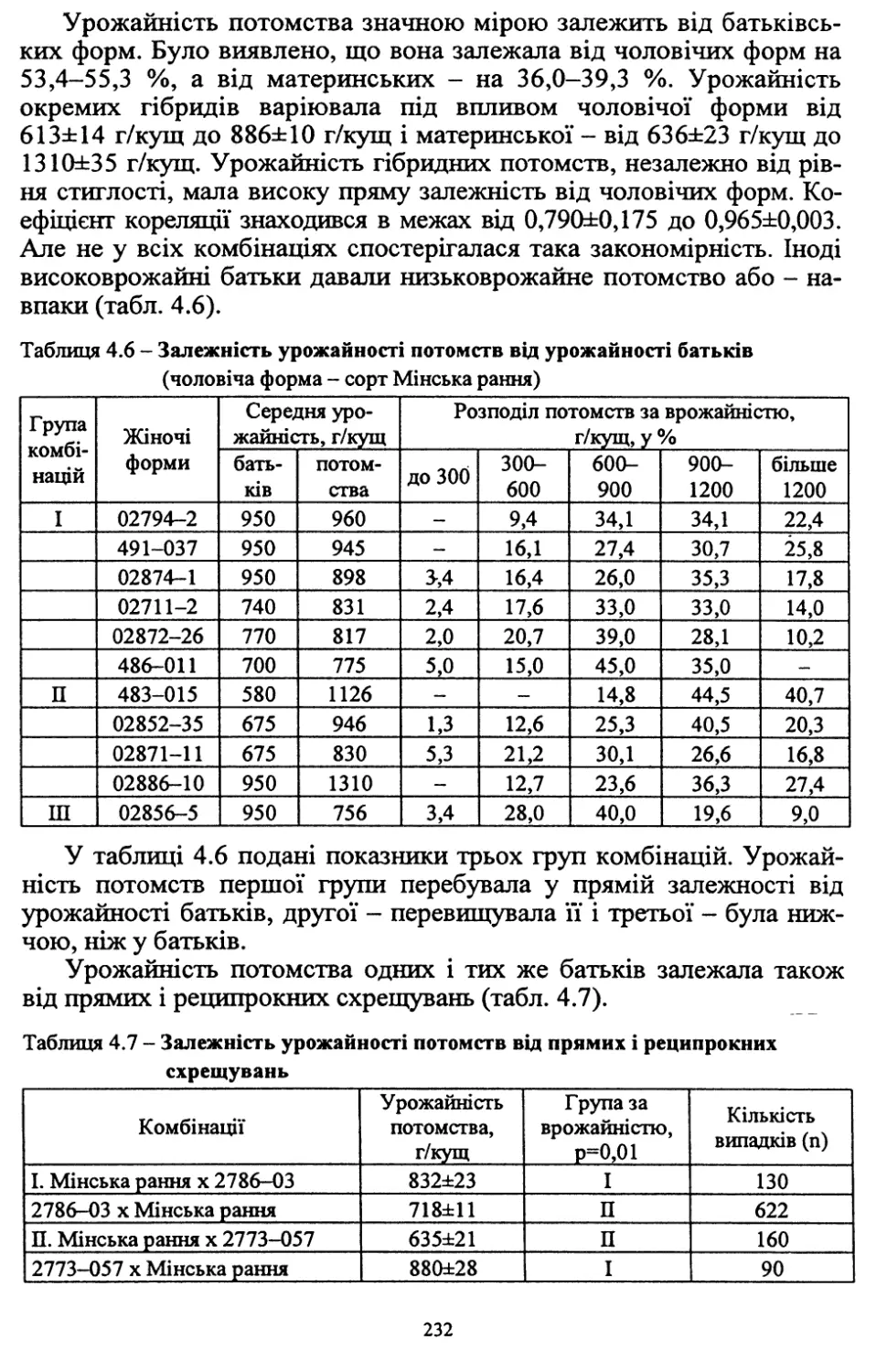
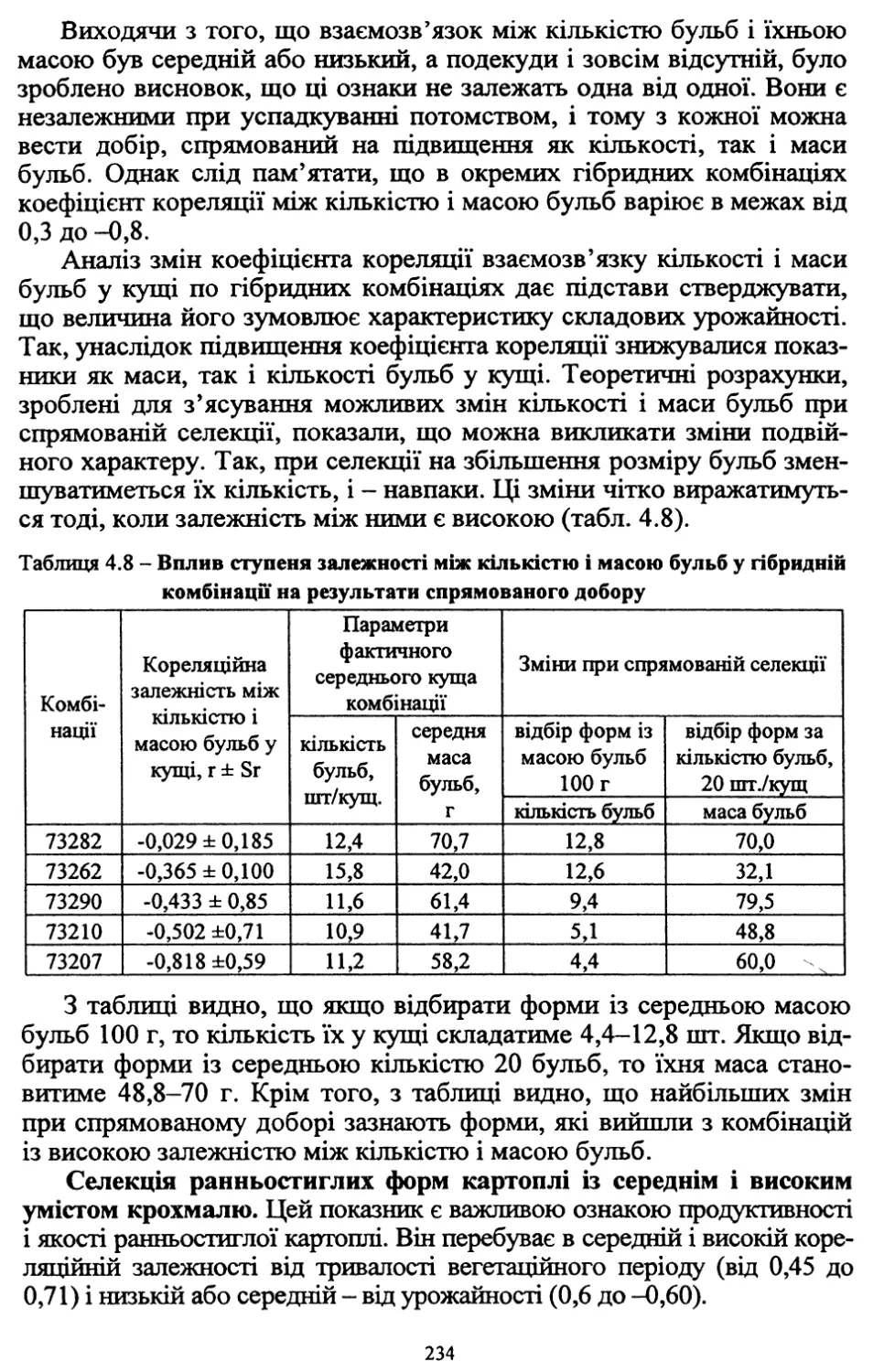
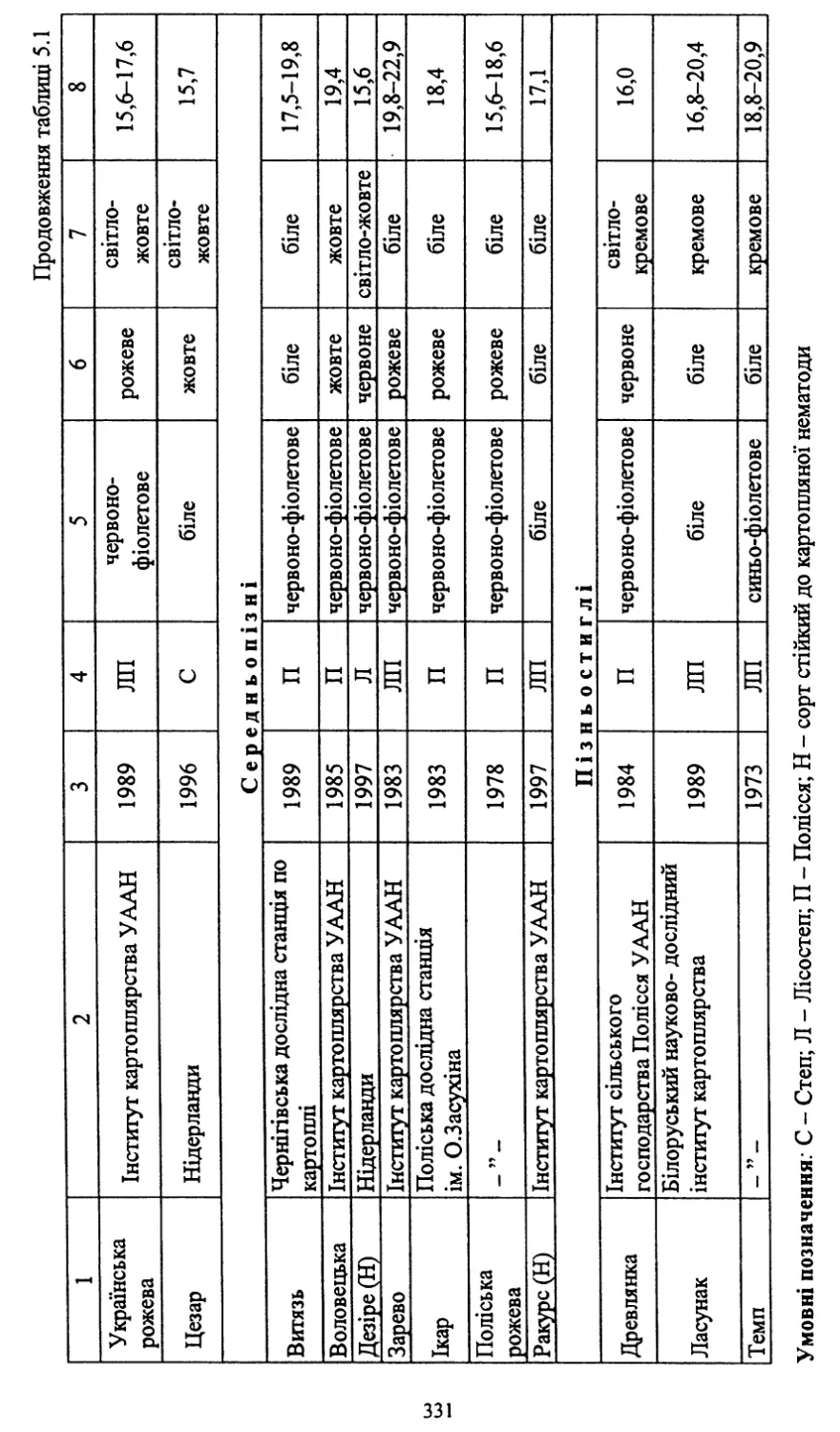
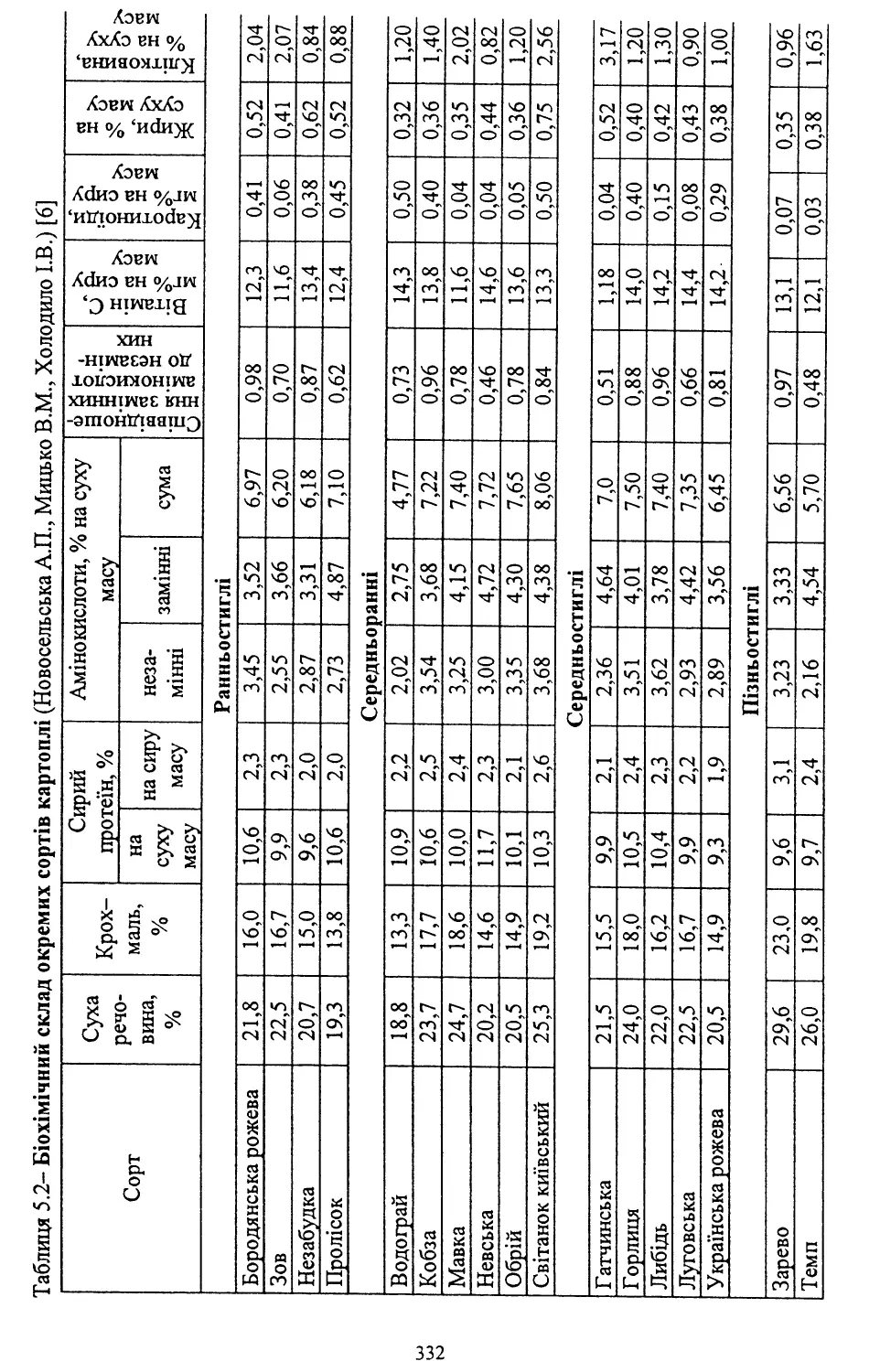
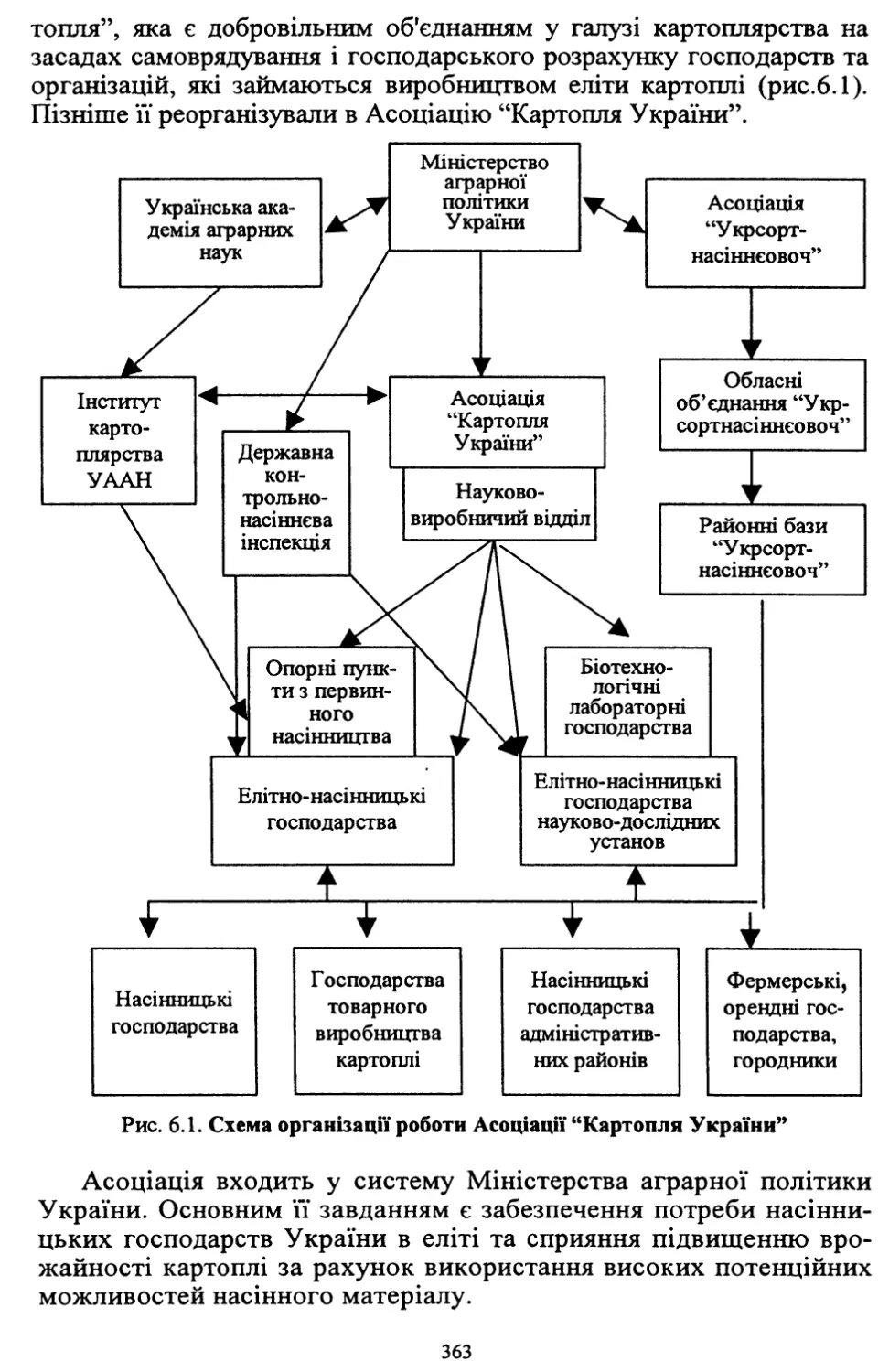
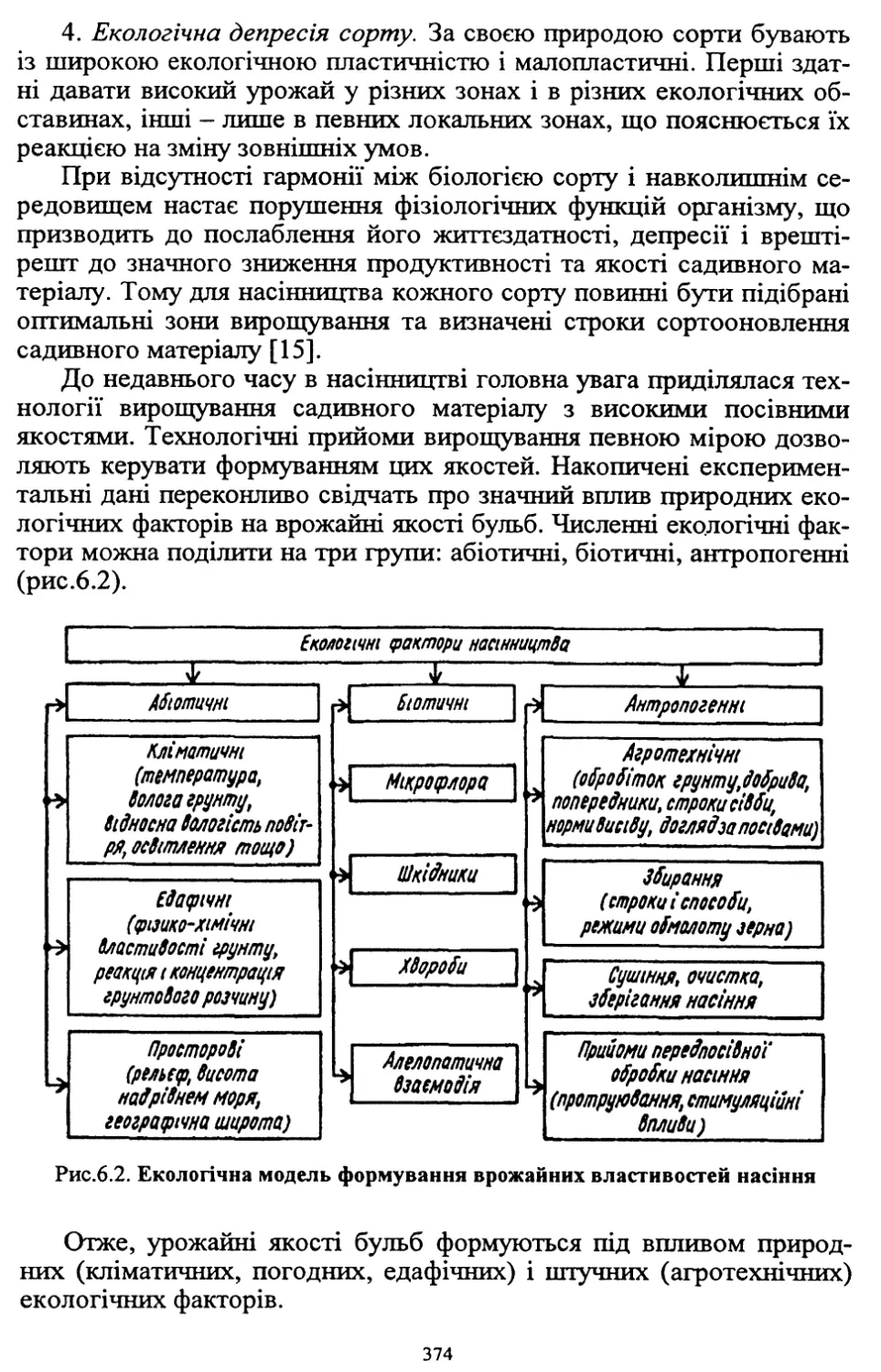
/

















































































































































































































































































































































































































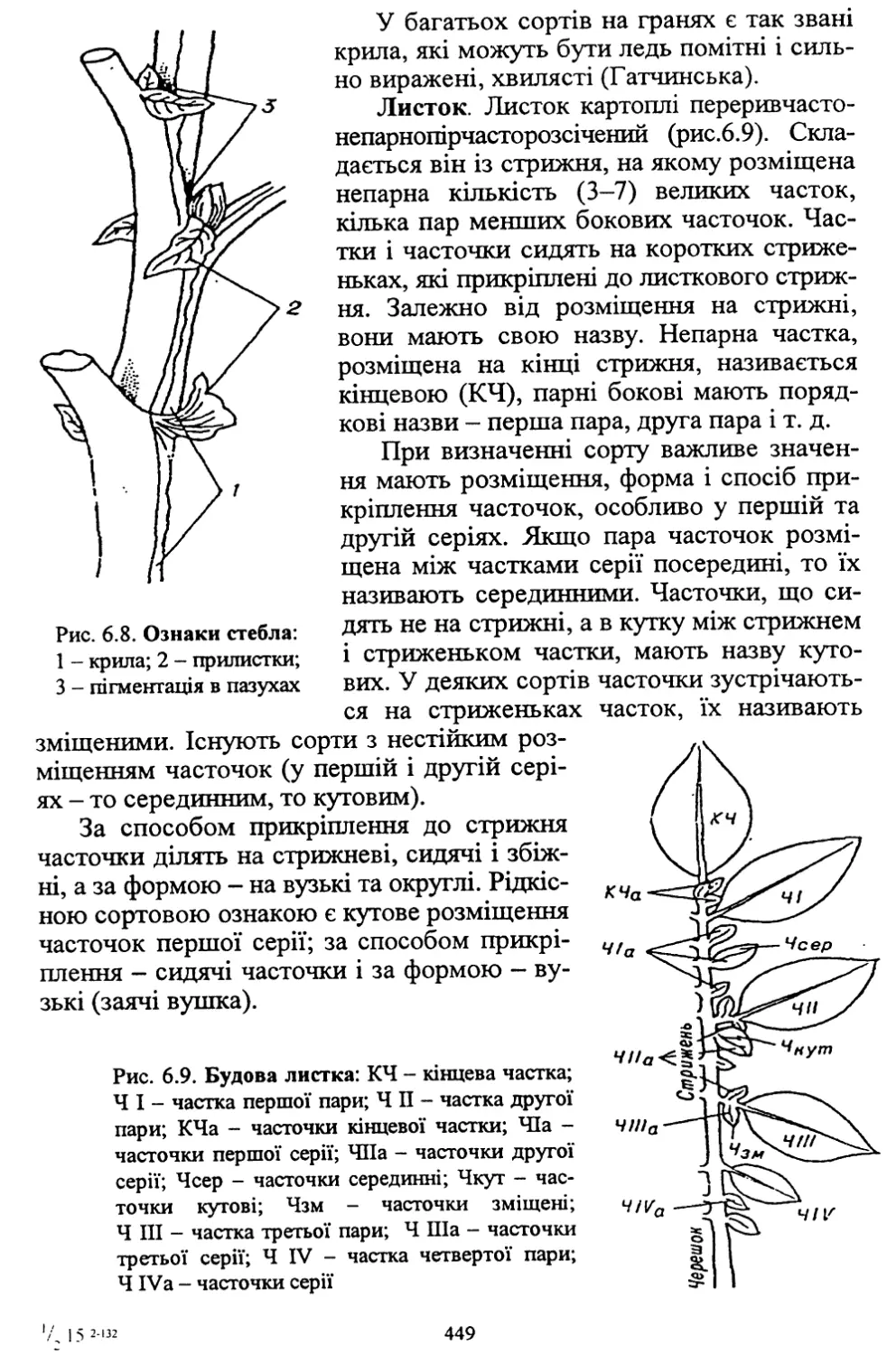
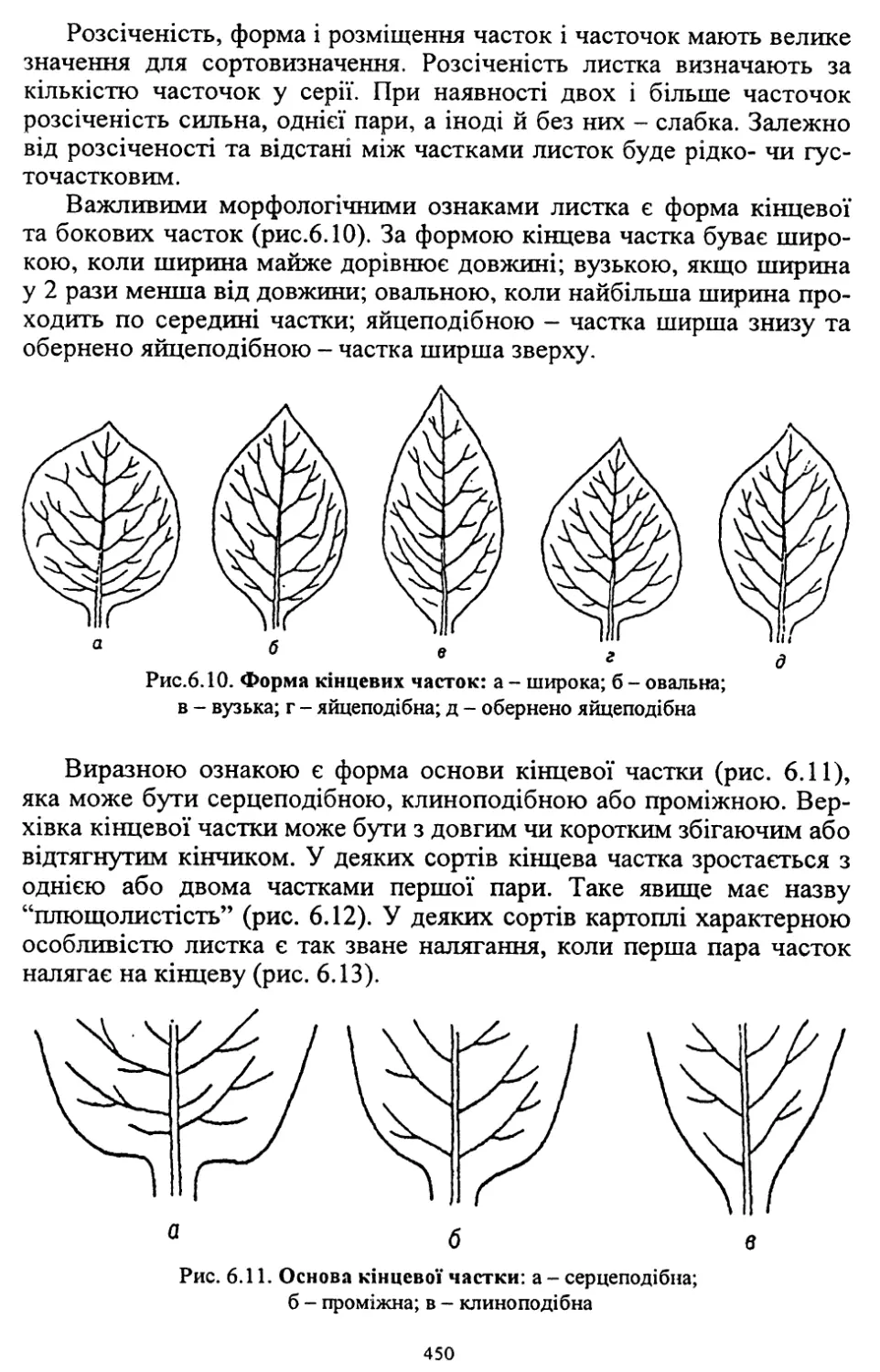
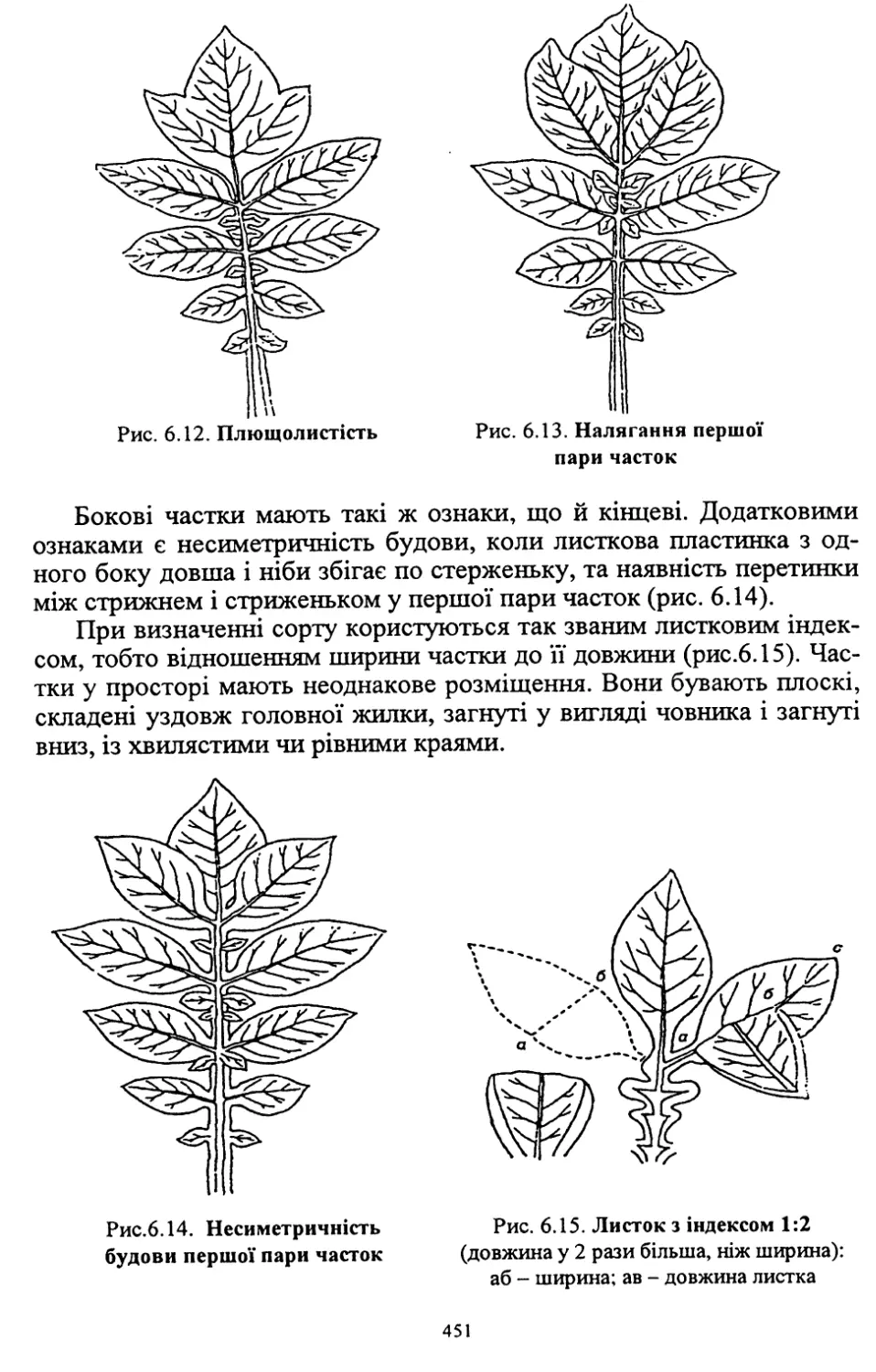
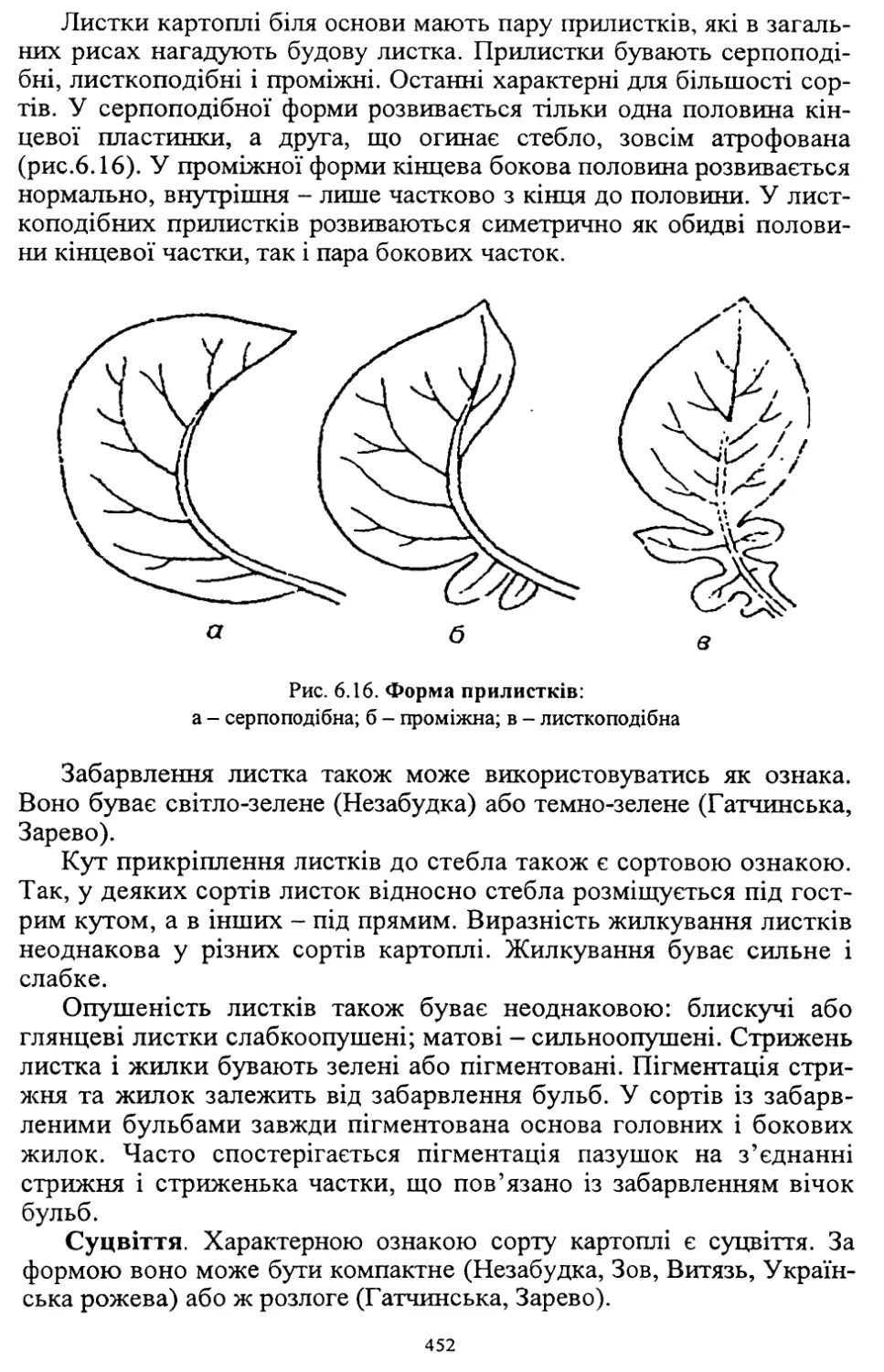











































































Author: Кононученко В.В. Молоцький М.Я.
Tags: монографія картопля аграрна політика видавництво біла церква картопляри урожаї бульби розвиток картоплярства
ISBN: 966-7417-32-8
Year: 2002




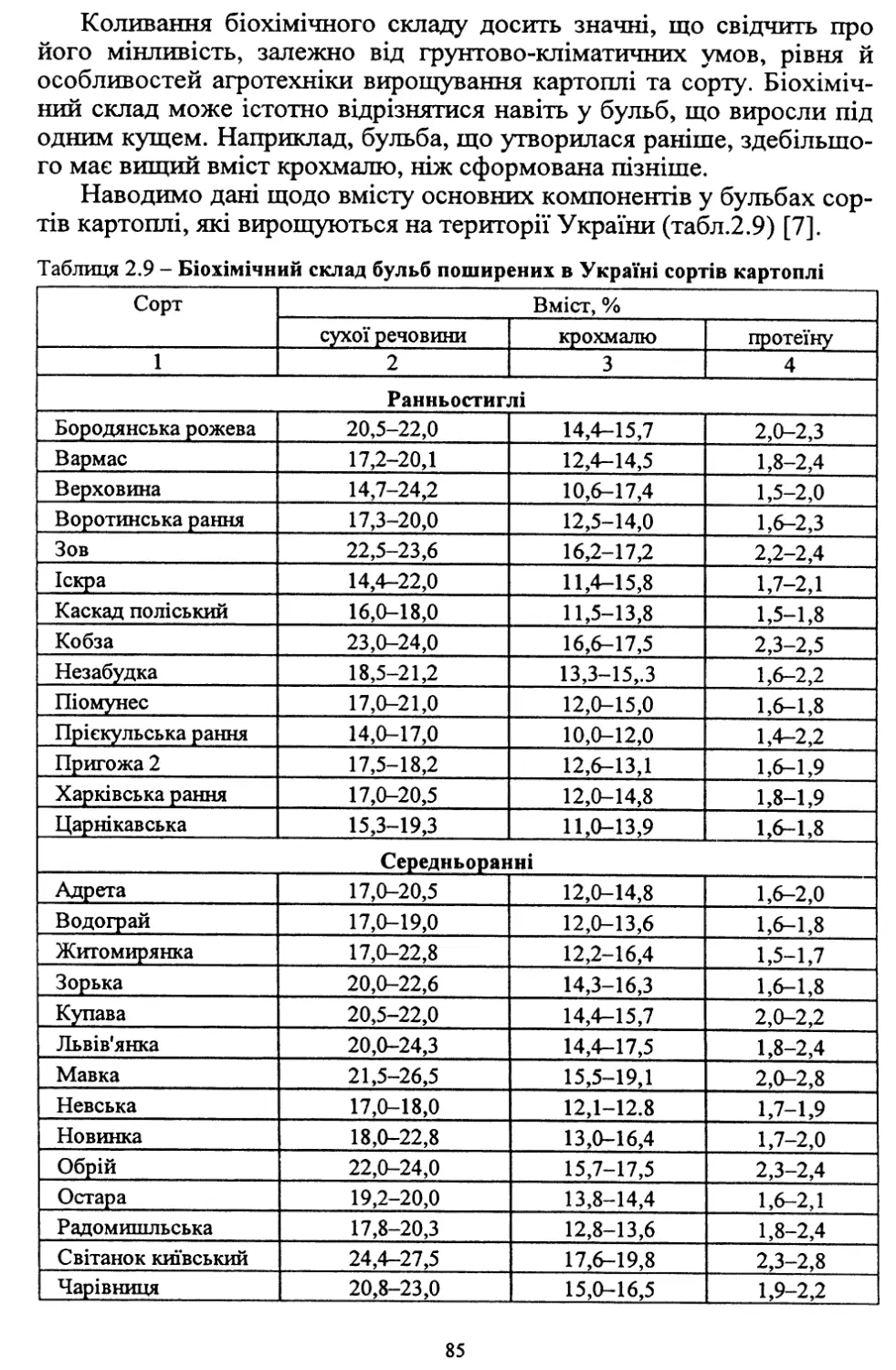
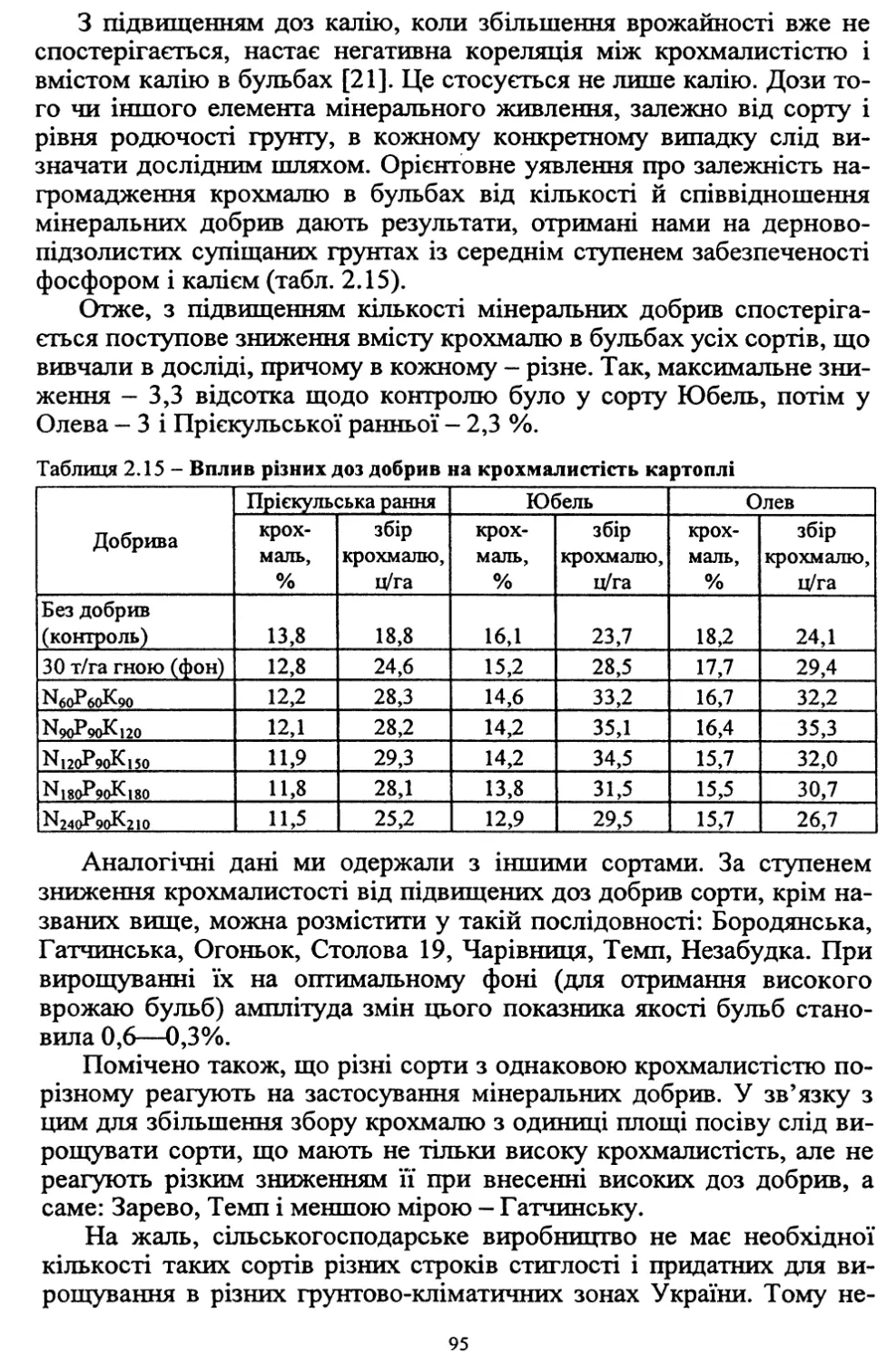
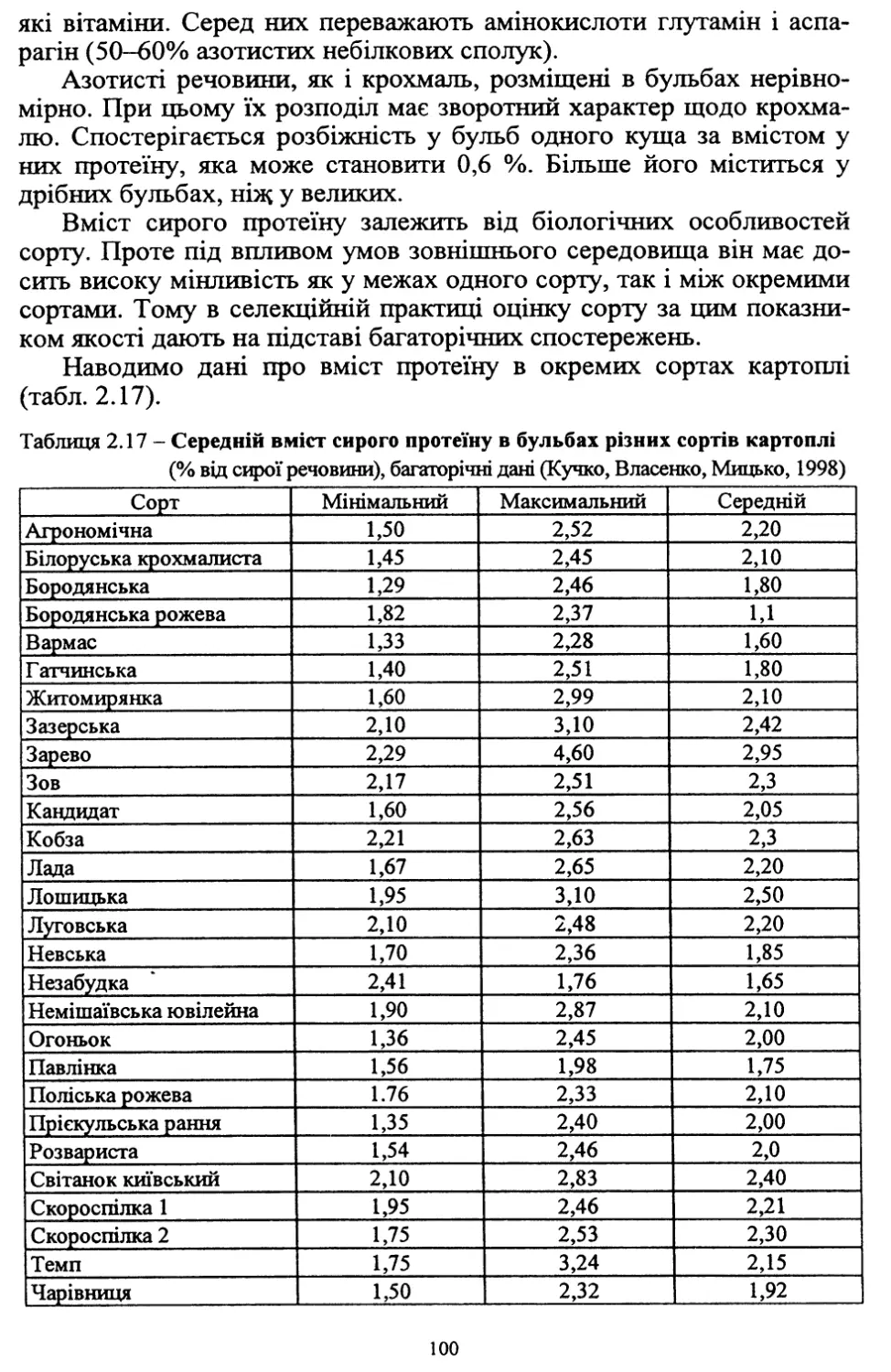
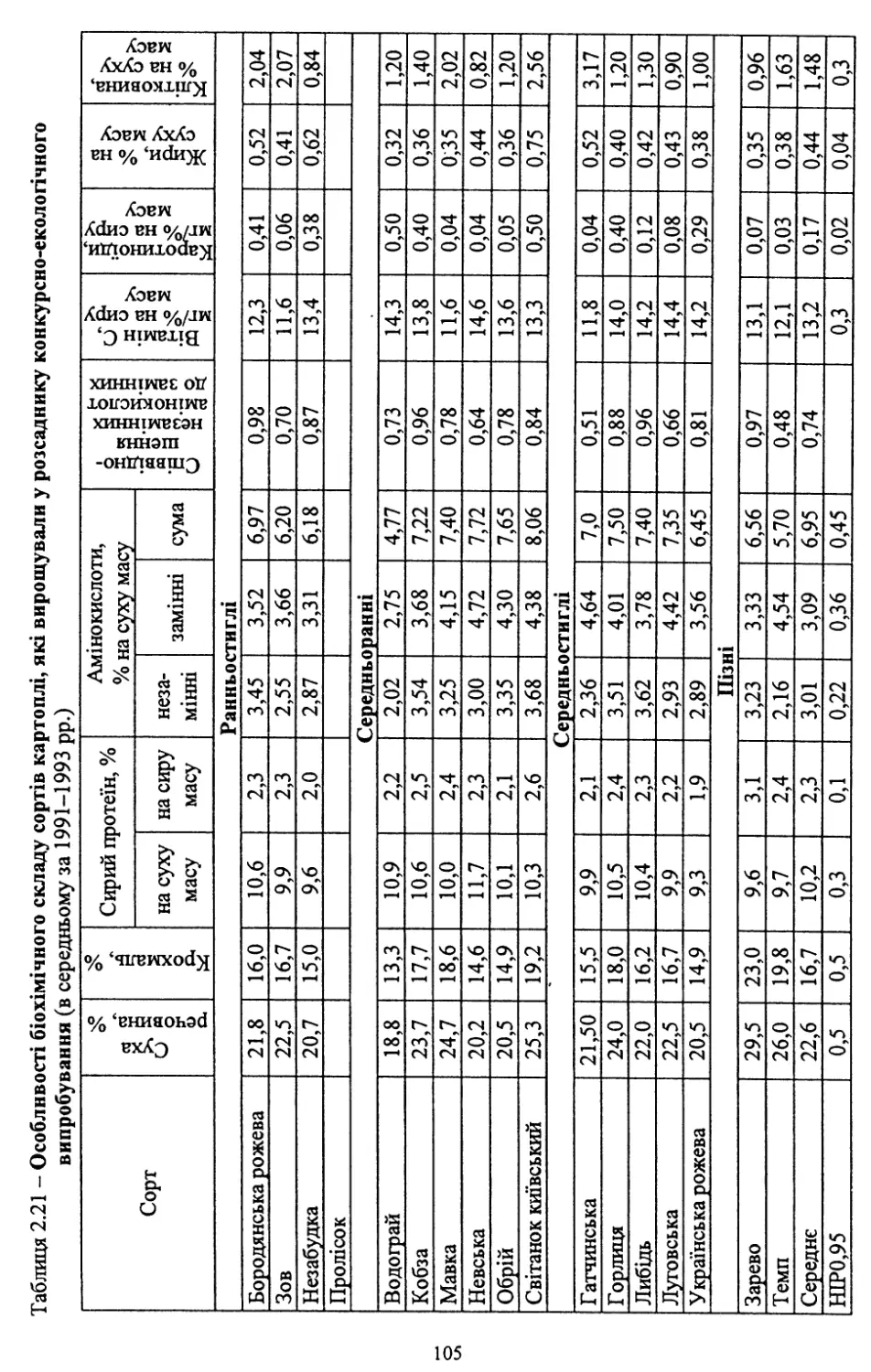
Text
МІНІСТЕРСТВО АГРАРНОЇ ПОЛІТИКИ УКРАЇНИ
УКРАЇНСЬКА АКАДЕМІЯ АГРАРНИХ НАУК
КАРТОПЛЯ
За редакцією
В.В. Кононученка, М.Я. Молоцького
Том перший
ББК 42.15
К 27
УДК 635.21:631.527/.53
Авторський колектив:
С.П. Васильківський, Ю.Я. Верменко, М.Ю. Власенко,
М.Д. Гончаров, Н.С. Кожушко, В.В. Кононученко,
М.Я. Молоцький, Т.М. Олійник, А.А. Осипчук,
А.А. Подгаєцький, В.І. Сидорчук, П.С. Теслюк,
І.І. Тимошенко
Картопля / За ред. В.В.Кононученка, М.Я.Молоцького - Біла
Церква, 2002.- Т. 1,— 536 с.
ISBN 966-7417-32-8
У першому томі висвітлено народногосподарське значення кар¬
топлі, її походження і розповсюдження та сучасний стан розвитку
картоплярства в Україні. Наведені ботанічна, морфологічна, фізіоло¬
гічна і біохімічна характеристики культури. Викладені сучасні уяв¬
лення з генетики картоплі та стратегія використання її генофонду.
Висвітлені новітні методи і основні напрями селекції картоплі: на
продуктивність, ранньостиглість, стійкість до хвороб і шкідників,
придатність до промислової переробки; наведено основні досягнення
селекції. Значна частина монографії відведена насінництву картоплі.
Розглядаються історія розвитку галузі в Україні, наукові та організа¬
ційні засади, розкриті причини виродження картоплі і заходи бороть¬
би з ним, наведені система насінництва і схеми відтворення еліти.
Приділена увага біотехнологічним методам одержання вихідного ма¬
теріалу, питанням репродукційного та внутрішньогосподарського на¬
сінництва, подані короткі характеристики сучасних сортів, занесених
до Реєстру сортів рослин України.
Розраховано на наукових співробітників, аспірантів, викладачів і
студентів вищих аграрних навчальних закладів, спеціалістів та картоп¬
лярів, які бажають отримати високі врожаї якісних бульб картоплі.
ББК 42.15
ISBN 966-7417-32-8
О Кононученко В.В.,
Молоцький М.Я., 2002
ВСТУП
За обсягами споживання та географічною поширеністю культура
картоплі займає одне з провідних місць у структурі продовольчої
продукції в Україні. Вона значною мірою використовується і як фу¬
раж у тваринництві. Виходячи з історичних, природно-кліматичних
та соціально-економічних умов, формування ринку картоплі має
виключно специфічні особливості, які зумовлюються рядом чин¬
ників, зокрема таких, як: ступінь забезпечення матеріально-технічної
бази виробництва, формування нових економічних взаємовідносин,
зміна структури виробництва і використання продукції, ціноутво-
реїшя та ін.
Результати аналітичної обробки статистичних даних щодо
картоплі свідчать про те, що в 90-х роках минулого століття по¬
чали виявлятися негативні тенденції в розвитку галузі картопляр¬
ства.
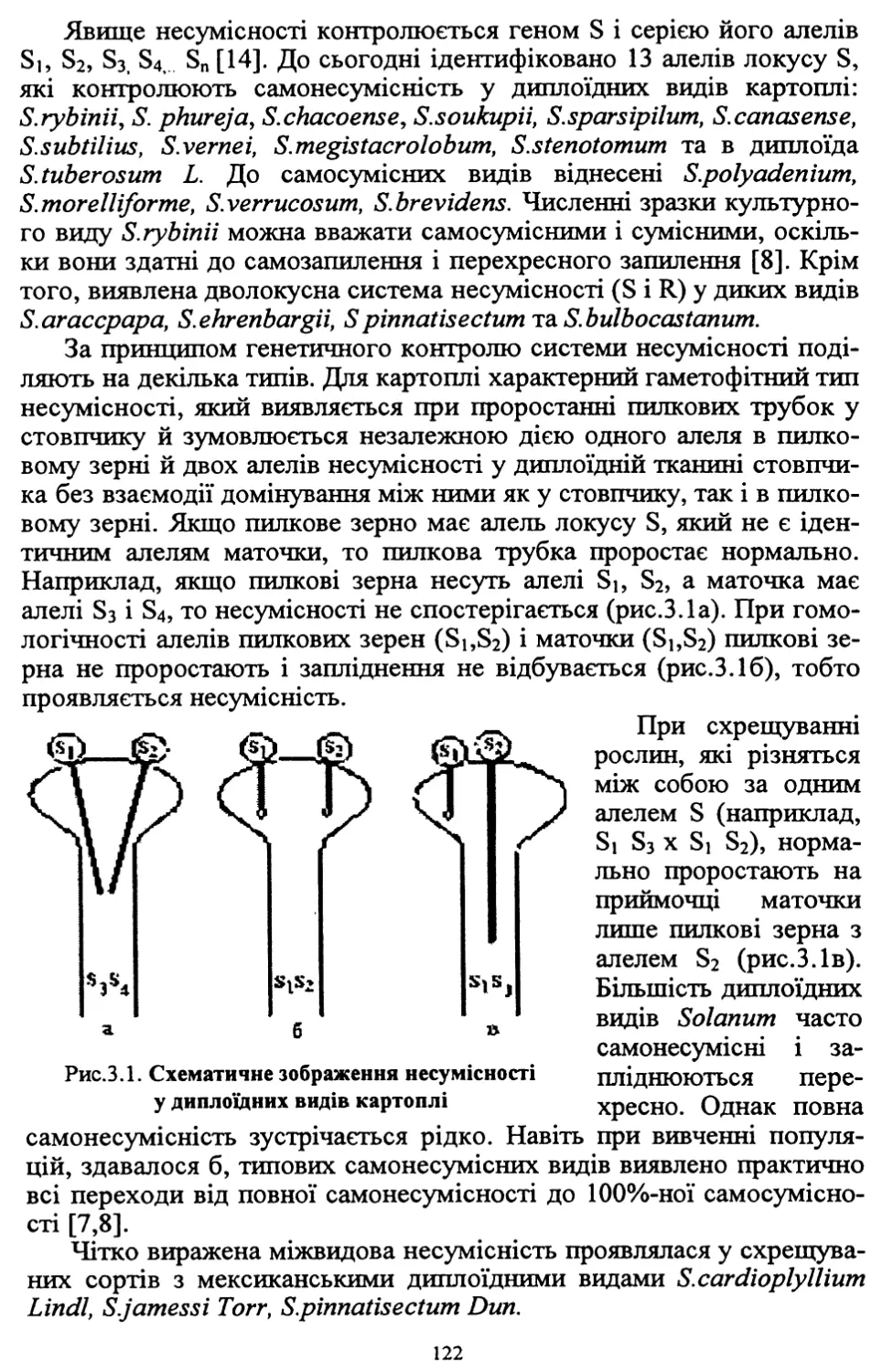
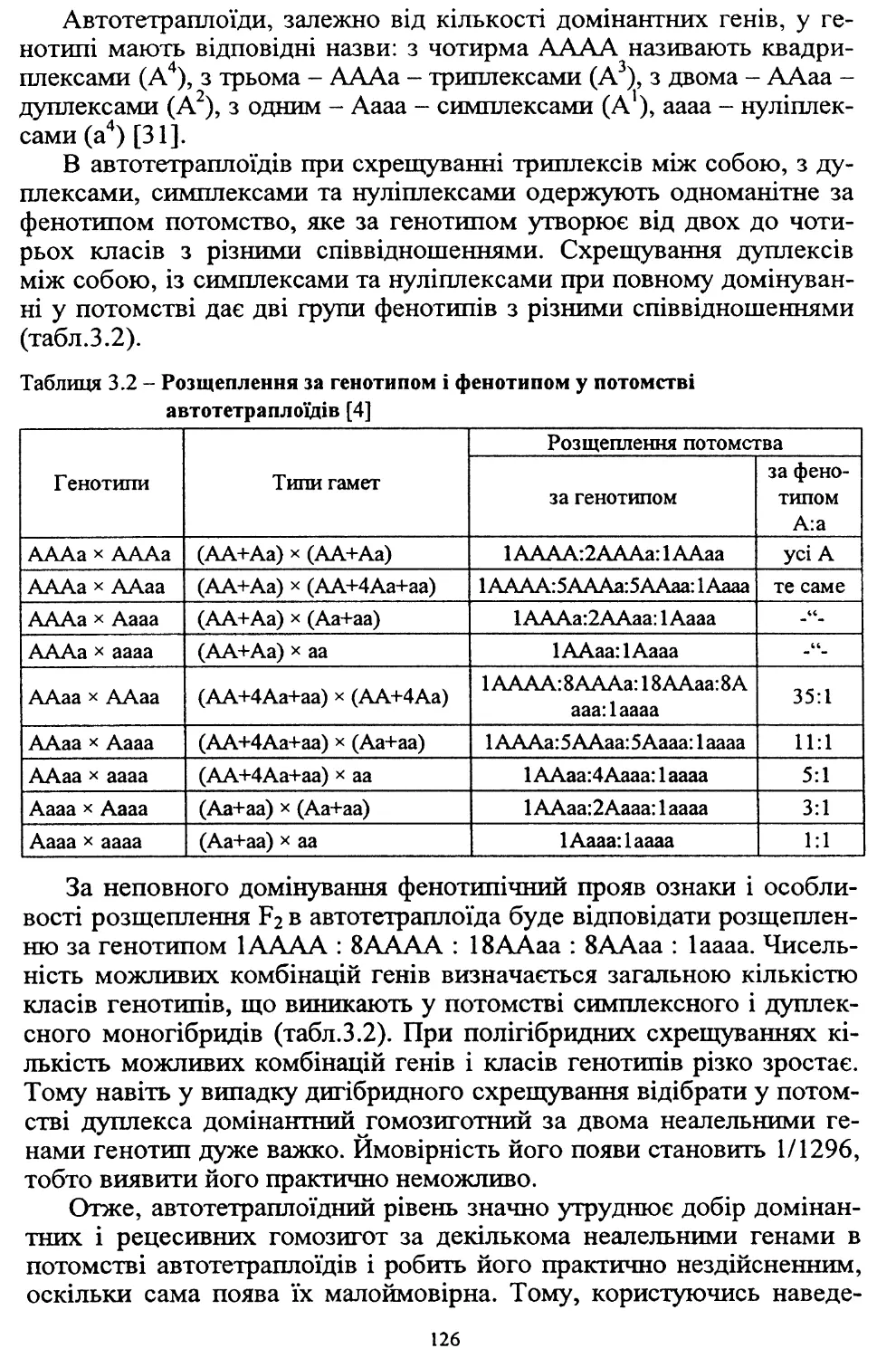
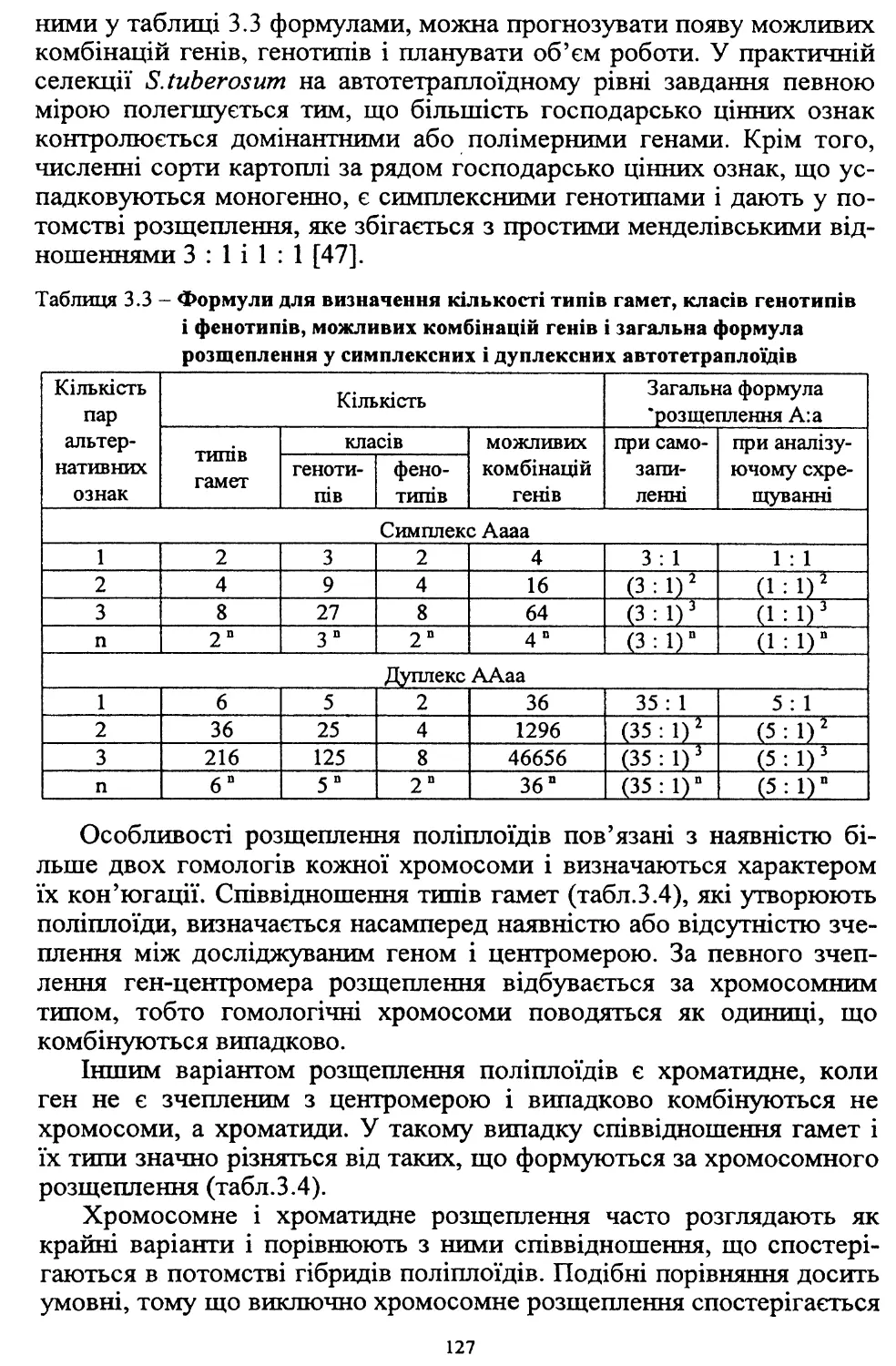
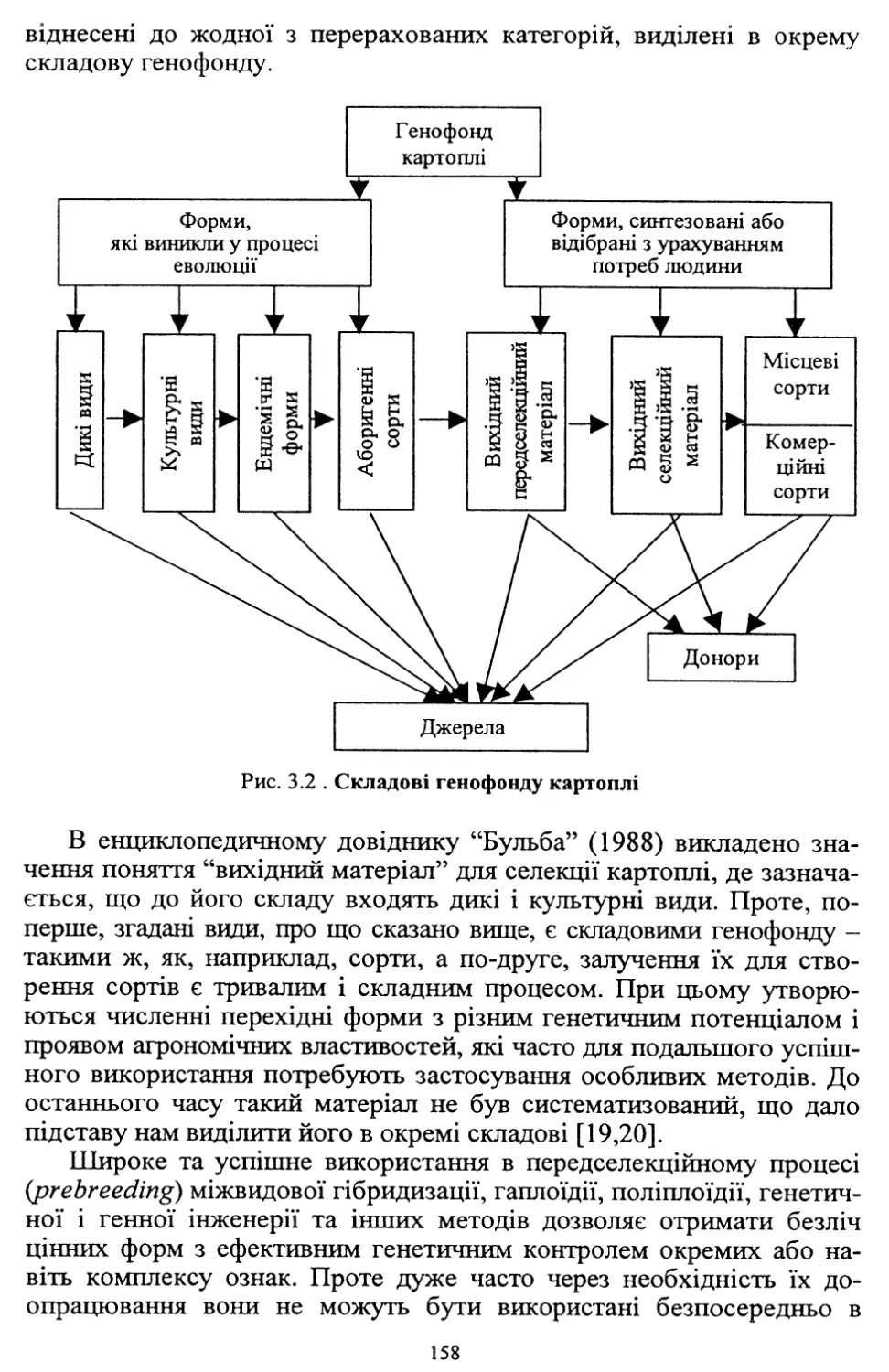
У зв’язку з високою трудо- та ресурсомісткістю виробництва, яке
при загальній енергетичній кризі в країні технологічно не забезпе¬
чується, площі насаджень картоплі в суспільному секторі госпо¬
дарств, які застосовували інтенсивні технології, значно скоротились.
Так, якщо в середньому за 1986-1990 рр. вони становили 477 тис. га,
то в 2000 р. - лише 30 тис. га, тобто в 16 разів менше. За названий
період в індивідуальному секторі господарств посіви культури зрос¬
ли з 996 тис. га до 1,5 млн га, або в 1,5 рази. У цілому по Україні
посівні площі під картоплею зросли незначно - з 1473 до 1551 тис.
га, тобто лише на 5 %.
Переміщення площ картоплі за останні роки в особисті підсобні
господарства і на дачні ділянки із середнім розміром 15-20 соток
призвело до деконцентрації посівів, розпорошування ресурсів, знач¬
ного зростання витрат виробництва. У той же час зростання рівня
самозабезпечення населення картоплею спричинює ряд негативних
наслідків, зокрема погіршення фітосанітарного стану умов вирощу¬
вання, різке зниження рівня механізації, втрату організованого ринку
та сировинної бази для переробної промисловості.
Тому значне скорочення товарного виробництва продовольчої
картоплі у спеціалізованих сільськогосподарських підприємствах та
концентрація посівів у господарствах населення потребують вирі¬
шення цілого комплексу надзвичайно складних і важливих проблем
як практичного, так і наукового рівня.
Досвід передових країн свідчить, що високопродуктивне картоп¬
лярство у них базується на досягненнях науково-технічного прогре¬
су. Основними напрямками його є використання інтенсивних сортів
та високоякісного насіннєвого матеріалу, прогресивних прийомів
агротехніки, застосування високоефективних пестицидів, високо¬
продуктивних тракторів, комбайнів, іншої сільськогосподарської
техніки та надійне матеріально-технічне забезпечення. Крім того, у
них відчутна економічна і нормативно-правова підтримка сільсько¬
господарського виробництва державою.
Поточна ситуація в картоплярстві нашої держави характери¬
зується недостатнім технологічним рівнем виробництва бульб, а час¬
то й відсутністю необхідних знань та ринкової інфраструктури щодо
забезпечення дрібних товаровиробників якісним насіннєвим ма¬
теріалом, засобами захисту рослин та добривами. Слід також надава¬
ти послуги з середньо- та дрібнооптової закупівлі й збуту вирощеної
продукції, інформаційно-навчального сервісу тощо. Загальновідомо,
що вирощування картоплі відзначається тісною залежністю агро¬
технічного і технологічного циклів. Приватний сектор на сьогодні не
має достатньої кількості засобів виробництва для виконання цього
технологічного процесу.
Дефіцит можливостей щодо поповнення техніки та інших матері¬
альних ресурсів призводить до того, що на практиці застосовуються
примітивні технології. В Україні фактично не виробляються машини
для забезпечення потреб галузі, а використовувана нині техніка є
здебільшого морально і фізично застарілою.
Розрахунки показують, що просте відтворення при виробництві
продовольчої картоплі досягається сьогодні при врожайності 150-
160 ц/га. Для того ж, щоб забезпечити розширене відтворення галузі,
тобто рентабельність більше 50%, необхідно вирощувати не менше
200-230 центнерів бульб з кожного гектара.
Заслуговує уваги ще одна причина зниження ефективності карто¬
плярства - його зональна переспеціалізація.
Унаслідок різкого скорочення товарного виробництва в госпо¬
дарствах Полісся, які раніше забезпечували потреби південних регі¬
онів країни в картоплі, відбулося значне розширення посівних площ
культури в зонах Лісостепу і Степу. Так, якщо в 1990 р. на Поліссі
розміщувались 59,2% посівів картоплі, у Лісостепу - 25,3, у Степу -
15,5 %, то в 2000 році - відповідно 49,3; 30,3; 20,4 %.
Для наукового забезпечення галузі розроблена науково-технічна
програма “Картоплярство”, над реалізацією якої працюють науковці
22 установ різного підпорядкування.
В Інституті картоплярства УААН проводяться дослідження з фор¬
мування, підтримання та вивчення генофонду картоплі України. У
цей час генетичні ресурси України нараховують 1060 зразків сортів
та міжсортових гібридів. На сьогодні колекція інституту налічує 325
зразків диких видів, 117 зразків культурного виду Solanum andigenum
та 57 зразків трьох диплоїдних видів картоплі.
4
За попередні 5 років (1996-2000 рр.) Інститутом картоплярства
УААН та установами-співвиконавцями створені й передані до Дер¬
жавного сортовипробування 27 нових сортів картоплі, з них Інститу¬
том картоплярства —11, Поліською дослідною станцією - 8, Інститу¬
том землеробства і тваринництва західного регіону - 2, Гірсько-Кар-
патською дослідною станцією - 1, Інститутом сільського господарс¬
тва Полісся - 1, Сумським державним аграрним університетом - 2,
Львівським державним аграрним університетом - 2. До Реєстру сор¬
тів рослин України з них уже занесені ранні сорти Повінь, Серпанок,
Поран; середньоранні Поліська 96 і Дубравка; середньостиглі Явір,
Віринея, Віра. За 1996-2000 рр. до Реєстру занесено у цілому 17 но¬
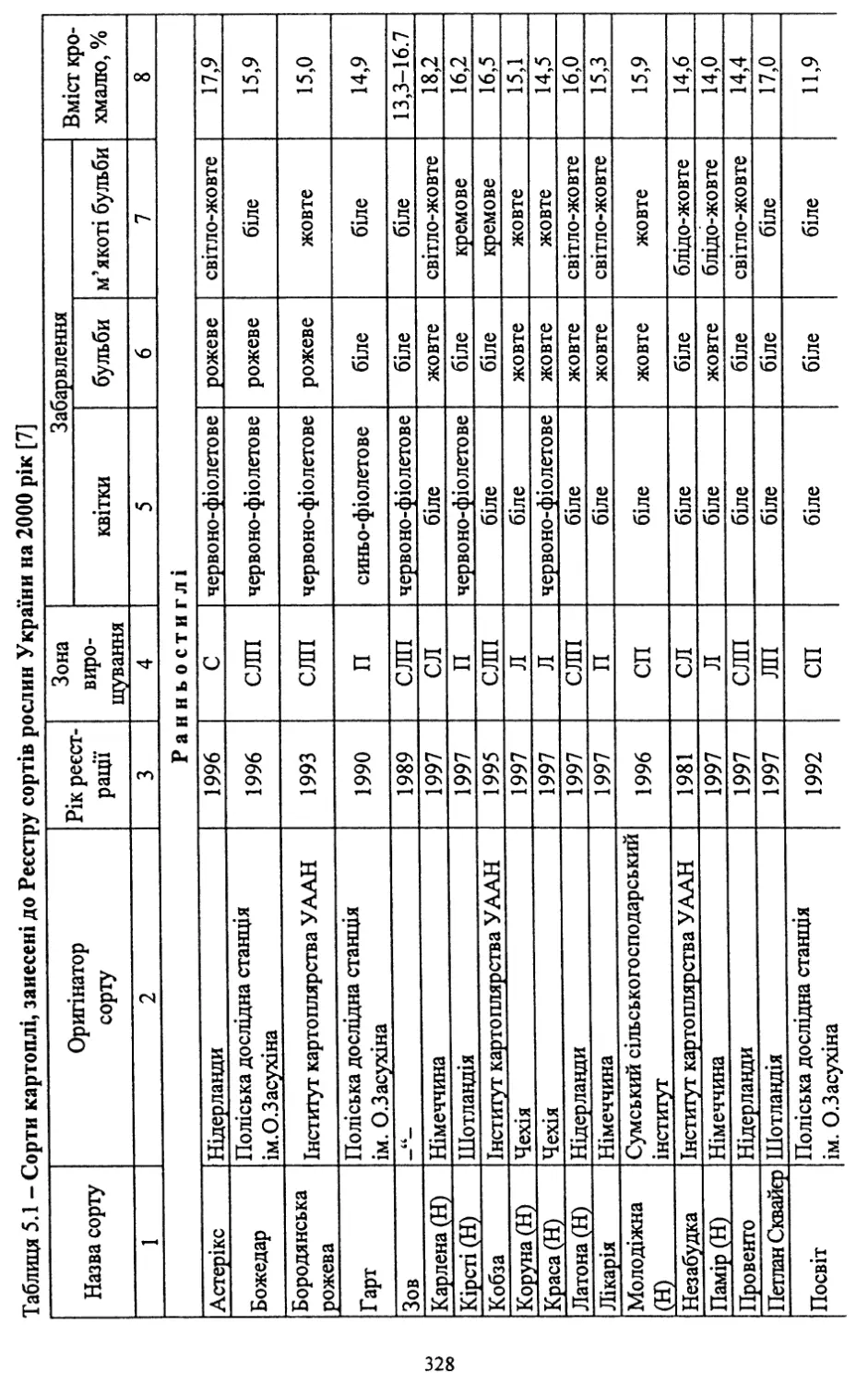
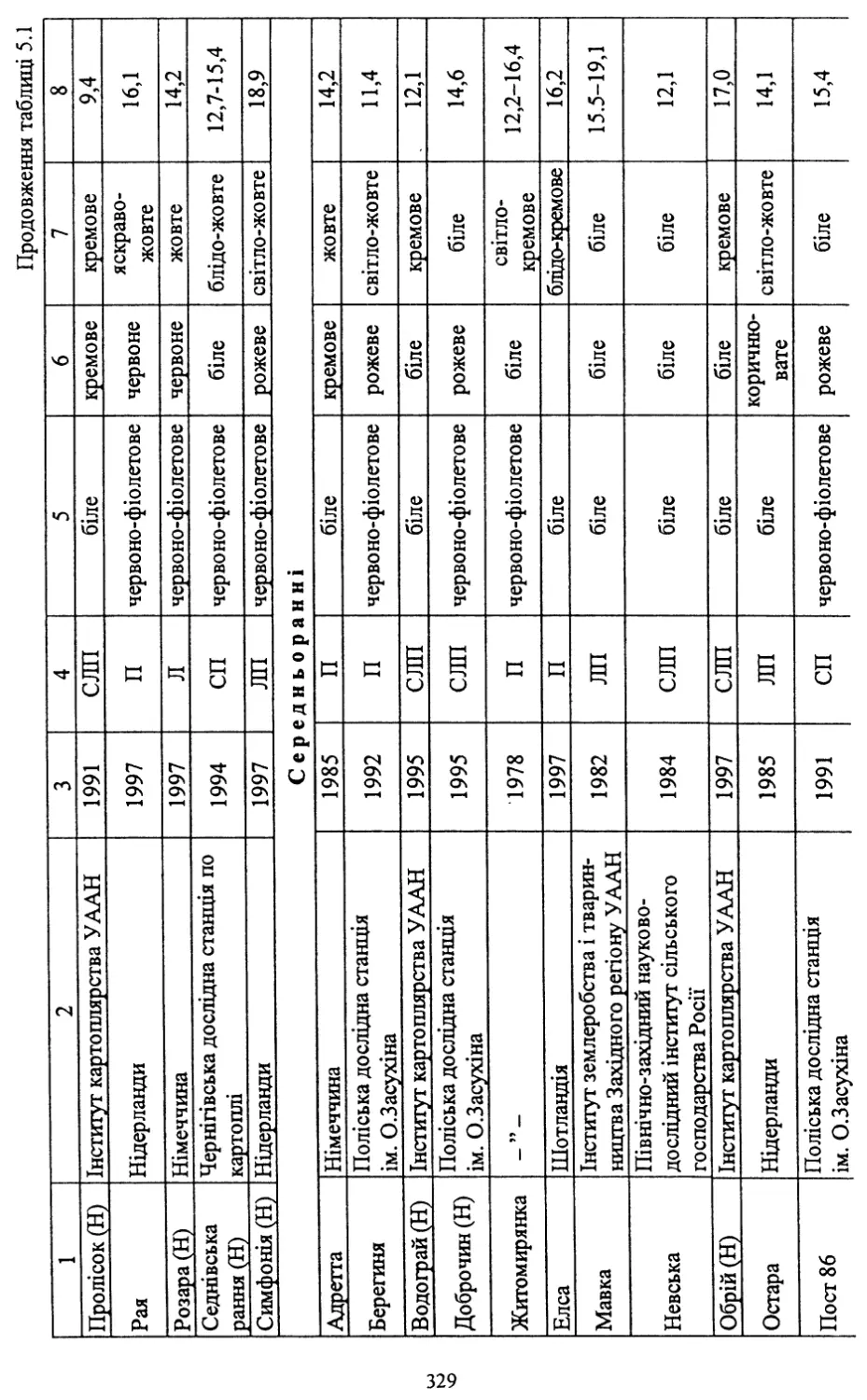
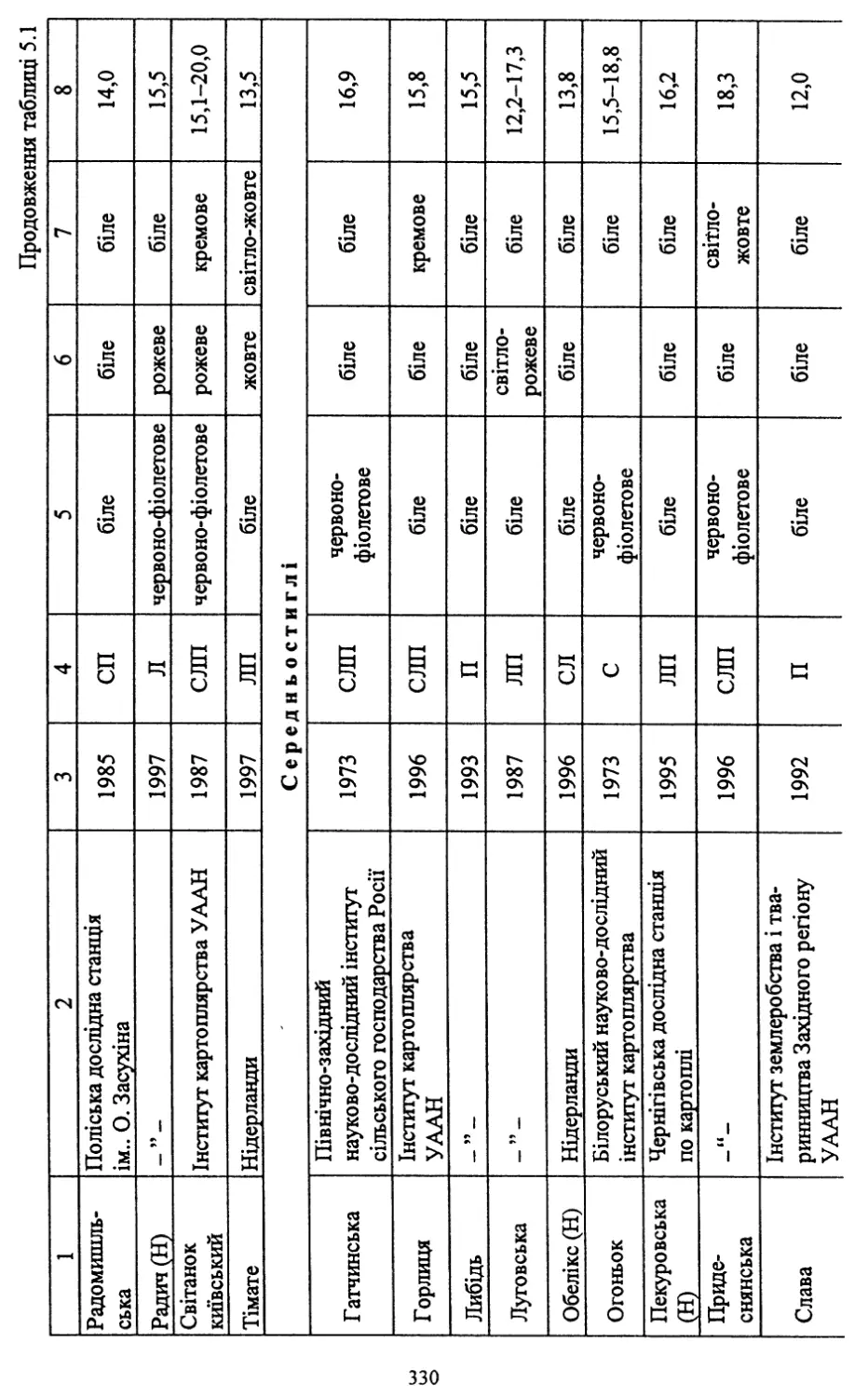
вих сортів. Із занесених до Реєстру сортів рослин України на 2000 рік
40 (54,8 %) є сортами української селекції.
Нові сорти відзначаються стійкістю проти раку картоплі, окремі з
них - проти картопляної нематоди (Обрій, Слов’янка, Поран, Полі¬
ська 96, Повінь, Західна, Дзвін, Молодіжна та ін.). Більшість із них
мають хороші смакові якості та високий уміст крохмалю. Частина
сортів придатна для переробки на картоплепродукти, що в даний час
має важливе значення для переробної промисловості.
Слід зазначити, що значна частина створених сортів може виро¬
щуватися в різних грунтово-кліматичних зонах України.
Помітними є й певні досягнення в селекції на стійкість проти та¬
кого шкодочинного захворювання, як фітофтороз. Із нових сортів
високою стійкістю проти нього відзначаються Явір, Віринея, Багря¬
на, Дубравка, Віра, Ракурс, Ольвія, Західна та Дзвін.
В Україні створено сорти, які мають відносну стійкість проти ко¬
лорадського жука (фактор витривалості та антибіотичної дії на ре¬
продуктивну функцію жука) - Зарево, Світанок київський, Волове¬
цька, Зов. Селекція на стійкість проти колорадського жука викорис¬
товує досягнення біотехнології - трансформацію генів стійкості про¬
ти шкідника в існуючі сорти картоплі. Розпочата така робота і в
Інституті картоплярства УААН.
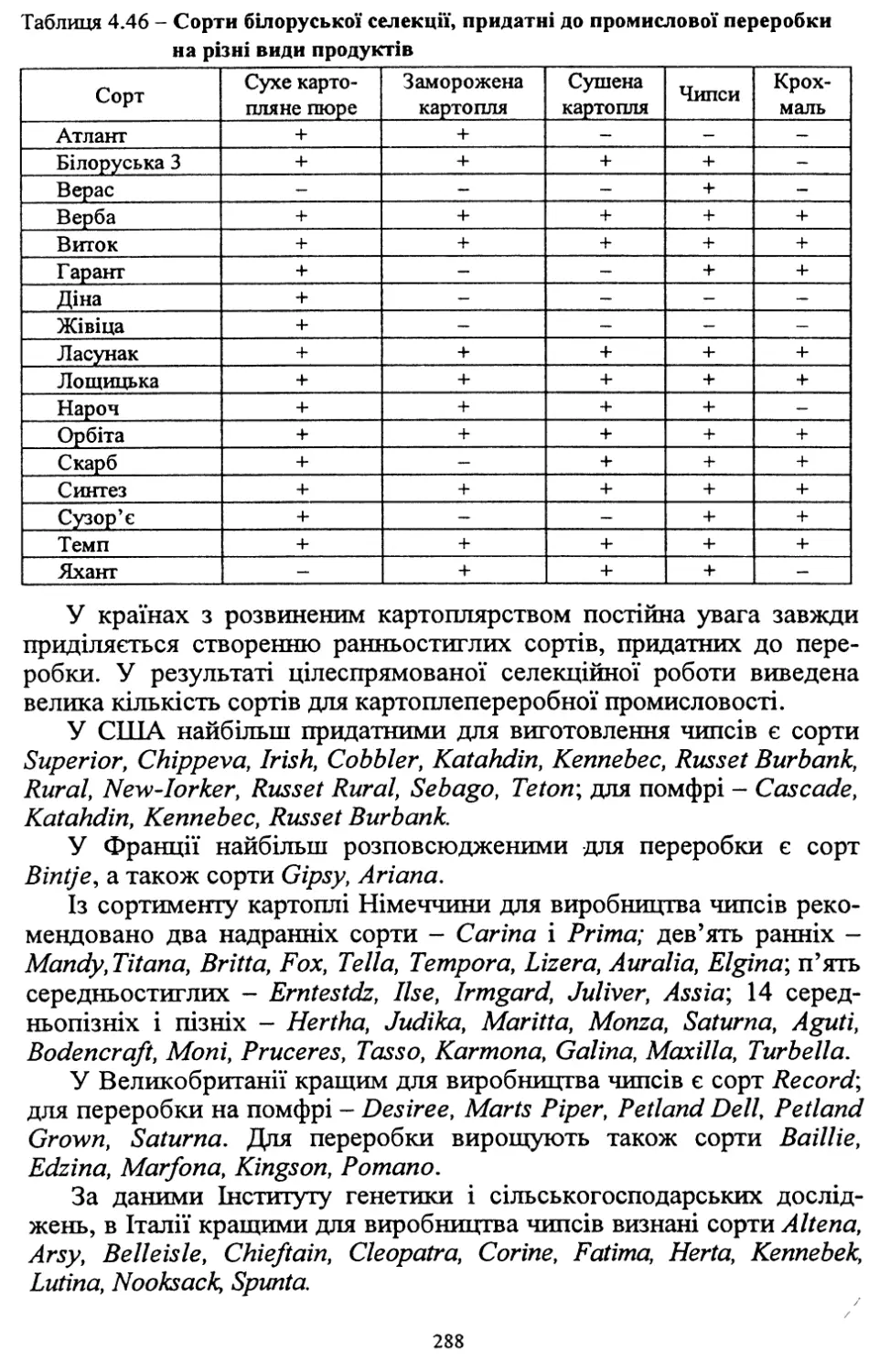
Деякі новостворені сорти є придатними для промислової пе¬
реробки. До початку 90-х років картопля в Україні була головним
чином продовольчою культурою і її використання обмежувалося
домашнім споживанням. Такий стан швидко змінився після появи
на ринку двох продуктів переробки картоплі - хрусткої (чипсів) і
картоплі гарнірної (фрі). Поширення цих картопляних продуктів
докорінно змінило споживання картоплі в багатьох країнах.
Подібне відбувається зараз і в Україні. У зв’язку з цим поглибле¬
на робота щодо створення сортів для картоплепереробної проми¬
словості.
Порівняльна оцінка нових вітчизняних сортів щодо їхньої при¬
датності до переробки показала перспективність таких, як Фантазія,
Зарево, Дзвін та ін.
5
Ринок продуктів, виготовлених з картоплі, швидко збільшується і
розширюється, і цей напрям у селекції сьогодні є одним із актуальних.
У галузі насінництва картоплі дослідження здійснюють 12 нау¬
ково-дослідних установ і вузів, зусилля і пошуки яких спрямовані на
розробку нових і вдосконалення існуючих способів одержання та
формування оригінального садивного матеріалу картоплі.
Зусиллями Інституту картоплярства УААН, Білоцерківського
ДАУ, Асоціації “Картопля України” удосконалена система насінниц¬
тва картоплі, яка більш повно враховує потреби в садивному матері¬
алі дрібного товаровиробника, опрацьовані науково обгрунтовані
норми садіння картоплі під запланований урожай.
Доведена рівноцінність як вихідного матеріалу в системі насін¬
ництва рослин мікро- і міні-бульб, одержаних біотехнологічним ме¬
тодом, а також макробульб першої бульбової репродукції від оздо¬
ровлених рослин. Цінним вихідним матеріалом є також бульби роз¬
міром менше ЗО мм, одержані від оздоровлених рослин, а також сор¬
тів, що відзначаються польовою стійкістю до вірусних хвороб.
Виявлена також рівноцінність міні-бульб, незалежно від способу
їх отримання, а саме: вирощених у культиваційних спорудах мето¬
дом гідропонної культури чи в польових умовах.
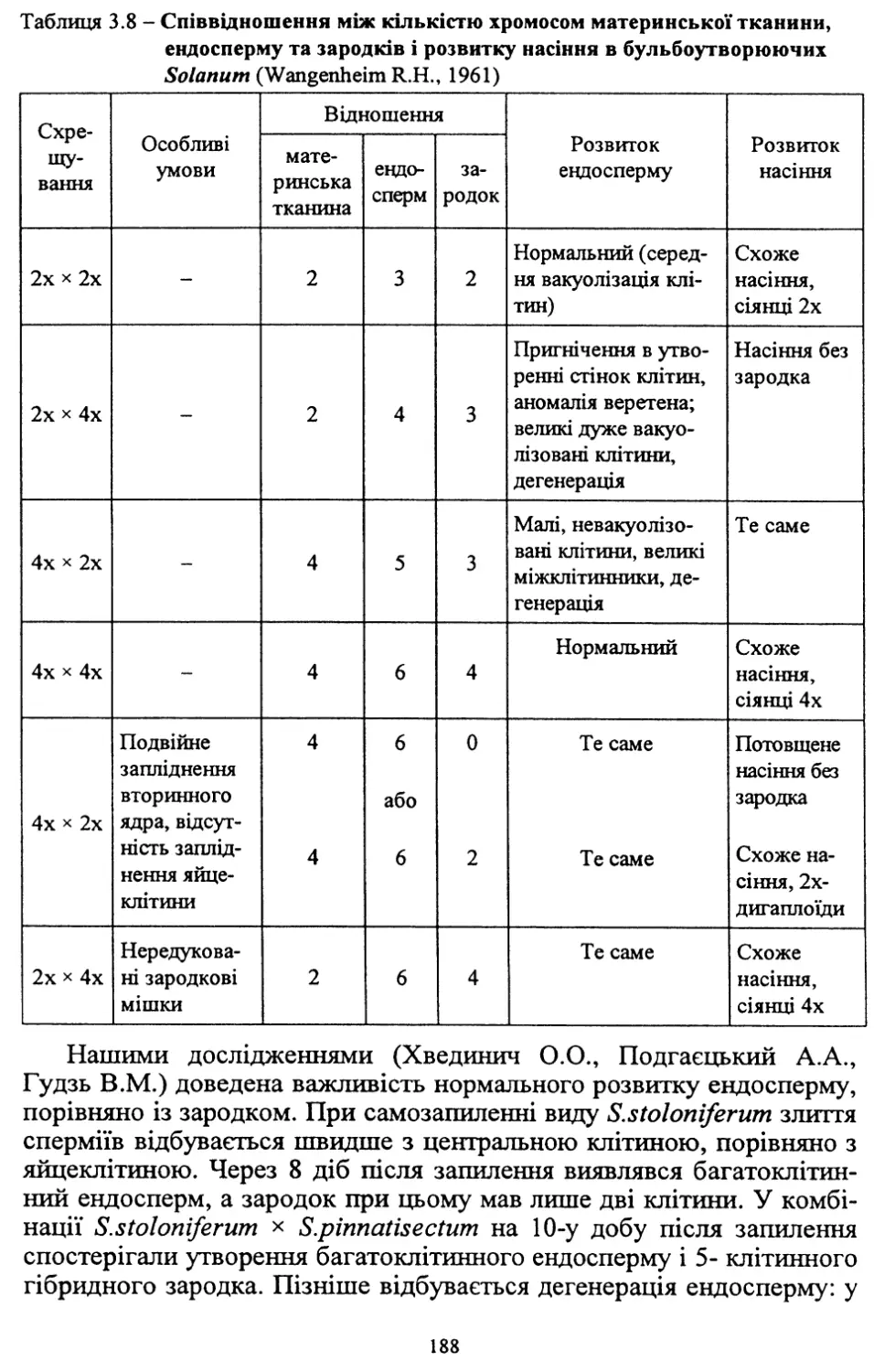
На основі аналізу взаємозв’язків цілісної системи рослина-господар-
паразит і механізму впливу на неї елементів технології оздоровлення та
клонального розмноження in vitro і in vivo доведено, що застосування
культури тканини забезпечує ефективне звільнення картоплі від основ¬
них вірусів, не викликаючи при цьому зміни сортотипових параметрів
фенотипу. Це підтверджують результати досліджень щодо збереження
сортової ідентичності картоплі у процесі оздоровлення від вірусної та
бактеріальної інфекції методом апікальної меристеми в поєднанні з тер-
мотепією при використанні сортів Божедар, Доброчин, Купава, Ольвія,
Слов’янка, Адретга, Серпанок та Віринея.
Проводиться значна робота з удосконалення технологічного про¬
цесу оздоровлення та прискореного розмноження оздоровлених рос¬
лин картоплі з метою оптимізації та підвищення його надійності при
використанні.
Результати досліджень у цьому напрямку використані для розробки
схем відтворення еліти на основі оздоровленого матеріалу. Такі схеми
(три- і чотирирічна) схвалені й рекомендовані для практичного викорис¬
тання в елітних господарствах Полісся і Лісостепу України. Досліджен¬
нями Інституту землеробства південного регіону в умовах зрошення до¬
ведена також доцільність застосування скороченої схеми польового ре¬
продукування оздоровленого матеріалу. Ці схеми є складовою частиною
науково-методичних рекомендацій з насінництва картоплі.
Використання скорочених схем відтворення еліти дозволяє на
один-два роки, порівняно з існуючою п’ятирічною, прискорити
впровадження у виробництво нових сортів, створених на основі ви-
6
сокорепродукційного насіннєвого матеріалу, на 30-50 ц/га підвищи¬
ти врожайність картоплі.
При визначенні економічно доцільних строків репродукування
еліти, сформованої як за п’ятирічною схемою на основі клонового
добору, так і за трирічною з використанням оздоровлених міні-
бульб, зі зниженням репродукції спостерігається зменшення врожай¬
ності бульб. Зниження врожайності значною мірою залежить від за¬
гальних фітосанітарних умов, здатності сорту протистояти вірусній
інфекції та дії негативних явищ природно-кліматичної зони.
На основі оздоровленого, у тому числі біотехнологічними методами,
вихідного матеріалу щорічно в Україні культивується 100-120 тис. Рос¬
лин in vitro і 250-300 тис. мікро- і міні-бульб. У розсадниках розмно¬
ження щорічно добирається близько 600 тис. клонів. Виробництву такої
кількості вихідного матеріалу сприяє робота з оздоровлення методом
термотерапії та культури меристеми значної кількості сортів, занесених
до Реєстру сортів рослин України та визнаних перспективними.
За 1996-2000 роки в елітгоспах України було вирощено 19 тис.
тонн супер-супереліти, 34 тис. тонн супереліти і 58,5 тис. тонн еліти,
реалізовано 32 тис. тонн елітних бульб. Насінницька робота в Украї¬
ні проводиться із 62 сортами картоплі.
У первинних ланках насінництва елітгоспів України сорти селек¬
ції Інституту картоплярства становлять 72,4 %, інших вітчизняних
науково-дослідних установ - 24,3, іноземні сорти - 3,3%. Частка но¬
вих і перспективних сортів складає 33,3 %.
Водночас усе гострішою проблемою стає підтримання сортових,
посівних та врожайних ознак вихідного матеріалу, які дуже часто
губляться вже на етапі елітного насінництва. На даний час обсяги
репродукційного насінництва не задовольняють вимог виробництва.
Відсутня інфраструктура, здатна забезпечити якісним насіннєвим
матеріалом високих репродукцій кращих сортів дрібних товаровиро¬
бників, фермерів та власників присадибних ділянок.
Значного вдосконалення потребують питання контролю, уніфіка¬
ції процесів та сертифікації насіння відповідно до сучасних правил
та вимог міжнародних організацій.
Технологічною основою виробництва картоплі залишається гре¬
бенева посадка з міжряддями 70 см. Вивчення питання використання
більш широких міжрядь в Інституті картоплярства та Білоцерківсь¬
кому ДАУ показало їх перспективність в умовах України. В останні
роки значно зросли площі під картоплею з міжряддями 75 см і біль¬
ше. Розроблена технологія вирощування картоплі із застосуванням
постійної технологічної колії із шириною 3,0 м на основі вітчизняно¬
го трактора ХТЗ-120.
Розробки останніх років з удосконалення способів боротьби з
бур’янами, систем удобрення, захисту картоплі від шкідників і хво¬
роб та режимів зберігання включені в технологічний регламент ви¬
7
рощування картоплі і с надійною основою високорентабельного кар¬
топлярства. Але відсутність оборотних коштів не дає можливості
картоплярам створити повне ресурсне забезпечення для одержання
високих урожаїв.
Підвищення попиту на картоплю, придатну для переробки, вима¬
гає глибшого вивчення усього комплексу факторів, що впливають на
якість вирощування і зберігання картоплі.
Вимоги покупця до товарного вигляду картоплі стали більш
жорсткими і специфічними. Для виробників картоплі стало ціл¬
ком очевидно, що садити картоплю, не маючи чіткого уявлення
про вимоги ринку - справа нерозумна і ризикована. Завдання сьо¬
годні полягає не в збільшенні валового виробництва продукту, а в
зменшенні площ під культуру на основі підвищення врожайності
та структурної перебудови картоплярства. Це зумовило потребу
відновлення спеціалізованого виробництва. У Київській, Черні¬
гівській, Житомирській, Тернопільській та інших областях ство¬
рені перші крупні зони спеціалізованого виробництва картоплі
для переробки на картоплю хрустку (чипси), картоплю гарнірну
(фрі). Швидко зростаюча мережа супермаркетів пред’являє під¬
вищені вимоги до якості та зовнішнього вигляду бульб, їх перед¬
продажної підготовки й упаковки.
На жаль, рівень розвитку ринкової інфраструктури як у цілому в
країні, так і в окремих регіонах не відповідає вимогам нормального
товарообігу. Для покращення ситуації в цьому плані необхідно при¬
скорити багатоцільове формування і розвиток сучасної інфраструк¬
тури ринку картоплі: організаційних і посередницьких виробничих
консорціумів, асоціацій, об’єднань, кооперативних господарств та
інших формувань.
У зв’язку із загальним реформуванням сільського господарства
України, постала необхідність самоорганізації в діяльності підпри¬
ємницьких структур, яка цінується так само високо, як і конкуренція,
з метою створення стабільного ринку з мінімальною участю і втру¬
чанням у нього держави.
Звичайно, остаточний вибір найбільш раціональних форм органі¬
зації ринку картоплі в Україні зробить саме життя. Та все ж стратегі¬
чно програшними будуть методи, зорієнтовані на суто державний
підхід. Якими б важкими і складними не були демократичні методи
за широкої участі сільськогосподарських виробників, ставку слід
робити саме на них.
Інституційною формою формування ринку картоплі в Україні по¬
винні бути класичні, демократичні, випробувані часом громадські ор¬
ганізації, товариства товаровиробників та переробних підприємств.
Можна з упевненістю стверджувати, що картоплярство України
перебуває в стадії переходу на ринковий шлях розвитку. Існуючі віт¬
чизняні наукові розробки дозволяють забезпечити інтенсифікацію
8
виробництва картоплі. У зв’язку з цим структурна перебудова картоп¬
лярства потребує відповідних інвестицій. Основними напрямками
інвестування мають бути:
- переробна промисловість;
- виробництво спеціалізованих сортів картоплі;
- розвиток насінництва;
- створення в регіонах вирощування картоплі агробізнесових
структур, які б займались забезпеченням товаровиробників насін¬
нєвим матеріалом, добривами, засобами захисту рослин та засобами
механізації, а також закупівлею й оптовою реалізацією вирощеної
продукції;
- виробництво спеціалізованої техніки й обслуговування нею
товаровиробників згідно з вимогами часу.
Досягти вирішення зазначених завдань можна за умови створен¬
ня^ знаходження спільних економічних інтересів ініціативних струк¬
тур - держави, наукових та дослідно-конструкторських організацій,
внутрішніх і зовнішніх інвесторів.
Запропоноване видання є монографією з культури картоплі і
складається з трьох томів. У перший том увійшли матеріали з
історії, стану та перспектив розвитку картоплярства в Україні, ви¬
кладені морфологічні, біологічні особливості картоплі та біохі¬
мічний склад її бульб. У цьому томі висвітлені питання генетики,
складові і характеристика генофонду картоплі, методи і результати
селекції, сортовий склад; приділена увага насінництву культури,
застосуванню методів біотехнології в селекційній та насінницькій
практиці. У другому томі докладно викладені матеріали із захисту
рослин від хвороб і шкідників, наведені малюнки та оригінальні
фотографії найбільш поширених і шкодочинних із них. У третьому
томі висвітлюються питання технології вирощування картоплі за
грунтово-кліматичними зонами України, в умовах зрошення, за
літнього садіння, наведені матеріали щодо зберігання бульб у
капітальних та пристосованих сховищах. Велика увага приділена
сучасній механізації виробничих процесів, питанням переробки
картоплі на картоплепродукти.
Монографія написана провідними вченими Інституту картопляр¬
ства УААН, Білоцерківського, Сумського та Львівського державних
аграрних університетів, Інституту зрошуваного землеробства УААН,
Асоціації “Картопля України”. У цьому виданні використано та уза¬
гальнено багаторічні експериментальні матеріали вітчизняних і за¬
кордонних дослідних установ та дослідників, а також досвід картоп¬
лярів України.
Видання призначене для наукових співробітників і аспірантів,
викладачів і студентів вищих аграрних навчальних закладів, спеціа¬
лістів з картоплярства та картоплярів, які бажають отримати високі
врожаї якісних бульб картоплі.
9
Редакція книги здійснена директором Інституту картоплярства
УААН, кандидатом технічних наук Кононученком Валерієм Ва¬
сильовичем, завідувачем кафедри селекції та насінництва БДАУ,
доктором сільськогосподарських наук, професором, заслуженим
діячем науки і техніки України Молоцьким Михайлом Якови¬
чем.
Окремі розділи і підрозділи першого тому написані наступними
науковцями:
Вступ - директором Інституту картоплярства УААН, к.т.н. Ко¬
нонученком В.В.
Розділ перший. Розвиток картоплярства в Україні - головою
Асоціації “Картопля України”, к. с.-г. н. Теслюком П.С.
Розділ другий. Морфологія, фізіологія та біохімія картоплі - за¬
відувачем кафедри ботаніки та фізіології рослин БДАУ, д. с.-г. н.,
професором Власенком М.Ю.
Розділ третій. Генетика картоплі (3.1 - 3.7) - деканом агро¬
номічного факультету БДАУ, д. с.-г. н., професором Васильківсь¬
ким С.П.
Генофонд картоплі, його складові, характеристика і стратегія ви¬
користання (3.8) - завідувачем лабораторії вихідного матеріалу Ін¬
ституту картоплярства УААН, д. с.-г. н. Подгаєцьким А.А.
Розділ четвертий. Селекція картоплі. 4.1. З історії розвитку се¬
лекції картоплі; 4.2. Генетичний потенціал картоплі; 4.4. Методи се¬
лекції картоплі; 4.5. Основні напрями та завдання селекції картоплі;
4.5.1. Селекція на вміст крохмалю і сирого протеїну; 4.7. Схема, ор¬
ганізація і техніка селекції; 4.8. Основні досягнення селекції кар¬
топлі за останні роки та їхнє народногосподарське значення - завіду¬
вачем відділу селекції Інституту картоплярства УААН, д. с.-г. н. Осип-
чуком А.А.; 4.3. Біологічний взаємозв’язок ознак картоплі і його
вплив на добір - завідувачем кафедри селекції та насінництва Львів¬
ського ДАУ, д. с.-г. н, професором Тимошенком І.І.; 4.5.2. Се¬
лекція картоплі на ранньостиглість; 4.5.3. Селекція картоплі на при¬
датність до механізованого виробництва; 4.5.5. Селекція на придат¬
ність до промислової переробки - завідувачем кафедри селекції та
насінництва, д. с.-г. н., професором Кожушко Н.С. і професором цієї
кафедри Сумського ДАУ, д. с.-г. н., заслуженим діячем науки і тех¬
ніки України Гончаровим М.Д.; 4.5.4. Селекція картоплі на стій¬
кість (імунітет) до хвороб і шкідників - директором Поліської дослід¬
ної станції ім. О.М. Засухіна ІК УААН, к. с.-г. н. Сидорчуком В.І.;
4.6. Вирощування картоплі з використанням ботанічного насіння;
4.6.1. Історія проблеми вирощування картоплі з ботанічного насіння;
4.6.2. Генетичні основи і підходи до створення сортів, придатних для
розмноження ботанічним насінням; 4.6.3. Технологія вирощування
картоплі з ботанічного насіння — завідувачем лабораторії вихідного
матеріалу Інституту картоплярства УААН, д. с.-г. н. Подгаєцьким А.А.
10
Розділ п’ятий. Сорти - головою Асоціації “Картопля України”,
к. с.-г. н. Теслюком П.С., д. с.-г. н., проф. Молоцьким М.Я. (БДАУ),
завідувачем відділу насінництва ПС УААН, к. с.-г. н. Верменком Ю.Я.
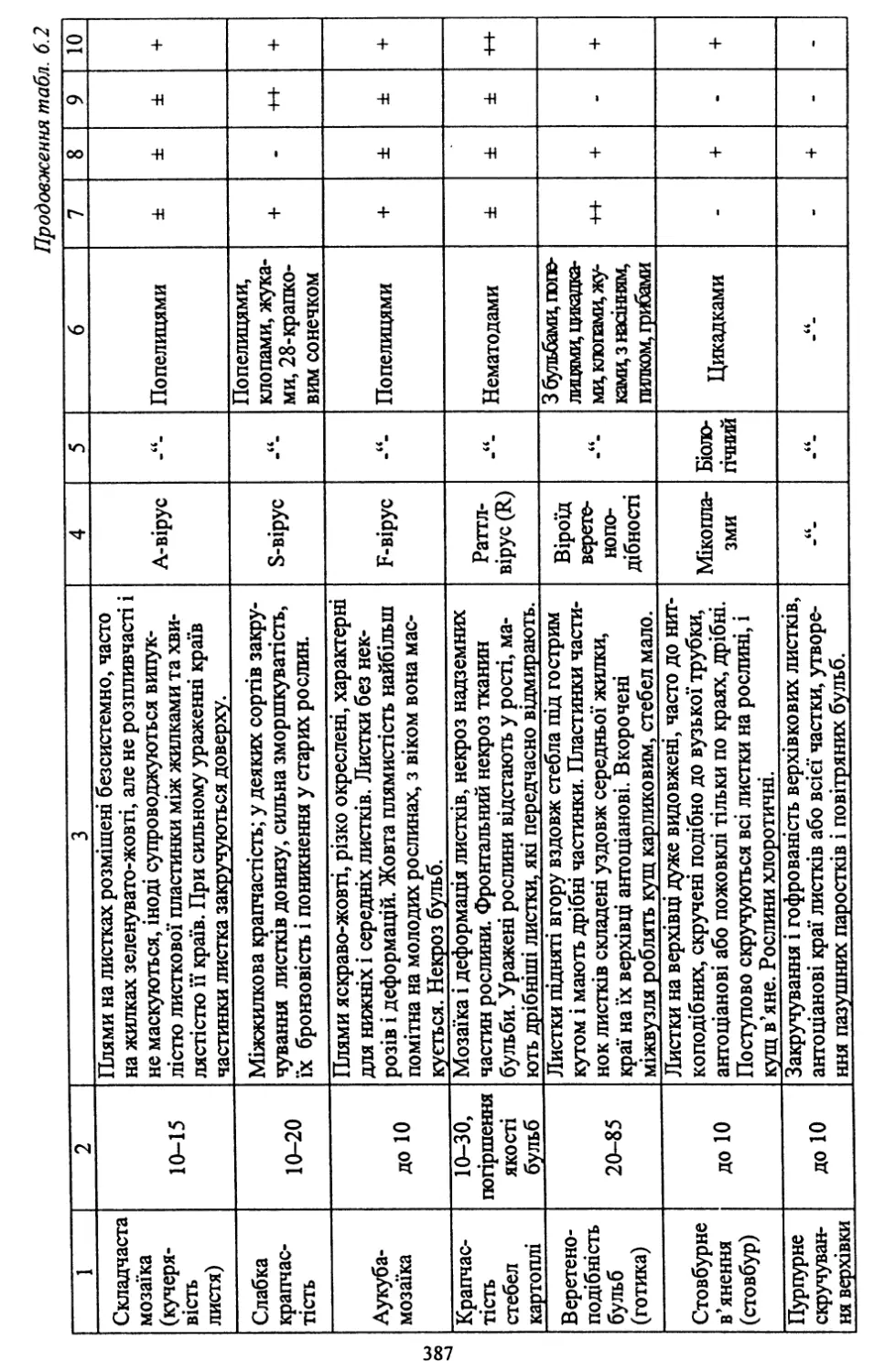
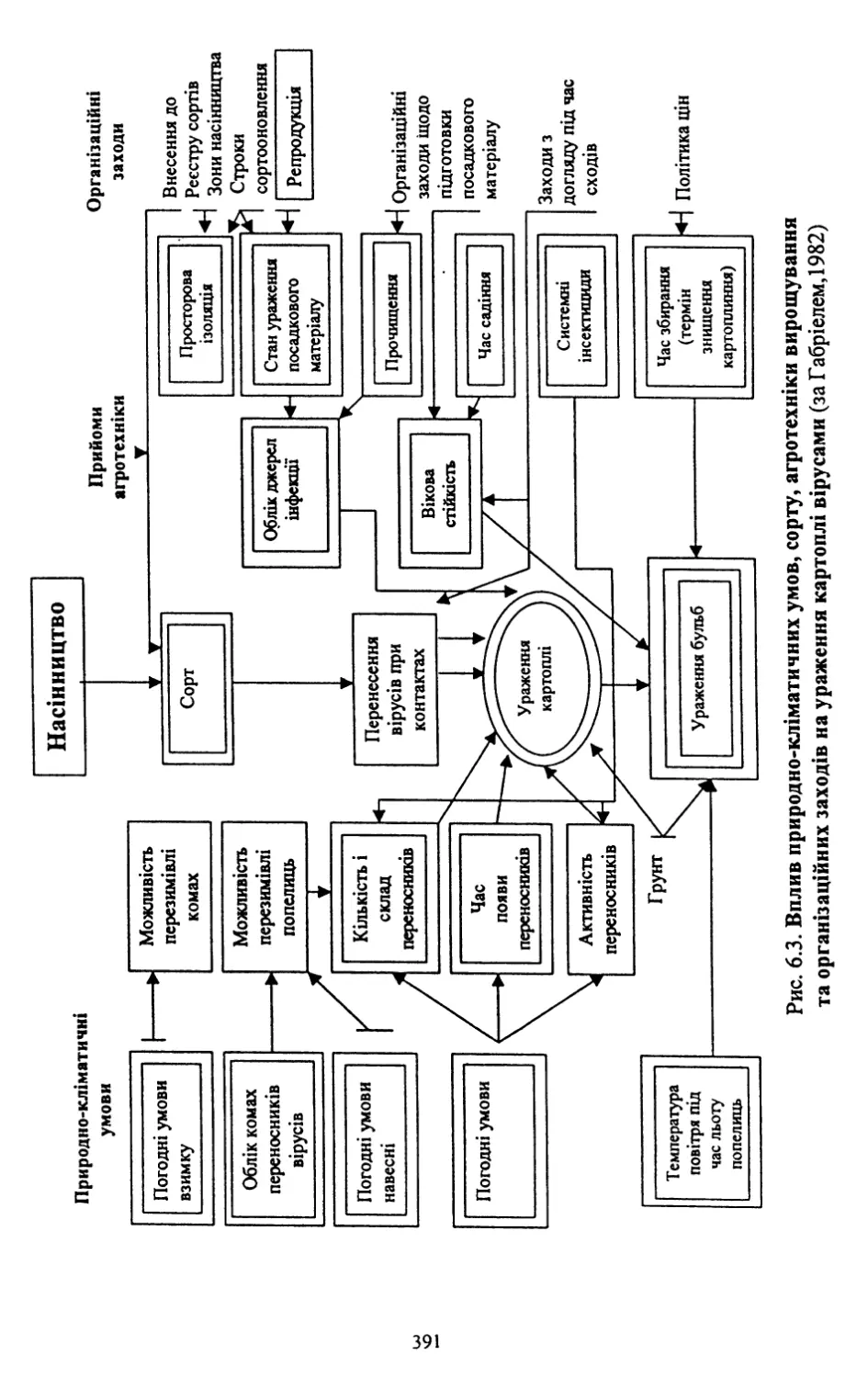
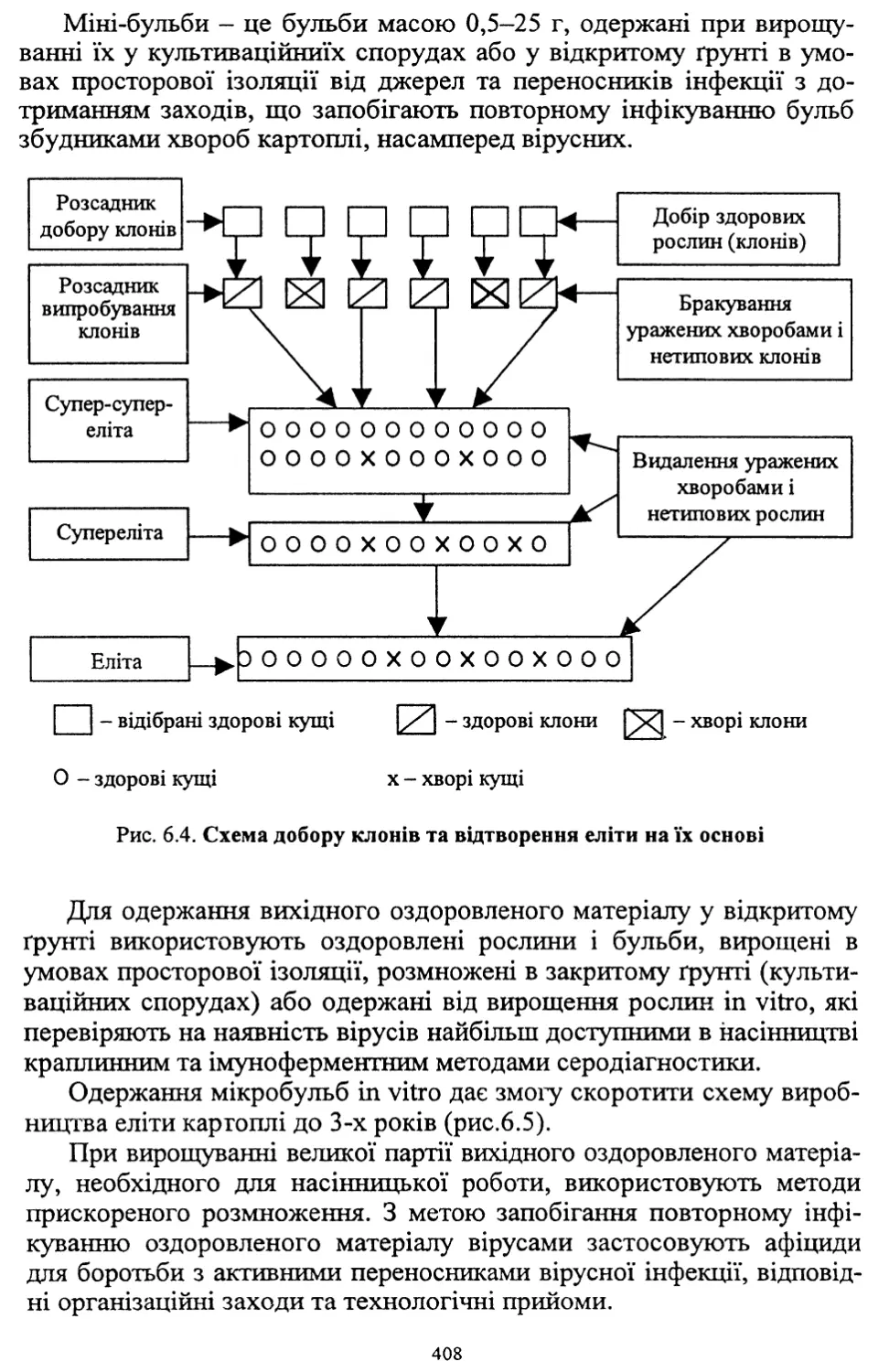
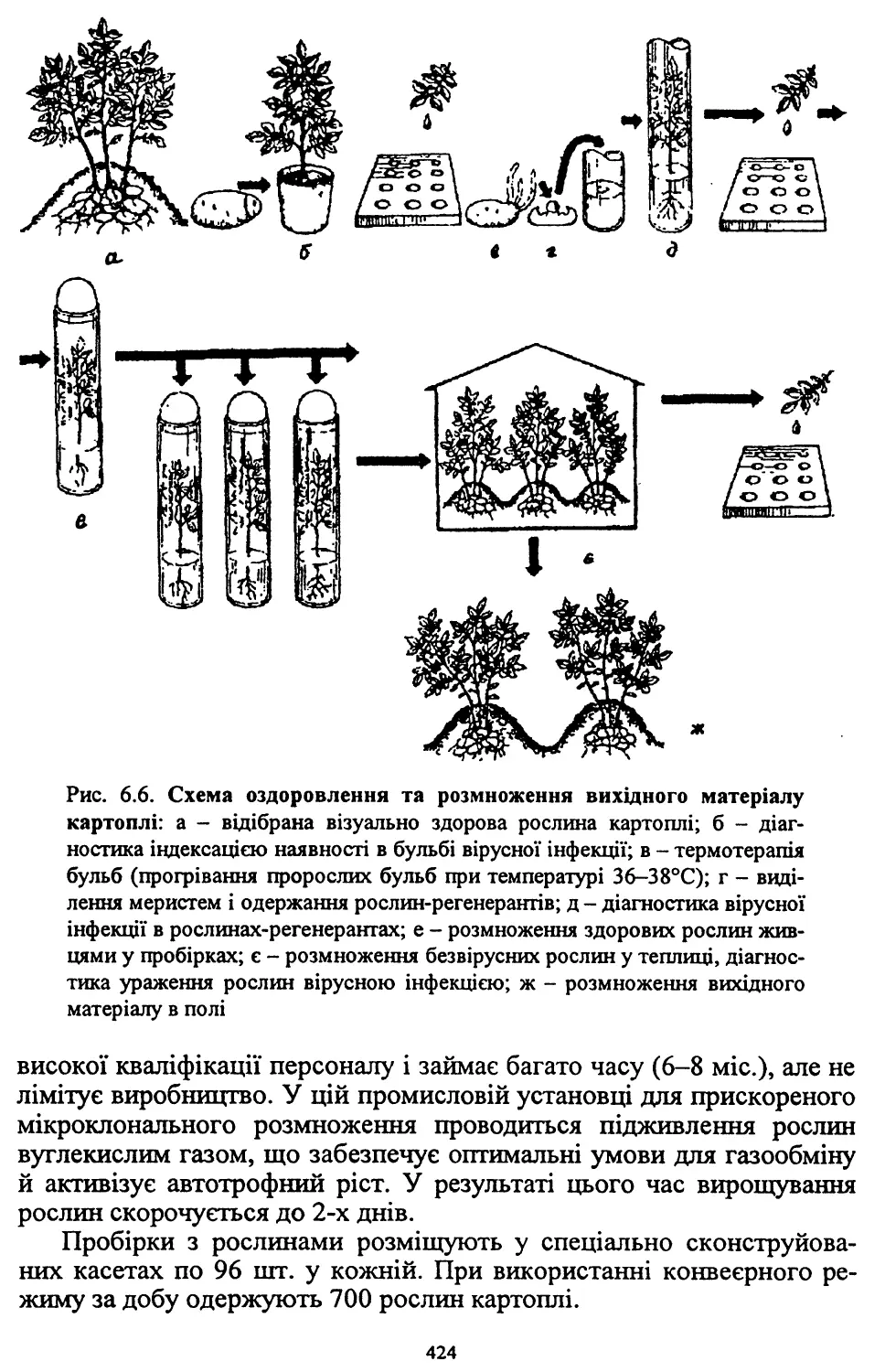
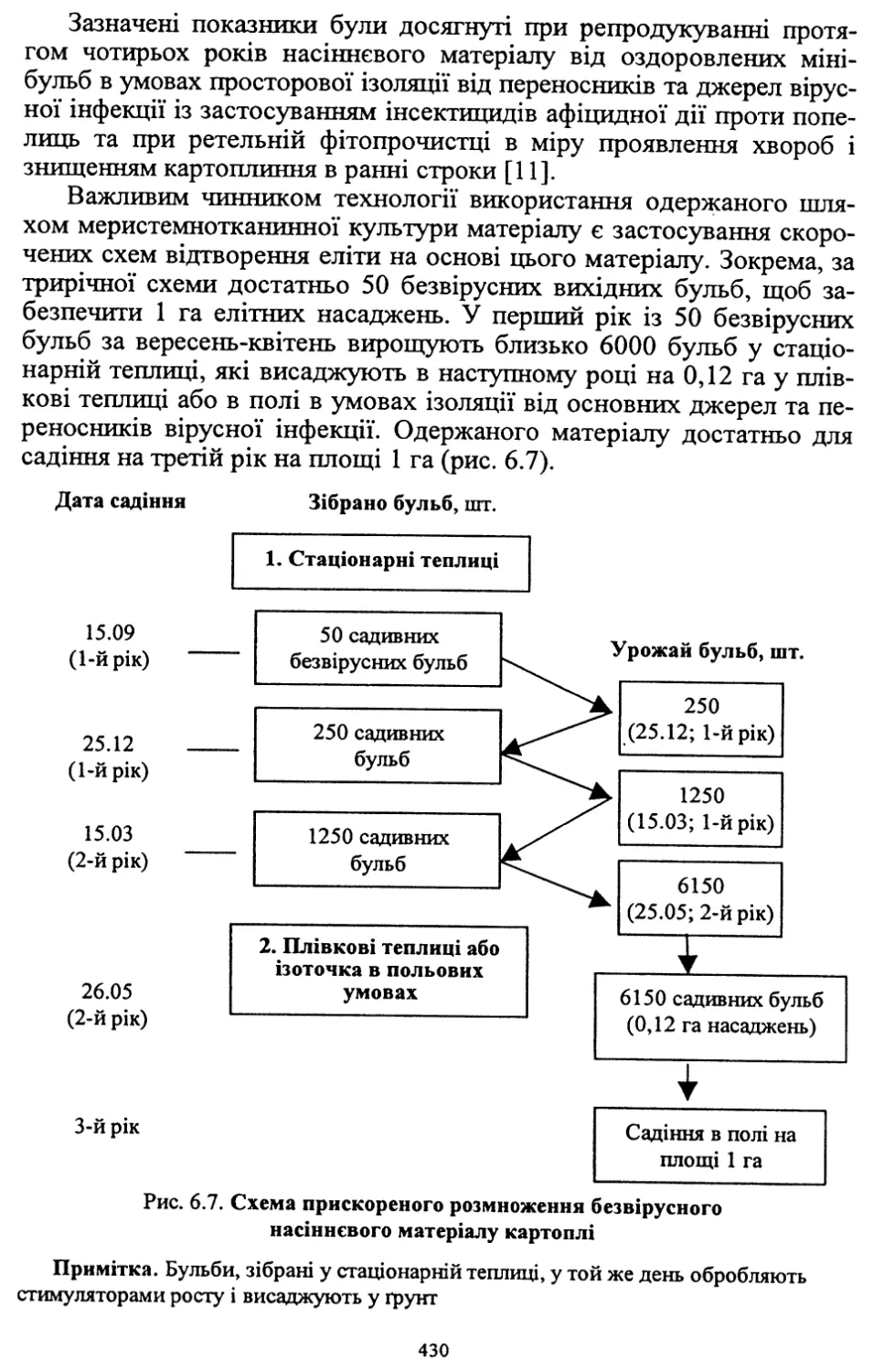
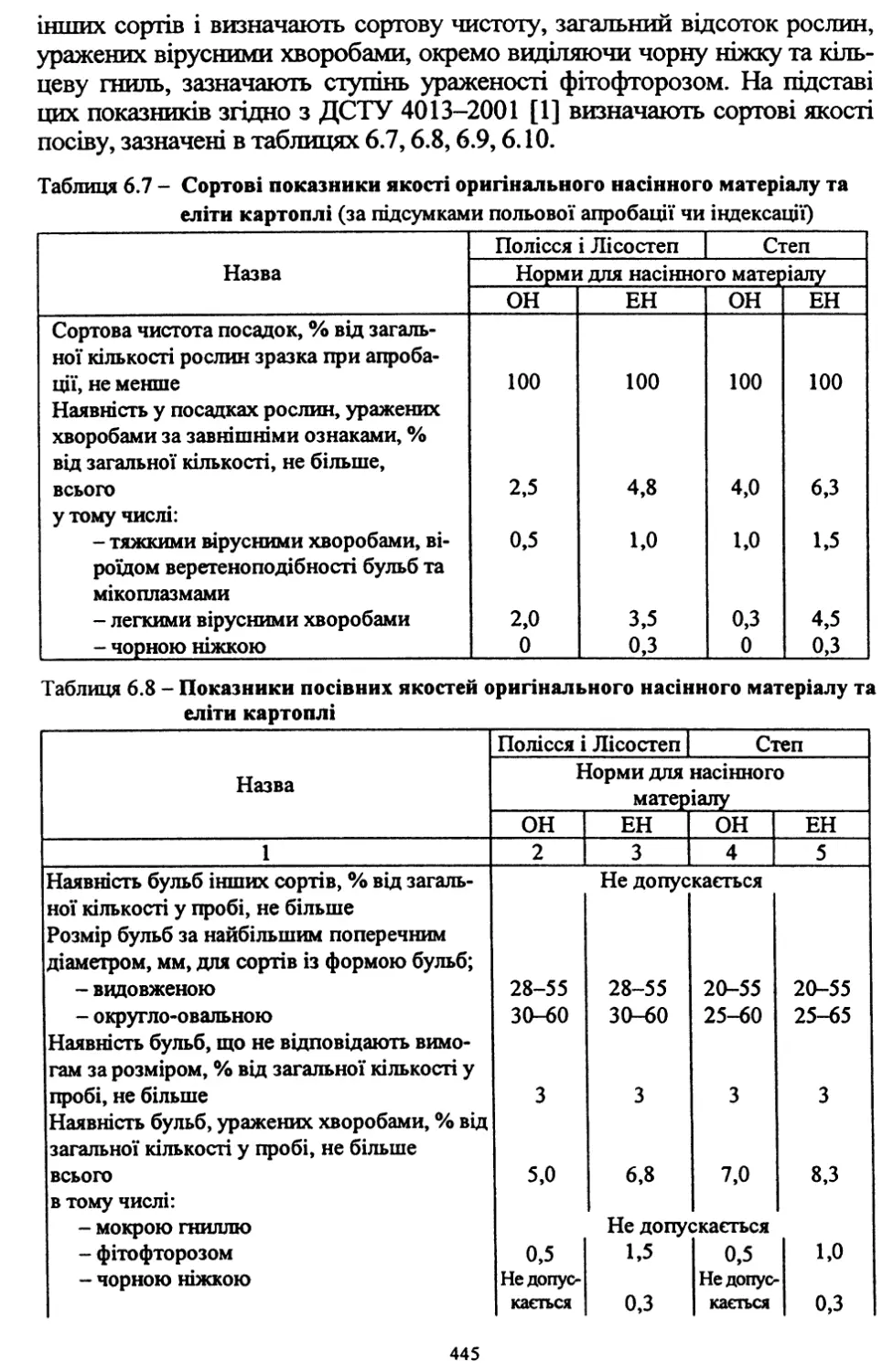
Розділ шостий. Насінництво картоплі (6.1, 6.2, 6.4, 6.8) - д. с.-г. н.,
проф. Молоцьким М.Я.; 6.3. Причини виродження картоплі і захо¬
ди боротьби з ним - к. с.-г. н. Верменком Ю.Я. і д. с.-г. н. Молоць¬
ким М.Я.; 6.5. Одержання вихідного матеріалу в насінництві карто¬
плі шляхом культури апікальної меристеми; 6.6 .Репродукційне та
внутрішньогосподарське насінництво - к. с.-г. н. Верменком Ю.Я.;
6.7. Апробація картоплі - к. с.-г. н. Верменком Ю.Я.
Розділ сьомий. Біотехнологія - завідуючою лабораторією біоте-
хнології та біохімії ПС УААН, к. б. н. Олійник Т.М.
п
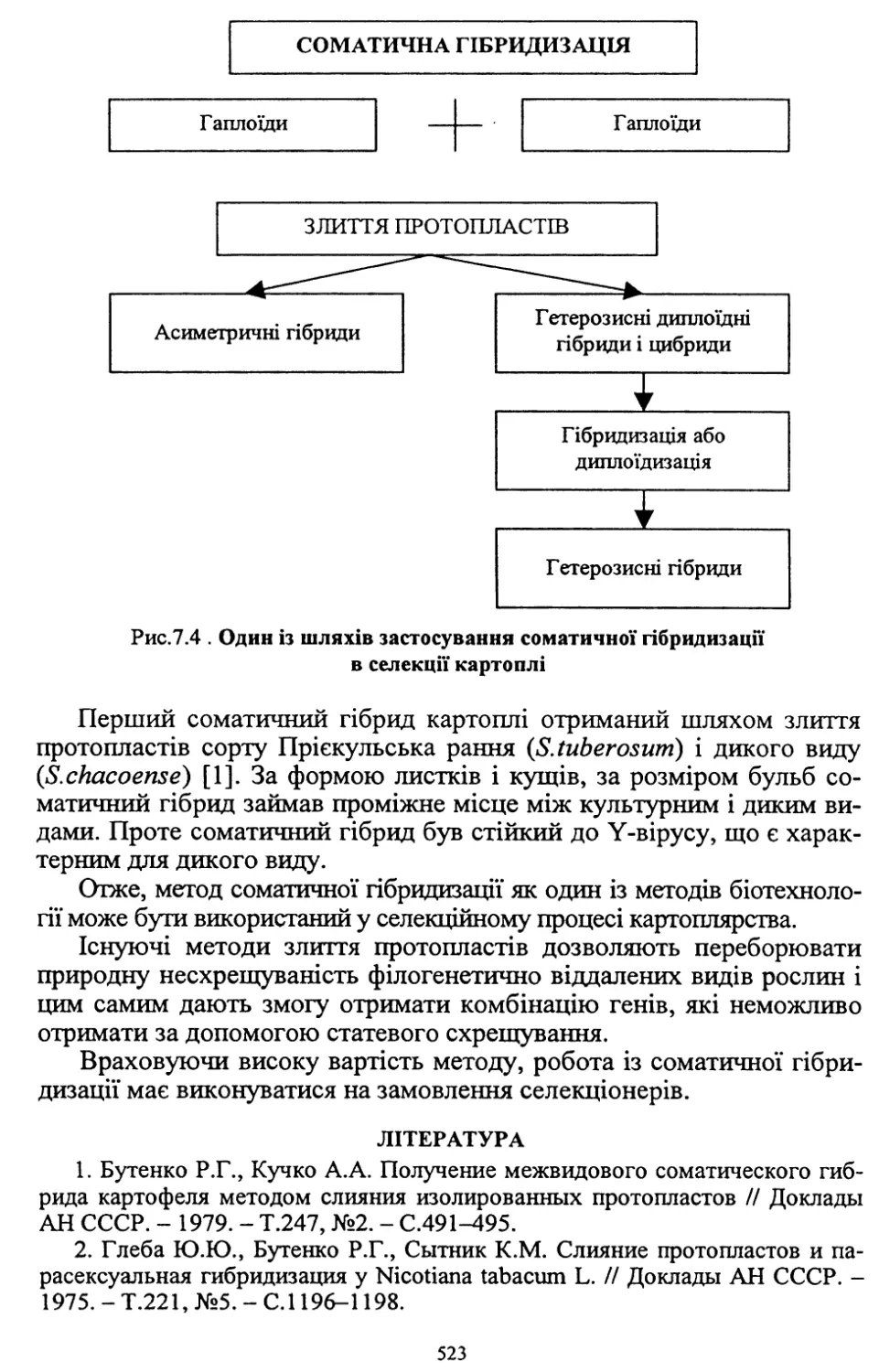
1. РОЗВИТОК КАРТОПЛЯРСТВА
В УКРАЇНІ
1.1. НАРОДНОГОСПОДАРСЬКЕ ЗНАЧЕННЯ
КАРТОПЛІ
Картопля займає важливе місце в сільськогосподарському вироб¬
ництві України. Будучи цінною продовольчою культурою, або дру¬
гим хлібом, як її називають у народі, картопля водночас є високока¬
лорійним кормом для худоби і однією з найголовніших технічних
культур, вирощуваних у республіці.
Вважають, що за універсальністю використання у різних галузях
народного господарства з картоплею не може зрівнятися жодна сіль¬
ськогосподарська культура.
Серед інших польових культур картопля займає одне з перших
місць за кількістю поживних речовин, яку можна одержати з одиниці
площі. Щоб забезпечити повну потребу в картоплі для продовольчих
і кормових цілей, для одержання насіння та переробки на картопле-
продукти, в Україні щорічно потрібно вирощувати 18-20 млн тонн
бульб. Площа під картоплею в республіці у 1995-2001 роках стано¬
вила понад 1,5 млн гектарів.
У бульбах картоплі міститься близько 26 % сухих речовин, із яких
80-85 % - це крохмаль і майже 3 % - білкові речовини. Її вуглеводи
є важливим джерелом енергії для людського організму, а білок за
якістю рівнозначний білку молока, яєць та яловичини, переважає бі¬
лок хлібних злаків, сої і бобів. Картопля є також своєрідним регуля¬
тором травлення, який підвищує засвоєння та біологічну цінність
інших продуктів. Цінність її як продукту харчування визначається
також високим умістом вітамінів, особливо протицинготної аскорбі¬
нової кислоти.
Калорійність картоплі дуже висока: при споживанні кілограма
бульб організм одержує близько 840 калорій. Споживання 300 г кар¬
топлі забезпечує майже 50 % добової потреби організму людини у
вітаміні С, 10-15 % - у вітаміні Ві і 15 % - у вітаміні РР.
Уміст різних поживних речовин у картоплі та її високі поживні і
смакові якості сприяють найрізноманітнішому застосуванню цієї
культури в кулінарії.
Відомо понад 1000 страв, які можна приготувати із бульб кар¬
топлі. Чимало з них стали національними в деяких країнах. Кар¬
топлю споживають печеною, вареною, тушкованою та смаженою.
Її використовують для приготування супів, борщів, гарнірів. Із
12
крохмалю готують різні дієтичні страви, фруктові та молочні ки¬
селі тощо.
Картопля є цінною кормовою культурою для всіх видів сільсько¬
господарських тварин. Гі бульби згодовують великій рогатій худобі,
використовують при відгодівлі свиней. Вона є цінним кормом і для
птиці.
Поживність 1 кг картоплі становить 0,3 кормової одиниці, 16 г
перетравного протеїну. З 1 га картоплі при врожаї 200 ц/га можна
одержати 6000 кормових одиниць. За кількістю кормових одиниць та
перетравного білка, які можна одержати з гектара посіву, картопля
поступається лише кукурудзі та цукровим бурякам і набагато пере¬
вищує ячмінь, овес, пшеницю, кормові буряки та інші культури. Се¬
редній урожай картоплі з одного гектара за кількістю кормових оди¬
ниць дорівнює врожаю вівса, зібраного з 2,2 га, ячменю - з 2 га або
вики - з 2,3 га.
Велике значення мають бульби картоплі як сировина для вироб¬
ництва крохмалю, клею, цукру, патоки, спирту, декстрину, глюкози
тощо. На заводах і фабриках із продуктів переробки картоплі вигото¬
вляють лаки, штучний шовк, парфуми, ліки та інші вироби. Картоп¬
ляний крохмаль, який виробляється в основному для продовольчих
цілей, використовується також у різних галузях промисловості: текс¬
тильній, консервній, м’ясо-молочній, паперовій, хімічній та ін.
Із тонни картоплі можна одержати 9,5 дкл спирту-сирцю або 140 кг
сухого крохмалю чи 100 кг декстрину. Із бульб картоплі виготовля¬
ють, крім того, сагову крупу, картопляну муку, сушену картоплю. У
1995-2001 роках в Україні значно зросло виробництво картоплепро¬
дуктів - чипсів, картоплі-фрі, картопляного пюре, мороженої і кон¬
сервованої картоплі та інших продуктів.
Крім важливої ролі, яку картопля відіграє як продовольча, кор¬
мова і технічна культура, вона має велике агротехнічне значення - є
хорошим попередником для зернових та інших культур. Після ран¬
ньої картоплі можна виростити непоганий урожай моркви, столових
буряків тощо.
В Україні картоплю вирощують в усіх грунтово-кліматичних зо¬
нах, однак найбільші площі вона займає в зоні Полісся.
Основним виробником картоплі у світі є Європа, частка якої ста¬
новить понад 50 % обсягів її виробництва. Найбільшими виробника¬
ми картоплі у світі є також Росія, Китай, Польща, Україна, СІЛА,
Німеччина, Білорусь.
1.2. СТАНОВЛЕННЯ І РОЗВИТОК
КАРТОПЛЯРСТВА В УКРАЇНІ
Картопля вважається чи не найголовнішим продуктом харчуван¬
ня в Україні. Почали її вирощувати тут понад 250 років тому.
13
У вирощуванні картоплі існувало кілька періодів, що пов’язано
як із поширенням інших сільськогосподарських культур, так і з на¬
буттям населенням знань про їх агротехніку.
Картопля є найпродуктивнішою сільськогосподарською культу¬
рою помірної зони. На території поліських і лісостепових областей
вона забезпечує найвищі врожаї і дає в півтора-два рази більше вуг¬
леводів з одиниці площі, ніж зернові культури. Крім того, картопля,
порівняно з іншими сільськогосподарськими культурами, має деякі
переваги з точки зору агроекономіки: вона добре росте в зоні Поліс¬
ся і Лісостепу на відносно бідних піщаних і супіщаних грунтах, які
менше придатні для зернових культур, а також дає хороші врожаї в
Степу при зрошенні.
Попит на картоплю як продукт харчування формувався поступо¬
во. Повільне поширення її на першому етапі пояснювалося незнан¬
ням основ агротехніки вирощування. Проте, населення поліської та
лісостепової зон України, маючи певні навички городництва, посту¬
пово долало ці труднощі, що й дало відповідний поштовх розвитку
культури. Наприкінці ХѴШ - на початку XIX ст. картопля стала для
нашого населення однією з основних сільськогосподарських куль¬
тур, серед яких важко знайти таку, котра б могла зрівнятися з нею за
універсальністю використання.
Поширення картоплі в Україні. Батьківщиною культурної кар¬
топлі є Південна Америка. Населення країн, які знаходяться на тери¬
торії гірського масиву Анд - від Чилі до Колумбії, знає цю рослину
дуже давно. Вважають, що зустріч первісної людини з дикою картоп¬
лею на території Південної Америки могла відбутися близько
14 тис. років тому, а, можливо, й раніше. Очевидно, спочатку первіс¬
на людина використовувала бульби дикої картоплі для харчування,
придумавши своєрідний спосіб її приготування - заморожування,
промивання та сушіння.
Вперше картоплю виявили у 1536-1537 рр. іспанські мореплавці
в індійському поселенні Сорокота на території сучасної Перу. Це
були учасники військової експедиції Гонсало де Касада. Через рік, у
1538 р., учасник другої експедиції Педро Сієза де Ліон у верхів’ї до¬
лини р. Каука, що на території сучасного Еквадору, також знайшов
м’ясисті бульби, які індійці називали “папа”. У написаній ним книзі,
яка вийшла у 1553 р. під назвою “Хроніка Перу”, читаємо: “Папа -
це особливий рід земляних горіхів, які, будучи звареними, стають
м’якими, як печений каштан; при цьому вони покриті шкіркою, яка
товстіша за шкірку трюфеля”.
Слово “картопля” (російською - “картофель”) походить від іта¬
лійського tartufo, що означає “трюфель”. Відомий із такою назвою
їстівний гриб, форма якого нагадує бульбу, росте в землі і є улюб¬
леним в італійців. Оскільки гриб відомий досить давно, то порів¬
няння його з картоплею було закономірним. З XVIII ст. слово “тар-
14
туфо” німецькою мовою з’явилось у сучасній формі Kartoffel, що й
стало основою російського варіанта назви “картофель”. Таким чи¬
ном, українська назва “картопля” запозичена з німецької та росій¬
ської мов.
В Україні, залежно від місцевості, слово “картопля” має чимало
синонімів, якими населення визначає однаково назву рослини і бу¬
льби, називаючи їх: картопелина, картоплина, картопля, картопа,
картофля, картофій, карток, картоха, картошка; надземну вегетати¬
вну масу - картоплиння, картофлиннє, картохлиннє, картошиннє;
площу, де вирощують картоплю, - картоплисько, картоплище, кар-
тофлисько; страви з картоплі - картопляник, картофляник, карто-
шник тощо.
У Європі, а пізніше й у Росії, картоплю називали також земляним
яблуком. На початку 1765 р. з ініціативи Державної медичної колегії
Сенат Росії видав указ “Про розведення і споживання земляних яб¬
лук, які звуться в деяких місцях “тартуфолями” або “картуфелями”».
Історики стверджують, що перша партія картоплі з Південної
Америки була завезена до Іспанії в 1565 р. Тоді ж з Іспанії картопля
потрапила до Італії. Проте в Іспанії, як і в інших країнах Європи, во¬
на спочатку вирощувалась не для споживання, а як декоративна або
лікарська рослина.
Одночасно з поширенням картоплі в Англії, Німеччині, Іспанії та
Франції ботаніки зробили опис рослини і подали її рисунок.
Так, у 1596 р. швейцарський ботанік Каспар Баухін у своїй праці
“Фітопінакс” описав рослину картоплі, вперше подав її латинську
назву {Solanum tuberosum). Ця назва існує й донині.
У 1601 р. австрійський ботанік Кароль Клузіус опублікував у книзі
“Історія рідкісних рослин” більш детальний опис картоплі. Вперше ко¬
льоровий малюнок рослини було виконано у 1588 р. Клузіусом із карто¬
плі, вирощеної ним у Віденському ботанічному саду.
У своєму каталозі рослин, виданому в 1596 р., та у “Травнику”,
який вийшов друком у 1597 р., англієць-ботанік Джон Джерарді та¬
кож подав опис картоплі і зробив рисунок стебла, квіток та бульб.
Професор Гамільтон знайшов у архіві однієї з лікарень Севільї
(Іспанія) матеріали, у яких зазначається, що в 1573 р. цей лікуваль¬
ний заклад купив на ринку для своїх хворих картоплю. Щоправда,
там не сказано, з якою саме метою здійснювали закупівлю - для лі¬
кування чи харчування.
У 1619 р. картопля входила до переліку страв англійського коро¬
лівського столу. В Англії відзначили заслуги в її розведенні адмірала
Вальтера Регеля. Хоч перші спроби пропаганди ним цієї культури не
відомі, він вважається поширювачем картоплі у цій країні. Вирости¬
вши картоплю на своєму городі, Регель приготував з неї якусь страву
з маслом та прянощами, але друзям адмірала вона не сподобалася,
тому що була із стебел та листків.
V
15
Не знайшла спочатку визнання картопля і у Франції. У народі на¬
віть стверджували, що вона отруйна. А французький парламент у
1630 р. спеціальним указом заборонив її поширення. У відомій “Ве¬
ликій енциклопедії-”, видання якої 1765 р. здійснили видатні францу¬
зькі вчені Дідро, Д’Аламбер та інші, повідомлялося, що картопля -
це груба їжа, придатна лише для невибагливих шлунків. У той же час
Путш повідомляв, що в 1616 р. картоплю подавали як делікатес до
королівського столу [9].
Ініціатор розведення картоплі у Франції аптекар Пармантьє у 1787 р.
подарував букет її квітів королю Людовику XVI. Останній приколов йо¬
го до свого одягу. Після цього в паризьких франтів стало модним носиш
квітку картоплі в петлиці. Селянам, що розводили картоплю біля фран¬
цузької столиці, за квітучу гілочку вони платили великі гроші.
Позитивно було оцінено значення картоплі в Німеччині. Прусь¬
кий король Фрідріх Вільгельм І у 1651 р. оголосив вирощування кар¬
топлі обов’язковим для всіх німців. Він навіть погрожував відрізува¬
ти носи і вуха тим, хто відмовлявся розводити цю рослину.
Так починалося вирощування картоплі по всій Європі. Однак
широкому її розповсюдженню у XVI-XVII ст. заважало невміння
використовувати бульби. Ті, хто починав вирощувати і використову¬
вати для харчування варені бульби, старалися їсти їх молодими. Але
в недостиглому вигляді вони мали гіркуватий присмак і містили чи¬
мало отруйної речовини (соланіну), що часто спричиняло отруєння
організму. Тому виникла думка, що картопля містить отруту, через
це й почали називати бульби “чортовим яблуком”.
Минуло ще чимало років, поки картопля стала другим хлібом.
Точних відомостей про рік завезення картоплі до Росії, куди входила
й територія України, поки що не знайдено. За даними Вільного еко¬
номічного товариства, вперше картопля була завезена в Росію з Гол-
ландії Петром І під час його подорожі по Європі - близько 1700 р. А
в 1736 р. картопля вже була серед інших культур на аптекарському
городі в Петербурзі. У 1741 р. страви з картоплі подавали в дуже ма¬
лій кількості на придворних банкетах.
У 1764 р. картоплю вже розводили під Ригою, в Естонії, у Новго¬
родській та Арзамаській провінціях, під Москвою.
Проте в Росії у ХѴШ ст. картоплю вважали розповсюджувачем
прокази, рослиною, що викликає дурощі в людини. Тоді її також на¬
зивали “німецьким овочем”, розведення якого принижувало націона¬
льну гідність. Існувала думка, що картоплю начебто висаджують для
того, щоб змінити віру [ 22 ].
Відомий російський історіограф картоплі В.Лехнович вважає, що
історію її поширення в Росії можна поділити на два періоди: перший -
до 1765 р., період стихійного проникнення культури торговими шля¬
хами; другий - після 1765 р., коли розпочалось організоване завезен¬
ня в країну насінного матеріалу [32].
іб
Якщо в 1736 р. у Петербурзі картоплю висаджували лише на ап¬
текарських городах, то вже через п’ять років бульби продавали на
ринках, щоправда, в дуже обмеженій кількості.
З різних архівних матеріалів відомо, що в 1791 р. картоплю ви¬
рощували у великій кількості на городах Вологди. Близько 1786 р.
вона вже була звичайною городньою культурою в Курську, а в 1796 р. -
польовою культурою в Казанській губернії.
Точно не з’ясовано, з якого року почали вирощувати картоплю на
території України. Відомо лише, що спочатку садили її на Лівобе¬
режжі, в Харківській та Полтавській губерніях, а з 1742 р. почали
вирощувати по всьому Правобережжю, тобто в Подільській, Волин¬
ській та Київській губерніях.
У 1765 р. Вільне економічне товариство надіслало всім губерна¬
торам анкети з питаннями про стан сільського господарства. В анке¬
ті, що надійшла з Київської губернії, повідомлялося, що тут картопля
з’явилася раніше, ніж її надіслали з Петербурга, і кияни вже виро¬
щують її на городах. Називали картоплю по-різному: земляні яблука,
тортофель, потетеси. Тоді ж, тобто у другій половині XVIII ст., за
пропозицією Медичної колегії була зроблена спроба організованого
завезення насінного матеріалу картоплі в Росію, у тому числі й в
Україну. З цією метою у 1765 р. Сенатом в Англії був закуплений
насінний матеріал, який розфасовували в бочки і відвантажували до
Росії. Усього було відправлено 464 пуди, з них у Малоросію - 4 боч¬
ки, у Слобідську Україну - 7 бочок по 5-6 пудів кожна. Проте з ра¬
порту губернатора цієї губернії Щербини за 1766 р. дізнаємося, що
ця картопля не була одержана - замерзла в дорозі [ 50 ].
На городах киян картопля була відома в 1764 р., хоча місцеві се¬
ляни її не вирощували. Дещо раніше почали вирощувати картоплю в
областях, які межують з Україною на північному заході. Так, існу¬
ють дані про те, що в Гродненській області (тепер Західна Білорусь)
картоплю почали розводити за Августа Ш, короля польського (1736—
1763 рр.) [7].
Широко була відома картопля в 1770 р. і в Закарпатській Україні,
яка тоді входила до складу Угорщини. З літературних джерел відомо,
що картопля в Галичині вперше з’явилась у 1780 р. у маєтку графа
Потоцького. Спочатку її вирощували в поміщицьких маєтках, а піз¬
ніше вона поширювалася і в селянські садиби. З кожним роком, за¬
лежно від зони, посіви картоплі збільшувалися, розширювалося її
використання.
Важливе значення для розширення посівів картоплі в Росії мало
заснування 21 жовтня 1765 р. Імператорського вільного економічно¬
го товариства, члени якого сприяли розведенню цієї культури.
Поширення картоплі в Україні пов’язують також із поселеннями
німців-колоністів. Відомо, наприклад, що в 1767 р. недалеко від Бор-
зни на Чернігівщині поселилися колоністи. У праці Е. Дракохруста,
17
де він посилається на історичні джерела, знаходимо, що в Борзнян-
ській економії в 1774 р. місцеве населення “знайшло смак у картоплі
і купувало в колоністів по 80 коп. за чверть. Бульби їли особливо у
великий піст” [21,56]. З-поміж інших культур картопля їм найкраще
вдавалась. Лишок врожаю бульб жителі обмінювали на жито та інші
злаки.
Поміщики засвоїли технологію винокуріння, для чого до картоплі
додавали третю частину житньої муки. При додаванні до картоплі поло¬
вини пшеничної муки пекли з картоплі досить хороший хліб [52].
Лікар А. Шафонський у книзі “Чернігівського намісництва топо¬
графічний опис 1783-1784 років” наводить дані про розведення кар¬
топлі в різних повітах країни на городах та полях. Він назвав карто¬
плю на той час звичайною для українців культурою [19, 56]. її роз¬
множували “звичайним паростком”.
З архівних матеріалів відомо, що у 80-х роках ХѴШ ст. у Ніжин¬
ському окрузі Чернігівського намісництва картопля на городах ро¬
дила майже щороку і давала задовільний урожай. Так було і в Ро-
менському окрузі. Проте весь урожай використовувався на харчу¬
вання. Досить високий урожай картоплі збирали в цей час у Харків¬
ській губернії. Так, 1795 р. на городах Чернігівського округу врожай
картоплі досягав “сам” двадцять і більше, а у Вовчанському окрузі -
“сам” двадцять п’ять [ 52,53 ].
У західних областях України (у маєтках Потоцьких) польова
культура картоплі була відома вже з 1780 р., а як городня культура
вона вирощувалась і раніше. Потім ці області стали основними райо¬
нами вирощування картоплі в Австро-Угорщині.
Як свідчать архівні матеріали, у Харківському намісництві на¬
прикінці XVIII ст. площа під картоплею швидко зростала. Так, у
1788 р. під нею було зайнято 18 десятин, а в 1790 р. - вже 195 деся¬
тин. У Катеринославському намісництві у 1794 р. на городах помі¬
щиків садили картоплю, а бульби використовували для харчування
[50,51].
У Таврійській губернії у 1796 р. бульби вирощували лише на де¬
яких городах. Місцеве населення висаджувало картоплю наприкінці
березня, а збирало на початку липня.
У Криму в 90-х роках XVIII ст. картоплю вирощували лише на
деяких городах росіян. Серед архівних матеріалів зустрічаються
дані про те, що в Катеринославській, Таврійській, частково
Херсонській і Чернігівській губерніях інтенсивне розведення кар¬
топлі в 1795-1804 рр. здійснювалося іноземними колоністами: нім¬
цями, болгарами, греками. У цих областях картоплею засаджували
близько 360 десятин [8,19].
Крім даних про обсяги виробництва картоплі в окремих райо¬
нах України в 1780-1800 рр., в архівах збереглися матеріали про її
врожайність. З них дізнаємося, що, наприклад, у Харківському на¬
18
місництві Білопільського округу картопля вирощувалась у поміщи¬
ків та селян, і врожай бував майже щорічно. В Охтирському окрузі
бульби використовували навіть на корм худобі та на продаж, коли
на удобрених грядках одержували високі врожаї. У Малоросійській
губернії, в Конотопському, Миргородському, Глухівському, Сос-
ницькому округах у 1798-1800 рр. на городах вирощувалось багато
картоплі [49].
Таким чином, наприкінці ХѴІП ст. картоплю вирощували на всій
території України.
Картоплярство у XIX та на початку XX ст. У першому десяти¬
річчі XIX ст. у поміщицьких і селянських господарствах картоплю
вирощували на невеликих площах, але з кожним роком вони зроста¬
ли. Картопля починала займати чільне місце як продовольча, кормо¬
ва і технічна культура, що використовувалася для переробки на
спирт. Вирощування бульб на спирт зробило картоплю для поміщи¬
ків джерелом прибутків.
Попит на картоплю формувався поступово. Українське населен¬
ня, маючи давній досвід вирощування городніх культур, поступово
засвоювало практику вирощування нової культури, вносячи власні
корективи до “закордонних” рекомендацій з агротехніки вирощуван¬
ня картоплі, що також сприяло її поширенню.
Розвиток картоплярства на початку XIX, як і в кінці ХѴШ ст., ба¬
зувався на вирощуванні сортів, завезених із європейських країн. Се¬
ляни розводили ті сорти, які були їм більше до вподоби [ 9 ]. Відомо,
наприклад, що в 1805-1813 рр. до Росії з-за кордону було завезено
насінний матеріал ряду сортів. Це були урядові закупівлі, але насіння
завозили також і окремі поміщики. Безумовно, частина насінного
матеріалу потрапляла й до українських губерній.
У 1805 р. площа під картоплею залишалася все ще незначною, її
висаджували в поміщицьких господарствах по 1-2 грядки, проте че¬
рез двадцять років, тобто у 1825 р., насадження картоплі помітно
зросли, і поступово вона посіла одне з перших місць серед польових
культур [ 32,33 ].
На розвитку картоплярства в Україні певною мірою позначилось
і те, що в різні періоди ХѴШ-ХІХ ст. окремі її території належали до
різних держав, які мали різний політичний і земельний устрій. Так,
наприклад, Східна Галичина була сільськогосподарською провін¬
цією Австро-Угорщини. У 40-х рр. Галичина виробляла 38 % зерна,
23 % - льону, 16 % - тютюну, 28 % - конопель і 38 % - картоплі. Зем¬
леробство мало вівсяно-картопляний напрям. У зв’язку з тим, що на
душу населення тут вироблялось лише 60 кг жига і пшениці, в хар¬
чуванні населення картопля займала помітне місце.
На початку XIX ст. у Східній Галичині, Північній Буковині та в
Закарпатті картопля стала найважливішою культурою - як продукт
харчування, корм для худоби та сировина для горілчаних підпри¬
19
ємств. Вона почала витісняти посіви зернових культур. За 1815-1840 рр.
збір бульб виріс майже у 18 разів. Картопляний напрям господарю¬
вання призвів до виснаження і без того бідних грунтів, а коли в 40-х
роках сталося масове ураження картоплі хворобами, - до бідувань
селян [ 2,54 ].
На поширення картоплі в західних губерніях Росії мало вплив її
вирощування та використання й на західному кордоні - у Польщі. На
початку XIX ст. картопля в Польщі займала значні площі, валовий
збір бульб був також досить високий. Так, на території Варшавсько¬
го князівства в 1810 р. було зібрано близько 50 тис. тонн бульб, а у
1827 р.- вже 115 тис. тонн [ 13,20 ].
Відомо, що в Європі у 1844-1845 рр. була досить поширеною та¬
ка хвороба, як фітофтороз, що призвело до значного скорочення
площ посіву картоплі. Проте у Польщі цієї хвороби не було, і в 1847 р.
площа під картоплею в королівстві займала 210 тис. га, а до 1870 р.
ще й збільшилась у два рази [14,41,46].
У 1845 р. на сусідній із Польщею території Волині картопля була
після хліба основним сільськогосподарським продуктом харчування
[47,48].
У 1845 р. Єсимонтовський описав поширення картоплі в Суз¬
дальському повіті Чернігівської губернії. За його свідченнями, 40
років тому в повіті картоплю висаджували лише в поміщицьких са¬
дибах, по одній-дві грядки. Через двадцять років почали садити ряд¬
ками під соху малими ділянками на овочевих городах, тому що ніхто
не вірив у можливість її вирощування на звичайному полі [ 56 ]. Він
писав: “Найбідніший селянин дбає про те, щоб зробити з неї запаси,
часто-густо, за браком іншої можливості, він поступається для неї
частиною площі, підготовленої для його улюблених конопель. Що
менший урожай і що більші злидні, то більше дбає селянин про те,
щоб посадити картоплю, купуючи її навесні за велику ціну на насін¬
ня, продаючи останній шматок конопляного прядива”. “Бульбу та
козу для бідних створено,” - говорили селяни повіту [ 57 ].
З архівних матеріалів відомо, що в 1844 р. в Чернігівській, Хар¬
ківській, Полтавській, Волинській та Київській губерніях посіви кар¬
топлі складали близько 150-200 м2 у середньому на душу сільського
населення [ 1, 35 ].
До 1847 р. розвиток картоплярства набрав відчутного розмаху. У
цей час бульби були підмогою в харчуванні. У деяких повітах картоп¬
ля майже повністю займала селянські городи, а частково висаджува¬
лась і в полі. Щорічно на Київщині площі під картоплею становили:
в Чигиринському повіті - 1000-2000 десятин, Таращанському - 600-
3000 і Лохвицькому - 900-1000 десятин.
В окремих літературних джерелах 1844 р. наводяться основні тех¬
нологічні вимоги щодо грунту для вирощування картоплі. У Харківсь¬
кій губернії, наприклад, були переконані, що “картофель можно иметь
20
только на огородней земле, т. е. хорошо прорыхленной и в низинных
местах”. Вважалось, що картопля погано родить на полях [20].
У 30-х роках XIX ст. картоплярство було дуже слабко розвинене
на землях Чорноморського війська, де картопля, незважаючи на
сприяння уряду, майже зовсім не вирощувалась [5, 40].
Відомо, що в Україні, як і в Росії, значну організаційну роботу
серед селян щодо агрономічної діяльності в 1870-1890 рр. проводи¬
ли губернські земства. Вони привертали увагу сільських господарів
до вирощування сіяних трав, розвитку садівництва та городництва,
догляду за худобою тощо. Але питанням насінництва, зокрема заве¬
зенням і пропагандою сортів різних культур, у тому числі й картоплі,
земства не займались [17].
У працях Вільного економічного товариства за 1843 р. знаходимо
відомості про те, що картопля все ширше починає вирощуватись для
винокуріння в Чернігівській та Могильовській губерніях [26].
Наприкінці XIX ст. вже існували певні висновки щодо агротехні¬
ки вирощування картоплі. У 1897 р., через 100 років від початку ви¬
рощування картоплі в Україні, С.Сутулов писав, що вона є однією з
найкорисніших рослин, яку доводилось вирощувати людині, проте
повної віддачі від картоплі не одержують лише тому, що не вміють її
вирощувати, особливо селяни.
З таблиці 1.1 видно, що наприкінці XIX ст. площі посівів картоплі
хоч і були незначними, але швидко розширювались. Причому, навіть у
південних областях (Херсонська і Катеринославська) вони збільшилися.
Найбільшими площі картоплі у відсотках до загальних посівів у 1885 р.
були у Київській та Катеринославській губерніях [21,57].
Таблиця 1.1 - Поширення картоплі в окремих губерніях України (1881-1895 рр.)
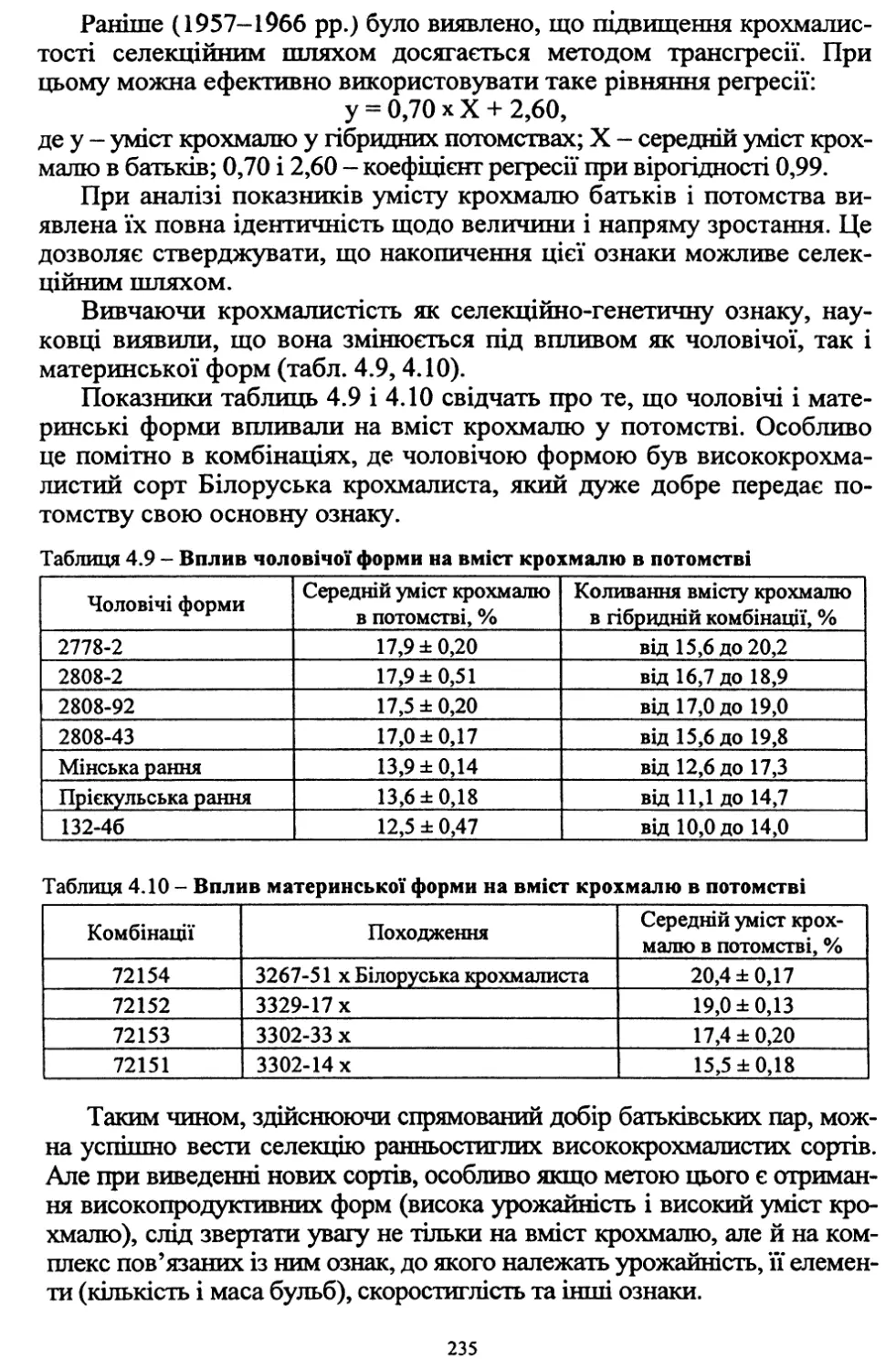
Губернія
Площа картоплі,тис. десятин
1885 р.
у процентах
до 1881 р.
1881р.
1885 р.
Чернігівська
21,7
45,3
346
Харківська
20,7
38,7
185
Полтавська
20,4
42,8
210
Херсонська
44,2
40,0
91
Катеринославська
29,2
38,2
131
Поліська
18,6
45,8
246
Київська
22,6
49,4
218
Волинська
47,7
78,7
165
Цікавим є порівняння площ посіву картоплі на півдні України в
Херсонській та Катеринославській губерніях у 1895 р. та через сто
років - у 1995 р. У 1885 р. у двох областях картопля висаджувалась
на площі 85 тис. га. А в 1995 р. в Херсонській, Кіровоградській, Оде¬
ській, Запорізькій, Дніпропетровській областях її вирощували на
площі 250 тис. га.
21
Подальшого розвитку картоплярство набуло в Україні на початку
XX ст. Так, у 1906-1910 рр. у структурі посівних площ ‘"хлібів” у селян
та поміщиків картопля займала в Чернігівській губернії - 8 %, Волинсь¬
кій - 4,9, Київській - 3,8, Харківській - 2,8, Херсонській - 1,2 %. В окре¬
мих районах вона значно перевищувала ці показники. Найменші площі у
структурі посівів сільськогосподарських культур були під картоплею в
Таврійській (0,7 %) та Катеринославській (1,7 %) губерніях.
У 1901-1905 рр. у Подільській губернії вирощували 51,3 тис. де¬
сятин картоплі, у Київській - 115,4 тис. десятин.
Врожаї у Чернігівській губернії становили 905-846 пудів з однієї
казенної десятини, у Волинській - 884-829, у Київській - 707-644, у
Харківській - 874-770 пудів.
У 1906-1910 рр. картоплею все активніше почали займатись як
продовольчою і кормовою культурою в поміщицьких маєтках. Ана¬
логічна робота проводилась і в селянських господарствах. У літера¬
турі того часу зустрічаємо запис: “Картофель - исключительный по
важности в крестьянском хозяйстве пищевой продукт, а затем - и как
незаменимое кормовое средство для всякого рода рогатого скота,
свиней и лошадей” [10].
Значно вплинуло на зростання посівів картоплі використання
бульб на винокуріння, в результаті чого у Європейській частині Росії
виникли відомі центри з виробництва спирту.
У 1910-1911 рр. у 50 губерніях Європейської частини Росії нара¬
ховувалось понад 2300 винокурних заводів, з них у чорноземній зоні
їх було 1129, а в нечорноземній - 1188. У середньому один завод пе¬
реробляв 62 тис. пудів картоплі на рік. Майже 60 % усієї переробле¬
ної картоплі припадало на чорноземні області, що й призвело до зро¬
стання виробництва картоплі в цій зоні.
На початку XX ст. кількість винокурних заводів на території су¬
часної України також різко зростала (табл.1.2).
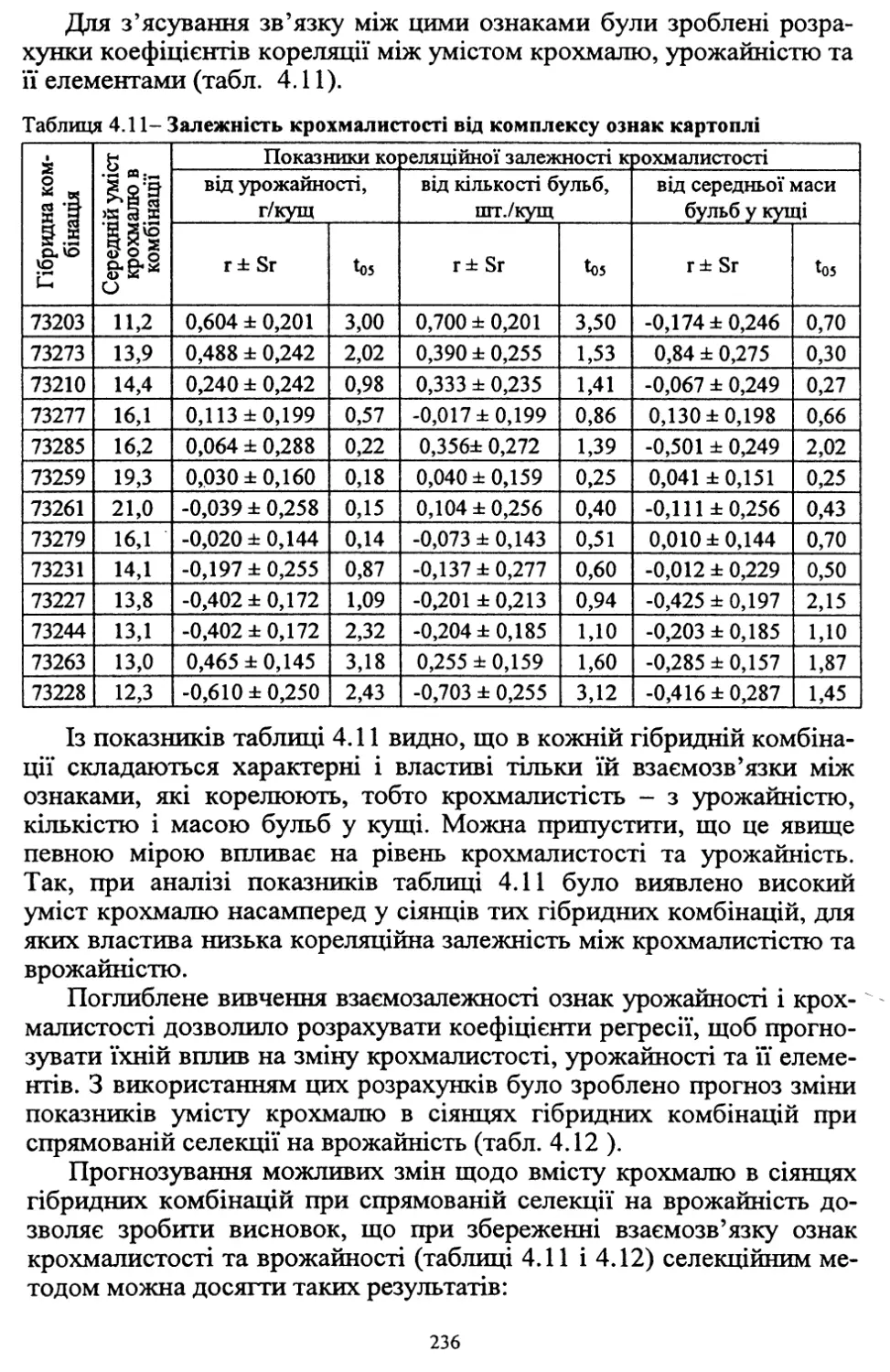
Таблиця 1.2 - Наявність винокурних заводів у різних губерніях України
Роки
Губернії
Подільська
Київська
Волинська
1868
7
19
5
1911
99
71
136
Кількість винокурних заводів збільшилась до 236 у Чернігівській,
65-у Полтавській, 51-у Харківській губерніях. У цей час на один
винокурний завод, залежно від зони, припадали різні обсяги перероб¬
лених бульб. У Чернігівській губернії, наприклад, переробили 84
тис. пудів, Київській - 73, Волинській - 58, Полтавській - 44, Хер¬
сонській - 4 тис. пудів картоплі. Важливо підкреслити, що навіть у
Полтавській та Херсонській областях, де в наш час під картоплею
зайняті незначні площі, бульби переробляли на спирт.
22
Таким чином, винокуріння стало значним фактором, який впли¬
вав не лише на розширення площ під картоплею, але й на підвищен¬
ня її врожайності.
На початку XX ст. зростає кількість заводів із переробки картоплі
на крохмаль. У 50 європейських губерніях Росії у 1908 р. нарахову¬
валось 205 таких заводів, у тому числі: у Харківській губернії - 1,
Полтавській - 2, Херсонській -1, Київській - 1, Волинській - 1 завод.
Слід зазначити, що в цей час на Півдні України з картоплі виробляли
також крохмаль. Досить потужним був Херсонський крохмальний
завод. Щорічно крохмалю вироблялось на суму 233 тис. руб, тоді як
у Росії один завод виробляв продукції на 54 тис. руб.
В інформації від поміщиків та селян Поділля зазначалося, що у
зв’язку із збільшенням кількості винокурних заводів картопля при
зростанні площ стає вигіднішою, ніж цукрові буряки [26].
Урожайність картоплі у 90-х роках XIX ст. була різною і залежа¬
ла від зони та господарства, однак, вищий урожай бульб збирали в
нечорноземних губерніях, де умови для вирощування картоплі були
кращі. Середню (за 1901-1910 рр.) врожайність картоплі по зонах
Росії подано в таблиці 1.3.
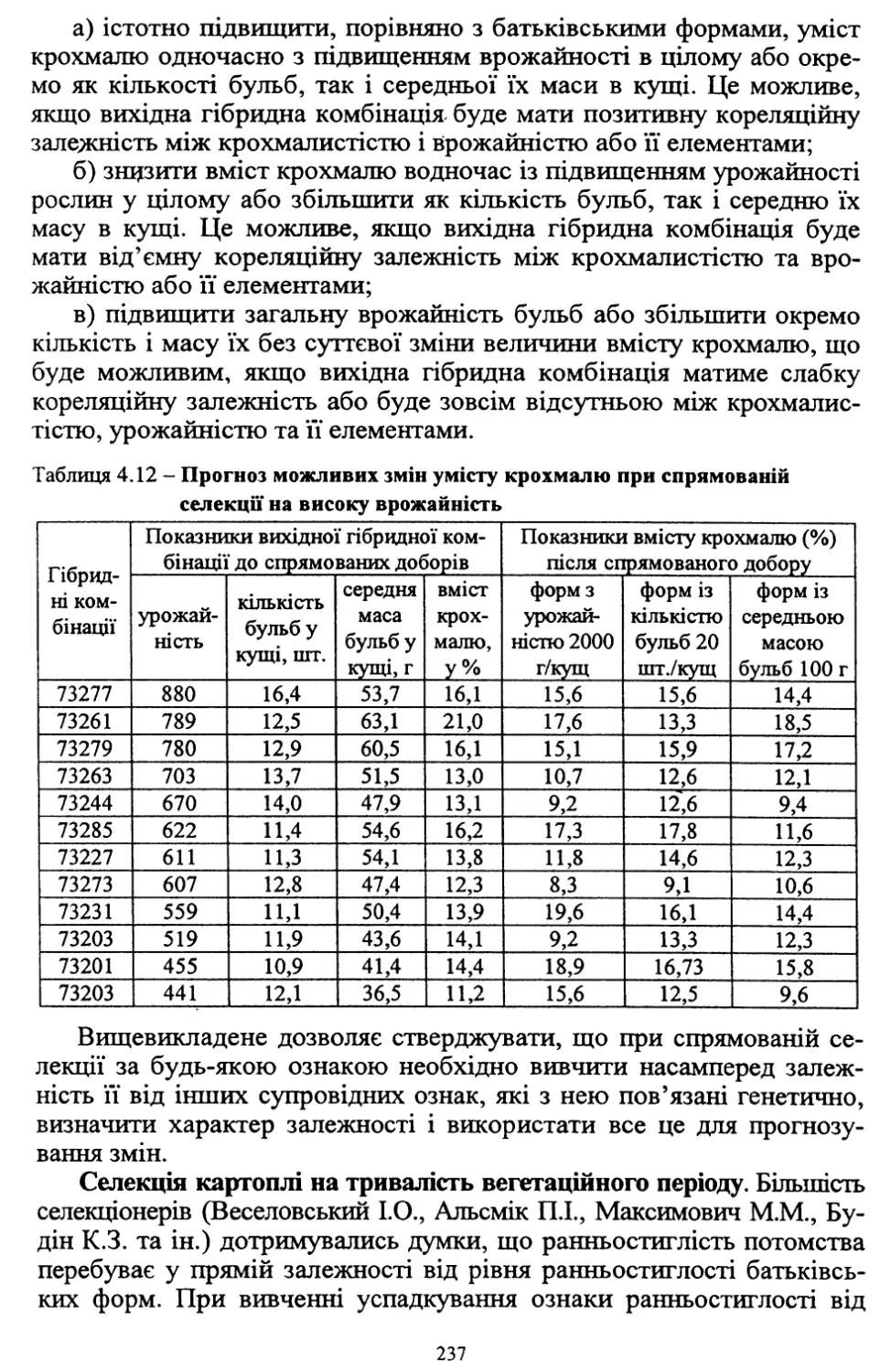
Таблиця 1.3 - Урожайність картоплі у різних господарствах, у пудах з десятини
Зона вирощування
Поміщики
Селяни
У Європейській частині Росії, у середньому
455
410
Чорноземна зона
442
393
Нечорноземна зона
466
427
У різних зонах України врожай бульб за цей період становив у
середньому (пудів із десятини):
новоросійські губернії (Херсонська, Таврійська, Катеринославсь¬
ка, Бесарабія) - 268;
південно-західні губернії (Подільська, Київська, Волинська) - 529;
малоросійські губернії (Харківська, Чернігівська, Полтавська) - 528.
Найнижчою врожайність за десятиріччя була в Херсонській гу¬
бернії - 237 пудів із десятини.
Про врожайність картоплі в 1906-1920 рр. свідчать також статис¬
тичні дані, зібрані від респондентів різних зон. Вищою врожайність,
як і раніше, залишалась у поміщицьких маєтках, де, як правило, ви¬
користовували кращий садивний матеріал та застосовували мінера¬
льні добрива.
Середню врожайність за 1906- 1910 рр. по губерніях наведено в
таблиці 1.4.
Причому, збір бульб значною мірою залежав від погодних умов,
вегетаційного періоду та агротехніки вирощування картоплі [ 25 ].
Сортовим посівам картоплі не надавали особливого значення і
використовували в основному різні місцеві сорти. Однак біля ви¬
23
нокурних заводів розводили такі сорти, як Алкоголь, Імператор,
рідше - Білий слон та Рання американка (очевидно, це була Рання .
рожева).
Таблиця 1.4 - Урожайність картоплі по губерніях у різних господарствах,
у пудах з казенної десятини
Губернія
Поміщики
Селяни
Подільська
870
685
Київська
784
711
Волинська
943
688
Херсонська
433
393
Чернігівська
815
789
Полтавська
824
739
За перше десятиріччя XX ст. площі під картоплею в різних губе¬
рніях України продовжували змінюватися. У 1910 р., порівняно з
1901 р., площі зросли у Волинській губернії на 34 %, Полтавській -
на 12 %, Подільській - на 18 %. Водночас після активного розши¬
рення площ наприкінці 90-х років XIX ст. в 1910 р. вони зменшилися
на півдні - на 18 % у Таврійській губернії. Не збільшилися площі в
Херсонській та Харківській губерніях. У Херсонській губернії в
1906-1910 рр. висаджували 43 тис. десятин картоплі.
Особливої уваги заслуговує практика вирощування картоплі на
Чернігівщині. На легких малородючих грунтах Чернігівщини, де на
початку XIX ст. зернові культури давали низькі врожаї, картопля по¬
повнювала господарство селянина продуктами харчування і кормом
для худоби.
Площі під картоплею на Чернігівщині, за даними С.П.Кульжин-
ського [50], швидко збільшувалися, спостерігалось і певне підви¬
щення врожайності бульб (табл.1.5).
Таблиця 1.5 - Посівні площі та врожайність картоплі на Чернігівщині
у 1881-1910 рр.
Рік
Площа посіву, тис. га
Урожайність, ц/га
1881
24
-
1892
46
57
1895
75
72
1900
108
76
1910
126
78
Отже, за 29 років посівні площі картоплі, як видно з таблиці, збіль¬
шились на 102 тис. га. Такого швидкого розширення посівів цієї
культури не спостерігалося в жодній іншій губернії України. Посту¬
пово зростала і врожайність картоплі, хоч вона і залишилась усе ще
досить низькою [ 11,29].
Причому, значна частина врожаю з України вивозилася на Кавказ
та в інші регіони Росії. Так, лише Харківська губернія відправила
24
залізницею у 1905-1909 рр. 1367 тис. пудів картоплі, Чернігівська -
1017, Полтавська - 263 тис. пудів. Найбільше вивозилось картоплі до
Одеської області, Криму, на Кавказ, Кубань, до Москви, Санкт-
Петербурга. Це були в основному сорти Імператор, Меркер, Вольт¬
ман, Канцлер та інші [ 26,36 ].
У 1913 р. посівна площа картоплі на території України складала
1080 тис. га. Цей показник прийнято за відліковий при визначенні та
оцінці площ картоплі в наступні десятиріччя. Валовий збір бульб до¬
сягав 8,5 млн тонн.
Картоплярство у 1920-1990 роках. Період післяреволюційної
розрухи та громадянська війна в Росії 1917-1922 рр. призвели до
значного скорочення посівів картоплі (табл. 1.6).
Таблиця 1.6 - Площі під картоплею у 1913-1921 рр.
1913 р.
1916 р.
1917 р.
1920 р.
1921 р.
100 %
83%
77%
52%
50%
Скоротились за цей час площі вирощування картоплі і в Україні.
Так, під картоплею було зайнято на одне господарство в 1916 р - 5,1,
а в 1922 р. - 3,2 десятини [55].
Щоб покращити становище, уряд почав вживати певних заходів.
У 1919 р., в Україні, наприклад, було створено місцеві губернські та
повітові комісії з насінництва, які повинні були займатися не лише
заготівлею, але й розподілом насінного матеріалу. Це значною мірою
стосувалось і картоплі [12,23, 38].
В архівних джерелах наявні відомості про те, що в 1919 р. із за¬
пасів Полтавської та Херсонської областей до 15 квітня 1919 р. було
видано наряд на садивний матеріал картоплі, який був украй потрі¬
бен у деяких повітах Чернігівської, Київської та Харківської губер¬
ній. Усього на насіння було виділено 26,0 тис. пудів бульб [24].
За статистичними даними [27], до 30-х років картопля більше ви¬
рощувалась на городах. Так, у 1926 р. площа під картоплею у різних
районах на городах і в польових посівах характеризувалась показни¬
ками, поданими в таблиці 1.7.
Таблиця 1.7 - Розподіл площі садіння картоплі по регіонах та місцях
вирощування, тис. га
Регіон
Город
Польові посіви
Разом
Правобережжя
184,0
182,0
366,0
Степ
133,0
86,0
219,0
Лівобережжя
88,0
81,0
169,0
Полісся
73,0
199,0
192,0
Валовий урожай картоплі в Україні у 1928 р. по зонах, за даними
ЦСУ, відображено в таблиці 1.8.
25
Таблиця 1.8 - Валовий збір картоплі у різних зонах, 1928 р.
Регіон
Урожай, тис. тонн
У процентах
Полісся
2275
25
Правобережний Лісостеп
4180
46
Лівобережний Лісостеп
ЗОЮ
29
У наступні роки площі під картоплею знову почали зростати.
Якщо в 1917 р. площу посіву картоплі на Київщині взяти за 100 %, то
у 1928 р. вона зросла на 70 %. Найбільше зросли площі в Переяслав¬
ському, Васильківському, Київському районах, які постачали карто¬
плю до Києва. Середній врожай бульб складав 90 ц/га. Однак, про
потенційний урожай картоплі сортів у різних районах України в
1925-1928 рр. за існуючої на той час технології можна судити з да¬
них, одержаних дослідними сільськогосподарськими станціями
[27,37] (табл. 1.9).
Таблиця 1.9 - Урожайність картоплі на дослідних станціях, 1925-1928 рр.
Дослідна станція
Область
Урожайність, ц/га
Носівська
Чернігівська
350
Уманська
Київська
102
Херсонська
Херсонська
64
Сумська
Сумська
174
Харківська
Харківська
217
Полтавська
Полтавська
206
Радомишльська
Житомирська
197
У 20-х роках XX ст. площі посіву картоплі в Україні були таки¬
ми: 1923 р. - 395 тис. десятин; 1925 р. - 439; 1926 р. - 470; 1928 р. -
662 тис. десятин, а з 1932 року вони постійно зростали (табл. 1.10)
[28,43].
Таблиця 1.10 - Площа посіву та врожайність картоплі в Україні, 1930-1935 рр.
Рік
Площа посіву картоплі,
тис. га
Середня
врожайність, ц/га
Валовий збір,
тис. тонн
1930
1199
74,7
8957
1935
1263
80,3
10909
Проте врожайність картоплі значною мірою змінювалась і зале¬
жала від грунтово-кліматичних умов [ 49 ]. Для порівняння можна
взяти врожайність за 1933 і 1935 роки в окремих областях (табл. 1.11).
Це офіційні матеріали перших років колективного господарювання, і
про їх об’єктивність важко судити.
У 1928 р. використали садивного матеріалу на десятину (пудів):
Полісся - 94,9, Правобережжя - 99,8, Лівобережжя - 94,8, Степ - 44,0.
У середньому висаджували на десятину 84,1 пудів насіннєвої карто¬
плі [ 15,43 ].
26
Таблиця 1.11 - Урожайність картоплі по областях України, 1933 і 1935 рр.
Область
Рік
Площа посіву,
Урожайність,
тис. га
ц/га
1933
210,3
79,2
Київська
1935
296,9
102,3
1933
162,2
75,0
Чернігівська
1935
215,7
109,9
1933
112,2
62,2
Харківська
1935
200,3
106,2
1933
60,1
59,4
Дніпропетровська
1935
104,8
34,7
На Носівській дослідній станції в 1936 р. на опідзолених чорно¬
земах при достатньому удобренні і високій агротехніці врожай досяг
562 ц/га.
Обстеження, проведені у 30-х роках XX ст. Київською дослідною
станцією, показали, що селяни здебільшого висаджували суміш сор¬
тів, які відрізнялись між собою різним забарвленням квіток і бульб.
Кущі достигали неодночасно. Сортові посіви займали лише одну чет¬
верту площ, але були значно врожайнішими.
Післявоєнний період (1945-1955 рр.) характеризувався інтенсив¬
ним розвитком галузі картоплярства (табл. 1.12). Проте за умов від¬
будови народного господарства картоплярство, як і сільське госпо¬
дарство в цілому, не одержувало достатньої кількості техніки, міне¬
ральних добрив і базувалося в основному на фізичній праці і живій
тягловій силі. У перші повоєнні роки певний вплив на розвиток кар¬
топлярства мала Постанова Ради Народних Комісарів від 14 березня
1945 р. “Про поліпшення насінництва картоплі в Українській РСР”.
Таблиця 1.12 - Виробництво картоплі у 1945-1955 рр.
Роки
Всього
У тому числі населенням
площа,
тис. га
валовий
збір,тис. т
урожай¬
ність, ц/га
площа,
тис. га
валовий
збір, тис.т
урожайність,
ц/га
1945
1833,5
13770
75,1
1254,5
11212
89,4
1946-1950
1802,8
17930
99,4
1246,9
4782
118,6
1951-1955
2040,8
16067
78,7
1182,6
14782
94,1
Як видно з таблиці, у цей період посівні площі картоплі щороку
збільшувалися, зростали валові збори, хоча врожайність була досить
низькою. У 1945 р. площа під картоплею досягала 89 %, а валовий
збір становив 66 % від довоєнного. Лише в 1951-1955 рр. загальна
площа посіву картоплі в колгоспах, радгоспах та у приватних госпо¬
дарствах досягла довоєнного рівня, хоча валовий збір бульб через
низькі врожаї був на 22 % нижчим за довоєнний.
У 60-ті роки XX ст. спостерігалася стабілізація виробництва картоп¬
лі в Україні, хоча її врожайність продовжувала залишатись низькою. З
27
року в рік збільшувалися посівні площі, які досягли максимуму в 1958—
1959 рр., однак у 1961-1965 рр. усі показники виробництва картоплі,
порівняно з попередньою п’ятирічкою, знизилися (табл. 1.13).
Таблиця 1.13 - Виробництво картоплі в господарствах усіх форм власності,
1956-1965 рр.
Категорія
господарств
1956-1960 рр.
1961-1965 рр.
площа,
тис.га
валовий
збір, тис.т
урожай¬
ність, ц/га
площа,
тис. га
валовий '
збір, тис.т
урожай¬
ність, ц/га
Усі категорії
господарств
2259,5
21110
93,4
2078,6
18557
89,3
у тому числі:
населення
1298,0
13621
104,9
12,76
12418
97,3
колгоспи
895,5
7063
78,9
706,9
5460
77,2
радгоспи та
інші держгоспи
65,9
426
64,6
85,3
680
71,3
Урожайність картоплі за десятирічку в колгоспах, радгоспах та
інших державних господарствах не перевищувала 80 ц/га, а в колгос¬
пах Ворошиловградської, Дніпропетровської, Донецької, Запорізь¬
кої, Миколаївської та Одеської областей - 40 ц/га (табл. 1.14).
Вищі врожаї картоплі в цей період у господарствах усіх категорій
було одержано в Чернівецькій, Чернігівській, Тернопільській, Львів¬
ській, Закарпатській та Вінницькій областях, проте і він не переви¬
щував 120 ц/га бульб.
Інтенсивний розвиток галузі картоплярства спостерігався після
1965 р. Народне господарство було відбудоване, і держава тепер мог¬
ла спрямувати значні матеріально-технічні ресурси в сільське гос¬
подарство, зокрема в картоплярство. Аналіз показує, що внаслідок
інтенсифікації виробництва картоплі за цей період зросли врожай¬
ність і рентабельність культури, знизилися затрати на одиницю про¬
дукції (табл. 1.15).
З підвищенням інтенсифікації вирощування картоплі з розрахун¬
ку на 1 крб додаткових виробничих затрат приріст продукції в серед¬
ньому становив 1,5 крб, а з розрахунку на 1 крб додаткових матеріа¬
льних витрат - 4 крб. Однак окупність затрат спостерігалася лише в
тих господарствах, де чітко виконувалася прийнята технологія ви¬
рощування картоплі, зокрема дотримання строків виконання усіх
робіт та їх максимальна механізація.
У середньому за 1966-1970 рр. питома вага картоплі в посівних
площах Полісся становила 9,4 %, передгірних і гірських районах Кар¬
пат - 7,7, Лісостепу — 2,9, північного і центрального Степу - 0,3, пів¬
денного Степу - 0,2 %, при середньому показнику по колгоспах
України -2,1 %.
28
Таблиця 1.14 - Посівна площа картоплі та врожайність по областях
у середньому за рік, 1961-1985 рр.
Посівна площа, тис. га
Урожайність, ц/га
Область
усі кате¬
горії гос¬
подарств
у ТОМ1
V числі
усі кате¬
у тому числі
кол¬
госпи
рад¬
госпи
горії
госпо¬
дарств
кол¬
госпи
рад¬
госпи
Вінницька
148,3
22,0
1,6
107
68
79
Волинська
96,7
46,6
3,1
98
75
83
Ворошиловградська
35,8
1,6
2,3
39
23
29
Дніпропетровська
55,0
5,4
3,1
48
39
46
Донецька
48,9
2,7
3,0
48
36
46
Житомирська
168,4
86,3
3,9
83
75
86
Закарпатська
36,4
15,0
1,3
107
91
81
Запорізька
32,9
2,6
1,3
42
33
39
Івано-Франківська
73,5
27,7
2,3
94
76
62
Київська
162,5
42,1
24,1
92
78
81
Кіровоградська
56,0
8,5
0,7
61
41
49
Кримська
11,5
1,7
1,3
75
63
53
Львівська
121,7
42,9
8,1
107
74
73
Миколаївська
30,8
2,0
1,2
35
21
43
Одеська
40,7
5,4
2,0
55
35
ЗО
Полтавська
81,2
29,2
1,7
66
60
74
Рівненська
87,4
42,3
4,1
95
73
79
Сумська
128,8
56,3
2,3
81
74
91
Тернопільська
106,2
53
2,2
106
88
91
Харківська
80,9
18,8
12,4
58
41
63
Херсонська
21,3
1,7
1,2
50
45
43
Хмельницька
117,4
44,1
1,8
116
84
88
Черкаська
87,9
12,8
0,8
99
82
89
Чернівецька
35,4
15,3
0,5
118
102
89
Чернігівська
213,2
120,7
7,0
109
93
88
Усього
по республіці
2078,6
706,9
95,3
88
77
71
Таблиця 1.15 - Ефективність інтенсифікації виробництва картоплі в колгоспах
Центрального Полісся (за Бапотою В.Г.)
Показник
Групи господарств за рівнем інтенсифікації
картоплярства
1
2
3
4
Виробничі затрати на 1 га, крб
552
653
735
974
Урожай, ц/га
97
128
147
196
Собівартість 1 ц, крб
6,0
5,1
5,0
4,9
Середня реалізаційна ціна 1 ц, крб
5,6
6,2
6,3
7,4
Рівень рентабельності, %
-7
+22
+26
+51
29
Досвід передових господарств свідчить, що при виробництві про¬
довольчої картоплі в основних картоплярських областях оптимальною
є така структура виробничих затрат: на насіння - 25-28 %, добрива -
12-15, пальне, амортизацію, автотранспорт - 15-18, на оплату пра¬
ці - 25-30 , інші витрати - 2-15 %.
Основним централізованим постачальником картоплі у 1966-
1970 рр. був державшій сектор. Так, питома вага колгоспів у прода¬
жу картоплі державі становила 71 %, радгоспів - 12, індивідуаль¬
ного сектора -17%. Однак у Київській, Донецькій та Харківській
областях, де були створені спеціалізовані трести радгоспів із забез¬
печення великих промислових центрів картоплею та іншою сільсь¬
когосподарською продукцією, питома вага радгоспів у продажу
державі картоплі була значною і становила по областях, відповід¬
но, 58, 74 і 73 %.
У 1976-1983 рр. інтенсифікація картоплярства в Україні тривала.
Процес удосконалення розміщення виробництва картоплі здійснюва¬
вся в напрямі концентрації її посівних площ у зонах із найсприятли¬
вішими природно-економічними умовами. Незважаючи на це, у ці¬
лому картоплярство залишалося трудомісткою галуззю.
Урожайність картоплі зростала незначною мірою і не була стабіль¬
ною по роках. Так, через несприятливі погодні умови вегетаційного
періоду в 1969 р. було зібрано лише 66 ц/га, у 1975-86, 1980 - 56, у
1981 і 1982 рр. - по 97 ц/га бульб.
Протягом 1966-1983 рр. у колгоспах навіть поліської зони вро¬
жайність бульб не перевищувала 121 ц/га (табл. 1.16).
Таблиця 1.16 - Динаміка врожайності картоплі в колгоспах різних зон, ц/га
Зона
1966-1970 рр.
1971-1975 рр.
1976-1980 рр.
1981-1983 рр.
Полісся
88
114
121
105
Лісостеп
79
90
91
91
Степ
58
56
59
57
У середньому по
УРСР
86
108
113
100
Така врожайність позначилась на економічних показниках вироб¬
ництва картоплі в цілому. Особливо низькими вони були в степовій
зоні (табл. 1.17).
У середньому в господарствах степових районів собівартість вироб¬
ництва 1 ц картоплі за наведені вище роки становила 17-21 крб. Такий
рівень витрат на вирощування бульб міг би виправдовуватися лише за
умов реалізації молодої картоплі не пізніше 15-28 серпня, коли закупі¬
вельні ціни за 1 ц бульб були вищими за 20-22 крб. А для цього потріб¬
но було вирощувати максимальну кількість ранніх сортів. Проте в 1981—
1983 рр. в цьому регіоні вони займали в середньому лише 17 % посівів.
Колгоспи і радгоспи південних областей до 1 вересня реалізовували не
більше 30-40 % бульб ранньою продукцією.
зо
Таблиця 1.17 - Економічна ефективність виробництва картоплі в колгоспах за
1976-1983 рр. (За Алексєєвим Л.О.)
Показник
Полісся
Лісостеп
Степ
1976-
1980 рр.
1981-
1983 рр.
1976-
1980 рр.
1981-
1983 рр.
1976-
1980 рр.
1981-
1983 рр.
Виробничі витрати на 1 га, крб
Затрати праці на 1 га, люд.-год
Трудомісткість 1 ц, люд.-год
Собівартість 1 ц, крб
Рівень рентабельності (+), зби¬
тковості (-), %
1122
326
2,69
8,32
+4
1329
299
2,87
11,31
+33
1040
417
4,60
10,67
-15
1311
396
4,38
13,34
+1
1282
306
5,41
21,90
-25
1005
299
5,35
17,03
-22
Незважаючи на деяке зростання врожайності картоплі в колгос¬
пах і радгоспах протягом 80-х років, собівартість її, як і в попередні
роки, загалом по Україні залишалася високою, а рентабельність ви¬
рощування - низькою.
Для прикладу візьмемо 1987 р., коли врожайність бульб у колгос¬
пах та радгоспах була однією з найвищих і складала 133 ц/га. Цього
року в зоні Полісся було зібрано 146 ц бульб з гектара, у Лісостепу -
126 ц, тоді як у зоні Степу - 71 ц/га.
Як і в попередні роки, у колгоспах і радгоспах Донецької області
від картоплярства в 1987 р. було одержано 5,7 млн крб збитків; Хар¬
ківської - 4,5; Ворошиловградської - 2,7; Дніпропетровської - 2,4
млн крб. Урожайність бульб у цих областях, відповідно, становила
90, 83, 68 та 72 ц з гектара. Збитковим було картоплярство і в госпо¬
дарствах Запорізької, Кіровоградської, Кримської, Миколаївської,
Одеської, Полтавської, Херсонської областей. Низькорентабельним
залишалось картоплярство в Сумській, Львівській та Київській обла¬
стях. Якщо в колгоспах і радгоспах Волинської області рентабель¬
ність вирощування картоплі складала 47,2 %, Івано-Франківської -
40,1, то в Сумській області вона не перевищувала 5 %, Львівській -
6,7, Київській - 10,8, Житомирській - 14,3, Тернопільській - 15,5 %.
Це можна пояснити і тим, що в ряді областей собівартість бульб була
досить висока. Якщо в господарствах Івано-Франківської області со¬
бівартість 1 ц бульб складала 9,9 крб, то в Чернігівській - 12,2,
Львівській - 13,5, Київській - 14,2, Рівненській - 15,5 крб. Проте в
окремих областях рентабельність картоплярства була високою, оскіль¬
ки господарства вирощували і продавали насінну картоплю, яка мала
вищу ціну, ніж продовольча.
Аналіз роботи господарств Чернігівської області за 1986-1987 рр.
показав, що, незважаючи на деяке зростання врожайності, собівар¬
тість картоплі збільшувалася, а рентабельність галузі знижувалась. У
1986 р. в області з площі майже 100 тис. гектарів зібрали по 160 ц/га
бульб, а в 1987 р. - по 145 ц/га. Так, якщо в середньому за 1981-1985 рр.
собівартість 1 ц картоплі складала 9,5 крб, то в 1986 р. - 10,81, у
1987 р. - 12,2 крб, рентабельність, відповідно, - 65,8, 39,8 і 24,2 %.
зі
Причиною такого явища було зниження в області на 14 % реаліза-
ційної ціни, у той час як по республіці вона зросла на 22 %.
Важливим фактором в одержанні належного врожаю картоплі є
внесення органічних та мінеральних добрив. Фактично без добрив
неможливі добрі врожаї бульб у поліській та лісостеповій зонах.
У середньому по Україні у 1976 та 1984 роках вносилося по 47,3-
56,7 тонн на гектар органічних та 259- 254 кг мінеральних добрив
(табл. 1.18).
Таблиця 1.18 - Внесення добрив під картоплю в окремих областях України
(колгоспи та радгоспи)
Область
Органічні, т/га
Мінеральні, кг/га
1976 р.
1984 р.
1976 р.
1984 р.
Вінницька
23,8
25,8
226
192
Волинська
57,0
77,5
298
292
Житомирська
48,1
54,9
277
270
Закарпатська
32,7
41,9
330
334
Івано-Франківська
28,7
35,8
236
279
Київська
54,2
60,9
293
289
Львівська
56,7
71,3
293
289
Полтавська
26,0
50,2
178
250
Рівненська
61,9
79.2
304
287
Сумська
56,4
65,5
285
262
Тернопільська
32,2
40,7
221
226
Хмельницька
33,5
39,9
185
184
Черкаська
26,9
38,0
205
215
Чернівецька
27,1
28,3
302
309
Чернігівська
65,7
73,7
249
219
Усього
47,3
56,7
259
254
З освоєнням нової системи насінництва щорічно чимала кількість
насінної картоплі із поліських областей завозилася до південних об¬
ластей. Основною організацією, через яку проводилось завезення
насіння, було об’єднання “Сортнасіннєовоч”. Найбільше насінного
матеріалу завозили в Донецьку та Харківську області (табл. 1.19).
Аналіз вирощування картоплі в Україні за десятиріччя (1980—
1990 рр.) показує, що в цей період було досягнуто деяких позитив¬
них результатів, проте врожайність бульб у колективних та держав¬
них господарствах у цілому залишалася невисокою - на рівні, що не
перевищував 126 ц/га.
У той же час у 1990 р., наприклад, як і в попередні роки, чимало
господарств збирали урожай бульби на рівні середніх світових дося¬
гнень - по 220-396 ц/га. Проведене групування господарств за вро¬
жайністю картоплі в 1990 р. показало, що 42 % господарств на 27 %
площі посіву картоплі зібрали менше 75 ц/га бульб, а ще 29 % госпо¬
дарств на 37 % площі виростили не більше 125 ц/га бульб.
32
Таблиця 1.19 - Завезення насінної картоплі в господарства південних областей
України з республіканського фонду, середньорічне
за 1971-1985 рр., тонн
Область
1971-1975
1976-1980
1981-1985
Ворошиловградська
7473
9442
11593
Дніпропетровська
13184
3704
18814
Донецька
17180
17972
26586
Запорізька
3627
4498
5296
Кіровоградська
4117
7925
5933
Кримська
6218
7173
9335
Миколаївська
2897
4907
5074
Одеська
6048
10869
12652
Полтавська
7187
8698
9539
Харківська
24063
28014
22235
Херсонська
2852
4368
7279
Черкаська
1832
3812
4625
Усього
96678
124999
138972
Кількість господарств, урожайність картоплі в яких за цей рік
була задовільною, становила лише 11 %. Вони мали врожайність по¬
над 176 ц/га лише на 13% площі від загальних посівів (табл. 1.20).
Таблиця 1.20 - Групування господарств (колгоспів і радгоспів) за врожайністю
картоплі
Урожайність
бульб, ц/га
Кількість
господарств
Площа посіву,
тис. га
У процентах
за кількістю
господарств
за посівною
площею
Усього
9252
423,9
100
100
До 75
3901
115,4
42
27
76-125
2655
155,0
29
37
126-175
1683
98,9
18
23
176-250
874
48,9
9
12
251-350
135
5,7
2
1
351 і більше
3
0,01
-
-
На той час у розвитку картоплярства України спостерігався ряд
негативних тенденцій: скорочення посівних площ під картоплею і
валового збору, зниження якості, втрати при зберіганні, низький рі¬
вень механізації та недосконалість вітчизняної техніки. Усе це не
лише спричинило труднощі в забезпеченні армії та населення міст
картоплею, але й у постачанні картоплі до державних установ гро¬
мадського і дитячого харчування, організацій. Особливо критичний
стан з виробництвом картоплі склався у 1990 році.
Картопля продовжувала залишатися однією з найбільш трудоміст¬
ких культур. На виробництво центнера бульб у колгоспах та радгос¬
пах у середньому по республіці втрачалося 2,2 люд.-год, а на частку
2 2'132
33
ручної праці в картоплярстві припадало близько 60 % затрат. Картоп¬
лярство як галузь стало збитковим.
Аналіз показує, що в переважній більшості картоплярських гос¬
подарств не могла застосовуватись передова спеціалізована техніка,
тому що в 50 %, або в 4500 господарствах, площа насаджень карто¬
плі становила до 25 гектарів і лише в 1460 господарствах (16 %) -
понад 100 га, на яких вирощувалося більше 40 % валового збору
бульб від загального виробництва їх суспільним сектором.
Крім того, давалася взнаки недосконалість механізмів, які вико¬
ристовувалися в картоплярстві. Із 56 назв спеціальних машин, що
були затверджені Системою машин на 1986-1990 рр., вітчизняною
промисловістю випускалося лише 23, з яких 9 були морально заста¬
рілими. Причому, більшість картоплезбиральних машин, що завози¬
лися в Україну, значно поступалися світовим стандартам і при зби¬
ранні рожаю, особливо на глинистих грунтах, травмували бульби.
Загальна кількість бульб, що зазнають механічного пошкодження
при використанні в господарствах сучасного збирального комплексу,
складає близько 55 %.
На основі багаторічної роботи науково-дослідних установ в
Україні розроблена і з 1985 року запроваджується в господарствах
ресурсозберігаюча технологія вирощування картоплі. Запропонована
технологія є сучасною і базується на використанні існуючої техніки.
Ця технологія виробництва картоплі розрахована на одержання 200-
300 ц/га. В основу її покладені заходи, які створюють оптимальні
умови для підвищення врожайності картоплі найбільш раціональни¬
ми методами.
Інтенсифікація картоплярства сприяла зростанню врожайності
цієї культури. Проте в наступні роки, коли увага до сільського гос¬
подарства ослабла, що проявилось зниженням рівня матеріально-
технічного постачання та інтенсифікації, знизились і темпи зростан¬
ня врожайності. Так, якщо за 10 років (1971- 1975 рр.) урожайність
картоплі в суспільному секторі, порівняно з 1966-1970 рр., зросла на
30 ц/га, то надалі вона не змінювалася. Зменшувалася різниця в
рівнях урожайності картоплі між індивідуальними господарствами і
суспільним сектором, і лише в 1986-1990 рр. колгоспи і радгоспи
досягли рівня врожайності приватних господарств (табл. 1.21).
Таблиця 1.21 - Динаміка врожайності картоплі в Україні
Категорія господарств
Роки
1966-1970
1971-1975
1976-1980
1981-1985
1986-1990
Усі категорії господарств,
у т.ч.
99,8
111
121
123
122
приватні господарства
108
113
126
130
120
колгоспи
86
109
113
110
127
радгоспи
80
101
106
105
123
34
У той час, коли інтенсивно розвивалося твариннищво і значно
зріс рівень добробуту народу, картопля, як основний продукт харчу¬
вання, відійшла на другий план. Відбулися зміни і в структурі спо¬
живання продуктів харчування.
За період з 1966 по 1990 рр. посівні площі під картоплею змен¬
шились на 680,3 тис. га, у тому числі у суспільному секторі - на
388,8 тис. га, у приватних господарствах - на 299,5 тис. га.
Найбільше скорочення площ під картоплею з 1960 по 1990 рік
відбулося в Житомирській, Київській, Сумській, Хмельницькій, Во¬
линській, Чернігівській та інших областях (табл. 1.22),
Таблиця 1.22 - Динаміка посівних площ картоплі в Україні (тис. га)
Категорія
господарств
Роки
1966-1970
1971-1975
1976-1980
1981-1985
1986-1990
Усі категорії гос¬
подарств,
у т.ч.
приватні гос¬
подарства
колгоспи
радгоспи
2033.3
1305.3
640.0
88.0
1888,9
1221,3
579,2
88,1
1695,0
1094,2
509,1
91,7
1628.4
1088,1
460.4
79,5
1472,9
996,2
404,8
71,9
У результаті переважаючих темпів скорочення посівних площ,
водночас із підвищенням урожайності відбулося поступове зменшен¬
ня середніх обсягів виробництва з 20-20,5 млн тонн у 1966-1980 рр. до
19,0 млн тонн у 1986-1990 рр. Але досягнутий рівень інтенсифі¬
кації галузі картоплярства на існуючій матеріально-технічній базі
та організації її використання не забезпечували щорічного висо¬
кого і стабільного врожаю. Якщо в 1979 р. урожайність картоплі
в колгоспах складала 137,6 ц/га, то в 1975, 1981, 1982 роках вона
знаходилася в межах 85-100, а в 1980 році становила лише 56 ц/га
(табл. 1.23, 1.24).
У цілому швидкі темпи збільшення грошово-матеріальних витрат
у розрахунку на 1 га картоплі, порівняно із підвищенням урожайнос¬
ті, зумовили зростання собівартості виробництва 1 ц продукції, а та¬
кож низьку загальну економічну ефективність її вирощування в су¬
спільному секторі. Так, у 1966-1990 рр. фактично відбувалося зви¬
чайне відтворення галузі.
Підвищення рівня рентабельності в 1981-1990 рр. до 15-20 %
було досягнуте не за рахунок різкого зростання врожайності, а за¬
вдяки підвищенню в цей період державних закупівельних цін. Проте
і за цих умов виробництво продовольчої картоплі залишалось мало¬
ефективним (табл. 1.25).
35
Таблиця 1.23 - Динаміка посівних площ картоплі по областях у приватному секторі
Область
Рік
1960
1970
1980
1990
Автономна Республіка Крим
2,2
3,8
2,9
2,8
Вінницька
25,2
23,4
17,0
12,6
Волинська
57,0
46,2
40,3
31,8
Дніпропетровська
12,5
8,5
7,8
4,8
Донецька
8,8
8,5
8,0
7,2
Житомирська
106,1
83,5
68,9
46,3
Закарпатська
18,2
13,3
11,7
8,8
Запорізька
5,4
2,9
2,8
2,1
Івано-Франківська
73,8
58,8
48,8
26,9
Київська
38,2
14,5
9.8
9,3
Кіровоградська
73,8
58,8
48,8
26,9
Луганська
9,0
8,0
8,3
8,2
Львівська
5,3
3,4
3,5
3,3
Миколаївська
59,4
34,4
22,1
16,5
Одеська
3,5
3,3
2,6
1,8
Полтавська
9,1
5,5
5,9
3,6
Рівненська
38,1
24,4
21,2
18,7
Сумська
50,4
38,4
39,2
28,3
Тернопільська
68,6
52,3
46,9
32,1
Харківська
64,8
48,6
31,5
22,0
Херсонська
42,9
15,1
13,0
10,1
Хмельницька
1,9
2,1
2,7
2,4
Черкаська
53,9
40,9
37,8
26,7
Чернівецька
14,3
12,6
10,5
8,3
Чернігівська
18,1
9,2
8,1
7,1
128,9
120,3
110,5
90,6
Усього
915,7
681,9
581,9
431,8
Таблиця 1.24 - Валовий збір картоплі в Україні, тис. тонн
Категорія
господарств
Роки
1966-1970
1971-1975
1976-1980
1981-1985
1986-1990
Всі категорії гос¬
подарств, у т.ч.:
приватні гос¬
подарства
колгоспи
радгоспи
20294
14086
5504
704
21019
13848
6286
885
20494
13775
5747
972
19998
14119
5945
834
17965
11937
5140
887
Прибутковість галузі картоплярства в колгоспах поліських та за¬
хідних областей становила 21,7 %. Такий її рівень не відповідав ви¬
могам розширеного відтворення галузі.
Збитковість виробництва продовольчої картоплі в колгоспах сте¬
пової зони (-2 %) є наслідком незначного рівня використання полив¬
них земель (не більше 40 %) та вирощування ранньої продукції, коли
використовувалися переважно середньостиглі сорти (до 60 %). Ці фак¬
36
тори зумовлюють низьку врожайність та високу собівартіть картоплі,
а також реалізацію молодої картоплі в більш пізні строки за нижчи¬
ми закупівельними цінами.
Таблиця 1.25 - Економічна ефективність виробництва картоплі в колгоспах
України (за Оверчуком П.В.)
Показник
Роки
1966-
1970
1971-
1975
1976-
1980
1981-
1985
1986-
1990
Урожайність, ц/га
86,0
108,3
112,5
104,6
126,2
Виробничі витрати на 1 га, крб
115,0
787
1094
1353
1749
Собівартість 1 ц, крб
5,98
7,28
10,8
12,33
12,58
Трудомісткість 1ц, люд.-год
5,22
3,74
3,10
2,92
2,32
Ціна реалізації 1 ц, крб
7Д5
8,08
9,35
16,31
15,78
Чистий прибуток на 1 га, крб
16
9
-6
132
112
Рівень рентабельності, %
9,8
3
-1,4
20,2
15,1
У цілому за 1986-1990 рр. в республіці лише в 14-18 % госпо¬
дарств рентабельніть картоплярства складала понад 45 %. У перева¬
жній більшості господарств ця галузь була збиткові. Розрахунки свід¬
чать, що у виробничих умовах колгоспів та радгоспів Полісся роз¬
ширеного відтворення галузі картоплярства можна досягти при вро¬
жайності не менше 165 ц/га, лісостепової і степової зон - 150 ц/га
при середній по Україні 160 ц/га. Таким чином, щоб досягти необ¬
хідної рентабельності, урожай бульб повинен зрости на 25-30 % в
основних зонах вирощування картоплі (табл. 1.26, 1.27).
Таблиця 1.26 - Розміщення виробництва картоплі в колгоспах і радгоспах
України, 1986-1990 рр.
Зона
Посівні площі
Валовий збір
тис. га
%
тис. га
%
Полісся і західні райони
86,8
77,3
6021
83,7
Лісостеп
81,9
16,4
915
12,7
Степ
31,8
6,3
257
3,6
Усього по Україні
500,5
100,0
7193
100,0
Таблиця 1.27 - Урожайність картоплі в різних природно-кліматнчних зонах
України, 1986-1990 рр.
Категорія господарств
Зона
Полісся
Лісостеп
Степ
Усі категорії господарств,
156,8
117,0
75,8
у тому числі:
приватні
157,0
118,0
75,2
колгоспи
155,4
113,4
67,2
радгоспи всіх систем
157,8
102,4
95,2
Якщо різниця в урожайності картоплі між окремими областями,
районами і господарствами однієї природно-економічної зони зумов-
37
лена переважно рівнем організації та інтенсивності виробництва, то
між зонами, головним чином,- природно-кліматичними умовами то¬
варного виробництва насінної картоплі.
Сучасний стаи картоплярства. У 1990-2000 рр. у галузі карто¬
плярства відбулися істотні зміни. Із колективних та державних гос¬
подарств посіви картоплі перемістились у приватні і фермерські гос¬
подарства, на городи і дачні ділянки (табл. 1.28).
Таблиця 1.28 - Показники виробництва картоплі в Україні у 1990-2000 рр.
Усі категорії господарств
У тому числі суспільний сектор
Рік
площа,
урожайність,
валовий збір,
площа,
урожайність,
валовий збір,
тис. га
ц/га
млн тонн
тис. га
ц/га
млн тонн
1990
1429
117
16,7
429
112
4,8
1992
1701
119
20,3
332
93
3,1
1994
1527
105
16,1
162
76
1,2
1996
1548
119
18,4
77
105
0,8
1998
1513
101
15,1
40
92
0,4
2000
1627
124
20,2
30
124
2,0
У 2000 р. незначні площі насінницьких посівів картоплі залиша¬
лися в державних господарствах науково-дослідних установ, вузів і
технікумів.
І хоча в приватному секторі збирали вищий урожай, ніж у колек¬
тивних та державних господарствах, він усе ще залишається дуже
низьким. Це відбувається при досить великих затратах праці, що
пов’язано в першу чергу з відсутністю малої механізації для невели¬
ких площ, а також недостатнім забезпеченням приватного сектора
здоровим насінним матеріалом сучасних високопродуктивних сортів.
Водночас при внесенні достатньої кількості добрив та при значно¬
му досвіді ведення власного господарства на селянських городах зби¬
рають не менше 200-300 центнерів бульб у перерахунку на гектар.
Найнижчі врожаї збирають дачники та міське населення на своїх горо¬
дах. Причини цього різні, але основна полягає в недостатньо удобре¬
них і погано оброблених ділянках, відсутності якісного садивного ма¬
теріалу та потрібних знань і навичок для вирощування картоплі.
Таким чином, хоч приватний сектор і став основним виробником
продовольчої картоплі, її вирощування є дуже затратним.
З урахуванням сучасного стану розвитку картоплярства, у держа¬
ві здійснюється ряд заходів, спрямованих на його стабільність. У пер¬
шу чергу це вдосконалення організації виробництва та реалізації
продовольчої і насінної картоплі господарствами усіх форм власнос¬
ті. У сучасному картоплярстві особливе значення має виробництво
висококласного насіння та забезпечення ним товаровиробників.
Картопля - одна з небагатьох культур, площі якої за десятиріччя
не зменшились. Причому, питома вага картоплі в структурі посівних
площ зросла (табл. 1.29).
38
Таблиця 1.29 - Питома вага картоплі у структурі посівних площ
Рік
1990 р.
1997 р.
1998 р.
1999 р.
2000 р.
Картопля у структурі
посівних площ, %
4,41
5,21
5,43
5,48
5,75
Оскільки виробництво картоплі зосереджене переважно на малих
площах, то при великих затратах ручної праці продуктивність її знач¬
ною мірою залежить від погодних умов відповідного року та природ-
но-економічної зони вирощування (табл 1.30, 1.31, 1.32).
Таблиця 1.30 - Динаміка виробництва картоплі в розрізі природно-економічних
зон
1998 р.
1999 р.
2000 р.
Зона
зібрана пло¬
ща, тис. га
валовий збір,
тис. т
урожайність,
ц/га
зібрана пло¬
ща, тис. га
валовий збір,
тис. т
урожайність,
ц/га
зібрана пло¬
ща, тис. га
валовий збір,
тис. т
урожайність,
ц/га
Степ
298,8
2206
73,8
309,1
2097
67,8
305,0
3078
100,0
Лісостеп
466,7
4491
96,2
471,5
3336
70,8
498,1
5754
115,5
Полісся
747,7
8708
116,5
770,5
7290
94,6
824.1
11417
138,5
У цілому
по Україні
1513,2
15405
101,8
1551,1
12723
82,0
1627,2
20349
124,4
Таблиця 1.31 - Виробництво картоплі в Україні в 1991-2000 рр.
Показник
Роки
1991-1995
1996-2000
у т.ч. 2000
Площа, тис. га
1566
1564
1627
Урожайність, ц/га
110,7
106,8
124,4
Валовий збір, тис. тонн
17333
16698
20249
Основними причинами невисокої врожайності картоплі і висо¬
кої собівартості вирощених бульб є недостатнє забезпечення усіх
товаровиробників насінним матеріалом сучасних інтенсивних сор¬
тів. Поки що відсутні і механізми та система заміни посадкового
матеріалу.
За останні роки значно скоротилося внесення органічних і міне¬
ральних добрив. Як правило, вносяться тільки органічні добрива, а
на значній площі картопля вирощується без добрив.
Забур’яненість посівів картоплі перевищує допустимі пороги
шкодочинності; значних втрат завдають колорадський жук та фіто¬
фтороз. У роздрібній торгівлі щорічно зростає кількість пестицидів,
які при застосуванні не забезпечують відповідної ефективності. При
такому стані картоплярі вимушені збільшувати норми застосування,
що негативно впливає на якість вирощеної картоплі. Відсутній і ді¬
йовий контроль за пестицидами, які реалізуються.
39
Таблиця 1.32 - Виробництво картоплі по областях у 1999-2000 рр.
Області
1999 р.
2000 р.
зібрана
площа,
тис. га
урожай¬
ність,
ц/га
валовий
збір,
тис. т
зібрана
площа,
тис. га
урожай¬
ність,
ц/га
валовий
збір,
тис. т
Автономна Республіка Крим
24,6
93,0
229,1
24,2
93,7
226,8
Вінницька
115,4
87,4
1008,5
121,5
116,6
1417,2
Волинська
72,6
131,1
952,0
78,8
151,5
1194,6
Дніпропетровська
53,3
69,0
367,4
50,6
149,0
754,4
Донецька
59,3
64,1
380,2
58,0
132,0
763,2
Житомирська
77,9
91,1
710.2
82,4
133,4
1099,0
Закарпатська
30,9
122,5
378,2
34,3
139,3
477,7
Запорізька
34,5
52,6
181,3
35,3
82,0
291,0
Івано-Франківська
52,8
117,8
621,6
62,3
135,6
845,0
Київська
94,8
70,6
669,2
98,2
138,0
1350,6
Кіровоградська
59,1
36,2
214,2
58,0
54,7
317,0
Луганська
45,7
75,5
345,6
47,8
75,0
358,7
Львівська
96,4
113,8
1096,7
105,9
143,0
1512,6
Миколаївська
29,4
50,3
148,0
26,5
27,3
72,4
Одеська
39,8
66,4
264,1
39,9
79,3
316,5
Полтавська
65,7
42,6
279,9
69,6
98,8
687,5
Рівненська
67,2
114,0
765,9
73,6
119,0
875,6
Сумська
72,2
47,6
343,6
77,5
120,0
928,9
Тернопільська
61,6
103,0
634,6
72,9
143,0
1040,3
Харківська
88,2
71,1
627,2
96,7
156,0
1511,0
Херсонська
22,5
80,2
180,8
22,7
129,7
294,7
Хмельницька
74,9
102,3
765,9
73,6
146,3
1076,5
Черкаська
80,4
59,9
481,6
86,7
108,0
938,0
Чернівецька
31,9
108,5
346,5
31,3
129,5
406,1
Чернігівська
100,0
73,0
730,5
98,9
151,0
1493,5
Усього
1551,1
82,0
12722,8
1627,2
124,4
20249,0
Слід зазначити, що до сьогодні ще не налагоджено випуск повно¬
го комплексу машин і механізмів для садіння, догляду, збирання та
післязбиральної доробки врожаю. Відсутня також малогабаритна
техніка та обладнання для вирощування картоплі на малих площах.
Не вистачає і сучасних картоплесховищ для зберігання картоплі.
Стабілізація виробництва та поліпшення якості продовольчої і
насінної картоплі можливі при широкому впровадженні наукових
розробок установ УААН та Міністерства аграрної політики України,
досвіду вітчизняних і зарубіжних виробників шляхом:
- навчання виробників продовольчої і насіннєвої картоплі на на¬
уково-практичних конференціях, семінарах і курсах;
- ознайомлення картоплярів із впровадженням нових прийомів
виробництва картоплі в наукових установах, державних дослідних
господарствах, навчально-дослідних господарствах вузів і безпосе¬
редньо у приватних господарствах;
40
- забезпечення товаровиробників іформаційними матеріалами,
у яких висвітлені рекомендації, нові розробки в картоплярстві
(преса, радіо, телебачення, видання окремих буклетів, брошур, ре¬
комендацій).
- безпосередньої участі вчених у запровадженні нових розробок.
Споживання картоплі. Картопля - одна з найбільш універсаль¬
них сільськогосподарських культур, а бульби - одне з важливих
джерел харчування людини і годівлі худоби. У світі вона займає
п’яте місце за кількістю енергії в харчуванні людини після пшениці,
кукурудзи, рису і ячменю. Проте не в усіх країнах вирощування кар¬
топлі має однакове значення для харчування. Основним продуктом
харчування вона є в Західній, Середній та Східній Європі. У півден¬
них регіонах Європи та в Азії, Африці і Південній Америці на харчо¬
ві цілі вона використовується значно менше. Мають місце також
традиції окремих народів та країн у вирощуванні і споживанні кар¬
топлі.
Слід зазначити, що використання картоплі на харчові цілі в роз¬
винених країнах Європи за останні роки зменшилось, але одночасно
зросла частка виробництва продуктів, виготовлених з картоплі.
Вважають, що більше 50 % світового виробництва картоплі ви¬
користовується до або ж після переробки для харчування, 30 - на
корм худобі, 3-4 % - для одержання крохмалю і спирту та приблизно
10 % - на насіння.
Структура використання картоплі в Україні за останніх 15 років
подається в таблиці 1.33. З даних таблиці видно, що до 25 % виро¬
щених бульб використовуються на насінні цілі. Це значно перевищує
світові показники і пояснюється низькою врожайністю цієї культури
в Україні.
Таблиця 1.33 - Структура використання картоплі в господарствах усіх
категорій, %
Використання картоплі
Роки
1986-1990
1991-1995
1997
1999
Загальне використання
100
100
100
100
у тому числі: на споживання
27,8
33
34,2
36,3
на насіння
20,9
22,8
23,8
25,9
на кормові цілі
25,8
26,6
27
23,0
інші виграти та втрати при збері¬
ганні
25,5
17,6
15,0
14,8
У 1981-1999 роках споживання картоплі в Україні на душу насе¬
лення при середньому 122-150 кг за рік становило у 1981 р. - 128;
1983 - 142; 1985 - 132; 1987 - 131; 1989 - 127; 1991 - 116; 1993 -
150; 1995 - 126; 1997 - 134; 1999 - 122 кг.
Раціональною нормою споживання картоплі на душу населення
на рік є 120 кг.
41
1.3. АГРОКЛІМАТИЧНА ХАРАКТЕРИСТИКА
ЗОН УКРАЇНИ ЗА ЇХ ПРИДАТНІСТЮ
ДЛЯ ВИРОЩУВАННЯ КАРТОПЛІ
Сільськогосподарська характеристика клімату складається з оці¬
нок температурних та світлових ресурсів, умов вологозабезпечення
вегетаційного періоду та його підперіодів, несприятливих явищ для
сільського господарства.
Оцінка термічних ресурсів потрібна не лише для вирішення
окремих технологічних питань, як, наприклад, строки садіння чи
збирання картоплі, але й для підрахунків забезпечення рослин
теплом при прогнозуванні кількісних та якісних показників уро¬
жаю. Це важливо також знати в різні міжфазні періоди, особливо
в період бульбоутворення і нагромадження врожаю. Здійснюється
оцінка за такими характеристиками: сумою середніх багаторічних
температур повітря вище 5°С (сума ефективних температур), 10°С
(сума активних температур) і 15°С. Для картоплі ще враховують
температуру понад 7°С, а також рівень температури грунту при
бульбоутворенні та нагромадженні врожаю, тривалість безмороз¬
ного періоду і т. ін.
Сума ефективних температур - це сума середніх добових темпе¬
ратур (понад 5°С), відрахована від біологічного мінімуму, при якому
розвиваються рослини картоплі. Для одержання повноцінних сходів
картоплі, наприклад, у період садіння-поява сходів потрібна сума
ефективних температур 295-305°С для ранньостиглих сортів та 367-
385°С - для середньопізніх.
Дані таблиці 1.34 свідчать про те, що в зоні Полісся і Лісостепу
України сума ефективних температур (понад 5°С) є достатньою для
проростання бульб картоплі, а в зоні Степу вона значно вища. Для
сільськогосподарської оцінки термічних ресурсів клімату викорис¬
товують також показник суми активних температур, який відобра¬
жає забезпечення теплом періоду активної вегетації сільськогоспо¬
дарських культур на певній території за рік чи певний час (вегета¬
ційний період, сезон, місяць). Сума активних температур склада¬
ється із середніх добових температур понад 10°С. Для повного
розвитку ранньостиглих та середньоранніх сортів картоплі вона
становить 1000-1400°С, для середньостиглих - 1300-1500, для піз¬
ньостиглих - 1400-1600°С. З даних, наведених у таблиці, видно,
що в поліській та лісостеповій зонах достигання ранньо- та серед¬
ньостиглих сортів може настати в липні, а в лісостеповій - у черв-
ні-липні.
В оцінці ролі сонячної фотосинтетичної активної радіації (ФАР)
розрізняють її двобічне використання: пряме, що визначає засвоєння
рослинами, і побічне - використання показників термічного режиму
і зволоження.
42
Таблиця 1.34 - Сума середніх добових температур повітря в різних зонах
України, °С
Місяць
Ефективні температури понад
5°С
Активні температури понад
10°С
Полісся
Лісостеп
Степ
Полісся
Лісостеп
Степ
Квітень
225
205
255
35
60
105
Травень
615
640
740
440
495
590
Червень
1130
1150
1310
940
1005
1160
Липень
1740
1740
1984
1520
1605
1835
Серпень
2245
2300
2625
2055
2160
2485
Вересень
2645
2720
3105
2445
2580
2925
В Українському науково-дослідному інституті метеорології ви¬
значено середньомісячні й річні значення сумарної ФАР на території
України (табл.1.35). Цей показник необхідно враховувати при ство¬
ренні умов на картопляному полі для активного нагромадження вро¬
жаю у якомога ранніші строки.
Таблиця 1.35 - Середньомісячні та річні значення сумарної ФАР, МДж/м2
Зона
Квітень
Травень
Червень
Липень
Серпень
Вересень
За рік
Полісся
213
289
305
318
172
201
2092
Лісостеп
222
301
322
326
289
201
2176
Степ
238
326
343
356
305
226
2372
Аналіз агрокліматичних ресурсів різних зон показує, що вони, за
винятком Степу, де картоплю бажано вирощувати при зрошенні,
сприятливі для одержання стабільних високих урожаїв якісних
бульб. Однак погодні умови вегетаційного періоду в деякі роки в
окремих районах обмежуються надмірною кількістю опадів, недо¬
статньою сумою активних температур та великою кількістю днів з
інтенсивною хмарністю (табл. 1.36, 1.37).
Таблиця 1.36 - Характеристика температурного режиму в різних зонах України
(середні багаторічні дані)
Показник
Полісся
Лісостеп
Степ
1
2
3
4
Середньомісячна температура повітря, °С
травень
14
14
15
червень
17
17
19
липень
19
19
22
серпень
17
18
21
Абсолютний мінімум температури січня, °С
-32...-38
-32...-38
-28...-41
Абсолютний максимум температури липня, °С
33-38
33-39
35-41
Середньорічні дані стійкого переходу добових
температур повітря:
весною через 5°С
через 10°С
через 15°С
7.04
27.04
30.05
4.04
24.04
20.05
43
Продовження табл. 1.36
1
2
3
4
восени: через 10°С
через 5°С
1.10
28.10
2.10
31.10
Імовірність заморозків (-3 °С) у повітрі, пер-
ша-третя декади квітня, %
55-20
50-10
55-10
Тривалість періоду з температурою повітря
понад 15 °С, днів
99-110
115-120
130
Імовірність прогрівання грушу на глибині 10
см у першій-другій декадах квітня до темпера¬
тури 5 °С, %
50-90
55-95
50-95
Середня кількість днів з максимальною темпе¬
ратурою повітря понад 25 °С (за червень-вере-
сень)
33
46
70
Таблиця 1.37 - Характеристика режиму зволоження різних зон України (середні
багаторічні дані)
Показник
Полісся
Лісостеп
Степ
Середньорічна сума опадів, мм
600
543
425
Кількість опадів у теплий період року (кві-
тень-жовтень), мм
400-500
350-400
165-290
травень
60
55
40
червень
80
70
55
липень
85
75
45
серпень
80
65
40
Імовірність бездощових періодів різної три¬
валості, %:
квітень-червень
10-20 днів
75
75
55
21 день і більше
25
25
45
липень-вересень
10-20 днів
75
70
40
21 день і більше
25
30
60
Середня кількість днів за червень-липень з
відносною вологістю повітря о 13-й год, %:
10-20
0
1
3
21-30
2
9
12
понад 30
60
50
43
Імовірність запасів продуктивної вологи в
шарі грушу 0-50 см у період бульбоутво¬
рення картоплі (пізньостиглі сорти), %
0-25 мм
26-50 мм
51-75 мм
76 і більше
10
35
40
15
5
35
45
15
44
За ступенем зволоження території атмосферними опадами та ін¬
тенсивністю випаровування води в Україні розрізняють три зони [З,
4, 5,6, 9].
1). Зона надмірного зволоження (Полісся), де надходження опа¬
дів у грунт перевищує їх витрати при стіканні та випаровуванні. За
середньобагаторічними даними тут випадає за рік 621-922 мм опа¬
дів, а випаровується 500-550 мм, гідротермічний коефіцієнт (ГТК)
досягає 1,3-1,6 (оптимальним вважається 1-1,4; при коефіцієнті 1,6
рослини пригнічуються від перезволоження). До цієї зони входять
північні райони Сумської, Чернігівської, Київської, Житомирської,
Хмельницької, Рівненської, Волинської, Тернопільської і Львівської
областей.
2). Зона нестійкого зволоження (Лісостеп). Надходження і ви¬
трати водного балансу в ній приблизно однакові. У середньому кіль¬
кість опадів за рік складає 541-645 мм, випаровується - 475-575 мм,
ГТК становить 1-1,3. До зони входять південні райони Тернопільсь¬
кої, Хмельницької, Львівської, Волинської, Рівненської, Житомирсь¬
кої, Чернігівської та Сумської областей, північні райони Вінницької,
Полтавської, Черкаської, Одеської та Кіровоградської областей, схід¬
на частина Івано-Франківської і північно-східна Харківської облас¬
тей, а також майже вся Чернівецька область, за винятком західної
частини, що входить у зону передгірних і гірських районів Карпат.
3). Зона недостатнього зволоження (Степ). Характеризується
тим, що витрати води перевищують її надходження з атмосферними
опадами у грунт. У зоні в середньому за рік випадає 406-514 мм опа¬
дів, а випаровується 300-450 мм, ГТК становить 0,4-1 (при ГТК 0,6
рослини пригнічуються посухою). До зони віднесена територія, що
лежить нижче південної межі Лісостепу, яка проходить по лінії Бал-
та-Кременчук-Чугуїв.
Є ще дві окремі зони - зона передгірних і гірських районів Криму
та зона передгірних і гірських районів Карпат.
В останні роки помітна тенденція до пом’якшення клімату на ве¬
ликих територіях земної кулі. Очевидно, це пов’язане із зростаючим
проявом так званого парникового ефекту, що є причиною збільшення
вмісту в атмосфері таких газів, як діоксид вуглецю (СОг), оксид вуг¬
лецю (СО), сірчистий ангідрид (SO3), оксид азоту (NO) та ін., а також
звичайного пилу. Україна не є винятком у цьому процесі. Розвиток
промисловості, урбанізація, розростання мегаполісів, інтенсифікація
сільського господарства поглиблюють процес парникового ефекту.
Відомо, що одна третина території з найбільш родючими грунта¬
ми знаходиться у нас в зоні недостатнього і нестійкого зволоження.
Посухи, які охоплюють половину території країни, спостерігаються
через 10—12 років, а 10 % їх - через 2-3 роки. За останні 30 років сім
із них відзначалися інтенсивними та обширними посухами (1972,
1975, 1979, 1981, 1990, 1994, 1999), а чотири - меншого ступеня
45
(1982, 1984, 1985, 1992 рр.)- Тобто, кожний третій рік був несприят¬
ливим з точки зору забезпечення рослин вологою.
Отже, тенденція до потепління клімату все більше заявляє про
себе, і з цим явищем не можна не рахуватися.
Світова наука, у тому числі й українська, обізнана з методами
подолання посухи, послаблення дії високих температур. Ці методи
передбачають виведення і впровадження у виробництво посухостій¬
ких сортів і гібридів; організацію зрошення, створення вітрозахис¬
них лісосмуг, впровадження відповідних сівозмін, нещадну боротьбу
з бур’янами, застосування технологій і сільськогосподарської техні¬
ки, спрямованих на накопичення і збереження вологи у грунті; про¬
гнозування погоди і своєчасне коригування технологій вирощування
сільськогосподарських рослин.
На жаль, нам доводиться констатувати, що довгострокові
прогнози часто-густо не справджуються. Так, за прогнозами
УДНДПТІ “Агроресурси” [7], літо 1999 року передбачалося
дощовим і холодним. Проте ми є свідками того, що воно було
посушливим і жарким. Тому будувати технологію, очевидно, слід
таким чином, щоб завжди залишалась можливість перебудови її в
необхідному напрямі, тобто робити її гнучкою. Наприклад, при
передбаченні нормального літа (опади і температура в межах
середньої багаторічної норми) картоплю в лісостеповій зоні садять
гребеневим способом або гладким із наступним проведенням од-
ного-двох підгортань. З настанням сухої погоди треба зменшити
висоту гребенів, здійснити мульчування грунту спеціальними бо¬
рінками, а підгортання не проводити зовсім. Відповідні зміни в
технології необхідно вносити по всіх культурах.
Помилки у прогнозі погоди на наступний рік призводять до по¬
милок у технологіях. Скажімо, 1998-й був сприятливим з точки зо¬
ру суми опадів та ефективних температур. За цих умов урожайність
картоплі в навчально-дослідному господарстві БДАУ при внесенні
50 т/га гною становила 167 ц/га, що вище, ніж без гною, на 36 ц/га;
внесення на фоні гною мінеральних добрив (азоту, фосфору і ка¬
лію) по 45 кг діючої речовини забезпечувало приріст бульб 72 ц/га,
а по 60 кг - 103 ц/га. У посушливому 1999 році при внесенні 50 т/га
гною зібрали по 124 ц/га бульб, що вище, ніж без добрив, лише на
19 ц/га; за внесення на фоні гною по 45 кг діючої речовини приріст
становив 9 ц/га, а на фоні гною + 60 кг урожайність, навпаки, зни¬
зилася на 15 ц/га.
Зрозуміло, що внесення високих норм добрив за низького вро¬
жаю призвело до значних збитків. Тому при вирішенні питання ви¬
бору певної технології і проведення окремих заходів, у тому числі з
метою найбільш ефективного використання добрив для формування
врожаю, важливо мати точний як довгостроковий, так і короткостро¬
ковий прогноз погоди.
46
1.4. ПЕРСПЕКТИВИ РОЗВИТКУ
КАРТОПЛЯРСТВА В УКРАЇНІ
Відповідно до науково обгрунтованих норм споживання та з ура¬
хуванням потреб у насінні, переробці, використанні на корм худобі й
інші цілі, в Україні щорічно потрібно виробляти близько 20 млн тонн
картоплі. На сьогодні середня площа посіву картоплі в товаровироб¬
ників усіх форм власності складає понад півтора мільйона гектарів, а
середня врожайність - не більше 130 ц з гектара. Втрати при збері¬
ганні бульб в окремі роки досягають 20% і більше Тому собівартість
вирощеної картоплі залишається усе ще досить високою.
Сучасні проблеми в галузі картоплярства мають вирішуватись з
урахуванням вітчизняних наукових розробок і досвіду зарубіжних
країн шляхом переведення галузі на інтенсивну основу при значному
підвищенні врожайності, скороченні посівних площ та раціонально¬
му використанні вирощеного врожаю. Цього можна досягти при
здійсненні у якомога коротші строки ряду організаційних, виробни¬
чих та економічних заходів, а саме:
- скорочення площ посівів картоплі до 1,0-1,2 млн га за рахунок
підвищення врожайності до 180-200 ц/га, а на зрошуваних землях
південних областей - їх розширення з метою вирощування ранньої
картоплі для власних потреб та забезпечення великих міст і промис¬
лових центрів ранньою продукцією у червні-липні. Цей процес має
відбуватися поступово, з урахуванням підвищення врожайності;
- забезпечення виробників продовольчої картоплі усіх форм влас¬
ності високоякісним насінним матеріалом, який має відповідати сві¬
товим стандартам сучасних високоврожайних сортів. Здійснення
цього заходу може забезпечити підвищення врожайності картоплі
щонайменше на 40-60 ц/га;
- селекцію картоплі необхідно спрямувати на створення високо¬
врожайних столових сортів різного призначення, стійких до раку,
вірусної інфекції, фітофторозу та картопляної нематоди, що дозво¬
лить зменшити необхідність завезення в Україну великими партіями
насіння сортів іноземної селекції;
- необхідно здійснити перехід на вирощування насінної картоплі
з якісними показниками, що відповідають міжнародній сертифікації.
Реалізація такого проекту має бути поступовою. Початком етапу має
стати сертифікація вихідного матеріалу;
- науково-дослідні установи-оригінатори сортів повинні вирощу¬
вати свій вихідний матеріал до супереліти з оцінкою за вищевказа¬
ними критеріями і забезпечувати ним на комерційній основі елітні
господарства;
- для потреб виробників продовольчої картоплі в Україні слід
вирощувати щорічно не менше 15-20 тис. тонн еліти. Для цього,
враховуючи набутий досвід, в різних областях (з метою зменшення
47
перевозок насіння) зони Полісся та Лісостепу потрібно залишити не
більше 25-30 спеціалізованих елітних господарств, які здатні забез¬
печити вирощування сертифікованого насінного матеріалу. Такими
мають стати кращі колективні, державні й фермерські господарства;
- вирощена еліта може завозитись безпосередньо в господарства
для внутрішньогосподарських потреб або в окремі господарства для
розмноження й наступної реалізації насіння першої-другої репроду¬
кцій. Використання насінного матеріалу має відбуватись лише на
комерційній основі з урахуванням рекомендацій наукових установ.
Виробництво елітного насіння потребує постійної державної фінан¬
сової підтримки;
- в умовах України є можливість щорічно вирощувати не менше
8-10 тис. тонн висококласного насінного матеріалу (за замовленням)
на експорт. Це можуть бути сорти як вітчизняної, так і зарубіжної
селекції;
- вирішення питань виробництва якісного і здорового насіння
картоплі не можливе без організації державної контрольно-
сертифіка-ційной служби. Тому створення такої служби є першочер¬
говим завданням у насінництві;
- для організації вирощування ранньої картоплі в потрібних для
країни обсягах у південних областях необхідно налагодити постійне
й організоване забезпечення їх відповідним садивним матеріалом.
Для цього потрібно організувати реальне випробування ранньостиг¬
лих сортів картоплі в південних областях та вирощування їх насіння
(першої-другої репродукції) у поліських насінницьких господарст¬
вах. Необхідно також забезпечити щорічне завезення у господарства
південних областей садивного матеріалу для виробництва власного
насіння шляхом літнього садіння з використанням його в наступному
році з метою одержання врожаю для продовольчих цілей. Можливе
часткове вирощування насіння на місці шляхом двоврожайної куль¬
тури;
- скорочення посівів картоплі в господарствах з великою пло¬
щею і збільшення їх у господарствах із площею 5-30 га та на го¬
родніх ділянках розміром 0,1-0,5 га вимагає перегляду концепції
забезпечення картоплярства спеціальною технікою. Виникає по¬
треба у дворядкових машинах. Тому, з метою зниження собівартос¬
ті врожаю картоплі та своєчасного проведення всіх робіт на посі¬
вах, для картоплярства потрібно налагодити виробництво вітчизня¬
ної техніки. Можливе придбання і зарубіжної техніки з необхідни¬
ми параметрами.
Одним із невирішених питань у картоплярстві є якість зібраного
врожаю, особливо на зв’язних грунтах лісостепової зони. Існуюча
збиральна техніка і та, що завозиться з Росії, призводить до механіч¬
ного травмування 30-50 % бульб. При цьому втрачаються товарні
якості врожаю. Тому метою і завданням імпорту зарубіжної чи виго¬
48
товлення власної збиральної техніки має бути забезпечення мінімаль¬
ного травмування бульб при їх збиранні;
- за наявності коштів господарства всіх форм власності можуть
бути забезпечені достатньою кількістю мінеральних добрив вітчиз¬
няного виробництва. їх сортимент дозволяє при раціональному ви¬
користанні збирати врожаї високоякісної продовольчої картоплі.
Причому, саме застосування добрив має забезпечити максимальну їх
віддачу. Це досягається за умови внесення добрив в оптимальні
строки, у рекомендованих співвідношеннях елементів живлення та
обов’язкового локального внесення у завчасно підготовлені гребені
при садінні;
- неабиякою проблемою залишається захист посівів картоплі від
шкідників та хвороб. Якщо стан боротьби з колорадським жуком
можна вважати задовільним, то за останні роки, особливо у приват¬
ному секторі, великими є втрати врожаю та його якості від фітофто¬
розу. З метою захисту картоплі, слід проводити 2-3-разове обприс¬
кування посівів проти шкідників і не менше 3-4 разів проти фіто¬
фторозу та інших хвороб.
Імпортовані та частково власні вітчизняні пестициди можуть за¬
безпечити реальний захист посівів, проте проблемою залишаються
рекомендації щодо внесення пестицидів, які розроблені переважно
на основі досліджень учених інших країн. Випробування засобів за¬
хисту, як це проводиться в останні роки в одній-двох наукових
установах, без широкої їх виробничої перевірки та визначення
фактичної ефективності в різних зонах України призводить часто до
завищення норм застосування, низької ефективності окремих із них,
значних затрат на їхнє використання. При цьому не завжди гаран¬
тується і безпечне споживання бульб.
Потрібно розробити науково обгрунтовані рекомендації щодо
використання пестицидів на городах, присадибних і дачних ділянках,
де між рядками картоплі або поряд вирощуються овочеві культури;
- при забезпеченні потреби країни в якісному насінному матеріа¬
лі та якісній продовольчій картоплі особливого значення слід нада¬
вати поліпшенню умов зберігання картоплі.
Для того, щоб садивний матеріал картоплі перед садінням відпо¬
відав міжнародним стандартам і давав максимальну віддачу, його
зберігання протягом осінньо-зимового періоду повинно відповідати
біологічним вимогам сортів. Існуючі картоплесховища у спеціалізо¬
ваних насінницьких господарствах, побудовані 10-20 років тому, не
здатні забезпечити підтримання заданих температур, необхідних для
зберігання насіння. Тому в усіх насінницьких господарствах для ви¬
рішення цієї проблеми потрібно реконструювати існуючі чи збудува¬
ти найближчим часом нові картоплесховища;
- реалізація насінної картоплі повинна здійснюватись в основно¬
му у весняний період. Організації, що займаються забезпеченням ви¬
49
робників садивним матеріалом (елітні господарства, асоціації, агро¬
фірми, об’єднання) повинні зберігати й реалізовувати лише
сертифіковане насіння.
Значні втрати під час зберігання продовольчої картоплі мають іс¬
нуючі у великих містах картоплесховища. Тому тут, як і в насінницт¬
ві, потрібна реконструкція діючих або спорудження спеціальних
сховищ для продовольчої картоплі. Без сучасних сховищ для кар¬
топлі галузь картоплярства не може стати рентабельною;
- потрібно розвивати виробництво картоплепродуктів. Пріоритет¬
ним у цій справі може бути виробництво швидкозаморожених на¬
півфабрикатів, хрусткої картоплі, сухого картопляного пюре, консер¬
вованої молодої картоплі та ін. Вирішення цього питання можливе
при вирощуванні придатних для переробки сортів, спорудженні ліній
з їхньої переробки, вивчення потреб ринку та його забезпечення;
- подальший розвиток картоплярства можливий лише при доско¬
налому й сучасному науковому та інформаційному забезпеченні га¬
лузі. Але вирішити поставлені завдання, на нашу думку, можна лише
після структурної реорганізації наукових установ і підвищення акту¬
альності досліджень;
- дослідження науково-дослідних установ з проблем картопляр¬
ства слід фінансувати з кількох джерел через державні бюджетні
кошти, кошти виробничо-торговельних об’єднань, фірм та асоціацій,
що займаються картоплярством, і власних коштів установ, одержа¬
них від реалізації наукових розробок. Окремі наукові установи мо¬
жуть входити в асоціації, фірми та інші об’єднання, як це має місце
за кордоном.
Основними мають бути дослідження на договірних умовах,
спрямовані на вирішення конкретних завдань. З цією метою дослі¬
дженнями з картоплярства в мінімальних обсягах повинні займатись
не всі науково-дослідні установи Полісся та Лісостепу, як це має міс¬
це останнім часом, а лише окремі з них в основних грунтово-клі¬
матичних зонах. План науково-дослідних робіт не обов’язково має
координуватися одним центром.
За рахунок бюджетних коштів доцільно проводити теоретичні до¬
слідження та селекцію. Усі прикладні дослідження, незалежно від місця
їх проведення, мають виконуватися за замовленням окремих організацій,
що створить здорову конкуренцію серед наукових установ.
За рахунок переважно комерційних надходжень має здійснюва¬
тись фінансування інформаційного забезпечення галузі;
- найважливішими проблемами в картоплярстві, які належить
вирішити науково-дослідним установам, є створення нових столових
сортів картоплі з показниками, що будуть перспективними у най¬
ближче десятиріччя; розробка зональних прийомів одержання здоро¬
вого насінного матеріалу та збереження високих потенційних мож¬
ливостей нових сортів у репродукціях; раціональне розміщення посі¬
50
вів картоплі в реорганізованих сільськогосподарських структурах, її
удобрення та захист; конструювання спеціальної техніки для механі¬
зації виробництва картоплі, збирання, зберігання тощо.
- налагодження сервісного забезпечення виробників насінної і
продовольчої картоплі сучасними матеріально-технічними засобами;
- організація реалізації, у тому числі фірмової, насінної та продо¬
вольчої картоплі з гарантією її якості в усіх регіонах України.
Ми зупинились лише на окремих проблемах перспективного роз¬
витку галузі. Вирішення зазначених проблем і завдань має бути мак¬
симально обмеженим у часі, що дозволить швидше наблизити картоп¬
лярство України до рівня світових досягнень, для чого можливості
країни є достатніми.
ЛІТЕРАТУРА
1. Абушев Н. Практическое руководство к усовершенствованию сельско¬
го хозяйства в Нечерноземной полосе.-Л., 1985.
2. Анфимов А.М. Кардовские планы Мекленбург-Стрелецких в конце
XIX - начале XX в. // Материалы по истории сельского хозяйства и кресть¬
янства СССР. Сб. 5.- М.: Изд-во АН СССР, 1962.
3. Бабицкий Б. А. Херсонская сельскохозяйственная опытная станция.-
Херсон, 1924.
4. Батыренко В.Г., Беликов А.Д. Сорта картофеля // Сводный отчет за
1925-1926 гг. Укр. сети по изучению сортов культурных растений при Все-
украинском обществе семеноводов. - Харьков, 1927.
5. Баранович А.Й. Хозяйство фольварка на юге Волыни в XVIII в. // Ма¬
териалы по истории земледелия СССР - М.: Изд-во АН СССР, 1952.
6. Благодаров В.О. Ранній вигін молодої картоплі- Харків: Держ-
сільгоспвидав, 1933.
7. Бобровский П. О. Гродненская губерния. Ч.І. - С.Пб.,1863.
8. Болотов А. Наказ для деревенского приказчика // Труды Вольного
экономического общества. Ч. XVI.-1770.
9. Веселовский Й.А. Картофель - Л.: Всесоюз. ин-т приклад, ботаники и
новых культур, 1930.
10. Волчанский Ф.М. Результаты опытов по культуре картофеля в Харь¬
ковской губернии за 1913 год. - Харьков, 1914.
11. Г аршин А. Г. Из практики опытных полей - Чернигов, 1909.
12. Гордеев Г.С. Сельское хозяйство в войне и революции - М.-Л.: Гос¬
издат, 1925.
13. Гохгут Г. Хозяйственное обозрение Киевской губернии // Журн. мин.-
ва гос. имущества. - Ч. XVII - 1845
14. Грачев Е. А. О мерах к улучшению и распространению огородниче¬
ства в России // Труды Вольного экономического общества. Ч. IV. - 1872.
15. Грінченко П.Л. Колективні досліди 1928 року - Суми, 1929.
16. Добриловський Ю.Д. Колективні (масові) досліди в селянських гос¬
подарствах Чернігівщини (1926-1927 рр.).- Чернігів, 1928.
17. Дорошенко В. З історії землеробства на Україні і податкова та
політико- і соціально-економічна діяльність - Львів-Київ, 1910.
51
18. Достижения сельскохозяйственных опытных станций.-Харьков: Рад.
селянин, 1928.
19. Дракохруст Е.Й. Борзенская экономия Кочубеев в Черниговском на¬
местничестве во второй половине XVIII в. // Материалы по истории сель¬
ского хозяйства и крестьянства СССР. Сб. 5.- М.: Изд-во АН СССР, 1962.
20. Есимонтовский П. Сельское хозяйство в Черниговской губернии-
Журн. мин-ва гос. имущества. -1844.-Ч. XI.
21. Еще некоторые примечания о картофеле // Экономический магазин. -
1937.-Ч.31.
22. Золотницкий Й.Ф. Наши садовые цветы, овощи и плоды, их истори¬
ческая роль в жизни и верованиях разных народов.- М., 1911.
23. Известия Народного Комиссариата Продовольствия Украины. - 1919.
-№ 3, 4.
24. Известия Народного Комиссариата Продовольствия Украины. -
1919.-№5, 6.
25. Картофель. Современное положение культуры картофеля й стои¬
мость его производства - Ч. I. Основные таблицы. - С.Пб., 1912.
26. Картофель. Современное положение культуры картофеля и стои¬
мость его производства -Ч. II. Стат. сведения - Петроград, 1914.
27. Картопля і її культура - 1932.
28. Крячко З.Ф. Распространение рака картофеля и борьба с ним на тер¬
ритории УССР.-К., 1957.
29. Кульжинський С.П. Як швидко розмножити нові сорти картоплі /
НКЗС УРСР. Носівська с.-г. дослідна станція. - Вип. 39.-К., 1924.
30. Кульжинський С.П. Питання культури картоплі на Україні // Укр. аг¬
роном. - 1928 - № 9.
31. Культура картоплі на Київщині.-К.: Вид-во Окрземвідділу, 1929.
32. Лехнович В.С. К истории культуры картофеля в России // Материалы
по истории земледелия в СССР. Сб. 2.- М.- Л.: Изд-во АН СССР, 1962.
33. Лялин П. Состояние сельского хозяйства Малороссийского края //
Журн. мин-ва гос. имущества. -1844.-Ч. X, разд. 2.
34. Машир М. Культура картоплі в посушливих районах-Харків:
Держсільгоспвидав, 1932.
35. Молоцький М. Землеробство та зміна клімату // Пропозиція - 2000-
№11гС. 38-39.
36. Островський М.Ф. Як одержувати високі врожаї картоплі - К.,1957.
37. Про картоплю - Харків, 1926.
38. Рабінович Я. Л. Культура картоплі // Із наслідків робіт Вінницької
досл. станції за 1926-1929 рр.- Вінниця, 1930.
39. Сільське господарство УРСР: Статист, зб.- К.: Статистика, 1969.
40. Сортовые посевы СССР за 1938 год - М.-Л.: Госполитиздат, 1939.
41. Статистические известия о Кавказской области и земель войска Чер¬
номорского. - ЖМВД, к. У., 1830.
42. Улащик И.Н. Изменение в хозяйстве крестьян Литвы и Западной Бе¬
лоруссии в связи с введением новых культур (картофеля).- М.: Изд-во АН
СССР, 1962.
43. Україна в цифрах - К., 1927.
44. Україна: Статист. щорічник.-Харків, 1929.
52
45. Фаворов О.М., Котов А.Ф. Літнє садіння картоплі в колгоспах і рад¬
госпах УРСР. - Одеса, 1949.
46. Франкфурт C.JL, Венгеровский А.М. Труды сети опытных полей
Всероссийского общества сахарозаводчиков. Опыты по культуре картофе¬
ля.-К., 1908.
47. Хозяйственные очерки Могилевской, Минской и Волынской губер¬
ний // Журн. мин-ва гос. имущества - 1845 - Ч. XVII.
48. Хозяин - 1896. - № 46.
49. Хозяйственное обозрение за 1859 г. // Журн. мин-ва гос. имущества. -
1851.-Ч. ХХХѴІІІ-І.
50. ЦГИАЛ.-Ф. 91, д. 542, л. 7,20, 34, 47,58,70, 79, 101. 112,123, 132. ГО.
155,167.
51. ЦГИАЛ.- Ф. 91, д. 537 (1794), № 6.
52. ЦГИАЛ.- Ф. 91, д. 442 (1794, 1796).
53. ЦГИАЛ.- Ф. 91, д. 525 (1794).
54. ЦГИАЛ.- Ф. 91, д. 542 (1795).
55. Целинский Б.А. Журн. мин-ва гос. имущества.- С.Пб., 1846.
56. Чаянов А.В. Економічні основи культури картоплі. - К.,1921.
57. Щафонский А. Черниговского наместничества топографические опи¬
сания-Киев, 1851.
58. Шадилов С. Аграрна реформа і прогрес сільського господарства - К.,
1918.
59. Щербанова В.Д. Звіт про селекційні роботи з картоплею за 1925-1929 рр.
/НКЗС УРСР. Носівська с.-г. дослідна станція - Вип. 122.- Носівка, 1930.
53
2. МОРФОЛОГІЯ, ФІЗІОЛОГІЯ
ТА БІОХІМІЯ КАРТОПЛІ
Картопля належить до родини пасльонових (Solanaceae) роду So¬
lanum. Батьківщина культурної картоплі - Південна Америка, де
росте більшість її видів. Культурна картопля цього континенту на¬
лежить до трьох серій - Tuberosa, Andigena, Acaulia.
Європейські культурні сорти походять від S. Tuberosym, але
останнім часом у селекційний процес залучаються й інші види (їх
понад 160) і форми картоплі. Картопля може розмножуватися веге¬
тативно і генеративно. Останній спосіб в Україні використовується
тільки в селекції. У деяких країнах, зокрема Китаї, його застосову¬
ють і для вирощування столової картоплі. Але основним способом
розмноження цієї культури є вегетативний.
2.1. МОРФОЛОГІЧНІ ОСОБЛИВОСТІ
Родина картоплі характеризується низкою морфологічних особ¬
ливостей.
Кущ. Залежно від будови кущів, розрізняють кілька морфолого-
біологічних типів рослин картоплі. Окремі властивості та ознаки ще не
дають цілісного уявлення про рослину. Організм рослини необхідно роз¬
глядати як єдину складну систему, де всі ознаки взаємозв’язані.
Перше уявлення про рослину щодо сортових ознак дає зовнішня
морфологічна форма, оскільки вона є відображенням фізіологічних
особливостей.
Доросла нормально розвинена рослина картоплі являє собою
кущ, що складається, як правило, з 4-8 стебел, розміщених паралель¬
но або під певним кутом. Кількість їх у кущі одного сорту корелює з
розмірами материнської бульби: чим більша насіннєва бульба, тим
більше стебел вона утворює.
За формою розрізняють сорти з прямостоячим і розлогим кущем,
за ступенем облистяності - слабо-, середньо- і сильнооблистяні.
Виділяють сорти стеблового і листового типу кущів.
Сорти стеблового типу мають велику кількість дрібних листків,
добре розподілених просторово; рослини листкового типу створю¬
ють біомасу за рахунок більших листків, які за фотосинтетичною
активністю поступаються листкам попереднього типу. За оптималь¬
них природних умов, вологи і температури габітус рослин стеблово¬
го типу сприятливіший для формування значної біомаси, розподіл
якої на початку вегетації в обох типів приблизно однаковий, але змі¬
54
нюється в онтогенезі. Перед закінченням вегетації на кущах листко¬
вого типу маса листків переважає над масою стебел (2:1); у рослин
стеблового типу ці величини майже однакові (1:1).
Кущі стеблового типу розвивають більше стебел і мають більшу масу
бульб при меншій кількості їх на одному кущі. Перевага надається сортам
із прискореним формуванням біомаси на початку вегетації, швидким пе¬
реходом до максимального росту і його закінченням через 80 днів після
появи сходів при ранньому формуванні бульб, що створює умови для
швидкого їх росту і отримання високого господарського урожаю.
Первинна продукція формується інтенсивніше, якщо фотосинте¬
тичний потенціал рослини достатній, що створюється великою кіль¬
кістю листків з високим умістом хлорофілу. Розміщення ярусів лист¬
ків на значній висоті у рослин стеблового типу створює умови для
доброго освітлення більшості їх, порівняно з рослинами листкового
типу, кущ яких відносно компактний. Тому стебловий тип рослин
більш придатний для середньо- і пізньостиглих сортів, а для ранньо¬
стиглих - листковий тип, що характеризується прискореними проце¬
сами формування біомаси, у тому числі бульб, на які припадає біль¬
ша частина всієї сухої біомаси врожаю.
Стебло. Стебла утворюються з паростків бульби. За формою во¬
ни бувають три- або чотиригранні; здебільшого зелені, інколи пігме¬
нтовані із синьо-фіолетовим або червоно-фіолетовим відтінком. На
ребрах граней стебел наявні різної товщини рівні або хвилясті відро¬
стки черешків листка, що називаються крилами. У багатьох сортів
вони відносно вузькі. Висота стебел коливається досить відчутно
(30-150 см) і досягає максимуму на час цвітіння рослин. Пізньостиг¬
лі сорти, як правило, мають вищі стебла, ніж ранні.
Стебла складаються з міжвузлів із потовщеними вузлами. У піз¬
ніх сортів їх більше, ніж у ранніх. Стебла часто галузяться у нижній
частині, а інколи й у верхній. За цією ознакою рослини картоплі по¬
діляють на дві групи - розгалужені і нерозгалужені.
Відомий білоруський селекціонер П.І.Альсмік [1] вважає, що з
галуженням сортів пов’язана їхня продуктивність, реакція на родю¬
чість грунту, стійкість до виродження. За цією ознакою усі сорти він
схематично поділяє на дві цілком відмінні групи: із розгалуженими в
нижньому ярусі стеблами і нерозгалуженими.
При вирощуванні на високому агрофоні нерозгалужені сорти на
початку філогенезу є більш продуктивними, ніж розгалужені, проте
швидко втрачають цю властивість. Розгалужені в нижньому ярусі
сорти довше зберігають властиву їм продуктивність.
Доведено, що найвищу продуктивність сорти картоплі мають у
перші 10-15 років після їх створення і включення у Державний ре¬
єстр сортів [2].
Дуже важливо, особливо для високопродуктивних сортів, уже на
початку їх філогенезу розробити сортову агротехніку, яка забезпечу-
55
вала б за цей період (10-15 років) максимальну врожайність. Якщо ж
сорт втрачає необхідну продуктивність, його виключають з Реєстру
сортів. У зв’язку з цим реакцію сортів на застосування добрив та ін¬
ші агротехнічні заходи слід вивчати ще до передачі сортів до Держав¬
них сортодільниць і сортостанцій з тим, щоб на період реєстрації
були відомі особливості технології їх вирощування.
Сортова технологія важлива ще й тому, що надлишкове або не-
збалансоване застосування добрив, особливо азотних, посилює виро¬
дження картоплі, насамперед сортів, схильних до ураження вірусни¬
ми хворобами.
Згідно з теорією П.І.Альсміка, існує чітко виражена залежність
між типом конституції рослини та її виродженням. Нерозгалужені
форми майже всі певною мірою вироджуються, незалежно від трива¬
лості їх вегетаційного періоду. Певні розгалужені форми (наприклад,
сорти Вольтман, Зарево, Світязь, Сілезія та ін.) майже не вироджу¬
ються. Вироджені рослини завжди прямостоячі і нерозгалужені, як¬
що навіть вони походять від сортів із розлогою формою куща і роз¬
галуженими стеблами.
Високою продуктивністю і достатньою життєздатністю виріз¬
няються сорти, що є своєрідною “конституційною популяцією”, по¬
єднуючи розгалужені і нерозгалужені форми (наприклад, сорти Гат¬
чинська, Коренєвська, Лорх, Луговська та ін.).
Отже, за конституцією окремого сорту можна прогнозувати його
поведінку у філогенезі, вивчати найбільш раціональну систему удоб¬
рення, технологію вирощування.
Знання конституції дуже важливе у селекційній роботі. Слід
пам’ятати, що при доборі продуктивних форм можливе значне погір¬
шення їх у наступних репродукціях, навіть при оптимальній агротех¬
ніці, що потребує застосування таких насінницьких та інших заходів,
які підтримували б продуктивність рослин на досить високому рівні.
Крім того, розгалужені форми в процесі філогенезу не будуть різко
погіршувати свою продуктивність.
Листок. Він є головним органом фотосинтезу, який формує ор¬
ганічну масу рослини.
Процесу фотосинтезу підпорядкована вся структура листка. За¬
вдяки дорсивентральній будові поверхня листка більша від його
об’єму. Цим забезпечується інтенсивний газообмін листка з атмос¬
ферою, здійснюється інтенсивне поглинання вуглекислого газу через
численні продихи, а також систему дихальних порожнин і міжклі¬
тинників. У більшості видів картоплі переважна кількість продихів
розміщена на нижньому боці листка, а менша - на верхньому, у де¬
яких на верхньому боці листка вони відсутні повністю. Кількість
продихів на 1 мм2 площі листка досить значна: 93-572 - на нижньо¬
му боці листка і 0-230 - на верхньому. Вуглекислий газ проникає в
листок через продихи до хлоропластів переважно завдяки дифузії за
56
градієнтом концентрації, який підтримується швидким зв’язуванням
вуглекислоти у процесі фотосинтезу. Це зв’язування тісно корелює з
кількістю ферменту 1,5-риболозодифосфаткарбокеилази, яка активі¬
зує роботу акцептора вуглекислого газу - рибулозо-1,5-дифосфату.
За нашими попередніми даними, сорти картоплі інтенсивного типу
характеризуються вищим умістом цього ферменту і ступенем розвит¬
ку провідної системи листків, ніж сорти невисокої продуктивності.
Успішному фотосинтезу сприяють також особливості структури
листка, що забезпечує максимальне поглинання сонячної радіації.
Епідерміс його прозорий, безбарвний, без хлоропластів, тому вільно
пропускає сонячні промені.
Вакуолі епідермальних клітин служать мініатюрними лінзами,
що концентрують світло. Клітини палісадної паренхіми розміщують¬
ся щільно, здебільшого в один або два ряди, торцем до поверхні епі¬
дермісу. Внаслідок цього промені ковзають поздовжніми стінками
клітини й освітлюють усю її порожнину.
Під палісадною паренхімою розміщуються клітини губчастої па¬
ренхіми, до яких доходить розсіяне світло. Але внаслідок пухкого їх
розміщення світло в міжклітинниках багаторазово відбивається від
клітинних стінок і поглинається майже повністю.
Достатньому освітленню сприяють також форма і розміщення
листків на стеблах.
При появі сходів картоплі листки завжди цілісні й невеликі. З ча¬
сом на рослині з’являються переривчасто-непарноперисторозсічені
листки, спочатку з однією парою бокових часток, потім - із двома.
Кількість їх в окремих сортів може досягати 6-7 пар. Між парними
частками є дрібніші утворення - часточки.
Листок складається із пластинок, черешка і стержня, на якому
окремо розміщені одна кінцева частка і одна-сім бокових. Непарна
частка, що розміщується на кінці стержня, називається кінцевою, а
парні бокові мають порядкові назви - перша, друга і т.д.
За характером прикріплення листки можуть бути черешковими і сидя¬
чими, а за формою - вузькими, округлими та проміжними. Розсіченість
листка залежить від кількості часток і часточок. Найбільша розсіченість
буває в середній частині стебла, ближче до його верхівки й основи ступінь
її зменшується. Біля основи листка є два прилистки різної форми (серпо¬
подібні, листоподібні, проміжні). Довжина листка - 15-25 см.
Серед загальних ознак листка як органа фотосинтезу особливе
значення має розмір, розміщення часток (рідке, густе), розсіченість
(кілька часток у серіях).
Поверхня листка буває блискуча або матова, опушена або не
опушена. Листки по стеблу розміщуються спірально - вліво і вгору
по рослині.
Основна частина листка густо пронизана провідними елементами
(жилками). Центральна жилка поділяє частки листка на дві рівні час¬
57
тини. Від неї під гострим кутом відходять вторинні жилки, які бага¬
торазово розгалужуються і пронизують усю паренхіму листка.
Квітка. Квітка в рослини картоплі п’ятипелюсткова. Вона скла¬
дається із чашечки з п’ятьма чашолистками, п’ятипелюсткового ві¬
ночка, п’яти тичинок із пиляками, що утворюють конусоподібну ко¬
лонку маточки. Маточка включає приймочку, стовпчик і зав’язь. За¬
барвлення віночка може бути білим, синім, синьо-фіолетовим, чер-
воно-фіолетовим із відтінками різної інтенсивності. Пиляки також
мають різне забарвлення і можуть бути лимонними, жовтими, зеле¬
нувато-жовтими, оранжевими.
У картоплі квітки зібрані в суцвіття, що розміщується на квітко¬
носі. Воно включає найчастіше 5-6 квіток.
Усі елементи квітки в комплексі із суцвіттям є однією з важливих
ознак сорту.
Плід. Плід у картоплі є двогніздовою багатонасінною ягодою
округлої або овальної форми. Насінини світло-жовтого кольору. Во¬
ни належать до категорії найдрібнішого насіння. Один грам вміщує в
середньому 1600 штук (трапляється від 1450 до 1750).
Багато сортів картоплі стерильні і не утворюють насіння.
Коренева система. Тип кореневої системи залежить від способу
розмноження картоплі. При садінні бульбами рослина утворює мич¬
кувату кореневу систему, а при садінні насінням - стрижневу.
У рослин, що розмножуються насінням, спершу утворюється
стрижневий корінь із численними корінчиками, а з часом із нижньої
підземної частини стебла розвиваються побічні корені.
У рослин, що розмножуються бульбами, у період проростання ві¬
чок навколо паростків утворюються потовщення з великою кількіс¬
тю горбиків початкових корінців, з яких виростає коренева система
мичкуватого типу. Незважаючи на те, що материнська бульба міс¬
тить велику кількість поживних речовин, картопля не сходить, якщо
не утворилася коренева система.
Приблизно протягом 20-25 діб ріст маси паростків і коренів від¬
бувається за рахунок поживних речовин материнської бульби.
Максимальна маса коренів наростає на 50-60-й день після садін¬
ня. Добре розвинена коренева система підвищує продуктивність
асиміляційної листкової поверхні.
Сорти картоплі різняться між собою довжиною і розміщенням
кореневої системи. Якщо вона більш горизонтальна і поверхнева, то
такі сорти менш стійкі до посухи.
Коренева система картоплі порівняно з іншими культурами розви¬
нена слабо. Співвідношення її маси до маси всієї рослини менше, ніж в
інших сільськогосподарських купыур. За даними Ф.А.Новікова [13],
вона становить тільки 3 % маси надземної частини і 8 % сухої маси
листків. Об’єм грунту, охопленого кореневою системою картоплі, в 1,4
рази менший від об’єму грушу кореневої системи ячменю і в 2,2 рази
58
- від цукрового буряка [12]. До того ж коренева система картоплі роз¬
міщується переважно у верхніх горизонтах грунту: на глибині - до 60
см, а в горизонтальному напрямку - до 50 см. Окремі корені можуть
проникати на глибину до 150 см і нижче.
Ступінь розвитку кореневої системи залежить від сортових особ¬
ливостей та умов вирощування картоплі. Так, у ранньостиглих сортів
вона розвинена менше, ніж у середньо- й пізньостиглих. Сорти, що
ростуть інтенсивно, характеризуються швидшим приростом коренів
у довжину: 2,5-3 см проти 1,9-2,5 см у сортів неінтенсивного типу.
За даними П.І.Альсміка [1], потужно розвинену кореневу систему
мають розгалужені сорти. Для типових представників нерозгалужено-
го типу більш характерні нетовсті корені з незначним горизонтальним
поширенням. Щодо проникнення вглиб, то особливих відмінностей
між розгалуженими і нерозгалуженими сортами здебільшого не спо¬
стерігається: до 70-75 % коренів у них проникає на глибину до 60 см.
Найбільшої маси і об’єму коренева система досягає в період цві¬
тіння рослин. У міру дозрівання бульб вона поступово відмирає.
Столони і бульби. Столони є видозміненими підземними стеб¬
лами, тому за своєю формою вони дуже схожі з надземними стебла¬
ми. Розвиваються столони після сходів картоплі з пазух лускатих
листочків підземної частини стебла. Вони ростуть спочатку в довжи¬
ну, утворюють кілька міжвузлів, потім верхівка їх потовщується і
започатковує бульбу. Тому бульбу можна розглядати як надмірно
потовщену частину столона. За своєю будовою вона також дуже на¬
гадує пагін. У молодих бульб є дрібні лускуваті листочки, у пазухах
яких закладається від 3 до 4 бруньок, що перебувають у стані спокою
й утворюють вічка.
При пробудженні бульб проростає лише одна (середня) брунька,
решта може проростати лише після пошкодження або видалення па¬
ростка. Брунька складається із зачаткових стебла і листочків, пазухових
бруньок і зачаткових корінців. Вічка на бульбі розміщені спірально; на
верхівці вони зближені внаслідок того, що бульба росте верхівкою.
За формою бульби бувають різними: округлими, округло-
овальними, видовжено-овальними і т. п., білого, червоного і синього
кольору з різними відтінками. М’якуш бульб також може бути білим,
жовтим, інколи кремовим або червоно-фіолетовим.
Паростки, що утворюються при проростанні бульб на світлі, короткі,
звичайно міцні. Вони мають буруватий відтінок, а колір, залежно від
сорту картоплі,- зелений, синій, синьо- або червоно-фіолетовий.
2.2. БІОЛОГІЧНІ ОСОБЛИВОСТІ
Вимоги до грунту. Біологічні особливості картоплі вимагають
пухких грунтів. Ці вимоги базуються на механічній дії бульб на
грунт під час їх росту. При врожайності бульб 0,5 кг на 1 кущ, тобто
59
250 ц/га, вони повинні вивільнити від грушу простір об’ємом 0,5 дм3,
що на 1 га посіву становить близько 20 м3 [4]. Корені картоплі погре¬
бують у десятки разів більше кисню, ніж інші сільськогосподарські
культури, що було доведено ще ЛБушнелем. За даними досліджень
цього автора, інтенсивність дихання бульб картоплі коливається в межах
7-12 мл 02 на 1 г сухої маси коренів, що у 5 разів вище за інтенсивність
дихання коренів соняшнику (цит. за Гончариком М.М. [4]).
При нестачі кисню в груші бульби погано утворюються і розви¬
ваються, затримуються у строках дозрівання. Вважають [4], що для
картоплі кращими грунтами, котрі забезпечують кореневу систему
киснем та чинять менший опір росту бульб, є легкі піщані грунти.
Проте існує думка, що найвища урожайність бульб забезпечується
вирощуванням картоплі на легкосуглинкових грунтах [9]; [6, 7].
Оптимальна щільність грунту при вирощуванні картоплі визна¬
чається його гранулометричним складом, вологістю. На дерново-
підзолистих супіщаних і чорноземних грунтах найкращі умови для
росту картоплі створюються при щільності 1,3-1,4 г/см3 [4]; [7], а на
дерново-підзолистих піщаних грунтах - 1-1,2 г/см3 [7, 8].
Вимоги до температури. Картопля належить до культур віднос¬
но холодостійких. Разом з тим вона дуже чутлива до дії негативних
температур. Бадилля ушкоджується при температурі мінус 1-1,5°С, і
за тривалої дії такої температури рослина гине. Окремі дикі види
картоплі можуть переносити приморозки до -6°С і навіть до -7-8°С.
Але отримати сорти, стійкі навіть до невисоких негативних темпера¬
тур, тобто відносно морозостійкі, шляхом гібридизації культурних
сортів із дикими морозостійкими видами не вдалося.
Від тривалої дії приморозків гине не тільки бадилля, але й бульби
картоплі. Наприклад, при мінус 6°С вони гинуть через 8 год, а при
мінус 9°С - через одну годину. Але за певних умов бульби здатні пе¬
реохолоджуватися і витримувати без ушкодження температуру до
мінус 7°С. Очевидно, цим пояснюється збереження життєдіяльності
бульб, що залишаються в грунті на зиму під снігом [4].
Температурним режимом регулюється проростання бульб, ріст і
розвиток рослин, їхня продуктивність.
Нормальне проростання бульб у польових умовах відбувається
тоді, коли грунт на глибині 10 см прогріється до температури 7-8°С.
Якщо картоплю посадити в холодний (нижче 6°С) і вологий грунт,
то сходи з’являться звичайно не раніше, ніж через 30-50 днів. За та¬
ких умов замість паростків і стебел можуть утворитися дочірні буль¬
бочки без появи сходів. Цей факт свідчить про те, що бульбоутво¬
рення можливе при досить низькій плюсовій температурі, при якій
ріст вегетативної маси не відбувається.
Повноцінний ріст молодих рослин спостерігається лише при по¬
яві коренів. Це є необхідною умовою і тоді, коли вони одержують
поживні речовини виключно з материнської бульби. Таке явище зу¬
60
мовлене передусім синтезом у кореневій системі цигокінінів та
пересуванням їх у точки росту паростків.
Температурні умови визначають не тільки швидкість проростан¬
ня, але й темпи подальшого росту та розвитку рослин. Існує певна
залежність між температурою грунту і появою сходів після садіння
картоплі. Так, при температурі грунту 11-12°С сходи з’являться че¬
рез 23 дні, 14-15°С - через 17-18, 18-25°С - через 12-13 і при 27-28°С -
через 16-17 днів [11]. В інших дослідженнях спостерігається подібна
закономірність, але тривалість періоду від садіння картоплі до її сходів
має мінливий характер. Зокрема при температурі грунту 10-12°С трива¬
лість періоду садіння-сходи становила 34, а при 20-22оС - 22 дні [15].
В умовах Білорусі при температурі грушу 9-13°С цей період тривав 25-
30 днів [5]. На дерново-підзолистих супіщаних грунтах Київської облас¬
ті в дослідах В.С.Куценка [7] при температурі грунту 5°С сходи з’я¬
вилися через 53, а при 8°С - через 31 день. Певні розбіжності цих
показників зумовлені фізіологічним станом бульб на період садіння,
кількісним співвідношенням між температурою і вологістю грунту,
його механічним складом та деякими іншими факторами.
Температура впливає на швидкість проростання бульб і строки
садіння картоплі. Через організаційні обставини в ряді господарств
на деяких полях картоплю садять у більш ранні строки, ніж передба¬
чено технологічною картою. У випадку перезволоження грунту це
призводить не тільки до затримання появи сходів, але й значного
зрідження їх, що зумовлюється загниванням бульб та ураженням па¬
ростків ризоктоніозом, чорною ніжкою і т.п. Зрідження сходів може
досягати 30-50% [6]. За низьких температур знижується доступність
і поглинання коренями із грунту фосфору, азоту та послаблення їх
метаболізму. Посилення фосфорного обміну на початку вегетації при
низьких температурах має важливе значення для підвищення стійкос¬
ті рослин до низьких температур грунту [4].
У дослідженнях з різними строками садіння та їх впливом на про¬
дуктивність картоплі тривалість періоду садіння-сходи, залежно від
погодних умов весни, при ранньому садінні (12-20 квітня) становила
33-40 днів, а при оптимальному - 20-22 [3]. При дуже ранньому
садінні зрідження сходів внаслідок загнивання бульб, ураження
паростків ризоктоніозом і частково чорною ніжкою було на 17,7—
20,6 % більшим, ніж при оптимальному садінні, а саме: 55-58 %, 31-
32 і 10-11 % відповідно від загальної кількості загиблих рослин. Ха¬
рактерно, що дуже пізнє садіння (20 травня) також призвело до поміт¬
ного зрідження посівів (табл. 2.1).
Таким чином, при ранньому чи пізньому садінні насадження кар¬
топлі або не досягають потрібної густоти стояння, або сходи з’яв¬
ляються занадто пізно. Це призводить до порушення оптимальних
температурних режимів проходження рослинами окремих фаз фено¬
логічного розвитку.
61
Таблиця 2.1 - Вплив строків садіния на схожість бульб картоплі сорту
Гатчинська
Строк
садіння
Розрахункова густота садіння, тис, бульб на 1 га
CQ
І
І
10 квітня
20 квітня
1 травня
10 травня
41,0
40.5
40.6
40.6
29.0
37.0
36,6
36,2
70,7
91.3
90,0
89.4
61,8
61,2
60,8
60,1
44.7
55.8
55.8
56,4
72.3
91.3
90.7
93.8
81,8
80,7
80.5
80.5
60,9
74.4
75,1
75.5
74,4
72.1
93.2
93,7
Найкращою температурою для проростання бульб вважається 18-
22°С. Зниження до 13-14°С подовжує період садіння-сходи на 2 дні,
до 11-12°С - на 5 і до 10-11°С - 6 днів. В інтервалі 11-18°С для заве¬
ршення появи сходів необхідною сумою ефективних температур є
243-316°С. Період від появи поодиноких сходів (10 %) до повних
(75 %) складає при цьому 3-4 дні із сумою температур до 45°С.
Позитивна дія зростаючих температур проявляється тільки до пев¬
ного періоду. У фазі садіння-сходи сума ефективних температур не
повинна перевищувати 565-593°С. Коли вона досягає 660-662°С,
рослини гинуть. Шляхом створення нових сортів можна підвищити
стійкість бульб під час проростання до високих температур. Трива¬
лість періоду сходи-бутонізація також залежить від температури.
Зниження її від оптимальної до 13-14°С подовжує цей період на 2-3
дні, а до 11-12°С - на 7 днів.
Середня тривалість періоду бутонізація-масове цвітіння становить
близько 14 днів при сумі середніх добових температур близько 230°С.
Кожен орган рослини картоплі для оптимального розвитку вимагає
певних температур. Максимальна маса листя утворюється при 12-14°С,
а стебел - при 18°С. Для фази цвітіння також характерні певні опти¬
мальні температури. При нічних 12°С формуються тільки квіткові бру¬
ньки, а при 18°С відбувається пишне цвітіння й утворення ягід. Денна
температура має менше значення для цвітіння. В цілому ці процеси най¬
більш активно відбуваються в інтервалі 18-25°С. Проте спостерігаються
певні відхилення від цього, залежно від географічних параметрів. Так, в
умовах Підмосков’я температурний оптимум для цвітіння і ягодоутво-
рення знаходиться в межах 18-21°С, Ставропольського краю - 18-23°С,
Казахстану - 24-25°С [8, 9, 10].
За вищих температур цвітіння слабшає, а при 27°С зовсім припи¬
няється.
Виявлено також особливості реакції картоплі на температурні
умови залежно від сорту. Так, ранньостиглі сорти розвиваються при
нижчій температурі, ніж пізньостиглі. При 16-17°С ранні сорти ра¬
ніше завершують розвиток асиміляційного апарату і починають буль¬
62
боутворення. Процес бульбоутворення, незалежно від сорту, най¬
більш інтенсивно відбувається при середньодобовій температурі 16-
18°С, при підвищенні її до 20°С він уповільнюється, а при 29°С і ви¬
ще при відсутності опадів повністю припиняється. Висока темпера¬
тура повітря і грунту призводить до теплового виродження картоплі і
внаслідок цього - до різкого зниження продуктивності [16, 14, 4].
Негативно впливають на ріст і розвиток картоплі понижені плю¬
сові температури. Наростання вегетативної маси майже повністю
припиняється при температурі, нижчій 7°С, а фотосинтез хоча й три¬
ває майже до заморозків, але відбувається досить повільно.
Важливе значення для росту картоплі має співвідношення денних
і нічних температур. Найвищі врожаї одержують при температурно¬
му режимі день-ніч 20°/10°С, нижчі - при 25°/15°С, а при 30°/20°С
рослини майже не утворюють бульб навіть через 130 днів.
Вивчення двох фотоперіодів (12 і 18 год) і чотирьох температур¬
них режимів (18/12, 22/16, 26/20, 30/24) показало, що високі темпе¬
ратури і тривалий фотоперіод, з одного боку, затримують експолі-
нійний розмір бульби, а з іншого - об’ємний розмір. Високі темпера¬
тури зменшують також загальний розмір бульб, а в поєднанні з дов¬
гим фотоперіодом - і наростання сухої маси. Фотоперіод не впливає
на загальний ріст бульби при низьких температурах [19].
Ранньостиглі сорти менше реагують на зміни температурного режи¬
му [19]. Бульбоутворення можна прискорити, якщо рослини, що виро¬
щувалися при 20°С, перенести у прохолодніші температурні умови, на¬
приклад 3-9°С. Нормально цей процес у картоплі відбувається при тем¬
пературі грушу 18-19°С, зокрема для ранньостиглих сортів найсприят¬
ливішою є температура 17°С, а середньостиглих - 19°С [11].
Поєднання високої температури і тривалого дня викликає пере¬
творення столонів на надземні пагони.
В умовах півдня України довгочасна дія високих температур (30-
40°С) у період формування бульб порушує нормальний обмін речовин,
знижує врожай і насіннєві якості бульб. При проростанні ці бульби
дають тонкі ниткоподібні паростки, з яких виростають ослаблені кущі.
При середньодобовій температурі 19-21 °С урожай містить 20 % таких
бульб, при 24°С - до 50, а при температурі понад 25°С - 75 %.
Найбільше впливає на втрату продуктивності бульб підвищена
температура в нічні години (табл. 2.2).
Таблиця 2.2 - Вплив температури грунту на утворення бадилля і бульб, г
(Лорх А.Г., 1968)
Температура
грунту,
°С
Ещ
зон
Ваі
эба
Лорх
бадилля
бульби
бадилля
бульби
бадилля
бульби
16-17
240
1480
280
1520
1050
900
19-22
790
770
820
810
420
1450
24-28
310
365
260
320
350
520
63
Рослини картоплі чутливі до різких коливань температури, зок¬
рема протягом вегетації вони часто призводять до ослаблення або
тимчасової зупинки бульбоутворення. Якщо ця зупинка тривала, від¬
бувається утворення багатошарового кіркового покриття, а новий
приток асимілятів прориває менш опробковілу ділянку вічка з утво¬
ренням на бульбах молодих бульбочок. Якщо зміни росту і гальму¬
вання бульбоутворення проходять часто, то спостерігається нерівно¬
мірний ріст частин бульби, що призводить до витягування тканин
біля вічок і утворення бульб неправильної форми.
Найсприятливішою для картоплі є місцевість з липневою ізотер¬
мою не вище 21 °С. У США врожай бульб 420 ц/га одержано при
опадах 300 мм і середньодобовій температурі 16,4°С а рекордний
урожай Канади - 1180 ц/га, відповідно, при 283 мм і 15°С.
Вимоги до світла. Рослини картоплі вимогливі до світла. При
його нестачі спостерігається незначне пожовтіння бадилля, видов¬
ження стебел рослин, слабке або повністю відсутнє цвітіння і внаслі¬
док цього - зменшення урожайності бульб.
Дослідження, проведені Пок’якалла, з тимчасовим затіненням
рослин в умовах Скандинавського півострова (цит. за Гончариком
М.М.) засвідчили, що нестача світла проявляється при 67 % повного
денного освітлення. Подальше зменшення освітленості до ЗО % ви¬
кликало гальмування синтезу сухої маси рослин на 38 %. При цьому
суха маса стебел збільшилася на 57 % (внаслідок етіоляції), а суха
маса бульб зменшилася на 80 %.
Для продуктивності картоплі важливе значення має не тільки
тривалість освітлення, але й якість світла - його спектральний склад,
час затінення рослин. У наших спеціальних вегетаційних дослідах із
ранньостиглим сортом картоплі Бородянська рожева при повному
денному освітленні урожайність бульб становила 807 г/кущ; уміст
сухих речовин у бульбах - 21,2 %; крохмалю - 15,5 %. При триго¬
динному полуденному затіненні рослин названі показники становили
відповідно 778 г/кущ; 21,0 % та 15,2 %.
При тригодинному ранковому затіненні урожайність бульб була
найменшою - 717 г/кущ; мінімальним був також уміст сухих речовин -
20,4 % та крохмалю - 14,8 %. Отже, ранкове затінення рослин було
більш негативним, ніж полуденне. Це пов’язано, можливо, насамперед із
тим, що вранці у спектрі сонячного проміння більше червоних проме¬
нів, ніж опівдні. Відомо, що червоні промені набагато ефективніше по¬
глинаються хлорофілом, ніж інші промені спектра. Крім того, вранці
більш сприятливою для картоплі є нижча температура повітря та грушу,
а вологість повітря вища, ніж опівдні. Все це сприяло кращому прохо¬
дженню процесу фотосинтезу, а отже - і вищій продуктивності рослин.
Під впливом різної тривалості освітлення та якості світла у рос¬
лин спостерігалися і морфологічні зміни. По-різному формувалася
площа листків, зокрема найбільш активно фотосинтезуючого четвер¬
64
того листка кожного пагона. При ранковому затіненні вона була най¬
більшою і становила 35 см2; при денному затіненні - 31 см ; а най¬
меншою - 29 см2 - при повному денному освітленні.
Залежно від тривалості освітлення та часу затінення рослин, змі¬
нювався і вміст хлорофілу в листках. Найменша кількість хлорофілу
спостерігалася в рослин, що не затінювалися, а найбільша - у рослин
із ранковим затіненням. Проте інтенсивність та продуктивність фо¬
тосинтезу була найвищою в рослин, що не затінювалися. Це свідчить
про те, що продуктивність фотосинтезу за різних умов освітлення не
завжди визначається кількістю хлорофілу в листках. Сказане підтвер¬
джується визначенням сухої маси листків четвертого, найбільш про¬
дуктивного ярусу листків. Так, у рослин, що отримували світло про¬
тягом світлового дня, маса сухих речовин становила 5,2 мг/см2; при
ранковому затіненні - 4,1 г/см2 і при денному - 4,3 г/см2. Така зале¬
жність фотосинтезу і визначала кінцеву врожайність бульб, яка по¬
вністю корелювала з інтенсивністю синтезу сухих речовин. Тобто,
найвища урожайність бульб спостерігалась у рослин без тимчасового
затінення, а мінімальна - у бульб із ранковим затіненням.
Вплив на картоплю чергування добових періодів освітлення не
можна розглядати ізольовано, без зв’язку з іншими одночасно дію¬
чими зовнішніми факторами. Є.М.Успенський у 1936 році, вивчаючи
реакцію картоплі на фотоперіоди залежно від температури, дійшов
висновку, що при 10-15°С рослини швидше дозрівають при довгому
дні, а при більш високій температурі - при короткому. Існує думка,
що тривалість фотоперіоду не є однаковою для окремих органів рос¬
лин [18, 10]. Для формування квітів картоплі потрібний довгий день,
а для формування бульб - короткий.
Є дослідні дані про сортові особливості вимог картоплі до фото¬
періоду та спектрального складу світла. У дослідах з різними сорта¬
ми картоплі при освітленні червоними променями кращу врожай¬
ність забезпечив сорт Берлінхінген, а при освітленні синьо-
фіолетовими - сорт Лорх [15].
Тому сорти, що мають вищу врожайність при освітленні довго¬
хвильовою радіацією, краще вирощувати у північних районах, а ті,
що сприймають короткохвильову сонячну радіацію, - у південних.
Вимогливість картоплі до води. Картопля є вибагливою до во¬
логості грунту і повітря, що пов’язане насамперед із формуванням
великої надземної маси та слабким розвитком кореневої системи. За
вегетаційний період рослини картоплі витрачають багато води, але
тільки незначна кількість її (менше 0,5 %) використовується на будо¬
ву їхнього тіла, решта - випаровується. Один гектар насаджень кар¬
топлі випаровує за вегетаційний період 2000-4000 м3 води, з яких на
рослини в середньому припадає 40, а на грунт - 60 %.
Коефіцієнт транспірації в картоплі невисокий (281-448), тобто на ут¬
ворення одиниці сухої речовини рослина витрачає 281-448 одиниць води.
з2132
65
Протягом доби максимум транспірації припадає на першу поло¬
вину дня, мінімум - на ніч.
Потреба рослин у воді під час онтогенезу є нерівномірною.
П.І.Альсмік [1] вважає, що картопля має три періоди витрати води.
Перший - це період від садіння до початку бутонізації, коли рослини
є маловимогливими до вологи і більше реагують на перезволоження.
У цей час оптимальна вологість грунту знаходиться на рівні 60 %
ПВ. Другий період охоплює початок бутонізації-кінець цвітіння, ко¬
ли рослини потребують найбільше води. Оптимальний рівень воло¬
гості у грунті - 80 % ПВ. Третій період - це кінець цвітіння-
збирання врожаю. Вимоги до вологості у цей період дещо зменшу¬
ються, оптимальна вологість грунту становить 80-70 % ПВ.
Важливе значення у стабілізації водозабезпечення рослин картоп¬
лі мають материнські молоді бульби, особливо в перший період рос¬
ту й розвитку. Проте і в подальшому материнська бульба відіграє
певну роль у цьому, оскільки вона є своєрідним резервуаром, здат¬
ним змінювати свій об’єм за рахунок води на 15-30 %. Нині на 1 га
насаджень витрачають 3,5—4,0 т картоплі. З такою масою бульб май¬
бутні рослини накопичують доступну вологу в кількості 2500-2700 кг.
Це - стартовий запас вологи для проростання і страховий фонд у най¬
більш напружені години доби. Таке ж значення має вода молодих
бульб. Звичайно, при нормальному водозабезпеченні вміст води в буль¬
бах відносно стійкий, проте під час посухи він значно коливається.
Наприклад, при зниженні вологості дерново-підзолистого грушу до 15 мм
кількість води в бульбах може зменшитися на 20 %, що спричинює їх
в’янення. Видалення бульб у посушливий період призводить до зни¬
ження вмісту вологи в листках на 3-4 абсолютні проценти, порівняно з
рослинами, у яких бульби не вилучалися. При в’яненні рослин і втраті
тургору виникають відхилення в обміні речовин.
У польових умовах забезпеченість рослин вологою тісно пов’я¬
зана з температурними умовами, переміщенням повітряних мас, ти¬
пом грунту, його станом, кількістю атмосферних опадів та ін.
Найбільш сприятливе співвідношення між кількістю опадів і су¬
мою температур за період вегетації виражається величиною гідротер¬
мічного коефіцієнта (ГТК). Він вираховується за формулою:
РГС = сума опадів 10
сума температур
Ці величини вказують на сприятливі умови вирощування карто¬
плі тільки при середній температурі повітря, близькій до 19°С. Коли
вона вища, то при такому ГТК ріст картоплі пригнічується. Наводи¬
мо дані (табл. 2.3) про врожай картоплі залежно від ГТК і темпера¬
тури повітря в період інтенсивного бульбоутворення [2].
Кількість опадів у період вегетації є важливим фактором урожаю.
Додатковий міліметр їх дає приріст урожаю від 0 до 440 кг/га.
бб
Результати досліджень залежності опадів і врожаю, проведені на
Волинській державній дослідній станції [17], подано у таблиці 2.4.
Таблиця 2.3 - Залежність урожайності картоплі від кліматичних умов
гтк
Температура повітря, °С
Урожайність, ц/га
0,5-0,7
19-22
100
0,5-0,7
17-19
120
0,7-1,0
22-23
70
0,7-1,0
19-22
100
0,7-1,0
17-19
140
1,0-1,5
22-23
110
1,0-1,5
17-19
190
1,5-1,8
19-22
100
1,5-1,8
17-19
220
1 >5-1,8
16
250
Таблиця 2.4 - Вплив кількості опадів за червень-серпень на врожайність
картоплі сорту Юбель
Кількість опадів, мм
Роки
Урожайність бульб,
ц/га
86-137
1961,1963,1967,1970
137-173
150-190
1962,1964, 1966,1971, 1974
210-215
204-286
1959, 1960,1958, 1965, 1969
196-217
У довготривалому стаціонарному досліді Інституту картоплярст¬
ва УААН виявлено, що для одержання максимально високих урожаїв
і ефективності добрив вирішальне значення для раннього сорту кар¬
топлі Прієкульська рання мали опади середини червня та початку
липня, а середньостиглого сорту Катюша - середини липня і початку
серпня (табл. 2.5).
Таблиця 2.5 - Залежність урожайності картоплі від кількості й розподілу опадів
Місяці
Кількість опадів, мм
Урожайність, ц/га
Прієкульська
рання
Катюша
Червень-липень
43,6
Липень-серпень
44,1
Сума
87,7
151
169
Червень-липень
112,3
Липень-серпень
136,0
Сума
248,3
329
252
Червень-липень
42,0
Липень-серпень
109,4
Сума
151,4
203
318
Червень-липень
158,0
Липень-серпень
183,0
Сума
341,0
381
217
67
Як видно з наведених даних, урожай цих сортів формувався по-
різному і залежав не тільки від кількості, але й від розподілу опадів
за періодами вегетації. Так, максимальний урожай бульб сорту Ка¬
тюша (318 ц/га) одержано при кількості опадів за червень-липень 42,
а за липень-серпень - 109,4, загальній сумі - 151,4 мм. Коли за чер-
вень-липень випало 158 мм опадів, то врожай становив 217 ц/га, тоб¬
то був на 99 ц нижчий, ніж у першому випадку. Щодо сорту Пріє¬
кульська рання помічена зворотна залежність: максимальний врожай
(381 ц/га) було одержано при великій кількості опадів (158 мм) у че-
рвні-липні і значно нижчий (лише 203 ц/га), коли за червень-липень
випало лише 42 мм опадів.
Результати наших багаторічних досліджень з різними сортами
картоплі і в різні за погодними умовами роки дозволяють зробити
висновок, що ефективність дії добрив значною мірою залежить не
лише від загальної продуктивності сорту, але й від його реакції на
погодні умови періоду вегетації рослин певної грунтово-кліматичної
зони. Висока віддача від застосування добрив спостерігається у ви¬
падку, коли максимальна потреба в поживних речовинах і надхо¬
дження їх у рослини збігається з оптимальним водно-повітряним та
температурним режимами.
ЛІТЕРАТУРА
Q7) Альсмик П.И., Амбросов A.JL, Вечер А.С. и др. Физиология картофе¬
ля- М.: Колос, 1979.
2. Андрюшина Н.А., Жемойц А.А., Клюквина Ю.Ф. Возделывание кар¬
тофеля при орошении.- М.: ВНИИТЭИСХ, 1975.
3. Власенко Н.Е. Условия высокой эффективности применения удобре¬
ний под продовольственный картофель, семенной картофель: Дис.... д-ра с.-х.
наук-Немишаево, 1983.
(4) Ганчарык М.М. Біялагічный і экалагічний асаблівасці бульбы //
Фізіялогія і біяхімія бульбы.- Минск: Ураджай, 1979 - С.19-44.
5. Дорожкин Н.А. Картофель - Минск: Ураджай, 1972.
б!)Куценко В.С. Місце картоплі в сівозміні // Картопля - К., 1978 - С.78-
102,
7.Жуценко В.С. Прогресивна агротехніка картоплі.- К., 1984.
® Кучко А.А., Власенко М.Ю., Мицько В.М. Фізіологія та біохімія кар¬
топлі. - К.: Довіра, 1998. - 335 с.
Кучко А.А., Мицько В.М. Потенційна продуктивність картоплі і ос¬
новні фактори її формування // Картоплярство - 1995 - Вип.26.- С.3-8.
10. Кучко А.А., Мицько В.М. Наш другий хліб // Дім, сад, город.- 1996-
№ 11.
11. Лорх А.Г. Динамика накопления урожая клубней.- М.: ОГИЗ,
1948.
12. Модестов А.П. Правда о корнях - М., 1932.
13. Новиков Ф.А. Водный режим картофельного растения //Картофель-
М., 1937.- С.88-89. м
14. Онищенко О.Й. Насінництво картоплі на Україні - К.: Урожай, 1966.
15. Писарев Б.А. Производство раннего картофеля - М.: Россельхозиз-
дат, 1986.
68
16. Руденко А.И. Климатическое вырождение картофеля и мероприятия
по борьбе с ним // Вопросы семеноводства картофеля и борьба с его вырож¬
дением-К., 1960-С.28-44.
17. Теслюк П.С., Клець СЛ. Влияние метеорологических условий веге¬
тационного периода на урожайность и качество картофеля // Картофелевод¬
ство- 1987.- № 18.- С.47-49.
(Пу Чайлахян NLX. Фитогормоны и фитотехника // Агрохимия - 1983 - № 12-
С. 105-110.
19. Dam F., Kooman P.L., Struik Р.С. Effects of temperature and photo-
period on early growth and final number of tubers in potato // Potato Research-
1996.-Vol. 39, № 1.- P.51-60.
2.3. ФІЗІОЛОГІЧНІ ОСОБЛИВОСТІ
Картопля належить до автотрофних рослин, а отже самостійно
синтезує всі необхідні для росту й розвитку органічні речовини. Цей
синтез розпочинається з утворення первинної органічної речовини,
яка в подальшому вступає до складних біохімічних реакцій з під¬
ключенням до них поглинутого з грунту азоту, фосфору, калію, ін¬
ших макро-, мікро- та ультрамікроелементів. І завершується синте¬
зом складних вуглеводів, білків, жирів, біологічно активних росто¬
вих речовин, фотосинтезуючих пігментів та інших необхідних для
живого організму сполук.
Первинна органічна речовина створюється у процесі фотосин¬
тезу. Фотосинтез - це поглинання рослиною вуглекислого газу і
відновлення його до вуглеводів. Здійснюється цей процес за учас¬
тю фотосинтезуючих пігментів, енергії світла, води й відповідних
ферментів.
Схематично фотосинтез можна зобразити формулою:
світло
6 СС>2 + 6 Н2О хлорофіл СвЯпОв + 6 О2.
глюкоза
Фотосинтез включає дві стадії: світлову і темнову.
Світлова стадія полягає у поглинанні фотосинтезуючими пігмен¬
тами квантів світла та перетворенні їх енергії на хімічну енергію
лабільних високоенергетичних сполук АТР і НАДР • Н2. Енергія цих
сполук використовується при темновій стадії фотосинтезу для віднов¬
лення вуглекислого газу до вуглеводів.
Утворення АТР і НАДР • Н2 включає два етапи: фотофізичний і
фотохімічний. При фотофізичному етапі енергія квантів світла пере¬
дається одному з Пі-електронів хлорофілу, а саме: електрону атому
магнію, що входить до складу цього пігменту.
Отримана електроном енергія використовується на фотохімічному
етапі для фотолізу води і фотосинтетичного фосфоршповання.
Фотоліз води, або реакція Хілла,- це розщеплення води, яке ін¬
дукується хлорофілом, окисненим під впливом світла.
69
Схематично фотоліз води можна зобразити так:
1. 2 Н20-*~2 ОЬГ + 2 ЇҐ + 2 є”
2. 2 ОН—2ЬҐ + 2е+20
3. 2 О -*Ю2 або 2 Н20—4 НҐ + 4 е“+ 02.
Кисень, що утворився при фотолізі води, виділяється в атмосфе¬
ру. Отже, кисень атмосфери - це кисень, що виділяється при розще¬
пленні води при світловій стадії фотосинтезу, а не той, що вивільню¬
ється від перетворення вуглекислого газу в темновій стадії фотосин¬
тезу, як вважали раніше.
Енергія квантів світла, що сприйнялась електроном хлорофілу,
витрачається не тільки для фотолізу води, але й на фотосинтетичне
фосфоршповання.
Фотосинтетичне фосфоршповання - це утворення АТР і НАДР • Н2.
Для утворення АТР електрон рухається по замкнутому електро¬
транспортному ланцюгу (ЕТЛ), на одній з ділянок якого віддає свою
енергію відповідному ферменту, наприклад АТРазі.
Цей фермент використовує передану енергію для приєднання не¬
органічного фосфору до молекули АДР, наслідком чого і є утворення
АТР, тобто:
АДр + Р неорганічну АТр
АТРаза
НАДР • Нг утворюється також за рахунок енергії електронів, що
рухаються по електротранспортному ланцюгу, але не замкнутому.
Тому для заповнення електронної “дірки" при цій схемі руху елект¬
ронів необхідне джерело додаткових електронів. Цим джерелом і є
фотоліз води. Розподіл електронів фотолізу води такий: з чотирьох
електронів і протонів, що утворилися при фотолізі, 2 протони і 2
електрони витрачаються на відновлення ферменту пластохінону, що
входить до ЕТЛ, 2 протони залишаються - на відновлення НАДР до
НАДР • Н2, а решта (2 електрони) - на заповнення електронної “дір¬
ки” в атомі магнію молекули хлорофілу, що бере участь у нецикліч-
ному фотосинтетичному фосфорилюванні
НАДР • Н2, що утворюється при нециклічному фотосинтетично¬
му фосфорилюванні, служить не тільки постачальником енергії для
темнової стадії фотосинтезу, але й протонів водню для відновлення
вуглекислого газу до вуглеводів. Тому цю сполуку називають також
“відновлювальною силою”.
ХІМІЗМ відновлення СС>2 до вуглеводів, або шлях вуглецю при
фотосинтезі, вперше детально вивчила група американських уче¬
них під керівництвом Кальвіна. Тому він також має назву циклу
Кальвіна.
Схематично цей цикл можна виразити так (за Лібертом):
70
,6С02
6[Cs]
РБФ
6[Сб] >12[Сз] ФГК
і
Гмф <-Фмр <- Фдіф<~12[Сз]
глзркоза ФГА+ДОАФ
42 НАДФ • Н2
^12 НАДФ
/12 АТФ
*12 АДФ
+ 12 Рнеорганічний
Ш!
крохмаль
Q2 ШО,
де ФГК - фосфогліцеринова кислота, ФГА - фосфогліцериновий альдегід,
ДОАФ - діоксіацетонфосфат, ФДІФ - фруктозодифосфат, ФМФ - фрукто-
зомонофосфат, ГМФ - глюкозомонофосфат, НАДР • Н2 - нікотинамідаде-
ніннуклеотид фосфат у відновленій формі, РБФ - рибулозобіфосфат.
Особливо важливим у циклі Кальвіна є те, що з 12 молекул тріоз
ФГА і ДОАФ лише дві витрачаються на синтез глюкози, а решта -
підключаються до регенерації акцептора вуглекислого газу - рибу-
лозобіфосфату. Завдяки цьому в процесі фотосинтезу відбувається
ланцюгова реакція регенерації акцептора, а отже, виключно інтен¬
сивний процес акцептування С02 і синтезу органічної речовини, не¬
зважаючи на те, що атмосфера містить лише 0,03 % вуглекислого газу.
Як видно із схеми шляху вуглецю при фотосинтезі, у циклі Каль¬
віна відбувається перетворення тріоз-тривуглецевих сполук, тому
рослини з таким циклом фотосинтезу називаються Сз-рослинами, а
шлях фотосинтезу в них — Сз-шляхом. Вважають, що близько 80 %
рослин належить до С3-рослин, у тому числі й картопля.
Картопля за вимогами до тривалості дня належить до кос¬
мополітів, тобто нейтральноденних культур. Проте існує точка зору,
що вона є коротко- або довгоденною рослиною [2], що пов’язане,
вочевидь, головним чином із сортовими особливостями. Різні сорти
неоднаково реагують і на спектральний склад світла. Зокрема, в до¬
слідах кращу врожайність забезпечував сорт Берліхінген при освітленні
червоно-хвильовими променями, а Лорх - синьо-фіолетовими [12]. То¬
му сорти, що мають вищу врожайність при освітленні довгохвильо¬
вою радіацією, краще вирощувати у північних районах, а ті, що
сприймають і короткохвильову сонячну радіацію,- в південних.
Мабуть, однією з причин невисокої продуктивності ряду сортів
картоплі на Півдні України є не тільки негативний вплив високих
позитивних температур на утворення бульб та посилене екологічне
виродження, але й негативне ставлення до синьо-фіолетового випро-
71
мінювання, що становить значно більший відсоток у загальному спе¬
ктрі світла південних районів, ніж північних.
Світло необхідне рослині для нормального розвитку структури
хлоропластів, утворення хлорофілу й високоенергетичних сполук
АТР та відновлювальної сили НАДР • Н2.
При підвищенні інтенсивності світла зростає й активність фото¬
синтезу, але тільки до певної межі. При інтенсивності світла, що за¬
безпечує максимум енергії, необхідної для світлової стадії фотосин¬
тезу, тобто утворення максимально можливої кількості АТР і НАДР •
Н2, спостерігається світлове насичення. Подальше її зростання не
призводить до збільшення активності фотосинтезу.
У картоплі, як і деяких інших світлолюбних культур, світлове на¬
сичення настає залежно від сорту при 60-90 тис. люкс за оптималь¬
них температур та вмісту С02 в атмосфері (рис. 2.1).
Рис. 2.1. Залежність фотосинтезу від інтенсивності світла
За оптимальних умов освітлення органічної речовини синтезу¬
ється значно більше, ніж витрачається на дихання, що сприяє інте¬
нсивному росту та розвитку рослин, підвищенню їх продуктивнос¬
ті. І навпаки, при значному зменшенні інтенсивності світла різко
сповільнюється синтез органічної речовини. Може виникнути та¬
кий стан, коли ці два процеси внаслідок дихання врівноважуються,
тоді настає компенсаційна точка. Під компенсаційною точкою (КТ)
розуміють інтенсивність освітлення, при якому поглинання С02 під
час фотосинтезу врівноважується виділеною С02 у процесі дихання
одними і тими самими фотосинтезуючими листками за однакових
умов.
У картоплі компенсаційна точка відмічається при освітленні
1200-2000 люксів. Коли воно менше, то розпад органічної речовини
рослин переважає над її створенням у процесі фотосинтезу і рослина
“простоює”, тобто не створює необхідних додаткових сполук, значно
уповільнює або повністю припиняє ріст і розвиток.
72
Це явище нерідко спостерігається у полуденні години ясних і
спекотних літніх днів, коли у листках закриті продихи і фотосинтез
гальмується, а інтенсивність дихання зростає. Саме тому добовий
перебіг фотосинтезу має вигляд двовершинної кривої (рис. 2.2). За
нашими даними, такий процес фотосинтезу часто спостерігається у
червні-липні, особливо за нестачі вологи в грунті та атмосфері.
Рис. 2.2. Добовий перебіг фотосинтезу в рослинах картоплі
Максимальна інтенсивність фотосинтезу характерна для Полісся
України вранці, з 8 до 12 години, коли найкращий спектральний
склад світла (велика кількість червоних променів), невисока темпе¬
ратура та достатня вологість повітря. Це сприяє оптимальному по¬
глинанню вуглекислого газу, швидкому перетворенню його на вуг¬
леводи, тому що максимально відкриті продихи листків. Другий пік
фотосинтезу, але значно менший, спостерігається з 17 до 19 години,
коли спадає денна спека, покращується спектральний склад світла,
підвищується відносна вологість повітря та зменшується інтенсив¬
ність транспірації.
Депресія фотосинтезу з падінням його активності нижче компен¬
саційної точки припадає на 13-16 годину внаслідок поєднання висо¬
кої інсоляції, температури повітря 20 °С і вище, денного дефіциту
вологи у ґрунті та відносної вологості повітря (нерідко нижче 40 %).
Ці фактори призводять не тільки до значного водного дефіциту, але
й до посиленого фотодихання.
Двовершинний перебіг фотосинтезу спостерігали в численних
дослідах за умов Харкова Ф. Я. Бузовер [1], Крайньої Півночі (Ігар¬
ка) М.М. Гончарик [2] та інші автори.
Інтенсивність фотосинтезу неоднакова у різних рослин, що
пов’язане зі структурою куща окремих сортів картоплі та умовами
вирощування.
73
Різна вона і у листків рослини залежно від їхнього віку та роз¬
міщення на стеблах. На кожному стеблі формується залежно від
сорту і умов вирощування 14-20 ярусів листків загальною площею
0,6-2,0 м2 на кущ. Звичайно найбільша кількість світла припадає на
листки двох верхніх ярусів, а нижні поступово поглинають все мен¬
ше енергії світла внаслідок взаємного затінення. Максимальна ін¬
тенсивність фотосинтезу, як правило, спостерігається у листків 3-4
ярусів зверху. Це пов’язане насамперед з тим, що вони отримують
достатню кількість енергії, характеризуються найвищим вмістом
елементів мінерального живлення і закінчили ріст, а отже фізіоло¬
гічно більш активні. Листки нижчих ярусів не тільки менше погли¬
нають сонячної радіації, але й старіші за віком. І чим нижче вони
розміщені, тим старіші, а отже мають невисоку активність фотоси¬
нтезу.
Для інтенсивності освітлення важливе значення має не тільки
розміщення листків на стеблах рослин, але й густота стояння рос¬
лин на одиниці площі посів. При загущених посівах спостерігаєть¬
ся не тільки взаємне затінення листків різних ярусів, але й однієї
рослини іншою внаслідок великої кількості листків і стебел. За да¬
ними М. М. Гончарика [2], при збільшенні загальної площі листків
понад 25 тис м2/га вони поглинають радіацію значно повільніше,
ніж до цього, а при площі робочої поверхні листків 40 тис м2/га во¬
но становить 60-80 %. Подальше збільшення площі листків у посі¬
вах картоплі малоефективне
Вважається, що залежно від сорту оптимальною площею листків
картоплі є 35-40 тис м2/га [3, 4, 8], тобто листковий індекс у посівах
картоплі має становити 3,5-4,0 м2/м2.
При більшому розвитку вегетативної маси картоплі нижні листки
рослин зазнають значної нестачі світлової енергії. Вони не тільки не
створюють додаткової органічної речовини, необхідної їм для ди¬
хання, а й перетворюються на непродуктивних споживачів речовини,
створеної високопродуктивними верхніми листками та інтенсивним
фотосинтезом. Ослаблене світло, що падає на затінені листки, при¬
зводить до підвищення вмісту в них хлорофілу, зменшення асиміля¬
ційного числа і товщини стовпчастої паренхіми. Таке листя здатне
до асиміляції при слабкому освітленні, проте рівень його активності
незначний і продуктивність рослин знижується. Вирощування карто¬
плі при слабкому освітленні, що супроводжується недостатнім за¬
безпеченням вологою та елементами живлення, призводить до фор¬
мування малоактивного фотосинтетичного апарату. Про вплив світла
на структуру й фотосинтетичну активність листків картоплі свідчать
дані таблиці 2.6.
Низка досліджень [2, 5, 6, 9 - 11, 13] показали, що обсяги і якість
врожаю до певного рівня знаходяться в тісній кореляції з розмірами
площі листків, тривалістю та інтенсивністю їх діяльності та відтоку
74
Таблиця 2.6 - Вплив інсоляції на структуру та фотосинтетичну активність
листків картоплі
Показник
Освітлення, %
100
40
10
Площа верхівкового листка, см2
21
32
28
Суха маса листка, мг/см2
4,6
4,1
3,0
Кількість клітин на листок, млн шт.
ІД
1,9
3,2
Кількість пластвд у полісадній клітині, шт.
67
85
98
Кількість хлорофілу, шт., в 1 хлоропласті
3,0-Ю9
3,5-109
4,1-Ю9
Інтенсивність фотосинтезу, мг С02/см2 за 1 год
0,54
0,49
0,37
асимілятів. У зв’язку з цим основне завдання рослинництва полягає в
тому, щоб засобами агротехніки, агрохімії, фізіології рослин і селек¬
ції формувати рослини та посіви, здатні до активного фотосинтезу і
найкраще використовувати його спочатку для оптимального росту
вегетативних, а потім - господарсько цінних репродуктивних та за¬
пасаючих органів.
Згідно з дослідженнями щодо формування густоти насаджень рос¬
лин картоплі [4], оптимальною густотою слід вважати таку, після
якої чиста продуктивність фотосинтезу, врожайність і відсоток това¬
рних бульб починають значно зменшуватися.
У дослідах, проведених нами з низкою сортів різної продуктив¬
ності, показано, що для низькопродуктивних сортів картоплі
(Юбель, Прієкульська рання) оптимальний листковий індекс зна¬
ходився в межах 2,5-3,1 м2/м , а врожайність бульб не перевищува¬
ла 200-280 ц/га, для високопродуктивних (Гатчинська, Луговська,
Незабудка і деяких інших) відповідно 37-43 мѴм2, тобто 37-43 тис./га
та 340-480 ц/га.
Для досягнення оптимального фотосинтезу, а отже продуктивнос¬
ті картоплі, необхідно досягти не тільки оптимального листкового
індексу рослин на певний період вегетації, наприклад, бутонізації -
цвітіння, а й на більш пізні строки.
Дуже важливо, щоб процес наростання площі листків інтенсивно
відбувався в ранні строки першої половини вегетації, яка характери¬
зується, як правило, більш сприятливими умовами інсоляції, тепло¬
вим та водним режимами ґрунту.
Сумарна фотосинтетична радіація для умов Києва розподіляється
за період вегетації рослин так (ккал/см2): квітень - 4,7; травень - 6,9;
червень - 7,7; липень - 7,7; серпень- 6,3; вересень - 4,3 [7] та ін.
У кінці травня - середині червня, крім того, більш сприятлива
температура грунту для утворення і росту бульб, ніж у липні - на
початку серпня, що сприяє формуванню вищого врожаю.
При вирощуванні картоплі в середніх географічних широтах оп¬
тимальним слід визнати такий графік наростання площі листків [10]:
через 20 днів після сходів - 5-6 тис. м2 /га; через 40 днів -20-25 і че-
75
рез 60 днів — 30—40 тис. м2/га. У південному Поліссі та на півночі
правобережного Лісостепу, залежно від сорту, рівня мінерального
живлення і погодних умов цей графік може зазнавати значних відхи¬
лень [3]. Зокрема, ранні й середньоранні сорти картоплі для утворен¬
ня врожаю бульб близько 280-300 ц/га через 20 днів після сходів по¬
винні мати площу листків не 5-6, а 8-10 тис. м2/га, а через 40 днів -
до 30—35 тис. м2/га. Середньостиглі й середньо-пізньостиглі сорти
наближаються до створеного А. О.Ничипоровичем графіка нарос¬
тання площі листків.
Для більш повного уявлення про вплив інтенсивності росту пло¬
щі листків і їх діяльності на утворення кінцевого врожаю бульб не¬
обхідно враховувати такі важливі показники фотосинтезу, як фото¬
синтетичний потенціал (ФП) і чиста продуктивність фотосинтезу.
Фотосинтетичний потенціал — це сумарна величина площі лис¬
тків у м2/га на кожну добу протягом усього вегетаційного періоду.
Він є одним із вирішальних факторів у визначенні врожайності,
тому що дає уявлення про те, яка фотосинтезуюча площа і протя¬
гом якого часу працювала на створення врожаю. Вважають, що чим
більший фотосинтетичний потенціал, тим вища врожайність, якщо
при цьому не сталося значного зменшення чистої продуктивності
фотосинтезу.
У наших дослідах [3] із вивчення впливу мінеральних добрив на
формування фотосинтетичного потенціалу і врожайність бульб у різ¬
них сортів картоплі доведено, що між цими показниками існує висо¬
ка корелятивна залежність. Зокрема, для сорту Юбель коефіцієнт
кореляції між фотосинтетичним потенціалом і врожайністю картоплі
становив 0,89; для Гагринської - 0,92.
Від рівня мінерального живлення залежить і чиста про¬
дуктивність фотосинтезу у різних сортів картоплі. При цьому най¬
вищий показник у дослідах був у варіанті без внесення органічних та
мінеральних добрив. З підвищенням рівня мінерального живлення
він закономірно зменшувався і досягнув мінімуму при внесенні мак¬
симальної кількості добрив.
У цілому простежується обернена залежність між фотосинтетич¬
ним потенціалом і чистою продуктивністю фотосинтезу. Виходячи з
їх величин стосовно конкретного сорту та рівня мінерального жив¬
лення рослин, оптимальним слід визнати таке поєднання цих показ¬
ників, яке забезпечує підвищення фотосинтетичного потенціалу, з
таким зменшенням чистої продуктивності фотосинтезу, при яких
одержують найвищу врожайність бульб без значного погіршення
їхньої якості. Для сорту Юбель найкращою виявилася чиста продук¬
тивність фотосинтезу 7,3 г/м2 за добу при середній величині фотоси¬
нтетичного потенціалу 1300238 м2/га за вегетацію. Саме за таких
умов у південному Поліссі України отримано максимальний урожай
сухої речовини. Відносно близькими виявились показники в інших
76
сортів картоплі (Бородянська, Гатчинська, Луговська), чиста продук¬
тивність фотосинтезу в яких знаходилась у межах 6,9-7,4, а фотоси¬
нтетичний потенціал 1,4-1,7 млн м2/га за вегетацію.
Інтенсивність та продуктивність фотосинтезу залежить не тільки
від сили освітлення рослин, яка значною мірою визначається габіту¬
сом куща та густотою насаджень картоплі, але й від фотоперіоду.
При довгому дні засвоєння рослинами фотосинтетично активної
радіації підвищується. З’ясовано, що при поглинанні енергії соняч¬
них променів картоплею енергетична цінність 1 г біомаси колива¬
ється в межах 14,03-16,29 кДж. Цей показник протягом вегетації
має високу позитивну кореляцію з використанням загальної радіа¬
ції (г=0,93). На 1 м насаджень картоплі припадає 14540 кДж (у
ранньостиглих сортів - 14608 кДж). Таким чином, картопля вико¬
ристовує в середньому 0,99% загальної сонячної і 2,2% фотосинте¬
тично активної радіації. На інтенсивність фотосинтезу, крім світла,
значно впливають температура, забезпеченість вологою, вуглекис¬
лим газом, елементами мінерального живлення та поєднання цих
зовнішніх факторів.
На формування врожаю бульб 500 ц/га, що становить половину
потенціальної продуктивності картоплі, потрібно асимілювати про¬
тягом вегетаційного періоду 30-40 т вуглекислого газу (в серед¬
ньому 300 кг на добу на площі 1 га посіву). Тому концентрація С02
у насадженнях картоплі має важливе значення: чим вона вища, тим
більша інтенсивність фотосинтезу. Відомо, що вміст С02 в атмос¬
фері становить у середньому 0,03%. Крива інтенсивності фотосин¬
тезу зростає пропорційно до росту концентрації вуглекислого газу
в атмосфері в 10 разів і більше залежно від температури й сили сві¬
тла. При нормальному вмісті С02 в повітрі (0,03%) інтенсивність
фотосинтезу становить у середньому 0,1-0,2 мг С02/1 см2 на годи¬
ну, а при зростанні її до 1 мг на 1 л повітря підвищується майже
вдвічі.
Концентрація вуглекислого газу в посівах картоплі значно нижча
від рівня його в атмосфері. Особливо це помітно, коли добре розви¬
нута вегетативна маса рослин (табл. 2.7).
Таблиця 2.7 - Вміст вуглекислого газу в посівах картоплі
Рівень розвитку
вегетативної маси картоплі
Вміст С02 в повітрі, (% об’єму)
22.ѴП
2.VIII
19. VIII
Високий
0,010
0,007
0,020
Середній
0,015
0,011
0,020
Слабкий
0,017
0,019
0,022
Дуже важливо підтримувати високий рівень вуглекислого газу в
насадженнях, тому що зниження його нижче 0,01% призводить до
значної депресії засвоєння С02 рослинами.
77
Концентрація С02 в посівах залежно від типу ґрунту, рівня удоб¬
рення й агротехніки коливається в межах 0,015- 0,040%. Важливим
джерелом цього газу є розпад у грунті його органічної речовини й
органічних добрив.
Мінімальна температура фотосинтезу картоплі 4-6°С. При під¬
вищенні її від 5 до 36°С інтенсивність фотосинтезу зростає на 0,7-2,3 мг
С02/см2 за годину на кожні 10°С, а при 40°С і вище вона різко спа¬
дає, що пов’язане насамперед із структурними змінами мембран хло¬
ропластів.
Депресія фотосинтезу настає і при порушенні водообміну, який
спричиняється зниженою вологістю грушу й повітря, високою кон¬
центрацією ґрунтового розчину, перегрівом рослин і посиленою
транспірацією листя. Фотосинтез картоплі досягає найбільшої вели¬
чини при частковому 5-10%-ному водному дефіциті листя. Швид¬
кість фіксації С02 зменшується як при збільшенні насичення водою,
так і в подальшому при зменшенні її. Сорти із слабкою водоутриму-
вальною здатністю тканин швидше піддаються депресії фотосинтезу.
Але якщо рослини картоплі в початковий період вегетації перенесли
водний дефіцит, то вони набувають досить високої водоутримуваль-
ної здатності листя і стійкості фотосинтезу до посухи. Це пов’язано з
тим, що листя, яке формується при нестачі вологи, за функціональ¬
ною активністю не поступається вирощеному при нормальній воло¬
гості грунту. Проте при дефіциті вологи зазнають сильної депресії
процеси росту і морфогенезу, ділення і росту клітин. Площа листка і
загальна кількість хлоропластів зменшується, що призводить до
зниження продуктивності рослин. Порушення процесу фотосинтезу
залежить від ступеня й періоду зневоднення. Коли воно довготрива¬
ле, руйнуються колоїдні структури клітин. Крім того, зменшення во¬
логозабезпеченості спричинює закривання продихів і, як наслідок,
припинення доступу вуглекислоти в рослину.
Умови забезпечення рослин мінеральним живленням впливають
на характер метаболічних процесів, співвідношення між фотосинте¬
зом і диханням. У зв’язку з цим вони є фактором, за допомогою яко¬
го можна впливати на фотосинтез. Висока забезпеченість азотним
живленням у перших фазах росту картоплі зумовлює направленість
фотосинтезу на ростові процеси. Із збільшенням доз азоту зростає
об’єм клітин мезофілу і кількість хлоропластів у клітині, а також
площа поверхні листкової пластинки і загальна площа листкової по¬
верхні насаджень картоплі на одиницю площі поля. Найшвидший і
найефективніший шлях формування листкової поверхні - це поси¬
лення азотного живлення в поєднанні з поливами (за потреби).
Збільшення врожаю під впливом азотних добрив відбувається в
основному за рахунок наростання листкової поверхні. Фотосинтети¬
чна активність у цих випадках істотно не змінюється. Водночас фос¬
форні й калійні добрива мають більший вплив на фотосинтез, але в
78
пізніх строках вегетації. Наприклад, посилене фосфорне живлення
під час бульбоутворення сприяє утворенню вуглеводів і їх відтоку в
бульби. Із зростанням дози добрив інтенсивність фотосинтезу на
одиницю листкової поверхні послаблюється, хоча продуктивність
рослин у цілому підвищується. Проте це відбувається тільки до пев¬
ної дози добрив, після якої вступають у дію інші фактори, параметри
яких знаходяться в несприятливому режимі. Внаслідок цього підви¬
щення врожаю не відбувається, а нерідко спостерігається навіть його
зниження. На інтенсивність фотосинтезу впливають також мікроеле¬
менти, особливо при позакореневому їх застосуванні під час масово¬
го цвітіння рослин.
Інтенсивність та продуктивність фотосинтезу регулюється не тіль¬
ки зовнішніми, але й внутрішніми факторами. Одним з них є кіль¬
кість глюкози як кінцевої сполуки циклу Кальвіна. Вона може нако¬
пичуватись у хлоропластах лише до певної концентрації, вище якої
фотосинтез різко уповільнюється або ж припиняється повністю. То¬
му для безперервного фотосинтезу обов’язковим є відтік чи видален¬
ня продуктів, що синтезуються в хлоропластах, до місць їх спожи¬
вання або запасаючих органів (бульб, коренів і т. п.).
У хлоропластах синтезується й концентрується не тільки глюкоза,
але й продукт її перетворення - первинний крохмаль. Він теж має ви¬
далятися з місць синтезу і прямувати до запасаючих органів. Ці проце¬
си потребують не тільки витрати метаболічної енергії, але й певних
перетворень, тому що є відповідні транспортні форми органічних ре¬
човин. Наприклад, вуглеводи і жири пересуваються по ситоподібних
трубках флоеми у формі сахарози, білки - у вигляді амінокислот.
З асимілюючих клітин до флоеми провідних судин листка вугле¬
води пересуваються у формі моноцукрів (глюкоза, фруктоза) через
живі клітини листка. У флоемі провідних судин вони перетворюють¬
ся на сахарозу, яка потім прямує до місць споживання чи запасання
по ситоподібних трубках флоеми листків і стебел рослини. Крохмаль
також гідролізується до глюкози, потім перетворюється на сахарозу.
У формі цієї сполуки він рухається від листків до бульб, і там знову
перетворюється до вторинного крохмалю.
Він і є основною запасною формою вуглеводів - головним джерелом
енергії та пластичних речовин при проростанні бульб картоплі.
Енергію для перетворення речовин та їх руху фотосинтезуючі ор¬
гани, головним чином листки, отримують за рахунок квантів*світла.
Нефотосинтезуючі органи, наприклад, корені, енергію для погли¬
нання елементів мінерального живлення, метаболізації цих елемен¬
тів, перетворення й руху органічних та мінеральних речовин отри¬
мують за рахунок дихання.
Дихання - це процес розщеплення органічних речовин, що су¬
проводжується перетворенням потенціальної енергії їх хімічних
зв’язків на доступну для організму енергію АТР. При диханні склад¬
79
ні органічні речовини окиснюються до простіших через ряд проміж¬
них сполук. Останні служать вихідним пластичним матеріалом для
синтезу нових речовин, необхідних організму та специфічних для
кожного виду рослин.
Спрощено дихання можна зобразити формулою:
С6Н1206+602 С02 + Н20 + 2870 кДж.
Дихання є джерелом енергії для нефотосинтезуючих органів і
пластичних речовин, а отже центральною ланкою енергетичного й
пластичного обміну у живій клітині. Воно тісно пов’язане із фотосин¬
тезом. У фотосинтезуючій клітині ці обидва процеси відбуваються
одночасно і мають багато спільного. Згідно з теорією В. І. Палладіна,
неодмінною складовою дихання, як і фотосинтезу, є вода. Саме ки¬
сень води витрачається на окиснення вуглецю дихального субстрату,
наприклад, глюкози, до вуглекислого газу. Молекулярний кисень
повітря, що бере участь у диханні, є кінцевим акцептором електронів
і протонів, наслідком чого є вода, що утворюється при диханні.
При окисненні глюкози гліколітичним шляхом утворюються такі
сполуки, як і при фотосинтезі: ФГК, ФГА, а також фосфоенолпіро-
виноградна (ФЕП) і піровиноградна кислоти та інші.
При окисненні глюкози пентозофосфатним шляхом утворюється
рибулозобіфосфат (РУБФ), рибоза, без якої неможливий синтез ДНК
і РНК, еритрозофосфат, що при взаємодії з ФЕП утворює шикимову
кислоту. Саме ця кислота є вихідною речовиною для синтезу сполук
ароматичного ряду - гормонів росту, хінонів та інших речовин, не¬
обхідних для росту й розвитку рослин.
Крім того, ФЕП є акцептором С02 при фотосинтезі у С4-рослин, а
РУБФ - у Сз-рослин, у тому числі й картоплі. Отже, дихання і фото¬
синтез - це два глибоко взаємопов’язані процеси, які є визначальни¬
ми для росту, розвитку, а отже й продуктивності рослин.
Дихання відіграє особливу роль при проростанні насіння, бульб
чи коренеплодів.
Для будови тіла, утворення коренів, стебел нової рослини енергія
і пластичні речовини надходять тільки за рахунок дихання бульб.
Дихання включає три послідовні етапи: деполімеризацію - роз¬
пад складних полімерних сполук на простіші, наприклад, гідроліз
крохмалю до глюкози; анаеробне або неповне окиснення продуктів
деполімеризації; аеробне чи повне окиснення дихального субстрату
до кінцевих продуктів - С02 і Н20.
На першому етапі виділяється лише незначна кількість тепла, що
розсіюється у навколишньому просторі. При анаеробному диханні,
якщо воно відбувається гліколітичним шляхом, утворюється чотири
молекули АТР: дві з них витрачаються на фосфоршповання фрукто¬
зи, а дві залишаються Утворюється також дві молекули НАД • Н2.
Кінцевим продуктом гліколізу є ПВК (піровиноградна кислота),
якою започатковується третій - аеробний етап дихання, що має назву
80
циклу Кребса. Це найбільш енергоємний етап, у якому утворюється
понад 40% енергії АТР, що витрачається на процеси росту, погли¬
нання, пересування та перетворення органічних і мінеральних спо¬
лук, а також цілий ряд проміжних сполук, що започатковують синтез
жирів, вуглеводів, амінокислот.
Органічні кислоти, що виникли у цьому циклі, наприклад, щав-
левооцтова, приєднуючи аміак, утворюють аспарагінову амінокисло¬
ту, а кетоглутарова - глутамінову. Ацетилкоензим А служить вихід¬
ною речовиною для синтезу жирних кислот, вуглеводів та ін.
Багато вчених вважають, що важливість циклу Кребса полягає
насамперед у підготовці пластичного матеріалу для синтетичних
процесів у молодих клітин під час росту паростків, пагонів, коренів,
тобто нефотосинтезуючих органів.
Звичайно, цей цикл має провідне значення при проростанні
бульб картоплі, формуванні у грунті коренів, молодих пагонів, які
не вийшли ще на денну поверхню і не приступили до фотосинте¬
зу. Аеробне дихання відіграє важливу роль при формуванні, осо¬
бливо рості, молодих бульб, які витрачають багато енергії на
розсування ґрунту внаслідок збільшення їхнього розміру. Вся ця
енергія утворюється тільки за рахунок дихання підземної частини
картоплі, тому що енергетичні сполуки АТР, АДР та інші, що
утворюються при фотосинтезі, не здатні пересуватися від листків
до коренів рослин.
Складовими енергії циклу Кребса, тобто молекул АТР, є утво¬
рені внаслідок окиснення ПВК 8 молекул НАД • Н2 та 2 молекули
ФАД • Н2.
При окисненні глюкози гліколітичним шляхом утворюється дві
молекули ФАД • Н2. Отже, при анаеробному та аеробному диханні
разом утворюється 8 молекул НАД • Н2 та 4 молекули ФАД • Н2. При
окисненні однієї молекули НАД • Н2 синтезується 3 молекули АТР,
при окисненні 8 - відповідно 24. При окисненні однієї молекули
ФАД • Н2 утворюється 2 молекули АТР, а з 4, відповідно - 8. Всього
це становить 24+8=32 молекули АТР. Крім того, при гліколізі утво¬
рюється 4 вільні молекули АТР та дві у циклі Кребса. Разом у про¬
цесі дихання утворюється 32 + 6 = 38 молекул АТР.
При гліколізі кожної молекули АТР під час дихання вивільню¬
ється 35 кДж енергії, що в сумі становить 35x38=1330 кДж. Відомо,
що у процесі дихання з одного грам-моля глюкози вивільнюється
2870 кДж енергії. У формі АТР запасається 1330 кДж, або
іззохіоов
2870
Ця енергія і витрачається на метаболічні процеси, а решта - 1540
кДж - виділяється у формі тепла і сприяє прискоренню біохімічних
реакцій та розсіюється у навколишньому просторі.
81
Дихання найбільш інтенсивно відбувається в листках молодих
рослин і послаблюється з їх старінням. Приблизно 20% органічних
речовин, утворених у процесі фотосинтезу і залишених на нічний
період у пластинці листка, витрачається на дихання. Інтенсивність
цього процесу коливається в межах 1,5-4,5 г С02 на добу на 1 м2
площі листка.
Залежно від інтенсивності дихання і швидкості фотосинтезу
встановлені три фази розвитку листя. Молоді листки у віці 4-7 тиж¬
нів, які містять 91-89% води, характеризуються високою інтенсивні¬
стю дихання і низькою - фотосинтезу. Цілком розвинуті яскраво-
зелені листки у віці 7-12 тижнів, які містять 88-85 % води, мають
слабке дихання і максимальну швидкість фотосинтезу. У старих, у
яких пожовтіло близько 1/4-3/4 площі, фотосинтез спадає, а дихання
залишається у середньому на одному рівні.
У стеблах картоплі при високих температурах і слабкому освіт¬
ленні дихання проходить неактивно і на балансі всього куща істот¬
но не позначається. При цьому в темряві воно проходить повільні¬
ше, ніж у листках, і в період старіння стає ще слабшим. В інтенси¬
вності дихання рослин різних сортів картоплі істотної різниці не
виявлено.
Дихання звичайно визначають за кількістю виділеного С02. Про¬
те рівень виділення і засвоєння його залежить від взаємодії двох
процесів: асиміляції та дисиміляції і фактично не відображає повніс¬
тю окислювальні процеси, оскільки частина вивільненого С02 знову
включається в синтез. У практичному картоплярстві доцільно вести
розмову про потребу рослин у кисні. Потреба рослин у кисні за
польових умов для вегетативної маси не лімітується, тому що його
цілком достатньо в повітрі. Щодо підземних органів, то ці частини
відчувають нестачу в кисні, особливо в районах з періодичним над¬
лишковим перезволоженням. У західних районах, на Поліссі спосте¬
рігаються часті випадки вимокання посівів картоплі, викликані витіс¬
ненням повітря з грушу водою.
Біологічною особливістю картоплі є висока чутливість до повіт¬
ряного режиму грунту. Це зумовлено значною потребою коренів,
столонів і бульб у кисні. При недостатньому забезпеченні киснем
затримується проростання бульб. Велику кількість кисню із грунто¬
вого повітря, в десятки разів більшу, ніж інші сільськогосподарські
культури, споживає коренева система картоплі. Інтенсивність дихан¬
ня коренів коливається в межах 700-1200 мг 02 на годину на 100 г
сухої маси. Зниження концентрації 02 до 4 % при температурі 13°С
призводить до зупинки дихання. Вищу потребу в кисні відчувають
бульби і столони. Максимальне споживання грунтового кисню при¬
падає на період найбільш інтенсивного приросту бульб.
Вміст повітря в грунті залежить від його шпаруватості, або пори¬
стості. На окультурених структурних грушах шпаруватість досягає
82
65 % об’єму грунту. Чим пухкіший грунт, тим більша його шпарува¬
тість і насиченість повітрям. У зв’язку з цим грунт необхідно утримува¬
ти в розпушеному стані з об’ємною масою не більше 1,0-1,2 г/см . В
умовах перезволоження при сильному ущільненні грунту вміст кис¬
ню може знизитися до 2 % і менше і тоді бульби задихаються і за¬
гнивають, а в грунті зростає вміст вуглекислого газу. Для нормаль¬
ного росту коренів концентрація вуглекислого газу має не перевищу¬
вати 1 %.
При ущільненні грунту колесами трактора із 3,3-4,3 до 7,5-8,3 кг/см3
урожай зменшується до 40 %, що неможливо компенсувати ні під¬
вищеними дозами добрив, ні зрошенням.
На ущільнених грунтах коренева система розвивається слабо,
столони розгалужуються й утворюються дрібні, часто деформовані
бульби. Це зумовлене тим, що для процесів, пов’язаних з перетво¬
ренням і переміщенням пластичних речовин бульб при проростанні
їх у грунті з утворенням коренів, паростків, столонів і бульб, потріб¬
ні великі затрати енергії. Вони забезпечуються окисненням органіч¬
них речовин і вимагають значних витрат кисню повітря в самому
грунті. Подібні процеси спостерігаються і в період бульбоутворення.
Крім того, при врожаї 0,5 кг на кущ бульбам потрібно перемістити
близько 50 см3 грунту, що на 1 га становить майже 20 м3. Це потре¬
бує також значних затрат енергії і кисню.
На ущільнених грунтах столони через особливість будови (відно¬
сно великі клітини) мають недостатню здатність розсовувати грунто¬
ві частинки і протидіяти їх механічному тиску. При нестачі кисню в
повітрі грунту бульби картоплі не тільки запізнюються із зав’язу¬
ванням, але й погано визрівають. При вологості грунту понад 80 %
повної вологоємності, в першу чергу на зв’язних грунтах, бульби
відчувають нестачу повітря і погано ростуть. У деяких випадках це
призводить до відкладання поживних речовин у повітряних бульбах,
які утворюються в пазухах нижніх листків.
На бідних, погано оструктурених грунтах дуже важливими є за¬
ходи, спрямовані на підвищення пористості грунту, зокрема застосу¬
вання органічних добрив і сидератів.
ЛІТЕРАТУРА
1. Бузовер Ф.Я. Водный режим картофеля // Исследования по физиоло¬
гии и биохимии растений - К., 1966 - Т.48 - С.81-88.
2. Вечер А.С., Гончарик М.Н. Физиология и биохимия картофеля-
Минск: Наука и техника, 1973.
3. Власенко Н.Е. Удобрение картофеля - М.: Агропромиздат, 1987.
4. Гончарик М.Н., Кручинина С.Г. Влияние густоты посадок на форми¬
рование и работу фотосинтетического аппарата растений // Физиолого¬
биохимические исследования растений- Минск: Наука и техника, 1965-
С.91-100.
83
5. Гончарик М.Н., Власенко Н.Е. Влияние форм минеральных удобрений
на фотосинтез и качество сахарной свеклы // Физиолого-биохимические ис¬
следования растений - Минск, 1968 - С.93-102.
6. Листопад Т.Г., Климов А.А., Иванов А.Ф. и др. Программирование
урожая //Тр. Волгоград. СХИ.- 1975- Т.5.- С. 140-150.
7. Молдау X., Росс Ю., Тооминг X., Уидла И. Географическое распреде¬
ление фотосинтетически активной радиации (ФАР) на территории Европей¬
ской части СССР // Фотосинтез и вопросы продуктивности растений.- М.:
Изд. АН СССР, 1963.- С. 149—158.
8. Ничипорович А.А. Задачи работ по изучению фотосинтетической дея¬
тельности // Фотосинтезирующие системы высокой продуктивности - М.,
1966.- С.7-50.
9. Ничипорович А.А. Основы фотосинтетической продуктивности расте¬
ний //Современные проблемы фотосинтеза - М., 1973.
10. Ничипорович А.А., Чмора С.Н. Об использовании солнечной радиа¬
ции на фотосинтез в посевах картофеля // Физиология растений.- 1958.-
Т.2.- Вып.4.
И. Ничипорович А.А. Фотосинтез и пути повышения продуктивности
растений // Программирование урожаев с.-х. культур.- К., 1976.- С. 9-15.
12. Писарев Б.А. Производство раннего картофеля.- М.: Россельхозиз-
дат, 1986.
13. Чаповская В.Г., Замараев А.Г. Фотосинтетический потенциал и про¬
дуктивность зерновых культур // Программирование урожаев сельскохозяй¬
ственных культур: Науч. тр. ВАСХНИЛ.- М.,1975 - С.232-235.
2.4. БІОХІМІЧНИЙ СКЛАД
Біохімічний склад бульб є одним із найважливіших показників
харчової цінності картоплі та її кулінарних властивостей. Основною
складовою частиною бульби є крохмаль. Інші органічні і мінеральні
речовини знаходяться в значно меншій кількості, проте відіграють
досить важливу роль, наприклад, протеїн, вітаміни, глікоалкалоїди,
зольні елементи, цукри та ін. (табл. 2.8).
Таблиця 2.8 - Біохімічний склад бульб картоплі (за зведеними даними різних
авторів)
Речовина
Вміст речовини, % до сирої маси
мінімальний
максимальний
середній
Вода
63,2
86,9
76,3
Суха речовина
13,1
36,8
23,7
Крохмаль
8,0
29,4
17,5
Цукри
0,1
8,0
1,0
Клітковина
0,2
3,5
1,0
Сирий протеїн
0,7
4,6
2,0
Жири
0,04
1,0
од
Зола
0,4
1,9
1,0
Органічні кислоти
0,1
1,0
0,6
84
Коливання біохімічного складу досить значні, що свідчить про
його мінливість, залежно від грунтово-кліматичних умов, рівня й
особливостей агротехніки вирощування картоплі та сорту. Біохіміч¬
ний склад може істотно відрізнятися навіть у бульб, що виросли під
одним кущем. Наприклад, бульба, що утворилася раніше, здебільшо¬
го має вищий вміст крохмалю, ніж сформована пізніше.
Наводимо дані щодо вмісту основних компонентів у бульбах сор¬
тів картоплі, які вирощуються на території України (табл.2.9) [7].
Таблиця 2.9 - Біохімічний склад бульб поширених в Україні сортів картоплі
Сорт
Вміст, %
сухої речовини
крохмалю
протеїну
1
2
3
4
Ранньостиглі
Бородянська рожева
20,5-22,0
14,4-15,7
2,0-2,3
Вармас
17,2-20,1
12,4-14,5
1,8-2,4
Верховина
14,7-24,2
10,6-17,4
1,5-2,0
Воротинська рання
17,3-20,0
12,5-14,0
1,6-2,3
Зов
22,5-23,6
16,2-17,2
2,2-2,4
Іскра
14,4-22,0
11,4-15,8
1,7-2,1
Каскад поліський
16,0-18,0
11,5-13,8
1,5-1,8
Кобза
23,0-24,0
16,6-17,5
2,3-2,5
Незабудка
18,5-21,2
13,3-15,.3
1,6-2,2
Піомунес
17,0-21,0
12,0-15,0
1,6-1,8
Прієкульська рання
14,0-17,0
10,0-12,0
1,4-2,2
Пригожа 2
17,5-18,2
12,6-13,1
1,6-1,9
Харківська рання
17,0-20,5
12,0-14,8
1,8-1,9
Царнікавська
15,3-19,3
11,0-13,9
1,6—1,8
Середньоранні
Адрета
17,0-20,5
12,0-14,8
1,6-2,0
Водограй
17,0-19,0
12,0-13,6
1,6-1,8
Житомирянка
17,0-22,8
12,2-16,4
1,5-1,7
Зорька
20,0-22,6
14,3-16,3
1,6-1,8
Купава
20,5-22,0
14,4-15,7
2,0-2,2
Львів’янка
20,0-24,3
14,4-17,5
1,8-2,4
Мавка
21,5-26,5
15,5-19,1
2,0-2,8
Невська
17,0-18,0
12,1-12.8
1,7-1,9
Новинка
18,0-22,8
13,0-16,4
1,7-2,0
Обрій
22,0-24,0
15,7-17,5
2,3-2,4
Остара
19,2-20,0
13,8-14,4
1,6-2,1
Радомишльська
17,8-20,3
12,8-13,6
1,8-2,4
Світанок київський
24,4-27,5
17,6-19,8
2,3-2,8
Чарівниця
20,8-23,0
15,0-16,5
1,9-2,2
85
Середньостиглі
Віхола
17,6-20,0
12,6-14,5
1,8-2,3
Г атчинська
19,0-25,5
13,7-18,4
1,6-2,3
Горлиця
20,6-22,4
14,8-16,1
1,7-2,1
Луговська
21,0-22,8
15,1-16,0
2,1-2,3
Огоньок
18,2-25,0
13,3-18,0
1,8-2,3
Радуга Полісся
19,0-20,6
13,5-14,8
1,7-1,9
Слов'янка
17,0-19,0
12,1-13,6
1,8-2,0
Юбель
20,0-26,4
14,2-19,0
1,6-2,0
Середньопізні
Воловецька
26,1-27,0
18,8-19,4
2,3-2.5
Гібридна 14
16,2-21,5
11,6-15,5
1,8-2,2
Зарево
26,0-32,0
18,7-24,0
2,5-3,2
Ікар
24,5-25,8
17,6-18,6
1,6-2,0
Лорх
18,6-23,6
17,0-20,6
1,9-2,3
Ольвія
22,5-24,0
16,2-17,5
2,0-2,3
Поліська рожева
21,6-25,9
15,6-18,6
2,0-2,4
Сулев
18,0-32,0
13,7-22,7
1,9-2,5
Пізньостиглі
Древлянка
21,8-23,0
15,8-16,5
2,0-2,2
Зарево
28,0-32,0
20,0-24,0
2,8-3,2
Комсомолець
27,2-29,2
19,6-21,0
2,4-2,5
Мета
17,6-21,5
14,8-15,5
1,7-2,0
Полонина
27,0-29,0
19,5-21,0
2,3-2,5
Темп
26,0-29,0
18,8-20,9
2,0-2,3
Вода. Бульби картоплі містять у середньому 75 відсотків води
(63-87). Вона відіграє велику роль як будівельний матеріал при біо¬
синтезі органічних сполук, а також як транспортний засіб і
температурний регулятор (шляхом транспірації).
Розрізняють вільну, гідратаційну і колоїдну форми води. Дві
останні структуровані й пов’язані різними колоїдними частками клі¬
тини, наприклад, білковими тілами, крохмалем тощо. Важливим є те,
що структурованість води у бульбах значно нижча, ніж у листках. Це
означає, що водоутримувальна здатність бульб низька і вони легко
втрачають воду. Відомо, що бульби можуть деякою мірою компен¬
сувати водний дефіцит листків. У період посухи вони втрачають тур¬
гор і віддають воду листкам, а інколи навіть повністю розсмокту¬
ються, тобто відбувається відтік не тільки води, але й органічної ре¬
човини на покриття витрат, пов’язаних з підвищенням дихання над¬
земної маси. Під час зберігання бульб кількість вільної води
зменшується, а зв’язаної (структурованої) - зростає.
З’ясовано, що здатність харчових продуктів до тривалого збері¬
гання визначається не кількісним вмістом води, а її станом, тобто
86
формою зв’язку із сухою речовиною продукту. Це стосується і при¬
датності картоплі до тривалого зберігання: чим більше в ній
зв’язаної форми води і менше вільної, тим менше вона загниває.
Сухі речовини становлять у середньому 25% маси бульби. Цей
показник якості нестабільний і, залежно від сортових особливостей,
ґрунтово-кліматичних та інших умов вирощування може коливатись
у межах 13-37%. Як правило, підвищеним вмістом сухих речовин
характеризуються пізньостиглі сорти, а ранньостиглі - низьким.
Відомо, що картопля з високим вмістом сухих речовин має біль¬
шу господарську цінність. Проте, в деяких країнах Західної Європи
цьому не надають особливого значення. У Великобританії, напри¬
клад, для консервування використовують молоді бульби з низьким
вмістом сухої речовини. З іншого боку, цей показник є основним при
використанні бульб для виготовлення напівфабрикатів. Високий
вміст сухих речовин не тільки забезпечує якість цих продуктів, але й
дозволяє скоротити витрати енергії у процесі сушіння. Особливо він
впливає на текстуру чипсів.
Нагромадження сухої речовини бульбами у процесі вегетації ро¬
слин зростає, але цей процес не має прямої залежності у зв’язку зі
зміною погодних умов. За посушливої і жаркої погоди, порівняно з
помірною, він підвищений, а за вологої і прохолодної - понижений.
Нагромадження сухої речовини в онтогенезі також нестабільне і не¬
рівномірне. Найінтенсивніше воно відбувається у початковий період:
для ранніх, середньоранніх і середньостиглих сортів - через 70-90
днів, а пізньостиглих - 80-100 днів від садіння картоплі. Це припа¬
дає звичайно на липень - першу половину серпня і збігається з акти¬
вним бульбоутворенням. Потім нагромадження сухої речовини від¬
бувається відносно рівномірно. Час настання максимуму визначаєть¬
ся передусім скоростиглістю сорту: в ультраранньостиглих сортів -
на 90-й день після садіння; дещо пізніше (на 100-й день) - у ранніх,
середньоранніх і середньостиглих сортів; у пізньостиглих - на 120—
130-й день після садіння. Вміст сухої речовини у наступний період,
як правило, істотно зменшується у всіх сортів. Проте і на цей процес
певний вплив справляють погодні умови. Тому в багатьох випадках
посушлива і тепла, а інколи жарка погода, призводить, з одного боку,
до скорочення вегетаційного періоду, з іншого - до максимального
вмісту сухої речовини в бульбах на період збирання врожаю. У пере¬
зволожені роки цей показник на час збирання врожаю завжди значно
нижчий, ніж у нормальні для конкретної грунтово-кліматичної зони.
Згідно з даними, отриманими нами на дерново-підзолистих супіща¬
них грунтах Полісся України, вміст сухих речовин у бульбах карто¬
плі в перезволожений рік становить: у сорту Прієкульська рання -
16,7%, Бородянська - 18,4; Юбель - 19,9; Олев - 22,3%. У сприятли¬
вий рік ці показники були значно вищими: відповідно, 19,2; 22,4;
24,9 і 26,3%.
87
Певне уявлення про мінливість вмісту сухих речовин у бульбах
картоплі залежно від року дає таблиця 2.10 [1].
Таблиця 2.10 - Мінливість вмісту сухих речовин у бульбах картоплі за роками
Сорт
Жовтень
Листопад
1993
1994
+ до 1993
1993
1994
+ до 1993
Витязь
22,4
24,7
+2,3
22,4
31,5
+9,1
Воловецька
21,9
28,1
+6,2
19,6
31,3
+11,7
Горлиця
18,4
28,2
+9,8
20,6
25,9
+5,3
Зарево
26,8
30,9
+4,1
28,8
34,1
+5,3
Ікар
24,4
28,3
+3,9
23,0
28,0
+5,0
Кобза
21,9
25,2
+3,3
19,6
27,2
+7,6
Либідь
25,3
28,0
+2,7
23,5
28,2
+4,7
Луговська
17,8
24,9
+7,1
22,5
24,8
+2,3
Обрій
19,5
26,2
+6,7
22,6
27,3
+4,7
Українська рожева
20,6
26,2
+5,5
20,2
26,9
+6,7
НІРо,5
2,3
2,4
2,5
2,4
Крохмаль - основна складова бульб картоплі (70-80%). У буль¬
бах він розподіляється нерівномірно. Найбільшою є кількість його в
тканинах камбіального кільця. Як у бік периферії, так і в напрямку
ядра бульби вміст крохмалю знижується. В цілому найбільша кіль¬
кість його нагромаджується у паренхімі кори, потім у зоні судинно-
волокнистих променів і найнижча - у серцевині бульби. В усіх тка¬
нинах основи бульби вміст крохмалю вищий, ніж у тканинах вер¬
хівкової частини (на 2-3%).
Основною умовою отримання високого виходу крохмалю є вибір
сорту. Як уже зазначалося, крохмалистість бульб залежить, як пра¬
вило, від тривалості вегетації. Але слід пам’ятати, що й у межах гру¬
пи стиглості, залежно від сорту, різниця в ній може становити 6-7 %.
Існує певна залежність між вмістом крохмалю в окремих бульбах
і їх розміром. Дрібні бульби містять його менше, проте у дуже вели¬
ких крохмалистість теж зменшується, особливо, якщо серцевина ду¬
плиста. Нашими дослідженнями доведено, що максимальна розбіж¬
ність за вмістом крохмалю в окремих бульбах одного сорту і навіть
фракції відносно однакового розміру може досягати 5-6 %.
У таблиці 2.11 наведено багаторічні дані амплітуди коливань
вмісту крохмалю і сухих речовин у бульбах висококрохмалистих со¬
ртів, що вирощувались у розсадниках лабораторії інтродукції Інсти¬
туту картоплярства УААН.
Відомо, що найціннішими є сорти, які характеризуються менши¬
ми показниками коливань вмісту крохмалю як між окремими буль¬
бами, так і в цілому за сортами, залежно від ґрунтово-кліматичних та
погодних умов.
Ціннішими є також сорти, в яких розбіжність вмісту крохмалю у
бульбах виражена меншою величиною.
88
Таблиця 2.11 - Максимальний і мінімальний вміст сухої речовини та крохмалю
в бульбах висококрохмалистих сортів картоплі
Сорт
Суха речовина,%
Крохмаль, %
мінімальний
максимальний
мінімальний
максимальний
Бекра
25,4
31,4
15,4
23,6
Берізка
20,1
32,2
17,7
22,0
Зарево
25,0
34,0
18,0
26,0
Кандидат
26,4
29,5
15,1
21,9
Лошицька
26,6
26,7
16,0
19,2
Лявоніха
24,2
30,5
16,3
21,6
Павлінка
29,0
33.3
18,9
22,2
Ритм
27 7
35,4
17,5
22,2
Світанок київський
24,0
29,0
17,0
22,0
Темп
20,5
28.8
17,1
24,6
Фекула
28,0
32,8
16,3
23,1
Форта
25,0
32,3
17,8
21,9
Важливо зазначити й те, що одного року одні сорти, а другого -
інші нагромаджують більше крохмалю, і тому різниця між ними мо¬
же збільшуватись або зменшуватись.
Підвищення крохмалистості картоплі є власне збільшенням фак¬
тичного врожаю цієї культури. З іншого боку, підвищення врожаю
картоплі призводить до більшого збору сухої речовини і крохмалю з
одиниці площі посіву.
Ріст вегетативної маси картоплі триває приблизно до закінчення
цвітіння, потім припиняється і настає період інтенсивного утворення
бульб. Лише невелика кількість продуктів фотосинтезу в цей час ви¬
трачається на підтримання життєздатності надземної частини росли¬
ни, більшість з них відтікає до бульб і там нагромаджується в основ¬
ному у формі крохмалю. Частина продуктів фотосинтезу спочатку
нагромаджується у листі у формі асиміляційного крохмалю, потім
уночі знову перетворюється у розчинні цукри, здебільшого сахарозу,
і транспортується до бульб. Крохмаль становить близько одного від¬
сотка сухої маси листків. Чим швидше рослина створює необхідну
вегетативну масу і чим довше буде функціонувати листковий апарат,
тим вища врожайність і крохмалистість бульб. Звідси випливає, що
всі фактори, які позитивно впливають на ріст рослин і їх життєдіяль¬
ність, будуть впливати й на вміст крохмалю у бульбах і врожай. Ці
фактори можна об’єднати в три основні групи: грунт і його родю¬
чість, погодні умови, агротехнічні заходи з вирощування картоплі.
Про вплив на крохмалистість типу ґрунту, рівня його родючості
та фізичного стану ми не маємо достатніх літературних відомостей.
У дослідах П. С. Теслюка [16] із сортом Смачна на дерново-
підзолистому супіщаному грунті вміст крохмалю в бульбах становив
16,0 %; чорноземі опідзоленому середньо-суглинковому - 17,2; а на
осушеному торфовищі - 15,2%.
89
Ще більш контрастні дані отримано в дослідах німецьких учених
(Hunnius, 1972). Крохмалистість картоплі, що вирощувалася на осу¬
шеному торфовищі, становила 13,9%; супіщаному ґрунті - 15,2 і суг¬
линковому- 17,2%.
Нашими вегетаційними дослідами з грунтом різного ступеня
окультуреності [4, 5] доведено, що максимальна крохмалистість
бульб картоплі сорту Немішаївська ювілейна - 17,8% спостерігалася
на дерново-підзолистому ґрунті, що містив гумусу 2,47%, рухомого
фосфору 10,3 мг/100 г ґрунту та обмінного калію 14,3 мг/100 г ґрун¬
ту. При гумусності того ж грунту 1,09%, 4,5 мг фосфору і 9,4 мг ка¬
лію крохмалистість була значно нижчою - 16,2%.
Нерівнозначний вплив типу ґрунту на крохмалистість картоплі
можна пояснити різними умовами повітряного й мінерального жив¬
лення рослин.
Найкращі умови для нагромадження крохмалю створюються на
легкосуглинкових ґрунтах. Це зумовлене тим, що ґрунти такого гра¬
нулометричного складу добре забезпечують кореневу систему рос¬
лини та молоді бульби киснем і разом з тим мають достатню ємність
поглинання. На важких глинистих ґрунтах, передусім перезволоже¬
них, холодних, і торфовищах вміст крохмалю знижується. Певне зна¬
чения має і стан ґрунту: недостатня шпаруватість його, як наслідок
замулювання і утворення ґрунтової кірки, уповільнює синтез крох¬
малю. За таких умов вміст його в бульбах знижується на 2-3 %. Ос¬
новною причиною зниження його на торфовищах є затримання онто¬
генетичного старіння рослин, внаслідок чого більшість пластичних
речовин втрачається на розвиток вегетативної маси, затримується
дозрівання бульб і синтез у них крохмалю. Спричиняє це здебільшо¬
го надлишок азоту в грунті. Доведено, що вміст легкодоступних
форм азоту, передусім нітратного, у торфовищах різко перевищує
вміст його у мінеральних ґрунтах. Кількість рухомих форм фосфору,
особливо калію, у цих ґрунтах, як правило, дуже мала. Тому тут спо¬
стерігається порушення необхідного співвідношення між основними
елементами мінерального живлення і, як наслідок, порушення опти¬
мального живлення рослин, їх розвитку.
При промисловій переробці на чипси, пюре, смажену картоплю
тощо потрібна сировина без ушкоджень, гнилизни, щоб легко очи¬
щувалася, з високим вмістом крохмалю.
Урожай з такою якістю бульб можна отримати не на всіх ґрунтах,
а на біологічно активних з належною структурою, що забезпечує
сприятливі умови для росту, живлення, бульбоутворення і нагрома¬
дження крохмалю. Механізоване збирання має проводитися без по¬
шкоджень бульб. У зв’язку з цим заслуговують на увагу рекоменда¬
ції щодо вирощування крохмалистої картоплі на промислову переро¬
бку в Німеччині, де після збирання зернових проводять ретельний
обробіток ґрунту з метою прискорення розкладання стерні і знтцен-
90
ня бур’янів. Потім рівномірно розкидають і приорюють гній 20—
25 т/га або дрібно порізану солому. Не рекомендують вносити перед
садінням картоплі великі дози органічних добрив, особливо свіжого
гною, тому що це підвищує ушкодження бульб паршею, зменшує
вміст крохмалю і затримує дозрівання врожаю. Не слід вносити в
грунт у рік вирощування картоплі рідкий гній через нестабільний
вміст у ньому хлору й азоту. Вапно вносять з такого розрахунку, щоб
pH на піщаних ґрунтах становила до 5,5; сильно оглеєних - 6,0 і су¬
піщаних - до 6,5. Із мінеральних добрив застосовують азотні - 120—
160, фосфорні - 90-150 і калійні - 120—200 кг/га діючої речовини.
Грунти мають містити 6-8 мг/ 100 г ґрунту або ж вносять по 40 кг/га
MgO. Густота садіння - 40-50 тис. кущів на 1 га, залежно від сорту і
садивного матеріалу. За 6-14 днів до збирання врожаю бадилля ви¬
даляють. Збирати врожай слід при температурі не нижче 10°С за су¬
хої погоди, щоб зменшити травмування бульб.
Нерівнозначність впливу географічної широти на крохмалистість
картоплі пояснюється зміною інтенсивності інсоляції, температури і
кількості атмосферних опадів. За даними О.Й.Оншценка [13], при
вирощуванні картоплі в північних областях України крохмалю в буль¬
бах нагромаджується на 2 менше, ніж у південних, а в західних -
менше, ніж у східних.
У Румунії протягом 10 років вивчали вплив ґрунтових і клімати¬
чних умов на вміст крохмалю в бульбах картоплі сортів Бінтьє, Ме-
гура, Меркур у 23 географічних точках. Виявлено, що кількість його
приблизно на 33 % забезпечується особливостями сорту картоплі, на
21 - сумою температур за вегетаційний період; на 18 - кількістю ат¬
мосферних опадів; на 15 - тривалістю сонячного освітлення; на 7 -
розміщенням над рівнем моря; на 5 - вмістом обмінного калію у
грунті та піску, глини і гумусу - відповідно, на 2,0; 0,5; 0,5 %. Кис¬
лотність ґрунту і вміст у ньому фосфору ніякого значення не мали.
Однак, слід зважати на те, що ці фактори в інших умовах можуть
мати інше співвідношення.
Дослідженнями лабораторії біохімії Всеросійського інституту
рослинництва ім. М.І.Вавилова виявлена чітка залежність між метео¬
рологічними факторами у період бульбоутворення та вмістом сухої
речовини і крохмалю в бульбах.
Найвищий вміст крохмалю (17-20%) у середньостиглих і в ранньо¬
стиглих (16-18%) сортів звичайно корелює з середньою температурою
за період цвітіння - початок відмирання бадилля 17-20 °С і гідротерміч¬
ним коефіцієнтом не більше 0,9-1,0 при сумі температур для ранньости¬
глих сортів не менше 300-350 °С, середньостиглих - 600-650°С [9]. При
недостатній кількості опадів на період інтенсивного росту бульб у них
міститься більше крохмалю, ніж при високій вологозабезпеченості. На¬
приклад, крохмалистість бульб картоплі сорту Прієкульська рання в до¬
слідах Українського науково-дослідного інституту землеробства УААН
91
в 1962 рощ становила 11,5 %, 1963-му - 14,7 і 1964-му - 10,3 %, а кіль¬
кість опадів у липні була, відповідно, 106,8; 31,3 і 177,4 мм. Вплив пого¬
ди особливо помітний, якщо порівняти крохмалистість бульб різних сор¬
тів. Так, у 1970 році на період інтенсивного бульбоутворення (липень-
серпень) у дослідному господарстві “Немішаєве” Інстшуіу картоплярс¬
тва УААН було 79,4 мм опадів, в 1974 р. за цей же період - 241,5 мм.
Крохмалистість бульб сортів Огоньок, Олев, Сулев становила відповід¬
но: 19,5; 23,1 і 19,5% та 13,1; 13,6 і 13,3 %. Аналогічні дані отримано в
Західному Поліссі України (табл. 2.12).
Таблиця 2.12 - Крохмалистість бульб картоплі при різній кількості опадів
за липень-серпень [17]
Рік
Сума опадів, мм
Вміст крохмалю, %
Смачна
Юбель
1961
136
16,6
16,9
1966
174
16,5
15,6
1968
223
16,5
16,5
1969
286
15,7
16,4
1972
329
16,7
11,5
Крохмаль є важливим нормованим показником якості сировини
для виробництва картоплепродуктів. Дослідження, проведені в Ін¬
ституті картоплярства УААН на 10 сортах [1], показали, що вміст
крохмалю і сухої речовини у бульбах змінюється за сортами у після¬
збиральний період. Згідно з вимогами до якості продукції, як сиро¬
вини для переробки, в бульбах має бути сухих речовин не нижче 22,
а крохмалю - 15-17%.
Як видно з наведених даних (табл. 2.13), в 1993 році різниця між
сортами за вмістом крохмалю в бульбах на час збирання врожаю бу¬
ла більша, ніж у 1994 році.
Таблиця 2.13 - Вміст крохмалю в бульбах різних сортів картоплі,
процент на сиру масу
Сорт
Жовтень
Листопад
1993 р.
1994 р.
+ до 1993 р.
1993 р.
1994 р.
+ до 1993 р.
Витязь
17,5
19,2
+ 1,7
16,2
20,7
+ 4,5
Воловецька
18,3
20,3
+2,0
17,3
20,9
+0,0
Горлиця
15,6
18,9
+3,3
16,0
16,4
+0,4
Зарево
23,9
21,5
-2,4
21.4
24,2
+ 2,8
Ікар
17,7
19,7
+2,0
16,2
21,9
+5.7
Кобза
17,6
16,5
-ІД
16,1
19,3
+3,2
Либідь
. 17,9
21,4
+3,5
14,8
22,0
+7,2
Луговська
13,5
17,1
+3,6
14,3
17,9
+3,6
Обрій
17,4
19,5
+2,1
17,3
17,3
-
Українська рожева
18,6
16,8
-1,8
17,1
18,1
+1,0
НСРо.5
2,9
2,6
92
У 1993 р. нижчим за норму (15%) був вміст крохмалю у Луговської,
а через місяць після збирання - у сортів Либідь, Луговська. У 1994 р. у
всіх сортів крохмалю було не менше 16,5 % сирої маси бульб.
Нагромадження крохмалю в бульбах відбувається нерівномірно:
найбільш інтенсивно - починаючи з 70-80-го дня після садіння кар¬
топлі, а максимально - залежно від сорту і погодних умов. Так, в ок¬
ремі сприятливі роки протягом 10 днів кількість крохмалю може зро¬
сти на 3,6-4,8 %.
На період інтенсивного бульбоутворення, який характеризується
також посиленим нагромадженням крохмалю, значний вплив має
температура повітря. Між цими показниками виявлена пряма залеж¬
ність, причому вищі коефіцієнти кореляції у пізніх сортів, слабші - у
ранньостиглих.
Разом з тим слід зазначити, що процеси синтезу і розпаду крох¬
малю не мають постійного температурного оптимуму. Він змінюєть¬
ся не тільки впродовж вегетації, але й протягом доби, залежно від
сорту.
Вплив температури на процес синтезу крохмалю і його нагрома¬
дження в бульбах дуже важливо враховувати в південному регіоні
України.
Особливого значення тут слід надавати строкам садіння і сорту
картоплі.
При весняному садінні в Одеській, Херсонській і Миколаївській
областях бульби дозрівають при високій температурі і пониженій
вологості повітря, що сприяє інтенсивному синтезу сахарози та кро¬
хмалю. Проте його більше нагромаджується в урожаї літнього садін¬
ня, тому що тривалість дня на період вегетації, температура повітря в
кінці літа - на початку осені більш сприятливі для біосинтезу крох¬
малю (табл. 2.14).
Таблиця 2.14 - Вплив строків садіння на вміст вуглеводів у бульбах картоплі [9]
Вуглеводи
Вміст вуглеводів, % на сиру речовину, при садінні
весняному літньому
весняному літньому
Одеська 24
Елла
Моноцукри
0,32
0,24
0,56
0,55
Сахароза
0,69
0,28
0,86
0,70
Крохмаль
14,4
16,8
15,2
18,4
Для максимального нагромадження крохмалю необхідне швидке
наростання листкового апарату і його тривале функціонування. Це
досягається шляхом пророщування і стимулювання росту паростків.
Численними дослідами доведено, що пророщуванням насіннєвого
матеріалу можна підвищити крохмалистість бульб на один відсоток.
Прискорення росту паростків підвищує крохмалистість на 0,5%.
Приріст виходу крохмалю від передсадивного пророщування бульб
93
досягає 5 ц/га. Садіння великими бульбами дозволяє розпочати зби¬
рання дозрілої картоплі на два тижні швидше, порівняно з садінням
дрібними бульбами.
Доведено, що максимум нагромадження сухих речовин і крохма¬
лю у більшості сортів настає раніше, ніж закінчується період вегета¬
ції. При подальшому перебуванні бульб на рослині кількість сухих
речовин, у тому числі й крохмалю, у них зменшується. Тому збирати
картоплю найвигідніше при їх максимальному вмісті. Винятком є
дуже пізні сорти, наприклад, Темп, Прикарпатський, в цих бульбах
процес акумуляції сухої речовини і крохмалю триває до кінця веге¬
тації. На період максимального вмісту сухої речовини в бульбах ба¬
дилля повністю ще не відмирає. Отже, строки збирання картоплі ма¬
ють визначатися не повним відмиранням бадилля, а максимальною
кількістю сухої речовини в бульбах. На цей період звичайно близько
80 % листя уже відмирає.
Серед агротехнічних прийомів вирощування картоплі найбільше
значення має правильне застосування добрив. Основні елементи мі¬
нерального живлення рослин не однаково впливають на синтез і на-
гомадження крохмалю.
В наших дослідженнях з вивчення впливу азоту, фосфору і калію
на якість продовольчої картоплі на дерново-підзолистих грунтах Пів¬
денного Полісся України застосування лише азотних добрив у дозі
60 кг/га діючої речовини призводило до зниження крохмалистості
бульб на 0,2-0,4 %.
Численними дослідами доведено, що кожен кілограм внесеного в
грунт азоту зменшує вміст крохмалю в бульбах у середньому на
0,007 %.
Є дослідні дані, які свідчать про те, що максимальні збори крох¬
малю з урожаєм бульб на легких грунтах одержують при вмісті в
шарі грунту 0-60 см 160 кг/га азоту [24].
Проте для отримання високого врожаю крохмалю з одиниці
площі посіву необхідне внесення певної кількості азоту, який акти¬
вує швидкий розвиток бадилля, особливо листкового апарату, чим і
забезпечується високий урожай.
Фосфорні добрива безпосередньо впливають на крохмалистість
бульб. При внесенні 90 кг/га Р205 вона підвищується на 0,8-1,0 відсотка
та прискорюється бульбоутворення на початку цвітіння рослин.
Калій сприяє синтезу крохмалю в листках і його переміщенню до
бульб. За внесенні під картоплю 120-160 кг/га цього елемента вміст
крохмалю в бульбах підвищується на 0,5-1,0%. При цьому слід
обов’язково враховувати форму калійного добрива. У дослідах Ін¬
ституту картоплярства УААН при внесенні калійного добрива у фор¬
мі сирих калійних добрив (каїніт) вміст крохмалю в бульбах стано¬
вив 12,4%, а збір його з 1 га - 31,5 ц; при внесенні сірчанокислого
калію - відповідно, 13,7 % і 37,1 ц.
94
З підвищенням доз калію, коли збільшення врожайності вже не
спостерігається, настає негативна кореляція між крохмалистістю і
вмістом калію в бульбах [21]. Це стосується не лише калію. Дози то¬
го чи іншого елемента мінерального живлення, залежно від сорту і
рівня родючості грунту, в кожному конкретному випадку слід ви¬
значати дослідним шляхом. Орієнтовне уявлення про залежність на¬
громадження крохмалю в бульбах від кількості й співвідношення
мінеральних добрив дають результати, отримані нами на дерново-
підзолистих супіщаних грунтах із середнім ступенем забезпеченості
фосфором і калієм (табл. 2.15).
Отже, з підвищенням кількості мінеральних добрив спостеріга¬
ється поступове зниження вмісту крохмалю в бульбах усіх сортів, що
вивчали в досліді, причому в кожному - різне. Так, максимальне зни¬
ження - 3,3 відсотка щодо контролю було у сорту Юбель, потім у
Олева — 3 і Прієкульської ранньої - 2,3 %.
Таблиця 2.15 - Вплив різних доз добрив на крохмалистість картоплі
Добрива
Прієкульська рання
Юбель
Олев
крох¬
маль,
%
збір
крохмалю,
ц/га
крох¬
маль,
%
збір
крохмалю,
ц/га
крох¬
маль,
%
збір
крохмалю,
ц/га
Без добрив
(контроль)
13,8
18,8
16,1
23,7
18,2
24,1
30 т/га гною (фон)
12,8
24,6
15,2
28,5
17,7
29,4
N60P 60К90
12,2
28,3
14,6
33,2
16,7
32,2
N90P90K120
12,1
28,2
14,2
35,1
16,4
35,3
N120P90K150
11,9
29,3
14,2
34,5
15,7
32,0
NisqP 90К180
11,8
28,1
13,8
31,5
15,5
30,7
N240P 90К210
11,5
25,2
12,9
29,5
15,7
26,7
Аналогічні дані ми одержали з іншими сортами. За ступенем
зниження крохмалистості від підвищених доз добрив сорти, крім на¬
званих вище, можна розмістити у такій послідовності: Бородянська,
Гатчинська, Огоньок, Столова 19, Чарівниця, Темп, Незабудка. При
вирощуванні їх на оптимальному фоні (для отримання високого
врожаю бульб) амплітуда змін цього показника якості бульб стано¬
вила 0,6—0,3%.
Помічено також, що різні сорти з однаковою крохмалистістю по-
різному реагують на застосування мінеральних добрив. У зв’язку з
цим для збільшення збору крохмалю з одиниці площі посіву слід ви¬
рощувати сорти, що мають не тільки високу крохмалистість, але не
реагують різким зниженням її при внесенні високих доз добрив, а
саме: Зарево, Темп і меншою мірою - Гатчинську.
На жаль, сільськогосподарське виробництво не має необхідної
кількості таких сортів різних строків стиглості і придатних для ви¬
рощування в різних грунтово-кліматичних зонах України. Тому не¬
95
обхідна ширша, поглиблена робота наших селекціонерів над ство¬
ренням сортів, що позитивно реагують приростом врожаю без суттє¬
вого зниження в бульбах найбільш цінних компонентів, у тому числі
й крохмалю, при вирощуванні їх на високих агрохімічних фонах.
М. І. Вавилов [2] ще півсторіччя тому писав, що хімізація земле¬
робства ставить на порядок денний питання про селекцію на чутли¬
вість до добрив. Нині це питання не тільки не втратило своєї актуа¬
льності, а й набуло ще більшого значення.
Мінеральне живлення відіграє дуже важливу роль у метаболізмі
рослинного організму, тому реакція сорту на застосування добрив
пов’язана з його стійкістю проти вірусних, грибних і бактеріальних
хвороб, що у свою чергу також впливає на вміст крохмалю.
У польових дослідах з різними нормами мінеральних добрив ми
виявили, що при вирощуванні картоплі сорту Олев протягом трьох
років на дерново-підзолистому супіщаному грунті без застосування
добрив ураженість рослин вірусними хворобами у післядії становила
20,2 %, а на оптимальному агрохімічному фоні - 30 т/га гною разом з
N6oP6oK9o - лише 10,1 %. У Прієкульської ранньої - відповідно 29,5 і
18,1 %.
Узагальнення багаторічних дослідів дає змогу зробити висновок,
що бульби рослин, уражених вірусами скручування листя, містять у
середньому на 3,2 % крохмалю менше, ніж від здорових рослин, а
уражених вірусами мозаїки, А, У, X - на 2,0-2,5 %.
Добрива не справляють прямої дії на збудника грибних хвороб,
зокрема фітофтори, але вони підвищують стійкість рослин проти неї
завдяки поліпшенню мінерального живлення. При цьому рослини
активно використовують вологу, посилюється процес фотосинтезу і
прискорюється накопичення урожаю до настання масового розвитку
фітофторозу. Це стосується насамперед ранньостиглих сортів. Своє¬
часне запобігання розвитку фітофторозу сприятливим поєднанням
мінерального живлення і хімічних засобів захисту картоплі забезпе¬
чує підвищення крохмалистості бульб на 0,5-1,5%, а врожай крох¬
малю на 6-9 ц/га.
Вплив густоти насаджень рослин на вміст крохмалю в бульбах
також визначається особливостями сорту й агрохімічним фоном, на
якому вирощується картопля. У дослідах з сортами Прієкульська
рання і Столова 19 на фоні 40 т/га ; торфогнойового компосту разом
з мінеральними добривами у розрахунку N90P90K120 і N120P90K180 вміст
крохмалю в бульбах Прієкульської ранньої при густості насаджень
50 тисяч бульб на 1 га при першій дозі мінеральних добрив становив
12,3%, другій - 12,0%. При густоті 80 тисяч - відповідно 11,8 і 12,8 %.
Для сорту Столова 19 крохмалистість бульб при зазначеній густоті й
дозах добрив була, відповідно, 13,4; 13,6 і 13,8%. Таким чином,
більш контрастні коливання крохмалистості бульб, залежно від умов
вирощування, були у Прієкульської ранньої. При цьому підвищений
96
агрохімічний фон був негативним для мінімальної і позитивним для
максимальної в досліді густоти садіння картоплі.
Щодо впливу строку садіння на крохмалистість бульб на підставі
наших багаторічних досліджень можна твердити, що запізнення са¬
діння картоплі на 20-30 днів призводить до зниження вмісту крох¬
малю на 0,2-1,2 %, причому більш відчутне воно на бідних агрохімі¬
чних фонах.
Слід звернути увагу й на те, що всі рекомендовані до вирощуван¬
ня сорти сильно реагують на нестачу магнію в ґрунті, особливо лег¬
кого гранулометричного складу (піщані, супіщані). Застосування ма¬
гнію під картоплю значною мірою нейтралізує негативний вплив
азоту на крохмалистість бульб [22].
Недостатня забезпеченість магнієм відчувається на мінеральних
ґрунтах Полісся України, особливо при підвищенні дози азотистих,
фосфорних і калійних добрив. Внесення у грунт 40 кг/га MgO під¬
вищує врожай крохмалю в середньому на 3 ц/га.
Слід також зазначити, що існує певна кореляція між вмістом су¬
хої речовини і крохмалю, яка полягає в тому, що різниця між цими
показниками є відносно постійною величиною. З підвищенням вміс¬
ту першої зростає вміст другого. На цьому ґрунтується прискорений
метод одночасного визначення крохмалю і сухої речовини за пито¬
мою масою бульб.
Наші дослідження показали, що в умовах Полісся України серед¬
ня величина цієї різниці, залежно від технології вирощування й біо¬
логічних властивостей сорту, коливається в межах 5,5-8,0; а в рес¬
публіці Білорусь [3] - у межах 4,76-6,48.
У зв’язку з цим слід пам’ятати, що визначення крохмалю на вагах
Парова, яке широко використовують у виробництві, не завжди дає
правильні результати. Доцільніше для практичних цілей використо¬
вувати такий показник, як питома вага бульб, не роблячи перерахун¬
ків на суху речовину і крохмаль. У зарубіжній практиці питома вага
як показник якості використовується давно, наприклад, при виготов¬
ленні чипсів.
Якісні параметри крохмалю картоплі (вміст амілази, глюкозо-6-
фосфату, розмір гранул) змінюються в процесі бульбоутворення. У
процесі росту бульби збільшується вміст глюкозо-6-фосфату; розмір
гранул крохмалю, а вміст амілази не змінюється [20].
Інші вуглеводи. При оцінці якостей бульб, особливо кулінарних,
значну увагу приділяють вмісту редукуючих цукрів і олігосахаридів.
Сумарна кількість їх у зрілих бульбах нового врожаю незначна: не
перевищує 0,5—1,3 % від сухої речовини. Це головним чином глю¬
коза, фруктоза і сахароза (цукор).
Кількість цукрів у бульбах при зберіганні збільшується. Досліда¬
ми Інституту картоплярства УААН показано, що накопичення цукрів
у бульбах окремих сортів картоплі залежить передусім від темпера-
4 2-П2
97
тури зберігання. Коли картоплю зберігали при 1-3°С, вміст їх, зале¬
жно від сорту, збільшувався в кінці строку зберігання у 3-5 і більше
разів. При 3-5°С кількість цукрів була в 2 рази меншою. В деяких
випадках при зберіганні картоплі при температурі, близькій до нуля,
вміст їх зростає до 7-8 % сирої маси бульб. Це пов’язане з тим, що
при низьких температурах посилюється гідроліз крохмалю, внаслі¬
док чого збільшується кількість цукрів.
Існує певна залежність між вмістом цукрів у бульбах і кліматич¬
ними умовами вирощування. Наприклад, за однакових умовах збері¬
гання, у бульбах з північних районів цукрів накопичується більше,
ніж у бульбах того самого сорту, але з південних районів. Кількість
цукрів і співвідношення сахарози до моноцукрів є досить стабільною
сортовою ознакою і впливає на лежкість бульб. Чим вищий процент
сахарози, тобто чим вище відношення сахарози до моноцукрів, тим
краще бульби зберігаються.
Бульби, що містять більше 0,25-0,50 % моносахарів (редукуючих
цукрів), вважаються малопридатними для виготовлення харчових
напівфабрикатів, тому що під час переробки сировини відбувається
прискорена реакція взаємодії редукуючих цукрів з вільними аміно¬
кислотами і утворюються меланіноподібні речовини темного кольо¬
ру. Внаслідок цього напівфабрикати мають непривабливий вигляд.
У дослідах з різними за біохімічним складом зразками сортів Рассет
Бербанк і Шеподі, які протягом 8-9 місяців зберігалися в різних темпе¬
ратурних режимах виявлено, що колір продуктів френч фрі більшою
мірою залежить від глюкози (коефіцієнт залежності г = 0,65-0,62), ніж
від фруктози, суми редукуючих цукрів, сахарози, суми всіх цукрів.
При вмісті глюкози понад 0,16 % на сиру масу в Рассет Бербанк і
понад 0,12 % в Шеподі затрати на одержання продукту приємного
кольору значно зростають [27]. Також втрата сировини на виробниц¬
тві крохмалю з картоплі залежать від вмісту цукрів. При 0,5 % цукрів
втрати становлять 2,8 %, а при 2,0 %—11,1 % [30].
З наведених даних (табл. 2.16) видно, що вміст цукрів у бульбах
деяких сортів картоплі значною мірою коливається [1]. Так, у сортів
Ікар, Обрій, Луговська, Українська рожева в жовтні 1993 р. вміст су¬
ми цукрів у бульбах був у 3-3,5 рази вищий, ніж у жовтні 1994-го.
При цьому особливо велика різниця спостерігалась за роками щодо
вмісту редукуючих цукрів, тоді як кількість нередукуючих (цукрози)
змінювалася в основному в 1,1-2 рази. Співвідношення їх, як вважа¬
ють, впливає на стійкість бульб при зберіганні у 1993 р. щодо всіх
сортів становило 0,16-0,17, лише у Кобзи - 3,09.
У бульбах врожаю 1994 р. разом зі зниженням вмісту цукрів спосте¬
рігалась і зміна співвідношення цукрів, яке у сортів Світанок київський,
Зарево, Витязь становило відповідно 9,3; 11,5; 12,6; в інших - 0,54-3,88.
Через місяць після збирання врожаю вміст цукрів у бульбах бі¬
льшості сортів у 1993 р. знизився, у 1994-му - підвищився або майже
98
не змінився. Найбільш стабільним він був у сорту Витязь, Кобза, За¬
рево, Воловецька.
У бульбах урожаю 1993 р. вміст редукуючих цукрів 0,5 % спо¬
стерігався лише у сортів Зарево, Кобза, Воловецька, Витязь. Через
місяць після збирання врожаю висока їх кількість збереглася лише в
Обрію, Горлиці (понад 0,5 %), Ікара, Світанку київського (0,48-0,50 %).
У бульбах усіх сортів урожаю 1994 р. кількість редукуючих цукрів
була значно нижчою за гранично допустиму (табл. 2.16).
Таблиця 2.16 - Вміст цукрів у бульбах картоплі, процент сирої маси [7]
Сорт
Жовтень
Листопад
1993
1994
1993
1994
сума цукрів
редукуючі
цукри
сума цукрів
редукуючі
цукри
сума цукрів
редукуючі
цукри
сума цукрів
редукуючі
цукри
Витязь
0,40
0,24
0,34
0,03
0,49
0,45
0,28
0,11
Воловецька
0,57
0,31
0,37
0,24
0,27
0,24
0,39
0,18
Горлиця
1,17
0,56
-
-
0,75
0,62
0,54
0,53
Зарево
0,56
0,25
0,38
0,05
0,30
0,09
0,21
0,04
Ікар
1,09
0,59
0,39
0,08
0,81
0,50
0,31
0,05
Кобза
0,30
0,09
0,09
0,03
0,38
0,28
0,11
0,04
Либідь
1,16
0,79
0,45
0,18
0,80
0,28
0,41
0,18
Луговська
1,64
1,02
0,33
0,12
0,98
0,49
0,67
0,31
Обрій
1,03
0,90
0,26
0,08
0,92
0,58
0,30
0,14
Світанок київський
0,87
0,72
0,27
0,03
0,76
0,48
0,29
0,09
Українська рожева
1,22
1,06
0,22
0,05
0,84
0,43
0,57
0,25
НІР 0.5
0,09
0,03
0,05
0,03
0,08
0,02
0,06
0,03
За певних несприятливих погодних умов спостерігається накопи¬
чення моноцукрів у пуповинній частині бульби, що призводить до
утворення її склоподібності. Така картопля має низькі кулінарні яко¬
сті та незадовільний смак.
Склоподібність сприяє утворенню гнилизни під час зберігання
бульб. Вміст редукуючих цукрів у пуповині склоподібних бульб ін¬
коли доходить до 8-9 %. Досліди свідчать, що розвиток склоподіб¬
ності зумовлюється вторинним ростом бульб наприкінці вегетацій¬
ного періоду. Він виникає під впливом зміни періоду тривалої посу¬
хи достатньою водозабезпеченістю і посилюється при зберіганні не¬
достиглих бульб, а також при пониженій температурі.
Азотисті речовини. Сукупність усіх азотистих сполук бульб
картоплі має назву “сирий протеїн”. Він включає суто білок і небіл-
кові азотисті речовини. Азот у сирому протеїні розподілений при¬
близно так: 60 % - білкового азоту і 40% - небілкові сполуки. До
складу азотовмісних небілкових речовин входять аміачні й нітратні
солі, алкалоїди, нуклеїнові кислоти, аміди, вільні амінокислоти, де¬
99
які вітаміни. Серед них переважають амінокислоти глутамін і аспа¬
рагін (50-60% азотистих небілкових сполук).
Азотисті речовини, як і крохмаль, розміщені в бульбах нерівно¬
мірно. При цьому їх розподіл має зворотний характер щодо крохма¬
лю. Спостерігається розбіжність у бульб одного куща за вмістом у
них протеїну, яка може становити 0,6 %. Більше його міститься у
дрібних бульбах, ніж У великих.
Вміст сирого протеїну залежить від біологічних особливостей
сорту. Проте під впливом умов зовнішнього середовища він має до¬
сить високу мінливість як у межах одного сорту, так і між окремими
сортами. Тому в селекційній практиці оцінку сорту за цим показни¬
ком якості дають на підставі багаторічних спостережень.
Наводимо дані про вміст протеїну в окремих сортах картоплі
(табл. 2.17).
Таблиця 2.17 - Середній вміст сирого протеїну в бульбах різних сортів картоплі
(% від сирої речовини), багаторічні дані (Кучко, Власенко, Мицько, 1998)
Сорт
Мінімальний
Максимальний
Середній
Агрономічна
1,50
2,52
2,20
Білоруська крохмалиста
1,45
2,45
2,10
Бородянська
1,29
2,46
1,80
Бородянська рожева
1,82
2,37
1,1
Вармас
1,33
2,28
1,60
Г атчинська
1,40
2,51
1,80
Жигомирянка
1,60
2,99
2,10
Зазерська
2,10
3,10
2,42
Зарево
2,29
4,60
2,95
Зов
2,17
2,51
2,3
Кандидат
1,60
2,56
2,05
Кобза
2,21
2,63
2,3
Лада
1,67
2,65
2,20
Лошицька
1,95
3,10
2,50
Луговська
2,10
2,48
2,20
Невська
1,70
2,36
1,85
Незабудка
2,41
1,76
1,65
Немішаївська ювілейна
1,90
2,87
2,10
Огоньок
1,36
2,45
2,00
Павлінка
1,56
1,98
1,75
Поліська рожева
1.76
2,33
2,10
Прієкульська рання
1,35
2,40
2,00
Розвариста
1,54
2,46
2,0
Світанок київський
2,10
2,83
2,40
Скороспілка 1
1,95
2,46
2,21
Скороспілка 2
1,75
2,53
2,30
Темп
1,75
3,24
2,15
Чарівниця
1,50
2,32
1,92
100
Високим вмістом сирого протеїну характеризуються окремі фор¬
ми диких і культурних південноамериканських видів картоплі:
S. demissum, S. Andigenum, S. schickii, horovitzii та деякі інші.
Важливою умовою підвищення кількості протеїну в бульбах є
достатня забезпеченість їх азотним живленням. Взаємозв’язок доз
азотних добрив з вмістом сирого протеїну досить високий. Вміст йо¬
го в рослинах картоплі свідчить про забезпеченість цієї культури
азотом [24, 5]. З підвищенням доз азотних добрив зростає і вміст си¬
рого протеїну в бульбах більшості сортів картоплі, про що свідчать
дослідні дані, одержані нами на дерново-підзолистих супіщаних ґрун¬
тах Південного Полісся України (табл. 2.18).
Таблиця 2.18 - Вміст сирого протеїну в бульбах картоплі залежно від рівня
азотного живлення, процент [7]
Сорт
Дози добрив, кг/га діючої речовини
N9oP9oK12o
N180P90K90
Бородянська
2,07
2,59
Г атчинська
1,75
2,75
Незабудка
2,82
2,84
Немішаївська біла
2,20
2,71
Огоньок
1,87
2,80
Прієкульська рання
2,03
2,33
Росинка
1,44
2,66
Темп
2,35
2,14
Чарівниця
1,75
2,39
Збільшення кількості протеїну в бульбах названих сортів, як ба¬
чимо, було досить різним: вміст у Темпу навіть дещо знизився при
вирощуванні рослин на досить високому фоні азотних добрив. Це дає
підстави вважати, що існує певний поріг або рівень забезпеченості
рослин азотом, перевищення якого негативно позначається навіть на
вмісті сирого протеїну в бульбах.
Це припущення підтверджується також взаємозв’язком рівня аг¬
рохімічного фону з вмістом білкового азоту в бульбах різних сортів
картоплі (табл. 2.19).
Підвищення кількості мінеральних добрив супроводжувалось по¬
ступовим зростанням вмісту сирого протеїну в бульбах і досягло мак¬
симуму в сорту Юбель із дозою азотних добрив N^o, а в Прієкульсь-
кої ранньої й Олева воно зростало до максимальних доз добрив, що
вивчались у досліді.
На відміну від сирого протеїну, уміст білка у бульбах цих трьох
сортів зростає лише до певного відносно невисокого рівня азотних
добрив — N90P90K120.
Ця норма добрив виявилася оптимальною і для якісного складу азо¬
тистих сполук бульб: частка білкового азоту в сирому протеїні була най¬
вищою в бульбах усіх сортів, що вивчались у досліді (табл. 2.20).
101
Таблиця 2.19 - Вплив мінеральних добрив на вміст білка і сирого протеїну
в бульбах картоплі, процент
Добрива
Прієкульська рання
Юбель
Олев
сирий
протеїн
білок
сирий
протеїн
білок
сирий
протеїн
білок
Без добрив (контроль)
1,78
0,86
2,0
1,05
1,77
0,89
Гній, ЗО т/га (фон)
1,86
0,91
2,08
1,14
2,07
1,10
І^боРбоКад
1,99
0,99
2,33
1,29
2,29
1,22
N90P90K120
2,26
1,33
2,40
1,40
2,44
1,36
N12oP9oK15o
2,28
1,05
2,73
1,35
2,48
1,33
N180P90K180
2,37
1,07
2,72
1,36
2,56
1,34
N240P90K210
2,46
1,11
2,79
1,32
2,69
1,33
Таблиця 2.20 - Вплив добрив на вміст білка в бульбах, процент до загальної
кількості азотистих сполук
Добрива
Прієкульська
рання
Юбель
Олев
Без добрив (контроль)
48,2
52,5
50,2
Гній, ЗО т/га (фон)
48,9
54,8
53,1
Neo? 60К90
49,7
55,4
53,2
N90P 90К120
58,8
58,3
54,0
N120P90K150
46,1
49,5
53,6
N180P90K180
45,1
50,0
50,0
N240P90K210
45,1
47,3
49,4
Серед інших факторів, що впливають на азотний обмін картоплі,
важливе місце належить погодним умовам під час вегетації.
Згідно з дослідами, проведеними в колишній Чехословаччині
[22], уміст загального азоту в бульбах в окремі роки в різних регіо¬
нах країни коливався в межах: у ранніх сортів - 0,366-0,390%; у
середньоранніх - 0,378-0,409; середньостиглих і пізніх - 0,361-
0,401%.
Відомо, що рослинні білки засвоюються організмом людини і
тварини гірше, ніж білки м’яса і молока. У дослідах із тваринами
рослинні білки, порівняно з білками молока, засвоювалися менше:
гороху - в 0,5; кукурудзи - в 0,55; пшениці - 0,65; овочів і картоплі -
в 0,7 рази. Отже, білки картоплі є більш повноцінними, ніж інших
рослин. Особливо вони багаті лізином і лейцином. Кількість інших
амінокислот відповідає потребі організму людини.
Сорти картоплі різняться між собою не тільки кількісним вміс¬
том, але й якісним складом білків. Основну частину їх (70-80 %)
становлять водорозчинні фракції, невелику - соле- і лугорозчинні
білки.
До складу білка входять усі біологічно незамінні амінокислоти,
тобто такі, що не синтезуються в людському організмі. Така аміно¬
102
кислота, як метіонін, що лімітує харчову цінність усіх рослинних
білків, не тільки є складовою чистого білка картоплі, але й небілко-
вих азотовмісних сполук.
Біологічна цінність білка визначається вмістом незамінних амі¬
нокислот і позначається індексом повноцінності (ЄАА). Еталоном
його є білок яйця курки. Якщо його взяти за 100 %, то у бульб картоп¬
лі індекс становить 60-92 %.
До незамінних амінокислот, що визначають індекс повноцінності
білка, належать: триптофан, фенілаланін, лізин, треонін, метіонін,
лейцин, ізолейцин, валін.
Щодобова потреба в них організму людини становить відпо¬
відно: 1,1; 4,4; 5,2; 3,5; 3,8; 9,1; 3,3; 3,8 грама на добу. До цих аміно¬
кислот відносять також цистин, гістидин і тирозин.
Важлива роль належить картоплі в дитячому та лікувально-
профілактичному харчуванні, оскільки в ній містяться всі незамінні
амінокислоти. Якщо ж однієї амінокислоти недостатньо або вона
зовсім не надходить до організму з продуктами харчування, то це
призводить до порушення обміну речовин і тяжких захворювань.
Амінокислоти виконують в організмі важливу пластичну і регу¬
ляторну функції. Так, серед речовин, які підвищують адаптацію
організму до гіпоксії (нестачі кисню), широко застосовують глута-
мінову кислоту. Вона посилює засвоєння кисню, активізує ряд фер¬
ментів.
Гамма-аміномасляна кислота, яка відсутня у багатьох рослинах,
але є у бульбах картоплі, гальмує передавання нервових збуджень у
синапсах нервової системи, посилює кровообіг у мозку і широко за¬
стосовується при лікуванні хвороб нервової системи, інсультах та
судорогах.
Незамінні амінокислоти, що містять сірку (метіонін, цистин і ци¬
стеїн), необхідні для росту й розвитку організму, підтримання маси
тіла і азотної рівноваги. Вони входять до складу білків, ферментів,
гормонів і визначають їх біологічну активність. Ці амінокислоти є
попередниками усіх сполук, які містять сірку в організмі, і джерелом
її в процесах детоксикації. Добова потреба в метіоніні у дітей до 2,5
років - 85, дорослих - 3 мг/кг маси тіла. Як показали наукові дослід¬
ження, при вживанні метіоніну хворими на атеросклероз судин серця
спостерігалося зниження частоти нападів стенокардії; покращення
скоротливої функції міокарда. Препарат корисний при анеміях, він
нормалізує обмінні процеси в печінці.
Цистин і цистеїн широко застосовують при радіаційних уражен¬
нях, отруєнні токсичними речовинами. Нестача в організмі цистину
призводить до затримки росту волосся, а також утворення інсуліну.
Якщо першому можна запобігти, додаючи до дієти еквімолярну кі¬
лькість метіоніну, то стимулювати утворення інсуліну можна тільки
введенням цистину.
103
Десять грамів картопляного білка може замінити 6-7 г білка
м’яса. За амінокислотним складом білок картоплі дуже близький до
жіночого молока, а за вмістом аргініну, гістидину та лізину, які від¬
сутні в багатьох рослинних продуктах, переважає його.
Дослідження [12, 6] показали, що у нових сортів картоплі, ство¬
рених в Інституті картоплярства УААН, за кількісним складом знач¬
ною мірою виділяється лізин, найбільше якого міститься у сортів
Бородянська рожева, Зов, Водограй, Світанок київський, Либідь, Гор¬
лиця, Кобза.
За сумою незамінних амінокислот найбільшим вмістом харак¬
теризуються сорти Світанок київський, Либідь, Кобза, Бородян¬
ська рожева, Горлиця, Обрій. Наведені дані (табл. 2.21) свідчать,
що в середньому вміст сирого протеїну в бульбах досліджуваних
сортів з розрахунку на суху масу становив 10,2 %, тобто був та¬
ким, як у зерні кукурудзи. Найбільше його на сиру масу містило¬
ся у сорті Зарево - 3,1 %, проте на суху масу він не переважав
інші сорти.
Усі агротехнічні заходи, що сприяють підвищенню протеїну в
бульбах, збільшують і вміст у них амінокислот [10].
Глікоалкалоїди. Це група глікозидів, у яких аглікон представле¬
ний алкалоїдом. Відомо ряд глікозидів пасльонових: а, (і, у-соланін і
а, р, у-чаконін, салакаулін, соласонін, саламандрин, демісин, лептин,
лептин-1, лептин-2 та ін.
Усі вони складаються з двох основних частин, що утворюються
при кислотному гідролізі цих речовин - аглікону (соланідину), який
є суто алкалоїдом, і одного або кількох простих цукрів (глюкози, рам¬
нози, галактози), що знаходяться в молекулі глікоалкалоїду у вигляді
глюкозидного ланцюжка, пов’язаного з агліконом.
Наприклад, а-соланін складається з соланідину, галактози, глю¬
кози і рамнози; р-соланін - з соланідину, галактози і глюкози; а
у-соланін - з соланідину і галактози.
Глікоалкалоїди, що містяться в бульбах реєстрованих сортів кар¬
топлі, являють собою суміш шести різних глікозидних сполук: а-, р-,
у-соланін, a-, J3-, у-чаконін.
а-соланін і а-чаконін є основними формами і становлять близько
95 % усієї суми глікоалкалоїдів групи соланіну. Крім названих алка¬
лоїдів, в окремих сортах культурного виду картоплі містяться демі¬
син, лептин, томатин, які властиві диким видам картоплі. Це пов’я¬
зане з використанням останніх як вихідного матеріалу при виведенні
стійких проти хвороб і шкідників сортів.
У рослині картоплі глікоалкалоїд розміщені вкрай нерівномірно:
найменше - в бульбі, найбільше - у паростках і квітках.
Абсолютний вміст глікоалкалоїдів у рослині безперервно збіль¬
шується під час вегетації (до припинення росту бадилля), незважаю¬
чи на те, що процентний вміст цих сполук зменшується.
104
Таблиця 2.21 - Особливості біохімічного складу сортів картоплі, які вирощували у розсаднику конкурсно-екологічного
випробування (в середньому за 1991-1993 рр.)
Сорт
Суха
речовина, %
Крохмаль, %
Сирий протеїн, %
Амінокислоти,
% на суху масу
Співвідно¬
шення
незамінних
амінокислот
до замінних
Вітамін С,
мг/% на сиру
масу
Каротиноїди,
мг/% на сиру
масу
Жири, % на
суху масу
Клітковина,
% на суху
масу
на суху
масу
на сиру
масу
неза¬
мінні
замінні
сума
Ранньостиглі
Бородянська рожева
21,8
16,0
10,6
2,3
3,45
3,52
6,97
0,98
12,3
0,41
0,52
2,04
Зов
22,5
16,7
9,9
2,3
2,55
3,66
6,20
0,70
11,6
0,06
0,41
2,07
Незабудка
20,7
15,0
9,6
2,0
2,87
3,31
6,18
0,87
13,4
0,38
0,62
0,84
Пролісок
Середньоранні
Водограй
18,8
13,3
10,9
2,2
2,02
2,75
4,77
0,73
14,3
0,50
0,32
1,20
Кобза
23,7
17,7
10,6
2,5
3,54
3,68
7,22
0,96
13,8
0,40
0,36
1,40
Мавка
24,7
18,6
10,0
2,4
3,25
4,15
7,40
0,78
11,6
0,04
0,35
2,02
Невська
20,2
14,6
11,7
2,3
3,00
4,72
7,72
0,64
14,6
0,04
0,44
0,82
Обрій
20,5
14,9
10,1
2,1
3,35
4,30
7,65
0,78
13,6
0,05
0,36
1,20
Світанок київський
25,3
19,2
10,3
2,6
3,68
4,38
8,06
0,84
13,3
0,50
0,75
2,56
Середньостиглі
Гатчинська
21,50
15,5
9,9
2,1
2,36
4,64
7,0
0,51
11,8
0,04
0,52
3,17
Горлиця
24,0
18,0
10,5
2,4
3,51
4,01
7,50
0,88
14,0
0,40
0,40
1,20
Либідь
22,0
16,2
10,4
2,3
3,62
3,78
7,40
0,96
14,2
0,12
0,42
1,30
Луговська
22,5
16,7
9,9
2,2
2,93
4,42
7,35
0,66
14,4
0,08
0,43
0,90
Українська рожева
20,5
14,9
9,3
1,9
2,89
3,56
6,45
0,81
14,2
0,29
0,38
1,00
Пізні
Зарево
29,5
23,0
9,6
3,1
3,23
3,33
6,56
0,97
13,1
0,07
0,35
0,96
Темп
26,0
19,8
9,7
2,4
2,16
4,54
5,70
0,48
12,1
0,03
0,38
1,63
Середнє
22,6
16,7
10,2
2,3
3,01
3,09
6,95
0,74
13,2
0,17
0,44
1,48
ШР0,95
0,5
0,5
0,3
0,1
0,22
0,36
0,45
0,3
0,02
0,04
0,3
Максимальна кількість глікоалкалоїдів міститься в молодих ор¬
ганах і з ростом їх вміст цих сполук знижується. Наприклад, кіль¬
кість глікоалкалоїдів як а-соланіну, так і а-чаконіну в листках на кі¬
нець періоду цвітіння рослин зменшується втричі порівняно з фазою
бутонізації [18].
Найбільше глікоалкалоїдів міститься у квітках і паростках, порів¬
нянно з бульбами - у кілька сот разів.
У бульбах ці сполуки сконцентровані в основному у вічках, потім
у шкірці, і найменше - в м’якуші (мг % на сиру масу):
ціла бульба -3-10; вічка -25-35;
шкірка (без вічок) - 15-20; м’якуш - 1—4.
Переважна кількість їх зосереджується у перших 10 рядках клі¬
тин бульби, розміщених під кірковим епідермісом. Вони містяться в
усіх клітинах бруньок, причому більшість - у клітинах пробуджених
бруньок, особливо ініціальному кільці - найбільш активній частині
апекса. З віддаленням від вічок вміст глікоалкалоїдів у тканинах буль¬
би різко зменшується.
Молоді зачаткові листочки, які оточують точку росту, містять
максимум цих сполук. Значна кількість їх зосереджена в молодих
листочках завдовжки близько 1 см: в усіх клітинах черешка і пласти¬
нки, крім клітин провідних судин. З формуванням листка вміст глі¬
коалкалоїдів у ньому знижується. Це стосується і стебел, що розви¬
ваються. Сформований листок містить незначну кількість цих спо¬
лук, як і стебла з закінченою диференціацією тканин. Розділ глікоал¬
калоїдів у клітині змінюється у міру її диференціації. У меристема-
тичній клітині вони локалізуються в протоплазмі, а в клітині, що
диференціюється, пересуваються до вакуолей.
Соланін і чаконін у незначній кількості розглядаються як норма¬
льні компоненти бульб картоплі, у підвищеній - вони погіршують її
якості. У бульбах виникає гіркий присмак, а при вживанні їх у їжу -
відчуття свербіння в горлі. Існує певна кореляція між вмістом гліко¬
алкалоїдів і органолептичними смаковими якостями вареної карто¬
плі: при дуже низькому (1-4 мг %) - смак варених бульб дещо гір¬
ший, ніж при середньому їх вмісті (5-10 мг %). Але підвищення глі¬
коалкалоїдів понад 25-30 мг % на сиру речовину значно погіршує
смак картоплі і стає небезпечним для здоров’я людини.
Доза цих сполук 0,2-0,4 г є летальною для людини. Бульби кар¬
топлі, що містять їх понад 20 мг%, не рекомендують вживати в їжу,
особливо, якщо картопля вариться в шкірці. Обчищені бульби менш
небезпечні, оскільки значна частина глікоалкалоїдів видаляється з
лушпинням, а також переходить у воду. Найбільшу небезпеку для
здоров’я людини має нерозчинний у воді соланін, зосереджений в
основі пророслих вічок.
Доведено токсичність підвищеного вмісту глікоалкалоїдів для
ссавців. При введенні в раціон великої рогатої худоби значної маси
106
бадилля картоплі спостерігається порушення координації рухів,
правцеві скорочення скелетної мускулатури, ціаноз слизових оболо¬
нок. При розтині виявляють запалення слизових оболонок шлунка,
кишечнику тощо. Глікоалкалоїди можуть справляти токсичний
вплив і на потомство сільськогосподарських тварин.
При варінні на пару кількість глікоалкалоїдів зменшується на 65,
у воді -на 80% (табл. 2.22). Відбувається це в основному за рахунок
термічного розпаду їх до простіших сполук і за рахунок вимивання із
бульб у воду.
Таблиця 2.22 - Вплив різних способів кулінарної обробки на вміст
глікоалкалоїдів у бульбах картоплі [7]
Спосіб варіння бульб
Вміст глікоалкалоїдів
мг % на сиру речовину
% до загального вмісту
Сирі
5,6
100
На пару
2,0
35,7
У воді
1,2
21,4
Сортові відмінності за вмістом глікоалкалоїдів можуть бути знач¬
ними. Наводимо дані досліджень щодо вмісту цих сполук у сортах
картоплі, вирощеної в дослідах Інституту картоплярства УААН (мг %
на сиру речовину):
Бородянська 3,2-14,7
Незабудка 2,0-8,3
Прієкульська рання 2,1-11,5
Гатчинська 2,4-7,5
Луговська 1,0-6,3
Зарево 3,0-9,1
Вміст глікоалкалоїдів у бульбах визначається не лише сортом. При
вирощуванні картоплі на сухому піщаному грунті рослини містять їх
більше, ніж на вологому грунті. З підвищенням рівня забезпечення
рослин елементами мінерального живлення при зрошенні біосинтез
глікоалкалоїдів у бульбах дещо посилюється. Підвищений вміст цих
сполук картоплі спостерігається, як правило, у північних районах, де
літо прохолодніше, дощове і її збирають недозрілою, а також у зонах
із жарким кліматом, що також пов’язано з гальмуванням дозрівання
бульб. Надлишкове накопичення глікоалкалоїдів відбувається і при
недостатньому зрошенні та значному нагріванні грушу. Отже, кліма¬
тичні умови істотно впливають на синтез цих сполук у бульбах окре¬
мих сортів. Мабуть, при затриманні дозрівання бульб на період зби¬
рання, незалежно від природи його, вміст глікоалкалоїдів вищий вна¬
слідок того, що в молодих бульбах завжди більше цих сполук.
Аналогічні дані отримано зарубіжними вченими, які твердять, що
вміст глікоалкалоїдів у картоплі залежить від генотипу і місця виро¬
щування рослин, але провідна роль належить їх генотипу [26].
107
Сприяють підвищенню вмісту в бульбах глікоалкалоїдів низькі
температури (до 60%), посуха (на 10-35%), значно меншою мірою -
нестача калію [29 ].
Рівень вмісту цих сполук змінюється під час зберігання картоплі,
особливо навесні, коли бульби починають проростати. Наприклад, в
одному з дослідів їх було (мг % на сиру речовину): 25 січня - 2,4; 12
березня - 8,4; 20 квітня - 9,6. Суттєвим фактором, що впливає на
вміст глікоалкалоїдів у бульбах, є світло. Так, при зберіганні картоп¬
лі сорту Бородянська в освітленому приміщенні з 3 лютого по 5 бе¬
резня кількість цих сполук змінювалася так (мг % на сиру речовину):
З лютого - 2,5; 9 лютого - 9,9; 15 лютого - 11,3; 24 лютого - 17,5; 16
березня - 22,6.
Різні сорти картоплі неоднаково накопичують глікоалкалоїди під
дією світла. Інтенсивність цього процесу залежить також від якості
світла. Так, бульби, що зберігаються при штучному освітленні нако¬
пичують їх більше, ніж за природного. Інтенсивніше синтезуються
глікоалкалоїди під пливом світла у молодих бульбах. Активність син¬
тезу цих сполук у рослинах картоплі може змінюватися також зале¬
жно від режиму освітлення в період онтогенезу.
Виявлено, що чим нижча температура зберігання, тим вищий
вміст глікоалкалоїдів при експозиції на світлі. Так, після зберігання
протягом 18 днів при температурі від 5 до 30°С з наступним витри¬
муванням на світлі протягом 48 годин кількість цих сполук станови¬
ла, відповідно, 21 і 3 мг на 100 г.
Глікоалкалоїди у підвищеній кількості синтезуються в місцях
пошкодження бульб при збиранні залежно від сорту, типу пошкод¬
ження, температури, тривалості зберігання картоплі. Найбільше їх
спостерігається у бульб з порізами, менше - з проколами, потім -
ударами при падінні і найменше - у бульб з подряпинами.
Таким чином, підвищення накопичення глікоалкалоїдів у бульбах
залежить від таких факторів: збирання недозрілої картоплі, освітлен¬
ня під час зберігання, механічних пошкоджень під час збирання
врожаю, транспортування і проростання бульб, періоду їх зберігання.
Тому доцільно визначати вміст глікоалкалоїдів у бульбах перед їх
споживанням.
Кількість глікоалкалоїдів значно вища у диких форм картоплі
(табл. 2.23).
Таблиця 2.23 - Вміст глікоалкалоїдів у культурної і дикої картоплі (мг % на суху
речовину) [7]
Вид
Бульби
Очистки
Паростки
Листки
Solanum tuberosum
10
260
5200
200
Solanum chacoense
60
2500
7600
5200
Solanum demissum
60
1100
8000
4500
Solanum poliadenium
80
190
8200
1200
108
У міжнародному центрі з картоплі (Перу) провели визначення
вмісту цих сполук у бульбах диких і культурних видів картоплі. Ви¬
вчали види S. juzepczukii, S. curtilobum, S. stenotonum, S. tuber os sum,
S. andigenum та дикий вид. Максимальний вміст глікоалкалоїдів (35-
120 мг% на сиру речовину) виявлено у S. асаиіе, дещо менше (15,7-
46,8 мг%) - у S. juzepczukii і S. cutrilobum (18,3-29,0 мг%). У решти
зразків він був нарівні S. tuberossum (не більше 6,5 мг%). Високий
рівень цих сполук виявлено в культурного сорту Лепар (до 50 мг%).
У складі глікоалкалоїдів знайдено а-чаконін, а-соланін, демісин, то-
матин, соламарин. У S. асаиіе переважали демісин (96 мг%) і тома-
тин; у S.curtilobum і S.stenotonum - Р-чаконін і а-соланін. Види з під¬
вищеною кількістю глікоалкалоїдів пропонують використовувати в
селекції при створенні сортів, стійких проти фітофтори [25].
Дослідження [28] показують, що включення в селекційний про¬
цес S. chacjense веде до підвищення вмісту глікоалкалоїдів у листках.
Глікоалкалоїди привертають увагу селекціонерів-картоплярів у
зв’язку з їх токсичною дією на бактеріальні й грибні збудники хвороб.
З’ясовано, що соланін гальмує ріст бактерій, а 0,1 і 0,5%-ний роз¬
чин його спричиняє відмирання протоплазми. Виявлено фунгіцидні
властивості глікоалкалоїдів щодо ряду патогенних грибів, зокрема
Microsporum audoine, Fusarium oxiporum, зооспор фітофтори Helmi-
tosporum carbanum.
Глікоалкалоїди а-соланін і а-чаконін мають більшу фунгіцидну
активність, ніж феноли (кофейна і хлорогенова кислоти). Останні у
концентрації 0,1 мг/мл не впливають на ріст фітофтори, тоді як глі¬
коалкалоїди пригнічують його майже на 70%. Зооспори фітофтори
при концентрації а-соланіну 5 мг/мл вимирали повністю.
Токсичність а-соланіну значною мірою визначається реакцією
середовища. Значення Д 50 при зміні pH від 5 до 8 коливається для
макроспор F. corculum у межах 2-11 мг/мл. Накопичення глікоалка¬
лоїдів у бульбах при їх позеленінні підвищує стійкість насіннєвого
матеріалу до захворювання.
Соланін, чаконін і соланідин інгібують радіальний ріст Alternaria
solani на картоплі. За ступенем інгібування перше місце посідає со¬
ланідин, потім - чаконін і соланін. При концентрації 500 мг/л сола¬
нін викликає 50% інгібірування росту після 96 годин інкубації при
24°С. Виявлено зростання чутливості листків до ушкодження і одно¬
часне зменшення концентрації глікоалкалоїдів у листках з їх ростом.
Наприклад, 30-денні листки містять їх 960 мг/кг і стійкі проти пара¬
зита, а старіючі 90-денні - лише 150 мг/кг і значно уражуються гри¬
бом. Незважаючи на те, що глікоалкалоїдам властива фунгітоксич-
ність, вони не є вирішальним фактором у фотоімунітеті картоплі.
Взагалі, після інфікування у бульбах нагромаджуються хлорогенова і
кофейна кислоти, скополін, а-соланін, а-чаконін, соланідин, рипш-
тин, фітуберин. Усі вони, за винятком ришитину і фітуберину, міс¬
109
тяться в шкірці бульби у кількостях, рівних або більших від тих, які
продукуються після інфікування патогеном очищеної картоплі. Хло¬
рогенова і кофейна кислоти знайдені в усіх частинах рослини як реак¬
ція на ушкодження бульб та інфекцію багатьма патогенами і непато-
генами, а також накопичуються або існують у деяких здорових тка¬
нинах у кількостях, які пригнічують мікроорганізми. Ушкодження
або інфекція в цілому призводять до швидкого окиснення цих спо¬
лук, продукти якого токсичні для багатьох мікроорганізмів. На ран¬
ніх стадіях інфекції після інокуляції бульб окремими расами фіто¬
фтори нагромаджуються поліфеноли. У тканинах, інокульованих не¬
сумісними расами фітофтори, або на зрізах - тільки хлорогенова ки¬
слота; а-салонін і а-чаконін локалізуються навколо ушкоджених
ділянок бульби. Високу токсичність до личинок колорадського жука
мають томатин, демісин - алкалоїди дикої картоплі.
Токсичність демісину і томатину зумовлена агліконами, водночас
цукри, що входять до молекул цих алкалоїдів, не справляють токсич¬
ної дії.
Стероїдні глікоалкалоїди використовуються як сировина для син¬
тезу різних гормонів. Щодо цього соланін і соласонін є зручними
вихідними сполуками.
Як сировину для глікоалкалоїдів можна використовувати парост¬
ки завдовжки 10-15 см і більше при одному обламуванні, або 3-
15 см при 2-3-х з одних і тих самих бульб. Сушать ці паростки тер¬
міново при 80°С без фіксування або при 40-60°С з попереднім фік¬
суванням сировини, чим забезпечується найбільша кількість глікоал¬
калоїдів. Зберігання їх протягом року в сухому приміщенні не при¬
зводить до втрати цих сполук, як у вологих умовах. Найбільш повно
соланідин вилучається метиловим, етиловим спиртами і ацетоном
при pH 12,0.
Вітаміни та інші речовини. У бульбах картоплі міститься чимало
вітамінів, зокрема С, Вь В2, В3, В6, РР, H, Р, каротиноїди, К (табл. 2.24).
Вітаміни відіграють важливу роль у підвищенні імунобіологічних
реакцій організму, які зумовлюють стійкість його до несприятливих
умов навколишнього середовища, що має суттєве значення в профі¬
лактиці інфекційних хвороб, відновленні порушених функцій органі¬
зму. Вони посилюють лікувальний ефект лікарських препаратів та
знижують їхню токсичну дію, запобігають професійним захворю¬
ванням та отруєнням. Вітаміни - це радіопротектори, тобто речови¬
ни, що знижують або послаблюють негативний вплив хемотоксикан-
тів та радіонуклідів на організм.
Вітамін С - аскорбінова кислота. Дія її багатогранна: підтримує
нормальний стан серцево-судинної системи, стабілізує майже всі фі¬
зіологічні процеси, сприяє холестериновому обміну, запобігає авіта¬
мінозам. За вмістом цієї кислоти картопля знаходиться на рівні
апельсинів і лимонів. Упровадження картоплі в Європу забезпечило
110
Таблиця 2.24 - Вміст вітамінів у бульбах картоплі, мг% на сиру масу
Вітамін
Орієнтовний вміст
Каротиноїди
0,0-0,46
В і (тіамін)
0,024-0,20
В2 (рибофлавін)
0,075-0,20
В3 (панготенова кислота)
0,20-0,32
В6 (ггіридоксин)
0,009-0,25
С (аскорбінова кислота)
5-35
РР (нікотинова кислота)
0,0008-0,0010
Н (біотин)
1,7-1,9
К (колахінони)
0,06-0,24
Р( цитрин)
25-40
успішну боротьбу з епідеміями цинги. Вважається, що споживання
300 г картоплі на добу забезпечує половинну потребу людини в цьо¬
му вітаміні. Знаходиться вітамін С у бульбах катоплі у формі аскор¬
бінової і дегідроаскорбінової кислоти (останньої не більше 20% від
загальної кількості). Вміст аскорбінової кислоти розподілений у бу¬
льбах нерівномірно: у верхівці в середньому 21%, пуповині - 17,2,
шкірці - 16,9, камбіальний частині - 17,7, серцевині - 16,1%. Най¬
вищий її вміст у період інтенсивного росту бульб, а потім дещо'зни-
жується. Основна втрата вітаміну відбувається в період зберігання -
з грудня по лютий (60%). Проте втрати значною мірою залежать від
умов зберігання. Наприклад, при низькій температурі (близько 0°С)
вітамін С майже повністю руйнується. Вміст аскорбінової кислоти
залежить також від сорту, агротехніки і особливо ґрунтово-
кліматичних умов. Як правило, на легких грунтах він вищий, ніж на
важких. Надлишок азоту і калію призводить до зниження вмісту ві¬
таміну С, а фосфору - до підвищення. Суха погода сприяє нагрома¬
дженню його, а волога і холодна - діє навпаки.
У період вегетації спостерігається коливання вмісту аскорбінової
кислоти, що пов’язане із змінами агрометеорологічних умов. Значні
опади, зниження температури повітря і похмура погода викликають
помітне зниження С- вітамінної активності в листках і молодих бу¬
льбах. З настанням теплої, ясної і помірно вологої погоди спостері¬
гається повторне підвищення вмісту вітаміну С в бульбах, особливо
у пізньостиглих сортів.
Вітамін Bj (тіамін) бере безпосередню участь в обміні вуглево¬
дів, білків і жирів; стимулює секреторну активність залоз, які беруть
участь у перетравлюванні їжі, і нормалізує рівень артеріального тис¬
ку. Вміст його в середньому становить 100 мкг% при коливанні від
24 до 200 мкг%. Дослідженнями Є. А. Ладигіної і В. П. Кирюхіна [8]
доведено, що при вирощуванні картоплі на супіщаному грунті вміст
його в різних сортах коливався в межах 66-131, на торф’яному ґрунті
- 62-154 мкг%. У міру дозрівання бульб вміст його зростає. 20-25% ві¬
111
таміну Ві знаходиться у зв язаному стані, а 75-80 - у вільному. Най¬
більша кількість його зосереджена навколо ситоподібних трубок і в
середній частині бульб. Більше його також у верхівковій частині, ніж
у пуповині. Вживання 300 г картоплі на добу компенсує 10-20% по¬
треби у вітаміні Bj.
Вітамін В2 регулює тканинний обмін речовин, сприяє утворенню
енергії в організмі, утворенню гемоглобіну, стимулює процеси росту
і заживления ран, сприяє роботі печінки, центральної і вегетативної
нервової системи. У сирих бульбах його міститься 40-100 мг%.
Споживання 300 г картоплі забезпечує 5-10% його добової потреби
для людини. Вміст вітаміну В2, як і Bj, різко змінюється за роками,
залежно від погоди.
Вітамін Вз бере участь у забезпеченні нормальних процесів кро¬
вотворення, синтезу білка, обміні нуклеїнових кислот.
Вітамін Вв регулює обмін вуглеводів і білків, стимулює утворен¬
ня крові, сприяє розвитку природного імунітету, нормалізує роботу
печінки.
Вітамін РР - необхідний для процесів біологічного окиснення,
запобігання запаленням; нормалізує вміст холестерину в крові.
Вітамін Н бере участь у жировому, вуглеводневому і пуринових
обмінах, процесах карбоксилювання, декарбоксилювання і дезаміну¬
вання амінокислот.
Вітамін Р сприяє зміцненню стінок капілярів, посилює виділення
жовчі.
Вітамін К бере участь у процесах згортання крові, утворення АТФ,
зменшує проникність стінок капілярів, стимулює утворення жовчі.
Проте вітаміни В2, В3, В4, РР, H, Р вивчені стосовно картоплі не¬
достатньо, а вітамін К почали недавно досліджувати, оскільки він
має велике профілактичне значення в районах, забруднених радіону¬
клідами. Виявлена різниця його вмісту в бульбах у сортовому розрізі.
У зв’язку з аварією на Чорнобильській АЕС в Інституті картоплярст¬
ва УААН значна увага приділяється також створенню сортів, що міс¬
тять каротиноїди, які відіграють роль радіопротекторів. Це сорти Не¬
забудка, Світанок київський, Кобза, Купава, Водограй, Обрій, Гор¬
лиця, Слов’янка, в яких вміст каротиноїдів коливається в межах
0,15-0,40 мг% на сиру масу. Вони створені шляхом залучення до се¬
лекційного процесу сортів з жовтим м’якушем, які містили 0,11-
0,15 мг% провітаміну А, а з білим - лише 0,014—0,053 [15].
Бульби містять клітковину та пектинові речовини, які відіграють
важливу роль у нормалізації процесів живлення. Поряд з каротиної¬
дами клітковина і пектин, маючи здатність утворювати комплексні
сполуки з радіонуклідами, важкими металами, різними токсичними
хімічними речовинами та холестерином, виводять їх зі шлунково-
кишкового тракту людини, що дає змогу зменшити їх шкідливий
вплив на організм та запобігти розвитку атеросклерозу [11].
112
Серед продуктів харчування картопля є основним джерелом ка¬
лію, який відіграє значну роль у нормалізації водного обміну та під¬
триманні роботи серця. В бульбах картоплі його міститься 1,5-2,0%
на суху масу, що значно більше, ніж у хлібі, рибі. При щоденному
вживанні до 500 г картоплі забезпечується добова потреба людини в
цьому елементі. Високий вміст калію визначає сечогінні властивості
картоплі, тому вона входить до дієтичних продуктів харчування лю¬
дей із захворюваннями серцево-судинної системи та нирок. Особли¬
во потребують калію м’язи серця (міокард). При зниженні рівня ка¬
лію в крові нижче 3 ммоль/к порушується ритм роботи серця, прові¬
дність судин.
Багато в картоплі також фосфору. Фосфатні солі беруть участь у
формуванні скелета, в біологічних процесах головного мозку та ін¬
ших органів, є акумуляторами енергії.
Залізо міститься в червоних кров’яних тільцях та як необхідна
складова частина більшості клітин організму входить до складу де¬
яких ферментів.
Кальцій у комплексі з фосфором є джерелом формування кісток і
зубів. Магній активізує фосфорний обмін, знижує кров’яний тиск,
бере участь у виведенні з кишкового тракту холестерину. Марганець
підвищує активність деяких ферментів.
При нестачі йоду розвивається хвороба щитоподібної залози. Йо¬
го органічні сполуки характеризуються антисклеротичною дією.
Але, основним є те, що в бульбах картоплі співвідношення мінераль¬
них елементів знаходиться у пропорції, близькій до потреб організму
людини. Наводимо орієнтовні дані середнього вмісту в бульбах еле¬
ментів мінерального живлення (табл. 2.25).
Таблиця 2.25 - Орієнтовний середній вміст у бульбах картоплі зольних
елементів, мг/100 г сухої маси
Елемент
Вміст
Елемент
Вміст
Калій
2000
Силіцій
10
Фосфор
300
Марганець
4
Натрій
50
Цинк
1,5
Кальцій
40
Алюміній
4
Магній
70
Миш’як
0,02
Сірка
150
Молібден
0,02
Хлор
300
Кобальт
0,006
Бром
0,6
Нікель
0,02
Залізо
6
Иод
0.02
Мідь
0,8
Фтор
од
Бор
0,7
Літій
0,03
Крім макро- й мікроелементів, у бульбах картоплі знайдено стеа¬
рини, ліпіди та органічні кислоти (кофейну, хлорогенову, лимонну,
щавлеву, яблучну та ін.). До складу ліпідів (жироподібні речовини) в
113
основному входять незамінні жирні кислоти - лінолева, ліноленова,
які беруть участь у біохімічних процесах обміну речовин, регулю¬
ванні проникності клітин, адсорбційних процесах протоплазми і не
синтезуються в організмі людини. Кількість жиру в бульбах у серед¬
ньому 0,15-0,16% на сиру масу, в деяких сортах - близько 1%. Ліпі¬
ди поліпшують смак бульб. При кулінарній обробці вміст їх знижу¬
ється в 1,5-2 рази.
У картоплі виявлено чимало фенольних сполук (тирозин, кофей¬
на, хлорогенова, абсцизова кислоти, скополін, скополетин та ін.).
Значна частина їх розглядається як похідні фенілпропану. Зосере¬
джені фенольні сполуки переважно в активно функціонуючих орга¬
нах (вічках, шкірці). Орієнтовний вміст їх у вічках 350-500; шкірці -
250—400; м’якуші - 15—45 мг% на суху масу. Фенольні речовини - це
ароматичні сполуки, як мають кислу реакцію і легко розчиняються в
лугах. Організм людей і тварин не здатний синтезувати їх. Вони
утворюються вищими рослинами й мікроорганізмами.
Хлорогенова, кофейна і корична кислоти є основними представ¬
никами фенольних сполук бульб картоплі. Нагромадження їх у буль¬
бах спостерігається при механічних пошкодженнях і ураженні хво¬
робами. Найбільша кількість хлорогенової кислоти утворюється, ко¬
ли середня температура повітря низька протягом трьох тижнів перед
збиранням картоплі. Наприклад, при середній температурі повітря
10-15°С цієї кислоти утворюється в пуповинній частині бульб в 1,5
рази більше, ніж при 20-25°С. Більше хлорогенової кислоти містить¬
ся також у повністю дозрілих бульбах. Фенольні сполуки відіграють
певну роль у стійкості бульб проти мокрої гнилі [7].
ЛІТЕРАТУРА
1. Буколова Т.П., Дуда В.В., Маленко І.М., Кравець В.С. Біохімічний
склад бульб і його вплив на якість картоплепродуктів // Картоплярство-
1977.- Вип. 27.-С. 153-160.
2. Вавилов Н.И. Генетика на службе социалистического земледелия //
Соц. растениеводство - 1932-№4 -С. 19-42.
3. Вечер А.С., Гончарик М.Н. Физиология и биохимия картофеля.-
Минск: Наука и техника, 1973.
4. Власенко Н.Е. Условия высокой эффективности применения удобре¬
ний под продовольственный и семенной картофель: Дис. ... д-ра с.-х. наук-
Немишаево, 1983.
5. Власенко Н.Е. Удобрение картофеля - М.: Агропромиздат, 1987.
6. Кучко А.А., Мицько В.М. Фізіологічні основи формування врожаю і
якості картоплі-К.: Довіра, 1997.
7. Кучко А.А., Власенко М.Ю., Мицько В.М. Фізіологія та біохімія кар¬
топлі-К.: Довіра, 1997.-335 с.
8. Ладыгина Е.А., Кирюхина В.П. Изменение содержания витаминов в
клубнях различных сортов картофеля при выращивании и хранении // Тех¬
нология производства картофеля - 1975 - Вып. 22 - С. 22-30.
114
9. Луковникова Г.А., Злотников М.Д. Влияние метеорологических усло¬
вий на содержание сухого вещества и крахмалистости картофеля // Вест,
с.-х. науки - 1968 - № 11- С. 26-29.
10. Мицько В.М., Мороз С.І., Таращенко Н.І. Вплив вермикомпосту на
біохімічний склад бульб И Картоплярство.- 1997.- Вип. 27.- С. 100-102.
11. Новосельська А.П., Осипчук А.А. Сорти картоплі, придатні для діє¬
тичного харчування // Картоплярство - 1994 - Вип. 25 - С. 75-77.
12. Новосельська А.П., Мицько В.М., Холодило І.В. Сортові особливості
біохімічного складу бульб картоплі // Картоплярство- 1995 - Вип. 26-
С. 42-45.
13. Онищенко О.Й. Насінництво картоплі на Україні - К.: Урожай, 1966.
14. Онищенко О.Й. Рання картопля - К.: Урожай, 1969.
15. Осипчук А.А. Створення картоплі з різним кольором м’якуша бульб
// Картоплярство - 1997 - Вип.27- С.56-69.
16. Теслюк П.С. Выращивание столового картофеля - К.: Урожай, 1976.-
97 с.
17. Теслюк П.С., Клець С.А. Влияние метеорологических условий веге¬
тационного периода на урожайность и качество картофеля И Картофелевод¬
ство.- 1987.- №> 18.- С.47-49.
18. Ahmed S., Mulier R. Seasonal changes and effect of nitrogen- and potash-
fertilization on the solanine parts of the potato piant // Z.pflan-zenermarh.-
Bodenk., 1979.-Vol. 142, № 2.- P.275-279.
19. Bodlaender V.B.A. Growth regulators in potato production - 1983.-P. 1-24.
20. Christensen D.H., Madsen M.H. Changes in potato starch quality during
growth // Potato Research - 1996 - Vol. 39, № 1.- P.43-50.
21. Marschuer H. und Kraus A. Beziehungen zwischen Kaliumgehalt und
Qualitat von Kartoffeln // Kartoffelbau.- 1980. - № 2.- P. 65-67.
22. Mica B., Vocal B. Vliv obsahu a kvality scroby na pevnost (texturu)
bramboravych hliz // Ros. Vyroba - 1986 - № 10 - P. 109-119.
23. Muresan S., Olteanu Q. Soiwri de cartof pentru industria amidoneclui //
Prod.veget. Hortic.- 1978.- Vol. 27, № 10.- P.l0-12.
24. Nitsch1 A., Klein K. Qualitat der Kartoffel in Abhandigkeit von der
Stikstofftdnung // Kartoffelbau - 1983. - Vol. 34, № 2 - P. 30-34.
25. Osmann S. et ali. Glycoalcaloid composition of wild and cultivated
tuberbearing Solanum species of potential value in potato breeding programs //
J. Agr. Food Chem.- 1978.- Vol. 26, № 5.- P.1246-1248.
26. Pamell A., Bhuva V.S., Binteliffe E.J. B. The glycoalkaloid content of
potato varieties // J. Nat. Inst Agr. Bot - 1984 - Vol.16, № З - P.535-541.
27. Pritchard M.K. and Adam L.R. Relationships between fry color and sugar
concentration in stored Russet Burbank and Shepody potatoes //Potato Joumal-
1994.- Vol. 71, № 1.- P.59-68.
28. Sanford L.L., Deahl K.L., Sinden S.L., Glycoalcaloid content in foliage of
hybrid and backcross populations from a S.tuberosum x S. chacoense // Potato
Journal.- 1994.- Vol. 71, № 4.- P.225-236.
29. Swaaij A.C. Effect of growth conditions on gycoalkaloid in potato tubers //
Potato Research - 1992 - Vol. 35, № 1- P.68-69.
30. Tucher Th. Entwicklungsmoglichkeiten der Kartoffel zur Starkegewinnung
// Kartoffelbau.- 1995.- № 7.- P.292-296.
115
3. ГЕНЕТИКА КАРТОПЛІ
У межах роду Solanum L. види утворюють безперервний поліпло-
їдний ряд, що вперше було виявлено [37] у результаті широкомасш¬
табних цитологічних досліджень. Поліплоїдний ряд включає диплої-
дні (2п=24), триплоїдні (2п=36), тетраплоїдні (2п=48), пентаплоїдні
(2п=60) і гексаплоїдні (2п=72) види. Основна кількість хромосом (х)
цього поліплоїдного ряду дорівнює 12.
В секції Tuberarium налічується близько 70% диплоїдних видів,
15% тетраплоїдних, 8% гексаплоїдних, а решту становлять триплоїд¬
ні й пентаплоїдні види [17].
Для генетичних досліджень картопля є надто трудомістким
об’єктом. Будь-який сорт цієї культури є гетерозиготним, якщо не за
всіма, то за більшістю ознак. При схрещуваннях відбувається розще¬
плення за багатьма генами, а відповідно й за ознаками, що значно
ускладнює гібридологічний аналіз. Однак, широке застосування від¬
даленої гібридизації із залученням диких і культурних диплоїдних
видів, використання поліплоїдії і гаплоїдії в останні десятиріччя сти¬
мулювало розвиток генетичних досліджень картоплі.
3.1. ГЕНЕТИКА ДИПЛОЇДНИХ
ВИДІВ КАРТОПЛІ
Диплоїдні види (2п=24) зустрічаються майже у всіх таксономіч¬
них серіях секції Tuberarium, за винятком Aculia, Subaculia, Inqae-
folia, Oxycarpa, Lonqipedicellata i Borealia [9].
Еволюція еукаріотів відбувається за рахунок регулярного чергу¬
вання гаплоїдної і диплоїдної фаз. Поява диплоїдів зумовлена злит¬
тям жіночої і чоловічої гаплоїдних гамет із збереженням об’єднаного
диплоїдного ядра в мільйонах клітинних вегетативних поколіннях
завдяки мітозу.
Збереження постійної кількості хромосом, характерної для кожного
виду, з покоління в покоління при статевому розмноженні зумовлюється
особливим типом поділу клітин - мейозом, у результаті якого кількість
хромосом у дочірніх клітинах зменшується наполовину.
Мейоз у диплоїдних видів. Огляд і детальний аналіз робіт до¬
слідників (Oppenheimer, 1933; Choudhuri, 1943; Lamm, 1945; Bains,
1951; Prakken, Swaminathan, 1952; Howard, 1961; Gilles, 1955; Buch,
1966 та ін.), які вивчали мейоз у диплоїдних видів і внутрішньо-
серійних міжвидових диплоїдних гібридів картоплі, зроблений
І.М.Яшиною, М.М.Букетовою та К.З.Будіним [45,6].
116
У переважній більшості робіт показано, що у диплоїдних видів
мейоз, як правило, перебігає нормально.
Суть нормального перебігу мейозу у диплоїдних видів полягає в
кон’югації гомологічних хромосом з утворенням у діакінезі профази-
I і метафазі-І 12 бівалентів. Однак іноді спостерігаються певні пору¬
шення нормального мейозу з різною частотою.
Зокрема, у культурних диплоїдів S.rybinii і S.phureja О.В.Іва-
новська [19] виявила, що в незначної частини клітин макро- і мікро¬
спор зустрічаються відхилення від норми. До 0,5 % у стадії спорад
зустрічаються діади, а в одному випадку виявлене формування двох
зародкових мішків.
При вивченні мікроспорогенезу у культурних ДИПЛОЇДІВ (іS. Ste-
notomum і S.rybinii) і дикого диплоїда S.verrucosum було виявлено
поряд з правильним мейозом у більшості рослин усіх трьох видів
чітко виражені хромосомні аберації різних типів. У рослин з аберація¬
ми хромосом спостерігалося зниження фертильності пилку. Крім
хромосомних аберацій, у диплоїдних видів виявлені в мейозі реститу¬
ційні ядра з соматичною кількістю хромосом, триваленти, а також ма¬
теринські клітини пилку з 11п+2і в діакінезі й 10ц+ 4і у метафазі-І.
Частота хромосомних аберацій у різних видів коливалася від 19 %
(у S. infundibuliforme) до 25,6 % (у S. stenotomum). У S. schikii виявле¬
но лише 53 % клітин з 12 бівалентами, а решта клітин крім бівален¬
тів мали по два і навіть по чотири уніваленти [45].
Іноді у диплоїдних видів у метафазі-П зустрічаються пластинки з
24 хромосомами. Поява тетраплоідів, що часто спостерігають дослідни¬
ки в потомстві від схрещування тетраплоідів з диплоїдами, може бути
зумовлена формуванням гамет з нередукованим числом хромосом.
У цілому нормальний перебіг мейозу, подібно як у диплоїдних
видів, виявлено у внутрішньосерійних і міжсерійних міжвидових ди¬
плоїдних гібридів. Наприклад, правильний мейоз у серії гібридів від
реципрокних схрещувань S.goniocalyx х S. rybinii, S. phureja х S. Ry-
binii, S. rybinii * S. stenotomum та інші спостерігав В.А.Рибін [37]. Всі
пластинки в метафазах-І і II мейозу мали по 12 елементів, в анафазах
спостерігалося правильне розходження хромосом, а в гібридів відмі¬
чалося добре утворення плодів.
За даними багатьох дослідників, у міжсерійних диплоїдних гіб¬
ридів у цілому описаний правильний мейоз. Однак зустрічалася не¬
значна кількість порушень, головним чином у діакінезі спостерігало¬
ся утворення унівалентів, відставання хромосом в анафазі-І, а на ста¬
дії спорад відмічалися пентади й гексади. В життєвому циклі організ¬
мів мейоз є унікальним явищем, окремі етапи, стадії і функції якого
контролюються особливими генними системами. Генетичний конт¬
роль мейозу остаточно доведено численними дослідженнями мей-
мутацій (мейотичних мутацій), більшість яких характеризується ці¬
лим спектром аномалій мейозу порівняно з нормальним [40]. Отже,
117
причиною появи унівалентів, відсутності або неповної кон’югації та
відставання хромосом в анафазі-І, формування гамет з нередукова-
ною кількістю хромосом, що спостерігаються в одногеномних дип¬
лоїдних видів, є мей-мутації, які, можливо, відіграли істотну роль у
виникненні поліплощних видів. Не виключено, що саме гетерозигот¬
ні (а, можливо, й гетерозисні) форми, які мають селективні переваги,
успішно розповсюджувалися при такому способі розмноження, коли
відсутня генетична рекомбінація за рахунок кросинговеру й комбі¬
нування хромосом при розходженні в редукційному поділі. А саме:
організми із заблокованим редукційним діленням, які продукують
здатні до самостійного розвитку диплоїдні гамети, могли бути під¬
хоплені природним добором і дати початок поліплоїдним формам.
Нередукований FDR-NCO- пилок формується у картоплі при по¬
єднанні двох порушень мейотичного ділення - десинапсису (ds) і
злиття веретен (fs) у фазах MII - А П [51]. В.Є. Подлісысих та ін. [35]
спостерігали у дигаплоїдних форм картоплі в мейотичному діленні
на стадії А І у мейоцитах злиття і триполярне розміщення веретен, а
також нерегулярне розходження хромосом в А П, зумовлене мутація¬
ми fs, tps та unr-A П відповідно. У дигаплоїдних форм з мутацією
iinr-A П на стадії М П окремі мейоцити замість двох метафазних пла¬
стинок формували одну, подібну до такої, що утворюється при злитті
веретен. Причому, на відміну від метафазної пластинки з мутацією fs
хромосоми розміщалися безладно в екваторіальній площині клітини.
Ці автори виявили, що злиття веретен з високим ступенем вірогідності
призводило до формування діади, замість тетради, і відповідно 2п - пил¬
ку. Для селекції картоплі з використанням маніпуляцій на диплощному
рівні найбільше значення мають FDR- і SDR- механізми (реституція
першого і другого мейотичного ділення) формування 2п-гамет.
Успадкування основних морфологічних ознак. Знання законо¬
мірностей успадкування морфологічних ознак як у диплоїдних, так і
поліплоїдних видів картоплі має не лише теоретичне значення. Як
відзначають окремі дослідники [27], на сьогодні картопля є одним із
найбільш зручних об’єктів клітинної біології та генетичної інженерії.
Це дало змогу суттєво розширити можливості генетичних маніпуля¬
цій з цією надзвичайно важливою сільськогосподарською культу¬
рою. Однак, навіть за умови найширшого застосування культури іп
vitro, польові дослідження залишаються обов’язковим і логічним
продовженням досліджень для одержання реальних результатів се¬
лекційного напряму.
Вступ України до Міжнародної спілки з правової охорони селек¬
ційних досягнень (UPOV) передбачає випробування сортів із засто¬
суванням особливих тестів на відмінність, однорідність і стабіль¬
ність (DUS- test). Цей тест грунтується на морфологічних маркерах,
які охоплюють певний набір якісних альтернативних і деяких кількі¬
сних ознак, за якими зручно ідентифікувати селекційні сорти. У цій
118
ситуації передбачається залучення до селекційного процесу джерел з
чітко вираженими ознаками, які б надавали сорту відмітних рис [5].
Це положення загострює необхідність вивчення генетики морфоло¬
гічних ознак. Крім того, у селекційній практиці морфологічними
ознаками користуються як генетичними маркерами при доборах. То¬
му часто постає необхідність ідентифікації генотипів за фенотипом.
Однак, до сьогодні генетика диплоїдних видів картоплі залишається
недостатньо вивченою.
Детальний огляд літератури з генетики диплоїдних видів картоп¬
лі зроблений Ю.П. Лаіггєвим [29].
Форма листка. При вивченні гібридів, одержаних від реципрокних
схрещувань диплоїдних видів S.rybinii, S.commersonii, S.emmeae,
S.Kesselb- rennerU О.К.Емме [43] вперше виявила, що у гібридів Fi домі¬
нує довгий листок виду S.emmeae над укороченим в інших видів. Як
правило, сильна розсіченість і вузькі частки листка домінують над мен¬
шою розсіченістю і крупнішими частками листка. Аналогічні дані були
одержані Чаудьюрі (цит. за [29]). Він виявив, що складний лист S.rybinii
домінує над простим листом S. simplicifolium, а в F2 спостерігається про¬
сте менделівське розщеплення за цією ознакою 3:1.
Л.І. Костіна [24], порівнюючи за морфологічними ознаками сіян¬
ці S.chilotanum з культурними диплоїдними видами S. stenotomum і
S.phureja та дикими видами S.leptostigma, S.molinae, S.mochicense і
S.chancoense, також спостерігала випадки домінування сильної роз¬
січеності листка над слабкою. Однак, вона зустрічала випадки, коли
у частини сіянців домінувала слабка розсіченість листка.
Форма віночка. За даними О.К.Емме [43], у диплоїдних гібридів
за участю S. еттеае в Fi формуються проміжні між зірчастою і ко¬
лесоподібною формою віночки, що вказує на неповне домінування
першої. І навпаки, у Fi Commersonii х var. Gandarae повністю домі¬
нує форма віночка останнього.
Ю.П.Лаїггєв [29] наводить дані одних дослідників про доміну¬
вання колесоподібного віночка над зірчастим у Fi і правильне
розщеплення у відношенні 3:1 в F2, а інших - про те, що це явище не
підтверджується.
На випадки домінування сильної розсіченості пелюсток віночка у
потомстві сіянців S.chilotanum вказує Л.І.Костіна [24].
Забарвлення віночка. Має широкий спектр кольорів- від білого
до темно-фіолетового, що зумовлюється наявністю пігментів. їх біо¬
синтез контролюється генами. У диплоїдних видів до сьогодні гени
забарвлення ще не ідентифіковані. Суперечливі дані про характер
успадкування забарвлення віночка вказують на складну природу ге¬
нетичного контролю цієї ознаки.
Однією із перших проб вивчення генетики забарвлення віночка у
диплоїдних видів були дослідження О.К.Емме ще в 1936 р. Вивчаю¬
чи характер успадкування цієї ознаки в потомстві 27 диплоїдних
119
міжвидових гібридів, вона виявила наявність низки факторів, які зу¬
мовлюють відповідне забарвлення: Ansp - ген антоціану на верхній
стороні віночка: An,nf - ген, що контролює наявність антоціану на
нижній стороні віночка; Сг - ген, що зумовлює забарвлення віночка
від червоно-фіолетового до червоно-малинового; Су - ген, який зу¬
мовлює синє і синьо-фіолетове забарвлення та Fi - ген, що детермі¬
нує наявність жовтих пігментів. Відсутність антоціану зумовлює біле
забарвлення віночка.
Крім того, було виявлено, що забарвлення віночка зумовлюється
взаємодією декількох неалельних генів, а ряд генів представлені
множинними алелями, що зумовлює широкий спектр інтенсивності
забарвлення внаслідок кумулятивної полімерії.
У потомстві гібридів, одержаних від схрещування S.verrucosum і
S.chacoense, Пропах (цит. за [29]) виявив наявність двох менделюю-
чих генів забарвлення віночка: An,nf - домінантного але ля синьо-
фіолетового забарвлення, рецесивний алель якого aninf зумовлює біле
забарвлення.
При схрещуванні клонів S.rybinii з синіми і S.rybinii з білими
віночками було виявлено [50] біфакторіальну природу забарвлення
віночка. Синє забарвлення віночка детермінується домінантним але-
лем Р, а червоне - полімерним геном R, який за наявності рецесивно¬
го гена р зумовлює червоне забарвлення віночка. Біле забарвлення
контролюється домінантним алелем Rpw у гомозиготному стані або в
присутності домінантного гена Р гомозиготним локусом г. Блакит¬
ний колір віночка є результатом взаємодії домінантних неалельних
генів P і Rpw Rpw.
Отже, у диплоїдних видів картоплі забарвлення віночка зумов¬
люється всіма відомими типами взаємодії генів, які контролюють
синтез пігментів. Рідше зустрічаються випадки простої міжалельної
взаємодії генів. У більшості ж випадків колір віночка є результатом
комплементарної, епістатичної та полімерної взаємодії генів. При¬
чому, як правило, синє забарвлення є домінантним відносно черво¬
ного й білого, а червоне домінує над білим.
Форма бульб. У диплоїдних видів має моногенну природу. Округ¬
ла форма домінує над видовженою.
Забарвлення бульб. У диплоїдних видів зустрічається біле, жов¬
те, рожеве, червоне, синьо-фіолетове забарвлення.
Жовте забарвлення бульб домінує над білим, і в потомстві гібри¬
дів між S.rybinii з S.goniocalyx спостерігається розщеплення у спів¬
відношенні 3:1 [43].
K.Dodds і D.Long [50] встановили, що забарвлення бульб у диплоід-
них видів контролюється трьома незалежними локусами - Р Д (з серією
множинних алелів) та І, з яких перші два діють плейотропно, зумовлю¬
ючи одночасно забарвлення бульби й віночка. Епістатичний рецесивний
алель і в гомозиготному стані зумовлює забарвлення бульб.
120
Забарвлення бульб у диплоїдних видів картоплі, як і в сортів тет-
раплоїдних культурних видів, перебуває в корелятивній залежності з
забарвленням паростків і віночка. Хоч у потомстві гібридів, одержа¬
них від схрещування тетраплоїдних сортів, спостерігаються інші
співвідношення фенотипів за спектром кольорів.
За забарвленням бульб, паростків і віночка у диплоїдних видів
картоплі було визначено [29] такі генотипи основних фенотипічних
класів (табл.3.1).
Таблиця 3.1 - Кореляція забарвлення бульб, паростків та квіток
Забарвлення
Можливий
генотип
бульб
паростків
віночка
синє
синє
синє
P-R-I-
червоне
червоне
червоне
рр R -1 -
біле
синє
синє
P-R-ii
червоне
червоне
рр R - іі
рожеве
рожеве
біле
рр RpwRpwI-
синє
синє
світло-синє
P-RpwRpwI-
Хімічна природа пігментів у диплоїдних видів картоплі була ви¬
явлена методом хроматографії англійськими дослідниками К.Дотсом
і Д.Лонгом ще в 1956 р.
Синє забарвлення бульб зумовлюється наявністю пеонідину й пе-
тунідину, а синій колір віночка - окрім пеонідину й петунідину, ще й
наявністю ціанідину. Червоний колір бульб проявляється в інших
генотипів за синтезу пігментів пеларгонідину і піонідину, тоді як та¬
кий самий колір віночка зумовлюють ціанідин і пеонідин (генотип
ppR -). Наявність у клітинах шкірки бульб лише пеонідину, за відсу¬
тності інших пігментів, фенотипічно проявляється рожевим забарв¬
ленням (генотип рр RpwRpwl - ), а у віночка - білим. Отже, дія генів R
і Rpw неоднозначна для бульб і віночка.
Самонесумісність і перехресна несумісність. Статевий процес
у рослин контролюється генетичними системами несумісності. Тер¬
мін “несумісність” означає нездатність пилкових трубок проникати
по тканині стовпчика й привносити до зародкового мішка гамети і
здійснювати запліднення. Однак, системи несумісності не слід ото¬
тожнювати з різними формами стерильності, коли нездатність утво¬
рювати насіння зумовлюється хромосомними або різного роду фізіо¬
логічними аномаліями.
У природі широко розповсюджене явище самонесумісності гер¬
мафродитних квіткових рослин, коли при самозапиленні не утворю¬
ється насіння. Цим самим у процесі еволюції створилися умови для
перехресного запилення й підтримання гетерозиготності.
Несумісність при самозапиленні називається самонесумісністю,
а та, що спостерігається при перехресному запиленні в межах виду
чи споріднених видів, - перехресною несумісністю.
121
Явище несумісності контролюється геном S і серією його алелів
Si, s2, S3> S4... sn [14]. До сьогодні ідентифіковано 13 алелів локусу S,
які контролюють самонесумісність у диплоїдних видів картоплі:
S.rybinii, S. phureja, S.chacoense, S.soukupii, S.sparsipilum, S.canasense,
S.subtilius, S.vernei, S.megistacrolobum, S.stenotomum та в диплоїда
S.tuberosum L. До самосумісних видів віднесені S.polyadenium,
S.morelliforme, S.verrucosum, S.brevidens. Численні зразки культурно¬
го виду S.rybinii можна вважати самосумісними і сумісними, оскіль¬
ки вони здатні до самозапилення і перехресного запилення [8]. Крім
того, виявлена дволокусна система несумісності (S і R) у диких видів
S.araccpapa, S.ehrenbargii, Spinnatisectum та S.bulbocastanum.
За принципом генетичного контролю системи несумісності поді¬
ляють на декілька типів. Для картоплі характерний гаметофітний тип
несумісності, який виявляється при проростанні пилкових трубок у
стовпчику й зумовлюється незалежною дією одного алеля в пилко¬
вому зерні й двох алелів несумісності у диплоїдній тканині стовпчи¬
ка без взаємодії домінування між ними як у стовпчику, так і в пилко¬
вому зерні. Якщо пилкове зерно має алель локусу S, який не є іден¬
тичним алелям маточки, то пилкова трубка проростає нормально.
Наприклад, якщо пилкові зерна несуть алелі Sb S2, а маточка має
алелі S3 і S4, то несумісності не спостерігається (рис.З.Іа). При гомо-
логічності алелів пилкових зерен (SbS2) і маточки (SbS2) пилкові зе¬
рна не проростають і запліднення не відбувається (рис.3.16), тобто
проявляється несумісність.
При схрещуванні
рослин, які різняться
між собою за одним
алелем S (наприклад,
Sj S3 х Si S2), норма¬
льно проростають на
приймочці маточки
лише пилкові зерна з
алелем S2 (рис.З.Ів).
Більшість диплоїдних
видів Solanum часто
самонесумісні і за¬
пліднюються пере¬
хресно. Однак повна
cff}
SlS,
Рис.3.1. Схематичне зображення несумісності
у диплоїдних видів картоплі
самонесумісність зустрічається рідко. Навіть при вивченні популя¬
цій, здавалося б, типових самонесумісних видів виявлено практично
всі переходи від повної самонесумісності до 100%-ної самосумісно-
сті [7,8].
Чітко виражена міжвидова несумісність проявлялася у схрещува¬
них сортів з мексиканськими диплоїдними видами S.cardioplyllium
Lindi, S.jamessi Torr, S.pinnatisectum Dun.
122
Перехресну сумісність і несумісність у межах виду або спорідне¬
них видів часто пов’язують із загальною гаметофітною системою
несумісності. При внутрішньовидових і міжвидових схрещуваннях у
ряду рослин, включаючи диплоїдні види картоплі, виявляють суміс¬
ність у таких комбінаціях: самонесумісний х самонесумісний, само-
сумісний х самосумісний, самосумісний х самонесумісний; несуміс¬
ність - у комбінації самонесумісний х самосумісний [29]. Так, перше
покоління від міжвидових схрещувань дикого самонесумісного
S.canasense і самосумісних форм S.rybinii, а також культурних само-
несумісних зразків S.phureja х S. stenotomum набувало здатності до
самосумісності [8].
Унікальні розміри серії алелів у локусі S у рослин вказують на те,
що алелям несумісності властива висока частота мутацій. У видів
Nikotiana, Trifolium pratense давно вже відома спонтанна мутація са-
монесумісних алелів до самосумісності (S -+-S2). У популяціях само-
несумісних видів Solanum іноді вдається виявити як спонтанні, так і
індуковані генні мутації самофертильності. Пилок мутантів Sf - типу
здатний проростати у будь-яких стовпчиках рослин з алелями несу¬
місності від Si до Sn.
Як спонтанні геномні мутації, так і експериментальна поліплоїдія
можуть порушувати самонесумісність гаметофітного типу до само¬
фертильності. Доказом цього є те, що спонтанні поліплоїдні види
картоплі (2П=36, 48, 60, 72), лише за невеликим винятком
(iS.tugerrense, 2П=48), самофертильні. Долати бар’єри несумісності
певною мірою дають змогу методи, спрямовані на подолання не¬
схрещуваності видів (див. Методи подолання несхрещуваності).
3.2. ГЕНЕТИКА ПОЛШЛОЇДНИХ
ВИДІВ КАРТОПЛІ
Поліплоїдія зіграла виключно важливу роль в еволюції роду
Solanum L. Поліплоїдія є геномною мутацією, зумовлена кратним
збільшенням кількості хромосом. Геном - це сукупність хромосом
гаплоїдного (гаметичного) набору (А), характерного для конкретного
біологічного виду. У соматичних клітинах диплоїдного виду міс¬
титься по два геноми (АА).
Поліплоїди за походженням поділяють на автополіплоїди й ало-
поліплоїди. Автополіплоїди виникають безпосередньо від диплоїдів
у результаті подвоєння наборів хромосом у клітинах соматичної тка¬
нини або злиття нередукованих гамет одного й того самого виду.
Прийнято розрізняти збалансовані поліплоїди з парною (4п, 6п і
т.д.) і непарною (3 п, 5 п, 7 п) кратністю хромосом.
У триплоідних (2п=36) і пентаплощних (2п=60) видів мейоз прохо¬
дить з відхиленнями від нормального, пилок стерильний, а тому вони не
утворюють плодів і насіння й розмножуються тільки вегетативно.
123
У тетраплоідів (2п=48) і гексаплоїдів (2п=72) мейоз проходить
нормально, пилок фертильний. Вони утворюють плоди й насіння і
можуть розмножуватися статевим шляхом.
Алополіплоїди (гібридні поліплоїди, або амфідиплоїди), на від¬
міну від автополіплоїдів, несуть набори хромосом двох і більше спо¬
ріднених видів, одержаних у результаті гібридизації з наступною по-
ліплоїдизацією. Справжні алополіплоїди можуть виникати лише при
гібридизації видів, хромосоми яких через істотні відмінності не зда¬
тні до кон’югації.
Алополіплоїди, які містять геноми різних видів з гомологічними
сегментами хромосом або навіть цілими хромосомами, називають
сегментними. Основними показниками, за якими можна визначати
тип поліплоїдії, є особливості мейозу (кон’югація хромосом й утво¬
рення бівалентів у профазі-І мейозу) і характер розщеплення.
Оскільки алополіплоїди виникають при гібридизації між генети¬
чно диференційованими видами, то первинний диплоїдний гібрид, з
якого формується поліплоїд, є стерильним. Подвоєння кількості хро¬
мосом (спонтанно чи штучно) у такого стерильного гібрида Fi
відновлює нормальну їх кон’югацію і певну фертильність. Хоча на¬
бори хромосом від двох різних видів не є гомологічними і не
кон’югують між собою, нормальна кон’югація відновлюється між
гомологічними хромосомами кожного виду після їх подвоєння, вна¬
слідок чого спостерігається повна фертильність.
Найширше розповсюдження мають сорти, що належать до тетра-
плоїдних видів. Тетраплоїди представлені двома типами - авто- й
алотетраплоїдами.
Автотетраплоїди (2п=48). На сьогодні більшість генетиків ви¬
знає, що з тетраплоїдних видів S. tuberosum і S. andigenum є автотет-
раплоїдами. Принципове значення для розуміння суті автотетраплої-
дії має вивчення поведінки хромосом у мейозі. В клітинах тетраплої-
да міститься чотири одиниці кожної хромосоми. Якщо у диплоїда
(2п=24) в профазі-І мейозу утворюється 12 бівалентів, то в автотет-
раплоїда (2п=48) може виникнути 12 квадривалентів - утворень, які
складаються з чотирьох гомологічних хромосом, що кон’югують між
собою. Однак, у момент кон’югації не всі хромосоми знаходять своїх
гомологів, тому, крім квадривалентів, можуть утворюватися трива-
ленти, біваленти та уніваленти. Середня кількість квадривалентів на
одну клітину, як правило, менша від максимально можливої. Квад-
ривалентна кон’югація є обов’язковою умовою утворення збалансо¬
ваних (диплоїдних) гамет. Через порушення кон’югації хромосом і
утворення три-, ди- та унівалентів можуть формуватися анеуплоїдні
спори, що призводить до зниження фертильності тетраплоїдів. Сте¬
рильність пилку проявляється сильніше, ніж яйцеклітин.
У селекції картоплі, особливо при застосуванні віддаленої гіб¬
ридизації, багато диких і культурних видів переводять на тетрапло-
124
їдний рівень, тобто створюють експериментальні тетраплоїди. Ус¬
пішне їх подальше використання в селекційному процесі значною
мірою залежить від знання особливостей тетрасомічного розщеп¬
лення та характеру успадкування селекційно і господарсько цінних
ознак.
Огляд результатів досліджень, проведених багатьма авторами,
особливостей розщеплення в автотетраплоїдного виду S.tuberosum
зроблений І.М.Яшиною і Н.П. Скляровою [47].
Характер розщеплення у поліплоїдів зумовлений наявністю у них
двох гомологів кожної пари хромосом і визначається особливостями
їх кон’югації. Так, у тетраплоїдів порівняно з диплоїдами при роз¬
щепленні в F2 спостерігається інше співвідношення домінантних і
рецесивних форм за простими менделюючими ознаками.
Диплоїд, гетерозиготний за однією парою алелів (Аа), у Fi утво¬
рює два типи гамет - А й а, тому в F2 при повному домінуванні дає
розщеплення за фенотипом 3:1, а за генотипом - ІАА : 2Аа : Іаа.
У автотетраплоїдних форм при схрещуванні рослин АААА х аааа
в Fj одержують гетерозиготи ААаа, в яких у результаті мейозу фор¬
муються диплоїдні мікро- і макроспори на основі рівновірогідного
парного розходження чотирьох гомологічних хромосом в анафазі-І,
що можна зобразити схематично:
аі ^ а2
Отже, у процесі гаметогенезу гібриди Fi (ААаа) можуть утворю¬
вати по три типи гамет у співвідношенні ІАА : 4Аа : Іаа. При рівно-
вірогідному злитті цих гамет у процесі запліднення в автотетраплої¬
дних рослин F2 можуть утворюватися п’ять груп генотипів: 1АААА :
8АААа : 18ААаа : 8Аааа : Іаааа, тобто:
ІАА
4 Аа
1 аа
1 АА
ІАААА
4 АААа
1 ААаа
4 Аа
4 АААа
16 ААаа
4 Аааа
1 аа
ІААаа
4 Аааа
1 аааа
У F2 за однією парою алелів у автотетраплоїдів спостерігається
розщеплення за фенотипом у відповідності 35А... : Іаааа.
У диплоїдних форм розщеплення в F2 за двома парами менделю-
ючих алелів дає співвідношення фенотипів 9 : 3 : 3 : 1, а в автотетра¬
плоїдів - 1225 : 35 : 35 : 1.
125
Автотетраплоїди, залежно від кількості домінантних генів, у ге¬
нотипі мають відповідні назви: з чотирма АААА називають квадри-
плексами (А4), з трьома - АААа - триплексами (А3), з двома - ААаа -
дуплексами (А2), з одним - Аааа - симплексами (А1), аааа - нуліплек-
сами (а4) [31].
В автотетраплоїдів при схрещуванні триплексів між собою, з ду¬
плексами, симплексами та нуліплексами одержують одноманітне за
фенотипом потомство, яке за генотипом утворює від двох до чоти¬
рьох класів з різними співвідношеннями. Схрещування дуплексів
між собою, із симплексами та нуліплексами при повному домінуван¬
ні у потомстві дає дві групи фенотипів з різними співвідношеннями
(табл.3.2).
Таблиця 3.2 - Розщеплення за генотипом і фенотипом у потомстві
автотетраплоїдів [4]
Розщеплення потомства
Генотипи
Типи гамет
за генотипом
за фено¬
типом
А:а
АААа х АААа
(АА+Аа) х (АА+Аа)
1 АААА:2 АААа: 1 ААаа
усі А
АААа х ААаа
(АА+Аа) х (АА+4Аа+аа)
1 АААА:5АААа:5ААаа: 1 Аааа
те саме
АААа х Аааа
(АА+Аа) х (Аа+аа)
1 АААа:2ААаа: 1 Аааа
АААа х аааа
(АА+Аа) х аа
1ААаа:1Аааа
tt
ААаа * ААаа
(АА+4Аа+аа) х (АА+4Аа)
1 АААА: 8 АААа: 18 ААаа: 8 А
ааа:Іаааа
35:1
ААаа х Аааа
(АА+4Аа+аа) х (Аа+аа)
1 АААа:5ААаа:5Аааа: Іаааа
11:1
ААаа х аааа
(АА+4Аа+аа) х аа
1 ААаа:4 Аааа: 1 аааа
5:1
Аааа х Аааа
(Аа+аа) х (Аа+аа)
1 ААаа:2Аааа: 1 аааа
3:1
Аааа х аааа
(Аа+аа) х аа
1 Аааа: Іаааа
1:1
За неповного домінування фенотипічний прояв ознаки і особли¬
вості розщеплення F2 в автотетраплоїда буде відповідати розщеплен¬
ню за генотипом ІАААА : 8АААА : 18ААаа : 8ААаа : Іаааа. Чисель¬
ність можливих комбінацій генів визначається загальною кількістю
класів генотипів, що виникають у потомстві симплексного і дуплек¬
сного моногібридів (табл.3.2). При полігібридних схрещуваннях кі¬
лькість можливих комбінацій генів і класів генотипів різко зростає.
Тому навіть у випадку дигібридного схрещування відібрати у потом¬
стві дуплекса домінантний ^гомозиготний за двома неалельними ге¬
нами генотип дуже важко. Ймовірність його появи становить 1/1296,
тобто виявити його практично неможливо.
Отже, автотетраплоїдний рівень значно утруднює добір домінан¬
тних і рецесивних гомозигот за декількома неалельними генами в
потомстві автотетраплоїдів і робить його практично нездійсненним,
оскільки сама поява їх малоймовірна. Тому, користуючись наведе¬
126
ними у таблиці 3.3 формулами, можна прогнозувати появу можливих
комбінацій генів, генотипів і планувати об’єм роботи. У практичній
селекції S.tuberosum на автотетраплоїдному рівні завдання певною
мірою полегшується тим, що більшість господарсько цінних ознак
контролюється домінантними або полімерними генами. Крім того,
численні сорти картоплі за рядом господарсько цінних ознак, що ус¬
падковуються моногенно, є симплексними генотипами і дають у по¬
томстві розщеплення, яке збігається з простими менделівськими від¬
ношеннями 3 : 1 і 1 : 1 [47].
Таблиця 3.3 - Формули для визначення кількості типів гамет, класів генотипів
і фенотипів, можливих комбінацій генів і загальна формула
розщеплення у симплексних і дуплексних автотетраплоїдів
Кількість
пар
альтер¬
нативних
ознак
Кількість
Загальна формула
‘розщеплення А: а
ТИПІВ
гамет
класів
можливих
комбінацій
генів
при само¬
запи¬
ленні
при аналізу¬
ючому схре¬
щуванні
геноти¬
пів
фено¬
типів
Симплекс Аааа
1
2
3
2
4
3 : 1
1 : 1
2
4
9
4
16
(3 : 1)2
(1 •' 1) 2
3
8
27
8
64
О : 1)3
(1 : 1) 1
п
2“
3“
2“
4°
(3:1)”
(1:1)”
Дуплекс ААаа
1
6
5
2
36
35 : 1
5 : 1
2
36
25
4
1296
(35: І)2
(5: І)2
3
216
125
8
46656
(35: І)1
(5: І)3
п
6“
5“
2“
36“
(35 : 1)”
(5:1)”
Особливості розщеплення поліплоїдів пов’язані з наявністю бі¬
льше двох гомологів кожної хромосоми і визначаються характером
їх кон’югації. Співвідношення типів гамет (табл.3.4), які утворюють
поліплоїди, визначається насамперед наявністю або відсутністю зче¬
плення між досліджуваним геном і центромерою. За певного зчеп¬
лення ген-центромера розщеплення відбувається за хромосомним
типом, тобто гомологічні хромосоми поводяться як одиниці, що
комбінуються випадково.
Іншим варіантом розщеплення поліплоїдів є хроматидне, коли
ген не є зчепленим з центромерою і випадково комбінуються не
хромосоми, а хроматиди. У такому випадку співвідношення гамет і
їх типи значно різняться від таких, що формуються за хромосомного
розщеплення (табл.3.4).
Хромосомне і хроматидне розщеплення часто розглядають як
крайні варіанти і порівнюють з ними співвідношення, що спостері¬
гаються в потомстві гібридів поліплоїдів. Подібні порівняння досить
умовні, тому що виключно хромосомне розщеплення спостерігається
127
дуже рідко за абсолютного зчеплення ген-центромера. Виключно
хроматидному розщепленню запобігає часткове зчеплення генів з
центромерами, вплив на особливості розщеплення у поліплоїдів по¬
ведінки гомологічних хромосом у мейозі (тип кон’югації - мульти-
валентна чи бівалентна) і характер розходження в анафазі-1.
Таблиця 3.4 - Співвідношення типів гамет у тетраплоідів [20]
Тетраплоїд
Генотип
Співвідношення типів гамет при розщепленні
хромосомному
хроматидному
Квадриплекс
АААА
АА
АА
Триплекс
АААа
1АА: ІАа
15АА : 12Аа : Іаа
Дуплекс
ААаа
1АА : 4Аа : ІАа
ЗАА : 8Аа : Заа
Симплекс
Аааа
ІАа: Іаа
ІАА : 2Аа : Іаа
Нуліплекс
аааа
аа
аа
Послідовність подій, внаслідок яких, наприклад, у тетраплоїда
триплекса (АААа) утворюються гамети аа, називається подвійною
редукцією. За хромосомного розщеплення поява такого типу гамет у
триплекса неможлива (див. табл.3.3).
Для здійснення подвійної редукції необхідна регулярна
кон’югація квадривалентами. Оскільки для центромери перший ме¬
йотичний поділ (анафаза-1) завжди редукційний, частота редукції за
маркером при першому діленні залежить від відстані між геном і
центромерою, а отже, від частоти кросинговеру на цій ділянці. У ме-
йозі-І кросоверні хроматиди можуть відійти до одного полюса. Лише
в такому випадку обидві хроматиди з рецесивними алелями можуть
потрапити в одну гамету при екваційному поділі в мейозі-ІІ, у ре¬
зультаті чого з’явиться гомозиготна гамета аа. Отже, редукція за ге¬
ном А/а проходить протягом двох послідовних поділів мейозу, звідси
й термін “подвійна редукція”.
Теоретичні розрахунки розщеплення у поліплоїдів, наведені в
таблиці 3.3, є ідеальними ситуаціями. Насправді ж параметри по¬
двійної редукції мінливі. Частота кросинговеру, характер кон’югації
і розходження хромосом знаходяться під генетичним контролем, а
отже варіюють залежно від генотипу та дії факторів середовища.
У сортів, що належать до автотетраплощного виду S. tuberosum,
хроматидне розщеплення було виявлено за генами D і R, які контролю¬
ють антоціанове забарвлення різних частин рослин, а також за геном Nx,
що детермінує летальну некротичну реакцію на вірус X [47].
Алотетраплощи (2п = 48). З алотетраплоїдів найперше викорис¬
товувалися і найбільш вивченими є S.stoloniferum, S.akaule, S.antipo-
viezii, S.ajuskoense та ін. Вважається, що вони виникли внаслідок
спонтанної гібридизації диплоїдних видів з наступним подвоєнням
кількості хромосом. Мейоз у них проходить правильно з утворенням
24 бівалентів, тип успадкування дисомічний.
128
У генетичному відношенні алотетраплоїди є функціональними
диплоїдами з парами повністю гомологічних хромосом кожного з
обох видів, об’єднаних в одному генотипі. Генотип гіпотетичного
алотетраплоїдного виду прийнято позначати формулою ААВВ. Оскі¬
льки у картоплі алотетраплоїди виникають внаслідок об’єднання ка¬
ріотипів генетично близьких видів, то користуються такою форму¬
лою: А1А1А2А2. Тому гетерозиготність в алотетраплоїдів може мати
декілька типів: Аіа]А2а2, Аіа]А2А2, Ajaia2a2, аіаіА2А2, aiajA2a2, а отже
утворювати різні типи гамет.
Для визначення типів гамет можна користуватися наступним
правилом: гомологічні хромосоми, які несуть алельні гени, потрап¬
ляють за умови перебігу нормального мейозу обов’язково в різні га¬
мети, а об’єднання різних алелів в одній гаметі здійснюється за зако¬
нами імовірності. Наприклад, для гетерозиготи AjaiA2a2 утворення
можливих типів гамет можна зобразити схематично:
Отже, цей генотип може формувати чотири типи гамет: Аіа2,
Аіа2, аі А2 й аі а2. Гетерозиготні алотетраплоїдні форми Аіа2Аіа2 при
самозапиленні або при схрещуванні між собою (АіаіА2а2 х АіаіА2а2)
дають розщеплення за фенотипом у співвідношенні 15:1, а при аналі¬
зуючому схрещуванні (AjaiA2a2 х а}аі а2а2) -3:1. Гетерозиготні фор¬
ми типу Аіаіа2а2 у першому випадку (Аіаіа2а2 х Аіаіа2а2) - 3:1, а в
другому (Аіаіа2а2 х а^а^) -1:1, відповідно.
Кількість типів гамет, фенотипів і генотипів, можливих комбіна¬
цій генів та співвідношення розщеплення в алотетраплоїдів можна
визначити за загальноприйнятими формулами (табл.3.5), а для полег¬
шення визначення кількості фенотипів і генотипів за кожним геном
користуються решіткою Пеннета. Однак при аналізі більше трьох
альтернативних пар ознак розписати можливі комбінації практично
неможливо, тому слід користуватися наведеними формулами.
Пентаплоїди (2п = 60) мають гібридне походження . До цієї гру¬
пи належать дикі види S. x semidemissum і S. x edinense, а також
культурний S. x curtilobum. Допускається, що вид S. x semidemissum є
результатом спонтанної гібридизації гексаплоїдного виду S.demissum
і диплоїдного S.verrucosum, причому гамета S.verrucosum була нере-
дукованою. Вид S. x edinense виник у результаті злиття редукованих
гамет тетраплоїдного виду S.andigenum і гексаплоїдного S.demissum.
Пентаплоїд S.curtilobum є результатом злиття нередукованої гамети
триплоїдного виду S. juzepczukii з нормальною гаметою S.andigenum
5 2*132
129
Таблиця 3.5 - Кількість типів гамет, фенотипів і генотипів, можливих
комбінацій генів і загальна формула розщеплення
алотетраплоїдів різних типів
Кількість
пар альтер¬
нативних
ознак
Кількість
Загальна формула роз¬
щеплення при
типів
гамет
класів
можливих
комбіна¬
цій генів
генотипи
феноти-
пи
самозапи¬
ленні
аналізуючо¬
му схрещу¬
ванні
Ai at а2 2
і2
1
2
3
2
4
3:1
1:1
2
4
9
4
16
(3:1)2
(1:1)2
3
8
27
8
64
(3:1)3
(1:1)3
n
2°
Зп
2 п
4 п
(3:1)”
(1:1)"
Аі а} А2 а2
1
4
9
2
16
15:1
3:1
2
16
81
4
256
(15:1)*
(3:1)2
3
64
729
8
4096
(15:1)
(3:1)3
16п
4 п
9 п
2 0
(15:1)”
. „(3:1)"
за спонтанної гібридизації. Пентаплоїди мають вищу фертильність,
порівняно з триплоїдами, якщо в гібридизації їх використовувати як
материнські рослини.
З погляду цитології, пентаплоїди вивчені відносно слабо. В основ¬
ному у цих видів проходить нормальна кон’югація, утворення біва¬
лентів з незначною кількістю унівалентів, тривалентів, квадривален¬
тів і пентавалентів [6].
Використання пентаплоїдів у схрещуваних з S.tuberosum є про¬
міжним етапом у селекції картоплі, яка проводиться на основі між¬
видової гібридизації з гексаплоїдами. У штучно одержаних пента-
плоїдних гібридів детально вивчена кон’югація хромосом, яка пере¬
бігає за типом 24 біваленти +12 унівалентів, тому формуються гаме¬
ти, не збалансовані за кількістю хромосом (переважно 24 - 36).
Гексаплоїди (2п=72), як правило, самосумісні і частіше пово¬
дяться як алополіплоїди. Найбільш відомий з семи гексаплоїдних
видів і широко використовується дикий мексиканський фітофторос¬
тійкий вид S.demissum. Цей вид є алогексаплоїдом і має високу цито¬
логічну збалансованість. Попарна аутосиндетична кон’югація хро¬
мосом зумовлює дисомічний тип успадкування ознак.
Алогексаплоїд у гетерозиготному стані АіаіА2а2А3а3 теоретично
може утворювати вісім типів гамет: А]А2А3> АіА2а3, А]а2А3, Аіа2а3,
аіА2А3, аіа2А3, аіА2а3, аіа2а3. За геном, локус якого міститься у всіх
трьох геномах, в алогексаплоїда спостерігається розщеплення за фе¬
нотипом у співвідношенні 63:1. Таке співвідношення відповідає
розщепленню диплоїдів за трьома полімерними генами. Однак, роз¬
щеплення в алогексаплоїда S. demissum не зустрічається взагалі або
130
зустрічається лише за поодинокими генами, спільними для всіх
трьох геномів, тому що у S.demissum два геноми подібні між собою,
а третій відрізняється від них, що можна виразити формулою
АААААіАі або ААААВВ. Автоалогексаплоїди такого типу характе¬
ризуються як диплоїдними, так і автотетраплоїдними співвідношен¬
нями розщеплення залежно від наявності домінантних генів у складі
обох або одного геному - диплоїдного чи автотетраплоїдного.
Відношення генотипних і фенотипічних класів у потомстві ви¬
значається ступенем гетерозиготності геномів.
3.3. УСПАДКУВАННЯ МОРФОЛОГІЧНИХ
І ГОСПОДАРСЬКО ЦІННИХ ОЗНАК
У S.TUBEROSUM
Основні господарсько цінні ознаки у картоплі контролюються
домінантними генами (забарвлення бульб, вічок, квіток, стійкість до
хвороб та ін.) або полігенами (вміст крохмалю, білка та ін.), тому
зростає ймовірність добору гетерозигот з домінантними ознаками.
За рядом ознак сорти картоплі в основному є симплексами, а за
окремими - можуть бути дуплексами і триплексами. Труднощі ви¬
вчення успадкування й розщеплення багатьох ознак пов’язані з авто-
тетраплоїдною природою S.tuberosum, тому що гени, які їх контро¬
люють, можуть бути в різному алельному стані. Тому при аналізі
кожної ознаки у формулі генотипу записується алельний стан гена і
можуть бути всі можливі співвідношення, характерні для розщеп¬
лення автотетраплоїдів.
Генетичний контроль забарвлення бульб і квіток. Найраніше
і досить добре вивчене успадкування забарвлення бульб, вічок і кві¬
ток, що зумовлено відносною (порівняно з іншими ознаками) легкіс¬
тю аналізу цих ознак і можливістю його проведення у рік вирощу¬
вання сіянців.
Огляд робіт з генетики успадкування забарвлення у S.tuberosum,
виконаних до 1937 р., був зроблений Т.ВЛсєєвою, пізніше, до 1945 р., -
М.Свамінатаном і Х.Говардом. Перші експеременти у цьому напрямі
були проведені у 1910 р. Е.Істом і Р. Саламаном [47].
Ще у 1910 р. Е.Іст виявив, що синє забарвлення бульб домінує
над червоним, кожне з них - над білим. Тоді ж Р.Саламан висловив
припущення, що забарвлення бульб контролюється трьома незалеж¬
ними генами: R- контролює червоне забарвлення, Р - синє, D - про¬
яв пігменту.
Пізніше Т.В. Асєєва і Н.В. ЬІіколаєва [1] доповнили цю гіпотезу,
розділивши гени, які контролюють антоціанову пігментацію, на гени
основи і гени прояву. Гєни основи P і R детермінують характер піг¬
ментів: Р зумовлює синьо-фіолетову, R- червоно-фіолетову пігмен¬
тацію. Характер розподілу пігментів залежить від наявності компле¬
131
ментарних генів прояву і може проявлятися в забарвленні бульб, кві¬
ток, вічок, різних частин стебла.
Пізніше, з нагромадженням експериментальних даних, ці гіпоте¬
зи були уточнені й доповнені. Сьогодні визнається, що забарвлення
контролюється двома групами комплементарних генів: генами осно¬
ви P і R і генами прояву E, F, D, S. Комплементарні гени P і R у при¬
сутності гена D (генотип Pp3Rr3Dd3) зумовлюють синьо-фіолетове, а
гена D (генотип Рр3 Dd3 г4) - синє забарвлення бульб. Червоне забар¬
влення бульб будуть мати рослини з генотипом р4 Rr3 Dd3. У рослин,
які у своєму генотипі мають лише один домінантний ген (Рр3 r4d4>
p4Rr3d4, р4 rr4Dd3) або всі рецесивні гени (рррр rrrr dddd), будуть білі
бульби.
Всі гени мають дублі, що підтверджує тетраплоїдну природу кар¬
топлі. Більшість сортів має симплексні генотипи за забарвленням
бульб. Наприклад, бульби червоного забарвлення мають генотипи:
рррр Rrrr Dddd; ppppRRrrDDdd; ppppRRRrDddd та ін., тобто від сим¬
плекса до квадриплекса за генами R і D. Враховуючи сутність понят¬
тя “доза гена”, можна пояснити причину різної інтенсивності забарв¬
лення. За значної громіздкості повних генетичних формул у автотет¬
раплоїдів застосовують їх у скороченому вигляді (Рр3 Rr3Dd3, або
P—R- D).
Генотипи сортів за генами забарвлення визначають методом гіб¬
ридологічного аналізу потомства за результатами аналізуючого
схрещування, тобто схрещування симплексів з повними рецесивами.
Плейотропна дія генів забарвлення, що зумовлює одночасно піг¬
ментацію різних частин рослини, детермінує чітку кореляційну за¬
лежність між забарвленням бульб, паростків і квіток. Ця залежність є
основою сортової систематики картоплі, якою користуються в селек¬
ційній і насінницькій роботі та генетичних дослідженнях.
В Україні сорти за забарвленням квіток і бульб поділяють на
п’ять груп.
І. Квітки білі, бульби білі або жовті: Віхола, Мавка, Незабудка та ін.
П. Квітки червоно-фіолетові, бульби білі: Житомирянка, Зов, Льві¬
в’янка та ін.
ПІ. Квітки червоно-фіолетові, бульби червоні чи рожеві: Зарево,
Ікар, Поліська рожева, Світанок київський.
VI. Квітки білі, бульби червоні або рожеві: Луговська.
V. Квітки синьо-фіолетові, бульби білі: Пригожа 2.
Забарвлення м’якуша бульби у картоплі буває біле, жовте, ро¬
жеве й синє. У селекційних сортів воно варіює від білого до світло-
жовтого.
В результаті численних схрещувань давно доведено (Salaman R.,
1910,1926; Fruwirth С., 1912; Okuno S., 1952; Schick R., 1956), що жов¬
те забарвлення м’якуша домінує над білим [46]. Ряд авторів того ча¬
су вважали, що жовте забарвлення контролюється одним домінант¬
132
ним геном, а біле - рецесивним алелем. Тепер визнано, що жовте
забарвлення зумовлюється дією полімерних генів, а інтенсивність
прояву його залежить від дози домінантних генів. Тому при схрещу¬
ванні сортів з світло-жовтим м’якушем у потомстві можуть появля¬
тися рослини з жовтим м’якушем. І навіть від схрещування сортів з
білим м’якушем іноді з’являються бульби із світло-жовтим м’я¬
кушем.
Червоне і синє забарвлення м’якуша бульби визначається ком¬
плементарною взаємодією неалельних генів С і У. Наявність в одно¬
му генотипі цих двох генів зумовлює утворення пігменту, а ген Z
пригнічує його синтез.
Форма бульб. Форма бульб значною мірою залежить від умов
вирощування картоплі. За формою вони бувають округлі (поздовж¬
ній і поперечний діаметри майже однакові); видовжені (поздовжній
діаметр більший за поперечний не менше ніж у 2,5 рази); овальні,
округло-овальні; видовжено-овальні та ін. Господарське значення
мають лише бульби округлої, овальної та видовженої форми.
Округла форма бульб домінує над округло-овальною і видовже¬
ною і контролюється полімерними генами. Роботи з вивчення гене¬
тики форми бульб нечисленні, в основному 30-50-х років XX століт¬
тя. Гени, що контролюють форму бульб, до даного часу ще не іден¬
тифіковані. Вважають, що переважна більшість сортів є гетерозигот¬
ними за цими генами, а тому як в інцухт-поколіннях, так і у
міжсортовій внутрішньовидовій гібридизації спостерігається поява
форм з округлими, округло-овальними, овальними, видовжено-
овальними бульбами. Крім того, частина генів контролю форми
бульб має проміжну дію.
Глибина вічок. Вічка на бульбі розміщені спірально. На верхній
частині бульби їх більше, на нижній - менше. В одних сортів вічок
на бульбі багато, в інших - мало. Розташовані вони поверхнево або
глибоко, по всій бульбі чи на верхівці. Отже кількість, характер роз¬
міщення і глибина вічок зумовлені генотипом сорту. Хоч до цього
часу відсутні чіткі дані про закономірності успадкування глибини
вічок.
Висловлювалися припущення про домінування глибини вічок і,
навпаки, що глибокі вічка є рецесивною ознакою. Зустрічаються
припущення, що дрібні поверхневі вічка є результатом комплемен¬
тарної взаємодії генів, інші допускають, що глибина вічок успадко¬
вується полігенно. Гени глибини вічок не ідентифіковані. Хоч
округла і округло-овальна форма бульб і поверхневі вічка без особ¬
ливих труднощів комбінуються з іншими господарсько цінними
ознаками [46].
Вміст крохмалю у бульбах варіює залежно від умов вирощуван¬
ня, однак зумовлюється генетично. Істотні спадкові відмінності за
цією ознакою спостерігаються у різних сортів.
133
Вивчаючи ступінь мінливості крохмалистості ЗО сортів і гібридів
картоплі залежно від умов вирощування, А.М. Кадичегов [21] ви¬
явив, що фенотипічна мінливість вмісту крохмалю зумовлена гено¬
типними відмінностями становила лише 24%.
Крохмалистість є полігенною ознакою й успадковується адитив¬
но, тобто розщеплення виражається безперервним варіаційним ря¬
дом, характерним для кількісних ознак, без дискретних переходів від
мінімального до максимального ступеня прояву. Така тенденція по¬
силюється при збільшенні кількості локусів, що контролюють полі-
генну ознаку. При цьому не завжди всі алелі однаково впливають на
прояв ознаки: дія одних алелів може бути сильнішою за дію інших
того самого чи іншого локусу [31]. Тому дія полігенів не завжди бу¬
де адитивною за домінування і взаємодії між локусами.
Доведено, що розщеплення за крохмалистістю потомства від са¬
мозапилення і схрещування селекційних сортів відбувається за ти¬
пом тетрасомії [4], яка властива для дуплексних генотипів (ААаа,
ААААВВЬЬ та ін.) з числовими відношеннями між класами геноти¬
пів варіаційного ряду в потомстві 1:8:18:8:1.
Найвищий вміст крохмалю (24 -26 %) у бульбах картоплі мали
генотипи, які несли по вісім домінантних полімерних генів
(ААААВВВВ). Генотипи ААААВВВЬ містили у бульбах 21-23 %
крохмалю, ААААВВЬЬ - 18-20 % (батьківські компоненти), ААА-
АВЬЬЬ — 14—17 %, ААААЬЬЬЬ - 12-14 %, що було найнижчим показ¬
ником у цьому варіаційному ряду. Отже, залежно від розподілу по¬
лімерних генів, у батьківських форм спостерігається неоднаковий
розподіл кількісних ознак у потомстві з появою позитивних транс¬
гресій (ААААВВВВ), які мають максимальний прояв ознаки, що має
важливе значення в селекції.
Вміст білка також є полігенною ознакою, визначається сумісною
дією багатьох неалельних генів, від кількості яких залежить ступінь
її прояву, хоч значною мірою модифікується, залежно від погодних
умов, типу грунтів, добрив, ураження хворобами та ін.
Скоростиглість - здатність рано зав’язувати бульби й інтенсив¬
но формувати урожай - зумовлена спадковими особливостями. Ско¬
ростиглість є полігенною ознакою [6] й успадковується рецесивно
[44,46]. Хоча були виявлені [11] сорти, в яких у потомстві від само¬
запилення домінувала ознака скоростиглості. Суперечливі дані вка¬
зують на складну природу генетичного контролю тривалості вегета¬
ційного періоду у картоплі. Ця ознака до нинішнього часу вивчена
недостатньо, полімерні гени, які її контролюють, не ідентифіковані.
Численними дослідженнями з вивчення гібридних популяцій за
скоростиглістю доведено, що розщеплення за тривалістю вегатацій-
ного періоду виражається асиметричною кривою, вершина якої змі¬
щена у бік пізньостиглості з втцепленням середньостиглих, серед¬
ньоранніх та ранньостиглих форм. Останні становлять близько 25 %
134
від усього потомства, одержаного від самозапилення ранньостиглих
сортів.
У практичній селекції на скоростиглість успішно застосовують
метод інцухту. При цьому в потомстві, одержаному від самозапилен¬
ня, виявляють гомозиготні форми, які використовують як вихідний
матеріал для гібридизації.
3.4. ГЕНЕТИКА СТІЙКОСТІ
ДО БІОТИЧНИХ ФАКТОРІВ
3.4.1. Стійкість картоплі до патогенів
Здатність протистояти хворобі може проявлятися у формі іму¬
нітету до зараження або у вигляді певного механізму стійкості,
який допомагає рослині уникати природного зараження й ослабляє
його дію.
Імунітетом є несприйнятливість рослин до хвороб у випадку
безпосереднього контакту із збудниками, здатними спричинити
хворобу, за наявністю сприятливих для зараження умов. Імунітет є
абсолютним станом, коли організм може бути або імунним, або не-
імунним. Ступінь же стійкості до хвороби різних генотипів рослин
часто коливається від майже повного імунітету до повної сприйня¬
тливості.
Основоположником генетичної теорії імунетету рослин є М.І. Ва¬
вилов. Він встановив зв’язок між генетичною різноманітністю видів
культурних рослин і їх стійкістю до хвороб, а також показав роль
генетичної диференціації паразитів у стійкості до них рослин.
Взаємозв’язок фітопатогенів і вищих рослин-живителів зумовлю¬
ється складною взаємодією їхніх ядерних і цитоплазматичних струк¬
тур на основі гомеостазу. В ході еволюції у рослин формуються за-
хисно-відновні системи, що захищають їх від генетичних і фізіологі¬
чних пошкоджень в умовах середовища, які постійно варіюють. Ці
системи забезпечують стабільність гетичних структур організму в
умовах дії несприятливих факторів середовища.
Походження й еволюція взаємозв’язків патогену і живителя гру¬
нтується на спонтанних мутаціях в обох компонентів взаємодії. По¬
пуляція патогенів, як і рослин-живителів, постійно знаходиться у
процесі мінливості.
Зміни в генетичній структурі фітопатогенів можуть бути наслід¬
ком різних біологічних процесів. Нові біотипи, штами і раси патоге¬
нів виникають за рекомбінацій як результат статевого процесу, му¬
тацій, гетерокаріозису й парасексуального циклу, а також внаслідок
трасформації, трансдукції, кон’югації та обміну екстрахромосомни-
ми факторами. Роль кожного з цих процесів у мінливості залежить
від особливостей розвитку представників різних груп патогенів.
135
Труднощі вивчення генетики імунітету у рослин полягають у то¬
му, що, на відміну від інших властивостей, стійкість чи сприйнятли¬
вість є результатом взаємодії двох геномів - паразита і рослини. Ці
труднощі ускладнюються ще й тим, що стійкість чи сприйнятливість
рослин залежить від екологічних умов, у яких взаємодіють паразит і
живитель. Ці умови можуть по-різному впливати на рослину, зміню¬
ючи її стійкість, і на паразита, змінюючи його вірулентність.
Поєднана еволюція рослин-живителів і їх патогенів у спільних
центрах походження приводить до виникнення комплементарних
генетичних систем у рослин і паразитів. Наявність у рослин і їх пара¬
зитів генів, що можуть взаємодіяти, експериментально підтвердив і
на підставі своїх робіт висунув гіпотезу “ген проти гена” фітопатолог
Н.Флор у 1956 р. За цією теорією кожному гену, який контролює
стійкість рослини, відповідає специфічний ген, що контролює віру¬
лентність патогену. Іншими словами, на кожний ген живителя, який
здатний мутувати так, щоб забезпечити живителю стійкість, припа¬
дає ген паразита, який мутує таким чином, щоб цю специфічну стій¬
кість подолати. Природний добір сприяв тим мутаціям живителя, які
захищали його від паразита, і тим мутаціям паразита, які долали цю
властивість живителя.
Гіпотеза Ван дер Планка “білок на білок” допускає, що білок па¬
тогену каталізує роботу гена рослини-живителя, який контролює син¬
тез комплементарного білка, зв’язаного з ним, і транспортується з
клітини паразита. При несумісності реакцій білок, що кодується ге¬
ном вірулентності, не сполімеризовується з білком рослини-
живителя і каталізує процеси, які позбавляють патоген елементів
живлення. Тому патоген не розвивається.
Відношення “ген проти гена” підтверджено для численних пар
живитель-паразит, куди входять рослини і збудники хвороб різних
систематичних груп: гриби, бактерії, віруси, комахи, рослини-
паразити. Гени стійкості часто утворюють алельні серії і зчеплені
блоки, а гени вірулентності, як правило, незалежні. Очевидно, не
тільки основні гени стійкості, а й їх модифікатори взаємодіють з мо¬
дифікаторами генів вірулентності за схемою “ген проти гена”. Тако¬
му ж принципу підпорядковується взаємодія малих генів стійкості і
малих генів патогенності.
Фітофторостійкість зумовлюється двома генетичними система¬
ми, одна з яких контролює реакцію надчутливості, тобто здатності
клітин рослини утворювати некроз у відповідь на проникнення пев¬
них рас патогену, а друга - польову стійкість, тобто здатність росли¬
ни протистояти проникненню збудника або обмежувати швидкість
інфекції й інтенсивність спороношення.
У селекції на стійкість проти фітофторозу враховуються як польо¬
вий, так і надчутливий типи стійкості. Однак при доборі перевага нада¬
ється тим генотипам, які поєднують обидва типи стійкості [33].
136
Надчутливість контролюється системою незалежних олігогенів
(major genes - великі, або основні гени), тобто гени, які проявлять
сильну фенотипічну дію. їх присутність у генотипі забезпечує імуні¬
тет до певних рас або групи рас фітофтори. При появі нових рас рос¬
лини уражуються . На відміну від цього типу, польова стійкість зу¬
мовлюється серією полімерних генів (minor genes - малі гени) з ади¬
тивним (кумулятивним) ефектом, різна кількість яких у генотипі за¬
безпечує різний рівень стійкості до всіх рас патогену.
За міжнародною номенклатурою генів стійкості і рас фітофтори
гени позначають символом R, наявність яких виявлена у гібридного
матеріалу, одержаного за участю S.demissum. Расову специфічність у
P .infestans вперше було виявлено ще в 1932 р. у Німеччині. За здат¬
ністю уражувати рослини, раси гриба поділяють на прості і складні.
За міжнародною класифікацією прості раси позначають однією циф¬
рою - 0; 1; 2;3; ... 10; 11 і так далі, а складні - кількома 1.2; 1.3; ...;
2.3; 2.4; ...; 1.2.3; 1.2.4; 2.3.4 і так далі.
Уніфікована міжнародна шкала наглядно ілюструє взаємовідно¬
шення “ген на ген” між генотипом живителя і расами P.infestans
(табл.3.6). Форми, які не мають домінантних генів (птг), уражуються
всіма расами. Ген К\ контролює стійкість до рас, у позначенні яких
відсутня цифра 1, тобто 2; 3; 4; 2.3; 2.4; і так далі. Ген R2 зумовлює
надчутливість до рас 1; 3; 4; 1.3; 1.4; 1.3.4, відповідно. Ураження
можливе, коли патоген позначений усіма цифрами, які збігаються з
індексами генів генотипу рослини. Наприклад, генотип RiR2 уражу¬
ється расами 1.2; 1.2.3; 1.2.3.4, але стійкий до всіх інших рас
(табл.3.6). З поданої схеми випливає, що для кожної раси є генотип-
живитель, на якому вона може проявити максимальну здатність рос¬
ту й спороношення.
Наведена схема наглядно демонструє наявність лише чоти¬
рьох основних генів стійкості, комбінації яких теоретично дають
16 генотипів за стійкістю відповідних їм 16 рас фітофтори. Ціл¬
ком очевидно, що ця схема не є завершеною, оскільки виявляють¬
ся нові R-гени у S.demissum, S. polyadenium, S.pinnatisectum,
S.stoloniferum та ін. Наявність додаткових генів R5 та R$ у
S.bulbocastanum і S.polytrichon, а також фактів виділення склад¬
них рас гриба 1.2.3.4.5; 1.2.3.4.5.6; 1.2.3.4.6 та ін. зумовлюють не¬
обхідність розширення схем в міру виявлення нових R-генів і рас
фітофтори. Проте вона залишається придатною для теоретичного
прогнозу одержання фітофторостійких форм картоплі із залучен¬
ням до гібридизації донорів R-генів.
Стійкість на основі надчутливості успадковується домінантно
за простою менделівською схемою. R-гени, локалізовані в різних
хромосомах, при самозапиленні і гібридизації незалежно комбі¬
нуються в потомстві й підлягають тетрасомічному типу успадку¬
вання.
137
Таблиця 3.6 - Міжнародна схема взаємозв’язків між генами надчутливості
S.demissum і расами P .infestant [10]
Генотип
Раса
0
1
2
3
4
1.2
1.3
1.4
2.3
2.4
3.4
1.2.3
1.2.4
1.3.4
2.3.4
1.2.3.4
г
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
R1
-
+
-
-
-
+
+
+
-
-
-
+
+
+
-
+
R2
-
-
+
-
-
+
-
-
+
+
-
+
+
-
+
+
R3
-
-
-
+
-
-
+
-
+
-
+
+
-
+
+
+
R4
-
-
-
-
+
-
-
+
-
+
+
-
+
+
+
+
R1R2
+
+
+
-
-
-
R1R3
+
+
-
+
-
+
R1R4
+
+
+
-
+
R2R3
-
-
-
-
-
-
-
-
+
-
-
+
-
-
+
+
R2R4
+
+
-
+
+
R3R4
+
+
+
RlR2R3
+
-
-
+
R1R2R4
+
-
-
+
R1R3R4
-
+
-
+
R2R3R4
+
+
R1R2R3R4
+
Примітка. - стійкий; + сприйнятливий
Генотипи з R-генами, як правило, позначаються не повними
формулами, а лише домінантними символами: Rb R2, Ri R2 і так да¬
лі. їх повні формули для сортів S.tuberosum, які є тетраплоїдними,
мають такий вигляд: гпт (нуліплекси), Rirrr (симплекси), RirR2r
(дуплекси) і т.д. (див. табл.3.4). При схрещуванні форм із різними
генотипами в потомстві може утворюватися різна кількість стійких
до фітофторозу рослин (табл. 3.7). Однак реальне співвідношення
форм, одержаних у практичній роботі, з багатьох причин часто не
тотожне з теоретично очікуваним, унаслідок чого збільшується кі¬
лькість нестійких генотипів.
Дія R-генів поширюється головним чином на листки, а бульби
можуть уражуватися. Виявлено, що всі сорти, надчутливі по бульбах,
є такими ж і по листках, а зворотна кореляція - надчутливість по ли¬
стках і надчутливість по бульбах - спостерігається значно рідше. Ген
Ri здебільшого зумовлює надчутливість як листків, так і бульб, а ге¬
ни R2, R3, R4 - надчутливість листків і лише в окремих випадках - од¬
ночасно й бульб.
Наявність і поява нових рас фітофтори значно ускладнює селек¬
цію стійких до неї сортів картоплі, а тому не розв’язує цієї проблеми
на основі надчутливості. У зв’язку з цим постає завдання поєднання
в одному сорті обох типів стійкості - надчутливості і польової стій¬
кості.
138
Таблиця 3.7 - Теоретично очікувані відношення в потомстві від схрещування
симплексних і дуплексних генотипів з різними R- генами [22]
Варіант
схрещування*
Можливий фенотип
Спів¬
відно¬
шення
феноти-
пів R:r
Rj х r
Rj + r
1:1
R2x r
R2 + r
1:1
Rj R2 x r
Rj R2 + Rjr + R2 r + r
3:1
R3R4 X г
R3 R4 + R3r + R» r + r
3:1
Rj x Rj
Ri R2 + Rjr + R2 r + r
3:1
Rj R2 x R,
3R, R2 + 3R,r + R2 r + r
7:1
Rj R2 X R3
RjR2R3 + Rj R2 + R2R3 + Rj R3 + Rj + R2 + R3 +r
7:1
Rj R2 X Rj R2
9 Rj R2 + 3R, + 3R2 +r
15:1
Rj R2 x R3R4
RjR2R3 R4 + RjR2R3 + RjR2R4 + R2R3 R4 + Rj R2 +
+ Rj R3 + Rj R4 + R2R3 + R2R4 + R3 R4 + Rj + R2 + R3 + R4 + Г
15:1
Rj x R1R2R3R4
3RjR2R3 R4 + 3RjR2R3 + 3RjR2R4 + 3R1R3R4 +
+ R2R3 R4 + 3Rj R2 + 3Rj R3 + 3Rj R4 + R2R3 + R2R4+ + 3Rj +
R2 + R3 + R4 + r
31:1
Примітка. * Позначення наведені лише за домінантними генами
Польова стійкість залежить від дії кількох захисних механізмів і
визначається одним із них або всіма разом. Контролюється багатьма
генами, успадковується рецесивно або за проміжним типом, ступінь
прояву модифікується великою кількістю факторів (температура,
вологість, добрива та ін.). Польова стійкість бульб є полігенною
ознакою і не корелює зі стійкістю листків. Тобто, обидві форми стій¬
кості контролюються різними системами полігенів.
Широкий розмах варіювання рівня стійкості у S.berthaultii,
S.chacoense, S.microdontum, S.vernei та ін. вказує на адитивну дію ма¬
лих генів [48]. Джерелами стійкості до фітофторозу є дикі й культур¬
ні родичі сортів. Обидва типи контролю стійкості виявлені у мекси¬
канських видів S.demissum, S.bulbocastanum, S.stoloniferum, S.polia-
denium, S.verrucosoum, S.papita, S.polytrichon та ін. [34].
Успадкування стійкості до раку картоплі. У збудника цієї хво¬
роби гриба Synchytrium endobioticum (Schilb.) Pers., починаючи з
1941 р., виявлено ряд агресивних біотипів. З двадцятих років XX ст.
вивченням успадкування стійкості проти раку картоплі займалося
багато авторів [10,22,44].
Здебільшого стійкість сортів і видів картоплі до раку базується
на реакції надчутливості рослини-живителя. Однак, у деяких сортів
відзначається стійкість до проникнення збудника. До найбільш по¬
ширеного у всіх країнах звичайного (далемського, або 1) біотипу D
стійкість контролюється трьома домінантними генами - X, Y і Z.
139
Ген X - основний, a Y i Z - комплементарні, які діють незалежно
від гена X. Комплементарні гени Y і Z зумовлюють стійкість до раку
лише у тому випадку, коли вони обидва наявні в одному геноти-
пі.Тому іноді в потомстві, одержаному від схрещування сприйнятли¬
вих сортів між собою (генотипи Yyyy X Zzzz), з’являються стійкі до
раку гібриди (Yy Zz). За відсутності комплементарних генів сорт,
який має ген X у симплексному стані (Хххх), теоретично дає розще¬
плення в інцухт-потомстві 3:1 (стійких : сприйнятливих). При схре¬
щуванні симплекса з нестійким сортом (Хххх х хххх) спостерігаєть¬
ся розщеплення у співвідношенні 50 % : 50 %, або 1:1 стійких і
нестійких сіянців. Отже, стійкість до раку картоплі, що
контролюється домінантним геном X, успадковується як проста мен-
делююча ознака. Характер розщеплення у потомстві гібридів,
одержаних від схрещування форм, які різняться за кількістю
домінантних генів, можна передбачити, користуючись таблицею 3.5.
Відомо, що успадкування стійкості до біотипу 2 має значно склад¬
нішу генетичну природу і базується не лише на домінантних, а й на
рецесивних генах. Часто сорти, стійкі до біотипу 2, характеризують¬
ся стійкістю до біотипу 1.
Існування більш агресивних біотипів раку (рахівський, буковинсь¬
кий, міжгірський) потребує пошуку нових джерел стійкості. До цих
біотипів виявлені стійкі форми у S.andigenum, S.acaule, S.vernei, S.car-
dyophyllum, S.stoloniferum, S.chacoense, S.curtilobum, S.maglia та ін.
Генетика стійкості до вірусів у картоплі може бути зумовлена
імунітетом, надчутливістю, польовою стійкістю і толерантністю.
Імунітет і надчутливість до вірусу X відомі у форм S.acule: імунітет
(ген Rx) домінує над чутливістю (Rxn)- ГЄН Rxs контролює утворення
некрозів з мозаїкою і домінує над сприйнятливістю (гх). Тобто, домі¬
нування цих генів можна виразити в такій послідовності: Rx > >
Rxs ^ Гх*
Наявність різних за патогенністю штамів виявлено у різних віру¬
сів, які уражують картоплю. Г. Корекхем (цит. за [15]) виділив чоти¬
ри групи штамів ХВК, які різняться за здатністю долати дію генів
стійкості картоплі Nx і Nb.
Імунітет виявлений лише до вірусів X, Y і А. У S.stoloniferum
стійкість до вірусу Y контролюється серією алелів одного домінант¬
ного гена: Ry > Ryn > Rys > гу, де Ry зумовлює імунітет, R^- надчутли¬
вість, Rys - некротичну реакцію з мозаїкою, гу - сприйнятливість.
Крім того, Ry діє плейотропно, контролюючи також імунітет до віру¬
су А.
Крім S.stoloniferum, стійкі до вірусу Y зразки виявлені у видів
S.vernei, S.chacoense, S. canas ense, S.commersoni та ін.
Тетраплоїдні гібриди, одержані на основі S.stoloniferum методом
багаторазового бекросу на S.tuberosum, як правило, є симплексними
(Ryryryry) формами за геном Ry. Розщеплення у їхньому потомстві за
140
стійкістю до вірусу Y відбувається за простим менделівським спів¬
відношенням 3:1 (стійкі : сприйнятливі) при самозапиленні і 1:1 —
при аналізуючому схрещуванні (Ryryryry х гугугугу),.
Надчутливі до вірусу Y форми виділені серед рослин диплоїдно-
го виду S.commersonii. у селекційних сортів реакція надчутливості до
штамів вірусів X, Y і А проявляється некрозом верхівки і контролю¬
ється домінантними генами N (Ny, Nx, Na, Nc, Ns), дія яких залежить
від пггамового складу вірусів і не забезпечує повного захисту від
них. Лише домінантний ген Na, який контролює надчутливість до
вірусу А, захищає картоплю від нього. Тому селекцію на стійкість
картоплі до вірусу А можна вести на основі S.tuberosum .
Значні перспективи у створенні стійких до вірусів сортів картоп¬
лі відкриваються із застосуванням методів генної інженерії та клі¬
тинної селекції [3]. Методами генетичної інженерії спеціалістами
фірм Galgene і Monsanto були одержані стійкі до вірусних хвороб
форми картоплі. В Інституті картоплярства УААН методом трансфор¬
мації рослин штамом Agrobacterium tumefaciens, що містить плазміду
з геном, який кодує білок оболонки Х-вірусу картоплі (стійкість до
Х-вірусу), і геном NPT-П (ознака канаміцинстійкості), одержані
трансгенні лінії, з яких 56 % мали високу польову стійкість до
Х-вірусу картоплі.
Нематодостійкість у картоплі базується на реакції надчутливос¬
ті і контролюється як олігогенами, так і полігенами. У S.andigenum
виявлено один домінантний ген Нь який проявляє повну пенетрант-
ність, тобто здатний пригнічувати утворення цист до мінімуму (від 0
до 10), порівняно з численною їх кількістю у сприйнятливого сорту.
Ген Ні контролює стійкість до патотипу Ro-І, ген Н2 (у
S.multidissectum Hawk) - до патотипу Ro-2. У S.spegazzinii виявлені
гени Fa, Fb, у S.vernei - гени Ѵь Ѵ2, Ѵ3, Ѵ4, у S.oplocense - гени Оь 02.
Стійкість до ряду рас нематоди успадковується полігенно.
Аналіз літератури з питань генетичного контролю стійкості про¬
ти патогенів приводить до висновку, що абсолютної межі між окре¬
мими типами стійкості немає. Стійкість рослин забезпечується лан¬
цюгом біохімічних реакцій, які, очевидно, не можуть контролювати¬
ся лише одним конкретним геном. Значна частина мінливості за
стійкістю до патогенів, що спостерігається у гібридів, як правило,
зумовлена генетичними причинами, а інша її частина - впливом умов
середовища. Природа функціонального аспекту геному стала зрозу¬
мілою після визнання в біології певного рівня елементарних струк¬
тур гена і відкриття регуляторних його систем. Функціональний про¬
яв гена здійснюється регуляторними системами і сумісною взаємоді¬
єю їх продуктів на різних етапах становлення надорганізмених сис¬
тем. У зв’язку з цим залежність між генотипом і фенотипом не є
однозначною і чіткою, тому не завжди можна точно пояснити при¬
чини варіабельності ступеня стійкості організмів спільного похо¬
141
дження. У процесі формотворення основну роль відіграють зміни прос¬
торово-часових закономірностей експресії генетичного матеріалу. Будь-
яке зрушення в цьому ланцюгу, випадання або, навпаки, включення в
нього нових реакцій дає різний ступінь стійкості рослин, тому що реалі¬
зація гена в ознаку проходить через серію надзвичайно складних біохі¬
мічних процесів, більшість з яких взаємозв’язані.
Саморегуляція ланцюгів метаболічних і морфогенетичних проце¬
сів через взаємодію продуктів генів на різних рівнях біологічної ор¬
ганізації, як реакція на ліміт або надміру факторів зовнішнього сере¬
довища, за своєю суттю є генетичним контролем адаптації. Регулю¬
вання цими потоками здійснюється за рахунок того, що в екосистемі
живитель-паразит “прагнуть” зберегти свій гомеостаз. Крім того,
гомеостаз можна розглядати як універсальну властивість саморегу¬
ляції живого в системі взаємозв’язків між організмами та між організ¬
мами і навколишнім середовищем. Оскільки будь-які прояви гомео¬
стазу генетично детерміновані, він притаманний усім сортам, але
різною мірою.
3.4.2. Стійкість до абіотичних факторів
Генотип, сформований у процесі природного чи штучного добо¬
ру, визначає спадкову пристосованість організму до умов середови¬
ща, за яких проходив його онтогенез. Фактори середовища різнома¬
нітні і мінливі в широких межах, і в процесі еволюції добір створює
спеціальні механізми адаптації.
Генотипна адаптація зумовлюється спадково детермінованими
змінами в генотипі, які забезпечують нормальне функціонування ор¬
ганізму або сорту за конкретних умов навколишнього середовища.
Стійкість культури або сорту до дії специфічних умов середовища
(екстремальних температур, посух, хімічних сполук та ін.) зумовлю¬
ється специфічною адаптивною здатністю. Рівень стійкості, а відпо¬
відно й її механізм, зумовлюються як інтенсивністю дії екстремаль¬
ного фактора, швидкістю його відхилення від норми, так і ступенем
адаптації рослин.
Сорт як відкрита біологічна система в польових умовах завжди
буде зазнавати дії регульованих абіотичних факторів. Тому генетич¬
ний захист продукційного процесу має розглядатися як здатність си¬
стеми включати в морфогенетичний процес будь-який продукт гена
або його комплексів, що призведуть до зміни характеру взаємодії
генів на різних рівнях морфогенетичних процесів, тобто як здатність
перевизначення генетичної організації конкретної ознаки. Труднощі
вивчення генетики адаптацій до абіотичних факторів полягають у
кількісному факторі виявлення ознак стійкості та в їх залежності від
зовнішніх умов середовища.
Пізні приморозки навесні пошкоджують листки і стебла картоп¬
лі, оскільки існуючі сорти не витримують тривалого зниження тем¬
142
ператури до -1° С. Серед видів картоплі S.bukasovii, S.canasense,
S.famatinae, S.vernei, S.curtilobum та інших виявлені зразки, здатні
витримувати приморозки до -7° С протягом 6-8 годин [6]. Такі зраз¬
ки можуть використовуватися як джерела стійкості до приморозків
при міжвидовій гібридизації.
Висока температура і сухість грунту можуть зумовлювати
в’янення, засихання листків і зупинку росту всієї рослини. Польова
жаростійкість, зумовлена високою інтенсивністю транспірації здоро¬
вих рослин за достатнього зволоження грунту, виявлена у диких ви¬
дів S.poliadenium, S.molinae та ін.
Виявлені стійкі до дії високих температур види картоплі, у яких
білок коагулює при температурі вище +58°С {S.bulbocastanum,
S.polyadenium, S.microdontum, S.chacoense та in.).
Несприятливі наслідки техногенного навантаження призводять
до підвищення концентрації в грунті та рослинах хімічних аналогів,
важких металів та ін., які зумовлюють зміни метаболізму рослинного
організму.
Генетичні дослідження рослин, стійких до солей важких металів,
показали, що толерантність до більшості з них контролюється полігенно
генами, які виявляють ефект домінування та адитивну дію [23].
Складність адаптивної системи рослин, яка забезпечує взаємо¬
зв’язок і взаємодію між процесами і функціями, що керують різними
генетичними системами, підтверджується численними генетичними,
фізіологічними і біохімічними дослідженнями. Стійкість, яка вини¬
кає при біохімічній та фізіологічній адаптації, зумовлює в цілому
фізіологічну реакцію на дію стресового фактора на рівні рослинного
організму.
Інформація в клітині передається за схемою ДНК <-> РНК —» бі¬
лок. Дійсно, холодостійкість або жаростійкість зумовлюється фізіо¬
логічними і біохімічними процесами, а генетичний контроль реалізу¬
ється за допомогою синтезу стресових білків. Під дією будь-якого
стресу дуже швидко змінюється експресія генів, індукується або ре¬
пресується синтез специфічних мРНК або їх трансляція, що призво¬
дить до зміни синтезу специфічних поліпептидів.
Внутрішньогеномна збалансованість організму виявляється в різ¬
них аспектах: через кореляційні зв’язки між ознаками, в основі яких
лежить явище зчепленого успадкування і плейотропії, та через зміни
дії генів-модифікаторів за нових умов середовища.
Отже, питання стійкості рослин картоплі до дії стресових фак¬
торів до сьогодні зовсім не вивчене. Гени, які контролюють певні
ланцюги цього складного явища, не ідентифіковані. Та й у най¬
ближчі десятиліття навряд чи це буде здійснено. Значні можливо¬
сті у вивченні стійкості до дії стресових факторів відкриваються з
інтенсивним розвитком культури картоплі in vitro, особливо клі¬
тинної селекції.
143
3.5. МУТАГЕНЕЗ У КАРТОПЛІ
У широкому розумінні мутації - це раптові спадкові зміни ознак
і властивостей організму, зумовлені змінами в генетичному апараті і
не пов’язані з рекомбінаціями.
Організм, у якого в результаті мутації виникла зміна певної озна¬
ки або властивості (кількох ознак), за якою він відрізняється від реш¬
ти особин, називається мутантом, а процес виникнення мутацій -
мутагенезом.
Мутації - це багатогранне загальнобіологічне явище, властиве
всім живим організмам. Вони є постачальниками нових генів для
природного і штучного добору.
За ступенем фенотипічного прояву розрізняють великі і малі му¬
тації. До великих (видимих, макромутацій) належать мутації, які
спричиняють чітко виражені зміни ознак організму. Наприклад, змі¬
ну тривалості вегетаційного періоду, розмірів і форми стебла, листя
та ін. До малих (мікромутацій) відносять мутації, у результаті яких
ознаки змінюються незначною мірою, тобто їх практично неможливо
виявити окомірно. Для їх виявлення застосовують біометричний ана¬
ліз мінливості окремих кількісних ознак, спеціальні прийоми (прово¬
каційні, аналізи на вміст і якість різних речовин та ін.).
За локалізацією в клітині мутації бувають ядерні і цитоплазма¬
тичні. Ядерні мутації пов’язані із змінами генетичного матеріалу
ядра. Оскільки при мейозі ядерні мутації, на відміну від цитоплазма¬
тичних, рекомбінують і успадковуються за законами Г. Менделя, то
розрізнити їх неважко.
Зміни ознак і властивостей організму, пов’язані зі змінами екст-
раядерних генів (плазмогенів), називають цитоплазматичними. Во¬
ни як і ядерні, стабільні і передаються з покоління в покоління.
Мутації можуть виникати в будь-яких клітинах і на різних етапах
розвитку організму. Мутації, що виникають у гаметах і клітинах, з
яких вони формуються, називають генеративними, а ті, що виника¬
ють у соматичних клітинах - соматичними. За природою зміни гене¬
тичного матеріалу генеративні й соматичні мутації відмінностей не
мають. Вони різняться лише особливостями успадкування і метода¬
ми їх виявлення.
Генеративні мутації успадковуються при статевому розмноженні
(через насіння), а соматичні - вегетативним способом (бруньками,
пагонами, бульбами), тобто частинами рослин, які сформувалися з
мутантних клітин. Особини, які мають ділянки мутантної тканини,
називають химерами, або мозаїками. Чим раніше в онтогенезі вини¬
кає мутація у соматичній клітині, тим більшою буде ділянка тканини
з даною мутацією.
За особливостями виявлення ознак і функцій організму розрізня¬
ють мутації домінантні і рецесивні. Домінантні мутації виявляються
144
в ознаках потомків першого (Мі) покоління, які одержали мутацію з
гаметою одного з батьків. Рецесивна мутація може бути виявлена
лише за умови, коли мутантний ген перейде в гомозиготний стан. У
диплоїдів це може відбутися, починаючи з М2.
За особливостями зміни ядерного генетичного матеріалу розріз¬
няють геномні мутації (зміна кількості хромосом, гетероплоїдія),
структурні мутації хромосом (делеції, нестача, дуплікації, інверсії,
транслокації) та генні (точкові).
Методи індукованого мутагенезу доповнюють усі інші розділи
вчення про вихідний матеріал, оскільки дають змогу залучити в селек¬
цію багатий різноманітний первинний матеріал у вигляді мутацій генів і
хромосом. Причому, самі мутації є, дійсно, лише вихідним матеріалом,
тому що, лише пройшовши через жорсткий добір, а в ряді випадків із
залученням до гібридизації, вони можуть дати початок новим сортам.
Перша публікація з експериментального мутагенезу картоплі з’яви¬
лася ще в 1935 р. у колишньому СРСР. Для ознайомлення з історією
цього питання відсилаємо читача до публікацій М.Д.Тарасенка,
К.П.Скворцова [42] та М.Д. Тарасенка [41], у яких наведено детальний
аналіз робіт з експериментального мутагенезу картоплі.
На сьогодні всі мутагенні фактори, що застосовуються для інду¬
кування мутацій у сільськогосподарських рослин, можна розділити
на три групи: фізичні, хімічні і біологічні.
Серед фізичних мутагенів найчастіше використовуються елект¬
ромагнітні і корпускулярні випромінювання. Іонізуючі у- і рентгенів¬
ські промені, проникаючи в клітину, надають електричний заряд
нейтральним атомам і молекулам. Вибиті з орбіти електрони приєд¬
нуються до інших атомів і молекул, що спричинює хімічні перетво¬
рення речовин, у які вони потрапляють.
Корпускулярні, або іонізуючі випромінювання (а-Р-частки, про¬
тони, нейтрони), проникаючи у клітини, спричинюють сильну іоні¬
зацію з утворенням вільних радикалів Н і ОН, що сприяє виникнен¬
ню органічних іонів, радикалів і перекисів у молекулах-мішенях.
При цьому їх реакція з ДНК може призводити до дегідроксилювання,
дезамінування основ, окиснення залишку цукру, розриву піриміди-
нового кільця та нуклеотидних ланцюгів. Високу мутагенну актив¬
ність мають і радіонукліди збР, 60CO,137Cs, 35S.
Нині відомі сотні хімічних речовин^що мають мутагенні власти¬
вості. Відкриті, а пізніше синтезовані Й.А.Рапопортом нові високо¬
ефективні органічні сполуки - супермутагени - індукують у сотні і
навіть у тисячі разів більше мутацій, порівняно з хімічними речови¬
нами неорганічної природи.
У мутаційній селекції на сьогодні найширше застосовуються спо¬
луки, що належать до таких класів:
алкілуючі ДНК сполуки - цей клас об’єднує велику групу
супермутагенів (етиленімін, диметилсульфат, нітрозометилсечо-
145
вина, нітрозоетилсечовина, ефіри сірчаної і сульфатної кислоти
та ін.);
аналоги азотистих основ, здатні включатися безпосередньо в ДНК
(азотиста кислота, штрозогуанідин, гідроксиламін, 5-бромулацил та ін.).
Мутагени біологічної природи - це очищені природні або синте¬
зовані ДНК і РНК, віруси, гормони, афлотоксини та ін.
Методи застосування фізичних мутагенів. Для опромінювання
об’єктів використовують медичні або промислові рентгенівські апа¬
рати і у-гармати (ГУП-Со60'50), якими зручно опромінювати пилок,
суцвіття й окремі квітки.
Існують спеціальні установки з великою потужністю випроміню¬
вання. Крім того, існують спеціальні у-поля, де рослини можна опро¬
мінювати протягом тривалого або короткого періоду у будь-якій фазі
росту і розвитку. Значно потужнішими є установки ГУБЕ-4000,
ГУПОС та ін.
Доза опромінювання обов’язково має контролюватися. Щодо оп¬
тимальної дози, то існують суперечливі погляди. На ефект прояву
дози впливає насамперед генотип, його фізіологічний стан. Крім то¬
го, доза може бути одноразовою або застосовуватися частинами, то¬
му в кожному конкретному випадку її вибирають індивідуально.
При обробці насіння картоплі гамма-випромінюванням найчас¬
тіше застосовують дозу в межах 150 Гр, а бульб - у межах 40-60 Гр.
Методи застосування хімічних мутагенів базуються на тому,
що мутаген має проникнути у клітину і вступити у взаємодію з її ге¬
нетичним матеріалом. Тому застосовується намочування об’єкта у
водному розчині мутагену, ін’єкція розчину в об’єкт за допомогою
медичного шприца та обробка в газовому середовищі.
Дозування хімічних мутагенів визначається концентрацією роз¬
чину, тривалістю дії та хімічною природою самого мутагену. У серед¬
ньому для всіх мутагенів експозиція замочування об’єктів варіює від
4 до 12 годин, а концентрація розчину - від 0,1% до 0,001%.
Об’єктом обробки мутагенами може бути будь-який орган роз¬
множення рослини, з яким найзручніше працювати в кожному конк¬
ретному випадку.
Найчастіше мутагенами обробляють бульби і насіння. Можна
працювати з окремими вічками і паростками. Для цього їх вирізають
до або після обробки мутагеном з частиною тканини бульб розміром
1 см для паростків і до 2,5 см - для вічок.
Опромінювати можна всю вегетуючу рослину, суцвіття, пилок.
Методи виявлення мутацій обов’язково потребують наявності
контрольного варіанта. Хоча сорти S.tuberosum є самозапильними,
перенесення пилку комахами не виключається. Тому серед зібраного
для обробки мутагенами насіння якась його кількість може мати гіб¬
ридне походження. Отже, поряд з обробленим мутагенами насінням
обов’язково слід висівати контроль - необроблене насіння.
146
Цілком можливо, що серед бульб, відібраних для обробки мута¬
генами, можуть траплятися химери. Для чистоти експерименту ба¬
жано кожну бульбу розрізати на дві частини. Тобто, використовують
метод половинок, де одна з половинок буде контролем, а інша - за¬
знаватиме дії мутагену.
Протягом вегетації рослини покоління Мі (сіянці чи бульбове по¬
томство) порівнюють із контрольними і позначають етикетками ті,
що за якою-небудь ознакою різняться від контрольних.
У рослин Мі морфологічні зміни здебільшого можуть проявитися
морфозами, хоч іноді можуть виникати домінантні мутації, які про¬
являються у першому поколінні. У переважної більшості мутації
рецесивні, тому виявити їх можна в М2- М3.
Кожну рослину Мі збирають окремо, вирощують М2 і М3 по
клонах.
Мутаційні зміни ознак. Численні дослідження з експеримента¬
льного мутагенезу [12, 38, 39] показали , що різні мутагенні фактори
індукують у картоплі мутаційні зміни забарвлення бульб і квіток,
пігментацію листкових пластинок, їх форми, кількості, розміри, а
іноді й нові форми з господарсько цінними ознаками. Однак, цей ме¬
тод у селекції картоплі ще не посів належного йому місця. Це можна
пояснити кількома причинами. По-перше, певними труднощами іден¬
тифікації мутацій у картоплі; по-друге, відсутністю чітких парамет¬
рів умов, які забезпечують відтворюваність (репрезентативність)
результатів; по-третє, досить низькою чистотністю появи форм з ба¬
жаними спадковими змінами, наявністю інших методів отримання
вихідного селекційного матеріалу.
Обробляючи хімічними мутагенами (ЕІ, ДМС, НЕС, і НМС) на¬
сіння від природного запилення сорту Агра, А.Сауліте [38] одержав
мутанти із зміненою шкіркою бульб, яка має високу стійкість до ме¬
ханічних пошкоджень, оскільки вона покрита товстим кірковим ша¬
ром. Крім того, бульби не уражуються паршею.
Обробкою насіння картоплі розчином НЕС 0,25-0,05%-ної кон¬
центрації індуковано і відібрано форми [12], які за врожайністю ві¬
рогідно перевищували контроль.
Аналіз численних експериментів з радіаційного мутагенезу зро¬
били М.Д. Тарасенко [41], Е.А. Сімаков та ін. [39], підтверджуючи
можливість одержання форм з підвищеною продуктивністю, умістом
крохмалю та білка порівняно з вихідними сортами. Крім того, дове¬
дена можливість зміни у мутантів кореляції між продуктивністю і
вмістом крохмалю в бульбах, що підвищує ефективність добору різ¬
них форм, а також можливість поєднання в одному генотипі корис¬
них спадкових змін за кількісними і морфологічними ознаками і ви¬
користання таких форм у гібридизації.
Мутаційний процес, як і рекомбіногенез у гібридизації, має імо¬
вірнісний характер, тому ефективність такої селекції ще низька. За-
147
порукою успішного використання індукованих мутантів є їх копітке
генетичне вивчення, тому що вони є новими формами із зміненими
генетичними системами, сформованими у вихідних сортів природ¬
ним і штучним добором у процесі селекції. Крім того, досить серйоз¬
ну трудність у мутаційній селекції картоплі становить утворення
анеуплоїдів і химер, особливо за обробки бульб мутагенами.
Анеуплоїди. Дія у-променів більш жорстка порівняно з іншими
видами опромінення і хімічними мутагенами, тому дає більше пору¬
шень структури хромосом і навіть їх кількості. Некратну гаплоїдно¬
му набору зміну кількості хромосом називають аиеуплоїдією.
Нестача хромосом у диплоїдних видів є, як правило, летальною
для генотипу (за насіннєвого розмноження), оскільки будь-яка хро¬
мосома необхідна для виживання організму. Часто це бувають моно-
сомики (2п-1) - генотипи, які втратили одну або нулісомики - дві
хромосоми.
Додаткові хромосоми до диплоїдного набору, особливо у трисо-
миків (2п+1) та тетрасомиків (2п+2), дають змогу вивчити їх генети¬
чне значення, оскільки життєздатність таких генотипів може бути
збережена. Хоча для селекційної роботи анеуплоїди не мають
практичного значення.
Химери - це організми, що несуть генетично неоднорідні ділян¬
ки тканин або клітин. При розмноженні насінням у картоплі така
мутація є нестійкою.
Мутагенез у картоплі вивчається, головним чином, на матеріалі
соматичних мутацій та за їх поведінкою при вегетативному розмно¬
женні. Генеративні мутації і статеве розмноження дають змогу про¬
вести глибокий генетичний аналіз індукованих змін. У картоплі та¬
кий аналіз утруднюється за високого рівня плоїдності, а отже - гете¬
розиготності та, частково, за самонесумісності у багатьох сортів. За
статевого розмноження цінні мутації можуть втрачатися і маскувати¬
ся комбінативною мінливістю, а нові мутантні ознаки важко виділя¬
ти при складному розщепленні у поліплоїдів. Окрім того, частина
мутантних генів елімінується у мейозі на стадії формування гамет та
раннього ембріогенезу.
З іншого боку, як твердив М.Д.Тарасенко [41], картопля є надзви¬
чайно зручним об’єктом для вивчення практичних питань мутагенезу,
оскільки за вегетативного розмноження легко зберігати цінні спадкові
зміни, особливо мутації, що виникли у соматичних тканинах.
Соматичний прояв мутації у тканині, де вона появилася, залежить
від стадії розвитку, на якій мутація виникла, від розміщення мутантних
клітин відносно інших тканин та від конкуренції між клітинами. Наяв¬
ність у картоплі трьох гістологічних шарів (Lb Ln, Ьш), які зберігають
певну самостійність, може призводити до формування генетично неод¬
норідних тканин. Залежно від того, на якому етапі і в якому шарі виник¬
ла мутація, можуть формуватися різного типу химери.
148
Периклинальні химери виникають у разі, коли внутрішні тка¬
нини оточені одним або кількома шарами клітин іншого генотипу.
Секторіальні химери - мутантна тканина утворює сектор, який
розташований від епідермісу до серцевини.
Мериклинальні химери - мутантна тканина утворює дискрет¬
ний шар (або шари) лише в одному секторі.
Міксохимери - нормальні і мутантні клітини хаотично розмі¬
щуються по всіх шарах. Міксохимери є нестабільними і в кінцевому
результаті можуть утворювати один із трьох типів стабільних химер,
названих вище.
Отже, при обробці мутагенами цілих бульб картоплі або за три¬
валого вегетативного розмноження можуть нагромаджуватися химер¬
ні тканини, які можна відділяти (вічка, пагони та ін.), розмножувати
і, порівнюючи з вихідними сортами, відбирати клони з господарсько
цінними ознаками і властивостями.
Вивчення біохімічної дії гена повністю базується на порівнянні
метаболічної активності нормальних і мутантних особин. Всі наші
знання про хімічну дію гена грунтуються на даних, одержаних при
вивченні мутантних фенотипів. Однак, дослідження дії гена, що ба¬
зуються виключно на спостереженнях і описі кінцевих результатів
мутації на рівні макрофенотипічних ознак, залишаються лише грубо
описовими. По суті, будь-яка спадково зумовлена фенотипічна зміна
є результатом зміни хімії гена і його впливу на біосинтез та обмін
речовин у клітині. Отже, для розуміння первинних наслідків генної
мутації слід вивчати первинні хімічні зміни, зумовлені мутантними
алелями в клітині.
Хоч і досягнуто значних успіхів молекулярної біології у вивченні
будови генів і складних механізмів регуляції їх експресії, до сьогодні
залишаються невідомими ключові події процесу становлення ознаки,
у якому взаємодіють системи генів. Цілком очевидно, що необхідний
пошук нових принципів і методів аналізу [13,25,26].
Одним із важливих шляхів у цьому напрямі є вивчення зв’язків
між фенотипічною мінливістю і генотипними особливостями біосин-
тетичних процесів із застосуванням молекулярних маркерів, тобто
методів електрофорезу білків. Повну інформацію про генетичну мін¬
ливість дають методи секвенування (визначення нуклеотидної послі¬
довності) генів клітинного ядра та полімеразної ланцюгової реакції
(RAPD).
3.6. РЕКОМБІНАЦІЯ ГЕНЕТИЧНОГО
МАТЕРІАЛУ
Рекомбінація - це перерозподіл генетичного матеріалу батьків¬
ських форм у потомстві, що призводить до появи нових генотипів,
тобто зумовлює спадкову комбінативну мінливість живих організмів.
149
Генетичною рекомбінацією може бути названий будь-який процес,
що забезпечує появу потомків з новим поєднанням двох або більшої
кількості спадкових факторів, за якими різнилися їхні батьківські
форми.
У вищих еукаріотів є три рівні рекомбінацій: міжхромосомні,
міжгенні і внутрішньогенні. У численних варіантах рекомбінацій
головна роль належить кросинговеру.
Реципрокні рекомбінації між генетичними варіантами є важли¬
вим джерелом генотипних комбінацій, які мають потенційно різну
селекційну цінність. У мейозі завдяки кросинговеру може виникати
величезна кількість комбінацій генів. Наприклад, у виду, що має ли¬
ше 1000 локусів, кожен з яких представлений серією із 10 алелів, за
рахунок рекомбінації теоретично може утворюватися Ю1000 різних
генотипів [18]. Основним джерелом генетичної варіабельності є му¬
тації генів і структурні зміни хромосом, які добре зберігаються міто¬
тичною системою за вегетативного розмноження картоплі.
У селекції картоплі особливо широкого застосування набула інт-
рогресивна гібридизація, тобто включення генів одного виду в ген¬
ний комплекс іншого шляхом віддаленої гібридизації. Однак, мінли¬
вість, що спостерігається при міжвидовій гібридизації, відображає
лише незначну частину її загальної потенційної можливості. Кіль¬
кість реальних рекомбінацій ознак обмежують елімінацію гамет і
зигот, плейотропію, групи зчеплення генів, дію репаративної систе¬
ми клітини та ін. Дійсно, загальновідоме явище інтерференції обме¬
жує частоту кросинговеру, оскільки гетерогенна структура хромати-
нової нитки базується на сильних електростатичних зв’язках гістонів
з ДНК. Крім того, частота рекомбінації в окремій зоні хромосоми
контролюється специфічними генами-регуляторами.
Частота рекомбінацій модифікується дією умов середовища та
генетичних факторів. Для підвищення частоти рекомбіногенезу за
ознаками урожайності і вмісту крохмалю у бульбах картоплі
Є.А.Сімаков та ін. (1991) застосовували у-опромінення бульб бать¬
ківських клонів і пилку в дозах 13-35 і 20 Гр, відповідно. Частота
рекомбінантів за ознаками продуктивності і вмісту крохмалю в ком¬
бінаціях схрещування опромінений х опромінений коливалася від 3,5 до
5 %, а в схрещуваннях неопромінений х опромінений - від 3 до 12 %.
У дослідах з іншими видами культур із застосуванням X-
променів доведено, що частота індукованих міжгенних і внутрішньо-
генних рекомбінацій залежить від дози й експозиції опромінення,
особливостей самої хромосоми або її сегментів. Як зазначала
Ш.Ауербах [2], більшість або ж усі мутагени здатні індукувати кро¬
синговер. На думку Й.А.Рапопорта [36], засобом підвищення частоти
рекомбінацій може бути мутокросинговер, тобто поєднання мутацій і
кросинговеру в одному комбінованому процесі. Дія мутагенів на гіб¬
риди Fi підвищує частоту кросинговеру між ділянками хромосом, які
150
в нормі ніколи між собою не обмінюються. У цілому проблема інду¬
кованого рекомбіногенезу має важливе значення, тому що метод
комбінативної селекції, яка базується на гібридизації і мейотичній
рекомбінації, залишиться в найближчому майбутньому основним в
управлінні спадковою мінливістю рослин.
Потужний потенціал генетичної інженерії, метою якої є конс¬
труювання організмів із запланованими властивостями, є розв’язання
питань такого рівня складності, який залишається на рівні обмеженої
маніпуляції окремими генами. Перенесення окремих генів за допо¬
могою методів генетичної трансформації дасть змогу розв’язувати
окремі питання поліпшення культурних сортів картоплі. Поява гене¬
тично модифікованих (трансгенних) сортів картоплі, стійких до ко¬
лорадського жука, свідчить, що генетична інженерія лише у поєд¬
нанні з традиційними методами селекції досягне нових успіхів у
створенні сортів картоплі, які б відповідали вимогам товаровиробни¬
ка і споживача.
Гетерозис як складне загальнобіологічне явище, що характери¬
зується перевагою гібридів Fi над батьківськими формами за однією
або кількома ознаками, виявлений і в картоплі.
Гетерозиготний стан багатьох локусів є неодмінною умовою ви¬
явлення гетерозису. Оскільки при насінному розмноженні гетерози¬
готи розщеплюються і ефект гетерозису виявляється лише в першо¬
му поколінні, то за вегетативного розмноження у картоплі він може
зберігатися у багатьох бульбових поколіннях.
Ефект гетерозису у картоплі виявлений у міжвидових схрещу¬
ваннях сортів з культурним тетраплоїдом Solanum andigenum, дигап-
лоїдними гібридами і диплоідними видами. У гібридного потомства
від схрещування S.tuberosum x S.andigenum, S.tuberosum х диплоїдні
гібриди (4х х 2х) виявлений гетерозис за врожайністю, що значно
рідше зустрічається у комбінаціях S.tuberosum x S.tuberosum.
Нині в селекції картоплі запропоновано декілька схем викорис¬
тання явища гетерозису.
Перша схема базується на використанні тетраплоїдної материн¬
ської форми S.tuberosum і диплоїдної форми (запилювач), яка фор¬
мує нередуковані 2х-гамети (FDR-гамети) у першому мейотичному
діленні. Такі форми виявлені у S.phureja, S.rybinii та у диплоїдних
гібридів за їх участю.
Друга схема базується на використанні двобічної статевої полі-
плоїдизації у диплоїдних видів. У цьому випадку підбираються як
компоненти схрещування диплоїдні міжвидові гібриди, здатні утво¬
рювати нередуковані гамети ( 2х FDR). Ефект гетерозису за такої
схеми схрещування (2xFDR х 2xFDR) залежить від ступеня спорід¬
неності або генетичних відмінностей цих компонентів схрещування.
Наступна схема базується на однобічний статевій поліплоїдизації
самонесумісних материнських диплоїдних форм (2xFDR х 4х).
151
Використання мейотичної поліплоїдії дає змогу зберегти рівень
гетерозиготності при переході з диплоїдного на тетраплоїдний рі¬
вень. За сучасними уявленнями, продуктивність гетерозисних гібри¬
дів можна умовно розділити на два компоненти - адитивний і неади-
тивний. Перший з них визначається загальною комбінаційною здат¬
ністю і практично не пов’язаний з гетерозиготністю. Другий, неади-
тивний, компонент є результатом взаємодії певних генів. Таким
чином, досягнення високого рівня гетерозиготності батьківських
форм і збереження її у гібридів слід розглядати лише як один із спо¬
собів одержання комбінацій певних генів, взаємодія яких забезпечує
гетерозис [49,16].
Цитоплазматична спадковість. Відкриття ДНК в органелах клі¬
тин (хлоропластах, мітохондріях) вказує на наявність позаядерної
генетичної інформації. Локалізовані в цитоплазмі одиниці спадково¬
сті, на відміну від ядерних генів, називають плазмогенами, а сукуп¬
ність цих одиниць - плазмоном, який відповідає генетичному мате¬
ріалу хромосомного набору - геному.
Геноми хлоропластів мають порівняно великі розміри - близько
140 тис. пар нуклеотидів. Сумарний вміст ДНК хлоропластів стано¬
вить кілька процентів від загального вмісту її у клітині.
Кожна мітохондрія містить кілька копій її геному. Оскільки в
клітині мітохондрій багато, то на одну клітину може припадати
значна кількість геномів даної органели. У рослин розміри ДНК
мітохондрій варіюють у широких межах. Є дані про те, що міто-
хондріальний геном рослин є не однією молекулою ДНК, а скла¬
дається з кількох різних за розміром молекул. Мітохондріальний
геном у рослин за розмірами однаковий із хлоропластним, тому
загальний обсяг генетичної інформації поза ядром, очевидно, до¬
сить великий.
Мітохондріальні гени кодують в основному дві групи ознак. По-
перше, це ознаки, пов’язані з діяльністю дихальних систем, по-друге,
ознаки стійкості до антибіотиків та інших клітинних отрут.
У картоплі цитоплазматична спадковість не вивчена, що пов’я¬
зано з трудністю проведення генетичного аналізу ознак, які контро¬
люються факторами цитоплазми. Дані аналізу реципрокних гібридів
Fi досить суперечливі. В одних випадках деякі ознаки і властивості
успадковуються по материнській лінії, в інших - це не підтверджу¬
ється. Отже, нагромаджені експериментальні дані лише вказують на
складну взаємодію геному і плазмону, які складають єдину генетич¬
ну систему клітини, реалізація якої через фенотип проходить у мін¬
ливих умовах середовища.
Цитоплазматична чоловіча стерильність у картоплі зумовлена
взаємодією плазмону і домінантними незалежними генами і проявля¬
ється морфологічно через деформованість і нерозтріскуваність пиля¬
ків, відсутність у пиляках пилку та частковість мікроспор.
152
Було виявлено, що нерозтріскуваність пиляків є результатом вза¬
ємодії між цитоплазматичним фактором Ins, знайденим у S.chacoense,
і п’ятьма домінантними незалежними In-генами у S.phureja,
S.kurtzianum, S.stenotomum. Також два домінантні гени-відновлювачі
фертильності виявлені у S.chacoense з плазмоном Ins [6]. Ці гени епі-
статичні відносно In-генів і зумовлюють формування життєздатного
пилку у рослин. Плазмон Ins існує у двох формах, які різняться за чу¬
тливістю до генів Іп, а плазмон Іпг - нечутливий. У свою чергу Іп-
гени функціонально неідентичні, тому що кожен з них взаємодіє з
певним геном-відновлювачем фертильності й однією з двох форм
плазмону.
Виявлений у деяких форм рослин S.chacoense рецесивний ген df
зумовлює деформованість квіток, а стерильність проявляє лише у
плазмонах деяких рослин (S.phureja і S.stenotomum).
Деформовані квітки зумовлює плазмон df у присутності рецеси¬
вних ядерних генів df df. Стерильність через нерозтріскування пиля¬
ків зумовлює плазмоген Ins у присутності домінантних генів Іп1, Іп2.
Плазмони Df, іпг, Р1г, 1ЬГ зумовлюють фертильність при взаємодії з
домінантними або рецесивними алелями ядерних генів.
Результати досліджень Діоне (цит. за [6]) показали, що чоловіча
стерильність у гібридів, одержаних від схрещування S. demissum х
S.tuberosum, зумовлена взаємодією цитоплазматичних факторів
S.demissum з ядерними генами S. tuberosum.
До сьогодні природа цитоплазматичних факторів або “плазмоге-
нів”, що зумовлюють цитоплазматичну чоловічу стерильність, поки
що не з’ясована. Тому їх відносять до цитоплазматичних спадкових
факторів невизначеної природи. Форми з ЦМС використовують як
для генетичних досліджень, так і для селекційної роботи.
3.7. ГАПЛОЇДІЯ У КАРТОПЛІ
Гаплоїди (від грец. haplos - одинарний, простий) - це організ¬
ми, в соматичних клітинах яких міститься половинна (гаметична)
кількість хромосом (п замість 2п). Вивчення гаплоїдії і гаплоїдів є
дуже важливим для аналізу цитогенетичної структури видів і шля¬
хів їх еволюції. Крім того, це є не менш важливим у пізнанні ряду
генетичних явищ взаємодії генів і ролі генотипного середовища,
ефекту дози гена, груп зчеплення ознак, асоційованості хромосом,
блоків генів та ін.
Термінологія. Залежно від рівня плоїдності вихідної особини у
поліплоїдному ряду, гаплоїди поділяють на дві основні групи:
1) моногаплоїди, або моноплоїди - це організми, які походять від
особин з диплоїдною (2 п) кількістю хромосом;
2) полігаплоїди: дигаплоїди - 2х, тригаплоїди - Зх, тетрагаплої-
ди - 4х - це організми, що походять від поліплоїдних особин.
153
Залежно від структури геному вихідного поліплоїда (автополіп-
лоїд чи алополіплоїд), полігаплоїди поділяються на автополігаплоїди
й алополігаплоїди [32].
Гаплоїди з кількістю хромосом, кратною основній кількості, нази¬
вають еугаплощами, а такі, що мають меншу або більшу їх кількість від
основної кількості - анеугаплоідами. Анеугаплоцда, у яких кількість
хромосом вища за основну кількість, називають гіпергаплощами.
В основу класифікації гаплоїдів покладено їхню генетичну кон¬
ституцію, що визначається ядром і цитоплазмою. За цією ознакою
гаплоїди поділяють на три типи.
1. Матроклинні (Мк) - розвиваються з редукованої яйцеклітини
або з клітин зародкового мішка, що функціонально її заміщують і які
мають ядро і цитоплазму материнського організму.
2. Андрогенні (Аг) - розвиваються з яйцеклітини або будь-якої
клітини зародкового мішка, у якій залишилася цитоплазма материн¬
ського організму, а хромосоми ядра замінені хромосомами, привне¬
сеними спермієм.
3. Андроклинні (Ак) - розвиваються з чоловічої гамети і мають
цитоплазму і ядро однієї особини. Такі гаплоїди отримують методом
культури пилку або пиляків in vitro.
Способи одержання гаплоїдів можна розподілити на дві групи:
виявлення спонтанних гаплоїдів та експериментальне одержання із
застосуванням різних методів їх індукції.
Спонтанна гаплоїдизація є звичайним способом деполіплоїдиза-
ції, який належить до апоміксису - розвитку зародка без запліднення
(гаплоїдний партеногенез, апогамія, апогаметія).
Дигаплоїд (n=2x=24) S. tuberosum на основі редукованого парте¬
ногенезу вперше був отриманий у колишньому СРСР О.В.Івановсь-
кою (цит. за [28]) шляхом запилення S.tuberosum (2п=48) пилком
S.rybinii (2іг=24). Цей метод одержання гаплоїдів S. tuberosum знай¬
шов широке застосування як один із шляхів подолання несхрещува¬
ності у віддаленій гібридизації картоплі. Шляхом запилення S. tube¬
rosum пилком S.vernei, S.simplisifolium, S.verrucosum, S.famatinae та
ін. у багатьох країнах світу були одержані гаплоїди.
Можливість одержання гаплоїдів з використанням гаплопро-
дюсера вперше була доведена на прикладі ячменю при схрещуванні
тетраплоїдів Hordeum bullosum (2n=28) і Hordeum vulgare (2n=14).
Запилення і запліднення тетраплоїдного H.bulbosum пилком диплої-
дного H.vulgare перебігали без порушень. Однак, у зародку, почина¬
ючи з третьої доби його розвитку, спостерігалася поступова еліміна¬
ція хромосом H.bulbosum і вже на 11-у добу 93,7% клітин зародка
мали гаплоїдну кількість хромосом (п=7). Гіпотези, які пояснюють
механізми елімінації хромосом, в основному зводяться до асинхрон-
ності, мітотичного циклу через відмінності між батьківськими вида¬
ми та аномалії веретена.
154
За віддалених схрещувань тетраплоїдів з диплоїдами у картоплі
іноді виникають гібриди з ознаками обох батьків, але мають вони
диплоїдний набір хромосом, а не три- або тетраплоїдний [ЗО]. У та¬
ких випадках зниження плоїдності може відбуватися внаслідок по¬
слідовної втрати окремих хромосом через анеуплоїдію до стабільно¬
го мінімуму, шляхом розвитку без запліднення редукованих (гаплоїд¬
них) яйцеклітин або синергід, що функціонально їх заміщують.
Багатьма дослідниками було виявлено, що при запиленні
S.tuberosum (2п=48) пилком деяких форм S.phureja (2п=24) виника¬
ють матроклинні гаплоїди. У таких запилювачів спостерігається зна¬
чна кількість пилкових трубок, які несуть одне реституційне ядро.
Тобто, замість двох гаплоїдних сперміїв, які містять по 12 хромосом
(п=12), пилкова трубка несе одне 24-хромосомне (2п=24) генератив¬
не ядро, яке не розділилося. За проникнення його в зародковий мі¬
шок можливе лише одинарне запліднення яйцеклітини або вторин¬
ного ядра. У першому випадку насіння не утворюється через нездат¬
ність ендосперму розвиватися без запліднення, у другому випадку
може виникати гексаплоїдний ендосперм і гаплоїдний зародок із не-
заплідненої яйцеклітини.
Успішними виявилися методи гібридизації S. tuberosum з S.phu¬
reja, S.stenototum, S.chacoense, S.vernei та інших видів для індукції
гаплопартеногенезу [34].
Можливим джерелом одержання гаплоїдів є поліембріонія - роз¬
виток зародків-близнюків, що зустрічається у багатьох культур, у
тому числі й картоплі. Однак, відібрати життєздатні дигаплоїдні сіян¬
ці серед виділених близнюкових сіянців у різних сортів і мутантів
картоплі дуже важко [42].
Для підвищення частоти утворення дигаплоїдів у картоплі вико¬
ристовують дію стресових факторів.
Іонізуюча радіація. Підвищений вихід дигаплоїдів у сортів S. tu¬
berosum та моногаплоїдів у диплоїдних видів (,S.phureja, S.kessel-
brenneri) після запилення опроміненим пилком (найчастіше у дозах
4-6 Гр) спостерігали Й.Букан, Н.А.Лебедєва [цит. за ЗО ].
Дія хімічних речовин. Витримування до 20 хв у водному розчи¬
ні (0,08 %) колхіцину, сахарози (10 %) і борної кислоти (0,006 %)
пилку S.multidissectum і S.torijense і запилення ним різних клонів
S.tuberosum значно підвищувало частоту утворення дигаплоїдів [41].
Це підтверджує, що нередуковані ядра пилку запилювачів є важли¬
вим фактором індукування гаплоїдії у картоплі.
Висока частота формування реституційних ядер у пилку картоплі
спостерігається при його обробці розчином закису азоту мутагенів та
ін., а запилення таким пилком сортів S.tuberosum стимулює появу
гаплоїдів.
Температурний шок. Доведено, що у багатьох видів рослин дія
високих (від 40 до 50°С) або низьких (від +4 до -3°С) температур
155
протягом 5-30 хв на пилок, зібраний для запилення, дає змогу інду¬
кувати гаплоїди. У картоплі помірна температура (у межах 20°С) в
період запилення і запліднення підвищує вихід дигаплоїдів.
Культура пиляків і мікроспор. В останні десятиріччя значного
розвитку набув перспективний спосіб одержання гаплоїдів - культи¬
вування ізольованих пиляків і мікроспор у культурі in vitro (див. роз¬
діл 9).
Гаплоїди як модельний об’єкт у генетиці картоплі. Вивчення
гаплоїдних рослин дає змогу визначити геномну структуру виду. Цей
напрям базується на вивченні мейозу в особин з редукованою кількіс¬
тю хромосом. Спорідненість окремих хромосом, яка проявляється
через їх нормальну кон’югацію у профазі- І мейозу, дає змогу судити
про походження геномів і окремих груп хромосом.
У гаплоїдів відносно легко виявити алелі, що знижують життє¬
здатність, напівлетальні та летальні, які нагромаджуються в популя¬
ціях поліплоїдів внаслідок мутацій і практично не піддаються конт¬
ролю, оскільки вкрай рідко зустрічаються в гомозиготному стані.
Форми з такими ознаками у гаплоїдів можуть бути відібрані в межах
одного покоління.
Гібридологічний аналіз у картоплі на дигаплоїдному рівні істот¬
но спрощується та полегшує вивчення закономірностей успадкуван¬
ня як незалежних, так і зчеплених ознак, а також добір форм із ко¬
рисними рецесивними генами. Дійсно, у випадку моногібридного
схрещування батьківських форм, що різняться за однією парою аль¬
тернативних ознак А і а, у диплоїдних зиготах можливе лише таке
сполучення генів - АА, Аа й аа, що значно простіше, ніж у тетрапло-
їдів (див. табл.3.2).
Гаплоїди *S.tuberosum, S.cmdigenum і їх диплоїдні гібриди можуть
дати генетикам і селекціонерам унікальну можливість створення
найбільш сприятливих генетичних комбінацій вихідного матеріалу.
3.8. ГЕНОФОНД КАРТОПЛІ, ЙОГО СКЛАДОВІ,
ХАРАКТЕРИСТИКА І СТРАТЕГІЯ
ВИКОРИСТАННЯ
3.8.1. Генофонд картоплі, його складові
і характеристика
Складові генофонду картоплі. Серед сільськогосподарських
культур картопля вигідно відрізняється надзвичайною різноманітніс¬
тю видів {species), підвидів (subspecies), груп різновидностей
(iconvarietas), різновидностей {varietas), форм {forma), зразків і сорто-
зразків (concultivar), сортів (cultivar), гібридів {hybrids). Ця особли¬
вість культури сприяла значному поширенню її в природних нішах
та при штучному розмноженні.
156
Названа таксономічна сукупність характеризується наявністю ве¬
ликого генетичного потенціалу з ефективним, еволюційно відшліфо¬
ваним контролем практично всіх ознак. Крім того, позитивним також
с те, що картопля досить легко адаптується при вирощуванні її в не¬
типових умовах зовнішнього середовища, що сприяє значному по¬
ширенню культури [29].
Усе різноманіття картоплі - це її генофонд (від грец. genos - рід,
походження і франц. fond - основа). Генофонд культури - це сукуп¬
ність усіх генів таксономічних її різновидностей, які характеризу¬
ються певною частотою. До останнього часу вважалося, що гено¬
фонд картоплі складається із сортименту культурних і аборигенних
сортів, ендемічних форм, культурних і диких видів (Букасов С.М.;
fiawkes J.G.; Костина Л.И.), [10].
Еволюція картоплі відбувалась і відбувається, з одного боку, в
напрямі природного добору форм з ефективним генетичним контро¬
лем ознак, які забезпечують їх збереження. При цьому відселектову-
ються високоефективні гени контролю, головним чином, стійкості
проти екстремальних факторів, у тому числі хвороб і шкідників, а
також генів, реалізація яких забезпечує оптимальне функціонування
органів розмноження. А тому в генетичних центрах походження кар¬
топлі має місце не лише видова різноманітність, а й відмінність за
домінантними, зокрема морфологічними ознаками [4]. З іншого бо¬
ку, М.І.Вавилов вважав, що "... селекція являє собою еволюцію,
спрямовану волею людини" [6, с.7]. А тому створення нових форм,
добір, що супроводжує селекційний процес, проводяться з ураху¬
ванням високого прояву тих властивостей, які задовольняли б люди¬
ну. Причому, біологічні (важливі для збереження форм картоплі) і
господарсько цінні якості часто не збігаються за вираженням. Ви¬
кладене стало підставою розділити генофонд картоплі на форми, які
утворилися у процесі еволюції, та синтезовані або відібрані з ураху¬
ванням потреб людини (рис. 3.2), [19].
До перших може бути віднесене природне різноманіття видів,
ендемічних форм, аборигенних сортів, яке формувалося під впливом
біотичних та абіотичних факторів: географічних, кліматичних, пато¬
генних та інших компонентів зовнішнього середовища.
За характеристикою продуктивності, кулінарних якостей тощо
види розділяють на дикі та культурні (культивовані) [10].
Види картоплі є основною таксономічною одиницею в система¬
тиці культури. Це сукупність близькоспоріднених організмів, що ха¬
рактеризуються тільки їм властивими морфоло-фізіологічними та
еколого-географічними особливостями, які заселяють певний ареал і
мають певний тип взаємозв’язків із середовищем існування. З ураху¬
ванням наявних ендемічних ареалів [12], форми, які утворилися в
таких локальних мікроцентрах, виділені як окремі складові генофон¬
ду картоплі. Значна кількість аборигенних сортів, що не можуть бути
157
віднесені до жодної з перерахованих категорій, виділені в окрему
складову генофонду.
В енциклопедичному довіднику “Бульба” (1988) викладено зна¬
чення поняття “вихідний матеріал” для селекції картоплі, де зазнача¬
ється, що до його складу входять дикі і культурні види. Проте, по-
перше, згадані види, про що сказано вище, є складовими генофонду -
такими ж, як, наприклад, сорти, а по-друге, залучення їх для ство¬
рення сортів є тривалим і складним процесом. При цьому утворю¬
ються численні перехідні форми з різним генетичним потенціалом і
проявом агрономічних властивостей, які часто для подальшого успіш¬
ного використання потребують застосування особливих методів. До
останнього часу такий матеріал не був систематизований, що дало
підставу нам виділити його в окремі складові [19,20].
Широке та успішне використання в передселекційному процесі
(prebreeding) міжвидової гібридизації, гаплоїдії, поліплоїдії, генетич¬
ної і генної інженерії та інших методів дозволяє отримати безліч
цінних форм з ефективним генетичним контролем окремих або на¬
віть комплексу ознак. Проте дуже часто через необхідність їх до¬
опрацювання вони не можуть бути використані безпосередньо в
158
практичній селекції. Зазначене стало підставою для виділення їх в
окрему складову генофонду картоплі - вихідний передселекційний
матеріал.
У процесі виведення сортів картоплі, з’ясування закономірностей
успадкування основних господарських ознак матеріалу внутрішньо¬
видового (у межах S.tuberosum) походження, створення донорів при
одержанні міжвидових гібридів із високим ступенем бекросування та
ітри інших дослідженнях втцеплюються форми, які через наявність,
як правило, однієї або кількох негативних ознак не можуть бути ре¬
комендовані виробництву. Проте вони характеризуються високим
ступенем прояву окремих або комплексом цінних властивостей, адап¬
товані до вирощування в конкретних еколого-патогенних умовах, здат¬
ні добре передавати певні ознаки нащадкам. Багато з них використо¬
вуються безпосередньо при виведенні сортів. Форми з таким генетич¬
ним контролем і проявом ознак та призначенням виділені нами в
окрему складову генофонду - вихідний селекційний матеріал [21].
Підтвердженням необхідності виділення нових складових гено¬
фонду картоплі з позицій обгрунтування методичних підходів до ро¬
боти із залученням співродичів культурних сортів у селекційну прак¬
тику може бути розроблена схема використання різноманіття картоп¬
лі при створенні вихідного селекційного матеріалу, як передостан¬
нього етапу в інтрогресії цінних генів конпролю агрономічних ознак
при виведенні нових сортів (рис. 3.3).
Комерційні сорти - загальновизнана складова генофонду картоп¬
лі, яка є результатом мобілізації генетичного потенціалу культури і
найбільшою мірою орієнтована на задоволення потреб людини.
З позицій ефективності генетичного контролю агрономічних влас¬
тивостей та шляхів практичного використання серед складових ге¬
нофонду можуть бути виділені або створені джерела чи донори
окремих, а інколи й комплексу ознак. Поява цих відносно нових у
селекційній практиці термінів часто зумовлює неоднозначність їх
трактування.
Згідно із твердженнями А.Ф.Мережко, “... джерелами можна
вважати виділені із світового різноманіття рослин дикі форми, які
мають бажаний рівень прояву тієї або іншої властивості, але не ви¬
вчені за генетичним контролем її” [17, с.7]. При цьому автор заува¬
жує, що відносно останнього термін є досить умовним, оскільки ві¬
дображає лише ступінь вивченості матеріалу. Погоджуючись із цим,
ми пропонуємо таке визначення категорії “джерело ознак картоплі”:
це - виділені із світового різноманіття дикі і культурні види картоплі
або створені на їх основі форми, які характеризуються високим фе-
нотипічний вираженням однієї або кількох властивостей та ефектив¬
ним генетичним їх контролем, але водночас із цінною ознакою пе¬
редають нащадкам і негативні. Тобто, обов’язковою вимогою до
джерел є їх дослідження в генетичному аспекті.
159
Експедиції
в генцентрм
походження
картоплі та
центри спорід¬
неної еволюції
рослин-
господарів і
патогенів для
збирання
зразків видів,
ендемічних
форм із висо¬
ким проявом
ознак або інтро¬
дукція їх із
світових
колекцій
Вивчення нор¬
ми реакції
складових ге¬
нофонду куль¬
тури на виро¬
щування в
певних еколо-
го- патогенних
умовах
Виділення
форм із висо¬
ким феноти-
пічним вира¬
женням гос¬
подарських
показників
Визначення
ефектив¬
ності гене¬
тичного
контролю
провідної
властивості,
гомозиго-
тизація
цінних
алелей,
виділення
та
створення
джерел
однієї або
групи ознак
І
Формуван¬
ня ознако-
вих колек¬
цій
Подолання
міжвидової
несуміснос¬
ті, визна¬
чення гене¬
тичного
контролю
провідних
ознак у Fj,
провідної та
інших ознак
у F2 і т.п.
Виявлення
оптимальних
шляхів інт-
рогресії цін¬
них генів Fi
у вихідний
передселек-
ційний
матеріал
Визначен¬
ня прояву
та успад¬
кування
ознак ком¬
понентами
бекросу-
вання
(сорти,
міжсортові
гібриди)
Виявлення
закономір¬
ностей
успадку¬
вання
провідних
та інших
ознак се¬
ред вихід¬
ного
селекцій¬
ного
матеріалу,
виділення
донорів,
позитивні
добори
Викорис¬
тання до¬
норів у
практич¬
ній селек¬
ції
Визначен¬
ня опти¬
мальних
схем схре¬
щувань,
донорів
вихідного
селекцій¬
ного
матеріалу
Збирання цінних зразків видів, форм, виділення
та створення джерел господарських ознак
Створення та вивчення вихідного
передселекційного матеріалу
Вивчення, створення та використання
вихідного селекційного матеріалу
Передселекційний процес
Рис. 3.3. Поетапне виконання досліджень з використанням генофонду картоплі при створенні
вихідного селекційного матеріалу та донорів окремих або кількох ознак
Уперше досить повне визначення категорії “донор” зробив
Т.Я.Зарубайло: “Донор (лат. donare - дарувати, жертвувати) - сорт
або лінія з високими показниками тієї або іншої ознаки, яку відносно
легко можна передавати іншим формам, але при цьому разом з нею
не передаються певні небажані ознаки, яких не можливо або важко
позбутись без одночасної втрати цінної ознаки”(1976, с.6). Вважає¬
мо, що донором може бути також форма, яка характеризується висо¬
ким проявом багатьох властивостей і їх ефективним генетичним конт¬
ролем, що є передумовою ймовірного створення з її участю багатьох
сортів із комплексом ознак.
Генетичні осередки (центри) походження картоплі. Перші, ор¬
ганізовані М.І. Вавиловим [9] експедиції в Мексику, Гватемалу, в
Колумбію на чолі з С.М. Букасовим (1925, 1926 рр.) і в Перу, Болі¬
вію, Чилі - С.В.Юзепчуком (1927, 1928 рр.), дозволили виявити знач¬
не різноманіття картоплі. Вже в той час результати експедицій до¬
зволили М.І.Вавилову зробити висновок про специфічний розвиток
землеробства Нового Світу на основі дикої флори та в інших матері¬
альних умовах, ніж у Старому Світі. А тому, наприклад, туземне на¬
селення Мексики вживало в їжу бульби диких видів картоплі. Під
час експедицій виявлено центри походження виду Solanum tuberosum
L. - Чилоанське вогнище (південна частина Чилі та острів Чилоє)
Андійського осередку. На відміну від перуанських, болівійських,
еквадорських видів картоплі, які зав'язують бульби лише при корот¬
кому дні, S.tuberosum здатний утворювати їх в умовах довгого фото¬
періоду, що, крім усього іншого, зумовило його поширення в євро¬
пейській культурі.
У цілому М.І.Вавилов [6] виділив два осередки походження кар¬
топлі - Північноамериканський і Центральноамериканський, вклю¬
чаючи Антильські острови, а також Південноамериканський (перуа-
но-еквадоро-бо лівійський). В останньому виділено два підосередки -
чилоанський і бразильсько-парагвайський.
Уперше вчення про осередки походження культурних рослин
М.І.Вавилов опублікував у “Трудах по прикладной ботанике и селек¬
ции" в 1926 р. (Т.16, вип.2). Пізніше (1931, 1940 рр. та ін.) учений
неодноразово повертався до проблеми, уточнюючи і розширюючи її.
Так, наприклад, у роботі “Учение о происхождении культурных рас¬
тений после Дарвина” (1940) ним були виділені Центральноамери¬
канський осередок із гірським Південноамериканським, Центрально¬
американським і Вест-індійським вогнищами та Андійським із влас¬
не Андійським, Чилоансысим, Баготанським вогнищами.
Спираючись на вчення М.І.Вавилова, П.М. Жуковський [12] виділяє
такі центри походження різноманіття картоплі: Центральноамерикансь¬
кий, Південноамериканський і Північноамериканський.
У Центральноамериканський осередок (генцентр) входять Мек¬
сика, Гватемала, Коста-Рика, Гондурас, Панама. Серед перерахова¬
161
них країн особливо слід виділити Мексику, яку окремі вчені (Hawkes
I.G.) вважають первинним осередком еволюції бульбоутворення у
видів. Саме тут виявлено найбільше різноманіття картоплі, у тому
числі і за рівнем плоїдності (ди-, три-, тетра-, гексаплоїди).
Незважаючи на те, що найбільш повний еволюційний розвиток
картоплі від ди- до гексаплоїдів відбувався в Мексиці, повною мірою
в культуру бульбоутворююча картопля увійшла на території Півден¬
ноамериканського осередку. Це в першу чергу стосується тетраплоїд-
ного S.andigenum і, особливо, S. tuberosum. У цьому осередку еволю¬
ціонували також ди- і триплоїдні культурні види: S.ajanhuiri, S.go-
niocalyx, S.phureja, S.stenotonum, S. x chaucha, S. x juzepczukii, S.curtilo-
bum та деякі інші.
Найбільш бідний на різноманіття картоплі Північноамери¬
канський осередок, де ростуть такі дикі види картоплі, як S.jamesii,
Sfondleri.
Крім генцентрів, П.М.Жуковський [12] виділяє мікроцентри по¬
ходження видів, що особливо стосується поліморфної секції бульбо-
утворюючої картоплі. Ця проблема ще не до кінця вивчена, але вва¬
жається, що S.tuberosum, S.papita, S.sanbucinum, S.clarum, S.maglia i
багато інших є /ендемічними для певних територій. Доведено, що ма¬
теріал із мікроцентрів може бути як гомозиготним, так і гетерозигот¬
ним.
Виділення осередків походження і різноманіття видів картоплі
має велике наукове і практичне значення. Шляхом визначення філо¬
генетичного зв’язку між ними можна з’ясувати еволюцію культури.
Цьому також сприятиме ідентифікація гібридогенних видів, які, як
правило, утворювалися при накладанні ендемічних географічних мік¬
роцентрів. Вивчення ступеня гомологічності геномів видів великою
мірою пояснює їх схрещуваність.
Розвиваючи ідею М.І.Вавилова [8] про те, що стійкі до шкідників
і збудників хвороб форми найчастіше зустрічаються в осередках їх
походження, П.М. Жуковський [12] створив теорію спорідненої ево¬
люції рослини-господаря і паразита, суть якої полягає в тому, що ви¬
хідний матеріал для селекції необхідно шукати в місцях сумісного
знаходження рослини-господаря і паразита. Осередки походження
культурних рослин характеризуються винятковою мінливістю існу¬
ючих там видів. Цьому сприяє виражене більше, ніж в інших місцях,
перехресне запилення навіть самозапильних форм. Крім того, умови
для росту рослин в осередках їх походження сприяють формотвор¬
чому процесу і значною мірою - прояву гетерозиготності. Ця теорія
має надзвичайно велике значення для селекції взагалі і селекції на
імунітет, зокрема. Вона дозволяє сконцентрувати пошук стійких
форм у мікроцентрах із значним поширенням патогенів, тобто вико¬
ристовувати природну еволюцію резистентних форм, утворених під
патогенним тиском збудників хвороб.
162
Яскравим прикладом спорідненої еволюції видів картоплі і пара¬
зита є взаємовідносини при сумісному існуванні збудника фі¬
тофторозу і бульбоутворюючих видів Мексики. Саме тут знаходить¬
ся центр расоутворення гриба і сприятливі умови для його по¬
ширення. Крім того, вперше статевий спосіб розмноження гриба ви¬
явлений саме в Мексиці (Gallegly М.Е.). Формотворчий процес диких
видів, ареал яких знаходиться в цьому регіоні, сприяв еволюціону¬
ванню стійких форм, пошук яких тут є надзвичайно перспективним.
Аналогічним є поширення в Аргентинських Андах картопляної не¬
матоди та еволюціонування там стійких видів S.andigenum, S.vernei,
S.kurtzianum та ін.
Існує припущення (Букасов С.М.), що осередки є одночасно
центрами домінантних ознак, тобто домінантні властивості форму¬
ються в генцентрах. Такий теоретичний підхід дозволяє визначити
види, перспективні для практичного селекційного використання за
якоюсь однією ознакою.
3.8.2.Систематика бульбоутворюючих видів
секції Petota D итоги роду Solanum L.
До останнього часу науково обгрунтованої остаточної системи кар¬
топлі немає, незважаючи на те, що перші спроби створити її були зроб¬
лені давно (Вальперс К.; Дюналь М.). Такий стан зумовлений багатьма
причинами, зокрема складністю секції Potata для систематиків (De
Candolle), різними підходами систематиків-картоплярів до виділення
видів як таксономічних одиниць у культурі, тобто встановлення діагно¬
зу, що відповідав би вимогам Міжнародного кодексу ботанічної номен¬
клатури 1975 р.; зникненням деяких видів у процесі діяльності людини;
відкриттям нових видів (тільки протягом 1980-1984 рр. було відкрито й
описано більше 30 нових видів картоплі) [11] тощо.
Крім видатного соланолога Г.Біттера [33], систематикою картоплі
займалися С.В.Юзепчук (1937), Л.Даннерт (1956), C.Ochroa (1958, 1964),
H.Brucher (1968) та ін. На сьогодні існують три найбільш повні й аргу¬
ментовані системи культури: С.М.Букасова (на ранньому періоді у спів¬
авторстві з С.ВЮзепчуком - 1959, 1971, 1978 рр.), американського бо¬
таніка Д.Коррелла [38] і англійського - Д. Хокса [39]. Певний внесок у
систематику виду S.andigenum зробив В.СЛехнович (1978), а південно¬
американських видів - Л.Є. Горбатенко [11].
Погляди С.М.Букасова і Д. Коррелла на таксономію виду і серії
майже завжди збігаються. На відміну від них, Д.Хокс вважає їх
більш значними комплексами. Наприклад, у Д.Хокса виділена одна
серія Comersoniana з п’ятьма видами, а у С.М.Букасова вона розділе¬
на на чотири дрібніші серії, до складу яких входить 20 видів.
Вважається, що система С.М.Букасова, порівняно із такою Д.Хокса,
характеризується більшою логічністю, простотою пояснення з позиції
диференційного ботаніко-географічного методу М.І.Вавилова [6].
163
Нині відомо більше 200 видів картоплі, об’єднаних у підсекцію
Potatoe (раніше Huperbasarthrum) секції Petota ( раніше Tuberarium)
(В.Агсу). В Україні, країнах СНД притримуються системи картоплі
С.М Букасова [4]. Наводимо перелік підсекцій, серій, видів, а також
загальноприйняті скорочення назви останніх (Huaman Z.,Ross R.).
Система диких і культурних видів картоплі
селекції Tuberarium (Dun.) (нині Petota Dumort.)
роду Solanum L.
Серія Berthaulitana Вик.
S.berthaultii Hawk. ber. 2n=24
Серія Transaeguatorialia Buk.
S.abancayense Qchoa. abn. 2n=24
S.alandiae Card. 2n=24
S.amabile Vargas aml. 2n=24
S. ambo sinum Ochoa amb. 2n=24
S.aracc-papa Juz. arp. 2n=24
S.brevicaule Bitt. brc. 2n=24
S.brevimucronatum brv. 2n=24
Hawk.
S.bukasovii Juz. buk. 2n=24
S.canasense Hawk. can. 2n=24
S.candollenum Berth end. 2n=24
S.catarthrum Juz. ctr. 2n=24
S. coelestipetalum Vargas. cop. 2n=24
S.doddsii Corr. dds. 2n=24
S.famatinae Bitt. et Wittm. fint. 2n=24
S.gourlayi Hawk. grl. 2n=24
S.kurtzianum Bitt. et ktz. 2n=24
Wittm.
S.incamayoense Okada. inm. 2n=24
S.lechnoviczii Hawk. lez. 2n=24
S.leptophyes Bitt. lph. 2n=24
S.liriuxiianum Card. et lm. 2n=24
Hawk.
S.multidissectum Hawk. mlt. 2n=24
S.multiinterruptum Bitt. mtp.2n=24
S.neohawkesii Ochoa nhk. 2n=24
S.neorosii Hawk. nrs. 2n=24
S.okadae Hawk. oka. 2n=24
S.oplocense Hawk. opi. 2n=24
S.parvicorollatum Lechn. 2n=24
S.ruiz-cevallosii Card. rzk. 2n=24
S.sandemanii Hawk. 2n=24
S.scadrifolum Ochoa 2n=24
S.setulosistylum Bitt. stl. 2n=24
S.soukupii Hawk. sou. 2n=24
S.sparsipilum (Bitt.) Juz.
et Buk. spl. 2n=24
S.spegazzinii Bitt. spg. 2n=24
S.sucrense Hawk. scr. 2n=24
S.vernei Bitt. et Wittm. vm. 2n=24
S.vidaurrei Card. vdr. 2n=24
Серія Piurana Hawk.
S.acroglossum Juz.
S.chiquidenum Ochoa
S.chomatophillum Bitt.
S.cuscoense Ochoa
S.hypacrarthum Bitt.
S.marinasense Vargas
S.pampasense Hawk.
S.paucissectum Ochoa pns. 2n=24
S.piurae Bitt . pur. 2n=24
S.immite Dun. imt
S.solisii Hawk.
S. blanco-galdosii Ochoa
Серія Cuneoalata Hawk.
S.infundibuliforme Phil. ifd. 2n=24
Серія Megistacroloba Card. et
Hawk.
S.ddichocremastrum Bitt. hst. 2n=24
S.hastiformum Corr. hst. 2n=24
S.megistacrolobum Bitt. mgs.2n=24
S.raphanifolium Card. et rap. 2n=24
Hawk.
S.sanctae-rosae Hawk. sct. 2n=24
Серія Lignicaulia Buk.
S.lignicaule Vargas lgl. 2n=24
Серія Ingaefolia Ochoa
S.jalcae Ochoa jlc. 2n=48
S.tuquerrense Hawk. tuq. 2n=24
acg. 2n=24
chq. 2n=24
chm. 2n=24
czc. 2n=24
2n=24
mm.2n=24
pam. 2n=24
2n=24
2n=24
2n=24
164
Серія Simpliciora Вик.
Серія Glabrescentia Buk.
S.gandarillasii Card. gnd. 2n=24
S.angustisectum (Hassl.)
S.gigantophyllum Bitt. gig.
2n=24
Buk.
2n=24
S.microdontum Bitt. mcd. 2n=24
S.boergeri Buk.
2n=24
S.simplicifolium Bitt. sim. 2n=24
S.chacoense Bitt.
2n=24
S.senturii Hawk.
S.cuevoanum Card.
2n-24
et Hjert. snt. 2n=24
S.dolichostigma Buk.
2n=24
S.virgultorum (Bitt.) Card. et
S.garciae Juz. et Buk.
2n=24
Hawk.
2n=24
S.gibberulosum Juz. et
Серія Alticola Buk.
Buk.
2n=24
S.boliviense Dun.
2n=24
S.horovitzii Buk.
2n=24
S.sogarandinum Ochoa
2n=24
S.emmeae Juz. et Buk.
2n=24
S.toralapanum Card. et Hawk. 2n=24
S.knappei Juz. et Buk.
2n=24
Серя^4caule Buk.
S.laplaticum Buk.
2n=24
S.acaule Bitt.
2n=48
S.parodii Juz. et Buk.
2n=24
S.aemulans Bitt. et Wittm.
2n=48
S.saltense Hawk.
2n=24
S.albicans (Ochoa) Buk.
2n=72
S.schickii Juz. et Buk.
2n=24
S.depexum Juz.
2n=48
S.subtilius Bitt.
2n=24
S.punae Juz.
2n=48
Серія Commersoniana
Buk.
S.schreiteri Buk.
2n=48
S. commersonii Dun.
2n=24,36
S.ujunense Card.
2n=48
S.enteriosense Buk.
Серія Conicibaccata Buk.
S.malmeanum Bitt.
S.acroscopicum Ochoa
S.pyriforme Buk.
2n=24
S.colombianum Dun.
2n=48
S.rionegrinum Buk.
2n=24
S.flahaultii Bitt.
Серія Collina Buk.
S.huancabambense Ochoa
2n=24
S.chacayense Ochoa
2n=24
S.laxissimum Bitt.
S.maglia Schlechtd.
2n=24,36
S.limbaniense Ochoa
S.medians Bitt
2n=24
S.moscopanum Hawk.
2n=72
S.mochiquense Ochoa
2n=24
S.pausijugum. Bitt.
S.pamiricum Perlova
S.santolallae Vargas
2n=24
S.weberbaueri Bitt.
2n=24
S.subpandurpatum Ochoa
Серія Tuberosa (Rydb.) Buk
S.tundalomense Ochoa
S.guaytecarum (Bitt.) Buk.
2n=48
S.urubambae Juz.
2n=24
S.leptostigma Juz.
2n=48
S.violaceimarmoratum
S.molinae Juz.
2n=48
Bitt.
2n=24
S.ochoanum Lechn.
2n=48
Серія Circaeifolia Hawk.
Серія Oxycarpa Buk.
S.capsicibaccatum Card.
2n=24
S.agrimonifolium Rydb.
2n=48
S.circaefolium Bitt.
S.oxycarpum Schiede.
2n=48
Серія Tarijense Corr.
Серія Verrucosa Buk.
S.tarijense Hawk.
2n=24
S.verrucosum Schlechtd.
2n=24
S. vallegrandense Card.
2n=24
Серія Demissa Buk
Серія Jungasensa Corr.
S.brachycarpum Corr.
2n=72
S.trigalense Card.
2n=24
S.demissum Lindi.
2n=72
S.yungasense Hawk.
2n=24
S.hougasii Corr.
2n=72
165
S.guerreroense Corr. 2n=72
S.iopetalum. (Bitt.) Hawk. 2n=72
S. semidemissum Juz. 2n=60
Серія Longipedicellata Buk.
S.ajuscoense Buk. 2n=48
S.antipoviczii Buk. 2n=48
S.coriaceifoliolatum 2n=48
Lechn.
S.fendleri A. Gray. 2n=48
S.hjertingii Hawk. 2n=48
S.malinchense Hawk. 2n=48
S. neoantipoviczii Buk. 2n=48
S .j orbiculatibaccatum 2n=4 8
Lecln. 2n=48
S.papita Rydb. 2n=48
S.polytrichon Rydb. 2n=A8
S.stoloniferum. Schlechtd. 2n=48
S.tlaxcalense Hawk. 2n=48
S.vallis-mexici Juz. 2n=36
Серія Borealia Corr.
S.wightianum Rydb. 2n=48
Серія Polyadenia Buk.
S.lesteri Hawk. 2n=24
S.polyadenium Greenm. 2n=24
Серія Pinnatisectum Buk
S. brachistotrichum (Bitt.)
Rydb. 2n=24
S.nayaritense (Bitt.) Rydb. 2n=24
S.jamesii Torr. 2n=24
S.pinnatisectum Dun. 2n=24
S. sambucinum Rydb. 2n=24
S.stenophyllidium Bitt. 2n=24
Серія Trifida Corr.
S.michoacanum (Bitt.)
Rydb. 2n=24
S.trifidum Corr. 2n=24
S.humectophilum Ochoa. 2n=24
Серія Cardiophylla Buk.
S.cardiophyllum Lindi. 2n=24
S.ehrenbergii (Bitt.) Rydb. 2n=24
Серія Bulbocasiana Rydb.
S.bulbocastanum Dun. 2n=24,36
Серія Clara Graham et
Dionne
S.clarum Corr. 2n=24
Серія Morelliformia Hawk.
S.morelliforme Bitt. et
Muench. 2n=24
Серія Andigena
S.ajanhuiri Juz. et Buk. 2n=24
S andigenum Juz et Buk. 2n=48
S.boyacense Juz. et Buk.
S.canarense Buk.
S.cardenasii Hawk
S.chocclo Buk. et Lechn.
S.ciec, al Buk. et Lechn.
S.chaucha Juz. et Buk. n=36
S.cuencanum Buk. 2n=36
S.goniocalyx Juz. et Buk.
S.kesselbrenneri Juz. et
Buk.
S.macmillanii Buk. et
Lechn.
S.mammillliferum Juz. et
Buk. n=36
S.multijugum Buk. et
Bavyko
S.phureja Juz. et Buk. 2n=24
S.rybinii Juz. et Buk. 2n=24
S.surimana Buk.
S.stenotomum Juz. et Buk. 2n=24
S. tenuifilamentum Juz. et
Buk. 2n=36
S.yabari Hawk.
Серія Subacaulia Buk.
S.juzepczukii Buk. 2n=36
S.curtilobum Juz. et Buk. 2n=60
Серія Chilotana Buk.
S.tuberosum ssp. 2n=48
chilotanum Buk. et Lechn.
S.tuberosum ssp.
europacum Buk. et Lechn. 2n=48
Дикі види. Дикі види картоплі - це види, які ростуть у малопри¬
датних для картоплі умовах і характеризуються наявністю дрібних і
несмачних бульб, що знаходяться на довгих столонах, відзначають-
166
ся, як правило, невеликим габітусом куща і в осередках походження
є бур’янами.
Детальний опис диких видів і їх селекційне значення наведені в
“Каталогах мировой коллекции ВИР” [14, 15].
Враховуючи те, що для виду S. tuberosum, а часто і для всіх куль¬
турних видів картоплі, не властивий ефективний генетичний конт¬
роль багатьох ознак, створення сортів із новим поєднанням ознак
шляхом гібридизації можливе лише при залученні в селекційну
практику диких видів.
Прояв ознак у диких видів дуже часто зумовлений їх ареалом, а
точніше - географічними, кліматичними та патогенними умовами,
що склалися в певному регіоні. Наприклад, Мексика є еколого-
географічним центром збудника фітофторозу. Тут склалися надзви¬
чайно сприятливі метеорологічні умови для існування гриба. Саме в
Мексиці вперше відкрито статевий спосіб розмноження збудника.
Існує думка, що в цьому регіоні є ще одна рослина, на якій патоген
підсилює свої фізіологічні властивості [14]. А тому більшість мекси¬
канських видів характеризується високою стійкістю проти гриба.
Особливо слід підкреслити, що значний ареал видів, наявність
їхніх ендемічних форм, грунтові і патогенні особливості мікрорегіо-
ну та інші причини зумовили поліморфізм видів за проявом і конт¬
ролем ознак. Навіть серед найбільш стійкого проти фітофторозу ви¬
ду S.bulbocastanum зустрічаються зразки з невисоким проявом цієї
властивості, зокрема по бульбах [31].
Через складність проведення міжвидових схрещувань, тривалість
процесу залучення диких видів у селекційну практику та інші причи¬
ни на сьогодні в сорти інтрогресовано гени лише окремих видів. При
створенні сортів картоплі найширше використовується дикий гекса-
плоїдний мексиканський вид S.demissum Lind, який поширений у
Мексиці та Гватемалі. У цілому вид характеризується стійкістю про¬
ти фітофторозу (надчутлива і польова), вірусів Y, L, різних патотипів
раку картоплі, цистоутворюючих нематод, стовбура, парші звичай¬
ної, кільцевої гнилі, ооспорозу. Наявність у рослинах алкалоїдів зу¬
мовлює певну стійкість проти колорадського жука, епіляхни, с лиз¬
нів. Недоліком цього виду слід назвати ураження альтернаріозом і
деякою мірою - ризоктоніозом. Бульби окремих зразків виду мають
високий уміст крохмалю (22-33%), білка - 2,5-5,4%. Вид характери¬
зується певною стійкістю проти заморозків, відносно легко схрещу¬
ється з дикими і культурними видами, селекційними сортами, особ¬
ливо при використанні його як материнської форми.
Меншою мірою в практичній селекції використовуються інші
дикі види картоплі, зокрема S.stoloniferum Schlechtd., 2п=48, який
широко розповсюджений у Мексиці. Окремі зразки його мають ефе¬
ктивний контроль стійкості (аж до імунітету) проти вірусів X, Y, А і
добре передають цю ознаку нащадкам. У цілому вони характеризу¬
167
ються високою резистентністю проти скручування листків, SBK,
МВК, фітофторозу, раку картоплі, парші звичайної, чорної ніжки,
альтернаріозу. У польових умовах вид стійкий проти картопляної і
стеблової нематод, персикової тлі, епіляхни, колорадського жука.
Бульби не уражуються ооспорозом і незначною мірою уражуються
ризокто¬
ніозом.
S.acaule Bitt., 2п=48. Має найбільший ареал серед усіх диких ви¬
дів - довжину 3000 км і по висоті до 5000 м над рівнем моря. Поши¬
рений у Перу, Аргентині, Лімі, Болівії. Є одним із найбільш морозо¬
стійких видів (витримує до —8° С при вологості повітря 85%) і добре
передає цю ознаку нащадкам. Стійкий проти звичайного і вірулент¬
них біотипів раку картоплі, парші звичайної і порошистої, ооспоро-
зу, альтернаріозу, вертициліуму, кільцевої і бурої гнилі, чорної ніж¬
ки, стеблової і картопляної нематод, МВК, SBK, вірусу скручування
листків. Імунний проти ХВК та надчутливий до УВК. Незначною
мірою пошкоджується епіляхною. Бульби мають підвищений уміст
крохмалю (до 23%) і білка (до 2,8%). Легко схрещується як з дикими
видами, так і з S. tuberosum.
S.chacoense Bitt., 2п=24. Поширений у Болівії, Парагваї, Арген¬
тині, Уругваї, Бразилії. Надзвичайно поліморфний вид. Його зразки
стійкі проти раку картоплі, альтернаріозу, парші звичайної і порошис¬
тої, чорної ніжки, кільцевої гнилі, сухої фузаріозної і мокрої гнилі,
вілту, УВК, АВК, SBK, МВК, вірусу скручування листків, стеблової і
картопляної нематод, а також проти шкідників (картопляного 28-
крапкового сонечка, колорадського жука, цикадки, картопляної молі,
епіляхни). За даними окремих дослідників, стійкий до фітофтори.
Бульби мають підвищений уміст крохмалю (16-29%) і білка (1,1-
3,3%). Відносно стійкий до заморозків, високої температури повітря
і засухи. Має цінність у селекції на врожайність.
S.vernei Bitt., 2п=24. Вид поширений в Аргентині, Болівії. Його
зразкам властива стійкість проти фітофтори (надчутлива і польова),
раку, картоплі, альтернаріозу, ризоктоніозу, парші звичайної, ооспо-
розу, чорної ніжки, ранньої сухої плямистості, мокрої бактеріальної
гнилі, ХВК, УВК, АВК, SBK, МВК, усіх відомих рас картопляної
нематоди, колорадського жука. Вид характеризується певною стійкіс¬
тю проти заморозків (до -4°С). Бульби мають підвищений уміст кро¬
хмалю і білка і низький - редукуючих цукрів.
S.spegazzinii Bitt., 2п=24. Вид поширений в Аргентині. Його зраз¬
ки характеризуються стійкістю до всіх видів нематод (картопляної,
стеблової, галової), патотипів раку картоплі, сухої фузаріозної гнилі,
до окремих шкідників. Витримує зниження температури. М’якуш
бульб не темніє. Вони мають незначний уміст редукуючих цукрів.
Інші дикі види ще меншою мірою використовуються у практич¬
ній селекції, а більша половини взагалі не залучені до цієї роботи,
168
хоч кожен із них має високий прояв властивостей, відсутніх у
S.tuberosum.
Наприклад, S.bulbocastanum Dun., 2n =24 характеризується дуже
високою стійкістю проти фітофторозу (Niderchauser Z.S.; Mills W.R.;
Лебедева Н.А.; Житлова Н.А.; Подгаєцький А.А. та ін.). Зразкам ви¬
ду властивий як олігогенний, так і полігенний тип контролю ознаки,
причому, з незначним проявом поліморфізму. За даними Д.Хермсена
і М.Раманна (Hermsen G., Ramanna М.), гібриди виду із S.acaule ( 9 ),
у яких ідентифіковано хромосоми S.bulbocastanum, характеризували¬
ся надзвичайно високою стійкістю проти фітофторозу. Крім того,
виду властива стійкість проти альтернаріозу, чорної ніжки, карто¬
пляної нематоди, ХВК, УВК, ризоктоніозу, колорадського жука та
інших шкідників. Разом з тим зразки виду уражуються ооспорозом,
слизнями, павутинним кліщем. Бульби мають підвищений уміст кро¬
хмалю (більше 20%) і білка (більше 3%). Враховуючи те, що еволю¬
ція виду відбувалась у сухих гірських районах на кам’янистих грун¬
тах, вид характеризується високою стійкістю до екстремальних умов
вирощування.
Оригінальний за формою листка вид S.pinnatisectum Dun2n=24
також росте в гірських районах Центральної Мексики на кам’я¬
нистих схилах, серед трав і кущів. Має стійкість проти фітофторозу
(r,Rx і полігени), чорної ніжки, УВК, АВК, LBK, колорадського жука,
епіляхни, стеблової і картопляної нематод, незначною мірою уражу¬
ється альтернаріозом. Бульби не уражуються ооспорозом і дуже ма¬
ло - ризоктоніозом. В окремі роки рослини проявляють низьку стій¬
кість проти альтернаріозу. Уміст крохмалю в бульбах - 28-37%, біл¬
ка-2,7-4,3%.
Вид S.polytrichon Rydb., 2п=48 поширений у північних районах
Центральної Мексики. Росте як на добре зволожених грунтах вулка¬
нічного походження, так і сухих кам’янистих, на відкритій місцевос¬
ті або в гірських ущелинах. Особливістю виду є те, що його насіння
не втрачає схожості навіть після 20-річного зберігання в кімнатних
умовах. Зразки виду характеризуються стійкістю проти фітофторозу
(r,Rx і полігена), УВК, АВК, чорної ніжки, колорадського жука, епі¬
ляхни, картопляної нематоди, альтернаріозу, ооспорозу. Завдяки силь¬
ному опушенню не заселяється попелицями. Середньостійкий проти
ризоктоніозу. Відносно стійкий до заморозків. Уміст крохмалю в бу¬
льбах - 22-26%, білка - 2,4-4,2%.
Культурні види картоплі. Це види, що мають значний габітус
куща, великі бульби на коротких столонах, хороші та задовільні сма¬
кові якості, які використовувались і використовуються для харчу¬
вання людини.
J.Hawkes (1979) вважає, що до культурної картоплі належать 4 прос¬
ті (S.stenotomum, S.phureja, S.goniocalyx, S.tuberosum) і 4 гібридогенні
(S. x chaucha, S. x ajanhuiri. S. x juzepczukii, S. x curtilobum) види. Ще
169
примітивнішу класифікацію пропонував K.Dodds (1962), який вважав
S.phureja, S.stenotomum, S. * chaucha, Ssp.andigena i Ssp.tuberosum гру¬
пами одного виду S.tuberosum. C.M. Букасов лише серед болівійських
культурних видів картоплі виділяє не менше восьми: S.phureja,
S.ajanhuiri, S.stenotomum, S.stenuifilamentum, S.chaucka, S.juzepczukii,
S.curtilobum, S.andigenum. За межами Болівії, вважається, існує ще шість
видів: S.goniocaliyx, S.mammiliferum, S.choeclo, S.canarense, S.cuencamun,
S.rybinii. Л.Е.Горбатенко [10] виділяє 11 культурних видів картоплі:
S.curtilobum, S.juzepczukii, S.ajanhuiri, S.goniocalyx, S.phureja, S.rybinii,
S.stenotomum, S.andigenum, S.higrothermicum. S.estradae, S.tuberosum.
Тобто, особливо значним є розходження вчених-систематиків картоплі
щодо культурних видів.
Для культурних видів характерні багатоярусне квітування, добре
зав’язування ягід від самозапилення, здатність зав’язувати бульби на
коротких столонах. Окремі види {S.phureja, S.rybinii та деякі інші)
мають короткий період спокою бульб, високий уміст крохмалю (до
26%), білка (до 5%), значну кількість каротину, вітаміну С, хороші
смакові якості та інші цінні агрономічні властивості, у тому числі
стійкість проти хвороб.
Характеристика окремих культурних видів картоплі S.tube¬
rosum L., 2п=48. Відносно систематики виду серед учених існують
діаметрально протилежні думки. Д.Хокс (Hawkes D.) вважає, що вид
S.tuberosum походить від високогірного S.andigenum після пере¬
несення останнього з Болівійських Анд в Чилі мігруючими індійсь¬
кими племенами з наступною адаптацією до умов довгого дня. С.М. Бу¬
касов і В.С. Лехнович (1971) вважають, що S.tuberosum походить від
місцевого дикого предка, який, імовірно, через деякі обставини (по¬
ширення фітофторозу) перестав існувати. Крім того, важко припус¬
тити можливість подолання видом S.andigenum природного бар’єра -
пустині Атакама, а також адаптації його в нетипових географічних
умовах (із території, розміщеної на 25-30° південної широти на 40-
50° південної широти).
Аборигенні чилійські сорти відзначаються високим умістом крох¬
малю, білка, аскорбінової кислоти, стійкі проти механічних пошко¬
джень, мають хороші смакові якості, підвищену продуктивність,
польову стійкість проти вірусних хвороб (Турулева Л.М.).
Селекційним сортам виду властива стійкість проти раку картоплі,
парші звичайної і порошистої, ХВК, УВК, АВК, LBK, МВК. Окремі
форми мають стійкість до нематод, у тому числі і блідої картопляної,
до фузаріозної гнилі, іржавості бульб, польову стійкость проти фіто¬
фторозу. У цілому це високоврожайні форми з хорошими смаковими
якостями бульб.
S.andigenum Juz.et Вик., 2п=48. Це надзвичайно поліморфний
вид (Лехнович В.С., 1978), серед якого виділені географічні форми і
підвиди, що мають особливі морфологічні і фізіологічні властивості.
170
В.С. Лехнович (1971) вважає, що майже за всіма ознаками кожен з
видів S.andigenum і S.tuberosum утворює свої ряди мінливості. Осно¬
вний ареал S.andigenum - Південна Америка (від Колумбії до Арген¬
тини) . В.С. Лехнович (1971) виділяє створений ареал виду - високо¬
гір’я Центральної Америки. Обидва ареали характеризуються корот¬
ким днем, що є основною особливістю виду.
Зважаючи на поліморфізм, серед зразків виду можливий пошук
джерел стійкості проти звичайного та агресивних біотипів раку кар¬
топлі, альтернаріозу, звичайної і порошистої парші, чорної ніжки,
ХВК, УВК, АВК, а окремих клонів - до LBK, золотистої цистоутво¬
рюючої картопляної нематоди (контролюється домінантним геном),
деяких клонів - до блідої та інших нематод. Зразки вторинного ареа¬
лу мають польову стійкість проти фітофтори. Окремі клони практич¬
но не пошкоджуються тлею, картопляною корівкою. Бульби містять
до 32% сухої речовини і до 4% білка, витримують заморозки до
-2°С. Як правило, представники виду мають високу врожайність,
хороші смакові і кулінарні якості бульб, в окремих випадках придат¬
ні для виготовлення чипсів.
S.rybinii Juz» et Buk.у 2n=24, 36. Ареал його - гірські райони Ко¬
лумбії. Місцеве населення серед виду виділило велику кількість сор¬
тів, які успішно вирощуються і до нинішнього часу. Виду властивий
короткий період спокою, а тому він використовується при виведенні
двоврожайних скоростиглих сортів. Бульби містять до 26% сухої ре¬
човини, у них високий уміст білка (до 4,5 %), вітаміну С. М’якуш їх
жовтий, не темніє в сирому вигляді. Кулінарні якості хороші. Виді¬
лені зразки стійкі проти раку картоплі, альтернаріозу, ооспорозу, ри¬
зоктоніозу, парші звичайної, порошистої, сріблястої, чорної ніжки,
кільцевої гнилі, ХВК, ВЛК, МВК, УВК, а також відзначаються від¬
носною польовою стійкістю проти фітофторозу.
S.phureja Juz, et Вик., 2п=24. Ареалом його є південь Перу і гірсь¬
кі райони Болівії. Росте переважно у вологій місцевості. Відрізняєть¬
ся від S.rybinii сильним антоціановим забарвленням бугристих, ова¬
льних бульб, переважно білим кольором м’якуша, рідким ягодоутво-
ренням, формою кінцевої частини листка (не збільшена). Для селек¬
ційної практики цінний як носій стійкості проти раку картоплі, аль¬
тернаріозу, парші звичайної, чорної ніжки, ризоктоніозу, кільцевої
гнилі, АВК, а окремі зразки - до SBK, МВК, ХВК, УВК. Дуже стій¬
кий проти вірусу скручування листків, зеленої персикової попелиці.
Може також бути використаний у селекції на стійкість проти нема¬
тод, у тому числі галової, і підвищений уміст сухої речовин, білка,
вітаміну С і низький - редукуючих цукрів. Добре витримує високі
температури.
Аборигенні сорти. Це сорти, які виділені серед диких культур¬
них видів місцевим населенням країн, віднесених до осередків похо¬
дження картоплі, і широко використовувались у далекому минулому
171
і знаходять своє застосування й сьогодні. Вважають [4], що комер¬
ційні сорти картоплі не лише Європи, але й Сполучених Штатів
Америки XIX, XX ст. є нащадками аборигенних сортів Чилі і Перу,
що підтверджує опис старих європейських сортів. Наприклад, вияв¬
лено, що сорт Sancisse аналогічний Conv. coraila і т.п.
Експедиції з пошуків картоплі в Південній Америці дозволили
виявити певну однорідність чилійських аборигенних сортів, що зу¬
мовило виділення їх в окремий вид - S.chilotanum. Крім того, на
островах Чилоє і в Арауканії збереглися стародавні індійські сорти
S.tuberosum. Частина з них до наших днів мають свою першу назву.
Незважаючи на те, що аборигенні чилійські сорти рідко викорис¬
товувались у практичній селекції США, країнах Європи, їхні окремі
цінні якості, такі, як урожайність, скоростиглість, стійкість проти
окремих вірусів та інші інтрогресовані у доробок селекціонерів. На¬
приклад, сорт Early Rose, виведений у СІЛА в 1961 р., є сіянцем від
самозапилення Garnet Chile. С.М.Букасов (1959) виділяє кілька груп
різновидів аборигенних чилійських сортів: var.chilotanum, var.ele¬
gans, var.villaroela, var.recurvatum, var.multibaccatum.
Значно ширшим таксономічним різноманіттям відзначаються абори¬
генні сорти Перу. Окремі з них належать до видів S.juzepczukii
S.curtilobum. Основна перевага їх - морозостійкість, що дозволяє виро¬
щувати картоплю на висоті до 4300 м над рівнем моря, хоча за врожай¬
ністю та іншими властивостями вони не можуть конкурувати з предста¬
вниками інших видів. Надзвичайно велике різноманіття серед перуансь¬
ких сортів має S.andigenum. Для практичного використання придатні
близько 1200 його форм. Порівняно із S.andigenum, менш урожайними є
сорти іншого культурного виду - S.stenotomum, яких нараховується
кілька десятків. Причому, вони характеризуються відносною стійкістю
проти хвороб, зокрема вірусних, хорошими смаковими якостями. Або¬
ригенні сорти інших видів ще менше поширені в Перу.
Значне розповсюдження мають стародавні сорти в Колумбії, особ¬
ливо в її гірських районах. Вони являють собою ди- і тетраплоїди.
Окремі з них за проявом високих якостей бульб, стійкістю проти низь¬
ких температур та доброю пристосованістю до несприятливих зов¬
нішніх умов (грунти, погода та ін.) мають певну цінність для практич¬
ної селекції (Горбатенко Л.Е., 1982). Найбільше поширення мають
сорти видів S.kesselbrenneri, S.rybinii, S.boyacense.
Особливістю аборигенних сортів Еквадору, порівняно з іншими
індійськими країнами Південної Америки, зокрема виду S.andi¬
genum, є великобульбовість, правильна їхня форма та ін. Ранні фор¬
ми віднесені до видів S. canar ense, S.kesselberenneri, S.cuencanum, де¬
якою мірою це стосується і виду S.goniocalyx.
Значна кількість аборигенних сортів Болівії є представниками ди-
і триплоїдних видів, зокрема S.ajanhuiri, S.juzepczukii, S.phureja,
S.chaucha, S.stenotomum, S.andigenum. Форми перших двох і остан¬
172
нього відзначаються морозостійкістю і використовуються для виго¬
товлення тунту (вид вимороженої сушеної картоплі). Сорти виду
S.phureja, як правило, мають досиггь високий уміст крохмалю, значно
різняться за забарвленням шкірки бульб. Сорти виду S.chaucha харак¬
теризуються глибоким заляганням вічок, рожевим забарвленням
бульб і специфічним смаком, який нагадує смак кукурудзи. Сорти
виду S.stenotomum мають різне забарвлення бульб, високий уміст
крохмалю, значну розварюваність.
Отже, аборигенні сорти, залежно від вимог до картоплі та умов
вирощування, відбиралися від різних видів, здебільшого культурних.
Широкий спектр розсіювання властивостей серед них (часто селек¬
ційно цінних) дозволяє вважати аборигенні сорти важливою складо¬
вою генофонду картоплі.
3.8.3. Вихідний передселекційний матеріал
(попередні дослідження та його створення)
Епоха міжвидової гібридизації в селекції картоплі зумовила не¬
обхідність визначення стратегічних і тактичних напрямів викорис¬
тання при створенні сортів перш за все диких і культурних видів, а
також інших складових генофонду картоплі. Доведено (Букасов С.М.),
що при схрещуванні комерційних сортів із дикими і навіть культур¬
ними видами практично не можливо отримати форми, які б за всіма
ознаками відповідали вимогам до сортів. Встановлена необхідність
виконання спектра робіт щодо зменшення прояву в матеріалі міжви¬
дового походження негативних властивостей і насичення його пози¬
тивними (Букасов С.М., Камераз А.Я.; Будін К.З.), [15].
Першим етапом щодо залучення співродичів культурних сортів у
практичну селекцію повинен бути збір зразків у місцях спорідненої
еволюції рослин-господарів і патогенів чи видів із високим проявом
інших агрономічних властивостей (біохімічний склад бульб тощо)
або інтродукція такого матеріалу від утримувачів світових колекцій.
Враховуючи те, що кожен регіон характеризується певною еко¬
логічною і пагогенною ситуацією, перша оцінка (фенотипічна) інт-
родукованого матеріалу має бути зроблена з урахуванням цих особ¬
ливостей. Попри те, що вид S.pinnatisectum вважається фітофторос¬
тійким, (Зотеева Н.М.), [12], фенотипічний прояв цієї властивості у
нього в умовах Полісся України має не дуже високий ступінь (Под-
гаєцький А.А., 1978, 1993).
Поліморфізм прояву ознак у межах видів [12, 21, 28] вимагає
проведення оцінки якомога більшої кількості зразків. Крім того, фе¬
нотипічний прояв ознаки часто не адекватний ефективності контро¬
лю її. Тому при виборі вихідної форми для залучення в міжвидову
гібридизацію необхідне визначення останнього. У своїх досліджен¬
нях з цією метою ми використовували аналіз прояву ознаки серед
нащадків від самозапилення [21,31].
173
Значний гетероалелізм контролю властивостей у картоплі (Яши¬
на И.М., Склярова Н.П.) зумовлює необхідність проведення в перед-
селекційному процесі ще одних досліджень - гомозиготизації цінних
алелів контролю ознак. Доведено, наприклад, що при фітофторостій-
кості листків або бульб гомозиготизація цінних алелів відбувається
до І4 [24,31], а сухої фузаріозної гнилі - до І3 -14 [32].
Труднощі отримання первинних (одержаних від безпосереднього
схрещування двох видів) міжвидових гібридів зумовлються генетич¬
ною та філогенетичною віддаленістю видів картоплі між собою.
Крім того, у нащадків від таких схрещувань відбувається особлива
алельна взаємодія генів, залучених у гібридизацію видів. Через це
уже на першому етапі безпосереднього створення вихідного перед-
селекційного матеріалу спостерігається зниження прояву властивос¬
тей, порівняно з кращою батьківською формою. Разом з тим навіть
серед цього матеріалу виявлено позитивний ступінь домінування і
високу позитивну частоту трансгресії [31].
При залученні в селекційну практику культурних видів картоплі,
особливо диких, отримати потомство з комплексом агрономічних
властивостей досить складно (Букасов С.М., Камераз А.Я.; Будин
К.З.), [25]. Для покращення якостей первинних міжвидових гібридів
проводять додаткове схрещування з іншим видом або видами (при
цьому утворюються вторинні міжвидові гібриди), бекросування (пов¬
торне схрещування із сортами), отримання генеративних поколінь
або чергування двох останніх. У процесі бекросування отримують
бекроси (за аналогією, як і при гібридизації, одержують гібриди), які
позначаються латинською буквою "В”. Ступінь біля "В" показує кі¬
лькість повторних, крім першого, схрещувань із сортами (В1, В , В3 і
т.п.). Сам термін “ступінь бекросування” вказує на це. На повтор¬
ність бульбового репродукування вказує індекс, що ставиться знизу (
Ві, В2, В3 і т.п.). Перше гібридне покоління позначається Fb а насту¬
пні - відповідно F2, F3 і т.д.
Тобто, передселекційний процес складається із збирання, виді¬
лення і створення форм, які залучаються в міжвидову гібридизацію і
власне етапів щодо створення вихідного передселекційного матеріа¬
лу. Цей матеріал отриманий при залученні у процес виведення сортів
їхніх співродичів або з використанням інших методів і характеризу¬
ється високим проявом ознак, відсутніх у S.tuberosum, ефективним їх
генетичним контролем, але поряд із провідною (рідше провідними)
властивістю (властивостями) передає потомству при схрещуванні
негативні ознаки, а тому, як правило, не може бути використаний
безпосередньо при створенні нових сортів.
Вихідний селекційний матеріал. Вихідний селекційний матері¬
ал - це форми, які отримані з використанням різних методів, що за¬
стосовуються безпосередньо при створенні нових сортів картоплі.
Він характеризується комплексом агрономічних властивостей, добре
174
передає їх потомству, а частка вищеплення форм із високим проявом
негативних ознак невелика, порівняно з вихідним передселекційним
матеріалом. Ці форми за більшістю своїх властивостей відповідають
вимогам до сортів, але через наявність, як правило, однієї або кіль¬
кох негативних не можуть бути рекомендовані для виробництва.
Вихідним селекційним матеріалом можуть бути аборигенні, міс¬
цеві, комерційні сорти, гібриди, отримані в селекційному процесі і за
проявом ознак залишені для подальшого використання при прове¬
денні генетичних досліджень (наприклад вивчення комбінаційної
здатності), у результаті досліджень із клітинної селекції або генетич¬
ної інженерії тощо.
Особливу цінність для практичної селекції як вихідний матеріал
мають форми, отримані у процесі бекросування первинних і вторин¬
них міжвидових гібридів. Значні успіхи в цьому відношенні досягну¬
ті в Інституті Макса Планка (Німеччина), Млоховському науково-
дослідному центрі (Польща), Всеросійському інституті рослинницт¬
ва (ВІР) ім. М.І.Вавилова (Росія), Інституті картоплярства УААН
(Україна).
З використанням вихідного селекційного матеріалу, створеного в
Інституті Макса Планка (назва багатьох його форм починається з
МРІ), виведена значна кількість сортів (Костина Л.И.). Гібриди цієї
установи відзначаються стійкістю проти цистоутворюючих нематод
(іноді навіть двох видів - золотистої і блідої), багатьох вірусів, фіто¬
фторозу, бактеріозів і т.п. Висока загальна і специфічна комбінацій¬
на здатність цього матеріалу дозволила отримати з його участю ви¬
соковрожайні пластичні сорти. Наприклад, із комбінації Вірулане х
МРІ 40 663/21 відібрано три сорти: Сулев, Олев, Ієгева Тальвік.
Гібриди Млоховського науково-дослідного центру Інституту се¬
лекції рослин і акліматизації мають, перш за все, високу стійкість
проти вірусів - одного, кількох або комплексу.
Починаючи з 20-х років минулого століття, успішно проводяться
дослідження щодо створення вихідного селекційного матеріалу із
залученням співродичів культурних сортів у Всеросійському (раніше
Всесоюзному) інституті рослинництва ім. М.І.Вавилова. Як генетич¬
ні джерела для одержання бекросів були використані форми фіто¬
фторостійких видів: S.stoloniferum, S.vernei, S.demissum, s.berthaultii,
S.simplicifolium, S.verrucosum, S.polytrichon, вірусостійких - S.gourliy,
S.vernei, S.stoloniferum, стійких проти нематоди - S.andigenum, S.kur-
tzianum, S.famatinae, S.vernei, S.megistacrolobum, S.simplicifolium, S.cho-
caense [i].
За даними працівників ВІР [1,2], у багатьох випадках створений
матеріал є донорами, зокрема стійкості (імунітету) проти вірусів (ча¬
сто кількох), фітофторозу (полігенний контроль), картопляної цисто¬
утворюючої нематоди G.rostochiensis (контроль Ні), раку картоплі, а
також відносної стійкості проти колорадського жука, несприятливих
175
екзогенних факторів (підвищеної температури, засухи тощо). Маючи
широку генетичну основу, а також функціонування нередукованих
гамет у залучених у дослідження вадів, створені донори відзначаються
високою врожайністю (до 8,2 кг/м2), крохмалистістю (до 22-23 %), гар¬
ною формою бульб, короткими столонами та іншими агрономічними
властивостями. В окремих форм гетерозис досягає 21%.
Використовуючи унікальний за стійкістю проти фітофторозу ди¬
кий мексиканський вид S.bulbocastanum, а як посередники - S.acaule,
S.demissum, а також культурні види S.phureja, S.andigenum, в Інсти¬
туті картоплярства УААН створено вихідний селекційний матеріал з
високим проявом багатьох агрономічних властивостей. Стійкість
проти фітофторозу у більшості створених форм вища, ніж у кращих
сортів-стандартів, у 2-5 разів (Подгаєцький А.А., Чередниченко Л.М.).
Численні бекроси багатовидових гібридів мають високу польову
стійкість проти найбільш поширених у зоні Полісся України вірусів,
а окремі, незважаючи на тривале бульбове репродукування, не мали
X, У, S, М у прихованому вигляді. Виділені бекроси, які зовсім або
майже не уражуються стебловою нематодою при випробуванні в
умовах штучного інфекційного фону. Багато бекросів мають стій¬
кість проти раку картоплі, чорної ніжки, кільцевої гнилі, золотистої
цистоутворюючої нематоди. Виділено форми з груповою і навіть
комплексною стійкістю проти фітофторозу, сухої фузаріозної гнилі,
альтернаріозу, чорної ніжки, картопляної нематоди, а також з висо¬
ким проявом інших агрономічних властивостей: частіше - урожай¬
ності, крохмалистості, багатобульбовості, рідше - скоростиглості,
великобульбовості [19].
Місцеві сорти. Ін^одукція картоплі з осередків походження у
країни Європи та інші здійснювалася двома способами - викорис¬
танням бульб або ботанічного насіння. З того часу вирощування кар¬
топлі з ботанічного насіння мало деяке поширення. Щоб упорядку¬
вати цю роботу, в 1933 р. була видана спеціальна постанова Нарком-
зему СРСР "Про масовий дослідно-господарський посів картоплі на¬
сінням". Вирощування картоплі з ботанічного насіння, мутаційні
зміни, що відбуваються постійно, та інші фактори зумовили появу
форм, які широко використовувались, і, можливо, дещо меншою мі¬
рою використовуються населенням при вирощуванні картоплі тепер.
Вони отримали назву “місцеві сорти”. Це - сукупність однакових за
походженням, господарськими властивостями і морфологічними
ознаками рослин картоплі, відібраних або створених під дією приро¬
дного (внаслідок природних мутацій та інших факторів) та найпрос¬
тішого штучного відбору (народна селекція) і які вирощуються в пе¬
вній місцевості.
Однією з основних особливостей місцевих сортів є висока адап¬
таційна здатність до вирощування в певних грунтово-метеоро-
логічних умовах. Практично основний принцип відбору таких форм
176
грунтувався на стабільності прояву основних господарських ознак
протягом тривалого часу вирощування без ретельного дотримання
насінницьких заходів. Іншою особливістю місцевих сортів є висока
стійкість до патогенного комплексу певного регіону, де вони мають
поширення. Ще одна особливість місцевих форм - задоволення ви¬
мог споживача до якості картоплі.
Результати досліджень, виконаних у лабораторії вихідного ма¬
теріалу Інституту картоплярства УААН, свідчать про стабільний по
роках прояв у багатьох місцевих сортів основних господарських
властивостей. Особливо це стосується років з екстремальним екзо¬
генним комплексом. Наприклад, у 1999 р., який мав саме таку ха¬
рактеристику, значна частина місцевих сортів колекції переважала
сорти-стандарти за врожайністю. Населення завжди було зацікав¬
лене в тому, щоб споживати картоплю у свіжому вигляді якомога
довший проміжок часу. Цього можна досягти, вирощуючи ранню
картоплю. А тому більшість із 49 місцевих сортів колекційного
розсадника Інституту картоплярства характеризується ранньостиг¬
лістю. Інша властивість, яка домінувала при відборі,- це хороші
смакові якості. Більшість місцевих сортів мають високий прояв цієї
ознаки. Тривале вирощування місцевих сортів без дотримання на¬
сінницьких заходів зумовило відбір форм із високою стійкістю
проти вірусних хвороб та тих, які мають значне поширення в пев¬
ній місцевості. З огляду на вищесказане, місцеві сорти є цінним
вихідним матеріалом для селекції картоплі, а отже - і складовою
генофонду культури.
Умовно місцеві сорти можна розподілити на три групи: віднос¬
но старі селекційні сорти, що втратили свою першу назву, часто
знеособлені; одержані з насіння від самозапилення або в резуль¬
таті схрещування любителями; сорти, які є природними мутанта¬
ми [ЗО].
Дуже часто в назві місцевих сортів зафіксований регіон їх виро¬
щування, наприклад: Полтавська рожева, Макарівський шедевр, Ро¬
жевий з Мілета, Малинківська сніжинка та багато інших.
Опис найбільш поширених в Україні місцевих сортів наведений у
науково-популярному альманасі "Картопля - другий хліб" (1995) і
книзі П.С.Теслюка, М.Я. Молоцького [ЗО].
Комерційні сорти. Сорти, які описані в науковій літературі із
згадуванням автора або установи-оригінатора і розмножуються для
виробничого вирощування чи використовуються для отримання си¬
ровини, є комерційними сортами. На сьогодні доробок селекціонерів
світу складає близько 5 тис. сортів, але, у зв’язку із зростаючими за¬
питами виробництва, кількість їх постійно збільшується.
Весь період селекції картоплі можна умовно поділити на два ета¬
пи. Перший, який тривав до 40-х років XX ст., відзначався тим, що
єдиним вихідним селекційним матеріалом при створенні сортів були
177
форми внутрішньовидового (у межах S.tuberosum) походження. При¬
чому, протягом цього періоду відбулися значні зміни в генетичному
потенціалі сортів. По-перше, навіть та незначна стійкість до деяких
збудників хвороб, шкідників, яка властива культурним сортам, на¬
прикінці XIX - на початку XX ст. здебільшого була втрачена через
цілеспрямований відбір високоврожайних, великобульбових з інши¬
ми господарськими властивостями сортів [20]. По-друге, зміни в со¬
ртименті сортів картоплі відбулися внаслідок сильного ураження і
випадання матеріалу в період епіфітотій фітофторозу, раку картоплі
[29]. Тобто, у цей період мало місце звуження генетичної основи ви¬
хідного селекційного матеріалу і зовсім не вирішувалася проблема
створення стійких сортів, тому що виду S.tuberosum не властивий
ефективний генетичний контроль резистентності проти багатьох
хвороб і шкідників (Букасов С.М.). Це дозволило окремим дослідни¬
кам стверджувати, що “...внутрішньовидова гібридизація уже не мог¬
ла дати нічого принципово нового. Селекція картоплі певною мірою
зайшла в тупик” (Камераз А.Я.).
Для другого етапу селекції характерним було залучення у прак¬
тичну роботу з виведення сортів картоплі диких і культурних видів.
Саме завдяки цьому вдалося повністю або значною мірою вирішити
проблему стійкості проти раку картоплі, картопляної нематоди, фі¬
тофторозу та інших збудників хвороб і шкідників, а також значно
розширити генетичну основу вихідного селекційного матеріалу
[22,29,40].
Окремі сорти широко залучались у схрещування, і за їх участю
були виведені нові, що дозволило виділити сорти-донори окремих
ознак. Наприклад, при схрещуванні із сортом Прієкульська рання
було отримано 37 сортів, і майже всі вони характеризуються скоро¬
стиглістю (Костина Л.И.). Численне потомство має сорт Рання роза
(Early Rose) - 816, а безпосередньо за його участю виведено 36 сор¬
тів, більшості з яких також властива скоростиглість.
Американський сорт Катадин лише в колишньому Радянському Со¬
юзі дав початок 30 сортам. Сказане такою ж мірою стосується і сорту
Цертифолія, за участю якого виведено 40 сортів (Костина Л.И.).
Деякі сорти мають значну морфологічну подібність. Це дозволи¬
ло виділити окремі з них у сортотипи (Костина Л.И.).
Добір при створенні нових сортів відбувався і відбувається в на¬
прямі збільшення висоти рослин, урожайності (не без впливу гетеро¬
зису), розсіченості листків, зміни контуру листка, підвищення крох¬
малистості, стійкості проти шкідників, хвороб, лімінації фіолетового
забарвлення поверхні бульб і їхнього м’якуша і т.п. (Костина Л.И.,
Жолудева З.П.).
Основні світові генобанки картоплі та їхнє значення. Залежно
від завдань, які вирішуються при створенні колекцій, їх умовно мож¬
на розділити на три категорії (Sawyr R.L.).
178
1. Прості. Головна мета при їх формуванні - це збільшення кількос¬
ті зібраних зразків. Опрацювання матеріалу, як правило, не проводиться.
2. Таксономічні. Крім поповнення колекцій, проводяться дослі¬
дження із систематизації генетичних ресурсів, але практичне вико¬
ристання інтродукованих форм відсутнє.
3. Практичні. Класифікований матеріал планомірно вивчається з
метою практичного селекційного використання.
Починаючи з перших організованих М.І.Вавиловим експедицій
працівників ВІРу С.М.Букасова, С.В.Юзепчука по збору співродичів
культурних сортів, поповнення колекцій цієї установи здійснюється і
до нинішнього часу. Тому вважається, що за обсягом та періодом
збору матеріалу колекція ВІРу є найбільшою у світі. На сьогодні во¬
на нараховує більше 5000 зразків диких і культурних видів у живому
вигляді [2]. Причому, там зібрана чи не найбагатша колекція сортів -
2500 (Будин К.З.). Завдяки цій установі всі наукові установи й орга¬
нізації колишнього Радянського Союзу мали необхідний для дослід¬
ження матеріал.
Крім збереження генетичних ресурсів картоплі, у ВІРІ прово¬
диться широке вивчення матеріалу із випробуванням його в умовах
штучного (Білоруський інститут захисту рослин - за стійкістю проти
раку картоплі, картопляної нематоди) та природного (Сахалінський
опорний пункт ВНДІФТ - за стійкістю проти фітофторозу) інфекцій¬
ного фону [1], а також з використанням лабораторних оцінок. А тому
каталоги, видруковані ВІРом, містять досить повну інформацію що¬
до прояву у зразків агрономічних властивостей.
Крім ВІРу, у світі існує ще кілька великих колекцій картоплі: На¬
ціональна колекція картоплі Шотландського науково-дослідного ін¬
ституту з проблем сільського господарства в Петландфілді (СРС)
(The commonwealth Potato collection), колекція міжрегіонального
проекту з картоплі (Вісконсин, CULA) (Hannemann R.E., 1979), все¬
світня колекція зародкової плазми Міжнародного центру з картоплі
(СІР) (Ochoa), аргентинська колекція картоплі Національного інсти¬
туту технології сільського господарства (INTA) (Ocada К.А.), чилій¬
ський генобанк картоплі у Вальдивії (Contereras А.Е. et al), голланд¬
сько-німецьке відділення генобанку картоплі у Брауншвейті - Німе¬
ччина (BGRC) (Van Sorst Е.М.).
Наступ цивілізації на первозданну природу, еволюція форм кар¬
топлі та інші фактори вносять корективи в існування різновидів цієї
культури. Деякі види зникли в природі, і сьогодні дослідники мо¬
жуть лише здогадуватися про їх існування за малюнками, описами
своїх попередників (Лехнович В.С.). Тому значення зібраного у сві¬
тових генобанках картоплі матеріалу надзвичайно велике не лише
для нас, але й прийдешніх поколінь. Під час наступних експедицій
не виявлені, наприклад, описані раніше чилійські види S.molinae,
S.leptostigm [4]. Інші перебувають під загрозою вимирання.
179
3.8.4. Стратегія і тактика використання
генофонду картоплі у практичній селекції
Раціональне використання генофонду картоплі. Однією з
найбільш складних проблем розробки положення щодо вико¬
ристання генофонду картоплі в селекційній практиці є визначення
стратегічних напрямів у створенні вихідного селекційного матеріалу,
донорів. Складність його зумовлюється значною тривалістю перед-
селекційного (за розробленою нами схемою 6-12 років) [19] і селек¬
ційного (згідно з прийнятою в Інституті картоплярства схемою виве¬
дення сортів - 10 років) (Методические рекомендации по проведе¬
нию исследований с картофелем, 1983) процесів, тобто необхідністю
передбачати властивості сортів на 16-22 роки.
Збільшення кількості видів, використаних при створенні вихід¬
ного селекційного матеріалу, розширює спектр стійкості його
проти хвороб і шкідників, підвищує гетерозиготність, і тим самим -
ймовірність вищеплення трансгресивних форм за багатьма озна¬
ками, у тому числі і полігенними (Skiebe K.; Bingham Е.Т.). У
зв’язку з цим в геномі деяких сортів присутні спадкові фактори не
менше шести диких і культурних видів (Stegemann H., Schnick D.), а
при виведенні канадського сорту Conestoga було використано сім
видів [29].
Генетичний потенціал стійкості проти картопляних нематод скла¬
дають 10 видів згідно із системою С.М.Букасова: S.andigenum,
S.kurtzianum, S.multidissectum, S.vernei, S.spegazzinii, S.gourlayi, S.oplo-
cense, S.sparsipilum, S.lignicaule, S.megistacrolobum (Mai W.F., Peter-
son L.C.; Langton F., Ross H.; Uhrig, ej.). Останнім часом одержані
сорти з генами чотирьох видів, які характеризуються стійкістю проти
цистоутворюючих нематод: S.andigenum, S.vernei, S.sparsipilum,
S.spegazzinii [29]. За участю S.gourlayi, S.oplocense створено вихідний
генетичний та селекційний матеріал також із значним рівнем прояву
ознаки, який успішно застосовується селекціонерами в практичній
роботі (Uhrig W.). Гібриди двох видів (S.multidissectum, S.kurtzianum)
використовуються як диференціатори при визначенні патотипів шкід¬
ника. Пошук джерел стійкості проти цистоутворюючих нематод роз¬
почався наприкінці 50-х - на початку 60-х років минулого століття
(Ellenby, с 1948; Mai U.F., Peterson L.L.). Перший неуражуваний па¬
разитом сорт СAntinema) був одержаний у 1961 році. Значне поши¬
рення останнім часом цистоутворюючих нематод стало передумо¬
вою для обов'язкового використання голландськими та німецькими
селекціонерами як одного із компонентів схрещувань стійких
форм, що зумовило залучення до створення вихідного селекційного
матеріалу широкого арсеналу генетичних ресурсів. Таким чином,
протягом майже 40 років більша частина видів, що відзначаються
стійкістю проти нематод (близько 80 %), залучена в селекційний
процес.
180
Подібне можна сказати і про використання генофонду картоплі
при створенні сортів, вихідного фітофторостійкого матеріалу, доно¬
рів ознаки. Вважають (Graham К.М.; Toxopeus H.J.; Black W.C.; Ка¬
мераз А .Я. та ін.), що висока стійкість проти фітофторозу властива
18-ти видам картоплі: S.demissum, S.bulbocastanum, S.oxycarpum,
S.verrucosum, S.pinnatisectum, S.stoloniferum, S.polyadenium, S.papita,
S.antipoviczii, S.hjertingii, S.trifidum, S.ambosinum, S.capsicibaccatum,
S .aeros copicum, S.vernei, S.berthaultii, S.simplicifolium, S.microdontum.
Починаючи з 30-х років, гени п’яти видів інтрогресовані в сорти.
Одержано перспективні гібриди ще із шести видами. Таким чином,
за 60 років більше 60 % фітофторостійких видів були залучені в до¬
слідження з виведення сортів картоплі.
Отже, за досить короткий історичний період більшість видів, які
відзначаються стійкістю проти фітофторозу, цистоутворюючих кар¬
топляних нематод, були використані при створенні нових, менш
уражуваних сортів. На нашу думку, при сучасних темпах залучення в
селекційну практику диких і культурних форм пройде зовсім небага¬
то часу, і всі види з ефективним генетичним контролем важливих
ознак будуть залучені в роботу. Враховуючи те, що через наявність
вищезгаданих причин надходження нових видів і зразків з ефектив¬
ними генами контролю основних господарських ознак з кожним ро¬
ком зменшується, необхідно розробити стратегію використання ге¬
нофонду культури. Уже зараз гострою є проблема раціонального ви¬
користання еволюційно відшліфованих ефективних факторів конт¬
ролю стійкості проти хвороб і шкідників для успішного проти¬
стояння людини з її можливостями виведення неуражуваних сортів
патогенам з їх пристосованістю до умов, що змінюються. Як зазна¬
чалося (Walker С.), мінливість збудників хвороб не дає змоги певно¬
му сорту залишатися таким протягом тривалого часу. А тому генети¬
кам і селекціонерам завжди потрібно мати резерв ефективних носіїв
спадковості контролю проти збудників хоча б найбільш шкідливих
та найпоширеніших хвороб і шкідників.
Раціональне використання джерел стійкості має зумовлюватись
еколого-патологічними характеристиками зони вирощування куль¬
тури, ступенем мінливості патогену, типами стійкості рослин-
господарів.
Через багато факторів зараженість грушу картопляними немато¬
дами може складати від поодиноких цист до 100-200 тис. личинок
або яєць на 100 см грунту, що є верхньою межею розмноження, після
чого вступають у дію механізми самообмеження популяції. Відповід¬
но до міжнародних правил, тільки сорти, які визначають швидкість
розмноження нематод (Pf/Pi<0,6), віднесені до стійких, і їх можна
вирощувати на заражених фітогельмінтозами площах. Проте Х.Росс
[29] вважає неправильним відносити до уражуваних сорти з
Pf/Pi<0,6-2,0. їх можна дозволити висаджувати на ділянках з невели¬
181
кою зараженістю (поодинокі цисти). Якщо за прогнозами у певному
регіоні після використання системи захисту не передбачається збіль¬
шення інфікованості грунту, то зовсім не обов'язковою є присутність
у геномі вирощуваних сортів ефективних носіїв спадковості стійкос¬
ті проти шкідника диких та культурних видів. У цьому й буде поля¬
гати ефективне використання генофонду картоплі.
Формотворчий процес збудника раку картоплі Sinchytrium endo-
bioticum (Scilb.) Percival незначний. У Західній Європі виявлено 10
біотипів патогену (Камераз А.Я. та ін.). В Україні у зоні Карпат іден¬
тифіковано три агресивні біотипи: рахівський, міжгірський, бу¬
ковинський, але значне поширення має лише звичайний - далемсь-
кий (“D” згідно з Неу А.). Враховуючи це, а також домінантний мо-
ногенний контроль стійкості проти патогену, можна стверджувати,
що присутності експресивного гена (генів) навіть у симплексній фор¬
мі достатньо для того, щоб сорт не уражувався грибом. А тому стра¬
тегія створення вихідного селекційного матеріалу, стійкого проти
раку картоплі, мало чим відрізняється від досліджень на нематодос-
тійкість.
Значно більшою мінливістю, порівняно з цистоутворюючими не¬
матодами, збудника раку картоплі характеризується Phytophtora
infestans (Mont.^ de Вагу. До 70-х років у популяції гриба нашої та
зарубіжних країн переважали відносно прості раси: 0,1,2,4, 1.2, 1.3,
1.4, 2.4, 3.4, 1.2.3, 1.2.4, 1.3.4, 1.2.3.4 (Федотова Т.И. та ін.). Протягом
наступного періоду популяція патогену зазнала змін у бік підвищен¬
ня вірулентності. У результаті цього були ідентифіковані складні
раси, включаючи 1.2.3.4.5.6.6+0.7.8.9.10.11 xyz (Чумакова А.И.). То¬
му раціональне використання генофонду картоплі в селекції на фі-
тофторостійкість повинно плануватись з урахуванням пластичності
гриба.
Стратегічний напрям у створенні фітофторостійкого вихідного
матеріалу в 20-30-х роках XX ст. базувався на широкому викорис¬
танні вертикального типу контролю ознаки. При пошуку джерел
стійкості також відбирались зразки з надчутливою реакцією на про¬
никнення патогену. Підвищення вірулентності Phytophtora infestans
стало причиною значного ураження фітофторозом форм, які після їх
створення вважались імунними проти гриба. І, навпаки, ті гібриди та
сорти, у геномі яких були присутні як домінантні, так і полігени, від¬
значалися певною стійкістю навіть після появи рас з більшою віру¬
лентністю, ніж набір R-генів створеної форми.
Втрата стійкості проти фітофторозу сортів картоплі, виведених
при використанні надчутливого типу контролю ознаки, внесла корек¬
тиви у стратегію і тактику створення неуражуваних сортів. Почина¬
ючи з 70-х років XX ст., використання лише вертикальної (Ван дер
Планк І.) стійкості розглядалось як безперспективний напрям дослі¬
джень (Shick R., Horfe А.). Не підтвердився прогноз про можливість
182
створення для патогену бар'єра шляхом поєднання в одному гено¬
типі 3—4 домінантних генів із виключенням при вирощуванні у ви¬
робничих умовах сортів, виведених на основі матеріалу з одним-
двома R-генами (Toxopeus H.J.). Значного ефекту в боротьбі з фі¬
тофторозом картоплі чекали від комбінування двох-трьох домінант¬
них генів у скоростиглих сортів картоплі (Zadina І.). Проте рання
поява хвороби, починаючи від повних сходів до квітування, а та¬
кож присутність у цей період високовірулентних рас збудника (Іва-
нюк В.Г., Константинович Г.П.) не дозволили при застосуванні та¬
кого підходу одержати позитивний практичний результат. Певної
уваги при створенні форм, здатних протягом тривалого періоду
протистояти інфекції, заслуговує думка про комбінування домінант¬
ного та рецесивного (полімерні гени) контролю стійкості (Букасов С.М.,
Камераз А.Я.). Особливо ефективне таке поєднання в ранніх форм
(Чумакова А.И.).
Отже, використання при створенні вихідного селекційного мате¬
ріалу складових генофонду картоплі повинно відбуватися з ураху¬
ванням патогенної ситуації регіону, де проводяться селекційні дослі¬
дження, визначення ефективності генетичного контролю ознак видів,
які залучаються в роботу, і можливості «прогресувати їх у вихідний
селекційний матеріал. Запропоновано ще один підхід раціонального
використання генофонду картоплі в селекційній практиці - залучен¬
ня в дослідження сортів, якщо це не вирішує проблеми, культурних
видів, а потім - і диких (Howard Н.).
Значення міжвидової гібридизації в селекції картоплі. До по¬
чатку 40-х років минулого століття селекція картоплі в основному
базувалася на використанні одного виду - S.tuberosum. Незважаючи
на те, що в його геномі виявлені гени контролю високої врожайності,
крохмалистості, ранньостиглості, стійкості проти парші звичайної,
раку картоплі, залізної плямистості бульб, надчутливості до ХВА,
АВК, правильної форми бульб, поверхневого залягання вічок, ком¬
пактності гнізда, високих кулінарних і технологічних якостей бульб
та деяких інших ознак (Ross Н.; Букасов С.М.; Камераз А.Я.), цей
вид характеризується наявністю й успадкуванням багатьох негати¬
вних властивостей, і перш за все - уражуваності багатьма хвороба¬
ми і шкідниками. Крім того, використання в селекційній практиці
лише виду S.tuberosum через залучення в схрещування близькоро-
динних форм значно звузило генетичну основу вихідного селекцій¬
ного матеріалу. Тому використання в селекційному процесі лише
невеликої кількості споріднених форм внутрішньовидового похо¬
дження охарактеризовано як консервативний шлях (Tucker І.). То¬
му в боротьбі із хворобами і шкідниками були здійснені спроби
Ивлити свіжу кровм у вихідний селекційний матеріал - сорти
(Hawkes I.Y.). Проте залучення в селекційну практику співродичів
культурних сортів не завжди було успішним. По-перше, у схрещу¬
183
вання іноді залучали неперевірений матеріал; по-друге, викорис¬
тання співродичів S.tuberosum при виведенні нових сортів не зав¬
жди було результативним.
Початок міжвидових схрещувань картоплі пов'язаний з іменем
Клотша (Klotzsch І.). Перший фітофторостійкий міжвидовий гібрид
S.maglia x S.tuberosum одержав Суттон (1896). Але незабаром гібрид
уразився хворобою, що дещо затримало поширення міжвидової гіб¬
ридизації в селекції картоплі. І тільки завдяки дослідженням
И.Бройлі (1921) шляхом схрещування сортів із видами S.edinense
(ймовірно S.demissum x S.tuberosum x S.commersonii) вдалося отри¬
мати фітофторостійкі гібриди, які були названі SW-клонами.
Продовжуючи дослідження в цьому напрямі, К.Мюллер (1925)
створив вихідні фітофторостійкі форми, які назвав Wilderassen. Вони
успішно використовувалися при виведенні нових сортів не лише в
Німеччині, але й в інших країнах світу: Канаді, Шотландії, США. За
неповних 20 років за участю цих гібридів було створено 41 сорт
(Rieman Y.H.). Незважаючи на те, що в період досліджень над вихід¬
ним матеріалом Й.Бройлі і К.Мюллера було відкрито не більше 30-ти
видів, результати залучення їх у селекційну практику свідчили про
перспективність проведення таких робіт.
У НДІКГ шляхом залучення в схрещування дикого мексиканського
виду картоплі S.demissum у 1931 р. був одержаний сіянець 8670 с/31,
який пізніше був районований під назвою Фітофторостійка (Пушка-
рев И.И.). Після цього міжвидова гібридизація стала основним методом
створення вихідного матеріалу та виведення нових сортів.
Використовуючи міжвидові гібриди, можна вирішувати не ли¬
ше практичні, але й теоретичні проблеми. Ефективний генетичний
контроль стійкості проти практично всіх збудників хвороб та шкід¬
ників дає змогу виявити взаємозв’язки між рослиною-господарем
та патогеном, з’ясувати закономірності успадкування цінних ознак.
Із використанням диких і культурних видів картоплі одержані ди¬
ференціатори для проведення популяційного аналізу збудників та
шкідників.
Міжнародна схема взаємовідносин між генами, які контролюють
надчутливість проти Phythophora infestans (Mont.) de Вагу, та расами
збудника базується на визначенні їх з використанням гомозиготних
ліній S.demissum (Black W. et al.). Пізніше (Malcolmson J., Black W.)
була доведена різна природа R-генів S.demissum та інших видів, зок¬
рема S.stoloniferum, що спонукало виділити гени цих видів в інші
схеми. При створенні рослин-диференціаторів для визначення складу
популяцій Globodera rostochiensis Woll та Globodera pallida Woll та¬
кож були використані дикі і культурні види, зокрема S.andigenum,
S.kurtzianum, S.vernei, S.multidisectum (Kort J. et al.). Як індикатори
вірусів Y, A, M використовуються види S.demissum, S.chacoense та
гібриди з ними (Букасов С.М., Камераз А.Я.).
184
Цитолого-генетичні та філогенетичні основи міжвидової не¬
схрещуваності картоплі. Міжвидова несумісність сільськогоспо¬
дарських культур, у тому числі картоплі, проявляється через несхре¬
щуваність батьківських форм, а також низьку життєздатність гібрид¬
ного насіння або повну його несхожість (Банникова В.П.). Наші до¬
слідження дозволяють назвати ще один фактор, який зумовлює
міжвидову несумісність - відсутність нормального вегетативного або
генеративного відновлення нащадків, що проявляється загибеллю
рослин у перший або другий рік вирощування, різким зниженням
загальної життєздатності одержаних форм, значними відхиленнями в
розвитку генеративних органів [20].
Незважаючи на те, що для більшості видів характерна наявність
фертильного пилку (Подгаєцький А.А.), значній кількості їх властива
само- або перехресна несумісність. У результаті вивчення міжвидо¬
вих гібридів і потомства від близькородинних схрещувань виявлений
контроль гаметофітної системи несумісності (Puschkamacht М.І.),
здебільшого аналогічний Nicotiana (East Е.М., Mangelsdorf А.І.). За¬
вдяки грунтовним дослідженням з вивчення запліднення численних
видів картоплі ідентифіковано 13 алелів гена S. Пізніше доведено
існування дволокусної системи самонесумісності, яка контролюється
ще одним геном - R. При цьому проростає лише той пилок, гени
якого відсутні у стовпчику маточки.
Перехресна несумісність при міжвидових схрещуваннях звичай¬
но зумовлюється загальною гаметофітною системою несумісності.
Остання спостерігається при гібридизації за схемою: самонесумісний
вид х самосумісний. При іншому комбінуванні - самосумісний х са¬
мо сумісний, самонесумісний х самонесумісний, самосумісний х са¬
монесумісний - відбувається нормальне проростання пилку. Зазначе¬
не явище виявлене при схрещуванні видів S.verrucosum, S.brevidens
(d) (AubertinN.; GrunP., Pandey K.).
У результаті досліджень з диплоїдними формами виділені акцеп-
торні клони, які сприймають пилок самосумісного виду, і неакцептор-
ні, які не запилюються ним. Враховуючи це, виділяють за сумісністю
диплоїдні форми з такими типами: самонесумісний неакцепторний
(наявні множинні алелі S-гена + гени, що контролюють неакцептор-
ність); самосумісний, неакцепторний (Sf + контроль неакцепт-
торності); самосумісний, акцепторний (Sf+гени акцепторності); са¬
монесумісний, акцепторний (множинні алелі S гена+фактори акцеп¬
торності (Gum Р., Aubertin N.).
Розщеплення y Fj на акцепторне і неакцепторне потомство відбува¬
ється за незначним винятком у співвідношенні 1 : 3 і 1 : 15, залежно від
характеристики материнської форми. Це дозволяє зробити висновок про
домінантний контроль ознаки, який здійснюється двома генами при
схрещуванні акцептор х неакцептор або множинними рецесивними
генами при схрещуванні акцептор х акцептор (Grun Р., Aubertin N.).
185
Зроблена спроба пояснити реалізацію гаметофітного контролю
несумісності (Levis D.), суть якої полягає в тому, що білки, які утво¬
рюються в пилковій трубці і приймочці маточки, складаються з двох
ланцюгів. При проростанні пилку відбувається з’єднання білка пил¬
кової трубки і приймочки маточки з утворенням ферменту, який
складається з чотирьох ланцюгів білка. У випадку формування одна¬
кового за будовою білка з’єднання ланцюгів не відбувається, і фер¬
мент не утворюється, що призводить до інгібування процесів пророс¬
тання пилку.
При міжвидових схрещуваннях виявлено, що відхилення у про¬
ростанні пилку спостерігається уже при контакті його з приймочкою.
Воно характеризується затримкою проростання пилкових зерен, тру¬
бок або зміною морфології останніх, що є проявом спорофітної не¬
сумісності (Банникова В.П.). Вважається (Hogenbohm N.G.), що в
родині Solanacae генетична система “стерильність/фертильність” має
більше поширення при віддалених схрещуваннях, порівняно з близь-
кородинними, і проявляється реалізацією додаткових, крім S-факто-
рів, генів. Механізм її прояву аналогічний описаним вище (Hermsen J.,
Sawiska E.).
Певною перешкодою для проростання пилку є взаємодія плазмо¬
ну і ядерних генів, що проявляється за типом цитоплазматичної чо¬
ловічої стерильності (Staub І.Е.). Залучення в міжвидову гібридиза¬
цію форм із рецесивними алелями стерильності плазмону (?) спри¬
чинює в певної частини потомства порушення функціонування чоло¬
вічих органів квіток. При цьому спостерігається нерозкриття
пиляків, тетрадна стерильність, деформація квіток (Abballa М.М,
Hermsen G.G.).
Проведення грунтовних досліджень у цьому напрямі дозволило
виявити ЦЧС серед сортів з участю S.phureja, S.maglia, S.vernei,
S.chacoense, а також у видів S.acaule, S.demisum, S.stoloniferum,
S.verrucusum (Dionne J.; Grun P.). Виявлена чутливість до ядерних
генів плазмону чилійських форм S.tuberrosum. А тому фертильність
пилку гібридів з участю видів картоплі вища, ніж у матеріалі з плаз¬
моном від S.tuberrosum (Staub І.Е.). Доведено, що ЦЧС контролюєть¬
ся плазмоном (мітохондріями, пластидами), через що визначити її
можна лише при отриманні гібридів. У деяких випадках (схрещуван¬
ня S.verrucosum x S.tuberrosum 2п=2х) чоловіча стерильність Fi про¬
являється світлим забарвленням пилку, викликаним лактофенольною
кислотою. Разом з тим спостерігається і нормальний мейоз, але ядра
мікроспор не діляться, хоча мікроспори не гинули, а формували пи¬
лок, який забезпечував нормальне запліднення. Це явище трактуєть¬
ся як результат взаємодії цитоплазми S.verrucosum і генів S.tu¬
berrosum.
Різноманіття картоплі являє собою поліплоїдний ряд від ди- до
гексаплоїдів. Більшість видів картоплі (близько 70%) - це диплоїди
186
(Будин К.З.). Залучення їх у схрещування за диплоідно-поліплоїдним
типом часто є причиною значних порушень, які не дозволяють сфор¬
мувати нормально розвинуте насіння. Вперше на це звернув увагу
А.Мюнтцинг (1934). Доведено, що відмирання зародка при схрещу¬
ванні форм із різним рівнем плоїдності не пов'язане з якісними від¬
мінностями геномів батьків. При гібридизації аутополіплоїдів із ви¬
хідними диплоїдними формами практично не вдається отримати нор¬
мально розвинуте насіння, що спостерігалось при схрещуванні
S.vernei (2п=48) х S.vernei (2n=24), S.acaule (2n=96) х S.acaule (2п=48) і
реципрокно (Wangenheim К.Н.) S.chacoense (2n=24) х S.chacoense
(2n=48) (Marks G.E.). Проте у деяких видів, наприклад S.cardi-
ophyllum, S.pinnatisectum, S.tarijense, в окремих випадках утворю¬
ються триплоїди. У комбінаціях генетично відмінних батьківських
форм, але з однаковим рівнем плоїдності, незважаючи на нормальне
запліднення, також спостерігалася загибель зародків, що можна
виявили при схрещуванні S.acaule (2n=48) х S.tuberosum (2п=48),
S.stoloniferum (2n=48) х S.tuberosum (2п=48) і реципрокно (Wangen¬
heim К.Н.).
В окремих випадках при інтерплоїдній гібридизації утворюється
незначна кількість нормально розвинутого насіння. Разом з тим, на¬
приклад у результаті схрещування аутотетраплоїда S.tuberosum із
диплоїдними культурними видами, утворене насіння, як правило,
мало тетраплоїдний зародок (Івановська Е.В.; Frandsen N.O.;
Hanneman R.E., Peloquin S .J.). Збалансованість у розвитку гібридного
зародка спостерігається за участі в заплідненні нередукованих гамет,
тобто в результаті мейотичної поліплоїдизації. Види S.acaule,
S.stoloniferum є функціональними диплоїдами, а тому схрещування їх
із диплоїдними видами відбувається успішно.
Причиною відсутності або поганого зав’язування насіння при ін-
терплоїдних схрещуваннях А.Мюнтцинг (1934) вважав відхилення у
співвідношенні між кількістю хромосом материнської тканини, ен¬
досперму і зародка, яке в нормі повинно бути 2:3:2.
Аналізуючи результати досліджень багатьох авторів, Р.Ванген-
хейм (Wangenheim R.H.) відзначив відсутність єдиної думки щодо
першочергового значення для нормального розвитку зародка спів¬
відношення між кількістю хромосом ендосперму і зародка або мате¬
ринською тканиною та ендоспермом. Зазначалося, що в інтерплоїд-
них схрещуваннях відхилення в розвитку насіннєвого зачатка спо¬
стерігається спочатку в ендоспермі, що потім негативно позначаєть¬
ся на розвитку гібридного зародка. Це доведено цитологічними
дослідженнями материнської тканини, ендосперму і зародка
(Wangenheim. R.H. et al.)* Узагальнені дані (табл. 3.8) свідчать, що
триплоїдний і гексаплоїдний ендосперм розвивається без відхилень.
А це при нормальному злитті яйцеклітини і спермію зумовлює утво¬
рення насіння, яке успішно проростає.
187
Таблиця 3.8 - Співвідношення між кількістю хромосом материнської тканини,
ендосперму та зародків і розвитку насіння в бульбоутворюхочих
Solanum (Wangenheim R.H., 1961)
Схре¬
щу¬
вання
Особливі
умови
Відношення
Розвиток
ендосперму
Розвиток
насіння
мате¬
ринська
тканина
ендо¬
сперм
за¬
родок
2х х 2х
-
2
3
2
Нормальний (серед¬
ня вакуолізація клі¬
тин)
Схоже
насіння,
сіянці 2х
2х х 4х
-
2
4
3
Пригнічення в утво¬
ренні стінок клітин,
аномалія веретена;
великі дуже вакуо-
лізовані клітини,
дегенерація
Насіння без
зародка
4х х 2х
-
4
5
3
Малі, невакуолізо-
вані клітини, великі
міжклітинники, де¬
генерація
Те саме
4х х 4х
-
4
6
4
Нормальний
Схоже
насіння,
сіянці 4х
4х х 2х
Подвійне
запліднення
вторинного
ядра, відсут¬
ність заплід¬
нення яйце¬
клітини
4
4
6
або
6
0
2
Те саме
Те саме
Потовщене
насіння без
зародка
Схоже на¬
сіння, 2х-
дигаплоїди
2х х 4х
Нередукова-
ні зародкові
мішки
2
6
4
Те саме
Схоже
насіння,
сіянці 4х
Нашими дослідженнями (Хвединич О.О., Подгаєцький А.А.,
Гудзь В.М.) доведена важливість нормального розвитку ендосперму,
порівняно із зародком. При самозапиленні виду S.stoloniferum злиття
сперміїв відбувається швидше з центральною клітиною, порівняно з
яйцеклітиною. Через 8 діб після запилення виявлявся багатоклітин¬
ний ендосперм, а зародок при цьому мав лише дві клітини. У комбі¬
нації S.stoloniferum x S.pinnatisectum на 10-у добу після запилення
спостерігали утворення багатоклітинного ендосперму і 5- клітинного
гібридного зародка. Пізніше відбувається дегенерація ендосперму: у
188
його клітинах призупиняється ділення, вони значною мірою вакуолі-
зуються, а шар цитоплазми розміщується біля оболонки. Надалі від¬
бувається повна дегенерація вмісту клітин, що є причиною загибелі
зародка.
Погоджуючись із гіпотезою Р.Вангенхейма, деякі дослідники
(Den Nijs E.F.JPeloguin S.J.; Jonston S.A., Hanneman R.E.; Ehlenfeld M.K.,
Hanneman R.E.) нормальний розвиток зародка пов’язують із рівнем
плоїдності ендосперму, кількість хромосом якого залежить від плоїд-
ності материнської і чоловічої форми. Для кожного з батьків визна¬
чається балансове число ендосперму (EBN). Однакове цифрове зна¬
чення EBN батьків зумовлює формування нормально функціонуючо¬
го ендосперму, що є умовою успішного розвитку зародка. При від¬
мінності EBN утворення нормального ендосперму не відбувається.
Рівень плоїдності видів не завжди відповідає балансовому числу ен¬
досперму. Наприклад, доведено, що у виду S.cardiophyllum (2п=2х=24)
EBN=1. Для подолання несхрещуваності його з диплоїдними видами
S.phureja, S.chacoense та іншими, для яких EBN=2, необхідною є по¬
передня поліплоїдизація S.cardiophyllum.
Значні труднощі міжвидової гібридизації зумовлені філогенетич¬
ною віддаленістю видів. Г.Д. Карпеченко (1935) вважав найбільш
правильним розділити схрещування на близькородинні певною мі¬
рою. Останні з позицій відмінності геномів батьківських форм (на¬
явність хромосом, здатних кон’югувати, утворення життєздатного
потомства, нормальне вегетативне і генеративне відтворення гібри¬
дів) розділяють на конгруентні та інконгруентні. До конгруентних
належать такі схрещування, при яких є подібність хромосом батьків¬
ських форм, що забезпечує отримання гібридів без зниження їхньої
життєздатності і порушень у розвитку генеративних органів. При
інконгруентних схрещуваннях батьки значно відрізняються кількіс¬
тю і, головне, - гомологічністю хромосом. У результаті таких схре¬
щувань утворюються гібриди з неправильним проходженням мейозу,
що призводить до часткової або повної їх стерильності, відхилень у
рості і розвитку.
До інконгруентних схрещувань здебільшого належать міжвидові
схрещування, батьківські форми яких значно філогенетично віддалені.
Це дозволяє стверджувати, що віддалена гібридизація включає геогра¬
фічну і філогенетичну віддаленість (Букасов С.М., Костина Л.И.).
Спонтанне утворення гібридогенних видів свідчить про значну
філогенетичну близькість форм, у результаті схрещування яких вони
отримані. Аналіз анатомо-морфологічних, імунно-хімічних власти¬
востей білків бульб дозволив виявити гібридогетгість: S.sambucinum=
S.pinnatisectum* S.cardiopyllum (Крашенинник Н.В.). Виявлений ге-
номний склад видів картоплі (Hawkes J.G.; Оглуздін А.С.) свідчить
про близькість походження лише деяких видів, що може пояснити
частий прояв міжвидової несумісності.
189
Наслідком філогенетичної віддаленості видів, що залучаються у
схрещування, є порушення росту і розвитку, а також генеративного
відтворення одержаних гібридів. Результати наших досліджень свід¬
чать, що аномалії геномної незбалансованості проявляються вже на
етапі проростання насіння. В окремих комбінаціях жодна насінина,
висіяна в чашку Петрі, не проросла. Частина наклюнутого насіння
(0-70 %) не дала сходів після висівання їх у спеціальні ящики. Деякі
гібриди (0-20%) випали в перший рік вирощування, інші - на дру-
гий-третій. Окремі з гібридів не утворили бульб, а більшість - ягід
від самозапилення [20].
З метою підвищення результативності міжвидових схрещувань, а
також часткового подолання несумісності при їх виконанні розроб¬
лені численні способи і методи покращання результативності міжви¬
дових схрещувань.
Основні методи і способи подолання міжвидової несхрещува¬
ності. Ефективність використання методів і способів подолання між¬
видової несхрещуваності різна як у цілому, так і залежно від схем
гібридизації. З урахуванням наявних у науковій літературі даних,
інтуїції дослідника, підхід до вибору їх повинен бути таким: спершу
необхідно забезпечити оптимальні умови для схрещування (Успен¬
ський Е.М.; Родионова З.В.; Букасов С.М., Камераз А.Я.; Подгаєць-
кий А.А.), [20] і лише після цього застосовувати методи і способи
подолання міжвидової несхрещуваності, починаючи із простих. Дані
багатьох дослідників, а також отримані нами свідчать, що на сучас¬
ному етапі розвитку науки при створенні вихідного передселекцій-
ного і вихідного селекційного матеріалу можна залучати практично
будь-який вид картоплі.
За типом дії факторів запропоноване таке групування методів по¬
долання міжвидової несхрещуваності (Банникова В.П.): механічні -
змивання виділень із несумісного рильця, укорочення довжини стовп¬
чика, запилення і запліднення in vitro, введення пилку всередину
зав’язі, виділення гібридних зародків у молодому віці та вирощуван¬
ня їх на штучних живильних середовищах; фізіологічні, до яких та¬
кож можна віднести і три останні з попереднього переліку - запи¬
лення молодих бутонів або майже зів’ялих квіток, схрещування на
рослинах наприкінці вегетаційного періоду, вплив на материнський
організм відносно низької температури, мікроелементів, фізіологічно
активних речовин, опромінення тощо, попередня пересадка зародків
або попереднє вегетативне зближення батьківських форм; генетичні -
амфідиплоїдія, поліплоїдія вихідних у схрещуванні форм із нижчим
рівнем плоїдності, метод посередника, схрещування на внутрішньо¬
видових гібридах.
Інші дослідники [20] групують методи і способи за спрямованіс¬
тю дії основних їх факторів: кратна зміна кількості хромосом (полі¬
плоїдія, гаплоїдія), “посередник”; затримка відтоку пластичних речо-
190
вин у бульби або обмін ними між батьківськими формами (трансплан¬
тація, декапітація стебел, стовпчика маточки, видалення столонів,
бульб); застосування фізіологічно активних речовин: використання
штучних живильних середовищ (вирощування насіння, ізольованих
зародків, соматична гібридизація, схрещування in vitro); інші типи
впливу на зав’язування гібридних ягід і насіння.
Поліплоїдія. Розрізняють два види поліплоїдизації - мітотичну і
мейотичну. Для подолання міжвидової несхрещуваності використо¬
вують обидва. Позитивний вплив мітотичної поліплоїдизації вихід¬
ної батьківської форми з нижчим рівнем плоїдності на результатив¬
ність міжвидових схрещувань уперше довів А.І.Мітін (Букасов С.М.,
Камераз А .Я.). Особливо широко стали проводитись дослідження в
цьому напрямі, починаючи з 1940 p. (Livermore J.R., Jonhston F.E.;
Swaminatan М.С.; Magoon M.H., Cooper D.C., Hougas R.W.; Лебедева H.A.;
Житлова H.A., 1971; Лаптев Ю.А. та ін.).
При застосуванні методу поліплоїдії слід враховувати те, що
окремі види {S.acaule, S.stoloniferum) за своєю природою є функціо¬
нальними диплоїдами, а тому при гібридизації із сортами ведуть себе
відповідно, що й зумовлює несхрещуваність у таких комбінаціях
(Яшина И.М.). Крім того, якісні зміни, які відбуваються в поліплоїд-
них формах, можуть коригувати результативність схрещування видів
(Карпеченко І.Д.), що також необхідно враховувати при використан¬
ні методу.
Застосування поліплоїдії дозволяє підвищити процент вдалих
схрещувань, кількість насіння в ягоді. Більшість дослідників вважа¬
ють за краще використовувати поліплоїдну форму як материнську
(Magoon М.Н., Cooper D.C., Hougas R.W.; Житлова Н.А. та ін.), але
існує й протилежна думка (Есипова З.И.).
Використанням методу поліплоїдії вдалося отримати позитивні
результати від залучення у схрещування із сортами найбільш важко-
схрещуваних філогенетичних віддалених видів, наприклад S.bulbo-
castanum (Лебедева Н.А.). Доведено, що дія поліплоїдії позитивно
позначається на отриманні потомства з одержаними первинними
міжвидовими гібридами (Камераз А.Я., Вавилова М.А., Житлова Н.А.,
Іванова В.Н.).
Після застосування інших методів подолання міжвидової не¬
схрещуваності утворюються триплоїдні форми, які успішно можна
залучити в подальшу роботу, використовуючи метод поліплоїдії, на¬
приклад у випадку з триплоїдними гібридами S.acaule х S.bulboca-
stanum (Hermsen J., Ramanna М.).
Найбільш ефективним способом отримання поліплоїдних форм
вважається використання колхіцину, зокрема обробка його розчином
точки росту у фазі сім'ядолей (Житлова Н.А.).
У поліплоїдів також спостерігається усунення само- і перехресної
несумісності (Swaminatan М.С.; Magoon М.Н. et al.). Разом з тим ме¬
191
тод мейотичної поліплоїдії через значне утворення великої кількості
химер, а також негативну реакцію окремих видів {S.demissum) на йо¬
го використання не здобув широкого застосування.
Мейотичне подвоєння хромосом може бути однобічним при
схрещуванні за схемою 4х х 2х або 2х х 4х або двобічними (2х х 2х).
Позитивним при використанні мейотичної поліплоїдії є те, що
при цьому не лише вдається отримати гібриди, але й збільшити
вищеплення гетерозисних форм. Урожайність отриманого таким
шляхом потомства не лише перевищує середню батьків, але й про¬
яв цієї ознаки у кращого з них (Mendibura А.О.). Причому, справе¬
дливість викладеного більшою мірою стосується затримки першого
ділення мейозу (FDR), порівняно з другим (SDR) (Мок D.N.,
Pelopuin S.J.).
Великий вплив на частоту мейотичної поліплоїдизації мають від¬
хилення від оптимального рівня факторів зовнішнього середовища,
особливо температури (Frandsen N.O.).
Гаплоїдія. Зменшення у два рази кількості хромосом сортів до¬
зволяє більш успішно проводити схрещування, перш за все з диплоїд-
ними видами. Зважаючи на низьку фертильність дигаплоїдів, перс¬
пективніше використовувати їх як материнські форми (Ross R.W.,
Pelopuin S.J., Hougas R.W.), проте існують повідомлення про успішне
застосування дигаплоїдів і як запилювачів (Брокш B.JL).
Використання методу гаплоїдії дозволяє не лише мати значне ягодо-
утворення, але й високу обнасіненість ягід (у середньому 37,3 пгг.). У
цілому за основними показниками результативності схрещування
метод дигаплоїдії має переваги над поліплоїдією (Будин К.З.).
Як стверджував С.М.Букасов (1940), негативні господарські вла¬
стивості при схрещуванні диких видів і їх гібридів із сортами можуть
успадковуватися не лише від співродичів, але й сортів. Отримання
дигаплоїдів, їх аналіз і відбір кращих дозволяє зменшити серед на¬
щадків вищеплення негативних ознак.
Найбільш простим і практично доступним методом отримання
дигаплоїдів сортів є міжвидова гібридизація з участю видів S.phureja,
S.kesselbrennei та деяких ін. Останнім часом набув поширення інший
метод - індукція гаплогенезу в культурі in vitro.
При багатьох позитивних сторонах використання методу гаплої¬
дії в міжвидовій гібридизації слід зазначити, що до останнього часу
не вдалося отримати гібриди між дигаплоїдами і філогенетично від¬
даленими видами, наприклад S.bulbocastanum, S.pinnatisectum (Собо¬
лева Е.И.).
Метод посередника. Видове різноманіття картоплі дозволяє ус¬
пішно використовувати для подолання міжвидової несхрещуваності
мічурінський метод, який називають посередником (Мічурін Й.В.).
Визначені основні критерії для вибору посередника (Букасов С.М.,
Лебедева Н.А.), хоча з деякими з них погодитися важко [20].
192
Враховуючи значне розсіювання серед F і властивостей, характер¬
них. лише для батьківської форми, основною вимогою до посередни¬
ка є наявність (бажано у високому ступені) ознаки, задля якої залуча¬
ється в селекційну практику основний вид. Причому, одержані ре¬
зультати (Dionne L.; Hermsen J., Ramanna М.; Оверчук B.I., Подгає-
цький А.А.) свідчать про можливість використання як посередника
виду, у якого провідна властивість іншого компонента схрещування
є слабо вираженою. При залученні в селекційну практику важкосхре-
щуваних із сортами видів доцільно використовувати як посередник
один (Подгаєцький А.А.) або кілька видів (Лебедева Н.А.; Hermsen Y.,
Ramanna М.; Оверчук B.I., Подгаєцький А.А.).
Позитивною стороною методу при його застосуванні є можли¬
вість залучити в селекцію практично будь-який вид і вже на першому
етапі (при отриманні первинних міжвидових гібридів) збагатити ге¬
ном потомства генами контролю властивостей, які відсутні в однієї з
батьківських форм. Крім того, ми вважаємо, що метод є одним із
найбільш "м’яких", тобто серед потомств, як правило, відсутні ано¬
малії, які зустрічаються при застосуванні інших методів.
Багатьма дослідниками доведено, що досить вдалим посередни¬
ком для численних видів є S.acaule (Dionne L.; Житлова Н.А.;
Hermsen J., Ramanna М.; Оверчук B.I., Подгаєцький А.А.). При залу¬
ченні в практичну селекцію виду S.stoloniferum доцільно використо¬
вувати як посередники S.acaule, S.demissum, S.rybinii, S.species,
S.simplicifolium, S.gourlayi, S.vernei і особливо S.chacoense, S.verru-
cossum, а виду S.jamesii - S.demissum [27].
Метод трансплантації є одним із найефективніших і багатогранних
при подоланні міжвидової несхрещуваності. Як підщепи при трансплан¬
тації широко використовуються томати (Успенський Е.М.), дикі види
картоплі, якщо на них прищеплюють культурний сорт, або культурний
сорт, якщо на нього прищеплюють дикий вид (Букасов С.М., Лебеде¬
ва Н.А.), паслін чорний, тютюн (Подгаєцький А.А., Чередниченко Л.М.).
Н.П.Кренке (1928) вважав, що в результаті трансплантації картоп¬
лі на небульбоутворюючі види відбувається затримка відтоку плас¬
тичних речовин із листків, що утворилися в результаті фотосинтезу, і
зменшення мінерального живлення, а іноді й водозабезпечення через
неповне відновлення провідної системи. Спільними із колишньою
лабораторією фізіології Інституту картоплярства дослідженнями
(Мицько В.М.) доведено, що вміст сухих речовин у листках прищепи
S.stoloniferum був на 0,3% вищий, ніж в інтактних рослин, а процент
цукру був у 2 рази більшим (1,1 проти 0,57).
Результати лабораторних досліджень свідчать, що використання
методу дозволило отримати гібридне насіння у 8-ми комбінаціях з 9-
ти, тоді як на інтактних рослинах зав’язування ягід та утворення на¬
сіння відбулося лише в одній комбінації - S.demissum х S.bulbo-
castanum (Подгаєцький А.А., Чередниченко Л.М.).
193
Крім зазначеного, використання методу сприяє значному поліп¬
шенню росту і розвитку підщеп. Наприклад, прищеплені рослини
видів S.demissum, S.bulbocastanum, S.ehrenbergii мали гілок у 3,7-5,6
разів більше, ніж інтактні, квіток - у 12,9-15,7, а ягодоутворення від
самозапилення у виду S.demissum було більшим у 18,8 разів (Подгає-
цький А.А., Чередниченко Л.М.).
Використання фізіологічно активних речовин. На думку бага¬
тьох дослідників (Dionne L.; Букасов С.М., Камераз А.Я.; Брокш B.J1.
та ін.), застосування для подолання міжвидової несхрещуваності фі¬
зіологічно активних речовин дозволяє підвищити результативність
гібридизації. Найбільш часто для поліпшення гібридизації викорис¬
товують регулятори росту: 2,4-Д-дихлорфеноксиоцгову кислоту;
2,4,5-Т-трихлорфеноксиоцтову кислоту; індолоцтову та оксилоцтову
кислоти (Пушкарев И.И.; Dionne L.; Fichnich О., Lubert G.), а також
азотисту основу - аденін (Брокш B.JL).
Вважається, що застосування фізіологічно активних речовин по¬
кращує зав’язування гібридного насіння (Букасов С.М., Камераз А.Я.,)
або запобігає опаданню ягід, що зав’язались (Dionne L.).
Успіх використання фізіологічно активних речовин залежить від
їх концентрації, строку і способу нанесення. Нами виявлено, що оп¬
тимальна концентрація 2,4,5-Т для забезпечення доброї обнасіненос-
ті ягід складає 25-35 мг/л. З підвищенням її збільшується кількість
партенокарпічних ягід [20].
Більш детальний розгляд наведених вище та інших методів подо¬
лання міжвидової несхрещуваності поданий у нашій оглядовій робо¬
ті [20].
Досягнення міжвидової гібридизації при виведенні сортів но¬
вого типу. Завдяки міжвидовій гібридизації вдалося створити сорти,
які характеризуються відносно високим проявом властивостей, від¬
сутніх у S.tuberosum, тобто сорти нового типу. Перший фітофторос¬
тійкий сорт картоплі з участю виду S.demissum одержали в Німеччині
(1934 p.) - Sandnudel [29], а через деякий час в СРСР (1937 р. ) - Фі¬
тофторостійка (Пушкарев И.И.).
Широке використання генофонду співробітниками ВІР дозволило
створити не лише цінний вихідний селекційний матеріал, але й сорти
картоплі з участю S.andigenum - Імандра, Сестра Імандри; S.boyacen-
се - Хібіни, Хібінська двоврожайна, Хібінська скороспілка; S.demi-
sum - Камераз 1; S.vallis-mexici - Пушкінська; декількох видів -
Дєтскосєльська, Волховська, Красносєльська, Уральська, Красно-
уфимська і багато інших (Букасов С.М.).
В Інституті картоплярства УААН основою для створення нових
високоінтенсивних і висококомплексних сортів, які відповідають
вимогам сучасного виробництва, став матеріал з участю видів
S.andigenum, S.leptostigma, S.comerrsonii, S.demisum (Терещенко O.I.).
У результаті використання цього та іншого вихідного селекційного
194
матеріалу виведені сорти Перлина, Смачна, Бородянська, Чарівниця,
Немішаївська біла, Росинка, Зарево, Лада, Луговська, Українська
рожева, Бородянська рожева, Обрій, Горлиця, Віринея. На основі
створеного в інституті вихідного селекційного матеріалу на Полісь¬
кій дослідній станції ім. Засухіна виведено такі сорти: Покра, Жито-
мирянка, Поліська рожева, Лепта, Пост 86, Веста.
Надзвичайно широка і багатогранна робота щодо створення вихід¬
ного селекційного матеріалу на основі міжвидової гібридизації не
лише зумовила велику популярність гібридів Інституту Макса План¬
ка (Німеччина), а й сприяла виведенню понад 60 сортів у Німеччині,
Нідерландах, Мексиці та інших країнах, зокрема Fatima, Sheriff,
Kirsty і багато інших (Костина. Л.И.), [29]. В Інституті картоплярства
УААН з участю гібрида МРІ 44.1066/10 виведено сорт Здвижанка.
Результатом широкого використання генофонду картоплі в селекційній
практиці Німеччини є інгрогресія цінних апелів співродичів диких і
культурних видів у комерційні сорти. Понад 83% їх мають гени S.de¬
missum, 26 % - гени інших видів, які часто доповнюють названий [29].
При виведенні мексиканських сортів широко використовувався
вид S.demissum. Високофітофторостійкі, з комплексом інших агро¬
номічних властивостей, у тому числі з пластичністю, сорти Мексики
є складними бекросами S.demissum: Аніта - В2 [(ДТ5 х ДТ4) х Т], Бер-
тита; Кончита - В2 [( ДТ5 х ДТ5) х ДТ2]; Дорита, Еленіта-В2 {([(ДТ3
х ДТ3) х ДТ4] х Т}, Ерендира -( ДТ х ДТ ) х Т; Грета - ДТ х Аквіла
(ступінь біля "Т" означає кількість повторних схрещувань із
S.tuberossum) (Toxopeus H.J).
При створенні висококрохмалистих (Терещенко О.І.; Альсмик П.И.),
високобілкових (Томчук Н.Г.), з польовою стійкістю проти фітофто¬
розу, нематодостійких, високогетерозисних, з високою загальною
комбінаційною здатністю (Landeo Y.A., Наплетал R.E.) батьківських
форм і сортів широко використовувався вид S.andigenum. На сьогодні
його інтрогресовано в більш як 200 сортів (Костина Л.И.).
Меншою мірою, порівняно з S.demissum, використовуються в
практичній селекції інші дикі види картоплі (Пушкарев И.И.; Будин
К.З.). Перший сорт із геном імунітету до ХВК - Сафір - був одержа¬
ний з участю S.acaule. Пізніше гени виду, як доповнення до інших,
були інтрогресовані ще в 4 сорти (Костина Л.И.). Дещо пізніше з
участю S.chacoense був створений сорт Hydra. Імунітет до УВК (ген
Ry) сортів Bison, Fanal, Cardia, Esta, Franzi, Heidrum, Wege та інших
зумовлений присутністю в їхньому геномі генів виду S.stoloniferum
(Ross Н.). Значні успіхи при використанні цього виду в селекційній
практиці отримані в Ленінградському СП (Котова К.А.).
У зв’язку з поширенням картопляної нематоди виникла потреба
виведення стійких проти патогену сортів. Крім виду S.andigenum, для
вирішення проблеми використані S.vernei, S.spegazzinii. За поперед¬
німи даними, обом видам властива стійкість не лише до патотипів
195
Globodera rostochiensis, але й G.pallida (Ross H., Hijsmann C.). Гени
контролю ознаки від S.vernei вже інтрогресовані в 35 сортів. Перший
нематодостійкий сорт із генами від S.spegazzinii - Cordia - виведе¬
ний у 1974 р. Згодом у геном ще 10 сортів були уведені гени цього
виду [29].
Підвищення вимог до сортів зумовило залучення до їх виведення
кількох видів. Найбільш часто зустрічається поєднання S.demissum і
S.andigenum, S.vernei і S.spegazzinii та ін. (Stegemann H., Schnick D.).
Позитивні результати від використання міжвидової гібридизації
при виведенні нових сортів характерні не лише для окремих із них, а
й у цілому. Інтрогресія факторів стійкості проти фітофторозу диких і
культурних видів у селекійні сорти дозволила підвищити польову
стійкість проти хвороб сортів, які є міжвидовими гібридами, порів¬
няно із внутрішньовидовими, на 1,36 балів за шестибальною шкалою
(Zadina J.). І такі приклади непоодинокі.
ЛІТЕРАТУРА
1. Будин К.З., Житлова Н.А., Соболева Т.И. Каталог мировой коллекции
ВИР. Вип. 477.-М., 1989. - 18 с.
2. Будин К.З. Генетические основы создания доноров картофеля. - С.Пб.:
ВИР, 1997.-39 с.
3. Паспорта доноров хозяйственно ценных признаков с.-х. культур, соз¬
данных сотрудниками ВИР и опытной сети в 1990 году/ Будин К.З., Пыжен-
ков В.И., Алимов Д.П. и др.- С.Пб.: ВИР, 1991.-25 с.
4. Букасов С.М. Принципы систематики картофеля // Труды по прикл.
ботанике, селекции и генетике - М., 1978. - Т.62, вып.1- С.3-35.
5. Вавилов Н.И. Мексика и Центральная Америка как основной центр
происхождения культурных растений Нового Света // Труды по прикладной
ботанике, селекции-1931 .-Т.26.-№3.
6. Вавилов Н.И. Ботанико-географические основы селекции // Теоретиче¬
ские основы селекции растений.- М. - Л., 1935. - Т.1- С. 17-74.
7. Вавилов Н.И. Учение о происхождении культурных растений после
Дарвина // Советская наука.-1940.-№2.
8. Вавилов Н.И. Иммунитет растений к инфекционным заболеваниям //
Избранные труды. В 5 т. - М. - Л., 1964 - Т.ІѴ.
9. Вавилов Н.И. Избранные произведения в двух томах. -Л.: Наука, 1967.
-Т.1 -424 с.
10. Горбатенко Л.Е. Южноамериканские виды картофеля (Petota Dumort.
род Solanum L.) и их интродукция в СССР: Автореф. дис... д-ра биол. наук:
06.01.05 /ВИР-Л., 1989.- 37с.
11. Горбатенко Л.Е. Конспект системы секции Petota Dumort. рода Sola¬
num L. из Южной Америки. Мобилизация мировых растительных ресурсов //
Труды по прикладной ботанике, селекции, генетике. - М., 1989. - Вып.126-
С.92-108.
12. Жуковский П.М. Культурные растения и их сородичи.- Л.: Колос,
1971.-752 с.
13. Картопля - другий хліб. - К.: Довіра, 1995. - Вип.1. - 282 с.
196
14. Каталог мировой коллекции ВИР: Виды картофеля Мексики и их
значение для селекции.,- Л., 1989. - Вып.439.- 88 с.
15. Каталог мировой коллекции ВИР: Южноамериканские виды кар¬
тофеля (секция Petota Dumort. рода Solanum L.). - JI., 1990.- Вып.569.-
398 с.
16. Межвидовые гибриды картофеля - генетические источники и доноры
устойчивости к патогенам.-JI.: ВИР, 1989. - Вып.477.-18 с.
17. Мережко А.Ф. Система генетического изучения исходного материала
для селекции растений: Методические указания.-Л.: ВИР, 1984.-70 с.
18. Мережко А.Ф. Проблема доноров в селекции растений- С.Пб.:
ВИР, 1994. - 27 с.
19. Подгаєцький А.А. Використання генофонду картоплі для інтрогресії
цінних генів при створенні вихідного селекційного матеріалу: Автореф.
дис...д-ра с.-г. наук: 60.01.05: Інститут землеробства.- К., 1993.- 44 с.
20. Подгаецкий А.А. Межвидовая несовместимость картофеля. Методьі и
способы ее преодоления: Методические рекомендации - К., 1993.- 99с.
21. Подгаєцький А.А. Використання генофонду картоплі в селекційній
практиці // Картоплярство.- К.: Урожай, 1995 - Вип.26. - С. 9-18.
22. Подгаєцький А А. Генофонд картоплі // Науково-популярний альма¬
нах "Картопля - другий хліб". - К.: Довіра, 1995 - Вип. II. - С. 109-112.
23. Подгаєцький А.А. Дослідження передселекційного етапу при виве¬
денні нових сортів картоплі // Тези доп. конф. молодих вчених. - Немішаєве:
ПСГ УААН-1996.-С.2.
24. Подгаецкий А.А. Использование сородичей культурного картофеля
для создания источников фитофтороустойчивости клубней // Цитология и
генетика-1997-Т31.-№ 6 - С. 37-45.
25. Подгаецкий А.А. Необходимость выделения на современном уровне
развития селекции картофеля предселекционного этапа // Современные тех¬
нологии, экономика и экология в промышленности, на транспорте и в сельс¬
ком хозяйстве: Сб. статей по материалам 6-й Междунар. науч.-метод, конф.
-Алушта, 1999-С.321-326.
26. Подгаєцький А.А., Чередниченко J1.M. Використання методу транс¬
плантації у передселекційному процесі // Картоплярство- К.: Урожай,
1995.- Вип.26.-С.33-38.
27. Подгаєцький А.А., Чередниченко Л.М. Використання методу посеред¬
ника при міжвидовій гібридизації картоплі // Картоплярство. - К.: Урожай,
1996.- Вип.27 - С.59-65.
28. Пушкарев И. И. Селекция картофеля на устойчивость к фитофторе-
К.: ГоссельхозиздатУССР, 1962.-115 с.
29. Росс X. Селекция картофеля. Проблемы и перспективы - М.: Агро-
промиздат, 1989.-184 с.
30. Теслюк П.С., Молоцький М.Я. Практичний порадник картопляра-
К., 1999.-268 с.
31. Чередниченко Л.М. Використання генофонду картоплі для створення
фітофторостійкого вихідного селекційного матеріалу: Автореф. дис. .. .канд.
с.- г. наук: 06.01-05 / Ін-т цукрових буряків - К., 2000.-20 с.
32. Чечітко І.П., Подгаєцький А.А. Створення генофонду стійкості проти
сухої фузаріозної гнилі // Вісн. аграр. науки-1998 - №10.- С. 23-26.
197
33. Bittner G. Solana nova vel minus cognita. // Fedde Repertorium Speci-
erum Novarum regni Vegetabilis: 1912—1914.-№ 10-13-566 p.
34. Brucher H. Problematisches zum Ursprung der Kulturkartoffel aus chiloe
// Z.Pflanzucht. - 1960.- №43.- S.241-265.
35. Arcy D.G. Solonaceae Studies II: Typification of subdivision of Sollanum
// Annals of the Missouri Botanical garden. - 1972. - 59 s.
36. Dannet L.S. Zur Sistamatik von Solanum tuberossum L. Die Kul-
turpflanze. - Berlin, 1956.-P.83-129.
37. Dodds K.S. Classification of cultivated potatoes. // Correi D.S.: The potato
and its wild relatives. - Texas Res/ Faund, 1962.-P.517-539.
38. Corell D. The Potato and its wild relatives. Section Tuberarium of genus
solanum.- Texas, 1962- 606 p.
39. Hawkes Jg. Biosystematies of the potato // The Potato Crop - London:
Chapman and Hali., 1978. -P.l5-69.
40. Hawkes J.G. Resept conce pts in the evolution of tuber bearing Solanums
rep. Plann. Conf. Exploration, Taxonomy and Maintenance of potato Germplasm.
III Inter. Pot. Center - Lima, 1979 - P.40-59.
41. Ochoa C. Determinacion sistematica y recuentos cromosomicos de las
popas indigenas. Cultivadas en el. Centro der Peru. - Lima, 1965. - P. 103-166.
42. Ochoa C. Recuentos cromosomicos y determinacion sistematica de papa
nativas cultivadas en el sur dei Peru. - Lima, 1964. - 41p.
4. СЕЛЕКЦІЯ КАРТОПЛІ
4.1. З ІСТОРІЇ РОЗВИТКУ СЕЛЕКЦІЇ
КАРТОПЛІ
В Україні селекція картоплі була започаткована професором
сільськогосподарського факультету Київського політехнічного ін¬
ституту М.К.Малюшицьким на Київській крайовій сільськогоспо¬
дарській дослідній станції (с.Халаєво Києво-Святошинського райо¬
ну). Тут він у 1914—1916 рр. зібрав першу в Україні колекцію картоп¬
лі, потім створив перший в Україні сорт під назвою Пиріжок Малю-
шицького. У 1923 р. розпочав роботу з поліпшення німецького сорту
Вольтман, бульби якого темніли при зберіганні, а інколи і в полі. Він
одержав клони сорту, які не мали цієї вади. У 1932-1937 рр. на Не-
мішаївській дослідній станції було виділено найурожайніший із них і
під сортовою назвою Вольтман 1177 районовано в Україні [3].
Селекційна робота з картоплі в Україні проводилася в різних нау-
ково-дослідних установах. При цьому селекціонери враховували
специфічні умови тієї чи іншої грунтово-кліматичної зони та вимоги
виробників цієї культури.
У 1929-1930 рр. на Носівській сільськогосподарській дослідній
станції селекцію картоплі розпочала В.Д.Щербачова.
З 1929 року селекція картоплі почала проводитись на Поліській
дослідній станції ім. О.М. Засухіна. Ця установа розташована в укра¬
їнському Поліссі на піщаних грунтах. Специфічні умови зони давали
змогу створювати сорти, пристосовані до них, й отримувати високо¬
якісний насінний матеріал. З одержаного від Київської сільського¬
сподарської дослідної станції селекційного матеріалу тут був ство¬
рений сорт картоплі Роза Полісся, а від Носівської сільськогоспо¬
дарської дослідної станції - Поліська 36. Були створені також сорти
Крепиш, Островська, Баранівська. Над створенням сортів картоплі
працювали відомі селекціонери М.Ф.Островський, І.М.Бодисько,
І.В.Карпович [3].
Основним напрямком селекції в той час було створення столових та
універсальних високоврожайних сортів, стійких проти вірусних хвороб,
кільцевої гнилі, фітофторозу, іржавої плямистості. Питання ракостійкос¬
ті тоді не було вирішене у зв’язку з відсутністю установи для оцінки се¬
лекційного матеріалу на стійкість проти цього захворювання [1].
Початковий період селекційної роботи відзначався тим, що в
ньому використовувався в основному метод внутрішньовидової гіб¬
ридизації в межах виду S.tuberosum.
199
Пізніше на Поліській дослідній станції, як і в інших селекційних
установах, широко почали використовувати як батьківські форми
сорти та гібриди міжвидового походження. Із насіння, одержаного на
Немішаївській дослідній станції від схрещування батьківських форм
міжвидового походження, у Поліській дослідній станції було створе¬
но середньоранній сорт Житомирянка та середньопізній Поліська
рожева. На станції були створені також інші цінні високопродуктив¬
ні сорти картоплі: Ікар, Малинчанка, Зов, Пост 86, Посвіт, Гарт, Бе¬
региня, Радич, Малич, Косень 95, Поран, Поліська 96, Дубравка, Те¬
терів та ін. З 1967 по 1982 рік селекцію картоплі на Поліській дослід¬
ній станції очолював заслужений агроном України, лауреат Держав¬
ної премії СРСР І.В.Карпович, а з 1982 року - кандидат сільсько¬
господарських наук В.І.Сидорчук.
У 1932 році роботу із селекції картоплі розпочали на дослідному
полі Всесоюзного науково-дослідного інституту ферментативної та
плодоягідної промисловості (с.Немішаєве Київської області). У 1933 р.
дослідне поле разом із полями спиртзаводів Головспирту було офор¬
млене в селекційно-насінницьку мережу цього ж інституту, на базі
якої в 1935 р. була створена Київська селекційно-дослідна станція
Всесоюзного НДІ спиртової промисловості (з 1944 року - Київська
сільськогосподарська дослідна станція Всесоюзного НДІ спиртової
промисловості, пізніше - Немішаївська дослідна станція Українсько¬
го НДІ землеробства). У 1968 році станція була реформована в Укра¬
їнський НДІ картопляного господарства (з 1992 року - Інститут кар¬
топлярства УААН). Нині він є центром із селекції картоплі в Україні.
Одним із головних завдань селекції дослідної станції в перший пері¬
од її роботи було створення сортів для спиртової промисловості. У
1932-1946 рр. під керівництвом Р.Д.Шехаєва були створені технічні
сорти картоплі: Стаханівська, Червоноспиртова, Рясна, Вольтман
1177. В основному в цей період у селекції використовувались внут¬
рішньовидова гібридизація в межах S.tuberosum та клоновий добір. З
1946 по 1973 рік селекцію картоплі на дослідній станції та в Україн¬
ському НДІ картопляного господарства очолював відомий селекціо¬
нер, заслужений агроном України, кандидат сільськогосподарських
наук О.І.Терещенко. Ним були створені високопродуктивні, з комп¬
лексом ознак сорти картоплі Катюша, Чотирьохсотая, Бородянська,
Смачна, Немішаївська ювілейна, Луговська, Українська рожева.
У наступні роки селекцію картоплі в Українському НДІ картоп¬
ляного господарства (пізніше Інститут картоплярства УААН) очо¬
лював заслужений працівник сільського господарства України, док¬
тор сільськогосподарських наук А.А.Осипчук. Разом із колективом
селекціонерів ним були створені сорти різних груп стиглості і госпо¬
дарського призначення, стійкі проти хвороб і шкідників та з іншими
цінними ознаками: Зарево, Воловецька, Віхола, Світанок київський,
Ромашка 8, Либідь, Бородянська рожева, Водограй, Обрій, Кобза,
200
Горлиця, Купава, Ракурс, Слов’янка, Повінь, Явір, Серпанок, Віри-
нея, Фантазія, Багряна, Поляна, Дніпрянка та ін. У зв’язку з поши¬
ренням в Україні хвороб і шкідників (рак, фітофтороз, зморшкувата,
смугаста мозаїки, закручування та скручування листків, готика, пар¬
ша звичайна, чорна ніжка, кільцева гниль, стеблова та картопляна
нематоди, колорадський жук) селекція картоплі в Інституті картоп¬
лярства спрямовується на створення сортів, стійких проти вказаних
патогенів.
У зв’язку з розвитком промисловості з переробки картоплі на
картоплепродукти з 90-х років в Інституті картоплярства ведеться
селекція сортів, придатних для зазначеного виду господарського ви¬
користання.
Над створенням нових сортів картоплі для півдня України у Все¬
союзному селекційно-генетичному інституті працювали О.М.Фаво-
ров та А.Ф.Котов. Слід зазначити, що для півдня України одним із
важливих завдань селекції картоплі є створення посухо- та жаростій¬
ких сортів. Тут були створені сорти Одеська 24 та Одеська 65, які
пристосовані до місцевих умов. Нині селекційна робота з картоплею
тут не проводиться.
У 1947 році була розпочата робота із селекції картоплі в Україн¬
ському НДІ овочівництва і картоплярства (східний Лісостеп Украї¬
ни) та його дослідній мережі. Методом внутрішньовидової гібриди¬
зації тут були створені посухостійкі сорти Харківська рання та Сія¬
нець 10/22 та ін. На Одеській овоче-картопляній дослідній станції
Українського НДІ овочівництва і картоплярства В.Г.Гойко методом
добору поліпшив сорт Богарна, який був районований на півдні
України під назвою Богарна одеська [3]. Український НДІ овочівниц¬
тва і картоплярства, крім селекції картоплі, проводив також коорди¬
націю робіт в Україні із зазначеного напряму досліджень. З 1968 року ці
функції передані Інституту картоплярства УААН.
Селекцією картоплі в західному регіоні України почали займа¬
тись після Великої Вітчизняної війни. У Науково-дослідному інсти¬
туті землеробства і тваринництва західного регіону України спочат¬
ку вивчили місцеві сорти, відібрані переважно на Львівщині, Івано-
Франківщині, Буковині. У 1956-1958 рр. було зібрано близько 300
зразків місцевих сортів. Окремі з них передавалися до державного
сортовипробування (сорт Трембіта). Своєрідним і особливим місце¬
вим зразком є сорт картоплі Карпатська, який було одержано від са¬
мозапилення форми, відібраної в горах. За участю сорту Карпатська
було створено ряд нових сортів, які відзначалися фітофторостійкіс-
тю, підвищеною і високою крохмалистістю та іншими цінними озна¬
ками [4]. Селекцією картоплі в цьому інституті займався доктор
сільськогосподарських наук, професор В.Г.Влох, пізніше - кандидат
сільськогосподарських наук Л.А.Ільчук. Тут були створені сорти
Мавка, Полонина, Слава, Віра, Оксамит 99.
201
У західному регіоні України селекція картоплі проводиться та¬
кож у Львівському державному аграрному університеті. Тривалий час
її очолював доктор сільськогосподарських наук, професор І.Д.Не-
чипорчук. Ним разом із колективом були створені високопродуктив¬
ні, з комплексом ознак сорти Львівська біла, Львівська синьовічкова,
Львівська рання, Гібридна 14, Прикарпатська, Тетянка, Нестерівська
та ін. На сьогодні роботу із селекції картоплі в цьому закладі очолює
доктор сільськогосподарських наук, професор І.І.Тимошенко, який
разом із колективом селекціонерів створив високопродуктивний,
стійкий проти фітофторозу сорт Західна. У селекції картоплі Львів¬
ського державного аграрного університету широко використовують
як батьківські форми сорти та гібриди, одержані від схрещування
S.tuberosum з дикими південноамериканськими видами [2]. У зв’язку
з цим тут створюються сорти, які відзначаються фітофторостійкістю.
З 1963 року селекція картоплі проводиться на Чернігівській до¬
слідній станції по картоплі (зона Полісся). У різні часи селекцією
картоплі тут займались заслужений агроном України Я.І.Прошко,
кандидати сільськогосподарських наук В.Д.Волков та В.М.Шолом.
Нині цю роботу очолює тут кандидат сільськогосподарських наук
О.Ф.Онищенко. На цій станції були створені сорти картоплі Радуга
Полісся, Витязь, Седнівська рання, Пекуровська, Придеснянська,
Чернігівська рання, Чернігівська 98.
У 60-х роках селекцію картоплі розпочали на Житомирській об¬
ласній державній сільськогосподарській дослідній станції (нині це
Інститут сільського господарства Полісся). Селекцію картоплі тут
проводили в різний час кандидати сільськогосподарських наук
В.М.Сергієнко та М.С.Литовченко. У цій установі були створені такі
сорти картоплі, як Древлянка, Рум’янка, Дзвін, Дарина.
У гірських умовах Карпат селекцію картоплі проводять на Гірсь-
ко-Карпатській дослідній станції. Селекціонером Д.В.Равликом тут
створені сорти картоплі Нижньоворітська та Свалявська. Слід зазна¬
чити, що гірські умови Карпат є унікальними для одержання ботаніч¬
ного насіння від схрещування і природного самозапилення, а також
для оцінки селекційного матеріалу на фітофторостійкість.
У Сумському державному аграрному університеті селекцію картоплі
розпочали у 80-х роках. Очолює її лауреат Державної премії СРСР, за¬
служений діяч науки і техніки України, доктор сільськогосподарських
наук, професор М.Д.Гончаров. Над селекцією картоплі тут працює та¬
кож доктор сільськогосподарських наук, професор Н.С.Кожушко. У цій
установі створено сорти картоплі Молодіжна, Ластівка та ін.
Таким чином, селекція картоплі на сьогодні проводиться в Інсти¬
туті картоплярства УААН, на Поліській дослідній станції ім. О.М. За¬
сухіна, в Інституті землеробства і тваринництва західного регіону
УААН, Інституті сільського господарства Полісся УААН, Сумсько¬
му та Львівському державних аграрних університетах, Гірсько-
202
Карпатській сільськогосподарській дослідній станції, Чернігівській
дослідній станції по картоплі.
ЛІТЕРАТУРА
1. Карпович І.В. Селекція картоплі на Поліській дослідній станції ім. О.М. За-
сухіна // Картоплярство. - К.: Урожай, 1970. - Вип. 1. - С.29-31.
2. Нечипорчук І.Д. Селекція картоплі в західних районах України // Кар¬
топлярство. -К.: Урожай, 1970. - Вип.1. - С.20-23.
3. Онищенко О.Й. Селекція картоплі в Україні. - К.: Вид-во УАСГН,
1960.- 114 с.
4. Фаворов О.М., Влох В.Г., Коптев М.Н., Ліорек С.Й. Досвід селекційної
роботи з картоплею в передгірській та гірській зонах українських Карпат //
Картоплярство. - К.: Урожай, 1970. - Вип. 1. - С. 13-19.
4.2. ГЕНЕТИЧНИЙ ПОТЕНЦІАЛ
КАРТОПЛІ
Методами селекції в сортах картоплі може бути поєднано понад
50 ознак [5], окремі з яких (стійкість до раку, картопляної нематоди,
деяких вірусів та ін.) контролюються основними генами. Генетичний
потенціал потомства в цьому випадку буде відзначатися високим
проявом наявних ознак.
Більшість ознак у картоплі (урожайність, уміст крохмалю і білка,
польова стійкість проти хвороб та ін.) контролюються полігенами і
під впливом умов зовнішнього середовища можуть змінюватися.
Максимальний прояв названих ознак відбувається при оптимальних
умовах вирощування.
Важливим показником для картоплі є потенційна врожайність.
Вона теоретично підрахована. Для країн Європи, у тому числі Украї¬
ни, вона становить близько 1000 ц/га. Фактична урожайність у дослі¬
дах називається польовою і може досягти 70-80% від потенційної.
Завдання полягає в тому, щоб максимально наблизити польову
продуктивність до потенційної. Цього можна досягти створенням
оптимальних умов для вирощування картоплі, а також створенням і
застосуванням сортів, які б відзначалися пластичністю і стабільністю
зазначеної ознаки при високих її показниках. Є багато прикладів
отримання високої врожайності в польових умовах. У Київській об¬
ласті урожайність сорту Гатчинська реєструвалась на рівні 744 ц/га,
а в Красноярскому краї сорту Бородянська рожева - 1087 ц/га . Уро¬
жайність сортів Водограй і Луговська в Інституті картоплярства до¬
сягла 640 ц/га, Світанку київського - 569 ц/га.
Складовою частиною продуктивності картоплі є вміст у бульбах
крохмалю. При сприятливих умовах уміст його у середньораннього сор¬
ту Світанок київський досягав 20-22% при 18% у звичайних умовах, а
середньопізнього сорту Зарево - 26-29% при 22% у звичайних умовах.
203
Вирішення практичних завдань із підвищення продуктивності
рослин здійснюється двома шляхами: по-перше, створенням бажаних
генотипів, по-друге, розробкою методів керування індивідуальним
розвитком. При вивченні впливу зовнішніх факторів з метою управ¬
ління розвитком організмів слід виявити ті умови, у яких найбільш
повно реалізується генотипний потенціал організму [2].
Велику роль у підвищенні продуктивності картоплі відіграє
управління ростом і розвитком рослин за допомогою відповідних
заходів агротехніки.
Визначення норми реакції селекційного матеріалу і сортів картоп¬
лі дає можливість розробити сортову науково обгрунтовану техноло¬
гію їх вирощування й одержувати потенційно можливі врожаї при
оптимальних затратах матеріалів і праці. Так, застосовуючи ряд аг¬
розаходів, у Львівській області отримали урожай сорту Тетянка
665 ц/га, а сорту Львів’янка -619 ц/га [1].
При застосуванні відповідних агрозаходів у 1942 р. під Кемерово
було одержано рекордний урожай сорту Лорх - 1331 ц/га [3].
Велика роль у використанні генетичного потенціалу сортів кар¬
топлі належить насінництву, же реалізує досягнення селекції шля¬
хом вирощування високоякісного насіння спеціальними прийомами.
Врожаї та валові збори сільськогосподарських культур підвищу¬
ються на 20-25% за рахунок використання високоякісного насіння
нових сортів [4].
ЛІТЕРАТУРА
1. Лтцак И.А. Генотипическая реакция новых фитофтороустойчивых сортов
и гибридов картофеля на условия питания // Селекция фитофтороустойчивых
сортов картофеля: Сб. науч. тр. Львовского СХИ. - Львов, 1983. - С.15Ф-161.
2. Лобашев М.Е. Генетика. - Л.: Изд-во Ленинград, ун-та, 1969. - 752 с.
3. Лорх А.Г. Экологическая пластичность картофеля. - М., 1968.
4. Молоцький М.Я., Васильківський С.П., Князюк В.І. Селекція та насін¬
ництво польових культур: Підручник. - К.: Вища шк., 1994 - 454 с.
5. Росс X. Селекция картофеля. Проблемы и перспективы / Перевод с
англ. В.А. Лебедева. - М.: Агропромиздат, 1989. - 183 с.
4.3. БЮЛОГШНИЙ ВЗАЄМОЗВ’ЯЗОК ОЗНАК
КАРТОПЛІ І ЙОГО ВПЛИВ НА ДОБІР
Питання про кореляційну мінливість ознак у живих організмів
здавна цікавить селекціонерів.
Кореляція (від лат. correlatio - співвідношення) - це взаємозв’язок
між окремими частинами та ознаками організму, який виявляється в то¬
му, що зміна однієї ознаки веде до корелятивної зміни іншої.
Після робіт багатьох дослідників, у яких певною мірою уже був
окреслений принцип кореляції у живих організмів, Ч.Дарвін присвя¬
204
тив цьому питанню спеціальні розділи у своїх працях “Происхожде¬
ние видов”, “Изменение животных и культурных растений в домаш¬
них условиях”, у яких він навів велику кількість прикладів кореляції
і вказав на їхнє значення в еволюції живих організмів.
Чимало дослідників констатують, що в багатьох організмів зу¬
стрічаються ознаки, взаємозв’язані між собою. У таких випадках при
визначенні однієї ознаки можна твердити про наявність й іншої. Зна¬
чення взаємозв’язку ознак сприяє добору потрібних форм і приско¬
рює процес селекційної роботи.
У практичній роботі спряженість може бути використана двояко -
залежно від зв’язку і його типу.
У першому випадку звертають увагу на величину коефіцієнта ко¬
реляції і його ознак і лише потім роблять висновок про практичну
цінність такої залежності. Прикладом цього є всі випадки чітко ви¬
раженої лінійної кореляції, яка вже давно використовується в селек¬
ційній роботі. У другому випадку величина коефіцієнта і навіть його
знак суттєвої ролі не відіграють, а тип зв’язку є прямо- або криволі¬
нійним.
У багатьох випадках продуктивність сільськогосподарських рос¬
лин зумовлюється численними кількісними ознаками, які є результа¬
том взаємодії спадкових особливостей рослин і комплексу умов зов¬
нішнього середовища. На величину кількісної ознаки можуть впливати
інші кількісні або якісні ознаки. Так, наприклад, анатомічна будова
тканини і вміст цукру в клітинному соці рослин можуть впливати на
їхню зимостійкість. Тому за кількісними ознаками, у тому числі і вро¬
жайністю рослин, можна говорити і про їхні якісні характеристики.
Наприклад, скоростиглість сіянців у молодому віці можна визна¬
чати за кількістю днів від садіння до цвітіння, оскільки дуже часто з
цвітінням рослин картоплі пов’язане їхнє бульбоутворення, а також
за врожаєм - при ранньому викопуванні.
Виходячи із зведених спостережень багатьох дослідників, дуже
простим і в той же час дуже надійним способом є визначення скоро¬
стиглості картоплі як у сіянців першого року, так і в бульбових ре¬
продукціях. Крім того, скоростиглість сіянців першого року в буль-
борепродукціях можна визначити за деякими морфологічними озна¬
ками, які змінюються з віком рослин. Відомо, що із скоростиглістю
рослин картоплі пов’язані швидке наростання розчленованості лист¬
ка, максимальна розсіченість у ранньому ярусі і мала кількість ярусів
монополія. І, навпаки, для пізньостиглих форм картоплі характерне
повільне наростання розчленованості листка, високе розміщення
листків із максимальною розсіченістю, велика кількість ярусів мо¬
нополія.
Виявлена багатьма вченими кореляція між різними ознаками кар¬
топлі являє практичний інтерес і може використовуватися в селек¬
ційній роботі, зокрема позитивна кореляція між умістом крохмалю в
205
бульбах сіянців першого року і при їх бульбовому розмноженні, яка
вказує на можливість добору висококрохмалистих форм за сіянцями
першого року, тощо.
Однак недостатньо вивченими в культурі картоплі є питання ко¬
реляційної мінливості в гібридних популяціях, одержуваних від
схрещування нових сортів картоплі, особливо тих, які виведені шля¬
хом міжвидової гібридизації, характер успадкування господарських і
біологічних ознак у таких гібридних популяціях та інше, знання яких
могло б сприяти поєднанню в одному сорті потрібних ознак і ство¬
ренню нових цінних сортів цієї культури.
Разом з тим вважаємо за доцільне продовжувати дослідження ко¬
реляції між різними ознаками, наявність якої могла б сприяти більш
ефективному добору сіянців картоплі і кращій оцінці їх у селекцій¬
ній роботі. Цілком очевидно, що кореляція не є постійною і в різних
популяціях є різною, тому використовувати її слід з урахуванням
особливостей кожної окремо взятої популяції.
Так, при першому бульбовому розмноженні сіянців ми вивчали
мінливість і кореляційну залежність між довжиною міжвузлів у ниж¬
ній частині стебла рослин та їхнім врожаєм, довжиною міжвузлів і
вмістом крохмалю в бульбах сіянців, довжиною міжвузлів і скоро¬
стиглістю рослин, довжиною міжвузлів і висотою рослин, товщиною
стебла у рослин та їхнім урожаєм, товщиною стебла і вмістом крох¬
малю в бульбах сіянців, товщиною стебла і скоростиглістю сіянців,
початком цвітіння сіянців та їх врожаєм, початком цвітіння сіянців і
вмістом крохмалю в їх бульбах, умістом крохмалю в бульбах сіянців
та їхнім урожаєм, скоростиглістю сіянців та їхнім урожаєм, скоро¬
стиглістю сіянців і вмістом крохмалю в їхніх бульбах, висотою рос¬
лин та їхнім урожаєм, врожаєм рослин і середньою масою товарної
бульби, врожаєм сіянців при першому бульбовому розмноженні і
врожаєм сіянців першого року, умістом крохмалю у рослин при
першому бульбовому розмноженні і вмістом крохмалю у сіянців
першого року.
Вивчення гібридних популяцій при першому бульбовому розмно¬
женні сіянців картоплі показало, що для більшості ознак суттєвої коре¬
ляції в їх мінливості не існує, за винятком окремих ознак, у мінливості
яких була виявлена більш-менш чітко виражена і математично доведена
кореляція. У мінливості окремих ознак спостерігалися певні відхилення
від закономірностей, характерних для сіянців першого року. Це стосува¬
лося в першу чергу зв’язку товщини стебла у рослин з їхнім урожаєм для
окремих гібридних популяцій, а також взаємозв’язку між початком цві¬
тіння рослин та їхнім урожаєм. Крім того, вивчали відмінності між гіб¬
ридними популяціями за ступенем мінливості ознак, щоб з’ясувати,
якою мірою досліджені популяції відрізняються між собою, а також ха¬
рактер мінливості кожної пари ознак у кожній гібридній популяції та
ступінь кореляційної залежності між ними.
206
Вивчаючи у різних популяцій залежність між довжиною міжвуз¬
лів у нижній частині стебла рослин та врожаєм сіянців при першому
бульбовому розмноженні, а також кореляцію для цих двох ознак,
закономірностей не виявили. Кожна популяція мала певні властивос¬
ті. Це стосується як середньої величини цих ознак, так і інтенсивнос¬
ті варіювання їх, а також ступеня взаємозв’язку між ними.
Вивчаючи кореляційну мінливість у гібридних популяціях картоп¬
лі за товщиною стебла в його нижній частині та врожаєм рослин при
першому бульбовому розмноженні сіянців, виявили, що, порівняно із
сіянцями першого року, у яких здебільшого спостерігалася значна
позитивна кореляція між цими ознаками, при першому бульбовому
розмноженні вона не зберігалася.
При вивченні кореляційної мінливості таких ознак, як початок
цвітіння гібридних рослин (кількість днів від садіння до початку цві¬
тіння) та їх урожай, при першому бульбовому розмноження сіянців у
всіх гібридних популяціях картоплі спостерігали закономірну незнач¬
ну від’ємну кореляцію. Слід зазначити, що між початком цвітіння
рослин і вмістом крохмалю у їхніх бульбах при першому бульбовому
розмноженні гібридних сіянців певної кореляційної залежності не
було виявлено. Щодо цих ознак у різних гібридних популяцій спо¬
стерігалася як позитивна, так і від’ємна кореляція, яка, однак, в обох
випадках математично не була доведена.
Значна увага була приділена вивченню кореляційної мінливості в
гібридних популяціях картоплі таких важливих господарсько корис¬
них ознак, як урожайність та крохмалистість бульб.
Одержані дані свідчать про те, що в популяціях, які вивчалися
при першому бульбовому розмноженні сіянців, як і в сіянців першо¬
го року, про що згадувалося раніше, не виявлялася будь-яка чітко
виражена кореляція. Причому, як і щодо інших ознак, на їхній
зв’язок не впливає ні середня величина врожаю, ні величина варіа¬
ційних коефіцієнтів.
Враховуючи те, що в селекції картоплі велике значення має знан¬
ня залежності між успадкуванням скоростиглості рослин та їхньою
врожайністю, вивченню саме цього питання було приділено значну
увагу, тобто характеру успадкування тривалості вегетаційного пері¬
оду (кількості днів від садіння бульб до початку природного відми¬
рання бадилля) і врожайності рослин у їх взаємозв’язку. Вивчаючи
кореляційну мінливість цих ознак у дев’яти гібридних популяцій
картоплі при першому бульбовому розмноженні сіянців, виявили
від’ємну кореляційну залежність. Однак у більшості випадків вона
була незначною і математично не доведена.
Значний інтерес являє і знання кореляційної мінливості таких
ознак, як скоростиглість рослин і вміст крохмалю в гібридних попу¬
ляціях картоплі, оскільки можливість поєднання короткого вегета¬
ційного періоду рослин з високим умістом крохмалю вже давно ці¬
207
кавить селекціонерів. Виявилося, що при суттєвій різниці інтенсив¬
ності варіювання досліджуваних ознак майже ніякої кореляції між
ними не існує. Величини кореляційного коефіцієнта для кількості
днів від садіння до природного відмирання картоплиння та вмісту
крохмалю в бульбах коливалися в різних гібридних популяцій від
-0,195 до +0,218. Це свідчить про те, що в цих популяціях довгий або
короткий вегетаційний період гібридних сіянцях здебільшого не
пов’язаний з високою або низькою крохмалистістю бульб.
Можна вважати, що використання кореляції між масою товарних
рослин і врожаєм рослин полегшить добір високоврожайних форм за
масою товарних бульб у гібридних популяціях картоплі при першо¬
му бульбовому розмноженні сіянців.
Таким чином, при першому бульбовому розмноженні сіянців у
досліджуваних гібридних популяціях картоплі добре виражена пози¬
тивна кореляція виявилася тільки між врожайністю рослин і масою
товарних бульб, а також між довжиною міжвузлів у нижній частині
стебла рослин та їхньою висотою.
Між товщиною стебла в нижній частині його та врожаєм рослин,
тривалістю вегетаційного періоду рослин та їх урожаєм, висотою
рослин і врожаєм, початком цвітіння гібридних рослин (кількістю
днів від садіння бульб до початку цвітіння рослин) і врожаєм, почат¬
ком цвітіння рослин і вмістом крохмалю в бульбах, довжиною між¬
вузлів у нижній частині стебла рослин і врожаєм, довжиною міжвуз¬
лів і тривалістю вегетаційного періоду рослин, товщиною стебла і
скоростиглістю рослин виявилася незначна позитивна або від’ємна
кореляція, що не дозволяє твердити про певну закономірність.
Але у взаємозв’язку між врожаєм рослин і крохмалистістю бульб,
товщиною стебла і вмістом крохмалю, довжиною міжвузлів у ниж¬
ній частині стебла рослин і вмістом крохмалю ніякої кореляції, по
суті, не було виявлено.
У селекції картоплі великої уваги заслуговує також питання ко¬
реляції врожаю бульб у сіянців картоплі першого року та їхнім уро¬
жаєм при першому бульбовому розмноженні. З цього приводу в літе¬
ратурі часто зустрічається думка про те, що між урожаєм сіянців
першого року та врожаєм їх при бульбовому розмноженні ніякої ко¬
реляції не існує, з чим, однак, не можна погодитися, оскільки коре¬
ляція повинна вивчатися з урахуванням умов вирощування сіянців у
першому і другому роках, їхньої урожайності в перші два роки і по¬
ходження сіянців. Вирішення цього питання повинно сприяти під¬
вищенню ефективності добору потрібних форм серед сіянців першо¬
го року, а отже - успішному виведенню нових урожайних сортів
картоплі.
Проведені дослідження свідчать про те, що в багатьох гібридних
популяціях картоплі врожайність рослин тісно пов’язана з їхньою
висотою і що в таких популяціях добір високоврожайних форм мож-
208
ливий за висотою рослин ще при першому бульбовому розмноженні
сіянців. В інших гібридних популяціях врожайність рослин пов’язана
з низькими стеблами. У таких гібридних форм, як правило, утворю¬
ються великі листки, що зумовлює велику асиміляційну поверхню
рослин.
Одночасно при першому бульбовому розмноженні сіянців вив¬
чають і залежність між ступенем ураження рослин фітофторозом та
їх урожаєм, а також умістом крохмалю. У досліджуваних популяціях
вивчали ступінь залежності ураження картоплі фітофторозом при
першій польовій оцінці та врожаєм її при першому бульбовому роз¬
множенні сіянців. При цьому проявилася чітко виражена і математи¬
чно доведена від’ємна кореляція, яка полягала в тому, що чим більше
гібридні рослини уражувалися фітофторозом, тим менший був їхній
врожай. Це спостерігалося, зокрема, у популяцій, одержаних від
схрещування сортів Львівська біла х Прієкульська рання (г±82 =
= - 0,415±0,095), Прієкульська рання х Львівська біла (г ± 82 = - 0,372 ±
± 0,102) та популяцій Фітофторостійка х Львівська біла (rt82 = - 0,445±
0,094).
З наведених даних видно, що в гібридних популяціях з великою
кількістю форм, нестійких до фітофторозу, величина врожаю знач¬
ною мірою залежить від польової стійкості гібридів до фітофторозу в
ранній період їх росту і розвитку, тобто тоді, коли ранні сорти най¬
більш інтенсивно накопичують урожай. Тому в таких популяціях
доцільно вести добір форм з інтенсивним накопиченням врожаю в
ранній період росту рослин для того, щоб до масової появи фітофто¬
розу рослини могли сформувати високий урожай бульб і нагромади¬
ти в них достатню кількість крохмалю.
При другому бульбовому розмноженні сіянців картоплі у біль¬
шості гібридних популяцій була виявлена позитивна кореляційна
залежність лише між висотою рослин та їхнім урожаєм. Щодо фіто¬
фторозу, то чим раніше гібриди уражувалися ним і чим вищий був
їхній ступінь ураження, тим більше знижувався врожай. Раннє ура¬
ження рослин фітофторозом призводить до зниження крохмалистості
бульб.
З метою визначення зв’язку між такими найважливішими госпо¬
дарсько корисними ознаками, як урожай і вміст крохмалю, нами бу¬
ла визначена кореляційна залежність у різних гібридних популяціях
картоплі, одержаних за участю сорту Львів’янка, який використову¬
вався як материнська форма (табл. 4.1). Відомо, що єдиної думки про
кореляційний зв’язок між урожайністю і крохмалистістю бульб у
рослин картоплі не існує. Ряд авторів вказують на від’ємну кореля¬
ційну залежність між високим умістом крохмалю і врожайністю
бульб. Інші виявили слабку позитивну або від’ємну кореляцію між
цими ознаками. Одержаний коефіцієнт кореляції для наведених гіб¬
ридних популяцій, який коливається від +0,02 (Львів’янка х Несте-
209
рівська) до -0,90 (Львів’янка х 10-116), свідчить про те, що врожай¬
ність і вміст крохмалю успадковуються незалежно один від одного
й одержані гібриди можуть бути як високо-, так і низькокрохмали-
стими.
Від’ємну кореляцію між врожайністю і крохмалистістю бульб, вияв¬
лену в перших трьох гібридних популяціях, а для схрещування сорту
Львів’янка х 10-116 і математично доведену, можна подолати шляхом
схрещування висококрохмалистих сортів із високоврожайними.
Таблиця 4.1 - Кореляційна залежність між урожайністю та вмістом крохмалю
Схрещування
n
X
г/кущ
min - шах
У,
%
min - шах
r
Sr
tr
Львів’янка х 301-125
21
1126
700-1555
17,5
12,4-21,9
-0,36
0,33
1,09
Львів’янках 10-116
17
1486
1116-2025
16,7
12,9-18,6
“0,90
0,18
5,00
Львів’янка х 294-248
23
1301
762-1840
17,6
15,1-20,0
-0,18
0,21
0,85
Львів’янка х Нестерівська
74
1220
820-1820
19,7
15,2-24,1
0,02
0,11
0,21
Львів’янка х 303-296
31
1309
910-1820
19,8
14,3-25,4
0,13
0,18
0,74
Вивчення кореляційної залежності ознак різних гібридних попу¬
ляцій картоплі, одержаних від міжвидових схрещувань, дозволяє
констатувати наступне. У сіянців першого року добре проявляється
позитивна кореляція в мінливості таких ознак картоплі, як товщина
стебла в нижній його частині і врожай, кількість бульб і врожай.
Незначна кореляційна залежність (позитивна) у сіянців першого
року виявляється між висотою розсади сіянців у період садіння її в
грунт і врожаєм, масою бульб, утворених із розсади перед висадкою
її в грунт, і врожаєм.
Незначна позитивна або від’ємна кореляція, яка не вкладається в
жодні закономірності, виявляється в сіянцях першого року для таких
ознак, як вміст крохмалю і врожай, довжина міжвузлів у нижній час¬
тині стебла і врожай, довжина міжвузлів і вміст крохмалю, товщина
стебла і вміст крохмалю, форма кінцевої частини листка і форма
бульб, кількість днів від садіння сіянців до початку їх цвітіння і
вміст крохмалю.
При першому бульбовому розмноженні сіянців картоплі добре
виявляється позитивна кореляція між умістом крохмалю сіянців
першого року і рослин першого бульбового розмноження їх, між
урожаєм рослин і масою товарних бульб, довжиною міжвузлів у ниж¬
ній частині стебла рослин і їхньою висотою. Як і в гібридних сіянців
першого року, при першому бульбовому розмноженні їх виявляється
незначна кореляція (позитивна або від’ємна) між мінливістю товщи¬
ни стебла в нижній його частині та врожаєм рослин картоплі, трива¬
лістю вегетаційного періоду (кількість днів від садіння бульб до по¬
чатку природного відмирання картоплиння) і їх урожаєм, висотою
рослин і врожаєм, початком цвітіння гібридних рослин (кількість
210
днів від садіння бульб до початку цвітіння рослин) і врожаєм, почат¬
ком цвітіння рослин і вмістом крохмалю, довжини міжвузлів у ни¬
жній частині стебла рослин і їх урожаєм, довжиною міжвузлів і
тривалістю вегетаційного періоду рослин, товщиною стебла і ско¬
ростиглістю рослин, товщиною стебла у сіянців першого року і
врожаєм рослин при їх першому бульбовому розмноженні, довжи¬
ною міжвузлів у нижній частині стебла у сіянців першого року і
врожаєм рослин при їх першому бульбовому розмноженні, урожа¬
єм сіянців першого року і врожаєм їх при першому бульбовому
розмноженні.
У взаємозв’язку врожайності рослин і крохмалистості бульб, то¬
вщини стебла і вмісту крохмалю, довжини міжвузлів і вмісту крох¬
малю при першому бульбовому розмноженні гібридних сіянців кар¬
топлі ніякої кореляції не було виявлено.
Між ступенем ураження рослин фітофторозом та їхнім урожаєм,
а також між ураженням рослин фітофторозом і нагромадженням
крохмалю в їхніх бульбах спостерігається чітко виражена від’ємна
кореляція.
У зв’язку з відсутністю чітко вираженої кореляційної залежності
в мінливості і крохмалистості сіянців у гібридних популяціях картоп¬
лі, врожайності і тривалості вегетаційного періоду рослин можна
вважати, що ці ознаки успадковуються гібридами картоплі незалеж¬
но одна від одної. Тому при міжсортових схрещуваннях сіянців кар¬
топлі можна поєднувати високу врожайність з високою крохмалистіс¬
тю бульб, високу врожайність із коротким періодом вегетації рослин,
високу крохмалистість бульб із коротким вегетаційним періодом ро¬
слин та високу врожайність із високим умістом крохмалю і коротким
вегетаційним періодом рослин. Завдяки цьому можна виводити нові
високоврожайні ранньостиглі сорти картоплі з високим або підви¬
щеним умістом крохмалю.
Наявність у гібридних популяціях картоплі значної позитивної
кореляційної мінливості товщини стебла і врожайності сіянців пер¬
шого року, а також кількості бульб у гнізді і врожайності свідчить
про те, що з великою вірогідністю можна відбирати високоврожайні
форми картоплі за товщиною стебла в нижній його частині і кількіс¬
тю бульб у гнізді.
Висока позитивна кореляція між умістом крохмалю в сіянців
першого року і бульбовим розмноженням повинна сприяти успішно¬
му добору серед гібридних популяцій висококрохмалистих форм із
сіянців першого року.
Незначна позитивна кореляція в гібридних популяціях картоплі
між урожаєм сіянців першого року і при їх бульбовому розмноженні,
що спостерігається в більшості випадків, свідчить про те, що в кар¬
топлі вибракування сіянців першого року за їхньою врожайністю
проводити не можна, оскільки високоврожайні за генотипом форми,
211
які не виявили високої врожайності на першому році життя, можуть
вибраковуватися. Тому перше вибракування гібридних сіянців кар¬
топлі за ознакою врожайності доцільно проводити при першому бу¬
льбовому розмноженні.
Цілком очевидно, що результати проведених досліджень будуть
становити певний інтерес для селекційної роботи, зокрема при об¬
грунтуванні напряму селекції і для підвищення ефективності добору
бажаних форм із міжсортових гібридних популяцій картоплі на сіян¬
цях першого року і при їх першому бульбовому розмноженні.
4.4. МЕТОДИ СЕЛЕКЦІЇ КАРТОПЛІ
Селекція картоплі, як і багатьох продовольчих культур, має дав¬
ню історію. У той час, коли людина почала вирощувати картоплю,
культурних сортів ще не було. Вирощували здебільшого популяції
різних видів, форм і зразків. Згодом для насіння почали відбирати
кращі рослини за їх розвитком, стійкістю проти хвороб, урожайністю
та іншими ознаками. Відбираючи й об’єднуючи бульби таких рос¬
лин, людина створювала популяції картоплі, які, на відміну від попе¬
редніх, характеризувались вищою продуктивністю і пристосованістю
до місцевих умов. З часом, відбираючи з популяції окремі рослини і
розмножуючи їх, люди створювали місцеві сорти.
Клонова селекція - найбільш давній метод створення сортів картоплі
в Європі. Він виник в середині 17 ст. унаслідок повної або часткової не¬
придатності д ля вирощування перших завезених у Європу сортів [6].
Нині метод клонової селекції застосовується як при створенні
нових сортів, так і в насінництві картоплі. Шляхом клонової селекції
можна, з одного боку, звільнити сорт від хвороб, що передаються
бульбами, а з іншого - відібрати господарсько цінні мутації [12].
Клоновий добір дає змогу істотно підвищити ефективність селе¬
кції картоплі, особливо на польову стійкість проти вірусів, продук¬
тивність і життєздатність, а іноді й адаптивність, фітофторостійкість,
скоростиглість, стійкість до механічних пошкоджень тощо [9].
На початку XX ст. Л.Бербанком методом клонової селекції був
створений сорт картоплі Рассет Бербанк, який і нині вирощується в
СІЛА. У наш час цим методом були створені сорти Вольтман 1177,
Скороспілка 1, Сніжинка, Богарна одеська, Баранівська, Сумська
поліпшена, Дачна та ін.
Перші зразки картоплі, завезені з Південної Америки в Європу,
належали до культурного виду S.tuberosum, на основі якого проводи¬
лася селекційна робота. Тривалий час селекція картоплі базувалася
на використанні насіння від природного самозапилення рослин.
Згодом почали схрещувати між собою різні сорти виду S.tube¬
rosum, оскільки одержання потомства від природного самозапилення
знижує його життєздатність.
212
Перші відомості про штучну гібридизацію у картоплі наводяться
в 1819 р. Путшем, а в 30-х роках цей метод широко застосовував ан¬
глійський селекціонер Петерсон [12]. Метод гібридизації є ефектив¬
нішим, порівняно із самозапиленням, оскільки в процесі запліднення
беруть участь статеві клітини двох батьківських форм, у результаті
чого розширюються можливості для поєднання багатьох ознак в од¬
ному генотипі. Розрізняють внутрішньовидову гібридизацію (у ме¬
жах одного виду S.tuberosum) і міжвидову (схрещування кількох ви¬
дів). При обох методах показники ознак потомства можна передба¬
чити на основі відомостей про батьківські пари (їхній фенотип і ге¬
нотип). Першим методом можна отримувати господарсько цінні
генотипи у Fi від простих схрещувань, а при міжвидовій (віддаленій)
гібридизації для їх одержання необхідно застосовувати бекросування
простих гібридів.
Методом внутрішньовидової гібридизації ботаніком-аматором де
Ври (Нідерланди) у 1905 р. був створений відомий сорт картоплі Бін-
тьє, який і зараз культивується в Західній Європі. У 1931 р. був райо¬
нований сорт Лорх, створений цим методом російським селекціоне¬
ром О.Г.Лорхом. Він набув значного поширення в Росії.
Методом внутрішньовидової гібридизації створено багато сортів
картоплі, зокрема такі, як: Ульянівська, Южанін, Передовик, Друж¬
ба, Волжанін, Волзька, Прієкульська рання, Фаленська, Сєдов, Сибі¬
ряк, Кореневська, Комсомолець, Калітинець, Новинка, Свердловська
та ін. В Україні цим методом створені сорти Островська, Бородянсь¬
ка, Роза Полісся, Крепиш, Стаханівська, Червоноспиртова та ін.
Слід зазначити, що в родоводі майже всіх сортів картоплі знахо¬
димо вид S.tuberosum. Цінність цього виду для селекції полягає в йо¬
го високій урожайності, підвищених крохмалистості і вмісту вітамі¬
ну С, хороших смакових якостях, стійкості проти раку і несприятли¬
вих умов зовнішнього середовища, добрій формі бульб та ін. [19].
При внутрішньовидовій гібридизації, як і міжсортовій з багатови¬
довим походженням, уже в першому генеративному поколінні прохо¬
дить розщеплення за господарськими, біологічними і морфологічними
ознаками. Це дає різноманітний матеріал для добору. Вегетативний
спосіб розмноження закріплює ознаки відібраних генотипів.
Незважаючи на результативність методу внутрішньовидової гіб¬
ридизації для селекції картоплі, він має і певну обмеженість. Створи¬
ти сорти з високою стійкістю проти фітофторозу, нематоди, коло¬
радського жука, комплексною опірністю проти хвороб і шкідників та
з високими показниками біохімічних ознак методом внутрішньови¬
дової гібридизації неможливо.
Міжвидова гібридизація застосовується для поєднання ознак
виду S.tuberosum з цінними ознаками диких та інших культурних
видів. На сьогодні цей метод є одним з основних у селекції картоплі.
Завдяки йому вдалося створити сорти з високою стійкістю проти фі¬
213
тофторозу, картопляної нематоди, вірусів, колорадського жука, з ви¬
соким умістом крохмалю та білка, високою врожайністю завдяки
гетерозису та з іншими ознаками.
Міжвидову гібридизацію у картоплі вперше застосував у 1850 р.
Клотш, який схрестив дикий вид S.demissum з S.tuberosum й одержав
у результаті цього морозостійкі гібриди [12].
У 1931 р. за участю виду S.demissum Пушкарьовим [13] вперше у
світовій практиці був створений фітофторостійкий сорт картоплі Фі¬
тофторостійка 8670.
Пізніше на Литовській держселекстанції за участю S.demissum
було одержано висококрохмалистий, стійкий проти фітофторозу
сорт картоплі Олев.
Методом міжвидової гібридизації створено сорти картоплі Мос¬
ковська, Імандра, Камераз, Агрономічна, Любимець, Веселовсыса,
Зауральська, Столова 19, Лошицька, Темп, Розвариста та ін.
В Інституті картоплярства УААН на багатовидовій основі ство¬
рено високостійкий проти фітофторозу з комплексом інших ознак
сорт Луговська, висококрохмалистий з підвищеним умістом білка та
відносною стійкістю проти колорадського жука сорт Зарево, високо¬
крохмалистий, стійкий проти картопляної нематоди сорт Обрій та ін.
На Поліській дослідній станції ім. О.М.Засухіна Інституту картоп¬
лярства УААН методом міжвидової гібридизації були створені сорти
Житомирянка, Поліська рожева та інші, які характеризуються віднос¬
ною стійкістю проти фітофторозу в комплексі з іншими цінними озна¬
ками. Завдяки методу міжвидової гібридизації у Львівському державно¬
му аграрному університеті створені сорти з високою стійкістю проти
фітофторозу - Гібридна 14, Прикарпатська, Нестерівська, Захід на та ін.
Сьогодні в усіх селекційних установах як батьківські форми ви¬
користовується вихідний матеріал, створений на міжвидовій основі.
Використання методу міжвидової гібридизації в селекції картоплі
має свої труднощі. По-перше, багато видів не схрещуються із селек¬
ційними сортами, а тому це обмежує їх використання в селекції.
Друга складність полягає в тому, що у диких видів є ряд негативних
ознак, які домінують у гібридів перших поколінь. До таких ознак
належать низька урожайність і дуже довгі столони - до 150 см [3].
Створити сорти з першого покоління міжвидових гібридів можна ли¬
ше в тих випадках, коли схрещуються між собою два культурні види [2].
У селекції з використанням диких видів можливі такі напрями:
перший - проведення повторних схрещувань міжвидових гібридів із
селекційними сортами і другий - одержання потомства від самоза¬
пилення міжвидових гібридів. Іноді для окультурювання міжвидових
гібридів проводять чергування зазначених прийомів.
На всіх етапах міжвидової гібридизації та використання міжви¬
дових гібридів вирішальне значення має культурний компонент,
оскільки він значною мірою впливає на формування потомства [3].
214
Мутагенез. У селекції картоплі можуть бути використані спон¬
танні та індуковані мутації.
Під час вегетативного розмноження протягом тривалого часу
можуть виникати як корисні, так і небажані мутації, які можуть бути
відібрані для селекції або вибраковані.
На підвищення інтенсивності мутаційної мінливості впливає висо¬
кий ступінь гетерозиготності генотипів, застосування активних методів
оздоровлення сортів від вірусних хвороб, а також клональне мікророзм-
ноження іп vitro при оздоровленні сортів і відтворення вихідного матері¬
алу, одержаного методами генної та клітинної інженерії [9].
Під дією хімічних і фізичних мутагенів можна прискорити одер¬
жання цінних мутацій у картоплі. Застосування названих мутагенів
на бульбах картоплі значно збільшує різноманітність рослин за мор¬
фологічними та господарсько цінними ознаками [10,15].
Обробляючи ботанічне насіння мутагенами, відбирали сіянці, які
перевищували вихідні форми за стійкістю проти хвороб, умістом
крохмалю та іншими ознаками [4,5].
Таким чином, мутагенез може бути використаний як допоміжний
метод селекції.
Гаплоїдія - це кратне зменшення кількості хромосом. У картоплі їх
отримують як способом культури пиляків і мікроспор, так і використан¬
ням партеногенезу при запиленні материнських форм пилком окремих
зразків S.phureja та інших видів. Селекція картоплі на диплощному рівні
може підвищити ефективність селекції традиційними методами. Дигап-
лоіди можуть використовуватись у різних напрямках селекції. Після до¬
бору на диплощному рівні їх переводять на тетраплощний. Дигаплощи
можуть бути використані як спосіб подолання несхрещуваності деяких
видів із культурними сортами та окремими видами картоплі.
У ряді країн цей метод використаний при створенні нових сортів
картоплі.
Метод гаплоїдії описаний в інших розділах книги.
Поліплоїдія - це кратне збільшення кількості хромосом. Розріз¬
няють мітотичне і мейотичне збільшення кількості хромосом.
При мітотичній поліплоїдії насіння чи інші частини рослин обро¬
бляють колхіцином. Мейотичне поєднання кількості хромосом вини¬
кає в результаті відхилення від нормалного мейозу й утворення не-
редукованих гамет. Поліплоїди можуть утворюватись при схрещу¬
ванні 4п х 2п та 2п х 2п.
З використанням мейотичної поліплоїдії в Північній Америці бу¬
ли створені сорти картоплі Jukon Yold, Ica і Sirena [14].
У Росії із застосуванням методу поліплоїдії був створений сорт
картоплі Гібрид 390.
Однією з важливих властивостей поліплоїдії є використання її
для ресинтезу дигаплоїдів у тетраплоїди і подолання несхрещуванос¬
ті видів (описано в інших розділах).
215
Біотехнологічні методи. Поруч із традиційними методами в се¬
лекції картоплі мають значення клітинна та генна інженерія, які пе¬
редбачають :
- використання гетерогенності соматичних клітин та
сомаклональної мінливості для відбору ліній із більш високими
показниками, ніж вихідні форми;
- селективний відбір у суспензійній культурі клітин на стійкість
до патогенів;
- соматичну гібридизацію шляхом злиття ізольованих протопластів;
- генетичну трансформацію (описано в інших розділах книги).
Використання зазначених методів має велике значення для по¬
ліпшення існуючих та створення нових сортів картоплі.
Використання гетерозису в селекції картоплі є важливим за¬
вданням практичної селекції. Успадкування гетерозису зумовлене
малими генами або побічними ефектами головних генів. їх спільна
дія може проявлятися адитивним (ЗКЗ) або неадитивним (СКЗ) спо¬
собом. У більшості випадків діють обидва фактори [14]. За допомо¬
гою ефекту гетерозису можна отримати генотипи з високим показ¬
никами ряду ознак.
Виявлена висока ефктивність методів статевої поліплоїдії, яка
дає змогу ввести в геном S.tuberosum зародкову плазму диплоїдних
видів і досягти гетерозисного ефекту за продуктивністю [16].
При схрещуванні диплоїдних форм із культурними диплоїдними
видами одержане потомство з високим гетерозисом за кількістю
бульб та іншими ознаками [1].
При схрещуванні батьківських форм, які несуть гени різних видів
картоплі, зростає гетерозисний ефект, що призводити» до одержання
більш урожайних гібридів [14].
Високий ступінь гетерозиготності, який досягався на основі бага¬
товидових схрещувань із сортами різного походження, введення на
певних етапах роботи помірного інбридингу дали змогу створити в
Інституті картоплярства сорти, які характеризуються високою врожай¬
ністю завдяки гетерозисному ефекту. Таким методом були створені
високоврожайні сорти Бородянська рожева, Обрій, Віринея та ін.
Високоврожайні сорти з гетерозисним ефектом за урожайністю
створені на основі використання батьківських форм різних груп
стиглості та походження: Водограй, Серпанок, Косень 95, Слов’ян¬
ка та ін.
Гетерозисним ефектом за врожайністю відзначається сорт Лу¬
говська, створений на багатовидовій основі.
Інцухт. Цей метод дає змогу виділити цінні форми рецесивного
порядку. При цьому негативні ознаки, які містить материнська рос¬
лина, у прихованому стані можуть бути виявлені і поступово вибра¬
ковані [7]. Питаннями використання в селекції картоплі інцухту в
різних його проявах займалось багато дослідників.
216
У результаті використання ягід, утворених на рослинах картоплі
від природного самозапилення, створені сорти Рання Роза, Красуня
Геброна, Костромська, Варба, Оссео, Вікторія Петерсона, Арран Чіф
та ін. [18]. Проте великого поширення цей метод селекції не набув.
Вплив інцухту позначається на картоплі протягом декількох по¬
колінь зниженням урожайності та інших ознак, аж до вищеплення
нежиттєздатних сіянців [3].
Виявлена депресія інцухт-ліній пояснюється наближенням їх до
гомозиготності [8].
Проте з проведеного дослідниками аналізу робіт випливає, що
застосування інцухту в селекції може мати певне значення.
Так, І.І. Пушкарьовим [12,13] помічено, що в потомстві від само¬
запилення простих міжвидових гібридів S.demissum, порівняно з бек-
росами Ві, отримують більше сіянців з раннім бульбоутворенням,
вищим урожаєм, великими бульбами та короткими столонами. Тому
при використанні виду S.demissum, залежно від батьківських компо¬
нентів і поставленого завдання, він рекомендує метод повторних
схрещувань гібридів S.demissum*S.tuberosum чергувати з отриман¬
ням потомства від самозапилення.
Виявлено, що при кожному повторному схрещуванні гібридів, які
мають польову стійкість проти фітофторозу, з S. tuberosum втрачаєть¬
ся половина факторів стійкості. Тому початковий рівень стійкості
може бути досягнутий їх самозапиленням [20].
Вважається, що внаслідок рецесивного характеру успадкування
стійкості проти колорадського жука гібридів Glabrescentia*S. tube¬
rosum для відбору більш стійких генотипів бекросування необхідно
чергувати з одержанням потомства від самозапилення [21].
У потомстві від самозапилення сортів картоплі можна відібрати
лінії, здатні виявляти гетерозис [17].
В Інституті картоплярства УААН унаслідок самозапилення бага¬
товидового гібрида до Із створено сорт картоплі Горлиця, висока
урожайність якого поєднується з комплексом інших цінних ознак.
Високий ступінь гетерозиготності, який досягнуто на основі багато¬
видових схрещувань із сортами різного походження, дав змогу створити
в цьому інституті сорти, що характеризуються високою врожайністю
завдяки проявленню в потомстві батьківських форм гетерозису [11].
ЛІТЕРАТУРА
1. Брокш В.Л., Корчемная Н.А. Качество клубней гибридов S.tuberosum с
культурными видами Южной Америки // Научные труды по прикладной
ботанике, генетике и селекции - Л., 1989. - T.122. - С.52-58.
2. Букасов С.М. Методы отдаленной гибридизации в селекции картофеля
// Материалы совещания по отдаленной гибридизации растений и животных.
-М., 1958.- Вып. 2.
3. Букасов С.М., Камераз А .Я. Основы селекции картофеля. - М.-Л.:
Сельхозгиз, 1959. - 528 с.
217
4. Волков В.Д., Данько Г.В. Вплив фізичних та хімічних мутагенів на
ріст і розвиток рослин картоплі // Картоплярство. - К.: Урожай, 1970. -
Вип.1. - С.51-56.
5. Волков В.Д., Данько Г.В. Використання мутагенів для одержання вихідно¬
го матеріалу в селекції картоплі // Картоплярство. -К.: Урожай, 1973. - Вип.4.
5. Лехнович В.С. Каталог-справочник мировой коллекции ВИР. Карто¬
фель / Под ред. акад. С.М. Букасова. - М., 1963.
6. Лобашев М.Е. Генетика. - Л.: Изд-во Ленинград, ун-та, 1969. - 752 с.
7. Максимович М.М. Методы селекции картофеля. - М.: Сельхозгиз, 1951.
8. Майщук З.М. Ефективність клонових доборів в селекції картоплі //
Картоплярство. - К.: Урожай, 1993. - Вип.24. - С.24-27.
9. Оверчук В.І., Піка М.А., Гладишко С.А. Вихідний матеріал для селек¬
ції картоплі // Сільськогосподарська інформація. - 1971. - № 2. - С.33-35.
10. Осипчук А.А. Селекція картоплі в умовах Полісся України: Дис....
д-ра с.-г. наук у формі наукової доповіді. - Харків, 1993. - 50 с.
11. Пуппсарев И.И. Селекция картофеля // Картофель. - М.: Сельхозгиз, 1937.
12. Пушкарев И.И. Новый фитофтороустойчивый сорт картофеля 8670. -
М.: Сельхозгиз, 1937. - 44 с.
13. Росс X. Селекция картофеля. Проблемы и перспективы / Пер. с англ.
В.А. Лебедева. - М.: Агропромиздат, 1989. - 183 с.
14. Сергієнко В.М., Литовченко М.С. Вплив хімічних мутагенів на по¬
ліпшення господарсько цінних ознак у картоплі // Картоплярство. - К.: Уро¬
жай, 1971, вип.2. - С.7-10.
15. Склярова Н.П., Петухов С.Н., Юдина Т.А. Использование полевой
полиплоидии в селекции картофеля // Селекция и биотехнология картофеля.
-М., 1990. -С.21-30.
16. Тринклер Ю.Г., Колачев В.Д., Матенкова Т.В. Поведение самоопы-
ленных линий при селекции картофеля, размноженного семенами // Докла¬
ды ВАСХНИЛ. - М., 1976. - №4. - С.7-8.
17. Уайтхед Т., Мак-Ингош У., Финдлей У. Картофель здоровый и больной /
Пер. с англ.; Под.ред. И.И. Пушкарева - М.: И-во иностран. лит. - 1955.
18. Филиппов А.С. Методы и схема селекции картофеля // Картофель /
Под.ред. Н.С.Бацанова. - М.: Колос, 1970. - С.124-134.
19. Яшина И., Попкова К., Ерохина С. и др. Генетические основы селекции
картофеля на фитофтороустойчивость // Картофель и овощи,- 1966. - №6.
20. Torka М. Resistenzziichtung gegen den Kartoffelkafer. Handb. der Pflan-
zenzuchtung, 2 Auflage, Bd.3 verlag P.Parey. - Berlin - Hamburg, 1958.
4.5. ОСНОВНІ НАПРЯМИ ТА ЗАВДАННЯ
СЕЛЕКЦІЇ КАРТОПЛІ
Україна відзначається різноманітністю грунтово-кліматичних
умов, які не завжди і не в усіх регіонах сприятливі для вирощування
картоплі. Часті епіфітотії фітофторозу, ураження вірусними хворо¬
бами і сильне виродження на півдні призводять до значного недобо¬
ру врожаю. Великої шкоди завдають парша звичайна, стеблова нема¬
тода та бактеріальні хвороби, колорадський жук. Надзвичайно не¬
218
безпечними є карантинні об’єкти - рак картоплі і золотиста цистоут-
ворююча картопляна нематода.
Картопля в Україні використовується як продукт харчування, си¬
ровина для переробної промисловості, корм для тварин. За кожним із
зазначених напрямів ставляться певні вимоги до сортів картоплі. Для
виробництва і споживання мають значення сорти різних груп стигло¬
сті. Велику роль у харчуванні населення картоплею у весняні і ран-
ньолітні строки відіграють скоростиглі сорти.
Основні завдання і напрями селекції картоплі випливають із вищеза¬
значених факторів, вимог виробників та споживачів цієї культури.
Важливим напрямом роботи є селекція на пластичність і стабіль¬
ність основних господарсько цінних ознак. Проте селекція картоплі в
якомусь одному напрямі не може мати вирішального значення при
створенні нових сортів.
Надзвичайно актуальне завдання селекції картоплі полягає в по¬
єднанні у сортах стійкості проти хвороб і шкідників з основними го¬
сподарсько цінними ознаками.
Вирішувати поставлені завдання можна завдяки цілеспрямованій
селекції, яка базується на використанні різноманітного вихідного
матеріалу, знанні генетичної природи батьківських пар, використан¬
ні ефективних методів оцінки та добору бажаних генотипів.
Бульби картоплі вищої якості повинні мати такі зовнішні ознаки:
середню величину, добру форму, дрібні вічка, відповідний колір
шкірки і м’якоті, відсутність пошкоджень, тріщин, позеленіння і ду-
пловатості, парші, придатність для миття. Внутрішні ознаки вклю¬
чають якість консистенції м’якоті, борошнистість, відсутність потем¬
ніння в сирому і вареному вигляді, хороший смак [27].
Велике значення має вміст сухої речовини, крохмалю, білка, цук¬
рів, вітамінів та ін.
4.5.1. Селекція на вміст крохмалю
і сирого протеїну
Відомо, що цінність картоплі в основному пов’язана з умістом у
бульбах крохмалю і білка - основних компонентів сухої речовини.
Сорти з підвищеним умістом крохмалю поживніші, ніж низько-
крохмалисті.
Більш придатні й економічно вигідніші для технічної переробки і
використання на картоплепродукти сорти з високим і підвищеним
умістом крохмалю в поєднанні з іншими біохімічними показниками
[1,2,4].
Крохмаль із картоплі добре засвоюється всіма сільськогосподар¬
ськими тваринами і характеризується винятково високою перетрав¬
ністю. Підвищення вмісту крохмалю у бульбах може здешевити
кормову одиницю, підвищити рентабельність використання картоплі
на кормові цілі [2].
219
Таким чином, підвищення в бульбах картоплі вмісту крохмалю
має велике народногосподарське значення.
Основною умовою нагромадження в бульбах крохмалю є прави¬
льне водне і мінеральне живлення рослин картоплі протягом усього
періоду їх росту і розвитку, дотримання елементів технології виро¬
щування, застосування ефективних заходів боротьби з хворобами і
шкідниками.
Одним із найефективніших заходів підвищення вмісту у бульбах
крохмалю є створення висококрохмалистих сортів.
Перші сорти з підвищеним і високим умістом крохмалю створені
методом внутрішньовидової гібридизації в межах виду S. tuberosum.
Значних успіхів у цьому напрямі досягнуто в Німеччині, де най¬
більш відомими були висококрохмалисті сорти Остботе, Парнасія,
Імператор, Меркер, Герой, Сілезія, Вольтман, Форан, Карнеа, Робус-
та, Фальке, Фламінгштерке, Гігант, Моніка, Росвіт, Тигр. Перші ви¬
сококрохмалисті сорти в Росії були створені в 20-х роках на Коренев-
ській селекційній станції - Кореневська і Радянська. На Петровській
державній селекційній станції був створений середньоранній високо-
крохмалистий сорт Октябрьонок. Потім у межах виду S .tuberosum
було створено ще ряд висококрохмалистих сортів [11].
На Немішаївській дослідній станції методом внутрішньовидової гіб¬
ридизації був створений висококрохмалистий сорт Червоноспиргова.
Тут також добором із сорту Вольтман був створений висококрохмалис¬
тий сорт Вольтман 1177, а на Литовській держселекстанції добором із
сорту Штерхерайхе - висококрохмалистий сорт Крохмалиста [6].
Найбільш повно завдання створення висококрохмалистих сортів
у поєднанні з іншими цінними ознаками можна вирішити на основі
міжвидової гібридизації [3].
С.М.Букасов вважає, що міжвидова гібридизація для підвищення
крохмалистості перспективна в тих випадках, коли висока крохмали¬
стість у перших поколіннях поєднується з відносно високою вро¬
жайністю, що можливе при використанні переважно великобульбо-
вих і більш урожайних видів, таких, як S.leptostigma або S.molinae.
На його думку, найбільш простим шляхом створення висококрохма¬
листих сортів є гібридизація з культурними видами S.andigenum і
S.curtilobum, які не мають негативних властивостей диких видів [6].
У результаті застосування міжвидової гібридизації створено такі
висококрохмалисті сорти, як Фітофторостійка 8670, Москвич, Олев,
Лошицька, Темп, Розвариста, Білоруська крохмалиста, Синтез [3,4,6].
В Україні на основі міжвидової гібридизації були створені такі
висококрохмалисті сорти: Смачна, Мавка, Зарево, Придеснянська,
Прикарпатська, Полонина, Ікар, Поліська рожева, Воловецька, Об¬
рій, Кобза та ін.
Як батьківські форми при селекції на крохмалистість використо¬
вували сорти та гібриди: Смачна, Мавка, Карпатська, Зарево, Білору¬
220
ська крохмалиста, Бекра, Світанок київський, 77.583/16, 1509с/55,
7692с/68, 498с/66, 1—35с/61, 1—53с/61, Білоруська 3, Леандер, Покра,
Перлина, Гранат, Поліська рожева та ін.
У селекції на крохмалистість використовувались наступні види:
S.andigenum, S.demissum, S.semidemissum, S.curtilobum, S.leptostigma,
S.commersonii, S.chacoense, S.gibberulosum та ін.
Використання в селекції вихідного матеріалу міжвидового поход¬
ження дало змогу створити сорти, у яких високий уміст крохмалю
поєднується зі стійкістю проти хвороб, шкідників та з іншими цін¬
ними ознаками [17].
Дослідженнями, проведеними у ВІРІ, доведено, що міжсортові
схрещування Робуста х Октябрьонок, Аквіла х Штеркерагіс та інші
мають значення при селекції на високу крохмалистість. Хорошими
вихідними формами при такій селекції можуть бути сорти Ердкрафт,
Хохпроцентіге й ін.
Висококрохмалисті гібриди можуть бути одержані при гібриди¬
зації багатьох видів серій Glabrescentia, Demissa, Longipedicellata,
Tuberosa, Andigena та інших, які вирізняються високою крохмалисті¬
стю бульб [12].
Крохмалистість має полігенний і здебільшого домінантний хара¬
ктер успадкування.
Чіткий фенотипічний ефект крохмалистості потомства проявля¬
ється лише при накопиченні великої кількості полігенів. Наявність у
батьків специфічної комбінаційної здатності вказує на комплемента¬
рну взаємодію полігенів [20].
Встановлена висока кореляційна залежність крохмалистості по¬
томства від проявлення цієї ознаки у батьківських форм.
У 34,6 комбінацій, які вивчалися в Республіці Білорусь, середня
крохмалистість потомства перевищувала середнє значення цього
показника у батьків, в іщних - виявлено проміжний характер успа¬
дкування та депресію. Остання характерна для комбінацій, одер¬
жаних від схрещування низькокрохмалистих батьківських форм із
висококрохмалистими, а також для пізньостиглих родин. Кореля¬
ційна залежність між середніми арифметичними показниками гіб¬
ридних родин і батьків коливалася від слабопозитивної до високо-
позитивної. Звідси випливає, що підбирати батьківські форми при
селекції на крохмалистість можна за їхнім фенотипом. Значною
мірою проявлення ознаки залежить від комбінаційної здатності ба¬
тьківських пар [4].
В успадкуванні вмісту крохмалю, крім домінантних, беруть
участь і рецесивні гени, оскільки були виявлені висококрохмалисті
гібриди і в потомстві від схрещування двох низькокрохмалистих ба¬
тьківських форм [25].
Полігенні ознаки не дають у потомстві чітких фенотипічних кла¬
сів, зумовлених проявом домінантних і рецесивних генів. Розщеплення
221
виражається безперервним варіаційним рядом форм, які поступово
переходять від максимального до мінімального прояву ознаки [14].
Щодо крохмалистості картоплі встановлено, що кожний сорт або
гібрид дає при самозапиленні варіаційний ряд форм, у більшої час¬
тини яких зберігається крохмалистість вихідної форми, а в гібрид¬
ному потомстві визначається середній показник крохмалистості обох
батьківських форм. Інші генотипи відхиляються як у бік підвищення
крохмалистості, так і в бік її зниження. Тому в селекції на крохмали¬
стість основна увага має бути приділена підбору генотипів у класах
високої крохмалистості [24].
У потомстві від самозапилення нагромадження крохмалю відбу¬
вається інтенсивніше, ніж при гібридизації, проте ефективність до¬
бору при цьому знижується через стерильніть і відсутність цвітіння у
більшості форм. Тому форми з високим умістом крохмалю частіше
отримують у гібридному потомстві.
Для поєднання високого вмісту крохмалю і врожайності потріб¬
но підбирати для гібридизації висококрохмалисті батьківські фор¬
ми, які відзначаються високою комбінаційною здатністю за вро¬
жайністю [9].
У групі врожайних родин спостерігалася тенденція до оберненої
залежності між урожайністю і крохмалистістю. У популяціях, де
врожайність і крохмалистість були підвищеними, встановлено неза¬
лежне їх успадкування, і процент генотипів, що поєднують високі
показники обох ознак, тут був вищим, ніж у попередній групі [4].
На цій підставі можна зробити висновок про можливість поєд¬
нання в одному генотипі високої крохмалистості й урожайністі.
В Україні створені такі сорти картоплі, зокрема, це Поліська ро¬
жева, Світанок київський, Обрій, Кобза, Українська рожева, Придес¬
нянська, Купава, Дзвін та ін.
Між умістом у бульбах крохмалю і ранньостиглістю, а також між
крохмалистістю і розміром бульби встановлена від’ємна кореляційна
залежність [22]. Це зумовлює трудність поєднання високої крохма¬
листості з ранньостиглістю та великобульбовістю.
Для поєднання високої крохмалистості і великобульбовості по¬
трібний великий обсяг селекційного матеріалу тих комбінацій, у
яких відсутня кореляційна залежність між названими ознаками [9].
У результаті цілеспрямованої роботи створено сорти картоплі, у
яких підвищена і висока крохмалистість поєднуються як із скоростиг¬
лістю (Кобза, Зов, Молодіжна, Обрій, Світанок київський, Купава,
Поліська 96), так і з великобульбовістю (Купава, Поліська 96).
Складовою частиною сухої речовини картоплі є білки. Вони не¬
обхідні як у продуктах харчування людини, так і в кормах для тва¬
рин. Біологічна цінність білка картоплі визначається вмістом у ньому
незамінних амінокислот, які в організмі тварин не утворюються. У бі¬
лку картоплі на незамінні амінокислоти припадає 48% його маси [8].
222
Білок і небілкові азотисті сполуки є складовими сирого протеїну. На
частку першого припадає 44-46% від маси сирого протеїну [19].
У бульбах більшості сортів картоплі міститься 1,5-2% сирого
протеїну [11]. Підвищення вмісту сирого протеїну важливе як для
столової, так і для кормової картоплі. Його вміст може змінюватися
під впливом умов середовища. Дією різних факторів можна збільшу¬
вати вміст сирого протеїну в картоплі.
На фоні оптимальних агротехнічних заходів для підвищення в
бульбах вмісту сирого протеїну велике значення має створення і
впровадження сортів із високим умістом насамперед чистого білка.
Сорти картоплі з підвищеним умістом сирого протеїну отримані
в Білоруському НДІ картоплярства: Білоруська крохмалиста - 2-2,2%,
Бекра - 2,4-2,7%, Лошицька - 2,4-3%.
На колишній Немішаївській дослідній станції на основі міжвидо¬
вої гібридизації були створені середньоранні сорти Смачна і Перли¬
на з підвищеним умістом сирого протеїну (2,2-2,8%), крохмалю (17-
18%), високими смаковими якостями та іншими цінними ознаками. В
Інституті картоплярства створені сорти Світанок київський і Зарево,
у яких високий уміст сирого протеїну (2,8-3,3%) поєднується з висо¬
кою крохмалистістю (20,4 і 22,8%) [16].
На Устимівській дослідній станції високим умістом білка відзна¬
чаються сорти Кобра (3,09%) та Немішаївська біла (2,95%) [16].
Багато південноамериканських диких і культурних видів картоп¬
лі значно перевищують за вмістом сирого протеїну селекційні сорти і
можуть бути цінним вихідним матеріалом для селекції на високі по¬
казники вказаної ознаки.
Так, у Білорусі при схрещуванні окремих зразків видів S.cha¬
coense, S.phureja із селекційними сортами отримали високоврожайні
гібриди з умістом у бульбах сирого протеїну понад 3% [10]. Від
схрещування форм S.phureja із сортами Бекра, Мінська рання, Весе-
ловська окремі гібриди мали 4,5% сирого протеїну [13]. Тут же при
схрещуванні ряду форм S.andigenum із селекційними сортами отри¬
мали гібриди з умістом сирого протеїну 3,0-3,1% [23]. При викорис¬
танні дикого виду S. demissum у бекросах отримані генотипи з підви¬
щеним умістом сирого протеїну - 2,8-4,2% [21].
В Інституті картоплярства при селекції на підвищений уміст си¬
рого протеїну широко використовують сорти Смачна, Перлина, Ло¬
шицька, Бекра, Зарево, Світанок київський, а також гібриди, одержа¬
ні за участю зразків видів S.demissum, S.andigenum, S. semidemissum
[16,17].
Успадкування білковості має полігенний характер, а тому підбір
батьківських пар при селекції на цю ознаку слід проводити за типом
насичуючих схрещувань [1].
У гібридному потомстві найчастіше проявляється домінування
ознаки низького вмісту сирого протеїну або проміжний характер ус¬
223
падкування з тенденцією наближення до батьківських форм із низь¬
ким його вмістом. При цьому вшцеплювались гібриди, які за назва¬
ною ознакою перевищували батьківські форми [13].
Характеристика селекційного матеріалу картоплі за вмістом білка
має враховувати мінливість цієї ознаки залежно від географічних, агро¬
технічних умов. З цією метою проводять відповідні дослідження.
В Інституті картоплярства виявлена висока та середня позитивна
залежність між умістом сирого протеїну у батьківських формах і по¬
томством, що свідчить про можливість їх підбору для гібридизації за
фенотипом. Виділено також у різних екологічних зонах комбінації
схрещувань Зарево х Божедар, Галина х Зарево, Либідь х Зарево, За¬
рево х 77.583/16, Зарево * Густо та інші, у потомстві яких вшцеплю-
ються генотипи з умістом сирого протеїну вище 3% [18].
Вважаєтся також, що при селекції на білковість слід проводити
оцінку комбінаційної здатності батьківських форм [7].
Між урожайністю і білковістю потомства виявлена слабка і сере-
дньопозитивна та слабка середньовід’ємна кореляційна залежність
[5]. Це вказує на те, що в ряді комбінацій спостерігається незалежне
успадкування зазначених ознак.
В Інституті картоплярства виявлена слабо- і середньопозитивна
кореляційна залежність між умістом сирого протеїну і врожайністю.
У цьому матеріалі окремі гібриди при врожайності бульб 320-435
ц/га містили в бульбах 2,5-3,3% сирого протеїну [16].
Наведені факти свідчать про можливість створення сортів, у яких
висока врожайність поєднувалася б з підвищеним і високим умістом
сирого протеїну.
Між умістом кромалю і білка в картоплі виявлений чіткий взаємо¬
зв’язок: сорти з підвищеним і високим умістом крохмалю мають та¬
кож високий рівень білка [26].
Між вказаними ознаками виявили середню позитивну кореляцій¬
ну залежність [1,15,16]. Це вказує на можливість створення сортів
картоплі з підвищеним умістом сирого протеїну і високим умістом
крохмалю.
Створені сорти з підвищеним та високим умістом сирого протеї¬
ну - Білоруська крохмалиста (2,0-2,2%), Бекра (2,4-2,7%), Лошицька
(2,4-3,0%), Зарево (2,8-3,3%), Світанок київський (2,8-3,0%) - поєд¬
нують цю ознаку з високою крохмалистістю та іншими цінними
ознаками [1,2,4,16,17].
ЛІТЕРАТУРА
1. Альсмик П.И. Селекция картофеля ка повышенное содержание сухих
веществ, крахмала и белка // Результаты исследований по селекции и семе¬
новодству картофеля. - М.: Колос, 1972. - С.15.
2. Альсмик П.И. Селекция крахмалистых сортов картофеля // Картофе¬
леводство. - Минск: Ураджай, 1974.- Вып.2. - С.3-11.
224
3. Альсмик П.И. Селекция картофеля в Белоруссии. - Минск: Ураджай,
1979. - 127с.
4. Альсмик П.И., Пискун Г.И. Селекция сортов картофеля с высоким
содержанием сухих веществ, пригодных для переработки // Картофель: се¬
лекция, семеноводство, технология возделывания. - Минск: Ураджай, 1988.
- С.70-85.
5. Анисимова Г.Л. О корреляционной зависимости между содержанием
протеина и урожаем в гибридах картофеля // Селекция и семеноводство кар¬
тофеля. - М., 1973. - Вып. XIV. - С.72-75.
6. Букасов С.М., Камераз А .Я. Основы селекции картофеля. - М.- Л.:
Сельхозгиз, 1959. - 528 с.
7. Букасов С. Исходный материал для селекции на белок // Картофель и
овощи. -1972. - №3. - С. 15.
8. Вечер А.С. Основные проблемы биохимических исследований
картофеля. - Минск: Ураджай, 1966. - С. 127-135.
9. Гончаров Н.Д. Селекция картофеля на скороспелость // Картофель /
Под.ред. Н.А. Дорожкина. - Минск: Ураджай, 1972. - С.33-48.
10. Демидко Я. Использование видов S.chacoense и S.phurja в селекции
картофеля на повышенное содержание белка // Научные труды БНИИКПО.
- Минск: Ураджай, 1979. - Вып.4. - С.8-12.
11. Иванченко Е.А., Иванченко Г.З. Основные направления селекции карто¬
феля //Картофель /Под.ред. Н.С. Бацанова. - М.: Колос, 1970. - С. 100-124.
12.Камераз А.Я. Селекция картофеля // Культурная флора СССР. Т.ХІ-
Л.: Колос, 1971. - С.430-445.
13.Купчина С.А. Использование видов S.phureja juz et Вик., S.rybinii juz
et Вик., S.kesselbrenneri Вик. и других при выведении сортов картофеля с
повышенным содержанием белка // Картофель / Под.ред. Н.А. Дорожкина. -
Минск: Ураджай, 1972. - С.70 -77.
14. Лобашев М.Е. Генетика. - Л.: И-во Ленинград, ун-та, 1969. - 752 с.
15. Онищенко О.Й., Поправко М.Й. Поєднання господарсько цінних
ознак у міжсортових гібридах картоплі // Картоплярство. - К.: Урожай,
1974. - Вип.5. - С.49-52.
16. Осипчук А.А. Селекція картоплі // Селекція і насінництво картоплі. -
К.: Урожай, 1988. - С.10-43.
17. Осипчук А.А. Селекція картоплі в умовах Полісся України: Дис...
д-ра с.-г. наук у формі наукової доповіді. - Харків, 1993. - 50 с.
18. Осипчук А.А. Створення та оцінка селекційного матеріалу картоплі
для різних грунтово-кліматичних зон: Автореф. дис... канд. с.-г. наук. - Харків,
1999.-19 с.
19.Прокошев С.М. Биохимия картофеля. - М., 1966. - С.127-135.
20. Росс X. Селекция картофеля: проблемы и перспективы / Пер. с англ.
В.А. Лебедева. - М.: Агропромиздат, 1989. - 183 с.
21. Семенова И.А. Использование вида S. demissum Lind. при селекции
картофеля на повышенное содержание сухих веществ // Картофель /
Под.ред. Н.А.Дорожкина. - Минск: Ураджай, 1972. - С.60-70.
22. Свижинский К. Создание исходного материала для селекции карто¬
феля в Польше И Картофель : селекция, семеноводство, технология возде¬
лывания. - Минск: Ураджай, 1988. - С. 135-150.
8 2132
225
23.Томчук Н.Г. Использование вида S.andigenum juz. et Вик. при селек¬
ции на белок // Картофель / Под.ред. Н.А. Дорожкина. - Минск: Ураджай,
1972. - С.77-80.
24. Яшина И.М., Першутина О А., Кирсанова Е.В. Генетика морфологи¬
ческих и хозяйственно ценных признаков картофеля // Г енетика картофеля.
- М.: Наука, 1973. - С.233-259.
25. Borger H., Huhnke W., КоЫег D., Schwanitz F., Sensbuch К Untersuchungen
uber die Ursachen der Zesitung vor Kulturpflanzen // Das Verhalten der Komponenten
des Sfariceertrages von kartoffeln.- Zuchter, 1956. - B.26. - S.363.
26.Hunnius W., Scheidt M., Munzert M. Der Eiweibgehaet der Kartrags -
und Wertmerkmalen und in seinen Sortenverhalten // Bayer. Zandu. Jb. 50. -
Sonderh, 1973. - S.120-133.
27. Howard W., Factor influencing the qualityof ware potatoes. I The genotype
// Pot. Ros. - 1974. - B. 17.- S.490-511.
4.5.2. Селекція картоплі на ранньостиглість
Народногосподарське значення культури ранньостиглої кар¬
топль Бажано, щоб ранньостигла картопля займала 30% посівних
площ цієї культури. Це дозволяє в кожному господарстві, яке вироб¬
ляє товарну картоплю, вирощувати не менше трьох сортів різної стиг¬
лості, тобто сортів з різними біологічними особливостями.
Відомо, що бульби при тривалому зберіганні втрачають не тільки
вологу і крохмаль, але й велику кількість вітамінів (Вь В2, С) та ін¬
ших корисних сполук. Особливо багато їх витрачається під час про¬
ростання або при несприятливих умовах зберігання [6]. Якщо порів¬
няти поживну цінність і смакові якості бульб, які довго зберігались,
із бульбами ранньостиглих сортів картоплі ранніх строків збирання,
то всі якісні показники будуть на користь останніх.
Виграти на виробництво картоплі минулорічного врожаю розпо¬
діляються таким чином: на виробництво - 44,4 %, на збереження -
15,6 %, втрати бульб при зберіганні - 40 %.
Виграти енергії для штучного охолодження бульб минулорічного
врожаю до червня вдвоє більші, ніж витрати на вирощування ранньої
картоплі.
Ранній урожай бульб має високі смакові і споживчі якості. Буль¬
би, зібрані через 60-70 днів після садіння, містять до 50 мг вітамі¬
ну С у 100 г бульбової маси. Такі бульби містять високоякісний бі¬
лок, біологічна цінність якого складає 80% від білка курячого яйця, а
також важливі мінеральні речовини, дуже потрібні людському орга¬
нізму.
Отже, вирощування ранньостиглої картоплі вирішує питання ра¬
ціонального харчування людей в умовах середньої кліматичної зони,
до якої належить територія України. Крім того, рання картопля
сприяє розширенню строків споживання якісної продукції від уро¬
жаю до урожаю.
226
В останні роки в Україні розвивається переробна промисловість,
широко стали виготовляти чипси та інші продукти. Кращою сирови¬
ною для цього є свіжозібрані бульби. Тому саме ранні сорти картоплі
сприятимуть забезпеченню переробної промисловості високоякісною
сировиною.
Ранньостигла картопля має важливе агротехнічне значення, оскіль¬
ки вона може з успіхом використовуватися як парозаймаюча культу¬
ра для посіву озимини. Після картоплі поле залишається чистим від
бур’янів і має високий агротехнічний фон, особливо якщо вносились
органічні добрива. Після картоплі добре ростуть ярі зернові. Після
такого попередника можна сіяти просо.
Ранні сорти картоплі можуть з успіхом використовуватись для
одержання екологічно чистої продукції. Дуже ранні посіви пророще¬
ними бульбами встигають дати повноцінний урожай до появи фіто¬
фторозу. Тому немає потреби обробляти їх отрутохімікатами.
Біологічні особливості ранньостиглої картоплі Великий інтерес у
селекції ранньостиглої картоплі викликають такі фактори, як урожай¬
ність і скоростиглість, їх взаємозв’язок, взаємодія і поєднання.
Урожайність картоплі завжди була предметом досліджень бага¬
тьох учених. Так, урожайність бульб у динаміці досліджували П.І. Аль-
смік, М.Н. Гончарик; М.М. Максимович; В.В. Арнаутов; Н.С. Баца-
нов; В.С. Шевелуха та ін. [1,2,3,5,7].
Урожайність як селекційну ознаку вивчали І.М.Яшина (1966),
М.Д.Гончаров (1981) та ін. [4,8].
Урожайність картоплі перебуває у прямій кореляційній залежно¬
сті від кількості і розміру бульб. У той же час ці ознаки не пов’язані
між собою.
При вивченні динаміки росту і накопичення врожаю було вияв¬
лено, що у процесі утворення урожайності бульб є два чітко вираже¬
ні періоди:
1-й - ріст і накопичення наземної маси (листкового апарату), роз¬
виток кореневої системи та утворення бульб;
2-й - збільшення маси бульб, утворених за рахунок використання
ресурсу накопиченої наземної і підземної маси коріння і листкового
апарату.
Як у перший, так і в другий період картопля по-різному реагує на
умови росту, які бувають інколи дуже контрастними. Так, у перший
період росту картопля терпимо реагує на нестачу вологи, тоді як у
другий потребує її у великій кількості. Отже, якщо умови першого
періоду будуть сприяти утворенню великої кількості бульб, а друго¬
го - збільшенню їх маси, то урожайність буде високою.
До сьогодні динаміку накопичення врожаю вивчали за результа¬
тами другого періоду, що є недостатнім. Тому необхідно проводити
дослідження протягом обох періодів росту і розвитку картоплі. Тоді в
селекційному процесі легше буде визначити і відібрати ранньостиглі
227
форми. При аналізі багаторічних показників урожайності було вияв¬
лено, що всі сорти картоплі свій потенціал при вирощуванні реалі¬
зують лише частково (табл. 4.2).
Таблиця 4.2 - Рівень реалізації потенційних можливостей картоплі
(середнє за 10 років)
Назва сорту
Рівень реалі¬
зації продук¬
тивності сор¬
ту, %
Параметри складових потенційної
врожайності
Фактична
маса вро¬
жаю бульб,
г/кущ
кількість
бульб,
шт./кущ
маса од¬
нієї буль¬
би, г
маса врожаю
бульб під
кущем, г
Прієкульська рання
60
18
74
1332
793
Білоруська рання
56
16
110
1760
990
Зорька
80
18
75
1350
1080
Дєтскосєльська
80
14
75
1050
840
Як видно з наведених даних, рівень реалізації потенційних можли¬
востей картоплі залежить від реакції сорту на зовнішні фактори. Це мож¬
на використати для оцінки сорту на екологічну пластичність. Чим ви¬
щий рівень реалізації продуктивності, тим вища його пластичність.
Важливою властивістю ранньостиглих форм картоплі є її здат¬
ність накопичувати ранній урожай. Відрізняють фізіологічну ранньо¬
стиглість і господарську. Фізіологічну ранньостиглість визначають
за тривалістю вегетації у днях. Вона досягає 95-100 днів від садіння.
За цей період рослини накопичують високу врожайність. Оцінюють
її методом пробного збирання врожаю по строках вегетації: на 60-,
70-, 80-, 90- і 100-й день від садіння. Дослідами за 10 років була
виявлена певна динаміка врожайності ранньостиглих форм картоплі
(табл. 4.3).
Таблиця 4.3 - Динаміка врожайності ранньостиглих форм картоплі, г/кущ
(середнє за 10 років)
Скоростиглість форм
картоплі
Урожайність бульб картоплі (г/кущ)
по строках вегетації, у днях
60
70
80
90
100
Дуже ранні
420
615
698
760
785
Ранні
405
690
770
863
882
Середньоранні
272
480
705
809
910
Як видно з таблиці, ранні форми давали товарну врожайність на
60-70-й день від садіння, тоді як середньоранні - на 90-100-й день.
Відомо, що на ріст і розвиток картоплі впливають погодні умови.
Суттєві зміни вони викликають у форм, які не відзначаються еколо¬
гічною пластичністю. Реакція рослин на погодні умови є сортовою
ознакою. Кожен сорт реагує на погоду відповідно до своїх властивос¬
тей. Так, виявлено, що форми картоплі з високим генетичним потен¬
ціалом продуктивності забезпечують більш високу відносну врожай¬
228
ність у різні за погодою роки. Тому можна стверджувати, що власти¬
вість генотипу давати відносно високу урожайність тісно пов’язана з
його екологічною пластичністю. Отже, відбираючи високоврожайні
форми, селекціонер з великою вірогідністю відбиратиме екологічно
пластичні форми.
При вивченні явища періодичності по роках формування складо¬
вих врожайності - кількості бульб і їх маси під кущем - була вияв¬
лена наступна закономірність: у вегетаційні періоди, коли зав’язу¬
валася відносно велика кількість бульб під кущем, вони були меншої
маси, а при меншій кількості мали більшу масу. Обгрунтувати це
явище можна тим, що в картопляної рослини існує зворотна кореля¬
ційна залежність між кількістю і масою бульб, яка коливається від 0
до -0,864. Виявлено, що у форм картоплі, які мають зворотну залеж¬
ність між компонентами його врожайності, існує виразна річна пері¬
одичність (табл. 4.4).
Як видно з таблиці 4.4, у даному випадку ступінь залежності ма¬
си і кількості бульб був зворотним у всіх сортів: високий - у ранньо¬
стиглих і нижчий - у пізньостиглих. Але це не вплинуло на річну
періодичність у формуванні кількості і накопиченні маси бульб у
кущі. Цю закономірність слід враховувати при відборах.
При вивченні реакції окремих компонентів, які визначають уро¬
жайність картоплі (кількість і маса бульб), було доведено, що кожен
із них окремо по-своєму реагував на зовнішні фактори.
Таблиця 4.4 - Ступінь впливу кореляційної залежності між кількістю бульб і їх
масою в кущі на річну періодичність структури врожаю картоплі
(середнє за 10 років)
Сорти
Ступінь коре¬
ляційної залежності
кількості і маси
бульб
Показники кількості
і маси бульб у кущі
r±Sr
*05
у роки з малою кіль¬
кістю бульб і під¬
вищеною їх масою
у роки з підвище¬
ною кількістю бульб
і малою їх масою
кількість
маса
кількість
маса
Прієкульська рання
-0,864±0,178
4,85
11,7
60,1
16,3
44,4
Білоруська рання
-0,834±0,199
4,27
10,9
82,2
15,6
56,0
Зорька
-0,729±0,365
1,99
13,4
64,2
16,2
52,0
Дєтскосєльська
-0,569±0,369
1,54
11,3
69,0
12,5
61,0
Лошицька
-0,573±0,368
1,55
11,8
67,0
13,1
52,0
Огоньок
-0,570±0,365
1,56
9,3
92,0
11,7
67,0
Темп
-0,434±0,370
1,17
6,9
109,0
8,7
83,0
Для виведення високоврожайних ранньостиглих форм картоплі
необхідно створювати і використовувати генотипи із наступними
характеристиками:
229
- форми, які забезпечують утворення максимальної кількості
бульб і накопичення їх високої маси, незалежно від характеру погод-
них умов і екологічних факторів;
- форми, які утворюють більше 20 штук бульб під кущем і нако¬
пичують 2,2-2,5 кг/кущ у звичайних умовах росту;
- форми, які не реагують або слабко реагують на несприятливі
фактори погоди (волога, тепло);
- форми, у яких відсутня або слабко виражена протилежна коре¬
ляційна залежність між кількістю і масою бульб у кущі.
Основною властивістю ранньостиглих форм є висока інтенсив¬
ність накопичення бульбової маси. Цей процес проходить у 2-3 рази
інтенсивніше, ніж у пізньостиглих. За один день вегетації в період із
60-го по 100-й день від садіння ранньостиглі форми накопичували за
добу до 25 г маси бульб на кущ, тоді як пізньостиглі - усього лише
10-15 г. За інтенсивністю накопичення маси врожаю бульб ранньо¬
стиглі сорти діляться на такі групи:
1-а - дуже рання; накопичується у середньому за добу 1 т/га і біль¬
ше; 2-а - рання - 0,8 до 1 т/га; 3-я - середньорання - 0,5-0,8 т/га; 4-а -
середньостигла - нижче 0,4 т/га.
За тривалістю (у днях) періоду бульбоутворення після 60-го дня
від садіння було виявлено такі групи форм: 1-а - з дуже коротким
періодом бульбоутворення (40-45 днів); 2-а - з коротким періодом
(45-50 днів); 3-я - із середнім періодом (50-55 днів); 4-а - з подов¬
женим періодом (55-60 днів).
Виходячи з вищезазначеного, можна стверджувати, що ранньо¬
стиглість залежить від двох процесів, які відбуваються незалежно
один від одного: інтенсивності бульбонакопичення і тривалості в
часі. Ці дві властивості мають різний генетичний контроль, тому успад¬
ковуються в потомстві по-різному.
Інтенсивність і тривалість періоду накопичення бульб у варіантах
визначають ступінь ранньостиглості і врожайності різних форм кар¬
топлі. Моделі скоростиглих форм відображені в таблиці 4.5.
Із таблиці видно, що високоврожайні форми виникають тоді, ко¬
ли висока інтенсивність поєднується з тривалим періодом накопи¬
чення бульб. З іншого боку, ранньостиглі і високоврожайні форми
виникають при наявності комплексу ознак, що забезпечують високу
інтенсивність бульбонакопичення за короткий період вегетації. Та¬
ким чином, доведено, що головною ознакою, яка забезпечуй ліоєд-
нання в одному генотипі скоростиглості і високої врожайності, є
властивість інтенсивного накопичення бульб. Тому саме ця ознака
повинна бути основним об’єктом вивчення селекціонерів.
Для практичної селекції з метою одержання цінних генотипів
слід шукати форми з оптимальним варіантом сполучення в ньому
високої інтенсивності і тривалості бульбонакопичення. При відборі
чоловічих форм потрібно мати форми, які мають хоча б одну з ви¬
230
щезгаданих ознак, щоб створити генотипи з оптимальним їх поєд¬
нанням.
Таблиця 4.5 - Вплив інтенсивності і тривалості бульбонакопичення
на врожайність і ступінь ранньостиглості картоплі
Група уро¬
жайності
Інтенсив¬
ність бульбо¬
утворення за
добу
Тривалість вегетації і періоду бульбонакопичення у днях
85-90 днів від
садіння, 40-45
днів бульбона¬
копичення
95-105 днів
від садіння,
45-50 днів
бульбона¬
копичення
105-115 днів
від садіння,
50-55 днів
бульбонако¬
пичення
115-125 днів
від садіння,
55-60 днів
бульбона¬
копичення
І
Дуже висока
- більше
1 т/га
дуже ранні
високоврожай¬
ні форми
ранні дуже
врожайні фор¬
ми
середньоранні
дуже вро¬
жайні форми
середньостиг¬
лі дуже вро¬
жайні форми
II
Висока - 0,8-
1,0 т/га
дуже ранні
середньовро-
жайні форми
ранні високо¬
врожайні фор¬
ми
середньоранні
високовро¬
жайні форми
середньостиг¬
лі дуже вро¬
жайні форми
т
Середня -
0,5-0,8 т/га
дуже ранні
низьковрожай¬
ні форми
ранні серед-
ньоврожайні
форми
середньоранні
врожайні
форми
середньостиг¬
лі високовро¬
жайні форми
IV
Низька -
нижче 0,5 т/га
дуже ранні
низьковрожай¬
ні форми
ранні низько¬
врожайні фор¬
ми
середньоранні
середньовро-
жайні форми
середньостиг¬
лі врожайні
форми
Селекція високопродуктивних ранньостиглих форм. Селекція
на продуктивність є одним із найскладніших завдань, що зумовлено
комплексністю цього показника. Складність цієї властивості полягає
в тому, що вона включає в себе більш прості елементи, кожен з яких
має свої біологічні, морфологічні та інші властивості і власний гене¬
тичний контроль. При аналізі показників існуючих сортів картоплі
доведено, що кожний із них по-своєму впливає на утворення вро¬
жаю. Вони залежать від різних варіантів складових урожаю, таких,
як кількість і маса бульб у кущі. Виявлено кілька варіантів складових
урожаю: 1-й - оптимальне поєднання складових урожайності (13 шт.
бульб/кущ із середньою масою кожної 65 г); 2-й - підвищена кіль¬
кість бульб із середньою їх масою (14 шт. і 65 г); 3-й - середня кіль¬
кість бульб із підвищеною їх масою (11 шт. і 77 г); 4-й - середня
кількість бульб із середньою масою (11 шт. і 59 г); 5-й - мала кіль¬
кість бульб із великою їх масою (8 шт. і 110 г).
Виявлено, що врожайність перебуває у прямій кореляційній за¬
лежності від кількості і маси бульб. Коефіцієнт кореляції за першим
фактором знаходився в межах від 0,236±0,0112 (t=l,93) до 0,767±0,77
(t=l,43); за другим - від 0,021±0,119 (t=0,17) до 0,898±0,052 (t=17,9).
Певної закономірності зміни коефіцієнтів кореляції під впливом
зовнішніх факторів виявити не вдалось, але доведено, що більшість
існуючих сортів відзначається дефіцитом кількості бульб. Тому ця
ознака потребує особливої уваги селекціонера.
231
Урожайність потомства значною мірою залежить від батьківсь¬
ких форм. Було виявлено, що вона залежала від чоловічих форм на
53,4-55,3 %, а від материнських - на 36,0-39,3 %. Урожайність
окремих гібридів варіювала під впливом чоловічої форми від
613±14 г/кущ до 886±10 г/кущ і материнської - від 636±23 г/кущ до
1310±35 г/кущ. Урожайність гібридних потомств, незалежно від рів¬
ня стиглості, мала високу пряму залежність від чоловічих форм. Ко¬
ефіцієнт кореляції знаходився в межах від 0,790±0,175 до 0,965±0,003.
Але не у всіх комбінаціях спостерігалася така закономірність. Іноді
високоврожайні батьки давали низьковрожайне потомство або - на¬
впаки (табл. 4.6).
Таблиця 4.6 - Залежність урожайності потомств від урожайності батьків
(чоловіча форма - сорт Мінська рання)
Група
комбі¬
націй
Жіночі
форми
Середня уро¬
жайність, г/кущ
Розподіл потомств за врожайністю,
г/кущ, у %
бать¬
ків
потом¬
ства
до 300
300-
600
600-
900
900-
1200
більше
1200
І
02794—2
950
960
-
9,4
34,1
34,1
22,4
491-037
950
945
-
16,1
27,4
30,7
25,8
02874-1
950
898
УА
16,4
26,0
35,3
17,8
02711-2
740
831
2,4
17,6
33,0
33,0
14,0
02872-26
770
817
2,0
20,7
39,0
28,1
10,2
486-011
700
775
5,0
15,0
45,0
35,0
-
П
483-015
580
1126
-
-
14,8
44,5
40,7
02852-35
675
946
1,3
12,6
25,3
40,5
20,3
02871-11
675
830
5,3
21,2
30,1
26,6
16,8
02886-10
950
1310
-
12,7
23,6
36,3
27,4
Ш
02856-5
950
756
3,4
28,0
40,0
19,6
9,0
У таблиці 4.6 подані показники трьох груп комбінацій. Урожай¬
ність потомств першої групи перебувала у прямій залежності від
урожайності батьків, другої - перевищувала її і третьої - була ниж¬
чою, ніж у батьків.
Урожайність потомства одних і тих же батьків залежала також
від прямих і реципрокних схрещувань (табл. 4.7).
Таблиця 4.7 - Залежність урожайності потомств від прямих і реципрокних
схрещувань
Комбінації
Урожайність
потомства,
г/кущ
Група за
врожайністю,
р=0,01
Кількість
випадків (п)
І. Мінська рання х 2786-03
832±23
І
130
2786-03 х Мінська рання
718±11
П
622
П. Мінська рання х 2773-057
635±21
П
160
2773-057 х Мінська рання
880±28
І
90
232
Важливим аспектом у селекції на врожайність ранньостиглих
форм є розкриття процесу формування кількості бульб у кущі і нако¬
пичення їхньої маси.
Кількість бульб у сіянців залежала як від чоловічої, так і від жіночої
форми батьків. Так, кількість бульб у кущі в потомстві під впливом чо¬
ловічих форм варіювала від 9,65±0,35 до 10,71±0,10 шт./кущ, жіночих
форм-від 7,05±0,12 до 13,58±0,13 шт./кущ.
Питання успадкування потомствами кількості бульб у кущі вив¬
чалося з метою цілеспрямованої селекції картоплі за цією ознакою.
Аналіз показників кількості бульб у потомстві, залежно від похо¬
дження гібридів, показав високу пряму залежність їх від батьків. Це
дозволило розрахувати коефіцієнти регресії і скласти наступне рів¬
няння:
К„ = 0,81 хКр+ 1,03,
де Кн - кількість бульб у кущі потомства; Кр - кількість бульб у кущі
батьків; 0,81 і 1,03 - коефіцієнти регресії вірогідності, коли Р = 0,99.
Використовуючи цю формулу, можна прогнозувати успадкування
кількості бульб у кущі гібридного потомства. Враховуючи прямий хара¬
ктер успадкування цієї ознаки, можна зробити висновок про можливість
поліпшення сортів за цією ознакою селекційними методами.
Маса бульб у кущі потомства залежала від походження. Була ви¬
явлена пряма залежність маси бульб потомств від батьків, що дозво¬
лило розрахувати коефіцієнти регресії і скласти наступне рівняння:
Вн = 0,68 х Вр + 14,1,
де Вн - маса бульб у кущі потомства; В! - маса бульб у кущі батьків;
0,68 — 14,1 — коефіцієнти регресії з вірогідністю 0,99 достовірні.
Таким чином, урожайність - це властивість, яка при спрямова¬
ній селекції може підвищуватися до певного високого рівня.
Важливим у теорії і практиці селекції є питання про взаємо¬
зв’язок складових урожайності та їхній взаємовплив і динаміку в
процесі спрямованої селекції. З урахуванням цього гібридні комбі¬
нації були розподілені на такі групи:
- перша група - комбінації, у яких урожайність високою мірою за¬
лежала від кількості і маси бульб у кущі. Сіянці не можуть бути вико¬
ристані для селекції на підвищення кількості бульб і їхньої маси;
- друга - комбінації, у яких урожайність високою мірою залежала
від кількості бульб і дещо менше - від їхньої маси. Сіянці можуть бути
використані для селекції тільки на підвищення маси бульб у кущі;
- третя - комбінації, у яких урожайність мала низьку або серед¬
ню залежність від кількості бульб і високу - від їхньої маси. Сіянці
цих комбінацій можуть бути використані для селекції тільки на під¬
вищення кількості бульб;
- четверта - комбінації, у яких урожайність мала середню або
низьку залежність від кількості і маси бульб. їх сіянці є найбільш
придатними для селекційної роботи.
233
Виходячи з того, що взаємозв’язок між кількістю бульб і їхньою
масою був середній або низький, а подекуди і зовсім відсутній, було
зроблено висновок, що ці ознаки не залежать одна від одної. Вони є
незалежними при успадкуванні потомством, і тому з кожної можна
вести добір, спрямований на підвищення як кількості, так і маси
бульб. Однак слід пам’ятати, що в окремих гібридних комбінаціях
коефіцієнт кореляції між кількістю і масою бульб варіює в межах від
0,3 до -0,8.
Аналіз змін коефіцієнта кореляції взаємозв’язку кількості і маси
бульб у кущі по гібридних комбінаціях дає підстави стверджувати,
що величина його зумовлює характеристику складових урожайності.
Так, унаслідок підвищення коефіцієнта кореляції знижувалися показ¬
ники як маси, так і кількості бульб у кущі. Теоретичні розрахунки,
зроблені для з’ясування можливих змін кількості і маси бульб при
спрямованій селекції, показали, що можна викликати зміни подвій¬
ного характеру. Так, при селекції на збільшення розміру бульб змен¬
шуватиметься їх кількість, і - навпаки. Ці зміни чітко виражатимуть¬
ся тоді, коли залежність між ними є високою (табл. 4.8).
Таблиця 4.8 - Вплив ступеня залежності між кількістю і масою бульб у гібридній
комбінації на результати спрямованого добору
Комбі¬
нації
Кореляційна
залежність між
кількістю і
масою бульб у
кущі, r ± Sr
Параметри
фактичного
середнього куща
комбінації
Зміни при спрямованій селекції
кількість
бульб,
пгг/кущ.
середня
маса
бульб,
г
відбір форм із
масою бульб
100 г
відбір форм за
кількістю бульб,
20 шт./кущ
кількість бульб
маса бульб
73282
-0,029 ±0,185
12,4
70,7
12,8
70,0
73262
-0,365 ±0,100
15,8
42,0
12,6
32,1
73290
-0,433 ± 0,85
11,6
61,4
9,4
79,5
73210
-0,502 ±0,71
10,9
41,7
5,1
48,8
73207
-0,818 ±0,59
11,2
58,2
4,4
60,0 ^
З таблиці видно, що якщо відбирати форми із середньою масою
бульб 100 г, то кількість їх у кущі складатиме 4,4-12,8 шт. Якщо від¬
бирати форми із середньою кількістю 20 бульб, то їхня маса стано¬
витиме 48,8-70 г. Крім того, з таблиці видно, що найбільших змін
при спрямованому доборі зазнають форми, які вийшли з комбінацій
із високою залежністю між кількістю і масою бульб.
Селекція ранньостиглих форм картоплі із середнім і високим
умістом крохмалю. Цей показник є важливою ознакою продуктивності
і якості ранньостиглої картоплі. Він перебуває в середній і високій коре¬
ляційній залежності від тривалості вегетшдйного періоду (від 0,45 до
0,71) і низькій або середній - від урожайності (0,6 до -0,60).
234
Раніше (1957-1966 рр.) було виявлено, що підвищення крохмалис¬
тості селекційним шляхом досягається методом трансгресії. При
цьому можна ефективно використовувати таке рівняння регресії:
у = 0,70 х X + 2,60,
де у - уміст крохмалю у гібридних потомствах; X - середній уміст крох¬
малю в батьків; 0,70 і 2,60 - коефіцієнт регресії при вірогідності 0,99.
При аналізі показників умісту крохмалю батьків і потомства ви¬
явлена їх повна ідентичність щодо величини і напряму зростання. Це
дозволяє стверджувати, що накопичення цієї ознаки можливе селек¬
ційним шляхом.
Вивчаючи крохмалистість як селекційно-генетичну ознаку, нау¬
ковці виявили, що вона змінюється під впливом як чоловічої, так і
материнської форм (табл. 4.9,4.10).
Показники таблиць 4.9 і 4.10 свідчать про те, що чоловічі і мате¬
ринські форми впливали на вміст крохмалю у потомстві. Особливо
це помітно в комбінаціях, де чоловічою формою був висококрохма-
листий сорт Білоруська крохмалиста, який дуже добре передає по¬
томству свою основну ознаку.
Таблиця 4.9 - Вплив чоловічої форми на вміст крохмалю в потомстві
Чоловічі форми
Середній уміст крохмалю
в потомстві, %
Коливання вмісту крохмалю
в гібридній комбінації, %
2778-2
17,9 ±0,20
від 15,6 до 20,2
2808-2
17,9 ±0,51
від 16,7 до 18,9
2808-92
17,5 ±0,20
від 17,0 до 19,0
2808-43
17,0 ±0,17
від 15,6 до 19,8
Мінська рання
13,9 ±0,14
від 12,6 до 17,3
Прієкульська рання
13,6 ±0,18
від 11,1 до 14,7
132-46
12,5 ± 0,47
від 10,0 до 14,0
Таблиця 4.10 - Вплив материнської форми на вміст крохмалю в потомстві
Комбінації
Походження
Середній уміст крох¬
малю в потомстві, %
72154
3267-51 х Білоруська крохмалиста
20,4 ±0,17
72152
3329-17 х
19,0 ±0,13
72153
3302-33 х
17,4 ±0,20
72151
3302-14 х
15,5 ±0,18
Таким чином, здійснюючи спрямований добір батьківських пар, мож¬
на успішно вести селекцію ранньостиглих висококрохмалистих сортів.
Але при виведенні нових сортів, особливо якщо метою цього є отриман¬
ня високопродуктивних форм (висока урожайність і високий уміст кро¬
хмалю), слід звертати увагу не тільки на вміст крохмалю, але й на ком¬
плекс пов’язаних із ним ознак, до якого належать урожайність, її елемен¬
ти (кількість і маса бульб), скоростиглість та інші ознаки.
235
Для з’ясування зв’язку між цими ознаками були зроблені розра¬
хунки коефіцієнтів кореляції між умістом крохмалю, урожайністю та
її елементами (табл. 4.11).
Таблиця 4.11- Залежність крохмалистості від комплексу ознак картоплі
Гібридна ком¬
бінація
Середній уміст
крохмалю в
комбінації
Показники ко]
реляційної залежності кі
рохмалистості
від урожайності,
г/кущ
від кількості бульб,
шт./кущ
від середньої маси
бульб у кущі
r ± Sr
*05
г± Sr
t<)5
г± Sr
t<)5
73203
11,2
0,604 ± 0,201
3,00
0,700 ±0,201
3,50
-0,174 ±0,246
0,70
73273
13,9
0,488 ± 0,242
2,02
0,390 ±0,255
1,53
0,84 ± 0,275
0,30
73210
14,4
0,240 ± 0,242
0,98
0,333 ± 0,235
1,41
-0,067 ± 0,249
0,27
73277
16,1
0,113 ±0,199
0,57
-0,017 ±0,199
0,86
0,130 ±0,198
0,66
73285
16,2
0,064 ± 0,288
0,22
0,356± 0,272
1,39
-0,501 ± 0,249
2,02
73259
19,3
0,030 ±0,160
0,18
0,040 ±0,159
0,25
0,041 ±0,151
0,25
73261
21,0
-0,039 ± 0,258
0,15
0,104 ±0,256
0,40
-0,111 ±0,256
0,43
73279
16,1
-0,020 ± 0,144
0,14
-0,073 ± 0,143
0,51
0,010 ±0,144
0,70
73231
14,1
-0,197 ±0,255
0,87
-0,137 ±0,277
0,60
-0,012 ± 0,229
0,50
73227
13,8
-0,402 ±0,172
1,09
-0,201 ±0,213
0,94
-0,425 ± 0,197
2,15
73244
13,1
-0,402 ±0,172
2,32
-0,204 ±0,185
1,10
-0,203 ±0,185
1,10
73263
13,0
0,465 ±0,145
3,18
0,255 ±0,159
1,60
-0,285 ±0,157
1,87
73228
12,3
-0,610 ±0,250
2,43
-0,703 ± 0,255
3,12
-0,416 ±0,287
1,45
Із показників таблиці 4.11 видно, що в кожній гібридній комбіна¬
ції складаються характерні і властиві тільки їй взаємозв’язки між
ознаками, які корелюють, тобто крохмалистість - з урожайністю,
кількістю і масою бульб у кущі. Можна припустити, що це явище
певною мірою впливає на рівень крохмалистості та урожайність.
Так, при аналізі показників таблиці 4.11 було виявлено високий
уміст крохмалю насамперед у сіянців тих гібридних комбінацій, для
яких властива низька кореляційна залежність між крохмалистістю та
врожайністю.
Поглиблене вивчення взаємозалежності ознак урожайності і крох-
малистості дозволило розрахувати коефіцієнти регресії, щоб прогно¬
зувати їхній вплив на зміну крохмалистості, урожайності та її елеме¬
нтів. З використанням цих розрахунків було зроблено прогноз зміни
показників умісту крохмалю в сіянцях гібридних комбінацій при
спрямованій селекції на врожайність (табл. 4.12 ).
Прогнозування можливих змін щодо вмісту крохмалю в сіянцях
гібридних комбінацій при спрямованій селекції на врожайність до¬
зволяє зробити висновок, що при збереженні взаємозв’язку ознак
крохмалистості та врожайності (таблиці 4.11 і 4.12) селекційним ме¬
тодом можна досягти таких результатів:
236
а) істотно підвищити, порівняно з батьківськими формами, уміст
крохмалю одночасно з підвищенням врожайності в цілому або окре¬
мо як кількості бульб, так і середньої їх маси в кущі. Це можливе,
якщо вихідна гібридна комбінація буде мати позитивну кореляційну
залежність між крохмалистістю і врожайністю або її елементами;
б) знцзити вміст крохмалю водночас із підвищенням урожайності
рослин у цілому або збільшити як кількість бульб, так і середню їх
масу в кущі. Це можливе, якщо вихідна гібридна комбінація буде
мати від’ємну кореляційну залежність між крохмалистістю та вро¬
жайністю або її елементами;
в) підвищити загальну врожайність бульб або збільшити окремо
кількість і масу їх без суттєвої зміни величини вмісту крохмалю, що
буде можливим, якщо вихідна гібридна комбінація матиме слабку
кореляційну залежність або буде зовсім відсутньою між крохмалис¬
тістю, урожайністю та її елементами.
Таблиця 4.12 - Прогноз можливих змін умісту крохмалю при спрямованій
селекції на високу врожайність
Гібрид¬
ні ком¬
бінації
Показники вихідної гібридної ком¬
бінації до спрямованих доборів
Показники вмісту крохмалю (%)
після спрямованого добору
урожай¬
ність
кількість
бульб у
кущі, шт.
середня
маса
бульб у
кущі, г
вміст
крох¬
малю,
у %
формз
урожай¬
ністю 2000
г/кущ
форм із
кількістю
бульб 20
шт./кущ
форм із
середньою
масою
бульб 100 г
73277
880
16,4
53,7
16,1
15,6
15,6
14,4
73261
789
12,5
63,1
21,0
17,6
13,3
18,5
73279
780
12,9
60,5
16,1
15,1
15,9
17,2
73263
703
13,7
51,5
13,0
10,7
12,6
12,1
73244
670
14,0
47,9
13,1
9,2
12,6
9,4
73285
622
11,4
54,6
16,2
17,3
17,8
11,6
73227
611
11,3
54,1
13,8
11,8
14,6
12,3
73273
607
12,8
47,4
12,3
8,3
9,1
10,6
73231
559
11,1
50,4
13,9
19,6
16,1
14,4
73203
519
11,9
43,6
14,1
9,2
13,3
12,3
73201
455
10,9
41,4
14,4
18,9
16,73
15,8
73203
441
12,1
36,5
11,2
15,6
12,5
9,6
Вищевикладене дозволяє стверджувати, що при спрямованій се¬
лекції за будь-якою ознакою необхідно вивчити насамперед залеж¬
ність її від інших супровідних ознак, які з нею пов’язані генетично,
визначити характер залежності і використати все це для прогнозу¬
вання змін.
Селекція картоплі на тривалість вегетаційного періоду. Більшість
селекціонерів (Веселовський І.О., Альсмік П.І., Максимович М.М., Бу-
дін К.З. та ін.) дотримувались думки, що ранньостиглість потомства
перебуває у прямій залежності від рівня ранньостиглості батьківсь¬
ких форм. При вивченні успадкування ознаки ранньостиглості від
237
різних батьків було виявлено, що підбір батьківських пар і типів
схрещування впливає на рівень ранньостиглості потомства. Напри¬
клад, від схрещування двох ранніх форм між собою одержували 65%
ранньостиглого потомства, ранніх із середньоранніми - 48,5%, серед¬
ньоранніх із ранніми - 41,8%, ранніх із пізніми - 34,4 і пізніх із ран¬
німи - 28,0%. Як бачимо, вихід ранньостиглого потомства спостері¬
гається здебільшого в ранньостиглих батьків, меншою мірою - у піз¬
ніх. Крім того, найбільш чіткою вираженою ранньостиглість була в
потомствах, отриманих від ранньостиглих батьків. У комбінаціях,
отриманих за участі середньоранніх або пізньостиглих батьківських
форм, типово ранньостиглих батьків, типово ранньостиглі форми
зустрічалися рідко, а найчастіше були зовсім відсутні.
Окремі батьківські форми значною мірою впливають на форму¬
вання ранньостиглих нащадків. Це такі сорти, як Прієкульська рання,
Рання роза, Кобблер, Еррен пілот, Епрон, Веселовська 2-4, Епікур,
Дайма, Фера, Бона, Майкопська, Розафолія, Мінська рання, Білору¬
ська рання (ряд складено у спадному порядку формування
ранньостиглого потомства).
Ранньостиглість є полігенною ознакою. На основі вивчення 29 гіб¬
ридних комбінацій із різними батьківськими формами було виявлено,
що тривалість вегетаційного періоду нащадків на 50% залежала від чо¬
ловічих форм і на 44% - від материнських (вірогідність - 0,99). Цей по¬
казник варіював під впливом чоловічих форм від 111±0,11 до 120±0,08
днів; материнських - від 109±0,14 до 125±0,74 днів.
Ранньостиглість значною мірою пов’язана також із тривалістю
вегетаційного періоду генотипу, тому в подальшому вивчали утво¬
рення форм із коротким періодом вегетації.
Аналіз показників потомств за тривалістю (у днях) вегетаційного пе¬
ріоду показав, що вони з вірогідності 0,99 розподілялися на 4 групи: І -
99-112, П -113-116, Ш - 117-120, IV - більше 121 дня (табл. 4.13).
З показників таблиці видно, що сіянці першої групи здебільшого
(40-60%) відзначалися дуже коротким періодом вегетації (до 100
днів), що має значення для відбору дуже ранньостиглих форм. У сіян¬
ців другої групи (40-70%) вегетаційний період тривав довше (110
днів), і тому вони мали велике значення для селекції ранньостиглих
форм. Третю групу склали сіянці від комбінацій пізньостиглих форм,
тому в них переважали генотипи середньораннього типу. У четверту
групу увійшли сіянці від комбінацій із пізньостиглими батьками, то¬
му це були здебільшого пізньостиглі форми. Кількість одержаних
ранньостиглих форм зумовлювалася тривалістю вегетаційного пері¬
оду сіянців окремих комбінацій. Так, у комбінаціях першої групи
було від 16 до 28,4 % дуже ранніх форм, у другій і третій групах їх
було значно менше, а в четвертій вони були зовсім відсутні. Отже,
виходячи з вищезазначеного, при селекції на скоростиглість слід
враховувати тривалість вегетаційного періду потомства.
238
Таблиця 4.13 - Тривалість вегетаційного періоду потомства (у днях) залежно від
походження (чоловіча форма - сорт Мінська рання)
Група
стиглості
материн¬
ської
Аооми
Походження материнської
форми
Розподіл (%) сіянців за групою ранньо¬
стиглості, у днях вегетації
99-112
113-116
117-120
більше
121
І
Прієкульська рання х Унікат
24,8
40,6
23,8
10,8
Бонву х Прієкульська рання
16,3
59,1
15,1
9,5
П
491-037 х середньорання
2,2
70,6
19,5
77
Прієкульська рання * Кобблер
9,2
42,5
36,0
12,3
Епікур х Прієкульська рання
5,7
60,0
28,5
5,8
Прієкульська рання * 1063
15,7
38,8
26,0
20,6
Бонімару х Прієкульська рання
-
47,0
40,6
12,4
Ш
К - 222 х Прієкульська рання
12,5
27,8
29,7
30,0
Розвариста х Прієкульська
рання
—
38,4
46,1
15,5
IV
486-011 х середньостигла
-
17,1
30,0
50,9
Бекра х пізньостигла
-
12,6
16,1
71,3
Ранньостиглість як селекційно-генетична ознака взаємозв’язана з
іншими ознаками (табл. 4.14 ).
Таблиця 4.14 - Залежність ранньостиглості від інших ознак картоплі
Ознаки, які корелюють
із ранньостиглістю
Показники
кореляції
t(>5
Урожайність, г/кущ
-0,127 ±0,120
1,58
Кількість бульб у кущі, шт.
+ 0,097 ±0,100
0,97
Середня маса бульб у кущі, г
-0,402 ±0,011
36,54
Уміст крохмалю, %
- 0,456 ± 0,009
50,67
Тривалість вегетації у днях
-0,709 ± 0,04
177,25
З таблиці видно, що ранньостиглість і врожайність мають низьку
протилежну залежність. Однак, у деяких комбінаціях вона буває ви¬
щою. Це виникає тоді, коли кущі мають більшу кількість бульб.
Скоростиглість і маса бульб перебувають у вираженій протилежній
залежності. Тому з’являються певні біологічні перепони, які вини¬
кають на шляху селекції форм, що поєднують скоростиглість та
врожайність і є досить великими за розміром.
Враховуючи те, що в цілому урожайність має низьку від’ємну
кореляційну залежність від ранньостиглості, виведення ранньостиг¬
лих високоврожайних форм є цілком важливим. При їх селекції не¬
обхідно надавати перевагу ранньостиглим формам, які мають досить
велику кількість бульб під кущем або багатобульбові у поєднанні з
великобульбовістю.
У селекції картоплі є певні труднощі в поєднанні ознак ранньо¬
стиглості і крохмалистості, між якими існує середня кореляційна за¬
лежність, яка іноді стає високою. Для подолання цього необхідно
239
враховувати й особливості кожної комбінації. Перспективними для
селекції таких форм є комбінації з участю типових ранньостиглих
форм у поєднанні з пізньостиглими висококрохмалистими батьками.
Серед сіянців таких комбінацій можливі випадки виникнення форм,
які поєднують ранньостиглість і крохмалистість.
Підсумки селекції картоплі на ранньостиглість. До Реєстру
сортів рослин України на 2001 рік внесено 27 скоростиглих сортів
картоплі, створених в Україні, у тому числі ранніх - 15, середньо¬
ранніх - 12. З них селекції Інституту картоплярства: ранні - Боро¬
дянська рожева, Кобза, Незабудка, Пролісок, Повінь, Серпанок; се¬
редньоранні - Водограй, Купава, Обрій, Світанок київський; Полісь¬
кої дослідної станції ім. О.М. Засухіна Інституту картоплярства: ран¬
ні - Божедар, Гарт, Зов, Поран, Посвіт, Косень 95; середньоранні -
Берегиня, Доброчин, Малич, Поліська 96, Пост 86, Радич, Дубравка;
Чернігівської дослідної станції по картоплі: ранні - Чернігівська
рання, Седнівська рання; Сумського державного аграрного універси¬
тету - ранній сорт Молодіжна; Інституту землеробства і тваринницт¬
ва західного регіону УААН - середньоранній сорт Мавка.
Ранні сорти створені методом статевої гібридизації з наступним
добором: Бородянська рожева (77.583/16 х Гідра), Кобза (Мануелла
х Мавка), Незабудка (Швальбе х Карла), Пролісок (Світанок київсь¬
кий х Омега), Повінь (76.198/175 х 79.533-38), Серпанок (Поля х Ро-
мано), Божедар (81.82-55 х Радомишльська), Зов (Поліська 36 х По¬
ліська рожева), Гарт (Сулев х Новинка), Посвіт (Гітте х С-227-73),
Косень 85 ( П.81.60/76 х Зов), Поран (Невська х Берегиня), Седнів¬
ська рання (ВіхолахГранола), Чернігівська рання (Гранола х Льві¬
в’янка), Молодіжна (Адретта х Кристал білоруський).
Переважна більшість сортів у ранні строки збирання і наприкінці веге¬
тації дають задовільний і високий врожай товарних бульб (табл. 4.15).
Таблиця 4.15 - Урожайність ранніх сортів картоплі української селекції
Сорт
Урожайність, ц/га
на 40-45-й день
після сходів
наприкінці
вегетації
Ранні
Божедар, Косень, Повінь
220-210
450-460
Бородянська рожева, Гарт, Зов, Кобза, Молодіжна,
Незабудка, Поран, Посвіт, Пролісок, Серпанок
205-200
400-450
Буян, Веста, Дніпрянка, Косовщинська
150-120
330-400
Середньоранні
Аграрна, Берегиня, Водограй, Доброчин, Купава,
Ластівка, Лілея Сумщини, Малич, Обрій, Поліська 96,
Світанок київський, Ювіляр 6070, Пост 86,
135-130
450-500
Дара, Немішаївська 100, Радич, Фантазія
120-110
350-400
Хорошими смаковими якостями відзначаються такі сорти, як Коб¬
за, Пролісок, Повінь, Зов, Молодіжна (табл. 4.16)
240
Таблиця 4.16 - Смакові якості ранніх сортів картоплі вітчизняної селекції
Сорт
Смак, балів
5-4,5
. 4,4—4,0
3,9-3,5
Кобза, Купава, Молодіжна, Обрій, Світанок
київський
+
Аграрна, Бородянська рожева, Буян, Веста,
Гарт, Дара, Дніпрянка, Доброчин, Дубравка,
Зов, Косовщинська, Ластівка, Лілея Сумщи¬
ни, Малич, Незабудка, Повінь, Поліська 96,
Пост 86, Радич, Серпанок, Фантазія
+
Водограй, Берегиня, Божедар, Косень, Посвіт,
Немішаївська 100
+
Примітки : 5-4,5 - відмінний; 4,4-4,0 - добрий; 3,9-3,5 - задовільний
Важливим показником у ранньостиглих сортів є стійкість бульб
до потемніння м’якоті. У таблиці 4.17 подані показники схильності
до потемніння сирої м’якоті бульб ранньостиглих сортів картоплі.
Таблиця 4.17 - Схильність сирої м’якоті бульб ранньостиглих сортів картоплі до
потемніння
Сорт
Потемніння, балів
9
8
7
6
5
4
Сортимент України
Кобза, Ластівка, Мавка, Молодіжна, Світанок
київський
+
Аграрна, Божедар, Бородянська рожева, Зов,
Косовщинська, Пролісок, Ювіляр 6070
+
Водограй, Доброчин, Незабудка, Пост 86, Сед-
нівська рання
+
Сортимент Білорусі
Гранат, Явар
+
Аксамит, Альтаїр, Добро, Пригожа 2
+
Росинка
+
Лазуріт
+
Сортимент Росії
Жуковська рання, Резерв
+
Волжанін
+
Бежицька, Брянська рання, Невська, Скороплід-
на, Удача
+
Прайса
+
Сортимент Німеччини
Каролін
+
Ацдра, Бербер, Делікат, Карла, Карлена, Лікарія,
Ліу, Серафима
+
Анелі, Аркула, Жаклін, Іветта, Каратон, Моллі,
Рікеа, Укама, Феліцигас
+
Адретта, Альвара, Карат, Коретта, Ліберія
+
Агаве
+
Примітки: 9 - дуже низька схильність; 8 - дуже низька до низької; 7 - низька;
6 - низька до середньої; 5 - середня; 4 - середня до високої;
З - висока; 2 - висока до дуже високої; 1 - дуже висока
241
З іноземних сортів рання група представлена в Реєстрі наступни¬
ми сортами: Джаерла, Латона, Симфонія, Юкама, Лікарія, Розара,
Карлена, Вінета, Колєт, Рікеа, Велокс, Маріс Бард, Коруна, Краса.
Високою врожайністю товарних бульб при збиранні в ранні
строки та наприкінці вегетації відзначалися сорти Джаєрла, Лікарія,
Розара, Карлена, Вінетта. Високим і підвищеним умістом крохмалю
характеризувалися сорти Симфонія, Карлена, Колет; смаковими яко¬
стями - Симфонія, Лікарія, Карлена.
Серед сортів іноземного походження високою врожайністю в
ранні строки збирання виділялися сорти Невська, Аргос, Альвара.
Усі сорти цієї групи вирізнялися низьким умістом крохмалю і подіб¬
ними смаковими якостями.
ЛІТЕРАТУРА
1. Альсмик П.И., Гончарте М.Н. Картофель и динамика его роста / Изд-
во АН БССР. Ин-т биологии. - Минск, 1933.-112 с.
2. Арнаутов В.В. Картофель. - М.: Сов. Россия, 1959 - 96 с.
3. Бацанов Н.С. Повышение продуктивности растения картофеля. - М.,
1969.-70 с.
4. Гончаров Н.Д. Селекция высококачественных и продуктивных сортов
картофеля интенсивного типа: Дис....д-ра с.-х. наук. - Самохваловичи,
1981.-183 с.
5. Максимович М.М. Методы селекции картофеля. - М.: Сельхозгиз,
1951.- 192 с.
6. Писарев Б А. Ранний картофель. - М.: Россельхозиздат, 1980 - 48 с.
7. Шевелуха В.С. Периодичность роста сельскохозяйственных растений
и пути ее регулирования. - Минск: Ураджай, 1977 - С.156-185.
8. Яшина И.М. Принципы генетических исследований при селекции на
повышенное содержание крахмала и устойчивость к фитофторе // Карто¬
фель. - Минск, 1966. - С.49-58.
4.5.3. Селекція картоплі на придатність
до механізованого виробництва
Якість і втрати картоплі при інтенсивному виробництві. З
переходом виробництва картоплі на промислову основу проблема
боротьби із втратами набула особливо важливого значення і актуаль¬
ності. Причиною втрат є передусім масове механічне пошкодження
бульб у процесі їх збирання, транспортування, закладання на збері¬
гання і затарювання [10]. У зв’язку з цим якість картоплі на сьогодні
є низькою: у Швейцарії виявляють 20,5% бульб із механічними трав¬
мами; у Франції - 46,7 %; Великобританії - від 22 до 35-44 %. У
Швеції при збиранні бульб травмується до 70%, при транспортуванні
і закладанні на зберігання - до 10 %, при сортуванні - 37 % значних і
25 % середніх травм, при навантаженні і затарюванні - 30 і 10 %. У
Польщі при збиранні травмується 25-40 % бульб, при транспорту¬
242
ванні - 10 %, при сортуванні - 10-15 %, а при закладанні на збері¬
гання близько 2/3 бульб виявилися пошкодженими [8].
Дані вітчизняних досліджень також вказують на значне травму¬
вання бульб. Так, М.Мосин і В.Толопилов (1967) довели, що при ме¬
ханізованому збиранні травмованість бульб становила 12-85 %. Ме¬
ханізоване збирання, перевезення самоскидом одночасно із сорту¬
ванням на КСП-15, за результатами досліджень, викликало в серед¬
ньому 21-24 % пошкоджень бульб. На ступінь травмування істотно
впливали строки і способи збирання картоплі. На Київській овоче-
картопляній дослідній станції розроблено конвеєрно-безперева-
лочний спосіб збирання картоплі, який включає лише 4 технологічні
операції замість 10-11. Механічна пошкоджуваність бульб при цьо¬
му знижувалася на 50 % з істотним підвищенням кулінарних власти¬
востей картоплі [7].
Втрати картоплі при механізованому збиранні значною мірою
зумовлюються грунтово-кліматичними умовами. Дослідження, про¬
ведені у Швейцарії, показали, що на легких грунтах травмування
бульб знижується у два рази, порівняно з цим показником при зби¬
ранні на важких грунтах. Надлишкове азотне живлення, підвищений
уміст вологи на торф’яно-болотних грунтах і пізні строки садіння
картоплі за низьких температур призводили до того, що бульби зби¬
ралися фізіологічно не дозрілими, внаслідок чого вони дуже травму¬
валися при збиранні [1].
При вивченні впливу температури повітря в період збирання на
стійкість бульб до механічного пошкодження виявлено, що близько
30% травм зумовлювалися збиранням при низькій температурі. Так,
при температурі 18-20°С бульби менше травмуються, ніж при 4-6°С
(Smittle D., Rice В.). Зниження температури з 18 до 10 °С збільшува¬
ло важкі травми м’якоті у 1,3 рази, зниження температури до 4 °С - у
2,2 рази (Сиим Я.М., Валуєва Т.). Сортування картоплі при темпера¬
турі 11 °С і нижче збільшувало кількість бульб із механічними по¬
шкодженнями, порівняно із травмованістю при температурі 16 °С і
вище (Grey Е., Paterson М., Leppack).
Збирання картоплі з дуже сухого грунту значно підвищує сту¬
пінь пошкоджуваності бульб (Pavasd, Holst J.). Роки з надмірною
вологістю також сприяли посиленню травмованості (Peterson С.,
Нааі С.).
Важлива роль у попередженні механічних пошкоджень належить
добривам: калійні і фосфорні добрива прискорюють дозрівання, під¬
вищують міцність шкірки і твердість м’якоті, що знижує травмуван¬
ня (Shaller R.); підвищені дози азоту подовжують термін достигання,
знижують уміст сухої речовини, підвищують схильність м’якоті
бульб до потемніння.
Спеціальними дослідами в країнах, що спеціалізуються з вироб¬
ництва картоплі, доведено, що ступінь пошкоджуваності бульб кар¬
243
топлі залежить і від сорту. Вивчення впливу окремих чинників на
фенотипічну мінливість стійкості бульб до механічних пошкоджень
показало, що частка впливу генотипу становила 78,8 %, середовища
- 8-7 %, сукупного впливу цих чинників - 12,5 %.
Стійкістю до травмування бульб характеризуються сорти селекції
Інституту картоплярства УААН: Луговська, Воловецька, Світанок
київський, Українська рожева (Кучко А.А., Куценко В.С., Осипчук АА.).
У Німеччині (Гросс-Люзевітц) більш стійкими до механічного трав¬
мування визнані сорти Аксилія, Астілла, Галина, Лібелла (Demel J.,
Specht А.); у Польщі - Вишеборський, Бінтьє, Санте (Kubicki К.); в
Австрії - Елла, Ганза, Дезіре, Остара (Gittle D.), у Чехії - Красава,
Сосна (Malina V.), у Голландії - Бінгьє (Sparks W.).
Різна чутливість картоплі до механічних пошкоджень поясню¬
ється високим умістом тирозину в клітинах бульби (Shaller R.), пи¬
томою вагою бульб, розміром крохмальних зерен (Hudson D.), міцніс¬
тю шкірки (Веселовский И.А. с соавт.; Валуєва Т.; De Maine М.).
Пошкоджені бульби різко знижують товарні та насінні якості,
призводять до великих втрат їх маси при зберіганні (18-25 %) [11] і
значних відходів при очищенні (Гусєв С.А., Сокол П.Ф., Burton W.,
Полищук С.Ф.). У Чехії, наприклад, втрати врожаю від травмування
бульб становлять близько 500 тис. тонн на рік (Malina V.), у Польщі
- 6,5 млн тонн (Whekowski S.), у штатах Вашингтон і Айдахо (США)
- 1 млн доларів щорічно (Thomtor R..). Пошкодження бульб при ін¬
тенсивному виробництві негативно впливає на лежкість, насінні та
споживчі якості. Так, наприклад, за наявності 1 % травмованих бульб
збільшувались відходи при очистці на 0,33-0,42 %, залежно від виду
травм, загальні втрати - на 0,98 %, у тому числі від гнилі - на 0,25 %,
знижувались насінні якості - на 5,8-8,6 ц/га.
Одним із можливих і важливих шляхів зниження втрат картоплі
від механічного пошкодження є створення і використання стійких
сортів [2,4,5]. У селекції при оцінці на стійкість картоплі до ме¬
ханічного пошкодження використовують різні методи: у Чехії - при¬
лад Instrom, що реєструє час і силу стискування бульби (Blahovek J.);
у Великобританії та Польщі - маятниковий метод (De Meine М.,
Jastzebski К.); у Росії - прилади типу ПДТ-С, ПСТ-ВИМ і ПКК-ВИМ
(Сафразбекян О.).
Кожний з описаних методів має свої переваги і недоліки, засто¬
совувався в різні роки, на різному бульбовому матеріалі. У зв’язку з
вищезазначеним, виникла необхідність розробити швидкий і надій¬
ний метод, що дозволяє виявляти перспективні зразки ще на ранніх
етапах селекції.
Ознаки та властивості, які характеризують придатність сор¬
ту до механізованого виробництва. Передумовою визначення кри¬
теріїв оцінки стійкості бульб до пошкоджень у селекційному процесі
було дослідження впливу погодних умов, агротехнічних і технологі¬
244
чних факторів не тільки на загальну травмованість, але й на окремі її
види у 14 сортів картоплі [9]. Виявлено, що загальна кількість травм
у різні роки змінювалася в 1,5-2,6 рази, подерта шкірка - у 2,5-4,5;
вириви - 1,2-2,4; тріщини - 1,1-5,5; потемніння м’якоті - 1,1-1,9
рази. З цього видно, що найбільш мобільними були травми у вигляді
подертої шкірки і тріщин. Особливо чітко простежувалося збільшен¬
ня як загальної кількості травмованих бульб, так і бульб із тріщина¬
ми в роки з високою зволоженістю повітря при його низькій темпе¬
ратурі. Загальне травмування бульб залежало на 43 % (г=0,656) від опа¬
дів і на 33 % (г=0,577) - від вологості повітря. Виявлена також
від’ємна залежність (г=-0,569) між травмуванням бульб у вигляді
тріщин від перепадів температури повітря. Травми у вигляді подер¬
тої шкірки зумовлювалися (г=0,666) в основному кількістю опадів за
вегетаційний період, меншою мірою (г=0,510) - вологістю повітря і
не залежали від його температури (г = 0,092).
На високому фоні добрив загальне травмування бульб збільшилося
на 16 %, у тому числі легких травм - на 10, важких - на 6 %. При цьому
травмованість підвищувалася за рахунок збільшення вмісту бульб із по¬
дертою шкіркою в 1,6-2,4 рази, із тріщинами - в 1,0-2,1, з виривами -
лише в 1,1 рази. Отже, мобільними травмами в даному випадку були
подерта шкірка і тріщини. Травми у вигляді виривів визначалися не
стільки умовами живлення, скільки сортовими особливостями.
Щодо впливу агротехнічних і технологічних чинників, а також
погодних умов на травмованість бульб можна зробити висновок, що
рівень травмування бульб від строків збирання зростає в 4,5 рази, від
видів післязбиральної доробки - у 4, від строку сортування - у 2,5
рази. Зміни за видами травм були такими: по подертій шкірці - ви¬
щими в 9,5 разів, по тріщинах - в 1,7, по потемнінню м’якоті від
ударів - в 1,5 рази. Пошкодження бульб у вигляді подертої шкірки
на 62 % визначалося дослідженими агротехнічними і погодними чин¬
никами, тріщини - на 18, вириви - на 11 і потемніння м’якоті - на 10 %.
Отже, кількість бульб із подертою шкіркою можна з успіхом регу¬
лювати агротехнічними заходами. Інші види травм, особливо вириви
і потемніння м’якоті, очевидно, більшою мірою зумовлювалися ге¬
нотипом.
Оцінка рівня травмування бульб показала, що при механізовано¬
му збиранні менше за інших пошкоджувалися сорти ранньої групи,
більше - пізньої [3,6]. З аналізу динаміки окремих -видів травм по
групах стиглості виявлено, що кількість бульб із подертою шкіркою і
потемнінням м’якоті від ударів показники збільшувалися з подов¬
женням тривалості вегетаційного періоду, а за наявністю травм із
виривами і тріщинами, навпаки, вони знижувалися від пізньої до
ранньої групи (табл. 4.18).
При цьому в середньоранніх і середньостиглих сортів, порівняно з
ранніми, кількість бульб із подертою шкіркою збільшувалася в 5 разів,
245
у середньопізніх - у 8, а в пізніх - у 50; з потемнінням м’якоті - від¬
повідно в 1,6; 2,4 і 5,8 рази. Травмованість за виривами від ранніх до
пізніх сортів знижувалася в 1,7; 1,5 і в 1,3 рази; за тріщинами - у 2,3 і
1,3 рази. Виняток становила травмованість за тріщинами в сортів се¬
редньоранньої і середньостиглої групи, де вона була найбільшою -
6,2 %. Отже, сорти з більш коротким вегетаційним періодом менш
стійкі до глибоких зовнішніх механічних пошкоджень, але стійкі до
легких зовнішніх травм і до потемніння м’якоті від ударів. Сорти з
тривалим періодом вегетації менш стійкі до внутрішніх і легких зов¬
нішніх пошкоджень, але більш стійкі до глибоких травм. Це зумов¬
лює необхідність цілеспрямованої селекції на поліпшення показни¬
ків травмованості бульб по групі ранніх сортів, тобто створення
форм, стійких до виривів і тріщин; по групі середньоранніх і серед¬
ньостиглих - до тріщин; по групі середньопізніх - до потемніння
м’якоті; по групі пізніх - до обдирання шкірки і потемніння м’якоті
від ударів.
Таблиця 4.18 - Вплив групи стиглості на травмування бульб
Група стиглості
Механічні пошкодження, %
всього
зовнішні
внутї
ЭІШНІ
подерта
шкірка
вириви
тріщини
потемніння
м’якоті
Ранні
9,8±0,52
0,2±0,03
3,5±0,15
4,0±0,30
2,1±0,12
Середньоранні,
середньостиглі
12,6±0,87
1,0±0,13
2,0±0,17
2,0±0,27
3,4±0,16
Середньопізні
14,7±0,79
5,7±0,20
2,3±0,20
1,7±0,17
5,0±0,34
Пізні
28,1±2,10
10,0±1,33
2,7±0,17
3,1±0,14
12,3±0,58
Прогнозуванню стійкості бульб до механічний пошкоджень при¬
діляється достатньо уваги як у вітчизняній, так і в зарубіжній літера¬
турі. Однак найчастіше досліджувався вплив якогось одного чинника
на травмованість бульб. Вивчення ступеня зв’язку травмованості як
за окремими ознаками і властивостями бульб, так і за їхнім сукупним
впливом дає змогу прогнозувати стійкість бульб до механічних
травм на ранніх етапах селекції.
Вплив фізичних і морфолого-анатомічних ознак і властивостей
бульб на їхню травмованість при комбайновому збиранні вивчався
на шести сортах картоплі. Уміст бульб із механічними пошкоджен¬
нями коливався від 22 до 44 %, питома вага досліджених сортів - від
1,0676 до 1,0993 г/см3; форма їх бульб - від округло-овальної (1,25-
1,35) до видовженої (1,46) і довгої (1,60); розміри бульб (найбільший
поперечний діаметр) - від 45 до 61 мм.
Виявлено, що пошкодження бульб при збиранні перебуває у пря¬
мій і високій (53,5 %) залежності від питомої ваги (у=-6018 + 10565х
- 4596х2; г=0,731); в міру подовження бульб збільшувалася інтенсив¬
ність їх пошкодження, а залежність між цими показниками складала
246
72,7 % (у=-65,98+103,7х-23,3х2, г=0,852). Збільшення розміру бульб
у 2 рази з ЗО до 60 мм спричинило підвищення травмування в 1,7 ра¬
зи, а пошкодженість бульб розміром 90 мм була вищою вже в 2,4 ра¬
зи. Травмування бульб на 93 % залежало від їхнього розміру (і=0,964,
у=33,3 - 0,93х + 0,015х2).
Існує пряма залежність між травмованістю бульб і товщиною
шкірки, яка складає 64,6 % (у=82,1 - 34,4х + 5,Ох2; г=0,803).
Вплив фізичних властивостей (питома вага) на окремі види ме¬
ханічних пошкоджень бульб. При підвищенні питомої ваги з 1,065 до
1,100 г/см3 кількість легких зовнішніх і внутрішніх травм збільшува¬
лася більш ніж у 3 рази, тяжких - зменшувалася в 1,5 рази. Стійкими
до виривів були бульби з питомою вагою більше 1,09 г/см3. Стійкість
до тріщин спостерігалася в бульб з найбільш низькою і найвищою
питомою вагою.
Найбільша залежність (74,8 %) була виявлена між питомою ва¬
гою і наявністю легких зовнішніх травм; висока (більше 60 %) - за
тяжкими травмами, у тому числі за виривами, а також за потемнін¬
ням м’якоті. Залежність між кількістю бульб із тріщинами та їхньою
питомою вагою складала 53,7 %.
Вплив товщини кіркового шару бульб на пошкоджуваність їх різ¬
ними видами травм. Збільшення товщини в 2 рази підвищувало кіль¬
кість бульб з виривами в 1,7 рази, з тріщинами - у 2,6, а з потемнін¬
ням м’якоті - у 8,9 рази. Ступінь зв’язку між видами травм і товщи¬
ною кіркового шару досягав 95 %.
Сукупний вплив питомої ваги, розміру і форми бульб, а також
товщини їхнього кіркового шару засвідчив наступне. У дрібних
бульб підвищення питомої ваги з 1,065 до 1,085 г/см3 призводило до
збільшення на 6,7 % загальної травмованості, більшою мірою за ра¬
хунок легких зовнішніх травм - на 11,1 % і значно меншою мірою -
за рахунок потемніння м’якоті (на 1,2 % уміст бульб із тріщинами і
виривами знижувався відповідно на 0,9 і 1,6 %). У бульб розміром
45-65 мм спостерігалась така ж тенденція, що й у дрібних бульб, але
з деякою різницею по травмованості за тріщинами і виривами. З під¬
вищенням питомої ваги кількість цих видів травм збільшувалася.
Травмованість великих бульб розміром понад 65 мм зростала по всіх
видах зовнішніх травм із підвищенням питомої ваги. Так, кількість
загальних пошкоджень із підвищенням питомої ваги на 0,025 г/см3
зростала на 33 %, у тому числі по легких травмах - на 20, по тяжких
- на 4 %. Травмованість у вигляді потемніння знизилася при цьому
на 10%.
Ступінь мінливості цих показників коливався в межах 59-89 %.
Винятком було потемніння м’якоті великих бульб, де кореляційне
відношення було низьким (г=0,223).
Вплив розміру бульб на вид пошкодження при збиранні. Дрібні
бульби мали найбільшу кількість легких травм, наполовину менше
247
було тяжких пошкоджень, а травм із потемнінням м’якоті - най¬
менше. У бульб середньої фракції кількість легких травм і травм із
потемнінням м’якоті була найбільшою і однаковою. Пошкодження
цих бульб тяжкими травмами було найменшим. Бульби великого
розміру були менше травмовані тяжкими пошкодженнями і потем¬
нінням м’якоті, але мали більше легких травм.
Вплив форми бульб на вид травмування. У межах дрібної фракції
картоплі видовженість бульб призводила до збільшення кількості
легких травм із високою залежністю - 71,3 %. Вплив подовження
бульб на потемніння м’якоті був невисоким - 32,5 %. Щодо серед¬
ньої фракції бульб залежність між видами травм і їх формою склада¬
ла близько 65 %. При цьому спостерігалася тенденція до зменшення
кількості бульб із потемнінням м’якоті і збільшення кількості тяж¬
ких травм у довших бульб. У бульб подовженої форми великої фрак¬
ції спостерігалася менша кількість потемнінь м’якоті і тяжких травм
при незначному підвищенні легкої травмованості.
Вплив кіркового шару на стійкість до різних видів травм. Чим
товща шкірка у дрібних бульб, тим більше вони мають легких травм
і потемніння м’якоті, але менше - тяжких травм. Залежність цих по¬
казників становила 70-94%. Залежність між зменшенням товщини
кіркового шару в бульб середньої фракції і зниженням кількості лег¬
ких пошкоджень становила 59,3 %, тяжких - 97,1 %, за потемнінням
м’якоті - 78,8 %. Ступінь зв’язку між кількістю тяжких травм і потем¬
нінням м’якоті та зменшенням товщини шкірки у великих бульб був
дуже високий і складав 88-86 %, а між збільшенням кількості бульб
із легкими пошкодженнями і зменшенням товщини кіркового шару -
77,7 %.
Для кожної залежності розроблені математичні моделі, викорис¬
тання яких дозволить з високою ефективністю проводити відбір
форм, стійких до механічних пошкоджень на ранніх етапах селек¬
ційного процесу.
Прогнозування селективного показника можна здійснювати та¬
кими рівняннями регресії, у яких ступінь пошкодження бульб визна¬
чається їхньою питомою вагою (Хі), індексом форми (Х2), розміром
бульб (Х3) і товщиною шкірки бульб (Х4):
1) у= -380,2 + 333,91Хі + 13,6Х2 + 0,51Х3 + 0,47X4, що на 68 %
пояснює зміни загальних травм (г=0,827);
2) у= -296,2 + 267,4Хі + 13,0Х2+ 0,001Х3 - 0,20Х4, що на 65 % по¬
казує зміни легких травм (г=0,807);
3) у= 183,0 + 170,8Х, - 2,8X2 + 0,1Х3 + 1,9Х4, що на 44 % відоб¬
ражає зміни тяжких травм (г=0,655);
4) у= 14,1 - 12,1Хі - 2,8X2 + 0,04Х3 - 0,5Х4, що на 41 % пояснює
зміни пошкоджень у вигляді тріщин (г=0,644);
5) у= 50,0 - 46,2 Хі - 0,50 Х2 + 0,04 Х3 + 0,20 Х4, - на 28 % пояс¬
нює зміни пошкоджень у вигляді виривів (г=0,534).
248
У селекційній практиці при оцінці форм на стійкість до механіч¬
них пошкоджень застосовують різні інструментальні методи, але час¬
тіше за все - прилад типу маятника. З урахуванням цього був прове¬
дений спеціальний дослід з порівняльною оцінкою цього методу із
фактичною травмованістю бульб при збиранні 26 сортозразків кар¬
топлі. Виявлено, що кількість травм у вигляді подертої шкірки при
збиранні корелює із стійкістю до обдирання шкірки в лабораторних
умовах на 29,5 %, кількість тріщин - на 32,9 %, виривів - на 74,7 %.
Тому найбільш адекватним показником інструментальної оцінки
стійкості бульб до механічних пошкоджень можна вважати динаміч¬
ну твердість м’якоті, тобто стійкість до виривів і тріщин.
Результати оцінки травмованості 20 сортів картоплі за допомо¬
гою приладів типу маятника і ПДТ-С показали, що зв’язок між дина¬
мічною твердістю і стійкістю до виривів був дуже високим (99,8 %), по
тріщинах - високим (52,3 %).
У зв’язку з цим динамічна твердість може слугувати показником
стійкості бульб до тяжких пошкоджень, починаючи вже з другого
бульбового покоління. Оскільки сприйнятливість бульб до потем¬
ніння м’якоті значною мірою корелює з питомою вагою, то відбір
стійких форм можна з успіхом проводити за цим показником. Трав¬
мування ж бульб у вигляді подертої шкірки певною мірою можна
регулювати агротехнічними і технологічними прийомами, тому до¬
бір форм на цьому етапі за ознакою міцності з’єднання шкірки з
м’якоттю бульб, очевидно, є не таким важливим.
При вивченні ступеня впливу різних властивостей і ознак бульб
на їх травмованість певний інтерес викликала залежність пошко¬
джень від структури кіркового шару.
Регресивний аналіз показав, що на 26,7 % твердість м’якоті бульб
середньостиглих сортів зумовлюється товщиною шкірки (г=0,517), у
пізніх - на 13, ранніх - на 5, середньоранніх - на 0,77 % (г=0,088), у
середньому - на 3,2 %. Крім загальної товщини шкірки, слід врахо¬
вувати ще й структуру тканини: щільніша структура через більшу
кількість клітин меншого розміру зумовлює вищу твердість м’якоті
бульб сорту Темп - 7,7 шт.уд./200 м; перидерма середньостиглого
сорту Лорх складалася з меншої кількості клітин, але більшого роз¬
міру. Динамічна твердість м’якоті бульб цього сорту становила 6,6
шт.уд. Найбільш пухка перидерма середньораннього сорту Дєтско-
сєльська зумовлена великими клітинами, твердість м’якоті складала
5,2 шт.уд ./200 м. Виявлено також позитивну кореляцію між твердіс¬
тю м’якоті бульб і вмістом сухої речовини, яка становила 79 %.
У розрізі груп стиглості зв’язок між твердістю м’якоті і питомою
вагою був таким: у ранніх сортів становив 99,9 %, середньоранніх -
75,0, середньостиглих - 92,3 і пізньостиглих - 43,5 %. Неоднакову стій¬
кість бульб до механічних пошкоджень слід пов’язувати не тільки з пи¬
томою вагою, умістом сухих речовин або крохмалю (тенденція та ж),
249
але й ураховувати якість крохмалю. Вплив розміру крохмальних зе¬
рен на твердість м’якоті становив 1-12 %, у тому числі вплив розмі¬
ру великих зерен на твердість м’якоті ранніх сортів - 20,1 %, серед¬
ньоранніх - 4,5, середньостиглих - 15,2 і пізньостиглих - 1,2 %.
Уміст великих крохмальних зерен також підвищувався з подов¬
женням вегетаційного періоду сортів і складав 27,5-32,5 %. Цей по¬
казник у ранніх і пізньостиглих сортів становив 12 %, у середньостиг¬
лих і середньоранніх - відповідно 5,5 і 4,1 %.
Залежність динамічної твердості від маси бульби виражалася на¬
ступними коефіцієнтами кореляції: у ранніх г=-0,344; середньоранніх
- г=-0,488; середньостиглих - г=-0,103; пізніх - г=0,226.
Індекс форми незначно (1,6-3%) впливав на динамічну твердість
бульб перших трьох груп стиглості. Вірогідним його вплив був у піз¬
ніх сортів (г=0,436).
Таким чином, вивчено вплив семи основних властивостей і ознак
бульб на їхню стійкість до механічних травм. Це дало змогу розробити
моделі прогнозу травмованості з урахуванням впливу якогось одного
чинника. Такі моделі доцільно застосовувати при доборі форм, стійких
до травм у першому бульбовому поколінні. У другому доцільно вико¬
ристовувати методи множинної кореляції і регресії. Для цього розроб¬
лені математичні моделі у вигляді множинних рівнянь регресії з ура¬
хуванням корелюючих чинників (табл.4.19).
Таблиця 4.19 - Математичні моделі для прогнозу динамічної твердості м’якоті
бульб
Група стиглості
дт,
У
Коефіцієнти регресії X змінні в УХ
загальний початок
відліку
суха речовина, Х|
*
Vо
І
VO
8
03
5
товщина кори, х3
форма бульби, Х4
розмір крохмаль¬
них зерен, х5
розмір великих
крохмальних зе¬
рен, х«
вміст великих кро¬
хмальних зерен, х7
R2
1
230
+0,10*1
■+0,005x2
+3,21x3
-1,91x4
+0,19x5
40,13x6
40,07x7
0,529
2
у=
12,05
+0,18*1
40,50x2
49,76x3
-0,01x4
+0,01x5
-038x6
+0,23 х 7
0,966
3
-335
■+031x1
40,007x2
-44,41x4
-0,08x4
-0,08x5
40,06x6
40,03x7
0,609
4
-11,72
+0,14x1
40,070x2
-60,10x3
-031x4
+031x5
40,11x6
4030x7
0,701
Середнє
У*
-6,54
-ЮД5ХІ
+0,004x2
+17,69x3
-0,13x4
40,13x5
40,05x6
-0,02x7
0,253
Примітки: 1 -ранні, 2 - середньоранні, 3 - середньостиглі, 4 - пізньостиглі
Як видно з таблиці, варіювання твердості бульб пов’язане з дією
хімічних властивостей і морфолого-анатомічних ознак на 53 %, по
середньостиглих сортах - на 61, по пізньостиглих - на 70, по серед¬
ньостиглих- на 96 %, у той час як математична модель для прогнозу¬
вання середньої динамічної твердості пояснює тільки 25 % змін цьо¬
го показника від чинників, що беруть у ньому участь.
250
Стійкість до механічних пошкоджень. Селекція картоплі на
стійкість бульб до механічйих пошкоджень багато в чому залежить
від вихідного матеріалу. Правильний підбір пар для схрещування
дозволяє отримувати потомство з високою стійкістю до травмування.
Отже, спрямована селекція може бути ефективною тільки після
попередньої оцінки вихідного матеріалу. Така робота була проведена
в 1978-1980 рр. на 633 сортах світової колекції в лабораторних умо¬
вах із застосуванням приладу типу маятника, а також доповнена ре¬
зультатами оцінки 60 сортів на ПДТ-С у 1986-1992 рр. Більшість
досліджених сортів (86 %) були стійкі до травмування. Однак висо¬
кою стійкістю характеризувалися лише 1,3 % сортів, 14 % сортів бу¬
ли досить стійкими до пошкоджень, понад 70 % мали середню або
відносну стійкість до травмування.
Під час досліджень сортів за їхньою стійкістю до різних видів
травм доведено, що більшість із них були чутливими до обдирання
шкірки (83 %). Бульби майже 80 % сортів темніли від ударів. Поло¬
вина сортів картоплі виявилася нестійкою до тріщин, а 40 % - до ви¬
ривів. Виявлена різна чутливість сортів до видів травм по групах
стиглості (табл. 4.20). Так, кількість виділених сортів, стійких до об¬
дирання шкірки, знижувалася від ранніх сортів до пізніх і складала
відповідно 93 і 87 %. Найбільша кількість (88-83 %) сортів, стійких
до потемніння м’якоті бульб від ударів, була виявлена в ранній і се¬
редньоранній групах; серед середньостиглих і пізніх сортів такими
виявились лише 71-74 %.
Таблиця 4.20 - Розподіл сортів за їхньою стійкістю до видів та характеру
пошкоджень, %
Вид пошкодження
Стійкість,
середня
У тому числі по групах стиглості
1
2
3
4
Подерта шкірка
83,4
93,0
91,1
62,2
87,3
Потемніння м’якоті
79,4
88,2
83,2
71,5
73,8
Колір некрозу
84,9
88,6
84,1
82,9
84,0
Розмір некрозу
73,4
89,7
82,3
60,1
63,5
Тріщини
56,5
58,3
51,0
58,4
58,6
Вириви
40,4
44,6
35,6
38,2
43,5
Примітки: 1 - ранні - 112 сортів; 2 - середньоранні - 165 сортів; 3 - середньо¬
стиглі та середньопізні - 184 сорти; 4 - пізні - 172 сорти
За стійкістю до тріщин з кожної групи стиглості виділено прак¬
тично однакову кількість сортів - 51-58 %, за стійкістю до виривів -
найменшу кількість (35-38 %), особливо в середньоранніх, серед¬
ньостиглих і середньопізніх групах.
Ступінь стійкості сортів колекції до пошкоджень узагальнено в
таблиці 4.21. Середній бал травмованості стійких сортів становив
7,8, перебуваючи в межах 6,4-9,0; нестійких сортів - 4,3 і коливався
в межах 1,9-5,5 балів.
251
Таблиця 4.21 - Стійкість сортів до окремих видів пошкоджень, бал
Вид пошкодження
Групи сортів
стійкі
нестійкі
середнє
коливання
середнє
коливання
Стійкість
7,8
6,4~9,0
4,3
1,9-5,5
У тому числі: подерта шкірка
8,8
7,5-9,0
6,1
1,0-9,0
потемніння м’якоті
7,7
6,0-9,0
4,9
3,0-6,0
тріщини
7,4
5,0-9,0
зд
1,0-6,0
вириви
7,4
6,0-9,0
3,1
1,0-4,0
Високостійкими до всіх видів пошкоджень були сорти Смєна,
Вельяміновська, Центифолія, Азалія, Амзель, Рясна, Електра, Маріє-
лла і Астілла; стійкими до обдирання шкірки - сорти Антінема, Крі-
ста, Калина, Адретта, Грата, Вісла і Яра; високостійкими до виривів
- Морена, Коретта, Луговська, Елла, Ласунак, Українська рожева,
Зарево, Мате, Фреско, Сарма; за стійкістю до тріщин - сорти Юлла,
Розвариста, Білоруська крохмалиста, Вінетта, Верба, Амальфія, Темп
і Юбель; стабільними до потемніння м’якоті бульб - Атцимба, Ли-
пенська, Тарпан, Пригожа 2, Ельвіра, Ректор, Таріста, Аякс, Білору¬
ська рання. Високочутливими до всіх видів травм виявилися сорти'
Принцеса, Бінова, Кастор і Кондор. Дуже нестійкими до виривів
м’якоті є сорти Корона і Біла роза, до тріщин - Гідра і Фріла, до по¬
темніння м’якоті від ударів - Ербіна і Мазур. Інші сорти за стійкістю
до пошкоджень займали проміжне місце.
У селекційний процес по створенню високоякісних сортів, стій¬
ких до механічних пошкоджень, було залучено 60 чоловічих форм, у
тому числі 36 сортів і 24 гібриди. Вихідний матеріал мав форми з
різною стійкістю до пошкоджень. Динамічна твердість м’якоті бульб
високостійких сортів і гібридів становила від 9 до 11,7 шт. уд., і та¬
ких форм було 20 %; понад 33 % - з досить твердою м’якоттю бульб
при коливанні від 8,9 до 7,0; 28 % стійких чоловічих форм мали твер¬
дість м’якоті від 6,9 до 6,0; 18 % сортів і гібридів мали відносну
стійкість до травмування - 5,9-5,0 шт. уд.
Для визначення характеру успадкування динамічної твердості були
взяті як високосгійкі чоловічі форми (11,7), так і відносно стійкі (4). У ви¬
вчених 89 гібридних комбінаціях динамічна твердість м’якоті бульб коли¬
валася від 3,6 до 11,3, що свідчить про тенденцію до зниження твердості
бульб у потомства. Взаємозв’язок цієї ознаки у вихідних форм і їхнього
потомства був невисокий (г=О,327±0Д01, t=3,24>toi=2,60). Однак при ви¬
вченні впливу окремо материнської і чоловічої форм було виявлено, що
твердість м’якоті бульб потомства залежала від них відповідно на 43 % і
на 61,7 %. У зв’язку з цим вивчався характер передачі стійкості за твердіс¬
тю м’якоті бульб потомства від батьків при різних типах схрещування
(табл. 4.22). Як видно з таблиці, твердість м’якоті бульб знижувалася у
формах від першого до четвертого типу схрещування: була більшою за
материнською формою і меншою - за чоловічою.
252
Таблиця 4.22 - Динамічна твердість м’якоті бульб батьків
Тип схрещувань
Батьківські форми
Середнє по
батьках
чоловіча
материнська
Високостійкий X високостійкий
10,41±0,31
6,95±0,25
10,02±0,21
Стійкий X стійкий
7,21±0Д4
7,57±0,35
7,28±0,13
Середньостійкий х середньостійкий
6,04±0,05
5,93±0,04
6,00±0,04
Слабостійкий х слабостійкий
4,97±0,08
4,81±0,13
4,87±0,08
Середнє
7,20±0,28
6,99±0,35
Динамічна твердість м’якоті бульб батьківських форм за типами
схрещувань була вірогідною (F=302,03>Fo5=2,79), а між материнсь¬
кими і чоловічими формами та між середніми значеннями взаємодії
типів схрещувань і їх батьківськими формами - неістотною.
Проведений багатофакторний дисперсійний аналіз дозволив
з’ясувати вплив типів схрещувань, батьківських форм і спільної дії
цих двох чинників на величину показника динамічної твердості
м’якоті бульб потомства. У результаті доведено значний вплив на
твердість бульб потомства типу схрещування (F=7,88 > F05=2,79);
вплив батьківських форм (F=2,33<F05=4,03), взаємодія типів схрещу¬
вання і батьківських форм (F=l,79<Fo5==2,79) були неістотними.
Аналіз значення показників динамічної твердості м’якоті бульб
гібридного потомства загалом виявив їхнє зниження, порівняно із
середнім значенням чоловічих форм. Діапазон розподілу індивідів
потомства за твердістю бульб був значно більшим, ніж у їх батьків¬
ських форм. Було також доведено, що твердість бульб гібридних
комбінацій була найбільшою в першому типі схрещування, най¬
меншою - у четвертому. Так, потомство від схрещування високо-
стійких батьківських форм між собою мало найбільшу твердість
м’якоті - 7,46±0,35. Цей показник у потомстві при схрещуванні між
собою форм із відносною твердістю м’якоті був найменшим і скла¬
дав 5,80±0,28. Твердість м’якоті бульб від схрещування стійкий х
стійкий була нижчою (6,74±0,24) за твердість бульб потомства від
схрещування високостійкий х високостійкий (7,46), але вищою, ніж
твердість бульб потомства (5,8), отриманого від відносно стійких
форм. Значення ж показника твердості бульб потомства від серед-
ньостійких батьківських форм виходило за рамки виявленої тенден¬
ції, що, очевидно, пов’язано з переважаючим впливом батьків над
типом схрещування. У середньому ж у комбінаціях схрещувань, де
брали участь високостійкі батьківські форми, частка таких гібридів
становила не менше 40 %.
Сказане вище є загальною тенденцією в характері передачі по¬
томству ознаки твердості м’якоті бульб за типом схрещувань. Сту¬
пінь цієї передачі батьківськими формами з додатковими проміжни¬
ми типами схрещувань відображено в таблиці 4.23.
253
Таблиця 4.23 - Залежність динамічної твердості бульб потомства від типу
схрещування і батьківських форм
Тип схрещування
Твердість м’якоті,
шт.удар. /200 г
Кореляційне відно¬
шення
Рівняння регресії,
у = а + Ьх + Ьх2
мати
бать¬
ко
потом¬
ство
t!±Sti
t
Високостійкий х
високостійкий
10,8
10,6
8,1
0,662 0,094
7,01
-21,0+4,86х-0,2х2
Високостійкий X
стійкий
10,7
8,4
7,7
0,666 0,093
7,11
12,3+5,0х-0,3х2
Середньостійкий
X високостійкий
6,3
9,0
7,7
0,637 0,091
7,00
-50,4+8,5х+0,1х2
Стійкий х серед¬
ньостійкий
8,0
6,0
7,4
0,502 0,074
6,82
84,1-23,1х+1,7х2
Середньостійкий
х середньостійкий
6,5
6,3
6,7
0,626 0,090
6,93
65,8-21,1х+2,0х2
Середньостійкий
X стійкий
6,2
8,4
6,4
0,647 0,092
6,99
/
/
-5,8+4,6х-0,4х2
Низькості ЙКИЙ X
високостійкий
5,0
9,3
6,7
0,252 0,061
4,13
-88,5+48,6х-5,0х2
Низькості ЙКИЙ X
низькостійкий
5,2
5,0
5,5
0,764 0,089
8,56
-111,6+48,6х-5,0х2
Виявлено, що залежність твердості бульб потомства від типу
схрещування і чоловічих форм коливалась у межах 25-76 %. При
цьому найвище кореляційне відношення між твердістю бульб батьків
і потомства виявлене при схрещуванні відносно стійкий * відносно
стійкий; високе кореляційне відношення - при схрещуванні між со¬
бою високостійких форм, високостійких і стійких. Отже, при пере¬
дачі потомству в цих типах схрещування ознака динамічної твердо¬
сті м’якоті бульб була від’ємною.
Твердість бульб потомства при схрещуванні відносно стійкий х ви-
сокостійкий від чоловічих форм залежала лише на 25 %. Потомство
при інших типах схрещування незначною мірою визначалося твердіс¬
тю бульб батьків. Отримані дані дозволяють зробити висновок про
те, що при відповідному підборі пар для схрещування можна отри¬
мувати потомство з високою твердістю м’якоті бульб. Розроблені
математичні моделі дають можливість за кожним типом схрещуван¬
ня прогнозувати значення твердості бульб потомства при певних
значеннях цього показника у батьківських формах.
Відібрано кращі батьківські пари для одержання потомства з ду¬
же твердою м’якоттю (17-11 шт. удар./200 г) - Темпора х 265-14N,
Кера х 305-27N, Супер х 305-27N, Кера х 4965-5, Лізера х 78573-
35N, Хейдрум х Ельзіна; із стійкою до виривів і тріщин м’якоттю
(10-8) - Сарма х Зарево, Г244 х Воловецька, Ельвіра х Білоруська З,
61.2107018 - 80N х 4938 - 45, Прем’єр х Ласунак, Кера х Світязянка,
Аркула х 305-27N, 61.2107018 - 80N х 3430-10, 1.198129-77N+S х
254
4952-20, Ельда х 1.37031-78N, Морена х Ласунак, Морена х Зарево,
Супер х Білоруська 3, 61.2107018 - 80N х Ласунак.
З 92 виділених у селекційних розсадниках стійких до пошкодження
гібридів практичний інтерес являли 28, у тому числі 32 % - високостійкі,
39 - досить стійкі і 29 % - стійкі. Велика частина цих гібридів зберігала
стійкість до пошкоджень, незалежно від умов вирощування.
Розроблені математичні моделі для прогнозування видів травм
наведено в таблиці 4.24.
Таблиця 4.24 - Математичні моделі прогнозування видів травм у картоплі
Ввди травм
Коефіцієнти регресії X змінні в YX
загальний
початок відліку, а
питома
вага, Хі
форма,
Х2
розмір,
Х3
шкір-
ка, Х4
R
Загальні
-380,20
+333,93
+13,61
+0,51
+0,47
0,685
Легкі
-296,23
+267,39
+ 13,03
+0,001
-0,16
0,645
Тяжкі
183,01
-170,86
-2,77
+0,10
+1,85
0,444
Вириви
50,02
-46,22
-0,52
+0,04
+0,19
0,286
Тріщини
14,08
-12,08
-2,81
+0,04
+0,51
0,415
Потемніння
-315,82
+282,01
+1,25
+0,39
-0,30
0,663
З використанням цих моделей було спрогнозовано рівень травмова¬
ності 14 сортів і гібридів картоплі, які оцінювались у 1995-1998 рр. у
розсаднику конкурсного сортовипробування. Максимальна загальна
травмованість бульб при механізованому вирощуванні досягала в серед¬
ньому 29 %, у тому числі за рахунок легких травм і потемніння - по 9 %,
тяжких травм - 7 %, виривів та тріщин - по 2 % (рис.4.1).
ЗО
*
ю
28
2J
29
29
29
ЗС
31
31
31
И3І
26
~ 77
"27
11
11
9
10
11
9
10
10
10
11
10
10.
ль
Сл\\
2
&
"Заплигі *
•Легкі -
S
U
•Потемнінні4
Tnad “■Ж"1 ■ Ьнрттн • О ■ Тріщиш
Рис. 4.1. Прогнозний максимальний вміст травмованих бульб
у сортах і гібридах картоплі
255
Доведено, що легких травм можна позбутися додержанням опти¬
мальних строків садіння і зберігання картоплі; тяжкі травми зале¬
жать від технічних факторів, тобто їх також можна виключити або
запобігти; оптимальна сукупність агротехнічних, технологічних і
технічних факторів практично може виключити потемніння бульб
від ударів. Останні травми у вигляді виривів та тріщин - це здебіль¬
шого прояв сортової ознаки. За цими травмами досліджені зразки
повністю можуть відповідати вимогам стандарту щодо продовольчої
картоплі. При нормі не більше 5 % таких травм зразки їх максималь¬
но можуть містити від 2 до 4 %, у тому числі сорт Злагода - 2, Гіб¬
рид 5 і сорт Ластівка -3, усі інші сорти і гібриди - по 4 %.
Підсумки сучасної селекції сортів картоплі, стійких до механіч¬
них пошкоджень. Світова практика картоплярства свідчить про постій¬
не зростання уваги до сортів, придатних до механізованого вирощуван¬
ня, збирання, транспортування, закладання на зберігання і затарювання.
Тому каталоги містять багато таких сортів (табл.4.25). і
Таблиця 4.25 - Стійкість сучасних сортів картоплі до механічних пошкоджень
Сорт
Потемніння, балів
9
8
7
6
5
4
Сортимент України
Божедар, Бородянська рожева, Водограй, Воловецька, Гор¬
лиця, Доброчин, Зов, Ластівка, Либідь, Луговська, Мавка,
Молодіжна, Пост 86, Првдеснянська, Світанок київський,
Українська рожева
+
Гібридна 14, Древлянка, Кобза, Незабудка, Пролісок, Седнів-
ська рання
+
Зарево
+
Сортимент Білорусі
Ласунак
+
Аксамит, Альтаїр, Атлант, Білоруська 3, Верас, Верба, Лазу-
ріт, Лошицька, Пригожа 2, Росінка, Сузор'є, Темп, Яхант
+
Архідея, Виток, Гранат, Дельфін, Діна, Добро, Живіця, На-
роч, Синтез, Скарб, Миловиця
+
Альпініст, Білоруська рання, Гарант, Огоньок, Орбіта
+
Сортимент Росії
Аспія, Бронницька, Резерв
+
Акросія, Жуковська рання, Заворовська, Москворецька, Нев¬
ська, Осінь, Прайса, Ресурс, Роднік, Скороплідна, Удача
+
Білоусівська, Брянська рання, Волжанін, Лук'янівська
+
Бежицька, Голубізна, Ефект
+
Сортимент Німеччини
Анеллі, Карат, Ханса
+
Альвара, Коретга, Ліу
+
Агаве, Адретта, Аркула, Бербер, Дезіре, Івера, Корал, Кара-
топ, Карлена, Каролін, Лікарія, Моллі, Кама
+
Делікат, Жаклін, Карата, Рікеа, Серафима
+
Феліцитас, Боденкрафт
+
256
Так, наприклад, у сортименті України майже 90% сортів високо-
стійкі до механічних пошкоджень, сортименті Білорусі - 80%, Росії -
70% (9-7 балів); натомість у Німеччині високостійких сортів близько
20%, а середньостійких - 70% (6-5 балів).
ЛІТЕРАТУРА
1. Власюк П.А., Власенко Н.Е., Мицко В.Н. Химический состав картофе¬
ля и пути улучшения его качества.- К.: Наук, думка, 1979.- 194 с.
2. Гончаров Н.Д. Оценка исходного материала картофеля при селекции
интенсивного типа сортов // Картофелеводство и плодоовощеводство: Науч.
тр. БНИИКПО.- Минск, 1976.- Вып.1.- С.3-7.
3. Гончаров Н.Д., Кожушко Н.С. Оценка пригодности исходного и селек¬
ционного материала картофеля к механизированной уборке и хранению //
Пути повышения продуктивности и качества с.-х. продукции в Сумской об¬
ласти: Материалы обл. науч.-практ. конф., 1990.- С.72-81.
4. Гончаров Н.Д., Кожушко Н.С. Оценка селекционного материала кар¬
тофеля на устойчивость к механическим повреждениям в интенсивных
условиях выращивания // Картофелеводство: Сб. науч. тр. / БНИИКПО.-
Минск, 1978.- Вып.З- С.9-11.
5. Гончаров М.Д., Кожушко Н.С. Підвищення продуктивності та якості
картоплі селекційними методами - Суми: Сумський СП, 1992.- 126 с.
6. Гончаров Н.Д., Кожушко Н.С. Травмируемость и лежкость клубней
при механизированной уборке картофеля // Картофелеводство и плодоово-
щеводство: Науч. тр. / БНИИКПО.- Минск, 1979.- Вып. 4 - С.41-48.
7. Довідник по зберіганню картоплі та овочів / С.Ф.Поліщук, М.М.Івакін,
Б.П.Федорець та ін.; За ред. С.Ф.Поліщука - К.: Урожай, 1986.- 280 с.
8. Картофелеводство за рубежом / В.Н.Киселев, В.И.Назаренко, И.П. Со¬
ломина и др. - М.: ВНИИТЭИ АПК, 1990- С.78-120.
9. Кожушко Н.С. Влияние видов механических повреждений на лежкость
картофеля // Картофелеводство и плодоовощеводство: Науч.тр. /БНИИКПО .-
Минск, 1976- Вып.1- С.60-63.
10. Кожушко Н.С., Кравченко И.В. Влияние травмированных клубней на
урожай картофеля // Картофель и овощи - 1976.- № 10 - С. 10.
11. Кожушко Н.С., Михальчик B.T. Хранение травмированных клубней //
Картофель и овощи.- 1975.- № 11- С. 14.
4.5.4. Селекція картоплі на стійкість (імунітет)
до хвороб і шкідників
Щорічні втрати врожаю у картоплярстві, що спричиняються хво¬
робами і шкідниками, становлять близько 20 %. Розмноження картоп¬
лі бульбами сприяє інтенсивній передачі збудників захворювань від
одного вегетаційного покоління іншому. Велика кількість різних па-
тогенів викликає необхідність виділити найважливіші з них, відповід¬
но до частоти появи, шкодочинності, можливостей хімічного захис¬
ту, здатності передаватися наступному поколінню, економічного
значення і витрат на селекційну роботу, спрямовану на створення
відповідної стійкості.
9 2-132
257
В Україні до найбільш поширених і шкідливих паразитів нале¬
жать фітофтороз, рак картоплі, ризоктонія, парша звичайна, кільцева
і мокра гниль, вірусні хвороби, картопляна і стеблова нематоди. Ви¬
рішальне значення в боротьбі з ними має створення стійких сортів
картоплі.
Один із засновників вчення про імунітет рослин М.І.Вавилов
вперше довів, що імунітет тісно пов’язаний із генетичною природою
рослин. Згідно з його теорією, стійкі форми слід шукати на території
співіснування рослини-господаря і паразита.
Розширення генетичної бази в новостворюваних сортах картоплі,
інтрогресія ефективних генів контролю стійкості до паразитів мож¬
ливі лише при широкому залученні в селекційну роботу культурних і
диких видів картоплі.
Важливим і актуальним завданням сучасної селекції картоплі є
поєднання в одному сорті стійкості до кількох захворювань. Труд¬
нощі цього завдання зумовлюються диференціацією видів паразитів
на численні патотипи, які виникають у різних умовах вирощування
картоплі.
Стійкість до хвороб і шкідників повинна поєднуватись із комплек¬
сом інших господарсько цінних ознак, зокрема з продуктивністю,
високими якісними показниками та ін.
Фітофтороз. Збудником хвороби є гриб Phytophthora infestans
de Вагу, який уражує практично всі органи рослини: листя, стебла,
квітки, ягоди, столони і бульби. Найбільша шкодочинність захворю¬
вання спостерігається в регіонах з помірною температурою та части¬
ми опадами протягом вегетаційного періоду.
Існує два типи стійкості до фітофторозу: надчутливість (вертикаль¬
на чи специфічна) і польова (горизонтальна чи загальна) стійкість.
Генотипи з надчутливістю характеризуються некротичною реакцією
заражених і сусідніх клітин. Надчутливість рослин зумовлюється
дією основних, незалежних генів, кожен із яких активізується пев-
ною расою гриба. Польова стійкість - це комплекс певних факторів,
що запобігають ураженню рослини патогеном, а її успадкування від¬
бувається завдяки дії малих генів. Обидва типи стійкості проявля¬
ються сумісно в диких видів картоплі S.demissum, S.bulbocastanum,
S.poliadenium, S.stoloniferum, S.vernei, S.verrucosum, у той час як не-
гібридизовані чисті форми S. tuberosum мають лише малі гени польо¬
вої стійкості.
Перші дослідження із селекції картоплі на стійкість до фіто¬
фтори були розпочаті в середині XIX ст. у зв’язку з катастрофіч¬
ною епіфітотією в Ірландії та швидким її поширенням по всій Захі¬
дній Європі.
Реальна перспектива створення фітофторостійких сортів картоплі
з’явилась із включенням у схрещування мексиканського гексаплоїд-
ного виду S.demissum, який, несучи головні гени надчутливості, за¬
258
безпечував повний захист гібридів-бекросів S.demissum х ssp.tube¬
rosum, але тільки в перші роки їх вирощування. Зазнаючи тиску генів
надчутливості, включених у гібриди картоплі, збудник починає фор¬
мувати нові раси. Так, уже в 1932 році в Німеччині було виявлено
сильне ураження гібридів, створених за участю S.demissum, які до
цього вважалися стійкими.
Численні дослідження, проведені в різних країнах, привели до
створення міжнародної системи “ген на ген”, у межах якої домінант¬
ні R-гени рослини-господаря відповідають певним генам вірулентно¬
сті патогену. Наприклад, генотип Rj уражується расою 1 або більш
складними - 1.2; 1.3; 1.2.3 і т.д., але не расами 2; 2.3; 3.4. Сорт кар¬
топлі з R-генами чи їх комбінацією залишається стійким доти, поки
не з’явиться спільна раса. На сьогодні відомо 11 R-генів.
У багатьох країнах (Німеччина, США, Великобританія, колишній
Радянський Союз) на основі використання виду S.demissum було
створено велику кількість сортів картоплі з R-генами: у Росії - Фіто¬
фторостійка 8670, Камераз, Столова 19, Гатчинська, Невська, в Біло¬
русії - Темп; в Україні - Поліська рожева, Зарево, Гібридна 14, Мав¬
ка; у Німеччині - Анко, Віргінія, Вертифолія, Пантер, Фортуна; у
США - Кенебек, Черокі, Емпайр; Великобританії - Оріон, Пентланд-
ас; у Польщі - Вулкан, Лава, Епока.
Проте селекція картоплі на надчутливість не вирішила проблему
фітофторозу. Наявність R-гена або їх комбінації в генотипі сорту
стимулює добір відповідних рас гриба. Недостатня їх роль особливо
помітна при порівнянні ступеня стійкості сортів з R-генами і без них.
Так, 64 % сортів із R-генами віднесено до групи з оцінкою від 1 бала
(висока) до 5 (середня). До цієї ж групи належить майже така ж кіль¬
кість сортів (62 %) без R-генів [3]. У зв’язку з цим в останні роки все
більше уваги приділяється польовій стійкості. Польова стійкість є
результатом дії кількох захисних механізмів: тривалого інкубаційно¬
го періоду, повільного поширення інфекції на рослині, зменшення
споруляції. У рослин із високою польовою стійкістю ураження на¬
стає значно пізніше і відбувається повільніше; спороутворення гриба
гальмується. Цей тип стійкості однаковою мірою захищає від усіх
рас фітофтори, проте в міру розвитку захворювання рослини почи¬
нають уражуватися з різною інтенсивністю.
На основі вивчення майже 500 сортів світового сортименту в ко¬
лишній Чехословаччині було виявлено, що близько 20 % з них тією
чи іншою мірою виділяються польовою стійкістю (Епока, Олев, Еве¬
рест, Фальке, Херкула, Сусанна, Уральська, Віргінія та ін.). Причо¬
му, у переважній більшості із них міжвидові гібриди отримані від
схрещування з дикими видами.
Дослідження, проведені І.М.Яшиною (1968), показали, що для
форм з R-генами і без них успадкування польової стійкості відбува¬
ється за одним типом і зумовлюється серією полімерних генів. У всіх
259
проаналізованих комбінаціях виділено гібриди, які перевищують на
2-2,5 бали стійкість батьківських компонентів.
Надаючи великого значення польовій стійкості, не слід недооці¬
нювати важливість поєднання її з R-генами в новостворюваних сор¬
тів картоплі. Без сумніву, польову стійкість значно частіше мають
рослини, які несуть R-гени.
Крім стійкості картоплиння до фітофторозу, важливе значення,
особливо в останній час, надається стійкості бульб. Як і для бадилля,
селекція на стійкість бульб базується на польовій стійкості, яка кон¬
тролюється малими генами.
Успадкування стійкості бульб у польових умовах вивчав Токсо-
пеус (Toxopeus H.J., 1961). Так, у потомстві від схрещування стійких
вихідних форм майже всі гібриди Fi мали стійкі чи слабоуражувані
бульби. При схрещуванні стійких і сприйнятливих форм виявлено
невелику кількість гібридів із сприйнятливими бульбами, але основ¬
на частина належала до стійких. Схрещування сприйнятливих форм
призводило до втцеплення лише одиноких високостійких за буль¬
бами генотипів. Отже, гібриди із стійкими до фітофтори бульбами
можна одержати при використанні будь-якого вихідного матеріалу,
але частота їх появи буде прямо пропорційною стійкості батьківсь¬
ких форм.
Узагальнюючи дані багатьох досліджень, при селекції на стій¬
кість до фітофтори доцільно використовувати як одну з батьківських
форм такі високостійкі до збудника сорти, як Гібридна 14, Прикар¬
патська, Карпатська, Літня Ш, Чепецька, Мімоза, Поліська рожева,
Ясень, Соната, Зубрьонок, Луговська, Бізон, Барака, Вулкан, Гунда,
Сафір, Сента, Атцимба, Флорита та ін.
Із досить значної кількості вихідного матеріалу в селекції на фі-
тофторостійкість велике значення мають складні міжвидові гібриди
картоплі, створені в лабораторії вихідного матеріалу Інституту кар¬
топлярства УААН. Причому, вони характеризуються наявністю ін¬
ших ознак, ступінь прояву яких значно вищий, ніж у сортів-
стандартів.
Результативність селекції на фітофторостійкість певною мірою
залежить і від досконалості методів оцінки.
Розроблено декілька методів оцінки польової стійкості бадилля і
бульб картоплі.
Візуальна оцінка в польових умовах на природному інфекційному
фоні. Облік проводять 3-4 рази, визначаючи середній і найвищий
бали ураження.
Лабораторно-польовий метод оцінки польової стійкості картоплі до
фітофтори з використанням мікрокамер. Порівняння індексів уражен¬
ня сортів картоплі, розрахованих на основі результатів обліку, із багато¬
річними характеристиками ураження фітофторою в польових умовах
підтверджує високу достовірність цього методу.
260
Відсутність чіткої кореляції між стійкістю бадилля і бульб кар¬
топлі потребує більше уваги в селекційній роботі до польової стійко¬
сті бульб. В Інституті картоплярства УААН протягом багатьох років
використовують методику випробування фітофторостійкості бульб
картоплі з використанням зараження суспензією зооспор цілих (не-
травмованих) бульб. Результати цих оцінок доповнюються випробу¬
ванням (до 1991 р.) на Сахаліні, де спостерігається відносно високий
і постійний тиск інфекційного фону і досить широкий спектр рас.
Отже, незважаючи на труднощі, які виникають при селекції кар¬
топлі на стійкість до фітофторозу,4 особливо при появі високовірулен-
тних та агресивних рас гриба, створення високостійких сортів є ціл¬
ком реальною справою. Для цього в арсеналі селекціонерів є різно¬
манітний вихідний матеріал з ефективним генетичним контролем
цієї ознаки та досконалими методами оцінки.
Рак картоплі. Рак картоплі, який викликається грибом Synchyt-
rium endobioticum (<Schilb), є одним із найбільш небезпечних каран¬
тинних захворювань, яке має досить широкий ареал в Україні, особ¬
ливо на присадибних ділянках.
Уже на початку XX ст. були виявлені стійкі до збудника сорти
картоплі, які залучались у схрещування (Дабер, Юбель та ін.). У
1941 р. проявилась епіфітотія раку картоплі, спричинивши появу аг¬
ресивних рас збудника й уразивши стійкі до того часу сорти. Тому в
багатьох європейських країнах було прийнято законодавчі акти, що
забороняли вирощування сортів картоплі, нестійких до раку.
У 1961-1962 рр. В.И.Яковлева [4] в гірськокарпатській зоні Украї¬
ни вперше виявила агресивні раси - міжгірську, рахівську і бис-
трицьку, а дещо пізніше (1976р.) - ясинську.
На сьогодні в різних країнах світу створено значну кількість ра¬
костійких до звичайного патогену сортів картоплі. Відомі також сор¬
ти картоплі, стійкі до кількох патотипів (Арго, Бланік, Ора, Сафір,
Тондра, Ультимус, Фонтана, Фортуна, Фрам, Хілла та Цайсиг). У
селекції картоплі найбільший інтерес викликають батьківські ком¬
поненти різного походження, які добре передають гібридному по¬
томству ракостійкість у поєднанні з іншими корисними ознаками.
На думку Мариса [7], стійкість до раку картоплі контролюється
одним домінантним геном, у той час як три комплементарні гени-
супресори обмежують дію першого. Цим можна пояснити появу
стійких генотипів у потомстві нестійких батьків. Для практичної се¬
лекції кращими джерелами стійкості до звичайного патотипу раку є
сорти Таборки, Дорідна, Прієкульська рання, Катюша, Олев, Агро¬
номічна, Камераз, Гіндебург, Лех, Катадін, які дають у потомстві від
самозапилення до 90-95 %, а при схрещуванні з нестійкими форма¬
ми - близько 80 % стійких рослин.
У селекції на стійкість до агресивних патогенів слід використо¬
вувати сорти Ора, Аполло, Хілла, Пандора, Тондра, Беті і Цейсиг, які
261
не уражуються звичайним і агресивним патотипами - міжгірським і
рахівським.
Дослідження, проведені В.І.Яковлевою і Л.П.Салтиковою (1966),
показали, що серед південноамериканських диких і культурних видів
картоплі можна виділити значну кількість форм, стійких до агресив¬
них патотипів раку картоплі. Так, зокрема, до міжгірського патотипу
стійкими виявилися окремі зразки таких видів: S.acaule, S.cardi-
ophyllum, S.pinnatisectum, S.vernei, S.chacoense, S.commersonii,
S.stoloniferum. Особливо перспективними у селекції на стійкість до
агресивних патотипів є різні форми S.andigenum. За даними тих же
авторів, із 365 зразків S.andigenum 74 % не уражувались міжгірським
патотипом. Деякі зразки були одночасно стійкими і до рахівського
біотипу.
В Україні випробування селекційного матеріалу картоплі на стій¬
кість до звичайного та агресивних патотипів раку проводиться лабора¬
торно-вегетаційним та польовим методами в Українській науково-
дослідній станції карантину рослин (с.Бояни Чернівецької області).
Отже, шляхом виведення стійких сортів картоплі можна обмежи¬
ти поширення і шкодочинність такого небезпечного захворювання,
як рак.
Ризоктоніоз. Захворювання викликається грибом Risoctonia
solami Kuhn. Базидіальна стадія Hupochus solami Prill утворюється на
уражених стеблах картоплі в середині літа під час вологої і теплої
погоди і свідчить про інтенсивний розвиток хвороби на підземних
органах рослин. Збудником ризоктоніозу є гетерокаріон, який у при¬
родній популяції складається з різних штамів.
Гриб уражує коріння, столони і молоді паростки, внаслідок чого
затримується поява сходів, рослини відстають у рості й утворюють
багато дрібних і деформованих бульб.
Нечисленні дослідження, проведені в різних країнах як у лабора¬
торних, так і польових: дослідах, не виявили достатньої відмінності у
стійкості сортів і диких видів до ризоктоніозу. Менш сприйнятливи¬
ми були сорти картоплі Юбель, Мажестік, Вольтман, Амзель, Фрю-
ка, Марієла, Форветс, Гієвонт, Меркур, рльба, Прокура та Спартан.
На наш погляд, зважаючи на те, щб стійкість до ризоктоніозу зу¬
мовлена дією багатьох факторів (різною здатністю сортів до віднов¬
лення коріння, еталонів і паростків; швидким розвитком на початко¬
вих фазах росту) спершу необхідно вивчити ці фактори, характер їх
успадкування і лише тоді розпочати селекцію на стійкість до ризок¬
тоніозу.
Суха (фузаріозна) гниль. Збудником її є гриб Fusarium var
coeruleum (iSacc.), головним чином рановий патоген. Захворювання
проявляється під час зимового зберігання картоплі. Оскільки інфек¬
ція залежить від наявності механічних пошкоджень на бульбах, то
стійкість до них значною мірою забезпечує захист від сухої гнилі.
262
Щік і Хопфе [8], узагальнивши дані багатьох досліджень, вказують
на велику кількість сортів картоплі з відносною стійкістю до
F. coeruleum - Арран банер, Епікур, Форан, Кінг, Едвард, Кенебек та ін.
Хермсен і Фенденіус повідомляють, що з 200 випробуваних форм
S.cmdigenum 72 були стійкими, а з 4 форм S.stenotonum - лише одна.
Серед складних міжвидових гібридів картоплі, створених в Ін¬
ституті картоплярства УААН за участю S.acaule, S.bulbocastanum,
S.demissum, S.phurejza i S.cmdigenum, виявлено окремі з них, що по¬
єднують високу стійкість до фітофтори на бадиллі і бульбах (8-9 ба¬
лів) і сухої гнилі (6-7 балів).
Мокра гниль і чорна ніжка. Збудником, який викликає ці захво¬
рювання картоплі, є бактерія Erwinia caratovora ssp. atroseptica.
У грунтовній праці Хана і Шюллера при оцінці стійкості сортів
картоплі до мокрої гнилі виявлена позитивна кореляція між проявом
мокрої гнилі на бульбах і чорною ніжкою на стеблах.
У 100 вивчених сортів найбільш стійкими були Сафір і Зікінген.
Учені вважають, що оцінки бульб за стійкістю лише до мокрої гнилі
достатньо, оскільки чорна ніжка розвивається услід за нею.
Лельбах [6] довів, що стійкість до мокрої гнилі контролюється
малими генами, проте вплив факторів навколишнього середовища
настільки великий, що сам він упевнений в успіхах селекції за цією
ознакою.
За повідомленням Т.В.Тимошенко (1994), серед сортів вітчизня¬
ного та зарубіжного походження виділено ряд зразків, які поєднують
стійкість до мокрої гнилі і фітофторозу і широко залучаються у
схрещування. Це сорти Ракурс, Либідь, Українська рожева, Брянець,
Лібелла, Санте та ін.
Кільцева гниль. Збудником захворювання є Corynebacterium
sepedonicum (S.et В). Стійкість до кільцевої гнилі успадковується
домінантно. У потомстві від самозапилення стійких сортів перева¬
жають стійкі форми, які складають від 55 до 85 %. Аналіз родоводу
ряду стійких форм (Вольтман, Мерімек, Пауль Крюгер) показує, що
вони походять від первинного схрещування Дабер х Ерсте фон Фре-
метдорф.
Щорічне вивчення стійкості перспективних гібридів картоплі при
штучному зараженні збудником кільцевої гнилі на Поліській дослід¬
ній станції ім. О. Засухіна дозволило виділити форми з відносною
стійкістю до захворювання (6-7 балів). Цінність їх у тому, що вони
поєднують стійкість бадилля і бульб картоплі до фітофторозу, парші
звичайної, стеблової нематоди та іржавої плямистості бульб.
Звичайна париш. Збудником парші звичайної є променисті гриби,
або актиноміцети Streptomyces scabies (Tяке) Wakstan et Henrice та ін.
Відомо, що в боротьбі із звичайною паршею найбільш радикаль¬
ним заходом є вирощування стійких сортів картоплі. Уже на початку
цього століття в Німеччині було створено стійкий до парші і рако¬
263
стійкий сорт Юбель. Беркнер уперше довів, що сорт Юбель є вихід¬
ним пунктом у селекції картоплі на стійкість до парші звичайної і
більшість резистентних до захворювання сортів мають його у своєму
походженні (Арніка, Гінденбург, Пепо, Пауль Вагнер та ін.).
Найбільший інтерес для використання в селекції мають сорти
картоплі, гомозиготні за стійкістю до парші, наприклад Гінденбург і
Острагіс. При схрещуванні цих сортів між собою можна отримати
досить стійке потомство, а при схрещуванні з нестійкими формами -
дещо меншу кількість стійких гібридів. З інших стійких до парші
сортів заслуговують на увагу Акерзеген і Юбель. Останній із них
гетерозиготний, оскільки дає при схрещуванні дуже розщеплене по¬
томство за стійкістю до збудника парші [1]. Наявні в даний час мате¬
ріали з вивчення генетики стійкості картоплі до парші звичайної вка¬
зують лише на те, що ця ознака успадковується домінантно.
А.Я.Камераз (1967), узагальнивши результати багатьох досліджень,
виділяє досить велику групу сортів картоплі, які придатні для викорис¬
тання в селекції на стійкість до актиномікозу (Акерзеген, Апта, Гінден¬
бург, Кардинал, Карнеа, Котнов, Таненберг, Юбель та ін).
У результаті випробування великої кількості вітчизняних і зару¬
біжних сортів картоплі на Поліській дослідній станції виділено гру¬
пу високостійких. Це, насамперед, Житомирянка, Новинка, Дарунок
поля, Полісянка, Поліська рожева, Промінь, Сафір, Фріла, Прикар¬
патська і Роза, які поєднують стійкість до парші з комплексом інших
господарсько цінних ознак і широко залучені в селекційну роботу.
Для селекції на стійкість до парші звичайної може бути залуче¬
ний різноманітний вихідний матеріал з деяких видів картоплі, перш
за все із серії Glabrescentia і Commersoniana.
Оцінка стійкості гібридного матеріалу проводиться на природ¬
ному інфекційному фоні. З цією метою використовують, як правило,
ділянку поля з pH грунтового розчину 6,8-7,0, створюючи тим самим
сприятливі умови для розвитку актиноміцетів. Крім того, інфекцій¬
ний фон створюється тривалою монокультурою картоплі, щорічним
внесенням свіжого соломистого гною і періодичним вапнуванням.
Селекція на стійкість до вірусних хвороб. У селекційній роботі
важливе місце належить виведенню сортів картоплі, стійких до вірус¬
них хвороб. Складність цієї проблеми пов’язана з наявністю багатьох
вірусних захворювань, труднощами виділення форм із комплексною
стійкістю до основних вірусів, а також із численними штамами, які
викликають різні симптоми на рослинах.
Найбільшу загрозу для картоплі в Україні являють віруси X, Y, S,
М та L, а також поєднання їх при змішаній інфекції.
У селекції на стійкість до вірусних хвороб вирішальне значення
має правильний підбір вихідного матеріалу, що грунтується на знан¬
ні генетичної природи цього типу стійкості та закономірностей його
успадкування.
264
У картоплі за реакцією на інфікування вірусами розрізняють чо¬
тири основні типи стійкості.
1. Імунітет, тобто найбільш високий ступінь стійкості. У рослин
із таким ступенем стійкості розмноження вірусу після його проник¬
нення гальмується настільки сильно, що не з’являються зовнішні
симптоми захворювання, і вірус не може бути виділений діагностич¬
ними методами.
2. Надчутливість - це некротична реакція рослин у відповідь на
проникнення вірусу, яка забезпечує повний захист і при якій на рос¬
линах у полі ознаки ураження зовсім не з’являються.
Х.Росс (1989) виділяє три різні типи надчутливості: локальну
надчутливість, крайню стійкість, системну надчутливість. При локаль¬
ній надчутливості розвивається некротизація верхівки пагона. Край¬
ня стійкість - це посилена локалізована надчутливість. Обидва типи
можна розрізнити за щепленням на уражений вірусом помідор. При
крайній стійкості симптоми зовсім не проявляються або на верхніх
листках розвивається крапчастий некроз. При обох типах стійкості
серологічні тести дають негативні результати.
Третій тип некротичної стійкості - системна надчутливість; вона
характеризується некротизацією більшої частини рослини при зара¬
женні в польових умовах.
3. Польова стійкість чи стійкість до зараження, як основна за¬
хисна реакція, викликається кожним вірусом і проявляється в кож¬
ному сорті, але різною мірою.
За повідомленням Баркера, сорти з польовою стійкістю містять мен¬
ше вірусів, ніж нестійкі, а також менше заселяються інфекційною тлею.
Порівняно з надчутливим типом стійкості, значення польової стійкості
до вірусних хвороб значно менше, і вони забезпечують достатній захист
лише при відносно нижчому інфекційному фоні.
4. Толерантність — це особливий ступінь терпимості до вірусів,
коли рослина уражена, але не знижує продуктивності і не проявляє
видимих ознак захворювання, будучи, однак, постійним резервато-
ром вірусу в латентній формі.
Вірус X. Селекція картоплі на стійкість до вірусу X базується на
таких її типах, як стійкість до зараження, локалізована надчутли¬
вість, крайня стійкість.
За класифікацією Х.Росса (1989), ген Rx контролює імунітет до
всіх штамів вірусу X, а ген Rxn - також надчутливість до всіх шта¬
мів. Ген Rx від чилійського сорту Villaroela через сіянець USDA
41956 було введено в американські сорти картоплі Атлангик, Шомо-
ні, Карлтон, Феліансе, Сако і Тава. У потомстві сіянця СПС 1673
S. andigenum було виділено ген Rxadg, який, крім того, стійкий до
двох патотипів картопляної нематоди.
Вірсема [10] довів, що стійкість контролюється одним домінант¬
ним геном й успадковується тетрасомічно. Цей зразок широко вико-
265
ристаний селекціонерами Голландії. Він поєднав стійкість до вірусу
X і картопляної нематоди (сорти Амарил, Превалент, Промінент).
Ген Rx від S. асаиіе включено в сорти картоплі Барбара, Мокі, Ната¬
лі і Сафір.
Надчутливість до вірусу X виявлена також у багатьох інших ди¬
ких видів: S.curtilobum, S.chacoense, S.juzepczukii та ін.
Польовою стійкістю до вірусу X відзначаються сорти Асока, Го-
ліаф, Веві, Леандер, Ехуд, Едзіна та Ердкрафт.
Вірус К Є одним з найбільш поширених і шкодочинних вірусів,
який викливає смугасту і зморшкувату мозаїку. Найбільш шкодо-
чинною зморшкувата мозаїка є при одночасному зараженні вірусами
X і Y. Наявність різних штамів вірусу значно ускладнює селекційну
роботу.
Селекція на стійкість до вірусу Y головним чином базується на
стійкості до зараження і надчутливості. В окремих сортів і деяких
гібридів картоплі, отриманих за участю S. phyreja і S. stenotonum, ви¬
сока стійкість до зараження пов’язана з некротичною реакцією, по¬
дібною до системної надчутливості. Така стійкість визначається ефек¬
том малих генів.
У селекції на некрогенну стійкість використовують основні гени N
та R. Гєни N керують некрогенною реакцією, що займає проміжне місце
між локалізованою і системною надчутливістю. Гени Ry контролюють
краще високу стійкість, забезпечуюючи повний захист рослин.
Висока стійкість до вірусу Y вперше була виявлена в дикого виду
S.stoloniferum, а характер її успадкування Росс [3] визначив як моно¬
мерно домінантний. Окремі генотипи з Ry проявляють найвищу
стійкість і до вірусу А, який близький до вірусу Y.
З нових сортів картоплі, які містять ген Ry, слід відзначити такі:
Коріне, Санте, Бобр, Піліца, Сан, Барбара, Бізон, Кардіа, Еста, Фа-
нал, Франзі, Пірола, Вега, Ресурс, Брда, Бзура.
Вірус L. Відомо, що єдиним типом стійкості до вірусу скручуван¬
ня листків є польова стійкість, виявлена в диких видів S. demissum,
S. chacoense, S. acaule, S. andigenum. Цей тип стійкості успадко¬
вується полігенно. Рівень стійкості потомства залежить головним
чином від ступеня стійкості обох батьківських форм. Так, при схре¬
щуванні двох форм стійких до вірусу скручування листків, у потомс¬
тві можна отримати 10-15 % стійких гібридів картоплі. У потомстві,
отриманому від кількох послідовних схрещувань стійких батьківсь¬
ких форм рівень стійкості значно підвищується. У такому випадку
спостерігається явище трансгресії, тобто поява серед гібридів фено-
типів, які перевищують за стійкістю батьківські форми.
Польова стійкість до вірусу скручування листків виявлена в до¬
сить великої групи сортів картоплі, зокрема в таких, як Аквіла, Апта,
Карла, їда, Кама, Катадін, Швальбе, Шпатц, Ора, Пірат, Уран, Сафір.
Окремі з них відносно стійкі і до мозаїчних вірусів.
266
Вірус S. Це один із найбільш поширених вірусів картоплі. У пе¬
реважній більшості сортів він перебуває в латентній формі. Стійкість
до вірусу S була виявлена в сорту Сако. На думку Береке [5], ця стій¬
кість успадковується рецесивно під контролем малих генів. При за¬
раженні потомства від самозапилення сорту Сако щепленням чи ме¬
ханічною інокуляцією всі 100 % рослин виявилися стійкими.
Надчутлива стійкість до вірусу S була виявлена в S. andigenum
(клон Р.І.258907). Її успадкування зумовлене дією одного домінант¬
ного гена. Клон Р.І.258907 є слабостійким до вірусів скручування
листків та Y, тому для схрещування необхідно підбирати батьківські
форми, стійкі до обох вірусів.
Вірус М. Цей вірус викликає в рослин картоплі скручування мо¬
лодого листя і мозаїку.
Сорти картоплі значно відрізняються за ступенем польової стій¬
кості. Із сортів S.tuberosum відносною стійкістю до вірусу М відзна¬
чаються Агрономічна, Берліхінген, Камераз, Світязь, Янтарна і Фі¬
тофторостійка. Польова стійкість до вірусу М виявлена в диких видів
S. tariense, S.commersonii і S.chacoense.
Домінантний ген Nm, який контролює надчутливість, було вияв¬
лено в клону EBS 1787 виду S. megistacrolobum.
Клон, як і його гібриди із сортами картоплі, відзначаються досить
високою стійкістю до зараження навіть при інокуляції щепленням.
Успадкування ознаки зумовлюється дією одного домінантного гена.
Для визначення стійкості сортів і гібридів картоплі до вірусних
хвороб використовують наступні методи:
1. Випробування сортів і гібридів картоплі в польових умовах, де
спостерігається сильний прояв окремого вірусного захворювання.
2. Зараження сіянців першого року різними штамами мозаїчних
вірусів шляхом втирання в листя вірусного соку з карборундом або
обприскування пістолетом-розпилювачем.
3. Щеплення випробовуваних рослин на попередньо заражені пе¬
вним вірусом (підщепа). Імунні форми симптомів не дають, а надчу¬
тливі утворюють некроз.
4. Посадка гібридів картоплі поряд із хворими рослинами з на¬
ступним обліком кількості уражених кущів, що дозволяє визначити
відносну стійкість до найбільш поширених вірусних хвороб.
Результати випробування визначають не тільки візуально, але й
шляхом зараження рослин-індикаторів, серологічними чи іншими
методами.
Картопляна нематода. На картоплі паразитують два види цис-
тоутворюючих нематод - Globodera rostochiensis і Globodera pallida,
які різняться між собою деякими морфологічними ознаками та ко¬
льором цист. Якщо цисти G.rostochiensis проходять фазу золотистого
кольору протягом кількох тижнів, то колір цист G.pallida залишаєть¬
ся білим чи блідо-жовтим до виходу личинок.
267
Дослідження, проведені в кількох областях України, показали,
що всі вивчені популяції були представлені виключно видом
G.rostochiensis і расою R01. Незважаючи на те, що цей шкідник є ка¬
рантинним об’єктом, ареал його з кожним роком збільшується. Так,
якщо перші вогнища картопляної нематоди в Україні були виявлені в
1958 році, то вже наприкінці 90-х років паразит практично поширив¬
ся по всій республіці.
Відомо, що картопляна нематода особливо шкодочинна на при¬
садибних ділянках, де часто відсутнє чергування культур, чи там, де
картопля повертається на попереднє місце вирощування через 2-
3 роки. З урахуванням ступеня зараження грунту (кількість личинок
на 100 см ) недобір урожаю нестійкого до глободерозу сорту карто¬
плі становитиме 11—43 %.
Як свідчить практика, для оздоровлення заражених ділянок недо¬
статньо застосування лише агротехнічних і хімічних прийомів.
Більш радикальним і екологічно безпечним заходом у боротьбі з
картопляною нематодою є створення і впровадження у виробництво
нематодостійких сортів картоплі.
При вивченні диких і культурних видів картоплі в середині 40-х
років було виявлено, що окремі форми S.cmdigenum і S.vernei мають
стійкість до глободерозу, що дозволило розпочати інтенсивну роботу
з виведення нематодостійких сортів картоплі - спершу у Великобри¬
танії, а згодом у Голландії, ФРН, Данії і СІЛА. Проте, вже в 1957 р. в
Шотландії була виявлена популяція, яка значною мірою уразила
стійкий до цього часу сорт картоплі. Агресивні патотипи були вияв¬
лені в Англії, Голландії, ФРН та Південній Америці.
Польові популяції можна поділити за допомогою рослин-
диференціаторів на п’ять патотипів G.rostochiensis і три G.pallida. У
1977 р. була запропонована класифікація, яка об’єднала 5 патотипів
золотистої картопляної нематоди і 3 блідої в одну систему, яка при¬
йнята за міжнародну (табл.4.26).
Таблиця 4.26 - Міжнародна система патотипів картопляної нематоди і зв’язок її
з національними номенклатурами
Номенклатура
Назва патотипів
Голландська
А В С-*-*-Д
Англійська
А-^В Е
Німецька
АВ-н^Е
Міжнародна система патотипів
Rol Ro2 Ro3 Ro4 Ro5 Раї Pa2 Pa3
Перші дослідження зі створення нематодостійких сортів картоплі
були проведені шляхом міжвидової гібридизації високостійких зраз¬
ків S. andigenum і S.vernei із сортами і гібридами S.tuberosum, цінни¬
ми в господарському відношенні. Дещо пізніше у схрещування було
включено третій вид S. spegazinii, гени стійкості якого було передано
268
ряду сортів картоплі. У нього було виділено три основні гени: Fa,
який контролює стійкість до Rol та Ro2; F6 - від Rol та Ro5; Fc - до
деяких патотипів обох видів картопляної нематоди.
На сьогодні асортимент нематодостійких сортів картоплі в Укра¬
їні та за її межами досить значний. Так, уже на початку 90-х років
понад 60% сортів картоплі у ФРН були стійкими до одного чи двох
патотипів глободерозу і займали близько 40 % площ, відведених під
насінні посіви.
В Україні селекція, спрямована на створення сортів, стійких до
картопляної нематоди, була розпочата в 70-х роках, спершу в
УНДПСГ, пізніше на Поліській дослідній станції ім. О.М. Засухіна і
Чернігівській дослідній станції по картоплі. У результаті було ство¬
рено перші нематодостійкі сорти картоплі Віхола (1986), Пролісок
(1991) і Берегиня (1992). Нині у державному Реєстрі сортів рослин
України майже половина сортів картоплі є нематодостійкими.
Численні матеріали досліджень свідчать про те, що стійкі до нема¬
тоди сорти і гібриди картоплі з ефективним генетичним контролем до
кількох патотипів є хорошими вихідними формами для подальшої робо¬
ти в цьому напрямі [2]. Так, сорти Гітте, Гідра та Міранда поєднують
стійкість до картопляної нематоди і вірусних хвороб. Комплексом таких
господарсько цінних ознак, як урожайність, стійкість до нематоди та
інших хвороб, високі якісні показники, відзначаються сорти Берегиня,
Пролісок, Доброчин, Поран, Турбела, Санте, Гранат, Водограй, Густо,
Бероліна, Обелікс, Карлена, Астерикс та ін.
Досить перспективним є включення у схрещування на стійкість
до глободерозу складних міжвидових гібридів картоплі з ефектив¬
ним генетичним контролем ознаки, які створені в лабораторії вихід¬
ного матеріалу Інституту картоплярства УААН. В окремих комбіна¬
ціях вихід сіянців, стійких до паразита, складав близько 67 %.
В Україні оцінка гібридів картоплі за стійкістю до картопляної
нематоди проводиться в Інституті захисту рослин УААН.
Отже, результативною селекція картоплі, спрямована на виве¬
дення нематодостійких сортів, може бути при залученні до схрещу¬
вання стійких сортів або складних міжвидових гібридів, які відзна¬
чаються комплексом інших господарсько цінних ознак.
Стеблова нематода. Стеблова нематода (Ditylenchus destructor
Torne) є досить поширеним в Україні захворюванням бульб, яке
спричиняє значні втрати врожаю.
Згідно з даними ряду дослідників, усі випробувані сорти картоплі
уражувалися стебловою нематодою, проте відрізнялися ступенем
ураження. Найбільш стійкими були сорти Катюша, Олев, Роза По¬
лісся, Сагітга, Спекула, Форан, Апта, Бородянська, Перлина, Жито-
мирянка, Цейсиг та ін.
З диких видів найбільший інтерес своєю стійкістю до стеблової
нематоди викликають певні форми S. andigenum і S.chacoence. Окре¬
269
мі з них відзначаються комплексною стійкістю до картопляної і
стеблової нематоди.
Випробування на стійкість до стеблової нематоди проводяться в
польових умовах, на провокаційному фоні, який створюється вне¬
сенням у лунки при посадці 40-50 г дрібно порізаних бульб, зараже¬
них паразитом (15-18 тис. нематод різного віку).
ЛІТЕРАТУРА
1. Букасов С.М., Камераз А .Я. Селекция и семеноводство картофеля-
Л.: Колос, 1972.- 510 с.
2. Подгаецкий А.А., Гайдук В.С. Исходный материал в селекции на
устойчивость к картофельной нематоде // Создание и использование исход¬
ного материала в селекции картофеля.- К., 1992.- С.91-108.
3. Росс X. Селекция картофеля: проблемы и перспективы. - М.: Агро-
промиздат, 1989 - 111 с.
4. Яковлева В.И. Ракоустойчивость картофеля и паразитическая измен¬
чивость возбудителя рака Synchytrium endobioticum: Автореф. дис. ... канд.
биол. наук - Л., 1963.- 24 с.
5. Baerecke M.Z. Uberempfindlichkeit gegen das S-virus der kartoffel in
einem bolivinnischen andigena-klon. Zuchter. 37. - 1967. - S.281-285.
6. Zelbach H. Schatzung genetische Parametr aus diallelen Krenzungen bei der
Najshaule-anfallegkeit der kartoffel. Arch. Zuchtungst. 8. - 1978. - S. 193-199.
7. Maris D. Races of the potato wart causing fundus Synchitrium endo¬
bioticum and some data on the inherifance of resistance to ruse // Euphytica. - 10.
-№3. 1961.-S.269.
8. Schir A., Hopfe A. Die Zuchtung der kartoffel.- Die kartoffel. Handbuch.
Bd. II. Berlin veb. Dtsch. Zandwirtshaft. 1962.- S.1462. /
9. Toxopeus H.J. On the inherifance of tuber resistance of s. tuberosum to Ph.
infestans in die field // Euphitica. 10 - № 3 - 1961- S.307.
10. Wiersema H.T. On vatbaarkeid voor het x-virus bif S. andigenum. T.O.
Plantenz - 64.- 1958.- S.215.
4.5.5. Селекція на придатність
до промислової переробки
Народногосподарське значення і перспективність селекції на
придатність до промислової переробки. Цілорічне забезпечення
населення свіжою картоплею пов’язане як з великими кількісними
втратами, так і зі зниженням її харчової цінності. Тому постає про¬
блема зміни структури її споживання із забезпеченням населення
широким і різноманітним асортиментом продуктів переробки.
Поживна цінність продуктів, виготовлених із свіжої картоплі в чет¬
вертому кварталі і спожитих у другому-третьому кварталі наступного
року, має таку ж якість, що і свіжа картопля цього періоду. При цьому
поживну цінність картоплепродуктів ще можна підвищити за рахунок
включення в них різних добавок - білків, жирів, вітаміну С, глютамінату
натрію, лимонної кислоти, сухого молока тощо [17].
270
Продукти переробки мають тривалий термін зберігання без за¬
стосування спеціальних технічних засобів; вигідні при транспорту¬
ванні, оскільки, наприклад, одна тонна сухого картопляного пюре
займає об’єм 0,18 м3 , тоді як така ж маса сирих бульб - 1,4 м3 [24];
вони значно полегшують працю робітників громадського харчування
і господинь; переробка картоплі в місцях її вирощування може бути
безвідходною при утилізації відходів при очистці і доочистці бульб;
картоплепродукти - це довгострокові страхові запаси.
Асортимент картоплепродуктів у світі зростає дуже швидко, і
нині вони представлені такими видами: сушені з тривалим терміном
зберігання (не менше 1 року) - сушена картопля (кубики), сухе кар¬
топляне пюре (пластівці, гранули, крупка); заморожені продукти не¬
тривалого зберігання (не менше трьох місяців) - помфрі, гарнірна
картопля, картопляні пиріжки, котлети, вареники, галушки тощо;
обсмажені продукти короткотермінового зберігання (не більше трьох
місяців) - хрустка картопля, або чипси, крекери, консервована ціла
картопля під назвою “Сервос” і “Гольдгар” або консервовані пластів¬
ці “Ганза”; маловідомі продукти - печена картопля, картопляний
крокет, хрусткий глазурований крокет, картопляні вафлі.
У США виробляють більше 100 найменувань картоплепродуктів. Ця
країна займає перше місце у світі за обсягом переробки картоплі. На кі¬
нець 80-х років у США цей показник досягав 60 % валового збору
бульб, у ФРН, Великобританії, Нідерландах, Франції - 30 % [20].
Україна, яка за валовим збором картоплі входить до числа семи
великих виробників картоплі у світі [4], за обсягом виробництва кар¬
топлепродуктів посідає лише 50-е місце [12]. За даними Інституту
аграрної економіки, в Україні на продовольчі цілі витрачається бли¬
зько 40 % внутрішнього споживання картоплі, а на переробку - лише
0,3 % (табл.4.27). Причому, частина підприємств, що спеціалізували¬
ся на виробництві сушених і консервованих продуктів, припинили
своє існування.
Таблиця 4.27 - Баланс споживання картоплі в Україні
Стаття
балансу
Маркетингові роки
1996-1997
1997-1998
1998-1999
1999-2000
тис. тонн
%
тис.тонн
%
тис.тонн
%
тис.тонн
%
Внутрішнє
споживання
16961,1
97,2
16295,0
97,0
15956,4
97,
2
13645,0
98,
2
у т.ч. продово¬
льство
6675,7
38,2
6692,1
39,8
6423,6
39,
1
5950,0
42,
8
Промислова
переробка
30,0
0,2
30,0
0,2
45,0
0,3
45,0
0,3
Натомість починають створюватись нові переробні підприємства
із залученням іноземних інвестицій, що спрямовані в основному на
виробництво чипсів (табл.4.28). За розрахунками споживання цього
271
продукту в Україні на душу населення за рік становить лише 0,9
грамів, у той час як у США ще в 1969 році цей показник досягав
2700, Англії - 1500, Голландії - 650, Бельгії, Швейцарії - по 250,
Франції, Італії, ФРН - по 200, Данії, Швеції - по 150 грамів [26,33].
Таблиця 4.28 - Виробництво продуктів харчування із картоплі в Україні,
тис. тонн
Картоплепродукти
1993 р.
1998 р.
Обсмажені, у т.ч.
1,1
8,1
чипси із свіжої картоплі
0,6
8,0
чипси формовані
0,5
0,1
Сушені, у т.ч.
3,0
0
сушена картопля
1,0
0
сухе картопляне пюре
2,0
0
Консервовані
0,6
0
Слід особливо підкреслити, що сировина для цих підприємств
вирощена з імпортованого посадкового матеріалу, якого в Україну в
1999 р. було завезено понад 4 тис. тонн з Нідерландів та Німеччини.
За прогнозом Інституту аграрної економіки попит на картоплю для
переробки буде постійно зростати, а імпорт садивного матеріалу мо¬
же досягти 7-10 тис. тонн.
Маючи досвід сучасного насінництва і виробництва та великий сор¬
товий потенціал картоплі для переробки, Україна вже до 2005 р. може в
два рази підвищити виробництво сушених продуктів та налагодити ви¬
готовлення картоплі фрі і напівфабрикату сухої картоплі (табл.4.29).
Таблиця 4.29- Прогноз виробництва картоплепродуктів в Україні,
тис. тонн за роками
Карто плепродукти
Роки
2000
2001
2002
2003
2004
2005
Чипси із свіжої картоплі
8,5
12,3
12,7
13,0
14,9
15,1
Картопля-фрі
1,0
4,0
4,0
4,0
4,0
4,0
Сухе картопляне пюре
1,0
4,0
4,0
4,0
4,0
4,0
Формовані продукти
0,9
2,0
2,8
3,6
4,0
Сирий напівфабрикат
5,0
14,0
20,0
20,0
20,0
20,0
Згідно з прогнозом Інституту картоплярства УААН щорічний
об’єм картоплі для переробки може становити близько 130 тис. тонн.
Це має бути сировина відповідної якості. Основні показники якості,
що характеризують придатність картоплі до переробки, визначають¬
ся сортом. Селекція таких сортів до недавнього часу зовсім не велась,
але технологічна оцінка існуючого асортименту картоплі постійно про¬
водилася відомим у колишньому СРСР науковим закладом - Білорусь¬
ким НДІ по переробці продуктів харчування з картоплі.
Випробування селекційного і вихідного матеріалу картоплі на
придатність до переробки вперше було розпочато в Білоруському
272
НДІ картоплярства і плодоовочівництва 1970 р., а селекція спеціаль¬
них сортів - з 1974 р. [15]. На сьогодні Сумський державний аграр¬
ний університет веде цілеспрямовану селекцію картоплі на придат¬
ність до переробки на харчові продукти.
Ознаки і властивості сорту, придатного до промислової пере¬
робки. Крім традиційних вимог до сорту, таких, як висока врожай¬
ність, стійкість до хвороб, шкідників і несприятливих умов вирощу¬
вання, лежкоздатність, придатність до механізованого виробництва,
стабільність якісних показників тощо, переробна промисловість ста¬
вить специфічні вимоги до картоплі.
Суха речовина - одна з основних ознак сорту картоплі при її пе¬
реробці. Уміст сухої речовини на 95 % (г=0,977) визначає вихід сухо¬
го картопляного пюре, на 81 % (г=0,903) - сушеної і на 78 %
(г=0,886) - хрусткої картоплі. Розрахунки на підставі даних таблиці
4.30 показують, що підвищення вмісту сухої речовини в бульбах на 1 %
може збільшити вихід сушених продуктів у середньому на 9 кг, чип¬
сів - на 14 кг з кожної тонни сировини; знизити вміст жиру в пластів¬
цях хрусткої картоплі на 1,62 % [ 7].
Таблиця 4.30 - Залежність виходу картоплепродуктів (%) від вмісту сухої
речовини
Продукти
Уміст сухої речовини, процент на сир;
у масу
18
20
22
24
26
28
ЗО
32
Сушена картопля
14,0
17,0
18,5
21,0
23,0
26,0
~
-
Сухе пюре
16,0
19,0
20,5
23,0
25,0
27,0
28,7
29,9
Чипси
32,9
35,8
37,7
39,3
41,5
43,5
44,2
47,2
Оптимальним умістом сухої речовини для виробництва сушених
продуктів є 24,6 % і більше, для чипсів - 20,6-24,5 %. Для чипсів із
висококрохмалистої речовини - 23,6-17,5 % [6, 5], але W.G. Burton і
В. Putz вважають, що це - 22,5-24,5% [25, 30], G.Adler і В.І, Шинка-
рев - 21-24 % [23, 21], В.А. Барановська - не нижче 22% [17]. При
виготовленні консервованих продуктів віддають перевагу картоплі з
низьким умістом сухої речовини - 17-19 % [ 24].
Крохмаль картоплі становить 70-80% сухої речовини [18]. Ще в
1935 p. K.Ratzack, а потім в 1971 p. G.Adler та в 1973 р. В.А. Бара¬
новська й інші встановили, що для переробної промисловості потріб¬
ні сорти не тільки з умістом крохмалю, але й з визначними його фі¬
зичними і хімічними властивостями, а саме: підвищена зернистість
(уміст великих і середніх крохмальних зерен, оптимальне співвідно¬
шення амілози і амілопектину [31,24,2]. Від цих властивостей крох¬
малю залежить нерозварюваність консистенції консервованої карто¬
плі чи розсипчастість пюре з гранул, крупки і пластівців.
Вміст крохмалю в картоплі та його структура пов’язані зі скоро¬
стиглістю: вона вища у пізніх сортів з більш тривалим періодом фо¬
тосинтезу (табл.4.31).
273
Таблиця 4.31- Уміст і якість крохмалю в бульбах картоплі у сортів різних груп
стиглості
Показник
Група стиглості
ранні
середньо¬
ранні
середньо¬
стиглі
пізні
Кількість сортів, пгг.
15
10
18
16
Уміст крохмалю, %
15,3±0,6
16,6±0,9
16,0±0,5
18,2±0,5
Зернистість, %
29,0±2,9
34,1±3,8
29,5±2,4
32,9±0,9
Великі крохмальні зерна, мк
50,0±1,1
52,4±1,7
49,4±1,2
53,6±1,3
Середні крохмальні зерна, мк
31,0±0,6
32,8±1,7
30,8±1,8.
32,9±0,9
Розмір крохмальних зерен - це сортова ознака (табл.4.32).
Таблиця 4.32 - Характеристика сортів картоплі за розміром крохмальних зерен
Сорт
Розмір зерен, мк
великі, 71 і >
середні, 70-31
дрібні, 30 і <
Лошицька
91,0±0,1
51,4±1,3
21,2±1,2
Сарма
80,0±2,8
50,4±1,5
24,6±1,2
Ескорт
77,3±1,2
49,6±1,4
25,4±1,9
Пост 86
77,0±0,1
52,3±1,2
19,6±2,0
Білоруська 5
76,4±1,4
57,8±1,3
16,9±2,2
Дєтськосєльська
76,0±2,9
60,2±2,3
27,8±0,8
Діамант
76,0±2,8
50,2±2,1
22,6±1,3
Пріор
75,8±1,7
54,2±3,3
23,2±2,4
Гібрид 12
75,7±1,3
44,0±1,3
23,0±1,2
Морена
75,0±1,6
52,0±1,4
24,4±1,8
Спадщина
75,0±1,3
56,6±2,1
22,6±1,4
Мавка
75,0±1,3
50,7±1,4
21,2±1,9
Зарево
74,7±1,3
52,4±1,4
26,4±1,8
Плай
74,0±1,2
50,2±1,3
23,0±1,2
Пролісок
73,7±0,7
47,1±2,7
22,0±1,1
Темп
73,0±0,1
53,6±1,4
26,6±1,4
Гібрид 6
72,7±0,9
53,2±1,4
22,4±1,2
Коретта
72,6±1,0
53,3±2,1
19,5±1,4
Українська рожева
72,5±1,1
58,4±1,4
25,8±1,5
Пригожа 2
72,2±0,8
56,8±1,4
22,2±2,3
Лорх
72,0±0,1
52,2±1,6
22,4±1,2
Білоруська рання
72,0±0,8
46,8±1,2
25,6±1,7
Світанок київський
71,5±0,3
50,2±2,3
22,8±1,4
Гібрид 22
71,5±0,3
54,4±1,3
21,2±2,1
Посвіт
71,5±0,4
47,8±2,2
19,0±1,3
Санте
71,0±0,1
46,2±1,4
25,2±1,6
За даними Сумського ДАУ, існує висока залежність текстури
м’якоті від умісту крохмалю (г=0,73 5-0,933): висока і середня - між
зернистістю крохмалю і консистенцією (г=0,800), борошнистістю
(г=0,729) та розварюваністю м’якоті бульб (г=0,622); середня - між
274
розміром крохмальних зерен і консистенцією (г=0,702), розварювані-
стю (г=0,624) та борошнистістю (г=0,518). Тому для виготовлення
кращого за якістю картопляного пюре доцільна переробка сортів з
умістом крохмалю понад 17 %, амілози - 20 %, а золи - менше 4 %
(табл.4.33); з розміром крохмальних зерен більше 50 мк і найбіль¬
шою кількістю великих (>71 мк) і середніх (70-30 мк) зерен [17,24,
6,5,31,19].
Таблиця 4.33- Хімічні показники сортів картоплі та якості пюре з крупки
Сорт
Крохмаль,
%
Амілоза, %
Зола, %
Консистенція
пюре
Оцінка
пюре, бал
Темп
24,8
17,2
3,5
розсипчасте
4,4
Лошицька
21,8
22,0
3,9
розсипчасте
4,3
Вілія
16,7
14,7
4,7
в‘язке
3,3
Янтарна
15,5
14,3
4,6
в‘язке
3,2
Форан
13,2
9,8
4,5
в‘язке
3,0
Примітка. Вміст амілози і золи в розрахунку на суху речовину картоплі
Редукуючі цукри справляють негативний вплив на якість картоп-
лепродуктів, насамперед на їхній колір і смак. Колір чипсів від уміс¬
ту цукрів залежить на 80 %, сухого картопляного пюре (гранули) -
на 83 % (табл.4.34).
Таблиця 4.34 - Залежність кольору продуктів (5-1 балів) від умісту цукрів
Продукт
Уміст цукрів, процент на сиру масу
r+Sr
t
<0,25
0,30-0,40
0,50-0,60
0,70>
Чипси
4,5
3,9
3,5
2,0
0,897±0,104
8,61
Сухе пюре
4,7
4,6
4,1
2,4
0,915±0,095
9,65
Аналіз наведених даних свідчить, що оптимальний уміст цукрів у
картоплі для переробки на чипси має бути не більше 0,25 %, на су¬
шені продукти - 0,6 %. G.Fdler і Y.W. Stricker допускають, що для
виготовлення пюре і картоплі фрі вміст редукуючих цукрів може ко¬
ливатися від 0,5 до 1,0 % на суху масу [24,32]. Однак слід зазначити,
що обсмажені продукти доброї якості, кольору і приємного смаку
можна отримати практично з будь-якого сорту картоплі шляхом від¬
повідної зміни температури і тривалості смаження, але це дуже
ускладнює процес виготовлення [27].
Важливим критерієм придатності сортів до промислової перероб¬
ки є нагромадження цукрів при зберіганні та їх ресинтез. Оцінка 280
сортів картоплі світової колекції показала, що більшість із них за
вмістом редукуючих цукрів при збиранні були придатні для виготов¬
лення сушених продуктів. Умовно придатних сортів залежно від
групи стиглості було 4-9 %, непридатних - 1-6 %. Природно, що
рівень цукрів підвищувався при зберіганні і знижувався при ресинте-
зі (табл. 4.35).
275
Таблиця 4.35 - Розподіл сортів колекції за вмістом редукуючих цукрів при
оцінці їх на придатність до переробки
Вміст
цукру
Придатність
ДО
переробки
Група стиглості
ранні
середньо¬
ранні
середньо¬
стиглі
пізні
<0,40
Придатні
92-11-65*
89-24—60
92-24-76
91-25-68
0,41-0,60
Умовно
придатні
4-73-35
9-52-40
6-46-22
8-53-32
0,61 і >
Непридатні
4-16-0
2-24-0
1-30-2
1-22-0
Примітка. Перша цифра - уміст цукрів при зберіганні, друга - після зберігання,
третя - після ресинтезу
У зв’язку з тим, що при виробництві чипсів вимоги до вмісту цу¬
крів більш жорсткі, при збиранні картоплі придатних сортів за цим
показником було на 7-15 % менше, ніж на сировину для сушених
продуктів. У процесі зберігання придатними для виготовлення чип¬
сів було лише 2-6 %, після ресинтезу - вже 23-26 % (табл.4.36 ).
Таблиця 4.36 - Розподіл сортів колекції за вмістом редукуючих цукрів при
оцінці придатності до переробки на чипси
Вміст
цукру
Придатність
до перероб¬
ки
Група стиглості
ранні
середньо¬
ранні
середньости¬
глі
пізні
0,10-0,30
Придатні
81-0-23
74-6-27
85-2-36
81-6-30
0,31-0,50
Умовно
придатні
15-39-69
22-42-67
112-42-60
16-37-64
0,51 і >
Непридатні
0-61-8
4-52-6
3-56-4
3-57-6
Однак кондиціювання під впливом підвищених температур, що
призводить до ресинтезу, веде до значних виробничих витрат, втрат
сухої речовини та збільшення кількості дрібних крохмальних зерен.
Тому більш ефективним є підбір сортів, стійких до накопичення цу¬
крів у процесі зберігання.
Дуже важливим критерієм придатності сортів картоплі до промис¬
лової переробки є оптимальне співвідношення вмісту сухої речовини
і редукуючих цукрів. Із 605 сортів світової колекції відібрані сорти з
умістом сухої речовини 27-28 % і редукуючих цукрів 0,12-0,24 %
(Зарево, Берізка, Верба), які придатні для приготування сухих карто-
плепродуктів; з умістом сухої речовини 25-26 % і редукуючих цук¬
рів 0,10-0,20 (Білоруська крохмалиста, Ексид, Ердкрафт, Ритм, До¬
нор, Фекула, Кинаво) - для виготовлення сухих і смажених продук¬
тів; з вмістом сухої речовини 23-24 % і редукуючих цукрів 0,11—0,28 %
(Форте, Надежда, Корделія, Ниса, Нарев, Оса, Беладона, Гитте, Гат¬
чинська, Лошицька, К-27, Хоорделінг) - придатні для приготування
картоплепродуктів усіх видів. При цьому була встановлена висока
від’ємна залежність між умістом сухої речовини і редукованих цук¬
рів (r=0,984; t=54,7).
276
Білки і вітаміни зумовлюють поживну цінність картоплепродук¬
тів. Тому бажано, щоб сировина містила якомога більше цих речо¬
вин, враховуючи ще й те, що при термічній обробці втрачається 30-
50 % вітаміну С. Найбільш придатними є сорти з умістом білка не
менше 2 %, вітаміну С - не менше 17 мг% [25]. Значний інтерес ста¬
новить залежність між умістом сухої речовини і вітаміну С в бульбах
(г =0,518±0,069, t=7,50 >to5=l,96). Однак реальність такого зв’язку
більшою мірою виявляється в межах одного сорту: Зоря Сумщини -
85 %, Ювіляр - 66, Слобожанка - 44, Молодіжна - 37 %.
Потемніння м’якоті бульб до і після варіння - це властивість, що
визначає товарну і споживчу якість картоплі як продукту безпосеред¬
нього вживання, так і сировини для промислової переробки. Потемніння
м’якоті обчищених свіжих бульб пов’язане з окисненням амінокислоти
тирозину поліфенолоксидазою, варених - залежить від співвідношення
хлорогенової і лимонної кислот [16]. Бульби, які мають схильність до
потемніння, змінюють природний колір на синювато-сірий або чорний.
Існує тісний зв’язок між потемнінням і кольором картоплепродукту
[9,10]. Так, колір чипсів на 83 % (г=0,915) залежить від ступеня потем¬
ніння як сирої, так і вареної м’якоті; колір сухого картопляного пюре -
на 84 і 85 % (г=0,919 і 0,923).
Для промислової переробки найбільш придатні сорти зі зниже¬
ною здатністю до потемніння, а за дев’ятибальною оцінкою - не ниж¬
че 6,6 балів [11].
Морфологічні ознаки, такі, як розмір і форма бульб, вічка і їхня
глибина в середньому на 12% визначають величину відходів при
очистці 204 сортів картоплі (F=6,72 > Fi0=3,98). Однак у межах
окремої групи стиглості частка їх впливу різна (табл.4.37).
Таблиця 4.37- Частка впливу ознак бульби на відходи при очистці картоплі, %
Морфологічні ознаки
По всіх
сортах
Група стиглості сортів
ранні
середньо¬
ранні
середньо¬
стиглі
пізні
Сукупний вплив
11,9
20,0
35,3
19,4
33,8
Маса
6,8
2,7
25,6
9,2
15,8
Форма
3,5
7,0
2,3
14,8
1,8
Вічка, кількість
0,6
9,3
6,8
0,0
2,4
Вічка, глибина
0,9
1,0
0,6
4,4
13,4
Так, найбільший сукупний вплив ознак на відходи був у серед¬
ньоранніх і пізніх сортів (34-35 %), однаковий - у ранніх і середньо¬
стиглих (19-20 %). Якщо відходи у ранніх сортів визначаються од¬
наковою мірою кількістю вічок і формою бульб, у середньоранніх -
масою бульби, у середньостиглих - формою, то у пізніх - масою і
глибиною вічок. Менше відходів при очистці (не більше 14 %) бульб
розміром не менше 50 мм, з індексом форми не більше 1,29, з кількіс¬
277
тю вічок і їх глибиною не більше, відповідно, 7,5 шт. і 1,3 мм [11].
Менші відходи зумовлюють більший вихід продукту при переробці.
Існує залежність між виходом сухого картопляного пюре і чипсів та
кількістю відходів у розрахунку на сухий продукт порівняно з необ-
чищеною картоплею. Збільшення відходів при механічній очистці
бульб на 1 % знижує вихід картопляного пюре на 9,5 кг, а чипсів - на
9,2 кг з кожної тонни сировини при переробці.
Колір м’якоті - це ознака сорту, вимоги до якої визначаються
традиціями тієї або іншої країни. Так, у більшості країн Європи пе¬
ревага віддається сортам із жовтою м’якоттю, у США, Англії, Канаді - з
білою. На думку білоруських учених, при переробці доцільно вико¬
ристовувати як білі, так і жовтом’якотні сорти [17].
Колір м’якоті бульб залежить від умісту каротиноїдів. Бі-
лом’якотна картопля містить від 0,014 до 0,053 мг% пігменту, жов-
том’якотна - 0,11—0,18 мг% [24]. За даними Сумського ДАУ, уміст
каротиноїдів у картоплі із жовтою м’якоттю становить 0,77 мг %, із
світло-жовтою - 0,71 із білою - 0,69 мг% на суху речовину.
Смак картоплі певною мірою впливає на якість продуктів пере¬
робки, але вона більше залежить від технології виготовлення і різних
добавок, ніж від особливостей сорту [25].
Скоростиглість - це властивість, якій все більше приділяє увагу
картоплепереробна промисловість. Доцільність переробки ранніх
сортів картоплі із середини липня до початку вересня вже у 80-х ро¬
ках обгрунтована вченими у Нідерландах і Німеччині. У Великобри¬
танії віддають перевагу раннім строкам садіння і збирання в липні-
серпні, що забезпечує 65-80 % бульб, придатних для переробки, при
загальній врожайності від 46 до 68 т/га [29].
Звичайно, що за якістю сировини і продуктів найбільш придат¬
ними для переробки є пізньостиглі сорти. Але середньоранні і серед¬
ньостиглі сорти дають цілком задовільну якість харчових продуктів,
що можна простежити за технологічною оцінкою середньопізнього
сорту Верба і середньоранніх сортів Пригожа 2 і Добро (табл.4.38).
Таблиця 4.38 - Технологічна оцінка за скоростиглістю сортів картоплі
Сорт
Якість сировини
для продуктів,
бали (1-9)
Органолептична
оцінка, бали (1-5)
Загальна оцінка, бали
(1-9)
сушені
смажені
пюре
суп
чипси
сухе
пюре
картопля
суха
хру¬
стка
Верба
8,1
6,0
4,8
3,1
4,2
9-8
6-5
7-8
Пригожа 2
4,5
3,5
2,7
4,4
3,6
4-3
7-6
6-5
Добро
4,5
4,5
2,8
4,2
3,5
4-3
7-6
6-5
Використання скоростиглих сортів і постачання сировини для
переробної промисловості в більш ранні строки - один із вагомих
278
факторів підвищення ефективності роботи підприємств і розванта¬
ження їх у найбільш напружений осінній період.
Придатність сорту до промислової переробки визначається не тільки
якістю сировини, але й якістю картоплепродуктів. Це зумовило необхід¬
ність створення і вдосконалення системи комплексної технологічної
оцінки селекційного матеріалу [56]. Система включає 28 змінних поло¬
жень якості за дев’ятибальною шкалою оцінок, у тому числі якість кар¬
топлі - 20 (хімічні показники - 8, морфоанатомічні - 5, технологічні - 7),
якість продуктів (сухе картопляне пюре, сушена і хрустка картопля) - 8.
Придатність селекційних зразків до переробки на ранніх етапах селекції
визначається за показниками якості бульб, а на завершальному - за ком¬
плексом технологічних показників картоплі і продуктів із неї. Виснув-
ки про придатність селекційного матеріалу до переробки базуються
на п’яти рівнях оцінки: 9,0-7,2 балів - найбільш придатні; 7,1-5,9 - при¬
датні; 5,8-4,6 - умовно придатні; 4,5-3,3 - малопридатні; 3,2 і нижче
балів - непридатні.
Комплексна оцінка показників якості сприяла визначенню пара¬
метрів сортів, придатних до переробки (табл. 4.39).
Таблиця 4.39 - Моделі сортів за їх ступенем придатності до переробки на
картоплепродукти
Показники якості
Придатність до переробки
най¬
більш
прида¬
тні
прида¬
тні
умовно
прида¬
тні
мало¬
при¬
датні
непри¬
датні
Морфолого-анатомічні:
розмір бульб, мм, не менше
60
50
45
40
35
форма бульби, індекс, не більше
1,0
1,3
1,4
1,5
1,6
вічка, шт., не більше
5
7
8
9
10
глибина вічок, мм, не більше
1,0
1,3
1,5
1,7
1,9
зернистість крохмалю, %, не менше
60
50
45
40
35
Хімічний уміст:
сухої речовини, %, не менше
31/25
27/22
25/21
22/19
20/17
цукрів, %, не більше
0,2/0,1
0,4/0,2
0,5/0,3
0,6/0,4
0,7/0,5
нагромадження цукрів при 6-7°С,
%, не більше
0,4/0,2
0,5/0,3
0,6/0,4
0,7/0,5
0,8/0,6
білка, %, не менше
2,7
2,4
2,1
1,8
1,5
вітаміну С, мг %, не менше
26
25
23
22
20
Технологічні*.
відходи при очистці, %, не більше
11
14
17
20
23
потемніння м’якоті, бал, не менше
8
7
6
5
4
період спокою, дні, не менше
200
180
160
140
120
Примітка. Чисельник - для сушених, знаменник - для чипсів
Взаємозв’язок мінливості ознак і властивостей, які визначають
придатність сорту до переробки, став основою для визначення на¬
279
прямів селекції на поліпшення показників якості картоплі різних
груп стиглості:
для групи ранніх - поліпшення смаку і текстури тканини, підви¬
щення стійкості до потемніння вареної м’якоті; підвищення вмісту
сухих речовин і зниження вмісту цукрів; зменшення відходів при
очистці за рахунок зменшення кількості вічок і відтворення округлої
форми бульби;
для групи середньоранніх і середньостиглих - поліпшення текс¬
тури вареної м’якоті (борошнистість, вологість); підвищення вмісту
сухих речовин; зменшення відходів при очистці за рахунок збіль¬
шення маси бульб округлої форми з малою кількістю вічок;
для групи середньопізніх і пізніх - поліпшення смаку, підвищен¬
ня стійкості до потемніння м’якоті після варіння; зниження вмісту
цукрів і відходів від очистки за рахунок збільшення маси бульб і
зменшення глибини вічок.
Особливості успадкування потомством ознак та властивостей
технологічної якості картоплі. Біохімічні властивості. Успад¬
кування вмісту сухої речовини має як домінантний [22], так і рецеси¬
вний характер [1]. За С.М.Букасовим (1959) і Б.Н.Дорожкіним
(1985), - це маловаріююча ознака. Результативність добору досяга¬
ється за використання гібридних комбінацій, які внаслідок розщеп¬
лення дають форми з підвищеним умістом сухої речовини [28]. На¬
громадження крохмалистості при селекції може здійснюватися ме¬
тодом трансгресії [3].
Оцінка 65 гібридних комбінацій показала, що при середньому
арифметичному значенні сухої речовини 25,07±0,23 максимальне -
становило 31,3, мінімальне - 19,6, найбільша втцепленість комбіна¬
цій з однаковим умістом сухої речовини - 20 %, найменша - 1,6%.
У 57 % потомства (r=0,755±0,082; t=9,20>toi=2,68) уміст сухої ре¬
човини зумовлений його рівнем у батьків.
Ця ознака в потомства на 78 % визначалася типом схрещувань і
на 22 % - батьками в кожному типі (F=55,42>F0i=3,78). Найвища за¬
лежність між умістом сухої речовини потомства і батьків виявлена у
таких комбінаціях: низький уміст х середній, високий х високий
(табл.4.40).
Таблиця 4.40 - Уміст сухої речовини у потомства від різного типу схрещувань
Комбінація
Суха речовина
Кореляційне
відношення
батьки
потомство
H±Sti
t
1 - Високий х високий
28,2 ± 0,73
26,2 ± 0,45
0,820 ±0,160
5,12
2 - Високий х середній
26,2 ± 0,35
24,5 ± 0,28
0,748 ± 0,296
2,52
3 - Середній х високий
25,9 ± 0,47
24,2 ± 0,29
0,749 ±0,177
4,23
4 - Середній х середній
23,9 ± 0,49
21,4 ±0,25
0,527 ± 0,200
2,63
5 - Низький х середній
21,8 ±0,49
20,3 ±0,14
0,865 ±0,132
6,55
280
Для кожного типу схрещувань розроблені рівняння регресії для
прогнозу ознаки в потомстві (у) при визначеному вмісті сухої речо¬
вини у батьків (х):
1) у=217,7-16,1х +0,33х2; 4) у=250,4-16,5х +0,29х2;
2) у=289,7-18,Ох +0,3х2; 5) у=218,3-20,Ох +0,5х2.
3) у = 135 - 10,5х + 0,24х2;
До батьківських пар, що мали в потомстві вміст сухої речовини
29-27%, належали: Форум * Сарма, Отрада х Зарево, Раритан х Во-
ловецька, 1.198129-77 N+8 х 2036-092; 27-26 % : Білоруська 3 * Гіб¬
рид 35, Білоруська 3 х Білоруська рання, 1.293112 N х 406-36 N, 76-
51 х Добро; 24-22 %: ВИР-377 х Зарево, Львів’янка х Зарево, При¬
гожа 2 х Білоруська 3, Супер х Білоруська 3, 1.445152- 74N х
1.371031-78N, Андра х 335-21, Супер х 305-27N; 23-22 %: Адретга
х Кристал білоруський, Адретга х Гібрид 22, 81716—18N х Добро,
Супер х Добро.
На генетичну зумовленість вмісту цукрів указують В.І.Байков і
Б.Путц. Рівень умісту редукуючих цукрів контролюється домінант¬
ними генами. Між ознаками батьків і потомства встановлена слабка
позитивна залежність.
Оцінка 700-ти гібридних комбінацій [10] виявила регресію взає¬
модії вмісту редукуючих цукрів батьківських форм (х) та їх потомст¬
ва (у): у = - 0,059+1,365х; r=0,542±0,101; t=5,0>to,=2,68.
Вивчення можливостей селекції картоплі на вміст цукрів прове¬
дено в гібридних комбінаціях від шести типів схрещування. Встано¬
влено рівень і вірогідність різниці середніх значень цього показника
у батьків (F=46,51) і в потомства (F=6,64). Уміст цукру в потомстві
залежав на 34% від типу схрещування і на 65 % - від батьківських
форм в окремому типі. Ступінь залежності показників умісту цукру в
потомства, одержаного різними типами схрещувань, був таким:
1) 98 % (г=0,993; t =14,47), високий х високий - у=1,3-10,4х+
+22,4х2. Батьківські пари, які мали у потомстві вміст цукрів не біль¬
ше 0,3 %: Форум х Зарево, 1.98129-77N+Sx2036-092,61.2107018—
-80N х Ласунак та ін.;
2) 98 % (г=0,992, t=22,22), низький х середній - у=0,22-0,15х+
+0,16х2. Кращі батьківські пари: Білоруська З х Гібрид 35, Ельда х
1.371031-78N, Андра х 335—21, Морене х Гібрид 35 та ін.;
3) 82 % (г=0,907, t=4,10), високий х високий - у= -2,0+-12,9х-16,9х2,
у Раменська х Добро, Добро х 81719-13N, Гібрид 22 х Дивосна;
4) 67 % (г=0,818, t=3,25), середній х низький - у= -2,3+24,6х-
59,2х2. Кращі батьківські пари: Отрада х Зарево, Кристал білорусь¬
кий х Ласунак, Пригожа 2 х Білоруська 3, Адретга х Кристал білору¬
ський, Морене х Зарево, Львів'янка х Зарево, Світанок київський х
Андра, 1.198129-77N+S х 4952-20, 80716-25N х 80897-47 та інші.
При типах схрещувань за вмістом цукру середній х середній та
низький х низький залежність показника в потомстві від батьків бу¬
281
ла, відповідно, середньою (50 %) і низькою (9 %), що свідчило про
незалежний характер успадкування ознаки. У першому випадку віді¬
брано 15 % малоцукристого потомства від батьків - Мавка х Гібрид
35, Астілла х 1.371031-78N, Гібрид 22 х Гібрид 35 та ін.; у другому -
50 %, батьки - Білоруська 3 * Білоруська рання, Прем’єр х Ласунак,
Морене х Ласунак, Кера х Віхола.
Висока від’ємна залежність між умістом сухої речовини і реду-
куючих цукрів у батьків (г=0,984) зберігалася в потомстві, але з
меншим впливом - від 33 до 58% (табл.4.41). Ця закономірність да¬
вала певну можливість вести контрольоване відтворення форм з ви¬
соким умістом сухої речовини і низьким - редукуючих цукрів. Сту¬
пінь варіювання цих показників у потомства був вищим, ніж у бать¬
ків. Якщо вимогам до сировини за біохімічним складом відповідали
80% комбінацій, то в потомстві - близько 59 %.
Таблиця 4.41- Вплив типів схрещувань на взаємозв'язок у комбінаціях вмісту
сухої речовини і цукрів
Тип схрещування
з урахуванням вмісту
сухої речовини
Уміст, %
Кореляційне
відношення
сухої
речовини
цукрів
ц±$ц
t
1 - Високий X високий
28 ± 0,7
0,24 ± 0,053
0,494 db 0,097
5,09
2 - Високий X середній
26 ± 0,3
0,19 ±0,003
0,571 ± 0,087
6,56
3 - Середній X високий
26 ± 0,4
0,23 ± 0,004
0,585 ± 0,064
9,14
4 - Середній X середній
24 ± 0,5
0,26 ± 0,025
0,425 ± 0,069
6,15
5 - Середній х низький
22 ± 0,5
0,24 ± 0,008
0,329 ±0,122
2,69
Морфологічні ознаки. Єдиної думки з приводу успадкування
морфологічних ознак не існує. Округла форма є домінуючою і має
проміжний характер, довга форма домінує над округлою [8].
При успадкуванні, на думку одних авторів, домінують глибокі ві¬
чка, на думку інших - мілкі. За даними А.Е.Зуйкова [8], залежність
кількості вічок від вихідних форм - середня, глибини вічок - слабка.
Оцінка 74-х гібридних комбінацій і 50-ти батьків за формою
бульб, кількістю і глибиною вічок показала, що чоловічі форми
впливали на успадкування всіх зазначених ознак більше, ніж жіночі.
Останні проявляли помітний ефект лише при успадкуванні форми
бульб - 25%. Статистично не доведена залежність між ступенем про¬
яву ознаки в батьків та їх потомства за кількістю вічок (г=0,013 від
чоловічих форм і г=0,063 від жіночих), за глибиною їх залягання
(г=0,220 і 0,504).
При вивченні гібридного матеріалу були виділені комбінації
схрещування батьків, які давали потомство з бульбами округлої фор¬
ми Астілла х 1.371031-78N, Пригожа 2 х Гібрид 22, 2031-0,23 х
78573-35N, Ельда х 1.371031-78N, 1.1981289-77N+S х 4938-45 та
ін.; з малою кількістю вічок (5-6): Пригожа 2 х Гібрид 22, Прем’єр х
282
Ласунак, Білоруська 3 х Гібрид 22, Берегиня * Гібрид 35, Морене х
Гібрид 35 та ін.; з неглибоким заляганням вічок (1,2-1,3 мм): Астіл-
ла х 1.371031—78N, Хейдрум х Добро, Морене х Зарево, Адретга х
Гібрид 22, Морене х Гібрид 35, 1.198129-77N+S х 4938--45 та Заре¬
во, Адретга х Гібрид 22, Морене х Гібрид 35, 1.198129-77N+S х
4938—45 та ін.
Технологічні ознаки. Ступінь потемніння м’якоті сирих і варе¬
них бульб гібридного потомства залежить від підбору батьківських
пар, що дає змогу ефективно селекціонувати цю ознаку. Між потем¬
нінням варених бульб у батьків і їх гібридним потомством є середній
позитивний зв’язок, потемнінням сирих бульб - сильний [8].
Регресія взаємодії м’якоті сирих бульб у 63 батьківських форм і
їх гібридних комбінацій була середньовірогідною (г=0,645±0,097;
t=6,58>; toi=2,68). Показник у потомстві був в 1,1 рази нижчим, а йо¬
го варіювання в 1,9 рази вищим, ніж у батьків. Ступінь залежності
між батьками і потомством знижувався в міру зменшення середнього
значення ознаки (табл.4.42).
Таблиця 4.42 - Вплив батьків на потемніння м’якоті сирих бульб потомства
Комбінації
Оцінка, бал
Кореляційне відношення
батьки
потомство
n±S\\
t
21
7 Л
7,6
0,667 ± 0,207
3,23
30
6,7
6,4
0,627 ± 0,178
3,51
17
6,2
5,5
0,404 ± 0,289
1,40
32
6,0
4,2
0,368 ± 0,199
1,81
Частка впливу на передачу ознаки від батьківських форм становить
61 %, від типу схрещувань - 39% (F=12,44>F0|=2,79). Сукупний вплив
цих факторів вивчено при чотирьох типах схрещувань (табл.4.43).
Таблиця 4.43 - Потемніння м’якоті сирих бульб потомства під впливом батьків
і типу схрещування
Тип схрещування за
якістю
Комбі¬
націй,
шт
Оцінка, бал
Кореляційне
відношення
батьки
потомство
Л±Бл
t
Висока х висока
19
7,5 ± 0,06
6,8 ± 0,22
0,665± 0,176
3,78
Середня х середня
23
6,5 ± 0,05
5,9 ±0,21
0,688 ±0,155
4,48
Середня х низька
12
5,8 ± 0,09
4,7 ± 0,23
0,407 ± 0,289
1,40
Низька х середня
9
5,6 ±0,10
5,1 ± 0,47
0,476 ± 0,332
1,43
Для прогнозування показників потемніння м’якоті в потомстві
від батьків пропонуються такі рівняння регресії: 1) при схрещуванні
батьків з нетемніючою м’якоттю - у = 226,4 - 60х + 4,їх (г=0,815;
t=3,78); 2) батьки із середніми показниками потемніння - у = -10,6 -
- 3,5х + 0,15х2 (r=0,829; t=4,44).
Кращі батьківські пари для одержання потомства з нетемніючою
м’якоттю: Прем’єр х Берегиня, Кристал білоруський х 78573-35N,
283
817116—18N х Добро, Дивосна х Гібрид 22, Морене х Ласунак, Мо¬
рене X Берегиня.
Якість картоплі в потомстві за показником стійкості до потем¬
ніння вареної м’якоті визначалася батьківськими формами на 57 %, а
типом схрещування - на 33 % (F=9,07>F0i=4,20). Частина взаємо¬
зв’язку та форма залежності потомства від батьків за цією ознакою
на фоні різних типів схрещування була такою:
1) високостійка х стійка: у = 21,7 - 3,6х + 0,24х2, детермінова¬
ність - 72 % (r=0,850; t=4,28>to5==2,09);
2) середньостійка х середньостійка: у = 20,2 - 3,6х + 0,20х2, детер¬
мінованість - 70 % (г=0,836; t=6,18>to5 =2,18);
3) високостійка х високостійка: у = 21,5 - 3,5х + 0,23х2, детермі¬
нованість - 69% (г=0,835; t=4,57>to5=2,06).
М’якоть, що не темніє при варінні, забезпечували батьки: Андра
х 335-21, 76-51 х Добро, Кера х Світязянка, 1.445152-74N х 4938-
45, ВИР-224 х Z-15, Гібрид 22 х Гібрид 35, Кристал білоруський х
Ласунак, Адретта х Гібрид 22.
Такий технологічний показник картоплі, як відходи при очистці
бульб у потомстві і батьків, досліджено на 57 комбінаціях. Виявлено,
що в потомстві рівень відходів від очистки бульб був вищим, ніж у
батьків. Встановлено вірогіднцй вплив батьків на цей показник
(г=0,714; t=7,59>to5==2,88). Кращі батьківські форми давали потомст¬
во з низьким рівнем відходів при очистці бульб. Так, 19 % гібридних
комбінацій дали потомство, у якого цей показник становив до 14 % -
Астілла х 1.371030-78N; Елла х 731030-78N; Супер х Добро; Море¬
не х Зарево; Світанок київський х Андра; 61.2107018-80N х 4938-45;
Кера х Віхола; Амалія х Віхола; 3031-023 х 78573-35N.
Найбільша кількість потомства (74%) характеризувалася серед¬
нім рівнем відходів - 15-16%, і лише у 7% потомства втрати досяга¬
ли більше 17%. Причиною великих відходів були дрібні бульби (40-
50 г), довга форма (1,67- 1,45), більша кількість вічок (10,6 шт.) при
глибокому їх заляганні (2,8 мм), як, наприклад, у гібридного потомс¬
тва Прем’єр х Гібрид 35.
Смак і текстура варених бульб та характер їх успадкування -
запорука споживчої якості картоплепродукту. Чим вищі такі ознаки в
батьків, тим більша ймовірність прояву їх у потомстві [13,14].
Дослідженнями 71 комбінації на смакові якості виявлено зни¬
ження середніх значень ознаки в потомстві, що є цілком припусти¬
мим. При цьому регресія взаємодії смаку батьків і потомства стано¬
вила 3,5%, але коефіцієнт кореляції був вірогідним на 99% (г=0,485;
t=5,16>toi=2,68).
Використовуючи закономірності успадкування потомством сма¬
ку, розробили рівняння регресії для різних типів Схрещування. При
цьому смакові якості потомства на 77% зумовлювалися батьківськи¬
ми формами і на 23 % (F=6,58) -типом схрещування. Для кожного
284
типу схрещування успадкування смакових якостей у потомства може
бути виражене такими залежностями:
1) у= -52,4 + 15,їх + 0,98х2, відмінний смак * добрий; детерміно¬
ваність - 66% (г=0,811; t=3,16>to5=2,13);
2) у = -172,7 - 52,2х + 4,06х2; добрий х добрий; 73 %; г=0,857;
3) у = -240,5 + 84,їх - 7,2х2; середній х відмінний; 74 %; г=0,860;
4) у = -125,4 + 43,2х - 3,6х2; середній х добрий; 79 %; г=0,889.
Із виділених кращих батьківських пар для одержання потомства з
високими смаковими якостями найбільший інтерес становили такі
комбінації: Адретга х Кристал білоруський, Астілла х 1.371031—
78N2, Пригожа 2 х Білоруська З, 612107018-80N х 4938-45.
Регресія взаємодії розварюваності бульб батьків і потомства до¬
рівнює 35% (r=0,588; t=5; 16>toi=2,68). Дисперсійний аналіз виявив
залежність розварюваності бульб потомства від батьківських форм
на 68 % і від типу схрещувань - на 32 % (F=7,41>F0i=4,20). Для кож¬
ного типу схрещувань ступінь генетичної детермінованості цієї
ознаки коливався від 56 до 91 %:
1 - сильна розварюваність х сильна; у = 531-13ІХ+ 8,2 х 2; детер¬
мінованість - 91 (г=0,955; t=5,43 tos=2.31);
2 - низька х сильна; у = 136,7 - 39,6х + 3,Ох2; 81 %; г=0,904;
3 - середня х середня; у = -54,7 - 18,6х + 1,42х; 67 %; г=0,822;
4 - сильна х середня; у = 580 - 165х + 11,8 х 2; 56 %; r =0,746.
Кращі батьки для селекції картоплі на підвищену розварюваність
бульб:
1) батьківська форма - сорт Зарево, материнська форма - Форум,
Морене, Сарма, Отрада, ВИР-377, ВИР-244;
2) батьківська форма - Білоруська 3, материнські - Супер, Ельві-
ра, Пригожа 2, Гібрид 22, Гібрид 35;
3) батьківська форма - Воловецька, материнські - Раритан, Гітте;
4) батьківська форма - Добро і 305-27М, материнська - Супер;
5) батьківська форма - Ласунак, материнська - 61.2107018-80N;
6) батьківська форма - Берегиня, материнська - Ласунак.
У спадкову ваність ознак і властивостей якості картоплі. Бі¬
льшість ознак і властивостей картоплі, які визначають її придатність
до переробки, мають кількісний характер, що свідчить про їх полі-
генний механізм контролю. Картопля - автотетраплоїд, що зумовлює
поліморфність більшості її ознак. Ця особливість вимагає значних
витрат при доборі в селекційній роботі.
Відомо, що мінливість при успадкуванні кількісних ознак безпе¬
речна і залежить не тільки від генотипу, але й від дії факторів зовніш¬
нього середовища. Останні можуть маскувати прояв генотипу. Тому,
щоб вичленити дію генотипу, визначали коефіцієнт успадковуванос-
ті, який є важливим елементом передбачення успішного добору. За
результатами аналізу якісних технологічних показників у потомства,
одержаного від 12 батьківських та 22 материнських форм, що були
285
об’єднані в короткі незамкнені діалельні цикли, було встановлено,
що рівень спадкоємності був високим у потомства від таких мате¬
ринських форм: Адретта, Львів’янка, Отрада, Сарма, Форум, Море¬
не, Раменська, Супер та ін.; батьківських: Ласунак, Дойна, Волове-
цька, Добро та ін.; за успадкуванням кулінарних властивостей і хо¬
роших смакових якостей - у потомства від материнських форм Ад¬
ретта, Отрада, Сарма, Форум, Морене, Віхола, Світанок київський та
ін., батьківських - Ласунак, Дойна, Добро, Білоруська рання та ін.
Результати сучасної селекції сортів картоплі, придатних до
промислової переробки. В Україні робота із селекції картоплі коор¬
динується Інститутом картоплярства УААН. За останні роки цією
науковою установою виведено більше 20 сортів картоплі, у тому чис¬
лі 13 - універсального призначення. До таких сортів із ранньої групи
стиглості належать Зов і Кобза; із середньоранніх - Обрій, Світанок
київський, Фантазія; із середньостиглих - Либідь, Українська роже¬
ва, Багряна, Лелека, Лілея; із середньопізніх - Зарево, Воловецька,
Ікар. Крім того, середньоранній сорт Свалявська виведений Гірсько-
Карпатською дослідною станцією Закарпатського інституту АПВ;
середньостиглі сорти Дзвін і Дарина - Інститутом сільського госпо¬
дарства Полісся; середньопізній сорт Тетерів - Поліською дослідною
станцією ім. О.М. Засухіна.
Столові сорти картоплі, які придатні до промислової переробки і
поєднують скоростиглість, хороші споживчі та технологічні якості,
виведено в Сумському ДАУ (табл. 4.44).
Таблиця 4.44 - Характеристика сортів картоплі, виведених у СДАУ, 9-1 балів
Сорт
Група
стиглості
Споживчі якості
Якість продуктів
тип
смак
ккал/кг
сухе
пюре
сушена
картоп¬
ля
чипси
Молодіжна
ранній
СВ
8,8
0,446
7,2
7,5
7,6
Косовщинська
« іі
АВ
7,0
0,424
6,9
7,1
7,1
Ластівка
середньо¬
ранній
С
7,9
0,483
7,3
7,5
7,8
Аграрна
11 СІ
В
7,4
0,461
7,3
7Д
7,8
Ювіляр 6070
tt U
В
7,1
0,424
7,1
7,0
7,3
Лілея Сумщини
11 14
С
7,4
0,404
7,0
7,0
7,2
Поліська
K <t
В
7,4
0,406
7,0
7,0
7,2
Злагода
11 ii
сд
7,2
0,499
7,3
7,5
7,7
Зоря Сумщини
It ii
сд
7,5
0,461
7Д
7,3
7,5
Використання універсальних сортів може забезпечити поліпшен¬
ня техніко-економічних показників роботи картоплепереробних під¬
приємств за рахунок підвищення виходу кінцевих продуктів
(табл.4.45). /
286
Для виробництва харчових, а найчастіше - технічних продуктів
іноді використовують суміш сортів. Різні сорти - це різна якість, яка
зумовлює неоднакову поведінку сировини в технологічному процесі
виготовлення продуктів. Особливо це стосується виробництва сухого
пюре і консервованої картоплі, оскільки навіть невелике збільшення
часу бланшування або варіння бульб може призвести до руйнування
клітин і в подальшому негативно вплинути на консистенцію готово¬
го продукту.
Таблиця 4.45 - Прогнозована ефективність виробництва харчових та технічних
продуктів
Сорт
Уміст
сухоі
речо¬
вини
Вихід продукту,
кг із 100 кг сировини
сушені
чипси
крохмаль
сухий,
20%-ний
крохмаль
сирий,
50%-ний
Умовний стандарт
Зов, Кобза, Дарина, Ювіляр 6070,
Тетерів, Свалявська
22
20
37
15
24
Молодіжна, Аграрна, Українська
рожева, Лілея, Ластівка
23
21
38
16
26
Фантазія, Ікар, Либідь, Лелека,
Лілея Сумщини
24
22
39
17
27
Воловецька, Обрій, Зоря Сумщи¬
ни, Дзвін
25
23
41
18
29
Світанок київський, Злагода
26
24
42
19
30
Зарево
29
27
44
22
35
Використання несортової картоплі при виготовленні чипсів при¬
зводить до погіршення зовнішнього вигляду пластівців за рахунок
різного їх кольору, а також до перевитрат олії. Так, якщо вміст жиру
в пластівцях сорту Злагода становить 40,8%, то у сортів Ластівка -
41,9, Зоря Сумщини - 42,8, Лілея Сумщини - 43,9, Молодіжна, Аг¬
рарна - 44,8 %, а у сортів Ювіляр 6070, Псільська і Косовщинська -
вже 45-46%. Слід відзначити, що при виготовленні чипсів заводсь¬
ким звичайним способом уміст жиру в пластівцях буде на 6-8%
меншим [30].
Отже, сортова чистота сировини має велике значення для проми¬
слової переробки, оскільки від неї залежить економіка підприємства,
якість продукту та його маркетинг.
Сучасною тенденцією в селекції картоплі в Білорусі водночас із
потребою подальшого підвищення врожайності гостро постало пи¬
тання про створення сортів, придатних до промислової переробки.
Тому із 27 сортів картоплі селекції Інституту картоплярства БААН
60% придатні до переробки на готові харчові продукти і напівфабри¬
кати, а також на крохмаль і спирт (табл.4.46).
287
Таблиця 4.46 - Сорти білоруської селекції, придатні до промислової переробки
на різні види продуктів
Сорт
Сухе карто¬
пляне пюре
Заморожена
картопля
Сушена
картопля
Чипси
Крох¬
маль
Атлант
+
+
-
-
-
Білоруська 3
+
+
+
+
-
Верас
-
-
-
+
-
Верба
+
+
+
+
+
Виток
+
+
+
+
+
Гарант
+
-
-
+
+
Діна
+
-
-
-
-
Жівіца
+
-
-
-
-
Ласунак
+
+
+
+
+
Лощицька
+
+
+
+
+
Нароч
+
+
+
+
-
Орбіта
+
+
+
+
+
Скарб
+
-
+
+
+
Синтез
+
+
+
+
+
Сузор’є
+
-
-
+
+
Темп
+
+
+
+
+
Яхант
-
+
+
+
-
У країнах з розвиненим картоплярством постійна увага завжди
приділяється створенню ранньостиглих сортів, придатних до пере¬
робки. У результаті цілеспрямованої селекційної роботи виведена
велика кількість сортів для картоплепереробної промисловості.
У США найбільш придатними для виготовлення чипсів є сорти
Superior, Chippeva, Irish, Cobbler, Katahdin, Kennebec, Russet Burbank,
Rural, New-Iorker, Russet Rural, Sebago, Teton\ для помфрі - Cascade,
Katahdin, Kennebec, Russet Burbank.
У Франції найбільш розповсюдженими для переробки є сорт
Bintje, а також сорти Gipsy, Агіапа.
Із сортименту картоплі Німеччини для виробництва чипсів реко¬
мендовано два надранніх сорти - Carina і Prima; дев’ять ранніх -
Mandy, Titana, Britta, Fox, Telia, Tempora, Lizera, Auralia, Elgina; п’ять
середньостиглих - Erntestdz, //se, Irmgard, Juliver, Assia; 14 серед¬
ньопізніх і пізніх - Hertha, Judika, Maritta, Monza, Saturna, Aguti,
Bodencraft, Moni, Pruceres, Tasso, Karmona, Galina, Maxilla, Turbella.
У Великобританії кращим для виробництва чипсів є сорт Record;
для переробки на помфрі - Desiree, Marts Piper, Petland Dell, Petland
Grown, Saturna, Для переробки вирощують також сорти Baillie,
Edzina, Marfona, Kingson, Pomano.
За даними Інституту генетики і сільськогосподарських дослід¬
жень, в Італії кращими для виробництва чипсів визнані сорти Altena,
Arsy, Belleisle, Chieftain, Cleopatra, Corine, Fatima, Herta, Kennebek,
Lutina, Nooksack, Spunta.
288
У Нідерландах з 17 вирощуваних ранніх сортів чотири рекомен¬
довано для переробки на чипси і катоплю фрі - Amason, Anosta,
Prior, Os tara; з 25 середньоранніх і пізніх сортів вісім придатні до
переробки — Agria, BintJe, Fijana, Hertha, Provita, Sante, Saturna.
ЛІТЕРАТУРА
1. Альсмик П.И. Селекция картофеля в БССР - Минск: Ураджай, 1979 - 127 с.
2. Барановская В.А., Шариков К.Е., Пшеничная Э.Е. Величина крахма¬
льных зерен как показатель качества картофеля //Вопросы товароведения и
технологии пищевых продуктов.- Минск: Вышейшая школа, 1972 - Вып.2-
С.124-128.
3. Гончаров Н.Д. Наследование крахмалистости при селекции на скоро¬
спелость // Картофель и овощи - 1966 - № 12.- С.73-75.
4. Гончаров М.Д., Кожушко Н.С., Адіб Абу Обайд. Стан і перспективи
розвитку галузі картоплярства //Вісник СДАУ.- Суми, 1999 - Вип.З- С.4-7.
5. Гончаров Н.Д., Кожушко Н.С., Кравченко И.В. К оценке пригодности сор¬
тов и сеянцев картофеля для промышленной переработки //Технология произ¬
водства картофеля: Науч.тр. /НИИКХ РСФСР - М., 1980 - Вып.37- С.58-64.
6. Гончаров Н.Д., Кожушко Н.С., Кравченко И.В. Технологическая оце¬
нка селекционного материала картофеля на пригодность к промышленной
переработке для пищевых полуфабрикатов: Метод, указания / БНИИКПО-
Минск, 1980.-28 с.
7. Гончаров М.Д., Кожушко Н.С., Оничко В.І. Експрес-метод для оцінки
вихідного та селекційного матеріалу картоплі // Картоплярство. - 1994-
Вип.26.- С.52-55.
8. Зуйков А.Е. Подбор исходного материала и оценка гибридных ком¬
бинаций для селекции картофеля на пригодность к промышленной перера¬
ботке на пищевые полуфабрикаты: Автореф. дис. ... канд. с.-х. наук / Бел-
НИИКХ.- Самохваловичи, 1988 - 23 с.
9. Кожушко Н.С. Моделювання якості при селекції картоплі // Вісник
СДАУ.- Суми, 1997.- Вип.І.-С.31-33.
10.Кожушко Н.С. Селекція картоплі на якість: Дис. ... д-ра с.-г. наук-
Харків, 1994 - 386 с.
11. Кожушко Н.С., Гончаров М.Д. Технологічна оцінка картоплі на при¬
датність до промислової переробки //Картоплярство.- 2000.- Вип.30.
12.Маленко І.М., Дуда В.В., Царенко М.І. Сучасний стан і перспективи
розвитку картоплепереробної галузі в Україні // Картоплярство- 1992-
Вип.29.- С.27-35.
ІЗ.Осипчук А.А. Селекція картоплі в умовах Полісся України: Автореф.
дис.... д-ра с.-г. наук / Інсішут рослинництва ім.В.Я.Юр’єва - Харків, 1993- 50 с.
14. Осипчук А.А., Положенець В.М., Мицько В.М. Комплексна оцінка в
селекції картоплі // Картоплярство - 1981.- Вип.12 - С. 12-16.
15. Оценка сортов и сеянцев картофеля на пригодность их для производ¬
ства сухого картофельного пюре / П.И. Альсмик, Н.Д.Гончаров,
Н.С.Кожушко, И.В.Кравченко // Картофелеводство и плодоовощеводство:
Науч.тр. / БНИИКПО - Минск, 1977.- Вып.2.- С.3-7.
16.Плешков Б.П. Биохимия сельскохозяйственных растений - М.: Агро-
промиздат, 1987 - С.423-434.
10 2.132
289
17. Проблемы картофеля: Материалы науч.-произв. конф. / Зап. отд.
ВАСХНИЛ.-Минск, 1974.- 142 с.
18. Прокошев С.М. Биохимия картофеля - М., 1947 - 226 с.
19. Пшеничная Э.Е., Трулль Л.А., Раптунович ГА. Определение некото¬
рых показателей, влияющих на качество сухого картофельного пюре // Воп¬
росы товароведения и технологии пищевых продуктов - Минск: Вышейшая
школа, 1973.- Вып.З.- С.61-65.
20. Соломина И.П., Суханова Р.С. Хранение и переработка картофеля в
местах производства: Обзор, информ. / ВНИИТЭИ АПК - М., 1992 - 48 с.
21.Шинкарев В.И. Изучение технологических свойств картофеля: Ме-
тод.указания / ВИР - М., 1988 - 135 с.
22. Яшина И.М. Наследуемость крахмалистости картофеля // Картофель
и овощи - 1976-№ 9- С. 13.
23.Adler G. Anforderung der Verorbeitungs industrie an den Rohstoff
Kartoffeln // Kartoffelbau.- 1977.- Bd.9.- S.24-25.
24. Adler G, Shiling M. Anforderugen an die Beschefenheit und Inhaltsstaffe
der Kartoffel fur Veredelungserzeugnisse // Land.Forseh.- Bd.26, № 2 - S.25-27
25.Burton W.G. Requirements of the urers of ware potatoes // Potato re-
search.- 1974.- № 4.- P.374-409.
26. Cleophas P. Th.M. Die Kartoffelverartende Iudustrie in Holland // Der
Kartoffelbau. - 1971.- Bd. 22, № 3.- S. 100-103.
27. Howard H.W. Factors influencing the quality of ware potatoes // Potato
Res. - 1974.- Vol.l7, № 4.-P.409-501.
28. Kaminski R. Phenotyre and genotipic correlationi of morphologial and
physiologial characters of potato // Jennet Piant- 1977 - Vol. 18, № 2 - P.l 25-133.
29.Nelson D.G. Jenkins PD Villison TC Processing potential of potato
cultivates at early harvests // Potato Res - 1988 - Vol.31, № 4 - P.633-642.
30. Putz B. Der zeitige Moglichketion zur Selektion von Verarbeitungssorten
durch den Zuchter // Kartoffelbau - 1985 - № 11- S.427- 431.
31. Ratzack K. Der Speisewert der Kartoffe - Berlin, 1935.
32. Strickerr H.W. Genalt an reduuzierenden Zuckeru in der Kartoffelknolle //
Der Kartoffelbau.- 1972.- Bd.23, №> 12.- S.298.
33.Thiers A. La consommtion de pomme de terre par habitant en hausse //La
pomme de terre fraucaise- 1970 - Vol. 32, № 336 - S.19-24.
4.6. ВИРОЩУВАННЯ КАРТОПЛІ
З ВИКОРИСТАННЯМ БОТАНІЧНОГО НАСІННЯ
4.6.1. Історія проблеми вирощування
картоплі з ботанічного насіння
До останнього часу серед учених немає єдиної думки щодо пер¬
шочергового типу розмноження картоплі - генеративного чи вегета¬
тивного. Вважається, що великий розмір квіток, яскраве забарвлення
їх віночка, наявність у них запаху свідчать про сприятливі умови для
відвідування їх комахами, що є підставою визнати основним спосо¬
бом розмноження картоплі генеративний. Додатково можна ще на^
290
звати у зв’язку з цим такі фактори, як зігнутість стовпчика, виступ
рильця над колонкою пиляка, багатонасінність ягід та ін. [14,16]. Ін¬
ші, базуючись на багатобульбовості, особливо диких видів, значній
довжині столонів, що зумовлює розповсюдження бульб на великій
площі, ідентичності матеріалу в поколіннях, твердять, що основним
способом розмноження картоплі є вегетативний.
По-іншому слід розглядати це питання, виходячи із сьогоднішніх
позицій щодо важливості біологічних і господарських властивостей
картоплі. По-перше, слід зазначити, що спрямованість селекційної
роботи не адекватна біологічним особливостям культури. Викорис¬
товуючи штучний добір, людина гіпертрофувала ті властивості кар¬
топлі, які їй необхідні, наприклад, розмір бульб, уміст крохмалю,
інших речовин тощо, хоча прояв їх не має великого значення для
збереження культури в цілому і навіть - навпаки. По-друге, значно
знизився прояв ознак, закладених у біології картоплі. Наприклад,
протягом усього періоду після інтродукції культури в Європу селек¬
ціонери пристосовували її для вирощування з використанням вегета¬
тивного розмноження [14]. А тому значна частина сортів втратила
здатність до ягодоутворення і навіть квітування. За даними Е.М. Ус¬
пенського [16], серед колекційного матеріалу Інституту картопляно¬
го господарства (Коренево) у середньому за 5 років серед 883 сортів
квітувало 86,8%, зав’язало ягоди від самозапилення 11,8% від квіту¬
ючих.
Вважається, що в Росії перші масові посіви ботанічного насіння
були проведені в 1767 р. на Камчатці і в Сибіру [4]. На початку XIX
ст. поширення культивування картоплі з використанням ботанічного
насіння відбувалося під егідою Вільного економічного товариства.
Вже в той час було встановлено, що іноді потомство від самозапи¬
лення перевершує батьківську форму за проявом основних господар¬
ських ознак. На думку деяких дослідників [4], імовірно, що саме та¬
ким чином у ті далекі часи були отримані перші місцеві сорти.
Враховуючи велику зацікавленість населення у використанні для
вирощування картоплі ботанічного насіння, Вільне економічне това¬
риство доручило П.А.Грачову описати агротехніку вирощування
картоплі з ботанічного насіння, що й було зроблено ним у 1867 р.
Проблема широкого використання істинного насіння для одер¬
жання продукції картоплі постійно цікавила вчених. Так, наприклад,
на Першому Всеросійському з’їзді генетиків у 1929 р. цьому пи¬
танню був присвячений виступ І.О.Веселовського. Численні спроби
виростити картоплю з ботанічного насіння здійснювалися і населен¬
ням. Щоб якось упорядкувати ці роботи, Наркомзем у 1933 р. видав
спеціальну постанову: “О массовом опытно-хозяйственном посеве
картофеля семенами”. Для втілення цієї постанови в життя “Сортна-
сіннєовоч” проводив заготівлю ботанічного насіння від сортів, які
рясно зав’язували ягоди від самозапилення. З цього часу значну ува¬
291
гу проблемі вирощування картоплі з використанням ботанічного на¬
сіння стали приділяти вчені. У газеті “Труд” за вересень 1933 р. дру¬
кується стаття академіка Р.В.Вільямса і професора В.А.Харченка, у
якій підтверджується надзвичайна важливість вирішення цього пи¬
тання для успішного задоволення потреб населення в картоплі. У цей
же період вперше С.М. Букасовим була висловлена думка, що ре¬
продукування картоплі істинним насінням дозволить значно знизити
загрозу ураження її хворобами.
Широкомасштабні дослідження з проблеми були виконані в 50-
60-х роках під керівництвом В. Едельштейна JL Лорхом, І. Таракано-
вим, А. Лахіним та ін. Ними, з метою механізації посіву насіння, за¬
пропоновано його дражування. Крім цього, була розроблена техно¬
логія вирощування картоплі з ботанічного насіння, яка передбачає
одержання в перший рік “сівку” (дрібних бульб масою 2-4 г), а на
другий рік від їх посадки - отримання повноцінного садивного буль¬
бового матеріалу. Дослідженнями академіка В.І.Едельпггейна та його
учнів доведене зменшення в матеріалі від ботанічного насіння, порів¬
няно з бульбовим репродукуванням, ураження рослин вірусними
хворобами. Залучення в роботу сортів Камераз, Епрон дозволило на
другий рік після посіву насіння одержати урожайність 20 т/га, що
свідчить про перспективність широкого застосування цього методу
отримання насіннєвого матеріалу у виробництві.
Вирощування картоплі з використанням ботанічного насіння
завжди викликало зацікавленість в Україні. Так, В.Д.Щербачова твер¬
дить про значно менше ураження вірусними хворобами сіянців, по¬
рівняно з бульбовим репродукуванням, і задовго до В.І.Едельпггейна
вона запропонувала дворічну схему вирощування насіннєвого мате¬
ріалу. Результати її досліджень свідчать, що картопля, вирощена з
насіння сорту Деодара, мала врожайність 343 ц/та, що на 12 % біль¬
ше, ніж із бульбового матеріалу.
Доцільним вирощувати картоплю з насіння вважав відомий кар¬
топляр професор І.ІІІушкарьов. Він і його аспірант Ю.Чердинов осо¬
бливо виділяли перспективність заходу в боротьбі з виродженням
картоплі. Позитивні практичні результати в цьому напрямі одержали
І.О.Веселовський, Г.Г.Лєпиліна, Е.Н.Сагалович, Е.А.Вовк, Ю.А. Ру¬
мянцев та багато інших.
Сьогодні майже в кожній країні, де займаються картоплярством,
проводяться дослідження або практично вирощується картопля з ви¬
користанням ботанічного насіння.
Позитивні та негативні сторони обох типів розмноження кар¬
топлі. Однією з основних переваг вегетативного способу розмно¬
ження картоплі є можливість відтворення всього комплексу агроно¬
мічних властивотей, який характерний для сорту, що є по суті веге¬
тативним потомком сіянців, тобто рослини, вирощеної з насіння.
Особливо велике значення це має при використанні бульб для пере-
292
робки на окремі напівфабрикати або глибокої переробки, наприклад
чипси і т.п.
Основних недоліків цього типу розмноження кілька. По-перше,
соковиті, багаті поживними речовинами бульби є хорошим субстра¬
том для численних шкідників і збудників хвороб. Накопиченню ін¬
фекції також сприяють оптимальні умови для її збереження під час
зимового зберігання бульб. Для оздоровлення картоплі застосовують
багато способів та методів. Останнім часом найбільшого поширення
набув спосіб звільнення від інфекції за допомогою термотерапії та
апікальної меристеми. Але при цьому матеріал не стає імунним. По¬
дальше вирощування його супроводжується повторним зараженням і
перезараженням. Тобто, великі витрати, які спрямовуються на оздо¬
ровлення і розмноження, як вважається (хоч не завжди так є), безві¬
русного матеріалу, дуже часто не окуплюються прибавкою врожаю
навіть у перші кілька років.
Крім цього, як справедливо вважають окремі вчені, у результаті
культивування обмеженої кількості клітин з використанням штучно¬
го живильного середовища відбуваються певні зміни і виникають
варіації. Оздоровлена рослина далеко не завжди є тотожною вихідній
формі. А тому метод не можна вважати надійним і ефективним.
По-друге, навіть незважаючи на використання насінницьких та
інших заходів при бульбовому репродукуванні відбувається старіння
сорту і його виродження. При цих явищах втрачається гетерозисний
ефект, який теоретично при вегетативному розмноженні повинен
існувати досить довго.
По-третє, близько третини зібраного врожаю використовується
на насінні цілі. А враховуючи втрати при зимовому зберіганні, яке
часто проходить не в оптимальних умовах, частка матеріалу, який
закладається як садивний, може досягати 50 % від зібраного врожаю.
По-четверте, велику масу насінної картоплі необхідно транспор¬
тувати з поля (при збиранні) і на поле (при садінні) та зберігати з до¬
триманням певних вимог.
По-п’яте, з кожним роком практично в усіх країнах світу спостеріга¬
ється подорожчання садивного матеріалу картоплі. За даними Департа¬
менту плодоовочівників Медисонського університету CLLLA, вартість
його на 1 акр зросла в 1989 р. порівняно з 1988 р. на 70 доларів.
Як позитивні сторони вирощування картоплі з використанням
ботанічного насіння можна зазначити наступні.
Насамперед, це мінімальні витрати на отримання насіння. З 1 га
батьківського розсадника можна отримати 4-6 т ягід, або 40 кг/га
чистого ботанічного насіння. Вартість 1 кг його становить 100 дола¬
рів США. Крім цього, потреба істинного насіння на 1 га дорівнює
100-120 г [2]. Тобто, насінням, отриманим з 1 га батьківського роз¬
садника, можна засіяти 333-400 га. Витрати на посів насіння станов¬
лять 5 % від загальних [12].
293
Важливим є те, що, як вважали видатні мікологи професор
А.А.Ячевський і професор Н.А.Наумов [4], що підтверджено і в наш
час (Jonce F.), через істинне насіння практично не передається вірус¬
на інфекція, відпадає потреба у застосуванні високовитратних насін¬
ницьких технологій і тестуванні садивного матеріалу.
Крім того, звільняється значна посівна площа, на якій за звичай¬
ної технології необхідно було б вирощувати садивний матеріал кар¬
топлі, а це становить при внутрішньогосподарському насінництві
30-35 % від загальної площі під картоплею.
Нарешті, ювенільні форми (з позицій онтогенезу картоплі) є
більш життєздатними, а тому більш урожайними, вони зазнають мен¬
шого впливу негативних зовнішніх факторів [14].
У цілому вважається [3], що найбільш доцільно вирощувати кар¬
топлю з ботанічного насіння в країнах із жарким кліматом.
Разом з тим сівба картоплі насінням має недоліки. Перш за все,
до сьогодні немає сортів або батьківських пар, які дають потомство
без розщеплення. Ті форми, які використовуються для отримання
ботанічного насіння, повинні майже не розщеплюватися за забарв¬
ленням бульб, їх формою і деякою мірою - за забарвленням квіток та
деякими іншими ознаками. Хоч окремі дослідники [14] не вважають
розщеплення при генеративному розмноженні перешкодою. Свою
думку вони обґрунтовують тим, що вшцеплення повного (в кожному
з наборів хромосом) рецесиву - надзвичайно рідке явище. Оскільки
негативні ознаки контролюються домінантними генами, то їх можна
зразу вибракувати. Розщеплення полігенно контрольованих ознак
має варіаційний ряд, який у середньому має такий прояв властивості,
як вихідна (і) форма (и).
По-друге, як правило, урожайність сіянців першого року нижча,
ніж при бульбовому репродукуванні. Проте, за даними НДПСГ Росій¬
ської Федерації, при оптимальних умовах і агротехніці сіянці можуть
утворювати до 1-3 кг бульб із куща [12].
Враховуючи те, що масштабні дослідження щодо вирощування
картоплі з ботанічного насіння проводяться лише в останні 10-15
років, не для всіх зон і регіонів відпрацьоване агротехнічне забезпе¬
чення виконання роботи.
4.6.2. Генетичні основи і підходи до створення
сортів, придатних для розмноження
ботанічним насінням
Генетичні і загальнобіологічні особливості картоплі. Культур¬
ні сорти мають у соматичних клітинах 48 хромосом, тобто є тетрап-
лоїдами. Чотири набори гомологічних хромосом зумовлюють значну
гетерозиготність сортів, що означає практичну неможливість отри¬
мання потомства, близького за проявом хоча б основних ознак. Крім
цього, більшість господарсько цінних ознак картоплі характеризу¬
294
ється полігенним контролем, а це означає, що важливе місце серед
потомства має варіаційний ряд різноякісних форм. Проте, з іншого
боку, якщо властивість контролюється домінантними генами, то зо¬
всім не обов’язково для отримання однорідного за ознакою потомст¬
ва мати повністю гомозиготну форму. За розщепленням триплекси
(АААа) характеризуються однаковою частотою домінантного по¬
томства, як і квадриплекси (АААА). Проте середнє вираження озна¬
ки потомства здебільшого близьке за значенням до батьківських
форм, тобто при такому генетичному контролі в середньому прояв
властивості достатній. Генетична відмінність картоплі від інших
сільськогосподарських культур зводиться до відсутності одноманіт¬
ності першого гібридного покоління, тому через розщеплення, яке
відбувається в Fj , неможливо отримати одноманітні форми. Це є
значною перешкодою при вирощуванні картоплі з ботанічного на¬
сіння.
Однорідне за проявом ознак генеративне потомство теоретично
можна отримати при схрещуванні двох гомозиготних форм. Одним із
найбільш поширених методів отримання їх є інбридинг. У картоплі
його використання значно ускладнене через значний прояв інбредної
депресії. Особливо це стосується сортів, що, як правило, мають вузь¬
ку генетичну основу і досить часто є близькородинними формами. І,
навпаки, вираження гетерозису має протилежну природу. Максима¬
льний його прояв відбувається, коли в локусі знаходяться чотири рі¬
зні алелі. Такий генетичний стан називається гетероалелізмом, або
тетраалелізмом. Тобто, через різку втрату життєздатності з кожним
наступним інцухтуванням практично не можна отримати гомозиготні
форми, а без них не вдається досягти гетероалелізму.
Дуже часто в картоплі внаслідок інбридингу порушується розви¬
ток генеративних органів. Саме через це здійснення процесу інцух-
тування є складним завданням.
До 30-40-х років XX ст. основним методом селекції картоплі бу¬
ла внутрішньовидова гібридизація, що різко звузила генетичну осно¬
ву сортів картоплі. Цьому сприяла також втрата цінних форм під час
епіфітотій раку картоплі та фітофторозу. Генетичною особливістю
картоплі є велика різноманітність диких і культурних видів. Залу¬
чення їх у селекційну практику значно розширило генетичну базу
культури, але одночасно, через вищеплення серед потомства диких
ознак, збільшило частоту селекційного браку. Тобто, відбулося збі¬
льшення гетерозиготності матеріалу, у зв’язку з інтрогресією генів
від співродичів культурних сортів, що ускладнило отримання близь¬
кого за проявом ознак генеративного потомства.
Селекція, як еволюція, що керується волею людини, завжди про¬
водилася у напрямі гіпертрофування одних ознак за рахунок інших.
Наприклад, коли в 20-30-х роках XX ст. вважалося можливим вирі¬
шити проблему захисту картоплі від фітофторозу за рахунок R-генів
295
(вертикальної стійкості), добори у цьому напрямі призвели до втрати
значної частини матеріалу з полігенною стійкістю. Спрямованість
селекції на отримання сортів високоврожайних, висококрохмалистих з
певним проявом інших ознак зумовила відсутність у певної частини
сортів квітування, ягодоутворення від самозапилення. Генетичною ос¬
новою останнього є порушення, що відбуваються при кон’югації хро¬
мосом у статевих клітинах. Встановлено, що при температурі 25-30°С
кількість моновалентів може досягати 48. Серед 50 залучених у дослі¬
дження сортів порушення спостерігаються на всіх стадіях розвитку
мікроспороцитів. При першому діленні, крім тетрад, формувалися діа¬
ди, тріади, що зумовлювало низьку фертильність пилку. Аналогічні
результати отримали ВА.Рибин, W.Ellison та багато інших.
Ріст і розвиток рослин картоплі з ботанічного насіння значно від¬
різняється від одержаних з бульб. Це стосується кореневої системи,
стеблостою, галуження стебел і т. п. Ймовірно, згадані відмінності
накладають відбиток на прояв господарських ознак. А тому здебіль¬
шого відсутня кореляція їх прояву серед сіянців першого року та бу¬
льбовими поколіннями, що не дозволяє проводити повний обсяг до¬
сліджень у рік посіву насіння.
У ягодах картоплі формується дрібне насіння. Маса однієї насі¬
нини в середньому становить близько 0,6 мг, що зумовлює труднощі
генеративного репродукування картоплі. При безпосередньому сіян¬
ні насіння в грунт навіть за умови обробітку його речовинами, що
стимулюють ріст, існує загроза, що значна частина його може не зі¬
йти. Більш ретельне спостереження за проростанням насіння можли¬
ве при розсадному способі, але, як свідчать наші дані та інших уче¬
них, кількість загиблих рослин за період проростання насіння зби¬
рання врожаю сіянців в окремих комбінаціях може досягати 50 %.
Це є значною перешкодою не лише при проведенні досліджень, але й
при вирощуванні картоплі з використанням ботанічного насіння.
Вимоги до сортів, придатних для вирощування картоплі з
використанням ботанічного насіння, та умови його отримання.
Використання генеративного способу розмноження картоплі перед¬
бачає вирощування популяцій, отриманих тим чи іншим шляхом.
Але для того, щоб мати насіння, потрібний батьківський розсадник
спеціальних форм. Вони мають забезпечувати високогетерозисні по¬
пуляції, які повинні характеризуватись такими властивостями: висо¬
кою польовою схожістю насіння, швидким ростом і розвитком рос¬
лин, скоростиглістю, стійкістю проти несприятливих метеорологіч¬
них і грунтових умов, хвороб, шкідників, високою врожайністю і
товарністю бульб, однорідністю і вирівняністю бульб та ще деякими
господарськими властивостями.
Важливими вимогами до сортів, придатних для генеративного
розмноження, є їхня ягодоутворювальна здатність, а з урахуванням
результатів наших досліджень - і обнасіненість ягід. Складність до¬
296
тримання цих вимог зумовлена втратою здатності сортів до генера¬
тивного відтворення, а тому в цьому напрямі має бути проведена пе¬
вна робота [15].
Безперечно, оптимальними будуть сорти, які здатні зав’язувати
велику кількість ягід (або з незначною зміною у прояві цієї ознаки) в
різних умовах. Проте, як свідчить селекційна практика, зав’язування
ягід також залежить і від багатьох агрометеорологічних факторів.
Важливим для ягодоутворення є наявність високофертильного
пилку. На його формування значною мірою впливають зовнішні
умови. Сприятливою є помірна вологість з невисокою температурою
повітря [19].
Враховуючи те, що заздалегідь заготовлений пилок не може бути
використаним (особливо при штучному запиленні) за 1-2 дні, постає
проблема його зберігання. Встановлено, що в сухому прохолодному
(з температурою не більше +5°С) місці пилок може зберігатися на¬
віть кілька місяців. Практично це досягається зберіганням пилку в
ексикаторах із вологовбирною речовиною, які розміщують у побуто¬
вих холодильниках. При цьому важливо, щоб не використаний за
період роботи пилок (день, половину дня) не зсипати разом із тим,
який знаходиться в холодильнику (Методические рекомендации по
проведению исследований с картофелем, 1983).
За впливом на розвиток і функціонування генеративної сфери
фактори зовнішнього середовища розміщуються таким чином: світ¬
ло, поливний режим, температура, вологість.
Багатьма дослідниками встановлено, що сприятливим для квіту¬
вання є червоний спектр, і, навпаки, сині промені діють негативно
[16]. Виявлена пригнічувальна дія синьої частини спектра на асимі¬
ляцію, що й зумовлює затримку і слабке квітування. У випадку за¬
безпечення рослини продуктами асиміляції розвиток генеративних
органів відбувається навіть у темноті. У рослин, які формують запа¬
сні органи (бульби, цибулини тощо), слід ураховувати інтенсивність
і спектр освітлення минулого року.
Важливий вплив на квітування має фотоперіодизм. Рослини, що
розвиваються в умовах довгого дня, швидше плодоносять при ціло¬
добовому освітленні, а рослини короткого дня не зможуть нормально
розвиватися в умовах довгого дня [16].
Значний вплив на квітування і ягодоутворення має поживний ре¬
жим грунту. Підживлення азотними добривами сприяє ягодоутво-
ренню і навіть збільшує масу насіння. Підвищення дози азоту, що
вносили в грунт, з 100 мг/100 г до 300 мг сприяло поліпшенню квіту¬
вання і проростанню пилку [23]. Збільшення дози азоту при піджив¬
ленні до 240 кг/га сприяло формуванню великого насіння. У вегета¬
ційних дослідах максимальне внесення азоту становить 800 кг/га. Ще
кращі результати можна отримати при збалансованому мінеральному
живленні або при збільшенні кількості підживлень з 3 до 6-ти [19].
297
Вивчали вплив високої температури (35-40°С) і високої вологос¬
ті на квітування сортів Смисловський, Альма, Вольтман в оранжереї
[16]. У результаті сорти, які у звичайних умовах добре квітують,
утворили на 15-ти кущах: перший - 4 квітки, другий - 7, а Вольтман
зовсім не мав квіток. Крім цього, в усіх трьох пилок був стерильним.
Результати дослідів узагальнив H.Klebs. Він встановив, що: 1) в
умовах інтенсивної асиміляції та при інтенсивному освітленні і за¬
безпеченні вологою та поживними речовинами рослини мають три¬
валий вегетативний ріст і розвиток; 2) при інтенсивній асиміляції та
освітленні, але обмеженому доступі води і мінеральних солей відбу¬
вається утворення квіток; 3) при середньому забезпеченні рослин
вологою і поживними речовинами напрям процесів (утворення кві¬
ток або подальший вегетативний ріст) повністю залежить від інтен¬
сивності освітлення.
Методи отримання форм, придатних для вирощування кар¬
топлі з ботанічного насіння. Першим етапом досліджень щодо ви¬
рощування картоплі з використанням ботанічного насіння є отри¬
мання форм, придатних для успішного використання цього способу.
Існує кілька підходів до вирішення проблеми. Перші способи ви¬
рощування картоплі з істинного насіння при використанні самозапи¬
лення проведені А.П.Герном. Урожайність цього матеріалу була ниж¬
чою, ніж у батьківських форм. Проте однорідність за забарвленням
квіток, бульб і скоростиглістю виявилася високою. В інших дослі¬
дах, які проводилися в Ленінградській області, потомство від само¬
запилення мало високу врожайність (у середньому близько 400 ц/га).
Виділені кращі в цьому відношенні сорти: Веселовська 2-4, Агроно¬
мічна, Анко, Ода, Г’євонт, Епрон, Гібридна 14, Амзель, Черокі, Ка-
мераз, Невська, Ізобіліє [4].; Швальбе, Богатир, Комсомолець, Етюд,
Адвіра, Видра, Г’євонт, Гідра, Маріелла, Ліра, Сотка, Умбра, Ураль¬
ська рання, Швальбе [17]. Порівнюючи результати, отримані різними
дослідниками, можна зробити висновок, що окремі сорти (Швальбе,
Г’євонт, Комсомолець, Анко та деякі інші) придатні для вирощуван¬
ня картоплі з ботанічного насіння в різних грунтово-кліматичних
умовах.
Отримання ботанічного насіння від самозапилення є найбільш де¬
шевим способом, порівняно із схрещуванням, а тому, незважаючи навіть
на дещо нижчу врожайність вирощених рослин, порівняно з гібридами,
цей метод, як вважають окремі дослідники, є перспективним.
Більш вирівняне за проявом морфологічних ознак потомство мож¬
на отримати в процесі інцухтування. Щодо депресивного впливу його
на прояв агрономічних властивостей, зокрема врожайності, серед уче¬
них немає єдиної думки. А.П.Герн [5] вважає, що врожайність самоза-
пилених ліній значно нижча, ніж у вихідних сортів. За даними
К.З.Будіна [1], вираження згаданої властивості різко знижується, по¬
чинаючи з Із, але здатність зав’язувати ягоди від самозапилення посту¬
298
пово втрачається з 11. На нашу думку, дещо протилежні результати до¬
сліджень можна пояснити генетичною характеристикою вихідного
матеріалу: у А.П.Герна використані культурні сорти, які деякою мірою
є близькородинними, а при використанні форм із широкою генетич¬
ною основою (бекроси багатовидових гібридів), з якими працював
К.З.Будін, негативний вплив інцухтування значно менший.
Щоб отримати насіння від гібридизації, необхідно виконати до¬
даткові роботи (кастрація, збір і нанесення пилку), що підвищує за¬
тратність методу. Але при використанні штучного схрещування, по¬
рівняно із самозапиленням, збільшується не лише врожайність
бульб, але й ягід [19].
Щоб зменшити негативний вплив інцухтування на прояв госпо¬
дарських ознак запропонована динамічна популяційна селекція
[11,14]. Селекція на гетерозис дуже часто базується на комбінаційній
здатності батьківських компонентів. Цей метод використовується
також і в селекції картоплі. Результати оцінки комбінаційної здатно¬
сті дозволяють повторно залучати в схрещування певні батьківські
форми для проведення додаткових доборів кращого потомства і за¬
кріплення прояву їх властивостей у бульбових поколіннях. Таку се¬
лекцію автори назвали статичною [11,14].
Створення форм картоплі, придатних для вирощування з викори¬
станням ботанічного насіння, неможливе в результаті статичної се¬
лекції. Для вирішення проблеми була запропонована динамічна по¬
пуляційна селекція. Вона враховує те, що картопля в основному є
самозапильною культурою, а тому високий урожай та інші господар¬
ські властивості мають зберігатися у F2. З іншого боку, культурні
сорти є складними гетерозиготами, тобто генетична константність
може бути досягнута лише протягом багатьох генерацій, що і стано¬
вить динамічний характер такої селекції. Перший етап нічим не від¬
різняється від класичних підходів при виведенні сортів, коли прово¬
диться схрещування батьківських форм. Вибракування і добір про¬
водять серед матеріалу 1-го бульбового покоління. Кращі форми
примусово самозапилюють для отримання F2 (автори ще позначають
такі популяції Іь що для даного випадку, мабуть, більш правильно).
Серед F2 здійснюють таку саму роботу й отримують F3 і т.д. У
зв’язку з тим, що сіянці від кожного примусового самозапилення
підтримуються через бульбове репродукування, їх на кожному з ета¬
пів легко залучати у схрещування й отримати нове Fi. Серед його
потомства також проводяться вибракування і добори в кожному на¬
ступному самозапиленні, як і в попередньому циклі. Тобто, відбува¬
ється чергування самозапильних ліній (F2-F5) і їх схрещування, що
дозволяє уникнути інбредної депресії і звести до мінімуму ефект
розщеплення.
При статевому розмноженні гетерозиготних організмів завжди
відбувається розщеплення. Враховуючи те, що культурні сорти є
299
складними гетерозиготами, це явище проявляється в них. Проблема
ще більш ускладнюється тим, що у створюваних форм повинно про¬
являтися понад 50 агрономічних властивостей [14]. Для часткової
гомозиготизації вихідних форм запропоновано ступінчастий добір з
метою їх подальшого дослідження. Суть його зводиться до поступо¬
вого добору форм із високим вираженням однієї ознаки. При наявно¬
сті достатньої кількості матеріалу з першою властивістю виконували
добір ще за однією ознакою. Відібрані форми схрещували між собою
і проводили добір за третьою ознакою і т.д.
При необхідності за такою схемою (або до названих властивос¬
тей) можна проводити і негативний добір. Обов’язковою умовою
успішного виконання ступінчастого добору є значна гетерозигот¬
ність вихідної популяції за найбільш можливою кількістю ознак, а в
цілому із загальної популяції за властивостями шляхом схрещування
їх потомства формуються окремі сіянці.
Теоретичні дослідження з проблем розробки по картоплі дозво¬
лили запропонувати ще один спосіб створення форм, придатних для
вирощування картоплі з використанням ботанічного насіння. Після
виявлення у деяких видів порушень мікроспорогенезу, які зумовлю¬
ють утворення нередукованих гамет (диплогамет), це явище почало
використовуватися практично, але теоретичне обгрунтування його
застосування було розроблене дещо пізніше. Встановлено, що 2п-га-
мети утворюються в результаті нерозходження хромосом в ІІ-му ді¬
ленні мейозу, що рівноцінно 1-му діленню. При цьому формуються
високогетерозисні гамети, які рівноцінні генотипу батьківської фор¬
ми. У цьому випадку спостерігається дисомічне успадкування, що
веде до високої гетерозисності, посилення міжалельного взаємо¬
впливу і, відповідно, до підвищення позитивного рівня прояву гос¬
подарських ознак.
На основі явища нередукованості гамет розроблено метод ство¬
рення гетерозисних гомозиготних самофертильних сортів, які прида¬
тні для вирощування картоплі з ботанічного насіння, тому що утво¬
рюють однорідне, з чітко вираженими агрономічними властивостями
потомство [13].
Розроблена схема отримання мейотичних поліплоїдів при ство¬
ренні гомозиготних, самофертильних сортів картоплі [1].
4п(С) х 2п(ДВ) = Fj4n + Fi3n +F12n(fl)
2п(Д)х2п(ДВ) = Р12п(МДГ)
4п(С) х F!2n(MДГ) = В4п(МП)
Р,2п(МДГ) х 4n (С) = В4п(МП),
де Д - дигаплоїдні форми, С - тетраплоїдні форми, ДВ - диплоїдні
види, МДГ - міжвидові дигаплоїдні гібриди, МП - мейотичні полі-
плоїди.
Реалізація цієї схеми дозволила створити сорти, придатні для ге¬
неративного розмноження, а саме: ВІР-3, ВЕР-6, ВІР-8 [1].
300
Відкриття у картоплі явища апоміксису дозволило дослідникам
прогнозувати використання його для отримання форм, придатних
для вирощування картоплі з ботанічного насіння. Вважається, що
потомство від таких форм було однаковим за проявом властивостей
батьків, зберігаючи при цьому всі переваги генеративного розмно¬
ження картоплі. Проте цей напрям досліджень лише розвивається.
Перспективним, на нашу думку, є пошук серед складних міжви¬
дових гібридів форм, придатних для генеративного розмноження
картоплі. Широка генетична основа такого матеріалу дозволяє отри¬
мувати із значною частотою гетерозисне потомство навіть від його
самозапилення. Нами виділені бекроси складних багатовидових гіб¬
ридів: 85.29ІСІ9; 85.119 с 2; 86.410 с 74; 90.672 с 24; 90.35 с 448; по¬
томство яких від самозапилення не дає розщеплення за кольором
бульб, а середнє популяційне значення врожайності знаходиться на
рівні сортів-стандартів [6].
Біологічні властивості насіння і стимуляція його проростан¬
ня. При вирощуванні картоплі з ботанічного насіння дуже важливо
знати його біологію, а також на цій основі розробити заходи подо¬
лання перешкод, нею зумовлених. Істинне насіння картоплі дуже
дрібне: маса 1000 насінин становить 0,6 г [5], або однієї - 0,6 мг [1].
Середня довжина насіння - 1,8-2,2 мм, або в три рази менша за на¬
сіння томатів.
Біологічною особливістю насіння є наявність твердої і сухої обо¬
лонки (насіннєвої шкірки), вологість якої становить 6,7 % [1]. Для
того, щоб насінина проросла, необхідно забезпечити її вологою, кі¬
лькість якої у 600 разів має перевищувати масу насінини. Крім цього,
оболонка насінини складається з великої кількості жирів і білків, які
погано пропускають воду до ендосперму зародка, тому для пророс¬
тання насіння потрібна велика кількість води.
Значний вплив на проростання насіння справляють умови його
зберігання. Встановлено, що зберігання істинного насіння картоплі в
рідкому азоті (-196°С) може бути досить тривалим, і що при цьому
навіть поліпшуються його посівні якості.
Доведено, що свіжозібране насіння має значно нижчу схожість,
порівняно з тим, яке зберігається 2 роки. Це можна пояснити періо¬
дом спокою насіння, який триває близько 20 місяців. Проте останнє
залеясить від багатьох факторів: повноти дозрівання ягід, технології
виділення насіння тощо. Позитивні результати дає ферментування
ягід протягом 24 годин перед виділенням із них насіння.
Широкі дослідження щодо впливу строку зберігання насіння на
його схожість проведені в Ленінградському СП [8]. Підтверджений
факт низької схожості насіння в перші 2-4 місяці після його виді¬
лення (32-69 %), а також енергії проростання (до 6 %). Уже на 7-й
місяць після виділення схожість насіння становила 92-96 %. Подібні
результати (91-99 %) отримані в період до 10 років. У цей час найбі-
301
лыною також була енергія проростання (46-94 %). Після 15 років
зберігання схожість і енергія проростання різко знижуються.
В цілому тривалість збереження доброї схожості насіння зумов¬
люється умовами та строком зберігання, способами отримання, по¬
ходженням. За даними багатьох дослідників [7, 10], таке насіння мо¬
же мати високе вираження схожості упродовж 10—13 років.
Враховуючи те, що в культурних сортів штучний добір відбував¬
ся не в напрямі посилення дії факторів, які б сприяли збереженню
форм при генеративному відношенні, посівні якості насіння в них
значно нижчі, ніж у їхніх родичів. А тому у диких видів, навіть при
зберіганні насіння в лабораторних умовах, життєздатність не знижу¬
ється 10-15 років, а, можливо, і довше.
Значний вплив на проростання істинного насіння картоплі справ¬
ляє стимуляція цього процесу. Виявлена позитивна дія коливань те¬
мператури на схожість та енергію проростання насіння, зміна темпе¬
ратури з 22-25°С вдень до +12°С вночі підвищує енергію проростан¬
ня насіння картоплі.
На температуру проростання насіння впливає і термін його збері¬
гання. Оптимальною для свіжозібраного насіння є температура 10—
12°С. Крім цього, свіжозібране насіння краще проростає на світлі, а
на те, яке зберігалося 2-3 роки, цей фактор не впливає .
Проморожування сухого насіння протягом 24 годин при темпера¬
турі -2-5 °С з наступним підвищенням її при намочуванні в розчині
гібереліну (0,005 %) до +5°С позитивно впливає на схожість насіння.
При вивченні дії на насіння гібереліну в концентрації 0,02-0,05 %
отримані різні дані. Більшість дослідників указують на позитивну
дію цього способу при проростанні насіння. Схожість та енергія
проростання при цьому підвищувалися на 20-25 %. У дослідженнях
К.З.Будіна [1] на 5-у добу не виявлено позитивної дії на проростання
ні гіберелінової кислоти, ні намочування в розчині солей (0,5 %
KNO3 + 0,5 % К3РО4). Проте, на 8-у добу кількість пророслого насін¬
ня у варіантах намочування у воді протягом 48 годин і замочування в
розчині гібереліну була приблизно однаковою (94 і 96 % відповідно).
Позитивний вплив на проростання насіння справляє застосування
розчину гібереліну після попереднього проморожування насіння до -
5°С протягом однієї доби (Вавилова М.А.; Лаптев Ю.П.). В інших
дослідженнях (Попов В.А.) значне підвищення схожості насіння до¬
сягається обробкою його кінетином (0,5 мг/л) у комплексі з індоліл-
оцговою кислотою (1 мг/л).
Встановлено, що насіння саме синтезує необхідні для нього ре¬
човини, але, наприклад, додаткове намочування його в розчині гібе¬
реліну позитивно впливає на розщеплення в ньому вуглеводів. Акти¬
візація ферментів, яка також відбувається під дією гібереліну, - це
лише перший етап змін, що відбуваються в насінні та рослинах. Се¬
ред усіх ферментів найбільш активізується оксидаза, яка посилює
302
активність росту. В інших роботах відзначається, що під час пророс¬
тання зародок сам продукує гіберелінову кислоту, яка активізує
алейроновий шар. Основним ферментом, що при цьому утворюється,
є а-амілаза, яка гідролізує ендосперм і забезпечує зародок поживни¬
ми речовинами. При додатковому додаванні гібереліну (до оптиму¬
му) підвищується швидкість цих процесів. На необхідність присут¬
ності гіберелінової кислоти для активізації алейронового шару вка¬
зують також інші дослідники. Оптимальною концентрацією речови¬
ни є більше 0,001, але менше 0,075 %, інші - пригнічують ріст
картоплі (СЕР, 1979).
Встановлено, що позитивний вплив гібереліну не припиняється
під час накльовування насіння, а продовжується в подальшому, на¬
приклад, при утворенні зелених паростків. При цьому також значно
знижується вплив негативних зовнішніх факторів. У дослідах, вико¬
наних у Китаї, доведена позитивна дія гібереліну (при намочуванні
насіння в його розчині) на вихід насіннєвої бульбової фракції. З 200
насінин за 3 роки одержано 10 т бульб при середній врожайності 262
ц/га, що на 40 ц/га більше, ніж у контролі. Аналогічні результати
отримали В.І.Разумов та Р.С.Лимар (1969).
Схожість та енергію проростання насіння можна посилити дією
іонізуючої радіації. Оптимальною дозою опромінення насіння є 300
рентген. Встановлено, що цей захід позитивно впливає також на роз¬
виток рослин. Застосування розчину радіоізотопів урацилу (Си) при
намочуванні насіння не тільки підвищило схожіть, але й збільшило
урожайність на 61 %.
В Індії вивчали вплив на схожість насіння і життєздатність сходів
хімічної скарифікації. На зріле насіння протягом 15 хвилин діяли
азотною кислотою концентрацією 0,1; 1,0; 5,0; 10,0 %, а потім до ро¬
ку з інтервалом в один місяць вивчали схожість і життєздатність
сходів. Результати досліджень свідчать про позитивний вплив ска¬
рифікації насіння протягом усього періоду. Максимальний прояв
ознак спостерігається при концентрації кислоти 0,1-1,0 % [21].
Для стимуляції проростання насіння також успішно можуть ви¬
користовуватися природні ростові речовини з торфу: хлоргумати,
нітрохлоргумати. Значний ефект може бути досягнутий при поєд¬
нанні гуматів, гідрохінону або гіберелової кислоти. Схожість і енер¬
гія проростання при цьому були на 22 % вищі, ніж у контролі.
Позитивний вплив на схожість насіння справляє сода (0,5 %), гі¬
дрохінон (0,1 %), бромистий калій (0,05 %) при різному комбінуван¬
ні або сік алоє і перманганат калію (5%) при намочуванні насіння
протягом 10 годин. В обох випадках схожість підвищилася на 10-15 %,
а у другому - на 3-5 днів прискорилося проростання насіння.
Особливості онтогенезу рослин картоплі, вирощених із ботаніч¬
ного насіння. Онтогенез рослин картоплі при вирощуванні з істинного
насіння значно відрізняється від бульбового репродукування, що необ¬
303
хідно враховувати при розробці технологій посіву картоплі насінням
(ПКН). Насамперед слід відзначити високу життєздатність сіянців і ще
кількох бульбових поколінь. Унаслідок цього підвищується стійкість
проти несприятливих зовнішніх факторів, чим і зумовлена рекомендація
застосування ПКН у зонах, не зовсім сприятливих для культури. Причин
цього явища є кілька. По-перше, молодий організм від злиття двох ста¬
тевих клітин має значно більші можливості для пристосування до спе¬
цифічних умов вирощування. По-друге, в матеріалі з насіння повністю
відсутній негативний вплив фізіологічного старіння бульб. По-третє,
відсутня дія загального старіння сортів. По-четверте, враховуючи те, що
через насіння практично не передається інфекція, зокрема вірусна, у
здорових рослин залишається більше енергії для боротьби з іншими не¬
сприятливими факторами.
Зважаючи на те, що сіянці мають одне стебло, продуктивність
його порівняно з бульбовим репродукуванням дуже велика. Звичай¬
ним є явище, коли сіянці дають урожайність з одного куща 1-3 кг.
Це досягається не лише вимогами згаданих вище факторів, але й
значним гілкуванням стебла.
На перших етапах сіянці ростуть і розвиваються значно повіль¬
ніше, ніж рослини з бульб. Це стосується всіх фаз. Але, якщо порів¬
няти габітус куща рослин, вирощених із насіння, і бульб наприкінці
вегетації, то практичної різниці між ними не виявимо.
Тривалість періоду вегетації рослин залежить від того, чи вони
виросли з насінини, чи з бульби. У ранніх сортів початок відмирання
бадилля настає приблизно на 70-й день після садіння, а в сіянців во¬
но залишається зеленим досить тривалий час. Через це бульби сіян¬
ців часто збирають фізіологічно молодими. Отже, сіянці значно від¬
стають у розвитку від рослин із бульб, як окремо по фазах, так і в
цілому за період вегетації, що слід ураховувати при виборі методу
посіву насіння в теплиці (розсадна культура) чи безпосередньо у
грунт. У зонах із коротким вегетаційним періодом застосування
останнього методу може призвести до загибелі рослин від ранніх
осінніх приморозків.
Істотні відмінності між сіянцями і рослинами з бульб спостері¬
гаються також за початком бульбоутворення. В останньому випадку
початок зав’язування їх в основному збігається в часі з початком бу¬
тонізації, а в сіянців - значно раніше бутонізації (майже на 50 днів).
На цьому навіть базується один із методів раннього виділення скоро¬
стиглих форм (за початком бульбоутворення у розсади).
За даними А.Мокроносова, бутонізація у вегетативно розмноже¬
них рослин картоплі настає через 20-23 дні після сходів. Перші бу¬
тони сіянців з’являються через 75-90, що й зумовлює тривалий зага¬
льний цикл їх розвитку.
Відомо, що висока температура (25-35°С) під час зав’язування
бульб призводить до їх фізіологічного старіння. А тому на півдні уже
304
в перший рік вирощування насінний матеріал стає малопридатним
для подальшого використання. А при вирощуванні сіянців їхній ріст
і розвиток відбуваються в оптимальних умовах, тому що надземна
частина формується при досить теплій погоді і довгому дні, а бульби -
при більш низькій температурі і короткому дні.
4.6.3. Технологія вирощування картоплі
з ботанічного насіння
Існує кілька способів вирощування картоплі з істинного насіння:
1 - прямий посів у грунт у гребні або без них; 2 - посів у гряди окре¬
мого розсадника і пересадка сіянців у поле (часто для цього викорис¬
товують парники, теплиці тощо); 3 - висадка бульб з окремого роз¬
садника в поле [23]; 4 - добір серед сіянців 1-го року клонів і виса¬
джування їх у наступному році.
Кожен із способів має позитивні і негативні сторони. Прямий по¬
сів насіння в грунт можна застосувати лише в регіонах, де кількість
днів із середньодобовою температурою більше 10°С становить 120.
Це пов’язано з тим, що період вегетації рослин від прямого посіву
насіння у грунт на 30-40 днів довший, ніж за використання розсад¬
ного способу. Крім цього, висіяне насіння дуже чутливе до нестачі
вологи у грунті, що може призвести до зрідженості насаджень кар¬
топлі. Рослини при прямому посіві насіння у грунт через подовження
періоду вегетації (епіфітотія хвороб, масова поява шкідників) не мо¬
жуть виділити енергію для захисту від дії екстремальних факторів.
Необхідність і складність боротьби з бур’янами (через низькі темпи
росту рослин з насіння на перших етапах росту і розвитку) також
ускладнюють поширення цього способу вирощування картоплі.
Розсадний спосіб, порівняно з іншими, знайшов найбільше по¬
ширення. В організаційному відношенні він близький до вирощуван¬
ня рису в країнах Азії або вирощування багатьох овочів у країнах
Європи. При розсадному способі відношення кількості висаджених
рослин до зібраних значно вище, ніж при прямому посіві у грунт.
Попереднє вирощування рослин в умовах закритого грунту дозволяє
їм адаптуватися до зовнішніх умов і при пересадці в поле вони здатні
відразу (або майже відразу) “включати в роботу” фотосинтезуваль-
ний апарат. При розсадному способі вирощування картоплі з ботаніч¬
ного насіШія усуваються недоліки, пов’язані із прямим його посівом:
скорочується вегетаційний період, висаджена в поле розсада має до¬
статню енергію для продовження активного росту навіть при не зо¬
всім сприятливих зовнішніх умовах. Водночас розсадний спосіб є
більш трудомістким, потребує більших витрат на основні засоби.
Проте його переваги є відчутнішими, ніж недоліки, а тому він знай¬
шов найбільше поширення, хоч деякі дослідники вважають ці пере¬
ваги тимчасовими, доки не буде вдосконалений комплекс заходів для
прямого посіву насіння у грунт.
305
Інші два способи вирощування картоплі з ботанічного насіння
через великі витрати не знайшли широкого застосування.
Порівняння таких трьох способів, як прямий посів у грунт, розсад¬
ний і бульбами з насіння в умовах жаркого клімату дослідної станції
університету штату Джорджія показало, що найбільш доцільним є
розсадний спосіб.
Аналогічні дослідження проведені в зоні помірного клімату
(штат Вашингтон), де тривалість вегетаційного періоду становить
200 днів. Одержані результати свідчать, що при розсадному способі
врожай був вищим на 20-50 % [20].
У Фінляндії вивчали два способи вирощування картоплі з ботаніч¬
ного насіння: прямий посів у грунт і розсадний метод. Розсаду виро¬
щували в теплиці і пересаджували в поле у віці 4-х тижнів. Загальний
період росту розсади становив 144 дні (у тому числі 28 днів у тепли¬
ці), а рослин від прямого посіву насіння в грунт - 124 дні. Середня
врожайність у першому випадку становила 3,38 кг/м2, а в другому -
2,33.
Аналогічні дослідження виконані в Казахському НДІ картопля¬
ного і овочевого господарства. Встановлено, що при прямому посіві
насіння в грунт сходи недружні, рослини розвиваються дуже повіль¬
но. Часто кількість отриманих рослин з висіяного насіння становить
10-11 %, а виживання сіянців було лише 5,2% .
У дослідах, виконаних в умовах Самаркандської області, уро¬
жайність рослин із розсади була на 17-89 ц/га вищою, ніж при пря¬
мому посіві насіння у грунт.
Уперше було запропоновано вирощувати розсаду у парниках,
проте цей спосіб через його трудомісткість не набув поширення. У
подальшому для вирощування розсади використовували пластмасові
лотки, контейнери, ящики тощо. У Канаді вважають, що найбільш
доцільно використовувати при вирощуванні картоплі із ботанічного
насіння пластикові контейнери квадратної або прямокутної форми.
Це дозволяє найбільш ефективно використовувати площу культива¬
ційних споруд, механізацію [18].
Останнім часом для вирощування розсади широко використову¬
ють касети, що дозволяє підвищити продуктивність, порівняно з гор¬
щиковою культурою, у 3-7 разів [9].
Враховуючи значні витрати на обігрів теплиць, запропоновано
вирощувати розсаду на відкритому повітрі. Найбільш доцільним ви¬
явився такий варіант: прямий посів насіння в теплиці з наступним
виносом паростків за її межі [18].
При вирощуванні розсади використовують різні грунтові суміші.
Найпоширенішою є наступна: дернова земля, перегній, торф, річко¬
вий пісок (в однаковій пропорції). Але, як зазначають інші автори,
використання такої суміші призводить до випадання або росту непов¬
ноцінної розсади - 20-50 %, паростків і молодих рослин. Крім цього,
306
такі субстрати є добрим середовищем розмноження і нагромадження
інфекції та шкідників, протягом експлуатації вони мають непостій¬
ний хімічний склад та фізичні властивості, і їхнє приготування по¬
требує значних витрат [9]. А тому останнім часом почали шукати
штучні субстрати, які б не мали недоліків природних. В Інституті
фізико-органічної хімії Білорусі створені штучні субстрати ИС-2В та
ИС-2С. Вирощені на них сіянці переважали у 2-3 рази за врожай¬
ністю ті, що росли на природних субстратах. Використання перших
дозволило збільшити виживання рослин, їх висоту, кількість листків,
а кількість бульб у кущі зросла залежно від особливостей популяції у
2-8 разів. Можливість повторного використання субстратів звільни¬
ла селекціонерів від необхідності заготовляти грунтові суміші. У
штучні субстрати рекомендується додавати добрива, що повільно
розчинюються, особлива увага звертається на їх дозування.
Встановлено, що температура вирощування розсади впливає на
подальший розвиток рослин. Багато дослідників рекомендують загар¬
тування її перед висаджуванням у поле. Цей захід забезпечує добре
приживлення рослин у полі навіть при перепаді температури.
Здебільшого розсаду вирощують із застосуванням пікірування.
Висаджують розсаду в поле з грудкою землі на корінні або без неї.
Перший спосіб особливо ефективний у засушливих і жарких зонах,
оскільки сприяє кращому приживленню розсади. В умовах відносно
низьких температур і високої відносної вологості різниця між обома
способами садіння практично відсутня.
При пікіруванні розсада дуже часто має столони і навіть маленькі
бульби. Дослідженнями Д.Т. Абдукаримова встановлено, що росли¬
ни, вирощені без пікірування, краще розвивались і утворювали біль¬
ше бульб, ніж із застосуванням названого заходу. Зрозуміло, що під
час пікірування відбувається часткове порушення кореневої системи,
що в подальшому негативно впливає на її ріст і розвиток. А тому пе¬
ресаджування розсади до початку утворення бульб забезпечує вищу
врожайність, ніж у пізніші строки.
Незважаючи на використовувані агротехнічні заходи, під час пе¬
ресаджування розсада зазнає шокової дії екзогенних факторів. Вико¬
ристання генотипів, стійких проти стресового фактора, є найбільш
надійним захистом від нього. Виявлена значна відмінність потомства
різних популяцій за реакцією на пересадку [23]. Перші автори вста¬
новили, що гібридні сіянці мають більш високу здатність до віднов¬
лення після пересаджування порівняно з матеріалом від самозапи¬
лення. Саме два фактори - висока схожість насіння і хороше прижи¬
вання розсади в полі - можуть забезпечити отримання врожаю до
332 ц/га. На контролі (бульбове розмноження) величина прояву
ознаки становила 234 ц/га.
Відпрацьовано також багато способів прямого посіву насіння у
грунт. У країнах Південної Америки, Новій Зеландії насіння сіють
307
разом із торфом або перегноєм. Для цього готують суміш у співвід¬
ношенні 1:12. Причому, торф повинен мати 100 %-ну вологість. До¬
зують суміш, використовуючи пластикові капсули об’ємом 0,25 л, у
які попадає в середньому по 5 насінин. Після дводенного знаходжен¬
ня насіння в капсулах їх висаджують на відстані 30 см одна від од¬
ної. Недоліком цього способу є великі затрати ручної праці. Значне
поширення набув спосіб гідровисіву насіння. Відомо, що в польових
умовах насіння проростає тривалий час, а зовнішні фактори для цьо¬
го часто не зовсім сприятливі. А тому, щоб зменшити негативний
вплив на проростання насіння температури і вологості, його перед
посівом пророщують, з іншого боку наклюнуте насіння потребує бе¬
режного поводження з ним, щоб не травмувалися первинні корінці. З
цією метою використовуються гідросівалки, а як середовище для
насіння використовують воду чи гель. Висіяне таким чином насіння
значно швидше проростає, що сприяє прискоренню проходження
фаз розвитку, зменшує негативний вплив зовнішнього середовища
[22]. Гідросівалками насіння висівають на глибину 2-3 см. У дослі¬
дах НДПСГ Російської Федерації застосування гідровисіву дозволило
отримати урожай на 10-30 % вищий, ніж при застосуванні розсадно¬
го способу [13]. Інший принцип, на якому можуть бути побудовані
гідросівалки,- використання стиснутого повітря. Одержані експери¬
ментальні дані свідчать, що продуктивність праці при використанні
таких сівалок зростає порівняно з базовим способом сівби у 10—
15 разів.
У Казахському НДІ картопляного і овочевого господарства для
посіву насіння використовують ручну селекційну однорядкову сівал¬
ку РС-1. При цьому запропоновано завантажувати в бункер разом з
насінням наповнювач.
Давню історію має дражування насіння. Уперше його застосував
у 1779 році на зерні вівса А.Болотов. Для цього він використав де¬
ревний попіл. Велику увагу цьому питанню приділяв академік
В.І.Едельпггейн. Він вважав, що дражуванням насіння можна вирі¬
шити ряд питань: збільшити запас поживних речовин, поліпшити
фізико-механічні властивості насіння, умови його проростання і роз¬
витку сходів, більш рівномірно проводити сівбу.
Пориста оболонка, яка утворюється в результаті дражування на¬
сіння, посилює контакт його з грунтом, активніше притягує вологу
до нього, добре забезпечує його поживними речовинами при пророс¬
танні.
Уперше дражування насіння картоплі застосували В.І.Едель-
штейн, Г.Г.Лєпиліна (1966). їх висновок: дражоване насіння має під¬
вищену схожість, через збільшення коефіцієнта тертя забезпечується
більш рівномірний посів.
Важливим фактором, який впливає на врожай та його складові, є
густота рослин. Оптимальною вважається 100 сіянців на 1 м2, хоч ця
308
величина значно змінюється залежно від схожості насіння та енергії
проростання. У дослідах СЕР найвища врожайність насінних бульб
була одержана при густоті сіянців 150 шт./м2 і мінімальній схожості
95 %. Встановлено, що розрідження не забезпечує підвищення уро¬
жаю бульб, а лише збільшує масу бадилля. Крім цього, підвищується
частка великих бульб.
В умовах 1986 року оптимальною була густота рослин 96 шт./м2.
При цьому отримано 976 бульб загальною масою 8,77 кг зимою в
теплиці і, відповідно, 782 і 104 кг - улітку в полі. Загущений посів
зменшує продуктивність рослин, а також знижує вміст крохмалю в
бульбах.
При повному забезпеченні вологою максимальна продуктивність
отримана при густоті сіянців 50-100 шт./м2. А в дослідах, виконаних
на дослідній станції університету штату Пжорджія (США), при гус¬
тоті висівання насіння 79, 131, 164 шт./м максимальний урожай був
отриманий в останньому варіанті.
Залежно від комплексу екзогенних факторів запропонована різна
норма висіву ботанічного насіння. Так, у дослідах ТСГА на 1 га ре¬
комендують висівати 5-10 кг насіння, у Горьківському СП - 2-2,5 кг. У
В’єтнамі при висіванні насіння в поле оптимальна густота становить
42 рослини на 1 м2, а при використанні розсади - 240 тис. рослин на
1 га [21].
В умовах теплиці з метою більш ефективного використання пло¬
щі при прямій сівбі, порівняно з горщиковою культурою, використо¬
вують загущений посів.
За даними СІР, у країнах Південної Америки при прямому посіві
насіння в грунт використовують малогабаритну сівалку з дисковими
і вакуумними висівними апаратами. Витрати насіння при цьому ста¬
новлять 800-10000 г/га, тоді як при оптимальному проведенні опера¬
ції ця величина може зменшитися до 150-200 г/га.
Оскільки розміри ботанічного насіння картоплі малі, важливим є
дотримання оптимальної глибини його посіву. Встановлено, що схо¬
жість насіння обернено пропорційна глибині загортання. Рекоменду¬
ється в полі висівати насіння на глибину 1,5-2 см, а при вирощуванні
розсади - 0,5-1 см. Вважається, що повільне проростання насіння
зумовлене погіршенням обміну повітря у грунті із збільшенням гли¬
бини посіву.
При вирощуванні картоплі з ботанічного насіння важливим захо¬
дом є боротьба з бур’янами. Значно нижчі темпи початкового росту
рослин з насіння порівняно з бульбами не дозволяють їм конкурува¬
ти з бур’янами за швидкістю росту. Проблема ускладнюється ще й
тим, що використання гербіцидів на таких площах обмежене через
негативну дію їх на молоді сіянці. А тому в багатьох випадках єди¬
ним методом боротьби з бур’янами на посівах картоплі з ботанічного
насіння є механічне їх знищення.
309
Після висадки розсади в поле, навіть у гребні, вона певний час
адаптується, а тому не росте і взагалі є малою. Через це, особливо на
перших порах, механізоване знищення бур’янів навіть у міжряддях
призводить до прикидання розсади, її травмування. Ще більшого не¬
гативного впливу при механізованому знищенні бур’янів зазнають
рослини від прямого посіву насіння в грунт. Виходячи з цього, в
обох випадках (а особливо в останньому) при догляді за посівами
використовується ручний обробіток.
Застосування гербіцидів навіть при бульбовому розмноженні ча¬
сто зумовлює зупинку росту рослин, негативно діє на них. Ще біль¬
шою мірою це стосується розсади або сіянців від прямого посіву у
грунт. У зв’язку з цим для боротьби з бур’янами при генеративному
розмноженні картоплі запропоновано використовувати зменшені нор¬
ми витрат препаратів. Дослідами, виконаними в СЕР, встановлена
можливість використання на матеріалі від ботанічного насіння зен-
кору зі зменшеною удвоє, порівняно з обробкою бульбового матеріа¬
лу, нормою.
В Угорщині встановлена висока ефективність застосування на
посівах картоплі з ботанічного насіння таких гербіцидів, як афалон
та арезин.
В умовах жаркого клімату Тифтонської дослідної станції універ¬
ситету Джорджія (СЕНА) на посівах картоплі з істинного насіння в
боротьбі з бур’янами використовують наступні засоби: перед садін¬
ням розсади - паракват, метабрамурон, через 20 днів після садіння -
метрибузин, напропамід. Для зменшення токсичної дії гербіцидів за
чотири дні до їх внесення в грунт рослини обробляли ціамінозидом.
В іншому штаті (Вашингтон) для боротьби з бур’янами викорис¬
товують такі гербіциди: метрибузин, дифенамід, напропамід, алах-
лор, пебулат, ЕРТС [20].
Звичайно, загущені посіви мають меншу забур’яненість, ніж
зріджені.
Враховуючи слабку, особливо на перших етапах росту і розвитку,
кореневу систему розсади або рослин від прямого посіву істинного
насіння у грунт, площі, де вони розміщуватимуться, мають бути
більш родючими, порівняно з бульбовим репродукуванням та добре
удобреними. Особливо позитивний вплив на рослини справляють
мінеральні добрива, зокрема азотні. У В’єтнамі під розсаду вносять
20-40 тонн гною і NPK у співвідношенні за діючою речовиною
100:27:100 кг/га.
В умовах жаркого клімату Тифтонської дослідної станції універ¬
ситету штату Джорджія (С1ЕІА) перед прямим посівом насіння у
грунт вносили NPK у дозах 44:39:112, а після посіву - дифенамід (4,5
кг/га) і додатково азот. В іншому штаті (Вашингтон) в аналогічних
умовах при прямому посіві у грунт вносилось 250:80:80 кг/га діючої
речовини NPK і цинкові добрива - 10 кг/га [20].
і
310 1
Інше співвідношення NPK застосовували при вирощуванні кар¬
топлі з ботанічного насіння у Фінляндії (університет Хельсинки) -
6:7:17 при внесенні 800 кг/га суміші.
Особливістю вирощування картоплі з використанням ботанічно¬
го насіння є отримання серед сіянців першого року порівняно дріб¬
них бульб, які не завжди можна використати для вирощування про¬
довольчої картоплі. А тому запропоновано кілька схем отримання
насіннєвого бульбового матеріалу з ботанічного насіння. Найбільш
поширені і вивчені дві - однорічна та дворічна.
Однорічна схема зводиться до вирощування товарної картоплі
через розсаду або прямим посівом насіння протягом одного року.
Дворічна забезпечує отримання товарного врожаю за два роки. Про¬
міжним (у перший рік) є одержання сівку.
При однорічній культурі насіння висівають взимку в теплиці.
Мінімальна густота стояння рослин забезпечує отримання максима¬
льного виходу бульб, які є сформованими до червня-липня. їх оброб¬
ляють стимуляторами і висаджують у полі з тим, щоб у жовтні зібра¬
ти врожай. Тобто, за цією схемою вже протягом одного року вдаєть¬
ся отримати товарний урожай.
Ще у 1932 р. В.Щербачова запропонувала використовувати для
товарного виробництва друге або третє бульбове покоління, оскільки в
умовах Полтавщини сіянці 1-го року утворюють дрібні бульби. Пізні¬
ше (1957 р.) вирощувати картоплю з ботанічного насіння за дворіч¬
ною схемою запропонували В.І.Едельпггейн і його учні.
Дворічна культура картоплі з ботанічного насіння передбачає
першого року отримати сівок (відносно дрібні бульби), які висаджу¬
ють на другий рік, щоб отримати товарний урожай.
У селекційних установах при вирощуванні картоплі з істинного
насіння використовується модифікація дворічної схеми - додається
ще один рік, а також розсадник (насіннєвий) [14]. З використанням
такої схеми на перший рік у насіннєвому розсаднику з бульб сортів,
гібридів, що мають добре ягодоутворення, отримують рослини і яго¬
ди з насінням. У наступному році з насіння вирощують рослини та
одержують сівок, і лише на третій - товарний урожай. Основними
показниками на кожному етапі є: у насіннєвому розсаднику - макси¬
мум ботанічного насіння, для розсадників сівку - максимальна кіль¬
кість бульб (а не їхня маса), для виробничих посівів - товарність уро¬
жаю.
У СІЛА протягом семи років вивчали ефективність двох схем виро¬
щування картоплі з ботанічного насіння - дворічної і однорічної. Одно¬
річну культуру отримували з розсади або після прямого посіву насіння у
грунт. Дворічна включала: перший рік - вирощування сівку розсадним
способом чи прямим посівом; другий рік - вирощування товарного вро¬
жаю із сівку. У результаті проведених досліджень зроблено висновок,
що в СІЛА більш ефективною, особливо для фермерів, є дворічна схема.
311
У південних районах однорічну культуру картоплі з ботанічного
насіння можна впроваджувати і без теплиць. У цих умовах можна за
один рік, висіваючи насіння рано навесні в поле, отримати сівок і
використати його в подальшому для вирощування товарної картоплі
(двоврожайна культура).
Численними дослідженнями встановлена необхідність дотриман¬
ня при вирощуванні картоплі з ботанічного насіння спеціальної агро¬
техніки, яка значною мірою визначається біологічними особливос¬
тями потомства. Грунт має бути добре окультуреним, з високим умі¬
стом поживних речовин, добре забезпечений вологою. Агротехніка
вирощування сівку значно впливає на врожай, товарність картоплі
при другому бульбовому поколінні.
На добре удобрених грунтах розмір сівка не впливає на товарний
урожай. Навіть найменша його фракція (менше 5 г) дозволяє отрима¬
ти урожай, який не поступається бульбовому при масі материнської
бульби 50-60 г.
Посадку невеликого за розмірами сівка (до 40 г) можна проводи¬
ти, використовуючи дещо вдосконалену сівалку для цибулі.
Таким чином, при доопрацюванні окремих питань вирощування
картоплі з використанням ботанічного насіння можна буде усунути
деякі моменти, що роблять галузь картоплярства високозатратною і
малоефективною.
ЛІТЕРАТУРА
1. Будин К.З. Перспективы семенного размножения картофеля // С.-х.
биология.- 1985. - № 5.- С.32-37.
2. Будин К.З., Киру С.Д. Приемы генеративного (семенного размноже¬
ния) картофеля //Бюл. ВИР - Л., 1990 - Вып.203 - С.3-8.
3. Будин К.З., Палеха С.Б., Киру С.Д. Выращивание картофеля из се¬
мян-Л., 1987- 11 с.
4. Веселовский И.А., Вовк Е.А. Оценка сортов картофеля при их ге¬
неративном размножении //Культура картофеля семенами.- Горький,
1983.- С.7.
5. Герн А.П. О самоопыленных линиях у картофеля //Доклады
ВАСХНИЛ- М., 1940 - Вып.7.- С.29.
6. Гордієнко В.С., Подгаєцький А.А. Аналіз складових генофонду кар¬
топлі за придатністю для вирощування з використанням ботанічного насіння
//Картоплярство - 2000 - Вип.30 - С.113-117.
7. Киру С.Д. Усовершенствование способов выращивания картофеля из
семян//Научн.-техн. бюл. ВИР-Л., 1986 -Вып. 165 -С.84-86.
8. Котова К.А. Технология выращивания сеянцев картофеля в горшеч¬
ной культуре и возможность браковки гибридов при селекции //Доклады
ВАСХНИЛ - М., 1989.- № 3 - С. 10—11.
9. Кучумов В.О. Выращивание сеянцев картофеля в кассетах //Селекция
и семеноводство - 1986 - № 6 - С.47-48.
10. Кучумов В.О. Схема производства картофеля на основе настоящих
гибридных семян //Селекция и семеноводство - 1990 - № 2 - С.41-43.
11. Румянцев Ю.А. Гетерозис при селекции картофеля, размножаемого
семенами: Автореф. дис. ... канд. с.-х. наук / НИИ картофельного хозяйст¬
ва.-М., 1989.- 22 с.
12. Склярова Н.П., Кучумов В.О. Влияние стимуляторов роста и микро¬
элементов на полевую всхожесть и урожайность гибридных популяций кар¬
тофеля при возделывании семенами // Селекция, семеноводство и
биотехнология картофеля - М.: НИИКХ, 1989 - С.57-64.
13.Тринклер Ю.Г. Большой цикл развития картофеля и возможность раз¬
множения его семенами: Автореф. дис.... д-ра биол. наук - М., 1975 - 35 с.
И.Тринклер Ю.Г. Динамическая популяционная селекция картофеля,
размножаемого семенами с помощью ступенчатого отбора: Сб. науч. тр.
Горьковского СХИ. - Горький, 1983 - С.8-21.
15. Тринклер Ю.Г., Румянцев Ю.А., Гувенков В.И. Создание межвидовых
гибридов для размножения семенами //Бюл. ВИР. Л., 1990 - Вып.203- С.8-12.
16. Успенский Е.М. Биология цветения картофеля //Работы НИИКХ.- М.,
1935.- 152 с.
17. Фаворов А.М. К истории вопроса о размножении картофеля семенами на
Украине // Культура картофеля семенами: Сб. науч. тр - Горький, 1983 - С.1.
18. De Jong H.Evalution and implementation of several bedding piant
technology components forthe raising of potato breeding seedlings // American
Potato Journal - 1989. -Vol. 66, № 2.- P.93-100.
19.Malagamba P. Potato production from true seed in tropical climates //
Hortscience- 1988.- Vol.23, № 3.- P.495-500.
20. Martin M.W. Cultural practices for using true seed in potato production
under temperate climates // Hortscience - 1988 - Vol.23, № 3.- P.505-510.
21. Mitra S., Kushari D. Response of seed during different storage periods //
Geobios.- 1987.- Vol. 14, № 6.- P.253-257.
22. Suhar R., Javorski A. Eugeneering need potato production from true seed
//American Potato Journal - 1986 - Vol.63, № 3 - P.l 31-140.
23. Wiersema S.J. A method of production seed tubers from true potato seed
//Potato research - 1986 - Vol. 29, № 29 - P.225-237.
4.7. СХЕМА, ОРГАНІЗАЦІЯ І ТЕХНІКА
СЕЛЕКЦІЇ
В Інституті картоплярства розроблена і прийнята схема селекції, згід¬
но з якою передбачається отримання, вивчення, розмноження і відбір
селекційного матеріалу відповідно до завдань із створення сортів.
У схему селекції входять такі складові: розсадник батьківських
форм, перший селекційний розсадник (сіянці й однобульбові), дру¬
гий селекційний розсадник, третій селекційний розсадник, основне
випробування, конкурсно-екологічне випробування і виробниче ви¬
пробування. Крім того, у спеціальних дослідах на інфекційних та
провокаційних фонах проводиться випробування селекційного мате¬
ріалу на стійкість проти хвороб і шкідників. У лабораторних умовах
селекційний матеріал оцінюється за біохімічними і смаковими якос¬
тями, стійкістю проти хвороб, лежкістю та іншими показниками.
313
На основі польових і лабораторних оцінок проводиться відбір се¬
лекційного матеріалу і передача кращих генотипів до державного
сортовипробування.
Розсадник батьківських форм. У ньому вирощують сорти, між¬
сортові та міжвидові гібриди, виділені за генотипом. Для розсадника
вибирають ділянку поблизу водоймища, яку удобрюють органічними
і мінеральними добривами. Набір вихідних батьківських форм і кіль¬
кість рослин кожної з них визначають об’єм і плани робіт. Площа
живлення рослин - 70x30x70 см.
У розсаднику проводять гібридизацію та збір ягід від самозапи¬
лення.
Батьківські форми вивчають за комплексом ознак, проводять фі-
топрочистку, для посадки в наступному році відбирають здоровий
матеріал.
Сіянці (генеративне покоління). їх одержують із гібридного на¬
сіння або від самозапилення вихідних батьківських форм. Вирощу¬
ють із розсади в польових умовах при площі живлення 70х30 см або
в горщиках, розміщених у теплиці чи відкритому грунті.
Відбір сіянців проводять за формою, забарвленням бульб, дов¬
жиною столонів, ураженням фітофторою, вірусними хворобами, пар¬
шею та іншими ознаками. За урожайністю проводять частковий ін¬
дивідуальний відбір гнізд із тих сіянців, які вирощувались у полі. У
такому разі їм присвоюється селекційний номер. Із невідібраних сіян¬
ців можуть набиратись однобульбові (по одній бульбі з куща й
об’єднуватись у межах комбінації).
Сіянці, вирощені горщиковою культурою, за урожайністю не ви¬
браковують. Тут відбирають по одній бульбі з куща, які об’єднують
у межах комбінації. Оцінку за врожайністю проводять у наступному
році.
Однобульбовки (перше бульбове покоління). У полі із площею
живлення 70x30x40 см вирощується матеріал, відібраний у поперед¬
ньому році із сіянців - по одній бульбі з куща. Кожен генотип пред¬
ставлений однією рослиною. Під час вегетації проводять вибраку¬
вання хворих рослин, мітять рослини з інтенсивним ростом бадилля.
Збирають урожай на початку відмирання бадилля, а при селекції
на ранньостиглість - на 70-80-й день після посадки. При цьому про¬
водять індивідуальний відбір за комплексом ознак: формою бульби,
кольором шкірки і м’якуша, ураженістю фітофторозом, паршею зви¬
чайною, вірусними хворобами, стебловою нематодою, умістом крох¬
малю, частково - за врожайністю. Відібраним генотипам присвою¬
ються селекційні номери. 1
Другий селекційний розсадник (перше або друге бульбове поко- 1
ліния). У ньому вирощують і вивчають матеріал, відібраний у поперед¬
ньому році за клоновим принципом: на сіянцях (перше бульбове поко¬
ління) і в розсаднику однобульбових (друге бульбове покоління).
314
Селекційні номери розміщують на одно-дворядкових ділянках по
5-20 рослин, залежно від наявності посадкового матеріалу. Через
кожні 25-100 селекційних номерів розміщують сорти-стандарти.
Площа живлення рослин - 70x28-30 см. У період вегетації прово¬
дять фенологічні спостереження, облік ураження хворобами, прочи¬
стки від хворих кущів і домішок. Позначають номери з інтенсивним
початковим ростом, роблять загальну оцінку за габітусом куща і
морфологічними ознаками. Для визначення скоростиглості прово¬
дять пробне викопування по 1-2 кущі з ділянки на 60-65-й день піс¬
ля посадки. Остаточне збирання врожаю проводять на початку від¬
мирання бадилля. Відбір виконують за комплексом ознак: урожайні¬
стю, кількістю і вирівняністю бульб у гнізді, глибиною вічок, ураже¬
ністю рослин і бульб хворобами, смаковими якостями, потемнінням
м’якуша, умістом крохмалю та ін.
Із числа відібраних номерів в Українську науково-дослідну стан¬
цію карантину рослин відсилається проба із 5 бульб для попередньо¬
го випробування ракостійкості першого року.
У третьому селекційному розсаднику (друге або третє буль¬
бове покоління) вирощують і вивчають селекційні номери на дво¬
рядкових ділянках по 80 кущів кожна. Через кожні 20-30 номерів
розміщують сорти-стандарти. Площа живлення рослин - 70x28-30 см.
У період вегетації проводять фенологічні спостереження, облік ура¬
ження рослин хворобами і шкідниками, видалення хворих рослин і
домішок.
Визначають польову стійкість проти фітофторозу - шляхом штуч¬
ного зараження листків і бульб у лабораторних умовах. Для визна¬
чення господарської стиглості на 60-65-й день після посадки прово¬
дять пробне викопування - по 4-6 кущів з ділянки. Збирають урожай
на початку відмирання бадилля. Відбір проводять за врожайністю,
вирівняністю і формою бульб, ураженістю та стійкістю проти хво¬
роб, умістом сухих речовин, смаковими якостями та іншими цінними
ознаками.
Восени зразки (по 10 бульб кожного відібраного селекційного
номера) відправляють в Українську науково-дослідну станцію каран¬
тину рослин для попереднього випробування ракостійкості другого
року, а також по три бульби в Інститут захисту рослин для поперед¬
нього випробування стійкості проти золотистої цистоутворюючої
картопляної нематоди першого року.
Основне випробування. У ньому селекційний матеріал вивча¬
ють один-два роки, порівнюючи із сортами-стандартами; ділянки -
дворядкові, по 40 кущів кожна, повторність - чотириразова.
На ділянках основного випробування проводяться фенологічні
спостереження, облік ураження хворобами, серологічні аналізи на
ураження вірусами, визначення стійкості листків проти фітофторозу
при штучному зараженні в лабораторних умовах.
315
Для визначення господарської стиглості проводять дві пробні коп¬
ки по 4 кущі з ділянки: першу - на 55-65-й день після посадки, другу -
через 14-15 днів після першої. Негативних відборів (прочистку) у
розсаднику випробування не проводять.
При остаточному відмиранні бадилля проводять повний збір
урожаю. Визначають загальну і товарну урожайність, товарність
бульб, середню масу товарної бульби. У лабораторних умовах визна¬
чають смакові якості (шляхом дегустації), стійкість проти фітофто¬
розу при штучному зараженні бульб її збудником, уміст сухої речо¬
вини та її компонентів.
Відбір і бракування селекційних номерів проводять на основі
узагальнених даних, отриманих при випробуванні та оцінках. Від
кожного відібраного номера для попереднього випробування стійко¬
сті проти золотистої цистоутворюючої картопляної нематоди друго¬
го року в Інститут захисту рослин відправляється зразок із 5 бульб.
Одночасно з основним випробуванням закладають розсадник
розмноження селекційних номерів (500 і більше кущів кожного). Тут
проводять фітопрочистки та інші заходи для підтримання рослин у
здоровому стані.
Конкурсно-екологічне випробування проводиться протягом 2-
3 років у різних грунтово-кліматичних зонах у порівнянні із сортами-
стандартами, які є наскрізними для всіх зон і певної, окремо взятої
зони.
Для першого конкурсно-екологічного випробування посадковий
матеріал селекційних номерів розсилається оригінатором із селек¬
ційного розмноження, а для стандартів - еліти чи супереліти одного
походження.
У другому і третьому конкурсно-екологічному випробуванні в
кожній точці використовується посадковий матеріал місцевої репро¬
дукції (метод накладання).
Ділянки в дослідах дворядкові, по 60 кущів кожна, повторність
дослідів чотириразова. Тут проводяться фенологічні спостереження,
облік ураження хворобами, серологічні аналізи на ураженість віру¬
сами, визначення стійкості проти фітофторозу при штучному зара¬
женні в лабораторних умовах. Фітопрочисток у досліді не проводять.
Для визначення господарської стиглості роблять два викопування
по 6 кущів із ділянки: перше - на 55-65-й день після посадки, друге -
через 14-15 днів після першого, остаточне - при повному відмиранні
бадилля. Після цього визначають загальну і товарну врожайність,
товарність бульб, середню масу товарної бульби. У лабораторних
умовах визначають смакові якості, стійкість проти фітофторозу та
фузаріозу (при штучному зараженні бульб їх збудниками), придат¬
ність для переробки, лежкість та проводять біохімічні аналізи.
У спеціальних дослідах проводять випробування селекційного мате¬
ріалу на стійкість проти вірусних хвороб, стеблової нематоди, парші
316
звичайної, кільцевої гнилі, чорної ніжки, іржавої плямистості, потемнін¬
ня м’якуша, вивчають елементи сортової агротехніки. В Інституті земле¬
робства південного регіону селекційний матеріал випробовують на при¬
датність до вирощування двоврожайною культурою.
Селекційні номери другого та третього конкурсно-екологічного
випробування продовжують вивчати на стійкість проти раку картоп¬
лі та золотистої цистоутворюючої картопляної нематоди.
Одночасно з конкурсно-екологічним проводять і виробниче ви¬
пробування селекційних номерів та розмноження із застосуванням
фітопрочисток, клонових доборів та інших насінницьких заходів.
За результатами основного випробування та проведених оцінок
кращі селекційні номери передаються в державне сортовипробування.
Гібридизація та отримання насіння. Схрещування проводять
на рослинах батьківського розсадника.
Квіти для одержання пилку збирають у ранковий час. їх просу¬
шують, а потім за допомогою вібраційних пристроїв з них витрушу¬
ють пилок. Його збирають у маленьку пробірку, яка закривається
пробкою. Протягом кількох днів пилок може зберігатися в побуто¬
вому холодильнику. Запилюють квіти батьківських форм на початку
розвитку або перед розкриванням. Старі квіти і молоді бутони обри¬
вають. Якщо квіти батьківських форм фертильні, схрещування вико¬
нують із застосуванням кастрації. У випадку, якщо пилок стерильний
або відсутній, кастрацію не проводять. У наступні дні проводять пов¬
торне запилення тих же квіток. Запилення найбільш доцільно прово¬
дити в похмуру погоду, у ранковий і вечірній час.
Проведення декапітації (запилення на зрізаних стеблах) значно
збільшує результативність гібридизації. Стебла при цьому поміща¬
ють у проточну воду або в посудини з водою, яку міняють через кож¬
ні 3-4 дні. Для попередження загнивання стебел застосовують слабкі
розчини азотнокислого срібла, марганцевокислого калію, антибіоти¬
ків або інших препаратів.
Наприкінці вегетації материнської форми ягоди збирають у мар¬
леві пакетики, які поміщають у прохолодне приміщення (близько
+ 15°С), де витримують до дозрівання. Видаляють насіння вручну за
допомогою скальпеля або подрібнювачем із наступним промиванням
у воді. Зберігають підсушене насіння в паперових пакетах, які помі¬
щають у побутовий холодильник.
Вирощування сіянців. Висів сухого або пророщеного насіння
проводять у посівні ящики, які виставляють у теплиці або парники.
Для підвищення схожості насіння та енергії проростання його намо¬
чують у слабких розчинах стимуляторів росту, а також діють холо¬
дом (—2...-3°С) протягом 1-2 діб або змінними температурами
(+3...+22°С).
Поживну суміш готують із дернової землі, перегною і піску у спів¬
відношенні 2:1:1. Ящики з висіяним насінням накривають філь¬
317
трувальним папером, який періодично зволожують. Після сходів па¬
пір знімають. Подальший догляд зводиться до розпушування між¬
рядь, прополювання від бур’янів та періодичного поливу.
У стадії 3-4 справжніх листочків проводять пікірування сіянців у
парники або теплицю без горщиків (для вирощування розсади на
польову культуру сіянців) або в горщики із сумішшю землі і торфу,
які ставлять у теплиці, парники або у відкритий грунт (на постійне
місце).
Догляд за сіянцями, які розпікіровані в горщики на постійне міс¬
це їх вирощування, включає розпушування, прополювання від
бур’янів, полив, боротьбу з колорадським жуком та попелицями, а в
теплиці - і з мухою-білокрилкою. За місяць до збирання бульб полив
сіянців припиняють. Під час збирання бульби кожного сіянця кла¬
дуть у горщики, після чого проводять їх відбір і бракування відпові¬
дно до схеми селекції.
Догляд за розпікірованими сіянцями на розсаду для польової
культури подібний до описаного вище. За тиждень до висадки в поле
розсаду сіянців загартовують. У другій половині травня, коли мине
загроза заморозків, розсаду сіянців у стадії 5-7 листочків висаджу¬
ють у полі в невисокі гребені. Догляд за польовою культурою сіянців
включає розпушування, прополювання від бур’янів, підгортання рос¬
лин, боротьбу з колорадським жуком, фітофторозом та ін.
Збирання бульб проводять на початку відмирання бадилля, відбір
і бракування здійснюють відповідно до схеми селекції.
Агротехнічні умови. Розсадники, передбачені схемою селекції,
розміщують у полях селекційної сівозміни, де попередниками є озима
пшениця чи жито. Після збирання попередника проводять лущення
стерні, а пізніше неглибоку оранку. Наприкінці липня - на початку
серпня висівають як сидерат жито з горохом, гірчицю або редьку олій¬
ну. Наприкінці жовтня - на початку листопада вносять гній, фосфорні
й калійні добрива і приорюють або придисковуюіь його разом із сиде¬
ратом. Азотні добрива вносять під весняну оранку. Рекомендується
вносити гній - по 40 т/ га, мінеральні добрива - N60-90 Рбо-9оК9о-ш-
Селекційний матеріал висаджують в оптимальні строки садиль¬
ною машиною або розкладанням бульб у попередньо нарізані бороз¬
ни з наступним їх закриванням тракторними загортачами.
Догляд за дослідами передбачає дворазове досходове розпушу¬
вання міжрядь з одночасним боронуванням сітчастою бороною і
дворазове післясходове розпушування міжрядь. Для боротьби з
бур’янами, хворобами і шкідниками використовують комплекс ре¬
комендованих заходів. Збирання врожаю сіянців, однобульбовок і
врожаю другого селекційного розсадника проводять вручну з викла¬
данням бульб у лунки. В інших розсадниках збирання проводять
картоплекопачем, а на великих площах селекційного розмноження -
комбайном.
318
Після збирання врожаю селекційний матеріал протягом 2-3 тиж¬
нів зберігається в сортувальному приміщенні або в кагатах. Після
проведення аналізів і перебирання його завозять у картоплесховище,
де підтримується постійна температура (+2.. ,+3°С).
4.8. ОСНОВНІ ДОСЯГНЕННЯ СЕЛЕКЦІЇ
КАРТОПЛІ ЗА ОСТАННІ РОКИ ТА ЇХНЄ
НАРОДНОГОСПОДАРСЬКЕ ЗНАЧЕННЯ
Українськими селекціонерами створено велику кількість сортів
картоплі, які відзначаються високими показниками господарсько
цінних ознак, груповою стійкістю проти хвороб і шкідників, придат¬
ністю для вирощування в різних грунтово-кліматичних зонах та ін.
Створені сорти відзначаються стійкістю до звичайного біотипу
раку картоплі, а окремі з них - і до агресивних. Значна їх частина
стійкі проти золотистої цистоутворюючої картопляної нематоди
(Пролісок, Повінь, Поран, Чернігівська рання, Молодіжна, Седнівсь-
ка рання, Водограй, Обрій, Берегиня, Доброчин, Поліська 96,
Слов’янка, Пекуровська, Західна, Дзвін). Використання цих сортів
дає змогу повністю обходитися без застосування хімічних препаратів
при захисті картоплі від зазначених патогенів.
Сорти картоплі Луговська, Явір, Віринея, Західна, Дубравка,
Ольвія, Дзвін, Ракурс, Мавка та інші відзначаються високою польо¬
вою стійкістю проти фітофторозу. Використання їх дозволяє значно
зменшити кількість хімічних обробок при захисті посівів від цього
захворювання.
Відносно стійкими проти парші звичайної є такі нові сорти, як
Либідь, Молодіжна, Поран, Бородянська рожева, Водограй, Обрій,
Явір, Віринея, Ракурс, Серпанок, Дубравка та ін. Ці сорти мають
значення для підвищення якості бульб продовольчої і насіннєвої
картоплі. Певних успіхів досягнуто в селекції на стійкість проти
бактеріальних хвороб і стеблової нематоди. Використання сортів з
відносною стійкістю проти зазначених патогенів дозволяє зменши¬
ти втрати врожаю і підвищити якість продовольчої і насіннєвої кар¬
топлі.
Переважна більшість створених українських сортів характеризу¬
ються польовою стійкістю проти вірусних хвороб, що є дуже важли¬
вим для виробництва продовольчої і насіннєвої картоплі. Викорис¬
тання сортів, стійких до патогенів, має велике народногосподарське і
природоохоронне значення.
Високі і хороші смакові якості мають сорти Повінь, Кобза, Зов,
Молодіжна, Світанок київський, Мавка, Обрій, Купава, Доброчин,
Малич, Поліська 96, Явір, Віринея, Придеснянська, Горлиця, Лугов¬
ська, Ольвія, Ракурс, Дзвін та ін. Для забезпечення населення висо¬
коякісною продукцією вони мають велике значення.
319
Вагомих успіхів досягнуто в селекції картоплі на високий уміст
крохмалю. Створено ряд сортів, які містять у бульбах 17-23% крох¬
малю (Кобза, Обрій, Купава, Світанок київський, Мавка, Поліська
96, Либідь, Українська рожева, Явір, Придеснянська, Зарево, Дзвін
та ін.). Використання цих сортів дає змогу підвищити ефективність
використання картоплі переробною промисловістю, а також при зго¬
довуванні як корму для худоби.
Ряд українських сортів картоплі, маючи підвищений і високий
уміст крохмалю (Зарево, Обрій, Зов, Світанок київський, Кобза та
ін.), відзначаються придатністю для переробки на картоплепродукти.
За останні роки, починаючи з 1996 року, до Реєстру сортів рос¬
лин України внесені такі сорти картоплі української селекції: ранні -
Божедар, Косень 95, Молодіжна, Повінь, Поран, Серпанок, Чернігів¬
ська рання; середньоранні - Водограй, Доброчин, Дубравка, Купава,
Малич, Обрій, Поліська 96, Радич; середньостиглі - Віринея, Віра,
Горлиця, Західна, Придеснянська, Слов’янка, Явір; середньопізні -
Дзвін, Ольвія, Ракурс.
Божедар. Ранній сорт столового призначення. Високоврожайний.
У бульбах міститься 13-14 % крохмалю; смакові якості задовільні і
хороші. Стійкий проти звичайного і чотирьох агресивних біотипів
раку. Придатний для вирощування двоврожайною культурою на пів¬
дні України.
Бульби рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М.Засухіна Інсти¬
туту картоплярства УААН.
Косень 95. Ранній сорт столового призначення. Високоврожай¬
ний. У бульбах міститься 14-15 % крохмалю. Смакові якості задові¬
льні і хороші. Стійкий до звичайного і двох агресивних біотипів ра¬
ку. Придатний для вирощування двоврожайною культурою на півдні
України.
Бульби рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М.Засухіна Інсти¬
туту картоплярства УААН.
Молодіжна. Ранній сорт столового та універсального призначен¬
ня. Високоврожайний. Крохмалистість підвищена - 15 % і вище,
смакові якості хороші. Стійкий проти звичайного біотипу раку і зо¬
лотистої цистоутворюючої картопляної нематоди, середньостійкий
проти парші звичайної.
Бульби і м’якуш кремові.
Створений у Сумському державному аграрному університеті та
Білоруському НДІ картоплярства.
Повінь. Ранній сорт столового призначення. Високоврожайний,
інтенсивного типу. У бульбах міститься 15-16 % крохмалю, має високі
смакові якості. Стійкий проти звичайного і двох агресивних патотипів
раку, а також золотистої цистоутворюючої картопляної нематоди.
320
Придатний для вирощування двоврожайною культурою на півдні
України.
Бульби рожеві, м’якуш кремовий.
Створений в Інституті картоплярства УААН.
Поран. Ранній сорт столового призначення. Високоврожайний. У
бульбах міститься 11-13 % крохмалю, смакові якості задовільні. Стій¬
кий проти звичайного біотипу раку та золотистої цистоутворюючої
картопляної нематоди, відносно стійкий до стеблової нематоди.
Бульби рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М.Засухіна Інсти¬
туту картоплярства УААН.
Серпанок. Ранній сорт столового призначення. Високоврожай¬
ний. У бульбах міститься 13,2 % крохмалю; смакові якості задовільні
і хороші. Стійкий до звичайного патотипу раку, відносно стійкий
проти кільцевої та мокрої бактеріальної гнилі, парші звичайної, вірус¬
них хвороб. Придатний для вирощування двоврожайною культурою
на півдні України.
Бульби рожеві, м’якуш кремовий.
Створений в Інституті картоплярства УААН.
Чернігівська рання. Ранній сорт столового призначення. У буль¬
бах міститься 13—14 % крохмалю, смакові якості задовільні і хороші.
Стійкий проти звичайного біотипу раку і золотистої цистоутворюю¬
чої картопляної нематоди.
Бульби білі, м’якуш кремовий.
Створений на Чернігівській дослідній станції по картоплі.
Водограй. Середньоранній сорт столового призначення, високо¬
врожайний, інтенсивного типу. У бульбах містяться 12,5-13,5 % крох¬
малю, компоненти лікувально-профілактичної дії. Смакові якості за¬
довільні. Стійкий проти звичайного та одного агресивного патотипу
раку та золотистої цистоутворюючої картопляної нематоди. Відносно
стійкий проти фітофторозу, парші звичайної, стеблової нематоди.
Бульби жовті, м’якуш світло-жовтий.
Створений в Інституті картоплярства УААН.
Доброчин. Середньоранній сорт столового призначення. Високо¬
врожайний, інтенсивного типу. У бульбах міститься 14,5-15,5 % крох¬
малю; смакові якості хороші. Стійкий до звичайного біотипу раку і
золотистої цистоутворюючої картопляної нематоди.
Бульби рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М. Засухіна Інсти¬
туту картоплярства УААН.
Дубравка. Середньоранній сорт столового призначення. Високо¬
врожайний, інтенсивного типу. У бульбах міститься 11-14 % крох¬
малю; смакові якості задовільні і хороші. Стійкий до звичайного біо¬
типу раку та відносно стійкий проти фітофторозу, парші звичайної і
стеблової нематоди.
її2132
321
Бульби світло-рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М. Засухіна Інсти¬
туту картоплярства УААН.
Купава. Середньоранній сорт столового призначення. Високо¬
врожайний. У бульбах міститься 17,5-18,5 % крохмалю, смакові яко¬
сті хороші. Стійкий проти звичайного біотипу раку; бульби відносно
стійкі проти фітофторозу.
Бульби рожеві, м’якуш жовтий.
Створений в Інституті картоплярства УААН.
Малич. Середньоранній сорт столового призначення. Високо¬
врожайний. У бульбах міститься 16-17% крохмалю; смакові якості
хороші. Стійкий проти звичайного біотипу раку та середньостійкий
проти фітофторозу.
Бульби білі, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М. Засухіна Інсти¬
туту картоплярства УААН.
Обрій. Середньоранній сорт столового та універсального при¬
значення. Високоврожайний, інтенсивного типу. У бульбах міс¬
титься 17,6-18,5% крохмалю; смакові якості хороші. Стійкий проти
звичайного біотипу раку та золотистої цистоутворюючої картопля¬
ної нематоди; відносно стійкий проти парші звичайної та інших
хвороб.
Бульби білі із забарвленими вічками, м’якуш білий.
Створений в Інституті картоплярства УААН.
Поліська 96. Середньоранній сорт столового призначення. Висо¬
коврожайний. У бульбах міститься 17,0-17,5%; смакові якості висо¬
кі. Стійкий до звичайного біотипу раку та золотистої цистоутворюю¬
чої картопляної нематоди; середньостійкий проти фітофторозу і
стеблової нематоди.
Бульби рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М. Засухіна Інсти¬
туту картоплярства УААН.
Радич. Середньоранній сорт столового призначення. У бульбах
міститься 15-15,5 % крохмалю; смакові якості задовільні і хороші.
Стійкий до звичайного біотипу раку.
Бульби рожеві, м’якуш білий.
Створений на Поліській дослідній станції ім.О.М. Засухіна Інсти¬
туту картоплярства УААН.
Горлиця. Середньостиглий сорт столового призначення. Висо¬
коврожайний, інтенсивного типу. У бульбах міститься 16,2-17,6 %
крохмалю; смакові якості хороші. Стійкий проти звичайного та од¬
ного агресивного біотипу раку; відносно стійкий проти фітофторозу,
мокрої бактеріальної гнилі та інших хвороб.
Бульби кремові, м’якуш жовтий.
Створений в Інституті картоплярства УААН.
322
Віринея. Середньостиглий сорт столового призначення. У буль¬
бах міститься 15,5 % крохмалю; має хороші смакові якості. Стійкий
проти звичайного біотипу раку, відносно стійкий проти фітофторозу,
кільцевої і мокрої бактеріальної гнилі, стеблової нематоди.
Бульби рожеві, м’якуш світло-жовтий.
Створений в Інституті картоплярства УААН.
Віра. Середньостиглий сорт столового призначення. Високовро¬
жайний. У бульбах міститься 15-16% крохмалю; смакові якості хо¬
роші. Стійкий до звичайного біотипу раку та відносно стійкий проти
фітофторозу.
Бульби білі, м’якуш білий.
Створений в Інституті землеробства і тваринництва західного ре¬
гіону УААН.
Західна. Середньостиглий сорт столового призначення. Високо¬
врожайний. У бульбах міститься 15-16% крохмалю; смакові якості
хороші. Стійкий проти звичайного біотипу раку і золотистої цистоу¬
творюючої картопляної нематоди; відносно стійкий проти фітофто¬
розу.
Бульби жовті, м’якуш кремовий.
Створений у Львівському державному аграрному університеті.
Придеснянська. Середньостиглий сорт столового та універсаль¬
ного призначення. Високоврожайний. У бульбах міститься 19-20%
крохмалю; смакові якості високі. Стійкий до звичайного біотипу ра¬
ку та відносно стійкий проти інших хвороб.
Бульби білі, м’якуш світло-жовтий.
Створений на Чернігівській дослідній станції по картоплі.
Слов’янка. Середньостиглий сорт столового призначення. Висо¬
коврожайний, інтенсивного типу. У бульбах міститься 12,9-13,5%
крохмалю; смакові якості задовільні. Стійкий проти звичайного біо¬
типу раку та золотистої цистоутворюючої картопляної нематоди;
відносно стійкий проти вірусних та інших хвороб.
Бульби рожеві, м’якуш кремовий.
Створений в Інституті картоплярства УААН.
Явір. Середньостиглий сорт столового призначення. Високовро¬
жайний. У бульбах міститься 17,6% крохмалю; смакові якості хоро¬
ші. Стійкий проти звичайного біотипу раку та відносно стійкий про¬
ти фітофторозу, парші звичайної, вірусних і бактеріальних хвороб.
Бульби жовті, м’якуш кремовий.
Створений в Інституті картоплярства УААН.
Дзвін. Середньопізній сорт універсального призначення Високо¬
врожайний. У бульбах міститься 17-19% крохмалю; смакові якості хо¬
роші. Стійкий проти звичайного біотипу раку та золотистої цистоутво¬
рюючої картопляної нематоди; відносно стійкий проти фітофторозу.
Бульби рожеві, м’якуш білий.
Створений в Інституті сільського господарства Полісся УААН.
323
Ольвія. Середньопізній сорт столового призначення. Високо¬
врожайний. У бульбах міститься 16,1% крохмалю; смакові якості
хороші. Стійкий проти звичайного біотипу раку; відносно стійкий
проти фітофторозу, кільцевої гнилі та інших хвороб.
Бульби та м’якуш білі.
Створений в Інституті картоплярства УААН.
Ракурс. Середньопізній сорт столового призначення. Високо¬
врожайний. У бульбах міститься 16,6 % крохмалю; смакові якості
хороші. Стійкий до звичайного біотипу раку; відносно стійкий проти
фітофторозу, парші звичайної, кільцевої і мокрої бактеріальної гнилі
та стеблової нематоди.
Бульби та м’якуш білі.
Створений в Інституті картоплярства УААН.
По зазначених сортах в Україні ведеться первинне насінництво.
Вони придатні для використання всіма товаровиробниками.
324
5. СОРТИ
Сортом картоплі вважають однорідну за морфобіологічними та
господарськими властивостями форму, одержану шляхом гібридиза¬
ції або при систематичному доборі, яка може за певних агротехніч¬
них умов вирощування відтворювати свої властивості в наступних
поколіннях.
Кожен сорт картоплі визначається за комплексом його власти¬
востей, а саме: урожайністю, вмістом крохмалю, білків та вітамі¬
нів, смаком, стійкістю проти раку, вірусних та інших хвороб і шкід¬
ників, придатністю до виготовлення картоплепродуктів, зберігання
тощо.
У сучасному землеробстві сорт виступає як самостійний засіб
підвищення врожайності і поряд з технологією має велике, а іноді й
вирішальне значення.
Знову реєстровані сорти переважають раніше вирощувані за ви¬
щою продуктивністю, кращою якістю продукції, стійкістю до хвороб
і шкідників.
Поєднати в одному сорті високу продуктивність з високою якіс¬
тю продукції дуже важко. Часто буває так, що з підвищенням уро¬
жайності знижується крохмалистість бульб картоплі. Щоб максима¬
льно використовувати можливості сорту, не погіршити якість продук¬
ції при підвищенні врожайності, слід правильно поєднувати застосу¬
вання органічних і мінеральних добрив з іншими прийоми, що до¬
кладно викладено у відповідних розділах.
Сьогодні у сільськогосподарському виробництві використовують
сорти екстенсивного та інтенсивного типу.
Сорти екстенсивного типу добре пристосовані до певної місцево¬
сті, менш вимогливі до умов середовища, їх можна вирощувати після
слабких попередників. Вони незначно реагують на внесення мінера¬
льних добрив, здатні максимально використовувати родючість ґрун¬
ту. Максимальна їх урожайність - 150-200 ц/га.
Сорти інтенсивного типу мають біологічні можливості щодо
підвищення врожайності при збільшенні норм внесення добрив і на
зрошенні. По суті їхня продуктивність мало залежить від природної
родючості ґрунту, а більше - від рівня агротехніки. Потенційна їх
урожайність - 500—800 ц/га. Особливість сортів цього типу - висока
вимогливість до агрофону, культури землеробства. При вирощуванні
після слабких попередників, за малих норм добрив, відсутності зро¬
шення в зоні нестійкого зволоження, низької культури землеробства
325
ці сорти різко знижують свої показники, іноді дають нижчий врожай,
ніж сорти екстенсивного типу [5].
На сучасному етапі розвитку світового сільськогосподарського
виробництва, коли відбувається його подальша інтенсифікація, до
сортів ставляться наступні вимоги: висока і стійка врожайність по
роках; стійкість до несприятливих умов вирощування, хвороб і шкід¬
ників; пристосованість до механізованого вирощування; висока
якість продукції.
За скоростиглістю, або за тривалістю вегетаційного періоду (кі¬
лькість днів від садіння до утворення товарного врожаю), сорти по¬
діляють на п’ять груп: ранньостиглі, середньоранні, середньостиглі,
середньопізні та пізньостиглі [8].
Ранньостиглі сорти мають вегетаційний період 80-100 днів.
Перші бульби утворюються на 10-15-й день після появи сходів,
на 55-60-й день уже можна одержати по 90-100 ц/га бульб. До
цієї групи належать столові сорти, які придатні для споживання
влітку.
Середньоранні достигають за 100-115 днів. Бульбоутворення по¬
чинається на 15—20-й день після сходів, господарський урожай одер¬
жують через 65-70 днів після садіння. До цієї групи належать здебі¬
льшого столові сорти.
Середньостиглі сорти мають вегетаційний період 115-125 днів.
Відчутний урожай нагромаджують через 60-75 днів. У групу входять
здебільшого столові сорти.
Середньопізні для повного достигання потребують 125-140 днів,
а для нагромадження господарсько відчутного врожаю - 75-80. Сю¬
ди відносять сорти різного господарського призначення.
Пізньостиглі сорти мають вегетаційний період понад 140 днів,
господарсько придатний урожай дають на 80-85-й день. До цієї гру¬
пи належать універсальні та технічні сорти.
Однак, наведені дані про тривалість вегетаційного періоду
орієнтовні і зумовлені якістю насіннєвого матеріалу, строками
садіння і доглядом за посівами. Дотримуючись рекомендацій,
можна строки наростання бульб дещо скоротити і одержати вро¬
жай через 40 днів.
За господарським призначенням усі сорти картоплі поділяють на
шість груп: столові, технічні (заводські), столово-технічні (столово-
заводські), кормові, універсальні, придатні для виготовлення напів¬
фабрикатів.
До групи сортів, які використовуються для приготування на¬
півфабрикатів (чипси, крекери, соломка, крупка тощо), належать
сорти з підвищеним умістом сухих речовин, у тому числі крохма¬
лю, та низьким - редукованих цукрів, з коротким періодом ресин-
тезу. М’якоть бульб не темніє ні в сирому, ні у вареному вигляді
[1,3].
326
5.1. ЗАГАЛЬНА ХАРАКТЕРИСТИКА
СОРТІВ
В Україні на городах, дачних ділянках, у кооперативних, фер¬
мерських та державних господарствах вирощується понад 100 сортів
рослин. Елітне насінництво ведеться з 50 сортами. Сорти української
селекції становлять 60 % (табл. 5.1).
Найбільш поширеними сортами у приватному та громадському
секторі є: ранньостиглі та середньоранні: Адретта, Бородянська ро¬
жева, Зов, Мавка, Світанок київський, Синьоглазка та інші; серед¬
ньостиглі: Гатчинська, Луговська; середньопізні та пізньостиглі: За¬
рево, Поліська рожева, Темп.
Серед сортів картоплі, які занесені до Реєстру сортів рослин
України на 2000 рік, найсмачнішими, оціненими від 4,5 до 5 ба¬
лів, є Світанок київський, Адретта, Кобза, Придеснянська, Ли-
бідь, Луговська, Поліська рожева, Мавка, Горлиця; від 4 до 4,5
балів - Бородянська рожева, Незабудка, Пролісок, Зов, Добро¬
чин, Пост-86, Обрій, Українська рожева, Слава, Зарево, Ікар та
інші.
Сорти значно різняться вмістом сухих речовин, крохмалю, проте¬
їну, цукрів, вітамінів та інших речовин (табл. 5.2). Наведені біохіміч¬
ні показники не є постійними, а залежать від зони і умов вирощуван¬
ня [6].
Столові сорти повинні мати в комплексі інших ознак високі сма¬
кові і кулінарні якості, підвищену стійкість проти іржавої плямисто¬
сті, потемніння м’якоті, бути без наростів, ростових тріщин, дуплу-
ватості, без ураження бульб різними хворобами, бути придатними
для механізованого очищення.
Сорти для технічної переробки (крохмаль, спирт та ін.) повинні
характеризуватися високим умістом та виходом крохмалю з одиниці
площі в поєднанні з іншими цінними ознаками.
Сорти, які за комплексом ознак придатні для використання на
столові цілі і для переробки, відносять до сортів універсального при¬
значення.
Для кормового сорту основною вимогою є високий вихід кормо¬
вих одиниць з гектара. Крохмалистість бульб має становити не ниж¬
че 17-18 %, уміст сирого протеїну - не менше 2,5-3 %. Спеціально
вирощувати кормові сорти доцільно лише тоді, коли за врожайності
понад 300 ц/га вони можуть дати вихід сухих речовин більший, ніж
сорти столового, універсального та промислового призначення і не
менше 100 ц/га кормових одиниць [4].
Сорти, які занесені до Реєстру, мають певний комплекс госпо¬
дарсько цінних ознак, різне господарське призначення і належать до
різних груп стиглості [9].
327
328
Таблиця 5.1 - Сорти картоплі, занесені до Реєстру сортів рослин України на 2000 рік [7]
Назва сорту
Оригінатор
сорту
Рік реєст¬
рації
Зона
виро¬
щування
Забарвлення
Вміст кро¬
хмалю, %
квітки
бульби
м’якоті бульби
1
2
3
4
5
6
7
8
Ранньостиглі
Астерікс
Нідерланди
1996
С
червоно-фіолетове
рожеве
світло-жовте
17,9
Божедар
Поліська дослідна станція
ім.О.Засухіна
1996
СЛП
червоно-фіолетове
рожеве
біле
15,9
Бородянська
рожева
Інститут картоплярства УААН
1993
СЛП
червоно-фіолетове
рожеве
жовте
15,0
Гарт
Поліська дослідна станція
ім. О.Засухіна
1990
п
синьо-фіолетове
біле
біле
14,9
Зов
1989
СЛП
червоно-фіолетове
біле
біле
13,3-16.7
Карлена (Н)
Німеччина
1997
СЛ
біле
жовте
світло-жовте
18,2
Кірсті (Н)
Шотландія
1997
п
червоно-фіолетове
біле
кремове
16,2
Кобза
Інститут картоплярства УААН
1995
СЛП
біле
біле
кремове
16,5
Коруна(Н)
Чехія
1997
Л
біле
жовте
жовте
15,1
Краса(Н)
Чехія
1997
Л
червоно-фіолетове
жовте
жовте
14,5
Латона (Н)
Нідерланди
1997
СЛП
біле
жовте
світло-жовте
16,0
Лікарія
Німеччина
1997
п
біле
жовте
світло-жовте
15,3
Молодіжна
(Н)
Сумський сільськогосподарський
інститут
1996
СП
біле
жовте
жовте
15,9
Незабудка
Інститут картоплярства УААН
1981
СЛ
біле
біле
блідо-жовте
14,6
Памір (Н)
Німеччина
1997
Л
біле
жовте
блідо-жовте
14,0
Провенто
Нідерланди
1997
СЛП
біле
біле
світло-жовте
14,4
Петлан Сквайєр
Шотландія
1997
ЛП
біле
біле
біле
17,0
Посвіт
Поліська дослідна станція
ім. О.Засухіна
1992
СП
біле
біле
біле
11,9
Продовження таблиці 5.1
1
2
3
4
5
6
7
8
Пролісок (Н)
Інститут картоплярства УААН
1991
ст
біле
кремове
кремове
9,4
Рая
Нідерланди
1997
П
червоно-фіолетове
червоне
яскраво-
жовте
16,1
Розара (Н)
Німеччина
1997
л
червоно-фіолетове
червоне
жовте
14,2
Седнівська
рання(Н)
Чернігівська дослідна станція по
картоплі
1994
СП
червоно-фіолетове
біле
блідо-жовте
12,7-15,4
Симфонія (Н)
Нідерланди
1997
ЛП
червоно-фіолетове
рожеве
світло-жовте
18,9
Середньоранні
Адретта
Німеччина
1985
П
біле
кремове
жовте
14,2
Берегиня
Поліська дослідна станція
ім. О.Засухіна
1992
п
червоно-фіолетове
рожеве
світло-жовте
11,4
Водограй (Н)
Інститут картоплярства УААН
1995
слп
біле
біле
кремове
12,1
Доброчин (Н)
Поліська дослідна станція
ім. О.Засухіна
1995
слп
червоно-фіолетове
рожеве
біле
14,6
Житомирянка
>>
1978
п
червоно-фіолетове
біле
світло-
кремове
12,2-16,4
Елса
Шотландія
1997
п
біле
блідо-кремове
16,2
Мавка
Інстиіут землеробства і тварин¬
ництва Західного регіону УААН
1982
лп
біле
біле
біле
15.5-19,1
Невська
Північно-західний науково-
дослідний інстиіут сільського
господарства Росії
1984
слп
біле
біле
біле
12,1
Обрій (Н)
Інститут картоплярства УААН
1997
слп
біле
біле
кремове
17,0
Остара
Нідерланди
1985
лп
біле
коричню¬
вате
світло-жовте
14,1
Пост 86
Поліська дослідна станція
ім. О.Засухіна
1991
СП
червоно-фіолетове
рожеве
біле
15,4
330
Продовження таблиці 5.1
1
2
3
4
5
6
7
8
Радомишль¬
ська
Поліська дослідна станція
ім.. 0. Засухіна
1985
СП
біле
біле
біле
14,0
Радич (Н)
»»
1997
Л
червоно-фіолетове
рожеве
біле
15,5
Світанок
київський
Інститут картоплярства УААН
1987
СЛП
червоно-фіолетове
рожеве
кремове
15,1-20,0
Тімате
Нідерланди
1997
ЛП
біле
жовте
світло-жовте
13,5
Середньостиглі
Гатчинська
Північно-західний
науково-дослідний інститут
сільського господарства Росії
1973
СЛП
червоно-
фіолетове
біле
біле
16,9
Горлиця
Інститут картоплярства
УААН
1996
СЛП
біле
біле
кремове
15,8
Либідь
1993
п
біле
біле
біле
15,5
Луговська
1987
ЛП
біле
світло-
рожеве
біле
12,2-17,3
Обелікс (Н)
Нідерланди
1996
СЛ
біле
біле
біле
13,8
Огоньок
Білоруський науково-дослідний
інститут картоплярства
1973
с
червоно-
фіолетове
біле
15,5-18,8
Пекуровська
(Н)
Чернігівська дослідна станція
по картоплі
1995
ЛП
біле
біле
біле
16,2
Приде-
снянська
а
1996
СЛП
червоно-
фіолетове
біле
світло-
жовте
18,3
Слава
Інститут землеробства і тва¬
ринництва Західного регіону
УААН
1992
п
біле
біле
біле
12,0
Продовження таблиці 5.1
1
2
3
4
5
6
7
8
Українська
рожева
Інстиіут картоплярства УААН
1989
ЛП
червоно-
фіолетове
рожеве
світло-
жовте
15,6-17,6
Цезар
Нідерланди
1996
С
біле
жовте
світло-
жовте
15,7
Середньопізні
Витязь
Чернігівська дослідна станція по
картоплі
1989
п
червоно-фіолетове
біле
біле
17,5-19,8
Воловецька
Інститут картоплярства УААН
1985
п
червоно-фіолетове
жовте
жовте
19,4
Дезіре (Н)
Нідерланди
1997
л
червоно-фіолетове
червоне
світло-жовте
15,6
Зарево
Інститут картоплярства УААН
1983
лп
червоно-фіолетове
рожеве
біле
19,8-22,9
Ікар
Поліська дослідна станція
ім. О.Засухіна
1983
п
червоно-фіолетове
рожеве
біле
18,4
Поліська
рожева
1978
п
червоно-фіолетове
рожеве
біле
15,6-18,6
Ракурс(Н)
Інститут картоплярства УААН
1997
лп
біле
біле
біле
17,1
Пізньостиглі
Древлянка
Інстиіут сільського
господарства Полісся УААН
1984
п
червоно-фіолетове
червоне
світло-
кремове
16,0
Ласунак
Білоруський науково- дослідний
інстиіут картоплярства
1989
лп
біле
біле
кремове
16,8-20,4
Темп
»
1973
лп
синьо-фіолетове
біле
кремове
18,8-20,9
Умовні позначення: С - Степ; J1 - Лісостеп; П - Полісся; Н - сорт стійкий до картопляної нематоди
Таблиця 5.2- Біохімічний склад окремих сортів картоплі (Новосельська А.П., Мицько В.М., Холодило І.В.) [6]
Сорт
Суха
речо¬
вина,
%
Крох¬
маль,
%
Сирий
протеїн, %
Амінокислоти, % на суху
масу
Співвідноше¬
ння замінних
амінокислот
до незамін¬
них
Вітамін С,
мг% на сиру
масу
Каротиноїди,
мг% на сиру
масу
Жири, % на
суху масу
Клітковина,
% на суху
масу
на
суху
масу
на сиру
масу
неза¬
мінні
замінні
сума
Ранньостиглі
Бородянська рожева
21,8
16,0
10,6
2,3
3,45
3,52
6,97
0,98
12,3
0,41
0,52
2,04
Зов
22,5
16,7
9,9
2,3
2,55
3,66
6,20
0,70
11,6
0,06
0,41
2,07
Незабудка
20,7
15,0
9,6
2,0
2,87
3,31
6,18
0,87
13,4
0,38
0,62
0,84
Пролісок
19,3
13,8
10,6
2,0
2,73
4,87
7,10
0,62
12,4
0,45
0,52
0,88
Середньоранні
Водограй
18,8
13,3
10,9
2,2
2,02
2,75
4,77
0,73
14,3
0,50
0,32
1,20
Кобза
23,7
17,7
10,6
2,5
3,54
3,68
7,22
0,96
13,8
0,40
0,36
1,40
Мавка
24,7
18,6
10,0
2,4
3,25
4Д5
7,40
0,78
11,6
0,04
0,35
2,02
Невська
20,2
14,6
11,7
2,3
3,00
4,72
7,72
0,46
14,6
0,04
0,44
0,82
Обрій
20,5
14,9
10,1
2,1
3,35
4,30
7,65
0,78
13,6
0,05
0,36
1,20
Світанок київський
25,3
19,2
10,3
2,6
3,68
4,38
8,06
0,84
13,3
0,50
0,75
2,56
Середньостиглі
Г атчинська
21,5
15,5
9,9
2,1
2,36
4,64
7,0
0,51
1,18
0,04
0,52
3,17
Горлиця
24,0
18,0
10,5
2,4
3,51
4,01
7,50
0,88
14,0
0,40
0,40
1,20
Либідь
22,0
16,2
10,4
2,3
3,62
3,78
7,40
0,96
14,2
0,15
0,42
1,30
Луговська
22,5
16,7
9,9
2,2
2,93
4,42
7,35
0,66
14,4
0,08
0,43
0,90
Українська рожева
20,5
14,9
9,3
1,9
2,89
3,56
6,45
0,81
14,2-
0,29
0,38
1,00
Пізньостиглі
Зарево
29,6
23,0
9,6
3,1
3,23
3,33
6,56
0,97
13,1
0,07
0,35
0,96
Темп
26,0
19,8
9,7
2,4
2,16
4,54
5,70
0,48
12,1
0,03
0,38
1,63
5.2. РАННІ СОРТИ
Астерікс. Виведений у Нідерландах.
Кущ малостебловий. Стебла досить товсті, з антоціановим забар¬
вленням. Листки великі, тверді, темно-зелені. Квіти червоно-фіоле-
тові.
Бульби великі, видовжені, рожеві. Шкірка лускувата, вічка мілкі.
М’якуш світло-жовтий, дещо твердий, не темніє після приготування.
Смакові якості хороші. Уміст крохмалю ( 17,9 %) і сухої речовини
високий і дуже високий.
Ракостійкий. Імунний до парші звичайної, чутливий до скручу¬
вання листків.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1996 року.
Божедар. Виведений Поліською дослідною станцією ім. О. Засу¬
хіна. Відзначається інтенсивним накопиченням урожаю, який при
відповідній технології вирощування на 40-45-й день після сходів
може досягти 240 ц/га бульб.
Кущ середньої висоти. Квіти червоно-фіолетові.
Бульби рожеві, округло-овальні, вічка середньозаглиблені. М’якуш
білий, після варіння не темніє. Смакові якості задовільні та хороші
(3,8 бала). Уміст крохмалю в бульбах - 13,7%.
Ракостійкий. Стійкість до фітофторозу невисока.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1996 року.
Бородянська рожева. Виведений Інститутом картоплярства
УААН. Високоврожайний. Потенційна врожайність на 40-45-й день
після сходів - 200-220 ц/га, а наприкінці вегетації - 500-600 ц/га.
Кущ прямостоячий, добре облиствлений. Стебла із слабким анто¬
ціановим забарвленням у пазухах листка та вздовж усього стебла.
Квіти рожеві.
Бульби рожеві, округлі. М’якуш жовтий, при розрізуванні не тем¬
ніє. Смакові якості хороші (4,1 бала). Уміст крохмалю в бульбах -
14-15%.
Ракостійкий. Відзначається високою стійкістю проти стеблової
нематоди, парші звичайної, бактеріальних хвороб, відносно стійкий
проти фітофторозу.
На легких грунтах, коли після засухи випадають сильні дощі, під
час вегетації можливе розтріскування бульб. Сорт не витримує пере-
зволоження грунту наприкінці вегетації, через що відбувається уду¬
шення бульб.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1993 року.
333
Велокс. Виведений у Німеччині. Потенційна врожайність - 450-
500 ц/га. Придатний для приготування фрі.
Кущ високий, добре облиствлений.
Бульби овальні, шкірка гладенька, жовта. Вічка поверхневі.
М’якуш - жовтий або світло-жовтий, при варінні не темніє. Уміст
крохмалю - 14,0%. Смакові якості відмінні. Бульби розварюються
слабко.
Ракостійкий. Стійкий до нематоди, відзначається високою стійкі¬
стю до гнилі.
Придатний для вирощування на всіх грунтах.
Рекомендується для вирощування в лісостеповій та степовій зо¬
нах.
Занесений до Реєстру сортів рослин України з 2001 року.
Вінета. Виведений у Німеччині. Високоврожайний, особливо при
садінні пророщеними бульбами.
Кущ добре розвинений, високий. Квіти білі.
Бульби округло-овальні, білі. Залягання вічок мілке. Шкірка
бульб дещо нерівна. М’якуш світло-жовтий. Смакові якості хороші.
При варінні бульби майже не темніють.
Ракостійкий. Стійкий до окремих вірусів, відносно стійкий до
парші звичайної, чорної ніжки, фітофторозу. Стійкий до картопляної
нематоди.
Рекомендується для вирощування в зонах Полісся та Степу.
Занесений до Реєстру сортів рослин України з 2001 року.
Гарт. Виведений Поліською дослідною станцією ім. О.Засухіна.
Кущ прямостоячий, високий. Листки зелені, матові. Квіти синьо-
фіолетові.
Бульби округлі, з тупою верхівкою, білі. Шкірка бульби дещо
лускувата. Вічка середньозаглиблені. М’якуш білий, при розрізу¬
ванні не темніє. Смакові якості хороші. Уміст крохмалю в бульбах -
14,9%.
Ракостійкий. Фітофторозом уражується середньою мірою, слабко
- макроспоріозом і сухою гниллю бульб.
Рекомендується для вирощування в зоні українського Полісся.
Занесений до Реєстру сортів рослин України з 1990 року.
Джаерла. Виведений у Нідерландах. Дуже ранній. Посухостійкий.
Кущ малостебловий. Стебла товсті, добре облиствлені. Листки
блідо-зелені, в пазухах світло-пурпурові, великі. Квіти білі. Сорт від¬
значається підвищеною енергією росту в початковий період.
Бульби великі, овальні, жовті. М’якуш світло-жовтий, не розва¬
рюється і не темніє при приготуванні страв. Смакові якості задовіль¬
ні. Уміст крохмалю - 15,3 %.
334
Ракостійкий. Стійкий до парші звичайної. Вважається досить
стійким до механічних пошкоджень. Відзначається польовою стійкіс¬
тю до фітофторозу.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1998 року.
Зов. Виведений на Поліській дослідній станції ім. О. Засухіна.
Досить урожайний. Потенційна врожайність на 40^5-й день після
сходів - 200 ц/га, наприкінці вегетації - 500 ц/га.
Кущ високий, дуже облиствлений, стебла слабопігментовані ан¬
тоціаном, нечисленні. Квіти червоно-фіолетові.
Бульби білі, округлі. Шкірка гладенька. Вічка середньозаглибле-
ні. М’якуш білий, при розрізуванні не темніє. Бульби переважно
крупні, середня маса їх - 90-130 г. Смакові якості хороші (4,1 бала).
Уміст крохмалю - 15-16 %.
Ракостійкий. Відносно стійкий до стеблової нематоди, парші
звичайної та вірусних хвороб. В окремі роки без належного захисту
може значно уражуватись фітофторозом.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1989 року.
Карлена. Виведений у Німеччині.
Кущ - від середнього до високого. Розвивається дуже швидко.
Картоплиння товсте. Квіти червоно-фіолетові.
Бульби округло-овальні, середнього розміру, гарної форми. Вічка
мілкі. Шкірка жовта, сітчаста. М’якуш світло-жовтий. Смакові якості
відмінні. Структура розвариста, майже не темніє, відзначається уні¬
версальною придатністю для переробки (чипси, пюре, сушена картоп¬
ля, картопля-фрі).
Ракостійкий. Стійкий до картопляної нематоди та окремих вірусів.
Слабко уражується фітофторозом, ризоктонією та чорною ніжкою.
Рекомендується для вирощування в лісостеповій та степовій зонах.
Занесений до Реєстру сортів рослин України з 1997 року.
Кобза. Виведений Інститутом картоплярства УААН. Урожайний.
Потенційна врожайність - до 500 ц/га, а ранньої картоплі на 40—45-й
день після сходів - 200-250 ц/га.
Кущ середньої висоти, багатостебловий. Квіти білі.
Бульби білі, округлі. На щільних грунтах при недостатньому
міжрядному обробітку часто деформуються. М’якуш кремовий.
Смакові якості хороші (4,5 бала). Уміст крохмалю - 17—18%. Збері¬
гається добре.
Ракостійкий. Стійкий до стеблової нематоди. Має середню стійкість
проти фітофторозу, парші звичайної, вірусних і бактеріальних хвороб.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1995 року.
335
Колєт. Виведений у Німеччині. При пророщуванні бульб може
давати дуже ранню продукцію. Добре реагує на зрошення.
Кущ добре розвинений. Квіти червоно-фіолетові.
Бульби видовжено-овальні, шкірка гладка, жовта, бульби дуже
вирівняні, вічка мілкі. М’якуш блідо-жовтий. Уміст крохмалю в буль¬
бах - 12,5-13,5 %. Бульби зберігаються добре. Смакові якості хоро¬
ші.
Ракостійкий і стійкий до картопляної нематоди та парші звичайної.
Рекомендується для вирощування в поліській та степовій зонах.
Занесений до Реєстру сортів рослин України з 2001 року.
Коруна. Виведений у Чехії. Характеризується дуже раннім нако¬
пиченням урожаю.
Кущ середньої висоти. Квіти білі.
Бульби округло-овальні, досить вирівняні в урожаї, жовті. Вічка
мілкі, середньозаглиблені. Смакові якості задовільні. Уміст крохма¬
лю в бульбах - 15,1 %.
Ракостійкий. Стійкий до картопляної нематоди і відносно стійкий
до окремих вірусів.
Рекомендований для вирощування в лісостеповій зоні.
Занесений до Реєстру сортів рослин України з 1997 року.
Косень 95. Сорт виведений Поліською дослідною станцією ім. О. За¬
сухіна. Потенційна врожайність - 400-450 ц/га.
Кущ прямостоячий, компактний, добре облиствлений. Листки се¬
реднього розміру. Квіти червоно-фіолетові.
Бульби овальні, рожеві. Вічка поверхневі. М’якуш білий. Смакові
якості задовільні (3,4-4,0 бали). Уміст крохмалю в бульбах - 14,4-
15,3 %. Розміщення бульб у гнізді компактне.
Ракостійкий. Стійкий проти парші звичайної, вірусних хвороб і
відносно стійкий проти стеблової нематоди.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1999 року.
Краса. Виведений у Чехії. Визначається інтенсивним накопичен¬
ням урожаю.
Кущ середньої висоти, прямостоячий. Листя світло-зелене. Квіти
червоно-фіолетові.
Бульби округлі, великі, вирівняні за масою і формою, жовті. Віч¬
ка мілкі. М’якуш жовтий. Смакові якості бульб хороші (4,0 бали).
Уміст крохмалю - 14,5%.
Ракостійкий. Стійкий до картопляної нематоди і середньостійкий
проти скручування листків, парші звичайної. Чутливий до фітофторозу.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1997 року.
336
Латона. Виведений у Нідерландах.
Кущ середньої висоти, стебла прямостоячі, блідо-пурпурові. Лис¬
тки досить великі, темно-зелені. Квіти білі.
Бульби великі, овальні, жовті. Шкірка лускувата. Вічка досить
мілкі. М’якуш блідо-жовтий, твердої текстури, не темніє після при¬
готування страв. Смакові якості задовільні. Уміст крохмалю в буль¬
бах-12-16%.
Ракостійкий. Стійкий до картопляної нематоди. Картоплиння чут¬
ливе до фітофторозу. Середньо уражується макроспоріозом і вірус¬
ними хворобами. Бульби не стійкі до парші звичайної, слабко ура¬
жуються сухою і кільцевою гниллю.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1997 року.
Лікарія. Виведений у Німеччині. Високоврожайний при макси¬
мальній кількості товарних бульб. Відзначається досить високою
пластичністю.
Кущ - від середнього до високого, розвивається швидко. Карто¬
плиння товсте. Квіти білі.
Бульби округло-овальні, від середнього до крупного розміру, гар¬
ної форми. Вічка мілкі. Шкірка жовта, сітчаста. М’якуш - від світло-
жовтого до жовтого. Смакові якості хороші. Бульби дещо розварю¬
ються, не темніють у сирому і вареному вигляді.
Ракостійкий. Стійкий до нематоди, середньою мірою уражується
фітофторозом, ризоктонією та чорною ніжкою.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1997 року.
Маріс Бард. Виведений у Шотландії. Вважається дуже раннім.
Кущ середній, добре облиствлений. Квіти червоно-фіолетові.
Бульби білі, видовжено-овальні, вічка середньозаглиблені. М’я¬
куш білий, після варіння не темніє. Текстура м’якуша тверда. Смако¬
ві якості хороші. Уміст крохмалю низький (10-12,8 %).
Нестійкий до фітофрозу і парші звичайної.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1999 року.
Молодіжна. Виведений у Сумському державному аграрному уні¬
верситеті. Відзначається раннім накопиченням урожаю.
Кущ високий, стебла численні, середньо облиствлені. Квіти білі.
Бульби округлі, жовті. М’якуш жовтий. Смакові якості бульб
відмінні (4,8 бала). Уміст крохмалю в бульбах - 15,0-17,0 %. Відзна¬
чається високою стійкістю бульб до механічних пошкоджень при
збиранні, придатністю до довготривалого зберігання.
Рекомендується для вирощування в зонах Полісся та Степу.
Занесений до Реєстру сортів рослин України з 1996 року.
337
Незабудка. Виведений Інститутом картоплярства УААН. Через
40-45 днів після сходів здатний нагромаджувати врожай 200-250 ц/га
бульб. Потенційна врожайність наприкінці вегетації - 400 ц/га.
Кущ середньої висоти, стебла мають мало гілок. Квіти білі.
Бульби білі, округло-овальні, шкірка гладенька. Вічка мілкі, по¬
верхневі. М’якуш світло-кремовий. Смакові якості хороші (4 бали).
Уміст крохмалю середній - 13,8 %. При хорошому врожаї утворює
багато великих бульб. Середня маса товарних бульб - 89—108 г.
Зберігається добре.
Ракостійкий. Має підвищену стійкість проти вірусних хвороб.
Рекомендується для вирощування в лісостеповій та степовій зонах.
Занесений до Реєстру сортів рослин України з 1996 року.
Повінь. Виведений в Інституті картоплярства УААН. Потенційна
врожайність - 450 ц/га, на 1-5 липня - 160-225 ц/га.
Кущ високий, облиствленість хороша. Квіти червоно-фіолетові.
І Бульби рожеві, округлі. М’якуш кремовий. Смакові якості хороші
(4,6 бали). Уміст крохмалю - 15,0-16,0 %. Середня маса товарних
бульб - 99—110 г.
Ракостійкий. Стійкий до картопляної нематоди і відносно стійкий
проти альтернаріозу та сухої фузаріозної гнилі.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 2000 року.
Поран. Виведений на Поліській дослідній станції ім. О. Засухіна.
Високоврожайний.
Кущ середньої висоти, напіврозлогий, стебла нечисленні, слабко-
гіллясті, зелені. Листки середнього розміру. Квіти червоно-фіолетові.
Бульби рожеві, овальні, з поверхневими вічками. М’якуш білий.
Смакові якості задовільні (3,2-3,6 бала). Уміст крохмалю в бульбах -
11,2-12,0%.
Стійкий до звичайного біотипу раку і картопляної нематоди та
відносно стійкий до парші звичайної.
Рекомендований для вирощування в зонах Полісся та Степу.
Занесений до Реєстру сортів рослин України з 2001 року.
Посвіт. Виведений на Поліській дослідній станції ім. О. Засухіна.
Потенційна врожайність середня: ранньої картоплі на 40-45-й день
після сходів - до 200 ц/га, а наприкінці вегетації - до 400 ц/га.
Кущ середньо облиствлений, стебла слабкогіллясті. Квіти білі.
Бульби білі, округлі. Шкірка гладенька, вічка середньозаглиблені.
М’якуш білий. Смакові якості задовільні (3,8 бала). Середня маса
товарної бульби - 97 г. Уміст крохмалю невисокий - 13,7 %.
Ракостійкий. Стійкість проти стеблової нематоди, фітофторозу,
вірусних і бактеріальних хвороб - середня.
338
Рекомендований для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1992 року.
Пролісок. Виведений Інститутом картоплярства УААН. Потен¬
ційна врожайність наприкінці вегетації - 400 ц/га, а на 40-45-й день
після сходів - близько 200 ц/га.
Кущ середньої висоти, стебла численні, мають багато гілок. Квіти
білі, цвітіння рясне.
Бульби кремові, овальні або округлі. Вічка мілкі, нечисленні. М’я¬
куш світло-жовтий, при розрізуванні не темніє. Смакові якості задо¬
вільні (4,1 ба- ла). Уміст крохмалю - 14,5 %. Середня маса товарних
бульб - 97 г.
Ракостійкий. Стійкий проти картопляної нематоди і відносно стій¬
кий проти кільцевої, мокрої бактеріальної та сухої фузаріозної гнилі.
Рекомендується для вирощування в усіх зонах України, особливо
на площах, уражених картопляною нематодою.
Занесений до Реєстру сортів рослин України з 1991 року.
Рікеа. Сорт виведений у Німеччині.
Надземна вегетативна маса розвивається добре, сходи дружні.
Квіти білі.
Бульби округло-овальні, гарної форми, із плоскими вічками, жовтого
кольору. М’якуш жовтий, при приготуванні не темніє. Смакові якості
хороші. Бульби не розварюються. Уміст крохмалю - 11,0-12,0 %.
Ракостійкий. Стійкий до картопляної нематоди, уражується
окремими вірусами.
Рекомендується для вирощування в зонах Полісся і Степу.
Занесений до Реєстру сортів рослин України з 2001 року.
Розара. Виведений у Німеччині. Урожайність ранньої картоплі - 100—
250 ц/га (через 45 днів після сходів), наприкінці вегетації - 400-450 ц/га.
Кущ напіврозлогий. Квіти червоно-фіолетові.
Бульби видовжено-овальні, червоні. Шкірка гладенька. Вічка міл¬
кі. М’якуш жовтий. Смак хороший та відмінний. Уміст крохмалю -
12,1-15,8 %. Середня маса товарних бульб - 81-115 г.
Ракостійкий. Стійкий до картопляної нематоди. Слабко уражу¬
ється фітофторозом та паршею звичайною.
Рекомендується для вирощування в зоні Лісостепу.
Занесений до Реєстру сортів рослин України з 1997 року.
Седнівська рання. Створений в ОНВП “Чернігівеліткартопля”.
Кущ середньорозвинутий, низький. Квіти червоно-фіолетові.
Бульби білі, округлі. Смакові якості задовільні (3,5-3,7 бала).
М’якуш блідо-жовтий. Уміст крохмалю в бульбах - 12,7-15,4 %.
Ракостійкий. Стійкий до картопляної нематоди і відносно стійкий
до вірусних хвороб. На уражених картопляною нематодою полях
339
здатний давати не тільки врожай бульб, а й очищати грунт від цист
нематоди.
Рекомендується для вирощування в зонах Полісся і Степу.
Занесений до Реєстру сортів рослин України з 1994 року.
Серпанок. Виведений в Інституті картоплярства УААН. Урожайність
на 40-45-й день після сходів -160 ц/га, наприкінці вегетації - 350 ц/га.
Кущ прямостоячий, невисокий, компактний, добре облиствлений.
Листки середньої величини. Квіти червоно-фіолетові.
Бульби овальні, рожеві. М’якуш кремовий. Смакові якості хороші
(4 бали). Уміст крохмалю - 13,0-14,0 %.
Ракостійкий. Стійкий проти парші і фузаріозу.
Рекомендується для вирощування в зонах Полісся та Степу. При¬
датний для двоврожайної культури на півдні України.
_ Занесений до Реєстру сортів рослин України з 2001 року.
Симфонія. Виведений у Нідерландах.
Кущ високий. Стебла товсті або середньої товщини, прямостоячі.
Пігментація темно- або дуже темно-пурпурова. Листки великі, тем¬
но-зелені. Квіти червоно-фіолетові.
Бульби рожеві, овальні. Шкірка лускувата. М’якуш світло-жовтий,
після приготування не темніє. Смакові якості хороші. Уміст крохмалю в
бульбах - 13,1-19,2%. Середня маса товарних бульб - 68-138 г.
Ракостійкий. Помірно чутливий до фітофторозу. Слабко уражу¬
ється паршею звичайною і макроспоріозом, нематодостійкий.
Рекомендується для вирощування в зонах Полісся і Степу.
Сорт занесений до Реєстру сортів рослин України з 1997 року.
Чернігівська рання. Виведений в ОНВП “Чернігівеліткартопля’’.
Кущ середній, середньооблиствлений. Квіти білі.
Бульби округло-овальні з мілкими вічками, білі. М’якуш кремо¬
вий. Уміст крохмалю - 12,3-14,7 %. Смакові якості хороші.
Ракостійкий і стійкий до картопляної нематоди. Чутливо реагує
на різання садивних бульб.
Рекомендується для вирощування в зонах Полісся і Степу.
Занесений до Реєстру сортів рослин України з 1999 року.
Юкама. Виведений у Нідерландах.
Кущ - від середнього до низького. Стебла прямі, зі слабким ангоціано-
вим забарвленням. Листки середнього розміру. Квіти червоно-фіолетові.
Бульби великі, видовжено-овальні, жовті з поверхневими вічками.
М’якуш світло-жовтий, при приготуванні страв не темніє. Уміст су¬
хих речовин середній.
Ракостійкий і стійкий до картопляної нематоди.
Рекомендується для вирощування в зоні Лісостепу.
Сорт занесений до Реєстру сортів рослин України з 1998 року.
340
5.3. СЕРЕДНЬОРАННІ СОРТИ
Адретта. Виведений у Німеччині. Потенційна врожайність на¬
прикінці вегетації - 400 ц/га.
Кущ високий, компактний; стебла прямі, зелені. Листки великі,
світло-зелені. Квіти білі.
Бульби округлі та округло-овальні. Шкірка жовта, сітчаста. Вічка
нечисленні, поверхневі. М’якуш жовтий, при розрізуванні і варінні не
темніє. Смакові якості відмінні (4,8 бала). Уміст крохмалю - 14,2 %.
Товарність бульб висока. Середня маса товарних бульб - 100-120 г.
Ракостійкий. Вірусними хворобами уражується слабко.
Здатний давати достатні врожаї при частковому затіненні наса¬
джень деревами. У дуже сухих умовах вирощувати не доцільно.
Рекомендується для вирощування в умовах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1985 року.
Альвара. Виведений у Німеччині.
Кущ добре розвинений; вегетативна маса активно наростає у по¬
чаткових фазах розвитку. Квіти червоно-фіолетові.
Бульби видовжено-овальні, гарної форми, червоні; шкірка гладень¬
ка. М’якуш жовтий, не змінює забарвлення. Смак хороший. Уміст
крохмалю в бульбах - 12,6-14,0%.
Стійкий до раку та картопляної нематоди. Бульби не пророста¬
ють при тривалому зберіганні.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 2001 року.
Берегиня. Виведений Поліською дослідною станцією ім. О. Засухіна.
Кущ компактний, середньої висоти. Квіти червоно-фіолетові.
Бульби рожеві, округло-овальні. Шкірка бульби лускувата, вічка по¬
верхневі. М’якуш світло-жовтий. Смакові якості задовільні (3,6 бала).
Уміст крохмалю середній - 13,6 %. Середня маса товарних бульб - 95 г.
Ракостійкий. Стійкий до картопляної нематоди і середньостійкий
проти фітофторозу, парші звичайної та вірусних хвороб.
Рекомендується для вирощування в зоні Полісся.
Сорт занесений до Реєстру сортів рослин України з 1992 року.
Водограй. Виведений Інститутом картоплярства УААН. Високо¬
врожайний, потенційна врожайність наприкінці вегетації - до 600 ц/га.
Кущ середньої висоти, стебла сильно облиствлені. Листки великі.
Квіти білі.
Бульби білі, округло-овальні. М’якуш кремовий. Смакові якості
задовільні (3,6 бала). Уміст крохмалю низький - 13,6 %. Середня ма¬
са товарних бульб - 95-120 г. Бульби відзначаються підвищеним
умістом вільних і незамінних амінокислот та пептидів. Сорт рекоме¬
ндований для дитячого і дієтичного харчування.
341
Ракостійкий. Стійкий проти картопляної нематоди та відносно
стійкий проти фітофторозу і парші звичайної. Має підвищену стій¬
кість проти стеблової нематоди.
Рекомендується для вирощування в зонах Полісся і Степу.
Занесений до Реєстру сортів рослин України з 1995 року.
Доброчин. Виведений на Поліській дослідній станції ім. О. Засу¬
хіна. Потенційна врожайність наприкінці вегетації - 350-400 ц/га.
Кущ прямостоячий, добре облиствлений. Квіти червоно-фіоле-
тові.
Бульби овальні, рожеві. Вічка поверхневі. М’якуш білий, не тем¬
ніє до і після варіння. Смакові якості хороші (4,3-4,5 бала). Уміст
крохмалю - 14,4-18,4%.
Ракостійкий. Стійкий до картопляної нематоди та середньостій¬
кий проти фітофторозу.
Рекомендується для вирощування в усіх зонах України.
Сорт занесений до Реєстру сортів рослин України з 1995 року.
Карін. Виведений у Чехії.
Кущ добре облиствлений. Квіти білі, ягід не утворюють.
Бульби видовжено-овальні, вирівняні за величиною та формоюю.
М’якуш жовтий. Шкірка жовта, вічка мілкі. Смакові якості хороші.
Уміст крохмалю у бульбах низький - 12,6%.
Ракостійкий та середньостійкий проти фітофторозу і парші зви¬
чайної. Сприйнятливий до скручування листя.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1998 року.
Купава. Виведений Інститутом картоплярства УААН. Урожай¬
ність на 40-45-й день після сходів - близько 150 ц/га, а наприкінці
вегетації - 450 ц/га.
Кущ прямостоячий, широкий, середньої висоти. Стебла гіллясті,
середньо облиствлені. Квіти червоно-фіолетові.
Бульби овальні, рожеві. М’якуш світло-жовтий. Смакові якості
хороші (4,5 бала). Уміст крохмалю в бульбах - 17,8 %. Середня маса
товарної бульби - 94 г.
Ракостійкий та стійкий проти стеблової нематоди, фітофторозу,
парші звичайної і вірусних хвороб.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1998 року.
Мавка. Виведений Інститутом землеробства і тваринництва за¬
хідного регіону УААН.
Кущ компактний, середньої висоти, має мало стебел. Вони гіллясті,
слабкооблиствлені. Листки великі, світло-зелені, матові. Квіти білі.
342
Бульби білі, овальні. Шкірка гладенька, вічка мілкі, середньоза-
глиблені. М’якуш білий. Смакові якості хороші. Уміст крохмалю в
бульбах - 15,5-19,1 %.
Ракостійкий. Фітофторозом уражується слабко, паршею і вірус¬
ними хворобами - середньою мірою.
Рекомендується для вирощування в лісостеповій та степовій зо¬
нах України.
Занесений до Реєстру сортів рослин України з 1982 року.
Малич. Сорт селекції Поліської дослідної станції ім.О. Засухіна.
Кущ середньої висоти, добре облиствлений. Квіти білі.
Бульби білі, округло-овальні. М’якуш білий. Заглиблення вічок
мілке. Смакові якості хороші та відмінні. Уміст крохмалю - 13,9—
18,9%.
Ракостійкий і відносно стійкий до стеблової нематоди, фітофто¬
розу, парші звичайної та бактеріальних хвороб.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1998 року.
Невська. Виведений Північно-Західним інститутом сільського
господарства Росії. Високоврожайний, потенційна врожайність на¬
прикінці вегетації - понад 600 ц/га.
Кущ добре облиствлений. Стебла зелені, заокруглені. Листки ве¬
ликі. Квіти білі.
Бульби білі, округлі. Шкірка гладенька, вічка поверхневі, блідо-
рожеві. М’якуш білий, при розрізуванні не темніє. Смакові якості
задовільні (3,1 бала). Уміст крохмалю низький - 12,1%. В урожаї пе¬
реважають великі бульби. Середня маса товарної бульби - до 150 г.
Ракостійкий. Фітофторозом та вірусними хворобами уражується
середньою мірою.
При садінні бульб у непрогрітий грунт або ж після кількаразово¬
го обламування паростків значна частина їх може не дати сходів.
Сорт рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1984 року.
Обелікс. Виведений у Нідерландах. Відзначається інтенсивним рос¬
том у початкових фазах і досить раннім бульбоутворенням.
Кущ середньої висоти. Стебла напівстоячі. Листки середні або
великі, зелені або темно-зелені. Квіти білі.
Бульби овальні, жовті. Шкірка здебільшого гладенька; вічка мілкі.
М’якуш блідо-жовтий. Текстура досить тверда, трохи темніє після при¬
готування. Смакові якості задовільні. Крохмалистість невисока - 13,8.
Ракостійкий. Імунний до парші звичайної та картопляної нематоди.
Рекомендований для вирощування в зонах Степу та Лісостепу.
Занесений до Реєстру сортів рослин України з 1996 року.
343
Обрій. Створений в Інституті картоплярства УААН. Потенційна
врожайність на 45-й день після сходів - 125 ц/га, наприкінці вегета¬
ції - 500 ц/га.
Кущ середньої висоти. Стебла гіллясті. Квіти білі.
Бульби білі із жовтим відтінком, короткоовальні. М’якуш білий.
Вічка мілкі. Смакові якості хороші (4,3-4,5 бала). Уміст крохмалю в
бульбах - 18-19 %.
Ракостійкий та стійкий до картопляної нематоди, парші звичай¬
ної і бактеріальних хвороб.
Рекомендований для вирощування в усіх зонах України.
Є одним із кращих сортів для вирощування на городах, де вияв¬
лено картопляну нематоду.
Занесений до Реєстру сортів рослин України з 1997 року.
Палма. Виведений у Німеччині. Придатний для виробництва на¬
півфабрикатів. Посухостійкий.
Кущ добре розвинений, листки великі. Рослини активно ростуть у
початкових фазах. Квіти білі.
Бульби жовті, видовжено-овальні, з мілкими вічками. М’якуш
жовтий, не темніє. Консистенція ніжна, при варінні не розсипається.
Смакові якості хороші; сорт відзначається приємним ароматом.
Стійкий до раку і ризоктонії.
Рекомендується для вирощування в зонах Полісся та Степу.
Занесений до Реєстру сортів рослин України з 2001 року.
Поліська 96. Виведений на Поліській дослідній станції ім. О. За¬
сухіна. Високоврожайний.
Кущ середній, добре облиствлений. Квіти червоно-фіолетові.
Бульби короткоовальні, рожеві, з поверхневими вічками. М’якуш
білий. Смакові якості хороші. Уміст крохмалю в бульбах - 17,4%.
Ракостійкий. Стійкий до картопляної нематоди та відносно стій¬
кий до фітофторозу.
Рекомендований для вирощування в зонах Полісся та Степу.
Занесений до Реєстру сортів рослин України з 2001 року.
Пост 86. Виведений Поліською дослідною станцією ім. О. Засу¬
хіна. Потенційна врожайність наприкінці вегетації - 450 ц/га.
Кущ прямостоячий, компактний. Кількість гілок на стеблах серед¬
ня. Квіти червоно-фіолетові.
Бульби рожеві, овальні. Шкірка лускувата, вічка численні, негли¬
бокі. М’якуш білий, не темніє до і після варіння. Смакові якості хо¬
роші (4,0 бали). Уміст крохмалю - 15,2 %. Середня маса товарних
бульб - 95 г.
Ракостійкий. Середньостійкий до парші звичайної, бактеріальних
хвороб та фітофторозу. У роки, сприятливі для розвитку хвороби,
344
може нею сильно уражуватися, має підвищену стійкість до вірусних
хвороб.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1991 року.
Радич. Виведений на Поліській дослідній станції ім. О. Засухіна.
Технологічна врожайність на 45-й день після сходів - 130 ц/га, а на¬
прикінці вегетації - 420 ц/га.
Кущ прямостоячий, компактний. Листки світло-зелені. Квіти чер-
воно-фіолетові.
Бульби округлі, рожеві. Вічка мілкі, нечисленні. М’якуш білий,
не темніє після приготування. Смакові якості хороші (4,0 бали).
Уміст крохмалю - 15,4 %.
Ракостійкий та відносно стійкий проти стеблової нематоди, фі¬
тофторозу і парші звичайної.
Рекомендований для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1997 року.
Санте. Виведений у Нідерландах. Інтенсивно накопичує вегета¬
тивну масу.
Кущ компактний, середньої висоти, добре облиствлений, стебел
має небагато. Листки великі, світло-зелені. Квіти білі.
Бульби овальні, блідо-жовті. Шкірка гладенька, вічка мілкі.
М’якуш блідо-жовтий, борошнистий, досить розсипчастий при ва¬
рінні. Смакові якості хороші (3,6-4,0 бали). Уміст крохмалю в буль¬
бах підвищений.
Ракостійкий та стійкий до картопляної нематоди і парші звичайної.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 2000 року.
Сатіна. Виведений у Німеччині. Придатний для виробництва каргоп-
лепродуктів. Урожайність на сортодільниці становила 310-340 ц/га.
Кущ високий; квіти білі.
Бульби білі. Середня маса - 90 г. М’якуш кремовий. Смакові яко¬
сті хороші (4,2 бала). Уміст крохмалю низький (10,7-12,4%).
Ракостійкий. Не стійкий до картопляної нематоди.
Рекомендований для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 2001 року.
Світанок київський. Виведений Інститутом картоплярства
УААН. Потенційна врожайність наприкінці вегетації - 450 ц/га.
Кущ невисокий, має багато стебел. Стебла округлі, мають мало
гілок. Листки невеликі, темно-зелені. Квіти червоно-фіолетові.
Бульби округлі, рожеві. Вічка поверхневі, нечисленні. М’якуш
кремовий, не темніє при розрізуванні та варінні. Смакові якості від¬
345
мінні (4,9 бала). Уміст крохмалю - 17-18%. Середня маса товарних
бульб - 90-130 г. За смаковими та кулінарними показниками є одним
із кращих сортів картоплі, що вирощуються в Україні.
Ракостійкий та відносно стійкий до стеблової нематоди, фітофто¬
розу і вірусних хвороб.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1987 року.
Солара. Виведений у Німеччині.
Кущ добре розвинений. Квіти білі.
Бульби овальні, вічка середньозаглиблені. Шкірка гладка, жовта.
М’якуш жовтий, розварюється слабко, при приготуванні страв не
темніє.
Ракостійкий та стійкий до парші звичайної, чорної ніжки й окре¬
мих вірусів. До вторинного росту бульб, внутрішніх дефектів, дуп¬
листості схильний слабко. Має довгі столони, а тому висаджувати
бульби слід досить мілко.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 2001 року.
Тімате. Виведений у Нідерландах. Рослини відзначаються інтен¬
сивним ростом у початкових фазах розвитку. Сорт належить до ви¬
соковрожайних.
Кущ добре облиствлений. Стебла нечисленні, прямостоячі до кін¬
ця вегетації, товсті, розгалужені помірно, мають світло-пурпурове
забарвлення. Листки досить великі, пониклі, темно-зелені. Квіти білі,
нечисленні.
Бульби видовжено-овальні, жовті. Смакові якості хороші. Уміст
крохмалю середній - 13,5 %.
Картоплиння досить чутливе до фітофторозу. Сорт імунний до
парші звичайної та стійкий до картопляної нематоди.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1999 року.
5.4. СЕРЕДНЬОСТИГЛІ СОРТИ
Віра. Створений в Інституті землеробства і тваринництва захід¬
ного регіону УААН. Потенційна врожайність - 300-450 ц/га.
Кущ середньої висоти. Квіти білі.
Бульби білі; вічка неглибокі. М’якуш білий. Смакові якості хо¬
роші (4,5 бала). Уміст крохмалю в бульбах - 17,1-18,0 %.
Ракостійкий. Відзначається високою стійкістю проти фітофторо¬
зу, вірусних хвороб та парші звичайної.
Рекомендований для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 2001 року.
346
Віринея. Виведений в Інституті картоплярства УААН.
Кущ прямостоячий, високий, добре облиствлений. Листки серед¬
нього розміру. Квіти червоно-фіолетові.
Бульби овальні, великі, рожеві. Середня маса товарної бульби -
117 г. М’якуш світло-жовтий. Смакові якості хороші (4,2 бала).
Уміст крохмалю - 15,5-15,8 %.
Ракостійкий. Високостійкий проти фітофторозу і кільцевої гнилі
га відносно стійкий проти мокрої гнилі.
Рекомендований для вирощування в зоні Полісся.
Занесений у Реєстр сортів рослин України з 2001 року.
Гатчинська. Виведений Північно-Західним науково-дослідним
інститутом сільського господарства Росії.
Кущ високий, добре облиствлений. Стебла товсті, пігментовані в
пазухах листків, ребристі. Листки великі. Квіти червоно-фіолетові.
Бульби білі, округло-овальні та плескаті із середньозаглибленими віч¬
ками. Шкірка бульб гладенька. М’якуш білий. Смакові якості хороші
(4,0 бали). Маса товарних бульб — до 140 г. Уміст крохмалю - 16,9 %.
Ракостійкий та відносно стійкий проти фітофторозу й вірусних
хвороб.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1973 року.
Горлиця. Сорт селекції Інституту картоплярства УААН.
Кущ високий, стебла потовщені, численні, гіллясті. Листки тем¬
но-зелені. Квіти білі.
Бульби білі, округлі, часто з висунутим столонним слідом. Шкір¬
ка гладенька, вічка нечисленні. М’якуш світло-жовтий. Смакові яко¬
сті хороші (4,5 бала). Уміст крохмалю в бульбах - 17 %.
Ракостійкий та відносно стійкий проти фітофторозу, основних ві¬
русних і бактеріальних хвороб.
Рекомендований для вирощування в усіх зонах України.
Занесено до Реєстру сортів рослин України з 1996 року.
Дубравка. Виведений на Поліській дослідній станції ім. О. Засу-
хіна. Високоврожайний.
Кущ високий, прямостоячий, середньо облиствлений, стебла ма¬
ють середню кількість гілок. Листки великі. Квіти білі.
Бульби округлі, світло-рожеві. Вічка середньозаглиблені. М’якуш
білий. Смакові якості задовільні і хороші. Уміст крохмалю в бульбах -
10,1-13,2%.
Ракостійкий та відносно стійкий проти фітофторозу і парші зви¬
чайної.
Рекомендований для вирощування в зонах Полісся та Степу.
Занесений до Реєстру сортів рослин України з 2001 року.
347
Західна. Виведений у Львівському державному аграрному уні¬
верситеті.
Кущ прямостоячий, середньої висоти. Листки зелені. Квіти чер-
воно-фіолетові.
Бульби округло-овальні, жовті, середнього розміру. М’якуш світло-
кремовий. Смакові якості задовільні. Уміст крохмалю в бульбах - 18 %.
Ракостійкий та стійкий до фітофторзу і картопляної нематоди.
Рекомендований для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1998 року.
Либідь. Виведений Інститутом картоплярства УААН. Потенцій¬
на врожайність наприкінці вегетації - 500 ц/га.
Кущ високий, добре облиствлений. Квіти і м’якуш бульб білі.
Смакові якості хороші (4 бали). Уміст крохмалю - 15,5%. Середня
маса товарної бульби - 120 г.
Ракостійкий та відносно стійкий проти фітофторозу, парші зви¬
чайної, кільцевої й мокрої гнилі та іржавої плямистості.
Рекомендований для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1993 року.
Луговська. Виведений Інститутом картоплярства УААН. Потен¬
ційна врожайність наприкінці вегетації - 600-650 ц/га.
Кущ компактний, середньооблиствлений. Стебла кутасті, мають
небагато гілок. Квіти білі. Наростання надземної вегетативної маси
на початку вегетації, порівняно з іншими сортами, уповільнене.
Бульби світло-рожеві, овальні, з дрібними рожевими вічками.
Шкірка гладенька. М’якуш білий, при розрізуванні не темніє. Сма¬
кові якості хороші (4,0-4,5 бала). Уміст крохмалю - 15-16 %. Серед¬
ня маса товарних бульб - 91-150 г. Зберігаються добре.
Ракостійкий. Відзначається високою польовою стійкістю проти фі¬
тофторозу, середньою - проти вірусних хвороб і стеблової нематоди.
Рекомендується для вирощування в зонах Лісостепу та Полісся
України.
Занесений до Реєстру сортів рослин України з 1987 року.
Пекуровська. Виведений Чернігівською дослідною станцією по
картоплі. Середньостиглий.
Кущ середньої висоти. Стебла гіллясті. Квіти білі.
Бульби овальні, білі, з мілкими вічками. М’якуш білий. Смакові
якості задовільні (3,7-4,2 бала). Уміст крохмалю в бульбах - 14,4—
17,8 %. Середня маса товарних бульб - 78-85 г.
Ракостійкий. Стійкий до картопляної нематоди та відносно стій¬
кий проти фітофторозу і вірусних хвороб.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1995 року.
348
Придеснянська. Виведений Чернігівською дослідною станцією
по картоплі.
Кущ середньої висоти, добре облиствлений. Квіти червоно-
фіолетові.
Бульби білі, округло-овальні на досить довгих столонах. Вічка
поверхневі. М’якуш світло-жовтий. Смакові якості досить хороші
(4,5-4,7 бала). Уміст крохмалю високий - 19,7-21,8 %. Середня маса
товарних бульб - 78-103 г.
Ракостійкий. Відзначається польовою стійкістю до фітофторозу,
вірусних хвороб і стеблової нематоди.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1996 року.
Слава. Виведений Інститутом землеробства і тваринництва за¬
хідного регіону УААН та Львівським державним аграрним універси¬
тетом. Потенційна врожайність наприкінці вегетації - 400 ц/га.
Кущ компактний, листки світло-зелені. Квіти білі.
Бульби білі, округлі. Шкірка гладенька, вічка поверхневі. М’якуш
білий, не темніє при розрізуванні і після варіння. Смакові якості хо¬
роші (4,0 бали). Уміст крохмалю невисокий - 12%. Середня маса то¬
варних бульб - 80-150 г.
Ракостійкий. Відзначається середньою стійкістю проти стеблової
нематоди, парші звичайної, вірусних хвороб та підвищеною - проти
фітофторозу.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1992 року.
Слов’янка. Виведений Інститутом картоплярства УААН. Потен¬
ційна врожайність - 570 ц/га.
Кущ прямостоячий, високий. Стебла слабогіллясті. Листки вели¬
кі, світло-зелені. Квіти червоно-фіолетові.
Бульби видовжено-овальні, великі, рожеві. Шкірка гладенька, вічка
мілкі. М’якуш кремовий. Смакові якості задовільні (3,7 бала). Уміст
крохмалю невисокий - 12,9%. Середня маса товарної бульби -118 г.
Ракостійкий. Стійкий проти картопляної нематоди. Має польову
стійкість проти фітофторозу, відносно стійкий проти фузаріозу і ві¬
русних хвороб. Бульби чутливі до ураження сухою та мокрою гнил¬
лю, особливо при надто дощовому чи сухому вегетаційному періоді.
Рекомендований для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1999 року.
Українська рожева. Виведений Інститутом картоплярства УААН.
Потенційна врожайність наприкінці вегетації - близько 500 ц/га.
Кущ високий. Стебла дуже гіллясті, численні, забарвлення стебел
антоціаном слабке, крапчасте. Листки середнього розміру, темно-
зелені. Квіти червоно-фіолетові.
349
Бульби короткоовальні, рожеві. Вічка дрібні, червоні. М’якуш
світло-рожевий, при розрізуванні не темніє. Смакові якості хороші
(4,1 бала). Уміст крохмалю - 17,6 %. Середня маса товарної бульби -
88-117 г.
Ракостійкий та відносно стійкий проти фузаріозу, макроспоріозу,
стеблової нематоди.
Рекомендується для вирощування в Лісостепу та на Поліссі України.
Занесений до Реєстру сортів рослин України з 1989 року.
Явір. Виведений Інститутом картоплярства УААН.
Кущ прямостоячий, компактний. Стебла не пігментовані. Листки
великі, матові.
Бульби білі, округлі. М’якуш кремовий. Смакові якості хороші
(3,9 бала). Уміст крохмалю в бульбах -16-18%. Товарність бульб у
врожаї висока.
Ракостійкий та відносно стійкий до фітофторозу, альтернаріозу,
мокрої гнилі, парші звичайної та вірусних хвороб.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 2000 року.
5.5. СЕРЕДНЬОПІЗНІ СОРТИ
Воловецька. Виведений Інститутом картоплярства УААН.
Кущ напіврозлогий, високий, має багато стебел. Стебла мають
мало гілок та крапчасте антоціанове забарвлення уздовж стебла. Силь-
нооблиствлений. Листки темно-зелені. Квіти червоно-фіолетові.
Бульби жовті, овальні. Шкірка сітчаста. Вічка середньозаглибле-
ні. М’якуш жовтий. Смакові якості хороші. Уміст крохмалю - 19,4%.
Ракостійкий. Відзначається польовою стійкістю проти фітофто¬
розу, вірусних хвороб, парші звичайної та бактеріальних хвороб.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1985 року.
Дезіре. Виведений у Шотландії. Посухостійкий.
Кущ добре облиствлений. Стебла численні, високі, товсті, з тем-
но-коричневим антоціановим забарвленням. Листки - від великих до
середніх, сіро-зелені. Квіти дають багато ягід.
Бульби видовжено-овальні, червоні. Шкірка гладенька. Вічка по¬
верхневі. М’якуш світло-жовтий. Бульби великі, малочутливі до внут¬
рішніх пошкоджень. Смакові якості хороші. Уміст сухої речовини і
крохмалю досить високий (15,6-21,5%).
Ракостійкий. Досить чутливий до вірусу скручування листя; іму¬
нний до парші.
Рекомендується для вирощування в зоні Лісостепу.
Занесений до Реєстру сортів рослин України з 1997 року.
350
Дзвін. Виведений в Інституті сільського господарства Полісся
УААН. Універсальний, придатний для переробки на чипси.
Кущ високий, прямостоячий. Квіти червоно-фіолетові.
Бульби овальні, рожеві, з мілкими вічками. Сорт відзначається
досконалою формою бульб. М’якуш світло-жовтий. Смакові якості
хороші (4,0-4,1 бала). Уміст крохмалю в бульбах - 18,0-18,6 %.
Ракостійкий. Стійкий проти картопляної нематоди та відносно
стійкий проти фітофторозу.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 2000 року.
Зарево. Виведений Інститутом картоплярства УААН. Потенційна
врожайність наприкінці вегетації - 400 ц/га.
Кущ низький. Стебла слабко пігментовані, численні. Квіти чер¬
воно-фіолетові.
Бульби рожеві, округлі. М’якуш білий, при розрізуванні не тем¬
ніє. Смакові якості хороші (4,4 бала). Уміст крохмалю досить висо¬
кий - 23%. Це один із найбільш крохмалистих сортів картоплі, по¬
ширених в Україні. Середня маса товарних бульб - 75-81 г. В окремі
роки на них можливі неглибокі тріщини.
Ракостійкий та відносно стійкий проти фітофторозу, парші зви¬
чайної. Іншими хворобами уражується середньою мірою.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1983 року.
Ікар. Виведений Поліською дослідною станцією ім. О. Засухіна.
Потенційна врожайність наприкінці вегетації - 400 ц/га.
Кущ компактний, середньої висоти. Стебла дуже гіллясті, в попе¬
речному розрізі кутасті. Листки середнього розміру, світло-зелені.
Бульби овальні, з тупою верхівкою і втиснутою пуповиною, ро¬
жеві. Вічка нечисленні, поверхневі. Смакові якості задовільні (3,9
бала). М’якуш білий, при розрізуванні не темніє. Уміст крохмалю -
18,4 %. Середня маса товарних бульб - 75 г.
Ракостійкий та відносно стійкий проти фітофторозу й вірусних
хвороб.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1983 року.
Ольвія. Виведений Інститутом картоплярства УААН. Потенцій¬
на врожайність наприкінці вегетації - 450 ц/га.
Кущ прямостоячий, компактний, високий. Стебла дуже гіллясті.
Листки середнього розміру, темно-зелені, матові. Квіти білі.
Бульби короткоовальні, білі. Шкірка гладенька, вічка мілкі. Сма¬
кові якості хороші (4,3 бала). Уміст крохмалю в бульбах - 16,1 %.
Середня маса товарної бульби - 108 г.
351
Ракостійкий. Відзначається високою польовою стійкістю проти
фітофторозу.
Рекомендований для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1999 року.
Пєтланд Делл. Виведений у Шотландії.
Кущ добре розвинений. Квіти білі.
Бульби білі, видовжено-овальні. Вічка середні. М’якуш світло-
кремовий. Смакові якості хороші. Розварюється слабко. Уміст сухих
речовин підвищений. До механічних пошкоджень бульби відносно
стійкі.
Ракостійкий та відносно стійкий до фітофторозу.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1999 року.
Поліська рожева. Виведений Поліською дослідною станцією
ім. О. Засухіна. Потенційна врожайність - 450 ц/га.
Кущ середньої висоти. Стебла за всією довжиною сильно забарв¬
лені антоціаном. Листки середнього розміру, темно-зелені. Квіти чер-
воно-фіолетові.
Бульби овальні, рожеві. Шкірка сітчаста. М’якуш білий, не темніє
до і після варіння. Смакові якості хороші (4,3 бала). Уміст крохмалю
- 15,6-18,6 %. Середня маса товарних бульб - 87-120 г.
Неракостійкий. Має підвищену стійкість проти фітофторозу.
Слабко уражується вірусними хворобами, паршею звичайною, чор¬
ною ніжкою та кільцевою гниллю.
Рекомендується для зони Полісся.
Занесений до Реєстру сортів рослин України з 1978 року.
Ракурс. Виведений Інститутом картоплярства УААН. Потенцій¬
на врожайність наприкінці вегетації - 450 ц/га.
Кущ прямостоячий, високий, компактний. Стебла гіллясті, серед-
ньооблиствлені. Квіти білі.
Бульби округло-овальні, білі. М’якуш білий, відносно стійкий до
потемніння. Смакові якості бульб хороші (4,0-4,5 бала). Уміст крох¬
малю в бульбах - 16-17 %. Середня маса товарної бульби - 80-92 г.
Ракостійкий та стійкий проти стеблової нематоди, фітофторозу і
парші звичайної.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1997 року.
Цезар. Виведений у Нідерландах.
Кущ високий, досить добре облиствлений. Стебла середньої ви¬
соти, з помітним антоціановим забарвленням. Листки середнього
розміру, зелені. Квіти білі.
352
Бульби великі, овальні, жовті. Шкірка гладенька, вічка поверхне¬
ві. М’якуш блідо-жовтий. Смакові якості задовільні. Уміст крохмалю
середній - 15,7%.
Ракостійкий та стійкий до картопляної нематоди. Резистентний
до скручування листків. Імунний до парші звичайної.
Рекомендується для вирощування в усіх зонах України.
Занесений до Реєстру сортів рослин України з 1996 року.
5.6. ІГОНІ СОРТИ
Древлянка. Виведений Інститутом сільського господарства По¬
лісся УААН.
Кущ прямостоячий, високий. Стебла гіллясті. Листки середнього
розміру, темно-зелені. Квіти червоно-фіолетові.
Бульби овальні, червоні. Шкірка сітчаста, вічка мілкі, численні.
М’якуш світло-кремовий. Смакові якості хороші. Уміст крохмалю в
бульбах - 16 %.
Ракостійкий. Має підвищену стійкість проти фітофторозу, серед¬
ню - проти парші звичайної, вірусних і бактеріальних хвороб.
Рекомендується для вирощування в зоні Полісся.
Занесений до Реєстру сортів рослин України з 1984 року.
Ласунак. Виведений Білоруським інститутом картоплярства і
плодоовочівництва. Потенційна врожайність - 400 ц/га.
Кущ прямостоячий, стебла нечисленні, мають мало гілок, дуже
облиствлені. Листки середнього розміру, темно-зелені. Квіти білі,
цвітіння рясне і тривале.
Бульби білі, овальні. Шкірка сітчаста. Вічка невеликі, поверхневі.
М’якуш кремовий. Смакові якості хороші (4,2-4,6 бала). Уміст кро¬
хмалю в бульбах - 16,8-20,0 %. Бульби великі, маса товарних бульб
- 90-120 г.
Ракостійкий. Фітофторозом і вірусними хворобами уражується
середньою мірою.
Рекомендується для вирощування в зонах Полісся та Лісостепу.
Занесений до Реєстру сортів рослин України з 1989 року.
Темп. Виведений Білоруським інститутом картоплярства і пло¬
доовочівництва. Високоврожайний. Потенційна врожайність напри¬
кінці вегетації - 400 ц/га.
Кущ високий. Стебла мають мало гілок, ребристі, забарвлені
антоціаном у пазухах листків і в основі стебла. Листки великі, тем-
но-зелені. Квіти блідо-синьо-фіолетові, наприкінці цвітіння - май¬
же білі.
Бульби округло-овальні, плескуваті, великі, білі з жовтим відтін¬
ком. Вічка неглибокі. Шкірка лускувата. М’якуш світло-жовтий.
12 2 ,32
353
Смакові якості досить хороші (4,8 бала). Уміст крохмалю в бульбах -
18,8-20,9 %. Маса товарних бульб - 115-180 г.
Ракостійкий та відносно стійкий проти фітофторозу і парші зви¬
чайної. Більш високі врожаї дає на зв’язних грунтах.
Рекомендується для вирощування в зонах Лісостепу і Полісся.
Занесений до Реєстру сортів рослин України з 1973 року.
ЛІТЕРАТУРА
1. Вітенко В.А., Молоцький М.Я. Насінництво картоплі. - К.: Урожай,
1977. - С.97-113.
2. Довідник картопляра. За редакцією Кучка А.А., Куценка В.С., Осипчу-
ка А.А., Батюти В.Г.- К.: Україна - 1991- С.10-19.
3. Зубченко О.О. Сорти і насінництво картоплі- К.: Урожай, 1970-
С.12-21.
4. Кучко А., Власенко М., Мицько В. Фізіологія та біохомія картоплі.-
К.: Довіра, 1998.-С.220-249.
5. Молоцький М.Я., Васильківський С.П., Князюк В.І. Селекція та
насінництво польових культур. - К.: Вища пік., 1994. - С. 53-60.
6. Новосельська А.П., Мицько В.М., Холодило І.В. Сортові особливості
біохімічного складу картоплі // Картоплярство. - К.: Урожай, 1995. -
Вип.26.
7. Реєстр сортів рослин України на 2000 рік. Ч.1.- К., 1999.- С.69-72.
8. Теслюк П.С., Молоцький М.Я. Практичний порадник картопляра - К.:
Кий, 1999.- С.57-96.
9. Теслюк П., Пасічник П., Верменко Ю., Панківська Ю. Сорти картоплі.
- К.: Агросвіт України, 2001.- 93 с.
6. НАСІННИЦТВО КАРТОПЛІ
6.1. ІСТОРІЯ РОЗВИТКУ НАСІННИЦТВА
На початку XX ст. селекційна і насінницька робота з картоплею в
Україні не провадилася. Сорти, що вирощувались у той час, мали
закордонне, переважно німецьке, походження. Вони були мало при¬
стосовані до місцевих умов і погано витримували посуху, швидко
вироджувались і давали низькі врожаї.
Сортовим посівам тоді не приділялось належної уваги, і селяни в
основному вирощували суміші різних сортів [9].
Вивченням урожайності завезених із-за кордону сортів і добором
кращих для розмноження займалися Полтавська, Сумська та Харків¬
ська науково-дослідні станції, Зміївське і Драбівське дослідні поля, а
також мережа дослідних полів Товариства цукрозаводчиків, Поділь¬
ського товариства сільського господарства, а пізніше дослідні поля з
культури картоплі в Харківській області [1]. У той час із завезених
сортів вивчались такі, як Імператор Ріхтера, Новий експорт, Сілезія,
Вольтман, Крюгер, Індустрія, Бісмарк, Витязь, Аграрія, Ісполін, Си¬
ній велетень, Юбель, Спаржевий та ін. [2, 10].
Найбільш урожайними вважалися сорти Вольтман і Крюгер. Во¬
ни та сорт Юбель ще до недавнього часу займали великі площі в по¬
ліських районах країни.
Планова робота з насінництва картоплі розпочалися в 1921 р. піс¬
ля виходу в світ урядового декрету “Про насінництво”, яким Народ¬
ний комісаріат землеробства зобов’язувався у найкоротші строки
налагодити організацію масового розмноження і розповсюдження
чистосортного насіння. Першим заходом у здійсненні цього декрету
була організація в 1921 р. “Держнасінкультури”. З того часу почало¬
ся порівняно швидке виявлення і розмноження в країні сортової кар¬
топлі.
З 1924 р. Українська сортовипробувальна мережа розгорнула ро¬
боту з випробування сортів картоплі, у якій брали участь такі дослід¬
ні установи: Харківська та Київська обласні дослідні станції, Полісь¬
ка, Чорторийська, Носівська, Уманська, Новозибківська, Чемерська,
Сумська, Полтавська, Вознесенська та Одеська дослідні станції, фі¬
ліал Дніпропетровської обласної дослідної станції «Надія» і При-
азовське дослідне поле [1].
Вивченням стану картоплярства займались також об'єднання
крохмальних заводів, “Укрспиртотрест” і сільськогосподарська коо¬
перація. Обстеженням посівів було виявлено, що більшість із них
355
являє собою суміш сортів із місцевими назвами (Винокурка, Фунтов-
ка, Американка, Фуксінка, Синя та ін.).
У 1925 р. на більшій частині посівів картопля була значною мі¬
рою вироджена, і тому потрібно було терміново впроваджувати не¬
обхідні заходи щодо її оздоровлення.
З різних сумішок виділялися більш цінні сорти, а також завози¬
лися кращі з півночі республіки, і цей садивний матеріал розмножу¬
вався. Одночасно ефективно працювала і мережа із сортовипробу¬
вання картоплі. У результаті сортовипробування були виділені кращі
сорти, серед яких можна назвати такі: Рання роза, Пам’ять Костіна,
Рожева з Мілет, Деодара, Парнасія, Вольтман та ін. [1].
З метою прискореного розмноження кращих сортів картоплі Но-
сівська сільськогосподарська дослідна станція розробила спосіб ви¬
рощування їх вічками [4,5,6]. Велику роботу з вирощування і роз¬
множення елітної картоплі провадила в той час Поліська дослідна
станція [7]. Питаннями розмноження кращих сортів картоплі і впро¬
вадженням їх у виробництво займалася також “Укрплодоспілка” .
Обстеження, проведені у 30-х роках Київською дослідною стан¬
цією, показали, що хоча сортові посіви на той час займали лише одну
четверту частину площ, але вони були значно врожайнішими.
Наприклад, сорт Вольтман в Україні забезпечував урожай бульб
на 325 пудів з десятини більше, ніж місцева селянська картопля.
Крім того, місцеві сорти містили менше крохмалю.
Завозилися також і нові сорти: Деодара, Крюгер, Лотос, Князів¬
ська корона та багато інших. Вирощувалися й ультраранні сорти, що
досягали стиглості через 45-50 днів. Це - Шеститижневий та Ранній
Маржелен. На жаль, нині ці сорти втрачені.
За даними А.Д. Бєлікова, керівника відділу селекції Українського
науково-дослідного комбінату картопляного господарства, у 30-х
роках районувалися також сорти Епікур, Бов’є, Джентельмен.
При невисоких урожаях у цілому по республіці особливо низь¬
кими вони були на півдні України, незважаючи на те, що сюди заво¬
зився насінний матеріал з північних районів. В посушливих районах
особливо врожайними вважалися сорти з рожевими бульбами. А то¬
му під виглядом Ранньої рози в 1932 р. сюди були завезені сорти
Епікур, Рожева з Мілет, Деодара та ін. Уже в той час було виявлено,
що в райони, де картопля швидко втрачає продуктивність, на насінні
цілі її доцільно завозити з північних областей.
У різних регіонах сортовий склад картоплі був дуже розмаї¬
тий.
Значну роботу з розповсюдження нових сортів на Чернігівщині (у
північній частіші лісостепової зони області) проводила Носівська
державна селекційна дослідна станція, заснована у червні 1911 р. До
її складу входили Лосинівське дослідне поле і Остерський опорний
пункт.
356
У 1930 р. на Чернігівщині вирощувались такі сорти картоплі: у зоні
Полісся - Рання роза, Вольтман, Крюгер, Рожева з Мілет, Альма; у зоні
Лісостепу - Епікур, Рання роза, Вольтман, Деодара, Бов’є, Альма.
У 30-х роках в Україні серед сортів картоплі були поширені так
звані “кучерявці”. їх виявили в насадженнях картоплі у селян (від 10
до 100 %). Це були змінені на вигляд сорти. На той час точно не зна¬
ли, чи це мутації, чи хвороби картоплі. Розрізняли “вперті кучерявці”
і “напівкучерявці”. “Напівкучерявці” - це середні на вигляд кущі між
нормальним сортом і “впертим кучерявцем”. “Кучерявці” давали до¬
сить дрібні бульби і низький урожай. При розмноженні “кучерявців”
бульбами потомствам передавались усі властиві бульбам ознаки. Бу¬
ли відомі кучерявці Ранньої рози, кучерявці Червоносотки, кучерявці
Народної. Очевидно, це були вироджені рослини картоплі, що до
певного часу зберігали продуктивність.
У статистичному довіднику за 1938 р. знаходимо склад сортових
насаджень картоплі в Україні та площі, зайняті під окремими сорта¬
ми. У колгоспах і радгоспах картоплю висаджували на площі 513,5
тис. га, у тому числі сортові насадження займали 35,2 %. Загальна
площа насаджень картоплі тоді в СРСР складала 3479,8 тис. га, а
сортових - 30,8 %.
Найбільшу площу серед сортових насаджень в Україні - 106,7
тис. га, або 59 %,- займав сорт Вольтман. По 10 %, або по 18 тис. га,
займали Рання роза та Елла. Сорт Деодара займав 4,3 % (7,7 тис. га),
Лорх - 3,2 % (5,7 тис. га). На площі 1460 га вирощували сорт Альма.
Сорт Рання роза найбільші площі займав у Миколаївській області,
Лорх - у Ворошиловградській, Альма - у Вінницькій.
У 1940 р. загальна площа насаджень картоплі в південних облас¬
тях складала: Одеській, Миколаївській, Запорізькій, Дніпропетров¬
ській, Ворошиловградській та в Криму - 172,3 тис. га, у тому числі
сортових насаджень - 77,2 тис. га. З них Рання роза займала 29,6 тис.
га; Епікур - 9,1; Кур’єр - 6,1; Елла - 11,2; Лорх - 17,2; Альма - 1,2;
Юбель - 1,9; Кобблер - 0,4; Епрон - 0,3; Майка - 0,1 тис. га.
У 1940 р. в Україні було районовано понад 20 сортів картоплі,
проте в переважній быльшості це були сорти зарубіжної селекції.
У перші повоєнні роки певний вплив на розвиток картоплярства
мала Постанова Ради Народних Комісарів від 14 березня 1945 р.
“Про поліпшення насінництва картоплі в Українській РСР”. У поста¬
нові, виходячи з існуючого стану справ, ставилось завдання довести у
1946 р. сортові насадження до 25—40 % від загальних площ картоплі
в республіці. Основою насінництва мала бути насінна ділянка, і вод¬
ночас рекомендувалось закладання в кожному господарстві розсад¬
ника поліпшення. Було також визначено завдання з вирощування
еліти картоплі в 10 наукових і навчальних установах, у тому числі у
Всесоюзному селекційно-генетичному інституті (Одеса); на селек¬
ційних овочево-картопляних дослідних станціях НКЗС УРСР-
357
Харківській, Одеській, Сталінській (Донецькій); Дніпропетровсько¬
му інституті зернового господарства та на Київській дослідній стан¬
ції ВХДІСПу НКХП.
На той час в Україні було районовано 19 сортів картоплі. Кра¬
щими серед них були Рання роза, Епікур, Кур’єр, Елла, Вольтман,
Червоноспиртовий, Парнасія, Карнеа, Альма, Лорх, Деодара та ін.
Насадженням картоплі в країні великої шкоди почав завдавати
завезений у роки війни рак. Тому насінницьку і селекційну роботу
було спрямовано на випробування зарубіжних та виведення вітчиз¬
няних сортів, стійких до цієї хвороби.
У 1956 р. в Україні почали створюватись обласні державні сіль¬
ськогосподарські дослідні станції, які, крім вирішення інших питань,
одержували завдання з виробництва та реалізації еліти картоплі для
сортооновлення та сортозаміни.
У 1958 р. в республіці було створено 884 районні насінницькі
колгоспні і радгоспні розсадники. Вихідний матеріал районованих і
перспективних сортів районний насінний розсадник одержував за
планом обласного управління сільського господарства від науково-
дослідних установ. Завдання з вирощування елітної картоплі одер¬
жували й науково-дослідні установи, розміщені в усіх природно-
кліматичних зонах республіки, у тому числі й на півдні.
До 1961 р. насінництво картоплі в усіх зонах України було орга¬
нізоване так, що в ньому брали участь елітні господарства, а в колгос¬
пах і радгоспах існували розсадники з розмноження еліти і насінні
ділянки. Така організація виробництва садивного матеріалу відіграла
позитивну роль у поширенні нових і дефіцитних сортів. Проте з ча¬
сом це стало гальмом у вирощуванні високоякісного садивного ма¬
теріалу в усіх районах, особливо південних. Це пояснюється швид¬
ким виродженням картоплі в південних областях через значне поши¬
рення вірусної інфекції та підвищену температуру в період форму¬
вання врожаю. Враховуючи це, з 1961 р. було прийнято такий
порядок виробництва насінної картоплі, при якому вирощування ви¬
соких репродукцій було сконцентроване у сприятливих грунтово-
кліматичних зонах. У насінницьких господарствах, розміщених у
поліських та західних регіонах, почали вирощувати садивну картоп¬
лю як для власних потреб, так і для вивезення в господарства пів¬
денних областей.
Постановою Ради Міністрів колишнього СРСР від 24 травня 1962 р.
№487 “Про заходи по поліпшенню насінництва картоплі в колгоспах
і радгоспах” було визначено порядок виробництва сортової насінної
картоплі (еліти) науково-дослідними установами і передачу її колгос¬
пам і радгоспам для сортооновлення. Союзним республікам було да¬
не завдання створити республіканські фонди сортової картоплі. На
виконання цієї постанови Рада Міністрів Української РСР прийняла
постанову, якою науково-дослідним установам і сільськогоспо¬
358
дарським вузам були доведені плани виробництва і продажу еліти: на
1963 р. -14,5 тис. тонн, на 1964 р. - 17,6 і на 1965р. - 17,6 тис. тонн; на¬
сінницьким господарствам - матеріалу високих репродукцій: на
1963 р. - 22,2 тис. тонн, на 1964 р. - 54,6 і на 1965 р. - 67,7 тис. тонн.
У 1965 р. уряд України ще раз розглянув питання виробництва і
забезпечення колгоспів і радгоспів насінною картоплею для сорто¬
оновлення. Науково-дослідні господарства одержали завдання збіль¬
шити виробництво і реалізацію елітної картоплі в 1967 р. до 36,8 тис.
тонн, а заготівлі до республіканського фонду - до 95 тис. тонн. Була
визначена і чітка система реалізації сортової насінної картоплі та
строки сортооновлення.
Слід зазначити, що значна кількість науково-дослідних установ
плани не виконували, а якість еліти була низькою. Елітна картопля,
вирощена в Лісостепу, за посівними якостями значно поступалася
перед елітою з Полісся. Тому господарства з більшим задоволенням
брали насінний матеріал для сортооновлення з поліських госпо¬
дарств.
Для виявлення кращих зон вирощування високоякісного насінно¬
го матеріалу в республіці була проведена велика робота з питань ви¬
значення ступеня виродження картоплі в різних природно-кліматич-
них умовах.
У результаті проведеного обліку (візуального) на сортодільницях
та в елітно-насінницьких господарствах дослідних установ було ви¬
явлено, що поширення вірусних хвороб зростало з північного заходу
на південний схід. Крім того, була виявлена закономірність нарос¬
тання чисельності персикової попелиці на посівах картоплі в тому ж
напрямку. При цьому не тільки збільшувалася кількість попелиць,
але й посилювалась інтенсивність їх розмноження, від чого залежить
ураженість картоплі вірусними хворобами [8].
Дослідники виявили поступове зниження насінних якостей кар¬
топлі з кожною наступною репродукцією, і чим нижчою була репро¬
дукція, тим швидше знижувалась її продуктивність. У зв’язку з цим
виникла потреба скорочення строків сортооновлення, а отже - збіль¬
шення виробництва здорового насіннєвого матеріалу. Учені багатьох
країн, у тому числі й вітчизняні, вирішуючи справу поліпшення на¬
сінництва картоплі, звільнення її від вірусів, дійшли висновку про
необхідність створити закриті райони (зони) товарного насінництва і
провести цілий комплекс відповідних заходів. Використовуючи віт¬
чизняний і зарубіжний досвід, Міністерство сільського господарства
Української РСР разом з ученими республіки розробили систему за¬
ходів щодо створення закритих районів і зон для товарного насінниц¬
тва безвірусної насінної картоплі для сортооновлення.
На початку 1968 р. Рада Міністрів України прийняла постанову
про організацію в республіці закритих районів товарного виробницт¬
ва сортової насінної картоплі в Житомирській, Рівненській та Черні¬
359
гівській областях. Такими районами були затверджені Овруцький,
Костопільський і Менський, як такі, що мають найсприятливіші
ірунтово-кліматичні, фітосанітарні умови для вирощування високо¬
якісного здорового насіннєвого матеріалу.
Було затверджено такий порядок виробництва і реалізації садив¬
ної картоплі: а) первинне насінництво і вирощування еліти проводи¬
ти у виділених у кожному районі одному-двох кращих господарст¬
вах, що є насінницькими господарствами першої групи. Вирощену
елітну картоплю мало реалізовувати об'єднання “Сортнасіннєовоч”;
б) розмножувати одержану від “Сортнасіннєовоч” еліту в насінниць¬
ких господарствах другої групи, до якої належить решта господарств
закритого району; в) контроль за вирощуванням елітної і сортової
картоплі, заготівлею і реалізацією її, відповідно до планів сортоонов¬
лення, здійснювати через новостворені в цих районах насіннєві бази
“Сортнасіннєовоч”.
У кожному господарстві, яке займалось вирощуванням еліти, бу¬
ло організовано опорний пункт науково-дослідних установ з первин¬
ного насінництва.
Але перед організацією закритих районів необхідно було провес¬
ти економічні розрахунки і здійснити практичні заходи, що дозволи¬
ли б всебічно оцінити придатність району для ведення насінницької
роботи з картоплею. Вченими республіки були розроблені відповідні
вимоги для визначення закритих районів і зон товарного насінництва
цієї культури. Ці вимоги передбачали:
- необхідність повного забезпечення насінною картоплею райо¬
нованих сортів для сортооновлення господарств своєї та інших облас¬
тей республіки;
- придатність ірунтово-кліматичних і фітосанітарних умов, а та¬
кож економічні можливості для ведення насінницької роботи з кар¬
топлею;
- відповідність структури посівних площ до завдань з вирощу¬
вання насінної картоплі;
- огляд земельних територій і всіх посівів як у колгоспах і радго¬
спах, так і на присадибних ділянках для виявлення придатних для
наступного використання насінного матеріалу;
- організаційні й матеріально-технічні заходи щодо організації
закритого району;
- наявність ґрунтово-кліматичних умов, сприятливих для вирощу¬
вання картоплі, і природних перепон (ліси, луки, пасовиська, болота,
ріки, озера) для захисту від попелиць, що можуть прилітати з-за меж
району. На межі з іншими районами, де немає природних перепон, необ¬
хідно було створювати лісові смуги або насаджувати захисні смуги кар¬
топлі здоровим насінним матеріалом.
Також було накреслено ряд інших заходів щодо захисту насінної
картоплі від ураження її вірусами.
360
Було визначено, що основою насінництва картоплі в плановій си¬
стемі сортооновлення є еліта науково-дослідних установ або насінна
картопля високих репродукцій, вирощена в закритих районах на ос¬
нові безвірусних клонів із застосуванням розроблених методів конт¬
ролю за її станом.
Положення про еліту картоплі передбачало вирощувати її тільки
методом клонового добору із застосуванням візуального і серологіч¬
ного контролю за її якістю. На той час була прийнята шестирічна
схема вирощування еліти: перший рік - добір клонів; другий - ви¬
пробування клонів першого року; третій - випробування клонів дру¬
гого року; четвертий - розмноження клонів (супер-супереліта); п'я¬
тий - супереліта; шостий рік - еліта. У деяких науково-дослідних
установах та елітно-насінницьких господарствах країни використо¬
вували п'ятирічну схему вирощування еліти. У цьому разі ланка ви¬
пробовування клонів другого року випадала.
Науково-дослідними установами і сільськогосподарськими орга¬
нами були розроблені і схвалені такі строки сортооновлення картоп¬
лі: у господарствах Полісся і західних районів республіки - один раз
у п'ять років на насінну площу; у господарствах центрального і схід¬
ного Лісостепу - один раз у три роки на насінну площу; у південних
областях - щороку на всю площу садіння картоплі; на присадибних
ділянках на Поліссі і в Лісостепу сортооновлення проводити, як і в
радгоспах та колгоспах, у південних областях - через кожні два ро¬
ки. Але це не виключало можливості зміни визначених строків у
зв’язку з тим, що з кожним роком поліпшувалася якість насіннєвого
матеріалу, що використовувався для сортооновлення, фітосанітарний
стан у районах виробництва продовольчої картоплі [3].
Пізніше, протягом 1971-1978 рр., було додатково організовано
ряд нових закритих районів з виробництва товарної насінної картоп¬
лі у поліських областях України, де також були виділені господарст¬
ва першої групи і створені опорні пункти з первинного насінництва.
Усього з 1978 р. було спеціалізовано 21 район і дві зони з насінницт¬
ва картоплі.
Наказом Міністерства сільського господарства від 17 січня 1978 р.
за № 16 “Про заходи по поліпшенню селекції і насінництва картоплі”
щорічне виробництво і реалізацію елітної та сортової картоплі пе¬
редбачалося довести в 1982-1985 рр. до 618 тис. тонн, у тому числі
75 тис. тонн еліти, з них у закритих районах і зонах - відповідно
590,5 і 48 тис. тонн.
У період організації і в перші роки діяльності закритих районів
проводилася велика робота з навчання кадрів картоплярів за участю
провідних учених науково-дослідних установ. У цей час широко
впроваджувалася механізація вирощування картоплі. У господарст¬
вах створювалися механізовані ланки по її вирощуванню з мінімаль¬
ними затратами праці. У закритих районах інтенсивно створювалася
361
матеріально-технічна база колгоспів і радгоспів з виробництва і збе¬
рігання картоплі та насінних баз “Сортнасіннєовоч”.
З насінної картоплі науково-дослідних установ, навчальних за¬
кладів і закритих районів створювали республіканський фонд насін¬
ної картоплі для сортооновлення. Заготівлю цієї картоплі і її реаліза¬
цію здійснювали організації “Сортнасіннєовоч”.
В обов'язки районної насінної бази “Сортнасіннєовоч” входило:
укладання договорів на виробництво і реалізацію насінної картоплі
(рис. 6.1); здійснення контролю за вирощуванням насінної картоплі в
господарствах; проведення польової апробації насінних посівів; від¬
бір проб насінних бульб для ґрунтового контролю в науково-дос¬
лідних установах.
Для проведення заготівель насінної картоплі в закритих районах і
науково-дослідних установах були організовані приймальні пункти,
підпорядковані насінній базі. Як правило, вони розташовувалися біля
транспортних магістралей.
Велику увагу приділяли затарюванню картоплі при перевезенні.
Найкращою тарою виявились контейнери. Тому організація “Сорт¬
насіннєовоч” перейшла на контейнерне перевезення картоплі в гос¬
подарства згідно із планом сортооновлення.
З організацією закритих районів вирощування товарної насінної
картоплі розпочалася спеціалізація окремих господарств, запрова¬
дження при вирощуванні насіння сучасної технології та спеціальних
насінницьких прийомів. Все це дало позитивні результати уже через
кілька років. У 1971-1973 рр. з вирощених у закритих районах і зо¬
нах 909,7 тис. тонн картоплі було реалізовано 295,3, з них для сорто¬
оновлення і сортозаміни - 124,8 тис. тонн.
У закритих районах картоплю вирощували на 56,3 тис. га, що
становило майже 20 % посівної площі цієї культури на Поліссі. Пло¬
щі під картоплею в господарствах дещо зросли, в окремих з них - до
500-600 га. До найбільших за виробництвом картоплі належали гос¬
подарства, у яких площа насаджень становила 15-20 % орних зе¬
мель. Таких господарств у закритих районах було близько 10 %.
З метою державної підтримки насінницьких, у першу чергу еліт¬
них, господарств було встановлено сортові надбавки до реалізаційної
ціни на насінний матеріал.
Виробництвом еліти картоплі у 1990-1995 рр. в Україні займало¬
ся понад 45 господарств закритих районів, науково-дослідних уста¬
нов та вузів. В окремих господарствах працювали спеціалісти опор¬
них пунктів по первинному насінництву, біотехнологічні лаборато¬
рії, відділи насінництва науково-дослідних установ. Первинне насін¬
ництво базувалося на клоновому доборі, рослинах, вирощених іп
vitro (у пробірках) та міні-бульбах.
У кінці 1991 р. була створена Українська науково-виробнича
асоціація з вирощування та реалізації еліти картоплі - “Укреліткар-
362
топля”, яка є добровільним об'єднанням у галузі картоплярства на
засадах самоврядування і господарського розрахунку господарств та
організацій, які займаються виробництвом еліти картоплі (рис.6.1).
Пізніше її реорганізували в Асоціацію “Картопля України”.
Рис. 6.1. Схема організації роботи Асоціації “Картопля України”
Асоціація входить у систему Міністерства аграрної політики
України. Основним її завданням є забезпечення потреби насінни¬
цьких господарств України в еліті та сприяння підвищенню вро¬
жайності картоплі за рахунок використання високих потенційних
можливостей насінного матеріалу.
363
Для виконання своїх завдань асоціація веде господарську, ко¬
мерційну, посередницьку і рекламно-інформаційну діяльність в Україні
та за її межами згідно з діючим законодавством, створює науково-
виробничі і господарські комерційні структури, встановлює творчі, еко¬
номічні та виробничі зв'язки з партнерами в країні і за кордоном.
З метою забезпечення виконання замовлень, виробництва здо¬
рової та високопродуктивної насінної картоплі асоціація через нау-
ково-виробничий відділ, лабораторії і спеціалістів господарств здій¬
снює в господарствах-учасниках організаційну, науково-методичну і
практичну роботу, а саме: сприяє в укладанні з членами асоціації до¬
говорів на реалізацію еліти з урахуванням потреб областей і госпо¬
дарств для сортооновлення та сортозаміни; разом із спеціалістами
господарств визначає обсяги первинного насінництва для виконання
замовлень на еліту; забезпечує членів асоціації вихідним матеріалом
поширених і перспективних сортів картоплі для організації елітного
насінництва; відпрацьовує в кожному господарстві особливості тех¬
нології виробництва здорового садивного матеріалу, виходячи з
ґрунтово-кліматичних і організаційних умов та прийнятої схеми ви¬
робництва еліти, виявляє резерви підвищення ефективності вироб¬
ництва еліти; запроваджує спеціальні насінницькі прийоми, спрямо¬
вані на зниження ураження картоплі вірусною інфекцією та збіль¬
шення садивної фракції бульб в урожаї; організовує внутрівідомчий
контроль за якістю садивного матеріалу в розсадниках первинного
насінництва, супер-супереліти, супереліти, еліти; організовує забез¬
печення елітою картоплі фермерів, селянські господарства та город¬
ників; організовує республіканські, обласні й районні практичні се¬
мінари по насінництву картоплі, навчання кадрів, забезпечує їх спе¬
ціальною інформацією.
В останні роки в картоплярстві відбулися значні зміни. Якщо на
початку 90-х років з 1,5 млн га дві третини площ знаходилися в дер¬
жавному і колективному секторах, то в приватному - лише одна тре¬
тина.
У 1997 р. фермери, селяни, городники і дачники вже вирощували
картоплю на площі 1476,5 тис. га, що складало понад 95 % від загаль¬
ної площі. На частку колективних і державних господарств припада¬
ло 70,8 тис. га, або близько 5 %. У цих господарствах у 1998 р. було
посаджено картоплі на площі 40 тис. га, а у приватному секторі - на
рівні 1997 року.
Таким чином, на сьогодні в картоплярстві переважають дрібні ви¬
робники картоплі. У 1997 р. у приватному секторі середня площа,
зайнята під картоплю, становила близько 0,20 га, а більшість їх ви¬
рощувала її на 0,02-0,05 га.
Підрахунки показують, що затрати праці на 1 га за звичайної механі¬
зованої технології складають 400-450, енергозберігаючої - 160 люд.-год,
а за ручної - понад 1000 люд.-год.
364
Але, незважаючи на величезні витрати праці та коштів (добрива,
пестициди, гербіциди тощо), урожаї не перевищують 100 ц/га, а в
багатьох регіонах - 40-60 ц/га. Головною причиною одержання
низьких урожаїв картоплі є використання для садіння вироджених
бульб.
З настанням в Україні перебудовчих процесів, особливо за еко¬
номічної кризи, існуюча система насінництва самоліквідувалася.
Дорожнеча змусила господарства Степу відмовитися від щоріч¬
ного завезення насінної картоплі, а Лісостепу - від сортооновлення.
З припиненням замовлення перестали вирощувати насінну картоп¬
лю в закритих районах. Картопля, як уже зазначалося, перейшла в
руки приватного сектора, який теж не став замовником.
Таким чином само ліквідувались і закриті райони з виробництва
товарної насінної картоплі. Постало питання про введення нової сис¬
теми насінництва картоплі.
Наукове забезпечення картоплярства, зокрема насінництва, в ми¬
нулі десятиріччя надавали Український науково-дослідний інститут
овочівництва і картоплі та його дослідні станції (Дніпропетровська,
Донецька, Київська, Сімферопольська і Сквирське дослідне поле);
Немішаївська дослідна станція, а пізніше УНДІКГ та його дослідні
станції (Чернігівська і Поліська); Всесоюзні інститути: селекції та
генетики (м.Одеса), землеробства (м.Київ), землеробства та тварин¬
ництва західного регіону (м.Львів); сільськогосподарські вузи -
Дніпропетровський, Львівський, Білоцерківський, Сумський, Хар¬
ківський та ін.; обласні державні сільськогосподарські станції - Во¬
линська, Житомирська, Чернігівська, Сумська та ін.
Вагомий внесок у подальший розвиток вітчизняного картоплярс¬
тва зробили відомі вчені-насіннєводи: у поліській зоні - С.П.Куль-
жинський, М.К.Малюшицький, О.О.Зубченко, А.П.Ящук, Я.І.Прош-
ко, М.Н.Стукало, П.С.Теслюк, О.Й.Оншценко, Ю.Я.Верменко, В.А.ВІ-
тенко, В.С.Свертока, М.Д.Гончаров, М.Г.Андрушко, Н.С.Кожушко,
Д.М.Остапенко; у лісостеповій зоні - А.Ф.Ілляшенко, І.І.Пушкарьов,
М.Я.Молоцький; у степовій зоні - Т.Д.Лисенко, О.М.Фаворов,
А.Ф.Котов, І.А.Лук’яненко, М.С.Бойко, М.Ф.Соколенко, І.П.Бугаєва;
у західному регіоні - І.Д.Нечипорчук, О.М.Фаворов, В.Т.Влох,
І.І.Тимошенко, М.С.Колотуха. За їх участю були розроблені методи
первинного і внутрішньогосподарського насінництва, строки сорто-
заміни і сортооновлення, обгрунтовані системи насінництва, техно¬
логії одержання високоякісних садивних бульб у різних грунтово-
кліматичних умовах, літні посадки і двоврожайна культура, біотех-
нологічні методи одержання вихідного матеріалу, створені закриті
райони з виробництва товарного садивного матеріалу, розроблена
технологія промислового різання садивних бульб та багато інших
заходів, які зумовили розвиток галузі в Україні і стали надбанням
світового картоплярства.
365
ЛІТЕРАТУРА
1. Батыренко В.Г., Беликов А.Д. Сорта картофеля // Сводный отчет за
1925-1926 гг. Укр. сети по изучению сортов культурных растений при Все-
украинском обществе семеноводов. - Харьков, 1927.
2. Вітенко В.А., Молоцький М.Я. Насінництво картоплі - К.: Урожай, 1977.
3. Волчанский Ф.М. Результаты опытов по культуре картофеля в
Харьковской губернии за 1913 год. - Харьков, 1914.
4. Козакевич В.В. Як вирощувати картоплю на Поліссі.- Вип.12/38. /
Поліська краєва с.-г. дослідна станція. - К., 1929.
5. Кульжинський С.П. Результаты опытов по изучению форсированного
размножения сортов картофеля. - Вып.59. - К.: Изд-во Носовской опытной
станции, 1928.
6. Кульжинский С.П. Питання культури картоплі на Україні // Укр. агро¬
ном.- 1928. -№9.
7. Кульжинський С.П. Як швидко розмножити нові сорта картоплі. -
Вип.39. / Носівська с.-г. дослідна станція. - К., 1924.
8. Онищенко В.В. Насінництво картоплі на Україні. - К.: Урожай, 1966.
9. Теслюк П.С., Щербенко О.В. Становлення і розвиток українського
картоплярства. - К., 1987.
10. Филипповский А.К. Результаты опытов по культуре картофеля за
1905-1911 гг.-К., 1912.
6.2. НАУКОВІ ЗАСАДИ НАСІННИЦТВА
Насінництво - це важлива ланка в організаційній структурі вироб¬
ництва картоплі. Насінництво - галузь рослинництва, що займається
розмноженням насіння, збереженням і поліпшенням його сортових,
посівних та врожайних якостей.
Вченими підраховано, а практикою доведено, що врожаї та ва¬
лові збори сільськогосподарських культур підвищуються на 20-25%
за рахунок використання високоякісного насіння нових реєстрованих
і перспективних сортів. Насіннєвий матеріал є, безперечно, одним із
найважливіших і незамінних чинників, через який із покоління в по¬
коління передаються генетичні властивості сортів.
Насінництво безпосередньо пов’язане із селекцією. У результаті
селекційної роботи створюються нові сорти. Насінництво ж реалізує
досягнення селекції шляхом впровадження у виробництво нових сор¬
тів і вирощування високоврожайного насіння.
В основі насінництва лежить генетика, однак урожайні якості на¬
сіннєвих бульб залежать не тільки від генетичної основи, а й від
умов формування, тобто умов розвитку материнської рослини, впли¬
ву на неї хвороб та інших чинників, тому при організації насінництва
слід ураховувати увесь їх комплекс.
Насінницька галузь повинна забезпечувати гнучкі форми органі¬
зації, які дають змогу швидко впроваджувати нові сорти у ви¬
робництво при збереженні їхніх спадкових властивостей і забезпе¬
ченні високої якості садивного матеріалу.
366
Знання закономірностей спадковості і мінливості, методів управ¬
ління онтогенетичним розвитком та використання позитивних моди¬
фікацій дозволяє практично реалізувати потенційну продуктивність
сортів [13].
У системі агротехнічних та організаційних заходів щодо під¬
вищення і забезпечення стабільності врожаїв картоплі провідне міс¬
це належить сортовому матеріалу, через який реалізуються потен¬
ційні можливості сорту, і, навпаки, найбільш високопродуктивний
сорт дає низький урожай при садінні низькоякісним насінним, тому
одним із об’єктів насінництва є сорт, який являє собою сукупність
рослин, створених шляхом селекції, що мають певні спадкові мор¬
фологічні, біологічні та господарсько цінні ознаки і властивості.
Сорт є одним із основних засобів сільськогосподарського вироб¬
ництва. Від його генотипу значною мірою залежать реалізація біопо¬
тенціалу поля, ефективність меліоративних і агротехнічних заходів,
особливо за несприятливих умов середовища.
Але біологічний потенціал поля визначається не лише генетич¬
ними особливостями сорту, а й екологічними ресурсами конкретної
ділянки і технологією вирощування.
За останні десятиріччя роль сорту в підвищенні врожайності кар¬
топлі зросла. За підрахунками спеціалістів, зростання врожайності у
світовій практиці картоплярства в цілому забезпечується однаковою
мірою за рахунок як агротехніки, так і впровадження нових, доско¬
наліших сортів. Однак потенційні можливості сорту можуть бути
реалізовані лише при високій якості насіннєвого матеріалу.
У виробничих умовах сорт поступово погіршується. Для під¬
тримання всіх його цінних біологічних властивостей на високому
рівні на всіх етапах насінництва застосовують спеціальні насінницькі
методи і заходи: добір типових здорових рослин у первинних ланках
насінництва при виробництві елітного насіння; вирощування рослин
за оптимальних агротехнічних умов, які сприяють формуванню ви¬
соковрожайних бульб; запобігання пошкодженню насаджень і бульб
хворобами та шкідниками; проведення сортових і фітопатологічних
прочисток; виділення для садіння найбільш повноцінних фракцій.
Одержаний за таких умов насіннєвий матеріал забезпечує збіль¬
шення врожайності на товарних насадженнях на 20-30 %. Ось чому
так важливо щороку забезпечувати на насінницьких насадженнях
комплекс агротехнічних, фітосанітарних і організаційних заходів,
спрямованих на одержання садивного матеріалу з високими врожай¬
ними якостями [14].
Сортові якості бульб залежать від генотипу самого насіння, тобто
якщо насіння належить до високопродуктивного сорту, то його потен¬
ційні можливості дають змогу вирощувати високий врожай, а якщо
до низькопродуктивного, то навіть при висіванні високопродуктив¬
ного насіння високий урожай одержати неможливо.
367
Під урожайними властивостями насіння розуміють здатність різ¬
ного насіння одного генотипу за однакових агротехнічних умов да¬
вати різні врожаї, а рослини, одержані з насіння з різними вро¬
жайними властивостями, можуть відрізнятися за рядом фенотипіч-
них і господарсько цінних ознак. Отже, врожайні якості бульб - це
сукупність властивостей і ознак насіння, здатних відповідно впли¬
вати на формування насаджень як фотосинтезуючої системи, їхню
структуру, ріст, розвиток, що в підсумку визначає біологічний і гос¬
подарський урожай.
Урожайні властивості бульб пов’язані з фенотипічною мінли¬
вістю, мають модифікаційний характер і є наслідком впливу на них
кліматичних і метеорологічних факторів, агротехніки, технології на¬
сінництва та збудників хвороб.
Першим із цих факторів є грунт. Встановлено, що різниця в уро¬
жайних властивостях бульб, вирощених на різних типах грунту, до¬
сягає значних величин. Кращими насінними властивостями відзна¬
чаються бульби, вирощені протягом кількох років на торфових і су¬
глинкових грунтах, порівняно з бульбами з піщаних або супіщаних.
Так, садивні бульби з торфових грунтів забезпечували приріст уро¬
жаю понад 70 %, порівняно з урожаєм від бульб із піщаних грунтів;
завезені із суглинкових грунтів підвищували врожай на 18-48 %, по¬
рівняно з урожаєм від бульб із піщаних. Добрі насінні властивості
формують бульби, вирощені в заплавах річок. Але за збалансованого
вмісту NPK різниця у формуванні врожайних якостей бульб на всіх
типах грунту спостерігається незначна. Це свідчить про те, що вплив
грунтових відмінностей на формування врожайних властивостей
бульб залежить здебільшого від рівня поживних речовин у них. Яко¬
гось специфічного впливу типу грунту не виявлено.
Значною мірою на формування врожайних властивостей бульб
впливає температура, особливо в період бульбоутворення. Найвро-
жайніші бульби формуються, коли середньодобова температура ста¬
новить 15-18°С. Як підвищення, так і зниження температури повітря,
особливо за незбалансованої вологості грунту в цей період погіршу¬
ють урожайні властивості бульб.
Негативний вплив на якості бульб має недостатня вологість грун¬
ту (нижче 50-60 % ППВ) і відносна вологість повітря (нижче 30 %).
Світло впливає на рослини у двох напрямах: як енергетичний фак¬
тор, воно визначає продуктивність фотосинтезу, а також виконує ре¬
гуляторну роль через різні системи.
З посівних якостей, залежно від фотоперіоду, змінюються насам¬
перед маса і показники сили росту бульб. Виявляється тенденція до
збільшення маси бульб під кущем при зменшенні тривалості дня, а
при його збільшенні картоплиння інтенсивніше набирає масу.
Кожна бульба має певні біологічні властивості, які визначають її
якість. Відмінності в якості можуть мати як морфологічний, так і фі¬
368
зіологічний характер. Навіть у межах одного сорту одна бульба (а в
межах бульби окремі вічка і паростки) біологічно відрізняються від
інших, зберігаючи загальні ознаки сорту.
Якість бульб є складним комплексом генетичних і фізіолого-
біохімічних властивостей. Останні можуть сильно змінюватися при
збереженні генотипу сорту.
Модифікаційні зміни, що акумулюються бульбами, значною мі¬
рою визначають і розвиток майбутнього покоління. Отже, різно¬
якісність бульб є проявом модифікаційної мінливості. Це явище по¬
ширене в рослинному світі. Воно виявляється в тому, що насіння од¬
нієї рослини або навіть колоса, волоті, качана, коробочки, куща нерів¬
нозначне за своїми морфологічними, анатомічними та фізіолого-
біохімічними властивостями.
Екологічна різноякісність бульб є наслідком взаємодії організ¬
му та умов зовнішнього середовища. Вона зумовлена вирощуванням
рослин на різних грунтах, дією метеорологічних та інших факторів,
які впливають на забезпечення бульб водою та метаболітами під час
їхнього формування, що призводить до зміни фізичних, морфологіч¬
них і фізіолого-біохімічних ознак та властивостей. Екологічна різно¬
якісність бульб не є спадковою, однак значною мірою впливає на
їхні урожайні якості.
Екологи вважають, що картопля, яка походить з Південної Аме¬
рики, де вологий і прохолодний клімат без сильної спеки та морозів
(середньодобова температура найтеплішого літнього місяця стано¬
вить 15°С) і переважають пухкі грунти, не має на більшій території її
розповсюдження подібних умов, що й призводить до її виродження.
В Україні умови вирощування картоплі погіршуються з півночі на
південь та із заходу на схід, що й зумовлює посилення її виродження
в цих напрямках.
Матрикальна (материнська) різноякісність є наслідком утво¬
рення бульб материнською рослиною в різні строки, за неоднакових
погодних умов і фітосанітарного стану.
Однією з причин материнської різноякісності є асинхронність ор-
ганоутворюючих процесів у різних рослин у кущі картоплі. Різноякіс¬
ність стебел у кущі і сформованих у них бульб є результатом неод¬
накового надходження поживних речовин, яке визначається розвит¬
ком кореневої системи.
У будь-якому організмі, передусім у рослин, які розмножуються
вегетативно, тканини й клітини генетично різноякісні. Особливо по¬
мітно виражене це явище в картоплі. Причину різноякісності тканин
бульби ряд учених вбачають у гібридному походженні сортів і сту¬
пені диференціації тканин і клітин під впливом умов зовнішнього
середовища.
Проте на причини виникнення різноякісності тканин і клітин
бульб картоплі є й інші погляди. Багато дослідників вважають, що
369
різноякісність у бульб пов’язана з різною глибиною спокою вічок
[27, 28, 29].
Ф.Н.Харитонович [26] вказує на факт одержання різноманітних
рослин під час вирощування їх із бруньок, узятих із різних частин
бульби. На його думку, це явище пов’язане із станом спокою. Стан
спокою властивий тканинам і клітинам бруньок материнської буль¬
би, тобто різноякісність бруньок бульби за періодом спокою переда¬
ється потомству в процесі вегетативного розмноження. Під час са¬
діння різних частин бульби одержують різні за біологічними власти¬
востями рослини, у тому числі і рослини, відмінні між собою пері¬
одом спокою їхніх клітин і тканин. Верхівкові бруньки бульби
мають більш слабкий стан спокою і тому починають проростати ра¬
ніше за бруньки, які розташовані біля основи (пуповини). Бруньки
середньої частини бульби займають проміжне положення. Різноякіс¬
ність бруньок бульби за періодом спокою передається потомству: із
бруньок верхівкової частини виростають бульби з коротким пері¬
одом спокою, з бруньок основи - з довгим, а із середніх бруньок - з
проміжним періодом спокою.
А.А.Горюнов [3] дійшов висновку, що різноякісність є наслідком
відмінностей в живленні бульб у різні періоди їхнього розвитку, а
також неоднорідності їх у кущі під час їхнього формування.
Цікаві досліди з вивчення цих факторів проведені В.Є.Космако-
вою і М.І.Прозоровою. Застосовуючи метод мічених атомів вуглецю,
вони помітили, що надходження продуктів асиміляції з листків до
різних бульб протягом вегетаційного періоду проходить неоднаково.
На початку бульбоутворення асимілянти більш активно надходять до
великих і середніх бульб. Наприкінці вегетації більша кількість ра¬
діоактивного вуглецю помічена у дрібних бульб. Автори також зафік¬
сували, що у випадку діткування бульб радіоактивний вуглець про¬
ходить транзитом через старі бульби і відкладається в молодих буль¬
бах. Крім того, продукти асиміляції СО2, які надходять у бульби,
розподіляються в них нерівномірно. Досліди показали, що вміст вуг¬
лецю наростає від пуповини до верхівки бульби. Звідси, на думку
авторів, причина різноякісності бульб у гнізді й генетична неоднорід¬
ність вічок окремої бульби. Насінні якості верхньої і нижньої частин
бульби або окремого вічка будуть залежати від умов, за яких прохо¬
дило їхнє формування.
І.З.Лазарєв [10] різноякісність бруньок вважає наслідком фізіоло¬
гічного стану організму і його життєдіяльності. Він пов’язував це
явище з локалізацією фізіологічно активних і поживних речовнсн у
внутрішніх тканинах верхівки й пуповини бульби.
Багато авторів зазначають, що залежно від розташування бруньок
на бульбі з них виростають рослини з більшим або меншим умістом
певних речовин. При цьому в потомстві виявляється перевага тих
речовин, які локалізувалися в певній частині бульби.
370
Так, О.Н.Савельєва [24] виявила в бульбах, вирощених із верхі¬
вок, знижений уміст крохмалю і підвищений аскорбінової кислоти,
порівняно з бульбами, вирощеними з пуповинок.
За даними С.І.Єфремова [4], верхівкові й пуповинні частини бульб,
відрізняючись між собою вмістом різних речовин і фізіологічними по¬
казниками, передають свої якості відповідним вічкам і рослинам, що
виросли з них. Це зумовлює відмінність стеблоутворюючих і бульбо-
утворювальних якостей вічок. Проведені автором досліди показали,
що після розрізання бульб на 3 рівні поперечні частини, кожна з них
утворює майже однакову кількість стебел, а на дві частини - верхівко¬
ві половинки утворюють їх більше, ніж пуповинні. У деяких випадках
цього не спостерігалося, навпаки, стеблоутворююча здатність вічок,
розташованих на пуповинній частині бульби, була значно вищою, ніж
верхівкових. Вічка середньої частини бульби за стеблоутворюючою
здатністю займали проміжне положення.
Після підрахунку кількості бульб у кущі з’ясувалося, що відно¬
шення кількості бульб у кущах від верхівок до кількості бульб у ку¬
щах від пуповинних частин бульби становило 100:93. Відношення
кількості бульб у кущах, вирощених з різних вічок, змінювалося від
сорту і за роками вирощування рослин. Усі ці факти свідчать про на¬
явність стеблоутворюючої різноякісності вічок.
Відомо, що рослини, одержані від різних бруньок, як за довжи¬
ною бульби, так і за їх походженням, неоднаково уражуються хворо¬
бами, зокрема вірусними.
У змістованій праці С.І.Радченка [22] наведено результати такого
досліду. Бульби розрізали на 6 частин і висаджували у грунт без про¬
рощування. Вирощені рослини аналізували за продуктивністю та
ураженістю вірусами. Виявилося, що рослини, одержані від серед¬
ньої частини бульби, були більше уражені вірусними хворобами.
Звідси випливає висновок про локальний характер розташування не
лише фізіологічно активних і поживних речовин, а й вірусів. Причо¬
му, від одного із шматочків середньої частини бульби в першому ве¬
гетативному поколінні одержано рослину з ознаками картопляної
мозаїки, а від іншого шматочка (теж середньої частини бульби) у
першому поколінні було одержано рослину без явних морфологічних
ознак хвороби, але в наступному вегетативному поколінні проявило¬
ся вірусне захворювання. Виявлено також, що рослини картоплі,
одержані від пуповинної частини вихідної бульби, протягом трьох
років не уражувалися вірусами, і, навпаки, одержані із верхівок,
уражувалися сильно. Дослідник пов’язує захворювання рослин вірус¬
ними хворобами з інактивацією ферментів вірусом, що сильно ре¬
продукується, або ж з утратою рослиною здатності до синтезу повно¬
го комплексу речовин типу ферментів з тієї ж причини. Оскільки фер¬
менти розташовані в окремих частинах бульби, то й локалізація віру¬
сів має певний характер.
371
Відомо, що здоровими і краще розвинутими є рослини, одержані
від бульб із добре розвинутими паростками на всій поверхні бульби.
Бульби з розвинутими верхівковими паростками часто схильні до
ураження готикою. Рослини, одержані від нижніх половинок, менше
уражуються вірусами X і S, порівняно з верхівковими, і дають біль¬
ший урожай, ніж цілі бульби.
Проводячи дослідження в різних грунтово-кліматичних умовах,
багато авторів дійшло протилежних висновків про продуктивні якос¬
ті верхівки, серединки і пуповинки бульби картоплі. Ще в XIX ст.
такі досліди були описані в працях В.А.Поггенполя, І.Стебута, а на
початку XX ст. - у праці І.ІЇІилова. Автори відзначають переваги
верхівкової частини бульби над середньою і пуповинною як за си¬
лою розвитку рослин, так і за кінцевим урожаєм.
Пізніше до аналогічних висновків прийшли В.В.Вінер і А.К.Фі-
ліповський, які вважали, що у верхівці бульби зосереджений більш
рухомий пластичний матеріал, тоді як у середній і пуповинній час¬
тинах клітини заповнені крохмалем, а тканини більше гіпертрофова-
ні. Цим автори пояснили більшу активність верхівки під час пророс¬
тання, а С.П.Кульжинський [9], посилаючись на дані Вольні, ствер¬
джував, що верхні половинки дають урожай у півтора рази більший,
ніж пуповинки однакової ваги, взяті з однієї й тієї ж бульби.
Детальне обгрунтування переваг верхівки бульби над іншими її
частинами міститься у праці Ф.А.Новикова [16]. Як зазначено у цій
роботі, верхівкове домінування не є фактором у поясненні їх пере¬
ваг над іншими частинами бульби. Різна продуктивність поясню¬
ється тим, що з бруньок верхівкової частини рослини розвиваються
швидше, ніж із бруньок пуповинних часток. Ці відмінності збері¬
гаються і в тому випадку, якщо усунути верхівкове домінування.
Прискорений розвиток рослин із бруньок верхівкової частини без¬
посередньо пов’язаний з особливостями локалізації поживних ре¬
човин у різних частинах бульби, ступенем інтенсивності основних
життєвих процесів, які по-різному проходять у різних частинах.
Верхівкова частина бульби містить багато вуглеводів, азотистих
речовин, зольних елементів, ауксинів, і в ній більш активними є окис-
нювальні ферменти. Аналогічні дані висвітлені в працях С.М.Про-
кошева [20,21].
Питання цінності різних частин бульби А.С. Кружилін [8] пов’я¬
зував із кліматичними і погодними умовами. На його думку, у зв’язку
із стадійною і генетичною різноякісністю тканин рослини, бруньки
нижньої половинки бульби є більш здоровими і врожайними, ніж
верхньої. Ці бруньки закладаються раніше, у прохолодний період
весни, з більш молодої тканини, знаходяться весь час у стані спокою,
і на них слабо діє висока літня температура. Бруньки верхньої поло¬
вини бульби утворюються влітку, до того ж із більш старої ткани¬
ни, вони більш активні, і тому під дією високої температури влітку
372
якісно змінюються, що призводить до швидкого їх старіння - вирод¬
ження.
Від верхніх половинок, на думку С.І. Радченка [22], одержують
скоростигліші форми, які дають більшу кількість стебел і бульб під
кущем. Але врожай від верхніх половинок буває вищим лише в ран¬
ній період збирання. При осінньому збиранні вищий урожай дають
більш пізньостиглі нижні половинки. Тому на півночі України за
умов короткого вегетаційного періоду кращі результати дають верх¬
ні половинки.
Узагальнюючи дані численних дослідів з вивчення верхівкових і пу-
повинних часток бульби, проведених різними авторами в різних грунто-
во-кліматичних районах, О.Й.Онищенко [17,18] дійшов висновку, що в
північних районах за врожайністю кращі результати одержують від вер¬
хівок бульб, у середній смузі - серединок, а на півдні - від пуповинних
частин бульб. У південних районах країни бруньки на верхівці бульби
закладаються під час високих температур, тому є більш виродженими, а
рослини, одержані від них, менш продуктивними.
Якщо проаналізувати численні праці зарубіжних дослідників за по¬
рівняльною цінністю і продуктивністю верхівок і пуповинок бульб, то
можна розділити їх на дві групи: європейські дослідники віддають пере¬
вагу верхівкам, а американські визначають усі частини бульб рівноцін¬
ними. Проте для одержання рівномірних кущів з однаковою кількістю
стебел вважають за потрібне різати бульби уздовж.
За результатами канадських дослідників, найбільш продуктив¬
ною частиною бульби є пуповина, найменше - середина, а верхівка
займає проміжне місце.
Нові високоврожайні сорти є важливим фактором інтенсифікації
сільського господарства, однак у процесі розмноження і ви¬
рощування у виробничих умовах якості сортів поступово погіршу¬
ються. Розглянемо основні причини погіршення сортових якостей.
1. Механічне засмічення. Воно може відбуватися в саджалках, ком¬
байнах, при збиранні і сортуванні, у кагатах тощо. Механічне засмі¬
чення іншими сортами призводить до вибраковування сортових по¬
сівів при апробації.
2. Поява соматичних мутантів - це безперервний біологічний
процес, який відбувається в рослинному світі в невеликих обсягах, а
нині, у зв’язку з аварією на Чорнобильській АЕС, в зоні ураження
може мати і практичні наслідки.
3. Зниження імунітету і збільшення захворюваності рослин. Кар¬
топля дуже швидко уражується хворобами, особливо новими расами
патогенів, тобто стійкі проти хвороб сорти через кілька років втра¬
чають свій імунітет. Отже, боротьба із хворобами обов’язкова на всіх
етапах насінницької роботи. Особливо велика робота здійснюється в
первинних ланках насінництва, де сорт можна повністю позбавити
від різних хвороб.
373
4. Екологічна депресія сорту. За своєю природою сорти бувають
із широкою екологічною пластичністю і малопластичні. Перші здат¬
ні давати високий урожай у різних зонах і в різних екологічних об¬
ставинах, інші - лише в певних локальних зонах, що пояснюється їх
реакцією на зміну зовнішніх умов.
При відсутності гармонії між біологією сорту і навколишнім се¬
редовищем настає порушення фізіологічних функцій організму, що
призводить до послаблення його життєздатності, депресії і врешті-
решт до значного зниження продуктивності та якості садивного ма¬
теріалу. Тому для насінництва кожного сорту повинні бути підібрані
оптимальні зони вирощування та визначені строки сортооновлення
садивного матеріалу [15].
До недавнього часу в насінництві головна увага приділялася тех¬
нології вирощування садивного матеріалу з високими посівними
якостями. Технологічні прийоми вирощування певною мірою дозво¬
ляють керувати формуванням цих якостей. Накопичені експеримен¬
тальні дані переконливо свідчать про значний вплив природних еко¬
логічних факторів на врожайні якості бульб. Численні екологічні фак¬
тори можна поділити на три групи: абіотичні, біотичні, антропогенні
(рис.6.2).
Екологічні (рактори насінництва
Абіотичні
Кліматичні
(температура,
Волога грунту,
Відносна Вологість повіт¬
ря, освітлення тощо)
Едафічні
(фізико-хімічм
властивості грунту,
реакція іконцентрація
грунтоВого розчину)
Просторові
(рте<р, Висота
найріВнем моря,
географічна широта)
біотичні
Мікрофлора
Шкідники
Алелопатична
Взаємодія
Антропогенні
Агротехнічні
(обробіток грунту,добриВа,
попередники, строки сіВби,
норми ВисіВу, догляд за посіВами)
Збирання
(строки і способи,
режими обмолоту зерна)
Сушіння\ очистка,
зберігання насіння
Прийоми передпосівної
обробки насіння
(протруювання, стимуляційні
ВплиВи)
Рис.6.2. Екологічна модель формування врожайних властивостей насіння
Отже, урожайні якості бульб формуються під впливом природ¬
них (кліматичних, погодних, едафічних) і штучних (агротехнічних)
екологічних факторів.
374
Завдання насінництва. Наявності хороших сортів недостатньо
для успішного ведення картоплярства. Необхідною умовою на всіх
етапах вирощування насіннєвого матеріалу є суворе дотримання ви¬
мог насінництва картоплі, які повинні забезпечити протягом трива¬
лого часу не тільки збереження, а й поліпшення цінних якостей сорту.
Недостатня увага до насінництва призводить до таких тяжких нас¬
лідків, як поширення на рослинах картоплі різних хвороб, швидке
зниження врожаю, вихід сорту в тираж [25].
Одним з основних завдань у насінництві картоплі є боротьба з
вірусними хворобами. Сильне поширення їх на насадженнях картоп¬
лі є найчастіше основною причиною низької якості насіннєвого ма¬
теріалу. Тому вирощування насінної картоплі, вільної від вірусної
інфекції, є головною передумовою забезпечення на належному рівні
якості насіннєвого матеріалу. Це найбільш складне завдання, однак
не єдине. Садивна картопля повинна бути повністю вільна також від
інших численних хвороб - грибкових і бактеріальних. Особливо не¬
допустиме зараження її небезпечними карантинними хворобами, та¬
кими, як рак картоплі, картопляна нематода та ін. [1,19,23,30,31,32].
Насіннєва картопля повинна мати належні продуктивні якості й
давати при відповідній агротехніці високі урожаї. Бульби повинні
бути в основному посадкового розміру, недопустимими є як дуже
дрібні, так і великі.
На всіх етапах насінництва слід створювати такі умови вирощу¬
вання, які б повністю виключали сортові домішки. Наявність їх мо¬
же зрештою призвести до втрати цінного сорту. Крім того, домішки
різних сортів можуть сприяти поширенню різних захворювань. Усі
ці завдання можуть бути вирішені лише при планомірному здійс¬
ненні продуманої і науково обґрунтованої системи насінництва.
Щоб зменшити можливість поширення вірусів, необхідна прос¬
торова ізоляція між первинними ланками, насіннєвими ділянками в
господарстві, а також від продовольчих посадок картоплі. Для по¬
передження змішаних вірусних інфекцій просторової ізоляції потріб¬
но дотримуватись і між посадками різних сортів картоплі. Своєчасне
виявлення і якомога раннє видалення хворих рослин із поля також
повинні сприяти запобіганню поширенню вірусів і збудників інших
хвороб.
Для того, щоб зменшити можливість передачі інфекції механіч¬
ним шляхом, при проведенні різних агротехнічних робіт у розсадни¬
ках необхідно дотримуватись ряду запобіжних засобів.
Поширення вірусів може здійснюватися різними комахами, особ¬
ливо безкрилими і крилатими попелицями. Роль останніх дуже знач¬
на, оскільки вони можуть переносити віруси на великі відстані.
Щоб успішно боротися з вірусами в районах поширення літаючої
попелиці, необхідно враховувати біологію попелиці та умови для її
розселення в певні роки, залежно від метеорологічних та інших особ¬
375
ливостей року з тим, щоб вибрати оптимальні строки для цих райо¬
нів садіння картоплі і провести обробку інсектицидами в періоди, що
найбільше сприяють знищенню попелиць. У кожній місцевості не¬
обхідно ретельно вивчити умови перезимівлі попелиці. Вірусна ін¬
фекція особливо сильно розповсюджується там, де попелиця, зокре¬
ма персикова, знаходить хороші умови для перезимівлі і де внаслі¬
док цього крилата попелиця може зустрітися на полях картоплі вже
ранньою весною. Це пов’язано з тим, що рослини картоплі в ранній
фазі росту більшою мірою схильні до зараження, ніж у пізніші стро¬
ки. Крилата попелиця, що залітає з інших місцевостей, зустрічається
на посадках картоплі значно пізніше, коли рослини вже більш дорос¬
лі і порівняно менше схильні до зараження.
У місцевостях з більш суворими зимами персикова попелиця
може зимувати на відкритому повітрі у стадії яйця, що було виявле¬
но на персику та абрикосі. У місцевостях з більш м’якими зимами
попелиця звичайно зимує на відкритому повітрі на зелених рослинах,
особливо на хрестоцвітих. Великі можливості для перезимівлі попе¬
лиці мають теплиці, постійні сховища і бурти, особливо з корене¬
плодами, де попелиця протягом усієї зими може харчуватися соко¬
витими частинами рослин. Навіть у тих місцях, де перезимівля тлі
неможлива через дуже холодну зиму і відсутність захищених місць
для її перезимівлі на рослинах, повної гарантії від зальоту попелиці
з інфекцією не може бути. Це трапляється іноді навіть і в гірських
місцевостях, якщо в сусідніх низинах є умови для раннього розвитку
крилатої персикової попелиці, яка з повітряними течіями може бути
занесена в більш високі місця розташування [11, 12, 25].
При вирощуванні насіннєвої картоплі необхідно здійснювати
основний комплекс агротехніки, який розроблений для отримання
високих урожаїв продовольчої картоплі в тій або іншій зоні країни.
Однак він повинен включати також деякі спеціальні заходи, спрямо¬
вані на отримання високоякісного насіннєвого матеріалу.
Ділянка насіннєвої картоплі повинна бути ізольована від можли¬
вих джерел інфекції. Її конфігурація також має істотне значення: вузь¬
кі ділянки більшою мірою вразливі для поширення інфекції, яка пе¬
реноситься крилатою попелицею. Особливості грунту, способи його
обробітку, співвідношення різних елементів живлення, методи під¬
готовки садивного матеріалу, строки і способи посадки і збирання -
ці та інші фактори мають важливе значення при вирощуванні насін¬
нєвої картоплі.
Строки посадки і збирання бульб можуть, наприклад, у деяких
умовах вирішальним чином впливати на якість насіннєвої картоплі.
Правильний вибір строку посадки дозволяє забезпечити розвиток
рослин у такий період, який не буде співпадати з максимальним льо¬
том попелиці. Важливе значення має при цьому передпосадкове
пророщування бульб, яке зменшує тривалість періодів між фазами,
376
прискорює ріст і розвиток рослин і дозволяє в більш ранні строки
зібрати повноцінний урожай до того, як рослинам найбільшою мі¬
рою загрожуватиме зараження вірусами внаслідок зростання кількос¬
ті переносників.
За даними багатьох дослідників у нас і за кордоном, збирання не¬
зрілої картоплі, особливо ранніх сортів, доцільне для отримання на¬
сінних бульб більш високої якості. При збиранні незрілих бульб зни¬
жується можливість їх інфікування, оскільки для того, щоб вірус із
місця первинної інфекції на листках дійшов до бульб, необхідний
іноді значний час.
Особливо ефективним збирання недозрілих бульб може бути там,
де небезпека зараження відносно невелика. У цьому випадку є під¬
стави передбачати, що значна частина рослин до часу раннього зби¬
рання ще вільна від інфекції. Позитивна роль цього заходу знижуєть¬
ся, якщо вірусна інфекція має масовий характер вже на початку рос¬
ту картоплиння. Причому, і при сильному поширенні вірусів зби¬
рання незрілої картоплі може якоюсь мірою сприяти збільшенню в
урожаї кількості здорових бульб.
Збирання незрілої картоплі перериває зв’язок між надземною час¬
тиною рослин і бульбами, що може певною мірою запобігти проник¬
ненню нової інфекції з листя в бульби. Цього ж можна досягти й ін¬
шими способами, наприклад обробкою бадилля картоплі певними
хімікаліями.
Природно, що збирання картоплі в більш пізні, звичайні для да¬
ної місцевості, строки дещо підвищує урожай бульб, однак при цьо¬
му знижуються їх насіннєві якості, які перебувають у прямому
зв’язку з тим, що після раннього збирання настають сприятливі умо¬
ви для інтенсивного льоту переносників вірусів. Насіннєва картопля,
вирощена при ранній весняній посадці від заздалегідь пророщених
бульб і при ранньому збиранні, має найбільші продуктивні властивос¬
ті при подальшому використанні їх як садивного матеріалу.
Ряд зарубіжних учених, позитивно оцінюючи ранню посадку, за¬
значають, що запізнення з нею посилює зараження рослин вірусами.
В інших випадках, відповідно до природно-історичних умов, велике
позитивне значення мають і правильно вибрані строки літніх поса¬
док [5, 6].
Раннє збирання врожаю бульб до появи фітофторозу або (у край¬
ньому випадку) при перших ознаках його є позитивним заходом у
боротьбі з ним. Правильному вибору часу початку збиральних робіт
може допомогти добре налагоджена система прогнозування відповід¬
ними станціями захисту рослин строків появи фітофторозу.
Крім своєчасної посадки і збирання картоплі, у районах поши¬
рення фітофторозу та інших хвороб, особливо при вирощуванні не¬
стійких до них сортів, необхідно застосовувати відповідні хімічні
заходи боротьби.
377
Із різних грибкових хвороб насінництву картоплі (іноді значною
мірою) завдає збитків ризоктоніоз. Причому, шкода, що заподіюєть¬
ся ним, пов’язана не тільки із втратою врожаю. У деяких випадках
зовнішні прояви цього захворювання на рослинах нагадують деякі
вірусні хвороби. Це дезорієнтує апробатора і нерідко приводить йо¬
го до помилкових висновків. Мали місце випадки бракування поса¬
док насіннєвої картоплі внаслідок сильного ураження їх нібито вірус¬
ними хворобами, у той час як вони були уражені ризоктоніозом.
Усі агротехнічні прийоми, що сприяють більш швидкій і друж¬
ній появі сходів і створюють сприятливі умови для подальшого рос¬
ту рослин, одночасно сприяють боротьбі з ризоктоніозом і зменшу¬
ють збитки від нього [2].
ЛІТЕРАТУРА
1. Амбросов А.Л. Вирусные болезни картофеля и методы выращивания
здорового семенного материала в Белорусской ССР: Автореф. дис. ... д-ра
наук. - Минск, 1965.
2. Верменко Ю.Я., Свертока В.Є. Відтворення еліти кароплі // Селекція і
насінництво картоплі. - К.: Урожай, 1988. - С. 151-156.
3. Горюнов А.Л. О разнокачественности частей клубня картофеля // Аг¬
робиология. - 1947 - №2.
4. Ефремов С.И. Некоторые вопросы биологии клубня картофеля (проб¬
ковый слой, глазки, влияние света) // Ученые записки Орловского гос. пед.
ин-та. - Вып.2. - Орел, 1963.
5. Картопля / Під ред. В.А. Вітенка, М.Ю. Власенка , В.С. Куценка. - К.:
Урожай, 1978. - 240 с.
6. Камераз А.Я. Селекция и семеноводство картофеля в связи с вирус¬
ными болезнями // Вирусные болезни с.-х. растений и меры борьбы с ними.
- М.: Изд-во МСХ СССР, 1960. - С.228-237.
7. Космакова В.Е., Прозорова М.И. Изучение вопроса генетической раз-
нокачественности клубней картофеля с помощью меченых атомов // Сооб¬
щение Дальневосточного филиала Сиб. отд. АН СССР им. В.Л. Комарова:
Сб.трудов. - Вып. 14. - Новосибирск, 1961.
8. Кружилин А.С. Биологические особенности орошаемых культур. -
М.: Сельхозгиз, 1954.
9. Кульжинский С.П. Результаты опытов по изучению форсированного
размножения картофеля. - К.: Изд-во Носовской опытной станции, 1928.
10. Лазарев 3.0. О молекулярно-структурной разнокачественности внут¬
ренней ткани апикальной и столоновой частей клубня картофеля // Биофи¬
зика. - Т.VI, вып.4. - М.: Изд-во АН СССР, 1961.
11. Молоцкий М.Я. Выращивание картофеля при малых нормах посадки.
-К.: Урожай, 1986.-240 с.
12. Молоцкий М.Я. Проблеми картоплярства: вибрані наукові праці. -
Біла Церква: НІВ “Мустанг”, 1996. - 332 с.
13. Молоцкий М.Я., Васильківський С.П., Князюк В.І. Генетика. - Біла
Церква, 1998. - 279 с.
14. Молоцький М.Я., Васильківський С.П., Князюк В.І. Селекція та
насінництво польових культур: Практикум. - К.: Вища шк., 1995. - 238 с.
378
15.Молоцький М.Я., Васильківський С.П., Князюк В.1. Селекція та
насінництво польових культур. - К.: Вища шк., 1994. - 454 с.
16. Новиков Ф.А. Физиолого-химические основы использования верхушек
клубней картофеля на^семенные цели: Дис. ... канд. с.-х. наук. - М., 1946.
17. Онищенко О.Й. Різноякісність бульб і виродження картоплі // Вісник
с.-г. науки. - 1958. -№ 1.
18. Онищенко О.Й. Виродження картоплі і його шкідливість // Насін¬
ництво картоплі на Україні. - К., 1966. - С.62-75.
19. Починок В .Я. Вплив умов вирощування та біологічних особливостей
сорту на ступінь ураження рослин картоплі вірусними хворобами // Картоп¬
лярство. - 1986. -№16. - С.29-31.
20. Прокошев С.М. Биохимия картофеля. - М. - Д.: Изд-во АН СССР, 1947.
21. Прокошев С.М. О раневых реакциях клубней картофеля // Биохимия. -
Т.ѴІІІ, вып. 2-3. - М., 1943.
22. Радченко С.И. Биология клубнеобразования у картофеля (значение
способов резки и посадки клубней) // Известия Естественнонаучного ин-та
им. П.Ф. Лесгафта. - Т.ХХѴІІ. - М.: Педиздат АН РСФСР, 1995.
23.Рожалин Л.В. Вырождение картофеля и несостоятельность теории за¬
разности мозаичных вирусных болезней картофеля // Вестник с.-х. науки.
Овощеводство и картофелеводство. - Вып. 1. - М.: Сельхозгиз, 1940.
24. Савельева О.Н. Биохимические особенности картофеля, выращенного из
различных участков картофеля: Автореф. дис.... канд. с.-г. наук. - М., 1955.
25.Теслюк П.С., Молоцький М.Я., Власенко М.Ю. Насінництво кар¬
топлі. - Біла Церква, 2000. - 200 с.
26. Харатинович Ф.Н. О разнокачественности почек картофельного клуб¬
ня в отношении периода покоя // Ученые записки Ленинабад, гос. пед. ин-та
им. Кирова. - Вып.1. - Ленинабад, 1948.
27. Чесноков П.Г. Исторический очерк развития знаний о вырождении
растений картофеля в СССР // Болезни вырождения картофеля в СССР. - Л.-
М., 1961.-С.5-14.
28. Чесноков П.Г. Диагностика вирусных заболеваний картофеля // Ви¬
русные болезни с.-х. растений и меры борьбы с ними. - М.: Изд-во МСХ
СССР, 1960. - С.207-228.
29. Чесноков Н.С., Михайлова В.Н. Качественное различие почек клуб¬
ней картофеля // Агробиология. - 1948. - №4.
30. Шмыгля В.А. Система мероприятий по защите картофеля от вирусных,
микоплазменных болезней // Болезни картофеля. - М.: Колос, 1980. - С.216-240.
31. Шмыгля В.А., Лодочник П.И. Некоторые вопросы развития безви-
русного семеноводства картофеля // Современные проблемы семеноводства
картофеля на безвирусной основе. - Владивосток, 1985. - С.73-76.
32.Ячевский А.А. Болезни и повреждения картофеля. - С.Пб.: Изд-во
Департамента земледелия, 1904.
6.3. ПРИЧИНИ ВИРОДЖЕННЯ КАРТОПЛІ
I ЗАХОДИ БОРОТЬБИ З НИМ
Для картоплі, як культури, що вегетативно розмножується, харак¬
терним є те, що при довготривалому беззмінному використанні на¬
сінного матеріалу спостерігається прогресуюче з роками зниження
379
врожаю. Зокрема, в Англії, починаючи з 1775 року, зниження вро¬
жаю картоплі набуло таких загрозливих масштабів, що уряд встано¬
вив премію за виявлення причин цього явища і розробку заходів бо¬
ротьби з ним. Було помічено, що зниження врожаю супроводжується
морфологічними змінами картопляної рослини: вона відстає в рості,
а листя її втрачає ознаки, типові для сорту (воно скручується, закру¬
чується, зморщується, змінюється забарвленість). У 1801 р. вже ви¬
словлювалась думка про те, що причиною є хвороба, яка поширю¬
ється комахами, зокрема попелицями. Розширенню досліджень з ви¬
вчення природи цього явища на початку XX ст. сприяв сильний спа¬
лах захворювання, відомого під назвою “скручування листя”. Тоді ж
була доведена інфекційність цього захворювання. У Росії захворю¬
вання картоплі такого типу було виявлене вперше в 1915 р. в Новго¬
родській губернії. У колишньому СРСР вивчення цього явища роз¬
почав А.А.Ячевський [34]. Робота проводилася на десяти дослідних
ділянках, розташованих у різних географічних зонах країни.
Стосовно загального терміна, що характеризував хворобливий
стан рослин картоплі, то він наприкінці XVIII ст. набув назву “виро¬
дження” (degeneration). У літературних джерелах цей стан називався
також “виснаження”, “старість”.
З вивченням причин погіршення насінних якостей картоплі на
початку XX ст. з’явилося кілька теорій, які по-різному пояснювали
це явище.
Теорія старіння. Було помічено, що нові сорти більш стійкі про¬
ти захворювань, ніж старі. Тому вчені дійшли висновку, що виро¬
дження є прогресуючим старінням рослин, які розмножуються веге¬
тативно і з часом втрачають свою життєздатність.
При цьому старіє не окрема рослина, а весь сорт. Автором цієї
теорії, зокрема у Франції, був такий відомий дослідник, як Перман-
тьє (1786). Цієї теорії з невеликими змінами притримувалися більше
100 років.
В Україні прихильник цієї теорії Г.М.Линник [10] вважав, що ви¬
родження картоплі є зовнішнім проявом її старіння і закономірним
процесом вікової мінливості, що закладено в самій природі рослин.
На його думку, вірус - не причина, а лише наслідок виродження і
старіння сорту.
Для одержання здорового садивного матеріалу науковці вважали
за потрібне здійснити насінну репродукцію. Хоч ця теорія не відпо¬
відала фактичним обставинам виродження картоплі, все ж вона віді¬
грала свою позитивну роль тим, що з насіння, одержаного від само¬
запилення і від схрещування, було створено велику кількість нових
сортів.
В Україні в 40-х рр. XX ст. вже були спроби вирощувати здорові
садивні бульби шляхом сіяння насіння картоплі, одержаного від са¬
мозапилення [29], а також за допомогою “омолодження”, тобто ско-
380
щування бадилля на картоплі весняного садіння і перенесення таким
чином бульбоутворення на осінній період, що дає можливість пізні¬
ше збирати молоді насінні бульби [10].
Теорія токсинів. Прихильниками цієї теорії були М.Г.Бондарєв
[3] та інші, які вважали, що причиною виродження картоплі на пів¬
дні є дія на рослину токсичних речовин маточної бульби. На їхню
думку, токсичні речовини нагромаджуються в бульбах під час їх фор¬
мування при високих температурах. Автори стверджують, що при
високих температурах у тканинах картоплі підвищується вміст от¬
руйних речовин, наприклад аміаку, який згубно впливає на живу
протоплазму клітини. Для запобігання виродженню вони пропону¬
ють висаджувати картоплю окремими вічками з невеликими шматоч¬
ками бульб або відокремленими від бульб паростками. Цим, очевид¬
но, обмежується згубна дія токсичних речовин маточної бульби на
рослину.
Теорія екологічного виродження. Прихильники її - Меркеншлагер
(1929), Циглер (1931), Т.ДЛисенко [11], О.МФаворов [28], Л.В.Рожалін
[22] та інші - головною причиною виродження картоплі вважали винят¬
ково несприятливі умови, що створюються при вирощуванні цієї куль¬
тури: невідповідність температурного і поживного режимів та інших
екологічних факторів, що призводять до глибоких патологічних змін, які
спадково через бульби передаються майбутній рослині.
Ще А.І.Стебут [25] вказував на те, що сорти картоплі при виро¬
щуванні їх в одних і тих же умовах, які цілком відповідають потре¬
бам цієї культури, зберігають свої якості досить довго. Якщо картоп¬
лю перенести в інші умови клімату, ґрунту, добрив, то й бульби її
через деякий час змінять свої якості відповідно до нових умов. Нерід¬
ко навіть у перший рік вирощування в нових умовах вони втрачають
свої первісні якості.
Культура картоплі на півдні недостатньо розвинена тому, що за¬
везений із північних областей насінний матеріал швидко втрачає свої
якості: урожайність різко зменшується, і на третій рік при весняному
садінні ледве повертає витрати садивного матеріалу.
Теорію екологічного виродження картоплі підтримували Ф.І.Нєм-
чин [18], Н.С.Чесноков [32], М.Ф.Соколенко [24], М.М.Балашов [2]
та інші дослідники, які дійшли висновку, що основним фактором,
який сприяє виродженню картоплі, є дія високої температури на кар¬
топляну рослину.
Часткове виродження картоплі в північних областях С.М.Букасов
пояснював наявністю в цій зоні періодів із високими температурами.
Виродження картоплі може статися навіть у тому випадку, коли
тривала дія високих температур припадає лише на кінець вегетації -
у період, близький до відмирання бадилля [30].
Л.В.Рожалін [22] зазначав, що виродження картоплі, зокрема
розвиток зморшкуватої мозаїки, готики, скручування листя та інші, є
381
порушенням співвідношення між вуглеводами та азотистими речо¬
винами в бік відносного збільшення останніх. Він поділяє всі форми
цього захворювання на дві групи: південне виродження і виродження
від неправильного кореневого живлення. В обох випадках відбува¬
ються морфологічні та фізіологічні зміни рослин. Подібні зміни в
морфології рослин, які відбуваються в результаті незбалансованості
фізіолого-біохімічних процесів у рослині, мають тимчасовий харак¬
тер: якщо бульби від таких рослин перенести у сприятливі умови, то
вже в перший рік, а найпізніше - на другий рік з них виростають
здорові рослини.
М.М. Максимович [12] вважав, що на насінні якості бульб впли¬
вають усі фактори зовнішніх умов, включаючи зберігання і збирання
врожаю.
Отже, прихильники екологічного виродження всі зміни, які від¬
буваються в картопляній рослині і призводять до її виродження,
пов’язують із впливом факторів, що не відповідають біологічним
потребам картоплі. Домінуючим із них є температурний фактор.
М.М.Муш [17] пояснює процес південного виродження картоплі
порушенням оптимального співвідношення між гетеротрофним і ав¬
тотрофним живленням. По суті, це є тією ж теорією токсинів.
Ниткоподібність паростків. А.І.Руденко [22] відносив ниткопо-
дібність паростків до особливої неінфекційної (кліматичної) форми
виродження картоплі, зумовленої занадто високими температурами
під час бульбоутворення. Бульби з ниткоподібними паростками да¬
ють дуже пізні і кволі сходи, рослини відстають у рості та розвитку
протягом усього вегетаційного періоду і, як правило, врожаю не да¬
ють. Дуже часто такі бульби взагалі не утворюють сходів.
Вплив умов вирощування на утворення ниткоподібних паростків
на бульбах вивчав на Дніпропетровській овочево-баштанній дослід¬
ній станції М.Я.Молоцький [13]. Картоплю завозили з Прієкульської
селекційної станції (Латвія) і вирощували літнім та весняним садін¬
ням (табл.6.1).
Таблиця 6.1 - Вплив умов вирощування на утворення ниткоподібних паростків
на бульбах картоплі, сорт Прієкульська рання
Репродукція
Кількість бульб із ниткоподіб¬
ними паростками, %
Одна літня
4
Дві літні
3-4
Три літні
6-8
Одна весняна
22
Дві весняні
54
Три весняні
60-61
Одна літня і одна весняна
28
Дві літні і одна весняна
42
382
Результати показали, що у Степу без зрошення за одну репродук¬
цію майже чверть від загальної кількості бульб мають ниткоподібні
паростки, за дві репродукції - понад половина їх, за три - понад 60 %.
У літньому садінні цей процес відбувається значно повільніше - за
три репродукції ниткоподібні паростки мають 3-8 % бульб. Але й за
літнього садіння відбуваються внутрішні зміни, які не проявляються
як зовнішні симптоми, проте при перенесенні культури у весняне
садіння прояв їх значно посилюється: одне літнє і одне весняне са¬
діння дають 28 % вироджених рослин (одна весняна - 22 %), дві літ¬
ні і одна весняна репродукції - 42 % їх.
Усі вищезгадані теорії споріднені і мають спільну рису: виро¬
дження є явищем фізіологічного, а не інфекційного характеру. Тому і
заходи боротьби з ним мають спільні риси. Пропонувалося перенес¬
ти бульбоутворення на більш сприятливий період року - осінь (літ¬
нім садінням), рано садити і рано збирати картоплю; застосовувати
двоврожайну культуру з використанням свіжозібраних бульб; виро¬
щувати здоровий садивний матеріал у гірських і північних районах;
здійснювати “омолодження” скошуванням картоплиння на весняних
насадженнях; садіння вічками або невеличкими шматочками бульб,
де міститься менша кількість токсинів тощо. Але, як показала прак¬
тика, ці методи виявились малоефективними: урожайність при цьому
підвищувалася повільно.
Вірусологічна теорія виродження. Уперше вірусні хвороби у
1890-1902 рр. почав вивчати російський вчений Д.І. Івановський. У
20-х роках XX ст. вірусні хвороби картоплі дослідили голландські
вчені Quanjer та ін. (1916) і Botjes (1920). З того часу цьому питанню
присвячується велика кількість досліджень як зарубіжних, так і віт¬
чизняних учених.
Прихильники вірусологічної теорії виродження в колишньому
Радянському Союзі М.С. Дунін [6, 7], B.J1. Рижков [21], К.С. Сухов
[26, 27], С.М. Букасов, А.Я. Камераз [4], М.К. Фом’юк [31], П.Г.Чес-
ноков [33] та ін. вважали, що виродження картоплі є патологічним
явищем, яке відбувається під дією вірусів. Екологічні умови, на їхню
думку, можуть прискорювати поширення вірусних захворювань і
певною мірою регулювати їх шкідливість.
У 1956 р. були опубліковані праці Gierer Schram і Frankel-Cjnrat,
у яких повідомлялося, що нуклеїнова кислота, виділена з часток ві¬
русу тютюнової мозаїки, інфекційна, тобто викликає при зараженні
нею рослин утворення некрозів і синтез вірусних частинок, які не
відрізняються від часток вірусу, властивих тютюновій мозаїці.
А.ЛАмбросов [1] зазначав, що в Білорусії шляхом створення
відповідних умов для росту і розвитку рослин картоплі не можливо
зберегти їхню продуктивність і знизити шкідливість вірусів. Навіть
оптимальні умови вирощування не запобігають вірусній інфекції.
При порушенні рівноваги “зовнішнє середовище - вірус” останні
383
інтенсивно розмножуються, що призводить до різкого зниження
продуктивності уражених рослин картоплі. За такого процесу поси¬
люється прояв зовнішніх ознак хвороб, тобто спостерігається незво-
ротне виродження.
Вірусні, віроїдні, мікоплазмові хвороби та шляхи поширення
їх. Картопля уражується близько 30-ма різноманітними вірусами,
мікоплазмами, а також віроїдом веретеноподібності бульб.
Багаті вуглеводами і водою картоплиння і бульби є сприятливим
середовищем для розвитку патогенів, включаючи і віруси. Вегетатив¬
не ж розмноження картоплі забезпечує можливість їх існування в
активному стані протягом тривалого часу в період вегетації у росли¬
нах і бульбах, а в період зберігання - у бульбах.
Патогени викликають хвороби, які призводять до зміни фізіоло¬
гічних процесів - дихання, фотосинтезу, утворення амінокислот, біл¬
ків, вуглеводів, нуклеїнових кислот, вітамінів та багатьох інших ре¬
човин. Подібні зміни супроводжуються зниженням не лише врожай¬
ності, а й товарних, смакових і насінних якостей бульб.
Для різних хвороб характерні різні патологічні зміни. Так, інфек¬
ціям мозаїчного типу властиве руйнування хлорофілу в паренхімі
листків і ростова деформація; при скручуванні листя і мікоплазмових
захворюваннях спостерігається ураження провідної системи - заку-
порення і некроз судин. В уражених рослин, як правило, посилюєть¬
ся транспірація, знижується вбирна здатність коріння. Внаслідок цих
змін рослини втрачають здатність ефективно використовувати воло¬
гу і мінеральне живлення.
Патологічний процес при вірусних інфекціях завершується міс¬
цевим і системним некрозом (відмиранням тканин). При цьому утво¬
рення бульб і нагромадження в них крохмалю майже повністю зупи¬
няється, а в деяких випадках відбувається гідроліз уже нагромадже¬
ного крохмалю.
Щодо механізму дії вірусу на рослину існує дві гіпотези. Прибіч¬
ники однієї з них вважають, що вірус, потрапивши у рослину, необ¬
хідний для себе “будівельний” матеріал бере з неї, що й призводить
до патологічних змін і навіть загибелі рослинних клітин, а все нові й
нові віруси атакують сотні інших здорових клітин. “Вірусна пожежа”
охоплює організм.
Згідно з другою гіпотезою, вірус, потрапивши в рослину, виділяє
отруйні речовини, які призводять до захворювання.
Найбільшої шкоди картоплі в районах розвинутого картоплярст¬
ва завдають віруси, які мають різко виражені інфекційні і хворобо¬
творні властивості. Вони є облігатними внутрішньоклітинними пара¬
зитами і здатні до відтворення лише в них.
Основною причиною, що впливає на ураження вірусом, є стій¬
кість рослин, яка сформувалась у філогенезі. Має значення інтенсив¬
ність накопичення вірусу в рослині. Стійкість рослин змінюється у
384
процесі їх розвитку: вони набувають стійкості в міру їх росту і роз¬
витку.
Віруси, що проникли у рослину, переміщуються в бульби, де й
нагромаджуються, знижуючи з кожним роком урожайність картоплі.
Інтенсивність перезараження рослин вірусами перебуває у прямій
залежності від стану здоров’я вихідного матеріалу, тривалості його
репродукування; фітосанітарних та природно-кліматичних умов ра¬
йону вирощування картоплі; біологічних особливостей сорту, агро¬
техніки, насінницьких заходів, що застосовуються; чисельності, ви¬
дового складу, часу появи, активності та інтенсивності льоту перенос¬
ників, кількості джерел інфекції та концентрації вірусів; фізіологіч¬
ного стану рослин на час активного льоту переносників, а також мож¬
ливості контактного перенесення патогенів.
При цьому умови, оптимальні для росту й розвитку картоплі, не
звільняють рослин від інфекції, а лише тимчасово послаблюють сту¬
пінь прояву симптомів хвороб та їхню шкодочинність. Тому при ре¬
продукуванні картоплі основним завданням є запобігання нагрома¬
дженню вірусів у рослині і бульбах.
Розрізняють первинну інфекцію, коли вірус проникає в рослину,
а згодом - у бульби. Так зване вторинне ураження спостерігається у
рослин, які виросли з уражених бульб. Потомство таких рослин ви¬
являється повністю ураженим вірусом.
При вторинному ураженні прояв вірусних хвороб на картоплинні
більш виразний, а зниження врожайності більш відчутне. Потрапля¬
ючи в рослину, більшість вірусів уражують паренхімні тканини лис¬
тків і стебел, а вірус скручування листків - і судини флоеми. В ура¬
жених клітинах утворюється велика кількість вірусних часток, жит¬
тєздатність клітин порушується, що призводить до виснаження, при¬
гнічення або ненормального посилення фізіологічних процесів, а при
тяжких ураженнях - до загибелі клітин і тканин.
Найбільше значення мають фітопатогенні віруси, які поділяються
на персистентні (стійкі) і неперсистенггні (нестійкі).
Фітопатогенні віруси самостійно у природі поширюватися не
можуть. Вони переносяться різними комахами, грибами, контактно
при стиканні хворих і здорових рослин, сільськогосподарськими
машинами і знаряддями, людиною, бульбами тощо.
Віруси мають антигенні властивості, на чому ґрунтується сероло¬
гічний метод їх діагностики.
У таблиці 6.2 наведено найбільш розповсюджені вірусні хвороби
картоплі, їх збудники, шляхи поширення та найефективніші способи
їх виявлення.
Щодо показників шкідочинності кожного вірусу, які одержані у
відповідних дослідах, то вони у виробничих умовах змінюються під
дією багатьох факторів. Потрібно враховувати і змішані інфекції, що
викликаються комплексом вірусів.
13 2 ,32
385
Таблиця 6.2 - Найбільш поширені та шкідливі вірусні, мікоплазмові і віроїдні хвороби картоплі та їх збудники
Хвороба
Зниження врожай¬
ності картоплі при
садінні ураженими
бульбами, %
Основні ознаки захворювання
Збудни¬
ки
Шляхи поширення збудника
Найефектив¬
ніший спосіб
виявлення
візуально
тест
спосіб передачі і поши¬
рення
можливість
контактного
ураження
серологічний
біологічний
1
2
3
4
5
6
7
8
9
10
Скручу¬
вання лис¬
тя
50(28-64)
Листки жорсткі, крихкі, листові частки згорнуті в
трубку або напівтрубку в будь-якій частині рос¬
лини або на всьому кущі. Сильно пригнічено ріст.
Некротизація розпочинається з найнижчого яру¬
су. Некроз флоеми листків.
L-вірус
Біоло¬
гічний
Попелицями,
клопами,
28-крапковим
сонечком
-
+++
-
+
Смугаста
мозаїка
(строката)
50(20-90)
Поздовжні чорні або бурі смужки, штриху в атий
некроз на стеблах, черешках і жилках з нижнього
боку листків. Рослина крихка. Дуже уражена рос¬
лина має оголене стебло з декількома листками на
верхівці.
Y-вірус
Меха¬
нічний
Попелицями,
клопами, жу¬
ками
+
++
±
+++
Зморшку¬
вата мо¬
заїка
40(20-70)
Листки мозаїчні, зморшкуваті, краї закручені
донизу, жилки бурі. Рослини крихкі, пригнічені.
Нижні листки засихають, але не відпадають і про¬
тягом тривалого часу висять на стеблі.
XY-
віруси
Попелицями,
жуками,
ґрунтовими
грибами
+
+
+
+
Закручу¬
вання лис¬
тя
25(9-48)
Хвилясті краї частинок листка загнуті доверху у
верхньому ярусі рослин. Складчастість і загальний
хлороз відсутні. У другій половині вегетації ознаки
хвороби послаблюються, іноді зникають зовсім.
М-вірус
tt
Попелицями,
клопами
++
±
++
+
Звичайна
мозаїка
10-28
На листках між жилками - зеленувато-жовті пля¬
ми, невеликі, розпливчасті, за жаркої сухої пого¬
ди маскуються і майже непомітні.
Х-вірус
Меха¬
нічний
Ґрунтовими
грибами, жука¬
ми, трипсами
++
++
4-
387
Продовження табл. 6.2
1
2
3
4
5
6
7
8
9
10
Складчаста
мозаїка
(кучеря¬
вість
листя)
10-15
Плями на листках розміщені безсистемно, часто
на жилках зеленувато-жовті, але не розпливчасті і
не маскуються, іноді супроводжуються випук¬
лістю листкової пластинки між жилками та хви¬
лястістю її країв. При сильному ураженні країв
частинки листка закручуються доверху.
А-вірус
Попелицями
±
±
±
+
Слабка
крапчас¬
тість
10-20
Міжжилкова крапчастість; у деяких сортів закру¬
чування листків донизу, сильна зморшкуватість,
їх бронзовість і поникнення у старих рослин.
S-вірус
Попелицями,
клопами, жука¬
ми, 28-крапко-
вим сонечком
+
-
++
+
Аукуб а-
мозаїка
до 10
Плями яскраво-жовті, різко окреслені, характерні
для нижніх і середніх листків. Листки без нек¬
розів і деформацій. Жовта плямистість найбільш
помітна на молодих рослинах, з віком вона мас¬
кується. Некроз бульб.
F-вірус
Попелицями
+
±
±
+
Крапчас¬
тість
стебел
картоплі
10-30,
погіршення
якості
бульб
Мозаїка і деформація листків, некроз надземних
частин рослини. Фронтальний некроз тканин
бульби. Уражені рослини відстають у рості, ма¬
ють дрібніші листки, які передчасно відмирають.
Ратгл-
вірус (R)
Нематодами
±
±
±
++
Веретено¬
подібність
бульб
(готика)
20-85
Листки підняті вгору вздовж стебла під гострим
кутом і мають дрібні частинки. Пластинки части¬
нок листків складені уздовж середньої жилки,
краї на їх верхівці антоціанові. Вкорочені
міжвузля роблять кущ карликовим, стебел мало.
Віроїд
верете¬
нопо¬
дібності
іі
3 бульбами, пош¬
линами, цикадка¬
ми, клопами, жу¬
ками, з насінням,
пилком, грибами
++
+
-
+
Стовбурне
в’янення
(стовбур)
до 10
Листки на верхівці дуже видовжені, часто до нит¬
коподібних, скручені подібно до вузької трубки,
антоціанові або пожовклі тільки по краях, дрібні.
Поступово скручуються всі листки на рослині, і
кущ в’яне. Рослини хлоротичні.
Мікопла-
зми
Біоло¬
гічний
Цикадками
-
+
-
+
Пурпурне
скручуван¬
ня верхівки
до 10
Закручування і гофрованість верхівкових листків,
антоціанові краї листків або всієї частки, утворе¬
ння пазушних паростків і повітряних бульб.
-
+
-
За О.Г.Зикіним [8], у разі, коли для конкретних умов і сортів по¬
казник шкідочинності для окремих вірусів не визначено, слід
користуватися середніми величинами: в умовах, сприятливих для
росту і розвитку рослин картоплі, для всіх латентних вірусів і
слабких форм вірусів така величина становить 10 %, для тяжких
форм - 40, а в несприятливих умовах - відповідно 20 і 80 %.
Згідно з дослідженнями, проведеними у Великобританії, при 10 %-
ній ураженості вихідних рослин вірусами втрати загального врожаю
становлять 5 %, а при 50 %-ній - 25 % [9]. У Польщі в середньому на
1% ураження вірусом Y і скручуванням листя припадає 0,5% втрат
урожаю [5].
Вірусологи вважають, що на перебіг вірусних хвороб картоплі
значною мірою впливають екологічні фактори. Негативний вплив їх
виявляється не тільки безпосереднім погіршенням насінних якостей
бульб, але й зменшенням опору рослин проти вірусних хвороб.
Уражені вірусами організми рослин активно борються із захво¬
рюванням. Кінець цієї боротьби залежить не тільки від внутрішніх
факторів, що зумовлюють імунність рослин, але й від різних факто¬
рів зовнішнього середовища.
К.С.Суховим [26] доведено, що при розвитку крилатої персикової попе¬
лиці, яка є найбільш активним переносником вірусів картоплі, вирішаль¬
ну роль відіграють екологічні фактори й умови живлення попелиці.
У ряді досліджень доведено вплив добрив на стійкість рослин
проти різних захворювань, у тому числі й вірусних. Як зазначав
В.Л.Рижков [20], не можна заперечувати й існування “депресії*” кар¬
топлі, незалежно від вірусних хвороб, але водночас не варто запере¬
чувати й те, що вірусні хвороби значною мірою інфекційні і за спри¬
ятливих умов швидко поширюються від хворих рослин до здорових.
Доведено, що ступінь ураження вірусними, як і грибковими та
бактеріальними хворобами, залежить головним чином від погодних
умов року вирощування картоплі, а також від умов зберігання і під¬
готовки бульб до садіння.
Факторами, що впливають на прояв зовнішніх симптомів хвороб та
концентрацію хвороботворних патогенів у рослинах, є температура, світ¬
ло, тривалість дня, вологість, удобрення і пора року. При цьому слід за¬
значити, що найбільший вплив на рослину і вірус вони проявляють у
взаємодії, проте експериментально доведена дія кожного з них.
Температура. Виявлено, що оптимальною температурою для рос¬
ту, розвитку і утворення бульб є 15-20°С. При температурному ре¬
жимі 16-28°С процес нагромадження вірусу в рослинах є найінтен-
сивнішим. Зміна температури протягом вегетації впливає на зміну
темпів нагромадження вірусів, внаслідок цього гірше чи краще про¬
являються симптоми хвороби на рослині. Щодо окремих вірусів, то
для інтенсивного нагромадження кожному з них відповідає різна тем¬
пература, хоч ця різниця в окремих випадках є незначною.
388
Так, для інтенсивного нагромадження і прояву характерних сим¬
птомів вірусу М оптимальною є температура близько 20°С, для віру¬
су Y - 15-20°С. При ураженні вірусом X більш чітко локальні некро¬
зи проявляються при температурі 16°С, ніж при 24°С. Навпаки, сис¬
темні некрози виразніші при 24°С, ніж при 16°С. При штучному за¬
раженні сіянців картоплі найбільш характерні симптоми вірусу X
проявилися при температурі 10-24°С. Вірус М проявляє зовнішні
ознаки більше на рослинах за температури 16°С, а типові і чіткі -
при 18-20°С.
У той же час, як свідчать спостереження, проведені польськими вче¬
ними, зовні ознаки хвороби під впливом вірусу М зникають на рослинах
у полі, коли температура протягом кількох днів буває вищою 24°С.
Світло. Доведено, що більша інтенсивність освітлення і трива¬
ліший день сприяють нагромадженню вірусів. При меншій інтенсив¬
ності освітлення прояв симптомів слабший у вірусів X, Y і L. Таку
закономірність щодо вірусу L спостерігаємо і при індексації бульб.
Інтенсивніший прояв симптомів вірусу S, як виняток, спостерігався
при затіненні нижніх листків картоплі.
Вологість. Тривала посуха впливає негативно на ріст і розвиток
рослин, викликає в’янення листя, що є подібним до симптомів, харак¬
терних для вірусу скручування листя; маскуються також ознаки мо¬
заїки. Надмірна вологість викликає некрози.
Удобрення. Норми добрив, які сприяють підвищенню врожайності,
також призводять до накопичення вірусів. Високі норми азоту, не збалан¬
совані за фосфором, маскують ознаки вірусу скручування листя, особли¬
во на молодих рослинах. За збільшених норм калію симптоми цієї хво¬
роби більш виразні. При нормальному удобренні фосфором і калієм, але
при надмірному - азотом ознаки хвороб маскуються.
Перенасичення рослин марганцем проявляється некрозами на стеб¬
лах і опаданням листя, подібно до симптомів, що викликає вірус Y.
Пора року. Залежно від пори року, на взаємозв’язок “рослина-
вірус” діє комплекс факторів. Ознаки хвороб проявляються сильніше
навесні і восени, слабкіше - літом. Жарка погода і підвищена інсо¬
ляція призводять до зниження концентрації вірусів і слабкого зовніш¬
нього прояву ознак.
Пора року впливає на прояв ознак вірусних хвороб та індексацію
бульб у теплицях: сильнішими вони є навесні і восени, ніж узимку та
влітку^ Винятком є вірус X, який проявляється сильніше взимку.
О.Й. Онищенко [19] зазначав, що для одержання здорового садив¬
ного матеріалу треба вирощувати картоплю в більш сприятливих
умовах, проводити боротьбу з рослинами-вірусоносіями і комахами-
переносниками. Зокрема, такий шлях організації насінництва загаль¬
ноприйнятий у країнах-експортерах насінної картоплі. Умовами, що
відповідають біологічним вимогам картоплі, є північні та гірські ра¬
йони України.
389
В умовах, сприятливих для вирощування, знищенням хворих рос¬
лин та ізоляцією від рослин-вірусоносіїв можна запобігти поширен¬
ню вірусів на здорові рослини, що є важливою умовою одержання
здорової насінної картоплі.
У несприятливих для вирощування картоплі умовах позбавитися ві¬
русних хвороб не можливо через велику кількість їхніх переносників.
Звичайно, в усіх випадках значна роль належить сорту, передусім
його стійкості до різноматнітних патогенів, а також реакції на умови
зовнішнього середовища та агротехнічні прийоми вирощування,
проведення насінницьких заходів (рис. 6.3).
Вивченням питань виродження картоплі (Молоцький М.Я.) [14] у
зоні Степу України (Дніпропетровська область), де умови для виро¬
щування картоплі є дуже складними (середньомісячна температура
липня становить близько 24°С, відносна вологість повітря знижу¬
ються до 45-30 %, а річна сума опадів складає 450—470 мм), було
доведено, що при вирощуванні її за весняного садіння, навіть в умо¬
вах зрошення, виродження значно зростає з кожною наступною ре¬
продукцією, а врожайність зменшується. Наприклад, у сорту Пріє-
кульська рання при вирощуванні за весняного садіння з кожною на¬
ступною репродукцією урожайність знижувалася на 20-22 %, а кіль¬
кість вироджених рослин у насадженнях після першої репродукції
складала 27 %, другої - 63, третьої - 91 %.
При вирощуванні картоплі в літньому садінні кількість виродже¬
них рослин зростала повільніше, а зниження врожайності було менш
помітним. Четверта літня репродукція була кращою за другу весня¬
ну. Але при використанні цього матеріалу у весняних посадках він
втрачав продуктивність за одну-дві репродукції.
Слід також зазначити, що поряд із симптомами вірусних хвороб спо¬
стерігається поява на бульбах у процесі їх пророщування ниткоподібних
паростків, що також призводило до втрати ними продуктивних властивос¬
тей. Але це явище не має ніякого відношення до вірусних хвороб.
Про позитивний вплив літнього садіння на бульби картоплі свід¬
чать дані Білоцерківської сортодільниці (Лісостеп), куди з півдня
надсилали зразки сортів на ґрунтоконтролю. Виявилося, що зразки
сортів Прієкульська рання і Вармас були уражені вірусними хворо¬
бами незначною мірою, дещо більше -Гатчинська. Південний зразок
сорту Прієкульська рання був більш стійким, ніж із поліської зони і
відрізнявся високою продуктивністю. Так, зразок другої літньої ре¬
продукції, що надійшов із Вознесенської сортодільниці в 1974 р., був
уражений вірусними хворобами на 8 %, а його врожайність станови¬
ла 228 ц/га; у зразка восьмої літньої репродукції, надісланого із Сім¬
феропольської сортодільниці в 1973 р., ці показники відповідно ста¬
новили 18 % і 257 ц/га. У 1972 р. зразок п’ятої літньої репродукції,
що надійшов із Херсонської сортодільниці, не мав зовнішніх ознак
вірусної інфекції і забезпечив урожайність 320 ц/га.
390
Природно-кліматичні
умови
Організаційні
заходи
Внесення до
Реєстру сортів
Зони насінництва
Строки
сортооновлення
Репродукція
Організаційні
заходи іцодо
підготовки
посадкового
матеріалу
Заходи з
догляду під час
сходів
Політика цін
Рис. 6.3. Вплив природно-кліматичних умов, сорту, агротехніки вирощування
та організаційних заходів на ураження картоплі вірусами (за Габріелем,1982)
Таким чином, можна стверджувати, що наявність вірусів у клі¬
тинному соці картопляної рослини ще не є причиною зниження вро¬
жайності бульб на півдні України. Тільки під впливом несприятли¬
вих умов вирощування приховані віруси спричиняють тяжкі вірусні
хвороби, і рослини стають неспроможними давати повноцінний вро¬
жай. Отже, першопричиною виродження картоплі на півдні України
є несприятливі умови, за яких ослаблені рослини швидше зазнають
дії вірусної інфекції.
Пізніше (у 1975-1985 рр.) була проведена серія дослідів з ви¬
вчення зниження урожайності картоплі при репродукуванні еліти в
умовах правобережного Лісостепу України в зоні помірного виро¬
дження (Молоцький М.Я., Тихоступ В.Ф.) [16], де середньомісячна
температура липня становить 21-22°С, а річна норма опадів - 490-
600 мм.
Було доведено, що в подальших від еліти репродукціях у клітин¬
ному соці рослин також зростає кількість вірусів, але їх прояв конк¬
ретними вірусними хворобами залежить від сорту та умов вирощу¬
вання. Дослідження свідчать, що за високого рівня агротехніки і
сприятливих погодних умов продуктивність сортів Чарівниця, Боро¬
дянська, Вармас у процесі репродукування в окремі роки навіть зрос¬
тала. Так, урожайність п’ятої-шостої репродукції сортів Бородянська
і Чарівниця у сприятливі роки становила понад 300 т/га, четвертої-
п’ятої репродукції сорту Вармас - 200-250 ц/га. Водночас урожай¬
ність сортів Прієкульська рання, Столова 19 уже в третій-четвертій
репродукціях опускалася нижче 200 ц/га, а в несприятливі роки у
сорту Прієкульська рання не перевищувала 100 ц/га. Проведені до¬
слідження переконливо доводять, що навіть у цій більш сприятливій
зоні основним фактором стійкості до виродження є генотип, а його
розвиток і прояв продуктивності залежать від технології та умов ви¬
рощування. Так, за сприятливих умов генотип здатний реалізувати
свої потенційні можливості та протистояти хворобам, у тому числі
вірусним, а пригнічений організм внаслідок екологічної депресії
швидше зазнає негативної дії вірусної інфекції.
Дослідження, проведені у 1991-1997 рр., у двох підзонах Лісо¬
степу (правобережній і центральній) показали, що в елітному матері¬
алі, одержаному шляхом клонового добору, присутня значна кіль¬
кість вірусів. І це залежить не лише від генотипу, але й від умов ви¬
рощування, включаючи технологію (Молоцький М.Я.) [14]. Так, се¬
ред еліти сорту Гатчинська рослин, інфікованих вірусом X, було
53 %, S - 27, М - 18 %, уражених вірусними хворобами - 7,6 %; у
п’ятій репродукції інфікованих рослин стало відповідно 84 %, 49, 55 % і
хворих — 41,1 %. У Світанку київського, відповідно, в елітних росли¬
нах - 26 %, 16, 14 і 7,1 %, а в п’ятій репродукції - 51%, 53, 45 і
30,4%; у Луговської - відповідно 21%; 13, 11 і 12,1 % та 58 %, 54, 29
і 48,5%.
392
Отже, найбільш інфікованою вірусами виявилась еліта сорту Гат¬
чинська, менше - Світанок київський і найменше - Луговська. Щодо
прояву вірусних хвороб, то розподіл сортів був таким (у спадному
порядку): Луговська, Гатчинська, Світанок київський. Ця тенденція
реакції сортів на вірусну інфекцію збереглася і в подальших репро¬
дукціях.
Урожайність від еліти сорту Гатчинська становила 364 ц/га, п'я¬
тої репродукції - 214, або 60%; Луговської - 338 і 213 ц/га, або 63%;
Світанку київського - 323 і 184 ц/га, або 57%.
Механізм зниження продуктивності рослин картоплі в наступних
від еліти репродукціях вивчався в дослідах, проведених в 1991-1993
рр. у центральному Лісостепу України [15].
Було виявлено, що в процесі репродукування еліти упродовж кіль¬
кох репродукцій погіршується схожість бульб та їхня стеблоутво-
рююча здатність. У середньому по п’яти сортах кількість стебел у
кущі зменшилася від першої до п’ятої репродукції на 1,9 пгг., що ста¬
новить 44%. Знижувалася також висота рослин і площа листкової
поверхні. Так, сорти Гатчинська, Невська, Луговська і Світанок київ¬
ський формували оптимальний листковий апарат (30-40 тис. м2/га)
до п’ятої-шостої репродукції, Зов - до четвертої, а Гарт - лише
на елітних насадженнях і в першій репродукції. Відповідною була
і врожайність: у перших чотирьох сортів вона становила понад
200 ц/га в п’ятій і навіть шостій репродукціях, а у сорту Гарт - ли¬
ше в першій.
Процес бульбоутворення, маса бульб під кущем і середня маса
бульби також залежали від репродукції та сорту. Так, кількість бульб
під кущем зменшилася від еліти до п’ятої репродукції у сортів Сві¬
танок київський на 20%, Луговська - 28, Гатчинська - 28%; маса
бульб під кущем - на 51%, 44 і 41%, середня маса бульби - на 38%,
23 і 46%, відповідно.
Проте слід зазначити, що в багатьох дослідженнях, які проводи¬
лись у цій зоні, у тому числі й наших, виявлено, що з кожною наступ¬
ною репродукцією, незважаючи на погодні умови, кількість рослин,
уражених вірусами, зростала, а продуктивність їх знижувалась. Але
це не суперечить висловленій раніше думці, а лише підтверджує, що
в Лісостепу, як і в Степу, рослини картоплі зазнають значної еколо¬
гічної депресії, яка є причиною їх виродження, тобто втрати здатнос¬
ті накопичувати врожай бульб.
Щодо вірусів, то у зв'язку із втратою рослинними організмами
стійкості внаслідок депресії, вони надзвичайно швидко розмножу¬
ються, проявляючись вірусними хворобами, які остаточно знижують
продуктивні властивості рослин.
В даному разі доцільно провести певну аналогію з організма¬
ми тварин і людей. Відомо, що тварини і люди тривалий час мо¬
жуть бути носіями збудників багатьох небезпечних хвороб, на-
393
приклад туберкульозу. Але в комфортних умовах (а для людини
ще й у сприятливих соціальних) хвороба в організмі не розвива¬
ється, і, навпаки, у дискомфортних і стресових ситуаціях швидко
розвивається й набуває масового поширення. Це пояснюється
тим, що в несприятливих умовах послаблюється імунітет організ¬
му, знижується резистентність, і патогени швидко розмножують¬
ся, спричиняючи захворювання. Отже, очевидно, що це є загаль-
нобіологічним явищем.
Таким чином, визнаючи шкодочинність вірусних хвороб, не мож¬
на нехтувати умовами вирощування картоплі, не шукаючи техноло¬
гічних рішень забезпечення її біологічних потреб.
Перш за все слід поліпшувати систему насінництва картоплі в
Україні і вдосконалювати технологію її вирощування.
Стратегія системи насінництва картоплі для півдня (зона сильно¬
го виродження) має ґрунтуватися на тому, що тут відтворення еліти
методом клонового добору в польових умовах є неможливим. Одер¬
жувати оригінальний садивний матеріал можна лише біотехнологіч¬
ним способом, що під силу висококваліфікованим спеціалістам з на-
уково-дослідних установ при наявності спеціального обладнан-
ня.Тому елітний і репродукційний матеріал у цю зону, як правило,
слід завозити з науково-дослідних установ та елітно-насінницьких
господарств Полісся та західного регіону. Завезений матеріал потріб¬
но розмножувати в насінницьких господарствах один-два роки у літ¬
ньому садінні або у двоврожайній культурі, одержані садивні бульби
слід використовувати для одноразового вирощування у весняному
садінні на продовольчі потреби.
Враховуючи складність зберігання завезених бульб для літнього
садіння (липень) і обмеженість стаціонарних сховищ із регульова¬
ною температурою повітря, першого року матеріал доцільно викорис¬
тати у двоврожайній культурі, а на другий - застосовувати обидва
варіанти.
У Криму для розмноження садивних бульб слід ширше вико¬
ристовувати високогірні райони, де картопля менше вироджу¬
ється.
У Лісостепу, який є зоною помірного виродження картоплі, еліту
картоплі можна відтворювати двома шляхами - біотехнологічним і
добором клонів та їх випробуванням у польових умовах. Не варто
виключати і завезення для розмноження супереліти та еліти з Поліс¬
ся і західного регіону. Добирати клони краще на оздоровлених від
вірусів насадженнях у науково-дослідних установах зони незначного
виродження, але можна й на здорових місцевих насадженнях. При
доборі та бракуванні в первинних ланках насінництва разом із візуаль¬
ним оглядом необхідно проводити обов’язкову серодіагностику віру¬
сів (X, M, S, Y). З цією метою можна також використати індексацію,
імуноферментний метод та ін.
394
На Поліссі і в Карпатах еліту можна створювати клоновим добо¬
ром і біотехнологічним методом із застосуванням усіх перерахова¬
них вище методів оцінки і контролю якості еліти [14].
Важливим фактором зниження шкодочинності вірусних хвороб є
створення і впровадженння вірусостійких сортів і таких, що менше
уражуються попелицями завдяки морфологічним особливостям (тов¬
ста кутикула, опушеність тощо).
Для боротьби з вірусною інфекцією вчені розробили і такі захо¬
ди, як відбір вірусних клонів за допомогою серодіагностики, індек¬
сація та інші методи, оздоровлення картоплі методом апікальної ме¬
ристеми в поєднанні з термотерапією, розмноження безвірусного
матеріалу в ізольованих умовах, боротьба з комахами-переносни-
ками вірусів, фітопатологічні прочистки тощо.
Не слід також нехтувати методами боротьби з виродженням кар¬
топлі, які розроблені екологами, про що вже згадувалось вище.
Але головним, на наш погляд, є створення шляхом технологічних
рішень сприятливих умов для вирощування картоплі, за яких росли¬
ни почувають себе комфортно, мають високу резистентність і здатні
чинити опір вірусній інфекції.
ЛІТЕРАТУРА
1. Амбросов А.Л. Вирусные болезни картофеля и методы выращивания
здорового семенного материала в Белорусской ССР: Автореф. дис. ... д-ра
с.-г. наук. - Минск, 1965.
2. Балашов Н.Н. Семеноводство картофеля на юге СССР. - М.: Сельхо-
зиздат, 1963.
3. Бондарев М.Т. Борьба с вырождением картофеля на юге СССР при
культуре его ростками. - M.J Росиздат, 1938.
4. Букасов С.М., Камераз А .Я. Основы селекции картофеля. - М.:
Сельхозгиз, 1959.
5. Габриэль В. Методические основы семеноводства картофеля // Кар¬
тофель: селекция, семеноводство, технология возделывания - Минск: Ура-
джай, 1988.-С. 172-204.
6. Дунин М.С. Вирусные болезни сельскохозяйственных растений. - М.:
Сельхозгиз, 1937.
7. Дунин М.С. О некоторых методах получения и применения антивоз-
будительных сывороток в фитопатологии и энтомологии // Вестник с.-х.
науки - 1958-№3.
8. Зыкин А.Г. Вирусные болезни картофеля - Л.: Колос, 1976.
9. Кучумов А. Организация и технология производства семенного кар¬
тофеля (зарубежный опыт) // Международный с.-х. журн. - 1970. - №5. -
С. 50-53.
Ю.Линник Г.Н. Причины вырождения картофеля // Картофель. -
1957. - №2.
11.Лысенко Т.Д. Заготовить больше верхушек картофеля. - М.:
Огизсельхозгиз, 1947.
12. Максимович М.М. Семеноводство картофеля. - М.: Сельхозгиз, 1951.
395
13. Молоцький М.Я. Виродження картоплі у південних районах України
та заходи боротьби з ним // Картопля, овочеві та баштанні культури: Респ.
міжвід. темат. наук. зб. - К., 1986. - Вип. 5. - С.78-84.
14. Молоцький М.Я. До питання виродження картоплі в степовій і лісо¬
степовій зонах України і заходи боротьби з ним // Картоплярство: Міжвід.
тем. наук. зб. - К.: Довіра, 1999 - С. 19-27.
15. Молоцький М.Я., Кривенко І.С. Строки сортопоновлення картоплі у
центральних районах Лісостепу України // Вісн. аграр. науки. - 1994. - №5.
- С.З 8-40.
16. Молоцкий М.Я., Тихоступ В.Ф. Строки сортообновления картофеля //
Картофель и овощи. - 1984. - №2. - С.9-10.
17.Муш Н.Н. О взаимодействии гетеротрофного и автотрофного питания
в онтогенезе картофеля // Физиология растений - Т.8, вып.2. - 1961.
18.Немчин Ф.И. Изучение и проверка методов борьбы с вырождением
картофеля // Мат. конф. по вопросам семеноводства картофеля. - М.: Изд-во
МСХ СССР, 1958.
19. Онищенко О.Й. Із історії теорій виродження картоплі // Вірусні хво¬
роби картоплі. - К.: Урожай, 1969. - С. 6-12.
20. Рыжков В.Л. Вирусные болезни растений. - М.: Сельхозгиз, 1935.
21. Рыжков В.Л. Фитопатогенные вирусы. - М.: Изд-во АН СССР, 1946.
22. Рожалин Л.В. Агротехника семенного картофеля // Картофель. -
1958.-№6.
23. Руденко А.И. Меры борьбы с климатическим типом вырождения кар¬
тофеля // Мат. конф. по вопросам семеноводства картофеля - М.: Изд-во
МСХ СССР, 1958.
24. Соколенко Н.Ф. Выращивание здорового семенного картофеля на
юге // Науч. тр. Всесоюз. ордена Трудового Красного Знамени селекционно¬
генетического института им.Т.Д. Лисенко. - Одесса, 1962.
25. Стебут И.А. Основы полевой культуры и меры к ее улучшению в Рос¬
сии. - М., 1882.
26. Сухов К.С. Вирусы - М.: Изд-во АН СССР, 1956.
27. Сухов К.С. Проблемы общей и сельскохозяйственной вирусологии //
Защита растений от вредителей и болезней, 1960. - №1.
28. Фаворов А.М. К истории вопроса о размножении картофеля семенами на
Украине // Культура картофеля семенами: Сб. науч. тр. - Г орький, 1983 .-С.1.
29. Фаворов А.М. Картопля в степу УРСР. - Харків, 1948.
30. Фаворов А.М., Котов А.Ф. Летняя посадка картофеля. - М.: Госсель-
хозиздат, 1952.
31. Фом’юк М.К. Розвиток готики картоплі залежно від умов вирощуван¬
ня // Питання насінництва картоплі та боротьби з її виродженням. - К.: Вид-
во УАСГН, 1960. - С.60-68.
32. Фом’юк М.К. - В кн.: Питання насінництва картоплі та боротьби з її
виродженням. - К.: Вид-во УАСГН, 1960. - ^8с.
33. Чесноков П.Г. Болезни вырождения картофеля в СССР. - М.-Л. -
Изд-во с.-х. лит-ры, журналов и плакатов, 1961.
34. Чесноков Н.С. Причины вырождения картофеля на юге и меры борь¬
бы с ним // Мат. конф. по семеноводству картофеля. - Л.: Изд-во ВАСХ-
НИЛ, 1958.
35.Ячевский А.А. Защита растений от вредителей, 4, 1927, с.62.
396
6.4. СИСТЕМА НАСІННИЦТВА КАРТОПЛІ
В УКРАЇНІ
Система насінництва - це комплекс взаємопов’язаних організа¬
ційних, наукових і агротехнічних заходів, спрямованих на забезпе¬
чення виробництва, реалізацію і використання насіннєвого матеріа¬
лу. Складається з ланок первинного, елітного і репродукційного на¬
сінництва, страхових фондів насіння та державних ресурсів.
Насінництво нерозривно пов’язане з селекцією і своєю організа¬
ційною структурою відображує рівень її розвитку й досягнуті ре¬
зультати. Важливим завданням насінництва є також забезпечення
своєчасної сортозаміни і сортооновлення.
Селекційно-насінницька робота в Україні ведеться на базі єдиної
централізованої системи. Вона об’єднує ланки з виведення (селек¬
ція), випробування і реєстрації нових сортів (Державне сортовипро¬
бування), масового їх розмноження, здійснення контролю за сорто¬
вими і посівними якостями насіння.
Перелік основних ланок, зв’язок між окремими виконавцями
можна подати у такому вигляді (табл.6.3).
Таблиця 6.3 - Система селекційно-насінницької роботи в Україні
Ланка системи
Завдання
Виконавці
Селекція
Виведення нових сортів та
їх первинне розмноження
Науково-дослідні інститути, вищі
навчальні заклади (ВНЗ), селекційно-
дослідні станції, селекціонери
Сорто¬
випробування
Випробування, оцінка і
реєстрація сортів
Державна комісія України по
випробуванню та охороні сортів
рослин
Насінництво
Розмноження сортового
матеріалу при одночасному
поліпшенні і підтриманні
його сортових якостей:
а) виробництво супереліти
та еліти
б) виробництво насіння
еліти і І репродукції
в) розмноження насіння
еліти і репродукційного
матеріалу для повного
забезпечення потреби
господарства для виробни¬
чих посівів
Науково-дослідні інсппуги, ВНЗ і
селекційно-дослідні станції-
оригінатори нових сортів
Дослідні станції, навчально-дослідні
господарства сільськогосподарських
ВНЗ, елітгоспи
Насінницькі господарства, ділянки
розмноження і насінні ділянки в
товарних господарствах
Сортовий і на¬
сіннєвий конт¬
роль
Контроль за сортовими
якостями садивного матеріалу
(апробація), перевірка його
посівних якостей
Державні насіннєві інспекції,
карантинна інспекція,
сільськогосподарські органи і
спеціалісти господарств
397
Принципи ведення насінництва визначені Законом України “Про
насіння” [2] і новою системою насінництва картоплі, яка розроблена
вченими і спеціалістами Міністерства аграрної політики України,
Української академії аграрних наук, Інституту картоплярства УААН,
Білоцерківського державного аграрного університету та асоціації
“Картопля України” і затверджена наказом Міністерства аграрної
політики України № 31 від 06.04.2000 р. [7].
Міністерство аграрної політики України забезпечує розвиток на¬
сінництва, спрямований на повне задоволення потреб виробників у
насіннєвому матеріалі, перевірку його якості; здійснює заходи щодо
державної підтримки насінництва, удосконалення економічного ме¬
ханізму взаємовідносин між виробниками і споживачами, веде і ви¬
дає Державний реєстр виробників насінного матеріалу.
Українська академія аграрних наук організовує вирощування і реалі¬
зацію вихідного, оригінального насіннєвого матеріалу та еліти, а також
розмноження і впровадження нових перспективних сортів. Вона вивчає
попит світового насіннєвого ринку, вступає у взаємовідносини з органа¬
ми насінництва інших держав, розробляє прогресивні технології виро¬
щування насіннєвого матеріалу, нормативні документи.
Державна комісія України по випробуванню та охороні сортів
рослин проводить випробування нових сортів та їх реєстрацію.
Українська державна насіннєва інспекція здійснює державний
контроль у насінництві. Спеціалісти інспекції контролюють дотри¬
мання суб’єктами насінництва методичних і технологічних вимог;
дають вказівки щодо усунення порушень у веденні насінництва;
ознайомлюють з документацією з насінництва; беруть участь у про¬
веденні апробації посівів; контролюють якість підготовленого до
реалізації насіннєвого матеріалу; за порушення вимог чинного зако¬
нодавства з насінництва накладають згідно із законодавством Украї¬
ни на винних осіб адміністративні стягнення; мають право забороня¬
ти реалізацію будь-якого насінного матеріалу, якщо під час перевір¬
ки виявлено невідповідність його якісних показників державним
стандартам України.
Державна насіннєва інспекція проводить роботу з насінництва у
взаємодії з органами виконавчої влади, Державним комітетом Укра-
ни з питань стандартизації, метрології та сертифікації і його терито¬
ріальними органами.
Державна карантинна інспекція здійснює карантинний контроль
за ввезенням, вивезенням і використанням насіння згідно з Законом
України “Про карантин рослин” [1].
Виробництво оригінального насіннєвого матеріалу і елітне на¬
сінництво зосереджується в науково-дослідних установах, елітно-
дослідних та елітно-насінницьких господарствах науково-дослідних
установ, навчальних господарствах вищих сільськогосподарських
навчальних закладів за умови одержання патенту за результатами
398
атестації та занесення до Державного реєстру виробників оригіналь¬
ного і елітного насіння.
Узгодженим рішенням Міністерства аграрної політики України
та Української академії аграрних наук право виробництва елітного
насіннєвого матеріалу картоплі може бути надане й іншим господар¬
ствам, незалежно від форми власності. Обов’язковою підставою для
цього має бути наявність угоди між елітгоспом та профільною нау-
ково-дослідною установою, яка сама виробляє оригінальний і еліт¬
ний матеріал, на проведення насінницьких робіт під науково-
методичним керівництвом та контролем з боку останньої.
Насінницькі господарства всіх форм власності за умови атестації
і отримання дозволу управлінь Автономної Республіки Крим і облас¬
тей вирощують репродукційне насіння і також заносяться до Держав¬
ного реєстру виробників насіннєвого матеріалу, який веде і видає
Міністерство аграрної політики України.
Реалізацію насіннєвої картоплі здійснюють безпосередньо його
виробники або посередницькі організації за їх дорученням.
Особи, що не мають дозволу на виробництво та використання на¬
сіннєвого матеріалу, можуть вирощувати його лише для власних по¬
треб без права реалізації.
Виробництво насінного матеріалу вищих категорій (оригіналь¬
ний насіннєвий матеріал, еліта) здійснюється в господарствах з най¬
більш сприятливими ґрунтово-кліматичними і фітосанітарними умо¬
вами для культури картоплі з достатнім матеріально-технічним за¬
безпеченням.
Юридичні і фізичні особи, що одержали дозвіл на виробництво
і використання насіння, мають право: на договірних умовах
купувати в інших виробників відповідні категорії насіння для на¬
ступного розмноження і реалізації; встановлювати договірні ціни
на вирощене насіння, що підлягає реалізації; звертатися до органів
державного управління і контролю в насінництві та до судових ор¬
ганів у разі порушення прав, наданих їм Законом України “Про на¬
сіння”.
Виробники насіння зобов’язані: укладати угоди на викори¬
стання сортів з власниками патентів на сорти, установами-
оригінаторами сортів, на які не видаються патенти, або з авторами
сортів; вирощувати насіння згідно з обсягом, зумовленим державним
контрактом; додержувати технологічних і методичних вимог насін¬
ництва щодо збереження сортової чистоти, біологічних і урожайних
властивостей сорту та посівних якостей насіння; гарантувати відпо¬
відність насіння, що підлягає реалізації, сортовій чистоті і посівним
якостям, зазначеним у документі на насіння; своєчасно здавати зраз¬
ки насіння для контролю якості до лабораторій Української держав¬
ної насіннєвої інспекції; вести по кожному сорту насінницьку доку¬
ментацію за встановленими формами і зберігати її протягом трьох
399
років; додержувати встановленого порядку маркування і затарюван-
ня насіння, що підлягає реалізації.
Застосування біотехнологічних методів розширює зону вироб¬
ництва оригінального і елітного насіння.
У господарствах, що вирощують елітний та сортовий насінний мате¬
ріал, за їхній рахунок карантинна служба проводить щорічне обстежен¬
ня насіннєвої картоплі з метою визначення карантинного стану. В цих
господарствах допускається вирощування насіннєвої картоплі лише тих
категорій, з виробництва яких вони спеціалізуються згідно з Державним
реєстром виробників насіннєвого матеріалу.
Завезення насіннєвого матеріалу картоплі зарубіжної селекції в
насінницькі господарства та включення їх у процес відтворення та
розмноження здійснюється за умови, що сорт занесений до Реєстру
сортів рослин України або визнаний перспективним. При цьому
обов’язковим є наявність сортових та карантинних документів, вида¬
них відповідними державними установами.
Внутрішньогосподарське насінництво для власних потреб вироб¬
ників продовольчої картоплі організовується при потребі в кожному
господарстві.
Термін поновлення (кількість поколінь відтворення) насіннєвого
матеріалу, що виробляється, визначають агропромислові формування
Автономної Республіки Крим та областей з урахуванням рекоменда¬
цій оригінатора сорту. Враховують ґрунтово-кліматичні та фітосані-
тарні умови зони вирощування, біологічні особливості сорту, насам¬
перед стійкість до виродження.
Сортооновлення - це заміна сортового садивного матеріалу в го¬
сподарствах бульбами цих самих сортів, але вищих репродукцій.
Сортооновлення в товарних господарствах проводиться елітою
або репродукційним насінням у строки, прийняті для конкретних
ґрунтово-кліматичних зон.
Строки сортооновлення, залежно від ґрунтово-кліматичних
умов України. У реєстрованих сортів протягом виробничого вико¬
ристання з часом можуть погіршуватися господарсько-біологічні
властивості, що зумовлюється механічним засміченням, збільшенням
ураження рослин хворобами, які передаються через бульби.
Урожайні властивості та сортові якості насіння також знижуються
внаслідок порушення правил ведення насінництва. Тому виникає
потреба проводити сортооновлення.
Цінність садивного матеріалу визначається його сортовими якостя¬
ми. Але помітну роль відіграють і ірунтово-кліматичні умови та агро¬
технічні прийоми вирощування, які впливають на майбутній врожай і
його якість. Садивні бульби є тим проміжним ланцюгом, через який
здійснюється зв’язок між попереднім і наступним поколіннями.
Індивідуальний розвиток материнських рослин відбувається в
конкретному середовищі, що складається на ділянках вирощування.
400
Середовище кожного поля характеризується рівнем родючості ґрун¬
ту, його хімічними і фізичними властивостями, фітопатологічним
станом, а також комплексом факторів, які можна назвати кліматом і
мікрокліматом (вологість ґрунту і повітря, освітленість, рух повітря
тощо), які по-різному складаються в насадженнях залежно від їх гус¬
тоти, напрямку рядків тощо.
Бульби з високими посівними і врожайними якостями формують¬
ся, як правило, на фоні високої агротехніки, при відповідній культурі
землеробства та застосуванні комплексу насінницьких прийомів. Це
дає змогу одержувати позитивні модифікації, які зберігаються в 1-2-х
поколіннях, тобто модифікації, спричинені умовами вирощування
материнських рослин, мають короткочасний характер. Ефект пози¬
тивних модифікацій можна використати при добре налагодженому
сортооновленні, високому рівні організації насінництва.
Модифікаційна мінливість не змінює генотип. Вона відображує
реакцію його на зміни зовнішніх умов, а реакція, у свою чергу, ви¬
значається адаптивним потенціалом сорту.
За агрокліматичною та ґрунтовою характеристикою, а також да¬
ними щодо умов для поширення вірусної інфекції, в Україні визна¬
чено окремі зони виродження картоплі. Таких природних зон існує
чотири: найменшого виродження - Карпати і Прикарпаття; слабкого
виродження - Полісся і західні області; помірного виродження - лі¬
состепові області; сильного виродження - степові області [6].
Зона найменшого виродження (Карпати і Прикарпаття). У зону
входять гірські й передгірні райони Закарпатської і Чернівецької об¬
ластей.
Передгірні та гірські райони Карпат відрізняються від інших су¬
мою опадів і температурним режимом. Висота гір коливається від
500 до 1500 м над рівнем моря. Середня температура липня знижу¬
ється від 17-18,5°С в передгір’ї до 12-15 °С у горах. Річна сума опа¬
дів, навпаки, збільшується із 700-850 мм в передгір’ї до 1000-1200
мм у горах. Ця зона має найкращі умови для вирощування насінної
картоплі. Проте в Закарпатській і Чернівецькій областях є низинні
райони (до 250 м над рівнем моря), умови яких не сприяють вирощу¬
ванню якісної насінної картоплі через високі температури та велику
кількість переносників вірусних хвороб.
Зона має різні ґрунти, більшість з яких придатні для вирощування
садивної картоплі.
Зона слабкого виродження картоплі - це Волинська, Житомир¬
ська, Івано-Франківська, Львівська, Рівненська і Чернігівська облас¬
ті, а також північні райони Вінницької, Київської, Сумської та Хме¬
льницької областей. Ґрунтово-кліматичні умови цієї зони найсприят¬
ливіші для вирощування садивної картоплі. Проте західні і східні
райони значно різняться між собою. У найтеплішому місяці (липні)
середня температура повітря на заході Полісся становить 17-18°С, а
401
в центральному районі і на сході вона вища 18-19°С. Максимальна
температура повітря в цей час у західних районах - 35-36°, а в інших
районах Полісся - 37-39 °С. Річна сума опадів - 550- 625 мм, але в
крайніх південно-західних районах вона місцями досягає 740-770 мм.
Найбільша річна сума опадів - 850-950, а найменша - 300-400 мм.
На Поліссі за період з квітня по липень випадає в середньому 230-
290 мм опадів, а в його крайній південно-західній частині - до 300-
340 мм. Із серпня по жовтень кількість опадів зменшується до 140-
170 мм, і лише на заході вона досягає 170-200 мм. О 13-й год віднос¬
на вологість повітря низька на всій території Полісся - 37-39 %. У цій
зоні найбільш поширені дерново-підзолисті ґрунти, які становлять
близько 70 % усієї території.
Зона помірного виродження картоплі. До неї входять Полтавсь¬
ка, Черкаська, південні райони Київської, Сумської, Хмельницької,
Вінницької, північні райони Кіровоградської та Харківської облас¬
тей. Ґрунтово-кліматичні умови тут є сприятливими для вирощуван¬
ня садивної картоплі. Але протягом вегетаційного періоду спостері¬
гаються різкі зміни метеорологічних факторів, які негативно впли¬
вають на розвиток рослин. Найвищі й одноманітні температури спо¬
стерігаються в липні і серпні. Середня липнева температура
становить 21-22°С, а в окремі дні липня й серпня максимальні тем¬
ператури повітря можуть досягати 36-39°С.
Кількість опадів за рік поступово зменшується у напрямку з пів¬
нічного заходу на південний схід від 600 до 490 мм, а за вегетацій¬
ний період - від 400 до 330 мм. Спостерігається значне коливання
відносної вологості повітря. Зниження її у весняно-літній період (до
30% і нижче) шкідливо позначається на розвитку картоплі, особливо
в період бульбоутворення.
У зоні поширені чорноземні і сірі лісові ґрунти, що утворилися
на карбонатних лісових материнських породах. Вони високородючі і
при достатній вологості забезпечують високі врожаї картоплі. На
Лівобережжі, в заплавах річок Недри, Супою, Трубежа, Ірпеня та
інших є значні масиви болотних ґрунтів. Після проведення належних
меліоративних робіт ці ґрунти з кислою реакцією ґрунтового розчину
використовують для вирощування насінного матеріалу.
Для вирощування картоплі більш придатні північні райони цих
областей.
Зона сильного виродження картоплі. До неї входять Запорізька,
Донецька, Луганська, Одеська, Дніпропетровська, Миколаївська,
Херсонська, південні райони Кіровоградської та Харківської облас¬
тей і Автономна Республіка Крим.
Кліматичні умови зони відзначаються великими коливанням вологи,
температури повітря та ґрунту за періодами і порою року. З півдня на
північ зони кількість опадів збільшується, безморозний період зменшу¬
ється, а температура знижується. Спостерігаються різкі коливання кіль¬
402
кості опадів - від 220 до 500 мм. За період квітень-липень тут випадає
від 170 до 200 мм, а за серпень-жовтень - до 110 мм.
Літній період характеризується високими температурами повітря
без значних змін. Середньомісячна температура липня (найтеплішо-
го місяця) - 20,5-24°С, максимальна температура - 30-42°С. Віднос¬
на вологість повітря (о 13-й год) є найменшою, порівняно з усією
територією України. У травні-серпні вона коливається в межах 40-
55, хоча окремі травневі дні бувають з вологістю меншою 30 %. Усі
зазначені метеорологічні показники зони негативно позначаються на
вирощуванні картоплі без зрошення.
Орієнтовні строки сортооновлення при використанні еліти як ви¬
хідного матеріалу для зони найменшого виродження - через 7-8 ро¬
ків, слабкого виродження - 5-7 років, помірного виродження - 3-5
років і зони сильного виродження - 1-2 роки.
Як зазначалося вище, врожай картоплі зумовлюється не лише
кількістю репродукцій, а ґрунтово-кліматичними, агротехнічними і
фітосанітарними умовами вирощування насіннєвого матеріалу.
Основними вимогами при сортооновленні мають бути стопро¬
центна або близька до неї сортова чистота, відсутність ураження хво¬
робами, високі посівні та врожайні якості. Встановлення єдиних
строків для різних сортів і для всіх господарств зони суперечить біо¬
логічним властивостям та мінливості сортів.
Це питання вивчалося у центральній частині Лісостепу протягом
3-х років (1991-1993 рр.) на чотирьох сортах: Невська, Зов, Гарт, Гат-
чинська [4, 5]. Дані про вплив репродукцій на врожайність картоплі
наведено в таблиці 6.4.
Таблиця 6.4 - Урожайність картоплі залежно від сорту і репродукції
Репродукція
Гатчинська
Невська
Гарт
Зов
ц/га
про¬
цент
ц/га
про¬
цент
ц/га
про¬
цент
ц/га
про¬
цент
Еліта
(контроль)
354
100
340
100
260
100
330
100
І
323
91
305
89
226
84
309
93
II
287
81
263
77
185
71
260
79
ПІ
246
70
226
66
151
58
218
66
IV
246
81
263
77
185
71
260
79
V
203
57
136
40
93
36
125
38
Аналіз даних свідчить про те, що незалежно від сорту з кожною
наступною репродукцією врожайність знижується. Порівняно з елі¬
тою, урожайність І репродукції знизилася на 21-35 ц/га, або на 7-
16%, II - відповідно на 66-77 і 19-30, III - на 108-114 і 30-42, ГѴ - на
140-178 і 40-55, V - на 151-205 ц/га і 43-64 %. Урожайність на кон¬
тролі (еліти) становила 354-260 ц/га, залежно від сорту.
403
Найменше зниження при репродукуванні еліти виявилося в сорту
Гатчинська. Порівняно з елітою, І репродукція його знизила врожай¬
ність на 9%, П - на 19, Ш - на ЗО, IV - на 40 і V - на 43%. Найбільше
зниження відбулося у сорту Гарт: І репродукція - на 16 %, П - на 29,
III - на 42, IV -на 55 і V - на 64 %.
Строки сортооновлення мають визначатися рівнем оргнанізації на¬
сінництва на основі даних апробації. За такого підходу до сортоонов¬
лення садивний матеріал одних і тих самих сортів можна використову¬
вати від двох до восьми років, залежно від умов вирощування.
Сортозаміна - це повна заміна на виробничих насадженнях од¬
ного реєстрованого сорту іншим. Здійснюється вона на основі даних
державного сортовипробування. Як правило, нові реєстровані сорти
істотно перевищують за врожайністю старі. Іноді новий сорт може
бути зареєстрованим, якщо його врожайність буде на рівні старого
сорту, однак з кращими показниками якості продукції (вміст крох¬
малю, білка, придатність до переробки) або стійкості до хвороб і
шкідників.
Швидка сортозаміна - надійний шлях підвищення врожайності.
Прискорене впровадження нових сортів у виробництво безпосеред¬
ньо залежить від роботи всіх ланок насінництва, і навпаки, повільне
здійснення сортозаміни - показник недосконалої організації насін¬
ництва.
Про вплив сорту на врожайність картоплі свідчать матеріали, на¬
ведені в таблиці 6.5.
Таблиця 6.5 - Потенційна врожайність сортів картоплі на різних етапах
сортозаміни
Роки
Сорти, що займали основні площі насаджень
Середня
врожай¬
ність,
ц/га
1
2
3
1921-1930
Імператор Ріхтера, Новий експорт, Сілезія, Вольтман,
Крюгер, Індустрія, Бісмарк, Витязь, Аграрія, Ісполін, Си¬
ній велетень, Юбель, Спаржевий, Меркер і місцеві сорти
240
1931-1940
Деодара, Крюгер, Лотос, Князівська корона, Шеститижне¬
вий, Ранній Маржелан, Рання роза, Епікур, Рожева з
Мілет, Вольтман, Апьма, Лорх, Епрон, Бов’є, Юбель, Коб-
блер, Народна, Стахановська, Рясна, Червоноспиртова,
Роза Полісся, Крепиш, Островська, Поліська 36 і місцеві
сорти
270
1941-1950
Рання роза, Епікур, Елла, Вольтман, Альма, Парнасія,
Карнеа, Лорх, Деодара, Кур’єр, Червоноспиртова, Рясна,
Стахановська, Гренцмарк, Народна, Роза Полісся, Крепиш,
Камераз, Кур’єр, Кобблер, Островська, Поліська 36 та
суміш сортів
305
404
Продовження табл. 6.5
1
2
3
1951-1960
Червоноспиртова, Стахановська, Катюша, Чотирисотка,
Камераз, Мажестік, Октябрьонок, Островська, Рання роза,
Роза Полісся, Елла, Юбель, Лорх, Альма, Поліська 36,
Баранівська, Прієкульська рання, Харківська рання, Агро¬
номічний, Парнасія, Кур’єр, Кобблер, Вольтман та інші
320
1961-1970
Смачна, Чарівниця, Немішаївська ювілейна, Катюша,
Островська, Українська рожева, Прієкульська рання,,Ста¬
хановська, Харківська рання, Юбель, Бородянська, Полі¬
ська 36, Альма, Елла, Рання роза, Камераз, Кур’єр, Львів¬
ська біла, Одеська 24, Пірмунес, Смисловська, Столова 19,
Сулев, Форан, Лорх, Вольтман, Краснооктябрська, Октяб¬
рьонок, Баранівська, Царнікавська, Агрономічний, Чоти¬
рисотка, Олев, Зазерська та інші
350
1971-1980
Баранівська, Білоруська рання, Бородянська, Вармас, Вер¬
ховина, Гатчинська, Гібридна 14, Іскра, Львівська біла,
Огоньок, Олев, Пірмунес, Прієкульська рання, Смачна,
Поліська 36, Харківська рання, Елла, Столова 19, Сулев,
Темп, Фаленська, Царнікавська, Чарівниця, Юбель, Лорх,
Одеська 24, Островська, Катюша, Камераз, Стахановська,
Роза Полісся, Малинова та інші
380
1981-1990
Воротинська рання, Зов, Іскра, Каскад поліський, Пірмунес,
Прієкульська рання, Пригожа 2, Адрета, Жигомирянка, Льві-
вянка, Мавка, Невська, Новинка, Радомишльська, Світанок
київський, Чарівниця, Віхола, Гатчинська, Луговська, Ого¬
ньок, Українська рожева, Витязь, Зарево, Ікар, Поліська роже¬
ва, Полонина, Ласунак, Темп, Древлянка та інші
410
1991-2000
Божедар, Бородянська рожева, Гарт, Зов, Кобза,
Молодіжна, Незабудка, Посвіт, Пролісок, Адрета, Водо¬
грай, Доброчин, Житомирянка, Мавка, Невська, Обрій,
Повінь, Радомишльська, Радич, Світанок київський,
Вітязь, Воловецька, Зарево, Ікар, Поліська рожева, Ракурс,
Древлянка, Ласунак, Темп, Купава, Либідь, Українська
рожева, Придеснянська, Слов’янка, Берегиня, Горлиця,
Ольвія, Слава, Західна та інші
460
Дані вказують на значні потенційні можливості нових сортів. За
минулі 80 років урожайність сортів селекції кінця XX століття зрос¬
ла більш як у два рази, порівняно з зарубіжними і місцевими сорта¬
ми, які вирощувалися на початку століття. Інша справа, що потенціал
сортів використовується погано, через що не реалізовується п’ята
частина їхніх можливостей.
Сортозаміна здійснюється в міру реєстрації нових сортів у най¬
стисліші строки.
Садивний матеріал нових реєстрованих сортів установою-оригінато-
ром передається іншим науково-дослідним установам, навчально-дос¬
лідним господарствам вузів для розмноження в зоні їх діяльності.
405
6.4.1. Оборот насіннєвого матеріалу
Насіннєвий матеріал надходить в оборот після його визнання
державними інспекційними і контрольними органами в насінництві,
що підтверджується відповідними документами встановленого зраз¬
ка, який належить до сорту, що за результатами державного випро¬
бування занесений до Реєстру сортів рослин України або визнаний
перспективним; за сортовою чистотою і посівними якостями відпо¬
відає вимогам відповідних державних стандартів України.
Норми сортових і посівних якостей насіннєвого матеріалу вста¬
новлюються державними стандартами України і визначаються мето¬
дом апробації, ґрунтового контролю і бульбового аналізу.
Обсяги апробації сортових посівів за сортами і репродукціями
визначають виробники насіннєвого матеріалу.
Апробацію сортових посівів здійснюють в порядку, визначеному
“Інструкцією по апробації сортових посівів” (1995 р.).
У випадку невідповідності посіву вимогам державних стандартів
України він вилучається з насіннєвих, а одержаний матеріал не вва¬
жається насіннєвим.
Визначення посівних якостей та аналіз бульб партії картоплі, що
підлягає реалізації, здійснюють спеціалісти Державної насіннєвої
інспекції, а насінного матеріалу для власних потреб виробника - сам
виробник та особи, уповноважені на те Державною насіннєвою ін¬
спекцією.
На насіннєвий матеріал, який відповідає вимогам державного
стандарту України, власник одержує документи про якість, а саме:
акт апробації та акт приймання посівів еліти, акт апробації репроду¬
кційних сортових посівів. Ці документи видаються в порядку, визна¬
ченому Міністерством аграрної політики України.
Насіннєвий матеріал за етапами насінництва підрозділяють на
такі категорії.
Оригінальний насіннєвий матеріал картоплі - бульби первин¬
них ланок насінництва, отримані в розсадниках розмноження супер-
супереліти та супереліти, які за сортовими та посівними якостями
відповідають вимогам цього стандарту.
Елітний насіннєвий матеріал картоплі - бульби, що отримані
від послідовного розмноження оригінальних бульб в елітно-насін-
ницьких та інших господарствах, занесених до Державного реєстру
виробників насіння, і які за сортовими якостями відповідають вимо¬
гам цього стандарту.
Репродукційний насіннєвий матеріал картоплі (перша-
четверта репродукція) - насіннєвий матеріал картоплі, одержаний
від послідовного пересіву бульб еліти і який за сортовими та посів¬
ними якостями відповідає вимогам державного стандарту.
Відтворення еліти. Відтворюють еліту на основі добору кращих
за продуктивністю здорових рослин з випробуванням їхнього вегета-
406
тивного потомства (клонів) або ж використовують як вихідний оздо¬
ровлений біотехнологічними методами матеріал без індивідуального
його випробування в потомстві.
На основі клонів еліту вирощують за такою схемою: 1-й рік - до¬
бір клонів; 2-й рік - розсадник випробування клонів; 3-й рік - розса¬
дник супер-супереліти; 4-й рік - розсадник супереліти; 5-й рік - роз¬
садник еліти.
На основі оздоровленого біотехнологічними методами вихідного
матеріалу (рослини і мікробульби in vitro, міні-бульби) відтворення
еліти може проводитися за три- або чотирирічною схемою.
Трирічна схема відтворення еліти
1-й рік - одержання вихідного матеріалу біотехнологічним мето¬
дом у лабораторних умовах, культиваційних спорудах та закладання
ним того ж року розсадника супер-супереліти в полі; 2-й рік - розсад¬
ник супереліти; 3-й рік - розсадник еліти.
Чотирирічна схема відтворення еліти
1-й рік - виробництво макро- і міні-бульб на основі культивуван¬
ня оздоровлених рослин у культиваційних спорудах або у відкрито¬
му ґрунті в умовах просторової ізоляції від переносників та джерел
вірусної інфекції; 2-й рік - розсадник супер-супереліти; 3-й рік - роз¬
садник супереліти; 4-й рік - розсадник еліти.
На півдні в умовах зрошення при двоврожайній культурі застосо¬
вують таку схему відтворення еліти.
1-й рік. Одержання міні-бульб у культиваційних спорудах на ос¬
нові мікробульб або рослин in vitro. Розсадник випробування (літнє
садіння міні-бульб).
2-й рік. Розсадник розмноження (весняне садіння бульбами з роз¬
садника випробування). Розсадник супер-супереліти (літнє садіння
свіжозібраними бульбами з розсадника розмноження).
3-й рік. Розсадник супереліти (весняне садіння бульбами з розсад¬
ника супер-супереліти). Розсадник еліти (літнє садіння свіжозібра¬
ними бульбами з розсадника супереліти).
Процес відтворення еліти на основі клонового добору та оздоров¬
лених рослин або мікробульб in vitro наведено на рис.6.4.
Одержання вихідного матеріалу біотехнологічними методами.
Для одержання вихідного оздоровленого насіннєвого матеріалу кар¬
топлі застосовують методи термотерапії і хіміотерапії різних моди¬
фікацій в поєднанні з культурою меристеми.
Оздоровлений таким чином вихідний насіннєвий матеріал, залежно
від схеми і методу одержання, розподіляють на вирощений:
- в умовах in vitro (рослини і мікробульби);
- в закритому ґрунті з рослин, мікробульб in vitro (міні- і макро-
бульби);
- у відкритому ґрунті (в польових умовах) з рослин, мікробульб
in vitro.
407
Міні-бульби - це бульби масою 0,5-25 г, одержані при вирощу¬
ванні їх у культиваційниїх спорудах або у відкритому ґрунті в умо¬
вах просторової ізоляції від джерел та переносників інфекції з до¬
триманням заходів, що запобігають повторному інфікуванню бульб
збудниками хвороб картоплі, насамперед вірусних.
Супер-супер-
еліта
Супереліта
Добір здорових
рослин (клонів)
Бракування
уражених хворобами і
нетипових клонів
ОООООООООООО
0000X000X000
0000X00X00X0
Еліта
Видалення уражених
хворобами і
нетипових рослин
Э00000Х00Х00Х000
II - відібрані здорові кущі И - здорові КЛОНИ |jxj - хворі клони
О - здорові кущі х - хворі кущі
Рис. 6.4. Схема добору клонів та відтворення еліти на їх основі
Для одержання вихідного оздоровленого матеріалу у відкритому
ґрунті використовують оздоровлені рослини і бульби, вирощені в
умовах просторової ізоляції, розмножені в закритому ґрунті (культи¬
ваційних спорудах) або одержані від вирощення рослин in vitro, які
перевіряють на наявність вірусів найбільш доступними в насінництві
краплинним та імуноферментним методами серодіагностики.
Одержання мікробульб in vitro дає змогу скоротити схему вироб¬
ництва еліти картоплі до 3-х років (рис.6.5).
При вирощуванні великої партії вихідного оздоровленого матеріа¬
лу, необхідного для насінницької роботи, використовують методи
прискореного розмноження. З метою запобігання повторному інфі¬
куванню оздоровленого матеріалу вірусами застосовують афіциди
для боротьби з активними переносниками вірусної інфекції, відповід¬
ні організаційні заходи та технологічні прийоми.
408
Рис. 6.5. Схема вирощування еліти картоплі з використанням
як вихідного матеріалу мікробульб та рослин in vitro
При створенні еліти клоновим добором, залежно від прийнятої
схеми, закладають відповідні розсадники.
Розсадник відтворення оздоровленого матеріалу та добору
клонів. У розсаднику висаджують матеріал, попередньо оздоровле¬
ний доборами, методом культури верхівкової меристеми або селек¬
ційний насіннєвий матеріал. Меристемний матеріал одержують з на-
уково-дослідних установ. Якщо безвірусний насіннєвий матеріал не
завозять щорічно, то в розсаднику висаджують здорові бульби за ре¬
зультатами серодіагностики.
Розсадники добору клонів закладають в умовах просторової ізо¬
ляції від резерваторів та переносників інфекції, зокрема попелиць.
Вони мають забезпечувати виконання завдання щодо відбору клонів
на заплановані обсяги еліти. При цьому враховують середню кіль¬
кість хворих кущів, що видаляються за період вегетації при фітопа-
тологічному прочищенні.
Усі рослини в розсаднику в період вегетації підлягають візуаль¬
ній оцінці. Проводять також оцінку на приховану вірусну інфекцію.
Після видалення всіх хворих рослин проводять добір клонів.
В окремих випадках добір клонів проводять у розсаднику випро¬
бування клонів, а по сортах, вперше занесених до Реєстру, - на селек¬
ційному матеріалі.
Для виробництва 100 т еліти за п’ятирічною схемою добирають
1000-1200 клонів.
Намічені для добору рослини мають бути добре розвинуті, типові
за морфологічною будовою, здорові за зовнішнім виглядом. Кіль¬
кість стебел у кущі характерна для сорту.
В усіх випадках остаточну оцінку проводять за врожайністю. Для
цього при збиранні врожаю бульби попередньо викладають з кожно-
409
го куща для оцінки і добору клону по гнізду. Добирають ті клони, де
всі бульби в кущі здорові, кількість товарних бульб характерна для
сорту, перехід від великих до дрібних звичайних типовий для основ¬
ної маси здорових рослин.
Відібрані від кожної рослини бульби збирають окремо, в паперо¬
ві чи в поліетиленові мішечки. У поліетиленових мішечках потрібно
зробити отвори діаметром до 1см для дихання бульб. Якщо поліети¬
ленові мішечки використовують повторно, їх дезінфікують 2 %-ним
розчином мідного купоросу або формаліну.
Взимку та на початку весни клони перевіряють методом індекса¬
ції. Перевіряють 1-2 бульби від клону. В теплицях за зовнішніми
симптомами визначають уражені хворобами рослини. Крім візуаль¬
ної оцінки, проводять серологічні аналізи для виявлення вірусів у
латентному стані.
Уражені вірусами рослини вибраковують. Вибраковують і відпо¬
відні їм клони.
Навесні перед садінням кожний клон оглядають при доброму
освітленні. При виявленні хоча б однієї ураженої бактеріальними чи
грибковими хворобами бульби увесь клон вибраковують.
При перегляді клонів поверхню столу і рукавиці періодично дез¬
інфікують, використовуючи 2 %-ний розчин мідного купоросу, фор¬
маліну або розчин вапна.
Здорові клони групують за кількістю бульб: 8-10, 11-12, 13-
14 і так далі, що значно полегшує розміщення їх у полі під час
садіння.
Усі дані про вихідний матеріал, який використовують для закла¬
дання розсадника відтворення оздоровленого матеріалу та добору
клонів, а також результати бракування за період зберігання та оцінки
під час вегетації заносять у спеціальний журнал.
Розсадник випробування клонів. У розсаднику висаджують ві¬
дібрані клони, розміщуючи їх один біля одного смугами, між якими
обов’язково залишаються поперечні доріжки завширшки не менше
1 м. При вирощуванні у розсадниках різних сортів їх висаджують на
відстані не менше 1,4 м.
Доріжки використовують також для проїзду сільськогосподарсь¬
ких машин, вивезення картоплиння під час прочисток та обробок
посівів пестицидами.
Клони висаджують шляхом розкладання бульб у попередньо
нарізані борозни з наступним загортанням їх з утворенням гре¬
бенів.
Схема садіння - 70x35-40 см. При проведенні серо діагностики
клони нумерують. Кілочки встановлюють через кожні 20 рядків і
нумерують порядковим номером ділянки (20, 40, 60, 80 і т.д.).
Спостереження за клонами та перше вибракування починають
при досягненні рослинами висоти 15-20 см; друге - на початку квіт¬
410
ня. Коли починає відмирати бадилля, проводять кінцеву оцінку та
вибракування клонів, після чого збирають урожай.
Щоб зменшити витрати на проведення серологічних аналізів, до¬
цільно спочатку дослідити середній зразок листків кожного клону,
потім перевірити всі кущі безвірусних клонів. Середній зразок фор¬
мують з 3-5 кущів клону. Це дає змогу вибраковувати максимальну
кількість уражених рослин - основного джерела інфекції при наступ¬
ному розмноженні відібраного вихідного матеріалу в процесі форму¬
вання еліти.
При вибракуванні видаляють усі клони, в яких хоча б на одній
рослині виявлено ознаки вірусних хвороб, чорної ніжки, кільцевої
гнилі, а також клони з пригніченими в рості рослинами.
Усі записи оцінки клонів та їх бракування ведуть у спеціальному
журналі.
Розсадник супер-супереліти. У розсаднику висаджують ретель¬
но перебрані бульби, вирощені в результаті випробування клонів,
або посадковий матеріал, одержаний біотехнологічними методами
(рослини та мікробульби від оздоровлених рослин). Садять ретельно
перебраними бульбами. Густота садіння становить 50-60 тис. бульб
на 1 га, залежно від розміру фракції, ширина доріжок між сортами -
1,4 м. Після садіння складають акт на закладання розсадника, зазна¬
чивши спосіб одержання вихідного матеріалу. Посіви супер-
супереліти прочищають 2-3 рази, починаючи тоді, коли рослини до¬
сягають висоти 15-20 см. На кожну прочистку складають акт.
Контрольний аналіз на приховану ураженість вірусами проводять
у фазі бутанізації-цвітіння рослин або в зимово-весняний період, ко¬
ристуючись методом індексації бульб. Для цього із розсадника по
кожному сорту беруть 50 рослин з 1 га, але не більше 200 рослин
(або бульб для індексації). Збирання проводять з попереднім зни¬
щенням картоплиння. Строки раннього збирання встановлюють за¬
лежно від властивостей сорту і зони вирощування.
Розсадник супереліти. Тут висаджують бульби, одержані в роз¬
саднику супер-супереліти. Сортове та фітопатологічне прочищення,
контрольний серологічний аналіз на приховану вірусну інфекцію, а
також оформлення документації, технологічні та насінницькі роботи
проводять так само, як у розсаднику супер-супереліти.
З насіннєвого матеріалу супереліти згідно з методикою відбира¬
ють бульби для ґрунтконтролю.
Розсадник еліти. Вирощують еліту з насіннєвого матеріалу, одер¬
жаного в розсаднику супереліти. Густота садіння - 55-60 тис. бульб
на 1 га. Тут застосовують комплекс технологічних і насінницьких
прийомів, рекомендованих для цієї зони, які сприяють одержанню
насіннєвого матеріалу, що відповідає вимогам державного стандарту
щодо сортових та посівних якостей еліти. Обов’язково проводять 2-3
сортофітопрочистки.
411
ЛІТЕРАТУРА
1. Про карантин рослин: Закон України №3348 - XII; ЗО червня 1993 р. //
Відомості Верховної Ради. - 1993. - №34. - С.352.
2. Постанова Верховної Ради України “Про введення в дію Закону Укра¬
їни “Про насіння”. - Київ, 23 грудня 1993 року.
3. Добірне насіння - золотий фонд України: Інструкція по апробації сор¬
тових посівів (затверджено Мінсільгосппродом України, протокол №11 від
14 березня 1994 р.) // Земля і люди України. - 1995. - №1. - С.22-25.
4. Молоцький М.Я., Кривенко І.С. До питання про сортооновлення кар¬
топлі у центральному Лісостепу України // Картоплярство: Міжвід. темат.
наук. зб. - К, 1994. - Вип.25. - С.72-74.
5. Строки сортооновлення картоплі у центральних районах Лісостепу
України // Вісн. аграр. науки. - 1994. - №5. - С.38-40.
6. Онищенко О.Й. Насінництво картоплі на Україні. - К: Урожай, 1966.
7. Про стабілізацію виробництва картоплі товаровиробниками агропро¬
мислового комплексу України в 2000 році: Наказ Міністерства аграрної по¬
літики. - №31. - К, 2000.
6.5. ОДЕРЖАННЯ ВИХІДНОГО МАТЕРІАЛУ
В НАСІННИЦТВІ КАРТОПЛІ ШЛЯХОМ
КУЛЬТУРИ АПІКАЛЬНОЇ МЕРИСТЕМИ
Особливістю сучасного насінництва картоплі є використання
вихідного матеріалу, оздоровленого шляхом термотерапії та культу¬
ри меристемної тканини, у поєднанні з клональним розмноженням.
Клональне розмноження оздоровленої картоплі покликане забез¬
печувати одержання вільного від хвороб вихідного матеріалу в обся¬
гах, достатніх для потреб первинних ланок насінництва. Застосову¬
ють при цьому як лабораторні методи (мікроживцювання в культурі
in vitro), так і методи прискореного розмноження безвірусних рос¬
лин, головним чином для одержання міні-бульб у культиваційних
спорудах або в польових умовах.
Мікроживцювання в культурі in vitro, або клональне мікро роз¬
множення, полягає в одержанні in vitro нестатевим шляхом здорових
рослин, генетично ідентичних вихідному сорту. У практичній роботі
клональне мікророзмноження здійснюють шляхом живцювання
рослин, одержаних із меристеми, у штучних умовах на живильному
середовищі, мінеральна основа якого модифікована з урахуванням
біологічних властивостей картоплі. Ці методи дозволяють досягти
дуже високих коефіцієнтів розмноження - 1 млн рослин за 120 днів
при клональному мікророзмноженні [48].
Регенеровані з меристеми рослини вільні від найбільш пошире¬
них і шкідливих збудників хвороб картоплі, передусім від вірусів.
Навіть при наявності вірусної інфекції в них, що знаходиться за ме¬
жею чутливості методів діагностики, такі бульби є цінним насіннє¬
412
вим матеріалом, оскільки під час їх репродукування синтез вірусного
білка в рослинах відбувається повільно, у результаті чого уповільню¬
ється процес накопичення вірусної інфекції [12, 49, 53].
У результаті цього оздоровлений активним способом насіннєвий
матеріал відзначається на перших етапах його використання підви¬
щеними продуктивними якостями [26].
У країнах із розвинутим картоплярством питанню забезпечення
насінницьких господарств оздоровленим матеріалом, який одержу¬
ють біотехнологічним методом, приділяється значна увага. Застосу¬
вання цього методу дозволяє прискорити більш ніж у два рази про¬
цес розмноження нових сортів і оздоровлювати при потребі ті, що
знаходяться у виробництві. Технологія розмноження постійно вдос¬
коналюється і має кілька модифікацій, основою яких є процес бага¬
торазової регенерації пробіркових рослин. У таких країнах, як Авст¬
ралія, Нідерланди, Франція практикують послідовне багаторазове
живцювання пагонів, одержаних із оздоровлених бульб, із проміж¬
ним укоріненням живців у ґрунті після кількох пасажів, що підвищує
їхню життєздатність. В інших країнах (Англія, Канада, СІЛА) поши¬
рена багаторазова регенерація (мікророзмноження) рослин у пробір¬
ках без проміжного укорінення рослин у ґрунті. Одержану в лабора¬
торії необхідну кількість живців укорінюють у теплиці і висаджують
у поле.
Значна увага в Англії та Франції приділяється також одержанню
мікробульб у пробірках, що дозволяє виробляти їх цілорічно.
Першою країною у світі, що перейшла на одержання всього са¬
дивного матеріалу картоплі методом культури тканини, була Да¬
нія. У країні з 1976 р. протягом 10 років здійснювалася програма,
спрямована на забезпечення всіх товарних посадок картоплі оздо¬
ровленим насінним матеріалом, що було викликане завезенням і
розповсюдженням у країні інфекції та забороною виходу у зв’язку з
цим на міжнародний ринок. По завершенні програми весь садивний
матеріал було приведено у відповідність до вимог європейського
ринку.
Метод культури тканини є основою процесу насінництва і в Ки¬
таї, а в Японії оздоровлення картоплі від вірусів цим методом здійс¬
нюється з 1964 р.
В Україні на сьогодні також проведена значна робота з одержан¬
ня оздоровленого матеріалу картоплі біотехнологічним методом. Зо¬
крема, в Інституті картоплярства УААН зосереджений генетичний
банк оздоровлених вітчизняних і закордонних сортів. На основі цьо¬
го матеріалу щорічно виробляється 1,0-1,2 млн шт. розсади, мікро-
та міні-бульб [43].
Раціональне використання оздоровленого матеріалу є важливим
чинником підтримання продуктивних якостей сорту в системі насін¬
ництва картоплі. У першу чергу це стосується способів прискореного
413
розмноження оздоровлених рослин та мікробульб in vitro та раціональ¬
них прийомів і методів формування вихідного матеріалу в елітному
насінництві.
Із прискорених методів розмноження найбільш доцільним щодо
одержання вихідного матеріалу в елітному насінництві вважається
мікророзмноження, яке має значний коефіцієнт розмноження. При¬
чому, при використанні цього методу значно меншою є вірогідність
повторного ураження матеріалу вірусами, порівняно з розмноженням
його стебловими живцями або бульбами, оскільки основна частина
операцій здійснюється в лабораторних стерильних умовах. Проте це
у свою чергу підвищує вартість одержаного насіннєвого матеріалу
[31,9].
Мікроживцювання рослин. Здебільшого цей метод застосову¬
ють при розмноженні перших здорових рослин, одержаних із верхів¬
кових меристем.
Розмножувати оздоровлені рослини можна на агаризованому
(твердому) середовищі у стерильних умовах або на рідкому середо¬
вищі в нестерильних умовах. Рослини з 5-8 листками пінцетом вий¬
мають із пробірок і на простерилізованій чашці Петрі гострим скаль¬
пелем або лезом безпечної бритви розрізають на живці, включаючи
частину стебла з одним листочком. На старому живильному середо¬
вищі можна залишити частину стебла з кореневою системою для пов¬
торного відрощування. Повторне використання живців з кореневою
системою і одним нижнім міжвузлям та пазуховою брунькою для
одержання нового рослинного організму дозволяє одержувати з них
рослини, що за своїм розвитком значно переважають рослини, одер¬
жані від знятих із них живців, за рахунок збільшення довжини стебел
і кількості міжвузлів.
Таке повторне живцювання маточної рослини можна здійснюва¬
ти тричі і одержати понад ЗО живців, тоді як при одноразовому жив¬
цюванні - лише п’ять. За три місяці з чотирьох вихідних рослин одер¬
жують тисячу рослин [43].
Відокремлені живці висаджують у пробірки із живильним сере¬
довищем на глибину міжвузля. Інструменти і чашку Петрі стерилі¬
зують перед живцюванням кожної рослини. Вирощують рослини із
живців при температурі 20-23°С, відносній вологості 70-80 %, освіт¬
ленні люмінесцентними лампами із силою світла 3-4 тис. лк і при
16-годинному світлоперіоді. На 3-4-й день після садіння в живців
починається ріст стебла і коріння. Через 12-15 днів рослини повніс¬
тю відростають і готові для повторного живцювання.
Для вирощування рослин із живців використовують модифікова¬
не середовище Мурасіге-Скуга. Застосовують також рідке живильне
середовище з мінеральною основою за Мурасіге-Скугом. На такому
середовищі живці відростають через 20-25 днів. При цьому для пов¬
торного живцювання використовують різні поживні середовища з
414
1-2 %-ним умістом цукру, а для висаджування в теплиці вміст його
необхідно збільшити до 4 %, освітлення забезпечувати упродовж
16 год, застосовуючи його періодично.
Після того, як із живців одержують необхідну кількість рослин у
пробірках, їх пересаджують у горщики або безпосередньо на добре
зволожений живильний субстрат у теплиці (торф, суміш торфу, піску
і ґрунту 3:1:1, удобрений зволожений ґрунт).
При пересаджуванні для доброго приживлення рослин, одержаних іп
vitro, що переносяться із штучного живильного середовища на субстрат,
необхідно запобігти травмуванню рослин, повністю очистити корені від
живильного середовища, укрити рослини на кілька днів (3-5) після пе¬
ресаджування склянками або плівкою, щоб забезпечити підвищену во¬
логість під укриттям. У перші дні приживлення необхідно також затіня¬
ти рослини, а надалі їм потрібне достатнє освітлення, що сприяє кращо¬
му розвитку листкової поверхні і стебла. Рослини із горщиків через
12-15 днів пересаджують у ґрунт теплиць. Такий спосіб розмноження
дозволяє упродовж 3—4 місяців одержати 2-3 тис. рослин, придатних
для пересаджування в поле, а за 8-10 місяців від однієї рослини можна
одержати близько 20 тис. бульб [ 6,43 ].
Метод розсади. Одним із способів розмноження рослин in vitro є
метод розмноження розсадою, який забезпечує прискорення їх росту
і розвитку, покращення умов адаптації, а в результаті цього - підви¬
щення їхньої продуктивності.
Для одержання розсади застосовують касети з комірками
об’ємом 65, 190 і 300 см3, заповненими торфоперегнійним субстра¬
том, а також спеціальні гідропонні установки “Мінівіт-2”. Оптималь¬
ним строком вирощування розсади із пробіркових рослин in vitro на
установці “Мінівіт-2” є 10 днів, у касетах - 20-25 днів. На установці
“Мінівіт-2” висаджені пробіркові рослини адаптуються протягом
кількох годин.
Застосування розсадного способу дозволяє на 25-30 % підвищи¬
ти урожай картоплі і на 15-17 днів прискорити дозрівання бульб.
Найбільш ефективний об’єм живлення розсади - 125-150 см3 [45].
Застосовують також живцювання рослин in vitro.
Живці укорінюють також у пастеризованих ґрунтових горщиках,
які розміщують на лотках. Живці обприскують водою, поміщають у
поліетиленовий пакет, який потім заклеюють і переносять у теплиці
під навіс із затіненням (до 60 %) і температурою вдень 24°С, уночі -
19°С (по 12 год). Через 10-14 днів плівку знімають, а коли більшість
живців дадуть паростки, їх переносять в умови затінення (до 30-50 %),
потім ці паростки живцюють. За кожні два тижні утворюється новий
паросток. Цю операцію повторюють необхідну кількість разів. Жив¬
ці укорінюють через 8-10 днів, а через 3-5 тижнів розвиваються мі¬
цні рослини заввишки 10—15 см. Перед садінням у поле укорінені
живці витримують 4-6 тижнів на повному освітленні, що сприяє
415
кращому розвитку листкової поверхні і стебла. Через 2-2,5 місяці
після висаджування їх у поле збирають урожай [29].
Для покращення приживання рослин in vitro в теплицях засто¬
совують іонітні субстрати, що мають збалансований склад мікро- і
макроелементів у вигляді катіонів і аніонів, які є добрим живильним
середовищем для укорінення рослин. Такий субстрат забезпечує не-
лімітоване саморегулююче мінеральне живлення рослин, що дозво¬
ляє значною мірою реалізувати продуктивність оздоровлених рос¬
лин. Маса бульб при вирощуванні рослин на таких субстратах збіль¬
шувалася на 41-55 %, а їх кількість - на 111-127 %, порівняно з кон¬
тролем, де як субстрат використовували торф. Кількість мінібульб
розміром 25-30 мм збільшувалася з 27 % у контролі до 56 % - у дос¬
лідному варіанті [46].
Найбільшу кількість бульб на такому субстраті одержують від жи¬
вців першого строку знімання з маточної рослини. Від укорінених жи¬
вців першого строку одержують 13 бульб з куща, третього - 5 штук.
При цьому середня маса бульб у першому випадку становила 20-25 г,
а від живців 2-го і 3-го строків знімання - відповідно 54 і 68 г [24].
З метою використання пробіркових рослин in vitro як вихідного
матеріалу в елітному насінництві, їх розмножують у культиваційних
спорудах у вересні-квітні, завдяки чому накопичується бульбовий
матеріал, достатній для закладання розсадника вихідного матеріалу.
Зокрема, при живцюванні 55 пробіркових рослин in vitro з верес¬
ня до квітня наступного року можна накопичити до 50000 бульб.
Накопиченого бульбового матеріалу вистачає для садіння на
площі близько 1 га. У цьому розсаднику матеріал оцінюють на наяв¬
ність вірусної інфекції з одночасним вибракуванням уражених рос¬
лин (табл. 6.6).
Підтримання у здоровому стані і розмноження оздоровленого ма¬
теріалу в подальшому здійснюється з дотриманням заходів, які попе¬
реджують інтенсивне вторинне ураження найбільш поширеними і
шкідливими патогенами картоплі, у першу чергу - вірусними [6, 24].
Висаджування в поле 50640 бульб на площі 1 га здійснюють
10.05. У період бутонізація-цвітіння проводять оцінку рослин на на¬
явність вірусів (візуально та методом серодіагностики).
Одержання мікробульб in vitro. Доведено, що при вирощуванні
рослин in vitro в теплицях вони можуть уражуватися їх вірусами,
оскільки при цьому не можливо повністю позбутися розповсюджу¬
вачів вірусів - попелиць і білокрилок. Крім того, у теплицях рослини
сильно перегріваються, що також негативно позначається на якості
насіннєвого матеріалу.
У дослідах Інституту картоплярства УААН з колекцією оздоров¬
лених сортозразків картоплі, що включає понад 150 сортів і гібридів,
виявлено, що 95% із них утворюють мікробульби за 2 місяці. В ін¬
ших бульбоутворення відбувається за триваліший період. Прискорити
416
Таблиця 6.6 - Використання оздоровлених рослин in vitro для формування
розсадника вихідного матеріалу в насінництві
Пробіркові рослини
Рослини, висаджені в
ґрунт
Збирання врожаю
втеплиці
дата
шт.
дата
шт.
дата
шт.
15.09
55
5.10
330
25.10
1980
1.11
1440
15.01
8640
1.11
440
20.11
2640
25.11
2440
15.02
14640
25.11
200
20.12
1200
20.12
720
15.03
4320
25.12
480
20.01 насту¬
пного року
2400
20.01
960
20.01
1440
1.04
8640
15.02
5760
20.02
2400
30.04
14400
3360(для
розсади)
Усього бульб
у теплиці
50640
Примітки: Для розрахунків узято: а) коефіцієнт розмноження при живцюванні
рослин у пробірках = 6; б) коефіцієнт розмноження при одержанні урожаю із пробір¬
кових рослин у теплиці - 6, у польових умовах - 8
цей процес можна шляхом оптимальної взаємодії основних факторів,
що стимулюють його: оптимального вмісту в поживному середовищі
вуглеводів і біологічно активних речовин, оптимальних величин фо¬
топеріоду і температури [42]. У зв’язку з цим застосовують розмно¬
ження оздоровленої картоплі мікробульбочками, одержаними у
пробірковій культурі з використанням спеціальних методів, які
індукують утворення мікробульб. Для індукції мікробульб у про-
бірочних рослин in vitro використовують низку чинників: збагачене
мінеральними елементами середовище, підвищений уміст (4-8 %)
цукру і кінетину (до 1 мг/л), скорочений фотоперіод на певному
етапі росту рослин, біологічно активні речовини. Оптимальне спів¬
відношення цих чинників дозволяє одержати повноцінні мікро-
бульби діаметром 0,7-0,8 см у 80-85 % висаджених живців. Такі
мікробульби забезпечують одержання рослин, які формують у се¬
редньому 10-12 бульб загальною масою 580-680 г [33, 37, 38].
Для збільшення розміру мікробульб до 9-11 мм рослини in vitro
культивують у пробірках розміром 20-200 мм за наявності в живильно¬
му середовищі 6 % сахарози та активованого вугілля і pH 4,7-5,6 [18].
Перевагою цього способу є також підвищена продуктивність мік¬
робульб у відкритому ґрунті, порівняно з розсадною культурою. При
застосуванні мікробульб для одержання врожаю в полі відпадає по-
|4 2-132
417
треба в теплицях, у 4—5 разів зростає інтенсивність використання об¬
ладнання на етапі розмноження в культурі in vitro. Мікробульби є ви¬
сокорентабельним і малооб’ємним садивним матеріалом [32, 39, 8].
Висота рослин, одержаних із мікробульб, менша, ніж звичайних
бульб середнього розміру, на 25-30 %. У структурі врожаю таких
рослин спостерігається збільшення фракції дрібних бульб: 25 г - 48-
53 % за кількістю і 10-17 % - за масою. У наступних бульбових ре¬
продукціях рослини не відрізняються від звичайних [37].
Елементи технології використання мікробульб. Після досяг¬
нення мікробульбами розміру 0,8-1,8 см і відмирання рослин їх вий¬
мають із пробірок, відмивають від агару в дистильованій воді і з ме¬
тою попередити висихання зберігають у колбах із застиглим від роз¬
плавлювання агаром, укритих щільним папером. Колби закривають
пробками з вати або целофановим папером і зберігають у холодиль¬
нику при температурі 2-4°С.
Після закінчення періоду спокою бульби зберігають при такій же
температурі, але з освітленням - для того, щоб попередити витягу¬
вання паростків. Перед садінням бульби упродовж 10-20 днів про¬
грівають при температурі 10-20°С. При потребі період спокою бульб
можна перервати.
Висаджують мікробульби в добре підготовлений грунт у поперед¬
ньо сформовані гребені на глибину 4-8 см. Максимальну врожай¬
ність із мікробульб одержують при густоті садіння 95 тис./га [25].
У технології вирощування картоплі важливе значення має забез¬
печеність вологою, особливо в перший період розвитку рослин, ре¬
тельне виконання всіх заходів, рекомендованих для насінницьких
посівів, включаючи захист від шкідників та хвороб [37].
Використання мікробульб як садивного матеріалу збільшує коефі¬
цієнт розмноження в 1,5-2,0 рази, порівняно з іншими способами са¬
діння (розсадою, укоріненням верхівкових живців або столонів). По¬
єднання цього методу із способом прискореного вирощування мікро¬
бульб дозволяє збільшити загальний вихід першого бульбового поко¬
ління в 3-4 рази, порівняно із загальноприйнятою технологією [20].
Разом з тим актуальною залишається проблема схожості мікро¬
бульб в екстремальних посушливих погодних умовах та догляду за
посівами в полі.
Одержання міні-бульб від оздоровлених рослин in vitro. В остан¬
ні роки досить широко в оригінальному насінництві як вихідний матері¬
ал використовують міні-бульби з масою менше 25 г. Міні-бульби одер¬
жують при культивуванні рослин in vitro як у культиваційних спорудах
на різних сипучих субстратах і в гідропонній культурі, так і в польових
умовах від рослин і мікробульб in vitro. Міні-бульби одержують також із
паросткових або стеблових живців від безвірусних бульб.
Застосування міні-бульб пов’язане також із тим, що за при¬
скореного розмноження рослин картоплі шляхом живцювання у про¬
418
бірках слабкою ланкою залишається перехід від культури на стериль¬
ному живильному середовищі до вирощування в ґрунті. Коренева
система, що утворюється в пробірці, майже повністю відмирає в ґрун¬
ті й розвивається нова, внаслідок чого рослини довго хворіють і від¬
стають у рості та розвитку. Використання міні-бульб дає можливість
уникнути впливу цього негативного чинника, оскільки в теплиці мо¬
жна підтримувати сприятливі умови не тільки для росту, але й розви¬
тку рослин. Це дозволяє вибракувати вже на початку репродукування
оздоровленого матеріалу максимальну кількість інфікованих рослин
- основного джерела вірусної інфекції при подальшому розмноженні
цієї партії оздоровленого насіннєвого матеріалу.
Мін-бульби мають перевагу і відносно мікробульб, висаджених у
ґрунт. При культивуванні мікробульб у польових умовах їх необхід¬
но висаджувати на порівняно легких за механічним складом ґрунтах.
При цьому слід ураховувати те, що при інтенсивних опадах ґрунт
ущільнюється, що не дозволяє одержати повноцінні сходи. В умовах
засухи також одержують зріджені сходи. Розвиток рослин від мікро¬
бульб розтягнутий у часі, що при інтенсивному льоті попелиць ство¬
рює загрозу значного їх інфікування вірусами. Зростає загроза ура¬
ження насаджень і фітофторозом.
Навіть міні-бульби розміром 10-20 мм здатні давати в польових
умовах урожай, що забезпечує одержання насіннєвих бульб від 80 до
280 тис. шт./га. При використанні як садивного матеріалу міні-бульб
розміром 20-30 мм одержують до 300 тис. шт./га, а 30-35 мм - до
400 тис. шт./га насіннєвих бульб [23].
Найкраще вирощувати міні-бульби в культиваційних спорудах,
тобто в контрольованих умовах, де є можливість створити оптималь¬
ні умови для рослин in vitro, попередити вторинне інфікування їх
різноманітними патогенами. Використання культиваційних споруд
дозволяє створити умови для накопичення протягом осінньо-
зимового періоду значної кількості міні-бульб і здійснити необхід¬
ний комплекс передпосадкових заходів, що також підвищує життє¬
здатність бульб у відкритому ґрунті. Водночас у тепличних умовах
створюються сприятливі умови не тільки для росту, але й для розви¬
тку рослин. Крім того, в інфікованих рослинах, не виявлених раніше,
відбувається інтенсивне нагромадження вірусів до концентрації, яка
дозволяє існуючими методами виявити їх у латентній формі.
У світовій науці й практиці нагромаджено чималий досвід ви¬
рощування насіннєвої картоплі на основі оздоровлених міні-бульб.
Так, у Нідерландах одержане бульбове потомство від рослин in vitro
прирівнюються до клонового матеріалу. З них вирощують супер-
супереліту. Загалом у країнах Західної Європи щорічно вирощують
приблизно 800 тис. шт. пробіркових рослин, в США і Канаді - 580 тис.
штук. Технологія одержання бульб у культиваційних спорудах достат¬
ньо відпрацьована. В Ірландії міні-бульби одержують при садінні мік-
419
ророслин із щільністю 400 шт./м2 у стерилізований торфокомпостний
субстрат. Кожна рослина за 90 днів дає приблизно 5 бульб діаметром
до 25 мм і масою близько 5 г. За рік можна отримати три генерації.
При садінні міні-бульб урожайність становить приблизно 42 т/га, у
тому числі - 32 т/га бульб розміром понад 30 мм [27].
У стерильних лабораторних умовах міні-бульби одержують та¬
кож шляхом живцювання паростків після тривалого пророщування
бульб. Для цього живці у великій кількості (до 500 на 1 м2) виса¬
джують на торф’яну грядку в теплиці. Використовують різноманітні
суміші хімічних регуляторів росту, що дозволяє контролювати темпи
розвитку рослин і стримувати формування бульб. Через десять тиж¬
нів кожна рослина картоплі дає до десяти бульб розміром від 5 до 15 мм,
тому з 1 м можна одержати до 2,5 тис. таких бульб. При вирощуван¬
ні чотирьох урожаїв на рік продуктивність 1 м становить 10 тис. мі-
ні-бульб. Оскільки за таким показником, як цінність садивного мате¬
ріалу, 15 кг міні-бульб еквівалентні тонні звичайного насіннєвого
матеріалу, достатню кількість бульб для садіння на одному гектарі
дають 5 м2 торф’яної грядки. Собівартість міні-бульб і звичайного
насіннєвого матеріалу суттєво не відрізняється [36].
Міні-бульби в теплицях одержують і з мікробульб in vitro, застосо¬
вуючи при цьому укорінення верхівкових та пазушних бруньок. Коефі¬
цієнт розмноження при такому способі становить 1:700 [40,36].
Для одержання міні-бульб у теплицях використовують також від¬
садки, верхівки і бокові пагони. Ефективне також багаторазове зби¬
рання бульб з-під рослини протягом вегетації. Підкопування бульб
починають тоді, коли під рослиною вже сформувалися 1-3 бульби
діаметром близько 3 см. Наступне збирання здійснюють через 15-20
днів після попередніх, у міру нагромадження бульб. За таких умов
коефіцієнт розмноження підвищується в 1,5-2 рази [37].
Водночас слід зазначити, що міні-бульби. одержані в теплицях
(5-10 шт. від однієї рослини, 150-300 шт./1 м ), відзначаються знач¬
ною неоднорідністю за масою. У структурі урожаю тепличних міні-
бульб найбільший відсоток становлять бульби з масою від 5 до 25 г
[2, 5]. Різниця міні-бульб за масою суттєво не впливає на їхні насінні
якості.
Результати імуноферментного аналізу рослин, одержаних від теп¬
личних міні-бульб масою 5-10 г, 10—20 г, 20-30 г, при польовому
випробуванні та їх візуальна оцінка не виявили суттєвих відміннос¬
тей залежно від маси. Ураженість рослин вірусами L і Y становила
1,2-2,5 %, вірусами X і М - 2,5-6,0 % [4].
У Російській Федерації міні-бульби, які використовуються в еліт¬
ному насінництві, повинні мати в найбільшому поперечному діамет¬
рі 1-3 см, що відповідає масі бульб від 5 до 25 г. Технологія одер¬
жання таких бульб дозволяє на 500 м2 теплиці виростити 180-200
тис. міні-бульб, використання яких у розсаднику супер-супереліти на
420
площі 4 га забезпечує виробництво еліти картоплі обсягом 1000 т за
трирічною скороченою схемою [4].
Необхідність висаджування у відкритому ґрунті значної кількості
оздоровлених бульб пов’язана з тим, що на невеликих ділянках здо¬
рові рослини порівняно швидко уражуються вірусами. Швидкість
ураження обернено пропорційна розміру ділянок, віддаленості
останніх від джерел інфекції, а також кількості й активності перенос¬
ників вірусів.
Так, у дослідах в умовах Пушкінської лабораторій ВІР здоровий
вихідний матеріал сорту Детскосєльська при садінні дворядковими
ділянками (по 40 бульб) вже після однорічного репродукування у
безпосередній близькості від хворих колекційних зразків картоплі
мав до 90 % інфікованих рослин; у середині поля площею 5 га серед
посівів товарної картоплі - відповідно 30-43 %, а серед здорових
клонів другого року випробування на площі 2 га - тільки 5-8 % рос¬
лин із латентною формою інфекції. Зовнішньо хворих рослин не бу¬
ло виявлено [21].
Незважаючи на те, що урожайність міні-бульб (3-5 г) під час ви¬
рощування їх у розсаднику першого року з використанням механізо¬
ваного збирання менша на 71,1%, а при ручному збиранні - на 63,8 %,
ніж урожайність макробульб (30-50 г) під час вирощування їх у роз¬
саднику першого року, при подальшому репродукуванні суттєвих
відмінностей у параметрах надземної і підземної частини куща не
спостерігалося [7].
Для одержання міні-бульб застосовують також живцювання ма¬
точних рослин, що вирощуються в теплицях із рослин in vitro. З од¬
нієї маточної рослини зрізають до 100 живців. Формування бульб на
живцях відбувається через 1-1,5 місяці після садіння. 3 1м2 теплич¬
ної площі одержують 500 і більше бульб розміром 1-3 см [27, 50].
Запропоновано також спосіб вирощування міні-бульб на торфо-
цеолітовому субстраті, для формування якого використовують цеоліт
(20 %) і торф (80%). Із стелажних теплиць на торфоцеолітовому суб¬
страті міні-бульби збирають у міру їх наростання. Вихід міні-бульб з
1 м2 становить 350-760 шт. Схема садіння на стелажах зимових теп¬
лиць - 15...20х40 см, а для сортів, що утворюють значну масу картоп¬
линня, - 15...25x40 см. Товщина шару субстрату - 10 см. Рослини
підгортають двічі, а після другого разу їх уздовж рядків закривають
світлонепроникною плівкою. У період бутонізації та на початку цві¬
тіння через кожні 2-3 дні гребені розмивають несильним струменем
води із шланга і збирають міні-бульби.
Використовують торфоцеолітовий субстрат і в спеціальних уста¬
новках, що забезпечують необхідний мікроклімат для одержання мі-
ні-бульб, які мають розмір 4,6x4,4x4,25 м і розміщуються в теплиці.
Конструкція установок являє собою циліндр, що обертається навколо
вертикальної осі, по його периметру розміщуються лотки з рослина¬
421
ми. Лотки - це ящики, що закриваються кришками і зсуваються до
центру. їх можна легко переставляти. У кришках є отвори для вихо¬
ду стебел. Більший коефіцієнт розмноження досягають у сортів, у
яких стебло в нижній частині краще галузиться.
З 1 м2 стелажних теплиць на торфоцеолітовому субстраті міні-
бульб збирають у міру їх наростання в 4-6,5 рази більше (350-760
шт.), ніж при садінні з максимальним загущенням (10x10 см) і одно¬
часним збиранням. Міні-бульби масою понад 10 г добре зберігають¬
ся - на 80 % і більше. Польова схожість міні-бульб з такою масою
перевищувала 80 %.
Використання сипучих субстратів із періодичним зволоженням
поживним розчином не тільки значно зменшує енергетичні затрати,
але й попереджує можливість поширення збудників вірусних хвороб,
а також віроїда веретеноподібності бульб, що є можливим при засто¬
суванні гідропоніки [19].
Використання міні-бульб в елітному насінництві при формуванні
вихідного матеріалу має певні особливості щодо агротехніки їх ви¬
рощування в розсадниках випробування в польових умовах. Перш за
все - це ретельний передсадивний обробіток ґрунту, який повинен
забезпечити достатнє розпушення на глибину оранки з наступною
культивацією та загущене садіння, враховуючи біологічні влас¬
тивості сорту.
Так, садіння міні-бульб двох фракцій за схемою 70x15 сприяло
зростанню врожайності сорту Жуковський ранній на 58-80%, сортів
Світанок київський і Росинка - відповідно на 30-32 і 28-49 %, порів¬
няно із схемою 70x35 см. Вихід стандартних насіннєвих бульб (30-
60 мм) за схеми садіння 70x15 зростає по сортах: Жуковський ранній
- у 2,9-3 рази, Світанок київський - в 1,8-2 , Росинка - у 2-3 рази,
порівняно із схемою 70x35 см [22].
Міні-бульби одержують також шляхом гідропонної культури.
Культивування гідропонним способом у проточній культурі рослин,
одержаних in vitro для промислового виробництва міні-бульб, є тех¬
нологічно контрольованим процесом. Це дозволяє одержувати пла¬
нову кількість міні-бульб необхідного розміру, активно керувати
технологічним режимом культивування і його автоматизацією, знач¬
но зменшити ймовірність ураження рослин фітопатогенними мікро¬
організмами, грибами, а також шкідниками, які розвиваються в сип¬
ких субстратах. Гідропонні бульби мають високу енергію пророс¬
тання та схожість, добру лежкість, стійкість до пошкоджень при
транспортуванні. Завдяки однорідності бульб за розміром їх виса¬
джування у ґрунт легко механізувати [29].
Виробництво міні-бульб за гідропонною технологією дозволяє
значно збільшити одержання вихідного оздоровленого матеріалу для
первинного насінництва. Так, на Чернігівській дослідній станції по
картоплі за вегетаційний період з однієї рослини на такій установці
422
одержують у середньому по сортах по 19 міні-бульб, а у сортів Сед-
нівська рання, Зов, Невська - від 60 до 115 міні-бульб. Міні-бульби
розміром 10-15 мм збирають через 3-5 днів. Багаторазове збирання
міні-бульб стимулює у рослин процес бульбоутворення [29].
На насінницькій станції “Анвек” у Франції в умовах гідропоніки
укорінення живців у горщиках дозволило прискорити розмноження
цінних сортозразків, підтримувати у здоровому стані насінницькі
насадження [34].
На сьогодні вже розроблені установки для прискореного розмно¬
ження рослин in vitro, а також налагоджено випуск у заводських умовах
спеціального устаткування для промислового виробництва гідропонних
міні-бульб від оздоровлених рослин картоплі. Розміщують такі гідро¬
понні установки в закритому приміщенні або в теплицях. Так, продукти¬
вність установки, розробленої Центром “Дока” (Москва-Зеленоград) за
3,5 місяці вегетації рослин складає близько 1650 міні-бульб з 1 ni із се¬
редньою масою 5 г. При культивуванні рослин in vitro на таких установ¬
ках забезпечується вільний доступ до кореневої зони рослин для пері¬
одичного збирання кондиційних міні-бульб.
Розроблені також установки для прискореного мікроклонального
розмноження рослин in vitro з метою подальшого культивування їх у
гідропонній культурі. Принцип побудови гідропонної установки і
установки прискореного мікроклонального розмноження модульний,
що дозволяє розміщувати їх у приміщенні з будь-якою заданою пло¬
щею. Управління технологічними процесами - автоматичне.
Одна з таких кондиціонерних установок, що виготовляється акці¬
онерним товариством “Дока”, має таку технологічну характеристику:
виробництво міні-бульб за одну вегетацію становить 200 тис. шт.; за
рік - 600 тис. шт.; виробництво пробірочних рослин за добу - 703 шт.;
кількість вегетацій за рік - три; енергоспоживання за добу - 90 квт./год.;
потреба води за добу: питної - 0,5 м3, технічної - 0,9 м3; сировина -
технічні мінеральні солі, витрати за вегетацію - 20 кг. Площа однієї
ізольованої кондиціонерної камери, де розміщується така установка,
становить 154 м2, корисна площа - 132 м2. Необхідна площа, де роз¬
міщується установка прискореного мікроклонального розмноження
рослин, становить 20 м2, приміщення для лабораторії - 35 м2, для
зберігання міні-бульб з автоматичним забезпеченням відповідного
режиму - 12 м2 [13].
Технологія виробництва оздоровлених міні-бульб картоплі включає
три основні етапи: 1-й етап - із апікальних меристем регенерують 10-15
оздоровлених рослин in vitro; 2-й етап - методом живцювання і культу¬
рою in vitro розмножують їх до необхідної кількості; 3-й етап - рослини,
одержані in vitro, висаджують для одержання міні-бульб (рис. 6.6).
Частину вирощених на другому етапі пробіркових рослин знову
використовують для живцювання. Цей процес може повторюватись
багаторазово, завдяки чому оздоровлення (1-й етап), хоч і потребує
423
б і г д
Рис. 6.6. Схема оздоровлення та розмноження вихідного матеріалу
картоплі: а - відібрана візуально здорова рослина картоплі; б - діаг¬
ностика індексацією наявності в бульбі вірусної інфекції; в - термотерапія
бульб (прогрівання пророслих бульб при температурі 36-38°С); г - виді¬
лення меристем і одержання рослин-регенерантів; д - діагностика вірусної
інфекції в рослинах-регенерантах; е - розмноження здорових рослин жив¬
цями у пробірках; є - розмноження безвірусних рослин у теплиці, діагнос¬
тика ураження рослин вірусною інфекцією; ж - розмноження вихідного
матеріалу в полі
високої кваліфікації персоналу і займає багато часу (6-8 міс.), але не
лімітує виробництво. У цій промисловій установці для прискореного
мікроклонального розмноження проводиться підживлення рослин
вуглекислим газом, що забезпечує оптимальні умови для газообміну
й активізує автотрофний ріст. У результаті цього час вирощування
рослин скорочується до 2-х днів.
Пробірки з рослинами розміщують у спеціально сконструйова¬
них касетах по 96 игг. у кожній. При використанні конвеєрного ре¬
жиму за добу одержують 700 рослин картоплі.
424
Для виробництва міні-бульб використовують гідропонний метод
вирощування рослин із періодичним притоком живильного розчину.
Із міні-бульб розміром 1,5-2 см, вирощених на гідропонній уста¬
новці, одержують типові для сорту кущі. Форма бульб, забарвлення
шкірки і м’якоті повністю відповідають сорту. Крохмалистість і
стійкість бульб проти фітофторозу також є типовими для сорту.
Урожай із куща від гідропонних міні-бульб складає 659-586 г, від
тепличних міні-бульб - 405 г [41].
Із збільшенням маси міні-бульб з 6-10 до 11-20 г зростала про¬
дуктивність щодо кількості бульб під кущем та їхня маса. Відповідно,
урожайність від гідропонних міні-бульб становила 465 і 587 г/кущ, від
тепличних міні-бульб - 388 і 485 г./кущ. Гідропонні міні-бульби масою
5-30 г без розподілу на фракції також сформували більш високий уро¬
жай (536 г/кущ), порівняно з тепличними бульбами (455 г/кущ). Кіль¬
кість бульб під кущем в обох випадках суттєво не відрізнялася [3].
Разом з тим при використанні міні-бульб у насінництві картоплі не¬
обхідно враховувати і зазначені особливості.
Міні-бульби, вирощені в гідропонних модулях, потрібно викорис¬
товувати диференційовано, залежно від строків їх збирання. Так, згід¬
но з дослідженнями, що були проведені в Росії, найбільшою про¬
дуктивністю відзначалися міні-бульби сорту Луговська, зняті в грудні
та січні з рослин, що культивувалися в гідропонних модулях протягом
52-84 днів. Міні-бульби, зняті в більш пізні строки з тих же рослин,
значно знижували продуктивність. Такі міні-бульби краще закладати в
холодильні камери на зберігання до наступного сезону садіння.
Диференційовано слід використовувати і міні-бульби, різні за ма¬
сою. Міні-бульби діаметром 8—10 мм (масою 4-5 г) обробляють спеціаль¬
ною живильною сумішшю, виготовленою на торф’яному субстраті, яка
містить всі необхідні макро- і мікроелемента. Цей прийом збільшує в
три рази вихід оздоровлених бульб з одиниці площі (з 87 до 329 тис.
пгг/га). Склад поживної суміші на 10 дм3 має бути такий: торфо¬
перегнійний компост - 5 кг; повільно діюче добриво (нітроамофоска з
умістом N10P25K25) - 0,2 кг; рідке комплексне добриво “Басфоліар-36-
екстра”, яке використовується для підживлення інтенсивних культур
(N - 10 %, Р - 10 %, Mg - 3%, Мп - 1%, Си - 0,2%, Во - 0,02%, Fe -
0,02%, Zn - 0,01%, Mo - 0,005%) - 0,2 л. Продуктивність міні-бульб у
цьому випадку підвищується за рахунок прискорення появи сходів на 3-
5 днів і підвищення їх польової схожості на 30-50 %. Рослини від таких
бульб мають більше стебел і більшу площу листкової поверхні. Урожай
одного куща сорту Луговська при використанні оброблених гідропон¬
них міні-бульб складала 258 г, на контролі - 178 г, а тепличних міні-
бульб - відповідно 362 г і 249 г [52].
Збільшення виходу крупніших фракцій (6—20 г) оздоровленого
матеріалу при використанні гідропонних міні-бульб можна досягти
завдяки загущеному садінню з площею живлення 70 х 15 см.
425
Висаджувати міні-бульби в полі необхідно відповідно до фракцій,
у добре оброблений ґрунт, дрібні - мілкіше, більші - дещо глибше.
При подальшому розмноженні насіннєвого матеріалу від бульб
різної маси різниці щодо продуктивності не виявлено, хоча в перший
рік їх випробування у відкритому ґрунті від менших за масою міні-
бульб одержують нижчий урожай.
Так, у дослідах, проведених на піщаних ґрунтах, урожай від гід¬
ропонних міні-бульб масою 6-10 г становив 465 г/кущ, масою 20 г -
587 г/кущ. Гідропонні міні-бульби без розподілу на фракції за масою
сформували кущ по 6,7 шт. бульб загальною масою 536 г/кущ [35].
Для зменшення втрат при довготривалому зберіганні міні-бульб
та підтримання їхніх продуктивних якостей зібрані гідропонні міні-
бульби промивають водою, а потім їх закладають у поліетиленові
ящики, пересипаючи прожареним річковим піском, а після цього за¬
кладають на зберігання в картоплесховище. Після зберігання повно¬
та сходів міні-бульб сорту Водограй була на 6% і сорту Луговська на
11% вищою, ніж при садінні непромитих міні-бульб, які зберігалися
без перешаровування піском. На продуктивні і насінні якості міні-
бульб ці фактори суттєво не впливали [30].
Репродукційні властивості насіннєвого матеріалу від гідропонних
міні-бульб значною мірою залежать і від сорту. Так, рослини сорту
Бородянська рожева 1—3-ої польової репродукції за візуальною оцін¬
кою були уражені вірусними хворобами на 0,3-0,5%, 4-5-ої репродук¬
цій - на 0,9—1,0%. У той же час рослини 1—3-ї польової репродукції
сорту Пост 86 були вільні від вірусів, а в 4-5-й репродукціях кіль¬
кість уражених рослин становила 0,8-1,0%. Проте в окремі роки
продуктивність і насіннєві якості репродукованого матеріалу різних
сортів перебували практично на одному рівні [30].
Гідропонні міні-бульби за своїми показниками, як вихідний на¬
сіннєвий матеріал, не відрізняються від міні-бульб, одержаних у
культиваційних спорудах та польових умовах. Так, дослідженнями,
проведеними в Інституті картоплярства УААН, доведена рівноцін¬
ність у насінництві картоплі одержаного вихідного матеріалу в теп¬
личних, гідропонних, польових від рослин in vitro, польових від мік¬
робульб in vitro.
Найбільша різниця в урожайності за три роки репродукування на¬
сіннєвого матеріалу від різних за походженням міні-бульб становила
по сорту Водограй 1,1 т/га, Бородянська рожева - 1,7 т/га, Світанок
київський - 2,4 т/га, Луговська - 2,3 т/га. Не виявлено також суттєвої
різниці щодо ураження насіннєвого матеріалу вірусами M і S різного
походження [11].
Застосовують метод гідропоніки і при вирощуванні мікробульб
in vitro у вегетаційних ємностях з різними наповнювачами (вулканіч¬
ний шлак, гравій), що виключає контакт рослин із ґрунтом, завдяки
чому подовжується період стійкості до ураження ґрунтовими хворо-
426
ботворними патогенами. Найбільш придатними є легкі субстрати з
малим діаметром частинок (5-10 мм) [16]. Модифікацією цього спо¬
собу є одержання розсади із рослин in vitro шляхом іонітопонної
культури. Іонообмінний субстрат за своїми властивостями близький
до ґрунтових умов. Маса міні-бульб, що одержують на цьому суб¬
страті, -20-68 г, кількість бульб — 5,4—13 шт. з однієї рослини [24].
Для одержання міні-бульб здійснюють культивування рослин та мік¬
робульб, вирощених in vitro, у польових умовах при дотриманні просто¬
рової ізоляції від джерел та переносників вірусної інфекції. При цьому
необхідно забезпечити достатні норми органічних та мінеральних доб¬
рив, оптимальний обробіток ґрунту та догляд за рослинами, ретельний
обробіток посівів проти шкідників та хвороб [44,48,17].
Одна рослина in vitro, висаджена в полі, формує під кущем 8,4-
10,8 бульб [48].
Одержання міні-бульб із паросткових і стеблових живців при
обмеженій площі живлення. Паросткові живці, одержані за звичай¬
ною методикою після довготривалого пророщування бульб, або
стеблові живці, одержані при живцюванні маточних рослин у фазі
бутонізації-початку цвітіння, що вирощуються в теплицях, виса¬
джують у ящики із живильним субстратом із площею живлення 6x6
см або 8x8 см) на глибину 1-1,5 см.
Стеблові живці - це частина стебла завдовжки близько 10 см з
одним листком (можна використовувати і пазушні пагони). їх виса¬
джують на глибину міжвузля.
Попередньо протягом шести годин для кращого укорінення стеб¬
лові живці бажано витримати в розчині гетероауксину (50 мг/л). За
такої технології від кожної рослини протягом двох місяців із парост¬
кового або стеблового живця утворюються одна-дві бульби масою до
10 г. При цьому кількість бульб з їм2 площі становила 300-500 шт.,
у той час як при звичайному вирощуванні з 1 м2 за 3-5 місяців одер¬
жують не більше 30-40 шт.
Для одержання паросткових живців замість маточних рослин ви¬
користовують оздоровлені бульби після довготривалого пророщу¬
вання. У такому разі потреба в тепличній площі скорочується удвічі,
оскільки немає необхідності вирощувати маточні рослини. Особливу
увагу при оцінці маточних бульб звертають на наявність чи відсут¬
ність у них вірусів. Зокрема, такі бульби можна проіндексувати.
Одержані з індексів рослини слід оцінити методом серодіагностики і
здійснювати добір живців для одержання міні-бульб тільки від віль¬
них від вірусів бульб.
Для одержання живців бульби закладають на пророщування. Перед
цим їх миють, обробляють проти грибкових інфекцій 1%-ним розчином
борної кислоти. Бульби вигримують при температурі 18-20°С і віднос¬
ній вологості 80-90 % до досягнення ними довжини 10-20 см і більше.
За 20-30 днів до садіння паростки обламують біля основи маточної буль¬
427
би і живцюють, розрізаючи на шматочки завдовжки 1,5 см за кількістю
зародкових бруньок. Живці розкладають на вологий фільтрувальний
папір у кювети, які покривають листком скла. Кювети час від часу про¬
вітрюють, знімаючи скло приблизно на 1 год. У такій вологій камері в
кімнатних умовах живці швидко проростають, на них з’являються зарод¬
кові корінці і молоді паростки. Через 2-4 дні живці висаджують безпо¬
середньо в закритий ґрунт або вирощують із них розсаду, яку в подаль¬
шому переносять у відкритий ґрунт.
Для одержання розсади живці висаджують у розсадні ящики або
горщики, заповнені живильною сумішшю (торфом і піском або дер¬
новою землею). Глибина садіння живців - 1 см, відстань між росли¬
нами у рядку і між рядками - 3-5 см. Полив при вигонці розсади по¬
мірний. Розсада через 15-20 днів придатна для висаджування у відкри¬
тий ґрунт або в плівково-марлеві теплиці. Для одержання міні-бульб
площа живлення має становити 10 х 10, 20 х 10, 30 х 10, 30 х 20 см, за¬
лежно від здатності сорту формувати масу картоплиння.
Для розсадної культури у відкритому ґрунті вибирають родючі
ділянки із сприятливим водно-повітряним режимом. Висаджують
60-70 тис. рослин на 1 га. Розпушують ґрунт неглибоко, оскільки
коренева система картоплі і столони залягають біля поверхні і є особ¬
ливо чутливими до пошкодження. Рослини підживлюють мінераль¬
ними добривами (1-1,5 ц/га нітрофоски або нітроамофоски) і 2-3
рази за сезон підгортають для кращого столоно- і бульбоутворення.
Використовуючи як посадковий матеріал паросткові живці, мож¬
на отримати коефіцієнт розмноження 1 : 1000 - 1 : 1500 і більше.
Маса бульб з одного куща досягає 500-2000 г, залежно від сорту та
умов вирощування [6].
Отже, підсумовуючи вшцевикладене, слід зазначити, що оздоров¬
лені міні-бульби є цінним вихідним матеріалом в елітному насінниц¬
тві. Про це, зокрема, свідчать дослідження, проведені як вітчизняни¬
ми, так і закордонними науково-дослідними установами.
Так, у результаті досліджень, проведених на Кам’янсько-Бузь-
кому опорному пункті первинного насінництва Інституту картопляр¬
ства УААН у Львівській області, доведено, що при репродукуванні
упродовж трьох років кількість рослин із зовнішніми ознаками ура¬
ження вірусами була практично однаковою на посадках картоплі з
насіннєвим матеріалом, сформованим на основі міні-бульб масою 1-
5 г, 6-10 г і 11-15 г. Не виявлено в межах сорту істотної різниці і в
урожайності репродукованого насіннєвого матеріалу.
Разом з тим для сортів, що вивчалися, а саме: Невська та Лугов¬
ська, які відзначаються толерантністю до вірусів, - бульбовий (від
здорових кущів) та клоновий добори є практично рівноцінними при
формуванні вихідного матеріалу на основі міні-бульб масою 1-5 г,
6—10 г і 11-15 г, одержаних від оздоровлених методом меристемної
тканини рослин in vitro. Різниця в ураженості рослин X, M, S віруса¬
428
ми картоплі між знеособленим і клоновим насіннєвим матеріалом
становила для сорту Невська 0,8-3,4%, для сорту Луговська - 1,6-
3,8%. Не виявлено також значної відмінності в ураженості вірусною
інфекцією потомства, залежно від маси вихідних міні-бульб. Різниця
в кількості уражених рослин на посадках потомства міні-бульб ма¬
сою 1-5 г та 11-15 г становила для сорту Невська 2,4% для знеособ¬
леного матеріалу і 3,3% - для клонового, для сорту Луговська - від¬
повідно, 2,9 % та 4,2 % [2].
Використовуючи в насінництві картоплі одержаний методом ме-
ристемнотканинної культури матеріал, зокрема, як вихідний для від¬
творення еліти, необхідно застосовувати заходи і методи, спрямовані
на підтримання його продуктивних якостей при одночасному отри¬
манні максимального економічного ефекту від його застосування.
На перших етапах розмноження меристемного матеріалу необ¬
хідно проводити контрольну оцінку його, щоб виключити нетипові
меристемні клони.
Слід також систематично оновлювати і вихідний матеріал на ос¬
нові введення в культуру і підтримання банку кращих вихідних лі¬
ній, ретельно перевірених на сортову типовість і наявність вірусної,
мікоплазмової і бактеріальної інфекції.
Одним із шляхів вирішення поставленого завдання є оцінка та
добір високопродуктивних оздоровлених ліній для мікроживцюван¬
ня і використання в оригінальному насінництві. Такі лінії дозволя¬
ють підвищити продуктивність рослин картоплі на 35-45%. Досяга¬
ють цього, вивчаючи протягом декількох років лінії нових оздоров¬
лених сортів при проведенні контролю за їхньою продуктивністю і
фенотипічністю. Для виробництва 5-10 тис. оздоровлених рослин іп
vitro достатньо мати 2-3 високопродуктивні лінії. Під лінією розу¬
міють потомство однієї оздоровленої рослини, розмноженої через
культуру in vitro [1].
При репродукції насіннєвого матеріалу, одержаного на основі
міні-бульб, особливої уваги потребують заходи, що попереджують
ураження рослин вірусами. Дотримання таких вимог дозволяє одер¬
жувати високоякісний насіннєвий матеріал у розсадниках елітного
насінництва.
Проведені з таким матеріалом в умовах Південного Полісся
України дослідження із сортами Водограй, Бородянська рожева, Сві¬
танок київський і Луговська засвідчили рівноцінність міні-бульб,
одержаних різними способами (тепличні, гідропонні, польові від рос¬
лин in vitro, міні-бульби in vitro), як вихідного матеріалу для відтво¬
рення еліти. У той: же час накопичення вірусної інфекції у процесі
репродукування насіннєвого матеріалу залежало від біологічних
особливостей сорту. Найменша кількість рослин з ознаками вірусних
хвороб була виявлена в сорту Світанок київський - 1,6-2,4%, най¬
більша - у сорту Луговська (2,8-3,4 %).
429
Зазначені показники були досягнуті при репродукуванні протя¬
гом чотирьох років насіннєвого матеріалу від оздоровлених міні-
бульб в умовах просторової ізоляції від переносників та джерел вірус¬
ної інфекції із застосуванням інсектицидів афіцидної дії проти попе¬
лиць та при ретельній фітопрочистці в міру проявлення хвороб і
знищенням картоплиння в ранні строки [11].
Важливим чинником технології використання одержаного шля¬
хом меристемнотканинної культури матеріалу є застосування скоро¬
чених схем відтворення еліти на основі цього матеріалу. Зокрема, за
трирічної схеми достатньо 50 безвірусних вихідних бульб, щоб за¬
безпечити 1 га елітних насаджень. У перший рік із 50 безвірусних
бульб за вересень-квітень вирощують близько 6000 бульб у стаціо¬
нарній теплиці, які висаджують в наступному році на 0,12 га у плів¬
кові теплиці або в полі в умовах ізоляції від основних джерел та пе¬
реносників вірусної інфекції. Одержаного матеріалу достатньо для
садіння на третій рік на площі 1 га (рис. 6.7).
Дата садіння
15.09
(1-й рік)
Зібрано бульб, шт.
25.12
(1-й рік)
15.03
(2-й рік)
26.05
(2-й рік)
3-й рік
Рис. 6.7. Схема прискореного розмноження безвірусного
насіннєвого матеріалу картоплі
Примітка. Бульби, зібрані у стаціонарній теплиці, у той же день обробляють
стимуляторами росту і висаджують у ґрунт
430
Оптимальні умови для вирощування картоплі у стаціонарних теп¬
лицях створюються в осінньо-зимовий і зимово-весняний періоди.
Вирощувати рослини в найбільш спекотні літні місяці (липень-
серпень), а також у зимовий період (грудень-січень) не доцільно. У
літніх теплицях повітряний обмін і зниження температури забезпе¬
чується за рахунок комбінованого плівково-марлевого покриття. У
середньому за один оборот у стандартній теплиці площею 1000 м2
одержують до 60 тис. бульб.
Як тепличний ґрунт можна використовувати чистий торф або су¬
міш торфу, піску і дернового ґрунту у співвідношенні 3:1:1. Поперед¬
ньо ґрунт ретельно перевіряють на відсутність карантинних об’єктів.
Рослини поливають в міру потреби. Основним заходом при вирощу¬
ванні вихідного насіннєвого матеріалу в закритому ґрунті є боротьба
з попелицями - переносниками вірусів. Найбільш ефективним афі-
цидом у боротьбі з ними є піримор у концентрації 0,05-1 %. Пері¬
одичність обробітку - один раз на 7-10 днів. Проте для попереджен¬
ня стійкості попелиць до одного афіциду, зокрема при довготрива¬
лому обприскуванні піримором, необхідно час від часу чергувати
його з обприскуванням афіцидом, який відрізняється хімічним скла¬
дом діючої речовини.
Позитивні результати у боротьбі з попелицями і білокрилкою
одержані при обприскуванні 0,4-0,5 %-ним амбушем; 0,5-0,6 %-ним
цимбушем. При цьому для знищення білокрилки необхідно провести
не менше чотирьох обприскувань через 3-4 дні.
Для попередження бурхливого наростання картоплиння і приско¬
рення бульбоутворення проводять обприскування посівів ретардан¬
тами. За підрахунками, проведеними за результатами досліджень у
Росії, при скороченій трирічній схемі насінництва за рахунок збіль¬
шення виходу кондиційних бульб елітної картоплі річний економіч¬
ний ефект у перерахунку на 100 га посадок еліти становить 150-200
тис. руб, або на 1 т еліти - 120-125 руб.
У випадках, якщо елітне господарство має наукову лабораторію і
в закритому ґрунті одержує оздоровлені вихідні бульби, то частину з
них реалізує іншим господарствам, а ті, що залишилися, використо¬
вує для виробництва елітної картоплі з наступною реалізацією. Ефек¬
тивність насінництва при цьому зростає. У такому разі при реалізації
З тис. т еліти і щорічному виробництві 1 млн шт. оздоровлених бульб
умовно-чистий прибуток на 1 т еліти становить 300 крб, окупність
витрат-2,18 руб [27].
Крім того, скорочені схеми відтворення еліти дозволяють значно
прискорити впровадження у виробництво нових сортів картоплі на
основі використання високоякісного насіннєвого матеріалу. Так, чо¬
тирирічна схема, за результатами досліджень, проведених на
Кам’янсько-Бузькому опорному пункті первинного насінництва кар¬
топлі Інституту картоплярства УААН, дозволяє на п’ятий рік при
431
використанні як садивного матеріалу 100 т еліти одержати додатко¬
во, порівняно із п’ятирічною схемою на основі клонового добору,
300 т першої репродукції. Використовуючи таку кількість садивного
матеріалу першої репродукції, можна одержати на п’ятий рік додат¬
ково 85-100 га насаджень нового сорту на основі високорепродук-
ційного насіннєвого матеріалу [10]. Аналогічні результати одержані
у дослідженнях, проведених із сортами Пролісок, Невська і Луговсь¬
ка в умовах північного сходу України. Щодо названих сортів вияв¬
лена рівноцінність еліти, одержаної на основі меристемного і клоно¬
вого матеріалу. При цьому доведена висока зворотна залежність між
продуктивністю і ураженістю рослин вірусною інфекцією [28].
У той же час щодо сорту Зов у дослідах Поліської дослідної ста¬
нції кращі результати були одержані на основі клонового добору,
проведеного у розсадниках, які закладалися оздоровленим матеріалом.
Урожайність еліти на основі клонових доборів становила 28 т/га, що
на 38-37% більше, ніж еліти, одержаної на основі міні-бульб, і рос¬
лин in vitro та міні-бульб in vitro. У сортів Гарт і Радомишльська
продуктивність еліти картоплі не залежала від вихідного матеріалу
та методів його формування [15].
Використання в насінництві вихідного матеріалу картоплі, одер¬
жаного шляхом меристемнотканинної культури, потребує чіткого до¬
тримання технологічного процесу його відтворення. Перш за все це
стосується використання кращих вихідних меристемних ліній, багато¬
разово і ретельно перевірених на сортову типовість і життєздатність,
на наявність вірусної, мікоплазмової і бактеріальної інфекції.
Важливо також систематично оновлювати вихідний меристемний
матеріал. Репродукуючи такий насіннєвий матеріал, особливу увагу
необхідно приділяти заходам, що попереджують інтенсивне уражен¬
ня рослин найбільш шкідливими та розповсюдженими патогенами
картоплі, у першу чергу - вірусами та мікоплазмами. Досягають цьо¬
го як організаційними, технологічними заходами і застосуванням
афіцидів, так і скорочуючи термін репродукування цієї партії мерис¬
темного матеріалу. Зокрема, застосовують прискорені методи роз¬
множення як вихідного, так і оригінального та елітного насіння.
Необхідно також зазначити, що при відтворенні еліти сортів кар¬
топлі з польовою стійкістю до вірусів застосовують комплекс загаль¬
ноприйнятих насінницьких заходів, у тому числі оцінку маточних
рослин з допомогою серологічної діагностики, коли немає потреби
використовувати меристемнотканинну культуру для одержання ви¬
сокопродуктивного вихідного матеріалу.
ЛІТЕРАТУРА
1. Адамова А.И. Оценка и отбор оздоровленных линий для семеноводс¬
тва новых и перспективных сортов картофеля // Новое в семеноводстве
картофеля.- Минск, 2000 - С.60-61.
432
2. Андрушко О.М. Продуктивність та якість насіннєвої картоплі, отри¬
маної на основі міні-бульб, залежно від прийомів формування вихідного
матеріалу: Автореф. дис. ... канд. с.-г. наук - К., 2000.- 20 с.
3. Анисимов Б.В., Сариев Г.М., Габель Б.В. Сравнительное изучение раз¬
личных по величине фракций миниклубней, полученных на основе теплич¬
ных и гидропонных технологий // Актуальные проблемы современного кар¬
тофелеводства- Минск, 1997 - С.91.
4. Анисимов В.В., Баранов А.Н^ Меньков А.Ф. Выращивание тепличных
миниклубней картофеля и их полевое испытание в условиях Брянской обл. //
Биология в картофелеводстве: Науч. тр. /НИИКХ- М., 1991- С.120-125.
5. Баранов А.Н. Оценка схем выращивания элитного картофеля из исход¬
ного материала, оздоровленного методом верхушечной меристемы: Авто¬
реф. дис. ... канд. с.-х. наук.- М., 1991- 21 с.
6. Безвирусное семеноводство картофеля: Рекомендации- М.: Агро-
промиздат, 1980 - 33 с.
7. Бердін С.І. Динаміка продуктивності картоплі залежно від репродук¬
ції насіння // Картоплярство - Вип.25- К.: Урожай, 1994,- С.51-53.
8. Биотехнологические методы получения и оценки оздоровленного
картофеля /Л.Н.Трофимец, В.В. Бойко, Т.В. Зейрук и др.- М.: Аіропромиз-
дат, 1998.-С. 17-18.
9. Блоцкая Ж.В. Вирусные болезни картофеля: Рекомендации - М.: Аг-
ропромиздат, 1980- С.ЗЗ.
10.Верменко Ю.Я., Андрушко О.М. Ефективність застосування міні-
бульб в насінництві картоплі // Вісник Львівського ДАУ.- № 5 - Львів,
1999 - С.153-156.
И.Верменко Ю.Я., Олійник В.П. Міні-бульби в насінництві картоплі //
Вісник ДАУ - Вип.Ю - Біла Церква, 2000- С.54-58.
12.Вызасте Э.Д. Влияние сокращения вегетационного периода и разных
методов отбора на качество семенного картофеля в Эстонской ССР: Авто¬
реф. дис. ... канд. с.-х. наук.- Таллин, 1970- 30 с.
13. Габель Б.В., Зайченко Е.С., Цоглин Л.Н. Индустриальное производст¬
во безвирусных миниклубней // Картофель и овощи.- 1993 - № 1- С.9-11.
14. Гидропонная установка для промышленного выращивания семенных
миниклубней картофеля /Б.В. Габель , О.С. Малик-Саркисов , Л.Н. Цоглин и
др. // Физиология растений.- 1989 - Т.38, вып.5.
15. Гончаренко О.П., Нечипоренко Г.Т. Технологический процесс вос¬
производства и ускоренного размножения оздоровленного исходного и эли¬
тного картофеля // Новое в семеноводстве картофеля.- Минск, 2000 - С.74-
75.
16. Деведжян А.Г., Майрапетян С.Х. Размножение безвирусного карто¬
феля в условиях гидропоники // Регуляция роста и развития картофеля - М.:
Наука, 1990.- С. 167-169.
17. Демченко Л.О. Продуктивність і якість насіннєвої картоплі в інтен¬
сифікації технологічного процесу при виробництві мікро- та міні-бульб: Ав¬
тореф. дис.... канд. с.-г. наук.- К., 1996 - 26 с.
18. Есипова З.И. Клубнеобразование in vitro как способ получения поса¬
дочного материала в первичном семеноводстве картофеля // Селекция и се¬
меноводство- 1987.-№ 3 - С.51-53.
433
19. Зернов В.Н., Петухов С.Н., Мусин С.М. Оздоровленные миниклубни
с торфоцеолитовых субстратов // Картофель и овощи - 1999 - № 6 - С.24.
20. Зубкевич О.Н., Сильванович Н.А. Ускоренный метод выращивания
микроклубней картофеля в пробирочной культуре // Эколого-экономические
основы усовершенствованной интегральной системы защиты растений от
вредителей, болезней и сорняков - Ч.ІІ - Минск, 1996 - С.17-18.
21. Зыкин А.Г. Вирусные болезни картофеля - JI.: Колос, 1976 - С. 118-120.
22. Иванова А.А. К вопросу получения семенного материала картофеля
из мини-клубней // Новое в семеноводстве картофеля.- Минск, 2000-
С.65-66.
23.Игонтов В.Г., Шакуров И.Ш. Изучение продуктивных качественных
показателей семенного картофеля в зависимости от методов получения су¬
пер-суперэлиты в условиях Среднего Поволжья // Актуальные проблемы
современного картофелеводства - Минск, 1997- С.97-98.
24. Изучение способов размножения оздоровленного исходного материа¬
ла картофеля с использованием современных биотехнологических средств /
А.И. У сков, Ю.П. Бойко, В.В. Бойко и др.// Новое в семеноводстве картофе-
ля.- Минск, 2000 - С.74.
25.Касян О .Я., Василинчук З.Д., Демченко J1.0. Вплив іустоти та глиби¬
ни садіння мікробульб на насінну продуктивність картоплі // Картоплярст¬
во.- Вип.25 - К.: Урожай, 1994.- С.48-51.
26. Киселев В.Н., Соломина И.П. Современные аспекты семеноводства
овощных культур и картофеля // Обзор МС “Агропроминформ”.- М., 1990-
С.3-12.
27. Князев В.А., Исаков А.И., Гавришова В.И. Перспективы использова¬
ния защищенного грунта в первичном семеноводстве картофеля // Селекция,
семеноводство и биотехнология картофеля.- 1989.- № С.75-79.
28.Кожушко Н.С., Дубовик В.И. Влияние методов оздоровления на про¬
дуктивность картофеля // Новое в семеноводстве картофеля - Минск, 2000-
С.50.
29.Колонтай Г.Н., Манойло Г.В., Гайструк Г.В. Способ получения исход¬
ного оздоровленного материала картофеля // Новое в семеноводстве карто¬
феля- Минск, 2000 - С.48-49.
30.Колонтай Г.Н., Мозоль J1.E. Влияние промывания миниклубней оздо¬
ровленного картофеля с гидропонных установок на их лежкость, продуктив¬
ность и семенные качества // Новое в семеноводстве картофеля - Минск,
2000.- С.67.
31. Культура ткани в семеноводстве картофеля / В.Н. Киселев, В.И. На¬
заренко, И.П. Соломина и др. // Картофелеводство за рубежом - М.: ВНИИ-
ТЭИ агропром, 1991- С.40-58.
32.Кучумов А.П., Князев В.А. Культура тканей и клеток в селекции и
семеноводстве картофеля - М.: ВНИИТЭИСХ, 1980 - С.34-36.
33. Майщук З.М. Клональне мікророзмноження картоплі in vitro - Львів,
1998.- C.95.
34. Макаров П.П. Картофелеводство Великобритании. - М.: ВНИИТЭИСХ,
1984.-С. 13.
35. Миниклубни: как лучше их использовать / Анисимов Б.В., Габель Б.В.,
Сариев Г.М., Шакуров И.Ж. // Картофель и овощи. - 1998. - № 6. - С.28.
434
36. Оздоровление и ускоренное размножение семенного картофеля: Ме-
тодич. рекомендации /Л.Н.Трофимец, Д.П.Остапенко, В.В. Бойко и др. - М.,
1985.- С.24-26.
37. Остапенко Д.П. Способи одержання вихідного матеріалу // Селекція і
насінництво. - К.: Урожай, 1998 - С.129-151.
38. Остапенко Д.П., Різник В.С. Бульбоутворення in vitro залежно від черен-
кування рослин // Картоплярство.- Вип.24- К.: Урожай, 1993- С.36-38.
39. Остапенко Д.П., Різник В.С., Костюк І.І. Використання мікробульб
при клональному мікророзмноженні картоплі // Картоплярство - Вип.25-
К.: Урожай, 1994.- С.26-28.
40. Пантелеймонов И.А. Размножение исходного материала картофеля //
Производство картофеля на индустриальной основе - М., 1988 - С.87-90.
41. Рассадина Г.В., Юрьева Н.Ю., Габель Б.В. Миниклубни с гидропон¬
ных установок // Картофель и овощи - 1994- № 2.- С. 10.
42. Різник В.С. Метод багаторазового знімання живців при клональному
мікророзмноженні картоплі // Картоплярство.- Вип.25.- К.: Урожай, 1994.-
С.43-45.
43. Різник В.С. Оздоровлення картоплі: проблеми і перспективи // Карто¬
плярство- Вип.27.- М., 1997 - С.23-24.
44. Різник В.С., Жила Е.Д. Вплив способів підготовки розсади на розви¬
ток і продуктивність оздоровлених рослин картоплі в польових умовах //
Картоплярство.-Вип.26.-К.: Урожай, 1995 - С.60-63.
45. Родькин О.И. Эффективность метода рассады в оригинальном семе¬
новодстве картофеля // Новое в семеноводстве картофеля.- Минск, 2000-
С.72-73.
46. Семенова З.А., Матусевич В.В. Использование ионитных субстратов
для получения миниклубней картофеля в условиях защищенного грунта //
Новое в семеноводстве картофеля - Минск, 2000.- С.59-60.
47. Технология и оборудование для промышленного производства безви-
русных миниклубней / О.С. Малик-Саркисов, Б.В. Габель, Л.П. Цоглин,
С.Л. Чернобровкин//Картофель и овощи - 1993.-№ 1.- С.9-11.
48. Трофимец Л.Н. Биотехнология в картофелеводстве.- М., 1989 - 45 с.
49. Трофимец Л.Н. Некоторые особенности инфекционного процесса при
заражении картофеля вирусами M, S, Y // Тр. НИИКХ.- Вып.8.- М., 1971-
С.244-251.
50. Трофимец Л.Н. Развитие биотехнологии в картофелеводстве // Селек¬
ция, семеноводство и биотехнология картофеля - М., 1989 - С. 100-105.
51. Цоглин Л.Н., Габель Б.В. Автоселекционные процессы при микро¬
клонировании растений. Фитофторное размножение картофеля // Физиоло¬
гия растений - 1994.- № З - С.436-440.
52. Шакуров И.Ж. Сравнительное изучение продуктивных и качествен¬
ных показателей супер-суперэлитного картофеля при использовании раз¬
ных по величине фракций тепличных и гидропонных миниклубней в усло¬
виях Среднего Поволжья: Автореф. дис. ... канд. с.-х. наук.- М., 1999-
22 с.
53.Шмыгля В.А. Биологическое обоснование, разработка и усовершенс¬
твование методов диагностики вирусов в селекции и семеноводстве карто¬
феля и томата.: Автореф. дис. ... д-ра биол. наук - Л., 1989 - 42 с.
435
6.6. РЕПРОДУКЦІЙНЕ ТА
ВНУТРІШНЬОГОСПОДАРСЬКЕ НАСІННИЦТВО
Виробніщгво товарного насінного матеріалу високих репродук¬
цій, що широко використовується для проведення сортозаміни та
сортооновлення, зосереджується у спеціалізованих насінницьких гос¬
подарствах різних форм власності. Вирощування і реалізацію репро¬
дукційного насінного матеріалу організовують асоціації й акціонерні
товариства. Для потреб окремого господарства насінництво здійсню¬
ється в ньому для всієї площі садіння.
Основою внутрішньогосподарського насінництва є насінні ділян¬
ки, які в господарстві займають не менше 30-35 % від загальної
площі садіння. Обсяг виробництва садивних бульб для власних по¬
треб визначають із розрахунку 4-4,5 т/га посівної площі.
На насінницьких посівах висаджують лише сортовий матеріал, що
підтверджується відповідними сортовими документами (акт апро¬
бації, атестат, сортове свідоцтво). У господарстві, як правило, ви¬
рощують 3-4 сорти різних строків достигання, занесені до Реєстру,
визначивши їх процентне співвідношення згідно із своїми завдан¬
нями.
Орієнтовні схеми внутрішньогосподарського насінництва такі:
господарства, що спеціалізуються з виробництва сортової репродук¬
ційної картоплі, щорічно завозять 28-30 тонн еліти на 100 га посівів і
дотримуються такої схеми розмноження еліти: 1-й рік - ділянка, роз¬
множення - 7,5 га, одержують першу репродукцію; 2-й рік - насінна
ділянка - 22,5 га, одержують другу репродукцію; 3-й рік - товарні
насінницькі посіви - 70 га, одержують третю репродукцію.
Господарства, які вирощують продовольчу картоплю, щорічно
завозять 10-12 тонн еліти або другої репродукції на 100 га посівів.
Дотримуються такої схеми внутрішньогосподарського насінництва:
1-й рік - ділянка розмноження - 2,5 га, одержують першу або третю
репродукцію; 2-й рік - ділянка випробувань - 7,5 га, одержують дру¬
гу або четверту репродукцію; 3-й рік - 25 га, одержують третю або
п’яту репродукцію; 4-й рік - товарні посіви - 65 га, з яких уся картоп¬
ля реалізується.
У господарства півдня України при необхідності завозять бульби
1—2-ої репродукцій.
Усі технологічні процеси: удобрення, підготовка ґрунту, садіння,
догляд, включаючи боротьбу із шкідниками та хворобами, - прово¬
дять в оптимальні строки, визначені рекомендаціями для госпо¬
дарств певної ґрунтово-кліматичної зони.
При вирощуванні на одному полі різних сортів і репродукцій
картоплі між ними залишають незайнятими смуги завширшки не мен¬
ше 1,4 м, запобігаючи цим змішуванню при садінні та збиранні на¬
сіннєвих бульб.
436
Надзвичайно важливе значення в поліпшенні продуктивних якостей
бульб мають прочистки насінницьких насаджень від хворих рослин та
домішок інших сортів. Це обов’язковий і найефективніший захід, який
дозволяє підтримувати чистосортні насадження у здоровому стані.
Прочистки у великих колективних господарствах проводять спе¬
ціально організовані ланки, за якими закріплюють відповідну площу
розсадників чи насінну ділянку. Проводять навчання членів ланки,
на посівах їх знайомлять з ознаками основних вірусних хвороб, на
окремих рослинах вивчають морфологічні ознаки основного сорту і
можливих домішок, що зустрічаються в насадженнях. Організовує
прочистки агроном-насінник, а там, де його немає, - головний агро¬
ном господарства, який несе повну відповідальність за якість та
строки виконання робіт.
У фермерських господарствах і на городах прочистки здійсню¬
ють безпосередньо господарі, які повинні самостійно вивчити апро¬
баційні характеристики сортів та симптоми поширених хвороб.
На насінницьких насадженнях за вегетаційний період рекоменду¬
ється проводити дві-три прочистки. Першу проводять при досягненні
рослинами висоти 15-20 см. У цей час найкраще проявляються озна¬
ки таких вірусних хвороб, як зморшкувата та смугаста мозаїка, скру¬
чування листя. Крім того, перша прочистка пов’язана з початком ма¬
сового льоту попелиць - основних переносників вірусів. Коли ж пе¬
ред цим насадження звільняються від уражених вірусами рослин,
тоді можна запобігти інтенсивному перенесенню вірусних хвороб із
хворих рослин на здорові. Саме тому перша прочистка проводиться у
стислі строки - за 3-4 робочі дні, що значно підвищує її ефектив¬
ність. Уражені кущі викопують, а картоплиння разом з маточними
бульбами вивозять з поля і знтцуть - спалюють чи закопують.
Другу прочистку проводять на початку цвітіння картоплі. У цей
час добре помітні сортові домішки та хворі рослини. Вона дещо
складніша від першої, тому що на цей час рослини мають велику ве¬
гетативну масу і молоді бульби, які також слід викопати і вивезти з
поля. Цю прочистку також проводять у стислі строки, оскільки за¬
тримка з її виконанням призводить до повторного зараження здоро¬
вих рослин вірусними хворобами. Крім того, після цвітіння дуже
важко видалити домішки інших сортів, які вже відцвіли.
У розсадниках часто доводиться проводити ще й третю прочист¬
ку, здебільшого незадовго до збирання врожаю, але обов’язково до
початку відмирання та скошування картоплиння. При цьому особли¬
во старанно прочищають насадження від рослин з ознаками в’яне¬
ння, що вказує на ураженість рослин кільцевою гниллю. Саме якісні
і своєчасні прочистки є основним заходом боротьби з цією хворо¬
бою.
Кількість домішок інших сортів та рослин, уражених вірусами,
бактеріальними чи грибковими хворобами, визначають взяттям проб
437
за методикою апробації. Після кожної прочистки складають акт, у
якому зазначають площу насаджень, номер поля, назву сорту, кіль¬
кість домішок та хворих рослин.
Насінницькі насадження всіх категорій після проведення прочис¬
ток мають відповідати сортовим вимогам державного стандарту що¬
до насіння.
Збирають бульби в ранні строки, попередньо видаливши бадилля
на початку його відмирання, при наявності під кущем не менше 70-
75% бульб насіннєвої фракції. Не раніше ніж через 14 днів після цьо¬
го викопують бульби. Спочатку їх витримують у тимчасових кагатах
до дозрівання, потім перебирають і закладають на постійне зберіган¬
ня. Урожай оприбутковують за масою по сортах і репродукціях.
Усі насінницькі посіви картоплі оформляють відповідними ети¬
кетками із зазначенням сорту і репродукції.
Контроль за якістю насіннєвої картоплі. Контроль за якістю
посівів бульб насіннєвої картоплі в усіх ланках елітного насінництва
здійснюється Державною насіннєвою інспекцією України при наяв¬
ності паспорта-патенту на право вирощування еліти. Інспекція орга¬
нізовує проведення грунтового контролю.
На основі результатів апробації елітні посіви приймаються спеці¬
альною комісією, створеною управлінням сільського господарства
облдержадміністрації. До її складу входять представники управління
сільського господарства, Державної насіннєвої інспекції, Державної
інспекції з карантину рослин, науково-дослідних установ, що здійс¬
нюють методичне керівництво виробником еліти картоплі. При не¬
обхідності залучаються ентомологи та фітопатологи. При прийманні
посівів комісія враховує правомірність використання запатентованих
сортів відповідно до Закону України “Про охорону прав на сорти
рослин” [1], а також результати ґрунтового контролю.
Науково-дослідні установи та елітно-насінницькі господарства
зобов’язані надсилати для ґрунтового контролю зразки супереліти
картоплі у визначені строки і за власний рахунок. Вартість витрат за
проведення контролю відшкодовують господарства-виробники еліти
згідно з рахунками установи, що проводить ґрунтоконтроль.
Посіви еліти господарств, які не подали зразки на ґрунтоконт¬
роль, не класифікуються як елітні.
Посівні якості бульб визначають згідно з існуючими нормами
відповідних державних стандартів.
Кожна партія насіннєвої картоплі, що реалізується, супроводжу¬
ється “Атестатом на супер-суперелітну, суперелітну, елітну картоп¬
лю” або “Свідоцтвом якості репродукційної сортової картоплі” та
карантинним сертифікатом.
Ці документи оформляються виробниками насіннєвої картоплі на
основі актів апробації, приймання еліти та аналізу насінних бульб
картоплі.
438
Супер-суперелітну, суперелітну, елітну картоплю з нижчими від
норм показниками, що передбачені державними стандартами, допус¬
кають до використання як репродукційне насіння згідно з нормами,
яким воно відповідає. Особи, що порушили технологічні вимоги ви¬
рощування, притягаються до адміністративної відповідальності згід¬
но із Законом України “Про насіння” [2].
Оцінка сортових і посівних якостей насіннєвої картоплі. Сор¬
тові якості визначають апробацією насінницьких посівів картоплі,
ґрунтоконтролем зразків супереліти, індексацією бульб у післязби¬
ральний період.
Насінницькі посіви картоплі повинні бути одного сорту, однієї
категорії розмноження за етапами насінництва і відповідати вимогам
державного стандарту.
З посівів насіннєвої картоплі з показниками якості, нижчими від
норм, визначених державним стандартом, допускається використо¬
вувати насіннєвий матеріал відповідно до категорії розмноження,
якій він фактично відповідає.
Посіви оригінального насіннєвого матеріалу вибраковують при
наявності рослин, уражених кільцевою і бурою бактеріальною гнил¬
лю, чорною ніжкою, еліти та першої і другої репродукції - при наяв¬
ності рослин, уражених кільцевою і бурою бактеріальною гниллю.
Вибраковуються посіви насіннєвого матеріалу всіх категорій за
етапами насінництва при ураженості рослин або при наявності каран¬
тинних об’єктів на полі згідно з переліком, затвердженим Держав¬
ною інспекцією з карантину рослин України.
Вимоги до посівних якостей. Бульби насінної картоплі за посів¬
ними якостями повинні відповідати вимогам державного стандарту.
Такі бульби повинні бути одного сорту, однієї категорії розмножен¬
ня за етапами насінництва, сухими, із затверділою шкіркою, мати
типову для даного сорту форму і забарвлення, бути непророслими
(при весняній реалізації допускаються бульби із паростками завдов¬
жки не більше 5 мм).
У насінному матеріалі не допускається наявність карантинних шкід¬
ників та їх личинок, а також карантинних хвороб відповідно до переліку,
затвердженого Державною інспекцією з карантину рослин України.
Бракуються партії садивного матеріалу, який зібраний з полів,
уражених картопляною нематодою.
Оригінальний садивний матеріал бракують при наявності бульб,
уражених стебловою нематодою, кільцевою та бурою бактеріальною
гниллю, мокрою гниллю, паршею порошистою; еліту - мокрою та
бурою бактеріальною гниллю, порошистою паршею, стебловою не¬
матодою; репродукційний матеріал - при ураженні мокрою гниллю,
кільцевою та бурою бактеріальною гниллю, порошистою паршею.
Усі категорії садивного матеріалу бракуються при наявності
бульб, уражених карантинними об’єктами, видалення яких при пере¬
439
биранні не гарантується, відповідно до переліку, затвердженого
Державною інспекцією з карантину рослин України.
Для всіх категорій садивного матеріалу розмір бульб при викорис¬
танні для власних потреб господарства не нормується.
Протруювання бульб садивного матеріалу та обробка інгібітора¬
ми чи стимуляторами не нормуються.
При застосуванні щодо партії насіннєвого матеріалу хімічних за¬
собів проти хвороб і шкідників, інгібіторів та стимуляторів росту в
атестаті або свідоцтві якості сортової картоплі зазначається назва
хімічного засобу і дата його застосування.
Визначення сортових якостей. Сортову чистоту та наявність
хвороб у посадках насіннєвої картоплі визначають апробацією поса¬
док згідно з чинними інструкціями.
Приймання посівів еліти здійснюють після апробації, враховую¬
чи результати ґрунтового контролю.
ЛІТЕРАТУРА
1. Закон України “Про охорону прав на сорти рослин” // Відомості Вер¬
ховної Ради України. - 1993. -№ 21. -С. 539-547.
2. Закон України “Про насіння”. №3690-ІІХ; 15 грудня 1993 р. // Відомо¬
сті Верховної Ради України. -1994. - №2. - С. 27.
6.7. АПРОБАЦІЯ КАРТОПЛІ
Серед заходів, спрямованих на підвищення врожайності картоп¬
лі, важливе значення в системі насінництва має контроль за сорто¬
вими і посівними якостями насіння. За умов перебудови економічних
відносин, з розвитком багатоукладних форм господарювання (інди¬
відуальні, фермерські господарства, орендні колективи) проблема
виробництва високоякісного садивного матеріалу стає ще гострі¬
шою, а контроль за його якістю залишається актуальним. Звичайно,
його форми змінюватимуться, однак цілі залишаються такими ж.
Ведення сучасного насінництва на науковій основі в Україні ґрун¬
тується на суворому дотриманні вимог, передбачених у трьох групах
взаємопов’язаних нормативно-технічних документів: державних і
міждержавних стандартах на сортові і посівні якості насіння, на ме¬
тоди визначення посівних якостей насіння та інструкції з апробації
сортових посівів.
В усіх ланках насінницької роботи - господарствах, науково-
дослідних установах - внутрішньогосподарський і державний конт¬
роль здійснюється постійно.
Внутрішньогосподарський контроль передбачає дотримання пра¬
вил розмноження сортового садивного матеріалу, ведення докумен¬
тації в межах господарства. Головне завдання контролю - забезпечи¬
ти правильну організацію комплексу заходів щодо виробництва сор¬
440
тового насіння, яке має відповідати вимогам стандарту, а також ви¬
ключати можливість механічного засмічення в період вегетації та
при збиранні, транспортуванні і зберіганні бульб.
Внутрішньогосподарський контроль здійснюється агрономічним
персоналом, фермерами та іншими господарями, які вирощують садив¬
ні бульби. Кожне господарство повинно гарантувати відповідність
сортових і посівних якостей показникам, зазначеним у документах.
Документи оформляються за єдиним зразком, згідно з вимогами
інструкції та стандартів, під методичним керівництвом і контролем
Державної насіннєвої інспекції, тобто внутрішньогосподарський кон¬
троль безпосередньо пов’язаний із державним.
Державний сортовий і насіннєвий контроль здійснюють Дер¬
жавні насіннєві інспекції. Сортовий контроль є науково обґрунтова¬
ною системою заходів, спрямованих на збереження типових для ре¬
єстрових сортів морфологічних, біологічних і господарсько цінних
ознак. Одним із основних видів сортового контролю є апробація [3].
Основне завдання апробації картоплі - визначення якості сортових
посівів. Шляхом польового обстеження визначаються достовірність
сорту, сортова чистота та ступінь ураження насаджень хворобами. Од¬
ним із завдань апробації є віднесення насінних посівів, що апробують¬
ся, на основі ознайомлення з насінницькою документацією до
відповідної категорії насіння за етапами його розмноження. Визнача¬
ються також конкретні заходи щодо поліпшення насінництва картоплі.
Польовій апробації підлягають усі насінницькі насадження, уро¬
жай з яких передбачено використовувати на насіння. Сортові наса¬
дження картоплі, урожай з яких не використовують на насінні цілі,
агрономи-апробатори оглядають без аналізу проб і на основі огляду
посіву та наявних сортових документів на висаджену картоплю
складають акт реєстрації сортових насаджень. Несортові насадження
не апробують і не реєструють.
До початку апробації, але не пізніше ніж за два тижні до неї ап¬
робатори проводять підготовчу роботу, а саме:
- визначають усі площі сортових насаджень, які підлягають ап¬
робації; перевіряють документацію, що характеризує якість виса¬
дженої насінної картоплі (атестат на елітну картоплю, свідоцтво на
сортову картоплю за умови завезення садивного матеріалу, акт апро¬
бації посівів, якщо для посадки використано садивний матеріал влас¬
ного врожаю, акт весняного аналізу бульб);
- знайомляться з актом закладки насінницької ділянки, актом
прочистки сортових насаджень картоплі;
- організовують додаткову прочистку, якщо вона була проведена
незадовільно. Прочистка має бути проведена, якщо видаленню під¬
лягає не більше 20% рослин. При наявності понад 20% кущів, що
підлягають викопуванню, насадження не прочищають, а вважають їх
несортовими. Після кожної прочистки складають акт;
441
- визначають, які агротехнічні заходи проведені господарством
на насадженнях насінної картоплі (строки садіння, способи підготов¬
ки садивного матеріалу, строки і види обробітку ґрунту і посадок,
види і дози добріш та ін.);
- на основі перевірки сортових документів і польового огляду
посадок реєструють усі посіви, що не підлягають апробації, запов¬
нюють акт їх реєстрації.
Насадження сортової картоплі на насінницьких ділянках після
проведення прочисток за сортовою чистотою і наявністю в посівах
рослин, уражених хворобами, мають відповідати вимогам державно¬
го стандарту. Якщо в господарстві немає документів, що засвідчують
сорт та його репродукцію, апробатор сам визначає сорт, апробує чи
реєструє його. Якщо виникає сумнів щодо правильності визначення
сорту, йому слід звернутися за допомогою до старшого апробатора
чи в науково-дослідну установу.
За вегетаційний період проводять дві-три сортові і фітопатологіч-
ні прочистки і після кожної складають акт. На сортових посівах під
час прочисток видаляють хворі і пригнічені рослини, а також доміш¬
ки інших сортів.
Кожну прочистку проводять у стислі строки за 3-4 дні, що знач¬
но підвищує її ефективність. Уражені кущі викопують з обов’язковим
вилученням бульб. Картоплиння разом з маточними бульбами виво¬
зять з поля і знищують (спалюють чи закопують).
Польову апробацію картоплі проводять під час цвітіння у присут¬
ності особи, яка відповідає за насінництво в господарстві. Апробацію
здійснюють методом проб по діагоналі поля. Пробою називають 20
рослин картоплі, які, не викопуючи, оглядають підряд на одному ря¬
дку, апробаційним зразком - кількість усіх оглянутих у пробах рос¬
лин (кущів) картоплі.
Враховуючи площу ділянки, яка підлягає апробації, апробатор
визначає кількість проб і рослин, які необхідно оглянути, і, врахову¬
ючи конфігурацію ділянки, визначає відстань між пробами.
На ділянці до 5 га оглядають 15 проб по 20 кущів, усього - 300 ро¬
слин; до 10 га - 20 проб по 20 кущів - 400 рослин; на площі до 15 га -
25 проб по 20 кущів - 500 рослин; понад 15 га на кожні наступні
5 га додатково оглядають ще по дві проби по 20 кущів у кожній. На¬
приклад: на ділянці площею 16-20 га слід взяти 27 проб, тобто по¬
трібно оглянути (25+2)х20= 540 кущів.
Висока якість апробації забезпечується рівномірністю розміщен¬
ня (огляду) проб на полі. Для цього ширину ділянки в метрах або
кількість рядків необхідно поділити на визначену кількість проб.
Одержане число визначатиме відстань між пробами по ширині поля
в метрах або в рядках. Відстань між пробами по довжині поля визна¬
чається діленням довжини поля на кількість проб. Наприклад: апро¬
бації підлягає насінницька ділянка площею 15 га. За нашими розрахун¬
442
ками необхідно взяти 25 проб. Ширина ділянки - 300 м, довжина - 500.
Якщо розділити ширину ділянки на кількість проб, то одержимо від¬
стань між пробами по ширині, тобто в даному випадку -12 м (300 : 25).
Отже, проби слід оглядати на кожному 17-му рядку. Після цього ви¬
значаємо відстань між пробами по довжині поля: 500:25=20 м. У ці
20 м входить і довжина рядка під рослинами, взятими як проба. При
схемі садіння 70x25 см довжина рядка в пробі (20 кущів) становити¬
ме 5 м (0,25 м х 20). Отже, кожна проба за довжиною ділянки буде
розміщена через 15 м (20-5). Тобто від кінця першої проби слід
пройти вперед по рядку 15 м і 17 рядків у бік до початку другої про¬
би і т. д. Таким чином, при огляді проб апробатор проходить по ла¬
маній діагоналі поля, що забезпечує рівномірність розміщення проб
на ділянці, яка підлягає апробації.
Закінчивши розрахунки, апробатор починає огляд проб, тобто
кожного куща в пробі. Ретельно оглядаючи кожну рослину, апроба¬
тор визначає основний сорт, домішки інших і наявність хвороб. При
наявності домішок інших сортів умовно позначає колір бульб і кві¬
тів: чер. (червоний), син. (синій), біл. (білий), рож. (рожевий) і т. д.
Результати обстеження кожної рослини у пробі (сорт, хвороба) апро¬
батор позначає умовними знаками (крапка, риска) і заносить до від¬
повідної графи журналу польового огляду картоплі, де всі записи
проводяться простим олівцем. Наприклад, оглянуті перші чотири
кущі позначають крапками, у результаті чого утворюється умовний
квадратик, наступні чотири кущі - рисками, якими з’єднують крапки
і одержують справжній квадрат jZT, наступні два кущі також позна¬
чають рисками, які з'єднують кути квадрата по діагоналі й утворю¬
ють “конверт” - ]Х[. Так само аналізують і позначають другий деся¬
ток кущів у пробі. Якщо в пробі не виявлено домішок інших сортів,
то в журналі в графу основного сорту буде записано два повні “кон¬
верти” - gc] [X] • При наявності однієї домішки один з них буде не¬
повним - [^] (23 > при Двох домішках - відповідно, [><] ; при
трьох - [^<] Q і т. д. Кожний кущ у пробі перший раз оглядають на
сортність, а другий - на наявність захворювання і дані огляду запи¬
сують у відповідній графі польового журналу. Уражені хворобами
кущі враховують, не зважаючи на те, чи є вони основним сортом чи
домішками.
Після закінчення огляду проб апробатор визначає повноту та гус¬
тоту насаджень, вирівняність посівів, наявність бур’янів, орієнтов¬
ний урожай та ступінь ураження насаджень фітофторозом. Усі ці да¬
ні записують у графі “Примітки” польового журналу.
Ураження фітофторозом визначають візуально. Слабким вважається
таке ураження, при якому на окремих кущах трапляються поодинокі
плями; середнім - якщо на всіх кущах листки уражені, але стебла ще
зелені; і сильним - листки і стебла уражені наполовину або більше.
Ураження посадок фітофторозом не є підставою для зниження категорії
443
насаджень, але робиться відповідна помітка в журналі польового огляду
і примітка у свідоцтві на сортову картоплю.
Вирівняність насаджень вважають достатньою, якщо картоплин¬
ня нормально розвинене і рослини змикаються в рядках, а кількість
рослин, що відстають у рості, не перевищує 10 %; середньою, якщо
25 % рослин відстають у рості, і поганою, якщо відстають у рості
понад 25 % рослин.
Повноту (зрідженість) посіву, виражену у відсотках, визначають, ви¬
ходячи із схеми садіння, тобто розрахункової густоти насадження. На¬
приклад, при схемі садіння 70*25 см розрахункова густота насадження
становить 57,1 тис. кущів на гектар. Проби добирають рівномірно по
всій площі. Для визначення фактичної густоти насаджень підраховують
загальну кількість кущів у кількох пробах і перераховують (у тисячах)
на гектар насаджень. Забур’яненість насадження визначають окомірно
(велика, середня, незначна), урожай - орієнтовно.
Сортову чистоту й ураженість рослин хворобами обчислюють у
відсотках. Так, для визначення сортової чистоти із загальної кількос¬
ті оглянутих кущів (100%) вираховують домішки з іншим забарвлен¬
ням бульб або квітів. Наприклад, із оглянутих 300 рослин виявлено З
рослини домішок, тобто (3*100) : 300 = 1 %. Отже, сортова чистота
такого посіву становить 100 % - 1 % = 99 %. Аналогічно визначають
відсоток хворих рослин по кожній хворобі окремо.
Згідно з одержаними даними насадження відносять до тієї чи ін¬
шої категорії за нормами стандарту на сортову картоплю.
Якщо домішок інших сортів і хворих рослин більше, ніж перед¬
бачено стандартом, урожай із цих насаджень визнають непридатним
для використання як садивного матеріалу і вибраковують.
Насадження одного й того ж сорту, для садіння яких використані
бульби різного походження, апробують окремо і на кожен з них
складають акт. Насадження одного й того ж походження, але виро¬
щені на різних полях, також апробують окремо.
Документація. Одним із відповідальних процесів при апробації
картоплі є оформлення документів, які засвідчують сортову якість
насінної картоплі.
За даними журналу польового огляду вираховують (у відсотках до
загальної кількості оглянутих кущів) сортову чистоту й ураженість рос¬
лин вірусними та іншими хворобами. На підставі цих даних, а також
документів про походження садивного матеріалу, даних про агротехніку
насінної картоплі складають акт апробації у трьох примірниках, один з
яких (обов’язково) залишається в господарстві. Акт апробації має два
розділи. Перший розділ заповнюють на підставі таких документів: атес¬
тата чи сортового свідоцтва на використаний садивний матеріал, або ж
акта апробації за минулий рік, актів про проведену сортову і фітосанітар-
ну прочистки. Другий розділ акта доповнюють даними з журналу
польового огляду. У другому пункті акта записують кількість домішок
444
інших сортів і визначають сортову чистоту, загальний відсоток рослин,
уражених вірусними хворобами, окремо виділяючи чорну ніжку та кіль¬
цеву гниль, зазначають ступінь ураженості фітофторозом. На підставі
цих показників згідно з ДСТУ 4013-2001 [1] визначають сортові якості
посіву, зазначені в таблицях 6.7,6.8,6.9,6.10.
Таблиця 6.7 - Сортові показники якості оригінального насінного матеріалу та
еліти картоплі (за підсумками польової апробації чи індексації)
Полісся і Лісостеп 1
Степ
Назва
Норми для насінного матеріалу
ОН
ЕН
ОН
ЕН
Сортова чистота посадок, % від загаль¬
ної кількості рослин зразка при апроба¬
ції, не менше
100
100
100
100
Наявність у посадках рослин, уражених
хворобами за завнішніми ознаками, %
від загальної кількості, не більше,
всього
2,5
4,8
4,0
6,3
у тому числі:
- тяжкими вірусними хворобами, ві-
0,5
1,0
1,0
1,5
роїдом веретеноподібності бульб та
мікоплазмами
- легкими вірусними хворобами
2,0
3,5
0,3
4,5
- чорною ніжкою
0
0,3
0
0,3
Таблиця 6.8 - Показники посівних якостей оригінального насінного матеріалу та
еліти картоплі
Назва
Полісся і
Лісостеп 1
Степ
Норми для насінного
матеріалу
ОН
ЕН
ОН
ЕН
1
2
3
4
5
Наявність бульб інших сортів, % від загаль¬
Не допускається
ної кількості у пробі, не більше
Розмір бульб за найбільшим поперечним
діаметром, мм, для сортів із формою бульб;
- видовженою
28-55
28-55
20-55
20-55
- округло-овальною
30-60
30-60
25-60
25-65
Наявність бульб, що не відповідають вимо¬
гам за розміром, % від загальної кількості у
пробі, не більше
3
3
3
3
Наявність бульб, уражених хворобами, % від
загальної кількості у пробі, не більше
всього
5,0
6,8
7,0
8,3
в тому числі:
- мокрою гниллю
Не допускається
- фітофторозом
0,5
1,5
0,5
1,0
- чорною ніжкою
Не допус¬
Не допус¬
кається
0,3
кається
0,3
445
Продовження табл. 6.8.
1
2
3
4
5
- сухою гниллю (фомоз, фузаріоз)
0,5
0,5
1,0
1,0
- ризоктоніозом при ураженості поверхні
бульб:
- від 1/8 до 1/4
1,0
1,5
0,5
1,0
- більше 1/4
Не допускається
- паршею звичайною і сріблястою (при
ураженості 'A поверхні бульби)
3,0
3,0
5,0
5,0
- паршею порош и стою
Не допускається
Наявність бульб, уражених шкідниками і
механічно, % від загальної кількості у пробі,
не більше:
всього
5,5
6,0
7,0
7,5
в тому числі:
- уражених стебловою нематодою
Не допускається
- уражених дротяником (більше трьох
ходів)
1,0
1,0
0,5
0,5
- уражених гризунами, хрущами, совками
(без ураження вічок)
1,0
1,0
1,0
1,0
- з механічними пошкодженнями ткани¬
ни (глибина більше 10 мм)
3,5
3,5
5,5
5,5
Наявність бульб, ушкоджених хімікатами, з
ознаками задухи; підморожених, з опіками,
виродливих, із наростами, роздавлених, рі¬
заних (менше Уг їхнього розміру), з обдер¬
Не допускається
тою шкіркою (більше 1А поверхні).
Наявність землі і сторонніх домішок, % від
маси бульб, не більше
2
2
2
2
Таблиця 6.9 - Сортові показники якості репродукційних посівів насінної
картоплі (за підсумками польової апробації)
Назва
Полісся і
Лісостеп 1
Степ
Норми для
эепродукцій
PH-1-2
PH-3-4
PH-1-2
PH-3-4
Сортова чистота посадок, % від загаль¬
ної кількості рослин зразка при апро¬
бації, не менше
97
95
97
95
Наявність у посадках рослин, уражених
хворобами за зовнішніми ознаками, %
від загальної кількості, не більше
всього
8,0
12,8
10,5
15,3
у тому числі:
- тяжкими вірусними хворобами,
віроідом веретеноподібності бульб і
мікоплазмами
1,5
2,0
2,5
3,5
- легкими вірусними хворобами
6,0
9,5
7,5
10,0
- чорною ніжкою
0,5
1,0
0,5
1,5
- кільцевою і бурою бактеріальною
Не допус¬
Не допус¬
гниллю
кається
0,3
кається
0,3
446
Таблиця 6.10 - Показники посівних якостей репродукційного насінного
матеріалу картоплі
Назва
Норми репродукцій
PH-1-2
PH-3-4
Наявність бульб інших сортів, % від загальної кількос¬
0,5
1,0
ті у пробі, не більше
Розмір бульб за найбільшим поперечним діаметром,
мм, для сортів із формою бульб:
- видовженою
28-55
28-55
- округло-овальною
30-60
30-60
Наявність бульб, що не відповідають вимогам за роз¬
міром, % від загальної кількості у пробі, не більше
5
7
Наявність бульб, уражених хворобами, % від загаль¬
ної кількості у пробі, не більше
7,8
13,0
у тому числі
- мокрою гниллю
Не допускається
- чорною ніжкою
0,3
0,5
- фітофторозом
2,0
4,0
- сухою гниллю (фомоз, фузаріоз)
1,0
1,5
- ризоктоніозом з ураженням поверхні бульб:
- від 1/8 до 1/4
1,5
2,0
- більше 1/4
Не допускається
- паршею звичайною і сріблястою (при ураженні
Ѵл поверхні бульби)
3,0
5,0
- паршею порошистою
Не допускається
Наявність бульб, уражених шкідниками і механічно,
% від загальної кількості у пробі, не більше
всього
8,0
10,5
у тому числі:
- стебловою нематодою
0,5
1,0
- дротяником
2,0
2,0
- хрущами, совками (без ушкодження вічок)
1,0
1,0
- з механічними пошкодженнями тканини (глиби¬
ною не більше 10 мм)
4,5
6,5
Наявність бульб з ознаками задухи, підморожених,
з опіками, виродливих, з наростами, роздавлених,
різаних (менше Ѵг їхнього розміру), з обдертою шкір¬
кою (більше 1А поверхні)
Не допускається
Наявність землі і сторонніх домішок, % від маси
9
о
бульб, не більше
z
447
В акти апробації вносять основні дані про агротехніку сортової
картоплі на підставі перевірки відповідних актів на проведені заходи
(внесення добрив, дати прочисток, густота насаджень та ін.).
Агроном-апробатор, використовуючи результати проведеної ап¬
робації, записує свої рекомендації щодо вирощування та зберігання
сортової картоплі. Акт підписують апробатор і особа, що відповідає
за насінництво картоплі в господарстві. Керівник господарства дає
гарантійне зобов’язання про виконання вказівок апробаторів і підпи¬
сує акт. Акт прочистки сортових посівів картоплі складають в одно¬
му примірнику і зберігають у господарстві. На сортові насадження,
які не підлягають апробації, складають акт реєстрації у двох примір¬
никах, один з яких залишається у господарстві.
Основні морфологічні ознаки сортів. При визначенні сортів
картоплі користуються морфологічними ознаками куща, стебла, лис¬
тка, квітки, бульби. Ці ознаки, залежно від умов вирощування (ґрунт,
погодні умови, агротехніка), можуть змінюватися. Мінливість їх не¬
однакова: деякі з них, такі, як забарвлення бульб, квіток, паростків,
форма листків та ступінь залистяності, залишаються постійними, то¬
ді як форма бульб, стебла, куща і його забарвлення можуть змінюва¬
тись і бути не зовсім типовими для сорту.
Кущ. Має відносно мало сортових ознах. Основною ознакою со¬
рту є загальний вигляд куща, тобто його габітус, який залежить як
від висоти і кількості стебел, так і від розміщення їх у просторі та
залистяності.
Кущі бувають високі (Луговська, Витязь), середні (Світанок київ¬
ський, Берегиня, Слава) і низькі (Зарево), а залежно від розташуван¬
ня стебел - прямостоячі, коли стебла розміщені майже паралельно
(Українська рожева, Зов, Зарево), напіврозлогі (Незабудка, Невська -
цей тип куща характерний для більшості сортів) і розлогі.
За кількістю стебел кущі поділяють на малостеблові (Гатчинська,
Ласунак) і багатостеблові (Незабудка, Світанок київський).
Характерною і відносно стійкою ознакою є залистяність куща,
яка залежить від кількості листків на стеблі. За цією ознакою сорти
поділяють на сильнозалистяні (Світанок київський, Зарево), серед-
ньозалистяні (Мавка, Поліська рожева, Українська рожева) і слабо-
залистяні. Більшість сортів мають середню залистяність.
Стебло. Характерними ознаками є розміщення стебла у прос¬
торі, наявність пігментації, ребристість, крилатість, галуження
(рис.6.8).
Стебло може бути прямим (Українська рожева), колінчастим
(Огоньок). В одних сортів стебла сильнопігментовані, крапчастопіг-
ментовані, пігментовані в основі стебла.
У більшості сортів стебла кутасті (Ласунак, Витязь, Українська
рожева, Зов) і лише в окремих сортів - округлі. Деякі сорти мають
гіллясті стебла (Берегиня, Пролісок).
448
Рис. 6.8. Ознаки стебла:
1 - крила; 2 - прилистки;
З - пігментація в пазухах
зміщеними. Існують сорти з нестійким роз¬
міщенням часточок (у першій і другій сері¬
ях - то серединним, то кутовим).
За способом прикріплення до стрижня
часточки ділять на стрижневі, сидячі і збіж¬
ні, а за формою - на вузькі та округлі. Рідкіс¬
ною сортовою ознакою є кутове розміщення
часточок першої серії; за способом прикрі¬
плення - сидячі часточки і за формою - ву¬
зькі (заячі вушка).
Рис. 6.9. Будова листка: КЧ - кінцева частка;
Ч І - частка першої пари; Ч П - частка другої
пари; КЧа - часточки кінцевої частки; ЧІа -
часточки першої серії; ЧІІа - часточки другої
серії; Чсер - часточки серединні; Чкут - час¬
точки кутові; Чзм - часточки зміщені;
Ч III - частка третьої пари; Ч ПІа - часточки
третьої серії; Ч IV - частка четвертої пари;
Ч IVa - часточки серії
У багатьох сортів на гранях є так звані
крила, які можуть бути ледь помітні і силь¬
но виражені, хвилясті (Гатчинська).
Листок. Листок картоплі переривчасто-
непарнопірчасторозсічений (рис.6.9). Скла¬
дається він із стрижня, на якому розміщена
непарна кількість (3-7) великих часток,
кілька пар менших бокових часточок. Час¬
тки і часточки сидять на коротких стриже-
ньках, які прикріплені до листкового стриж¬
ня. Залежно від розміщення на стрижні,
вони мають свою назву. Непарна частка,
розміщена на кінці стрижня, називається
кінцевою (КЧ), парні бокові мають поряд¬
кові назви - перша пара, друга пара і т. д.
При визначенні сорту важливе значен¬
ня мають розміщення, форма і спосіб при¬
кріплення часточок, особливо у першій та
другій серіях. Якщо пара часточок розмі¬
щена між частками серії посередині, то їх
називають серединними. Часточки, що си¬
дять не на стрижні, а в кутку між стрижнем
і стриженьком частки, мають назву куто¬
вих. У деяких сортів часточки зустрічають¬
ся на стриженьках часток, їх називають
'/, 15 2-132
449
Розсіченість, форма і розміщення часток і часточок мають велике
значення для сортовизначення. Розсіченість листка визначають за
кількістю часточок у серії. При наявності двох і більше часточок
розсіченість сильна, однієї пари, а іноді й без них - слабка. Залежно
від розсіченості та відстані між частками листок буде рідко- чи гус-
точастковим.
Важливими морфологічними ознаками листка є форма кінцевої
та бокових часток (рис.6.10). За формою кінцева частка буває широ¬
кою, коли ширина майже дорівнює довжині; вузькою, якщо ширина
у 2 рази менша від довжини; овальною, коли найбільша ширина про¬
ходить по середині частки; яйцеподібною - частка ширша знизу та
обернено яйцеподібною - частка ширша зверху.
Рис.6.10. Форма кінцевих часток: а - широка; б - овальна;
в - вузька; г - яйцеподібна; д - обернено яйцеподібна
Виразною ознакою є форма основи кінцевої частки (рис. 6.11),
яка може бути серцеподібною, клиноподібною або проміжною. Вер¬
хівка кінцевої частки може бути з довгим чи коротким збігаючим або
відтягнутим кінчиком. У деяких сортів кінцева частка зростається з
однією або двома частками першої пари. Таке явище має назву
“плющолистість” (рис. 6.12). У деяких сортів картоплі характерною
особливістю листка є так зване налягання, коли перша пара часток
налягає на кінцеву (рис. 6.13).
Рис. 6.11. Основа кінцевої частки: а - серцеподібна;
б - проміжна; в - клиноподібна
450
Рис. 6.12. Плющолистість
Рис. 6.13. Налягання першої
пари часток
Бокові частки мають такі ж ознаки, що й кінцеві. Додатковими
ознаками є несиметричність будови, коли листкова пластинка з од¬
ного боку довша і ніби збігає по стерженьку, та наявність перетинки
між стрижнем і стриженьком у першої пари часток (рис. 6.14).
При визначенні сорту користуються так званим листковим індек¬
сом, тобто відношенням ширини частки до її довжини (рис.6.15). Час¬
тки у просторі мають неоднакове розміщення. Вони бувають плоскі,
складені уздовж головної жилки, загнуті у вигляді човника і загнуті
вниз, із хвилястими чи рівними краями.
Рис.6.14. Несиметричність
будови першої пари часток
Рис. 6.15. Листок з індексом 1:2
(довжина у 2 рази більша, ніж ширина):
аб - ширина; ав - довжина листка
451
Листки картоплі біля основи мають пару прилистків, які в загаль¬
них рисах нагадують будову листка. Прилистки бувають серпоподі¬
бні, листкоподібні і проміжні. Останні характерні для більшості сор¬
тів. У серпоподібної форми розвивається тільки одна половина кін¬
цевої пластинки, а друга, що огинає стебло, зовсім атрофована
(рис.6.16). У проміжної форми кінцева бокова половина розвивається
нормально, внутрішня - лише частково з кінця до половини. У лист-
коподібних прилистків розвиваються симетрично як обидві полови¬
ни кінцевої частки, так і пара бокових часток.
Рис. 6.16. Форма прилистків:
а - серпоподібна; б - проміжна; в - листкоподібна
Забарвлення листка також може використовуватись як ознака.
Воно буває світло-зелене (Незабудка) або темно-зелене (Гатчинська,
Зарево).
Кут прикріплення листків до стебла також є сортовою ознакою.
Так, у деяких сортів листок відносно стебла розміщується під гост¬
рим кутом, а в інших - під прямим. Виразність жилкування листків
неоднакова у різних сортів картоплі. Жилкування буває сильне і
слабке.
Опушеність листків також буває неоднаковою: блискучі або
глянцеві листки слабкоопушені; матові - сильноопушені. Стрижень
листка і жилки бувають зелені або пігментовані. Пігментація стри¬
жня та жилок залежить від забарвлення бульб. У сортів із забарв¬
леними бульбами завжди пігментована основа головних і бокових
жилок. Часто спостерігається пігментація пазушок на з’єднанні
стрижня і стриженька частки, що пов’язано із забарвленням вічок
бульб.
Суцвіття. Характерною ознакою сорту картоплі є суцвіття. За
формою воно може бути компактне (Незабудка, Зов, Витязь, Україн¬
ська рожева) або ж розлоге (Гатчинська, Зарево).
452
Квітконоси бувають довгі (Берегиня, Гатчинська) і короткі (Не¬
забудка, Посвіт). Деякі сорти мають пігментовані квітконоси і квіт¬
коніжки (Зарево), а інші - зелені квітконоси (Зов, Ласунак, Неза¬
будка). В окремих випадках пігментація чітко виражена тільки в
розгалужених завитках та в місці з’єднання верхньої частини квіт¬
коніжки з нижньою (рис.6.17), що має вигляд кільця (Поліська
рожева).
У деяких сортів на розгалужених суцвіттях зустрічаються верхові
листочки (рис.6.18), що є також розпізнавальною ознакою (Зарево,
Житомирянка).
2 - на з’єднанні квітконіжки
Квітка. Квітка картоплі складається з чашечки з п’ятьма чашо¬
листками, п'ятипелюсткового зрощеного віночка, п’яти пиляків та
маточки із стовпчиком, приймочкою і зав’яззю (рис. 6.19).
Найбільш виразною ознакою квітки є чашечка, що характеризу¬
ється пігментацією, опушенням, формою і довжиною чашолистків.
Чашечка може бути слабкоопушена і сильноопушена без пігментації
(Незабудка, Світанок київський), суцільно пігментована лише в ос¬
нові (Зарево). Чашолистки бувають листоподібні, шилоподібні та
ланцетоподібні (рис.6.20).
Віночок квітки має деякі особливі ознаки. Найхарактернішою з
них є забарвлення. Воно буває білим (Незабудка, Мавка, Ласунак,
Слава), червоно-фіолетовим (Берегиня, Зов, Гатчинська, Світанок
київський) і синьо-фіолетовим (Темп, Гарт).
Розмір віночка не є характерною ознакою, але інколи також може
бути використаний для визначення сорту. Як правило, великий віно¬
чок характерний для сортів, які цвітуть рясно.
2
Рис. 6.17. Пігментація:
1 - на розгалуженнях квітконоса;
Рис.6.18. Суцвіття з верховими
листочками
7 + 15 2'132
453
Рис. 6.19. Будова квітки:
1- віночок; 2 - пиляки, зі¬
брані в колонку; 3 - зірка
Рис. 6.20. Чашечка квітки:
а - з шилоподібними чашолист¬
ками; б - з листоподібними
чашолистками
“Махровість” віночка також є ознакою для визначення сортів
(рис.6.21); вона буває внутрішньою та зовнішньою.
Велике значення для визначення сорту має забарвлення, розмір та
форма пиляків. За забарвленням вони бувають оранжеві, жовті та
жовто-зелені. Колонка, в яку зібрані пиляки, може бути правильної
(циліндрична чи конусоподібна) і неправильної форми (рис.6.22).
Зав’язь квітки також має свої сортові ознаки. Вона буває овальною
або грушоподібною, а на розрізі забарвленою (у сортів, що мають чер¬
воні бульби) або ж не забарвленою (у сортів з білими бульбами).
Плід картоплі — багатонасінна, двогнізда ягода. Утворення ягід
буває рясним (Зарево), середнім або не відбувається (Незабудка).
Бульби. Характерними ознаками бульб є форма, глибина залягання
вічок, забарвлення і характер шкірки, забарвлення м’якоті і паростка.
За формою бульби бувають округлі (Зарево, Світанок київський,
Гарт, Радич, Либідь, Кобза, Посвіт, Зов), округло-овальні (Берегиня,
а
б
Рис. 6.21. Махровість квітки:
а - зовнішня; б - внутрішня
Рис. 6.22. Форма колонки пиляків:
а - правильна (конічна);
б - неправильна (потворна)
Незабудка, Темп, Водограй, Доброчин, Малич), овальні (Українська
рожева, Купава, Ікар, Поліська рожева), видовжено-овальні (Сло¬
в’янка), довгі (Дезіре).
Залягання вічок у різних сортів буває неглибоким (Луговська,
Світанок київський, Берегиня, Адретта, Незабудка), середнім (Зов,
Посвіт) і глибоким (Обрій).
Шкірка може бути гладенька (Луговська, Незабудка, Зов), сітчас¬
та (Витязь), лускувата (Українська рожева).
Забарвлення бульб буває рожеве (Берегиня, Купава, Древлянка,
Зарево, Доброчин, Радич, Українська рожева), біле (Кобза, Гарт, Зов,
Водограй, Гатчинська, Ласунак), синьо-фіолетове. Більшість сортів
мають червоне і біле забарвлення бульб, але зустрічається і жовте чи
кремове (Адретта, Пролісок, Західна).
Забарвлення м’якоті буває білим (Луговська, Невська, Божедар,
Гарт, Радич, Обрій, Косень 95), жовтим (Бородянська рожева, Воло-
вецька) і кремовим (Незабудка, Житомирянка, Слов’янка, Чернігів¬
ська рання, Водограй, Кобза).
Паростки за забарвленням бувають червоно-фіолетовими (Мавка,
Невська) або синьо-фіолетовими (Темп, Гарт). Переважна більшість
сортів має червоно-фіолетові паростки бульб.
Порядок визначення сортів. Найкращим строком визначення
сортів є період цвітіння першого ярусу суцвіття.
Квіти беруть із суцвіть, у яких розпустилися не менше 2-3 квіток
у слабкоцвітих сортів і не менше 3-4 квіток - у рясноцвітих. При
визначенні кольору квіток потрібно використовувати квіти, які тіль¬
ки розпустились, а для визначення оптимальних ознак слід користу¬
ватись квітами, які вже повністю закінчили свій ріст і ознаки у яких
чітко виражені.
Найбільш типові листки для сорту розміщені у середній частині
куща (це 5-7-й лист від суцвіття). Ці листки в цей час закінчили ріст,
і ознаки у них чітко виражені.
Визначають сорт не за однією, а за сукупністю ознак. При цьому
основну увагу слід звертати на основні ознаки куща, стебла, листка,
квітки.
В усіх сортів картоплі існує кореляція між забарвленням бульб, па¬
ростків і квітів. За ознаками забарвлення бульб і квіток сорти картоплі
України для зручності їх визначення можна поділиш на п'ять таких груп:
I - квіти білі, бульби білі або жовті (Адретта, Кобза, Обрій, Ли-
бідь, Горлиця, Ласунак, Мавка, Невська, Незабудка, Посвіт, Водо¬
грай, Слава);
II - квіти червоно-фіолетові, бульби білі (Придеснянська, Воло-
вецька, Гатчинська, Зов);
III - квіти червоно-фіолетові, бульби червоні або рожеві (Радич,
Косень 95, Зарево, Берегиня, Поліська рожева, Українська рожева,
Божедар, Слов’янка, Повінь);
455
ГѴ - квіти білі, бульби червоні або рожеві (Луговська);
V - квіти синьо-фіолетові, бульби білі (Гарт, Темп).
Ґрунтовий контроль еліти проводить Українська державна насін¬
нєва інспекція. Контролю підлягає еліта за зразками супереліти,
які надсилають з усіх елітних господарств країни. Для здійснення
ґрунтового контролю в господарствах, які вирощують еліту, восени
від супереліти відбирають по кожному сорту середню пробу відпові¬
дно до ДСТУ 4014-2001. Із відібраної картоплі для середнього зраз¬
ка беруть 300 бульб. Кожен зразок пломбують і супроводжують ети¬
кеткою, на якій зазначають господарство, сорт, масу партії, дату від¬
бору, прізвище й посаду особи, що проводили цю роботу в
господарстві. Елітгосп за свій рахунок у визначені строки відправляє
зразки на адресу інспекції.
Весь зразок (300 бульб) аналізують за методикою бульбового
аналізу на наявність зовнішніх симптомів захворювань (без розрізу¬
вання бульб), потім 50 бульб використовують для лабораторного
аналізу (у зимовий час проводять індексацію бульб), а інші бульби,
незалежно від ураження їх хворобами (крім гнилі), весною висаджу¬
ють у польових умовах на спеціально відведеній для цього ділянці,
яка ізольована (не менше 500 м) від інших посадок картоплі і приса¬
дибних ділянок. Під час вегетації проводиться оцінка рослин на ура¬
женість вірусними, бактеріальними і грибковими хворобами.
Результати ґрунтового контролю еліти по кожному господарству
у визначені строки щорічно надсилають у Міністерство аграрної по¬
літики України, елітним господарствам, обласним управлінням сіль¬
ського господарства.
У випадку, якщо різниця між показниками ураженості посівів у
ґрунтоконтролі і польової оцінки під час приймання посівів еліти
становить понад 20%, проводять післязбиральну індексацію бульб,
яку проводять згідно з існуючою методикою.
Вимоги до сортових якостей еліти за результатами індексації ви¬
значають за показниками, зазначеними у державному стандарті.
Документом, що засвідчує сортові якості оригінального і репро¬
дукційного насіннєвого матеріалу, є акт апробації сортових посівів
картоплі.
Документами, що засвідчують сортові якості еліти, є акт апроба¬
ції сортових посівів картоплі, акт приймання посівів еліти, результа¬
ти ґрунтоконтролю еліти за надісланими зразками супереліти, ре¬
зультати індексації бульб у післязбиральний період.
Визначення посівних якостей. Посівні якості насінної картоплі
визначають шляхом відбору проб бульб від партії насіннєвого мате¬
ріалу картоплі та їх аналізу згідно з ДСТУ 4014-2001, у якому пе¬
редбачено певний порядок проведення аналізу.
Відбір проб неупакованого матеріалу. Виїмки насіннєвого ма¬
теріалу в кількості 8-10 бульб кожна у необхідній для об’єднаної
456
проби проводять у місцях зберігання перед відвантаженням або під
час контрольної перевірки за схемою (рис.6.23).
Рис.6.23. Схема відбору та аналізу проб під час визначення
посівних якостей насіннєвого матеріалу картоплі
Відбір проб з упакованого насіннєвого матеріалу. Виїмки для
об’єднаної проби відбирають від однієї партії або певної її частини з
розшитих мішків або сіток. При відборі проби із середнього і ниж¬
нього шарів мішка шар бульб, що знаходиться вище, висипають. За¬
лежно від обсягу партії кількість мішків і виїмок з них змінюється
(табл. 6.11).
Таблиця 6.11 - Кількість мішків і виїмок з них, що відбираються для формуван¬
ня проби за різної величини партії насіннєвого матеріалу
Кількість мішків, сіток,
ящиків у партії
Кількість мішків, сіток, з яких від¬
бирають виїмки
Кількість виїмок 3
мішка, сітки та їх
розміщення
До 100 включно
10
3 (з верху, середи¬
ни і низу мішка)
Від 101 до 200 включно
15
те саме
Від 201 до 400 включно
20
*
Від 401 до 600 включно
25
і»
Понад 601
25 і додатково по одному мішку,
сітці, ящику від кожних 100 наступ¬
них після заповнених або неповних
мішків, сіток або ящиків у партії
И
457
У кожному ящиковому піддоні (контейнері) виїмки відбирають із
верхнього, середнього і нижнього шарів бульб. Залежно від обсягу пар¬
тії кількість ящикових піддонів і виїмок з них змінюється (табл. 6.12).
Таблиця 6.12 - Кількість ящикових піддонів і виїмок з них, що відбираються
для формування проби за різної величини партії насіннєвого ма¬
теріалу
Кількість ящикових під¬
донів у партії
Кількість ящикових піддо¬
нів, з яких відбирають виїм¬
ки
Кількість виїмок з ящи¬
кового піддону та їх
розміщення
До 10 включно
3
3 (з верхнього, серед¬
нього і нижнього шарів
бульб у піддоні)
Від 11 до 30 включно
4
те саме
Від 31 до 50 включно
6
2 (з верхнього і ниж¬
нього шарів бульб у
піддоні)
Від 51 до 100 включно
8
II
Понад 101
8 і додатково по одному
ящиковому піддону від кож¬
них 50 наступних повних
або неповних ящикових
піддонів у партії
II
Виїмки з мішків, сіток, ящикових піддонів, що знаходяться в
транспортних засобах (у вагонах, автофургонах та ін.), допускається
відбирати з тих, які розміщені в доступних місцях без перекладання.
Від партії незатареної картоплі виїмки відбирають рівномірно з
різних місць шару насипу. Кількість поверхневих і глибинних проб
повинна бути рівнозначною.
Виїмки в кількості ЗО шт. від партії незатареної картоплі з пло¬
щею поверхні насипу до 20 м2 беруть рівномірно по поверхні насипу
і зверху - 15 пгг.; на глибині 50 см - 15 шт. (рис.6.24, схема А). Якщо
поверхня насипу більша, то на кожних додаткових 10 м2 площі повер¬
хні насипу беруть 6 виїмок, зверху - 3 пгг.; на глибині 50 см - 3 шт.
(рис.6.24, схема Б). Усі виїмки складають в одну об’єднану пробу.
Кожна виїмка складається з 30 бульб.
S=20 м2
S=30 м2
0 0 0 0 0
0 О О 0 О 0
0 0 0 0 0
0 0 0 0 0 0
0 0 0 0 0
0 О 0 0 0 0
Схема А Схема Б
Рис.6.24. Схеми взяття виїмок насіннєвого матеріалу від партії
незатареної картоплі
458
Виїмки відбирають руками, дерев’яними совковими лопатами
або дерев’яними, вкритими гумою, совками у відра, щільні мішки
або ящики із суцільними дном і стінками, не допускаючи втрат землі
і сторонніх домішок.
Об’єднану пробу формують із усіх відібраних виїмок, по черзі
висипаючи їх на чисту гладеньку поверхню (брезент, клейонку, плів¬
ку, фанеру та ін.), не допускаючи втрат землі та інших сторонніх до¬
мішок.
Виділення середніх проб. Висипавши бульби об’єднаної проби
на рівну поверхню, рівномірно розгортають їх у формі квадрата і
підряд з будь-якої сторони відділяють 250 бульб. Це й буде середня
проба, яку зважують.
Від дрібних партій (до 1 тонни картоплі) допускається відбирати
середню пробу обсягом 250 бульб.
У масу середньої проби включають масу землі та інших сторон¬
ніх домішок, що обсипалися з бульб під час виділення середньої
проби.
Залишок об’єднаної проби після виділення середньої проби зважу¬
ють і повертають у партію насіннєвого матеріалу, звідки вона була взята.
Масу бульб об’єднаної проби, що залишилася після формування серед¬
ньої проби, додають до маси середньої проби і використовують у підра¬
хунках маси наявної вільної землі і сторонніх домішок.
Визначення посівних якостей. Аналіз здійснюється в такій по¬
слідовності: визначення кількості землі і сторонніх домішок; визна¬
чення розміру бульб; кількості бульб із зовнішними ознаками ура¬
ження хворобами і пошкодженнями; кількості бульб, уражених
хворобами у прихованій формі (кільцева гниль, бура бактеріальна
гниль, чорна ніжка, стеблова нематода, фітофтороз) і бульби інших
сортів.
Визначення наявності землі та інших сторонніх домішок. По¬
слідовно визначають масу вільної землі і сторонніх домішок; масу
землі, що налипла на поверхні бульб.
Середню пробу зважують у тарі і визначають її масу, віднімаючи
масу тари від результатів зважування.
Бульби перекладають із мішків або ящиків на чисту поверхню
або брезент і підраховують. Вільну землю, що залишилася в мішку
або ящику, і сторонні домішки збирають і зважують. Масу наявної
вільної землі і сторонніх домішок (разом) обчислюють у відсотках
від маси об’єднаної проби.
Середню пробу (після виділення вільної землі і сторонніх домі¬
шок) зважують і відмивають від землі водою. За незначної забрудне¬
ності допускається обтирання бульб ганчірками. Обмиті або обтерті
бульби кладуть на решета або в корзину і витримують 5-10 хв для
того, щоб з поверхні бульб стекла вода. Потім чисті бульби зважу¬
ють із похибкою ±0,01 кг. Для визначення маси чистих бульб від їх
459
маси віднімають масу води, що залишилася на поверхні бульб, яка
умовно становить один відсоток маси відмитої картоплі.
Масу землі, що налипла на поверхні бульб, визначають за різни¬
цею маси проби до і після відмивання і обчислюють у відсотках до
маси об’єднаної проби.
Землю і сторонні домішки, що залишилися в транспортних засо¬
бах після вивантаження картоплі, збирають і зважують. Обчислюють
їх масу у відсотках від маси картоплі.
Загальну кількість землі і сторонніх домішок обчислюють у від¬
сотках, підсумовуючи результати, одержані відповідно до маси зем¬
лі, що налипла на поверхні бульб, і маси землі та сторонніх домішок,
що залишилися в транспортних засобах.
Визначення розміру бульб. У бульб об’єднаної проби вимірю¬
ють штангенциркулем найбільший поперечний діаметр з похибкою
±1 мм або пропускають через квадратні калібри решіт із розміром от¬
ворів 28 і 55 мм для сортів із видовженою формою бульб; ЗО і 60 мм -
для сортів з округло-овальною формою бульб і сортують їх на дві
фракції: бульби, розміри яких відповідають вимогам ДСТУ 4013 [1];
бульби, розміри яких не відповідають вимогами ДСТУ 4013 [1].
Підраховують кількість бульб, що не відповідають вимогам
ДСТУ 4013 [1] за розмірами, і цю кількість віднімають від загальної
кількості в об’єднаній пробі. Результат виражають у відсотках до
загальної кількості бульб в об’єднаній пробі.
Визначення наявності бульб із зовнішніми ознаками ураження
хворобами, пошкодженнями та інших сортів. В об’єднаній пробі піс¬
ля видалення землі і домішок за зовнішніми ознаками виділяють бульби
з дефектами в такій послідовності: з ознаками задухи, підмерзлі, з опі¬
ками, потворні, із вторинними бульбами (дітками) і наростами, що легко
обламуються; уражені хворобами (мокрою гниллю, кільцевою і бурою
бактеріальною гниллю, чорною ніжкою, фітофторозом, стебловою не¬
матодою, сухою гниллю, ризоктоніозом, паршею звичайною, срібляс¬
тою, порошистою); з ознаками відмирання вічок; пошкоджені механіч¬
но, шкідниками, гризунами. Результати обліку виражають у відсотках до
загальної кількості бульб в об’єднаній пробі.
Глибину механічних пошкоджень бульб картоплі і пошкоджень
шкідниками і гризунами визначають методом послідовного зрізуван¬
ня пошкоджень м’якоті бульб картопляним ножем, що забезпечує
зріз висотою 1,5 мм. Довжину механічних пошкоджень вимірюють
лінійкою з похибкою ± 1 мм.
Бульби вважаються пошкодженими, якщо глибина пошкодження
м’якоті є більшою ніж 4,5 мм. Площу пошкоджень і уражень визна¬
чають візуально.
Визначення наявності бульб інших сортів проводять одночасно з
визначенням зовнішніх дефектів, відбираючи окремо бульби з нети¬
повим забарвленням шкірки і нетипові за формою. Належність до
460
даного сорту нетипових за формою бульб визначають згідно з Реєст¬
ром сортів рослин України.
Визначення наявності бульб, уражених хворобами у прихова¬
ній формі (кільцева гниль, бура бактеріальна гниль, чорна ніжка,
стеблова нематода, фітофтороз). Із різних місць об’єднаної проби
після відокремлення бульб, уражених за зовнішніми ознаками мок¬
рою, кільцевою або бурою бактеріальною гниллю, стебловою нема¬
тодою, фомозом, фузаріозом і фітофторозом, відбирають без вибору
не менше 200 бульб, розрізують їх уздовж ножем через місце при¬
кріплення до столону й оглядають м’якоть бульби. На місці прояв¬
лення кільцевої гнилі зрізують тонкий шар покривної тканини.
Одночасно із хворобами у прихованій формі на розрізі м’якоті
визначають наявність бульб, що мають забарвлення, відмінне від ос¬
новного сорту. Результати аналізу виражають у відсотках до загаль¬
ної кількості бульб в об’єднаній пробі.
Результати аналізу переносять на всю партію і відображають в
акті аналізу бульб насіннєвої картоплі.
Правила обробки результатів аналізу. Бульби з наявними де¬
фектами підраховують по кожному виду ураження хворобами чи
пошкодженнями, що перераховані в ДСТУ 4013 [1]. Наявність хво¬
рих і ушкоджених бульб, а також бульб інших сортів обчислюють у
відсотках від кількості проаналізованих бульб із точністю до першо¬
го десяткового знака.
Під час підрахунку бульб, уражених хворобами, що мають зов¬
нішню і приховану форму прояву (кільцева і бура бактеріальна
гниль, чорна ніжка, стеблова нематода, фітофтороз), підсумовують
процентний уміст виявлених хворих бульб.
За наявності на одній бульбі кількох видів хвороб або пошко¬
джень враховують одне найбільш шкодочинне; дефектів, забороне¬
них ДСТУ 4013 [1] або таких, що мають допуски, та пошкодження
або хвороби, для яких у названому стандарті є допуски, враховують
один вид дефекту, пошкодження або хвороби, залежно від ступеня їх
шкідливості.
За ступенем шкідливості хвороби і пошкодження розподіляють у
спадаючій послідовності: мокра гниль, кільцева гниль, бура бактері¬
альна гниль, чорна ніжка, стеблова нематода, фітофтороз, суха гниль
(фомоз, фузаріоз), ризоктоніоз, парша звичайна, парша срібляста,
парша порошиста, пошкодження сільськогосподарськими шкідника¬
ми, механічні пошкодження, інші хвороби.
Правила оформлення результатів аналізу. Результати аналізу
переносять на контрольовану партію насіння картоплі і записують в
акт аналізу насіннєвих бульб.
Документом, що засвідчує посівні якості оригінального садивно¬
го матеріалу, еліти або репродукційного садивного матеріалу, є акт
аналізу насіннєвих бульб картоплі.
461
Визначення посівних якостей сортового садивного матеріалу
картоплі здійснюється згідно з вимогами ДСТУ 4013-2001.
ЛІТЕРАТУРА
1. ДСТУ 4013-2001. Сортові та посівні якості картоплі насінної. Технічні
умови. - К.: Держстандарт України, 2001.
2. ДСТУ 4014-2001. Картопля насіннєва. Відбір проб і методики визна¬
чення посівних якостей. - К.: Держстандарт України, 2001.
3. Інструкція по апробації сортових посівів // Земля і люди України. -
1995.-№1.
6.8. ПРИСКОРЕНЕ РОЗМНОЖЕННЯ
КАРТОПЛІ
Найчастіше картоплю розмножують вегетативним способом, пе¬
реважно цілими бульбами. При загальноприйнятих способах підго¬
товки садивного матеріалу проростають лише 3-4 бруньки, розташо¬
вані на верхівковій і середній частинах бульби. Якщо врахувати, що
на бульбі є в середньому 6-12 вічок і в кожному з них по 3-5 бру¬
ньок, то коефіцієнт використання закладених бруньок дуже низький і
не перевищує 16-17%. Обробка бульб різними фізіологічно актив¬
ними речовинами або витримування їх у приміщенні з регламентова¬
ним вмістом кисню (5%) сприяють проростанню значно більшої кі¬
лькості бруньок. У дослідах вдавалося одержати на бульбах окремих
сортів по 26-35 паростків. Проте, під дією наявної в бульбах системи
“регулятори-інгібітори”, яка регулює пробудження бруньок і ріст
паростків, нормально розвивалося лише 6-8 стебел. Спостереження
за ростом і розвитком рослин з такою кількістю стебел показали, що
в урожаї переважали дрібні бульби. Здебільшого загальна кількість
бульб була такою самою, як у кущах із 3-4 стеблами. Раціональне
використання кущів із 6-8 стеблами потребує посиленого живлення
та оптимального режиму вологості й аерації грунту. Разом з тим ідея
підвищення коефіцієнта використання закладених на бульбі бруньок
заслуговує на увагу.
Відомі кілька способів прискореного розмноження картоплі: зе¬
леними паростками, частинами куща, зеленими живцями, етіольова-
ними паростками, різаними бульбами та насінням.
Крім того, у селекційному процесі і насінницькій практиці відомо
ще кілька способів, зокрема: повторне садіння маточних бульб, вий¬
нятих з-під куща, повторне садіння зелених кущів після раннього
збирання, розмноження апікальною (верхівковою) меристемою,
живцюванням та ін.
Розмноження картоплі зеленими паростками. За 20-25 днів до
садіння бульби пророщують на світлі і висаджують в утеплені гряд¬
ки, парники або горщечки в теплиці. Великі бульби ріжуть навпіл
462
упоперек і кладуть зрізом донизу, щільно одна біля одної, присипа¬
ють сумішшю перегною або торфу із землею шаром 3—4 см. Після
цього поливають теплою (15-18°С) водою, укривають поліетилено¬
вою плівкою, щоб підтримувати температуру на рівні 14-17°С. Після
появи сходів плівку на день знімають. Коли паростки досягнуть 10-
15 см, бульби виймають з ґрунту, обламують з них паростки, які ра¬
зом з корінням укладають у корзини або ящики, накривають вологою
мішковиною і вивозять у поле для садіння. Садити найкраще у по¬
хмуру погоду в добре підготовлений ґрунт з одночасним поливан¬
ням. Бульби знову висаджують у теплі грядки і через 10-15 днів
удруге обламують паростки. За 3-4 обламування з кожної бульби
можна одержати 25-30 паростків. Останній раз паростки не обламу¬
ють, а розрізають бульби на частини і висаджують у ґрунт.
Розмножуючи таким чином цінний безвірусний матеріал картоп¬
лі, можна одержувати по 25-30 рослин з однієї бульби. Урожай з ко¬
жної рослини становить 400-500 г, а всього з однієї материнської
бульби може бути одержано по 12 кг картоплі. При цьому коефіцієнт
використання бруньок зростає у 4-7 разів.
Розмноження картоплі частинами куща і зеленими живцями.
Цей метод полягає в тому, що в польових умовах вибирають добре
розвинені кущі з чотирма і більше стеблами заввишки 10-15 см до
початку бутонізації і, відокремивши кожне стебло разом з кореневою
системою, пересаджують їх на нове місце. Пересаджені рослини по¬
ливають і ретельно доглядають. Таким способом можна з одного
одержати 2-3 повноцінні кущі.
Розмножувати картоплю зеленими живцями можна у теплицях
або парниках, де їх висаджують у легкий ґрунт, добре поливають і
накривають у перші 2-3 дні поліетиленовими ковпаками. Після уко¬
рінення їх висаджують у спеціальні грядки або безпосередньо в поле.
З однієї рослини можна взяти без шкоди для основного куща 6-7
живців. Цей спосіб розмноження трудомісткий і потребує певних
навиків.
Розмноження картоплі етіольованими паростками. При під¬
вищеній температурі у сховищах картопля напровесні інтенсивно
проростає, пронизуючи товщу бульб великими етіольованими па¬
ростками. Як правило, їх обламують і викидають, не знаючи про те,
що це цінний матеріал для одержання розсади. Якщо цим займатися
спеціально, то картоплю слід закладати у темному приміщенні у во¬
логе середовище (торф, тирса) шаром 2-3 бульби і витримувати до
тих пір, доки не утворяться паростки завдовжки 3-5 см. Ці паростки
разом з корінцями обламують і висаджують в утеплені грядки з та¬
ким розрахунком, щоб верхівка паростка виходила з ґрунту не більш
як на 1 см. При досягненні рослинами висоти 10-15 см їх висаджу¬
ють у ґрунт. Паростки обламують кілька разів, а в останній раз буль¬
бу розрізають на частини за кількістю утворених паростків і теж ви-
463
саджуютъ в утеплені грядки. Таким способом з однієї бульби можна
одержати 20-25 рослин.
Використання різаних бульб картоплі для садіння. Садивна нор¬
ма, яка досягає 40-45 ц/га бульб, стає перешкодою у виробництві деше¬
вої картоплі. При розрахунку садивної норми виходять з необхідності
садіння на гектар 50-60 тис. бульб середньою масою 50-80 г. Для одер¬
жання бульб такої маси рекомендується проводиш загущене садіння,
раннє збирання, пізнє літнє садіння тощо. Проте ці заходи не завжди
дають бажані результати. Часто, залежно від метеорологічних умов, що
склалися, і рівня агротехніки в урожаї садивної продукції одержують
значну кількість бульб масою 150-200 і більше грамів. Такі бульби ви¬
користовують, як правило, на господарські потреби, хоч з агрономічного
погляду це невірно. Адже великі бульби є більш якісним садивним ма¬
теріалом як за врожайністю, так і за життєвістю потомства.
Надійним шляхом, який дає змогу використати переваги великих
бульб і водночас зменшити садивну норму, є садіння картоплі части¬
нами великих бульб.
Питання зменшення садивної норми картоплі не нове. Перші дослі¬
дження з різання бульб проводилися ще в XIX столітті [12-14]. Вони
показали, що різання бульб дозволяє вдвоє зменшити садивну норму,
разом з тим від половинок одержують врожай вищий, ніж від цілих
бульб середнього розміру, а інколи навіть близький до врожаїв від цілих
бульб, у два рази більших за масою, ніж половинки.
На початку XX ст. фундаментальні дослідження з різання садив¬
них бульб були проведені І. Шиповим [15], В.В. Вінером [1] і С.П.
Кульжинським [2], у яких було обґрунтовано методи різання бульб
перед садінням, вивчені строки і способи, показана цінність різних
частин бульби картоплі.
Автори допускали можливість різання великих, середніх і навіть
дрібних бульб, віддаючи перевагу верхівкам. Застосування для са¬
діння верхівок бульб давало змогу, не знижуючи врожайності, знач¬
но зменшити садивну норму і використовувати дві третини бульби
на господарські потреби.
Детально вивчивши фізіологічні і біохімічні процеси, які прохо¬
дять у бульбах та в окремих їх частинах, Ф.А. Новіков [10, 11] дій¬
шов висновку про перевагу верхівкової частини бульб над серед¬
ньою і пуповинною, розробив технологію зрізування верхівок з
бульб, зберігання і садіння їх, довів можливість використання верхі¬
вок для садіння. Ці праці були взяті за основу для розробки рекомен¬
дацій з використання верхівок бульб у виробничих умовах.
Н.С. Ничипоренко [8, 9], проводячи дослідження в аналогічних
умовах, довів, що різання бульб на поздовжні половинки збільшило
кількість пророслих вічок від 48,1 до 67,9%, тобто майже на 20% від
їх загальної кількості на бульбі, при цьому кількість рослин, ураже¬
них грибковими хворобами, не збільшилася. Різання, проведене без¬
464
посередньо перед садінням, не зменшує густини насадження і дозво¬
ляє одержати здорові рослини.
У колишньому Радянському Союзі різання картоплі особливо
поширилося під час Другої світової війни.
Науково-дослідну роботу в той час вели у напрямі вивчення фізіо¬
логічних особливостей різних частин бульби, обґрунтовували пере¬
ваги верхівок перед іншими частинами бульби, вивчали строки заго¬
тівлі верхівок і способи їх зберігання.
Досягнення науки й практики у вивченні і впровадженні у вироб¬
ництво оптимальних строків різання і способів зберігання верхівок,
безумовно, незаперечні. Проте ці процеси були дуже громіздкими. У
післявоєнні роки, як тільки послабла напруженість з садивним мате¬
ріалом, різання як агрозахід втратило свою актуальність і стало мало
застосовуватися.
Питання щодо скорочення садивної норми картоплі вивчалося і
вивчається й тепер у багатьох країнах світу.
Слід відзначити, що в країнах Західної Європи за умов вологого
клімату не виключається можливість перезараження різаних бульб
грибковими хворобами, тому в цій кліматичній зоні даний захід за¬
стосовують обережно, за нестачі садивного матеріалу або для при¬
скореного розмноження нового цінного сорту.
Велика увага приділялася захисту різаної картоплі від грибкових
і бактеріальних хвороб у СІЛА. Основним методом вважали ствот
рення механічного бар’єра, що захищає шматочок від зневоднення і
зараження мікроорганізмами. Вивчали різні сипкі матеріали і дуети
(сірчаний порошок, гіпс, вапно, мелену цеглу тощо), а також хімічні
речовини, здатні створити біологічний бар’єр (агроміцин, семесан-
бел, кахетин та ін.). Результатом проведених досліджень були реко¬
мендації, застосування яких давало змогу одержати задовільний
врожай від зниженої в 2-3 рази садивної норми. Разом з цим поліп¬
шувалися товарні показники врожаю і збільшувалася середня маса
товарної бульби, що є одним з важливих факторів оцінки якості про¬
довольчої картоплі.
Таким чином, більшість наукових праць характеризує різання
бульб як вигідний агрозахід, який дозволяє не лише зекономити са¬
дивний матеріал, а й збільшити середню масу бульб в урожаї та їхню
товарність [4].
Аналізуючи питання про можливість зменшення норми садіння
картоплі за рахунок різання бульб, інколи стикаємося з суперечли¬
вими висновками численних авторів, які проводили свої досліди в
різні роки і в різних ґрунтово-кліматичних умовах.
Невдачі в одержанні високого врожаю від різання бульб, на нашу
думку, зумовлюються кількома причинами. По-перше, незадовіль¬
ними умовами зберігання завчасно нарізаного садивного матеріалу
(невідповідність вологості повітря, температури, газового складу).
465
По-друге, відсутністю сприятливих умов у ґрунті в момент садіння
неопробковілих шматочків бульб, у зв’язку з чим вони уражуються
гнильними мікроорганізмами (надлишок або нестача вологи в ґрунті,
висока або низька температура, погана аерація тощо). По-третє, не¬
дотриманням запобіжних заходів під час різання картоплі, що при¬
зводить до перезараження шматочків грибковими і бактеріальними
хворобами (відсутність дезінфекції приміщення, тари, різальних апа¬
ратів машин або ножів). По-четверте, ігноруванням сортових відмін¬
ностей під час різання бульб у весняні і літні строки (відсутністю
паспортизації сортів за їх реакцією на різання бульб у певні строки).
І, нарешті, низьким рівнем механізації трудомістких процесів,
пов’язаних з різанням, зберіганням нарізаного матеріалу, садінням
його в ґрунт (відсутністю спеціальних машин).
Внаслідок згаданих причин урожайність картоплі від різаних
бульб залежить від багатьох випадкових факторів і буває високою за
сприятливих умов, і, навпаки, низькою під час порушення або відсут¬
ності хоч би одного з факторів.
У зв’язку з цим різання садивних бульб розглядають як у сільсь¬
когосподарській науці, так і в практиці у нас в країні як вимушений
захід і користуються ним лише тоді, коли відчувається гострий дефі¬
цит у садивному матеріалі.
Разом з тим питання забезпечення садивним матеріалом залиша¬
ється не повністю вирішеним. Мається на увазі зростаюча норма са¬
діння, необхідність використання великих садивних бульб, потреба
швидкого розмноження цінних сортів. Тому питання різання садив¬
них бульб залишається й надалі актуальним як для сільськогоспо¬
дарської науки, так і для практики особливо.
Стосовно сучасних умов різаний садивний матеріал картоплі має
відповідати наступним вимогам:
не поступатися за врожайністю і товарними якостями цілим са¬
дивним бульбам і не сприяти поширенню вірусних та інших хвороб
картоплі;
мати на зрізах надійний кірковий шар, який запобігав би зневод¬
ненню і ураженню шматочків мікроорганізмами.
Виконання цих вимог дає змогу одержувати високоякісний на¬
сіннєвий матеріал, у 2-3 рази зменшувати садивну норму, швидше і
краще використати нові високоврожайні сорти і на цій основі підви¬
щити врожайність картоплі.
Технологія різання садивних бульб. Строки різання бульб за¬
лежать від ґрунтово-кліматичних умов і строку садіння. Перед садін¬
ням різати садивні бульби можна лише тоді, коли температура та во¬
логість ґрунту дозволяють утворитись на шматочку бульби задовіль¬
ній рановій перидермі на зрізах у ґрунті. Якщо для цього умов немає,
різати слід завчасно, за 10-12 днів до садіння. У день садіння різати
можна лише бульби, пророщені на світлі.
466
Як уже зазначалося, всі частини бульби придатні для садіння.
Якщо бульби ріжуть вручну на дві частини, їх слід розрізати уздовж.
Великі бульби розрізають на 4-6 частин, спочатку навпіл, а потім
кожну половинку впоперек ще два-три рази залежно від розміру
бульби та кількості вічок. Машини зарубіжного виробництва ріжуть
бульби на 2, 3 і 4 частини тільки вздовж, внаслідок чого одержують
видовжені шматочки.
Доведено, що мінімальною масою шматочка бульби для одер¬
жання повноцінного врожаю є 25-30 г (табл. 6.13).
Таблиця 6.13 - Вплив різання садивних бульб та їхньої маси на врожайність і
якість картоплі
Садивні
бульби, їхня
маса, г
Норма
садіння,
ц/га
Урожайність, ц/га
Середня
Кількість
бульб під
кущем,
шт.
Коефіці¬
зага¬
льна
чиста
товар¬
ність,
процент
маса
бульби,
г
єнт
розмно¬
ження
Цілі,
80-100
45
256
211
97
121
5,9
6
50-60
26
248
222
97
132
5Д
10
Половинки,
40-50
22
246
224
97
153
4,2
11
25-30
13
224
211
98
146
4,5
15
Третинки,
27-33
12
236
222
98
161
4,3
17
Четвертинки,
20-25
11
200
189
97
182
3,8
18
Для різання більш придатні сорти, бульби яких округлої чи округло-
овальної форми, і вічка рівномірно проростають як на верхівці, так і
на пуповинній частині. Виявлено, що краще реагують на різання
бульб ранньостиглі сорти з невисоким вмістом крохмалю.
Слід пам’ятати, що різати можна лише здоровий садивний мате¬
ріал. Якщо картопля погано збереглася і в партії нараховується 10-
15% бульб, уражених грибковими і бактеріальними хворобами чи
нематодою, її різати не рекомендується. Садивні бульби слід різати
гострими ножами в захищеному від сонця та вітру приміщенні. У
таких умовах швидше відбувається суберинізація зрізів, що сприяє
захисту шматочків від зневоднення і хвороб.
За добу до різання бульби переносять у приміщення з температу¬
рою 15-20°С, щоб вони угрілись і в них активізувалися фізіологічні
процеси. Треба мати на увазі, що чим швидше утвориться ранова пе¬
ридерма (кірка) на різаних бульбах, тим менша ймовірність уражен¬
ня їх хворобами. Дотримання оптимального режиму є запорукою ви¬
сокої продуктивності садивного матеріалу. Оптимальною температу¬
рою для процесу регенерації кіркової тканини є 15-20°С. Проте, для
інтенсифікації ранових реакцій можливе підвищення її до 25-30°С.
467
Температура, вища за 30°С і нижча за 15°С, призводить до послаб¬
лення ранових реакцій і, відповідно, до погіршення якості опробко-
вілих шматочків бульб.
Оптимальною вологістю повітря слід вважати 94-96%. При низь¬
кій вологості шматочки бульб втрачають багато води і знижують
здатність до проростання.
Щоб не переносити інфекцію під час різання бульб, ножі і шматочки
бульб систематично дезінфікують знезаражуючими речовинами.
За даними Сквирської селекційно-дослідної станції [4], для при¬
скорення ранових реакцій різані бульби обробляють ферментами, що
містяться в екстракті ячмінного солоду 5-10 %-ної концентрації, які
сприяють швидкому перетворенню крохмалю в редуковані цукри і
звільненню верхнього шару клітин від крохмальних зерен. Підви¬
щенню інтенсивності дихання - необхідної умови для посилення ра¬
нових реакцій - сприяє бурштинова кислота, яку додають у розчин
солоду в концентрації 0,00063-0,0025% [3]. Про позитивний вплив
солодо-бурштинової суміші на утворення ранової перидерми свід¬
чать дані, наведені на рис. 6.25.
вода (контроль)
— солод 5%
282
24 години
48 годин
72 години
Рис. 6.25. Товщина суберинового шару і перидермальних клітин
залежно від обробки зрізів фізіологічно активними речовинами, мк
468
%
З інших вивчених ди- і трикарбонових кислот позитивно впли¬
вають на відкладення суберину цисаконітова, яблучна і фумарова, а
на утворення перидерми - лимонна і цисаконітова. Однак, у суміші
з солодом вони поступаються дії бурштинової кислоти.
У циклі Кребса бурштинова кислота перетворюється в фумарову,
яблучну і щавлевооцгову кислоти, що призводить до утворення АТФ.
Отримана в надлишку щавлевооцгова кислота і АТФ використовуються
в новому колі циклу Кребса, сприяючи різкому посиленню процесу ди¬
хання. Утворені при цьому фумарова, кетоглутарова, щавлевооцгова і
піровиноградна кислоти можуть перетворюватися в замінні амінокисло¬
ти, інкрустуючі речовини та інші важливі продукти метаболізму.
Проведені досліди показали, що внаслідок обробки шматочків
бульб солодо-бурштиновою сумішшю у прирановій зоні збільшуєть¬
ся вміст нуклеїнових кис¬
лот, амінокислот, аскор¬
бінової кислоти і розчин¬
ного білка, що служить
основою для прискорення
ранових реакцій (рис.
6.26). Показником сприят¬
ливого впливу на регене¬
раційні процеси солодо-
бурпггинової суміші слугує
рівень інтенсивності ди¬
хання цілих і різаних бульб
протягом зимово-весня¬
ного періоду (рис. 6.27).
Обробка нею зрізів сприяє
різкому посиленню тка¬
нинного дихання і більш
швидкій нормалізації про¬
цесу при завершенні рано¬
вих реакцій.
Нанесення на зрізи бульб
деяких похідних фенолу
також сприяє прискоренню
регенерації захисних тка¬
нин. Кращі результати
отримані від розчинів хі¬
нону і гідрохінону (0,001-
0,0001 %), резорцину і пі¬
рогалолу (0,005-0,001 %).
ііталтік С
год.
%
1?
1Р
0£
0/5
0,4
0,2
0
24 48 72
у*
загальник
білок
водорозчинним
білок
год. 24 48 72
Рис. 6.26. Вплив різання та обробки бульб
солодо-бурштиновою сумішшю на їхній
хімічний склад
469
g
&
о
&
К
и
2
о
U
/
/А \
'И
Л
/
а
w
ѵ
W
\\
ІЛ
\
Очень
Цілі бульби
Різані бульби завочені в водопровідній воді
—«-Різані бульби, оброблені солодом, 5% і
бурштиновою кислотою, 0,0025%
Лютий Березень
години 2 24 48 72 96 2 24 48 72 96 2 24 48 72 96
Рис.6.27. Інтенсивність дихання різаних бульб при обробці їх
солодо-бурштиновоко сумішшю
Проведені дослідження щодо обробки зрізів бульб вітамінами і
вітаміноподібними речовинами підтвердили позитивний вплив на
відкладення суберину розчинів нікотинової кислоти (0,01-0,005 %),
інозиту (0,005-0,0005 %) і ціанокобаламіну (0,0001 %); на утворення
перидермальних клітин - піридоксину (0,005-0,0005 %), рибофлаві¬
ну (0,005-0,0005 %) і тіаміну (0,001-0,0005 %). Найбільш ефектив¬
ний вплив на утворення ранової перидерми справляє 0,5-1,0 %-ний
розчин аскорбінової кислоти.
Обробка частин бульб розчинами деяких макро- і мікроелементів
прискорювала процеси загоювання поранень. Задовільні результати
отримані при використанні розчинів кобальту (СоС12 • 6Н20) - 0,1 мг/л,
молібдену (NH4)6Mo7024 * 4Н20 - 0,54 мг/л і міді в суміші з бором
(ChS04 + Н3ВО3) - 0,05+0,15 мг/л.
Виявлено, що обробка різаних бульб розчинами регуляторів росту
рослин - гібереліну (0,01-0,0025 %) і гетероауксину (0,005-0,00125 %)
прискорює регенераційні процеси. Однак, їх можна включити до
складу суміші для обробки різаних бульб лише у весняний період,
оскільки вони виводять їх із стану спокою і сприяють активації рос¬
тових процесів.
470
Польовими дослідами виявлено, що завчасна обробка різаних
бульб фізіологічно активними речовинами приводить до стабільного
підвищення врожайності картоплі. З органічних кислот у суміші з
солодом найвища прибавка урожаю була отримана від бурштинової
кислоти. У середньому за 5 років вона становила 33 ц/га, або 13,1 %
при рівні врожайності на контролі 249 ц/га (рис. 6.28).
Нанесення на зрізи бульб кобальту і суміші міді з бором забезпе¬
чило підвищення врожайності в середньому за 4 роки, відповідно, на
18 і 20 ц/га, або 7 і 8 % до контролю (267 ц/га).
Фенольні сполуки сприяли підвищенню урожаю на 11-14 %, що
однак виявилося в аналогічних умовах нижче на 4-7 % у порівнянні з
солодо-бурштиновою сумішшю, тому їх застосування в чистому ви¬
гляді недоцільне.
Рис. 6.28. Вплив обробки різаних бульб біологічно активними
речовинами на врожайність картоплі, ц/га: А - обробка органічни¬
ми кислотами; 1 вода (контроль); 2 - солод + фумарова кислота;
З - солод + лимонна кислота; 4 - солод + бурштинова кислота; В - феноль¬
ними сполуками; 1 вода (контроль); 2 - солод + хінон; 3 - солод + са¬
ліцилова кислота; 4 -солод + бурштинова кислота (другий контроль);
С - обробка макро- і мікроелементами; 1 вода (контроль); 2 - цинк;
З - бор; 4 - залізо; 5 мідь + бор: 6 - кобальт; 7 - молібден
Кращими строками різання садивних бульб при обробці їх фізіо¬
логічно активними речовинами потрібно вважати жовтень - першу
декаду березня. У цей час, завдяки фізіологічному стану бульб, про¬
ходить швидке загоювання ранових поверхонь, а підвищення вро¬
жайності, порівняно з пізніми строками, становить 25-51 ц/га при
рівні на контролі 275 ц/га (табл. 6.14).
На практиці різані бульби ще обробляють деревним попелом,
який містить у собі дезінфекційні речовини та мікроелементи, що
сприяє кращому загоюванню ран.
471
Таблиця 6.14 - Вплив на врожайність картоплі строку різання бульб і обробки
зрізів солодо-бурштнновою сумішшю, ц/га
Строк різання
Необ
роблені
Оброблені
Надбавка
врожайності від
солодо-бурштинової
суміші
урожай¬
ність
у процен¬
тах до
контролю
урожай¬
ність
у процен¬
тах до
контролю
ц/га
%
Жовтень
267
116
300
109
33
11
Грудень
253
109
304
110
51
17
Січень
262
113
306
111
44
14
Лютий
280
121
326
118
46
14
Березень (1 дек)
277
120
306
111
29
9
Березень (2 дек)
234
101
272
99
38
14
Квітень (1 дек)
210
91
243
88
33
14
Квітень (в день
садіння)
231
100
275
100
44
16
Для підвищення інтенсивності нагромадження в рановій пери-
дермі соланіну, який має високі фунгітоксичні властивості, різані
бульби під час пророщування опромінюють люмінесцентними лам¬
пами (40 Вт) протягом 10-15 днів. Тару, машини і приміщення дез-
інфікують 2 %-ним розчином формаліну. Використовувати формалін
як знезаражуючу речовину для бульб, не рекомендується, тому що
він має інгібуючі властивості, що викликає гальмування ранового
процесу та зниження схожості шматочків бульб.
Знезаражування ножів, машин, тари, приміщення, видалення
бульб і їх шматочків, уражених різними хворобами, сприяють оздо¬
ровленню картоплі (табл. 6.15).
Таблиця 6.15 - Ураженість бульб і рослин хворобами залежно від способу
підготовки садивного матеріалу
Садивний матеріал
Ураженість, %
рослин
вірусами
бульб
фітофторозом
гнилями
паршею звичайною
Цілі бульби
60,0
4,9
1,9
21,9
Опробковілі
шматочки бульб
45,0
1,8
2,0
17,0
Вплив сорту на швидкість утворення ранової перидерми на
зрізах садивних бульб та врожайність картоплі. Інститутом моле¬
кулярної біології та генетики разом з Білоцерківським державним
аграрним університетом були синтезовані, підібрані концентрації і
випробувані суміші нових біостимуляторів, які прискорюють утво¬
рення на зрізах та механічно пошкоджених бульбах надійного кірко¬
вого шару за значно коротший строк, ніж існуючі стимулятори [6].
Кращим виявився нуклеїновий біостимулятор - аномальний нуклео-
зид 6-азацитидин у суміші з н-пропіловим ефіром галової кислоти.
472
Аномальний нуклеозид 6-азацитидин має широкий спектр біоло¬
гічної активності з регуляторними властивостями. У низьких кон¬
центраціях (0,001—0,000001 %) він стимулює білково-нуклеїновий
обмін у живих організмах і рослинах, тим самим прискорюючи про¬
цес регенерації кіркової тканини.
Н-пропілгалат має антибактеріальні та антивірусні властивості,
додатково захищає ранову поверхню різаних бульб, завдяки чому
сприяє поліпшенню умов їх зберігання до висаджування в полі.
Сумісне застосування цих біостимуляторів, з одного боку, при¬
скорює процеси регенерації живої кіркової тканини, а з іншого - за¬
хищає ранову поверхню від проникнення гнильних та інших мікро¬
організмів, що приводить до взаємного підсилення ефекту прискоре¬
ного утворення суберину і ранової перидерми.
Щоб виявити строки утворення ранової перидерми на зрізах садив¬
них бульб, оброблених сумішшю біостимуляторів, у нових сортів, зане¬
сених до Реєстру сортів рослин України, протягом 1996-1998 рр. були
проведені досліди в лабораторії кафедри селекції та насінництва і в
навчально-дослідному господарстві БДАУ [7]. Вивчали 10 сортів кар¬
топлі: два ранньостиглі (Бородянська рожева, Кобза), чотири серед¬
ньоранні (Берегиня, Водограй, Обрій, Світанок київський), два
середньостиглі (Горлиця, Либідь) і два середньопізні (Дезіре, Ракурс).
Бульби масою 80-150 г різали, залежно від маси, на 2, 3 і 4 час¬
тини із середньою масою 35-45 г. За контроль брали різані без оброб¬
ки та цілі бульби.
Як свідчать дані досліджень, розмір листкової поверхні рослин за¬
лежить від сорту та способу підготовки садивного матеріалу. З подов¬
женням періоду вегетації сортів площа листкової поверхні зростає. Так,
по цілих бульбах вона підвищилася від ранньостиглих до середньопізніх
на 4,6 тис. м2/га, по різаних бульбах без обробки - на 5,5 тис. м2/га і по
різаних бульбах, оброблених біостимулятором, - на 4,7 тис. м2/га.
Стосовно способу підготовки садивного матеріалу, то спостеріга¬
ється наступна закономірність. Порівняно з цілими, у різаних бульб
без обробки площа листкової поверхні скоротилася на 9,3 тис.мѴга, а
в оброблених - лише на 0,1 тис. м2/га. Це означає, що обробка різа¬
них бульб біостимуляторами дозволяє сформувати такий самий фо¬
тосинтетичний апарат, як і від цілих бульб.
Порівняльні дані щодо стеблоутворювальної здатності цілих
бульб, різаних без обробки та різаних і оброблених біостимулятора¬
ми, показують, що при однаковій масі цілих і різаних садивних
бульб (30-50 г) на їх стеблоутворювальну здатність певною мірою
вплинув спосіб підготовки садивного матеріалу. За роки досліджень
цілі бульби в середньому по сортах утворили 4,1 стебла на кущ, тоді
як різані без обробки - лише 3,5 стебла, а оброблені - 4,0.
Обробка різаних бульб біостимуляторами сприяє пробудженню
сплячих бруньок, з яких розвивається певна кількість додаткових
16М32
473
стебел, що в подальшому практично наближує цей показник до рівня
цілих бульб.
Важливим показником роботи фотосинтетичного апарату рослин
є величина фотосинтетичного потенціалу (ФП) та чиста продуктив¬
ність фотосинтезу (ЧПФ).
Дослідження показали, що фотосинтетичний потенціал рослин
картоплі залежав певною мірою від сортових особливостей та спосо¬
бів підготовки садивного матеріалу. У ранньостиглих та середньо¬
ранніх сортів при використанні для садіння цілих бульб ФП становив
у середньому 2.7; у середньостиглих сортів - 2,8-2,9; у середньопіз¬
ніх - 3,0 млн м -добу/га. При садінні різаними необробленими буль¬
бами величина ФП зменшувалася на 20-23%, порівняно з цілими бу¬
льбами, і становила, відповідно, 2,2; 2,3 і 2,4 7 млн м2-добу/га.
При використанні для садіння оброблених біостимуляторами рі¬
заних бульб ФП становив, відповідно, 2,7; 2,8; 3,0 7 млн м2 -добу/га,
тобто був на рівні ФП насаджень, де висаджували цілі бульби.
Важливим показником формування врожаю картоплі є чиста продук¬
тивність фотосинтезу, яка обернено пропорційно змінювалася стосовно
ФП. Так, при використанні цілих бульб у ранньостиглих і середньоран¬
ніх сортів цей показник становив у середньому 5,5 г/м^добу, у серед¬
ньостиглих - 5,4; у середньопізніх - 5,2 г/м2/добу. Аналогічні показ¬
ники, а у деяких сортів і вищі, отримані і при використанні для са¬
діння оброблених біостимуляторами різаних бульб. Рослини, отри¬
мані з різаних необроблених бульб, мали на 15% нижчий показник
ЧПФ, порівняно з цілими.
Таким чином, можна зробити висновок, що ФП насаджень карто¬
плі підвищується із збільшенням тривалості періоду вегетації рос¬
лин, а ЧПФ обернено пропорційна до ФП. Застосування цілих і різа¬
них, оброблених біостимуляторами, садивних бульб забезпечує фак¬
тично однакові величини ФП і ЧПФ, що підтверджують і показники
врожайності бульб.
При збиранні картоплі було виявлено, що врожайність залежить
від генотипу та способу підготовки садивного матеріалу.
Спостерігається тенденція до незначного збільшення валового
врожаю із збільшенням груп скоростиглості незалежно від способу
підготовки садивного матеріалу. Так, різниця в урожайності між се¬
редньопізньою групою і ранньостиглою була: при садінні цілими
бульбами - 21 ц/га; різаними бульбами без обробки - 21 ц/га і різа¬
ними бульбами, обробленими сумішшю біостимуляторів, - 13 ц/га.
За роки досліджень із груп ранньостиглих сортів при садінні цілими
бульбами найвищу врожайність мав сорт Кобза - 218 ц/га; із серед¬
ньоранніх - Світанок київський (220 ц/га); середньостиглих - Горли¬
ця (223 ц/га) і середньопізніх - Дезіре (237 ц/га).
Аналогічна закономірність спостерігається на ділянках, де вико¬
ристовували як садивний матеріал різані бульби без обробки та об¬
474
роблені сумішшю біостимуляторів. При використанні різаних бульб
без обробки їх біостимуляторами, порівняно з цыими, валова вро¬
жайність незалежно від сорту значно знизилася. Ця різниця була в
межах 39-59 ц/га. Слід зазначити, що найбільша різниця в урожайності
була у сортів Кобза (59 ц/га), І орлиця 55 (ц/га) та Дезіре (53 ц/га). Об¬
робка різаних бульб сумішшю біостимуляторів сприяла одержанню
врожаю на рівні з цілими бульбами ( рис. 6.29).
Аналізуючи отримані результати за чистою врожайністю (за мі¬
нусом використаного садивного матеріалу ), бачимо, що різниця в
урожаї від цілих бульб і різаних без обробки дещо скоротилася, але
була значною, а за обробки сумішшю біостимуляторів вона зросла до
5-19 ц/га. Таким чином, можна зробити висновок, що використання
різаних бульб без обробки їх біостимуляторами, порівняно з цілими
бу'льбами, знижує як валову, так і чисту врожайність. При викорис¬
танні різаних бу'льб, оброблених біостимулятором, вдається отрима¬
ти однакову валову і навіть вищу чисту врожайність.
Сорт - F(9.90)=5.11; р<.0000; НІР05=9,7 ц/га
Способи - F(2 90)=175,46; р< 0000. НІН05=6,4 ц/га
о
X
-5
Г0
ї
О
>1
X
190
180
170
160
150
-о- Бородянська рожева
--о- Кобза
-о- Ьерегиия
-л- Водограй
Обрій*
-* Світанок київський
Горлиця
Либідь
•••+•• Дезіре*
Ракурс*
Цілі бульби Різані оброблені
Різані без обробки
Пр. - дані за 1997-1998 p.p.
НІР 05 (способи)=6,4 ц/га, F(2,90,1=175,46<0,000
Рис. 6.29 Вплив сортів та способів підготовки садивних бульб
на врожайність картоплі (середнє за 1996 - 1998 рр )
Проведені багаторічні лабораторні і польові досліди дозволяють
запропонувати технологію виготовлення опробковілих шматочків
бульб (табл. 6.16),
475
Таблиця 6.16 - Основні показники технології приготування опробковілих
частин бульб картоплі
Показник
Оптимальний
Допустимий
Маса частин бульби
(г) при врожайності
до 200 ц/га
понад 200 ц/га
25-30
35-50
20-40
40-70
Напрям розрізу:
на 2 частини
на 3 частини
на 4 частини
на 8 частин
У будь-якому напрямку
Уздовж довгої осі
Уздовж і впоперек
те саме
уздовж або впоперек
зрізати верхівку, а іншу
частину - навпіл
уздовж довгої осі
те саме
Температура, °С
20-25
12-30
Вологість повітря,
процент
94-96
85-100
Різання в день
садіння
При вологості ґрунту 85-100 %
ПВ і температурі 10-12°С
При вологості грунту 70%
ПВ, температурі 7- 8°С
Різання за 3-4 дні до
садіння
Зберігання шматочків бульб при
активній вентиляції
Зберігання шматочків
бульб з перешаруванням
перегноєм, торфом, піском
Завчасні строки
різання
При циклічності параметрів:
І цикл (3 дні) - вологість повіт¬
ря 95 100 %, температура 20°С,
безперервна подача повітря зі
швидкістю 0,14 м/с;
П цикл (7 днів) - вологість 90-
95 %, температура 5,5°С, по-дача
повітря через кожні 45 хв, по 15 хв
зі швидкістю 0,14 м/с;
Ш цикл (весь період зберіган¬
ня) - вологість 90-95 %, темпе¬
ратура 4,5-5,4°С, подача повітря
через кожні 55 хв протягом 5 хв
зі швйдкістю 0,14 м/с;
IV цикл починається за 3 дні до
вивантаження різаної картоплі.
Температуру підвищують до 9-
10°С, вологість до 85-90%, венти¬
ляцію здійснюють по 30 хв кожної
години, швидкість потоку 0,14 м/с
Зберігання шматочків
бульб з перешаруванням
перегноєм, торфом, піском;
в ящиках і на стелажах
тонким шаром
Обробка різаних
бульб біологічно
активними
речовинами
Біостимулятори: аномальний
нуклеозид 6-азацитидин+
Н-пропілгалат (0,0001-0,0002 %)
1.Солод (5 %)+бурштинова
кислота (0,0025 %) протягом
зимово-весняного періоду.
2.Солод (5 %)+бурштинова
кислота (0,00063%)+гібе-
релін (0,0025 %) або гете-
ро-ауксин (0,0125 %) -
лише навесні
Обробка ножів
різальних машин і
бульб знезаражуючи¬
ми речовинами
Саліцилова кислота - 0,05 %,
перманганат калію 0,1 %
За діючими
рекомендаціями
476
Деякі питання технології вирощування картоплі з різаних
садивних бульб. Строк садіння картоплі шматочками бульб має ду¬
же велике значення. Розпочинають цю роботу, коли земля на глибині
10-12 см прогріється до 6-8°С. Глибина садіння різаними бульбами
залежить від типу ґрунту, способу і строку. При садінні шматочками
глибина садіння буває завжди дещо меншою (на 2-3 см). Оптимальна
глибина загортання різаних бульб - 5-10 см. Слід пам’ятати про необ¬
хідність дотримання відповідної густини садіння картоплі. Схема са¬
діння шматочками бульб становить 70x20 і 70x25 см (табл. 6.17).
Таблиця 6.17 — Залежність урожайності картоплі від густоти садіння різаними
бульбами
Схема
садіння, см
Висаджено
бульб,
тис. шт./га
Кількість рослин
на 1 га, тис.шт.
Кількість
стебел,
тис. шт./га
Урожайність,
ц/га
70x20
71
65,9
211
265,2
70x25
60
53,1
155
213,0
70x30
47
46,8
140
193,9
70x35
41
40,0
112
183,0
Практика показала, що найкращі результати одержують, коли ви¬
саджують в лунку одну половинку, одну-дві третинки чи дві четвер¬
тинки бульби (табл. 6.18).
Таблиця 6.18 - Урожайність картоплі залежно від садивного матеріалу
Садивний матеріал, г
Висаджено
на 1 га, ц
Урожайність, ц/га
валова
чиста
Цілі бульби, 80-100
45,0
191
146
Одна половинка, 40-50
22,5
174
151
Дві половинки
45,0
190
145
Одна третинка, 27-33
15,0
163
148
Дві третинки
30,0
185
155
Три третинки
45,0
195
150
Одна четвертинка, 20-25
11,3
153
142
Дві четвертинки
22,6
182
159
Три четвертинки
34,0
188
154
Чотири четвертинки
45,0
196
151
Шматочки бульб перед садінням слід обов’язково проростити.
Передсадивне пророщення сприяє прискоренню появи сходів, на¬
стання фази бутонізації, цвітіння; інтенсивному росту картоплиння і
коренів; збільшенню поверхні листків; нагромадженню врожаю
бульб та поліпшенню їхніх товарних якостей [5].
ЛІТЕРАТУРА
1. А.С. 1540058 СССР МКИ Ао /43/54, Ао /С1/00. Состав для обработки
резаных клубней картофеля /М.Я.Молоцкий и др. - № 4180074/30-15; Заяв¬
лено 12.01.87; Опубл. 30 янв. 1990. Бюл. №4.-2 с.
477
2. А.С. 316419 СССР; МКИ Ао/п 21/00. Состав для обработки срезов
семенного картофеля /М.Я. Молоцкий. - № 1472445; Заяв. 04.09.70; Опубл.
05.08.71. Бюл. № 30- 4 с.
3. Винер В.В. Кулыура картофеля в Северной и Центрально-Чернозем¬
ной полосе России - СПб, 1905.
4. Кульжинський С.П. Питання культури картоплі на Україні // Укр. аг¬
роном.- 1928 - № 9.
5. Молоцкий М.Я. Выращивание картофеля при малых нормах посад-
ки.- К.: Урожай, 1986.- 240 с.
6. Молоцкий М.Я., Тихоступ В.Ф. Сроки сортообновления картофеля
//Картофель и овощи. - 1984. - № 2. - С.9-10.
7. Молоцький М.Я. Виродження картоплі в степовій і лісостеповій зонах
України та заходи боротьби з ним // Картоплярство: Міжвід. темат. наук. зб.
-1999. -№ 29. - С. 19-27.
8. Молоцький М.Я. Прискорене розмноження картоплі - К.: Урожай,
1972.-198 с.
9. Молоцький М.Я., Кривенко І.С. Строки сортооновлення картоплі у
центральних районах Лісостепу України // Вісн. аграр. науки. - 1994. - № 5.
- С.38-40.
10. Молоцький М.Я., Поліпшайко Ю.М. Вплив сорту на швидкість утво¬
рення ранової перидерми на зрізах садивних бульб та врожайність картоплі
//Вісн. Білоцерків. держ. аграр. ун-ту: 36. наук, праць.- Вип. 8, ч.З- Біла
Церква, 1999.-С.121-126.
11. Муш Н.Н. О взаимодействии гетеротрофного и автотрофного питания
в онтогенезе картофеля // Физиология растений. -Т.8, вып. 2. - М., 1961.
12.Насырова Т. Новые способы посадки картофеля - Ташкент: Госиздат
УзССР, 1962.-39 с.
13.Немчин Ф.И. Изучение и проверка методов борьбы с вырождением
картофеля // Материалы конф. по вопросам семеноводства картофеля. - М.:
Изд-во МСХ СССР, 1958.
14.Ничипоренко Н.С. Влияние гнездовой посадки долями клубня и орга-
но-минеральных удобрений на урожай картофеля: Автореф. дис... . канд.
с.-х. наук. - М., 1956.
15. Новиков Ф.А. Физиолого-химические основы использования верхушек
клубней картофеля на^семенные цели: Дис.... канд. с.-х. наук. - М., 1946.
16. Онищенко О.И. Із історії теорій виродження картоплі // Вірусні хво¬
роби картоплі. - К.: Урожай, 1969. - С.6- 12.
17.Поггенполь В.А. Картофель, его культура, сохранение, болезни и сор¬
та.-СПб, 1879.
18.Путш К.Е. Описание картофеля с подробным изложением оного, раз¬
ных пород и способов ведения и употребления в хозяйстве. - СПб, 1821-
185 с.
19.Рожалин Л.В. Агротехника семенного картофеля // Картофель. -
1958.-№6.
20. Руденко А.И. Меры борьбы с климатическим типом вырождения кар¬
тофеля // Материалы конф. по вопросам семеноводства картофеля. - М.:
Изд-во МСХ СССР, 1958.
21. Рыжков B.JI. Вирусные болезни растений. - М.: Сельхозгиз, 1935.
478
22. Рыжков В.JI. Фитопатогенные вирусы. - М.: Изд. АН СССР, 1946.
23. Соколенко Н.Ф. Выращивание здорового семенного картофеля на
юге И Науч. труды Всесоюз. Ордена Трудового Красного Знамени селек-
цион.-генетич. ин-та. - Одесса, 1962.
24. Стебут И.А. Основы полевой культуры и меры к ее улучшению в Рос¬
сии. - М., 1882.-Т.1, №1.-483 с.
25. Сухов К.С. Вирусы. - М.: Изд-во АН СССР, 1956.
26. Сухов К.С. Проблемы общей и сельскохозяйственной вирусологии //
Защита растений от вредителей и болезней. - 1960. - № 1.
27. Фаворов А.М. К истории вопроса о размножении картофеля семенами
на Украине // Культура картофеля семенами: Сб. науч.тр - Горький, 1983. -
С.7-8.
28. Фаворов А.М. Картопля в Степу УРСР. - Харків, 1948.
29. Фаворов А.М., Котов А.Ф. Летняя посадка картофеля. - М.: Госсель-
хозиздат, 1952 - 110 с.
30. Фомюк М.К. Питання насінництва картоплі та боротьби з її виро¬
дженням. - К.: Вид-во УАСГН, 1960. - 68 с.
31. Хранение верхушек клубней картофеля // Сад, огород. - 1947. - № 2.
32. Чесноков Н.С. Причины вырождения картофеля на юге и меры борь¬
бы с ним // Материалы конф. по семеноводству картофеля. -Л.: Изд-во
ВАСХНИЛ, 1958.
33. Чесноков П.Г. Болезни вырождения картофеля в СССР. - М.-Л.:
Изд-во с.-х. лит-ры, журн. и плакат., 1961- 319 с.
34. Ячевский А.А. Защита растений от вредителей. - М., 1927. - С.62.
479
7. БІОТЕХНОЛОГІЯ
Завдяки досягненням біотехнології створені умови для цілеспрямо¬
ваного і науково обгрунтованого впровадження біологічних процесів у
виробництво, переробку та використання сировини. Крім відомих тех¬
нологічних процесів, сучасна біотехнологія включає в себе нові техноло¬
гії в сільському господарстві, зокрема в селекційному процесі.
Історія селекції свідчить про її великі досягнення у створенні нових
сортів рослин. І сьогодні актуальним залишається завдання створення в
порівняно короткі строки високоякісних сортів, стійких проти негатив¬
ної дії зовнішнього середовища та з іншими цінними ознаками [31,60].
Впровадження досягнень біотехнології в генетико-селекційний
процес сприяє пошуку нових перспективних напрямів, насамперед
таких, як:
розробка нових технологій селекційного процесу на основі вда¬
лого поєднання традиційних методів селекції і досягнень клітинної
та генетичної інженерії;
удосконалення методів клітинної інженерії рослин, придатних
для використання у створенні нових сортів;
створення на основі генетичної інженерії не тільки нових форм рос¬
лин із бажаними ознаками, але й принципово нових селекційних форм.
Методи біотехнологічних досліджень набули широкого викорис¬
тання в багатьох країнах світу, але в Україні на сучасному етапі в
біотехнологічній науці відчутно відстає розробка нових універсаль¬
них узагальнюючих методик та положень, що стримує розвиток цьо¬
го експериментального напряму для цілей генетики і селекції.
7.1. КЛІТИННА ІНЖЕНЕРІЯ КАРТОПЛІ
На сьогодні картопля є одним із найбільш зручних об’єктів клі¬
тинної та генетичної інженерії. Це дало змогу значно розширити мож¬
ливості генетичних маніпуляцій з цією надзвичайно важливою сіль¬
ськогосподарською культурою.
Використання клітинної інженерії для селекційних потреб перед¬
бачає різні напрями:
сомаклональна мінливість як один із напрямів широко викорис¬
товується в різних наукових центрах світу і, безумовно, має перс¬
пективу для збільшення фонду гермоплазми картоплі;
одна із основних цілей клітинної селекції картоплі - підвищення
стійкості до різних збудників хвороб. Цей метод дозволяє проводити
спрямований добір клітин з використанням селективних факторів;
480
для селекціонерів і генетиків досить цікавими є методи культури
зародків, запліднення in vitro, які дозволяють отримувати гібридні
рослини, що є недосяжним при схрещуванні природним шляхом. Ці
методи дають змогу переборювати період спокою і прискорювати
пророщування насіння;
культури пиляків і мікроспор знайшли застосування при масо¬
вому одержанні гаплоїдних рослин;
метод соматичної гібридизації є насамперед перспективним для
одержання віддалених гібридів картоплі і досягнення тетраплоїдного
рівня при злшті двох диплоїдних ліній. При цьому за допомогою со¬
матичної гібридизації можна комбінувати ознаки, що успадковують¬
ся полігенно [47];
у насінництві картоплі перспективним є швидке клональне мік-
ророзмноження цінних і рідкісних форм та оздоровлення рослин від
вірусів.
У цілому з клітинної інженерії проводяться різнобічні дослід¬
ження, що у свою чергу зумовлює необхідність пошуку нових мето¬
дів і підходів для збільшення генотипного різноманіття вихідного та
селекційного матеріалу [123].
Сомаклональна мінливість. Історія та сучасний стан розвит¬
ку. Феномен сомаклональної мінливості (варіабельності), виявлений
близько ЗО років тому, розглядався спочатку як артефакт у культурі
рослинних тканин, і тільки в 1981 році було зроблено аргументова¬
ний висновок, що дивергентні варіанти, які виникли серед різнома¬
ніття ідентичних клонів, можуть бути корисним джерелом нових
ознак та змін у рослин [111, 38, 48, 39].
Успішний розвиток методу культури тканин та клітин вищих ро¬
слин започаткували праці французького дослідника П.Вайта [163,
164], який довів наступне: якщо кінчики культивованих корінців пе¬
ріодично пересаджувати на свіже поживне середовище, то вони мо¬
жуть рости і культивуватися тривалий час.
З роками стрімко збільшується кількість рослин, тканини яких
вирощують in vitro [80, 94, 122, 124]. Список, наведений у класичній
монографії з культури тканин Р.Готре [9], уже на той час включав
142 види вищих рослин.
Найбільш важливою подією періоду 60-70-х років XX ст. була роз¬
робка професором Нотангемського університету (Великобританія)
Е.Коккінгом [99] методу отримання ізольованих протопластів із тканин
корінця та плодів томатів шляхом обробки їх сумішшю пектолітичних
та целюлітичних ферментів, що виділяли із культуральної рідини грибів.
У подальшому продовжувався швидкий розвиток техніки in vitro,
вивчення біології культивованих об’єктів та створених на їх основі
технологій.
На території колишнього СРСР перші сомаклональні варіанти
(тютюну) були одержані Р. Бутенко із співавторами [77].
481
Після публікації у 1984 р. австралійськими дослідниками [78]
статті про сомаклональну мінливість як джерело генетичного різно¬
маніття у створенні нових форм рослин, кількість праць, виконаних з
різними видами рослин, різко збільшилася [10, 44, 51, 65, 77, 78, 115,
123, 138, 154, 157].
Вивчення сомаклональних варіантів показало, що від прототипу
вони відрізняються не лише моногенними якісними ознаками, але й
полігенними кількісними, зокрема інтенсивністю росту, продуктив¬
ністю, толерантністю до несприятливих факторів середовища [39].
Спостерігалися випадки прояву сомаклональних варіантів, що
поєднують ознаки, які неможливо або дуже важко поєднати в одному
генотипі традиційним селекційним шляхом. Л.Кучеренко [26] у своїх
дослідженнях виділила із сомаклональних варіантів, утворених з ка-
лусної культури рису, рослини, що характеризувалися швидким до¬
зріванням та довгозернятковістю. На їх основі створений новий сорт
рису Біоріза.
Американський дослідник Д.Еванс [89] довів, що використання
сомаклональних варіантів може удвічі прискорити процес створення
нових сортів, навіть тих, що розмножуються насінням. Учені отри¬
мали й вивчили генетичні сомаклональні варіанти томатів та виділи¬
ли ряд форм з такими цінними господарськими ознаками, як колір і
форма плодів, форма куща, плодоніжка, що легко обламується.
Значних успіхів у вивченні сомаклональної мінливості культури
льону маслянистого досяг канадський учений G. Rowland [136]. Основну
кількість сомаклональних ліній він одержав із сорту McGregor. Мінли¬
вість щодо врожаю насіння, тривалості періоду до початку цвітіння, ви¬
соти рослин і маси 1000 насінин була вищою, ніж селекційних ліній цьо¬
го самого сорту при одночасному випробуванні в польових умовах. У
сомаклональних ліній варіації стабільні і мають генетичну основу. Лінія
F86343 виявила стабільно більшу врожайність, ніж сорт McGregor, і бу¬
ла включена до сортовипробування 1992 року.
Нині роботи з виявлення сомаклональної мінливості у важливих
сільськогосподарських культур ведуться у всьому світі.
Серед рослин-регенерантів знайдені варіанти з відхиленням від
батьківських форм як за морфологічними, так й іншими інтеграль¬
ними ознаками - біохімічними параметрами [74, 102] стійкості про¬
ти: патогенів [3, 72, 83, 95, 98, 118, 133, 143, 151, 153], стресів [51,
100], а також за строками стиглості врожаю [65, 89, 139, 143].
Природа сомаклональних змін. Третє десятиріччя в клітинній
біології рослин не стихають суперечки про природу сомаклональної
мінливості і можливості її практичного використання. Залежно від
інтересів дослідників, сомаклональну мінливість розглядають як до¬
ступне джерело генетичного різноманіття, загрозу стабільності гено¬
типів, що підтримуються в ізольованій культурі, або як модель дивер¬
генції генів у популяції [8, 37, 43, 90, 104, 110].
482
Причини сомаклональної варіабельності досить широко обгово¬
рюються в науковій літературі [62, 81, 116, 159], і це явище може
бути використане для потреб практичної селекції [27, 29, 35, 48, 141].
Культура in vitro створює можливість експериментальної дії без¬
посередньо на соматичну клітину, а властивість тотипотентності
(здатності до регенерації рослин із культивованих клітин) дозволяє
одержати змінені форми із змінених клітин [11].
Виявлено, що в процесі культивування клітин і тканин часто спо¬
стерігаються генетичні зміни [114, 123, 155], більшість з яких прояв¬
ляються як спадкові мутації у потомстві регенерованих рослин. Це
явище, що називається сомаклональною мінливістю, може бути ви¬
значене як генетична мінливість, яка нагромаджується у процесі
штучного культивування тканин [111].
Генетична природа і механізми виникнення сомаклональної мін¬
ливості ще недостатньо вивчені. Але для управління сомаклональ¬
ною варіабельністю важливо зрозуміти її причини та розмах.
Основними причинами виникнення сомаклональних варіантів є
насамперед генетична гетерогенність соматичних клітин вихідного
експланта, а також генетична та епігенетична мінливість, індукована
умовами культивування.
При отриманні первинних культур, вибираючи експланти та умо¬
ви культивування, можна збільшити чи зменшити гетерогенність пер¬
винних культур, але повністю виключити її, очевидно, неможливо.
Значна варіабельність серед клітинних популяцій спостерігається
за різними ознаками: ростом, морфологією, потребою в компонентах
середовища, відношенням до стресової дії тощо.
Природа мінливості може бути різною. Найчастіше зустрічається
фізіологічна мінливість, що входить у норму реакції певного геноти¬
пу. Вона визначає високу адаптивність культур і особливо про¬
являється при зміні умов культивування [14, 24, 25].
Епігенетична варіабельність визначається функціональною ак¬
тивністю генів і проявляється при різних рівнях диференціювання
[120]. Визначити цей тип досить важко, оскільки частота епігене¬
тичних змін може дати той же порядок, що й мутаційних, але на
відміну від останніх вона не підвищується при дії мутагеном [91,
117].
Генетична мінливість, викликана мутаціями, успадковується не
тільки в ряді клітинних поколінь, але і в потомстві регенеранта. Без¬
умовно, що цей тип мінливості має найбільше значення, оскільки
саме він створює нові генотипи. Тому дуже важливо мати чіткі кри¬
терії визначення генетичної мінливості.
Варіабельність популяцій соматичних клітин виявляється на різ¬
них рівнях, і розмах її може бути різним [5, 27, 44]. На геномному і
хромосомному рівнях виділені культури з різним ступенем плоїднос-
ті та із транслокаціями [64]. Виникнення їх пов’язане з порушенням
483
мітотичного апарату. Така мінливість характерна не лише для ядер¬
ного геному, але й мітохондріального. Розподіл кільцевих фрагмен¬
тів різних розмірів у клітинних суспензіях дає значно більший роз¬
мах, ніж в інтактних рослин [86]. Зміни на генному рівні чітко вияв¬
ляються при використанні селективних систем [165]. Реалізація цих
змін у цілій рослині дає сомаклональні варіації [89]. Але правильні¬
ше називати сомаклональними варіантами не тільки зміни рослин,
але і клітинних ліній, оскільки і ті й інші є результатом генетичних
змін соматичних клітин.
У соматичних тканинах рослин нерідко присутні клітини із змі¬
неною анеуплоїдною кількістю хромосом. Особливо часто химеризм
і міксоплоїдія з високою частотою анеуплоїдних клітин зустрічаєть¬
ся в апоміктичних рослин, що розмножуються вегетативно [7]. Так, у
картоплі [4] може відбуватися значне нагромадження “вегетативних”
мутацій. При цьому, індукуючи in vivo чи in vitro утворення адвентив¬
них пагонів, можна одержати нові форми рослин [82, 148]. Таким
чином, існує значна ймовірність одержання нових варіантів із клітин
з уже існуючими генетичними змінами через культуру поодиноких
клітин та протопластів.
Численні цитологічні дослідження свідчать, що варіабельність,
індукована умовами культивування in vitro, пов’язана з дійсними ге¬
нетичними змінами [23, 25, 52].
Такі хромосомні зміни, як розриви, ацентричні і дицентричні
фрагменти, кільцеві хромосоми, реципрокні і негомологічні трансло-
кації, делеції та інверсії, які спостерігалися у процесі диференцію¬
вання у рослин, індукуються з високою частотою при культивуванні
in vitro [81, 111].
Причина, що зумовлює підвищену частоту хромосомних абера¬
цій, була в основному з’ясована Г.Бензіоном та іншими [69]. Автори
вважають, що реплікація гетерохроматину в мітотичному циклі
культивованих клітин може інколи відбуватися так пізно, що форму¬
вання місточків і подальше розщеплення хромосом настає в анафазі.
Це може призвести до циклу розрив-злиття місточків з такими на¬
ступними генетичними ефектами, як делеції і хромосомний обмін.
Точкові мутації можуть дати початок мутантам, які поводяться
відповідно до класичного закону Менделя. Генетичний аналіз рос¬
лин, одержаних із культури тканин різних видів рису, пшениці, ку¬
курудзи, латуку, тютюну і ріпаку, підтверджують появу точкових
мутацій. Більшість із цих мутантів алельні іншим відомим мутантам.
У багатьох випадках є вагомі докази того, що такі мутації виникають
протягом культивування тканин, оскільки сестринські рослини, ре¬
генеровані з тієї самої калусної тканини, не були мутантами.
Очевидно, частина змін серед регенерантів гороху (рослини з
хлорофільною недостатністю і без воскового нальоту) зумовлена точ¬
ковими генними мутаціями. Про те, що виникнення таких мутацій
484
(у частотах, порівняно з тими, які спостерігають при іонізуючому
опроміненні) можливе в умовах культури тканин, відомо із дослід¬
жень на кукурудзі, томатах, рисі [14, 39, 89, 128]. Один такий мутант,
у якого ідентифікована мутація на рівні нуклеотидів, одержаний се¬
ред регенерантів кукурудзи і виявлений шляхом скринінгу на ізози¬
ми локусу алкогольдегідрогенази (Аоіп) [72].
Транспозиція генетичних елементів для пояснення змін у сомак-
лонах пшениці була запропонована П.Ларкіним [99].
Запропоновано декілька механізмів, що пояснюють природу со¬
маклональної мінливості, які П.Ларкін і В.Скофкрофт [111, 112] по¬
діляють на такі можливі категорії: грубі каріологічні зміни; критич¬
ні, непомітні при цитологічному аналізі, хромосомні перебудови; рух
мобільних генетичних елементів; ампліфікація і редукція генів; со¬
матичний кросинговер та обмін сестринських хроматид; криптичні
елімінації вірусів.
Одним із основних джерел появи фенотипічних варіантів є різні
каріологічні зміни та перебудови, але з’ясувати, які з них мають фе-
нотипічний ефект і будуть успадковуватись як стабільні мутації ге¬
нів, часто досить складно. Як грубі, так і тонкі хромосомні зміни
(дрібні делеції, дуплікації, транслокації, інверсії) можуть спричиняти
істотні фенотипічні зміни не тільки в первинних регенерантах, але і в
їхньому потомстві.
Ампліфікація генів - один із способів забезпечення клітини ве¬
ликою кількістю специфічного генного продукту. Перше повідом¬
лення про ампліфікацію генів у рослин, індуковану in vitro, зробив
Г.Донн [87]. Було відібрано суспензійну лінію люцерни у 20-100 ра¬
зів більш стійку проти гербіциду (фосфінотрицину), порівняно з ди¬
ким видом. Цей гербіцид є конкуруючим інгібітором глутамінсисте-
тази (ГС), і стійкі клітини мають підвищений у 3-7 разів рівень цього
ферменту.
Використовуючи специфічний ГС-зонд, автори довели, що стій¬
кість є результатом 4-11-разової ампліфікації ГС-гена, яка приз¬
водила відповідно до 8-разового підвищення рівня РІЖ.
Сомаклональну мінливість можна простежити на молекулярному
рівні, оцінюючи тонку перебудову ядерної ДНК [39, 72]. Так, 2 із 12
протоклонів картоплі виявилися дефектними за 252 рибосомними
РНК генами [109]. Ці зміни, які не проявлялися фенотипічно, збері¬
галися у вегетативного потомства.
Причина такої мінливості досі залишається нерозкритою. Мож¬
ливо, в ній певну роль відіграють мобільні генетичні елементи, їх
ампліфікація та переміщення в нові ділянки геному. Про це свідчать
молекулярно-генетичні дослідження, проведені останнім часом на
регенерантах льону [70].
Крім сомаклональної варіабельності, пов’язаної з успадкуванням
перебудови геному, спостерігали фенотипічні зміни, що визначають
485
як “епігенетичні”. Вони можуть стабільно передаватися дочірнім клі¬
тинам, але не проявлятися в регенерованих рослинах або їхньому
статевому потомстві [70].
Залежність виникнення фенотипічних варіантів серед рослин-
регенерантів від експресії генів, яку оцінювали за ступенем мети¬
лування ДНК розкрита в роботі Х.Лерца і П.Броуна [114]. їх поява,
очевидно, пов’язана із змінами характеру метилування ДНК у куль¬
тивованих in vitro клітин. Зміни рівня метилування ДНК, виявлені у
рослин-регенерантів кукурудзи, і, що особливо цікаво, у К2-рослин
(за ступенем метилування значно відрізнялися 25% рослин).
Проходження рослинами стадії неорганізованого росту in vitro
може також індукувати модифікації цитоплазматичних генетичних
детермінант і відповідно проявлятися зміною ознак, що успадкову¬
ються по материнській лінії. Сомаклональна мінливість може прояв¬
лятися перебудовою мітохондріона. Перші зміни мітохондріона ви¬
явили серед сомаклонів кукурудзи, які були одержані з калусної тка¬
нини, індукованої із недозрілих зародків [105]. Рослини, що несли
Т-тип (Техаська) цитоплазми, мали чоловічий тип стерильності і бу¬
ли високосприйнятливими до Helminthosporium maydis раси Т і ток¬
сину цього гриба. Рослини з N-типом (нормальна) цитоплазми є чо-
ловічостерильними і стійкими проти N.maydis. Серед рослин кукуру¬
дзи, одержаних із калусної тканини з Т-цитоплазмою, пересаджених
як у присутності токсину [95], так і без нього [71,72], виявлені чоло-
вічофертильні токсиностійкі рослини. Ця ознака успадковувалася по
материнській лінії. Подібні зміни мтДНК, які спостерігали при куль¬
тивуванні in vitro, ніколи не виявляли під час звичайного розмно¬
ження кукурудзи насінням.
Перебудову мтДНК також виявлено в популяції протоклонів кар¬
топлі [106]. Виділено дві варіантні форми із зміненою мтДНК. Серед 47
протоклонів у 5 клонів були зміни в послідовності високомолекулярної
мтДНК, дві рослини мали додаткові низькомолекулярні форми мтДНК.
Відносно невелика кількість повідомлень є про зміни у сомакло¬
нальних варіантів пластидного геному. Виникнення альбіносних со¬
маклональних варіантів - явище досить часте [13, 40, 85, 127, 132].
Сомаклональна варіабельність супроводжується також змінами
функціональної активності фотосинтезу і його первинних реакцій.
Так, одержана в дослідах В.Моргуна та В.Логвиненко [33] сомакло¬
нальна лінія №6 пшениці сорту Зоря переважає вихідну форму не
лише за інтенсивністю фотосинтезу і його світлових реакцій, але й
виявляє зниження оксигенної активності РБФКО на фоні підвищеної
карбоксил азної.
Крім того, ця лінія може бути використана як донор ознаки ви¬
сокої загальної продуктивності і зимостійкості. Отже, автори роб¬
лять висновки, що за допомогою культури тканини in vitro можна до¬
сягти збільшення фотосинтетичної продуктивності рослин. Це у
486
свою чергу вказує на ймовірність генетичної регуляції внутрішньо¬
клітинних процесів у напрямі зниження витрат на виробництво оди¬
ниці біомаси рослини.
За оцінкою А.Жученка [94], важливим механізмом генотипної
мінливості в культурі in vitro є мітотичний кросинговер. Оскільки в
культурі соматичних клітин у процесі утворення калусу відбувається
велика кількість мітозів, зазначена частота мітотичного кросинговеру
має досить відчутне практичне значення. Воно особливо важливе у тет-
раплощної картоплі, яка має значний запас прихованої варіабельності у
формі рецесивних генів і можливість утворення різних типів гетерозигот
(Аааа, ААаа, АААа). Збільшення чи зменшення гомо- чи гетерозиготно¬
сті (зміни Аааа -*-Аааа-*~аааа чи Аааа -*~Аааа —►АААа) призводить до
збільшення генотипного різноманіття соматичних клітин in vitro [19].
Ідеальним варіантом мінливості окремої соматичної клітини є
одночасне збільшення гетерозиготності за генами, що контролюють
урожайність, і гомозиготності за адитивними генами, які контролю¬
ють крохмалистість або польову стійкість проти фітофтори. Імовір¬
ність змін однієї й тієї ж соматичної клітини за декількома кількіс¬
ними ознаками дуже мала. Тому результати експериментів показу¬
ють, що відібрані рослини-регенеранти характеризуються в основ¬
ному якоюсь однією позитивно зміненою ознакою [56].
Проте слід враховувати, що в процесі утворення калусу зміни мо¬
жуть відбуватись послідовно і накопичуватися поступово. Тому теоре¬
тично можлива поява варіантів, які б несли кілька позитивних змін.
Для розширення спектра сомаклональних варіантів використо¬
вують як спонтанний, так і індукований мутаційний процес. Визна¬
чення темпу мутації за окремими локусами (порядку 10 на клітину в
одному поколінні) дозволяє накопичувати зміни клітини при довго¬
тривалому культивуванні і виділяти їх потім, використовуючи різні
селективні системи [165].
Інший шлях збільшення мінливості - це індукований мутагенез
[44, 56].
Якщо ж використовувати звичайні умови, то в результаті неста¬
більності геному, зворотних мутацій, рекомбінацій, хромосомних пе¬
ребудов знову виникають гетерогенні клітинні популяції. Популя¬
ційні відносини на клітинному рівні дуже складні і маловивчені.
Очевидно, що клітини, які мають селективну перевагу, більш корот¬
кий клітинний цикл, і у котрих відсутня легфаза повинні витіснити
своїх конкурентів, що менш активно проліферують [53]. Цей процес
дійсно спостерігається на початку дестабілізуючого добору, коли в
кожній наступній регенерації збільшується частка клітин дикого ти¬
пу. Але повної заміни маркера не відбувається.
Таким чином, умови культивування, за яких виникає перебудова,
можуть змінити регуляцію експресії блоку генів, а саме: ген, який
попередньо експресувався, може бути репресований, а “мовчазний”
487
ген може почати експресуватися. Здатність генерувати такі зміни в
культурі може виявитися цінним інструментом при аналізі цих ген¬
них блоків. Оскільки деякі цінні білки кодуються мультигенними
блоками (наприклад, гліадини, зеїни, глютеїни, гордеїни, амілази), то
це може мати важливе значення для селекції.
Успадковані цитоплазматичні мутації можна виявити в таких со-
маклонів:
а) морфологічні зміни у латуку, що успадковуються по материн¬
ській лінії [147];
б) стійкі проти Т-токсину, які мають чоловічу фертильність і змі¬
ни у профілі рестрикції мт ДНК у Т-ЦЧС кукурудзи [72, 95, 157];
в) зі змінами спектра рестрикції мтДНК у томатів [89];
г) зі змінами спектра рестрикції мтДНК у картоплі [106].
Важливість генів, локалізованих у цитоплазмі, стимулює подаль¬
ші дослідження цього явища.
Часто менделівське розщеплення мутантних генів спостерігаєть¬
ся в другому поколінні. Проте зафіксовані випадки появи форм у по¬
колінні сомаклонів, що не розщеплюються [88, 99, 147].
Найбільш цікавою є передача модифікованого гена в покоління
шляхом статевого розмноження, однак ще необхідно з’ясувати меха¬
нізми виникнення цих гомозиготних мутацій. Деякі з них можуть
бути результатом гемізиготності, яка існує в рослині або виникає в
культурі. Інші можуть бути суто гомозиготними мутаціями. В обох
випадках вони виникають у деяких видів і дозволяють вести добір
рецесивних мутацій.
Часто постає питання: сомаклональні варіант - це результат му¬
тацій попередніх чи зовсім нових локусів? Поки що немає доказів, що це
мутують нові локуси. Деякі генетичні зміни, які рідко виникають спон¬
танно, досить часто з’являються в процесі культивування, тому виникає
думка, що сомаклональна мінливість “створює” нові гени.
На сьогодні вже доведено, що сомаклональна мінливість має більш
високу частоту, ніж мінливість, яка виникає при широкому використанні
хімічних і фізичних мутагенів. Переваги її полягають у тому, що для
добору бажаних мутантів достатньо проаналізувати незначну кількість
рослин, проте більше мутацій може бути сконцентровано тільки в по¬
одиноких сомаклонах [48, 93, 99]. Щоб з’ясувати, чи індукуються нові
категорії мутацій, необхідно провести, з одного боку, паралельний гене¬
тичний аналіз потомства сомаклонів, з другого - рослин, відібраних хі¬
мічними або фізичними мутагенами.
Отже, шляхи створення генетичного різноманіття клітинних по¬
пуляцій розроблені досить широко. Після конструювання і виділення
нових генотипів постає завдання стабілізації та збереження цих цін¬
них форм з метою залучення їх у селекційний процес.
Регенерація рослин із клітин протопластів. Добір сомаклонів
у картоплі та їх аналіз. Одержання нових сортів картоплі здебіль¬
488
шого пов’язане з виділенням специфічних ліній та подальшою їх оцін¬
кою. Класичні методи селекції сьогодні неможливо замінити мето¬
дами in vitro, хоча їх розумне поєднання відкриває зовсім нові мож¬
ливості для селекційно-генетичних досліджень.
Культура протопластів, калусна і суспензійна культури картоплі
мають високий ступінь ядерної нестабільності, що призводить до
полі- та анеуплоїдії і хромосомних змін. На ступінь нестабільності
впливають як генетичні, так і культуральні фактори.
Картопля - полісоматична культура, тому попередні генетичні
зміни в тканинах переходять у культуру in vitro [130].
Перше повідомлення про індукцію калусу із паренхімної тканини
бульб з’явилося в 1951 році [152]. Після цієї праці дослідники на¬
магалися індукувати пагоноутворення із калусу різного походження,
проте тільки з 1975 року [66, 67, 108, 134, 149] розпочалися широкі до¬
слідження щодо одержання рослин-регенерантів із калусу [1, 2, 15, 49,
50, 57,60,73,119, 129, 160]. Перші роботи були спрямовані на огггиміза-
цію умов індукції та культивування калусу, але основні зусилля - на ін¬
дукцію пагоноутворення в культурі калусної тканини. Основними лімі¬
туючими факторами виявилися специфічна реакція генотипу і співвід¬
ношення між фітогормонами ауксинової і цитокінінової дії.
Усі без винятку дослідники відзначають необхідність значного
перевищення цитокінінів над ауксинами в середовищі для регенера¬
ції, а як інші фактори - необхідність різної інтенсивності освітлення
і тривалості світлового дня [2, 57, 59, 79]. Деякі вчені також спосте¬
рігали залежність інтенсивності регенераційних процесів від пори
року [49, 57]. У ролі експлантів використовували диски бульб, лист¬
кові та стеблові експланти, сегменти кореня і гіпокотиля.
Аналіз одержаних різними дослідниками результатів з індукції та
культивування калусу і регенерації рослин дозволяє відзначити, що
єдиної технології, у тому числі поживного середовища та інших
умов, не існує. Є лише загальна умова: на першому етапі (індукції та
культивування калусу) переважають ауксини над цитокінінами, а на
другому (регенерації ) - навпаки. Цей загальний принцип дотриму¬
ється при забезпеченні достатнього енергетичного запасу (сахарози)
і наявності органічних добавок (вітамінів, гідролізату казеїну).
Морфологічний аналіз рослин-регенерантів свідчить про значне
різноманіття морфології листків і стебла, кольору бульб і квітів [27,
41, 59, 61, 75, 79, 104, 107, 154], характеру апікального домінування і
ступеня розвитку кореневої системи та низькорослості [16, 15]. Час¬
тота морфологічної мінливості у регенерантів перебувала в межах
від Одо 7% [107].
Уперше регенерація рослин із поодиноких клітин суспензійної
культури дигаплоїдних ліній картоплі була продемонстрована
М.Бенке [66]. Активні дослідження на початку 70-х років у багатьох
провідних біологічних лабораторіях світу дозволили розв’язати ряд
489
складних методичних проблем культури протопластів картоплі, ре¬
генерацію з них рослин та детально вивчити рослини-регенеранти.
Перше повідомлення про одержання рослин із протопластів було
зроблене відомим індійським ученим М. Упадхія [158]. У своїй ро¬
боті він детально описав методику виділення і культивування прото¬
пластів мезофілу листка картоплі до одержання регенерантів, не
акцентуючи уваги на їхній характеристиці.
Майже одночасно із дослідженнями М.Упадхія, А.Кучко із спів¬
авторами розробили свою оригінальну методику культивування про¬
топластів картоплі та регенерації з них рослин [12, 28]. Уже тоді ці
дослідники виявили значну мінливість серед рослин-регенерантів із
протопластів. Але розглядали це явище швидше як виняток, ніж за¬
кономірність. Тобто, автори ще перебували у полоні давнього гене¬
тичного міфу про клональну одноманітність серед рослин-
регенерантів у культурі in vitro і не змогли дати правильної оцінки
цьому явищу. Але це не завадило Дж.Шепарду та Р.Тоттену [146],
П.Ларкіну та В. Скоукрофту [111] виявити і дати визначення та роз¬
виток цьому напряму - сомаклональній мінливості. Д.Шепард і
Р.Тоттен одержали і дослідили велику групу рослин картоплі із про¬
топластів (близько 10000). Культивуючи протопласти сорту Рассет
Бербанк, вони теж зіткнулися з несподіваним спалахом мінливості.
Подальші експерименти Д.Шепард а із співавторами [144, 145] під¬
твердили ці спостереження.
Сомаклони також були одержані Р.Нельсоном зі співавторами
[125] із S.brevidens, Т. Барсбі та іншими [63] із S.brevidens, S.tube¬
rosum, S.bernandezianum ( дикі диплоїдні види, стійкі проти скручу¬
вання листя).
М.Джоне із співавторами [103] одержали калуси з 35 % прото¬
пластів сортів Maris, Piper, Desiree і в 50—80% випадків - регенера¬
цію пагонів. Мінливість включала:
1) виражені аберантні рослини;
2) більш чи менш помітні морфологічні відхилення габітусу ку¬
ща, форми листків і бульб, кольору листків та квітів, якості шкірки
та кольору м’якоті, компактності гнізда і т.д.;
3) фізіологічні зміни - час цвітіння, тривалість вегетаційного пе¬
ріоду;
4) зміни стійкості проти Globodera pallida, Alternaria solani,
Phytophthora infestans (листки), парші звичайної, приморозків, а та¬
кож умісту редукуючих цукрів. Кількість різних відхилень морфоло¬
гічних ознак в окремих протоклонів варіювала від 1 до 16. Частота
змін залежала від тривалості культивування калусу.
При вивченні ліній сомаклональних варіантів картоплі виявлено, що
від прототипу вони відрізняються не тільки моногенними якісними змі¬
нами, але й полігенними — інтенсивністю росту, продуктивністю, толе¬
рантністю до несприятливих факторів середовища та хвороб.
490
Із 500 протоклонів, перевірених на стійкість проти альтернаріозу,
5 були стійкі, порівняно з вихідним сортом, у тому числі 4 мали
польову стійкість [118].
Д.Шепард із співавторами [143], аналізуючи 800 протоклонів
картоплі на стійкість проти збудника фітофторозу, виявив, що майже
2% ліній мали вищу стійкість, ніж вихідний сорт Рассет Бербанк.
А.Каселз зі співавторами [75] із культури експлантів та клітин
картоплі одержали цілий ряд фенотипічних варіацій за стеблом. Се¬
ред них були значні відхилення та хлорофілдефектні типи. Подібні
аномалії спостерігала Л.Хромова [156].
С.Аустін із співавторами [61] виявили значні відхилення в сома-
клональних лініях картоплі порівняно зі стандартом за деякими
ознаками морфології листків, висотою рослини та життєздатністю.
При цьому розмах варіації регенерантів у межах одного калусу був
таким, як і між усім калусним матеріалом.
Однак досліди показали, що частота появи цінних форм у куль¬
турі тканини без застосування селективних факторів досить низька.
Так, в Ірландії за допомогою культури тканин планували отримати
на основі старого сорту Голден Уандер новий, з підвищеною стійкіс¬
тю проти вірусів. Регенеранти відбирали за фенотипом. Із 586 реге¬
нерантів 473 виявилися вільними від вірусів у першому бульбовому
поколінні, із 85 відібраних ліній 36 мали польову стійкість проти ві¬
русу Y, а 27 - до механічної інокуляції цим вірусом. Проти вірусу X
стійкими виявилось 17 ліній, 12 були стійкими проти обох вірусів. У
другому поколінні стійкими були 8 ліній: 6 - проти вірусу Y; 2 -
проти вірусу X. З комплексною стійкістю ліній не виявлено. У всіх
поколіннях спостерігали химерність за ознакою кольору шкірки [75].
При вивченні причини сомаклональної мінливості картоплі пер¬
шою визнано змінену кількість хромосом [96, 103]. Детальне дослід¬
ження регенерантів показало, що рослини, одержані із протопластів
(протоклони), мають значно більшу варіабельність за морфологічни¬
ми ознаками, ніж ті, що регенерують безпосередньо із експлантів
рослини (сомаклони). Так, від 5 до 70% рослин першого типу залеж¬
но від сорту і умов регенерації були еуплоїдами (2п-4х-48), а грубі
аберанти - анеуплоїдами з високою кількістю хромосом (88-95). І
навпаки, в середньому близько 90% рослин, регенерованих із екс-
плантів, були ауплоїдами. Проте в обох випадках рослини з нормаль¬
ною кількістю хромосом відрізнялися від батьківських сортів. Для
прикладу: регенеранти із живців червонобульбового сорту Дезіре
мали бульби білого кольору. У регенерантів сорту Фюртифолд, що
має строкаті бульби, вони також були білі. У наступних поколіннях у
частини регенерантів колір бульб повертався до дикого типу.
Дані цитологічного аналізу [18] свідчать, що серед рослин-
регенерангів сорту Лорх майже половина були регулярними тетраплої-
дами, а решта - анеу- і міксоплоідами. У протоклонів сорту Українська
491
рожева практично не спостерігали мінливості рівня площності, порівня¬
но з вихідним генотипом (2п-4х-48). Вихід нормальних за фенотипом
регенерантів був дуже низьким (20%), а серед протоклонів Pg-438 25%
їх були ідентичними за фенотипом, а 92% - за каріотипом.
Технологія регенерації в культурі тканини і протопластів кар¬
топлі добре відпрацьована. Сомаклони і протоклони одержані в сор¬
тів Бард, Биліна, Гатчинська, Годцен Уандер, Дезіре, Зарево, Зміна,
Маріс, Міранда, Прієкульська рання, Рассет Бербанк, Резерв, Столо¬
ва 19, Ульяновська та ін. У результаті їх вивчення виділені форми з
важливими господарськими показниками, які є селекційною цінніс-
тю [11, 45, 135].
Мінливість серед калусних культур не обмежувалася тільки
морфологічною і змінами кількості успадкованого матеріалу (хро¬
мосом і ДНК). Деякі дослідники вивчали також білкову мінливість,
що виникала в калусних культурах та рослинах-регенерантах [16,
30, 34, 101, 150].
С.Мусін [34] у своїх дослідженнях з вивчення сомаклональної
мінливості серед регенерантів із калусних тканин дійшов висновку,
що сомаклональна мінливість “виражається як незначна флуктуація
біля білкового “портрета” вихідного генотипу”. Водночас інші до¬
слідники твердять про значну мінливість серед білкових спектрів у
сомаклонів картоплі [16].
При вивченні електрофоретичних спектрів білків листків вихід¬
них рослин, калусних ліній і регенерантів не виявлено швидкорухо-
мих білків у калу сів. Рослини-регенеранти також відрізнялися від
вихідних рослин кількістю та інтенсивністю спектрів. Крім того, бу¬
ли показані кількісні зміни в амінокислотному складі загального біл¬
ка при однаковому якісному його складі в регенерантів і вихідних
рослин [30].
Т.Долбік зі співавторами [16] спостерігали помітні зміни в білко¬
вих спектрах не тільки регенерантів порівняно з вихідними рослина¬
ми, але й між різними фенотипічними лініями рослин-регенерантів.
Крім морфологічної, цитологічної та білкової мінливості вияв¬
лено регенеранти з різними господарсько цінними показниками.
М.Меулеманс [121] одержав 300 ліній регенерантів, із яких 2 бу¬
ли стійкі проти Phytophthora infestans.
Детальний аналіз мінливості морфологічних і господарсько цін¬
них показників був проведений JI. Хромовою [54]; В. Сидоровим
[45]; Н.Юр’євою [58]; А.Кучком [27]; Т.Олійник [35] та іншими ав¬
торами (табл. 7.1).
На думку дослідників [50], значні варіаційні зміни характерні для
регенерантів, отриманих із експлантів первинного та тривало пасо-
ваного калусу. У сортів Биліна, Любимець, Смєна, Ульяновська та
інших спостерігали різноманітний тип змін, а саме: підвищення стар¬
тової інтенсивності росту, збільшення габітусу куща, прискорення
492
фаз розвитку, підвищення рівня врожаю, крохмалистості, горизонта¬
льної стійкості проти фітофторозу. Зміни цих цінних ознак in vitro
дають змогу використовувати метод сомаклональної варіабельності
для доопрацювання сортів і вихідних форм у кількох різних напря¬
мах. А саме: зміни, що спостерігають у регенерантів у темпах розви¬
тку, особливо в перший період росту (стартова інтенсивність), та в
збільшенні габітусу куща (без зниження інтенсивності бульбоутво¬
рення) мають значення для підвищення здатності сорту активно про¬
тистояти бур’янам (один із параметрів придатності сорту до промис¬
лової технології вирощування).
Таблиця 7.1 - Сомаклональні варіанти картоплі (S.tuberosum) за важливими
господарськими ознаками
Змінені характеристики
Літературне джерело
1
2
Ізольовані протопласти
Стійкість проти Alternaria solani і Phytophthora infestans,
форма бульб, урожай, строки дозрівання, фотоперіод,
морфологія рослин
Matem et al., 1978
Shepard et al., 1980
Secor, Shepard, 1981
Secor, 1984
Форма бульб, каріотип
Рівень плоїдності, морфологія, строкатолистість, хлорофіл-
дефектність
Wenzel et al., 1979
Ramulu et al.,1983,
1984
Морфологічні та кількісні ознаки, каріотип, стійкість проти
хвороб
Сидоров и др., 1985
Калусна і суспензійна культури
Форма і колір бульб, морфологія листків
Harten et al., 1981
Морфологія листків і бульб, урожайність, уміст крохмалю,
стійкість проти фітофторозу, кільцевої гнилі
Хромова и др., 1983,
1988
Стійкість проти вірусів X та Y
Cassels, 1987
Морфологія, продуктивність, уміст крохмалю, стійкість
проти фітофторозу
Юрьева, 1987
Збільшена кількість стебел та бульб, низькорослість,
урожайність
Schenk et al., 1988
Морфологія листків, стебел, бульб і квітів
Kostrica, 1988
Морфологія, біохімічні параметри
Звездина, 1989
Морфологія, продуктивність
Рассадина и др., 1990
Морфологія, урожайність, уміст крохмалю, стійкість проти
фітофторозу
Кучко, 1992
Морфологія та фенотипічні ознаки
Лопухин, 1994
Стійкість проти Vertillium dahliae
Sebastiani et al., 1994
Стійкість проти PH.ifestans
Рассадина, 1994
Нетемніючий сорт White-Baron із сомаклонального варіанта
сорту Irish Cobbler
Arihara et al., 1995
Толерантність до хлориду натрію
Burgutin et al., 1996
Морфологія, фізіологія, продуктивність
Thieme et al., 1996
Рівень плоїдності
Mozafari et al., 1997
493
В.Сидоров і співавтори [44] одержали й дослідили більше 320
рослин картоплі сорту Зарево, що регенерували із незалежних клі¬
тинних клонів (протоклони). Приблизно таку ж кількість рослин ви¬
бракували через аномальну морфологію, повільний ріст або відсут¬
ність нормального ризогенезу. Мінливість у популяції протоклонів
картоплі за морфологічними та господарсько цінними ознаками ви¬
вчали в польових умовах. Морфологічні зміни спостерігалися за: то¬
вщиною, пігментацією, облистяністю, вираженістю крил на ребрах
стебла; розсіченістю, кольором, опушеністю, жилкуванням та фор¬
мою часток листка і кінцевою часткою. Лише 20% протоклонів сорту
Зарево були морфологічно ідентичними з вихідною формою. Серед
господарсько цінних ознак спостерігались зміни за урожайністю,
кількістю бульб у кущі, середньою масою однієї бульби, умістом су¬
хої речовини, крохмалю та сирого протеїну.
Значну, хоч і меншу, ніж у сорту Зарево, варіабельність багатьох
ознак спостерігали також серед рослин-регенерантів, одержаних із
мезофільних протопластів дигаплоїда сорту Гатчинська 859Г. Зокре¬
ма, це стосується дещо інших каріотипових змін: близько 55% рос¬
лин виявилися тетраплоїдами, решта - дигаплоїдами і поодинокі -
анеуплоїдами. Відібрані також хлорофілдефектні клітинні лінії, спон¬
танна частота творення яких становила 2 х Ю'5.
Вивчали також спонтанну варіабельність серед протоклонів ди¬
кого виду картоплі Solanum pinnatisectum (2п-2х-24), який характе¬
ризується особливо високою регенераційною здатністю. Усі дослід¬
жені рослини-регенеранти мали типову диплоїдну кількість хромо¬
сом і нормальну морфологію.
Висока спонтанна варіабельність у тетраплоїдної картоплі може
бути зумовлена складною гібридною природою цього виду з прита¬
манним йому високим рівнем гетерозиготності, який підтримувався
завдяки вегетативному способу розмноження [27].
Дослідження Н.Юр’євої [58] дозволили виділити в популяції
сомаклонів сорту Любимець три форми, які перевищують вихідну за
стійкістю проти фітофторозу, підвищеною продуктивністю та досить
високим умістом крохмалю. Серед регенерантів сорту Смєна за ком¬
плексом господарсько цінних ознак виділили дві форми. Одержані
результати дозволяють припустити, що потенційний, найбільш віро¬
гідно видовий діапазон варіювання ознаки, має в кожному сорті пев¬
ний фіксований рівень, який за допомогою сомаклональної варіабель¬
ності можна довести до максимальних значень і таким чином вирі¬
шувати завдання сортополіпшення.
Г.Шенк із співавторами спостерігали появу лінії, що виникла при
регенерації в калусній культурі сорту Аркула [137]. Ця лінія відріз¬
нялася від вихідного сорту більшою кількістю стебел та бульб, низь¬
корослістю та нижчою врожайністю. Такі ознаки лінія зберігала про¬
тягом 8 років вегетативного розмноження.
494
В. Солодовниченко [46] відзначав, що при культивуванні in vitro
зазнають змін як якісні, так і кількісні ознаки. Частота сомаклональ-
них варіантів залежить від виду експланта, вихідного генотипу, умов
культивування. Ядерні і цитоплазматичні гени однаково відповіда¬
льні за їх виникнення. Сомаклональні зміни успадковуються, але в
деяких випадках передаються тільки мітозом і є потенційно зворот¬
ними при регенерації і переході до мейозу. Змінюватися можуть од¬
ночасно кілька ознак, які успадковуються незалежно.
Значні варіаційні відмінності характерні також для регенерантів,
похідних експлантів через первинний та тривалопасований калус
[55]. Із багатьох типів змін, що спостерігали у сортів Биліна, Смєна,
Любимець, Ульяновська та ін., особливу увагу привертають приско¬
рення стартової інтенсивності росту, збільшення фаз розвитку, під¬
вищення рівня врожайності, крохмалистості та горизонтальної стій¬
кості проти фітофторозу.
У дослідженнях П.Костріка із співавторами [107] вивчені основні
морфологічні, цитологічні та агрономічні властивості регенерантів
картоплі із калусів стеблового, листкового і бульбового походження.
У 8 вивчених сортів частота мінливості серед регенерантів становила
від 0 до 7%.
У групі калусів стеблового походження лише регенеранти сорту
Apta характеризувалися 100%-ною морфологічною мінливістю. Де¬
які з них мали інший рівень плоїдності, пов’язаний, як правило, із
змінами всього габітусу регенерантів. У регенерантів сорту Apta та
інших сортів виявлені сомаклони із зміненою стійкістю проти пато-
генів. У тетраплоїдних генотипів зустрічались зміни кольору шкірки
бульб та квітів, а також морфології листків. Деякі з них були химер¬
ної природи.
Широкий спектр мінливості в культурі in vitro, що спостерігали
багато авторів, пояснюється не тільки гетерозиготністю соматичних
клітин картоплі. Важливою причиною її є умови культивування іп
vitro: можлива стимулювальна дія середовища і стимуляторів росту
та найбільш вірогідно - виникнення у процесі поділу соматичних
клітин різних хромосомних перебудов (делеції, транслокації, інвер¬
сії, заміщення), соматичний кросинговер, що призводить до рекомбі¬
нації та нових поєднань генів [20].
Т.Олійник [35] вивчала варіабельність сомаклональних ліній сор¬
тів селекції Інституту картоплярства УААН. Проведені багаторічні і
різнопланові дослідження дали змогу розробити умови, що за¬
безпечують регенерацію рослин у суспензійній культурі картоплі
(рис.7.1). Одержані рослини-регенеранти оцінювали в польових умо¬
вах за комплексом господарсько цінних ознак з урахуванням морфо¬
логії і стійкості проти грибкових та бактеріальних хвороб. Аналіз
частоти виникнення в популяціях сомаклонів ліній, які перевищують
за продуктивністю вихідну форму, показує, що в сомаклонів сортів
495
Бородянська рожева, Зарево, Кобза, Луговська, Незабудка такі лінії
зустрічаються значно частіше, ніж у Світанку київського,
Львів’янки, Проліску.
Рис.7.1. Схема одержання сомаклонів картоплі
496
Так, наприклад, у каліклонів сорту Зарево із 300 випробуваних у
полі 9 були за продуктивністю, вмістом у бульбах сухої речовини,
сирого протеїну вищими за вихідну форму. Щодо стійкості цих реге¬
нерантів проти мокрої гнилі, то 3 із них мали 7-8 балів стійкості (конт¬
роль - 6 балів). Із суспензійної культури сорту Зарево було відібрано
300 генотипів. Ці рослини відрізнялися від вихідної форми за фено¬
типом (міцно розвинені рослини із вкороченим або нормальним між¬
вузлям, міцним стеблом, великим листям та добре розвиненою коре¬
невою системою). Найбільш продуктивними з позитивними відхи¬
леннями за господарсько цінними ознаками були форми з 5-го по 20-
й пасаж калусу.
Помічено, що частота появи позитивних відхилень за продуктив¬
ністю вища при одержанні регенерантів після 2-3-х місяців культи¬
вування на регенераційних середовищах [21].
У дослідженнях А.Кучка [27] серед протоклонів сорту Зарево
спостерігали велике різноманіття морфологічних і основних госпо¬
дарсько цінних показників як у бік їхнього збільшення, так і знижен¬
ня. Частка протоклонів, що відрізнялися за всіма вивченими ознака¬
ми (близько 30) від сорту Зарево, становила 93,3%. У сорту Гатчин-
ська цей показник не перевищував 60%, хоч за деякими ознаками
(урожайність, якість бульб) і для протоклонів цього сорту характер¬
ний високий коефіцієнт варіації (20%). Отримані зміни стабільно
спостерігались при вегетативному розмноженні.
Використовуючи метод культури тканин, японські дослідники в
1995 році [60] отримали сорт картоплі White-Baron (Білий барон) з
нетемніючого сомаклонального варіанта сорту Irish Cobbler (Айриш
коблер).
З використанням методу сомаклональної мінливості співробітни¬
ки Інституту картоплярства УААН отримали сорт Ольвія, який з
1999 року занесено до Реєстру сортів рослин України [126].
Отже, одержані експериментальні дані про сомаклональну мінли¬
вість з метою селекції дозволяють сьогодні досить точно деталізувати
програму робіт у напрямі поліпшення сортів та вихідного матеріалу за
тими ознаками, варіабельність яких у культурі досить висока.
Клітинна селекції картоплі. Добір змінених форм. В останні
роки більш широкого застосування набуває метод клітинної селекції
(рис. 7.2). Перспективність цього методу доведена для томатів, лю¬
церни, ячменю, картоплі і т.д. Найбільш ефективна вона у тих куль¬
тур, для яких розроблені методи масової регенерації рослин із клітин
та протопластів. Форми, стійкі до стресових факторів, одержують за
допомогою простої селективної системи, що дозволяє добирати відпові¬
дні генотипи, оскільки в умовах in vitro виживають поодинокі, але спад¬
ково стійкі до стресорів генотипи [17]. Якщо селекція проводиться на
стійкість до патотоксину, то використовуватися можуть спонтанні й ін¬
дуковані мутанти, що виникають у клітинній культурі [6].
497
Рис.7.2. Схема етапів проведення досліджень з клітинної селекції
498
Вперше можливість добору стійких рослин на клітинному рівні з
використанням хімічного аналога токсину довів американський до¬
слідник П.Карлсон [76]. Для виділення форм тютюну, стійких проти
бактеріальної рябухи, ним був використаний аналог токсину, що
продукується Pseudomonas tabaci - метіонін сульфоксимін. Після
додаткового тестування відібрали три калуси: один одержаний від
клітин, два - від протопластів. Із них були регенеровані диплоїдні
рослини, стійкі проти патогену.
У дослідженнях М.Бенке [68] з Інституту Макса Планка (ФРН)
вивчалася можливість одержання фітофторостійких форм картоплі
при відборі калусів на середовищі, яке містило культуральний фільт¬
рат Р.infestans. На основі проведених досліджень зроблено висновок,
що рослини, одержані із відібраних у селективних умовах калусів,
мають загальну (горизонтальну) стійкість.
Відбір стійких проти фузаріозу форм люцерни на клітинному рів¬
ні був проведений К.Хартмен та Д.Лінг зі співавторами [98, 113] із
університету штату Невада (США) серед клонів з високим регенера¬
ційним потенціалом. Стійкі проти культурального фільтрату лінії
були одержані і при прямому доборі. Ці регенеранти мали нормальну
кількість хромосом (2п-32), що свідчить про перевагу такого добору.
Форужі Вехр та інші [92] вивчали також реакцію генотипів кар¬
топлі на фільтрат Р.infestans. При розміщенні на поживне середови¬
ще з культуральним фільтратом патогену двох дигаплоїдних калус¬
них ліній з різною польовою стійкістю виявили, що калуси рослин із
високою стійкістю через чотири тижні некротизувались, а ріст калу¬
сів сприйнятливих рослин уповільнювався. Перша реакція відповідає
надчутливості, друга — аналогічна тій, яка проявлялась у калусів
сприйнятливих форм у дослідах М.Бенке [68]. У тканині бульб стій¬
кого клону вже в перші три доби після дії культурального фільтрату
спостерігалося накопичення фітоаклексинів (100 мг на 1 г сирої ма¬
си), у той же час як у його калусах не було виявлено акумуляції цих
речовин. Очевидно, механізм стійкості, пов’язаний з нагромаджен¬
ням фітоалексинів, не має для калусів ніякого значення.
Дослідження показали, що добір in vitro на стійкість, який визна¬
чається надчутливістю (некрозами), не може бути ефективним, оскі¬
льки стійкі клітини гинуть.
Система добору на стійкість проти Ervinia atroseptica вивчена в
дослідах Т. Олійник [35]. Для добору резистентних калусів викори¬
стовували комплекс екзоферментів (СФ), що продукуються збудни¬
ком E.atroseptica.
Як показали результати досліджень, зміни на рівні перших па¬
гонів візуально спостерігалися у всіх сортів. Третя частина одержа¬
них рослин-регенерантів складалася із фенотипічно нормально роз¬
винених пробіркових рослин. Поряд з цим з’являлися регенеранти з
очевидними відхиленнями від норми: рослини з дуже вкороченим
499
міжвузлям, розеткові, багатостеблові або гіллясті з послабленим вер¬
хівковим домінуванням; з редукованою різною мірою листковою
пластинкою; частковою хлорофілдефектністю; повні альбіносні та
низькорослі рослини. Найбільше відхилень від норми давали лінії
сорту Незабудка. Цікаво те, що на одному й тому ж калусному екс-
планті, де нерідко відбувалась регенерація низькорослих чи альбінос-
них рослин, водночас з’являлися цілком життєздатні регенеранти з
нормальним стеблом та зеленим кольором.
Унаслідок проведеної роботи були одержані рослини-регенеран-
ти з різною стійкістю проти мокрої гнилі.
У дослідженнях Е. Мазіна [31, 32] розроблений спосіб реге¬
нерації конюшини, люцерни і картоплі з клітин суспензійних куль¬
тур. У результаті проведення добору клітинних ліній конюшини,
стійких проти культуральної рідини збудника склеротиніозу, одер¬
жані рослини-регенеранти та оцінені на стійкість проти патогену при
штучному зараженні. Проведено добір клітинних ліній, стійких про¬
ти культурального фільтрату збудника фітофторозу.
Особливої уваги заслуговує питання про можливість клітинної
селекції картоплі на стійкість проти грибкової та бактеріальної ін¬
фекції [21, 42]. Вона можлива в тих випадках, коли доведено, що фак¬
тором патогенності збудника є токсин і його наявність викликає по¬
вний спектр симптомів цього захворювання. Цій умові повністю від¬
повідає досить незначна кількість збудників, оскільки в більшості
випадків поряд з токсинами факторами патогенності є також екзофер-
менти, що продукуються патогенами. У лабораторії клітинної селек¬
ції Інституту картопляного господарства РАСГН проводили дослід¬
ження щодо отримання форм картоплі з підвищеною стійкістю проти
кільцевої гнилі методом культури тканин. Як селективний фактор
використовували токсин, що продукується бактеріями С. sepedo-
пісит. Всі отримані рослини-регенеранти були розподілені за стійкі¬
стю на 4 групи: високостійкі, толерантні, із стійкістю на рівні вихід¬
ної, нестійкі.
Таким чином, отримано генетичне різноманіття сомаклональних
варіантів з різною стійкістю проти патогенів. І хоч вимоги селекціо¬
нерів значно випереджають реальні можливості їх забезпечення, все
ж одержані рослини-регенеранти мають склад генів, який може зни¬
зити швидкість пристосування збудників.
Приклади досягнення позитивних практичних результатів свід¬
чать про необхідність більш ефективного впровадження різних при¬
йомів клітинної інженерії у практичну селекцію.
ЛІТЕРАТУРА
1. Аветисов В.А. Индуцированный морфогенез картофеля в культуре in
vitro // Культивирование клеток растений и биотехнология.- Кишинев: Шти-
инца, 1983. - С. 110.
500
2. Аветисов В.А., Мелик-Саркисов О.С. Генотипические особенности
морфогенеза в каллусных культурах различных сортов картофеля // С.-х.
биол.-1985.-№3. - С.67-70.
3. Аникеева Н.В. Полевая оценка сомаклонов пшеницы на устойчивость
к корневой гнили / Саратовская гос. с.-х. акад. - Саратов, 1994 - С.90-94.
4. Асеева Т.В. Вегетативные мутанты картофеля // Генетика-1968. -
Т.4, №3. - С. 145-164.
5. Атанасов Атанас. Биотехнология в растениеводстве. - М., 1993.
6. Биотехнология и проблемы селекции растений на устойчивость / Н.Н.
Гусева, О.С. Афанасенко, Р.И. Высоцкая и др. // Защита растений-1990. -
№8. -С.3-5.
7. Бреславец Л.П. Полиплоидия в природе и опыте. - М.: Из-во АН
СССР, 1963. -364 с.
8. Бугаенко Л.А., Бугара А.М. Клеточные технологии в селекции эфи¬
ромасличных растений // Тезисы докл. междунар. конф. "Агробиотехноло¬
гии растений и животных” (29-30 мая 1997 г., Киев). - К., 1997. - С.84.
9. Бутенко Р.Г. Культура изолированных тканей и физиология морфоге¬
неза растений. - М.: Наука, 1964. - 272 с.
10. Бутенко Р.Г. Технология in vitro в сельском хозяйстве // С.-х. биол. -
1983. - №5. - С.3-7.
11. Бутенко Р.Г. Морфогенез in vitro как важнейший этап клеточной и генной
инженерии растений // Новые методы биотехнологии растений: Тезисы П Рос.
симпозиума (18-20 мая 1993 г., Пущино).-Пущино, 1993. - С.118.
12. Бутенко Р.Г., Кучко А.А., Витенко В.А. Получение и культивирова¬
ние изолированных протопластов из мезофилла листа Solanum tuberosum и
Solanum chacoense // Физиология растений. - 1977. - Т.24. - С.660-665.
13. Гострицкий С А., Багрова А.М., Ежова ТА. Обнаружение и цитогенетичес¬
кий анализ изменчивости, возникающей при регенерации растений из культуры
ткани полевого гороха // Докл. АН СССР. -1985 - Т.283, №4. - С. 1007-1011.
14. Давоян Э.И. Мутагенез в культуре ткани риса и получение на его основе
нового исходного материала И Г енетика. - 1983. -Т. 19, №10-С.1714-1719.
15. Долбік Г.М., Бердзічавец Л.П. Калусаутварэнне і морфогенез у куль¬
туры тканах бульбы // Весці АН БССР, Сер.біял.наук. - 1986. - №3- С.22-
26 (Білорус, мовою).
16. Долбик Г.М., Быкова Л.Н., Юшкевич А.Т. Сомаклональная изменчи¬
вость картофеля in vitro // Тез. докл. Всесоюз. конф. по генет. сомат. клет. в
культ. - М., 1989. - С.43.
17. Долгих И.Ю., Ларина С.Н., Шамина З.Б. Толерантность отобранных
in vitro растений кукурузы к засолению и низким температурам // Материа¬
лы научн.конф. "Актуальные проблемы биотехнологии в растениеводстве,
животноводстве и ветеринарии". - М., 1996 - С.82.
18. Жила Э.Д., Кучко А.А., Сидоров В.А. Хромосомная изменчивость
протоклонов картофеля //Цитология и генетика. - 1987.-Т.21. - С. 105-108.
19.Жученко А.А. Экологическая генетика культурных растений. - Ки¬
шинев, 1980.-587 с.
20.3вездина Т.Н. Изучение сомаклональной вариабельности некоторых
сортов картофеля // Селекция, агротехника и орошение овощных культур. -
1989. - С.15-19.
501
21. Клеточная селекция на устойчивость к кольцевой гнили картофеля:
оптимизация оценки устойчивости сомаклональных линий картофеля к ко¬
льцевой гнили / Г.В. Рассадина, Л.М. Хромова, Р.Г. Бутенко, Н.Г. Иванова //
Биология культивированных клеток и биотехнология растений / АН
СССР.Ин-т физиологии растений. - М., 1991. - С. 133-137.
22. Кунах В.А. Изменчивость числа хромосом в онтогенезе высших рас¬
тений И Цитология и генетика. - 1978. - Т. 12, №2. - С.160-173.
23. Кунах В.А. Геномная изменчивость соматических клеток растений и
факторы, регулирующие этот процесс // Цитология и генетика. - 1980. -
Т.14, №1.- С. 73-81.
24. Кунах В.А. Получение клеточных культур-продуцентов биологически
активных веществ и пути регуляции их продуктивности // Тез.докл. между-
нар.конф. "Пути решения проблем и перспективы развития биотехнологии в
декоративном садоводстве и плодоводстве” (25-26 сент.1997. Ялта) - Ялта,
1997.- С.6
25. Кунах В.А., Алпатова Л.К. Роль фитогормонов в изменчивости числа
хромосом в культуре тканей Haplopappus // Докл. АН СССР. - 1979. - Т.245,
№4. - С.967-970.
26. Кучеренко Л.А. Индуцированный морфогенез в культуре тканей риса
и его использование для создания исходного селекционного материала //
Культура клеток растений и биотехнология. - М.: Наука, 1986. - С.211-213.
27.Кучко А.А. Разработка и применение биотехнологических методов
создания ‘исходного селекционного материала картофеля: Дис... д-ра с.-х.
наук в форме научного доклада. - К., 1992. - 42 с.
28.Кучко А.А., Бутенко Р.Г. Культура изолированных протопластов
S.tuberosum L и S.chacoense Butt // Использование тканевых культур в селек¬
ции растений: Тезисы докл. на междунар. совещании в Оломоуце, ЧССР. -
Оломоуц,1976. - 29с.
29. Кучко А.А.,Олійник Т.М. Сомаклональна мінливість у картоплі //
Картоплярство. - Вип.28. - 1998. - С.28-36.
30. Леонова Н.С., Солоненко Л.П. Биохимические различия у картофеля
на разных уровнях дифференциации в культуре тканей // Использование
клеточных технологий в селекции картофеля. - М., 1987. - С.89-94.
31.Мазин В.В. Клеточная селекция клевера лугового на устойчивость к
возбудителям корневых гнилей (Sclerotinia trifoliorum, Fusarium sp.sp.) //
Молекулярные и генетические механизмы взаимодействия микроорганиз¬
мов с растениями. - Пущино, 1989. - С.164-167.
32.Мазин В.В. Биологические технологии получения генотипов расте¬
ний, устойчивых к биотическим и абиотическим стрессам // Материалы
научн. конф."Актуальные проблемы биотехнологии в растениеводстве, жи¬
вотноводстве и ветеринарии". - М., 1996. - С.82.
33. Моргун В.В, Логвиненко В.Ф. Мутационная селекция пшеницы - К.:
Наукова думка, 1995 - 628 с.
34. Мусин С.М. Полиморфизм белков и возможности его использования
в идентификации генотипов картофеля в культуре in vitro: Автореф.... канд.
биол. наук. - М.: МГУ, 1989. - 19 с.
35. Олійник Т.М. Клітинна селекція картоплі на стійкість проти Erwinia
atroseptica: Автореф.... канд. с.-г. наук-К. 1995.-24 с.
502
36.Осипчук А.А. Селекція картоплі в умовах Полісся України: Дис....
д-ра с.-г. наук у формі наукової доповіді. - Харків, 1993.-50 с.
37. Перспективы применения тканевых культур в селекции плодовых и
ягодных растений /В.М. Тюленев, А.В. Будаговский, Р.П. Евсеева, Н.В. Со¬
ловых // Материалы научн.конф. "Актуальные проблемы биотехнологии в
растениеводстве, животноводстве и ветеринарии".- М., 1996.- С.82.
38. Пирузян Э.С. Основы генетической инженерии растений. - М.: Наука,
1988.-302 с.
39. Получение сомаклональных и трансгенных клеток растений пшеницы
Triticum aestivum, устойчивых к ультрафиолетовому излучению / Р.Г. Бутен¬
ко, Л.Г. Копертех, Л.А. Коппель, П.В. Лапшин // Тезисы докл. Междунар.
конф. “Молекулярно-генетические маркеры растений” (11-15 ноября, 1996 г.,
Киев).-К., 1996.-С.44.
40. Получение сомаклональных линий у злаков (Triticum aestivum L. И
Hordeum vulgare L.) / A.K. Гапоненко, Н.И. Маликова, Г.Н. Охрименко, А.А.
Созинов // Докл. АН СССР. - 1985. -Т.283, №6. - С. 1471-1475.
41. Попухин Н.И. Индукция фенотипической и генетической изменчивос¬
ти в зависимости от методов получения сомаклональной вариации в культу¬
ре in vitro и использование ее в селекции картофеля. - Новосибирск, 1994.-
С. 19-21.
42. Рассадина Г.В. Клеточная инженерия для сортоулучшения картофеля
/ НПО по картофелеводству РАСХН. - М., 1994. - №4.- С.22.
43. Редько В.В. Особливості онтогенезу та формування продуктивності
цукрових буряків і соняшнику. - К.: Укр ІНТЕІ, 1994.-140 с.
44. Сидоров В.А. Биотехнология растений. - К.: Наукова думка, 1990 - 280 с.
45. Сидоров В.А., Кучко А.А. Культура изолированных протопластов для по¬
лучения новых форм картофеля // Науч. тр. НИИКХ. - М., 1984. - С. 108-114.
46. Солодовниченко В.Д. Сомаклональная изменчивость как способ увели¬
чения формообразовательного процесса // Прогресс в селекции озимой пшеницы
как фактор интенсификации производства зерна. - Мироновка, 1988. - С.34-38.
47. Сомаклональные вариации среди растений, полученных из протокло¬
нов картофеля / В.А. Сидоров, А.А. Кучко, П.П. Гайдук, Ю.Ю. Глеба // Докл.
АН СССР. - 1985. - Т.281, №3. - С.704-707.
48. Сомаклональная изменчивость / З.Витанов, М. Влахова, П.Денчев и
др. // Биотехнология и биохимия. - 1990. - Т.4, №1. - С.8-13.
49. Суриков И.М. Регенерация побегов из первичного клубневого каллу¬
са у различных сортов картофеля // Культивирование клеток растений и био¬
технология. - Кишинев: Штиинца,1983. - С.92-93.
50.Трофимец Л.Н., Волкова Т.В., Миренкова Н.Н. Метод культуры тка¬
ней в картофелеводстве / Тканевые и клеточные культуры в селекции расте¬
ний. -М.: Колос, 1979. - С. 123-128.
51. Тучин С.В. Экспрессия и наследование термодинамических парамет¬
ров воды у засухоустойчивых сомаклонов пшеницы // Генетика. - 1994-
№30.-С. 160.
52. Тучин С.В. Влияние условий культивирования изолированных тканей
на частоту сомаклональной изменчивости пшеницы // Материалы научн.
конф. "Актуальные проблемы биотехнологии в растениеводстве, животно¬
водстве и ветеринарии." - М., 1996. - С.82.
503
53. Фролова Л.В., Шамина З.Б. Цитогенетическая характеристика куль¬
туры тканей растений из семейства бобовых // Цитология и генетика. - 1974.
-Т.8, №5. - С.413-418.
54. Хромова Л.М. Каллусо- и морфогенез в культуре тканей картофеля //
Исследования по клеточной селекции картофеля: Науч. тр- М., 1984. -
С.81-88.
55. Хромова Л.М. Система ранних тестов для скрининга сомаклонов кар¬
тофеля // Биотехнология культивируемых клеток и биотехнология: Тезисы
междунар. конф. - Новосибирск, 1988. - С.255-256.
56. Хромова Л.М., Седнина Г.В. Условия получения регенерантов в куль¬
туре ткани картофеля // Культивирование клеток растений и биотехноло¬
гия- Кишинев: Штиинца, 1983. - С.93-94.
57. Цилосани Г.М. Морфогенез в культуре каллусной ткани картофеля//
Бюл. ВНИИ растениеводства. - 1980 - Т.98. - С.64-65.
58. Юрьева Н.О. Изучение различий в вариационных потенциях геноти¬
пов сортов картофеля // Использование клеточных технологий в селекции
картофеля: Науч. тр. - М., 1987. - С.6-10.
59. .Ahloowalia B.S. Piant regeneration from callus culture in potato //
Euphytica. -1982.- Vol.31.- P.755- 759.
60. White-baron - a non -browning somaclonal variant of danshakuimo (Irish-
cobbler) / Arihara A., Kita Т., Igarashi S., Goto M.,Irikura Y. // American potato
joumal. - 1995. - Vol.287, №11. - P.701-705.
61.Austin S., Cassels A.C. Variation between plants regenerated from
individual calli produced from separated potato stem callus cells // Piant Sci. -
1983.- Vol.31. -P.107-114.
62.Barabas Z. Biotechnology: a way to increase the efficiency of piant
breeding // Economic Bull.Eur. - 1986. - Vol.38, №1. - P.100-102.
63.Somatic hybridization in the genes Solanum: Solanum tuberosum and
Solanum brevidens / Barsby T.L., Shepard J.F., Kemble R.J.,Wong I.R. // Piant
Cell Rep. - 1984. - Vol.3. - P.165-169.
64. Bayliss M.W. Chromosomal variation in piant tissues in culture // Int. Rev.
Cytol. - 1980. - Vol.llA, suppi. - P.l 13-114.
65.Bebeli Р., Kaltsikes P. Superior yielding R3 families in hexaploid triticale
through tissue culture // J.Genet. and Breed. - 1995. - Vol.49, №2. - P.l05—110.
66.Behnke M. Culturen isolierter Zellen von einigen dihaploiden Solanum
tuberosum - klonen und ihre Regeneration // Z.Pflanzenphysioi. - 1976. - Vol.78.
- P.177-181.
67. Behnke M. Selection of potato callus for resistance in culture filtrates of
Phytophthora infestans and regeneration of resistant plants // Theor. and
Appi.Genet. - 1979. - Vol.55, №2. - P.69-71.
68. Behnke M. General resistance to late blidht of Solanum tuberosum plants
regenerated from callus resistant to culture filtrates of Phytophthora infestans //
Ibid. - 1980. - Vol.56. - P.151-152.
69. Benzion G., Philips R.L., Rines H.W. Cell culture and somatic cell genetics of
plants / Ed.I.K. Vasil. - New York: Acad. press, 1986. - Vol.3 - P. 156-162.
70. Binns A., Meins F.J. Evidence that habituation of tobacco pith cell
division-promoting factors is heritable and potentially reversible // Proc. Nat.
Acad. Sci. USA. - 1973. - Vol.70. - P.2660-2662.
504
71.Brettell R.I.S., Goddard B.V.D., Ingram D.S. Selection of Tms-cytoplasm
maize tissue cultures resistant to Drechslera maydis T - toxin // Maydica. - 1979.
- Vol.24.-P.203-213.
72.Brettell R.I.S., Thomas E., Ingram D.S. Reversion of Texas malesterile
cytoplasm maize in culture to give fertile, T-toxin resistant plants // Theor. and
Appi. Genet. - 1980. - Vol.58, №2. - P.55-58.
73.Burgutin A.B., Butenko R.G., Kaurov B.A., Iddagoda N. In vitro selection
of potato for tolerance to sodium - chloride // Russian joumal of piant physiology.
-1996. - Vol.43, №4. - P.34-38.
74. Cai Т., Ejeta G., Butler L. Screening for grain / polyphenol variants from
high-tannin sorghum somaclones // Theor. and Appi. Genet. - 1995. - Vol.90,
№2.-P.211-220.
75.Cassels A.C., Tarrell G., Coleman M.C. Somaclonal variation as a source
of novel virus resistance in potato character improvement // Abst.of Conf.of
Papers and Poster of the oth Trienn Conf.of the Eur. Ass. Potato Res. 23-31 July.
- 1987.-423 p.
76. Carlson P.S. Methionine sulfoximine-resistant mutants of tobacco // Ibid. -
1973. - Vol.180. - P. 1366-1368.
77.Chaleff R.S., Keil R.L. Genetic and physiological variability among
cultured cells and regenerated plants of Nicotiana tabacum // Мої. and Gen.
Genet-1981. - Vol.181. -P.254-258.
78. Chaleff R.S. Ray T.B. Herbicide-resistant mutants from tobacco cell
culture // Science. - 1984. - Vol.223. - P. 1148-1151.
79. Chapman H.W. Potato tissue culture // Amer. Potato Jour. - 1955. -
Vol.32. -P.207-210.
80. Cocking E.C. A method for the isolation of piant protoplasts and vacuoles
// Nature. - 1960. - Vol.187. - P.962-963.
81.Cooper D.B., Sears R.G., Lookhart G.L., Lones B.L.Heritable somaclonal
variation in gliadin proteins of wheat plants derived from immature embryo callus
culture // Ibid. - 1986. - Vol.71, №6. - P. 784 -790.
82. D'Amato F. Nuclear cytology of tissue cultures // Piant Tissue and Cell
Culture Application to Crop Improvement: Proc. Int Symp. Olomouc. (Czecho-
slovakia, Sept. 24-29,1984). - Prague, 1984. - P.295-303.
83.Dan Yinghui.,Stephens C. The development of asparagus somaclones with
high levels of resistance to Fusarium oxysporum f.sp.asparagi and F.proliferatum
// Piant Disease. - 1995. - Vol.79, №9. - P.923-927.
84. Davies P.A.,Pallotta M.A.,Ryan S.A. et.al. Somaclonal variation in wheat:
genetic and cytogenetic characterizeation of alcoholde - hydroginase I mutants //
Theor.Appl.Genet. - 1986. - Vol.72. - P.644-653.
85.Deambrodio E.,Dale P.I. EfFect of 2,4-D on the freguency of regenerated
plants in barley (Hordeum vulgare) cultivar “Akka” and genetic variability
between them // Cereal Res. Commun. - 1980. - Vol.8. - P.417-424.
86. Diversity of Vicia faba circular mtDNA in whole plants and suspension
cultures TAG / Negruk V.L., Eisner G.J., Redichkina T.D., et al - 1986 - Vol.72.-
P.541-547.
87.Donn G., Tischer E., Smith J.A., Goodman H.M. Herbicide - resistant
alfalfa cells: an example of gene amplification in plants // J. Мої. Appi. Genet. -
1984. - Vol.2, №6. - P.621-635.
J 7 2-132
505
88. Evans D.A., Sharp W.R. Single gene mutations in tomato plants
regenerated from tissue culture // Science. - 1983. - Vol.221. - P.949-951.
89. Evans D.A., Sharp W.R., Bravo J.E. Cell culture methods for crop
improvernent //Handbook of piant cell culture. - New York, London: Macmillan,
1984. - Vol.2. - P.47-68.
90. Evans D.A., Sharp W.R. Applications of somaclonal variation. //
Biotechnology. - 1986. - Vol.4, №6. - P.528.
91. Flick C.E. Isolation of mutants from cell culture / Handbook of Piant Cell
Culture // Technigues for Propagation and Breeding. - New York: Macmilan,
1983. - Vol.l. - P.393-441.
92. Foroughi-Wehr B., Stolle K. Resistenz selection in vitro am Bcispieldes
systems kartoffel (Phytophthora infestans Mont.) de Barry // Nachr. Dt.
Pflanzenschutz. -1985. -Bd.71- S. 115-211.
93. Fukui Y., Fukushima М., Ono H. Fertilization and cleavage of bovine
follicular oocytes in rabbit reproductive tracts afler maturation in vitro // J. Exp.
Zool. -1983. - Vol.226. - P.137-142.
94. Gautheret R.J. La culture des tissus vegetsux. - Paris, 1959. - 864 p.
95. Gengenbach B.G., Green C.E., Donovan C.M. Inheritance of selected
pathotoxin resistance in maize plants regenerated from cell cultures // Proc. Nat.
Acad / Sci. USA. - 1977. - Vol.74, №11. - P.5113-5117.
96. Gill B.S., Kam-Morgan L.N. W., Shepard J.F. Cytogenetic and phenotipic
variation in mesophyle cell-derived tetraploid potatoes // S.Heredity. - 1987-
Vol.78,№l.-P. 15-20.
97. Harten A.M.V., Bouter H., Broertjes C. In vitro adventicious bud techniques
for vegetative propagation and mutation breeding of potato (Solanum tuberosum L.).2.
Significance for mutation breeding. //Euphytica. - 1981. - Vol.30. -P.l-8.
98. Hartman C.L., McCoy T.Y., Knous T.R. Selection of alfalfa (M.sativa)
cell lines and regeneration of plants resistant to the toxin (s) produced by
F.oxysporum f.sp.medicaginis // Piant Sci.Lett. - 1984. - Vol.34. - P.l 83-194.
99. Heritable somaclonal variation in wheat / Larkin P .J., Ryan S.A., Brettell
R.I.S., Scowcrorft W.R. // Theor. Genet, and Appi. Genet. - 1984. - Vol.67, №5. -
P.443-455.
100. Hiroshi., Amagai Hiroshi., Kikuta Isao., Kubota Mitsuru., Oosawa Katsuji.
Selection of somaclonal variants with low-temperature germinability in melon
(Cucumis melo L.) // Piant Cell Repts. -1995. - Vol.14, №11.- P.684-688.
101. Allicchio R., Antonioli С., Grasiani L. et al. Isozyme variation leaf-callus
regenerated plants of solanum tuberosum // Piant Sci. - 1987. - Vol.53. - P.81-86.
102. Jacobsen E. Isolation, characterization and resistant cell line of dihaploid
potato // J.Plant Physiol. - 1986. - Vol.123. - P.307-315.
103. Jones M.G.K., Bright S.W.S., Nelson R.S., Foulger D., Creissen G.P.,
Angela Каф I.G.Ooms. Variations in plants regenerated from protoplasts and
complex explants of potato // Proc. 6 Intem Protoplast Symp., Basel, Ciba-
Geigy, poster proc. - 1983. - P.l50-151.
104. Каф A. Somaclonal variation as a tool for crop improvement //
Euphytica. -1995. - Vol.85. - P.295-302.
105. Kemble R.J., Flavell RJB., Brettell R.I.S. Mitochondrial DNA analyses of
fertile and sterile maize plants derived from tissue culture with the Texas male
sterile cytoplasm // Theor. and Appl.Genet. - 1982. - Vol.62, №3. - P.213-217.
506
106. Kemble R.J., Shepard J.F. Cytoplasmic DNA variation in a potato
protoclonal population // Ibid. - 1984. - Vol.69, №2. - P.211-216.
107. Kostrica P., Opatmy Z., Fricova E. et al. Somaclonal variation of callus-
derived potato regenerants // Genet, slecht. - 1988. - Vol.24. - P.245-256.
108. Lam S.L. Shoot formation in potato tuber discs in tissue cultures // Amer.
Potato J. - 1975. - Vol.52. - P.103-106.
109. Landsmann J., Uhrig K. Somaclonal variation in Solanum tuberosum
detected at the molecular level // Theor. and Appi. Genet. - 1985. - Vol.71, №3.
- P.500-505.
110. Larkin P.J. II Iowa State J.Res. - 1987. - Vol.61. - P.393-434.
111. Larkin P.J., Scowcroft W.R Somaclonal variation - a novel source of
variability from cell culture for piant improvement // Theor. Appi. Genet. - 1981.
- Vol.60. -P. 197-214.
112. Larkin P.J., Scowcroft W.R. Somaclonal variation: a new option for piant
improvement // Piant improvement and somatic cell genetics. - Orlando etc.:
Acad. press, 1983. - P.159-178.
113. Ling D.H., Vidhyaseharan P., Borromeo E.C.et al. In vitro screening of
rice germplasm for resistance to brown spot disease using phytotoxin // Theor.
And Appi. Genet. - 1985. - Vol.71, №1 - P. 133-135.
114. Lorz H., Brown P.T.H. Variability in tissue culture derived plants -
possible origins; advantages and drawbacks // Genetic Manipulation in Piant
Breeding. -Berlin; New York: Walter de Gruyter Со. - 1986. - P.513-534.
115. Lyssine overproducer mutants wih an altered dihydrodipicolinate
synthase from protoplast culture of nicotiana sylvestris (Spegazzini and Comes.) /
Negrutin I., Cattoir-Reynearts A., Verbruggen I., Jacobs M. // Piant. - 1984. -
Vol.68.-P. 11-20.
116. Maddock S.E., Lancaster V.A., Risiott Ret.al. Piant regeneration from
cultured immature embryos and inflorescences of 25 cultivars of wheat ( Triticum
aestivum) // J. Exp. Bot. - 1983. - Vol.34, №.144. - P.915-926.
117. Maliga P., Xuan L.T., Dix P.J., Cseplo A. Antibiotic resistance in
Nicotiana // Piant Cell Cultures: Results and Perspectives. - Amsterdam: Elsevier,
1980.-P.161-167.
118. Matem U., Strobel G., Shepard J.F. Reaction to phytotoxins in a potato
population derived from mesophyll protoplasts // Proc. Nat. Acad. Sci. USA. -
1978. - Vol.75, № 10. - P.4935-4939.
119. Maroti М., Rudolf J., Bognar J.et al. In vitro piant leaves from potato
shoot segments // Acta Bot. Acad. Sci.Hung. - 1982. - Vol.28. - P. 127-132.
120. Meins F.J., Binns A. Epigenetic variation of cultured'somatic cells:
Evidence for gradual changes in the requirement for factors promoting cell di-
vision // Proc. Nat. Acad. Sci. USA. - 1977. - Vol.77. - P.2928-2932.
121. Meulemans M. Regeneration et selection de variants de pomme de terre
du cultivar Bintje resistants a Phytophthora infestans (Mont.) de Вагу // Ann.
Gembloux. - 1984.- Vol.90. - P.35-38.
122. Morel G. Biochemistry of morphogenesis of piant shoots Proc.IV.
Intemat // Congr. Biochem. - 1959. - Vol.l. - P.221-222.
123. Mozafari J., Wolyn D., Alikhan S. Chromosome doubling via tuber disc
culture in dihaploid potato as determined by confocal microscopy // Piant cell
reports. -1997. - Vol.5. - P.329-333.
507
124. Murashige Т., Scoog F. A revised medium for rapid growth and
bioassays with tobacco tissue cultures // Physiol. Piant. - 1962. - Vol.15, №13. -
P.473-497.
125. Nelson R.S. Genetics of horizontal resistance to piant disease // Ann.Rev.
Phytopath. - 1978. - Vol.16, №3. - P.359-378.
126. Oleinik T. Biotechnological methats for developing pest-resis-tant potato
lines // Bulletin OEPP / EPPO Bulletin 28,000-000.- 1999.
127. Oono K. In vitro methods applied to rice // Piant tissue culture. Methods
and applicati ons in agri culture. - New York: Acad. press, 1981. - P.273-298.
128. Oono K. Genetic variability in rice plants regenerated from cell culture //
Cell and tissue culture technigues for cereal crop improvement. - Beijing (Chine):
Sci. press, 1983. -P.95-104.
129. Ouraishi A., Rossignol-Bancilhon L., Noreran K. Effect de 1'origine du frug-
ment sur la callogeneses in vitro // Ann. amelior. piant. - 1979. - Vol.29. - P.639-663.
130. Pijnacker L.P.,Sree R.K. Somaclonal variation in potato a korryotypis
evaluation // Acta.bot.neerl. - 1990. - Vol.39, №2. - P.163-199.
131. Ramulu K.S., Dijkhuis P., Roest S. Phenotypic variations and ploidy
level of plants regenerated from protoplasts of tetraploid potato // TAG. - 1983. -
Vol.65, №4. - P.329-338.
132. Ramulu K.S., Dijkhuis P., Roest S. Genetic instability in protoclones of
potato (Solanum tuberosum L.cv."Bintje"): new types of variation after vegetative
propagation // Ibid. - 1984. - Vol.68, №6. - P.515-519.
133. Rines H.W., McCoy T.I. Tissue culture initiation and piant regeneration
in hexaploid species of oats // Crop Sci. - 1981. - Vol.21. - P.837-842.
134 . Roest S.,Bokelmann G.S. Vegetative propagation of Solanum tuberosum
L. in vitro//Potato Res - 1976 - Vol.19- P.173-178.
135. Ross H. Potato breeding - Problems and Perspektives. - Berlin and
Hamburg: Verlag Paul Parley, 1986. - 183 c.
136. Rowland G.G. Flax research // Univ. of Saskatchewan Crop Dev. Center.
-1993.-43 p.
137. Schenk G., Moli A., Kleinhenpel D. Morphophysiologische Untersu-
chungen an einer Mutante der Kartoffelsorte Arkula // Arch. Zuchtungsforsch. -
1988. - Vol.18. - P.275-284.
138. Scowcroft W.R. The analysis of somaclonal mutants // Piant. Genetics.,
Ed. Alan R. Liss.,1985. - P.799-815.
139. Scowcroft W.R., Ryan S.A. Tissue culture and piant breeding // Piant
Cell Culture Technology - Oxford: Blackwell Sci. Pubi., 1986. - P.67-95.
140. Sebastiani L., Lenzi A., Pugliesi С., Fambrini M. Somaclonal variation
for Resistance to verticillium-dahliae in potato (Solanum-tuberosum L) plants
egenerated from callus // Euphytica. - 1994. - Vol.80, №1-2. - P.5-11.
141. Secor G.A. The potato protoplast system // Proc. ND .Acad. Sci. - 1984.
-Vol.38.-P.801.
142. Secor G.A., Shepard J.F. Variability of protoplast-derived potato clones
// Crop Sci. - 1981. - Vol.21. - P.l02-105.
143. Shepard J.F. Abscisic acid - enhanced initiation in protoplast - derived
calli of potato // Piant Sci. Lett. - 1980. - Vol.18. - P.334.
144. Shepard J.F. Protoplasts as souruces of disease resistance in plants //
Ann. Rev. Phytopath. - 1981. - Vol.19. - P.l45-166.
508
145. Shepard J.F. The regeneration of potato plants from leaf cell protoplasts
// Scientific Am. - 1982. - Vol.246. - P.l 12-121.
146. Shepard J.F., Totten I.R.E. Mesophyll cell protoplasts of potato.
Isolation, proliferation and piant regeneration // Piant Phys. - 1977. - Vol.60. -
P.313-316.
147. Sibi M. Variability in plants regenerated from tissue culture // Ed. E.D.
Earle, Y. Demarley. - Paris, 1982. - P.228-244.
148. Skene K.G. M.,Barlass M. Studies on the fragmented shoot apex of
grapevine // J. Exp. Bot. - 1983. - Vol. 34, №147. - P. 1271-1280.
149. Skirvin R.M., Lam S.L., Janick J. Plantlet formation from potato callus in
vitro // Hort-Sci. - 1975. - Vol. 10. - P.413.
150. Smith D.B. Variation in the electroforetic band pattem of tuber proteins
from somaclones of pototo // J.Agric. Sci. - 1986. - Vol. 106. - P.427-428.
151. Soulie O., Roustan J., Fallot J. Early in vitro selection of entypinetolerant
plantlets. Application to screening of Vitis vinifera cv. Ugni blanc somaclones//
Inst. National Polytechnique. Lab. de Biotechnologie et Amelioration des Plantes,
Toulouse. Vitis. -1993. - Vol.32, №4 - P.243-244.
152. Steward F.C., Carlin S.M. A tissue culture from potato tubenthe
synergistic action of 2,4-D and coconut wilk // Science. - 1951- Vol.lll. -
P.518-520.
153. Thanutong P.,Furusawa I.,Gamamoto M. Resistant tobacco plants from
protoplast-derived calluses selected for their resistance to Pseudomonas and
Altemaria toxins//Theor and Appi. Genet - 1983 - Vol.66. -P.209-215.
154. Thieme R, Griess H. Somaclonal variation of haulm growth, earliness
and yield in potato // Potato research. - 1996. - Vol.3. - P.355-365.
155. Tibok A., Blackhall N., Power J., Davey M. Optimized piant re¬
generation from callus derived from seedling hypocotyls of Eucalyptus
urophylla // Plan Genetic Manipulation Group. - 1995. - Vol.110, №1. -
P.139-145.
156. Transisent cycloheximide resistance in a tobacco cell line / Maliga P., Lazar
P., Svab Z., Nagy F. // Мої. and Gen. Genet. - 1976. - Vol. 149. - P.267-271.
157. Umbeck P.F., Gendenbach B.G. Reversion of male sterile T-cytoplasm
maize (Zea mays) to male fertility in tissue culture // Crop. Sci. - 1983. - Vol.23.
- P.584-588.
158. Upadhya M.D. Isolation and culture of mesophyll protoplasts of potato
(Solanum tuberosum L.) // Potato Res. - 1975. - Vol.l 8. - P.438-445.
159. Vasil I.K. Advantages of embryogenic cell cultures of Graminea // Nuci.
Techn. and in vitro Cult Piant. Improv. Proc. Int. Symp. - Vienna, 1986. - P.71-75.
160. Webb K.J.,Henshaw G.G. Morphogenesis in leaflet discs of potato
(Solanum tuberosum subsp.tuberosum) // Piant Tiss. Cult. Tokyo: Jap. Ass. Piant
Tiss.Cult, 1982.-P. 157-158.
161. Wenzel G., Schieder О., Przewozny T. et al. Comparison of single cell
culture derived Solanum tuberosum L. plants and a model for their application in
breeding programs // Theor. Appi. Genet. - 1979. - Vol.55. - P.49-55.
162. Wheeler V.A., Evans N.E., Foulger D., Webb K.J., Karp A., Frankiin J.,
Bright S.W.J. Shoot regeneration from expiant cultures of fourteen potato culti-
vars, and studies of the cytology and morphology of regenerated plants. -1985. -
P.361-434.
509
163. White P.R. Piant tissue cultures a preliminary report of results obtained
in the culturing of certain piant meristems // Arch. exptl. Zellforsch. - 1932. -
Vol.12. -P.602-620.
164. White P.R. Potentially unlimited growth of excised tomato root tips in a
liquid medium // Piant. Physiol. - 1934. - Vol.12. - P.777-791.
165. Widholm J.M. Selection and characterization a Daucus carota L.cell line
resistant to four amino acid analogues // J. Exp. Bot. - 1978. - Vol.29. - P.l 111-
1116.
7.2. ЕКСПЕРИМЕНТАЛЬНИЙ АНДРОГЕНЕЗ
Одержання гаплоїдних рослин становить великий інтерес для
практичної селекції і генетики. В основі виникнення гаплоїдів ле¬
жать фізіологічні процеси, пов’язані із змінами нормального шляху
розвитку чоловічих і жіночих статевих клітин (андрогенез, гіноге¬
нез). Ці явища мало вивчені на клітинному і молекулярному рівнях і
регулюються в основному емпірично - підбором оптимальних умов
для індукції аномального процесу і реалізації індукованого стану, що
завершується появою калусної тканини гаметичного походження або
андрогенних і гіногенних ембріоїдів.
Спосіб одержання гаплоїдів шляхом гібридизації з гаплопродю-
серами, як і внутрішньовидове запилення, міжвидова та міжродова
гібридизація, обробка пилку фізичними і хімічними мутагенами, не
вирішують проблеми масового одержання гаплоїдів.
Найбільш сучасним і перспективним способом одержання гаплої¬
дів є андрогенез. При культивуванні пиляків у культурі in vitro час¬
тина молодих мікроспор, не здатна до нормального перетворення в
зрілий пилок, редиференціюсться, багаторазово ділиться й утворює
калус або емріоїди. У гаплоїдному калусі можна індукувати утво¬
рення de novo бруньок чи ембріоїдів, а з них у подальшому одержу¬
вати гаплоїдні рослини [13, 14,48].
Проте багато аспектів цієї проблеми ще маловивчені, а експе¬
риментальна гаплоїдія для багатьох сільськогосподарських культур,
у тому числі і для картоплі, сьогодні ще не привела до реальних успі¬
хів. Однією з причин є відсутність швидких, надійних та ефективних
методів одержання гаплоїдів.
Культурна картопля Solanum tuberosum - генетично складний
вид, аутотетраплоїд. У тетраплоїдів розщеплення в поколіннях знач¬
но складніше, ніж у дигаплоїдів, тому практично неможливо отрима¬
ти гомозиготні форми, що характеризувалися б домінантними гена¬
ми. Лише переведення картоплі на дигаплоїдний рівень 2п-24 допо¬
може досягти високого ступеня гомозиготності в короткий термін.
Особливий інтерес становить отримання за допомогою андроге¬
незу 12-хромосомних моногаплоїдних рослин картоплі [38]. Це дасть
змогу більш глибоко вивчити як морфологію окремих хромосом, так
510
і локалізацію в них генів, що контролюють різні ознаки та власти¬
вості картоплі.
Культура пиляків і мікроспор. У середині 60-х років С.Гуха та
С.Магешварі [28-30] вперше спостерігали виникнення гаплоїдних
рослин у культурі пиляків дурману. Вони екпериментально довели,
що можна різними способами стимулювати мітотичні ділення в мік¬
роспорах або незрілих пилкових зернах з утворенням гаплоїдних ка¬
лусів чи ембріоїдів.
Відкриття можливості використання культури пиляків для отри¬
мання гаплоїдів викликало великий інтерес у всьому світі. Метод
культури пиляків розроблений для майже 200 видів рослин. Проте
слід зазначити, що андрогенез властивий не всім видам і має певні
труднощі: калуси чи зародки виникають із невеликої кількості пил¬
кових зерен; часто виникають рослини-альбіноси; крім гаплоїдів,
виникають рослини з різним рівнем плоїдності; у багатьох рослин
через поліплоїдний характер батьківських геномів гаплоїди можуть
мати не тільки один набір генів.
Подальший розвиток ці дослідження отримали в роботі
Дж.Буржена та Дж. Ніча [17]. їм вдалося на простому поживному
середовищі в культурі пиляків тютюну отримати велику кількість
гаплоїдних рослин. їхні дослідження підтвердили японські дослід¬
ники [53], які шляхом культивування пиляків також отримують гап¬
лоїдні рослини тютюну.
Перше успішне отримання рослин із пиляків в однодольних рос¬
лин здійснене у рису [58].
При культивуванні ізольованих пиляків існують чотири основні
шляхи розвитку мікроспор.
Прямий ембріогенез мікроспор. Ембріоїди виникають усередині
пиляка із мікроспор чи пилкових зерен. Не проходячи періоду спо¬
кою, ембріоїди проростають і утворюють гаплоїдні рослини.
Морфогенез через калусну тканину. При культивуванні пиляків
мікроспори і молоді пилкові зерна редиференціюються, діляться, ут¬
ворюють калусну тканину, в якій індукується органогенез (ембріоге¬
нез) і формуються рослини. Проте, при формуванні калусної тканини
із соматичних клітин пиляків можуть утворюватися рослини з різним
рівнем плоїдності.
Утворення гаплоїдного калусу з обмеженою морфогенетичною
здатністю. При індукуванні росту гаплоїдного калусу не можна вик¬
ликати його диференціацію.
Утворення диференційованих ембріоїдів для отримання калу¬
су та наступне індукування органогенезу. Очевидно, що метод
культури пиляків у майбутньому може відігравати важливу роль
при виведенні нових форм рослин. Поки що для картоплі немає
системи, яка б продукувала гомозиготні лінії в кількості, необ¬
хідній для практичної селекції. Проте останнім часом широко об¬
511
говорюються результати досліджень щодо використання дигапло-
їдів у селекції.
Аналіз факторів, що впливають на калусо- і ембріогенез: ста¬
дія розвитку мікроспор, умови вирощування рослин-донорів,
передобробка бутонів, живильні середовища. У результаті інтен¬
сивних досліджень, що проводилися в багатьох країнах, стали відомі
фактори, які впливають на реакцію пиляків у культурі in vitro. Про¬
цес ізолювання і культивування пиляків у стерильній культурі може
приводити до дедиференціювання або до розвитку пилкового зерна
шляхом ембріогенезу.
Багато дослідників підкреслюють залежність індукції андрогене¬
зу від генотипних особливостей рослин [2, 40, 59, 75, 78].
В.Сухановим [3] було виявлено, що на ефективність ембріогенезу
і калусогенезу впливають як сортові особливості, так і рівень плоїд-
ності вихідних форм. Ефективність калусогенезу пшениці сорту Са¬
ратовська 36 у 5 разів перевищувала Саратовську 42, а тетраплоїдна
кукурудза майже в 40 разів -диплоїдну. За даними У.Ірікури [4,37],
найбільший вихід гаплоїдних ембріоїдів отримано у диплоїдного
гібрида S. tuberosum х S.verrucosum.
Значний вплив на індукцію гаплоїдних ембріоїдів із мікроспор та
пилкових зерен справляє фізіологічний стан рослини-донора. Важ¬
ливими факторами є вік рослини, сезонність, різні обробки.
Найбільш сприятливим є вирощування рослин-донорів за опти¬
мальних для виду кліматичних умов [71]. Більш пізнє цвітіння у
зв’язку з несприятливими кліматичними умовами чи, наприклад, зи¬
мою в теплиці, знижує кількість ембріогенних пиляків [13].
Важливим фактором для індукції ембріогенезу є час після почат¬
ку цвітіння. Максимум гаплоїдних проростків спостерігається через
3-6 днів після початку цвітіння [71]. Пиляки, взяті з рослин напри¬
кінці цвітіння, як правило, не утворювали ні ембріоїдів, ні рослин.
Поряд з умовами, що впливають на фізіологічний стан рослини-
донора, ряд авторів указують на стимулювальну дію стресових фак¬
торів безпосередньо на репродуктивну сферу. Позитивний вплив на
стимуляцію ембріогенезу справляє попередня обробка квіткових бу¬
тонів і пиляків низькими позитивними температурами, опроміненням
[1, 31, 42], абсцизовою кислотою; витримування в умовах аноксії,
зниження атмосферного тиску [33,34] центрифугування [65].
Щодо S. tuberosum з успіхом була застосована обробка бутонів
пониженими температурами (48 годин при 6°С), опромінення гамма-
квантами 60Со і обробка мутагенами як неспецифічний шок-індуктор
[62, 65]. При цьому було досягнуте триразове збільшення виходу ем¬
бріогенних структур.
Серед факторів, що впливають на андрогенез, є підбір оптималь¬
ної стадії мікроспорогенезу. Детальне вивчення розвитку мікроспор
дурману, тютюну і петунії дослідив К.Ендвілд [8]. Ним доведене ут¬
512
ворення гаплоїдних рослин із молодих мікроспор; диплоїдних рос¬
лин - із більш старих мікроспор, що перебували у стадії реплікації
ДНК перед першим мітозом пилку; диплоїдних рослин - після злиття
двох ядер у мікроспорофітах після першого мітозу пилку і тетрапло-
їдних рослин при злитті ядер після реплікації ДНК у генеративному
ядрі перед другим мітозом. Автором зроблений висновок, що саме
стадія мікроспорогенезу є вирішальною в андрогенезі пиляків.
Утворення зародка при культивуванні пиляків картоплі спостері¬
гали при експлантації у фазі мікроспор, інколи - у фазі молодого пил¬
кового зерна [7,9].
Важливим фактором, що впливає на хід розвитку мікроспор, є по¬
живне середовище. Для культивування пиляків використовують різні
прописи середовищ: Уайта [17], Мурасіге і Скуга [52], Гамборга і
Евелега [26], Еріксона [24], Ніча [60] і №6 [19]. Середовища засто¬
совують без змін або в модифікаціях. Використовують різні співвід¬
ношення фітогормонів і стимулювальних факторів, добавки рослин¬
них екстрактів, амінокислот, цукрів.
При культивуванні пиляків картоплі застосовують мінеральний
склад середовища за Мурасіге і Скуга з використанням ауксинів у
концентраціях від 0,01 до 10 мг/л. Із цитокінінів - кінентин і 6-бен-
зиламінопурин від 0,2 до 2 мг/л. Стимулювальними в розвитку емб¬
ріоїдів картоплі є добавки кокосового молока та картопляного екст¬
ракту [50, 69, 74].
Моногаплоїди та їх використання. Накопичені дані про особ¬
ливості андрогенних рослин свідчать, що в багатьох випадках спо¬
стерігається значне морфологічне різноманіття рослин-регенерантів
та мінливість їхньої плоїдності.
Серед рослин, отриманих із пилку шляхом ембріогенезу, частіше
зустрічаються гаплоїдні, тоді як рослини, отримані через калусну
тканину, були еуплоїдами, інколи анеуплоїдами на гаплоїдному,
триплоїдному чи тетраплоїдному рівнях. Це пов’язано зі швидкою
поліплоїдизацією в калусній тканині. Дорослі гаплоїдні рослини в
багатьох видів можна легко відрізнити від диплоїдних. Вони, як пра¬
вило, менші за розмірами, з більш вузьким листям. Листя має менші
за розмірами продихи.
Характерною особливістю андрогенних рослин пшениці, ячменю
та рису є високий відсоток регенерантів, дефектних за синтезом хло¬
рофілу [66, 77].
Рослини, отримані при культивуванні пиляків картоплі [9] через
калусну тканину, мали варіабельну кількість хромосом від 18 до 80,
часто зустрічалися химерні форми. Рослини, отримані ембріогенним
шляхом, як правило, були завжди дигаплоїдами (2п-24) або монопло-
їдами для диких диплоїдних видів. Можна припустити, що різні рівні
плоїдності можуть бути результатом поліплоїдизації, типової для
клітинних популяцій in vitro, або пояснюються соматичним поход¬
513
женням калусу. На морфологічні та фізіологічні характеристики
отриманих рослин впливав рівень плоїдності. Рослини з кількістю
хромосом, яка дорівнювала вихідному сорту, були морфологічно
ідентичні, а ди- і триплоїди (як еуплощи, так і анеуплоїди) значно
відрізнялися від вихідної форми. Ці відмінності стосувалися інтенсив¬
ності розвитку габітусу куща, форми листків, квітконоса та суцвіття,
розміру та кольору бульб.
У подальшому інтенсивне вдосконалення методу дозволило
отримати андрогенні рослини в культурі пиляків різних видів рослин
(табл. 7.2).
Таблиця 7.2 - Приклади використання методу культури пиляків для різних
видів рослин
Вид
Результат
Літературне джерело
1
2
3
Anemone canadensis
РОСЛИНИ
Johansson L.JEriksson Т.,
A. multifida
1977
Arabidopsis thaliana
Keathley D.N.,Scholl R.L., 1982
Arachis hupogaea
Mroginski LA., Femandez A. 1979
Beta vulgaris
Rogozinska J.N., Goska M., Kuzdo
wicz 0., 1977
Lux H., Herrmann L., Wetzel C.,1990
B. manerorhiza
Rogozinska J.N., Goska M., 1982
Brassica campestris
Keller WA. et al., 1975
Bromus inermis
Niizeki M., Kita F., 1974
Capsicum annuum (2n-36)
Kuo J.S. et al., 1972
Chrysarithenum indiocum
(2n-72)
Watanabe K. Et al .,1972
Ch. Japonense (2n-54); (2n-72)
Wata-iabe K. Et al .,1972
Ch. boreale (2n-18)
Watanabe K. et al .,1972
Coriandrum satinum
Егорова E.A., Резникова С.А., 1982
Datura metel
Narayanaswamy S., Chandy Z.P., 1971
D. stramonium
Maheshwari S.C. et al., 1982
Festuca arundinacea
Zenkteler M., Misiura F.,1975
Fragaria ananassa
Laneri U, Damiano C., 1980
Heliantus mollis
Nenova N., Ivanov P., Christoy U., 1998
Hevea brasiliensis
Chen Z., et al., 1982
Hordeum vulgare
ClaphamD., 1972
Davies P..Morton S.,1998
Hyoscyamus niger
WemickeW., Kohlenbach H.W., 1977
Ipomoea batatas
Sehgal C.B., 1978
Litchi chinensis
Fu L.,TarigD. ,1983
Lycopersicum esculentum
Zagorska N.A., et al.,1982
Medicago sativa
Thomas P. T., Thomas H., 1973
Nicotiana affinis
Maheshwari S.C.,1980
514
Продовження таблиці 7.2.
1
2
3
Orysa perennis
Heszky L., Pauk J., 1975
NguyenManh Don., 1997
Paeonia hybrida
Ono K.Jiarashima S., 1981
Paraver somniferum
ElbemB., 1984
Pennisetum typhoideum
Bui Dang Duc, Pemes J., 1982
Petunia parodii
Gupta P.P., 1982
Glycine max
Croughan S, Yao S., Zhoo G., 1996
Sal via sclarea
Русин H.B. и др., 1997
Secale cereale
Шкаврон A.C. и др., 1992
Solanum bulbocastanum
Irikura Y., 1975
Маруненко И., 1986
S.chacoense
Кучко А.А., Маруненко И.М., 1983;
Калинин А.В., 1987
S. demissum
Irikura Y., 1975
S. ducam ara
Zenkteler М., 1973
S.fendleri
Inikura Y., 1975
S.hjertingii
Irikura Y., 1975
S.hougasii
Irikura Y., 1975
S. infundibuliforum
Irikura Y., 1975
S. megistaerolotum
Irikura Y., 1975
S. melongena
Isouard G.,1979
S. michoacanum
Irikura Y., 1975
S. microdontum
Mix G., 1982
S. multidissectum
Irikura Y., 1975
S. nigrum
Irikura Y.,1975
S. pennellii
Irikura Y.,1975
S. phureja
Irikura Y.,1975
S. stenotomum
Irikura Y. ,1975
S. stoloniferum
Irikura Y.,1975
S. surattense
Sihha S. et al., 1979
S. tuberosum 4n
Irikura Y., 1975
S.tuberosum 2n
Irikura Y., 1975; Маруненко И., 1986
S.torvum
Jaiswal V., Narayan P., 1981
S.verrucosum
Weatherhead M., Henshaw G., 1979
Triticale
Soziniv A., et al., 1981
Machii, Mizuno et al., 1998
Vitis vinifera
Gresshoff Р. M., Doy C.K., 1974
Zea mays
Пиралов Г.Р., Абраимова O.E., 1997
Одержання експериментальним шляхом моноплоїдів культурних
сортів і диких видів відкриває нові перспективи в селекційній роботі.
При цьому полегшується перенесення бажаних генів від диких видів
у культурні сорти. Елімінування небажаних ознак диких видів про¬
ходить значно швидше, ніж при гібридизації.
515
Використання спрямовано розроблених селекційних схем дозво¬
ляє поєднати в одному геномі стійкість до патогенів з високою уро¬
жайністю.
ЛІТЕРАТУРА
1. Атанасов А.И., Бутенко Р.Г. Культура изолированных пыльников сахарной
свеклы // Физиол. и биохимия растений. - 1980. - Т. 12, № 1. - С. 49-56.
2. Егорова Н.А., Резникова С.А. Исследование культуры изолированных
пыльников кориандра в связи с индукцией андрогенеза // Физиология расте¬
ний. -1982. - Т.29, вып. 1. - С. 142-148.
3. Использование культуры пыльников для получения гаплоидов / В.М.
Суханов, В.П. Клочков, С.С.Хохлов, В.С.Тырнов // Культура клеток расте¬
ний : Труды II Всесоюз. конф. - К.: Наукова думка, 1978 - С.312.
4. Использование культуры тканей и органов в селекции растений и про¬
изводстве посадочного материала /Г. Лейке, П.Лабес, К.Эртель,
М.Петерсдорф. - М.:Колос, 1980. - С.22-23.
5. Калинин А.В. Оптимизация условий получения андрогенных гаплои¬
дов у картофеля //Науч. труды. - М., 1987. - С. 94-97.
6. Кучко А.А.,Маруненко И.М. Культура изолированных пыльников кар-
тофеля//С.-х. биология. -1983. - №5. - С. 16-20.
7. Кучумов А.П., Князев В.А. Культура тканей и клеток в селекции и се¬
меноводстве картофеля. - М.,1980. - С.21-25.
8. Левенко Б.А. Применение культуры изолированных клеток, тканей и
органов растений в генетике и селекции // Общая генетика. - М., 1978. -
С.166.
9. Маруненко И.М. Условия, способствующие развитию микроспор по
спорофитному пути, и получение гаплоидов в культуре пыльников картофе¬
ля: Автореф. дис.... канд. биол. наук. - М.,1986. - 24 с.
10. Пиралов Г.Р., Абраимова О.Е. Влияние биологических особенностей
исходного материала и состава питательных сред на каллусогенез и регене¬
рацию в культуре незрелых зародышей кукурузы // Физиология и биохимия
культ, растений. - 1997. - Т.29, №1. - С. 44-50.
11. Русин Л.В., Бугара А.М., Бугаенко Л.А. Использование метода культуры
изолированных зародышей для получения межвидовых гибридов шалфея // Фи¬
зиология и биохимия культ, растений. - 1997. -Т.29, №2. - С. 121-128.
12. Харченко П.Н., Кучеренко П.А. Получение растений риса из пыльни¬
ков // Докл. ВАСХНИЛ. - 1977. - №4. - С. 15-16.
13. Шамина З.Б. Андрогенез и получение гаплоидов в культуре пыльников и
микроспор // Культура клеток растений. - М.: Наука, 1981. - С. 124-131.
14. Шамина З.Б., Берндт Б. К проблеме получения гаплоидов картофеля
на основе андрогенеза // Исследования по клеточной селекции картофеля. -
М.,1984. - С.37-55.
15. Шкаврон А.С., Рабинович С.В., Сидоров В.А. Культивирование пыльни¬
ков ржи: эффективность использования глюкозы как источника углеродов // Фи¬
зиология и биохимия культ, растений. - 1992. - Т.24, №5. - С. 511-515.
16. Anther culture techniques of rubber tree and sugarcane / Chen Z., Quian
C., Xu X., Deng Z.//Plant Tissue Cult., Tokyoand Lfrt Yamanaka, July 11-
16,1982. -Tokyo, 1982. -P.533-534.
516
17. Bourgin J.P., Nitsch J.P. Obtention de Nocotiana haploides a partir
d'etamines in vitro // Ann. physiol. veg. -1967. - Vol.9, №4. - P.377-382.
18. Bui D., Pernes J. Pearl millet microspore cultire: potentialities and
regeneration of whole plants // Biol. Cell. - 1982. - Vol.45, №2. - P.l00.
19. Chin Ching C. The N6 Medium and ist applications to anther culture of
cereal crops // Ичуан сюэбао. - 1977. - Vol.4, №4. - P.302-310.
20. Clapham D. Haploid Hordeum plants from anthers in vitro // Z. Pflan-
zenzucht. -1973. - Vol.69. - P.l42-155.
21. Croughan S.,Yao S.,Zhoo G.Pollen culture and antherculture of soybeans
// In vitro Cell. and Dev.Biol.Anim. - 1996. -Vol. 32, №3, Pt2. - P.85A.
22. Davies P., Morton S. A comparison of barley isolated microspore and
anther culture density // Piant Cell Repts. - 1998. - Vol. 17, №3. - P.206-210.
23. Elbem Bettina. Antherenkultur und Regeneration haploider Pflanzen von
Mehn (Papaver somniferum) // Landbauforsch. Volken.rode-1984- Vol.34,
№1.- P.63-66.
24. Eriksson T. Studies on the prowth requirements and growthmeasurements
of cell cultures of Haplopappus gracilis // Physiol. Piant -1965. - Vol. 186, №4. -
P.976-993.
25. Fu L., Tang D. Индукция регенерантов в культуре пыльников Litchi
chinensis // Ичуан сюэбао, Acta genet, sin. - 1983. - Vol.10, №5. - P. 369-374.
26. Gamborg O.L.JEveleigh D.E. Culture methods and detection ofglucanases in
cultures of wheat and barley // CanJ.Biochem. - 1968. - Vol.46, №5. - P.417-421.
27. Gresshoff P.M., Doy C.H. Derivation of a haploid cell line from Vitis vinifera
and the importance of the stage of meiotic development of anthers for haplois culture of
this genera // Z. Pflanzenphysiol. - 1974. - Vol.73. - P.l32-141.
28. Guha S., Maheshwari S. In vitro production of embryos from anthers of
Datura // Nature (London). - 1964. - Vol.204, №4957. - P.497.
29. Guha S., Maheshwari S. Cell division and differentiation of embryos in the
pollen grains of Datura in vitro // Nature (London) -1966 - Vol.212, №5057-
P.97-98.
30. Guha S., Maheshwari S. Development of embryoods from pollen grains of
Datura in vitro // Phytomorphology. - 1967 - Vol.17- P.454-461.
31. Gupta P. Genesis of microspore-derived triploid Petunias // Theor. and
Appi. Genet.- 1982.- Vol.61, №4.- P.327-331.
32. Heszky L., Pauk I. Induction ofjiaploid rise plants of different origin in
anther culture // Riso.- 1975. - Vol.24 - P.l97-294.
33. Imamura J., Harada H. Effects of abscisic acid and water strees on the
embryo and plantlet formation in anther culture of Nicotiana tabacum cv. Samsun
//Zeitschrift Pflanzenphysiologie. - 1980 - Vol.100 - P.285.
34. Imamura J., Harada H. Stimulatory effects of reduced atmospheris
pressure on pollen embryogenesis // Naturwissenshaften. - 1980. - Vol.67. -
P.357-358.
35. Induction of haploidy from pollen grains in angiosperms - the current
status / Maheshwari S., Tyagi A., Malhotra K., Sopory S // Theor. Appi. Genet.-
1980.-Vol.58. - P.193-206.
36. Investigations on the anther culture in vitro of Nicotiana tabacum L. and
Capsicum annuum L / Kuo J., Wang Y., Chien N. et al // Acta Bot., Sinica-
1973.-Vol.15. - P.37-50.
517
37. Irikura Y. Induction of haploid plants by anther culture in tuber-bearing
species and interspecific hybrids of Solanum // Potato Res.- 1975. - Vol.18. -
P.133-140.
38. Irikura Y., Sakaguchi S. Induction of 12-chromosome plants from anther
culture in a tuberous Solanum // Potato Res. - 1972. - Vol. 15, №2. - P. 170.
39. Isouard G., Rsquin C., Demarly Y. Obtention de plantes haploides et
diploides par culture in vitro d’antheres d'Aubergine (Solanum melongena L.) //
C.r.Acad. sci.- 1979. - Vol.288D, №12. - P.987-989.
40. Jacobsen E., Sopory S. The influence and possible recombination of
genotypes on the production of microspore embryoids in anther cultures of
Solanum tuberosum and dihaploids // Theoret. Appi. Genet-1978- Vol.52-
P. 119—123.
41. Jaiswal V.,Narayan P. Induction of pollen embryoids in Solanum torvam
Swartz // Curr.Sci (India). - 1981. - Vol.50. - P.998-999.
42. Johansson L. Induced embryogenesis in tetraploid and dihaploid anther
cultures of Solanum tuberosum // Abetraet of Conference Papers of the 9 Tri-
ennial Conference of the EAPR- 1984. - P.50-51.
43. Johansson L., Eriksson T. Induced embrio formation in anther cultures of
several Anemone species // Physiol. Piant - 1977 - Vol.40, №3. - P. 172-174.
44. Keathley D.,Scholl R. Culture of Arabidopsis thaliana anthers on liguid
medium // Z.Pflanzenphysiol- 1982. - Vol.103, №3. - P.199-212.
45. Keller W., Rajhathy Т., Lacapra J. In vitro production of plants from
pollen in Brassica campestris // Can.J.Genet., Cytol - 1975. - Vol.17. - P.667.
46. Laneri U.,Damiano C. Strawberry anther culture. - Appi. cult. in vitro
amelior. piant, potageres // Reun. EUCAPPLA sect. Legum. - Versaclles, 1980. -
P.l 50-155.
47. Lux H., Herrmann L., Wetzel C. Production of haploid sudar beet (Beta
vulgaris L.) by culturing unpollinated ovules // Piant Breed-1990- Vol. 104,
№3.-P. 177-183.
48. Maheshwari S., Rashid A., Tyagi A. Haploids from pollen grains-
retrospect and prospect. // Amer.J.Bot. - 1982. - Vol.69, №5. - P.865-879.
49. Mix G. Anther culture of Solanum tuberosum L. and some wild species. -
Piant Tissue Gult. 1982, Proc. 5 Int. Congr. Piant. Tissue snd Cell Cuit. Tokyo
and Lake Yamanaka, July 11-16,1982. - Tokyo, 1982. - P.573-574.
50. Monohaploid plants from anthera of dihaploid genotype of Solanum
tuberosum / Foroughi-Wehr B., Wilson H.M., Mix G., Gaul H. // Euphytica.-
1977.-Vol.26.-P.361.
51. Mroginski L.,Femandez A. Cultivo in vitro de antheras de especies de
Arachis (Leguminosea) // Oleagimaux-1979. - Vol.34, №5. - P.243-248.
52. Murachige Т., Skoog F. A revised medium for rapid growth and bioassays
with tobacco tissue cultures //Physiol. Plant-1962. Vol.16. №13. - P.473-479.
53. Nakata К., Tanaka M. Differentiation of embryoides from developing
germ cell in anther culture of tobacco // Jap.J. Genet - 1968 - Vol.48. - P.65-71.
54. Narayanaswamy S., Chandy L. In vitro induction of haploid and triploid
androgenic embryoids and plantlets in Datura metel // Ann.Bot.- 1971- Vol.35-
P.535.
55. Nenova N, Ivanov P., Christoy M. Anther culture regeneration breen Heliantus
mollis (M-020) and its hybrid combinations. - 1998. - Vol. 12, №2. -P.58-60.
518
56. Nguyen M.Nong nghiep cong nghiep thuc pham // Adr.and Food Ind. -
1997. -№2. - P.78-80.
57. Niizeki M., Kita F. Studies on piant cell and tissue culture. In vitro
induction of callus from anther culture of forage crops // J.Fac> Agrie.Hokkaido
Univ.-1974. -Vol.57, №3. - P.293-300. .
58. Niizeki M., Oono K. Induction of haploid rice piant anther culture // Proc.
Jap.Acad. - 1968. - Vol.44. - Р.55Ф-557.
59. Nitsch C. Experimental androgenesis in Nicotiana // Phytomorphology. -
1969. - Vol.19. - P.3 89-404.
60. Nitsch C. Pollen culture - a new technigue for mass production of haploid
and homozygous plants // K.J.Kasha (ed) Haploids in higher plants.1974. -
P.123-135.
61. Ono K., Harashima S. Induction of haploid callus from isolated
microspores of peony in vitro // Piant and Cell Physiol. - 1981. - Vol.22, №2. -
P.337-341.
62. Przewdzny Т., Schider O., Wenzel G. Induced mutants from dihaploid
potatoes after pollen mother cell treatment // Theoret.Appl>Genet.- 1980.-
Vol.58.-P.145.
63. Rogozinska J., Goska M., Kuzdowiez O. Induction of plants from antheres
of Beta vulgaris cultured in vitro // Acta Soc. Bot. Pol.-1977. - Vol.46, №3. -
P.471-479.
64. Rogozinska J., Goska M. Attempts to induce haploids in anther cultures of
sugar, fodder and wild species of beet // Acta Soc.bot.pol.-1982. - Vol.51, №1. -
P.91- 105.
65. Sangwan-Norreel B. Androgenic stimulating factors in the anther and
isolated pollen grain culture of Datura innoxia Mill // J.Exp.Bot.- 1977.-
Vol.28.-P.843- 852.
66. Screening wheat genotypes for high callus induction and regeneration
capability from anther and immature embryo cultures /Machii H., Mizuno H. et al.
//Piant Cell. Fissue and organ Cult. - 1998. - Vol.53, №1. - C.67-74.
67. Sehgal С.Дгога S., Narang K. Production of pollen embryoids in anther
cultures of Sambucus nigra L// Curr. Sci (India).- 1982-Vol.52, №2- P. 104-105.
68. Sinha S., Roy R., Jha K.Callus formation and choot bud differentiation in
anther culture of Solanum surratense // Can.J.Bot.-1979. - Vol.576, №22. -
P.2524-2527.
69. Sopory S., Rogan P. Induction of pollen divisions and embryoid formation
in anther cultures of some dihaploid clones of Solanum tuberosum //
Z.Pflanzenphysiol. - 1976. - Vol.80, №1. - P.77-80.
70. Sozinov A., Lukjanjuk S., Ignatova S. Anther cultivation and induction of
haploid plants in triticale // Z.Pflanzenzucht. - 1981. - Vol.86. - P.272-285.
71. Sunderland N. Anther culture: a Progress report // Sci Prog-1971. -
Vol.59. -P.527-549.
72. Thomas P., Thomas H. Welsch Piant Breed.Sta.Rept. - 1972. - P. 125-131.
73. Watanabe K.,Nishii V., Tanaka R. Anatomical observations the high
freduency callus formation from anther culture of Chrysanthemum //Jap.J.,
Genet.-1972. - Vol.47, №4. - P.249-255.
74. Weatherhead M.,Hehshaw G. The induction of embrioids in frec pollen
culture of potatoes // Z.Pflanzenphysiol. - 1979. - Vol.94. - P.441-447.
519
75. Wenzel G.,Hofbnann F., Thomas E.Increased induction and chomosome
doubling of androgenetic haploid rye // Theoret.Fppl.Genet-1977. -Vol.51, №2.
- P.81-86.
76. Wemicke W., Kohlenbach H. Versuche zur Kultur isolierter Microsporen von
Nicotiana undHyoscyamus // Z.Pflanzenphysiol.- 1977. - Vol.81. - P.330-340.
77. White P. The cultivation of animal and piant cells. - New York, 1954. -
P.239.
78. Woo S.,Mok T. Anther culture of Orysa sativa L., and Oryza perensis
M.hybrids // Abstr. IV Inter. Congr.Calgsry-1978. - P.54.
79. Zenkteler M. In vitro development of embryos and seedlings from pollen grains
of Solanum dulcamara // -Z. Pflanzenphysiol. - 1973. - Vol.69. - P. 189-192.
80. Zenkteler M., Misiura E.,PonitkaA. Induction of androgenetic embryoids
in the in vitro ciltured anthers of sereal species // Experientia - 1975. - Vol.ll,
№3. -P.289-340.
7.3. СОМАТИЧНА ГІБРИДИЗАЦІЯ
Традиційна селекція широко використовує статеве схрещування,
при якому жіночі і чоловічі статеві клітини беруться від різних сор¬
тів і навіть від видів. Статева гібридизація обмежена можливістю
використання як вихідних форм тільки споріднених видів рослин,
тому в селекції не завжди вдається отримати бажані статеві гібриди.
Подолання статевого бар’єра стало можливим, коли вдалося отрима¬
ти за допомогою методів біотехнології гібриди потрібного типу
шляхом злиття соматичних клітин. У природних умовах злиттю со¬
матичних клітин перешкоджають целюлозно-пектинові стінки. Дією
спеціальних ферментів (пектинази і целюлази) пошкоджуються стін¬
ки клітин. Рослинні клітини, що втратили зовнішню стінку, дістали
назву протопластів.
Сьогодні протопласти виділяють із рослинних тканин і клітин,
що культивуються in vitro. Ензиматичний метод дає змогу ізолювати
протопласти із будь-якого органа чи тканини багатьох рослин. Для
отримання великої кількості гомогенних і гомоплоїдних протоплас¬
тів часто використовують мезофіл листка [4, 5, 19, 20].
Методи ізолювання протопластів мають відповідати двом основ¬
ним вимогам: мацерації клітинної стінки без пошкодження прото¬
пласта та забезпечення стабільності протопластів шляхом підбору
оптимального іонно-осмотичного складу середовища.
Розробка способів ензиматичного виділення протопластів рослин
і відкриття можливості злиття ізольованих протопластів визначили
принципово новий напрям під назвою “соматична гібридизація”. Со¬
матична гібридизація - це процес злиття в одну клітину двох (або
більше) протопластів соматичних клітин рослин. Розрізняють такі
етапи соматичної гібридизації: виділення протопластів, злиття, реге¬
нерація клітинної стінки, злиття ядер, розмноження гібридних клі¬
тин, регенерація рослин (рис. 7.3).
520
Оскільки самозлиття про¬
топластів у суспензії спосте¬
рігається вкрай рідко, дослід¬
ники стали застосовувати
різні речовини, які приско¬
рюють цей процес. Індукова¬
не злиття протопластів уперше
вдалося отримати в 1970 ро¬
ці в лабораторії Е.Кокінга [16].
Як індуктор злиття був вико¬
ристаний нітрат натрію. Від¬
соток злитих протопластів
був невисокий. Спроби вико¬
ристовувати для злиття кон-
кавалін, позитивно заряджені
синтетичні фосполіпіди та
інші речовини були малоус¬
пішними. Ю. Глєба з дослід¬
никами запропонували як
індуктор злиття використо¬
вувати розчини з високим
значенням pH (9-11) і висо¬
кою концентрацією іонів
кальцію (100-300 мм) [2, 7, 9,
10, 12]. При цьому протопла¬
сти попередньо аглютинують
за допомогою концентрова¬
них розчинів поліетилен глі¬
колю (ПЕГ). Методику “ПЕГ
- високе pH - висока концен¬
трація кальцію” у різних модифікаціях використовують в більшості
лабораторій світу і тепер. Використовуючи цю методику, вдається
залучити до злиття 10-50% протопластів [6].
У 1981 році з’явилися роботи [18, 21], в яких описаний принци¬
пово новий метод, коли для аглютинації і злиття протопластів вико¬
ристовують електричне поле. Протопласти розміщують у дуже неод¬
норідному змінному полі. Під дією електричного поля протопласти
утворюють містки із кількох клітин між електродами. Накладання
поодинокого імпульсу (15В, 50 мсек) призводить до злиття клітин.
Злиття за допомогою електричного поля можливе для протопластів
різного тканинного та видового походження. Проте ця методика по¬
ки що малоефективна.
Частота утворення гібридних клітин дуже низька, тому слід ство¬
рювати умови для їх розвитку, тобто селективні умови. При гібриди¬
зації соматичних клітин рослин існують три типи селективних сис¬
Злнггя
Формування клітинної
стінки, злиття ядер
Утворення
кялусу
Морфогенез
та регенерація
рослин
Рис. 7.3. Схема процесу соматичної
гібридизації
521
тем: візуальна ідентифікація гібридних клітин; механічний добір та
використання селективних середовищ, де протопласти партнерів не
здатні до поділу [З, 13].
Метою індукованого злиття ізольованих протопластів є утворен¬
ня гібридних клітин з наступним утворенням клітинних ліній та ам-
фіплоїдних рослин.
Слід зазначити, що соматична гібридизація за допомогою злит¬
тя протопластів дає змогу по-новому комбінувати батьківські гени
у вищих рослин [13, 18]. Але тут потрібно підкреслити, що вона
можлива лише тоді, коли відпрацьована методика регенерації рос¬
лин із протопластів. Використання соматичної гібридизації відкри¬
ває можливість перенесення не тільки частини ядерної генетичної
інформації від одного виду до іншого, а й цитоплазматичних генів
від однієї лінії до іншої та заміщення цитоплазми. Дослідженнями
визначено, що при соматичній гібридизації ядерні гени можуть ус¬
падковуватись як двобатьківські, так і однобатьківські. Частка по-
заядерних детермінант (пластома, локалізованого у хлоропластах, і
мітохондріона, локалізованого в мітохондріях) соматичних гібри¬
дів відмінна від статевих. У соматичних гібридах незалежно від
того, відбувається елімінація ядерних хромосом чи ні, позаядерні
генетичні детермінанти успадковуються як двобатьківські. Можли¬
ва елімінація або сегрегація цитоплазматичних детермінант, що
може проходити незалежно для мітохондріона і пластома. У ре¬
зультаті таких змін у гібридних клітинах може сформуватися циб-
рид (цитоплазматичний гібрид). Цитоплазматичні гібриди були де¬
тально вивчені А. Бургутіним [8].
Перший соматичний гібрид був створений П. Карлсоном із спів¬
робітниками [13]. Це був міжвидовий гібрид Nicotiana glauka +
N. langsdorffii.
Методами соматичної гібридизації були отримані внутрішньови¬
дові (тютюн, дурман, петунія), міжвидові (тютюн, петунія, морква,
дурман, картопля), міжродові (томати + картопля, арабідопсис +
турнепс, дурман + беладона) гібриди. Список одержаних гібридних
комбінацій і літературних джерел висвітлений в книзі Ю. Глєби та
К. Ситника [3].
Метод соматичної гібридизації насамперед перспективний для
отримання віддалених гібридів картоплі і досягнення тетраплоїдного
рівня при злитті двох дигаплоїдних ліній. При цьому за допомогою
соматичної гібридизації можна комбінувати ознаки, що успадкову¬
ються полігенно (рис.7.4).
Дослідження щодо соматичної гібридизації картоплі ведеться до¬
сить обмежено. Міжвидові соматичні гібриди картоплі отримані між
видами S.tuberosum + L.esculentum chl (yg) [15]; S.tuberosum + L.escu-
lentum chl (giva) [18]; S .tuberosum + L.esculentum chl; S.tuberosum +
N.tabacum.mtr (yg) [12]; S.tuberosum + S.chacoence [1].
522
Рис.7.4 . Один із шляхів застосування соматичної гібридизації
в селекції картоплі
Перший соматичний гібрид картоплі отриманий шляхом злиття
протопластів сорту Прієкульська рання (,S. tuberosum) і дикого виду
(іS.chacoense) [1]. За формою листків і кущів, за розміром бульб со¬
матичний гібрид займав проміжне місце між культурним і диким ви¬
дами. Проте соматичний гібрид був стійкий до Y-вірусу, що є харак¬
терним для дикого виду.
Отже, метод соматичної гібридизації як один із методів біотехноло¬
гії може бути використаний у селекційному процесі картоплярства.
Існуючі методи злиття протопластів дозволяють переборювати
природну несхрещуваність філогенетично віддалених видів рослин і
цим самим дають змогу отримати комбінацію генів, які неможливо
отримати за допомогою статевого схрещування.
Враховуючи високу вартість методу, робота із соматичної гібри¬
дизації має виконуватися на замовлення селекціонерів.
ЛІТЕРАТУРА
1. Бутенко Р.Г., Кучко А.А. Получение межвидового соматического гиб¬
рида картофеля методом слияния изолированных протопластов // Доклады
АН СССР. - 1979. - Т.247, №2. - С.491-495.
2. Глеба Ю.Ю., Бутенко Р.Г., Сытник К.М. Слияние протопластов и па-
расексуальная гибридизация у Nicotiana tabacum L. // Доклады АН СССР. -
1975. - Т.221, №5. - С.1196-1198.
523
3. Глеба Ю.Ю., Сытник К.М. Клеточная инженерия растений.- К.: Наук,
думка, 1984. - 160 с.
4. Игнатова J1.K. Получение протопластов из С3- и С4- растений и их фо-
тосинтетическая активность // Физиология растений. - 1980. - Т.27, №1. -
С.80-85.
5. Культура изолированных протопластов / Ю.Ю. Глеба, Л.Г. Швыдкая,
Р.Г. Бутенко, К.М. Сытник // Физиология растений. - 1974. - Т.21, №3. -
С.598-605.
6. Кучко А.А. Гибридизация соматических клеток растений методом
слияния изолированных протопластов // Культура клеток растений и биотех¬
нология : Сб. науч. тр. - М.: Наука, 1986 - С. 144-159.
7. Межтрибные гибридные клеточные линии Atropa beladonna + Nicotiana
chinensis, полученные слиянием изолированных протопластов / В.П. Момот,
А.Н.Околот, М.В.Скаржынская и др. // Доклады АН СССР. - 1982. - Т.267,
№4. - С.959-962.
8. Некоторые особенности растений, возникших из слившихся протоплас¬
тов мутантов табака Nicotiana tabacum L / А.Б. Бургутин, Р.Г. Бутенко,
Л.В.Фролова, Ю.Ю.Глеба // Физиология и биохимия культурных растений. -
1977.- Вып.9, №3. - С.291-300.
9. Парасексуальные цитоплазматические гибриды (цибриды) Nicotiana
tabacum L.+N. debneyi, полученные слиянием протопластов / Ю.Ю. Глеба,
Н.М. Пивень, И.К. Комарницкий, К.М.Сытник // Доклады АН СССР. - 1978.
- Т.240, №5.- С. 1223-1226.
10. Скаржинська М.В., Череп Н.Н., Глеба Ю.Ю. Соматическая гибриди¬
зация и получение клеточных линий и растений картофеля + табак // Цито¬
логия и генетика. - 1982. - Т. 16, №6. - С.42-48.
11. Скаржинська М.В., Череп М.Н., Глеба Ю.Ю. Отримання та вивчення
гібридних клітинних клонів Nicotiana tabacum +Solanum tuberosum та
Nicotiana tabacum +S.sucrense // Укр. ботан. журн. - 1983. - Т.40, №1. -
С.64-68.
12. Ультраструктурний анализ слияния клеток человека с протопластами
высших растений / В.А. Сидоров, Ю.Ю. Глеба, Л.Д. Кривохатская, К.М. Сы¬
тник // Доклады АН СССР. - 1978.- Т.240, №1. - С.211-212.
13. Carlson P.S., Smith Н.Н. Dearing R.D. Parasexual interspecific piant
hybridization // Proc. Nat. Acad. Aci. USA - 1972. - Vol.69, №10. - P.2292-2294.
14. Genetic transfer in plants through interspecific protoplast fusijn / Shepard
J., Bidney D., Barsby Т., Kemble R // Science. - 1983. - Vol.219, № 4595. -
P.683-688.
15. Melchers G., Sacristan M., Holder A. Somatic hybrid plants of potato and
tomato regmerated from fused protoplasts // Galsberg Res. Commun. -1978. -
Vol.43.- P. 203-218.
16. Melchers G.. The first decennium of somatic hybridization // Proc, 5th.
Intem. congr. piant tissue and cell cult (Tokyo, July 11-16, 1982). - Tokyo, 1982.
-P.l 3-18.
17. Power J.B., Cummins S.E., Cocking E.C. Fusion of isolated piant
protoplasts // Nature. - 1970. - Vol.225, №5374. - P.l016-1018.
18. Scheurich P., Zimmermann U., Schnabl H. Electrically stimulated fusion of
different piant cell protoplasts // Piant Physiol. - 1981. - Vol.67, №4. - P.849-853.
524
19. Takebe J., Otsuki V., Aoki S. Isolation of tobacco mesophyli calls in intact
and active state //Piant Cell Physiol. - 1968. - Vol.9, №1- P.l 15-124.
20. Upadhya M.D. Isolation and culture of mesophyli protoplasts of potato
(Solanum tuberosum L.) // Potato Research. - 1975. - Vol.18, №3. - P.438-445.
21. Zimmerman U., Scheurich P. High frequency fusion of piant protoplasts
by electric fields // Planta. - 1981. - Vol. 151, №4. - P. 26-32.
7.4. ГЕНЕТИЧНА ІНЖЕНЕРІЯ
Генетична інженерія є основним розділом сучасної біотехнології.
Методологія напряму базується на конструюванні, виділенні, іден¬
тифікації та перенесенні генів і тісно пов’язана з проведенням
молекулярно-біологічних досліджень.
Генетична інженерія складається із генної та геномної інженерії.
Генна інженерія використовує методи in vitro або in vivo, забезпечує
введення в геном реципієнтної клітини одного чи кількох чужорід¬
них генів або ж створює в геномі нові регуляторні зв’язки. При цьо¬
му вид не змінюється, але набуває не властивих йому раніше ознак.
При геномній інженерії відбуваються більш глибокі зміни організму
аж до створення нових штучних генетичних програм.
Метою створення трансгенних рослин-організмів є виділення по¬
трібних генів із чужої ДНК і вбудовування їх у молекулу ДНК даної
рослини. Цей процес дуже складний.
Найбільш розповсюджений спосіб переносу чужих генів у спадко¬
вий апарат рослини - за допомогою хвороботворної для рослин бактерії
Agrobacterium tumefaciens (у буквальному перекладі з латині - польова
бактерія, що викликає пухлини). A. tumefaciens - це грунтова бактерія із
родини Rhizobiaceae, вона здатна поселятися на ранових поверхнях рос¬
лин, переносиш частину своєї Ті-плазміди (кільцеві молекули ДНК, які
стабільно передаються потомству бактеріальних клітин незалежно від
хромосомної ДНК) у геном рослинних клітин.
Для конструювання штучної плазміди використовують ферменти
рестриктази, що розділяють довгу молекулу ДНК на окремі діляноч¬
ки, які набувають “липкості” на кінцях, що дає змогу їм вклеюватися
в розрізану такими ж рестриктазами чужу ДНК [8]. Створення штуч¬
ної плазміди та процес трансформації продемонстровано на рисунку
7.5. Потрібний ген вклеюють за допомогою рестриктаз у кільцеву
молекулу ДНК бактерії, так звану плазміду. Ця ж плазміда несе ген
стійкості до антибіотика. Ті бактеріальні клітини, які приймуть у
свій генетичний апарат “прооперовані” плазміди, отримають, крім
нового корисного гена, ще й стійкість до антибіотика. Клітини, які
вдало сприйняли плазміду, ростуть і розмножуються, а всі інші ги¬
нуть [8]. Для генної інженерії спеціально виведений штам агробакте-
рії, що не викликає пухлин, але має здатність вносити свою ДНК у
рослинну клітину.
525
Й Введення
плазм ід в
бактеріальну
^ клітину
са>
ТРАНСФОРМАЦЇЯ
Рис. 7.5. Створення штучної плазміди та процес
трансформації
Перші експерименти з генетичної інженерії з використанням аг-
робактерій проводили шляхом простого нанесення культури бакте¬
рій на ранові поверхні рослин. Пухлини, що утворювалися, перено¬
сили на безгормональні поживні середовища, які містили антибіоти¬
ки для елімінації бактерій. Цей спосіб дуже зручний у роботі, оскіль¬
ки передбачав використання тільки онкогенних штамів агробактерій.
Проте значну кількість у тканинах пухлин становили нетрасформо-
вані клітини, що росли за рахунок фітогормонів, які виділялися транс¬
формантами.
У подальшому для отримання трансгенних рослин був розробле¬
ний порівняно простий та зручний метод “листкових дисків” [9]. Цей
метод із незначними модифікаціями з успіхом застосовується щодо
різних сільськогосподарських культур і сьогодні.
У тих випадках, коли важко регенерувати цілі рослини із окре¬
мих клітин, для трансформації використовують A.rhizogenes. Росли¬
ни багатьох видів можуть регенерувати із культури кореня [28, 32].
Але використання A.rhizogenes для трансформації обмежене з тієї
причини, що регенеровані із “кошлатих коренів” рослини часто
морфологічно змінені [42].
Методи отримання трансгенних рослин без використання агробак-
теріальних векторів отримали назву “пряме перенесення генів” [18].
Ця група методів має ряд переваг: по-перше, виключається необхід¬
ність введення клонової ДНК до складу плазмідної ДНК, по-друге,
прямий метод переносу придатний для клітин, однодольних, які ма¬
лоефективні в плазмідній системі A. tumefaciens.
526
Оптимізація умов проникнення плазмідної ДІЖ у протопласти
рослин привела до розробки методу електропорації. Метод грунту¬
ється на використанні тимчасових (тривалістю від кількох десятків
мікросекунд до кількох десятків мілісекунд) імпульсів електричного
струму високої напруги (від 200 до 1500 В/см) [25]. Цей метод до¬
зволив отримувати генетичну трансформацію з ефективністю від
0,01 до 0,1% від кількості протопластів, що використовувалися в дос¬
ліді, а в деяких випадках ефективність трансформації зростала до
10% [17].
Найбільшим недоліком методу електропорації є необхідність ро¬
боти з протопластами, що обмежує його застосування щодо багатьох
сільськогосподарських культур.
Один із способів поліпшення процесу трансформації є спосіб мі-
кроін’єкції ДНК у рослинну клітину. Перші успішні експерименти із
трансформації протопластів тютюну шляхом мікроін’єкцій ДНК
отримали П. Мельников [5] та Т. Пастернак [6] зі співавторами.
Найбільш вартісним і трудомістким є спосіб генетичної трансфор¬
мації рослин із використанням обладнання під назвою “shortgun” -
“дробовик”. Суть методу полягає в тому, що на частинки металу (воль¬
фраму чи золота) осаджується ДНК. Ці частини розганяються в об¬
ладнанні, і ними обстрілюють рослинні тканини [31]. Частинки осі¬
дають у рослинній клітині, а потім проводять тестування на “тран-
зієнтну” активність перенесених генів. Така активність була виявле¬
на після обстрілу тканин кукурудзи [27], пшениці [2, 47], ячменю
[14], сої [39, 44], тютюну і баклажанів [36].
За допомогою трансгенних методів, які розроблені практично для
усіх корисних організмів, можна змінювати організми в бажаному
напрямі для вирішення наступних завдань селекції.
Стійкість до гербіцидів. Сьогодні уже отримані і проходять
польове випробовування трансгенні рослини, що несуть різні госпо¬
дарсько цінні гени, серед яких насамперед, судячи з активності лабо¬
раторій, зайнятих вирішенням цієї проблеми, необхідно виділити
гени стійкості до гербіцидів.
Серед генів, що визначають стійкість до гербіцидів, вже кпоновані
гени стійкості до таких гербіцидів, як гліфосат (комерційна назва
Roundup) [22, 41], фосфінотрицин (комерційна назва BASTA) [20],
хлорсульфуронові та імідазолінові гербіциди [34]. З використанням
цих генів уже отримані трансгенні рослини у різних сільськогоспо¬
дарських видів, зокрема кукурудзи, сої, льону, бавовнику та ін.
Шляхом введення мутантного аго А гена Е.соїі отримані транс¬
генні рослини картоплі, стійкі проти гербіциду гліфосату [3].
Стійкість проти комах. Уже досить давно відома бактерія
Bacillus thuringiensis, що продукує білок, який дуже токсичний для
багатьох видів комах. Виявилося, що вмощування гена цього білка в
геном рослини дає змогу отримати трансгенні рослини, які не поїда¬
527
ються комахами. Водночас практичне застосування генноінженерних
методів щодо створення стійких проти комах рослин потребує вели¬
кої роботи з підбору необхідних штамів B.thuringiensis і створення
генноінженерних конструкцій, що дають найбільший ефект [43].
Для отримання трансгенних рослин, стійких проти комах, розроб¬
лені два основні підходи. Перший включає використання кодуючих
послідовностей дельта-ендотоксину B.thuringiensis. Другий - вико¬
ристання рослинних генів-інгібіторів ферментів або лектини [43].
Сьогодні фірми “Monsanto” і “Ciba - Geigy” вже пропонують для
практичного використання трансгенні форми томатів, бавовнику,
кукурудзи і картоплі, стійкі проти комах, у тому числі і проти коло¬
радського жука.
Стійкість проти вірусів. Одним із підходів у боротьбі з вірус¬
ними хворобами є клонування і вмонтовування в геном рослин гена
синтезу білка оболонки (coat - protein) вірусів. Активний синтез та¬
кого білка, що дуже подібний до РІЖ вірусу, не дає змоги останній
активно реплікуватися в клітині господаря, що й приводить до до¬
сить високої стійкості такої трансгенної рослини проти вірусів [23,
24, 48]. Використовуючи такий підхід, отримали стійкі до Х-вірусу
трансгенні рослини картоплі [1, 10].
У дослідженнях канадських учених [33] стійкість до вірусів
трансгенних рослин досягається в результаті експресії білка, реплі-
каз, протеаз, а також нетрансльованих послідовностей чи псевдо-
справжніх РНК, сателітів та ін. Уже отримані трансгенні рослини
картоплі, стійкі до вірусів S, X, Y та скручування листя. Подальші
дослідження ведуться з комбінування стратегії з включенням у дос¬
лідження вірусів із різних таксономічних груп.
До інших підходів у боротьбі з вірусною інфекцією можна віднес¬
ти повідомлення про отримання стійкості проти вірусів при вмонто¬
вуванні в геном рослин генів, що кодують специфічні авгтитіла, які
пізнають вірусні білки [45]; послідовність ДНК, котрою кодуються
нетрансльовані послідовності РНК вірусів [43] або ж мутантного ге¬
на білка рг17, залученого в рух лютеовірусів між клітинами [40].
Стійкість проти грибкових та бактеріальних хвороб. Активно
ведуться роботи з клонування генів, що беруть участь у захисті рос¬
лин проти грибкових інфекцій [30, 37]. Передусім це ген хітинази із
квасолі. Наступним кроком є вмонтовування гена хітинази рису су¬
місно з геном глюконази люцерни [21] і отримання рослин тютюну,
стійких проти інфекції. Трансформація люцерни конструкцією з ге¬
ном глюконази показала, що рослини мали підвищену стійкість про¬
ти Phytophthora megasperma f. sp. medicaginus [13].
Нові підходи застосовують при отриманні трансгенних рослин із
геном дефензину, що несе стійкість до грибкових та бактеріальних
хвороб [7]. Так, у різних видів рослин були відкриті багаті залишка¬
ми цистеїну короткі пептиди, що мають антимікробні властивості і
528
названі дефензинами [4]. Найбільша концентрація дефензинів синте¬
зується у проростаючому насінні. Діють вони проти грибів, а в де¬
яких проявляється інсектицидна активність.
Китайські біотехнологи створили повністю синтетичні гени для
захисту рослин від бактеріальних хвороб. При трансформації картоп¬
лі отримано трансгени, стійкі до бактеріального вілту картоплі [11].
Іншим підходом для створення рослин, стійких проти грибкових
захворювань, є використання генів, що беруть участь у синтезі феніл-
пропанових сполук, здебільшого фітоалексинів [19]. Так, у рослину
тютюну був уведений ген стинбенсингази із винограду [16]. Цей фер¬
мент бере участь у синтезі фітоалексинарезерватолу. Отримані
трансгенні рослини тютюну мали підвищену стійкість проти грибко¬
вих патогенів Botrytis cinerea.
Учені із компанії “ Monsanto” [15] розробили спосіб, який дозво¬
ляє отримати трансгенні рослини, стійкі як проти бактеріальної, так і
грибкової інфекцій. У рослини картоплі був перенесений ген грибко¬
вого походження, що кодує синтез ферменту, який здійснює реакцію
окиснення глюкози з утворенням перекису водню. Отримані рослини
виявилися майже повністю стійкими до м’якої гнилі, збудником якої
є бактерії з роду Erwinia, мали стійкість проти фітофторозу.
Стійкість до абіотичних стресів. Велика увага приділяється
створенню рослин, стійких до абіотичних стресів [12]. Важливе зна¬
чення в розвитку стресу має накопичення окиснювальних радикалів.
Одним із підходів до створення рослин, більш стійких до таких стре¬
сів, є перенесення генів супероксиддисмутази. Так, одержані транс¬
генні рослини тютюну [26] з підвищеним умістом супероксидцисму-
тази виявили більш підвищену стійкість до гербіцидів окиснюваль-
ної дії і до озону. Трансгенні рослини люцерни, що несуть додатко¬
вий ген супероксиддисмутази із N.plumbaginifolia, були більш
стійкими до водного дефіциту, і протягом трирічних польових ви¬
пробувань їхня продуктивність була значно вищою, ніж нетрансфор-
мованих контрольних рослин [49].
Пошук підходів до створення холодостійких рослин привів до
розробки методів, що базуються на зміні співвідношення насичених і
ненасичених жирних кислот у мембранах рослинних клітин. У ре¬
зультаті переносу цього гена в тютюн були створені рослини, більш
стійкі до понижених температур [29, 50].
Регуляція строків дозрівання. Генноінженерний підхід був ви¬
користаний для створення трансгенних томатів. У цьому випадку був
виключений один із генів синтезу етилену - фітогормон, що регулює
дозрівання плодів. Одержані плоди у таких трансгенних рослин мо¬
жуть зберігатися практично необмежено довго, аж до примусової
обробки етиленом [35]. Розроблений метод також цілком може бути
використаний у регуляції строків зберігання зібраних плодів і для
інших видів рослин [38].
529
Слід зазначити, що основні проблеми в цій галузі досліджень
пов’язані з виділенням генів та створенням генетичних конструкцій.
Існуючі способи генетичної трансформації вищих рослин забезпе¬
чують сьогодні можливість одержання трансформантів майже у всіх
господарсько цінних видів.
ЛІТЕРАТУРА
1. Болвелл П., Диксон Р. Биотехнология растений: культура клеток. - М.:
Агропромиздат, 1989. - 280 с.
2. Генетическая трансформация с помощью высокоскоростной механи¬
ческой инъекции ДНК и характеристика канамицинустойчивых клеточных
линий озимой пшеницы /Н.В.Сидорова, В.А.Колесников, А.А.Амилов и др.
//Тез. док. I Всесоюз. симпозиума “Новые методы биотехнологии расте¬
ний”.- Пущино,1991. - С.79.
3. Карбаль Эскаланте Бьенвенидо. Получение трансгенных растений
картофеля (Solanum tuberosum L.), устойчивых к гербициду глифосату путем
введения мутантного аго А гена Е.соіі: Автореф. дис. ... канд.биол. наук. -
М.:ВНИИ с.-х.биотехнологии, 1998. - 21 с.
4. Лети Джое, Регунатан Пилат, Калашникова Е.А. Разработка методов
клеточной селекции пшеницы на устойчивость к Septoria nodorum
//Молекулярная генетика и биотехнология: Материалы междунар. конф.
(Минск, 6-8 апреля 1998 г.) - Минск, 1998. - С.221-222.
5. Микр о инъекции ДНК в клетки высших растений /П.В.Мельников, Т.П.
Пастернак, Ю.Ю. Глеба, К.М. Сытник // Доклады АН СССР - 1985 - №10. -
С.69-71.
6. Пастернак Т.П., Мельников П.В. Генетическая трансформация клеток
высших растений с помощью микроинъекции ДНК // Изв. АН СССР. - 1986.
- №2. - С.314-316.
7. Получение трансгенных растений рапса и томата, экспрессирующих
ген дефензина редьки /Е.В. Парашина, Л.А. Сердобинский, Е.Г. Калле и др.
// Физиол.раст. - 2000. - Т.47, №3. - С.471-478.
8. Созинов А.А. Трансгенные растения: как это делается // Наука и
жизнь.- 1998. - №10. - С.24-25.
9. A simple and general method for transfearing genes into plants /. R.Horsch,
J.Fry, N. Hoffinan et al // Science. - 1985. - Vol.227, №4692. - P.1231-1299.
10. Barchend G. Ergebnisse der PVY-Resistenz-testung von transgenen
Kartoffelgenotypen nach Anbau im Freiland: //Bioi/Bundensanst. Land-und
Forstwirt. - Berlin-Dahlem. - 1998. - №357. - P.l 13.
11. Behnke M. Selection of potato callus for resistance in culture filtrates of
Phytophthora infestans and regeneration of resistant plants // Theor. and
Appl.Genet. - 1979. - Vol.55, №2. - P.69-71.
12. Bohnert H.,Nelson D., Jensen R. Adaptation to enviromental stresses //
Piant Cell.- 1995.- Vol.7, №7. - P. 1099-1111.
13. Constitutive expression of an inducible b-l,3-glucanase in alfalfa reduces
disease severity caused by the oomycete pathogen Phytophthora egasperma
f.sp.medicaginis, but does not reduce disease severity of chitin-containing flingi /.
S.Masoud, Q. Zhu, C. Lamb, R. Dixon // Transgenic Res. - 1996. -Vol.5. -
P.313-323.
530
14. Delivery of foreign genes to intact barley cells by high-velocity
microprojectiles /R.Mendel, B. Mulier, J. Schulze et al // Theor. Appi.Genet. -
1989. - Vol.78, №1. - P.31-34.
15. Disease resistance conferred by expression of a gene encoding H202 -
generating glucose oxidase in transgenic potato plants /. G.Wu, B. Shortt, E.
Lawrence et al // Piant Cell. - 1995. - Vol.7, №9. - P.1357-1368.
16. Disease resistance results from foreign phytoalexin expression in novel
plant/R.Hain, H. Reif, E. Krause et al. //Nature. - 1993.- Vol.361. - P. 153-156.
17. Direct gene transfer to plants /J.Paszcowski, R. Shillito, M.Saul et al. //
The EMBO J. - 1984. - Vol.3, №12. - P.2717-2772.
18. Direct gene transfer. State of the art and future potential /I.Potrykus,
R.Shillito, M.Saul, J.Paszcowski // Piant Mol.Biol.Rep~ 1985 - Vol.3 - №3.
P.l 17-128.
19. . Dixon R., Paiva N. Stress-indued phenylpropanoid metabolism // Piant
Cell. - 1995. - Vol.7, №7. - P.l085-1097.
20. Engineering herbicide resistance in plants by expression of a detoxifying
enzyme /M.De Block, J. Botterman, M.Vandewiele et al. // EMBOJ. - 1987. -
Vol.6, №9. - P.2513-2518.
21. Enhanced protection against fimgal attack by constitutive co-expression of
chitinase and glucanase genes in trangenic tobacco /Q.Zhu, E. Maher, S. Masoud
et al. // Biotechnology. - 1994. - Vol.12, №8. - P.807-812.
22. Experession in plants of a mutant Salmonella gene from Salmonella
typhimurium confers tolerance to glyphosate /l.Comai, D. Facciotti, W. Hiatt et al.
//Nature. - 1985. - Vol.317, №6039. - P.741-744.
23. Expression of alfalfa mosaic virus coat protein confers cross protection in
transgenic tobacco and tomato plants /N.Tumer, T. Stone, Y.Muskopf et al. //
EMBO J.- 1987. - Vol.6, №5. - P. 1181-1188.
24. Expression of alfalfa mosaic virus RNA 4 in transgenic plants confers
virus resistance / L.Loesch-Fries, D.Merlo, T.Zinnen et al. // EMBO J - 1987-
Vol.6, №7. - P.l845-1852.
25. Fromm M., Tayor L., Walbot V. Expression of genes transferred into
monocot and dicot piant cells by electroporation // Proc. Nati. Acad. Sci.USA. -
1985. - Vol.82, №14. - P. 5824-5825.
26. Genetic engineering of oxidative stress resistance in higher plants/
D.Herouart, C. Bowler, H. Willekens et al. // Phil. Trans.R.Soc. Lond.B - 1993 -
Vol.342. -P.235-240.
27. Genetic transformation of maize cells by particle bombardment / T.Klein, L.Kom-
stein, J. Sanford, M.Fromm // Piant Physiol. -1989. - Vol.91, №1. - P.440-444.
28. Genetic transformation of potato with binary vector in Agrobacterium
rhizogenes / O.Melik-Sarkisov, V.Avetisov, G.Sobolkova, A.Gartel // Abstracts
VII th International Congress of Piant Tissue and Cell Culture. Amsterdam, June
24-29, 1990. - Amsterdam, 1990. - P.69.
29. Genetically engineered altemation in the chilling sensitivity of plants
/N.Murata, O.Ishizaki-Nishizawa, S. Higashi et al. // Nature-1992. - Vol.356,
№6371 - P. 710-713.
30. Glutatione ahd fungal elicitor regulation of a piant defence gene promoter
in electroporated protoplasts / M.Dron, S. Clouse, R.Dixon et al. // Proc. Nat.
Acad.Sci. USA.- 1988. - Vol. 85, №18. - P.6738-6742.
531
31. High velocity microprojectiles for delivering nucleic acids into living
cells / T.Klein, E.Wolf, R.Wu, J.Sanford // Nature - 1987 - Vol.327, №6743 -
P.70- 73.
32. Introduction of foreign genes into Pharbitic nil calli using a vector derived
from Agrobacterium pTi /T.Araki, H.Hirano, S.Naito, Y.Komeda // Piant Cell
Repts. - 1989.- Vol.8, №5.- P.259-262.
33. Kawchuk L., Prufer D. Molecular to potato viruses // Can. J. Piant Pathol.
- 1999. - Vol.21, №3. -P.231-247.
34. Li Z., Hayashimoto A.,Murai N. A sulfonylurea herbicide resistance gene
from Arabidopsis thaliana as a new selectable marker for production of fertile
transgenic plants // Piant Physiol. - 1992. - Vol. 100, №2. - P.662-668.
35. Molecular characherization, enzyme expression, and fruit ripening traits
/ J.Reed Andrew et al. // J. Agr. and Food Chem. - 1995.- Vol.43, №7. -
C.1954-1962.
36. Morikawa H., Lida A., Yamada G. Transient expression of foreign genes
in piant cells and tissues obtained by a simple biolistic devise ( particle-gun) //
Appl.Microbiol. Biotechnol. - 1989. - Vol.31, №3. - P.320-322.
37. Optimizing the expression of chimaeric gene in piant cells/J.Taylor, J.
Jones, S. Sondler et al. // Мої. Gen.Genet. - 1987. - Vol.210, №3. - P.572-577.
38. Reversible inhibition of tomato fruit senescence by antisense RNA /
P.Oeller, L.Min-Wong, L.Taylor et al. // Science. - 1991. - Vol.254, №5030. -
P.437-439.
39. Soybean genetic engineering - commercial production of transgenic
plants/P.Christou, D.Mc Cabe, B.Martinell, W. Swain // Trends in Biotechnology.
- 1990. - Vol.8, №6. - P. 145-151.
40. Таске E., Salamini F., Rohde W. Genetic engineering of potato for broad-
spectrum protection agains virus infection // Nature Biotechnology. - 1996-
Vol.14, №11. - P.1597-1601.
41. Targeting herbicide-resistsnt enzyme from Escherichia coli to chloroplasts
of higher plants /. G.Gioppa-della, S. Bacur, M. Taylor et al. // Biotechnology.-
1987. -Vol.5, №4. - P.579-584.
42. Tepfer D. Transformation of several species of higher plants by
Agrobacterium rhizogenes: sexual transmission of the transformed genjtype and
phenotype // Cell. - 1984. - Vol.37, №5. - P.959-967.
43 Transdenicplants for insect resistance / Jouanin Lise, Bonade В ottino
Micchel et al. //Piant Sci. - 1998.- Vol.131, №1. - C. 1-11.
44. Transformation of cereal cell cultures by microprojectile bombardment /
D.Chen, N. Batty, J. Evans, P. Dale // Abstracts VII th International Congress of
Piant Tissue and Cell Culture. Amsterdam, June 24-29, 1990. - P.50.
45. Transgenic piant expressing a functional singlechain Fv antibody are
specifically protected from virus attack / P.Tayladoraki, E.Benvenuto, S.Trinca et
al. // Nature. - 1993.- Vol.366, №6454. - P.469-472.
46. Transgenic piant virus resistance mediated by untranslatable sense RNAs:
Expression, regulation,and fate of nonessential RNAs / H.Smith, S.Swaney, T.
Parks et al. // Piant Cell. - 1994. - Vol.6, №10. - P.l441-1453.
47. Transient expression of foreign genes in rice, wheat and soybean cells
following particle bombardment / Y .Wang, T.Klein, M.Fromm et al. // Piant.
Мої. Biol. -1988. - Vol. 11, №4. - P.433-439.
532
48. Virus tolerance piant growth, and field performance of transgenic tomato
plants, expressing coat protein from tomato mosaic virus / R.Nelson, S. McCormic,
X.Delannay et al. // Biotechnology. - 1988 - Vol.6, №3. - P.403-409.
49. Water-deficit tolerance and field perfomance of transgenic alfalfa
overexpressing superoxide dismutase / B.McKersie, S. Bowley, E.Harjanto, O.
Leprince // Piant Physiol. - 1996. - Vol.l 11, №4. - P. 1177-1181.
50. Wolter F., Schmidt R., Heinz E. Chilling sensitivity of Arabidopsis
thaliana with genetically engineering membrane lipids // EMBO J - 1992 - Vol.l 1-
P.4685-4692.
ЗМІСТ
вступ з
1. РОЗВИТОК КАРТОПЛЯРСТВА В УКРАЇНІ 12
1.1. Народногосподарське значення картоплі 12
1.2. Становлення і розвиток картоплярства в Україні 13
1.3.Агрокліматична характеристика зон України за їх придатністю
для вирощування картоплі 42
1.4. Перспективи розвитку картоплярства в Україні 47
2. МОРФОЛОГІЯ, ФІЗІОЛОГІЯ ТА БІОХІМІЯ КАРТОПЛІ 54
2.1. Морфологічні особливості 54
2.2.Біологічні особливості 59
2.3.Фізіологічні особливості 69
2.4.Біохімічний склад 84
3. ГЕНЕТИКА КАРТОПЛІ 116
3.1. Генетика диплоїдних видів картоплі 116
3.2.Генетика поліплоїдних видів картоплі 123
3.3.Успадкування морфологічних і господарсько цінних ознак у
S. tuberosum 131
3.4.Генетика стійкості до біотичних факторів 135
3.4.1. Стійкість картоплі до патогенів 135
3.4.2. Стійкість до абіотичних факторів 142
3.5.Мутагенез у картоплі 144
3.6.Рекомбінація генетичного матеріалу 149
3.7.Гаплоїдія у картоплі 153
3.8.Генофонд картоплі, його складові, характеристика і стратегія
використання 156
3.8.1 .Генофонд картоплі, його складові і характеристика 156
3.8.2.Систематика бульбоутворюючих видів секції Petota
Dumort роду Solanum L 163
3.8.3.Вихідний передселекційний матеріал (попередні
дослідження та його створення) 173
3.8.4.Стратегія і тактика використання генофонду картоплі у
практичній селекції 180
4. СЕЛЕКЦІЯ КАРТОПЛІ 199
4.1.3 історії розвитку селекції картоплі 199
4.2. Генетичний потенціал картоплі 203
4.3.Біологічний взаємозв’язок ознак картоплі і його вплив на добір 204
4.4.Методи селекції картоплі 212
4.5.Основні напрями та завдання селекції картоплі 218
4.5.1 .Селекція на вміст крохмалю і сирого протеїну 219
4.5.2.Селекція картоплі на ранньостиглість 226
534
4.5.3.Селекція картоплі на придатність до механізованого
виробництва 242
4.5.4. Селекція картоплі на стійкість (імунітет) до хвороб і
шкідників 257
4.5.5. Селекція на придатність до промислової переробки 270
4.6. Вирощування картоплі з використанням ботанічного насіння 290
4.6.1. Історія проблеми вирощування картоплі з ботанічного
насіння 290
4.6.2. Генетичні основи і підходи до створення сортів, придат¬
них для розмноження ботанічним насінням 294
4.6.3. Технологія вирощування картоплі з ботанічного насіння 305
4.7. Схема, організація і техніка селеьсції 313
4.8. Основні досягнення селекції картоплі за останні роки та їхнє
народногосподарське значення 319
5. СОРТИ 325
5.1.Загальна характеристика сортів ,. 327
5.2.Ранні сорти 333
5.3.Середньоранні сорти 341
5.4. Середньостиглі сорти 346
5.5.Середньопізні сорти 350
5.6.Пізні сорти 553
6. НАСІННИЦТВО КАРТОПЛІ 35^
6.1.Історія розвитку насінництва *. 355
6.2.Наукові засади насінництва ^66
6.3.Причини виродження картоплі і заходи боротьби з ним 379
6.4.Система насінництва картоплі в Україні 397
6,4.1 .Оборот насіннєвого матеріалу 406
6.5. Одержання вихідного матеріалу в насінництві картоплі
шляхом культури апікальної меристеми 412
6.6.Репродукційне та внутрішньогосподарське насінництво 436
6.7. Апробація картоплі 440
6.8.Прискорене розмноження картоплі 462
7. БІОТЕХНОЛОГІЯ 480
7.1.Клітинна інженерія картоплі 480
7.2. Експериментальний андрогенез 510
7.3.Соматична гібридизація 520
7.4. Г енетична інженерія 525
535
Наукове видання
КАРТОПЛЯ
За редакцією
КОНОНУЧЕНКА Валерія Васильовича
МОЛОЦЬКОГО Михайла Яковича
Редактори: С.О. Заїка, Л.В. Міщенко
Технічний редактор: С.І. Сидоренко
Комп'ютерний набір: Т.О. Іштванік, Н.П. Граждан
Здано до набору 22.04.2001. Підписано до друку 20.05.2002.
Формат 60x90V16. Папір офсетний №1. Гарнітура Times New Roman.
Друк офсетний. Ум. друк. арк. 31,2. Зам. 2-132. Тираж 1000 прим.
Сектор оперативної поліграфії РВІКВ БДАУ.
09117, м. Біла Церква, Соборна пл. 8/1, тел. 3-11-01.
Віддруковано на ВАТ “Білоцерківська книжкова фабрика”,
м. Біла Церква, вул. Леся Курбаса, 4.