здесь совершенно отсутствуют.
Таким образом, мы видим, что гипотеза Стеншиб о происхождении покровных костей в черепе Acipenseridae путем распадения крупных окостенений головы первичных ископаемых Saurichthyidae не согласуется с только что изложенными сравнительно-анатомическими и сравнительноэмбриологическими фактами.
Вместе с тем отметим, что палеонтологи, создавшие эту гипотезу, исходят при этом из другой гипотезы, именно из предположения, что первичной стадией в эволюции скелета позвоночных был слизистый хрящ, который в течение филогенеза превратился в кость, в свою очередь превратившуюся в дальнейшем путем дегенерации в обыкновенный хрящ; поэтому все формы современных рыб с хрящевым скелетом, т. е. круглоротые, акуловые, химеры и осетровые, рассматриваются ими как дегенерировавшие формы.
Палеонтологи говорят также о том, что череп предков Acipenseridae первично состоял из эндохондральных костей и превратился в хрящевой череп вторично лишь впоследствии. Исследования В. А. Ракова1 показали, что в хрящевом черепе различных осетровых (Acipenser giildenstadti, Acipenser ruthenus, Acipenser stellatus, Pseudo-scaphirhinchus kaufmanni и Huso huso) имеются окостенения перихондрального характера. Окостенения эти встречаются только у взрослых форм и притом наиболее резко выражены лишь у очень старых экземпляров. Они располагаются, как правило, около отверстий выхода сосудов и нервов и вариируют в величине, форме и числе. В. А. Раков гомо-логизирует их на основании их расположения в черепе с окостенениями, имеющимися в черепе Crossop
terygii, Hoiostei и Teleostei и находит: в орбитальной области—orbitospheno-ideum (у Acipenser giildenstadti) и alisphenoideum (у Acipenser giildenstadti, Acipenser. stellatus, Acipenser ruthenus), в слуховой области—prooticum и opistoticurri (у Acipenser giildenstadti и Pseudoscaphirhynchus kaufmanni) и в затылочной области—occipitalia lateralia (у всех исследованных им форм). Это могло бы навести нас на мысль, что у предков осетровых рыб некогда имелось большое число впоследствии редуцирующихся хрящевых окостенений. Однако Раков рассматривает указанные окостенения как орименты черепных костей, закладывающихся в онтогенезе осетровых на
1 R a k о w W. A., Zur Frage der Verknocherungen des Knorpelschadels der Acipenseridae, Zool. Jahrb. (Anat.), Bd. 57, 1933.
232
ГЛАВА VIII
очень поздних стадиях развития. При изучении истории развития Huso huso ему удалось показать, что окостенения эти с возрастом увеличиваются в размерах, вытесняя хрящ иногда настолько, что, например, occipitalia на некотором участке затылочной области черепа представляют собой всю толщу стенки черепа и выходят в полость его, не имея под собой хрящевой подстилки. Вместе с теммы не имеем абсолютно никаких доказательств того, что, кроме этих немногих костей, у предков Acipenseridae существовали еще и другие кости, образовавшие полностью костный осевой череп этих животных. Повидимому, и у ископаемого хрящевого Chondrosteus череп был еще хрящевым.
Если бы мы приняли вторичное усиление хрящеобразования в черепе Chondrostei и его происхождение от вторично изменившегося черепа палео-нисцид (Ватсон), то нам пришлось бы также разъяснить, почему костный осевой череп палеонисцидного типа, т. е. тропибазальный череп с миодомом и тонкой глазничной перегородкой, мог вторично приобрести признаки примитивного пла-тибазального селахоидного черепа без миодома, т. е. вторично приобрести признаки, которыми не обладали их палеонис-цидные предки. Этот факт остается непонятным, если стоять на точке зрения гипотезы дегенерации.
ЭВОЛЮЦИЯ HOLOSTEOIDEI PRIMITIVI
По вопросу о путях эволюции костных рыб я держусь взгляда, что: 1) Actinopterygii, т. е. Palaeoniscoidei, Holostei и Teleostei в широком смысле слова, а также Crossopterygii и Dipnoi образуют одну большую группу—Holosteoidei и что 2) Palaeoniscoidei во многих отношениях более примитивны, чем Crossopterygii (А. Н. Северцов, 1923).
Сравнительно морфологические исследования показывают нам, что эти группы рыб (Holostei + Teleostei-}-Crossopterygii-]-Dipnoi) в целом ряде важных признаков отличаются от хрящевых ганоидов (Chondrosteoidei mihi) и что именно теми признаками, которыми они отличаются от этих последних, они сходны друг с другом.
Так, кожный скелет Holosteoidei эволюирует в ином направлении, чем у Chondrostei. Плакоидные чешуи эласмобранхийных предков этих форм сохранились у наиболее примитивных представителей современных Actinopterygii (Lepidosteidae) и Crossopterygii (Polypterus), что является признаком их примитивной организации. На поверхности чешуй и костей этих рыб образовалась особая остеоидная ткань, так называемый космин, вследствие чего эти образования утолщаются и образуют плотный и непроницаемый панцырь, защищающий тело животного. Продольные гребни чешуй и покровных костей головы здесь редуцировались на всех тех костях, которые не имеют отношения к каналам боковой линии.1 У Holosteoidei развиты некоторые кости, отсутствующие у Chondrostei (Acipenseridae, Polyodontidae). В жаберной крышке, например, развивается ргаеорег-culum, в котором располагается канал боковой линии, идущий к нижней челюсти. Здесь имеются также парные, несущие зубы, сошники, отсутствующие у- Chondrostei (А. Н. Северцов, 1926). У Chondrostei имеется лишь одна птеригоиднаякость, у остальных же Actinopterygii, так же как и у Crossopterygii, их имеется целых три: ento-,ecto-n metapterygoideum. Число rostralia у Chondrostei гораздо больше, чем у Holosteoidei, причем у этих последних совершенно отсутствуют basirostralia. На нижней челю
1 В костях, имеющих отношение к этому каналу, они сохранились лишь в эмбриональном состоянии, принимая участие в построении костного канала боковой линии, как, например, у Amia, Lepidosteus, Polypterus.
эволюция oscteichthyes
233
сти Holosteoidei, кроме dentale, имеющейся и у Chondrostei, развиваются еще и другие покровные кости, несущие зубы, как, например, spleniale, coronoideum, angulare и т. д. Praemaxillare имеет у Holosteoidei совершенно другое положение, чем у Chondrostei (Acipenseridae и Polyodontidae): у Chondrostei оно сидит на palatoquadratum (рис. 68 А, р. mx., р. qu'.), у Holosteoidei (Holostei, Palaeoniscidae, Crossopterygii),—на передней части (этмоидальном отделе) осевого черепа (рис. 68 В, р. mx., р. antor.).
Мы видим, таким образом, что
в отношении костного скелета Chon7 drosteoidei отличаются от Holosteoidei целым рядом морфологически важных признаков и что именно эти признаки сближают между собой Holostei (L е pi-do s t е u s, Amia ит. д.), Р а-laeoniscidae и Crossopterygii, образуя единую группу (Holosteoidei mihi).
Этот вывод подтверждается сравнением других характерных для представителей обеих этих групп (Chondrosteoidei и Holosteoidei) признаков. Если мы сравним нейрокраниум Holosteoidei и Chondrosteoidei, то увидим, что эти образования отличаются друг от друга как по своему строению у взрослых форм, так и по характеру своего эмбрионального развития.
Так, у всех Holosteoidei отсутствует базальный угол (processus palatobasalis medialis), а также processus palatobasalis lateralis; rostrum развит слабо или его вовсе нет. Трабекулы прирастают не к нижней поверхности парахорда-лий, как у селахий и осетровых рыб, а к их передним концам.
Все это относится к черепу взрослых форм; но и общая конфигурация эмбрионального осевого черепа, почти одинаковая у Holostei и у Crossopterygii, существенно отлична от конфигу-
Рис. 68 А. Сагиттальный разрез через переднюю часть головы довольно крупного малька Acipenser ruthenus.
Рис. 68 В. Соответствующий сагиттальный разрез через переднюю часть головы малька Amia calva.
d. pmx.—премаксилярный зуб; N,—носовая ямка; р. antor.—processus antorbitalis осевого черепа (заштрихован); р. mx.—praemaxilla; р. qu.— небноквадратный хрящ (пунктир). На обоих рисунках обратить внимание на соотношение между praemaxilla и нёбнокваДратным хрящом (верхней челюстью). У малька стерляди (рис. 68 А) видно, что praemaxilla со своим зубом (d. pmx.) сидит на нёбноквадратном хряще; у малька Amia, так же как и у всех остальных Holosteoidei (включая Palaeoniscidae и Crossopterygii), ргае-maxilla со своим зубом сидит на осевом черепе, на processus antorbitalis (из А. Н. Северцова, 1923).
рации нейрокраниума у Chon-
drostei и у акул. У Holosteoidei ростральные концы нёбноквадратного хряща лежат далеко друг от друга, так что symphysis palatoquadrati, столь характерный для Chondrostei и Elasmobranchii, здесь не образуется. У всех Holosteoidei имеется квадранторбитальное сочленение верхней челюсти с черепом, т. е. у них palatoquadratum сочленяется с processus antorbitalis черепа (рис. 69 D, a. antor.). Как мы это видели выше, это сочленение хорошо развито у Chlamydoselachus (рис. 69 A, a. antor.) и, вероятно, существовало
234
ГЛАВА VIII
также у Cladodontidae; у остальных акул (рис. 69 В) и у Chondrosteoidei (рис. 69 С) оно исчезло. Прежде я считал, что у Holosteoidei не имелось палатобазального сочленения верхней челюсти с черепом, но теперь я склоняюсь ж мысли, что такая связь в несколько измененной форме у них все же суще-
рм
Рис. 69 А—D2. Ряд рисунков, изображающих эволюцию сочленений между нёбноквадратным хрящом и осевым черепом. А—череп Chlamydoselachus anguineus; В— череп Laemargus borealis; С—череп Pseudoscaphirhynchus kaufmanni; D—череп малька Amia calva; Dx—череп малька Amia calva на несколько более поздней стадии развития; D2—череп взрослой Amia calva (рис. А, С, D из А. Н. Северцова, 1931; рис. В из А. Н. Северцова и Н. Н. Дислера, 1924; рис. Dx—из Шмальгаузена; рис. D2—из Гудрича). Рис-D и Dt сильно увеличены.
a. anto.—квадранторбитальное сочленение; a. pl. Ь.—палатобазальное сочленение; р. pl. Ь.—processus palatobasalis осевого черепа; ph. m.—pharyngomandibulare; р. mtpt.—processus metapterygoideus; p. orb.—proc, orbitalis нёбноквадратного хряща; p. qu.—нёбноквадратный хрящ. На рис. 69 А мы видим, что у Chlamydoselachus (по нашему мнению, также и у Cladodus) развиты оба сочленения верхней челюсти; квадранторбитальное (a. anto.) и палатобазальное (a. pl. Ь.); у большинства селахий и у всех Chondrosteoidei сохранилось палатобазальное сочленение между processus palatobasalis черепа и processus orbitalis нёбноквадратного хряща; у Laemargus (В) и у Scaphirhynchus (С) имеется также и pharyngomandibulare, которое сохранилось еще у этих форм. У малька Amia (D) мы еще видим хорошо сохранившееся квадранторбитальное сочленение (a. anto.) примитивного селахоидного черепа; на несколько более поздней стадии (О>) мы находим ясно выраженный processus metapterygoideus, который соответствует, по нашему мнению, сдвинувшемуся каудально processus orbitalis*(pnc. 69 A, B,p.orb.) акул. Поэтому мы думаем, что у Holosteoidei (D), повидимому, сохранились и развились дальше оба сочленения верхней челюсти с осевым черепом, существовавшие у их селахоидных прародительских форм (А).
ствовала; вполне возможно, что processus metapterygoideus (рис. 69 Dn р. mtpt.) хрящевых ганоидов и костистых рыб1 соответствует несколько сдвинувшемуся назад processus orbitalis селахий; в таком слу
1 У кистеперых processus metapterygoideus рудиментарен (Sewertzoff A. N. Der Ursprung der Quadrupeda, Palaeont. Zschr., 1926.)
ЭВОЛЮЦИЯ OSTEICHTHYEC
235
чае метаптеригоидное сочленение Holosteoidei будет соответствовать палатобазальному сочленению предков хрящевых ганоидов. У Holosteoidei, вероятно, сохранились и достигли дальнейшего развития оба сочленения типа Chlamydoselachus, т. е.'у них имелась весьма прочная связь между верхней челюстью и черепом; у хрящевых ганоидов сохранилось только переднее сочленение.
Таким образом, мы действительно имеем перед собою ряд признаков, относящихся к биологически важным органам1—признаков, общих для Palaeoniscidae, Holostei, Teleostei и Crossopterygii, и мы считаем поэтому, что имеем основания для объединения всех этих рыб в одну систематическую группу Holosteoidei, которую мы вправе противопоставить хрящевым ганоидам (Chondrosteoidei).
Мы только что отметили, что у потомков первичных Osteichthyes развились два различных способа прикрепления верхней челюсти к осевому черепу, а именно: палатобазальный (Chondrosteoidei) и палатобазаль-ный+квадранторбитальный (Holosteoidei) тип сочленения, и мы считаем теперь своевременным поставить вопрос о том, из какой анцестральной формы челюстного аппарата могли развиться оба эти типа?
Интересно, что исходную форму челюсти мы встречаем у низших акул. У Chlamydoselachus, например, верхняя челюсть (palatoquadratum) связана с осевым черепом в двух пунктах, а именно: processus orbitalis palatoquadrati сочленен здесь с processus palatobasalis lateralis глазничного отдела черепа, а передняя часть palatoquadratum соединена посредством соединительной ткани с нижней поверхностью processus antorbitalis cranii (рис. 69 А). Первая из этих связей (рис. 69 А, р. pl. Ь.) полностью соответствует палатобазальному сочленению у акул (Laemargus, Spinax, Acan-thias etc.) и подвешиванию верхней челюсти у Acipenseridae (рис. 69 С). Вторая связь соответствует квадранторбитальному сочленению, имеющемуся у Holostei (Amia; рис. 69 D), у Teleostei, Palaeoniscidae, Saurich-thyidae и у Crossopterygii.
Е. Стеншиб, которому удалось наблюдать вполне сохранившийся череп > ископаемой акулы Cladodus, нашел, как он мне сообщил об этом, что череп этот необычайно похож на череп современного Chlamydoselachus, и мы можем поэтому думать, что и обе связи верхней челюсти с осевым черепом были у Cladodus такими же, каковы они сейчас у Chlamydoselachus; это приводит нас к выводу, что оба типа сочленений, наблюдаемые нами у Chondrostei и у Holosteoidei, а именно, палатобазальное и квадранторбитальное, развились из типа сочленений Chlamydoselachus-Cladodus; сочленение, существовавшее у примитивных Osteichthyes, по всей вероятности, было построено по типу сочленения Cladodus.
У предков Chondrostei и у большинства селахий нёбная часть верхней челюсти сделалась короче, анторбитальное сочленение исчезло и сохранилось только палатобазальное сочленение.1 2 У Holosteoidei, наоборот, сохранились и развились в прогрессивном направлении оба сочленения верхней челюсти.
Гипотеза эта, объясняющая нам происхождение обоих типов сочленения верхней челюсти (т. е. сочленений Chondrosteoidei и Holosteoidei) из сочленения типа Chlamydoselachus-Cladodus (рис. 69 А—В—С, А—D— Dj—D2), где у одной и той же формы встречаются оба вида сочленения, находится в полном соответствии со взглядом, по которому конечности Osteichthyes развились из плавников типа Cladodus.
1 Мы считаем, например, способ подвешивания верхней челюсти (квадранторбитальное или палатобазальное причленение верхней челюсти) важным биологическим признаком, так как здесь способ сочленения определяет собою способ захватывания пищи.
2 Как, например, у Acanthias и у Spinax.
236
ГЛАВА VIII
При обсуждении перечисленных признаков, различия между Holosteoidei и Chondrostei мы намеренно оставили в стороне такой важный признак, как строение их плавников, хотя именно на этом признаке прежние авторы главным образом и строили свою классификацию Osteichthyes.
Современная теория конечностей, о которой мы уже упоминали выше, исходит из двух основных фактов: во-первых, из того, что плавники всех известных нам форм закладываются в виде широкой (эврибазальной), горизонтально лежащей боковой складки (рис. 39 А—Е) и во-вторых, из того, что закладки плавниковых мускулов (дорзальные и вентральные мускульные почки, или мускульные пучки) представляют собою образования метамерные, причем метамерия их соответствует метамерии туловищных миотомов. Это относится не только к плавникам Selachii и Actinopte-rygii, но также и к закладкам плавников Crossopterygii (Polypterus) и Dipnoi. В. Заленский1 показал, что у Ceratodus1 2 закладываются в течение развития типичные метамерные мускульные отростки, из которых впоследствии образуются столь же типичные мускульные ночки, являющиеся в свою очередь материалом для построения мускулов плавника. Из этого наблюдения необходимо следует, что у предков Ceratodus, у которого во взрослом состоянии имеются типичные бисериальные архиптеригии, были развиты эврибазальные плавники с сегментированной мускулатурой.
Вместе с тем, мы имеем все основания считать, что не только мускулы плавника были первично сегментированы, но и что скелет примитивных плавников (а также скелет плавников предков Dipnoi) первоначально состоял из метамерно расположенных плавниковых лучей; лучи эти сидели на длинном, сегментированном и прилежащем к стенке тела метаптеригии.
Такая примитивно метамерная структура скелета плавников встречается в почти неизмененном виде у ископаемого Cladodus [грудные (рис.46) и брюшные плавники], у современного Pleuracanthus (брюшные плавники), у мальков и у взрослых осетровых рыб [брюшные плавники (рис. 41 В)] и у ископаемых Palaeoniscidae [брюшные плавники (рис. 42)]. Мы имеем полное основание считать, что и грудные плавники эласмобранхий и Osteichthyes также были первоначально построены по этому типу: во всяком случае, все известные нам формы плавников этих рыб эврибазальны, причем и у них хрящевые (или костные) лучи сидят на унисериальном, прилегающем к стенке тела и отчасти еще сегментированном архиптеригии.
Теперь возникает вопрос, как из этих плавников эврибазального типа, столь характерных для Chondrostei, Hoiostei, Palaeoniscoidei и Teleostei, мог развиться стенобазальный плавник типа Crossopterygii и Dipnoi?
На этот вопрос эмбриологическое исследование дает нам исчерпывающий ответ (рис. 39 А—Е). У Ceratodus на ранних стадиях развития широкая складка плавника прилегает всей своей длиной к боковой стенке тела; позже, однако, на заднем конце этой складки образуется incisura metapterygoi-dea (рис. 39 Е и70Ь), которая, углубляясь по мере своего развития, внедряется между метаптеригиумом и стенкой тела и отделяет их таким образом друг от друга. У Elasmobranchii (рис. 70 В) эта выемка не образуется вовсе, у хрящевых и у костных ганоидов она выражена настолько слабо [эври-базальный плавник (рис. 70 F, G)], что у них лишь самая незначительная дистальная час!Ь метаптеригия оказывается отделенной от мускульной стенки туловища, и у этих рыб лишь одно основание плавника делается несколько уже; у Crossopterygii [Polypterus, Sauripterus и др. (рис. 70, J, К)] incisura metapterygoidea (оооо°) становится уже столь глубокой, что весь метаптеригий оказывается отделенным от боковой стенки тела, при
1Salensky W., Sur le developpement de I’ichtyopterygie des poissons ganoides etdipnoides, Ann. Mus. zool. Acad., St. Petersburg, t. 3, 1898.
2 Как известно, архиптеригиальная гипотеза Гегенбаура была построена именно на основании строения плавников взрослого Ceratodus.
ЭВОЛЮЦИЯ OSTEICHTHYES
237
чем основание плавника превращается в сравнительно тонкий стержень, на котором сидит широкая пластинка плавника (стенобазальный плавник). Таким образом возникает первично унисериальный архиптеригий, сво-
Рис. 70 А—М. Схема филогенетического развития грудных плавников рыб.
А—гипотетическая прародительская форма, из которой развились грудные плавники. Плавники эврибазальны и состоят из довольно значительного числа членистых плавниковых лучей; базальные членики этих лучей (на рисунке заштрихованы) прилегают к мускульной стенке тела. Сегментальное расположение лучей соответствует сегментации мускульных почек и образующихся из них мускульных пучков конечности. Из этой исходной формы А развились эврибазальные плавники акул (В), у которых из базальных члеников плавниковых лучей развился метаптеригий и плечевой пояс, а из проксимальных члеников путем срастания—проптеригий и мезоптеригий. Плавники Ichtyotomi (С), развившиеся из той же самой прародительской формы А,более примитивны, чем плавники акул, в том отношении, что у них метаптеригий еще сохранил свое первоначальное расчленение (сегментацию) и что у них еще не образовался ни проптеригий, ни мезоптеригий. Из примитивного плавника акул (В) развился плавник скатов (D). Резкое вторичное изменение первичной формы грудного плавника (А) мы находим yPleuropterygii (Е), у которых образовался типичный бисериальный архиптеригий. Из примитивного плавника эласмобранхий, у которых уже образовался плечевой пояс и у которых передние базальные членики путем срастания уже образовали метаптеригий, но у которых еще не имеется ни проптеригия, ни мезоптеригия, развились плавники Chondrosteoidei (F—Acipenser) и Holosteoidei (G—Amia). Эти последние плавники унисериальны и эврибазальны; incisura metapterygoidea у них только намечается. Из примитивных унисериальных эврибазальных плавников (у) развились стенобазальные плавники (z) кистеперых (J—Sauripterus, К—Polypterus), двоякодышащих (L.—Ceratodus) и предков четвероногих (М—Quadrupeda). У всех этих последних форм (J, К, L, М) образовалась глубокая метаптеригоидная выемка, изображенная на рисунках рядом маленьких белых кружочков, г.—плавниковые лучи; г. р.—проксимальные членики плавниковых лучей. На всех рисунках метаптеригий заштрихован (из А. Н. Северцова, 1926).
бодно подвижный у своего основания (А. Н. Северцов, 1926); из этого типа плавников развился тип бисериального плавника, характерный для кистеперых и двоякодышающих рыб (рис. 70 L).
233
ГЛАВА VIII
ЭВОЛЮЦИЯ HOLOSTEOIDEI ACTINOPTERYGII И HOLOSTEOIDEI CROSSOPTERYGII
Мы видим, что ход эмбрионального развития различных плавников рыб определенно указывает нам на то, что бисериальный архиптергий Cros-sopterigii и Dipnoi не является первичным образованием, но представляет собой вторичное приобретение эврибазальных предков кистеперых и двоякодышащих рыб.
Ведущие нектонный образ жизни Chondrostei,-Holostei, Palaeoniscoidei и Teleostei сохранили в своих плавниках этот первичный эврибазаль-ный план строения в более или менее неизмененном виде (рис. 70 F, G); наоборот, Crossopterygii и Dipnoi, перешедшие к ползанию на дне водоемов, к бентонному образу жизни, приобрели свободно подвижные стенобазальные плавники (рис. 70 К, L), служащие им не только для плавания, но или речного дна.
Поэтому мы можем разделить всех Holosteoidei по строению их грудных плавников на две большие группы, именно на более или менее консервативных Actinopterygii (Palaeoniscoidei, Holostei и Teleostei), построенных, как и их предки—примитивные Selachii и Osteichthyes, приспособительно к нек-тонному образу жизни, и на более прогрессивных Crossopterygii и Dipnoi, перешедших к новому, бентонно-ползающему образу жизни.
Существенным различием между Actinopterygii с одной стороны, и Crossopterygii и Dipnoi—с другой, является наличие у первых дорзального плавате-
льного пузыря (рис. 71 А), тогда как у вторых развит пузырь, открывающийся в кишечник с вентральной его стороны (рис. 71 В).
Прежними авторами делались попытки вывести дорзальный (т. е. открывающийся в кишечник дорзально) плавательный пузырь из вентрального (т. е. открывающегося в кишечник вентрально) плавательного пузыря типа кистеперых рыб. При этом обычно принималось, что лежащий первоначально вентрально плавательный пузырь (легкие) сдвинулся сначала в латеральном направлении, а потом уже занял дорзальное положение. Однако более новые исследования кровеносных сосудов и нервов плавательного пузыря Actinopterygii и легких кистеперых рыб показали, что эту гипотезу сдвигания приходится оставить, что дорзальный плавательный пузырь и вентральное легкое произошли независимо друг от друга и что эти два морфологических образования нельзя выводить одно из другого.
Мы знаем, что вентральные легкие имеются также и у Quadrupeda; кроме того, в настоящее время считается общепринятым, что парные легкие примитивных амфибий произошли путем преобразования непарных легких примитивных кистеперых рыб. Изучение развития легких у амфибий (А.
и для передвижения по поверхности морского
Рис. 71 А. Дорзальный плавательный пузырь, характерный для всех актиноптеригий.
int.—кишечный тракт; v.d.—спинной плавательный пузырь (из Нормана).
/Ztf.
4* V. UV.
Рис. 71 В. Вентральный плавательный пузырь, характерный для всей группы Holosteoidei crossopterygii.
int.—кишечный тракт; v. v.—парный брюшной плавательный пузырь (из Нормана).
ЭВОЛЮЦИЯ OSTEICHTHYES
239
Гётте,1 М. Е, Макушок1 2) показало, что они представляют собою видоизмененные задние жаберные мешки (более точно, повидимому,—видоизмененные вентральные части жаберных мешков), наружные отверстия которых заросли eiiie у пятижаберных акул, Энтодермальные выпячивания жаберного отдела кишечника преобразовались здесь в легкие.
Новейшие исследования В. В. Васнецова3 4 и Л. Неймайера1 показали, что не только легкие, но и плавательный пузырь также образуется из задних (вероятно, из шестой пары) жаберных мешков, но не из вентральных, а из дорзальных их частей.
Мы знаем, что у Chondrostei, Hoiostei, Teleostei и, вероятно, у Palaeoniscoidei имеется дорзальный плавательный пузырь и что его, наоборот, нет у Crossopterygii, Dipneusta и Quadrupeda, у которых имеется вентральное легкое. Спрашивается: каким путем и из какой прародительской формы могли развиться оба эти типа кишечных выростов, легкие и плавательный пузырь?
Можно было бы предположить, что у Osteichthyes primitivi, от которых происходят все упомянутые выше формы животных, некогда имелось два плавательных пузыря, вентральный и дорзальный, и что у одной ветви их потомков (Crossopterygii и Dipneusta) редуцировался дорзальный плавательный пузырь и сохранился, превратившись в легкие, вентральный, тогда как у другой (Chondrosteoidei и Actinopterygii), эволюция которой шла противоположным путем, сохранился один лишь дорзальный плавательный пузырь. Одно время я придерживался этой гипотезы, теперь, однако, мне кажется, что другое (опять-таки чисто гипотетическое) предположение, пожалуй, больше согласуется с известными нам фактами.
Можно предположить, что у Osteichthyes primitivi вообще не существовало определенно выраженного плавательного пузыря и что у них лишь едва закрылись те жаберные мешки, из которых впоследствии у их потомков развились как плавательный пузырь, так и легкие; мешки эти представляли собою сильно разросшиеся парные энтодермальные выпячивания жаберного отдела кишечника. У Chondrosteoidei развился дорзальный плавательный пузырь, у Holosteoidei—соответствующие ему выпячивания жаберных мешков сначала оставались неизмененными, а затем (уже у предков Holosteoidei actinopterygii) из дорзальных отделов этих выпячиваний развился дорзальный плавательный пузырь, а у предков дипной и кистеперых рыб из вентральных отделов—вентральные легкие. Но если это так, то повидимому, дорзальное положение плавательного пузыря у Chondrosteoidei и у Holosteoidei actinopterygii приходится рассматривать как параллелизм в развитии этих органов.
Это последнее предположение кажется нам во всяком случае более обоснованным, чем взгляд, по которому у Osteichthyes primitivi были одновременно развиты оба типа плавательных пузырей. Но как бы то ни было, мы не можем придавать большого значения этим гипотезам, ибо убеждены в том, что обе они имеют вообще смысл лишь как одна из возможных эволюционных схем.
Как известно, Гудрич высказал мнение, что Polypterus и Calamoichthys относятся вовсе не к кистеперым рыбам, но представляют собой сильно
1 G о е 11 е A., Uber den Ursprung der Lungen. Zool. Jahrb. (Anat.), Bd. 21, 1905.
“Makuschok M. E., Zur Frage uber die phylogenetische Entwicklung der Lungen bei den Wirbeltieren, s. I, II, III, IV. Anat. Anz. Bd. 39, 42, 46; 1911, 1912, 1914. Id. Uber genetische Beziehungen zwischen Schwimmblase und Lungen. Anat. Anz. Bd. 44, 1913.
8 Wasnetzoff W., Zur Frage iiber die Morphologie der Schwimmblase. Anat. Anz., Bd. 66, 1928.
4 Neumayer L., Die Entwick lung des Darms von Acipenser. Acta zool., Bd. 11, 1930..
240
ГЛАВА VIII
видоизменившихся палеонисцид.1 Мы затрудняемся присоединиться к этому взгляду. Нам кажется прежде всего, что развитие грудных плавников у Polypterus,1 2 * где имеется ясно выраженная incisura metapterygoidea, вследствие чего плавники их из эврибазальных становятся стенобазальными, решительно говорит против этой точки зрения. Вместе с тем мы знаем теперь, что такой же точно стенобазальный унисериальный архиптеригий имелся и у некоторых типичных ископаемых кистеперых, например, у Sauripterus® и у Eustenoptheron. С другой стороны, строение грудного плавника Polypterus, который по Бёджету (1. с.) происходит из унисериаль-ного архиптеригия, но делается тем не менее стенобазальным, заставляет нас относить Polypterus к группе кистеперых рыб. Присутствие же у Polypterus вентрального легкого не позволяет нам включить его в группу актиноптеригий, все представители которой характеризуются дорзальным плавательным пузырем.4 Этому последнему признаку, т. е. признаку дорзального или вентрального положения плавательного пузыря, мы придаем большое морфологическое значение, так как здесь речь идет не о количественном видоизменении, но об изменении плана строения животного. Вследствие всего вышесказанного, мы придерживаемся общепринятой точки зрения, по которой Polypterus не относится к палеонисци-дам, но представляет собою кистеперую рыбу.
Эволюцию двоякодышащих рыб мы разберем здесь лишь в самых кратких чертах.
На основании сказанного выше представляется вероятным, что уже на весьма ранних стадиях эволюции Holosteoidei от них ответвилась группа форм, характеризовавшаяся стенобазальными плавниками и вентральным плавательным пузырем. Группа эта перешла к жизни в пресных водах; мы можем обозначить ее именем Holosteoidei crossopterygii. От этих'стенобазальных Holosteoidei crossopterygii произошли только что описанные нами кистеперые рыбы, вероятно предки наземных позвоночных и двоякодышащие рыбы. Все же остальные Holosteoidei должны сохранить за собой название Holosteoidei actinopterygii. В пользу того взгляда, что предки всех двоякодышащих рыб действительно отделились от общего ствола Holosteoidei crossopterygii на очень раннейстадии эволюции этих последних, говорят следующие факты: 1) строение древнейших ископаемых двоякодышащих (Dipterus и родственных ему форм), характеризовавшихся весьма примитивно построенным черепом, состоявшим из небольших индиферентных покровных костей, 2) организация современного Neoceratodus с его весьма примитивно построенным позвоночником, хрящевым черепом, хрящевым висцеральным скелетом и т. д.
Предки двоякодышащих очень рано специализировались. Подобно Holocephali среди эласмобранхий, они приспособились к определенному роду пищи, недоступной для других форм—к ракообразным и моллюскам (т. е. к животным с твердой скорлупой); при этом их нёбноквадратный хрящ плотно сросся с осевым черепом, а мелкие зубы, сидящие как на верхней, так и на нижней челюстях (а также сошниковые зубы), частично срослись между собой, образуя большие зубные пластинки, приспособлен
1 Goodrich Е. S., Polypterus a Palaeoniscid? Palaeobiologica, vol. I, 1928,
2 В u d g e t t J. S., On the structure of the larval Polypterus. Trans, of the Zool, Soc., vol. 16, 1902.
Kerr J. G., The development of Polypterus senegalus. Budgett Memor. vol., 1907.
8 Grego ry W. K., Present Status of the Porblem of the Origin of Tetrapoda. New Jork Acad. Annals, vol. 26, 1915.
4 Если бы было найдено, что палеонисциды имели вентральный плавательный пузырь, то это несомненно могло бы послужить основанием для того, чтобы объединить палеонисцид в одну группу с кистеперыми рыбами и противопоставить их всем остальным акт ин о-птеригиям.
ЭВОЛЮЦИЯ OSTEICHTHYES
241
ные к прокусыванию и размельчению твердой скорлупы ракообразных и моллюсков;.
Мы имеем все основания предполагать, что именно эта специализация дипной, т. е. их приспособленность к определенному роду пищи и определенному образу жизни (жизни в болотистых- водах с гниющими водорослями, что вызвало у них образование легочного дыхания), явилась причиной того, что современные нам двоякодышащие могли удержать в своем строении многие черты примитивной организации предков, сохранивши их при этом в столь мало измененной форме. Подобного же рода явления мы наблюдаем у форм, приспособившихся, как и дипнои, к постоянным и своеобразным условиям существования: вспомним, например, бесчерепных позвоночных (ланцетника), круглоротых, Holocephali и многих других.
Crossopterygii в собственном смысле этого слова, т. е. Osteolepidoti, Coelacanthini и Polypterini, достигли своего полного расцвета в позднем палеозое. В настоящее время все они, за исключением Polypterini, вымерли. Однако в высокой степени вероятно, что именно от какой-то очень примитивной ветви кистеперых произошли предки всех наземных позвоночных (Tetrapoda). Ниже'мы будем говорить об этом более подробно.
Мы не будем останавливаться на дальнейшей эволюции Actinopterygii; отметим только, что Гудрич (1909) разделяет современных актиноптеригий на три большие группы: Amioidei, Lepidosteoidei и Teleostei. Среди них наибольшее количество признаков примитивного строения ископаемых предков сохранилось у Lepidosteoidei; так, мы находим у них ясно выраженные ромбические космоидные чешуи, образующие на поверхности тела плотный панцырь, а также покровные кости и остатки плакоидных чешуй, т. е. наиболее примитивные формы чешуй. Amioidei имеют уже типичные циклоидные чешуи типа Teleostei. Как у Amia, так и у Lepidosteoidei сохранились типичные метаптеригии в грудных плавниках, на которых сидят плавниковые лучи, спиральный клапан в кишечнике и conus arteriosus, т. е. целый ряд примитивных анцестральных признаков, большей частью исчезнувших у костистых рыб.
Что касается костистых рыб, то они представляют собою прогрессивную, чрезвычайно богатую формами группу, современные представители которой приспособились к самым разнообразным условиям существования в морских и пресных водах. Большинство костистых рыб сохранило нектонный образ жизни своих предков, причем одни формы приспособились к жизни на поверхности вод, другие (абиссальные формы)—к жизни на огромных глубинах. Многие Teleostei приспособились к прибрежной жизни, или к жизни среди скал, многие стали бентонными животными, лежащими или ползающими по дну. Некоторые из Teleostei, ведущие прибрежный образ жизни, приобрели способность к передвижению по суше в поисках добычи. Нам хотелось бы обратить внимание читателя на то, что морфологические и физиологические приспособления к чрезвычайно разнообразным условиям среды вызывают глубокие изменения в типичной организации костистых рыб и что изменения Эти в огромном числе случаев совершенно не похожи на те, которые имели место в свое время у предков костистых рыб, когда эти последние в свою очередь переходили к специализированному образу жизни.
Прежде чем перейти к описанию эволюции Quadrupeda, мы даем в наглядной форме в виде прилагаемой здесь таблицы (табл. 3) характерные особенности главных групп Osteichthyes. •
Из рассмотрения этой сводной таблицы прежде всего явствует, что у Chondrosteoidei (Chondrosteus, Acipenseridae, Polyodontidae) встречаются многие морфологически важные признаки селахий, вследствие чего они стоят более близко к примитивным эласмобранхиям, чем какая-либо Морфологические закономерности 16
Таблица 3
Примитивные Elasmobranchii (Cladodus, Chlamydoselachus) Chondrosteoidei Holosteoidei actinopterygii Holosteoidei crossopterygii Dipneusta
1. Эврибазальные грудные плавники. 1. Эврибазальные грудные плавники. 1. Эврибазальные грудные плавники. 1. Стенобазальные грудные плавники. 1. Стенобазальные грудные плавники, бисериальный архиптеригий. 2. Метаптеригий грудных плавников мало или вовсе не сегментирован. 3. Плакоидные чешуи отсутствуют. 4. Костные чешуи не имеют продольного гребня. 5 Эпителиальный канал боковой линии отсутствует, вследствие чего отсутствует и его костный канал.
2. Сегментированный метаптеригий. 3. Имеются только плакоидные чешуи. 4. Костные чешуи (покровные кости) отсутствуют.. 5. Эпителиальные каналы органов боковой линии проходят под плакоидной чешуей (медиально от нее). 2. В метаптеригии грудных плавников следы первичной сегментации; метаптеригий брюшных плавников ясно сегментирован. 3. Плакоидные чешуи отсутствуют. 4. Имеются метамерные ромбические костные чешуи с продольным гребнем на поверхности (покровные кости черепа также снабжены продольным гребнем) 5. Эпителиальный канал органов боковой линии проходит снаружи (латерально) от костей, внутри костных каналов, образованных костными чешуями (покровными костями с продольным гребнем). 6. Осевой череп хрящевой, селахоидного типа; базальный угол, processus palatobasales laterales и rostrum хорошо развиты. 7. Носовые ямки лежат далекодруг от друга; истинное septum nasale отсутствует. 8- Имеется symphysis palatoquadrati. 9. Развито только articulatio palatobasalis. 10. Имеется дорзальный плавательный пузырь. 11. Praeoperculum и парный сошник, несущий зубы, отсутствуют. 12. На несущих зубы костях ротовой полости ^развиты многочисленные отдельно сидящие зубы. овапиям (А. Н. Северцов, 192 лучом. 2. Метаптеригий грудных плавников не сегментирован, метаптеригий брюшных плавников сегментирован у наиболее примитивных форм (Palaeoniscidae). 3. Существуют рудиментарные плакоидные чешуи. 4. Костные чешуи и покровные кости головы не имеют продольного гребня. 5. Эпителиальный канал органов боковой линии проходит в голове внутри костного канала, который образован у личинок костями с продольным гребнем на поверхности. 2. Метаптеригий грудных плавников не сегментирован.1 3. Существуют рудиментарные плакоидные чешуи. 4. Костные чешуи и покровные кости головы не имеют продольного гребня. 5. Эпителиальный канал органов боковой линии проходит в голове внутри костного канала, который образован у личинок костями с продольным гребнем на поверхности.
6. Осевой хрящевой череп типично селахоидный с базальным углом; имеются processus palatobasales laterales и rostrum. 7 . Носовые ямки лежат далеко друг от друга; истинное septum nasale отсутствует. 8. Имеется symphysis palatoquadrati. 9. Развиты оба типа подвесочного аппарата (подвешивание palatoquadratum к черепу): artieulatio palatobasalis и articulatio quadrantorbitalis. 10. Плавательный пузырь отсутствует. 1 Согласно моим исслед теригием, но сильно развитым 6. Осевой череп голостеоид-ного (не селахоидного) типа; имеются processus palatobasales laterales; rostrum отсутствует или слабо выражен. 7. Носовые ямки лежат вплотную друг около друга; имеется высокое и тонкое septum nasale. 8. Symphysis palatoquadrati отсутствует; partes palatinae нёбноквадратных хрящей отделены друг от друга. 9. Articulatio palatobasalis сохранилось, articulatio quadrantorbitalis верхней челюсти хорошо развито. 10. Имеется дорзальный плавательный пузырь. 11. Praeoperculum и парные сошники, несущие зубы, хорошо развиты. 12. На несущих зубы костях ротовой полости развиты многочисленные отдельно сидящие зубы. 6>, медианная ось грудных илав 6. Осевой череп голостео-идного (не селахоидного)типа; processus palatobasales laterales и rostrum отсутствуют. 7. Носовые ямки лежат вплотную друг около друга; имеется высокое тонкое septum nasale. 8. Symphysis palatoquadrati отсутствует; partes palatinae palatoquadrati отделены друг от друга. 9. Articulatio palatobasalis рудиментарно, articulatio quadrantorbitalis верхней челюсти хорошо развито. 10, Имеется вентральный плавательный пузырь (или легкие). 11. Praeoperculum и парные сошники, несущие зубы, хорошо развиты. 12. На несущих зубы костях ротовой полости развиты многочисленные отдельно си-1 дящие зубы, ников кистеперых и двоякоды 6. Осевой череп голосте-оидного типа. 7. Носовые дмки тесно прилегают друг к другу, существует топкое septum nasale. 8. Palatoquadratum срастается с осевым черепом (ау-тостилия), вследствие этого: 9. никакого сочленения верхней челюсти с черепом не имеется. 10. Имеется вентральный плавательный пузырь (или легкие). 11. Существуют парные сошники. 12. Все зубы превратились в крупные, сложные зубные пластинки. ш.ащих рыо является не метан-
242 ГЛАВА VIII
ЭВОЛЮЦИЯ OSTEICHTHYES
243
иная группа, принадлежащая к Osteichthyes. Это обстоятельство, а также, строение костного скелета этих форм, позволяет нам рассматривать группу Chondrosteoidei как наиболее примитивную группу среди Osteichthyes и противопоставлять им всех остальных Osteichthyes. Мы видим, с другой стороны, что Actinopterygii (Amioidei, Lepidosteoidei, Teleostei) и Crossopterygii (Polypterini, Coelacanthini, Osteolepidoti) сходны между собой в столь многих отношениях, что мы имеем полное право объединить их всех в одну большую группу, названную нами Holosteoidei, в систематическом отношении вполне равноценную Chondrosteoidei; к этой последней группе, вероятно, относятся и Dipnoi.
Мы можем, таким образом, классифицировать Osteichthyes следующим образом:
Osteichthyes
I. Chondrosteoidei (Chondrosteidae, Acipenseridae, Polyodontidae).
II. 1. Holosteoidei actinopterygii (Palaeoniscoidei, Lepidosteoidei, Amioi-dei, Teleostei).
2 A. Holosteoidei crossopterygii (Osteolepidoti, Coelacanthini, Polypterini).
2 B. Dipneusta.
Наш взгляд на филогенетические соотношения между всеми этими группами представлен на родословном древе (схема XV). В эту схему, однако, мы не поместили палеонисцид (Palaeoniscoidei): они должны были бы находиться где-то поблизости от Holosteoidei actinopterygii, но их филогенетические отношения к остальным группам высших актиноптеригий, по нашему мнению, еще недостаточно выяснены; поэтому мы пока не можем яснее определить их положение в общей схеме родословного древа.
ПРОИСХОЖДЕНИЕ QUADRUPEDA
Взгляды различных авторов относительно происхождения наземных позвоночных (Quadrupeda, Tetrapoda) и, в особенности, относительно происхождения стегоцефалов в настоящее время все еще в значительной степени расходятся. Многие зоологи и сравнительные анатомы (Р. Сэмон, Т. Браус, К. Боас, М. Фюрбрингер, Е. Гудрич и др.) придерживаются того мнения, что примитивные амфибии, т. е. стегоцефалы, происходят от двоякодышащих рыб, большинство же палеонтологов (Т. Осборн, В. Грегори, А. Смите Вудвард, Муди, Д. Ватсон и др.) склоняется скорее к тому, чтобы производить их от кистеперых рыб. Однако все они согласны между собой в том, что никакие другие группы рыб, т. е. ни эласмобранхий, ни хрящевые, ни костные ганоиды, ни костистые рыбы отнюдь не могут рассматриваться как непосредственные предки Tetrapoda.
При этом особенно отмечается, что между современными двоякодышащими (Neoceratodus) и современными амфибиями существует большое сходство, которое можно объяснить только общностью их происхождения. Так, например, и у двоякодышащих рыб, и у амфибий имеется аутостилистический череп; у двоякодышащих, как и у амфибий, мы находим хоаны, соединяющие носовые полости с ротовой полостью этих животных;1 у двоякоды
1 В моей работе о происхождении Quadrupeda я довольно подробно разобрал вопрос о происхождении органов обоняния у двоякодышащих и у Quadrupeda. Мне представляется невероятным, чтобы обонятельный орган амфибий, в котором мы находим как наружные носовые отверстия (ноздри), лежащие на наружной поверхности передней части головы, так и внутренние, открывающиеся в ротовую полость (хоаны), мог бы произойти из обонятельного органа двоякодышащих, у которых ноздри лежат не на наружной поверхности рыла, а внутрь от верхней губы (см. Северцов А. Н., Palaont. Zschr., Bd. 8, 1926).
16*
244
ГЛАВА VIII
шащих, так же как и у амфибий, крыша больших полушарий головного мозга состоит из нервной ткани, а мозг имеет, в общем, амфибийный характер; conus arteriosus у Dipnoi построен по тому же принципу, что и conus arteriosus амфибий и, наконец, существует большое'сходство в гистологическом строении тканей Dipnoi и Urodela.
Большинство этих признаков отсутствует у кистеперых рыб: так, череп Crossopterygii гиостиличен, крыша больших полушарий их головного мозга состоит не из нервной, а из эпителиальной ткани, у них совсем нет хоан и т. д.
Против гипотезы о происхождении наземных позвоночных от двоякодышащих рыб говорят следующие факты: все известные нам формы двоякодышащих рыб так сильно специализованы, что нам представляется крайне невероятным, чтобы амфибии (стегоцефалы) могли произойти от них. Возьмем хотя бы такой важный признак, как строение их челюстного аппарата. Мы знаем, что даже у наиболее примитивных из всех известных нам ископаемых двоякодышащих (Dipterus, Phaneropleuron и т. п.) были развиты очень сложные зубы, состоящие из сросшихся между собой отдельных маленьких зубов, и что у них quadratum был, по всей вероятности, уже сращен с черепом (аутостилия). Трудно представить себе, чтобы из этих сильно специализованных зубов могли развиться многочисленные зубы амфибийного типа, столь характерные для стегоцефалов. Поэтому нам приходится откинуть взгляд, что примитивные Tetrapoda, т. е. предки стегоцефалов, могли произойти от известных нам двоякодышащих. Самое большее, что можно было бы утверждать, это то, что предки Tetrapoda ответвились от предков известных нам Dipneusta еще на очень ранней стадии эволюции этих последних. Челюсти этих предшественников двоякодышащих рыб были снабжены многочисленными отдельными зубами. Вместе с тем эти животные примыкали по всем своим другим морфологическим признакам к очень примитивным Holosteoidei.
Возвращаясь к гипотезе о происхождении стегоцефалов от кистеперых рыб, мы должны отметить, что эта гипотеза лучше освещает нам ход эволюции некоторых существенных органов стегоцефалов, чем гипотеза о происхождении их от двоякодышащих рыб. Так, например, нам довольно трудно провести сравнение между костями черепа примитивных стегоцефалов с костями черепа наиболее древних из известных нам двоякодышащих, хотя Грегори1 и удалось доказать, что положение костей черепа у Diplop-terus (Crossopterygii, Osteolepidoti) и у Trimerorachis (Stegocephalia) очень сходно: разница между этими двумя формами черепа состоит главным образом в отсутствии у стегоцефалов некоторых костей жаберной крышки (operculum, interoperculum). Для всех же остальных покровных костей черепа Crossopterygii имеются гомологи в черепе стегоцефалов.
У тех же Crossopterygii (Sauripterus) Грегори находит и своеобразную форму плавников (грудных плавников), являющуюся по форме и по положению своих скелетных элементов переходной ступенью между типичными плавниками рыб и типичной конечностью стегоцефалов (рис. 70 J, Sauripterus). То, что у Crossopterygii имеются одновременно как нижние, плевральные, так и верхние, септальные ребра, между тем как у Dipnoi имеются лишь одни нижние ребра—несомненно тоже говорит в пользу гипотезы кроссоптеригиального происхождения наземных позвоночных.
И, однако, если мы примем во внимание всю организацию известных нам форм кистеперых рыб, то увидим, как трудно нам предста
1GregoryW. К., Present status of the Problem of the origin of Tetrapoda. New York Acad. Annals, vol. 26, 1915.
ЭВОЛЮЦИЯ OSTEICHTHYES
245
вить себе, чтобы Quadrupeda могли произойти от какого-нибудь известного нам представителя кистеперых рыб.
Мы знаем, что у амфибий (и у стегоцефалов) задняя конечность состоит из большего числа скелетных элементов, чем передняя конечность этих форм. Вместе с тем у нас имеется основание предполагать, что конечность предков Quadrupeda (И. И. Шмальгаузен)1 некогда была не пяти-, а, вероятно, семилучевой: по всей вероятности, у них был хорошо развит praepollex s. praehallux и postminimus, рудименты которых сохранились у их потомков, т. е. у примитивных амфибий, вплоть до настоящего времени. Мы должны были бы поэтому признать, что брюшные плавники рыбообразных предков амфибий были в свое время столь же развиты (если не больше), как и их грудные плавники и, по меньшей мере, столь же диференцированы. Однако, мы видим, что даже у наиболее примитивных из известных нам кистеперых рыб брюшные плавники диференцированы гораздо слабее, чем, например, грудные плавники ископаемого Sauripterus, от которого, по Грегори, произошли стегоцефалы.
Нам приходится поэтому предположить, что примитивные Quadrupeda произошли от какой-то палеонтологически нам еще неизвестной формы кистеперых рыб—формы, у которой имелись одинаково хорошо диферен-цированные грудные и брюшные плавники, обладавшие, по меньшей мере семью лучами. Другими словами, они произошли от какой-то гораздо более примитивной формы кистеперых рыб, чем те* которые нам сейчас непосредственно известны, и невольно напрашивается мысль, не стоит ли эта нам еще неведомая форма Crossopterygii очень близко к Dipnoi?
Обе только что разобранные нами гипотезы о присхождении Quadrupeda указывают, что последние некогда произошли от какой-то, пока нам еще точно неизвестной, но очень примитивной формы, близко стоящей к общим стенобазальным предкам Crossopterygii и Dipneusta: назовем ли мы когда-нибудь эту прародительскую форму именем Crossopterygii или именем Dipnoi—в настоящее время еще трудно решить, да и вряд ли это может иметь существенное значение.
Гораздо интереснее то, что известные нам потомки примитивных Dipnoi (Dipteridae), которые приобрели в течение своего филогенеза легкие, приспособленные для воздушного дыхания, и сильно выраженные стенобазальные конечности, предназначенные для поддерживания туловища, все же не превратились в наземных позвоночных: по всей вероятности, препятствием к этому превращению для них явилась специализация их ротового скелета.
Кроме только что упомянутых видоизменений, в организации вероятных гипотетических предков Quadrupeda произошел еще ряд других перемен, приведших в конечном итоге к тому, что типичные рыбы превратились в настоящих амфибий: плавники, приспособленные столь же для плавания, сколь и для ползания по дну мелководных водоемов, превратились в настоящие лапы с типичными stylopodium, zeugopodium и семилучевым autc-podium, а их мускулы, в соответствии с новоприобретенной функцией, диференцировались и усилились: из туловищных миотомов развилась новая, уже вторичная, мускулатура, соединяющая плечевой пояс конечностей с туловищем (А. Н. Северцов, 1908). Повысилась также подвижность головы (articulatio cranio-vertebrale), образовался типичный таз и крестец (sacrum); редуцировались непарные плавники; вентральный плавательный пузырь превратился в типичные легкие с гортанью и трахеями и, наконец, развились новые воздушные пути через хоаны, что имело
ЧПмальгаузен И. И., Развитие конечностей амфибий и их значение в вопросе о происхождении конечностей наземных позвоночных. Москва, 1915.
246
ГЛАВЫ V—VIII
огромное, значение как для функции дыхания, так и для функции обоняния.
Все эти перемены сделали возможным для предков наземных позвоночных, бывших до сей поры типичными водными животными, пребывание на суше, пребывание сначала лишь факультативное, во время пересыхания водоемов, а затем все более и более постоянное, когда они окончательно приспособились к жизни на поверхности земли.
На этом мы можем закончить наш сжатый очерк филогенетического развития позвоночных животных. Результаты, к которым мы пришли, представлены нами на схеме XV.
ОБЩАЯ СВОДКА ГЛАВ
ОБ ЭВОЛЮЦИИ НИЗШИХ ПОЗВОНОЧНЫХ
(V —VIII)
1. В качестве исходной формы, от которой произошли позвоночные животные, мы принимаем Acrania primitiva, организация которых может быть нами восстановлена только на основании сравнительно морфологических данных. Acrania primitiva представляют собой весьма примитивных бесчерепных сегментированных Chordata с довольно большим числом (около 20) жаберных щелей, свободно открывающихся в наружную среду (отсут-.ствие г!ерибранхиальной полости), со слабо диференцированным головным мозгом, с глазками Гессе. Органы высших чувств у Acrania primitiva не были развиты, их питание было пассивным (эндостиль, околоротовое скелетное кольцо и cirri).
2. От этих Acrania primitiva ответвились Branchiostomatidae (Amphio-xus), перешедшие к подземно-подводному образу жизни и остановившиеся, подобно многим другим специализованным группам животных, на низкой ступени своего филогенетического развития (едва намечающийся головной мозг и органы высших чувств). У Branchiostomatidae число жаберных щелей вторично сильно возросло; у них развилась жаберная решетка и rostrum, появилась защищающая жабры атриальная складка и перибран-хиальная полость, образовавшаяся из ее стенок; их питание осталось таким же пассивным, каким оно было у Acrania primitiva.
3. Другая ветвь потомков гипотетических Acrania primitiva—Protocraniata—развивалась в прогрессивном направлении, и мы можем, на основании сравнительно анатомических и сравнительно эмбриологических данных, представить себе довольно детально основные черты ее строения. У этих гипотетических Protocraniata уже имелся головной мозг, орименты парных глаз с глазными мускулами, орименты слуховых пузырьков и носовых ямок; имелся слабо развитый череп и диплоспондильно расположенные верхние дуги позвоночника. Ряд жаберных щелей и лежащих между ними нерасчлененных жаберных дуг доходил у них до рта, снабженного околоротовым жаберным кольцом. Повидимому, у них было также развито мешковидное венозное сердце. Их питание было еще пассивным (эндостиль), но в их пищеварительном тракте из мешковидной печени предков уже развилась трубчатая печень типа Craniata. Protocraniata по своему образу жизни были, по всей вероятности, нектонными животными.
4. От Protocraniata произошли две линии потомков: одна, ведущая к вымершим Ostracodermi и ныне живущим Cyclostomata, другая, ведущая к предкам современных рыб. У представителей первой линии потомков, у гипотетических Entobranchiata, образовались большие складчатые энто-дермические жаберные мешки и сложно построенная жаберная решетка, доходившая до самого ротового кольца, что привело к интенсификации функции дыхания. В их слуховом органе развились два полукружных канала, а на дорзальной поверхности головы образовалась назогипофизарная
ЭВОЛЮЦИЯ низших позвоночных
247
полость, в которую открывались с одной стороны сближенные между собой обонятельные ямки, а с другой—канал гипофиза; назогипофизарная полость открывалась в свою очередь на спинной поверхности головы посредством медиального назогипофизарного отверстия.1 Питание Entobranchiata было первоначально пассивным (стадия AmmocoetesB филогенезе энтобранхиат), но все потомки Entobranchiata перешли к активному питанию.
Схема XV. Схематическое изображение родословного древа низших позвоночных (см. также стр. 213)..
На этой диаграмме в виде обычной схемы родословного древа представлены вероятные, по нашему мнению, филогенетические соотношения между различными родоначальными группами низших позвоночных. Описанные нами в тексте промежуточные гипотетические формы (Protocraniata, Entobranchiata, Ectobranchiata и т. д.) также вошли в это древо. Черными кружками обозначены вымершие формы животных, светлыми кружками обозначены группы, дожившие до нашего времени. Одним восклицательным знаком (!) обозначены группы, не обнаруживающие признаков вымирания, знаком минус (—) обозначены группы, близкие к вымиранию. Буквой S изображены идиоадаптивные или специализованные группы, двумя восклицательными знаками (!!)—'Прогрессивные современные группы. Связь между Anaspida и Ectobranchiata я лично считаю возможной, но спорной; поэтому на линии, идущей от Ectobranchiata к Anaspida имеется вопросительный знак (?).• Palaeoniscidae совсем не вошли в наше родословное древо; их надо было бы поместить поблизости от Holosteoidei actinopterygii.
5. У одной линии этих потомков, а именно у предков Ostracodermi, развился твердый костный панцырь. Число жаберных щелей у них редуцировалось до 10.1 2 Из костных чешуй, окружающих рот, образовался ротовой скелет, и животные эти перешли к активному способу питания. Голов
1 Назогипофизарное отверстие прежде считалось за непарное носовое отверстие круглоротых.
2 Среди них имелись две функционирующие пребранхиальные жаберные щели.
248
ГЛАВЫ V—VIII
ной мозг и органы высших чувств у них были, невидимому, хорошо развиты: так, в слуховом органе имелись два полукружных канала, глаза были крупны и хорошо развиты, обонятельные органы были сближены наподобие того, что мы наблюдаем у ныне живущих круглоротых, и открывались непарным дорзальным отверстием в ту же назогипофи-зарную полость, в которую открывался и гипофиз. Большинство членов этой крупной группы панцырных Entobranchiata перешло к бентонному образу жизни, причем их широкое тело сплющилось в дорзо-вентральном направлении. Они вымерли еще в палеозое.
6. У другой ветви потомков Entobranchiata, а именно у Protocyclo-stomata, костного скелета в виде панцыря не образовалось. Передние жаберные щели их закрылись, а передние части жаберной решетки превратились в сложно построенные части сосущего ротового аппарата, скелет которого был снабжен роговыми зубами. От Protocyclostomata произошли две ныне живущие группы круглоротых, миноги и миксины (Petromy-zontes и Myxinoidea), сохранившие вплоть до нашего времени многочисленные следы примитивного строения своих отдаленных предков.
7. В другую сторону от Protocraniata ответвилась прогрессивная ветвь более высокоорганизованных форм (Ectobranchiata), ведущая к современным рыбам. У гипотетических Ectobranchiata хрящевые жаберные дуги расчленились на четыре типичных отдела; ряд этих дуг, точно так же как и ряд лежащих между ними жаберных щелей, достигал рта. На наружной стороне жаберных дуг развились эктодермальные жаберные нити, превратившиеся впоследствии в эктодермальные жаберные лепестки. Таким образом, у этих форм также образовался более совершенный дыхательный аппарат, но результат этот был достигнут иным путем, чем у Entobranchiata.
8. У Ectobranchiata в течение филогенетического развития на туловище и в ротовой полости развились многочисленные плакоидные чешуи (зубы). У потомков Ectobranchiata (у Protognathostomata) передние жаберные щели закрылись таким же путем, как и у Entobranchiata, а из передних частей расчлененных жаберных дуг развилась челюсть хватательного типа, а также губные хрящи и подъязычный аппарат. Мелкие плакоидные зубы, сидящие на челюсти, развились в прогрессивном направлении и превратились в интегральные части ротового скелета, приспособленного для кусания. Весьма вероятно, что у Protognathostomata не только сама челюсть, т. е. вторая преджаберная висцеральная дуга, но и задний губной хрящ, т. е. третья преджаберная висцеральная дута, были снабжены зубами и выполняли функцию кусания и захватывания пищи. Таким образом, у первичных предков челюстных рыб, у Protognathostomata, переход от пассивного питания предков к активному осуществился иным путем, чем у Entobranchiata. Органы движения Protognathostomata также развились в прогрессивном направлении, а именно, у них из непарной плавниковой складки предков образовался сначала протоцеркальный хвостовой плавник и непарные плавники с хрящевыми плавниковыми лучами, а затем уже из горизонтальных боковых складок туловища развились эврибазальные парные плавники с метамерной мускулатурой и метамерно расположенными сегментальными плавниковыми лучами. Весьма возможно, что уже Protognathostomata обладали весьма просто построенными парными плавниками, но утверждать это с полной определенностью мы не можем.
9. От Protognathostomata—этих еще очень примитивных предков рыб— произошла группа примитивных Chondrichtyes selachoidei, которые, также будучи еще весьма примитивными животными, напоминают нам во многих отношениях некоторых из низших палеозойских Elasmobranchii, а именно Cladodontidae. Формы эти обладали хорошо развитыми челюстями, снабженными крепкими зубами; palatoquadratum у них был соединен с осе
ЭВОЛЮЦИЯ низших позвоночных
249
вым черепом- в двух пунктах—в глазничной (articulatio palatobasalis) и в анторбитальной (articulatio quadrantorbitalis) областях. Их осевой череп был построен по типу осевого черепа Cladodus (и Chlamydoselachus). Их хвостовой плавник был эпицеркален, а парные плавники были весьма примитивны, с еще сегментированным метаптеригием, причем на каждом сегменте этого метаптеригия сидело по одному хрящевому плавниковому лучу (radius).
10. От этих примитивных селахоидных форм (Chondrichthyes selachoidei) произошли родоначальники всех известных нам Elasmobranchii, Pleuropterygii, Pleuracanthodei, Holocephali, Squaloidei и Batoidei; по всей вероятности, Acanthoidei и Arthrodira также представляют собой сильно видоизменившиеся боковые ветви этого ствола. Некоторые представители этой крупной группы примитивных рыб в большей или меньшей степени специализировались в течение своего эволюционного развития (например скаты, Holocephali), другие сохранили первичные черты своей организации вплоть до наших дней (Notidanidae). В общем, современные Elasmobranchii представляют собой и поныне процветающуюв и богатую формами группу рыб, сохранившую со времени палеозоя основные черты своего строения.
11. В ином направлении развивались происшедшие также от Chondrichthyes selachoidei предки всех современных Osteichthyes—Osteichthyes primitivi. Эти гипотетические предки высших рыб и всех Quadrupeda могут быть охарактеризованы следующим образом. Их тело было покрыто мелкими плакоидными чешуями, под которыми находились сравнительно крупные, метамерно расположенные костные чешуи ромбической формы с продольным гребнем на поверхности; первоначально эти гребенчатые чешуи имели на голове такое же правильное расположение, как и на туловище, но впоследствии некоторые из них срослись друг с другом, иные увеличились в объеме, иные, наконец, совсем редуцировались. Таким образом образовались типичные покровные кости черепа Osteichthyes. Важным биологическим изменением было развитие снабженных зубами костей ротового скелета (praemaxillae, maxillae, dentalia, palatina, pterygoidei и т. д.), явившихся прочной основой для расположенных в ротовой полости зубов. Роговые лучи плавников (ceratotrychia) постепенно заместились костными лучами (lepidotrychia).
Дыхательный аппарат также претерпел весьма важное изменение: свободно открывающиеся наружу жаберные щели Chondrichthyes (села-хоидный тип) прикрылись у Osteichthyes primitivi сидящей на гиоидной дуге кожной складкой—жаберной крышкой. Из костных чешуй этой области развились оперкулярные кости и radii membranae branchiostegiae. Таким образом, изменился аппарат дыхания; число жаберных щелей уменьшилось до пяти. Из задней пары не открывшихся жаберных щелей > образовался плавательный пузырь, взявший на себя функцию гидроста-; тического аппарата.
[ 12. От Osteichthyes primitivi произошли две ветви рыб с костным ске-
i летом, а именно: предки хрящевых ганоидов (Chondrosteoidei) и предки j всех высших рыб и Quadrupeda (Holosteoidei primitivi). Chondrosteoi-j dei сохранили в своем строении многие примитивные признаки своих : селахоидных предков. Следы эти заметны в общей структуре их хрящевого ' скелета, т. е. в строении хрящевого черепа с rostrum и характерными для : селахий отростками, в широко расставленных обонятельных капсулах, в строении их челюстного аппарата (symphysis palatoquadrati), в строении позвоночника и примитивных непарных и парных плавников. Строение их костного скелета также осталось еще весьма примитивным. В настоящее время эта примитивная группа представлена лишь осетровыми рыбами (Acipenseridae) и Polyodontidae.
-250
ГЛАВЫ V—VIII
13. Другая-ветвь потомков Osteichthyes primitivi, а именно Holosteoidei primitivi, наоборот, развилась в основном в прогрессивном направлении и дала целый ряд потомков. Для Holosteoidei характерны следующие морфологические признаки: их обонятельные капсулы сближены, и между ними находится типичное septum nasale; у них имеется квадранторбитальное и, вероятно, палатобазальное (метаптеригоидное) сочленение верхней челюсти с осевым черепом; symphysis palatoquadrati отсутствует; rostrum редуцирован. У Holosteoidei имелись парные плавники первоначально эврибазального типа (типа Actinopterygii). Весьма характерным признаком этой группы является также прогрессивное окостенение хрящевого скелета у высших Holosteoidei.
Эволюция потомков Holosteoidei также пошла в двух направлениях: одна линия (Holosteoidei actinopterygii) повела к образованию широко распространенных нектонных Actinopterygii, другая (Holosteoidei crossopterygii)—к образованию бентонных Crossopterygii и Dipnoi, а также предков Quadrupeda.
14. У всфс Actinopterygii первоначально имелись парные грудные плавники с широким основанием и слабо развитой incisura metapterygoidea. Эти плавники имели сначала горизонтальное расположение, но у высших форм краниальный край их повернулся в дорзальном направлении, так что сам плавник занял по отношению к оси тела косое положение, получив возможность прикладываться к боковой стенке туловища. У Actinopterygii имеется дорзальный плавательный пузырь. К этой группе относятся Palaeo-niscoidei, Lepidosteoidei, Amioidei и вся многочисленная группа костистых рыб (Teleostei).
15. У второй ветви рыб, происшедших от Holosteoidei primitivi, а именно у Holosteoidei crossopterygii, сильно развилась incisura metapterygoidea, так что грудные плавники этих животных стали стенобазальными. У них имеется вентральный плавательный пузырь (легкие).
От этих бентонных пресноводных форм произошли Crossopterygii, Dipneusta и группа рыбообразных предков Quadrupeda. От последних, в свою очередь, начался новый ряд прогрессивных форм.
ГЛАВА IX
ОБЩИЙ ХОД ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ
ЭВОЛЮЦИЯ И ПРИСПОСОБЛЕНИЕ
Прежде чем перейти к обсуждению общих закономерностей филогенеза, попытаемся на основании того, что нам теперь известно относительно филогенетического развития крупных групп животного царства, построить очень общую схему хода эволюции животного мира. Такая схема, конечно в весьма упрощенной форме, даст нам возможность окинуть общим взглядом ход эволюции и позволит нам расчленить этот исключительно сложный процесс на отдельные, следующие друг за другом и поэтому более доступные нашему анализу стадии или фазы.
Разбирая какую-либо крупную группу животных с морфологической точки зрения, мы неизбежно приходим к выводу, что ее филогенетическое развитие в общем и целом представляет собою приспособительный процесс и что ее биологический прогресс определяется в конечном итоге филогенетическим развитием отдельных приспособлений. При этом как приспособления мы обозначаем такие изменения строения и функций органов, которые оказываются биологически полезными животным при изменениях, происходящих в окружающей их среде.
В предыдущих главах этой книги (главы V—VIII) мы привели эволюцию целого ряда подобного рода приспособительных изменений. Сюда относится, например, эволюция жаберного аппарата и его кровеносных сосудов у энто- и эктобранхиат, т. е. увеличение дыхательной поверхности этих форм путем образования эктодермальных жаберных складок или эктодермальных жаберных нитей и лепестков; сюда относится эволюция плакоидных чешуй эласмобранхий, служащих защитой при нападении врагов, эволюция роговых зубов предков миног и миксин, а также плакоидных зубов примитивных челюстных позвоночных, в обоих случаях представляющая собой развитие приспособлений к активному захватыванию пищи; сюда относится, наконец, эволюция различных форм парных и непарных плавников рыб и парных конечностей наземных позвоночных, являющихся полезными приспособлениями этих форм к различным способам передвижения в воде и на суше.
Мы не станем задерживать внимание читателя перечислением всех многочисленных и крайне разнообразных форм приспособлений животных к различным условиям обитания:1 подавляющее большинство филогенети
1 См. книги Plate L, Selektionsprinzip und'Probleme der Artbildung, 1913, и Allgemeine Zoologie, Bd. 1, 1922 и 2, 1924, а также книгу Hesse R., Tiergeographie auf okologischer Grund lage, 1924, в которых оба автора приводят большое число примеров самых различных приспособлений и дают им подробный анализ.
254
ГЛАВА IX
ческих изменений строения животных форм представляет собой именно подобного рода приспособления, и поэтому нам приходится скорее ставить вопрос о том, какие изменения в организме животного не являются приспособлениями, чем наоборот.
Изучая морфологически и физиологически органы животных, мы видим, что их легко можно подразделить на две большие категории, а именно на пассивные и на активные органы или признаки. К активным относятся те органы, которые активно функционируют в течение жизни животного, все равно будет ли это происходить в течение онтогенеза или во взрослом состоянии, будет ли это происходить часто или редко. Сюда относятся, например, все мускулы и все железистые клетки организма, вся нервная система и органы чувств, выделительные и половые органы и т. д. Мы знаем, что функции всех этих регулярно и постоянно действующих органов полезны либо самой особи как таковой, либо, как это относится, например, к функции половых органов, тому виду, к которому данная особь принадлежит.
Другую категорию образуют так называемые пассивные части организма: разнообразная окраска кожи и внутренних органов, многие части кожного скелета, например чешуи и покровные кости, различные шипы и выросты этого скелета, его так называемая скульптура и пр. Мы можем с полной определенностью сказать, что огромное большинство пассивных органов и частей полезны животному уже одним лишь фактом своего присутствия. Так, полезны, например, все виды покровительственных окрасок и форм, панцырь бентонных морских животных(панцырь Ostracodermi и ракообразных, раковины моллюсков), все многочисленные органы защиты животных, как, например, шипы рыб и иглокожих, различного рода стрекательные капсулы и т. д.
Большинство пассивных органов, в особенности же те, которые имеют более или менее сложное строение, безусловно необходимо тем формам, у которых они встречаются. Однако среди них попадаются и такие органы, которым мы при современном состоянии нашего знания никак не можем приписать какое-либо биологическое значение. Правда, при этом нам всегда приходится помнить о том, цто наши сведения относительно биологического значения того или иного признака все еще крайне недостаточны и что поэтому нам всегда приходится быть очень осторожными, когда мы обозначаем какой-либо признак или орган животного как «индиферентный». Укажем, например, читателю на весьма интересное исследование Р. Гессе, касающееся биологического значения относительной величины ушной раковины у различных млекопитающих, живущих в различных широтах. Гессе удалось показать, что этот признак, который прежде всегда относили к числу «инди-ферентных», имеет большое биологическое значение в качестве регулятора теплоты тела животного и что поэтому он является полезным тому животному, у которого находится.
И тем не менее, имеется не мало признаков, которым мы при теперешнем состоянии наших знаний все же никак не можем приписать какое-либо определенное биологическое значение и которые, повидимому, являются действительно индиферентными признаками. К числу таких признаков относится, например, признак окраски внутренних органов животных с непрозрачными тканями: у нас не имеется никаких оснований считать, что красный цвет крови позвоночных, или окраска их мускулов и костей, имеют какое-либо биологическое значение для организма.1 То же самое
1 То обстоятельство, что у животных, подверженных постоянному действию яркого света,образуется во внутренних органах пигмент, предохраняющий их от световых лучей, указывает нам, как будто, на то, что в некоторых случаях окраска внутренных органов животных не является столь индиферентным признаком, как это нам теперь кажется.
ОБЩИЙ ХОД ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ 255
можно сказать и о различных особенностях в строении наружной скульптуры чешуй у многих рыб, например, у костных ганоидов, различной у различных особей, о многочисленных индивидуальных и видовых вариациях формы и окраски у рыб, амфцбий, рептилий, птиц и млекопитающих, биологическое значение которых нам все еще пока неизвестно.
Мы не станем задерживать внимание читателя перечислением всех инди-ферентных признаков животных, отметим только одно, а именно, что все они являются не более как признаками, т. е. незначительными; морфологически простыми особенностями, которыми особи, принадлежащие к одному и тому же виду или к близким между собой видам, отличаются друг от друга.
Даже среди органов пассивных нам не удалось найти индиферентных органов, т. е. бесполезных образований, состоящих из функционально или морфологически координированных тканей—образований, прошедших более или менее сложную и длительную эволюцию и не бывших при этом когда-либо полезными тому животному, у которого они развились.1 Поэтому мы считаем, что хотя «индиферентные» образования и попадаются нам относительно часто, однако они, как таковые (т. е. постоянно сохраняя свой индиферентный характер), обычно уже далее не эволюируют.1 2 В общем эволюция органов представляет собой приспособительный процесс.
ВЗАИМООТНОШЕНИЯ МЕЖДУ ОКРУЖАЮЩЕЙ СРЕДОЙ И ЭКТОСОМАТИ-ЧЕСКИМИ И ЭНТОСОМАТИЧЕСКИМИ ОРГАНАМИ
Мы пришли к выводу, что все активные и большинство пассивных органов животных эволюируют в качестве полезных образований, т. е. как .приспособительные признаки. Теперь мы можем поставить вопрос о том, как эти приспособительные признаки образуются и как они развиваются в дальнейшем. Чтобы ближе подойти к разрешению этой проблемы, нам придется сначала сделать некоторую предварительную классификацию приспособлений.
Как известно, попытки классификации приспособлений делались не раз. Так, Плате 3 дает следующую классификацию главных типов приспособлений животных.
1 Само собой разумеется, мы не имеем здесь в виду рудиментарных органов животных, несомненно бесполезных в настоящее время тем животным, у которых они встречаются, но в свое время полезных их предкам (см. ниже главы XII, XV).
2 Исключением из этого правила, повидимому, являются некоторые из так называемых «эксцессивных» органов, которые в одних случаях бывают индиферентными, а в других—прямо-таки вредными для тех животных, у которых они встречаются. К первым относятся, например, многие из украшающих перьев птиц, тех перьев, которым мы никак не можем приписать определенного полового значения и которые зачастую имеют чрезмерную длину и чрезвычайно своеобразную форму и цвет. Ко вторым относятся такие образования, как огромные ветвистые рога вымерших гигантских оленей, клыки мамонтов и т. п., т. е. такие образования, которые, по мнению некоторых палеонтологов, послужили непосредственной причиной вымирания этих форм. К сожалению, биология птиц с сильно выраженными эксцессивными украшающими перьями нам известна слишком мало для того, чтобы иметь возможность утверждать с определенностью, что перья эти действительно лишены всякого биологического значения. Отметим только, что главное различие между эксцессивными и нормальными органами, т. е. органами, имеющими определенное биологическое значение, в огромном большинстве случаев зависит от чрезмерного роста эксцессивных органов,—обстоя-ятельство, отнюдь не предполагающее предшествующую сложную эволюцию этих органов и, может быть, легко объяснимое какими-либо ненормальностями во внутренней секреции животных. Во всяком случае, в настоящее время все эти образования нам не достаточно известны для того, чтобы составить себе более или менее обоснованное представление о филогенезе и о биологическом значении эксцессивных органов.
3 Р 1 a t е L. Selektionsprinzip und Probleme der Artbildung, 1913; см. также П л а-те Л., Эволюционная теория, М., 1928.
256
ГЛАВА IX
Он рассматривает как приспособления:
I. Единство индивидуальной организации, или коррелятивную целесообразность. Этим термином Плате обозначает гармоничную приспособленность всех органов сложного организма друг к другу, благодаря которой функции одного органа, например почек, целесообразно координированы с деятельностью целого ряда других органов (например, кровеносной системы, других желез организма и т. д.). Только полная координация всех частей организма и делает его жизнеспособным.
П. Целесообразность строения, или внутреннюю целесообразность — приспособление строения одних органов к строению других: расположение чувствующих клеток в органах чувств, расположене костных перекладин в длинных костях, благодаря чему получается способность к наибольшему сопротивлению при наименьшем весе, присутствие клапанов в сердце и сосудах позвоночных, определяющих направление тока крови, и многие другие особенности животных.
III. Наружную целесообразность, под которой Плате подразумевает все приспособления тела животных к условиям окружающей живой и неживой природы, т. е. приспособления к жизни в воде и на суше, близ берега или в открытом море, на поверхности или в абиссальных глубинах, при свободном образе жизни или при паразитическом и т. д.
IV. Рефлекторную и инстинктивную целесообразность при нормальных условиях жизни, т. е. приспособительные реакции организма на простые или сложные раздражения, исходящие из внешней среды или зависящие от изменений состояния собственного тела (выделение желудочного сока при введении пищи, сокращение зрачка при усилении светового раздражения, половые инстинкты самцов и самок животных и т. д.).
V. Санаторную; или регуляторную целесообразность: деятельность лейкоцитов при бактериальных болезнях, выделение антитоксинов, аглютининов и преципитинов, регенерация утраченных частей организма и т. д.
VI. Функциональную целесообразность: способность активных органов усиливаться при усиленном функционировании, ослабевать при отсутствии деятельности.
VII. П р и с п о с о б л е н и я к поддержанию жизни вида: желточные и белковые железы, совокупительные аппараты, обеспечивающие внутреннее оплодотворение, органы, служащие для откладывания яиц в защищенных местах.
VIII. Эмбриональную целесообразность, обнимающую бесчисленные приспособления эмбрионов к условиям эмбриональной жизни (амнион, аллантоис, приспособления к перевариванию желтка и т. д.) и к гармоничному развитию органов взрослого животного.
Я позволил себе привести классификацию Плате, во-первых, чтобы дать понятие о той точке зрения, с которой обычно рассматриваются приспособления, во-вторых, чтобы напомнить читателю о том громадном разнообразии целесообразных изменений, которые произошли в организации животных в течение эволюционного процесса.
Г. Ф. Осборн в своей книге о происхождении и эволюции жизни1 дает такой обзор целесообразных изменений организмов.
1. Приспособления, находящиеся в соотношении с окружающей средо й—дыхательные, обонятельные, зри
'Osborn Н. F., The origin and evolution of life on the theory of action, reaction and interaction of energy, 1918.
ОБЩИЙ ХОД ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ
257
тельные, слуховые, статические функции и органы, координированные и соотносительные (correlative) с изменениями света, тепла, влажности и сухости, вызванными миграциями индивидуума или изменениями среды.
II. Приспособления, представляющие собой внутренние корреляции: корреляции и координации роста «функций внутренних органов вследствие внутренней секреции,действия энзимов и нервной системы.
III. Приспособления к питанию: неорганическими веществами, бактериями,протофитами, водорослями одноклеточными животными, высшими растениями (травоядное питание), высшими животными (хищный образ жизни); паразитарное питание—на поверхности или внутри растений и животных.
IV. Приспособления к индивидуальному состязанию и подбору.
А. Подбор, касающийся вариаций, ректиградаций, мутаций, происхождения и развития единичных признаков, пропорций и т. д.
В. Подбор, касающийся всех органов размножения, первичных и вторичных.
V. Приспособления к расовому состязанию и подбору, касающиеся главным образом всех органов движения, нападения и защиты, внутреннего и внешнего скелета, а также скорости размножения.
Если мы сопоставим между собой классификацию приспособительных изменений организмов Осборна с приведенной нами выше классификацией Л. Плате, то нам станет совершенно ясна та точка зрения, с которой эти приспособления рассматриваются авторами. Внимание авторов обращено главным оразом на целесообразность приспособлений и на соотношения между одними органами и функциями животного и другими органами и функциями того же животного или животных другого пола в различные эпохи их жизни, или же на соотношение между органами и функциями животных и различными сторонами окружающей сре-д ы. Осборн, несколько расширяя понятия приспособления, вводит в свой обзор и способы, посредством которых совершается эволюция.
Но к вопросу о целесообразных изменениях можно подойти и с другой точки зрения, а именно, поставив вопрос о том, как различные приспособительные изменения отражаются на дальнейшей эволюции животных.
Обычно под именем приспособления обозначаются такие особенности в строении какого либо органа, которые находятся в целесообразно-функциональном или целесообразно-биологическом соотношении с какой-либо особенностью среды, окружающей данное животное, или с какой-либо особенностью в строении какого-нибудь другого органа того же животного.
Здесь мы не будем подробно останавливаться на том, что собственно h следует подразумевать под словом внешняя среда: этим термином мы обозначаем всю совокупность факторов окружающего мира, факторов, стоящих i в каком бы то ни было отношении к живому организму (особи, i виду), т. е. тех факторов, изменение которых способно вызвать морфологи-; ческие или физиологические изменения в интересующих нас организмах. ‘ Условия внешней среды могут быть подразделены следующим образом: 1) условия неорганической среды, т. е. условия температуры, сухости и влажности почвы и воздуха, условия атмосферного давления и : Морфологические закономерности 17
258
ГЛАВА IX
давления воды, условия строение почвы, силы освещения и т. д. и 2) условия органической среды или биологические условия, т. е. отношения данного организма к окружающим его животным или растительным организмам (пища, условия обитания, различные способы защиты от врагов и т. п.). Далее мы различаем условия отношений данной животной формы к животным'того же вида (спаривание, потомство, сотрудничество, конкуренция и т. д) от условий его отношения к животным, принадлежащим к другим видам (комменсалы, паразиты, враги, добыча и т. д.).
Если какая-либо животная форма просуществовала в течение определенного периода времени в определенных условиях внешней среды и приспособилась к ним, то мы обозначим ее органы и признаки, стоящие в непосредственных, функциональных или (при пассивных органах) биологических соотношениях с условиями этой среды, как э к то-сом а т и ч е с к и е1 органы и признаки. Эктосоматическим органам и признакам мы противопоставим те органы и признаки, которые не стоят в непосредственной функциональной связи с условиями внешнего мира, и назовем эти последние эктосоматическими органами и признаками.
В качестве примеров эктосоматических органов позвоночных животных можно привести кожу с ее придатками (волосы, перья), кожный скелет, органы кожного чувства и органы высших чувств, зубы, когти, копыта, ногти и рога, плавники рыб, конечности наземных четвероногих, кишечный канал с его железами1 2 и т. д. Каждый из этих эктосоматических органов стоит в совершенно определенном функциональном соотношении с о д н им й*л и с нескол ькими фа к-jro_P а м и внешней с редПя, н о н е со в с ею в н е ш н е~ й с р е д о й, в з я три в цело м. Так, глаза ж:ивотных~стоят в определенном функциональном соотношении с нормальными условиями освещения места его обитания: они изменяют свое строение в связи с изменениями в силе освещения (ср., например, строение глаза удневных и у ночных птиц). Кожа наземных позвоночных стоит в совершенно определенных функциональных соотношениях с климатом (температура, влажность и т. д.); строение их зубов—с пищей, которую употребляют данные животные (ср., например, строение зубов у травоядных и всеядных млекопитающих, а также у питающихся исключительно мясной пищей хищников).3
В качестве примеров эктосоматических органов животных можно привести сердце со всеми отходящими от него большими кровеносными сосудами (за исключением капиллярных сосудов кожи и кишечного тракта, которые несомненно относятся к эктосоматическим органам), почки, центральную нервную систему, железы внутренней секреции и многое другое. При этом подчеркнем, что энтосоматические органы также являются приспособленными органами, но что их «приспособленность» относится не к условиям внешней среды, но к строению и функциям эктосоматических органов и к строению и функциям других эктосоматических органов. Здесь, однако, нам уже приходится иметь дело с функциональными корреляциями или координациями между энто- и эктосоматическими органами, о которых речь будет впереди.
Чтобы облегчить читателю понимание функциональных соотношений между внешней средой и органами животного, живущего в условиях этой среды, например какого-либо наземного позвоночного животного, приведем следующую схему (схема XX), в которой примем в качестве основной предпосылки, что условия внешней среды остаются
1 Я заимствовал этот термин из превосходных неврологических работ И. Джонсона.
2 Ибо пищу животного можно рассматривать как одну из важнейших сторон его внешней среды.
3 Здесь мы могли бы увеличить число наших примеров до бесконечности.
ОБЩИЙ ход филогенетического развития
259
константными, т. е. что в ней не происходит никаких заметных и длительных изменений в определенном направлении. Тогда мы можем представить себе следующим образом взаимоотношение между организмом, и внешней средой:
в с ।----Условия окружающей среды
till
\ { "1...этосоматические органы
I I I I
? { 5 f----Знтосоматичесш органы izj порядка
1111
у ю ю -— Знтосоматичесние органы порядка
Организм а:
Схема XX. Объяснение в тексте.
В нашей схеме мы принимаем, что буквы А, В, С, D, Е.... символически изображают различные условия той внешней среды, в которой обитает данная животная форма X, причем, например, А обозначает собою условия освещения, В—температурные условия, С—строение и свойства почвы, D—пищу, которую принимает животное, и т. д. Буквы h, i, k, 1, m... символически обозначают собою эктосоматические органы формы X, причем h представляет собою органы, находящиеся под непосредственным воздействием светового раздражения (глаза и хроматофоры кожи), i—части организма, функционально реагирующие на колебания температуры или вообще стоящие в каком-либо соотношении с этими колебаниями (капиллярные сосуды кожи, подкожный жир, такие кожные придатки, как волосы, перья, пух и т. д.), к—органы движения, связанные со средой, в которой передвигаются животные (конечности с их скелетом и мускулами, кожа подошвы, когти, копыта и т. п.), 1—органы питания, функционально связанные с видом пищи, присущей форме X (челюсти и зубы, язык, слюнные железы, эпителий и железы кишечного тракта и т. п.). Буквы р, q, г, s, t... символизируют в нашей схеме те из энтосоматических органов формы X, которые стоят в непосредственной функциональной зависимости от эктосоматических органов h, i, k, 1, гп... Таким образом р соответствует зрительным нервам и зрительным центрам, q—вазомоторным нервам и сосудам кожи, г—нервам и сосудам конечностей и т. д. Буквы u, v, w, х, у... символически изображают другие энтосоматические Органы, а именно те, которые находятся в коррелятивной связи с первой группой энтосоматических органов—р, q, г, s, t. Взаимоотношения всех этих органов между собой изображено на нашей схеме при помощи стрелок. Мы обращаем особенное внимание читателя на то, что здесь эти взаимоотношения до крайности упрощены и что в действительности они являются бесконечно более запутанными и сложными.
Говоря о функциональных соотношениях, или о функциональных координациях между экто- и энтосоматическими органами, мы имеем в виду физиологические, но отнюдь не филогенетические соотношения. При этом мы считаем, что, например, нормальное строение и нормальная функция зрительных нервов и зрительных центров совершенно необходимы для нормальной зрительной функции ретины, а для отправления нормальной функции конечностей (ходьба) безусловно необходимы нормальное строение и нормальная функция мускулов, нервов и сосудов как самих конечностей, так и поясов конечностей. Именно этого рода соотношения мы и обозначаем термином «функциональные корреляции».
Итак, мы принимаем, что все органы и признаки животных подразделяются на две большие групп ыз .1) на эктосоматические органы и признаки, 17*
260
ГЛАВА IX
находящиеся под непосредственным воздействием условий внешней среды, и 2) на энтосоматические органы и признаки, не стоящие в непосредственных отношениях с внешней средой, но морфологическими функционально связанные с эктосоматическими или другими энтосо-матическими органами или признаками.
Мы установили, кроме того, что каждая категория экто-соматических органов или признаков находится в функциональном отношении не со всей суммой условий внешней среды, но лишь с одной какой-либо определенной категорией этих условий.
ВЛИЯНИЕ ИЗМЕНЕНИЙ ВНЕШНЕЙ СРЕДЫ НА ЭКТОСОМАТИЧЕСКИЕ И ЭНТОСОМАТИЧЕСКИЕ ОРГАНЫ
Теперь нам приходится поставить вопрос о том, каким образом воздействуют изменения, происходящие в условиях органической и неорганической внешней среды, сначала на эктосоматические органы, находящиеся под непосредственным воздействием этих условий, а затем на энтосоматические органы, связанные с данными эктосоматическими органами.
Филогенетические и экологические исследования совершенно недвусмысленно указывают нам на то, что изменения известных условий внешней среды вызывают те или иные изменения в органах, находящихся в состоянии функциональной зависимости именно от этих сторон окружающей среды (теория онтогенетической адаптивной эволюции). Возьмем, например, какой-нибудь определенный вид животных, живущий в течение долгого времени в известных условиях окружающей среды и вполне приспособившийся к ним, и представим себе, что какая-либо одна из сторон этой среды начинает длительно изменяться в том или ином определенном направлении; представим себе, например, что постоянно, но постепенно начинает понижаться интенсивность освещения. Это может случиться, если какая-нибудь форма наземных животных начнет переселяться в темную пещеру или постепенно переходить к подземно роющему образу жизни. Первый случай мы имеем у пещерных амфибий и рыб, второй—у гимнофионов и подземных млекопитающих (кротов, Spalax, Chrysochloris и т. п.). Мы наблюдаем при этом, что животное отвечает на изменение среды изменениями в тех именно органах, которые находятся в функциональной зависимости от света. У протея редуцируются глаза и пигментация кожи, аналогичные изменения встречаются у постоянно живущих в пещерных водах рыб.
Другие примеры того, что одностороннее изменение внешней среды вызывает соответствующее изменение только в очень н е м н о г и х экто-коматических органах, мьГ встречаем"у многих рептилий. Так, например, весьма вероятно, что предки гекконов перешли в течение своего филогенеза к такому образу жизни, который был связан с лазанием по отвесным скалам; для этого их конечности должны были соответственным образом изменить свое строение. И мы видим, что изменение какого-либо одного условия в обычной среде обитания, в данном случае переход от движения по горизонтальной поверхности к движению по отвесным, порою почти вер-тикальнььм утесам и скалам, вызвал соответствующие структурные изменения в строении конечностей гекконов. У гекконов пальцы на передних и задних лапках становятся широкими и плоскими, причем у них на вентральной поверхности образуются сложно построенные складки с рядами
ОБЩИЙ ХОД ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ 261
роговых коротких волосков. Все же другие органы гекконов по сравнению с соответствующими органами остальных Lacertilia сохранили весьма примитивное строение.
У амфибий и рептилий, перешедших к подземно роющему образу жизни, изменения, происшедшие в эктосоматических органах, более многочисленны, чем, например, у амфибий, живущих постоянно в пещерах (протей и подобные ему формы). Так, у гимнофионов переход к подземно роющему образу жизни вызвал редукцию глаз; редукция глаз имеет место и у животных, населяющих пещерные воды. С другой стороны, с жизнью в темноте связано прогрессивное развитие органов обоняния и осязания (щупалец). Иногда наблюдается и прогрессивное развитие кожных желез. К роющему образу жизни приспособлены изменения в форме головы, а также целый ряд изменений в черепных костях, которые у роющих амфибий (гимнофионов) становятся крепче и толще, чем у нероющих амфибий. Различные изменения в строении туловищных чешуй этих форм также должны рассматриваться как приспособления к роющему образу жизни. Таким образом, в строении тела гимнофионов произошел ряд изменений, из которых одни вызваны приспособлением к жизни в темноте (изменения в органах зрения, обоняния и осязания), а другие— к новому способу передвижения этих форм (изменения в форме головы и черепных костей, изменения в кожных железах и чешуях).1
Все эти примеры, число которых при желании можно было бы значительно умножить, с полной ясностью указывают нам на то, что изменения во внешней среде вызыв а ют соотвТт-с т в у ю'щиё и з“м ёТПГн йТ лИ ш ь~TTVejOi ,KJC_o..C D маги ч.е-с к и х о р г а нах ж иво т и о г о, . кото р ы е - находятся в непосредственном функцио нал ь ном от н о ш е-н и и ~к~~и~з~м~е н~и в ш й м с я сторонам этойсреды.
FT аналогичным результатам приходйлГмы и при изучении эволюции некоторых бентонных рыб, например скатов. Здесь изменения во внешней среде состояли в том, что нектонные предки скатов, перешедшие от плавающего образа жизни к лежанию на дне, вступили в определенные функциональные отношения с почвенными условиями обитаемых ими водоемов, т. е. с донной флорой и донной фауной. Филогенетические исследования показали нам, что изменения в эктосоматических органах вызвали коррелятивные изменения в тех эктосоматических органах, которые с ними функционально связаны (функциональная корреляция).
Эмбриологические исследования определенно говорят о том, что скаты произошли от веретенообразных акул (рис. 72). Широкая и плоская форма тела скатов произошла оттого, что грудные плавники их акулоподобных предков сильно разрослись в каудальном и ростральном направлениях, так что их propterygium и mesopterygium постепенно превратились в длинные палочковидные хрящи. Число хрящевых плавниковых лучей при этом возросло, сами же лучи удлинились (рис. 139). Это были те первичные изме
1 Мы имеем все основания предполагать, что удлинение тела (увеличение количества позвонков) и дегенерация конечностей имели место у гимнофионов еще до того, как эти формы перешли к подземному образу жизни (ср. главу X). Ход изменений, происшедших в строении тела предков гимнофионов, можно представить себе следующим образом. Предки гимнофионов (стегоцефалы) были животными, передвигавшимися на четырех лапках и напоминавшими своей общей формой саламандр; тело их было пок-крыто чешуями.Впоследствии они перешли к извивающемуся, ползающему способу передвижения, причем тело их удлинилось, а конечности начали редуцироваться, и животные постепенно приобрели змееобразную форму тела. Еще позже формы эти, которые теперь стали легко укрываться от врагов в трещинах и щелях земной поверхности, постепенно перешли к чисто подземному (роющему) образу жизни и приобрели свойственные ему особенности (упомянутые нами изменения в коже, в черепе и в щупальцах, редукция глаз).
262
ГЛАВА IX
нения, которые впоследствии повлекли за собою основное изменение в форме тела скатов. Другой ряд первичных изменений (приспособлений к бентонному образу жизни) появился в ротовом аппарате этих форм: первично острые зубы акулообразных предков скатов превратились в плоские мостовидные зубы, приспособленные для перетирания раковин, ползающих по морскому дну различных ракообразных, которыми питаются ныне живущие скаты.
В связи с первичными изменениями адаптивного характера эктосома-тических органов развивается целый.ряд координированных с ними филогенетических изменений, которые сами по себе не имеют никакого отношения к изменениям во внешней среде, но которые находятся в непосредствен-
Рис. 72 А. А'. В. С. D. Четыре последовательных стадии развития наружной формы тела Trygon pastinaca.
Abd.—брюшные плавники; br.j—br.5—жаберные щели 1—5; pect.—грудные плавники; spir.^ брызгальце. На рис. А и Д'1 зародыш имеет еще акулообразную форму; грудные плавники лежат позади последней жаберной щели (Ьг.д) На рис. В мы видим, что грудные плавники выросли спереди и почти прикрывают собою жаберные щели. На рис. С и D мы видим, что грудные плавники разрослись над вентрально сдвинувшимися жаберными щелями (Ьг.г—Ьг.д) и что их передний край лежит непосредственно позади глаз. На рис. D зародыш уже принял типичную форму ската: на стадиях А, В, С видно, как форма тела зародыша постепенно становится все более плоской и более широкой (из А. Н. Северпова, 1931).
ной связи с изменениями в только что упомянутых нами эктосоматических органах. Так, увеличение груднЬх плавников скатов имеет следствием более прочное прикрепление плечевого пояса к позвоночнику. Плечевой пояс соединяется теперь с позвоночником при помощи плотных соединительнотканных связок, что в свою очередь влечет за собой новые изменения во внутреннем скелете: в области причленения плечевого пояса к позвоночнику отдельные позвонки срослись между собою, так что в этом месте образовался очень прочный, негнущийся отрезок осевого скелета (рис. 139, Sc. г). У нектонных акул, у которых грудные плавники не увеличены и плечевой пояс лежит свободно, позвонки в этом месте соединены между собою подвижно. Разрастание передней части грудного плавника (propterygium) в ростральном направлении вызывает смещение жаберных щелей на вентральную сторону тела и обусловливает дорзальное положение спиракулярной щели (рис. 72, br. 1—Ьг. 5, spir). Эмбриологические исследования ясно показывают нам, каким путем осуществляется это смещение щелей и как тесно оно действительно зависит от разрастания передней части грудного плавника (propterygium) скатов.
На этом примере прекрасно видно, как.первичные адаптивные измене-ния в.эктосоматических органах вызывают коррелятивные филогенетические изменения^ тех: именноэнтосоматических органах, кбтррыелщэтими. эктосоматическими органами функционально свазаны. Указанные нами морфологические изменения во внутренних энтосоматических органах,
ОБЩИЙ ХОД ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ
263
собственно, не имеют непосредственного отношения к изменениям, происходящим во внешней среде, окружающей эволюирующее животное, но они необходимы для отправления нормальной функции адаптивно изменившихся эктосоматических органов и ими каузально обусловлены. Мы обозначаем этого рода изменения именем филетических корреляций или координаций.
Только что описанные нами филогенетические изменения, имеющие место в эктосоматических и энтосоматических органах в условиях изменяющейся среды, могут быть символически представлены при помощи схемы XXI, являющейся некоторым видоизменением нашей предыдущей схемы.
/ в с' о
Организм х
...... Условия окру тающей среды
m.......Эктосоматические органы
........Энтоеоматические органы № порядка
у -•*-------кЭнтосоматические органы дю лирника
Схема XXI. Объяснение в тексте.
Примем, что изменяется один или несколько факторов окружающей среды, что, например, фактор А (освещение) превращается в А', или что фактор С (свойство почвы) превращается в С'; тогда соответственно изменится строение тех органов, которые находятся в каком-либо отношении к этим изменившимся условиям среды. Иными словами, возвращаясь к приведенным нами выше примерам, если изменятся условия освещения в месте обитания протея—изменится строение его глаз, если изменится место обитания (свойство почвы) предков гекконов при их переселении на отвесные утесы—изменится строение их конечностей.
Если эти изменения в эктосоматических органах будут незначительны, то они не вызовут заметных морфологических изменений в органах, функционально с ними связанных, но если первичные изменения эктосоматических органов будут значительны и будут кроме того носить длительный характер, то они породят соответствующие филетические корреляции в структуре, форме и величине тех энтосоматических органов, которые с ними функционально связаны. Так, изменения в органах hx и кх (схема XXI) обусловливают коррелятивные изменения в органах рх и гх, т. е. в зрительных центрах в одном случае и в скелете, нервах и мускулах конечностей—в другом.
На приведенных нами примерах мы видим, что коррелятивные морфологические изменения в одних энтосоматических органах вызывают со своей стороны новые коррелятивные изменения (корреляции второго порядка) в других энтосоматических органах: так, разрастание плавников у скатов (первичное эктосоматическое изменение) вызывает соединение плечевого пояса конечностей с позвоночником (филетическая корреляция первого порядка), а это последнее, в свою очередь, вызывает срастание грудных позвонков между собой (филетическая корреляция второго порядка). Прогрессивное развитие птичьих ног (первичное эктосоматическое изменение) вызывает увеличение размеров таза (филетическая корреляция первого порядка), что обусловливает собою развитие synsacrum (филетическая корреляция второго порядка). [На схеме филетические корреляции второго порядка обозначены превращением букв и и w в буквы их и wj.
264
ГЛАВА IX
В действительности соотношения между первичными и коррелятивными морфологическими видоизменениями гораздо сложнее, чем мы их изобразили на схемах XX и XXI, так как каждый эктосома-тический орган функционально связан не с одним, но со многими энтосо-матическими органами и так как функциональные корреляции между различными энтосоматическими органами фактически гораздо многочисленнее, чем это нами представлено на тех же схемах (см. схему XXI, стрелки, идущие от h1 и к1). Сейчас мы не будем вдаваться в дальнейшее обсуждение всех этих соотношений, ибо впоследствии мы снова вернемся к этому вопросу (в главе XIII о координациях).
Краткости ради мы обозначим морфологические адаптации в эктосо-матических органах как первичные, или проталлактические изменения (проталлакс ы),1 а вторичные и третичные морфологические изменения в энтосоматических органах как филетико—морфологические корреляции, или дейталлакс ы.1 2
В предыдущем, базируясь на фактах филогенеза, мы попытались дать общую схему эволюции животных. При этом мы пришли к выводу, что эволюция эта в общем является эктогенетиче-ской эволюцией, т. е. что она обусловливается изменениями во внешней среде, окружающей животных. Филогенетическое развитие организма можно сравнить с мозаикой в том отношении, что од новременно и равномерно изменяются не .все части организм а, *н сГ л 'и;щ ь те ёгоэктосоматич ё с к и и части, кот о ры е н а ходя т с я в функциональных соотношениях с изменениями, происходящими в окружающей среде в данный период.
Интенсивность изменений эволюирующих организмов стоит в известном количественном соотношении с интенсивностью и длительностью изменений внешней среды, т. е. сильные и длительные изменения внешней среды вызывают в организме более значительные изменения, чем слабые и непродолжительные. Мы знаем, что все глубокие изменения, происходящие в неорганической среде (изменение климата, орографические изменения и т. д.) точно так же как и все резкие изменения в органической среде (значительное изменение вегетации, среди которой живет данная животная форма, изменения той части фауны, с которой она находится в непосредственных соотношениях), обычно совершаются очень медленно. Даже в тех случаях, когда интересующий нас вид животных находится на пути активного переселения из одного места обитания в другое, даже в этих случаях переселение вида (не отдельных особей) происходит лишь очень медленно и очень постепенно.
Из этого следует, что незначительные, но идущие все время в одном определенном направлении изменения окружающей среды, когда они непрерывно следуют друг за другом, медленно суммируются и вызывают соответствующие изменения в эктосоматических органах животного. При этом очень глубокие и очень длительные изменения во внешней среде обычно обусловливают соответственные изменения в доволы10. зна-чительном числё эктосдматичёских органов и в еще более значительном числе энтосоматических органов. Наблюдения показывают нам, что это имеет место тогда, когда животные переходят пассивным3 или актив-
1 От тсротс? — первый и — изменяю.
2 От йебтеро? — второй и аДДаааФ — изменяю.
3 Т. е. либо путем изменения во флоре (например, при смене леса степью, или пустыней, или наоборот), либо путем изменения климата (ледниковые периоды и т. п.), либо путем изменения почвенного рельефа (орографические процессы), причем животное пребывает на своем старом месте обитания (пассивное изменение окружающей среды).
ОБЩИЙ ХОД ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ
265
ним1 путем в новую среду, во многих отношениях резко отличающуюся от прежней.
Как мы это только что отметили, изменения условий окружающей среды обычно происходят лишь очень медленно и очень постепенно; поэтому большей частью случается так, что при переселении из__однр.го места в другое животное выбирает сначала такие стации, которые, отли-чаются от старого места обитания лишь очень немногими признаками. Животное продолжает свое дальнейшее странствование (миграцию) лишь тогда, когда оно оказывается достаточно приспособленным к новым условиям. Таким образом изменение функций у мигрирующего вида происходит с еще большей медленностью и еще большей постепенностью, че'м изменения во внешней среде .^Приспособления к новой пище, к новому способу передвижения, т. е. к новым условиям почвы, к иной добыче, которую надо поймать, или к иному врагу, которого надо научиться избегать, развиваются н е о д н о н н о, но так, что каждое из них
соответствует, отдельным., друг за другом следующим изменениям , окружающей среды в строгой последовательности, одно пос ле д“руттггтгг~——- .......
‘“"' Именпо поэтому мы и можем сравнить эволюцию новых эктосоматических признаков с мозаикой: отдельные эктосоматические признаки часто развиваются независимо друг от друга. Но более часто случается, однако, так, что первично изменяющийся эктосоматический орган изменяет собой не только ход эволюции функционально с ним связанных энтосоматических органов (см. схему XXI), но вместе с тем влияет и на эволюцию некоторых других эктосоматических органов, так что филетические корреляции возникают также и между различными эктосомати-ческими органами.
выводы
Мы можем резюмировать наши взгляды на общий ход филогенетического развития многоклеточных животных в следующих немногих положениях.
1. В подавляющем большинстве случаев филогенетические изменения организации животных являются приспособлениями к изменениям окружающей среды, в которой существуют и эволюируют данные животные формы.
2. Изменения в окружающей среде обычно протекают таким образом, что не все условия этой среды меняются одновременно: при изменении одних сторон этой среды другие ее стороны остаются в течение долгого времени неизменными.
3. Для эволюции животных имеют значение лишь те изменения окружающей среды, которые протекают медленно и непрерывно в определенном направлении и влияние которых на организацию животных может постоянно суммироваться.
4. Органы животных могут быть нами подразделены на две большие группы, а именно на: А) эктосоматические органы, находящиеся в непосредственных и определенных физиологических и биологических соотношениях с условиями'окружающей среды и В) эн.то-соматические органы, не имеющие непосредственных физиологических соотношений с окружающей средой.
5. Каждый эктосоматический орган функционально связан с одним или несколькими энтосоматическими органами (функциональная коорди
1 Под активными изменениями окружающей среды мы подразумеваем такие ее изменения, когда само животное вследствие каких-либо причин меняет место своего обитания (миграция, перенос потоками вод, течением рек и т. п. самих животных, либо их яиц или личинок).
266
ГЛАВА IX
нация). Эктосоматический орган приспособлен к условиям окружающей среды, энтосоматический орган—к функциям эктосоматических органов, так что в каждом организме имеются двоякого рода приспособления: внешние и внутренние адаптации.
б. Эволюция животных представляет собою эктогенетический процесс, иными словами, адаптивные изменения функций и строения эктосоматических органов' обусловлены изменениями, имеющими место в неорганической и органической среде, окружающей эволюирующее животное.
7. В общем, эволюция любой группы животных протекает по следующей схеме: известные стороны той внешней среды, в которой обитает данная животная форма, изменяются в некотором определенном направлении; те эктосоматичес. кие органы, которые имеют непосредственное отношение к изменяющимся сторонам окружающей среды, приспособляются к изменившимся условиям этой среды (проталлаксисы); эктосоматические органы, находящиеся в функциональном соотношении с изменившимися эктосоматическими органами, приспособляются со своей стороны к наступившим изменениям в структуре и функциях эктосоматических органов, в результате чего образуются новые координации (координации первого, второго и т. д. порядка) между эктосоматическими и эктосоматическими органами, с одной стороны, и между различными функционально между собою связанными энтосо-матическими органами, с другой. Схематически этот процесс может быть изображен следующим образом:
Изменения тех энтосоматических органов, которые находятся в функциональном соотношении с изменившимися эктосоматическими органами (координации первого, второго и т. д. порядка).
Изменения тех эктосома- ’ тических органов, которые находятся в функциональном соотношении с этими сторонами окружающей среды.
Изменения некоторых сторон окружающей среды.
ГЛАВА X
ОСНОВНЫЕ НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
БИОЛОГИЧЕСКИЙ ПРОГРЕСС И ГЛАВНЫЕ ПУТИ, КОТОРЫМИ ОН ДОСТИГАЕТСЯ
В предыдущей главе мы сделали попытку дать очень общую схему типичного хода филогенетического развития животных. При построении этой схемы мы исходили главым образом из результатов филогенетических и экологических исследований, намеренно оставляя в стороне результаты целого ряда не менее важных дисциплин. Отметим в особенности, что изложенное нами в этой главе под рубрикой « Выводы», должно рассматриваться только как голая схема и что основная задача настоящей и следующих глав этой книги заключается в том, чтобы в эту схему вложить конкретное содержание.
Начнем со много обсуждавшегося, но доныне все еще очень темного вопроса относительно направления прогрессивной эволюции животных. Мы не будем входить в историю полемики, возникшей вокруг этой проблемы,1 но чтобы иллюстрировать степень притиворечия во взглядах авторов, занимавшихся этим вопросом, отметим, что одни ученые держатся того мнения, что прогрессивной эволюции, как таковой, вообще нет и быть не может, ибо прогресс совпадает с приспособлением, и амеба, например, так же хорошо приспособлена к условиям окружающей ее среды, как тигр или человек к условиям окружающего его мира (Шауинсланд, 1909), другие же, наоборот, считают, что в природе существует непреложный, так сказать механически действующий закон прогрессивного усовершенствования животного мира, так что вообще всякий новый признак, независимо от его приспособленности к окружающим условиям, является прогрессивным. Это различие во взглядах зависит в значительной степени от смешения биологической и морфологической сторон эволюционного процесса.
Вдумываясь в характер приспособительной эволюции, мы находим два различных типа прогресса, а именно, прогресс биологический и прогресс морфофизиологический. Остановимся более подробно на разборе этих двух чрезвычайно важных понятий.
Если мы видим, что какая-либо группа животных находит обильный корм, быстро и свободно размножается, мало истребляется врагами и благодаря этому сильно увеличивается численно, если повышается долговечность особей вида и этот вид в течение последующей своей эволюции расселяется и захватывает новые ареалы распространения; если, наконец, данный вид при этом процессе численного увеличения и расселения рас
1 В работах В. Франца можно найти подробное изложение вопроса о филогенетическом «усовершенствовании» организмов (см. Franz V., Die Vervollkomnjinung in der iebenden Natur, Jena, 1920, а также в его книге Geschichteder Organismen, Jena, 1924).
268
ГЛАВА X
падается на более мелкие таксономические группы, т. е. на целый ряд разновидностей, которые стремятся в свою очередь превратиться в новые виды, то ясно, что данная группа животных изменяется в прогрессивном направлении, и этот прогресс имеет биологический характер. Непосредственное наблюдение учит нас тому, что при благоприятных условиях окружающей среды, т. е. если климатические условия подходящи, если имеется в достаточном количестве пища, если в привычных местах обитания животного находится лишь немного опасных врагов, то число особей данного вида возрастает с каждым новым поколением. Так, например, нам зачастую приходится видеть, что какая-либо форма рыб, обычно лишь изредка попадающаяся в тех или иных реках, при особо благоприятных условиях существования (в условиях искусственной охраны) довольно скоро становится обычным представителем нашей ихтиологической фауны, или же, что какая-нибудь редкая птица с течением времени начинает вдруг усиленно размножаться. Напомним читателю о голубях, которые в некоторых городах, как, например, в Венеции или в Падуе, не только не истребляются населением, но которых там даже прикармливают, или же о воробьях и галках, обычных обитателях наших деревень.
Подвижные свободно живущие млекопитающие, попадая в благоприятные для них условия существования, также размножаются с чрезвычайной быстротой и достигают необычайной многочисленности в короткое время. Так, например, Колумбом на остров С. Доминго было привезено несколько голов рогатого скота, которые одичали и размножились так быстро, что через 27 лет стада в 4000—6000 голов были здесь довольно обыкновенны. Позднее рогатый скот был перевезен с острова С. Доминго в Мексику и другие части Америки, и в 1567 г., т. е. через 65 лет после завоевания Мексики, испанцы вывезли 64350 голов из Мексики и 35 444 головы с острова С. Доминго, что указывает на то, какое множество этих животных существовало здесь, так как пойманные и убитые составляли, конечно, только небольшую часть всего количества. Мы знаем также, с какой быстротой размножались кролики, вывезенные в Австралию, и как они сделались настоящим бедствием для этой страны. Эти примеры касаются животных, вывезенных и охраняемых человеком, но мы имеем основание думать, что в благоприятных условиях животные размножаются с той же быстротой и в диком состоянии, и мы знаем примеры такого размножения. Отметим, что здесь мы вообще имеем в виду не случайное, не неповторяющееся или лишь периодически повторяющееся повышение числа особей известного вида в известной местности,1 но именно постоянное, из года в год повторяющееся прогрессивное увеличение Числа особей, образующих данный вид. Мы можем с уверенностью сказать, что незначительное само по себе, но непрерывно возрастающее увеличение числа особей, образующих данный вид, является первым признаком биологически прогрессивной эволюции этого вида.
За последнее время С. А. Северцов опубликовал ряд работ, касающихся условий борьбы за существование и ее значения для биологического прогресса. В этих работах он подробно разбирает вопросы, связанные с численностью вида.
Согласно С. А. Северцову, границы местообитания каждого вида . определяются положительными и отрицательными моментами. Каждый
1 Так, например, довольно часто встречающееся у насекомых, птиц и т. д. спорадическое увеличение числа особей в течение только одного какого-нибудь лета ни в коем случве не может быть рассматриваемо как признак прогрессивной эволюции (см. С. А. Северцов, 1931).
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ 239 вид приспособлен к определенным условиям температуры, влажности, содержания солей в воде и почве и другим биотическим и абиотическим факторам. Только в условиях своей стации .он находит наибольшие возможности добывать пищу, спасаться от врагов и непогоды и т. д. В смежных стациях эти возможности меньше. Отдельные особи вида выходят за границы своего местообитания, но не могут в них закрепиться, так как смертность выше естественного прироста. Для хищников вне их стации смертность возрастатет, так как они там не могут поймать достаточного количества добычи, а для растительноядных увеличиваются трудности укрыться от хищников.
Но и в пределах своего собственного местообитания каждый вид подвергается весьма интенсивной гибели, которая покрывается естественным приростом. С. А. Северцов различает гибель индивидуальную и массовую, вызванную стихийными причинами или эпидемическими болезнями. Случаи массовой гибели для каждого вида периодически повторяются, и численность вида не остается постоянной, но волнообразно колеблется в весьма широких пределах.
Величина смертности в единицу времени изменяется с возрастом животных. Новорожденные обычно обладают наибольшим коэфициентом гибели. По мере того, как они развиваются и приближаются к строению взрослых, смертность падает. У взрослых особей она может считаться постоянной. Когда же наступает старческое ослабление организма, вероятность .гибели опять возрастает. Мы можем принять, что отмирание закончено, когда от поколения осталась одна особь. Тогда время отмирания окажется предельной продолжительностью жизни особи в данных природных условиях борьбы за существование. Так как вид ежегодно размножается, то эта же величина равна предельному числу поколений, которое может существовать одновременно. Таким образом долговечность характеризует и плотность населения.
Одинаковая смертность наблюдается при разнообразных приспособлениях к условиям среды. Чем меньше смертность в борьбе за существование, тем, очевидно, выше значение приспособлений. В пределах одной и той же группы более крупные виды долговечнее. Хорошо защищенный дикообраз доживает до 20 лет, а мышь всего до 2.5—3 лет. Олень крупнее косули и живет до 25—30 лет, косуля—до 16, но хотя косуля крупнее дикообраза, она менее долговечна, чем дикообраз. У млекопитающих живородных и питающих детенышей молоком матери до возраста половой зрелости гибнет (косуля) менее 50%. У яйцекладущих, но заботящихся о потомстве птиц равной или большей долговечности (глухари, тетерева) догибает в среднем около 80—85%. У рептилий смертность выше, чем Ф птиц, так как, хотя они откладывают яйца подобные яйцам птиц, но у них нет заботы о потомстве. О смертности амфибий у нас нет данных. У рыб смертность очень велика. На 100 тысяч икринок у воблы достигает промыслового возраста всего 10—15 рыбок, у лососевых, обладающих большим количеством желтка и инстинктом закапывать икру в гравий, выживает 30—40 особей на 100 тысяч.
До сих пор мы говорили об индивидуальной гибели особей в борьбе за существование, но от индивидуальной гибели С. А. Северцов отличает гибель массовую от стихийных причин и заразных болезней (эпизоотий).
Массовая смертность от стихийных причин описана Дарвином и многими другими биологами после него. С. А. Северцов приводит много примеров этой гибели в специальной работе. Суровые зимы, глубокие снега, гололедицы, иногда отсутствие снега, когда земля глубоко промерзает, слишком большие засухи и, наконец, катастрофы вроде ураганов и мятелей, губят животных в огромном количестве; эти причины действуют или непосредственно, или косвенно, вызывая неурожаи кормов и голодовки. Такие
270
ГЛАВА X
отклонения от обычных климатических условий повторяются с известной периодичностью. При стихийных депрессиях гибель каждой отдельной особи зависит от случая. Поэтому, чем больше была численность населения перед падежом, тем больше особей имеет шансы выжить. Следовательно, плодовитость вида должна быть такова, чтобы в промежуток между депрессиями он успевал размножиться. В этом надо видеть основную причину той плодовитости, которая так часто удивляла наблюдателей.
Громадное значение имеет массовая гибель от заразных болезней (эпизоотий).
Наиболее важным с экологической и эволюционной точки зрения отличием климатических депрессий от эпизоотий является то, что последние связаны с плотностью населения, создающей условия для переноса заразы. Заразные начала всегда имеются в природе, но обычно дают очень незначительную смертность. Эта смертность достигает большой величины в период, когда имеют место эпидемии.
По законам Фарра и Вильсона вероятность заражения и смерти при эпидемии прямо пропорциональны плотности населения. Так как в нормальные годы естественный прирост стада выше смертности, то каждый вид периодическидостигаеттакойчисленности, прикоторой неизбежно начнется эпидемия. Эпизоотия продолжается до тех пор, пока существует контакт между здоровыми и больными особями, когда же этот контакт прерывается, эпидемия затухает. Указанная связь между плотностью населения и эпидемическими болезнями устанавливает для каждого вида некоторую минимальную и некоторую максимальную плотность населения. От минимума до максимума вид нарастает в численности, и длительность периода нарастания опеределяется плодовитостью вида и коэфициентами индивидуальной смертности.
Очень важно отметить противоречивость описанных моментов борьбы за существование. Понижение индивидуальной смертности или повышение рождаемости способствуют учащению эпидемий. Понижение рождаемости и повышение индивидуальной смертности снижает их частоту. Снижение рождаемости или повышение смертности выше известного предела должно повести к вымиранию вида от периодических климатических депрессий, которые губят животных назависимо от плотности населения. Чем вид плодовитее, тем легче он восстанавливает свою численность после такой депрессии, но зато тем чаще повторяются эпидемии.
Постоянное повышение численности наступает в результате приобретения видом новых адаптивных изменений. Когда в результате прогрессивной эволюции он приобретает новые приспособления, то происходит снижение смертности и увеличение долговечности особей, которое отражается на всей жизни населения. После каждой депрессии вид быстрее возрастает в числе, и плотность населения, при которой возникает эпидемия, будет достигаться чаще. Учащение эпидемий усиливает отбор иммунных особей, что повышает начальную плотность населения после эпизоотии и еще более учащает ритм эпидемий, а вместе с тем вызывает более частое повторение высоких плотностей населения, которое усиливает конкуренцию между животными и стимулирует их расселение.
Естественным следствием постепенного и постоянного повышения численности особей, образующих данный вид, является столь же постепенное и постоянное увеличение ареала его распространения. Если число особей данного вида постоянно увеличивается в той области, где этот вид обитает, и население местности становится, таким образом, очень густым, то животные начинают постепенно расселяться в соседние районы; при этом они останавливаются на таких местностях, которые наиболее соответствуют их привычкам и их строению. В результате ареал распространения данного вида постепенно рас
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
271
ширяется, и это обстоятельство мы рассматриваем как второй характерный признак биологического прогресса.
При изменении исходного места обитания, что связано с расселением данного вида, различные представители этого вида приходят в соотношения с несколько иными, отличными от первоначальных и большей частью неблагоприятными условиями окружающей среды; к этим новым условиям существования, т. е. к несколько иному климату, к несколько инойпище, к другим врагам и другому рельефу почвы, животные постепенно приспособляются путем незначительных адаптивных изменений в своих привычках, в своих инстинктах, в строении своих энтосоматических органов. В результате этого процесса первоначально единый вид начинает постепенно распадаться сначала на подвиды, а позднее на дочерние виды.’ Давно известно, и отмечено еще Дарвином, что широко распространенные, богатые особями виды обычно распадаются на большое число подвидов, и этот общий факт, т. е. именно то, что вследствие увеличения числа особей и расширения ареала распространения э в о л ю и р у ю щ и й вид распадается сначала на подвиды, а поздне е— на дочерние виды, мы считаем третьим характерным признаком биологического прогресса животных.
Биологический прогресс характеризуется, таким образом, следующими признаками: 1) численным увеличением особей данной систематической группы, 2) прогрессирующим расселением, т. е. захватом новых ареалов обитания, 3) распадением, по мере того как животные попадают в новые условия существования, на подчиненные систематические единицы (разновидности, группы и т. д.).
С. А. Северцов указывает еще на один факт, являющийся, по его мнению, признаком биологического прогресса. Иногда в силу резких различий в условиях смежных местообитаний вид не может расселиться и остается в' прежней стации. В этом случае при повышении долговечности усиливается конкуренция, приводящая к снижению плодовитости. Более плодовитые самки при усилившейся конкуренции оказываются в невыгодных условиях сравнительно с менее плодовитыми, так как сильнее истощаются на произведение потомства. У млекопитающих и птиц, кроме -того, родители тратят много сил на выкармливание детенышей. Снижение плодовитости и повышение долговечности приводят к уменьшению колебаний численности, что является, по мнению С. А. Северцова, признаком биологического прогресса.1 2
Теперь читателю, мне кажется, стало совершенно ясным то содержание, которое мы вкладываем в наше понятие «биологический или экологический прогресс»: оно равнозначуще понятию «победы в борьбе за существование» и обозначает собою то, что потомки какого-либо биологически прогрессивного вида занимают доминирующее положение в природе по сравнению с представителями родственной им группы, не прошедшей подобного рода эволюции.
1 Само собой разумеется, что истинный ход дивергентного филогенетического развития животных далеко не так прост, как это может здесь показаться и, конечно, он не сводится исключительно к изменениям в географических условиях существования. Мы высказываем лишь то общее положение, что дивергентное филогенетическое развитие находится в связи с изменением условий внешней среды и с увеличением общего числа особей, образующих данный вид.
2 С. А. Северцов, 1936.
272
ГЛАВА X
Еще лучше мы поймем этот процесс, если сравним его с процессом прямо ему противоположным, а именно с биологически регрессивной эволюцией животных. В последнем случае можно видеть, что у регрессирующей группы число особей, составляющих данный вид, с- течением времени постепенно падает, что ареал распространения прежде обычно и широко расселенной формы с течением времени постепенно суживается, т. е. что местности, населенные данным видом, все более и более принимают вид островков, отделенных друг от друга все большими и большими пространствами, что, наконец, число видов, образующих данный род, т. е. число подчиненных таксономических групп, постепенно уменьшается. Все эти признаки характеризуют собою вымирание животных.
Биологически прогрессивная эволюция приводит к образованию крупных систематических групп, состоящих из большого числа подчиненных групп. Биологически регрессивная эволюция какой-либо группы животных ведет к вымиранию этой группы, от которой в лучшем случае остаются в живых лишь немногие географически далеко отстоящие друг от друга реликтовые формы.
Таким образом, мы видим, что каждому виду животных представляются две возможности дальнейшего существования. Во первых, он может продолжать существовать, изменяясь морфологически с течением времени очень незначительно, и тогда, вообще говоря, дальнейшей эволюции не происходит (статические или константные формы). Палеонтологические исследования показывают нам, что такие случаи действительно встречаются, хотя и относительно редко: многие простейшие, некоторые брахиоподы (например Ligula) и многие другие формы пережили целые геологические эпохи, совершенно не изменяясь морфологически. Во вторых, и это случается гораздо чаще, животные с течением веков меняются в своих функциях и в своем строении, т. е. огромное большинство животных эволюирует в том или в другом направлении. Мы только что отметили, что эта эволюция может итти в двух направлениях, а именно: либо в направлении биологического прогресса, приводящего к биологическому расцвету эволюирующего вида, либо в направлении биологического регресса, приводящего в конечном итоге к вымиранию данной группы животных.1
Филогенетические исследования привели нас к выводу, что существуют по меньшей мере четыре общих направления биологического прогресса,2 т. е. четыре различных способа морфофизиологических изменений, которые одинаково приводят к биологическому расцвету видов, а именно к увеличению общей численности вида, к его прогрессирующему расселению, к его постепенному распадению на новые подчиненные систематические группы.
Мы можем следующим образом обозначить эти- четыре направления эволюции:
1. Направление морфофизиологического прогресса, или направление ароморфоза.
2. Направление морфофизиологического приспособления (адаптация в узком смысле этого слова), или направление идиоадаптации.
3. Направление эмбрионального приспособления, или направление ценогенеза.
4. Направление морфофизиологической дегенерации.
1 Нет особенной нужды напоминать читателю о том, что и многие статические формы в свое время также вымерли.
’Северцов А. Н., Главные направления эволюционного процесса, Москва, 1925; 2-е изд., 1934.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
273
С биологической точки зрения все эти четыре направления равноценны, но с логической точки зрения, по сравнению с биологическим прогрессом, эти четыре способа имеют подчиненное значение. Таким образом, и морфофизиологический прогресс, понятие которого так часто смешивалось.с понятием биологического прогресса, на самом деле представляет собой лишь частный случай биологического прогресса, лишь один из способов, которыми этот последний достигается.
Разберем теперь каждое из этих направлений эволюции в отдельности и начнем с направления ароморфозов, т. е. с морфофизиологического прогресса.
МОРФОФИЗИОЛОГИЧЕСКИЙ ПРОГРЕСС ИЛИ АРОМОРФОЗ
Напомним читателю о том, что все физиологические изменения, с которыми мы будем иметь дело в этом и в следующих отделах настоящей главы, являются прежде всего приспособлениями, т. е. такими изменениями организма, которые имеют известное биологическое значение для данной эволюирующей формы, иными словами, которые ей так или иначе полезны. Мы займемся при этом не теми мелкими изменениями структуры, которые генетики обозначают как наследственные вариации или мутации, но теми крупными изменениями в строении и функциях органов, которые имеют биологическое значение для вида. Эти изменения образуются главным образом путем постепенного суммирования упомянутых выше элементарных вариаций.
Филогенетические исследования показали нам, что различные приспособления организмов далеко не равноценны с биологической и филогенетической точек зрения. Нашей непосредственной задачей является показать, каким образом различные формы приспособлений могут влиять и определять^обою различные направления биологического прогресса для того, чтобы можно было классифицировать эти приспособления с такой проспективной точки зрения.
Мы начнем с группы не особенно многочисленных, но весьма важных приспособлений, ведущих к повышению морфофизиологического уровня организации животных, приспособлений, которые мы назвали «аромор-фозами».1 Под именем «ароморфозов» мы объединяем такие измен е-ния организации и функций животных, которые, имея общее значение, поднимают энергию жизнедеятельности организма животных. Выражением «общее значение» мы хотим сказать, что изменения эти оказываются полезными животному не только при некоторых не особенно благоприятных изменениях внешней среды, но также и при многих неблагоприятных ее изменениях.1 2
1 Ароморфоз от aipw—поднимаю и цоруЛ—'форма; изменения формы.
2 Э. Геккелем была высказана мысль, которую разделяет, между прочим, и В. Франц (F г a n z V., Die Vervollkommnung inder lebenden Natur, 1920), что прогресс организмов в природе происходит вследствие взаимодействия двух факторов: централизации и диференциации. С первого взгляда можно было бы предположить, что «закон совершенствований» Франца соответствует нашему понятию «ароморфоз». Однако это не.так: эти понятия нетождественны. Самое характерное для ароморфозов—это подъем жизнедеятельности активных,органов животного. Между тем мы знаем, что существует много случаев централизации пассивных органов и частей пассивных органов, например срастание покровных костей и т. п. Плате (1928) следующим образом определяет совершенствование ('Vervollkommnung): «Биологическое совершенствование»,— говорит он,—«заключается в гармоническом повышении численности, сложности и функциональной деятельности приспособлений». Такой процесс скорее соответствовал бы нашему понятию ароморфоза. Для ароморфоза в нашем смысле этого слова характерно повышение деят ел ьности «активных» приспособлений, (ср. статью Плате Л., fiber Vervollkommnung, Anpassung und die Unterscheidung von niederen und hoheren Tieren, Zoolog. Jahrb., Bd. 45, 1928. В этой статье он дает подробный критический обзор интересующей нас проблемы).
Морфологические закономерности 18
274
ГЛАВА X
До сих пор мы давали лишь абстрактное определение морфофизиологических прогрессивных изменений организма (ароморфозов). Попробуем теперь вложить конкретное содержание в это понятие. Для этой цели приведем в качестве примера несколько наиболее значительных ароморфозов, имевших место в течение филогенетического развития позвоночных животных.
Прекрасным примером ароморфоза служат явления концентрации органов, когда органы, первоначально раздельные, соединяются друг с другом в целях повышения функции. На рис. 73 нами приведен один из случаев такой концентрации. Здесь изображено строение центральной нервной системы обычного мотыля (Chironomus plumosus). Мы видим
Рис. 73. Строение центральной нервной системы Chironomus plumosus.
Типичная брюшная нервная цепочка насекомых, состоящая из ряда отдельных ганглиев (п^ —n9); С— головной ганглий (из А. Ланга).
Рис. 74. Строение центральной нервной системы Sarcophaga carnaria.
Отдельные ганглии брюшной нервной цепочки слиты в один очень крупный ганглий (n); С—головной ганглий (из А. Ланга).
типичную брюшную нервную цепочку, состоящую из девяти отдельных ганглиев (гц—п9). На рис. 74, изображающем строение нервной системы у одного из представителей высших насекомых (Sarcophaga carnaria), мы видим, что отдельные ганглии брюшной нервной цепочки слились в один очень крупный ганглий (п). При этом изменении несомненно происходит повышение специфической функции нервной ткани и в связи с этим повышение интенсивности, разнообразия и сложности функции центральной нервной системы, т. е. подъем общей энергии жизнедеятельности всего организма, что собственно и является главным признаком ароморфоза.
Другим примером филогенетического развития, идущего путем ароморфоза, является эволюция сердца позвоночных, несомненно повысившая энергию жизнедеятельности активных органов этих животных и обусловившая у них возможность прогрессивной эволюции центральной нервной системы и органов чувств. Эволюция эта, хорошо известная на основании сравнительно анатомических и эмбриологических данных, изображена нами на рис. 75. На рис. 75 А схематически представлено двухкамерное сердце рыб: венозная кровь проникает в цельное, непод-разделенное предсердие, а оттуда в желудочек, из которого течет в аорту и затем в жаберные сосуды, где она окисляется и откуда, окисленная, разносится по всем органам. У амфибий (рис. 75 В) предсердие уже частич-
направления филогенетического развития животных
275
но разделилось на две половины, причем правая половина оста паев венозной, а левая сделалась артериальной: в желудочек проникает таким образом смешанная кровь, причем к голове идет преимущественно артериальная^ часть крови, к вегетативным органам—преимущественно венозная. У рептилий (рис. 75, С) мы имеем уже полное разделение предсердия, но еще не полное—желудочка: здесь левая половина сердца почти целиком артериальная, правая—венозная. Наконец, у птиц и млекопитающих (рис.75, D) обе перегородки сердца становятся полными, так что у них вся левая половина сердца артериальная, вся правая—венозная; у этих животных кровь уже совсем не смешивается, и ихактивные органы и органы растительной жизни получают вполне окисленную кровь, т. е. у них
Л в с о
Рис. 75. Схематическое изображение эволюции сердца позвоночных животных; происходящей путем ароморфоза.
А—двухкамерное сердце рыб; В—трехкамерное сердце амфибий; предсердие (atr.) неполной перегородкой подразделяется на две части; С—четырехкамерное сердце рептилий: предсердие целиком разделено на две части перегородкой; желудочек (vt.) еще разделен не вполне; D—четырехкамерное сердце птиц и млекопитающих; и предсердие и желудочек вполне разделены сплошной перегородкой на правую п левую половины.
а—аорта; air.—предсердие; s—перегородка; vt—желудочек (из Кингсли).
происходит наиболее интенсивный обмен веществ и наиболее интенсивная жизнедеятельность. Таким образом, мы видим, что вся эволюция сердца позвоночных животных имеет совершенно определенное направление: она направлена к тому, чтобы доставить всем органам тела животного возможно большее количество совершенно окисленной крови, и это вполне достигается у птиц и млекопитающих. Мы можем легко представить себе, что степень окисления крови при дальнейшем усовершенствовании органов дыхания, т. е. при более совершенном развитии в них капиллярной системы, может еще повыситься, но нам трудно представить себе возможность дальнейшего прогресса в самом сердце и в главнейших сосудах, отходящих от него.
Характеризуя в общих чертах процесс прогрессивной эволюции, мы говорили до сих пор об изменениях в отдельных органах; теперь можно поставить вопрос о том, в какой мере перечисленные нами признаки прогрессивной эволюции мы в праве переносить на целые организмы. Законность такого вопроса обусловливается тем фактом, что прогрессивные изменения весьма часто сопровождаются регрессивными, т. е. что при прогрессивной эволюции одних частей организма другие части этого организма редуцируются, а в некоторых случаях и атрофируются. По этому поводу надо принять в соображение следующее: если некоторые части организма изменяются в прогрессивном направлении, а остальные 18®
276
ГЛАВА X
части организма не изменяются (остаются статическими), то мы имеем полное право говорить об общем прогрессивном изменении или подъеме организации; степень этого подъема, конечно, зависит от того, насколько велики происшедшие изменения и какие части организма они захватили. Если некоторые части организма изменились прогрессивно, а другие— регрессивно, то вопрос о том, в каком направлении произошло общее изменение организации, может быть разрешен только на основании сопоставления прогрессивных и регрессивных изменений друг с другом. Если изменение при этом произошло путем субституции органов (см. ниже), т. е. если некоторые части организма атрофировались, но были замещены другими частями с тою же или аналогичной функцией, то обычно мы имеем интенсификацию функций, и тогда общее изменение организма как целого идет в сторону прогресса. Такие случаи мы имеем при замещении спинной струны низших позвоночных хрящевыми позвонками, при замещении хрящевых позвонков костными и т. п. Если .же общее изменение произошло таким образом, что одни органы изменились прогрессивно, а другие (с совершенно иной или несколько иной функцией) редуцировались, то вопрос о направлении изменения всего организма можно решить, только учитывая интенсификацию функций и повышение их сложности и разнообразия в организме как в целом или, что то же, учитывая происшедший при этом общий подъем жизнедеятельности организма. Так, сопоставляя потерю конечностей у змей (регрессивное изменение) с удлинением тела и изменениями в челюстном аппарате (прогрессивные изменения), мы, по всей вероятности, должны будем признать, что если организация змей не понизилась, то и не повысилась: общая энергия жизнедеятельности этих животных осталась приблизительно на том же уровне. Сопоставляя прогрессивное развитие ног с редукцией крыльев у страусов, т. е. сопоставляя изменения, которые привели к утрате способности к полету, но повысили способность к передвижению по суше (выработался быстрый бег), мы, вероятно, должны будем признать наличие общего понижения организации. Конечно, при такой оценке приходится быть очень осторожным, ибо здесь легко ввести субъективный элемент в рассуждение.
ЭВОЛЮЦИЯ ПОЗВОНОЧНЫХ С ТОЧКИ ЗРЕНИЯ ТЕОРИИ АРОМОРФОЗА
В предыдущем мы разобрали вопрос о билогическом и морфофизиологическом прогрессе лишь в очень общей форме. Теперь нам надо сделать попытку применить эти соображения к вопросу об эволюции позвоночных и определить, благодаря каким изменениям о р г а низа ц и и и у них совершался процесс прогрессивной эволюции. Само собой разумеется, что главное внимание нам придется обратить на те изменения, которые мы можем охарактеризовать как прогрессивные изменения морфофизиологического типа, т. е. как ароморфозы.
В первой части нашей книги мы попытались реконструировать на основании сравнительно морфологического иселедования строение общих предков энтобранхиат и эктобранхиат, т. е. протокраниат. Попробуем теперь дать биологическую оценку тем особенностям в структуре этих гипотетических форм, которыми они отличались от своих предков, первичных бесчерепных позвоночных.Главные отличия бесчерепных предков позвоночных (Acrania primitiva) от протокраниат заключались, ка-к об этом уже было сказано выше, во-первых, в том, что у протокраниат из трубчатого сердца бесчерепных развилось мешковидное двухкамерное мускулистое сердце, во-вторых, в том, что у них из мешковидной печени типа ланцетника образовалась состоящая из многочисленных железистых протоков
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
277
печень типа краниат, наконец, в том, что у них развилась центральная нервная система и органы высших чувств по типу краниат. Весьма вероятно, что зачатки черепа и верхних дуг позвонков уже были развиты у протокраниат. Ротовой скелет, за исключением ротового кольца, невидимому, еще не существовал, так что питание протокраниат было вполне пассивным. Их многочисленные жаберные щели также имели весьма простое строение; в них еще не образовалось ни эктодермальных, ни энтодер-мальных нитей, или лепестков, так что и в этом отношении протокраниаты еще целиком сохранили первичную организацию своих бесчерепных предков (Acrania primitiva).
Если мы спросим себя, в чем же собственно заключался биологический прогресс в филетическом развитии первичных предков всех позвоночных, то мы увидим, что у них развивались прогрессивно те именно органы, от функционирования которых непосредственно зависел подъем общей жизнедеятельности организма животных. Так, в результате прогрессивного развития сердца, появилась некоторая интенсификация кровообращения, т. е. окисление крови в жабрах повысилось, что привело в свою очередь к усилению обмена веществ во всех тканях организма (а р о м о р ф о з). Прогрессивное развитие печени обусловило улучшение питания тканей, что, конечно, также способствовало подъему жизненной энергии животного (ароморфоз). Прогрессивное развитие мозга и органов высших чувств привело к той же цели: развитие органов высших чувств позволило животным легче находить места с более богатой и более питательной пищей; вместе с тем несколько повысилась их способность противостоять нападению врагов и, таким образом, борьба за существование облегчилась (ароморфоз). Все эти изменения в нервной системе и органах чувств внесли громадное разнообразие в общую жизнедеятельность протокраниат и, расширив область их восприятия, позволили им отвечать на раздражения, исходящие из внешней среды более сложными и более координированными действиями, чем это имело место у Acrania primitiva. Таким образом, если изменения в органах кровообращения и питания привели к интенсификации жизненных процессов у протокраниат, то прогрессивное изменение в органах высших чувств и в центральной нервной системе вызвало диферен-циацию и усложнение в действиях и поведении животных.
Весьма примечательно, что все только что перечисленные нами функции, обусловившие, как это мы только что видели, общий подъем жизнедеятельности у протокраниат, повысились также (хотя и разными путями) и у потомков протокраниат, т. е. у энто- и эктобранхиат. При превращении протокраниат в энтобранхиаты совершилось весьма важное изменение в их органах дыхания: развились энтодермические жабры и появились жаберные мешки, вследствие чего общая поверхность дыхания у этих форм сильно увеличилась (ароморфоз). Совершенно ясно, что здесь мы имеем перед собою интенсификацию функции дыхания, т. е. резкое повышение общей энергии жизнедеятельности этих в общем еще чрезвычайно примитивных форм1—первичных предков современных круглоротых и ископаемых Ostracodermi.
При постепенном превращении энтобранхиат в ныне живущих круглоротых, с одной стороны, и в вымерших Ostracodermi, с другой, в их жаберном аппарате произошел ряд параллельных изменений: самые передние жаберные щели закрылись, и животные перешли от пассивного питания к активному. Но переход этот в обеих группах осуществился различными путями: в то время как у панцырных Osteostraci из пластинок кожного скелета образовался своеобразный ротовой аппарат, у циклостом из передних жаберных дужек образовался сложно построенный сосущий ротовой скелет и развились эктодермальные роговые зубы.
С биологической точки зрения оба эти ряда изменений имели перво
278
ГЛАВА X
степенную важность, ибо протокраниатам при их пассивном питании приходилось довольствоваться лишь взвешенными и плавающими в воде органическими частицами, которые попадали в их ротовую или бранхиальную полость вместе стоком воды, служащей им для дыхания, и проталкивались далее в кишечник действием ресничатого эпителия эндостиля. Таким образом, выбора наиболее подходящей пищи у них не было; к тому же вряд ли можно думать, чтобы маленькие, взвешенные в воде организмы были вообще очень питательны. При активном способе питания животные, снабженные органами высших чувств, выбирают для себя ту пищу, которая им наиболее подходит и которую они легче всего находят в достаточном количестве в месте своего обитания. Выгода этого последнего способа питания очевидна. Поэтому мы легко можем себе представить, какой подъем общей энергии жизнедеятельности вызвал этот переход к активному питанию у потомков примитивных энтобранхиат.
Что касается циклостом, то они перешли к очень высокому в качественном отношении способу питания, а именно к питанию посредством сосания крови других позвоночных, т. е. к очень высокостоящему в смысле питательности и усвояемости роду животной пищи.
Циклостомы, сохраняя весьма примитивные черты своей организации, дожили до нашего времени, да и теперь не являются вымирающими животными. Между тем большая часть особенностей их организации представляется неизмененным или очень мало измененным пережитком весьма отдаленной от нас эпохи,—эпохи, во всяком случае предшествовавшей силурийским временам. Этот парадоксальный факт переживания объясняется, по всей вероятности, тем, что высокое совершенство способа питания циклостом компенсировало все биологические недочеты других черт их организации (по сравнению с остальными водными позвоночными, претерпевшими весьма сложную прогрессивную эволюцию) и позволило им выжить, сравнительно мало изменяясь, в той же среде, в которой происходила эволюция другой ветви потомков протокраниат, а именно рыб. С другой стороны, приспособление к одному определенному роду пищи, т. е. специализация, не позволила их организации эволюировать дальше в прогрессивном направлении; насколько мы можем судить, от круглоротых не произошло ни одной ветви животных с более высокой организацией. Достигнув определенной высоты, они остановились в своем эволюционном развитии.
Замечательно, что вторая ветвь потомков протокраниат (см. родословное древо на схеме XV), давшая начало челюстным позвоночным, также претерпела ряд морфологических изменений, которые с билогической точки зрения протекали аналогично изменениям, имевшим место у энтобранхиат и круглоротых. Так, у эктобранхиат развились эктодермальные жаберные нити (либо лепестки), а их жаберные дуги распались на ряд палочковидных жаберных элементов. Вследствие этого у них, с одной стороны, увеличилась дыхательная поверхность, а с другой, возросла подвижность жабр, в результате чего интенсифицировался процесс окисления крови и произошел общий подъем энергии жезнедеятельности (ароморфоз).
Важное изменение, имевшее место в организации эктобранхиат при переходе их в Protognathostomata, произошло в их способе питания; зак-лючалосьоно, также как и у предков круглоротых и Ostracodermi, в переходе от пассивного питания к активному. Этот переход осуществился благодаря превращению самых передних висцеральных дуг в части жаберного скелета: передние дуги изменили свою функцию и превратились в губные хрящи и в элементы челюстной и гиоидной дуги. Плакоидные чешуи этого отдела увеличились в размерах, изменили свое расположение и превратились в зубы. Таким образом возник ротовой аппарат кусательного или хватательного типа. Все эти изменения привели к такому же результату, как
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
279
и только что описанные изменения в передних частях висцерального аппарата у предков круглоротых иostracodermi, а именно к общему повышению энергии жизнедеятельности первичных челюстных позвоночных (Protognathostomata). Поэтому к а ж д о е из этих изменений мы должны рассматривать как прогрессивное изменение строения, или ароморфоз. Кроме того, как мы это только что отметили, у Protognathostomata развились плакоидные чешуи с острыми шипами, покрывающими всю поверхность тела животного (орган защиты).
Другим крупным изменением, имевшим место при превращении эктобранхиат в протогнатостом, была интенсификация функции движения, т. е. прогрессивное развитие у них хвостового гетероцеркального плавника. Дальнейшим и весьма важным этапом эволюции челюстных позвоночных было развитие у них парных плавников, т. е. органов ориентировки при движении, иначе говоря, органов, изменяющих направление движения. Каждому, конечно, понятно биологическое значение этих органов: развитие сильного хвостового плавника, являющегося у рыб главным органом движения, позволило этим формам двигаться в воде с гораздо большей быстротой, чем это могли делать их предки, а развитие парных и непарных плавников позволило им лучше регулировать свое движение при плавании и лучше ориентироваться в пространстве. В результате этих изменений Protognathostomata стали лучшими пловцами, чем их еще не обладавшие этими органами предки, и стали лучше и легче, чем эти последние, преследовать и ловить свою добычу. Все это привело к лучшему питанию и, как следствие этого, к новому подъему жизнедеятельности всех органов и тканей (ароморфоз). Отметим еще, что с функциональной точки зрения развитие рыбьего хвоста и парных плавников имеет особый характер: эти органы по своему строению не являются органами специализованными, приспособленными к одному какому-нибудь виду движений (как, например, конечности лазающих или копытных млекопитающих), но органами очень общего характера, повышающими и быстроту и разнообразие движений в той среде, в которой животное обитает.
Мы думаем, что читателю ясен общий характер изменений, имевших место при постепенном превращении эктобранхиальных предков рыб в формы, которые мы обозначили как Chondrichthyes selachoidei, т. е. акулоподобные хрящевые рыбы. Каждое из этих изменений носило универсальный характерв том смысле, что все они представляли собой приспособления, полезные не при одном каком-либо определенном условии существования данной формы, но при самых различных условиях. При этом каждое из перечисленных изменений усиливало в той или иной степени интенсивность активных функций животного и вызывало, таким образом, некоторый подъем в общей энергии его жизнедеятельности; каждое из них являлось, следовательно, новым ароморфозом.
От Chondrichthyes selachoidei произошли, с одной стороны, ископаемые эласмобранхий и современные акулы и скаты, с другой—примитивные Osteichthyes. Ископаемые эласмобранхий в общем остановились в своем прогрессивном развитии и у них, повидимому, не образовалось ни одного нового, более или менее значительного ароморфоза.1 Вероятно, поэтому
1 К сожалению, многие важные черты организации ископаемых эласмобранхий, например строение их центральной нервной системы, до сих пор остаются нам совершенно неизвестными. Мы знаем, однако, что у современных эласмобранхий имеется в общем довольно высоко организованный мозг (prosencephalon, cerebellum, corpora restiformia), что, конечно, не может рассматриваться иначе как ароморфоз, но мы не знаем, были ли эти признаки развиты уже у отдаленных предков этих животных (что, судя по общему строению черепа ископаемых эласмобранхий, далеко не представляется нам невозможным), или же они являются лишь новоприобретением современных нам форм? Тот же вопрос мы можем поставить себе и относительно высокоразвитой мочеполовой системы эласмобранхий.
280
ГЛАВА X
вымерла эта примитивная группа рыб. Непосредственные же потомки ископаемых эласмобранхий—современные акулы и скаты—представляют собой богатую формами и в общем процветающую группу рыб, хотя современные акуловые рыбы и отличаются от своих давно вымерших предков лишь сравнительно второстепенными признаками и до сих пор остались формами с весьма низкой организацией, живыми пережитками палеозойских времен. Перед нами встает вопрос, благодаря каким признакам прогрессивного характера эти эласмобранхий, живя в той же обстановке, в которой жила сильно прогрессировавшая группа костных рыб (Osteichthyes), и в частности, доминирующая в настоящее время группа костистых (Teleostei), выдержали конкуренцию с этими формами, а не вымерли подобно многим другим группам примитивных рыб.
Рассматривая организацию современных эласмобранхий с этой точки зрения, мы находим в некоторых системах органов черты прогресса: например, их головной мозг, сохраняя черты примитивного строения, велик и сложно построен, органы чувств хорошо развиты ит. д. Но в общем эти изменения не слишком велики и не могут никоим образом сравниться с той суммой изменений, через которую прошли современные костистые рыбы. Строго говоря, у эласмобранхий не появилось ни одного сколько-нибудь заметного ароморфоза. Лишь в одной системе органов, а именно в органах размножения, а также в способе эмбрионального развития мы находим весьма большие изменения прогрессивного характера. У эласмобранхий на очень ранней стадии эволюции развилось внутреннее оплодотворение и целый ряд приспособлений, обеспечивающих жизнь и благополучное развитие зародыша: плотная скорлупа яиц с покровительственной окраской и с приспособлениями для прикрепления яиц к подводным предметам (рис. 87); громадное количество желтка, обеспечивающее питание зародыша в течение всего периода эмбрионального развития, эмбриональные органы кровообращения для питания зародыша желтком и т. д. Благодаря всем этим эмбриональным приспособлениям зародыш хорошо защищен во время онтогенеза и выходит из яйца не в виде маленькой слабой личинки, как у большинства даже высших рыб,а в виде довольно крупной молодой рыбки, обладающей всеми главными особенностями взрослого животного и'не подвергающейся опасности со стороны большинства врагов, от которых терпят мальки других рыб.
Биологический факт, что у эласмобранхий выработались приспособления, защищающие молодь в течение самого раннего периода развития, т. е. в тот именно период, когда происходит наибольшее истребление особей у других рыб, как мне кажется, был чрезвычайно важен, и, может быть, именно прогрессивное развитие этих сторон организации акуловых рыб имело преимущественное значение в парадоксальном факте процветания весьма примитивной группы рыб в той же обстановке и в той же биологической среде, в которой живут гораздо более высоко организованные группы рыб. Если это толкование верно, а у нас нет пока оснований в этом сомневаться, то мы имеем здесь новое подтверждение общего высказанного нами в начале этой главьгположения, что морфофизиологический прогресс не есть единственный путь прогресса, но лишь частный случай более общего биологического прогресса, который может совершаться различными путями и который в данном случае, как мы это увидим ниже, произошел путем ценогенеза.
У другой ветви потомков Chondrichthyes selachoidei—у Osteichthyes primitivi—появился ряд признаков, имевших большое проспективное значение для всей дальнейшей эволюции позвоночных и в значительной степени повысивших энергию жизнедеятельности этих форм. Наиболее важным из этих прогрессивных признаков, конечно, является развитие костного скелета.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
281
Метамерно расположенные костные чешуи заменили плакоидные зубы и покрыли все тело животного, причем в области головы они образовали сплошной и крепкий панцырь (костный череп, рис. 54). Как известно, появление внутреннего костного скелета имело большое значение. Кость является гораздо более резистентным образованием, чем хрящ, и поэтому, даже при частичной замене массивного хрящевого скелета костным, общая масса подвижных скелетных частей становится меньше, а деятельность мускулов, движущих этими частями, соответственно интенсивнее. В результате животные становятся более ловкими и подвижными, чем их снабженные хрящевым скелетом предки. Этот подъем в деятельности туловищной мускулатуры мы, конечно, не можем рассматривать иначе как ароморфоз.
Образование некоторых костей, между прочим костей ротового скелета (maxilla, praemaxilla, pterygoideum, dentale etc.), а также усложнение структуры челюстной мускулатуры, имели следствием усиление деятельности ротового аппарата, что до известной степени, конечно, повысило питание животных и должно поэтому также рассматриваться как ароморфоз.
Важные изменения произошли кроме того и в дыхательном аппарате первичных Osteichthyes: у них образовалась костная жаберная крышка (ср. Воскобойников, 1932) и прогрессивно развилась жаберная мускулатура. Не вдаваясь в подробности, отметим, что здесь мы имеем перед собою более высокий тип жаберного дыхания, чем это имело место у предков первичных Osteichthyes.
Наконец, у них образовался из одной из задних пар жаберных мешков гидростатический аппарат—зачатки плавательного пузыря,—что также, конечно, является крупным ароморфозом. Вернее сказать, у них появились зачатки двух плавательных пузырей: дорзального и вентрального. Это позволило рыбе свободно и без всяких усилий пребывать на любой глубине.
При постепенном превращении Osteichthyes primitivi в Holosteoidei primitivi произошло важное изменение в способе прикрепления челюстной дуги к осевому черепу. У близко родственных селахиям первичных Osteichthyes небноквадратный хрящ с сидящим на нем praemaxillare сочленялся с нёбнобазальным отростком (processus palatobasalis) глазничного отдела осевого черепа; у Chondrosteoidei эта связь с черепом сохранилась и удержался кроме того столь характерный для селахий symphysis palatoquadrati; но у прогрессивной ветви потомков Osteichthyes primitivi, т. е. у Holosteoidei primitivi, прогрессивно развилось также и сочленение между нёбноквадратным хрящом и несколько сдвинутым вперед processus antorbitalis черепа, так называемое articulatio antorbi-talis: praemaxilla вступила, таким образом, в связь с ростральным концом осевого черепа, в результате чего образовался новый, более прочный тип соединения верхней челюсти с осевым черепом. В непосредственной связи с этими изменениями ротового скелета стоит диференцировка челюстной мускулатуры.
Физиологическое значение всех этих* изменений еще не исследовано с достаточной подробностью, но смысл их уже теперь нам ясен: результатом этих изменений явилось, во-первых, то, что прикрепление верхней челюсти к черепу стало прочнее, во-вторых, то, что челюстная мускулатура стала сильнее, и, в-третьих, то, что функция хватания ротового аппарата стала значительно интенсивнее.
Выше мы уже разобрали вопрос о том, какое биологическое значение имеет усиление деятельности ротового аппарата. Поэтому нам совершенно понятно, что и в данном случае мы имеем дело с приспособлением очень общего характера, приспособлением,—ведущим к усилению функции пита
282
ГЛАВА X
ния и, следовательно, к подъему общей жизнедеятельности организма, иными словами, здесь мы снова имеем дело с ароморфозом.
Первая линия потомков примитивных Holosteoidei, а именно Holosteoidei actinopterygii (см. схему XV), продолжала свое водное существование и прогрессивно развивалась дальше применительно к этому образу жизни. В общем можно сказать, что у представителей этой группы животных в прогрессивном направлении эволюировали главным образом органы движения. Уже на ранних стадиях их филогенеза у лучеперых рыб появляется плавательный пузырь, т. е. гидростатический орган, позволяющий животным регулировать свое положение в воде, что имело для них первостепенное значение с биологической точки зрения. Главный орган поступательного движения рыб, хвостовой плавник, имевший у предков гетероцеркальную форму, становится у потомков (актиноптеригий) гомоцеркальным. Парные грудные плавники меняют свое положение и до известной степени свою функцию: так, у Osteichthyes primitivi (а также у примитивных палеонисцид) грудные плавники имели горизонтальное положение и широкое основание (плавники эврибазального типа); при постепенном превращении этих форм в высшие актиноптеригии (Holosteoidei actinopterygii) положение грудных плавников изменилось таким образом, что их передний край переместился в дорзальном направлении, а та сторона свободного плавника, которая прежде была его нижней поверхностью, стала его боковой стороной, причем первичные плавниковые лучи редуцировались, но лепидотрихии развились прогрессивно и плавниковая мускулатура усложнилась. Все эти изменения постепенно увеличили подвижность грудных плавников, которые с течением времени превратились в весьма активные органы, направляющие движение этих форм. Благодаря всему этому движение костистых рыб в воде стало более быстрым и более разнообразным, чем оно было до сих пор, так что потомки стали во всех отношениях лучшими пловцами, чем были их предки. Поэтому мы рассматриваем как аромор фозы все те биологически важные изменения в строении органов движения, которые произошли у потомков примитивных актиноптеригий.
В известном соотношении с только что описанным усилением подвижности стоят и те изменения, которые имели место в строении чешуйчатого покрова актиноптеригий. У первичных Osteichthyes чешуи были очень толсты (ганоидные чешуи); они образовали плотный и при движении мало гибкий костный панцырь на теле рыбы. У актиноптеригий развились тонкие и легко подвижные циклоидные или ктеноидные чешуи. Вместе с тем процесс окостенения внутреннего скелета во всех его частях пошел дальше, так что у актиноптеригий массивные хрящевые массы, из которых состоял скелет предков, окончательно заменились менее массивными и более резистентными костями. Мы видим, таким образом, что все только что коротко перечисленные нами изменения происходили в направлении усиления подвижности и общего подъема жизнедеятельности высших рыб: весь филогенез этих форм следовал, таким образом, по пути ароморфозов.
Вторая линия потомков Holosteoidei primitivi—Holosteoidei crossopterygii (см. схему XV) не пошла далеко по пути прогрессивной эволюции в водной среде и, поскольку мы можем судить, не выдержала конкуренции с более приспособленными (в биологическом отношении) к подвижной и свободной жизни в открытой воде актиноптеригиями: большая часть Holosteoidei crossopterygii вымерла в мезозое и только немногие формы дожили до нашей эпохи. Но особенности, которые, быть может, служили препятствием к прогрессивной эволюции кроссоптеригий в качестве типично водных животных, а именно вентральный плавательный пузырь (рис. 71 В), служивший при их береговом образе жизни добавочным
направления филогенетического развития животных
283
органом воздушного дыхания (легким), и плавники с мясистой лопастью, сильно выраженной метаптеригоидной впадиной и хорошо развитым скелетом проксимального отдела (рис. 70К) были признаками, которые позволили им пойти по другому пути прогрессивной эволюции, а именно превратиться в наземных животных.
От этих примитивных кистепёрых форм ответвилась новая прогрессивная ветвь животных, а именно дышащие воздухом наземные позвоночные, или Quadrupeda. Ароморфозы, имевшие место при постепенном превращении Holosteoidei crossopterygii в Quadrupeda, т. е. при переходе позвоночных из воды на сушу, касались целого ряда органов, и мы имеем все основания для того, чтобы предполагать, что по крайней мере некоторые из них произошли в то время, когда предки амфибий еще не оставили водную среду.
Нам представляется весьма вероятным, что кистеперые предки наземных позвоночных были обитателями мелких водоемов. В жаркое время года, когда, вследствие частичного высыхания этих водоемов, условия для жаберного дыхания становились неблагоприятными, животные эти стали пользоваться своим вентральным плавательным пузырем в качестве добавочного органа дыхания (для воздушного дыхания). Мы знаем, что и современные нам двоякодышащие (Ceratodus, Protopterus) и кистеперые (Polypterus) рыбы обычно, когда вода начинает портиться в водоемах вследствие жары или загнивания водорослей, выплывают на поверхность воды и заглатывают воздух. Они ползают по дну, опираясь на свои сильные, мясистые стенобазальные плавники и ловко пробираются между густыми зарослями водяных растений. Весьма вероятно поэтому, что и те первичные кистеперые (типа Sauripterus), от которых произошли примитивные стегоцефалы, были животными, еще жившими в воде, но уже часто выплывавшими на поверхность и уже дышавшие воздухом. Вероятно, они могли уже свободно передвигаться по дну мелких водоемов при помощи своих лопатовидных стенобазальных плавников.
Ясно, что только такие животные могли покинуть воду и приспособиться к жизни на земле, у которых уже имелись известные предпосылки к переходу насушу. Таких предпосылок у интересующих нас форм было две: способность к факультативному воздушному дыханию и способность к поступательному ползающему движению. Когда потомки этих форм стали постепенно проводить на суше все большее и большее время и находить себе там пропитание, у них естественно развились новые адаптации приспособительно к новым условиям существования.
Изменения в конечности, происшедшие при этом превращении предков наземных позвоночных, заключались главным образом в том, что из стенобазальных плавников постепенно образовалась целая система рычагов (stylo-, zeugo-, meta- и autopodium примитивных Tetrapoda). При этом особенное значение имело то, что на проксимальном конце stylopodium (humerus, femur) образовалось шаровидное сочленение, давшее возможность животному в значительной степени разнообразить движение своих конечностей. Усиление же общей подвижности конечностей несомненно вызвало общий подъем жизнедеятельности организма и поэтому, конечно, не может нами рассматриваться иначе как ароморфоз.
Аналогичный характер носит и другое изменение конечностей, касающееся мускулатуры. У рыбообразных предков Tetrapoda мускулатура конечностей была, невидимому, слабее развита и менее диференцирована, чем у первичных амфибий, происшедших от этих форм. При переходе к более или менее постоянному наземному образу жизни мускулатура конечностей диференцировалась, причем отдельные мускулы стали сильнее, ибо при передвижении на суше в воздушной среде их функция стала
284
ГЛАВА X
интенсивнее, чем при плавании в воде или при поддерживании тяжести тела при ползании по дну водоемов.
Из других изменений, имевших место в органах движения, отметим лишь прогрессивное развитие грудного и брюшного поясов, а также образование крестца, развившегося как новое приспособление для поддержания тяжести задней части тела (ароморфозы).
Кроме изменений в органах движения, происшедших при превращении Holosteoidei crossopterygii в Quadrupeda, произошли еще важные изменения в органах дыхания и кровообращения этих форм. Из плавательного пузыря предков у потомков развились крупные парные легочные мешки с большей поверхностью дыхания. Вместе с тем образовалась перегородка в сердце (septum atriorum), разделяющая первоначальносплошное предсердие на две части (рис. 75В), что является приспособлением органа к ставшему постоянным воздушному дыханию. Сердечные мускулы стали при этом также сильнее.1 Оба эти приспособления являются, конечно, ароморфозами.
Прогрессивные изменения мы находим также в органах высших чувств: в глазу, который, не утратив характерной для рыб способности видеть вблизи, приспособился и к зрению вдаль, в ухе (развитие слуховых косточек и прогрессивное изменение внутреннего уха) и в органах обоняния (развитие хоан и якобсонова органа). Все эти прогрессивные изменения в органах высших чувств, само собой разумеется, не приходится рассматривать иначе как ароморфозы.
Древнейшими наземными позвоночными являются принадлежащие к классу амфибий стегоцефалы, следы которых найдены в верхнем девоне, а хорошо сохранившиеся скелеты—в нижних слоях каменноугольных отложений.
Мы знаем, что от первичных стегоцефалов произошли две линии потомков: одна, ведущая к современным амфибиям, другая—к рептилиям. Современные амфибии подразделяются на три группы: на хвостатых (Urodela), бесхвостых (Anura) и безногих (Gymnophiona). Наименее измененную сравнительно со стегоцефалами группу представляют собой хвостатые амфибии, которые и по общему облику и по внутренней организации сохранили в относительно неизмененном виде многие черты своих палеозойских предков.1 2 Группу эту мы можем считать несомненно адаптивной. Гимнофионы пошли в своей эволюции по пути специализации: они приспособились к подземному образу жизни и представляют собой во многих отношениях очень примитивную группу амфибий; тело их удлинилось и приобрело змеевидный облик, конечности вполне атрофированы во взрослом состоянии, глаза редуцировались и т. п.
Что касается бесхвостых амфибий, то у них мы находим целый ряд признаков прогрессивной эволюции, т. е. целый ряд ароморфозов, хотя и должны признать, что эта прогрессивная эволюция не пошла особенно далеко и что бесхвостые амфибии, в сущности, невысоко поднялись над общим уровнем высших хвостатых. Как на признаки такой прогрессивной эволюции Anura мы можем указать на изменения в строении скелета и мускулатуры конечностей, благодаря которым лягушки и жабы сделались животными гораздо более подвижными, чем хвостатые амфибии, на прогрессивные изменения в мозгу и органах высших чувств и т. д. Напомним также, что у многих высших бесхвостых амфибий уже наблюдается тенденция порвать последнюю связь с водной средой и перейти к размножению на суше, при этом животные то носят яйца и детенышей на
1 Hesse R., Die Grosse des Herzens bei den Wirbeltieren, Handb. d. norm. u. path. Physiol., Berlin, 1931.
2 Мы не упоминаем здесь о вторичных явлениях неотении, имеющих место у некоторых хвостатых амфибий.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
285
спине (некоторые Hylidae, Arthroleptis seychellensis), то, как, например, Rhinoderma darwini, таскают их в глоточных мешках, где происходит все развитие зародыша, то, наконец, приклеивают яйца, окруженные массой пенистой слизи, к листьям растений (Hylodes martiniensis).
Второй ветвью потомков, ответвившихся от общего ствола стегоцефалов, были, как мы это уже указали выше, первичные рептилии. Аромор-фозы, обусловившие подъем организации и общей жизнедеятельности у рептилий, для нас более понятны, чем ароморфозы, имевшие место при эволюции амфибий.
Мы только что отметили, что по современным взглядам предками древнейших рептилий (котилозавров) являются стегоцефалы. Как это уже было сказано, стегоцефалы по своему образу жизни были тесно связаны с водой, причем одной из основных причин этой связи с водной средой является их способ размножения, так как стегоцефалы откладывали свои яйца вводу, где их дышавшие жабрами личинки развивались до метаморфоза, после которого они выходили на сушу. Такой способ развития, сохранившийся и у большинства современных хвостатых и бесхвостых амфибий, является последним остатком водного происхождения наземных позвоночных. Но этот способ размножения в те эпохи, когда стегоцефалы являлись единственными наземными или, точнее, земноводными позвоночными,1 ограничивал их распространение берегами водных бассейнов, так как, ввиду указанного способа размножения и по другим особенностям своей организации, эти неуклюжие и медленно двигавшиеся на суше животные не Могли удаляться далеко от берегов озер, рек и прочих водоемов; другими словами, в эти отдаленные от нас эпохи все промежутки суши между водными бассейнами еще не были заселены позвоночными.
Заселение суши в собственном смысле этого слова было осуществлено рептилиями, и основными факторами их эволюции были такие изменения способа размножения, которые позволили им удаляться далеко от водных бассейнов и освободили их от необходимости периодически возвращаться в воду, т. е. делаться в период размножения водными животными и проводить период личиночного развития в водной среде.
Таким образом, изменение в способе эмбрионального развития было для наземных позвоночных основным условием их расселения, давшим им возможность покинуть берега водных бассейнов, у которых жили их земноводные предки, и углубиться внутрь суши.
Но ни изменение в способе эмбрионального развития первичных рептилий, ни ряд других не менее важных изменений, например ороговение их кожи, отнюдь не носило характера ароморфозов. Мы не можем рассматривать как ароморфозы ороговение кожи или даже такие полезные для животных эмбриональные приспособления, как увеличение желточной массы, меробластию, образование амниона и аллантоиса, развитие яичной скорлупы и т. д., ибо, хотя все эти изменения с биологической точки зрения и носили прогрессивный характер, так как все они имели следствием расширение ареала распространения первичных рептилий, тем не менее они отнюдь не вызывали общего подъема жизнедеятельности организма, что является главным характерным признаком ароморфоза. Все эти явления принадлежат к группе ценоге-незов и идиоадаптаций, о которых мы подробно будем говорить ниже.
Первичные рептилии были животными, которые ни в один из периодов своей индивидуальной Жизни не были связаны с водой, и мы можем поэтому принять, что их органы дыхания—легкие—с самого начала своей эволю
1 В начале каменноугольной эпохи, но, может быть, и в конце девона.
286
ГЛАВА X
ции развивались в прогрессивном направлении. Правда, в настоящее время мы пока еще ничего не знаем относительно строения легких и легочных альвеол у вымерших примитивных рептилий (котилозавров, аномо-донтов), но факт, что у всех этих форм уже имелись грудные ребра, указывает нам на то, что процесс дыхания у этих животных происходил интенсивнее, чем у их амфибиеобразных предков. Об этом же мы можем заключить и из того, что даже у наиболее низко стоящих современных нам рептилий, например ySphenodon, или у такой примитивной ящерицы как геккон, легкие развиты лучше, чем у амфибий. Кроме того, подвижность грудной клетки (на что указывает нам строение грудных ребер и межреберной мускулатуры), а, стало быть, и функция дыхания у рептилий стала интенсивнее, чем у амфибий (и у стегоцефалов). Мы можем поэтому принять, что окисление крови у первичных рептилий происходило более совершенным путем, чем у их стегоцефалообразных предков, вследствие чего у них повысился обмен веществ и произошел общий подъем жизнедеятельности всех активных функций. Все это, конечно, нам нельзя рассматривать иначе, как ароморфоз.
В тесной связи с повышением окисления крови стояли прогрессивные изменения, происшедшие в сердце и кровеносных сосудах примитивных рептилий: преобразование трехкамерного сердца типа амфибий в четырехкамерное сердце типа рептилий (образование сплошной перегородки между предсердиями и еще не сплошной перегородки между желудочками, рис. 75 С) и прогресивное изменение в артериальных сосудах. Все эти изменения мы также рассматриваем как ароморфоз.
Из изменений внутреннего скелета отметим развитие одного затылочного бугра и видоизменение двух первых шейных позвонков, превратившихся в атлас и эпистрофей, а также развитие шейного отдела позвоночника. У амфибий, так же как у рыб, нет шейного отдела тела, и голова их неподвижна или мало подвижна по отношению к туловищу; благодаря развитию атласа и эпистрофея и сочленению их с одним затылочным бугром голова рептилий сделалась способной свободно поворачиваться на шее. У амфибий пояс передней конечности лежит непосредственно позади головы, а шеи нет; у рептилий шея образуется благодаря сдвиганию пояса передней конечности спереди назад, что непосредственно можно видеть у зародышей во время эмбрионального развития.1 Вследствие этого сдвигания у них образовался подвижный передний отдел позвоночника— шея, в котором ребра укоротились, мускулатура диференцировалась и образовалось особое шейное нервное сплетение (plexus cervicalis).
Функциональное значение этих изменений весьма важно: голова, т. е. главный орган захватывания пищи (челюстной аппарат), нападения и защиты (зубы) и восприятия внешних впечатлений (органы высших чувств), приобрела способность к разнообразным движениям и поворотам относительно туловища, и благодаря этому разнообразие действий животных было повышено. Таким образом, эти последние изменения являются несомненными ароморфоза ми.
К прогрессивным с морфофизиологической точки зрения изменениям позвоночника надо далее отнести более совершенное окостенение позвонков, усовершенствование в способе их сочленения и образование крестца из двух позвонков, что связано с прогрессивным развитием задней конечности. И передние и задние конечности первичных рептилий развились прогрессивно по сравнению с конечностями стегоцефалов, что выразилось прежде всего в удлинении костей stylo- и zeugopodium, а также в увели-
1 S е wertzo f f A. N., Studien uber die Entwicklung der Nerven, Muskeln und des Skeletts der Extremitaten der Tetrapoda, Бюлл. О-ва испытателей природы, М., 1908.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ 287 чении числа фаланг и в срастании проксимальных элементов плюсны.1 Благодаря всем этим изменениям, а также благодаря изменению строения поясов конечностей1 2 способность к передвижению по суше значительно повысилась, и животные перешли от ползания к хождению и даже к беганию, причем они уже перестали касаться • земли своим брюхом. Все эти прогрессивные изменения в органах движения являются ароморфоза м и, ибо они ведут, как и описанное выше увеличение подвижности головы, к постоянному улучшению питания (более легкая ловля лучше усвояемой добычи), что в свою очередь приводит к дальнейшему подъему жизнедеятельности описываемых форм.
В черепе мы также можем констатировать некоторые прогрессивные изменения, но ввиду большой сложности этого вопроса я на нем останавливаться не буду.
Значительно изменилась в прогрессивном направлении мускулатура, так что почти в каждом ее отделе мы находим более сложное строение мускулов: отметим усложнение строения челюстной мускулатуры, развитие мускулатуры языка, диференцировку шейной мускулатуры и, в особенности, как это явствует из работ американских палеонтологов, мускулатуры конечностей.3 •
Важное значение при эволюции рептилий имели прогрессивные изменения головного мозга, в котором наблюдается сильное развитие мозжечка, повидимому связанное с прогрессивными изменениями в органах движения (в мускулатуре туловища и конечностей), а также прогрессивные изменения в органах высших чувств—в глазу, в органе обоняния и в ухе (дифе-ренциация среднего уха и улитки внутреннего уха).
Этих отрывочных замечаний, мне кажется, достаточно для того, чтобы охарактеризовать в общих чертах направление прогрессивных изменений в организации рептилий, так как в мои задачи не входит разбирать вопрос об их филогенезе.
Резюмируя сказанное, мы видим, что ароморфозами, т. е. чисто прогрессивными изменениями, повысившими общую энергию жизнедеятельности и активность рептилий, могут считаться изменения в сердце и кровеносных сосудах, увеличение дыхательной поверхности легких (повышение интенсивности процессов окисления в тканях организма) и изменения в органах движения (развитие подвижной шеи, прогрессивные изменения в мускулатуре, изменения в скелете и мускулатуре конечностей и т. д.). В результате животные сделались, во-первых, более подвижными, во-вторых (и это особенно важно), движения частей их тела по отношению друг к другу сделались более разнообразными. Наконец, к прогрессивным изменениям, повысившим сложность функций организма, т. е. к аромор-фозам, мы должны отнести изменения в нервной системе и органах чувств.
Эти прогрессивные изменения организации рептилий имели своим последствием их решительную победу в борьбе за существование и захват всех свободных мест на суше. Низшие рептилии, а именно котилозавры и аномодонты, известны нам со второй половины каменноугольной эпохи,4 но уже в триасе мы находим все главные группы их хорошо диференциро-
1 Образование astragalus, calcaneus и tarsale proximale у ящериц и у черепах.
2 Не останавливаясь на этом вопросе подробно, упомяну только о том, что у рептилий грудина сочленяется с ребрами и что вследствие этого образовалась более прочная, чем у амфибий, связь между поясом передней конечности и туловищем.
3 См. Gregory М. С. и Camp С. L., Studies in comparative Myology and Osteology, Bull, of Amer. Mus. of natural Hist., vol. 38, 1918.
Romer A. S., Locomotor Apparatus of Reptiles, ibid., vol. 46, 1922.
4 Отделение рептилий от стегоцефалов произошло раньше, как предполагают— в начале каменноугольного периода.
288
ГЛАВА X
ванными.1 Другими словами, группа рептилий в течение пермского и триасового периодов распалась на громадное число форм, которые захватили все свободные места на суше и расселились по всей поверхности земли, причем различные группы приспособились к самым различным обстановкам и условиям, так что мы среди рептилий встречаем животных лесостепи, степи, пустыни, болот, гор и т. д.; некоторые рептилии переходят к роющему образу жизни и становятся животными подземными, другие, как вся группа птерозавров, превращаются в животных летающих, причем передние лапы их делаются крыльями. Наконец, целый ряд форм, принадлежащих к самым различным группам, возвращается вторично в водную среду, из которой некогда вышли их предки, и становятся морскими животными (ихтиозавры, плезиозавры, талаттозавры, многие черепахи и т. д.). Все эти приспособления к различным местопребываниям и средам сопровождаются крупными изменениями целого ряда органов, которые носят главным образом чисто адаптивный характер и о которых мы подробно будем говорить ниже, разбирая вопрос об идиоадаптациях.
Мы можем считать установленным, что при превращении рептилий в птиц имела место значительная интенсификация энергии жизнедеятельности этих форм. К этому привело: во-первых, прогрессивное развитие легких и, главное, особых легочных мешков, столь характерных для птиц, в результате чего кровь стала окисляться более энергично (возрастание общего количества бронхов и легочных альвеол, усиление вентиляции легких при помощи легочных мешков); во-вторых, образование сплошных перегородок между обоими желудочками, т. е. развитие четырехкамерного сердца (рис. 75 D); в-тре'тьих, более интенсивная деятельность сердечной мускулатуры. Все эти изменения, само собой разумеется, вызвали повышение обмена веществ и обусловили дальнейшую интенсификацию всей мускулатуры, в особенности же мускулатуры крыльев и задних конечностей (полет, хождение и прыгание на задних ногах). На ряду с этим происходило дальнейшее прогрессивное развитие мозга и органов высших чувств.
Необходимо также упомянуть о развитии пуха, поддерживающего постоянную температуру тела, и перьев, делающих возможным летательное движение (ароморфоз). Образование клюва и развитие весьма подвижной шеи, равно как и прогрессивное развитие органов пищеварения, конечно, способствовало улучшению питания и, стало быть, повышению жизнедеятельности всех активных органов птиц. В общем вся сумма только что перечисленных нами изменений не может нами рассматриваться иначе как ряд последовательных ароморфо-з о в, высоко поднявших общую организацию птиц над уровнем организации их рептилийных предков.
Все вышеперечисленные изменения протекали, по всей вероятности, не одновременно, но в известной последовательности. Мне кажется, что изменения в органах дыхания и кровообращения наступили раньше других и что именно они сделали возможным дальнейшее прогрессивное развитие нервной системы, органов высших чувств, мускулатуры и скелета.
Особый интерес представляет для нас та группа ароморфозов, которая привела к образованию и прогрессивному развитию млекопитающих. Само собой разумеется, я не буду подробно останавливаться на эволюции всей группы млекопитающих и ограничусь только немногими замечаниями относительно общего характера их прогрессивной эволюции.
1 По Бройли (1923): Cotylosauria, Anomodontia, Rhynchocephalia, Thalattosauria, Palaeohatteriidae, Brontosauridae, Kadaliasauridae, Squamata, Paterosauridae, Ichthyo-sauria, Sauropterygia, Placodontia, Mososauria, Testudinata, Parasuchia, Crocodilia, Dinosauria, Pterosauria.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
289
Млекопитающие произошли от весьма примитивных рептилий, по всей вероятности от форм, близких к Theriodontia, от которых они отделились очень давно, повидимому, еще в течение пермского периода. У нас есть основание предполагать, что в течение всей мезозойской эры потомки этих первичных млекопитающих существовали в виде мелких форм, от которых до нас дошло сравнительно мало ископаемых остатков. Во всяком случае, в течение мезозоя эволюция их шла крайне медленно, и фауна млекопитающих триаса, юры и мела никоим образом не могла конкурировать с крайне богатой и разнообразной рептилийной фауной этих времен. Наоборот, с начала эоцена эта эволюция пошла гораздо более быстрым темпом, так что в продолжение сравнительно весьма короткой третичной эры1 развилось громадное число весьма разнообразных форм млекопитающих.
Повидимому, и в этом ряду прогрессивная эволюция началась с изменений, возникших в органах дыхания, в органах кровообращения и в ротовом аппарате рептилийных предков млекопитающих. Прогресс в органах дыхания сводился здесь, как это было и у птиц, к увеличению общих размеров легких, т. е. к количественному увеличению легочных альвеол и мелких бронхов (ароморфоз). Из многочисленных изменений, имевших место в кровеносной системе, отметим лишь образование четырехкамерного сердца с вполне разделенным предсердием и желудочком (рис. 75 D). В качестве прогрессивных изменений ротового аппарата укажем на образование жевательных зубов гетеродонтного типа, на замену многочисленных косточек нижней челюсти одной костью (dentale) и на непосредственную связь dentale с височною костью черепа. Все эти изменения, в результате которых появился лучший по сравнению с прежним способ захватывания и пережевывания пищи, привели к дальнейшему улучшению питания и, в свою очередь, вызвали, подобно только что описанным изменениям в органах дыхания и кровообращения, новый подъем в общей энергии жизнедеятельности примитивных млекопитающих. Все они, таким образом, являются ароморфозами. В качестве прогрессивного признака млекопитающих следует также отметить развитие у них волосяного покрова, делающего животных мало чувствительными к переменам температуры.
Трудно решить, является ли развитие млечных желез у млекопитающих также ароморфозом. Его несомненно придется признать таковым, если будет установлено, что питание детенышей молоком подымает их общую жизнедеятельность и делает их крепче и сильнее.
Относительно целого ряда этих изменений мы, к сожалению, не знаем, в каком порядке и в какие эпохи они появились, так как они касаются органов, которые не сохранились палеонтологически. Это тем более вызывает наше сожаление, что восстановление порядка и времени появления прогрессивных признаков, может быть разъяснило бы нам парадоксальный ход эволюции млекопитающих. Появившись, как мы это только что указали, в очень давние времена (в перми), млекопитающие очень долго, в течение всего мезозоя, не эволюируют прогрессивно, а затем внезапно начинают сильно изменяться и дают начало большому числу сильно отличающихся друг от друга групп. Объяснить это явление только тем, что, вследствие' вымирания мезозойских рептилий в конце мелового периода, освободились свободные места в природе, едва ли 1 * 3
1 По подсчету Осборна, который берет, по всей вероятности, слишком низкие
цифры, мезозойская эра продолжалась около 9 000 000 лет, третичная—около
3 000 000 лет, т. е. втрое короче По современным данным, как указал мне В. И. Вернадский, эти цифры, вероятно, должны быть гораздо более значительными, а именно 220 000 000 лет для мезозоя и 80 000 000 лет для третичного периода.
Морфологические закономерности
19
290
ГЛАВА X
возможно, так как свободное место само по себе не есть фактор, обусловливающий прогрессивную эволюцию данной группы животных. Невольно является предположение, что причиной прогрессивной эволюции млекопитающих в начале третичного периода было появление у них каких-то прогрессивных признаков, повысивших их организацию сравнительно с их мезозойскими предками и давших им победу в борьбе за существование, которой они в предшествующие периоды не могли достигнуть. Весьма интересно было бы на основании сравнительно морфологического анализа попытаться выяснить вопрос о том, в чем состояли эти признаки, и это составило бы благодарную задачу исследования для специалистов маммологов Л
Весьма важным прогрессивным изменением, имевшим место при превращении рептилий в млекопитающих, мы считаем увеличение общих размеров мозга, в частности мозжечка и, главное, больших полушарий, другими словами, развитие высших психических способностей млекопитающих, при помощи которых они получили возможность приспосабливаться к самым разнообразным изменениям внешней среды, не изменяя при этом своего строения. Действительно, мы хорошо знаем, что млекопитающие очень редко приспособляются к быстро наступающим переменам в окружающей среде (новые враги, новая добыча и т. п.) путем медленного изменения своих органов и функций; гораздо чаще это происходит путем быстрого изменения прежних привычек и навыков и образования новых, приспособленных к новым условиям среды. Здесь впервые
1 Такой анализ порядка появления прогрессивных признаков в классе млекопитающих недавно сделал С. А. Северцов (С. А. Северцов, 1936). Сопоставляя систематическое положение и признаки современных представителей класса с родословным древом млекопитающих, как оно установлено палеонтологами (Romer, 1933), он приходит к выводу, что млекопитающие приобрели свои основные прогрессивные особенности (ароморфозы) еще в триасе, но до конца мезозоя оставались яйцеродными. Эти ароморфозы, превратившие их из рептилий в млекопитающих, позволили им выдержать конкуренцию с прогрессивными мезозойскими рептилиями: они обеспечили млекопитающим достаточную подвижность и выносливость, чтобы конкурировать с динозаврами. Живородность и выкармливание детенышей молоком матери развились у сумчатых и плацентарных независимо друг от друга в конце меловой эпохи. Это вызвало громадное усиление конкуренции, которое было непосредственной причиной широкого развития класса в третичную эпоху. Таким образом, С. А. Северцов считает, что мы имеем два цикла адаптивной радиации млекопитающих: первый—в триасе—был вызван ароморфозами, второй—в третичную эпоху— эмбриональными приспособлениями.
Повышение выживаемости детенышей млекопитающих сравнительно с рептилиями, на ряду с усилением общей жизнедеятельности благодаря приобретению теплокровности и другим ароморфозам, определили более высокий коэфициент элиминации класса, установившийся в третичную эпоху. С. А. Северцов обращает внимание на то, что повышение долговечности в ряду млекопитающих идет медленнее, чем в ряду птиц и рептилий. Это означает большую интенсивность борьбы за существование у млекопитающих и объясняет быстрые темпы эволюции класса. Образование главных ветвей рептилий началось в триасе, и они достигли расцвета в юре и нижнем мелу приблизительно через 150—200 миллионов лет. Третичное развитие млекопитающих началось в палеоцене и достигло развития в миоцене, т. е. через 20—25 миллионов лет. Гораздо более короткий срок, в течение которого образовались водные, наземные и воздушные формы, может быть объяснен только большей интенсивностью борьбы и отбора. Вто же время С. А. Северцов отмечает, что имеет огромное значение не только интенсивность отбора, но и его направление. Так, грызуны и насекомоядные, наиболее плодовитые и наименее долговечные представители млекопитающих, в то же время сохраняют наибольшее число примитивных признаков. Это объясняется тем, что их эволюция пошла в направлении количественных приспособлений и что значительное повышение плодовитости компенсировало частую массовую гибель от климатических депрессий, после которых они легко восстанавливали свою численность. Прогрессивные же формы пошли по пути повышения стойкости особи. Это направление отбора привело их к прогрессивной эволюции, несмотря на меньшую плодовитость и смертность.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
291
выступает на сцену новый и весьма важный фактор адаптивной эволюции позвоночных животных, именно его психика. Поэтому мы и рассматриваем прогрессивное развитие мозга, и в особенности его больших полушарий, у млекопитающих (отчасти и у птиц, также обладающих высокоорганизованным мозгом) как весьма важный ароморфоз.
Так как вопрос о развитии этого ароморфоза (высших психических способностей) имеет первостепенное значение в эволюции млекопитающих,, то я позволю себе остановиться на нем несколько подробнее.
В одной из своих предшествующих работ,1 рассматривая различные способы приспособления животных к изменениям в окружающей среде, я подробно остановился на «способе приспособления посредством изменения поведения (behaviour) животных без изменения их организации», иными словами, на вопросе о значении психики как фактора биологически прогрессивной эволюции животных.
Как известно, у животных существуют три основных типа психической деятельности: рефлекторная, инстинктивная и деятельность, которую мы условно обозначим как деятельность «разумного типа». Термином «рефлекс» обычно обозначаются правильно повторяющиеся, однообразные приспособительные реакции организма на специфические раздражения окружающей среды. Рефлекторная деятельность является наследственной, т. е. молодое животное начинает производить те же рефлекторные действия, которые производили их родители, без всякого предварительного обучения, вполне правильно. Точно так же, как и рефлекторная деятельность, инстинктивная деятельность является целесообразной, наследственной и до известной степени машинообразной, но отличается от рефлекторной большей сложностью. Здесь мы обычно имеем длинный ряд сложных действий, являющихся ответом на определенное внешнее раздражение. Инстинкты, так же как и рефлексы, являются приспособлениями к медленно протекающим изменениям внешней среды, и притом во многих случаях приспособлениями очень сложными и биологически вполне стойкими, т. е. повторяющимися у каждой особи неизменно из поколения в поколение с замечательной правильностью. В инстинктах наследственна не способность к действиям, но самые действия со всеми их типичными чертами.
Деятельность так называемого «разумного порядка» является таюке целесообразной и приспособительной, но в отличие от рефлексов и инстинктов не наследственной и не машинообразной. Здесь наследственной является способность к деятельности данного типа, но не самые действия, и животные являются наследственно весьма различными в этом отношении: одни способны к сложным действиям, другие к весьма элементарным, но сами действия их не предопределены наследственно и в индивидуальной жизни не являются готовыми, как рефлексы и инстинкты: для производства определенного действия требуется определенная выучка. Они не являются и «машинообразными»: за определенными раздражениями могут следовать весьма разнообразные действия..
В различных группах животных преимущественное значение имеет либо тот, либо другой тип психической деятельности: у членистоногих преимущественное значение приобрел инстинкт, у птиц и особенно у млекопитающих—психика так называемого «разумного порядка».
Инстинкты и рефлексы, как и все наследственные приспособления, эволюируют крайне медленно и постепенно, посредством суммирова
> Северцов А. Н., Эволюция и психика, М., 1922.
19*
292
ГЛАВА X
ния мелких наследственных вариаций инстинктов. Это—приспособление к медленно протекающим изменениям внешней среды. Наоборот, в психических свойствах организма, которые мы относим к категории «разумных», наследственной является только известная высота психики и способность к определенным действиям (эволюция этой способности, как и все наследственные процессы, идет, само собою разумеется, также чрезвычайно медленно и постепенно), но сами действия не предопределены наследственно. Они могут быть крайне разнообразны. Каждая особь выучивается им заново, и все они являются быстрым ответом организма на быстрые, но большей частью незначительные изменения в экологических условиях окружающей среды (появление нового врага, уменьшение обычной добычи и т. д.).
Низшая форма «разумной» деятельности подходит под тип условных рефлексов. Как известно, условный рефлекс отличается от обычного типа рефлексов тем, что он не наследственен и что он приобретается животным в необычайно короткий срок. В естественной обстановке часто устанавливаются целесообразные условные рефлексы, имеющие приспособительный характер. Мы знаем, например, что дикие птицы и млекопитающие, живущие на необитаемых островах, при первом появлении человека ведут себя как ручные, не боятся его и не убегают. При повторном появлении человека, после того как они испытали на себе опасности, проистекающие от его присутствия, они начинают пугаться и убегать; установился новый условный рефлекс.
Близко к условным рефлексам стоит способность домашних животных к различного рода дрессировке, т. е. к приобретению новых навыков. Вместе с тем является несомненным, что и дикие животные вырабатывают под влиянием изменений внешней среды новые способы действия и новые привычки приспособительного характера. Это становится особенно заметным, когда высшим млекопитающим приходится сталкиваться в своей борьбе за существование с человеком. Так, например, Рузвельт приводит наблюдение, что африканские слоны изменили свое поведение с тех пор, как за ними стали охотиться охотники с дальнобойными винтовками; они перестали пастись в открытой местности и стали держаться в лесу, где отыскать их гораздо труднее и где дальнобойное оружие не представляет преимуществ. Охота за ними стала гораздо труднее, и истребление приостановилось. Это изменение у слонов произошло очень быстро, в течение. одного поколения, так что о наследственном изменении инстинкта здесь говорить нельзя. Аналогичные изменения в повадках произошли у канадских бизонов: они тоже под влиянием преследования из степных животных сделались лесными и тоже в короткое время. Наоборот, африканские носороги, гораздо более тупые, не приобрели этой привычки и поэтому усиленно истребляются. В этом отношении интересен также пример «контроблав» оленей. Когда в данной местности произведено несколько облав, то обычное поведение оленей изменяется, и они, вместо того, чтобы бежать от шума, производимого загонщиками, на стоящих тихо и спрятанных охотников, бегут на шум, т. е. на загонщиков, прорываются через их линию итаким образом уходят. Это становится настолько постоянным, что стрелкам приходиться становится позади загонщиков, и тогда олени нарываются на них.
С биологической точки зрения, т. е. с точки зрения приспособляемости, мы имеем здесь фактор чрезвычайной важности, биологическое значение которого до сих пор не было достаточно оценено. Значение психической деятельности «разумного порядка» состоит в том, что она весьма значительно повышает пластичность млекопитающих по отношению к быстрым изменениям среды. При быстрых изменениях внешних условий животное отвечает не измене-
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ 293 нием своей организации, которое требует для своего осуществления огромных периодов времени (в геологическом ^смысле слова), но быстрым изменением своего поведения, и поэтому в очень большом числе случаев оно может приспособиться к новым условиям весьма быстро.
Итак, психическую деятельность животных можно подразделить на две большие группы:
I. Наследственные приспособления к очень медленным изменениям условий среды—наследственные изменения поведения без изменения строения (рефлексы и инстинкты).
II. Ненаследственные приспособления к сравнительно быстрым, но не очень значительным изменениям среды—ненаследственные изменения поведения «разумного типа».
Развитие наследственно не фиксированных действий шло прогрессивно в ряду млекопитающих. Приспособления посредством изменения поведения в течение индивидуальной жизни имеют громадное биологическое значение, ибо позволяют высшим млекопитающим быстро приспособляться к изменениям, вносимым в их жизнь другими животными и человеком.
Наибольшее значение приспособление этого типа имело при эволюции человека. Мы можем с полной уверенностью сказать, что развитие психики «разумного типа» повысило в исключительной степени способность непосредственного предка человека и самого человека к приспособлению и что именно благодаря этой способности человек и занял свое доминирующее положение в ряду млекопитающих. Он может приспособляться в чрезвычайно короткое с эволюционной точки зрения время решительно ко всяким изменениям в условиях существования, ибо над наследственной приспособляемостью организма у него появилась огромная надстройка индивидуальной приспособляемости поведения.
ВЫВОДЫ ОБ АРОМОРФОЗАХ
Подводя итоги всему сказанному об ароморфозах, а также сопоставляя между софой перечисленные нами признаки морфофизиологической прогрессивной эволюции, мы видим, что прогрессивная эволюция каждой крупной систематической группы позвоночных характеризуется развитием определенного комплекса очень общих по своему физиологическому характеру признаков. Прежде всего мы встречаем ряд изменений, значение которых заключается в том, что они способствуют общему подъему энергии жизнедеятельности животных, у которых они появляются; сюда относятся: переход от пассивного питания к активному, изменения в пищеварительных железах, приспособления к более совершенному захватыванию пищи (зубы и изменение в челюстном аппарате), повышение дыхательной функции (все изменения в жабрах и легких) и т. д. Затем следуют прогрессивные изменения активных проявлений жизнедеятельности, куда в первую очередь относится повышение подвижности животных (усиление мускулов, диференцировка их, замена хрящевого скелета костным, развитие и совершенствование хвостового плавника, развитие парных плавников и т. д.); при этом характерно то, что, благодаря разобранным нами морфологическим изменениям, повышаются и интенсивность и разнообразие движений. Сюда же относятся прогрессивные изменения в органах нападения (зубы и т. д.) и защиты (чешуи, панцыри). Наконец, отметим повышение разнообразия и сложности реакции животных на раздражения, получаемые из внешней среды, и прогрессивное развитие психики. Я не останавливался специально на изменениях головного мозга и органов высших чувств, но должен здесь отметить,
294
ГЛАВА X
что во всем ряду рассмотренных нами прогрессивно изменявшихся форм мы видим прогрессивное изменение этих органов, выражающееся частью в увеличении количества нервного вещества, частью в усложнении структуры мозга и органов чувств.
Не вдаваясь в подробности, особенно подчеркиваю одну особенность всех перечисленных нами изменений, а именно, их универсальный характер. Эти изменения строения самых различных органов ( и, само собой разумеется, зависящие от них изменения их функций) не суть приспособления к каким-либо специальным условиям жизни, т. е. приспособления к жизни на определенной глубине, или на дне, или к питанию особой пищей и т. д., но изменения очень общего характера. При этом мы видим, что изменения такого универсального типа оказываются весьма стойкими: они сохраняются в своих основных чертах неизменными и при дальнейшей прогрессивной, иногда очень долгой эволю-ц и и животных. Иначе говоря, всякий ароморфоз, раз появившись, остается у потомков данной формы в качестве весьма постоянного признака. Так, например, четырехкамерное сердце, однажды образовавшись у реп-тилийных предков птиц, наследуется всеми представителями этой богатой родами и видами группы; совершенно то же мы можем сказать и относительно сердца, легких и мозга всех млекопитающих. Гетеродонтные зубы, унаследованные млекопитающими от тех же рептилийных предков, сохранились у огромного большинства их потомков. У всех потомков первичных гнатостом, эволюировавших в направлении прогрессивного изменения, сохранился тип хватательных челюстей; у всех рыб сохраняется основной тип строения плавников; у прогрессивных наземных позвоночных—основные черты пятипалой конечности и т. д.
Отмеченные выше факты указывают нам на причину, почему признаки, которые мы обозначаем как ароморфозы, обыкновенно бывают так постоянны: эти признаки имеют столь большое биологическое значение, что они являются полезными при весьма разнообразных условиях жизни. В тех случаях, когда они перестают быть полезными, они начинают изменяться: так, йапример, гетеродонтные зубы первичных млекопитающих у предков китообразных становятся однородными, а у современных китов редуцируются совершенно. Подобный случай встречается, однако, крайне редко, обычно же ароморфозы наследуются всеми потомками данной животной формы и становятся характерным признаком крупных систематических групп животного царства, другими словами, ароморфозы наследуются всеми дивергентно развивающимися потомками той формы, у которой они впервые образуются.
Вдумываясь глубже в вопрос об ароморфозах, мы не можем не заметить, что ароморфозы обычно представляют собою изменения в тех системах органов, которые прямо или косвенно связаны, с обменом веществ в организме. Так, ароморфозами являются все прогрессивные изменения в органах дыхания, кровообращения и питания; в последнем случае ими являются, кроме изменений, имеющих место в кишечнике и его железах, также и все изменения прогрессивного характера, происходящие в строении зубов, челюстей и челюстной мускулатуры. Ясно, что все прогрессивные изменения в органах движения, нападения и защиты, в органах высших чувств и в мозгу животных имеют, хотя и косвенное, но постоянное влияние наспособ их питания (отыскание и ловля добычи, нахождение свежей и обильной
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
295
растительной пищи), т. е. на качество и количество потребляемой ими пищи; таким'образом, все они косвенным путем повышают энергию жизнедеятельности эволюирующих форм. То же относится и к изменениям, имеющим место в строении жаберных дуг, жаберной мускулатуры, жаберных лепестков у рыб и торакальных мускулов у высших Quadrupeda (грудные ребра и межреберные мускулы), которые косвенным образом тоже влияют, конечно, на весь процесс дыхания этих животных. Общее же значение почек для действия обмена веществ понятно само собой.
Итак, мы можем сказать, что всякая интенсификация функций органов кровообращения, дыхания и питания, произошла ли она прямым или косвенным путем, является ароморфозом. Если же мы встречаемся с прогрессивными изменениями в энтосоматических органах, с изменениями, стоящими в прямой функциональной зависимо-jjc т и от особых, частных условий окружающей среды и не имеющих влияния на органы обмена веществ, то, как мы это сейчас увидим, мы будем иметь дело с идиоадаптациями, т. е. с изменениями, не ведущими к повышению уровня общей жизнедеятельности животного.
Таким образом я назвал «ароморфозами» изменения универсального характ.ера, благодаря которым организация животных поднимается на более высокую ступень и которые дают возможность дальнейшего. прогрессивного изменения, и назвал «идиоадаптациями» все изменения приспособительного характера, все приспособления к строго определенным условиям среды, которые не повышают общей энергии жизнедеятельности живо т н ы х.1
Лишь в редких случаях удается понять, почему та или другая животная форма пошла по тому или иному пути эволюции. Причина того, что в одних случаях прогрессивно развиваются органы кровообращения, питания и дыхания (ароморфозы), а в других появляются только идиоадап-тации, остается до сих пор для нас мало понятной.
ИДИОАДАПТАЦИИ ИЛИ ЧАСТНЫЕ ПРИСПОСОБЛЕНИЯ
Вторым направлением биологического прогресса является направление морфофизиологического приспособления (адаптации в узком смысле этого слова) или направление идиоадаптации. Как это нами было сказано, идиоадаптации отличаются от а р о-морфозов тем, что при них не происходит какого бы то ни было усиления или подъема общей энергии жизнедеятельности организма, но эволюирующие формы, изменяясь вместе с изменениями среды, лучше приспособляются к определенным условиям окружающей
1 Отмечаем, что мы перечислили очень крупные изменения организации, которыми отличаются друг от друга большие группы животных. Дальнейшей задачей эволюционной морфологии является распространение этого исследования и на отличия более мелких групп, т. е. на роды, виды и т. д. Поэтому мы подчеркиваем, что наше исследование является только первым приближением к решению поставленной нами задачи,
296
ГЛАВА X
среды, чем предки. Иными словами, животные, вступившие на этот путь биологического прогресса, в противоположность животным, эволюи-рующим путем ароморфозов, не поднимаются на более высокую ступень организации: в этом отношении они стоят не выше и не ниже своих предков. Это коренное различие между обоими типами биологически-прогрес-сивной эволюции станет понятным читателю из следующих примеров.
Мы знаем, что черепахи, филогенетические соотношения которых с другими рептилиями в настоящее время до известной степени выяснены благодаря открытию триасовой черепахи (Triassochelys dux), повидимому, очень рано, еще в пермскую эпоху, отделились от котилозавров и приобрели характерные черты своей своеобразной организации: вследствие срастания костных щитков кожи с ребрами и остистыми отростками позвонков у них образовался плотный костный спинной щит, а благодаря аналогичному срастанию костей пояса передней конечности (episternum, interclaviculae) и элементов кожного скелета развился брюшной щит. Оба эти щита вместе образовали прочный панцырь, в который могут в случае нужды спрятаться голова и конечности. Кроме этих крупных изменений^ произошло и довольно большое число менее значительных изменений в других частях скелета: редуцировались зубы и заменились роговыми челюстями, число фаланг в конечностях сократилось и т. д. Эти изменения оказались весьма стойкими, и черепахи в виде довольно обильной формами группы дожили до нашего времени и не выказывают никакой склонности к вымиранию: их существует и в настоящее время около 200 видов.
Обыкновенно говорят, будто черепахи представляют собой весьма специализованную группу животных, но в сущности довольно трудно указать у них признаки специализации. Специализованными мы называем животных, приспособившихся к определенным, однообразным и постоянным условиям существования, например животных подземных: гимнофионов среди амфибий, кротов и слепышей среди млекопитающих; или животных, ведущих исключительно древесный образ жизни, как хамелеоны среди рептилий, ленивцы между млекопитающими; специализованными являются рыбы с чисто донным образом жизни (скаты, камбалы и т. д.); далее животные, приспособившиеся к одному только роду пищи (сосущие кровь насекомые) и т. д. Если же мы обратимся к черепахам, то увидим, что они представляют собой животных с разнообразным образом жизни: мы знаем чисто наземных черепах, которые живут в сухих местностях (Testudo graeca); знаем болотные формы, как наша Emys lutaria, знаем пресноводные (Clemmis) и, наконец, чисто пелагические формы (Sphargis, Chelone); одни из них питаются только растительной пищей, другие—различными видами животной пищи.
Таким образом, черепахи при своей эволюции приспособились к весьма разнообразным условиям существования и при этом организация их сильно менялась приспособительно к тем или иным условиям среды: например, у пелагических Dermochelidae (Sphargidae) редуцировался костный панцырь и удлинились пальцы передних конечностей, вследствие чего они превратились в приспособленные к плаванию ласты. Интересно при этом, как показал Долло, что редукция костного панцыря произошла дважды: береговые предки Sphargidae, снабженные плотным панцырем, перешли к пелагическому образу жизни и потеряли при этом панцырь, затем снова сделалйсь береговыми животными, причем у них развился новый костный панцырь, который при вторичном переходе к пелагическому образу жизни снова редуцировался. Однако, несмотря на изменения адаптивного характера, общая высота организации черепах в течение их весьма долгой эволюции не п о в ы с и л а с ь, и они остались примитивными рептилиями: эволюция у них шла по пути идиоадаптации, а не по пути ароморфоза.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
297
То же, что мы находим у черепах, мы видим и у змей; змеи произошли сравнительно поздно (по всей вероятности, в течение мелового периода) от общего корня с ящерицами и образовали богатую формами группу с очень широким географическим распространением. От ящериц они отличаются главным образом способом питания и передвижения. Как известно, змеи—хищные рептилии, глотающие свою добычу целиком, не разрывая ее на части, и в связи с этим способом питания у них развивались своеобразные изменения в челюстном аппарате,1 благодаря которым ротовое отверстие способно растягиваться весьма значительно. Кроме того у них атрофировались обе пары конечностей и сильно увеличилось число позвонков, благодаря чему удлинилось тело. С ползающим способом передвижения связаны характерные изменения строения позвонков, брюшных чешуй и туловищной мускулатуры. Характерная для змей редукция одного легкого и некоторые другие изменения во внутренних органах стоят, повидимому, в связи с удлинением тела. Здесь мы имеем ряд крупных изменений, которыми организация змей отличается от организации ящериц, но, несмотря на это, сравнивая организацию ящериц (Lacertilia) с организацией змей (Ophidia), мы не имеем основания сказать, что последние изменились в прогрессивном направлении сравнительно со своими ящерицеобразными предками, так как организация змей не выше организации ящериц, хотя и сильно от нее отличается. Змеи перешли к другому способу передвижения, стали питаться иначе, чем их ящерицеобразные предки, но мы не можем сказать, что у них от этого повысилась общая энергия жизнедеятельности или что их организация усложнилась, как это мы ясно видим, например, при сравнении рептилий с амфибиями1 2 или птиц с рептилиями. Змеи, таким образом, в своей эволюции пошли по пути идиоадаптации.
Пример идиоадаптивной эволюции мы находим и наблюдая изменения, происшедшие в организации скатов по сравнению с организацией их акулоподобных предков. Сравнительно анатомические и, в особенности, сравнительно эмбриологические исследования (рис. 72) ясно указывают нам на то, что скаты произошли от акул, перешедших к бентонному об-расу жизни. Но значительные изменения организации, имевшие место при этом переходе, т. е. общее дорзовентральное сплющивание тела, черезвычайное разрастание грудных плавников, редукция хвостового плавника, вентральное положение жаберных щелей и дорзальное положение увеличенного брызгальца, никоим образом не могут быть рассматриваемы как ароморфозы; они являются лишь простыми приспособлениями скатов к бентонному образу жизни, ибо, хотя общая организация этих форм по сравнению с организацией их акуловых предков и изменилась очень сильно, однако она нисколько не стала от этого выше.
Совершенно то же можно сказать, сравнивая хамелеонов и гекконов
1 Удлинение quadratum (рис. 134 В, 135 А и В) и squamosum, свободное сочленение этих костей друг с другом и с другими костями челюстного аппарата и т. д.
2 Здесь можно сделать возражение, что нам за недостатком палеонтологического материала (который, действительно, и редок и отрывочен) приходится в данном случае сравнивать современных змей с современными же ящерицами, которые безусловно не могут считаться предками Ophidia; это возражение, конечно, вполне основательно. Тем не менее, я думаю, что сделанный нами вывод правилен, так как несомненно характерные черты Lacertilia выработались уже в очень древние времена, как предполагают, еще в течение пермского периода. У триасовой Palaeiguana, например, уже развиты все главные признаки Lacertilia, так что мы имеем полное основание думать, что тип этот со своими характерными признаками выработался уже давно, во всяком случае еще задолго до того времени, как от него отделились предки змей. К тому же выводу мы приходим и при сравнении скелета Dolichosauria и Mososauria со скелетом Lacertilia. Насколько мы можем судить, Lacertilia представляют собой группу рептилий, которая, несмотря на целый ряд изменений идиоадаптивного характера, не пошла по пути прогрессивной эволюции.
298
ГЛАВА X
с типичными ящерицами или сравнивая гимнофионов с типичными хвостатыми амфибиями; то же относится и к огромному большинству костистых рыб, как, например, к Pleuronectidae, Gobiidae, Lophiidae, к рыбам, населяющим коралловые рифы, словом, ко всем формам, приспособленным к узко определенным условиям существования.
Большинство приспособительных из м е не.н и й эк т о с о м а т "и чес к и х "о Р г а но в я~в~л‘яПед'с'я* .„.и д_и о а д а-^тацйяИГК идиоадаптациям относятся, например, все бесчисленные 1 измененитгтп?р а с к и животных, соответствующие той среде, в которой они обитают, в том числе все покровительственные и защитные окраски. Мы не можем, например, считать подъемом организации, т. е. подъемом общей энергии жизнедеятельности животного, когда при переселении какой-либо формы из одного места в другое приспособительно меняется ее окраска: так, организация полосатой зебры нисколько не выше организации однотонно окрашенной лошади Пржевальского, и изменние в окраске льва по сравнению с тигром нисколько не подняло высоты его организации. Мы знаем, что очень многие пелагические животные имеют прозрачное тело, делающее их невидимыми в водной среде; тот же признак мы находим и у многих свободно плавающих личинок рыб; биологическое значение этих особенностей настолько очевидно, что на нем нечего останавливаться. Напомним еще белую окраску брюшной стороны тела, очень распространенную у рыб, держащихся близ поверхности воды; окраска эта, благодаря своему сходству по цвету с пеной гребней волны, выгодна для обладателей ее как средство защиты от хищных рыб, подплывающих к ним снизу из глубины.
У рыб, живущих близ дна или среди растений, цвет верхней стороны тела близок к окраске среды, причем часто рисунок, т. е. расположение полос и пятен разных цветов и оттенков, приспособлен к тому, чтобы сделать животное мало заметным. Во многих случаях этот способ защиты еще более усовершенствован, и окраска меняется приспособительно под влиянием нервной системы; например, камбалы меняют свой цвет, делая его вполне подобным цвету того дна, на котором они в данное время находятся.
Мы напомнили читателю некоторые примеры приспособительной окраски у рыб, так как мы до сих пор говорили главным образом о них, но мы знаем, что во всякой группе живущих на свету животных мы найдем бесчисленные примеры защитных и маскирующих окрасок и рисунков; не опасаясь сделать большой ошибки, мы можем сказать, что большая часть окрасок наземных животных является приспособительной и что все изменения этого рода являются идиоадаптациями, ибо совершенно понятно, что различная окраска и различное расположение пятен на спине у вальдшнепов, перепелок, куропаток, дроф и рябчиков, делающих этих животных настолько похожими на окружающую их среду, что они становятся в ней почти невидимыми для своих обычных врагов,1 нисколько не повышают общей высоты их организации по сравнению с организацией родственных им форм, лишенных этой покровительственной окраски. То же можно сказать и о разнообразнейших формах мимикрии у многочисленных насекомых, например у бабочек (Callima paralecta, Siderone strigosa), внутрен
1 В молодости мне прищлось быть свидетелем того, как один охотник подошел невзначай вплотную к мирно дремавшей в степной траве дрофе, не замечая ее; он обратил внимание на дрофу только тогда, когда она, поднявшись и громко хлопая крыльями, стала от него убегать. Он был так ошеломлен этим, что схватился за ружье лишь тогда, когда дрофа находилась вне выстрела. Замечу при этом, что мы в это время находились в степи специально для охоты за дрофами и что охотник, о котором шла речь, был опытным собирателем птиц и страстным охотником; он был препаратором моего отца, орнитолога Н. А. Северцова.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
299
няя поверхность крыла которых во всех подробностях подражает окраске и форме листа тех растений, на который бабочка обычно садится (рис. 76). То же наблюдается и у прямокрылых, например у «блуждающего листа» (Phyllium scythe, рис. 77), зеленая окраска которого и форма крыльев в совершенстве имитирует оторвавшийся от ветки лист.
Выше я уже указывал на некоторые случаи, когда форма тела является адаптацией к той обстановке, в которой живет животное, как, например, плоская форма тела скатов (рис. 133), служащая приспособлением к донному образу жизни. Аналогичным примером мимикрии может служить плоская форма тела, камбал (рис. 78). Тот и другой случаи
интересны как примеры того, насколько значительные изменения организации мо-гут быть вызваны приспособлением к определенному образу жизни. По поводу камбаловых рыб отмечу, что для хордовых животных би-
Рис. 76. Бабочки, имитирующие формой своего тела древесный лист.
A—Callima paralecta во время полета; а—та же бабочка, сидящая со сложенными крыльями на ветке (из Уоллеса); В—Siderone strigosa во время полета; b—та же бабочка, сидящая со сложенными крыльями на ветке (из К. Штерне).
Рис. 77. Phyllium scythe $> (из Гертвига).
латеральная симметрия тела является одним из очень стойких и древних признаков организации. Благодаря переходу к донному образу жизни камбалы изменились именно в этом признаке, причем произошли коренные преобразования всего строения: изменилось строение черепа, рот сделался асимметричным, один глаз переселился с одной стороны тела на другую и т. д. (рис. 78).
Из адаптивных изменений строения рыб упомяну еще об удивительных листовидных выростах кожи на теле некоторых морских коньков (Phyl-lopteryx equus), делающих их похожими как раз на те водоросли, среди которых они живут; о своеобразных жгутовидных гибких лучах непарного плавника так называемого «морского чорта» (Lophius, Chirolophius), служащих приманкой для добычи, когда эти хищные рыбы лежат на дне (рис. 79 г1( г2, г3, ...).
300
ГЛАВА X
Рис. 78. Камбала (Bothae maximus). (Из Смита).
Рис. 79. Морской 4opT(Lophius piscatorius) со своеобразным приманочным аппаратом на голове и на спине, образованным видоизмененными лучами переднего спинного плавника (гт—г6).
а—анальный плавник; с—хвостовый плавник; da—неизмененный задний спинной плавник; pct— грудной плавник; vt—брюшной плавник (из А. Н. Северцова, 1927).
Рис. 80. Gobius capito.
а—анальный плавник; ап—анальное отверстие; с—хвостовой плавник; di, d2—передний и задний спинные плавники; pct—грудной плавник; vt—сросшиеся между собой брюшные плавники, образующие крепкую присоску, с помощью которой Gobius удерживается на скалах во время сильного прибоя (из А. Н. Северцова, 1927).
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
301
Рис. 81 А. Тригла. Грудные плавники (pct) с сильно измененными передними лучами (rdj, rd2, rd3), при помощи которых тригла может ползать по дну; В— эти же плавники триглы, изображенные снизу (из А. Н. Северцова, 1927).
Интересен пример превращения брюшных плавников в присоски, при помоши которых некоторые рыбы (Gobiidae, Lepadogaster и др.), живущие в области прибоя, прикрепляются к прибрежным скалам (рис. 80, vt). В качестве примеров адаптивных изменений у рыб можно привести и изменение грудных плавников у тригл, у которых три передних луча этих плавников превратились в свободные и гибкие, напоминающие пальцы, при,-датки (рис. 81, rd1; rd2, rd3), при помощи которых они ползают по дну и ощупывают добычу. В качестве очень своеобразного приспособления к образу жизни на поверхности воды упомянем глаза четырехглазки (Anableps tetraphtalmus) (рис. 82); рыба эта плавает на поверхности воды так, что верхняя часть ее глаз находится в воздухе, а нижняя в воде, и соответственно этому роговица и зрачок глаза разделены горизонтальной перегородкой на два отдела — верхний и нижний (собственно говоря, в каждом глазу находится по два зрачка), причем верхний приспособлен к видению в воздухе, а нижний—к видению в воде; аккомодация обеих половин глаза различна.
Все изменения этого рода являются идиоадаптациями, ибо совершенно понятно, что различная окраска и различное расположение пятен на теле животных, точно так же как и различные изменения формы их тела, нисколько не повышают общей энергии их жизнеде’ятельности, равно как и высоты их организации по сравнению с организацией родственных им форм.
К этой же категории идиоадапта-ций относятся ц все многочисленные изменения, происходящие в о р г а-,'н~а"х~~Д"в и ж е н и я^животных, т. е. все пр испосо б лё ния" "к” оп'рё дёлён ному ЗТОду движения' в отличной^ от пред-т1Ж^еТ®7’Здё"сь7пЬмЖд''только что упомянутых нами приспособлений к движению по дну у различных рыб, например у тригл, прекрасными при
мерами идиоадаптаций служат различные случаи «адаптивной радиации», приводимые Осборном.
Путем сопоставления громадного палеонтологического и зоологического материала Осборн пришел к важному выводу, что во всякой крупной группе животных, попавшей в новые условия существования, потомки исходной формы приспособляются к этим условиям, и те органы их, которые имеют непосредственное отношение к новым особенностям внешней среды, тоже приспособляются к ним и изменяются в соответственных направлениях. В результате этого оказывается, что один и тот же орган предков у потомков, перешедших к разным условиям существования, принимает весьма разнообразные формы и что строение его делается весьма различным у разных потомков исходной формы. Мы знаем, например, что предки всех млекопитающих первоначально были типично стопоходящими пятипалыми животными (рис. 83 А); некоторые из их потомков, переселившихся в среду, где было важно быстрое передвижение по более или менее ровной и твердой почве, приспособились к бегающему способу передвиже
302
ГЛАВА X
ния, и у них сначала развилась пальцеходность (конечности опираются на землю концами пальцев), а затем краевые пальцы редуцировались, кости запястья и предплюсны значительно удлинились и когти превратились в копыта, приспособленные к очень быстрому бегу. У парнокопытных, т. е. ужирафф, оленей, быков, антилоп, верблюдов и т. д., вес тела покоится на третьем и четвертом пальцах, у непарнокопытных, как, например, у лошадей и вымерших Litopterna,— на третьем пальце, причем краевые пальцы (I, II, V—у парнопалых, I, II, IV и V—у непарнопалых) редуцированы отчасти или совсем (рис. 83 G). У других потомков первичных пятипалых стопоходящих млекопитающих, именно у тех, которые перешли к полуводному образу жизни, конечности изменились в другом направлении и приспособились к плаванию, превратившись в ласты типа
Рис. 82. Четырехглазка (Anableps tetraphthalmus). (Из Никольского).
тюленей и моржей. При переходе к чисто водному образу жизни они превратились в ласты китов (рис. 83 F), кашалотов и дельфинов, совершенно непригодные к передвижению на суше, но прекрасно приспособленные к плаванию. При переходе к жизни на деревьях стопоходящая пятипалая конечность превращается в конечность лазающего типа, причем когти делаются ногтями, а первый палец руки и ноги приобретает способность противополагаться другим пальцам [хватательная конечность лемуров и обезьян (рис. 83 Е)]; другой тип лазающей конечности развился у ленивцев (рис. 83 D). При переходе от лазанья по деревьям к прыжкам с дерева на дерево, а затем к скольжению по воздуху (парашют белок-летяг), наконец—к летанию, конечности того же исходного типа снова изменяются и превращаются в крылья летучей мыши, летучих собак (рис. 83 С) и т. д.
При переходе к роющему образу жизни конечности приспособляются к рытью почвы: пальцы становятся толстыми, они снабжены большими когтями и т. д.При этом у тех животных, которые роют рыхлую почву, как например, у кротов (также Condylura, Ctenomis), передняя лапа расширяется на подобие лопаты и на ней развивается шестой палец; у форм, которые разрывают твердую почву, передняя лапа сужена, а третий и чет
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
303
вертый пальцы увеличены и служат для рытья. Аналогичные изменения мы находим у муравьедов, у которых передние лапы служат для взламывания твердых куч, в которых живут термиты. Сходные приспособления конечностей к разрыванию почвы мы находим и у американских броненосцев.
Подобного же рода адаптивную, или, как мы это теперь называем, «и ди о ад а пт ив н у ю» радиацию, мы имеем и у рептилий; весьма примечательно, что у них, так же как и у млекопитающих, дивергентная эволюция началась после того, как у их предков развились те прогрессивные признаки строения, которые поднимают организацию рептилий на более высокую ступень по сравнению с амфибиями (развитие типичных для рептилий легких и сердца, образование длинной шеи, развитие грудных ребер и т. д.), т. е. после появления целого ряда ароморфозов. При этом
Рис. 83. Примеры идиоадаптивной радиации передней конечности млекопитающих.
А—типичная стопоходящая конечность носухи (Nasus socialis); В—типичная роющая конечность крота; С-—летательная конечность летучей собаки; D-—лазательная конечность ленивцев; Е— хватательная конечность обезьяны; F—плавательная конечность^кита; G—пальцеходная конечность лошади. Н—humerus; с—carpalia; ш—metacarpalia; г—radius; u—ulna; 1—V—пальцы 1—V (из А. Н.
Северцова, 1934).
все дивергирующие ветви, отделившиеся от общего ствола рептилий, сохраняют свои типично рептилийные признаки, но каждая из них, кроме того, приобретает еще и ряд новых признаков, являющихся приспособлениями к самым разнообразным условиям среды, в которой приходится жить этим новым формам. Так, например, одна линия потомков первичных рептилий приспособилась к жизни в воде (стегозавры, талаттозавры, плезиозавры), другая—к жизни в болотах и реках (крокодилы и их родичи), третья—к жизни на утесах и отвесных скалах (многие ящерицы и гекконы), четвертая—к воздушной жизни (птерозавры), пятая, как например динозавры,—к жизни на суше, на берегу мелких водоемов и т. д. Конечности всех этих форм являются прекрасно приспособленными к различного рода движениям в различной среде, в которой обитают данные животные. Так же как и у млекопитающих, в одних случаях конечности эти превращаются в лопасти для плавания, в других—в крылья для полета, в третьих—в конечности резко выраженного дигитиградного типа, приспособленные для быстрого бега и т. д. В то же самое время и ротовой
304
ГЛАВА X
аппарат рептилий приспособился к разнообразной пище и к различному способу защиты от многочисленных врагов, с которыми им приходится иметь дело. Все эти изменения в конечностях только Что упомянутых нами форм являются приспособлениями к движению животных в определенной обстановке, в условиях среды, отличающихся от тех условий, в которых жили и двигались их близкие и дальние предки. Мы не считаем, что превращения конечностей предков крота в роющие конечности его потомков, или конечностей типа ящерицы в хватательные конечности хамелеона, или же стопоходящих конечностей хищника, живущего на берегу моря, в лопасти тюленей, моржей, морских львов и т. п. повлекло бы за собою повышение организации перечисленных нами форм, короче говоря, мы не думаем, чтобы эти изменения были ароморфозами: все они являются идиоадаптациями, т. е. приспособлениями к иному, но не более совершенному образу жизни. Сами животные, конечно, при этом меняются более или менее сильно, но общая энергия их жизнедеятельности не повышается и общее строение их не делается ни более сложным, ни более совершенным.
Мы привели немногие примеры адаптаций, главным образом у рыб, рептилий и млекопитающих, причем выбрали наиболее резкие примеры, касающиеся сравнительно немногих органов. Примеры эти можно было бы умножить почти до бесконечности, так как идиоадаптации существуют во всех органах как наружных, так и внутренних, имеющих отношение к наружной среде в широком смысле этого слова, и встречаются у всех решительно групп животных: наземных и подземных, водных, летающих и т. д. Мы имеем огромное число приспособлений к условиям неорганической среды—холоду и теплу, условиям освещения, сухости и влажности и т. д.—и еще большее количество приспособлений к условиям биологической среды—к особенностям растений, среди которых животные живут, к растительной или животной пище, которой они питаются, к врагам, с которыми им приходится бороться или которых приходится избегать, к добыче, которую приходится преследовать, и т. п.,—другими словами, вся организация животных в любой период эволюции являе<т-ся адаптивной. При этом идиоадаптации эти бывают весьма различными в зависимости от условий среды, к которой приспособляется животное; мы имеем приспособления к средам с разнообразными и переменными условиями, в которых живут подвижные животные с более или менее сложной организацией, и целый ряд переходов в приспособлении к средам с однообразными и постоянными условиями. В последних случаях, как, например, у животных, приспособленных к подземному образу жизни (гимнофионы, кроты, слепыши), или к жизни на отвесных скалах (гекконы), или к жизни на высоких деревьях (хамелеоны, ленивцы), или как у животных, приспособленных к какому-либо одному роду добываемой пищи, мы встречаемся уже со специализацией, т. е. с приспособлениями к узким, определенными не вари-ирующим условиям среды. Эти специализации, о которых мы будем подробно говорить ниже, мы можем рассматривать как частные случаи идиоадаптации.
СПЕЦИАЛИЗАЦИЯ КАК ЧАСТНЫЙ СЛУЧАЙ ИДИОАДАПТАЦИИ
Особым видом идиоадаптаций являются так называемые специализации свободно живущих Metazoa, примеры которых легко можно найти среди любой группы беспозвоночных и позвоночных животных. Особенно характерные примеры специализации обычно встречаются среди таких форм, которые приспособились к каким-либо не особенно сложным, но
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
305
очень постоянным условиям окружающей среды. Подобные формы стоят обычно как бы в стороне от своих ближайших! сородичей, эволюирующих в направлении общих идиоадаптаций. Сюда откосятся животные, приспособившиеся, например, к жизни в глубоких пещерах, к жизни в земле, к жизни на отвесных, недоступных другим животным скалах, к жизни на тонких ветвях высоких деревьев и т. д.
Среди млекопитающих типично специализованными формами являются ленивцы (Choloepus, Bradypus, рис. 84), приспособившиеся к древесной жизни на высоких ветвях. Как известно, ленивцы живут в густых лесах Южной Америки; большей частью они висят неподвижно, ухва-
Рис. 84. Ленивец (Bradypus tridactylus). (Из Брэма.)
тившись своими сильными и длинными конечностями, снабженными большими изогнутыми когтями, за наиболее высокие ветви, обычно недоступные даже для хорошо лазающих хищников; когда им приходится передвигаться, что случается довольно редко, они двигаются очень медленно и незаметно; кроме того, они защищены от врагов покровительственной окраской своего волосяного покрова. Так как ленивцы питаются почками и плодами деревьев, на которых живут, то обычно они совсем не спускаются на землю и пребывают в течение всей своей жизни в этом недоступном для их врагов месте обитания. Вся организация ленивцев: своеобразие строения их органов движения, свойства и окраска меха, общий вид, напоминающий собой повиснувшие кучки мха, привычка висеть неподвижно или двигаться очень медленно, способность Морфологические закономерности 20
306
ГЛАВА X
выдерживать длительную голодовку,—все это вместе ^взятое является рядом приспособлений животного к его особому, защищенному от врагов месту обитания. У нас имеются все основания предполагать, что ленивцы представляют собой последние пережитки некогда богатой представителями фауны Edentata, в свое время населявшей Южную Америку и сохранившейся вплоть до наших дней исключительно благодаря специализации, т. е. благодаря идиоадаптивным изменениям своей, организации, приспособленной к постоянным условиям своеобразного, недоступного врагам места обитания. Отметим, что ленивцы, на ряду с только что перечисленными нами признаками специализации, сохранили и целый ряд признаков весьма примитивного строения.
Разбирая характер только что упомянутых нами приспособительных видоизменений ленивцев, мы узнаем в них типичные черты идиоадапта-
Рис. 85. Геккон (Gecko verticellatus), ползающий по вертикальной стеклянной поверхности (по фотографии Коха).
ций, т. е. приспособлений к строго определенным условиям своеобразнойсреды, позволяющих животным безопасно существовать именно в этой среде. Общее строение организма, несмотря на приобретение этих новых признаков, не становится от этого выше— наоборот, в организации ленивцев встречаются, как мы об этом только что упоминали, многие анцестральные признаки строения их предков—примитивных третичных Edentata. Сохранение анцестральных признаков можно приписать только постоянству условий древесной среды, которая после переселения потомков третичных Edentata все время пребывала неизменной. Резко выраженные идиоадаптивные признаки ленивцев (конечности, когти, волосяной покров) позволили им дожить вплоть до настоящего времени, в то время как большинство родственных им форм среди неполнозубых погибло в борьбе за существование.
Весьма вероятно, что и многие другие современные нам неполнозубые (муравьеды, броненосцы, ящеры, трубкозуб) сохранились до настоящего времени только благодаря специализации.
направления филогенетического развития животных
307
В качестве примеров специализации среди рептилий можно привести гекконов (рис. 85) и хамелеонов (рис. 86). Как известно, гекконы приспособились к жизни на отвесных утесах и скалах, где они стали недоступны обычным врагам ящериц; при этом у них развился приспособительно к этим новым условиям жизни ряд идиоадаптаций: покровительственная окраска и плоская форма тела, присоски на конечностях и т. д., но вместе с тем у них сохранился и ряд примитивных анцестральных признаков (амфицельные позвонки, строение кожи и т. д.). Хамелеоны (рис. 86) великолепно приспособились к жизни на высоких и тонких ветвях, недоступных их врагам. Соответственно этому у них развились следующие идиоадаптации: весьма характерные, напоминающие собой клешню, хватательные конечности, покровительственно меняющаяся, окраска, своеобразно приспособленная к месту обитания форма тела, чрезвычайно подвижные глаза, быстро выбрасывающийся, снабженный приспособитель-
Рис. 86. Хамелеон (Chamaeleo vulgaris). (Из Брэма.)
ным аппаратом язык1 и медленность движений. На ряду с этим у них сохранились также и некоторые черты примитивного строения предков, исчезнувшие у других видов ящериц. Такими чертами являются строение чешуй, короткая шея, положение plexus brachialis и другие менее существенные признаки.
Мы легко могли бы привести еще ряд примеров специализации животных, но нам кажется, что и сказанного вполне достаточно, чтобы сделать понятным для читателя различие, существующее между идиоадаптацией и специализацией. Под именем .щдиоадаптаций мы подразумеваем такие морфологические изменения организкцщцкйвотных, с помощью которых данный вид приспособляется к разнообразным рд сложным изменениям окружающей среды, что позволяет ему сохранитьсяТГббрьбе-за'сущёствб" ваниеГНсГесли какой-либо вид животных переселится в среду с п он о-явными условиями, где он вместе с тем будет защищен от большинства врагов, угрожающих его сородичам, то обычно" достаточно будет развиться лишь'ТГёТГТГо "г""и м_ ка-ч е с тве р н:о~;зТнГачи т е л ь н ы м ид ио-ада пт'а'Дй я м (с пе ц и а л и з а ц и я), чтобы обеспечить ему“победу
1 S е we г t z о f f S. A., Zur Entwicklungsgeschichte cpr Zunge bei den Reptilien, Acta Zool. Bd. 10, 1929.
2 J*
308
ГЛАВА X
в борьбе за существование. При этом данный вид сохраняет, как общее правило, в неизменном состоянии многие черты примитивного строения своих предков. Эти черты первичного строения исчезают у огромного большинства сородичей специализованного вида в тех сложных и разнообразных условиях окружающей среды, в которых им приходится жить, и заменяются новыми идиоадаптациями (редукция анцестральных признаков). Мы видим, таким образом, что специализация является лишь особого рода разновидностью обычных идиоадаптаций и что главное отличие между ними заключается лишь в различии внешних условий существования, в которых приходится жить животным, ив степени изменения организации и функций самых животных. •
СООТНОШЕНИЯ МЕЖДУ АРОМОРФОЗАМИ И ИДИОАДАПТАЦИЯМИ
Если мы теперь, после того как нами установлено различие между эволюцией животных, идущей путем ароморфозов и путем идиоадаптаций, обратимся снова к ходу эволюционного процесса у низших позвоночных, изображенного на нашем родословном древе на схеме XV, и сравним его с диаграммой, изображающей общий ход эволюции (схема XXII), то мы не сможем не заметить, что появление в данной группе животных одного или нескольких ароморфозных признаков, хотя и обусловливает собою некоторый подъем их организации, но что эти формы, поднявшись на определенную высоту строения и функций, не идут по пути прогресса далее, пока у них не разовьется новый ароморфоз или группа таковых. Между появлениями двух групп прогрессивных изменений организаций, т. е. между двумя ароморфозами, может пройти весьма значительный промежуток времени, так что процесс прогрессивной эволюции идет в большинстве случаев как бы уступами, причем периоды под ъ.е м а организации чередуется с периодами, когда морфофизиологической прогрессивной эволюции не происходит вовсе.
Периоды, в течение которых не происходит морфофизиологической прогрессивной эволюции в данной группе животных, не означают остановки эволюционного процесса вообще. Тщательно всматриваясь в ход этого процесса, мы видим, что в промежутках между периодами ароморфозов организация животных может изменяться очень сильно и что изменения некоторых форм весьма велики количественно, но что все эти изменения качественно носят характер идиоадаптаций. Периоды ароморфоза представляют собой как бы узловые точки эволюционного процесса, после которых начинается процесс усиленной идиоадап-тивной радиации, когда данная прогрессивная группа распадается на большое число дочерних групп, характеризующихся адаптивными признаками.
Анализ примеров адаптивной радиации указывает нам на то, что вслед за периодом ароморфоза, когда развиваются те морфофизиологические признаки, которые подымают общую жизнедеятельность данных форм, следует период идиоадаптив-ного филогенетического развития, во время которого потомки ароморфозно изменившейся прародительской формы расширяют свой географический ареал и- приспосабливаются к наступающим при этом новым и разнообразным условиям
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
309
окружающей среды. Во время этого периода дивергентной адаптации все ароморфозные признаки животных остаются без изменения (они становятся признаками высших систематических групп: семейств, отрядов, классов), но нар яду с ними появляются новые приспособительные признаки,1 и только в отдельных, очень немногих, случаях ароморфозные признаки исчезают совсем (редукция зубов у китов и т. д.).
Таким образом, обычно вслед за периодом аромор-фозов следует не новый период ароморфозов, но период дивергентной идиоад aji т а ц и и.
Это общее правило можно было бы подтвердить многими случаями, взятыми из филогенеза многоклеточных животных. Не повторяя только что приведенных нами примеров млекопитающих и рептилий, у которых период идиоадаптивной радиации наступил после периода накопления целого ряда ароморфозов, упомянем еще об идиоадаптивной радиации, сменившей период ароморфоза и имевшей место в группе эласмобранхий. Так, рассматривая филогенетическое развитие потомков Chondrichthyes selachoidei, мы можем с полной уверенностью сказать, что от этой группы примитивных рыб отделились, по крайней мере, две ветви, из которых одна (Osteichthyes) пошла по пути прогрессивной эволюции,1 2 а другая остановилась в своем прогрессивном развитии и, сохраняя весьма примитивную организацию, вполне благополучно дожила до нашего времени (Elasmobranchii). Эта остановка на’ пути прогрессивной эволюции отнюдь не сопровождалась остановкой эволюционного процесса вообще: элас-мобранхии в течение всего своего существования, т. е. с силура до наших дней, продолжали изменяться и дали начало большому числу форм, из которых некоторые более или менее специализировались, приспособившись к определенным, в общем весьма разнообразным, условиям жизни, другие же не проявили признаков узкой специализации. Представителями специализованных форм среди эласмобранхий могут считаться скаты, организация которых испытала сильные и в количественном и в качественном отношениях изменения в связи с донным образом жизни. Спе-циализованными формами являются также Holocephali с их приросшим к осевому черепу нёбноквадратным хрящом, измененными зубами и т. д.
Но особенно интересным для нас в данном случае являются те акуловые рыбы, которые в общем не специализировались, но тем не менее сохранили в сравнительно неизмененном виде целый ряд признаков своей примитивной организации3 и вместе с тем живут в той же обстановке, в какой обитают и высшие рыбы (Teleostei), и ведут тот же образ жизни, что и эти последние. Здесь мы имеем типичный пример того, что морфофизиологический прогресс, или ароморфоз, является лишь частным случаем общебиологического прогресса: акуловые рыбы остановились на пути ароморфоза, но приобрели некоторые биологические особенности (главными из них я считаю указанные выше особенности, касающиеся их способа развития), которые позволили им выдержать конкуренцию с гораздо более высокими формами рыб. Но и здесь эволюционный процесс не остановился; группа акуловых рыб в течение своего существования распалась на целый ряд более или менее различающихся между собой форм (идиоадаптивная радиация). Для того чтобы представить себе, насколько в некоторых случаях различия в организации,
1 Лопасти у ихтиозавров и китов, крылья у птерозавров и летучих мышей и т. д.
2 Точнее, некоторые члены которой эволюировали прогрессивно.
3 Вероятно, акуловые рыбы и скаты сохранили примитивные признаки в своей организации в значительной степени благодаря развившимся у них ценогенезам, но различия между отдельными группами акул и скатов имеют характер идиоадаптации.
310
ГЛАВА X
а, следовательно, и эволюционные изменения акуловых рыб, велики^ достаточно вспомнить строение хотя бы таких форм, как пила-рыба (Pris-tis antiquorum) или молот-рыба (Sphyrna zygaena) и т. д.
Итак, мы пришли к выводу, что обычно вслед за периодом ароморфозов следует период дивергентной эволюции (адаптивной радиации—по терминологии Осборна, идиоадаптивной радиации—по нашей терминологии). При этом ареал распространения животных расширяется, и они начинают приспособляться к новым, часто весьма разнообразным и своеобразным условиям окружающей среды. Это приспособление осуществляется путем идиоадаптации, и мы можем сказать совершенно определенно, что здесь мы имеем перед собою обычнейший и наиболее распространенный тип филогенетического изменения животных. Само собою разумеется, все идиоадаптивные изменения эктосоматических органов являются биологически (экологически) прогрессивными изменениями, обеспечивающими виду, у которого они являются, увеличение численности индивидуумов, больший ареал распространения и образование новых соподчиненных систематических групп у потомоков.
Таким образом, в прогрессивной эволюции животных всегда встречаются' периоды ароморфозов, и эти периоды сменяются периодами идиоадаптации.
Теперь мы дожны поставить вопрос: встречаются ли идиоадаптации также и во время периодов ароморфоза у ароморфозно изменяющихся животных? Иными словами, встречаются ли идиоадаптации одновременно с ароморфозами?
Я думаю, что на этот вопрос нам придется дать утвердительный ответ, ибо нам не известно ни одной животной формы ни в один из периодов ее филогенетического существования, когда у нее не было бы тех или иных идиоадаптаций. Быть может, скорее нам следует изменить самую постановку вопроса и спросить себя: какого рода идиоадаптации могут развиваться у животных во время периода ароморфоза?
Мы знаем, что главным признаком ароморфозов является повышение активных функций эволюирующих форм или, что то же, подъем общей энергии жизнедеятельности организма. Поэтому в течение периода ароморфоза Moryf развиваться только такие идиоадаптации, которые не препятствуют, так сказать, не стоят на пути развития активных функций данной ф о р м ы: все же иные типы идиоадаптации препятствовали бы параллельно идущему образованию ароморфозов и отклоняли бы животное от пути морфофизиологического прогресса.
Представляется довольно вероятным, что такого рода факты действительно имели место в истории развития видов. Так, когда в эволюции некоторых форм млекопитающих появились идиоадаптации, позволившие нм перейти к малоподвижному образу жизни, как это, например, имело место у лазающих предков ленивцев, которые, благодаря некоторым особенностям строения конечностей, стали очень медленно передвигаться по веткам, то мы смело можем сказать, что такого рода идиоадаптации несомненно препятствовали этим формам эволюировать дальше в направлении морфофизиологического прогресса.
То же, конечно, относится и к изменениям, происшедшим в строении тела, и, в особенности, в строении конечностей у хамелеонов (рис. 86), также приспособившихся к медленному передвижению по очень тонким веткам, среди густой листвы. Морфологические и физиологические изменения этих форм, точно так же как и выработавшаяся у них привычка (инстинкт) неопределенно долгое время спокойно держаться в одном и том же положении на одном и том же месте, конечно, для них полезны, тем более, что они при этом защищены от врагов, благодаря своей способности
направления филогенетического развития животных
311
приспособительно менять окраску и пребывать на таких высоких и тонких ветках, доступ к которым для огромного большинства хищников совершенно невозможен. Однако, к филогенетическому подъему жизненной энергии хамелеоны, повидимому, уже более неспособны. То же можно сказать и о многочисленных роющих животных (амфисбены, гимнофионы и т. д.).
Мы видим, таким образом, что многие специальные идиоадаптации, действительно, могут парализовать дальнейшую эволюцию ароморфозов, если они не находятся с ними в соответствии. Но нам хорошо известен целый ряд идиоадаптаций, которые никоим образом не препятствуют направлению морфофизиологического прогресса в эволюции. Так, например, помимо различного рода покровительственных окрасок и форм, отнюдь не препятствующих развитию ароморфозов, некоторые идиоадап-тивные изменения органов движения лазающих животных (обезьян, гекконов и т. п.) не только не парализуют интенсивности движения животных, у которых они появляются, но, наоборот, в значительной степени поднимают ее. Мы это наблюдаем особенно часто при таких идиоадаптивных изменениях конечностей, когда общее число возможных движений органа увеличивается (см. ниже принцип расширения функций Плате), как это бывает, например, при развитии хватающей конечности у обезьян, длинной шеи у рептилий и у птиц, своеобразных плавников у кистеперых и двоякодышащих рыб и т. д.
Эколого-эволюционные исследования показывают нам, что направление идиоадаптации является самым распространенным направлением эволюции; мы можем с полной уверенностью сказать, что огромное большинство животных выдержало борьбу за существование только благодаря идиоадаптациям.
Сводя воедино все сказанное, можно отметить следующее: 1) п 'е р и о д ароморфозов характеризуется тем, что в: это время развиваются такие морфофизиологические изменения организации, при которых интенсивность активных функций повышается, и такие идиоадаптации, которые этому повышению не препятствуют; 2)пе риод идиоадапта-ц и й характеризуется тем, что в это время развиваются одни только идиоадаптации; эти идиоадаптации изменяют строение животных, но не поднимают интенсивности их жизнедеятельности, так что общая высота жизнедеятельности и общая сложность строения организма животных вэ том случае у потомков остается натом же уровне, на котором она была у предков.
ЭМБРИОНАЛЬНЫЕ ПРИСПОСОБЛЕНИЯ ИЛИ ЦЕНОГЕНЕЗЫ
Третье направление биологически прогрессивной эволюции мы обозначили как направление ценогенеза или эмбриональных приспособлений организма. Ценогенеза ми мы называем такие эмбриональные или личиночные приспособления, которые развиваются в организме в течение онтогенеза, а затем исчезают, но которые непосредственно полезны развивающемуся организму. Сюда относятся также все случаи более раннего начала функционирования эмбриональных органов, например, функционирования сердца у зародышей: сердце начинает работать еще во время эмбрионального развития, что имеет большое значение для самого процесса онтогенетического развития.
312
ГЛАВА X
Мы употребляем здесь слово ценогенез в том смысле, в котором первоначально употреблял его Э. Геккель, когда вводил этот термин в лите-
ратуру, т. е. мы принимаем, что ценогенезы являются полезными эмбриональными приспособлениями. Как известно, позднейшие исследователи (Менерт, Кейбель и др.) обозначали термином «ценогенез» вообще все уклонения анцестрального хода эмбрио-
нального развития животных, даже такие, которые не имели для зародыша
Рис. 87. Яйцо акулы со стекловидной оболочкой и с длинными нитями оболочки, при помощи которых оно прикрепляется к тонким ветвям коралла (из Нила, 1936).
никакого биолого-физиологического значения. Так, они называли ценогенезами различные явления гетерохронии и гетеротопии, явления, важные для образования органов взрослых животных, но не имеющие никакого значения для жизни самих зародышей. Ценогенезами считались также всякие увеличения или уменьшения величины эмбриональных закладок (см. ниже явления положительного или отрицательного архаллаксиса), безразличные как таковые для жизни зародышей, но имеющие важное значение для прогрессивного развития взрослых форм. С нашей точки зрения все эти явления и признаки не являются эмбриональными приспособлениями организма и, стало быть, не являются ценогенезами.1
Мы знаем, что в каждом многоклеточном организме в течение его эмбрионального развития обязательно появляются различные приспособления к специфическим условиям его зародышевой или личиночной жизни, т. е. истинные ценогенезы. Приведем некоторые хорошо известные примеры таких ценогенезов.
Несомненными ценогенезами являются различные виды яйцевых оболочек, защищающие зародышей от различного рода механических повреждений, как, например, всем известные известковые оболочки яиц рептилий и птиц, или стекловидные оболочки яиц акул с их длинными нитями,
при помощи которых они прикрепляются к тонким стеблям подводных растений (рис. 87); ценоге-незом является яичный желток (рис. 88 и 89, dt), представляющий собой совершенно готовую пищу для зародышей и личинок во время их эмбрионального развития и избавляющий их от опасных и часто бесплодных поисков подходящей пищи; ценогенезом
является амнион рептилий, птиц (рис. 88, ат) и млекопитающих, защищающий зародышей этих животных от толчков и ударов, которые иногда прихо-
дится выдерживать материнскому организму; ценогенезом является аллан-
1 Сам Геккель причислял к числу ценогенезов явления гетерохронии и гетеротопии: он считал, что все эти «исказители палингенезов» являются «эмбриональными приспособлениями». Однако теперь, когда гетерохронии и гетеротопии стали лучше известны, оказалось, что большинство из них не имеет никакого физиологического или биологического значения для самих эмбрионов: таким образом, они не могут считаться ценогенезами в геккелевском смысле этого слова. Поэтому нам думается, что, во избежание недоразумений, лучше было бы обозначить их каким-либо другим термином (архаллаксис, девиация и т. д.; см. ниже, гл. XIV).
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
313
тоис тех же животных с его богатым сплетением эмбриональных кровеносных сосудов (рис. 88, al), а также плацента млекопитающих, являющаяся, как и аллантоис, органом эмбрионального дыхания животных; таким же органом личиночного дыхания и, стало быть, таким же ценогене-
Рис. 88. Эмбрион цыпленка с тремя зародышевыми придатками.
ат—амнион; al—аллантоис; dt—желточный мешок (из Кольмана).
Рис. 89. Эмбрион гимнофиона (Ichtyophis glutinosus) с наружными жабрами и кожными органами чувств (из Сарацина).
зом являются и длинные наружные жаберные нити, встречающиеся у зародышей селахий (рис. 72 А) и у личинок амфибий (рис. 89); ценогене-зом является, наконец, твердая оболочка хризолид и различного рода эмбриональные кровеносные сосуды зародышей рыб, рептилий, птиц и млекопитающих.
Очень интересные эмбриональные приспособления найдены С. Г. Кры-жановским1 при исследовании им органов дыхания у личинок различных
Рис. 90. Личинка Hemichromis вскоре после вылупления.
Стадия максимального развития дыхательных сосудистых сетей в преданальном и анальном плав’ никах. Рисунок с живого малька, с. о.—органы приклеивания; d. cuv.—ductus cuvteri; md.—arcus mandibularis aortae; sp.—spiraculum; v. c. i.—vena caudaUs inferior; v. si.—vena subintestinalis (из Крыжановского, 1933).
рыб. На рис. 90 изображена личинка Hemichromis вскоре после вылупления из яйца; мы видим, что ее анальный и преданальный плавники сплошь пронизаны густой сетью кровеносных сосудов, совершенно отсутствующих во взрослом состоянии. Крыжановский пришел к выводу, что оба эти плавника у эмбриона Hemichromis являются органами дыхания личинок
’Крыжановский С. Г., Органы дыхания личинок рыб (Teleostomi) и п'ев-добранхия, Труды лаб. гвол. морф. Акад. Наук СССР, т. 1, ЮЧ9.
314
ГЛАВА X
сот. ' hu(d1
Рис. 91. Голова личинки Amia (реконструкция по сагиттальным срезам). Сосудистая сеть в жаберной крышке, образованная гиоидной дугой аорты.
com.—conassura hyoideo-mandibularis; с. тес. меккелев хрящ; hy(d)—arcus hyoideus aortae, pars dorsalis; hy (v)—arcus hyoideus aortae, pars ventralls; md—arcus mandibularis aortae (из Кры-жанивского, 1933).
' в тот период, когда у них не развились еще настоящие жабры. По мере развития этих последних кровеносные сосуды в обоих плавниках постепенно облитерируются. Еще более интересным примером в этом отношении является развитие эмбриональных кровеносных сосудов у Amia. На рис. 91 изображена ранняя стадия развития головы личинки Amia, у которой жаберная крышка покрыта густою сетью дыхательных кровеносных сосудов, отходящих от гиоидной дуги аорты. Таким образом, у Amia жаберная крышка является органом личиночного дыхания. Эта сосудистая сеть на жаберной крышке по мере развития у Amia настоящих жабр постепенно редуцируется.
Внимательно изучая все эти эмбриональные приспособления, мы видим, что каждое из них усиливает главные функции развивающегося организма и является поэтому совершенно необходимым для того животного, у которого оно встречается.
Общее число известных нам це-ногенезов весьма велико. Если мы попытаемся классифицировать их по группам, то увидим, что одну группу образуют ценогенезы, являющиеся защитой зародышей и личинок от врагов, особенно опасных для маленьких, слабых организмов, лишенных возможности активно защищаться в этот ранний период их развития. К этой группе относятся все виды приспособительных окрасок зародышей: своеобразная пигментация яиц и личинок амфибий, соответствующая окраске окружающей среды, прозрачность неразличимых в воде яиц и личинок ланцетника и целого ряда других беспозвоночных и позвоночных животных (в том числе многих костистых рыб), зеленая окраска живущих на листьях личинок насекомых (гусениц), слож
ная расцветка гусениц бабочек, живущих на стволах и ветвях кустарников и деревьев, все случаи окрасок, подражающих разнообразным неодушевленным предметам и столь характерных для личинок различных насекомых, наконец, все твердые, не пропускающие дневного света оболочки яиц насекомых, известковые скорлупы яиц многих рептилий и всех птиц и т. д.
Не надо при этом забывать, что все эти часто сложнейшие защитные приспособления зародышей и личинок образуются и функционируют именно в те периоды жизни и только в те периоды жизни, когда они бывают ‘ •особенно нужны, и что они начинают редуцироваться, когда надобность в них исчезает. Так, например, обычная специфически покровительственная окраска скорлупы птичьего яйца держится лишь то время, пока зародыш питается желтком этого яйца; к концу этого периода, ко времени развития яйцевого .зуба, она исчезает вместе со скорлупой яйца и заменяется другими эмбриональными приспособлениями (эмбриональными инстинктами птенцовых и выводковых птиц).
Вторую группу эмбриональных приспособлений представляют собою те ценогенезы, которые предназначены для питания развива ю-
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
315
щи х с я зародышей и личинок. Сюда относятся: желток голобластических и, в особенности, меробластических яиц, а также различные эмбриональные органы, например кровеносные сосуды, служащие для абсорбции этого желтка в меробластических яйцах (желточные сосуды); сюда же можно причислить и различные органы личиночного питания, например роговые челюсти личинок бесхвостых амфибий (головастиков) и многие другие.
Т ретью группу ценогенезов составляют эмбриональные или личиночные органы дыхания, например длинные наружные жаберные нити зародышей селахий (рис. 72 А) и личинок амфибий (рис. 89), многочисленные сосудистые сплетения у эмбрионов костистых рыб, сосудистая система аллантоиса рептилий, птиц (рис. 88) и млекопитающих, плацента млекопитающих и т. д.
Упомянем, наконец, об эмбриональных почках позвоночных (головных почках), служащих в качестве экскреторных органов для зародышей.
Разделяя ценогенезы на группы, мы в основу своей классификации положили функциональный принцип, т. е. подразделили ценогенезы на группы с точки зрения их функций: защиты, питания, дыхания.
Б. С. Матвеев (1936)1 классифицирует ценогенезы, которые он называет эмбриоадаптациями, с иной точки зрения, а именно, с точки зрения их происхождения. Он различает пять групп эмбриональных приспособлений.
1. Приспособления к зародышевой жизни, полученные через материнский организм и связанные с эволюцией материнского организма, а не зародыша: сюда относятся, например, слизистые оболочки икринок рыб и амфибий,известковые оболочки яиц рептилий и птиц, плацента млекопитающих, желток яйца и т. д.
2. Приспособления зародышей, связанные с эволюцией самих зародышей и выработавшиеся в течение эволюции онтогенеза путем преобразования уже имеющихся у зародышей органов (примером могут служить различные типы желточных мешков, обусловливающие различные формы движения мальков костистых рыб).
3. Новые провизорные органы, развивающиеся самостоятельно у заро-дыша и не сохраняющиеся у взрослых форм; сюда относятся, например, различного рода присоски, развивающиеся у мальков рыб и личинок амфибий и исчезающие у взрослых животных, а также амнион, аллантоис и т. н.
4. Регулируемые гетерохронии, т. е. временные ускорения и временные замедления в развитии органов зародышей; сюда относится ускоренное развитие плавников у многих рыб, мальки которых приспособйлись к пелагическому образу жизни (например, у многих пелагических рыб брюшные и грудные плавники развиты очень сильно, но затем они приостанавливаются в развитии и у взрослых приобретают нормальную величину).
5. Древние палингенетические признаки, которые сохраняются у зародыша, потому что органы имеют у него специфическую функцию, нужную организму в течение зародышевой жизни и не сохраняющуюся у взрослых форм.
Итак, мы видим, что главные функции организма у эмбрионов и личинок выполняются особыми ценогенетическими, т. е. развивающимися у зародышей, органами, и что эти органы позднее редуцируются. Все эти часто весьма сложно построенные ценогенетические органы развиваются в течение эмбрионального или личиночного периода жизни животного и начинают атрофироваться либо у зародышей и личинок, либо уже у молодых животных, так что у взрослых форм от них не остается ни малейшего
1 Матвеев Б. С. Современные задачи эволюционной морфологии. Изв. Ак. Наук СССР, Серия биолог. № 5, 1936.
316
ГЛАВА X
следа. Биологическое значение этих чисто эмбриональных, исчезающих у взрослых форм приспособлений весьма велико: они дают возможность зародышам и позже молодым личинкам перенести без особого урона наиболее опасный период их индивидуального существования, тот период, когда молодые животные еще малы и беззащитны. С этой точки зрения понятно, почему развитие каждого нового эмбрионального признака так важно для жизни вида: оно позволяет достигнуть взрослого состояния большему числу особей, чем это имело место у предшествовавших поколений. Другими’ словами, каждое новое эмбриональное приспособление обеспечивает выживание большего числа особей, чем это имело место у предков, у которых это приспособление еще не было развито; таким образом увеличивается общее число взрослых индивидуумов, образующих данный вид.
В самом деле, представим себе существование определенного вида животных в определенной местности, причем среднее количество индивидуумов здесь остается неизменным, так что данный вид экологически не развивается дальше, но и не вымирает; если у зародышей данного вида разовьется какое-либо важное эмбриональное приспособление (ценогенез), вследствие которого деятельность одной из главных функций зародыша, например функции питания, дыхания или защиты от врагов, повысится, то увеличится число зародышей или личинок, доживающих до взрослого состояния, а тем самым и число взрослых особей, населяющих данную область. В результате имеет место перенаселенность данной формы в данном географическом ареале, что влечет за собой расселение отдельных особей данного вида по новым ареалам. Возникают новые идиоадаптации, соответствующие новым условиям существования, что в свою очередь ведет к образованию новых дочерних видов. Причины этих биологически прогрессивных изменений лежат не в изменении строения взрослых предков данной формы, но в только что упомянутых эмбриональных приспособлениях (ценогенезах) потомков, приспособлениях, исчезающих у взрослых животных, но делающих возможным значительное увеличение их числа.
Можно принять, что большое количество примитивно построенных форм, например примитивные акулы, вероятно, многие примитивные рептилии (например Sphenodon) смогли дожить до наших дней главным образом благодаря своим эмбриональным приспособлениям.
МОРФОФИЗИОЛОГИЧЕСКИЙ РЕГРЕСС ИЛИ ОБЩАЯ ДЕГЕНЕРАЦИЯ
Результаты филогенетических, биологических и систематических исследований приводят нас к выводу, что общее регрессивное направление эволюции часто ведет в конечном итоге к биологическому (экологическому) прогрессу вида. Мы знаем, что целые группы дегенерировавших форм, например все оболочники, ленточные черви, усоногие раки и многие другие, увеличили в течение филогенеза число особей, составляющих данный вид, расширили свой ареал распространения и распались на соподчиненные систематические категории (дочерние. виды).
Следует различать «общую дегенераци ю», т. е. общий регрессивный процесс, обусловливающий филогенетическое развитие животного, как, например, общая дегенерация, наблюдающаяся у сидячих форм и у эктопаразитов, и «частичную дегенераци ю», или редукцию органов—явление, встречающееся при ароморфозах и идиоадаптациях; об этой «частичной дегенерации» речь будет впереди в главах, посвященных теории филэмбриогенеза и филогенетической редукции органов. В настоящий момент мы имеем в виду лишь общую дегенерацию.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
317
При регрессивной эволюции какой-либо животной формы мы находим, что одна система органов развивается прогрессивно, в то время как другая редуцируется. При этом обычно редуцируются органы, несущие активные функции, а прогрессивно развиваются пассивные, но важные для животного органы или системы органов, так что мы можем сказать, что общая дегенерация является противоположностью морфофизиологического прогресса (ароморфоза). Так, например, у многих дегенерировавших форм более или менее редуцированными (у сильно дегенерирующих форм целиком редуцированными) являются органы движения, т. е. вся мускульная система и те части скелета, которые непосредственно служат для движения (скелет конечностей). При этом обычно редуцируется также
и центральная нервная система со всеми нервами и органами чувств, а также органы активного питания, и, наоборот, прогрессивно развивается половая система. У таких форм функция движения и активного расселения переносит-
0V
Рис. 92. Асцидия. Схематизированный продольный разрез.
ап—анальное отверстие; atr.—околожаберная или перибранхиальная полость; Ьг.—жаберные отверстия; еп.—эндостиль; h—сердце; G-—нервный узел; int— кишечник; ov.—яичники; ts.—семенники (из Боаса).
Рис. 93. Схема строения одного из представителей стебельчатых усоногих (Lepas), у которого правая сторона раковины и находящаяся под нею мантия удалены.
с—carina; cd—цементная железа, открывающаяся у основания первой антенны; cf—усоножки; m — мускул, замыкающий scuta; 1—печеночная часть кишечника; od— яйцевод; ov—яичники; р—penis; sc—scutum; te—tergum; ts—семенники; vd.— vas deferens (из Клауса, по Гертвигу).
ся со взрослых форм на свободно плавающие личинки, у которых функция движения интенсифицируется. В иных случаях, например у многих паразитов, распространение вида происходит при помощи свободно движущегося хозяина паразитирующего животного.
Вероятными причинами общей дегенерации животных являются, во-первых, переход от свободно подвижного образа жизни к сидячему или неподвижному образу жизни и, во-вторых, переход от самостоятельного образа жизни к паразитизму.
Исследования экологии сидячих форм, например многих кишечнополостных (Hydrozoa, Anthozoa, Madreporaria), большей части оболочников (рис. 92), усоногих раков (рис. 93) и паразитических форм [лен
318
ГЛАВА X
точных червей (рис. 94), некоторых паразитических ракоооразных (рис. 95 ) и др.], с несомненностью говорят о том, что все эти группы животных развиваются в направлении биологическогопрогресса: здесь отдельные виды представлены большим, числом индивидуумов, а крупные систематические группы—большим числом родов, видов и т. п., имеющих широкий ареал распространения. Мы можем считать достоверно установленным, что многие сидячие (сессильные) дегенерировавшие формы принадлежат к числу наиболее цветущих групп животного мира.
Можно считать также установленным, что сидячие и паразитические группы животных произошли от свободно двигающихся и самостоятельно питающихся форм. Мы знаем, что дегенерация сидячих форм выражается главным образом в редукции органов движения, а также в атрофии всех тех органов, которые служат для отыскания и преследования добычи. Многие формы, как, например, обо-
Рис. 94. Caryophyllaeus mutabilis. D—желточники; h—семенники; ov—яичники; р—конуляционный сосочек; ut—матка; vd—семенной проток; vs—семенной пузырь; w—выделительные каналы. Обратить внимание на отсутствие пищеварительной н нервной системы и на чрезвычайное развитие мочеполовой системы у этой типичной энтопаразитической формы (из Боб-рецкого).
лочники (Tunicata), которые, вероятно, перешли от активного способа питания к пассивному, не обладают, как это видно из рис. 92, органами, приспособленными для захватывания и удержания добы-
v 3J
Рис. 95. Sacculina carcini, прикрепившаяся к нижней стороне брюшка обыкновенного краба (Carcinus maenas).
а—антенна краба;''d—задний проход кра-ба; m—отверстие раковины; о—глаза краба; г—корневидные отростки саккулины, внедряющиеся во внутренности хозяина, оставляя свободной жаберную область; s— стебелек (по Делажу из Гертвига).
чи; к тому же они утратили и значительную часть центральной нервной системы, а также органы высших чувств.
У морских уточек (рис. 93) питание также происходит пассивно при помощи ритмических движений усоножек (рис. 93, cf.), пригоняющих к ротовому отверстию вместе с током свежей воды взвешенные в ней мелкие организмы, служащие пищей для этих сидячих стебельчатых форм.
То же можно сказать и о некоторых энтопаразитах, у которых редукция пошла дальше, чем у сидячих форм. У эктопаразитов атрофированными оказываются не только органы движения и органы захватывания пищи и добычи, но также и весь пищеварительный тракт со всеми его придатками, как это, например, имеет место у гвоздичников среди ленточных червей (рис. 94), паразитирующих в кишечном канале карповых рыб, или у саккулин среди усоногих раков, паразитирующих на нижней стороне хвоста
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
319-
обыкновенного краба (рис. 95). Питание этих форм происходит эндосмотиче-ским путем при помощи всей кожной поверхности тела животного. Все эти регрессивные изменения, столь резко упростившие строение перечисленных выше форм, компенсируются целым рядом новых прогрессивных изменений (иногда весьма сложного характера) в половой системе животного, а также многими эмбриональными изменениями (ценогенезами) зародышей и личинок.
Причина всех этих на первый взгляд столь поразительных изменений до известной степени нам понятна. Мы знаем, например, что во многих случаях функция защиты, в особенности, когда у животного много опасных врагов, имеет первостепенное значение для жизни вида; в подобных случаях у животных развивается прочный наружный скелет, защищающий его от нападения врагов. Понятно, что для преследуемого животного резистентность наружного скелета и связанная с этим толщина его имеют огромное значение. Однако увеличение толщины наружного скелета влечет за собой увеличение общего веса тела животного, вследствие чего (ceteris paribus) уменьшается его подвижность, так что животное с течением времени начинает передвигаться все медленнее и все с большим и большим трудом. Если тяжесть наружного скелета с течением эволюции будет постоянно увеличиваться, то можно легко представить себе, что наступит, в конце концов, момент, когда животное из медленно двигавшегося превратится сначала в факультативно, а затем и в постоянно сидячую форму. При этом у него разовьются адаптации к пассивному питанию. Весьма вероятно, что многие моллюски (Lamellibranchiata) и асцидии пошли именно по этому пути и что у их предков переход от свободно подвижного к сидячему образу жизни был вызван именно недостаточностью активных органов защиты и связанным с этим прогрессивным развитием тяжелого наружного скелета.
Наше толкование, однако, не годится для объяснения сидячего образа жизни кишечнополостных животных, у которых, как известно, имеются стрекательные капсулы, являющиеся прекрасными органами защиты от нападения врагов. Существование большой группы морских анемон указывает нам на то, что капсулы эти являются вполне достаточной защитой даже для совершенно голых форм, и тем не менее морские анемоны все же перешли к сидячему образу жизни. Таким образом, стимулом к переходу к неподвижному образу жизни здесь является не образование тяжелого наружного скелета; сидячий образ жизни в данном случае представляет, невидимому, какие-то другие, особые преимущества, о настоящей природе которых мы пока еще судить не можем.
Не менее трудным является вопрос о стимуле, обусловливающем собою переход животных к паразитическому образу жизни. С известной долей вероятности можно предположить, что предки энтопаразитов были некогда эктопаразитами, вернее сказать, факультативными эктопаразитами, т. е. мелкими хищниками, питавшимися соками более крупных животных, к которым им удавалось тем или иным способом прикрепиться. В этих случаях стимулом для перехода к эктопаразитизму, вероятно, явилась, во-первых, возможность получить постоянный источник питания, во-вторых, возможность иметь в лице хозяина постоянную защиту от врагов. Мы хорошо знаем, что всякий паразит, поскольку он не вызывает смертельного заболевания своего хозяина, защищен от врагов в такой же мере, как и его хозяин, значительно превосходящий его в силе, крепости и размерах.
Результатом паразитизма и сидячего образа жизни явилось то, что органы, служащие для отыскивания пищи и для свободного передвижения животных в пространстве, стали ненужными и вследствие этого дегенерировали. Наши исследования над редукцией органов, как мы это увидим.
320
ГЛАВА X
ниже, подтверждают этот вывод и еще раз показывают, что биологически ненужные органы, т. е. органы, которые уже не являются приспособлениями к изменившимся условиям среды, всегда редуцируются. Это правило целиком .подтверждается теми явлениями, которые мы наблюдаем у паразитов и у сидячих форм: степень редукции обусловлена общим числом ставших ненужными органов и в некоторых случаях является весьма значительной. Вообще можно считать установленным, что у дегенерировавших форм редукция активных органов вполне компенсируется биологически прогрессивными изменениями, наступившими в других органах и в первую очередь—в половых органах и в органах зашиты; она компенсируется также развитием значительного числа биологически важных ценоге незов у эмбрионов и у личинок и увеличением численности потомства.
Выше мы отметили, что основной характер эволюции дегенерировавших форм заключается в атрофии некоторых активных органов и даже целых систем органов; именно этот признак прежде всего бросается в глаза наблюдателю. Более детальное изучение явлений редукции приводит нас, однако, к мысли, что атрофия активных органов у паразитов и у сидячих форм является не самостоятельным процессом, а ,лишь результатом прогрессивного развития целого ряда других органов, функции которых заменили собою функции вышеупомянутых активных органов и сделали излишним, таким образом, само их существование. Это относится как к эволюции органов пассивной защиты и половых органов (большое число потомков, новые приспособления к их защите), так и к эволюции личиночных органов (ценогенезы). Самое характерное для эволюции дегенерировавших форм заключается, конечно, в прогрессивном развитии у них только что упомянутых нами органов, т. е. половой системы и различного рода ценогенезов; ибо именно это и обусловило их победу в борьбе за существование. С равным правом мы могли бы поэтому заменить наш подзаголовок: «Направление общей дегенерации» подзаголовком «Направление прогрессивной эволюции половых и пищеварительных органов, органов пассивной защиты и т. д.»; мы сохраняем, однако, ходячее название потому, что оно короче и удобнее, а также потому, что мы употребляли его уже и раньше.1
О НАПРАВЛЕНИЯХ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНОГО МИРА
В предыдущем мы описали четыре различных направления биологически (экологически) прогрессивной эволюции животных, а именно, направление ароморфозов, направление идиоадаптации, направление ценогенезов и направление общей дегенерации.
В приведенных нами примерах мы старались показать, что в течение морфофизиологической прогрессивной эволюции, т. е. в течение эволюции путем ароморфозов, строение и функции эволюирующих животных усложняются, а интенсивность функций активных органов повышается. Мы показали, что у животных, развивающихся путем идиоадаптаций, появляются многочисленные приспособления к изменившимся условиям окружающей среды, приспособления, часто весьма значительно изменяющие общее строение и функции их органов, но не подымающие организацию измененных этим путем животных на более высокую ступень и не
1Северцов А. Н., Главные направления эволюционного процесса (прогресс, регресс и адаптация), изд. А. В. Думнова, Москва, 1925, 2-е изд., Биомедгиз, 1934.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
321
повышающие общей энергии их жизнедеятельности. Мы видели, что при эволюции путем эмбриональных приспособлений (ценогенезов) у животных изменяются различным образом строение и функции зародышей и личинок, вследствие чего значительно возрастает количество взрослых особей, но не изменяется их строение. Мы видели, наконец, что при эволюции, протекающей в направлении общей дегенерации, у животных изменяются прогрессивно только некоторые системы органов, в то время как большинство активно функционирующих органов или систем органов атрофируется вследствие перехода животных к паразитическому или сидячему образу жизни.
На основании разобранного нами фактического материала нам удалось обособить понятие «морфофизиологический прогресс» («ароморфоз») от понятия «идиоадаптации»; при этом мы пришли к выводу, что существуют не два направления биолого-экологического прогресса (прогресс и регресс), как это прежде думали, но целых четыре. Естественно теперь поставить вопрос о том, что же, собственно, дает нам это новое представление об общих путях филогенеза?
Расчленение понятий ароморфозов, идиоадаптаций и эмбриональных изменений (ценогенезов) помогает нам прежде всего понять, почему очень многие группы весьма примитивных животных не только благополучно дожили до наших дней, но и сейчас не обнаруживают признаков вымирания. Это обстоятельство было необъяснимо с точки зрения прежнего представления об эволюции, когда считалось, что существуют лишь прогрессивная и регрессивная эволюция, причем каждое новое приспособление рассматривалось как прогрессивное изменение организма. С этой точки зрения факт переживания примитивных животных форм, этих живых остатков фауны древнейших геологических эпох, действительно является по меньшей мере парадоксальным.
Обычно принято думать, что древние примитивные формы потому только дожили до наших дней, что они постоянно существовали в неизме-няющихся условиях внешней среды. Однако целый ряд примеров указывает нам на то, что взгляд этот в корне неправилен: о ч ен ь часто случается так, что п р и м итивные формы живут ц т е х ж е г е о г'р а ф ических и эко "логических условиях внешней среды, в каких живут их прогрессив но изменившиеся потомки. Вспомним, например, современных круглоротых или примитивных акул, живущих в тех же условиях среды, что и происшедшие от селахоидных предков костистые рыбы, или примитивных ракообразных, пребывающих в тех же водных бассейнах, что и высоко организованные десятиногие раки и т. п.
Если мы сравним нашу современную фауну с фауной давно прошедших геологических эпох, то увидим, что большинство крупных животных групп развивалось по типу идиоадаптаций и что эти группы—по своей организации и но своим функциям—стоят и теперь не выше, чем их давно вымершие предки. Современная нам фаун а—по общей высоте своей организации и по общим признакам характеризующих ее форм— состоит из форм, принадлежащих к самым разнообразным геологическим эпохам. Так, в любом Морском бассейне с относительно богатой фауной одновременно встречаются представители самых разнообразных геологических эпох: костистые рыбы, крабы и омары—представители современной морфофизиологически прогрессивной фауны, акулы—сравнительно очень мало изменившиеся представители силурийской фауны, плеченогие (Brachiopoda), кораллы, медузы и черви—поныне процветающие пережитки бесконечно отдаленной от нас докэмбрийской фауны; наконец, бесчисленные простейшие—живые Морфологические закономерности 21
322
ГЛАВА X
реликты еще более отдаленного, примордиального (протерозойного) периода жизни .нашей планеты.
Парадоксальный факт одновременного существования и биологического процветания форм, принадлежащих к столь различным геологическим эпохам, может быть нами- понят только в том случае, если мы станем на точку зрения обоснованных выше закономерностей направления эволюционного процесса, иначе говоря, если мы примем, что биологически прогрессивная эволюция может следовать различными путями: и путём ароморфозов, и-путем идиоадаптаций, и путем ценогенезов, и путем общей дегенерации. Ибо, хотя морфофизиологические различия между всеми этими типами филогенетического развития и очень велики, однако мы считаем, что с биологической (экологической) точки зрения все они равноценны и в равной мере ведут к биологическому процветанию видов.
Мы только что упоминали о том, что наша современная фауна носит • гетерогенный характер, т. е.что она состоит как из примитивных, так и из прогрессивных форм, и указали на некоторые примеры выживания этих примитивных форм. Однако, едва ли многие читатели имеют представление о том, насколько это явление оказывается распространенным и закономерным в природе. Филогенетические исследования последних 25 ( лет позволяют привести ряд дополнительных примеров выживания древнейших низкоорганизованных форм, обусловливающих гетерогенное строение современной фауны.
Мы имеем основания считать, что все многоклеточные животные (Metazoa) произошли в свое время от одноклеточных простейших (Protozoa), точнее говоря, от некоторых палеозойских, несомненно еще докембрийских форм, близких к жгутиковым.1 Когда произошло это отделение предков Metazoa от Protozoa, мы точно сказать не может, но вполне определенно знаем, что оно произошло задолго до кембрийского периода, так как в кембрии мы встречаем уже довольно богатую фауну многоклеточных животных. Между тем одноклеточные и, в частности, жгутиковые живут и поныне, сохраняя при этом черты строения своей первичной организации; об этом говорит сравнение строения раковинок кембрийских простейших с раковинками современных Protozoa (рис. 96). Можно считать установленным, что весь тип одноклеточных представляет собою биологически процветающий пережиток какой-то бесконечно отдаленной от нас фауны.
Об исключительной древности этого типа животных можно судить хотя бы по тому, что уже в кембрии существовала довольно высоко организованная, богатая различными группами фауна, насчитывающая среди своих представителей как простейших, так и кишечнополостных, как моллюсков, так и иглокожих и членистоногих животных. Если, как это весьма вероятно, все эти формы произошли от одноклеточных, то приходится признать, что их эволюция происходила в течение весьма длинного (в геологическом смысле) периода, или, иными словами, что общие предки всех Protozoa и Metazoa жили в период времени, геологически по меньшей мере столь же отдаленный от кембрия, как кембрий отдален от нас.
С другой стороны, Сравнивая палеозойских одноклеточных с современными, а мы можем это сделать на основании сохранившихся скелетов ископаемых радиолярий (рис. 96) и корненожек, мы видим, что изменения, которые они претерпели за этот длинный промежуток времени с кембрийских времен и до нашей эры, не особенно велики, так как уже в ту отдаленную эпоху существовали отряды, находимые нами и в современной фауне.
1 См. Plate L., Allgemeine Zoologie, Bd. I, 1922; также H eid er К., Phylogenie der Wirbellosen, Kultur der Gegenwart, Bd. 4, Abstammungslehre, 1914.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
323
Рис. 96. Два вида радиоларий. А—ископаемая (докембрийская) Stanosphaera; В—соврем менная Spumellaria spheroidea (из М. В.
Павловой).
Мы приходим, таким образом, к выводу, что некоторая часть Protozoa в своем филогенетическом развитии пошла по пути ароморфоза и что от этой линии простейших произошли все Metazoa. Другая же и, вероятно, большая часть Protozoa в своей эволюции пошла по пути идиоадаптаций, сохранив общие черты строения простейших и изменившись сравнительно мало; она распалась на громадное число одноклеточных форм, хорошо приспособившихся к различным условиям окружающей среды.1
К тому же выводу приводит нас и исследование целого ряда других типов Metazoa, в первую очередь : кишечнополостных. Большинство авторов считает, что как Bilateralia, так и
Coelenterata происходят от одного общего корня,1 2 т. е. от некоторой гипотетической, ближе неизвестной нам гастреадной формы, тело которой состоит из двух слоев эпителиальных клеток (эктодермы и энтодермы); заключающих в себе гастреальную полость (архентерон), которая открывается наружу посредством бластопорного отверстия.
Организация современных кишечнополостных в общем сохранила этот тип строения и остановилась на стадии, недалеко ушедшей от первичного типа. Можно считать, что животные типа пресноводных гидр еще довольно близки к тем формам, от которых ответвились в одну сторону предки кишечнополостных, в другую—предки билатерально симметричных животных.
Вообще Coelenterata недалеко ушли вперед по пути прогрессивного развития (по пути ароморфоза), и среди них встречается много форм, которые по своему строению стоят не выше, чем примитивные Hydrozoa, хотя приходится признать, что с морфологической точки зрения организация высших Hydrozoa, в особенности Scyphozoa и Anthozoa, является гораздо более сложной, чем организация примитивных гидрополипов. Что же касается до гребневиков (Ctenophora), то новейшие авторы склоняются к тому, чтобы производить их от личиночных форм Anthozoa.
Мы знаем, что главные группы кишечнополостных животных отделились друг от друга в чрезвычайно отдаленные от нас времена. Уже в кембрийских отложениях встречаются отчетливые отпечатки медуз (Peytoia nathorsti) и других кишечнополостных, в остальных же палеозойских
1 Вместе с большинством исследователей, работающих над этими вопросами (ср. К. Рейдер, Л.Плате, В. Франц и др.) я принимаю точку зрения монофилетической теории филогенеза. Вопрос же о том, произошли ли все типы Metazoa, например кишечнополостные и губки, от одного единственного протозойного корня или нет, я оставляю пока открытым.
2 Гейдер предполагает, что губки (Spongia) произошли от одного корня с кишечнополостными; но так как другие авторы признают, что губки эволюпровали независимо от остальных Metazoa, т. е. принимают дифилетическое происхождение животного царства, и так как вопрос о филогении губок до сих пор еще темен, то я оставляю его пока в стороне. Во всяком случае, едва ли могут быть споры относительно того, что они произошли от одноклеточных предков, и в таком случае мы должны признать, что губки некоторое время эволюировали в прогрессивном направлении, но что затем эта прогрессивная эволюция остановилась и они пошли ио пути идиоадаптации и дожили до нашего времени, сохранив очень примитивную организацию. Эволюция в направлении идиоадаптации началась уже очень давно, ибо губки известны еще с кембрия,
21*
324
ГЛАВА X
слоях представлены все главные группы кишечнополостных, за исключением Ctenophora.
Сравнивая палеозойских кишечнополостных с современными нам целентератами, мы видим, что, несмотря на крупные морфологические изменения, происшедшие в их форме и строении за чрезвычайно долгий период их существования, их организация в общем стала не выше, чем она была, и что поэтому все изменения, которые им пришлось за это время претерпеть, произошли у них не в направлении ароморфозов, но в направлении идиоадаптаций, а во многих случаях даже в направлении общей дегенерации.
Подводя итоги только что сказанному, мы можем следующим образом представить общий ход филогенетического развития этой группы животных. В начале своей эволюции кишечнополостные, ответвившись от предков Bilateralia, пошли по пути прогрессивного развития: их организация постепенно стала усложняться, и они распались на ряд подчиненных групп. За этим геологически кратким, периодом ароморфозов последовал очень длительный период идиоадаптаций, в течение которого кишечнополостные, приспособляясь к различным условиям окружающей среды, во многих случаях сильно изменили свое строение и распались на целый ряд соподчиненных систематических групп: видов, родов, семейств и даже отрядов. В течение этого доныне длящегося периода их организация в общем 'не стала выше. У нас, по крайней мере, нет никаких оснований предполагать, что от известных нам кишечнополостных со времен силура произошли какие бы то ни было высшие группы животных. Единственная же прогрессивная группа, ответвившаяся от общих предков современных и палеонтологически известных нам первичных Coelenterata, а именно, предки Bilateralia, отделилась от них гораздо раньше, вероятно еще в докембрийский период.
Можно думать, что предки Bilateralia развились из форм близких к гребневикам (Ctenophora) еще в то время, когда эти последние находились на ранних стадиях своего филогенетического развития. В настоящее время у нас еще нет возможности, за отсутствием достаточного палеонтологического материала, правильно судить о ходе развития примитивных Bilateralia, ибо уже в древнейших кембрийских слоях встречается целый ряд вполне диференцированных представителей этого типа животных; среди них попадаются и Brachiopoda, и Echinodermata, и Annelidae, и Arthro-poda и даже моллюски. Ясно поэтому, что первичная диференциация всех этих форм началась еще задолго до кембрия. Мы принуждены, таким образом, строить филогенез Bilateralia только на основании сравнительно анатомических и сравнительно эмбриологических данных, но мы не можем при этом не отметить, что в течение последних двадцати пяти лет все эти проблемы были еще раз переисследованы целым рядом авторов, так что многие вопросы, касающиеся истории развития Bilateralia, стали для нас яснее, чем они были для исследователей предшествующего поколения.1 Здесь, конечно, мы не можем вдаваться в подробное рассмотрение этой исключительно трудной проблемы, и поэтому я остановлюсь на развитии только некоторых типов этих животных.
Как известно, двусторонне симметричные животные (Bilateralia) распадаются на две главные группы, а именно—на первичноротых (Protostomia), у которых ротовое отверстие развивается непосредственно из бластопора или первичного рта зародыша, и на вторичноротых (Deuterostomia),
1 В нашем изложении этого вопроса мы придерживаемся главным образом упомянутой нами выше мастерски написанной сводки К. Гейдера, 1914.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
325
у которых ротовое отверстие взрослого животного является новообразованием, развивающимся независимо от бластопора. Из первичноротых нам особенно интересны кольчатые черви, или а н н е л и д ы. Я не буду рассматривать сложного вопроса о происхождении аннелид и ограничусь указанием на то, что в среднекембрийских отложениях мы уже встречаем некоторые формы аннелид (Canadia spinosa, Worthenella cambria), близких, поскольку мы можем судить, к современным формам. Сравнивая эти формы и остатки других палеозойских аннелид с современными формами, мы приходим к заключению, что основные черты организации, кольчатых червей выработались уже в течение палеозоя, а может быть и еще раньше, и что организация их за весь длинный промежуток времени, протекший с кембрийского периода, не претерпела заметных прогрессивных изменений (ароморфозов). Ввиду . того, что аннелиды в настоящее время представляют собой процветающую и богатую формами группу, ясно, что эволюция их пошла по пути идиоадаптаций.
С точки зрения занимающих нас вопросов о направлениях эволюционного процесса весьма интересным является факт, что от аннелид произошли три крупнейшие группы членистоногих животных, а именно—ракообразные (Crustacea), группа Antennata, куда относятся Peripatus, тысяченожки и вся громадная группа насекомых, и, наконец, паукообразные (Arachno-morpha).1 •
Предки Articulata (Crustacea) ответвились от общего ствола аннелид на той стадии их филогенетического развития, когда эти последние уже приобрели большую часть характерных для аннелид (и даже для щетинконогих червей) признаков, а именно, когда их тело, покрытое хитином, уже было сегментировано, параподии образовались, а нервная система лежала на брюшной стороне тела под кишечником и уже состояла из брюшной нервной цепочки с двумя крупными парами подглоточных и надглоточных ганглиев. Эти парные ганглии соединялись между собою боковыми окологлоточными комиссурами; в это время уже имелся лежащий над кишечником спинной кровеносный сосуд, по которому кровяной поток протекал в ростральном направлении; имелись типично построенные нефридии, функционировавшие в качестве органов выделения и т. д. На основании сопоставления этих анцестральных признаков мы можем сказать, что Articulata происходят от некоторой, пока еще ближе неизвестной нам группы кольчатых червей, напоминающей но своим признакам щетинконогих червей (Chaetopoda).
От этих щетинконогоподобных предков произошли предки ракообразных, а от примитивных ракообразных—предки паукообразных (промежуточными формами между этими двумя группами являются Palaeostraci) и, по всей вероятности, предки тысяченожек и насекомых; Peripatus рассматривается обычно как форма, промежуточная между Crustacea и Antennata.
Сравнивая с точки зрения направления эволюции строение этих трех t главнейших групп членистоногих (Crustacea, Antennata и Arachnomorpha), мы видим перед собою группы, которые в общем развивались в направлении морфофизиологического прогресса (ароморфоза): у всех представителей потомков аннелид,
1 Мы не касаемся вопроса о том, развились ли Antennata и Arachnoidea из какой-либо определенной группы ракообразных (Crustacea), например из группы листоногих раков (Phyllopoda), как это предполагает Гатчек, или же они произошли независимо друг от друга от различных групп аннелид. Разрешение этой трудной проблемы не имеет значения для наших целей.
326
ГЛАВА X
т. е. у всех Arthropoda общее строение и диференциация органов стала сложнее, чем у их аннелидоподобных • предков; вместе с тем функции активных органов у них заметно интенсифицировались. ’
Различные группы Arthropoda в настоящее время стоят на различных ступенях филогенетического' развития: на верхних ступенях лестницы, как конечный результат прогрессивного развития путем ароморфозов, стоят высшие ракообразные (Decapoda), т. е. наиболее высоко организованные представители водных членистоногих, а также пауки и насекомые— * высшие представители фауны беспозвоночных на суше.
Таким образом, от общего ствола первичных, вероятно, еще докембрийских аннелид, произошли четыре линии потомков, развивавшихся в различных эволюционных направлениях: одна из этих линий шла в направлении идиоадаптаций и привела к образованию ныне живущих аннелид, которые, хотя и отличаются во многих отношениях от своих вымерших предков, однако по своей общей организации и по энергии своей жизнедеятельности стоят немногим выше, чем эти предки; три другие линии потомков были прогрессивными линиями: одна из них ведет к наиболее высокоорганизованным водным беспозвоночным, а именно, к современным омарам, крабам и ракам, две другие—к высшим представителям наземных членистоногих: к паукам и насекомым.
Не останавливаясь на трудной проблеме филогенеза моллюсков, отметим попутно, что среди них головоногие в своем развитии несомненно пошли по путчи ароморфозов.
Перейдем теперь к группе вторичноротых (Deuterostomia) и отметим прежде всего, что весь этот ствол, т. е. Chaetognata (Sagitta), Enteropneusta (Balanoglossus и родственные ему формы), Echinodermata и Chordata (асцидии и сальпы, Branchiostomatidae и позвоночные), развивался моно-филетически и находится в состоянии лишь отдаленного родства с группой первичноротых (Protostomia). Мы отмечаем это в связи со старой гипотезой Семпера-Дорна (1875), производящей позвоночных от аннелид, т. е. от первичноротых. Все новейшие исследования приводят к выводу, что эта гипотеза неверна и что все новейшие попытки возродить ее (Дельсман)1 являются несостоятельными. Мы имеем основания утверждать, что Deuterostomia после их отделения от остальных Bilateralia вступили на самостоятельный путь филогенетического развития.
Гипотетические предки всех Deuterostomia (Dipleurula, по К. Гейдеру) представляли собой билатерально симметричные формы, снабженные апикальным органом и тремя парами целомических мешков; ротовое и анальное отверстия открывались у них на вентральной стороне тела. Эти Dipleurula во многих отношениях напоминали собою личинок иглокожих—Bipinnaria. Первичные иглокожие после своего отделения от этих гипотетических предков перешли к прикрепленному (сидячему) образу жизни и приобрели пятилучевую радиальную симметрию тела. Организация потомков этих примитивных форм достигла впоследствии более высокой степени развития; многие из них затем вторично перешли к свободно подвижному образу жизни. На этом, однако, прогрессивная эволюция иглокожих приостановилась, и они вступили на путь идиоадаптаций; при этом они распались на большое число самых разнообразных форм, из которых одни благополучно дожили до наших дней, а другие вымерли.
Исходя из вышеизложенного, мы имеем возможность установить два главных периода в эволюции иглокожих. Первый период, период прогрессивных морфологических изменений, во время которого их организация
1 Подробное обсуждение совершенно необоснованной гипотезы Дельсмана мы находим в работе Д. Третьякова (1929, 1. с.).
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
327
постепенно усложнялась: из билатерально симметричного их тело стало радиально пятилучевым; у них развился весьма сложно построенный известковый скелет, а также ряд других общих признаков, характерных для всех Echinodermata. Это—и ериод ароморфозов. После этого прогрессивного периода эволюции в филогенезе иглокожих наступил второй, ид ио ад апт ивны й период. Все вновь возникшие с этого времени, часто весьма сложные морфологические изменения иглокожих, уже не выходят из этого новоприобретенного радиального типа строения (морские ежи, морские звезды, морские лилии, голотурии). Ийыми словами, ни один из многочисленных потомков этих форм уже больше не поднялся над общим уровнем, на котором стояли их предки и не дал начала какой-либо новой прогрессивной группе, вышедшей за пределы этого типа строения.
Филогенез позвоночных и Branchiostomatidae был нами подробно описан в гл. V—VIII этой книги, но мы совсем не касались вопроса об эволюции низших хордовых животных — оболочников (Tunicata). Между тем филогенез этих форм является во многих отношениях столь интересным, что мы не можем не остановиться на нем несколько подробнее.
Изучение онтогенеза асцидий ясно указывает, что эволюция их предков в течение долгого времени шла в направлении морфофизиологического прогресса. Их массивная нервная трубка (нейроцель) постепенно расширялась в своем переднем отрезке, принимая форму мозгового пузыря, их осевой скелет (chorda dorsalis) уже был хорошо развит, а мускулатура сегментирована; были также хорошо выражены и органы высших чувств, уже существовал слуховой пузырек и снабженный хрусталиком глаз. Мы имеем все основания думать, что предки современных туникат представляли собою в этой стадии своего филогенетического развития свободно плавающих нектонных животных, передвигавшихся в воде при помощи длинного хвостового плавника.
Позднее направление их эволюции изменилось, и предки асцидий перешли ксидячемуобразу жизни, причем общее строение их тела значительно упростилось: хорошо развитая центральная нервная система редуцировалась до единственного небольшого Ьанглия (рис. 92 G), лежащего дорзально от кишечника; сегментальная мускулатура атрофировалась целиком; то же можно сказать и относительно органов высших чувств, хорды и длинного хвостового плавника. Наоборот, прогрессивно развились целлюлозная мантия, служащая защитой для внутренних органов, сложный жаберный аппарат (рис. 92, Ьг) и органы размножения (рис. 92, ov., ts).
Мы видим, таким образом, что в течение эволюции большинство активных органов асцидий редуцировалось и что следствием этого явилось значительное упрощение в строении современных асцидий по сравнению со строением их свободно живших предков. Произошел ряд регрессивных изменений, вследствие чего организация современных асцидий значительно снизилась. Связь между этим снижением и сидячим образом жизни понятна само собою и дальнейших пояснений не требует.
Сальпы во время своей эволюции претерпели ряд аналогичных изменений с той лишь разницей, что они перешли не к сидячему, но к пелагическому (пассивно-подвижному) образу жизни.
Таким образом, в эволюции оболочников можно установить две следующие стадии развития: 1) период прогрессивной эволюции, т. е. ряд следующих друг за другом ароморфозов; в течение этого длительного периода организация предков Tunicata поднялась на довольно зачительную высоту (хорошо развитая центральная нервная система, снабженные хрусталиком глаза, слуховой пузырек, хорошо развитая мускулатура, по всей вероятности активное питание и т. д.)
328
ГЛАВА X
и 2) период общей дегенерации, когда большинство активных органов резко, порой даже целиком, атрофировалось, а организация в общем и целом упростилась.
Мы несколько задержались на истории развития оболочников, потому что здесь мы имеем дело с очень большой группой животных, целым подтипом, развитие которого протекало в направлении общей дегенерации, и также потому, что в данном случае для нас понятна причина, заставившая эти формы пойти именно по этому, а не по другому пути эволюции. Причина эта лежит в переходе от . активного образа жизни свободно подвижных нектонных животных к пассивно-подвижному (сальпы) или к неподвижно-сидячему (асцидии) образу жизни.
ВЗАИМООТНОШЕНИЯ МЕЖДУ РАЗЛИЧНЫМИ НАПРАВЛЕНИЯМИ ЭВОЛЮЦИИ! животных
В предыдущем мы дали краткий обзор эволюции некоторых типов животного царства и показали, что установленные нами направления биологически прогрессивной эволюции, т. е. направления ароморфозов, идиоадаптаций, ценогенезов и общей дегенерации, имеют очень широкое распространение. У нас имеются все основания предполагать, что эволюция каждого типа как такового, точно так же как и эволюция каждой подчиненной этому типу группы (класса и т. д.) в течение определенных периодов времени непременно шла по одному из указанных нами направлений. Подробный анализ каждого отдельного случая указывает на то, что внутри каждой крупной группы животных всегда находятся формы, стоящие на различных ступенях прогрессивной (ароморфозной), идиоадаптивной или дегенеративной эволюции, или же, наконец, эволюции, происходящей путем эмбриональных изменений (ценогенезов).
Рассматривая животный мир как целое, с точки зрения высоты организации, мы видим, что многие крупные группы животных, например все типы, развились в течение первых стадий своего филогенеза путем ароморфоза, после чего для них обычно наступал период идиоадаптаций или, в некоторых случаях, период общего регресса (дегенерации). Часто случается так, что какая-нибудь определенная группа животных сначала эволюирует прогрессивно и поднимается в своем развитии все выше и выше по пути ароморфозов, затем направление ее эволюции меняется, и она начинает развиваться путем идиоадаптаций; при этом организация ее уже более не повышается. Эта вторая линия развития часто распадается на два новых направления, причем одна линия, не изменяя общей высоты своего строения, продолжает эволюировать идиоадаптивно вплоть до наших дней, а для другой наступает новый период ароморфозов. Так возникает ряд более высоко стоящих по своему строению и функциям прогрессивных форм. Эта схема филогенетического развития повторяется у простейших, у губок, у кишечнополостных, у аннелид и у многих других групп животных.
Рассмотрим несколько примеров такого эволюционного пути. От первичных одноклеточных отделилась прогрессивная ветвь многоклеточных, но примитивная идиоадаптативная ветвь одноклеточных продолжает существовать в виде громадного числа форм и в настоящее время. От кишечнополостных, а именно от форм, близких к Ctenophora, отделилась прогрессивная ветвь Bilateralia, но гребневики и остальные группы кишечнополостных существуют и поныне, частью в виде адаптивных, частью в виде регрессивных (вследствие сидячего образа жизни) форм, и организация их в общем не повысилась. От аннелид, которые живут и теперь, в давно прошедшие времена отделилась прогрессивная группа ракообразных,
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
329
а ст примитивных ракообразных—прогрессивные высшие ракообразные, Antennata и Arachnoidea. Тот же тип эволюции мы находим и в остальных группах двусторонне симметричных животных.
Аналогичную картину мы наблюдаем и у низших позвоночных животных. Так, от общего ствола всех позвоночных, а именно от первичных бесчерепных (Acrania primitiva), отделилась прогрессивная ветвь протокраниат, от которых, в свою очередь, произошли все рыбы; но боковая ветвь, ответвившаяся еще в палеозойские времена от общего корня бесчерепных, т. е. branchiostomatidae, пошла по пути идиоадаптации и в виде весьма примитивной формы (ланцетник) благополучно дожила до нашего времени. От протокраниат, кроме указанной прогрессивной ветви, отделилась еще другая линия потомков, которая развивалась сначала в прогрессивном направлении (Entobranchiata—Protocyclostomata), но затем также пошла по пути идиоадаптации и специализации и вследствие этого отчасти сохранилась и по сию пору в виде процветающей группы круглоротых (миноги и миксины).
Что касается первичных челюстных, то и от них также отделилась ветвь, развивавшаяся частью путем идиоадаптации, частью путем ценогенеза; современные представители этой ветви (акулы и скаты) сохранили и теперь все примитивные черты своего строения, образуя в то же время многочисленную и богатую формами группу. Одна линия потомков первичных рыб с костным скелетом, а именно осетровые рыбы, также дожила до нашего времени, благодаря тому, что пошла в своей эволюции по пути идиоадаптации; по той же причине осетровые и поныне сохранили многие примитивнейшие черты своего строения, и организация их мало повысилась с древнейших времен. Между тем другая, чисто прогрессивная линия потомков первичных Osteichthyes, Holosteoidei actinopterygii, значительно изменившись, привела к образованию самой крупной и наиболее процветающей сейчас группы костистых рыб. От нее ответвились еще две линии потомков, частью вымерших, а частью развившихся в направлении идиоадаптаций и сохранивших поэтому многие примитивные черты своей организации; я имею в виду современные формы ильных рыб (Amia) и панцырных ujyK(Lepidosteus). Наконец, если мы обратимся к Holosteoidei crossopterygii, то увидим, что и здесь, кроме происшедшей от них высоко прогрессивной группы наземных позвоночных (Quadrupeda), мы имеем еще и линию двоякодышащих и линию кистеперых, эволюировавших в направлении идиоадаптации и специализации и сохранивших, таким образом, наравне со многими чертами адаптивного характера также и многие признаки примитивной анцестральной организации.
Такое же чередование различных направлений эволюционного процесса, начинающихся с периода прогрессивной морфофизиологической эволюции (ароморфоза), мы имели случай наблюдать и тогда, когда вкратце разбирали вопрос о ходе эволюции наземных позвоночных. Так, от прогрессивного ствола вымерших стегоцефалов ответвились две идиоадаптив-ные ветви потомков (одна из них пошла по пути специализации), сохранивших многие черты своей примитивной организации вплоть до наших дней: это—хвостатые и безногие амфибии (гимнофионы), и одна прогрессивная ветвь—бесхвостые амфибии. В эволюции огромной группы рептилий, как это мы видели выше, после длинного периода ароморфозов и ценогенезов также наступает длинный период идиоадаптивной радиации, и только две группы рептилий, та, от которой произошли птицы, и та, от которой произошли млекопитающие, оказались чисто прогрессивными группами.
Чередование различных направлений эволюции и ход эволюционного процесса можно представить в очень упрощенном виде посредством, прилагаемой здесь схемы.
330
ГЛАВА X
Схема XXII показывает нам, что на некоторой стадии эволюционного процесса у данной формы животных А развиваются признаки прогрессивного характера, т. е. происходит ароморфоз, и благодаря этому повышение высоты организации; этот подъем организации на схеме обозначен некоторым подъемом линии а4—а2, символизирующей ход эволюции данной группы. Вслед за этим подъемом организации данная группа начинает приспособляться к различным условиям той физико-химической и биологической среды, в которой она живет и с которой ей приходится сталкиваться в результате расселения, т. е., другими словами, у нее появляются адаптации Ь, Ь' и т. д. к различным условиям
Схема XXII. Схема эволюционного процесса (объяснение в тексте).
среды, причем данная группа, попадая в различные условия, распадается на большее или меньшее число систематически подчиненных групп, из которых каждая приспособлена к определенному комплексу условий среды. Этот период жизни потомков нашей группы мы можем обозначить как период идиоадаптаций. Факт, что потомки данной исходной формы распадаются на целый ряд форм, мы выражаем на схеме многократным дихотомическим разделением линии, символизирующей эволюцию данной группы, причем получается знакомая нам форма родословного древа. Но чтобы обозначить, что при этом процессе изменения не происходит подъема организации (ароморфоза), мы располагаем на схеме эти идиоадаптации на горизонтальной плоскости (Q), причем отклонения вправо и влево от прямой линии, проведенной пунктиром, обозначают степень изменений адаптивного характера, сильные же уклонения в сторону (ч, s) обозначают специализации.1
Вслед за периодом идиоадаптивной эволюции а2...а3 может произойти ряд прогрессивных изменений (ароморфозов), которые на схеме символизированы новым подъемом линии а3...а4 над горизонтальной плоскостью Q, и затем новый период идиоадаптации (а4...а8), который происходит уже в плоскости R, но при этом часть потомков Ьх—Ь2 продолжает изменяться адаптивно и не повышается в своем строении (это выражено на схеме тем, что они остаются в той же плоскости Q) и в виде сравнительно примитивной группы доживают до настоящего времени. Выше мы привели целый ряд примеров такой эволюции, когда потомки прогрессивной группы (а5) и потомки примитивной группы (Ь2) существуют одновременно.
1 Выше мы уже упомянули о том, что идиоадаптации могут имееть место также и в течение периодов ароморфоза (а,—аа, а3—а4); в это же время могут возникнуть и новые боковые линии; мы не изобразили их здесь, чтобы не усложнять нашей схемы.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
331
На нашей схеме представлен графически и третий тип эволюции, а именно регрессивная эволюция, или эволюция путем общей дегенерации, которая, согласно принятой нами схеме обозначений, представлена понижением линии, изображающей ход эволюции данной группы, т. е. понижением линии г1...г2; после этого понижения, которое обозначает, что организация данной группы животных стала менее сложной, происходит снова ряд приспособлений (идиоадаптации г2—г3, плоскость Р) к различным условиям, в которые попадает измененная группа животных, например к условиям окружающей среды, характерной для паразитов или для сидячих, форм.
Направление эмбриональных приспособлений, или ценогенезов, которое должно было бы быть выражено линиями, проецированными все на ту же горизонтальную плоскость (ибо строение взрослых особей осталось на прежнем уровне развития), на нашей схеме не изображено вовсе. Мы это сделали намеренно для того, чтобы излишне не усложнять этой и без того достаточно сложной схемы.
Наша схема могла бы изобразить, например, ход эволюции хордат. В этом случае буква А изображала бы билатерально симметричного предка высоко стоящих хордат, линия гг... г2—ход эволюциии их дегенерировавших потомков—туникат, линии aj ... а2 и а3 ... а4 символизировали бы ход прогрессивной эволюции потомков свободно живущих хордат. Факт остановки форм в своем развитии на относительно примитивной стадии эволюции и перехода их к идиоадаптивным изменениям, как уже сказано, изображен нами графически тем, что родословное древо (bL—b2) проецировано на горизонтальную плоскость Q. Поднимающаяся вверх линия а3 ... а4 символизировала бы в этом случае прогрессивный филогенез краниат.1
Внимательно рассматривая нашу схему и сравнивая ее со схемой XV, представляющей родословное древо низших позвоночных, мы видим, что периоды ароморфоза сравнительно редки. По всей вероятности, они были также и относительно коротки; именно это может служить причиной, почему палеонтологически мы находим так мало переходных форм между большими группами животных, в частности между большими группами позвоночных.
Другой причиной редкости переходных форм между крупными систематическими группами является, по всей вероятности, тот факт, что они эволюировали монофилетически.
ИЗМЕНЕНИЯ ОБЩИХ НАПРАВЛЕНИЙ ЭВОЛЮЦИИ И ПРИЧИНЫ ПЕРЕХОДА ЖИВОТНЫХ ОТ ОДНОГО НАПРАВЛЕНИЯ К ДРУГОМУ
Филогенетические исследования показали нам, что общее направление эволюции любой группы животных может меняться не раз в течение геологических эпох. Часто за периодом ароморфозов следует период идиоадаптивной эволюции, когда прогрессивная форма, именно благодаря подъему своей организации, начинает усиленно размножаться, когда она увеличивается численно и вследствие этого расселяется, т. е. захватывает новые для нее места жительства. Потомки этой прогрессивной формы попадают в новые условия, в новую биологическую и физико-химическую обстановку и начинают приспособляться к этим условиям:
1 Мы подчеркиваем еще раз, что приведенная нами схема крайне упрощена: периоды ароморфозов (линии а,... а2, а3... а4) собственно должны были бы быть изображены в виде пологой лестницы с очень низкими ступенями, ибо весьма вероятно, что отдельные подъемы организации эволюирующих форм отделялись друг от друга периодами идиоадаптаций. Схема наша, однако, имеет только одну цель—сделать возможно более наглядной основную мысль теории эволюционных направлений, а для этого она мне кажется достаточной даже и в своем настоящем виде.
332
ГЛАВА X
наступает период идиоадаптивной эволюции или же период ценогенеза, которые могут длиться очень долго, пока не наступит новый период ароморфоза или же пока не появятся условия, которые заставят потомков данной группы изменить направление своего филогенетического развития и пойти по направлению специализации или по направлению регресса.
Установив четыре главных направления эволюционного процесса, посредством которых достигается биологический прогресс: морфофизиологическую прогрессивную эволюцию (ароморфоз), идиоадап-тивную эволюцию, частным случаем которой является специализация, эволюцию путем эмбриональных изменений или ценогенезов и регрессивную эволюцию или общую дегенерацию, совершенно естественно поставить вопрос о том, каким образом каждый из.этих типов морфофизиологических изменений отражается на дальнейшем ходе эволюции данной группы, т. е.в какой мере каждый из названных типов эволюции способен переходить в другие типы.
Разбирая приведенные выше многочисленные примеры различных направлений эволюции, мы, между прочим, находим и примеры того, что идио-адаптивная эволюция данной группы нередко сменяется прогрессивной, т. е., другими словами, что адаптация сама по себе отнюдь не препятствует возникновению ароморфозов; с другой стороны, мы видели также, что и периоды ароморфоза сменяются периодами идиоадаптаций. Вообще, повидимому, оба эти типа эволюции свободно переходят друг в друга. Вместе с тем идиоадаптивный тип эволюционного процесса способен перейти, во-первых, в тип регрессивной эволюции, во-вторых, что бывает еще чаще, в тип специализации. Мы находим много примеров такого перехода. Наобо1 рот, я не нашел примеров того, чтобы группа специализованных животных или группа животных, вступивших на путь общего регресса или общей дегенерации, пошли по пути прогрессивной эволюции. Во всяком случае такие переходы очень редки и представляются чем-то исключительным. Насколько мы можем судить, специализация, а тем более дегенерация, служат препятствием к появлению прогрессивных изменений у животных и вступлению их на путь ароморфоза.
Нам представляется весьма интересным разобрать вопрос о причинах-изменений направлений эволюционного процесса, т. е. выяснить факторы, обусловливающие переход какого-либо вида животных с одного пути эволюции на другой. Другими словами, чрезвычайно интересно поставить вопрос о том, почему животные, изменявшиеся в течение одной эпохи в идиоадаптивном направлении, в следующую эпоху переходят на путь прогрессивной эволюции (ароморфозов) или на путь ценогенеза, в других же случаях—на путь специализации или на путь дегенеративных изменений.
Отмечаю, что в данном случае я не ставлю вопроса о причинах эволюционных изменений вообще, а касаюсь только причин перемен направления эволюционного процесса, т. е. до известной степени суживаю задачу. Но и в этой форме вопрос настолько сложен, что последующие замечания скорее могут служить лишь указанием на то, в каком направлении можно искать решения этой проблемы, чем самым решением ее.
Основным положением, из которого нам приходится исходить, является положение, что перемены в условиях существования в случаях, когда они не ведут к непосредственному вымиранию данной группы животных, вызывают изменения в строении и функциях этих животных, благодаря чему они выживают при новых измененных условиях. Но эти перемены в строе
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
333
нии животных могут произойти либо в том же направлении, в каком они шли раньше, т. е. могут появиться новые идиоадаптации вместо старых (если данная группа животных эволюировала в направлении идиоадаптации), либо они могут произойти в ином направлении, т. е. животные могут в одних случаях пойти по пути прогрессивной эволюции (по пути ароморфоза), в,других—по пути ценогенеза, в третьих—по пути дегенерации.
В сущности очень трудно решить, почему именно происходит то, а не другое изменение в направлении филогенеза эволюирующих животных. A priori можно было бы думать, что общий характер изменений в направлении эволюции зависит от общего характера изменений в условиях окружающей среды. Однако вопрос разрешается далеко не так просто, как это кажется на' первый взгляд. Хотя в природе и встречаются случаи, когда резкие изменения условий окружающей среды вызывают сходные изменения в направлении эволюции данных форм, тем не менее гораздо чаще встречаются такие случаи (примеры которых мы приводим ниже), когда аналогичные изменения в окружающей среде вызывают различного рода изменения в направлении эволюции тех животных форм, на которых эти изменения воздействуют. Здесь, следовательно, направление эволюции не зависит непосредственно от изменений, происходящих в окружающей средё, а связано еще с какими-то иными факторами. Чтобы облегчить читателю понимание этой трудной проблемы причины изменения эволюционных направлений в филогенезе животных, я приведу несколько относительно простых примеров, которые помогут понять причину того, почему вообще могут изменяться направления эволюции животных.
Рассмотрим для начала переход от адаптации к специализации. Мы знаем, что среди позвоночных довольно большое число животных приспособилось к подземному образу жизни и благодаря этому строение их изменилось в направлении специализации. Среди млекопитающих, приспособившихся почти исключительно к подземному образу жизни, мы можем назвать: из насекомоядных—наших кротов и капского Chrysochloris, из грызнунов—слепышей и из сумчатых—австралийских сумчатых кротов (Notoryctes); среди рептилий мы имеем роющих змей (Typhlopidae) и различных Amphisbenidae, среди амфибий—гимнофионов. У всех этих животных имеется, несмотря на все несходство их организации, ряд общих черт: для всех их характерна большая или меньшая редукция глаз, часто скрытых под кожей, а в некоторых случаях прикрытых костями; у всех изменено строение черепа, форма которого приспособлена к рытью и к продвиганию в земле. У роющих млекопитающих конечности хорошо развиты, и на коротких и широких передних лапах, приспособленных к разрыванию почвы, имеются длинные и крепкие когти; роющие змеи, ящерицы и гимнофионы все утратили как передние, так и задние конечности и приобрели змеевидную форму тела.
Если мы спросим, какие изменения внешней среды могли вызвать переход к подземному образу жизни животных, ближайшие родичи которых не специализованы и живут на поверхности земли, то следующие предположения являются наиболее вероятными: переход к подземному образу жизни был вызван либо необходимостью спасаться от хищников, преследовавших и истреблявших данную форму в ее прежнем месте обитания и лишенных возможности последовать за нею в ее новую подземную среду, либо большей легкостью добывания пищи под землей, чем на поверхности. Разберем каждое из этих предположений в отдельности.
Прежде всего поставим вопрос о том, при каких условиях наземные животные, предки перечисленных нами млекопитающих, могли при преследовании врагами перейти к подземному образу жизни. Первой
334
ГЛАВА X
предпосылкой такого перехода является предположение, что эти животные до времени перехода к подземному образу жизни были хотя бы в незначительной степени роющими животными: мы знаем, что многие насекомоядные и грызуны живут в вырытых ими норах, выводя в них детенышей. пряча запасы пищи и. спасаясь от опасностей. Если мы вспомним землероек, сусликов, хомяков, сурков и многих других, то предположение, что предки кротов, слепышей и им подобных животных и прежде вели этот , образ жизни и рыли норы в земле, является вполне вероятным.
Мы предположили, что одним из факторов, определяющих переход к подземному образу жизни, могло быть появление в местности, населенной предками наших роющих млекопитающих, истребляющего их хищника. Весьма вероятно, что переход к подземному образу жизни происходил только в тех случаях, когда существовало большое несоответствие между силами хищника и силами преследуемого животного. Если хищник настолько превосходил преследуемое им животное в силе, резвости, чуткости и т. п., что непосредственная борьба с ним, бегство от него и прятание, т. е. обычные методы спасения, не приводили к цели, и животное усиленно истреблялось, то единственным средством выживания для преследуемого вида оставался уход в такое место обитания, куда враг не мог следовать за ним. Весьма возможно, что у роющего животного развился инстинкт по возможности постоянного пребывания в вырытых им норах и отыскивания там той пищи, которую оно могло найти под землей. Те формы, у которых этот инстинкт стал развиваться, выживали, те, у которых он не развился, погибали в борьбе за существование. Но подземный образ жизни, в свою очередь, должен был оказаться стимулом к целому ряду вторичных изменений, носящих характер специализации, т. е. изменений в строении глаз, органов обоняния, черепа и конечностей, о которых мы уже упоминали выше.
Совершенно то же мы можем сказать и о живущих под землею амфибиях й рептилиях, а также о гекконах, обычно держащихся на отвесных скалах, где их уже больше не может преследовать огромное большинство их обычных врагов. То же относится и к хамелеонам, живущим на таких тоненьких веточках, которые не могут выдержать тяжести тела превышающих их по размерам хищников,1 а также к целому ряду других специализировавшихся в этом направлении форм.
Таким образом, переход к постоянному подземному образу жизни у упомянутых нами грызунов и насекомоядных совершился лишь тогда, когда появилась необходимая для этого предпосылка (привычка в случае надобности рыть норки и т. п.), с одной стороны, и когда наступило резкое несоответствие между силами преследующего хищника и преследуемой добычей, с другой.
Если же такого резкого несоответствия между силами хищника и силами преследуемой им добычи не существует, т. е. если хищник только немногим превосходит в своих силах добычу, так что при преследовании некоторый процент особей будет спасаться, сохраняя при этом надземный образ жизни, то, конечно, существует большая вероятность, что преследуемые . животные останутся в своих старых местах обитания и будут приспособляться к ухудшившимся условиям существования другими путями.
Как мы отмечали выше, другая причина перехода животных в иную, сильно отличающуюся от прежней среду заключается в избытке пищи в этой новой среде и в возможности добывать ее более легким путем, чем в прежнем месте обитания. Предпосылка к этому лежит, само
1 От хищных птиц, налетающих сверху, хамелеоны защищены 'своеобразной, меняющей цвет покровительственной окраской.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
335
собой разумеется, так же как и в предшествующих случаях, в организации предков данной формы: если строение и привычки животного позволяют .ему порою покидать свое обычное место обитания и переходить на некоторое время в новую среду, то закрепление такого перехода вполне возможно, в противном случае, т. е. если этого нет, переход вообще не может иметь места. Второе условие перехода заключается в количественном соотношении между питанием животных в старой и новой среде: если разница между количеством пищи и легкостью ее добывания в старой и новой среде очень значительна, то существует большая вероятность того, что животные переселятся в эту новую среду, если же этого нет, например, если разница между количеством пищи на поверхности земли и под ней невелика, то более вероятным является то, что животные останутся в старых местах своего обитания и будут приспособляться к ухудшившимся условиям существования каким-то другим образом.
Вполне возможно, что именно избыток находящейся в водных бассейнах пищи был причиною перехода некоторых наземных млекопитающих, например предков тюленей, моржей, морских львов и др., с суши в воду. Предпосылка к этому явлению лежала в способности некоторых млекопитающих плавать в случае необходимости, т. е. в общем строении их тела, в строении их конечностей, а также в их психических способностях, так как, если психические способности нашего наземного млекопитающего были не достаточно развиты для того, чтобы позволить ему понять, что рыба может годиться в пищу, то оно не сумеет использовать в этом отношении свою способность плавать. Но если животное станет постоянно, из поколения в поколение, голодать на суше и если при плавании ему случайно, без особых затруднений, удастся поймать рыбу, или какую-нибудь иную добычу (ракообразное), то весьма возможно, что спорадическое плавание превратится с течением времени в постоянное пребывание в воде и что наступит некоторое определенное изменение в направлении эволюции нашей формы.
В приведенных случаях мы видели, как некоторые животные формы, вследствие определенных и притом значительных изменений в условиях своего существования, переходят от пути идиоадаптации на путь специализации. Весьма возможно, что аналогичный процесс, имевший место у других форм, например у ракообразных, привел этих последних к паразитизму. Предпосылка к этому переходу могла состоять в том, что какое-либо свободно живущее ракообразное, которое прежде охотилось за какой-либо свободно плавающей добычей, получило привычку спорадически прикрепляться или присасываться к этой добыче, к другому, большему по размерам животному и питаться его соками. Этот более совершенный способ питания привел к тому, что полезная привычка мало-по-малу стала постоянным признаком, а животное постепенно превратилось в эктопаразита и стало эволюировать дальше именно в этом направлении.
Таких примеров, перехода от свободного образа жизни, т. е. от филети-ческого развития в направлении идиоадаптации, к экто-, а затем и к энто-.паразитизму, т. е. к развитию в направлении общей дегенерации, а также примеров перехода от свободно-подвижного (развитие в направлении идиоадаптации) к сидячему образу жизни (развитие в направлении специализации) можно было бы привести очень много. Стимулом к подобного рода переходам являются во всех этих случаях те крупные преимущества, которые животные встречают в условиях новой среды (большее количество пищи, большая защита от врагов), а основной предпосылкой—те особенности в их организации, которые делают возможным этот переход. Именно этим путем очень многие животные формы перешли с пути идиоадаптаций на путь дегенерации.
336
ГЛАВА X
Мы особенно подчеркиваем тот факт, что здесь мы говорим о переходе животных от одного эволюционного направления к другому лишь очень кратко и очень схематично, останавливаясь на этом процессе лишь постольку, поскольку это необходимо для того, чтобы сделать ход наших мыслей понятным читателю. В действительности же процесс этот протекает неизмеримо сложнее, чем это нами представлено: в каждом отдельном случае имеется налицо не одна,, но много предпосылок, необходимых для осуществления такого перехода, существует не один, но много переходов между различными стадиями филогенетического развития эволюирующих форм.
Во многих случаях перехода эволюирующих животных от одного эволюционного направления к другому причину этого явления приходится искать не столько в изменениях экологических условий окружающей среды (враги, добыча), сколько в корреляции между интенсивностью вызывающих изменения факторов и интенсивностью активных функций эволюирующего животного, т. е. тех функций, которые имеют непоср ед ственное отношение к этим факторам.
Значение этих количественных соотношений станет для нас более ясным, если мы допустим существование несколько иных отношений между данными животными и изменениями среды, чем те, с которыми мы имели дело до сих пор. Представим себе, что мы имеем перед собой ту же форму животных, которую мы брали выше в качестве примера, т. е. грызунов или насекомоядных, и что влияние изменяющего фактора (преследующего хищника) также осталось неизменным, вернее, осталось влиянием того же рода, каким оно было раньше, но что количественное соотношение между активными функциями преследуемой добычи и преследующего хищника стало несколько иным, чем оно было раньше. Предположим, например, что в данной местности появился новый хищник, преследующий упомянутое нами в качестве примера млекопитающее, но допустим, что этот хищник только в незначительной степени резвее, сильнее и умнее преследуемого им животного. Как отразится это обстоятельство на направлении дальнейшей эволюции потомков преследуемой формы?
Если между силами хищника и силами преследуемой им добычи существует лишь незначительная разница, так что более сильные из преследуемых особей все же могут спастись от врага либо активной защитой, либо бегством, либо тем, что они сумеют укрыться от него в естественно защищенных местах, и если при этом погибнут только наиболее слабые, наименее быстро двигающиеся особи, то переход к специализации, скажем, к подземному образу жизни, для этих форм уже не является безусловной необходимостью. Тогда среди преследуемого вида будут выживать наиболее сильные, наиболее ловкие, наиболее быстро бегающие особи, и от них произойдет новая раса потомков, которым уже не будет нужды переселяться под землю, но которые будут эво-люировать в прогрессивном направлении на поверхности земли.
У животных вообще, и в особенности у млекопитающих, средства защиты от врагов весьма разнообразны; поэтому потомки преследуемой формы могут эволюировать в самых разнообразных направлениях, и появление каждого защитного признака будет зависеть от всей организации и всех функций, инстинктов и привычек преследуемого животного в период наступления изменения, т. е. в тот именно период его эволюции, когда появляется новый враг. Может случиться, например, что животные уже обладают покровительственной окраской, до известной степени защищающей их от врагов; тогда при появлении нового врага эта покровительственная окраска может развиться сильнее и оказаться
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
337
достаточной для защиты от преследователя, если острота зрения этого преследователя не будет очень велика. В этом случае к прежде существовавшим идиоадаптациям прибавится новая идиоадаптация, и общее направление эволюции не изменится. Но может произойти и иное. Если есть задатки для этого в организации животного, то могут выработаться и какие-нибудь другие защитные особенности идиоадаптивного свойства, например могут развиться пахучие железы, делающие данное животное противным для преследующего врага, или его волосы (щетины) могут превратиться в колючие иглы, как это имеет место у ежей и у дикообразов. Во всех этих и подобных им случаях развиваются новые идиоадаптации, но общее направление эволюции не меняется.
Но может оказаться, что хищник настолько зорок, что покровительственная окраска преследуемой добычи уже не достигает цели, или же, что у этой добычи не имеется в данный период эволюции никаких предпосылок для образования пассивных органов защиты, иначе говоря, никаких зачатков новых защитных приспособлений: зловонных желез, игл, шипов и т. п. Тогда среди преследуемого вида смогут избегнуть уничтожения лишь такие особи, которые окажутся в состоянии противостоять врагу активной защитой, или же такие, которые сумеют избавиться от него бегством, иными словами, из представителей преследуемрй группы выживать будут наиболее резвые особи, наиболее зоркие и сторожкие, которые будут во-время замечать опасность, наиболее выносливые, которые будут лучше выдерживать преследование и т. д. Потомки таких особей образуют новую расу. Другими словами, животные в этом случае вступят на путь прогрессивных изменений, причем общая интенсивность их жизнедеятельности повысится. Эти прогрессивные изменения могут выразиться различно в зависимости от соотношения между организациями преследователей и добычи; в некоторых случаях прогрессивные изменения произойдут в органах высших чувств: глазах, органах слуха или обоняния, и животные станут более чуткими; в других случаях—в органах движения, и произойдет повышение резвости; в третьих—в сердце и легких, причем повысится выносливость, и т. д. Примеры таких прогрессивных изменений приведены выше, и мы на них не будем останавливаться. Во всяком случае для преследуемой формы наступит новый период ароморфозов. Направление эволюции при этом, конечно, изменится: животные с пути идиоадаптации перейдут на путь ароморфозов.
Мы можем себе представить, однако, что прогрессивная эволюция не остановится на этой первой фазе, но что и хищники, своим преследованием вызвавшие прогрессивные изменения в строении добычи, станут приспособляться к измененному строению преследуемых животных и, изменившись прогрессивно, в свою очередь вызовут новые прогрессивные изменения в организации последних. Тогда прогрессивное развитие хищника пойдет параллельно прогрессивной эволюции добычи.1
1 Отметим, что здесь, разбирая вопрос о различных взаимоотношениях между хищником и добычей, мы имеем дело только с такими филетическими изменениями органов, которые определяют собою те или иные изменения в направлении эволюции рассматриваемых животных: переход от направления ароморфозов к направлению идиоадаптации или обратно, наступление периода новых ароморфозов, переход к специализации и т. д. Сюда никак не могут относиться те случаи, о которых мы говорили выше, когда высшие млекопитающие спасаются от появившегося врага (человека) не путем наследственного, медленного, прогрессивного изменения своих органов (или своих привычек и инстинктов), но путем быстрого изменения своего поведения (behaviour) в результате действия психики «разумного порядка» (упомянутые нами примеры «контроблав» оленей, переход слонов и бизонов от степного образа жизни к лесному и т. д.). Само собою разумеется, что в этих последних случаях никакие изменения в направлении эволюции не могут иметь места.
Морфологические закономерности 22
338
ГЛАВА X
В разобранном нами сравнительно простом случае, когда изменяющим фактором служит появление нового хищника (случай непосредственной борьбы за существование в смысле Дарвина) или нехватка в пище, направление, в котором пойдет эволюционный процесс, зависит от сложно вариирующих соотношений между изменяющим фактором (хищник) и организацией изменяющегося животного (добыча). По всей вероятности, это положение, разобранное нами для одного рода отношений между изменяющим фактором и изменяющимся животным, можно до известной степени обобщить и сказать, что биологически важная перемена в условиях существования (среде) данного вида животных является стимулом к изменению его организации; характер же изменения среды и количественное и качественное соотношение между изменением среды и строением и функциями изменяющегося организма определяют направление, в котором пойдет эволюция изменяющегося вида в данную эпоху.
Мы видели, как один и тот же стимул (преследование добычи более сильным хищником) в зависимости от степени его интенсивности меняет направление эволюции преследуемого вида тем или другим образом и направляет его филогенетическое развитие в одних случаях по пути ароморфоза, в других—по пути идиоадаптации, в третьих—по пути специализации.
Мы пришли к выводу, что, насколько мы можем судить в разобранных нами случаях, н а п_р ajs л е н и е эволюции о пр еде л я лось во-первы х, сТр оением и функциями органов ж и в о т.п о г о, .под вергающегося дейст викг^тгзм е-"~ня ю щ_и х с я условий, во-вторых, измен ё“н и ё м^в'З е-ш и е й среды и, в-третьих, с о о т н о ш е н й е м между ~эГггр"ЯТГт ером и интенсивностью изменения среды й с тр'о ением изменяющегося животного.
Значение характера организации данной группы животных в данный период их эволюции едва ли было в достаточной мере оценено зоологами эволюционистами, а между тем его необходимо постоянно учитывать при рассмотрении вопроса о направлениях эволюции. Напомним для примера разобранный нами в начале этой книги вопрос о переходе предков позвоночных от пассивного питания к активному. Мы видели, что переход этот произошел совершенно независимо друг от друга в двух группах низших черепных позвоночных, а именно, у предков круглоротых и у предков челюстных позвоночных (гнатостом). Если мы допустим, что этот переход совершился под влиянием аналогичных условий (что довольно вероятно), то мы должны будем все-таки признать, что результаты были весьма различны: в одном случае из передних жаберных дуг развился сосущий ротовой аппарат круглоротых, в другом—хватательные челюсти рыб. .
Сравнительно анатомическое исследование с полной определенностью показывает нам, что различия в направлениях эволюции ротового скелета в том и другом случаях зависели от различий в направлениях эволюции жаберного аппарата в предшествовавший период, в частности, от строения жаберных дуг. Мы знаем, что у предков круглоротых, т. е. у Entobranchiata, жаберные дуги остались нерасчлененными и образовали жаберную решетку, а у предков всех рыб. т. е. у Ectobranchiata, они расчленились каждая на четыре отдела. Именно это расчленение, связанное с развитием наружных жабр, и дало возможность передним жаберным
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ 339
дивергентных направлениях.
Рис. 97. Opisthoproctus soleatus. Глубоководная рыба с очень крупными телескопическими глазами (из Хуна).
дугам предков гнатостом превратиться в челюсти и подвесочный аппарат, а развитие ротового скелета этого типа сделало возможной всю дальнейшую прогрессивную эволюцию рыб и затем наземных позвоночных. Как известно, у круглоротых образовался сосущий рот, и этот сосущий ротовой аппарат привел к специализации на определенном роде пищи, что, по всей вероятности, послужило причиной остановки их дальнейшей эволюции, между тем как челюсти оказались органами настолько пластичными, что этот тип ротового скелета сохранился при всех дальнейших прогрессивных изменениях потомков первичных рыб.
На этом примере, при сравнении направлений эволюции у круглоротых и у рыб, мы видим насколько велико значение организации животного в тот период, когда у него наступают изменения, для всей его последующей эволюции. Таких примеров мы могли бы привести очень много (эволюция зубов, эволюция пятипалой конечности), но я на них останавливаться не буду и только отмечу, что значение организации как фактора, определяющего направление последующей эволюции животных (на которое, как я уже говорил, до сих пор недостаточно обращали внимания), особенно бросается в глаза именно при детальном изучении филогенеза и, в частности, при исследовании групп, развивавшихся в
Мы признали, что вторым фактором, изменяющим строение животных организмов, является изменение внешней среды, но при этом подчеркнули, что направление изменений зависит от соотношения между интенсивностью и скоростью изменения среды и ор
ганизацией и функциями изменяющегося животного. Так как это соотношение чрезвычайно важно, то для выяснения его дополним сказанное нами еще некоторыми примерами.
Представим себе, что мы имеем некоторое изменение физико-химической среды, например ослабление интенсивности освещения, при котором живет животное. Это изменение среды отражается в первую очередь на глазах (а также на расцветке кожи и ее придатков), так что в этом случае мы видим определенное соотношение между характером изменения среды и изменением организма. Но направление, в котором пойдет эволюция глаз (т. е. филогенетическая реакция организма) может быть весьма различно в зависимости от соотношения между интенсивностью изменения среды и строением животного. В некоторых случаях мы находим, что при постепенном переходе в темноту глаза развиваются прогрессивно и приспособляются к тому, чтобы видеть при плохом освещении или почти при полной темноте. Прогрессивное развитие глаз выразится в том, что будут постепенно увеличиваться размеры глаз, повышаться чувствительность ретины и расширяться зрачок, так что животное сможет видеть даже при слабом освещении. В этом направлении эволюировали, например, глаза ночных птиц (сов, филинов, сычей), глаза таких ночных млекопитающих, как Nycticebus, Tarsius и многие другие, а также глаза многих глубоководных рыб (рис. 97). Все эти формы характеризуются большими глазами, их зрачки могут сильно расширяться, что позволяет даже самым слабым лучам достигать ретины. Наоборот, у других форм, живущих также в тем-
340
ГЛАВА X
ноте, как, например, у упомянутых нами выше подземных млекопитающих (кротов, слепышей и т. п.), рептилий и амфибий (амфисбены и гимно-фионы), а также у многих пещерных животных и у глубоководных рыб, живущих в полной темноте (рис. 98), глаза в большей или меньшей степени
редуцировались, а в некоторых случаях и совсем исчезли.
Для нас является совершенно ясным, что во всех этих случаях исходной формой эволюции были нормальные глаза, приспособленные к видению при обычных для наземных и водных животных условиях освещения, и что стимулом к их изменению было уменьшение количества яркости света. Между тем эволюция в обеих группах перечисленных нами форм пошла в разных направлениях: в одних случаях в сторону прогрессивной эволюции, т. е. в сторону большей чувствительности глаз к свету, в других случаях—в сторону регрессивной эволюции, т. е. в сторону редукции глаз, доходящей до полной атрофии и слепоты. Едва ли мы можем сомневаться в том, что здесь различные направления эволюции зависели именно
е н и я между интенсивностью изменения среды (переходом в большую или меньшую степень темноты) и организацией животного.1 Переход в среду со слабым, даже значительно более слабым освещением ведет к прогрессивному развитию глаз; переход в совершенно темную среду2 ведет к атрофии
от количественного соотнош
Рис. 98. Onirodes glomerosus. Глубоководная рыба глаз.
с редуцированными, скрытыми под кожей глазами К аналогичным резуль-(из Элькока). татам мы придем, если
будем сравнивать изменения, вызванные у животных с различной организацией такими факто-
рами, как, например, понижение температуры, которое приводит у насекомых к тому, что у них в одних случаях перезимовывают в защищенных от холода местах взрослые формы (причем вырабатываются более или менее сложные приспособления для зимовки), в других же случаях все взрослые особи погибают с наступлением холодов, а перезимовывают
только личинки.
Приведенные выше примеры прямой борьбы за существование между хищником и добычей, а также примеры обильной пищи, находящейся в иной, но все же доступной данному животному среде, показывают, что при изменении биологической среды количественная разница в интенсивности действующего на определенную форму стимула может изменить направление эволюции. Это общее правило, повидимому, распространяется также и на случаи изменения неорганической среды.
Можно было бы привести довольно много примеров, как один и тот же изменяющийся фактор, в зависимости от того, медленно или быстро
* Мы употребляем выражение «организация животного», а не просто «строение глаз», так как отношения здесь гораздо более сложны, чем это кажется на первый взгляд, и направление эволюции определяется не только отношением изменяющего фактора к органам зрения, но и строением других органов, в первую очередь органов чувств, т. е. органов осязания, обоняния и слуха.
а Во многих случаях полная редукция глаз делается возможной благодаря прогрес-сивному развитию других органов чувств, например органов осязания или обоняния, замещающих в темноте функцию органов зрения.
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
341
наступает изменение среды, интенсивно оно или нет, определяет у близких форм различные направления эволюционного процесса (идиоа-даптацию или специализацию, ароморфоз, ценогенез или общую дегенерацию), но нам кажется, что и сказанного вполне достаточно для того, чтобы сделать нашу мысль совершенно понятной читателю. Отметим только еще раз, что результаты современных филогенетических исследований совершенно недвусмысленно говорят нам о том, что изменения эволюционного направления происходили в течение филогенеза не редко. В животном мире направление идиоадаптации встречается, повидимому, наиболее часто; довольно часто встречается также и переход от этого направления к направлению ароморфозов, а также и обратный переход, т. е. переход от направления ароморфозов к направлению идиоадаптации. То же самое мы можем сказать и относительно перехода от направления идиоадаптаций к направлению ценогенезов, а также относительно перехода от направления ароморфозов к направлению эмбриональных приспособлений (ценогенезов). Часто встречаются и случаи перехода от направления идиоадаптаций к направлению специализации. Наоборот, мы не знаем случаев перехода от направления специализации к направлению ароморфозов, точно так же как не знаем и случаев, когда животное, раз вступив на путь дегенерации, сошло бы с него и стало бы развиваться дальше в направлении ароморфозов. Повидимому, в этих последних случаях переход от одного направления эволюции к другому, если в принципе и не невозможен, то во всяком случае чем-то сильно затруднен.
Мы, конечно, никоим образом не можем утверждать, что изменение эволюционных направлений всегда происходит именно так, как мы это только что описали, но сказанного, думается нам, совершенно достаточно для того, чтобы показать, что здесь мы имеем дело с необычайно сложной проблемой, разрешить которую можно лишь путем целого ряда специальных эколого-филогенетических исследований. Наша задача была гораздо скромнее: мы лично имели в виду лишь указать читателю на то, что здесь перед исследователем открывается огромное поле интереснейших и еще почти никем не затронутых проблем и что проблемы эти далеко не так неразрешимы, как это кажется на первый взгляд.
Нам хотелось также наметить и возможный путь к их разрешению. Путь этот, как мы видели, заключается в критическом и по возможности детальном сопоставлении данных филогенетического исследования, т. е. данных сравнительной анатомии, палеонтологии и сравнительной эмбриологии, с данными физиологии и биологии (экологии) исследуемых групп животных. Те выводы, которые высказаны здесь, имеют, конечно, значение лишь предварительных положений, с которыми можно приступить к дальнейшему, более подробному исследованию. С этими оговорками я позволю себе в форме кратких тезисов резюмировать эти выводы.
ВЫВОДЫ
I. Биологический прогресс какой-либо группы животных или успешное выживание ее в борьбе за существование характеризуется, во-первых, увеличением числа составляющих ее особей, во-вторых, расширением занимаемой ею географической площади (расселение) и, в-третьих, распадением данной группы животных на большее или меньшее число подчиненных систематических групп. Понятием, противоположным биологическому прогрессу, является понятие вымирания, характеризующееся обратными признаками, т. е. уменьшением числа особей, сужением ареала распространения и уменьшением числа подчиненных систематических групп.
11. Биологический прогресс достигается четырьмя различными, но биологически равноценными направлениями эволюционного процесса:
342
ГЛАВА X
путем морфофизиологического прогресса (ароморфоз), путем приспособления в узком смысле этого слова (идиоадаптация), частным случаем которого является специализация, путем эмбриональных приспособлений, исчезающих во взрослом состоянии (ценогенез), и путем морфофизиологического регресса (дегенерация).
а) Морфофизиологический прогресс, или ароморфоз, характеризуется усложнением и диференцировкой организации животных и усложнением, диференцировкой и интенсификацией функций их активных органов (органов дыхания, кровообращения, питания и движения, а также центральной нервной системы и органов чувств), в результате чего происходит общий подъем энергии жизнедеятельности животных. Изменения строения эволюирующих в направлении ароморфоза животных носят общий характер и не представляют собой приспособлений к каким-либо определенным и специальным условиям и особенностям окружающей среды.
Ь) Приспособление в узком смысле слова, или идиоадаптация, характеризуется тем, что здесь также происходят полезные изменения в строении животных, которые соответствуют определенным изменениям внешней среды, но сама организация животных при этом н е усложняется по сравнению с организацией их предков, и подъема общей энергии жизнедеятельности, имеющего место при ароморфозе, здесь не происходит. Частным случаем идиоадаптации является специализация, когда одни признаки эволюирующих животных изменяются резко приспособительно, а другие, наоборот, остаются примитивными.
с) При эмбриональных приспособлениях, или ценоге незах полезные приспособления развиваются только у зародышей и личинок, у взрослых форм эти приспособления исчезают бесследно, так что взрослые особи обычно сохраняют строение своих предков. Но благодаря ценогенетическим изменениям, происходящим в строении зародышей и личинок, число доживающих до взрослого состояния особей становится больше, чем оно было у предков, и достигнутое таким образом увеличение общего количества взрослых особей ведет, как мы это отметили выше (I), к победе в борьбе за существование.
d) При морфофизиологическом регрессе, или общей дегенерации, достигнутые в течение предшествующих периодов сложность организации и интенсивность проявления жизнедеятельности понижаются, активные органы или целые системы активных органов редуцируются, но развиваются 'прогрессивно половая система и пассивные органы защиты, вследствие чего также достигается победа в борьбе за существование (биологический прогресс).
III. Целый ряд крупных групп животного царства, многие простейшие, губки, кишечнополостные, свободно живущие плоские, круглые и кольчатые черви, иглокожие и многие другие в начале своей эволюции развивались в прогрессивном направлении или в направлении ароморфозов. Достигнув известной высоты организации, потомки их, оставив путь ароморфозов, пошли по пути идиоадаптации, причем за громадный промежуток времени с палеозоя до настоящей эпохи их организация изменялась, но не повышалась и сохранила вплоть до настоящего времени целый ряд примитивных признаков своих давно вымерших предков.
IV. Ракообразные, паукообразные, насекомые и позвоночные животные пошли в общем по пути прогрессивной эволюции (ароморфоза). Но и здесь многие группы вступили на путь идиоадаптаций и по своей организации остались на более низких ступенях эволюции, чем их потомки и родичи. Наша современная фауна представляет собой, таким образом,
НАПРАВЛЕНИЯ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНЫХ
343
весьма сложную систему, состоящую из множества форм, по характеру своей организации и высоте строения принадлежащих к очень различным и часто весьма отдаленным от нас геологическим эпохам.
V. Выживание некоторых форм с очень примитивным строением, например многих акуловых рыб (возможно, и некоторых рептилий—примитивных ящериц, гаттерий), объясняется главным образом тем, что у них развился целый ряд эмбриональных приспособлений (крупный желток, эмбриональное кровообращение, яйцевые оболочки и т. д.). Эти ценогенезы были настолько выгодны, что благодаря им не требовалось больших изменений формы и строения взрослых организмов для их выживания. Поэтому здесь общая организация взрослых особей изменилась сравнительно мало в течение геологических периодов (филетически статические формы во взрослом состоянии).
VI. Направление общей дегенерации было, повидимому, весьма выгодным для многих форм, ибо некоторые крупные систематические группы, достигнув при своей эволюции известной высоты, перешли с пути идиоадаптаций на путь дегенерации. Сюда мы должны отнести многих сидячих кишечнополостных, паразитических плоских и круглых червей, весь подтип оболочников, усоногих раков и многих других.
VII. Любая группа животных в течение геологических эпох может изменять (и притом не раз) направление своей эволюции. Повидимому, переход с пути мбрфофйзиологической прогрессивной эволюции (ароморфоза) на путь идиоадаптации, специализации и ценогенеза, а также-переход с пути идиоадаптации на путь ароморфоза, точно так же как переход от идиоадаптивной эволюции к эволюции дегенеративной и к специализации, совершается беспрепятственно. Наоборот, специализация и дегенерация являются, повидимому, препятствиями к переходу животных на путь прогрессивной эволюции (ароморфоза).
- VIII. Все эволюционные изменения животных происходят в прямой или косвенной зависимости от изменений окружающей среды (э к т о г е-н е з). Изменение направления эволюционного процесса, т. е. переход от одного направления к другому, по всей вероятности, зависит: I) от характера и высоты организации изменяющегося животного в период, когда наступает изменение внешней среды, вызывающее соответствующее изменение животного, 2) от характера, интенсивности и скорости изменения этой внешней среды и, наконец, 3) от количественного соотношения между изменениями среды и организацией и функциями животного. Последний фактор особенно интересен, так как при одном и том же качественном изменении окружающей среды и одной и той же организации изменяющегося организма животные могут эволюировать в различных направлениях в зависимости от различной скорости, с которой наступает изменение среды, и от различной интенсивности его.
ГЛАВА XI
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ METAZOA
ВВЕДЕНИЕ
В предыдущей главе мы разобрали вопрос об общих направлениях эволюции и пришли к выводу, что приспособления эволюирующих животных к изменяющимся условиям существования осуществляются четырьмя различными способами: путем ароморфозов, путем идиоадаптаций, путем ценогенезов и путем общей дегенерации. Но перечисленные общие направления эволюции определяют собой эволюционные изменения организма, взятого как биологическое целое, ими обыкновенно характеризуются крупные систематические группы животных. Эти общие приспособления животных к изменяющимся условиям среды достигаются посредством частных филогенетических изменений строения и функций отдельных органов. Именно этими частными приспособительными изменениями отдельных органов и отдельных функций мы и займемся в настоящей главе; кроме того, так как нам удалось установить довольно значительное число типов, или «принципов» таких частных изменений, мы попытаемся внести некоторый порядок и систематическую ясность в бесчисленное разнообразие филогенетических приспособлений животных.
Во многих учебниках сравнительной анатомии мы можем найти попытки классифицировать филогенетические изменения органов животных, но обычно эта классификация проведена в них с иной точки зрения, чем предлагаемая нами, и имеет иную цель.1 В этих учебниках филогенетические изменения распределен по системам органов (органы пищеварения, органы дыхания, органы кровообращения, скелетная система, мочеполовая система и т. д.) и по систематическим группам (рыбы, амфибии, рептилии и т. д.). Мы же постараемся систематизировать приспособления животных к изменившимся условиям среды, руководствуясь несколько иными соображениями, а именно, исходя из «принципов» эволюции, т. е. распределяя их соответственно различным типам или способам филогенетического развития. Поэтому мы начнем с вопроса о том, что собственно надо понимать под словами «принципы или типы филогенетических изменений органов».
Отметим прежде всего, что проблему эволюции различных типов (modi) приспособлений мы рассматриваем с биологической точки зрения и что поэтому мы переносим центр тяжести с анатомических изменений органов на их физиологическую сущность, т. е. оцениваем каждое морфологическое изменение органа как с г> о с об развития акт и в н б й фу н к ц и и ил и п ассивног'о «Три хгпо«-
1 Классификация в учебниках проведена главным образом с точки зрения педагогики, с целью наибольшей легкости обозрения материала при преподавании.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 345
собления в полезном для всего организма „н а-~ц р а вл с н и и.
"~с этой точки зрения морфологическое изменение какого-либо органа имеет для нас значение лишь постольку, поскольку оно является усовершенствованием функции этого органа и поскольку обусловливает собою выживание организма в борьбе за существование и биотогическое процветание вида. Для данного вида безразлично, выполняется ли данная функция у потомков тем же органом, что и у предков, или же она выполняется каким-либо другим органом, лишь бы биологически, т. е. с точки зрения выживания в борьбе за существование, она выполнялась лучше, чем у предков. Этим, собственно, и отличается наша точка зрения от точки зрения специалистов физиологов: они исследуют функции органов как таковые, мы же рассматриваем их как средство, с помощью которого виды сохраняются и выживают в борьбе за существование. Биологически образованный морфолог имеет, конечно, право говорить об изменениях в строении органов (и в последующем мы сами будет широко пользоваться этим правом), но при этом надо постоянно иметь в виду, что изменяющиеся органы являются лишь орудиями, благодаря которым у потомков данной формы образуются биологически важные активные или пассивные приспособления.
Насколько мне известно, эта точка зрения впервые была выдвинута А. Дорном,1 когда им был установлен принцип смены функций. Из аналогичной установки исходил впоследствии и Л. Плате,1 2 когда он впервые сформулировал свой принцип расширения функций. Наконец, подобного же рода воззрений придерживается в своих работах Р. Гессе.3
В 1875 г. А. Дорном был открыт принцип смены функций, о котором мы будем говорить ниже. Сейчас отметим лишь вкратце, что каждый орган по Дорну обычно имеет не одну, но несколько функций, из которых одна является наиболее „развитой и рассматривается как главная функция органа, по отношению к которой все остальные его функции являются второстепенными. Во время происходящих в течение эволюции перемен в условиях существования и в окружающей среде отношения между функциями меняются, и часто случается так, что у потомков одна из второстепенных функций предков становится главной. При этом изменяется и общая функция и все строение эволюирующего органа.
В 1886 г. Н. Клейненберг4 в своей работе о происхождении аннелид установил второй принцип (тип) филогенетического развития органов, а именно, принцип субституции органов. Сущность этого принципа заключается в следующем: в течение филогенеза часто случается, что какой-нибудь.щрган с„ определенной функцией замещается у потомков другим органом с той >ке функщеи, причем орган этот расположен на том же месте,' что и у предков; так, например, в течение филогенеза скелета позвоночных хрящ замещается костью.
В 1912 г. Л. Плате был установлен важный принцип расширения функций, возбуждающий наше особое внимание уже тем, ’чтоГэтот принцип дает объяснение существованию большого числа возможных направлений эволюции у высоко стоящих животных. Плате сформулировал в 1924 г. также приинцип интенсификации функций.5 6
1 Dohrn A., Der Ursprung der Wirbeltiere und das Prinzip des Funktionswechsels, Leipzig, 1875.
2 P 1 a te L.', «Deszendenztheorie» в Handworterbuch d. Natur, Jena, 1912.
3 Hesse R-, Die Okologie der Tiere, ihre Wege und Ziele, Речь на X интернациональном конгрессе зоологов в Будапеште (1926). Ср. также его «Tiergeographie auf okologischer Grundlage» и целый ряд его специальных экологических работ.
4 Kleinenberg N., Die Entstehung des Annelid aus der Larve von Lopado-
rhynchus, Ztschr. f. wissensch. Zoologie, 1886.
6 Plate L., Allgemeine Zoologie, Jena, 1924.
346
ГЛАВА XI
Д. М. Федотов в своем исследовании об иглокожих1 установил принцип физиологической субституции, по которому орган предков замещается у потомков другим органом, несущим ту же функцию.
Если мы спросим себя, какое значение имеют только что перечисленные нами принципы, то нам придется ответить, что, в сущности, они все являются типами тех способов, по которым идет филогенетическое изменение организации животных. Так, изучая. филогенез какого-нибудь органа, мы видим, что в течение эволюции он развивался по типу дорновской смены функций, т. е. что в данном случае общая функция анцестрального органа закономерно изменялась у потомков именно этим путем. В другом случае филогенетическое изменение происходит по типу клейненберговской субституции: это значит, что здесь общая функция орТанТТ<ачёстещ{нб~^Т”йзменилась у потомков, но что она изменилась у них количественно, будучи выполнена структурно изменившимся органом.
Возникает и другой, не менее важный вопрос: достаточно ли перечисленных пяти принципов1 2 для того, чтобы объяснить все известные нам случаи филогенетических изменений животных, или же существуют еще и иные, до сих пор неизвестные нам типы филогенетического развития органов?
Систематическое изучение этого вопроса у позвоночных животных показало нам, что возможность установления новых типов филогенеза далеко не исключается.
В основе всех частных типов филогенетического изменения органов лежит м у л ь т и ф у н к ц и о н а л ь н о с т ь органа, т. е. то свойство организма, по которому каждая часть его, начиная от простейшей клетки и кончая сложнейшим органом, обладает не одной, но многими функциями, причем одна является главной функцией и употребляется очень часто, а остальные являются второстепенными функциями и обычно пускаются в ход более или менее редко.
Изучение примитивного одноклеточного животного или индиферентной эмбриональной клетки многоклеточного животного, например эпителиальной мезенхимной клетки, говорит о том, что клетки имеют не одну, но несколько необходимых для их жизни элементарных активных и пассивных функций. Клетка может перемещаться в пространстве (функция движения), воспринимать раздражение и реагировать на него (функция раздражимости), питаться и выделять различные секреты и экскреты (функция обмена веществ, функция выделения) и т. д. Мы знаем, что все эти элементарные функции одинаково жизненно необходимы для организма и что приостановление хотя бы одной из них неминуемо ведет к смерти клетки или одноклеточного животного.
Конечно, все то, что относится к единичным клеткам, приложимо и к органам животных. У тех и других мы различаем главные и второстепенные функции. Если главная функция органа изменяется количественно, то при соответственном изменении строения других частей органа, связанных с этой функцией, происходит изменение в сторону усиления или ослабления функции органа и соответствующее изменение его строения, причем общая функция органа качественно остается одной и той же. Если же главная функция органа ослабевает и на ее место становится одна из второстепенных функций, то общая функция органа становится качественно другой.
1 Федотов Д. М. О некоторых случаях морфологической и физиологической субституции. Труды второго съезда зоол., анат. и гист., Москва, 1927.
2 А. Д о р н (1875), Н. Клейненберг (1886), Плате 0912 и 1924), Д. М. Федотов, 1927.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 347
Изучая филогенетические изменения органов, мы нашли, что усиление и ослабление функций органов происходит различными способами, так что мы находим различные типы усиления и ослабления функций, т. е., говоря иными словами, различные типы, при которых эволюирующая функция данного органа не изменяется качественно, но изменяется количественно.
Точно так же и качественное изменение функций органов происходит очень различными способами, так что и здесь мы находим целый ряд вторичных типов изменений функции эволюирующего органа.
Соответственно этому мы подразделяем различные типы (принципы) прогрессивных филогенетических изменений органов на две большие категории. К первой категории филогенетических изменений мы относим те типы (принципы) эволюционных изменений, при которых г л а в-фу,.н к ,ц ия. органов остается качественно н е и з м е н н о й,_н о у с и л и в а е т с я количестве н и о. Сюда относятся: ТУ принцип интенсификации функций Плате, 2) принцип фиксации фаз А. Н. Северцова'7 3) принцип выпадения промежуточных функции~А7 Н. Северцова, 4) принцип субституции„органов Клейнен-берга, 5) принцип физиологической субституции Федотова, б) принцип уменьшения числа функций С. А. Северцова.
Ко второй категории прогрессивных филогенетических изменений мы относим те типы (принципы) эволюционных изменений, при которых главная функция эволюирующего органа изменяется качественно. Сюда относятся: 1) принцип расширения функций Плате, 2) принцип ..смены функций Дорна, 3) принцип'субституции функций А. Н. Северцова, 4) принцип активаций функций А. Н. Северцова, 5) принцип иммобилизации функций А. Н. Северцова, б) принцип сими-ляции функций А. Н. Северцова, 7) принцип филогенетического разделения органов или функций А. Н. Северцова.
Обзор типов или принципов, при которых эволюирующая функция органа изменяется лишь количественно, мы начнем с принципа интенсификации функций. Этот важный принцип, весьма кратко сформулированный Плате в 1924 г., хотя и известен многим исследователям, однако никем еще подробно не разбирался.
ПРИНЦИП ИНТЕНСИФИКАЦИИ ФУНКЦИЙ
Первый тип адаптивного изменения органов животных мы можем обозначить как «тип интенсификации функций» активных органов; для пассивных органов и признаков это будет «тип интенсификации полезности органа». Принцип этот принимался implicate уже Дорном, когда он устанавливал свой принцип смены функций (1875), в качестве же самостоятельного типа развития он был впервые выдвинут Л. Плате.1 Плате в этом случае поступил правильно, ибо мы знаем о существовании многих случаев интенсификации функций, имевших место также и тогда, когда смены функции в собственном смысле этого слова не происходило вовсе. Так как этот принцип имеет очень большое значение (я лично считаю его одним из основных принципов эволюции), то мы остановимся на нем несколько подробнее.
Мы различаем два вида интенсификации функций: ^интенсификация функций клеток и тканей и 2) интенсификация функций органов. Начнем с первого типа, т. е. с интенсификации функций клеток и тканей.
1 Термин «интенсификация функций» введен мною. Плате называет этот тип филогенетического изменения органов «физиологическим принципом усиления функций» (Allgemeine Zoologie und Abstammungslehre, Bd. 2, S. 163, 1924).
348
ГЛАВА XI
1. Интенсификация функций клеток и тканей
Говоря о мультифункциональности, мы упомянули о том, что всякая животная клетка, будет ли это индиферентная эмбриональная клетка тела Metazoa или единичная клетка Protozoa, обладает несколькими элементарными жизненными функциями. Каждая из этих функций в течение филогенеза может интенсифицироваться и соответственно этому строение клетки может частично изменяться.
В качестве простого примера возьмем интенсификацию функции защиты у корненожек. Тело простейших корненожек—Amoeba proteus
Рис. 99. Amoeba proteus (из Дбфлейна).
(рис. 99) покрыто на поверхности лишь слоем эктоплазмы. При дальнейшей эволюции начинается образование более плотной кутикулы. На следующей стадии филогенеза, представленной на рис. 100 (Difflugia), появляется примитивная раковина, образованная либо из песчинок кварца или известняка, либо из осколков панцырей диатомей, захватываемых при помощи псевдоподий внутрь и экскретируемых Difflugia на поверхность. Наконец, на рис. 101 мы видим образование уже настоящего наружного скелета в виде сплошной кремнеземной раковины, выделяемой протоплазмой Euglypha. Здесь большая прочность раковинки сочетается с большой ее легкостью.
Но в единичной клетке, например в клетке тех же Protozoa, может усилиться и функция движения, и
тогда в ней образуются сократимые волоконца или же реснички (инфузории), либЬ функция питания (образование питательных вакуоль и глотки у инфузорий), либо функция выделения (выделительные вакуоли), либо функция опоры клетки (обра-
зование внутреннего скелета у радиолярий) и т. д. Словом, каждая из отдельных функций Protozoa может интенсифицироваться в течение фило-
генеза.
Мы знаем,что при размножении Protozoa делением клетка дает начало двум дочерним клеткам и каждая дочерняя клетка продолжает существовать далее как самостоятельный и изолированный организм. У Metazoa дочерние клетки оосперма после деления не расходятся, и вследствие этого у многоклеточных возникает новый тип интенсификации функций, не встречающийся у Protozoa, а именно тип интенсификации функций путем увеличения числа клеток. Мы считаем его наиболее важным способом интенсификации функций.
У всех Metazoa и, в особенности, у всех позвоночных специализо-ванные тканевые клетки, т. е. те клетки, у которых интенсифицируется та или иная из первичных функций и соответственно этому меняется строение, не живут изолированной жизнью и обычно еще до специализации усиленно размножаются. Поэтому при интенсификации какой-либо определенной функции эта интенсификация происходит не в одной, но о д н о-временново многих связанных между собой клетках. Интенсификация органа будет тем значительнее, чем
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 349
большее число одновременно изменяющихся клеток будет ею захвачено.
Специализация клеток многоклеточных организмов, которую мы наблюдаем при всяком онтогенетическом развитии, заключается в том, что в зародышевых клетках, из которых разовьется впоследствии определенная ткань, одна из нормальных функций начинает постепенно гипертрофироваться и делается главной фунцией клетки, в то время как остальные функции зародышевой клетки сохраняются в тех пределах, в которых они являются необходимыми для жизни клеток.1
Прекрасным примером интенсификации функции тканей путем прогрессивных гистологических изменений является филогенез мышечной ткани.
Рис. 100. Первичная, образованная из отдельных песчинок раковина Difflugia (из Делажа).
Рис. 101. Сплошная кремнеземная раковина Euglypha (из Делажа).
У зародышевой мезенхимной клетки, превращающейся в мускульную клетку, интенсифицируется функция движения и соответственно этому меняется ее форма и строение: клетка удлиняется и принимает вид веретена, в ней появляются сократимые волоконца, соответствующие миофанам простейших, и индиферентная мезенхимная клетка превращается в гладкое мускульное волокно. Из этих гладких мускульных волокон в течение филогенеза постепенно образуются поперечнополосатые волокна. Рис. 102 иллюстрирует нам ход этого процесса. Здесь мы видим (рис. 102 А) мышечную клетку паразитического червя, в которой образовались гладкие мышечные волоконца; часть клетки, в которой лежит ядро, еще состоит из индиферентной протоплазмы. Это—первая ступень интенсификации двигательной функции клетки. Следующая стадия повышения двигательной функции изображена на рис. 102 В, где показаны два гладких мускульных волокна, взятых из кишечного канала саламандры. Мы видим, что индиферентная протоплазма здесь уже совсем исчезла и мезенхимная клетка превратилась в гладкие мускульные волоконца; ядро клетки лежит в середине волокна. На рис. 102 С представлены попереч
1 В этом именно и заключается главное отличие дорновской смены функций от разбираемого нами типа интенсификации функций: при смене функций Дорна функция, которая прежде была главной, не сохраняется, но сначала ослабевает, а затем и вовсе исчезает.
350
ГЛАВА XI
нополосатые волокна из мускулатуры сердца млекопитающих. Здесь интенсификация мускульного сокращения достигла своих высших пределов. Отметим, что при всех этих превращениях остальные нормальные функции индиферентной клетки, т. е. способность питаться, воспринимать раздраженней реагировать.на него, способность выделять продукты обмена и т. д., сохраняются. То же наблюдается и при развитии клеток органов чувства, в которых гипертрофируется способность реагировать на определенный вид раздражения, вследствие чего все строение этих клеток соответственно изменяется (образование чувствующих волосков или ресничек, образование палочек и колбочек в клетках ретины и т. д.). В тех эпителиальных клетках, которые в течение онтогенеза превращаются
Рис. 102 А, В, С. Три последовательные филогенетические стадии интенсификации двигательной функции (сократимости) мускульных клеток.
А —мышечная клетка паразитического червя, в которой образовались гладкие мышечные волоконца (f );часть клетки, в которой лежит ядро (п) еще состоит из индиферентной протоплазмы (частичная интенсификация двигательной функпии). В—следующая стадия интенсификации двигательной функции мускульной ткани; в гладких мускульных волокнах (взятых из кишечного канала саламандры) индиферентная протоплазма исчезла совсем и мезенхимная клетка целиком превратилась в гладкие мускульные волокна; ядро клетки (п) лежит в середине волокна. С—поперечнополосатые волокна из мускульной стенки сердца млекопитающих; здесь интенсификация двигательной функции мускульной ткани достигла высших пределов; fi—поперечнополосатые мускульные волоконца; п—ядра мускульных клеток (из Маурера).
в железистые клетки, гипертрофируется секреторная функция и соответственно этому меняется строение.
Подобное же усиленное развитие одной из нормальных функций клеток находим мы и при образовании пассивных гистологических приспособлений в тканевых клетках; так например, способность зародышевых мезенхимных клеток до известной степени противостоять растяжению интенсифицируется путем образования соединительнотканных фибрилл: этим именно путем в теле многоклеточных животных происходит образование различных видов соединительной ткани.
В предыдущем мы рассматривали интенсификацию функций и изменение гистологического строения клеток, посредством которого она дости
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 351
гается, лишь с онтогенетической точки зрения. Однако мы имеем все основания предполагать, что и в филогенезе процесс этот осуществлялся тем же путем. Само собой разумеется, что при современном состоянии наших знаний мы еще не в состоянии проверить правильность этого предположения палеонтологическим исследованием. Мы считаем, однако, весьма вероятным, что в очень отдаленную от нас эпоху в клеточных колониях, которые, как нам думается, представляли собою предков известных нам примитивных Metazoa, некоторые из первичных функций известных групп клеток постепенно интенсифицировались, так что в одних группах клеток интенсифицировалась функция движения (из этих клеток впоследствии образовались предшественники мускульных клеток), в других—функция выделения (предшественники железистых клеток) и т. д. Т а к и м образом, образование различного рода тканей в теле Metazoa обусловлено происходившей в течение филогенеза интенсификацией отдельных первичных функций в индиферентных клетках тела предков многоклеточных животных.
II. Интенсификация функций органов
Прогрессивное развитие не слишком сложно построенных органов также покоится на принципе интенсификации функций. Мы знаем, что в очень многих случаях прогрессивной эволюции мускулов, нервов, желез и органов высших чувств филогенетические изменения заключаются именно, в том, что у потомков данной формы определенный орган функционирует более интенсивно, чем у предков. Эта интенсификация функций может происходить двумя различными путями: во-первых, может измениться прогрессивно строение всех клеток ткани, из которой состоит эволюирующий орган, в результате чего усилится функция органа. Так например, мы вправе считать, что во многих мускулах примитивных позвоночных животных гладкие мускульные волокна превратились в более интенсивно сокращающиеся поперечнополосатые волокна, ибо мы знаем, что мускулы кишечного тракта в его заднем и среднем отделе состоят из гладких мускульных клеток, в то время как в жаберном отделе кишечника висцеральная мускулатура бранхиального аппарата состоит из поперечнополосатых волокон. Происхождение и положение переднего отдела кишечной мускулатуры, а также его иннервация висцерально двигательными нервами заставляют час думать, что первоначально отдел этот также состоял из гладкой мускулатуры и что изменение его гистологического строения явилось результатом интенсификации функций.
Во-вторых, интенсификация функций может быть достигнута увеличением числа компонентов, из которых состоит данный орган. Так, интенсивность секреторной функции железистого органа достигается увеличением общего числа секреторных клеток, или же увеличением количества секреторных долек. Мы можем принять с большой долей вероятности, что, например, деятельность заушных желез жвачных животных усиливается в течение филогенеза именно этим путем; то же мы можем сказать и относительно интенсификации функций больших млечных желез млекопитающих, которые, как известно, развились из маленьких трубчатых желез. Аналогичным образом увеличивается и дыхательная поверхность легких при прогрессивном развитий легких наземных позвоночных, где в течение филогенеза число легочных альвеол постепенно увеличивается путем размножения клеток легочного эпителия.
352
ГЛАВА XI
Можно было бы думать, что в только что приведенных случаях гистологической интенсификации мы имеем перед собою скорее примеры не интенсификации, но смены функций. Однако мы думаем, что правильнее рассматривать их именно как случаи интенсификации, ибо здесь первичные функции клетки не исчезают, как это бывает с анцестральной главной функцией при развитии по типу смены функций А. Дорна, но сохраняются, как важные функции хрящевой, мускульной, нервной и т. п. тканей.
В очень многих случаях главная функция органа (см. ниже «принцип смены функции» А. Дорна) усиливается путем интенсификации функций отдельных клеток этого органа или же путем усиленного размножения этих клеток. Мы знаем, что в течение филогенеза грудные мускулы птиц стали гораздо более сильными, чем грудные мускулы их рептилеобразных предков. Это зависит от того, что у птиц значительно возросло общее число мускульных волокон. Мы знаем об этом потому, что то же произошло и у ныне живущих форм. У птиц интенсивность полета в значительной степени обусловлена развитием грудной мускулатуры; развитие же грудной мускулатуры стоит в тесной связи с развитием киля грудины, к которому она прикрепляется. По наблюдениям Ббкера,1 своего наиболее сильного развития crista sterni достигает у птиц с порхающим полетом (Schwirrflug), т. е. с полетом, связанным с наиболее частым движением крыльев. Ббкер дает схему, изображенную на рис. 103, иллюстрирующую постепенное увеличение crista sterni у различных птиц, начиная с козодоя (2) и кончая колибри (4 и 5), т. е. у птиц с постепенно усиливающимся движением крыльев. Мы имеем здесь яркий пример интенсификации функций (в данном случае функции полета) путем увеличения путем увеличения числа му
скульных волокон в грудной мускулатуре птиц и связанным с ним увеличением размеров киля грудины).
Этот же тип интенсификации функций путем одностороннего или разностороннего роста органов играет большую роль также и при адаптивном изменении пассивных органов животных. Мы знаем, например, какое большое значение имеют некоторые скелетные образования животных в качестве органов пассивной защиты. У низших позвоночных к такого рода органам относятся плакоидные чешуи и первичный череп селахий, костные чешуи и покровные кости Osteichthyes ит.д. Мы видим, как по мере филогенетического усиления функции защиты все эти образования постепенно становятся крупнее и толще, т. е. как они растут либо односторонне, либо во все стороны, что сводится к размножению клеток. Мы могли бы привести много примеров усиления защитной функции у рыб путем образования крупных чешуй или покровных костей, но мы ограничимся лишь тем, что отметим попутно образование большого, крепкого и толстого панцыря, состоящего из сросшихся между собой чешуй, у многих ископаемых рыб и образование большого головного панцыря у осетровых. Наблюдая увеличение разме-
} 2 2 4 5
Рис. 103. Схематическое изображение crista sterni у различных птиц с постепенно усиливающимся движением крыльев (сравнительно-анатомический ряд).
1—грудина ласточки; 2—грудина козодоя; 3—грудина стрижа; 4—грудина колибри Agyrtia apez; 5—грудина колибри Chryosolmpis elotus (из Ббкера, 1935).
числа компонентов органа (в данном случае
i В б k е г Н., Vergleichende biologische Anatomie der Wirbeltiere, Jena, 1935.
О ПРИНЦИПАХ или ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 353 ров чешуй у рыб, мы часто видим, что они разрастаются на тех именно частях тела, где они прикрывают собою хрящевой скелет животного и где они поэтому бывают неподвижны. Так, например, у осетровых рыб хрящевой плечевой пояс конечностей прикрывается двумя костями, cleitrum и clavicula, которые по своей форме и строению являются разросшимися чешуями. Здесь мы имеем перед собою две типичные туловищные чешуи, которые вследствие своего определенного отношения к хрящу плечевого пояса сильно увеличились в размерах. У высших Osteichthyes обе эти кости уже потеряли свое сходство с чешуями и превратились в обычные покровные кости.
Тот же принцип усиления функции органа путем одностороннего роста (местный процесс клеточного размножения) мы находим и в тех скелетных частях организма, которые служат для прикрепления мускулов или для причленения подвижных скелетных образований. Функция опоры или сопротивления этих частей усиливается благодаря образованию различных выростов (костные гребни в черепе у различных млекопитающих, отростки на черепе у селахий и Osteichthyes), т. е. путем одностороннего роста отдельных частей черепа.
Несколько иное значение имеют такие сами по себе пассивные, но подвижные и функционально связанные с мускулами скелетные образования, как, например, части скелета свободных конечностей, кости ротового скелета и жаберного-аппарата позвоночных и т. п. Их односторонний рост часто обусловливает собою интенсификацию активной функции всего органа, частями которого они являются. Так, например, интенсивность двигательной функции задних конечностей бесхвостых амфибий усиливается благодаря удлинению проксимальных тарсальных косточек, так называемых astragalus и calcaneus (tibiale и fibulare, рис. 105). Сравнивая между собой рис. 104 и 105, мы видим, насколько длиннее эти кости у лягушки, чем у саламандры.
То же самое мы находим и у пальцеходящих млекопитающих, у которых как metatarsalia, так и metacarpalia сильно выросли в длину.
К этому же типу интенсификации функций близко примыкает интенсификация путем увеличения общего числа повторяющихся в теле Metazoa органов.
Нам представляется весьма вероятным, что сегментация тела метамерно построенных Metazoa произошла именно этим путем. Можно думать, что первоначально туловищная мускулатура первичных хордат состояла из двух еще не сегментированных боковых мускульных тяжей, и что такая важная способность животного, как способность одновременно сгибать отдельные части тела в различных направлениях, т. е. производить змееобразные движения, образовалась вследствие того, что туловищная мускулатура распалась на отдельные мускульные сегменты (миотомы), из которых каждый иннервировался особой парой спинных нервов. Мы знаем, что у многих животных, например среди рыб—у угрей, у морских игл, среди амфибий—у гимнофионов, у Amphiuma и Menopoma, среди рептилий—у безногих ящериц и у змей, усиление гибкости тела достигается именно путем увеличения числа туловищных сегментов. Нам известно также, что у птиц подвижность головы является следствием увеличения числа шейных позвонков, а у рыб и хвостатых амфибий усиление функции метанефроса достигается, путем увеличения числа нефридий. Сюда же относятся все многочисленные случаи увеличения числа фаланг на пальцах передних и задних конечностей у морских рептилий, например на пальцах ихтиозавров, у которых число фаланг сильно возросло по сравнению с примитивными рептилиями. Если мы сравним между собою конечности вымерших морских рептилий, изображенные Морфологические закономерности 23
354
ГЛАВА XI
на рис. 106 А, Ви С, что увидим, что у них постепенно увеличивается число фаланг, начиная от Mososaurus и кончая Ichtyosaurus; при этом вследствие расщепления краевых пальцев у них образуются еще и добавочные пальцы также с большим числом фаланг.1 У ихтиозавров конечность, как известно, превратилась в гибкую лопасть. Вполне понятно поэтому, что появление добавочных пальцев расширяет эту лопасть, а увеличение числа фаланг повышает ее эластичность, вследствие чего повышается полезность ее функции.
Прекрасным примером интенсификации функций путем умножения органов является также развитие глазной мускулатуры у позвоночных животных. Еще со времени фан Вайя1 2 стало известно, что глазные мускулы позвоночных развиваются из передних миотомов головы, и мы имеем все основания думать, что эти передние миотомы, которые первоначально
были морфологически и функционально идентичны всем остальным!
Рис. 105. Скелет задней конечности Rana esculenta. Обратить внимание на относительно крупные размеры tibiale и fibulare (из Видерсгейма по Гауппу). tib.—tibiale; fib.—fibulare; tars. I — tarsale I; tars П+Ш—tarsale II+III; e—centrale;
p. h.—praehallux; I—V—пальцы 1—5.
Рис. 104. Скелет задней конечности Sa-lamandra maculosa. Обратить внимание на относительно незначительные размеры tibiale и fibulare (из Вершлюса, 1912). t.—tibia; f.—fibula; int.—intermedium; ‘ tib.>— tibiale; fib.—: fibulare; c.—centrale; tars. I4-II—tarsale 14*11; tars. V—tarsale V; met. I —met. V—metatarsale I—V; I—V—фаланги пальцев 1—5; for. art.—отверстие для сосуда.
каудально от них расположенным сегментам, изменили в течение филогенеза свою функцию (принцип смены функций А. Дорна) и превратились в глазные мускулы. При этом мы находим, что из первого миотома образуется не один глазной мускул, как из второго или из третьего миотома, но целых четыре (m. т. recti internus, superior, inferior и т. obliquus superior, которые все иннервйруются n. ocuiomoto-
1 У котилозавров формула фаланг была 2 3 4 5 3, у Eurypterygius communis число фаланг значительно возросло. Кроме того у Eyrypterygius communis имелось девять (вместо пяти) пальцев.
2 Van W i j h e J. W., Ober die Mesodermsegmente und die Entwicklung der Nerven des Selachierkopfes, Amsterdam, 1882. Ср. также A. H. Северцов, 1898 и Б. С, Матвеев, 1915.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 355
rias). Благодаря увеличению числа дериватов этого миотома их функция, т. е. приведение в движение глазного яблока, сильно повышается.
Весьма вероятно, что способы, которыми осуществляется интенсификация функций органов, разнообразнее и многочисленнее, чем нам это удалось показать на только что приведенных примерах, и что подробный анализ фактов филогенетической интенсификации функций выявил бы нам еще и другие способы интенсификации, в особенности, если бы мы занялись изучением многочисленных случаев интенсификации функций у беспозвоночных животных. Мы ограничимся, однако, лишь вышеприведенными примерами в надежде, что нам все же удалось показать огромное значение
Рис. 106. Интенсификация функции движения путем увеличения числа органов (фаланг) у примитивных рептилий.
А—передняя конечность Mososaurus; В—задняя конечность Taumatosaurus; С—передняя конечность Ichtyosaurus (из Вертлюса, по Циттелю, 1927).
этого принципа. Мы увидим ниже, что этот принцип входит в качестве главного действующего начала во все остальные принципы прогрессивного филогенетического развития животных.
Мы должны здесь упомянуть о противоположном принципе филогенетического развития, а именно о принципе филогенетического ослабления функций. Сущность его можно вкратце выразить следующим образом: каждая функция как клетки, так и органа может в течение эволюции сделаться более слабой. При регрессивной эволюции органа могут целиком исчезнуть как большинство его функций, так и весь орган или же те или иные части органов. При регрессивной эволюции клетки каждая из первичных, т. е. из необходимых для жизни клетки функций, может ослабеть до известного минимума, но не исчезнуть совсем. Ослабление функций может итти теми же путями, 23*
356
ГЛАВА XI
что и интенсификация функций, а именно путем регрессивных гистологических изменений, путем уменьшения размеров органов и путем уменьшения числа органов. Во избежание повторений здесь мы не будем приводить примеров регрессивных процессов в филогенезе животных, ибо в главе XV этой книги мы займемся подробным рассмотрением вопроса о редукции органов.
Мы можем следующим образом резюмировать наши выводы относительно прогрессивного филогенетического развития путем интенсификации функций организма.
I. Каждая индиферентная клетка Protozoa или зародыша Metazoa мультифункциональна, т. е. обладает многими первичными жизненно необходимыми функциями: подвижностью, раздражимостью, способностью ассимилировать пищу, способностью выделять секреты и экскреты и т. д. У Protozoa и у клеточных комплексов, происходящих из индиферентных зародышевых клеток, одна из первичных функций может интенсифицироваться и сделаться главной функцией клетки или ткани, Образованной этими клетками. Интенсификация главной функции в клетке осуществляется путем соответствующего этой функции изменения в строении клетки, т. е. путем образования тех или иных органелл: сократимых волокон, опорной субстанции, нервных и соединительнотканных фибрилл, выделительных вакуолей ит. д. Остальные первичные функции сохраняются в этих односторонне специализованных клетках в той степени, в которой они необходимы для жизни клетки, но для этого рода функций обычно не развивается никаких специальных органелл. Благодаря такой односторонней интенсификации одной из нормальных первичных функций клеток осуществляется гистологическая диференциров-ка тканей животного организма. У нас имеются все основания считать, что филогенетическое развитие тканей произошло именно этим путем, т. е. путем интенсификации тех или иных первичных функций клеток.
II. По тому же принципу идет и прогрессивное филогенетическое развитие просто построенных органов животных. При этом либо усиливается главная функция отдельных гистологических составных частей органа (превращение гладких мускульных волокон в поперечнополосатые, более совершенное строение клеток органов чувств и т. д.), либо увеличивается число этих составных частей. Сюда же относится и изменение формы пассивных и активных частей организма (односторонний рост), которое ведет к интенсификации общей функции органа.
Перейдем теперь к следующему типу филогенетических изменений, к типу, в котором также происходят лишь количественные изменения главной функции эволюирующего органа и который мы обозначаем как «принцип фиксации фаз».
ПРИНЦИП ФИКСАЦИИ ФАЗ
При исследовании изменений функций органов, происходящих при филогенетическом развитии животных, нам часто встречаются случаи, когда функция каког о-л ибо сло'жно построенного органа состоит из единичных, повторяющихся и следующих друг за другом фаз. Чаще всего это явление наблюдается при изучении процесса ходьбы у наземных позвоночных.
Мы знаем, например, что примитивные наземные позвоночные были типично стопоходящими животными, чрезвычайно медленно передвигавшимися в пространстве, наполовину ступая, наполовину ползая на брюхе: среди ископаемых—это стегоцефалы, среди ныне живущих—четвероногие наземные хвостатые амфибии, например саламандры. У происшедших от стегоцефалов рептилий конечности удлинились (интенсификация функций движения), вследствие чего животные смогли перейти к ходячему
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 357 и бегающему способу передвижения. При этом очень многие формы остались, наподобие своих предков, вполне стопоходящими животными и касались поверхности земли в состоянии покоя и при некоторых фазах движения всей ладонной и подошвенной поверхностью своих конечностей (ринхоцефалы, наземные и болотные черепахи, крокодилы и многие ящерицы).
Но у некоторых других ящериц переход к дигитиградному, т. е. к паль-цеходному движению, уже осуществился: при беге и при прыжках, т. е. при более быстром передвижении в пространстве, их tarsus и metatarsus, точно так же как carpus и metacarpus уже отделяются от почвы, так что конечности этих форм касаются земли лишь вентральной поверхностью своих пальцев. Так как при всяком поступательном движении главная работа приходится на долю задних конечностей, то мы обычно находим, что изменение способа передвижения морфологически выражается преимущественно в том, что у быстро бегающих форм более или менее резко удлиняются именно средние metatarsalia (Phrynocephalus, Calotes, Chlamydosau-rus, Draco volans и т. д.).
Среди примитивных рептилий типично стопоходящими формами были тяжеловесные, неуклюжие, медленно передвигающиеся тероморфы, такими же типично стопоходящими формами были и их потомки, первичные плацентарные млекопитающие (Insectivora, Creodontia, первичные хищники), такими же остались и многие современные представители этой группы.
Мы хорошо знаем, однако, что даже при очень медленном движении типично стопоходящего медведя, точно так же как и типично стопоходящего человека, бывают фазы, когда пятка целиком отделяется от поверхности земли и когда тело животного опирается только на вентральную поверхность пальцев; таким образом, в этом стопоходном движении бывают пальцеход-ные фазы. При быстром движении (беге) стопоходящего млекопитающего мы можем наблюдать, как это я наблюдал у выхухоли, у землеройки и у некоторых других форм, что животное касается земли только вентральной поверхностью своих пальцев и переходит, таким образом, в о время бега от стопоходного к пальцеходному образу передвижения. Такой переход от одного рода движения к другому можно легко наблюдать и при собственном движении, чередуя бег и медленную ходьбу: человек при медленной ходьбе является стопоходящим, при быстром беге—пальцеходящим животным.
Наблюдая бег любого стопоходящего животного, в том числе, конечно, и человека, мы видим, что здесь некоторая определенная, при медленном движении периодически повторяющаяся фаза его делается главно йсоставной частью движения. Так, человек при беге и при танце становится в той же степени пальцеходящим животным, в какой обычно является собака или кошка, а если мы захотим продолжать наше сравнение и дальше, то мы можем сказать, что, например, танцовщица классической школы, когда ей приходится танцевать, опираясь лишь на конец носка, вполне уподобляется в своем движении типично копытному животному, или говоря более вежливым языком, легкой газели. Мы знаем, что очень многие хищники, например собаки, кошки и т. п., перешли в течение своего филогенеза от стопоходящей формы движения к пальцеходящей, так что у них этот вид движения, представляющий собою у предков лишь переходящую фазу обычной ходьбы, становится уже постоянным признаком вида. Даже находясь в состоянии покоя, животные эти опираются всей тяжестью тела только на вентральную поверхность пальцев.
Крайний тип этого рода движения мы находим у парнокопытных и непарнокопытных, где животные касаются поверхности земли лишь превратившимися в копыта когтями.
358
ГЛАВА XI
Рис. 107 А, В, С, D. Четыре последовательных фазы движения конечностей стопоходящего млекопитающего (бурого медведя) при ходьбе. Фотография из фильма Ufa, из которого взяты только наиболее характерные фазы движения лап медведя. Все промежуточные фазы опущены.
Вышеизложенное можно иллюстрировать фотографиями, приведенными на рис. 107, 108, 109 и ПО. На рис. 107 А, В, С и D переданы последовательные фазы движения при ходьбе типично стопоходящего млекопитающего, именно бурого медведя..На фотографии 107 А мы видим, что три лапы медведя (обе передние и левая задняя) касаются земли всей своей ладонной поверхностью (стопоходная фаза движения); правая задняя лапа только начинает приподниматься—плантарная поверхность пальцев касается земли (полупальцеходная фаза движения конечности). На фотографии 107 В представлена следующая по времени фаза движения медвежьих лап. Левая передняя и левая задняя лапы находятся в прежнем положении; правая передняя начинает приподниматься (полупальцеходная фаза); правая задняя лапа поднялась несколько выше. На фотографии 107 С левая передняя и левая задняя—в прежнем положении, правая передняя приподнята, а правая задняя имеет вполне дигитиградное положение.
На фотографии 107 D правая передняя лапа уже опустилась совсем, так что теперь обе передние лапы касаются земли своими ладонями; левая задняя лапа, так же как и йа предыдущей фотографии, имеет вполне стопоходное положение, правая—вполне пальцеходное.
Если мы проследим по этим фотографиям следующие друг за другом различные фазы движения каждой конечности, то увидим, что в каждой из них можно ясно различить следующие отдельные положения: 1) вполне стопоходное, 2) полу-пальцеходное (поднятие пятки), 3) пальцеходное, .4) движение конечности в воздухе (шаг), 5) пальцеходное (опускание лапы на землю), б) полупальцеходное
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 359
положение, 7) стопоходное положение и т. д. Таким образом, мы можем воочию наблюдать, как у типично стопоходящего животного при ходьбе все время чередуются стопоходные и паль-цеходные фазы движения.
То же самое мы видим, наблюдая за движением ноги человека и человекообразной обезьяны (шимпанзе). Так, на рис. 108 А, В, С и D совершенно отчетливо видно, как у человека и у обезьяны при каждом шаге чередуются стопоходная фаза, характерная, как это было и у медведя, для стадии покоя, с пальцеходной фазой движения.
Фотографии на рис. 109 А, В, С и D показывают нам движение тигра при медленной ходьбе. Здесь мы можем наблюдать, как пальцеходное положение конечностей, которое у медведя было лишь одной из фаз движения (фазой 2-й, 3-й, 5-й и б-й), переходит в нормальное состояние как во время покоя, так и во время движения (рис. 109 А—D). Внимательно рассматривая эти фотографии, мы видим, что и при типично пальцеходном движении можно различить две друг за другом следующие фазы: при одной животное касается земли вентральной поверхностью своих фаланг (то же в состоянии покоя), при другой—оно опирается на землю только концами пальцев.
Это последнее положение делается нормальным с о-стояниемкак при покое, так и при движении (ходьба и бег) у копытных млекопитающих. Так, антилопа касается земли при ходьбе уже только концами фаланг пальцев (рис. ПО А—D); то же можно наблюдать и у стоящей позади нее зебры.
Чтобы совершенно точно изобразить изменения положения конечностей, происходящие при фиксации фаз движения млеко-
Рис. 108 А, В, С, D. Четыре последовательных фазы движения двуногого стопоходящего млекопитающего. На фотографии изображены различные фазы движения ног человека и трех шимпанзе (из фильма Ufa).
360
ГЛАВА XI
Рис. 109 А, В, С, D. Четыре последовательных фазы движения пальцеходящего млекопитающего. На фотографиях изображено положение лап тигра в различных фазах движения, (из фильма Ufa).
питающих, мы приводим еще один добавочный рисунок, а именно рис. 111 А, В, С. На рис. 111 А изображено движение (4 фазы движения: 1, 2, 3, 4) костей задней конечности медведя (стопоходное движение с пальцеходными фазами), на рис. 111 В—движение конечности тигра(чисто дигитиградное— пальцеходное — движение), а на рис. Ill С — движение лошади (унгулиградное движение). Здесь мы можем видеть гораздо более отчетливо, чем на фотографии, как фиксируется пальцеходная фаза движения у Felidae и у Perisso-dactyla.
Биологическое значение тех изменений, которые имели место при переходе от стопоходного движения к пальцеходному и от этого последнего к движению на концах копыт (унгулигр адному движению) нам совершенно понятно: животное постепенно приобретает возможность все более и более быстрого движения на поверхности земли, ибо к двучленной системе рычагов стопоходящей конечности (stylopodium + zeugopodium) прибавляется еще третий член, а именно, carpus + metacarpus для передней конечности и tarsus+metatarsus для задней. Таким образом, длина всей системы рычагов сильно возрастает. У примитивных стопоходящих живбтных это удлинение системы рычагов было лишь ф а-культативным явлением (см. схему на рис. 111), у их дигитиградных потомков оно превратилось в постоянное состояние, и конечности этих последних форм превратились из нормально двухрычаго-вого в трехрычаговый орган движения.
В предыдущем мы рассматривали лишь те изменения, которые происходили в скелетных частях конечностей. Само собой разумеется, однако, что параллельно с изменениями во взаимоотношении частей скелета имеют место также и изменения в мускулатуре, особенно в тех мускулах, которые не
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 361
посредственно связаны с новым, третьим членом упомянутой нами системы рычагов, а именно с саг-pus-j-metacarpus или с tarsus+me-tatarsus.
Только что установленную нами закономерность можно вкратце сформулировать следующими словами: эволюция неко-
торых функций иногда происходит таким образом, что сначала новая функция появляется только как одна из периодически повторяющихся фаз общей функции данного органа, в течение же дальнейшей эволюции этапре-ходящая фаза общей функции закрепляется и превращается в постоянное, нормальное состояние органа как во время его функционирования, так и во время покоя; общий характер функции остается при этом без изменения, т. е. таким же, каким он был у предков, сама же функция интенсифицируется.
Изучая филогенетические изменения функции движения у наземных позвоночных, мы можем встретить, кроме вышеперечисленных, немало и других примеров принципа фиксации фаз. Мы уже упоминали о том, что ящерообразные рептилии при быстром движении—беге и прыжках — переходят от стопоходного рода движения конечностей к пальцеход-ному; мы можем добавить к этому, что и динозавры происходят от стопоходящих форм, так что и у них также в свое время произошел переход от стопоходного движения к пальцеходному. Судя по строению конечностей, можно сказать, что у тяжеловесных Atiantosauridae переход этот осуществился не целиком и что эти звери остановились в своем развитии на полу-пальцеходной стадии движения.
Рис. ПО А, В, С, D. Четыре последовательных фазы движения конечностей непарнокопытного млекопитающего (антилопы) при медленной ходьбе (из фильма Ufa).
•362
ГЛАВА XI
Подобная полупальцеходная форма конечностей наравне с чисто диги-тиградной формой встречается среди ископаемых рептилий у двуногих Theropoda. Такие же формы как Compsognathus и Struthiomimus
Рис. 111. А, В, С. Схематическое изображение четырех последовательных фаз (1—4) движения задней конечности различных млекопитающих при ходьбе.
А—движение конечности медведя: чередование стопоходной (1) и пальцеходной (2) фаз; В—тигр: пальцеходное движение; С—лошадь: унгулиградное движение (из А. Н. Северцова, 1931).
были уже вполне пальцеходящими животными, и совершенно то же мы можем сказать относительно стегозавров и цератопсид.
На основании общего сходства в строении скелета конечностей только что перечисленных нами ископаемых можно заключить, что у этих реп-талий преобразование скелета конечностей шло по тому же типу, что и у
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 363 млекопитающих, т. е. по принципу фиксации дигитиградной фазы движения, именно той фазы движения, которая у предков была лишь одной из ритмически повторяющихся фаз, когда движение факультативно становилось более быстрым.
Мы хорошо знаем, что все птицы по существу пальцеходны и что при передвижении по земле они касаются субстрата только вентральной поверхностью своих пальцев. Так как птицы вне всякого сомнения происходят от стопоходящих рептилий, то нам приходится признать, что и здесь также переход от стопоходящего движения к дигитиградному произошел по принципу фиксации фаз, причем весьма вероятно, что этот переход осуществился не у непосредственных, но, наоборот, у еще очень отдаленных рептилеобразных предков птиц.
Известно, что и хвостатые амфибии являются типично стопоходящими животными; у бесхвостых амфибий, однако (судя по наблюдениям, которые я производил над движениями нашей Rana arvalis), произошла очень своеобразная комбинация стопоходного и пальцеходного движений. Передние конечности Rana, повидимому, остаются всегда стопоходящими, т. е. лягушка как при медленном, так и при быстром движении ставит на землю передние лапки всей их пальмарной поверхностью вниз.1 Что же касается задних конечностей, то в этом отношении лягушка в состоянии покоя, подобно стопоходящим животным, касается почвы всею подошвенною поверхностью задних лапок, включая и отдел calcaneus—astragalus; при медленном движении, например, когда лягушка взбирается на более высокую плоскость и при этом поднимает одну лапку за другой, пятка задних конечностей сначала приподнимается, так что животное в этот момент касается субстрата лишь вентральной поверхностью своих пальцев (дигити-градная фаза движения), и только затем уже лапка совсем отделяется от земли. После этого то же движение, т. е. приподнимание сначала пятки, а вслед за нею и пальцев, повторяет и вторая лапка. Припрыжке сначала приподымаются пятки (astragalus и calcaneus) обеих задних лапок одновременно и отделяются от почвы, затем быстрым движением отделяются от почвы кончики пальцев, так что здесь мы собственно наблюдаем то же движение, что и примедленой ходьбе, с той лишь разницей, что при прыжке это движение происходит в обеих лапках одновременно и, конечно, очень быстро.
Таким образом, мы видим, что лягушка в состоянии покоя является вполне стопоходящим животным; во время передвижения по суше ее передние конечности остаются стопоходящими и только задние, да и то лишь в момент прыжка, делаются частично пальцеходящими, т. е. у лягушки дигитиградная фаза движения фиксируется только в задних конечностях и только во время очень быстрого движения (прыжка).
Весьма вероятно, что принцип фиксации фаз играл немаловажную роль и при эволюции движения плавников у рыб, но процесс этого движения еще не настолько хорошо изучен, чтобы мы могли позволить себе высказаться более определенно по этому вопросу.
Впрочем, по последним исследованиям моего ученика А< Н. Дружинина,2 этот принцип несомненно играл весьма существенную роль при превращении плавника рыбообразного предка Quadrupeda в наземную конечность. В основу происхождения пятипалой конечности наземных позвоночных Дружинин в своей схеме (рис. 112) кладет двуосную конечность ископа-
1 Относительно прыжка, вследствие большой быстроты движения передних лапок, я не могу утверждать этого наверное.
’Дружинин А. Н., Общее в строении конечностей Dipnoi и Quadrupeda, Труды лаборатории эволюц. морф. Акад. Наук СССР, Т. I вып. 1, 1933. Д р у-ж и н и н А. Н., Новые данные об исходном положении конечностей Tetrapoda, Труды 2-го съезда зоол., анат. и гистол., 1925.
364
ГЛАВА XI
Рис. 112 А, В, С. Фиксация фаз движения плавника при его превращении в наземную конечность.
А и В—различные фазы движения плавника Ceratodus в момент плавания (А) и в момент опоры (В); С—фиксированное как при плавании, так и при опоре положение конечностей аксолотля,
А—Ceratodus во время плавания: грудной плавник приподнят преаксиальным краем вверх и обращен кнаружи своей первично вертикальной поверхностью; все скелетные элементы лежат в одной плоскости (A-l). Брюшной плавник опущен преаксиальным краем вниз и обращен кнаружи своей первично дорзальной стороной. Все элементы его скелета лежат в одной плоскости (Аа).
В—Ceratodus .во время опоры (при лежании на дне). Грудной плавник скручен в своем основании так» что его преаксиальный край Опущен вниз и первично дорзальная сторона обращена кнаружи. Скелет плавника не лежит в одной плоскости (Вх), но скручен между 1-м и 2-м члениками оси (*): в дистальной части он обращен первично дорзальной поверхностью кнаружи и преаксиальным краем вниз, в проксимальной части 1-й членик оси лежит горизонтально; преаксиальный край плавника обращен кнаружи, а первично дорзальная сторона занимает строго дорзальное положение. Брюшной плавник (Ва) занимает то же положение, что и в предыдущей фазе (А2).
С—Siredon pisciformis. Передняя лапка аксолотля остается постоянно скрученной в локтевом суставе (•) как во время плавания, так и во время опоры (Cj). Предплечье и кисть обращены своим преаксиальным краем вниз и первично дорзальной поверхностью кнаружи, плечо же лежит горизонтально: оно обращено преаксиальным краем кнаружи и первично дорзальной поверхностью вверх. Задняя лапка (С2) целиком обращена преаксиальным краем вниз и первично дорзальной поверхностью кнаружи. Первично вентральная поверхность скелета передней конечности обозначена пунктиром; первично дорзальная поверхность скелета передней конечности оставлена белой; рг—преаксиальный край конечности; ♦—место скручивания передней конечности (из А. Н. Дружинина. 1933).
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИИ ОРГАНОВ 365 емых кистеперых рыб, т. е. конечность типа Saurypterus. Одноосный плавник современных двоякодышащих рыб проходит эту стадию двуос-ности в течение своего эмбрионального развития, а затем уклоняется в сторону, как современная боковая ветвь, от линии, идущей от Crossopterygii к Quadrupeda.1 Однако по своим функциям плавники Ceratodus могут сравниться с конечностями наземных позвоночных и весьма близки к плавникам рыбообразных предков четвероногих. Последние несомненно были донными животными; их плавники, в особенности грудные, выполняли две функции: плавательную и опорную при стоянии и, вероятно, ползании по дну водоемов.
Среди современных донных рыб можно указать на ряд форм (цератод, налим, щука и др.), которые в момент стояния на дне, т. е. в момент «опорной фазы» функции плавника, скручивают грудной плавник так, что преаксиальный край его (рис. 112, рг), обычно поднятый при плавании дорзально (рис. 112 А и Ах), опускается вентрально [за исключением самой проксимальной своей части (рис. 112 В, BJ, а смотрящая кнаружи первично вентральная поверхность плавника (рис. 112 А, заштрихованная) обращается при этом внутрь (рис. 112В, В^. В этом случае кнаружи смотрит уже первично дорзальная поверхность плавника (на рисунках оставленная белой), обычно прижатая при плавании к телу рыбы (рис. 112 А). Любопытно, что у цератода, как это видно на рис. 112 В,, подобное скручивание грудного плавника совершается в суставе между первым и вторым члениками главной оси архиптеригия, т. е. в том месте, которое соответствует локтевому суставу четвероногих.
Что касается брюшных плавников цератода, то они во всех фазах своей функции сохраняют одно и тоже положение: они обращены преакси-альным краем вентрально и первично дорзальной стороной кнаружи (ср. рис. 112 Аа и В2).
Положение, свойственное грудными брюшным плавникам цератода в «опорной фазе» их функции, следует признать также характерным и для рыбообразных предков четвероногих. Интересно, что у всех Quadrupeda в течение эмбрионального развития мы наблюдаем во всех классах явное скручивание передней конечности, локализованное в локтевом суставе. От этого сустава в дистальном направлении вся конечность обращена преаксиальным краем вниз и первично дорзальной стороной кнаружи (рис. 112 С, Сх). Отличие скрученной передней лапы четвероногих от скрученного плавника их рыбообразных предков заключается лишь в том, что передняя конечность Quadrupeda, в основном несущая опорную функцию, сохраняет навсегда то скрученное состояние (фиксация фаз ы), которое для плавника предка было лишь временной, преходящей фазой его функции. Задняя конечность Quadrupeda, не испытывающая скручивания, как и брюшной плавник цератода, целиком опущена вниз своим преаксиальным краем и обращена кнаружи своей первично дорзальной стороной (рис. 112 С, С2).
Мне кажется, что мы привели достаточно примеров принципа фиксации фаз для того, чтобы сделать его понятным читателю и чтобы показать, насколько значительную роль играл этот тип филогенетических изменений органов при эволюции органов движения позвоночных животных. Нам приходится только еще раз подчеркнуть, что в действительности филогенетическое развитие шло и здесь гораздо более сложным путем, чем это было нами только что описано.
Отметим еще, что этот принцип развития, точно так же, впрочем, как и некоторые другие принципы, не действует в одиночку при эволюции сложно построенных органов. Мы только что видели, что при эволюции
1 Holmgren N., On the origin of the tetrapod limb, Acta Zoologica, 14, 1933.
366
ГЛАВА XI
дигитиградного движения у млекопитающих изменения в конечностях состоят не только в том, что расположение скелетных частей относительно друг друга становится не таким, каким оно было прежде, но и в том, что некоторые кости при этом удлиняются (это имеет место, например, при развитии пальцеходного движения у хищников, у которых удлиняются средние metatarsalia и средние metacarpalia), другие дегенерируют (редукция боковых пальцев у некоторых форм), третьи срастаются между собою (образование os canon у Artiodactyla) и т. д. Таким образом в филогенезе на ряду с принципом фиксации фаз обычно действуют и другие принципы. Так, например, мы можем сказать, что при удлинении metacarpalia и metatarsalia и при их срастании в одну os canon, кроме принципа фиксации фаз, действуют еще и принцип интенсификации и принцип иммобилизации (см. ниже).
В общем, можно думать, что введенный нами тип филогенетических изменений органов не имеет столь широкого распространения при развитии филогенетических изменений, как, например, принцип интенсификации или принцип смены функций, ибо он вступает в действие только при эволюции таких органов, у которых функция состоит из периодически повторяющихся фаз, т. е. в первую очередь при эволюции органов движения. При эволюции же органов с пассивной функцией или таких органов, которые функционируют не периодически, но постоянно, этот принцип не имеет никакого применения. Однако, так как в природе, кроме органов движения, существует еще значителное количество и других периодически функционирующих органов, то принцип фиксации фаз, как один из принципов, объясняющих нам пути филогенетического развития органов, безусловно не лишен известного значения.
ПРИНЦИП ВЫПАДЕНИЯ ПРОМЕЖУТОЧНЫХ ФУНКЦИЙ
Тип выпадения промежуточных функций также является одним из тех типов филогенетического развития органов, при которых эволюирующая функция органа изменяется только количественно. Так же как и только что разобранный нами принцип фиксации фаз, этот тип эволюции органов не имеет особенно широкого распространения. Тип филогенетического изменения органов по принципу выпадения промежуточных функций был мною впервые изучен при исследовании эволюции брюшных плавников рыб, и поэтому, чтобы яснее представить себе его сущность, мы попытаемся в самых коротких чертах разобрать ход филогенеза этих органов.
На основании сравнительно эмбриологических исследований1 мне удалось установить, что гипотетическая исходная форма скелета брюшных плавников всех рыб представляла собою ряд расчлененных хрящевых или костных лучей, лежащих в кожных складках плавника. К дистальным концам этих палочковидных лучей (рис. 113 А, г) прикреплялись роговые упругие цератотрихии (рис. 113 A, crtr), проксимальным же своим концом каждый такой луч причленялся к одно- или двухчленному базальному элементу (рис. 113 A, bs). Ряд этих basalia, числом равный числу палочковидных лучей, и представлял собою первичный основной скелет плавника. Из такого рода плавников позднее развились плавники типа Cladodus (рис. 113 В); в них передние базальные элементы срослись друг с другом, образуя хрящевой треугольный таз (базиптеригий, рис. 113 В, Bspt), на котором сидели базиптеригиальные лучи (рис. 113 В, г); задние же basalia сохранили свое членистое строение и образовали метаптеригий (рис. 113 В, Mtpt), на котором сидели метаптеригиальные лучи. По этому же примитивному типу были построены и плавники Hybodus (рис. 113 С), по нему же построены и плавники современных высших акул.
1 S е we г t z о f f A. N., Evolution der Bauchflossen der Fische, Zool. Jahrb., Bd. 58, 1934.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИИ ОРГАНОВ 367
»
Из плавников примитивных акул произошли плавники первичных рыб с костным скелетом (рис. 113 D); при этом первичный тип плавника
Рис. 113 А—L. Предполагаемый филогенез брюшных плавников рыб;
д__гипотетическая прародительская форма брюшного плавника эласмобранхий: виден ряд.
плавниковых лучей и basalia proxlmalian distalia; к концам лучей причленяются цератотрихии. В—брюшной плавник CladodusjBHfleH базиптеригий и сегментированный метаптеригий с сидящими, на них лучами; и базиптеригиальные и метаптеригиальные лучи хорошо развиты и имеют поперечное положение по отношению к оси тела. С—брюшной плавник Hybodus; базиптеригий имеет поперечное положение, видны следы его срастания с basalia; метаптеригий еще сегментирован--на своем дистальном конце; базиптеригиальные и метаптеригиальные лучи хорошо развиты и имеют поперечное расположение. D—исходная форма брюшного nnaBHHKaOsteichthyes primi-tivi; строение плавников напоминает то, что мы видели на рис. А и В, но цератотрихии здесь земещены лепидотрихиями. Е— брюшной плавник Acipenseridae; базиптеригий имеет поперечное положение по отношению к оси тела, метаптеригий сегментирован, бази- и метаптери-гиальне лучи хорошо развиты, лепидотрихии имеются. F—брюшной плавник Birgeria; брюшной плавник занимает поперечное по отношению к оси тела положение (|), метаптеригий сегментирован на своем дистальном конце; бази- и метаптеригиальные лучи хорошо развиты, лепидотрихии имеются, Ft—плавниковый луч, базиптеригий (заштрихован) и расчлененный ле-пидотрихий палеонисцид. G—брюшной плавник Holostei (Amia); базиптеригий имеет продольное по отношению к оси тела положение (*-); метаптеригий и базиптеригиальные лучи рудиментарны, матаптеригиальные лучи совершенно исчезли; лепидотрихии имеются. Gi—лепидотрохии, рудиментарный плавниковый луч и базиптеригий (заштрихован) Holostei. Н—•брюшной плавник Teleostei (Esox); базиптеригий имеет то же положение, от метаптеригия остался лишь незначительный рудимент, плавниковые лучи исчезли.—лепидотрихии и базиптеригий одного из представителей высших Teleostei. На рис. F1S Gt и Ht виден процесс выпадения промежуточной функции, выполняемой плавниковыми лучами. J—брюшной плавник высших Teleostei;, матаптеригий совершенно исчез; в качестве новообразования появились костные отростки бази-птеригия. К и L—брюшные плавники Crossopterygii; брюшной плавник имеет тоже положение по отношению к оси тела, что и у Holostei; р. il.—processus iliacus хорошо развит у мальков. К—брюшной плавник Polypteridae; метаптеригий совершенно редуцировался, имеются четыре базиптеригиальных луча; L—брюшной плавник Eusthenopteron; базиптеригиальные лучи полностью редуцированы, сохранился метаптеригий с тремя метаптеригиальными лучами, bs—basalia; ВэрЪ--базиптеригий; crtr.—цератотрихии; Iptr.—лепидотрихии; Mtpt.—метаптеригий; mtr.—мета-
птеригиальные лучи (у Eusthenopteron); г—плавниковые лучи (из А. Н. Северцова, 1934).
остался тот же, но произошло важное изменение: у Osteichthyes primitivi прогрессивно развилась активная функция плавника, роговые цератотрихии у них были замещены костными, происшедшими из костных чешуй,
368
ГЛАВА XI
лепидотрихиями. Каждый лепидотрихий состоит из двух рядов костных члеников и кончается проксимальной вилкой, охватывающей дистальный конец плавникового луча.
Из такого рода лучей развились плавники осетровых рыб (рис. 113 Е) и некоторых палеонисцид (Birgeria, Saurichthys, рис. 113 F). И у тех и у других передние базалии плавника слились между собой и образовали треугольную пластинку (рис. 113 Е и F, Bspt), на которой сидят 4—5 передних лучей со своими лепидотрихиями. Оси этих лучей и лепидотрихий направлены латерально по отношению к продольной оси тела, а базипте-ригий занимает поперечное положение. Интересно, что метаптеригий {рис. 113 Ей F, Mtpt) еще сохранил следы метамерного членистого строения.
В дальнейшем весьма важные изменения произошли в положении скелетных частей плавника у всех потомков палеонисцид. Мы видели, что у палеонисцид и у хрящевых ганоидов хрящевые лучи (рис. 113 Е и F, г) сидели на наружном латеральном крае бази- и метаптеригия; у костных же ганоидов лучи развиваются на заднем крае базиптеригия, причем число их постепенно убывает. Метаптеригий в течение последующей эволюции также постепенно редуцируется и сидящие на нем лучи исчезают. В общем, у всех потомков палеонисцид, т. е. у костных ганоидов (рис. 113 G), у костистых рыб (рис. 113 Н и J) и у предков кистеперых рыб (рис. 113 К), произошло очень важное изменение в положении пояса брюшного плавника: он повернулся так, что ось его из поперечной сделалась продольной и сидевшие на нем лучи оказались направленными не латерально по отношению к продольной оси тела, но параллельно ей: лучи и лепидотрихии базиптеригия, а также рудиментарный метаптеригий этих рыб направлены не в сторону, а назад.
У большинства кистеперых рыб, например у ископаемого Eusthenopte-гоп (рис. 113L), редуцировались базиптеригиальные плавниковые лучи, т. е. лучи, сидевшие непосредственно на поясе конечностей, но зато сохранился членистый метаптеригий (Mtpt) с сидящими на нем лучами (mtr) и лепидотрихиями (Iptr). Исключение представляют Polypteridae, у которых редуцировался целиком метаптеригий, но сохранились базиптеригиальные лучи (рис. 113 К, Bspt, г).
Наиболее интересные изменения произошли у Holostei и у костистых рыб, происшедших от Holostei. У низших костистых рыб (G и Н) редуцировалась вся дистальная часть метаптеригия и сильно редуцировались лучи, сидящие на поясе; у высших же костистых рыб (рис. 113 J) все эти элементы полностью редуцировались, так что лепидотрихии, являющиеся активными элементами скелета брюшных плавников, теперь причленяются прямо к каудальному краю пояса. Первоначально у рыб с костным скелетом (Palaeoniscidae, Acipenseridae) подвижными частями скелета были и лепидотрихии и лучи; у костистых же рыб подвижны одни лепидотрихии, к которым прикрепляются мускулы, двигающие свободную лопасть плавника. Таким образом, можно сказать, что у предков костистых рыб активным органом движения брюшного плавника была поддерживаемая лепидотрихиями гибкая лопасть плавника и что опорой этой активной части органа был базиптеригий; палочковидные же плавниковые лучи (г—г) поддерживали лепидотрихии и несли (несут и теперь у осетровых рыб) промежуточную функцию*движения брюшного плавника, т. е. эти лучи с их пассивной функцией расширения опорной поверхности плавника были промежуточной, или передаточной
О ПРИНЦИПАХ или ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 369
частью этого органа. При дальнейшей эволюции у костных ганоидов (рис. 113 G и Gx) и у низших костистых рыб (рис. 113 Н и плавниковые лучи в значительной степени редуцировались, а у высших костистых рыб (рис. 113J) и совсем исчезли, и активная часть луча, т. е. лопасть плавника (лепидотрихии) вступила в непосредственное сочленение с опорной частью плавника, т. е. произошло выпадение промежуточной функции лучей и уси
ление главной функции подвижной лопасти плавника (л е-пидотрихий), в результате чего получился выигрыш в интенсивности движения плавника. Этот-то именно тип филогенетических изменений органов мы и называем «типом или принципом выпадения промежуточных функций».
Чтобы читателю стало ясным, как все это в действительности произошло, мы даем два добавочных рисунка: рис. 114 А ирис. 114 В. Рисунки эти представляют собой реконструкции строения брюшных плавников у стерляди (рис. 114 А) и у щуки (рис. 114 В).
На этих рисунках мы видим, что у стерляди к базиптеригию (рис. 114 A, Bspt) причленяются два плавниковых луча (рис. 114 А, г, г'), которые в свою
Рис. 114 А—реконструкция по поперечным срезам скелета и мускулатуры базального отдела брюшного плавника Acipenser ruthenus.
очередь соединены посредством тонкой связки (lig.) с дорзальными и вентральными лепидо-трихиями (рис. 114 A, Iptr. d, Iptr. v). Дорзальные и вентральные мускулы плавника (рис. 114 A, dm, vm) прикрепляются к концам лепидотрихий и к концам плавниковых лучей. У щуки имеется базиптеригий (рис. 114 В, Bspt), рудиментарный
Рис. 114 В—реконструкция по поперечным срезам скелета и мускулатуры базального отдела брюшного плавника Esox lucius.
Bspt—базиптеригий; dm—дорзальная мускулатура плавника; dors—дорзальная сторона плавника; 1g — соединительнотканная связка,служащая для прикрепления мускулов; lig — связка, соединяющая оба лепидотрихия одной пары; Iptr—крючкообразный конец лепидотрихия; Iptrj—проксимальный конец лепидотрихия соседней пары; Iptr. d.—дорзальный лепидотрихий; Iptr. v.—вентральный лепидотрихий; Mtpt—рудимент метаптеригия у щуки; г—плавниковые лучи стерляди; rm—туловищная мускулатура; vm—вентральная мускулатура плавника; ventr. — вентральная сторона плавника (из А. Н.
Северцова, 1934).
метаптеригий (рис. 114 В, Mtpt)
и совершенно не имеется плавниковых лучей. Дорзальные и вентральные лепидотрихии (рис. 114 В, Iptr. d, Iptr. v) причленяются к бази-птеригиуму и соединены посредством плотной связки (рис. 114 В, lig.)
с сильно разросшимися дорзальными и вентральными мускулами (рис. 114 В, dm, vm). Мы видим здесь, как в связи с исчезновением плавниковых лучей’ усилилась двигательная функция лепидотрихий или, что то же, двигательная функция лопасти плавника. *
Если мы захотим схематически изобразить эволюцию брюшных плав-
Морфологические закономерности
24
370
ГЛАВА XI
ников костистых рыб, то нам придется представить ее в виде следующей таблицы:
Орган
Функция
1. Предки рыб.
Плавники состояли из ряда базалий (basalia proxima-lia и distalia), к которым причленялись хрящевые плавниковые лучи.
2. Акуловые, осетровые и палеонисцид ы.
Путем срастания передних базалий развился сначала хрящевой, а затем костный пояс брюшного плавника (базиптеригий), к которому причленяются базиптери-гиальные лучи; эти лучи и цератотрихии (у костных рыб—лепидотрихии) были направлены поперечно к оси тела.
3. Костные ганоиды и костистые рыбы
Функция вполне пассив-)> ная: поддержание равнове-) сия задней части тела.
Главная функция еще пассивная, но к ней прибавилась второстепенная активная функция изменения направления движения тела рыбы в вертикальном направлении путем поднятия и опускания переднего и заднего края плавника.
Горизонтальный поворот базиптеригия,. ось которого . из поперечной делается продольной и занимает положение параллельное продольной оси тела (латеральный край базиптеригия, несущий базиптеригиальные лучи, становится каудальным); метаптеригий и метаптери-гиальные лучи редуцируются в значительной степени (костные ганоиды и низшие Teleostei) или вполне (высшие Teleostei); лепидотрихии причленяются непосредственно к базиптеригию и направлены кау-дально. '
Усиление активной функции лепидотрихий путем выпадения промежуточной пассивной функции базиптери-гиальных лучей.
Другим прекрасным примером типа (принципа) выпадения промежуточных функций у позвоночных является эволюция quadratum, менявшей в течение филогенеза несколько раз свою функцию. Так, у предков рыб (рис. 115 А) челюстная дуга состояла из четырех элементов, из которых два средние, epimandibulare и ceratomandibulare (рис. 115 А, ер. т., сг. т.), соответствующие epibranchiale и ceratobranchiale, несли жаберные лепестки и служили в качестве органов дыхания. У акуловых и у низших рыб с костным скелетом квадратная кость представлена задним отделом хрящевой верхней челюсти (рис. 115 В, pl. qu.) и несет зубы; она сочленена с нижней челюстью, функция ее—кусание.
У высших рыб с костным скелетом функция нёбноквадратного хряща изменилась: его задний участок превратился в самостоятельную квадратную кость (рис. 115 С, qu.) и', оставаясь в связи с остальными окостенениями верхней челюсти, утратил свою прежнюю функцию кусания и стал служить только в качестве промежуточного элемента между нижней челюстью и черепом.
У рептилийных предков млекопитающих произошло новое филогенетическое изменение по типу выпадения промежуточной функции, а именно quadratum и articulate (рис. 115 D, qu., art.) вышли из состава челюстной дуги, так что нижняя челюсть (теперь dentale) стала непосредственно причленяться к височной области черепа. На этой стадии эволюции предков млекопитающих произошло выпадение промежуточной функции этих костей, т. е. функции поддерживания нижней челюсти: мы знаем, что у териодонтов (рис. 115 Е) нижняя челюсть уже непосредственно причленяется к черепу. Интересно, что quadratum и articulate при этом не редуцировались, подобно остальным костям нижней челюсти (spleniale, coronoideum), но снова изменили свою функцию и превратились в слуховые косточки [рис. 115 F, mal. (art.), inc. (qu.)].
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 371
Orb
С D
Рис. 115 А, В, С, D, Е, F. Схема филогенеза квадратной кости.
А—схематическое изображение висцерального аппарата гипотетических предков акуловых рыб. В—схематическое изображение висцерального аппарата акул и низших рыб с. костным скелетом, у которых сильно увеличились epimandibulare(palatoquadratum)H ceratomandibulare (cart. Meckeli). Ha palatoquadratum и на меккелевом хряще из плакоидных чешуй развились зубы, причем оба хряща превратились в верхнюю и нижнюю челюсть с кусательной функцией. С—схематическое изображение висцерального аппарата высших рыб с костным скелетом, у которых произошло окостнение хряща в задней части palatoquadratum, причем эта часть (quadratum), изменив свою функцию (кусание и сочленение с нижней челюстью), превратилась в кость с промежуточной функцией поддерживания нижней челюсти. Эту же функцию квадратная кость сохраняет у амфибий и у рептилий (D и Е). D—схематическое изображение черепа и висцерального аппарата амфибий. В—схематическое изображение нижней челюсти Cynognathus, у которого quadratum вышла из ряда челюстных костей и начала редуцироваться; то же произошло и в нижней челюсти carticulare. Здесь мы имеем перед собой выпадение промежуточной функции этих двух костей, т. е. функции поддерживания нижней челюсти; последняя у териодонтов причленяется уже непосредственно к черепу. F--схематическое изображение нижней челюсти зародыша млекопитающего (Tatusia), у которого, как и у териодонтов, она образована только одною dentale; quadratum и articulate вошли в полость среднего уха и превратились в слуховые косточки: наковальню и молоточек.
a. pb. 1—подъязычная дуга; a. pb. 2—челюстная дуга; a. pb. 4, a. pb. 3—второй и третий губные хрящи; art.—articulare; ang.—angulare, bi—первая жаберная щель; рЬ.г—pb2—рудиментарные преджаберные щели 1—3; ст. т.—ceratomandibulare; cr. h.—ceratohyale; с._ т.—меккелев хрящ dnt.—dentale; ер. h.—epihyale; ent. pt.—entopterygoideum; hm.—hyomanbidulare; nd.— hyoideum; inc. (qu.)—incus (quadrature^; jug.—jugale; mt. pt.—metapterygoideumf mal (art.)— maleus (articulare); orb.—глазница; pal.—palatinum, pl. qu.—palatoquadratum; qu.—quadratum; qu,. j.—quadratojugale; sqm.—squamosum. На всех рисунках quadratum обозначена косой штриховкой, a articulare—пунктиром (из А. Н. Северцова, 1933). >
24*
372
ГЛАВА XI
Если мы захотим схематически представить себе эволюцию квадратной кости, то нам придется изобразить ее в следующем виде:
Орган
Функция
Предки рыб.
’ Челюстная дуга состоит из че-.' тырех элементов; pharyngo-, epi-, cerato-и hypomandibulare, которые соответствуют pharyngo-, epi-, cerato-и hypobranchiale жаберных дуг.
Epi-и ceratomandibulare служат поддержкой жаберных лепестков при дыхательном движении.
Акуловые и низшие рыбы.
Челюстная дуга состоит из меккелева хряща и palatoquadratum. Quadratum представлено задним нижним концом палатоквадрат-. ного хряща.
Quadratum служит для кусания и сочленения верхней челюсти с нижней (смена функций).
Амфибии, боль- ( Quadratum—самостоятельная шая часть низших 5 кость между височной областью рептилий. ( черепа и нижней челюстью.
Млекопитающие
Quadratum и articulare вышли из состава челюстных костей.
Quadratum прикрепляет нижнюю челюсть к черепу (промежуточная функция quadratum).
Выпадение промежуточной функции quadratum и смена фун-кции. Превращение quadratum и articulare в слуховые косточки.
Таким образом мы видим, что челюстная дуга и, в частности, quadratum в течение своей эволюции несколько раз изменяли свою функцию: дыхательная функция сменилась функцией кусания, затем квадратная кость снова изменила свою функцию и долгое время была частью, поддерживавшей нижнюю челюсть (промежуточная функция), затем функция нижней челюсти усилилась, и quadratum (и articulare) выпали из цепи челюстных костей и стали редуцироваться; но произошла новая смена функций, и они превратились в слуховые косточки, молоточек и наковальню.
Итак, мы имеем здесь типичный случай выпадения промежуточной функции: скелетные элементы, несшие функцию поддерживания нижней челюсти и связи ее с осевым черепом, вышли из состава челюстных костей, и нижняя челюсть стала непосредственно причленяться к осевому черепу, что привело к интенсификации ее функции.
Но здесь мы имеем интересное отличие этого случая выпадения промежуточной функции от случая, разобранного нами на плавниках рыб. В брюшных плавниках Hoiostei и Teleostei плавниковые лучи редуцировались, в челюстном же аппарате квадратная кость начала также редуцироваться и, сильно уменьшившись в размерах, утратила свою опорную функцию, но затем вступила в функциональное соотношение с лежащим по соседству слуховым аппаратом и превратилась в слуховую косточку.
Примеров выпадения промежуточной функции можно привести довольно много. Не разбирая этих случаев в отдельности, мы можем указать еще на выпадение карпальных и тарсальных элементов в пятипалых конечностях амфибий, например на редукцию дистальных тарсалий в задней дане лягушки, а также на редукцию episternum и clavicula, выпадающих как промежуточный элемент в течение филогенеза в плечевом поясе конечностей у млекопитающих.
Относительно этого последнего случая напомним, что у ископаемых рептилий (тероморф, ринхоцефалов и др.) связь . грудины с плечевым поясом конечностей осуществлялась через clavicula, episternum и сога-coideum. Все упомянутые кости прочно соединялись между собой, что ограничивало свободу движения конечностей. Подобного же рода прочное
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 373
соединение существует еще и сейчас у примитивных млекопитающих (Monotremata), но уже у Marsupialia исчезает связь ключицы с грудиной через episternum, оставляя следы лишь в эмбриональном состоянии, где коракоидный отросток направлен к грудине. У Monodelphia и у ряда грызунов связь через praeclavicula (episternum) также сохраняется лишь в течение эмбрионального развития. У более высоко стоящих форм связь грудины с поясом конечностей сохраняется уже только через ключицу. Затем, благодаря редукции ключиц, порывается непосредственная связь грудины с поясом конечностей, и у лошадей, коров и собак наблюдается уже полное исчезновение ключицы. Мы видим, что и здесь также выпадение clavicula, episternum и coracoidem и связанной с ними промежуточной функции связи грудины] с плечевым поясом ведет к усилению функции движения бегающей конечности.
Наконец, еще одним примером развития, происходящего по типу выпадения промежуточной функции, может служить выпадение функции conus arteriosus;-этот случай имеет место при превращении сердца предков Quadrupeda в сердце амфибий и затем рептилий с редуцированным conus arteriosus.
Резюмируя все сказанное, мы должны отметить, что во всех разобранных нами случаях выпадение промежуточной функции всегда приводит к усилению главной функции органа.
Теперь мы перейдем к одному из важнейших и самых распространенных типов филогенетического изменения органов, а именно к принципу субституции органов Н. Клейненберга.
ПРИНЦИП СУБСТИТУЦИИ ОРГАНОВ Н. КЛЕЙНЕНБЕРГА
Н. Клейненбергом в 1886 г. был впервые введен в морфологическую науку важный принцип субституции органов. На основании своих исследований над аннелидами Клейненберг пришел к выводу, что во многих случаях филетическое развитие органа происходит таким образом, что определенный орган предков замещается у потомков другим органом, выполняющим ту же функцию, что и орган предков.
Так как точная формулировка этого принципа для нас очень важна, то я позволю себе привести соответствующую цитату из работы Клейненберга.1 «Под общим термином „замена или субституция органов" я объединяю все те случаи, когда старые органы разрушаются в течение филогенетического и онтогенетического развития и замещаются новыми органа ми, все равно, будут ли эти новые органы по своим первичным соотношениям им равноценны или нет. Ту часть, которая является предпосылкой или стимулом для новообразования, я называю промежуточным или посредствующим органом, само же новообразование —субституирующим органом» (стр. 216). «Глотка аннелид образуется из других частей тела аннелид путем исключительно сложного процесса субституции. Стомодеум у многих аннелид является личиночным органом, который у взрослого червя исчезает и замещается дефинитивной глоткой... хотя субституция стомо-деума дефинитивной глоткой и очень совершенна, однако генетические соотношения обоих органов гораздо ближе, чем соотношения следующих друг за другом нервных образований».1 2 «Таким образом дефинитив
1Kleinenberg N., Die Entstehung des Annelids aus der Larve von Lopa-dorhynchus, Zschr. f. wiss. Zool., Bd. 44, стр. 216 и 221, 1886.
2 Субституции в только что перечисленных органах описаны Клейненбергом очень подробно на предыдущих страницах его работы.
374
ГЛАВА XI
ная глотка является дериватом стомодеума; она находится в прямой морфологической зависимости от размножения и диференцировки некоторых клеток стомодеума. Строение парных зачатков приводит нас к выводу, что глотка произошла путем преобразования, замыкания и слияния двух добавочных желез стомодеума; в этом случае образование дефинитивной части тела совершается через посредство смены функций и субституции органов... Здесь промежуточные органы и субституирующие органы отчасти гомологичны. Замена органа или ткани вполне гомологичными им органами или тканями встречается очень часто. Примером этого является у аннелид субституция личиночной эктодермы эпидермисом взрослого животного».
«Достаточно лишь переглядеть развитие других форм, чтобы найти новые примеры субституции органов. Так, скелет позвоночных дает нам пример комбинации смены функций и субституции органов. Здесь первичный элемент скелетной системы, хорда, образуется путем прямого превращения из ранее существовавшего на этом месте энтодермального отрезка кишечного канала, т. е. образуется путем смены функций. Хорда является ’ единственным осевым скелетом низших хордат и появляется в онтогенезе всех без исключения настоящих позвоночных животных как первый зачаток скелетной системы. Но далее из хорды ничего более. не происходит, из ее клеток не развивается уже никакой другой ткани, никакого другого органа, и поэтому ее физиологическое значение падает все более и более', пока она у высших позвоночных не разрушится и не превратится в ничтожные остатки. На ее место выступают новые образования, генетически ей не равноценные и не гомологичные—части дефинитивного скелета: здесь вступает в силу типичная субституция. Эти части развиваются в зависимости от мускульной деятельности, которая определяется в свою очередь существованием хорды. Хорда, таким образом, здесь действует как промежуточный орган при образовании скелета. Хотя этот скелет теперь уже не содержит ни одной из своих прежних составных частей, его строение делается понятным, благодаря предшествующему развитию хорды. Наконец, можно подвести под понятие субституции замечательные явления, сопровождающие гистологические превращения хрящевых частей скелета, ибо здесь хрящевая ткань не превращается непосредственно в костную, но сначала растворяется, а затем уже вытесняется остеобластами, проникающими в нее снаружи. При этом замещается лишь ткань, сам же орган сохраняется как таковой. И как много еще других случаев субституции находим мы в выделительной системе, в аппарате кровообращения и не знаю еще в чем!..» (1. с., стр. 222)
На этих примерах, приводимых самим Клейненбергом, мы хорошо видим, что, собственно, он подразумевает под понятием «субституция» или «смена» органов: «орган, вполне развитый и хорошо функционирующий у предков данной формы, дегенерирует у ее потомков и замещается другим органом, занимающим его место и выполняющим его функци ю». В огромном большинстве примеров, приводимых Клейненбергом, субституирующий орган возникает на том же м е с т е, где находился анцестральный, теперь исчезнувший орган. При этом Клейненберг различает два вида субституции: 1) орган замещается гомологичным образованием1 и 2) орган замещается негомологичным образованием. Первый случай, по его мнению, встречается гораздо чаще, чем последний.
Субституция органов Клейненберга играет важную роль в филогенезе.
1 Отметим, что термин «гомологичный» употреблен здесь Клейненбергом в необычном и, по моему мнению, в неправильном смысле этого слова.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 375
Этот принцип широко распространен в животном мире, и мы легко можем найти целый ряд примеров, иллюстрирующих этот тип филогенетического развития органов. Мы знаем, например, что у стегоцефалов condyli occi-pitales образованы двумя выростами occipitalia. У примитивных рептилий, происшедших от этих стегоцефалов, мы находим только один condylus occipitalis tripartitus, образованный из выростов exoccipitalia и из вклинившегося между ними выроста basioccipitale (Dicynodon и т. п., а среди современных рептилий—черепахи); у происшедших от рептилий птиц этот condylus образован только basioccipitale. Таким образом, оба condyli occipitales здесь замещены .................
У селахий дорзальный отрезок жаберной артерии типичной жаберной дуги происходит из части первичного кровеносного сосуда этой дуги, т. е. из arcus aortae primarius (рис. 116 А), а у Osteichthyes (начиная от хрящевых ганоидов) этот сосуд, занимающий, кстати сказать, совершенно то же положение, что и у селахий, замещается новообразованием, а именно сосудом, вновь возникшим из связи сосудистых петель, идущих к жаберным нитям (рис. 116 В, ab'),1 Этот случай интересен втом отношении, что первичный, возникший из аортальной дуги ствол arteria branchialis не дегенерирует, но превращается в непарный ствол vena branchialis communis, т. е. меняет свою функцию. Таким образом, здесь, как увидим ниже, мы имеем перед собою особый случай совместного действия принципа субституции органов Клейненберга и принципа смены функций Дорна.
У Polypterus maxilla замещается suborbitale. У рептилий вентральный отрезок мандибулярной дуги, меккелев хрящ, покрыт довольно значительным числом покровных костей (dentale, spleniale, coronoideum, goniale, angulare, supraangulare), у млекопитающих же все эти кости замещаются однрй сильно разросшейся dentale, образующей собою всю костную нижнюю челюсть.
Мы обращаем особое внимание читателя на то, что во всех этих случаях клейненберговской субституции функция замещающего органа остается той же, что и функция замещенного органа.
Мы можем привести и такие случаи, когда замещающий орган состоит
condylus basioccipitalis.
одним
Рис. 116. А—схематизированная графическая реконструкция (по поперечным разрезам) сосудов 1-й жаберной дуги эмбрионов акулы (Pristiurus mela-nostotnus).
a. a. d_arcus aortae dorsalis; a. b.—arteria branchialis; ao.
d.—aorta dorsalis; ao. v.—aorta ventralis;b—место, где первичная жаберная дуга прерывается; v. b. а.—vena branchialis anterior; v.b. p.—vena branchialis posterior.
В—схематизированная графическая реконструкция (по поперечным разрезам) сосудов'1-й жаберной дуги эмбриона Acipenser ruthenus.
ab' — дорзальный отдел arteria branchialis, развившийся как новообразование.Остальные обозначенияте же, что и на рис. А. Перерыв ь находится более вентрально, чем у акул (рис. А), поэтому парные* вены (v. b. a., v. Ь. р.) слабее развиты.
аал.
аал.
v.b.p.
ab’
v.b.p.
1 Se we г t z of f A. N., Die Entwicklung der Kiemen und Kietnenbogenge-fasse der Fische, Zschr. f. wiss. Zool., 1923.
376
ГЛАВА XI
Рис. 117. Схема филогенетического развития хордовых оболочек и позвоночника.
А—первая стадия: пс.—клетки хорды; sh. 1—первичная оболочка хорды; sk. 1.—мезодермальная ткань. В—вторая стадия (круглоротые, хрящевые галоиды): центральные клет» ки хорды вакуолизированы, периферические эпителиальные клетки (пс. ер.) принимают правильное расположение; sh.2-— вторичная оболочка хорды. В мезодермальной ткани закладываются дорзальные (d. а.) и вентральные (v. а.) дуги позвоночника. С—третья стадия: хрящ проникает сквозь первичную оболочку хорды во вторую ее оболочку (Holocephali, Selachii, Dipnoi).’D—четвертая стадия: хрящ обрастает хорду снаружи от оболочек, которые постепенно исчезают (хрящевые галоиды, костистые рыбы, амфибии, амниоты). (Позвонки А—D взяты, из хвостового отдела позвоночника). Е—пятая, более поздняя стадия развития туловищного позвонка: хорда (пс) сильно редуцирована; отдельные хрящи срослись между собою, образуя цельный хрящевой позвонок, в котором мы различаем тело позвонка (с), верхние дуги (п. а.), остистый отросток (n. sp.), поперечные отростки (tip.) и сочленовные отростки (art.) F—схема строения грудного позвонка млекопитающего: 1—тело позвонка, 2—верхние дуги, 3—спинномозговой канал, 4—processus articularis, 5—processus transversus (diapophysis), 6—processus spino-sus, 7—processus maxillaris, 8—сочленовная поверхность дли capitulum, 9—сочленовная поверхность для tuberculum реб-- ра, 10—ребро.
На рисунках A—F изображен постепенный ход субституции первичного осевого скелета (А. хорда пс) сначала хрящевыми (В, С, D, Е), а затем и костными (F) позвонками Здесь мы видим, как первичный хрящевой позвонок замещается костью (F); замещающие части (тело позвонка и костные дуги позвонка) в этом случае принимают форму и положение замещенных частей (хорда, хрящевые дуги позвонка);
(из Видерсгейма).
из совершенно другой ткани, а иногда даже и происходит из совершенно другого зародышевого листа, чем замещенный орган. К этой категории явлений относятся случаи, приводимые самим Клейненбер-гом, а именно случай замещения хорды дефинитивными позвонками, а также и многие другие случаи гистологической субституции, как например, случай замещения хряща костью путем эндохондрального окостенения.
Для иллюстрации случая замещения хорды костными позвонками нами приведен рис. 117 А—F. Здесь мы имеем весь ход субституции первичного осевого скелета (хорды) сначала хрящевым (рис. 117, А—Е), а затем костным веществом (F). На рисунке видно, как появляются хрящевые дуги позвонков (рис. 117 В, С, D, V. a, d. а.), как постепенно хрящ обрастает хорду сначала снаружи от оболочек, которые постепенно распадаются и исчезают, и как он образует тело позвонка. Хорда при этом сильно редуцируется (рис. 117 А— Е, пс), а отдельные хрящи сливаются между собою, превращаясь в сплошной хрящ (рис. 117, Е), в котором можно различить тело позвонка, верхние дуги, остистый отросток, поперечные и сочленовные отростки. На рис. 116 F видно, как хрящевые дуги позвонков замещаются костью. Замещающие части (костное тело позвонка и костные дуги) в данном случае
занимают место замещенных частей (хорды, хрящевых дуг позвонка). Если мы обратимся к беспозвоночным, то здесь прекрасный пример
Клейненберговской субституции мы находим у пиявок, у которых кровеносная система, производная первичной полости тела, морфологически
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 377
и физиологически замещается лакунарной системой, возникающей из вторичной полости, тела. Повидимому, филогенетически этот процесс шел следующим путем. У малощетинковых предков пиявок существовала типичная вторичная полость тела, а также настоящая кровеносная система, состоящая из спинного и брюшного продольных сосудов, которые были соединены друг с другом кольцевыми сосудами; у.древних примитивных пиявок в связи с развитием соединительной ткани
и мускулатуры вторичная полость тела начала распадаться на участки и продольные каналы. При этом у таких примитивных форм, как например, Acanthobdellidae, редукция вторичной полости тела ушла еще недалеко вперед, полость тела еще значительна и в ней еще находятся спинной и брюшной кровеносные сосуды. У более высоко стоящих Ichthyobdellidae вторичная полость тела в связи с дальнейшим разраста-
нием паренхимы и мускулатуры превращается в систему лакун или каналов, из которых главнейшими являются спинная, брюшная и две боковые лакуны, причем весьма интересно, что здесь в спинной и брюшной лакуне еще лежат спинной и брюшной кровеносные сосуды. Наконец, у челюстных пиявок (Gnathobdellidae) происходит дальнейшее развитие сложной лакунарной системы и полное вытеснение ею настоящей кровеносной системы, так что последняя совершенно редуцируется. У конской пиявки, например, лакунарная система с ее тончайшими капиллярами обслуживает все системы органов и совершенно неотличима от настоящей
Рис. 118 А, В. Схема субституции вторичной полости тела офиур (А) третичной полостью тела (бурсами) у Euryalae (В).
Кровеносной системы КОЛЬ- а—вторичная полость тела; b—бурса; с—кишечник; d — стенка тела (из Д. М. Федотова, 1927), чатых червей.
Весьма интересный пример клейненберговской субституции наблюдал Д. М. Федотов в группе офиур Euryalae. В то время как у типичных офиур (Ophiurae) имеется вторичная полость тела (рис. 118 А, а), в которой помещается кишечник (рис. 118 А, с) и бурсы, или половые мешки [органы эктодермального происхождения (рис. 118 А, Ь)], у офиур группы Euryalae бурсы (рис. 118 В, Ь) начинают разрастаться, соединяются друг
с другом и, вытесняя вторичную полость тела, становятся сами дефинитивной, третичной полостью тела. Вторичная же полость тела (рис. 118 В, а} не редуцируется целиком, но низводится до степени узких щелей.
Интересно, что процесс субституции вторичной полости тела третичной еще не закончен; он продолжается в группе Euryalae и в настоящее время. У них же наблюдается и процесс рекапитуляции. Так, во время онтогенеза представители группы Euryalae проходят стадию офиуры с еще обширной вторичной полостью тела, в которую вдаются не сообщающиеся бурсы, т. е. здесь воспроизводятся отношения, характерные для взрослых типичных офиур.
378
ГЛАВА X
Особый тип эмбриональной субституции выдвинут В. М. Шимкевичем под именем меторизиса.1 Во многих случаях мы видим, что дериваты определенного зародышевого листка вытесняются и замещаются дериватами другого зародышевого листка. Так, мы имеем все основания предположить,- что, например, энтодерма ль ный эпителий переднего конца кишечного канала был вытеснен эктодермальным эпителием ротовой полости, так что задняя граница эктодермальной выстилки ротовой полости в течение филогенеза передвинулась у позвоночных назад в каудальном направлении. Шимкевич приводит довольно много случаев подобного рода сдвигов и совершенно ясно, что этот процесс меторизиса подпадает под рубрику субституций Клейненберга.
Приведем еще несколько дополнительных примеров субституции органов. Мы знаем, например, что у селахий краевая часть плавника усажена эластическими роговыми лучами; у Osteichthyes эти роговые лучи замещаются костными лепидотрихиями, представляющими собою метаморфозированные чешуи. Следы ранее существовавших здесь роговых лучей сохранились в этих плавниках в виде рудиментарных актино-трихий плавников Osteichthyes, гомологичных роговым лучам. Далее, у непосредственных предков птиц, как известно, край челюсти был усажен зубами. У современных птиц зубы атрофировались, а их функцию стал выполнять развившийся на их месте острый край рогового клюва. Подобного же рода субституцию мезодермальных зубов острым, развившимся из роговой ткани краем челюсти, мы находим у черепах.
Во всех приведенных нами случаях (а мы можем легко умножить их число) мы видим, что замещающий орган потомков имеет такое же функциональное значение, как и замещенный орган предков. В огромном большинстве случаев мы можем, кроме того, отметить, что при этом функция органа интенсифицируется в течение филогенеза у потомков вследствие субституции. Так, мы можем с полной определенностью сказать, что замена хорды хрящевыми позвонками, вытеснение хрящевого скелета костным, замена мезонефроса метанефросом, замена роговых лучей плавника костными лепидотрихиями и тому подобные явления ведут к усилению функции данного органа.
В большинстве случаев субституции замещающий орган образуется натом же месте тела животного, на котором у предков лежал замещенный орган (хрящевые позвонки образуются на том же месте, где лежала хорда, dentale—на том же месте, где лежали многочисленные покровные кости нижней челюсти, лепидотрихии—на месте церато-трихий и т. д.). Но бывают и такие случаи, когда замещающий орган, хотя он и несет ту же функцию, что и замещенный им орган, образуется в другом месте тела животного.
Так, у предков актиноптеригий брюшные плавники занимали абдоминальное положение, а их мускулы и нервы развивались из задних миото-мов и из задних спинных корешков нервов. Мы знаем, однако, что у современных актиноптеригий с их торакальными и югулярными брюшными плавниками мускулатура и нервы сдвинувшегося вперед брюшного плавника развиваются совсем из других миотомов и из других (кпереди лежащих) нервных сегментов, чем это имеет место у предков. Все эти многочисленные случаи субституции можно рассматривать как примеры уже иного рода субституции, ибо здесь органы (мускулы и нервное сплетение брюшного плавника), выполняющие у потомков ту же функцию,
1Schimkewitsch W. М., Die Methorisis als embryologisches Prinzip, Zool. Anz., Bd. 33, 1908.
Schimkewitsch W. M. Les phSnomenes de Methorisis chez les Vert6br6s, IX Korgr. Intern. Zool., 1914.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 379
которую они выполняли у предков, лежат в другом месте тела животного (в торакальном и югулярном отделе) и происходят из других эмбриональных зачатков (передние миотомы и передние спинномозговые нервы). Аналогичный случай представляет собой замещение мезонефроса Anamnia метанефросом млекопитающих: функция остается той же, но выполняется она новым органом, развившимся в другом месте тела животного.
К этому же типу субституции можно было бы, вероятно, свести и многие другие случаи преобразования метамерных органов. Этот тип филогенетического изменения органов представляет собою известного рода переход между типичными случаями субституции органов в смысле Клейненберга и случаями «субституции функций», о которых речь будет впереди.
Итак, мы различаем двоякого рода субституции органов: 1. когда замещающий орган развивается на том же месте тела животного, на котором находился у предков замещенный орган, и 2. когда замещающий орган развивается на другом месте тела животного. В обоих случаях замещающий орган выполняет туже функцию, что и замещенный орган, и в обоих случаях замещенный орган редуцируется.
ФИЗИОЛОГИЧЕСКАЯ СУБСТИТУЦИЯ Д. М. ФЕДОТОВА
В 1925 г. Д. М. Федотов очень коротко сформулировал новый принцип филогенетического развития, назвавши его принципом «физиологической субституции». При физиологической субституции один орган
Рис. 119 А, В. Схема физиологической субституции бурсы Stegophiura nodosa (А, b) вторичной полостью тела Ophionotus hexactis (В, а), принявшей на себя выводковую функцию бурсы.
а—вторичная полость тела; b—бурса; с—кишечник; d—стенка тела; е—личинки офиур (из Д. М. Федотова, 1927).
замещает одну из функций другого органа, морфологически ему уподобляясь; субституируемый же орган при этом не редуцируется. В качестве примера физиологической субституции Федотов приводит развитие личинок у некоторых типичных живородящих офиур. Обычно у живородящих офиур, например у Stegophiura nodosa, выводковыми полостями служат бурсы (рис. 119 А, Ь), имеющиеся в количестве пяти пар мешков, замкнутых со стороны полости тела и сообщающихся с наружной средой. Все развитие личинок (рис. 119 А, е), как это видно из рисунка, происходит у таких форм в бурсах. При физиологической же субституции у других форм живородящих офиур, например у Ophionotus hexactis, происходит следующее: личинки вынашиваются
380
ГЛАВА XI
не в бурсах, которые все время остаются свободными от молоди, но во вторичной полости тела (рис. 119 В, а); при этом личинки (рис. 119 В, е) размещаются здесь между стенками тела и кишечником (рис. 119 В, с). Таким образом вторичная полость тела (а) принимает на себя одну из функций бурсы, аим.енно функцию выводковой полости. Сами же бурсы, продолжая нести остальные свои функции (дыхательную, половую), целиком сохраняются. Федотов предполагает, что такой тип субституции мог возникнуть филогенетически недавно. . .
ПРИНЦИП УМЕНЬШЕНИЯ ЧИСЛА ФУНКЦИЙ
Ниже мы увидим, что при усложнении строения органов число функций каждого органа может увеличиваться путем приобретения новых второстепенных функций. Но именно у высокоорганизованных форм мы нахо-, дим часто случаи, когда общее число функций какого-нибудь органа уменьшается у потомков по сравнению с его числом у предков. На эти случаи обратил мое внимание С. А. Северцов, и здесь, повидимому, мы имеем перед собою тип филогенетического развития, представляющий собою противоположность принципу расширения функций Плате, о котором сейчас будет речь.
Нам известно, например, что и парно- и непарнокопытные животные происходят от примитивных пятипалых млекопитающих. Нам известно также, что конечности, снабженные пятью пальцами, могут производить большее число движений и более сложные движения, чем конечности парно- или непарнокопытных. У этих последних основная функция конечностей, т. е. ходьба или бег, настолько усилилась, что она, так сказать, подавила собою целый ряд второстепенных функций.
То же самое можно сказать и относительно функций конечностей водных млекопитающих: китов, морских коров, тюленей, моржей и т. п., т. е. таких животных, у которых конечности превратились в приспособленные для плавания ласты. То же, конечно, относится и к укороченной шее кита, который не имеет возможности делать все те многочисленные движения головою, которые были вполне доступны его длинношеим предкам. Так, кит или дельфин уже не могут, например, счищать зубами паразитов с поверхности своей кожи, что легко могли делать их наземные предки, обладавшие длинной шеей.
Можно было бы привести еще целый ряд примеров уменьшения числа функций, но и сказанного, мне кажется, достаточно, чтобы мы имели право утверждать, что часто при филогенетическом усилении главной функции какого-либо органа общее числофункций этого органа у потомков уменьшается.
Из этого принципа, так же как и из принципа расширения функций Плате, можно вывести следующее заключение о возможностях дальнейшей эволюции органа. Если расширение функций увеличивает число возможных эволюционных направлений для данного органа, то уменьшение числа функций какого-либо органа, конечно, уменьшает число возможных эволюционных направлений этого органа. Это значит, что данной форме животных будет все труднее и труднее приспособляться к изменяющимся условиям среды. Число возможных направлений приспособления конечности непарнокопытных животных в течение эволюции стало, безусловно, гораздо меньше, чем число возможных направлений приспособления конечности у их пятипалых предков.
Этим принципом мы заканчиваем рассмотрение тех типов филогенетических изменений органов, при которых главная функция органов изменяется у потомков лишь количественно. Обратимся теперь
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 381
к другой группе принципов (типов) эволюции органов, а именно к той, в которой главная функция эволюирующего органа изменяется у потомков качественно.
Свой обзор этих типов эволюции мы начнем с широко распространенного принципа расширения функций Плате.
Рис. 120. Брюшной плавник и копуляционный орган Rhinobatis productus (из Ф. Даниэля, 1928).
ПРИНЦИП РАСШИРЕНИЯ ФУНКЦИЙ
Принцип расширения функций был впервые сформулирован Л. Плате. В своей книге «Abstammungslehre» (1912, 1925) Плате говорит, что «когда какой-нибудь орган достигает в течение своей филогенетической диферен-цировки известной высоты жизнедеятельности и сложности, то он нередко принимает на себя новые задачи. Этот принцип расширения функций имеет большое значение, ибо он объясняет нам, почему орган с течением времени иногда соверешенно меняет свою функцию, причем первоначально второстепенная функция становится главной функцией». В качестве примера расширения функций Плате приводит в первую очередь случай расширения функций жабр у La-mellibranchiata, которые у этих форм, помимо своей основной функции дыхания, стали выполнять еще целый ряд важных побочных функций,' а именно они пригоняют вместе с током воды частички пищи к ротовому отверстию и служат, кроме того, у самок в качестве выводковой полости. У водяных блох длинные антенны являются не только органом чувств, но служат также, и даже главным образом, в качестве органа движения. У обыкновенного речного рака первая пара абдоминальных ножек является вместе, с тем и органом совокупления, а у акул, как это видно из рис. 120, в качестве копулятивного органа служат выросты брюшных плавников.1 У летающих рыб грудные плавники превращаются в своеобразные парашюты (рис. 121); у птиц клюв служит не только для еды, но также и в качестве орудий при самых разнообразных действиях: при постройке гнезда, при чистке и смазывании перьев, при нападении на врага и защите от него и т. д.
Кроме случаев, приводимых Плате, можно найти еще прекрасные примеры расширения функций при исследовании филогенетического развития парных конечностей у позвоночных. Так, например, мы знаем, что парные плавники представляют собой у акул гибкие горизонтальные пластинки, расположенные по бокам тела животного. Основание плавника, которым он прикрепляется к стенке тела, имеет у акул и у хрящевых ганоидов вид широкой пластинки (эврибазальный тип плавника); в проксимальной части каждого грудного плавника лежат крупные базальные хрящи, к которым причленяются многочисленные хрящевые лучи (рис. 122), а к концам этих лучей причленяются, в свою очередь, многочисленные роговые лучи плавника. По обеим сторонам плавникового скелета, дорзально и вентрально, лежат m. т. pterygiales, состоящие из отдельных, метамерно расположенных мускульных пучков. Функция
1 Пожалуй, эти случаи следует скорее рассматривать как примеры смены функций по принципу А. Дорна.
382
ГЛАВА XI
таких плавников, подробно изученная И. И. Шмальгаузеном,1 целиком пассивна: плавники эти значительно увеличивают нижнюю поверхность туловища и препятствуют опусканию тяжелой верхней части тела при плавании; они служат также в качестве органа равновесия, регулирующего нормальное положение тела в воде. У высших рыб, начиная от костных ганоидов и кончая костистыми рыбами, мы находим различные степени перехода от пассивной функции грудных плавников к активной их функции. Здесь имеется прежде всего целый ряд переходов от эвриба-зального типа грудных плавников селахий и хрящевых ганоидов к стенобазальному типу этих плавников у высших рыб (А. Н. Северцов, 1926): основание плавника постепенно становится все уже и вследствие этого сам плавник все подвижнее. У высших рыб (костные ганоиды, костистые рыбы) изменилось, кроме того, и само положение грудных плавников: мы находим, что у этих форм передний конец основания плавника приподнялся кверху, а сам плавник сдвинулся и вместо горизонтального принял косое по отношению к оси тела положение. Вследствие того, что основание плавника стало короче, а сам плавник сделался косым, значи-
Рис. 121. Приспособленные, кроме плавания, также и для взлета грудные плавники Exocoetus rondeletti (из Г. Ббкера, 1935).
тельно возросло разнообразие возможных движений грудного плавника по отношению к месту его прикрепления, т. е. к плечевому поясу, и мы действительно видим (см. Шмальгаузен), что грудные плавники костных ганоидов и костистых рыб производят самые разнообразные движения как при повороте тела во все стороны, так и при его поступательном движении.
Наивысшей степени подвижности грудные плавники достигают у кистеперых (Polypterus и вымершие Crossopterygii) и у двоякодышащих рыб, у которых сужение основания плавника достигает предельных размеров, а связь основания с плечевым поясом осуществляется у Polypterus при помощи сросшихся между собою проксимальных концов постаксиальных и преаксиальных лучей (рис. 123), а у Ceratodus—при помощи единственного базального членика плавниковой оси (рис. 112 В). Здесь плавник служит уже не только для плавания и для изменения направления движения, но и в качестве опорного органа при покое и при движении (ползании по дну водоемов); мы знаем, например, что Ceratodus может ползать при помощи своих грудных плавников взад и вперед по дну аквариума.
Сравнительное изучение движений у рыб говорит о том, что все морфологические изменения в скелете и мускулатуре грудных плавников ведут
1 См. Шмальгаузен И. И., О функциональном значении плавников у рыб, Русск. зоол. журнал, т. 1, 1916.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 383'
к повышению количества возможных движений этих плавников. Из эври-базального плавника типа селахий, обладающего лишь пассивной функцией, постепенно образовался активный мультифункциональный грудной плавник высших рыб. Плавник этот сохранил свою главную функцию (направлять движение тела при плавании), но вместе с тем приобрел и целый ряд новых побочных функций (у двоякодышащих и кистеперых—опорную функцию, функцию ползания и т. д.). Здесь мы ясно видим, что в течение филогенеза наряду с главной функцией произошло и значительное расширение побочных функций грудных плавников.
При переходе предков наземных позвоночных из водной среды к жизни на суше функция их конечностей резко изменилась: они приспособились к ползанию, к ходьбе и к бегу, а также стали служить в качестве опоры для несения тяжести тела животного при его передвижении по суше. При этом изменились как способ сочленения свободных конечностей с пле
Рис. 122. Эврибазальный плавник акул с ограниченным числом движений и пассивной функцией поддерживания передней части тела при плавании.
Рис. 123. Стенобазальный плавник кистеперых, приспособленный к разнообразным движениям в связи с приобретением значительного числа побочных функций.
чевым поясом, так и строение костного скелета самих конечностей и их мускулатура. На проксимальных концах плечевой и бедренной кости развились круглые сочленовные головки, благодаря чему первичные наземные позвоночные стали производить своими свободными конечностями такие движения, которые были недоступны их рыбообразным предкам. Сами конечности постепенно превратились в трехчленную систему рычагов, а строение их мускулатуры стало гораздо более сложным, чем у их .кистеперых предков. Вследствие всего этого они смогли постепенно приспособиться сначала к ползанию по земле, а затем и к настоящей ходьбе: здесь мы имеем дело с типичным случаем смены функций.
Но наряду с этим сложным комплексом движений, необходимых для поступательного передвижения животных по суше, у них появилась возможность (именно благодаря только что перечисленным нами изменениям в строении скелета конечностей) производить целый ряд также и других движений, к которым прибегают иногда даже очень примитивно построенные позвоночные в случае опасности или в случае крайней нужды. Напомним читателю, например, об известном опыте с лягушкою, которой касаются палочкою, смоченной в крепкой кислоте: если коснутся ею правой стороны туловища животного, то лягушка тотчас же начнет счищать с кожи кислоту своею правою задней лапкой, если же эту лапку отрезать, или отшнуровать, то она счищает кислоту с кожи левою задней лапкой, производя при этом целый ряд совершенно непривычных для нее движений. Строение сочленений и мускулатуры конечности лягушки делают, конечно, все эти «неестественные» для наблюдателя движения^
.384
ГЛАВА XI
вполне возможными, но совершенно несомненно, что лягушке в своей обычной жизни приходится прибегать к такого рода движениям лишь в исключительно редких случаях.
Таким образом, мы видим, что эволюция конечностей позвоночных протекает в направлении усовершенствования главной их функции, т. е функции поступательного движения по суше (ползание, ходьба, бегание), но что при этом далеко не исключена возможность производить целый ряд и других движений, уже не имеющих никакого отношения к передвижению тела в пространстве. В случае нужды животные обычно широко пользуются этой возможностью. Так, млекопитающие пользуются при необходимости своими конечностями и для плавания, и для рытья почвы, и для защиты •от нападения, и для того, чтобы вскарабкаться на дерево, и для того, чтобы почесаться, и для многих других потребностей. Само собой разумеется, что способность использовать в случае нужды все возможное разнообразие этих движений покоится на прогрессивном развитии центральной нервной системы, которое делает возможным возникновение ассоциаций между известными движениями и известными раздражениями, исходящими из внешнего мира.
Как это уже было отмечено Л. Плате (и как это мы увидим ниже), •принцип расширения функций является чрезвычайно важной предпосылкой при эволюции путем смены функций по А. Дорну. Вместе с тем мы видим, что и само по себе расширение функции может быть чрезвычайно полезно животному, ибо для существования и благоденствия вида весьма важно, чтобы отдельные его представители могли в случае крайней нужды спастись от опасности непривычными в обычной обстановке действиями (в нашем примере—непривычными движениями).
ПРИНЦИП СМЕНЫ ФУНКЦИЙ А. ДОРНА
Теперь мы займемся чрезвычайно важным и до сих пор, к сожалению, еще мало оцененным принципом, а именно принципом смены функций А. Дорна. По нашему мнению, дорновский принцип эволюции является одним из важнейших принципов, разъясняющих нам многие случаи филогенетического развития, и мы можем лишь поражаться таланту автора, который сумел его сформулировать еще в 1875 г.
Принцип Дорна говорит о том, что каждый орган имеет нормально не одну, а несколько функций, причем одна из них является главной, а другие—второстепенными.
В течение эволюции часто случается так, что вследствие изменившихся условий окружающей среды у данной животной формы одна из второстепенных функций органа делается его главной функцией и соответственно этому меняется все строение органа.
Сам Дорн следующим образом выразил сущность своего принципа: «Вследствие смены функций, носителем которой является данный орган, происходит изменение и самого органа. Функция органа представляет собою равнодействующую многих компонентов, из которых один является главной или первичной функцией органа, в то время как другие представляют собою его побочные или второстепенные функции. Ослабление главной функции и усиление одной из второстепенных функций меняет общую функцию органа: второстепенная функция постепенно становится главной функцией, общая функция делается иной, и следствием этого процесса является перестройка •всего органа».
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 385
желудочной мускулатуры (второ-
Рис. сгей
124. Изменение формы конечноречного рака в связи со сменой функций.
1—усик первой пары; 2—жало; 3 и 4—первая и вторая челюсть; 5—7—челюстные ножки; 8— ходильная ножка; 9—абдоминальная ножка (из Бобрецкого).
Дорн приводит в своей работе целый ряд случаев смены функций. Он говорит, например, об изменениях, происходящих при филогенетическом развитии желудка у птиц, об изменениях, имеющих место при развитии ротового аппарата у рака и у многих других. При этом он утверждает, что желудок имеет две функции, а именно функцию выделения желудочного сока, доставляемого желудочными железами (главная функция) и функцию перетирания пищевой массы действием степенная функция). При эволюции желудка птиц в одном из участков этого желудка усилилась выделительная функция желез, и этот участок превратился в особый—секреторный—отдел желудка, во втором участке усилилась первично второстепенная функция желудка, т. е. функция перетирания пищи, и стала его главной функцией, секреторная же функция этого участка атрофировалась совсем, и он превратился в чисто мускульный желудок, приспособленный для механического перетирания проглоченных зерен.
Ротовой аппарат ракообразных, по Дорну, сначала образуется как типично вильчатая конечность Schi-zopoda, т. е. как чисто плавательный аппарат. Но с течением эволюции шипы и иглы, сидящие на тех конечностях, которые были расположены в непосредственной близости от ротового отверстия, принимают все большее и большее участие в захватывании добычи, и постепенно эта второстепенная функция усиливается до такой степени, что передние конечности ракообразных превращаются в типичный ротовой аппарат. На рис. 124 мы видим типичную ходильную ножку речного рака (8) и различные формы, которые она принимает, превращаясь при смене функций в усик(1), в различные части челюстного аппарата (2, 3, 4), в клешню и т. д.
Некоторые другие случаи, приводимые Дорном в качестве примеров смены функций (мы имеем в виду его утверждение, что органы совокупления позвоночных произошли путем смены функций из жабр), в настоящее время уже потеряли для нас свое значение, ибо выставленная им и поддержанная его последователями гипотеза, касающаяся происхождения этих органов, не выдержала критики позднейших исследований. Однако ценность и общее значение принципа смены функций, конечно, не были поколеблены этим неудачным с нашей современной точки зрения выбором примеров, вполне соответствующим уровню знаний восьмидесятых годов прошлого столетия. Теперь, после более че.м пятидесятилетнего периода Морфологические закономерности 25
386
ГЛАВА XI
в области исследования филогенеза животных,'мы видим, что принципом смены функций можно объяснить гораздо ббльшее число случаев превращения органов, чем это думали прежде. В особенности же принцип смены функций приложим в тех случаях, когда эволюирующие формы, изменяя
Рис. 125. Плавун (Chironectes minimus). (Из Брема.)
люции рептилии и млекопитающих.
Рис. 126. Морские львы (Emetopias california-cus). (Из фильма Ufa).
место своего обитания, переходят в новые условия среды, сильно разнящиеся от тех старых условий, в которых жили их предки, например когда наземные животные вторично переходят в водную среду.
Мы знаем, что такое явление неоднократно имело место в течение эво-“ предки талаттозавров, морских черепах, ихтиозавров и плезиозавров некогда были типично наземными формами, которые лишь впоследствии вторично стали обитателями морей;1 то же самое произошло и с предками тюленей, морских коров и. китов. При этом изменении среды произошли, конечно, и значительные изменения в строении и функциях целого ряда органов этих животных.
Хотя во многих случаях мы и не можем с точностью установить родословную этих форм, однако мы можем позволить себе, на основании палеонтологических, экологических и срав
нительно анатомических исследований, восстановить в общих чертах филогенетический путь их развития. Для этого нам приходится несколько более подробно остановиться на вопросе о том, как именно происходило изменение функций при переселении наземных позвоночных в воду.
1 Ср. Abel О., Die Eroberungsziige der Wirbeitiere in die Meere der Vorzeit, Jena, 1924.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 387
Мы знаем, что типично наземному млекопитающему вода требуется для двух целей: во-первых, для утоления жажды и, во-вторых, для купания. Мы знаем также, что очень многие млекопитающие умеют плавать и что они обычно утилизируют эту способность в случае нужды. Плавают лошади, олени, медведи, собаки, тшдзы и многие другие животные. Не раз наблюдалось, что благородный олень плыл в случае нужды на расстоянии 20 км, а северный олень, как известно, плавает еще лучше его; известно также, что лошади могут переплывать очень широкие реки. Мне самому пришлось однажды видеть волка, переплывавшего в широком месте Дон.
Способность плавать у млекопитающих, связанных биологически с водой, развивалась прогрессивно. Это наблюдается, например, у северных
Рис. 127. Гигантский кит (Balaenoptera musculus L., из Чулока).
медведей, у плавунов (Chironectes minimus, рис. 125) и у многих других животных, которым приходилось либо добывать себе необходимую пищу в воде, либо бросаться туда, спасаясь от врагов.1 У всех этих форм развивалась прогрессивно функция плавания, т. е. та именно функция конечностей, которая имелась у них и раньше, но которая при прежней наземной жизни употреблялась лишь в случае крайней необходимости и играла поэтому лишь второстепенную роль. У всех этих животных между пальцами прогрессивно развились плавательные перепонки.1 2
Чем сильнее выступает у животных на первый план вторичная функция плавания, тем резче перестраивается в этом направлении строение его
1 Возможно, что это случилось у некоторых прибрежных хищников, умеющих при случае поймать в реке какое-либо водное животное, как это делают, например, дикие кошки: весьма вероятно, что у них, в случае недостатка пищи на суше, главной добычей делается именно водное животное (в данном случае рыба). Можно также предположить, что и у травоядных морских млекопитающих, например у предков морских коров (Manatidae, Halicoridae), главным стимулом для перехода в воду был именно недостаток растительной пищи на суше и поиски этой пищи в воде (водоросли).
2 Зачатки этих перепонок часто встречаются и у наземных млекопитающих.
25*
388
ГЛАВА XI
конечностей: передние конечности северных медведей, выдр, Lutrinae (рис. 125), ластоногих (рис. 126), сирен и, наконец, китов (рис. 127) представляют собой различные ступени преобразования конечностей, приспособленных для движения по суше, в конечности, приспособленные для движения в воде, т. е. в плавательные лопасти.1
К аналогичным выводам приходим мы и при изучении перехода наземных рептилий к водному образу жизни (нотозавры, плезиозавры), а также при изучении перехода летающих водных птиц к постоянной жизни в море (пингвины). Своеобразные изменения, происшедшие в передней конечности пингвинов, изображены на рис. 128.
Во всех этих изменениях мы ясно видим, как именно происходила здесь смена функций. Первоначально конечности всех млекопитающих были приспособлены для поддерживания тяжести тела животного и для его передви
Рис. 128. Изменения, происшедшие в крыле пингвина в связи со сменой функций при переходе от полета к плаванию (из И. И. Шмальгаузена, по Осборну, 1935).
жения по твердой поверхности (ходьба и бег). В этом и заключа
Рис. 129. Крот снизу. Обратить внимание на роющие конечности (из Г. Кокера, 1935).
лась главная функция конечностей. У береговых млекопитающих к этой главной функции присоединилась еще и второстепенная, а именно, функция плавания, которая сначала пускалась в ход относительно очень редко. При изменении образа жизни этих животных эта второстепенная функция конечностей постепенно превратилась в главную, а прежняя главная функция (поддерживание тяжести тела, ходьба) превратилась сначала в редко употребляемую второстепенную функцию (приблизительно на этой стадии эволюции остановились моржи и тюлени), а затем редуцировалась совсем (киты).1 2
1 Само собой разумеется, все эти современные нам формы не связаны друг с другом непосредственными филогенетическими связями. Все они являются конечными стадиями развития различных филогенетических рядов, исходящих от аналогичного состояния и развивающихся в одном и том же направлении (переход к плавательному движению в воде).
2 Здесь мы говорим только об эволюции передних конечностей, так как задние конечности ластоногих и китообразных развивались в дивергентных направлениях: у первых они превратились в ласты, у последних, т. е. у китов, у которых образовался горизонтальный хвостовой плавник, они исчезли совсем.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 389
Вообще говоря, конечности позвоночных и, в особенности, млекопитающих дают нам целый ряд прекрасных примеров смены функций, ибо здесь, исходя из типично построенной пятипалой конечности стопоходящих насекомоядных, конечности развиваются в самых разнообразных направлениях. Мы имеем в виду подробно разобранную нами выше адаптивную радиацию Осборна. Мы хорошо знаем, что из примитивной пятипалой конечности первичных млекопитающих (рис. 83 А) произошли и лазящие конечности лемуров и обезьян с отходящим в сторону большим пальцем (рис. 83 F), и приспособленные к висению на ветках конечности ленивцев с их огромными когтями (рис. 83 Е), и роющие конечности кротов и землероек (рис. 83 С; ср. рис. 129), и приспособленные к полету конечности летучих мышей и летучих собак(рис. 83D), и, наконец, только что подробно описанные нами конечности водных млекопитающих1 (рис. 83 G, рис. 124, 125, 126 и 127).
Во всех этих случаях мы находим в органах животных признаки, которые могли образоваться только путем смены функций, иными словами, мы находим у исходных форм, т. е. у форм с пятипалой стопоходящей конечностью, те второстепенные функции, которые могут впоследствии стать главной функцией. На этих примерах выступает особенно ясно тот фундаментальный факт, который лежит в основе принципа смены функций, а именно, что почти всякий более или менее сложно построенный орган является носителем целого ряда функций; как это уже было нами сказано выше,1 2 по существу он мультифункционале н. Даже такие высокоспециализированные органы, какими являются, например, конечности лошади, и те нормально служат не только для ходьбы и бега, но при случае являются орудием нападения и защиты; лошадь пользуется ими и при плавании и при рытье почвы. То же самое можно сказать и о конечностях других копытных. Felidae пользуется своими конечностями в качестве орудия при борьбе, как органом движения на суше (ходьба, бег, прыжок) и в воде (плавание), как органом цепляющимся, лазающим и т. д. Крылья крупных птиц служат для полета, но в случае надобности птица ими пользуется как органами нападения и защиты: орлы наносят крыльями сильные удары и ударами сбрасывают при случае добычу со скал. Ноги птиц служат не только для передвижения по земле, но и для обхватывания веток при сидении на них, для схватывания и перенесения добычи при полете, для нападения и защиты.
Почти все эктосоматические органы позвоночных по существу мульти-функциональны, и лишь немногие из них, как, например, млечные железы и органы совокупления, имеют только одну единственную функцию.
Интересный пример развития, происходящего по типу смены функций, наблюдается в филогенезе яйцеклада у пчел и у ос. По неопубликованным данным А. А. Махотина, у предков современных насекомых все тело было сегментировано, причем каждый сегмент, наподобие того, что мы видим у многоножек, нес одну пару конечностей (рис. 130 А). Втечение эволюции насекомых конечности на брюшке редуцировались, сохранившись лишь в виде зачатков у эмбрионов, но конечности восьмого и девятого абдоминального сегмента превратились в придатки, окружающие половые отверстия: у самок—в яйцеклад (рис. 130 В), у самцов—в так называемые парамеры, т. е. органы, при помощи которых самец захватывает и удерживает самку. В дальнейшем у высших перепончатокрылых (Hymenoptera acu-leata) яйцеклад постепенно превратился в жало, служащее, для защиты и нападения. У многих Aculeata, например у осы, жало сохраняет еще функ
1 Мы не упоминаем здесь о конечностях копытных, так как они развились не путем смены функций.
2 См. выше принцип интенсификации функций.
390
ГЛАВА ХГ
цию яйцеклада, но у пчел (по крайней мере у рабочих пчел, рис. 130 С) эта функция уже теряется и сохраняется лишь одна защитная функция.
Таким образом, в течение своей эволюции восьмая и девятая пара абдоминальных конечностей
насекомых дважды изменяла свою функцию: сначала ходильная конечность превратилась в яйце клад, а .затем яйцеклад превратился в жало.
В предыдущем мы уже упоминали о том, что принцип интенсификации функций лежит в основе целого ряда типов (принципов) эволюции органов; между прочим, лежит он и в основе принципа смены функций. Чтобы наглядно изобразить переход от первичной мультифункциональности
Рис. 130 A, С. Схема изменений, происшедших в филогенезе конечностей высших перепончатокрылых в связи со сменой функции, имевшей место при превращении абдоминальной конечности сначала в яйцеклад, а затем в жало.
А—абдоминальные конечности гипотетического предка насекомых (основная ходильная функция). В—яйцеклад стрекозы (Aeschna): половая функция—откладка яиц (первая смена функции). С—жало рабочей пчелы (Apis mellifica): защитная функция (вторая смена функций). Т8—Т9—тергиты 8-го и 9-го абдоминальных сегментов; соха изображена сеткой; subcoxa изображена штриховкой; stilus—черной краской; telopodit (дистальная часть ножки)—двойным пунктиром (из А. А. Махотина).
Интенсификац.и я: одна из этих функций интенсифицируется и становится главной функ-< ц и е й органа, остальные функции (второстепенные функции) сохраняются целиком
Смена функций: вторичная функция интенсифицируется и становится < новой главной функцией органа; бывшая главная функция органа редуцируется.
органа к интенсификации функций и далее к смене функций, мы приводим следующую таблицу:
Первичная мультифункциональность к леток и органов: ряд равноценных функций клеток и клеточных комплексов (подвижность, раздражимость, ассимиляция, секреторная функция, функция опоры и т. д.)
Мы говорили до сих пор о значении смены функций, имея в виду лишь активные органы животного. Теперь нам надо поставить вопрос о том; применим ли этот принцип также и в отношении пассивных органов? Нам думается, что на этот вопрос следует ответить утвердительным образом, если только мы будем говорить не о принципе смены функций, а о принципе изменения полезности пассивных экто-
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИИ ОРГАНОВ 391 соматических органов и признаков при изменении условий окружающей среды. Ради краткости и принимая только что упомянутую нами оговорку, мы будем называть его «принципом смены пассивных функций».
Поясним это следующим примером. Мы уже говорили о том, что покровные кости и, в особенности, чешуи рыб с функциональной точки зрения являются пассивными органами. Так, плакоидные чешуи эласмобранхий в общем представляют собою одновременно органы пассивной и активной защиты этих рыб: дентинные пластинки чешуй образуют, правда, несовершенный, но все же довольно крепкий панцырь (пассивная защита), а сидящие на этих пластинках зубы легко могут поранить любого из нападающих на них врагов (активная защита). Мы знаем, однако, что именно этот последний признак (побочная функция активной защиты), совершенно второстепенный для большинства эласмобранхий, в некоторых случаях становится их главным признаком (в смысле Дорна—главной функцией). У Spinaceidae, например, плакоидные чешуи, лежащие впереди от дорзальных плавников, сильно увеличились в размерах и превратились в длинные и крепкие шипы, служащие в качестве органов активной защиты животного. То же самое можно сказать и относительно развившихся из обычных плакоидных чешуй зубов, сидящих по бокам чрезвычайно удлинившегося рыла у Pristiurus и у Pristiophorus; то же относится и к хвостовым шипам Trygon, Raja и Myliobatis californicus (рис. 133): в этих последних случаях органы, первоначально служащие лишь для активной защиты животных, превратились в настоящее орудие нападения. Сюда же можно причислить и превращение плакоидных чешуй, сидевших по краям ротового отверстия у предков эласмобранхий, в типичные, приспособленные для кусания зубы.
К этой же категории пассивных адаптивных изменений принадлежат и все изменения, наблюдаемые в форме и окраске животных: мы имеем в виду защитные формы и покровительственные окраски. Мы знаем, что многие млекопитающие, изначально обитавшие в лесах или джунглях, приобрели желтую окраску, испещренную пятнами (ягуары, пантеры, оцелоты и т. п.) или полосами (тигр) шкуры. Мы знаем также, что когда такие формы переселяются в пустыню или в степь, то с течением времени изменяется их окраска приспособительно к одноцветножелтой среде, и их потомки становятся сами равномерно желтыми. Таким образом, у,них один из признаков окраски предков, а именно черные пятна и полосы, исчезает совсем, а другой (желтый фон) делается главным признаком.
Весьма вероятно, что многие адаптивные изменения формы и окраски, например, изменения цветных пятен кожи и кожных придатков, происходят по принципу смены функций при переходе животных из одной среды в другую. При этом следует отметить, что животные, для которых приспособительная окраска кожи и приспособительный ее рисунок имеют важное биологическое значение, могут переселиться только в такую среду, общий тон которой, хотя бы в некоторых цветовых отношениях, соответствует общей окраске их кожи. Во всякой другой среде, к которой они не подходили бы по своей окраске, они слишком резко бросались бы в глаза своим врагам, и это, конечно, создало бы для них чрезвычайно неблагоприятные условия существования. Очень многие приспособительные изменения в окраске животных (вспомним, например, как различна бывает расцветка болотных и древесных лягушек) становятся понятными нам, если только мы станем на эту точку зрения.
Мы видим, таким образом, что принцип смены функций А. Дорна распространяется (правда, в несколько видоизмененной форме) также и на пассивные органы и на пассивные признаки многих животных.
392
ГЛАВА XI
СМЕНА ФУНКЦИЙ, РАСШИРЕНИЕ ФУНКЦИЙ И ЧИСЛО ВОЗМОЖНЫХ НАПРАВЛЕНИЙ В ЭВОЛЮЦИИ ЖИВОТНЫХ
В литературе, посвященной проблемам эволюции, часто поднимался вопрос относительно числа возможных направлений, в которых может итти филогенетическое развитие той или иной формы животных. Сторонники ортогенетической теории филогенеза и, в особенности, сторонники аутогенетического ортогенеза обычно принимают, что направление прогрессивной эволюции каждой животной формы предопределено организацией этой формы и что поэтому данная форма может эволюировать лишь в одном, много в двух или в трех филогенетических направлениях. Сторонники эктогенеза полагают, наоборот, что каждой животной форме предоставляется возможность развиваться в самых разнообразных эволюционных направлениях.
Тщательный анализ принципа смены функций дает нам, повидимому, возможность несколько ближе подойти к разрешению этого сложного и трудного вопроса. Мы видели, что в случаях, когда эволюция какого-либо органа идет в направлении смены функций, число возможных эволюционных направлений в развитии органа в известной степени определяется числом его функций, т. е. совокупностью его главной и всех его второстепенных функций, ибо, как мы это уже установили выше, каждая из второстепенных функций при известном изменении условий окружающей среды может стать главной функцией. Это положение действительно для каждого активно функционирующего органа и для каждого пассивного приспособления. Оно действительно и для каждой отдельной части организма, но для животного, взятого как целое, число возможных филогенетических направлений, в которых оно может эволюировать, конечно, еще значительнее, ибо одно и то же животное может приспособиться к различным условиям окружающей среды путем смены функций в различных своих органах.
Возьмем, например, случай, когда млекопитающее, постоянно живущее на берегу какого-нибудь водоема, начинает постепенно, вследствие недостатка в пище на суше, переходить к постоянной жизни в воде, иными словами, случай, когда конечности бегающего типа превращаются в конечности плавающего типа. Главной функцией (схема XXIП,Н) конечностей наземного хищника является, как известно, ходьба или бег, но кроме того его конечности обладают целым рядом второстепенных функций (п), которые употребляются лишь в сравнительно редких случаях. Для млекопитающих характерны следующие второстепенные функции конечностей: функция рытья передними лапами (nJ, функция захватывания и передвижения пищи передними лапами (п2), функция царапанья передними лапами (п3), функции плавательных движений при плавании (п4) и т. д. Символически это можно представить при помощи схемы XXIII (А—Е).
’Представим себе, что у формы X в случае А изначально главной функцией конечностей будет Н, а пх, п2, п3 и п4 будут только что перечисленные нами второстепенные их функции. В случае В прежняя главная функция редуцируется, что мы обозначаем, помещая букву Н в скобки и сопровождая ее символом редукции г,—(Н)г, а побочная функция пх развивается прогрессивно и делается вторичной главной функцией конечности— лхН. В случаях С, D и Е прогрессивно развиваются та или иная из второстепенных функций: п2, п3ип4; последние превращаются при этом в главные функции (п2Н, п3Н, п4Н). Таким образом, данный орган в данном случае имеет возможность развиваться в пяти различных направлениях: в направлении прогрессивного развития первичной главной функции (случай А: интенсификация функций) и в направлениях прогрессивного развития каждой из четырех второстепенных функций (случаи В, С, D и Е: смена функций), где каждая из этих функций может сделаться главной функцией у потомков.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 393
Но этот же вид, переселившись в какую-нибудь другую местность, может, конечно, эволюировать и в другом направлении и приспособиться к новым условиям окружающей среды не изменением органов движения, но, например, изменением своей окраски (т. е. приобретая покровительственную окраску волосяного покрова) или же переходом к другому роду питания. При этом будут использованы эволюционные возможности, заложенные в других мультифункциональных органах, т. е. в окраске и расцветке волосяного покрова, в строении зубов, в структуре челюстного а аппарата и т. д. Так как каждое животное обладает большим числом эктосоматических органов и так как большинство этих органов по существу своему мультифункционально, то и число возможных адаптивных направлений в эволюции, осуществляемых путем смены функций (число функций эктосоматического органа, умноженное на число эктосоматических органов), может быть относительно очень высоким.
В предыдущем мы уже отметили главное значение принципа расширения функций, заключающееся в том, что этот принцип увели-
Схема XXIII. Объяснение в тексте.
чивает число направлений, в которых может последовать филогенетическое развитие потомков эволюирующей формы, т. е. что он умножает число возможных эволюционных направлений.
Выше нами было установлено, что это число, согласно дорновскому принципу смены функций, определяется совокупностью главной функции плюс всех второстепенных функций каждого органа. Если число второстепенных функций увеличится, то этим самым и в - той же мере увеличится и число новых потенциально возможных направлений в эволюции данной формы. Мультифункциональность каждого органа свидетельствует о том, что множественность эволюционных и адаптивных направлений мыслима не только для всего организма, взятого в его целом, но и для каждого его органа, взятого в отдельности. Принцип Плате, т. е. принцип расширения функций, показывает нам, что вместе с усложнением организации более высокостоящих форм увеличивается и число возможных эволюционных направлений для данной животной группы, иначе говоря, увеличивается ее способность к приспособлению. Все это рассуждение станет более понятным, если мы внимательно рассмотрим следующую схему (схема XXIV).
Здесь мы видим, что число второстепенных функций увеличилось, возросши с четырех (на схеме XXIII п* п2, п3 и п4) до пяти (схема XXIV, пх, п2, п3, п4, п5) и что, соответственно этому, число возможных направлений.
394
ГЛАВА XI
в филогенетическом развитии данной группы увеличилось с пяти до шести •(схема XXIV, направления Н, nv п2, п3, п4 и п5). Таким образом, данный орган сделался более гибким в смысле приспособления к условиям окружающей среды. Так как это относится ко всем органам, которые приспособляются путем смены функций, то отсюда можно заключить, что именно более высоко организованные животные, те, у которых число второстепенных функций больше, лучше и легче приспособляются к неблагоприятным изменениям в условиях окружающей среды, чем низко организованные . Все это решительно говорит против ортогенетической точки зрения, утверждающей, что каждая форма может развиваться лишь в од-
ном единственном предопределенном для нее направлении, а также против известной теории Д. Роза,1 согласно которой более высоко организованные животные менее способны к приспособлению, чем низко организованные. В этом отношении принцип Плате имеет большое теоретическое значение для правильного понимания общего хода эволюционного процесса.
ПРИНЦИП СУБСТИТУЦИИ ФУНКЦИЙ
В некоторой связи с описанным нами принципом субституции органов Н. Клейненберга стоит еще другой принцип или тип филогенетического развития, а именно принцип, который мы обозначаем термином «субституция функций» и который мы можем коротко охарактеризовать в следующих выражениях; функция органа предков, э в о л roil р у го щ е г о согласно этому типу филогенетического развития, замещается (субституируется) у потомков другой функцией, биологически ей •равноценной, но выполняемой другим органом, расположенным на другом месте тела животного и развившимся из другого зародышевого зачатка.
Мы лучше всего представим себе сущность и значение этого принципа, если обратимся к разбору изученного мною случая удлинения тела и редукции конечностей у безногих ящериц.1 2
1 Daniele Rosa, La Reduzione progressiva della Variabilita e i sui rapporti •coll’Estinzione e coll Origine delle Specie, Torino, 1899.
2 Cm. S ewertzoff A. N., Studien liber die Reduktion der Organe der Wirbel-tiere, Zool. Jahrb. (Anat.), Bd. 31, 1931.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 395
Мы знаем, что у огромного большинства ящериц поступательное движение тела осуществляется путем перемещения их конечностей. Тело животного опирается при этом на лапки, так что брюхо типичных ящериц, например Agama (рис. 131 А), ни при ходьбе, ни при беге не касается земли. У некоторых ящериц.можно еще, кроме того, наблюдать, как их туловище изгибается при ходьбе или при беге то вправо, то влево.
Изучая развитие ящериц, мы можем наблюдать во многих группах, что число туловищных сегментов в течение онтогенеза увеличивается, так что тело животного постепенно становится все длиннее (рис. 131 В). При этом увеличении числа позвонков и соответствующих им мускульных
Рис. 131 А, В, С, D. Схематическое изображение перехода от ходьбы к ползанию у рептилий (субституция функций).
А—Agama,—пример формы, бегающей на четырех лапках; В—Ablepharus, тело которого стало длиннее и брюхо которого при ходьбе касается земли (переход от ходьбы к ползанию); С—Seps—ползающая форма с еще существующими рудиментарными конечностями; D—Vipera—ползающая форма, конечности которой полностью редуцировались (из А. Н. Северцова, 1931).
сегментов, спинномозговых нервов и т. п. задние конечности вместе с тазовым поясом отодвигаются все более и более назад в каудальном направлении, так что крестец оказывается образованным уже не из туловищных позвонков, как это было у предков, но из лежащих каудально от анцестрального крестца хвостовых позвонков. Параллельно сдвиганию назад задних брюшных конечностей и таза идет и изменение в нервном сплетении задних конечностей: оно образуется теперь уже из ветвей спинномозговых нервов, лежащих более каудально.
У предков безногих ящериц редукция свободных конечностей шла одновременно с удлинением отрезка туловища, расположенного между
396
ГЛАВА XI
передними и задними конечностями. При этом сами конечности постепенно становились все меньше и меньше, конечные фаланги пальцев у них редуцировались в дистопроксимальном направлении, а краевые пальцы исчезли совсем. На этой стадии эволюции остановились многие ящерицы, например, Seps chalcides (рис.- 131 С). Вместе с увеличением числа сегментов и удлинением туловища изменяется и сама форма движения животного, ибо тело его приобретает способность извиваться при ходьбе и беге гораздо сильнее, чем это бывает, например, у ящериц с относительно коротким туловищем и нормально развитыми конечностями. С другой стороны, по мере того как туловище его удлинялось, а конечности укорачивались, животное начало касаться земли при ходьбе брюхом1 все в большей и большей степени, т. е. постепенно переходило к ползающему движению тела.
Непосредственное наблюдение показало мне, что и такие формы, у которых, как, например, у Seps, конечности, пояс конечностей, а также соответствующие мускулы и нервы только уменьшились в размерах, но-еще вполне сохранились, также перешли к чисто ползающему движению: их конечности плотно прилегают к туловищу и не имеют никакого функционального значения при ползании.
У многих ящериц и тот и другой ряд филогенетических изменений идет гораздо дальше: с одной стороны, число мускульных сегментов у них значительно возрастает, причем туловище становится очень длинным, с другой стороны, конечности совершенно атрофируются, так что животное уже целиком переходит к ползающему способу передвижения и приобретает вполне змееобразный вид. На этой стадии эволюции остановились, например, наши обычные медянки (Anguis fragilis), а также желтопузики (Ophisaurus). Настоящие змеи (рис. 131 D), у которых приспособление к ползающему движению пошло еще далее, чем у безногих ящериц, представляют собою конечный член этого эволюционного ряда.
Здесь мы имеем перед собою типичный случай субституции функций: анцестральная функция (ходьба на лапках) постепенно замещается иной функцией (ползанием на брюхе), выполняемой совершен но иными органами (позвоночником, ребрами, туловищной мускулатурой).
Так как нам часто приходится встречаться с этим типом эволюции и так как принцип этот делает для нас понятным многие явления филогенеза, в том числе, например, многие случаи редукции органов, которые оставались темными до сих пор, то мы позволим себе остановиться на нем несколько подробнее. Особенного интереса в этом смысле, пожалуй, заслуживает процесс филогенетического увеличения числа туловищных сегментов у безногих ящериц, представляющий собой необходимую предпосылку для перехода этих животных от бегающего движения к ползающему.
На схеме XXV схематически представлены позвонки с ребрами и конечности исследованных нами ящериц. Длина конечнЪстей здесь нанесена пропорционально по отношению к общей длине тела данного животного. В качестве исходной формы для нашей схемы мы взяли Scincus officinalis, форму с неудлиненным телом и с нормально развитыми конечностями. Шейный отдел позвоночника состоит здесь из шести позвонков, из коих три задних несут на себе короткие ребра. Позади него лежит грудной отдел позвоночника (позвонки 7—20), характеризуемый длинными ребрами (ребра 9—13 являются стернальными ребрами); далее назад расположен поясничный отдел позвоночника (позвонки 21—26) с укороченными ребрами, затем идет крестец (сросшиеся между собой позвонки 27 и 28) и, наконец, хвостовой отдел позвоночника, причем на схеме изображена
1 Я наблюдал это у быстро двигавшегося Eumeces.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 397 лишь его передняя часть (позвонки 29—38). У нас имеются все основания предполагать, что позвоночник прародительской формы, от которой, вероятно, произошли исследованные нами безногие ящерицы, имел приблизительно только что описанное нами строение.1
Если мы сравним между собой строение позвоночников изображенного нами на схеме XXV ряда форм (Scincus, Eumeces, Chalcides, Lygoso-ma, Thyphlosaurus и т. д.), то мы увидим, что кроме постепенного увеличения общего количества позвонков, отдельные перестройки и изменения произошли и в различных. отделах позвоночников. Изменения эти относятся главным образом к шейному и поясничному отделам позвоночника и касаются также положения крестцовых позвонков.
Обращаясь к шейным позвонкам ряда Scincus—Anguis, мы видим, что у тех форм, у которых число туловищных сегментов увеличилось не слишком резко, например у Eumeces, Lygosoma, Chalcides и т. д., шейный отдел позвоночника состоит из трех позвонков,1 2 не имеющих ребер (атлас, эпистрофей и третий шейный позвонок), и из трех позвонков, снабженных короткими ребрами (позвонки 4—6). Затем следуют два позвонка с длинными ребрами, которые, однако, не соединяются с грудиной. Аналогичное строение мы находим и у Seps, у которого, однако, число позвонков уже сильно возросло (61 предкрестцовый позвонок). У форм же, где редукция передних конечностей пошла еще дальше и где общее количество позвонков сильно увеличилось, мы находим, что число задних шейных позвонков с короткими ребрами сделалось еще меньше. Так, у Ophiodes имеются три передних шейных позвонка без ребер и только два шейных позвонка с короткими ребрами, шестой позвонок уже несет длинное грудное ребро; у Ophisaurus и у Anguis три передних шейных позвонка также не имеют ребер, но, начиная с четвертого позвонка и далее назад, начинают развиваться длинные грудные ребра. Иными словами, четвертый, пятый и шестой шейные позвонки превратились в типичные туловищные позвонки, а шейный отдел позвоночника укоротился на три позвонка.
У Scincus поясничный отдел позвоночника состоит из шести позвонков. Мы видим, что у форм с увеличенным числом позвонков этот отдел постепенно укорачивается, т. е. состоит из постепенно уменьшающегося числа позвонков: у Chalcides ocellatus—из пяти, у Eumeces schneideri и Lygosoma punctulatum—из четырех, у Chalcides sepoides—из трех, у Ophiomorus, Ophiodes, Ophisaurus и Anguis—из двух, у Typhlosaurus и Seps—из одного позвонка с укороченным поясничным ребром.
Рассматривая положение поясничных позвонков у форм, изображенных на схеме XXV, мы видим, что у форм с резко увеличенным числом позвонков, т. е. у Typhlosaurus, Ophiomorus, Seps, Ophiodes, Ophisaurus и Anguis, все те позвонки, которые у предков с малым числом позвонков (для сравнения возьмем Scincus, для которого характерны 26 предкрестцовых позвонков) были поясничными позвонками, превратились в течение эволюции в типичные туловищные позвонки с длинными (грудными) ребрами.
1 Само собою разумеется, что на нашей диаграмме мы имеем перед собой не филогенетический, но лишь сравнительно анатомический ряд: каждая из представленных нами форм (Eumeces, Lygosoma, Chalcides и т. д.) является конечным членом самостоятельного филогенетического ряда, но все эти ряды эволюировали в одном и том же направлении, а именно в направлении постепенного умножения числа туловищных сегментов и т. п. Таким образом, все они в такой же мере иллюстрируют нам следующие друг за другом стадии филогенетического процесса перестройки скелетных сегментов туловища безногих ящериц, в какой, например, сравнительно анатомические ряды конечностей современных парно- и непарнокопытных животных иллюстрируют эволюцию конечностей предков этих форм.
2 В шейном отделе позвоночника Ophiomorus мы находим безреберные атлас и эпистрофей, вслед за которыми идут три позвонка с короткими ребрами.
398
ГЛАВА XI
Обращаясь к положению крестца, т. е. обоих крестцовых позвонков, мы можем твердо установить тот факт, что крестец в течение эволюции форм со все увеличивающимся числом предкрестцовых позвонков постепенно продвигается все далее и далее назад. Так, у Scincus крестец образован позвонками 27 и 28, у Eumeces—позвонками 31 и 32 , у Lygoso-ша—позвонками 36 и 37, у Chalcides ocellatus—позвонками 40 и 41, у Chalcides sepoides—позвонками 49 и 50 и т. д., вплоть до Ophiodes, обладающего среди исследованных мною форм наибольшим числом позвонков, у которого крестец образован позвонками 66 и 67. У Ophisaurus и у Anguis крестец состоит из одного позвонка (у Ophisaurus из позвонка 58, а у Anguis из позвонка 63).
Если мы предположим (а это предположение весьма вероятно), что Ophiodes или Seps например, произошли от форм, у которых общее количество позвонков равнялось общему их количеству у семейства Scincoidae с их вполне развитыми конечностями (Scincus имеет 26, a Eumeces 30 предкрестцовых позвонков), то нам придется далее предположить, что крестцовые позвонки развились из позвонков, которые у отдаленных предков этих форм были типично хвостовыми позвонками или, иными словами, что анцестральные дистальные хвостовые позвонки без ребер, но с хорошо выраженными нижними дугами, превратились в крестцовые позвонки с расширенными processus transversi и т. д, Сравнивая между собою позвоночники таких форм как Scincus и Ophiodes, или как Ophisaurus и Anguis (схема XXV), нам приходится далее предположить, что оба крестцовых позвонка в течение эволюции превратились сначала в поясничные, а затем в грудные позвонки.
Мы приходим, таким образом, к выводу, что у предков безногих ящериц с большим числом пресакральных позвонков (например у Ophiodes, Ophisaurus, Seps, Anguis) целый ряд хвостовых позвонке в (например у Seps с 29-го по 61-й хвостовой позвонок его сцинкообразных предков) сначала превратились в поясничные позвонки с короткими ребрами, а затем в туловищные позвонки с типичными длинными грудными ребрами (ребрами с расширенными ’ концами).
Итак, мы очертили вкратце те крупные изменения в скелетных элементах позвоночника, которые имели место в течение эволюции безногих ящериц и которые коснулись как передней части позвоночника (его шейного отдела), так и его задней части (поясничные, крестцовые и хвостовые позвонки). Не вдаваясь в подробности, добавим к сказанному, что и сегментальная туловищная мускулатура также изменилась в течение этой эволюции: резче выразилась сегментация отдельных мускулов, например m. intercostalis externus и т. intercostalis inferior, увеличилось общее число мускульных сегментов. Изменения произошли также и в грудной области: у таких форм, как Ophisaurus и Anguis грудинные ребра потеряли связь с грудиной и превратились в типичные торакальные ребра.
Относительно всех прочих изменений, происшедших в течение эволюции безногих ящериц, отметим лишь следующие: одно легкое у них удлинилось, другое укоротилось;1 отверстие клоаки в ряду Scincus—Anguis постепенно перемещалось спереди назад, так что оно всегда лежало позади таза; соответственно этому удлинился и кишечный тракт. Нервное сплетение задних конечностей у форм с увеличенным числом позвонков также сильно сдвинулось в каудальном направлении и его положение соответствует положению крестца; поэтому у ящериц с удлиненным телом оно обра-
1 У змей одно легкое атрофируется вполне, так что от него остается лишь очень маленький рудимент, другое, наоборот, сильно увеличивается в размерах (см. И. И. Шмальгаузен, 1905).
Scincus officinalis 12 3 4 5 6 7 В 9 Ю 11 12 13 14 15 16 17 15 19 20 21 222324252627282930 31 32333433363738 394041 42434445464748 4950 5152 5354555657585960 61626364656667686970 4 727374757677787960
Eumeces Schneider/
Lygosoma punctulatum
Chalcides ocellatus \ \ mi/Z 111111111111111111 ,J'wWUnnninnrrrnfn"'rTg
Chvlcides sepoides ”,wvwrrfffrrrrnrnrrnrrfrrnrnFfrrrrr|y
Typhlosaurus \\ \u/Z ГН 1111111111111111 H 1111 H 11111 ""wmmmfflimimmmimmm'’
*“ ''^тп1ш1шшшт1ттштшмтгв—...................
"''штштшшг1шм1шшш11ш11тт[г'в—..............
.....
*'’''м,ш1[т[шшт1тт1[1П[тт1т1П1т1Г11!т''‘......
Схема XXV. На этой диаграмме представлено сегментальное строение позвоночника и его придатков у исследованных мною ящериц
Изученные формы располагаются на диаграмме в по- * .
Рядке возрастания числа предкрестцовых позвонков: I позвонок, I I грудные ребра;
Sclncua — 26, Ешпесез—30, Lygosoma— 35 и т. д. Отно- короткие шейные ребра; | |
знтельная длина конечностей изображена толстой I I короткие поясничные ребра; Й| I крестцовые позвонки.
Горизонтальной чертой ——— ’ * IM I
^... грудинные ребра; 1/ I
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 399' зуется из тех нервов, которые у их предков с коротким туловищем принадлежали к хвостовому отделу позвоночника.
Таким образом, мы видим, что изученный нами процесс приспособления-предков безногих ящериц к ползающему образу передвижения, т. е. субституция движения посредством перемещения конечностей на движение путем изгибания сильно удлиненного туловища, является по существу очень сложным процессом, складывающимся из целого ряда строго координированных изменений, происходящих во многих функционально связанных между собою органах. Первичное филогенетическое изменение заключалось в увеличении числа туловищных сегментов:1 после этого, как мы-это только что показали, стали адаптивно изменяться очень многие органы: позвонки (включая крестец), ребра, мускулы туловища, легкие, кишечный канал и клоака. Конечным результатом этого длинного ряда изменений координированных между собою органов было образование у упомянутых выше животных нового органа движени я—г и б-кого, удлиненного туловища, заменившего собою парные конечности.
Чтобы выяснить механизм этих координированных сегментальных перестроек, я поручил своему ученику В. Ф. Еремееву исследовать вариации, происходящие в скелетных элементах и мускулах заднего отдела тела Ophisaurus apus. Хотя число исследованных им экземпляров-и не было особенно велико (у него было только 50 желтопузиков), однако это исследование все же дает нам возможность бросить взгляд на самый-ход преобразования скелетных элементов; оно позволяет нам также судить о вариационном материале, лежащем в основе филогенетического изменения (см. схему XXVI на стр. 400).
На горизонтальных рядах I—XVI схемы XXVI представлены различные типы вариаций, замеченные в положении крестца и задних конечностей (Е), а также вариаций, встречающихся в ребрах (А, В, С), в processus tfansversi (F, G, H, К) ив некоторых туловищных сегментах (I, J) желтопузика. Цифры 53—62, помещенные над верхним горизонтальным рядом, обозначают сериальное положение соответствующих сегментов (считая от затылка назад). В левом вертикальном ряду помечено, в каком процентном отношении к числу исследованных экземпляров встречается данная вариация. Все вариации расположены сериально по отношению к положению крестца или ileum.
В первых трех горизонтальных рядах (ряды I, II, III) крестцовый позвонок", к которому во всех исследованных случаях причленяется ileum, и, стало быть, вся задняя конечность, располагается в 57-м сегменте. Главные отличия между этими тремя случаями касаются строения и положения поясничных и торакальных ребер, а также m. т. intercostales externi et ventrales; поясничный отдел представлен либо двумя (I ряд) поясничными позвонками, либо одним. Горизонтальные ряды IV—VII изображают случаи, когда крестец образован 58-м сегментом; здесь также мы имеем в поясничном отделе либо два позвонка с короткими ребрами (IV, V), либо один (VI—VII ряд). Это положение крестца встречается чаще, чем случаи первого типа (случаи I—III), а именно в 11 случаях (22%). Случаи, когда крестец образован 59-м позвонком, встречаются еще чаще (33 случая, или 66%). В 18 случаях (36%) поясничный отдел состоит из двух позвонков с короткими ребрами, а в 15 случаях (30%) он состоит из одного поясничного позвонка. Наконец, в нескольких еди-
1 Здесь мы не будем останавливаться на обсуждении вопроса об увеличении числа хвостовых сегментов, которое иногда, как, например, при эволюции Ophisaurus, протекает параллельно увеличению числа предкрестцовых позвонков.
400
ГЛАВА XI
ничных случаях (в трех случаях, или 6%) крестец образован позвонком 60-м; поясничный отдел состоит при этом из двух позвонков с короткими ребрами (позвонки 58 и 59).
количество тлетчватп: окоем моров В %
S3 50 , 60 67 52
У V у
V у ' Ш
у V у "
у w V v
у V W
у W v *
V у V й"
V у V
у 1/
у U V «
V W W V W
\ у \1 *
у W V v ™
\ v у v ”
V < V if *”
В- укороченное ребро
А- нормальное туловищноеребро(грудноеребра»
С- очень короткое ребро
, О- отросток после Он его предкрестцотго v позвонка (приросшее ребро)
fsc
и,[ б - крестцовый позвонок (sc) с ileum (Ц)
б- вильчатый pr transverse; (1Йтип) )
6- вильчатый pr transversus (гётил)
V В- pr tгаniversus
= 1-т intercostalis eiternus
J- т, intercostalis трепал(s. ventral/s)
\[ К-pr. transferees ZIPпрейкреапцивого
позвонка
• Г. G а И из гвостоаой часто тела1
Схема XXVI. Объяснение в тексте.
Нам представляется чрезвычайно вероятным, что здесь мы имеем дело с теми наследственными вариациями, которые «являются исходным материалом для филогенетических изменений элементов позвоночника. Факт, что эти вариации появляются у относительно молодых зародышей (это мы наблюдали попутно при исследовании развития таза), говорит, конечно, в пользу того, что они н е являются соматическими изменениями.
Обычно принято думать, что преобразование всего органа или части органа происходит очень медленно и очень постепенно путем суммирования очень мелких наследственных вариаций в величине и строении органов, и весьма вероятно, что в большинстве случаев дело именно так и обстоит. Очень медленная прогрессивная эволюция некоторых скелетных частей у предков семейства Equidae, например эволюция их зубов или эволюция их metatarsalia III и metacarpalia III, о которой мы говорили в главе II, свидетельствует о том, что здесь эволюция происходила именно путем суммирования мельчайших количественных вариаций. Можно было бы думать, что и в случае преобразования элементов позвоночника, т. е. при превращении хвостовых позвонков в крестцовые или при превращении коротких поясничных ребер в длинные грудные, это превращение
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 401
осуществляется крайне медленно и крайне постепенно путем суммирования очень незначительных количественных наследственных вариаций. Однако произведенное В. Ф. Еремеевым исследование вариаций в элементах позвоночника желтопузика совершенно ясно говорит о том, что здесь эволюция шла иными путями или, правильнее говоря, исходила из иного вариационного материала. Мы видим, что преобразование позвон-ков здесь происходит, так сказать, скачками (сальтациями), так что, например, хвостовой позвонок 58 (схема XXVI, ряды 1—III) внезапно превращается в крестцовый позвонок 58 (схема XXVI, ряды IV—VII); то же происходит и при превращении коротких поясничных ребер в длинные грудные ребра (схема XXVI, ряды I—XVI, сегменты 55, 56, 57).
Мы видим, что в этих случаях отбор оперирует не с мельчайшими ко личественными изменениями, а исходит из довольно значительных спонтанных вариаций, т. е. из внезапных превращений одного органа в другой. Эти вариации можно сравнить с тем, что в ботанике принято называть «Sports». Обычно принято думать, что «Sports» в природе встречаются настолько редко, что для эволюции они не могут иметь никакого значения. Здесь мы, однако, видим, что только что описанные нами скачки встречаются относительно часто. Даже в незначительном числе исследованных выше случаев их было найдено довольно много.
Чрезвычайно вероятно, что змеи также развились только что описанным нами образом из четвероногих ящероподобных предков. Но удлинение туловища, увеличение числа позвонков и атрофия конечностей у них пошла еще дальше, чем у безногих ящериц; к тому же у них развились и новые приспособления к змееобразному роду движения (мускулы брюшных чешуй и т. п.).
Таким образом, изучение филогенетического развития органа движения у безногих ящериц и у змей ясно указывает нам на то, что процесс этот слагается из трех главных фаз: 1. Увеличение числа позвонков, соответствующее ему удлинение туловища и усиление его гибкости (рис. 131 В), а также смещение задних конечностей в каудальном направлении; 2. Переход к ползающему роду передвижения и более или менее резкая редукция конечностей (рис. 131 С, Seps); 3. Дальнейшее удлинение туловища (Anguis, Ophisaurus, змеи) и полная редукция конечностей (рис. 131 D).
Для нас совершенно ясно биологическое значение всех этих коротко нами описанных изменений: ящерица с ее торчащими в сторону конечностями1 легко и быстро передвигается по открытой местности, например по песку; но это именно положение ее конечностей затрудняет движение животного, когда ему приходится быстро пробираться сквозь кустарники или густую траву. С другой стороны, мне приходилось часто наблюдать движение медянки, которая медленно передвигается по открытому песчаному пространству и гораздо быстрее ползает среди густой травы. Удлинение тела у безногих ящериц и у предков змей является приспособлением этих животных для передвижения в тесноте, например среди травянистых зарослей, или же для того, чтобы можно было легко пролезть в узкие щели среди скал или в сухие трещины в земле и т. п. При этом общая масса тела животного увеличивается, но ширина туловища уменьшается.
Мы видим, что во всех этих случаях способ передвижения предков (ходьба и бег посредством передвижения конечностей) замещается у потомков совершенно иного рода движением, а именно ползанием
1 Мы знаем, что у ящерицы humerus и femur расположены перпендикулярно к длинной оси тела, что конечно, увеличивает ширину туловища животного.
Морфологические закономерности 26
402
ГЛАВА XI
на брюхе посредством змеинообразного движения длинных ребер, т. е. движением, лучше приспособленным к новым условиям существования.
Это явление отнюдь не идентично явлению субституции органов # Клейненберга, так как при клейненберговской субституции один орган животного замещается другим органом с тою же функцией (см. выше), вытесняющим анцестральный орган. Кроме того, в случаях клейненберговской субституции замещаемый орган большей частью локально связан с замещенным органом. В приведенных же нами случаях (ползание на ребрах) субституируюшая функция ползания ничем не похожа на вытесненную анцестральную функцию ходьбы при помощи конечностей. Общее значение новой функции, т. е. поступательное продвижение тела по земле, остается при этом тем же, но характер движения становится совершенно иным. Поэтому, описанные нами случаи не могут быть сведены к принципу субституции органов Клейненберга и не могут быть им объяснены.
Не могут эти случаи быть обт яснены и посредством принципа, выдвинутого Д. М. Федотовым, ибо общая функция органа (ходьба) не остается при этом неизменной, но замещается другой функцией (ползанием): ползание на брюхе и ходьба (или бег) на лапках не могут рассматриваться как одна и та же функция. У Федотова же один орган замещается другим органом стой же самой функцией.
Можно было бы, пожалуй, думать, что только что приведенные нами случаи подчиняются принципу Дорна и могут рассматриваться как примеры смены функций. Но нам кажется, что такое толкование в равной степени не верно. Дорн, формулируя свой принцип, особенно подчеркивал, что. при смене функций главная функция эволюирующего органа замещается второстепенной функцией того же органа.1 В рассмотренных же только что случаях мы видели, что функция поступательного движения тела у безногих ящериц осуществляется не посредством преобразования функций конечностей, но посредством изменений, наступающих всовершенно других органах (в позвоночнике и в ребрах) этих животных.
Таким образом, мы видим, что во всех перечисленных нами случаях филогенетическое изменение функций движения животных не может быть объяснено ни с точки зрения принципа смены функций Дорна, ни с точки зрения принципа субституции органов Клейненберга, ни с точки зрения физиологической субституции Федотова, и что нам приходится для объяснения этого явления прибегнуть к новому принципу эволюции, именно к тому, который нами обозначен выше как «принцип субституции функций».
Отметим еще, что в упомянутых нами случаях замещения нервного сплетения новым сплетением, образовавшимся из более каудально или более рострально расположенных спинномозговых нервов, функция, макроскопическое строение и гистологическая структура самого сплетения остались теми же, какими они были у предков, но местоположение этого образования в теле животного стало в течение филогенеза иным.
Эти примеры являют собою ряд переходов от случаев типичной субституции органов в смысле Клейненберга к явлениям субституции функций, какими они представляются нам на основании изучения изменения функций движения тела животных при превращении четвероногих ящериц в типично безногих ящериц и змей. В одних случаях мы видим, что в зачатке, из которого развивается орган, функция и строение остаются сходными (гомодинамия), и что меняется только местоположение органа (нервное сплетение), в других случаях изменяется не только место возникновения
1 «Преобразование органа осуществляется через последовательность функций, носителем которых остается один и тот же орган». А. Дорн, 1. с., стр. 60, 1875.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 403 замещенного органа, но также и строение этого последнего, и только функция остается прежней; при этом новый орган развивается из того же эмбрионального зачатка, что и замещенный им орган. В третьей категории относящихся сюда случаев ц зачаток органа и его общее строение остаются неизмененными, но меняется место возникновения замещенного органа, многие особенности его строения и до известной степени сама его функция (торакальные и югулярные плавники высших костистых рыб).
В сущности, с логической точки зрения мы можем говорить только ободном общем принципе субституции, все же перечисленные нами случаи, как-то: случаи, подчиняющиеся принципу Клейненберга, случаи, подчиняющиеся принципу физиологической субституции и, наконец, случаи, подчиняющиеся принципу субституции функций, можно рассматривать как явления, соподчиненные этому общему принципу.
Выше мы видели, что принцип субституции функций является действующим принципом при эволюции целого ряда животных форм, именно при эволюции безногих ящериц и предков змей. Нам могут указать, что во всех этих случаях мы имеем дело с рядом чрезвычайно сходных филогенетических изменений и спросить нас при этом: годится ли этот принцип только для случаев преобразования метамерных органов, или он имеет более широкое распространение?
На этот вопрос мы можем дать утвердительный ответ. Мы знаем, например, что при переходе предков водных млекопитающих в водную среду предки китов утратили свой волосяной покров. У наземных млекопитающих главная функция волосяного покрова заключалась в том, чтобы защищать тело животного от холода; при постоянном же пребывании животного в воде волосы уже не могут выполнять этой функции. При переходе к водному образу жизни функцию защиты внутренних органов от холода принимает на себя слой подкожного жира, который в соответствии с этой задачей становится гораздо более толстым, чем он .был у наземных предков. Мы видим, что здесь общая функция защиты тела от низкой температуры морской воды переходит к другому органу, расположенному в другом месте тела животного, к органу, иначе построенному и первоначально выполняющему иную функцию. Само собой разумеется, что эта общая функция выполняется жировым слоем кожи иным образом, чем она выполняется эпидермальными, лежащими на поверхности кожи волосами.
Мы знаем также, что у акул (рис. 132) только один хвостовой плавник служит для поступательного движения тела в воде, парные же плавники в этом движении не принимают никакого участия и являются, согласно Шмальгаузену, только пассивными органами, регулирующими движение. У скатов (рис. 133), которые произошли от акул, грудные плавники развились прогрессивно и взяли на себя функцию передвижения тела: эти рыбы плавают при помощи волнообразного движения своих больших грудных плавников; что же касается хвостового плавника, то он, наоборот, утратил у них свою первичную локомоторную функцию, край его редуцировался, и у Raja, Trygon и Myliobatis хвост превратился в орган защиты.
Здесь мы снова имеем перед собой случай субституции функции: локомоторная функция хвостового плавника замещается функцией грудных плавников, т. е. функцией других органов, иначе построенных и иначе функционирующих и притом расположенных в ином месте тела животного. В самом же хвостовом плавнике произошла смена функций: из органа движения он превратился в орган защиты.
У большинства млекопитающих зубы, лапы и, в особенности, когти обычно служат в качестве орудия защиты и нападения; так было, по всей вероятности, и у более или менее отдаленных предков парнокопытных 26*
404
ГЛАВА XI
животных. Но у современных представителей этой группы можно наблюдать своеобразный случай субституции функций: функция защиты и нападения переходит у них к совершенно другим органам, а именно к выростам черепных костей (рога).
У наземных млекопитающих конечности, и, в особенности, задние конечности, служат, как известно, для поступательного передвижения по земле; у ластоногих, которые перешли целиком к водному образу жизни, задние конечности изменяют свою функцию и превращаются в ласты, приспо-
Рис. 132. Cotorhynus maximus, у которого хвостовой плавник несет функцию поступательного движения (из Ф. Даниэля, 1928).
поступательного движения перешла у них к
грудные плавники, соответственно изменившие свою форму (из Ф. Даниэля, 1928).
собленные для плавания; здесь мы имеем перед собою типичный пример смены функций по Дорну. Но если мы обратимся к китам, то увидим, что у них произошло явление совершенно другого порядка: у предков китов, у которых задние конечности также служили для передвижения тела, эти конечности в течение филогенеза постепенно атрофировались, и функция совершенно другому органу, именно к хвосту, на котором образовался горизонтально расположенный плавник. Здесь мы снова имеем перед собою типичный пример субституции функций: функция поступательного передвижения тела в новой среде перешла к совершенно другому органу, который прежде для этой цели никогда не служил.
Чрезвычайно интересный случай субституции функций мы находим еще у слепых, перешедших к подземному образу жизни гимнофионов.
У нас имеются все основания предполагать, что предки этих животных, совершенно также, как и все остальные хвостатые амфибии, ориентировались в пространстве при поисках пищи и в других своих движениях главным образом чувством зрения. При переходе к подземному образу жизни, т. е. при переходе в полную темноту, глаза их, конечно, уже не могли служить для ориентировки в пространстве, и функция зрения заменилась у этих животных прогрессивно развившейся функцией обоняния. Так, мы знаем, что у гимнофионов носовые ямки имеют гораздо более сложное
О ПРИНЦИПАХ НЛП ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 405
строение, чем у хвостатых амфибий; кроме того, у них образовался еще очень своеобразный якобсонов орган, соединяющийся посредством сместившихся слезных каналов с наружной средой,1 а также совершенно особого рода щупальцы, которые служат не только для осязания, но и для восприятия обонятельных раздражений.1 2 Таким образом, при переходе в темноту угимнофионов функция зрения заместилась функцией обоняния. В новой среде .активная функция органа предков (глаз) замещается у потомков другой функцией, столь же активной, но выполняемой совсем другим органом (носовые ямки, щупальцы).
Другой пример такого рода субституции функций мы находим снова у амфибий. Чрезвычайно вероятно, что предки современных хвостатых и бесхвостых амфибий были некогда снабжены хорошо развитым чешуйчатым панцырем и что этот панцырь служил им в качестве органа пассивной защиты. Как известно, все современные амфибии в настоящее время голы, и функция защиты у них переходит к совершенно другому органу, а именно к ядовитым железам кожи (Urodela, Anura, Gymnophiona); в этих железах прогрессивно развилась активная функция выделения и выбрызгивания ядовитого секрета. Здесь, стало быть, функция пассивной защиты, выполняемая панцырем, замещается другой функцией (в данном случае активной функцией) другого органа (кожных желез). Эта новая функция, однако, биологически вполне равноценна замещенной старой функции предков.
Все эти примеры, а число их можно легко умножить, показывают нам, что принцип субституции функций играет далеко не второстепенную роль в филогенезе животных.
В случае подробно разобранного нами примера субституции функций у безногих ящериц анцестральная активная функция (ходьба) замещается у потомков активной и притом аналогичной функцией (ползанием); в случае субституции функции защиты у амфибий пассивная функция предков замещается у потомков совершенно не похожей на нее, но биологически ей равноценной активной функцией. Поэтому, резюмируя все сказанное выше, мы можем теперь следующим образом сформулировать установленный щами принцип; под именем принципа субституции функций мы понимаем такой процесс в филогенезе, когда функция органа предков замещается у потомков функцией другого и иначе построенного органа, расположенного в другом месте тела животного; замещаю ш а я функция потомков обычно имеет лишь общее сходство с замещенной функцией предков; она биологически ей равноценна, но никогда ей не тождественна.
Мы видим, таким образом, что существует целый ряд крайне постепенных переходов между принципом субституции органов Клейненберга, когда анцестральная функция замещается у потомков идентичной функцией другого органа, развившегося из другого эмбрионального зачатка, и крайними случаями субституции функций А. Н. Северцова, когда анцестральная функция замещается у потомков совершенно другой, но биологически ей равноценной функцией.
Мы приходим, таким образом, к выводу, что значительное число филогенетических изменений структуры и функций органов, которые на пер
1 S a г a s i n Р. u. F., Ergebnisse naturwissenschaftlicher Forschungen auf Ceylon in den Jahren 1884—1886. Bd. 2. Zur Entwicklungsgeschichte und Anatomie der ceylonischen Blindwiihle Ichthyophis glutinosus, 1890.
2 Laubmann W., Uber die Morphogenese von Oehirn und Geruchsorgan der Gymnophionen, Zschr. f. Anat u. Entwicklungsgesch., Bd. 84, 1927.
406
ГЛАВА XI
вый взгляд имеют очень мало сходства между собой, происходят, тем не менее, путем субституции функций. Что же касается общего биологического зйачения этого принципа, то мы будем подробно говорить о нем в следующей главе этой книги.
ПРИНЦИП АКТИВАЦИИ ФУНКЦИЙ
Кроме перечисленных нами выше принципов, или типов филогенетического изменения органов, мы можем выставить еще несколько других принципов, которые, по нашему мнению, играют далеко немаловажную роль в эволюции органов животных.
Так, например, довольно часто встречаются случаи, когда орган, который у предков данной формы не выполнял никакой активной функции, но был полезен как пассивное образование, превращается у потомков в активно функционирующий орган.
Рис. 134. А—череп Sphenodon punctatus; В—череп удава (Boa constrictor). (Из Шим-кевича, по Боасу).
Рис. 135 А, В. Череп ядовитых змей. Общие обозначения для рис. 134 и 135:
Di.—m. depressor mandibulae; Fr.—frontale; g.—ядовитая железа; Ju.—jugale; Mx.—maxilla; Na.—nasale; Ol.—occipitale laterale; os.—supraoccipitale; Pa.—parietale; Pal.—pa-latinum; Pfwl—postfrontale 1; Pf.2—postfrontale 2; pmx.—praemaxilla; Prf.—praefrontale; Pt.—pterygoideum; Q—quadratum; Qj—qua-dratojugale; sq.—squamosum; Tr.—transversum (из Гильцгеймера).
Нам хорошо известно, что змеи и ящерицы произошли от общих предков. У этих предков неподвижная maxilla была плотно вклинена между другими костями переднего отдела черепа наподобие того, что мы видим у Palaeohatteria и у Sphenodon (рис. 134 А). У неядовитых змей (рис. 134 В) связь maxilla с другими черепными костями стала более свободной, а у ядовитых змей (Crotalus) челюстные кости, несущие ядовитые зубы, превратились в свободно подвижные косточки, связанные с praefrontalia хорошо развитым сочленением (рис. 135 А, В). Когда у этих змей открывается рот, то под действием m. т. sphenopterygoidei верхняя челюсть поворачивается таким образом, что ядовитые зубы, сидящие на ней, обращаются своими верхушками вниз и вперед (рис. 135 В); когда у них рот закрывается, то ядовитые зубы автоматически вкладываются обратно в пасть и плотно прилегают к небу (рис. 135 А). Мы видим, таким образом, что челюстные кости, бывшие у предков ядовитых змей пассивными и неподвижными органами, служив
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 407
шими лишь в качестве опоры для челюстных зубов, превращаются у потомков, благодаря изменениям, происшедшим в их отношениях к соседним костям, в свободно подвижный орган, столь же активно функционирующий, как и нижние челюсти.
В качестве другого примера превращения пассивно неподвижного состояния в состояние активно подвижное можно привести переход от монимостилии примитивных рептилий к стрептостилии высших форм. Мы знаем, что у примитивных рептилий (Palaeohatteria, Sphenodon, рис. 134 А) квадратная кость (Q), с которой сочленяется нижняя челюсть, неподвижна и плотно вклинена между squamosum, jugale и другими костями; у некоторых современных форм (крокодилы, а также Sphenodon, рис. 134 А) это первичное состояние сохранилось. У Lacertilia сочленение между квадратной костью и нижней челюстью также сохранилось, но на проксимальном конце нижней челюсти образовался сустав, так что квадратная кость из неподвижной стала подвижной. У змей подвижность квадратной кости стала еще значительнее (см. рис. 135 А, В): она соединена теперь лишь свободной связью с squamosum (sq), так что обе эти кости (quadratum и squamosum) образуют двухчленный рычаг, поддерживающий нижнюю челюсть.
Эволюция позвоночника и его придатков у предков безногих ящериц сопровождалась многими изменениями в этих органах, которые также, вероятно, были подчинены принципу активации функций. Мы знаем, например, что постсакральный позвонок у типичных ящериц не несет на себе свободных ребер. У происшедших от этих форм безногих змееобразных ящериц (Anguis, Ophisaurus) из этих бывших постсакральных сегментов образовались типичные грудные позвонки с подвижными торакальными ребрами и реберными мускулами: здесь мы имеем перед собой образование нового активного органа и новой активной функции, которых не было у предков безногих ящериц.
Прекрасным примером активации функций является строение таза у Pelobates fuscus; как известно, у лягушек подвздошные кости (ilea) плотно соединены с поперечными отростками крестца, так что таз является образованием неподвижным по отношению к крестцу. У Pelobates fuscus processus transversi крестца стали очень широкими, и таз, вернее подвздошные кости, получили возможность свободного относительно них движения. Таким образом, таз из образования неподвижного стал свободно подвижной скелетной частью, и эта подвижность целиком используется животным, когда Pelobates приходится закапываться в землю своими задними лапками. Так же как и в упомянутом выше случае, здесь пассивная функция органа (таза) замещается активной его функцией.
Мы надеемся, что приведенные нами примеры ясно показывают читателю, что собственно мы подразумеваем под принципом активации функций. Мы видим, что органы, которые не обладали у предков активной функцией, приобретают таковую у потомков. Здесь мы, конечно, не можем говорить, как это было в случаях смены функций или в случаях субституции функций, об изменении анцестральной функции, но мы можем говорить об образовании новой активной функции там, где ее прежде никогда не было.
Пассивные функции (опора для зубов, защита внутренних органов покровными костями и т. п.) не могут рассматриваться как функции в том же смысле этого слова, как, например, функция какого-нибудь мускула или функция какой-нибудь подвижной части скелета. Главное отличие только что описанного нами принципа активации функций от принципа расширения функций Плате заключается в том, что при расширении функции при прогрессивном развитии органа приобре-
408
ГЛАВА XI
т а ю т с я новые вторичные функции, старая же главная функция, пассивна она или активна, остается при этом неизменной. В случае же активации функций орган приобретает новую активную главную функцию. Мы думаем, что значение этого отличия совершенно понятно читателю и что поэтому мы имеем право рассматривать этот принцип как особый тип филогенетического изменения органов.
ПЕРЕХОД ОТ АКТИВНОЙ ФУНКЦИИ К ПАССИВНОЙ (ИММОБИЛИЗАЦИЯ)
Гораздо более широкое распространение, чем принцип активации функций, имеет противоположный ему принцип, который мы можем назвать, исходя из тех случаев, где он всего яснее выражен, принципом иммобилизации органов. Принцип активации касается тех случаев, когда какая-нибудь пассивная часть организма делается в течение филогенеза активной; принцип иммобилизации, наоборот, относится к случаям, когда орган, который упредков действовал активно, превращается у потомков в пассивную часть организма, полезную ему лишь одним своим
Рис. 137. Срастание (иммобилизация) palatoquadratum (pq.) с черепом у химер (Chimaera monstrosa); (из К. Гегенбаура, 1898).
Рис. 136. Подвижное сочленение palatoquadratum (pq.) с черепом у акул Hexanchus (из К. Гегенбаура, 1898).
присутствием и положением. Последнее чаще всего наблюдается при срастании прежде свободно подвижных скелетных частей.
Мы знаем, например, что palatoquadratum (рис. 136) подвижно соединен с черепом у примитивных эласмобранхий и что для его движения служат определенные мускулы. У Holocephali и у Dipneusta palatoquadratum срастается с черепом (рис. 137) и делается пассивной частью хрящевого черепа, служащей лишь в качестве опоры для расположенных на ней костей (pterygoidea) и зубов.
Аналогичные случаи срастания свободно подвижных частей,которые при этом теряют свою активную функцию, мы находим также при срастании первично свободных позвонков, образующих крестец у рептилий, птиц и млекопитающих. То же имеет место и при срастании туловищных позвонков у черепах; то же происходит при развитии tarsale proximale (astragalus+f i-bulare) (см. рис.210 В и рис. 199) у ящериц и черепах; при развитии tarsome-tatarsus у птиц; при срастании элементов os canon у парнокопытных и т. д. Мы видим таким образом, что существует очень' много случаев, когда свободно подвижные части скелета превращаются в массивные опорные элементы скелета позвоночных животных.
О ПРИНЦИПАХ ПЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 409
Но мы отнюдь не можем сказать, что все известные нам случаи срастания скелетных элементов могут быть сведены к этому принципу иммобилизации функций. Наоборот, существует немало случаев, когда скелетные части, которые первично не были подвижны и, следовательно, не функционировали активно, срастаются, тем не менее, между собой; это имеет место, например, у многих черепных костей (dermosupraoccipitalia etc.) осетровых рыб (А. Н. Северцов),1 у зубов Ceratodus (Семен).1 2 В этих случаях, конечно, никакого превращения активной функции в пассивную не происходит, самое большее, о чем мы можем здесь говорить, это об интенсификации пассивной функции.
Нам могут заметить, что с логической точки зрения изложенный нами только что принцип (иммобилизация) в некотором отношении близко стоит к дорновскому принципу смены функций. В перечисленных нами случаях иммобилизации при срастании palatoquadratum с черепом у Holocephali и у Dipnoi, при срастании поясничных и хвостовых позвонков с крестцом у предков птиц и т. п. упомянутые скелетные части имели кроме активной функции (движение посредством двигавших их мускулов) еще и пассивную функцию. Первая (активная функция) редуцируется, а вторая (пассивная опорная функция) при срастании интенсифицируется (смена функций).
Разница между приведенными нами выше типичными случаями смены функций и случаями, которые мы обозначали под именем иммобилизации, заключается главным образом в том, что в типичных случаях смены функций по Дорну главная активная функция предков замещается у потомков второстепенной, но такой же активной функцией; в случае же иммобилизации активная функция предков (движение) замещается у потомков пассивной, однако полезной организму функцией, которую по настоящему едва ли можно даже назвать функцией. Эта разница, мне кажется, дает нам право выделить многочисленные случаи иммобилизации в особую группу филогенетических изменений животных. Впрочем, по существу вопроса, довольно безразлично, будем ли мы эту группу изменений рассматривать как особую группу, подчиненную принципу смены функций, или будем, как это мы делаем сейчас, считать ее за самостоятельный тип филогенетического изменения органов.
СИМИЛЯЦИЯ ФУНКЦИЙ
Изучая условия, предшествующие редукции конечностей у безногих ящериц, мы видели, что у этих форм число туловищных сегментов сильно возросло и что при этом произошли значительные изменения в ребрах, обусловившие переход этих животных к ползающему движению тела. При этом поясничные, крестцовые и целый ряд хвостовых позвонков превратились в типичные грудные позвонки с хорошо развитыми грудными ребрами.
У типичных ящериц позвонки, расположенные впереди от крестцового отдела позвоночника (поясничные позвонки 4 и 5), несут на себе короткие ребра, резко отличающиеся как от грудных, так и от крестцовых ребер, сросшихся с крестцовыми позвонками. При удлинении туловища безногих ящериц мы видели, что число поясничных позвонков постепенно уменьшилось с шести до одного, т. е., иначе говоря, что ребра поясничных позвонков превратились в грудные ребра (схема XXV). Тот же процесс мы снова находим в шейном отделе позвоночника, где задние шейные ребра делаются совершенно похожими на типичные грудные ребра. Мы видели, что
1 Sewer tzoff A. N., Studies on the bony skull of Fishes, Quart. Journ. of Microsc. Science, 1926.
2 S e m о n R., Die Zahnentwicklung des Ceratodes Foisteri, Zool.Forschungsrci-sen in Australien u. dem Malayschen Archipet.. Jena, 190k
410
ГЛАВА XI
у безногих ящериц и стернальные ребра делаются вполне подобными остальным грудным ребрам (Ophisaurus, Anguis). И, наконец, мы нашли, что у форм с сильно выраженным удлинением тела оба крестцовых позвонка, "а также целый ряд хвостовых позвонков становятся совершенно похожими на типичные грудные позвонки и что на них развиваются типичные грудные ребра.
Очень многие части скелета, которые у предков по своей форме и функциям были совершенно непохожи друг на друга (шейные и грудные позвонки, стернальные ребра и каудально от них лежащие ребра, крестцовые и хвостовые позвонки и т. д) стали в течение филогенетического развития крайне схожими образованиями. Мы обозначаем этот тип филогенетических изменений как принцип или тип «симиляции» органов или функций. Если различным образом построенные, различно функционирующие органы сделаются по своей функции подобными, то это сходство функций вызовет сходство в строении данных органов.
Я пришел к выводу, что этот принцип действует в очень большом числе случаев эволюционных изменений. Так, мы должны были бы безусловно принять, что сходство сегментов тела у всех Ophidia филогенетически произошло путем симиляции функций. Тот же принцип оказался действующим при эволюции сегментов тела гимнофионов, при эволюции таких форм, как Amphiuma и Menopoma среди амфибий, при эволюции угрей и других рыб с сильно удлинившейся формой тела. Чрезвычайно вероятно, что мы могли бы найти много примеров этого типа филогенетического развития также и среди беспозвоночных, например, среди червей (аннелид) и многих других форм.
ПРИНЦИП РАЗДЕЛЕНИЯ ОРГАНОВ И ФУНКЦИЙ
Наконец, мы должны упомянуть еще о некоторых случаях филогенетических изменений, играющих известную роль при эволюции органов позвоночных животных: мы имеем в виду разделение функций и органов. Часто случалось так, что какой-нибудь первично целостный орган, какая-нибудь скелетная часть, мускул или нерв, расщеплялся в течение филогенеза и таким образом из него получается несколько более или менее самостоятельных органов, несущих различную функцию.
Как мы это отметили уже выше, чрезвычайно вероятно, что хрящевые жаберные дуги представляли собой у предков первичных челюстных рыб цельные, гибкие хрящевые палочки наподобие того, что мы имеем теперь у циклостом. Мы знаем, однако, что в течение филогенеза они распались на отдельные части (pharyngobranchialia, epibranchialia, ceratobranchialia, hypobranchialia). При этом функции различных частей развивались в различных направлениях. Так, например, функций pharyngobranchiale отличается от функций epi- и ceratobranchiale.
Мы знаем также, что первично единые висцеральные мускульные тяжи жаберной мускулатуры расщепились в течение филогенеза на отдельные, самостоятельные мускулы: m. m. constrictores, interarcuales, adductores arcuum и т. д.; функция каждого из этих мускулов становится при этом несколько иной, чем была функция первичного мускульного тяжа. Поэтому мы. действительно, можем сказать, что филогенетическое разделение органа ведет к изменению его функции.
Пример принципа разделения органов представляет собою разделение первично непрерывной плавниковой складки (рис. 138 А), характерной для предков современных рыб, на отдельные непарные плавники. Это разделение сопровождалось разделением функций: задняя часть плавниковой
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 411
складки превратилась в хвостовой плавник (рис. 138, В и С), взявший на себя роль главного органа движения, дорзальная часть складки превратилась в спинные плавники, а нижняя ее часть—в анальный плавник, исполняющий, так же как и спинные плавники, роль руля, направляющего движение рыбы. Такой же путь филогенетического распадения на отдельные части прошли и первично непрерывные парные боковые кожные
Рис. 138 А, В, С. Три стадии образования парных и непарных плавников. Филогенетическое разделение непрерывной плавниковой складки примитивных предков рыб (А) на отдельные плавники (В и С) у потомков (из И. И. Шмальгаузена, 1935).
складки, распавшиеся впоследствии на парные’грудные и брюшные плавники (рис. 138 С).
ВЫВОДЫ
Так как существует довольно значительное число типов филогенетического изменения органов, то естественно возникает вопрос, который из этих типов или принципов мы должны рассматривать как первичный путь изменения органов.
Выше мы уже отметили, что в основе всех перечисленных нами принципов лежат два основных свойства органов Metazoa: во-первых, основная способность каждой из известных нам функций меняться в течение филогенеза количественно; во-вторых, общее свойство живого организма, которое состоит в том, что каждый орган, точно так же как и каждая часть органа, начиная от слабо диференцированной клетки и кончая сложнейшими образованиями, по существу своему мультифункциональны, т. е. обладают несколькими активными или пассивными функциями. Эти два основных свойства организма позволяют всем его клеткам и всем его клеточным комплексам (органам), во-первых, изменять свою анцестральную функцию (свою главную функцию) количественно, т. е. усиливать ее в течение филогенеза или ослаблять,1 а во-вторых, изменять ее качественно. Мы видели, что оба эти свойства оказались действующими
1 Так как здесь мы интересуемся главным образом принципами прогрессивных филогенетических изменений органов, то в последующем бы будем говорить только об интенсификации.
412
ГЛАВА XI
при эволюции гистологического строения клетки и однородных клеточных комплексов (тканей). При превращении индиферентной мезенхимной клетки в гладкое мускульное волокно и при дальнейшем превращении этого последнего в поперечнополосатое, волокно действует интенсификация функций; при превращении эпителиальной клетки в железистую клетку, наоборот, вступает в действие качественное изменение функций.
Нас могут спросить, не следует ли считать за самостоятельный тип филогенетического изменения частей организма только что упомянутую нами мультифункциональность клетки, ткани или органа? Я лично думаю, что нет. Мультифункциональность представляет собою именно первичное общее свой ст в’о организма, от которого зависят, как мы это видели выше, многие типы филогенетических изменений органов (интенсификация, смена функций и т. д.). Это свойство лежит в основе всех типов эволюции.
Такой же точно вопрос могут нам поставить и относительно повсеместно встречающегося явления диференциации клеток, тканей и органов. Под диференциацией обычно понимают такой филогенетический процесс, в результате которого части организма (т. е. клетки или органы), бывшие у предков подобными друг другу, в течение эволюции делаются несхожими между собой. В предыдущем мы видели, что в большинстве случаев филогенетически^ изменений органов эволюирующие части становятся непохожи друг на друга, иными словами, диференцируются. Можно сказать, что дифференциация является необходимым и общим последствием описанных нами выше типов филогенетичестких изменений и что поэтому ее нельзя рассматривать как особый тип, или принцип филогенеза, равноценный другим его типам. Но здесь я не буду рассматривать этой трудной проблемы эволюции.
Если мы обратимся к более сложным случаям филогенетического изменения органов, то мы увидим, что все перечисленные нами типы свободно укладываются в следующие две большие группы:
I. К первой группе принципов относятся все те типы филогенетических изменений органов, при которых анцестральная главная функция эволюирующего органа остается у потомков качественно такой же, какой она была у предков, но количественно изменяется, т. е. интенсифицируется (количественные филогенетические изменения функций).
II . Ко второй группе принципов относятся все те типы филогенетических изменений органов, при которых анцестральная функция филогенетически изменяющегося органа замещается у потомков новой функцией (качественные филогенетические изменения функций).
Если мы спросим, какой собственно признак является общим всем исследованным нами типам филогенетических изменений органов, то нам придется ответить, что наиболее широкое значение имеет принцип интенсификации функций, входящий как общая составная часть во все перечисленные нами типы изменения органов. Оставляя в стороне интенсификацию функций в собственном смысле этого слова, мы увидим, что интенсификация одной из анцестраль-ных функций происходит при всех остальных типах положительных филог е‘н етических изменений орган» в. Разница между двумя упомянутыми нами группами (между группой I и II) заключается лишь в том, что в одних слу
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 413
чаях (I) эта интенсификация касается главной функции анцестрального органа, а в других (II)—одной из его второстепенных функций или функций другого органа.
При субституции органа в смысле Клейненберга замещение анцестрального органа другим органом потому выгодно для данной животной формы, что новый замещающий орган лучше выполняет данную функцию, чем старый замещенный орган. Так, опорная функция в организме лучше выполняется хрящевыми и особенно костными позвонками, чем она выполнялась у предков замещенною ими хордою: главная функция (в данном случае функция опоры) остается у потомков неизменной, но она значительно усиливается.
В случаях физиологической субституции Федотова главная функция замещенного органа предков также не меняется, но она выполняется у потомков другим органом, расположенным в другом месте тела животного и, главное, органом, лучше выполняющим ее, чем орган предков (интенсификация).
Мы видели, что в типе филогенетических изменений, который мы обозначили как принцип фиксации фаз, при переходе от стопоходного движения к пальцеходному, а от пальцеходного к копытному, самый способ движения (ходьба, бег на четырех лапах) остается неизменным, но движение количественно усиливается: пальцеходное млекопитающее может легче и быстрее двигаться, чем его стопоходящий предок. Мы имеем перед собою и в этом случае интенсификацию анцестральной функции, но не имеем никакого ее качественного изменения.
То же относится и к уменьшению числа функций какого-нибудь органа. При укорочении, например, шеи у предков кита, что вызвало уменьшение подвижности головы, число возможных функций рта, т. е. челюстей, сильно уменьшилось; однако это не коснулось главной функции рта, которая осталась прежней и даже усилилась (интенсификация).
Наконец, в случаях выпадения промежуточной функции главная функция также остается качественно неизмененной и изменяется лишь количественно. Например, при выпадении quadratum и articulare из состава челюстных костей у млекопитающих нижняя челюсть стала непосредственно причленяться к черепу, причем функция ее интенсифицировалась. Также интенсифицировалась и функция дижения брюшных плавников при выпадении плавниковых лучей и при непосредственном причленении лопасти плавника (лепидотрихий) к брюшному поясу конечностей (базиптеригию).
Таким образом, все перечисленные нами типы филогенетических изменений образуют особую группу, характеризующуюся тем, что г л а в н ая функция эволюирующего органа остается у потом к о в такой же, какой она была упредков, но интенсифицируется в течение филогенеза.
Другую группу образуют, как мы уже об этом упоминали выше, те типы филогенезов, где функция эволюирующего органа (его главная функция) изменяется у потомков качественно. К этой второй группе относится прежде всего принцип или тип расширения функций Плате, при котором главная функция эволюирующего органа остается почти неизменной, но увеличивается число его второстепенных функций, так что общая функция всего органа делается качественно несколько иною, чем она была у предков.
При смене функций Дорна мы видим, что анцестральная главная функция, например функция ходьбы и функция поддерживания тяжести тела у предков кита, у потомков редуцируется вполне и замещается другою функцией (функцией плавания). В этом и в подобных этому случаях мы имеем перед собою полное изменение анцестралыюй функции эволюирующего органа.
414
ГЛАВА XI
Такое же изменение анцестральной функции находим мы и в тех случаях, которые нами обозначены как принцип симиляции. Если, например, какой-нибудь типичный поясничный позвонок с короткими ребрами превращается у безногих ящериц в грудной позвонок с длинными ребрами или если два анцестральных хвостовых позвонка превращаются у потомков в крестцовые позвонки, то функции этих грудных и крестцовых позвонков будут уже иными, чем они были у предков. Здесь мы имеем перед собою типичные случаи качественного изменения функции (но не смены функций в смысле Дорна), и поэтому мы должны причислить их ко второй из упомянутых нами выше групп.
К этой же категории могут быть безусловно отнесены и случаи активации функций. Как известно, при этом типе филогенетического раз-
вития органы, которые у предков имели лишь пассивное значение, становятся у потомков активными. Верхняя челюсть становится подвижной у ядовитых змей; то же самое можно сказать и о symplecticum хрящевых ганоидов и о клюве попугаев. Противоположное явление мы встречаем при принципе иммобилизации функций: здесь активная функция предков замещается у потомков пассивною функциею, и таким образом анцестральная функция органа резко меняется.
К этой же группе принципов относится и тип филогенетического изменения органов, который мы обозначили как принцип субституции функций. Здесь анцестральная функция замещается у потомков другой функцией другого органа, функцией, которая для данной животной формы при изменившихся условиях существования имеет то же биологическое значение, что и функция органов предков (бег на четырех лапках замещается ползанием на брюхе у безногих ящериц, зрение замещается осязанием и обонянием у некоторых гимнофионов и т. д.). Само собой разумеется, что все эти случаи также относятся к нашей второй группе принципов, точно также как и случаи филогенетического разделения органов.
Поэтому мы можем установить на основании изложенных нами соображений следующее подразделение типов филогенетического изменения
органов:
I. Анцестральная функция эво-люирующего органа сохраняется у потомков качественно той же, но усиливается количественно.
Типы
1. Интенсификация.
2. Субституция органов Н. Клейненберга.
3. Физиологическая субституция Д. М. Федотова.
4. Фиксация фаз.
5. Уменьшение числа функций.
6. Выпадение промежуточных функций.
II. Анцестральная функция эво-люирующего органа изменяется у потомков качественно.
1. Расширение функций Л. Плате.
2. Смена функций А. Дорна.
3. Субституция функций.
4. Активация функций.
5. Иммобилизация.
6. Симиляция функций.
7. Филогенетическое разделение органов или функций.
Из сказанного мы видим, что все перечисленные нами типы прогрессивного филогенетического изменения органов животных покоятся на принципе интенсификации функций и на мультифункциональности органов.
О ПРИНЦИПАХ ИЛИ ТИПАХ ФИЛОГЕНЕТИЧЕСКИХ ИЗМЕНЕНИЙ ОРГАНОВ 415
Нам могли бы, конечно, поставить вопрос, стоит ли в таком случае устанавливать и перечислять различные типы филогенетического развития органов и не проще ли было бы просто сказать, что в основе всех филогенетических изменений лежат интенсификация функций и мультифункциональность органов. Мы думаем, что это не так. Задача филогенетика заключается не в том, чтобы свести все известные ему филогенетические изменения к общим основным принципам, но, конечно, в том, чтобы проследить во всех деталях и объяснить весь путь филогенетических изменений органов. Именно для этого ему и нужны перечисленные нами типы. Если мы, описывая превращение приспособленных для ходьбы конечностей стопоходящего млекопитающего в конечности кита или в конечности парнокопытного, для объяснения этого явления просто скажем, что оно представляет собою комбинацию интенсификации функций и мультифункциональности органов, то этим мы не сделаем более понятным самого процесса превращения органа; но если нам удастся свести первый случай к типу смены функций, а второй к типу фиксации фаз, то, конечно, оба данные процесса филогенетического изменения органов будут объяснены в гораздо большей мере. Порою нам удается проследить пути филогенетического превращения органов вплоть до мельчайших деталей.
На этом мы закончим обсуждение проблемы о типах или принципах филогенетических изменений органов. Нам остается лишь еще раз подчеркнуть, что мы никоим образом не думаем, чтобы перечисленными нами, типами ограничивалось все разнообразие случаев филогенетического изменения органов; наоборот, весьма вероятно, что дальнейшее углубление и расширение филогенетических исследований откроет нам еще целый ряд новых принципов эволюции. Здесь перед нами лежит новое и широкое поле исследований, изучение которого едва лишь намечается.
ГЛАВА XII
ОСНОВЫ МОРФОЛОГИЧЕСКОЙ ТЕОРИИ РЕДУКЦИИ ОРГАНОВ
ОСНОВНЫЕ ПРЕДПОСЫЛКИ РЕДУКЦИИ ОРГАНОВ
Разбирая в предыдущей главе отдельные принципы или типы филогенетических изменений органов, мы касались главным образом вопроса о прогрессивных преобразованиях строения и функций органов. Мы знаем, однако, что в течение эволюции часто встречаются и случаи регрессивного развития органов.1 Нам надо прежде всего попытаться ответить на вопрос о том, когда, т.е. при каких условиях, органы начинают редуцироваться?
Наши исследования показали, что орган начинает атрофироваться тогда, когда он перестает быть биологически полезным животному. Так, изучение редукции конечностей у безногих ящериц показало, что парные конечности начинают редуцироваться, когда животные переходят к ползающему образу жизни и перестают нуждаться в конечностях для поступательного движения. Глаза роющих животных и животных, обитающих в пещерах (Gym-nophiona, Ampiiisbaenidae, роющие насекомоядные и грызуны, пещерные амфибии и рыбы), только тогда начинают дегенерировать, когда эти животные переходят в темную среду, где они уже не могут пользоваться глазами. У очень многих рыб редукция чешуй начинается только тогда, когда эти формы переходят к бентонному образу жизни, т. е. кбгда начинают проводить большую часть времени на морском дне. Так, например, мы видим, что у Batoidei чешуи редуцируются на брюшной стороне тела, т. е. на той стороне, которая прилежит ко дну и не подвержена нападению врагов. У Pleuronectidae, например, у Pleuronectes flesus, которая лежит на дне на правой либо на левой стороне, редукция чешуй начинается на той стороне тела, которая обращена ко дну. Само собой разумеется, что мы говорим не о превращениях, происходящих у отдельного организма, но об эволюционных преобразованиях, протекающих у ряда поколений под воздействием естественного отбора.
Из этих примеров, число которых может быть увеличено до бесконечности, можно сделать вывод, что обычно атрофируются только ненужные органы. Но если это так, то возникает новый вопрос, а именно, когда, т. е. при каких обстоятельствах, органы становятся ненужными их обладателю?
Рассматривая различные типы редукции органов, мы видим, что это может происходить в двух случаях: во-первых, когда животное переходит в такую новую среду, в которой данный орган ему перестает быть нужным, во-вторых,
1 Ср. в главе X раздел о направлении общей морфофизиологической деградации.
ОСНОВЫ МОРФОЛОГИЧЕСКОЙ ТЕОРИИ РЕДУКЦИИ ОРГАНОВ
417
когда функция данного органа замещается функцией како й-л ибо его части или функцией какого-либо др у г ого органа.
РЕДУКЦИЯ ВСЛЕДСТВИЕ ИЗМЕНЕНИЯ УСЛОВИЙ ОКРУЖАЮЩЕЙ СРЕДЫ
Примеры дегенерации, имеющей место вследствие простого изменения условий окружающей среды, мы находим главным образом во всех многочисленных случаях редукции частей наружного скелета позвоночных. Сюда, например, относятся упомянутая нами редукция чешуй на брюшной стороне тела скатов, частичная редукция чешуй на нижней стороне тела у камбалы и многие другие случаи редукции чешуй у рыб. Совершенно понятно, что привычка постоянно лежать на брюхе на дне моря делает ненужным существование чешуй на брюшной стороне тела: чешуи, вообще говоря, служат для защиты, а часть тела, обращенная ко дну, не нуждается в этой защите, так как она с этой стороны не испытывает нападения. В этих случаях функция пассивной защиты теряет свое значение у чешуй ине замещается никакой другой функцией, ибо при особых условиях существования камбал, скатов и т. п. она становится ненужной.
Я думаю, что мы можем удовольствоваться теми немногими случаями, которые мы привели в качестве примеров редукции органов, ставших ненужными вследствие изменения условий окружающей среды, и что мы можем предоставить самому читателю отыскание дальнейших примеров этого рода.
РЕДУКЦИЯ ВСЛЕДСТВИЕ СУБСТИТУЦИИ
Второй тип редукции, т. е. замена у потомков какой-нибудь функции предков новой, лучше приспособленной к изменившимся условиям существования функцией, встречается гораздо чаще, чем простая редукция анцестральной функции, наступившая вследствие изменившихся условий существования, при-которых старая функция становится излишней у потомков.
Наши исследования привели нас к выводу, что именно этот тип филогенетических изменений органов лежит в основе большинства случаев филогенетической редукции органов. Чтобы избавить читателя от необходимости возвращаться к изложенному нами в предыдущей главе обзору различных случаев субституции, мы приведем здесь очень коротко все наиболее существенное, относящееся к этим случаям.
Мы видели, что существуют различные виды субституции. Н. Клейнен-. берг открыл тип (принцип) субституции органов, который можно формулировать следующим образом: во многих случаях филогенетического развития «орган предков» замещается (субституируется) у потомков новым органом, расположенным на месте анцестрального органа и выполняющим туже функцию, что и он, но развившимся из другого эмбрионального зачатка. Замещенный орган при этом редуцируется.1 Мы привели много случаев также и другого рода субституции органов:
1 Исключением из этого правила является описанный нами выше случай замещения arcus aortae primarius вторичной артерией жаберной дури у осетровых рыб (рис. 116). Здесь перед нами редко встречающийся случай совместного действия принципа субституции органов Клейненберга и принципа смены функцией А. Дорна: замещенный орган ко времени своей филогенетической субституции используется организмом в несколько ином, чем у предков, направлении, именно, как vena branchialis communis, и не становится поэтому, как в большинстве аналогичных случаев, ненужным органом. Поэтому он и не редуцируется. Такие редко наблюдаемые случаи лучше других показывают нам, что первичным явлением в процессе редукции следует считать именно наступившую бесполезность бывшего прежде необходимым анцестрального органа.
Морфологические закономерности 27
418
ГЛАВА XII
в этих случаях анцестральный орган замещается у потомков новым органом, расположенным в ином месте тела животного и развившимся из иного эмбрионального зачатка, но выполняющим ту же функцию, которую выполнял и орган предков. Наконец, мне (А. Н. Северцов, 1928) удалось установить третий тип субституции, который мы можем обозначить как субституцию функций. Этот тип субституции в свою очередь подразделяется на несколько подтипов. Во всех случаях субституции функций «орган предков» замещается у потомков другим органом, расположенным в другом месте тела животного и развившимся из другого, чем у предков, эмбрионального зачатка, но функция замещающего органа у потомков может быть различной в различных случаях. Замещенный орган при этом также редуцируется.
В случаях Клейненберга функция замещающего органа идентична функции замещенного органа (см. таблицу 4); замещающий орган потомков при этом развивается либо в том же самом (1-й вид субституции), либо в другом (2-й -вид субституции) месте тела животного. В случаях «субституции функций» А. Н. Северцова анцестральная функция замещается не идентичной, но всегда несколько иной, чем у предков, функцией: при этом замещающая функция может быть похожа на замещенную функцию (аналогична ей, 1-й модус1), или не аналогична ей, но «того же рода» (2-й модус1 2), или даже совсем другого рода, но «биологически ей равноценна (3-й модус3), т. е. столь же полезна организму, как и замещенная ею функция.
Во всех этих случаях замещенная (субституируемая) функция и ее орган редуцируются. Другими словами, субституция органов и функций является предпосылкой редукции замещенных функций и их органов.
При исследовании различных случаев субституции мы находим целый ряд постепенных переходов от типичных случаев субституции органов Клейненберга до крайних случаев субституции функций А. Н. Северцова, в которых «функция предков» замещается у потомков совершенно иной, абсолютно непохожей на нее функцией. Единственным условием этой субституции является биологическая равноценность функций: замещающая функция во всяком случае должна быть столь же полезна потомкам, сколь была полезна предкам замещенная ею функция.
Только что указанные нами переходы между различными видами субституции наглядно изображены на табл. 4 (стр. 419).
Эта таблица показывает нам, что между первым видом субституции органов Н. Клейненберга, где «орган предков» замещается у потомков органом, развившимся из другого эмбрионального зачатка, но занимающим то же положение и выполняющим ту же функцию, что и анцестральный орган, и крайними случаями субституции функций (3-й модус субституции функций А. Н. Северцова) имеется целый ряд переходов. Самый ход эволюции во всех этих случаях одинаков: замещающая функция и орган, выполняющий эту функцию, во всех этих случаях развиваются прогрессивно; поэтому старые, замещенные функции и выполняющие эти функции органы теряют свое, биологическое значение и редуцируются.
Мы видим, что принцип субституции можно положить в основу очень многих случаев редукции органов.
1 Субституция функции ходьбы на четырех лапах функцией ползания на брюхе у безногих ящериц.
2 Субституция функции зрения у гимнофионов совокупностью функции обоняния и осязания (орган обоняния+щупальцы).
s Субституция функции движения, позвонков и ребер у предков черепах защит-: ной функцией спинного щита у черепах.
ОСНОВЫ МОРФОЛОГИЧЕСКОЙ ТЕОРИИ РЕДУКЦИИ ОРГАНОВ
419
Таблица 4
Вид субституции Замещенный орган предков Замещающий орган потомков
Эмбриональный зачаток Местоположение Функция
(по сравнению с замещенным органом предков)
I ' Первый вид субституции органа Клейненберга 1 редуцируется другой то же та же
I Второй вид субститу- I ции органа ч Клейненберга1 2 редуцируется другой другое та же
ь ' Субституция функций (А. Н. Северцов) 1-й модус редуцируется другой другое аналогичная
2-й модус редуцируется другой другое того же вида, но не аналогичная
3-й модус редуцируется другой другое совершенно другого вида, но биологически равноценная
Вполне вероятно, что именно этот принцип был действующим началом при редукции целых систем органов, имевшей место в эволюции многочисленных сидячих и паразитических форм. Мы знаем, например, что присутствие состоящего из целлюлозы защитного органа, мантии, является характернейшим признаком всех туникат (асцидий и сальп). История развития этих животных вполне ясно указывает нам на то, что как сидячие асцидии, так и свободно плавающие сальпы произошли от свободно подвижных хордат с хорошо развитой мускулатурой и органами движения, с хорошо развитой центральной нервной системой и органами высших чувств. По аналогии с тем, что мы нашли у черепах,3 можно думать, что прогрессивное (идиоадаптивное) изменение организации, определившее общее направление эволюции предков туникат, состояло в том, что пассивная функция кожи интенсифицировалась, благодаря выделению плотной целлюлозной мантии (tunica) и что преимущество прогрессивного развития этой функции защиты во много раз превысило полезность функции движения. Функция движения у этих животных первоначально ослабела и в конце концов исчезла вовсе, вследствие развития тяжелой мантии: это произошло потому, что функция движения потеряла свое биологическое значение и вследствие усиления пассивной функции защиты, начала атрофироваться. Здесь, так же как и в случае развития панцыря у черепах, анцестральные
1 Субституция хорды хрящевыми позвонками.
2 Субституция задних спинномозговых нервов, иннервировавших сдвинувшийся вперед брюшной плавник, передними спинномозговыми нервами у югулярных аканте птеригий.
3 Прогрессивное развитие спинного панцыря=срастание позвонков и редукция спинной мускулатуры.
27*
420
ГЛАВА XII
активные функции и их органы1 заменились совершенно иной, но биологически им равноценною пассивной функцией,1 2 причем органы, соответствующие активной функции, тоже начали редуцироваться. Эти два противостоящих процесса (резкое развитие органов защиты, с одной стороны, редукция органов движения, органов чувств и нервной системы, с другой) позволили свободно подвижным предкам туникат перейти к совершенно другому образу жизни, т. е. к сидячему образу жизни асцидий и к пассивно подвижному образу жизни сальп.
Весьма вероятно, что принцип субституции функций был действующим принципом и в других случаях общей редукции органов, имевшей, место у энтопаразитов, например у паразитических циррипедий (Rhizo-cephala) и цестод. Мы знаем, что у этих животных редуцировались не только органы движения и органы чувств, но также и весь кишечный тракт со всеми его придатками. Потеря всех этих органов возмещается интенсивным развитием половой системы взрослых особей и различного рода эмбриональных приспособлений личинок.
В дальнейшее и подробное обсуждение этой проблемы мы, однако, входить не станем, так как надеемся, что и сказанного совершенно достаточно для того, чтобы сделать нашу мысль вполне понятной читателю.
Мы приходим к выводу, что редукция органов или систем органов, существовавших у предков, часто осуществляется у потомков путем замены ряда органов другими органами с другими функциями, лучше приспособленными к изменившимся условиям существования (субституция органов и функций). При этом функция предков замещается у потомков либо той же функцией (Клей-ненберг, субституция 1-го и 2-го рода), либо иной, но похожей на нее функцией (Северцов, 1-й модус), либо отдаленно на нее похожей функцией (Северцов, 2-й модус), либо, наконец, совершенно на нее не похожей, но равноценной ей функцией (Северцов, 3-й модус).
Сводя все сказанное нами выше, мы можем следующим образом представить себе общий ход филогенетических изменений, ведущих к редукции органов.
1. Атрофия важных органов животного обычно связана со значительными изменениями, происшедшими в условиях существования и в образе жизни предков данной формы животных. Так, предки безногих ящериц перешли от жизни в открытой местности к жизни в густой траве, предки слепых подземных амфибий и рептилий—от жизни на поверхности земли, открытой для солнечного света, к жизни под землею в полной темноте и т. д.
2. Некоторые органы животных при этом приспособились к изменившимся условиям существования и соответственно изменили свое строение (позвонки и ребра безногих ящериц, органы обоняния гимнофионов, панцырь черепах и т. п.).
3. Благодаря прогрессивному развитию новых приспособлений старые замещенные ими приспособления теряют свое биологическое значение и начинают редуцироваться, следуя правилам отрицательного филэм-бриогенёза.3 Во всех исследованных нами случаях редукции органов первичное изменение организма заключается в образовании новых приспособлений, т. е. в прогрессивном изменении организации, которая делает возможным последующую редукцию других органов.
Итак, мы видим, что принцип филогенетической субституции функции и органов играет большую роль в процессе филетической редукции органов. Естественно поэтому поставить вопрос, не играют ли какой-нибудь
1 Мускулатура, нервная система и органы высших чувств предков туникат,
2 Пассивная функция защиты мантии туникат.
s См. ниже, процессы рудиментации и афанизии.
ОСНОВЫ МОРФОЛОГИЧЕСКОЙ ТЕОРИИ РЕДУКЦИИ ОРГАНОВ 421
роли в явлениях редукции также и другие принципы (типы) филогенетических изменений органов?
На этот вопрос нам придется дать положительный ответ.
РЕДУКЦИЯ ВСЛЕДСТВИЕ СМЕНЫ ФУНКЦИЙ
Я нашел, что процессы редукции имеют место также и при других типах (принципах) филетического изменения органов. Мы знаем, например, что при смене функций прежняя главная функция изменяющегося органа постепенно слабеет и становится второстепенной. Можно думать, что при этом части органа, связанные с этой функцией, будут атрофироваться и сделаются рудиментарными. Это предположение подтверждается целым рядом фактов.
Конечности наземных млекопитающих, приспособленные для ходьбы по суше, превратились путем смены функций в лопасти водных млекопитающих (ср. предыдущую главу). Согласно Дорну, это случилось потому, что у предков ластоногих, сирен и китов, живших по берегам водоемов, прогрессивно развилась второстепенная функция плавания и, сделавшись сначала равноценной главной функции (ходьбе) (Lutra, Ursus maritimus), стала впоследствии сама главной функцией конечностей (Carnivora pinnipedia). У тех форм, у которых функция плавания сделалась главной функцией, части конечностей, прежде служившие только для ходьбы (прежняя главная функция), начинают дегенерировать. Когда же прежняя главная функция (ходьба) полностью атрофируется, то вместе с ней редуцируются и все те части органа, которые были связаны с функцией ходьбы, например все осо-бености кожи, подошвенной поверхности конечностей, ногтей и когтей. Как известно, когти ластоногих и сирен превращаются в незначительные рудименты, а в некоторых случаях (Manatusinunguis) совершенно редуцируются. У китов и у китообразных они также исчезают совсем. То же самое целиком относится и к анцестральным пальцевым и подошвенным подушечкам наземных млекопитающих, которые бесследно исчезают у типично водных форм.
На этих примерах мы видим, что и при эволюции, которая происходит путем смены функций, отдельные части эволюирующих органов могут атрофироваться. При смене функций одна из второстепенных функций эволюирующего органа развивается прогрессивно, а прежняя главная функция органа утрачивает свое значение и ослабевает. Части эволюирующих органов, находившихся в функциональном соотношении с прежней главной функцией, становятся при этом рудиментарными,и, наконец, целиком исчезают.
РЕДУКЦИЯ ВСЛЕДСТВИЕ УМЕНЬШЕНИЯ ЧИСЛА ФУНКЦИЙ
Разрабатывая проблему редукции органов, я пришел к выводу, что эволюция, происходящая по типу уменьшения числа функций, также ведет к редукции сделавшихся ненужными частей., Мы уже видели, что при эволюции органа, у которого имеется одна главная функция и ряд второстепенных, общее число функций может уменьшиться. Это мы наблюдаем, например, при эволюции пятипалых конечностей предков непарно- и парнокопытных млекопитающих. Как известно, у этих животных функция поддерживания тяжести тела (главная функция) падает главным образом на три средних пальца (II, III, IV) конечности, крайние же пальцы (I, V) имеют в этом отношении лишь второстепенное значение. При прогрессивном развитии главной функции она концентрируется у предков парнокопытных на III и IV луче конечности, так что лучи I, II и V эту функ-
422
ГЛАВА XII
цию поддерживания тяжести тела утрачивают совсем. У предков непарнокопытных функция эта концентрируется только на одном III пальце, так что здесь лучи I, 11, IV и V утрачивают ее в такой же мере, как и лучи I, II и V у парнокопытных млекопитающих. Во всех этих случаях мы видим, что пальцы, утратившие свою главную функцию и переставшие касаться земли как при покое, так и при движении, начинают постепенно редуцироваться все больше и больше, пока, наконец, не превращаются в незначительные рудименты (грифелевидные косточки) или же не исчезают совсем. Уменьшение числа функций у предков семейства Equidae можно представить себе следующим образом (схема XXVII):
Функция поддерживания
I II III IV V
2 3 3 3 3
Пятипалые предки Equidae
Функция поддерживания
О г III г О 0 0 3 0 0
Equidae
Схема XXVII. «Функция поддерживания» обозначает главную функцию несения тяжести тела животного.
Латинские цифры I, II, III, IV и V обозначают metacarpalia или matatarsalia I—V лучей; арабские цифры 2, 3,3,3, 3 обозначают формулу фаланг предков Equidae; г, г обозначают рудиментарные metacarpalia или metatarsalia II и IV у Equidae (грифелевидные-косточки); 0, 0, 0,0—исчезнувшие пальцы Equidae; 111,3—третье matacarpale или meta tarsale и фаланги III пальца конечностей Equidae.
Мы обращаем внимание читателя на то, что эволюцию, происходящую путем уменьшения числа функций, никак не следует смешивать с эволюцией, происходящей путем субституции органов или субституции функций. Мы видели, что при эволюции, происходящей путем субституции органов или путем субституции функций, у потомков эволюирующей формы всегда образуется новый орган, который либо принимает на себя функцию замещенного им органа, либо вырабатывает новую функцию, аналогичную или не аналогичную замещенной анцестральной функции, но во всяком случае всегда ей биологически равноценную. При эволюции, происходящей путем уменьшения числа функций, у потомков не происходит замещения главной функции предков какой-нибудь новой функцией, наоборот, здесь имеет место гипертрофия прежней главной функции, выполняемой теперь определенной частью эволюирующего органа, что делает излишним существование остальных его частей. Эти части, сделавшись бесполезными, начинают редуцироваться. Таким' образом, сильное развитие одних частей органа (III и IV луча у парнокопытных, или одного III луча у непарнокопытных млекопитающих) и перенесение всей функции органа на одни эти части ведет к дегенерации остальных частей этого органа.
Мы видим, следовательно, что по меньшей мере четыре типа (принципа) филогенетического изменения органов сопровождаются процессом редукции органов: тип субституции органов, тип субституции функций, тип смены функций и тип уменьшения числа функций. При субституции органов, также как и при субституции функций, орган предков (и его функция) замещается у потомков другим органом (с тою же или другой, но равноценной замещенной функции функцией), сам же орган, сделавшись биологически бесполезным, редуцируется. При смене функций (Дорн) главная функция эволюирующего органа замещается у потомков одной из второстепенных функций этого органа; эта второстепенная функция
ОСНОВЫ МОРФОЛОГИЧЕСКОЙ ТЕОРИИ РЕДУКЦИИ ОРГАНОВ
423
и соответствующие ей части органа развиваются прогрессивно, а прежняя главная функция и части органа, функционально с ней связанные, редуцируются. При уменьшении числа функций та функция, которая у предков выполнялась несколькими частями эволюирующего органа, у потомков выполняется уже только одной сильно развившейся частью этого органа; остальные же, более не функционирующие части органа, редуцируются совсем.
ВЫВОДЫ
Достигнутые нами результаты могут быть вкратце сформулированы следующим образом.
1. Органы (или части органов) редуцируются во всех тех случаях, когда в течение эволюции они перестают быть необходимыми организму и теряют, таким образом, свое биологическое значение.
2. Бывают случаи, когда органы перестают быть нужными и редуцируются только вследствие изменившихся условий существования: активная или пассивная функция органа теряет свое биологическое значение, и орган начинает дегенерировать. Подобные случаи, однако, встречаются, повидимому, не очень часто.
3. Гораздо более распространенными являются случаи, когда редукция органа предков происходит у потомков вследствие прогрессивной эволюции другого органа или вследствие прогрессивного развития некоторых частей старого органа, благодаря чему функция анцестрального органа утрачивает свое биологическое значение. Такого рода случаи встречаются при эволюции, идущей по типам (принципам) субституции органов и функций, смены функций и уменьшения числа функций.
4. При всех этих типах филогенетического изменения орган предков замещается у потомков другим органом или некоторыми частями того же органа, лучше приспособленными к изменившимся условиям существования, чем анцестральный орган. Замещенный анцестральный орган при этом атрофируется.
5. В различных случаях субституции органов или функций орган предков замещается у потомков другим органом, расположенным на том же месте и выполняющим ту же функцию (1-й случай Клейне нберга); или он замещается другим органом, расположенным в другом месте тела, но выполняющим ту же функцию (2-й случай Клейненберга); или же он замещается другим органом, расположенном в другом месте тела животного и выполняющим другую (аналогичную или не аналогичную) функцию (А. Н. Северцов). Замещенный орган редуци p-у ется во всех этих случаях.
6. При смене функций А. Дорна главная функция органа предков замещается у потомков одной из прогрессивно развившихся второстепенных функций того же органа; эта функция постепенно становится главной функцией, а прежняя главная функция редуцируется; при этом редуцируются те части органа, которые были функционально связаны с прежней главной функцией.
7. При эволюции, идущей по типу уменьшения числа функций, функция, выполняемая у предков несколькими частями эволюирующего органа, выполняется у потомков меньшим числом этих частей или даже какой-нибудь одной сильно развившейся частью органа; остальные части этого органа становятся функционально бесполезными и исчезают.
424
ГЛАВА XII
Различные типы редукции органов наглядно изображены нами на трех следующих ниже диаграммах (схема XXVIII, I, II, III). Внимательно рассмотрев их, читатель лучше поймет различие между этими типами редукции, чем из длинных и подробных описаний.
Смена "7 функции
Уменьшение числа функции
Схема XXVIIL
Схема XXVIII. I. В верхнем ряду мы видим у предков функционирующий (левая сторона схемы) орган X с функцией Рис ориментом новой функции—(or) у органа у. У потомков (правая сторона схемы) орган X замещается органом у, а функция F замещается биологически равноценной ей функцией F,. При этом орган X у потомков редуцировался, превратившись в нефункционирующий рудимент Хг.
II. В среднем ряду представлен, орган предков (Хх), имеющий главную функцию (Hf) и три второстепенных' функции (Nfx, Nf2, Nf3). У потомков в этом органе (Х2) второстепенная фунция (Nft) стала главной функцией (Hfx), прежняя главная функция Hf редуцировалась [r(Hf)], а соответствующая ей часть органа атрофировалась.
III. В нижнем ряду мы видим, что главная функция (Hf) органа предка (X') выполняется тремя частями органа О, О, О с соответствующими им функциями (fv f2 и f3); у потомков в органе X" функция 1г становится главной функцией [Fj(Hf)], а функции f2 и f3 и соответствующие им части органов редуцируются.
ГЛАВА XIИ
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ
ИСТОРИЧЕСКИЙ ОБЗОР
В предыдущих главах этой книги, излагающих основную часть морфобиологической теории филогенеза, мы имели дело главным образом с эволюцией эктосоматических органов, стоящих в непосредственных соотношениях с окружающей средой, и с приспособлениями этих органов. Теперь мы должны рассмотреть вопрос о соотношениях между эктосоматическими и энтосоматическими органами, иными словами, вопрос об эволюции внутренних органов и об их филетических корреляциях с эктосоматическими органами. При этом мы должны заранее оговориться: проблема закономерностей эволюции внутренних органов так сложна и так мало разработана, что о том, чтобы разрешить ее в настоящее время целиком, еще не может быть и речи. Поэтому все то, что читатель найдет в настоящей главе, должно им восприниматься скорее как постановка вопроса и как введение в проблему коррелятивных соотношений органов, чем как попытка окончательного разрешения ее.
Наш взгляд на эволюцию эктосоматических органов был уже изложен в общих чертах в главе IX этой книги. Мы признали при этом, что первона-
чально изменяются эктосоматические органы, приспособляясь к изменениям в окружающей среде, и что лишь затем к этим изменениям строения и функций эктосоматических органов приспособляются функционально и морфологически координированные с ними внутренние энтосо-матические орган ы.1 Мы признали также, что изменения строе-
ния и функций эктосоматических органов являются, в общем, приспособлением этих органов к изменениям органической и неорганической среды, происходящим в течение геологических эпох. Таким образом филетические
изменения энтосоматических органов тоже представляют собой приспособления, но уже приспособления к только что упомянутым нами изменениям эктосоматических органов (филетические корреляции или коорди-
нации).
Мы не будем задерживаться на истории возникновения понятия корреляций, ибо этот вопрос в свое время был подробно изложен Е. Радлем.2 Отметим лишь самым кратким образом следующее. 1 2
1 Это отнюдь не означает, что эволюция внутренних органов по времени происходит позднее, чем изменение строения эктосоматических органов, т. е. позднее их приспособления к уже изменившимся условиям окружающей среды. Мы лишь хотим подчеркнуть, что эволюция эктосоматических органов всегда давала тон для эволюции энтосоматических органов. ,
2 См. Radi Е., Uber die Bedeutung des Prinzips von der Korrelation in der Bio-logie, Biol. Zentralbl., Bd. 21, 1901. Из собственных построений Радля, касающихся корреляций, я не мог извлечь для себя ничего положительного, ибо он стоит на резко антиэволюционной точке зрения, т. е. на точке зрения, прямо противоположной моей-
426
ГЛАВА XIII
Как известно, понятие о корреляциях было впервые введено в морфологию Кювье, который считал, что два органа находятся в коррелятивных соотношениях друг с другом в тех случаях, когда они постоянно сосуществуют у тех или иных животных форм; это константное сосуществование он обозначает как «correlation des formes». Современные биометрики, работающие над корреляциями индивидуальных вариаций животных, придают термину «корреляция» приблизительно тот же смысл, что и Кювье. В аналогичном смысле употребляли этот термин Дарвин и его ближайшие последователи, с той лишь разницей, что в понятие о корреляциях, данное Кювье, они ввели историческую точку зрения. Дарвин подчеркивает, что все части организма связаны между собой; однако в некоторых отдельных случаях эта связь частей так слаба, что ее едва можно заметить; поэтому одна какая-нибудь часть организма может исчезнуть целиком без того, чтобы остальные его части от этого заметно изменились. Зато во многих других случаях он находит, что при изменении одной части организма одновременно изменяет свое строение и другая его часть. Такого рода корреляции хорошо известны у домашних животных. Именно они, эти мелкие коррелятивные вариации, представляют собой тот материал, из которого с течением времени развиваются крупные филетические коррелятивные изменения. Здесь в понятие Кювье о статической корреляции форм вносится исторический момент филогенетических изменений.
В настоящее время различают несколько видов корреляций; правильнее говоря, слово «корреляция» теперь употребляется в различных смыслах. Подразделение корреляций дается Л. Плате.1 Он следующим образом определяет понятие корреляции: «Под понятием корреляции в биологии подразумевают состояние взаимоотношения или зависимости между частями организма. При этом различают косвенную или фи лети ческу ю корреляцию и настоящую индивидуальную корреляцию». Интересующую нас филетическую корреляцию Плате определяет следующими словами: «Если в течение филогенеза изменяется тот или иной орган какого-нибудь вида, то это очень часто влияет на другие органы». Он приводит также несколько примеров филетическйх корреляций: «Если резцы становятся очень велики, то увеличивается также и межчелюстная кость и если лобные кости несут на себе рога, то они становятся больше и толще. У жвачных животных образование рогов идет параллельно с потерей клыков; когда в связи с удлинением туловища из ящериц развиваются змеи, то у них редуцируются лапки, пояс конечностей и т. д.» Плате подчеркивает, что мы не можем обойтись без понятия о филетических корреляциях, хотя нам постоянно возражают, что мы не можем дать экспериментального доказательства предполагаемого нами взаимного соотношения частей. «При этом существует большая опасность», говорит он, «что филетические изменения, которые возникли-совершенно независимо друг от друга, будут относиться за счет внутренних коррелятивных влияний».
При индивидуальных корреляциях, говорит он далее, «части организма зависят друг от друга, безразлично, являются ли они будущими, или уже готовыми органами, или же наследственными единицами зародышевой плазмы». Здесь в старое понятие корреляций (сосуществование частей) вносится новый, каузальный момент, чуждый прежним авторам (Кювье, Дарвин и т. д.), рассматривавшим корреляции лишь с точки зрения постоянного сосуществования частей.
Согласно Плате, существуют две главные формы Корреляций, которые он обозначает как функциональные и как идиоплазм а-
'Plate L., Vererbungslehre und Deszendenztheorie; in Festschrift R, Hertr wigs, Bd. 2, 1910.
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИИ ИЛИ КООРДИНАЦИИ ОРГАНОВ 427 тические корреляции; первая форма корреляций соответствует взаимоотношениям частей развивающегося или взрослого организма; вторая касается взаимоотношений находящихся в зародышевой плазме единиц наследственности. В качестве примеров функциональной корреляции Плате приводит взаимоотношения между различными железами внутренней секреции (щитовидной, поджелудочной, половой, зобной), с одной стороны, и органами, находящимися под их влиянием (скелет, вторичные половые признаки и т. д.), с другой.
В случаях, когда существование таких соотношений не может быть доказано экспериментальным путем, но когда корреляции все же имеются на лицо, исследователю приходится прибегнуть к «детерминативной, или идиоплазматической корреляции», т. е. признать существование определенных соотношений наследственных единиц. На основании своих исследований Плате различает следующие виды идиоплазматических корреляций:
«1. Корреляции вследствие плейотропных возбудителей, когда один и тот же фактор приводит к выявлению нескольких признаков.
2. Корреляции вследствие плейотропных условных факторов (Kondi-tionaifaktoren), когда необходимо присутствие того же фактора для того, чтобы определенные наследственные единицы могли выявить в организме свойственные им признаки.
3. Корреляции вследствие ложного аллеломорфизма, когда определенные детерминанты отталкивают друг друга if вселяются в различные половые клетки той же особи, хотя они и не принадлежат к одной аллеломорфной паре».
Таким образом, корреляции вторичных половых признаков находят свое объяснение в тех случаях, когда они не вызываются функциональными корреляциями. Эти идиоплазматические корреляции были подробно исследованы Плате генетическими способами, т. е. путем скрещивания специально для этих целей подобранных форм, и приходится признать, что для изучения индивидуальных корреляций эти исследования имели большое значение.
Мы видим, следовательно, что термин «корреляция» употреблялся: 1) для обозначения постоянных условий сосуществования между различными органами любой животной формы (корреляции Кювье и Дарвина, филетические корреляции Плате, корреляции индивидуальных вариаций при биометрических исследованиях); 2) для обозначения каузальных соотношений между эмбриональными органами и органами взрослых форм, развитие которых зависит от развития этих эмбриональных органов; 3) для обозначения соотношений между единицами наследственности и детерминированными ими признаками взрослых форм (индивидуальные корреляции Плате и корреляции физиологов, занимавшихся вопросами развития). В этом последнем смысле термин «корреляция» обычно употребляется только новейшими исследователями.
С. Бехер1 в своей работе стоит именно на этой точке зрения и дает следующее подразделение корреляций. Существуют прежде всего псевдокорреляции, которые, по его мнению, более или менее совпадают с понятием филетических корреляций Плате и которые Бехер обозначает как «непрямую зависимость между двумя частями тела, которая большей частью объясняется их общей зависимостью от некоторой третьей его части». Этим псевдокорреляциям противополагаются истинные корреляции, или как он их называет «э у ко р р е ляции». «Эукорреляция», говорит он, «есть истинная, непосредственная и прямая зависимость,
1 Becher S.; Untersuchungen liber nicht funktionelle Korrelation, Zool. Jahrb., Abt. f. allg. Zool. u. Physiol., Bd. 31, 1911—1912.
428
ГЛАВА XIII
основанная на том, что определенное влияние исходит тем или иным образом из одной какой-нибудь части организма и действует на какую-нибудь другую его части». В специальной части своей работы Бехер рассматривает различные эукорреляции между частями кожного скелета безногих голотурий (синаптид) и приходит к подтверждению теории наследственной мнемы Сэмона.
Подробная сводка корреляций’ принятых в смысле функциональных корреляций Плате (эукорреляций Бехера), дана Б. Дюркеном.1 Этот автор утверждает, что «истинные корреляции являются взаимными, прямыми, вызванными формативными раздражениями, влияниями морфологически характеризуемых компонентов». В качестве примеров таких корреляций Дюркен приводит свои опыты с различного рода экстирпациями у головастиков. Если у головастика на ранних стадиях развития экстирпиро-вать заднюю конечность, то можно увидеть, что в спинном мозгу, в среднем мозгу и в больших полушариях переднего мозга, а также в ретине соответствующей стороны тела наступают далеко идущие явления дегенерации. Отсюда он заключает, что развитие задней конечности находится в коррелятивной зависймости от нервной системы. Если у личинки лягушки экстирпировать глаз, то в результате этого также наступают далеко идущие явления дегенерации (задержки в развитии): подвергаются воздействию не только зрительные центры в среднем мозгу, но также и нервы и нервные центры конечностей-в спинном мозгу, так что в развитии конечностей наступают резко выраженные задержки. Из этих опытов можно вывести заключение, что стимулы к развитию конечностей у амфибий исходят от центральной нервной системы.
Если у эмбрионов зародышей Rana fusca или Hyla arborea разрушить зачаток глазного бокала,1 2 то у них не развивается хрусталик. Развитие хрусталика связано, таким образом, с развитием глазного бокала, вернее, с сохранением контакта глазного бокала с эпителием кожи, из которого развивается хрусталик. Если слуховой пузырек амфибий на очень ранних стадиях развития этого последнего, когда еще нет никаких следов хрящевой капсулы, пересадить на какое-нибудь другое место тела зародыша (Д. Филатов),3 то хрящевая слуховая капсула образуется здесь за счет находящейся на этом месте эмбриональной мезенхимы. При этом Филатов нашел, что когда хрящевой эмбриональный слуховой пузырек находится поблизости от трабекул, то от зачатка этих трабекул исходит целый поток эмбриональных мезенхимных клеток, направляющихся к слуховому пузырьку и образующих вокруг него слуховую капсулу. Интересно, что трабекула той стороны тела, куда был пересажен пузырек, становится тоньше, чем трабекула противоположной стороны.
Таким образом, исследование физиологии развития позволяет нам бросить взгляд на причинную связь, существующую между различными процессами онтогенетического развития органов: мы видим, что образование органа зависит от некоторого морфофизиологического процесса (возбудителя развития), разыгрывающегося в ранее заложившихся эмбриональных органах. Удивительные опыты Шпемана и его школы показывают нам, каких больших результатов можно достигнуть в этой области и как правильна была в своей основе мысль Ру, утверждавшего, что онтогенез основан отчасти на самостоятельной, отчасти на зависимой (коррелятивной) диференциации. Механика развития, невидимому, действительно стоит на пути к тому, чтобы перебросить мост через бездну, разделяющую работу исследователей лад строением насле
1 Diirken В., Einfiihrung in die Experimentalzoologie, Berlin, 1919, 192Э.
2Y Rana esculenta хрусталик развивается независимо от глазного бокала.
3 F i 1 a t о w D., Aktievierung des Mesenschyms durch eine Ohrblase und einen FremdkOrper bei Amphibien, Arch. f. Entwick., Bd. 110, 1927, ibid., Bd. 122, 1930.
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 429
ственных единиц и работу над признаками взрослых форм, составляющих предмет исследования теоретиков-менделистов.
Приходится все же отметить, что, несмотря на этот блестящий, по нашему мнению, результат, механика развития и, в особенности, учение о функциональных корреляциях (эукорреляциях) до сих пор еще очень мало дали для разрешения проблемы эволюции внутренних органов. Это зависит главным образом от того, что в этой дисциплине, так же, впрочем, как и в физиологии человека, в с а м ой постановке проблемы отсутствует исторический момент. Механика развития интересуется причинной связью между следующими друг за другом процессами развития какой-нибудь одной определенной животной формы, и она заключает из своих опытов, что, например, у Rana arvaiis развитие хрусталика зависит от развития глазного бокала. Вопрос же о том, как могла развиться в филогенезе амфибий эта причинная связь между предшествующим и последующим процессами в онтогенезе лягушек, совершенно не ставится. Некоторые замечания Дюркена в его книге об эволюции1 заставляют нас все же надеяться, что наступит время, когда механика развития вплоЖую подойдет к вопросам эволюции; к сожалению, надо сказать, что до сих пор ею_в этом отно-• шении сделано очень немного.
У нас вопросом о корреляциях, принятых в прежнем значении этого слова (Кювье, Дарвин), т. е. рассматриваемых как корреляции фг.летиче- . ские, занялся Б. Домбровский.1 2 Автор попытался проанализировать некоторые корреляционные факты на сложном, но хорошо ему известном по личному исследованию примере корреляций между различными слуховыми косточками у Quadrupeda. Результаты, к которым он пришел, он резюмирует в виде нескольких «правил», на обсуждении которых мы здесь останавливаться не будем; при этом он дает следующую классификацию корреляций, которые разделяет на три группы. К первой группе, по его мнению, относятся «тектонические корреляции, обусловливающие собой состояние аппарата и вследствие этого характеризующие крупные систематические группы животных—классы, подклассы и отчасти отряды. Корреляции второй группы находятся внутри отдельных аппаратов и обусловливают собой их строение и форму (но не их состояние)». Домбровский обозначает их термином «конструктивных корреляций»; они характеризуют собой малые систематические группы— отряды, семейства, виды и роды. Наконец, имеются еще «элементарные корреляции», наблюдаемые у индивидуумов внутри отдельных родов.
Из сопоставления результатов, к которым пришли только что упо- * мянутые нами авторы, занимавшиеся вопросами корреляции, мы видим прежде всего, что те различные значения, которые вкладываются ими в понятие «корреляция», легко могут привести к недоразумениям. Устанавливается разграничение между филетическими и индивидуальными корреляциями, между псевдо-и эукорреляциями (Бехер), или истинными корреляциями (Дюркен), наконец, между тектоническими, конструктивными и элементарными корреляциями (Домбровский). Поэтому, может быть, было бы удобнее совершенно оставить выражение «корреляция», принятое в его прежнем значении (Кювье, Дарвин), и заменить его каким-нибудь новым термином. Дюркен, со своей стороны, предложил для этого термин «конъюнкция», мне же кажется более подходящим
1 Эйгкеп В., Allgemeine Abstammungslehre, Berlin, 1924.
2 Dombrowski , Ein Versuch der Analyse eimger Korrelationen, Zschr. Anat. u. Entwicklungsgesch., Bd. 79, 1926,
430
ГЛАВА XIII
выражение «координация», однажды употребленное в этом смысле Дарвином; ниже я буду пользоваться именно этим термином.
«Координациями», или «филетическими корреляциями» мы будем обозначать постоянное сосуществование двух или нескольких органов одного и того же организма. Критерием координации мы принимаем, таким образом, признак константного сосуществования. При этом мы различаем две большие группы координаций: первую мы назовем группой «м о р ф о ф и-зиологических координаций», вторую—группой «т о п о-графических координаций».
Мы утверждаем, что два или несколько органов бывают «морфофизио-логически» координированы, когда функция одного органа необходима для функционирования другого или других органов; в этих случаях мы можем говорить о морфофизиологической цепи координаций. Примеры подобного рода координаций мы найдем в функциональных и морфологических соотношениях между двиг^ельными нервными центрами спинного мозга позвоночного животного ^отходящими от этих центров нервами конечностей; в соотношениях между этими нервами и мускулами, которые они иннервируют; в соотношениях между этими мускулами и теми скелетными частями, которые они приводят в движение. Все эти органы, вместе взятые, представляют собой координационную пару или координационную цепь.
Мы обращаем особенное внимание читателя на то, что морфофизиологическими координациями мы обозначаем только такие соотношения между органами, при которых функция одного органа необходимо нужна для функции другого органа. В случаях же, когда такого ясного функционального соотношения между органами наблюдать не приходится (например в соотношениях между костными носовыми раковинами у птиц и мускулами их задних конечностей), мы, конечно, не можем говорить о каких бы то ни было морфофизиологических координациях. Только что упомянутые нами образования (носовые раковины и мускулы конечностей) являются членами различных координационных цепей, между которыми не существует никаких непосредственных, доступных нашему наблюдению связей.
Термином топографические координации (филетические корреляции) мы обозначаем соотношения органов, связанных между собой не функционально, но лишь топографически, т. е. местом своего расположения в организме. Примером топографических координаций являются нервные стволы, сосуды и неподвижные скелетные части, расположенные друг около друга; сюда же относятся соотношения между мозгом и окружающей его черепной коробкой, между обонятельным эпителием и скелетом носовой капсулы и многие другие. Коротко говоря, всякие соотношения между частями организма, расположенными в ближайшем соседстве друг от друга, но не связанными между собой функционально, будут представлять собой топографические корреляции.
Отметим, между прочим, что выражение «морфофизиологическая координация» (это относится также и к выражению «топографическая координация») представляет собой не историческое, но статическое понятие: у каждого животного в любой период его филогенетического развития все его органы находятся в такого рода координационных соотношениях, и физиологи вместе с морфологами могут судить по состоянию современных форм о состоянии морфофизиологических связей, существовавших между членами координационной цепи. С историчес-ской,т. е. с филогенетической, точки зрения мы обязаны поставить вопрос
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 43» о том, как могли образоваться, развиваться и изменяться в течение эволюции эти координационные пары и цепи, т. е. о том, как развиваются из координаций предков новые координации у потомков.1
МОРФОФИЗИОЛОГИЧЕСКИЕ КООРДИНАЦИИ
Обратимся теперь к вопросу о филогенетических изменениях морфофизиологических координационных соотношений позвоночных животных. Чтобы несколько ограничить нашу задачу, мы здесь займемся не всеми координационными соотношениями вообще, но главным образом теми, которые существуют между эктосоматическими органами и связанными с ними энтосоматиче-скими органами позвоночных. Кроме того, среди великого множества филогенетических координационных изменений мы выберем лишь те, функция которых нам вполне понятна; поэтому в первую голову мы остановимся на координациях, существующих в органах движения.
Разрабатывая вопрос о филетических изменениях координационных соотношений между экто- и энтосоматическими органами, нам пришлось сопоставить между собой громадное число подобного рода изменений; для приведения в известный порядок этого фактического материала надо было прежде всего попытаться классифицировать имевшиеся в нашем распоряжении данные. Мы настойчиво подчеркиваем, однако, что подразделение имевшегося в нашем распоряжении материала отнюдь не является классификацией всех существующих филетических координаций, число которых действительно огромно, но лишь подразделением типичных,
1 Уже после того как рукопись этой книги была закончена, вышло новое издание «Основ сравнительной анатомии» Шмальгаузёна (1935), где между прочим, разбирается вопрос и о «соотносительном развитии органов». И. И. Шмаль-гаузен принимает разделение на собственно корреляции и координации. Собственно корреляции он разделяет на генетические (в смысле Л. Плате), на морфогенетические и на функциональные. Морфогенетические корреляции основаны на эмбриональных формативных зависимостях, а функциональные —на зависимостях в дефинитивных функциях. Что касается координаций, то И. И. Шмальгаузен принимает мое деление на топографические и морфофизиологические координации, но предпочитает для последних название динами ческих координаций. Кроме того, он выделяет еще особую категорию «биологических координаций». Под последними И. И. Шмальгаузен понимает относительное постоянство соотношений между органами, не связанными топографической ил» непосредственной функциональной зависимостью, но обнаруживающими ясную связь с определенной биологической обстановкой, определяющей функции данных органов. Многие примеры корреляций, приводимые Ж. Кювье, относятся, в сущности, к этой именно группе зависимостей. Таковы соотношения между формой и структурой зубов млекопитающих, с одной стороны, и формой челюстного сочленения и развитием челюстной мускулатуры, с другой. Эти соотношения определяются через посредство факторов внешней среды—в данном случае через род пищи. Биологическая координация между длиной передних конечностей и длиной шеи у травоядных копытных определяется способом добывания пищи. Зависимость между удлинением тела и редукцией конечностей у змей определяется способом передвижения животного по земле. Зависимость между образованием хвостового плавника, ластов-и развитием веретеновидной формы тела у водных рептилий и млекопитающих определяется способом передвижения в водной среде. В особенности при специализации животного обнаруживаются многие биологические координации, как, например, связь между хватательными лапами, цепким хвостом, подвижностью глаз и длинным языком у хамелеонов, связь, которая определяется местообитанием и способом питания этих животных. Примером обратной биологической координации может служить зависимость между редукцией волосяного покрова и развитием слоя подкожного жира у китообразных. Оба процесса, являющиеся примерами субституции функций, связаны между собой не непосредственно, а только тем, что животные обитают в водной среде, определяющей эти изменения.
432
ГЛАВА XIIl
-по нашему мнению, изменений координационных соотношений.1 При этом мы займемся только координациями между экто- и эктосоматическими органами и пока оставим в стороне координации, существующие между различными - эктосоматическими органами.
Итак, обратимся к вопросу о морфофизиологических координациях. В некоторых случаях мы находим, что филетические изменения морфофизиологических координационных соотношений между органами имеют чисто количественный характер.Так, например, мы знаем, что у млекопитающих поверхностный жевательный мускул, m. temporalis, отходит от височной ямки и прикрепляется к processus coronoideus; его функция заключается в поднятии нижней челюсти. Если этот мускул развит слабо, то боковые и дорзальные стенки черепа бывают гладкими (например, у симплицидентатных грызунов, у селенодонтных копытных и у Hominidae); если же на первый план выступает движение нижней челюсти вверх и вниз, как, например, у хищных и насекомоядных млекопитающих, то работа m. temporalis усиливается, и от него отделяется поверхностный слой, отходящий расширенным основанием от височной поверхности черепа. Образование этого нового мускула вызывает развитие костных гребней на темени (crista saggitalis) и на затылке (crista occipitalis). Здесь мы видим, что функция мускула не изменяется, но интенсифицируется, а это обуслов-вливает собою количественное изменение в скелете: поверхность костей, от которых отходит m. temporalis, в течение филогенеза увеличивается. Мы знаем, что образование гребней на височной и затылочных костях, вследствие интенсификации функции m. temporalis, происходит у многих ископаемых и современных млекопитающих. М. Вебер утверждает, что эти гребни являются палеонтологическим мерилом образования и развития височных мускулов (m. temporalis).1 2
Здесь мы имеем хороший пример того, как при количественном изменении функции эктосоматического органа (в нашем случае функции нижней челюсти) функция координированного с ним органа (m. temporalis—кости черепа) остается прежней, но изменяется количественно в том же самом направлени и.
Случай количественного изменения, происходящего в обратном направлении, т. е. в сторону ослабления функции , мы находим при изучении плечевой мускулатуры бескилевых птиц. Мы знаем, что у килевых птиц сильное развитие грудных мышц (m. т. pectorales major et minor) связано с сильным развитием киля на грудине (carina sterni). Бескилевые птицы, наоборот, утратили способность к полету, и крылья их более или менее редуцировались. У страусов, например, крылья стали очень малы, но способности к движению они не потеряли; грудные мускулы этих птиц очень ослабли, и мы видим, что у них carina sterni целиком редуцировалась. Функция грудных мускулов (m. т. pectorales) осталась прежней, но ослабление этих мускулов привело к редукции (уменьшение поверхности прикрепления), т. е.,к количественно отрицательному изменению в костях.
Так как к этим примерам можно было бы легко прибавить любое количество других аналогичных случаев, то, мы можем выставить следующее положение: Если анцестральная функция эктосо-
1 Домбровский (I. с.) уже попытался дать подобного рода классификацию филетических координаций; он основывался при этом на обработке одного весьма сложного случая координаций. Мы пошли другим путем, а именно, исходя из простейших известных нам случаев, пытались перейти ко все более и более сложным фактам. Трудно сказать, приложимы ли ко всем случаям координаций «правила» Домбровского, являющиеся основной предпосылкой для его классификации корреляций и представляющие собой обобщение процессов, наблюдаемых им при филогенезе слуховых косточек.
2 Weber М., Die Saugetiere, Jena, 1927.
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКЙХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 433
магического органа остается прежней, но с течением филогенеза усиливается или ослабляется, то у потомков сохраняются те же морфофизиологические координации с функционально с ними связанными э н-
тосоматическими органами, что и у предков, но функции этих последних органов будут при этом количественно изменяться в том же направлении, что и функции эктосоматических
органов, т. е. будут усиливаться или ослабляться. Этот процесс мы можем обозначить как количественное изменение координационных соотношений.
В тех случаях, когда в течение филогенеза происходят качественные изменения функций эктосоматических органов, мы обычно находим, что качественно изменились и координационные соотношения этих органов, причем степень самого изменения бывает весьма различна. Поясним это следующими примерами.
Мы знаем, что скаты происходят от акулообразных предков, у которых грудные плавники были в общем построены по тому же типу, что и у современных акул. У акул поступательное движение тела осуществляется действием хвостового плавника; грудные плавники служат при этом лишь в качестве руля для изменения направления движения тела сверху вниз и снизу вверх. У скатов же грудные плавники изменили свою функцию1, поскольку они стали органами активного движения. Эти формы, как известно, плавают при помо-
рие. 139. Raja batis. Вид1 сверху.
щи волнообразных движений своих чрезвычайно разросшихся грудных плавников (рис. 72d, 133и 139);
г—плавниковые лучи; sc.—scapula; sc. г.—ска. пулярная часть плечевого пояса скатов, связанная с позвоночником (из Гудрича, по Оуену).
в этих плавниках сильно увеличи-
лось число лучей (рис. 139, г) и мускульных пучков, что позволило им соответственным образом изменить свою функцию.
Соответственно этому изменились у скатов и соотношения между плечевым поясом и позвоночником. У акул и, повидимому, у акулообразных предков скатов плечевой пояс не имеет непосредственного отно-
шения к позвоночнику, у скатов же мы видим, что скапулярная часть плечевого пояса (рис. 139, sc. г.) тесно связана с прилегающей к ней частью позвоночника и что отдельные позвонки этой части позвоночника
1 Субституция функций: функция хвостового плавника заместилась функцией Грудных плавников.
Морфологические закономерности 28
434
ГЛАВА XIII
плотно срослись между собой, образуя сплошную трубку. Мы видим, таким образом, что здесь в связи с изменением функции плавника произошло некоторое, правда не очень значительное, изменение морфофизиологических координаций функционально связанных между собой частей. Совершенно ясно, что функциональные отношения плечевого пояса к позвоночнику и функциональные соотношения между сросшимися позвонками у скатов будут иными, чем у акул. Очевидно также, что эти изменения плечевого пояса и позвоночника вызваны изменениями, происшедшими в грудных плавниках, так как функция плавников, ставших у скатов органами движения, требует более прочного причленения плечевого пояса к позво'ночнику, чем это имеет место у акул, у которых эти плавники являются лишь рулевым органом.
Другой пример качественного изменения функции эктосома-тического органа и координированного изменения функции связанного с ним энтосоматического органа мы находим при изучении эволюции органов движения у бесхвостых амфибий.
У хвостатых амфибий задние конечности служат главным образом для поддерживания тяжести задней части тела, а передняя часть тела поддерживается передними лапками. Поэтому у них подвздошная кость имеет почти вертикальное положение, параллельное направлению силы тяжести. У этих животных поступательное движение осуществляется так, что с каждым движением задней лапки связано продвижение тела вперед на один шаг. Повидимому, при медленном движении хвостатых амфибий горизонтальные компоненты этого движения не влияют на положение подвздошной кости.
У примитивных стегоцефалов, от которых происходят предки бесхвостых амфибий, подвздошная кость имеет то же положение, что и у хвостатых амфибий. Мы можем поэтому принять, что стегоцефалы двигались очень медленно. У бесхвостых амфибий подвздошная кость стала гораздо длиннее и приняла совершенно иное положение чем у хвостатых амфибий: она лежит не вертикально, а изогнулась вперед, принимая почти горизонтальное положение, параллельное уростилю. Чрезвычайно вероятно, что это изменение в положении подвздошной кости и, следовательно, в положении всего таза, а также изменение в длине этой кости связано с изменением направления и интенсивности движения задних конечностей у бесхвостых амфибий. Предки бесхвостых амфибий, т. е. стегоцефалы, передвигались, так же как и современные хвостатые амфибии, посредством ходьбы, т. е. каждая их лапа передвигалась одна после другой. При превращении стегоцефалов в бесхвостых амфибий у них изменился самый тип движения тела: бесхвостые амфибии передвигаются путем чередующегося движения задних лапок только в исключительных случаях, например, когда они медленно взбираются на кочки. Обычно же у них при скачках обе задние лапки одновременно двигаются в направлении спереди назад, причем туловище их продвигается вперед; направление движения при этом совпадает с направлением продольной оси тела и с положением обеих подвздошных костей. Поэтому я и считаю, что изменение строения и положения таза у бесхвостых амфибий было, связано с изменением движения их задних конечностей, т. е. с переходом от медленной ходьбы к быстрым прыжкам.
В этом случае изменения координационных соотношений мы видим, что функция эктосоматического органа, т. е. функция задних конечностей предков бесхвостых амфибий, являющаяся основной функцией для всего процесса движения, очень сильно изменилась в течение филогенеза в к а-чественномотношении по сравнению с анцестральным состоянием, наблюдаемым у стегоцефалов и у хвостатых амфибий; животные от медленной ходьбы посредством чередующихся движений задних конечностей перешли к движению скачками, т. е. к движению интенсивного
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 435
одновременного прыжка обеими задними конечностями. Таким образом, координационные соотношения между тазом и позвоночником (крестцом) также изменились качественно: подвздошная кость удлинилась и получила другое местоположение и другую функцию: она перестала служить органом для'поддерживания тяжести тела и стала по преимуществу органом, приспособленным для скачкообразного движения вперед.
В разобранном нами случае изменилась функция и положение таза, но его отношение к бедру и к крестцу осталось неизменным. .
Теперь приведем случай, когда изменяются также и отношения частей друг к другу, т. е. когда устанавливаются новые взаимоотношения между членами координационной цепи. Очень интересное видоизменение такого рода координаций мы.наблюдаем, изучая эволюцию челюстной мускулатуры у Acipenseridae.
Мы видели, излагая филогенез Protognathostomata и Chondrichthyes selachoidei, что из мускульного тяжа, из которого у селахий развивается челюстная мускулатура (рис. 37, см. стр. 183), образуется много мускулов. Так, из средней части мускульного тяжа развивается m. adductor mandibu-lae (рис. 37, add. m.), из переднего его отдела развивается m. levator labiae superioris (рис. 37, 1. 1. s.) и из дорзальной его части—m. levator palatoquadrati (рис. 37, 1. р. qu), который нас теперь особенно интересует.1 У взрослых акул этот мускул отходит от посторбитального отдела черепа и прикрепляется к верхнему краю нёбноквадратной кости. У хрящевых ганоидов мускульный тяж челюстной мускулатуры закладывается совершенно так же, как и у селахий, но из его дорзо-каудального отдела [рис. 140, М. Md'. (р. h.)] развивается мускул, который, отходя от посторбитального отдела черепа, идет не к нёбноквадратной кости (m. levator palatoquadrati у селахий), но к hyomandibular е, ккоторому он и прикрепляется: это и есть М. protractor hyomandibularis [рис. 140, М. Md' (р. h.), рис. 141, р. h.]. Сравнительно анатомическое исследование иннервации этих мускулов приводит нас к выводу, что этот мускул хрящевых ганоидов (m. protactor hyomandibularis) гомологичен т. levator palatoquadrati эласмобранхий (Б. Феттер1 2, А. Лутер3). У хрящевых ганоидов функция этого мускула (m. levator palatoquadrati=m. protractor hyomandibularis) также изменилась: вместо того чтобы поднимать нёбноквадратную кость, он тянет вперед, hyomandibulare, т. е. совершенно другую часть скелета (рис. 141). Изменение места прикрепления мускулов, о которых идет речь, здесь непосредственно связано с изменением движения частей ротового скелета.
Вопрос этот детально выяснен в только что упомянутой нами работе А. Лутера (1913). При сокращении m. protractor hyomandibularis, sym-plecticum и обе связанные с ним челюсти сдвигаются вперед и опускаются вниз, а челюстной аппарат хоботообразно выдвигается. Как известно, этот далеко выдвигающийся ротовой аппарат Acipenseridae имеет большое биологическое значение при захватывании добычи: осетры зарывают свое рыло в ил и захватывают челюстями находящихся в нем бентонных животных, которые и служат им пищей.
Мы имеем здесь типичный случай смены функций (А. Дорн): ротовой аппарат кусающего типа эласмобранхий изменил свою функцию и превра-
1 Северцов А. Н., Этюды об эволюции низших позвоночных, ч. III, Русский архив анат., гист^ и эмбр., 1925.
Sewertzoff A. N., The head skeleton and muscles of Acipenser ruthenus, Acta Zoologica, Bd. 9, 1928.
2 V e t t e r B., Untersuchungen zur vergleichenden Anatomie der Kiemen- und Kiefermuskulatur der Fische, t. II, Jen. Zschr. f. Naturwiss., Bd. 12, 1878.
2 Luther A., Uber die vom N. trigeminus versorgte Muskulatur der Ganoiden und Dipneusten, Acta Soc. Scien. Fennicae, Bd. 41, 1913.
28*
436
ГЛАВА XIII
Рис. 140. Графическая реконструкция нервов, мускульных зачатков и частей скелета головы зародыша Acipenser ruthenus (обозначения—при рис. 141).
Рис. 141. Графическая реконструкция мозга, нервов, мускулов и скелетных частей головы несколько более позднего зародыша Acipenser ruthenus.
Общие обозначения для рисунков 140 и 141. а. br. 1—первая жаберная дуга; a. h.—гиоидная дуга А. па.—па. adductor mandibulae articularis; au—слуховая капсула: с. br.x—ceratobranchiale 1; ch.— хорда; с. М.—меккелев хрящ; спа. h.—артериальная гиоидная комиссура; С. п—зачаток т. constrictor, cr. h.—ceratohyale; h. m.—hyomandibulare; i. a. v.1} i. a. v2—зачатки m. m. interarcuales ventrales 1 и 2; 1Ь1Э lb8, lb3 —m. m. levatores arc. branchlalium 1, 2 и 3; lb4-6-4-tp зачатки—па. па. levatores arc. branchialium 4, 5 и m. trapezius; M. Md' (p. h.)=p. h.—верхняя часть зачатка челюстной мускулатуры, соответствующая na. protractor hyomandibularis; М. Md" (А. па.)—вентральная часть зачатка челюстной мускулатуры, соответствующая m. adductor mandibulae articularis; М. Md"' передняя часть зачатка челюстной мускулатуры, соответствующая М. adductor mandibulae symphysialis; М. Н. (сп)—па. consrictor hyoideus, М. Н' (В. Ь.)—m. retractor hyoideus; N.—носовая ямка; ос.—глаз; pq.—palatoquadratum; R. h.—m. retractor hyoideus; sh.—stylohyale; spir.—брыз-гальце; sy.—synaplecticum; tr.—trabeculae cranii; gV—ганглий n. trigemini; g VII—ганглий n. facialis; g IX—ганглий n. glossopharyngei; g X—ганглий n. vagi; III—n. oculonaotorius; Vj—r. ophthalmicus n. trigemini; Vx—r. maxillaris n. trigemini; V8—r. naandibularis n. trigemini; VII hm— truncus hyomandibularis n. facialis; VIIt—r. ophthalmicus n. facialis; VII2—r. buccalis n. facialis;
VII3—r. palatinus n. facialis; VIII—n. acusticus (из A. H. Северцова, 1928).
еволюция ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИИ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 437
Рие. 142. Мускулы, нервы и скелет головы Rhina squatina.
Нт,—hyomandibulare; pq.—palatoquadratum; M. sp"—m, levator palatoquadrati, часть которого (M. sp.) огибает брызгальце (spir.) и прикрепляется к hyomandibulare (Нт) (из А. Лутера, 1909).
тился в вытянутый беззубый ротовой аппарат осетровых рыб. Исследование А. Лутера1 совершенно ясно показало нам, каким путем происходит этот переход от одной функции к другой. У типичных селахий m. adductor palatoquadrati служит лишь для движения (приведения) верхней челюсти; у некоторых сплюснутых бетоннных форм [Pristiophorus, Rhina squatina (рис. 42), скаты] часть волокон этого мускула (рис. 142, М. sp")также прикрепляется к hyomandibulare (рис. 142, Нт). Лутер это расширение функции m. adductor palatoquadrati у бентонных форм эласмобранхий ставит в связь с изменением функции, имеющим место у хрящевых Ганоидов. Довольно вероятно, что у непосредственных прелков'осетровых рыб произошло подобное же изменение места прикрепленияТп. adductor palatoquadrati (одновременное прикрепление как к нёбноквадратной кости, так и к hyomandibulare) и что впоследствии из этого состояния развилась мускулатура типа Chondrostei (превращение m. adductor palatoquadrati в т. protractor hyomandibularis). Согласно Лутеру, у осетровых рыб изменилась вся функция челюстного аппарата: из прочно связанного с черепом хватательного аппарата типа Elasmobranchii развился очень подвижный, также хватательный, сильно высовывающийся вниз рот.
Если мы примем, что новая функция челюстного аппарата (яс. е. новый способ захватывания пищи) вызвала полное изменение в строении мускулатуры челюстного и гиоидного подвесочного аппарата (а что это действительно так,
в этом нам теперь не приходится сомневаться), то мы можем с уверенностью утверждать, что резкое изменение функции эктосоматиче-ского органа, например челюстей, функционально обусловливает собою крупное изменение в функции энтосоматического • органа, т. е. в данном случае превращение одного мускула в другой: m. levator palatoquadrati в т. protractor hyomandibularis. В этом случае мы видим, что качественное изменение функции экто с оматичес к о г о органа вызывает крупное качественное изменение в функции координированных с ним энтосоматических органов (hyomandibulare, m. protractor). Интересно, что здесь старая корреляция (m. levator palatoquadrati—palatoquadratum) замещается новой.
Еще более значительное изменение коррелятивных соотношений мы находим при изучении эволюции слуховых косточек среднего уха млекопитающих. Как мы уже об этом упоминали (излагая в главе о принци-
1 L u t h е г A., Untersuchungen Uber die vom N. trigeminus innervierte Musku-5 latur der Selachier (Hale und Rochen), Acta Soc. Scient. Fennicae, Bd. 36, 19U9S
438
ГЛАВА XIII
пах филогенетического изменения органов тип выпадения промежуточных функций), большинство исследователей, занимавшихся этим вопросом, приходит к выводу, что слуховые косточки млекопитающих развились из частей двух передних висцеральных дуг (мандибулярной и гиоидной). Этот взгляд подтверждается и новейшими эмбриологическими (Вершлюс,1 Шмальгаузен1 2) и палеонтологическими (Брум3) исследованиями. Как известно, у млекопитающих в среднем ухе имеются три слуховые косточки, образующие цепь, идущую от барабанной перепонки к fenestra ovalis: молоточек, наковальня и стремя, соединенные друг с другом сочленениями; молоточек прилегает к барабанной перепонке, а стремя замыкает fenestra ovalis. Эти косточки передают вызванные воздухом колебания натянутой барабанной перепонки эндолимфатической жидкости внутреннего уха. Упомянутые исследования показали, что внутренний член этой цепи, т. е. стремя, принадлежит гиоидной дуге и соответствует pharyngohyale рыб, что средний член этой цепи, т. е. наковальня, соответствует квадратной кости рыб и низших Tetrapoda и что, наконец, наружный член цепи, т. е. молоточек, соответствует articulare рептилий. Стремя (pharyngohyale) уже у рептилий функционирует в качестве слуховой косточки (columella рептилий), но превращение articulare и quadratum в элементы среднего уха, т. е. настоящая смена функций, происходит только при превращении рептилийных предков млекопитающих в типичных млекопитающих.
Мы знаем, что у рептилий нижняя челюсть первоначально подвешивалась на квадратной кости: большая квадратная кость была подвижно или неподвижно сочленена с височной областью черепа, ее дистальный конец сочленялся с проксимальным концом (articulare) нижней челюсти, состоявшей из нескольких костей (dentale, angulare, articulare, coronoideum). Мы можем сказать, что квадратная кость с самого начала находилась в известных координационных соотношениях с нижней челюстью; об этом мы заключаем потому, что эта кость изменяет свое строение тогда, когда изменяется питание животных, т. е. изменяется строение челюсти и зубного аппарата. У современных форм квадратная кость либо соединяется с черепом подвижно (стрептостилия), как это бывает у ящериц и змей, либо плотно вклинена между костями черепа, что делает ее совершенно неподвижной (монимостилия крокодилов и черепах), либо срастается с черепом (аутостилия), что наблюдается у амфибий и двоякодышащих (Dipneusta). При всех этих переменах квадратная кость неизменно сохраняет свое отношение к нижней челюсти, являясь посредствующим звеном, при помощи которого нижняя челюсть сочленяется с осевым черепом.
Исследования Брума показывают, что квадратная кость у некоторых тероморф, а именно у Cynognathus, была сильно редуцирована и потеряла свою функцию (рис. 115 Е); при этом dentale оказалась настолько увеличенной, что она достигала заднего конца края нижней челюсти и на ней образовался новый сочленовый бугорок—condylus mandibularis. Нижняя челюсть теперь сочленяется с осевым черепом не посредством articulare, как у всех остальных рептилий, но посредством отростка dentale (procesus
1 См. Versluys J., в отделе «Скелет» в Vergleichende Anatomie der Wirbeltiere, Jhle, v. Kampen, Nierstratz u. Versluys, 1927 г., а также Versluys J., Die mittlere und aussere Ohrsphare usw., Zool. Jahrb., 1898; Versluys J., Entwicklung der Columella auris bei den Lacertilieren, Zool. Jahrb., Bd. 19, 1903. Ср. также работы этого автора, посвященные вопросу о стрептостилии,—вопросу, в котором он по праву является наиболее крупными авторитетом.
2 Шмальгаузен И. И., Der Suspensorialapparat der Fische und das Problem der Gehdrknochelchen, Anat. Anz., Bd.59, 1923. Его же «О строении подвесочного аппарата у наземных позвоночных, Труды 1-го съезда зоологов, анатомор и гистологов СССР, Ленинград, 1923.
3 Broom R., On the origin of the Mammalian auditory ossicles, Trans, south. Afric. phylos. Soc., Proc. Zool. Soc., London, 1912.
еволюция ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 439 condyloideus dentale). Quadratum, articulare и angulare при этом уменьшаются в размерах и теряют свою прежнюю функцию (начало рудиментации и выпадение промежуточной функции поддерживания нижней челюсти). Однако эти кости (quadratum, articulare) у потомков Therocephalia, т. е. у первичных млекопитающих, не редуцировались целиком, а, как мы это только что указали, лишь уменьшились в размерах; при этом они вошли в состав близлежащего среднего уха и превратились в слуховые косточки—молоточек и наковальню; прежнее сочленение, существовавшее между ними (articulatio quadrato-articularis)1 также сохранилось (рис. 115, F). Мы не можем определенно сказать, как-произошла эта смена функций и как возникла связь со старыми слуховыми косточками (collumella s. pharyngohyale) рептилий, но общий ход морфологических изменений, происшедших в этом отделе черепа, ясен.
Мы видим, что изменение строения и функции эктосоматического органа, т. е. нижней челюсти и в особенности dentale, вызвало очень значительные изменения в тех частях черепа, которые были морфологически и функционально с ней связаны,—в articulare и quadratum. Старые координации (височная часть черепа—квадратная кость—articulare—нижняя челюсть) при этом полностью исчезли и заменились новыми, прежде не существовавшими координациями (барабанная перепонка—articulare— quadratum—наковальня—овальное окошечко слуховой капсулы). Функция всех этих элементов при этом резко изменилась, ибо соответственные скелетные элементы вместо старой функции подвешивания нижней челюсти приспособились теперь для новой функции передачи звуковых волн. От старой анцестральной связи между элементами координационной цепи сохранилось лишь сочленение между молоточком и наковальней, т. е. сочленение articulare—quadratum.
Мы видим, таким образом, что в данном случае изменение функции эктосоматического органа сопровождалось очень резким изменением координированных с ним органов; прежние координации при этом перестали существовать и заменились новыми координациями с частями другого органа, а именно с частями среднего уха.1 2
Резюмируя все только что нами изложенное относительно филетических изменений морфофизиологических координаций, мы можем сказать следующее. На каждой стадии эволюции любой животной формы каждый эктосоматйческий .орган функционально связан с целым рядом энтосоматических органов, функция которых совершенно необходима для нормального функционирования этого органа. Эту морфологическую и функциональную связь между энто- и эктосоматическими органами (а также между различными энтосоматическими органами там, где таковые имеются) мы обозначаем как морфофизиологическую координацию органов (координационная цепь). При филетическом изменении строения и функции одного какого-либо члена такой координационной цепи, напримрр, при изменении эктосоматического конечного члена этой цепи, изменяются также и морфофизиологические соотношения между ним и следующим за ним членом цепи,
1 Сочленение между молоточком и наковальней.
2 Мы не можем сказать определенно, как именно развилось у первичных млекопитающих это изменение функции quadratum и articulare. Вполне возможно, что мы имеем здесь перед собою два независимых друг от друга процесса: во-первых, выключение quadratum и articulare из старой координационной цепи и начало фх редукции, во-вторых (после того как эти кости потеряли отношение к челюстному аппарату и резко уменьшились в размерах), возникновение новых морфофизиологических отношений к среднему уху. Мы подчеркиваем, что здесь для краткости мы изложили этот процесс лишь в самых общих чертах: при более подробном обсуждении только что изложенных изменений мы должны были бы принять во внимание также и изменения в форме и взаимном расположении интересующих нас скелетных элементов, точно так же как и изменения среднего уха и лежащих в этой области мускулов, нервов и т. п.
440
ГЛАВА XIII
точно так же как изменяются соотношения и между всеми остальными ее членами. Таким образом, изменение в конечном эктосоматическом члене координационной цепи вызывает изменение во всех остальных членах этой цепи, т. е. в совершенно определенном ряду внутренних энтосоматических органов данной животной формы. Те же органы животного, которые к данной координационной цепи не принадлежат, изменением в эктосоматическом органе не будут затронуты вовсе или будут затронуты очень мало.
Мы можем установить следующие виды филетических изменений членов координационной цепи:
1. При количественном изменении, т. е. при усилении или ослаблении функции эктосоматического органа, функции координированных с ним частей (органов) в течение филогенеза качественно не изменяются, а лишь усиливаются или ослабляются, так что координационные соотношения между отдельными членами координационной цепи в общем остаются неизменными.
2. Если же функция эктосоматического органа в течение филогенеза изменится качественно, то качественно изменятся и функционально связанные с ним энтосоматические органы, так что взаимоотношения между членами координационной цепи также становятся иными. При этом надо различать следующие случаи:
а) отношения частей остаются прежними, но местоположение частей по отношению друг к другу изменяется, так что общая функция органа становится несколькой иной (пример—взаимоотношение между конечностями и тазом у хвостатых и бесхвостых амфибий);
Ь) между членами координационной цепи частично возникают новые взаимоотношения, так что функция всего органа становится иной (пример—изменение функции ротового аппарата осетровых, вследствие изменения места прикрепления m. aductor palatoquadrati = m. protractor hyomandibularis);
с) вследствие выпадения некоторых членов координационной цепи и замены старых координационных соотношений новыми наступает полная смена функции (пример—изменение координационных соотношений и функции при превращении quadratum и articulare в слуховые, косточки у млекопитающих).
ТОПОГРАФИЧЕСКИЕ КООРДИНАЦИИ
Кроме только что описанных нами морфофизиологических координаций существует еще другой вид соотношений между органами, отличающийся от первых тем, что органы, находящиеся в подобного рода соотношениях, связаны между собою не функционально, но по положению. Этот вид филетических корреляций Дюркен обозначает термином «конъюнкции», мы же предложили называть их «топографическими координациями».
У всех позвоночных животных подобного рода топографические соотношения существуют между величиной и наружной формой мозга, с одной стороны, и величиной и формой охватывающей его черепной коробки, с другой; между величиной сосудов и нервов и величиной соответствующих им каналов в черепе и т. д.
Мы должны теперь поставить вопрос о том, как изменяются в течение филогенеза взаимоотношения между членами подобного рода топографических координационных цепей (мозг—череп; нерв—нервный канал в скелете и т. п.) при изменении величины и формы одного какого-нибудь члена этой цепи?
Мы знаем, что у птиц и млекопитающих мозг резко увеличился и сильно диференцировался по сравнению с мозгом их рептилиеобразных предков и что соответственно этому увеличилась в размерах и их хрящевая или
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 441 костная черепная коробка. При этом, само собой разумеется, степень увеличения мозговой коробки соответствует степени увеличения мозга. С другой стороны, мы знаем, однако, что оба эти органа, т. е. мозг и череп, не стоят по отношению друг к другу в непосредственных функциональных отношениях: главная функция мозга, т. е. координация нервных процессов, не зависит от главной функции черепа, которая заключается в защите мозга и органов чувств от внешних механических повреждений. Другая функция черепа заключается в том, что он является местом прикрепления мускулов головы: мозг и череп являются функционально независимыми друг от друга органами, хотя биологически череп совершенно необходим мозгу, как орган защиты.
Эмбриологические исследования показывают нам, как возникает и как образуется этот параллелизм при филогенетическом развитии мозга и черепа у птиц и млекопитающих. Сравнение хода эмбрионального развития этих органов у рептилий и у происходящих от рептилий птиц и млекопитающих показывает нам, что в двух последних группах животных мозг закладывается раньше и развивается быстрее, чем у рептилий, так что все его главные части являются уже вполне диференци-рованными и относительно большими на тех стадиях развития животных, когда череп начинает только развиваться. Мы можем сказать вместе с Менертом,1 что здесь мозг обнаруживает ускоренное, а череп несколько замедленное развитие.
Как известно, именно Менерт выставил положение, что все филогенетически прогрессивные органы имеют ускоренное развитие (акцелерация), а все филогенетически регрессивные органы—замедленное развитие (ретардация). Мои личные исследования привели меня, однако,к другому выводу. Я нашел (см. нижеследующие главы), что дегенерирующие органы (например редуцирующиеся задние конечности у Seps и у Ophisaurus) закладываются на той же стадии развития, что и нормальные органы (например, конечности Ascalabotes), и очень часто бывает так, что прогрессивные органы животных (например, челюсти Belone) закладываются поздно и развиваются очень медленно. Поэтому мы никак не можем сказать, что положение Менерта имеет универсальное значение.
Скорее можно думать, что акцелерация и ретардация в онтогенетическом развитии животных имеют не то значение, которое им приписывает Менерт: оба эти процесса являются способами изменения анцестрального хода эмбрионального развития, являются приспособлениями, развивающимися в тех случаях, когда возникают новые топографические координации. В только что описанном нами случае (мозг и череп высших амниот) ретардация в развитии черепа является средством, при помощи которого форма и размеры черепа приспособляются к прогрессивному увеличению охватываемого им мозга. У рыб и у амфибий эти органы растут гармонично и в своем росте идут, так сказать, равномерным шагом. У птиц же и у млекопитающих мозг диференцируется очень рано, в то время как его защитная оболочка, черепная коробка, начинает развиваться только ко времени развития конечных стадий мозга, и лишь после этого начинается одновременный гармоничный рост этих двух топографически координированных органов; ретардация в развитии черепа является, таким образом, филогенетически важным приспособлением—способом изменения топографической координации между филе-тически увеличивающимся мозгом и приспособляющимся к этому увеличению черепом.
Можно привести еще много примеров топографических координаций
1 Mehnert Е., 1. с.
442
ГЛАВА XIII
органов животных. Например, если у какого-нибудь четвероногого животного сильно развиваются конечности, то части спинного мозга,соответствующие plexus lumbalis и plexus brachialis, становятся толще, настолько толще, что здесь образуются intumescentia cervicalis (рис. 143, int. cer.) и intumescentia lumbalis
sm rhomb.
mt. cer,
int. Lum.
и
Рис. 143. Головной мозг Gallus domesticus.
int cer.—intumescentia cervicalis; int. him.—intumescentia lumbalis; sin. rhomb.—sinus rhomboidalis (из Вершлюса, по Вагнеру, 1927).
спинной
которым они проходят И
(рис. 143 int. lumb.). У птиц в intumescentia lumbalis развивается sinus rhomboidalis (рис. 143, sin. rhomb.). Среди млекопитающих intumescentia lumbalis очень сильно развита у кенгуру в связи с развитием его задних конечностей. Среди ископаемых рептилий у многих двуногих динозавров она была так велика, что в том месте, где она помещалась, полость канала спинного мозга была иногда больших размеров, чем полость черепа. Вообще говоря, внутренняя полость костного, состоящего из позвоночных дуг канала спинного мозга, бывает сильно расширена у тех форм, у которых хорошо развиты in-tumescentiae lumbalis и cervicalis. Таким образом, здесь мы имеем другой пример топографической координации между рпинным мозгом и костными дугами позвонков.
Мы знаем также много случаев, когда нервы и кровеносные сосуды, проходящие через хрящевые и костные каналы черепа и крестца, становятся с течением филогенеза более толстыми. В таких случаях параллельно с увеличением объема нервов и кровеносных сосудов идет и расширение нервных каналов в скелете,1 и оба эти филетические процесса мы не можем рассматривать иначе как топографические координации.
В приведенных нами выше случаях в обоих взаимно зависящих друг от друга органах филогенетические изменения идут в одном и том же н а п р а в л е н и и; мы можем привести, однако, случаи, когда изменения в соответственных двух органах идут в противоположных направлениях. В моих «Этюдах по теории эволюции» (1912) я обозначил такого рода координации (филетические корреляции) термином «обратные координации». Поэтому мы различаем два типа топографических координаций: 1) тип «соответственных координаций», при которых структура и функция энтосоматических органов изменяется в том же направлении, что и структура и функция энтосоматических органов (мозг и череп, сосуды и каналы, по т. д.) и 2) тип «обратных коорди
1 Исследование истории развития этих образований показывает нам, что у высших рыб и у Quadrupeda нервные и кровеносные каналы образуются в черепе гораздо позднее, чем нервы и сосуды: здесь мы имеем дело с акцелерацией развития нервов и кровеносных сосудов (гетерохрония), подобной той, которую мы наблюдали в случае координации между мозгом и черепом у высших амниот.
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 443
наций», при которых один орган развивается в прогрессивном направлении, а другой, топографически координированный с ним, наоборот, соответственно редуцируется.1 В качестве примеров таких обратных координаций мы можем привести многочисленные случаи субституции органов (Клейненберг).
Мы знаем, например, что у осетровых рыб в костной крышке черепа имеются медиально расположенные кости, вклиненные между парными черепными костями; я обозначаю их как mediodorsalia.1 2 Кости эти явно стоят на пути к редукции; обычно они бывают небольших размеров и у высших рыб исчезают без остатка. Но и у Acipenseridae процесс их редукции уже определяется довольно четко. Так, у Pseudoscaphirhynchus Kaufmanni rnediodorsale posterius, лежащая между обеими frontalia, имеет еще довольно значительные размеры. У Acipenser ruthenus medio-dorsaie posterius охвачена с боков обеими frontalia и как бы вытеснена ими, так что мы можем сказать, что прогрессивное развитие frontalia ведет к регрессивному развитию (редукции) rnediodorsale posterius. Когда же rnediodorsale posterius исчезает совершенно, как это наблюдается среди осетровых у Huso huso, то frontalia занимают ее место, так что редукция rnediodorsale posterius идет параллельно прогрессивному развитию frontalia. Интересно, что rnediodorsale posterius закладывается в онтогенезе оче н ь поздно; весьма вероятно, что эта поздняя закладка rnediodorsale, так же как и в выше приведенных случаях (мозг и череп у птиц и млекопитающих), координирована с прогрессивным развитием frontalia (ранняя закладка frontalia).
У примитивных позвоночных череп построен по платибазальному типу. У костистых рыб, у рептилий и у птиц развивается тропибазальный тип черепа; trabeculae cranii при этом сближаются, так что дно черепа суживается в глазничной области, а между глазами образуется тонкая межглазничная перегородка (septum interorbitale). Положение мозга при этом сильно меняется: у рыб с платибазальным черепом мозг с окружающей его частью черепа лежит между глазами, тогда как у форм с тро-пибазальным черепом глаза отделены друг от друга тонкой перегородкой, а мозг расположен дорзально от нее. Чрезвычайно вероятно, что образование этой перегородки и вообще тропибазального черепа коррелятивно связано с прогрессивным развитием глаз. Глаза высших рыб и рептилий резко увеличились в размерах по сравнению с глазами их предков; поэтому трабекулы оказались сближенными, а расстояние между обоими глазами сильно уменьшилось; увеличение глаз имело, таким образом, следствием сближение и даже частичное срастание лежащих между ними частей черепа (узкое parasphenoideum).3
Утончение septum nasale, наблюдаемое у высших рыб, может также служить примером обратной топографической координации. У селахий и у Acipenseridae носовые ямки расположены далеко друг от друга и носовая перегородка у них очень толста; у Hoiostei (Amia), Crossopterygii и Teleostei носовые ямки увеличились в размерах и septum nasale превратилось в тонкую перегородку.
В только что приведенных случаях отношения между органами, находящимися в состоянии обратной координации, ясны: увеличение одного органа ясно координировано с уменьшением другого органа. В литературе
1 Повидимому, «обратные координации» появляются только в случаях топографических координаций.
2 Sewertzoff A. N., Studies on the bony skull of Fisches, Quart. Journ. of mic> roscop. science, vol. 70, 1926.
3 Sewertzoff A. N., The head skeleton and muscles of Acipenser ruthenus, Acta Zool., Bd. 9, стр. 279—299, 1928.
444
ГЛАВА XIII
мы находим утверждение, что не только расположенные по соседству органы, но и органы, так далеко отстоящие друг от друга, как, например, передние и задние конечности кенгуру, также находятся между собою в подобного рода координационных соотношениях. Я лично в данном случае воздержался бы от столь определённого утверждения, так как здесь отсутствует та характерная гетерохрония в эмбриональных зачатках, которая свойственна топографическим координациям, где обычно изменяется темп онтогенетического развития проталлактического прогрессивно изменяющегося органа.
Очень интересный случай гетерохронии при топографической координации мы находим, изучая филогенетическое развитие конечностей у парнокопытных животных. Мы знаем, что при этом прогрессивно развиваются III и IV пальцы конечностей и дегенерируют II и V луч. По аналогии с тем, что происходит у безногих ящериц (см. ниже), можно было бы ожидать, что и здесь сначала перестанут закладываться конечные фаланги редуцирующихся лучей, затем постепенно перестанут закладываться следующие фаланги, пока, наконец, не исчезнут metacarpalia и metatarsalia. Мы видим, однако, что, например, у оленей дело обстоит иначе, а именно, у них сохраняются фаланги II и Vпальца, а метаэлементы большей частью, или даже целиком, редуцируются. В некоторых случаях у парнокопытных редуцируется средняя часть малой берцовой кости (Сушкина-Попова),1 ибоковые лучи (II и V) закладываются позднее, чем средние лучи (III, IV). Мы видим, таким образом, что редукция пальцевых лучей происходит здесь иным путем, чем у исследованных нами рептилий. У парнокопытных средние лучи конечностей закладываются несколько раньше, чем их краевые лучи; гетерохрония в закладке средних и боковых лучей приводит к прогрессивному развитию средних лучей и к дегенерации боковых лучей, утративших свою функцию и ставших бесполезными для организма.
Итак, новое приспособление частей организма к изменившимся условиям существования достигается часто Образованием гетерохроний в морфогенезе соответствующих органов. Гетерохрония является способом образования топографических координаций, т. е. новых приспособлений частей друг к другу. При этом мы видим, что первично изменяющиеся органы закладываются в онтогенезе раньше, а приспособляющиеся к ним (координированные с ними) органы образуются в онтогенезе позже. Так, первично прогрессивно эволюирующий мозг (а также глаза и носовые ямки) закладывается раньше, а приспособляющиеся к мозгу части (череп и т. д.) позже.
Мне кажется, что читателю теперь стала ясна разница, существующая между нашими взглядами на акцелерацию и ретардацию и воззрениями Менерта. По Менерту, акцелерация, наблюдаемая в онтогенезе, всегда связана с прогрессивной эволюцией данного органа, ретардация всегда связана с его редукцией: оба эти процесса являются, так сказать, способами прогресса и регресса. Что касается меня, то я пришел к выводу, что ретардация во времени закладки органа является способом координации прогрессивно развивающегося органа с топографически связанными с ним частями организма.
Мы видим, таким образом, что редукция может происходить и при топографической координации: поздно закладывающиеся части организма могут настолько уменьшиться в размерах, что перестают быть объектами подбора (Плате) и, сделавшись ненужными организму элементами, начи-
1 Сушкина-Попова, Н. Н. Онтогенез скелета конечностей iaus и Bos, Билл, Нос. о-ва исп. природы, 1914.
ЭВОЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 445 нают редуцироваться (боковые лучи в конечностях парнокопытных, medio-dorsalia cranii и т. п).1
Можно сказать, что гетерохрония не непосредственно связана с прогрессивной и регрессивной эволюцией животных: она является средством, при помощи которого устанавливаются новые, биологически важные координации между прогрессивно развивающимися органами и остальными (не прогрессивно развивающимися) частями организма. Имеет ли гетерохрония еще и какое-либо иное значение (что никоим образом не исключается)—этого мы еще пока не знаем.
Итак, мы видим, что существуют два типа топографический координаций: соответственный или гармонический тип, когда оба члена координационной пары филогенетически развиваются у потомков в одном и том же направлении, и обратный тип, когда один член топографически координированной пары разивается прогрессивно, а другой— регрессивно.
О СПОСОБАХ ИЗМЕНЕНИЯ КООРДИНАЦИОННЫХ СООТНОШЕНИЙ
До сих пор мы занимались описанием филетических корреляций или, по нашей терминологии, филетических координаций органов и не ставили вопроса о том, как происходят в течение филогенеза изменения координационных соотношений.
Некоторые указания на разрешение этого вопроса можно найти при изучении меристических вариаций у сегментированных животных. Здесь мы будем говорить только о морфофизиологических координациях, а именно о вариациях в положении задних конечностей у позвоночных животных. По Июне1 2 первый крестцовый позвонок у ящерицы может принадлежать и 27-’му, и 28-му, и 29-му, и 30-му, и даже 31-му сегменту тела;3 таким образом, его положение сильно вариирует. Но во всех случаях индивидуаль-* ного изменения местоположений крестца подвзошная кость всегда сочленяется с обоими крестцовыми позвонками, так что мы можем сказать, что изменение положения всего таза и задних конечностей стоит в полной гармонии с изменением положения обоих крестцовых позвонков. Мы наблюдаем здесь вполне константную корреляцию между вариациями в положении крестца и таза.
Изучая вариации в положении состоящего из единственного позвонка крестца желтопузика, В. Ф.Еремеев пришел к тому же результату:сериаль-ное положение крестцового позвонка вариирует довольно значительно. Так, у различных экземпляров желтопузика крестцовый позвонок принадлежит то 57-му, то 58-му, то 59-му, то, наконец, 60-му сегменту туловища, но во всех случаях он сочленяется с подвздошной костью, так что мы и здесь можем сказать, что смещение таза и задних конечностей всегда соответствует смещению крестцового позвонка. Мы находим также, что и положение клоакального отверстия изменяется соответственно смещению таза, так что это отверстие вместе с ограничивающими его измененными чешуями при всех вариациях в положении таза неизменно лежит позади подвздошной кости. Координация между задними конечностями, тазом, крестцовым позвонком, клоакальным отверстием и клоакальными чешуями при всех смещениях этих образований, будут ли они происходить в краниальном или
1 Повидимому, в случае уменьшения числа функций редукция органов происходит именно этим путем, и этим отличается от типичной редукции, имеющей место при субституции органов и функций.
2 Kiihne К., Ober die Variationen der Wirbelsaule, des Brustkorbes und des Extremitatenplexus bei Lacerta muralis Dum. u. Bibr. und Lacerta muralis Jaqu. Mor-phoL, Jahrb., 1915.
3 В 1,5% случаев крестцовый позвонок принадлежит 27-му сегменту тела, в 49% случаев—28-му, в 20,4% случаев—30-му и в 0,8% случаев—31-му сегменту.
446
ГЛАВА XIII
в каудальном направлении, остается неизменно постоянной. Во всех этих случаях взаимоотношения между членами морфофизиологической координационной цепи остаются одинаковыми, несмотря на все изменения,происходящие в сериальном местоположении компонентов. Мы видим, что здесь происходит как бы перекрещивание двух морфофизиологических координационных цепей; одна из этих цепей образована свободными конечностями, тазом и крестцом, другая—поясничным, крестцовым и хвостовым позвонками; место перекрёста лежит в области крестцового позвонка.
Выше мы видели, что у некоторых ящериц в течение филогенеза происходит известное смещение задних конечностей в каудальном направлении. Весьма вероятно, что только что упомянутые индивидуальные вариации в положении крестцового позвонка и таза указывают нам на путь, которым шли все эти смещения. Вполне возможно, что они происходили посредством суммирования идущих в одном и том же направлении вариаций. Ясно, что координационные соотношения при этих вариациях сильно изменяются. Если у ящерицы (Июне) или у желтопузика таз передвинется на один сегмент вперед (Lacerta А., В.) или на один сегмент назад (Ophisaurus А, В),
Lacerta
........................А 27 28 S S 311
В 27 S S 30 31
Ophisaurus
А 57 58 S 60 61
В 57 58 59 S 61
то морфофизиологические соотношения между лежащими один подле другого позвонками изменятся очень сильно. У ящерицы поясничный позвонок превратится в крестцовый, а крестцовый в хвостовой, у желтопузика же превращение позвонков пойдет в обратном направлении. При этих значительных изменениях строения и функции позвонков отношение таза (подвздошной кости) к позвоночнику остается таким же, каким оно было у предков.
Мы приходим к несколько иному результату, наблюдая соотношения между позвонками и ребрами у ящериц.- Возвращаясь к схеме XXV, мы видим, что у безногих ящериц параллельно с каудально направленным смещением крестца происходят некоторые изменения и в ребрах; короткие ребра поясничных позвонков превратились в течение филогенеза в длинные ребра. Обращаясь, однако, к индивидуальным вариациям, встречающимся, например, у желтопузика (схема XXVI), мы видим, что координации между ребрами и крестцом не имеют того постоянного характера, который они имеют в соотношениях между крестцом и подвздошной костью. В некоторых случаях у .желтопузика между последним длинным ребром и крестцом имеются два коротких поясничных ребра (ряды I, IV, V, VIII— XI, XV, XVI), в других случаях—только один поясничный позвонок (ряды II, III, VI, VII, XII—XIV.) То же самое можно сказать и относительно взаимоотношений между мускулами и крестцовой костью: здесь мы не можем установить в вариациях прочной и постоянной координации. Во всяком случае, мы можем принять, что эти вариации представляют собой тот материал, из которого образуются изменения координационных соотношений между органами.
В свое время Уоллес2 высказал мысль, что новые координации (филетические корреляции) образуются постепенно, так что сначала образуется, скажем, один какой-нибудь биологический признак, т. е. изме-
1 S—крестцовый позвонок.
’Уоллес А. Р., Дарвинизм, Москва, 1898.
5В0ЛЮЦИЯ ФИЛЕТИЧЕСКИХ КОРРЕЛЯЦИЙ ИЛИ КООРДИНАЦИЙ ОРГАНОВ 447 няется, один какой-нибудь эктосоматический орган, приспособляясь к новым условиям существования, и только затем у его более или менее отдаленных потомков изменяется функционально связанный с ним внутренний (эктосоматический) орган, приспособляясь к новому строению и новой функции измененного. эктосоматического органа. Однако исследование индивидуальных вариаций эктосоматических и энтосоматических органов скорее указывает на то, что этот взгляд не вполне правилен; в ряде случаев (крестец—таз ящериц) мы видим, что новые координации устанавливаются уже при индивидуальных вариациях и что в действительности не существует никаких периодов дисгармонии между функциями экто-и энтосоматических органов.
В других случаях, а именно там,1 где соотношения между энто-'и эктосоматическими органами не так тесны (поясничные позвонки—крестец), мы усматриваем различные возможности для дальнейшей эволюции: именно здесь открывается возможность изменения старых (анцесдраль-ных) координаций и установления новых соотношений.
Мы хотели бы еще раз подчеркнуть, что приведенные нами рассуждения отнюдь не должны рассматриваться как полноценное разрешение координационной проблемы; это не более как попытка указать на направление, в котором можно искать способы ея разрешения. Мы думаем, что детальное изучение изменчивости (вариаций) координационных цепей, состоящих из экто-и энтосоматических органов, должно помочь разрешению этой проблемы. Изучение координационных вариаций указывает, повидимому, на то, что изменения в соотношениях между экто- и энтосома-тическими органами эволюирующей в течение филогенеза формы следуют не одно после другого, но что совокупность их является заранее предопределенной наличностью тех индивидуальных вариаций, из которых развиваются крупные филетические изменения органов. Это положение имеет значение как для количественных изменений координаций, при которых анцестральные координационные соотношения в общем остаются такими же, какими они были прежде, так и для качественных изменений эктосоматических органов, при которых происходят более или менее значительные изменения прежних координационных взаимоотношений и устанавливаются новые координационные связи.
выводы
Все изложенное нами относительно филетических координаций органов-мы можем резюмировать в следующих положениях.
1. Мы различаем два вида координаций, а именно: а) координации, существующие между функционально связанными между собою органами, или так называемые морфофизиологические координации и Ь) координации топографические, при которых координированные органы связаны между собой не функционально, но лишь по своему положению; топографические координации имеют только общебиологическое значение. Отношения между морфофизиологически и топографически координированными органами, между двумя членами координационной цепи, могут изменяться в течение эволюции различным образом.
2. При количественных изменениях, происшедших в одном из членов морфофизиологической координационной цепи, другой член цепи изменяется также количественно и притом в том же самом направлении, что и первый член цепи (интенсификация функций или ослабление функций), так что координационные соотношеия между двумя данными органами остаются теми же.
3. При качественном изменении одного из членов морфофизиологической координационной цепи координационные соотноше
448
ГЛАВА XIII
ния между членами цепи изменяются также качественно. При этом мы можем различать следующие случаи:
а) взаимоотношение между координированными частями остается прежним, но общее положение частей изменяется, так что общая функция эволюирующего органа становится несколько иной, чем она была прежде;
Ь) частично между членами координационной цепи устанавливаются новые соотношения, так что функция эволюирующего органа становится в общем иной;
с) некоторые члены координационной цепи выпадают и заменяются членами другой координационной цепи, вследствие чего в органе происходит полное качественное изменение функции.
4. Изменение координационных соотношений в течение филогенеза происходит, вероятно, путем суммирования постоянных и спорадических вариаций координационных соотношений между органами.
5. Изменение топографических координационных соотношений между органами происходит путем акцелераций (ускорений) или путем ретардаций (замедлений) хода морфогенеза одного из членов координационной пары органов или координационной цепи органов. Первично прогрессирующий орган закладывается при этом более рано, приспособляющийся к нему, т. е. координированный с ним орган,—более поздно.
ОБЩАЯ СВОДКА ГЛАВ IX—XIII О ЗАКОНОМЕРНОСТЯХ ФИЛОГЕНЕЗА
В предыдущем1 мы довольно подробно обсуждали вопрос об о б щ е м направлении адаптивной (биологически-прогрессивной) эволюции; мы говорили при этом об ароморфозах, идиоадаптациях, ценогенезах и дегенерации. Всеми этими способами эволюирующий организм, рассматриваемый как морфологическое целое, приспособляется к различным изменениям окружающей среды. Эти общие изменения строения совершаются посредством чрезвычайно разнообразных частных приспособительных изменений строения и функции отдельных органов. Мы изучили различные типы или принципы этих частных изменений и классифицировали их (принцип интенсификации, принцип фиксации фаз, принцип смены функций, принцип субституции органов и функций ит. д.).1 2 При этом мы нашли, что некоторые типы изменений, такие, например, как субституция, смена функций, уменьшение числа функций, всегда приводят у потомков форм, развивающихся в биологически-прогрессивном направлении, к редукции замещенных и ставших ненужными органов. Выдвинутая нами морфологическая теория редукции3 является необходимым логическим следствием только что упомянутой теории типов филогенетических изменений.
Наконец, мы попытались установить взаимоотношения, существующие между адаптивными изменениями эктосоматических органор и теми изменениями, которые имеют место в органах энтосоматических. При этом мы пришли к выводу, что прогрессивные и регрессивные изменения в эктосоматических органах вызывают соответствующие адаптивные изменения в функционально и топографически связанных с ними энтосоматических органах. В подобных случаях первично изменяющиеся эктосоматические органы действуют на координированные с ними внутренние органы наподобие вновь изменяющихся условий внешней среды и изменяют ход эволюции этих органов.
Направление изменения внутренних органов в общем зависит о т характера первичных изменений эктосоматических органов: при количественных изменениях функций эктосоматических или проталлактических органов морфофизиологически координированные с ними части изменяются лишь количественно; при качественном изменении функций проталлактических органов функции и строение координированных с ними органов изменяется также и качественно.
Только что перечисленные отдельные части нашей работы—части, касающиеся направления эволюции, принципов (типов) филогенеза, редукции органов и изменения координационных соотношений—образуют
1 Глава X.
2 Глава XI.
8 Глава XII.
Морфологические закономерности
29
450 ОБЩАЯ СВОДКА ГЛАВ IX—X11I
все вместе одну общую теорию, объясняющую ход адаптивной эволюции внутренних и внешних органов животных: мы можем обозначить ее как морфобиологическую теорию хода прогрессивной и регрессивной эволюции животных.
Эта теория показывает нам, что эволюция, взятая в целом, является в своей сущности приспособительным процессом: наружные, т. е. эктосоматические органы путем общих и частных изменений своей структуры и своих функций (проталлаксы) приспособляются к изменениям, ' происходящим в условиях окружающей среды; внутренние, т. е. энтосоматические органы, в свою очередь, приспособляются к изменившимся наружным органам (дейталлаксы)—и таким путем изменяется все строение эволюирующего организма.
Но теория морфобиологического хода эволюции не касается вопроса о том, как и когда в течение индивидуальной жизни происходят описанные выше изменения строения и функций эволюирующего организма. Ответ на этот вопрос дает нам теория филэмбриогенеза, изложению которой посвящены следующие главы этой книги.
ГЛАВА XIV
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА1
ОЧЕРК ИСТОРИИ БИОГЕНЕТИЧЕСКОГО ЗАКОНА И ЕГО КРИТИКИ
Когда у морфологов идет речь об отношении между филогенезом’и онтогенезом животных, то обычно имеются в виду факты рекапитуляции (повторения) анцестральных признаков в онтогенезе потомков, иными словами, биогенетический закон Э. Геккеля и Ф. Мюллера. Ниже мы увидим, что эти отношения гораздо сложнее, чем они в свое время представлялись Э. Геккелю. Однако, с чисто исторической точки зрения, нам кажется естественным начать наш обзор соотношений между филогенезом и онтогенезом именно с теории рекапитуляции в той ее форме, какой она представлялась Геккелю.
История биогенетического закона, или история соотношений между филогенезом и онтогенезом, в общих чертах известна каждому биологу; поэтому мы ограничимся лишь кратким историческим введением, которое в то же время позволит читателю уяснить себе и нашу личную точку зрения на этот сложный вопрос.
Отношение между эмбриональным развитием и высотой организации взрослого животного стало обращать на себя внимание исследователей еще в начале прошлого столетия. Так, И. Меккель1 2 уже в 1821 г. высказал мысль, что последовательные ступени в эмбриональном развитии высших животных и человека (с начала развития и кончая взрослым состоянием) соответствуют постоянным признакам строения в восходящем ряду животных форм. Он говорит также о том, что одно и то же животное, если судить о нем по положению его главных органов на различных стадиях развития, принадлежит с систематической точки зрения не только к различным родам, но иногда и к различным классам животного царства. Из этого явствует, что Меккель в своем понимании морфологических процессов уже довольно близко подошел к так называемому закону рекапитуляции анцестральных признаков.
Несколько позже, а именно в 1828 г., вопросом об отношении между эмбриональными признаками животных и признаками взрослых форм заинтересовался К. Бэр; тогда же им был впервые формулирован его знаменитый закон, который гласит: «1) то, что является общим для большой группы животных, образуется в зародыше раньше, чем частное; 2) из более общего в соотношении форм образуется менее общее и так далее, пока, наконец, не образуется самое специальное; 3) каждый зародыш определенной животной формы, вместо того чтобы проходить стадии других определенных животных форм, все более отдаляется от них; 4) по существу
1 Здесь теория филэмбриогенеза, по сравнению с моими работами 1912, 1927 и 1931 гг., разработана более подробно и значительно дополнена.
2 Meckel J. F. System der vergleichenden Anatomie, Halle, 1821.
454
ГЛАВА XJV
зародыш высшей животной формы никогда не бывает похож на другую животную форму, но лишь на ее зародыш».1
Два первые положения заключают в себе то, что я называю собственно законом К. Б э р а, т. е. обобщение, к которому он пришел на основании своих эмбриональных наблюдений. Два последних положения носят полемический характер и направлены против только что приведенного мнения Меккеля, с которым К. Бэр был несогласен. Как известно, К. Бэр в течение всей своей жизни был решительным противником как меккелевской точки зрения, так и биогенетического закона Мюллера-Геккеля.
Позже, а именно в 1876 г., К. Бэр в одной из своих речей дал более конкретную формулировку открытому им закону. Он говорит, что «в зародыше позвоночного животного сначала закладываются признаки класса, к которому этот зародыш принадлежит; только после того, как эти признаки, например признаки птицы, образуются, начинают развиваться признаки водяной или наземной птицы, еще позже появляются, скажем, куриные признаки (сем. Gallinae), еще позже образуются признаки рода Gallus, а затем вида Gallus domesticus и, наконец, позже всего появляются индивидуальные признаки данного цыпленка».
В законе К. Бэра установлены два весьма важных общих факта: во-первых, факт, что признаки взрослого животного закладываются у зародыша в определенной закономерной последовательности; во-вторых, факт, что эмбриональное образование этих признаков протекает в направлении обратном той последовательности, в которой по степени общности располагаются те систематические категории, к которым принадлежит данное животное.
Теоретическое значение открытого К. Бэром закона весьма велико. К сожалению, однако, его обобщения были мало использованы позднейшими исследователями и мало разработаны. Этот закон принято приводить с уважением, но он никогда еще не служил в качестве рабочей теории для систематика или эмбриолога, каковой, по нашему мнению, он мог бы стать.
Наоборот, основная мысль Меккеля была быстро воспринята эволюционистами дарвиновской школы и разработана дальше. Сам Дарвин вполне оценил значение эмбриологических исследований для понимания эволюции вымерших предков современных животных, и в своем «Происхождении видов» уже говорит об отношении между эмбриональным развитием высших форм и строением их примитивно построенных предков. Он принимает, что сходство в строении зародышей таких животных, которые во взрослом состоянии резко отличаются друг от друга, указывает на общность их происхождения. Но он подчеркивает также, что различия в эмбриональном развитии животных не являются непременным доказательством того, что данные формы животных происходят от различных предков, ибо многие стадии часто являются как бы скомканными в течение развития, а некоторые, вследствие различных условий окружающей среды, сильно изменяются. Таким образом, Дарвин уже в 1859 г. предвосхищает не только основную мысль закона рекапитуляций, но и значение геккелевских ценогенезов.1 2
Вопрос о зависимости между эмбриональным и филогенетическим развитием животных был снова выдвинут в 1864 г. Ф. Мюллером, и именно ему принадлежит заслуга, что он в своей краткой статье «За Дар
1 v. Baer К., Uber Entwicklungsgeschichte der Tiere, Beobachtung u. Reflexion., Bd. 1, стр. 224, 1828.
2 «Embryology rises greatly in interest, when we thus look at the embryo as a picture, more or less obscured, of the common parent-form of each great class ot animals». Цитировано по Дарвину, «Origin of species etc., гл. XIII, стр. 450, 1880.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
455
вина»1 ясно и точно формулировал так называемый закон рекапитуляций и пояснил его целым рядом прекрасно подобранных примеров. Он говорит следующее (1. с., стр. 76): «В короткий срок, исчисляемый немногими неделями или месяцами, меняющиеся формы зародышей и личинок дают нам более или менее полную, более или менее верную картину изменений, через которые прошел вид в течение бесчисленных столетий, пока он не достиг настоящего своего состояния». Здесь закон рекапитуляций формулирован ясно. Далее он говорит: «Историческое свидетельство, сохранившееся в развитии, понемногу стирается, в то время как развитие идет .по все более прямому пути от яйца к окончательному строению животного; это свидетельство часто искажается борьбой за существование, которую должны вести свободно живущие личинки». В этой фразе выражен закон ценогенезов Геккеля. Мы видим, таким образом, что так называемый биогенетический закон Э. Геккеля, т. е. метод эмбриолого-филогенетического исследования (ср. главу III), был сформулирован Ф. Мюллером еще в 1864 г.
В конце его работы мы находим мысль, на которую ученые мало обратили внимания,1 2 но которая, по нашему мнению, имеет огромное значение для понимания хода филогенетического развития. Именно Ф. Мюллер впервые принял, что изменение взрослых форм при эволюции происходит не только путем суммирования тех мелких вариаций, которые встречаются увзрослых форм (то, о чем говорили Дарвин, Геккель и Вейсман), но и путем постепенного изменения хода эмбрионального или личиночного развития. При этом он считает, что потомки определенной формы животных, эво-люирующей в определенном направлении, достигают конечного состояния (состояния взрослой формы), либо «отклоняясь рано или поздно» от анцестрального хода онтогенеза предков (т. е. что у них появляется иной ход, иное течение онтогенеза, чем то, по которому протекало индивидуальное развитие предков), либо «выполняя данный путь без уклонения в сторону, с тем чтобы вместо остановки, пойти даль-ш е». «Таким образом потомки достигают нового конечного состояния, либо отклоняясь рано или поздно в сторону от пути развития предковой формы, либо, выполняя данный путь без отклонения в сторону, не останавливаются на состоянии родителя, но продвигаются дальше. Первый случай, повидимому, преобладал там, где потомство общих предков образует круг форм, стоящих в главных чертах на одинаковой ступени, подобно всем Amphipoda, всем крабам или всем птицам. Мы приходим, напротив, ко второму случаю прогрессивного развития, когда пытаемся произвести от общей родоначальной формы животных, среди которых одни сходны с молодыми стадиями других. В первом случае история развития потомков будет совпадать с историей развития предков только до момента расхождения их путей, так что о строении предков во взрослом состоянии она нам ничего не говорит. Во втором случае потомки проходят все развитие предков, и, поскольку происхождение данного вида основывается на этом втором способе продвижения вперед, историческое развитие вида будет отражаться в истории его индивидуального развития». (1. с., стр. 75, 76).
Ниже мы увидим, что эти мысли Ф. Мюллера могут быть разработаны дальше и глубже.
Э. Геккель3 принял закон рекапитуляций Мюллера, дал ему имя «Био
1 М ii 1 1 е г F., Fiir Darwin, Leipzig, 1864; Мюллер Ф., За Дарвина, изд. «Классики естествознания», Медгиз, 1932.
2 В этом повинен и я в моих работах 1912 и 1927 г.
3 Haeckel Е., Generelle Morphologie, 1866.
456
ГЛАВА ХЛ'
генетического закона», разработал его дальше и применил к изучению филогенеза животных. Главная же историческая заслуга Геккеля заключается, по нашему мнению, в том, что он построил превосходное родословное древо животного мира.
В своей «Общей морфологии» он дает следующее определение «основного биогенетического закона»: «История развития организма распадается на две близко родственные и тесно связанные между собой ветви: онтогения, или история развития органических индивидуумов, и филогения, или история развития органических родов. Онтогения является быстрой и краткой рекапитуляцией филогении, обусловленной физиологическими функциями наследственности (размножение) и приспособления (питание). Органическая особь повторяет во время быстрого и короткого течения своего индивидуального развития важнейшие из тех изменений формы, через которые прошли ее предки во время медленного и длительного течения их палеонтологического развития по законам наследственности и приспособления».
Повторения филогенетических изменений формы в онтогении Геккель обозначает именем палингенезов; при этом он особенно подчеркивает то обстоятельство, что рекапитуляции не являются полными повторениями филогении, но, наоборот, в очень многих случаях повторениями «укороченными и искаженными». «Палингенетическими процессами, или эмбриональными повторениями, мы называем все те явления в индивидуальной истории развития, которые неизменно передаются консервативной наследственностью из поколения в поколение и которые позволяют нам заключать о соответствующем процессе филогенетического развития взрослых предков. Ценогенетическими процессами, или эмбриональными нарушениями, мы называем все те процессы в эмбриональном развитии, которые не могут быть сведены к наследственности от древнейших предковых форм, но которые скорее всего произошли лишь позже путем приспособления зародышей или молодых особей к определенным условиям эмбрионального развития. Эти ценогенетические явления представляют собой чуждые примеси, которые не позволяют нам делать никаких непосредственных заключений относительно соответствующих процессов в филогенезе предков, но наоборот, искажают и скрывают от нас познание этих процессов».1
Геккель попытался классифицировать эмбриональные приспособления или ценогенезы, и его работа в этом отношении имела большое влияние на работы позднейших исследователей (Оппель, Кейбель и многие другие). Геккель говорит, что ценогенезы большей частью можно свести к сдвиганиям местоположения органов и к различным по времени закладкам их: ценогенезы первого рода он называет гетеротопиями, ценогенезы второго рода—гетерохрониями. При этом он приводит примеры этих двух категорий.
Даже из этих кратких цитат, мне кажется, становится ясной та основная точка зрения, с которой Геккель рассматривает соотношение между филогенезом и онтогенезом. Для него биогенетический закон имеет главным образом чисто методологическое значение; он рассматривает его как средство реконструировать филогенез животных на основе эмбриологических наблюдений над индивидуальным развитием особи. Поэтому такое исключительное значение он придает палингенезам, и поэтому он рассматривает ценогенезы как неприятные для исследователя «искажения» и «фальсификации».
Биологическое значение и палингенетических рекапитуляций и ценогенетических «примесей», интересует его мало, и в этом отношении он ограничивается лишь довольно неопределенными и общими пояснениями,
J Haeckel Е., Anthropogenie, Bd. I, 1803.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
457
что палингенезы обусловлены консервативной наследственностью, а цено-генезы—приспособлением. Согласно с этим он думает, что «филогенез является механической причиной онтогенеза».
Более основательно и подробно рассматривается вопрос об отношениях между онтогенезом и филогенезом (с геккелевской точки зрения) А. Вейсманом.1 По мнению Вейсмана эволюция взрослых животных протекает таким образом, что у эволюирующих животных форм появляются во взрослом или почти взрослом состоянии мелкие наследственные изменения строения органов и что эти изменения (наследственные вариации в смысле Дарвина) суммируются в течение поколений путем естественного отбора и превращаются в крупные изменения строения органов. В течение этого эволюционного процесса происходит сдвигание во времени закладок эволюирующих орг а, нов на все более и более ранние стадии онтогенеза, вследствие чего ход онтогенеза потомков данной формы вторично меняется. Новый орган, заложив-шийся у предков во взрослом состоянии, закладывается у потомков сначала у молодых животных, а позднее уже у зародышей. Таким образом, закладка органа механически сдвигается на все более молодые стадии эмбрионального развития, в результате чего происходит повторение признаков взрослых предков у зародышей их иначе построенных потомков. Как говорил Геккель, филогенез является «causa efficiens» онтогенеза: последний, согласно этой точке зрения, является процессом вторичного характера, обусловленным филогенезом взрослых органов предков.
В доказательство правильности своего взгляда Вейсман ссылается на наблюдения Вюртембергера,1 2 которому, как известно, удалось констатировать такое сдвигание времени онтогенетического образования нового признака (бугорки на раковинах аммонитов) на все более ранние стадии онтогенеза. Изменения, которые с течением времени образуются в спирально завитых раковинах, впервые появляются на последнем завитке, с тем чтобы в последующем продолжиться на предыдущем завитке, а затем и на самых молодых завитках; в то же время нередко и на последнем завитке образуется снова новый признак. Так, например, если мы проследим филогенез бугорков на раковинах аммонитов, то увидим, что эти бугорки постепенно передвигаются назад, сначала с последнего завитка раковины на предпоследний, позднее с предпоследнего на предыдущий, с него еще на следующий и т. д. Одновременно на последнем завитке бугорки начинают превращаться в шипы. Таким образом, новый филетический признак появляется впервые у взрослого животного (последний завиток на раковине), но затем, по мере появления у взрослых особей все новых и новых признаков, постепенно передвигается в онтогенезе назад, перемещаясь на все более молодые стадии, так что в истории вида все время имеет место отодвигание новоприобретенных признаков взрослых особей на все более ранние стадии онтогенеза.
Надо сказать, что Вейсман едва ли дает правильное толкование наблюдениям Вюртембергера, принимая, что здесь происходило сдвигание признака на более раннюю стадию развития и признавая это явление общим законом эволюции.
Обозначая камеры аммонита цифрами 1, 2, 3, 4, 5 и т. д., а появляющиеся на них бугорки буквами b1? b2, Ь3 и т. д., мы можем себе представить, что на одной из стадий филогенетической эволюции аммонита на камере 5 появляется бугорок (bj, на последующей стадии его филогенетического развития такой же бугорок (Ь2) появляется на камере 4, на еще более позд
1 Weis mann A., Vortrage iiber Deszendenztheorie, Bd. 2, 5, 27 G. Fischer, Jena, 1902.
2 Wil r te mberger L., Studien iiber die Stammesgeschichte der Ammoniten, Leipzig, 1880.
458
ГЛАВА XIV
ней филогенетической стадии подобный же бугорок (Ь3) появляется уже на камере 3 и т. д. Таким образом, мы имеем здесь не сдвигание бугорков, но новообразование новых бугорков (bu b2 и т. д.) на лежащих более проксимально камерах. Наглядно мы можем представить себе этот процесс в виде следующей схемы:
I 1 2 3 4 5
b2 bj
II 1 2 3 4 5.
b3 Ь2 Ьх
III 1 2 3 4 5
Мы видим, что, в сущности, мы не имеем здесь сдвигания признака b на более раннюю стадию развития, но лишь появление нового аналогичного признака (Ь2, Ь3) на другом месте тела потомков данного животного. Этот процесс мы можем сравнить с изменением строения и формы позвонков и ребер у таких метамерных животных, как змеи, желтопузики и т. п. при филогенетическом метамерном преобразовании их строения. Как мы это увидим ниже, это—не общий процесс развития, но один из специальных видов архаллаксиса, т. е. это не общее правило эволюции, но частный случай, проявляющийся лишь при одном из исследованных нами способов (модусов) филэмбриогенеза.
В своих дальнейших работах Вейсман проверил высказанную им точку зрения, изучая развитие рисунка у гусениц, и снова пришел к выводу, что более древние филетические признаки постепенно вытесняются в онтогенезе, передвигаясь на все более молодые стадии, пока они, наконец, не исчезнут совсем. Так Вейсман придал форму разработанной теории мысли Геккеля, что палингенетическая рекапитуляция представляет собою повторение признаков взрослых предков.
Уясним себе еще раз основную точку зрения, на которой стоял как сам Геккель, так и его последователи и противники.
Основная задача Геккеля состояла в том, чтобы восстановить филогенез главных групп (Phyla) животного царства и построить правильное родословное древо современных животных. И в своей «Общей морфологии», точно так же как и в своей «Систематической филогении», он такое древо в основных чертах действительно построил. Как общее направление геккелевских исследований, так и добытые им результаты подверглись резкой критике со стороны его многочисленных противников; не раз говорилось, что его метод в корне неправилен, что построенное им родословное древо животного царства фантастично. Теперь, когда со времени появления на свет его «Общей морфологии» прошло 70 лет (1866—1936), когда жизненная задача Геккеля и ее значение для эволюционизма лежит перед нами как завершенное целое, теперь, кажется мне, можно лучше оценить его труды, чем это удавалось сделать как его страстным сторонникам, так и его не менее страстным, часто увлеченным полемикой противникам.
Как известно, эволюционная теория была создана впервые Дарвином в 1859 г. В своей книге «Происхождение видов» Дарвин сумел подобрать самые убедительные для его современников доказательства правильности теории эволюции и ввести в биологические науки историческую точку зрения.
Мы имеем в виду именно эволюционную теорию, но не дарвинизм, т. е. не теорию естественного отбора. В «Происхождении видов» и в своих последующих трудах Дарвином были даны убедительные доказательства в пользу эволюции животных и растений, но нигде им не был прослежен ход эволюционного процесса, нигде не была прослежена история
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
459
изменений организации предков современных животных в предшествующие геологические эпохи. Разрешением этой важнейшей проблемы занялись сторонники и последователи Дарвина, и если бы им не удалось этого сделать, то эволюционная теория осталась бы голой схемой филогенетического развития, лишенной конкретного содержания.
Еще Ламарк в свое время признал необходимость исторических, т. е. чисто филогенетических исследований. Как известно, он первый попытался построить филогенетическое родословное древо животного царства в своей «Философии зоологии» (1809). Осуществлена эта задача была Геккелем в его «Общей морфологии», «Естественной истории творения» и «Систематической филогении». Само собой разумеется, что при попытках Геккеля восстановить ход филогенеза животных было допущено много ошибок и было высказано много не очень обоснованных гипотез: при тогдашнем уровне знания это и не могло быть иначе.1 Непреходящая историческая заслуга Геккеля заключается в том, что он эту стоявшую в то время перед теорией эволюции проблему ясно осознал и пытался разрешить.
Выделяя значение геккелевских исследований, мы не имеем намерения преуменьшить значения трудов остальных крупных филогенетиков после-дарвиновского периода (Гегенбаура, Гё'ксли и многих других), но мы все же подчеркиваем, что незабываемая заслуга Геккеля заключается в том, что он очень скоро (1866) после появления «Происхождения видов» (1859) сумел дать стройную и связную картину филогенеза животных. Только теперь, после более чем полувекового промежутка, протекшего со времени появления его главнейших трудов, мы можем вполне оценить действительное значение его родословного древа, и мы с удивлением видим, что бесчисленные последующие исследования систематиков, палеонтологов, анатомов и эмбриологов внесли мало существенных изменений в нарисованную им картину. Филогенез животных в его общих чертах, поскольку мы можем судить о нем на основании наших современных знаний, был правильно намечен Геккелем в его первых же трудах.
Мы видим, таким образом, что основной задачей Геккеля были собственно филогенетические исследования, т. е. построение родословного древа животного царства, и что закон рекапитуляций Ф. Мюллера, так называемый биогенетический закон, имел для него значение лишь как один из способов восстановления общей картины филогенеза. Поэтому так интересны были для него все «палингенезы» и так неприятны «ценогенезы», как досадные «добавления и искажения», на которые приходится неизбежно наталкиваться исследователю при восстановлении истории филогенетического развития предков. Для Геккеля был важен лишь вопрос о том, достаточным ли является в том или ином случае общее количество палингенезов для того, чтобы можно было восстановить ход истории развития предков данной животной формы в предшествующие геологические эпохи. Весьма примечательным является то, что такое понимание биогенетического закона самим Геккелем не принималось во внимание не только его последователями, но также и его критиками и противниками.
Как известно, биогенетический закон в последнее двадцатилетие прошлого столетия подвергся резкой критике со стороны целого ряда ученых. При этом сами критики стояли на той же точке зрения, что и Геккель, т. е. мало интересовались закономерностью соотношений между онтогенезом и филогенезом, но зато очень оживленно обсуждали вопрос о том, достаточно ли часто происходят рекапитуляции для того, чтобы на основании
1 Сам Геккель в последующих изданиях своих трудов исправил, согласно более поздним исследованиям, многие свои ошибки.
460
ГЛАВА XIV
их можно было исследовать филогенез предков, или ценогенезы настолько подавляют и искажают палингенетические признаки, что делают подобное исследование немыслимым. Так, Гегенбаур1 в своей полемике с Дорном относительно теории метамерии головы высказал мнение, что метод эмбриологического исследования для изучения филогенеза не приемлем, именно из-за ценогенезов, которые не позволяют вскрыть анцестраль-ные признаки, что может быть сделано лишь путем сравнительно анатомического исследования над взрослыми животными.
Оппель1 2 и Кейбель3 занялись вопросом о сдвигах, происходящих во времени закдадки (гетерохронии) различных органов и признаков, и эти исследования позволили Кейбелю выполнить свои «Нормальные таблицы» развития позвоночных животных, содержащие массу фактов относительно последовательности во времени закладки органов различных позвоночных. Оба эти исследователя приходят к отрицанию биогенетического закона как метода филогенетического исследования. Многочисленные сдвиги во времени закладок органов и признаков (гетерохронии), по их мнению, так велики, что'они решительно не позволяют думать о восстановлении отдельных стадий филогенетического развития.
Менерт4 попытался в своих работах классифицировать сдвиги, наблюдаемые во времени появления закладок при развитии органов, и поставить в связь эти гетерохронии с процессами прогрессивного и регрессивного развития органов. Он принимал, что «всякая особенность, приобретенная во время индивидуальной филогении ряда предков,—независимо от того, приводит ли она к прогрессу или регрессу—всегда связана с изменением всего процесса развития, начиная от первой закладки и кончая полной остановкой развития» («Биомеханика»). Эти изменения процесса развития заключаются, с одной стороны, «в ретардациях развития-органов, т. е.: 1) в запаздывании в закладке органов, 2) в замедлении гистологической диференцировки, 3) в замедлении всего процесса роста», а с другой стороны «в акце ле рации развития, т. е.: 1) в ранней закладке, 2) в быстрой смене первых гистологических стадий и в раннем наступлении последующего развития тканей и 3) в быстром росте и, вследствие этого, в раннем увеличении размеров». В конце книги Менерт дает формулировку своего «основного закона органогенеза»: «Скорость процесса онтогенетического развития (роста) органа пропорциональна достигнутой им высоте развития. Она повышается всякий раз с повышением и падает с понижением однажды достигнутой высоты развития».
Мы видим, что Менерт сделал весьма интересную попытку каузально связать различные гетерохронии (в смысле Геккеля) с процессами прогрессивной и регрессивной эволюции органов взрослых животных. Теория рекапитуляции (как метод филогенетического исследования) принимается им с некоторыми ограничениями: по его мнению, «онтогенез является измененным повторением филогенеза».
Мы воздержимся здесь от подробного обсуждения критики биогенетического закона со стороны Гарбовского,5 6 который целиком отвергает его состоятельность. Критика Гарбовского представляет некоторый интерес
1 Gegenbaur К., Die Metamerie des Kopfes und die Wirbeltheorie des Kopf-skelettes, Morph. Jahrb., 13, 1888.
2 О p p e 1 A., Vergleichung des Entwicklungsgrades der Organe zu verschiedenen Entwicklungszeiten bei Wirbeltieren, G. Fischer, Jena, 1891.
3 К e i b e 1 F., Das biogenetische Grundgesetz und die Coenogenese, Erg. d. Anatom. u. Entwicklungsgeschichte, Bd. 8, 1898.
4 Mehnert E., Kainogenesis als Ausdruck differenter phylogenetischer Ener-gien, G. Fischer, 1897.
Idem, Biomechanik, erschlossen aus dem Prinzip der Organogenese, G. Fischer, 1898.
6 Garbowsky Tad., Morphogenetische Studien. Als Beitrag zur Methodologie zoologischer Forschung, G.,Fischer, Jena, 1903.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
461
лишь в том отношении, что ему удалось собрать и сопоставить между собой большое число случаев, говорящих против факта существования рекапитуляции анцестральных признаков. Случаи эти относятся главным образом к первым стадиям эмбрионального развития и, согласно Гарбовскому; решительным образом говорят как против гипотезы гастреи Геккеля, так и против правильности теории гомологии первичных зародышевых листков. Мне кажется, что отрицание теории рекапитуляций на основании изучения только первых стадий онтогенеза, тех именно, которые никак не могут подвергнуться контролю путем- палеонтологических исследований, в корне не верно, ибо испытание этой теории с методологической точки зрения возможно лишь путем сравнения результатов, полученных палеонтологическим и сравнительно эмбриологическим исследованием.
С точки зрения теории наследственности, биогенетический закон подвергся подробному обсуждению со стороны Эмери1 и 0. Гертвига.1 2 Эмери пришел к выводу, что рекапитуляция анцестральных признаков имеет, так сказать, лишь «метафорическое» значение, но что биогенетический закон может тем не менее найти применение при исследовании филогенеза. О. Гертвиг отрицает существование реальной рекапитуляции, так как, согласно его мнению, всякая животная форма, например любое млекопитающее, содержит в своей зародышевой плазме наследственные зачатки (по современной терминологии—гены) именно данной формы млекопитающих, но не зачатки досилурийских простейших, из которых эти млекопитающие филетически развились. Это последнее положение, само собой разумеется, верно. Он говорит далее, что нам приходится оставить как неправильное выражение «повторение форм вымерших предков», заменив его выражением «повторение форм, закономерных для органического развития и идущих от простого к сложному».
В своих собственных исследованиях, например в его теории целома3 и в теории происхождения зубов и покровных костей, О. Гертвиг широко использовал биогенетический закон, и от результатов, полученных этим путем, впоследствии не отказался. Мы можем поэтому предположить, что Гертвиг в свое время признавал значение биогенетического закона как метода филогенетического исследования.
Посмотрим теперь, каково было поведение противников биогенетического закона, когда они принимались за собственные филогенетические исследования. Мы видим, например, что Гегенбаур, который, как мы это уже отметили выше, отнюдь не может считаться сторонником теории рекапитуляции, в своем учебнике сравнительной анатомии неоднократно приводит примеры повторения анцестральных признаков у потомков.4 Он сравнивает, например, задние конечности зародыша птицы с задними конечностями рептилий (ящерицы) и т. д.
Большинство морфологов интересующего нас периода, т. е. конца XIX и начала XX века, применяли в своих филогенетических исследованиях закон рекапитуляции в качестве одного из основных методов исследования, т. е. применяли его именно в том смысле, в каком его применял сам Геккель; при этом они нашли много новых прекрасных примеров рекапитуляции.
1 Emery С., Gedanken zur Deszendenz und Vererbungstheorie, Homologie ti. Atavismus usw. Biol. Zentralbl., Bd. 16, 1896.
2 H er t wi g O., Zelle und Gewebe, Jena, 1893; Idem., Allgemeine Biologie, Jena, 1906; Idem., Handb. der Entwicklungslehre, Bd. I u. Ill, 2. Jena, 1906.
3 H e r t w i g O. u. R., Die Coelomtheorie, Jena, 1881.
4 Gegenbaur K., Vergleichende Anatomie der Wirbeitiere, Bd. I, стр. 578— 579, 1898.
462
ГЛАВА XJV
Т. Н. Монтгомери1 целиком отвергает закон рекапитуляции («the геса-pitulations-hypothesis is scientifically untenable») и утверждает, что все построения, основанные на применении этой гипотезы, не могут дать ничего хорошего для исследования филогенеза.
Мы привели некоторые примеры воззрений различных авторов на отношения между филогенезом и онтогенезом и думаем, что даже из этого далеко не полного обзора можно увидеть, какое огромное влияние оказал Геккель на ход мыслей своих современников. Для Геккеля, как мы это неоднократно подчеркивали, биогенетический закон всегда был лишь способом, при помощи которого он пытался реконструировать филогенез предков, т. е. был одним из методов филогенетического исследования. Именно с этой точки зрения морфологи и занимались тогда так много гетерохрониями и ценогенезами, т. е. всеми теми посторонними- «примесями», которые искажают и скрывают палингенетические повторения признаков.
Теперь, сохраняя историческую последовательность, я позволю себе высказать свое личное мнение относительно проблемы рекапитуляций. Когда я впервые занялся проблемой о соотношениях между филогенезом и онтогенезом, передо мною прежде всего встал вопрос о том, действи-
тельно ли повторение анцестральных признаков в онтогенезе современных потомков происходит настолько часто, что этим повторением можно пользоваться в качестве метода филогенетического исследования? Мне казалось, что этот вопрос можно было бы разрешить, если бы мне удалось сравнить результаты, полученные путем онтогенетических исследований над скелетом позвоночных, с результатами, полученными путем палеонтологических исследований над скелетом вымерших предков этих же животных, или, по крайней мере, над формами, близко стоящими к этим последним. Поэтому я в период между 1908 и 1934 г. предпринял целый ряд работ, касающихся исследования развития различных частей скелета позвоночных животных, работ, часть которых я произвел лично, а часть поручил проделать своим ученикам (см. список литературы, помещенный в начале книги). До первого издания этой книги я не опубликовывал общей сводки этих результатов, но для меня лично они уже тогда были решающими. Я убедился в том, что повторение признаков предков в онтогенезе потомков происходит достаточно закономерно и достаточно часто, чтобы позволить нам с успехом пользоваться эмбриологическим методом исследования для филогенетических целей. Ноя нашел, кроме того, что имеется ряд случаев (и встречаются они довольно част о), когда рекапитуляция анцестральных признаков не имеет места, причем это вовсе не зависит, как это думали прежде, от присутствия тех или иных ценогенезов (Геккель). Ниже мы увидим, что теория филэмбриогенеза2 нам объясняет как причины повторения, так и причины неповторения признаков предков в онтогенезе потомков.
РЕКАПИТУЛЯЦИЯ ПРИЗНАКОВ ОРГАНОВ И ТКАНЕЙ
Мы только что употребили выражение: «в онтогенезе потомков повторяются признаки предков». Так как это выражение несколько неопределенно, то нам кажется необходимым пояснить более подробно, чте
1 Montgomery Т. N., The analysis of racial Descent in animals, New York, 1906.
8 Северцов A. H., Этюды по теории эволюции, Киев, 1912, II изд. Берлин 1922, а также: «Uber die Beziehungen zwischen der Ontogenese und der Phylogenese der Tiere», Jen. Zschr., Bd. 63, 1927.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
463
мы, собственно говоря, под этим выражением подразумеваем. Zeugopodium современных взрослых Anura состоит из одной кости (рис. 144,T+F), в то время как у личинок (головастиков) мы различаем в этом отделе конечности два длинных хрящевых скелетных образования—radius и ulna или tibia и fibula (рис. 145, Т и F), которые только в течение дальнейшего развития сливаются между собой в один цельный элемент. Мы знаем, с другой стороны, что у стегоцефалов, от которых происходят современные амфибии, и в том числе Anura, в zeugopodium также имеются две отдельные кости или, точнее, два отдельных хряща, одетых костью (рис. 146, F и Т). Наличие
двух раздельных хрящевых элементов zeugopodium в конечности современного головастика мы и обозначаем как повторение (рекапитуляцию) анцестрального состояния скелета zeugopodium у первичных Anura.
Само собою разумеется, что скелетные элементы головастика, о которых только что шла речь, отличаются от соответствующих им скелетных частей взрослых стегоцефалов. Первые гораздо меньше вторых, имеют иное гистологическое строение, несколько иную форму и т. д. Но в двух отношениях они вполне походят на кости zeugopodium стегоцефалов, а именно:
145. Скелет задней головастика
Рис. 146. Скелет * задней конечности взрослого стегоце* фала Trematops milleric самостоятельными большой и малой берцовыми костями.
F—малая берцовая кость; Fm — бедро; Т—большая берцовая кость (из Вершлюса, 1927).
конечности pachipus.
Рис. 144. Скелет задней взрослого Bombinator
T+F—сросшиеся между собой большая и малая берцовые кости; f—fibulare; t—tibiale (из Вершлюса, по Штейнеру, 1927).
Рис. конечности Pelobates fuscus с раздельно закладывающимися большой и малой берцовыми костями.
Em—бедро; F—малая берцовая кость; f—fibulare: Т— большая берцовая кость; t—tibiale.
они имеют то же расположение по отношению к другим скелетным элементам конечности (к humerus и femur или к костям carpus и tarsus), что и у стегоцефалов, и они, так же как и у стегоцефилов являются элементами, разделенными друг от друга. Таким образом, в этих двух признаках данные части скелета конечностей бесхвостых амфибий совершенно подобны соответствующим частям скелета их стегоцефалоподобных предков. Поэтому мы можем с полным правом сказать, что признак «существования отдельных, не сросшихся друг с другом элементов zeugopodium» рекапитулируется в онтогенезе современных бесхвостых амфибий.
Мы могли бы привести очень много примеров такой рекапитуляции признаков. Так, например, в онтогенезе акул и хрящевых ганоидов
464
ГЛАВА XIV
(А. Н. Северцов, 1895, 1928) повторяется анцестральное состояние затылочного отдела черепа, который у предков этих форм состоял из отдельных, не сросшихся между собой дуг позвонков, расположенных непосредственно позади блуждающего нерва. Здесь повторяются «признак раздельного существования верхних дуг позвонков» и «признак срастания этих последних»; не повторяется вовсе «признак размеров дуг позвонков» и только частично повторяется «признак формы верхних дуг позвонков».
Изучая развитие плюсны у ящериц и черепах (А. Н. Северцов, 1908), мы видим, что -astragalus и fibulare (calcaneus) закладываются у этих форм как отдельные элементы (рис. 147, а и f) и только впоследствии, на более поздних стадиях развития, срастаются в одно цельное tarsale proximale (рис. 148, tr. рг.). Так как мы в то же время знаем, что у примитивных ископаемых рептилий (Ophiocodon mirus, Procolophon, Stereosternum), эти tarsalia существовали как раздельные элементы (рис. 149, a, cal.) то мы можем утверждать, что здесь также происходит рекапитуляция двух признаков, а именно: «признака самостоятельной закладки» и «признака срастания». Во всяком случае, можно сказать, что здесь мы имеем рекапитуляцию эмбрионального состояния предков, и это будет совершенно правильно; но так как признаки взрослых животных часто закладываются в онтогенезе очень рано (А. Н. Северцов, 1927), то в этих случаях повторение эмбриональных признаков предков является в то же самое время и повторением признаков взрослых предков. Это можно было бы символически выразить, соединив знаком равенства следующие три выражения: «эмбриональный признак современного животного= эмбриональный признак вымершего предка=признак взрослого предка».
В качестве характерного примера соответствия результатов палеонтологических исследований эмбриологическим приведем еще эволюцию скелета конечности лошади такой, какой она нам представляется на основании данных палеонтологии и эмбриологии. Сравнивая филогенез пальцев предков лошади (Mesohyppus, Miohippus, Meryhippus, Hipparion, Equus) с эмбриональным развитием конечности современной лошади, мы отмечаем, что у шестинедельного эмбриона имеются еще три пальца, причем каждый из боковых пальцев несет три фаланги. Интересно при этом, что III палец уже сильно увеличен, а боковые пальцы уже начинают превращаться в так называемые грифелевидные косточки. Фаланги боковых пальцев можно видеть еще у пятимесячного эмбриона лошади, причем конечная фаланга уже отличается по своей форме от остальных. Срастание фаланг при превращении боковых пальцев в грифелевидные косточки происходит очень поздно. Здесь развитие конечности, начиная с трехпалой стадии, рекапитулирует ход эволюции скелета конечности и ее фаланг очень полно. Четырехпалая стадия эволюции, повидимому, не рекапитулируется вовсе.
Мы привели только немногие примеры рекапитуляции признаков для того, чтобы сделать понятным для читателя смысл и значение этого выражения, но мы должны подчеркнуть, что подобных случаев повторения признаков предков в онтогенезе потомков встречается так много, что мы имеем полную возможность использовать их для восстановления хода филогенеза различных групп животных. Рекапитуляция признаков является не исключением, но правилом, постоянно повторяющимся при одном из типов филогетического изменения.органов, а именно, как мы это увидим ниже, при анаболии, т. е. при изменении органов, идущем по типу надставок конечных стадий онтогенеза. Ниже мы будем говорить также о случаях, которые являются исключениями из этого правила и которые, будучи исключенями, все же вполне закономерны.
Теперь же мне хочется остановить внимание читателя на одном весьма важном обстоятельстве, которому, как мне кажется, до сих пор не было
Рис. 147. Ранняя стадия развития задней конечности геккона Ascalabotes fascicularis, на которой ясно видна раздельная закладка astragalus (tibiale) и calcaneus (fibulare).
а—astragalus (tibiale); с—centrale; F—малая берцовая кость; f—fibulare; Fm—бедро; T—большая берцовая кость; ti—14—tarsalia distalia I—IV; I—V — metatarsalia I — V; 1, 2, 3 -г фаланги 1, 2, 3 (из A. H. Северцова, 1908).
Рис. 149. Скелет задней конечности взрослой ископаемой рептилии Ophiocodon mirus с вполне раздельными tibiale"(astragalus) и fibulare (calcaneus).
а—astragalus (tibiale); cal.—calcaneus (fibulare); cj— c2—centralia I, II; t,— t6—tarsalia distalia I—V;
I— V— metaiarsalia I—V (из Вершлюса, 1927).
Рис. 148. Стадия развития скелета задней конечности Ascalabotes fascicularis, на которой видно срастание fibulare и tibiale водно цельное tarsale proximale.
F — малая берцовая кость; Т — большая берцовая кость; ti—14—tarsalia distalia I—IV; tr. pr.—tarsale proximale; I—V—metatarsalia I—V (пз A. H. Северцова, 1908).
466
ГЛАВА XIV
уделено достаточного внимания. Я имею ввиду тот факт, что рекапитуляции в смысле биогенетического закона Мюллера-Геккеля в такой же мере существуют при г и с т о генетическом развитии тканей, как и при онтогенетическом развитии органов, иными словами, я считаю, что в течение гистогенетического развития какой-либо ткани современного животного появляются признаки, характерные для клеток данной ткани предков этого животного.
Так как нам в настоящее время еще недостаточно известно строение тканей ископаемых животных, то при изучении гистологических рекапитуляций нам приходится пользоваться главным образом данными сравнительной гистологии и сравнительного гистогенеза, а именно, сравнивать гистогенез специализованных элементов тканей той или иной группы более высоко стоящих животных со строением тех же тканей у более низко стоящих форм, филогенетически близких к предкам данной группы, иначе говоря, приходится пользоваться тем же методом, которым работают сравнительные анатомы и эмбриологи-филогенетики, когда они не могут пользоваться палеозоологическим материалом для своих выводов.1
Рис. 150. Поперечный разрез через однослойный эпидермис зародыша человека на ранней стадии развития, epid,—клетки цилиндрического эпителия эпидермиса; сог.—corium (из Вершлюса, 1927).
Рис. 151. Поперечный разрез через эпидермис зародыша человека на несколько более поздней стадии развития, когда эпидермис состоит из двух слоев эпителия: нижнего—цилиндрического и верхнего—плоского (из Вершлюса, 1927).
В качестве примеров гистогенетических рекапитуляций мы могли бы привести гистогенез эпителиальной ткани (эпидермиса), гистогенез крови, гистогенез униполярных нервных клеток, гистогенез ткани электрических органов некоторых скатов и гистогенез эпителия, выстилающего пищевод у высших позвоночных (А. Н. Северцов, 1935).1 2 Разберем самый простой из этих примеров, а именно, гистогенез эпителиальной ткани высших млекопитающих.
Изучая развитие кожи высших позвоночных, например человека, мы видим, что на ранних стадиях онтогенеза эпидермис зародыша человека, как это видно из рис. 150, состоит из однослойного цилиндрического эпителия, который путем деления своих клеток на более поздних стадиях развития превращается сначала в двуслойный (рис. 151), а затем и в много
1 Понятно, что для филогенетических целей сравнение мускулатуры позвоночных С мускулатурой, например, насекомых (как это иногда делают гистологи), т. е. сравнение органов или тканей, принадлежащих к двум очень высоким и далеким друг от друга типам животного царства, не может дать ничего. В лучшем случае оно может раскрыть лишь некоторые параллелизмы между эволюцией этих двух типов животных. Напротив, сравнительно гистогенетическое исследование тканей млекопитающих, рептилий, хвостатых амфибий, кистеперых рыб, акуловых рыб и бесчерепных позвоночных могло бы дать интересные с филогенетической точки зрения результаты для некоторых тканей. К сожалению, такое систематическое исследование гистологами еще не проделано.
2 С е в е р ц о в А. Н., Морфологическая и гистологическая рекапитуляция, Доклады Академии наук СССР, т. III, 1935.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
467
слойный однородный эпителий зародыша. В течение дальнейшего развития происходит изменение гистологического строения поверхностного слоя эпидермиса: клетки его становятся плоскими и, ороговевая, теряют ядра. У взрослого человека мы имеем многослойный специализованный эпидермис, состоящий из ряда вполне диференцированных слоев: stratum corneum, stratum lucidum, stratum granulosum и stratum malpigii (рис. 152).
Перед нами встает вопрос, не представляют ли собой различные стадии гистогенеза эпидермиса человека рекапитуляции признаков, характерных для его более близких и более отдаленных предков? Чтобы ответить на это нам, за отсутствием палеогистологических данных, приходится обратиться к строению эпителиальной ткани в ряду примитивных родичей высших позвоночных. При этом мы находим, что у ланцетника во взрослом состоянии имеется однослойный эпидермис (рис. 153). Мы можем предположить, что стадия однослойного эпителия в онтогенезе эпидермиса заро-
Рис. 152. Поперечный разрез через кожу взрослого человека, состоящую из многослойного специализованного эпидермиса, в котором виден ряд диференцировав-шихся слоев.
str. cor.—stratum согцеит; str. luc.—stratum lucidum; str. gran.—stratum granulosufri; str. mal.— stratum malpigii (из Бема и Давыдова).
дыша человека является гистогенетической рекапитуляцией, т. е. повторением в гистогенезе высшего млекопитающего строения однослойного эпи-телия, характерного для очень далеких бесчерепных предков высших позвоночных. Стадию гистогенетического развития эпидермиса при превращении однослойного эпидермиса в многослойный без ороговения поверхностного слоя клеток мы можем сравнить со строением эпидермиса костистых рыб, состоящего, как это видно из рисунка 154, из многих слоев камбиальных,т.е. индиферентных эпидермических клеток. Такимобразом, в стадии онтогенеза эпидермиса человеческого зародыша, изображенного на рис. 151, мы можем видеть, как в строении клеток, так и в их расположении, рекапитуляцию строения эпидермиса рыбообразных предков млекопитающих [колбовидным и слизистым клеткам, находящимся в эпидермисе рыб (рис. 154) филогенетического значения мы, конечно, придавать не можем]. Стадию начального ороговения поверхностного слоя эпидермиса мы находим в коже Salamandra maculosa (рис. 155). Соответствующая стадия в онтогенезе эпидермиса зародыша человека является, повидимому, рекапитуляцией строения эпидермиса амфибийных предков млекопитающих.
30*
'468
ГЛАВА XIV
Таким образом, с довольно большой долей вероятности можно предположить, что эпителиальный покров тела высших млекопитающих, в частности человека, прошел последовательно в своем филогенезе стадию инди-ферентного однослойного эпителия, стадию индиферентного многослойного эпителия, стадию многослойного эпителия с поверхностным, просто по-
Рис. 153. Поперечный резрез через эпидермис взрослого ланцетника, состоящий из одного слоя цилиндрических эпителиальных клеток (из Рабля).
Рис. 154. Поперечный разрез через многослойный эпидермис кожи костистой рыбы Barbus fluviatilis, состоящий из камбиальных эпидермических клеток, между которыми расположены колбовидные и слизистые клетки (из Вершлюса, по Мауеру, 1927).
в данном случае проявляется не только
Рис. 155. Поперечный разрез через многослойный эпидермис кожи Salamandra maculosa, на котором видны первые следы поверхностного ороговения эпителиальных клеток (из Рабля).
строенным роговым слоем и, наконец, стадию сложно построенного эпителия высших позвоночных.
Отметим при этом, что рекапитуляция анцестральных признаков во взаимных отношениях клеток ткани, но и в форме и в строении самих эпителиальных клеток.
Интересный пример гистоге-нетической рекапитуляции, имеющей место в изолированных клетках, мы можем наблюдать в красных кровяных тельцах млекопитающих. Как известно, у большинства млекопитающих (в том числе и у человека) существенные отличия красных кровяных телец сводятся к появлению двух признаков, а именно к накоплению в плазме гемоглобина и к потере клеткой ядра; при чем эти признаки появляют
ся при развитии в определенном порядке: сначала накопляется в теле инди-ферентной клетки гемоглобин, а затем исчезает ядро. Сравнивая гистогенез эритроцитов у высших млекопитающих с гистогенезом эритроцитов у низших позвоночных, мы видим, что признак присутствия ядра на ранних стадиях гистогенеза и потеря его на самых поздних представляет собой гистогенетическую рекапитуляцию филогенетического развития крови высших млекопитающих. То же можно сказать и относительно развития гемоглобина в крови, причем филогенетически гемоглобиновый пигмент появился в эритроцитах позвоночных гораздо раньше, чем стало исчезать
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
469
ядро. Так, у наиболее древних черепных позвоночных, т. е. у круглоротых и у акуловых рыб, эритроциты уже содержат гемоглобин, у ланцетника же кровь бесцветна; таким образом, образование гемоглобина, повидимому, произошло при превращении бесчерепных предков позвоночных в протокраниат, т. е. еще в досилурские времена.
Типичную гистогенетическую рекапитуляцию анцестральных признаков мы можем наблюдать при изучении образования в черепных и спинномозговых ганглиях эмбрионов всех высших позвоночных униполярных нервных клеток, развивающихся, как известно, из типично биполярных ганглиозных клеток. Исследуя спинные корешки сегментальных нервов у ланцетника, можно ясно видеть, что его спинномозговые ганглии состоят из типичных биполярных ганглионарных клеток, вполне похожих на клетки в спинномозговых ганглиях у зародышей высших позвоночных животных. У некоторых акуловых рыб мы также имеем в спинномозговых ганглиях типичные биполярные клетки, по существу вполне подобные клеткам спинных корешков ланцетника. У костистых рыб в спинномозговых ганглиях на ряду с большинством биполярных клеток попадаются уже редкие униполярные клетки. И только у более высоко стоящих форм биполярные клетки исчезают совсем. Все это показывает нам, что превращение биполярных ганглионарных клеток в униполярные происходило в филогенезе очень медленно и постепенно.
Наконец, гистогенез электрических и псевдоэлектрических органов у скатов (Torpedo, Raja) представляет собой великолепный пример рекапитуляции исторического хода эволюции этих органов. Не менее типичен в этом отношении и гистогенез эпителия пищевода высших позвоночных животных.
Все изложенные нами факты хорошо известны как морфологам, так и гистологам, но последние до сих пор как-то мало обращали внимания на их филогенетическое значение, которое нам хотелось бы здесь особенно выделить. Наши немногие примеры гистогенетической рекапитуляции ясно указывают на то, что в онтогенезе тканей потомков повторяются не только сами анцестральные признаки, но также и порядок их появления в ряду предков. Анцестральными признаками в этих примерах являются: биполярное строение ганглионарных клеток спинномозговых узлов, из которого развивается униполярное строение этих клеток у высших позвоночных; строение булавовидных клеток электрических органов, дающих электрические пластинки; содержащие ядра клетки кровеобразующих органов, из которых развиваются безъядерные кровяные тельца высших млекопитающих; однослойный эпителий зародышей высших млекопитающих, дающий начало многослойному эпидермису взрослых форм; мерцательный эпителий пищевода высших млекопитающих, из которого развивается выстилающий пищевод эпителий взрослых форм. Все эти анцестральные стадии являются необходимыми для дальнейшего развития более высоких видов ткани.
Изучение эволюции гистогенеза тканей наводит нас на мысль, что эта эволюция шла по типу анаболии, о котором мы будем говорить ниже, с тем основным различием, однако, что здесь надставки образуются не в группах эмбриональных клеток, но в самих клетках. Порою же эта эволюция шла по типу девиации.
Может показаться странным, что рекапитуляция выявляется в отдельных признаках тканей и органов, а не захватывает собою целые, органы или системы органов. Здесь нам кажется уместным напомнить читателю о тех выводах, к которым приходит по этому поводу Осборн1 на основании обширнейшего палеонтологического- и зоологического материала.
1 О s Ь о г n Н. F., Origin of single characters as observed in fossil and living animals and plants, American Naturalist, 1915.
470
ГЛАВА XIV
Осборн рядом прекрасно' подобранных примеров показывает, что эволюция животных в общем напоминает «мозаичную работу» и по существу своему является не чем иным, как эволюцией отдельных признаков (single characters). При эволюции различные признаки животных наследуются как отдельно друг от друга, так и целыми группами, т. е. эволюируют в известной локальной или функциональной корреляции по отношению друг к другу. В предыдущей главе мы привели ряд примеров подобного рода корреляций (морфофизиологические и топографические координации), показывающих, что отдельные признаки связаны друг с другом коррелятивными связями и не могут быть рассматриваемы независимо от организма. Осборновские же концепции для нас имеют значение постольку, поскольку они делают понятным тот факт, что рекапитуляция признаков идет тем же путем, что и их прогрессивная эволюция, т. е. что рекапитулируются отдельные признаки (single characters).
ОЧЕРК ИСТОРИИ ТЕОРИИ ФИЛЭМБРИОГЕНЕЗА
Выше мы видели, что непосредственйые ученики и последователи Геккеля, а также современные ему критики, стояли на его точке зрения, т. е. принимали, так же как и сам Геккель, что главное значение теории рекапитуляций или биогенетического закона, заключается в том, чтобы пользоваться им как методом при реконструкции хода филогенеза взрослых предков современных животных и что этот филогенез взрослых предков является действующею причиною («causa efficiens») онтогенеза потомков. Это последнее мнение Геккеля принималось слепо огромным большинством его современников и последователей, противоположный же ему взгляд Ф. Мюллера мало принимался во внимание или даже был вовсе забыт.
Как уже было сказано выше, Ф. Мюллер еще в 1864 г. высказал мнение, что эволюция органов взрослых животных происходит путем постепенного изменения хода онтогенеза их потомков. При этом он считал, что онтогенез животных имеет совершенно иное значение, чем то, которое ему приписывал Геккель. Онтогенез, по его мнению, служит не только для построения взрослого организма, но является важнейшим средством для филогенетических изменений потомков данной животной формы. Исследование- онтогенеза интересно не только тем, что оно нам открывает те или иные палингенезы, но и тем, что оно вскрывает закономерности в филогенетическом изменении органов.
Мнение Ф. Мюллера, что филетические изменения строения взрослых оробей обусловлены изменениями хода онтогенеза их предков, не было принято ни самим Геккелем, ни большинством его многочисленных сторонников. Тем не менее, ряд исследователей, независимо от Ф. Мюллера, пришел к тому же выводу, что и он. Так, в том же году, когда появился труд Мюллера, была опубликована работа знаменитого гистолога и эмбриолога А. Колликера1 под названием: «О дарвиновской теории творения», в которой он излагает свою гипотезу гетерогенного развития животных. Основная мысль этой гипотезы состоит в том, что «под влиянием общего закона развития животные производят из своих зародышей иные, т. е.. уклоняющиеся от них, формы».
Таким образом, Колликер принимает, что существует некоторый общий, имманентный закон развития, по которому типичный для каждого данного вида ход онтогенеза изменяется в некоторые определенные момен
1 К о 1 1 i k е г A. Uber die Darwinsche SchOpfungstheorie, Ztschr. f. wissensch. Zoologie, Bd. 14, 1864. ' '
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
471
ты, и вследствие этого появляются новые формы животных, уклоняющиеся в своей организации от родительских форм. При этом он подчеркивает, что эволюция происходит «скачками». Главная его мысль заключается в том, что «в основе происхождения органического мира лежит большой план развития», поэтому он отвергает дарвиновский принцип естественного отбора и принцип полезных вариаций. У Колликера мы не находим разработанной теории, к тому же он иллюстрирует свои воззрения лишь очень немногими примерами, взятыми преимущественно из эмбриологии кишечнополостных, примерами, которые, по его мнению, говорят в пользу выставленной им гипотезы. Для нас имеет значение лишь его основная мысль, что эволюция взрослых животных покоится на изменениях, происходящих в ходе онтогенеза этих животных.
Те же мысли встречаем мы и у Копа.1 Коп, на основании анатомосистематических исследований, пришел к выводу, что новые признаки у животных развиваются в течение эволюции, во-первых, путем естественного отбора, во-вторых, путем «акцелераций и ретардаций» роста изменяющихся животных форм. При этом он принимал, что эволюция родов и еще более высоких групп животных идет независимо от эволюции видов и что она обусловливается именно вышеупомянутыми замедлениями и ускорениями в развитии признаков.
Коп полагал, что признаки высших групп (родов, семейств и т. д.) образуются иначе, чем признаки видов, а именно, аутогенетически, путем изменения хода онтогенеза, т. е. путем ускорений и замедлений в индивидуальном развитии животного. В настоящее время, при современном уровне наших экологических знаний, нам уже невозможно принять этот аутогенетический принцип. Сравнивая признаки видов с признаками родов или с признаками других более высоких систематических групп, мы не видим никакого качественного различия между этими двумя типами признаков, но видим различия в смысле общности и количества: оба эти типа признаков являются, по существу, по своему происхождению признаками приспособительными (ср. главу IV этой книги).
Как мы об этом подробно говорили выше, нам представляется чрезвычайно невероятным, чтобы приспособления животных к различным изменениям внешней среды могли образоваться аутогенетически, т. е. независимо от влияния этой среды. Видовые признаки носят, как мы знаем, тот же характер, что и родовые, и отличаются от этих последних только степенью своей общности и интенсивностью своего развития. Поэтому мы не имеем никаких оснований предполагать эктогенетическое происхождение признаков мелких систематических групп и аутогенетическое происхождение признаков крупных систематических групп. Так как среди признаков мелких систематических групп (видов,, подвидов) встречается больше индиферентных признаков, чем среди признаков крупных систематических групп, приспособительный характер которых больше бросается в глаза (ср. главу IV), то аутогенетическую гипотезу, если уже принимать, то следовало бы принимать скорее для мелких систематических групп, чем для крупных.
Что касается моего личного мнения (как это уже отмечено в главах II — IV, я стою на позиции дарвинизма, защищающего эктогенетическую точку зрения), то мне кажется, что трудно постулировать различные принципы при объяснении образования признаков элементарных и более высоких групп. И во всяком случае, различия между обоими родами признаков должны были бы быть более точно определены, чем это делают Коп и его единомышленники..
1 С о р е Е. D., Essays on the Ciprinoid fishes of Pensylvania, Trans, amer. Philos. Soc., 1866., I d e m., The origin of the fittest, London, 1887, I d e m., Origin of the genera, Proc, of Philadelph. Acad, of sc., 1868.
472
ГЛАВА XIV
Разрабатывая мысль о том, что крупные группы животных развиваются посредством некоторого внутреннего (имманентного) эволюционного принципа, Коп приходит к гипотезе батмизма, носящей идеалистический характер. Согласно этой гипотезе, различные роды одного и того же семейства образуются вследствие воздействия некоторой внутренней направляющей силы, которая (путем акцелераций и ретардаций) изменяет в определенном направлении животные формы. Здесь мы не будем снова останавливаться на разборе гипотезы аутогенетического эволюционного принципа (батмизма), так как она не представляет для нас интереса, но отметим, что у Копа мы встречаем мюллеровскую концепцию относительно роли онтогенеза в филогенезе животных, ибо Коп считает, что фйле-тические изменения взрослых животных обусловлены изменениями, происходящими в ходе их онтогенеза. Являются ли эти изменения акцелера-циями или ретардациями, для нас не имеет существенного значения.
А. Седжвик1 опубликовал в 1910 г. короткую статью, касающуюся взаимоотношений между эмбриональным развитием животных и их эволюцией. Он решительно отвергает биогенетический закон и считает, что эмбриональное развитие не удлиняется от того, что к нему в течение филогенеза постепенно добавляются конечные стадии.1 2 Он принимает, что эволюция признаков взрослых форм происходит только путем изменений общего хода их онтогенеза.3 Ясно, что Седжвик пришел к тем же мыслям, которые были сформулированы Ф. Мюллером еще в 1866 г.: теория Седжвика соответствует так называемым «отклонениям» (девиациям) Ф. Мюллера.4 *
В том же 1910 г. мной была опубликована речь, произнесенная на XII съезде русских естествоиспытателей ц врачей,6 где я в очень сжатой форме впервые высказал свои основные мысли относительно взаимоотношений между филогенезом и онтогенезом животных. Эти же мысли, в виде более или менее разработанной теории, были затем положены в основу отдельной книги, которая вышла из печати в 1912 г.6
Так как мы в последующем будем довольно подробно излагать теорию филэмбриогенеза, то здесь мы ограничимся лишь указаниями на то, что я, так же как и Ф. Мюллер, взгляды которого остались мною в то время незамеченными, пришел к выводам: 1) что эволюция признаков взрослых животных происходит вследствие изменения хода онтогенеза их предков и 2) что биогенетический закон, поскольку он касается рекапитуляции отдельных анцестральных признаков, должен быть признан правильным; иными словами, я считал, что встречается много случаев, когда отдельные признаки предков повторяются в онтогенезе потомков, но что трудно говорить о рекапитуляции целых организмов.
1 Sedgwick A., The influence of Darwin on the Study of animal embryology в книге «Darwin and modern science» ed. by A. Seward, Cambridge, 1910.
2 «Из этого, повидимому, следует, что на заре жизни жизненный цикл должен был быть либо в действительности, либо в потенции,по крайней мере столь же длительным, как и в настоящее время, и что способность живой материи проходить через целый ряд стадий, в течение которых она приобретает все новые и новые свойства, является первичным признаком этой материи». Сэджвик, к счастью, ввел в это положение выражение «в потенции», так как предположение, что докэмбрийская клеточная колония, похожая на колонию Volvox, от которой, по всей вероятности, произошли все Metazoa и,следовательно, позвоночные и млекопитающие, жила столь же долго, как, например, человек, и что ее эмбриональное развитие, как и у человека, длилось девять месяцев, как мне кажется, является слишком смелым.
8 «Факты,повидимому, указывают нам не на то, что некоторая стадия прибавляется К концу жизненного цикла,но на то, что некоторые из стадий этого цикла изменяются».
4 Имя Ф. Мюллера Седжвиком не упоминается.
8 Северцов А. Н., Эмбриология и эволюция, Дневник XII съезда русских естествоиспытателей и врачей, 1910.
"Северцов А. Н., Этюды по теории эволюции. Индивидуальное развитие и эволюция, Киев, 1912; 2-е издание, Берлин, Госиздат, 1922.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
473
А. Нэф в своих методологических работах по систематической морфологии1 также занимается теорией рекапитуляций. Он также считает, что изменения в филогенезе взрослых животных являются результатом изменений, происходящих в онтогенезе предков. Он принимает, что существует модификация трех различных возможностей эмбриональных изменений: 1) когда максимум изменений падает на начальные стадии развития,1 2 2) когда максимум изменений падает на средние стадии развития и 3) когда максимум отклонений падает на конечные стадии развития. При этом различия в ходе развития нарастают с увеличением темпа развития. Отсюда его закон терминального изменения, который он формулирует следующими словами: «Стадии являются тем более консервативными, чем раньше они появляются в онтогенетическом ряду, и тем более прогрессивными, чем позднее они появляются».
Мы видим, таким образом, что по своей основной мысли взгляды Нэфа совпадают со взглядами Ф. Мюллера.
В 1922 г. появилась критическая статья Гарстанга,3 посвященная разбору биогенетического закона, по своему содержанию тесно примыкающая к только что изложенным взглядам Седжвика. Гарстанг считает, что «филогенез является не чем иным, как рядом онтогенезов, идущих по линии филетических модификаций. Филогенез представлен рядом следующих друг за другом взрослых структур, но зиготы последовательных онтогенезов прошли через параллельные модификации своих плазматических и ядерных структур, которые обусловили последовательность онтогенетических изменений формы. Филетическая последовательность взрослых форм является продуктом филетической последовательности онтогенезов. Онтогенез не повторяет филогенеза, но творит его. Жизненный цикл протекает не путем прибавления новых взрослых стадий к конечным старым взрослым стадиям, но путем дальнейшей диференцировки органов или частей органов. Древние признаки взрослых форм (adult characters) элиминируются из их онтогенеза в тех случаях, когда они не являются необходимыми в качестве временной базы для новых признаков. Так как каждый индивидуум, изменяющий свою форму в течение своего жизненного цикла, представляет собой эволюирую-щую и функционирующую единицу,то различные изменения его личиночных или взрослых фаз вызывают координированные изменения в его пассивных или формативных жизненных фазах (эмбриональные стадии, личиночные стадии, стадии метаморфоза). В известных случаях онтогенез имеет тенденцию повторять в нормальных условиях жизни те же последовательные изменения формы, через которые прошли и предки, но он является в то же время способным и к самостоятельным изменениям в каждый период своего жизненного цикла: в положительном направлении—путем развития приспособлений личиночных и взрослых стадий к изменяющимся условиям различного их образа жизни, в отрицательном направлении—путем аббревиации, т. е. ускорения, идущего до полного исчезновения их формативных процессов».
Мы видим, таким образом, что Гарстанг стоит на той же почве, что и Седжвик, и что он в общем не продвинулся в этом направлении вперед по сравнению со своим предшественником.
В. Франц, написавший не так давно обширную сводку филогенеза живот
1 Naef A., Studien zur generellen Morphologie der Mollusken, t. II, Ergebn. u. Forschr. d. Zool., Bd., 3, 1913.
Idem. Die individuelle Entwicklung organischer Formen als Urkundeihrer Stammes-geschichte, Jena, 1917.
2 С p. Северцов A. H., Этюды по теории эволюции, Киев, 1912.
3 Garstang W., The theory of Recapitulation. A critical Restatement of the Biogenetic Law, Journ. of the Linnean Soc. of London, Zoology, 35, 1922.
474
ГЛАВА XIV
ных1 и мнение которого поэтому нам интересно, также стоит на той точке зрения, что филетические изменения строения взрослых форм обусловлены изменениями, имевшими место в течение хода онтогенеза их предков.1 2 Эти филетические изменения онтогенеза происходят следующими четырьмя биометаболическими способами (модусами): 1) путем пролонгации (удлинения), 2) путем аббревиации (укорочения), 3) путем увеличивающейся и 4) на известных стадиях достигающей своего кульминационного пункта девиации (отклонения) онтогенеза. В этих четырех различных случаях наблюдаются следующие различные соотношения между данным организмом и его предком. «В первом случае молодая стадия тождественна взрослой стадии предка (теоретически говоря, в действительности же только подобна, но приблизительно равна по величине); во втором случае взрослая стадия тождественна с молодой стадией предков (теоретически говоря, в действительности, опять таки, только подобна, но приблизительно равна по величине); в третьем случае молодая стадия в первую очередь подобна молодой стадии предков: она всегда более похожа на взрослую стадию предков, чем на собственную взрослую стадию; в четвертом случае молодая стадия непохожа ни на одну из стадий предков, так что взрослое состояние более похоже на предков, чем уклоняющаяся молодая стадия».
В последнее время вопрос о соотношении между онтогенезом и филогенезом был затронут в книге де Беера.3 * В общем точка зрения этого автора довольно близко совпадает с точкой зрения Седжвика и Гарстанга. В своем предисловии де Беер говорит, что в данном случае его побудило к писанию разочарование в ценности биогенетического закона. Он отрицает сдвигание признаков взрослых предков на все более и более ранние стадии эмбрионального развития потомков (т. е. отрицает рекапитуляцию), но принимает, что существует некоторая «репетиция» анцестральных признаков в онгогенезе потомков.1 В сущности, автор является противником не рекапитуляции как таковой, но геккель-вейсмановской теории, полагающей, что признаки взрослых предков сдвигаются на ранние стадии онтогенеза потомков. Он говорит далее о том, что «репетиции» всегда являются признаком родства между предками и потомками, того родства, которое часто ускользает от исследователя при изучении им взрослых форм, и в этом собственно и заключается главное значение эмбриологии для истолкования эволюции.
Мы видим, таким образом, что де Беер признает методологическое значение своих «репетиций», которые, как мы это установили, стоят в центре геккелевского направления филогенетического исследования. Он приходит в 1930 г. к тому взгляду, которого мы придерживаемся с 1910 г.,5
1 Franz V., Geschichte der Organismen, Jena, 1924.
2 Franz V., Zur Kennzeichnung der allgemeinen Entwicklungsrichtungen der Organismenreihe, Ztschr. ind. Abst. Vererbl., 36, 1924.
3 De Beer G. R., Embryology and Evolution, Oxford, 1930.
i «Не существует рекапитуляций в геккелевском смысле этого слова, т. е. ускоренного повторения взрослых стадий, но существуют «репетиции». Признаки, появляющиеся у предков, имеют тенденцию появляться и у потомков, и какие бы стадии предков эти признаки не «представляли», трудно избежать впечатления, что энтузиазм сторонников рекапитуляции основан на этом очевидном факте повторений (репетиций). Если бы только сторонники рекапитуляции перестали утверждать, что то, что повторяется, представляет собой взрослое состояние предков, то у нас не было бы причин не соглашаться с ними».
5 См. Северцов А. Н., Эмбриология и эволюция, Москва, 1910. «Репетиции» признаков де Беера тождественны с моими «рекапитуляциями признаков конечных стадий морфогенеза, весьма сходных с признаками взрослых». Де Беер принимает вместе с К. ф. Бэром, что в онтогенезе постоянно рекапитулируются только эмбриональные признаки. Но это именно было нами сказано уже давно (1910, 1912). Значение рекапитуляций для исследования филогенеза заключается, как это было отмечено выше, в том, что эмбриональные признаки потомков, сходные с признаками взрослых предков, повторяются в онтогенезе потомков. Отметим попутно,
ТЕОРИЯ филэмбриогенеза
475
и мы думаем, что если бы он произвел на основании собственных исследований сравнение результатов палеонтологического и эмбриологического изучения скелетной системы, то он, вероятно, оценил бы еще выше методологическое значение репетиций для изучения филогенеза.
В отношении толкования хода эволюции де Беер стоит на той же точке зрения, что и Ф. Мюллер, Северцов, Седжвик, Нэф и др., т. е. он полагает, что эволюция взрослых животных происходит путем изменения хода онтогенеза их предков («Phylogeny is therefore due to modified ontogeny»). Но при этом он считает, что изменение темпа онтогенеза (т. е. гетерохронии, наблюдаемые в эмбриональном развитии органов) имеет даень большое филогенетическое значение. «Новые признаки могут появиться на всех стадиях онтогенеза, и появление их может быть ускорено или задержано гетерохронией, так что в последующих онтогениях они могут образоваться позднее или раньше. Признаки, существовавшие на ранних стадиях онтогенеза (следовательно, в случаях, когда они не могли быть очень специализованными), играли большую роль при эволюции, идущей путем педоморфоза; они внесли значительные изменения в строение потомков, не нанеся ущерба их пластичности».
Де Беер приходит,, таким образом, к выводу, что крупные систематические группы животного царства (семейства, отряды, классы, даже типы) развивались путем педоморфоза, т. е. путем эмбриональной девиации и неотении. При этом он склоняется к гипотезе Гарстанга, предполагающего, что самые крупные систематические группы (Phyla) развились путем неотении из личиночных форм, т. е., что, например, все хордовые развились из неотенических личинок иглокожих.
Другой тип филогенеза де Беер обозначает именем «геронтоморфоза». Он говорит, что «признаки, которые появляются на поздних стадиях онтогенеза, играют большую роль при эволюции, следующей путем геронтоморфоза» и вносят незначительные изменения в строение эволюирующих форм, потерявших пластичность.
Мелкие группы (расы, подвиды, виды, роды, самое большее—семейства) происходили путем образования вариаций у взрослых через гиперморфоз и акцелерацию.
Из этого, повидимому, следует, что, по де Бееру, существует два различных типа эволюции: один для крупных и другой для мелких систематических единиц, и в этом пункте его теория напоминает собой теорию Копа. С точки зрения де Беера, можно думать, что новый класс образуется не путем постепенного преобразования анцестрального вида в род, рада в семейство и т. д., но таким путем, что представители более примитивного класса, например рептилий, посредством девиации и неотении превращаются в представителей более высокого класса, т. е. в птиц или млекопитающих. Мысль во всяком случае интересная и стоящая того, чтобы для ее проверки произвести соответствующую детальную сравнительно анатомическую и палеонтологическую работу.* 1
что автор, повидимому, не всегда правильно понимает цитируемую им мою работу 1927 г.: он путает понятие «архаллаксиса» (см. de Beer, 1. с., стр. 11) с принципом девиации («Abbiren» Ф. Мюллера); как это читатель увидит ниже, я целиком принимаю принцип девиации и резко отличаю его от «изменений, происходящих в первых закладках, т. е. от архаллаксисов».
1 Можно было бы поставить вопрос, действительно ли главные признаки классов развиваются путем неотении и девиации? Факт, что в признаках крупных групп имеется много повторений анцестральных признаков (по терминологии де Беера—«репетиций», по нашему—«рекапитуляций»), повидимому, указывает на то, что если не все, то несомненно многие признаки крупных групп (например, классов) развиваются путем надставки конечных стадий («overstepping» де Беера), т. е. путем его «геронтоморфоза». Так, мы могли бы сказать, что, например, строение хвоста птицы с сидящими на нем с обеих сторон перьями весьма напоминает собой длинный хвост археоптерикса, а также, может быть, и хвост еще более отдаленного рентилеобразного предка птиц.
476
ГЛАВА XIV
На этом мы закончим наш критический обзор взглядов де Беера; подчеркнем только еще раз, что в основном они совпадают со взглядами Ф. Мюллера (1863) и с моими (1910, 1912, 1927, 1931).
Из рассмотренных нами работ мы видим, что на ряду с направлением Э. Геккеля, который видит в биогенетическом законе только метод исследования (реконструкции) хода филогенеза предков, существует и другое направление, восходящее к Ф. Мюллеру и ставящее своей задачей изучение закономерностей в соотношениях между филогенезом и онтогенезом. Основной мыслью этого последнего направления является положение, что изменение строения взрослых форм (филогенезы в смысле Геккеля) обусловлены изменениями, имеющими место в онтогенезе предков.
Эту точку зрения мы находим в особенности в работах более новых исследователей: Нэфа (1913, 1917), Седжвика (1910), Гарстанга (1922), Франца (1927), Северцова (1910, 1912, 1927), Матвеева (1929, 1930, 1932), де Беера (1930) и Крыжановского. Ее же встречаем мы и в одной из последних работ Э. Наука.1
Этот автор в оценке исследованного им процесса «окольного развития» исходит из той же основной мысли, что и мы, а именно, что «эволюция органов взрослых потомков обусловлена изменениями хода онтогенеза (морфогенеза) этих органов у предков» (стр. 67). Он отмечает при этом, что его взгляд разделяется также О. Бейтом (1911 и 1920) и Некелем (1901), что мною было в свое время упущено из виду. Любопытно, что Ф. Мюллер, являющийся основоположником этой теории, Э. Наукой при этом также не упоминается.
В фактической части своей работы Наук, исходя из собственных наблюдений над развитием различных органов позвоночных животных, исследует процесс окольного развития («umwegige Entwicklung»). Под этим несколько необычным названием он понимает следующее: «Если бы могло случиться так, что в результате диференциации данная индивидуальная форма уже более или менее сформировалась бы и если бы в течение дальнейшего хода онтогенеза появились какие-нибудь привходящие влияния, которые эту форму изменили бы, то могло бы случиться, что эти привходящие воздействия из такой более или менее готовой формы, т. е. формы, похожей на постоянное состояние, сделали бы форму вовсе на это состояние не похожую. Если же затем эти воздействия прекратятся, то первоначально существовавшая форма может снова восстановиться, и тогда начальное и конечное состояние данной формы будут более похожи друг на друга, чем на состояние, находящееся между ними, т. е. на промежуточное состояние данной формы». «Такой окольный процесс развития в действительности имеет место». Автор приводит в дальнейшем целый ряд подробно исследованных случаев, подтверждающих его положение.
Мы не входим в подробное обсуждение результатов, полученных Нау-ком, но не можем не отметить, что дальнейшая разработка заинтересовавших его проблем эмбриональной корреляции может привести к очень интересным выводам.
что эволюция крестца (synsacrum), птичьих ног, скелета крыльев, затылочных позвонков и целого ряда других признаков, характерных для класса птиц и прошедших в своем развитии через рептилеобразные стадии, шла путем анаболии, т. е. надставки конечных стадий, иными словами, путем «геронтоморфоза» де Беера. Из этого следует, что признаки классов могут образоваться путем «overstepping», т. е. путем «геронтоморфоза»;
1 Nauck, ~Е. Th., Uber umwegige Entwicklungsuntersuchungen iiber eine ontogenetische Entwicklungsweise und ihre Beziehungen zur Phylogenese, Morph. Jahrb., Bd. 66, 1931.
ТЕОРИЯ ФИЛ8 МБРИОГЕНЕЗ А
477
Что касается лично меня, то я работал над проблемой закономерностей соотношений между онтогенезом и филогенезом еще с 1908 г.; при этом я пытался, в отличие от большинства перечисленных мною авторов, подойти к разрешению этого вопроса несколько иным путем. Перечисленные мною ученые, каждый по своему, пробовали разрешить проблему соотношений между онтогенезом и филогенезом животных, исходя из критического обсуждения отдельных случаев, взятых ими из огромной эмбриологической литературы. Мне же казалось более целесообразным собрать необходимые для меня факты путем собственных планомерно продуманных сравнительно эмбриологических исследований, проведенных над специально собранным и подходящим для этого материалом.1 При этом я работал сначала над прогрессивным (А. Н. Северцов, 1912 и 1927), а затем над регрессивным (А. Н. Северцов, 1931) развитием органов.
Мне представляется чрезвычайно ценным,что и мои ученики также пошли путем личного исследования. Так, дальнейшее развитие теории филэмбриогенеза имело место в работах Б. С. Матвеева (1929, 1930, 1932) и С. Г. Крыжановского (1933, 1938). Матвеев принимает вместе со мной, что существуют различные типы сохранения анцестральных признаков в онтогенезе потомков и так же, как и я, считает, что при изменении конечных стадий эмбрионального развития, т. е. при анаболии, происходит полная рекапитуляция признаков взрослых предков, что при отклонениях на средних стадиях развития, т. е. при девиации, происходит частичная рекапитуляция (зародыш потомка повторяет зародыш предка) и что при эмбриональных изменениях, происходящих на начальных стадиях развития, т. е. при архаллаксисах, никакой рекапитуляции признаков предков в онтогенезе потомков не происходит.
Интересно, что Матвеев, сопоставляя только что перечисленные модусы филэмбриогенеза с изложенными нами принципами или типами филогенетических изменений органов, указывает на ряд случаев; когда анцестральный признак сохраняется у высших форм в эмбриональном развитии потому, что орган, которому принадлежит данный признак, получил новую функцию, т. е. благодаря смене функций. Такого рода рекапитуляцию он называет «функциональной рекапитуляцией».
Изучая развитие передних позвонков у карповых рыб, Матвеев отметил особые случаи появления анцестральных признаков в онтогенезе потомков, случаи, которые он обозначает как «вторичные рекапц-т у л я ц и и». Причиной появления такого рода рекапитуляций является гетерохрония: при сдвигании времени закладки органа на более ранние стадии развития самый процесс развития идет более полно, вследствие чего в онтогенезе потомков выявляются такие анцестральные признаки, которые обычно выпадают из онтогенеза родственных этим потомкам форм. Примером таких «вторичных рекапитуляций» будет появление у Cyprinidae диплоспондилии в позвонках, связанных с веберовским аппаратом.
В своих более поздних работах (1930, 1932) Матвеев на основании филэмбриологического изучения волос, чешуй и перьев исследует с точки зрения теории филэмбриогенеза различные случаи выпадения эмбриональных стадий и разбирает значение этого выпадения в изменении органов в течение филогенеза. Так, тип развития, который я называю отрицательной анаболией (см. ниже) и который заключается в выпадении конечных, стадий эмбрионального развития, он называет «аббревиацией» и объясняет ею различные случаи неотении, эпистазы и фета-лизации. Случаи выпадения средних стадий развития предков из онтоге
1 Таким же путем работал и Коп (1. с.), но у него была другая цель, чем у меня: для него важно было изучить происхождение родов и крупных систематических групп, теория же соотношений между онтогенезом и филогенезом его не интересовала.
478
ГЛАВА XIV
неза потомков, например случай выпадения хрящевых стадий из онтогенеза костных элементов, что ведет к ускорению развития, Матвеев называет «а к ц е л е р а ц и е й». Матвеев также принимает тот тип отрицательного архаллаксиса, который я называю «отрицательным архаллаксисом через выпадение»; при этом он указывает на случаи, когда при целом ряде наслаивающихся друг на друга надставок конечных стадий, начальные стадии постепенно стираются из онтогенеза потомков.
ФИЛЭМБРИОГЕНЕЗ И ПЕРИОДЫ ЖИЗНИ
Теория филэмбриогенеза пытается разрешить вопрос о том, как происходят и в какой период индивидуальной жизни возникают те изменения органов, которые ведут к филогенетическому преобразованию строения взрослого организма. Она пытается разрешить проблему модусов прогрессивного и регрессивного филогенетического изменения органов взрослых животных. При этом, как мы это уже отметили выше, теория филэмбриогенеза исходит из мысли, что прогрессивное филогенетическое развитие органов взрослых животных происходит путем изменения хода их онтогенеза (Ф. Мюллер). Попытаемся теперь изложить несколько подробнее те заключения,к которым мы пришли по поводу этого вопроса на основании наших собственных исследований.
Но прежде чем перейти к изложению полученных нами результатов, нам необходимо сказать несколько слов относительно периодов жизни многоклеточных животных.
Жизнь Metazoa, как известно, распадается на два главных периода: на период индивидуального развития (онтогенез)1 и на период половой зрелости, т. е. взрослого состояния. Мы знаем, что центр тяжести жизни вида падает именно на взрослое состояние (я не касаюсь здесь явлений педогенеза и смены поколений), ибо именно в течение этого периода появляется и воспитывается молодое поколение, те потомки, которыми поддерживается в течение геологических эпох жизнь вида. Период индивидуального развития, или онтогенеза, рассматриваемый с этой точки зрения, является подготовительным этапом к периоду половой зрелости; он характеризуется крупнейшими изменениями в строении и величине органов, происходящими в организме в это время.
Онтогенез в свою очередь распадается на два периода, а именно на период морфогенеза и на период роста. Первый период начинается для каждого органа в момент его закладки у зародыша или у личинки, кончается же он той стадией развития, когда молодое животное уже приобрело большинство признаков взрослой формы и отличается от этой последней лишь своей незначительной величиной и некоторыми мелкими морфологическими признаками. Период морфогенеза характеризуется весьма значительными преобразованиями формы, морфологического строения и гистологической структуры эмбриональных органов зародыша или личинки и весьма интенсивным, очень часто ди-ференциальным ростом всех частей органов.
Хорошим примером морфогенеза является развитие позвоночника какого-нибудь позвоночного животного, например акулы. Здесь на ранних стадиях развития зародыша из спинной стенки эмбрионального кишечного тракта образуется хорда, первоначально имеющая чисто эпителиальное строение, которое впоследствии постепенно изменяется благодаря вакуолизации клеток хорды; кроме того, за счет эпителия хорды образуются хордовые оболочки. Из перихордальной ткани развиваются диплоспон-
1 Мы расширяем здесь понятие онтогенеза, ибо вводим в него и период молодости.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
479
дильные верхние и нижние дуги и тела позвонков, которые частично вытесняют хордовую ткань. Позднее наступает объизвествление хряща позвоночного столба. На этом заканчивается морфогенез, состоящий из целого ряда морфологических и гистологических изменений формы и строения различных составных частей позвоночника рыб.
Итак, отдельные составные части развивающегося органа образуются из более или менее индиферентных тканей зародыша. При этом мы особенно подчеркиваем, что отдельные органы, а также составные части органов, образуются не одновременно, но в известной закономерной последовательности друг после друга. Так, первой закладывается эпителиальная хорда, затем появляются дуги позвонков, еще позднее закладываются тела позвонков.
Если мы станем рассматривать развитие парных плавников, то увидим, что начало морфогенеза, т. е. образование плавниковых складок, падает на значительно более поздний период общего развития зародыша, чем начало морфогенеза позвоночного столба. Еще более поздно начинается внедрение в эти складки мускульных почек, из которых впоследствии образуется плавниковая мускулатура, и только после всего этого начинает закладываться хрящевой скелет плавника.
Плакоидные чешуи акул закладываются на еще более поздних стадиях развития, когда морфогенез большинства органов уже заканчивается. Такую же позднюю закладку наблюдаем мы и при образовании волос у млекопитающих, контурных перьев у птиц, рогов у млекопитающих и многих других органов.
Отметим, что во всех случаях, когда мы будем говорить о конце периода морфогенеза, мы должны будем иметь в виду, что речь идет о морфогенезе того или иного органа, но отнюдь не об общем состоянии всех остальных органов.
В течение второго периода онтогенеза, т. е. в период роста, форма и строение различных частей тела молодого животного изменяются обычно очень мало, зато продолжается интенсивно рост, вследствие чего к концу этого периода молодыми животными достигается нормальная, типичная для половозрелой формы величина. В течение периода роста большинство частей тела животного растет гармонично, ив этом лежит момент существенного отличия периода роста от периода морфогенеза.
Взрослое состояние или период половой зрелости, отличается, по сравнению с обоими предшествующими периодами (периодом морфогенеза и периодом роста), своим относительным постоянством, развитием половых органов и вторичных половых признаков. В течение этого периода строение взрослого животного изменяется в общем очень мало: обычно животное более не растет или, если и растет в некоторых отдельных случаях, то лишь очень медленно.
Итак, мы различаем следующие периоды жизни:
I. Период индивидуального развития, или онтогенеза, который распадается на:
1. Период морфогенеза, характеризующийся: а) очень резкими изменениями в форме и строении органов и Ь) очень интенсивным, часто диференциальным ростом.
2. Период роста, 'характеризующийся: а) немногими и незначительными морфогенетическими изменениями и Ь) более или менее гармоничным увеличением размеров, т. е. интенсивным ростом отдельных частей тела животного.
II. Период половой зрелости, или взрослое состояние, характеризующееся: а) отсутствием морфогенетических изменений, Ь) отсутствием роста или крайне медленным ростом и с) размножением вида половым или бесполым путем.
480
ГЛАВА XIV
После этих предварительных замечаний перейдем к изложению теории филэмбриогенеза.
ФИЛЭМБРИОГЕНЕЗ И СПОСОБЫ (МОДУСЫ), КОТОРЫМИ ОН ОСУЩЕСТВЛЯЕТСЯ
Основная мысль теории филэмбриогенеза, как мы об этом уже упоминали выше, заключается в том, что эволюционные изменения состоят в изменении хода онтогенетического развития, т. е. в изменении тех процессов, совокупность которых образует собою онтогенез многоклеточных животных. Мы знаем, что таких процессов в основном имеется четыре: 1) общий рост, 2) частный или неравномерный рост, 3) гистогенез и 4) странствование или переселение клеток.
Чтобы разобрать каждый из этих процессов в отдельности, нам придется коснуться самым кратким образом вопроса о том, как происходит сам онтогенез.
Известно, что эмбриональное развитие (онтогенез) всякого многоклеточного животного начинается с размножения дериватов оплодотворенной яйцеклетки посредством дихотомического деления бластомеров, которое сначала идет равномерно и гармонично, причем число клеток умножается в геометрической прогрессии со знаменателем 2 по формуле 1, 2, 4, 8, 16 и т. д. (общий рос т). При этом клетки остаются индиферентными и образуют шаровидный клеточный комплекс (зародыш).
Несколько позднее темп деления некоторых клеток ускоряется, т. е. одни клетки начинают делиться быстрее других, и таким путем образуются зачатки эмбриональных органов, еще состоящих из индиферентных клеток. У зародышей позвоночных животных таким образом развивается выпячивание первичного кишечного канала, зачаток хорды и мезодермы, нервная трубка и т. д. Здесь мы имеем перед собою процесс частного или неравномерного роста сначала бластомеров, а позднее— отдельных зачатков зародыша.
Итак, мы можем сказать, что при всяком процессе развития происходит: 1) общий или равномерный рост, т. е. увеличение общего числа клеток всего зародыша и 2) частный или неравномерный рост, т. е. рост отдельных зачатков органа, иначе говоря, увеличение числа клеток этих зачатков.
Для нас особенное значение имеет этот последний (частный) рост зачатков органов, приводящий к диференцировке и изменению формы и к усложнению строения многоклеточного организма. Отметим, что в зародыше одного и того же животного этот частный неравномерный рост происходит в разных зачатках с разной скоростью и что начало развития зачатков имеет место на различных стадиях общего развития организма: одни зачатки образуются раньше, другие позднее.
Мы говорили до сих пор о зачатках органов, состоящих из индиферентных, т. е. эмбриологически подобных оосперму клеток. На более поздних стадиях развития начинается процесс гистогенеза, т. е. гистологическая специализация клеток различных зачатков зародыша. Отдель-,ные группы клеток каждого зачатка начинают неравномерно расти (удлинение клеток, образование отростков) и в них появляются специальные органеллы, т. е. мускульные волоконца, нервные волокна; мезенхимные клетки скелетных зачатков выделяют межклеточное вещество и т. д. Эти гистогенетические изменения происходят в каждой клетке и захватывают одновременно большое число клеток данного зачатка. Так, клетки мышечного сегмента все вытягиваются в длину и в них развиваются мускульные волоконца, клетки спинного и головного мозга разделяются на чисто нервные клетки с плазматическими отростками и осевым цилиндром и на опорные клетки ит. д. После этой морфологической (рост, изменение формы клеток) и гистогенетической (образование .межклеточного
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
481
вещества, волоконец внутри клетки и т. п.) диференцировки клетки становятся способными выполнять свои специальные функции внутри тела животного: опорную и защитную функцию в скелетных клетках, сократительную функцию в мускульных, нервную функцию в мозгу и т. д.
Мы видим, таким образом, что нормальное эмбриональное развитие многоклеточного организма состоит из трех главных процессов.
1. Общий рост, или равномерное размножение клеток оосперма. Этот процесс в некоторых органах (кожа, скелетная ткань) продолжается до взрослого состояния, а иногда и всю жизнь, в других органах (центральная нервная система) он заканчивается в течение эмбрионального периода жизни.
2. Частный, или неравномерный рост отдельных групп клеток зародыша, который приводит к развитию отдельных зачатков органа, т. е. к морфологической диференцировке организма.
3. Г и с т о г е н е з, или гистологическая диференцировка организма.
К этим трем процессам онтогенетического развития прибавляется на более поздних стадиях еще четвертый процесс, а именно:
4 Процесс странствования и переселения клеток, когда группы клеток, переместившись на новое место в теле зародыша, дают начало новому зачатку и претерпевают в конце своего развития крупные гистологические изменения.
Хотя этот последний процесс встречается очень часто и является, наравне с общим и частным (неравномерным) ростом дериватов яйцеклетки и гистогенезом, одним из самых распространенных способов филэмбриогенеза, однако он не настолько известен, как упомянутые выше первые три способа, и нуждается поэтому в некотором дальнейшем пояснении.
Непосредственно наблюдать процесс переселения клеток мне удалось при изучении развития зачатков мускулатуры в передних конечностях Lacertilia.1 Мускулатура свободных конечностей, как известно, образуется путем активного выселения клеток из так называемых мускульных почек вентральных отростков миотомов, лежащих в области свободной конечности. При этом из каждой мускульной почки в зачаток свободной конечности направляются два потока мезенхимных клеток: дорзальный, из которого образуется зачаток дорзальной мускулатуры свободной конечности, и вентральный, дающий начало ее вентральному зачатку. Рис. 156 показывает нам одну из стадий этого процесса. Мы видим здесь, как из девятой дорзальной почки (рис. 156, ms9) выселяется густой поток клеток, идущих в зачаток передней конечности и дающих зачаток дорзальной мускулатуры (рис. 156, dm) этой конечности.
Нам представляется чрезвычайно вероятным, что своеобразный процесс эмбрионального развития, изученный Б. С. Матвеевым при исследовании им развития скелетных элементов (позвонков рыб и конечностей млекопитающих) и названный им «способом заимствования частей»,1 2
1 Sewertzoff A. N., Studien liber die Entwicklung der Nerven, Muskeln u. des Skeletes der Extremitaten der Tetrapoda, Бюлл. о-ва испытателей природы, Москва, 1908.
2 Б. С. Матвеев в своей работе «О принципе заимствования при развитии органов» (Бюлл. Научно-исслед. института зоологии МГУ, 1935) утверждает, что: «в ряду гомо-динамных органов существует общая для всего ряда закономерность развития. В случае появления новообразования в отдельных сегментах материал для этих новообразований заимствуется в течение эмбрионального развития из соседних частей. В случае же появления какого-нибудь нового этапа в морфогенезе гомодинамного ряда органов процесс диференциации в той или иной степени захватывает весь ряд сериальных органов».
Примером процесса заимствования частей для Матвеева являются изменения, происходящие в передних туловищных позвонках у костистых рыб, имеющих так называемый веберов аппарат. У мальков таких рыб над передними позвонками развивается хрящевая крыша, защищающая веберов аппарат от механических повреждений и со-Морфологические закономерности 31
482
ГЛАВА XI
процесс, о котором он умозаключал лишь косвенно, осуществляется именно путем указанного переселения или странствования клеток.
Все перечисленные нами изменения, не регулирующиеся впоследствии, всегда отражаются на строении взрослых животных, происходящих из зародышей, претерпевших такие изменения. Эти изменения в числе клеток зародыша и в форме зачатков его органов, приводящие к изменению величины и формы взрослых органов, т. е. филогенетические изменения
Рис. 156. Поперечный разрез через заднюю часть зачатка передней конечности молодого Ascalabotes fascicularis (при большом увеличении), иллюстрирующий процесс «переселения клеток».
msg—девятая дорзальная мускульная почка, из которой выселяется поток клеток, направляющийся в зачаток передней конечности, где эти клетки образуют зачаток дорзальной мускулатуры конечности (dm); еп—нерв конечности; mes—мезенхимная ткань (из А. Н. Северцова, 1908).
трясений. При исследовании развития этого образования Матвеев нашел, что верхние и нижние дуги I и II позвонков, лежащие под этой крышей, по объему значительно меньше, чем дуги III, IV и следующих позвонков. Это навело его на мысль, что образование хрящевой крыши произошло за счет мезенхимного материала, заимствованного из лежащих под нею дуг позвоночника. Для проверки этого предположения он подсчитал у мальков костистых рыб на разрезах, взятых из этой области, объем верхних дуг первых и последующих позвонков и крыши веберова аппарата. При этом оказалось, что объем крыши и лежащих под ней редуцированных дуг приблизительно равен объему нормальных позвонков, что является подтверждением гипотезы заимствования.
Затем Матвеев изучал способ редукции боковых пальцев конечности парнопалых копытных; при этом оказалось, что редукция в этом случае происходит путем заимствования скелетогенного материала от боковых пальцев (II и V) двумя средними пальцами (III и IV), развивающимися, как оказывается, прогрессивно именно за счет краевых пальцев.
К сходным результатам пришли, независимо от Матвеева, Боголюбский (О типах хвостов овец и их развитии, Тр. ла5. эвол. морфол. Акад, наук СССР, 1934) и Шмальгаузен (Розвиток деяких расовихознак укурей, Труды Инет. зоол. и биол. Ак. Наук УССР, т. 1, 1934). Боголюбский описывает явление заимствования клеток при развитии хвоста у овец, Шмальгаузен отмечает еще более интересный случай у мохноногих кур мощные зачатки перьев при развитии заимствуют мезодермальные клетки из закладок скелета.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
483'
хода онтогенеза, происходят на различных стадиях индивидуального развития животного. Прежде всего необходимо отметить изменения, которые происходят на поздних стадиях развития, в конце периода морфогенеза или в начале периода роста. При этом онтогенетическое развитие данного зачатка удлиняется. Этот способ филогенетического изменения органов мы называем способом надставки стадий, илианабо лие й. Если изменения происходят на средних стадиях эмбрионального развития, мы их называем девиациями хода онтогенеза. И, наконец, если они происходят в самом начале эмбрионального развития органа и изменяют весь ход эмбриогенеза и строение взрослых органов, мы называем их начальными изменениями, или архаллаксисами. Эти три способа положительных приспособительных изменений органов приводят к увеличению (филогенетическому росту) органов или к их прогрессивной адаптации.
Трем способам положительного изменения филогенеза соответствуют три способа отрицательных филогенетических изменений хода онтогенеза, а именно: отрицательная анаболия (уменьшение величины зачатков на поздних стадиях), отрицательная девиация (уменьшение величины зачатков на средних стадиях эмбрионального развития) и- отрицательный архаллаксис (уменьшение величины зачатков на ранних стадиях эмбрионального развития). Продолжаясь в течение многих поколений, эти отрицательные изменения хода онтогенеза, как мы это увидим ниже, приводят к редукции органов у взрослых животных и, в конце концов, к их полному исчезновению.
Кроме перечисленных способов (модусов) филогенетического изменения хода онтогенеза, мы имеем еще и другие способы, а именно, изменение самого темпа онтогенетического развития органов. Так, у некоторых форм зачатки определенных органов закладываются у потомков раньше, чем у предков, у других форм они закладываются позже, чем у предков. И те и другие изменения отражаются на коррелятивных соотношениях данных органов с соседними органами, точно так же как они отражаются и на строении взрослых органов и на времени, когда они начинают функционировать.
Наконец, мы должны отметить еще один способ филогенетического изменения хода онтогенеза, а именно способ периодического изменения хода эмбрионального развития органов.
Разберем теперь все эти способы (модусы) филэмбриогенеза по возможности подробнее и приведем конкретные примеры хода эволюционного процесса.
НАДСТАВКА КОНЕЧНЫХ СТАДИЙ РАЗВИТИЯ ИЛИ АНАБОЛИЯ1
Мы остановимся прежде всего на изложении одного из наиболее распространенных модусов филэмбриогенеза, а именно на модусе анаболии, т. е. того способа филетического изменения органов, который происходит путем надставки конечных стадий онтогенеза.
В качестве типичного примера анаболии разберем развитие экспес-сивных органов позвоночных животных и остановимся в особенности на развитии эксцессивных органов у костистых рыб.
Изучая сравнительно эмбриологическим путем эксцессивные органы рыб, мы нашли, что они в своем развитии никогда не останавливаются на той стадии онтогенеза, на которой обычно завершается развитие соответ-
1 От ’алгафоЛт)—удлинение.
31»
484
ГЛАВА XIV
Рис. 157. Belone acus—взрослая форма.
На рисунке видно, что верхняя (о) и нижняя (и) челюсти сильно вытянуты в длину, а—анальный плавник: ап—анальное отверстие; с—хвостовой плавник; d2—задний спинной плавник; п—носовая ямка; net—грудной плавник; г. Ьг.—radii membranae branchiostegiae; vt—брюшной плавник (из А. Н. Северцова, 1927).
На рисунке видно, что нижняя челюсть (U) вытянута в длину так же, как и у Belone, и что верхняя челюсть (О) значительно короче нижней. N—носовая ямка (из А. Н. Северцова, 1927).
Рис. 159. Belone acus. Малек 10 мм длины.
а --анальный плавник; ап—анальное отверстие; с—хвостовой плавник; d2—задний спинной плавник; п—носовая ямка; о—верхняя челюсть; ра.—преданальный плавник; pct—грудной плавник;
и—нижняя челюсть (из А. Н. Северцова, 1927).
Рис. 160. Belone acus. Малек 21 мм длины.
и—носовая ямка; о—верхняя челюсть; os—ротовое отверстие; pct—грудной плавник; и—нижняя челюсть (из А. Н. Северцова, 1927).
/7
Рис, 161. Belone acus. Малек 9,1 см длины.
те же, что на рис. 157, 159 и 160, (из А. Н. Северцова 1927).
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
485
ствующих органов у их ближайших сородичей и предков, но что они всегда развиваются далее в том же или в несколько ином направлении, т. е. что здесь имеет место известная надставка стадий морфогенеза.
Возьмем, например, развитие Belone acus. Как известно, Belone отличается от своих сородичей строением челюстного аппарата (рис. 157).
Рис. 162. Atherina hepsetus. Малек 12 мм длины.
а—анальный плавник; ап—анальное отверстие; с—хвостовой плавник; d,—d2—зачаток дорзального плавника; п—носовая ямка; os—ротовое отверстие; pct—грудной плавник; v—вентральная плавниковая складка (из А. Н. Северцова, 1927).
У нее как верхняя, так и нижняя челюсть сильно вытянуты в длину, так что обе они вместе образуют длинное прямое рыло. У Hemiramphus
Рис. 163. Взрослая Atherina hepsetus.
Обозначения те же, что на рис. 162. Кроме того: ор—жаберная крышка; vt—брюшной плавник (из А. Н. Северцова, 1927).
(рис. 158) верхняя челюсть коротка, а нижняя вытянута, каку Belone. Изучая историю развития Belone, я нашел, что у эмбрионов в 10 мм длины еще не удлинены ни верхняя, ни нижняя челюсть (рис. 159); вернее говоря, нижняя челюсть лишь слегка выступает вперед по сравнению с верхней челюстью. Это состояние у большинства других Scombresocidae сохраняется также и у взрослых животных, но у Belone, наблюдая его дальнейшее развитие, мы видим, что нижняя челюсть начинает сильно расти в длину, так что у малька в 21 мм длиною она уже во много раз превосходит верхнюю челюсть (рис. 160). Еще позднее, т. е. у мальков в 9,1 см длиною, начинает удлиняться и верхняя челюсть, но она растет гораздо медленнее, чем нижняя челюсть (рис. 161). Аналогичное состояние, т. е.',сравнительно короткую верхнюю челюсть и сильно удлиненную нижнюю челюсть, мы находим у взрослого Hemiramphus (ср. рис. 158).
Мы видим, таким образом, что мальки Belone в отношении развития своих челюстей отличаются от большинства Percesoces: эти последние
486
ГЛАВА XIV
остаются и во взрослом состоянии на той стадии развития Belone, которая нами изображена на рис. 159; у Belone же в течение ее эмбрионального развития прибавляются, по крайней мере, еще две новые стадии, а именно, стадия Hemiramphus (рис. 161) и стадия взрослой Belone (рис. 157).
Для наглядного сравнения развития экцессивного органа у Belone с ходом нормального развития того же органа у какого-нибудь типичного представителя семейства Percesoces, например, у Atherina hepsetus, мы приводим следующую схему (схема XXIX), где символически изображено развитие верхней и нижней челюсти у. Belone и у Atherina hepsetus (см. рис. 162 и 163).
Atherina: верхняя челюсть: ах а2 а3 а4 . . . . А
нижняя челюсть: Ь4 Ь2 Ь3 Ь4 . . . . В
Belone: стадия I верхняя челюсть: а4 а2 а3 а4
нижняя челюсть: bt b2 bs b4
Belone: стадия II верхняя челюсть: а, а2 а3 а4
нижняя челюсть: b, b2 ba b4 b5 Ь,
Beione: стадия III верхняя челюсть: а, а2 а3 а4 as as
нижняя челюсть: bj b2 b3 b4 b5 Ьв Ь, Ь8
Belone: стадия IV верхняя челюсть: а, а2 а3 а4 а5 а3 а, а8 а9... .А'
нижняя челюсть: bx b2 b3 b4 b5 Ь3 Ь, Ь8 Ь9 .... В'
Схема XXIX. Объяснение в тексте.
Буквы ах, а2, а3, а4 символически изображают следующие друг за другом стадии морфогенеза верхней челюсти, буквы Ьх, Ь2, Ь3, Ь4—стадии развития нижней челюсти. У Atherina hepsetus последние стадии развития, г. е. стадии а4 и Ь4, сходны со взрослым состоянием (А, В) и отличаются
Рис. 164. Lophius piscatorius. Малек S мм длины.
с.—хвостовой отдел плавниковой складки; сЦ—d2—части плавниковой складки, соответствующие переднему (di—d°i) и заднему (d2) спинному плавнику; 115 1‘8 , 23—личиночные нитеобразные выросты брюшного плавника; md—нижняя челюсть; тх—верхняя челюсть; pct—грудной плавник; гх—г5— лучи переднего спинного плавника; v—вентральная плавниковая складка; vt—брюшной плавник (из А. Н, Северцова, 1927).
от этого последнего лишь своей малой величиной. У Belone мальки на самой молодой стадии, т. е. на стадии I (ах, а2, а3, а4, Ьх, Ь2, Ь3, Ь4), напоминают собой мальков Atherina на тех же стадиях развития (рис. 162), но затем развитие челюстей у Belone удлиняется на несколько стадий: в стадии II у Belone развитие нижней челюсти удлиняется на две стадии: на
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
487
be и Ьв; в стадии III развитие нижней челюсти удлиняется еще на две стадии—на Ь7 и Ь8; при этом начинает удлиняться и верхняя челюсть: к ней также прибавляются две стадии: а5 и ав; в стадии IV развитие верхней челюсти Belone снова удлиняется на три стадии: на а7, а8, а9—А', а развитие нижней—на стадию Ь9—В'.
Мы обращаем особое внимание читателя на то, что эти удлинения (надставки стадий) относятся только к челюстям Belone и что остальные части ея тела развиваются гармонично, т. е. так же, например, как они развиваются у Atherina hepsetus. .
В качестве другого примера анаболии, т. е. развития путем надставок конечных стадий, можно привести морфогенез некоторых органов морского чорта (Lophius piscatorius), в особенности его грудных плавников.
Как известно, у костистых рыб грудные плавники имеют в начале своего развития горизонтальное положение (т. е. такое же, как и у взрослых
Рис. 165. Малек Lophius piscatorius 12 мм длины. А—основание грудного плавника и fissura branchialis (f. br.) несколько более молодого малька Lophius piscatorius.
а—анальный плавник; ап.—анальное отверстие; Ьг.—membrana branchiostegia; с—хвостовой плавник; <1,—d\—передний спинной плавник; d2—задний спинной плавник; Л Ьг.—бранхиальная щель; Г—боковая поверхность грудного плавника; m—вентральная поверхность грудного плавника; md—нижняя челюсть; mx—верхняя челюсть; п—наружные носовые отверстия; ор—operculum; pct— грудной плавник; —передний свободный луч переднего спинного плавника; г.»—гв—остальные лучи переднего спинного плавника; го—костные лучи переднего спинного плавника; vt—брюшной плавник (из А. Н. Северцова, 1927).
акул), так что первично вентральная поверхность плавника обращена книзу. Позднее происходит изменение положения этих плавников, а именно, первично вентральная поверхность плавника поворачивается наружу, т. е. делается боковой поверхностью, а первично дорзальная поверхность плавника поворачивается внутрь и прилегает к боковой стенке тела. Это положение грудных плавников является типичным для большинства взрослых костистых рыб (рис. 163 и 157), оно типично также и для молодых мальков морского чорта (рис. 164 и 165). Но у большинства костистых рыб
488
ГЛАВ'А XIV
передние конечности остаются в этом положении в течение всей жизни особи, у морского же чорта изменение в положении плавников идет дальше: грудные плавники поворачиваются своим дорзальным краем вентрально (рис. 166 А и 166 В), так что они вторично принимают то же положение, что и у акул (рис. 167 А, 167 В). Это изменение положения грудного плавника видно еще яснее на рисунках 168 А и В, на которых изображены спереди молодой малек (рис. 168 А) и взрослый Lophius (рис. 168 В). Пунктирной линией (II, рис. 168 А) изображено положение, которое занимали эти плавники на стадии, иллюстрируемой рис. 166.
Рис. 166. А—малек Lophius piscatorius 3.7 см длины; В— та же личинка с брюшной стороны.
Обозначения те же, что и на рис. 165. Кроме того: 1'—боковая поверхность грудного плавника; пт—медиальная поверхность грудного плавника; s—верхний край грудного плавника; pl.—плавниковая лопасть (из А. Н. Северцова, 1927).
Таким образом, мы видим, что развитие плавников морского чорта не остановилось, как это бывает у других костистых рыб, на той стадии развития, когда плавники имеют горизонтальное положение (ср. рис. 165 с рис. 157 и 163), но что здесь к этой конечной стадии развития остальных Teleostei прибавилась еще новая стадия, а именно стадия вентрального поворота грудных плавников (надставка конечной стадии).
Итак, у большинства Teleostei (Belone, Atherina и т. д.) грудные плавники проходят в своем развитии через следующие стадии:
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
489
1. Горизонтальное положение (вентральная поверхность плавника обращена книзу).
2. Вертикальное положение (первично вентральная поверхность плавника обращена вбок).
У Lophius piscatorius мы наблюдаем три стадии:
1. Горизонтальное положение (вентральная поверхность плавника обращена книзу.
2. Вертикальное положение (первично вентральная поверхность плавника обращена вбок).
А
Рис. 167. А—Lophius piscatorius. Взрослый экземпляр. Вид сбоку. В—тот же экземпляр Lophius piscatorius. Вид снизу.
Обозначения те же9 что и на рис. 165 и 166.
На рисунке видно, что та поверхность грудного «
плавника, которая на предыдущей стадии была боковой поверхностью (1', рис. 166), стала теперь ”
вентральной поверхностью плавника, а та поверхность грудного плавника, которая па предыдущей стадии была медиальной поверхностью плавника (т',рис. 166),стала теперь его дорзальной поверхностью (из А. Н. Северцова, 1927).
3. Вторично горизонтальное положение (первично вентральная поверхность плавника вторично обращена книзу).
Весьма интересны также соотношения между грудными плавниками и жаберной крышкой у того же Lophius. На ранних стадиях развития основание плавников лежит непосредственно позади заднего края жаберной крышки (membrana branchiostegia), так что оно до известной степени покрыто этой последней (рис. 164). Оперкулярное отверстие представляет собою, как и у большинства костистых рыб, заметную щель. На несколько более поздней стадии развития membrana branchiostegia начинает срастаться (снизу вверх) с боковой стенкой тела (рис. 165 А)„
490
ГЛАВА XIV
так что оперкулярное отверстие суживается, и, 'Наконец, принимает вид маленькой щели, охватываемой основанием плавника (рис. 165, 166, 167, f. br.). Мы видим, что здесь к типичному ходу морфогенеза оперкулярной щели, характерному для большинства костистых рыб, прибавилась еще одна новая стадия развития, а именно стадия частичного
срастания.
Наконец, если мы возьмем еще один признак Lophius, а именно форму его тела, то.увидим, что и здесь в течение онтогенеза имеет место надставка конечных стадий. Так, у мальков морского чорта, как и у большинства нектонных костистых рыб, голова сжата с боков ( рис. 164, 165, 166), так
oct
pct
что у них дорзо-вентральный’ поперечник тела длиннее, чем латеральный (справа налево). В течение дальнейшего развития латеральный поперечник становится большим, чем дорзо-вентральный, и вся фор-
Рис. 168. А—личинка Lophius piscatorius на той же стадии развития, что и на рис. 165. Вид спереди. Пунктирной линией нанесено положение грудных плавников в начальной стадии их поворота (ср. рис. 166). В—Lophius piscatorius, взрослый экземпляр. Вид спереди.
Обозначения те же, что и на предыдущих рисунках. Кроме того: ос—глаза, I—положение грудных плавников на ранней стадии развития; 1'—боковая поверхность этого плавника; II—положение грудных плавников в начале поворота; 1"—боковая поверхность плавника загибается вниз. У взрослого Lophius боковая поверхность грудного плавника повернулась вниз (1") и превратилась в вентральную поверхность плавника (из А. Н. Северцова, 1927).
ма передней части тела становится другая: она становится плоской и широкой, вследствие приспособления к жизни на дне (рис. 167,168). Таким образом, к характерной для большинства пелагических форм конечной стадии морфогенеза (передняя часть тела не широкая, но высокая) прибавляется новая стадия, а именно стадия расширения и уплощения передней части тела.
Как известно, для Centriscus nScombresox является характерным сильное удлинение передней части головы, а именно ее преназального отдела. Мы находим, что у молодых мальков этих рыб, которые, в общем, имеют уже признаки взрослой особи, преназальный отдел головы еще короток; разрастание преназальной части происходит у них в течение периода роста.
То же самое можем мы сказать и относительно своеобразной формы и положения головы у Lbphobranchii, например у Syngnathus и у Hippocampus. У ранних зародышей морского конька голова является непосредственным продолжением туловища; преназальный отдел головы (рис. 169 ра) у них очень короток, так что вся голова зародыша, как это видно на рис. 169, принимает мопсообразный вид. У более поздних личинок Hippocampus преназальный отдел сильно разрастается в длину, а голова сгибается книзу. Вследствие этого получается своеобразное сходство
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
491
с головой лошади, которое так бросается в глаза у взрослой формы (рис. 170)
и которому морской конек обязан своим названием.
Таким образом, мы видим, что у Lophobranchu, как и в предыдущих случаях, к конечной стадии развития, характерной для морфогенеза
Рис. 169. Малек Hippocampus на очень ранней стадии развития.А—его хвостовой плавник в несколько увеличенном виде, а—анальный плавник; с—хвостовой плавник; d—
большинства костистых рыб, прибавляются две новые стадии, а именно: 1)стадия удлинения преназального отдела головы и 2) стадия резкого вентрально направленного сгибания отдела тела, лежащего позади грудных плавников.
Интересный пример надставки конечных стадий (анаболии) дает нам развитие грудных плавников у триглы (рис. 171, 172, 173). Как известно, у взрослой триглы три нижних краевых луча грудных плавников превратились в свободные членистые пальцеобразные придатки, при помощи которых
Рис. 170. Hippocampus guttulatus, взрослый экземпляр (из А. Н. Северцова, 1927).
спинной плавник; ра—преназальный отдел передней части головы; pct — грудной плавник; ш— борозда, отделяю.щая верхнюю челюсть от лобной части головы; и—нижняя челюсть (из А. Н.
Северцова, 1927).
эти рыбы ползают по дну моря. Уже у довольно поздних мальков триглы, как это видно на рис. 171, все лучи грудного плавника еще соединены между собой, так же как и у других костистых рыб, общей перепонкой. У еще более поздних мальков часть этой перепонки, находящаяся между только что упомянутыми тремя нижними лучами, начинает все более и более отставать в росте, так что концы этих трех лучей свободно выдаются вперед (рис. 172); этот процесс продолжается в течение всегс периода роста,
492
ГЛАВА XIV
в результате чего образуются три свободно подвижных, несвязанных между собой перепонкой плавниковых луча (рис. 173). При этом, так же как и у морского чорта, происходит поворот этих трех плавниковых лучей, правда, в сторону, противоположную той, в которую поворачивались плавники у Lophius. Таким
образом, и здесь также имеет место надставка конечных стадий морфогенеза.
В предыдущем мы привели целый ряд примеров анаболии, причем надставка конечных стадий касалась лишь развития внешних признаков, или, вернее,комплексов внешних признаков у костистых рыб, и мы легко могли бы умножить число этих примеров. Мы находим их также и при изучении развития внутре н -них органов.Напомним здесь хотя бы о много-
Рис. 171. Передняя часть тела довольно крупного малька триглы (Trigla nigripunctata), у которого все лучи грудного рлавника еще соединены между собой общей перепонкой.
pct.—грудной плавник; vt.—брюшной плавник (из А. Н. Северцова, 1927).
численных случаях срастания различных скелетных элементов у позвоночных животных.
Так, указанное нами выше различие в строении tarsus какой-нибудь примитивной формы рептилий, например Procolophon, Ophiocodon minis (рис. 149), и tarsus какой-ни-будь ящерицы, например Бите- /С, \\\\
cesschneideri(pnc. 194 В), заключается в том, что к стадии развития конечностей, через которые прошли предки ящериц и которые привели к образованию самостоятельных tarsalia proximalia (astragalus, calcaneus, cen-trale), прибавилась новая стадия развития, а именно стадия срастания этих элементов. С другой стороны, у предков ящериц tarsalia distalia I и II были самостоятельными элемен
Рис. 172. Передняя часть тела еще более крупного малька Trigla nigripunctata, у которого уже наметилось отделение трех плавниковых лучей (rd,—rd3) грудного плавника от общей перепонки.
pct—грудной плавник; rd,, rd2, rd:;—три передних плавниковых луча; vt—брюшной плавник (из А. Н. Северцова, 1927).
тами, такими, какими они были у Stereosternum tumidum, Proco-lopohon trigoniceps и Ophiocodon minis (рис. 149). У современных ящериц к их эмбриональному развитию прибавилась еще одна новая стадия, а именно,
стадия срастания: tarsalia distalia I и II срастаются с проксимальными концами metatarsalia I и II.
Изучая эволюцию гистологических признаков, мы и здесь находим различные случаи надставок конечных стадий. Так, при образовании первых окостенений хрящевого скелета скелетные
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
493
части, состоящие первоначально из хряща, например дуги позвонков у Dipneusta или длинные кости конечностей у амфибий, начинают впоследствии обхватываться на свбей поверхности тонкой костной оболочкой, которая постепенно становится все толще и толще и, наконец, вытесняет весь хрящ.
Мы видим, таким образом, что под понятием надставки конечных стадий, или анаболии, подразумевается не только «дальнейший шаг» в общем ходе эволюции (Weiterschreiten, по выражению Ф. Мюллера или overstepping, по выражению де Беера). Правда, в некоторых случаях эта надставка заключается лишь в продолжающемся в определенном направлении росте, зато в других случаях мы находим, что орган, уже достигший взрослого состояния предков, развивается дальше не в том направлении, в каком он развивался у предков, а вступает на новый путь эволюции. Это именно мы находим при изучении поворотов грудных плавников у морского чорта,
Рис. 173. Взрослая тригла (Trigla nigripunctata), у которой завершилось преобразование трех передних лучей грудного плавника в свободные пальцеобразные придатки этого плавника.
pct—грудной плавник; rdlt rda, rd3—три свободных плавниковых луча (из А. Н. Северцова, 1927).
при сгибании головы у морского конька, при развитии трех нижних свободных лучей грудных плавников у триглы, при срастании элементов плюсны у рептилий, при надставках новых гистологических структур и т. д.
Мы пришли, таким образом, к выводу, что во всех приведенных нами случаях, как и во многих других, филогенетическое изменение органов заключается в том, что в конце периода морфогенеза к конечным стадиям развития эволюирующего органа (который в это время уже вполне подобен взрослому органу предков и отличается от него лишь своей незначительной величиной) прибавляются новые стадии онтогенеза. «Новизна» эта в некоторых случаях заключается лишь в наступлении более интенсивного роста (в относительном увеличении размеров органа), в других случаях—в изменении онтогенетического направления развития органа (изменение формы или направления роста, изменение гистологической структуры, срастание отдельных элементов, разделение цельного зачатка на отдельные отделы и т. д.).
Мы подчеркиваем, что, по нашему мнению, эти новые надставки конечных стадий образуются не внезапно, не «скачками», как это думал Кол
494
ГЛАВА XIV
ликер, но путем суммирования мелких нерегулирующихся эмбриональных вариаций. Схематически, взяв в качестве примера эволюцию челюстей у Belone, мы можем прёдставить себе следующим образом ход эволюции, идущей путем анаболии (схема XXX):
Период морфогенеза Период роста Взрослое состояние
(а b . с Cj c2 c3 c4 c5 С — С Г рудные плавники
f g h hj h2 h3 h4 hs- Н — Н Верхняя челюсть
( TH n 0 O3 О-S O5 О — О Нижняя челюсть
( a b c C1 c2 c3 c4 c5 С — С Г рудные плавники (константные)
< f g 11 hj h2 h3 h4 h5 Н — Н Верхняя челюсть (константная)
1 TH n 0 I P Pi P2 Рз Pl Р — Р Нижняя челюсть (удлиненная)
a b c Ci Cg C3 C5 С — С Грудные плавники (константные)
< f g h 1 ij . i3 I — I Верхняя челюсть (удлиненная)
m n о p q 4i q2 qs Q — Q Нижняя челюсть (очень длинная)
( a b c Ci C2 c3 C4 Сд С — С Грудные плавники) (константные)
] f g h i k lq k8 ks К — К Верхняя челюсть (очень длинная)
| m и o P 4 qi Чз Рз Q — Q Нижняя челюсть (очень длинная)
Схема XXX; Объяснение в тексте;
Ряд а—Ь—с изображает собою морфогенез органа, который в течение эволюции предков не изменился или, вернее сказать, изменился очень мало, например грудные плавники Belone. Морфогенез этого органа заканчивается одновременно с морфогенезом других не сильно видоизмененных органов.1 Ряд Ci—с5 изображает те стадии роста грудных плавников, в течение которых этот орган растет очень сильно, но морфологически не изменяется. Ряд f—g—h изображает стадии морфогенеза верхней челюсти предков Belone (хг); буквы h,—h5 изображают стадии роста, а Н—Н взрослое состояние верхней челюсти формы xv Ряд m—п—о— Oi....o5 изображает последовательные стадии морфогенеза и роста органа О (нижней челюсти) у той же анцестральной формы хт.
У отдельных потомков формы xv т.е. у формы хп, орган С не изменился; поэтому стадии его морфогенеза (а—b—с) и роста (q—с5) остались такими же, как и у формы xv Не изменился также у формы хп и орган Н, т. е. строение верхней челюсти (f—g—h—hj h5—H—Н).'3ато строение нижней челюсти у формы хп уже стало изменяться: к обычному морфогенезу у этой последней (т—п—о) в течение времени Xj—хп прибавилась новая стадия р, так что ее морфогенез, носивший прежде характер m—п—о, теперь принимает вид m—п—о—р. У взрослого животного короткая нижняя челюсть предков (О) превращается в несколько более длинную нижнюю челюсть Р..
На следующей стадии развития челюстного аппарата у Belone, т. е. на стадии xq, орган С (грудные плавники) остается неизменным, но эволюция нижней челюсти (Р) уходит уже далеко вперед: к предшествующему мор
1 Морфогенезы других гармонически развивающихся органов, например морфо-
генез глаз, хвоста и т. п., простоты ради нами в эту схему не введены.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
495
фогенезу этого органа ш—п—о—р прибавляется еще одна стадия, а именно-стадия роста в длину—q, вследствие чего орган Р у формы xq превращается в Q (еще более длинная нижняя челюсть). В это же время и орган Н, т. е. верхняя челюсть, также начинает изменяться в том же направлении, что-и нижняя, а именно, к ее предшествующему морфогенезу, имевшему вид f—g—h, прибавляется новая стадия i, вследствие чего взрослый орган Н—Н (стадии Xi—хп) превращается у потомков xq во взрослый орган I— I. Стадия хп приблизительно соответствует стадии Hemiramphus (рис. 158) в филогенезе сем. Belonidae.
Наконец, в еще более поздней стадии филогенетического развития, предков Belone, а именно в стадии xz, к морфогенезу верхней челюсти, имевшему вид!—g—h—i, снова прибавляется новая конечная стадия к, так что морфогенез этого органа принимает вид f—g—h—i—k, а сама челюсть превращается в орган К—К, т. е. верхняя челюсть по длине становится, равной нижней челюсти. Орган С (грудные плавники) и Q (нижняя челюсть) остаются при этом неизменными (стадия Belone в филогенезе Belone, рис. 157). •
Таким образом, на этой схеме наглядно представлена и наблюденная нами на фактическом материале надставка конечных стадий и удлинение морфогенеза экцессивно развивающихся органов. Если бы мы захотели, то могли бы здесь вместо верхней и нижней челюстей Belone вставить грудные плавники Lophius или tarsalia proximalia ящериц. Схема в принципе осталась бы неизменной.
При эволюции признаков, происходящей по принципу анаболии, анцестральные признаки рекапитулируются («репетируются» по выражению де Беера), и приведенная выше схема наглядно показывает нам, как именно эта рекапиту ля ц и я происходит. Если мы обратимся к развитию челюстей у костистых рыб, например у Atherina, то увидим, что ее челюсти в течение морфогенеза претерпевают целый ряд изменений и достигают, наконец, стадии, изображенной на рисунке 162. Дальнейшее развитие челюсти заключается здесь в гармоническом росте всех частей, но форма и положение самих челюстей остаются в течение всего периода роста неизменными и во всем подобными форме и положению челюстей взрослой рыбы (рис. 163).
Наблюдая развитие челюстей у Belone, мы твердо установили, что у этой рыбы челюсти в своем развитии проходят через те же стадии морфогенеза, что и у Atherina, достигая при этом стадий, обозначенных нами на схеме XXX буквами h и о (рис. 159); но на этом развитие челюстей не останавливается, они развиваются дальше и растут в длину. В таких случаях говорят, что здесь рекапитулируется состояние взрослого предка. Это, однако, неверно, и если мы обратимся к нашей схеме, то нам станет ясно, что здесь повторяется не состояние (не признаки) взрослых предков, снабженных короткой челюстью, но лишь последняя стадия морфогенеза челюстей (h, о) этих предков, т. е. одна из поздних стадий их онтогенеза (ср. xlt xn, xq, xz). Эта последняя стадия морфогенеза, т. е. стадия h для верхней челюсти и стадия о для нижней челюсти, была в отношении признаков формы, положения и относительной величины похожа на взрослое состояние предков; поэтому всегда и казалось возможным, исходя из этой стадии, заключать о состоянии взрослого предка.
Мы видим, таким образом, что в онтогенезе потомков повторяются не признаки взрослых предков современных форм (как это утверждали Геккель и Вейсман), но лишь конечные стадии их морфогенеза (ср. К. Бэр), обычно по многим признакам (форма, положение,
496
ГЛАВА XIV
относительная величина и иногда гистологическое строение) похожие на взрослое состояние предков.
То, что мы говорили относительно стадий h и о, относится, конечно, и к предшествующим стадиям морфогенеза, а именно к стадиям g, f, и, m, которые у более отдаленных предков данной формы в свою очередь являлись конечными стадиями морфогенеза этих предков, весьма похожими на их взрослое состояние.
Исследование развития грудных плавников Lophius приводит нас к аналогичным выводам. У всех костистых рыб первые зачатки грудных плавников имеют приблизительно горизонтальное положение, т. е. такое же положение, как у эмбрионов и у взрослых акул, у хрящевых ганоидов и у взрослых палеонисцид, от которых произошли костистые рыбы. На более поздних стадиях развития положение грудных плавников изменяется, ибо передний край свободного плавника поднимается, так что первично дорзальная поверхность плавника прилегает к боковой стенке тела, а первично вентральная поверхность оказывается обращенной наружу (рис. 164—168). У большинства костистых рыб именно это положение плавника характеризует конечную стадию морфогенеза, сохраняющуюся и у взрослой формы без изменения. Мы обозначим, краткости ради, это положение грудных плавников как положение «вертикальное».
У Lophius piscatorius развитие грудных плавников проходит через обе только что упомянутые нами стадии, т. е. через стадию горизонтального и стадию вертикального положения плавника. В дальнейшем развитии к этим двум стадиям, как мы это видели выше, добавляется еще одна новая конечная стадия, л именно стадия, когда грудные плавники снова поворачиваются в дорзо-вентральном направлении и принимают опять свое первоначальное положение, т. е. то положение, которое является типичным для акул, хрящевых ганоидов и палеонисцид и которое мы здесь обозначаем как «вторично горизонтальное положение». Последовательные изменения в положении грудных плавников у вышеупомянутых нами рыб приведено на следующей схеме (схема XXXI).
Положение грудных плавников
Первично горизонтальное—>то же —>тоже—>тоже —>то же
Chondrostei Морфогенез Период роста взрослое
состояние
Первично горизонтальное —> вер-
Большинство тикальное —>то >ке—>то же —>тоже
костистых '-----------------——————- '------------------ 4--v'
рыб Морфогенез Период роста Взрослое
состояние
Lophius
Первично горизонтальное—вертикальное —> вторично горизонтальное > то же —> то же
Морфогенез Период роста Взрослое состояние
Схема XXXI. Объяснение в тексте.
Мы видим, таким образом, что у хрящевых ганоидов (и у эласмобранхий) признак «горизонтального положения» грудных плавников является характерным для конечной стадии морфогенеза, для стадий молодости, т. е. для периода роста, и для взрослого состояния. У большинства костистых рыб этот признак сохраняется только в течение морфогенеза и уже в конце этого периода заменяется признаком «вертикального положения» грудных плавников. Здесь мы имеем перед собой рекапитуляцию эмбрионального признака (К. Бэр), который
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
497
Рис. 174. Поперечный разрез через зуб первой генерации (Cyprinus carpio) при большом увеличении (из Васнецова, по Грибу, 1930).
является, однако, характерным также и для взрослых предков (Ф. Мюллер, Геккель), следовательно, рекапитуляцию в смысле биогенетического закона, т. е. повторение состояния, характерного как для малька, так и для взрослого предка Lophius.
У морского чорта (схема XXXI) мы имеем для грудных плавников повторение состояния, типичного для группы Chondrostei, т. е. «первично горизонтальное положение» этих плавников, являющееся рекапитуляцией (репетицией) № 1. Затем у него появляется то состояние плавников, которое является характерным для большинства костистых рыб, т. е. непосредственных предков сем. Lophiidae, а именно, вертикальное положение грудных плавников; это будет рекапитуляция (репетиция) №2. В обоих случаях мы имеем перед собой повторение конечной стадии морфогенеза, которая представляла собой для данного признака полное подобие состояния взрослого предка. Наконец, грудные плавники Lophius принимают «вторично горизонтальное» положение, т. е. в конце морфогенеза Lophius наступает состояние, характерное для взрослых форм.
Для нас совершенно ясно, почему эти признаки повторяются в онтогенезе потомков. При эволюции, происходящей путем надставки конечных стадий (анаболии), они являются необходимой предпосылкой онтогенетического развития нового состояния, ибо эволюция является не изменением признаков взрослых предков, но изменением хода их онтогенеза.
В заключение отметим еще особый способ анаболии, впервые отмеченный В. В. Васнецовым (1936)1 при исследовании им развития глоточных зубов у различных родов семейства карповых рыб.
Как известно, у карповых глоточные зубы сменяются ежегодно, а в первый год жизни этих рыб сменяют друг друга даже две генерации зубов. Наиболее полно процесс анаболии может быть прослежен на глоточных зубах сазана (Cyprinus carpio), имеющих наиболее сложное раз
витие. Первая генерация зубов сазана имеет простую конусовидную форму, рекапитулирующую обычный тип зубов костистых рыб (рис. 174). Вторая генерация первого года с самого начала своего появления уже имеет более сложную форму: коронка наклонена под углом к шейке и на вершине загнута крючком, внутренний край ее снабжен двумя зазубренными валиками (рис. 175 А—В). Именно такой формы зуб мы находим у многих примитивных форм карповых рыб во взрослом состоянии, например у взрослой шемаи (рис. 175 G, Н). Далее происходит усиленное разрастание одного из валиков внутреннего края, приводящее к форме зубов, изображенной на рис. 175 С. Эта форма зубов имеется во взрослом состоянии у пескаря (Gobio gobio) (рис. 175 J). Дальнейшее утолщение всего зуба приводит затем к форме, изображенной на рис. 175 D, близко напоминающий форму зубов взрослого усача (Barbus; рис. 175 К). К концу своего развития зуб второй генерации претерпевает еще некоторые изменения (рис. 175 Е): на утолщенной его коронке появляется грань, выделяющая более плоскую жевательную площадку (рис. 175 Е, а); крючок на вершине уменьшается в размерах. На этом развитие зубов второй генерации заканчивается.
Генерация следующего года уже при первом своем появлении имеет
1 Васнецов В. В., Пути филогенетических исследований костистых рыб. Известия Акад. Наук СССР, 1936.
Морфологические закономерности
32
468
ГЛАВА XIV
форму, сильно отличающуюся от формы, наблюдаемой на последней стадии генерации предыдущего года: зуб имеет теперь форму бокала с плоской полулунной жевательной площадкой. Далее зуб утолщается еще более и приобретает форму, свойственную взрослому сазану (рис. 175 F). Таким образом, появление новых признаков происходит, с одной стороны, у одного органа постепенным изменением на последовательных стадиях развития, как в случае типичной анаболии, с другой,—сменой органа другой его генерацией, имеющей уже новый признак.
В данном случае мы действительно имеем перед собою анаболию особого рода, так как здесь последовательное появление новых признаков происходит не у одного, но у нескольких сменяющих друг друга органов.
Здесь, в филогенезе более мелких систематических групп (родов) расхождение признаков путем анаболии играет первенствующую роль и
Рис. 175 А—К. Развитие глоточных зубов сазана, сопоставленное с формой глоточных зубов взрослых рыб других родов семейства карповых.
А—Е—последовательные стадии развития второй генерации глоточных зубов сазана. F—зуб взрослого сазана (следующая генерация); G—зуб малька шемаи; Н—зуб взрослой шемаи; J—зуб взрослого пескаря; К—зуб взрослого усача; а—шейка зуба; Ь—коронка зуба; с—валик коронки зуба;
d—жевательная площадка зуба (из В. В. Васнецова, 1936).
проявляется с большей яркостью, как это указывают, между прочим, и разобранные нами выше примеры развития семейств Belonidae и Tri-glidae.
В заключение остановимся на вопросе о вторичной анабол им. Как мы увидим ниже (см. ниже—вторичный архаллаксис), филогенетическое развитие органов не всегда происходит в течение эволюции при помощи только одного какого-либо модуса филэмбриогенеза. Наоборот, часто случается так, что филогенетический процесс, начавшийся одним способом филэмбриогенеза, естественно продолжается другим, т. е. что в эволюции органов имеет место комбинация двух способов филогенетического развития. Примером такой комбинации и вместе с тем примером вторичной анаболии может служить ранняя закладка и развитие крупных чешуй туловища осетровых рыб, т. е. чешуй непарного дорзального ряда и чешуй верхнего и нижнего боковых рядов (жучки). Весьма вероятно, что костные чешуи осетровых рыб возникли как новообразования путем архаллаксиса. На более поздних стадиях онтогенеза они растут в определенном направлении, на них появляются в определенном порядке характерные бугорки (скульптура чешуй) и они сильно увеличиваются в размерах. На основании изучения дальнейшего развития и появления рекапитуляций мы вправе признать, что, заложившись способом архаллаксиса, они в своем дальнейшем развитии эволюировали по способу надставок стадий, т. е. анаболии. Эту анаболию, сменившую собой архал-
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
499
лаксис, мы и называем вторичной анаболией. Таким образом, вторичная а набо лия обычно имеет место тогда, когда, орган, развившийся у предков путель а р х а л л а к с и с а, начинает развиваться у потомков путем ан а.б о л и и.
ЗАКОН к. Э. БЭРА
Теория анаболии делает’для нас понятным процесс рекапитуляции признаков, но, кроме того, она объясняет нам и открытый К. Бэром закон, касающийся последовательности в появлении и развитии признаков крупных и мелких систематических групп животного царства.
Закон К. Бэра устанавливает два важных общих факта: во-первых, то, что признаки взрослого животного закладываются у зародыша не одновременно, но в известной последовательности, во-вторых, то, что последовательность в образовании признаков взрослого животного соответствует последовательности систематических категорий, к которым данное животное принадлежит, последовательности по убывающей степени общности.1
При обсуждении различными авторами закона К. Бэра очень мало внимания было уделено тому факту, что общие систематические
признаки, т. е. признаки типов и классов, являются в то же время признаками филогенетически более старыми, чем признаки менее общих систематических групп, т. е. признаки семейств, родов, видов и т. д., и что поэтому обобщение К. Бэра имеет не только таксономическое, но и филогенетическое значение.
Вместе с тем для нас ясно, что более общие систематические признаки образуются филогенетически раньше, чем менее общие признаки: что, например, признаки, которые отличают современного человека (Ното sapiens) от Homo primigenius, образуются в филогенезе позднее, чем признаки, отличающие вообще род Ното; что признаки рода Ното являются более молодыми, чем признаки, характерные для всех Anthropomorpha; что признаки Catarrhina являются более древними, чем признаки Anthropomorpha и, наконец, что признаки Anthropoidea являются еще более древними, чем признаки Catarrhina и т. д.
Поэтому, мы можем формулировать закон К. Бэра следующим образом: последовательность появления в онтогенезе характерных признаков взрослого животного соответствует последовательности появления этих признаков в филогенезе пред-ков этого животного.
Мы особенно подчеркиваем, что последовательность появления признаков, о которой сейчас идет речь (К. Бэр), никоим образом не следует смешивать с последовательностью анцестральных признаков, появляющихся в онтогенезе при явлениях рекапитуляции (Мюллер, Геккель). Закон К. Бэра показывает нам,’ в какой последовательности закладываются сохранившиеся по сию пору у взрослого животного признаки его предков; наоборот, закон рекапитуляции показывает нам, в какой последовательности закладываются анцестральные признаки, н е-
1 «1) У зародыша общие признаки крупной животной группы образуются раньше, чем частные; 2) из более общих структур возникают все более частные, до тех пор пока не появятся наиболее специальные признаки» (К. v. Baer, 1828).
32*
500
ГЛАВА
когда существовавшие у взрослых предков данного животного и замещенные у него другими признаками.
Как можем, мы объяснить закон К. Бэра с точки зрения теории филэмбриогенеза? Чтобы иметь возможность ответить на этот вопрос, попробуем сначала разрешить две следующие проблемы:
1. Почему признаки взрослых животных развиваются не одновременно, но один после другого, в строго определенной последовательности?
2. Почему признаки систематически крупных групп, т. е. филогене-
тически древние признаки, появляются на ранних стадиях онтогенеза, а признаки систематически малых групп, т. е. филогенетически новые признаки,—на поздних стадиях онтогенеза?
Анализируя первый из этих вопросов, я пришел к мысли, что опреде-
ленная последовательность в появлении признаков взрослого животного
зависит от того, развиваются ли эти признаки путем надставки конечных стадий или нет. Пересматривая известные мне случаи эволюции животных, происходящей путем надставки конечных стадий (анаболии),
я нахожу, что указанная последовательность в закладке признаков взрослых форм представляет собою естественное следствие эволюции, происходящей по типу анаболии.
Эту мысль сделает нам более ясной следующая схема (схема XXXII), изображающая в уже знакомой нам форме эволюцию упомянутых выше органов С, Н и О (грудные плавники Lophius, верхняя и нижняя челюсти Belone и т. п.), происходящую путем анаболии.
I II III IV V VI VII VIII IX
( а b c Cl c2 C3 c4 C5 C I
Х1 f g h hi h2 h3 h4 h5 H > . . . .... 1
m n 0 °1 o2 O3 o4 °5 0 1
1 а b c Cl ^2 C3 c4 C5 c 1
хп f g h i h k is i4 J . . . .... 2
1 m n 0 p q qi q2 q3 Q 1
( а b c Cl C2 C3 c4 C5 c )
хд f g h i ki k2 k3 &4 к > . . . 3
1 m n 0 p q r s Si s J
: J :----->- : К : I
. . q,_______у . g. ! Эволюция взрослых органов С, Н и О на
’ 4 ’ I стадиях филогенеза xt, xn , xq
хп xq J
С : Н: О :
*1
Схема XXXII. Объяснение в тексте.
Мы принимаем, что буквы С, Н и О изображают, как и на предыдущей схеме, строение упомянутых трех органов взрослой животной формы х на одной из ранних стадий ее филогенеза (Xj). Как и прежде, буквами а—b—с, f—g—h, m—n—о мы обозначаем стадии морфогенеза, буквами Cj—с5, йг—h5, Oj—о5—стадии роста этих органов. Здесь можно было бы, конечно, привести онтогенезы и других эволюирующих и статических органов, но мы не сделали этого, чтобы не усложнять схемы.
На более поздней стадии эволюции животной формы х1;т. е. на стадии хп, орган С остается без перемен (хп , С); такие, не изменяющиеся в течение филогенеза органы мы будем впредь обозначать как константные или статические орган ы.1 Орган Н изменялся путем анаболии:
1 Конечно, статичность мы понимаем условно, так как в силу закона корреляции изменения, происходящие в других органах, не могут не повлиять и на данный орган.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
501
в конце его морфогенеза к его развитию прибавилась стадия i, так что Нпревратилсяв J. Орган О в течение промежутка времени х4—хп изменился более значительно, чем орган Н: к его морфогенезу постепенно прибавились стадии р и q, вследствие чего у взрослой формы он превратился сначала в Р_, а затем в Q.1
Предположим теперь, что эволюция органов С, Н и О, идущая путем анаболии, пошла и дальше этим же путем и что xq представляет собой гораздо более позднюю стадию эволюции потомков х, чем х2. При этом онтогенез и строение взрослого органа С остались без перемен, но к морфогенезу органа J (по сравнению с филогенетической стадией. хп) прибавилась новая стадия k (f—g—h—i—к}—k2—k3—k4), вследствие чего J превратилось в К. К морфогенезу органа Q (стадия хп) прибавились за это время две новые стадии: ths, вследствие чего у взрослого животного орган Q превратился в S. Таким образом, в течение промежутка времени х4—xq органы О и Н превратились в видоизмененные органы К и S, тогда как орган С совершенно не изменился.
Вышеприведенная схема в наглядной форме иллюстрирует нам отношения, существующие между эволюцией органов (филогенезом) и их индивидуальным развитием (онтогенезом) в тех случаях, когда эта эволюция идет путем анаболии (надставки конечных стадий). Следуя этой схеме, мы могли бы легко представить себе, что, например, буква С изображает хвостовой плавник предков Lophius, который в своей эволюции очень рано остановился на стадии гомоцеркии, характерной для большинства костистых рыб. Тогда ряд Н—»J—»К (схема XXXII) изобразил бы различные стадии частичного срастания оперкулярной щели (рис. 164, 165, 165 А), а ряд О—>Q—>S—различные стадии эволюции грудных плавников (срастание radialia, различные степени вентрального вращения плавника и т. д.). Те же ряды Н—»J—»К и О—>Q—>S могли бы нам дать представление о развитии челюсти у Belone и т. д.
Мы приходим, таким образом, к выводу, что определенная последовательность в онтогенетическом развитии различных органов зависит от количества прибавленных в морфогенезе признаков, т. е. от степени удлинения морфогенеза. Так, у органа С, который образовался у очень отдаленных предков х и который с тех пор в течение очень долгого периода времени (х4—хп —xq) совсем не изменялся (константный признак), морфогенез (а—b—с) остается таким же, каким он был у предков (х4), т. е. относительно очень коротким; если же другой какой-нибудь орган того же животного, например орган Н, в течение того же промежутка времени изменялся путем анаболии и к его морфогенезу прибавлялось несколько конечных стадий (I, к), то этот морфогенез станет у потомков длиннее и дефинитивное состояние такого органа (К) будет достигаться онтогенетически позже, чем это имело место у первого органа.
Если,- наконец, какой-нибудь третий орган (О) в течение того же промежутка времени изменится путем анаболии очень сильно и к его анце-
1 Мы не останавливаемся здесь подробно на обсуждении того, что различные органы одного и того же животного развиваются с различной скоростью, так как факт этот нам кажется и без того достаточно общеизвестным. В качестве одного из примеров различия в темпе эволюции можно привести филогенез челюстей у той же Belone. Сравнивая между собой развитие челюстей у Hemiramphus, Scombresox и Belone, мы ясно видим, что нижняя челюсть этих рыб филогенетически эволюирует быстрее, чем их верхняя челюсть. К тому же выводу приводит нас и исследование других одинаково построенных у предков органов, например передних и задних конечностей бесхвостных амфибий: задние конечности Anura филогенетически развивались более быстрым темпом, чем их передние конечности.
502
ГЛАВА XIV
стральному морфогенезу (ш—п—о) прибавится много конечных стадий (прибавка стадий р—q—г—s), то морфогенез такого органа удлинится еще больше и его дефинитивное состояние (S) наступит у потомков еще позднее, чем у нашего второго органа. Мы видим, таким образом, что последовательность в образовании дефинитивных (взрослых) признаков различных 'органов какой-нибудь эволюирующей животной формы зависит от различной длины морфогенезов этих органов, т. е. от относительного числа надставок новых конечных стадий к морфогенезам этих
Х,(СНО)
(С1Р)Хп, Xn2(CJQ)
Схема ХХХШ. Объяснение в тексте.
органов.
Этим, как мне кажется, мы даем удовлетворительный ответ на первый из поставленных нами вопросов.
Попробуем теперь подойти к разрешению второй проблемы, а именно, вопроса о том, почему признаки, образующиеся в онттогенезе рано, являются в то же время признаками, характерными для крупных систематических групп животных, а признаки, образующиеся в онтогенезе поздно, являются в то же время признаками, характерными для мелких систематических групп.
По нашему мнению этот факт является необходимым следствием дивергентной монофилетической эволюции крупных и мелких групп животного царства и притом эволюции, идущей путем анаболии. Чтобы пояснить нашу мысль, обратимся к нижеследующей диаграмме (схема XXXIII).
Как мы видим, на нашей схеме дано обычное изображение монофилетического родословного древа. Примем, что от какой-нибудь родоначальной формы X} (ср. диагр. XXXII) произошли две дочерние формы, Хщ и Хп2, развивающиеся в течение своей дальнейшей эволюции в дивергентном направлении, и что от каждой из этих форм ответвились в течение этой эволюции по два внучатных вида (Xq1; Xq2, и Xq3, XqJ.1 Первичная анцестральная форма Хг характеризовалась при этом уже известными нам признаками С, Н и О. Примем вместе с тем, что орган С (или группа органов) в течение всей последующей эволюции совсем не изменялся (константный орган) и что органы Н и О изменялись с различной интенсивностью.
У потомков формы Х1; т. е. у Хщ и Хп2, органы Н и О, развивающиеся путем анаболии, изменялись в несколько отличных друг от друга направлениях, а именно: Н превратился в I у Хпх и он же превратился в J у Хп3; О превратился в Р у Хщ и в Q у Хп2 (схема XXXIII). У отдаленных потомков Хп1; т. е. у Xqx и Xq2, органы С и I остались неизмененными (константные органы), но орган Р изменился снова, превратившись у формы Xq4 в R, а у формы Xq2 в S; у потомков Хп2, т. е. у Xq3 и Xq4, органы С и J сохранились неизменными (константные органы), но орган Q изменился, превратившись у формы Xq3 в Т, а у формы Xq4 в U.
Рассматривая диаграмму XXXIII, мы видим, что константные признаки, т. е. признаки, неизменяющиеся в течение очень дли-
1 Само собою разумеется, мы должны принять, что формы Хх, Xhj, Хп2, и Xqu Xq2, Xq3, Xq4 были отделены друг от друга геологически длинными промежутками времени.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
503
тельного периода времени (С), характеризуют собой всех потомков родоначальной формы Хн т. е. XqI; Xq2, Xq3 и Xq4: эти именно признаки закладываются у потомков формы X, онтогенетически рано. Признаки, мало изменившиеся в теченгк эволюции, закладываются несколько позднее:
• Наконец, признаки, сильно изменившиеся в течение эволюции, онтогенетически закладываются всего позднее:
( R
I и
Первые, т. е. константные признаки, характеризуют собою всех потомков исходной формы Хх; вторые, т. е. мало изменившиеся признаки, принадлежат к числу признаков, характеризующих крупные систематические группы; наконец, третьи, т. е. сильно изменившиеся признаки, характеризуют мелкие систематические группы потомков исходной формы Х4 (схема XXXIII).
Все это, само собой разумеется, касается только тех признаков, которые образуются путем надставки конечных стадий.
Таким образом, гомоцеркальное строение хвостового плавника является характерным для всего семейства Scombresocidae, ибо оно образовалось еще у их общих предков и не изменилось в течение всей последующей эволюции этого семейства (С—филогенетически константный признак); вместе с тем, онтогенетически развитие этого признака заканчивается рано. Строение и положение челюстей у Belone, Scombresox, Hemiramphus и т. д., т. е. у представителей отдельных родов, принадлежащих к этому семейству, изменяются более или менее сильно (R, S, Т, U), и признаки эти (поскольку мы можем об этом судить, изучая развитие их у Belone) образуются онтогенетически поздно, т. е. опять-таки следуя закону К. Бэра.
Некоторые признаки резко изменяются в течение эволюции, и тогда онтогенез их удлиняется вследствие надставки конечных стадий; другие признаки в течение того же промежутка времени н е изменяются совсем и их онтогенетическое развитие не удлиняется; этим именно и обусловливается открытый К. Бэром закон онтогенетически раннего образования общих (древних или филогенетически постоянных) признаков и онтогенетически позднего образования менее общих (филогенетически новых или анаболически измененных) признаков. Мы видим, таким образом, что закон К. Бэра является необходимым следствием теории анаболии.
Итак, результатом эволюции, происходящей путем надставки конечных стадий морфогенеза, является, с одной стороны, факт рекапитуляции исчезнувших анцестральных признаков (Ф. Мюллер), а с другой,— факт закономерной последовательности развития признаков взрослых животных (К. Э. Бэр). Оба эти общих факта объясняются одной и той же теорией анаболии.
Мы этим, конечно, не хотим сказать того, что все факты, касающиеся определенной последовательности в развитии различных органов животного, например, факты, что некоторые органы закладываются раньше, а другие позднее, сводятся целиком к закону К. Бэра. Наоборот, мы подчеркиваем, что только те признаки, которые эволюируют по типу анаболии, подчинены этому закону. Ниже мы увидим, что существует
504
ГЛАВА XIV
целый ряд других признаков, развивающихся иным путем и не подчиняющихся в своем онтогенетическом развитии бэровскому закону.
Мы уже упоминали, что онтогенетическая рекапитуляция исчезнувших у современных взрослых форм признаков объяснялась А. Вейсманом фактом значительного сдвигания признаков взрослого животного назад, на все более ранние стадии онтогенеза. Вейсман1 считал, что это сдвигание признаков взрослых предков на все более ранние стадии онтогенеза потомков является важным процессом эволюции, протекающим, так сказать, «механически». Но в этом приходится весьма усомниться: существует много случаев сдвигания во времени закладок эмбриональных признаков, протекающих как в направлении ускорения, так и в направлении замедления их развития (Менерт,1 2 де Беер 3).
Хорошие примеры такого «сдвигания» мы находим в различных случаях развития неотенических форм, у которых, как известно, образование и созревание половых органов происходит на сравнительно ранних стадиях развития животного. Подобные же сдвиги в образовании и функционировании органа на эмбриональных или личиночных стадиях его развития мы находим и при развитии других органов, например при раннем развитии сердца и кровеносных сосудов у птиц и млекопитающих, у которых эти органы образуются и начинают функционировать гораздо раньше, чем у рыб и амфибий. Во многих случаях мы имеем возможность наблюдать, что эти сдвиги во времени закладки органов являются в сущности приспособлением этих органов к особенным условиям их развития. Это имеет, например, место при образовании сердца у высших амниот. Его раннее развитие является здесь приспособлением к раннему функционированию органов дыхания и питания зародыша (желток и питательные соки материнского организма).
Трудно представить себе, чтобы подобные «сдвигания» признаков взрослых предков на ранние стадии онтогенеза потомков представляли собою действительно «общий механический закон», как это думает Вейсман, ибо в очень многих тщательно изученных нами случаях развития сильно измененных органов, где, казалось бы, легче всего ожидать присутствия таких сдвигов, мы не находим ни малейших их следов. Мы видим, например (см. нижеследующую главу), что редуцированные конечности безногих ящериц закладываются на тех же стадиях развития, что и конечности обычных ящериц, у которых, как известно, они развиты вполне нормально. Мы видим, что эксцессивно развитые челюсти Belone закладываются на тех же стадиях развития, на которых закладываются нормальные челюсти других костистых рыб, например челюсти Atherina, что до известной стадии онтогенеза они развиваются таким же темпом, как и челюсти других костистых рыб и только значительно позже начинают расти весьма интенсивно (ср. рис. 159—161).
У очень многих из исследованных нами эксцессивных органов различия в величине и в форме зависят не от того, что у зародыша сильно измененный орган закладывается онтогенетически рано, а мало измененный орган закладывается онтогенетически поздно, но от того, что их диферен-циальный рост идет различным темпом в конце морфогенеза. Вообще приходится сказать, что сдвигания первых зачатков органа на более ранние стадии развития, которого мы вправе ожидать при образовании эксцессивных органов, как правило, не происхо
1 Weismann A., Vortrage fiber Deszendenztheorie, Jena, 1902.
2 Mehnert E., Kainogenesis als Ausdruck differenter phylogenetischer Ener-gien, Jena, 1897.
Idem., Biomechanik, erschlossen aus dem Prinzip der Organogenese, Jena, 1898.
3 De B'eer G. R., Embryology and Evolution, Oxford, 1930.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
505
дит вовсе, поэтому нам вообще нельзя говорить о существовании какого-либо «общего закона» применительно к таким сдвигам.
Ни Вейсману, ни Геккелю не пришло в голову попытаться объяснить, т. е. свести к какой-нибудь более общей закономерности эволюции, закон К. Бэра, касающийся, соотношений между онтогенезом животных и их положением в системе животного мира. Как мы видели выше, это объяснение возможно найти в теории анаболии, из которой установленный эмпирически, но до сих пор необъяснимый закон К. Бэра неизбежно вытекает.
Во избежание недоразумений, я считаю нужным более точно определить свою точку зрения в отношении вейсмановской теории. Вейсман, как это уже было упомянуто выше, принимает, что сдвигание закладок прогрессивных признаков на все более ранние стадии онтогенеза является необходимым, общим процессом эволюции и что поэтому все прогрессивные признаки животных в течение эволюции закладываются постепенно все раньше и раньше. Я же, наоборот, нахожу, что большей частью новые прогрессивные признаки закладываются у потомков в то же время, т. е. на тех же стадиях развития, что и у предков, и что различные сдвиги во времени появления признаков, различные гетерохронии, которые действительно происходят в онтогенезе довольно часто, зависят каждый раз от особых причин. Эти сдвиги обычно представляют собой приспособления к определенным условиям жизни животного: в некоторых случаях это будут приспособления к условиям эмбриональной жизни (ценогенезы), как, например, в случае ранней закладки и ускоренного развития сердца и кровеносных сосудов у зародышей птиц и млекопитающих (приспособления к условиям дыхания и питания зародыша на относительно ранних стадиях его развития); в других случаях, как это мы видели, рассматривая топографические координации органов, это будет способ, с помощью которого различные эктосоматические и энтосоматические органы взрослого животного приспособлются друг к другу.
Общий ход мыслей Вейсмана, т. е. основная причина, почему он пришел к своей теории «сдвигания признаков», нам совершенно понятна. Если к органу, находящемуся во взрослом состоянии, прибавятся новые-стадии развития, то время, в течение которого этот готовый к функционированию орган сможет действительно функционировать, естественно-укоротится, так как каждая новая стадия потребует для своего развития некоторого промежутка времени; чтобы этого не случилось, каждая новая стадия должна предыдущую стадию несколько отодвинуть назад во времени. По Вейсману, такое сдвигание стадий происходит главным образом у органов, прошедших в своем развитии через длинную и интенсивную эволюцию, т. е. у таких органов, к эволюции которых в течение филогенеза прибавилось много новых стадий, вследствие чего эти органы и должны закладываться у животных онтогенетически рано; те же органы, которые в течение филогенеза изменялись мало, должны, по Вейсману-Геккелю, закладываться относительно поздно. Мы видим, однако, что это противоречит известному нам, установленному К. Бэром, общему факту, что филогенетически древние (т. е. константные) признаки закладываются у животных онтогенетически рано, а филогенетически новые (т. е. филогенетически сильно измененные) признаки закладываются у них онтогенетически поздно. Так как закон J{. Бэра является не гипотезой, а представляет собой обобщение большого числа число эмпирических данных, то такое противоречие для теории сдвиганий является фатальным; поэтому мы думаем, что эту теорию Вейсмана в настоящее. время приходится оставить.
Что касается до теории.анаболии, то она, как мы это уже видели, на такое противоречие не наталкивается.
503
ГЛАВА XIV
Для полной ясности подчеркнем еще раз, что мы отклоняем лишь «гипотезу сдвиганий» Вейсмана, но не тот его взгляд, по которому новые признаки могут появляться у животных также и во взрослом состоянии. Как это сейчас будет видно, мы имеем все основания думать, что это последнее обстоятельство не стоит в противоречии с теорией анаболии.
Мы знаем, что завершение процесса морфогенеза может иметь место на самых различных стадиях развития организма, рассматриваемого в данном случае как сумма составляющих его признаков. Теория анаболии принимает, что филогенетически важные изменения органов происходят именно в конце морфогенеза, т. е. на тех стадиях, когда развивающийся орган еще является достаточно пластичным, чтобы иметь возможность изменить свою структуру, форму и т. п.
Если морфогенез какого-нибудь органа животных завершается по каким-либо причинам много позднее, чем морфогенез остальных его органов,—тех, которые к этому времени уже вполне завершили свое развитие, а в некоторых случаях также и свой рост,—то к морфогенезу данного органа может прибавиться еще новая стадия развития в то время, когда само животное уже достигло половозрелого состояния; тогда может получиться впечатление, что филогенически изменяется взрослый орган. Мы знаем, что такие случаи, действительно, иногда имеют место: многие признаки, например рога у парнокопытных, дефинитивные черты лица у человека и многие другие, образуются очень поздно. Однако здесь мы имеем дело не с образованием нового признака у взрослой особи, но лишь с очень поздним, по сравнению с морфогенезами остальных органов, завершением морфогенеза данного органа.
МОДУС ДЕВИАЦИИ, ИЛИ ОТКЛОНЕНИЕ (АБЕРРАЦИЯ) НАПРАВЛЕНИЯ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Ф. Мюллер считал, что, кроме надставок стадий в конце онтогенеза, существует еще другой род соотношений между онтогенезом и филогенезом животных, а именно, такое отношение, когда эмбриональное развитие какого-нибудь органа принимает у потомков данной формы на средних стадиях онтогенеза несколько иное, чем у предков, направление, иначе говоря, когда оно «отклоняется» («abirrt») от старого эмбрионального пути, в результате чего данный орган принимает у взрослых потомков несколько иной вид, чем тот, который он имел у предков. А. Нэф называет такой род филогенетического развития «аберрацией на средних стадиях онтогенеза». Мы предпочитаем название, данное этому процессу В. Францем—«девиация на средних стадиях развития».
Прекрасным примером развития, происходящего путем девиации на средних стадиях онтогенеза, является развитие обонятельных органов у костистых рыб.
Как известно, носовые ямки у всех Osteichthyes закладываются у зародыша в передней части головы и имеют в начале вид обонятельных плакод или парных неглубоких ямок, состоящих из утолщенного эктодермального эпителия. Затем на дне этих обонятельных ямок образуются эпителиальные складки .(рис. 176 А, В и 178 Вх), которые в течение дальнейшего развития принимают радиальное расположение, различное для различных видов рыб (рис. 176 D, 177,178G). Параллельно с изменением дна ямки изменяются и ее края (рис. 178 А, В, t, t), а именно,'противоположные края начинают расти друг другу навстречу, образуя своим срастанием поперечный мост (рис. 178 С, Т), перекрывающий обонятельную ямку.
Образование и развитие этих складок у Acipenser ruthenus показано на рис. 176 А, В, С и D. Мы видим, что сначала на дне ямки образуется незначительное число слегка расходящихся -складок (рис. 176 А и В), благодаря которым увеличивается поверхность обонятельного эпителия.
Рис. 176. А—D. Развитие обонятельной ямки Acipenser ruthenus.
А-ранняя стадия развития обонятельной ямки стерляди, на которой впервые появляются складки слизистого обонятельного эпителия (реконструкция по сагиттальным разрезам); В—несколько более поздняя стадия развития обонятельной ямки; С—стадия, на которой обонятельные складки начинают приобретать радиальное расположение благодаря образованию новых складок по обеим сторонам от первичных обонятельных складок; а—место, где складки еще не начали развиваться; D—обонятельная ямка взрослого Acipenser ruthenus с радиально расположенными складками (из А.Н. Северцова, 1931).
Рис. 177. Обонятельная ямка Onchorhynchus keta с радиально расположенными складками слизистого обонятельного эпителия.
На рисунке видно,что радиальное расположение складок происходит, так же как и у стерляди, путем образования новых складок по обеим сторонам от первичного ряда обонятельных складок; а—место, где складки еше не образовались.Ср.рис. 176, С (изА.Н. Северцова, 1931).
Рис. 179. А—D. Четыре стадии развития обонятельной ямки Belone acus.
А—стадия 13 мм; здесь изображена обонятельная ямка, в которой еще не началось образование обонятельных складок; В—стадия 38 мм! на дне обонятельной ямки образовался вырост слизистого обонятельного эпителия а; С—стадия 82 мм; вырост а сильно увеличился в размерах, отделился от дна обонятельной ямки и изменил свою форму; D—взрослый экземпляр длиной в 40 см; вырост а сильно разросся и принял форму гриба, сидящего на относительно тонкой' ножке. На этом рисунке мы видим, что здесь не развиваются выросты t—t (ср. рис. 178 А, В), из которых образуется поперечный мостик Т у Sargus (рис. 178 С);
о—край глаза (из А. И. Северцова, 1931).
Морфологические закономерности, стр. 507.
Рис. 178 А—G. Развитие обонятельной ямки у Sargus vulgaris.
А—ранняя стадия развития (малекг9-ммУдлины) обонятельной ямки, когда на.ее слизистом обонятельном эпителии еще не образовалось ни одной складки; t, t—выросты краев обонятельного эпителия, из которого впоследствии образуется поперечный мостик; о—край глаза. В—несколько более поздняя стадия развития (малек 11 мм длины, где выросты t, t приблизились друг к другу. В<—та же стадия развития (малек 11 мм); края обонятельной ямки срезаны таким образом, чтобы можно было увидеть дно ямки, где образовалась первая обонятельная складка (f). С—немного более поздняя стадия (малек 11,5 мм длины), на которой уже образовался поперечный мостик Т путем срастания выростов t,t. D—стадия 12,5 мм дл., на которой по обе стороны от первичной складки f развиваются две новых складки Гг, f2. Е—ещеболее поздняястадия (малек 14 мм длины), на которой развитие обонятельной ямки продвинулось вперед по сравнению с предыдущей стадией: здесь первичная обонятельная складка f путем появившегося в ней углубления J делится на две части; на рисунке видно, как вся эта группа складок (f, f, ft, f2) подымается co дна обонятельной ямки; F— стадия 19 мм, на которой развились новые обонятельные складки Г3 и f4. G-— обонятельная ямка взрослого Sargus; здесь видно радиальное расположение складок слизистого обонятельного эпителия. На рисунках В, Е, F, G- срезаны края обонятельной ямки, а также поперечный мостик Т (из А. Н. Северцова, 1931).
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
507
Несколько позднее начинают образовываться складки на дне ямки по обеим сторонам от первоначальных складок, придавая обонятельной ямке полукруглый вид (рис. 176 С), а затем, постепенно, благодаря образованию все новых и новых складок, обонятельный орган принимает тот вид, который он имеет у взрослых рыб: складки имеют теперь радиальное расположение (рис. 176 D). Это радиальное расположение характерно также и для примитивных костистых рыб (рис. 177). На рис. 178 А, В и С показано образование поперечного моста в обонятельных ямках у костистой рыбы (Sargus vulgaris); отметим, что у осетровых (Acipenser ruthenus) образование этого моста идет таким же путем. У -взрослых стерлядей (рис. 176 D), так же как и у Onchorhynchus keta (рис. 177), все эпителиальные складки выходят из средней круглой и гладкой части дна и направляются от этого центра к краям почти круглой обонятельной ямки.
У многих костистых рыб мы находим незначительные видоизменения этого радиального расположения эпителиальных складок. На ранних стадиях развития обонятельных ямок у Sargus со дна ямки поднимается только одна невысокая складка (рис. 178 В1; f); на следующей стадии по обеим сторонам этой первой складки развиваются еще две боковые складки (рис. 178 D, f, и f2), так что все это образование принимает в этот момент форму заостренного конца гарпуна. На еще более поздней стадии развития мы видим, что по бокам от этих трех первичных складок образуется еще по одной новой складке (рис. 178 F, f3 и f4), причем все они исходят от гладкого, лишенного складок среднего поля. Наконец, у взрослого Sargus (рис. 178 G) все складки принимают обычное радиальное расположение.
Разница в строении носовых ямок у взрослого Sargus и взрослого Acipenser состоит в том, что, как это видно из сравнения рис. 178 G и 176 D, средняя часть дна носовой ямки у стерляди имеет круглую форму, тогда как у Sargus она сильно удлинена. Кроме того, у Sargus складки прямо поднимаются со дна носовой ямки вверх. Но в общем ход всего развития носовых ямок у обеих этих форм в принципе одинаков.
У Belone, Hemiramphus и Scombresox мы наблюдаем сильные уклонения от этого типа строения носового аппарата. Прежде всего, у этих последних форм не образуется никакого моста, перекрывающего носовые ямки; поэтому эти ямки у взрослых рыб открываются наружу не при помощи двух носовых отверстий, отделенных друг от друга мостом (nares anteriores et posteriores), а, подобно тому как это имеет место у зародышей других рыб лишь на ранних стадиях развития, при помощи одного единственного большого отверстия (рис. 158, 179 А—D). Первый зачаток носовой ямки у Belone (рис. 179 А) похож на первичный зачаток этих ямок у стерляди и у других костистых рыб, например у Sargus (рис. 178 А). На несколько более поздней стадии развития (рис. 179 В) у Belone со дна носовой ямки начинает подниматься вырост (а), который мы можем сравнить с первой складкой, образовавшейся на том же месте у Sargus (рис. 178 Вг), но дальнейшее развитие этого органа идет уже совершенно другим путем, чем у большинства костистых рыб. Так, у Belone по сторонам этой первоначальной складки уже не образуется никаких боковых складок, зато сама она сильно увеличивается в размерах (рис. 179 С), образуя на дне обонятельной ямки большой грибообразный вырост (рис. 179 D, а). Образование перекрывающего носовую ямку поперечного моста также не имеет места. Если мы сравним рис. 178 А—G (Sargus) с рисунком 179 А—D (Belone), то увидим,, что у Belone выпал из развития весь процесс образования поперечного моста, и в этом отношении у него носовые ямки остановились в своем развитии на эмбриональной стадии открытых ямок.
Кроме-Belone, этот же тип строения обонятельного органа мы находим
508
ГЛАВА XIV
и у взрослых Hemiramphus и Scombresox, и мы имеем все основания предположить, что развитие обонятельного органа у этих форм происходило тем же путем, что и у Belone.
Если мы поставим вопрос о том, как мог развиться этот особый тип носовых образований у всех Scombresocidae, то нам придется представить себе две различные возможности. Мы могли бы, например, думать, что это происходит путем надставки конечных стадий; но тогда мы должны были бы a priori приняты, что у очень отдаленных предков Scombresocidae на той стадии их филогенеза, когда у них обонятельный орган был еще представлен лишь открытой ямкой без поперечного мостика и лишь с одной эпителиальной складкой на дне, у них произошла надставка конечных стадий (более интенсивный рост донной складки, отсутствие образования мостика), в результате чего было достигнуто то состояние носового органа, которое мы имеем сейчас у взрослых экземпляров Belone и Hemiramphus.
С такой гипотезой мы, конечно, не можем считаться всерьез, так как тогда нам пришлось бы implicite принять, что предки Scombresocidae отделились от предков остальных костистых рыб на такой стадии развития этих последних, когда у них еще были развиты примитивные открытые носовые ямки с первичной складкой на дне. Так как не только большинство костистых рыб (и в том числе формы, родственные Scombresocidae), но и все хрящевые и костные ганоиды имеют многочисленные складки на дне носовых ямок, а также поперечный мостик, перекрывающий их, то это предположение приходится оставить, а вместе с ним оставить и попытку применить теорию анаболии к истолкованию только что описанных случаев.
Но если это так, то для нас остается лишь предположение, что все Scombresocidae произошли от костистых рыб с типично построенными носовыми органами, т. е. с органами обоняния, снабженными многочисленными складками и поперечным мостиком. При этом гораздо более вероятным является предположение, что у предков Scombresocidae ход онтогенетического развития начал меняться (отклоняться), начиная со средних стадий развития (девиация): первичная складка обонятельного эпителия стала расти у потомков более интенсивно, чем у предков, и вместе с тем стала менять свою форму. Рост вторичных складок и отростков t, t, при помощи которых происходит образование поперечного мостика (рис. 178, А, В и С; Т) задерживается, а таким образом и общий ход онтогенеза и состояние взрослого органа постепенно изменяются. В настоящее время я пока еще не могу сказать точно, как именно происходила редукция этого поперечного мостика и боковых складок эпителия, расположенных на дне носовых ямок, так как у исследованных мной современных форм даже в эмбриональном состоянии не сохранилось никаких рудиментов этих образований.
Согласно принятому нами способу буквенного обозначения, мы можем следующим образом символически представить протекающий по типу девиации филогенез грибообразного отростка, находящегося на дне носовой ямки у Belone (схема XXXIV).
I а1 . а2 а3 а4 а5 ае А
II Я1 а2 а3 Я1 а4 л11 35 я111 ав AIV
ш а1 а2 аз я11 а4 „ш 35 aIV ав Av
IV Э1 а2 аз аш а4 ар nv AVI
Схема XXXIV. Объяснение в тексте.
Мы принимаем здесь, что ряд а,—а2—а3—а4—а5—а6—А представляет собою онтогенетическое развитие первичной обонятельной складки у предков Belone (стадия филогенеза I). На следующей стадии филогенеза (стадия
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
509
Рис. 180. Схема филогенетического развития костных и роговых чешуй позвоночных животных.
А, В, С, D, Е, F—стадии развития плакоидной чешуи Heptanchus cinereus; G-, Н, К, L, М—стадии развития роговых чешуй Lacerta muralis (из Б. С. Матвеева, 1932).
II) развитие эпителия дна носовой ямки примерно до стадии а3 идет так же,как оно шло у предков (I), но, начиная со стадии а4, некоторая определенная часть дна носовой ямки (первичная складка) растет более интенсивно, чем она росла у предков(а1 *—а1/—-а”1—AIV), и этот процесс более сильного роста и изменения формы (рис. 179 В, С, D) усиливается в течение филогенеза все больше и больше (филогенетические стадии III и- IV, а1/—а1*1—а1}"—Av, а1*1—а1^—а^—AVI), в результате чего общая форма органа у взрослого животного в течение эволюции (А— AIV—Av—AVI) сильно изменяется.
Но теперь мы имеем право спросить себя, не происходит ли здесь в течение онтогенеза все же некоторой рекапитуляции (репетиции) признаков предков? Мы можем принять, что в начальной стадии развития обонятельного органа у Belone стадии онтогенеза а„ аа, а3 повторяют собою эволюцию общих предков всех рыб (включая, конечно, и эласмобранхий), протекавшей по типу надставки конечных стадий; начиная же со стадии а1 (схема XXXIV), мы уже не можем констатировать в онтогенезе разбираемого нами признака рекапитуляций признаков, схожих с признаками взрослых предков. Тем не менее является довольно вероятным, что интенсивный рост и изменение формы выроста а (рис. 179 С, D, а) все же является своего рода рекапитуляцией, а именно, повторением зародышевого состояния предков, причем последующее эмбриональное развитие и взрослое состояние потомка изменено. Здесь мы имеем перед собою рекапитуляцию чисто эмбриональных признаков в смысле К. Бэра.
Хороший пример девиации дает нам филогенетическое развитие роговых чешуй у рептилий. По наблюдениям Матвеева,1 начальные стадии развития роговых чешуй рептилий (Lacerta muralis) (рис. 180, G, Н, К) чрезвычайно близко напоминают собой начальные стадии развития плакоидных чешуй акуловых рыб (рис. 180 А, В, С): и в том и в другом случае образование и первоначальное развитие чешуй идет в Направлении уплотнения слоя эпидермиса и скопления под ним соединительной ткани. После этого, однако, развитие чешуй рептилий
1 Матвеев Б. С., Об эволюции кожных покровов позвоночных путем эмбрио-
нальных изменений, Зоологический журнал, 1932.
510
ГЛАВА XIV
резко уклоняется от пути развития чешуй низших водных позвоночных. В то время как чешуя рыб развивается в направлении сложных преобразований в соединительнотканном сосочке (рис. 180 D), постепенно превращаясь в плакоидный зуб (рис. 180 Е, F), чешуи рептилий филогенетически изменяются путем прогрессивного развития эпидермальной части чешуи: постепенно ороговевая. именно эта, эпидермальная, часть и дает нам типичную роговую чешую рептилий (рис. 180 L, М).
Таким образом, в развитии чешуй рептилий рекапитулируются лишь ранние эмбриональные стадии развития чешуй рыб, т. е. стадии, изображенные на рис. 180 А, В и С, после чего развитие чешуй рептилий отклоняется в своем пути (девиация) от пути развития чешуй рыб, и дальнейшее развитие костных чешуй низших водных позвоночных в онтогенезе рептилий уже не повторяется.
Прекрасным примером эволюции, идущей по пути девиации, является развитие спиракулярного мешка, превращающегося в течение филогенеза в среднее ухо наземных позвоночных. Как известно, ранние стадии развития среднего уха высших позвоночных чрезвычайно сходны с ранними стадиями развития спиракулярного мешка рыб. Но, начиная со средних стадий морфогенеза, развитие этих двух образований уже идет совершенно различными путями.
Мы имеем также полное основание думать, что многие случаи срастания частей скелета, найример срастание элементов carpalia и элементов tarsalia между собой, имеющее место у амфибий и у .рептилий, а также срастание отдельных позвонков крестца у птиц и у млекопитающих, произошло по способу (модусу) девиации.
Разбирая различные способы (модусы) филогенетического изменения строения органов, мы установили, что по окончании периода морфогенеза, т. е. в начале периода роста, молодое животное становится похожим на взрослое и что дальнейшее его развитие и изменение сводится к приблизительно «гармоничному» росту. Однако все это может считаться правильным лишь до известной степени, так как существует много животных, у которых именно в течение периода роста меняются более или менее значительно относительные пропорции органов и органокомплексов. Так, например, мы знаем, что у ребенка относительные размеры рук, ног, туловища и головы являются иными, чем они бывают у взрослого человека. Мы можем сказать, что морфогенез во многих случаях заканчивается не сразу, но что некоторые признаки (преимущественно относительные пропорции тела) могут изменяться и позже, т. е. в течение периода роста организма.1 В некоторых случаях эти изменения происходят путем надставки конечных стадий, но в большинстве случаев они совершаются посредством только что описанного нами способа девиации.1 2
АРХАЛЛАКСИС, ИЛИ ИЗМЕНЕНИЕ ПЕРВЫХ ЗАЧАТКОВ
Исследуя способы, при помощи которых органы взрослых животных изменяются в течение филогенеза в прогрессивном направлении, мы пришли к выводу, что это происходит не только путем надставки конечных стадий морфогенеза (анаболия), не только путем изменения средних стадий развития (девиация), но также и иным способом, а именно, изменением начальных стадий морфогенеза, способом, названным нами а р х а л-л а к с и с о м.
1 Увеличение ширины тела Sargus (см. Sewertzoff A. N. Beziehungen zwi-schen Ontogenese und Phylogenese, Jen. Ztschr., 1927).
2 Мы можем принять, что именно этим путем совершалась эволюция пропорций тела и эволюция черт лица у человека. Еще Больк отметил, что ребенок похож не на своих взрослых антропоидных предков, но на молодых обезьян. См. Bolk L., Das Problem der Menschenwerdung, Jena, 1926.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
5П
Для пояснения того, что собственно представляет собой архаллаксис^
обратимся к нижеследующей схеме (схема XXXV).
I а ь С Cl с2 сз С4 С5 С
п а' Ь' с' c'j С 2 С з С'4 с'з С'
Ш а" Ь" с" с 1 С 2 С 3 С4 с 5 С"
IV а'" Ь'" с"' c'i С 2 3 с 4 сД С'"
V а"" Ь"" с"" с'," С 2 3 С 4 с'б” С'"
N ап ь1 сп с? pH р?4 V 2 С 3 L 4 pH С 5 С”
Схема XXXV. Объяснение в тексте.
Представим себе, что у нас имеется какой-нибудь орган, характеризуемый признаком С, и что этот орган филогенетически развивается в течение многих поколений (I, II, III, IV, V...). Тогда, согласно принятому нами обозначению, буквы а—b—с—сг..с5—С будут символически изображать морфогенез (а—b—с) и период ростд (сг—с5) данного органа у какого-нибудь очень отдаленного предка (I) современного животного. Филогенетическое изменение в случае архаллаксиса заключается в том, что у потомков исходной формы I, т. е. у формы II, начнет изменяться в определенном направлении первый эмбриональный зачаток нашего органа (а), т. е. что этот зачаток при закладке будет, например, несколько большего размера, чем он был у предка I, или несколько иной формы, или, наконец, он появится на несколько ином месте. Все изменения этого рода мы обозначим индексом а'. При этом мы принимаем, что первичное изменение зачатка а' сохраняется в течение всех последующих стадий развития вплоть до взрослого состояния, так что ряд а—b—с—с1....с5 превращается в ряд а'—Ь'—с'—с/...c'g, а взрослое состояние органа С—в С'.
Продолжая наше рассуждение, мы можем себе представить, что наследующей стадии филогенеза изменение зачатка а' продолжалось все в том же направлении, так что у потомков, т. е. у формы III, зачаток а'превратился в зачаток а", а взрослый орган С'—в С".
Мы можем принять далее, что этот процесс продолжал итти все в том же направлении и на следующих стадиях филогенеза, т. е. на стадии IV и на стадии V, и что зачаток а" превратился с течением времени в зачаток а'", а затем и в а"" и, наконец, по истечении очень долгого промежутка времени в зачаток а”. Соответственно этому изменялось и взрослое состояние органа С, так что орган С" постепенно превратился в С'", затем в С"" и, наконец, в С”.
Дать непосредственное доказательство существования этого способа эволюции органов, мне кажется, очень трудно; тем не менее мы постараемся показать, что многие признаки животных не могут развиваться никаким иным путем, кроме этого.
Когда этот вопрос встал передо мною впервые,1 мне казалось несомненным, что число метамерно повторяющихся эмбриональных органов, из которых каждый в своем дальнейшем развитии проходит через сложный процесс диференциации, может изменяться только путем изменения первых закладок этих органов или путем архаллаксиса. Число метамерных органов уже потому одному фиксируется на относительно ранних стадиях развития животного, что для всего своего даль
1 Северцов А. Н., Эмбриология и эволюция, Дневник XII съезда русских, естествоиспытателей и врачей, Москва, 1910.
512
ГЛАВА XIV
нейшего развития и последующей диференцировки эти органы требуют известного промежутка времени.
Туловищные сегменты позвоночных, как известно, представляют собой систему метамерных, т. е. повторяющихся по всей длине тела животного, эмбриональных зачатков, претерпевающих в течение своего онтогенеза целый ряд изменений, пока они не достигнут конечной стадии морфогенеза. В туловище взрослых позвоночных животных мы находим не одну, но несколько координированных между собой систем сегментально (метамерно) расположенных органов. Примером этого служат миотомы, позвонки, ребра, сегменты нервной системы, т. е. дорзальные и вентральные нервные корешки или спинномозговые нервы, симпатические нервные корешки и ганглии, сегменты выделительной системы, т. е. мезонефрические воронки и канальцы, мальпигиевы тельца и т. д. Сейчас, однако, мы будем говорить только о мезодермальных сегментах и об их дериватах.
Мы знаем, что мезодермальные сегменты у черепных позвоночных образуются прежде всего в передней части головы; позднее сегментация мезодермы идет в двух противоположных направлениях: вперед (рострально) и назад (каудально), так что в туловище самые задние сегменты мезодермы появляются всего позднее. Мезодермальные сегменты или сомиты лежат в начале своего развития по бокам от нервной трубки и от хорды. После их отделения от висцеральной мезодермы наружная стенка каждого сомита распадается в мезенхиму; из вентромедиальной части сомита, т. е. из склеротома, образуются сегментальные части скелета туловища (дуги позвонков, тела позвонков и т. д.), из дорзомедиальной стенки сомита развиваются миотомы тела животного. Мы видим, таким, образом, что каждый сомит, после того как он образовался, проходит сложный путь гистологической и морфологической диференцировки. Предпосылкой к тому, что у взрослого животного образуется определенное число позвонков, ребер и мускульных сегментов, является факт, что у зародыша этого животного на ранних стадиях его развития закладывается такое же число индиферентных мезодермальных сегментов или сомитов.
Мы знаем, что общее число сегментов в теле того или иного рода позвоночных животных может считаться постоянным только в известных границах. Правда, у млекопитающих и у бесхвостых амфибий мы не наблюдаем особенно заметных вариаций в этом отношении, но у рептилий, птиц и рыб можно констатировать значительные колебания в общем числе сегментов тела животного. Мы можем сказать с полной определенностью, что, например, у рептилий это число сильно изменялось в течение филогенеза, причем изменялось в обоих возможных направлениях, а именно, как в направлении увеличения общего числа сегментов, так и в направлении его уменьшения.
На основании сравнительно анатомических и палеонтологических данных мы можем определенно сказать, что у предков современных змей число туловищных сегментов сильно увеличилось в течение филогенеза. У примитивных рептилий число позвонков было не очень велико; например, в позвоночнике у Palaeohatteria имелось приблизительно 80 позвонков, из которых 50 приходилось на хвостовой отдел. У современного Sphenodon зто число еще меньше: у него имеется всего 61 позвонок. У крокодилов мы находим 2&предхвостовых и от 34 до 40 хвостовых позвонков. У ящериц число позвонков непостоянно: оно меньше у короткохвостых форм и естественно больше у длиннохвостых (от 70 до 90). У долихозауров число позвонков было больше, чем у аутозауров; например у Platecarpus их было более ПО, у Mesosaurus—120. У змей, родственных Pythonomorpha, их число стало еще более значительным, достигая 250 и даже 300 позвонков. Судя по положению клоаки и рудиментов задней конечности, мы можем сказать,
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
513
что увеличение числа позвонков касалось только туловищного отдела тела;
число хвостовых позвонков у них не увеличилось.
Чтобы получить представление о том, как именно могло произойти увеличение общего количества позвонков, ребер и мио-
томов у змеи, мы сравнили развитие мезодермальных сегментов, особенно в отношении их числа, у геккона.(Ascalabotes fascicularis) с их развитием у ужа (Tropidono-tus natrix).
На рис. 181 изображен зародыш Ascalabotes fascicularis, а на рис. 182—зародыш Tropidonotus, находящиеся, судя по общей степени развития их органов, например мозга, слуховых пузырьков, глаз, жаберных щелей и т. д., приблизительно на одной и той же стадии развития. При очень тща-
Рис. 181. Зародыш Ascalabotes fascicularis на стадии, когда у него развит© 28 туловищных сомитов (из А. Н. Северцова, 1927).
тельном сравнении можно даже сказать, что зародыш ужа является
несколько более молодым, чем зародыш геккона. Но если мы сравним
Рис. 182. Зародыш Tropidonotus natrix на стадии, когда у него развито 34 туловищных сомита (из А, Н. Северцова, 1927).
между собой число сомитов у этих двух форм, то увидим, что у более молодого зародыша ужа (Tropidonotus) это число равняется 34 сомитам, тогда как у зародыша геккона (Ascalabotes) оно равно всего 28 сомитам. Таким образом процесс сегментации ме-
зодермы у Tropidonotus с самого начала идет быстрее, чем у Ascalabotes.
Это ускорение процесса сегментации туловищной мезодермы продолжает быть резко выраженным и на следующих стадиях развития. Так,
на рис. 183 и 184 изображены голова и передняя часть туловища несколько
Рис. 183. Зародыш Ascalabotes fascicularis на стадии 42 туловищных сомитов (из А. Н. Северцова, 1927).
Рис. 184. Зародыш Tropidonotus natrix на стадии 142 туловщиных сомитов (из А. Н. Северцова, 1927).
более поздних зародышей Ascalabotes и Tropidonotus, причем взятый нами зародыш геккона снова немного старше зародыша ужа; но в то время как у зародыша геккона на этой стадии развития имеется только 42 миотома, у зародыша ужа имеется уже целых 142 миотома. Процесс увеличения числа мезодермальных сегментов у зародышей обоих видов заканчивается Морфологические закономерности 33
514
ГЛАВА XIV
на стадиях, изображенных на рис. 185 и 186; у Ascalabotes к этому времени образовалось 63 сегмента, у Tropidonotus—250 сегментов. У взрослого Ascalabotes имеется от 60 до 65 туловищных сегментов (позвонков), у взрослого Tropidonotus их имеется до 250 и даже до 260. Мы можем сказать, что у этих двух форм образование дефинитивного числа туловищных сегментов (позвонков, ребер, миотомов, спинномозговых нервов) заканчивается на сравнительно ранних стадиях эмбрионального развития, т. е. на таких стадиях, когда развитие других органов (конечностей, головы ит. д.) является еще далеко незаконченным (см. рис. 185 и 186).
Совершенно ясно, что здесь филогенетическое изменение (в данном случае весьма значительное увеличений числа туловищных сегментов) может заключаться лишь в изменении хода развития (морфогенеза) на очень ранних стадиях, и мы знаем,
Рис. 185. Зародыш Ascalabotes fasci-cularis на стадии 63 туловищных сомитов (из А. Н. Северцова, 1927).
Рис. 186. Зародыш Tropidonotus natrix на стадии 250 туловищных сомитов (из А. Н. Северцова, 1927).
в чем именно это изменение состоит: у предков змей в свое время имело место резкое ускорение (акцелерация) развития мезодермальных сегментов и спинномозговых нервов на самых ранних стадиях развития этих органов. Противоположное предположение, а именно, что здесь эволюция, может быть, совершалась по типу анаболии или девиации, совершенно неприемлемо, ибо если бы это было так, то мы должны были бы ожидать, что первоначальное развитие сегментов как у Ascalabotes, так и у Tropidonotus шло одинаковым темпом и что акцелерация могла наступить только на поздних стадиях развития, в течение периода роста. Но приведенные нами эмбриологические факты говорят против этого предположения самым решительным образом.
Итак, мы можем принять, что увеличение числа метамерно повторяющихся органов во многих случаях происходит путем этого модуса филогенетического изменения органов, т. е. путем архаллаксиса.
Возвращаясь к нашей схеме (схема XXXV), мы видим, что ход онтогенетического развития разбираемого нами признака (т. е. признака увеличенного числа туловищных сегментов в змеином теле) соответствует п о-с л е д н е й стадии нашего гипотетического филогенеза, т. е. стадии N; начало же увеличения числа сегментов падает именно на первые стадии морфогенеза сомитов, и процесс этот заканчивается гораздо раньше, чем морфогенез других органов ужа (рис. 186).
Другой пример развития органов, совершающегося по типу изменения первых стадий морфогенеза, или по типу архаллаксиса, мы находим при образовании торакального и югулярного положения брюшных плавников
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
515
у высших костистых рыб. Как известно, брюшные плавники у примитивных костистых рыб (Salmonidae, Clupeidae) имеют абдоминальное положение и лежат близ анального отверстия (рис. 187, vt). У некоторых форм, как, например, у Atherina hepsetus, эти плавники (рис. 163, 188, vt) сдвинулись вперед и лежат недалеко от грудных плавников (pct), несколько позади от них. У Acanthopterygii1 брюшные плавники находятся либо непосредственно позади, как, например, у Sargus (рис. 189, vt), либо даже, как, например,.у Lophius (рис. 164, vt) и у Gobius (рис. 190, vt), непосред-
Рис. 187. Малек Salmo trutta (21 мм длиной), у которого брюшной плавник расположен вблизи анального отверстая.
' а—анальный плавник; ап—анальное отверстие; с—хвостовой плавник; d,—спинной плавник; н—носовые ямки; ра—преанальный плавник; pct—грудной плавник; vt—брюшной плавник (из А. Н. Север-пова, 1927).
ственно впереди от грудных плавников (pct), так что тазовый пояс у этих форм сочленяется с плечевым поясом (cleitrum). Подобное положение брюшных плавников обычно обозначается как торакальное или югулярное. Интересно, что брюшные плавники из области задних спинномозговых нервов, которыми они иннервируютсжу всех Pisces abdominales, перешли в область передних спинномозговых нфвов. Так, у Lophius грудные плавники иннервируются IV, V и VI парой спинномозговых корешков, а брюшные плавники—VII, VIII, IX, X парой; у Gobius к грудным плавникам
Рис. 188. Малек Atherina hepsetes (18.5 мм длиной), у которого брюшной плавник расположен несколько позади от грудных плавников.
а—анальный плавник; ап—анальное отверстие; с—хвостовой плавник; dt—d2—передний и задний спинные плавники; п—носовое отверстие; pct—грудной плавник; vt—брюшной плавник (из А. Н. Северцова, 1927).
идут I и II пара спинномозговых нервов, а к брюшным—III и IV пара, и т. д. Мы видим, таким образом, что факт сдвигания брюшного плавника в ростральном направлении отражается не только на скелете этих плавников, но и на их мускулатуре и на их иннервации.
Исследуя эмбриональное развитие брюшных плавников у Gobius и у Lophius, мы находим, что эти плавники у них закладываются в той же области, в которой они лежат впоследствии и у взрослой рыбы, и что они, таким образом, не изменяют своего положения в течение онтогенеза. Если мы допустим, что изменение положения брюшных плавников совершалось по типу надставки конечных стадий или по типу девиации, то мы должны были бы ожидать,
1 За исключением сем. Percesocidae.
33*
516
ГЛАВА
XIV
что брюшные плавники будут закладываться в абдоминальной области тела животного и что они в течение онтогенеза будут постепенно продвигаться в ростральном направлении. Такие случаи, вообще говоря, мы знаем. Так, например, мы наблюдали подобный процесс в некоторых случаях изменения положения конечностей. Я сам, изучая эмбриональное развитие гекконов (Ascalabotes fascicularis),1 нашел, что у них передние конечности закладываются впереди от того места, где они впоследствии находятся, у взрослого животного и что в образова-
ние. 189. Малек Sargus vulgaris (10 мм длиной). Вид снизу.
У малька Sargus брюшные плавники (vt) расположены непосредственно позади грудных плавников (pct); ап—анальное отверстие; pct—грудной плавник; vt—брюшной плавник (из А. Н. Северцова, 1927).
нии нервного сплетения конечностей геккона принимают участие задние шейные нервы. На более поздних стадиях развития ветви, идущие от шейных нервов к конечностям, редуцируются, а сами конечности сдвигаются назад. Здесь перед нами типичный пример развития, идущего путем анаболии, принципиально отличный от^разбираемого нами случая сдвигания брюшных плавников в ростральном направлении у Acanthopterygii, ибо в последнем случае конечности не передвигаются в течение онтогенеза, как это имеет место у геккона, а закладываются на том именно месте, на котором они находятся
Рис. 190. Малек Gobius capito (7,5 мм длиной), у которого брюшные плавники (vt) расположены впереди грудных плавников (pct)
ап—анальное отверстие; pct—грудной плавник; vt—брюшной плавник (из А. Н. Северцова, 1927).
у взрослого животного и иннервируются теми нервами, которые здесь лежат. Поэтому мы имеем все основания принять, что изменения положения брюшных плавников происходят в данном случае не путем анаболии, но путем архаллаксиса, т. е. путем изменений, происходящих на первых стадиях морфогенеза этого органа.
Для пояснения этого процесса мы приводим нижеследующую диаграмму (схема XXXVI). Следуя этой схеме, мы можем представить себе, что у предков тех рыб, которые имеют теперь торакально расположенные брюшные плавники, эти плавники, как и у примитивных Teleostei, были распо
’ Севе рцов, А. Н., 1908.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
517
ложены абдоминально и иннервировались 18—20 парами спинномозговых нервов; грудные же плавники иннервировались 2—4 парами нервов и лежали напротив этих нервов. Первое филетически важное изменение в положении брюшных плавников (схема XXXVI, стадия филогенеза II) заключалось, по нашему мнению, в том, что эмбриональная закладка этих плавников впервые появилась несколько впереди от того места, где она обычно появлялась у предков, т, е. приблизительно в районе расположения 17—19 пары спинномозговых нервов. Мы знаем, что
Схема XXXVI. Объяснение в тексте.
подобные вариации в местоположении конечностей происходят не только у рыб, но и у Quadrupeda (см. субституцию функций в главе XI). Это новое местоположение брюшных плавников сохранилось и у взрослых форм.
На следующей стадии филогенеза (схема XXXVi, стадия III) этот процесс сдвигания пошел еще дальше и брюшные плавники стали уже закладываться не в районе 17—19 пары спинномозговых нервов, но в районе расположения 16—18 пары, передвинувшись, таким образом, еще более вперед. На схеме XXXVI мы ясно видим (стадии филогенеза III —XIV), как это смещение эмбриональных закладок брюшных плавников, постоянно идущее в определенном направлении, т. е. сзади вперед, в конце концов может привести (и действительно приводит) к торакальному положению брюшных плавников.
Все перечисленные нами выше архаллаксисы мы можем рассматривать как первичные архаллаксисы.
Но есть другая группа архаллаксисов, которую мне пришлось впервые наблюдать при исследовании эволюции брюшных плавников рыб (1934).1
1 Sewertzoff A. N., Evolution der Bauchflossen der Fische, Zook Jairrb., Bd. 58, 1934.
518
ГЛАВА XIV
Исследуя филогенетическое развитие брюшных плавников Acipenseridae, (см. принцип выпадения промежуточной функции, гл. XI) я нашел, что оно происходит путем надставки стадий, т .е. путем анаболии. Здесь в онтогенезе брюшного плавника последовательно сменяются: 1) стадия метамерной закладки basalia (рис. 191 А), 2) стадия охрящевения
Рис. 191 А. Передний зачаток скелета брюшного плавника Acipenser ruthenus. bd+bp—общий мезенхимный зачаток basalia proximalia и basalia distalia; pnv—контур брюшного плавника: гх—г9—первые зачатки плавниковых лучей 1—9.
Рис. 191 В. Несколько более поздняя стадия закладки брюшного плавника Acipenser ruthenus.
bdx—bd,—basalia distalia 1—9; bpx—bp,—basalia proximalia 1—9; Iptr—лепидотрихии; pnv.—контур брюшного плавника; rx—г,—плавниковые лучи 1—9.
Рис. 191 С. Горизонтальный разрез через еще более позднюю стадию закладки скелета брюшного плавника Acipenser ruthenus (срастание basalia).
bd,—bd4—срастающиеся между собой basalia distalia; bd6—несросшееся basale; bp,—bp,—срастаю-щиеся между собой и с basalia distalia basalia proximalia; rx—r5—плавниковые лучи 1—5 (из A. H. Северцова, 1934).
basalia (рис. 191 В), 3) стадия их срастания (рис. 191 С), 4) стадия поворота оси базиптеригия и т. д. Исследуя эволюцию брюшных плавников Holostei . и Teleostei, несомненно происшедших из плавников типа Acipenseridae, я установил, что в онтогенезе этих групп первые три стадии развития брюшных плавников Acipenseridae полностью выпадают и вместо них происходит образование цельного базиптеригиального зачатка в виде сплошного,
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
519
сначала мезенхимного, а затем хрящевого стержня (рис. 192 А). Позднее образуется рудиментарный метаптеригий и еще позднее—рудиментарные плавниковые лучи (рис. 192 В).
Ясно, что мы имеем здесь в образовании нового цельного базиптеригия типичный положительный архаллаксис (ибо он ведет к прогрессивному развитию органа), но это—архаллаксис особого рода, так как он замещает собою первичный способ развития плавника, который шел путем анаболии. Эту группу архаллаксисов мы назвали вторичными архаллаксис а м и. Вторичные архаллаксисы обычно имеют место тогда, когда орган, развивавшийся у предков путем анаболии, начинает развиваться
у потомков путем архаллаксиса. При этом ряд начальных стадий развития (стадий анцестральной рекапитуляции) выпадает, онтогенез укорачивается (акцелерация), и образуется новый, несколько иной зачаток того же органа. В данном случае в результате развития этого зачатка конечное состояние
Рис. 192 А. Горизонтальный разрез через закладку плавника Amia (очень ранняя стадия развития).
Bspt—прохондральный зачаток базиптеригия; Bspt*—каудальный мезенхимный край базиптеригия; mtpt—мезенхимный зачаток метаптеригия; pnv—очертание плавниковой складки (из А. Н. Северцова, 1934).
Рис. 192 В. Брюшной плавник несколько более позднего зародыша Amia.
Bspt—хрящевой стержень базиптеригия; mtpti, mtpta, mtpt3—1-й, 2-й и 3-й членики метаптеригия; pnv—контур плавниковой лопасти; г*2—мезенхимная закладка рудиментарного плавникового луча (из А. Н. Северцова, 1934).
достигается быстрее, чем у предков, и брюшной плавник потомков начинает функционировать раньше, чем он функционировал у предков.
Таким образом, мы различаем две категории положительных архаллаксисов: первичные и вторичные арха л л а к с и с ы.
Мы принимаем, что изменения первых стадий развития, обусловливающих собою изменение всего дальнейшего хода морфогенеза животных, т. е архаллаксисы, развиваются постепенно в течение эволюции. Весьма вероятно, что они происходят путем суммирования нерегулиру-ющихся эмбриональных вариаций.
Как известно, эмбриональные вариации могут быть подразделены на две категории.
1. Эмбриональные вариации, выравнивающиеся в течение дальнейшего развития и не оказывающие, таким образом, никакого влияния на
520
ГЛАВА XIV
строение взрослого органа;1 такие вариации обычно обозначаются как регулирующиеся вариации хода эмбрионального развития.
2. Эмбриональные вариации, имеющие определенное влияние на строение образующихся из этих зачатков органов. Этот последний тип вариаций мы обозначаем как нерегулирующиеся эмбриональные вариаций.1 2
Из исследований К. Петера,3 произведенных над зародышами различных беспозвоночных животных, мы знаем, что зародыши вариируют по тем же законам, что и взрослые животные. Многие эмбриональные вариации являются нерегулирующимися вариациями и имеют определенное влияние на строение соответствующих органов взрослых животных, а поэтому мы можем принять, что эмбриональные вариации до известной степени наследственны.
Оба эти вида эмбриональных вариаций одинаково важны для эволюции органов животных. Так, нерегулирующиеся вариации, встречающиеся на первых стадиях развития, являются именно теми изменениями строения органов, которыми начинаются архаллаксисы. Мы можем, например, легко себе представить, что смещение в ростральном направлении брюшных плавников, происходившее в течение филогенеза у предков Gobius, Lophius и других Acanthopterygii, было обусловлено тем, что у этих форм постепенно накоплялись (суммировались) мелкие, нерегулирующиеся наследственные эмбриональные вариации в положении и иннервации брюшных плавников (схема XXXVI; I, II, III, IV и т. д.). Мы знаем, что такого рода вариации часто происходят у рыб, и поэтому принимаем, что нерегулирующиеся эмбриональные вариации, имеющие место в начальных стадиях развития, дают материал для эволюции признаков взрослых животных, происходящей путем архаллаксиса.
Весьма вероятно, что и регулирующиеся эмбриональные вариации имеют свое филогенетическое значение. Мы хорошо знаем, что существуют чисто эмбриональные органы, редуцирующиеся у взрослых животных, которые образуются у зародышей и личинок в течение филогенеза. Геккель, как известно, обозначил эти органы, имеющие громадное биологическое значение для тех форм, у которых они встречаются, именем ценогенезов. Не входя в подробное обсуждение вопроса о происхождении ценогенезов, отметим все же, что все эти образования происходят, по нашему мнению, путем суммирования естественным отбором регулирующихся эмбриональных вариаций. Мы принимаем, что в течение филогенеза накопляются те регулирующиеся эмбриональные вариации, которые имеют функциональное значение для животного в течение его эмбриональной жизни, и что из них именно развиваются те ценогенетические или провизорные органы (признаки), которые исчезают у взрослого животного.
Рассматривая эволюцию, происходящую путем анаболии, мы пришли к выводу, что рекапитуляция анцестральных признаков является следствием именно этого способа (модуса) филогенетического развития. Мы видели, что при этом повторяются не признаки взрос
1 У различных индивидуумов одного и того же вида эмбриональные зачатки различны; взрослые органы одинаковы.
2 У различных индивидуумов одного и того же вида эмбриональные зачатки различны; взрослые органы также различны.
’Peter С., Experimentelle Untersuchungen iiber die individuelle Variation in der tierischen Entwicklung, Arch. f. Entw. Meeh., Bd. 27, 1907.
Idem. Neue experimentelle Untersuchungen iiber die Grosse der Variabilitat usw., Arch. f. Entw. Meeh., Bd. 31, 1911.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
521
л ы х предков, но лишь конечные стадии морфогенеза анцестральных признаков, похожих по всему своему характеру на признаки взрослого животного. Мы уже тогда отметили, что при этом типе эволюции морфогенез потомков удлиняется в течение филогенеза путем постепенной надставки конечных стадий, и мы видели, что рекапитуляция происходит только тогда, когда имеет место эта надставка конечных стадий.
При эволюции, идущей путем архаллаксиса, не имеет места ни надставка конечных стадий, ни удлинение морфогенеза; при архаллаксисе мы не находим никаких рекапитуляций анцестральных признаков у потомков. Признаки, развивающиеся этйм способом, не подчиняются и рассмотренному нами выше закону К. Бэра.
Выше мы пришли к выводу, что торакальное и югулярное положение плавников у акантоптеригий образовалось путем архаллаксиса. Я изучал развитие таких плавников у некоторых акантоптеригий и нашел, что, например, у Sargus и у Gobius брюшные плавники имеют торакальное расположение уже на очень ранних стадиях развития (см. рис. 189 и 190). Таким образом, этот новый характерный признак образуется чрезвычайно рано. С другой стороны, мы знаем, что присутствие чешуй является характерным признаком для всех костистых рыб, представляя собой, таким образом, общий признак для целой группы форм, к которой принадлежат также и акантоптеригий. Непосредственные наблюдения показали, однако, что у акантоптеригий чешуи закладываются гораздо позднее, чем торакальные брюшные плавники. Мы видим, таким образом, что признак, характеризующий мелкую систематическую группу (акан-топтеригии), закладывается в онтогенезе животного раньше, чем признак, характеризующий крупную систематическую группу (Teleostei), к которой эта мелкая группа принадлежит. Таким образом, здесь мы имеем перед собою исключение из закона К. Бэра.
К аналогичному результату придем мы и тогда, когда будем сравнивать образование торакальных брюшных плавников с образованием какого-нибудь другого признака, характерного для всех костистых рыб, например с образованием гомоцеркального хвостового плавника. Наблюдение показало нам, что признак «гомоцеркальная форма плавника» (признак отряда Teleostei) образуется позже, чем признак «торакальное положение брюшного плавника» (признак подотряда Acanthopterygii) (А. Н. Северцов, 1927). То же самое находим мы и в целом ряде других случаев. Из всех этих примеров можно сделать вывод, что все признаки, образующиеся путем первичного изменения первых стадий развития, т. е. путем архаллаксиса, в своем развитии не подчиняются закону К. Бэра, т. е. что здесь древние (общие) признаки не образуются в онтогенезе раньше новых (менее общих) признаков. Эти новые признаки в тех случаях, когда эволюция идет по типу архаллаксиса, развиваются в то же время, что и более древние признаки, а иногда даже и раньше их.
Таким образо?л, как это видит читатель, выводы, к которым мы здесь приходим, сильно отличаются от ходячих воззрений на биогенетический закон и на закон К; Бэра.
Обычно принимают вместе с Геккелем, что закон рекапитуляций (биогенетический закон) действительно является основным законом филогенеза, т. е., что все новые признаки взрослых предков первично сдвигаются на ранние стадии онтогенеза и рекапитулируются у потомков и что эти признаки впоследствии вторично затемняются и вытесняются»
522
ГЛАВА XIV
эмбриональными приспособлениями (ценогенезами). Закон К. Бэра при этом рассматривается как общераспространенный закон.
По нашему мнению биогенетический закон имеет несколько иное значение, чем то, которое ему обычно приписывается авторами: рекапи-
туляция признаков.взрослых предков, т. е. признаков последних стадий морфогенеза, похожих на признаки взрослых предков и могущих служить целям филогенетического исследования, имеет место лишь при одном из. трех способов (модусов) филогенетического исследования, именно при эволюции, идущей путем анаболии, т. е. при надставке конечных стадий морфогенеза. При этом типе эволюции повторение признаков предков происходит всегда, и здесь мы, действительно, с полным правом можем говорить о законе рекапитуляций. При двух других
типах эволюции, т. е. при архаллаксис. е и при эволюции, происходящей путем девиации на средних стадиях онтогенеза, рекапитуляция признаков взрослых предков у потомков не.происходит вовсе. Правда, при девиации рекапитулируются ранние эмбриональные признаки предков, но для целей филогенетического исследования они обычно не имеют никакого значения.
Рекапитуляция у потомков анцестральных признаков, сходных с признаками взрослых предков, не является, как это часто принято говорить, каким-то «капризным и своенравным» процессом эволюции, имеющим место в одних случаях и не имеющим его в других; рекапитуля-ц и я—э то такой процесс, который всегда происходит при одном определенном способе эволюции (анаболия) и никогда не происходит при других ее способах. Затемнение рекапитуляций анцестральных признаков различного рода ценогенетическими процессами, по моему мнению, сильно переоценивалось. Обычно, когда исследователь не находит ожидаемых им рекапитуляций, он говорит, что здесь, по всей вероятности, рекапитуляции «затемнены» или «нарушены» ценогенезами. При этом большей частью эти «затемняющие» эмбриональные приспособления не исследуются вовсе. Часто говорят о ценогенетических акцелерациях, ретардациях и т. д., совершенно не входя в ближайшее рассмотрение этих процессов с биологической точки зрения, что, конечно, было бы крайне желательно, а в иных случаях даже более того—совершенно необходимо.
Мы знаем, что и сильно выраженные ценогенетические признаки, например образование большего количества питательного желтка, очень мало, а иногда и вовсе не отражаются на многих процессах эмбрионального развития, сопровождаемых рекапитуляциями. Если мы сравним развитие хорды и центральной нервной системы, сегментацию мезодермы (включая и образование головных сегментов) и пр. в полилецитальных и олиголе-нитальных яйцах позвоночных, то мы увидим, что все эти процессы в общем весьма мало нарушаются от присутствия большого количества желтка. Даже образование таких сложно построенных эмбриональных органов, какими являются, например, амнион и аллантоис у птиц, рептилий и мле- • копитающих, или присутствие длинных нитей наружных жабр у эласмобранхий ит. п., очень мало влияет на развитие соседних органов. Мы, конечно, не отрицаем влияния ценогенезов на ход эмбрионального развития организма, но думаем, что их «нарушающее» действие весьма переоценено авторами и что для правильного понимания этого процесса нужно •тщательно изучать ценогенезы в каждом отдельном случае.
Мне думается, что вся проблема вторичных приспособительных измене
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
523
ний, имеющих место в ходе анцестрального онтогенеза, еще недостаточно созрела и для своего окончательного разрешения настоятельно требует дальнейшего специального изучения и исследования.
ПЕРИОДИЧЕСКОЕ ИЗМЕНЕНИЕ ХОДА РАЗВИТИЯ ОРГАНОВ
Кроме разобранных нами модусов филэмбриогенеза нам нужно еще отметить так называемые периодические изменения строения органа, т. е. такой способ его развития, когда эволюирующий орган доходит до известной стадии развития, до известного конечного результата, после чего весь орган или какой-либо участок его отпадает; затем, в соответствующий период жизни животного, из остатка того же зачатка, из которого развивался орган, он начинает развиваться наново. Процесс этот повторяется из года в год вплоть до взрослого состояния.
Этот модус, редко встречающийся у позвоночных животных, но очень распространенный среди членистоногих, мы обозначили как «способ периодического изменения хода развития органо в».
Самым простым примером развития органов по этому способу является процесс линьки у высших позвоночных, чрезвычайно напоминающий собою периодическую линьку у членистоногих, а именно у ракообразных и у насекомых.
Как известно, у ракообразных и насекомых весь наружный хитиновый скелет образуется за счет эпидермиса, как выделение и преобразование его наружного слоя. При каждой линьке этот твердый скелетный панцырь со всеми своими добавочными образованиями, т. е. со скульптурой наружного покрова и хитиновыми частями органов высших чувств, отпадает и заменяется новым слоем хитина, который образуется из того же эпидермического зачатка и который дает начало всем отпавшим в течение первой линьки органам. При этом хитиновый панцырь сильно увеличивается в размерах соответственно росту мягких частей тела животного и пропорционально этому делается толще. Таким образом, период смены панцыря есть в то же время период роста всего тела и всех его частей (конечностей, клешней, усиков, ротовых органов и т. д.). Здесь мы имеем эмбриональный зачаток органа, потенция которого изменяется периодически каждой весной, (после линьки) и из которого скачками развиваются все новые и новые слои хитина с новой скульптурой и с новыми придаточными частями органов чувств. Мы имеем основание предположить, что у членистоногих этот модус является очень древним модусом развития, так как здесь мы не находим, или, вернее, мало находим надставок стадий: слой хитина после линьки мало растет в толщину.
При линьке рептилий мы находим, что в средней части мальпигиева слоя эпидермиса под новообразующимся роговым слоем развивается система упругих волосков, которые отделяют сбрасывающийся наружный слой эпидермиса от внутреннего остающегося слоя. У змей весь наружный слой отделяется при этом целиком, так что линяющая змея в буквальном смысле слова выползает из старой кожи, остающейся в виде сплошного прозрачного футляра. Таким образом при линьке змей внутри эпидермиса каждой весной происходит сложное гистологическое изменение клеток, определяющее рост тела змеи в течение следующего года.
Как известно, у гекконов на брюшной поверхности лапок находятся поперечные складки кожи, покрытые снаружи целой массой тонких упругих волосков. Прикладывая лапы к гладкой поверхности и выдавливая воздух, геккон как бы прилипает к поверхности. Этим объясняется то, что он может свободно держаться и бегать по вертикально стоящим скалам или по потолку комнат. Упомянутые упругие волоски геккона заменяются новыми при каждой линьке. Это показывает нам, что роговые воЛоски
524
ГЛАВА XIV
могут быть использованы как приспособления не только при сбрасывании кожи, но и при бегании.
Итак, мы видим: 1) что линька, т. е. смена кожи, происходит периодически каждый год и 2) что при линьке всегда происходят сложные гисто-генетические изменения, правильно повторяющиеся через определенные промежутки времени.
Несколько иначе происходит линька у птиц. Здесь каждый год отпадает не весь эпидермис кожи, а только одни перья. Исключение составляют пингвины, у которых при линьке сходит наружный слой эпидермиса и которые в этом отношении несколько напоминают своих реп-тилийных предков. У куриных перья на крыльях развиваются очень рано, так что маховые перья заканчивают свое развитие еще у очень молодой птицы. При первой линьке, т. е. через год после вылупления, на крыльях куриных развиваются длинные и широкие перья, соответствующие по своей величине перьям взрослой птицы, так что перья первой линьки по величине отличаются от птенцовых перьев; другими словами, только при первой линьке перья достигают значительной величины, приближающейся к величине перьев взрослой птицы. Если мы вспомним, что это увеличение размеров касается роста всех частей пера,т.е. бородок, крючочков ит.д., то мы придем к выводу, что здесь происходят довольно сложные морфологические изменения в строении перьев.
Еще более сложные изменения происходят при смене рогов у оленей. Известно, что детеныши оленя родятся безрогими и что только с конца первого года жизни у них на лобных костях начинают развиваться рога в виде прямых костных отростков, еще покрытых кожей с волосами и не имеющих разветвлений (см. рис. 193 А). На следующий год у основания этого костного стержня, между ним и лобной костью, образуется полость (рис. 193 В, х), выполненная тканью паренхимы надкостницы и кровеносными сосудами. Полость эта постепенно увеличивается в размерах и, наконец, первичный костный стержень рога отваливается (рис. 193 С), а над образовавшейся ранкой разрастается кожа головы и закрывает ранку (рис. 193 D). Весной следующего года на том же месте из надкостницы вырастает новый стержень (рис. 193 Е), сначала паренхимный, пронизанный кровеносными сосудами, отходящими от артериального сосудистого кольца, окружающего основание рога (в свою очередь отходящего от arteria temporalis головы оленя), а затем костный. Но этот отросток не остается прямым стержнем, подобно отростку первого поколения, но раздваивается (что, повидимому, определяется направлением роста артериальных сосудов рога1) и образует так называемый глазной отросток (рис. 193 FhG,z) и продолжение стержня рога (рис. 193 F и G, у). Еще через год рога оленя снова отпадают и через некоторое время из того же зачатка вырастают снова, причем этот новый отросток, так называемый нижний отросток, вырастает от конца стержня. При следующей смене образуется третий отросток и т. д. В общем каждый новый отросток образуется путем дихотомического деления конца стержня. Так идет дело до конца седьмого года, когда у оленя вырастает полный рог с семью отростками. На этом процесс развития рогов у оленя заканчивается.
Мы видим, таким образом, что онтогенетическое развитие зачатка второго поколения рогов идет в несколько другом направлении, чем развитие рогов первого поколения той же особи, и что это изменение направления развития повторяется и в каждом следующем году, благодаря чему образуются новые дополнительные костные отростки рогов оленей при каждой периодической смене рогов. Мы здесь, действительно, имеем особый способ
1 Rhumbler L., Der Arterienverlauf auf des Zehnerkolbenstange von Cervus elafus L. und sein Einfluss auf die Geweihform, Ztschr. f. wissen. Zool., B. 115, 1916.
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
525
эволюции, при котором на некоторой стадии развития появляется зачаток нового органа (дополнительный отросток), который некоторое время растет в определенном направлении; затем рог с одним, двумя, тремя и т. д.
Рис. 193 А—G. Схема развития рогов у оленей.
А—развитие первого рога. В и С—сбрасывание первого рога; в стадии В происходит сбрасывание кожи и начинается резорбция кости (образование полости х); в стадии С стержень первого рога (у) отпадает. D и Е—разрастание кожи и заживление раны. F—начало развития второго рога (глазного отростка z). G—завершение развития глазного отростка z и новое сбрасывание кожи.
сог.—corium; epid.—эпидермис; fr.—лобная кость; х—полость, образовавшаяся между первым рогом и лобной костью при сбрасывании первого рога; у—стержень рога первого поколения; z—рог второго поколения или глазной отросток (из И. И. Шмальгаузена, по Нитше, 1935). *
отростками отпадает почти полностью, а из ничтожной оставшейся части (остатка основания рога) вырастает новый зачаток, подобный прежним зачаткам, но с новой потенцией роста на конце.
526
ГЛАВА XIV
Рост и смену рогов у оленей мы можем выразить при помощи нижеследующей диаграммы (схема XXXVII).
I а — а,— а, — О
/Ь
II а!—а1 — а,—а, — О /Ь /с
III а2 а2 а2 а2 а2 О
/Ь /С yd
IV а3 а3 а3 а3 а3 — а3 • О
Схема XXXVII. Объяснение в тексте.
В нашей схеме буквой а мы обозначаем еще мезенхимный зачаток рога; тогда at будет обозначать выросший из него прямой стержень, отваливающийся при следующей смене рогов, что мы обозначаем буквой О. Таким образом, первая строка нашей схемы иллюстрирует рост рогов первой смены. На второй год рост стержня изображен несколько измененным: зачаток at обладает теперь уже двойной потенцией, он производит глазной отросток b и растет дальше. Следующее поколение рогов (III) имеет еще новую потенцию: кроме продолжения роста стержня рога он дает еще средний отросток с. На четвертый год (IV), помимо роста стержня, глазного и среднего отростка, появляется еще третий отросток d; к концу года и этот стержень отваливается, что мы снова обозначаем через букву О.
Таким образом, мы видим, что рост рогов является процессом, аналогичным линьке других животных. Действительно, несмотря на все разнообразие животных, у которых происходят филэмбриологические изменения, протекающие по этому способу развития, в самих процессах гистогенети-ческого изменения существуют общие черты, которые позволяют сказать, что во всех этих случаях мы имеем дело с одним и тем же способом филэмб-риогенетических изменений. Во-первых, все рассмотренные нами признаки относятся к наружным покровам в широком смысле этого слова; во-вторых, во всех рассмотренных случаях периоды роста зачатков органов отделены друг от друга одинаковыми и притом довольно значительными промежутками времени; в-третьих, в течение каждого периода роста происходят сложные гистогеыетические изменения, причем скорость этих гистогенетических процессов со временем возрастает, и, наконец, в-четвертых, закономерно изменяется при последовательных процессах роста и направление гистогенетических изменений (потенция и направление роста).
С другой стороны, между различными случаями описанного нами периодического изменения органов имеются и более или менее существенные отличия. Так, в описанных нами примерах периодической линьки у рептилий мы имели перед собою случаипериодического изменения органов, когда каждая смена органа подготовляется изменением его гистологического строения (образование роговых волосков в эпидермисе), и весь орган сбрасывается как одно целое. При линьке у птиц отпадает не весь орган, но лишь некоторые сложно построенные части его (перья); при этом в некоторых случаях, например у куриных, второе поколение перьев несколько отличается от первого. В случае же смены рогов у оленей мы имеем при каждой смене изменение как в потенции, так и в направлении роста.
Таким образом, здесь мы имеем перед собой некоторый, как нам кажется, особый способ изменения хода филэмбриогенеза, который еще нуждается в сложном и трудном дополнительном анализе.
Правда, можно было бы предположить, что разобранный нами случай смены рогов у оленей представляет собой лишь своеобразную комбинацию
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
527‘
двух известных нам способов филэмбриогенеза, а именно архаллаксиса и анаболии. Архаллаксисом является образование нового зачатка с определенной потенцией роста (схема XXXVII, мезенхимные зачатки рога аъ а2, а3...); растут же эти зачатки по типу анаболии, причем при каждом новом' периоде роста интенсивность роста и его направление меняются. Но, с другой стороны, мы не можем не признать, что имеются-весьма существенные отличия между обыкновенными анаболиями (см. схема XXIX) и анаболиями, происходящими при эволюции рогов оленей. В этом последнем случае развитие зачатков 2-го, 3-го и последующих годов идет иначе, чем развитие зачатка 1-го года, т. е. потенция этого зачатка в течение жизни животного несколько раз меняется. Вместе с тем существует также и отличие между обыкновенными архаллаксисами (см. диагр. XXXV) и архаллаксисами, имеющими место при развитии путем периодического изменения хода развития органов. Так, при эволюции позвонков у безногих рептилий, происходящей, как это мы знаем, путем архаллаксиса, мутации позвонков происходили у филогенетически последующих поколений (диагр. XXXV, I, II, III, IV, V ряды), т. е. у разных особей в ряду предков—потомков. В разобранном же нами случае они происходят у одной и той же особи в одном и том же органе в последующие периоды жизни этой особи.
В настоящее время мы затрудняемся сказать, насколько полно мы перечислили случаи, подходящие под этот тип филэмбриогенетических изменений органов. Например, мы не знаем еще, можно ли сюда отнести явления смены зубов. У многих примитивных форм, например у рыб,, зубы сменяются в течение всей жизни по мере изнашивания. В. Васнецов,1 изучивший развитие и смену зубов у карповых рыб, рассматривает это явление как пример анаболии особого рода, так как здесь последовательное появление новых признаков происходит не у одного органа, но у нескольких сменяющих друг друга органов. Что касается до млекопитающих, то у них происходит только одна смена зубов и притом, невидимому, независимо от их пригодности или непригодности к выполнению функции жевания.
ГЕТЕРОХРОНИЯ И ФИЛОГЕНЕЗ
В предыдущем мы рассмотрели вопрос о некоторых способах, которыми осуществляются изменения хода онтогенеза животных, т. е. вопрос о модусах филогенетического изменения органов взрослых животных. Мы упомянули вскользь об основных процессах онтогенеза, об общем и частном росте (неравномерное деление клеток) и о гистогенезе. Затем мы рассмотрели более детально вопрос о переселении клеток и о периодическом изменении хода развития органов и, наконец, очень подробно остановились на вопросе об архаллаксисе, девиации и анаболии, ибо эти три последних модуса филэмбриогенеза являются наиболее важными способами прогрессивной эволюции органов. Обратимся теперь к гетерохрониям, т. е. к вопросу о более поздних закладках и более медленном развитии и о более ранних закладках и более быстром развитии органов.
В настоящее время вопрос об изменении темпа онтогенетического развития, т. е. вопрос о гетерохронии, рассматриваемой как самостоятельный способ филогенетического изменения' взрослых органов, выступил на первый план. Уже Менерт высказал предположение, что онтогенетические акцелерации и ретардации являются признаками прогрессивной и регрессивной эволюции, а в послед
1 Васнецов В. В., 1. с.
528
ГЛАВА XIV
нее время Гарстангиде Беер приписывают гетерохрониям очень большое филогенетическое значение. Что касается меня лично, то я не придаю чрезмерного значения вопросу о темпе онтогенеза, но я согласен с тем, что во многих случаях ускорение и замедление в онтогенетическом развитии частей органов могут привести к некоторым изменениям структуры органов взрослых форм.
Мы знаем, что при развитии тех или иных органов очень часто встречаются мелкие вариации во времени закладки и в темпах морфогенеза этих органов. Мы можем заключить, что некоторые из этих мелких вариаций, являющихся наследственными, представляют собой тот материал, из которого медленно (путем суммирования) образуются те значительные изменения во времени закладки и в быстроте онтогенетического развития органов, т. е. те акцелерации и ретардации, о которых только что была речь. Мы знаем вместе с тем, что значительные изменения хода морфогенеза, особенно если в результате появляются изменения в признаках взрослых форм, не лишены биологического значения. Поэтому является чрезвычайно вероятным, что втех случаях, когда филогенетически развиваются более или менее значительные гетерохронии, они являются полезными для данного организма во время его онтогенеза или во взрослом состоянии. Естественно возникает вопрос: в каких случаях гетерохрониям можно приписать биологическое значение и какие приспособления могли развиться этим путем? Иначе говоря, нам приходится поставить вопрос о функциональном значении гетерохронии.
Начнем с проблемы акцелерации, и спросим себя, когда ускоренное эмбриональное развитие какого-нибудь органа может быть полезно организму, у которого закладывается этот орган?
Ранняя закладка и быстрое развитие какого-либо органа ведут обычно к тому, что морфогенез этого органа завершается раньше, чем морфогенез остальных (не акцелерированных) органов, так что данный орган становится способным к функционированию раньше, чем другие органы животного. Непосредственное наблюдение приводит нас к выводу, что завершение морфогенеза того или иного органа, т. е. его готовность к функционированию, наступает обычно тогда, когда по условиям существования того животного, у которого данный орган имеется, он делается ему необходимым. Так, например, развитие ротового аппарата у рыб обычно заканчивается тогда, когда запасы питательного желтка исчерпываются мальком, и животное принуждено перейти к самостоятельному питанию. В этом случае функционирование ротового аппарата является необходимой предпосылкой для дальнейшего развития организма. Мы можем представить себе, что если бы условия существования у какой-нибудь формы изменились таким образом, что потомки этой формы стали бы потреблять желток раньше, чем его предки, то ускорение (акцелерация) в развитии ротового скелета и его мускулатуры у потомков было бы средством для того, чтобы спасти данную форму от вымирания. Для такой формы, конечно, безразлично, образуется ли у нее дефинитивный ротовой аппарат более быстрым темпом, чем прежде, или развиваются вновь какие-нибудь временные органы, которые впоследствии будут заменены дефинитивными образованиями. Достаточно того, чтобы ее ротовой аппарат мог обеспечить развивающееся животное подходящим и достаточным питанием.
В этом именно и лежит, по нашему мнению, причина того своеобразного явления, что у головастиков очень рано,—в то время, когда они еще живут в воде,—образуются особые губные хрящи с роговой оболочкой и с особой мускулатурой. Хрящи эти очень скоро после метаморфоза, т. е. после переселения животных из воды на сушу, исчезают и заменяются дефинитив-
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
529
ними челюстями со своей специальной мускулатурой. Здесь мы имеем перед собою образование нового ценогенетического органа.
Весьма вероятно, что образование дефинитивного ротового аппарата, т.е. его готовность функционировать, завершается у наземных животных (например, у рептилий) тогда, когда эти животные начинают в нем нуждаться, т. е. ко времени, когда желток зародыша уже потреблен и наступает время для перехода к самостоятельному питанию. Вспомним, с другой стороны, о времени образования челюстей и зубов у ящериц, ядовитых змей и т. п. Весьма вероятно, что и развитие молочных зубов у млекопитающих (первое поколение зубов) стоит в связи с прекращением питания материнским молоком и с переходом молодых животных к самостоятельному питанию. Здесь мы также имеем перед собой пример ускоренного развития ротового аппарата. Быть может, многие случаи раннего завершения развития целого ряда органов, между прочим, органов движения, а также некоторые инстинкты выводковых птиц, в сущности, сводятся к адаптивной акцеле-рации. В этих случаях развиваются органы, функционирующие у молодых и у взрослых животных. То же можно сказать и о завершении развития органов высших чувств, а также о целом ряде других эктосоматических органов.
Явление акцелерации мне пришлось наблюдать при превращении брюшных плавников типа Acipenseridae в брюшные плавники типа Holostei и Teleostei.1 Как мы это уже отметили выше, у Holostei порядок закладки элементов брюшного пояса конечностей изменился в течение эволюции, и базиптеригий стал у них закладываться онтогенетически раньше, чем он закладывался у предков, и развиваться быстрее, чем у них. Это произошло потому, что в закладке базиптеригия выпали первые стадии развития [закладка дискретных базалий, их охрящевение и срастание (рис. 191 А, В и С)], характерные для онтогенеза брюшных плавников осетровых, и базиптеригий у Amia стал закладываться сразу в виде сплошного мезен-химатозного (а затем хрящевого) стержня (рис. 192 А и В). Для нас важно то, что в результате этой акцелерации у Holostei и у Teleostei брюшные плавники начинают функционировать раньше, чем у предков, и функционируют лучше, чем у них.
Явления акцелерации наблюдались также Б. С. Матвеевым1 2 и С. В. Емельяновым3 при изучении развития позвоночника многих костистых рыб (Cyprinidae, Cobitis, Gasterosteus, Atherina, Gobius и др.), у которых прямо из соединительной ткани, минуя хрящевые стадии, развиваются костные тела, верхние и нижние дуги позвонков и ребра. При этом соединительнотканные окостенения более гибки и более тонки, чем у предков, вследствие чего мальки этих костистых рыб раньше превращаются в маленьких рыбок, чем это имеет место у мальков с хрящевым скелетом, что выгодно для жизни вида.
Таким образом, завершение морфогенеза того или иного органа зависит от того, в какой именно период жизни животного, в зависимости от общих условий его существования, требуется функционирование данного органа.
Если при изменившихся условиях существования завершение морфогенеза, т. е. начало функционирования органа, должно начаться раньше, чем-это было у предков данного животного, то развитие акцелераций
1 Sewertzoff A. N., 1934, 1. с.
2 М a t v е i е v В. S., Die Entwicklung der vorderen Wirbel und des Weberschen Apparates bei Cypriniden, Zool. Jahrb., Bd. 51, 1929.
3Емельянов С. В., Развитие ребер и соотношение их с позвоночником, ч. IV, Русск. зоол. журн., т. 8, вып. 2, 1928.
Морфологические закономерности 34
530
ГЛАВА XIV
является одним из способов приспособления животного к изменившимся условиям существования.
Как известно, у птиц сердце и кровеносные сосуды развиваются раньше, чем у рыб и у амфибий. Весьма вероятно, что это ускоренное развитие является приспособлением к специфическим условиям эмбриональной жизни этих форм, т. е. приспособлением к зародышевому питанию при помощи очень большого количества желтка и к зародышевому дыханию при помощи аллантоиса.
В общем, мы можем принять, что акцелерация, т; е. ранняя закладка органа, имеет место тогда, когда прогрессивное развитие органа требует для своего завершения относительно большего промежутка времени, чем это было необходимо предкам. Последнее случается обычно у тех органов, которые эволюируют путем надставки конечных стадий. Так, мозг птиц и млекопитающих развивается быстрее, чем мозг рептилий, от которых млекопитающие и птицы происходят. В то же время мозг птиц и млекопитающих относительно гораздо больше и более диференцирован, чем мозг рептилий. Поэтому к конечной стадии развития анцестрального рептилийного мозга должен прибавиться целый ряд стадий, из которых каждая требует для своего течения некоторого промежутка времени, так что если бы развитие здесь началось на той же стадии, что и у рептилий, то его завершение, т. е. начало функционирования мозга, наступило бы значительно позднее, чем у рептилийных предков высших амниот; само собою разумеется, это было бы невыгодно для рассматриваемых нами форм. Но мы видим, что такое запоздалое завершение морфогенеза мозга в действительности не имеет места именно благодаря его ускоренному развитию.
Все примеры акцелераций, приведенные нами, делают для нас понятным значение этого процесса в филогенезе: акцелерация является приспособлением для осуществления своевременного функционирования органа, потребного организму. Функция эволюирующего органа при этом не изменяется, но в некоторых отдельных случаях, когда изменение условий существования требует, чтобы данный орган начал функционировать раньше, чем у предков, выступает акцелерация как средство для своевременного начала функционирования эволюирующего органа. Мы видим, таким образом, что филогенетическая акцелерация является одним из видов приспособления—адаптивным изменением организма.
Трудно свести все количественные изменения функций и строения органов к такого рода изменениям хода морфогенеза, но во многих случаях именно благодаря акцелерациям достигается интенсификация функций и приспособление к раннему наступлению взрослого состояния, т. е. к самостоятельному, независимому от эмбриональных приспособлений образу жизни.
Мы думаем, что с той же точки зрения нам следует подходить и к вопросу о неотении, т. е. рассматривать явления неотении как приспособления животных к особым условиям существования. Так, например, если мы обратимся к одному из наилучше изученных явлений неотении, а именно, к факту неотении, наблюдаемому у Perennibranchiata, то увидим, что здесь неотения является средством к обратному переходу предков наземных хвостатых амфибий в воду, причем переход этот осуществляется совершенно другим путем> чем у водных рептилий и млекопитающих.
Индивидуальная жизнь у предков Perennibranchiata разделялась на два периода, а именно на период молодости, который проводился личинкой в воде, и на период взрослого состояния, которое половозрелое животное проводило на суше. Какое-то, ближе нам еще неизвестное, изменение в условиях существования этих животных (может быть конкуренция с другими наземными Urodela за добычу, служившую пищей взрослым формам),
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
531
привело наземных предков Perennibranchiata к обратному переходу в воду. Переход этот достигался тем, что время наступления метаморфоза, т. е. период редукции личиночных органов, приспособленных к водному образу жизни, стал задерживаться все более и более, так что период жизни, проводимый личинкою в воде? постепенно удлинялся, а период жизни на суше становился все короче. Мы не знаем, ускорялось ли при этом развитие половых органов, т. е. происходила ли здесь акцелерация, но знаем, что наступило такое время, когда ретардация метаморфоза сделалась настолько значительной, что половая зрелость стала наступать у еще не метаморфозированных животных, и они, еще будучи личинками, стали тем не менее размножаться в воде. Именно на такой стадии филогенеза находится аксолотль (Siredon). Аксолотль размножается, как известно, в воде, но у некоторых экземпляров метаморфоз все же имеет место,-и животное превращается в амблистому с признаками наземных сала-мандрит. Другие Perennibranchiata (Menobranchus, Proteus, Siren), невидимому, прошли эту стадию эволюции и в настоящее время у них уже не наступает превращения в наземную форму. Мы видим, таким образом, что у Perennibranchiata неотения, т. е. появление гетерохронии в развитии половых органов и органов водной жизни (жабры, хвост в виде плавника), есть приспособление к переходу из наземной среды обратно в водную.
У водных млекопитающих этот переход был достигнут другим способом, а именно тем, что строение их органов движения сильно изменилось путем смены функций: наземные конечности превратились в ласты и т. д. Их органы дыхания и половые органы не претерпели принципиальных изменений в строении и функции: разница между наземными и водными.млекопитающими заключается в том, что последние приобрели способность глубоко нырять и долго оставаться под водою.
Что касается вопроса об онтогенетической ретардации, то здесь нам придется сказать лишь несколько слов. Отметим прежде всего, что типичным примером запоздалой закладки органов (ретардация) является закладка половых органов у млекопитающих. Половые органы млекопитающих достигают своего полного развития лишь у вполне взрослых форм. Здесь замедление в развитии и самих половых органов и вторичных половых признаков вполне понятно, так как функционирование этих органов (т. е. производство на свет потомства и заботы о нем родителей) должно наступить в тот период индивидуальной жизни особи, когда по сумме биологических условий родители достигают полной силы и способны прокормить и защитить подрастающих детенышей. В этих случаях сдвигание развития половых органов и вторичных половых признаков на более поздние стадии развития является признаком биологически полезным для любого данного вида. Это касается не только млекопитающих, но и всех форм, у которых выработалась забота родителей о детях (например у птиц, у многих рептилий, амфибий и рыб). Но, кроме этого примера, случаев биологически понятной ретардации чрезвычайно мало. Мы можем еще указать на позднюю закладку зубов мудрости у человека (биологически очень мало понятную) и на позднюю закладку эндохондральных костей (orbitosphe-noideum) в черепе осетровых рыб.
Гипотеза Менерта, утверждавшего, что поздняя закладка органа есть свидетельство его редукции, едва ли выдерживает критику. Во-первых, в очень многих случаех, описанных самим Менертом, ему пришлось наблюдать лишь некоторую разновременность в закладке органов, причем совершенно неизвестно, происходит ли здесь действительно более поздняя закладка органов, вследствие ретардации редуцирующихся зачатков, или же это лишь кажется таковым, вследствие некоторой акцелерации в развитий соседних зачатков. Во-вторых, эта гипотеза не подтверждается
34*
532
ГЛАВА XIV
изучением тех многочисленных случаев редукции, которые были подробно прослежены мною. Не отрицая целиком наблюдений Менерта, мы все же думаем, что они нуждаются в пересмотре с точки зрения теории филэмбриогенеза, еще неизвестной в то время, ^<огда Менерт писал свои работы.
ТЕМП ЭВОЛЮЦИИ ПРИ РАЗЛИЧНЫХ'МОДУСАХ ФИЛЭМБРИОГЕНЕЗА
Мы изложили некоторые типы изменений хода морфогенеза органов, ведущих к прогрессивной адаптивной эволюции этих органов (архаллаксис, девиация, анаболия, гетерохрония и пр.). Теперь нам надо постараться ответить на вопрос, почему существует вообще не один, но несколько видов такого рода изменений органов? Ибо легко можно было бы себе представить, что все существующие филетические изменения органов могут произойти, следуя одному какому-нибудь типу развития, например путем девиации, к чему склоняется де Беер, или путем анаболии, как это думал Геккель и другие.
Чтобы найти правильный ответ на поставленный нами вопрос, постараемся более точно определить разницу между характером изменений, происходящих при различных модусах эволюции; иными словами, попробуем сравнить результаты эволюции, следующей по пути анаболии или девиации, с результатами эволюции, происходящей путем архаллаксиса.
Проводя такое сравнение, мы увидим, что исходные стадии развития, т. е. те стадии, с которых начинается эволюция органа, во всех этих случаях различны. При развитии способом надставки, конечных стадий эволюция органа начинается с той стадии, на которой основные черты строения взрослого животного уже вполне определились, при девиации— с той стадии, когда значительное число частей состоит еще из относительно индиферентной эмбриональной ткани; при архаллаксисе зачатки органов образуются из совершенно индиферентной эмбриональной ткани.
Мы уже раньше отметили, что надставка конечных стадий (анаболия) представляет собою путь постепенного преобразования органа. Это зависит от того, что в данном случае эволюция исходит из такой начальной стадии развития, когда орган является уже во многих отношениях вполне готовым образованием. Правда, все его части еще могут расти и далее, и некоторые из них все еще состоят из тканей вполне способных к дальнейшему превращению, но проспективные его потенции гораздо ограниченнее, чем на ранних стадиях развития. Относительная интенсивность роста каждой из многочисленных его частей еще может возрасти, и поэтому у данного органа существует много возможностей изменения формы. При этом и гистологические преобразования могут протекать несколько иным путем, чем у предков.
Число возможных направлений эволюции соответствует в этих случаях числу образовавшихся частей органов, и так как число это на поздних, стадиях развития очень велико, то и число возможных эволюционных направлений, в которых может развиваться данный орган, также велико. Но темп роста на поздних стадиях морфогенеза является весьма замедленным по сравнению С ранними стадиями развития. Поэтому мы можем принять, что наследственные вариации относительной интенсивности роста, из которых в данном случае исходит филогенетическое развитие органа, здесь будут гораздо меньшими, чем вариации, из которых исходит, например, эволюция, идущая путем архаллаксиса.
Мы пришли, таким образом, к выводу, что эволюция, происходящая путем анаболии, протекает очень медленно. Палеонтологические исследования, повидимому, подтверждают это предположение. Обычно мы не представляем себе той огромной мед
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
533
ленности, с которой совершается эволюционный процесс. Попробуем следующими примерами сделать это явление более наглядным для читателя. Если мы примем вместе с Осборном,1 что минимальная продолжительность третичной эпохи равнялась трем миллионам'лет (в последнее время считается, что. эта эра длилась гораздо дольше, а именно от 30 до 60 миллионов лет) и если мы примем в соображение, что предки современных лошадей, например Eohippus, существовали уже в палеоцене, то мы можем приблизительно вычислить темп, которым происходила эволюция размеров современной лошади.1 2 Мы знаем, что. высота Eohippus колебалась между 25 и 51 см (у Eohippus resartus высота в плечах равнялась 51 см) и знаем, что высота плеч у современной лошади равняется приблизительно полутора метрам. Таким образом, мы можем принять, что в течение третичной эпохи, т. е. в период времени от верхнего палеоцена до верхнего плиоцена, потомки Eohippus выросли приблизительно на один метр. Так как эволюция лошади (от Eohippus до Equus) длилась около двух с половиной миллионов лет, то.ежегодный прирост высоты лошади должен приблизительно равняться 1/25 000 см, т. е. величине, равной 40 микромиллиметрам или 0,04 мм в столетие. Не забудем при этом, что мы взяли минимальную цифру, рассчитанную для продолжительности третичной эпохи. Если, что весьма вероятно, продолжительность этой эпохи действительно исчисляется 30—60 миллионами лет, то темп эволюции лошади становится еще более замедленным. Даже если мы примем, что в течение известного периода времени эволюция вовсе не двигалась вперед (период остановки), а в течение другого периода она шла более быстрым темпом, чем обычно, то и тогда ход эволюционного процесса все же остается крайне медленным.
Ввиду всего сказанного, нам кажется вероятным, что эволюция, происходящая посредством анаболии, шла путем очень медленных, но многосторонних филогенетических изменений. Этим способом животные могли приспособляться лишь к таким условиям существования, которые сами изменялись очень медленно и постепенно.
Но величина изменений животного организма зависит не столько от темпа изменений в окружающей среде, сколько от длительности этих изменений, идущих все время в одном и том же направлении. Если продолжительность периода изменений велика (как это имеет место в случае эволюции лошади), то изменения, происходящие в строении органов животных, являются, в конце концов, очень значительными.
Ясно, что эволюция, протекающая путем девиации, занимает в смысле темпа среднее положение между двумя крайними путями эволюции, т. е. между анаболией и архаллаксисом; поэтому мы, в целях большей наглядности изложения, перейдем теперь к изложению вопроса о темпе эволюции, происходящей путем архаллаксиса.
В свое время мы пришли к выводу, что при архаллаксисе первые зачатки эволюирующих органов начинают изменяться с момента своего появления. При этом изменению подвержены как величина, так и строение, как местоположение, так и направление роста эволюирующего органа. В рассмотренном нами случае увеличения числа туловищных сомитов, спинномозговых нервов и пр. у змей (Tropidonotus) мы видели, что хвостовой отдел туловищной мезодермы сегментируется наново. Образуются новые сомиты, не существовавшие прежде у предков. Каждый из этих новых сомитов обладает способностью образовать мускулы, позвонки, ребра ит. п. Из заднего отдела новообразовавшегося спинного мозга вырастают новые
1 Osborn, The Origin and Evolution of Life, London, 1918.
2 Ниже приводимые цифровые данные взяты из работ известного нашего геолога А. П. Павлова.
534
ГЛАВА XIV
спинномозговые нервы, иннервирующие эти новые мезодермические сегменты. Все это—новообразования, отсутствовавшие у предков.
При исследовании меристических вариаций позвонков и ребер у желтопузика мы отметили, что здесь изменения происходят скачками (сальтации). Позвонок, который у предков желтопузика был хвостовым позвонком, закладывается у потомков с самого начала своего развития как крестцовый позвонок. То же можем мы сказать относительно коротких поясничных ребер предков, которые у потомков закладываются с самого начала как длинные торакальные ребра. Тоже относится и к крестцовым позвонкам, превратившимся в поясничные, и т.'д. Таким образом, мы видим, во-первых, что здесь имеются перед нами не постепенные превращения, но относительно крупные скачкообразные изменения (сальтации), и, во-вторых, что здесь происходит образование новых органов, отсутствовавших у предков.
Нам, конечно, можно было бы возразить, что в сущности здесь не происходит новообразований в строгом смысле этого слова, так как преобразовавшийся позвонок превращается не в какой-либо другой орган, но лишь в иначе построенный и. иначе расположенный позвонок того же рептилийного типа. Однако у нас имеются основания принимать, что путем архаллаксиса происходит не только перестройка органов, но и действительное их новообразование.
Мы знаем, например, что у предков эласмобранхий на известной стадии филогенеза существовали плакоидные чешуи. В очень многих местах кожи у них имелись эмалевые органы, которых прежде у них не было и которые, таким образом, являются новообразованиями. Под этими эмалевыми органами образовались скопления мезенхимных клеток cutis, в которых развивался дентин эмалевых зубов. Из эмалевых органов развилась эмалевая покрышка плакоидных зубов. У некоторых потомков эласмобранхий, т. е. у предков Osteichthyes, в глубоких слоях cutis, на определенных местах, а именно напротив миосепт, образовались довольно крупные первичные костные пластинки чешуй. Мы знаем, что они лежали под сосудистой сетью кожных капилляров, кнутри от плакоидных чешуй. Таким образом, они явились новообразованиями.
Только что сказанное относится также к происхождению первых зачатков хрящевого скелета (дуг позвонков и т. п.), к образованию эндохондральных окостенений черепа и туловищного скелета и ко многим другим образованиям, относительно которых мы не можем утверждать с определенностью, что они развились путем преобразования из ранее существовавших частей. Здесь можно было бы возразить, что, например, покровные кости развились путем локального преобразования из уже ранее существовавшей там мезенхимы, но при этом следует принять во внимание, что части костей, состоящие из извести, происходят не путем преобразования, а путем выделения, т. е. являются также новообразованиями в собственном смысле этого слова.
Итак, мы приходим к выводу, что филогенетические новообразования происходят именно путем архаллаксиса.
Выше мы видели, что всякое удлинение туловища безногих ящериц происходит при помощи сальтаций, причем каждый скачок (мутация) соответствует преобразованию одного позвонка в иначе построенный позвонок. Если мы примем, что какая-нибудь форма .с очень длинным туловищем, например Seps с его 61 предкрестцовыми позвонками, произошел от какой-нибудь сцинкообразной формы, имевшей всего лишь 26 позвонков, то мы примем вместе с тем, что крестец переместился здесь на рас
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
535
стояние 35 сегментов; иными словами, у предков Seps сначала 27-й и 28-й позвонки, затем позвонки 28-й и 29-й, затем позвонки ЗО-й и 31-й и т. д. превратились в крестцовые позвонки. Преобразование каждого из этих позвонков соответствовало, как это уже было сказано, одному шагу вперед (одной сальтации) в эволюционном процессе; в общем таких шагов (сальтаций) было сделано 35.
Мы могли бы совершенно произвольно принять, что период в 1000 лет является достаточно большим промежутком времени для наступления и для фиксации такого, рода мутаций, т. е. для превращения мутации в постоянный видовой признак.1 Если это так, то превращение короткотуловищных сцинкообразных прародительских форм в длинных сепсов С сильно редуцированными конечностями продолжалось только 35 000 лет. Этот промежуток времени в сравнении с теми двумя с половиной миллионами лет (а вернее, с теми 30—60 миллионами лет), в течение которых длилась эволюция лошади, конечно, может рассматриваться нами как промежуток времени чрезвычайно короткий.
Мы говорили до сих пор о новообразованиях и о меристических вариациях, происходивших (и те и другие) путем архаллаксиса. Но кроме того существует очень большое число вариаций в величине и в форме органов, которые также произошли этим же путем. Мне не- пришлось, однако, исслед'овать вопроса о tomJ являются ли эти последние вариации настолько значительными, чтобы их можно было рассматривать как настоящие сальтации, поэтому мои выводы относительно темпа эволюции, происходящей путем архаллаксиса, следует рассматривать лишь как гипотезу. Вероятно, эта гипотеза верна для меристических органов.1 2 3 Но вопрос о том, всегда ли происходящая путем архаллаксиса прогрессивная эволюция величины, формы и строения других органов совершается быстро или нет, можно разрешить только путем последующих исследований.
Выше мы упомянули о том, что девиация занимает промежуточное положение между архаллаксисом и анаболией. Поэтому можно предположить с большой дозой вероятности, что чем раньше наступит изменение хода анцестрального онтогенеза, тем быстрее будет итти филогенетическое изменение строения эволюирующего органа, а чем позднее оно наступит, тем медленнее будет протекать эволюция. Но пока я воздерживаюсь от более подробного обсуждения этой, ближе мною не исследованной, проблемы.
выводы
Результаты, к которым мы пришли в этой главе, могут быть кратко формулированы в виде следующих положений:
1. Эволюция формы, величины, строения и положения органов взрослых животных является непосредственным следствием происходящих в течение филогенеза изменений в ходе онтогенеза этих органов.
2. Индивидуальное развитие (онтогенез) у всех многоклеточных животных распадается на два периода:. на период морфогенеза и на период роста.
3. Существуют четыре основных процесса, происходящих в ходе индивидуального развития организма: 1) общий равномерный рост, 2) частный, или неравномерный рост, когда при размножении дериватов яйцеклетки неравномерно размножающиеся клетки остаются вместе, образуя группу клеток, связанных друг с другом, т. е. зачаток органа или ткани, 3) гисто
1 Мы знаем, что у домашних животных для образования хорошо выраженной расы требуется гораздо более короткий промежуток времени.
2 У позвоночных животных меристическими органами являются ребра, зубы,
позвонки, фаланги пятипалой конечности, членики плавниковых лучей, спинномозговые нервы, сегментальные мускулы, чешуи и др.
536
ГЛАВА XIV
генез, т. е. изменения клеток, связанные с превращением их в специализо-ванные клетки с одной главной функцией и 4) странствование, или переселение клеток, когда группа клеток, переместившись на новое место в теле зародыша, дает начало новому зачатку и претерпевает в конце своего развития значительные гистологические изменения.
4. Филогенетические изменения признаков взрослых животных (признаков формы, строения, величины, местоположения и т. д.) происходят на различных стадиях индивидуального развития животного (на ранних, средних и Поздних стадиях онтогенеза) и совершаются при прогрессивной эволюции животных различными способами или модусами: 1) путем положительной анаболии, 2) путем положительной девиации, 3) путем положительного архаллаксиса, 4) путем периодического изменения хода развития органов, 5) путем гетерохронии, т. е. путем более ранней закладки (акцелерации) или более поздней закладки (ретардации) органов.
5. При анаболии к конечной стадии морфогенеза эволюирующего органа прибавляется новая стадия развития, так что в каждом случае прогрессивного изменения признака данного взрослого органа его морфогенез удлиняется на одну стадию за счет периода роста. Вследствие этого в каждом случае филетического преобразования органа удерживается в онтогенезе потомков конечная стадия морфогенеза предков, и это является необходимой предпосылкой для образования данного признак’а у потомков.
6. Конечные стадии морфогенеза предков современных животных, из которых каждая в своих главных чертах напоминает анцестральное взрослое состояние, повторяются у потомков в той же последовательности, в которой они появляются у предков, и в своих общих чертах похожи на взрослое состояние предков. Таким образом, закон рекапитуляций Мюллера-Геккеля (биогенетический закон) является непосредственным и необходимым следствием эволюции, происходящей по типу надставки конеч-’ ных стадий.
7. В законе К. Бэра заключаются два важных обобщения: 1) признаки современных животных закладываются и развиваются в онтогенезе этих животных не одновременно, но в определенной последовательности; 2) эта последовательность соответствует убывающей степени общности данного признака. Признаки крупных систематических групп (филогенетически древние признаки) образуются онтогенетически раньше, чем признаки мелких систематических групп (филогенетически новые признаки). Этот закон также является необходимым следствием эволюции, происходящей путем анаболии. Те признаки предков, которые образуются на ранних стадиях филогенеза и которые в течение последующих геологических эпох более не изменяются (древние константные признаки), закладываются у современных форм на тех же стадиях онтогенеза, что и у предков, т. е. закладываются рано. Признаки тех органов предков, которые в течение эволюции сильно изменились и морфогенез которых вследствие этого удлинился, закладываются в онтогенезе потомков тем позднее, чем больше будут изменения, произошедшие в органах. Древние неизмененные анцестральные признаки при дивергентной монофилетической эволюции переходят ко всем потомкам, происшедшим от прародительской формы, обладавшей этими признаками, и становятся у них признаками крупных систематических групп. Новые, т. е. относительно поздно образовавшиеся признаки, переходят только к потомкам тех форм, которые, вследствие недостаточного времени, дивергировали лишь в незначительной степени,, т. е. переходят к мелким систематическим группам.
8. Иногда начало филогенетического изменения анцестрального хода онтогенеза имеет место на средних стадиях онтогенеза, и это изменение также ведет к изменению строения взрослого органа. Этот модус филогенетического изменения органов мы обозначили именем девиации. При эво
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
537
люции, происходящей путем девиации, продолжительность морфогенеза не увеличивается, и происходит рекапитуляция только эмбриональных признаков предков данной формы.
9. Существует еще один очень важный модус изменения эволюции признаков взрослых животных, а именно изменение первых зачатков закладывающегося органа, или архаллаксис. Архаллаксис осуществляется путем суммирования нерегулирующихся наследственных вариаций величины, формы и местоположения первого эмбрионального зачатка органа, ведущих к изменению признаков взрослых органов эволюирующей животной формы. В этих случаях филогенетические изменения органа не являются преобразованиями ранее существовавшей структуры, но представляют собою новообразования (новая форма, новое местоположение и строение органа, возникновение ориментов совершенно новых органов и т. д.).
10. При этом типе эволюции не происходит никакого удлинения морфогенеза, так же как и никакого повторения анцестральных признаков. Закон К. Бэра относительно онтогенетической последовательности в образовании признаков крупных и мелких систематических групп у современных животных, также не приложим к органам, развивающимся путем архаллаксиса.
11. Таким образом, закон Мюллера-Геккеля и закон К. Бэра являются действительными только для таких признаков, которые развиваются по типу анаболии..
12. Периодическое изменение хода развития органов заключается в том, что развивающийся орган доходит в своем развитии до известной стадии, после чего он отпадает, а на его месте из тогб же зачатка развивается в соответствующий период жизни животного такой же, но идущий в своем развитии несколько дальше орган, который снова отпадает, причем процесс этот продолжается до достижения животным взрослого состояния.
13. Изменения в темпе онтогенетического развития (гетерохрония) обычно приводят к образованию эмбриональных приспособлений. Ускорение (акцелерация) онтогенетического развития какого-нибудь органа позволяет этому органу начать функционировать раньше, чем это было возможно у предков данной животной формы. Неотения может быть рассматриваема как специальный случай гетерохронии; она является способом, при помощи которого взрослые потомки эволюирующей формы приспособляются к тем условиям, в которых живут их зародыши и личинки и которые давно уже оставили их предки.
14. Темп, которым происходит филогенетическое изменение, невидимому, зависит оттого модуса, путем которого идет это изменение, т. е. от анаболии, архаллаксиса или девиации. Весьма вероятно, что эволюция, происходящая при помощи анаболии, является способом очень медленного преобразования древнего органа, а архаллаксис—способом относительно быстрой эволюции органа, при котором формируются все новообразования. Девиации, повидимому, принадлежит среднее место между этими двумя крайними модусами.
ГЛАВА XV
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ а ТИПЫ РЕДУКЦИИ ОРГАНОВ
Мы говорили до сих пор о прогрессивной эволюции органов и рассматривали те способы, которыми она осуществляется, главным образом способы анаболии, архаллаксиса и девиации. Нам остается теперь рассмотреть три отрицательных модуса филэмбриогенеза, а именно: отрицательную анаболию, отрицательный архаллаксис и отрицательную девиацию. Так как эти модусы тесно связаны с явлениями филогенетической редукции органов, на которых они и были впервые изучены, то нам придется обратиться к проблеме регрессивной эволюции или редукции органов и установить различные типы этой редукции.
Как известно, в мире животных регрессивные изменения органов и даже целых систем органов чрезвычайно распространены, и мы можем смело сказать, что нам неизвестна ни одна группа более или менее высоко организованных животных, у которой в течение эволюции не редуцировался бы целиком или отчасти тот или иной когда-то вполне развитый орган. Мы знаем, что существуют даже крупные группы животных (паразитические или сидячие формы), наиболее характерным признаком которых является именно редукция большинства активных органов. К таким группам относятся, например, оболочники, ленточные черви, усоногие и другие формы.
Исследование различных случаев редукции органов показало мне,1 что существуют два основных типа редукции, которые по своему характеру и по своему течению резко отличаются друг от друга, а именно рудиментация органов и полная редукция или а ф а-низия органов.
Дегенерация органов, происходящая по типу рудиментации, характеризуется тем, что редуцирующийся орган ни на одной из стадий своего существования не бывает полезен тому животному, у которого он находится, вследствие чего его структура с самого начала является упрощенной, а сам он постепенно становится все меньше и меньше, пока, наконец, не превратится в рудимент, иногда очень долго сохраняющийся у потомков данной формы животных.
Наоборот, при афанизии редуцирующийся орган на некоторых стадиях своего развития (обычно на эмбриональных стадиях или у молодых особей) бывает полезен тому животному, которому он принадлежит, и поэтому он обычно не претерпевает никаких изменений в начале своего морфогенеза. Он развивается в течение некоторого времени совершенно нормально, т. е. так же, как и не редуцирующийся орган, зато позднее редуцируется целиком очень быстро,
1 Sewertzoff A. N., Studien uber die Reduktion der Organe der Wirbeltiere, .Zool. Jahrb., Bd. 53, 1931.
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
539
не оставляя после себя никаких следов в виде того или иного рудимента.
Характерные признаки этих двух типов редукции органов, а также различия между ними, станут понятны читателю, если он обратится к тем примерам редукции, описание которых мы даем ниже, ибо эти примеры помогут ему лучше уяснить себе сущность этого процесса, чем те короткие определения, которые мы только что привели.
РУДИМЕНТАЦИЯ ОРГАНОВ
Основной предпосылкой для рудиментации хорошо развитого и нормально функционирующего органа является обычно тот факт, что известный орган, вследствие изменения некоторых условий существования, перестает быть полезным потомкам данной формы. Это обыкновенно происходит в тех случаях, когда орган предков замещается у потомков каким-нибудь новым органом, лучше приспособленным к изменившимся условиям существования; иными словами, когда на месте старого органа появляется другой, вновь развившийся и лучше приспособленный к новым условиям существования орган, и когда животное вследствие этого теряет нужду в старом органе; последний начинает при этом дегенерировать и редуцируется путем рудиментации.
Во всех этих случаях редукция осуществляется либо путем выпадения отдельных частей редуцирующегося органа, которые тогда не закладываются вовсе, либо путем общего (а иногда частичного) уменьшения всего органа. Оба эти процесса можно легко иллюстрировать примерами, взятыми из нашего исследования редукции конечностей безногих ящериц, о котором мы уже упоминали выше, рассматривая принцип субституции функций.
На рис. 194 А и 194 В представлены изображения нередуцированных конечностей Eumeces schneideri, на рис. 195—203 изображен ряд более или менее редуцированных конечностей различных безногих ящериц, наглядно показывающий ход процесса рудиментации конечностей у этих животных.
У Eumeces schneideri, как видно на рис. 194 А, скелет передней конечности состоит из плечевой кости (на рисунке не помещенной), из лучевой (R) и локтевой (U) костей, из девяти канонических carpalia, т. е. из radiale (г), рудиментарного intermedium (i), ulnare (u), pisiforme (p), cen-trale (с), четырех carpalia distalia (cx, c2, c3, c4), пяти metacarpalia (I—V) и пяти пальцев.
Формула фаланг у ящерицы имеет следующий вид:1
2. з. 4. 5. з.
I. II. III. IV. V.
Задняя конечность Eumeces schneideri, изображенная на рис. 194 В, состоит из бедра (на рисунке не помещенного), из большой берцовой (Т) и малой берцовой (F) костей, из большого tarsale proximale (t. рг.) и из двух tarsalia distalia (t3, t4). Кроме того имеется пять metatarsalia и пять пальцев. Формула фаланг задней конечности ящерицы имеет следующий вид:
2. 3. 4. 5. 4.
I. II. III. IV. V.
В задней конечности имеется, стало быть, одна лишняя фаланга на V пальце.
1 Во всех этих формулах арабскими цифрами обозначено число фаланг, римскими— metacarpalia и metatarsalia.
540
ГЛАВА XV
На рис. 195 изображена передняя конечность Lygosoma punctulatum. По своему строению, как это видно из сравнения рисунков 195 и 194 А, она вполне подобна передней конечности Eumeces schneideri, но редукция здесь уже началась. Мы видим, что в IV пальце имеются уже не пять, а только четыре фаланги; конечная фаланга на этом пальце не образуется вовсе. С другой стороны, сама конечность стала несколько меньше по величине. Задняя конечность еще не редуцируется и состоит из канонических элементов.
На рис. 196 изображена передняя конечность Chalcides sepoides. Здесь редукция обозначилась несколько яснее: IV палец, так же как и у Lygosoma,
Рис. 194 В. Скелет задней конечности Eumeces schneideri.
F—fibula; T—tibia; t. pr.—tarsale proximate; ta—14—tarsalia distalia 3—4; L— V—metatarsalia I—V; 1—5—фаланги 1—5 (иг A. H. Северцова, 1931).
Рис. 194 А. Скелет передней конечности Eumeces schneideri.
С—centrale; cj—С4—carpalia distalia 1—4; i—intermedium; p—pisiforme; Pi—radius; r—radiale; U—ulna; u—ulnare; I—V— metacarpalia I—V; 1—5—фаланги 1—5 (из A. H. Северцова, 1931).
состоит из четырех фаланг (выпадение пятой фаланги), а от элементов I пальца осталось лишь metacarpale (I), ибо обе фаланги I пальца совершенно исчезли. Задняя конечность еще имеет вполне нормальный вид.
На рис. 197 А и 197 В мы видим конечности Ophiomorus tridactylus, которые дегенерировали еще более значительно. В передней конечности (рис. 197 А) I палец, тот самый, который еще у Chalcides sepoides был пред-ставлентолько одним metacarpale I, исчез совершенно; кроме того мы видим, что все фаланги V пальца (1, 2, 3) тоже дегенерировали, так что от этого пальца осталось только одно сильно укороченное metacarpale. Уменьшилось число фаланг и в остальных пальцах: II палец здесь имеет только две фаланги, III—три, IV—четыре. На каждом пальце редуцировалась дистальная фаланга, так что формула фаланг здесь вместо обычного вида:
2. 3. 4. 5. 3.
L Т ПТ ГТ \Г
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
541
имеет следующий вид:
о. 2. з. 4. о.
и. и. 111. IV. V.
В запястье исчезло centrale carpi (рис. 194 А).
В задней конечности Ophiomorus tridactylus полностью атрофировался I пален и все фаланги V пальца, a metatarsale V сильно укоротилось (рис. 197В, V). Редуцировалось также и tarsale distale Ш(ср. рис. 194 В,t3).
Рис. 195. Скелет передней конечности Lygosoma punctulatum.
Обозначения те же, что и на рис. 194 А (из А. Н. Северцова, 1931).
Рис. 196. Скелет передней конечности Chalcides sepoides.
Обозначения те же, что и на рис. 194 А (из А. Н. Северцова, 1931).
Рис. 197 В. Скелет задней конечности Ophiomorus tridactylus.
Обозначения те же, что и на рие. 194 В (из А. Н. Северцова, 1931).
Рис. 197А. Скелет передней конечности Ophiomorus tridactylus.
Обозначения те же, что и на рис. 194 А (из А. Н. Северцова, 1931).
Формула фаланг в задней конечности Ophiomorus tridactylus такова же, как и в передней, т. е. имеет следующий вид:
0. 2,- 3. 4. 0.
0. II. HI. IV. V.
У Seps tridactylus (рис. 198и 199) редукция конечностей пошла несколько дальше, чем у Ophiomorus. Здесь на четвертых пальцах передней и задней конечности атрофировалось еще по одной фаланге, так что формула фаланг для обеих конечностей у сепса принимает следующий вид:
0. 2. 3. 3. 0.
о: 1Тг~тн. tv., v.
В запястье сохранилось лишь одно centrale (С) но оно уже представлено хрящом (редукция); в предплюсне имеется два tarsale—t3 и t4.
542
ГЛАВА XV
Мы видим, таким образом, что редуцирующаяся конечность в начале остается еше в течение некоторого периода времени пятилучевой (Lygo-soma). У Chalcides sepoides она впервые становится четырехлучевой (передняя конечность), у Ophiomorus и Seps делается уже трехлучевой.
У Ophiodes striatus (рис. 200) мы находим впервые двулучевую конечность. У этой формы передняя конечность вообще совершенно редуцировалась, сохранился лишь рудимент задней конечности. Он состоит из бедренной кости (на рис. 200 не представленной), из большой и малой берцовых костей, из предплюсны и остатков плюсны. Стопа построена более сложно, чем это можно бьТЛо бы ожидать: она состоит из трех прог ксимальных элементов, которые можно истолковать как tibiale (t), fibu-
Рис. 198. Скелет передней конечности Seps tri-dactylus.
Обозначения те же, что и на рисунке 194 А (из А. Н. Северцова, 1931).
Рис. 199. Скелет задней конечности Seps tri'da^ ctylus.
Обозначения те же, что и на рис. 194 В (из А. Н. Северцова, 1931).
Рис. 200. Скелет задней конечности Ophiodes striatus.
С —centrale; f— fibulare; t— tibiale. Остальные обозначения те же, что и на рисунке 194 В (из А. Н Северцова, 1931).
lare (f), centrale tarsi (С), из одного дистального элемента (вероятно—tarsale distale 4), из двух metatarsalia (вероятно metatarsale III и metatarsale IV) и из остатков первой фаланги IV пальца.1 Формула фаланг двулучевой конечности Ophiodes имеет, таким образом, следующий вид:
о. о. о.
1. о.
0. 0. Ш. IV. 0.
1 Весьма странно, что здесь, в предплюсне Ophiodes, мы не находим, как это обычно бывает у ящериц, одного цельного tarsale proximale (tibiale + centrale + fibulare), но имеем три независимых друг от друга элемента: t, f и С. Невероятно, чтобы tarsale proximale в процессе своей редукции снова распался на свои первичные компоненты, (tibiale, centrale, fibulare). Скорее можно думать, что редукция конечностей у Ophiodes началась очень давно, еще на той стадии филогенеза, когда у предков ящериц эти элементы не срослись между собой.
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
54?
Итак, мы перешли от пятипалой конечности типичной ящерицы через, четырех- и трехлучевые формы к двулучевым формам (Ophiodes). Пойдем теперь еще дальше.
Задняя конечность Ophisaurus является примером однолучевой конечности. Наши исследования показали нам, что скелет задней конечности, желтопузика (рис. 201, А, В, С, D) обычно состоит из бедра (Fm), из части,
Рис. 201 А, В, С, D. Скелет четырех рудиментарных задних конечностей Ophisaurus apus (взрослые формы).
Fm.—бедро; T + F—tibia+fibula; tr.—тарзальная часть; Dig. IV—рудимент четвертого пальца (из А. Н. Северцова, 1931).
соответствующей zeugopodium (T-J-F, рис. 201 А, В, С, D), из одного хряща предплюсны (tr), а также из остатков одного пальцевого луча — Dig. IV, вероятно, metatarsale IV. В некоторых случаях конечности желто
пузика редуцированы еще сильнее, так что они состоят только из трех.
следующих друг за другом скелетных элементов Передняя конечность желтопузика (рис. 202), а также задние конечности Anguis (рис. 203) и Typhlosaurus представляют собой примеры еще более сильной редукции скелета конечностей безногих ящериц. У этих последних форм исчезает целиком весь acropodium, basi-podium и zeugopodium, так что у желтопузика остается лишь небольшая часть stylopodium и проксимальная часть плеча, а у Anguis и Typhlosaurus—лишь проксимальная часть бедра.
Мы изложили только что ход редукции конечностей у безногих ящериц таким, каким он нам представляется на основании данных сравнительно анатомического исследования, и в заключение можем сказать, что, в общем, чем больше увеличивается число туловищных сегментов у этих форм (см. диагр. XXIV), тем более и более редуцируются у них конечности. Эти конечности состоят из постепенно уменьшающегося числа скелетных элементов. У таких
(рис. 201 D).
Рис. 202. Плечевой пояс и-рудимент передней конечности Ophisaurus apus (70 см длины).
форм как Anguis и Typhlosaurus скелет задней конечности состоит, как это мы только что видели, из остатков одного единственного элемента, а именно бедра, и чем ббльшее число туло-
cl. —clavicula; сог. —coracoide-um; Н.—рудиментарный humerus; sc.—scapula; ssc.—supra* scapulare (из A. H. Северцова, 1931).
вищных сегментов имеет животное, тем меньшей величины у него руди-мент бедренной кости.
М. Фюрбрингер,1 изучавший в свое время ре’дукцию конечностей срав
нительно анатомическим путем, пришел к выводу, что редукция этих органов является функцией месторасположения их частей. По его мнению,
1 Fiirbringer М., Die Knochen und Muskelnder Extremitaten bei den schlar-genahnlichen Sauriern, Leipzig, 1870.
544
ГЛАВА XV
редукция конечностей у ящериц происходит в дистопроксимальном и пре-аксиально-постаксиальном направлении. Это значит, что прежде всего редуцируются конечные фаланги пальцев, затем редуцируются metacarpalia s. metatarsalia и т. д., пока, наконец, не начнут редуцироваться дистальные концы humerus (s. femur).
Я изучал процесс рудиментации конечностей не только сравнительно анатомическим, но и эмбриологическим путем, и это последнее исследование привело меня к несколько иным, чем у Фюрбрингера, взглядам на ход редукции этих органов. Я исследовал развитие скелета конечностей у трех различных форм: у ящериц с вполне развитой конечностью (Ascalabotes fascicularis),1 у ящериц с довольно сильно редуцированной конечностью (Seps chalcides) и, наконец, у ящериц с очень сильно редуцированной конечностью (Ophisaurus apus).1 2
il.—ileum; isch.—ischium; Fm. —рудимент femur; pb—pubis (из A. H. Северцова, 1931).
Рис. 204. Зародыш Ascalabotes fascicularis на стадии развития, когда у него только что заложилась передняя конечность, е—зачаток передней конечности (из А. Н. Северцова, 1931).
Здесь я коротко приведу результаты этих исследований.
В предыдущем мы отметили, что при рудиментации, вследствие исчезновения целого ряда частей редуцирующегося органа* его строение упрощается, причем сам орган в течение филогенетической редукции сильно уменьшается в размерах и превращается в совершенно, незначительный рудимент, как это, например, мы видим, изучая заднюю конечность желтопузика. Это уменьшение общих размеров органа происходит путем уменьшения первого зачатка редуцирующегося органа.
На рисунках 204, 205 и 206 изображены три зародыша, а именно зародыш Ascalabotes fascicularis (рис. 204), зародыш Seps chalcides (рис. 205) и зародыш Ophisaurus apus (рис. 206), которые, судя по степени развития их органов (мозга, глаз, слуховых пузырьков, жабр и т. д.), находятся на одной и той же стадии развития. У всех этих зародышей передние конечности только что заложияись и, как видно из сравнения этих трех рисун-
1 Sewertzoff A. N., Studien uber die Entwicklung der Muskeln, Nerven und des Skeletts der Extremitaten der niederen Tetrapoda, Бюлл. о-ва испытателей природы* Москва, 1908.
2 Sewertzoff A. N., Studien liber die Reduktionder Organe der Wirbeltiere, Zool. Jahrb., Bd. 53, 1931.
ФПЛОГЕНЕТИЧЕОКАЯ РЕДУКЦИЯ ОРГАНОВ животных,
' 545
ков, имеют одинаковое положение и одинаковую форму. Но мы видим, что величина зачатков конечностей у всех трех зародышей различна: так, зачаток конечностей у Ascalabotes (рис. 204, е) значительно больших
Рис. 205. Зародыш Seps chalcides на той же стадии развития, что и зародыш, изображенный на рис. 204.
е—зачаток передней конечности (из А. Н. Северцова, 1931).
размеров, чем зачаток конечностей у Seps, а зачаток конечностей у Seps (рис. 205, е) почти вдвое больше, чем зачаток конечностей у Ophisaurus1 (рис. 206, е). Эта. разница в размерах зачатков конечностей сохраняется е
Рис. 206. Зародыш Ophisaurus apus на той же стадии развития, что и зародыши, изображенные на рис. 204 и 205.
с—зачаток передней конечности. А—разница в величине зачатков передней конечности на ранних стадиях развития у Ascalabotes, Seps и Ophisaurus. Ср. рис. 204, .205 и 206 (из А. Н. Север-
в течение всего эмбрионального развития, равно как и в течение всего периода роста, вплоть до взрослого состояния, так что мы можем с полным правом сказать, что раз н и ц а м еж д у величиной конеч-
1 Следует обратить внимание на то, что сами зародыши Seps и Ophisaurus больше, чем зародыш Ascalabotes.
Морфологические закономерности
35
546
ГЛАВА XV
ностей у взрослого Seps и взрослого Ascalabotes является непосредственным следствием разницы в величине, существующей между их зачатками.
Различия между развивающимися конечностями геккона и сепса видны при сравнейии рисунков 207 А—Е и 208 А—Е. Рисунки эти показывают нам следующие друг за другом стадии развития задних конечностей геккона (рис. 207 А—Е) и сепса (рис. 208 А—Е) при одинаковом увеличении. Мы видим, что разница в величине между этими двумя развивающимися зачатками конечностей остается приблизительно одинаковой во все время развития, так что малая величина конечностей сепса1 зависит от малых размеров зачатков этих конечностей.
Рис. 207 А—Е. Пять последовательных стадий развития наружной формы задней конечности у Ascalabotes fascicularis.
е—зачаток задней конечности (из А. Н. Северцова, 1931).
Исследуя прогрессивную эволюцию органов, мы в свое время видели,, что эта эволюция во многих случаях осуществляется путем прогрессивного изменения первых зачатков эволюирующих органов: зачатки таких органов в течение филогенеза постепенно увеличиваются в размерах, меняют свою форму, свое положение и т. д. Благодаря этим изменениям меняются в том же направлении и происходящие из этих зачатков органы. Некоторые из этих органов становятся больше, иные меняют свою форму,
1 Я не мог достать полную серию зародышей желтопузика (Ophisaurus apus) и вследствие этого не имел возможности проследить ход процесса рудиментации очень сильно дегенерировавшей конечности этой формы. Тем не менее, исследование даже неполной серии развития задней конечности желтопузика и изучение конечностей взрослых форм ясно указывают на то, что и здесь уменьшение размеров конечностей взрослых животных зависит от уменьшения величины первых зачатков этих органов...
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
547
иные—свое положение и т. д. Такое прогрессивное изменение первых зачатков эволюирующих органов мы обозначили термином «положительного архаллаксиса» и поэтому регрессивные изменения первых зачатков редуцирующихся органов, ведущие к постоянно прогрессирующему уменьшению их размеров, мы можем обозначить термином «отрицательный архаллаксис». Таким образом, положительному архалла-ксису, путем которого осуществляется прогрессивная эволюция многих органов, соответствует при' рудиментации «отрицательный архаллаксис», ведущий к уменьшению дегенерирующих органов и к их конечному превращению в незначительный рудимент.
Итак, мы видим, что и при редукции органов главную роль играет изменение хода эмбриональ-
Рис. 208 А—Е. Пять последовательных стадий развития наружной формы задней конечности Seps chalcides.
е—зачаток задней конечности (из А. Н. Северцова, 1931).
ного развития, т. е. филэмбриогенез, в данном случае—отрицательный филэмбриоген е з.
Отрицательный архаллаксис, как это мы только что видели, объясняет нам процесс постепенного уменьшения величины рудиментов органов. Но как можно объяснить наблюдаемое нами при сравнительно анатомическом исследовании постепенное упрощение строения редуцирующегося органа? Чтобы ответить на этот вопрос, мы должны несколько подробнее рассмотреть ход эмбрионального развития конечностей рептилий. Как известно, отдельные скелетные элементы конечностей рептилий закладываются в определенной закономерной последовательности. Эта последовательность изображена нами на прилагаемой ниже схеме (схема XXXVIII), в которой нами приводится порядок появления хрящевых и скелетных элементов в передних конечностях геккона.
Рассматривая эту схему, мы видим, что на самой ранней из исследованных нами стадий развития заложились humerus, radius и ulna. Судя по состоянию тканей, первым заложился humerus, затем ulna и только после них radius. На этой же стадии развития появилась и прохондраль-ная закладка четвертого пальцевого луча.
Здесь мы также видим, что в порядке появления закладок хрящевых скелетных частей господствуют два общих направления: 1) проксимо-дисталь-
35*
548
ГЛАВА XV
Стадия I палец II палец III палец IV палец V палеи
1 Н. R. U. — — v4 —
2 Н. R. U. U. — __ C3(V8) с4 v4 V5
3 Н. R. U. U. V1 V2 С3 III. 1 с4 IV. 1 (с5) v5
4 Н. R. U. г. u. (С) V1 (c2)(II) C3III. 1. с4 IV. 1 с3 V3
5 Н. R. U. г. u. С. (Сх)(1) са II с3Ш. 1 с4 IV. 1.2. c5V
6 Н. R. U. г. u. i. С. Ci I. 1.2 с2 II 1. 2 с3 111. 1.2.3 с4 IV. 1.2.3.4. c5V. 1.2
7 Н. R. U. г. и. 1. С. Cjl. 1.2.3 с2П 1.2.3 с3Ш. 1,2.3.4 с4 IV. 1.2.3.4.5 с, V. 1.2.3
Схема XXXVIII. Развитие передней конечности Ascalabotes fascicularis.
ci, С2, с3, с4, сз—carpalia distalia i—5; С—centrale carpi; i—intermedium (y Ascalabotes рудиментарное); H—humerus; R—radius; r—radiale carpi; U—ulna; u—ulnare carpi; vj—v6—прохондраль-вые зачатки I—V пальцев; I—V—metacarpalla I—V; 1, 2, 3, 4, 5—первая, вторая, третья, четвертая и пятая фаланги пальцев.
ное направление (humerus—»ulna—»radius—»carpalia distalia—»metacar-palia—»фаланги 1,2, Зит. д.) и 2) локте-лучевое направление. Это последнее направление выражено не вполне ясно, так как здесь первым закладывается IV палец, затем V и III, затем II и I. Вместе с тем мы можем наблюдать, что первыми развиваются проксимальные и вообще постаксиальные (локтевые) элементы скелета конечностей ящериц, затем появляются дистальные элементы, т. е. конечные фаланги пальцев, и последней— конечная пятая фаланга IV пальца (схема XXXVIII).
На рис. 209 А—D изображены четыре последовательные стадии развития скелетных элементов передней конечности Seps chalcides, а на рис. 210 А и В—две стадии развития задней конечности той же формы. Сравнение развития скелета конечностей сепса и геккона показало, что у сепса в его редуцированной конечности наблюдается такая же последовательность в образовании скелетных элементов, как и в нормально развитой конечности Ascalabotes.
Э. Менерт является родоначальником широко распространенного в свое время взгляда, принимавшего, что редуцированные органы имеют замедленное развитие, т. е. что они закладываются позднее, чем нормальные органы, и развиваются медленнее, чем они (ретардация). Однако наши наблюдения над развитием конечностей у безногих ящериц показали, что это не соответствует истинному положению вещей. Мы видели, что первые зачатки нормальной конечности Ascalabotes, сильно редуцированной конечности Seps и совершенно рудиментарной конечности Ophisaurus (рис. 204, 205, 206) закладываются на одной и той же стадии развития, т. е. в одно и то же время, и что здесь, таким образом, не наблюдается никакого запаздывания в закладке редуцирующегося органа. То же можно сказать и относительно развития отдельных скелетных элементов. Мы видим, например, что сильно редуцированное metacarpale V (resp. metatarsale V) у Seps закладывается совершенно так же, как и у Ascalabotes, одновременно с еще недегенерировавшим metacarpale III (s. metatarsale 111) (рис. 209 В, V и III), а не позже его, как этого можно было бы ожидать на основании утверждения Менерта. Рудиментация проявляется здесь главным образом в первичном уменьшении величины первого зачатка органа, а не в ретардации его закладки.
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
549
Последовательность во времени образования скелетных элементов конечности одинакова у Seps и у Ascalabotes (ср. схему XXXVIII). Прежде всех закладываются проксимальные элементы конечности (рис. 209 А), затем закладываются дистальные (рис. 209 В—D и рис. 210 А—В), а пост-
Рис. 209 А—D. Четыре последовательных стадии развития передней конечности Seps chalcides.
С—centrale; с2—с4—carpalia distalia 2—4; Dig. IV—зачаток четвертого пальца; Н—humerus; pi— pisiforme; R.—radius; r.—radiale; U.—ulna; u.—ulnare; II—V—metacarpalia II—V; 1—3—фаланги 1—3 (из Д. H. Северцова, 1931).
аксиальные части скелета образуются раньше, чем преаксиальные. Редукция же заключается в том, что здесь, в конечностях Seps, некоторые элементы не закладываются вовсе. Они не редуцируются во время онтогенеза,1 а вообще не закладываются, так что здесь мы можем говорить именно о выпадении эмбриональной закладки органа.
1 Как, например, мускулы черепахи, хвостовой плавник у личинок бесхвостых амфибий и т. д. (афанизия).
550
ГЛАВА XV
Чтобы лучше понять ход редукции органов, происходящий путем рудиментации, было важно проследить эмбриональное развитие конечности, еще более редуцированной, чем конечность Seps. К счастью, мне
Рис. 210 А—В. Две последовательных стадии развития задней конечности Seps chalcides А—astragalus; Fe—femur; F—fibula; f—fibulare; T—tibia; tg—14—tarsalia distalia 3—4; II—V— metatarsalia II—V; 1—1-я фаланга (из A. H. Северцова, 1931).
удалось получить для этого соответствующий материал. Как видно из рисунков 201 А—D, задняя конечность желтопузика является однолучевой конечностью, но состоит обычно из большего числа скелетных элементов, чем это прежде думали. На самой ранней из исследованных нами стадий
Рис. 211. Позвоночник, тазовый пояс и задняя конечность зародыша Ophisaurus apus на очень ранней стадии развития.
с. 1.—поясничные ребра; с.th—грудные ребра; Dig. IV—четвертый палец (metatarsale IV);E—зачаток конечности; F.—fibula; Fm.—femur; il.—ileum; isch.—ischium; pb.—pubis; S—крестцовый позвонок; T—tibia; v. c.—хвостовой позвонок; v. 1.—поясничный позвонок; v. th.—грудной позвонок (из А. Н. Северцова, 1931).
задняя конечность зародыша желтопузика имела форму маленького тупого бугорка (рис. 211, Е); в еще хрящевом тазу мы видим его составные части: ileum, ischium, pubis;1 ileum сочленяется с крестцовым позвонком.
1 Здесь таз представлен одним цельным неразделенным на части хрящом. Так как на более поздней стадии развития ileum, ischium и pubis являются самостоятельными костями, то вполне вероятно, что эти элементы онтогенетически закладываются как самостоятельные хрящи.
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
551
В свободной конечности зачаток femur (рис. 211,Рт)еще сравнительно большой величины.1 Zeugopodium представлен сильно редуцированной tibia (рис. 211, Т) и менее редуцированной fibula (рис. 211, F); возле них находятся остатки одного пальца; по всей вероятности, это metatarsale IV (dig. IV). Мы видим, что у желтопузика охрящевение идет совершенно тем же путем, что у Seps и у Ascalabotes, т. е. образуются хрящевые зачатки •stylopodium (Fm) и zeugopodium (F, Т); в плюсне еще не развилось никаких охрящевений, но уже образовался пальцевой луч.
Таким образом, при рудиментации уменьшается не только величина всей конечности, но и величина отдельных ее частей, причем дистальные скелетные зачатки (F, Т) уменьшаются в большей степени, чем проксимальные скелетные зачатки (Fm).
На следующей исследованной нами стадии развития задней конечности желтопузика (рис. 212) мы видим, что zeugopodium (T-J-F) состоит
Рис. 212. Позвоночник, тазовый пояс и задняя конечность зародыша Ophisaurus apus (более поздняя стадия развития).
Обозначения те же, что и на рис. 211. Кроме того: А—astragalus (tibio-centrale); f—fibulare-Иаг-sale distale IV (?) (из A. H. Северцова, 1931).
уже из одного единственного крупного хряща. Возможно, что этот хрящ соответствует сросшимся между собой tibia и fibula предшествующей стадии. Дистально от zeugopodium расположены два маленьких tarsalia, вероятно fibulare (рис. 212, f) и astragalus (рис. 212, A). Fibulare (f) непосредственно связано с пальцем (dig. IV), который мы видели на предыдущей стадии. Представляется вполне вероятным, что этот скелетный зачаток (dig. IV) соответствует ерошимся между собой metatarsale IV и tarsale distale IV (ср. рис. 200, Ophiodes).1 2
Если мы сравним только что описанную нами позднюю стадию развития скелета задней конечности желтопузика со строением скелета взрослого животного, то увидим, что изменения, происшедшие в этой конечности в течение ее дальнейшего развития, повидимому, состояли лишь в том, что оба tarsalia proximalia, т. е. fibulare и astragalus (рис. 212, f и А),
1 Само собой разумеется, он меньше, чем, например, femur зародыша ящерицы на соответствующей стадии развития.
2 Настоящее толкование скелетных элементов tarsus и metatarsus желтопузика я даю с известной оговоркой, так как Строение эмбриональной задней конечности этого животного сильно вариирует; поэтому без детального исследования этих вариаций трудно дать исчерпывающую гомологизацию отдельных скелетных частей; для наших целей, однако, мы можем довольствоваться приведенной нами интерпретацией этих элементов скелета.
552
ГЛАВА XV
срослись между собой, образовав одно цельное tarsale proximale, типичное для всех ящериц (ср. рис. 194 В, 197 В, 199, t. рг.).
Мы можем, таким образом, изобразить ход морфогенеза задней конечности желтопузика, согласно принятому нами выше обозначению, следующими тремя формулами: ’
IV IV IV
A. f. A. +f.
Т. F. Т. + F. Т. + F.
Fm. Fm. Fm.
X—— —-Х
1 2 3
Мне не удалось проследить шаг за шагом развитие передней конечности желтопузика. Можно лишь сказать, что у него (рис. 202) проксимальный конец (проксимальный эпифиз) humerus закладывается на относительно поздней стадии развития. Средние и дистальные части humerus и все скелетные элементы, расположенные дистально от плеча, не образуются вовсе.
Сравнивая развитие сильно редуцированной задней конечности желтопузика с развитием этой конечности у Seps и у Ascalabotes, мы видим, что, несмотря на редукцию, общий ход эмбрионального развития остается у всех этих трех форм одинаковым: развитие идет так же, как и у нередуцированных форм, в проке им о-ди стальном и по стакс и а л ь но-п р е а к с и а л ь н о м (ульно-радиальном или фибула-тибиальном) направлении, т. е. проксимальные элементы конечностей закладываются раньше, чем дистальные, ит. д. Интересно, что даже такое уклонение от этого правила, как, например, факт, что элементы tarsus и carpus образуются у ящериц несколько позднее, чем метатарсальные и метакарпальные элементы, проявляется в сильно редуцированной конечности желтопузика так же, как и в нормальной конечности Ascalabotes.
Это решительно говорит против взгляда, утверждающего, что ход онтогенетического развития, т. е. последовательность в закладке скелетных элементов зависит от прогрессивного или регрессивного направления эволюции этих образований (Э. Менерт). Согласно этому взгляду, в сильно редуцированных конечностях должны были бы происходить очень значительные гетерохронии. Мы видим, однако, что этого нет. У желтопузика наблюдается та же последовательность в закладке скелетных частей, что и у Seps и у Ascalabotes (схема XXXVIII). Таким образом, результаты, полученные нами при исследовании развития конечностей Seps, находят здесь новое подтверждение: 1) отрицательный архаллаксис привел к уменьшению величины как всей конечности, так и отдельных скелетных элементов входящих в состав ее частей, 2) редукция остальных скелетных элементов обусловлена тем, что эти элементы совсем не закладываются в онтогенезе.
Сравнительно анатомическое исследование показало нам, что в исчезновении скелетных элементов существует определенная последовательность. Расположив исследованные нами формы в порядке увеличения числа их позвонков (схема XXIV), мы видим, например, что у Lygosoma punctu-lAtum, имеющей 35 предкрестцовых позвонков, нехватает пятой фаланги IV пальца, у Chalcides sepoides, имеющего 48 предкрестцовых позвонков, отсутствуют, кроме пятой фаланги IV пальца, еще первая и вторая фаланга I пальца, у Ophiomorus, имеющего 52 предкрестцовых позвонка, исчезли уже не только пятая фаланга IV пальца и четвертая и третья фаланги III
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ 553
и II пальцев, но и все фаланги V пальца, а также все фаланги и metacarpaie I пальца; наконец, у Ophiodes,- имеющего 65 предкрестцовых позвонков, редуцировались metatarsalia I и II и все фаланги Г и II пальца задней конечности, кроме того все фаланги III пальца, вторая, третья, четвертая и пятая фаланги IV пальца, а также все фаланги и metatarsale V пальца. Таким образом, как на это указал еще М. Фюрбрингер, скелетные элементы конечностей безногих ящериц редуцируются в определенной последовательности. Эмбриональные же исследования нам показали, что р е-дукция эта зависит от отсутствия эмбриональных закладок этих скелетных частей.
У Lygosoma punctulatum первой выпадает пятая фаланга IV пальца, т. е., как это было только что сказано, она не закладывается вовсе в течение эмбрионального развития. Таким образом, первым исчезает тот скелетный’ элемент, который в нередуцированной конечности ящерицы закладывается’ позже всех. У Chalcides sepoides выпадают пятая фаланга IV пальца и, кроме того, первая и вторая фаланги I пальца. Мы видели, что у Ascalabotes (нормальная конечность) I палец закладывается позже всех остальных. Мы видели также, что у форм, у которых редукция пошла еще дальше, выпадают конечные фаланги Пи III пальца, а также все фаланги V пальца, т. е. что они у них эмбрионально совсем не закладываются. Иными словами, отрицательные изменения хода онтогенеза скелетных элементов (отсутствие их закладки) происходят в дисто-проксимальном направлении.
М. Фюрбрингер считал, что последовательность в редукции скелетных элементов конечностей ящериц является функцией местоположения этих элементов. Сравнительно эмбриологическое исследование привело меня к выводу, что эта последовательность является функцией времени эмбриологической закладки этих элементов. Скелетные части, закладывающиеся позднее всех, т. е. конечные фаланги пятипалой конечности, редуцируются первыми, иначе говоря, первыми перестают закладываться. Следующими начинают выпадать те скелетные части,которые закладываются несколько раньше их, т. е. предпоследние фаланги и т. д. Элементы преаксиальной стороны конечности (лучевая сторона) онтогенетически закладываются несколько позднее, чем элементы постаксиальной стороны. Поэтому они и редуцируются несколько раньше, чем элементы постаксиальной стороны (I палец редуцируется быстрее, чем V и т. д.).
Чем раньше какой-нибудь скелетный элемент закладывается в онтогенезе, тем позднее редуцируется он в филогенезе. Развитие конечностей у Seps и у желтопузика потдверждает это правило. Выше мы видели, что у желтопузика в задней конечности сохранился лишь один единственный палец, metatarsale IV, который в нередуцированной конечности закладывается раньше всех остальных metatarsalia (см. Ascalabotes на схеме XXXVIII, ср. Seps в рис. 210 А). Сильно уменьшившиеся остатки элементов, лежащих проксимально от metatarsus, также сохранились в онтогенезе задних конечностей желтопузика. В передних конечностях желтопузика сохранилась лишь проксимальная часть humerus, т. е. именно та часть скелета, которая в свободной конечности закладывается раньше всех остальных, скелетных частей. Все же более поздно закладывающиеся скелетные элементы редуцировались.
Мы видели, что многие органы эволюируют путем изменения их первых зачатков, и назвали этот путь филогенетического развития органов^ архаллаксисом. Возникновение филогенетически важного изменения органов, т. е. время наступления архаллаксиса, зависит от момента образования данного органа или его частей. Если закладка органа
554
ГЛАВА XV
происходит на ранних стадиях эмбрионального развития животного, то и изменение этого зачатка также наступает рано. Если же эволюирующая часть органа, как, например, прохондральная закладка какой-нибудь дистальной фаланги, образуется относительно поздно, то и ход онтогенеза начинает изменяться также поздно. Мы видели, что развитие (филогенетическое новообразование) дистальных фаланг происходит на поздних стадиях онтогенеза1 и что выпадение конечных стадий (в данном случае отсутствие закладки) начинается на последних стадиях и переходит онтогенетически на более ранние стадии, т. е. на более проксимально лежащие фаланги.1 2
Раньше я был склонен рассматривать этот процесс как отрицательную анаболию, теперь же я думаю, что правильнее было бы толковать его как отрицательный архаллаксис. Если конечные фаланги безногих ящериц, о которых идет речь, вследствие медленного роста или вследствие каких-либо других причин, изменяются в состоянии эмбриональных закладок на более поздних стадиях развития, то мы имеем право говорить об отрицательной анаболии; если же зачатки какой-либо скелетной части с самого начала отсутствуют (дистальные фаланги, затем проксимальные фаланги, элементы metapodium и т. д.) или если эти зачатки -с самого начала возникают как относительно малые образования (все скелетные части безногих ящериц), то мы должны этот вид редукции обозначить как отрицательный архаллаксис. При этом мы различаем двоякого рода архаллаксисы: 1) отрицательный архаллаксис, выражающийся в уменьшении пер’вых зачатков, и 2) отрицательный архаллаксис, выражающийся в выпадении первых зачатков.
Мы можем, следовательно, сказать, что рудиментация органов всегда осуществляется путем отрицательного архаллаксиса.
При этом дегенерирующие органы постепенно превращаются во все более и более уменьшающиеся рудименты со все более и более упрощенной структурой, очень долго сохраняющиеся у потомков, но все же в конце концов целиком исчезающие. Именно поэтому мы и обозначили этот, тип дегенерации органов термином рудиментация.
Если мы теперь спросим, является ли рудиментация широко распространенным в природе процессом, то на этот вопрос нам придется дать утвердительный ответ. Во всех тех случаях, когда мы находим у животных рудименты каких-либо органов, т.,е. совершенно бесполезные, более или менее уменьшенные и вторично упрощенные части, мы можем уверенно сказать, что здесь соответствующие органы предков редуцировались путем рудиментации.
Нам известно, что число встречающихся у многоклеточных животных рудиментов, вообще говоря, огромно. Напомним читателю о более или менее рудиментарных глазах у животных, постоянно пребывающих в темноте [глаза роющих амфибий (Gymnophiona), рептилий и млекопитающих], о глазах пещерных животных (рыбы, хвостатые амфибии, раки, насекомые), о глазах некоторых глубоководных рыб, о редукции плакоидных чешуй на брюшной стороне тела у скатов и костных чешуй у многих костистых рыб, о редукции пальцев на ногах у парно- и непарнокопытных животных, о редукции задних конечностей у китообразных и о многих других примерах рудиментации органов. Все это говорит о том, что рудиментация является весьма распространенным модусом редукции органов.
1 Проксимо-дистальное направление прогрессивной эволюции этих образований.
2 Дисто-проксимальное направление рудиментации соответствующих частей.
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ 555
ПОЛНАЯ ДЕГЕНЕРАЦИЯ ОРГАНОВ ИЛИ АФАНИЗИЯ
Обратимся теперь к описанию второго типа редукции органа, того процесса, который мы обозначили как активную редукцию или афанизию органов.1
Процесс афанизии легче всего понять на примере дегенерации спинной мускулатуры у черепах, которая специально для этой цели была мною исследована. Мы знаем, что у черепах спинные позвонки плотно срастаются между собой и с ребрами, ребра же, кроме того, прочно срастаются с внутренней поверхностью сагарах, так что мускулы, которые у предков черепах были расположены между подвижно соединенными между собой позвонками и ребрами и двигали эти последние, потеряли у потомков при развитии спинного панцыря свою функцию и утратили вследствие этого свое физиологическое значение. Поэтому у взрослых1 черепах оказалась редуцированной вся спинная мускулатура, т. е. все интервертебральные и интеркостальные мускулы туловищной области.
У зародышей черепахи туловищные сомиты и происходящие из них миотомы закладываются совершенно так же, как они закладываются у форм со свободно подвижными позвонками, например у ящериц или у крокодилов. В дальнейшем туловищные миотомы развиваются также совершенно нормально: из них образуются зачатки мускулатуры конечностей и развиваются лежащие между ребрами mm. intercostales.
Сагиттальный разрез такой интеркостальной мускулатуры, лежащей между двумя ребрами, изображен на рис. 213 (Emys lutharia). Здесь мы видим, что вполне развитые мускульные волокна М (на этой стадии еще без поперечной полосатости) идут от одного ребра (С) к другому. Одна ветвь спинномозгового нерва (spj проходит дорзально от этих мускулов, другая (sp2)—вентрально от них, а третья (sp3) находится в середине мускульного зачатка, между мускульными волокнами. На этой стадии редукция между-реберной мускулатуры еще не наступила.
Разрез, изображенный на рис. 214, проходящий через эмбриональную межреберную мускулатуру, принадлежит несколько более позднему зародышу другой черепахи (Chelydra serpentina), у которого редукция межреберной мускулатуры уже началась. Мы видим, что лежащий между ребрами С и С мускульный слой М' состоит из небольшого числа мускульных волокон, идущих между ' ветвями спинномозговых нервов (spj и (sp2). Существенное различие между мускульным зачатком этой более поздней стадии и стадии, изображенной на рис. 213, заключается в том, что здесь развивается большое число капиллярных сосудов (рис. 214, Сар.), проходящих между мускульными волокнами и образующих сеть кровеносных сосудов, пронизывающую мускульную ткань. Лучше, чем на сагиттальном разрезе, эта капиллярная сеть (Сар.) видна на поперечном разрезе, проходящем через редуцирующийся зачаток межреберной мускулатуры, изображенной на рис. 215; здесь мы видим очень большое число эмбриональных капилляров, лежащих между мускульными волокнами.
На рис. 216 редукция межреберной мускулатуры уже пошла далеко вперед. Здесь от довольно значительной мускульной массы, изображенной на рис. 213, остался лишь маленький мускульный пучок (рис. 216, М'), густо пронизанный капиллярными сосудами (Сар). Этот мускульный пучок сохраняет, однако, свое характерное положение между двумя ветвями спинномозговых нервов (spx и sp2).
На рис. 217 и этот последний остаток вполне развитого у более молодых зародышей миотома редуцировался целиком, так что между только что
1 Б своих прежних работах (см. Изв. Акад. Наук СССР, 1930) я обозначил этот тип редукции органов термином «аннигиляция». Теперь слово «афанизия» (от a'fmicu— исчезновение) кажется мне лучше передающим значение этого процесса.
556
ГЛАВА XV
упомянутыми нами ветвями спинномозговых нервов (sp, и sp2) уже не видно никаких 'мускульных волокон. Очень интересно, что и капиллярные сосуды, столь сильно развитые в редуцирующемся мускульном зачатке (рис. 215, 216), на этой более поздней стадии развития также совершенно исчезают. Повидимому, развитие этих сосудов стоит в известной связи с редукцией миотомов: они развиваются, когда редукция начинается, и исчезают, когда редукция приходит к концу.
Повидимому, мускульные волокна при редукции превращаются в инди-ферентную соединительную ткань. На рис. 218, где изображен при более сильном увеличении сагиттальный разрез через последние остатки уже почти целиком редуцированного миотома, видно, как конец мускульного волокна М' постепенно переходит в соединительнотканные волокна (с. f., с. Г). Такая картина, встречающаяся, между прочим, довольно часто, приводит к мысли, что редукция туловищной мускулатуры черепах заключается в превращении мускульных волокон в тонкие индиферентные соединительнотканные волокна.1
Мы видим, таким образом, что здесь перед нами также имеется один из случаев редукции органов, но что эта редукция происходит совершенно другим путем, чем происходила рудиментация конечностей в разобран
ном нами выше примере безногих
При рудиментации
1. Эмбриональный зачаток дегенерирующего органа уменьшается в размерах, начиная с самых ранних стадий развития.
2. Строение дегенерирующего органа постепенно упрощается в течение следующих друг за другом поколений из-за отсутствия образования конечных стадий развития частей данного органа. Уменьшенный и структурно упрощенный рудимент сохраняется у взрослых животных в течение очень многих поколений.
3. Редуцирующийся орган бесполезен животному во все периоды жизни животного.
ящериц.
При афанизии
1. Эмбриональный зачаток дегенерирующего органа закладывается не-уменьшенным, и его эмбриональное развитие вплоть до известной стадии протекает совершенно нормально.
2. Начиная с известных стадий развития, дегенерирующий эмбриональный орган начинает уменьшаться в размерах и в течение короткого времени редуцируется целиком, так что у взрослого животного от него не остается никаких рудиментов.
3. Редуцирующийся орган полезен животному либо в течение эмбрионального периода, либо в период молодости.1 2
Из этого сопоставления характерных черт рудиментации и афанизии явствует, что существуют два различных вида редукции органов: I) рудиментация, которая совершается путем параллельного действия уменьшения величины зачатков редуцирующегося органа в начальных стадиях морфогенеза (отрицательный архаллаксис
1 Мы позволим себе выразить эту мысль лишь с известной оговоркой, так как окончательное решение этого вопроса можно получить только путем гистологического исследования специально для гистологических (гистогенетических) целей зафиксированного материала. К сожалению, для такого детального исследования'материал, имевшийся в нашем распоряжении, оказался недостаточным.
2 Мы видели, что у черепах первые зачатки мускулатуры конечностей взрослых животных происходят из миотомов и что у них редукция (афанизия) только тогда вступает в силу, когда эти зачатки отделяются от миотомов и переселяются в конечности.
Рис. 213. Сагиттальный разрез через туловищный миотом зародыша черепахи (Emys lutharia).
G — поперечный разрез через хрящевое ребро; Ер — эпидермис наружной поверхности тела; М— зачаток межреберной мускулатуры; Mes—мезенхима; spt, зр2 —ветви спинномозговых нервов соответственных сегментов; зр5—ветвь тех же нервов, проходящая внутри мускулов (из А. Н. Северцова, 1931).
Рис. 214. Сагиттальный разрез через туловищный миотом черепахи (Chelydra serpentina) на несколько более поздней стадии развития (11мм), когда межреберная мускулатура начинает редуцироваться.
Обозначения те же, что и на рис. 213. Кроме того: Сар. -капиллярные сосуды; М'—редуцирующаяся межреберная мускулатура (из А. Н. Северцова, 1931).
Морфологические закономерности, стр. 557
М1
Рис. 215. Поперечный разрез через редуцирующуюся межреберную мускулатуру зародыша черепахи на той же стадии развития, что и на рис. 214.
Обратить внимание на увеличившееся число капиллярных сосудов (Сар.'. Обозначения те же, что и на предыдущих рисунках (из А. II. Северцова, 1931).
Л"
Рис. 216. Сагиттальный разрез через несколько более поздний зародыш черепахи (Emys lutharia) на стадии развития, когда межреберная мускулатура еще более редуцировалась.
Обозначения тс же, что и па предыдущих рисунках (иа Л. II. Северцова, 1931).
Рис. 217. Сагиттальный разрез через спинную область еще более позднего зародыша черепахи (Chelydra serpentina) на стадии развития, когда межреберная мускулатура уже полностью редуцировалась (ср. положение ветвей спинных нервов spx и sp2 с их положением на рис. 213).
С—ребро; Mes. d.— плотная соединительная ткань; Mes. I. рыхлая соединительная (из А.. Н. Северцова, 1931).
Рис. 218. Часть сагиттального разреза (при более сильном увеличении) через остаток редуцировавшейся межреберной мускулатуры Chelydra serpentina.
М'. -мускульные волокна межреберных мускулов; с. Г., с. Г'.—соединительнотканные волокна (из А. И. Северцова, 1931).
Рис, 219. Сагиттальный разрез через переднюю часть головы малька Acipenserruthenus. На рисунке виден еще вполне развитой премаксплярпый зуб,
d. ртх. — премаксплярпый зуб; р,—полость пульпы этого зуба; р. ц,— пёбноквадратвый хрящ; р. их,-ргаешахШа; х.—задняя стенка ауба; у.- передняя стенка зуба (из А. Н. Северцова, 1931'.
Морфологические закономерности, стр. 557
Рис. 220. Сагиттальный разрез через переднюю часть головы несколько более позднего малька Acipenser ruthenus. Редукция премаксилярпого зуба пошла дальше, чем на предыдущей стадии; задняя стенка зуба (х) резорбирована.
d. pmx. — премаксплярный зуб; mes. — мезенхима; pmx. — praemaxilla; р. — полость пульпы; p.q.—нёбноквадратный хрящ; у,—передняя стопка зуба (из А. Н. Северцова, 1931).
Рис. 221. Сагиттальный разрез через переднюю часть головы еще более позднего малька Acipenser ruthenus (32 мм длины). Конечная стадия редукции премаксилярпого зуба.
d; pmx,—отпадающая верхушка премаксилярпого зуба; pmx.—praemaxilla; р. q.—нёбпоквадрат-ный хрящ; х.— задняя стенка вуба; у. — передняя стенка зуба (из А. И. Северцова, 1931).
филогенетическая редукция органов животных
557
через уменьшение) и выпадения конечных стадий онтогенеза (отрицательный архаллаксис через выпадение) и 2) афанизия, при которой отрицательный архаллаксис вообще не имеет места. При афанизии морфогенез известного органа, в нашем случае морфогенез туловищной мускулатуры черепах, не достигает своего нормального завершения, ибо в нем выпадают его конечные стадии. При этом мы особенно подчеркиваем, что здесь редукция конечных стадий происходит совершенно не тем путем, каким происходила редукция конечных стадий, наблюдаемая при рудиментации. При рудиментации отдельные части редуцирующегося органа вообще не закладываются, при афанизии же за л оживши еся части органа постепенно исчезают.
Мы рассмотрели процесс афанизии только на одном примере этого типа редукции органов и должны поэтому поставить вопрос о том, много ли нам известно вообще таких примеров? Я лично думаю, что этот тип редукции органов очень распространен в животном мире. Все органы, полезные животному в течение периода онтогенеза или в течение периода молодости и функционирующие у него в течение этих периодов, но теряющие свое значение у взрослых форм и исчезающие у них, обычно редуцируются именно по этому способу редукции.
Мы знаем, что у личинок всех бесхвостых амфибий1 имеется типичный хвостовой плавник, совершенно подобный хвостовому плавнику личинок хвостатых амфибий. Этот хвостовой плавник функционирует у головастиков в течение их жизни в воде, но исчезает, когда они переходят к жизни на суше. Как известно, к этому времени хорда, мускулы и все остальные ткани хвоста головастика разрушаются фагоцитами, так что весь его хвост оказывается целиком редуцированным. Хвостовой плавник личинок бесхвостых амфибий унаследован ими от их хвостатых предков,1 2 но у взрослых форм он замещается уростилем, лучше приспособленным для прыгающих движений, совершаемых анурами на суше. И мы видим, что этот хвостовой плавник, так же как и межреберная мускулатура черепах, развивается сначала совершенно нормально (т. е. так же, как и у хвостатых амфибий, и, поскольку мы можем судить, у предков бесхвостых амфибий) и функционирует довольно долгое время; когда же при новом образе жизни на суше и новом способе передвижения он перестает быть полезным, то быстро редуцируется. Здесь мы имеем типичный пример афанизии: нормальная закладка, нормальное развитие и функционирование до известной стадии развития, затем дегенерация и полная редукция без сохранения рудиментов.
Примером афанизии является также редукция зубов у некоторых рыб, например у Acipenser ruthenus. На рис. 219—221 изображен процесс редукции зубов у Acipenser ruthenus. У мальков стерляди зубы обычно развиваются нормально (рис. 219, d. pmx.) и функционируют в течение некоторого периода. Затем наступают изменения. На рис. 220 мы видим, что в полости пульпы образуется большое отверстие, которое, повидимому, происходит оттого, что задняя дентиновая стенка зуба начинает постепенно разрушаться близлежащими мезенхимными клетками (рис. 220, х., mes.). На более поздних стадиях редукции зубов разрушается также и передняя стенка зуба (рис. 221, у), так что от самого зуба остается лишь одна верхушка (рис. 221, d. pmxv), уже не связанная с премаксиллярной костью, на которой сидел зуб (рис. 221, pmx). Верхушка эта вскоре совсем отпадает.
1 Напомним еще о хвосте личинок асцидий, который также редуцируется этим путем.
2 Как известно, у хвостатых амфибий хвостовой плавник существует и функционирует как у молодых, так и у взрослых форм; у бесхвостых амфибий он существует только в течение личиночных стадий.
558
ГЛАВА XV
Здесь перед нами имеется типичный случай афанизии. Орган, хорошо развитый у предков Acipenseridae, развивается у современных осетровых до известной стадии совершенно нормально, функционирует у мальков и затем редуцируется в течение очень короткого времени, не оставляя никаких рудиментов у взрослого животного.
Я имел случай наблюдать целый ряд афанизии, исследуя развитие жаберных сосудов у рыб.1 Многие кровеносные сосуды, например комиссуры, лежащие между venae branchiales posteriores и arcus aortae branchiales dorsales в эмбриональных жаберных дугах селахий, редуцируются именно этим путем. То же можно сказать и относительно редукции жаберных нитей у личинок селахий, а также относительно редукции многик кровеносных сосудов, образующихся в течение онтогенеза у рыб, функционирующих некоторое время, а затем бесследно исчезающих.
Итак, во всех известных нам случаях редукция органов животных происходит путем ряда закономерных отрицательных изменений хода морфогенеза этих органов, т. е. путем отрицательного филэмбриогенеза. Таким образом, мы видим, что как прогрессивная, так и регрессивная эволюция животных происходит при помощи филэмбриогенеза.
В предыдущей главе нашей книги мы изложили ход эволюции органов, развивающихся в прогрессивном и идиоадаптивном направлении, т. е. изложили теорию позитивного филэмбриогенеза. Основная мысль этой теории заключается в том, что ход морфогенеза эволюирующих .органов животных может изменяться различными способами (модусами) и что эти изменения вызывают собою различного рода положительные преобразования в строении органов взрослых форм. При этом мы знаем три способа, которыми происходят прогрессивные (положительные) изменения органов: архал-л а к с и с (изменения на первых стадиях развития), девиация (изменения на средних стадиях развития) и анаболия (изменения на конечных стадиях развития).
При изучении явлений регрессивной эволюции мы пришли к выводу, что редукцию органов обусловливают модусы отрицательных изменений хода морфогенеза, соответствующие модусам положительных изменений, а именно: отрицательный архаллаксис (выпадение первых зачатков органа или уменьшение размеров первых зачатков органа), который приводит к рудиментации органов, и своеобразная отрицательная анаболия (выпадение конечных стадий), которая приводит к афанизии дегенерирующих органов.
Впрочем, я не вполне уверен, что при афанизии имеет место именно отрицательная анаболия. Возможно, что здесь скорее наблюдается отрицательная девиация. Действительно, мы видим, что при афанизии орган (зубы осетровых рыб, спинные мускулы у черепах и т. д.) развивается до более или менее поздней стадии эмбрионального или личиночного развития, а затем быстро редуцируется. Можно думать, что редукция зубов осетровых рыб произошла здесь либо по модусу «своеобразной отрицательной анаболии», как я это думал раньше,1 2 либо, как мне это кажется теперь, по модусу отрицательной девиации на более поздних стадиях онтогенеза. Редукция же спинных мускулов у черепах шла по модусу отрицательной девиации на средних стадиях развития.
Изложенные здесь результаты являются второй частью теории филэмбриогенеза. С теорией положительного филэмбриогенеза, выясняющей
1 Sewer tzoff A. N., Die Entwicklung der Kiemen und Kiemenbogengefasse der Fische, Zschr. f. wiss. Zool., 1923.
2 CM.Seweftzoff A.N.MorphologischeGesetzmassigkeitenderEvolution. Jena, 1931.
ФИЛОГЕНЕТИЧЕСКАЯ РЕДУКЦИЯ ОРГАНОВ ЖИВОТНЫХ
554'
нам ход прогрессивной эволюции органов, непосредственно связана теория отрицательного филэмбриогенеза, объясняющая ход регрессивной эволюции, в которой мы находим те же типы филогенетического изменения, органов, но со знаком минус. Теперь мы действительно имеем право сказать, что теория филэмбриогенеза способна объяснить нам ход развития органов как при прогрессивной, так и при регрессивной эволюции этих животных.1,
ВЫВОДЫ
Сопоставляя результаты, к которым мы пришли в этой главе, мы можем сделать следующие выводы:
1. Существуют два различных способа редукции органов, обозначенные нами терминами: рудиментация и афанизия.
2. При рудиментации редуцирующийся орган в ряду следующих друг за другом поколений постепенно уменьшается в размерах, и его строение вторично упрощается так, что он превращается, в конце концов, в незначительный рудимент. Этот процесс обычно длится филогенетически очень долго, пока, наконец, последний рудимент органа не-исчезнет совсем.
3. Уменьшение размеров редуцирующегося путем рудиментации органа вызвано тем, что его первая закладка в течение следующих друг за другом, поколений постепенно становится все меньше и меньше. Уменьшенные размеры сохраняются в течение всего онтогенеза, так что и взрослый орган животного с течением поколений становится все меньше и меньше, пока, наконец, он не превратится в незначительный рудимент. Этот процесс мы обозначили термином отрицательного архаллаксиса через уменьшение. При отрицательном архаллаксисе через уменьшение ретардация не имеет места ни при закладке рудимента, ни при его развитии.
4. Параллельно с отрицательным архаллаксйсом через уменьшение при рудиментации органов происходит также и процесс выпадения конечных стадий, т. е. отрицательный архадТлаксис через выпадение. В редуцирующемся органе те его части, которые у предков данной формы закладываются в онтогенезе позднее всех, у потомков, у которых началась рудиментация, не закладываются вовсе, т. е. у них происходит выпадение или незакладка конечных стадий редуцирующегося органа. Существует определенная последовательность в выпадении отдельных, частей органа: первыми исчезают (не закладываются) те части органов, которые в соответствующих нередуцирующихся органах предков закладываются позже всех, вторыми редуцируются те части органов, которые у предков закладываются предпоследними и т. д. Последовательность выпадения частей редуцирующегося органа в филогенезе соответствует обратной последовательности появления этих частей в онтогенезе нереду-цирующегося органа предков данной формы.
Таким образом мы видим, что рудиментация происходит путем комбинации двух видов отрицательного архаллаксиса: первый ведет к уменьшению размеров рудимента, второй—к закономерному упрощению строения дегенерирующего органа.
1 Я отнюдь не берусь утверждать, что эта морфологическая теория дает нам окончательное и всестороннее объяснение хода эволюции животных. Ее роль заключается главным образом в том, что фи л эмбриогенез объясняет нам значение фактора времени в образовании эмбриональных закладок органа для эволюции органов взрослых животных. Вопрос, почему те или иные филогенетически важные изменения органов появляются именно на ранних, на средних или на поздних стадиях развития морфогенеза, по моему мнению, лежит вне компетенции морфологии и должен, повидимому, решаться исследователями, занимающимися проблемой наследственностиj.
560 ГЛАВА XV
5. Другой характер носит второй тип редукции органов, а именно полная дегенерация органов или афанизия. Органы, редуцирующиеся по этому второму типу, закладываются совершенно нормально (не уменьшенные первые зачатки) и некоторое время развиваются дальше так же, как и нередуцирующиеся органы. Это нормальное развитие продолжается до тех пор, пока данный орган по общему состоянию развивающегося организма или по общим условиям окружающей среды не перестает быть полезным животному. Когда этот момент наступает, начинается дегенерация органа, которая обычно протекает очень быстро и ведет к полному его исчезновению. Отрицательный архаллаксис при афанизии не имеет места, но орган дегенерирует путем своеобразной отрицательной анаболии, т. е. путем выпадения конечных стадий, или путем отрицательной девиации на более поздних стадиях онтогенеза.
ГЛАВА XVI
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА METAZOA
Как мы видели выше, модусы онтогенетических изменений, посредством которых происходит эволюция органов взрослых животных, многочисленны и в достаточной мере разнообразны. Мы можем сказать, что любой морфологический признак взрослого животного может изменяться на любой стадии своего онтогенетического развития в положительном и отрицательном направлении, т. е. что организация любого животного с эво-люцонной точки зрения очень пластична. '
Теперь нам нужно попытаться разрешить вопрос о том, как возникли модусы филэмбриогенеза, какие из них являются с эволюционной точки зрения первичными и какие—вторичными, иначе говоря, как возник и как эволюировал онтогенез многоклеточных животных. Нам надо поставить вопрос о филогенезе онтогенеза Metazoa, т. е. установить, каким путем осуществляется эволюция онтогенеза многоклеточных животных.
Отметим, что на этот вопрос мы при данном состоянии наших сведений можем ответить только более или менее вероятной гипотезой. Гипотеза эта следующая. Мы имеем полное основание предполагать, что многоклеточные животные произошли от одноклеточных форм, живших в очень отдаленную от нас геологическую эпоху. Сравнительно морфологическое исследование показало, что у Protozoa нет эмбрионального развития в том смысле, в каком мы его находим у многоклеточных, происшедших от них. У многоклеточных животных онтогенез происходит путем повторного деления оплодотворенной яйцеклетки и ее потомков, бластомеров, другими словами, онтогенез обнимает собой весь цикл делений и гистогенетических изменений, происходящих от времени конъюгации яйца и сперматозоида до взрослого состояния, т. е. до периода развития новых половых продуктов (яиц или сперматозоидов). У Protozoa этому циклу соответствует цикл делений потомков одной особи после конъюгации до нового периода конъюгации (М. Гартман); Очевидно, что изменения, происходящие в теле одноклеточных при развитии в нем органелл (ресничек, сократимых фибрилл и т. д.), соответствуют не онтогенезу Metazoa, но их гистогенезу, когда в теле каждой данной клетки образуются сократимые фибриллы, реснички и т. п.
Правда, между гистогенезом многоклеточных и гистогенезом одноклеточных имеется существенное различие: у высших одноклеточных дифе-ренцировка органелл (гистогенез) происходит или до окончания деления материнской клетки, или тотчас по окончании его. В очень многих случаях новообразования органелл не происходит вовсе, а органеллы материнской клетки просто распределяются между дочерними клетками (мионемы Vorticella и других инфузорий, реснички, пищеварительные и выдели-Морфологические закономерности 36
562
ГЛАВА XVI
тельные вакуоли и т. д.).1 В других случаях органеллы одной из дочерних клеток образуются заново (перистом стентора, пластинки скелета у Euglypha и т. п.) и заканчивают свое развитие очень быстро, так что эта дочерняя клетка скоро делается подобной своей отделившейся сестре. Подчеркнем в особенности, что такое новообразование происходит после каждого деления клеток, так что между двумя диференцировками органелл нет поколения индиферентных клеток, как это имеет место у Metazoa. То же происходит и в низших колониях одноклеточных, например у Рап-dorina, у которой каждая жгутиковая клетка колонии, делясь при бесполом размножении, дает начало двум подобным ей жгутиковым клеткам. Однако при половом размножении Pandorina между первым делением гаметоциты и образованием жгутиковой колонии существует несколько поколений индиферентных амебообразных клеток (рис. 222).
Обращаясь к гистогенезу типичных Metazoa, который мы свободна можем приравнять .к образованию органелл у одноклеточных, мы находим, что типичная специализация клеток любой ткани происходит у них н е тотчас после деления яйцеклетки надвое, как у Protozoa, но через большое число делений потомков яйцеклетки. У большинства многоклеточных последовательные стадии развития зародыша (морула, бластула, гаструла ит. д.) состоят из неспециализованных эмбриональных клеток, которые только в конце морфогенеза начинают специализоваться гистологически в разных направлениях. Во всех клетках данной ткани происходит гипертрофия одной функции и соответственное изменение строения (гистогенез), т. е. образование мускульных или нервных волоконец, развитие межклеточного вещества, хряща и кости и т. д.
Таким образом, мы видим, что изменения строения одной из дочерних клеток, образующихся в результате деления одноклеточного организма, мы можем приравнять к гистогенетиче-ским изменениям отдельных клеток эмбриона многоклеточного животного (например, к превращению мезенхимной кЯетки в мускульную или эктодермической клетки в нервную), но отнюдь не к онтогенезу многоклеточного животного.
Итак, мы принимаем, что у одноклеточных нет эмбрионального развития, что материнская клетка дает после деления начало не зародышу, которому надо проделать ряд сложных превращений, прежде чем он превратится во взрослое животное, а сразу двум взрослым дочерним клеткам, начинающим самостоятельное существование.
Если мы примем, что многоклеточные животные произошли от одноклеточных и что у этих первичных одноклеточных периода эмбрионального развития (онтогенеза) не было, то необходимым следствием из этого будет предположение, что онтогенез многоклеточных, т. е. весь сложный ряд морфологических превращений от оплодотворенной яйцеклетки (оосперма) до сложно построенного взрослого животного (человека, слона, собаки и т. д.), развился в течение эволюции многоклеточных форм, и перед нами вст&ет вопрос о том, как произошла эта эволюция онтогенеза.
Если бы мы знали онтогенез первичных ископаемых многоклеточных, происшедших от Protozoa, то нам было бы легко дать ответ на этот вопрос. Но палеонтология не сохранила нам не только онтогенеза предков Metazoa или переходных форм между Protozoa и Metazoa, но и взрослых первичных многоклеточных, и нам приходится ограничиваться лишь сравнительно
1 Отметим, что у инфузорий макронуклеус, который мы тоже можем считать за органеллу, образуется заново в обеих дочерних клетках.
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА METAZOA
563
анатомическими данными. К счастью, среди современных Protozoa имеется ряд колониальных форм жгутиковых, которые в своей эволюции остановились на стадиях, близких к вымершим переходным формам между Protozoa и Metazoa, что дает нам возможность восстановить приблизительное строение и развитие анцестральных форм первичных многоклеточных.
Все филогенетики, занимавшиеся этим вопросом, начиная с Э. Геккеля (1866) и кончая К. Рейдером (1918), признают, что некоторые колониальные формы жгутиковых подобны вымершим анцестральный формам первичных многоклеточных. Такими колониями являются, например, Pandorina morum и Vertvox globator. Колония из 16 жгути-♦
Рис. 222. Развитие Pandorina morum.
1—свободно плавающая колония из 16 клеток. 2—колония, в которой из каждой нлетки бесполым путем образовалась такая же дочерняя колония. 3—половая колония, из которой выходят отдельные половые клетки (гаметы). 4 и 5—конъюгация этих гамет, дающая зиготы. 6—только что образовавшаяся зигота. 7—выросшая зигота. 8—превращение плазмы зиготы с оболочкой в 9-жгути-ковую клетку без оболочки. 10—из последней путем повторного деления (начального онтогенеза) образуется молодая 16-клеточная колония (морула), превращающаяся путем роста во взрослую колонию (1) (из Принцгейма).
ковых клеток Pandorina изображена на рис. 222, 1. Бывают колонии в 32 и 64 клетки. Каждая из клеток колонии способна делиться на 2, 4, 8 и 16 клеток и дать начало новой колонии, подобной материнской (рис. 222, 1, 2). Здесь мы имеем нечто аналогичное дроблению яйца у многоклеточного животного, причем дочерние клетки не расходятся, а образуют новый многоклеточный (16-клеточный) взрослый организм. Аналогия с эмбрионом Metazoa, называемым морулой, ясна: каждая клетка проходит определенный ряд стадий, в данном случае четыре стадии, прежде чем произвести новый многоклеточный организм. Мы видим, что самый онтогенез здесь очень короток и что он происходит путем надставок стадий (анабо-
Зс*
564
ГЛАВА XVI
л и и): из одноклеточной стадии произошла двуклеточная (первая надставка), из двуклеточной—четырехклеточная стадия (вторая надставка) и т. д:
I II III IV стадии
2 '4 8 16 клеток.
Рис. 223. 1. Колония Volvox globator, состоящая из очень многих соматических клеток (som.) и немногих половых клеток (g), из которых образуются яйцеклетки и сперматозоиды. В соматических клетках произошла ди-ференциация на наружные жгутиковые и внутренние мезенхимные клетки; 2 и 3 — последовательные стадии развития оплодотворенной яйцеклетки Volvox (из Штейна).
Известно, что Pandorina размножается и половым путем, причем каждая клетка колонии выходит из общей связи и делается свободной (рис. 222, 3). Эти клетки конъюгируют друг с другом, и такая гамета (рис. 222, 4—9), соответствующая оосперму Metazoa, «снова делится на 2, 4, 8, 16 клеток и дает новую молодую колонию 4рис. 222, 10), которая растет и чере§ некоторое время превращается во взрослую колонию (рис. 222, 1).
Колония Volvox (рис. 223) представляет собой полый шар и состоит из гораздо большего числа клеток, чем колония Pandorina: в ней бывает до 22 000 клеток со жгутиками, соединен-, ных друг с другом протоплазматиче-’ скими перемычками. Диференцировки клеток пошла далее, чем у Pandorina. Здесь не все клетки способны делиться и давать начало новым колониям. В весенних колониях только восемь клеток колонии, более крупные, чем остальные, делятся дихотомически и дают начало новым колониям. После их выхода остальная часть весенней колонии, т. е. вся оставшаяся масса клеток, отмирает. Мы здесь имеем начало разделения тела многоклеточного организма на зародышевые клетки и на сому.1
Онтогенез у бесполых особей Volvox принципиально таков же, как у Pandorina, т. е. каждая из восьми круп- ных клеток колонии делится надвое, начетверо и т. д.; из нее получается однослойная многоклеточная пластинка (рис. 223, 2), из которой развивается однослойный шар (рис. 223, 3.), т. е. молодая колония. Ее клетки размножаются делением и диференцируются на мелкие жгутиковые и крупные зачатковые клетки, которые либо размножаются, как
указано, бесполым путем, либо дают начало гаметам, сперматозоидам и яйцеклеткам. После конъюгации начинается новый цикл развития, т. е. дробления, заключающий много стадий соответственно большему числу клеток.
Итак, мы видим, что в колониях одноклеточных существует очень простой онтогенез, происшедший путем последовательных надставок стадий; онтогенез этот очень короток
1 Как известно, этот вопрос связан с вопросом о потенциально неопределенной продолжительности жизни одноклеточных. В ряду поколений Protozoa нет старения и нет естественной смерти. Точно так же нет естественной смерти от старения и у таких форм, как колония Pandorina, где каждая клетка способна воспроизвести целый организм (новую колонию). У Volvox тело разделено на немногие половые клетки (рис. 223, g), способные дать начало новым колониям, и на громадное число соматических—стареющих и умирающих—клеток (рис. 223, som), т. е. здесь переход от Protozoa к Metazoa еще более ясен, чем у Pandorina.
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА METAZOA
565
у Pandorina, гораздо длиннее у Vol vox, но в обоих случаях он протекает по одному и тому же типу: сначала ряд надставок до стадии шарообразной многоклеточной колонии, затем стадия гистологической диференцировки; в общем—это типичная анаболия.
У низших многоклеточных животных (рис. 224) эмбриональное развитие удлинилось, т. е. к стадии шаровидной многоклеточной колонии прибавился ряд новых стадий. Примером такого онтогенеза может служить эмбриональное развитие низшего кишечнополостного животного. Одноклеточная исходная, стадия представлена здесь оплодотворенной яйце-
Рис. 224. Онтогенез очень примитивного многоклеточного животного, а именно гидроида (кишечнополостные).
1—зрелое яйцо. 2—оплодотворенное яйцо, прикрепленное ножкой к телу материнского организма. 3—начало дробления яйцеклетки. 4—однослойная бластула: из бластодермы в полость бластулы выселяются отдельные клетки, из которых образуется энтодерма зародыша (образование энтодермы будущей гаструлы путем миграции клеток). 5—внутри бластодермы образовался слой энтодермы (ent), остаток эктодермы превратился в эктодерму (ect) зародыша и образовалась наружная эмбриональная оболочка (ch). 6—зародыш выползает из оболочки и переходит к свободному образу жизни. 7— зародыш ползает по дну бассейна. 8—у этого зародыша развиваются состоящие из двух слоев (эктодермы и энтодермы) щупальцы (t). 9—молодая гидра с щупальцами и ртом (бластпором Ьр). 10—взрослая гидра. Отметим, что у гидры к стадиям онтогенеза колониальной формы (1—3) прибавились стадии 4—10, так что онтогенез соответственно этому значительно удлинился (схемы па И. Делажу).
клеткой (рис. 224, 1,2). Эта клетка многократно делится (рис. 224, 3) и из нее получается сначала шарообразный клеточный зародыш (морула), а затем зародыш с однослойной стенкой и полостью внутри (бластула, рис. 224, 4). Стадии дробления приблизительно соответствуют стадиям развития колонии одноклеточных (Pandorina, Volvox), но затем внутрь полости бластулы выселяются отдельные клетки бластодермы, которые образуют второй, внутренний одноклеточный слой. Зародыш из однослойного превратился в двуслойный (рис. 224, 5—9)—в гаструлу с отверстием (первичным ртом, или бластопором). Одновременно в этой гаструле появляются полые пальцевидные выросты вокруг бластопора (рис. 224, 8, 9), и гаструла превращается в маленькую гидру с эктодермой, эндотермой, гастральной полостью, бластбпором и щупальцами. Этим морфогенез гидры заканчивается (рис. 224, 10) и к нему прибавляется только стадия роста и гистологической диференцировки, т. е. стадия образования стрекательных клеток на щупальцах, нервно-мышечных клеток и т. д.
566
ГЛАВА XVI
Мы видим, таким образом, что онтогенез этой примитивной формы многоклеточного животного удлинился несколькими новыми стадиями, прибавленными к дефинитивному состоянию колониальных форм Protozoa. Еще боль-
* St ОШ
Рис. 225.. Онтогенез более высокоорганизованного кишечнополостного животного, а именно ктенофоры.
1—бластула: бластодерма, состоящая из макромеров (та) и микромеров (mi); рЫ—псевдобластопор; 2—гаструла, в которой мы видим состоящую нз микромеров эктодерму (ect) и образованную макромерами энтодерму (ent); кроме того, виден зачаток мезодермы (mes), отделившийся от энтодермы путем врастания части околоротовой эктодермы в полость архентерона (arch); образовался stomodaeum (stom). 3 и 4—более поздние стадии развития, на которых видно разрастание мезодермы в поперечном направлении; 5—дальнейшее развитие залоснившихся на предыдущих стадиях зачатков; на аборальном конце тела образовался орган чувств—статоцист (stc.); 6—та же стадия развития, что ина рис. 5; вид с аборального полюса; видна крестообразная форма зачатков мезодермы (mes) и восемь рядов мерцательных пластинок (ра); 7 й 8—поперечные разрезы через основание щупалец на двух последовательных стадиях, предшествующих образованию гастроваскулярных каналов; tt—парные щупальцы; cn. tt—соответствующие им впячивания энтодермы; ш—впячивания энтодермы, из которых образуются гастроваскулярные каналы; dv—дивертикул в основании щупалец; ра—ряды радиальных мерцательных пластинок (схемы по И. Делажу).
шее удлинение онтогенеза путем новых надставок мы находим у более высокой формы кишечнополостных, например у ктенофоры, развитие которой представлено на рис. 225.
Ранние стадии развития ктенофоры (рис. 225) приблизительно соответствуют тем, которые мы видели у гидры (рис. 224), но к ним добавляются
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА METAZOA
567
новые надставки. Так, в конце онтогенетического развития кте-нофоры в кишечнике развиваются полые радиальные выросты, превращающиеся в радиальные каналы взрослого животного (рис. 225, 8), образуются новые радиальные полосы двигательных пластинок (рис.'225, 6), развиваются два длинных щупальца, усаженных книдобластами (рис. 225, 7 и 8, tt), на аборальном конце тела появляется особый орган чувств (статоцист, рис. 225, 5 stc) и т. д. Мы видим, что эмбриональное развитие удлинилось на несколько добавочных стадий (надставок). На рис. 226 изобра-
жено строение взрослого животного.
Отметим, что ряд других новых признаков, наблюдаемых нами при развитии ктенофор, повидимому, произошел не путем надставок, но путем вторичных изменений филогенеза.
Чтобы проверить установленное нами положение, т. е. что э в о л ю-ц и я признаков происходила путем удлинения онтогенеза (надставкою стадий), мы приводим эмбриональное развитие более высоко стоящего билатерально симметричного животного, а именно примитивного хордового Balano-glossus, представителя Hemichordata. Ранние стадии развития—яйцо, дробление, образование бластулы и гаструлы (рис. 227, 1—5)—в общем подобны тому, что мы видели у гидры и ктенофоры. На более поздних стадиях мы видим, что от энтодермы отшнуровываются полые выросты, соответствующие гастроваскулярным каналам ктенофор, которые обра
Ь—рот; stom.—глотка (stonaodaeum); inf.—глоточная воронка; cn. ph.—глоточные каналы; stc.—орган чувств (статоцист); cn. аг., cn. ir., vs. mr—отделы системы гастроваскулярных каналов; tt—щупальцы; g. tt—полость, в которой лежит основание щупалец (из И. Делажа).
зуют пять парных мешков, зачатки стенок общей полости тела (рис. 227, 6—16, coel. 1, coel. 2, coel. 3), лежащей между кишечным каналом (int)
и наружной стенкой тела.
Из передней дорзальной стенки кишечного канала вырастает полый мешковидный вырост (рис. 227, 12—16, ch), который по своему гистологи
ческому строению может быть приравнен к спинной струне хордовых жи-
вотных. Над кишечником и зачатком спинной струны развивается зачаток нервной трубки. На рис. 228 схематически представлен передний отдел тела взрослого Balanoglossus, на котором видно взаимное отношение зачатков хорды (ch) и нервной системы (n, n. col). В передней части кишечного канала образуются метамерно расположенные отверстия (рис. 227 и 228, brt, br2), которыми передний отдел кишечного канала сообщается с наружной средой (водой). Положение этих жаберных щелей и их сосудов хорошо видно на рис. 228. Образование всех этих органов, т. е. отделов полости тела (рис. 227, coel. 1, 2, 3), хорды (ch), нервной системы (n. col.), жаберных щелей и их сосудов, происходит надставками, появляющимися в конце морфогенеза и удлиняющими время его развития. Мы видим, что к концу морфогенеза к нескольким системам органов приба
вилось по нескольку стадий развития.
У взрослого Balanoglossus перечисленные нами органы (центральная нервная система, хорда) остаются на стадии ориментов, у потомков вымер-
568
ГЛАВА XVI
ших гемихордат они путем дальнейших надставок проделали долгую и сложную эволюцию. Так, у ланцетника оримент спинной струны совсем отделился от энтодермы, начал сильно расти в ростральном и каудальном направлениях и образовал длинный и плотный упругий клеточный тяж, лежащий непосредственно под центральной нервной трубкой и представляющий собою осевой скелет тела животного. Спинной отдел туловищной части целомических мешков также претерпел сложные преобразования:
Рис. 227. Онтогенез Balanoglossus.
1—стадия бластулы; bld—бластодерма. 2 и 3—образование гаструлыгес!—эктодерма;еп!—энтодерма; Ыр.—бластопор. 4 и 5—зарастание бластопора. 6—образование переднего непарного целомического мешка (coel. 1), т. е. полости тела хоботка личинки и взрослого животного. 7 и 8—образование второй и третьей пары целомических мешков (coel. 2, coel. 3), т. е. полостей тела воротничка и туловища. 9 и 10—отшнуровывание этих полостей от архентерона (int.) 11 и 12—стадия, на которой появляется porus buccalis и зачатки хорды (ch) и рта (о); виде дорзальной стороны(11)и сбоку(12).< 13 и 14—более поздняя стадия развития (вид с дорзальной стороны и сбоку), когда хорда представлена уже полым выростом (ch) и когда образовалась первая жаберная щель ,(Ьг). 15 и 16—еще более поздняя стадия развития (вид с дорзальнойстороныисбоку),когда образуется вторая жаберная щель (Ьг2), анальное отверстие (ан), присоска (р) и нервный ствол (n. col.) (схемы по И. Делажу).
он распался на ряд дорзальных сомитов, из которых образуются главные органы движения—парные миотомы, изгибающие тело животного. Воротниковый отдел нервной системы разросся назад, дав начало спинному мозгу и спинномозговым нервам, и вперед (головной мозг и жаберные нервы). Главным образом путем ряда надставок развился мозг и связанный с ним череп и органы высших чувств с их скелетом и т. д.
Мы видим, что здесь произошел очень длинный ряд анаболий в отдельных сложных органах потомков первичных хордовых. Для каждой системы органов шла самостоятельная эволюция, которая путем целого
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА METAZOA
569'
ряда надставок стадий усложняла эволюцию организма как целого и удлиняла его эмбриональное развитие. Зачатки отдельных систем органов эволюировали более или менее самостоятельно, и в зависимости от того, какой зачаток в данную эпоху эволюировал прогрессивно путем ряда надставок, у одной и той же группы предков получились различные ряды потомков, ибо у одних прогрессивно развивалась одна система органов (например, органы движения), у других—другая (например, органы защиты), у третьих—третья и т. д. Постепенное удлинение онтогенеза высших форм является свидетельством их эволюции как процесса индиви-
дуального развития.
Естественным выводом из предыдущих фактов и соображений является
положение, что надставка стадий (анаболия) есть действительно первичный тип эволюции многоклеточных животных, тот тип, благодаря которому у них мало-по-малу развился особый начальный
период жизн и—о н -т о г е н е з или эмбриональное развитие. Этот период жизни не существовал у их одноклеточных предков, был очень короток у первичных колониальных форм и постепенно удлинялся, по мере осложнения организации, у высших многоклеточных.
Выше мы показали, что у одноклеточных нет онтогенеза в собственном смысле этого слова и что он начал постепенно эволюировать у предков современных многоклеточных, причем самое изменение этого онтогенеза путем надставок было сп
Рис. 228. Сагиттальный разрез через переднюю-часть тела взрослого Balanoglossus. Изображена передняя часть кишечного канала и диафрагма между воротником и хоботом.
Ьг—--жаберные щели; сое—кардиальный мешок; ch—хорда; n. col —нервный ствол воротника; п—его продолжение в туловище; prt. trp—peritoneum хобота; prt. col-peritoneum воротника; vd—спинной кровеносный сосуд; V. vnt—брюшной кровеносный сосуд; v. Ьг —жаберные сосуды, (из Делажа).
особом эволюции многокле-
есть.
точных животных. Изменение хода онтогенеза
средство усложнения организации Metazoa.
Мы нашли, что эволюция многоклеточных происходит путем изменения хода онтогенеза, но что онтогенез этот изменяется не толькр путем надставок (анаболий, удлинения онтогенеза), но и иными путями, а имен-
но: положительными и отрицательными изменениями первых зачатков органов и систем органов (архаллаксисы) и положительными и отрицательными девиациями хода развития этих зачатков на любой стадии их развития. Кроме того, происходят изменения темпа развития отдельных_ зачатков (ускорение и замедление развития). Все эти последние (вторичные) изменения хода первичного онтогенеза изменяют строение взрослых органов, но в общем не .удлиняют онтогенез.
В настоящее время, после ’детальной разработки теории филэмбрио
генеза, мы имеем полное основание принять, что этими путями вторичных изменений происходили очень крупные перемены в’строении органов.
и систем органов.
Как только у потомков одноклеточных началось эмбриональное раз-
570
ГЛАВА XVI
питие, к первичному типу эволюции прибавился целый ряд вторичных способов, которые появились филогенетически очень раной привели к большим изменениям организации взрослых форм многоклеточных. Первым из этих вторичных изменений было, повидимому, разделение всех клеток колонии на половые и соматические. Путем архаллаксиса в клеточной колонии (рис. 223) от массы соматических клеток отделились половые клетки, которые в свою очередь диференцировались на зачатковые клетки сперматозоидов и на яйцеклетки. Это разделение клеток на половые и соматические сохранилось у всех потомков первичных колониальных форм Metazoa до высших форм включительно. С течением времени у бойее высоких форм многоклеточных путем архаллаксиса от зачатка энтодермы отделился зачаток среднего листка (мезодермы, рис. 225, mes), претерпевший у их потомков очень сложную дальнейшую эволюцию и давший начало всем мезодермическим органам: мускулатуре, внутреннему скелету, нефридиальной и кровеносной системам и другим органам высших Metazoa. Таким образом, путем архаллаксиса возникли очень крупные и существенные признаки, изменившие весь тип строения организмов потомков данных форм без удлинения их онтогенеза.
Не менее существенны были изменения, происшедшие по типу девиа-ц и и и ускорения закладки зачатков органов и систем органов, словом, всех вторичных способов образования органов взрослых животных.
Мы пришли к выводу, что первичный способ эволюции органов взрослых животных есть способ надставки стадий (анаболия), следствием которого является рекапитуляция признаков предков в онтогенезе потомков. В течение эволюции к этому первичному способу прибавился ряд вторичных способов, а именно положительные и отрицательные архаллаксисы и положительные и отрицательные девиации и гетерохронии. Эти вторичные изменения хода онтогенеза меняют ход первичной эволюции и приводят к редукции последствий первичного способа, т. е. к редукции рекапитуляций.
Эволюция онтогенеза Metazoa и развитие вторичных типов филогенеза органов в очень упрощенном и сокращенном виде представлены на нижеследующей диаграмме (схема XXXIX). На схеме XXXIX, 1 изображено размножение свободно живущих (точнее—свободно живших в докэмбрий-ском времени) одноклеточных, от которых (схема XXXIX, 2) произошли первые колониальные формы (первая надставка) и у которых уже имело место разделение клеток тела на половые и соматические (первичный архаллаксис). На схеме XXXIX, 3 представлена эволюция низших кишечнополостных (вторая, третья, четвертая надставки). На схеме XXXIX, 4 представлена эволюция высших кишечнополостных, у которых путем вторичного архаллаксиса развилась новая система органов—мезодерма. Наконец, у потомков этих форм, первичных Bilateralia (схема XXXIX, 5), снова появились новые надставки (образование вторичного рта, нейроцеля, органов высших чувств и т. д.), т. е. вторичная анаболия, и первые девиации, т. е. появился ряд новых стадий развития и новых дефинитивных состояний сложного организма.
Мы можем следующим образом резюмировать все сказанное в настоящей главе.
1. У одноклеточных предков Metazoa не было периода эмбрионального развития: после каждого размножения делением у дочерних клеток наступало взрослое состояние и, новый период размножения делением.
2. Начало эволюции многоклеточных состояло в образовании колоний одноклеточных, происшедших дихотомическим делением материнской клетки, причем дочерние клетки не расходились в разные стороны, а оставались вместе, образуя двух-, четырех-, восьми- и шестнадцатиклеточную
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА METAZOA
571
колонию (Pandorina), каждая из клеток которой была способна таким же дихотомическим делением дать начало такой же новой колонии.
3. Первая диференцировка в колониях, состоящих из большого числа клеток, заключалась в разделении всех клеток колонии на половые (с неопределенной потенциальной продолжительностью жизни) и на соматические.
4. Первые стадии эволюции колрний одноклеточных состояли в увеличении числа клеток колонии путем дихотомического деления до образо-
Схема XXXIX. Гипотетический ход эволюции онтогенеза Metazoa bilateralia.
1—размножение свободно живущих одноклеточных животных; дочерние клетки после каждого деления расходятся и начинают жить самостоятельной жизнью. 2—онтогенез колонии одноклеточных (по Volvox); материнская клетка многократно делится, но дочерние клетки не расходятся и образуют шаровидную многоклеточную колонию, в которой происходит диференциация клеток на половые (черные), способные к дальнейшему делению, и соматические (светлые), умирающие естественной смертью. Из подобного простого онтогенеза произошел более сложный онтогенез низших Metazoa (Hydra). 3—дробление оплодотворенной яйцеклетки гидры, которое приводит к образованию шаровидного многоклеточного зародыша (морула), из которого образуется впоследствии бластула и затем гаструла, в принципе похожая на взрослое животное. Черным цветом обозначены первичные половые клетки, но гистогенез не представлен на схеме. Мы видим, что к стадии онтогенеза колониальных форм (2) прибавились стадии бластулы и гаструлы (в действительности прибавилось ббльшее число стадий). 4—эволюция онтогенеза первичного двустороннесимметричного животного, у которого, кроме энтодермы и эктодермы, путем архаллаксиса развилась еще и мезодерма, отделившаяся от слоя энтодермы. Здесь показано, как значительно появление вторичных способов эволюции (архаллаксис) изменяет первичный ход онтогенеза и строение взрослого организма (число стадий на схеме 4 то же, что и на схеме 3, так что здесь ойтогенез не удлинился). Крестообразная штриховка обозначает мезодерму. 5—онтогенез высшего двусторонне-симметричного многоклеточного животного, у которого эмбриональное развитие еще удлинилось путем новых надставок.
вания многоклеточной колонии. Pandorina и Volvox остановились на этой стадии эволюции.
5. У низших кишечнополостных прибавились еще две стадии эмбрионального развития: к стадии морулы прибавилась стадия полого шара с однорядной стенкой (бластула) и стадия двуслойного бокала со щупальцами вокруг бластопора (гаструла).
572
ГЛАВА XVI
6. У высших кишечнополостных (путем архаллаксиса) развилась мезодерма и прибавился путем надставок ряд новых признаков: stomodaeum, билатерально симметричное строение тела, статоцисты и т. д., т. е. их эмбриональное развитие удлинилось еще на несколько конечных стадий.
7. Таким образом, у потомков примитивных Metazoa к концу онтогенеза прибавляются все новые и новые конечные стадии и тем самым удлиняется первичный онтогенез. У Protozoa онтогенеза в собственном смысле этого слова, т. е. онтогенеза путем деления клеток, не было. Колониальные формы одноклеточных остановились на стадии морулы. Coelenterata дошли до стадии гаструлы и т. д. Мы видим, что онтогенез есть процесс, развившийся в течение филогенеза путем надставок конечных стадий и приведший к усложнению строения Metazoa. Этим путем развились первичные рекапитуляции признаков предков.
8. Этот первичный способ эволюции в течение времени изменился вторично, причем способов вторичного изменения хода онтогенеза было несколько: архаллаксис, девиация, гетерохрония и т. д. Эти вторичные способы эволюции начали действовать очень рано, с самого начала образования колоний одноклеточных, и привели к весьма крупным и существенным изменениям строения органов взрослых многоклеточных. Мы видели, что первые диференцировки в клетках произошли уже у колониальных форм (диференцировка колоний на половые и соматические клетки);этим же путем вторичного изменения появились новые системы органов (мёзодерма И т. д.).
9. Главный вывод, к которому мы приходим на основании изложенного ряда фактов и гипотез, тот, что у одноклеточных не было онтогенеза, но что у их многоклеточных потомков постепенно развился эмбриональный период жизни сомы. Развитие онтогенеза есть средство, которым была достигнута эволюция новых, более высоких форм жизни.
ГЛАВА XVII
СООТНОШЕНИЯ МЕЖДУ ПРИНЦИПАМИ ИЛИ ТИПАМИ ЭВОЛЮЦИИ И МОДУСАМИ ФИЛЭМБРИОГЕНЕЗА
В главе об отношениях между филогенезом и онтогенезом мы в общей форме разобрали главные способы изменения хода онтогенеза (анаболия, девиация, архаллаксис и т. д.) и указали их значение для изменений филогенеза (теория филэмбриогенеза). В главе о принципах эволюции нами изложены результаты наших исследований относительно различных типов (принципов) филогенетических изменений органов (принцип интенсификации, фиксации фаз и т. д.), причем мы не разбирали отношений этих типов к теории филэмбриогенеза. Теперь нам надо ближе заняться . этим последним вопросом и разобрать, в каких отношениях стоят различные способы филэмбриогенезов к известным нам типам филогенетических изменений. Другими словами, нам надо определить, какими способами филэмбриогенетических изменений (анаболией, девиацией, архаллаксисом, гетерохронией и т. д.) осуществляется интенсификация, смена функций органов и т. д.
Начнем с основного типа филогенетического изменения органов, а именно, с принципа интенсификации функций. Как это уже было установлено нами, существует интенсификация функций клеток и тканей и интенсификация функций органов. Во многих случаях интенсификация функций тканей происходит путем анаболии. Так, мы имеем полное основание думать, что висцеральные поперечнополосатые мускулы (жаберная и челюстная мускулатура) произошли от гладких мускулов. Мезодермическая стенка кишечного канала на большей части своего протяжения превращается в гладкие мускульные волокна. Передняя часть стенок кишечной трубки проходит стадию гладких мышечных волокон, но затем эти волокна превращаются в поперечнополосатые, вследствие чего энергия их сокращения усиливается. Эти изменения строения клеток мускульной ткани произошли на поздних стадиях их развития и поэтому мы их относим к способу надставки стадий.
В иных случаях гистологические изменения произошли на очень ранних стадиях развития данного органа, и нам приходится признать, что они произошли по типу архаллаксиса. С другой стороны, приведенный нами пример эволюции обонятельных ямок у костистых рыб показывает, что в некоторых случаях интенсификация может происходить и по типу девиации функций. Несомненно, что у предков Belone преобразование обонятельных ямок произошло в эволюции на стадии онтогенеза, когда у них только начиналось развитие складок эпидермиса.
1 Как это было уже сказано выше (см. стр. 5.1), настоящая глава представляет первую попытку автора связать теорию принципов эволюции с теорией модусов филэмбриогенеза; она должна поэтому рассматриваться лишь как конспект задуманных им в этом направлении трудов. (Прим, ред.)
574
ГЛАВА XVII
•
Мы предполагаем, что развитие грибообразного выроста^ т. е. увеличение обонятельной поверхности, повысило обонятельную функцию. У большинства костистых рыб это увеличение произошло иным путем, т. е. образованием многочисленных складок обонятельного эпителия. И у тех и у других результат был один и тот же: усиление обонятельной функции.
Отметим, что кроме указанных способов усиление функции происходит посредством частичного роста, т. е. ускоренного развития клеток эволюирующих органов. Этот фактор ведет к относительному увеличению размеров данного органа. В качестве примеров можно привести увеличение частей головного мозга высших позвоночных, задних ног бесхвостых амфибий и многие другие случаи неравномерного роста. Гетерохрония, т. е. более ранняя закладка частей, также приводит к большему росту (величине) органов. Так, более ранняя закладка чешуй верхнего, среднего и двух боковых рядов у Acipenser позволяет им сильнее вырасти.
Резюмируя все сказанное, мы приходим к выводу, что интенсификация функций происходит всеми тремя главными способами филогенетического изменения органов, т. е. анаболией, девиацией и архаллаксисом, а также путем неравномерного роста органов и гетерохронией.
Далее нам приходится поставить вопрос о том, каким, способом эмбриональных изменений происходит эволюция по типу фиксации фаз. Мы подробно разобрали этот принцип на примере прогрессивной эволюции конечностей млекопитающих, т. е. на примере перехода стопоходящих млекопитающих в пальцеходящие и копытные. Основное морфологическое •изменение, происшедшее при этом, состояло, как мы это видели выше, в прогрессивном развитии metacarpus и metatarsus, которое привело к переходу от стопоходности к пальцеходности. Metacarpus и metatarsus сильно выросли в длину, т. е. в этих отделах конечности произошел усиленный частичный рост клеток хряща. Мы видели, что при эволюции копытных произошла редукция боковых пальцев конечностей: у непарнопалых редуцировались I и V, а затем II и IV пальцы, а у парнопалых I, II, V пальцы. Как произошла эта редукция? Мы можем сказать, что в общем она произошла по типу отрицательного филэмбриогенеза, причем, повидимому, при редукции скелета (по исследованиям Матвеева) здесь большую роль играл процесс заимствования эмбриональной ткани редуцирующегося органа (пальца) соседними прогрессирующими частями скелета. Так как это заимствование происходит на очень ранних стадиях развития, в мезенхимных закладках, то, вероятно, можно обозначить процесс редукции боковых пальцев как вид о т р и ц а-тельного архаллаксиса.
Отметим, что это исследование позволяет нам углубить наше понимание теории архаллаксиса, недавно высказанной и до сих пор еще мало разработанной. Не вдаваясь в обсуждение этого вопроса, мы переходим к разбору принципа смены функций А. Дорна.
Мы уже отметили связь принципа смены функций с другим, более основным принципом, а именно с мультифункциональностью, т. е. с тем общим фактом, что всякий сложный орган имеет несколько функций, между которыми мы различаем одну главную и несколько второстепенных или добавочных. Так, в конечности млекопитающих главной функцией является хождение (бегание), второстепенной—прыгание, рытье земли, плавание, лазание по деревьям и т. д. Мы знаем, что в органах движения большинства других позвоночных существует аналогичная мультифункциональность с преобладанием главной функции (плавательная функция у рыб, полет у птиц, хождение у рептилий и т. д.) и факультативным действием добавочных функций.
Принцип смены функций, как известно, заключается в том, что
СООТНОШЕНИЕ МЕЖДУ ПРИНЦИПАМИ И ФИЛЭМБРИОГЕНЕЗАМИ
575
в соответствии с изменением образа жизни животного (часто при переходе в другую среду обитания) одна из второстепенных функций эволюирует прогрессивно, т. е. усиливается, а прежняя главная функция регрессирует и может вполне редуцироваться (функция хождения передних конечностей птиц, китообразных и т. д.). Здесь мы имеем два параллельно идущих процесса: 1) процесс прогрессивного изменения частей органа, связанных с прогрессивно развивающейся второстепенной функцией, и 2) процесс редукции частей органа, служивших прежней главной функции.
Мы видим, что здесь действуют два знакомых нам типа изменений, а именно интенсификация и редукция органов или частей органов. В качестве примера эволюции, происходящей по' типу смены функций, мы взяли случай превращения конечностей наземных млекопитающих в ласты водных млекопитающих (моржей, тюленей и,.наконец, китообразных). В этом случае второстепенная для наземных хищных млекопитающих функция плавания усилилась, а прежняя главная функция—хождение—редуцировалась. Мы с большой вероятностью можем принять, что изменение мускулов конечности произошло Путем анаболии, а изменение числа фаланг (как у ихтиозавров, у которых произошло аналогичное изменение в передней конечности)—путем архаллаксиса. Кроме этих прогрессивных изменений, в конечности китообразных произошла редукция частей, которые у их предков служили для функции хождения, т. е. бугорков на концах пальцев и когтей. На основании наших исследований о редукции конечностей у рептилий мы можем считать, что это, вероятно, произошло путем рудиментации, т. е. отрицательного архаллаксиса. Мы знаем, что у китообразных редуцировались задние конечности и от них остались рудименты скелета, и можно предположить, что редукция бесполезных частей передней конечности происходит по этому же способу.
Мы видели, что в очень многих случаях при эволюции произошло уменьшение числа функций. В этих случаях происходило: 1) усиление главной функции и перенос ее деятельности на часть органа и 2) редукция второстепенных функций всего органа. Усиление главной функции происходило, как это нами уже разобрано выше, всеми главными способами филэмбриогенеза; редукция сделавшихся ненужными частей шла путем рудиментации, так как палеонтологическое исследование конечностей предков копытных показало, что у них существовали рудиментарные пальцы, не доходившие до земли. По всей вероятности, происходило и заимствование тканей (мезенхимы) боковых пальцев соседними средними пальцами. Существование рудиментов пальцев показывает, что это заимствование происходило очень медленно.
Перейдем теперь к принципу субституции функций, при которой функция органов предков замещается аналогичной или совершенно другой, но биологически ей равноценной функцией. Мы разобрали этот процесс на примере субституции функции хождения и бегания при помощи четырех лап у рептилий (предков безногих Lacertilia и Ophidia) функцией ползания удлинившегося туловища на брюхе (безногие ящерицы и змеи). Первичным изменением в этом случае является увеличение числа позвонков туловища, которое происходит путем положительного архаллаксиса, т. е. новообразования зачатков позвонков позади крестца и постепенного превращения крестцовых позвонков в поясничные, развития новых крестцовых позвонков позади прежнего крестца, образования задних конечностей в области нового крестца и новообразования последних хвостовых позвонков.1
1 Мы не упоминаем о постепенном сдвигании заднепроходного отверстия, о новообразовании окружающих его чешуй, задних почечных' сегментов и т д.
576
ГЛАВА XVII
Наши исследования показали, что все эти изменения и, в частности, целый ряд новообразований (развитие поясничных ребер, преобразование поясничных и шейных ребер в туловищные ребра, преобразование крестцовых позвонков, развитие новых хвостовых позвонков, развитие новой мускулатуры и скелета задних конечностей, преобразование туловищных мускулов и нервных сплетений и т. д.) происходило по типу вторичного архаллаксиса и что этим путем произошла новая функция, а именно ползание животного на брюхе, т. е. на концах туловищных ребер. На некоторой стадии этой прогрессивной эволюции позвоночника задние и передние конечности стали ненужны для функций передвижения тела, и путем отрицательного архаллаксиса начался процесс их рудиментации, который привел к окончательной редукции и даже исчезновению конечностей у змей.
Мы видим, что весь этот очень сложный процесс прогрессивной эволюции ряда органов туловища и редукции другого ряда органов (конечностей -с их скелетом, нервами, сосудами и мускулатурой) произошел посредством положительного и отрицательного архаллакс[иса, т. е. аналогичными путями. Но мы можем естественно поставить вопрос о том, всегда ли субституция функций происходит по этому типу.
Выше было разобрано, что у гимнофионов, при переходе их к подземному образу жизни, функция зрения заместилась функцией обоняния. Мы не знаем точно, каким путем происходил этот тип субституции функций, но можем предположить, что при переселении предков гимнофионов в подземную среду прогрессивное развитие органа обоняния и усиление обонятельной функции произошло не посредством архаллаксиса, но путем надставки стадий, или анаболии. К сожалению, мы еще не можем сказать, каким образом развилась на щупальцах обонятельная функция и каким образом развились сами щупальцы, т. е. новый орган гимнофионов; здесь нужны дополнительные эмбриологические исследования.
В некоторых случаях активная функция предков замещается пассивной, но равноценной функцией потомков. Так, у черепах (а также у асцидий и у сальп) активная функция движения органов тела замещается пассивной функцией защиты (панцырь черепах и усоногих, мантия туникат), совсем иной, конечно, чем функция движения, но при измененных условиях жизни потомков биологически ей равноценной.
Разберем этот тип изменения на исследованном нами случае изменения организации черепах. Мы должны прежде всего поставить вопрос о том, как произошло у этих животных прогрессивное изменение, т. е. развитие защитного органа (сагарах и plastron). Мы знаем, что у предков черепах существовали все составные части этих органов, т. е. позвонки ребра и орименты брюшного скелета: грудина, ключица, episternum и т. д. В чем состояло новое изменение, приведшее к образованию панцыря? Мы можем принять, что первичное изменение состояло в прогрессивном изменении частей рогового скелета и разрастании костных ребер (костных чешуй), которое ограничило подвижность. Вследствие этого понижения подвижности произошло изменение направления эмбрионального развития частей скелета—спинные позвонки начали срастаться друг с другом. Это произошло, повидимому, по способу а н а бол и и. Мы думаем, что анаболия и была первичным изменением направления онтогенетического развития, которое привело корреллятивно к срастанию остальных частей спинного и брюшного скелета.
Нам теперь надо разобрать, как происходит эволюция по типу активации функций. В качестве примера активации функций мы разобрали случай, когда пассивная функция опоры (поддерживание зубов, место прикрепления мускулов) челюстной кости низших рептилий превратилась в активную функцию движения этой кости у змей. У предков змей эта кость «была неподвижна, у более примитивных змей (Boidae) она приобрела неко
СООТНОШЕНИЕ МЕЖДУ ПРИНЦИПАМИ И ФИЛЭМБРИОГЕНЕЗАМИ
577
торую подвижность, наконец, у ядовитых змей на ней развились ядовитые зубы и, благодаря сочленению, она стала поворачиваться, причем при открывании рта ядовитые зубы поворачиваются вперед, при закрывании— прилегают к небу. Изменения в эмбриональном развитии челюстной кости состояли в том, что она укоротилась и на ней развился длинный ядовитый зуб с каналами внутри; этот один зуб развился прогрессивно, а другие, задние зубы редуцировались, и между этой костью и невральным черепом развилось сочленение; кроме того, образовалось прогрессивное сочленение между maxilla и transversum.
Мы должны признать, что основным способом, которым произошли эти изменения, была девиация в развитии maxilla, которое пошло в другом направлении, чем у предков. Весьма вероятно, что при этом процессе имели место и другие способы эволюции. Мы можем предположить, что редукция задних зубов (у ядовитых змей) произошла посредством отрицательного архаллаксиса, а образование новых сочленений произошло путем положительного архаллаксиса.
В качестве примера принципа симиляциимы взяли преобразование позвонков и ребер у змей. При преобразовании позвонков и ребер у предков змей ребра различных областей позвоночника (шейные, поясничные и крестцовые) превратились в типично туловищные ребра, причем грудина редуцировалась и вентральные концы ребер, соединявшие грудные позвонки с грудиной, атрофировались. В результате этих процессов ребра большинства туловищных позвонков сделались похожи друг на друга. Аналогичный процесс уподобления (симиляции) произошел и в метамер-ных мускулах позвоночника. Эмбриологическое исследование показало нам, что эти изменения произошли по способу (модусу) архаллаксиса, т. е. по тому же способу изменений, по которому шло изменение туловищных позвонков.
В качестве примера принципа разделения органов мы привели расчленение жаберных дуг на сегменты. Можно думать, что это изменение произошло по способу архаллаксиса. Очень вероятно, что и распадение мускульных зачатков (например в конечностях Quadrupeda) произошло по тому же способу.
В заключение остановимся на вопросе о том, путем каких филэмбриогенезов осуществлялась эволюция органов, идущая по типу выпаде-ния промежуточных функций.
Как мы это уже отметили выше, по этому типу происходит детально исследованное нами развитие брюшных плавников Holostei и Teleostei, которые несомненно произошли от форм, близких по строению своих плавников к типу осетровых рыб.
Как это нам уже известно (см. главу XI), при превращении плавников осетровых рыб в плавники типа Holostei и Teleostei произошло, связанное с редукцией метаптеригия и плавниковых лучей, выпадение функции этих частей и усиление функции лепидотрихий (лопасти плавника). Следствием этого явился выигрыш в интенсивности движения самого плавника.
Мы нашли, что скелет плавников осетровых рыб проходит в своем онтогенезе следующие стадии развития: 1) стадию образования сегментальных мезенхимных зачатков базальных элементов, закладывающихся одновременно с плавниковыми лучами; 2) стадию охрящевения дискретных basalia proximalia и distalia; 3) стадию срастания дискретных базальных элементов в один цельный basipterygium.
При эволюции брюшных плавников Holostei и Teleostei эти три стадии развития выпали из онтогенеза этих форм. Здесь базиптеригий обычно закладывается в виде сплошного хрящевого стержня, лежащего парал-Морфологические закономерности 37
578
ГЛАВА XVII
лельно оси тела; несколько позднее закладывается рудиментарный метаптеригий, позднее—лепидотрихии и еще позднее—рудиментарные бази-птеригиальные плавниковые лучи.
Мы можем принять с большой дозой вероятия, что у Hoiostei порядок закладки элементов брюшного пояса изменился в течение эволюции и что у них базиптеригий онтогенетически стал закладываться раньше, чем он закладывался у предков (акцелерация).1 Если это так (а что это действительно так, в этом нам не приходится сомневаться), то здесь выпадение промежуточных стадий произошло путем отрицательного архаллаксиса.
Эмбриологические исследования показали нам далее, что после только что упомянутого периода отрицательного архаллаксиса последовал период положительного архаллаксиса, когда вместо исчезнувших стадий развития скелета брюшного плавника стал сразу закладываться сначала мезенхимный, а затем хрящевой зачаток базиптеригия, рудиментарных лучей и рудиментарного метаптеригия нового голостеоидного типа. Здесь мы несомненно имеем дело со вторичным а р х а л-лаксисом, благодаря которому старый тип эволюции сразу замещается новым, характерным для высших рыб типом.
Что касается закладки плавниковых складок и лежащих в них лепидотрихий, то она происходит у костных ганоидов и у костистых рыб так же, как она происходит у осетровых. У осетровых лучи, сидящие на базипте-ригии, причленяются к латеральному краю базиптеригия; так же они причленяются на ранних стадиях развития у Amia и у низших костистых рыб (Salmo, Osmerus, Cyprinidae), затем при постепенном разрастании заднего края базиптеригия они, а с ними вместе и лепидотрихии, оказываются сидящими на каудальном крае базиптеригия. Здесь филогенетическое развитие идет по способу анаболии. Таким образом, нам приходится принять, что эволюция брюшных плавников рыб идет двумя способамифилэмбриогенеза: по способу архаллаксиса для базиптеригия и по способу анаболии для лепидотрихий и плавниковых складок. Дальнейшая эволюция брюшных плавников (перекрест их передних концов у костных ганоидов и костистых рыб) также происходит по способу анаболии.
Все это показывает нам, что принципиальной разницы между всеми этими способами эволюции, в сущности, не существует, что процесс развития, начавшийся по одному способу (архаллаксис); может продолжаться и закончиться по другому (анаболия). Все это—с п о с о б ы, которыми достигается конечный результат, т. е. функционирующее состояние органа взрослого животного и его прогрессивное развитие.
Резюмируя рее сказанное о способах (модусах) филэмбриогенеза, которыми совершается эволюция различных типов филогенетического изменения органов, мы находим, что в развитии каждого типа участвуют различные способы положительного и отрицательного филэмбриогенезов.
1 Обратная гипотеза, а именно предположение, что здесь произошла ретардация в закладке рудиментарных лучей, нам представляется невероятной. При исследовании явлений рудиментации у безногих ящериц (А. Н. Северцов, 1931) мы нашли, что редуцированные конечности закладываются у них в то же время, что и гомологичные им нередуцированные части скелета, так что нам пришлось вообще отказаться от гипотезы ретардации Менерта.
ГЛАВА XVIII
ОБЩИЕ ВЫВОДЫ
МОРФОБИОЛОГИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
Основная задача наших исследований заключалась в попытке выяснить морфологические закономерности филогенетического развития животных на основании данных филогенеза. Устанавливая эти закономерности, мы исходили из собственных исследований эволюции низших позвоночных, изложенных во второй, специальной части этой книги. Добытые в этих исследованиях результаты мы всегда старались проверять на возможно более обширном фактическом материале, взятом из сравнительно анатомической, эмбриологической и палеонтологической литературы. Здесь мы, конечно, не будем снова излагать результатов наших филогенетических исследований. Мы лишь напомним читателю о двух важнейших выводах, к которым мы пришли в течение многолетней работы и которые мы разделяем с большинством современных морфологов, работающих над различными вопросами филогенеза. Выводы эти заключаются, во-первых, в том, что эволюция животных шла монофилетическим путем, и, во-вторых, в том, что эта эволюция представляет собой э к т о г е н е-тический процесс.
В начале третьей части нашей книги мы дали упрощенную схему хода адаптивной эктогенетической Эволюции. При этом мы исходили из предпосылки, что весь эволюционный путь является в общем процессом приспособления, т. е. что все органы многоклеточных животных постоянно изменяются вследствие приспособления к изменяющимся условиям существования.
Органы многоклеточных животных по своим отношениям к окружающей среде могут быть подразделены на две большие группы: на органы эктосоматические (наружные), стоящие по своим функциям в определенных и непосредственных отношениях к условиям окружающей среды, и на органы энтосомагические (внутренние), функции которых находятся в связи с функциями эктосоматических и других энтосоматических органов.
Каждый эктосоматический орган находится в совершенно определенных функциональных соотношениях с определенной группой условий окружающей среды. Если какие-нибудь из этих условий меняются, если, например, меняется климат или меняется сила освещения в окружающей среде, то изменяются только те эктосоматические органы, которые были приспособлены именно к этим условиям среды: образуются новые приспособления, касающиеся тех эктосоматических органов, которые находились в определенных соотношениях именно с данной группой условий внешней среды. Те же эктосоматические органы, которые связаны с другой, неизменившейся группой условий существования, изменяются мало или не изменяются совсем.
Функции каждой группы эктосоматических органов связаны с опре-
37*
580
ГЛАВА XVIII
деленным числом функций энтосоматических органов; эти последние органы приспособляются к изменяющемуся строению и изменяющимся функциям эктосоматических органов. Таким образом возникают новые филетические корреляции или, по нашей терминологии,—новые координации между определенной группой эктосоматических органов и связанной с ними группой энтосоматических органов, с одной стороны, и между этими изменившимися эктосоматическими органами и другими, функционально связанными с ними, энтосоматиче- ъ скими органами (вторичные или третичные координации, или координационные цепи)—с другой.
Итак, каждая группа изменений, происшедших в условиях окружающей среды, вызывает известное число первичных изменений в эктосоматических органах (проталлаксы) и некоторое число вторичных изменений в связанных с этими последними внутренних (энтосоматических) органах (дейталлаксы или координации первого, второго и следующих порядков). Таким образом, общий ход эктогенетической адаптивной эволюции можно представить себе в виде следующего схематического процесса:
Окружающая среда
Организм
Изменение в определенной группе условий су- < шествования.
Приспособление тех эктосоматических органов, которые функционально связаны с этими условиями существования (проталлаксы, или первичные адаптивные изменения)
Приспособление (адаптация) тех энтосоматических органов, которые функционально или топографически связаны с изменившимися эктосоматическими органами (дейталлаксы, или координации 1-, 2-, 3-го и т. д. порядков).
Исходя из этой схемы, мы пытались изучать закономерности хода филогенеза, и эти исследования привели нас к построению двух общих теорий, а именно: 1) морфобиологической теории хода эволюционного процесса, устанавливающей общие и частные закономерности филогенеза, и 2) теории филэмбриогенеза, устанавливающей зависимость филогенетического изменения органов от фактора времени, т. е. от тех периодов индивидуальной жизни (в частности от периода онтогенеза), когда возникают и развиваются филогенетические изменения строения эволюирующих органов.
Разрабатывая морфобиологическую теорию эволюции, мы поставили перед собой задачу—определить возможные направления хода эволюционного процесса и пришли к выводу, что эволюция может итти только в двух основных направлениях, а именно, в биологически (экологически) прогрессивном направлении (победа в борьбе за существование) и в направлении биологически регрессивном (вымирание).
Биологически прогрессивное направление эволюции характеризуется следующими признаками: 1) число особей данной группы животных постепенно увеличивается; 2) ареал распространения данной группы животных, вследствие этого увеличения, постепенно расширяется; 3) данная группа животных постепенно распадается на все большее число подчиненных систематических групп (подвиды, новые виды, роды и т. д.). При биологически регрессивной эволюции: 1) число особей, образующих данную группу животных, постепенно уменьшается, 2) ареал распространения данной группы животных съуживается, постепенно распадаясь на ряд небольших островных ареалов, отделе иных друг от друга все большими пространствами
ОБЩИЕ ВЫВОДЫ
581
и 3) число подчиненных систематических единиц в данной группе животных (подвиды, новые дочерние виды, роды и т. д.) постепенно уменьшается.
Биологически прогрессивная эволюция ведет к биологическому расцвету животных, биологически регрессивная эволюция ведет к в ы-мирами ю.1
Исследование филогенеза различных групп животных показало нам, что в природе существуют четыре общих направления, по которым может следовать биологически прогрессивная эволюция и которые ведут к биологическому расцвету данной группы животных.1 2
Первым из этих направлений является направление морфофизиологического прогресса или путь ароморфоза, т. е. то направление эволюции, при котором в организации эволюирующего животного происходят изменения, повышающие общую жизнедеятельность его активных функций. Эти изменения ведут к прямому или косвенному подъему функций активных органов (органов движения, дыхания, питания, кровообращения и выделения, центральной нервной системы и органов внешних чувств); они поднимают общую организацию животного на более высокую ступень в прямом смысле этого слова.
Вторым направлением эволюции является направление идиоадаптации, т. е. то направление эволюции, которое идет путем приспособления эктосоматических органов к строго ограниченным изменениям, происходящим в условиях окружающей среды. При этом организация животного не повышается, т. е. не происходит усложнения органов, и их функция в общем не интенсифицируется, иначе говоря, подъема общей энергии жизнедеятельности, имеющего места при ароморфозе, здесь не существует. Частным случаем идиоадаптации является специализация, при которой данные животные формы переходят в особую, отделенную от общей борьбы за существование, константную среду, к которой они и приспособляются путем изменения немногих органов. Остальные органы, находящиеся вне зависимости от действия этой среды, при этом не изменяются.
Третьим направлением эволюции является направление ц е н о-генеза, т. е. направление эмбриональных приспособлений; при этом образуется одно или несколько эмбриональных приспособлений, исчезающих у взрослых особей, так что организация взрослых форм остается неизменной. Образование эмбриональных приспособлений приводит к тому, что у эволюирующих форм все большее число молодых особей достигает взрослого состояния, вследствие чего число взрослых особей данного вида постепенно увеличивается (биологический прогресс).
Четвертым направлением является направление морфофизиологического регресса или общей дегенерации, т. е. такой эволюции животных, при которой некоторые пассивные органы эволюирующей животной формы (органы пассивной защиты, органы размножения) развиваются в прогрессивном направлении, между тем как ее активные органы становятся ненужными и постепенно редуцируются.
Эти четыре направления филогенеза являются равноценными в том отношении, что все они ведут к биологическому прогрессу, т. е. к победе в борьбе за существование. При любом из этих четырех путей эволюции число особей эволюирующего вида растет, ареал распространения расши
1 В этой книге мы занимаемся только вопросом о биологически прогрессивной эволюции животных, понимаемой в том смысле, в каком мы только что о ней говорили; важной и интересной проблемы вымирания крупных групп животных мы не касались здесь вовсе.
2 Мы особенно настаиваем на том, что здесь мы говорим только об о б щ и х направлениях эволюции, т. е.о тех направлениях, при которых эволюирующий организм изменяется как единое биологическое целое.
582
ГЛАВА XVIII
ряется и вид распадается на новые подчиненные систематические группы (подвиды, виды, роды и т. д.).
Эволюция во многих случаях может изменять свое направление. Так, группа животных, эволюировавшая до известной поры в направлении ароморфоза, может перейти на путь идиоадаптации (адаптивная радиация Осборна), и, наоборот, в группе животных, эволюировавших идиоадаптивно, могут возникнуть те или иные ароморфозы. То же мы можем сказать и относительно перехода от идиоадаптивного направления эволюции к направлению эмбриональных приспособлений, а также относительно перехода от направления ценогенеза к эволюции путем ароморфозов. Все эти направления могут беспрепятственно переходить одно в другое. Иначе обстоит дело со специализацией и дегенерацией: здесь не наблюдается обратимости. Эволюция с пути идиоадаптаций и с пути эмбриональных приспособлений может перейти на путь специализации и деградации, но обратный переход, невидимому, не имеет места. И если, как это было только что сказано, четыре перечисленных направления биологически прогрессивной эволюции являются биологически равноценными, то их никак нельзя признать таковыми в смысле их проспективного значения. Несомненно, возможность биологически регрессивной эволюции или вымирания в большей степени грозит тем формам, которые уже вступили на путь специализации и дегенерации, чем тем формам, которые пошли по пути ароморфозов или идиоадаптаций.
Изменение направления эволюционного процесса, т. е. переход от одного направления эволюции к другому, по всей вероятности, зависит: 1) от характера и высоты организации изменяющегося животного в период, когда наступает изменение внешней среды, вызывающее соответствующее изменение животного, 2) от характера, интенсивности и скорости изменения этой внешней среды и, наконец, 3) от количественного соотношения между изменениями среды и организацией и функциями животного. Последний фактор особенно интересен, так как при одном и том же качественном изменении окружающей среды и одной и той же организации изменяющегося животного животные могут эволюировать в различных направлениях в зависимости от различной скорости, с которой наступает изменение среды, и от различной интенсивности этого изменения.
Только что изложенные нами четыре направления эволюции представляют собой лишь очень общие пути биологического прогресса. Филогенетические изменения, имеющие место внутри каждого из этих направлений, происходят посредством особых более или менее частных изменений строения и функции отдельных органов или отдельных групп органов эволюирующих организмов, приспособляющихся к измененным условиям окружающей среды. Мы попытались анализировать более подробно различные виды или типы этих частных изменений, имеющих место при биологически прогрессивной эволюции, иными словами, мы попытались сделать шаг вперед на том пути, на который вступили до нас наши предшественники (А. Дорн, Н. Клейненберг, Л. Плате). При этом мы исходили, так же как и они, из функциональной точки зрения.
Эти исследования показали нам, что существуют еще несколько типов (или принципов) частных филогенетических изменений органов, не известных упомянутым нами авторам. Такими типами являются: 1) фиксация фаз, 2) субституция функций, 3) активации функций, 4) иммобилизация функций, 5)симиляция функций, 6) уменьшение числа функций, 7) разделение функций или органов, 8) выпадение промежуточных функций.
ОБЩИЕ ВЫВОДЫ
583
Все эти частные адаптивные филогенетические изменения органов можно разделить на две большие группы.
К первой группе относятся те филогенетические изменения органов, при которых функция эволюирующего органа изменяется количественно, оставаясь качественно неизменной.
Ко второй группе относятся те филогенетические изменения органов, при которых функция эволюирующего органа изменяется качественно, т. е. делается иной, чем она была прежде.
К первой группе относятся прежде всего: 1) все бесчисленные случаи усиления (интенсификации) и ослабления активных, и пассивных функций эволюирующих органов; 2) весьма многочисленные случаи субституции органов, когда функция остается прежней,но выполняется другим органом, расположенным в том же или в другом месте тела животного (Н. Клейненберг); сюда относятся также случаи физиологической субституции Федотова; 3) случаи фиксации фаз (А. Н. Северцов), когда одна из периодически повторяющихся фаз какой-либо определенной функции эволюирующего органа фиксируется и заменяет собой другие фазы этой функции; 4) случаи уменьшения числа функций (С. А. Северцов), когда ряд второстепенных функций' данного органа замещается усилившейся главной функцией этого органа; 5) случаи выпадения промежуточной функции органа, в результате чего главная функция этого органа усиливается (А. Н.Северцов).
Ко второй группе относятся:!) случаи расширения функций Л. Плате, когда вследствие возникновения новых побочных функций число функций эволюирующего органа увеличивается, и в результате этого его общая функция становится несколько иной, чем она была прежде; 2) случаи смены функций А. Дорна, когда главная функция анцестрального органа замещается одной из его второстепенных функций, вследствие чего общая функция эволюирующего органа изменяется; 3) случаи субституции функций А. Н. Северцова, при которых анцестральная функция органа замещается аналогичной или неаналогичной, но биологически ей равноценной функцией другого органа; 4) случаи активации функций А. Н. Северцова, когда пассивная анцестральная функция эволюирующего органа замещается у потомков активной функцией, т. е. становится совершенно иной, чем она была прежде; 5) случаи иммобилизации функций А. Н. Северцова, когда активная функция эволюирующего органа замещается у потомков пассивной функцией, т. е. также становится качественно иной, чем прежде; 6) случаи симиляциифункций А. Н. Северцова, когда анцестральный орган, делаясь похожим на другой орган, теряет свою старую функцию и приобретает новую; 7) случаи филогенетического разделения органов А. Н. Северцова, когда функции отдельных частей органов изменяются у потомков и делаются качественно отличными от общей функции разделившегося анцестрального органа предков.
Наша классификация адаптивных филогенетических изменений органов в настоящее время, конечно, еще несовершенна, так как она далеко не охватывает всех бесчисленных целесообразных изменений, имевших место в течение эволюции в строении Metazoa. Тем не менее, мне кажется, что уже теперь она вносит известный закономерный распорядок в великое разнообразие филогенетических изменений органов и позволяет наилучше представить себе закономерную последовательность в развитии этих изменений.
Детальное ознакомление с типами (принципами) филогенетического изменения органов привело нас к выводу, что теория принципов позволяет нам бросить взгляд также и на проблему редукции органов. Мы попытались поэтому разработать морфологическую теорию редукции органов животных. При этом выяснилось, что тот или иной орган
584
ГЛАВА XVIII
тогда начинает редуцироваться у животного, когда, вследствие изменившихся условий существования, он перестает ему быть необходимым или полезным, т. е. когда он утрачивает свое биологическое значение.
В некоторых случаях бесполезность органа вызывается непосредственно изменениями, происшедшими в условиях окружающей среды (например редукция органов защиты на тех местах тела животного, которые перестали подвергаться нападению врага и т. п.). Гораздо чаще, однако, орган становится бесполезным главным образом потому, что у потомков эволюи-рующей формы его функцию берет на себя другой орган, лучше приспособленный к изменившимся условиям существования. Это, несомненно, имеет место при следующих типах (принципах) филогенетических изменений органов: при субституции органов Клейненберга и субституции функций А. Н. Северцова, т. е. в большинстве случаев субституции, когда замещаемый орган предков становится ненужным потомкам и поэтому редуцируется; при смене функций Дорна, когда главная функция анцестрального органа замещается одной из второстепенных функций этого органа и когда части органа, связанные с прежней главной функцией, становятся ненужными и редуцируются. При уменьшении числа функций (С. А. Северцов) значение главной функции гипостазируется, иона одна принимает на себя общую функцию всего органа, те же части органа, которые у предков данной формы принимали участие в выполнении общей функции, перестали функционировать и редуцировались. При выпадении промежуточной функции и связанном с этим усилении главной функции органа, части органа, выполняющие эту промежуточную функцию, становятся ненужными и также редуцируются.
Мы видим, следовательно, что во всех этих случаях определяющий момент эволюции лежит не в регрессивных, но в прогрессивных адаптивных изменениях, имеющих место в органах эволюирующего животного и делающих биологически бесполезным существование анцестрального органа.
Таким образом, изучение различных типов (принципов) филогенетического изменения органов дает нам биологическое объяснение явлениям редукции. Это же биологическое толкование позволяет нам понять и причину существования двух видов редукции. Если орган, стоящий на пути редукции, не бывает полезен животному ни в один из периодов его существования, то он редуцируется путем рудиментации. При этом редуцирующийся орган уменьшается в размерах филогенетически постепенно и его строение вторично упрощается, так что в конце концов он превращается в незначительный рудимент. Процесс этот длится геологически очень долго. В тех случаях, когда орган имеет биологическое значение для животного в период его эмбрионального развития или в период молодости и лишь впоследствии перестает быть ему полезным, он редуцируется путем афанизии. Редукция в этом случае протекает быстро и ведет к полному исчезновению органа.
В только что изложенной нами части морфологической теории эволюции мы обсуждали главным образом вопрос об эктосоматических органах, стоящих в непосредственных соотношениях с окружающей средой, и лишь попутно коснулись проблемы эволюции тесно связанных с ними внутренних (энтосоматических) органов. Вторая часть этой теории посвящена проблеме закономерностей эволюции этих внутренних органов.
Мы пришли к выводу, что изменения, происходящие в энтосоматических органах (дейталлаксы), большей частью находятся, в связи с изменениями, происшедшими в эктосоматических органах (проталлаксы). Существуют, однако, случаи, когда первичное филогенетическое изменение исходит не из эктосоматических, но из энтосоматических органов и влияет
ОБЩИЕ ВЫВОДЫ
58&
на другие энтосоматические, а также и на эктосоматические органы. Это наблюдается, например, при многих явлениях ароморфоза. И в том и в другом случае развиваются филетические корреляции или, по нашей терминологии, координации между различными первично и вторично изменяющимися органами. Мы попытались произвести некоторое предварительное разделение этих филетических координаций. Мы нашли, что филетические координации можно разделить на две большие группы.
I. Морфофизиологические координации1, к которым относятся цепи функционально связанных между с,обою органов или частей органов,—те цепи, в которых каждое звено необходимо для правильного функционирования следующего звена.
II. Топографические координации, при которых органы не связаны между собою функционально.
Мы поставили также вопрос о том, как идет изменение в тех координационных цепях, в которых функция и строение одного из членов цепи, обычно исходного проталлактического ее члена, начинают филогенетически изменяться. При этом мы нашли, что при морфофизиологических координациях наблюдаются различные случаи изменения соотношений между членами координационной цепи, а именно:
1. Если функция эктосоматического члена координационной цепи изменяется количественно, то соотношение между остальными членами цепи большей частью остается неизменным, но интенсивность их функций обычно изменяется в том же направлении, в каком она изменяется в исходном эктосоматическом члене цепи (проталлактическая интенсификация—коррелятивная интенсификация).
2. При качественном изменении функции одного из членов координационной цепи различают следующие случаи:
а) функции остальных членов координационной цепи изменяются также качественно, но не слишком резко, и морфологические соотношения между ними остаются прежними;
Ь) функции остальных членов координационной цепи изменяются очень резко и в соответственной мере изменяются и морфологические соотношения между ними, так что устанавливаются новые корреляционные соотношения между членами координационной цепи.
Мы видим, таким образом, что при морфофизиологических координациях функциональные соотношения координированных органов претерпевают и количественные и качественные изменения.
При топографических координациях обычно изменяется темп онтогенетического развития проталлактического прогрессивно изменяющегося органа. Большей частью данный орган начинает закладываться у потомков раньше, чем он закладывался у предков, и развивающийся онтогенетически позднее координированный с ним орган приспособляется в своем развитии к форме и величине этого первично эволюирующего органа.
Теория направлений эволюции, теория типов (принципов) филогенеза, теория редукции органов и теория изменения координационных соотношений внутренних органов образуют все вместе одно внутренне связанное между собой целое—морфобиологическую теорию, объясняющую ход адаптивной эволюции органов взрослых животных.
Но теория морфобиологического хода эволюции не касается, однако, вопроса о том, как и когда в течение индивидуальной жизни происходят описанные выше филогенетические изменения строения и функций эволюирующего организма. Ответ на этот вопрос дает теория филэмбриогенеза.
586
ГЛАВА XVIII
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
Теория филэмбриогенеза пытается разрешить вопрос о том, как происходят и в какой период индивидуальной-жизни возникают те изменения органов животного, которые ведут к преобразованию строения взрослого организма, иными словами, она пытается разрешить проблему модусов прогрессивного и регрессивного филогенетического изменения органов взрослых животных.
Обычно принято думать, что филетические изменения начинаются с мелких наследственных вариаций (мутаций), возникающих в органах взрослых животных. Наши исследования привели к совершенно иному выводу, а именно, ко взгляду, высказанному впервые еще Ф. Мюллером (1864), что филогенетические изменения строения взрослых органов происходят путем изменения хода эмбрионального развития этих органов. Филогенез является, таким образом, функцией онтогенеза.
В основе онтогенетического развития органов лежат четыре процесса, а именно: 1) общий или равномерный рост, т. е. увеличение общего числа клеток всего зародыша, 2) частный или неравномерный рост, т. е. рост зачатков органа, иначе увеличение числа клеток этих зачатков, приводящее к морфологической диференциации, т. е. к изменению формы и усложнению строения многоклеточного организма, 3) гистогенез или гистологическая диференциация организма и 4) процесс странствования и переселения клеток, когда группы клеток, переселившись на новое место в теле зародыша, дают начало новому зачатку органа.
Все эти нерегулирующиеся изменения, происходящие в числе клеток зародыша и в форме зачатков его органов, т. е. все эти филогенетические изменения хода онтогенеза, имеющие столь важное значение для эволюции органов взрослых животных, могут возникнуть в самые различные периоды эмбриональной жизни организма, и это дает нам возможность установить различные модусы в изменении хода онтогенеза животных.
Эти изменения возникают иногда в самом конце периода морфогенеза эволюирующего органа, так что в этих случаях онтогенетическое развитие делает шаг вперед по сравнению с онтогенезом предков. При этом модусе к анцестральной конечной стадии органа прибавляется новая конечная стадия, что приводит к образованию нового строения взрослого органа. Этот процесс мы обозначаем как надставку конечной стадии или анаболию.1 При этом модусе филогенетического изменения конечная стадия морфогенеза данного органа предка, похожая на взрослое состояние большинством признаков формы и строения (но не величиной), сохраняется у потомков и, являясь исходной стадией нового дефинитивного состояния, развивается у них далее; как таковая она постоянно повторяется у потомков (рекапитуляция по Ф. Мюллеру и Геккелю, репетиция поде Бееру). Таким образом, анаболия объясняет нам своеобразное явление рекапитуляции анцестральных признаков, похожих на признаки взрослых предков. Она объясняет также открытый К. Бэром общий факт закономерной последовательности в образовании (закладке) признаков больших и малых систематических категорий, к которым данная животная форма принадлежит.
По закону К. Э. Бэра признаки крупных систематических групп (типов, классов, отрядов) закладываются онтогенетически рано, признаки мелких систематических групп (семейств, родов, видов и т. д.) заклады
1 Завершение морфогенеза органа иногда наступает на очень поздних стадиях развития животного; так, например, дефинитивное установление общих пропорций тела и черт лица у человека, образование некоторых половых признаков и т. д. (герон-томорфозис де Беера) завершаются очень поздно. Мы рассматриваем эти случаи как особые случаи анаболии.
ОБЩИЕ ВЫВОДЫ
587
ваются онтогенетически поздно. Иными словами, признаки животных появляются в их онтогенезе тем раньше, чем они филетически древнее, и тем позднее, чем они филетически моложе. Выше мы видели, что этот закон К. Бэра представляет собой необходимое следствие теории надставки конечных стадий онтогенеза. Анцестральные признаки органа, образующиеся у предков и остающиеся неизменными у потомков (древние константные признаки), закладываются у потомков на тех же стадиях онтогонеза, на которых они закладывались у предков, т. е. на ранних стадиях развития. Наоборот, те признаки органа, которые сильно изменяются у потомков путём надставок новых конечных стадий, закладываются у них соответственно позже. Древние константные признаки являются характерными признаками для всех потомков данной прародительской формы и рассматриваются нами как признаки крупных систематических категорий (классов, отрядов и т. д.). Новые признаки являются характерными именно для мелких подчиненных систематических групп (видов, пород и т. д.).
Теория сдвигания признаков на ранние стадии развития (теория Геккеля-Вейсмана) не дает нам никакого объяснения этой удивительной закономерности, так хорошо объясняемой явлением анаболии. Можно сказать, что все признаки, развившиеся путем надставки конечных стадий, подчинены закону К. Бэра, те же признаки, которые развиваются другими путями, этому закону не подлежат.
В некоторых случаях признаки взрослых животных могут эволюировать путем изменения хода морфогенеза через уклонение (девиацию) развития на средних стадиях онтогенеза (Ф. Мюллер). При этом модусе эволюции направление онтогенетического развития начинает изменяться на средних стадиях развития, так что взрослое состояние органа является измененным в адаптивном направлении: таким путем возникают новые приспособления взрослого органа. Здесь ход морфогенеза изменяется иначе, чем при анаболии: морфогенез не удлиняется.
При этом модусе эволюции также происходит рекапитуляция анцестральных признаков, но повторяются только одни ранние стадии морфогенеза (К. Бэр). Рекапитуляция этого рода не позволяет нам делать никаких заключений относительно строения непосредственных предков данной формы, а поэтому она не имеет палингенетического значения в смысле Э. Геккеля, и признаки, развившиеся этим путем, не подчинены только что упомянутому нами закону К. Бэра.
Филетически важные изменения хода морфогенеза могут наступить и на первых стадиях онтогенеза животного (А.* Н. Северцов, 1912). Эти изменения, возникающие в первых зачатках органа (архаллаксисы), влияют на все последующее течение эмбрионального развития органа, а также и на его взрослое состояние. При этом модусе эволюции не существуют ни рекапитуляции анцестральных признаков, ни удлинения морфогенеза. Признаки, эволюирующие по этому типу развития, не подчиняются закону К. Бэра.
Мы должны еще раз подчеркнуть, что ни закон рекапиту-л а ц и и, ни закон К. Э. Бэра не являются общими законами: оба эти закона действительны в полной мере лишь для тех органов, которые развиваются путем надставки конечных стадий, или анаболии. Рекапитуляция анцестральных эмбриональных признаков (К. Бэр) имеет место и при эволюции путем девиации. При архаллаксисе не происходит ни рекапитуляций, ни закономерной последовательности в образовании древних и новых признаков.
588
ГЛАВА XVIII
В последнее время мы установили еще один более редко встречающийся модус филэмбриогенеза, а именно модус периодического изменения хода развития органов. Он имеет место тогда, когда развивающийся орган доходит в своем онтогенезе до известной стадии развития, после чего отпадает, а на его месте из того же зачатка развивается в соответствующий период жизни животного такой же орган, который снова в определенный период жизни отпадает и т. д.; этот процесс продолжается до достижения животным взрослого состояния.
Не надо думать, что филогенетическое развитие органов всегда происходит в течение эволюции только одним каким-либо модусом филэмбриогенеза. Наоборот, часто случается так, что в филогенетическом развитии органов мы имеем комбинацию двух способов филэмбриогенеза, т. е. что эволюционный процесс, начавшийся у предков путем анаболии, естественно продолжается у потомков путем архаллаксиса, или наоборот. В этом случае мы имеем дело с в т о р и Фн ым архаллакси-сом. Вторичная анабо ли я будет иметь место тогда, когда орган, развившийся у предков путем архаллаксиса, начнет развиваться у потомков путем анаболии.
Кроме морфологических вариаций формы, строения, величины и местоположения эмбриональных зачатков, существуют еще изменения в темпе онтогенетического развития (гетерохронии). Весьма вероятно, что и эти вариации в некоторых случаях являются способом приспособительного изменения органов. Они происходят тогда, когда потомкам бывает полезно завершить морфогенез более быстро, чем это имело место у предков (ускорение развития). Ретардация в закладке органов, повидимому, является способом координировать топографические соотношения между органами. Неотении, которые также должны рассматриваться как гетерохронии, могут быть истолкованы как приспособления к значительным изменениям в условиях существования эволюирующего вида. В общем, в филогенезе животных гетерохронии играют лишь второстепенную роль: сами по себе они не ведут к изменениям в строении и функции органов.
Все филогенетически важные (т. е. ведущие к изменению строения взрослых огранов) изменения в ходе морфогенеза развиваются путем суммирования нерегулирующихся наследственных вариаций.
Весьма вероятно, что вновь образующиеся и позднее исчезающие эмбриональные приспособления (амнион, аллантоис и т. п.) развиваются в течение эволюции путем, суммирования регулирующихся, т. е. исчезающих во время онтогенеза вариаций. Другую группу эмбриональных приспособлений образуют те органы, которые, развиваясь у предков далее и превращаясь у них во взрослые органы, существуют и бывают полезны у потомков только во время личиночной жизни, позднее же у взрослых форм теряют свое биологическое значение и редуцируются.
Коротко очерченную здесь теорию соотношений между онтогенезом и филогенезом животных мы назвали теорией филэмбриогенеза. Эта теория была разработана мною на основании исследования прогрессивно-адаптивной эволюции органов, поэтому у меня возникал вопрос, применима ли она также к явлениям регрессивной эволюции (редукции) и годится ли она для объяснения этих последних.
При исследовании явлений редукции нам удалось установить, что существуют два главных типа редукции органов, которые мы обозначили терминами: «рудиментация» и «афанизия». При рудиментации первый зачаток редуцирующегося органа уменьшается с самого начала закладки, вследствие чего орган взрослого животного становится все меньше и меньше. Этот процесс вполне соответствует процессу архаллаксиса (положительный архаллаксис) у прогрессивных органов, и эта
общие ВЫВОДЫ
589
дает нам основание обозначить его как «отрицательный архаллаксис, происходящий путем уменьшения». Органы, редуцирующиеся по типу «отрицательного архаллаксиса через уменьшение», уменьшаются постепенно, превращаясь, в конце концов, в незначительный рудимент.
Параллельно с уменьшением редуцирующегося рудимента идет также и вторичн’ое упрощение его строения. Те части, которые закладывались в органе предков онтогенетически позднее других, у потомков не закладываются вовсе (отрицательный архаллаксис, происходящий путем выпадения первого зачатка органа или части , органа). Последовательность в выпадении частей органа при филогенетической рудиментации соответствует обратной последовательности развития этих частей в нередуцирующихся органах. Поэтому «отрицательный архаллаксис через выпадение» целиком соответствует положительному архаллаксису, который, как мы знаем, ведет к прогрессивной эволюции органов. Процесс рудиментации длится очень продолжительное время, так что маленькие рудименты органов сохраняются у животных в течение целых геологических эпох. «Отрицательный архаллаксис через выпадение зачатков», повидимому, всегда приводит к полному исчезновению и этих рудиментов.
Второй тип редукции органов мы обозначили именем афанизии. Редуцирующиеся по этому типу органы закладываются совершенно нормально и до известной стадии развиваются тоже нормально, т. е. также, каки у форм, у которых они не редуцируются. Но на более поздних стадиях их развитие останавливается, и они начинают уменьшаться и исчезать на наших глазах (фагоцитоз, превращение в индиферентную ткань и т. д.), не оставляя рудиментов. При этом мы наблюдаем, что в течение известного периода они являются полезными для животного и иногда в это время еще функционируют, позднее же утрачивают свое биологическое значение. Самым характерным в этом модусе дегенерации является то, что данный орган в начале своего развития не уменьшается, т. е. что здесь отрицательный архаллаксис не имеет места; к тому времени, когда орган утрачивает свое значение, он постепенно исчезает путем отрицательной анаболии или путем отрицательной девиации, не оставляя после себя никаких рудиментов. Повидимому, здесь в каждом случае к определенному времени развивается и начинает функционировать особый «редукционный аппарат» (фагоциты, капиллярные кровеносные сосуды и т. п.).
Оба эти типа редукции можно вкратце охарактеризовать следующим образом:
Рудиментация
С 1. Отрицательный архаллаксис через уменьшение
| первых зачатков.
4 2. Отрицательный архаллаксис через выпадение
| первых зачатков (орган не функционирует ни
V на одной из стадий развития).
(
Афанизия . . .
I. Отрицательный архаллаксис не имеет места.
2. Имеет место либо отрицательная девиация на средних стадиях развития, либо своеобразная отрицательная анаболия (орган или его зачаток функционирует на ранних стадиях развития).
Мы видим, таким образом, что теория филэмбриогенеза значительно расширяется. Теперь она объясняет нам не только процесс п р от р е с-сивной эволюции органов, но также и различные типы
590
ГЛАВА XVIII
регрессивной эволюции, или дегенерации органов.
Наконец, возникает вопрос о том, какое значение имеют эти различные способы филэмбриогенетических изменений органов. Почему одни признаки животных развиваются путем архаллаксиса, другие—путем девиации, третьи—путем анаболии, или надставки конечных стадий?
Изучение этого вопроса показало нам следующее. .
1. При архаллаксисе эволюция многих органов идет скачками, т. е. путем суммирования довольно крупных наследственных изменений. Это—путь возникновения. н о в ых, не существовавших у предков признаков и ориментов органов и это—способ быстрых (в геологическом смысле этого термина) изменений уже существующих органов (ускоренный темп эволюции, приспособление к сравнительно быстро изменяющимся условиям окружающей среды).
2. При надставке конечных стадий, или анаболии, суммируются мелкие наследственные вариации, а поэтому приспособление здесь может итти в самых разнообразных направлениях, но всегда чрезвычайно медленным путем (адаптация к медленно изменяющимся условиям окружающей среды).
3. Модус девиации занимает среднее положение между этими двумя крайними типами изменения темпа адаптации.
Мы приходим, следовательно, к выводу, что прогрессивная и регрессивная эволюция строения и функций взрослых органов происходит путем изменения хода морфогенеза этих органов, изменений, которые могут иметь место в начальных, в средних стадиях развития и в самом конце морфогенеза органа, проще говоря, прогрессивная и регрессивная эволюция происходит путем изменений, возникающих на любой стадии развития органов животного.
Нам могли бы, быть может, заметить, что если это так, то принятое нами разделение отдельных модусов эволюции, т. е. архаллаксиса, девиации, анаболии и др., в сущности излишне и что, в конце концов, можно было бы просто ограничиться положением, что изменения хода онтогенеза, приводящие к филогенетическим изменениям в строении взрослых органов, могут возникнуть на любой стадии индивидуального развития этих органов. Однако мы видим, что дело обстоит совсем не так: повторение или неповторение анцестральных признаков, факт, что некоторые признаки подчиняются закону К. Э. Бэра, а другие ему не подчиняются, различные способы редукции органов (рудиментация и афанизия) и, наконец, самый темп эволюции зависят от того, как именно идет эволюция, т. е. протекает ли она по тому или по иному модусу прогрессивного и регрессивного развития органов.
Наконец, мы попытались разрешить вопрос о том, как возникли в течение эволюции эти различные модусы филэмбриогенеза, какие из них являются первичными и какие вторичными, иначе говоря, как возник и как эволюировал онтогенез?
Мы установили, что у Protozoa нет онтогенеза в том смысле, как мы его понимаем для Metazoa, ибо те изменения, которые происходят при индивидуальном развитии Protozoa, соответствуют не онтогенезу Metazoa, а их гистогенезу. Онтогенез (и при том очень короткий) впервые возникает лишь с появлением колониальных жгутиковых форм и осуществляется путем анаболии, которая, таким образом, является первичным типом филэмбриогенеза. С течением эволюции онтогенез Metazoa все более и более удлиняется путем надставки конечных стадий.
На ряду с этим первичным способом эволюции, т. е. с постепенным удлинением онтогенеза путем анаболии, мы находим в онтогенезе колониальных форм жгутиковых и первые архаллаксисы (диференцировка клеток колоний на соматические и половые, образование мезодермы).
ОБЩИЕ ВЫВОДЫ
59 Г
Позднее у билатерально симметричных Metazoa в онтогенезе появляются первые девиации. Таким образом, архаллаксисы и девиации являются вторичными способами изменения хода онтогенеза. Отметим при этом, что ни один из этих двух последних способов не удлиняет онтогенез Metazoa.
Итак, онтогенез (или эмбриональное развитие) не существовал у Protozoa, был очень короток у колониальных форм и постепенно удлинялся по мере филогенетического развития и усложнения организации Metazoa. Развитие онтогенеза является, таким образом, средством, при помощи которого была достигнута эволюция более высоких форм жизни.
В заключение мы попытались разобрать, в каких соотношениях с известными нам типами филогенетического изменения органов находятся различные модусы филэмбриогенеза, т. е. попытались определить, какими способами филэмбриогенеза осуществляется тот или иной принцип (тип) филогенетического изменения органов. Основной вывод, к которому мы пришли, заключается, повидимому, в том, что развитие каждого принципа (типа) филогенетического изменения органов редко осуществляется при помощи одного какого-нибудь модуса филэмбриогенеза; наоборот, большей частью случается так, что процесс развития, начавшийся по одному способу филэмбриогенеза, продолжается и заканчивается по другому или даже по другим его способам. Чаще всего в различных типах изменения органов нам приходится наблюдать комбинации двух модусов филэмбриогенеза: архаллаксиса и анаболии, но бывает и так, что один какой-нибудь принцип эволюции, например широко распространенный принцип интенсификации функции, осуществляется в различных случаях при помощи почти всех известных нам способов филэмбриогенеза, т. е. и анаболией, и архал-лаксисом, и девиацией, и гетерохронией (акцелерацией).
Морфобиологическая теория хода эволюции и теория филэмбриогенеза тесно связаны друг с другом и последняя дополняет первую: м о р-фобиологиче.ская теория хода эволюции животных раскрывает нам общие и частные закономерности прогрессивной и регрессивной эволюцииэкто-иэнтосоматическихорганов взрослых животных, теория филэмбриогенеза дополняет ее, разъясняя нам тот путь, которым возникают изменения в органах взрослого животного в течение его индивидуальной жизни, в частности—в течение его онтогенеза.
ИМЕННОЙ УКАЗАТЕЛЬ
А
Абель, О.—76, 78 , 83, 104, 124—126, 386.
Б
Бабак.—97, 98.
Балабай, П. П.—145.
Беер де—78, 474 —476, 495, 504, 528, 532„ 586.
Бёджет, 14. С.—240.
Белобородова, О. В.—155.
Бем. А. А.—467*.
Берг, Л. С. —111.
Бергман—96.
Бехер, С,—427—429.
Боас, 14,—211,212, 243, 317*
Бобренкий, Н. В.—318*, 385*
Бовери, Ф.—77, 133, 141
Боголюбский, С. Н.—482
Бокер, Г,—352*, 382*, 388*.
Больк, Л,—510.
Брайд Мак—136*.
Брандес, Л.—97.
Браус, Т.—243.
Бридж, Т.—209, 212, 229.
Бройли, Ф,—288.
Брум, Р.—438.
Брэм, А. Е.—305*, 310*, 338*
Бэр, К. Э.—81, 453 , 454 , 474 , 495, 496, 499, 500, 503—505, 509, 521, 522, 536, 537, 586, 587, 590.
Бэтсон, В. 75.
Бюффон, Л.—81, 82.
В
Вагнер—442*
Вай фан, И,—72, 99, 133, 354.
Васнецов, В. В, —193, 214, 239, 497,
498*, 527
Ватсон, Д.—76 , 222 , 232 , 243.
Вебер, М.—432
Вейденрейх, Ф.—83—87, 94.
Вейсман, А,—73, 74, 82, 455, 457, 458, 495, 504, 505, 587
Вейт, О,—79, 476.
Вернадский, В. 14.—289.
Вершлюс, И. (Versluys, J)—79, 438, 455*, 456*, 463*.
Видерсгейм, Р. —188*, 37о*.
Виллистон, С. В.—76.
Вильсон, Е.—74.
Вирхов—78.
Воскобойников, М. М. —185, 205, 206, 281.
Вудвард, А. С.—76, 212, 215, 243.
Вюртенбергер, Л.—357.
Г
Газе, А.—198.
Гадов, Г.—ПО, 111
Гарбовский, ф.—460, 461
Гарстанг, В.—473—476, 528.
Гартман, М.—561
Гаупп, Е.—205, 354*
Гегенбаур, К,—72, 189, 214, 236, 408*, 459—461
Гейдер, К.—322—324, 326, 563.
Геккель, Э.—72, 73, 79, 82, 106—108, 207, 208,210,211,222, 273,312,453—462, 470, 476, 497, 499, 505, 520, 521, 532, 536, 537, 563, 586, 587.
Гексли, Т.—72, 459.
Гербст—74.
Гертвиг, О.—72 , 239*, 318*, 461.
Гертвиг, Р.—72.
Гессе, Р.—80, 96, 98, 133, 135, 141, 253, 254, 284, 345.
Гетте, А.—172, 239.
Гильцгеймер—406*
Годри, А.—72, 74.
Гольдшмидт, Р. —133.
Гольмгрен, Н,—79, 149, 154, 155, 157, 162, 164, 165, 169, 218, 365.
Грегори В. К,—76 , 216—218 , 240, 243— 245, 287
Грегори, М. С. (см. Кэмп)—287.
Грей Аза—72
Гудрич, Е. С,—78, 104, 168, 169, 209, 210, 212, 234*. 239, 241, 243.
Гукер—72.
Гюнтер—96.
Д
Давыдов, М. М.—467*.
Дакке—75, 111, 115.
Даниэль—381*, 404*
Дарвин, Ч,—71—73, 75—78, 80—82, 209, 271, 426, 427, 429, 454, 455, 458, 459.
Делаж, И.—318*, 349*, 565*, 566*, 567*, 568*, 569*
Дельсмап, Г.—326.
Джонстон, И.—79, 142.
Дин Башворд—209, 214.
автора.
* Звездочкой обозначены номера тех
страниц, где имеется рисунок упомянутого
ИМЕННОЙ УКАЗАТЕЛЬ
Дислер, Н. Н,—175, 176*, 234*.
Долло, Л,—73. 74, 76, 78, 296.
Домбровский Б.—429, 432
Дорн, А,—72, 73, 326, 345—347, 349, 352, 354, 375, 381, 384, 385, 391, 402, 413, 414,417,421—423, 435, 460, 574, 582 — 584.
Дофлейн—348*
Дри-ш, Г,—74, 75, 84.
Дружинин, А. Н.—295, 296*.
Дюбуа, Е.—78, 79.
Дюркен, Е..—428, 429.
Е
Емельянов, С. В. —192, 220, 529.
Ермаков, Н. В.—92.
Еремеева, Е. Ф.—188.
Еремеев, В. Ф.—399, 401, 445.
3
Заварзин, А. А.—181 Заленский,. В. В.—72, 236.
К
Каммерер—86, 87,
Каптерев, П. Н.—92.
Карпентер—72.
Кейбель, Ф.—312, 436, 460.
Кемп, С. (см. Грегори, М.)—287
Кез—76.
Керр, Г.—78, 187*, 240.
Кингсли —275*
Киэр, И,—76, 139, 153, 156—159, 167, 170, 208.
Клейненберг, Н,—73, 345—347, 373—379, 394, 402, 405, 413, 414, 417—419, 423, 443, 582—584.
Ковалевский, А. О.—72.
Ковалевский, В. О.—72, 74, 78.
Колликер, А.—470, 471, 493, 494.
Коллиндж, В. Е.—209.
Кольман—313*
Коп, Е. Д,—72, 74, 76, 81, 214—216, 218, 222, 471, 472, 475, 477
Корренс, К.—74.
Крыжановский, С. Г. —187, 188*, 214, 313, 314*, 476, 477
Купфер, К.—187*
Кювье, Ж.—71, 74, 75, 426, 427, 429, 431
Кюне, К.—445.
Л
Лайель, Ч,-72
Ламарк, Ж.—71, 73, 76, 81—83, 85, 94, 459.
Ланг—274*
Ланге, Н. О,—226, 229.
Лаубман, В. 405.
Лейбниц—81.
Лехе—85.
Лотси, Ж.—111, 112.
Лутер, А. 435, 437
Лэнкестер Рай—72.
М
Мак Брайд—136*
Макушок, М. Е.—239.
Маркус, Г.—79.
Марш—72, 74.
Морфологические закономерности
Матвеев, Б. С.—134, 141, 315, 354, 476, 477, 481, 482, 509, 529, 574.
Маурер, Ф.—350*, 468*.
Махотин, А. А.—389, 390*
Машковцев, А. А.—134, 147.
Меккель, И. 453, 454.
Мендель, Г.—74, 75, 85.
Менерт, Е,—312, 441, 460, 502, 527, 531, 532 , 548, 552, 578.
Мечников, И. И.—72.
Монтгомери, Т.—462.
Морган, Т.—74.
Моров, Т.—172.
Муди—243.
Мюллер, И.—207, 210, 222.
Мюллер, Ф,—72, 73, 106, .107, 453—455, 459, 470, 472, 473, 475, 476, 478, 497, 499, 504, 506, 536, 537, 586, 587.
Н
Наук, Э.—476.
Неймайер, Л.—79, 193, 239.
Неф, А, —116, 473, 475, 476, 506.
Нил, Г,—79, 141, 142.
Нитше—525*.
Норман—238*.
О
Оппель, А.—456, 460.
Осборн, Г.—72, 73, 76, 118, 209—212 243 256, 257, 289, 301, 388*, 389, 469’ 470’ 533, 582
Оуэн, Р,—71.
П
Павлова, М. В.—323*
Павлов, А. П.—533.
Паркер, В. 137*.
Паттен, В.—149, 157, 169.
Паули—75, 83.
Петер, К.—520.
Плате, Л.—82, 116, 118, 124, 141 253 273, 322, 345, 347, 381, 384, 393 394’ 407, 413, 414, 426—428, 583.
Прайс, Г,—140, 145, 155.
Принцгейм—563*.
Р
Рабль, К,—72, 468*.
Радль, Е.—75, 425.
Раков, В. А.—231.
Рейгерт—187*.
Ромене, А.—72.
Ромер, А.—287
Роза Даниэль—394.
Рындзюнский, А. Г,—220.
Ру, В,—74, 84, 87, 89, 97, 428.
Рузвельт—292
РумбЛр, Л.—524.
С
Саве-Сбдербсрг, П,—216—218.
Сарацин П. и Ф.—313*, 405.
Северцов, С. А,—268, 269, 271, 290, 307 347, 380, 583, 584.
Седжвик, А.—472—476.
Семон, Р.—84, 187, 243, 409, 428.
Семпер, К.—96, 326.
Сент-Илер Ж.—73.
38
594
ИМЕННОЙ УКАЗАТЕЛЬ
Серебровский, П. В.—95.
Соболев, Д. Н,—111, 115.
Спенсер, Г.—73.
Стеншиб, Ё.—76, 138, 139, 145, 154, 156, 158*, 159, 160*, 161*—165, 168—170, 208, 215—217, 221, 222, 231, 235.
Сушкина—Попова, Н. Н,—444.
Т
Третьяков, Д. К, —132, 143, 155 , 326.
Трэквер, Р,—76, 169, 212, 214, 215, 217, 222.
У
Уоллес, А. Р,—73, 82, 446.
Ф
Фан Вай—72, 99. 133, 354.
Федотов, Д. М.—346, 347, 379, 402, 413, 414, 583.
Феттер, Б.—435.
Филатов, Д. Н.—428.
Фишер, Р,—102.
Фохт, К.—72.
Франсе—75 , 83.
Франц, В, —136, 267, 273, 347, 474, 476.
Фриз де, Г.—74.
Фульд, Э.—00.
Фюрбрингер, М.—72, НО, 111, 243, 543, 544. 553.
X
Хун—339*.
ц
Циттель, К.—74, 355*.
Ч
Чермак, Н.—74.
Чулок—387*
Ш
Шауинсланд, Г,—267.
Шепельман—97
Шимкевич В. М.—378, 406*
Шмальгаузен, И. И.—186, 192 , 245, 382 398, 403, 431, 438, 482.
Шпеман, Г,—428.
Штандфус—102.
Штейнман, Г,—111, 112, 114, 115.
Штейнер, Г.—463*.
Штейн—564* ,
Штерне—299*.
Э
Эвклид—71.
Эджеворт—78.
Эдмонстон—96.
Эймер, Ф. 113, 114.
Эллис, Е.—78.
Элькок—340*.
Эмери, К.—461.
ПРЕДМЕТНЫЙ
А
Аббераци я —506.
Аббревиация—477.
Адаптивные признаки крупных систематических групп—118—119.
Адаптивные признаки мелких систематических групп—119—120.
Адаптивная (идиоадаптивная) радиация-73, 118, 301—304, 308, 389, 582.
Активация функций—347, 406—408, 414, 415, 576, 582, 583.
Активные органы—254.
Актиноптеригии (см. Actinopterygii) — .215, 218, 282.
Акуловые рыбы (см. Elasmobranchii) — 191, 194, 195, 207, 217
Акулы—192, 195 , 206 , 219, 220, 279, 329, 381, 403.
Акцелерация—441, 442, 444, 448, 471, 472, 475, 478, 528—530, 536, 537, 578, 591.
Амфибии бесхвостые (см. Anura)—218, 284, 329, 434, 463, 549.
Амфибии хвостатые (см. Urodela)—218, 329, 434, 530, 531.
Аммониты—457, 458.
Анаболия (надставка стадий)—469, 477, 483—499, 516, 522, 527, 532, 533, 536, 537, 568—576, 578, 586—588, 590, 591.
— ее связь с законом К. Э. Бэра— 499—505.
Анаболия вторичная—498, 499, 588.
Анаболия отрицательная—483, 538, 558, 560, 589.
Аннелиды (см. Annelidae, Кольчатые черви)—325, 328, 329, 373, 410.
Аномодонты—286, 287
Антропоидные обезьяны—78.
Ароморфоз—272—295, 321, 323—332, 337. 338, 341—343, 581, 582. — примеры—274, 275. — соотношения е идиоадаптациями — 308—311 — соотношения со специализациями— 310—311.
Ароморфозы при эволюции амфибий—284.
» » Chondrichthyes selachoidei и эласмобранхий—279, 280.
» >> Holosteoidei actinopterygii и Holosteoidei crossopterygii—282, 283.
» » круглоротых—278.
» » млекопитающих—288—293.
УКАЗАТЕЛЬ1
Ароморфозы при эволюции Osteichthyes и Holosteoidei primitivi—280, 281 s- » первичных бесчерепных и энтобранхиат—277, 278.
первичных Quadrupeda—283, 284.
» » птиц—288.
» » рептилий—285—288.
» » эктобранхиат и первичных челюстных—278, 279.
Архаллаксис—477, 483, 510—523, 527, 532—537, 570—578, 587, 590, 591 — отношение к биогенетическому закону и закону К. Э. Бэра—521, 522.
Архаллаксис вторичный—454, 519, 576, 578, 588.
Архаллаксис отрицательный—483, 538, 589. Архаллаксис отрицательный через выпадение—478, 549, 552—554, 557, 559, 589.
» » через уменьшение—544—548, 554, 557, 559, 589.
Архетип—71
Асцидии—327, 328.
Аутогенез—81,82, 113, 471.
Афанизия—538, 554—550, 584, 588, 589.
Б
Батмизм—81, 472
Белуга (см. Huso huso)—225—227.
Бесчерепные позвоночные (см. Acrania primitiva)—132—138, 142.
Бесхвостые амфибии (см. Anura)—218, 284, 329, 434, 463, 549.
Биогенетический закон—453—456, 459— 462 , 470 , 472 —474 , 497, 521, 522, 536.
— его определение—456.
— его история и критика—453—462. Биологические координации—431 Биологически прогрессивная эволюция (биологический прогресс)—267— 273, 580, 581
— определение—267, 268.
— характерные признаки—271 Биологически регрессивная эволюция (регресс)—272, 581
Бранхиомерия примитивных ПОЗВОНОЧНЫХ 143, 145, 146.
Бранхиомерия круглоротых—163—167.
В
Вариации индивидуальные (в скелете и мускулатуре желтопузика) -399, 400, 445, 446, 447.
Звездочкой обозначены номера тех страниц, где имеется рисунок.
3»*
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Вариации координационные—445, 446, 447. » » меристические—445, 534, 535.
Вариации (эмбриональные) нерегулирую-щиеся—519 , 520 , 588.
» » регулирующиеся—519, 588. Вторичноротые (см. Deuterostomia)—326. Вторичные анаболии—498, 499, 588. Вторичные архаллаксисы—454, 519, 576, 578, 588.
Вторичные рекапитуляции—477 Выживание примитивных форм—321, 322. Вымирание—272 , 580, 581.
Выпадение промежуточной функции—347, 366—373, 413, 577, 578, 582—584.
Г
Ганоиды (см. Ganoidei)—192.
Гекконы (см. Ascalabotes fascicularis)—260, 286, 297, 304, 306.
Генетические корреляции—431. Геронтоморфоз—405, 406, 586.
Гетерохрония—312, 442, 444, 456, 460, 505, 536, 537, 570, 572, 574, 588, 591
Гетерохрония и филогенез—527—532.
Гетеротопия—312, 456.
Гидра—565, 566.
Гимнофионы Сем. Gymnophiona)—260, 261, 234, 329, 333, 404, 405. 410.
Гиппарион (см. Hypparion)—125.
Гипотеза дегенеративной эволюции хрящевых ганоидов—214, 215, 221, 222
— ее критика—221—235.
Гипотеза «сдвигов»—504—506.
Гистогенез—480, 481, 536, 561, 562, 586. Гистогенетическая рекапитуляция—466— 469.
Гнатостомы (см. Gnathostomata)—168, 177, 178, 180, 181, 189, 191, 218.
Гнатостомы первичные (см. Protognatho-stomata)—185, 186, 191
Гребневики (см, Ктенофоры)—324, 329. Губки (см. Spongia)—-323, 328.
Д
Даман—105.
Двоякодышащие (см. Дипнои, Dipnoi, Dip-neusta)—186, 192, 195, 199, 211, 212, 217, 218, 233, 329.
Девиация—475, 477, 505—511, 523, 533, 534, 536—538, 571, 573, 574, 578, 591, 592.
Девиация отрицательная—483, 538, 560, 589. Дегенерация (общая)—272, 316—320, 321, 324, 328, 335, 342, 343, 582, 583.
Дегенеративная эволюция хрящевых ганоидов—214, 215, 221, 222.
Дейталлаксы—264, 581, 585.
Дельфины—115, 302, 380.
Дивергентное развитие (дивергентная эволюция)—1 16—1 18, 120—123, 124— 128.
Динозавры—77, 303, 361, 442.
Дипнои (см. Двоякодышащие, Dipnoi, Dip-neusta)—172 , 203 , 210.
Диференциация—353, 412.
Е
Ехидна—77.
Ж Желтопчзик (см. Ophisaurus apus)—399, 401, 543—546, 550—553.
3
Заимствование эмбриональной ткани (см. Странствование клеток)—574, 575. Закон К. Э. Бэра—499—505, 521, 522, 536, 537
— его связь с анаболией—500—504, 586, 587
Закон органогенеза Менерта—460.
Закон терминальных изменений Нэфа — 473.
Заселение суши—285.
Змеи—297, 401, 406, 407.
И
Иглокожие (см. Echinodermata)—326, 327. Игуанодоны—115.
Идиоадаптации—272, 295—311, 321, 328 — 330, 332, 335, 337, 338, 341—343, 581, 582.
— определение—295.
— примеры—296—304.
— соотношение с ароморфозами— 307—311
— соотношение со специализациями— 304.
Идиоадаптивная эволюция у аннелид—325, 326.
» » у губок—323.
» » у змей—297
» » у иглокожих—326, 327.
» » у кишечнополостных—324.
» » у позвоночных—329.
» » у простейших—323.
» » у скатов—297
» » у черепах—296.
Идиоплазматические корреляции—427 Изменение направлений эволюции—323— 341, 342, 343.
— причины и предпосылки—331—341, 343.
Иммобилизация—347, 408, 409, 414, 415, 582, 583.
Индивидуальные вариации—399, 400, 445, 446.
Индивидуальные корреляции—426. «Индиферентные признаки»—4 19. Интенсификация функций—347—356. Интенсификация функций клеток—347,348. » >> органов—348, 351—356.
» » тканей—347, 349—351
Интенсификация путем изменения строения клеток—351.
» » роста—352, 353.
» » увеличения числа компонентов— 351,352.
» » умножения органов—353, 354. Ихтиозавры (см. Ichthyosaurus)—77, 115, 288.
Ихтиостегиды (см. ichthyostegalia) — 218. К
Казуары—110.
Камбалы (см. Pleuronectidae)—87, 300*, 416.
Кашалоты—115, 302.
Киви — ПО.
ПРЕДМЕТНЫЙ. УКАЗАТЕЛЬ
Кистеперые (см. Кроссоптеригии, Crossopterygii)—100. 1.72, 197, 199, 203, 204, 207, 210, 212, 217, 218, 243, 244, 245, 283, 329, 368.
— отношение к Quadrupeda—243— 246, 329.
Киты —118, 302, 380, 386—388, 421 Кишечнополостные (см. Coelenterata)—323, 324, 565, 566.
Классификация гетерохроний—460.
Классификация корреляций—(по Бехеру) 427.
» » (по Домбровскому)—429.
» » (по Плате)—426—427
» » (по А. Н. Северцову)—430.
» » (по И. И. Шмальгаузену)—431.
Классификация модусов филэмбриогенеза 480—483.
Классификация Ostracodermi—162, 163.
» » Osteichthyes—243.
Классификация принципов (типов) эволюции—347.
Классификация приспособлений (по Осборну)—256, 257
» » (по Плате)—255, 256.
Классификация рыб—207—218.
Классификация ценогенезов—314—315.
Кольчатые черви (см. Аннелиды, Anneli-dae)—96, 137, 325.
Конвергентное монофилетическое развитие:—111.
Конечности:
— идиоадаптивная радиация конечностей у млекопитающих и рептилий— 301—304.
происхождение конечностей—186— 189, 236, 237, 244—245.
— редукция конечностей у безногих ящериц—539—554.
стенобазальные плавники—236— 238, 240, 383*
— строение парных плавников у Chondrichthyes selachoidei—192, 193.
— » у Pleuracanthodei—194—195.
— » у Protognathostomata—186—191
— строение конечностей у первичных Quadrupeda—244—245.
— . фиксация фаз движения конечно-— ‘ стей у бесхвостых амфибий—363. — » у млекопитающих—357—361.
— » у рептилий—357, 361—363.
— фиксация фаз движения плавника у Ceratodus—363—365.
— филогенез брюшных плавников— 366—370.
— филогенез грудных плавников—236-
237
— эврибазальные плавники —192, 236 —238, 381—385.
Конъюнкции—429, 470.
Координации биологические—431.
» » обратные—442 —445.
» » соответственные—442, 445.
Координационные вариации—446, 447.
Копытные—118.
Корненожки—323, 348.
Корреляции генетические—431
» » идиоплазматические—427
» » индивидуальные—426.
» » конструктивные—429.
Корреляции морфогенетические—431 » » тектонические—429.
» » функциональные—259, 261, 265, 427—429, 431
» » элементарные—429.
Корреляции (координации) морфофизиологические—430—440, 447, 470, 585. » » топографические—430, 440—445, 447, 470, 585.
Корреляции (координации) филетические— 123, 124, 12 8, 263, 425—449.
— исторический обзор—425—429.
— способы изменений—445—447
Костные ганоиды (см. Holosteoidei)—100, 197, 199, 202, 203, 209, 212, 219, 222, 223, 368—370, 382.
— двойное сочленение нижней челюсти—234, 235.
— строение скелета—232, 233.
Костистые рыбы (см. Teleostei)—96, 197, 199, 203, 205, 210, 212, 214, 222, 223, 329, 368—370, 382.
Котилозавры—285—287, 354.
Кроссоптеригии (см. Кистеперые, Crossopterygii)—209, 214, 215, 218, 222, 282
Кроты—118, 260, 302, 333, г>88*.
Круглоротые (см. Циклостомы, Cyclostomata)—133—135, 138—143, 146, 147, 149, 154, 155, 159, 161, 164 — 170, 172, 174, 175, 191, 208, 217, 277, 278, 329.
Ктенофоры—566, 507
Л
Ламарки зм —82 —83.
Ланцетник (см. Amphioxus)—133—138, 146, 177, 32.4.
Лемуры—118, 302, 389.
Ленивцы—118, 302, 305, 306, 389.
Летучие мыши—118, 302, 389.
» » рыбы—381, 382.
» » собаки 118, 302 389.
Линька —524—526.
М
Мегатерии—115.
Меланизм—96.
Меристические вариации—445, 534, 535. » » органы—535.
Меторизис—378.
Миксины (см. Myxinoidea)—139, 147, 148, 153 — 156, 159, 162, 167—169, 171, 180, 217, 329.
Мимикрия—298, 299.
Миноги (см. Petromyzontes)—139, 143, 147—156, 159, 162, 164—169, 171, 175, 180, 217, 329.
Млекопитающие—77, 288—293.
Модусы филэмбриогенеза (см. анаболия, архаллаксис, гетерохрония, гистогенез, девиация, общий рост, отрицательная анаболия, отрицательный архаллаксис, отрицательная девиация, периодическое изменение хода развития органов, странствование или переселение клеток, частный рост)—480—560.
Мезозавры—77
Моллюски—155t
предметный указатель
Мо ни мости ли я —4 07
Монофилетическая теория—109, ПО, 116— 118, 123, 124, 128.
— ее значение—116 —118.
Моржи—118, 380.
Морские коньки (см. Hippocampus)—490, 491.
Морские коровы—118, 380.
Морской чорт—(см. Lophius)—488, 490, 497
Морфобиологическая теория эволюции — 253—449, 579—585.
Морфогенетические корреляции—431 Морфологическая теория редукции органов—416—424, 583—584.
Морфофизиологические корнеляции—430— 440, 447, 470.
Z/юрфофизиологический прогресс (см. апо-морфоз)—267, 272—295, 321—332, 337, 338, 342, 343, 581, 582.
Морфофпзиологический регресс (см. дегенерация)—272, 316—320 , 328, 342, 343, 581, 582.
Мультифункциональность—346, 389, 390, 392—394, 412, 579.
Муравьеды—303, 306.
Мутации—534, 535.
Н
Нанду—110.
Направления филогенетического развития животных—267—343, 580, 581
— изменение этих направлений—331 —
343.
— стимулы изменения направления— 335—338.
— зависимость изменения направления от интенсивности стимула— 334—338, 343.
— » от характера организации жи-
— » вотного—336—338, 339, 343.
— от соотношения между этими двумя факторами—336—338, 340, 343. Необратимость эволюционного процесса— 73.
Неовитализм—8I Неодарвинизм—73.
Неоламаркизм—73, 81—102.
Неотения—475, 477, 530, 531, 537, 588.
Нерегулирующиеся эмбриональные вариа-ции—520, 521, 589.
Носорог—105.
О
Обезьяны—118, 302, 389.
Оболочники» (см. Tunicata)—317 318, 327, 328, 419.
Общий рост—480, 481, 535, 586.
Окольное развитие—476.
Онтогенез—478, 480, 535, 561—572, 590.
— его эволюция у Metazoa—561, 572, 590, 591
Органы активные—254.
» индиферентные—254.
» пассивные—91, 93, 94, 100, 254.
» эксцессивные—255, 483—486.
Органы эктосоматические—258—266, 425, 431—434 , 437 , 439 , 440 , 447—449, 579, 580, 584, 585.
— определение—258.
— соотношение с константной внешней средой—258—260.
— » » с изменяющейся внешней средой—260—264.
Органы энтосоматические—258, 259, 262— 266, 425, 431, 433, 434, 437, 439, 440, 447—449, 579, 580, 584, 585. — определение—258.
— соотношение с эктосоматическими органами—259, 262, 263, 425, 131 — 440, 447
Ортогенез—81, 392, 394.
Осетровые рыбы (см. Acipenseridae)--189, 200—202, 204, 206, 207, 214, 220— 223, 329.
Отрицательная анаболия—483, 538, 558, 560, 589.
Отрицательный архаллаксис через выпадение—478, 549, 552—554, 557, 559, 589.
» » через уменьшение—544—548, 554, 557, 559, 589.
Отрицательная девиация—483, 538, 560, 589.
Офиуры—377, 379, 380.
П
Палеаспиды (см. Palaeaspidae)—163.
Палеобиология—78.
Палеонисциды (см. Palaeoniscidae)—197, 199, 217, 218, 221, 368, 370.
Палеонтологический метод филогенетического исследования—103—104.
Палингенезы—456, 457, 459.
Пальцеходящие животные—357, 359.
«Панцырные рыбы»—156 —171, 207, 208.
— строение —157—162.
— классификация—162, 163.
—• соотношения с круглоротыми —167— 171
Параллелизмы—ИЗ, 128, 181, 197, 466.
Парнокопытные—302.
Пассивные органы—91, 93, 94, 100, 254.
Пассивное питание—133, 146, 149.
Педоморфоз—475.
Первичные бесчерепные (см. Acrania primitiva)—132—135, 246, 329.
— нервная система—133.
— пищеварительный тракт—133.
—• скелет—133—134.
— выделительная система—134.
«Первичные истинные ганоиды»—210.
Первичные костные рыбы (см. Osteichthyes primitivi)— Г97—207, 212, 213, 249, 280,281.
— костные чешуи и покровные кости черепа—197—204.
— жаберный аппарат—205, 20о.
Первичные челюстные (см. Protognathostomata)—180—191, 248, 278, 279. — наружный скелет—180, 181
— органы движения—186—191
— ротовой аппарат—183—185.
Периоды жизни—478—479.
Периодическое изменение хода развития органов—523—527, 537, 588.
Пескоройка (см. Ammocoetes)—143—146, 149, 153, 162, 164, 165, 169, 175.
Пингвины—388.
Питание пассивное—133, 146, 149.
ПРЕДМЕТНЫЙ 'УКАЗА ТЕЛЬ
Питекантропы—77, 78.
Пиявки—376, 377
Плезиозавры—77 288 , 303 , 386, 388.
Покровительственная окраска—298.
Полифилетическая теория—110—116. — ее несостоятельность—114—116.
Принципы, или типы филогенетического изменения органов—344—415.
Принцип (тип) .активации функций—347, 406—408, 414, 415, 576, 582, 583. » » выпадения промежуточной функции 347 366—373, 413, 577, 578, 582 — 584.
» » иммобилизации—347, 408, 409, 414, 415, 582, 583.
» » интенсификации функций—345, 347—356, 412, 414, 573, 583.
» » ослабления функций—355.
» » разделения органов и функций— 347, 410, 411, 414, 577, 582, 583.
» » расширения функций—345, 347, 380, 381—384, 393, 394, 407, 413, 583.
» » симиляции—347, 409, 410, 414, 577, 582, 583.
» » смены функций—345, 347, 349, 352 , 354, 372 , 381, 384—394 , 402 — 404, 409, 413, 417, 421—423, 435, 574, 575, 583, 584.
» » субституции органов—345, 347, 3'73—379, 402 , 403, 413, 414, 417— 420, 422, 423, 583, 584.
» » субституции функций—347, 394— 406, 414, 417—420, 433, 575, 576, 582—584.
» i> уменьшения числа функций—380, 381, 413, 414, 421—423, 582 —584.
» » физиологической субституции—346, 347, 379, 380, 402, 413, 414, 583.
» » фиксации фаз—347, 356—366, 413, 574, 582, 583.
Принцип неупражнения органов—83. — его несостоятельность—92—94.
Принцип упражнения органов—83. — его несостоятельность—88 —92.
Принцип целесообразной реакции—85—88. — критика—94—102.
Приспособительная дивергентная эволюция—120—123.
Приспособление (адаптации)—253—257, 265.
— классификация—255—257
— определение—257.
Приспособления специальные (специали-заци и)—304—307 частные (идиоадаптации)—295—304 эмбриональные (ценогенезы)—311 — 316.
Прогресс биологический—267, 268, 271, 273, 580, 581
— пути, которыми он достигается— 272.
Прогресс морфофизиологический (см. ано-морфоз)—267, 272—295, 321—332, 342, 343, 581, 582.
Происхождение конечностей—186—189, 236 , 237, 244—245.
Происхождение костного черепа—200—204, 230.
• Проталлаксы—264, 580, 584.
Протей—260, 261.
Протогнатостомы (см. первичные челю-— стные, Protognathostomata)—180— 191, 248, 278, 279.
Протокраниаты (см. Protocraniata)—138— 148, 154, 171, 175, 177, 179, 180, 246, 276, 277, 329.
— вкутрений скелет—139, 140.
— кожный скелет—139.
— мочеполовая система—147.
— мускулатура—141
— органы высших чувств и нервная система—141 —143.
— органы дыхания—140, 143—145. Протоциклостомы (см. Protocyclostomata 153—155, 164, 171, 175.
переход от пассивного питания к активному—155, 156.
Псевдокорреляции—427, 429. Пте родакти ли —7 7 Птерозавры—288, 303.
Р
Радиация адаптивная—73, 118 , 301—304 308, 389, 582.
Радиолярии—323.
Разделение органов и функций—347, 410, 411, 414 , 577, 582 , 583.
Рамфоринхи—77
Расширение функций—345, 347, 380—384, 393, 394, 407, 413, 583.
Регулирующиеся эмбриональные вариации—520, 589.
Редукция органов—316, 320, 416—424, 444, 538—560.
— ее предпосылки—416, 417. Редукция вследствие изменений условий внешней среды—417.
» вследствие субституции—417—420, 423.
вследствие смены функций—421,423. вследствие уменьшения числа функций—421 —424.
Рекапитуляция признаков—106, 107, 188, 462—470, 474, 522, 586, 587.
Рекапитуляция при анаболии—495—497, 520—521, 536, 586, 587
» при девиации—509, 537, 587. Рекапитуляция «вторичная»—477
» гистогенетическая—466—469.
» функциональная—477 Репетиция—474, 475, 586.
Ретардация—441, 444, 448, 471, 472, 531, 536, 548, 578.
Ри нхоцефалы —357
Родословное древо—72, 73, 77, 79, 207, 209, 211, 213, 214, 216—218, 459. Рост общий—480, 481, 535, 586.
» частный—480, 481, 535, 586.
Рудиментация—538, 539—554, 556, 559, 584, 588, 589.
Рудиментарные органы—106.
С
Саламандры—356.
Сальпы—327 , 328.
Сальтации—534, 535.
Селахии (см. Selachoidei)—172, 174—176, 182, 187, 193, 205—207, 211, 220, 222, 375.
600
ПРЕДМЕТНЫЙ УКАЗА ТЕЛЬ
Севрюга—226.
Симиляция функций—347, 409, 410, 414, 577 582 583.
Скаты—(см. Batoidci)—191, 192, 195, 261, 262*,279, 297, 329,403, 417, 433*, 434.
Слепыши—118, 333.
Смена зубов—497—498.
Смена рогов—524—526.
Смена функций—345, 347, 349, 352 , 354, 372 , 381, 384—394, 402 —404 , 409, 413, 417, 421—423, 435, 574, 583, 584.
Соответственные координации—442, 445. Специализация—136, 137, 192, 196, 296,
304—307, 309, 349, 581, 582.
Специализации у гимнофионов—284. » у двоякодышащих—240, 241, 244.
» у эласмобранхий 192, 196.
Сравнительно анатомический метод—104— 106.
Сравнительно анатомические ряды—104, 105.
Сравнительно эмбриологический метод— 106—108.
Стегозавры—303, 362.
Стегоцефалы—76, 77, 218, 219, 261, 283— 285, 329, 356, 375, 434.
— происхождение—244, 245.
Стенобазальные плавники—236—238, 240, 383*.
Стерлядь (см. Acipenser ruthenus)—188, 199, 201, 226, 227.
Стопоходящие животные—356—359.
Странствование или переселение клеток— 481, 482, 536, 586.
Страусы—ПО, 432
Стрептостилия—407.
Субституция органов—345, 347, 373—379, 402, 403, 413, 414, 417—420, 422, 423, 583, 584.
Субституция органов 1-го рода—375—378, 419.
Субституция органов 2-го рода—378, 379, 419.
Субституция функций—347, 394—406, 414, 417—420, 433, 575, 576, 582, 584.
Т
Талаттозавры—115, 288, 303.
Тапиры—105.
Тектонические корреляции—429.
Темп эволюции—264—265, 501, 532 —535, 590.
Теория бранхиомерии головы—143, 145, 146, 163—167.
Теория естественного подбора—71, 72, 82.
Теория морфобиологическая—253—449.
Теория монофилии —109, 110, 116—118, 123, 124, 128.
Теория направлений эволюции—267—343.
Теория полифилии—110—116.
Теория редукции органов—416—424, 538— 560.
Теория строения зародышевой плазмы—74.
Теория типов (принципов) филогенеза— 344—415.
Теория адаптивной эктогенетической эволюции—253—266 579, 580.
Теория филэмбриогенеза—453—537.
Териодонты—77.
Тероморфы—2S2.
Тетракленодоны—124.
Топографические корреляции (координации). —430, 440—445, 447, 470, 585.
Тригла—301 *, 492*, 493*
Тройной параллелизм—108.
Тюлени—118.
У
Унисериальный архиптеригий—236,237, 240-
Уменьшение числа функций—380, 381, 413, 414, 421—423, 582—584.
Усоногие—317, 318.
Утконос—77
Ф
Физиологическая субституция—346, 347, 379, 380, 402, 413, 414, 583.
Фиксация фаз движения конечностей у бесхвостых амфибий—363.
» » у млекопитающих—357—361.
» » у рептилий—357, 361—363.
Фиксация фаз движения плавника у Ceratodus—363—365.
Филетические корреляции (координации)— 124 , 263, 425—427, 430—448, 585.
Филогенез брюшных плавников—366—370.
» грудных плавников—236 , 237.
» плавательного пузыря—238—239.
» плечевого пояса млекопитающих
372 —373.
слуховых косточек—370—372, 437—439.
сочленений нижней челюсти с черепом—233—235.
» челюстной мускулатуры—435—437.
» яйцеклада у пчел—389, 390.
Филогенетическое древо—72, 73, 77, 79, 207—214, 216—218, 459.
Филогенетическое исследование—72—78.
— его задачи—79, 80.
— его методы—103—108.
Филогенетические ряды—104.
Функции: активация их—347, 406 —408, 414, 415, 576, 582,. 583.
— выпадение их—347, 366—373, 413, 577, 578, 582—584.
— иммобилизация их—347, 408, 409, 414, 415, 582, 583.
— интенсификация их—345, 347 —366, 412, 414, 574, 583.
— ослабление их—355.
— разделение их—347, 410, 411, 414, 582, 583.
— расширение их—345,347,381—384, 393, 394, 407, 413, 583.
— симиляция—347, 409, 410, 414, 577,
582, 583.
— смена их—345, 347, 349, 352 , 354, 372, 381, 384—394, 402—404, 409, 413, 417, 421—423, 574, 575 , 583, 584.
— субституция их—347, 394—406, 414, 417—420 , 433, 575, 576, 582 —584.
— уменьшение их числа 380, 381, 413, 414, 421—423, 582—584.
— фиксации их фаз—347, 356—366, 413, 574, 582, 583.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
60S
Функциональные корреляции—259, 261, * 265, 427—429, 431.
Функциональные рекапитуляции —477.
X
Хамелеоны—297, 304, 307
Характеристика главных групп Osteich-tyes—241—243, 284, 434.
Хвостатые амфибии (см. Urodela)—86, 218, 329, 434, 530, 531.
Химеры—211.
Хрящевые ганоиды (см. Chondrosteoidei)— 172, 182 , 207 , 209—212 , 214 , 216— 222
— несостоятельность гипотезы об их дегенеративной эволюции—222—
232.
— происхождение и систематическое положение—209—219.
— строение костного черепа—223—232.
— черты сходства с селахиями и элас-мобранхиями и отличия от кост-стных ганоидов, кистеперых и костистых рыб—219—221.
ц
Целесообразная реакция—85—88, 94—102.
Ценогенезы—106, 272 , 311—316, 331, 456, 457, 520, 522, 581
— классификация—314.
— определение—311, 312, 456.
— примеры—312—314.
Циклостомы (см. Круглоротые, Cyclostomata)—135, 138, 139, 141, 143, 149, 160, 164 — 168, 170 — 172, 175, 177, 180, 208, 214.
Ч
Челюстные рыбы—(см. Гнатостомы, Gna-thostomata)—141, 170.
Чередование различных направлений эволюции •—328—331.
Череп: двойное сочленение нижней челюсти с черепом—234—235.
— покровные костн черепа у Osteichthyes—197—207.
— происхождение костного черепа— 200—204, 230.
— развитие челюстей у протогнато-
стом—181 —185.
— строение висцерального черепа у
Cephalaspidae—158—159.
— » у Chondrichthyes selachoidei —192.
— » у Protocyclostomata—155.
— строение костного черепа у хрящевых ганоидов—221—232.
— строение осевого черепа у Chondrichthyes selachoidei—192.
— » у Protocraniata—139—140.
— строение черепа у змей—406.
— » у Holosteoidei primitivi—232 —235.
— » у эктобранхиат—177—178.
— » у энтобранхиат—148—149, 153.
— филогенез сочленений нижней челюсти с черепом—233—235.
— » слуховых косточек—370 — 372, 437—439.
Черепные позвоночные (см. Craniata)—132.
Черепахи—288, 296, 357, 375, 464, 554— 556, 576.
Э
Эволюционное учение—71—80.
— задачи—72, 79, 80.
— влияние на развитие различных дисциплин—72—76 , 78.
Эволюция и приспособление—253—255.
Эволюция (филогенез):
Acrania primitiva —132—135, 246, 329.
аннелид—325, 328.
безногих ящериц—395—398.
Bi lateralia—324—325.
Branchiostomatidae—136, 246, 329. Coelenterata (кишечнополостных)— 323, 324, 329.
Chondrychthyes selachoidei—191, 196, 249.
Chondrosteoidei (хрящевых ганоидов)—218—221
Deuterostomia—326—327
Dipnoi (двоякодышащих)—240— 241, 243—244, 245, 250.
Echinodermata (иглокожих)—326— 327.
Ectobranchiata—172—180, 248.
Elasmobranchii—279, 280, 309—310.
Entobranchiata —148—153, 248.
Equidae—124—127, 380, 421—422,.
464.
Holosteoidei actinopterygii—238 — 243, 250, 329.
Holosteoidei crossopterygii—238 — 243, 250, 329.
Holosteoidei primitivi —232—237, 250.
низших позвоночных—131—250.
оболочников (Tunicata)—327 —328. онтогенеза Metazoa—561—572, 590, 591.'
Osteichthyes—197—207, 249.
Ostracodermi—156—161.
Protocraniata—138—148, 246, 329.
Protocyclostomata—153—156, 248. Protognathostomata—180—191, 248, Protostomia—325, 326.
Protozoa—322 —323, 328.
Quadrupeda—243—246, 329. человека—77—78.
членистоног. (Articulata)—325 —326. фигйгтических корреляций—425— 448.
Эврибазальные плавники —192, 236—238, 381—383*
Эктобранхиаты (см. Ectobranchiata)—172, 174—181, 248, 277, 278.
— мочеполовая система —179.
— органы дыхания—172 —177.
— пассивное питание—177.
— строение черепа—177—178.
Эктогенетическая адаптивная эволюция— 253—266 , 579, 580.
Эктопаразиты—319. '
Эктосоматические органы—258—266, 425, 431, 432, 434, 437, 439, 440, 447— 449, 579, 580, 584, 585.
— воздействие изменяющейся внешней среды—260 —264.
— » константной внешней среды—258—• 260.
— определение—258.
002
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Эксцессивные органы—255, 483—486. Эласмобранхий (см. Elasmobranchii)—138, 145, 162, 182, 184, 188, 191—197, 206, 207, 212, 218—221, 279, 280, 309 , 391, 408.
Эмбриональные вариации нерегулирую-шиеся—520, 521, 589.
» » регулирующиеся—520, 589. Эмбриональная субститу ни я —378. Эктобранхиаты (см. Entobranchiata)—81, 151,' 153, 154, 156, 157, 169—171, 173, 177—179, 246, 277—278.
— органы дыхания—148—149.
— органы чувств—150—151.
— скелет—149, 153.
Эктопаразиты—318, 319.
Энтосоматические опганы—253, 259, #32— 266, 425, 431, 433, 434 , 437, 439, 440, 447—449, 579, 580, 584 , 585. — взаимодействие с актосоматическими органами (см. Функциональные корреляции)—259, 262, 263, 425, 431—440, 447,
Эукорреляции—427, 428, 429.
Я
Ящерицы (см. Lacertilia)—297, 396, 408, 409, 539, 542—544, 548.
Ящерицы безногие—394, 3^5, 397, 398, 401, 416, 543, 548, 552, 554.
INDEX1
A
Ablepharus—395*
Acanthias vulgaris —175* 235.
Acanthobdellidae—377
Acanthodii (Acanthoidei) 192, 249.
Acanthopterygii—93, 515, 516, 520. 521
Acipenseridae (см. Осетровые) —199—202, 207, 215, 218—222, 226, 230—233, 249, 367, 368, 435, 443, 518, 529, 558.
Acipenser nudiventris—220.
Acipenser giildenstadti—220, 222,223*, 226, 227*, 231
Acipenser ruthenus (см. Стерлядь)—187*, 188*, 190* 198*—202, 220, 222, 224, 226—228*, 231, 233*, 241, 243, 369*, 375*, 436*, 443, 506, 518*, 557
Acipenser stellatus—199*, 200, 222—224*, 227*, 231
Acrania primitiva —132 —135, 140, 142, 146, 147, 246, 276, 329.
Actinistia—215.
Actinopterygii—209, 215, 217, 220, 232, 236, 238, 239, 241, 243, 250.
Aculeata—389.
Aepiornis—110.
Aeschna—390.
Aga ma—395*.
Agnathi—163.
Alytes obstetricans—86.
Amblystoma sp.—86.
Amia calva —187*, 188*, 203*, 204, 207, 222, 230, 232—235, 237*, 241, 314*, 367*, 443, 519*, 529, 578.
Amioidei—241, 243, 250.
Ammocoetes (см. Пескоройка)—141, 144*, 145, 149* —153, 247
Amoeba proteus—348*
Amphioxus lanceolatus (см. Ланцетник)— 132, 134, 136, 140, 246.
Amphisbenidae—333, 416.
Amphitima—353, 410.
Anableps tetraphtalmus—302*.
Anamnia—134.
Anaspida—161—163, 167, 170, 191
Ananthrodira—217
Anchitherium—125, 126.
Ancylopsetta quadrocellata—96.
Anguis fragilis—396—398 , 401, 410 , 542, 544*
Annelidae—324.
Anseriformes—111.
Antennata—325, 326, 329.
Antiarchi—76, 162, 217, 218.
Anthozoa—317, 324.
Anthracosauria—218.
Anthropoidea—499.
Anthropomorpha—499.
Anura (см. Бесхвостые амфибии) —118, 284, 463, 501
Apoda—118.
Arachnoidea—325, 329.
Arachnomorpha—325, 326.
Archaehippus—125, 126.
Ardeiformes—111
Arthrodira—76, 162, 192,217,218.
Arthroleptis—285.
Arthropoda—324, 326.
Artiodactyla—366.
Articulata—325.
Arvicolae—121
Ascalabotes fascicularis (см. Гекконы)—441, 465*, 482, 513*, 514*, 516, 544*—546*, 548, 549, 551—553.
Asteresthes stomias—96.
Atherina hepsetus—485*—488, 495, 515*, 529.
Atlantosauridae—361
В
Balaenoptera musculus—322* Balanoglossus—326, 567*—569* Barbus fluviatilis—468, 497, 498. Batrachomorpha—218.
Batoidei (см. Скаты)—194, 249, 416.
Bdellostomatidae—140, 143. 154, 155,
168.
Bdellostoma stouti —140, 143, 145, 146, 155, 168.
Belone acus —186, 484*—488, 494, 495, 500—504, 507—509.
Belonidae—495 , 498.
Belonorhynchidae—215.
Benneciaspis —161
Bilateralia—323—326, 329.
Bipinnaria—326.
Birgeria—368.
Boa constrictor—406*.
Bobasatraniidae—215.
Boidae—576.
Bombinator pachipus—463*.
Boneilia—120.
Boreaspis—161.
Bothae maximus—300*.
1 Звездочкой обозначены номера тех страниц, где имеется рисунок.
604
INDEX
Bradypus tridactylus—305*.
Brachiopoda—322, 324.
Branchiostomatidae—132, 133, 135—138, 140, 147, 246, 326, 327, 329.
C
Calamoichthys—199, 203, 239.
Callima paralecta—299*
Calotes—357
Cani.s—96.
Canadia spiriosa—325.
Carcinus maenas—318*
Caryophyllaeus mutabilis—318*
Carnivora pinnipedia—421.
Catarrhyna—499.
Catopteridae—215.
Cephalaspidae—140, 156—162, 165, 167— 169, 207.
Cephalaspis murchisoni —157*.
Cephalaspidomorphi—163, 167
Ceratodus—187, 206, 236, 237, 283, 364*, 365, 382, 409.
Certhia—121.
Chaetognatha—326.
Chaetopoda—325.
Chalcides sepoides—397, 398, 540 —542 , 552, 553.
Chalcides ocellatus—397, 398.
Chamaeleo vulgaris (см. Хамелеон)—307*.
Chelone—296.
Chelydra serpentina—555.
Chimaera monstrosa—408*
Choronectes minimus—386*, 387.
Chironomus plumosus—274*
Chlamydosaurus—357
Chlamydoselachi i—191
Chlamydoselachus—192, 193*, 195, 206, 207, 220, 233, 234*, 235, 249.
Choanata—218.
Choloepus—305.
Chordata—132, 246, 326.
Chondrichthyes selachoidei—191 —194, 197, 207, 249, 280, 309, 435.
Chrondostei (см. Хрящевые ганоиды)—207, 209, 214, 219, 221, 232 , 233, 235, 236, 238, 239, 437, 497
Chondrosteoidei (см. Хрящевые ганоиды)— 91, 187, 191, 207, 212, 217—222, 233, 234, 235, 237,239, 241, 243, 249, 281.
Chondrosteus—209, 232, 241.
Chrysochloris—118, 260, 333.
Cladistia—215.
Cladodontidae—191, 207, 234, 249.
Cladodus—193, 194*, 206, 220, 235, 236, 249, 366, 367.
Cladocera—92.
Clemmis—296.
Cobitis—529.
Coelacantini (Coelacanthidae)—210,241,243.
Coelenterata(CM. Кишечнополостные)—323, 324, 572
Coelolepidae—169.
Colymbriformes—111.
Compsognathus—362.
Condylarthra—124.
Condylura—302.
Cotorhynus maximus—404*.
Craniata—133, 134, 246.
Creodontia—357.
Crossopterygii (см. Кистепсрые)—91, 131, 200, 207. 212, 215, 219, 220, 231-233, 235—239,241,243—245, 365, 367, 382, 443.
Crotalus—406.
Crustacea—325, 326.
Crypturiformes—110.
Ctenomis—302
Ctenophora1—324, 328.
Cyclostomata (см. Круглоротые) —132 139, 141, 145, 148 — 151, 159—170, 173, ISO, 246.
Cynognathus—371, 438.
Cyprinidae—529, 578.
Cyprinus Carpio—497.
D
Decapoda—326.
Deltatherium—77
Dermochelidae—296.
Deuterostomia—183, 325, 326.
Dicynodon—375.
Difflugia—348, 349*.
Dinornis—110.
Diplopterus—244.
Dipleurula—326.
Dipnoi (cm. Dipneusta, Дипнои, Двоякодышащие)—78, 220, 232, 236, 238, 243— 245 250 409
Dipneusta—207, 219, 239, 243—245, 250, 408, 438.
Dipterus—244.
Dipteridae—245.
Dolichosauria—297.
Dorypteridae—215.
Draco volans—357
Drepanaspidae—168.
Dryopitheci—77.
E
Echinodermata (см. Иглокожие)—324 , 326, 327.
Ectobranchiata (см. Эктобранхиаты) —147, 172 — 180, 248, 338.
Edentata—306.
Elasmobranchii (см. Эласмобранхий)—139, 186, 191 — 196, 217, 233, 236, 248, 309, 437.
Emetopias californicus—386*.
Emys—296, 555.
Enteropneusta—140, 326.
Entobranchiata—(см. Энтобранхиаты) —147 — 151, 153, 246—248, 329, 338.
Eohippus—124, 125, 126, 533.
Enihippus—125, 126, 533.
Equidae—118, 124, 127, 128, 400, 422
Equus—125, 127, 464.
Esox lucius—369*
Euglypha—348, 349*
Eumeces schneideri—396—398, 539, 540*.
Euryalae—377
Eustenoptheron—240, 367*, 368.
Eutetrapoda—218.
Exocoetus rondoletti—382*.
F
Felidae—360, 389.
INDEX
605
G
Galliformes—110.
Gallus domesticus—442*, 454.
Ganoidei—209.
Gecko verticellatus—303*.
Gnathobdellidae—377
Gnathostomata—145, 152, ISO. 181, 218.
Gnathostomata apterygiata—191.
Gnathostomi—163.
Gobiidae—298, 301.
Gobio gobio—497
Gobius capito—300*, 515, 516*, 520, 521.
Gruiformes—110, 111
Gymnophiona—284, 405, 416, 554.
H
Halicoridae—387.
Haplistia—215.
Hem ichordata—567.
Hemichromis—313*.
Hemiramphus—484*, 486, 495, 501, 503, 507, 508.
Heptanchus—140, 155, 188*, 195.
Hesperornis—110.
Hesperornitidae—111.
Heterostraci—162, 163, 168—170.
Hexanchus—155, 176, 182, 195, 408*.
Hypparion—125, 464.
Hippidium—125.
Hyppocampus—490, 491*
Hyppoglossoides platesoides—96.
Hippoglossus hippoglossus—96.
Hoelaspis—161
Holocephali (см. Химеры)—176, 192, 194— 196, 206, 240, 241, 249, 309, 408, 409.
Holostei (см. Костные ганоиды)—91, 207, 209 , 219, 232 —239 , 368, 372 , 443, 518, 529 577.
Holosteoidei—217, 219, 220,221,232—240, 243, 244, 250.
Holosteoidei mihi—233.
Holosteoidei actinopterygii—212, 238—243, 250, 282, 329.
Holosteoidei crossopterygii—212, 238—243, 250, 232, 283, 284, 329.
Holosteoidei primitivi—212, 218, 219, 250, 281, 282.
Hominidae—77, 432
Homo primigenius—499.
Homo sapiens—499.
Huso huso(CM. Белуга)—222,225*—229,443.
Hybodus—366, 367*
Hydrozoa—317, 323, 324.
Hyla arborea—428.
Hylidae—285.
Hylodes martiniensis—285.
Hymenoptera aculeata—389.
Hypohippus—125.
Hyracotheria—124.
Hyrax—105.
I
Ichthyobdellidae—377.
Ichthyophis glutinosus—313*.
Ichthyosaurus—354, 355*.
Ichthyostegalia—218.
Ichthyotomi —191, 237.
Iguanodon —127
I nsectivora—357.
К
Kiaeraspis—158*—161 *.
Kalohippus—125.
L
Lacerta—446, 509.
Lacerti'lia (см. Ящерицы)—261, 297, 407 481, 575.
Laemargus borealis—176*, 182, 220, 234, 235.
Lamellibranchiata—319, 381.
Lepadogaster—301.
Lepas—317*.
Lepidosteoidei—241, 243, 250.
Lepidosteus—199, 203, 204, 207, 219, 222, 230, 232, 233, 329.
Lepus—120.
Lophiidae—298.
Lophius piscatorius (см. Морской чорт)— 299*, 300*, 486*, 490*, 492, 495—497, 500, 501, 515, 520.
Lophobranchii—490, 491.
Loricata—114.
Lutra—421.
Lutrinae—388.
Lygosoma punctulatum—397, 398, 540, 541 *, 542, 552.
M
Madreporaria—317.
Manatidae—387
Manatus inunguis—421.
Marsijfobranchii—217.
Marsupialia—373.
Menobranchus—531.
Menopoma—353, 410.
Meryhippus—125, 464.
Mesohippus—125, 464.
Metazoa—322, 323, 351, 353, 356, 561—564, 569—572, 583, 590, 597.
Miohippus —125, 464.
Monodeiphia—373.
Monodon—91
Moniezia—84.
Monotremata—373.
Mososauria—297.’
Mososattrus—354, 355, 512.
Mui titubercu lata—77.
Mustelus —182, 220.
Myliobatis californicus—391, 403, 404*
Myxinoidea (см. Миксины)—149, 150, 153, 156, 162, 163.
N
Nasus socialis—303*
Neoceratodus—240, 243.
Neohipparion—125.
Neohippus—125.
Notharctus—77.
Notidanidae—191, 220, 249.
Notoryctes—333.
Nycticebus—339.
О
Onchorhynchus keta—507.
Onirodes glomerosus—340*.
Ophidia—297, 410, 575.
Ophiocodon mirus—464, 465*, 492
Ophiodes—397, 398 , 542*, 543, 551, 553.
INDEX
Ophiomorus tridactylus—397, 540—542, 552.
Ophionotus hexactis—379*.
Ophisaurus apus (см. Желтопузик)—395-— 401, 407, 410, 446, 543*—546, 548, 550*, 551*.
Opisthoproctus soleatus—339*.
Orohippus—124, 126.
Osmerus—578.
Ospiidae—215
Osteichthyes —155, 185, 186, 191—194, 197— 207, 214, 218—221, 236, 243, 279, 309, 352, 353, 378, 506, 534.
Osteichthyes primitivi—197, 198, 200—206, 213, 219, 221, 222, 235, 238, 239, 249, 250, 279—281, 329, 367
Osteolepidoti—241, 243, 244.
Osteostraci—76, 132, 138, 139, 141, 143, 145, 148, 151, 153, 155, 157, 159, 161 — 163, 167—170, 177, 180, 197, 217.
Ostracodermi—132, 148, 153, 156, 157, 161— 163, 165, 168—173, 181, 247, 278, 279.
P
Palaeaspidae—165, 167, 170.
Palaeohatteria—406, 407, 512.
Palaeiguana—297
Palaeoniscidae (см. Палеонисциды)—207, 215 , 219 , 233, 235, 236 , 368.
Palaeoniscoidei —191, 206 , 207, 219, 232, 236, 238, 239, 243, 250.
Palaeospondyloidea—162, 163.
Palaeostraci—325.
Pandorina morum—562—565, 571.*
Parahippus —125.
Paralichthys californicus—96.
Parus—120, 121
Parasemionotidae—215.
Pelobates fuscus—97, 407, 463*.
Percesoces—486.
Perennibranchiata—530, 531.
Peripatus—325.
Perleididae—215.
Petromyzon—134, 150, 151 *
Petromyzontes (см. Миноги)—156, 248.
Petromyzontia—162, 163.
Peytoia nathorsti—324.
Phacochoerus—84.
Phaenacodontidae—54.
Phaneropleuron—244.
Phanerorhynchidae—215.
Phoenicurus phoenicurus—121.
Phoenicurus ochrurus—121 Pholidopleuridae—215. Phrynocephalus—357.
Phyllium scythe—299*
Phylopoda—325.
Phyllopteryx equus—299.
Pisces abdominales—515.
Pithecanthropus—78.
Placodermi—207.
Plagiostomi —191, 194, 195.
Planorbis multiformis—118.
Platecarpus—512.
Platysomatidae (Platysomidae)—215.
Plesiohippus —125.
Pleuracan thodei (Pleuracanthodii)—191, 192, 194, 196 , 249.
Pleuracanthus—188*, 220 , 236.
Pleuronectus flesus—87, 93, 416.
Pleuronectus platessa—87
Pleuronectidae (см. Камбалы)—298, 416.
Pleuropterygii —191, 194, 237, 249.
Pliohippus —125.
Polacanthus —114, 115.
Polyodon—209, 222, 229, 230*, 231*
Polyodontidae—207, 215, 2 18, 232, 233, 248.
Polypteridae—243, 367, 368.
Polypterini—217, 241, 243.
Polypterus —187*, 199, 203, 204, 207, 210, 232, 236, 237*, 239, 240, 283, 375, 382.
Pristiophorus—391, 437.
Pristis antiquorum—309'.
Pristiurus melanostomus—173*, 178*, 183*, 184, 375*, 391.
Procolophon trigoniceps—464, 492.
Proganoides—207, 211.
Propliopitheci—77
Proselachii —143.
Proteus anguineus—86, 531.
Protocraniata (см. Протокраниаты)—131 — 132, 138—148, 246, 248.
Protocyclostomata (см. Протоциклостомы) — 153, 154, 169, 171, 248, 329.
Protognathostomata (см. Первичные челюстные)—180, 181, 191, 248, 435.
Protognathostomata apterygiata—191.
Protohippus—125.
Protopterus—283.
Protostomia—325 , 326.
Prototheria—77
Protozoa—322 , 323 , 348 , 356, 561—554, 566, 572, 590, 591
Pseudoscaphirhvnchus kaufmanni—176, 221, 222, 226, 229, 231,234*, 443.
Pteraspidae—169.
Pteraspidomorphi —163, 167.
Pteraspis—158*
Pythonomorpha—512.
Q
Quadrupeda—132, 155, 212, 237—239,241, 243—245, 249, 250, 283, 284, 329, 363, 365, 517, 577
R
Rana arvalis—97, 363.
Rana esculenta—354*.
Rana fusca—97, 428.
Raja—391, 403, 469.
Raja batis—433*
Ratitae—110, 111.
Reptiliomorpha—218.
Rhina squatina—437*.
Rhinobatis productus—381*.
Rhinoderma darwini—285.
Rhinoceros—105.
Rhipidistia—215.
S
Sacculina carcini—318*
Sagitta—326.
Salamandra maculosa—354*, 457, 468.
Sarcophaga carnaria—274*
Sargus vulgaris—507 , 515 , 516*, 521.
Saurichthys—216, 368.
Saurichtyidae—215, 222, 231, 235.
Sauripterus—76, 236, 237, 240, 244, 245, 283, 365.
Salmonidac—515.
INDEX
Salmo truta—515*, 578.
Schizopoda—324 , 385.
Scyncus officinalis—396 —398.
Scolopax rusticola—120.
Scombresocidae—485, 503, 508.
Scombresox—490, 501, 503, 507, 508.
Scyllium—187*, 220.
Scymnus lichia—195*.
Scyphozoa—324.
Selachii (см. Селахии)—236, 238.
Selachoidei—209.
Seps chalcides—395—398, 401, 534, 535, 544—553.'
Seps tridactylus—541, 542.
Siderone strigosa—299*.
Sinanthropus pekinensis—78.
Siredon mexicanus aut.—86, 531.
Siredon pisciformis—364.
Spalacidae—118.
Spalax—260.
Spatularia—207.
Sphargidae—296.
Sphargys—296.
Sphenodon—286, 406*, 407, 512.
Sphyrna zygaena—309.
Spinaceidae—220, 321.
Spinax—235.
Spongia—323.
Spumelaria spheroidea—323*.
Squaloidei—194, 195, 249.
Stanosphaera—323 *
Stegocephalia (см. Стегоцефалы)—244.
Stegophiura nodosa—379*.
Stereosternum—464, 492.
Struthiomimus—362.
Sturiones—207, 211
Syngnathus—490.
T
Tarsius—339.
Tatusia—371
Taumatosaurus—355 *
Teleostei (см. Костистые рыбы)—204, 212, 219, 230—232, 235—236, 238, 239, 241, 243, 250, 280, 309, 367, 372, 443, 488, 518, 521, 529, 577.
Teleosteidei—209.
Teleostorni—209, 215.
Tetraclaenodon—124.’
Tetraclaenodontidae—126.
Tetrapoda (cm. Quadrupeda)—217, 243—-244, 283.
Testudo graeca—296.
Thelodus—169, 180.
Therocephalia—128, 439.
Theropoda—362.
Theriodontia—289.
Thyphlosaurus—397, 543.
Tremataspis—158*
Tremataspidae—161
Trematops milleri—463*.
Triassochelis dux—296.
Triceratops—127.
Triglidae—498.
Trigla nigripunctata (см. Тригла)—492 *fc 493*.
Trimerorachis—244.
Tropidonotus natrix—513* 514*, 533.
Trygon—262 , 391, 403.
Tunicata (см. Оболочники)—318 , 327 , 328»
Typhlopidae—333.
Typhlosaurus—397, 543.
U
Urodela (см. Хвостатые амфибии)—244, 284.
405, 530.
Ursus spelaeus—78.
Vipera—395*.
Volvox—472, 563, 564*, 565, 571.
Vorticella—561.
W Worthenella Cambria—325.
X Xistreuris liolepis—95.
Z Zalambdalestes—77.
ОГЛАВЛЕНИЕ
Стр.
От редакции......................................................... 3
Алексей Николаевич Северцов (биографический очерк).................. 9
Предисловие А. Н. Северцова ....................................... 55
Список трудов А. Н. Северцова и его учеников....................... 59
Часть I
ВВЕДЕНИЕ
Глава I. Эволюционное учение после Дарвина и задачи филогенетического исследования............................................................. 71
Глава II. Неоламаркизм................................................... 81
Аутогенез и эктогенез.............................................. 81
Ламаркизм и неоламаркизм .......................................... 82
Принцип целесообразной реакции..................................... 85
Несостоятельность принципа упражнения для объяснения прогрессивной эволюции . 88
Несостоятельность принципа неупражнения для объяснения регрессивной эволюции ... 92
Критика принципа целесообразной реакции............................ 94
Глава III. Методы филогенетического исследования.......................... ЮЗ
Палеонтологический метод филогенетического исследования............. 103
Сравнительно анатомический метод филогенетического исследования . 104
Сравнительно эмбриологический метод филогенетического исследования 106
Глава IV Теория монофилетической эволюции................................. 109
Монофилетическая теория........................................... 109
Полифилетическая теория............................................. 110
Значение монофилетической теории.................................. 116
Адаптивные признаки крупных систематических групп................. 118
Адаптивные признаки мелких систематических групп.................. 119
Приспособительно-дивергентная эволюция............................ 120
Первичные приспособления и корреляции............................. 123
Выводы.............................................................. 128
Часть II
ЭВОЛЮЦИЯ НИЗШИХ ПОЗВОНОЧНЫХ
Глава V Эволюция Acrania и Protocraniata................................ 131
Acrania primitiva................................................. 132
Эволюция Branchiostomatidae....................................... 136
Эволюция Protocraniata ........................................... 138
Глава VI. Эволюция Entobranchiata....................................... 148
Строение Entobranchiata........................................... 148
Эволюция Protocyclostomata...................................... 153
Эволюция Ostracodermi............................................. 156
Взаимоотношения между Cyclostomata и Ostracodermi................. 161
Выводы ........................................................... 171
Глава VII. Эволюция предков челюстных позвоночных....................... 172
Эволюция характерных признаков Ectobranchiata..................... 172
Эволюция Protognathostomata....................................... 180
Эволюция Chondrichthyes selachoidei и Elasmobranchii.............. 191
Глава VIII. Эволюция Osteichthyes....................................... 197
Общая характеристика Osteichthyes................................. 197
Эволюция Chondrosteoidei и вопрос о классификации рыб............. 207
ОГЛАВЛЕНИЕ 609
О покровных костях черепа Chondrosteoidei ...............'........ 221
Эволюция Holosteoidei primitivi ................................ 232
Эволюция Holosteoidei actinopterygii и Holosteoidei crossopterygii . . . 238
Происхождение Quadrupeda..............................•........... 243
Общая сводка глав об эволюции низших позвоночных (V—VIII) .... 246
Часть Ill
ЗАКОНОМЕРНОСТИ ФИЛОГЕНЕЗА
Глава IX. Общий ход филогенетического развития.......................... 253
Эволюция й приспособление......................................... 253
Взаимоотношения между окружающей средой и эктосоматическими и энтосоматическими органами . . . ............................... 255
Влияние изменений внешней среды на эктосоматические и энтосомати-ческие органы..................................................... 260
Выводы............................................................ 265
Глава X. Основные направления филогенетического развития животных . . 267
Биологический прогресс и главные пути, которыми он достигается . . . 267
Морфофизиологический прогресс или ароморфоз . ..........• ... 273
Эволюция позвоночных с точки зрения теории ароморфоза .... ... 276
Выводы об ароморфозах............................................ 293
Идиоадаптации или частные приспособления.......................... 295
Специализация как частный случай идиоадаптации.................... 304
Соотношения между ароморфозами и идиоадаптациями.................. 308
Эмбриональные приспособления - или ценогенезы..................... 311
Морфофизиологический регресс или общая дегенерация................ 316
О направлениях филогенетического развития животного мира.......... 320
Взаимоотношения между различными направлениями эволюции животных ............................................................. 328
Изменения общих направлений эволюции и причины перехода животных от одного направления к другому .............................. 331
Выводы...................................................... ... 341
Глава XI. О принципах или типах филогенетических изменений органов Metazoa................................................................. 344
Введение..............................................• . ... 344
Принцип интенсификации функций.................................... 347
I Интенсификация функций клеток и тканей.......................... 348
II Интенсификация функций органов...............................' 351
Принцип фиксации фаз.............................................. 356
Принцип выпадения промежуточных функций........................... 366
Принцип субституции органов Н. Клейненберга...................... 373
Физиологическая субституция Д. М. Федотова........................ 379
Принцип уменьшения числа функций.................................. 380
Принцип расширения функций........................................ 381
Принцип смены функций А. Дорна.................................... 384
Смена функций, расширение функций и число возможных направлений в эволюции животных............................................... 392
Принцип субституции функций ..................................... 394
Принцип активации функций ........................................ 406
Переход от активной функции к пассивной (иммобилизация)........... 408
Симиляции функций................................................. 409
Принцип разделения органов и функций.............................. 410
Выводы........................................................... 411
Глава XII. Основы морфологической теории редукции органов .............. 416
Основные предпосылки редукции органов.............................. 416
Редукция вследствие изменения условий окружающей среды............. 417
. Редукция вследствие субституции................................... 417
Редукция вследствие смены функций ................................ 421
Редукция вследствие уменьшения числа функций ..................... 421
Выводы............................................................ 423
Глава XIII. Эволюция филетических корреляций или координаций органов . 425
Исторический обзор . . . •....................................... 425
Морфофизиологические координации................................... 431
Топографические координации........................................ 440
О способах изменения координационных соотношений ......... 445
Выводы............................................................. 447
Общая сводка глав IX—Х111 о закономерностях (Ьилогенеза............ 449
610
ОГЛАВЛЕНИЕ
Часть IV
О СООТНОШЕНИИ МЕЖДУ ОНТОГЕНЕЗОМ И ФИЛОГЕНЕЗОМ ЖИВОТНЫХ
Глава XIV. Теория филэмбриогенеза..................................... 453
Очерк истории биогенетического закона и его критики............ . 453
Рекапитуляция признаков органов и тканей ......................... 462
Очерк истории теории филэмбриогенеза.............................. 470
Филэмбриогенез и периоды жизни...........................• . . . . 478
Филэмбриогенез и способы (модусы), которыми он осуществляется . . . 480
Надставка конечных стадий развития или анаболия................... 483
Закон К. Э. Бэра............................................. . . 499
Модус девиации или отклонение (аберрация) направления эмбрионального развития................... •................................ 506
Архаллаксис или изменение первых зачатков......................... 510
Периодическое изменение хода развития органов..................... 523
Гетерохрония и филогенез........................................ 527
Темп эволюции при различных модусах филэмбриогенеза............... 532
Выводы..................................................... . 535
Глава XV. Филогенетическая редукция органов животных •................. 538
Типы редукции органов ............................................ 538
Рудиментация органов.............................................. 539
Полная дегенерация органов или афанизия........................... 555
Выводы............................................................ 559
Глава XVI. Эволюция онтогенеза Metazoa................................ 561
Глава XVII. Соотношения между принципами или типами эволюции и модусами филэмбриогенеза................................................. 573
Глава XVIII. Общие выводы........................................• . . 579
Морфобиологическая теория эволюции .... 579
Теория филэмбриогенеза............................................ 586
Именной указатель................................................ 592
Предметный указатель............................................ 595
Index.....................•.................................. . . 603
ОПЕЧАТКИ
Стр. Строка Напечатано Следует
19 3 св. А. Доном А. Дорном
24 15 св. „Переписку с друзьями" переписку с друзьями
128 4 св. приспособления приспособления к
142 подпись брахиомера бранхиомера
145 под рис. 6, стр. 13 сн. 16 сн. (А.р'о. — 1 А.рЬ.4; (A.pb. 1 — А.рЬ.4;
147 149 (нонпарель) подпись под рис. 10, стр. 2 сн. подпись эндотермальные энтодермальные
354 под рис. 12 1 св. медиально что увидим медиально то увидим
371 подпись под рис. 115, стр. 5 сн. 10 сн. 4 и 8 сн. 1 сн. 2 св. nd. морфологически ему уподобляясь личинок личинки личинки hd. морфологически ему не уподобляясь молоди молодь молодь
(Морфологические закономерности
Алексей Николаевич Северцов
Николай Алексеевич Северцов с сыном Алексеем Николаевичем
A. H. Северцов в студенческие годы
А. Н. Северцов в 1891 г.
А. Н. Северцов в бытность профессором в Киеве
A. H. Северцов, профессор Московского университета, в 1’928 г.
A. H. Северцов в 1935 г.