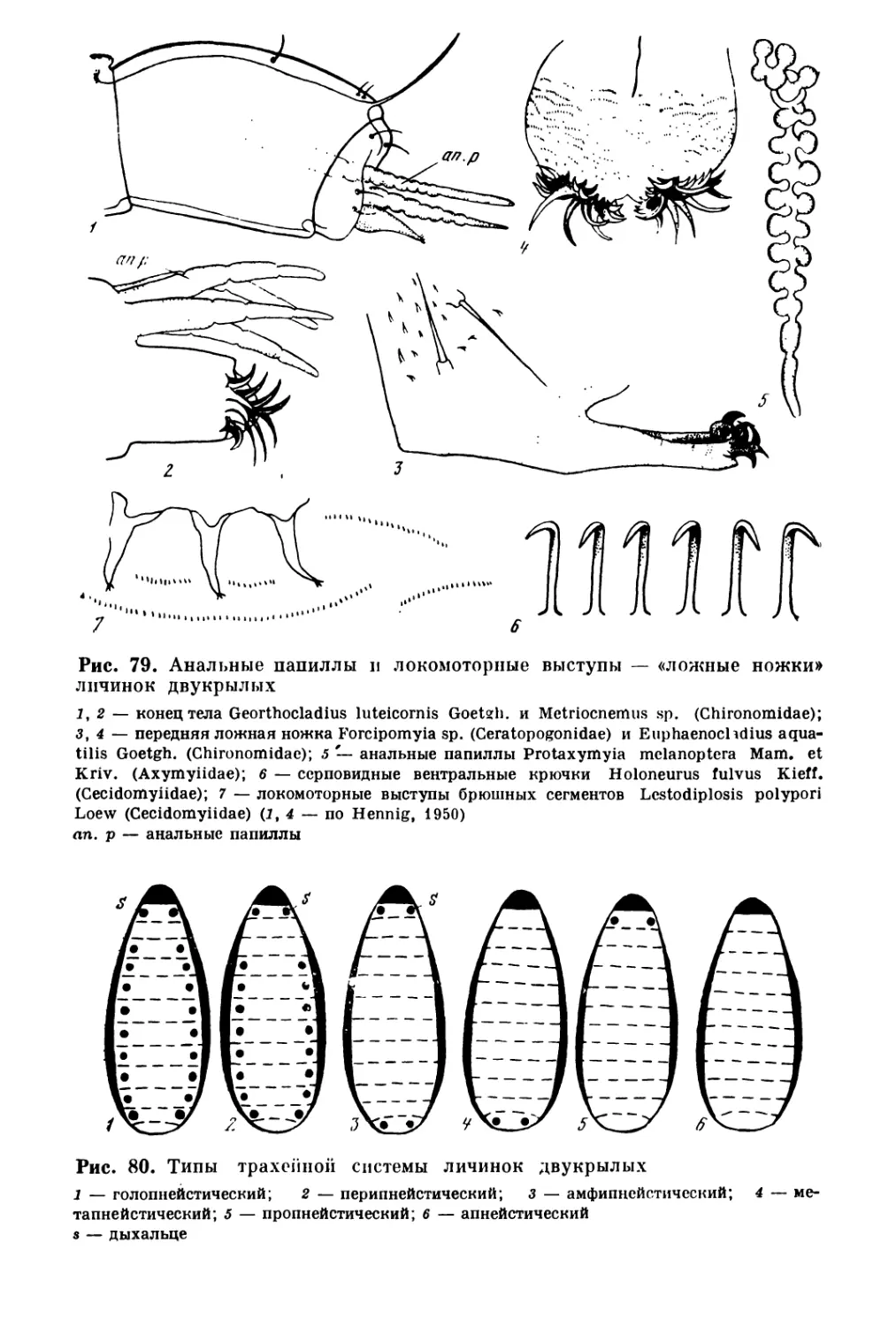
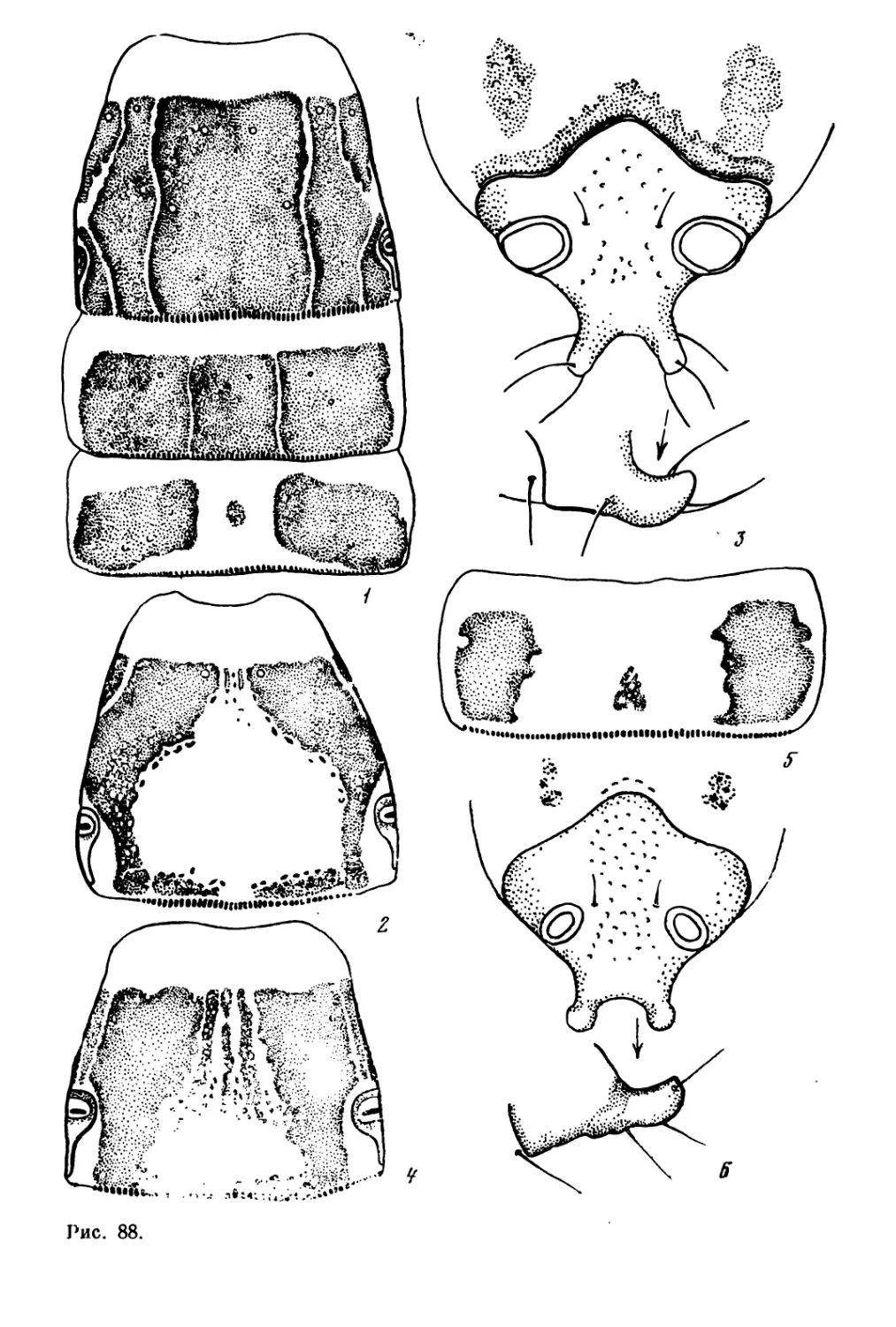
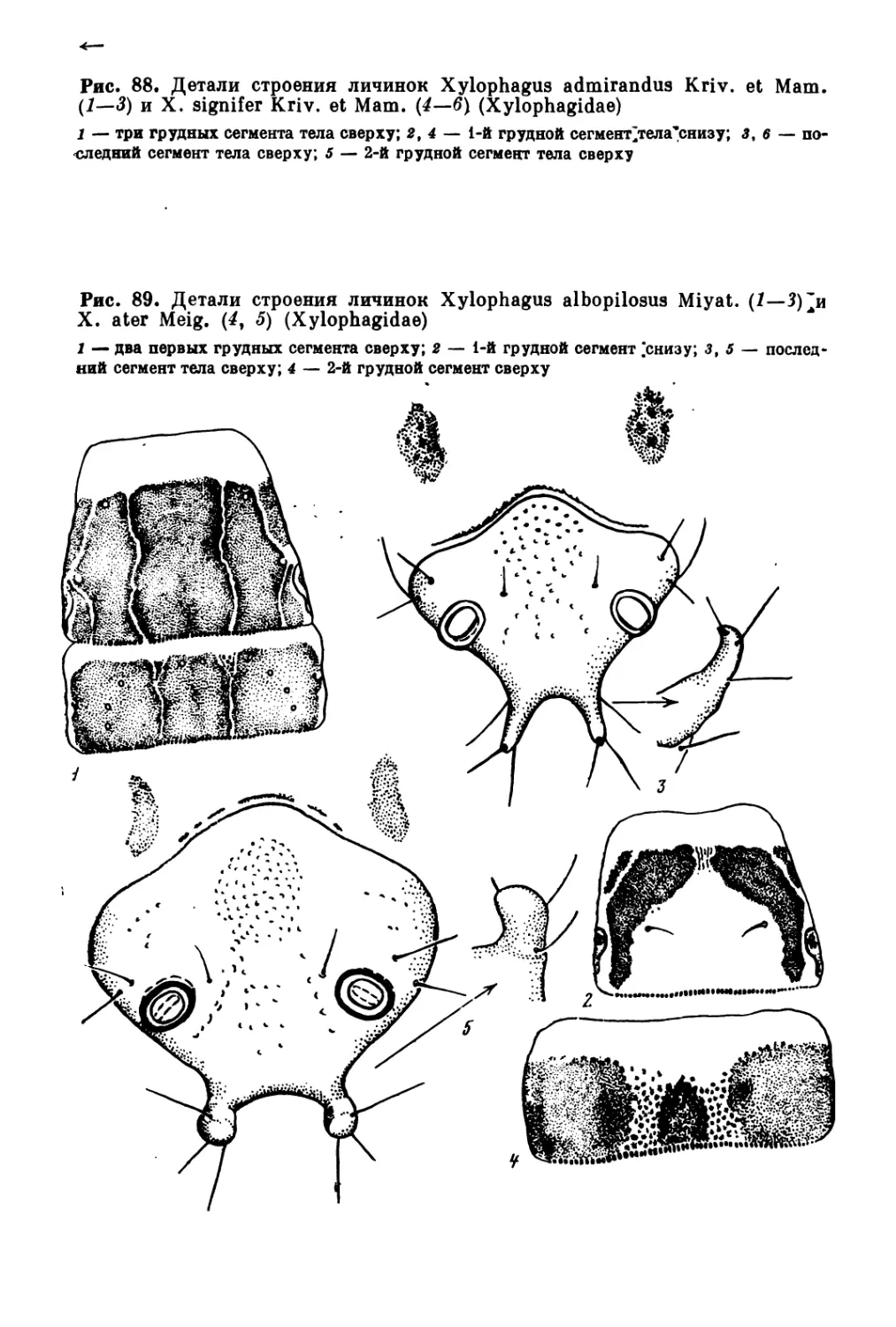
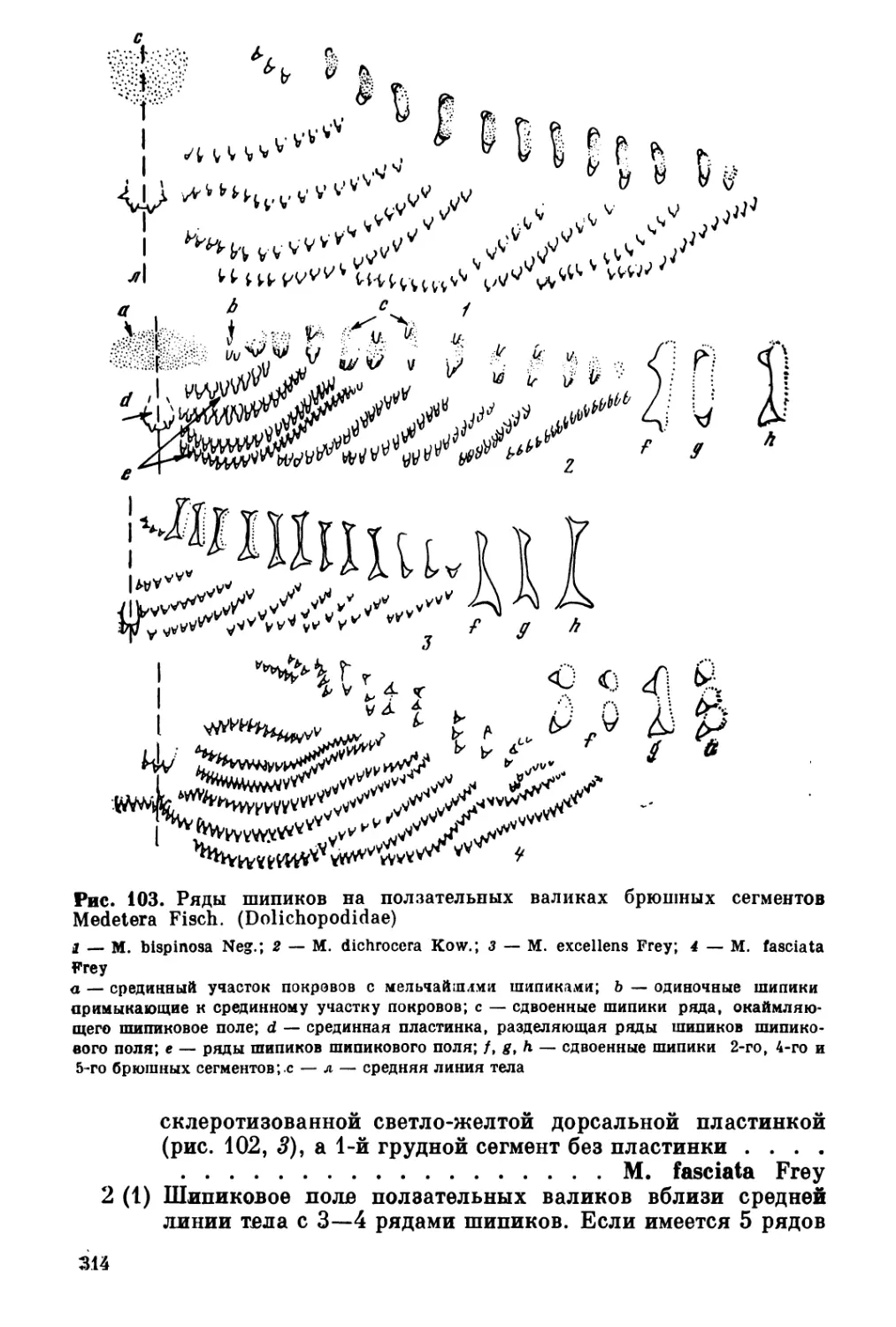
/
































































































































































































































































































































































































Author: Мамаев Б.М. Кривошеина Н.П. Потоцкая В.А.
Tags: insecta hexapoda насекомые энтомология зоология
Year: 1977




Text
Б.М.МАМАЕВ, Н.П.КРИВОШЕИНА
В.А.ПОТОЦКАЯ
Определитель
личинок
ХИЩНЫХ насекомых-
энтомофагов
стволовых вредителей
АКАДЕМИЯ НАУК СССР
Институт эволюционной морфологии
и экологии животных им. А. Н. Северцова
Б. М. МАМАЕВ, Н.П.КРИВ0ШЕ11НА,
В. А. ПОТОЦКАЯ
ОПРЕДЕЛИТЕЛЬ
ЛИЧИНОК
хищных НАСЕКОМЫХ-
ЭНТОМОФАГОВ
СТВОЛОВЫХ ВРЕДИТЕЛЕЙ
ИЗДАТЕЛЬСТВО «НАУКА»
Москва 1977
УДК 595.7
Мамаев Б. М., Кривошеина Н. П., Потоц¬
кая В. А. Определитель личинок хищных насекомых — эн-
томофагов стволовых вредителей. М., «Наука», 1977, 392 с.
Монография представляет собой обзор материалов по мор¬
фологии и экологии личинок 20 семейств насекомых, относящих¬
ся к отрядам жесткокрылых, верблюдок и двукрылых. В осно¬
ву положены оригинальные материалы авторов по биологии
наиболее массовых видов, которые относятся к облигатным или
факультативным хищникам и примыкающим к ним трофическим
группам. На современном уровне рассмотрены закономерности
смены комплексов насекомых в разлагающейся древесине,
разработаны основы классификации пищевых режимов ксп-
лобионтов, охарактеризованы взаимоотношения хищных насе¬
комых с другими компонентами ксилофильных сообществ. Со¬
ставлены определительные таблицы, охватывающие основные
труппы хищных насекомых-ксилобионтов фауны СССР.
Предназначена для специалистов в области общей экологии
и биогеоценологии, энтомологов широкого профиля и работни¬
ков по защите леса и древесины.
Илл. 117, библ. 428 назв.
Ответственный редактор
доктор биологических наук
Ф. Н. ПРАВДИН
^1008 182
М ^^02)-77 502-77, кн. 2 © Издательство «Наука», 1977 г.
ВВЕДЕНИЕ
В современный период развития науки о защите растений
основное внимание уделяется вопросу о замене сильных, устойчи¬
вых и отрицательно влияющих на окружающую среду ядохими¬
катов комплексными интегрированными истребительными систе¬
мами мероприятий, которые предусматривают широкое приме¬
нение биологических средств ограничения численности вредителей.
Именно по этой причине в последнее время резко интенсифи¬
цировалось изучение видового состава и биологии энтомофагов
и энтомопатогенных организмов.
Сложность возникающих при этом проблем усугубляется раз¬
норечивостью представлений о значении отдельных экологических
групп энтомофагов. Несмотря на успешный опыт применения, на¬
пример, хищных жуков против червецов — вредителей цитрусов,
хищных муравьев — для борьбы с хвое-листогрызущими вреди¬
телями, хищных клещиков и хищных личинок галлиц — против
растительноядных клещиков и тлей в закрытом грунте, энтомо-
фаги-хищники еще не привлекли к себе того внимания, которого
они заслуживают. Особенно слабо остаются разработанными воп¬
росы применения комплекса энтомофагов для ограничения чис¬
ленности отдельных групп вредных насекомых.
В лесном хозяйстве в этом отношении наблюдаются значитель¬
ные положительные сдвиги. Уже сравнительно давно привлекли
внимание и стали интенсивно изучаться отдельные виды хищных
насекомых, например пестряки — истребители короедов. В пос¬
леднее время привлекли внимание мухи-зеленушки и копьехвост-
ки, личинки которых обычны в короедных сообществах. Были
предприняты попытки изучить весь комплекс энтомофагов,
ограничивающих численность короедов.
Тем не менее дальнейший прогресс исследований в этом нанрав-*
ленни оказался практически невозможным из-за отсутствия необ¬
ходимой справочной и теоретической базы. До сих пор нет исчер¬
пывающих данных о видовом составе насекомых, сопутствующих
короедам и другим стволовым вредителям; для многих насеко¬
мых отсутствуют достоверные сведения о режимах питания и на¬
правлениях пищевой специализации; по существу достаточно хо¬
рошо известна только имагинальная фаза развития этих насе^
комых, личинки же многих групп остаются неизученными. Между
тем именно личинки в большинстве случаев являются энтомо-
фагами, уничтожающими стволовых вредителей. Более того, срав¬
нительно немногочисленные и иногда поверхностные наблюдения,
опубликованные много десятилетий назад, до сих пор широко
цитируются многими авторами без какого-либо критического пере-
“€мотра и добавления собственных оригинальных материалов. Ме¬
жду тем основной задачей изучения ксилофильпых, как, впрочем,
и всех других групп насекомых, в текущий период является не
«обобщение» (в который уже раз) материалов других авторов, а их
пересмотр на основе собственных данных и наблюдений. Все из¬
ложенное свидетельствует о назревшей необходимости публикации
•обзора хищных ксилофильных личинок насекомых — естествен¬
ных врагов стволовых вредителей.
Настоящий обзор подготовлен в лаборатории экологии сооб¬
ществ наземных беспозвоночных Института эволюционной мор-
^фологии и экологии животных им. А. Н. Северцова АН СССР глав¬
ным образом на основе оригинальных коллекционных материалов.
Была поставлена задача охватить исследованиями все основные
лесорастительные ассоциации в различных зоогеографических ре¬
гионах в основном к югу от 55-й параллели северной широты, где
лесные фитоценозы на территории СССР особенно разнообразны.
В 1958—1976 гг. были проведены экспедиции в Закарпатье,
в Крым и на Кавказ, в центральные районы Европейской части
СССР, в горные, тугайные и пустынные леса Средней Азии, в юж¬
ные районы Средней Сибири, в Забайкалье, Приамурье и Южное
Приморье, а также на Курильские острова.
Материал ^ был собран в следующих точках Советского Союза:
в Закарпатской области в ельниках на высоте более 1000 м, в сме-
шанных‘елово-буковых лесах с участием пихты (окрестности г. Ра-
:хова и пос. Квасы) и в буковых лесах около г. Хуста; на Северном
Кавказе в Краснодарском крае в дубовых лесах около села Ново-
Прохладное, в смешанных лесах с участием дуба, бука, граба,
осины, сосны в Убинском лесничестве (ст. Убинская), в горных
пихтовых лесах с участием бука на территории Кавказского за-
ловедника (пос. Гузерипль), в районе г. Красная Поляна в пой¬
менных лесах по р. Мзымта и в пихтовых лесах урочища «Мед¬
вежьи Ворота»; в Европейской части СССР — в Московской
области (пос. Павловская Слобода, Звенигород, Успенское, Пешки,
Красная Пахра, Левинская и др.) в ельниках, сосняках, березня¬
ках, осинниках, смешанных лесах с участием дуба, липы и клена;
в Тульской области в дубравах Тульских засек (пос. Ярцево)
с примесью березы, липы, осины, ели; в Ярославской области
в ельниках с участием осины, ольхи и березы (пос. Волково);
в дубравах Воронежской области (Теллермановское лесничество);
1в древостоях со значительным разнообразием пород в Архангель¬
* В тексте видовых характеристик в дальнейшем будут приводиться только
самые общие указания на район сбора материала.
ской (пос, Обозерская), Вологодской (пос. Кадниковский) и не¬
которых других областях; в Средней Азии в горных орехово¬
плодовых лесах заповедника Сары-Челек и в урочищах Каракала
и Ипайкала Копетдага, в пойменных турангово-лоховых лесах
по рекам Амударья (окрестности г. Дейнау), Мургаб (пос. Кзыл-
Байдак), Или (пос. Чилик), в саксауловых лесопосадках и защит¬
ных лоховых лесопосадках Туркменской ССР (Марыйская область,
пос. Акибай), в естественных саксауловых лесах Центральных
Каракумов (колодец Донгусбурун); в Сибири в лиственничниках
и березняках, а также в пойменных лесах с преобладанием ивы и
тополя в Тувинской АССР (пос. Иштии-Хем), в хвойных (ель, кедр,
лиственница) и мелколиственных (береза, осина) лесах Забай¬
калья (пос. Бабушкин и Таежный); в Приамурье и в Приморье
в разнообразнейших смешанных лесах с участием в древостоях бо¬
лее чем 20 основных древесных пород, на территории заповедников
«Кедровая Падь», «Комарово-Заповедное» (Приморский край),
Хехцирском и Хинганском (Хабаровский край); в наиболее разно¬
образных по составу пород островных лесах Курильской гряды
(о. Кунашир).
В результате были собраны оригинальные материалы более
чем из 40 древесных пород В связи с тем, что не всегда по отмер¬
шему стволу или пню можно было определить до вида изучаемую
древесную породу, в тексте нередко используются лишь родовые
названия деревьев. Ниже приводится список тех древесных пород,
которые нами в той или иной степени были изучены: хвойные —
ель европейская, или обыкновенная (Picea excelsa Link.), кедр
корейский (Pinus koraiensis Sieb. et Zucc.), кедр сибирский (Р. si-
birica Mayr.), лиственница европейская (Larix decidua Mill.),
лиственница сибирская (L. sibirica Ldb.), пихта европейская, или
белая (Abies alba IVnil.), пихта кавказская (А. nordmanniana Stev.),
пихта цельнолистная ( А. holophylla Maxim.), сосна обыкновенная
(Pinus silvestris L.); лиственные — бархат амурский (Phellodend-
ron amurense Rupr.), бархат сахалинский (Ph. sachalinensis Sarg.),
береза бородавчатая (Betula verrucosa Ehrn.) и береза пушистая
(В. pubescens Ehrh.), бук восточный (Fagus orientalis Lipsky),
бук европейский (F. silvatica L.), вяз обыкновенный (Ulmus
laevis Pali.), вяз шершавый (U. scabra Mill.), граб обыкновенный
(Carpinus betulus L.), дуб черешчатый (Quercus roDur L.), дуб мон¬
гольский (Q. mongolica Fisch.), ива (Salix cinerea L. и S. alba L.),
ильм долинный (Ulmus propinqua Koidz.), липа амурская (Tilia
amurensis Rupr.), липа мелколистная (Т. cordata Mill.), лох узко¬
листный (Elaeagnus angustifolia L.), маакия амурская (Maackia
amurensis Rupr. et Maxim.), ольха пушистая (Alnus hirsuta Turcz.),
ольха серая (А. incana Moench.), ольха клейкая (А. glutinosa
Gaertn.), ольха японская (А. japonica Sieb. et Zucc.), орех грец¬
^ В дальнейшем изложении в видовых характеристиках будут указываться
русские названия древесных пород.
кий (Juglans regia L.)» орех маньчжурский (J. manshurica Maxim.),
осина (Populus tremula L.), тополь лавролистный (Р. laurifolia
Lab.), тополь Максимовича (Р. maximowiczii Henry), туранга,
или тополь разнолистный (Р. diversifolia[Schrenk.), черемуха Маа-
ка (Padus maackii Кот.), чозения крупночешуйчатая (Chosenia
macrolepis Кот.), ясень маньчжурский (Fraxinus mandshurica
Rupr.).
В процессе исследования ксилофильных сообществ на каждой
отдельной древесной породе был определен видовой состав соответ-
ствуюпщх энтомокомплексов, намечены ассоциации индикаторных
видов — ксилофагов и энтомофагов, изучены их пищевые связи
и значение в ксилофильных ассоциациях, особенности развития*
формирование приспособлений к обитанию в древесине, приуро¬
ченность к породам и зонам ствола и другие вопросы. В результате
были охарактеризованы энтомокомплексы, слагающиеся под ко¬
рой и в древесине большинства основных древесных пород в различ¬
ных типах леса — от хвойных лесов таежной зоны до саксауловых
лесов пустынь.
Основное внимание уделялось обработке материалов по личин¬
кам насекомых. Разнообразие и значительная степень новизны этих
материалов позволили внести вклад не только в систематику личи¬
нок, но и значительно расширить представления по эволюционной
морфологии двукрылых и жесткокрылых насекомых, подготовить
обобщения по их экологии и биоценологии. Достаточно отметить»
что в результате проведенных исследований были впервые полу¬
чены данные о личинках 46 видов жесткокрылых, относящихся
к 9 семействам, и 205 видов двукрылых из 31 семейства.
Новые материалы частично были опубликованы в статьях, при¬
веденных’ в списке литературы, частично — в монографиях и те¬
матических сборниках: «Определитель личинок двукрылых насе¬
комых — обитателей древесины» (М., «Наука», 1967), «Роль на¬
секомых в лесных биогеоценозах Приморья» (Владивосток, 1972),
«Энтомологические исследования на Дальнем Востоке» (Ново¬
сибирск, «Наука», 1973), «Насекомые —разруп.ители древесины
в лесных биоценозах Южного Приморья» (М., «Наука», 1974),
«Насекомые как компоненты биогеоценоза саксаулового леса»
(М., «Наука», 1975), «Эволюционная морфология личинок насеко¬
мых» (М., «Наука», 1976).
Весь изученный личиночный материал хранится в коллекции
Института эволюционной морфологии и экологии животных им.
А. Н. Северцова АН СССР. Эта коллекция создана в Лаборатории
экологии сообществ наземных беспозвоночных в результате 18-
летних регулярных экспедиционных работ Б. М. Мамаева, II. П.
Кривошеиной, В. А. Потоцкой. В последние годы значительные ма¬
териалы, в том числе и но энтомофагам, были собраны сотрудни¬
ками лаборатории Е. Б. Антоновой, Т. В. Гусаковой, М. Л. Да¬
нилевским, А. И. Зайцевым, А. В. Комнанцевым, М. А. Кравчен¬
ко, Н. Б. Никитским.
Методика собирания таких материалов опубликована ранее
(Кривошеина, Мамаев, 1967).
Публикуемый определитель включает теоретическую часть,
где на основании новых материалов характеризуются этапы сук¬
цессии беспозвоночных в древесине, освещаются основы экологии
энтомофагов, предлагается подробная классификация их пищевых
режимов, выделяются индикаторные виды, приуроченные к после¬
довательным стадиям разрушения древесины, и т. д. Во второй
специальной части освещаются общие вопросы сравнительной и
эволюционной морфологии личинок, приводятся иллюстрирован¬
ные определительные ааблицы и сведения по элементарной эко¬
логии отдельных видов.
При пользовании систематической частью монографии необхо¬
димо учитывать, что она рассчитана на определение только тех
групп и видов, которые относятся к постоянным компонентам
ксплофильных сообществ. Наиболее подробно в монографии оха¬
рактеризованы семейства ксилофильных насекомых, имеющие в
своем составе активных массовых хищников или зоонекрофагов.
В этом случае в определительных таблицах семейств и родов в кон¬
це тезы с названием таксона дается ссылка на страницу, где при¬
водится его дифференциальный диагноз и сведения по экологии.
Почти все рисунки, включенные в монографию, выполнены ав¬
торами, при использовании иллюстраций из печатных работ дру¬
гих исследователей приводится соответствующая ссылка на источ¬
ник.
В определении имагинальных материалов неоценимую помощь
оказали сотрудники Зоологического института АН СССР и других
научных учреждений: А. А. Штакельберг, О. Л. Крыжановский,
Г. С. Медведев, Л. Н. Медведев, О. П. Негробов. Рисунки выпол¬
нены Н. П. Кривошеиной, В. А. Потоцкой, А. И. Зайцевым, Г. И.
Козловой и Г. Б. Титковой, при оформлении рукописи к печати
помогали Н. В. Крылова и Т. Н. Кондрашова. Приносим самую
глубокую благодарность всем энтомологам, кто своим трудом спо¬
собствовал выходу в свет публикуемой монографии.
Глава 1
ОСНОВЫ ФОРМИРОВАНИЯ СООБЩЕСТВ
КСИЛОФИЛЬНЫХ НАСЕКОМЫХ
Хищные несекомые, обитающие в древесине, тесно связаны с
определенными группами ксилофагов и сапроксилофагов. Поэтому
их всестороннее изучение предполагает предварительное ознаком¬
ление с некоторыми общими основами формирования и сукцессии
ксилофильных энтомокомплексов, которые рассматриваются ниже
применительно к задачам публикуемой монографий.
ОБЩАЯ ХАРАКТЕРИСТИКА ЭТАПОВ СУКЦЕССИИ
КСИЛОФИЛЬНЫХ НАСЕКОМЫХ
В каждом лесном биогеоценозе древесина относится к числу ос¬
новных сред обитания животных, представленных в ней почти ис¬
ключительно беспозвоночными. Как среда обитания она имеет це¬
лый ряд особенностей. В первую очередь следует отметить, что дре¬
весина практически целиком состоит из органических веществ и по¬
этому представляет собой мощный источник энергии. Эти органи¬
ческие вещества, преимущественно клетчатка и лигнин, весьма
устойчивы к разрушению, в связи с чем разлагающиеся стволы
деревьев являются долговременным источником энергии, сущест¬
вующим в качестве конкретного объекта от нескольких лет до нес¬
кольких десятков лет. Это обстоятельство очень важно, так как
в древесине нередко развиваются такие группы насекомых, жиз¬
ненный цикл которых измеряется несколькими годами.
В отличие от миогих других сред обитания лесных биогеоце¬
нозов, которые связаны друг с другом постепенными переходами,,
древесина представлена четко ограниченными монолитами — от¬
дельными стволами, масса которых может достигать нескольких
тонн. Поэтому и сообщество, формирующееся в таких объектах,
пространственно четко отграничено от других сообществ леснога
биогеоценоза.
Одним из основных свойств древесины как среды обитания
является также закономерное изменение во времени таких ее ка¬
честв, как твердость и химический состав. Поскольку эти качест¬
ва определяют ее доступность как источника питания для различ¬
ных групп беспозвоночных, видовой состав зоокомплексов, фор¬
8
мирующихся иа последовательных этапах разложения древесины,
различен. Изучение закономерных изменений видового состава
сообщества ксилофильных беспозвоночных в зависимости от стадии
разрушения древесины представляет собой одну из интереснейших
проблем лесной биогеоценологии.
На протяжении последнего столетия ксилофильные сообщества
привлекали внимание многих исследователей. Крупные работы по
сукцессии ксилофильных беспозвоночных выполнены в Средней и
Северной Европе. Однако только один автор — Шимичек (Sclii-
mitschek, 1952—1953) имел в качестве объекта как пни, так и раз¬
лагающиеся стволы деревьев и мог характеризовать сукцессию
беспозвоночных в древесине в полном объеме, что и было им сде¬
лано на примере бука и пихты. В работах Крогеруса (Krogerus,
1927), Дерксена (Derksen, 1941) и Вянцковского (Wi^Qkowski,
1957) исследовались сообщества, формировавшиеся в пнях соот¬
ветственно пихты, бука и сосны, поэтому, несмотря на больнюй
объем полученных данных, характеристика сукцессии, приведен¬
ная этими авторами, не могла быть всесторонней. Аналогичное ис¬
следование по сукцессии беспозвоночных в пнях сосны выполнено
в Англии Уоллесом (Wallace, 1953). Общие соображения о законо¬
мерностях формирования сообществ в разлагающейся древесине
имеются в монографии Элтона (Elton, 1966), написанной на осно¬
ве наблюдений, также проведенных в Англии.
Кроме этих работ, в которых сукцессия ксилофильных беспоз¬
воночных была основным предметом обсуждения, по европейским
материалам опубликованы крупные фаунистические обзоры дре-
воядных насекомых. В их числе в первую очередь следует назвать
монографии Сааласа (Saalas, 1917, 1923), Палма (Palm, 1951, 1959),
Вите (Vite, 1952) и Гикина (Hickin, 1963). Эти обзоры содержат
важные фактические материалы, привлечение которых помогает
глубже познать экологию основных ксилофильных групп и видов.
Главным образом на основе перечисленных выше работ
Б. М. Мамаевым (1960а, б, в, 1961) были сформулированы общие
закономерности сукцессии беспозвоночных в разлагающейся дре¬
весине, причем впервые был применен зоологический подход к ха¬
рактеристике отдельных этапов сукцессии ксилофильных сооб¬
ществ. Это прежде всего выразилось в том, что стадии разрушения
древесины, определявшиеся ранее по внешним признакам (твер¬
дость древесины, плотность прилегания коры, наличие в древе¬
сине полостей и т. д.), стали характеризоваться в соответствии
с видовым составом населяющих древесину беспозвоночных и полу¬
чили названия, созвучные с латинскими названиями доминирую¬
щих групп. В частности, в первоначальном варианте для коры бы¬
ли выделены ипидная, церамбицидная, формицидно-пирохроидная
и лумбрицидная стадии разложения; для древесины — церам¬
бицидная, луканидная, формицидная и лумбрицидная стадии.
В дальнейшем была разработана также и классификация режимов
питания личинок насекомых-ксилофагов (Мамаев, 1966).
Эти зоологические представления об основных закономерностях
сукцессии’ ксилофильных сообществ, каждое из которых вносит
свой вполне определенный вклад в процессы естественного разру¬
шения древесины, в целом соответствовали материалам, извест¬
ным к тому времени из Северной Америки по работам Блэкмена
и Стейжа (Blackman, Stage, 1918, 1924) и Сейвели (Savely, 1939).
Интересно отметить, что при сравнении палеарктических и не¬
арктических комплексов насекомых прослеживался определен¬
ный параллелизм в формировании ксилофильных сообш,еств в раз¬
личных подобластях Голарктики (Мамаев, 1961).
Тем не менее пока еще весьма немногочисленные данные о сук¬
цессии беспозвоночных в древесине в тропических областях —
Западной Африке (Eidmann, 1943) и Южной Америке (Вескег^
1955), где широко представлены амброзийные ксиломицетофаги и
такие ксилофаги, как термиты, свидетельствовали о необходимо¬
сти уточнения общей схемы сукцессии, составленной в основном
по материалам из умеренной климатической зоны. Эти уточнения
тем более стали очевидными в процессе проведения наших иссле¬
дований в Южном Приморье, где в связи с влажным климатом в
участием в формировании ксилофильных сообществ некоторых
групп насекомых тропического происхождения сам характер сук¬
цессии беспозвоночных в древесине несколько изменился и имеет
сходство с аналогичными процессами, протекающими во влажных
тропиках (Мамаев, 1972 г).
По европейским материалам, первую (церамбицидную) стадик>
разрушения древесины осуществляет сообщество несекомых, со¬
стоящее из представителей семейств короедов (древесинников)^
сверлильщиков, дровосеков, златок, рогохвостов, горбаток и те¬
нелюбов (Мамаев, 19606). Дальнейшее изучение пищевых режимов
ксилофагов (Мамаев, 1966) показало, однако, что в составе этого
сообщества имеются группы, резко отличающиеся по особенно¬
стям питания. Особый интерес в этом отношении представляют так
называемые амброзийные ксиломицетофаги — короеды-древе¬
синники и сверлильщики. Личинки этих насекомых проклады¬
вают ходы в еще свежей древесине, выталкивая из ходов нагры¬
зенные опилки наружу. На стенках образующихся галерей на¬
чинает развиваться мицелий симбиотических грибов, который п
служит основным источником питания личинок. Амброзийные кси¬
ломицетофаги первыми поселяются в мертвой еще свежей древе¬
сине, несколько опережая дровосеков и златок. Уже эти факты да¬
вали основание для выделения особой стадии разрушения древе¬
сины, осуществляемой амброзийными ксиломицетофагами.
В тропических лесах с их влажным климатом эта группа ксило¬
фагов получает широкое развитие. В ее состав, кроме сверлиль¬
щиков и короедов-древесинников, входят многочисленные виды
плоскоходов и слоников. Именно амброзийные ксиломицетофаги в
тропических странах в силу их массовости относятся к числу эко¬
номически наиболее важных вредителей заготовляемой древесины.
40
в 1Ржном Приморье, как и во всей Палеарктике, сверлиль¬
щики й короеды-древесинники обычны в древесине самых различ¬
ных древесных пород на ранних этапах ее разложения. В рассмат¬
риваемый комплекс подключаются такие группы, как слоники
Sipalinus gigas F., а также плоскоходы из рода Crassotarsus, кото¬
рым сопутствуют львинки рода Xylopachygaster. Это — фауни-
стические элементы тропических лесов Южной и Юго-Восточной
Азии, которые в Южном Приморье вместе с транспалеарктически-
ми видами формируют смешанное по своему составу лимексило-
нидное сообщество. Сверлильщики рода Elateroides в тропические
леса не проникают, в Евразии — это представители палеаркти-
ческой фауны, которые в Южном Приморье нередко даже в одном
объекте (пне или стволе) встречаются вместе с Sipalinus gigas F.,
распространенным в тропиках Юго-Восточной Азии с северной
границей ареала в Южном Приморье и на Японских островах.
Таким образом, тот факт, что по крайней мере представители
четырех семейств насекомых (короеды, слоники, плоскоходы, свер¬
лильщики) осуществляют разрушение древесины на ранних эта¬
пах ее разложения — до проникновения в древесину усачей и
златок, свидетельствует о целесообразности подразделения ранее
выделявшейся единой церамбицидной стадии разрушения дре¬
весины на две: первую — лимексилонидную и вторую — церам-
бицидную
Ко времени наступления второй (церамбицидной) стадии разру¬
шения древесины групповой состав развивающихся здесь насе¬
комых резко изменяется: доминирующее положение в комплексах
разрушителей занимают личинки жуков-дровосеков, златок, тене¬
любов, рогохвостов и некоторых других насекомых. Преобладаю¬
щее значение в этих ассоциациях как в европейско-кавказском
и сибирском, так и в дальневосточном районах имеют одни и те же
или близкие виды и группы насекомых, что свидетельствует о срав¬
нительно слабом влиянии фаунистических комплексов Юго-Во¬
сточной Азии на церамбицидный энтомокомплекс Южного При¬
морья.
Изменения, которые наступают в ксилофильных сообществах
на третьей (луканидной) стадии разрушения древесины, в зоогео-
графическом аспекте весьма сложны и интересны.
Виды и группы насекомых, широко распространенные в Па¬
леарктике и формирующие комплекс разрушителей древесины на
луканидной стадии, в первую очередь Sinodendron cylindricum
L., Ceruchus chrysomelinus Hochw., Dorcus parallelopipedus L.,
до Приморья не доходят. Они замещаются в Приморье и Уссурий¬
ском крае дальневосточными видами рогачей — Prismognathus
subaeneus Motsch. и Eurytrachelus rubrofemoratus Woll., которые
в Восточную Сибирь не проникают и по своим родственным свя¬
зям относятся к группам тропического происхождения. Важное
место в комплексе разрушителей древесины на этой стадии как
в Южном Приморье, так и в Юго-Восточной Азии занимают круп¬
11
ные личинки чернотелок трибы Misolampini. На Дальнем Йосто-
ке — это представители родов Misolampidius и Stenophanes^
в Индии — виды близкого рода Setenis. Несомненно, что /это еще
один компонент фауны тропической Азии в дальневосточных кси-
лофильных сообществах.
Особый интерес представляет тот факт, что на Дальнем Востоке
нет рогачей, которые были бы специализированы к обитанию в бу¬
рых гнилях древесины, и по этой причине не возникает сооб¬
ществ, аналогичных таежным европейско-сибирским энтомоком-
плексам, лидером которых является рогач Ceruchus chrysoiuelinus.
Hochw. В Южном Приморье на луканидной стадии разрушения
древесины обитает реликтовый таракан Cryptocercus relictus В.-
Bien. из подсемейства Panesthiinae, ведущий полусоциальный об¬
раз жизни. В тропиках южной Индии тараканы из упомянутого
подсемейства занимают ведущее положение в комплексах разру¬
шителей древесины. Наш дальневосточный вид — несомненный
элемент южноазиатских ксилофильных сообществ, не встречаю¬
щийся ни в Вост. Сибири, ни на Японских островах (Мамаев, 1975).
Изучение особенностей сукцессии беспозвоночных в Южном
Приморье и анализ материалов, собранных во время полевых работ
в тропических лесах южной Индии, привели к необходимости уточ¬
нения последовательности стадий разрушения не только соб¬
ственно древесины, но и коры. Б. М. Мамаевым (1961) третья ста¬
дия разру'шения коры была названа формицидно-пирохропдной.
При этом имелось в виду то обстоятельство, что в случае, если кора
не заселяется муравьями, подкоровая зона становится местом раз¬
вития личинок жуков-плоскотелок. Последующие наблюдения
показали, однако, что разрушение подкоровой зоны личинками
плоскотелок не исключает последующего поселения под корой му¬
равьев. После же муравьев личинки плоскотелок под корой уже
не встречаются. Таким образом, для большей точности следует раз¬
личать самостоятельные стадии разрушения коры: третью — пиро-
хроидную и четвертую — формицидную.
Следовательно, сукцессия беспозвоночных как в коре, так и
в древесине по мере их разрушения осуществляется в пять стадий^
которые для коры получили названия сколитидной, церамбицпд-
ной, пирохроидной, формицидной и лумбрицидной, для древе¬
сины — лимексилонидной, церамбицидной, луканидной, форми¬
цидной и лумбрицидной. Каждая из этих стадий характеризуется
индикаторной ассоциацией видов, которая может существенно от¬
личаться в различных регионах Палеарктики.
НАСЕКОМЫЕ КАК ИНДИКАТОРЫ СТАДИЙ РАЗРУШЕНИЯ
КОРЫ И ДРЕВЕСИНЫ
Вопрос о наиболее рациональных способах диагно?1^ки стадий
естественного разрушения древесины относится к числу важней¬
ших в комплексе исследований по экологии ксилофильных сооб-
12
1деств.\ Подходы к решению этого вопроса могут быть самыми раз¬
личными. Наиболее обычный способ выделения стадий разрушения
таких объектов, как разлагающиеся в естественных условиях пни
или стволы,— использование внешних признаков разрушения.
К числу \зтих признаков относятся цвет и прочность древесины,
плотность\прилегания коры, количество трухи под корой и в дре¬
весине и т1 п. Некоторыми исследователями были сделаны попыт¬
ки привлешть данные об удельной плотности древесины или о ее
химическом составе. Иногда стадии разрушения коры и древеси¬
ны выделялись независимо, некоторые ИчС исследователи предпо¬
читали рассматривать особенности разрушающегося объекта
(ствола, пня) в целом.
Этот круг вопросов, включая соответствующую литературу*
был проанализирован ранее (Мамаев, 19606, 1961), причем в ка¬
честве одного из методов выделения стадий естественного разруше¬
ния древесины был рассмотрен зоологический. Принцип этого ме¬
тода определения стадии разрушения древесины аналогичен тем
подходам, которые используются в индикационной геоботанике»
микологии, почвенной зоологии и т. д. Индикация особенностей
различных сообществ организмов в этом случае ведется по видово¬
му составу этих организмов и их количественным соотношениям.
Таким образом, ксилофильные сообщества зоолог может характе¬
ризовать на основе видового разнообразия беспозвоночных и осо¬
бенностей их сукцессии, т. е. смены видовых комплексов но мере
разрушения древесины. Особое значение при этом имеет выделение
отдельных индикационных таксонов и их сочетаний, что позволит
уточнить особенности процессов естественного разрушения дре¬
весины как в различных регионах (географический аспект), так
и в различных лесорастительных условиях (экологический
аспект).
Было бы неправильным рассматривать зоологический подход
к определению стадии разрушения древесины в качестве универ¬
сального, позволяющего оценить степень разложения каждого кон¬
кретного анализируемого объекта — ствола или пня. Достаточно
сказать, что в некоторых ситуациях беспозвоночные могут и не за¬
селить слишком сухую или, наоборот, переувлажненную древеси¬
ну, они избегают также древесины, перенасыщенной мицелием
грибов, и т. д. Поэтому представление о составе ксилофильных эп-
томокомплексов и их сукцессии в исследуемом регионе склады¬
вается на основе сопоставления результатов анализов многих
конкретных объектов и представляет собой определенную абст¬
ракцию. Особое значение имеет анализ таких объектов, для кото¬
рых характерен кульминационный период развития той или иной
стадии разрушения древесины. В составе именно этих энтомоком-
плексов следует искать виды или более крупные таксономические
группы, имеющие индикационное значение.
Ниже излагаются конкретные материалы, характеризующие
индикаторные видовые ассоциации, слагающиеся на пяти после¬
13
довательных этапах естественного разрушения коры и древесины
(Мамаев, 1974 г). /
Сколитидная стадия разрушения коры. Сколитидный комплекс
беспозвоночных, представленных главным образом насетомыми,
повсеместно хорошо выражен на хвойных породах. На лиотвенных
породах массовое поселение короедов и сопутствующих зидов на¬
блюдается значительно реже; тем не менее следует отметить доста¬
точно стабильное прохождение этой стадии на ильме и дубе в цент¬
ре Европейской части СССР, на грабе и буке на Кавказе. В Юж¬
ном Приморье в составе сколитидного комплекса на лиственных по¬
родах нередко доминируют слоники. Из изложенного следует,
что основными индикаторами сколитидной стадии разрушения дре¬
весины являются все представители семейства короедов, поселяю¬
щиеся под корой или в ее толще, а также подкорные виды слони¬
ков (рис. 1, i, 2).
Из короедов особенно активны представители родов Polygrap-
hus, Hylurgops, Hyla^es, Dryocoetes, Scolytus, Ips, Taphrorychus,
Orthotomicus и др. Слоники на хвойных представлены родами
Hylobius, Pissodes, Nyphades, на лиственных — Magdalis, Tracho-
des, Acicnemis и др. В частности, на Кавказе кору пихты весь¬
ма активно разрушают личинки слоника Pissodes picea 111., кото¬
рые нередко доминируют в сколитидном энтомокомплексе; в Юж¬
ном Приморье под корой пихты многочисленны личинки Nyphades
variegatus Roel., под корой ольхи, бука и граба па Северном Кав¬
казе наряду с короедами обычны личинки различных видов Trach-
odes, в Южном Приморье массовым разрушителем луба ольхи
является слоник Acicnemis pallida Pasc.
За основными разрушителями коры на сколитидной стадии уст¬
ремляются многочисленные сапрофаги, некрофаги и хищники,
которые также характерны для формирующегося здесь энтомо-
комплекса.
Среди хищных насекомых индикаторными родами из жуков-
пестряков является Thanasimus, из двукрылых — зеленушки
рода Medetera. Личинки этих насекомых уничтожают преимущест¬
венно личинок короедов. Характерны для этой стадии также ли¬
чинки и имаго жуков-карапузиков из родов Plegaderus, ^aroma-
lus, Cylister и др. Среди жуков-блестянок к числу обычных спут¬
ников короедов относятся некоторые представители родов Еригаеа
и Rhizophagus. Из узкотелок короедам на ильме сопутствует Aulo-
nium trisulcum Geoffr. В Южном Приморье весьма важным ком¬
понентом сколитидного энтомокомплекса являются личинки щи¬
товидки Trogossita japonica Rtt.
В ряде случаев очень тесно связаны с короедами двукрылые-
копьехвостки. Такой широко распространенный в Палеарктике
вид, как Lonchaea collini Наск., весьма характерен для сколитид¬
ной стадии разрушения коры хвойных деревьев. Важными инди¬
каторами сколитидной стадии разрушения коры являются львинки
рода Zabrachia, особенно вид Z.minutissima Zett., сопутствующий
14
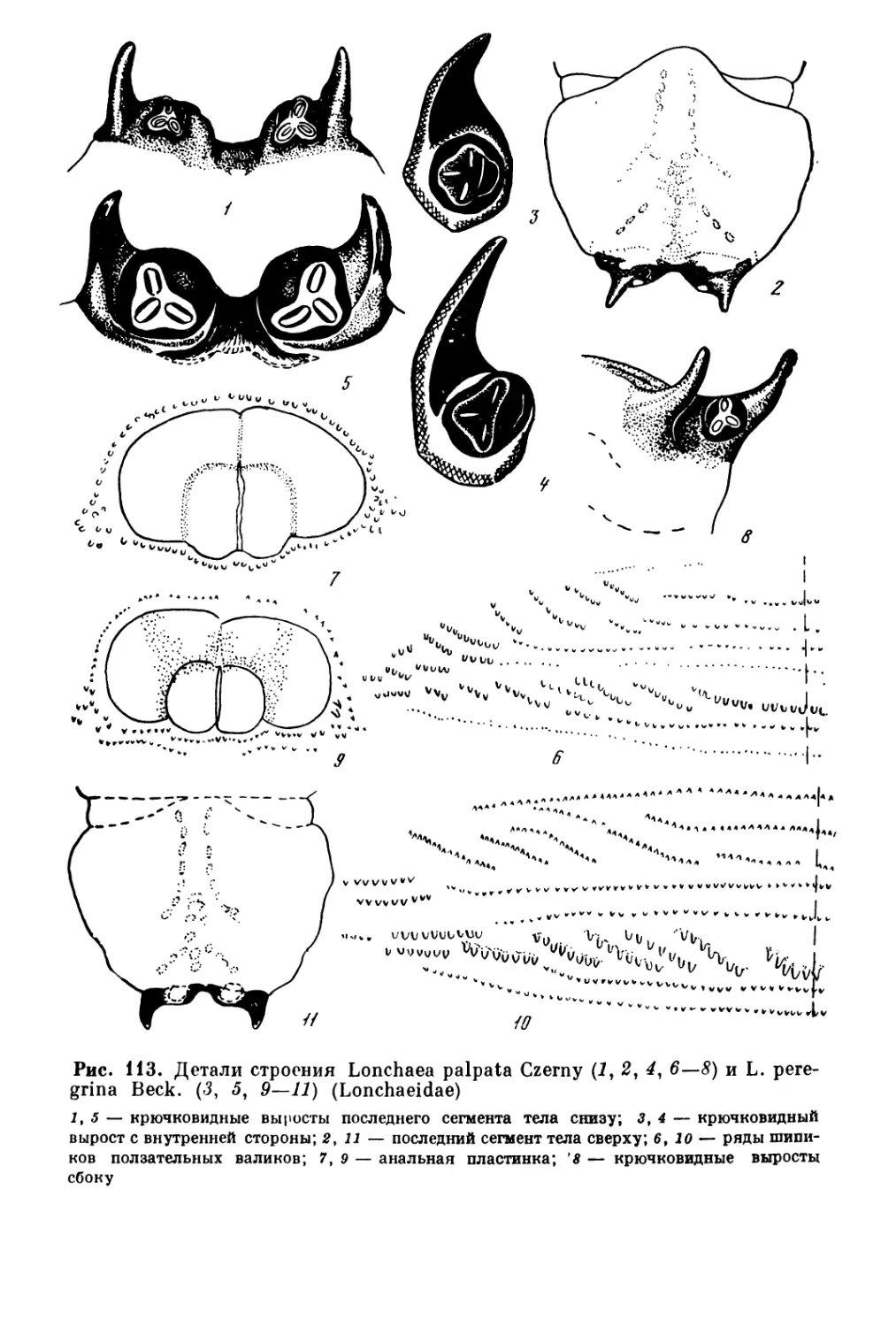
Рис. 1. Личинки жесткокрылых па сколптпдпоп стадии разрушения коры
1 — короед Scolytus multistiiatus Marsch. под корей глl^a; 2 — cj:cim Fif^4c4s piiii L.
под корой пихты кавказской (фото Б. М. Мамаева)
короедам па хвойных породах по всей территории И^еарк-
тики. /
Таким образом, в типичном случае разрушение коры на сколи-
тидной стадии осуществляется личинками короедов и слоников,
которым сопутствуют другие, перечисленные выше видй насеко¬
мых. Когда же по тем или иным причинам перечисленны^выше раз¬
рушители коры оказываются неактивными, следует кон^атировать
выпадение сколитидной стадии разрушения. В этом cjw4ae под ко¬
рой формируется н е п о л н о ч л е н н ы й сколитидный энтомо-
комплекс, который обычно бывает представлен личицками жуков-
блестянок и некоторых двукрылых (рис. 2, 1—2), г^ллиц и неко¬
торых круглошовных мух. В этом варианте процессы разрушения
коры насекомыми по существу начинаются со следующей — це-
рамбицидной стадии разрушения.
Церамбицидная стадия разрушения коры. Как показывает на¬
звание стадии, индикаторной группой церамбицидного энтомоком-
плекса являются жуки-дровосеки. Однако в экологическом отно¬
шении представители этого семейства необычайно разнообразны —
некоторые виды дровосеков способны развиваться в жизнеспособ¬
ных деревьях, некоторые, наоборот, предпочитают сильно разру¬
шенную кору и древесину. Поэтому сам факт обнаружения под ко¬
рой личинок дровосеков еще не свидетельствует о наступлении
церамбицидной стадии ее разрушения, необходимо уточнить систе¬
матическое положение видов и характер слагающегося под корой
сообщества.
В пределах Палеарктики можно назвать несколько широко
распространенных родов дровосеков, характерных именно для
церамбицидной стадии разрушения коры (рис. 3, 2). К их числу
относится прежде всего род Rhagium, а также роды Mesosa, Р1а-
gionotus, Pyrrhidium, Morimus, Xylotrechus и некоторые другие.
Личинки дровосеков из родов Monochamus и Acanthocinus харак¬
терны для наиболее ранних этапов прохождения стадии, а виды
рода Tetropium нередко поселяются в коре одновременно с ко¬
роедами.
Второй группой, индикаторной для церамбицидной стадии раз¬
рушения коры, являются златки. В этом отношении особое поло¬
жение занимает род Chrysobothris, в первую очередь вид Ch. affi¬
nis F. (рис. 3, 2) — широко распространенный полифаг. Сходная
экология характерна для златок рода Lampra и некоторых видов
Agrilus, которые также составляют основу церамбицидного энто-
мокомплекса подкорных насекомых. На хвойных, в частности на
лиственнице, высокой численности достигают личинки златки Ме-
lanophila acuminata Deg. Из сопутствующих хищных двукрылых
характерны личинки подкорных видов рода Xylophagus, которые,
однако, могут встречаться также на сколитидной стадии разру¬
шения коры, а также ктырей из родов Pogonosoma и Choerades.
Сходные экологические связи характерны для пестряка Clerus
mutillarius F. и для щелкунов из рода Stenagostiis и Denticollis,
10*
Рис. 2. Подкорные личинки двукрылых
1 — ксиломиида Xylornya czekanovskii Pl. под корой чозснии; 2 — стволосдка Ху1о-
p‘iagus sp. (слева) и копьехвостки Lonchaca sp. под корой липы (фото Б. М. Мамаева)
Рис. 3. Лпчппкм vic(‘CTKOKi)bi;ibix на ио1)амГтцидпо11 стадии разрушения коры
1 — дрово(Ч‘Ь' Hha^i[iiiin inquisitor L. под норой пихты ьавиа.чской; 2 — златка Chiysoboth-
ris affinis l\ под Kopoiijmu.i (фото Г.. ЛЦМамасна)
лйчинкл которых обычны под корой на церамбицидной стадии ее
разложе^я.
Таким юбразом, церамбицидная стадия разрушения коры лег¬
ко опреде^ется по наличию личинок соответствующих дровосе¬
ков и злато^. В случае отсутствия этих личинок характеризуемая
стадия може^ выпадать.
Пирохрои^ная стадия разрушения коры. Соответствующий эн-
томокомплекс ^формируется под корой тогда, когда она несколько
отслаивается. В его состав входят насекомые, получившие назва¬
ние «плоскотелок», поскольку обитание в щелевидных ходах и по¬
лостях отразилось на их внешнем облике (рис. 4, 1, 2). Одно из
индикаторных семейств жесткокрылых — плоскотелки (Cucuji-
dae) сохранило это название. Его наиболее характерный род —
Cucujus представлен в подкорных сообществах повсеместно в Па-
леарктике главным образом видом Cucujus haematodes Er.— весь¬
ма важным индикатором пирохроидной стадии разрушения коры.
Одновременно под корой в Европейской части СССР и на Кавказе
обычны личинки огнецветок Pyrochroa coccinea L. и Schizotus
pectinicornis L., а в Южном Приморье, особенно же на о. Куна-
шир — виды из рода Pseudopyrochroa. Широко в Палеарктике
распространен вид Boros schneideri Pz., личинки которого встре¬
чаются под корой как лиственных, так и хвойных пород
(рис. 4, 2).
Следующие две стадии разрушения — формицидная и лумбри-
цидная будут охарактеризованы ниже при рассмотрении энтомо-
комплексов, формирующихся в толихе древесины, так как для
этих стадий характерна общность видового состава беспозвоночных,
заселяющих кору и древесину.
Лимексилонидная стадия разрушения древесины. Эта стадия
особенно характерна для тропических стран, где находятся цент¬
ры видового разнообразия разрушителей древесины из группы амб-
розийных ксиломицетофагов. В Палеарктике она выражена в наи¬
большей степени во влажных лесах Южного Приморья.
Индикаторами лимексилонидной стадии разрушения древеси¬
ны повсеместно в Палеарктике служат два вида сверлильщиков —
Elateroides dermestoides L. на лиственных породах и Е. flabelli-
cornis Schn.— на хвойных. Эти виды образуют массовые поселе¬
ния в свежеотмерших стволах и пнях (рис. 5, 1). Их личинкам не¬
редко сопутствуют виды болотниц из рода Libnotes. В этот же ли-
мексилонидный комплекс видов входят короеды-древесинники
из родов Xyleborus, Trypodendron, Anisandrus, которые нередко
встречаются совместно со сверлильщиками. На Кавказе в дубо¬
вых пнях нередок Platypus cylindrus Hbst. Еще более обогащается
лимексилонидный комплекс на Дальнем Востоке за счет включения
в его состав некоторых групп жуков тропического происхождения,
например слоника Sipalinus gigas F. (рис. 5, 2), плоскохода Crasso-
tarsus sp. и двукрылых — львинок Xylopachygaster mamaevi
Kriv., а также некоторых других насекомых.
19
Piic. 4. Личинки жесткокрылых па nupoxpoiiAHoii стадии разрушения коры
J — трухляк Pytho depressus L. пор корой сосны обыкновенной; 2 — лжечсрнотелка
Boros schneideri Pz. под корой ясеня (фото Б. М. Мамаева)
Рис. 5. Личинки насекомых па лимексилопидиой стадии разрушения древе¬
сины
1 — сверлильщик Elateroides dermcstoidcs L. и болотница Libnotes sp. (в центре) в древе¬
сине липы; 2 — слоник Sipalinus gigas F. в древесине кедра корейского (фото Б. М. Ма¬
маева)
в целом лимексилонидный комплекс часто выпадает, й разру¬
шение древесины начинается с церамбицидной стадии. ^
Церамбицидная стадия разрушения древесины. Личинки жу-
ков-дровосеков представляют собой наиболее характерную со¬
ставную часть ксилофильных сообществ и чрезвычайно мощный
фактор разрушения древесины.
Древесину хвойных пород в Европейской части СССР разру¬
шают личинки Toxotus cursor L., Spondylis buprestoides L., Crio-
cephalus rusticus L.; na Кавказе к ним присоединяются Ergates
faber L. и Hylotrupes bajulus L.; в Закарпатье — Tragosoma dep-
sarium L. В Южном Приморье наиболее активны такие развиваю¬
щиеся хвойных породах виды дровосеков, как Megasemum
quadricostulatum Кг., Leptura variicornis Dalm., Sachalinobia
koltzei lleyd. и представители рода Asemum. Для лиственных по¬
род индикаторными видами могут служить представители родов
Strangaila и Leptura, особенно вид S. quadrifasciata L., на Кавказе
высокой активностью отличаются также личинки Megopis scabri-
cornis Scop. и Toxotus mirabilis Motscli., в Южном Приморье —
Callipogon relictus Sem., Strangalia thoracica Cr. и S. arcuata Pz.
(рис. 6, i). Все эти роды и виды могут рассматриваться в качестве
наиболее типичных для церамбицидного энтомокомплекса, фор¬
мирующегося в древесине.
Среди златок, которые также входят в упомянутый энтомо-
комплекс (рис. 6, 2), следует специально отметить крупный вид
Buprestis mariana L.— разрушителя древесины сосны, а также
представителей родов Dicerca, Eurythyrea и Ancylocheira. Весь¬
ма активен, например, вид А. araratica Mars, в иазрушении дре¬
весины пихты в условиях Северного Кавказа. ^
Семейство тенелюбов в полном составе также должно быть приз¬
нано индикаторным для церамбицидного энтомокомплекса разру¬
шителей древесины, хотя следует подчеркнуть их приуроченность
к более поздним этапам этого комплекса. В Европейской части
СССР и на Кавказе особенно активны виды рода Melandrya,
а также родов Orchesia, Hypulus и Dircaea, развивающиеся в дре¬
весине, прочность которой уже незначительна. В Южном Приморье,
наряду с видами Melandrya, заметную роль в ксилофильных энто-
мокомплексах играют Phloetrya bellicosa Lew. и Enchodes orienta¬
lis Nik. Такой вид тенелюбов, как Serropalpus barbatus Schall.,
относится к числу важнейших разрушителей твердой древесины
хвойных пород, он распространен во всей Палеарктике. Из дру¬
гих разрушителей древесины, достигающих высокой численности
и широко распространенных в лесах Палеарктики, необходимо от¬
метить таких узконадкрылок, как Ditylus laevis F. и Calopus ser-
raticornis L., которых в отличие от других представителей этого
-семейства следует считать и^^дикаторными для церамбицидной ста¬
дии разрушения древесины.
На церамбицидной стадии в комплекс разрушителей древе¬
сины входят все рогохвосты — представители отряда перепончато-
22
Рис. 6. Личинки жесткокрылых на церамбпцпдноп стадии разрушения дре¬
весины
1 — дровосек Pyrrhidium sanguineum L. в древесине дуба (на окукливании); 2 — златка*
Eurythyrea oxiana Sem. в древесине туранги (фото Б. М. Мамаева)
з^рылых (рис. 8, 2), Из них древесину хвойных пород заселяют
представители родов Urocerus, Sirex, Paururus, лиственные породы
предпочитаются видами Tremex и особенно Xiphydria.
Из хищных насекомых с дровосеками и златками тесно связаны
ктыри из родов Laphria и Choerades, личинки которых нередко
достигают высокой численности, в частности на хвойных поро¬
дах.
Луканидная стадия разрушения древесины. Основу луканид-
ного комплекса видов составляют представители двух родствен¬
ных семейств жесткокрылых — рогачей (рис. 7, 1) и пластинчато¬
усых жуков. Что касается рогачей, то эта тропическая группа на¬
секомых сравнительно бедно представлена на материковой части
Палеарктики. Тщ не менее почти все виды рогачей лесной зоны
нашли благоприятные условия для своего развития и, как прави¬
ло, достигают здесь высокой численности.
Виды рогачей подразделяются па две группы — разрушителей
светлых и разрушителей бурых гнилей древесины. В Европейской
части СССР светлые гнили древесины активно разрушают в основ¬
ном два вида рогачей — Sinodendron cylindricum L. и Dorcus ра-
rallelopipedus L. Из них первый вид имеет обширный ареал, про¬
двигается далеко на восток Палеарктики и активен в северных ши¬
ротах. Второй вид широко распространен на Карпатах и Кавказе,
однако на север не выходит из зоны широколиственных лесов.
Бурые гнили древесины в таежной зоне разрушает рогач Ceruchus
chrysomelinus Hochw. На Кавказе его замещает другой вид того
же семейства — Aesalus ulanowskyi Gglb. На Дальнем Востоке
рогачи участвуют только в разрушении светлых гнилей древеси¬
ны. Особенно активен в этом регионе рогач Prismognathus subaene-
us Motsch. Все упомянутые виды рогачей имеют первостепенное
индикационное значение. Им сопутствуют некоторые виды пла¬
стинчатоусых жуков. В светлых гнилях развиваются, например,
виды родов Trichius, Lasiotrichius и почти все представители рода
Gnorimus. Из изученных нами видов только G. bartelsi Fald. свя¬
зан в своем развитии с бурыми гнилями древесины. В состав групп
насекомых, типичных для лукапидной стадии разрушения древе¬
сины, следует включить ксилофильных чернотелок. К их числу
относятся достигающий высокой численности на Кавказе Helops
coeruleus 1.. и обычный в Южном Приморье Misolampidius tenty-
rioides Sols. К луканидному энтомокомплексу принадлежат почти
все виды щелкунов рода Ampedus, а также плоскотелки рода Pro¬
stomis, развивающиеся в бурых гнилях древесины на Кавказе
и в Южном Приморье.
На Дальнем Востоке, где рогачи не принимают участия в раз¬
рушении бурых гнилей древесины, наступление луканидной
стадии отмечается после поселения в древесине таракана Crypto-
cercus relictus B.-Bien., плоскотелки Prostomis mordax Rtt., a так¬
же жуков Istrisia rufobrunnea Lew. и Nematoplus semenovi Nik.
Все эти виды могут рассматриваться как индикаторы начала
.24
Рис. 7. Личинки насекомых па луканидпоп стадии ралрупюпия дрсвсспны
I _ рогач Prismognathus subacncus Motsch. в дрсвесине1орсха маньчжурского: 2 —
долгоножка Phoroctenia vittata Mg. в древесине ильма долинного (фото Б. М. Мамаева)
периода формирования луканидного энтомокомплекса разрушителей
бурых гнилей древесины.
Двукрылые насекомые сопутствуют рогачам главным образом
в энтомокомплексах разрушителей светлых гнилей древесины.
К их числу относятся в первую очередь личинки комаров-долго-
ножек (рис. 7, 2) из родов Phoroctenia, Dictenidia, Tanyptera и
болотниц из рода Epiphragma. Большинство видов ксилофильных
галлиц также развивается на луканидной стадии разрушения
древесины. То же следует отметить и в отношении мух-пятнокры-
лок из родов Clusia, Paraclusia и Clusiodes. На Дальнем Востоке в
характеризуемое сообщество активно включается Hesperinus
rohdendorfi Kriv. et Мат., личинки которого встречаются повсе¬
местно. С бурыми гнилями древесины связаны главным образом
комарики из семейства детритниц, принадлежащие к родам Tricho-
sia, Caenosciara, Scythropochroa и Epidapus.
Лукани;^кая стадия разрушения древесины так же, как и це-
рамбицидная, относится к числу основных.
Формицидная стадия разрушения древесины. Если рассмотрен¬
ные выше энтомокомплексы, формирующиеся в коре или древеси¬
не, имели различный видовой состав, то при поселении в стволе или
пне муравьев такие различия утрачиваются — муравьи колонизи¬
руют объект в целом и, как правило, лишают возможностей раз¬
вития все другие группы насекомых. Гнездо муравьев в древесине
конструируется как на основе уже имеющихЛ в древесине ходов,
проложенных личинками насекомых, так и в результате дополни¬
тельного механического разрушения древесины самими муравьями.
Муравьи могут заселять кору и древесину на всех стадиях ее
разложения, однако наибольшую активность как разрушители
субстрата проявляют представители родов Lasius, Camponotus,
Crematogaster, которые Могут служить индикаторными группами.
В затененных сырых участках леса формицидная стадия обычно
выпадает, так как муравьи в этих условиях представлены в основ¬
ном видами рода Myrmica, которые обычно сооружают в древе¬
сине небольшие локальные гнезда.
Лумбрицидная стадия разрушения древесины. Начало этой ста¬
дии отмечается по значительному появлению в древесине дождевых
червей, энхитреид, кивсяков, мокриц, моллюсков, т. е. групп,
двойственных главным образом почве и лесной подстилке. В этот
же период в древесине возрастает численность коллембол и кле-
щей-орибатид, древесина превращается в составную часть почвы
д сукцессия беспозвоночных в ней завершается.
ХИЩНЫЕ НАСЕКОМЫЕ
КАК КОМПОНЕНТЫ КСИЛОФИЛЬНЫХ СООБЩЕСТВ
Изучение значения хищных насекомых в ксилофильных сооб¬
ществах имеет не только научную, но и практическую ценность.
Хотя в большинстве опубликованных работ отсутствует строгая
26
математическая оценка эффективности влияния ксилофильных
хищников на вредные энтомокомнлексы, многие авторы считают^
что деятельность энтомофагов может в ряде случаев оказать ре¬
шающее воздействие на процессы ограничения численности ство¬
ловых вредителей (Schimitschek, 1929; Куренцов, 1941; Шиперо-
вич, 1954; Зиновьев, 1957; Morge, 1967а, и др.)« Это мнение заслу¬
живает самого внимательного изучения и конкретизации.
Экологические особенности хищных ксилофильных насекомых.
Хищные ксилофильные насекомые обладают целым рядом общих
экологических параметров. Прежде всего следует отметить, что
почти все они отличаются широтой трофических связей, так как
один и тот же вид хищника может питаться несколькими видами
вредителей, зачастую относящихся к разным семействам. Так^
например, Н. 3. Харитонова (1972) отмечает, что жертвами му-
равьежука Thanasimus formicarius L. являются более 25 видов
насекомых. Чернотелка Hypophloeus fraxini Kug. нападает на
6 видов короедов и 2 вида слоников. Карапузик Cylister oblongum
F. уничтожает по крайней мере 6 видов короедов и 2 вида слони¬
ков, а стафилиниды рода Placusa — 18 видов ксилофагов. Из дан¬
ных Н. Г. Коломийцаи Д. А. Богдановой (1973), по лесам Западной
Сибири, где ими были исследованы хищники и паразиты 15 видов
короедов, следует, что более 28 видов хищных жуков нападали в
среднем на 3—5 видов других насекомых, иногда же в их рационе
насчитывалось по 7—10 видов. В тех же условиях из 9 видов хищ¬
ных мух полифагия была отмечена для 7 видов. Эта закономерность
прослеживается не только тогда, когда хищники питаются корое¬
дами, но и на примере других "групп стволовых вредителей, один
и тот же вид хищника, как правило, нападает на несколько видов
дровосеков, златок, слоников и т. д. Эти данные подтверждаются
материалами работ многих авторов (Померанцев, 1902 а, б, в;
1903а, б, в; 1904; Saalas, 1917; Гусев, 1928; Буковский, 1940; Ни-
китюк, 1951; Nuorteva, 1956; Зиновьев, 1957; Земкова, 1965; Тара¬
сова, 1965; Плешанов, 1966; Кривошепна, Мамаев, 1972а; Козак,
1972; Коломиец, Богданова, 1973, и др.). .
Несмотря на то, что полифагия широко распространена среди
хищных ксилофильных насекомых, следует учитывать и то об¬
стоятельство, что в природе существуют экологические факторы,,
ограничивающие ее реализацию и нередко превращающие потен¬
циальных полифагов в олпго- и даже монофагов. К числу таких
факторов относится преяде всего местообитание хищника. Так,
личинки двукрылых, относящиеся к роду Xylopliagus Meig.,
могут питаться многими насекомыми, но круг жертв у разных ви¬
дов ограничивается микростацией хищника. Для развивающихся
под корой личинок X. cinctus Deg. и X. compeditus Wied. — это
личинки и куколки насекомых нодкорового комплекса, в то вре¬
мя как для живущих в толще древесины X. ater Meig. пищей^
естественно, служат личинки и куколки насекомых, обитающих
в толще древесины. Другим фактором, ограничивающим полифа-
27
ГИЮ, являются продолжительность и сроки развития насекомых.
Так, несмотря на то, что личинки рода Medetera способны питать¬
ся личинками и куколками многих видов короедов и слоников,
в природе круг их жертв ограничивается точной синхронизацией
сроков развития как хищника, так и жертвы.
К числу экологических особенностей, характерных для хищ¬
ных ксилофильных насекомых в целом, относится их повышенная
чувствительность к неблагоприятным факторам среды по сравнению
€ ксилофагами. По данным В. М. Яновского (1968), при переходе
от среднегорья к неблагоприятным условиям высокогорья числен¬
ность хищных насекомых резко падает, причем численность хищ¬
ников снижается значительно интенсивнее, чем плотность популя-
дии короедов.
Таким образом, важное значение при формировании комплек¬
сов энтомофагов имеет не столько их пищевая специализация,
сколько многие другие особенности экологии хищника и жертвы.
IlpiiiiepoM этому может служить формирование комплексов энто¬
мофагов короедов, златок и дровосеков. Многие авторы отмечали
тот факт, что в регуляции численности короедов основная роль
принадлежит хищным насекомым, та%-как паразиты в тех же ус¬
ловиях достигают достаточно высокой численности значительно
реже. В регуляции численности дровосеков и златок, напротив,
паразиты имеют большее значение, чем хищники. Это явление
объясняется тем, что яйца, личинки и куколки короедов распо¬
лагаются в общем гнезде, в которое хищники свободно проникают
ло единой системе ходов. Личинки же дровосеков и златок разви¬
ваются разобщенно и поэтому становятся для хищников более труд¬
нодоступными.
Ксилофаги, обладающие сходной экологией и сходным жизнен¬
ным циклом, имеют и близкие по составу комплексы энтомофагов,
что подтверждается, в частности, на короедах или на примере ли¬
ственничной златки и тонкоусого елового дровосека (Паньшин,
1964; Исаев, Яновский, 1966; Харитонова, 1972; Коломиец, Бог¬
данова, 1973, и др.). .
Как уже было показано выше, древесина в процессе разложения
проходит несколько стадий, каждая из которых характеризуется
определенным комплексом ксилофильных насекомых. Поскольку
начальные стадии разрушения древесины привлекали основное
внимание специалистов, существующие энтомологические публи¬
кации отечественных и зарубежных авторов посвящены комплек¬
сам, формирующимся на сколитидной стадии разрушения коры,
значительно слабее изучена церамбицидная стадия разрушения
коры и древесины; что же касается других стадий, то о них в ли¬
тературе имеются лишь отрывочные сведения.
Сколитидный комплекс хищных ксилофильных насекомых. Ли¬
тература, посвященная хищникам короедов, исключительно об¬
ширна и дать исчерпывающий обзор ее в данной работе не пред¬
ставляется возможным. Число фундаментальных обзорных работ,
28
в которых можно найти данные о хищных ксилофильных насеко¬
мых сколитидного комплекса, невелико. Из иностранных публи¬
каций подобного рода следует прежде всего отметить двутомную
монографию Сааласа (Saalas, 1917, 1923), посвященную ксило-
фильным насекомым — обитателям ели, монографию Эшериха
{Escherich, 1923), публикацию Нуортева (Nuorteva, 1956), содер¬
жащую обзор хищных насекомых, обитающих в ходах Hylurgops
palliatus Gyll. на ели и сосне, и монографию Палма (Palm, 1951,
1959), в которой имеются общие характеристики насекомых, оби¬
тающих на различных лиственных породах деревьев в Швеции. Из
специальных источников следует упомянуть монографию Н. 3. Ха¬
ритоновой (1972), посвященную энтомофагам короедов хвойных
пород, и А. А. Гирица (1975), где дана характеристика видов, со¬
путствующих короеду-типографу. Последние работы важны в том
отношении, что в них приведена обширная библиография.
Прежде чем рассматривать конкретные материалы о значении
отдельных групп хищников в сколитидном сообществе, необходимо
подчеркнуть нестабильность слагающегося здесь энтомокомплекса,
подверженного быстрым сукцессионным изменениям. Качествен¬
ный и количественный состав его компонентов меняется в зависимо¬
сти от длительности развития короедов, а также от тех изменений
среды обитания, которые вызывают сами обитатели. В этом плане
прежде всего следует упомянуть статью А. И. Куренцова (1936),
автор различал па Дальнем Востоке шесть этапов развития ско¬
литидного энтомокомплекса. На первом этапе происходит лёт
короедов и вбуравливание их по^ кору. Хищные насекомые в этот
период уничтожают имаго короедов на коре заселяемых стволов.
На втором этапе происходит вселение короедов под кору. Построй¬
ка маточных ходов закончена, идет откладка яиц и появляются ли¬
чинки короедов первого возраста. Для этого этапа характерно на¬
личие в ходах короедов хищников Librodor christophi Rtt., Ну-
pophloeus fraxini Kug., мелких стафилинид (очевидно представи¬
телей рода Placusa) и Lyctocoris sp., реже встречается Platysoma
deplanatum Gyll. Для третьего этапа — фазы расцвета характер¬
но развитие личиночной стадии короедов и их окукливание. Коли¬
чество хищных насекомых по сравнению с предыдущей стадией
увеличивается, к имевшимся видам прибавляются новые — кара¬
пузики Cylister lineare Er., Paromalus parallelepipedus Hbst.,
Hypophloeus linearis F., a также различные стафилипиды, питаю¬
щиеся в основном личинками короедов, появляются насекомые,
питающиеся всякого рода остатками. На четвертом этапе появляют¬
ся взрослые короеды нового поколения, а личинки дровосеков и
златок уходят в древесину. Число видов хищных насекомых
уменьшается: становятся редкими или совсем исчезают Platysoma
deplanatum Gyll., Hypophloeus fraxini Kug., Librodor christophi
Rtt. Луб разрушается, появляются плесневые грибки и связанные
с ними клопы, стафилипиды и личинки других насекомых. На пя¬
том этапе из ходов вылетает больше половины короедов молодого
29
поколения, остальные, продолжая дополнительное питание, ухо¬
дят в заболонь. Все хищники короедов, за исключением Рагота-
lus parallelepipedus Hbst., оставляют ходы. Остаются личинки
жуков-щелкунов и жуки, связанные с плесенью. Шестой этап —
отпадение коры. Сколитидный комплекс прекращает свое сущест¬
вование.
В. М. Яновский (1968) для ксилофильного сообщества, связан¬
ного с короедом Ips subelongatus Motsch. в лиственничниках Ту¬
винской АССР, дает несколько иную схему его динамики и выде¬
ляет три основных этапа. На первом этапе в период откладки яиц
короедом в ходах последнего преобладают стафилиниды — фа¬
культативные хищники, уничтожающие яйца. Все виды паразитов
на этом этапе отсутствуют. Второй этап характеризуется появле¬
нием личинок короедов младших и средних возрастов. В это вре¬
мя плотность энтомофагов снижается за счет уменьшения числен¬
ности стафилинид, но одновременно увеличивается участие таких
эффективных хищников, как личинки жуков рода Thanasimus,
Hypophloeus, Cylister, личинки клопов рода Scoloposcelis, и т. д.
В этот же период появляются паразиты. На третьем этапе личинки
Ips subelongatus Motsch. средних и старших возрастов, куколки
и молодые жуки подвергаются ^иболее активному воздействию
хищников и паразитов, число которых на этом этапе резко возра¬
стает.
Публикации о хищных насекомых — врагах короедов, в основ¬
ном представляют собой списки видов, найденных в ходах различ¬
ных короедов, и сообщения об элементарной биологии отдельных
видов с предположениями об их возможной эффективности как
энтомофагов (Perris, 1863; Померанцев, 1902а, б, в; 1903а, б, в;
1904; Kleine, 1908, 1909а, Ь; 1944а,Ь; Гусев, 1928; Буковский,
1940; Никитюк, 1951, 1957; Зиновьев, 1957, 1959; Земкова, 1965;
Тарасова, 1965; Beaver, 1966Ь, 1967; Плешанов, 1966; Юрченко,
1967; Козак, 1972, 1974; Коломиец, Богданова, 1973; Богданова,
Коломиец, 1974; Арефин, 1974а, и мп. др.). Значительно меньшее
число работ посвящено динамике сколитидиого комплекса в целом
(Куренцов, 1936; Исаев, Яновский, 1966; Японский, 1968).
В настоящее время в фаунд СССР в ходах короедов, согласно
литературным данным, отмечено около 160 видов жуков, около
30 видов двукрылых и несколько видов верблюдок н полужестко¬
крылых. Следует, однако, подчеркнуть, что далеко не все насе¬
комые, встречающиеся в ходах короедов, являются облигатными
хищниками. Многие нз них питаются различными органическими
остатками или являются факультативными хищниками.
Недостаточная изученность питания насекомых, встречаю¬
щихся в ходах короедов, приводит к тому, что нередко один и тот
же вид характеризуется различными авторами то как хищник,
то как сапрофаг. Так, часто встречающийся в ходах короедов има¬
го Silvanus bidentatus F., по данным В. И. Гусева (1928) и А. С.
Плешанова (1966), является хищником, а по данным Н. Б. Никит-
30
€Кого (1971) — сапрофагом. Имаго Bitoma crenata F. по одним
сведениям — хищник, уничтожающий яйца, личинок и неокрашен¬
ных жуков короедов (Saalas, 1917; Mokrzecki, 1933; Плешанов,
1966; Филиппенкова, 1970), а по другим — факультативный хищ¬
ник (Погориляк, 1963), факультативный мицетофаг (Никитский,
1971) или некрофаг (Гусев, 1928). Личинок Pytho depressus L.
одни авторы считают сапрофагами (Saalas, 1923; Криволуцкая,
1965), а другие — хищниками, уничтожающими личинок, куколок
и молодых жуков ребристого рагия и серого длинноусого усача
(Гусев, 1928; Машнина, 1960; Богданова, Коломиец, 1974), а Н. 3.
Харитонова (1972) считает личинок этого вида наряду с личинками
мух наиболее массовыми и эффективными хищниками, уничтожаю¬
щими куколок и молодых жуков короедов. Личинок Zabrachia
minutissima Zett. Г. О. Криволуцкая (1965) относит к хищникам,
Д. А. Тарасова (1968) и II. 3. Харитонова (1972) считают их фа¬
культативными зоофагами, а Г. А. Зиновьев (1958) — мусорщи¬
ками.
Современные авторы обычно делят энтомофагов, связанных с
короедами, в том числе и хищников, на три группы в зависимости
от их хозяйственного значения (Буковский, 1940; Харитонова,
1972; Коломиец, Богданова, 1973; Богданова, Коломиец, 1974).
К первой группе относят наиболее массовых и эффективных энто¬
мофагов, образующих популяции высокой плотности и являю¬
щихся, по мнению указанных авторов, возможными высокоэф¬
фективными факторами биологического контроля вредителей.
Виды, входящие в эту группу, иногда называют «хозяйственно¬
ценными». Ко второй группе относят виды, которые обычно не
являются массовыми и не образуют популяций высокой плотно¬
сти, но иногда, при еще недостаточно выясненных условиях, могут
существенно влиять на численность стволовых вредителей.
Н. 3. Харитонова (1972) называет эту группу группой второстепен¬
ного значения. Наконец, все остальные энтомофаги составляют
третью группу. Значение их в истреблении короедов несуществен¬
но. Совершенно недостаточное внимание пока еще привлекает
проблема использования не единичных видов, а всего комплекса
энтомофагов, куда могли бы войти не конкурирующие друг с дру¬
гом представители, независимо от их принадлежности к одной
из трех групп.
Состав группы «хозяйственно-ценных» энтомофагов у разных
авторов довольно существенно отличается. Так, Н. 3. Харитонова
(1972) включает в эту группу 7 видов хищников, Н. Г. Коломиеп,
и Д. А. Богданова (1973) — также 7, В. С. Арефин (1974а) —
4 вида, В. Т. Козак (1972) —14. Обобщая накопленные материалы
на данном этапе изучения энтомофагов, можно выделить группу
наиболее эффективных и широко распространенных хищных на¬
секомых, истребляющих короедов. К ним относятся такие жестко¬
крылые, как представители рода Thanasimus, ряд видов Нуро-
phloeus, все связанные с короедами виды рода Cylister, а также
31
Rhizophagus dispar Pk., Nudobius lentus Grav., некоторые виды ро¬
да Placusa, а из двукрылых насекомых — личинки представите¬
лей рода Medetera и некоторые виды Lonchaea.
Некоторые авторы (Зиновьев, 1957; Никитский, 1971) делят на¬
секомых, истребляющих короедов, на три группы по особенностям
питания: яйцеедов, хищников, нападающих на личинок и куко¬
лок или уничтожающих взрослых короедов. Такое деление являет¬
ся в значительной мере условным, так как обычно хищники (как
имаго, так и личинки) питаются не одной, а несколькими фазами
развития жертвы, отдавая предпочтение одной из них.
Значение хищных ксилофильных насекомых в снижении чис¬
ленности ксилофагов сколитидного комплекса изучено еще край¬
не недостаточно. Несмотря на то, что эта группа энтомофагов при¬
влекла в последнее время пристальное внимание исследователей
разных стран, конкретные сведения о влиянии хищных насекомых
на численность ксилофагов весьма невелики.
Некоторые представления об эффективности отдельных видов
ксилофильных хи'цников были получены экспериментально.
Г. А. Зиновьев (1957, 1959) и А. И. Никитюк (1951) определяли в
лабораторных условиях прожорливость ^ некоторых видов дву¬
крылых и жуков. По данным %А. Зиновьева (1957), средняя про¬
жорливость личинок рода Medetera была стабильной и для Mede¬
tera pinicola Kow. и М. signaticornis Loew составляла 0,4 (в ка¬
честве корма были использованы личинки и куколки гравера и
полиграфа). У личинок рода Lonchaea на таком же корме прожор¬
ливость сильно варьировала в зависимости от вида и колебалась
от 0,2 до 0,54 (Зиновьев, 1957). При выкармливании личинками
короеда Polygraphus subopacus Thoms. прожорливость личинок
чернотелки Hypophloeus linearis F. была равна 0,5, личинок Н. fra¬
xini Kug.— 0,8, а взрослых личинок Thanasimus formicarius L.—
1,6 (Зиновьев, 1959). В. С. Арефин (19746) установил, что в лабо¬
раторных условиях среднесуточная прожорливость личинок Tha¬
nasimus substriatus Gebl. равна 0,94—1,28 (при кормлении личин¬
ками и куколками Ips sexdentatus Boern.).
По данным А. И. Никитюка (1951), прожорливость имаго чер¬
нотелки Hypophloeus fraxini Kug. при питании яйцами ко-
роеда-типографа колебалась от 0,2 до 8,7, а при питании яйцами
короеда-халькографа — от 5,8 до 9,0. Для Cylister oblongum F.
эти цифры составили соответственно 9,0—13 при выкармливании
яйцами типографа и 10,0—12,0 — при использовании яиц халь-
кографа. Следует, однако, иметь в виду, что из-за высокой чув¬
ствительности хищных ксилофильных насекомых к абиотическим
условиям прожорливость в лаборатории может сильно отличаться
от таковой в природе; данных же об эффективности энтомофагов,
основанных на полевых наблюдениях, очень мало.
^ Среднее за время эксперимента число экземпляров жертвы, уничтожае¬
мое хищником, в пересчете на один день.
32
в некоторых работах имеются отрывочные сведения о значении
того или иного вида хищника. А. И. Воронцов (1949) отметил, что
имаго чернотелок Hypophloeus pini Pz. уничтожили 56% всех от¬
ложенных вершинным короедом яиц и около 20% его личинок,
а имаго карапузиков Cylister oblongum F. истребили 42% яйце¬
кладок большого соснового лубоеда. Известно, что при наиболее
благоприятных для деятельности хиш,ника условиях от теорети¬
чески предполагаемой численности потомства большого соснового
лубоеда выживало лишь 3,8% личинок, все остальные личинки
были уничтожены личинками Thanasimus formicarius L. (Ворон¬
цов, Харитонова, 1971). В. П. Малахова (1902) подчеркивает тот
факт, что существует прямая связь между количеством имаго Pla¬
cusa tachyporoides W^ltl и числом пустующих яйцевых колы¬
белек короеда-стенографа. В некоторых короедных гнездах хищ¬
ником было уничтожено до 40% яиц. По данным Р. И, Земковоп
(1965), в темнохвойных лесах Западного Саяна жуками Rhizop-
hagus puncticollis С. Sahlb. было уничтожено 90% молодого поко¬
ления смолевки и 80% молодого поколения короеда-автографа.
Лозовский (Losowski, 1922, цит. по Зиновьеву, 1957) отмечает,
что личинки Lonchaea palposa L. уничтожают до 60% личинок
короедов, Нуортева (Nuorteva, 1956) считает, что личинки Medetera
уничтожают 32% потомства короедов в одном гнезде. В монографии
Н. 3. Харитоновой (1972) имеются данные о том, что в условиях
Брянского лесного массива хищные насекомые на хвойных породах
уничтожают от 4 до 13% яиц и до 33% молодых личинок короедов.
В последнее время делаются попытки оценить роль хищных
насекомых с учетом состояния популяции жертвы. Бивер (Beaver,
1967), изучая влияние различных факторов на плотность попу¬
ляции Scolytus scolytus L., приходит к выводу, что интенсивность
влияния энтомофагов на численность личинок короеда трех по¬
следних возрастов прямо пропорциональна плотности, если она
низка, и обратно пропорциональна, если плотность короедов высо¬
ка (в этом случае значение хищников становится несущественным).
Б. Н. Огибин (1974), изучавший факторы, регулировавшие плот¬
ность популяции короеда Ips typographus L., приходит к сход¬
ным выводам и отмечает, что деятельность личинок Thanasimus
formicarius L., Т. rufipes Brahm. и Medetera signaticornis Loew
была наиболее эффективна при низкой плотности личинок типо¬
графа. В. М. Яновский (1971, 1974а, б), детально исследовавший
роль энтомофагов в динамике численности короеда Ips subelonga-
tus Motsch., установил, что наиболее ощутимо влияние энтомофагов
проявляется в разреженных и повышенно плотных популяциях
ксилофагов, а также на заключительных этапах развития очага
стволовых вредителей — фазе рассеивания и депрессии.
Церамбицидный комплекс хищных ксилофильных насекомых.
Имеющиеся в литературе сведения о хищных ксилофильных на¬
секомых церамбицидного комплекса крайне скудны. В. И. Гусев
(1928) отмечает, что личинки Pytho depressus L. поедают личинок
2 в. м. Мамаев и др. 33
дровоееков из родов Rhagium и Acanthocinus; о нападении личи¬
нок Pytho depressus L. на личинок Rhagium и Asemum сообщает
Р. И. Земкова (1965). В качестве активных хищников на церамби-
цидпой стадии разрушения древесины отмечены личинки щелку¬
нов Adelocera conspersa Gyll. и Stenagostus undulatus Deg. (Ма¬
маев, Данилевский, 1975).
Невелико и число видов двукрылых, личинки которых питают¬
ся за счет личинок дровосеков на церамбицидной стадии разруше¬
ния древесины. Д. А. Богданова, Н. Г. Коломиец (1974) сообщают,
что личинки ктырей Choerades gilvus L. нападают на личинок дро¬
восеков Monochamus urussovi Fisch. и Rhagium inquisitor L.
H. П. Кривошеиной и Б. М. Мамаевым (1973, 1975) опубликованы
данные по экологии личинок нескольких видов ктырей. Личинки
Choerades gilvus L., предпочитающие хвойные породы, зарегист¬
рированы как хищники, уничтожающие личинок Criocephalus
rusticus L. Было установлено также, что личинки Andreno-
soma atrum L. уничтожают личинок домового дровосека Hylot-
rupes bajulus L. Обитающие под корой дубовых колод личинки
Pogonosoma maroccanum F. уничтожают личинок дровосеков,
в частности Pyrrhidium sanguineum L., а личинки Pogonosoma
lugens Loew развиваются за счет личинок Xylotrechus grumi Sem.
Мы не обнаружили в литературе сведений об эффективности
отдельных видов хищных ксилофильных насекомых, обитающих
в древесине на церамбицидной стадии ее разложения.
^Подводя итог приведенным выше материалам, следует подчерк¬
нуть, что имаго и личинки хищных ксилофильных насекомых иг¬
рают определенную роль на всех этапах развития ксилофильных
сообществ, оказывая влияние на численность насекомых-ксило-
флгов. В основном это влияние заключается в уничтожении пре-
имагинальных фаз развития стволовых вредителей. В перспекти¬
ве — длительный период кропотливых количественных иссле¬
дований динамики ксилофильных сообществ, результаты которых
позволят надежнее оценить значение тех или иных групп энтомо-
фагов, истребляющих насекомых-ксилобионтов.
Глава 2
ЗАКОНОМЕРНОСТИ
ЭКОЛОГИЧЕСКОЙ СПЕЦИАЛИЗАЦИИ
КСИЛОФИЛЬНЫХ НАСЕКОМЫХ
До последнего времени экология ксилофильных насекомых бы¬
ла достаточно хорошо исследована только для группы стволовых
вредителей. Большинство сопутствующих видов в отношении тро¬
фических связей оставалось неясным, а тем более не были охарак¬
теризованы основные направления их специализации. Хотя со¬
бранные нами материалы также не являются исчерпывающими, они
дают возможность в общих чертах рассмотреть этот круг вопро¬
сов.
ОСНОВНЫЕ НАПРАВЛЕНИЯ СПЕЦИАЛИЗАЦИИ
КСИЛОФИЛЬНЫХ НАСЕКОМЫХ
Основными направлениями экологической специализации для кси¬
лофильных насекомых являются следующие: специализация к раз¬
витию на определенных группах древесных пород; специализация
к развитию в различных зонах ствола — подкоровой зоне и тол¬
ще древесины; специализация к развитию в определенных типах
гнили древесины; специализация к питанию определенным пище¬
вым субстратом, а для хи1цников — определенным кругом жертв.
Ниже все упомянутые направления будут последовательно рас¬
смотрены, главным образом на примере зоо- и некрофагов приме¬
нительно к фауне Палеарктики, причем последнее направление,
как особо важное для публикуемой монографии, будет выделено
в самостоятельный раздел.
Специализация к развитию на определенных группах древес¬
ных пород, В настоящее время накоплены факты, свидетельствую¬
щие не только о специфике энтомокомплексов, формируюшихся
на хвойных или лиственных породах, но и о специализации насе¬
комых в отношении отдельных групп лиственных пород или одной
породы.
Различия в качественном составе насекомых-ксилобионтов
хвойных и лиственных пород кажутся вполне закономерными. Их
кора и древесина различаются по плотности, строению проводящих
систем, а главное — по наличию в древесине специфических смо¬
ляных ходов. Смола играет определенную защитную роль, так как
содержит вещества, ядовитые для большинства насекомых. Боль¬
2^ 35
шое значение имеют также физические свойства смолы, ее липкость
и вязкость. Возможно, что в историческом плане на состав компле¬
ксов насекомых-ксилобионтов оказал определенное влияние вре¬
менной фактор, т. е. большая древность голосеменных растений по
сравнению с покрытосеменными. Следует принимать во внимание
также условия произрастания хвойных, их преимущественное
распространение в областях с более прохладным климатом.
Если оценить соотношение в ксилофильных сообществах двух
основных отрядов насекомых — жесткокрылых и двукрылых, то
создается вполне определенное впечатление, что жесткокрылые в
древесине хвойных пород представлены не менее обильно, чем на
лиственных, тогда как комплексы двукрылых на хвойных выгля¬
дят значительно беднее. На хвойных, например, не зарегистриро¬
ваны представители некоторых семейств этого отряда, таких, как
Pachyneuridae, Hesperinidae, Otitidae, Xylomyidae, Tephritidae,
Chloropidae и др., которые связаны исключительно с корой и дре¬
весиной лиственных деревьев.
Если сосредоточить внимание на различиях, можно констати¬
ровать, что комплексы ксилофильных насекомых, формирующиеся
на хвойных или лиственных породах, нередко представлены на
этих породах близкими таксонами — родами или видами. Среди
жесткокрылых в семействе карапузиков (Histeridae) в пределах
ксилофильного рода PIegaderu|| два вида — Р. vulneratus Pz.
и Р. saucius Ег. специализированы к развитию на хвойных, тогда
как Р. caesus Hbst. предпочитает лиственные деревья. Личинки ви-
jfoB рода Hololepta под корой хвойных не встречаются. Все пред¬
ставители этого семейства—хищники, поэтому у них, казалось бы,
нет экологической основы для специализации в отношении раз¬
личных древесных пород.
Другим хорошим примером служат хищные жесткокрылые из
семейства пестряков (Cleridae). В частности, род Thanasimus,
представленный в лесной зоне тремя видами, специализирован
к развитию на хвойных породах и лишь в виде исключения может
встречаться на лиственных. С другой стороны, палеарктические
виды близкого рода Clerus развиваются только на лиственных по¬
родах.
В семействе плоскотелок (Cucujidae) большинство видов при¬
урочено к лиственным породам. Такая же специализация харак¬
терна для рода Brontes, тогда как близкий род Dendrophagus раз¬
вивается под корой хвойных. Представители небольшого семейства
огнецветок (Pyrochroldae), являющиеся сапроксилофагами, спе¬
циализированы к развитию на лиственных породах, тогда как ви¬
ды рода Pytho из близкого семейства трухляков (Pythidae) — на
хвойных.
Что же касается истинных ксилофагов, примером которых мо¬
гут служить такие крупные семейства, как дровосеки (Ceramby-
cidae) или короеды (Scolytidae), то в большинстве случаев виды,
развивающиеся на ранних стадиях разрушения древесины, встре¬
36
чаются либо только на хвойных, либо только на лиственных поро¬
дах, а иногда приурочены к еще более узкому кругу древесных по¬
род или к одной породе. В числе слоников имеются такие роды,
как Hylobius и Pissodes, представители которых развиваются
только на хвойных. В составе семейства ложнослоников (Anthri-
biclae) все ксилофильные палеарктические виды развиваются иск¬
лючительно на лиственных породах и т. д.
Не менее показательны примеры из отряда двукрылых. Резко
различен на хвойных и лиственных породах видовой состав льви-
TIOK (семейство Stratiomyidae). Для хвойных пород характерен один
пшроко распространенный в Палеарктике вид — Zabrachia minu¬
tissima Zett. На лиственных породах развивается целый комплекс
ЕНДОВ — представителей таких родов львинок, как Neopachygas-
ter, Aspidacantha, Craspedometopon и др. Из лиственных деревьев
личинки львинок предпочитают тополь, осину, ильм, маакию
и сравнительно редко встречаются на других породах.
Существенные различия в видовом составе обитателей листвен¬
ных и хвойных пород намечаются также у личинок копьехвосток
(Lonchaeidae). Довольно четко очерчивается небольшой комплекс
видов этого семейства, связанный с хвойными породами. Такая
специализация в Палеарктике характерна для видов группы Lon-
chaea corticis (L. zetterstedti Beck., L. seitneri Hend., L. bruggeri
Morge, L. collini Hack.). Более многочисленна и разнообразна
группа видов, развивающихся на лиственных породах, из которых
личинки копьехвосток, как и львинок, предпочитают тополь и
осину. Личинки львинок и копьехвосток проявляют себя в основ¬
ном как некрофаги, иногда как хищники. Тем не менее приурочен¬
ность отдельных видов к разным породам деревьев проявляется
у них достаточно четко.
Целый комплекс хищных видов зеленушек (Dolichopodidae)
из рода Medetera (М. dichrocera Kow., М. fasciata Frey, М. striata
]\^r., М. signaticornis Loew и др.) характерен только для хвойных
пород. На лиственных этот род представлен другими видами (М.
bispinosa Neg., М. stackelbergiana Neg.), несколько отличающи¬
мися от первых по образу жизни. В хвойных породах личинки зе¬
ленушек приурочены в основном к ходам лубоедов, в лиственных
они предпочитают в первую очередь заболонников.
Имеющиеся в настоящее время данные позволяют говорить
также об определенной приуроченности к хвойным и лиственным
породам облигатных хищников — личинок различных видов рода
Xylophagus (Xylophagidae). Только под корой хвойных деревьев
встречаются личинки X. sachalinensis Р1. и X. cinctus Deg., под
корой лиственных — X. admirandus Kriv. et Мат. Только для
лиственных пород характерна определенная группа видов, раз¬
вивающихся в толще древесины (X. albopilosus Miyat., X. signi¬
fer Kriv. et Мат., X. ater Meig.).
Не менее четкая специализация в отношении древесных пород
прослеживается у облигатных хищников — личинок ктырей подсе¬
37
мейства Laphriinae (Asilidae). Представители рода Andrenosoma
Rond. зарегистрированы только в хвойных, а рода Pogonosoma
Rond.— только в лиственных породах. Только из хвойных пород
известны личинки Choerades igneus Meig., предпочитают хвойные
породы личинки Ch. gilvus L., Laphria gibbosa L. и др. Наоборот,
только в лиственных породах встречаются личинки Choerade^
amurensis Herm., Ch. fimbriatus Meig., Ch. fulvus Meig., Laphria
varia Loew и др.
Кроме специализации в отношении таких групп древесных
пород, как хвойные и лиственные, на палеарктических материалах
можно иллюстрировать более узкую приуроченность некоторых
видов к развитию на одной породе. Такая специализация возни¬
кает главным образом по отношению к специфическим эндемичным
породам.
В долинных лесах Дальнего Востока произрастает маакия
амурская — представитель древесных бобовых (Leguminosae), до-
стигаюш,ая иногда высоты 25 м. Она является дальневосточным
реликтом, распространенным в Приморском крае и на юге Амур¬
ской области. Для маакии характерен мощный слой луба, издаю-
ш,его при гниении специфический запах. Произрастание маакнп
на преувлажненных участках почвы приводит к сохранению по¬
вышенной влажности древ^ны на всех этапах разложения. По¬
этому маакия привлекает Тчень разнообразных насекомых и в
первую очередь двукрылых.
В Приморском крае состав двукрылых — обитателей маакии
очень своеобразен и значительно отличается от такового на дру¬
гих породах. На маакии обнаружен целый ряд интересных моно¬
типных родов и видов двукрылых, .известных только из Южного
Приморья или оказавшихся новыми для науки. Среди новых ви¬
дов заслуживают внимания Pergratospes holoptica Kriv. et Мат.
(Cramptonomyiidae), Maackiana laminiformis Kriv. (Stratiomyi-
dae), Lonchaea ussuriense Kov., L. mamaevi Kov. (Lonchaeidae).
Только в маакии обнаружены личинки Metalimnobia mendax AI.
(Limoniidae), Acanthoneura amurensis Portsch. (Tephritidae),
Graptomyza alabeta Seguy (Syrphidae), Xylomya moiwana Mats.
(Xylomyidae). Преобладаюш,ее большинство обитаюш,их в маакии
двукрылых являются представителями фауны восточной Пале-
арктики, причем большинство видов имеют хорошо прослеживаю-
ш,иеся генетические связи с группами, распространенными в Юго-
Восточной Азии. Этот перечень специфических для маакии видов
насекомых можно иллюстрировать также характерным примером
из отряда жесткокрылых. В частности, в древесине маакии были
обнаружены личинки вида из рода Tenerus (Cleridae), который
характерен именно для фауны Юго-Восточной Азии.
Специфические энтомокомплексы разрушителей коры и древе¬
сины слагаются также на таких породах, как черемуха Маака,
амурский и сахалинский бархат, тополь Максимовича, орех маньч¬
журский, тисс и др.
38
Завершая рассмотрение вопроса о специализации ксилофиль-
ных насекомых в отношении различных древесных пород, следует
отметить, что это направление специализации не всеобъемлющее.
Как среди жесткокрылых, так и среди двукрылых имеются грун-
пы (роды и виды), которые с известным безразличием относятся
к породе заселяемого ими дерева, даже на тех стадиях разложения
древесины, когда специфика той или иной породы еще не утрачена.
Дровосек-ксилофаг Prionus coriarius L. развивается в корнях как
лиственных, так и хвойных деревьев. Личинки-сапроксилофаги
лжечеюнотелки Boros schneideri Pz. (Boridae) зарегистрированы
под кброй всех широко распространенных хвойных и многих ли¬
ственных деревьев. Щелкун Stenagostus undulatus Deg., личинки
которого хищничают под корой, также не специализирован в от¬
ношении заселяемых пород. Среди двукрылых также имеются не¬
специализированные виды — это в первую очередь представите¬
ли некрофагов и хищников. Среди некрофагов к их числу относят¬
ся личинки Lonchaea sylvatica Bel. (Lonchaeidae) — обычного в
Палеарктике вида, встречающегося под корой самых разнообраз¬
ных пород. Из хищников следует отметить личинок некоторых ви¬
дов стволоедок рода Xylophagus (Xylophagidae), например X. mat-
sumurae Miyat. и X. compeditus Wied., развивающихся под корой
как хвойных, так и лиственных деревьев.
Отсутствие избирательности по отношению к породам наблю¬
дается у хищных личинок некоторых видов Medetera Fisch., на¬
пример М. impigra СоП. Среди Laphriinae (Asilidae) имеются от¬
дельные виды, не проявляющие избирательного отношения к
заселяемой древесной породе, как, например, Laphria ephippium
F., L. flava L. и др.
Насекомые-ксилобионты независимо от того, специализирова¬
ны они к развитию на определенных породах или пет, обычно чет¬
ко приурочены к определенным зонам ствола, подразделяясь в пер¬
вую очередь на подкорников и обитателей собственно древесины,
среди которых отмечаются группы, осваивающие всю толщу дре¬
весины или заселяющие только заболонь. Кроме того, важной ос¬
новой для различных направлений топической специализации яв¬
ляется разнообразие возникающих в разлагающейся древесине
дополнительных микробиотопов. В процессе разложения коры и
древесины в них возникают специфические среды обитания (запол¬
ненные древесной трухой и соком дупла; натеки сока на стволах;
скопления смолы). На стволах и пнях деревьев развиваются раз¬
личные грибы, в основном трутовики, мицелий которых пронизы¬
вает древесину.
В зависимости от микростации обитания насекомые, связан¬
ные в своем развитии с древесиной, могут быть подразделены
на ряд групп:
насекомые-ксило5ионты — обитатели коры и древесины на
различных стадиях разрушения, соответственно делятся на две
подгруппы — обитателей коры и обитателей древесины;
насекомые-резиниколы — обитатели натеков смолы на стволах
различных хвойных деревьев;
насекомые-латициколы — обитатели натеков сока, образую-
щихся в трещинах и отслоениях коры и пустотах стволов;
насекомые-мицетобионты — обитатели мицелия и плодовых тел
различных древесных грибов;
насекомые-детритовионты — обитатели древесной трухи, скап¬
ливающейся в дуплах и у основания стволов.
Наибольшее разнообразие как видового состава, так и приспо¬
собительных особенностей отмечается у обитателей коры и д])ове-
сины. Направления специализации этих обитателей рассматри¬
ваются нами более подробно.
Специализация к развитию в коре. В толще разлагающейся ко¬
ры и под корой формируется очень разнообразный в качественном
отношении комплекс насекомых-ксилобионтов. В этом комплексе
основными агентами разрушения коры являются личинки жестко¬
крылых, в первую очередь короедов, дровосеков и златок, которые
начинают развиваться на самых ранних этапах отмирания дерева.
Двукрылые насекомые заселяют кору отмерших или частично
усохших деревьев и относительно редки на начальных этапах ее
разложения, так как сюда способны проникать лишь немногие
их группы. В большинстве случаев двукрылые выступают в роли
вторичных поселенцев, сопутствующих первичным ксилофильным
насекомым.
В нашу задачу не входит подробное рассмотрение комплекса
жесткокрылых — подкорников из группы стволовых вредителей.
Необходимо отметить, однако, что именно личинки короедов, дро¬
восеков, а в южных районах — златок осуществляют первичное
разрушение луба и открывают путь в подкоровую зону другим
весьма многочисленным группам насекомых, включая хищников.
Среди стволовых вредителей многие виды специализированы
к развитию в толще луба. К их числу относятся почти все виды ко¬
роедов, за исключением группы так называемых короедов-древесин-
ников, слоники из родов Pissodes, Hylobius, Trachodes и др., очень
многие виды златок из родов Agrilus, Chrysobothris, Lampra и
др., а также значительная группа дровосеков, в составе которой
имеются такие обычные и массовые представители, как Rhagium,
Tetropium, Saperda, Acanthocinus, Plagionotus и др.
Четкой специализацией к жизни под корой при питании пре¬
имущественно разлагающимся лубом и погибшими личинками
насекомых отличаются представители таких семейств жесткокры¬
лых, как плоскотелки (Cucujidae), огнецветки (Pyrochroidae),
трухляки (Pythidae) и некоторые другие сходные по экологии
группы. В составе названных семейств имеются виды с различным
характером питания, в том числе облигатные хищники и зоонекро-
фаги.
Несмотря на разнообразие видового состава двукрылых —
обитателей коры, можно с достоверностью констатировать, что
40
лишь отдельные виды двукрылых сйособны интенсивно перераба¬
тывать луб. Среди них в первую очередь следует назвать личинок
пестрокрылок рода Acanthoneura (Tephritidae), встречающихся
в больших количествах под корой маакии на начальных этапах
ее разложения. В несколько побуревших волокнах луба обычны
личинки болотниц Libnotes kariyana Alex. (Limoniidae), ко¬
торые интенсивно перерабатывают лубяные волокна в труху. Пре¬
обладающая часть личинок двукрылых-подкорников обитает в
толще лубяных волокон и на границе коры и древесины, однако
они не относятся к энергичным разрушителям коры.
Среди этих двукрылых можно назвать много семейств, которые
встречаются только в зоне коры или преобладают в ней. Предста¬
вители таких семейств, как Trichoceridae из длинноусых двукры¬
лых, а из короткоусых круглошовных Megamerinidae, Caloba-
tidae, Strongylophthalmyiidae, Pallopteridae, Otitidae и др., раз¬
виваются в стволах только под корой. Большое число семейств
длинноусых двукрылых представлено в основном, за исключением
специфических родов и видов, обитателями коры. Так, из ксило-
фильных групп гнильниц (Scatopsidae) под корой развиваются
многочисленные виды рода Scatopse, из мокрецов (Ceratopogoni-
dae) — виды родов Culicoides, Dasyhelea и др. С корой связана
большая группа родов галлиц (Cecidomyiidae): Peromyia, Wyat-
tella, Camptomyia, Winnertzia и др. Преимущественно в коре раз¬
виваются ксилофильные группы прямошовных короткоусых дву¬
крылых, таких, как львинки подсемейства Pachygasterinae (Stra-
tiomyidae) и большинство видов стволоедок (Xylophagidae). В толще
коры развивается большая часть ксилофильных видов копье-
хвосток рода Lonchaea (Lonchaeidae), журчалок из родов Hammer-
schmidtia, Graptomyza, Brachyopa (Syrphidae) и др.
Особую группу подкорников формируют личинки жесткокры¬
лых и двукрылых, которые приурочены к ходам стволовых вреди¬
телей, хотя и не всегда по характеру питания относятся к обли¬
гатным хищникам. Так, в ходах короедов имеется целый ряд та¬
ких видов, к числу которых относятся жесткокрылые Aulonium
trisulcum Geoffr. (Colydiidae), многие Epuraea (Nitidulidae), ряд
видов Rhizophagus (Nitidulidae), представители рода Thanasi-
mus (Cleridae). Из двукрылых примером таких групп могут слу¬
жить хищные личинки Medetera (Dolichopodidae), Pogonosoma
(Asilidae) и некоторые другие.
Также специфическую в экологическом отношении группу об¬
разуют обитатели щелевидных пространств, возникающих под
корой в результате ее отслоения. Здесь нередко встречаются боль-
Н1ие скопления своеобразных личинок двукрылых рода Forcipo-
myia (Ceratopogonidae), а из жесткокрылых — также не менее
своеобразные подвижные личинки Brontes, Silvanus (Cucujidae)
и некоторые другие.
Небольшую, но экологически своеобразную группу насекомых
составляют виды, поселяющиеся под поверхностным корковым
41
слоем, т. е. на наружной поверхности луба таких пород, как бере¬
за, ольха, черемуха и др. В этот комплекс из жесткокрылых вхо¬
дят некоторые виды Rhizophagus (Nitidulidae) и Laemophloeus
(Cucujidae), а из двукрылых — несколько видов галлиц из родов
Camptomyia, Winnertzia (Cecidomyiidae), детритниц (Sciaridae)
и некоторые круглошовные мухи.
Специализация к развитию в толще древесины. Закономерности
специализации ксилофильных насекомых к жизни в различных
слагающихся в толще древесины условиях весьма многообразны.
Характер этой специализации зависит от таких факторов, как
влажность древесины, продолжительность ее разрушения, осо¬
бенности воздействия на нее дереворазрушающих грибов и т. д.
Это многообразие слагающихся в древесине условий отражено
в простейшей схеме:
дрквесина
сухая
»умеренно
влажная
^переувлаж¬
ненная
на ранних
стадиях
разрушения
на поздних
стадиях
разрушения
*■ зона
заболони
► вся толща
древесины
разрушенная
► по типу свет¬
лых гнилей
разрун1енная
^ по типу бурых
гнилей
В этой схеме обозначены те специфические среды обитания, ко¬
торые возникают в древесине в различных ситуациях. Наиболее
обычна специализация в шести разных направлениях:
1) к жизни в заболони умеренно влажной древесины на ранних
стадиях разрушения;
2) к использованию всей толщи умеренно влажной древесины
па ранних стадиях разрушения;
3) к развитию в светлых гнилях умеренно влажной древесины
па поздних стадиях разрушения;
4) к развитию в бурых гнилях умеренно влажной древесины на
поздних стадиях разрушения;
5) к развитию в сухой древесине;
6) к развитию в переувлажненной древесине.
Необходимо в первую очередь отметить, что специализация к
жизни в толще древесины па ранних стадиях ее разрущения воз¬
можна почти исключительно у тех насекомых, которые обладают
крепкими мандибулами и способны такую древесину разрушать.
Поэтому это направление специализации в большей степени харак¬
терно для представителей отряда жесткокрылых и тех перепон¬
чатокрылых, которые консолидировались в группу рогохвосто¬
образных. Личинки двукрылых не в состоянии самостоятельно
42
проделывать ходы в толще древесины. Они способны проникать
в нее лишь по ходам различных других ксилофильных насекомых.
Зона заболони на ранних этапах разрушения древесины при¬
влекает многочисленных насекомых, причем первопоселенцами в
этой зоне обычно бывают личинки сверлильщиков (Lymexyloni-
clae), короедов-древесипников (Scolytidae), плоскоходов (Platy-
podidae) и некоторых слоников, например Sipalinus gigas F.
(Curculionidae). Личинки этих жесткокрылых прокладывают в забо¬
лони систему главным образом концентрических ходов, освобож¬
даемых от нагрызенных опилок. Па стенках этих ходов развива¬
ются симбиотические грибы и туда устремляются многочисленные
сопутствующие виды. В их числе следует упомянуть из жестко¬
крылых личинок чернотелок из рода Hypophloeus (Tenebrionidae)
II узкотелок из рода Colydium (Golydiidae). Из двукрылых в ходах
обычны некоторые представители родов Odinia (Odiniidae), Lib-
notes (Limoniidae). Личинки упомянутых насекомых — хищни¬
ки и мицетофаги. Болотницы рода Libnotes в целом характерны
для сильно увлажненных субстратов, обычны в натеках сока, на¬
пример на ильме (L. undulata Mats.), могут встречаться и в доста¬
точно увлажненной коре (L. kariyana Alex.). Личинки одного
вида этого рода (L. ladogensis Lacksch.) приспособились к обита¬
нию во влажных ходах жуков-сверлильщиков. По ходам других
насекомых в толщу древесины способны проникать отдельные
представители львинок (Stratiomyidae). Так, личинки Xylopachy-
gaster mamaevi Kriv. развиваются в толще древесины в ходах
слоника Sipalinus gigas F. и плоскохода Grassotarsus sp. Ходы ли¬
чинок этих жуков богаты жидкими продуктами разложения дре¬
весины, а также мицелием грибов, что создает благоприятные ус¬
ловия для развития личинок львинок. По ходам жуков, в основ¬
ном короедов-древесипников и сверлильщиков, в толщу древе¬
сины проникают личинки различных видов журчалок из рода
Brachyopa, в первую очередь В. conica Pz. (Syrphidae).
Все упомянутые выше виды насекомых характерны именно
для заболони стволов в первые два года их отмирания и разру¬
шения.
Несколько позднее в заболони появляются личинки ±;екоторых
дровосеков и златок. Интенсивное разрушение ими древесины при¬
водит к тому, что в этот процесс вовлекаются все более и более
глубокие слои древесины до тех пор, пока не заселяется вся ее
толща. Однако такие жесткокрылые, как горбатки (Mordellidae),
некоторые тенелюбы (Melandryidae), например Hypulus quercinus
Guens., а из перепончатокрылых — такой рогохвост, как Xiphy-
dria prolongata Geoffr., заселяя стволы старых деревьев, специа¬
лизированы в основном к развитию только в заболони этих ство¬
лов. Из двукрылых-ксилофагов к развитию в заболони в ранний
период ее разложения специализированы личинки Pergratospes
holoptica Kriv. et Маш. (Cramptonomyiidae), Bolitophila aperta
Lund. (Bolitophilidae), некоторые галлицы (Cecidomyiidae).
43
Толща древесины разрушается главным образом личинками
жесткокрылых из семейств дровосеков (Cerambycidae) и златок
(Buprestidae), некоторыми тенелюбами, например Serropalpus
barbatus Schall., и видами рода Xylita (Melandryidae), а также
перепончатокрылыми из семейства рогохвостов (Siricidae). Инте¬
ресную, хотя и не очень многочисленную, группу образуют дву¬
крылые, для хищных личинок которых ходы дровосеков и златок
являются постоянным местообитанием. Из них в первую очередь
следует упомянуть личинок большинства видов ктырей из подсе¬
мейства Laphriinae в частности, представителей родов Choerades,
Laphria, Andrenosoma и др.
Древесина лиственных и хвойных пород на заключительных
этапах разложения постепенно теряет специфические свойства.
В различных условиях в зависимости от направления процессов
разложения происходит изменение в соотношении основных ком¬
понентов древесины (клетчатки и лигнина). В зависимости от вида
гриба происходит ускоренное разрушение одного из этих основ¬
ных веществ. При сохранении клетчатки и уменьшении содержа¬
ния лигнина формируются светлые гнили древесины, при более
интенсивном разрушении клетчатки — бурые гнили. Светлые гни¬
ли при различных условиях влажности также приобретают раз¬
личные оттенки. Более сухая древесина имеет светлый, слегка се¬
роватый цвет, сильно увлажненная древесина темнеет, приобре¬
тая почти черный оттенок (черненые гнили). Видовой состав на¬
секомых, развивающихся в светлых (включая черненые) и бурых
^нилях, существенно различается. Насекомые в целом предпочи¬
тают светлые и черненые гнили и значительно реже встречаются
в бурых гнилях.
Четкая специализация к развитию в светлых гнилях древеси¬
ны, включая черненые, отмечается у таких видов рогачей, как
Dorcus parallelopipedus L. и Sinodendron cylindricum L., a na Даль¬
нем Востоке — Prismognathus subaeneus Motsch. и Eurylrachelus
rubrofemoratus WolL, у пластинчатоусых жуков из рода Trichius,
у большинства видов узконадкрылок (Oedemeridae), у чернотелок
из триб Helopini и Misolampini (Tenebrionidae), тенелюбов (Melan¬
dryidae) и других жесткокрылых. Из двукрылых специализирова¬
ны к развитию в этих условиях представители семейств Hesperi-
nidae и Pachyneuridae, гребенчатоусых комаров-долгоножек из
подсемейства Ctenophorinae, отдельные группы мохнатокрылок на¬
пример виды рода Trichomyia (Trichomyiidae), гнильниц из рода
Ectaetia ((Scatopsidae), виды из семейств детритниц (Sciari-
dae) и галлиц (Cecidomyiidae) и т. д. Характерными обитателями
толщи древесины являются личинки пятнокрылок (Clusiidae).
В более рыхлой, преимущественно трухлявой древесине развива¬
ются личинки комаров-толстоножек из родов Bibio и Dilophus
(Bibionidae), плециид — Plecia nigra Lund. (Pleciidae), некото¬
рых видов Tipula (Tipulidae), например Т. stenostyla Sav. Для
этих условий характерны также виды рода Elephanlomyia (Limo-
44
niidae). В толще светлых гнилей древесины обитают также личин¬
ки-хищники таких видов, как Xylophagus ater Meig., X. admiran¬
dus Kriv. et Mam., X. albopilosus Miyat. (Xylophagidae), a также
ктырей рода Mesoleptogaster (Asilidae). Здесь развиваются неко¬
торые представители семейств зеленушек (Dolichopodidae), копье-
хвосток (Lonchaeidae) и других высших двукрылых (Cyclorrhapha).
К развитию в чрезвычайно специфических бурых гнилях дре¬
весины специализированы рогачи из родов Ceruchus и Aesalus
(Lucanidae), различные щелкуны преимущественно из рода Ат-
pedus (Elateridae), плоскотелки рода Prostomis (Cucujidae), а на
Дальнем Востоке — Istrisia rufobrunnea Lew. (Salpingidae), Nema-
toplus semenovi Nik. (Nematoplidae) в сообществе с тараканами
Cryptocercus relictus B.-Bien. Из двукрылых в бурых гнилях
встречаются личинки различных видов детритниц (Sciaridae),
представители туманниц рода Hyperoscelis (Hyperoscelididae),
а также хищники — отдельные виды ктырей из рода Choerades
(Asilidae) и стволоедок из рода Xylophagus (Xylophagidae).
ХАРАКТЕРИСТИКА ПИЩЕВЫХ РЕЖИМОВ
КСИЛОФИЛЬНЫХ НАСЕКОМЫХ
Классификация пищевых режимов насекомых наталкивается
на значительные трудности, обусловленные недостаточностью дос¬
товерных данных. Даже в крупных сводках по этому вопросу
(Brues, 1946, и др.) не приводится общей схемы соподчинения раз¬
личных типов питания. Р. Шовен (1953) характеризует режимы
питания также без попытки найти их соподчинение и выделяет
в качестве равнозначных совершенно неравноценные как по объе¬
му, так и по степени специализации трофические группы.
Такой эмпирический подход, хотя и нуждается в некоторой
критике, тем не менее пока единственно реальный, так как объем
накопленных сведений о пищевых режимах тех или иных насе¬
комых, особенно их личинок, еще крайне недостаточен. Это за¬
ставляет нас характеризовать трофические связи насекомых-кси-
лобионтов (следует еще раз подчеркнуть, что объектом нашего
исследования были их личинки), в первую очередь такими обще¬
принятыми терминами, как фито- и сапрофагия, зоо- и некрофа-
гия. Однако даже эта терминология в применении к ксилобион-
там нуждается в пояснении, в частности в уточнении таких поня¬
тий, как ксилофагия и мицетофагия.
Древесина представляет собой специфический субстрат, тре¬
бующий наличия у насекомых определенных приспособлений для
его усвоения. Основную часть древесины (от 40 до 60% сухого ве¬
са) составляет клетчатка, принадлежащая к наиболее стойким хи¬
мическим веществам. Помимо клетчатки, в древесине содержатся
другие углеводы, в том числе лигнин (от 18 до 38%). Имеются по¬
лисахариды в виде гемицеллюлоз, из них пентозаны составляют от
45
6 до 23% и гексозаны — от 2 до 14%. В древесине содержатся крах¬
мал (0—5,9%) и белок (1,1—2,3%).
Личинки ксилобионтов усваивают в первую очередь наиболее
доступные вещества (полисахариды, крахмал, белки). Истинные
потребители древесины способны перерабатывать клетчатку. Это
достигается различными способами: при помощи специальных
пищеварительных соков, содержащих целлюлазу, или главным
образом при помощи симбиотических организмов (простейших и
бактерий), живущих в кишечнике ксилобионтов. Наконец, целый
ряд насекомых-ксилобионтов питается не древесиной, а различ¬
ными поселяющимися в ней организмами, в первую очередь гри¬
бами.
Лишь немногие ксилофильные насекомые, заселяющие кору и
древесину живых деревьев или в начальные периоды их отмирания,
могут быть отнесены к истинным фитофагам. По мере дальнейше¬
го разложения коры и древесины в питании усиливаются элементы
сапрофагци (в широком смысле), но переходы между тем и другим
режимом питания установить в достаточной мере сложно. Только
насекомых, питающихся сильно разложившимися потерявшими
структуру остатками коры и древесины, можно считать истин¬
ными сапрофагами. Поэтому вполне обоснованно выделение тако¬
го самостоятельного режима питания, как ксилофагия, для харак¬
теристики трофических связей насекомых, питающихся отмершей,
но еще не разложившейся, сохраняющей свою структуру, древе¬
синой, т. е. занимающих промежуточное положение между ис¬
тинными фитофагами и сапрофагами.
К фитофагам \ таким образом, мы относим личинок насекомых,
питающихся живыми растительными тканями или их соками под
корой или в древесине отмирающих деревьев. Учитывая особен¬
ности питания личинок-ксилобионтов, поселяющихся в заключи¬
тельный период отмирания деревьев, видимо, следует ввести
промежуточный термин — фитоксилофагия.
К сапрофагам принадлежат личинки насекомых, питающиеся
сильно разложившимися растительными остатками, в том числе
утратившей структуру древесиной. Учитывая особенности пита¬
ния личинок-ксилобионтов, поселяющихся на заключительных
стадиях разрушения древесины, для их трофической характерис¬
тики, видимо, следует использовать термин сапроксилофагия.
Собственно ксилофагов также можно подразделить на истин¬
ных ксклофагов — потребителей собственно древесины (эуксило-
фагов) и флоеофагов — потребителей коры (Schedl, 1958).
Основываясь на изложенных выше материалах, с учетом ха¬
рактеристики пищевых режимов ксилобионтов, разработанной
Б. М. Мамаевым (1966), можно выделить пять трофических групп
насекомых-ксилофагов:
^ По-^ебители тканей низших растений — грибов обычно обозначаются
особым термином — мицетофагп, который нами также широко исполь¬
зуется.
46
1. Фитоксилофаги. Личи:^ки питаются по границе отмирания
тканей усыхающего дерева, хготребляя как живую, так и отмер¬
шую кору и древесину. JB качестве примера могут быть упомянуты
некоторые дровосеки, особенно роды Aeolesthes и СегатЬух, злат¬
ки, в частности род Asilus, некоторые слоники, например
Cryptorrhynchus lapathi L.
2. Амброзийные ксиломицеШфаги. Личинки обитают в ходах,
свободных от трухи, на стенках которых развиваются [симбиоти¬
ческие грибки. Личинки питаются в основном грибками. Древе¬
сина может заглатываться, но не является основной пищей, так
как слабо переваривается. Насекомые этой группы либо самостоя¬
тельно заносят в древесину споры специфических грибов, либо
сопутствуют таким насекомым. Сюда принадлежат жесткокрылые —
первопоселенцы в толще древесины из семейств сверлильщи¬
ков (Lymexylonidae), короедов (Scolytidae), плоскоходов (Platypo-
didae), биология которых неоднократно описывалась в литературе.
Из двукрылых к группе, экологически сходной с амброзийными
ксиломицетофагами, относятся личинки аксимиид Mesaxymyia и
Protaxymyia (Axymyiidae), самостоятельно проделывающие ходы
в толще древесины и имеющие приспособления для освобождения
ходов от трухи. К этой же группе относятся своеобразные личин¬
ки журчалок рода Temnostoma, обладающие специальными скреб¬
ками для прокладывания ходов.
К числу спутников амброзийных ксиломицетофагов принадле¬
жат личинки журчалок из родов Brachyopa и Hammerschmidtia
(Syrphidae), болотниц рода Libnotes (Limoniidae), львинок рода
Xylopachygaster (Stratiomyidae). Личинки журчалок Hammer-
schmidtia и большинства видов Brachyopa проникают в толщу ко¬
ры на самых начальных этапах ее заселения короедами. Личинки
Brachyopa conica Pz. встречаются как в толще коры, так и в тол¬
ще древесины, где они довольно обычны в ходах жуков-сверлиль-
щиков. Постоянно сопутствуют личинкам сверлильщиков также
личинки Libnotes, не способные проделывать ходы самостоятель¬
но. Очень близки по образу жизни к личинкам Brachyopa conica
Pz. личинки львинок Xylopachygaster mamaevi Kriv.
3. Ферментативные ксилофаги. Личинки развиваются в све¬
жей коре и древесине, в ходах, забитых буровой трухой. Они об¬
ладают наиболее полным набором ферментов, способны перевари¬
вать клетчатку (Parkin, 1940; Schlottke, 1948, Deschamps, 1953,
и др.) и не заносят в древесину споры симбиотических грибов при
откладке яиц. К этой группе относится большинство представите¬
лей семейств дровосеков (Cerambycidae), златок (Buprestidae),
точильщиков (Anobiidae) и ряд других жесткокрылых. Среди
двукрылых имеются лишь единичные представители этой трофи¬
ческой группы, относящиеся к потребителям коры. Сюда относят¬
ся, например, личинки пестрокрылок из рода Acanthoneura: А.
pteropleuralis Hend. и А. amurensis Portsch. (Tephritidae), актив¬
но перерабатывающие почти свежие лубяные волокна.
47
4. Деструктивные ксиломицетофаги. Личмки прокладывают
ходы, забитые буровой трухой, и заглатыв^т кору или древеси¬
ну, пронизанную мицелием грибов и уж^ определенным образом
ими разрушенную. Представители этой /руппы либо заносят спо¬
ры грибов в древесину в период откла^и яиц (рогохвосты, корое¬
ды), либо у них возникает трофическая связь с определенными ви¬
дами грибов — дереворазрушителей (тенелюбы, горбатки, узко-
падкрылки и др.). /
В качестве характерного примера можно привести рогохвостов
подсемейства Siricinae (Siricidae). В основании яйцеклада самок
этих насекомых имеется карман, набитый спорами и фрагментами
грибного мицелия. Во время откладки яиц эти споры заносятся в
ствол дерева и дают начало мицелию, который пронизывает дре¬
весину (в зоне, где грызет личинка), обогащая ее питательными
веществами. Грибы принадле;кат к родам Stereum, Amblystereum,
Polyporus, Daedalea, Daldinia, в том числе к видам, встречающим¬
ся в природе и вне связи с рогохвостами. В отношении некоторых
жуков-д1Л)восеков (Cerambycidae) и златок известно, что их личин¬
ки приспособлены к питанию древесиной, предварительно перера¬
ботанной дереворазрушающими грибами. Если этих личинок пе¬
ресадить в древесину, из которой экстрагированы продукты ее раз¬
рушения грибами, их рост и развитие задерживаются.
5. Сапроксилофаги. Личинки питаются сильно разрушенными
(грибами и насекомыми) корой и древесиной (сукцессия грибов
уже завершается, обильны остатки отмершего мицелия). Для пи¬
щеварительной системы личинок, перерабатывающих такую дре¬
весину, характерны особые «бродильные камеры» —• расширения
кишечника (Areekul, 1957), где концентрируются экстрацеллюлар-
ные внутрикишечные симбионты (Wiedemann, 1930).
Из жесткокрылых типичными представителями сапроксилофа-
гов являются рогачи (Lucanidae) и пластинчатоусые жуки (Scara-
Laeidae), кишечник которых весьма объемист, особенно в задних
отделах. Среди двукрылых личинки-сапроксилофаги составляют
довольно обширную группу. К ней относятся представители це¬
лого ряда семейств длинноусых двукрылых —• таких, как Hespe-
rinidae, Pachyneuridae, Cramptonomyiidae, а также комаров-дол-
гоножек подсемейства Ctenophorinae, развивающихся в достаточ¬
но плотной, сохранившей свою структуру древесине. Типичными
представителями сапроксилофагов являются личинки комаров-
толстоножек (Bibionidae) и плециид (Pleciidae), заселяющие преи¬
мущественно сильно разрушенные кору и древесину.
Весьма важный раздел трофической характеристики ксило-
фильных насекомых составляет классификация пищевых режимов
плотоядных групп. Как уже отмечалось, среди насекомых, питаю¬
щихся 8а счет животных организмов, выделяются две крупные
группы —• зоофагов и некрофагов. Зоофаги естественным образом
подразделяются на паразитов и хищников. Насекомые-паразиты
экологически весьма своеобразны. Их ксилофильные группы изу¬
48
чены недостаточно, и мы еще не считаем возможным приводить
даже их самую общук? хара1<теркстику. Поэтому в дальнейшем из¬
ложении основное BihiMaHHe будет уделено хищным личинкам
насекомых.
Хищники характеризуется тем, что ведут свободный подвиж¬
ный образ жизни; питаясь, они убивают свою жертву. В литера¬
туре применительно в основном к хищникам и паразитам исполь¬
зуются такие определения, как «облигатный» или «факультатив¬
ный». Облигатность питания — это сохранение определенными
группами насекомых в течение всего периода развития стабильных
пищевых связей. Факультативность питания характеризуется пе¬
риодическим изменением его режима в процессе развития ли¬
чинки.
Наибольший интерес как энтомофаги имеют облигатные хищ¬
ники.
В отряде жесткокрылых имеются крупные семейства, ксило-
фильпые личинки которых относятся исключительно к облигатным
хищникам. Среди них. привлекает внимание разнообразнейшее в
экологическом отношении семейсв'во карапузиков (Histeridae),
представленное в фауне Палеарктики почти 300 видами. Все кси-
лофильные виды этого семейства — хищники, уничтожающие ли¬
чинок других насекомых, среди которых в первую очередь следует
назвать двукрылых. Также исключительно хищниками являются
немногие виды близкого семейства Synteliidae (Мамаев, 1975)
К облигатным хищникам относятся все виды семейства пестря¬
ков (Cleridae). Эти жесткокрылые представлены в Палеарктике
более чем 50 видами, которые развиваются как под корой, так и в
толще древесины. Они специализировались главным образом
к питанию личинками жесткокрылых-ксилофагов. Предполагают,
что только виды рода Necrobia от хищничества перешли к пита¬
нию мясными продуктами. Близкое к пестрякам семейство Тго-
gossitidae имеет аналогичное направление пищевой специализации.
Огромное семейство коротконадкрылых жуков (Staphylinidae),
насчитывающее 20000 видов, представлено в самых различных
средах обитания, в том числе под корой и в древесине. Некоторые
ксилофильные роды этого семейства имеют в своем составе только
облигатных хищников. Есть облигатные хищники и в составе се¬
мейства щелкунов (Elateridae), в частности, к ним принадлежат все
виды рода Adelocera, Stenagostus, Denticollis и др.
В отряде двукрылых облигатное хищничество развито не ме¬
нее широко. В числе хищников в первую очередь следует назвать
личинок представителей рода Xylophagus (семейство Xylophagidae).
Все виды этого рода, единственного в семействе, развиваются под
корой и в древесине и относятся к категории почти всеядных хищ¬
ников (Кривошеина, Мамаев, 1966, 1972а). Они охотно питаются
личинками короедов, усачей, комаров-долгоножек, галлиц, гриб¬
ных комариков и даже личинками щелкунов, имеющими твердые
покровы. Были зарегистрированы даже случаи питания дождевы¬
49
ми червями и отмечены факты каннибализма. Личинки стволоедок
прокалывают покровы жертвы, выделяя внутрь их тела пищева¬
рительные соки, а затем высасывают содер>^<имое так, что остается
почти пустая шкурка. В литературе имеется указания на способ¬
ность личинок отдельных видов рода Xylophagus питаться мерт¬
выми остатками насекомых (Зиновьев, 1957), однако нами явле¬
ния некрофагии не были зарегистрированы. Преимущественно
с древесиной связано развитие одного из подсемейств ктырей —
Laphriinae. Все известные в настоящее время личинки Laphriinae
ведут активный образ жизни облигатных хищников и связаны в
первую очередь со стволовыми вредителями — личинками златок
и дровосеков (Кривошеина, Мамаев, 1967, 1973, 1975; Кривошеи-
па, 1974 г.). В толще относительно сильно разложившейся древеси¬
ны развиваются личинки ктырей Mesoleptogaster eous Lehr, пи¬
тающиеся различными представителями группы сапроксилофагов
(Pachyneura, Hesperinus и др.). Облигатными хищниками являют¬
ся личинки зеленушек рода Medetera, развивающиеся в поселе¬
ниях короедов. Образ жизни личинок других родов этого семей¬
ства изучен относительно слабо. Ксилобионтами, помимо Medete¬
ra, являются личинки Systenus, развивающиеся в натеках сока и
во влажной трухе дупел. Можно предполагать, что они также яв¬
ляются хищниками.
Облигатными хищниками среди других прямошовных коротко¬
усых двукрылых являются личинки Therevidae и все изученные
в настоящее время личинки Scenopinidae, а также бекасниц из ро¬
дов Rhagio и Chrysopilus (Rhagionidae). Но в древесине представи¬
тели этих семейств встречаются редко.
Значительно сложнее установить факты факультативного хищ¬
ничества.
В принципе факультативным хищником при наличии подходя¬
щей жертвы может стать любой ксилофаг. Основной лимитирую¬
щий фактор развития ксилофагов — это дефицит белковых соеди¬
нений. Поэтому достаточно широко известны факты нападения
личинок ксилофагов друг на друга. Это выражается в явлениях
каннибализма или в уничтожении крупным видом более мелкого.
Финский энтомолог М. Нуортева (Nuorteva, 1962) наблюдал, на¬
пример, массовое уничтожение личинками дровосека Acanthoci-
nus aedilis L. личинок короеда. Могут нападать друг на друга
также личинки златок.
Многие виды насекомых, сопутствующих стволовым вредите¬
лям, уничтожают их яйца, личинок и куколок. Известны случаи,
когда на куколок дровосеков нападали крупные личинки плоско-
телок из рода Сиси jus. Такое же поведение характерно для трух¬
ляков рода Pytho. Личинки щелкунов рода Ampedus могут дол¬
гое время существовать без животной пищи, однако в куколок не
превращаются (Долин, 1964а). Они охотно нападают на личинок
и куколок ксилофагов, проявляя свои наклонности к факульта¬
тивному хищничеству.
50
Факультативное хищничество в природных условиях чрезвы¬
чайно трудно отдифференцировать от некрофагии, т. е. от пита¬
ния уже погибшими животными, так как практически невозмож¬
но определить, напали ли личинки-пожиратели на живую личин¬
ку жертвы или нашли и поедают ее после гибели.
Некрофагия как пищевая специализация насекомых изучена
далеко не достаточно. Данные последних лет (Кривошеина, 1969,
1974а, б) показывают, что способность к некрофагии может про¬
являться у целого ряда групп насекомых, исходно питающихся
растительной пищей. В то же время пока трудно назвать группы
насекомых-ксилобионтов, которые являлись бы только некрофага-
ми. Видимо, некрофагия почти всегда будет комплексироваться
с какими-либо другими типами питания, и пищевой режим таких
личинок адекватно можно будет выразить такими терминами, как
«сапронекрофагия», «зоонекрофагия», «сапромицетонекрофагия»
и т. п.
Очень многие личинки жесткокрылых из различных семейств
охотно поедают полуразложившцеся остатки погибших особей,
восполняя таким образом дефицит белковых веществ в древесине.
В эксперименте добавление в субстрат мертвых личинок насеко¬
мых способствовало ускоренному росту личинок пластинчатоусых
жуков (Scarabaeidae), дровосеков (Cerambycidae), плоскотелок
(Cucujidae), трухляков (Pythidae), блестянок (Nitidulidae) и др.
Особенно охотно поедают погибших насекомых личинки дву¬
крылых. Так, например, личинки большинства ксилофильных ви¬
дов двукрылых из семейств львинок (Stratiomyidae) и ксиломиид
(Xylomyidae) приурочены к лиственным породам, где на началь¬
ных этапах своего развития концентрируются на участках, обиль¬
но смачиваемых бродящим соком и другими жидкими продуктами
разложения коры и древесины. Поэтому характер питания личи¬
нок в этот период может быть охарактеризован как сапроксило-
фагия. Однако на заключительных этапах развития перед окук¬
ливанием нередко можно наблюдать питание рассматриваемых
личинок погибшими личинками других насекомых. Очевидно,
подобные данные привели к ошибочным заключениям о хищниче¬
стве личинок ксиломиид (Malloch, 1917).
Наблюдения в лаборатории показали, что некрофагия способ¬
ствует ускорению и завершению развития. У некоторых видов,
развивающихся на хвойных породах (например, Zabrachia minutis¬
sima Zett.), некрофагия становится основой пищевого режима.
Определяя особенности пищевой специализации данного вида,
правильнее будет говорить о некрофагии с элементами сапромице-
тофагии.
У отдельных изученных к настоящему времени групп двукры¬
лых некрофагия может быть выражена в более определенной фор¬
ме и наблюдаются переходы от некрофагии к хищничеству. Так,
У мух-копьехвосток рода Lonchaea можно наблюдать все переходы
от сапрофагии к некрофагии, а у отдельных видов — к хищниче¬
51
ству. Хотя пищевые связи группы исследомны далеко не пол¬
ностью, уже сейчас можно утверждать, что для многих видов, ли¬
чинки которых ранее характеризовались как сапроксилофаги,
некрофагия — необходимый элемент питания (например, у личи¬
нок L. sylvatica BeL). У личинок целого ряда видов (L. bruggeri
Morge, L. collini Наск., L. zetterstedti Beck. и др.), развиваю¬
щихся на хвойных, зарегистрировано хищничество (Morge, 1963,
1967а; Гаприндашвили и др., 1967). Личинок копьехвосток тем
не менее нельзя рассматривать в качестве облигатных хищников.
Личинки не способны нападать на крупных личинок других насе¬
комых. Они питаются преимущественно куколками и личинками
короедов, имеющими небольшие размеры и мягкие, нежные пок¬
ровы; обычно нападают на живых, но поврежденных личинок. Не¬
совершенен и сам способ уничтожения жертв, подробно рассмот¬
ренный в работе Г. А. Зиновьева (1957). Очевидно, в рассматри¬
ваемой группе хищничество находится в фазе становления, поэто¬
му у личинок копьехвосток еще не выработались специфические
морфологические приспособления, характерные для многих грунц
облигатных хищников.
Глава 3
СИСТЕМАТИЧЕСКИЙ ОБЗОР ЛИЧИНОК
Специфичность древесины как среды обитания обусловила фор¬
мирование морфологически и экологически своеобразного комплек¬
са насекомых-ксилобионтов. Однако кроме истинных ксилобион-
тов, развитие которых проходит в древесине, представители от¬
дельных отрядов насекомых, в целом для древесины нетипичных,
могут с достаточным постоянством встречаться под корой или в
толще сильно разрушенных стволов и пней. В некоторых случаях
истинными ксилобионтами стала лишь небольшая, как правило,
обособленная группа видов того или иного отряда.
ГРУППЫ КСИЛОБИОНТНЫХ НАСЕКОМЫХ
В монографии рассматриваются только наиболее типичные кси-
лобионты из отрядов жесткокрылых, верблюдок и двукрылых,
так как именно в этих группах широко представлены ксилофиль-
ные хищные личинки. В целом же из насекомых с полным превра¬
щением в древесине можно обнаружить представителей 7 отрядов.
Отряд Coleoptera — Жесткокрылые, или жуки. Характеризу¬
ется в основном активными личинками с хорошо развитой голов¬
ной капсулой и функционирующими грудными ногами. В очень
редких случаях головная капсула уменьшается в размерах, теряет
склеротизацию, модифицируется, но никогда не исчезает полностью.
Ноги в большинстве случаев состоят из 4 отделов (голень и лапка
сливаются в голенелапку). Среди ксилобионтов много энтомофагов.
Отряд Raphidioptera — Верблюдки (рис. 8, 7). Личинки верб¬
людок развиваются в лесной подстилке, но некоторые их виды
стали истинными ксилобионтами. Ноги личинок состоят из 5 от¬
делов (голень и 1-члениковая лапка не сливаются). По этому при¬
знаку и по общему габитусу они напоминают личинок жужелиц,
но не имеют церок. Кроме того, для личинок верблюдок характе¬
рен специфический коричневато-фиолетовый рисунок. Облигат¬
ные неспециализированные хищники.
Отряд Neuroptera — Сетчатокрылые, В древесине во влажных
местообитаниях встречаются личинки видов рода Osmylus. Они
являются облигатными хищниками. Охотятся под отслоившейся
корой и в полостях в толще древесины. Хорошо отличаются силь¬
но удлиненными игловидными мандибулами.
53
Piic. 8. Личинки насекомых, встречающиеся в древесине, из отрядов
верблюдок, чешуекрылых и перепончатокрылых
1 — Raphidla sp. (Raphldloptera); 2 — Urocerus gigas L.; 3 — Rhyssa persuasorla L.
(Hymenoptera); 4 — Cossus cossus L. (Lepidoptera)
Отряд Mecoptera — Скорпионницы. В древесине их личинки
встречаются редко. Это прежде всего белые С-образные личинки-
Boreus, характеризующиеся резким удлинением и утолщением
3-й пары ног. Они развиваются под мхом, в том числе на древес¬
ных стволах и пнях. Личинки Bittacus sp. иногда заползают в хо¬
ды насекомых, где, по-видимому, хищничают или поедают погиб¬
ших беспозвоночных. Этих гусеницеобразных личинок легко отли¬
чить по наличию на теле ветвистых выростов и ложных ножек.
Отряд Lepidoptera — Чешуекрылые, или бабочки (рис. 8, 4).
В ослабленных стволах развиваются крупные гусеницы древоточ¬
цев и стеклянниц. В сильно разложившейся древесине обитают гу¬
сеницы некоторых молей. Этих гусениц легко отличить по наличию
54
брюшных ног (до 5 пар), подошва которых снабжена упорядочен¬
но расположенными крючками. Фитоксилофаги и ксиломицето-
фаги, лишь немногие виды способны потреблять животную пиш,у.
Отряд Hymenoptera —, Перепончатокрылые. В древесине пред¬
ставлены большой группой энтомофагов, исключительно парази¬
тов. В древесине же сооружают гнезда и выкармливают личинок
некоторые пчелы и осы. Личинки ксилофильных перепончатокры¬
лых имеют С-образное безногое тело (рис. 8, 5), мягкие покро¬
вы и слабо склеротизованную голову со слабо развитым ротовым
аппаратом. Такие перепончатокрылые-ксилобионты, как рого¬
хвостообразные, характеризуются цилиндрической личинкой, от-
личающ,ейся хорошо развитой головой, наличием рудиментар¬
ных ног и развитием опорного шипа на конце тела (рис. 8, 2). Сле¬
дует подчеркнуть, что последний сегмент тела этих личинок поделен
дорсальной продольной бороздкой и не несет многочисленных во¬
лосков. В сильно разрушенной древесине нередко окукливаются
падающие с деревьев личинки пилильш^иков, которые имеют 6—
8 пар брюшных ног.
Отряд Diptera — Двукрылые, или комары и мухи. Представле¬
ны в древесине крупной группой сапроксилофагов, некрофагов
и энтомофагов, как хиш,ников, так и паразитов. Личинки их ли¬
шены грудных ног и в большинстве случаев имеют редуцирован¬
ную, по крайней мере в задних отделах, головную капсулу. У
личинок высших двукрылых головная капсула полностью исчеза¬
ет. Форма их тела разнообразна, но, за редким исключением, не
бывает С-образной. Дыхальца на конце тела бывают особенно
крупными, расположенными на особом стигмальном поле.
Изложенные выше характеристики показывают, что личинки
представителей различных отрядов, встречаюш,ихся в древесине,
по комплексу признаков легко различимы.
ВВЕДЕНИЕ В МОРФОЛОГИЮ ЛИЧИНОК НАСЕКОМЫХ
Как и взрослые насекомые, личинки имеют 3 пары грудных
ног — это их основной отличительный признак. Иногда ноги
отсутствуют.
Большинство личинок насекомых обладает явственно расчле¬
ненным на сегменты телом (рис. 9, i), причем 3 первых после го¬
ловы сегмента составляют грудной отдел (передне-, средне-, зад-
негрудь).
Брюшко личинок состоит из 7—10 сегментов. Последний
сегмент часто снабжен различными выростами или шипиками.
В строгой морфологической трактовке этот сегмент считается пред¬
последним, так как за последний сегмент морфологи принимают
различные рудименты на конце тела (анальную подпорку, плас¬
тинку и т. п.).
Голова личинок (рис. 9, 2) имеет различную ориентацию от¬
носительно туловиш,а. Она может быть направлена ротовыми час-
55
Рис. 9. Общий вид (i), голова сверху (2), нижняя губа и левая максилла
снизу (5), мандибулы (4—5), ноги \7—9) и урогомфы (10) личинок жестко¬
крылых
1,8 — Nudoblus lentus Grav. (Staphyllnldae); 2 — Gucujus caucasicus Motsch. (Gucujida^;
3 (слева — нижняя губа, lb\ справа — максилла, тх), в — Grynocharls oblonga L. (Lopho-
caterldae); 4 — Adelocera conspersa Gyll. (Blaterldae); 5 — Malthodes sp. (Cantharidae);
7 — Dromius sp. (Garabidae); 9 — Plagionotus arcuatus L. (Gerambycidae); lo — Pogono-
cerus thoracicus Flsch. (Pyrochroidae) (9 — no Мамаеву, Данилевскому, 1975)
о. ped — подталкиватель; abd — брюшко; ant — усики; cap — голова; cer — церии;
cl — клипеус; cx — тазик; crd — кардо; fem — бедро; /г — лоб; inb—-впязив^ие; гь—
нижняя губа; 1Ьг — верхняя губа; m — ментум; та — мала; md — мандибула; тх —
максилла; ос1 — глазок; р. 1Ь — нижнегубной щупик; р, тх — нижнечелюстной щупик;
ped — ноги; рг — простека; ргт — прементум; ге — ретинакулум; в. ver — теменной
шов; в. /г — ЛЪбный шов; st — стипес; iar — лапка; th — грудь; tib — голень; tibt — голе-
нелапка; tr — вертлуг; ung — коготок; иг — урогомфы
тями вперед (прогнатные формы) или вниз (гипогнатные формы).
Головная капсула разграничена швами — непарным средин¬
ным теменным и отходящими от него лобными, которые ограничи¬
вают лобный треугольник. К лобному треугольнику присоединя¬
ется наличник и верхняя губа, по бокам от которых располагают¬
ся мандибулы. На боковых сторонах головы находятся точечные
глазки, а у личинок насекомых с неполным превращением — фасе¬
точные глаза и прикрепляются усики. На нижней поверхпостп
головы различают срединную и боковые лопасти, которые носят
названия нижней губы и нижних челюстей (максилл). У личинок
некоторых групп насекомых (высшие двукрылые) головная кап¬
сула полностью не развивается.
Грудные сегменты у многих групп личинок внешне почти не
отличаются от брюшных, особенно когда грудные ноги и зачатки
крыльев отсутствуют. Вполне развитые грудные ноги у Holometa-
bola состоят из 5 отделов: тазика, вертлуга, бедра, голени и 1-чле-
никовой лапки (рис. 9, 7). Однако у многих личинок ноги бывают
сильно укорочены, голень может сливаться с лапкой, и нога под¬
разделяется на 4 отдела: тазик, вертлуг, бедро и голенелапку
(рис. 9, 8), Ноги, как правило, заканчиваются коготками.
Брюшные сегменты обычно слабо отличаются друг от друга,
за исключением 1—2 последних, часто снабженных придатками.
Эти придатки могут иметь самое различное строение. Различают
два основных типа придатков: хвостовые нити, или церки (рис.
9, 2), и прочные опорные отростки (рис. 9, 10), которые обычно
называют урогомфами.
Покровы тела скрытноживущих личинок обычно гладкие или
в мелких волосках и шипиках. Открытоживущие личинки, нао¬
борот, как правило, покрыты волосками, иногда очень густыми,
или причудливыми выростами, шипами или шипиками. Интен¬
сивность окраски тела также значительно ниже у скрытноживу¬
щих форм, тогда как обитатели растений обычно окрашены либо
в зеленоватые и бурые маскирующие тона, либо, наоборот, обла¬
дают очень яркой отпугивающей окраской.
Все перечисленные особенности строения и окраски личинок
насекомых придают им вполне определенный облик. Поэтому даже
при чисто поверхностном осмотре легко отличить друг от друга
личинок представителей различных отрядов, а иногда и семейств
насекомых.
ОТРЯД COLEOPTERA-
ЖЕСТКОКРЫЛЫЕ, ИЛИ ЖУКИ
В Палеарктике достаточно тесные связи с древесиной имеют
представители более чем 60 семейств жесткокрылых. Из подотря¬
да Archostemata в древесине развиваются чрезвычайно интерес¬
ные в эволюционном отношении древнянки (Cupedidae). Хотя в
57
состав другого подотряда жесткокрылых — Adephaga входят преи¬
мущественно почвенные и подстилочные формы, он представлен
в древесине также истинными ксилобионтами — семейством нан-
цирников (Rhysodidae). Основные же группы ксилофильных жест¬
кокрылых относятся к подотряду Polyphaga.
В составе этого подотряда имеются крупные надсемейства,
обязанные своим становлением древесине как среде обитания.
К их числу относятся, например, пестряковидные (Cleroidea),
капюшонниковые (Bostrychoidea), а также многочисленные отдель¬
ные семейства, представленные истинными ксилобионтами, осо¬
бенно в группе Heteromera.
Жесткокрылые как энтомофаги уже привлекли внимание спе¬
циалистов по защите леса, так как их отдельные виды, особенно
связанные с короедами, достигают высокой численности и могут
существенно влиять на динамику популяции вредителей.
СРАВНИТЕЛЬНАЯ МОРФОЛОГИЯ ЛИЧИНОК ЖЕСТКОКРЫЛЫХ
^Црчинки жесткокрылых отличаются весьма разнообразной
формой тела (рис. 10—22), но всегда имеют достаточно хорошо раз¬
витую голову и в большинстве случаев функционирующие грудные
ноги.
Их тело состоит из 3 грудных и 10 брюшных сегментов, однако
последние сегменты могут редуцироваться или видоизменяться,
так что хорошо развитых, четких брюшных сегментов — 9,
а иногда даже 8. Известны единичные случаи, когда тело личинок
приобретает дополнительную сегментацию, тогда большинство
брюшных сегментов бывает поделено складкой на две неравные
части, как это наблюдается у личинок некоторых щелкунов (Саг-
diophorinae) из семейства Elateridae (рис. 19, 1) и древоедов (Ме-
lasinae) из семейства Eucnemidae. У личинок некоторых пестря¬
ков, например Orthopleura sanguinicollis L. (Cleridae), тело силь¬
но удлинено и также со следами вторичной сегментации.
Покровы тела личинок отличаются по степени склеротизал,ии,
окраске и наличию различных кутикулярных структур.
Тело личинок, за исключением головы, в большинстве случаев
имеет слабо склеротизованные бесцветные покровы, в результате
чего их окраска зависит от цвета жирового тела. Личинкам жест¬
кокрылых присуща вся гамма цветовых оттенков, однако подав¬
ляющее большинство из них — белые, что в целом свойственно
скрытнообитающим формам. С другой стороны, в отряде имеются
семейства, например щелкуны (Elateridae), для личинок которых
характерна интенсивная пигментация покровов — большинство
их личинок бурые, некоторым личинкам свойствен почти черный
цвет. Ярко-красными бывают личинки пестряков Thanasimus
(Cleridae), причем интересно, что личинки одного и того же вида
демонстрируют всю гамму окрасок от красной до сине-фиолето-
58
вой. у личинок этой группы семейств иногда наблюдается мрамор¬
ная расцветка (Tillus elongatus L., Lepidopteryxsquamulosa Gebl.).
У многих личинок на дорсальной стороне грудных сегментов,
особенно на переднегруди, бывают развиты темные склеротизо-
ванные пятна. Личинки разных возрастов даже одного и того же
вида могут существенно отличаться как по интенсивности окрас¬
ки, так и в редких случаях — по цвету.
Тело большинства ксилофильных личинок жесткокрылых вы¬
глядит гладким — па сегментах имеется лишь небольшое число
малозаметных волосков и щетинок. Однако, особенно у хищных
форм, встречаются личинки, которые имеют бархатистое тело бла¬
годаря густому покрову коротких волосков. Иногда цвет этих во¬
лосков определяет окраску всей личинки. На теле личинок могут
быть развиты крепкие опорные щетинки со всеми переходными
формами, которые различные авторы называют либо щетинками,
либо волосками. На сегментах тела (вооружение последнего сег¬
мента будет рассмотрено особо) могут быть развиты различные ку-
тикулярные опорные образования. К их числу в первую очередь
следует отнести поля шипиков, которые обычно имеются на дор¬
сальной, реже также на вентральной и очень редко — на боковых
сторонах. Такие поля шипиков имеются, например, у личинок уз-
котелок рода Colydium (Colydiidae; рис. 16, i), но особенно они
типичны для сапроксилофагов из семейств Zopheridae (рис. 14, 2),
Sienotrachelidae, Eucnemidae и др. Кроме шипиков, в ряде слу¬
чаев развиваются бугорковидные опорные образования, ряды
зубцов или поперечный склеротизованный гребень. Поля бугор¬
ков на дорсальной стороне тела особенно типичны для видов Ери-
гаеа (Nitidulidae; рис. 18, 4). Поперечный склеротизованный гре¬
бень в виде тонкой линии развит на дорсальной и вентральной сто¬
ронах личинок Cucujus (Cucujidae; рис. 18, i), Pytho (Pythidae;
рис. 16, 2) и т. д. У обитателей сыпучих субстратов, таких, как
древесная труха в дуплах, тело идеально гладкое (некоторые Е1а-
teridae, Alleculidae и Tenebrionidae).
Головная капсула у личинок жуков в большинстве случаев
хорошо развита, так как обеспечивает размещение и прикрепле¬
ние мускулатуры, приводящей в движение мощные верхние че¬
люсти-мандибулы и другие ротовые придатки. Лишь у представи¬
телей немногих групп (Trixagidae, Eucnemidae, Lycidae, Cerylo-
nidae) головная капсула уменьшена в размерах или видоизменена.
У личинок Cerylon, например, она утратила склеротизацию и под¬
верглась весьма сильной редукции.
На дорсальной стороне головной капсулы, как правило, мож¬
но различить линии, по которым головная капсула растрескива¬
ется при линьках: это срединный (теменной) и симметрично расхо¬
дящиеся к углам лобные швы (рис. 9, 2), Лобные швы ограничива¬
ют лобный треугольник (лоб). Головные швы хорошо развиты у
хищных личинок из семейств стафилинид (Staphylinidae), малашек
(Melyridae) и др. У пестряков (Cleridae) и темнотелок (Trogossiti-
59
dae) полушария головы позади точки слияния лобных швов спая¬
ны прямолинейным швом или треугольным склеритом. У личинок
карапузиков (Histeridae) головные швы едва намечены. Иногда
теменной шов отсутствует, и лобный треугольник заканчивается
на заднем крае головной капсулы. Лобная пластинка может
иметь и трапециевидную форму.
Задний край головной капсулы ограничивает затылочное от¬
верстие, а ее боковые части могут сливаться на вентральной по¬
верхности, где в этом случае имеется срединный продольный шов,
но чаш,е с вентральной стороны головы имеется выемка для раз-
меш;ения ротовых органов. В заднем отделе головной капсулы с
вентральной стороны обычно имеется склерит, соединяюш,ий края
боковых частей головной капсулы в монолитное образование —
это гуларная, или горловая, пластинка. По названию этой плас¬
тинки обозначаются и соответствующие швы.
С дорсальной стороны лобный треугольник переходит в спаян¬
ную с ним трапециевидную пластинку — наличник, или клипеус.
Граница между лбом и наличником в большинстве случаев явст¬
венная, хотя и не представляет собой истинного шва. К свобод¬
ному краю наличника подсоединяется подвижная верхняя губа,
котор#1Я в большинстве случаев имеет округло-сердцевидную фор¬
му. Ее внутренняя поверхность несет многочисленные шипики,
щетинки и другие кутикулярные структуры и носит название эпи-
фаринкса, или надглоточника. Личинки жужелиц (Garabidae),
щелкунов (Elateridae), многих карапузиков (Histeridae) не имеют
верхней губы, передний край их головной капсулы преобразован
в склеротизованный зубчатый выступ — назале.
Мандибулы, или верхние челюсти (рис. 9, 4—6),— важнейшая
часть ротового аппарата личинок жесткокрылых. Обычно они
направлены вершинами навстречу друг другу, лишь у немногих
личинок, например у представителей семейства древоедов (Еиспе-
midae), жвалы развернуты вершинами наружу. Внутренняя по¬
верхность типичных грызущих мандибул в основании образует
покрытую ребрами илп бугорками жевательную, или моларную,
поверхность. Апикальная доля молы может формировать так назы¬
ваемый премоларный зубец. Вторичное расширение основания ман¬
дибул некоторых преимущественно хищных личинок жесткокрылых
называют псевдомолой. На внутреннем крае мандибулы в ее средин¬
ной части может быть развит склеротизованный зубец — ретина-
кулум (рис. 9, 5). Ретинакулум хорошо выражен у личинок мно¬
гих хищников, таких, как пестряки (Cleridae), темнотелки (Тго-
gossitidae), многие мягкотелки (Cantharidae) и др. Если зубец не
склеротизован или сочленен с поверхностью мандибулы или если на
внутреннем крае мандибулы имеется группа щетинок, то такие обра¬
зования носят название простеки (рис. 9, 6). Иногда одновременно
бывают развиты и ретинакулум и простека. Вершина мандибулы
может быть простой или несет 2 или несколько зубцов. Колющие
мандибулы с простой вершиной характерны для хищных личппок
60
из семейств мягкотелок (Cantharidae), светляков (Lampyridae) и
близких групп (Lycidae), у которых па мандибулах имеется внутрен¬
ний или наружный канал для всасывания разжиженной пищи при
внекишечном пищеварении.
Максиллы, или нижние челюсти (рис. 9, 5), отличаются слож¬
ным строением. В их основании расположен обособленный скле¬
рит — кардо. Иногда кардо поделен на две части. К кардо причле-
няется крупная пластинка, основание которой примерно до уров¬
ня нижнечелюстных щупиков составляет особый отдел — стипес.
У некоторых личинок, например у карапузиков (Histeridae), сти¬
пес подразделяется на основную (собственно стипес) и придаточ¬
ную (костипес) части. Соотношение размеров кардо и стипеса мо¬
жет быть различным. Обычно они лишь незначительно отличаются
по величине, иногда же, как, например, у темнотелок (Trogossi-
tidae), кардо значительно меньше стипеса. Концевая часть рас¬
сматриваемой пластинки нередко делится на две лопасти: внут¬
реннюю — лацинию U наружную — галеа. Такое подразделение
достаточно хорошо выражено лишь у немногих групп, среди ко¬
торых, например, пилильщики (Byrrhidae), жужелицы (Carabi-
dae) и некоторые другие. В большинстве же случаев концевая
часть максилл не дифференцирована и имеет название жеватель¬
ной лопасти, или малы. Жевательная лопасть на вершине по внут¬
реннему краю усажена характерными кинжаловидными или обыч¬
ного типа щетинками и может нести пальцевидные придатки. На
месте лацинии нередко сохраняются один или несколько склеро-
тизованных зубцов. Наличие или отсутствие этих зубцов является
важным диагностическим признаком. Нижнечелюстные щупики
обычно состоят из трех члеников и сидят на щупиконосном бугор¬
ке, который нередко выглядит как самостоятельный членик,
у хищных личинок жужелиц (Carabidae) и некоторых стафилинид
(Staphylinidae) они 4-члениковые. На конце щупиков имеются
различные сенсорные придатки.
Нижняя губа подразделяется на три отдела: субментум, мен-
тум и прементум (рис. 9, 5). Иногда эти отделы сливаются друг с
другом или с другими склеритами вентральной части головы. Ес¬
ли сливаются субментум с ментумом, то в результате возникает
единый отдел — постментум. Субментум может сливаться также
с гуларным склеритом. Прементум заканчивается язычком, или
лигулой, который иногда недоразвивается. У личинок жесткокры¬
лых между основаниями максилл и нижней губы нередко имеется
склерит — сочленовная доля (рис. 9, 3). Активно хищничающие
личинки таких семейств, как мягкотелки (Cantharidae), щелкуны
(Elateridae), темнотелки (Trogossitidae) и др., имеют малоподвиж¬
ные, спаянные друг с другом вентральные ротовые части. Внут¬
ренняя сторона нижней губы имеет сложное строение, покрыта
волосками и нередко несет различные склеротизованные струк¬
туры. Она называется гипофаринксом, или подглоточником, а ее
склеротизованная часть — гипофарингеальной склеромой. Пос¬
61
ледняя особенно хорошо развита у личинок, питающихся твердой
пищей, у специализированных хищных форм отсутствует. Нижне¬
губные щупики, как правило, 2-члениковые, реже 1- или 3-члени-
ковые (в некоторых работах их первым члеником считают щупико¬
носный бугорок).
Усики у большинства личинок жесткокрылых хорошо развиты,
у хищных форм их длина может превышать длину головы. Они си¬
дят на усиконосном бугорке у наружных углов лобного треуголь¬
ника. Обычно усики состоят из трех члеников, но у личинок жуже¬
лиц (Carabidae) бывают 4-члениковыми, а у некоторых полувод-
ных жесткокрылых (Helodidae) выглядят многочлениковыми (рис.
14, 3) из-за многократного вторичного подразделения их 3-го чле¬
ника на кольца. У личинок слоников (Gurculionidae) усики, на¬
оборот, сильно редуцированы и представлены нечленистым бугор¬
ком. Предпоследний членик нормально развитых усиков обычно
значительно толще последнего, несет на вершине одну, реже не
сколько конусовидных сенсилл. Наибольшее число сенсилл — 4,
отмечено у личинок некоторых темнотелок (Trogossitidae). На
конце усиков также имеются различные мелкие сенсорные придат¬
ки. У хищных личинок Gleridae и близких групп усики сидят на
перепончатом основании и могут глубоко втягиваться в головную
капсулу. У основания усиков на головной капсуле располагаются
глазки. Развитый глазок состоит из пигментного пятна, которое
расположено под умеренно выпуклой линзой. Максимальное чис¬
ло глазков — по 6 с каждой стороны головы. Такой признак, как
наличие или отсутствие глазков, а также их число, как правило,
констацтен в пределах рода. У представителей отдельных семейств,
например у плоскотелок (Cucujidae), число глазков варьирует от
О до 6 (с каждой стороны головы). У фиксированных в спирте ли¬
чинок пигментное пятно может отслаиваться от линзы, в связи:
с чем число глазков не всегда служит надежным диагностическим
признаком.
Грудной отдел у подвижных личинок жесткокрылых, как пра¬
вило, хорошо обособлен, так как несет функционирующие конеч¬
ности. У представителей таких семейств, как дровосеки (Geramby-
cidae), златки (Buprestidae) и некоторые другие, личинки имеют
укрупненную хорошо развитую переднегрудь (рис. 22, 2), на ко¬
торой размещены опорно-фиксирующие приспособления, а сред¬
не- и заднегрудь слабее отличается от брюшных сегментов. На дор¬
сальной стороне переднегруди нередко бывает развит щиток (рис.
12, J; 13, 5; 20, 1—3), разделенный продольной светлой или тем**
ной линией, по которой происходит растрескивание шкурки при
линьках. Такой щиток наиболее характерен именно для хищных
личинок семейств карапузиков (Histeridae), пестряков (Gleridae),
темнотелок (Trogossitidae) и т. д. У личинок пластинчатоусых
(Scarabaeidae), слоников (Gurculionidae) и некоторых других групп
грудные сегменты, включая переднегрудь, с дорсальной стороны
поделены несколькими глубокими поперечными складками.
«2
Ноги (рис. 9, 7—9) у личинок жесткокрылых могут быть хо¬
рошо развитыми, рудиментарными или полностью редуцирован¬
ными. Хорошо развитая нога состоит чаш,е всего из 4 отделов (рис.
9, 8): тазика, вертлуга, бедра и голенеланки. У личинок жужелиц
(Carabidae) и других представителей подотряда Adephaga голень
и лапка обособлены в виде самостоятельных члеников (рис. 9, 7),
что соответствует более примитивному состоянию конечностей
жесткокрылых. Такую состояш,ую из 5 отделов конечность имеют,
например, личинки древнянок (Cupedidae). Лапка или голенелап-
ка заканчивается 1, реже 2 коготками. Коготки могут быть прос¬
тыми или снабженными зубчиком. У личинок, интенсивно пере-
двигаюш;ихся в субстрате, нередко передняя пара ног развита бо¬
лее сильно. К их числу относятся пыльцееды (Alleculidae) и мно¬
гие чернотелки (Tenebrionidae). Наоборот, у личинок, которые
прокладывают ходы в основном с помош.ью челюстей (Buprestidae,
Cerambycidae и др.), ноги сильно укороченные, не функционирую¬
щие, а во многих случаях полностью редуцированные.
Части ног, особенно бедро и голенелапка, бывают вооружены
шипиками и опорными ш,етинками. У личинок пестряков (Cleri-
dae) и близких семейств на конце голенеланки развиты длинные
ш;етинки, крючковидно загнутые на конце и, очевидно, имеюш,ие
значение зацепок для подтягивания тела. Иногда, например у
некоторых личинок пыльцеедов (Alleculidae), на вертлугах и бед¬
рах имеются своеобразные, сильно измененные шаровидные или
конусовидные ш,етинки. Тазики ног нередко несут опорные пло¬
щадки, помогающие движению.
Брюшко личинок насчитывает до 10 сегментов, однако в ре¬
зультате специализации концевого отдела их число может сокра¬
щаться до 8 видимых сегментов. Только 8 явственных брюшных
сегментов из ксилофильных хищников имеют, например, личинки
водолюбов рода Сегсуоп (Hydrophilidae), 1—7-й брюшные сегмен¬
ты обычно более или менее одинаковы. Они могут нести следы
склеротизации как на дорсальной, так и на вентральной сторо¬
нах, но в большинстве случаев их покровы равномерно склеро-
тизованы, причем степень склеротизации, как правило, неве¬
лика.
Брюшные сегменты могут иметь различные локомоторные обра¬
зования. У личинок дровосеков (Cerambycidae), крупных тене¬
любов (Melandryidae), темнотелок (Trogossitidae) и некоторых дру¬
гих групп брюшные сегменты, за исключением последних, снаб¬
жены дорсальными и вентральными двигательными мозолями
(рис. 13, 7), которые имеют морщинистую или гранулированную
поверхность. У большинства личинок узкокрылок (Oedemeridae)
на вентральной стороне брюшных сегментов развиты «ложные»
ножки (рис. 12, 4),
На боковых сторонах брюшных сегментов многих личинок жест¬
кокрылых, в том числе имеющих плотные покровы, хорошо выра¬
жена стерно-плевральная складка. Личинки некоторых пыльце-
вз
едов (Alleculidae), однако, такой складки не имеют, покровы их
сегмента образуют единое склеротизованное кольцо. Существует
большая группа личинок, у которых в соответствии с особенно¬
стями их передвижения в субстрате брюшные сегменты (особенно
с дорсальной стороны) поделены многочисленными поперечнымп
складками. Этот признак наиболее характерен для некоторых
С-образных личинок, у которых при передвижении опорную на¬
грузку несут брюшные тергиты. К их числу относятся личинки
бронзовок (семейство Scarabaeidae; рис. И, 5), капюшонников
(Bostrychidae; рис. И, 4), точильщиков (Anobiidae; рис. И, 6),
слоников (Curculionidae; рис. 22, 3) и др.
Предпоследний и особенно последний брюшные сегменты, как
правило, несу! различные онорно-фиксирующие образования.
Простейшими из этих образований являются ноля или ряды ши-
пиков с вентральной стороны или 1—2 крупных опорных дорсаль¬
ных шипа. Такие шипы характерны для личинок бугротелок (Zop-
heridae; рис. 14, 2), некоторых тенелюбов (Melandryidae; рис. 13,
2), чернотелок (Tenebrionidae; рис. 17, 2) и др. У некоторых личи¬
нок на конце тела развиваются шагренированные или ребристые
склеротизованные пластинки. Однако наибольшее значение для
диагностики личинок имеют парные концевые придатки на дорсо-
каудальной части 9-го брд)шного сегмента — церки и склеротизо¬
ванные выросты — урогомфы. Различия между этими опорными
образованиями скорее функционального порядка — считается,
что церки являются чувствующими придатками, а урогохмфы не¬
сут только опорную нагрузку. Однако очевидно, что и церкам,
и урогомфам присущи обе функции, только в первом случае чув¬
ствующая развита в большей степени, чем во втором. Церки ли¬
чинок жуков чаще всего состоят из 2 члеников. Наиболее прими¬
тивные церки — 4-члениковые, как это наблюдается, например,
у личинок представителей семейства Synteliidae. Нередко церки
1-члениковые, но тогда они, как правило, сочленены с 9-м брюш¬
ным тергитом. Урогомфы могут быть простыми, как, например,
у личинок огнецветок (Pyrochroidae; рис. 16, 5), разветвленными,
как у многих личинок щелкунов (Elateridae; рис. 19, 4), снабжен¬
ными боковыми шинами (Peltidae; рис. 13, 3) пли иметь в основа¬
нии склеротизованную площадку — такую, как у представителей
семейств группы Cleroidea (рис. 20, 1—3).
Между крайними вариантами опорных структур 9-го брюш¬
ного сегмента существуют все переходные ступени, поэтому в ря¬
де конкретных случаев трудно однозначно отнести их либо к цер¬
кам, либо к урогомфам. Следует подчеркнуть, однако, что урогом¬
фы не бывают членистыми и, как правило, отличаются сильной
склеротизацией. Вместе с видоизмененным 9-м брюшным сегмен¬
том урогомфы нередко обеспечивают не столько поступательное,
сколько возвратное движение по ходам и полостям. У личинок
огнецветок (Pyrochroidae), плоскотелок (Cucujidae) и других плос¬
котелых форм обслуживающая урогомфы мускулатура расположе-
64
lia внутри 8-го брюшного сегмента, размеры которого нередко
cHJrbHO увеличиваются.
Конусовидный или цилиндрический выступ на конце тела с
вентральной стороны в большинстве случаев представляет собой ви¬
доизмененный 10-й брюшной сегмент. Независимо от особенностей
его происхождения у разных групп личинок он обозначается как
подталкиватель, или анальная подпорка. У личинок стафилинид
(Staphylinidae; рис. 12, 3), некоторых плоскотелок (Gucujidae;
рис. 18, 2) и др. анальная подпорка длинная, конусовидная.
У мягкотелок (Cantharidae) она короткая, более или менее цилинд¬
рическая.
Дыхальца у личинок жесткокрылых расположены на средне-
груди и 1—8-м брюшных сегментах. Личинки, развивающиеся
в жидких средах, имеют существенные отклонения от этого основ¬
ного типа, так как на конце тела нередко возникают дыхальце¬
носные выросты (рис. 21, i), камеры и иные образования.
Приводимая ниже определительная таблица имеет прикладной
характер: она базируется в основном на признаках внешнего обли¬
ка личинок, без учета филогенетических отношений между рас¬
сматриваемыми группами. Она составлена по палеарктическим
материалам и учитывает только признаки личинок, относящихся
к ксилофильным группам. Обитатели подстилки и плодовых тел
грибов включены в тех случаях, когда они встречаются также и в
древесине.
Таблица для определения семейств отряда
Coleoptera — Жесткокрылые
1 (106) Ноги имеются — вполне развитые, функционирующие
или по крайней мере в виде отчетливых рудиментов. Ес¬
ли ноги полностью редуцированы (некоторые Cerambyci-
dae), то первые 6 или 7 брюшных сегментов с дорсальной
и вентральной сторон снабжены парными двигательными
мозолями (рис. 10, i), разделенными продольным вдав-
лением.
2 (7) Ноги состоят из 5 отделов (тазик, вертлуг, бедро, го¬
лень, 1-члениковая лапка) и заканчиваются 1—2 когот¬
ками.
3 (4) На конце тела имеется непарный склеротизованный от¬
росток (рис. 10, 2). Моларная часть мандибул хорошо
развита. В древесине. Сапроксилофаги
Cupedidae — Древнянки
4 (3) На конце тела придатки отсутствуют или имеются пар¬
ные церки.
5 (6) На конце тела имеются церки (рис. 10, 5), с вентральной
стороны — подталкиватель. Вентральные ротовые час¬
ти сильно вытянутые. Почвообитающие формы, некото¬
рые виды развиваются под отслоившейся корой или в
3 в. м. ]\1амаев и др. 65
Рис. 10. Общий вид личинок жесткокрылых семейств Cerambycidae (/)»
Gupedidae (2), Garabidae (3) и Rhysodidae (d)
г — Mesosa myops Dalm; 2 — Cupes sp.; 3 — Trechus sp.; 4 — Rhysodes comes Rtt. (2 —no
Boving, Craighead, 1931; 5 — no Шаровой, 1958)
толще сильно разложившейся древесины. Хищники
Garabidae — Жужелицы
6 (5) На конце тела церки и подталкиватель не развиты (рис.
10, 4). Вентральные ротовые части втянутые. Образуют
скопления в сильно разложившейся древесине. Пита¬
ние не изучено Rhysodidae — Панцирники
7 (2) Ноги состоят не более чем из 4 отделов (тазик, вертлуг,
бедро, голенеланка), заканчиваются 1 коготком или ко¬
готки редуцируются.
6в
8 (21) Тело короткое, толстое, согнутое С-образно, на конце
закругленное, без церок, ур^гомф, дорсальных крюч¬
ков или шиников.
9 (10) Личинки зеленовато-бурые, с хорошо склеротизованны-
ми покровами (рис. И, 1). Переднегрудной щит хорошо
выражен. Почвенные формы. Под отставшей корой,
Сапрофаги Byrrhidae — Пилюльщики
10 (9) Личинки белые или грязновато-белые. Переднегрудной
щит не выражен.
И (20) Ноги всегда хорошо развиты.
12 (15) Усики 4-члениковые. Личинки крупные, длиной до
100 мм. Во влажной древесине.
13 (14) Тергиты брюшных сегментов гладкие. На конце тела
имеются парные выпуклые мозоли (рис. И, 2), В древе¬
сине. Сапроксилофаги Lucanidae — Рогачи
14 (13) Тергиты брюшных сегментов разделены поперечными
складками (рис. И, 3). На конце тела нет парных вы¬
пуклых мозолей. В сильно разложившейся древесине
и древесной трухе. Сапроксилофаги
Scarabaeidae — Жуки пластинчатоусые
15 (12) Усики укороченные, состоят не более чем из 3 члеников.
Личинки мелкие, длиной до 15 мм. В сухой древесине.
16 (19) Голова частично втянута в переднегрудь (рис. И, 5).
Усики состоят из 2—3 члеников. На брюшных тергитах
нет поперечных рядов шиников.
17 (18) Последняя пара дыхалец (на 8-м брюшном сегменте)
значительно крупнее остальных (рис. И, 5). В древеси¬
не. Ксилофаги Lyctidae — Древогрызы
18 (17) Последняя пара дыхалец не крупнее остальных (рис. И,
4). В древесине. Ксилофаги
Bostrychidae — Капюшонники
19 (16) Голова свободная (рис. И, 6). Усики короткие, неяв¬
ственные, 1—2-члениковые. Брюшные тергиты с попе¬
речными рядами шипиков. Под корой и в древесине.
Ксилофаги
Anobiidae — Точильщики
20 (И) Ноги короткие, рудиментарные. В древесине, реже под
корой Anthribidae — Ложнослоники
21 (8) Тело различной формы, никогда не бывает согнутым
С-образно, нередко на конце с церками, урогомфами,
дорсальными крючками или шипиками.
22 (31) На 9-м брюшном сегменте имеются 2—4-члениковые или
1-члениковые, но подвижно сочлененные церки (рис. 12,
1-3).
23 (24) Церки состоят из 4 члеников. Внешне личинки весьма
сходны с личинками некоторых карапузиков (рис. 12»
1). Единственный род Syntelia Westw., представленный
3* 67
Рис. и. Общий вид С-образных личппок жесткокрылых
1 — ByiThus sp. (Byrrhidae); 2 — Lucanus (crvus L. (Lucanidae); 3 — Cetoiiia aurata L.
(Scarabaeidac); 4 — Stcphanopachys substriatus Pk. (Bostrychidac); 5 — Lyctus brunneus
Steph. (Lyctidae); 6 — Anobium punctatum Deg. (Anobiidae)
В фауне СССР видом S. histeroides Lew. (о. Кунашир).
Облигатные всеядные хищники, питающиеся в основном
личинками двукрылых . . .Synteliidae —Чернопузики
24 (23) Церки состоят из 1—2 члеников.
25 (26) Тело сильно расширенное и уплощенное, мокрицеоб¬
разное. Тергиты с крупными боковыми выростами. На
68
Рис. 12. Общий вид личинок жесткокрылых с истинными церками или вен¬
тральными локомоторпыми выростами
1 — Hololepta plana Sulz.; 2 — Plepadenis vulneratus Pz. (Histeridae); 3 — Placusa comp¬
lanata Er. (Staphylinidae); 4 — Galopus serraticornis L. (Oedemeridae)
падали, иногда встречаются в трухлявой древесине . .
Silphidae — Мертвоеды
26 (25) Тело существенно не расширенное. Крупных боковых
выростов на тергитах нет.
27 (28) Брюшные тергиты разделены поперечными складками
(рис. 12, i, 2). Лереднеспипка обычно такой же длины
69
или длиннее средне- и заднеспинки, вместе взятых.
Анальная подпорка очень короткая
Histeridae — Карапузики (стр. 86)
28 (27) Брюшные тергиты без поперечных складок (рис. 12, 3).
Переднеспинка короче средне- и заднеспинки, вместе
взятых. Анальная подпорка длинная, конусовидная.
29 (30) Усики длиннее головы. Тело в длинных простых щетин¬
ках. Подстилочные формы, встречающиеся на поверх¬
ности древесины и в трещинах. Сапромицетофаги . . .
. Scaphidiidae — Челновидки
30 (29) Усики короче головы, если длиннее (Trigonurus Miils.),
то голова и тело несут крупные булавовидные щетинки
Staphylinidae — Стафилиниды (стр. 97)
31 (22) На 9-м брюшном сегменте членистых или 1-члениковых
сочлененных церок нет. Опорные приспособления на
конце тела могут быть представлены несочлененными
урогомфами, крючками, шипиками и т. п.
32 (33) На вентральной стороне средних брюшных сегментов
имеются 2—4 пары длинных локомоторных выростов,
подошва которых несет шипики (рис. 12, 4). В древеси¬
не. Ксилофаги и сапроксилофаги
Oedemeridae — Узконадкрылки
33 (32) На вентральной стороне средних брюшных сегментов
парные локомоторные выросты отсутствуют.
34 (39) На дорсальной и обычно также на вентральной сторонах
брюшных сегментов имеются умеренно выпуклые мор¬
щинистые или гранулированные двигательные мозоли
(рис. 10, 1; 13, 1) Личинки умеренно длинные, мя¬
систые, белые. Опорные структуры на конце тела не вы¬
ражены или представлены 2 мелкими дорсальными или
1 концевым шипиками, реже — 2 загнутыми вверх крюч-
ками (Xylita Рк., Zilora Muls. из Melandryidae) ^ или
сложными урогомфами — раздваивающимися на конце
или снабженными боковыми шипами (Calitys Thoms.^
Thymalus Latr. из Peltidae).
35 (36) Продольный срединный киль (эндокарина) между лоб¬
ными швами на головной капсуле хорошо выражен.
Анальная щель в большинстве случаев тре^хлучевая.
Под корой и в древесине. Ксилофаги
Cerambycidae — Дровосеки
^ У некоторых Melandryidae двигательные мозоли слабо развиты, а у
Xylita Рк.— не выражены и функционально замещены поперечным рядом:
шипиков, который в отличие от сходных личинок других семейств имеется
только на средне- и заднегруди и двух первых брюшных сегментах
(рис. 13, 2).
2 Сюда попадает также личинка Synchita Hellw. (рис. 14, 1) — рода, отно¬
сящегося к Colydiidae (стр. 176) и характеризующегося аберрантными:
признаками.
70
Рис. 13. Общий вид личинок жесткокрылых с двигательными мозолями или
поперечными рядами шипиков
1 — Melandrya dubia Schall.; 2 — Xylita livida Sahlb. (Melandryidae); 3 — Thymalus
limbatus F. (Peltidae)
36 (35) Продольный срединный киль (эндокарина) между лоб¬
ными швами не выраихен. Анальная щель поперечная или
многолучевая.
37 (38) На конце тела имеются сложные урогомфы (рис. 13, 3)
или 2 шипика, но тогда боковые стороны головной кап¬
сулы густо усажены волосками. В разрушенной древе¬
сине, реже в грибах и под корой. Сапроксилофаги (ли¬
чинки Calitys scabra Thunb. считаются хищниками и,
возможно, относятся к семейству Trogossitidae) ....
Peltidae — Щитовидки
71
38 (37) На конце тела сложные урогомфы отсутствуют; опор¬
ные структуры не выражены (рис. 13, 1) или представ¬
лены 2 крючками или 2 шипиками, но тогда боковые сто¬
роны головной капсулы почти голые Под корой, в дре¬
весине, в плодовых телах грибов. Ксилофаги, сапрокси-
лофаги и мицетофаги
• • • Melandryidae — Тенелюбы
39 (34) На дорсальной и вентральной сторонах брюшных сег¬
ментов двигательные мозоли не выражены а если на
средних брюшных сегментах имеются (некоторые С1е-
ridae и Trogossitidae), то урогомфы простые, с хорошо
развитой склеротизованной плош,адкой в основании.
40 (41) Усики удлиненные (рис. 14, 5), так как их 3-й членик
вторично поделен на большое число колец. Обитатели
микроводоемов в дуплах деревьев
Helodidae — Трясинники
41 (40) Усики не удлиненные, их 3-й членик цельный.
42 (43) Тело (рис. 15, 1) покрыто, особенно в каудальной
части, густыми длинными нередко яркоокрашенными
волосками. Некрофаги. Ксилофильные виды живут под
корой и питаются сухими погибшими насекомыми . . .
Dermestidae — Кожееды
43 (42) Тело обычно с очень слабым покровом из волосков.
44 (45) Переднегрудь личинок сильно вздута и капюшенообраз-
но прикрывает голову (рис. 15, 5). На конце тела име¬
ется непарный склеротизованный вырост или куполовид¬
ная опорная плош,адка. В древесине. Амброзийные кси-
ломицетофаги и ксилофаги
Lymexylonidae — Сверлильш,ики
45 (44) Переднегрудь личинок не вздута.
46 (93) На конце тела имеются различные опорные структуры:
урогомфы, загнутые крючки, опорный вырост, шипики
и т. п. или последний сегмент закругленный, куполо¬
видный или уплош,енный, покрытый длинными волоска¬
ми (Alleculidae, некоторые Elateridae, включая Cardio-
phorinae, и Tenebrionidae).
47 (48) Ноги представлены нефункционирующими мясистыми
придатками. На конце тела имеется непарный склероти¬
зованный вырост (рис. 15, 2). В древесине. Ксилофаги
Mordellidae — Горбатки
48 (47) Ноги вполне развитые, функционирующие, иногда уко¬
роченные, но в этом случае на конце тела нет непарного
склеротизованного выроста.
Сюда попадают также сильно отклоняющиеся по форме личинки Tcncrus
Cast. (Cloridao, стр. 117).
У Zophoridac и Stenotrachclidae имеются слабо выраженные двигательные
мозоли. Однако эти мозоли нетипичны, так как покрыты овалом или полем
крупных ппшиков (рис. 14, 2).
72
Рис. 14. Общий вид морфологически уклоняющихся личинок жесткокрылых
1 — Synchita separanda Rtt. (Colydiidae); 2 — Phellopsis amurensis Heyd. (Zopheridae);
8 — Prionocyphon serricomis Muli. (Helodldae) (3 — no Стригановой, 1961)
49 (60) В основании урогомф имеются 1 или 2 симметричных от¬
четливых кармановидных впячивания со склеротизован-
ной поверхностью (рис. 9, 10). Если такие впячивания
не выражены, то на дорсальной стороне брюшных сег¬
ментов имеются развитые склеротизованные зубцы (рис.
16, 1), расположенные беспорядочно (Colydium F.) или
образующие группы, упорядоченные в поперечный ряд
(Dechomus J.-Duv., Colydiidae).
50 (55) В основании урогомф 1 кармановидное впячивание
(рис. 16, 2) или впячивания не выражены.
73
Рис. 15. Общий вид морфологически уклоняющихся личинок жесткокрылых
(продолжение)
1 — Megatoma undata L. (Dermestidae); 2 — Tomoxia blguttata Gyll. (Mordellidae);
3 — Elateroides dennestoides L. (Lymexylonidae)
51 (52) Ha дорсальной стороне 9-го брюшного сегмента в осно¬
вании урогомф имеется поперечный ряд крепких шипор.
(рис. 16, 2). Личинки крупные, плоские, длиной до 30 мм
Pythidae — Трухляки (стр. 183)
74
Рис. 16. Общий вид плоскотелых личинок жесткокрылых
1 — Colydlum elonga tum F. (Colydiidae); 2 — Pytho depressus L. (Pythidae); 3 — Pyrochroa
coccinea L. (Pyrochroldae); 4 — Boros schneideri Pz. (Boridae)
52 (51) Ha дорсальной стороне 9-го брюшного сегмента попе¬
речный ряд шипов отсутствует. Личинки мелкие, дли¬
ной до 10 мм.
53 (54) Урогомфы расш;еплены на вершине на 2 равновеликих
острия или клещевидные с 3—4 длинными внутренними
75
игловидными шипами. В толще коры и древесины. Сап¬
роксилофаги Salpingidae
54 (53) Урогомфы в виде простых загнутых кверху крючков ^
Colydiidae — Узкотелки (стр. 176)
55 (50) В основании урогомф 2 кармановидных впячивания
(рис. 9, 10),
56 (59) 9-й брюшной сегмент увеличен, его длина в 1,2—2 раза
больше длины 8-го брюшного сегмента (рис. 16, 5, 4),
Урогомфы не изогнуты на дорсальную сторону.
57 (58) На конце тела с вентральной стороны имеется дуговид¬
ный ряд шипиков. Выступ между урогомфами не выра¬
жен (рис. 16, 3), а если имеется, то трапециевидный.
В основном под корой. Сапроксилофаги
Pyrochroidae — Огнецветки
58 (57) На конце тела с вентральной стороны ряд шипиков от¬
сутствует. Выступ между урогомфами длинный, треу¬
гольный (рис. 16, 4). Подкорные сапроксилофаги с эле¬
ментами некрофагии
Boridae — Лжечернотелки
59 (56) 9-й брюшной сегмент не увеличен, такой же длины, как
и предшествующие. Урогомфы резко изогнуты на дор¬
сальную сторону
Pedilidae — Лжебыстрянки (стр. 186)
60 (49) Впячивания в основании урогомф и крупные склероти¬
зованные зубцы на дорсальной стороне брюшных сег¬
ментов отсутствуют.
61 (62) Голова сильно уменьшена. Мандибулы преобразованы
в острые стилеты, снабженные внутренним каналом.
Хищники. Нередко встречаются скоплениями в древе¬
сине Lycidae — Яркокрылки
62 (61) Голова вполне развита, а если уменьшена (Trixagidae),
то мандибулы не видоизменены в острые стилеты с внут¬
ренним каналом.
63 (76) На конце тела развиты 2 мелких шипика или 1—2 прос¬
тых, загнутых кверху крючка. Если последний сегмент
закругленный (куполовидный — Alleculidae и Uloma
Cast. из Tenebrionidae, стр. 188) или уплощенный (Ну-
pophloeus F.), то верхняя губа закругленная, четко от¬
делена от клинеуса. Тело умеренно длинное, без ложной
сегментации.
64 (65) На конце тела имеются 2 мелких шипика. Ноги сильно
укорочены. Голова уменьшена, ротовые органы видоиз¬
менены. Сапроксилофаги Trixagidae
65 (64) На конце тела имеются 2 крючка или последний сегмент
^ Видимо, сюда попадают личинки Aderidae (Hayashi, 1972), которые отли¬
чаются сильно разобщенными в основании опорными крючками 9-го
брюшного сегмента.
76
закру/лен. Ноги длинные. Голова крупная, с типичными
грызущими ротовыми органами.
66 (69) На дорсальной стороне брюшных сегментов имеются вы¬
пуклости, напоминающие двигательные мозоли, покры¬
тые овалами или полями крепких шипиков, которые упо¬
рядочены в поперечный ряд только на переднем крае
сегмента.
67 (68) На дорсальной стороне 9-го брюшного сегмента, кроме
мелких округлых, имеются также крупные продольно
вытянутые пятна (рис. 14, 2), В бурых гнилях древеси¬
ны. Сапроксилофаги .... Zopheridae — Бугротелки
68 (67) На дорсальной стороне 9-го брюшного сегмента имеются
мелкие округлые склеротизованные пятна. В древеси¬
не, разрушенной грибом Fomes fomentarius. Сапро¬
ксилофаги Stenoirachelidae — Коготники
69 (66) На дорсальной стороне брюшных сегментов нет покры¬
тых шипиками выпуклостей; если шипики имеются (не¬
которые Tenebrionidae), то они образуют один попереч¬
ный ряд.
70 (75) Покровы тела личинок хорошо склеротизованы, желто¬
ватые, желто-бурые или бурые. Урогомфы, если имеют¬
ся, склеротизованы полностью или почти полностью.
71 (74) Последний сегмент тела вооружен крючками, шипиком
или уплощен и закруглен (рис. 17, 1—4),
72 (73) Теменной шов хорошо развит. Мандибулы симметрич¬
ные Tenebrionidae — Чернотелки (стр. 188).
73 (72) Теменного шва нет. Мандибулы асимметричные. Мицето-
фаги с элементами сапронекрофагии, встречаются под
корой на грибах и плесени
Mycetophagidae — Грибоеды
74 (71) Последний сегмент тела не вооружен и не уплощен, глад¬
кий, куполовидный. В дуплах и полостях в древесной
трухе. Сапронекрофаги
Alleculidae — Пыльцееды
75 (70) Покровы тела личинок белые, мягкие, слабо склероти¬
зованные. Урогомфы длинные, склеротизованы только
в вершинной части. В основном педобионты, встречаются
в сильно разрушенной древесине. Сапроксилофаги ^
Anthicidae — Быстрянки
76 (63) На конце тела развиты прямые длинные направленные
назад простые урогомфы или сложные урогомфы, снаб¬
женные зубцами, выростами или же имеющие в основа¬
нии склеротизованную площадку ^ (рис. 18—20). Если
^ Сюда попадают также личинки Anaspis Geoffr. с неясным систематическим
положением.
2 Если площадка слабо склеротизована, то личинки очень длинные, со сле¬
дами ложной сегментации (некоторые Gleridae, стр. 117).
77
Рис. 17. Общий вид проволочникообразных личинок жесткокрылых
1 — Hypophloeus linearis Р.; 2 — Bius thoracicus Р.; 3 — Uloma culinarie F. (Tenebrio-
nidae); 4 — Mycetophagus sp. (Mycetophagidae)J;P, 3 — no Бызовой, [Келейниковой, 1964)
• последний сегмент закругленный (уплощенный — Cant¬
haridae, Lampyridae или куполовидный — некоторые
Elateridae), то верхняя губа сливается с клипеусом, не¬
редко образуя зубчатое назале. Тело ^южет быть вто¬
рично сегментированным.
77 (78) 8-й брюшной сегмент немного длиннее 7-го (рис. 18,1),
у светлых личинок более интенсивно склеротизован. Ес-
78
Рис. 18. Общий вид подкорных личинок жесткокрылых
J — Cucujus haematodes Er.; 2 — Brontes planatus L, (Cucujidae); 3 — Epuraea unicolor
01.; 4 — Garpophilus hemipterus L. (Nitldulidae) *
ЛИ различия в длине этих сегментов несущественные
(DendrophagusSchoenh., Brontes Fabr.), то имеется длин¬
ная конусовидная анальная подпорка и светлые направ¬
ленные назад урогомфы (рис. 18, 2)
Cucujidae — Плоскотелки (стр. 166)
78 (77) 7-й и 8-й брюшные сегменты существенно не отличаются
79
по длине и степени склеротизации. Анальная подпорка
короткая или не выражена.
79 (82) На дорсальной стороне 9-го брюшного сегмента перед
урогомфами имеются парные склеротизованные бугорки,
выступы или зубчики (рис. 18, 5, 4).
80 (81) Конец тела с вентральной стороны с дуговидным рядом
шипиков. Под корой. Сапроксилофаги
Elacatidae
81 (80) Конец тела с вентральной стороны без дуговидного ряда
шипиков. У крупных форм дыхальца обычно расположе¬
ны на склеротизованных бугорках (рис. 18, 5), у мел¬
ких — дорсальная сторона сегментов бугорчатая или
несет поперечные ряды волосков
Nitidulidae — Блестянки (стр. 157)
82 (79) На дорсальной стороне 9-го брюшного сегмента перед
урогомфами нет парных бугорков, выступов или зуб¬
чиков
83 (84) 9-й брюшной сегмент снабжен сложными, обычно рас¬
щепленными на вершине урогомфами (рис. 19, 4), или
без урогомф, куполовидный, реже — уплощенный, с
3 закругленными зубцами. Тело может быть вторично
сегментированным (рис. 19, 7) ^
Elateridae — Щелкуны (стр. 140)
84 (83) 9-й брюшной сегмент всегда снабжен простыми, не рас¬
щепленными на вершине урогомфами, никогда не быва¬
ет невооруженным, куполовидным или уплощенным.
85 (86) Урогомфы не склеротизованы, тупые, палочковидные,
направленные каудально (рис. 19, 3). В сильно разру¬
шенной древесине. Сапроксилофаги
Cephaloidae — Головачи
86 (85) Урогомфы склеротизованы целиком или в вершинной
части, обычно загнутые вверх, заостренные или на кон¬
це с зубчиком (рис. 20, 1—3).
87 (88) Склеротизованная площадка на дорсальной стороне
9-го брюшного сегмента разделена на две части попереч¬
ной несклеротизованной полосой (рис. 20, i). Между
урогомфами имеется треугольный выступ. Под корой и в
древесине представлен один род — Grynocharis Thoms.
Неспециализированные облигатные хищники
Lophocateridae
88 (87) Склеротизованная площадка на дорсальной стороне
9-го брюшного сегмента единая, иногда слабо склеро-
1 У личинок Tillus elongatus L. и близких видов Cleridae (стр. 117) бугорки
расположены не перед урогомфами, а в их основании на наружной стороне.
У личинок одного из неидентифицированных видов Opilo Latr. перед уро¬
гомфами имеются 2 сближенных зубцевидных выступа.
2 Сюда попадают личинки Drapetes Redt. (рис. 19, 2), положение этого рода
нуждается в дальнейшем уточнении.
80
Рис. 19. Общий вид личинок жесткокрылых из семейств Elateridae (2, 4),
Trixagidac (2) и Gephaloidac (3)
1 - Cardiophorus sp.; 2 - Drapetes biguttatus Pili.; 3 — Cephaloon variabilis Motsch.;
4 — Adelocera conspersa Gyll.
Рис. 20. Общий вид личинок жесткокрылых с крепкими урогомфами
1 — Grynocharis oblonga L. (Lophocateridae); 2 — Thanasimus formlcarius |L. (Clerldae);
8 — Trogossita Japonica Rtt. (Trogossitidae)
тизована. Между урогомфами треугольный выступ не
выражен (рис. 20,2,3).
89 (90) Теменной шов длинный, хорошо выраженный. Эндока-
рина не выражена. В основном подстилочные формы,
встречаются под корой, реже в древесине. Облигатные
хищники Melyridae — Малашки
82
90 (89) Теменной шов отсутствует (полушария головы в темен¬
ной тасти спаяны треугольным склеритом) или очень
короткий. Эндокарина хорошо развита
91 (92) Стипес и кардо максилл одинаковой длины. Тергит 9-го
брюшного сегмента без железистых отверстий ....
Cleridae — Пестряки (стр. 117)
92 (91) Стипес максилл значительно длиннее кардо. Тергит
9-го брюшного сегмента с 2 парами небольших железис¬
тых отверстий
Trogossitidae — Темнотелки (стр. 133)
93 (46) На конце тела опорные структуры отсутствуют. Послед¬
ний сегмент тела не куполовидный а если уплош,ен и
закруглен (Cantharidae, Lampyridae), то голый или в
очень коротких волосках.
94 (95) Последний сегмент тела уплощен и вытянут в треуголь¬
ную дыхательную трубку (рис. 21, 2). В натеках сока
на стволах. Сапрофаги , . . Nosodendridae — Сочноеды
95 (94) Последний сегмент тела не вытянут в дыхательную
трубку
96 (99) Жвалы серповидные с простой вершиной (рис. 9, 5) и с
желобком или внутренним каналом, всегда хорошо раз¬
витые.
97 (98) Голова втянута в переднегрудь. Лобные швы хорошо
развиты. Жвалы с внутренним каналом. Подстилочные
хищники, проникающие в сильно разрушенную древе¬
сину Lampyridae — Светляки
98 (97) Голова не втянута в переднегрудь (рис. 21, 1), Лобные
швы не развиты. Подстилочные хищники, проникающие
в сильно разрушенную древесину
Cantharidae — Мягкотелки
99 (96) Жвалы не серповидные, с расширенным основанием, без
желобка или внутреннего канала, иногда сильно реду¬
цированные.
100 (103) Ротовые органы заметно, а иногда и значительно редуци¬
рованы. Тело короткое и широкое, с боковыми и тер-
гальными почковидными выпячиваниями (рис. 21, 3),
101 (102) Ротовые органы сильно редуцированы и видоизменены.
Личинки белые, длиной 1—3 мм. Подкорные и подсти¬
лочные формы. Сапромицетофаги Cerylonidae
102 (101) Ротовые органы заметно редуцированы, однако все их
составные части достаточно хорошо выражены. Личин¬
ки окрашены в серовато-голубые или другие тона. Под¬
корные и подстилочные мицетофаги, характер питания
подлежит уточнению
Endomychidae — Плеснелюбы
^ Личинки Nematoplus l.oc. (Nematoplidae), у которых последний сегмент
тела напоминает куполовидный, отличаются асимметричной головной кап¬
сулой.
83
Рис. 21. Общий вид личинок жесткокрылых из семейств Cantharidae (1),
Nosodendridae (2), Cerylonidae (3), Nematoplidae (4)
1 — Cantharis fusca L.; 2 — Nosodendron fasciculare 01; 5 — Cerylon sp. 4 — Nematop-
lus semenovi Nik.
103 (100) Ротовые органы хорошо развиты, типичного грызущего
типа. Тело удлиненное, его покровы без боковых и тер-
гальных почковидных выпячиваний.
104 (105) Голова асимметричная. Усики намного короче головной
капсулы. Анальная подпорка короткая (рис. 21, 4), втя¬
нутая в 9-й брюшной сегмент. Сапроксилофаги ....
Nematoplidae — Буроеды
105 (104) Голова симметричная. Усики почти такой же длины, как
головная капсула. Анальная подпорка длинная, кону¬
совидная, не втянутая, или лишь незначительно втяну¬
тая в 9-й брюшной сегмент. Под отслоившейся корой.
Сапромицетофаги . . . Silvanidae — Лжеплоскотелки
84
Рис. 22. Общий вид личинок жесткокрылых с редуцированными ногами
1 — Buprestis mariana L. (Buprestidae); 2 — Otho spondyloides Germ. (Eucnemidae);
3 — Hylobius abietis L. (Gurculionidae)
106 (1) Ноги ПОЛНОСТЬЮ отсутствуют. Брюшные сегменты тела
без парных двигательных мозолей, разделенных про¬
дольным вдавлением.
107 (112) Голова направлена ротовыми органами вперед. Тело
прямое.
108 (111) Переднегрудь сильно расширена, если не расширена, то
ротовые органы видоизменены, мандибулы неявственны
или направлены вершинами наружу.
85
109 (110) Мандибулы нормальные. Переднегрудь всегда сильно
расширена, с одной срединной или 2 расходящимися
линейчатыми бороздками и различными опорными струк¬
турами (рис. 22, 1), Под корой и в древесине на ранних
стадиях разложения. Ксилофаги
Buprestidae — Златки
110 (109) Мандибулы видоизменены, неявственные или направлен¬
ные вершинами наружу. Переднегрудь не расширена
(рис. 22, 2) или сильно расширена, но тогда без линей¬
чатых бороздок, снабжена округлыми или Т-образными
склеротизованными мозолями. В древесине на различ¬
ных стадиях разложения. Бсиломицетофаги и canporfa-
ги Eucnemidae — Древоеды
111 (108) Переднегрудь не расширена. Мандибулы хорошо разви¬
ты, колюш,ие. В мокрой подстилке и переувлажненной
древесине встречаются виды рода Сегсуоп Leach. Хищ¬
ники Hydrophilidae — Водолюбы
112 (107) Голова направлена ротовыми органами вниз. Тело обыч¬
но С-образное (рис. 22, 5). Под корой и в древесине на
ранних стадиях разложения. Амброзийные и деструктив¬
ные ксиломицетофаги и ксилофаги
Curculionidae — Слоники ^
СЕМЕЙСТВО HISTERIDAE - КАРАПУЗИКИ
Семейство карапузиков насчитывает в настоящее время в фау¬
не СССР около 230 видов (Крыжановский, 1965а) и разделяется
на 10 подсемейств (Wenzel, 1944). Личинки карапузиков встреча¬
ются в различных местообитаниях, обычно вместе с имаго, и яв¬
ляются хищниками (Perris, 1876; Померанцев, 19036; Saalas, 1917;
Гусев, 1928; Рейхардт, 1941 и др.). Эти данные подтверждаются
более поздними исследованиями (Lindner, 1967; Харитонова, 1972)
и нашими наблюдениями.
Экология. В соответствии с экологическими особенностями ка¬
рапузиков можно разделить на 5 основных биологических групп
(Рейхардт, 1941).
1. Обитатели падали, навоза и разлагающихся растительных
остатков. Сюда относятся некоторые виды рода Acritus., большин¬
ство Histerini и почти все Sarpinini.
2. Обитатели гнезд и нор животных. В эту группу входят виды
из родов Pholioxenus, Dendrophilus, Dendrophilopsis, большинст¬
во видов рода Gnathoncus и др.
3. Обитатели пещер — небольшая группа мелких видов.
^ Здесь для удобства определения приняты в объеме, предложенном Кроу-
соном (Crowson, 1955), т. е. включают короедов (Scolytidae) иплоскоходов
(Platypodidac).
86
4. Мирмекофилы и термитофилы — почти все представители
подсемейства Hetaeriinae.
5. Обитатели ходов насекомых-ксилофагов. Ксилофильные ли¬
чинки этого семейства представляют собой обширную и разнооб¬
разную группу, включающую карапузиков из подсемейства Teret-
rinae, некоторые роды подсемейства Abraeinae (Plegaderus, Acri-
tus), часть Dendrophilinae (Paromalus) и почти всех Platysomini.
Их личинки — облигатные хищники, одпако узкой пищевой спе¬
циализации у них не наблюдается: все опи являются полифагами
и питаются яйцами, личинками и куколками многих ксилофагов,
особенно различных короедов. Помимо короедов, пищей им слу¬
жат также преимагинальные стадии развития капюшонников
(Bostrychidae), древогрызов (Lyctidae), дровосеков (Cerambyci-
dae), грибовиков (Cisidae), слоников (Curculionidae) и различных
двукрылых насекомых. Виды к&рапузиков, обитающие в ходах на¬
секомых-ксилофагов, встречаются, как правило, па сколитидной
стадии разложения древесины в тех же микростациях, что и има¬
го. Они не обладают приуроченностью к отдельным породам, но
в ряде случаев специализированы к развитию либо на лиственных,
либо на хвойных деревьях. Так, представители родов Cylister и
Paromalus развиваются па хвойных, а родов Acritus, Hololepta —
на лиственных породах. Виды рода Platysoma встречаются боль¬
шей частью под корой лиственных пород и довольно редко — под
корой хвойных. Довольно четко приуроченность к хвойным или
лиственным породам деревьев прослеживается и у видов рода Ple¬
gaderus.
Литературные данные о морфологии личинок карапузиков-кси-
лобионтов очень скудны. Некоторые сведения о строении личинок
родов Hololepta, Paromalus и Hister содержатся в монографии Бе-
винга и Крайгхеда (Boving, Craighead, 1931). В работах Перри
(Perris, 1877), Сааласа (Saalas, 1917, 1923) и Линднера (Lindner,
1967) имеются описания личинок отдельных видов карапузиков
и составлены таблицы для их определения.
Afорфология. Тело удлиненное (рис. 12, i, 2), дорсовентральпо
уплощенное или более или менее цилиндрическое. Голова и перед-
неспинка склеротизованы и пигментированы гораздо сильнее, чем
средне- и заднеспинка. Брюшко мягкое, лишь с отдельными слабо
склеротизованными пластинками.
Голова, как правило, сильно дорсовентрально уплощенная,
иногда явственно расширяющаяся в задней части (Hololepta), но
чаще с более или менее параллельными боковыми сторонами. Швы
го^ювной капсулы видны только у личинок 1-го возраста, у взрос¬
лых личинок они почти незаметны или обозначены вдавлениями.
По одному глазку с каждой стороны головы или глазки отсутству¬
ют. Верхняя губа, наличник и лоб слиты. Передний край головы
несет зубчатое, в очень редких случаях пластинчатое назале. Уси¬
ки (рис. 23, i, 5) 3-члениковые, их 2-й членик несет на наружном
крае около вершины 2—3 чувствующих выроста (сенсиллы).
87
Рис. 23. Детали строения Acritus minutus Hbst. (1—3), Cylister oblongum b.
(4—6), C. lineare Er. (7), Hister cadaverinus Hoffm. (8—11) (Histeridae)
1,5,8— усик; 2 — церк; з — прементум, нижняя челюсть; 4, 9 — нижняя челюсть;
6, 10 — прементум; 7 — назале; ii — нога
crd — кардо; cost — костипес; сх — тазик; fem — бедро; та — мала; р. 1Ь — нижнегуб¬
ной щупик; ргт — прементум; р. тх — нижнечелюстной щупик; tibt — голенелапка;
tr — вертлуг; ung — коготок
Мандибулы сильно склеротизованные, серповидно изогнутые,
с пучком щетинок у основания внутреннего края. Ретинакулум
имеется (за исключением рода Teretrius, у личинок которого внут¬
ренний край мандибул гладкий). Иногда на внутреннем крае ман¬
дибул имеется 1 дополнительный зубчик, расположенный несколь¬
ко выше ретинакулума. Максиллы (рис. 23, 9) состоят из кардо.
88
стипеса, нижнечелюстного щупика и малы. Кардо маленький,
обычно треугольный, сросшийся со стипесом, но отграниченный от
него глубоким вдавлением. Стипес сильно удлиненный, обычно с
параллельными боковыми сторонами, реже (Hister) расширяющий¬
ся к вершине. К стинесу причленяется короткий костипес, несу¬
щий на вершине малу, имеющую вид членика. Нижнечелюстные
щупики 3- или 4-члениковые. Субментум и ментум нижней губы
слились с головной капсулой. Прементум сильно варьирует по
форме и величине. Нижнегубные щупики 2- или 3-члениковые,
расположены на явственных щупиконосцах. Язычок отсутствует.
Ноги (рис. 23, 11) 4-члениковые.
Брюшко из 10 сегментов, из них 1—8-й разделены межсегмент¬
ными валиками. Тергиты и стерниты брюшка несут многочислен¬
ные мелкие склеротизованные пластинки (Hololepta, Platysoma),
либо их склеротизация крайне слабая, практически не выражена
(Plegaderus, Acritus). Тергиты и стерниты брюшка часто несут
покрытые мелкими шипиками мозоли. Иногда такие мозоли быва¬
ют и на межсегментных валиках (Hololepta). Реже шипиконосные
мозоли отсутствуют (Plegaderus, Acritus). 9-й брюшной сегмент с
2-члениковыми церками. Церки либо расположены на мясистых
выростах заднего края этого сегмента, широко расставленные,
с цилиндрическими склеротизованными члениками (рис. 24, i, 5),
(Hololepta, Cylister, Paromalus, Hister, Gnathoncus, Saprinus),
либо мясистые, конусовидные, соприкасающиеся основаниями
(Acritus, Plegaderus) (рис. 25, 7). Анальная подпорка имеет вид
короткой мясистой трубки, расположенной на вентральной
стороне 9-го брюшного сегмента.
^Таблица для определения родов семейства Histeridae
1 (2) Внутренний край жвал гладкий, без зубца
. Teretrius Er.
2 (1) Внутренний край жвал с явственным зубцом.
3 (12) Церки длинные, с цилиндрическими склеротизован¬
ными члениками. Основания церок не соприкасаются
(рис. 24, i, 5). Нижнечелюстные щупики 3-члениковые
(рис. 23, 4, 9), нижнегубные — 2-члениковые (рис. 23,10, 6)^
Прементум явственно короче стипеса. 1-й членик усиков
явственно длиннее 2-го (рис. 23, 5).
4 (5) Голова явственно расширена в задней трети. Назале с 1
двураздельным выступом (рис. 24, 2, 4), Теменной шов у
. основания головной капсулы явственно выражен. 9-й
брюшной стерпит несет по бокам по одному цилиндриче¬
скому, склеротизованному выросту (рис. 24, 1,5). Шипико-
носные мозоли на межсегментных валиках имеются...
Hololepta Рк. (стр. 94).
5 (4) Голова не расширена в задней трети. Назале с 2 зубчаты¬
ми выростами (рис. 23, 7). 9-й брюшной стерпит несет по
Рис. 24. Детали строения Hololepta amurensis Rtt. (i—5) и Н. plana Sulz.
(4, 6) (Histeridae)
1,5 — конец брюшка снизу; 2,4 — назале; 3 — срединный склерит [переднегруди 1
сег — церк; VIII — восьмой стернит
бокам только маленькие бугорки. Шипиконосные мозоли
на межсегментных валиках отсутствуют. Теменной шов не
выражен.
6 (7) Головная капсула с сильными вздутиями позади усиков.
Переднеспинка с глубокими крючковидными вдавлениями.
Коготок очень длинный, примерно равный по длине голе-
нелапке (рис. 23, 11). Стипес явственно расширяется к
вершине (рис. 23, 9). Членики нижнегубных щупиков
одинаковой длины (рис. 23, 10) . . . .Hister L. (стр. 94)
7 (6) Головная капсула без вздутия позади усиков. Передне¬
спинка без крючковидных вдавлений. Коготок в 3—4 раза
короче голенелапки. Стипес не расширяется к вершине
(рис. 23, 4). 2-й членик нижнегубных щупиков явственно
длиннее 1-го.
8 (И) Голова слегка поперечная или ее длина и ширина одина¬
ковы. Прементум явственно расширяется кпереди (рис.
23, 6). Стипес слегка изогнут внутрь (рис. 23, 4). 1-й чле¬
ник усиков в основании не изогнут (рис. 23, 5).
9 (10) Голова слегка поперечная. Передняя полоса шипиков на
1-м брюшном тергите по бокам крючковидно загнута
назад Cylister Coom. (стр. 92)
90
Рис. 25. Детали строения Paromalus flavicomis Ilbst. (/), Р. parallelepipo-
dus Hbst. (2—4), Plegaderus saucius Er. (J, 6) и P. vulneratus Pz. {7—У)
(Histeridae)
1,4 — прементум; 2 — нижняя челюсть; з, 9 — усик; 5 — церк; 6 — нижняя челюсть,
прементум; 7 — церки; 8 — назале
10 (9) Голова равной длины и ширины. Передняя полоса шипи-
ков на 1-м брюшном тергите прямая, без крючковидных
изгибов Platysoma Leach. (стр. 95)
И (8) Голова явственно удлиненная. Прементум с параллельными
боковыми сторонами и резким расширением в основании
(рис. 25, 1,4), Стипес не изогнут (рис. 25, 2), 1-й членик
усиков в основании изогнут (рис. 25, 5)
Paromalus Ег. (стр. 95)
12 (3) Церки короткие, конусообразные, мясистые, основания их
соприкасаются (рис. 25, 7). Нижнечелюстные щупики 4-
члениковые, нижнегубные — 3-члениковые. Прементум
очень длинный, почти равный по длине стипесу (рис.
25, 6). 1-й членик усиков явственно короче 2-го (рис. 25, 9).
91
13 (14) Прементум слабо расширяется в задней трети, затем слег¬
ка сужается (рис. 23, 3). 1-й членик церок чуть больше в
длину, чем в ширину, с параллельными боковыми сторо¬
нами (рис. 23, 2) Acritus Lee. (стр. 92)
14 (13) Прементум сильно расширен в задней трети, затем резко
сужается (рис. 25, 6). 1-й членик церок явственно больше
в длину, чем в ширину, сужается к вершине (рис. 25, 5)
Plegaderus Ег. (стр. 96)
Род Acritus Lee.
Назале с 2 двухзубцовыми выступами на переднем крае. Кос-
типес поперечный. В наших сборах представлен единственным
видом — А. minutus Hbst.
А. minutus Hbst. Длина личинки последнего возраста около
3 мм. По А. Н. Рейхардту (1941), встречается в трухе и под корой
дуба, бука, тополя, реже березы. Отмечен в ходах Xyleborus
monographus F. Серия личинок этого вида найдена нами 13 октяб¬
ря 1958 г. в Воронежской области под корой осины.
Род Cylister Coom.
1-й членик нижнечелюстных щупиков немного короче 2-го,
2-й членик в 2,5 раза короче 3-го. Нижнегубные ш,упики длиннее
прементума. 1-й членик церок втрое толш,е 2-го.
Таблица для определения видов рода Cylister Coom.
1 (2) Дорсальная сторона головной капсулы матовая, густо шаг¬
ренирована С. angustatum Hoffm.
2 (1) Дорсальная сторона головной капсулы блестяш,ая, с ред¬
кими тонкими морщинками.
3 (4) 1-й членик нижнегубных щупиков в 1,5 раза короче 2-го
(рис. 23, б) С. oblongum F.
4 (3) 1-й членик нижнегубных щупиков вдвое короче 2-го.
С. lineare Ег.
С. angustatum Hoffm. Длина личинки последнего возраста
около 8 мм. Вид четко приурочен к хройным породам, хотя Саль-
берг (Sahlberg, 1899, цит. по Saalas, 1917) указывал на то, что он
встречается также в ходах короедов под корой дуба. По данным
Д. Померанцева (19036), обитает под корой еловых пней в ходах
короедов Orthotomicus laricis F., Hylurgorps palliatus Gyll.,
Dryocoetes autographus Ratz., Crypturgus pusillus Gyll., a также
под корой еловых стволов в ходах Ips typographus L. В. И. Гу¬
сев (1928), проводивший наблюдения в сосновых посадках, отме¬
чает, что личинки карапузика поедают личинок и куколок коро-
еда-типографа, а Клайне (Kleine, 1909а) считает его естественным
врагом Hylastes opacus Er. А. С. Плешанов (1966) отмечал его
Я2
Ii Прибайкалье под корой лиственницы, где этот вид (стадия раз¬
вития не указана) нападал на личинок и куколок Ips subelongatus
Alolsch. По данным Н. 3. Харитоновой (1972), карапузик встре¬
чается на сосне обыкновенной совместно с С. oblongum F. в
лодах Blastophagus minor Hart.
Вид отмечен также в ходах Orthotomicus proximus Eichh..
Д. А. Богданова и Н. Г. Коломиец (1974) отмечали личинок этого
вида под корой сосны в ходах смолевки Pissodes piniphilus Hbst.
Личинки С. angustatum поедали яйца личинок и куколок смолев¬
ки. Нами вид был найден в Московской области под корой сосны
обыкновенной в ходах Blastophagus piniperda L., и в Ярославской
области под корой пней ели европейской и сосны обыкновенной.
С. lineare Er. Длина личинок последнего возраста около 9 мм.
Типичный обитатель хвойных пород встречается в ходах корое¬
дов на сосне, ели, лиственнице и кедре. В. И. Гусев (1928) впер¬
вые указал на тот факт, что личинки этого вида уничтожают ли¬
чинок и куколок короедов, в ходах которых они живут, а именно:
Blastophagus piniperda L., Trypodendron lineatum 01., Pityogenes
chalcographus L., Ips duplicatus Sahlb., I. sexdentatus Boern.,
I. amitinus Eichh. и Orthotomicus laricis F. По данным А. С. Пле¬
шакова (1966), вид (стадия развития не указана) встречается в
Прибайкалье в ходах Ips subelongatus Motsch. под корой лист¬
венницы и уничтожает личинок и куколок короеда. С. lineare
относится к ценным энтомофагам короедного комплекса (Харито¬
нова, 1972). В предгорьях Енисейского кряжа карапузик отмечен
на сосне и кедре в ходах короедов Ips typographus L. и Orthoto¬
micus proximus Eichh., в ходах смолевок Pissodes pini L. и Pis¬
sodes piniphilus Hbst., a также в ходах дровосеков Acanthocinus
aedilis L. и Rhagium inquisitor L. Вид может играть существен¬
ную роль в снижении численности короедов и смолевок (Богда¬
нова, Коломиец, 1974). Нами серия личинок С. lineare была най¬
дена в Тувинской АССР (Иштии-Хем) под корой лиственницы
сибирской и в Убинском лесничестве Краснодарского края под
корой сосны обыкновенной в ходах Ips acuminatus Gyll.
С. oblongum F. Длина личинки последнего возраста 10—И мм.
Вид приурочен к хвойным породам деревьев, отмечен на сосне,
кедре, ели и лиственнице. Самки карапузика откладывали яйца
в маточные ходы I. typographus L., а вылупляющиеся личинки
поедали яйца и личинок короеда (Померанцев, 19036). Вид от¬
мечен также в ходах короедов Orihotomicus laricis F., Ips ty¬
pographus L., Blastophagus piniperda L., Dryocoetes autographus
Ratz. (Померанцев, 19036). В. И. Гусев (128) указывает, что в
сосновых посадках в Ульяновской области жуки и личинки хищ¬
ника уничтожали яйца, личинок, куколок и молодых слабоокра-
тенных жуков Ips sexdentatus Boern. и I. acuminatus Gyll.
Личинки карапузика обнаружены в ходах дровосека Spondylis
buprestoides L. По данным А. С. Плешанова (1966), вид (стадия
развития не указана) встречается под корой лиственницы в
93
ходах Ips subelongatiis Molscli. и поедает личинок и куколок это¬
го короеда. Н. 3. Харитонова (1972) указывает, что в условиях
Брянской области карапузик образует популяции значительной
плотности в комлевой части сосен. Максимальное количество ли¬
чинок на 1 м^ подкорной поверхности достигало 150 экз. По мне¬
нию Н. 3. Харитоновой (1972), этот вид является особенно ак¬
тивным, способным в природе влиять на динамику численности
вредителей. В предгорьях Енисейского кряжа Д. А. Богданова
и Н. Г. Коломиец (1974) обнаружили единичные экземпляры
С. oblongum на кедре и установили, что личинки питаются яйца¬
ми, личинками, куколками и слабоокрашенными жуками Ips sex-
dentatus Boern. и Ips typographus L., однако из-за незначитель¬
ной численности вид в этом районе существенного значения как
энтомофаг не имеет. Нами серии личинок карапузика были най¬
дены в Убинском лесничестве Краснодарского края под корой
сосны обыкновенной, в Тувинской АССР (Иштии-Хем) под
корой лиственницы сибирской и в Приморском крае (Комарово-
Заповедное) в ходах Ips sexdentatus Boern. под корой кедра
корейского.
Род Hister L.
В наших сборах представлен единственным видом Н. cada¬
verinus Hoffm. Длина личинки последнего возраста около
14 мм. Костипес несет на вентральной стороне, немного апи-
кальнее места причленения малы, несколько коротких щети¬
нок. 1-й членик нижнечелюстных щупиков равен 2-му, 3-й — в
1,5 раза длиннее 2-го. 1-й членик усика сужается в средней части
(рис. 23, 8). Прементум резко расширен спереди (рис. 23, 10).
По данным Линднера (Lindner, 1967), вид встречается на падали,
в гниющих растительных остатках и навозе. Нами его личинки
были обнаружены во влажной древесной трухе в дупле грецкого
ореха в урочище Кара-Кала (Копет-Даг).
Род Hololepta Рк.
Прементум постепенно расширяется кпереди. Членики ниж¬
нечелюстных щупиков удлиняются по мере приближения к вер¬
шине. 2-й членик церок слегка длиннее 1-го.
Таблица для определения видов рода Hololepta Рк.
1 (2) Полосы шипиков на 8-м брюшном стерпите смыкаются, об¬
разуя овал (рис. 24, 5). Срединный выступ назале узкий,
расширяется к вершине (рис. 24, 4). Центральный склерит
переднегруди с 2 темными пятнами . . . .Н. plana Sulz.
2 (1) Полосы шипиков на 8-м брюшном стерпите параллельные
(рис. 24, 7). Срединный выступ назале широкий, с парал¬
лельными боковыми сторонами (рис. 24, 2). Центральный
94
склерит переднегруди с 3 темными пятнами, причем средин¬
ное пятно, длинное и узкое, занимает почти всю длину скле¬
рита (рис. 24,5) Н. amurensis Rtt.
Н. amurensis Rtt. Длина личинки последнего возраста около
18 мм. Личинки найдены нами в Приморском крае (Комарово-
Заповедное) под корой амурского бархата и маакии амурской,
где питались в основном личинками различных двукрылых.
Н. plana Sulz. Длина личинки последнего возраста около
15 мм. Д. Померанцев (19036) обнаружил этот вид под корой
осины. Нами личинки были найдены в Убинском лесничестве
Краснодарского края и в .Тувинской АССР (Итии-Хем) под ко¬
рой тополя. Вид был также отмечен в Приморском крае (Комаро-
во-Заповедное)под корой ильма долинного, ивы, липы, ореха мань¬
чжурского, бархата амурского и чозении.
Род Paromalus Ег.
Нижнегубные щупики по длине не превышают прементум.
Таблица для определения видов рода Paromalus Ег.
1 (2) Расширенная часть прементума поперечная — ее ширина
больше длины (рис. 25, 4) .. . parallelepipedus Hbst.
2 (1) Расширенная часть прементума почти квадратная (рис. 25,7)
Р. flavicomis Hbst.
Р. flavicomis Hbst, Развивается под корой сосны, кедра и ели.
Отмечен в ходах короедов Ips sexdentatus Boern., I. typographus L.
и Orthotomicus sutiiralis (iyH. (Померанцев, 19036; Saalas, 1917).
B. C. Арефин (1974а) обнаружил вид также в ходах короедов
(Polygraphus proximus Blandf., Pityogenes chalcographus L., Ips
acuminatus Gyll.).
P. parallelepipedus Hbst. Длина личинки последнего возраста
около 7 мм. Личинки и жуки отмечены под корой сосны, ели,
кедра. Они встречаются в ходах Ips sexdentatus Boern., I. acumi¬
natus Oyll. и питаются их личинками и куколками (Escherich,
1923; Гусев, 1928), обнаружены также в ходах короедов Hylurgops
pallialus Gyll., Blastophagus piniperda L., Crypturgus pusillus
Gyll., Orthotomicus longicollis Gyll. и 0. laricis F. (Гусев, 1928;
Nuorleva, 1956). Д. A. Богданова и Н. Г. Коломиец (1974) заре¬
гистрировали жуков и личинок этого вида в ходах короеда Ips
typographus L. В коллекции лаборатории имеются личинки этого
вида, найденные в Убинском лесничестве Краснодарского края под
корой сосны обыкновенной в ходах, покинутых Ips acuminatus
Gyll.
Род Platysoma Leach.
Род морфологически очень близок роду Cylister. 1-й членик
церок в 4 раза толще 2-го.
95
Таблица для определения видов рода Platysoma Leacb.
1 (2) 3-й членик усиков вдвое короче 2-го, 3-й членик нижнечелю¬
стных щупиков в 1,5 раза короче 4-го, 1-й членик нижие-
губпых щупиков в 1,5 раза короче 2-го
Р. deplanatum Gyll.
2 (1) 3-й членик усиков в 1,5 раза короче 2-го, 3-й членик нижне¬
челюстных щупиков вдвое короче 4-го, 1-й членик нижне¬
губных щупиков вдвое короче 2-го
Р. compressum Hbst.
Р. compressum Hbst. Длина личинки последнего возраста около
12 мм. Вид встречается под корой лиственных пород деревьев.
Д. Померанцев (19036) находил этот вид в дубовых пнях. Нами
серии личинок этого вида были собраны в Тувинской АССР (Иштии-
Хем) под корой березы и в 1{раснодарском крае — в Красной
Поляне под корой бука, в Убинском лесничестве — под корой
дуба черешчатого и осины. Питается личинками двукрылых.
Р. deplanatum GylL Длина личинки последнего возраста око¬
ло 10 мм. Д. Померанцев (19036) в изобилии находил этот вид под
корой дуба, поврежденного короедом Scolytus intricatus Ratz.
По данным Сааласа (Saalas, 1917), вид встречается также под ко¬
рой ели, в ходах короеда Ips typographus L. А. С. Плешанов
(1966) обнаружил его в ходах ips subeloiigatus Motsch. под корой
лиственницы, где этот вид уничтожал личинок и куколок короеда.
Нами этот вид также неоднократно отмечался под корой лист¬
венницы сибирской в Тувинской АССР. Помимо этого, вид отме¬
чен нами в Приморском крае (Комарово-Заповедное) под корой
кедра корейского и ильма долинного, в Ярославской области под
корой осины и в Тульской области под корой дуба черешчатого.
Род Plegaderus Ег.
Назале с одним нерасчлененным сильно выступающим вперед
пластинчатым выступом в средней части переднего края. Кости-
пес удлиненный. В наших сборах представлен видами Р. vulnera¬
tus Pz. и Р. saucius Er.
Таблица для определения видов рода Plegaderus Ег.
1 (2) 1-й членик усиков в 1,5 раза длиннее 3-го . .Р. saucius Ег.
2 (1) 1-й членик усиков в 2 раза длиннее 3-го (рис. 25,9) ....
Р. vulneratus Pz.
Р. saucius Er. Вид развивается под корой сосны и ели в ходах
короедов Blastophagus piniperda L. и Ips typographus L. и явля¬
ется естественным врагом этих вредителей (Kleine, 1909, цит. но
Saalas, 1917). Отмечен также в ходах короедов Orthotomicus lon-
gicollis Gyll. и О. suturalis Gyll. (Рейхардт, 1941). Личинки этого
вида найдены нами под корой сосны обыкновенной в Убинском
лесничестве Краснодарского края.
96
Р, vulneratus Pz, Длина личинки последнего возраста 6 мм.
Имаго и личинки встречаются под корой хвойных пород дере¬
вьев. Они отмечены под корой сосны, кедра, пихты, ели и лист¬
венницы. Имеются Случайные находки под корой осины, ивы и
бука. Саалас (Saalas, 1917) указывает, что чаще всего этот вид
встречался в ходах Ips typographus L. По Клейне (Kleine, 1909,
цит. по Saalas, 1917), является хищником, уничтожающим корое¬
дов Crypturgus pusillus Gy]l. По данным В. И. Гусева (1928), вид
развивается в ходах Ips sexdentatus Boern., где жуки и личинки
карапузика уничтожают молодых личинок короеда. А. С. Пле-
шанов (1966) отмечает, что -карапузик (стадия развития карапу¬
зика не указана) поедает яйца Ips subelongatus Motsch. На севере
СССР часто встречается также в] ходах короедов Crypturgus
pusillus Gyll., С. cinereus Hbst. и Blastophagus piiiiperda L. Кроме
того, Р. vulneratus отмечен как энтомофаг еще для 15 видов ко¬
роедов (Рейхардт, 1941; Гусев, 1928). Серия личинок этого вида
была найдена нами в Московской области под корой сосны обык¬
новенной в ходах Blastophagus piniperda L. и под корой ели ев¬
ропейской в ходах Hylastes opacus Er.
СЕМЕЙСТВО STAPHYLINIDAE - СТАФИЛИНИДЫ
Стафилиниды -- обширное и богатое видами семейство.
В Советском Союзе их насчитывается более 2000 видов, относя¬
щихся к 175 родам (Киршенблат, 1965). Объем и таксономический
ранг отдельных групп стафилинид трактуется различными авто¬
рами по-разному. Достаточно сказать, что согласно данным Сме¬
таны (Smetana, 1958), чья точка зрения является наиболее рацио¬
нальной, в фауне средней Европы насчитывается И подсемейств,
а согласно Лозе (Lohse, 1964) — 22 подсемейства.
Экология, Личинки стафилинид встречаются в самых различ¬
ных местообитаниях — в почве, подстилке, под камнями, в раз¬
личных гниющих органических остатках, разлагающейся траве,
наносах водорослей, навозе, компосте, грибах, гнездах птиц, но¬
рах грызунов, ходах кротов, муравейниках и термитниках. Реже
они встречаются на цветках, особенно в соцветиях сложноцвет¬
ных. Личинки старших возрастов некоторых видов рода Aleo-
chara паразитируют в пупариях двукрылых.
По типу питания личинки стафилинид делятся на 2 неравные
группы. Большинство их является хищниками (Paederinae,
Staphylininae, многие Aleocharinae), некоторые питаются трупамц
животных (Omalium). Гораздо меньшее количество видов относится
к группе с растительноядными личинками. Так, личинки рода
Bledius питаются диатомовыми водорослями, личинки Carpeli-
mus, Oxytelus, Platystethus — всевозможными растительными
остатками.
В разлагающейся древесине развивается богатый и обильный
видами комплекс стафилинид, принадлежащих почти ко всем под--
4 Б. М. Мамаев и др. 97'^
семействам этого семейства. Очень часто их личинок можно об¬
наружить под корой как сухостойных, так и ветровальных дере¬
вьев, в ходах короедов и дровосеков, в толще луба, между лубом
и древесиной, а также, хотя и гораздо реже, в толще влажной,
'Сшившей древесины. Личинки отдают явное предпочтение бурым
^гнилям, и в древесине, пораженной светлой гнилью, в:^тречаются
гораздо реже и в значительно меньших количествах. В некоторых
случаях у стафилинид отмечается приуроченность к хвойным или
лиственным породам. Однако чаще всего отдельные виды ксило-
фильных стафилинид встречаются как на лиственных, так и на
хвойных породах, отдавая, однако, заметное предпочтение либо
тем, либо другим. Состав энтомокомплекса ксилофильных стафи¬
линид зависит от стадии разложения древесины: виды родов Р1а-
tjusa, Phloeonomus, Nudobius и Zeteotomus заселяют древесину
на сколитидной стадии разложения, причем личинки встречаются
Ь тех же микростациях, что и имаго, в то время как большинство
Представителей родов Gabrius, Quedius, Olisthaerus встречается
на более поздних стадиях разрушения древесины, когда вредители
уже покинули ее. Питание личинок стафилинид-ксилобионтов до
сих пор изучено крайне слабо. Вследствие этого нередко один и
тот же вид одни авторы относят к хищникам, а другие — к сапро-
фагам. Так, Бегнелл *(BagrialI, 1906) и Шеерпельц (Scheerpeltz,
1925) считают, что личинка Acrulia inflata Gyll. является хищ¬
ником Trypodendron domesticum L., в то время как по данным
Сааласа (Saalas, 1917) и Стила (Steel, 1970) они поедают различ¬
ные остатки под корой или в ходах короедов. Все же следует
подчеркнуть, что для личинок стафилинид — обитателей древе¬
сины, как и вообще для личинок этого семейства, сапрофагия —
редкое явление. К сапрофагам относятся, по нашим наблюдениям,
личинки Phloeonomus, а к сапроксилофагам — личинки Trigonu-
rus, Siagonium, а также, по данным Полиана (Paulian, 1941), ли¬
чинки Leptochirus и др. Что же касается хищных личинок стафи¬
линид, то все они являются широкими полифагами и приурочен¬
ности к определенному виду жертвы не наблюдается. Эта особен¬
ность наиболее наглядно проявляется у личинок стафилинид, свя¬
занных с короедами. Для всех известных видов стафилинид —
обитателей ходов короедов отмечена связь с несколькими видами
жертв. Так, Nudobius lentus Grav. отмечен в ходах более чем
10 видов короедов. Placusa depressa Markl. — по крайней мере
6 видов, Р. tachyporoides Waltl. — 4 и т. д. Личинки стафилинид,
ве связанных с короедами, также не проявляют узкой пищевой
специализации.
Литературные данные по морфологии ксилофильных личинок
Стафилинид довольно скудны. Описания отдельных родов и видов
таких личинок можно найти в работах Сааласа (Saalas 1917, 1923),
Ферхеффа (Verhoeff, 1919), Полиана (Paulian, 1941), Касуле (Ка-
sule, 1966, 1968), Стила (Steel, 1970) и В. А. Потоцкой (1967,
19736). В этих работах, помимо описаний, имеются таблицы для
определения личинок стафилинид, включающие ксилофильные
виды. Обзор личинок стафилинид-ксилобионтов дан в статье
В. А. Потоцкой (1976).
Морфология, Личинки стафилинид делятся на 2 большие,
сильно отличающтеся друг от друга группы — стафилиноморфы
и алеохароморфы. К первой группе относятся личинки подсемейств
Paederinae и Staphylininae, ко второй — личинки Oxytelinae,
Habrocerinae, Tachyporinae и Aleochariiiae.
Стафилиноморфные личинки (рис. 9,1), Голова гипогнатическая
головная капсула сильно склеротизована, всегда с шейным пере¬
хватом. С каждой стороны головы от1до 6 глазков, в редких слу¬
чаях глазки отсутствуют. Верхняя губа, наличник и лоб слиты.
Передний край головной капсулы образует зубчатое назале,
Эпикраниальный, лобные, горловой и гипостомальные швы явст¬
венные (рис. 26, i, 2). Гула отсутствует. Усики (рис. 26, 1) 4-чле-
никовые, расположены на усиковом бугорке. Членики усиков
склеротизованные, их 3-й членик косо срезан у вершины и несет
на срезе 1 крупную и 1 или несколько мелких сенсилл и мутовку
ш,етинок, 4-й членик с несколькими узкими, длинными чувствую¬
щими выростами и мутовкой щетинок на вершине.
Мандибулы (рис. 96, i) сильно склеротизованные, серповидные,
с простой вершиной, как правило, с гладким внутренним краем,
в очень редких случаях их внутренний край в средней части за¬
зубрен. Никогда не бывает ни молы, ни простеки. Максиллы
(рис. 26, 2) состоят из короткого кардо, длинного стипеса, малень¬
кой галеи и 3-или 4-членикового нижнечелюстного щупика. Га л еа
имеет вид членика и причленяется к вершине стипеса. Лациния
отсутствует. Нижнечелюстной щупик расположен на щупико-
носце, который часто хорошо развит и имеет вид склеротизован-
ного полукольца. Субментум склеротизованный, ментум пере¬
пончатый, втяжной, прементум склеротизованный, трапециевид¬
ный. Нижнегубные щупики 2- или 3-члениковые, расположены
на сильно развитых часто опушенных щупиконосцах. Язычок
удлиненный, чаще всего конусовидный, как правило, почти пол¬
ностью склеротизованный, реже (у Paederinae) мясистый, опушен¬
ный, со склеротизованным полукольцом у основания.
Ноги 4-члениковые, состоят из массивного тазика, короткого
вертлуга, длинного бедра и длинной голенелапки, оканчивающей¬
ся коготком. Голенеланка с крупными шипиками, иногда расши¬
рена в вершинной части. Внутренняя поверхность голенелапкц
первой пары ног часто несет более мелкие шипики. Эти шипики
бывают простыми или раздвоенными на вершине и образуют ряды
или поля, иногда они покрывают всю внутреннюю поверхность
голенелапки, образуя так называемую тибиальную щетку (рис.
28, 9). Коготок слабоизогнутый, всегда одновершинный. Строение
коготка различно. У Staphylininae он постепенно расширяется к
основанию и несет 2—3 шипика в средней части. У Paederinae
коготок тонкий, и 2 шипика расположены на резко расширенном
4* 99
^ис. 26. Схема строения головы стафилиноморфных (1, 2) и алеохароморф-
ных (5, 4) личинок стафилинид (Staphylinidae)
ifS — голова сверху; 2,4 — голова снизу
ani — усик; crd — кардо; gal — галеа; gul — горло; lac — лациния; Ibr — верхняя губа;
m — ментум; mi — М1нднбула; п — назале; ос1 — глазок; р. 1Ь — нижнегубной щупик;
р. ттс — нижяечелюзгной щупик; ргт — прементум; s. fr — лобный шов; з. gul —
^уларный шов; з. ver — теменной шов; зт — субментум; 3t — стипес
1рсновании (рис. 30, 9). Брюшко состоит из 10 сегментов. Первые
8 сегментов сходны по строению. Тергиты и стерниты состоят из
2 пластинок. 9-й брюшной сегмент уже предшествующих и силь¬
нее склеротизован, с одной склеротизованной пластинкой как
на дорсальной, так и на вентральной сторонах. К нему подвижно
причленяются церки, которые чаще всего бывают 2- или 3-чле-
•никовыми, в очень редких случаях ложночлениковыми или узло¬
ватыми. Анальная подпорка представляет собой более или менее
удлиненную трубку, несущую на дорсальной стороне обычно 2,
а на вентральной — 4—6 продольных рядов щетинок. Ее вер¬
шина вооружена втяжными мясистыми выростами, покрытыми
многочисленными хитиновыми крючками.
1Q0
Алеохароморфные личинки (рис. 12, 3). Как правило, мелкие
формы (длина личинок последнего возраста редко достигает 1 см).
Голова 6^3 шейного перехвата. Гула имеется. Верхняя губа (рис.
26, 5) имеется, она отделена от остальной части головной капсулы
четким швом. Наличник, как правило, сливается с головной кап¬
сулой. Усики 3-члениковые (рис. 26, 3) (только у личинок рода
Tachinus усики неявственно 4-члениковые).
Мандибулы (рис. 26, 3; 27, 5, 7) часто с предвершинными зуб¬
цами, нередко с сильно развитой молой, иногда имеется простека.
Максиллы (рис. 26, 4) состоят из маленького треугольного кардо.
Стипес сливается с лацинией, которая всегда хорошо развита,
3-члениковый щупик расположен на явственном нцупиконосце.
Галеа чаш;е всего отсутствует, иногда представлена лопастью или
перистым образованием, находяш,имся на наружном крае лаци-
нии. Строение лацинии сильно варьирует. Она может быть силь¬
но вытянутой, четырехугольной, треугольной. Жевательный край
лацинии несет шипики, ш;етинки, зубцы, пучки нцетинок. Субмен-
тум и ментум в виде склеротизованных пластинок, иногда слив¬
шихся, но чаш,е разделенных четким швом. Прементум трапецие¬
видный. Язычок чаш,е всего конусовидный, реже поперечный,
иногда сливается с прементумом. В редких случаях язычок вооб¬
ще отсутствует.
Ноги всегда без тибиальной щетки. Нередко тергиты и стерни-
ты брюшка представлены каждый цельной неразделенной посере¬
дине пластинкой. Церки 1- или 2-члениковые. У паразитических
личинок Aleochara и некоторых мирмекофилов церки отсутствуют.
Таблица для определения родрв семейства Staphylioidae
1 (32) Верхняя губа имеется (рис. 26, 5). Галеа отсутствует, либо
значительно реже, рудиментарна (рис. 27, 2,6), расположе¬
на на вершине лацинии, которая всегда хорошо развита.
Усики 3-члениковые (рис. 26, 3) (исключение — род Tachi¬
nus, у которого усики 4-члениковые). Шейный перехват
не развит.
2 (5) Язычок нижней губы поперечный (рис. 27, 4). Мандибулы
с резко выраженной молой (рис. 27, 5), либо с лопастью
близ вершины, совкообразные. Простека отсутствует.
3 (4) Тело вальковатое, слабосклеротизованное. На голове и
теле имеются очень крупные булавовидные, опушенные
щетинки. 1-й и 2-й членики усиков покрыты многочислен¬
ными щетинками и волосками (рис. 27, 1). Рудимент галеи
в виде продольно исчерченной лопасти (рис. 27, 2). Лациния
значительно короче нижнечелюстного щупика, доходит
лишь до верхней трети его 2-го членика. Церки в 3 раза
длиннее анальной подпорки, 1-й членик церок вдвое длин¬
нее 2-го. По 6 глазков с каждой стороны головы. Сапро-
ксилофаги Trigonurus Muls.
101
Рис. 27. Детали строения Trigonurus asiaticus Reiche (1—3), Siagonium vit¬
tatum Fauv. (4—6), Phloeonomus pusillus Grav. (7), Tachinus bipustulatus
F. (8), Olisthaerus substriatus Gyll. (P, 10) и Bolitochara obliqua Er. (11)
(Staphylinidae)
1 — усик; 2, 6, 11 — вершина лацинии; 3, 7 — мандибула; 4 — прементум; 5, 10 —
конец брюшка сверху; в, 9 — максилла.
о. ped — анальная подпорка; свг — церк; gal — галеа; lac — лациния; рг — проств-
ка (7 — по VAFhoeff» 1919; 11 — по Paulian, 1941)
4 (3) Тело сильно уплощено, его склеротизация явственная.
Сложные булавовидные щетинки отсутствуют. 1-й членик
усиков гладкий, 2-й только с мутовкой предвершинных
ще-Цнок. Рудимент галеи имеет вид отграниченного зубца
(рис> 27, 6), Лациния почти равна по длине нижнечелюст¬
ному щупику. Церки лишь немного длиннее анальной
подпорки, 1-й членик церок в 5 раз длиннее 2-го (рис.
27, 5). По 4 глазка с каждой стороны головы. Сапроксило-
фаги Siagonium Kirby
5 (2) Язычок нижней губы удлиненный, самое большее равной
длины и ширины. Мандибулы с предвершинным зубцом,
либо с лопастью в средней части режущего края, часто
с простекой (рис. 27, 7; 32, 7),
6 (13) Церки 1-члениковые. Мандибулы с простекой (рис. 27, 7;
32, 7).
7 (8) Мандибулы с крупной веерообразной простекой (рис. 27, 7).
Язычок нижней губы широкий. Церки примерно равны
по длине анальной подпорке. Сапрофаги
Phloeonomus Неег.
8 (7) Мандибулы с тонкой короткой простекой (рис. 32, 7). Язы¬
чок нижней губы узкий, конусовидный (рис. 29,1). Церки
в 1,5—2 раза длиннее анальной подпорки.
9 (10) Наличник имеется (рис. 32, 6). Передний край верхней
губы с 4 шипиками и глубокой вырезкой в средней части
(рис. 32, 6). Лациния очень узкая и длинная, с цельной
вершиной, несет в верхней части жевательного края
продольный ряд из 5 мелких коротких шипиков, а в его
нижней части — 1 крупный короткий тупой шипик. Сен¬
силла 2-го членика усиков не доходит до вершины 3-го
членика. 2-й членик усиков в 1,5 раза длиннее 1-го.
Хищники Xylostiba Gglb. (стр. 116)
10 (9) Наличник отсутствует (рис. 29, 2). Передний край верх¬
ней губы прямой или слабо выемчатый, без шипиков.
И (12) Вершина лацинии раздвоена (рис. 29, 1). Мандибулы асим¬
метричные, предвершинный зубец на левой мандибуле
развит значительно сильнее, чем на правой. Ментум в
виде узкой поперечной пластинки (рис. 29, 1). Длина го¬
ловы немного больше ширины (рис. 29, 2). Жевательный
край лацинии несет мелкие шипики в вершинной части
и 1—2 крупных шипа в нижней половине (рис. 29, 1)
Hapalaraea Thoms. (стр. 112)
12 (И) Вершина лацинии не раздвоена. Мандибулы симметрич¬
ные. Ментум трапециевидный. Голова округлая, длина ее
равна ширине. Жевательный край лацинии только с 2
крупными шипами на режущем крае. Сапрофаг
Acrulia Thoms.
13 (6) Церки 2-члениковые. Мандибулы без простеки.
14 (19) По 5—6 глазков с каждой стороны головы. На 8-м брюш¬
103
ном тергите никогда пе бывает железы, а на анальной
подпорке — крючков.
15 (16) Тело сильно уплощено, с явственной склеротизацией.
Лациния очень короткая, треугольная, не длиннее 1-га
членика нижнечелюстных щупиков, с пучком длинных
тонких щетинок на внутренней поверхности (рис. 27, 5).
1-й членик церок в 5 раз длиннее 2-го (рис. 27, 10), Под
сильно сгнившей корой хвойных деревьев. Сапрофаги
Olisthaerus Неег.
16 (15) Тело выпуклое, слабо склеротизованное. Лациния четы¬
рехугольная (рис. 27, 5), равная по длине, по крайней
мере, 1-му и 2-му членикам нижнечелюстных щупиков
вместе взятым, без пучка длинных щетинок на внутрен¬
ней поверхности. 1-й членик церок в 1,5 раза длиннее
2-го или равен ему.
17 (18) Жевательный край лацинии образует угол с ее внутрен¬
ним краем, несет шипы и зубцы, причем 2 нижних шипа
очень крупные, с массивными основаниями (рис. 27, 8).
Верхняя губа на переднем крае со слабо выраженной
трехдольчатой лопастью. Язычок нижней губы длинный,
его длина более чем в 2 раза больше ширины. 1-й членик
церок в 1,5 раза длиннее 2-го. Усики 4-члениковые. Пи¬
тание не изучено Tachinus Grav.
18 (17) Жевательный край лацинии плавно, не образуя угла,
переходит во внутренний край, несет ряд шипиков оди¬
наковой величины. Передний край верхней губы слабо
выпуклый. Язычок нижней губы равной длины и шири¬
ны. 1-й членик церок равен по длине 2-му. Усики 3-чле-
никовые. Питание не изучено . .Bryocharis Boisd. et Lac.
19 (14) Не более 1 глазка с каждой стороны головы. 8-й брюшной
тергит часто с крупной железой (рис. 28, 2; 30, 7), Аналь¬
ная подпорка обычно несет на выступах хитиновые крюч¬
ки (рис. 28, 5).
20 (31) Верхняя губа на переднем крае с лопастями, зубчиками,
папиллами или щетинконосными бугорками (рис. 28,4\
30.6); иногда папиллы трудно различимы (Placusa; рис.
31.6), но тогда усики длинные и тонкие (рис. 31,2).
21 (22) Верхняя губа на переднем крае с лопастями (рис. 28, ^).
8-й брюшной тергит без железы. Анальная подпорка с
хитиновыми крючками (рис. 28, 5)
Atheta Thoms. (стр. 110)
22 (21) Верхняя губа на переднем крае с зубчиками, папиллами
или щетинконосными бугорками (рис. 28,7; 30,6).
23 (30) Передний край верхней губы прямой, с зубчиками или
слабо различимыми папиллами в средней части (рис.
30,6; 31,6).
24 (25) Усики длинные, тонкие (рис. 31, 2). Мандибулы с несколь¬
кими мелкими зубчиками у вершины (рис. 31, 4). Лациния
104
Рис. 28. Детали строения Anomognathus cuspidata Er. (1, 2), Atheta amicula
Steph. («3—6), Atrecus affinis Pk. (7—9) и A. pilicornis Pk. (10—12) (Staphy-
linidae)
J, 4 — верхняя губа; 2,3 — конец брюшка сверху; 5, 8, 12 — усик; 5, 7, 11 — максилла;
9 — вершина голенелапки, коготок; 10 — назале
i — тибиальная щетка (7, 5 — по Paulian, 1941)
с крупными листовидными зубцами в нижней части же¬
вательного края (рис. 31,5). Язычок короткий, равен по
длине 1-му членику нижнегубных щупиков (рис. 31,5)
Placusa Ег. (стр. 114)
25 (24) Усики короткие, массивные (рис. 30, 5). Мандибулы с
предвершинным зубцом, часто с зубчиками базальнее его.
Лациния с более или менее крупными шипами на жева¬
тельном крае (рис. 27, 11), крупные листовидные зубцы на
жевательном крае отсутствуют. Язычок длинный, длин¬
нее 1-го членика нижнегубных ш,упиков.
26 (27) 8-й брюшной тергит без железы. Передний край верхней
губы в средней части с 4 папиллами. Лациния несет на
жевательном крае около 20 тонких шипиков, увеличиваю-
ш,ихся по мере приближения к основанию. Питание не
изучено Cyphaea Fau^v.
27 (26) 8-й брюшной тергит с крупной железой (рис. 30,^7). Пе¬
редний край верхней губы в средней части с 4 зубчиками
(рис. 30, б). Шипы или зубцы в апикальной части жева¬
тельного края лацинии мелкие, в базальной — крупные
(рис. 27, 11),
28 (29) Сенсилла усиков длинная, доходит до мутовки щетинок на
3-м членике (рис. 30, 5). Лациния с зубчиками на вершине
и с длинными зубцами в базальной части жевательнога
края, короткая, доходит до вершины 2-го членика ниж¬
нечелюстного щупика. Анальная подпорка с 4 крючками
(рис. 30, 7) Leptusa Кг. (стр. 112)
29 (28) Сенсилла усиков короткая, доходит только до вершины
2-го членика. Лациния с короткими шипиками на вер¬
шине и длинными шипами в базальной части жевательно-
10G
Рис. 30. Детали строения Gabrius expectatus Smet. (2, 2), G. splendidulus
Grav. (5), G. trossmus Nordm. (4), Leptusa angusta Aub6 (5—7) и Medon
laticeps Ramb. (5, P) (Staphylinidae)
1, 7 — конец брюшка сверху; «—4 — назале снизу; 5 — усик; в — верхняя губа; tf —
«азале сверху; 9 — коготок (5—7 — по Paullan, 1941)
ГО края, удлиненная, лишь слегка короче нижнечелюст¬
ного щупика (рис. 27,11), Анальная подпорка без крюч¬
ков. Питание не изучено Bolitochara Мапп.
30 (23) Передний край верхней губы выпуклый, с 2 щетинконос¬
ными бугорками по бокам (рис. 28,1). Мандибулы с пред¬
вершинным зубцом и гладким внутренним краем ....
Anomognathus Sols. (стр. 109)
31 (20) Верхняя губа с округлым, гладким передним краем. 8-й
брюшной тергит с железой. Мандибулы зазубрены базаль-
нее предвершинного зубца. Лациния с мелкими зубчиками
в верхней части и с толстыми крупными шипами в нижней
части жевательного края. Питание не изучено ....
РЫоеорога Ег.
32 (1) Верхняя губа, наличник и лоб слиты в единую лобную
пластинку, передний край которой образует зубчатое
назале (рис. 26,1; 28,10). Галеа имеется, члениковидная
107
Рис. 31. Детали строения Placusa complanata Er, (i—б) и Nudobius lentus
Grav. (7—P) (Staphylinidae)
1 — конец брюшка сверху; 2 — усик; S — верпшна! лацинии; 4 — мандибула; 5^ 9 ^
прементум; в — верхняя губа; 7 — навале сверху; S — субментум
(рис. 28, 7, ii). Лациния редуцирована до нескольких ще¬
тинок в вершинной части стипеса. Усики 4-члениковые
(рис. 28, S, 12). Шейный перехват имеется.
33 (34) Основания коготков расширены, несут 2 шипика (рис.
30, 9). По 6 глазков с каждой стороны головы. Глазки соб¬
раны в кольцо. Внутренний край мандибул мелко зазуб¬
рен. Назале 6-зубцовое, отграниченное от лобной пластин¬
ки (рис. 30, S) Medon Steph. (стр. ИЗ)
34 (33) Коготки у основания не расширены, с шипиками у сере¬
дины (рис. 28, 9; 32, 4). Не более 4 глазков с каждой сто¬
роны головы. Внутренний край мандибул гладкий.
35 (40) Боковые стороны головы параллельные. 1 глазок с каж¬
дой стороны головы или глазки редуцированы. Коготки с
2 шипиками, расположенными один позади другого (рис.
28, 9; 32, 9). Крупные щетинки на тергитах, анальной под¬
порке, урогомфах простые.
108
36 (37) Навале 5-зубцовое (рис. 32,11), Голенелапка расширена в
вершинной части, с венцом крупных шипов над коготком.
Тибиальнад ш,етка отсутствует (рис. 32, 9). Галеа очень
маленькая, тонкая, малозаметная (рс. 32, 8). Прементум
расширяется к основанию (рис. 32, 10)
Zeteotomus Jacq. (стр. 116)
37 (36) Навале 11-вубцовое (рис. 28, 10\ 31, 7). Голенелапка не
расширена в вершинной части. Тибиальная ш,етка име¬
ется (рис. 28, 9), Галеа хорошо раввита (рис. 28, 7, 11).
Прементум суживается к основанию (рис. 31, 9),
38 (39) 1-й и 2-й боковые вубцы навале широко расставлены, 1-й
вубец острый, 2-й нритунлеп (рис. 28, 10). Субментум с
перехватом у основания. Длина личинок последнего вов-
раста около 6 мм Airecus Jacq. (стр. 110)
39 (38) 1-й и 2-й боковые зубцы навале сближены, притуплены
(рис. 31, 7). Субментум без перехвата у основания (рис.
31, 8). Длина личинок последнего возраста около 9 мм.
Nudobius Thoms. (стр. ИЗ)
40 (35) Боковые стороны головы округлые. По 4 главка с каждой
стороны головы. Коготки с 3 шипиками, расположен¬
ными мутовкой в средней части коготка (рис. 32, 4). Круп¬
ные щетинки на тергитах, анальной подпорке, урогом-
фах часто сложные.
41 (42) Церки короче анальной подпорки, мясистые (рис. 32,2,5)
Тибиальная ш,етка имеется, состоит ив раздвоенных ши-
пиков (рис. 32, 5, 4). Нижнечелюстные ш,упики 3-членико-
вые, нижнегубные — 2-члениковые. Навале 9-зубцовое
(рис. 32, i) Quedius Steph. (стр. 114)
42 (41) Церки не короче анальной подпорки (рис. 30,1). Тибиаль¬
ная щетка отсутствует. Нижнечелюстные щупики 4-чле-
никовые, нижнегубные — 3-члениковые. Навале 7-зуб-
цовое Gabrius Curt. (стр. 111)
Род Anomognaihus Sols.
8-й брюшной тергит с крупной железой (рис. 28,2), церки ко¬
роткие, с массивными члениками. Лациния очень короткая, мас¬
сивная, с мелкими зубчиками на жевательном крае.
А. cuspidata Er. Вид, по имеюпц1мся данным, предпочитает
лиственные породы. Саалас (Saalas, 1923) находил его под
корой осины в ходах Cryphalus grothii Hagel, а также под корой
березы, ясеня, ольхи и дуба, в хвойных породах встречается ред¬
ко. Перри (Perris, 1863, цит. по Saalas, 1923) отметил личинок и
куколок этого вида под корой сосны в ходах Orthotomicus laricis
F., а Саалас (Saalas, 1923) — под корой ели в ходах Ips typo-
graphus L. и Orthotomicus suturalis Gyll. В Убинском лесничестве
Краснодарского края личинки заселяли толщу бурого луба, дуба
черешчатого и липы. Хищники. Питаются коллемболами и дру¬
гими мелкими членистоногими. Трофические связи изучены слабо.
109
Род Atheta Thoms.
Внутренний край мандибул зазубрен ниже предвершинного
зубца. Церки длиннее анальной подпорки. 1-й членик церок ко¬
роче 2-го (рис. 28, 3).
Таблица для определения видов рода Atheta Thoms.
1 (2) Нижнечелюстной щупик вдвое длиннее лацинии. Сенсилла
усиков расположена на стебельке А. aequata Ег.
2 (1) Нижнечелюстной щупик самое большее в 1,5 раза длиннее
лацинии (рис. 28, 6). Сенсилла усиков сидячая, без стебель¬
ка (рис. 28, 5) А. amicula Steph.
А, aequata Er, Вид отмечен на ели в ходах Dryocoetes auto¬
graphus Ratz., Hylurgops glabratus Zett., Polygraphus punctifrons
Thoms. и других короедов, а также под корой сосны, осины (Sa-
alas, 1917) и березы (Paulian, 1941). Нами личинки А. aequata Ег.
были найдены в Вологодской области под корой ели европейской
в лежащем на земле стволе, покинутом короедами. Личинки хищ¬
ники. Трофические связи слабо изучены.
А. amicula Steph, Личинки найдены нами в Бухарской области
под корой колоды туранги. Хищники. Трофические связи слабо
изучены.
Род Atrecus Jacq.
Личинки этого рода сходны по строению с личинками рода
Nudobius, отличаются от них более короткой тибиальной щеткой,
состоящей из 5—6 шипиков, наличием четко выраженных глазков
и более мелкими размерами — около 6 мм.
Таблица для определения видов рода Atrecus Jacq.
1 (2) 1-й членик усиков в 2 раза короче 2-го, 4-й — в 1,5 р^за
короче 3-го (рис. 28, 12). Кардо более чем в 2 раза короче
стипеса (рис. 28, /i) А. pilicornis Рк.
2 (1) 1-й членик усиков более чем в 4 раза короче 2-го, 4-й в
2 раза короче 3-го (рис. 28, 8). Кардо более чем в 5 раз
короче стипеса (рис. 28, 7) А. affinis Рк.
А. affinis Рк. Вид приурочен к хвойным породам, на листвен¬
ных встречается лишь в виде редкого исключения. По данным
Сааласа (Saalas 1917), чаще всего встречается под корой в старых
пнях сосны с легко отслаивающейся сильно разложившейся корой.
Полиан (Paulian, 1941) зарегистрировал личинок под корой граба.
Намп личинки отмечены под корой сосны обыкновенной и пихты
кавказской (Убинская, Гузерипль, Краснодарский край). Хищ¬
ники. Питаются мелкими личинками насекомых, в частности, ли¬
чинками двукрылых.
110
А. pilicornis Рк. Вид приурочен к хвойным породам. В Евро¬
пе личинки встречаются под корой ели, реже — под корой сосны
в старых лежащих на земле, сильно разложившихся стволах, под
легко отстающей корой в почерневшем лубе (Saalas, 1917). В При¬
морском крае (Комарово-Заповедное) личинки найдены в колодах
кедра корейского и пихты цельнолистной в условиях, сходных
с описанными выше. Личинки — хищники, питаются мелкими
личинками насекомых, в частности, личинками двукрылых.
Род GaLrius Ciirt.
Внутренние боковые зубцы назале несут 2 маленьких шиппка
на внутренней поверхности. На наружном крае 1-го членика це-
рок имеется продольный ряд из 3—5 щетинок (рис. 30,1).
Таблица для определения видов рода Gabrius Curt.
1 (4) Из 2 маленьких шипиков на внутреннем боковом зубце
назале один расположен у основания зубца, близ внутрен¬
него края, а другой — на внешнем крае зубца (рис. 30,2, 3).
2 (3) Внутренний боковой зубец назале тупой (рис. 30, 3). Язычок
нижней губы явственно короче последнего членика нижне-
губного щупика G. splendidulus Grav.
3 (2) Внутренний боковой зубец назале острый (рис. 30, 2). Язы¬
чок нижней губы одинаковой длины с последним члеником
нижнегубных щупиков G. expecfaius Smet.
4 (1) Оба маленьких шипика на внутреннем боковом зубце назале
расположены близ внутреннего края зубца (рис. 30, 4)
G. trossulus Nordm.
G. expectaius Smet. Дорсальная сторона анальной подпорки с
2 четкими рядами из 3 сложных щетинок каждый (рис. 30, i).
Личинки в Вологодской области заселяли подкоровую зону
лежащих на земле стволов ели европейской с легко отслаиваю¬
щейся корой, сильно перегнившим бурым лубом и твердой дре¬
весиной. Хищники, питаются мелкими личинками насекомых,
в частности, личинками двукрылых.
G, splendidulus Grav, По данным Сааласа (Saalas, 1917), вид живет
в почве, под мхом, в подстилке. Однако он часто встречается под
корой разложившихся стволов и пней ели. Д. Померанцев (lS02a)
находил этот вид под корой осины, дуба, бука, ясеня и ильма.
Нами личинки были найдены в Тульской области, под корой
березы и в Московской области под корой ели европейской. Хищ¬
ник, питается мелкими личинками насекомых.
G, trosbulus Nordm, Дорсальная сторона анальной подпорки
с поперечным рядом из 4 коротких щетинок у основания. Продоль¬
ные ряды щетинок на анальной подпорке состоят из простых и
сложных щетинок. Личинки были найдены нами в Московской,
Вологодской областях в лиственном компосте, в лесной подстилке,
111
а также под корой лежащих на земле стволов ели европейской
с легко отслаивающейся корой, сильно перегнившим бурым лу¬
бом и твердой древесиной. Хищники. Питаются мелкими личин¬
ками насекомых.
Род Hapalaraea Thoms.
Передний край верхней губы прямой или слабо выемчатый.
2-й членик усиков в 1,5—2 раза длиннее 1-го.
Таблица для определения вищов рода Hapalaraea Thoms.
1 (2) Боковые стороны головы почти параллельны (рис. 29, 2),
2-й членик усиков в 1,5 раза длиннее 1-го. Лациния корот¬
кая, не доходит до вершины 2-го членика нижнечелюстных
щупиков. Шипы на жевательном крае лацинии расположе¬
ны примерно на равных расстояниях друг от друга, 2 ниж¬
них шипа крупнее остальных (рис. 29, 1). Режущий край
мандибул не зазубрен. Церки в 1,5 раза длиннее аналь¬
ной подпорки. Коготок в 2 раза короче голенелапки . . .
Н. ioptera Steph.
2 (1) Боковые стороны головы округлые. 2-й членик усиков в
2 раза длиннее 1-го. Лациния доходит до 2-го членика ниж¬
нечелюстных щупиков. Жевательный край лацинии с про¬
дольным рядом шипов у вершины и одним крупным шипом,
расположенным на значительном расстоянии от этого ряда,
у основания жевательного края. Режущий край мандибул
зазубрен ниже предвершинного зубца. Церки в 2 раза
длиннее анальной подпорки. Коготок в 4 раза короче голе¬
нелапки Н. pygmaea Рк.
Я. ioptera Steph. Вид живет под корой сосны, ели и бука
(Saalas, 1917). По данным Стила (Steel, 1970), личинки встречают¬
ся под корой отмерших деревьев, склонны к каннибализму и пред¬
положительно питаются мелкими насекомыми и другими живу¬
щими под корой членистоногими.
Н. ру^таеа Рк. Личинки, по-видимому, селятся под корой от¬
мерших деревьев. В лабораторных условиях они питались свеже¬
убитыми насекомыми (Steel, 1970).
Род Leptusa Кг.
1-й членик церок в 4 раза длиннее 2-го и чуть толще его.
Личинка известна для единственного вида — L. angusta Aube.
L. angusta Aube. Вид отмечен на осине, ели и сосне. Охотнее
всего заселяет лежащие деревья и пни, реже — сухостойные де¬
ревья (Saalas, 1917). Зарегистрирован в ходах короедов Ips ty-
pographus L., Pityogenes chalcographus L., Crypturgus pusillus
Gyll. и других видов.
112
Род Medon Steph.
Нижнегубные щупики 2-члениковые, нижнечелюстные —
3-члениковые. Средние зубцы назале широкие и плоские
(рис. 30, 8), внутренние боковые — треугольные, наружные —
редуцированные.
Л/, laticeps Ramb. В Приморском крае (Комарово-Заповедное)
отмечен нами под корой ильма, липы, тополя Максимовича, ясеня
маньчжурского, кедра корейского и пихты цельнолистной (По¬
тоцкая, 1973а). Несколько реже он встречается в толще влажной,
расслаивающейся древесины липы, ясеня и пихты. Иногда вид оби¬
тает в сухой бурой трухе, оставшейся от гниения луба. Питаются
мелкими личинками насекомых.
Род Nudobius Thoms.
Личинки сходны по строению с личинками рода Atrecus, но
отличаются от них более длинной тибиальной щеткой, состоящей
из 7—9 простых шипиков, редуцированными глазками и более
крупными размерами — 8—9 мм.
Таблица для определения видов рода Nudobius Thoms.
1 (2) 3-й боковой зубец назале равен 4-му (рис. 31, 7). Язычок
нижней губы короче 3-го членика нижнегубных щупиков
(рис. 31, Р) N. lenius Grav.
2 (1) 3-й боковой зубец назале больше 4-го. Язычок нижней гу¬
бы такой же длины, как и 3-й членик нижнегубных щу¬
пиков N. umbratus Motsch.
N. lentus Grav. Вид типичен для сколитидной стадии разложе¬
ния древесины и отмечен в ходах короедов — Orthotomicus pro¬
ximus Eichh., О. laricis F., О. suturalis Gyll., Ips typographus L.,
Pityogenes chalcographus L., Dendroctoiius micans Kiig., Hylur-
gops palliatus Gyll., H. glabratus Zett., Polygraphus polygraphus
L., Blastophagus piniperda L., B. minor Hart. Dryocoetes auto¬
graphus Ratz., a также в ходах дровосеков — Tetropium, Mono-
chamus, Rhagium и слоников — Pissodes cembrae Motsch. и P. pi-
niphilus Hbst. По литературным данным и нашим наблюдениям,
личинки встречаются под корой хвойных деревьев — сосны обык¬
новенной и ели европейской в свежих или покинутых ходах, реже
под отставшей корой, где нет ходов. В редких случаях их можно
найти под корой старых, сильно разложившихся стволов осины
(Saalas, 1917; Померанцев, 1902а; Харитонова, 1972). В Примор¬
ском крае (Комарово-Западное) личинки, как правило, встреча¬
лись под корой кедра корейского, реже — под корой пихты цель¬
нолистной в ходах короедов Ips sexdentatus Boern. и дровосеков.
Личинки неоднократно отмечались как хищники, поедающие
яйца, личинок и куколок короедов (Saalas, 1917; Померанцев,
ИЗ
1902а; Гусев, 1928; Буковский, 1940; Зиновьев, 1959; Харито¬
нова, 1972; Потоцкая, 1973а, и др.).
N. umbratus Motsch. Серия личинок этого вида была собрана
в Убинском лесничестве Краснодарского края под корой сосны
обыкновенной и пихты кавказской в старых и заселенных ходах
короедов, а также под корой, где не было ходов. Личинки пита¬
ются личинками и, возможно, яйцами короедов, а также другими
мелкими личинками насекомых.
Род Placusa Ег.
8-й брюшной тергит без железы (рис. 31,2). 2-й членик церок
явственно длиннее и тоньше 1-го.
Р. complanata Er, Личинки встречаются в ходах короедов хвой¬
ных пород. Нами серия личинок этого вида была найдена в Убин¬
ском лесничестве Краснодарского края под корой сосны обык¬
новенной в ходах Ips sexdentatus Boern., ]. acuminatus Gyll.^
Blastophagus piniperda L., B. minor Hart., Orthotomicus laricis F.^
0. suturalis Gyll. В литературе имеются данные о том, что этот
вид встречается также в ходах Ips typographus L. (Харитонова^
1972). Личинки — хип^ники, питаются яйцами и, возможно^
первыми возрастами личинок короедов, а также коллемболами.
Род Quedius Steph.
В наших материалах род представлен тремя видами. В древе¬
сине отмечены личинки четырех видов.
Таблица для опред.елеБия видов рода Quedius Steph.
1 (2) Срединный зубец назале почти равен по длине примыкаю-
ш,им зубцам (рис. 32,1), Церки чуть короче анальной под¬
порки (рис. 32,2) Q. plagiatus Mnnh.
2 (1) Срединный зубец назале явственно короче и уже п]римыкаю-
щих зубцов. Длина церок составляет максимум длины
анальной подпорки (рис. 32, 5).
3 (4) 1-й членик церок в 2—2,5 раза длиннее 2-го. Из 6 раз¬
двоенных шипиков, составляющих тибиальную щетку, 4-й
расположен явственно проксимальнее остальных шипиков
(рис. 32, 5) Q. scitus Grav.
4 (3) 1-й членик церок в 4—5 раз длиннее 2-го (рис. 32,5).
5 (6) 2-й членик нижнечелюстных щупиков длиннее 3-го. Тиби-
альная щетка из 9 раздвоенных шипиков (рис. 32, 4). Дор¬
сальная сторона анальной подпорки с 2 продольными ряда¬
ми коротких сложных щетинок, по 9 щетинок в каждом
(рис. 32, 5) Q. xanthopus Ег.
6 (5) 2-й членик нижнечелюстных щупиков короче 3-го. Тибиаль-
ная щетка состоит из 7 раздвоенных шипиков. Дорсальная
114
Рис. 32. Детали строения Quedius plagiatus Mnnh. (2, 2), Q. scitus Grav.
(3), Q. xanthopus Er. (4, 5), Xylostiba monilicomis Gyll. (6, 7) и Zeteotomus
scripticollis Hochh. (8—11) (Staphylinidae)
1, 11 — назале; 2, 5 конец брюшка сверху; 5, 4, 9 — вершина голенелапки; в —
верхняя губа; наличник; 7 — мандибула; 8 — максилла; ю — прементум (6, 7 — по
Steel, 1970)
сторона анальной подпорки с 2 продольными рядами про¬
стых и сложных щетинок, по 6 щетинок в каждом ряду . .
Q. picipes Mnnh.
Q. picipes Mnnh, Субментум с основанием в виде длинной язы¬
ковидной пластинки, морщинистый, начиная от верхней пары
щетинок. Кончик языковидной пластинки нежно шагренирован.
1-й членик урогомф расширяется к вершине. Личинки найдены
нами в Кодрах Молдавской ССР, в сильно разложившихся пнях
бука и граба обыкновенного. Хищники.
Q. plagiatus Mnnh, Субментум морщинистый в верхней поло¬
вине, с основанием в виде короткого треугольника. 1-й членик
церок цилиндрический. По данным Сааласа (Saalas, 1917), типич¬
115
ный обитатель хвойных пород — сосны и ели. Личинки и имага
часто встречаются в ходах Ips typographus L. и дровосеков, как
в сухостойных деревьях, так и в лежащих на земле стволах, их
можно также обнаружить и в стволах, уже покинутых короеда¬
ми. Питается личинками короедов или других жуков, в чьих хо¬
дах обитает. Нами серия личинок этого вида была собрана в Во¬
логодской области под корой ели европейской.
Q. plagiatus var. ater J. Sahlb. В Приморском крае (Комарово-
Зановедное) личинки найдены во влажном буром лубе самых раз¬
личных хвойных и лиственных пород (Потоцкая, 1973а, 1976).
Хищники питаются мелкими личинками насекомых.
Q. scitus Grai\ Серия личинок найдена в дубовой колоде, по¬
раженной бурой гнилью (Paulian, 1941). Характер питания тре¬
бует уточнения.
Q. xanthopus Er. Субментум морщинистый в средней части,,
с основанием в виде короткого языка. 1-й членик урогомф цилинд¬
рический. По данным Сааласа (Saalas, 1917), вид встречается под
корой старых, сильно разложившихся стволов ели и реже — оси¬
ны. Нами личинки были найдены в июле 1958 г. в Тульской об¬
ласти в бурой гнили древесины дуба. Хищники. Трофические
связи изучены слабо.
Род Xylosliba Gglb.
До сих пор этот род считался подродом рода Phloeonomus
Неег. Стил (Steel, 1970) выделяет Xylostiba в самостоятельный
род. Своеобразное строение верхней губы и четкое развитие на¬
личника делают подобное предположение вполне вероятным.
Личинка известна только для одного вида — X. monilicornis
Gyll.
X. monilicornis Gyll. Вид обычно встречается под корой су¬
хостойных елей и других хвойных деревьев, часто — в ходах ко¬
роедов (Saalas, 1917; Steel, 1970). По данным Лузе (Luze, цит. по
Saalas, 1917), встречается также под корой бука и клёна. Личин¬
ки всех возрастов встречаются в мае — июне. Личинки — хищ¬
ники (Steel, 1970).
Род Zeteotomus Jacq.
В наших сборах представлен единственным видом.
Z. scripticollis Hochh. Серия личинок этого вида была найдена
нами в Краснодарском крае (Убинское лесничество, пос. Гузе-
рипль) под корой граба обыкновенного и бука восточного в ходах
короеда Taphrorychus bicolor Hbst., личинками которого они в
ьсновном питались.
116
СЕМЕЙСТВО CLERIDAE - ПЕСТРЯКИ
Пестряки относятся к группе жесткокрылых, систематика
которых достаточно хорошо разработана не только для имаги-
нальной, но и для личиночной фазы развития. Причины этога
заключаются как в значительном биологическом своеобразии се¬
мейства, привлекавшем внимание специалистов, так и в перспек¬
тивности использования некоторых массовых видов пестряков в.
системе биологических способов борьбы со стволовыми вредите¬
лями.
В числе работ, посвященных общей характеристике семейства,
следует указать публикацию Винклера (Winkler, 1961), содер¬
жащую уточненный перечень родов мировой фауны, и сводку по
фауне СССР, опубликованную В. А. Рихтер (1961). Последняя
работа представляет особый интерес, так как в ней имеются так¬
же некоторые сведения по личинкам. Наиболее фундаментальной
ревизией личинок пестряков является публикация Бевинга и
Чемплейна (Boving, Champlain, 1920), включающая 16 родов
североамериканской фауны, половина которых представлена
также и в Палеарктике. Личинки рода Tenerus, наряду с некото¬
рыми другими родами, распространенными в фауне южной
Азии, охарактеризованы Гарднером (Gardner, 1937). Личинки ос¬
новных европейских представителей семейства описаны как r
некоторых работах начала текущего столетия (Haulbert, Betis,
1905, цит. по Boving, Champlain, 1920), так и более детально Ларе-
соном (Larsson, 1938) и Эмденом (Eniden, 1943). Если к этому
добавить выполненные около столетия назад, но не утратившие
своего значения работы Перри (Perris, 1854, 1855, 1877),
то можно констатировать, что для подавляющего большинства ро¬
дов, распространенных в Палеарктике, характеристики личинок
в той или иной степени уже известны.
Экология, Все представители семейства относятся к облигат¬
ным хищникам, причем такой способ питания характерен как для
личинок, так и для имаго. В отношении видов рода Necrobia
предполагается, что они могут развиваться в богатых жирами
портящихся продуктах как животного, так и растительного
происхождения (Рихтер, 1961). Так, N. rufipes Deg. в массе раз¬
множается на складах, где ее многочисленные личинки, прокла¬
дывая ходы в копченостях и других продуктах, могут наносить
серьезный вред, этот же вид в зависимости от ситуации может
быть и хищником, и мицетофагом. Литературные сведения о том,
что личинки некоторых видов пестряков на ранних стадиях раз¬
вития могут быть эктопаразитами (Boving, Champlain, 1920),
нуждаются в проверке.
Личинки ксилофильных видов семейства отличаются опреде¬
ленной специализацией в отношении объектов питания, хотя r
эксперименте поедают личинок любых насекомых. Можно наме¬
тить 3 основных направления пищевой специализации.
ИТ
Первым, важнейшим направлением является специализация
к питанию главным образом личинками и куколками короедов.
В качестве наиболее типичных представителей этой экологической
группы в Палеарктике могут быть названы только виды рода
Thanasimus и отчасти Clerus. Именно в этой экологической груп¬
пе имеются виды, высокая численность которых свидетельствует
об их возможном значении как важнейших истребителей корое¬
дов.
Вторым, пожалуй, наиболее распространенным направлением
специализации является питание личинками насекомых — разру¬
шителей подсохшей древесины, таких, как точильш;ики (Anobi-
idae), капюшонники (Bostrychidae) и древогрызы (Lyctidae).
В Палеарктике это прежде всего представители подсемейства Til-
linae (Tillus, Tilloidea, Denops). Их тонкие и длинные личинки
преследуют свои жертвы — личинок названных выше групп
жесткокрылых по их ходам. Численность их, однако, не на¬
столько высока, чтобы говорить о суш;ественном практическом
значении. К этой же экологической группе относятся некоторые
палеарктические Korynetinae. В мировой же фауне пестряки —
истребители точильш;иков, капюшонников и древогрызов особен¬
но многочисленны в тропических странах. Видимо, в качестве
обособленной подгруппы этой экологической группы следует вы¬
делять виды, например Orthopleura sanguinicollis L., связанные с
такими сверлильш;иками, как корабельный (Lymexylon navale L.).
К весьма многочисленной третьей экологической группе отно¬
сятся пестряки, развиваюш;иеся в основном за счет личинок дро¬
восеков и златок. Наиболее типичным представителем группы
является род Clerus. Вид С. mutillarius F. развивается под корой
таких усохших лиственных деревьев, которые с высокой плотно¬
стью заселены златками и дровосеками. Такие же трофические
связи имеют из встречающихся в Палеарктике некоторые виды
родов Tenerus и Trichodes. Что касается последнего рода, то в
его составе есть виды, личинки которых предпочтительно пита¬
ются личинками пчел, причем в роде есть не только виды, унич¬
тожающие диких пчел, устраивающих соты в древесине, но и ви¬
ды, вредные для пчеловодства. Некоторые виды Trichodes счи¬
таются истребителями кубышек саранчовых. Личинки видов из
рода Tenerus встречаются не только под корой, но и в древесине,
в качестве их жертв зарегистрированы златки, дровосеки и ко¬
роеды.
Три основных направления пищевой специализации пестря¬
ков служат предпосылкой для их приуроченности к определенным
местообитаниям. Группа видов, развивающаяся за счет короедов,
относится к подкорным насекомым. Поскольку в лесах умеренно¬
го климатического пояса короеды особенно многочисленны па
хвойных породах, такие пестряки, как представители рода Tha¬
nasimus, также предпочитают хвойные. На лиственных эти виды
встречаются значительно реже. Виды, преследующие личинок
118
точильщиков, капюшонников и древогрызов, относятся к обита¬
телям толщи древесины, так как среди объектов их питания пре¬
обладают именно такие формы. Наиболее широкой экологической
валентностью отличаются пестряки, уничтожающие личинок зла¬
ток и) дровосеков, в их рацион, кроме предпочитаемых объектов
питания, входят также короеды и слоники. Они встречаются как
под корой, так и в толще древесины.
Все экологические группы пестряков предпочитают сухую
или умеренно влажную древесину, в переувлажненной древесине
не встречаются.
Морфология. Личинки разнообразного габитуса —- от очень
длинных и топких, цилиндрических (рис. 33, 1\ 34, i) до умеренно
длинных, вальковатых, уплощенных (рис. 33,-2; 34,5), но всегда
с мягкими покровами и хорошо развитыми урогомфами на конце
тела. Цвет личинок также разнообразен: они могут быть белы¬
ми, розоватыми, бледно-красными, фиолетовыми, а некоторые
имеют на светлом теле темный рисунок.
Голова личинок (рис. 35, 1—4) прогнатическая, уплощенная,
слегка втянутая в переднегрудь. Она может быть вытянутой или^
наоборот, поперечной, ее боковые стороны обычно слегка выпуклые,
но могут быть и параллельными. Лобные швы обычно хорошо раз¬
виты, сливаются в заднем отделе головной капсулы. Полушария
головы позади точки слияния (или сближения) лобных швов^
обычно спаяны треугольным склеритом, реже имеется короткий
теменной шов. У изученных нами личинок Tenerus лобные швы
не выражены, а лобный отдел головы прослеживается в виде про¬
дольной трапециевидной полосы. Эндокарина хорошо развита.
Иногда на лобном треугольнике развиты сильно склеротизован-
ные выступы (рис. 35, 2). У большинства личинок — по 5 глазкоа
с каждой стороны головы, однако в семействе имеются также все
варианты редукции глазков вплоть до их полного исчезновения.
Усики 3-члениковые, прикреплены слабосклеротизованной ба¬
зальной мембраной, благодаря чему могут глубоко, нередко це¬
ликом, втягиваться внутрь головной капсулы. У большинства
изученных личинок 1-й членик усиков в 2-—4 раза длиннее 2-го;
на конце 2-го членика имеются 1 конусовидная сенсилла и сме¬
щенный к краю палочковидный 3-й членик.
Мандибулы массивные, треугольные, с неразделенной верши¬
ной, в большинстве случаев с ретинакулумом, с неразвитой мо-
ларной частью и слабо выраженной простекой. Верхняя губа
отделена от клипеуса, песет волосовидные и кинжаловидные
щетинки. Нижняя губа (рис. 36, i—4) состоит из 3 хорошо отдиф¬
ференцированных частей: субментума, ментума и прементума.
Ментум обычно с несколькими волосками, прементум несет 2-члени-
ковые нижнегубные щупики. Кардо и стипес максилл представ¬
ляют собой поперечные лентовидные склериты, соединенные мем¬
бранами, их размеры примерно одинаковые. Жевательная доля
обычно двулопастная, иногда поделена на 2 части поперечной
11»'
Рис. 33. Общий вид личинок пестряков (Cleridae)
1 — Denops longicollis F.-W.; 2 — Trichodes spectabilis Kr.; 3 — Til-
lus elongatus L.
Рис. 34. Общий вид личинок Orthopleura sanguinicollis L. (/),
Tarsostenus univittatus Rossi (2), Tenerus sp. (3) (Cleridae)
J \ у .
Рис. 35. Голова личинок пестряков (Cleridae)
J — Denops longicollis F.-W.; 2 — Orthopleura sangulnicollis L., 3 — Clerus mutilla-
rius F., 4 — Trlchodes spectabilis Kr.
складкой, ее апикальный отдел покрыт волосками, у некоторых
видов несет также несколько кинжаловидных щетинок. В средней
части жевательной доли ближе к ее внутреннему краю в глубине
складки имеется длинный пальцевидный вырост, несущий на
конце 1—2 крепкие щетинки. Нижнечелюстные щупики 3-члени-
ковые.
Степень склеротизации тергита переднегруди может быть раз¬
личной. Он может быть склеротизован почти полностью или
только в передней половине. Медиальная линия, делящая передне¬
грудной щит на 2 половины, может быть светлой или, наобо¬
рот, темной, так как в последнем случае она представлена внут¬
ренним склеротизованным килем. На средне- и заднеспинке мо¬
гут быть развиты разобщенные склеротизованные пятна —- по 2
на каждом сегменте. Ноги заканчиваются простым слабоизогну¬
тым коготком, на вершине голенелапки у большинства видов
имеются длинные крючковидные щетинки. Брюшные сегменты
нередко имеют парные, иногда гранулированные мозоли. 9-й
брюшной сегмент обычно с дорсальной склеротизованной площад-
121
Рис. 36. Нижняя губа и правая максилла снизу и вершина максиллы с внут¬
ренней стороны личинок пестряков (Cleridae)
1 —'Тепегиз’зр.; 2 — Thanaslmus substriatus Gebl.; з — Tillua elongatus L.; 4 — Tricho¬
des sp.
crd — Kapio; m — ментум; mi — мала; prm — прементум, st — стипес
кой, на заднем крае которой расположены сальносклеротизован-
ные урогомфы (рис. 37,1—6), Передний край площадки без скле-
ротизованного бортика. Анальный сегмент покрыт многочислен¬
ными щетинками. Форма дыхалец существенно варьирует.
Таблица для определения родов сеиейства Cleridae
1 (4) Глазков нет. Лобные швы соединяются в задней трети
головной капсулы (рис. 35, 2) или неявственны, тогда лоб¬
ный отдел представлен расширяющейся книзу полосой.
122
2 (3) Тело тонкое и длинное (рис. 34,1), длина средних сегмен¬
тов в 2,6—3,0 раза превышает ширину. Голова (рис. 35, 2)
в^редней части с крупным сильносклеротизованным выс-
"^упающим килем. Лобные швы четкие
Orthopleura Spin. (стр. 128)
3 (2) Тело (рис. 34, 3) короткое, утолщенное, длина средних
сегаентов в 3,5—4,0 раза меньше ширины. Голова без
выступа. Лобные швы неявственные, лобный отдел пред¬
ставлен расширяющейся книзу полосой
Tenerus Cast. (стр. 129)
4 (1) Глазки имеются. Лобные швы явственные, соединяются в
задней четверти головной капсулы (рис. 35, 2, 5).
5 (14) Тело взрослых личинок покрыто густыми длинными во¬
лосками (рис. 33, 2). На 9-м брюшном сегменте волоски по¬
крывают его переднебоковые стороны и нередко образуют
спутанные ряды вдоль переднего края склеротизованной
площадки.
6 (13) С каждой стороны головы по 5 глазков. Урогомфы загну¬
тые вверх, постепенно заостряющиеся или толстые, более
или менее цилиндрические, на конце с острым, направ¬
ленным вверх шипом (рис. 37,2—6).
7 (10) Переднегрудной щит полностью или по крайней мере спе¬
реди разделен светлой срединной линией. Длинные щетин¬
ки на 2-м членике усика отсутствуют.
8 (9) Урогомфы (вид сверху) расходящиеся, в их основании
иногда имеется впячивание покровов (рис. 37, 4—6). Кар-
до и стипес с немногими краевыми щетинками
Opilo Latr. (стр. 126)
9 (8) Урогомфы (вид сверху) параллельные, торчащие вверх,
разобщенные, в основании без каких-либо впячиваний
покровов. Кардо и стипес с небольшими группами щетинок
в средней части (рис. 36, ^) . . .Trichodes Hbst. (стр. 132)
10 (7) Переднегрудной тергит с темным срединным килем, без
следов светлой линии. Длинные щетинки на 2-м членике
усика имеются.
И (12) 9-й сегмент с широким поясом волосков вдоль переднего
края склеротизованной площадки. Позади глазков име¬
ется густой продольный ряд из 8—12 щетинок (рис. 35, 3),
Щетинки на мептуме смещены к краям склерита ....
Clerus F. (стр. 12/i>
12 (11) 9-й сегмент вдоль переднего края склеротизованной пло¬
щадки без полного пояска волосков или с отдельными ред¬
кими волосками. Позади глазков имеется ряд из 4—6 раз¬
реженных щетинок. Щетинки на ментуме удалены от края
склерита Thanasimus Latr. (стр. 130)
13 (6) С каждой стороны головы по 2 глазка. Урогомфы
(рис.37, 1) цилиндрические, на конце с бугорком ....
Korynetes Hbst. (стр. 125)
123
14 (5) Тело личинок (рис. 33, 1,3\ 3i,2—3) почти полностью ли¬
шено волосяного покрова. На 9-м брюшном сегменте во¬
лоски имеются только на боковых сторонах около осно¬
вания урогомф, на передне-боковые углы не заходят.
15 (16) С каждой стороны головы по 4 глазка. 6-й и 7-й брюшные
сегменты с хорошо выраженными дорсальными двигатель¬
ными мозолями (рис. 34, 2) . . Tarsostenus Spin. (стр. 129)
16 (15) С каждой стороны головы не более 3 глазков. Сегменты
тела без четких дорсальных мозолей.
17 (18) С каждой стороны головы по 3 глазка. В основании уро-
гомф (рис. 33, 5) наружный бугорок имеется. Тело обычно
с фиолетовым рисунком Tillus 01. (стр. 131)
18 (17) С каждой стороны головы по 1 глазку или глазки неявст¬
венны. В основании урогомф наружный бугорок редуци¬
рован. Тело одноцветное, белое.
19 (20) Склеротизованная площадка 9-го брюшного сегмента с 2
отверстиями для пор. На конце передней голенелапки 3
крючковидные щетинки. Темный срединный киль на пе¬
реднегрудном щите значительно короче половины его
длины. Поперечные склериты на заднем крае переднегру-
ди штриховидные Tilloidea Cast. (стр. 132)
20 (19) Склеротизованная площадка 9-го брюшного сегмента без
отверстий для пор. На конце передней голенелапки 4—5
крючковидных щетинок. Темный срединный киль на
переднегрудном щите не короче половины его длины. По¬
перечные склериты на заднем крае переднегруди ленто¬
видные, слегка изогнутые . . . Denops F.-W. (стр. 125)
Род Clerus F.
Личинки крупные, розовато-красные, тело их густо покрыто
волосками, последний сегмент снабжен хорошо склеро-
тизованной площадкой и сильносклеротизованными загнутыми
кверху урогомфами. Переднегрудь с дорсальной стороны почти
полностью склеротизована, темный срединный киль не короче
общей длины переднегрудного щита. На средне- и заднегруди
имеются по 2 склеротизованных пятна. Брюшные сегменты со
слабогранулированными двигательными мозолями. В межсегмент¬
ном пространстве имеются выпуклые валики, густо покрытые во¬
лосками. Голенелапка на конце с 2 крючковидными щетинками.
Длина и ширина головной капсулы (рис. 35, 3) примерно одина¬
ковы. Лобные швы слегка извилистые, четкие. Эндокарина дохо¬
дит до середины лобного треугольника.
Таблица для определения видов рода Clerus F*
1 (2) Верх в длинных (как торчащих, так и прилегающих) волос¬
ках. Дорсальные мозоли средних брюшных сегментов поде¬
лены на 2 части срединной полоской волосков. Низ в спу-
124
тайных, большей частью прилегающих волосках. Вент-
jja^bHan поверхность 9-го брюшного сегмента отчетливо
склеротизована, между основаниями урогомф на значи¬
тельном протяжении без волосков . . .С. mutillarius F.
2 (1) Верх в коротких торчащих волосках. Дорсальные мозоли
средних брюшных сегментов в виде единой голой полоски,
без волосков в средней части. Низ в коротких торчащих во¬
лосках. Вентральная поверхность 9-го брюшного сегмента
слабо склеротизована, покрыта волосками
.. С. dealbatus Кг.
С, dealbatus Kraatz. В литературе имеются указания на раз¬
витие этого вида в ходах короеда Ips sexdentatus Boern. (Рихтер,
1961). Нами большая серия личинок была собрана 22 сентября
1964 г. в заповеднике Комарово-Заповедное Приморского края
под корой ильма долинного, заселенного сравнительно крупным
короедом Scolytus jacobsoni Spess.
C. mutillarius F. Серия личинок была собрана в 1958 г. в Тел-
лермановском лесничестве Воронежской области и в 1970 г.
в Убинском лесничестве Краснодарского края главным образом
под корой дуба, где они уничтожали личинок дровосека Pyrrhi-
dium sanguineum L.
Род Dehops F.-W.
Личинки (рис. 33,1) белые, длинные и тонкие, с хорошо разви¬
тыми крючковидными урогомфами, от основания которых от¬
ходят 2 продольных киля, ограничивающих вогнутую площадку.
Переднегрудь отчетливо склеротизована только в передней по¬
ловине. По задней границе передне- и среднегруди имеются пар¬
ные поперечные запятовидные склериты. Сегменты тела сильно
удлинены, с увеличенными межсегментными промежутками.
Длина головной капсулы (рис. 35,7) несколько превышает ее ши¬
рину. Лобные швы дуговидно выгнутые. Эндокарина доходит
до половины лобного треугольника.
D, longicollis F~W,, известный из южной Европы, Крыма и
Закавказья, обнаружен в Туркмении (Акибай), где его личинки
питались капюшонниками Xylogenes dilatatus Rtt., поселяющи¬
мися на усохшем тамариксе. В тугаях Копет-Дага (Ипайкала)
пестряк развивался в усохших ветвях лоха, также разрушаемых
личинками капюшонников Enneadesmus scopini Furs., и в усох¬
шей виноградной лозе, где пищей его личинкам служили личин¬
ки древогрыза Lyctus sp.
Род Korynetes Hbst.
В роде насчитывается около 15 видов, один из которых —
К. coeruleus Deg. широко распространен в Палеарктике. Данные
о личинках этого вида имеются у Ларссона (Larsson, 1938) и Эм-
125
дена (Emden, 1943). Личинки средних размеров, тело густо по¬
крыто волосками. Последний сегмент с хорошо склеротизованной
площадкой и толстыми цилиндрическими урогомфами (рис. 37, 1),
которые к концу не утончаются, снабжены бугорком. Передне-
грудь с дорсальной стороны полностью склеротизована, с редкими
волосками и светлой срединной линией. Пятна на средне- и зад-
негруди имеются. Голенелапка на конце с длинными неявственно
крючковидными щетинками. Длина и ширина головной капсулы
примерно одинаковы. Лобные швы слегка вогнуты к средней ли¬
нии, в дистальной трети с незначительным изломом. Эндокарина
простирается на 7з длины лобного треугольника.
Описывается по шкурке личинки, обнаруженной вместе с жу¬
ком (Краснодарский край, село Ново-Прохладное) в сухой за¬
болони комля усохшего дуба. Древесина дуба была заселена
личинками точильщика.
Род ОрПо Latr.
Кроме европейских видов (Кетпег, 1913; Larsson, 1938; Em¬
den, 1943, и др.), описаны личинки некоторых представителей
индо-малайской фауны (Gardner, 1937). Личинки розовато-крас-
ные, их тело густо покрыто волосками. Урогомфы (рис. 37, 4—6)
равномерно утолщенные или утончающиеся, на вершине пере¬
ходящие в шипик. Переднегрудь с дорсальной стороны почти
полностью склеротизована, с темным срединным килем, который
лишь частично перекрывает светлую срединную линию. На средне-
и заднегруди имеются слабо выраженные небольшие склеротизо-
ванные пятна. Брюшные сегменты с очень слабо выраженными
дорсальными двигательными мозолями, с вентральной стороны, по
крайней мере у некоторых видов, с небольшой склеротизованной
пластинкой, в центре которой имеются 2 темных пятна. Голене-
лапка на конце с пучком щетинок, в том числе крючковидных, или
с немногими щетинками. Длина и ширина головной капсулы при¬
мерно одинаковы. Лобные швы прямые, в дистальной трети слегка
изогнутые. Эндокарина короткая, не заходит за середину лобного
треугольника.
Таблица для определения видов рода Opilo Latr.
1 (6) Урогомфы на конце с вентральным округлым расширением.
2 (5) Урогомфы на конце с очень густым пучком длинных волос¬
ков, между ними имеется глубокое впячивание покровов
(рис. 37,4, 5). Голенелапка с густым пучком щетинок.
Рис. 37. 9-й брюшной сегмент личинок пестряков (Cleridae)
; — Korynetes coeruleus Deg.; 2 — Orthopleura sanguinicollis L.; 5 — Tillus elongatue
L.; 4 — Opilo mollis L., 5 — Opilo sp., 6 — O. carinatus Lew. (?)
126
3 (4) Впячивание между урогомфами длинное, конусовидное (рис.
37, 4), На склеротнзованной площадке 9-го брюшного сег¬
мента нет 2 зубцевидных выступов О. mollis L.
4 (3) Впячивание между урогомфами неглубокое, чашевидное
(рис. 37, 5). На склеротизованной площадке 9-го брюшного
сегмента имеется 2 зубцевидных выступа . . . Opilo sp.
5 (2) Урогомфы на конце примерно с 10 редкими длинными волос¬
ками, между ними имеется лишь точечное впячивание по¬
кровов (рис. 37,5). Голенелапка на конце с 2—4 волосками
О. carinatus Lew.(?)
6 (1) Урогомфы к концу постепенно сужаются, без округлого
расширения О. domesticus Sturm.
О, carinatus Lew. (?). Регистрация этого вида в фауне СССР
нуждается в проверке. Личинка была обнаружена в заболони
черемухи Маака в Комарово-Заповедное Приморского края в
мае 1969 г. В заболони были обнаружены также личинки ксило¬
фагов из родов Melandrya sp. (Melandryidae) и Lissodema sp.
(Salpingidae).
0. domesticus Sturm. B. A. Рихтер (1961) обобщила материалы
по экологии этого вида, указав, что по имеющимся сведениям его
личинки на некоторых фруктовых деревьях преследуют личинок
дровосеков рода Clytus. Имеются данные, свидетельствующие о
важном значении личинок пестряка в уничтожении личинок до¬
мового дровосека (Hylotrupes bajulus L.), а также об их питании
личинками точильщиков.
О, mollis L, Небольшая серия личинок этого вида была собрана
в июне 1966 г. в Красной Поляне Краснодарского края в дре¬
весине вишни, разрушаемой точильщиками, личинки которых
истреблялись пестряком.
Opilo sp. Имаго характеризуемого вида получено из личинки,
найденной в Убинском лесничестве Краснодарского края в июле
1970 г. в необычных условиях — в сильно разрушенной древеси¬
не клена, где обитали личинки рогача Dorcus parallelopipedus L.,
чернотелки Helops coeruleus L.
Род Orthopleura Spin.
Личинки (рис. 34, 1) очень длинные, тонкие, червеобразные,
белого цвета. Последний сегмент тела без дорсальной склероти¬
зованной площадки, с разобщенными, короткими, загнутыми квер¬
ху урогомфами (рис. 37, 2). Переднегрудь с крупным дорсальным
щитом, разделенным светлой срединной линией, средне- изадне-
грудь — с крупными склеротизованными пятнами. Длина грудных
сегментов постепенно увеличивается. Брюшные сегменты чрезвы¬
чайно сильно удлинены (их длина в 3,5—4,0 раза больше попереч¬
ника). Каждый сегмент в передней трети с 2 двигательными
дорсальными мозолями, в задней части — с вентральными мозоле¬
видными выпуклостями. Последний, 9-й брюшной сегмент корот¬
128
кий, покрыт волосками, особенно длинными в основании урогомф.
Голенелапка на конце с 1—2 крючковидными щетинками, с вент¬
ральной стороны — с короткими торчащими волосками. Длина и
ширина головной капсулы (рис. 35, 2) примерно одинаковы. Лоб¬
ные швы хорошо выражены, в мелких изломах. Позади точки их
слияния полушария головы спаяны удлиненным склеритом. Бо¬
ковые стороны полушарий выпуклые. Лоб с крупным, склероти-
зованным, сильно выступающим бугром, который включает в себя
и эндокарину. На полушариях головы имеется по 1 склеротизо-
ванному зубцу.
О, sanguinicollis L. Значительная серия личинок и 2 имаго
были добыты в сентябре 1959 г. в Теллермановском лесничестве
Воронежской области из сухобочины перестойного дуба, плотно
заселенной личинками корабельного сверлилы (Lymexylon navale
L.).
Род Tarsostenus Spin.
Личинки белые, червеобразные, с длинным и тонким телом
(рис. 34, 2), почти лишенным покрова из волосков, и довольно
хорошо развитой склеротизованной площадкой на 9-м брюшнол*
сегменте, который несет склеротизованные загнутые кверху уро-
гомфы. Переднегрудь с дорсальной стороны почти полностью
склеротиссвана, средне- и заднегрудь — без следов склеротиза-
ции. На задних брюшных сегментах двигательные мозоли доста^
точно хорошо выражены (рис. 34, 2), Ширина головной капсулы
несколько больше ее длины. Лобные швы четкие. Эндокарина длин¬
ная, ее длина составляет 7з длины лобного треугольника.
Т, univittatus Rossi, Личинки истребляют личинок капюшон-
ников и древогрызов в сухой древесине. Описание личинок состав¬
лено по литературным источникам.
Род Tenerus Cast.
Личинки (рис. 34, 3) белые, имеют толстое цилиндрическое ко¬
роткое тело, почти полностью лишенное покрова из волосков,
и небольшие короткие, направленные вперед, зубцевидные урогом-
фы, расноло/кенпые па небольшом 9-м брюшном тергите, который
не несет следов склеротизации. Переднегрудь с дорсальной сто¬
роны склеротизована очень слабо, средне- и заднегрудь — без
следов склеротизации. Брюшные сегменты, кроме последних,
с дорсальными парными гладкими выпуклостями. Голенелапка с
обычными щетинками, среди которых нет крючковидных. Голова
округлая, ее длина и ширина примерно одинаковы.
В Приморском крае (Комарово-Заповедное) личинки Tenerus
sp. связаны с эндемичной породой — маакией амурской, они
обитают в толще древесины, где преследуют личинок дровосе¬
ков — Aglaophis colobotheoides Bat. и Strangalia arcuata Pz.
5 Б. ЛГ. ЛГамаев и др. 129
Род Thanasimus Lafr.
В энтомологической литературе существует целый ряд публи¬
каций, посвященных личинкам видов из данного рода. Многократ¬
но описаны личинки Th. formicarius L. Внешне хорошо отличаю¬
щиеся от них личинки Th. rufipes Brahm., характеризуемые
Сааласом (Saalas, 1917), по всей вероятности, принадлежат предпо¬
следнему возрасту личинок Th. formicarius L. (Emden, 1943). Это
Предположение подтверждается и нашими наблюдениями. Ли¬
чинки североамериканских видов Th. dubius F. и Th. repandus
Ноги, описаны Бевингом и Чемплином (Boving, Champlein, 1920).
Описание личинок Th. lewisi Jacobs. имеется в японской литера¬
туре (Kawada, 1959). Из Индии описаны личинки Th. himalayen-
sis Stebb. (Gardner, 1937). Нами собраны личинки Th. formica¬
rius L. и Th. substriatus Gebl., методом исключения определена
также личинка третьего вида фауны СССР — Th. rufipes Brahm.
Личиночные признаки Thanasimus Latr. (рис. 20, 2) такие же, как
Clerus Fabr. (см. выше). Основные отличия сводятся к тому, что
у личинок Thanasimus Latr. заметно беднее волосяной покров,
особенно на голове и 9-м брюшном сегменте. Есть различия и в
хетотаксии нижней губы (рис. 36, 2), они использованы в опреде¬
лительной таблице родов.
Таблица для определения видов рода Thanasimus Latr.
1 (4) Передний край склеротизованной площадки 9-го брюшного
сегмента четкий,, между ним и краем сегмента расположено
самое большее несколько волосков. Боковые стороны 8-го
брюшного сегмента покрыты более короткими и редкими во¬
лосками по сравнению с 9-м сегментом.
2 (3) Тело с дорсальной стороны более или менее равномерно по¬
крыто волосками. Урогомфы (вид сбоку) прямые, в вершинной
трети загнутые кверху, менее чем в 1,5 раза короче склеро¬
тизованной площадки ТЬ. substriatus Gebl.
3 (2) Тело, особенно 8-й и средние брюшные сегменты, с большими
голыми срединными участками. Урогомфы (вид сбоку)
постепенно загибаются в основании и резко в вершинной
трети, более чем в 1,5 раза короче склеротизованной пло¬
щадки ТЬ. formicarius L.
4 (1) Передний край склеротизованной площадки 9-го брюшного
сегмента размытый, между ним и краем сегмента имеется
1—2 спутанных ряда волосков. Боковые стороны 8-го брюш¬
ного сегмента покрыты такими же длинными и густыми во¬
лосками, как и 9-го ТЬ. rufipes Brahm.
Th, formicarius L, Отличительные признаки личинок, упоми-
1наемые в определительной таблице, требуют проверки на массо¬
вом материале. В нашем распоряжении была серия личинок, со¬
бранная в Финляндии осенью 1971 г. под корой сосновых пней.
130
Биология вида хорошо изучена, так какой был объектом исследо¬
вания многих энтомологов. Из последних отечественных работ
наиболее полной является публикация А. И. Воронцова и
Н. 3. Харитоновой (1971). Личинки пестряка всегда встречаются
под корой в ходах различных короедов. Вид предпочитает сосну^
однако, по литературным данным, встречается также под корой
других как хвойных, так и лиственных пород. Питается преиму¬
щественно личинками короедов как наиболее доступной пипдей^
однако, по-видимому, не имеет какой-либо пищевой специализа¬
ции и нападает на личинок любых других подкорных насекомых.
Считается полезным энтомофагом, имеющим первостепенное зна¬
чение в комплексе хищных насекомых, истребляющих короедов.
Подсчеты, произведенные Н. 3. Харитоновой (1972) в Бряпскон
лесничестве, показали, что личинки муравьежука достигают сред¬
ней численности свыше 100 экз. на 1 м^ коры при максимуме в
333 экз., на отдельных деревьях при сплошном учете обнаружи¬
вали свыше 250 личинок.
Th, rufipes Brahm, Как уже отмечалось, в литературе после
работы Сааласа (Saalas, 1917) личинки вида характеризовались
по признакам экземпляров 2-го возраста. Возможно, что к этому
виду относили личинок ранних возрастов других видов, поэтому
нет уверенности, что опубликованные в литературе данные по его
биологии полностью безупречны. Нами личинки пестряка были
собраны в 1961 г. в Ярославской области из-под коры пней ели
обыкновенной и в 1970 г. в Краснодарском крае (Убинская) из-
под коры пихты кавказской. Питались они личинками короедов.
Th. substriatus GebL Большие серии личинок собраны нами в
1967—1969 гг. в Приморском крае (Комарово-Заповедное) из-под
коры кедра корейского. Они отличаются сравнительно густым, но
коротким волосяным покровом и удлиненными урогомфами.
Тщательно выполненная работа В. С. Арефина (1974 б) содержит
экспериментальные данные, показывающие, что личинкам этого
вида пестряков достаточно для завершения развития 50—60 личи¬
нок шестизубого короеда. Они развиваются только на хвой¬
ных породах. В остальном биология рассматриваемого вида весь¬
ма сходна с таковой Th. formicarius L.
Род Tillus 01.
В составе рода числится немногим более 40 видов, хорошие
описания опубликованы для личинок типового вида — Т. elon-
gatus L. и 3 видов, распространенных в Индии (Gardner, 1937),
В наших материалах точно идентифицированы только личинки
Т. elongaius L. Длинные тонкие личинки (рис. 33, 5), грязно-
белые, с фиолетовым рисунком из продольных полос. Урогомфы
тонкие, широко расставленные, у основания с крупным наруж¬
ным зубцом. Дорсальная площадка 9-го брюшного сегмента хо¬
рошо склеротизована (рис. 37, 3), Переднегрудь с дорсальной
5* 131
стороны почти п0Л110СТ1>ю склеротизовапа, срединная линия свет¬
лая, в задней трети — с просвечивающим темным гребнем. На
средне- и заднегруди склериты рудиментарны и слабо выделяются
по цвету. Брюшные сегменты с длинными межсегментными участ¬
ками и слабо развитыми дорсальными мозолями, имеющими вид
€кладки. Голенелапка с несколькими концевыми крючковидными
щетинками. Ширина головной капсулы несколько больше ее
длины. Лобные швы слегка извилистые. Эндокарина доходит до
середины лобного треугольника.
Наиболее часто личинки пестряка встречаются в древесине,
заселенной точильщиками. В лесах обычными жертвами пестряка
служат лнчинки Ptilinus pectinicornis L., в сельских деревянных
аданиях — Anobium punctatum Deg.
Род Tilloidea Cast.
Объем рода, выделенного из рода Tillus, точно не установлен.
В наших материалах имеются личинки типового вида — Т. uni-
fasciata F., что делает возможным составление родовой харак¬
теристики. Комплекс признаков личинки оказался очень сход¬
ным с родом Denops F.-W. Основные отличия заключаются в иной
форме заднего края головной капсулы, сближающей рассматри¬
ваемый род с родом Tillus, а также в более слабом вооружении
голенелапки, на которой отсутствуют крепкие торчащие щетин¬
ки, типичные для Denops.
Род Trichodes Hbst.
Ксилофильными видами, уничтожающими стволовых вредите¬
лей, являются, видимо, лишь немногие представители рода, кото¬
рый характеризуется на примере Т. spectaoilis Кг. и некоторых
других еще не определенных видов. Личинки (рис. 33, 2) умеренно
длинные, толстые, уплощенные, розоватые, с 4 ярко-розовыми
пятнами на каждом брюшном сегменте. Конец тела с короткими
разобщенными загнутыми вверх урогомфами. Тело, нередко вклю¬
чая голову, очень густо покрыто волосками. Переднегрудь с дор¬
сальной стороны на значительном протяжении сильно склероти-
зована, со светлой срединной линией и иногда просвечивающим
темным склеротизованным гребнем. Средне- и заднегрудь (каж¬
дая) с 2 небольшими темными склеротизованными пятнами.
Брюшные сегменты широкие и короткие, без явственных двига¬
тельных мозолей. Плевральные складки большие, хорошо раз¬
витые. Голенелапка на конце с многочисленными щетинками,
среди которых могут быть и крючковидные. Ширина головной
капсулы явственно больше ее длины. Лобные швы отчетливые,
слабо извитые. Длина эндокарины варьирует, она может сос¬
тавлять до V3 длины лобного треугольника.
132
т. spectabilis Кг. Тело личинок густо покрыто волосками,
включая голову. На сегментах, начиная со среднегруди, имеются
4 ярких малиновых пятна, образующих поперечный ряд (на груд¬
ных сегментах иногда отчетливы только 2 срединных).
Личинки пестряка развивались под корой усыхающих деревьев
лоха (Акибай Туркменской ССР), где были собраны весной 1970 г.
Они уничтожали личинок дровосека Xylotrechus namanganensis
Sem., а также златок Chrysobothris affinis F. nCratomerus elaeag-
m Richt. В тугаях Амударьи под г. Дейнау личинки были найде¬
ны в ходах дровосека Turanium sp. также в усохших ветвях
лоха.
СЕМЕЙСТВО TROaOSSITlDAE — ТЕМНОТЕЛКИ
Кроусон (Crowson, 1964), сравнив как имагинальные, так и
личиночные признаки, обосновал необходимость выделения са¬
мостоятельных семейств Peltidae и Trogossitidae. Этот же автор
(Crowson, 1970) п&сл€ изучения личинок Grynocharis выделил са¬
мостоятельное семейство Lophocateridae.
Экология. Все представители семейства относятся к облигат¬
ным хищникам. Только в отношении Tenebroides mauritanicus L.
имеются наблюдения, показывающие, что при отсутствии животной
пищи этот вид, встречающийся также на зерновых складах,
может питаться зародышами зерен, нанося тем самым вред.
Как большинство ксилофильных хищных насекомых, предста¬
вители семейства Trogossitidae мало специализированы в отноше¬
нии объектов питания. Видимо, здесь ведущее значение имеет
соотношение размеров хищников и их жертв. Так, сравнительно
мелкие личинки Nemosoma хищничают за счет также относительно
мелких личинок короедов; более крупные личинки Trogossitan
Melambia питаются в основном личинками златок и дровосеков.
Виды семейства Trogossitidae отличаются определенной специа¬
лизацией в отношении заселяемых ими древесных пород. Это вы¬
ражается в том, что виды, встречающиеся на хвойных породах,
как правило, не встречаются на лиственных, и наоборот. Значи¬
тельная часть изученных видов семейства развивается под корой
на начальных стадиях ее разрушения, когда здесь поселяются
короеды или златки и дровосеки. Преследуя жертву по ее ходам,
хищники нередко проникают и в толщу древесины. Имеющиеся
в нашем распоряжении наблюдения показывают, что высокое
численности в заселенных объектах могут достигать личинки
Nemosoma, Trogossita и Melambia. В комплексе с другими энто-
мофагами они вносят существенный вклад в процессы регуляции
численности стволовых вредителей.
Морфология. Личинки (рис. 20, 5; 38, 2—2) длинные, мясистые,
слегка уплощенные, с мягкими покровами и склеротизованными
урогомфами на конце тела. Они могут быть одноцветными —
белыми, серовато-белыми или розоватыми или имеют на сег¬
133
ментах тела темный мраморный рисунок. Голова личинок упло¬
щенная, как правило, удлиненная, реже ее ширина превышает
длину. Лобные швы отчетливые, они сливаются в заднем отделе
головной капсулы. Полушария головы позади точки слияния
лобных швов либо на некотором протяжении спаяны, образуя
короткий теменной шов, либо сразу же расходятся, а пространст¬
во между ними занимает треугольный склерит. Эндокарина хо¬
рошо развита. Глазков — 5, они либо представлены пигментными
пятнами, либо имеют также и четкие линзы. Усики 3-члениковые,
могут глубоко втягиваться в головную капсулу. На апикальной
площадке 2-го членика усиков расположены конусовидные сен¬
силлы и смещенный на край площадки небольшой 3-й членик.
Мандибулы с двузубчатой вершиной, моларная часть не раз¬
вита. Простека имеется чаще всего в виде пучка игловидных вы¬
ростов. Верхняя губа отделена от клипеуса. Нижняя губа имеет
вид узкой склеротизованной полоски. Прементум хорошо отгра¬
ничен, несет 2-члениковые нижнегубные щупики. Кардо значи¬
тельно короче стипеса (рис. 39, 1—5), что является характерней¬
шим признаком личинок рассматриваемого семейства. Жеватель¬
ная доля без склеротизованного зубца, покрыта волосками
и толстыми кинжаловидными щетинками. Нижнечелюстные
щупики 3-члениковые, нередко сильно вытянутые.
Тергит переднегруди почти полностью склеротизован, на сред¬
не- и заднегруди могут быть дорсальные склеротизованные пятна —
по 2 пятна на каждом тергите. Ноги короткие, 4-члениковые,
заканчиваются простым слабо изогнутым коготком, в основании
которого имеется щетинка. В апикальной части голенелапки
имеются длинные щетинки, простые или на конце крючковидно
загибающиеся. Брюшные сегменты с боковыми, дорсальными и
вентральными мозолями, из них дорсальные в некоторых случаях
снабжены овалом из гранул. Урогомфы сильносклеротизованные,
длинные. Ветви урогомф (рис. 38, 3—5) либо плавно изогнутые
вверх, постепенно утончающиеся и заостряющиеся, либо равно¬
мерно утолщенные, на конце с дорсальным шипом. На каждой
из ветвей обычно имеются 1—2 длинные щетинки, иногда так/ке
дополнительно многочисленные короткие щетинки. Анальный
сегмент обычно с рядом мелких шипиков и щетинок.
Таблица для определения родов семейства Trogossitidae
1 (4) Урогомфы (рис. 38,5, 5) к вершине слегка утолщаются и за¬
канчиваются острым, направленным вверх зубцом.
2 (3) Личинка одноцветная, серовато-белая. Стипес с предвер¬
шинной боковой группой щетинок. На голенелапке в осно¬
вании коготка щетинки короткие, некрючковидные ....
Tenebroides Pili, et Mitt. (стр. 139)
3 (2) Сегменты тела светлые, с темным неправильным рисунком.
Стипес максимум с 2—3 предвершинными боковыми щетин-
134
Рис. 38. Общий вид {1, 2) и 9-й брюпшой сегмент (5—5) личинок темнотелок
(Trogossitidae)
1 — Melambia tekkensis Коп.; 2 — Neittosoma elongatum L.; [5 — Lepidopteryx squamulo-
sa Gebl.; 4 — Trogossita coerulea L., 5 — Tenebroides maurltanicus L«
нами (рис. 39, i). Ha голенелапке в основании коготка
имеются 2 очень длинные крючковидные щетинки ....
Lepidopteryx Норе (стр. 136)
4 (1) Урогомфы (рис. 38,4) к вершине равномерно утончаются и
заостряются, загнуты вверх.
135
Рис. 39. Нижняя губа и левая максилла личинок темнотелок (Trogossi-
tidae) снизу
1 — Lepidopteryx squamulosa Gebl.; 2 — Melambia t3kkensis Коп.; 3 — Nemosoma
cornutum Sturm.
crd — кардо; m — ментум; та — мала; prm — прементум; st — стипес
5 (6) Голова поперечная (рис. 38,1). Полушария головы позади
точки слияния лобных швов спаяны треугольным склери¬
том Melambia Erich. (стр. 137)
6 (5) Голова вытянута в длину (рис. 20, 5; 38, 2). Полушария го¬
ловы позади точки слияния лобных швов на некотором
протяжении спаяны по прямой линии.
7 (8) Отростки урогомф (рис. 20,3) параллельные, с 1-—2 крупными
и несколькими микроскопическими волосками. Последний
членик нижнечелюстных щупиков значительно короче
предпоследнего. Боковые стороны стипеса в апикальной
половине покрыты многочисленными волосками .....
Trogossita 01. (стр. 139)
8 (7) Отростки урогомф сходящиеся (рис. 38, 2), несут по
1 крупной и некоторое количество умеренно длинных щети¬
нок. Последний членик нижнечелюстных щупиков значи¬
тельно длиннее предпоследнего. Боковые стороны стипеса все¬
го с 2—4 волосками (рис. 39, 3) . . Nemosoma Latr. (стр. 137).
Род Lepidopteryx Норе
Представлен в наших материалах видом L. squamulosa Gebl.
Его личинка хорошо отличается от известных личинок других
палеарктических родов семейства по наличию неправильного
темного рисунка на светлых сегментах. Голова удлиненная, силь-
136
носклеротизованная, с почти прямыми лобными швами. Полуша¬
рия позади точки слияния лобных швов спаяны треугольным
склеритом. Эндокарина хорошо выражена на протяжении более
половины длины лобного треугольника. Урогомфы темные, их
боковые стороны покрыты многочисленными длинными волоска¬
ми. Передний край склеротизованной площадки 9-го брюшного
сегмента с приподнятым бортиком (рис. 38, 3).
Личинки развиваются под корой, нередко заходят и в толщу
древесины. Они были обнаружены, например, в Приморском крае
(Комарово-Заповедное) в древесине ольховского стволика, засе¬
ленного личинками Melandrya sp., и в стволе клена маньчжурско¬
го с многочисленными личинками рогохвоста Тгетех fuscicornis F.
В Хабаровском крае (Хинганский заповедник) личинки развива¬
лись под корой дуба, где питались личинками дровосека Mesosa
myops Dalm.
Род Melambia Erich.
В литературе опубликовано хорошее описание личинок М. саг-
doni Lew. (Gardner, 1931), которые были обнаружены в Индии
в стволе Grewia sp., где развивались личинки дровосеков, златок
и слоников.
М, tekkensis Коп. Личинки белые. Тергиты средне- и заднегру-
ди слабосклеротизованы (рис. 38,1). Площадка в основании уро-
гомф светлая, без склеротизованного бортика по переднему краю.
На конце 2-го членика усиков имеется 4 конусовидные сенсиллы.
На жевательной лопасти максилл (рис. 39, 2) расположены
3—4 кинжаловидные и многочисленные волосовидные щетинки,
а также пальцевидный вырост.
В засушливых районах Средней Азии (Акибай Туркменской
ССР и др.) личинки М. tekkensis развиваются в отмирающих фрук¬
товых деревьях итуранге. Это — облигатные подкорные хищники,
питающиеся личинками златок и дровосеков. Они были отме¬
чены под корой абрикоса и алычи, но особенно высокой численно¬
сти достигали в усыхающих деревьях лоха, кора и древесина
которых были плотно заселены личинками дровосека Xylotrechus
namanganensis Sem.
Род Nemosoma Latr.
Определительная таблица этих видов по макроскопическим
признакам и характеристика рода по личинкам опубликованы
Н. Б. Никитским (1974). Личинки (рис. 38,2) розовато-белые. Все
три грудных тергита со склеротизованными пятнами. Урогомфы
резко загнуты кверху и нередко сближаются своими вершинами.
Склеротизованного бортика по переднему краю площадки, распо¬
137
ложенной в основании урогомф, нет. Полушария головы сливают¬
ся по прямому шву. Эндокарина длинная, доходит почти до кли-
пеуса. На 2-м членике усика 1 конусовидная сенсилла. Крючко¬
видные щетинки па голенелапке не выражены. Длина личинок
до 10—12 мм.
Таблица для определения видов рода Nemosoma Latr.
1 (2) Пятна на тергитах средне- и заднегруди бледные, значитель¬
но слабее окрашены, чем переднегрудной тергит. Нижняя
губа в основании прементума слабо склеротизована. Личин¬
ки белые N. caucasicum Мёл.
2 (1) Пятна на тергитах средне- и заднегруди почти так же интен¬
сивно окрашены, как переднегрудной тергит. Склеротизация
нижней губы в основании прементума по своей интенсивности
не отличается от склеротизации других ее участков. Личинки
розоватые.
3 (4) Между урогомфами имеется до 10 волосков, большинство
которых заметно длиннее урогомф. 3-й членик нижнечелюст¬
ных Ш.УПИКОВ в 2 раза длиннее 2-го. Внутренний край жева¬
тельной лопасти в апикальной трети без четкой вырезки.
Длина конусовидной сенсиллы 2-го членика усиков состав¬
ляет не менее половины длины 3-го членика
N. cornutum Sturm.
4 (3) Между урогомфами имеется 5—6 коротких волосков, как
правило, не достигающих своими вершинами конца урогомф.
3-й членик нижнечелюстных щупиков в 1,5 раза длиннее
2-го. Внутренний край жевательной лопасти в апикальной
трети с четкой вырезкой. Длина конусовидной сенсиллы 2-го
членика усиков составляет около трети длины 3-го члени¬
ка N. elongatum L.
N, caucasicum Мёп, Личинки данного вида в Краснодарском
крае (Убинское лесничество) развиваются под корой ясеня, гру¬
ши и бука в ходах различных короедов, личинками которых пи¬
таются.
N. cornutum Sturm. Личинки — активные хищники. Они встре¬
чались в Краснодарском крае (Красная Поляна) под корой граба,
бука и реже — дуба в ходах двухцветного (Taphrorychus bicolor
Hbst.) и некоторых других видов короедов, а также среди личинок
слоников Trachodes sp., которые служили им пищей.
N. elongatum L. В большинстве случаев вид развивается под
корой хвойных деревьев (ель, сосна, кедр, туя и др.) на различ¬
ных видах короедов. Обнаружен нами в Краснодарском крае
(Красная Поляна). На лиственных породах, среди которых упо¬
минаются в первую очередь бук и дуб, вид развивается как на ко-
роедах-заболонниках, так и на короедах-древесинниках (Xylebo-
rus saxeseni Ratz., Trypodendron domesticum L.).
138
Род Tenebroides Pili, et Mitt.
Эти личинки отличаются серовато-белым цветом, наличием на
всех трех грудных тергитах четких склеротизованных участков
и несколько расширенными к вершине выростами урогомф. Голова
личинок продолговатая, ее полушария позади точки слияния лоб¬
ных швов спаяны треугольным склеритом. Лобные швы прямые.
Эндокарина хорошо развита, почти достигает передней четверти
лба.
Таблица для определения видов рода
Tenebroides Pili, et Mitt.
1 (2) Склеротизованная площадка 9-го брюшного тергита по пе¬
реднему краю со склеротизованным бортиком. В основании
отростков урогомф снизу нет длинных волосков. Конусовид¬
ная сенсилла 2-го членика усиков короче окружаюш,их ее
волосков т. mauritanicus L.
2 (1) Склеротизованная площадка 9-го брюшного тергита по пе¬
реднему краю с нечеткой границей, без склеротизованного
бортика. В основании отростков урогомф снизу по 2 длинных
щетинки. Конусовидная сенсилла 2-го членика усиков длин¬
нее окружающих ее волосков . . .Т. turkestanicus Ball.
Т. mauritanicus L. В естественных биотопах личинки встречают¬
ся обычно поодиночке под корой лиственных деревьев в ходах са¬
мых различных насекомых, главным образом личинок златок и дро¬
восеков. В частности, нами личинки были обнаружены в Красно¬
дарском крае (Убинская) под корой дуба в ходах дровосека Pyrr¬
hi dium sanguineum L.
Т. turkestanicus Ball. Личинки были обнаружены в Акибае
Туркменской ССР под корой усыхающих деревьев лоха, заселен¬
ных главным образом личинками дровосека Xylotrechus grumi
Sem.
Род Trogossita 01.
Цвет личинок различен. Склеротизованные участки развиты
с дорсальной стороны на всех грудных сегментах (рис. 20, 3), Эндо¬
карина достигает лишь середины лобного треугольника. На 2-м
членике усика имеется только 1 конусовидная сенсилла. На же¬
вательной лопасти максилл развит очень густой покров из разно¬
образных волосков. Двигательные мозоли на брюшных сегментах
гранулированные. На вентральной стороне сегментов имеются
сильно извитые волоски. Площадка в основании урогомф хорошо
склеротизована, без бортика или с неполным бортиком по перед¬
нему краю (рис. 38, 4). Крючковидные щетинки на ногах имеются.
139
Таблица для определения видов рода Trogossita 01.
1 (2) Личинки белые. Гранулы на двигательных мозолях хорошо
выражены только на средних брюшных сегментах, на боковых
сторонах этих сегментов — по 2—3 длинных волоска. На
выростах урогомф по 1 длинному волоску. Склеротизованная
площадка 9-го брюшного сегмента по бокам всего с 2—3 длин¬
ными волосками . . .Т. coerulea L.
2 (1) Личинки сероватые. Гранулы на двигательных мозолях хо¬
рошо выражены на всех брюшных сегментах, кроме 2 послед¬
них; на боковых сторонах этих сегментов — порядка 10 длин¬
ных волосков. На выростах урогомф по 2 длинных волоска.
Склеротизованная плош;адка 9-го брюшного сегмента по бокам
с 7 длинными волосками Т. japonica Rtt.
Т, coerulea L. Материалы по экологии вида крайне скудны.
Согласно Эмдену (Emden, 1943), личинки развиваются под корой
сосны. По-видимому, они хищничают за счет личинок других раз¬
вивающихся здесь насекомых.
Т, japonica Rtt. В Приморском крае, где изучалась экология
вида (Мамаев, 1971 г.), его личинки обычны под корой усыхающего
кедра корейского в ходах крупного шестизубчатого (Ips sexdenta-
tus Boern.) и других видов короедов. В число жертв этого хищника
следует включить также личинок различных дровосеков (Tetro-
pium, Acanthocinus и др.) и некоторых слоников (Lamyrus). На
одном крупном кедре удавалось собрать 70—150 личинок япон¬
ской щитовидки. Вид развивается также под корой пихты цельно¬
листной, но на этой древесной породе встречается значительно
реже.
СЕМЕЙСТВО ELATERIDAE - ЩЕЛКУНЫ
Щелкуны представляют собой крупное семейство, насчитываю¬
щее в пределах СССР свыше 500 видов (Гурьева, 1965).
Личинки щелкунов (так называемые проволочники) чаще всего
встречаются в почве и подстилке, причем значительное число поч¬
вообитающих форм является серьезными вредителями сельскохо¬
зяйственных, лесных и садовых культур. Развитие многих видов
протекает под корой и в толще разлагающейся древесины. Ли¬
чинки щелкунов, как правило, всеядны, но в спектре питания
обычно преобладает какой-нибудь один из пищевых источников.
Ксилофильные личинки щелкунов, развивающиеся под корой и в
толще разлагающейся древесины, в большинстве случаев являют¬
ся облигатными хищниками (Alaus, Adelocera, Calambus, Нуро-
ganus, Sienagostus, Ludius), реже всеядными видами, предпочи¬
тающими хищничество (Melauolus), или хищниками с элементами
некросапрофагии (Ampedus).
В большинстве случаев хищные личинки щелкунов ]ie специа¬
лизированы в отношении объектов питания, среди которых обыч-
140
пой пище{^ являются личинки златок и дровосеков, а в более силь¬
но разрушенной древесине — личинки рогачей, узконадкрылок,
древоедов и тех двукрылых, которые поселяются колониями (на¬
пример, Sciaridae). Четко выраженной специализации к обитанию
в определенных породах также не отхмечено, хотя тенденция к та¬
кой специализации имеется. Так, некоторые виды рода Adelocera
предпочитают хвойные породы, тогда как личинки Ludius встреч
чаются в дуплах только лиственных деревьев. Большинство видов
щелкунов развивается в толш,е древесины; нередко личинки тех
же видов выходят и в подкоровую зону. Чаще всего под корой
встречаются личинки видов рода Stenagostus. К настоящему вре¬
мени нет данных о возможной специализации личинок щелкунов
к жизни в различных типах гнили древесины. Особенно обычны
в бурых гнилях личинки Ampedus и Melanotus.
Данные по морфологии личинок щелкунов, в том числе и по
морфологии ксилофильных форм, имеются в работах Ф. Эмдена
(F. Emden, 1945) и X. Эмдена (Н. Emden, 1956). Наиболее полные
сведения содержатся в монографиях В. Г. Долина (19(>4а) и
А. И. Черепанова (1965). Нами используется терминология, при¬
нятая В. Г. Долиным, и опубликованные им материалы.
Морфология. Тело дорсовентрально уплощенное или цилин¬
дрическое, с явственно выраженной склеротизацией, у личинок
Cardiophorinae — несклеротизированное, за исключением головы
и переднегруди, червеобразное, с дополнительной (ложной) сег¬
ментацией.
Голова прогнатическая, хорошо склеротизованная, дорсовен¬
трально уплощенная, обычно примерно одинаковой длины и ши¬
рины, только у Cardiophorinae длина головной капсулы заметно
превышает ширину. Теменной шов либо очень короткий, как, на¬
пример, у Agrypnini и Agriotini, либо отсутствует (Cardiophorinae,
Athouini, Corymbitini). Лобные швы явственные, четко ограничи¬
вают расположенную между ними копьевидную лобную пластин¬
ку, основание которой сильно варьирует по форме. В средней час¬
ти переднего края лобной пластинки расположено назале. Как
правило, наличник слит с лобной пластинкой, но у представителей
подсемейств Cryptohypninae и Cardiophorinae шов между налич¬
ником и лобной пластинкой имеется. Гипокраниальный шов может
быть хорошо выражен в задней части головной капсулы или неяв¬
ствен. Гуларная пластинка развита не у всех форм. С каждой сто¬
роны головы по одному глазку или глазки отсутствуют. Усики
3-члениковые, прикреплены кнаружи от места причленения жвал.
1-й членик самый крупный, 2-й несет на вершине 1 или несколько
сенсилл, 3-й членик маленький, иногда еле заметный. У Cardio¬
phorinae самый крупный членик усиков —2-й, а сами усики прикре¬
пляются сверху, перед местом причленения жвал.
Мандибулы сильносклеротизованные, треугольные, серповид¬
но изогнутые. Их режущий край либо гладкий, либо с крупным
зубцом (ретинакулумом) в средней части. У личинок некоторых ви¬
141
дов имеется добавочный вершинный зубец. В нижней ^асти внут¬
реннего края мандибул развита щетка коротких волоскс^в. У Cardio-
phorinae мандибулы двулопастные, их дорсальная Лопасть, как
правило, снабжена зубцами, вентральная — гладкая. Максиллы
и нижняя губа образуют единую подвижную пластинку. Максиллы
состоят из небольшого, иногда двураздельного кардо и длинного,
Массивного стипеса. На вершине стипеса, около наружного края,
расположен 4-члениковый нижнечелюстной ш;упик, обычно 2-чле-
ииковая или гораздо реже — 1-члениковая галеа и нечленистая
лациния, внутренний край которой снабжен густой ш;еткой волос¬
ков. Нижняя губа состоит из удлиненного, массивного ментума
и короткого прементума, несущего 2-члениковые нижнегубные
Щупики.
Грудные сегменты хорошо склеротизованы. Наиболее крупным
из них является переднегрудь. Средне- и заднегрудь обычно оди¬
наковой длины. У личинок Cardiophorinae грудные сегменты при¬
мерно одинаковой величины, причем хорошо склеротизована толь¬
ко переднегрудь, а средне- и заднегрудь склеротизованы очень
слабо. Ноги 4-члениковые, все три пары ног сходны по размерам
и вооружению.
Брюшко состоит из 10 сегментов. Первые 8 сходны по строению
и размерам и несут каждый по паре дыхалец. У личинок Cardio¬
phorinae первые 7 сегментов брюшка имеют ложную сегментацию,
в результате чего брюшко кажется состоящим из 23 сегментов. 9-й
сегмент брюшка щелкунов бывает конусовидным или уплощенным,
в последнем случае он может быть либо цельным, либо несущим
ветвистые или неветвистые урогомфы. Анальная подпорка смеще¬
на на вентральную поверхность 9-го брюшного сегмента, она мо¬
жет быть простой или снабженной по бокам 2 шипиками (Adeloce-
га) или 2 мягкими серповидными долями (Cardiophorinae).
Таблица для определения родов семейства Elateridae
1 (14) 9-й брюшной сегмент уплощен. Урогомфы имеются
(рис. 40, 2, 3; 44, 7, 4).
2 (5) Анальная подпорка с 2 склеротизованными крючками
(рис. 40, 4, 8).
3 (4) Дорсальная сторона головной капсулы с 2 продольными
килями, усаженными щетинками. Анальная подпорка, по¬
мимо крючков, несет крючковидные бугорки (рис. 40, 8).
Дыхальца треугольные Alaus Esch. (стр. 146)
4 (3) Дорсальная сторона головной капсулы без килей. Крюч¬
ковидные бугорки па анальной подпорке отсутствуют
(рис. 40,4). Дыхальца овальные . . Adelocera Latr. (стр. 144)
5 (2) Анальная подпорка без склеротизованных крючков.
6 (11) Назале 3-зубцовое. Щетинки на последних сегментах тела
личинок всегда короче, чем ширина тела.
142
7 (8) Наружные ветви урогомф с округлой вершиной (рис. 43,
ii, J2) Stenagostus Thoms. (стр. 156)
8 (7) Наружные ветви урогомф с крючковидной вершиной
(рис. 44, i, 4).
9 (10) Дыхальца на 8-м брюшном сегменте расположены в его
передней трети Aihous Esch. (стр. 153)
10 (9) Дыхальца на 8-м брюшном сегменте брюшка расположены
в его средней части Denticollis Pili. (стр. 153)
И (6) Назале 1-зубцовое (рис. 45,5). Щетинки на последних сег¬
ментах тела личинок очень густые и длинные, их длина
не уступает ширине тела.
12 (13) Наружные ветви урогомф редуцированы, имеют вид не¬
большого бугорка, внутренние ветви широкие с округлыми
вершинами (рис. 44, 2) .. . Calambus Thoms. (стр. 153)
13 (12) Наружные ветви урогомф равны по длине внутренним,
или чуть короче их, цилиндрические, внутренние ветви
удлиненные, конические, с коготком на вершине (рис. 44, 5)
Hypoganus Kiesw. (стр. 155)
14 (1) 9-й брюшной сегмент брюшка куполовидный или конусо¬
видный (рис. 42, i, 4), если уплош,ен, то только в задней
половине (рис. 43, 10\ 44, 6). Урогомфы отсутствуют.
15 (26) Тело склеротизовано, без дополнительной сегментации.
Мандибулы серповидные, со срединным зубцом.
16 (17) Вершина 9-го брюшного сегмента уплош,ена, оканчивается
3 зубцами (рис. 43,10\ 44, 6) . . Melanotus Esch. (стр. 155)
17 (16) 9-й брюшной сегмент куполовидный или конусообразный,
вершина его не уплош,ена.
18 (19) 9-й брюшной сегмент с округлой вершиной и гладкими по¬
кровами (рис. 42, S) Ludius Latr. (стр. 155)
19 (18) 9-й брюшной сегмент несет на вершине 1—3 шипа или бу¬
горок (рис. 40, i; 42, ^), покрыт грубыми точками или мор-
ш,инистостью.
20 (25) 9-й брюшной сегмент с грубой пунктировкой (рис. 42,
1.2),
21 (22) 9-й брюшной сегмент постепенно суживается к вершине
(рис. 42, i, 4, 7). Назале 1-зубцовое. Мускульные вдавления
и боковые бороздки на тергитах средне-, заднегруди и
брюшка имеются Ampedus Dej. (стр. 146)
22 (21) Задняя половина 9-го брюшного сегмента резко сужена.
Назале 3-зубцовое. Мускульные вдавления и боковые бо¬
роздки отсутствуют.
23 (24) Вершина 9-го брюшного сегмента с 1 загнутым вниз ши¬
пом. Питание не изучено Procraerus Redtb.
24 (23) Вершина 9-го брюшного сегмента с 3 шипами. Питание не
изучено Megapenthes Kiesw.
25 (20) 9-й брюшной сегмент с морщинистой поверхностью и бу¬
горком на вершине (рис. 40, 1), Всеяден, в основном фи¬
тофаг Ectinus Esch.
143
/
26 (15) Тело с дополнительной (ложной) сегментацией,/за исклю¬
чением головы и переднегруди, не скле/ротизовано
(рис. 19, 1). Мандибулы двулопастные . . . /
Cardiophonis Esch. (стр. 153)
Род Adelocera Latr.
Площадка 9-го брюшного сегмента с 2 продольными параллель¬
ными бороздками, между которыми расположены 1—2 пары круп¬
ных щетинок. Вершинная треть задней части лобной пластинки
сильно сужена, часто сосцевидно вытянута (рис. 40, 5).
Таблица для определения видов рода Adelocera Latr»
1 (4) Площадка 9-го брюшного сегмента с 1 парой крупных ще¬
тинок в центре. Вырезка между урогомфами маленькая
(рис. 40,5), по крайней мере вдвое уже ширины урогомфы.
Задний край лобной пластинки широко округлый, ее вер¬
шинная треть постепенно суживается.
2 (3) Вырезка между урогомфами удлиненно-овальная
(рис. 40, 3). Площадка 9-го брюшного сегмента гладкая . .
А. conspersa Gyll.
3 (2) Вырезка между урогомфами поперечно-овальная. Площад¬
ка 9-го брюшного сегмента в редких мелких склеротизован-
ных бугорках А. fasciata L.
4 (1) Площадка 9-го брюшного сегмента с 2 парами крупных
щетинок (рис. АО, 2, 7). Вырезка между урогомфами круп¬
ная, глубокая, ширина ее по крайней мере равна ширине
урогомфы. Вершина задней лопасти лобной пластинки
сосцевидно вытянутая или треугольная (рис. 40, 5).
5 (()) Площадка 9-го брюшного сегмента, помимо 2 пар крупных
щетинок, несет многочисленные мелкие щетинки (рис. 40, 7).
Вершина задней лопасти лобной пластинки треугольная
А. quercea Hbst.
6 (5) Мелкие щети1[ки на площадке 9-го брюшного сегмента от¬
сутствуют (рис. 40, 6). Вершина задней лопасти лобной
пластинки сосцевидно вытянута (рис. 40, 5).
7 (8) Ширина вырезки между урогомфами равна ширине урогом¬
фы (рис. 40, 6) А. punctata Hbst.
8 (7) Ширина вырезки между урогомфами примерно вдвое шире
урогомфы (рис. 40, 2).
9 (10) Основание вырезки между урогомфами с небольшой насеч¬
кой. Зубцы по бокам площадки 9-го брюшного сегмента не
искривлены А. lepidoptera Pz.
10 (9) Основание вырезки между урогомфами равномерно округ¬
лое. Зубцы по бокам 9-го брюшного сегмента крючковидно
изогнутые (рис. 40, 2) А. altaica Cand.
144
Рис. 40. Детали строения Ectinus aterrimus L. (2), Adelocera altaica Cand.
(2), A. conspcrsa Gyll (<3), A. fasciata L. (4), A. punctata Hbst. (J, 6), A. quer¬
cea Hbst. (7), Alaus parreysii Stev. (<^) (Elateridae)
2—3, 6, 7 — 9-Й сегмент брюшка сверху; 4, 8 — конец брюшка сбоку; 5 — лобная плас¬
тинка
а. ped — анальная подпорка (J, 4, 5—8 — по Долину, 1964а; 2,3 — по Черепано¬
ву, 1965)
А. altaica Cand. Личинки развиваются в разлагающейся древе¬
сине пней хвойных пород. Хищники, уничтожающие личинок
различных жесткокрылых.
А. conspersa Gyll. Личинки развиваются в разлагающейся дре¬
весине и под корой гнилых пней и упавших деревьев хвойных по¬
род. Уничтожают личинок стволовых вредителей.
145
А. fasciata L. Личинки] развиваются в разлагающейся древеси¬
не и под корой хвойных пород. Питаются личинками стволовых
вредителей.
А. lepidoptera Pz, Личинки развиваются под корой и в разла¬
гающейся древесине лиственных пород. Хищники.
А. punctata Hbst. Личинки развиваются под корой и в разлагаю¬
щейся древесине сосны. Хищники.
А. quercea Hbst, Личинка найдена в разлагающейся древесине
дуба. Хищник.
Род Aiaus Esch.
Площадка на 9-ом брюшном сегменте неокаймленная, с 2 про¬
дольными бороздками, густо усажена склеротизованными щетин¬
коносными бугорками. Задняя лопасть лобной пластинки удли¬
ненная, с округлым задним краем. В Советском Союзе 1 вид.
А, parreysii Stev. В сосновых пнях. Хищник.
Род Ampedus Dej.
Сегменты тела с резкими мускульными вдавлениями и боковыми
продольными бороздками, отходящими от их наружного края.
Последний сегмент шлемовидный, заканчивается шипиком или
бородавкой.
Таблица для определения видов рода Ampedus Dej. i
1 (2) Стерниты, плейриты и боковые части тергитов за боковой
продольной бороздкой на сегментах брюшка гладкие, пол¬
ностью лишены пунктировки. Вершина 9-го брюшного сег¬
мента с плоской бородавкой (рис. 41, 5; 42, 7)
А. megerlei Lac.
2 (1) Стерниты, плейриты и боковые части тергитов брюшка яв¬
ственно пунктированы. Базальная часть стернитов брюш¬
ка с пояском крупных грубых точек, иногда прерванных
посредине. Вершина 9-го брюшного сегмента со склероти-
зованным шипом.
3 (8) Пунктировка тергитов, начиная со среднегрудного, ниже
пояска грубых точек равномерная и мелкая, точки на вер¬
шине тергитов не более чем в 1,5—2 раза меньше точек
в средней части. Поясок крупных точек в базальной части
стернитов брюшка однорядный и посредине прерванный.
Пунктировка 9-го брюшного сегмента иногда несколько
крупнее, чем предыдущих (рис. 41, 4\ 42, 6).
4 (5) На вентральной поверхности 9-го брюшного сегмента у
вершины анальной подпорки всего одна пара развитых
щетинок (рис. 43, 2) А. nigerrimus Lac.
^ По В. Г. Долину (1964а).
146
Рис. 41. Детали строения Ampedus balteatus L. (i), А. cinnabarinus Esch.
(2, 5), A. erythrogonus Miill. (4, 5), A. megerlei Lac. (6)^ A. nigrinus Hbst.
(7), A. ochropterus Germ. (S, 9)^ A. pomonae Steph. (JO), A. pomorum Hbst.
(ii), A. sanguineus L. (i2), A. sanguinolentus Schrk. (13, 14) и A. satrapa
Kiews. (15) (Elateridae)
Jt 3, 5, 7, JOt 22, 14 — мускульное вдавление и дыхальце; 2,4,8,11 — 2-й сегмент брюш¬
ка сверху; 6 — 8-й сегмент брюшка снизу; 9 — плейриты средне- и заднегруди; J5 —
среднегрудь сверху; 15 — 2-й сегмент брюшка сбоку (по Долину, 1964а)
5 (4) Вентральная поверхность 9-го брюшного сегмента у вер¬
шины подпорки с двумя парами развитых щетинок
(рис. 43, 5).
6 (7) Боковая продольная бороздка на 3—7 тергитах брюшка
только в 1,5 раза длиннее расстояния от ее конца до пояс¬
ка тонкой пунктировки на границе лентовидного окаймле¬
ния. Придыхальцевая щетинка всегда расположена против
середины дыхальца. Пунктировка тергита переднегруди
много мельче, чем на последующих сегментах (рис. 41,4,5)
А. erythrogonus Miill.
7 (6) Боковая продольная бороздка на 3—7-м тергитах брюшка
вдвое длиннее расстояния от ее конца до пояска тонкой
147
пунктировки на границе лентовидного окаймл^шя. При-
дыхальцевая щетинка на всех или части сегментов брюшка
расположена против конца дыхальца. Пунктировка терги-
та переднсгруди грубая, лишь вдвое мельче, чем на после¬
дующих (рис. 41,7) А. nigrinus Hbst.
8 (3) Пунктировка тергитов груди и брюшка неравномерная,
точки на вершине тергитов во много раз меньше крупных
точек в средней части. 9-й брюшной сегмент значительно
мельче пунктирован, чем предыдущие (рис. 43, 8).
9 (12) Плейриты средне- и заднегруди явственно пунктированы
только в передней половине. Плейриты брюшных сегмен¬
тов редко рассеянно пунктированы или гладкие, в базаль¬
ной части лишь со следами крупных точек.
10 (11) Тергит среднегруди в средней части по обе стороны от сре¬
динной линии с 2—5 крупными точками, приближающими¬
ся по размерам к точкам в базальном пояске, остальная по¬
верхность тергита очень мелко редко пунктирована. Мус¬
кульные вдавления поперечно-овальные, поперечная дли¬
на их втрое больше продольной (рис. 41, 13, 14) ....
А. sanguinolentus Schrk.
И (10) Тергит среднегруди по обе стороны срединной линии с 8—10
крупными точками, не меньшими, чем в базальном пояс¬
ке. Мускульные вдавления поперечно вытянутые, отноше¬
ние поперечного диаметра к продольному больше 4
(рис. Ai, 10) А. pomonae Steph.
12 ( 9) Плейриты средне- и заднегруди по всей длине грубо пунк¬
тированы (рис. 41, 9). Плейриты брюшных сегментов с яв¬
ственной грубой пунктировкой в базальной части.
13 (24) Мускульные вдавления распространены в сторону, за уро¬
вень боковой продольной бороздки примерно на ширину
вдавления у бороздки, но не менее чем на двойную тол¬
щину самой бороздки (рис. 41, 1, 12).
14 (19) Мускульные вдавления распространены в сторону, за пре¬
делы боковой продольной бороздки на ширину вдавления
у бороздки (рис. 41, i). 9-й брюшной сегмент при рассматри¬
вании сбоку лишь слегка выпуклый в вершинной трети
(рис. 43, 1). Задняя лопасть лобной пластинки вдвое или
почти вдвое длиннее ширины.
15 (16) Мускульные вдавления на 9-м брюшном сегменте не мень¬
ше или едва меньше, чем на предыдущем, и доходят до
средней пары продольных бороздок, на предпоследнем
брюшном сегменте лишь едва не доходят до срединной
линии А. praeustus F.
16 (15) Мускульные вдавления на 9-м брюшном сегменте брюшка
почти вдвое уже и короче, чем на предпоследнем, и на поло¬
вину длины доходят до средней пары продольных бороз¬
док, на предпоследнем брюшном сегменте обычно на поло¬
вину не доходят до средней линии (рис. 42, i, 2).
148
Рис. 42. Два последних сегмента брюшка личинок щелкунов сверху (Е1а-
teridae)
1 — Ampedus aethiopa Lac.; 2 — А. balteatus L.; 3 — A. cinnabarinus Esch.; 4 — A. ele-
gantulus Schonh.; 5 — A. elongatulua F.; 6 — A. erythrogonua Miill.; 7 — A. megerlel
Lac.; 8 — Ludiua ferrugineua L. (no Долину, 1964а)
17 (18) 9-й брюшной сегмент в 1,5—1,7 раза длиннее ширины в пе¬
редней трети. Переднегрудной сегмент почти квадратный,
не длиннее ширины. Отношение длины тела к ширине око¬
ло 10 (рис. 42, i) А. aethiops Lac.
18 (17) 9-й брюшной сегмент вдвое длиннее ширины. Переднегруд¬
ной сегмент спереди заметно сужен. Отношение длины тела
к ширине не менее 12 (рис. 42, 2) . . .А. balteatus L.
19 (14) Мускульные вдавления распространены в сторону за пре¬
делы продольной бороздки на двойную ширину бороздки и
сильно расширены в боковой наружной части (рис. 41, 5,12),
9-й брюшной сегмент при рассматривании сбоку с замет¬
ным вздутием в передней трети (рис. 43, 7). Задняя лопасть
лобной пластинки не более чем в 1,5—1,7 раза длиннее
ширины.
20 (21) Мускульные вдавления на 9-м брюшном сегменте не мень¬
ше, чем на предыдущем, и доходят до срединной пары бо¬
роздок, вдавления на предпоследнем сегменте почти или
сомкнутые у срединной линии (рис. 43, 6). Крупные точки
в средней части тергитов передних брюшных сегментов
крупнее точек в базальном пояске . . .А. sanguineus L.
21 (20) Мускульные вдавления на 9-м брюшном сегменте почти
вдвое короче и уже, чем на предыдущем, и на половину
длины не доходят до средних бороздок (рис. 42, 3). Вдавле¬
ния на предпоследнем сегменте значительно не доходят до
срединной линии.
22 (23) Мускульные вдавления на тергитах 2—7-го брюшных тер¬
гитов довольно широкие, поперечная длина их в 3—3,5
раза больше ширины, не доходят до срединной линии более
чем на половину своей поперечной длины (рис. 41, 2) . .
А. cinnabarinus Esch.
23 (22) Мускульные вдавления 2—7-го брюшных тергитов сильно
поперечно вытянутые, в 5 и более раз превышают ширину
и не доходят до срединной линии не более чем на V3 попе¬
речной длины (рис. 41, S) . . . .А. ochropterus Germ.
24 (13) Наружный край мускульных вдавлений ограничен боко¬
вой продольной бороздкой. Иногда вдавление переходит
бороздку, но не более чем на толщину самой бороздки
(рис. 41,75). Пунктировка тергитов груди и брюшка густая
и грубая, расстояния между точками в средней части тер¬
гитов равны или меньше размеров точек. Мускульные
вдавления широколентовидные, обычно значительно шире
длины дыхалец или равны ей.
25 (26) 9-й брюшной сегмент уплощенно-цилиндрический, на вер¬
шине широко округлен и с маленьким шиником (рис. 42, 4)
А. eleganiulus Schonh.
26 (25) 9-й брюшной сегмент удлиненно-конический (рис. 43, 9),
27 (28) Боковые части тергитов брюшка за боковой продольной
бороздкой так же грубо и рельефно пунктированы, как
средняя часть тергитов (рис. 41, 15) . . А. satrapa Kiesw.
28 (27) Точки на боковой части тергитов за боковой продольной
бороздкой много мельче, чем па прилегающих срединных
частях тергита, края их очень слабо склеротизованы.
29 (32) 9-й брюшной сегмент от середины к вершине остро-коничес¬
кий, с прямолинейными боковыми сторонами (рис. 43, 5).
30 (31) Мускульные вдавления сильно поперечно вытянутые, ме¬
нее чем на половину своей длины не доходят до срединной
линии (рис. 41,11), а на первом и предпоследнем брюшных
сегментах почти смыкаются. Мускульные вдавления на 9-м
150
Рис. 43. Конец брюшка Ampedus balteatus L. (i), А. nigerrimus Lac. (2),
A. nigrinus Hbst. (5), A. nigroflavus Goeze (4), A. pomorum Hbst. (5), A. san¬
guineus L. (6, 7), A. sanguinolentus Schrk. (5), A. satrapa Kiesw. (9), Melano-
tus rufipes Hbst. (10), Stenagostus undulatus Deg. (ii), S. villosus Geoffr.
(12) (Elateridae)
J, 7 — 9-Й сегмент брюшка сбоку; 2, 3 — 9-й сегмент брюшка снизу; 4, 6, S, 9 — 8-й в
9-й сегменты брюшка сверху; S, JO — 12 — 9-й сегмент брюшка сверху (по Долину, 19в4а>
Рис. 44. 9-й сегмент брюшка личинок щелкунов сверху (Elateridae)
1 — Athous rufiventris Eschz.; 2 — Calambus bipustulatus L.; 3 — Denticollis linearis L.;
4 — D. rubens Pili.; 5 — Hypoganus cinctus Pk.; 6 — Melanotus crassicollis Er. (J, 3 —
no Черепанову, 1965; 2, 4—6 no Долину, 1964)
сегменте не меньше или незначительно меньше, чем на пред¬
последнем А. pomorum Hbst.
31 (30) Мускульные вдавления распространяются только на поло¬
вину длины до срединной линии. Мускульные вдавления
па 9-м сегменте вдвое короче и уже, чем на предпоследнем
А, ochrinulus Rtt.
32 (29) Боковые стороны 9-го брюшного сегмента выпукло округ¬
ленные (рис. 42, 5; 43, 4),
33 (34) 9-й брюшной сегмент очень короткий и уплощенный, всего
в 1,5 раза длиннее ширины. Мускульные вдавления широ¬
152
колентовидные, немного не доходят до срединной линии,
на 9-м брюшном сегменте не меньше, чем на предпослед¬
нем, и почти доходят до средней пары бороздок. Концевой
шип мощный и длинный (рис. 43, 4) . .А. nigroflavus Goeze.
34 (33) 9-й брюшной сегмент почти вдвое длиннее ширины, на вер¬
шине с коротеньким шиником, длина которого не превы¬
шает ширину основания. Мускульные вдавления узколен¬
товидные, почти на половину не доходят до срединной ли¬
нии. На 9-м брюшном сегменте почти вдвое меньше, чем
па предпоследнем (рис. 42, 5) А. elongatulus F.
Личинки рода Ampedus — хищники с элементами некросапро-
фагии.
Род Athous Esch.
В древесине развиваются 2 вида этого рода — А. mutilatus
Rosenh. и А. rufiventris Eschz.
Таблица для определения вддов рода Athous Esch.
1 (2) Боковые бороздки на площадке 9-го брюшного сегмента
параллельные, срединные бороздки смыкаются (рис. 44, 1)
А. rufiventris Esch.
2 (1) Боковые бороздки на площадке 9-го сегмента брюшка рас¬
ходятся к основанию пластинки, срединные бороздки не смы¬
каются А. mutilatus Rosenh.
А, mutilatus Rosenh. Личинки развиваются в пнях. Хищник.
А. rufiventris Esch. Личинки развиваются в разлагающейся
древесине березы. Хищник.
Род Calambus Thoms.
В древесине зарегистрирован единственный вид.
С. bipustulatus L. Личинки развиваются в разлагающейся дре¬
весине широколиственных пород, особенно дуба, уничтожают мел¬
ких личинок и куколок разрушителей сухой древесины (точиль¬
щиков, капюшонников и др.).
Род Cardiophorus Esch.
В древесине зарегистрирован единственный вид.
С. ruficollis L. Личинки развиваются в сосновых пнях, реже
Б почве и подстилке хвойных лесов. Хищник с элементами некро-
сапрофагии.
Род Denticollis Pili.
Тело уплощенное, сверху темноокрашенное. Назале 3-зубчатое.
Кроме 5 основных, имеется дополнительная проксимальная пара
лобных щетинок. Внутренние ветви урогомф удлинены и заост¬
рены.
153
Рис. 45. Лобная пластинка щелкунов (1—6) (Elateridae)
1 — Denticollis linearis L.,— D. rubens Pili.; 3 — Hypoganus cinctus Pk.; 4 — Stenagos-
tus undulatus Deg.; 5 — S. villosus Geoffr. (J — no Черепанову, 1965; 2—5 — no Долину,
1964а)]
Таблица для определедия видов рода Denticollis Pili.
1 (2) Площадка 9-го брюшного сегмента без продольных светлых
пятен (рис. 44, 5). Задняя лопасть лобной пластинки расшире¬
на в средней части (рис. 45, 7) D. linearis L.
2 (1) Площадка 9-го брюшного сегмента с 2 продольными светлы¬
ми пятнами (рис. 44, 4), Задняя лопасть лобной пластинки не
расширяется в средней части (рис. 45, 2).
3 (4) Площадка 9-го брюшного сегмента слабо поперечная. Вырез¬
ка поперечно-овальная, незамкнутая (рис. 44, 4), Задняя
лопасть лобной пластинки более чем в 3 раза уже передней
части лобной пластинки (рис. 45, 2) . . .D. rubens Pili.
4 (3) Площадка 9-го брюшного сегмента слабо удлиненная. Вы¬
резка округлая, замкнутая. Задняя лопасть лобной пластин¬
ки в 2,5 раза уже передней части лобной пластинки ....
D. varians Germ.
D, linearis L, Личинки развиваются в разлагающейся древеси¬
не и под корой хвойных пород. Хищник с элементами некрофагии.
D, rubens PilL Личинки развиваются в разлагающейся древе¬
сине и под корой широколиственных пород. Активные хищники.
D, varians Germ. Личинки развиваются в разлагающейся дре¬
весине упавших деревьев и в пнях. Хищники.
154
Род Hypoganus Kiesw.
Килевидпая кайма на тергитах брюшка сомкнутая. Глубина
вырезки между урогомфами в 2—2,5 раза меньше длины плош;адки
9-го брюшного сегмента.
Таблица для определения видов рода Hypoganus Kiesw.
1 (2) Каждый из тергитов брюшка с 3 четкими светло-желтыми пят¬
нами. Щетинки па площадке 9-го брюшного сегмента отсут¬
ствуют (рис. 44, 5). Задняя лопасть лобной пластинки широко
ромбовидная (рис. 45, 5) Н. cinctus Рк.
2 (1) Каждый из тергитов брюшка с 3 продольными полосами не¬
правильной формы. Площадка 9-го брюшного сегмента с 2 па¬
рами щетинок. Задняя лопасть лобной пластинки с почти па¬
раллельными боковыми сторонами и широко закругленной
вершиной Н. stepano vi Denis.
Н. cinctus Рк. Личинки отмечены в сухой древесине березы
и ивы. Уничтожают мелких личинок короедов и капюшонников.
Я. stepanovi Denis. Встречаются в трухлявых пнях и почве.
Питание не изучено.
Род Ludius Latr.
Назале поперечное, 3-зубчатое, 2-й членик усиков с 6 чувст¬
вующими выростами. Тергиты груди и брюшка с 2 парами малень¬
ких удлиненных пятен у переднего края.
L. ferrugineus L. Личинки развиваются в дуплах главным обра¬
зом широколиственных древесных пород, заселенных личинками
бронзовок, которыми, видимо, питаются.
Род Melanotus Esch.
Крупные бурые личинки со слегка уплощенным телом. Назале
клиновидное. Мускульные вдавления хорошо выраженные.
Таблица для определения видов рода Melanotus Esch.
1 (2) Площадка 9-го брюшного сегмента поперечно-морщинистая,
без пунктировки, ее концевой зубец узкий и длинный
(рис. 43, 70) М. rufipesHbst.
2 (1) Площадка 9-го брюшного сегмента с пунктировкой, ее кон¬
цевой зубец короткий и широкий (рис. 44, 6).
3 (4) Мускульные вдавления на 9-м брюшном сегменте представ¬
лены 2 пятнышками, не доходящими до внутренних бороздок,
или полностью отсутствуют. Основание 9-го брюшного терги-
та в редкой пунктировке М. tenebrosus Ег.
4 (3) Мускульные вдавления на 9-м брюшном сегменте доходят до
155
внутренних бороздок. Весь тергит 9-го брюшного сегмента
густо и грубо пунктирован (рис. 44, 6) . .М. crassicollis Ег.
М. crassicollis Er. Личинки развиваются в почве, на юге иногда
и пнях. Хищник.
Л/, rufipes Hbst. Личинки развиваются в разлагающейся дре¬
весине различных пород, очень часто в трухлявых сосновых пнях,
иногда в почве и подстилке. Хищник, уничтожающий личинок
стволовых вредителей и других ксилофильных насекомых. Может
развиваться как некросапрофаг.
ЛУ. tenebrosus Er, Личинки развиваются в почве и подстилке,
иногда в разлагающейся древесине. Хищник с элементами некро-
сапрофагии.
Род Stenagostus Thoms.
Верх тела темно-коричневый. Тергиты груди и брюшка в гру¬
бой пунктировке. Боковые стороны площадки 9-го брюшного сег¬
мента несут по 3 крупных бугорка. Наружные ветви урогомф ци¬
линдрические, внутренние уплощенные, изогнутые вовнутрь.
Таблица для определснля видов рода Stenagostus Thoms.
1 (4) Площадка 9-го брюшного сегмента полностью покрыта круп¬
ной пунктировкой (рис. 43, 22). Задняя лопасть лобной пла¬
стинки на вершине обрублена (рис. 45, 5).
2 (3) Площадка 9-го брюшного сегмента несколько больше в дли¬
ну, чем в ширину, с 3 узкими, продольными овальными пят¬
нами в средней части. Вырезка поперечно-овальная; устье
вырезки узкое, не шире наружной ветви урогомфы ....
S. rufus Deg.
3 (2) Площадка 9-го брюшного сегмента явственно продолговатая,
без пятен. Вырезка округлая (рис. 43, 12), устье вырезки ши¬
рокое, явственно шире наружной ветви урогомфы ....
S. villosus Geoffr.
4 (1) Грубая пунктировка имеется только в основной части пло¬
щадки 9-го брюшного сегмента (рис. 43, 22). Задняя лопасть
лобной пластинки на вершине округлая (рис. 45, 4) .
S. undulatus Deg.
S. rufus Deg, Личинки развиваются в сосновых пнях. Питаются
личинками других насекомых.
S. undulatus Deg. Личинки развиваются под корой, реже в дре¬
весине самых различных древесных пород, предпочитая хвойные.
Активные хищники, уничтожающие всех доступных личинок насе¬
комых, в том числе дровосеков и златок.
S. villosus Geoffr, Длина личинки последнего возраста около
40 мм. Назале поперечное. Личинки развиваются в разлагающей¬
ся древесине лиственных пород. Хищник.
156
СЕМЕЙСТВО NITIDULIDAE — БЛЕСТЯНКИ
Небольшое семейство, насчитывающее в фауне СССР около 230
видов. Относительно объема семейства в настоящее время нет еди¬
ной точки зрения. Так, род Rhizophagus многие авторы выделяют
в самостоятельное семейство Rhizophagidae. Этот вопрос пока еще
нельзя считать окончательно решенным, поэтому в данной работе
род Rhizophagus рассматривается в составе семейства Nitidiilidae.
В литературе, посвященной преимагинальным фазам развития
блестянок, имеются как общие очерки строения личинок этого се¬
мейства (Boving, Craighead, 1931; Peterson, 1951), так и работы,
в которых морфология личинок разработана более подробно (Bo¬
ving, Rozen, 1962). Детальные описания личинок отдельных видов
блестянок, сведения об их экологии и определительные таблицы
имеются в монографии Сааласа (Saalas, 1917, 1923) и в статье Бе-
винга и Розена (Boving, Rozen, 1962). В целом группа, однако,
остается еще слабо изученной.
Экология, Личинки блестянок встречаются в самых различных
местообитаниях: в цветках, бутонах и почках (AmarLus, Brachyp-
terus, Meligethes), в грибах, в том чпсле и трутовиках (Pallodes,
Cyllodes), в гниющих растительных остатках (Stelidota), на сухих
трупах (Omosita), в вытекающем бродящем соке деревьев (Сгур-
tarcha, Soronia), в запасах растительных продуктов (некоторые
виды рода Carpophilus). Личинки многих родов и видов развивают¬
ся в разлагающейся древесине. Чаще всего они заселяют подкоро-
вую зону (большинство видов Glischrochilus, а также представи¬
тели родов Librodor, Cryptarchips и Ipidia), многие виды таких
родов, как Еригаеа и особенно Rhizophagus, встречаются в ходах
различных короедов.
Личинки блестянок не обнаруживают строгой приуроченности
к отдельным породам деревьев, но часто имеет место приурочен¬
ность либо к хвойным, либо к лиственным породам.
Среди личинок блестянок отмечены практически все типы пи¬
тания, за исключением паразитизма. Так, личинки некоторых ви¬
дов Meligethes, Brachypterolus и Carpophihis являются фитофагами,
личинки Cryptarcha и, по-видимому, Soronia — сапрофагами, ли¬
чинки Omosita — некрофагами. Обитающие под корой личинки
Glischrochilus, Librodor, Cryptarchips являются сапросилофагами
с элементами пекрофагии. Имеющиеся в литературе данные о хищ¬
ничестве личинок Glischrochilus quadripunctatus L. (Гусев, 1928,
Харитонова, 1972) нашими наблюдениями не подтверждаются.
Наконец, среди представителей родов Pityophagus, Еригаеа
и Rhizophagus встречаются в основном сапроксилофаги с элемен¬
тами хйщничества, которое особенно отчетливо выражено у личи¬
нок Rhizophagus. Специализации к определенному кругу жертв
у личинок этих родов не обнаруживается. Единственное известное
исключение — личинки Rhizophagus grandis Gyll., которые, воз¬
157
можно, являются монофагами, истребляющими короеда Dendro-
ctonus micans Kug.
Морфология, Тело личинок, как правило, уплощенное, реже
вальковатое. Головная капсула расширена в задней части (Glisch-
rochilus, Librodor, Cryptarcha и др.), реже с равномерно округлы¬
ми боковыми сторонами (Еригаеа, Rhizophagus). Лобные швы до¬
ходят почти до заднего края головы и имеют лировидную или
(у Еригаеа и Rhizophagus) удлиненно-лировидную форму. Усики
3-члениковые, расположены на крупных мясистых усиковых бу¬
горках, 2-й членик усиков несет на вершине с вентральной сторо¬
ны удлиненную сенсиллу. Верхняя губа обособлена. Наличник не
отделен ото лба.
Мандибулы с сильно развитой моларной частью, несущей ряды
склеротизованных гранул, и узкой вершинной частью, с несколь¬
кими субапикальными зубцами и часто раздвоенной вершиной.
Простека имеется. В большинстве случаев она несет склеротизо-
ванные зубцы. У представителей рода Rhizophagus простека про¬
стая. Гипостомальная выемка (выемка на вентральной стороне
головной капсулы, в которой расположены нижние челюсти
и нижняя губа) трапециевидная, конусовидная или с параллель¬
ными боковыми сторонами. Максиллы состоят из удлиненного или
треугольного кардо, стипеса и округлой или (у Rhizophagus) клю¬
вовидной малы, несущей короткие щетинки. Нижнечелюстные щу¬
пики 4- или (у Rhizophagus) 3-члениковые. Нижняя губа состоит
из разделенного на 2 части перегибом постментума и прементума,
несущего 1-члениковые щупики (у личинок Rhizophagus щупики
2-члениковые). Язычок слабо развит.
Переднеспинка склеротнзована сильнее, чем остальные сегмен¬
ты тела. Ноги короткие, с небольшим сильно изогнутым коготком.
Сегменты брюшка либо гладкие, несклеротизованные (Glischrochi-
lus, Librodor, Ipidia, Pityophagus и др.), либо несут на не всегда
явственно выраженных склеритах многочисленные бугорки. Ино¬
гда боковые стороны стернитов брюшка вытянуты в длинные мя¬
систые выросты (Cryptarcha и др.). Дыхальца расположены на
крупных склеротизованных бугорках (Librodor, Ipidia, Cryptarcha
и др.), либо находятся па уровне тела (Pityophagus, Carpophilus).
9-й брюшной сегмент снабжен урогомфами и отличается по строе¬
нию от остальных сегментов брюшка. Нередко тергит 9-го брюш¬
ного сегмента представляет сильно склеротизованную овальную
пластинку, в средней части которой находятся склеротизованные
бугорки или зубцы. Реже (у Еригаеа и Rliizophagus) степень скле-
ротизации 9-го брюшного сегмента не отличается от склеротизации
остальных сегментов тела. Урогомфы сильно варьируют, но строе¬
нию они бывают неразветвленными (Glischrochilus, Ipidia, Pityopha¬
gus), с раздвоенными вершинами (Librodor, Cryptarchips), с до¬
бавочными бугорками у основания (Еригаеа) п трехзубцовыми
(Rhizophagus). Анальная подпорка короткая, мясистая, цилин¬
дрическая, часто с рядом крючков на вершине.
158
Таблица для определения родов семейства Nitidulidae
1 (2) Гипостомальная выемка трапециевидная, от ее задних уг¬
лов по направлению к задним углам головной капсулы от¬
ходят явственные кили (рис. 46, 6), Жевательная лопасть
максилл клювовидная Rhizophagus Hbsl. (стр. 164)
2 (1) Гипостомальная выемка конусовидная или с почти парал-
Рис. 46. Общий вид и детали строения личинок Glischrochilus quadripuncta-
tus L. (1—4) и Rhizophagus bipustulatus F. {5, 6)
1,5 — общий] вид сверху;- 2 — площадка 9-го брюшного тергита и урогомфы сверху;
3 — площадка 9-го брюшного сегмента и урогомфы сбоку; 4, 6 — вентральная сторона
головной клпсулы
inv — гипостомальная выемка
159
Рис. 47. 9-й брюшной тергпт Carpophilus hemipterus L. (7, 2), Cryptarcha
sp. (5, 4) Librodor quadriguttatiis F. (5, 6) и Pityophagus ferrugineus L. (7, 8)
J, 3,5,7 — вид сверху; 2, 4, 6, 8 — вид сбоку
лельными боковыми сторонами. Вентральная поверхность
головной капсулы гладкая, без килей. Жевательная ло¬
пасть максилл округлая (рис. 46, 4).
3 (14) Брюшные тергиты гладкие, без бугорков.
4 (И) Дыхальца брюшных сегментов расположены на склероти-
зованных бугорках (рис. 46, 1).
5 (8) Вершины урогомф простые (рис. 46, 2).
6 (7) Урогомфы с крупным бугорком в вершинной трети внут¬
реннего края (рис. 46, 2), расходяш,иеся, со слабо загнуты¬
ми кверху вершинами. Бугорки на 9-м брюшном тергите
плоские (рис. 46, 5). Сапроксилофаг с элементами некро-
фагии Glischrochilus Мигг.
7 (6) Урогомфы с гладким внутренним краем, параллельные,
их вершины изогнуты кверху почти под прямым углом.
Бугорки на 9-м брюшном сегменте в виде высоких при¬
тупленных конусов. Сапроксилофаг с элементами некро-
фагии Ipidia Ег.
8 (5) Вершины урогомф двуветвистые (рис. 47, 5, 6).
9 (10) Зубцы на вершине урогомф одинаковой величины. Сапро¬
ксилофаг с элементами некрофагии .... Librodor Rtt.
10 (9) Внутренние зубцы на вершине урогомф явственно короче
наружных. Сапроксилофаг с элементами некрофагии . .
Cryptarchips Rtt.
И (4) Дыхальца брюшных сегментов не возвышаются над по¬
верхностью тела.
160
12 (13) Боковые стороны склеротизованной площадки 9-го брюш¬
ного тергита с очень длинными крупными щетинками, на¬
много превосходящими по длине урогомфы (рис. 47,1), Уро-
гомфы изогнуты навстречу друг другу, вырезка между
урогомфами трапециевидная. Бугорки 9-го брюшного тер¬
гита в виде зубцов, слабо приподнятых над поверхностью
тергита (рис. 47,2). Сапрофаг с элементами некрофагии
Carpophilus Leach
13 (12) Боковые стороны склеротизованной площадки 9-го брюш
ного тергита с мелкими щетинками. Урогомфы параллель
ны продольной оси тела (рис. 47, 7), вырезка между урогом
фами глубокая, дугообразная. Бугорки 9-го брюшного тер
гита в виде притупленных конусов, образующих с поверх
ностью тергита угол около 60° (рис. 47, S)
Pityophagus Shuck. (стр. 164)
14 (3) Все или по крайней мере 5-й — 8-й брюшные тергиты несут
склеротизованные бугорки.
15 (16) Боковые стороны брюшных сегментов вытянуты в длинные
мясистые отростки. 5-й —8-й брюшные тергиты несут по
4 щетинконосных бугорка, 2 срединных бугорка — трехвер¬
шинные, 2 боковых — простые. Боковые стороны 9-го брюш¬
ного сегмента гладкие (рис. 47,5), урогомфы без бугорков
у основания (47, 4), Сапрофаг . . . .Cryptarcha Shuck,
16 (15) Боковые стороны брюшных сегментов без мясистых вырос¬
тов. 1-Й — 8-Й брюшные тергиты несут по 6—8 продольных
рядов склеротизованных бугорков. Боковые стороны 9-го
брюшного тергита с зубцами (рис. 48, 4), урогомфы с бугор¬
ками у основания (рис. 48,5, S) ...Еригаеа Ег. (стр. 161)
Род Epuraea Ег.
Тергит 9-го брюшного сегмента слабо склеротизован, несет мно¬
гочисленные бугорки и по 3—4 зубца на боковых сторонах. В сред¬
ней части тергита расположены 2 крупных склеротизованных зуб¬
ца — срединные зубцы, а несколько выше их и ближе к средней
линии тела находятся 2 склеротизованных зубца меньших разме¬
ров — околосрединные зубцы (рис. 48, 4), Тергиты 1-го —8-го
брюшных сегментов с продольными рядами бугорков (исключение
Е. angusUiIa Sturm., у которого бугорки образуют один попереч¬
ный ряд на заднем крае тергитов). Отсчет рядов начинается от
средней линии тергита.
Таблица для определения видов рода Epuraea Er.
1 (2) Площадка 9-го брюшного тергита с 2 очень крупными дву¬
вершинными зубцами (рис. 48,5) . . .Е. pusilla 111.
2 (1) Площадка 9-го брюшного сегмента с простыми зубцами
(рис. 48,5,6).
6 Б. М. Мамаев и др. 161
Рис. 48. 9-й брюпшой тергит личинок Еригаеа angustula Sturm. (i), Е. lae-
viuscula Gyll. (2), E. longula Er. (5, 4), E. pusilla 111. (5), E. quadrimaculata
Rtt. (5), E. planulata Er. (?) (7), E. rubronotata Rtt. (<^), E. unicolor 01. (9)
1,4— вид сверху; 5, 5 — 9 — вид сбоку (J, г — по Saalas, 1917)
3 (4) Продольные ряды бугорков на брюшных тергитах отсут¬
ствуют, они несут на заднем крае по 3 бугорка с каждой
стороны (рис. 48,7) Е. angustula Sturm.
4 (3) Брюшные тергиты с 4 продольными рядами бугорков с
каждой стороны
5 (8) 1-й и 3-й ряды бугорков на тергитах состоят из 2 бугорков.
6 (7) Бугорки 2-го и 4-го рядов на брюшных тергитах одинако¬
вой величины. Урогомфы короткие, массивные, срединные
зубцы на плош;адке 9-го брюшного тергита слабо приподня¬
ты над поверхностью тела (рис. 48, 2) .Е. laeviuscula Gyll.
7 (6) Бугорки 2-го ряда на брюшных тергитах самые крупные,
гораздо крупнее бугорков 4-го ряда. ^ рогомфы длинные
и тонкие. Срединные зубцы на площадке 9-го брюшного
тергита образуют прямой угол с поверхностью тела . . .
Е. thoracica Tourn.
Ъ (5) 1-й и 3-й ряды бугорков на тергитах состоят из 3—4 бугор¬
ков каждый.
9 (14) Урогомфы резко изогнуты кверху, образуют угол с поверх¬
ностью 9-го брюшного тергита (рис. 48, 7, 9). 9-й брюшной
тергит без поперечного ряда бугорков у основания.
162
10 (И) Урогомфы крючковидные, образуют прямой угол с поверх¬
ностью 9-го брюшного тергита. Срединные зубцы 9-го брюш¬
ного тергита не изогнуты назад, околосрединные зубцы
очень маленькие, втрое короче срединных (рис. 48,7). Пе-
реднеспинка без бугорков Е. planulata Ег. (?)
И (10) Урогомфы прямые, образуют тупой угол с поверхностью
9-го брюшного тергита. Срединные зубцы 9-го брюшного
тергита изогнуты назад, околосрединные зубцы крупные,
лишь немного ниже срединных (рис. 48, 6, 9), Переднеспин-
ка покрыта склеротизованными бугорками.
12 (13) Дорсальная поверхность урогомф с крупным бугорком
в средней части (рис. 48,9) Е. unicolor 0L
13 (12) Дорсальная поверхность урогомф без бугорка в средней
части (рис. 48,6) Е. quadrimaculata Rtt.
14 (9) Урогомфы плавно изогнуты кверху, не образуют угла
с поверхностью 9-го брюшного тергита (рис. 48, 5, S). 9-й
брюшной тергит с поперечным рядом бугорков у основа¬
ния (48, 4).
15 (16) Околосрединные зубцы на 9-м брюшном сегменте очень
мелкие, почти невидимые сбоку (рис. 48, 3). Вырезка меж¬
ду урогомфами глубокая, отношение длины вырезки к ши¬
рине 2 : 2,5 (рис. 48, ^) Е. longula Er.
16 (15) Околосрединные зубцы хорошо развиты, явственно разли¬
чимы при рассмотрении сбоку (рис. 48, 8). Вырезка между
урогомфами мелкая, отношение длины вырезки к ширине
1 : 2,5 Е. rubronotaia Rtt.
Е. angustula Sturm. В Европе встречается под корой ели, сос¬
ны и березы. Отмечен в ходах короедов Trypodendron lineatum 01.
и Dryocoetes hectographus Rtt. По данным Сааласа (Saalas, 1917),
является хищником, уничтожаюн;им короедов.
Е, laeviuscula GylL По данным Сааласа (Saalas, 1917), является
врагом Т. lineatum 01. Отмечен под корой ели и сосны.
Е. longula Er, Личинки найдены в Убинском лесничестве Крас¬
нодарского края под корой бука восточного, в Московской области
под корой сосны обыкновенной. Питание не изучено.
Е, planulata £'г.(?). Серия личинок найдена в Хабаровском
крае (Хинганский заповедник) под корой пихты. Питание не изу¬
чено.
Е, pusilla 111. Серия личинок найдена в Московской области
под корой сосны обыкновенной в ходах короеда Blastophagus pini-
perda L.
E, quadrimaculata Rtt. Серия личинок найдена в Хабаровском
крае (Хинганский заповедник) под корой дуба, сильно пораженной
плесенью. Питание не изучено.
Е. rubronotata Rtt. Серия личинок найдена в Хабаровском крае
(Хинганский заповедник) под корой пихты цельнолистной в ходах
короедов Orthotomicus suturalis Gyll.
6* 163
Е, thoracica Tourn, Отмечен под корой сосны и ели в ходах ко¬
роедов Ips typographus L. Реже встречаются в ходах Dendroctonus
micans Kug., Polygraphus polygraphus L. и Blastophagus pini-
perda L. Уничтожают яйца и личинок короедов.
Е, unicolor 01. Личинки найдены под корой ели и березы в Мос¬
ковской области и под корой бука в Убинском лесничестве Красно¬
дарского края. Питание не изучено.
Род Pityophagus Shuck.
Бугорки 9-го брюшного тергита сдвинуты к переднему краю
'склеротизованной площадки. Мандибулы с мягкой простекой, по-
^крытой поперечными рядами стекловидных зубчиков.
Р. ferrugineus L. Саалас (Saalas, 1917) находил личинок этого
Ьида под корой елового пня, заселенного короедами Orthotomicus
laricis F., Ips duplicatus Sahlb., Hylurgops palliatus Gyll. H. 3. Ха¬
ритонова (1972), ссылаясь на других авторов, указывает, что ли¬
чинки этого вида уничтожают личинок слоника Hylobius abietis L.
и короедов Blastophagus minor Hart., В. piniperda L., Hylastes
ater Pk. и Ips typographus L. Несколько личинок найдены в Мос¬
ковской области в ходах короедов Hylurgops palliatus Gyll.
Род Rhizophagus Hbst.
Тело вальковатое (рис. 46, 5). Мандибулы с простой, незубчатой
цростекой, нижнечелюстные щупики 3-члениковые, нижнегуб¬
ные — 2-члениковые.
Таблица для определения видов рода Rhizophagus Hbst.
1 (2) 9-й брюшной тергит с 4 бугорками в средней части
(рис. 49,1), Срединные зубцы урогомф загнуты на дорсаль¬
ную поверхность (рис. 49, 2) Rh. aeneus Richt.
2 (1) 9-й брюшной тергит с 2 бугорками в средней части (рис. 49,
4, 6), Срединные зубцы урогомф не загнуты на дорсальную
поверхность (рис. 49, 5, 5, 7).
= 3 (8) Брюшные тергиты несут по 2 поперечпых ряда явственных
мелких бугорков. Выемка между урогомфами открытая,
расширяющаяся в задней части (рис. 49, 6, 8).
4 (5) Брюшные тергиты с 2 поперечными рядами щетинок рав¬
ной длины (рис. 49, 8). Срединный зубец урогомф конусо¬
видный, с мелким добавочным притупленным конусом . .
Rh. grandis Gyll.
5 (4) Щетинки переднего поперечного ряда на брюшных терги-
тах гораздо короче, чем щетинки заднего ряда. Срединный
зубец урогомф крючковидный (рис. 49, 7).
6 (7) Вершины срединных зубцов урогомф резко загнуты кни¬
зу, а сами зубцы слегка изогнуты навстречу друг другу
(рис. 49, б, 7) Rh. dispar Pk.
164
Рис. 49. 9-й брюпшой тергит и урогомфы Rhizophagus aeneus Richt. (Z, 2),
Rh. bipustulatus F. (5), Rh. depressus F. (4, 5), Rh. dispar Pk. {6, 7), Rh.
grandis Gyll. (5), Rh. nitidulus F. (9)
1, 4, 6, 8, 9 — ВИД сверху; 2, 3, S, 7 — вид сбоку (8 — по Saalas, 1917)|
7 (6) Вершины срединных зубцов урогомф изогнуты самое боль¬
шее под прямым углом, а сами зубцы параллельны продоль¬
ной оси тела (рис. 49, .9) Rh. nitidulus F.
8 (3) Брюшные тергиты почти гладкие, бугорки на них почти
не выражены. Выемка между урогомфами полузамкнутая,
суживаюш;аяся в задней части (рис. 49, 4).
9 (10) Выемка между урогомфами удлиненно-овальная (рис. 49, 4).
Срединный зубец урогомф с маленьким добавочным кону¬
сом на вершине, изогнут почти под прямым углом к поверх¬
ности тела, намного мельче срединных бугорков 9-го брюш¬
ного тергита (рис. 49, 5) Rh. depressus F.
10 (9) Выемка между урогомфами примерно равной длины и ши¬
рины. Срединный зубец урогомф в виде притупленного
конуса, слабо приподнят над поверхностью тела, значи¬
тельно крупнее срединных бугорков 9-го брюшного терги¬
та (рис. 49, 5) Rh. bipustulatus F.
Rh, aeneus Richt. Серия личинок этого вида была найдена в
Красной Поляне Краснодарского края в коре вишни. Питание
не изучено.
Rh. bipustulatus F. Приурочен к лиственным породам деревьев.
Встречается как под корой, так и, хотя и значительно реже, в вы-
1^5
текающем древесном соке. Д. Померанцев (19026) отмечал этот вид
на дубе, буке и липе. В коллекциях лаборатории представлен се¬
риями личинок из-под коры березы (Тульская обл.), бука (Крас¬
нодарский край), дуба и березы (Московская область).
Rh. depressus F. Встречается под корой как хвойных, так и лис¬
твенных пород. По данным В. И. Гусева (1928), личинки — хищ¬
ники. Они встречаются в ходах Ips acuminatus Gyll., I. sexdenta-
tus Boern., где уничтожают личинок короедов. Отмечены также в хо¬
дах Hylurgops palliatus GylL, Blastophagus piniperda L., B. mi¬
nor Hart., Trypodendron lineatum 01., Dendroctonus micans Kug.
и др. Более поздние наблюдения других исследователей подтвер¬
дили эти данные. В Убинском лесничестве Краснодарского края
личинки найдены под корой сосны в ходах Ips sexdentatus Boern.
и I. acuminatus Gyll.
Rh, dispar Pk, Встречается под корой березы, осины, ели, сос¬
ны, в бродящем березовом соке и на нижней поверхности древес¬
ных грибов. Личинки встречались в ходах короедов Hylurgops
palliatus Gyll., Polygraphus subopacus Thoms., Dendroctonus mi¬
cans Kug., Crypturgus hispidulus Thoms., Hylurgops glabratus
Zett., Orthotomicus laricis F., Trypodendron lineatum 01., a также
под корой деревьев и пней, в которых ходы короедов отсутствова¬
ли (Saalas, 1917; Nuorteva, 1956). Саалас (Saalas, 1917) подчерки¬
вает несомненную связь личинок этого вида с различными видами
короедов. В коллекциях лаборатории имеется несколько личинок,
найденных под корой пихты кавказской и бука восточного (Крас¬
нодарский край, село Ново-Прохладное).
Rh. granSis Gyll, По данным Сааласа (Saalas, 1917), вид строго
приурочен к ходам Dendroctonus micans Kug. Личинки также встре¬
чаются исключительно в ходах этого короеда и питаются его ли¬
чинками. В Грузии проводятся опыты по искусственному расселе¬
нию Rh. grandis Gyll. (Кобахидзе, Супаташвили, 1967).
Rh. nitidulus F. Серия личинок этого вида найдена в древесине
ольхи (Красная Поляна, Краснодарский край). Одна личинка най¬
дена в Московской области в березе, в ходах, покинутых Trypo¬
dendron signatum F.
СЕМЕЙСТВО сисиJIDAE -ПЛОСКОТЕЛКИ
Семейство принимается нами в несколько меньшем объеме по
сравнению с тем, какой был определен О. Л. Крыжановским
(1965в), так как семейство Silvanidae рассматривается в качестве
самостоятельного.
Несмотря на то, что личинкам плоскотелок посвящено неболь¬
шое число публикаций, их личиночные характеристики на уровне
)одов могут считаться сравнительно хорошо разработанными.
3 работе Перри (Perris, 1877) содержатся сведения о личинках
представителей родов Prostomis Latr. (с видом Р. mandibularis F.),
166
Brontes (с видом В. planatus L.), Dendrophagus (с видом D. crena-
tus Pk.), Laemophloeus (c видом L. testaceus F.) и Silvanus (c видом
S. unidentatus F. и др.). В этой же работе приводятся сведения,
ранее имевшиеся в публикациях с описанием личинок Cucujus
liaematodes Er., Pediacus dermestoides F. и других плоскотелок.
Весьма важный вклад в описание личинок плоскотелок был вне¬
сен Сааласом (Saalas, 1917), который изучил 7 видов, из них —
Pediacus fuscus Er., Laemophloeus abietis Wank., L. alternans Er.
были описаны им впервые. Бевинг и Крайхед (Boving, Craighead,
1931) хорошо иллюстрировали морфологические особенности ли¬
чинок 13 родов плоскотелок, однако никаких суш,ественных добав¬
лений к списку ксилофильных представителей семейства по их
материалам сделать уже не удается. Из работ, посвяш,енных япон¬
ской фауне, заслуживает внимания публикация Хаяси (Hayashi,
1969). В списке индийских видов плоскотелок, личинки которых
известны, фигурируют главным образом синантропные формы
(Gardner, 1948).
Экология, Плоскотелки возникли как подкорные формы —
большинство видов сохраняет эти экологические связи. Биологи¬
чески сильно уклонившаяся группа, иногда выделяемая в само¬
стоятельное семейство Prostomidae, специализировалась к разви¬
тию в толще сильно сгнившей древесины на луканидной стадии
ее разложения. В составе семейства имеются также нексилофиль-
ные виды — обитатели складов, гнезд общественных насекомых
п т. д.
Плоскотелки-ксилобионты отдают явное предпочтение листвен¬
ным породам. Тем пе менее виды, массовые под корой лиственных
деревьев, единично встречаются и под корой хвойных. У предста¬
вителей рода Laemophloeus такая специализация выражена более
четко: L. alternans Er. и L. abietis Wank. встречаются на хвойных,
а L. monilis F., L. muticus F., L. testaceus F.— na лиственных по¬
родах; L. muticus F. среди лиственных отдает явное предпочтение
березе; L. ferrugineus Steph.— широко распространенный обита¬
тель складов с запасами зерна и других продуктов, в естественных
условиях встречается под корой лиственных деревьев.
В отношении питания личинок различных видов данных в ли¬
тературе почти не приводится. По мнению Палма (Palm, 1951,
1959), суммировавшего собственные материалы и литературные
сведения, представители родов Brontes, Cucujus, Dendrophagus,
Pediacus и многие виды Laemophloeus имеют тенденцию к хищни¬
честву, Наши материалы, однако, показывают, что личинки Bron¬
tes и Dendrophagus относятся к сапромицетофагам и не проявляют
себя как хищники. Виды рода Laemophloeus, как L. monilis F.
и L. muticus F., а также представители рода Pediacus — в основ¬
ном сапромицетофаги, а некоторые — почти исключительно мице-
тофаги.
Морфология. Личинки плоскотелок подразделяются на 2 мор-
фо-экологических типа. К первому из них относятся истинные
167
Рис. 50. Общий вид личинок плоскотелок (Cucujidae)
1 — Prostomis mordax Rtt.; 2 — Pediacus depressus Hbst.; 3 — Laemophloeus muticus F.
плоскотелки, личинки которых очень сильно уплощены (рис. 18,1\
50,7—5), имеют плоскую голову, в большинстве случаев короткие
усики и сильносклеротивованные урогомфы или покрытые бугор¬
ками выросты на конце тела. Эти выросты сильной мускулатурой
8-го брюшного сегмента могут приводиться в вертикальное поло¬
жение и, упираясь в свод хода или полости, обеспечивают как по¬
ступательное, так и возвратное движение личинок. Анальная под¬
порка у этих личинок весьма слабо развита; по краю анального от¬
168
верстия могут быть развиты опорные шипики. В целом личинки
первого морфо-экологического типа заселяют узкие (по сравнению
с размерами личинок) подкорные пространства, в которых они мед¬
ленно передвигаются.
Ко второму морфо-экологическому типу (рис. 18, 2) относятся
личинки с умеренно уплощенным телом, включая голову, длин¬
ными усиками, хорошо развитой длинной конусовидной подпор¬
кой; урогомфы длинные, на конце заостренные, слабосклероти-
зованные. По краю анального отверстия шипиков нет. Такие ли¬
чинки заселяют крупные полости, образующиеся при отслоении
подсыхающей коры. Они очень подвижны.
Цвет личинок разнообразен — от беловатого до различных от¬
тенков желто-коричневого.
Голова личинок (рис. 9,2) прогнатическая, уплощенная или со¬
вершенно плоская, почти не втянутая в переднегрудь. Как прави¬
ло, она округлой формы, реже — поперечная, с угловидно оття¬
нутыми заднебоковыми отделами. Лобные швы хорошо выражены
только у крупных хорошо склеротизованных личинок, у которых
имеется также короткий теменной шов (Cucujus). У прочих личи¬
нок как лобные, так и теменной швы слабо выражены. У личинок
Laemophloeus лобный отдел представляет собой трапециевидный
склерит. Личинки крупных видов плоскотелок имеют по 6 глазков
с каждой стороны головы. Глазки располагаются в 2 неправиль¬
ных поперечных ряда (4 -f* 2). У мелких личинок глазки каждой
стороны могут группироваться в 2 или 1 крупное пигментное пят¬
но. V некоторых видов глазки отсутствуют. Усики 3-члениковые,
прикрепляются на усиконосном бугорке. Соотношение длины
члеников может быть различным. На конце 2-го членика распола¬
гается одна конусовидная сенсилла, 3-й членик несет на вершине
группу щетинок.
Мандибулы в большинстве случаев симметричные, реже асим¬
метричные (Prostomis), с хорошо развитой моларной частью
и сложной обычно 3—4-зубчатой вершиной. Простека, как прави¬
ло, хорошо развита, представлена внутренним срединным пере¬
пончатым выростом, реже (у видов с асимметричными мандибула¬
ми) отсутствует, но тогда левая жвала в основании с крупным зуб¬
цом. Основание мандибул нередко покрыто пучками щетинок.
Верхняя губа отделена от клипеуса, вооружена крепкими щетин¬
ками. Нижняя губа состоит из субментума, ментума и пременту-
ма, несущего 2-члениковые нижнегубные щупики. Кардо и стипес
максилл хорошо развиты. Жевательная доля обычно заостренная,
на конце и по внутреннему краю покрытая различной формы ще¬
тинками, реже (у Laemophlaeus) она на конце более или менее за¬
кругленная. Нижнечелюстные щупики 3-члениковые.
Грудные сегменты склеротизованы с такой же интенсивностью,
как и брюшные, за исключением 8-го, который может быть сильно
удлиненным и значительно сильнее склеротизованным, особенно
на участках, примыкающих к урогомфам. Ноги короткие, направ¬
169
лены в стороны. Бедра и голенелапки приблизительно одинако¬
вой длины; голенелапки часто несут крупные шипики, заканчи¬
ваются коготком. Брюшные сегменты могут нести поперечные
опорные гребни и редкие длинные щетинки. Урогомфы различного
строения (рис. 51—53).
Первая пара дыхалец наиболее крупная, расположена на бу¬
горковидных выступах между 1-м и 2-м грудными сегментами.
Таблица для определения родов семейства Cucujidae
1 (4) Последний брюшной сегмент преобразован в длинный ко*
нусовидный подталкиватель (рис. 18, 2).
2 (3) Выросты 8-го брюшного сегмента тонкие, пальцевидные
(рис. 18, 2). Усики, включая последний членик, покрыты
многочисленными микроскопическими волосками. Кону¬
совидная сенсилла, расположенная на конце 2-го членика
усика, сидит на отчетливом цилиндрическом выступе. Под
отслоившейся корой преимущественно лиственных пород.
Сапромицетофаги с элементами некрофагии . .Brontes F.
3 (2) Выросты 8-го брюшного сегмента толстые, конусовидные.
Усики, особенно последний членик, почти не несут микро¬
скопических волосков. Конусовидная сенсилла, располо¬
женная на конце 2-го членика усика, сидит непосредственно
на его поверхности. Под отслоившейся корой преимущест¬
венно хвойных пород. Сапромицетофаги с элементами не¬
крофагии Dendrophagus Schoenh.
4 (1) Последний брюшной сегмент не образует конусовидного
подталкивателя (рис. 50, 1—3).
5 (6) Голова в 1,5 раза шире переднегруди (рис. 50, i), асимме¬
тричная, с более сильно развитым правым полушарием.
Глазки отсутствуют. Урогомфы бугорковидные. В бурых
гнилях древесины. Сапроксилофаги . . . Prostomis Latr.
6 (5) Голова не шире или незначительно шире переднегруди,
симметричная (рис. 18, 7; 50, 2, 3), Глазки имеются. Уро¬
гомфы шипообразные.
7 (8) Задне-боковые части головной капсулы угловидно оття¬
нуты (рис. 18, 1). Сегменты тела с поперечным опорным
гребнем. 8-й брюшной сегмент с 2 опорными выростами.
В основании каждой урогомфы имеется двувершинный зу¬
бец (рис. 51,1—4) Сиси jus F. (стр. 171).
8 (7) Задне-боковые части головной капсулы плавно закругле¬
ны. Сегменты тела без поперечного опорного гребня. 8-й
брюшной сегмент без опорных выростов. Урогомфы без
зубца в основании.
9 (10) Урогомфы расположены на длинном стержневидном осно¬
вании (рис. 53,1—2) Pediacus Shuck. (стр. 175)
10 (9) Урогомфы расположены на широком поперечном основа¬
нии (рис. 52,1—4) LaemophloeusSteph. (стр. 173)
170
Род Cucujus F.
В Европейской части СССР представлен 2 видами, один из ко¬
торых (С. haematodes Ег.) широко распрострапен в Палеарктике.
В фауне Кавказа и Дальнего Востока дополнительно имеется по
1 виду. Бевипг и Крайхед (Boving, Craighead, 1931), а затем Петер¬
сон (Peterson, 1951) опубликовали данные о личинках С. clavi-
pes F. В монографии о личинках японских насекомых (Kawada,
1959) имеются рисунки урогомф С. opacus Law. и С. mniszechii
Grouv. Личинки обычных видов — С. haematodes Ег. и С. cinnaba-
rinus Scop. детально еще не изучались. Иллюстрации к личинке
С. cinnabarinus Scop., опубликованные М. С. Гиляровым (1964а),
в действительности относятся к американскому виду С. clavipes F.
(Boving, Craighead, 1931). Вид С. caucasicus Motsch. восстанавли¬
вается из синонимов по результатам изучения личиночных приз¬
наков, которые свидетельствуют о его самостоятельности.
Личинки выделяются своими размерами (длина тела до 30 мм),
буроватым цветом очень плоского тела и наличием длинных ост¬
рых своеобразных урогомф (рис. 51,1 — 4), 8-й брюшной сегмент,
где размепцена мускулатура этих урогомф, несет различные опор¬
ные образования и более интенсивно склеротизован.
Таблица для определения вицов рода Cucujus
1(4) Выемка между ветвями урогомф полукруглая (рис. 51, i, 2).
Дорсальный и вентральный поперечные гребни 8-го брюш¬
ного сегмента на боковых сторонах заканчиваются на разных
уровнях. Дорсальная светлая срединная линия выражена на
всех или почти на всех сегментах.
2 (3) Дорсальная срединная светлая линия явственна на всех сег¬
ментах. Наиболее крупный боковой опорный бугорок 8-го брюш¬
ного сегмента вытянут в острие (рис. 51,1), Под корой различ¬
ных лиственных пород. Сапронекрофаг . .С. opacus Lew.
3 (2) Дорсальная срединная светлая линия на средних сегментах
неявственна. Наиболее крупный боковой опорный бугорок
закруглен, не вытянут в острие (рис. 51,2), Под корой пихты
кавказской. Сапронекрофаг С. caucasicus Motsch.
4 (1) Выемка между ветвями урогомф полуовальная (рис. 51,3,4).
Дорсальный и вентральный поперечные гребни 8-го брюшно¬
го сегмента на боковых сторонах заканчиваются на одном
уровне. Дорсальная светлая срединная линия выражена на
грудных, 1-м и 3-м брюшных сегментах. На остальных сег¬
ментах она темная.
5 (6) 2-й членик усиков явственно длиннее 1-го. Опорный бугорок,
расположенный перед треугольными опушенными выростами
8-го брюшного сегмента, вытянут и закруглен (рис. 51, 5) . .
С. cinnabarinus Scop.
171
Рис. 51. 9-й брюшной сегмент плоскотелок рода Cucujus F. (Cucujidae)
J — С. opacus Lew.; 2 — С. caucasicus Motsch.; 3 — С. cinnabarinus Scop.; 4 — C. liaema-
todes Er.
6 (5) 1-Й И 2-Й членики усиков примерно одинаковой длины. Опор¬
ный бугорок, расположенный перед треугольными опушен¬
ными выростами 8-го брюшного сегмента, короткий, более
или менее треугольный (рис. 51,4) . . .С. haematodes Ег.
С, cinnabarinus Scop. Наблюдения за экологией этого вида
и литературные данные (Saalas, 1917; Palm, 1940, 1959; Lundblad,
1943, и др.) свидетельствуют о том, что его личинки могут разви¬
ваться под корой как хвойных (например, ели), так и лиственных
деревьев, причем явно предпочитают лиственные: в южных дубра¬
вах — дуб, а на севере — осину. Они питаются гниюш;им лубом^
однако для их развития, видимо, обязательно определенное коли-
172
\чество животной пищи. Как некрофаги они уничтожают остатки
)^ругих личинок насекомых, а иногда нападают на живых куколок
или мелких личинок. В наших материалах имеется значитель¬
ная серия личинок, собранных в Теллермановском лесничестве
Воронежской области под корой дуба, а также несколько личинок,
обнаруженных в Закарпатье (окрестности г. г. Хуста и Рахова)
под корой ели и клена.
С. haematodes Er, Несмотря на то, что этот вид весьма широко
распространен в Палеарктике, литературные сведения о его био¬
логии чрезвычайно скудны. Наши материалы свидетельствуют
о его более или менее четкой специализации к развитию под корой
самых различных, но исключительно лиственных пород. По харак¬
теру питания сходен с предыдуш,им видом. В собранной нами кол¬
лекции представлены серии личинок из средней полосы Европей¬
ской части СССР, с Кавказа, Саяна и из Приморского края. Предпо¬
читаемыми породами являются осина и тополь, ива и чозения,
а также дуб.
Род Laemophloeus Steph.
Личинки (рис. 50, 3) белые, у некоторых видов желтоватые или
розоватые, с сильно уплоп^енным телом и хорошо развитыми уро-
гомфами, ветви которых сидят на широком основании (рис. 53,2,2),
Голова округлая. Мандибулы симметричные, разнообразны по
строению: в их основной части у крупных видов имеется сложная
простека, состоящая из загнутого внутрь зубца, в основании ко¬
торого расположен такой же дополнительный зубец и длинный
сочлененный саблевидный придаток; у мелких видов эти два зубца
преобразуются в бугристую жевательную поверхность, однако
сочлененный придаток в основании этой поверхности также со¬
храняется. Лобная часть головы сложно устроена: она слабее
склеротизована и представляет собой длинную срединную непра¬
вильно-трапециевидную полосу, ограниченную склеротизованны-
ми утолщениями, от которых обособляются светлые лобные швы,
расходящиеся к основаниям усиков.
Таблица для определения видов рода Laemophloeus Steph.
1 (4) Ширина 8-го брюшного сегмента почти такая же, как его
длина (рис. 52,3,4), Боковые стороны урогомф бугристые.
3-й членик усика значительно короче 2-го.
2 (3) Боковые стороны урогомф с 3—4 бугорками (рис. 52, 3).
На дне выемки между урогомфами бугорок отсутствует.
Под корой лиственных пород. Сапронекромицетофаг . .
L. monilis F.
3 (2) Боковые стороны урогомф с 1 крупным бугорком в основа¬
нии и неявственным — в средней части (рис. 52,4). На дне
выемки между урогомфами с вентральной стороны имеется
173
Рис. 52. Последние сегменты тела и урогомфы плоскотелок рода Laemophlo-
eus Steph. (Cucujidae)
1 — L. alternans Er; 2 — L. testaceus F.; 3 — L. monilis F., 4 — L. muticus F.
бугорок. Под корой лиственных пород. Сапронекромице-
тофаг L. muticus F.
4 (1) Ширина 8-го брюшного сегмента в 1,2—1,8 раз меньше его
длины (рис. 52,7,2). Боковые стороны урогомф гладкие.
3-й членик усика не короче 2-го.
5 (10) 8-й брюшной сегмент от основания урогомф плавно рас¬
ширяется (рис. 52, 1). Дно выемки между урогомфами
в мелких зубчиках.
6 (7) Дно выемки между урогомфами широко закругленное.
Глазки явственные. Под корой лиственных деревьев. Са-
промицетофаг L. ferrugineus Steph.
7 (6) Дно выемки между урогомфами треугольное (рис. 52,1)
или треугольно-копьевидное. Глазки редуцированы.
8 (9) Выемка между урогомфами треугольная (рис. 52, 1) . , ,
L. alternans Ег.
9 (8) Выемка между урогомфами треугольно-копьевидная . .
L. abietis Wank.
10 (5) 8-й брюшной сегмент от основания урогомф резко расши¬
ряется (рис. 52,2). Дно выемки между урогомфами гладкое.
Под корой лиственных деревьев. Сапромицето фаг ....
L. testaceus F.
174
\ L. abietis Wank- В Финляндии личинки этого вида развиваются
\10д корой елей, усыхающих в болотистой местности, особенно
после пожаров. Они встречаются в ходах короедов Ips typograp-
hus L., Polygraphus polygraphus L., Pityogenes chalcographus L.
и др. Предположительно отнесены к хищникам, уничтожающим
личинок короедов (Saalas, 1917; Palm, 1959).
//. alternans Er, Встречается совместно с предыдущим видом,
с которым сходен по многим экологическим параметрам.
Род Pediacus Shuck.
Личинки представителей рода желтовато-кремовые, имеют
сильно утолщенное тело и характерные вильчатые хорошо склеро-
тизованные урогомфы (рис. 50, 2), Длина 8-го брюшного сегмента
в 2—3 раза больше длины каждого из предшествующих брюшных
сегментов. Голова округлая. Мандибулы более или менее симмет^
ричные, с хорошо развитой зубцевидной простекой. Одно глазко-
вое пятно с каждой стороны головы или глазки редуцированы.
Ноги хорошо развиты. Урогомфы с крючковидно загнутыми вверх
концами, частично покрыты микроскопическими зубчиками, в их
средней части у основания ветвей имеется 4 (иногда 6) бугорка»
снабженных щетинкой. Задне-боковые части 8-го брюшного сег¬
мента с выемками, поверхность которых может быть гладкой или
несет поля шипиков.
Таблица для определенил видов рода Pediacus Shuck.
1 (4) Поверхность боковых выемок заднего края 8-го брюшного
сегмента гладкая или с небольшим полем бугорков. В средин¬
ной части стержневидного основания урогомф имеется 4 ще¬
тинки, дополнительной пары длинных щетинок нет (рис.
53,2).
2 (3) Глазки явственны. Глубина выемки между урогомфами в
2 раза больше поперечника их стержневидного основания
(рис. 53, 2). 4 длинные щетинки, имеющиеся в основании вет¬
вей урогомф, расположены примерно на одном уровне . .
Р. dermestoides F.
3 (2) Глазки неявственны. Глубина выемки между урогомфами
в среднем в 2,3 раза больше поперечника их стержневидного
основания. Внутренняя пара длинных щетинок,. имеющихся
в основании ветвей урогомф, сдвинута к их вершине . . .
Р. depressus Hbst.
4 (1) Поверхность боковых выемок заднего края 8-го брюшного
сегмента покрыта большим полем острых шипиков. В средин¬
ной части стержневидного основания урогомф, кроме обыч¬
ных 4 щетинок, имеется дополнительная пара длинных щети¬
нок, расположенных с дорсальной стороны па крупных бу¬
горках (рис. 53, 7) Pediacus sp. (? luscus Er.)
175
Рис. 53. 8-й брюпшой сегмент и урогомфы плоскотелок рода Pediacus Shuck.
(Cucujidae)
i — Р. sp. (? fuscus Er.); 2 — P. dermestoides F.
P. depressus Hbst. В наших материалах имеются личинки, соб¬
ранные под корой дубовой колоды в Теллермановском лесничестве
Воронежской области и в районе г. Хуста Закарпатской области.
Палм (Palm, 1959) предполагает, что личинки вида — факульта¬
тивные хищники с элементами сапронекрофагии.
Р, dermestoides F. Питание точно не прослежено: видимо, у под¬
корных форм это преимущественно некро- и сапрофагия, у обита¬
телей бурых гнилей — сапро- и мицетофагия.
Pediacus sp, (? fuscus Er.). Личинки обитали в лиственничниках
Тувинской АССР в окрестностях пос. Иштии-Хем под корой тонко¬
мерных вывороченных лиственниц.
СЕМЕЙСТВО COLYDIIDAE -УЗКОТЕЛКИ
О личинках узкотелок в энтомологической литературе опубли-
1«овано сравнительно много исследований. Несмотря на несовер¬
шенство описаний, одной из основных работ является монография
Перри (Perris, 1877), в которой описаны такие интересные личин¬
ки, как Endophloeus spinosulus Latr. и Colobicus emargina tus Latr.,
развивавшиеся под корой дуба, а также Langelandia anoph-
thalma Aube., обнаруженные в сильно сгнившей древесине. В этой
-176
работе имеется перечень видов, личинки которых были описа¬
ны более столетия назад. К их числу относятся Bitoma crenata F.,
Synchita juglandis Helw., Aulonium sulcatum 01., Colydium casta¬
neum Hbst., C. elongatum F., C. filiforme F., a также некоторые
другие, относящиеся к выделяемому в настоящее время семейству
Cerylonidae. Из работ, выполненных на высоком уровне, следует
упомянуть обобщающую работу Бевинга и Крайхсда (Boving,
Craighead, 1931), где иллюстрированы детали строения личинок
Bitoma crenata F. (идентификация вида, вероятно, неточна), пред¬
ставителей родов Synchita и Aulonium, а также некоторых северо¬
американских представителей семейства. В монографии Сааласа
(Saalas, 1923) переописаны личинки Bitoma crenata F. и очень
подробно охарактеризованы личинки Lado jelskii Wank. Исчерпы¬
вающие описания личинок Langelandia anophthalma Aube и Anom-
matus duodecimstriatus Miill. опубликованы Дажо (Dajoz, 1968).
H. Б. Никитским (1972) изучены личинки Aulonium ruficorne 01.
Выполненная в Японии работа Хаяси (Hayashi, 1972) важна в том
отношении, что в ней охарактеризованы своеобразные личинки
японской фауны.
Экология, В соответствии с образом жизни личинок узкотелок
можно подразделить на несколько экологических групп. К первой
группе относятся обитатели толщи сильно разрушенной грибами
древесины на луканидной стадии ее разложения. По характеру
питания эти личинки относятся к сапроксилофагам. Типичным
представителем группы является обычный на Кавказе вид Decho-
mus sulcicollis Germ., относящийся к наиболее обычным разруши¬
телям светлых гнилей древесины лиственных пород, но иногда
встречающийся также в древесине хвойных (пихта, тисс). Вместе
с многочисленными личинками в древесине встречаются также
и имаго. Сходный образ жизни описан в литературе для Langelan¬
dia anophthalma Aube (Dajoz, 1968).
Вторую экологическую группу составляют обитатели коры на
ранних стадиях ее разрушения. Массовое поселение личинок Sync¬
hita было отмечено нами осенью 1959 г. в коре кленового пня
в Теллермановском лесничестве Воронежской области. Палм
(Palm, 1959) находил личинок этого вида под корой ольхи, а также
в наружных слоях древесины стволов, разрушенных сверлильщи¬
ками. Отмечены случаи нахождения личинок в плодовых телах
грибов-трутовиков.
Третья экологическая группа включает виды, тесно связанные
с короедами — заболонниками и древесинниками. В ее состав вхо¬
дят представители родов Aulonium и Lado, развивающиеся под
корой, и рода Colydium, обитающие в ходах короедов и толще дре¬
весины.
Четвертая экологическая группа личинок узкотелок отличает¬
ся существенными особенностями биологии. Ее представители так¬
же очень часто заселяют подкоровую зону на сколитидной стадии
разрушения, однако уже тогда, когда короеды в основном закон-
177
ЧИЛИ свое развитие, а кора неплотно прилегает к древесине. В этЬт
период под корой имеются разнообразные источники питаниш—
живые личинки короедов, погибшие личинки, мицелий гриоов,.
экскременты и т. д. Если учесть, что личинки Bitoma crenata F.
и других представителей этой группы встречаются также под ко¬
рой деревьев и в отсутствие короедов и других насекомых, их сле¬
дует считать ксиломицетофагами с элементами некрофагии. Неко¬
торые исследователи (Palm, 1951) публикуют точные указания на
факты хищничества личинок В. crenata, уничтожавших личинок
и куколок короедов, однако хищничество едва ли можно считать
основным способом питания этого вида.
Приведенные материалы показывают, что лишь немногие пред¬
ставители узкотелок в фазе личинки могут быть отнесены к полез¬
ным энтомофагам, способным влиять на численность короедов.
Морфология. Личинки узкотелок (рис. 14, i; 16, i; 54, 7,2) отли¬
чаются большим разнообразием. Среди них есть белые мясистые
формы, весьма напоминающие внешне личинок дровосеков
(рис. 14, i). Однако большинство личинок светло-желтой или бу¬
роватой окраски, с цилиндрическим или слабоуплощенным телом
(рис. 54, i, 2), Для всех известных личинок семейства характерна
наличие на конце тела хорошо развитых крючков или шипиков.
Голова личинок поперечная. Лобные швы в большинстве слу¬
чаев расходятся уже у заднего края головной капсулы, ограничи¬
вая лобную часть копьевидной или булавовидной формы. Рисунок,
образуемый лобными швами, характерен для рода, однако этот
признак, особенно на препаратах, воспринимается с трудом и по
этой причине в дальнейшем не используется. Иногда бывает раз¬
вит короткий теменной шов. Клипеус обычного строения, между
лбом и клипеусом в некоторых случаях имеется более или менее
явственный шов. Глазков 5 или они отсутствуют. Усики 3-члени-
ковые, сидят на крупном члениковидном усиконосном бугорке.
Их 2-й членик несет конусовидную сенсиллу и несколько волос¬
ков, 3-й членик снабжен несколькими волосками, из которых один
особенно длинный.
Мандибулы обычно слегка асимметричные, с 2—3-зубчатой вер¬
шиной и хорошо развитой моларной частью. Верхняя губа закруг¬
ленная. Нижняя губа состоит из субментума, сливающегося с гу-
ларной пластинкой, ментума и прементума, несущего 2-членико-
вые нижнегубные щупики и заканчивающегося язычком. Послед¬
ний иногда покрыт крупными острыми шипами. Гипофарингеаль-
ная склерома достаточно интенсивно склеротизована лишь у неко¬
торых групп. Максиллы с хорошо выраженным небольшим кардо
и длинным стипесом, слившимся с жевательной лопастью. Между
кардо, стипесом и нижней губой имеется крупная сочленовная до¬
ля, которая некоторыми авторами рассматривается как часть сти-
песа. Жевательная лопасть разнообразного строения. Она может
иметь округло-вытянутую форму или к вершине расширяться.
Внутренний угол жевательной лопасти у некоторых личинок снаб-
178
/Аен одним или несколькими склеротизованными зубцами, иногда
н^ет сильносклеротизованный изолированный шип. Поверхность
жевательной лопасти по внутреннему краю покрыта рядами кин-
их'аловидных и волосовидных щетинок. Нижнечелюстные щупики
3-члепиковые.
Грудные сегменты без склеротизованных пятен. Ноги обычного
строения, снабжены группами шипов и волосков и заканчиваются
простыми слабо изогнутыми коготками. Брюшные сегменты с мяг¬
кими покровами, тогда они нередко несут парные двигательные мо¬
золи. Если покровы сегментов склеротизованные, то обычно они
желтоватые, на них имеются различные опорные структуры —
волоски, шипики, нередко формирующие поперечную полоску,
поперечный валик и т. д. 9-й, а иногда также 8-й брюшные сегмен¬
ты с дорсальной стороны склеротизованы значительно сильнее,
иногда с округлыми рыжеватыми пятнами; 9-й сегмент несет заг¬
нутые вверх склеротизованные крючки, между которыми нередко
имеется кармановидное впячивание, или мелкие шипики. На вен¬
тральной стороне последнего сегмента у переднего края анального
отверстия может быть развит поперечный ряд опорных шипов.
Таблица для определения^родов семейства Colydiidae
1 (2) Тело белое, включая последний сегмент. На конце тела
2 маленьких сближенных шипика (рис. 14, 2)
Synchita Hellw.
2 (1) Тело желтоватое, последний сегмент иногда бурый. На
конце тела 2 длинных, раздвинутых, загнутых вверх крюч¬
ка (рис. 16,2; 54, 1—2),
3 (12) Между урогомфами на заднем крае сегмента имеется явст¬
венное, сильносклеротизованное кармановидное впячива¬
ние (рис. 54,2).
4 (5) Два последних сегмента тела с дорсальной стороны склеро-
тизованы значительно сильнее остальных (рис. 54,2), пос¬
ледний — с вогнутой крупнобугристой площадкой . . .
Aulonium Ег. (стр. 181)
5 (4) Более интенсивно с дорсальной стороны склеротизован
только последний сегмент тела или склеротизованы лишь
зубцы урогомф; поверхность сегмента не бугристая.
6 (9) Тергит 9-го брюшного сегмента у переднего края с попереч¬
ным склеротизованным пояском. Кармановидное впячива¬
ние между урогомфами (вид сверху) треугольной формы.
По переднему краю анального отверстия проходит попереч¬
ный ряд опорных шипов.
7 (8) Тергит 9-го брюшного сегмента склеротизован так же ин¬
тенсивно, как урогомфы. Бедра с группой крепких шипов.
Последний членик усиков явственно длиннее предпослед¬
него Bitoma Hbst.
179
Рис. 54. Общий вид (i, 2) и 2-й п 6-й брюшные сегменты сверху личинок узко-
телок (Colydiidae)
1 —Aulonium trisulcum Geoffr.; 2,4 — Dechomus sulcicollis Germ.; 8 — Colydium elon-
gatum F.
8 (7) Тергит 9-Г0 брюшного сегмента склеротизован значительно
слабее урогомф. Бедра с немногими светлыми волосками,
без шипов. Последний членик усиков примерно такой же
длины, как предпоследний Lado Wank. (стр. 182)
9 (6) Тергит 9-го брюшного сегмента у переднего края не несет
склеротизованных структур в виде пояска. Кармановид¬
ное впячивание между урогомфами пальцевидное. Ряд ши-
пиков по переднему краю анального отверстия не выражен.
180
10(11) у основания урогомф имеется поле бугорков. Округлые
\ рыжеватые пятна имеются на двух последних сегментах
тела Colobicus Latr.
11 (10) У основания урогомф поле бугорков не выражено. Округ¬
лые рыжеватые пятна имеются только на последнем брюш¬
ном сегменте Endophloeus Ег.
12 (3) Между урогомфами на заднем крае сегмента нет кармано¬
видных впячиваний (рис. 16, 1\ 54, 2).
13 (16) Глазков нет. Тергиты 6—8-го брюшных сегментов светлые,
слабосклеротизованные, покрытые редкими волосками
(рис. 54, 4).
14 (15) Брюшные сегменты с дорсальной стороны с поперечными
рядами крепких шипов (рис. 54, 4), Язычок на конце в длин¬
ных шипиках. Жевательная лопасть на внутреннем крае
в апикальной части несет склеротизованный бугорок и от¬
дельно стояш,ий крепкий шип . . . .Dechomus J.-Duv.
15 (14) Брюшные сегменты с дорсальной стороны не несут попе¬
речных рядов шипов. Язычок на конце в мелких структу¬
рах. Жевательная лопасть на внутреннем крае без выде-
ляюш,егося шипа и бугорка Langelandia Aiibe.
16(13) Глазки имеются. Тергиты 6—9-го брюшных сегментов жел¬
товатые, покрытые, кроме волосков, склеротизованными
зубцами, несущими щетинку (рис. 54,5)
Colydium F. (стр. 182).
Род Aulonium Ег.
Личинки более или менее цилиндрические, с умеренно пиг¬
ментированными желтоватыми покровами и 2 крупными разоб¬
щенными крючками на конце тела. С дорсальной стороны сегмен¬
тов, начиная со среднегруди, имеется поперечное опорное ребро.
Тергиты 8-го и 9-го брюшных сегментов склеротизованы зна¬
чительно интенсивнее. Поверхность кармановидного впячива-
ния (между опорными крючками) покрыта волосками. Перед
анальным отверстием имеется поперечный ряд шипиков. 5 глаз¬
ков (3 2) с каждой стороны головы.
Таблица для определения видов рода Aulonium Ег.
1 (2) Площадка 9-го брюшного сегмента целиком крупнобугорча¬
тая. Дорсальная поверхность 8-го брюшного сегмента в свет¬
лых поперечных морщинах. На 7-м брюшном сегменте имеет¬
ся поперечная пигментированная полоса
А. trisulcum Geoffr.
2 (1) Площадка 9-го брюшного сегмента крупнобугристая только
на боковых сторонах. Дорсальная поверхность 8-го брюш¬
ного сегмента в светлых точках. На 7-м брюшном сегменте нет
поперечной пигментированной полосы . . . .А. ruficorne 0L
181
А. ruficorne 01. На Северном Кавказе личинки этого вида встре¬
чаются под корой сосны в ходах 6 различных видов короедов (ш-
китский, 1972), главным образом вершинного (Ips acuminatus
Gyll.). Видимо, он предпочитает сосну, о чем свидетельствуют и
литературные источники. В период завершения развития короеда
численность всех фаз развития узкотелки превышает 5 экз. в рас¬
чете на 1 дм^ коры. Личинки поедают преимуш;ественно взрослых
личинок короедов и отродившихся неокрашенных жуков. В по¬
кинутых короедами ходах они заканчивают развитие за счет раз¬
личных органических остатков и мицелия грибов.
А. trisulcum Geoffr, Сведений об образе жизни этого вида недо¬
статочно. Д. К. Померанцев (1902в) указывает его в перечне вра¬
гов короедов. Нами личинки были найдены осенью 1958 г. в Тел-
лермановском лесничестве Воронежской области под корой вяза
шершавого в ходах, покинутых короедами, с обильно развив¬
шимся мицелием грибов. Не исключено, что сапромицетофагия —
основной способ питания этого вида.
Род Colydium F.
Личинки слабоуплощенные, со светлыми умеренно склероти-
зованными покровами. 8-й и 9-й брюшные сегменты склероти-
зованы значительно интенсивнее. В передней четверти брюшных
сегментов имеется дорсальное поперечное опорное ребро, а в сред¬
ней части — волоски и неупорядоченные склеротизованные зубцы,
каждый из которых несет также ш;етинку. Все тело покрыто ред¬
кими длинными волосками.
С. elongatum F, Личинки этого вида развиваются в ходах коро-
едов-древесинников, сверлильщиков и других так называемых
амброзийных насекомых. Тот факт, что личинки этого вида живут
в свежих ходах, заселенных личинками короедов, приводит к выво¬
ду об их возможном факультативном хиш;ничестве. Можно пред¬
положить, однако, что одним из источников питания личинок яв¬
ляется мицелий симбиотических грибов, разрастающийся на стен¬
ках ходов амброзийных насекомых. Во всяком случае пока нет
оснований относить личинок Colydium к облигатным хищникам.
Серия личинок добыта нами летом 1970 г. в Убинском лесничестве
Краснодарского края, из ходов короедов-древесинников в ольхе.
Род Lado Wank.
Личинки мелкие, со слабосклеротизованными покровами. 9-й
брюшной сегмент с поперечным опорным ребром, его остальная
поверхность, кроме опорных крючков, не склеротизована. Карма¬
новидное впячивание неглубокое, с немногочисленными щетин¬
ками. Перед анальным отверстием проходит полоска мелких ши-
пиков. С каждой стороны головы по 5 глазков. Конусовидная сен¬
силла на 2-м членике усиков хорошо развита.
182
L. jelskii Wank. Согласно Сааласу (Saalas, 1923), личинки это¬
го вида встречаются на ели в ходах короедов Ips typographus L.,
Polygraphus polygraphus L. и P. subopacus Thoms. Видимо, яв¬
ляются факультативными хищниками или сапромицетофагамп.
Точных наблюдений над образом жизни не опубликовано.
СЕМЕЙСТВО PYTHIDAE — ТРУХЛЯКИ
При определении объема семейства нами принята концепция
авторов, выделяющих Salpingidae в качестве самостоятельного так¬
сона. Таким образом трухляки в фауне СССР оказываются пред¬
ставленными только родом Pytho F. На материковой части Пале-
арктики — три широко распространенных вида: Р. abieticola
J. Sahlb., Р. depressus L. и Р. kolwensis С. Sahlb. Подробные опи¬
сания их личинок и таблицу для их определения опубликовал
Саалас (Saalas, 1923). Бураковский (Burakowski, 1962) зарегист¬
рировал Р. kolwensis С. Sahlb. в Средней Европе и описал все
стадии развития этого вида. Хорошо описаны Хаяси личинки
представителей родов — Р. nivalis Lew. и Р. jezoensis Кбпо (На-
yashi, 1969). Бевинг и Крайхед (Boving, Craighead, 1931) описали
личинку Р. niger КЬу, а Петерсон (Peterson, 1951) — личинку
близкого вида Lecontia discicollis Lee.
Экология, Личинки трухляков — типичные подкорники, спе¬
циализировавшиеся к развитию только на хвойных породах. Они
предпочитают поваленные стволы деревьев, влажность которых
более высока, а на сухостое поселяются только в комлевой ча¬
сти; обычны они также и в пнях. Предпочитаемой породой трух¬
ляков является сосна, но они охотно заселяют также ель, кедр,
пихту и лиственницу. Личинки ранних возрастов обитают в хо¬
дах короедов, более крупные личинки выходят в сильно разру¬
шенные участки луба, где завершают развитие.
Мнения различных авторов о пищевых связях личинок трухля¬
ков весьма противоречивы. Саалас (Saalas, 1923), Бураковский
(Burakowski, 1962) и другие авторы считают, что они питаются гнию¬
щим лубом, частичками древесины, экскрементами других насекомых,
заглатывая их вместе с развившейся здесь микробной флорой.
Ряд авторов отмечают, что личинки трухляков проявляют себя как
хищники, уничтожающие личинок дровосеков (Гусев, 1928; Бог¬
данова, Коломиец, 1974, и др.). Н. 3. Харитонова (1972) считает
личинок трухляков весьма эффективными истребителями коро¬
едов. Наши наблюдения показывают, что трухляки относятся
к всеядным насекомым, развитие которых ускоряется при нали¬
чии животной пищи, будь то погибшее беспозвоночное или живая
личинка, которую трухляки способны одолеть. Видимо, сле¬
дует ожидать, что мелкие личинки трухляков нападают па личи¬
нок короедов, однако в сроках поселения они сильно отстают от
кульминационного периода развития сколитидного энтомоком-
183
плекса и по этой причине существенного влияния на численность
короедов не оказывают. Для взрослых личинок характерна сапро-
некрофагия также с элементами хищничества.
Морфология. Личинки (рис. 16, 2) длинные, с плоским парал-
лельнюсторонним желтым или бурым телом, снабженным на конце
сложными урогомфами. Голова округлая, с неявственным темен¬
ным и хорошо выраженными лировидными лобными швами. Ман¬
дибулы более или менее симметричные, со сложной вершиной и
хорошо развитой молой. Усики умеренно длинные, 3-члениковые,
сидят на усиконосном бугорке. Их 2-й членик с 1 короткой конусо¬
видной сенсиллой. 5 глазков с каждой стороны головы. Рото¬
вые части интенсивно склеротизованы. Жевательная лопасть па¬
раллельносторонняя, с крупным трехвершинным субапикальным
зубцом. Ее внутренняя и апикальная части с рядами коротких
щетинок. Сочленовная доля с шагренированной поверхностью.
Гипофарингеальная склерома весьма сильно склеротизована. Ноги
с вентральной стороны в густых крепких шипиках. Каждый брюш¬
ной сегмент в передней четверти с дорсальным поперечным скле-
ротизованным ребром, которое в средней части либо сильно изги¬
бается и вместе с продольным ребром образует 2 угловатых вы¬
ступа, либо соединяется двумя продольными склеротизованными
линиями, не образуя такого изгиба. Урогомфы (рис. 55, i, 2)
с внутренними зубцами. По переднему краю анального сегмента
проходит поперечный ряд шипов.
Род Pytho F.
Единственный палеарктический род семейства, характеристика
которого приведена выше.
Таблица для определения видов рода Pytho F.
1 (4) На тергитах средних брюшных сегментов 2 срединные про¬
дольные склеротизованные линии, отходящие от попереч¬
ного опорного ребра, длинные, не образуют треугольного из¬
гиба; крепкие щетинки расположены по бокам от этих линий.
Ширина устья кармановидного впячивания около 1/3 общей
ширины 9-го брюшного сегмента (рис. 55, 1).
2 (3) В основании урогомф имеется крупный внутренний зубец.
Шипики на дорсальной стороне 9-го брюшного сегмента не
образуют правильного поперечного ряда
Р. abieticola J. Sahlb.
3 (2) В основании урогомф имеется мелкий внутренний зубец
(рис. 55, 1). Шипики на дорсальной стороне 9-го брюшного
сегмента образуют правильный поперечный дуговидный ряд
Р. depressus L.
4 (1) На тергитах средних брюшных сегментов 2 срединные про¬
дольные склеротизованные линии короткие, вместе с по¬
184
Рис. 55. 9-й брюшной сегмент {1—3,5) и конец урогомфы сбоку (4, 6) ли¬
чинок трухляков (Pythidae) и лжебыстрянок (Pedilidae)
1 — Pytho depressus L.; ^ — P. kolwensis Sahlb.; 3, 4 — Agnathus decoratus Genn.; 5, tf —
Pedilus sp.
перечным опорным ребром образуют 2 треугольных изгиба,
крепкие щетинки расположены позади этих изгибов. Ширина
устья кармановидного впячивания не более ^/в общей ши¬
рины 9-го брюшного сегмента (рис. 55, 2)
Р. kolwensis С. Sahlb.
Р, abieticola /. Sahlb. Личинки по комплексу признаков сходны
с Р. depressus L., но имеют в основании урогомф крупный внут¬
ренний зубец. Заселяет в болотистых местностях нависшие над
землей еловые стволы со сравнительно плотно прилегающей ко¬
рой. Поселяются после короедов Pityogenes chalcographus L. и не¬
которых видов Polygraphus и Hylastes. Сапронекрофаг с элемен¬
тами зоофагии. Л. Н. Медведев (1965) считает возможной идентич¬
ность Р. abieticola J. Sahlb. и Р. niger КЬу. М. С. Гиляров
(1964в) приводит рисунки деталей строения личинок Pytho sp.,
которые в действительности принадлежат Р. niger КЬу (Boving,
185
Craighead, 1931). Предполагаемая Л. Н. Медведевым (1965) сино¬
нимика требует дополнительной проверки. Признаки личинок при¬
водятся по Сааласу (Saalas, 1924).
Р. depressus L, Отмечен на всех обычных хвойных породах.
Согласно Сааласу (Saalas, 1923), предпочитает стволы, диаметр
которых для ветровала составляет более 13, для сухостоя — более
15 см; минимальный диаметр заселяемых пней — 8 см. Следует за
короедами Hylurgops glabratus zett., Ips typographus L., Dryocoetes
hectographus Rtt., Pityogenes chalcographiis L. и др. В отношении
характера питания личинок в литературе нет единого мнения. По-
видимому, данные об их первостепенном значении как энтомофа-
гов (Харитонова, 1972) несколько преувеличены или имеют ло¬
кальное значение. Наиболее адекватно режим питания трухляков
можно охарактеризовать как сапронекрофагию с элементами хищ¬
ничества. Нами личинки были собраны в Комарово-Заповедном
Приморского края под корой кедра в сентябре 1968 г.
Р. kolwensis С, Sahlb. По стациям обитания и характеру пи¬
тания существенно не отличается от предыдущего вида. Саалас
(Saalas, 1923) находил его личинок в ассоциации с теми же видами
короедов, причем личинки всех трех видов трухляков нередко
обитали совместно. Согласно Бураковскому (Burakowski, 1962),
вид распространен до Приморья включительно. Не исключено, что
очень близкий японский вид Р. nivalis Lew. окажется его сино¬
нимом.
Нами большие серии личинок собраны в Хинганском заповед¬
нике Хабаровского края под корой кедра в июне 1975 г.
СЕМЕЙСТВО PED1LIDAE - ЛЖЕБЫСТРЯНКИ
Статус данного семейства еще неясен, так как большинством
специалистов оно рассматривается как подсемейство семейства
быстрянок (Anthicidae). Нами несколько изменен и его объем,
так как сюда включается также род Agnathus, ранее относившийся
к семейству Lagriidae или Conoiiotidae.
Экология, Биология семейства (в принятом нами объеме) изу¬
чена крайне слабо. Образ жизни Pedilus sp. изучен нами впервые;
морфология личинок Agnathus decoratus Germ. также ранее не бы¬
ла изучена. Личинки этих лжебыстрянок могут быть охарактери¬
зованы как ксилофильные формы, обитающие под корой (Agnat-
hus) или в толще сильно разрушенной древесины (Pedilus). В на¬
стоящее время еще неясны пути экологической специализации
семейства. Следует отметить, однако, что личинки Agnathus deco¬
ratus Germ. связаны с короедами.
Морфология. Личинки буровато-желтого цвета, умеренно уп¬
лощенные, на конце с 2 загнутыми вверх крючками и 2 карманами
между урогомфами (рис. 55, 5). Голова поперечная. Теменной шов
отсутствует, лобные швы образуют лировидную фигуру. Глазки
186
хорошо выражены. Усики 3-члениковые, с конусовидной сенсил¬
лой на 2-м членике. Верхняя губа широкоовальная. Мандибулы
с хорошо развитой моларной частью. Максиллы хорошо вы¬
ражены. Кардо значительно короче стнпеса. Жевательная доля
максилл параллельно-сторонняя, на конце закругленная, по
внутреннему краю с рядом кинжаловидных щетинок и в апикаль¬
ной части с 3 зубцами — 1 острым крупным и 2 более мелкими.
Кроме кинжаловидных, на поверхности жевательной доли мак¬
силл имеются длинные игловидные ш,етинки. Нижнечелюстные
ш,упики 3-члениковые. Нижняя губа с хорошо склеротизованноп
гипофарингеальной склеромой, 2 пучками гипофарингеальных во¬
лосков и широким мясистым язычком, несуш,им 2 длинные щетин¬
ки. Нижнегубные щупики 2-члениковые. Грудные сегменты, как
и брюшные, слабо склеротизованы, в редких волосках. Ноги
обычного строения. Брюшные сегменты с дорсальной стороны по
переднему краю с опорным ребром или поперечным рядом мелких
зубчиков, 9-й брюшной сегмент (рис. 55, 3, 5) с широко расстав¬
ленными выростами урогомф, между которыми имеются 2 глубо¬
ких кармана. Их поверхность сильно склеротизована, в попереч¬
ной штриховке, несет редкие короткие щетинки.
Таблица для определения родов семейства Pedilidae
1 (2) Задний край 9-го брюшного сегмента (рис. 55, 5, 6) между
урогомфами выпуклый, его тергит более или менее гладкий.
В основании урогомф имеется внутренний зубец. Сенсилла
на 2-м членике усика очень крупная, значительно толще 3-го
членика. В древесине. Сапроксилофаги
Pedilus Fisch.
2 (1) Задний край 9-го брюшного сегмента (рис. 55, 5, 4) между
урогомфами ровный, его тергит в многочисленных пластинках,
каждая из которых несет волосок. Сенсилла на 2-м членике
усика небольшая, тоньше 3-го членика
Agnathus Germ. (стр. 187)
Род Agnathus Germ.
Глазков 5, линзы и пигментные пятна хорошо выражены. Уси¬
ки сидят на бугорке, их 2-й членик вдвое длиннее 1-го, несет 1 ко¬
нусовидную сенсиллу и тонкий 3-й членик, который снабжен на
конце 4 щетинковидными сенсиллами. Клипеус с прямым краем.
Верхняя губа с 2 щетинками в средней части и 10 щетинками по
краю. Мандибулы симметричные, с 2 зубцами на вершине и 3
дополнительными на режущем крае. Моларная часть с попереч¬
ной ребристостью. Дорсальная поверхность 9-го сегмента в мел¬
ких склеротизованных пластинках, снабженных короткой щетин¬
кой.
А, decoratus Germ, Специализирован к развитию под корой
ольхи во влажных поймах рек. Отмечен в ходах короеда Ху1е-
187
bonis pfeili Ratz. Предположительно сапронекрофаг, с элементами
зоофагии. Нами найден в районе г. Красная Поляна Краснодар¬
ского края.
СЕМЕЙСТВО TENEBRIONLDAE - ЧЕРНОТЕЛКИ
Крупное семейство, насчитывающее в фауне СССР более
1000 видов. Представляет собой характерный элемент пустынной
энтомофауны. В лесной зоне немногочисленно, но именно с древеси¬
ной экологически связаны наиболее примитивные группы семей¬
ства.
Экология. Личинки чернотелок в основном развиваются в поч¬
ве и лесной подстилке. Ряд видов чернотелок вредит запасам про¬
дуктов. Наконец, личинки многих видов этого семейства развива¬
ются в разлагающейся древесине, под корой, в ходах короедов и в
древесных грибах.
Среди личинок чернотелок отмечается как сапро- и фитофагия,
так и хищничество. Многие личинки-фитофаги относятся к числу
серьезных вредителей сельскохозяйственных культур. Сапрокси-
лофагия — наиболее обычный тип питания личинок чернотелок,
развивающихся в разлагающейся древесине. Примером этому
могут служить личинки Upis ceramboides L., Helops coeruleus L.,
Misolampidius tentyrioides Sols., виды рода Uloma и др. Хищни¬
чество — редкое явление среди личинок чернотелок. Наиболее
четко оно проявляется у личинок представителей рода НурорЫо-
eus, которые обитают в ходах короедов. Многие авторы неодно¬
кратно отмечали существенную роль личинок Hypophloeus в ист¬
реблении яиц, личинок и 1^уколок этих вредителей (Гусев, 1928;
Никитюк, 1951; Зиновьев, 1959; Богданова, Коломиец, 1974;
Никитский, 1976).
Большинство ксилофильных личинок чернотелок входят в лу-
канидный комплекс разрушителей древесины. Они развиваются
главным образом в светлых и черненых гнилях древесины лист¬
венных древесных пород. Хищные формы (личинки Hypophloeus)
специализированы к развитию под корой во время ее разрушения
короедами (сколитидная стадия), имеются также виды, прони¬
кающие в древесину по ходам короедов-древесинников. Некоторые
виды этого рода, например Н. linearis F. и Н. longulus Gyll.,
встречаются только на хвойных, некоторые — не специализиро¬
ваны к развитию на определенной группе древесных пород.
Морфологически неотличимые личинки Uloma culinaris L. и
и. perroudi Muls. хорошо отличаются экологически, так как первый
вид развивается в древесине лиственных, а второй — хвойных
пород. Такие виды, как Neatus picipes Hbst., Bius thoracicus F.,
Tenebrio sp., заселяют в древесине щели и полости, заполненные
трухой, а монолитную древесину не заселяют. У Bius thoracicus F.
предпочитаемой породой является береза.
188
Рис. 56. Детали строения Gryphaeus cornutus F.—W. (/), Helops coeruleus
L. (2), Misolampidius tentyrioides Sols. (5), Palorus subdepressus Woll. (4),
Tenebrio molitor L. (5), Uloma culinaris L. (6), Upis ceramboides L. (7, S)
{Tenebrionidae)
1—5, 7 — конец брюшка сверху; 4,6 — конец брюшка [сбоку; 8 — 1-й сегмент брюшка
сверху (J—2f 6—8 — по Бызовой, Келейниковой, 1964; 3 — по Келейниковой, Мамаеву,
1972; 4 — по Emden, 1948; 5 — Emden, 1947).
Морфология, Тело длинное, цилиндрическое или уплощенное
снизу (рис. 17, 1—3\ 57, 1—3).
Голова гиногнатическая, с округлыми боковыми сторонами.
Теменной и лобные швы явственные. С каждой стороны головы от 1
до 5 глазков. Гуларная пластинка хорошо развита. Усики 3-чле-
никовые. Они сидят на массивном, мясистом, несклеротизованном
бугорке, легко втягивающемся в головную капсулу, который не¬
которые авторы считают самостоятельным члеником (соответ¬
ственно усики считаются 4-члениковыми). 2-й членик усиков не¬
сет на вершине сенсорное поле в виде незамкнутого полукольца,
jfn6o, гораздо реже, крупную коническую сенсиллу, иногда рав¬
ную по длине 3-му членику, который снабжен длинной концевой
щетинкой.
189
Мандибулы массивные, силыюсклеротизованные, с теркой на
жевательной поверхности молы и обычно с явственными предвер¬
шинными зубцами. Наружный край мандибул у основания часто
несет щетинки или шипики. Верхняя губа поперечная, с округ¬
лым передним краем, явственно отделенная от наличника, несет
многочисленные краевые и дискальные щетинки. Наличник имеет¬
ся. Максиллы состоят из кардо, стипеса, жевательной лопасти (ма¬
лы) и трехчленикового нижнечелюстного щупика. Жевательная
лопасть обычно округлая, редко прямоугольная, часто слегка под¬
разделенная на галею и лацинию. Внутренний край жевательной
лопасти несет 2 продолышх ряда шипиков или крепких щетинок.
Нижняя губа состоит из прементума, ментума и субментума. Язы¬
чок чаще всего небольшой, конический, с 2 вершинными щетин¬
ками, редко (у рода Hypophloeiis) крупный, округлый, доходящий
до средины 2-го членика пижнегубных щупиков; иногда язычок
отсутствует. Нижнегубные щупики 2-члепиковые.
У многих почвенных чернотелок 1-я пара ног крупнее и отли¬
чается по вооружению от 2-й и 3-й. Коготок обычно простой, изо¬
гнутый, сильно сулчивающийся к вершине, очень редко дву вер¬
шинный. Форма и строение 9-го брюшного сегмента сильно варь¬
ируют (рис. 56, 1—7): он бывает округлым или заостренным,
может оканчиваться одним или 2 вершинными выростами. 9-й брюш¬
ной сегмент несет многочисленные щетинки или шипики, нередка
и то и другое одновременно. На его вентральной стороне распо¬
ложен подталкиватель. Он обычно имеет 2 лопасти, иногда же при¬
обретает форму полукруглой складки, а у Hypophloeus и Bolito-
phagus — мягкой трубки.
Таблица для определения родов семейства TenebriOnidae
1 (6) Передние ноги значительно крупнее средних и задних
и отличаются более сильным вооружением.
2 (3) Личинки беловатые. Тергиты грудных и первых брюшных
сегментов со складками (рис. 50, 8), Вершина 9-го брюш¬
ного сегмента с 2 широко расставленными шипами и много¬
численными щетинками (рис. 56, 7). В древесине и сильна
разложившейся коре. Сапроксилофаг Upis F.
3 (2) Личинки желтые. На тергитах грудных и первых брюш¬
ных сегментов нет складчатых участков. Вершина 9-го
брюшного сегхмента с 2 сближенными шипиками.
4 (5) На боковых сторонах 9-го сегмента брюшка, помимо ще¬
тинок, имеется по 2 мелких шипика (рис. 56, 5). В трухе
в дуплах или полостях в древесине. Санронекрофаг . .
Tenebrio L.
5 (4) На боковых сторонах 9-го брюшного сегмента имеются
только щетинки. В древесной трухе в дуплах, щелях и по¬
лостях. Санронекрофаг Neatus Lee.
190
Рис. 57. Обпщй вид личинок чернотелок (Tenebrionidae)
I — Helops coeruleus 2 — Hypophloeus longulus Gyll.; 3 — H. fraxini Kug.
6 (1) Передние ноги не отличаются по строению от средних и
задних.
7 (8) 9-й брюшной сегмент слегка уплощен, пшроко закруглен,
с острым краем, покрыт длинными щетинками (рис. 57,
2, 5). Язычок крупный, округлый, доходит примерно до
середины 2-го членика нижнегубных щупиков (рис. 58,
2, 5) Hypophloeus F. (стр. 193)
8 (7) 9-й брюшной сегмент вооружен опорными крючками или
шипиком (рис. 56, 1—6),
191
9 (10) 9-й брюшной сегмент куполовидный, покрыт грубыми точ¬
ками, с 1 маленьким шипиком (рис. 56, 6), В разлагающейся
древесине. Сапроксилофаг Uloma Cast.
10 (9) 9-й брюшной сегмент на конце с 2 выростами.
И (12) Выросты 9-го брюшного сегмента с массивными широкими
основаниями (рис. 56, i). Тергиты с сильносклеротизо-
ванными волнистыми гребнями близ переднего края. Го¬
лова и тергиты позади гребней с крупными плоскодонными
ямками и щетинконосными бугорками. В светлой древе¬
сине. Сапроксилофаг Cryphaeus Klug.
12 (И) 9-й брюшной сегмент с 2 коническими изогнутыми кверху
крючками.
13 (14) Выросты 9-го брюшного сегмента загибаются на спинную
поверхность (рис. 56, 2\ 57, 1). Тергиты 7-го и 8-го брюш¬
ных сегментов с крупными плоскодонными ямками. Зад¬
ний край 8-го брюшного тергита несет двувершинное воз¬
вышение. В разлагающейся древесине. Сапроксилофаг
Helops F.
14 (13) Выросты 9-го брюшного сегмента не загибаются на его
спинную сторону. Тергиты 7-го и 8-го брюшных сегментов
без плоскодонных ямок.
15 (16) Выросты 9-го брюшного сегмента лишь слегка изогнуты
кверху (рис. 56, 4), Щетинконосные бугорки на наруж¬
ных сторонах выростов отсутствуют. Под корой и в трух¬
лявой древесине. Сапромицетофаг . . . .Palorus Muls.
16 (15) Выросты 9-го брюшного сегмента изогнуты кверху почти
под прямым углом.
17 (18) Жевательная доля максилл округлая. Выросты 9-го
брюшного сегмента несут на внешней стороне по 1 малень¬
кому щетинконосному бугорку. Длина до 15 мм. В трухе
р полостях и дуплах. Сапронекрофаг . . . .Bius Muls.
18 (17) Жевательная доля максилл прямоугольная. Выросты 9-го
брюшного сегмента без щетинконосных бугорков на внеш¬
ней стороне. Длина до 30 мм.
19 (20) Голень с несколькими крупными щетинками в средней
части, вертлуг с 4 щетинками. 9-й брюшной сегмент к вер¬
шине резко суживается, несет небольшое количество длин¬
ных щетинок. Выросты его сближены (рис. 56, 3). В сильно
сгнившей древесине. Сапроксплофаг
Misolampidiiis Sols.
20 (19) Голень целиком покрыта щетинками, вертлуг с много¬
численными беспорядочно расположенными щетинками.
9-й брюшной сегмент слабо суживается к вершине, несет
большое количество щетинок среднего размера. Выросты
его широко расставлены. В сильно сгнившей древесине.
Сапроксилофаг Stenophanes Sols.
192
Род Hypophloeus F.
В отечественной литературе определительная таблица видов
этого рода была опубликована Ю. Б. Бызовой и С. И. Келейни-
ковой (1964). Н. Б. Никитский (1976) обработал большее число
видов, но не использовал для диагностики ряда точных, требую¬
щих микроскопического исследования признаков. Нам также не
удалось подготовить полную определительную таблицу видов
этого рода, личиночная таксономия которого нуждается в даль¬
нейшей разработке.
Таблица для опредслония видов рода Hypophloeus F.
1 (2) Среднегрудь, заднегрудь и 4 первых тергита брюшка несут в
задней трети по одному поперечному ряду из 4 склеротпзо-
ванных бугорков. Длина теменного шва составляет 1/3
длины головы. 2-й членик усиков в 4—5 раз длиннее 1-го
(рис. 58, 6). 2-й членик нижнечелюстных 1цупиков самый
длинный (рис. 58,7) И. unicolor Pili.
2 (1) Тергиты без склеротизованных бугорков. Длина теменного
шва составляет 1/5—1/6 длины головы. 2-й членик усиков
в 2,5—3 раза длиннее 1-го (рис. 58, 5, 4). Членики нижнечелю¬
стных щупиков равной длины или же их 3-й членик самый
длинный.
3 (4) Тело сверху сильно склеротизовано, темно-коричневое,
с явственной белой пунктировкой, особенно четко выражен¬
ной на 5—7-м тергитах брюшка (рис. 17, 1), 2-й членик ниж¬
негубных шуников явственно короче 1-го (рис. 58,5). 2-й
членик усиков более или менее цилиндрический (рис. 58. 4)
Н. linearis F.
4 (3) Тело сверху золотисто-желтое. 1-й и 2-й членик ни^ьнегуб-
ных щупиков одинаковой длины (рис. 58, 2), 3-й членик уси¬
ков расширяется к вершине (рис. 58, 3).
5 (6) Сегменты тела с длинными дорсальными щетинками по зад¬
нему краю (рис. 57, 5). 3-й членик нижнечелюстных щупи¬
ков слегка длиннее второго (рис. 58, 1). 2-й членик нижне-
губных щупиков явственно уже 1-го (рис. 58, 2) ....
Н. fraxini Kiig.
6 (5) Сегменты тела без длинных дорсальных щетинок (рис. 57, 2).
3-й членик нижнечелюстных щупиков в 1,5 раза длиннее
2-го. Членики нижнегубных щупиков одинаковой iiinj)ni ы
«Н. longulus (iyll.
Н. fraxini Kug. Вид отмечен на сосне, ели, кедре, ильме, буке,
грабе, вязе, ясене и дубе в ходах 16 видов короедов, его личипки
уничтожают личинок и куколок короедов (Гусев, 1928; Ники:юк,
1951). Н. Б. Никитский (1976) отмечает, что личинки этого вида
могут также поедать неокрашенных жуков. В. Буковский (19^()
находил его как в ходах, заселенных короедами, так и в нокинуилх
7 Б. м. 1\1амаев и др. 193
Рис. 58. Детали строения Hypophloeus fraxini Kug. (1—3), Н. linearis F.
{4, 5), Н. unicolor Pili. (6, 7) (Tenebrionidae)
1,7 — вершина максиллы; 2,5 — прементум; з, 4, б — усик
lig — язычок; та — мала; р. 1Ь — нижнегубной щупик; р. тх — нижнечелюстной щупик
ходах. Серия личинок была найдена в Комарово-Заповедном под
корой кедра корейского, а также под корой сосны обыкновенной
в Убинском лесничестве Краснодарского края.
Н. lineans F, Вид приурочен к хвойным породам, был отмечен
на сосне, ели, кедре, пихте в ходах более чем 20 видов короедов,
его личинки — активные хищники, поедающие яйца, личинок и
куколок короеда. Д. А. Богданова и Н. Г. Коломиец (1974) от¬
мечают, что в районе Енисейского кряжа жуки и личинки этого
вида встречаются в ходах короедов с июня до конца августа.
2 личинки этого вида найдены в Московской области под корой
сосны обыкновенной и под корой ели европейской.
Н. longulus GylL Встречается с июня по сентябрь на хвойных
породах в ходах 7 видов короедов. По данным В. И. Гусева
(1928), личинки — хищники, уничтожающие личинок короедов.
194
Н. Б. Никитский (1976) считает вид факультативным хищни¬
ком, личинки которого питаются, при отсутствии короедов, гриба¬
ми, растущими в ходах. Серия личинок этого вида найдена в Мо¬
сковской области под корой сосны обыкновенной.
Н. unicolor Pili. Встречается на ясене, березе, буке, ели и сос¬
не в ходах 7 видов короедов (Погориляк, 1963; Balazy, Michal-
sky, 1960). Палм (Palm, 1959) указывал на тот факт, что Н. uni¬
color встречается также под гнилой, отстающей, пронизанной гри¬
бным мицелием корой хвойных деревьев, а также бука, реже бе¬
резы, иногда в трутовиках. Нами вид обнаружен в колодах пих¬
ты, вяза, ильма, вишни, дуба. По данным Н. Б. Никитского
(1976), вид является факультативным хищником; в условиях опы¬
та личинка, питаясь грибами, может завершить полный цикл
развития. 2 личинки этого вида найдены в Красной Поляне
Краснодарского края в древесине вяза и ольхи, еще 2 личинки —
под корой сосны обыкновенной и бука в Убинском лесничестве
Краснодарского края.
ОТРЯД RАРН1D10PTERA - ВЕРБЛЮДКИ
Отряд 3 фауне СССР представлен 2 небольн ими семействами,
почти совершенно не изученными.
Экология. Личинки верблюдок заселяют различные местооби¬
тания. По данным Аспока (Aspock et al., 1974Ь), личинки всех
известных в Центральной Европе видов, за исключением почвен¬
ного вида Raphidia flavipes Stein., развиваются под корой и в коре
дерев1)ев и реже кустарников. Личинок можно также встретить
в гнездах птиц, древесных грибах, покинутых муравейниках,
шишках, галлах и других подобных средах, которые, возможно,
являются временными убежищами. Следует подчеркнуть, что ли¬
чинки верблюдок встречаются почти исключительно в усыхающих
деревьях, в отмерших стволах только в очень редких случаях на¬
ходят единичные экземпляры. Личинки верблюдок являются все¬
ядными хищниками (Wichman, 1957; Aspock et al., 1974Ь) и, по
накопленным данным, питаются личинками короедов и пилиль¬
щиков, гусеницами бабочек и другими членистоногими с мягкими
покровами. Активность личинок повышается в темноте (W yniger,
19Г)9). Некоторые авторы считают, личинки верблюдок уходят
для питания в почву (Wichman, 19о7)или в крону деревьев (Met-
zger, 1960). Мецгер (Metzger, 1960) даже считает крону основным
местом питания личинок, где они во время вегетационного периода
проводят большую часть времени. Пребывание личинок под корой,
по мнению этого автора, служит укрытием в дневные часы и убе¬
жищем в зимний период (защита от неблагоприятных климатг-
ческих условий). По данным Аспока (Aspock et al., 1974Ь), посе¬
ление личинок в стволе, наоборот, не зависит от погодных услов! й
195
и времени года, а лереход личинок подкорных видов верблюдок
в почту является редким и слу 1айным.
Развитие личинок длится по крайней мере год, а иногда до¬
стигает почти двух лет и включает около 10 линек.
Морфология наиболее полно дана в работе Аспока и соавторов
(Агроск et al., 1974а). Личинки веретеновидные. Голова и перед-
негрудь резко уп.иощенные, одноцветные, сильносклеротизован-
ные. Среднегрудь, заднегрудь и брюшко мясистые, с характерным
мраморным рисунком на тергитах. Ноги 5-члениковые, с 2 когот¬
ками. Брюшко состоит из 10 сегментов, веретеновидное. Наиболь¬
шая ширина его приходится на 3—4-й брюшные сегменты. 10-й
брюшной сегмент служит подталкивателем. Церки отсутствуют.
Таблица для опрпд('Л(»ния с(^м(41ств отряда
Raphidioptcra—Верблюдки^
1 (2) С ка>1х’дой стороны головы по 4 глазка. Срединный рисунок
на тергитах брюшка не отделен от боковых рисунков. Тер-
гиты 1—8-го брюшных сегментов с узко прямоугольным
или трапециевидным светлым пятном в задней части ....
Inocellidae — Малоглазки (стр. 200)
2 (1) С каждой стороны головы по 7 глазков. Срединный рисунок
па тергитах брюшка отделен от боковых рисунков. Тергиты
1—8-го брюшных сегментов иные
Raphidiidae — Верблюдки настоящие (стр. 196)
СЕМЕЙСТВО RAPHIDIIDAE —ВЕРБЛЮДКИ НАСТОЯЩИЕ
В Европе по последним данным насчитывается 13 видов этого
семейства (Aspock, 1974а).
Экология. Личинки встречаются как в хвойных, так и в лист¬
венных породах деревьев и, как правило, обнаруживают резко
выраженное предпочтение тех или других. Эта приуроченность,
однако, не является абсолютной, особенно у видов, развивающих¬
ся в хвойных породах, особенно если учесть изменчивость эколо¬
гических связей в пределах ареала. Например, Raphidia nigricol-
lis Alb. в средней Европе развивается почти исключительно на
лиственных деревьях, на хвойных встречается только в порядке
исключения. В горных областях юго-восточной Европы вид оди¬
наково часто встречается на соснах, елях и лиственных деревьях.
Обычно имеет место и явное преобладание вида на какой-нибудь
определенной породе. Так, личинки Raphidia major Burm. встре¬
чаются в 10 лиственных породах, но около 60% популяции со¬
средоточено под корой груши (Aspock et al., 1974b).
Морфология. Голова чуть расширяется к основанию, с яв¬
ственным шейным перехватом. Лобный и теменные швы явствен¬
ные. На вентральной стороне от переднего до заднего края головной
капсулы тянутся 2 параллельных шва. Усики 4-члениковые, 2-й
196
членик усиков очень маленький. Верхняя губа и наличник име¬
ются. Жвалы сильносклеротизованные, с зубцами на режущем
крае. Нижние челюсти состоят из короткого кардо, массивного
стипеса. Нижнечелюстной щупик 4-члениковый, расположен на
небольшом шупиконосце, имеющем вид склеротизованного полу¬
кольца. Мала не полностью отделена от стипеса, склеротизована
в нижней части, верхняя ее половина мясистая, несет короткие
щетинки.
Нижняя губа состоит из двураздельного склеротизованного
субментума, перепончатого ментума и двураздельного склеро-
тизованного прементума, от которого отходят 3-члениковые ниж¬
негубные щупики. Язычок мясистый, в виде тупого конуса. Сле¬
дует отметить, что гомологизация частей нижней губы дается в из¬
вестной степени условно, так как для окончательного решения это¬
го вопроса требуются специальные исследования.
Род Raphidia L.
Таблица для определения видов рода Raphidia L.
1 (2) Срединный рисунок на 8-м брюшном тергите с прямым
задним краем (без выступов) . . . .R. nigricollis Alb.
2 (1) Срединный рисунок на 8-м брюшном тергите с 2 выступами
на заднем крае.
3 (8) Задние выступы срединного рисунка 3-го брюшного тер-
гита доходят до заднего края сегмента, боковые стороны
их параллельны (рис. 59, 5; 60, 2, 4),
4 (5) Средне- и заднеспинка с отчетливой светлой срединной по¬
лосой R. ratzeburgi Brauer
5 (4) Средне- и заднеспинка без светлой срединной полосы.
6 (7) Срединный рисунок на 3-м брюшном тергите без светлой
срединной полосы (рис. 59, 5) .... R. notata F.
7 (6) Срединный рисунок на 3-м брюшном тергите со светлой,
в большинстве случаев сплошной, реже прерывистой,
срединной полосой (рис. 60, 4) .R. xanthostigma Schum.
8 ^3) Задние выступы срединного рисунка на 3-м брюшном тер¬
гите не доходят до заднего края сегмента (рис. 59, 5; 60, 5),
если же доходят, то в задней части слабо выражены и гра¬
ница между срединным рисунком и задними выступами
резко выражена (рис. 59, 4; 60, 1).
9 (10) Срединный рисунок на 3-м брюшном тергите связан с
боковым рисунком широкой пигментированной зоной
(рис. 59, 4) R. maior Burm.
10 (9) Срединный рисунок на 3-м брюшном тергите не связан
или очень слабо связан с боковым рисунком (рис. 60, 1,3),
И (12) Внутренний край бокового рисунка на 3-м брюшном тер¬
гите с крупным выступающим пятном (рис. 59, 5) . . .
R. cognata Ramb.
197
12 (И) Внутренний край бокового рисунка на 3-м брюшном тер-
гите без пятна (рис. 60, 1, 3).
13 (14) Средне- и заднеснинка со светлой срединной полосой
R. ulrikae Asp.
14 (13) Средне- и заднеснинка без светлой срединной полосы
R. ophiopsis L.
R. cognata Ramb. Специфическим признаком вида является X-
образная форма бокового рисунка. Личинки найдены только на
лиственных породах деревьев. Явно предпочитают грушу. Помимо
этого, найдены на яблоне, клене и дубе.
R, maior Вигт, В большинстве случаев срединный рисунок об¬
разует в задней трети остроугольные выступы (рис. 59, 4), Личинки
и куколки этого вида были найдены на лиственных деревьях, яб¬
лоне и особенно груше. Гораздо меньшее количество экземпляров
отмечено на дубе. Личинки найдены также под корой миндаля,
боярышника и единично на тополе, сливе, иве и грецком орехе.
R. nigricollis Alb, Характерным признаком вида является тем¬
ная полоска на боковых сторонах брюшных сегментов и наличие
широкой желтой полосы, отделяюш,ей срединный рисунок от бо¬
кового. Личинки этого вида в Средней Европе приурочены к ли¬
ственным породам деревьев и встречаются чаще всего под корой
сливы, реже под корой яблони, клена и дуба. Единичные находки
были сделаны на каштане и тополе черном. Одна личинка найде¬
на под корой сосны. В горах юго-восточной Европы вид встре¬
чается одинаково часто на соснах, елях и лиственных породах.
R. notata F. Срединный рисунок отделен от бокового широкой
непрерывной светлой полосой (рис. 59, 5). В задней трети боко¬
вого края срединного рисунка расположено округлое светлое пят¬
но, иногда открытое спереди. Чаще всего личинки встречались под
корой ели и сосны, реже — под корой лиственницы, еще реже —
пихты. Из лиственных пород наибольшее количество личинок от¬
мечено под корой яблони и груши. На ольхе, дубе, каштане, кле¬
не, сливе, березе и бузине находки единичны.
R. ophiopsis L. Брюшко позади срединного рисунка с 2 слабо
пигментированными полями, являющимися продолжением зад¬
них выступов срединного рисунка. Личинки развиваются под ко¬
рой сосны.
R, ratzeburgi Brauer. Срединный рисунок брюшных тергитов
со светлой срединной полосой (рис. 60, 2), вследствие чего на дор¬
сальной стороне личинки образуется светлая полоса, идущая по
всей длине тела. Личинки приурочены в основном к хвойным по¬
родам деревьев. Чаще всего они встречаются под корой сосны, реже
ели, лиственницы и пихты. Из лиственных пород (яблоня, дуб,
груша, клен и каштан) личинки были отмечены в незначительных
количествах.
R. ulrikae Asp. Брюшко позади срединного рисунка желтое.
Личинки найдены под корой сосны и единично под корой дуба.
R, xanthostigma Schum, Желтая или желто-коричневая полоса
Рис. 59. 3-й сегмент брюшка
личинок Raphidioptera (сверху)
1 — Inocellia braueri Alb.;
2 — 1. crassicomis Schum.;
3 — Raphidia cognata Ramb.;
4 — R. maior Burm.;
5 — R. notata F. (oo Aspock et al.,
1974a)
Рис. 60. 3-Й сегмент брюшка личинок Raphidioptera (сверху)
1 — Raphidia ophiopsis L.; 2 — R. ratzeburgi Brauer; 5 — R. ulrikae Asr.; 4 — R. xan-
thostigma Schum. (no Aspdck et al., 1974a)
между срединными и боковыми рисунками нечеткая, у многих
экземпляров вообще отсутствует. Личинки предпочитают листвен¬
ные породы и чан1е всего встречаются под корой груши и яблони.
Незначительное число экземпляров было отмечено на вязе и дубе.
Единичные находки были сделаны под корой сосны.
СЕМЕЙСТВО INOCELLIDAE — МАЛОГЛЛЗКИ
Небольшое семейство, представленное в Европе 2 видами.
Личинки встречаются под корой как хвойных, так и листвен¬
ных деревьев. Согласно литературным данным, для личинок
Inocellidae характерно наличие 4 глазков с каждой стороны го¬
ловы (Aspock et al., 1974b).
Род Inocellia Schneid.
Единственный род, представители которого встречаются зна¬
чительно реже Raphidiidae.
Таблица для определения видов рода Inocellia Schncid.
1 (2) Тергит 3-го брюшного сегмента, помимо заднего светлого
пятна, несет светлые пятна также и по бокам срединного
рисунка (рис. 59, 2) I. erassicornis Schum.
2 (1) Тергит 3-го брюшного сегмента только с крупным светлым
трапециевидным пятном в задней части (рис. 59, 7) . . .
I. braueri Alb.
/. braueri Alb. Личинка темная, почти черная. Вид не выка¬
зывает явного предпочтения определенным породам, личинки жи¬
вут как на хвойных, так и на лиственных деревьях, но все же на
лиственных они встречаются несколько чаш,е.
L crassicornis Schum, Личинка светлая, красновато-коричне¬
вая. Вид приурочен к хвойным породам, чаш,е всего встречается
под корой сосны, гораздо реже — под корой лиственницы. Еди¬
ничные находки сделаны под корой ели и груши.
ОТРЯД DIPTER А - ДВУКРЫЛЫЕ,
ИЛИ КОМАРЫ И МУХИ
Среди насекомых отряд двукрылых по обилию и разнообразию
представителей занимает одно из первых мест. В настояш,ее время
насчитывается приблизительно 80 ООО видов этого отряда. На тер¬
ритории нашей страны распространено не менее 120 семейств дву¬
крылых (Штакельберг, Нарчук, 1969; Кривошеина, Мамаев,
1970; Кривошеина, 1971в, 19726; Родендорф, Калугина, 1974).
Большинство современных двукрылых (кроме реликтового семей¬
ства Nymphomyiidae) объединяется в 3 крупные подотряда; Ne-
200
matocera — длинноусые двукрылые, Brachycera — Orthorrhap-
ha — короткоусые прямошовные двукрылые и Brachycera —
Cyclorrhapha — короткоусые круглошовные двукрылые. Такое
деление на подотряды широко используется в мировой литера¬
туре систематиками, хотя недостаточно при рассмотрении эволю¬
ционных вопросов. Требованиям филогенетики в большей мере
соответствует более дробная система деления двукрылых на ин¬
фраотряды, предложенная Б. Б. Родендорфом (1964).
СРАВНИТЕЛЬНАЯ МОРФОЛОГИЯ ЛИЧИНОК ДВУКРЫЛЫХ
Личинки двукрылых очень разнообразны по строению. Для
всех них характерна общая черта — отсутствие развитых члени¬
стых истинных грудных ног, однако имеются формы с хорошо раз¬
витыми мягкими выростами («ложными ножками»). Для личинок,
помимо головы или в случае ее редукции головного сегмента,
характерны 3 грудных сегмента и не более 10 (обычно 9) брюшных
сегментов.
Форма тела личинок двукрылых исключительно разнообразна.
Удлиненное и слегка С-образно изогнутое тело (рис. 61) характер¬
но для личинок различных групп бибионоидных двукрылых (Р1е-
ciidae, Pachyneuridae, Cramptonomyiidae и др.), комаров-звон-
цов (Chironomidae), мокрецов (Ceratopogonidae), детритниц (Sci-
aridae) и др. Удлиненное прямое тело (рис. 61, 4; 62, 5; 63, 7, 3)
характерно для личинок болотниц (Limoniidae), многих галлиц
Рис. 61. Общий вид личинок двукрылых сбоку
1^— Plecia nigra Lund (Pleciidae); 2 — Pergratospes holoptlca Kriv. et Mam. (Cramptono¬
myiidae); S — Dilophus febrilis Loew (Bibionidae); 4 — Epiphragma ocellaris L. (Limoni¬
idae)
201
Рис. 62. Общий вид личинок двукрылЫхХсбоКу (i, сверху (2, 3)
1 — Sylvicola fenestralis Scop. (Anisopodidae); 2 — Mycetobia sp. (Mycetobiidae);
3 — Trichopteromyia modesta Will. (Cecidomyiidae); 4 —^Thereva sp. (Therevidae)
Рис. 63. Общий вид личинок двукрылых сбоку и сверху
1 — Trichomyia sp. (Trichomyiidae); 2 — Mesaxymyial kerteszi Duda (Axymyiidae);
3 — Symmerus annulatus Meig. (Ditomyiidae) — сверху
(Cecidomyiidae), отдельных родов бабочниц (Psychodidae), дито-
миид (Ditomyiidae) и др. У многих личинок, например, у личинок
бекасниц (Rhagionidae), зеленушек (Dolichopodidae), стволоедок
(Xylophagidae), у большинства круглошовных двукрылых тело
заметно сужено на переднем конце (рис. 64, 1, 2), Для личинок
отдельных семейств, например слепней (Tabanidae), характерна
веретеновидная форма тела (рис. 64, 5). Сильно удлиненное, зме-
202
Рис. 64. Общий вид личинок двукрылых сбоку
1 — Xylophagus ater Meig. (Xylophagidae); 2 — Medetera fasciata Frey (Dolichopodidae):
3 — Tabanus karybenthinus Szil. (Tabanidae); 4 — Choerades igneus Meig. (Asilidae)
евидное тело (рис. 62, i, 2, 4) имеют личинки длинноусых двукры¬
лых (Anisopodidae, Mycetobiidae, отдельных групп мокрецов —
Ceratopogonidae), а также короткоусых двукрылых: лжектырей
(Therevidae), сценопинид (Scenopinidae) и др.
Среди личинок двукрылых сравнительно много групп, обла¬
дающих дорсовентрально уплощенным телом (рис. 65—67). Это —
личинки отдельных родов гнильниц (Scatopsidae), бабочниц
203
Рис. 65. Общш"! вид личинок двукрылых сверху
1 — Scatopse subnitens Verr. (Scatopsidae); 2 — Parepldosis ulmicorticis Mam. (Cecido-
myiidae); 3 — Brachyopa bicolor Fll. (Syrphidae)
(Psychodidae), галлиц (Cecidomyiidae) ив длинноусых двукрылых,
острокрылок (Lonchopteridae), грибных мушек (Platypezidae),
настоящих мух (Muscidae) из круглошовных двукрылых и др.
Личинки отдельных групп характеризуются сильно утолш,енным
вальковатым телом (рис. 63, 2\ 68, 5; 69, 2; 70, 1). Своеобразный
вид внешнему облику личинок придают различные выросты тела
(рис. 67, 2, 3).
У представителей отряда наблюдаются все переходы от вполне
развитой до целиком редуцированной головной капсулы. В за¬
висимости от степени редукции обычно выделяют три типа голов¬
ной капсулы (Вгаиег, 1883): хорошо развитая массивная (Еисер-
hala), частично редуцированная (Hemicephala) и полностью реду¬
цированная (Acephala). Подробно различные типы строения разви¬
той головной капсулы проанализированы в работе Н. П. Кри-
вошеиной (1969).
204
Рис. 66. Общий вид личинок двукрылых сверху
1 — Neopachygaster intermedia Kriv. (Stratiomyidae); 2 — Psychoda phalae noides L'
(Psychodidae); 3 — Solva marginata Meig. (Xylomyidae) {2 — no Satchell, 1947)
1. Группа Eucephala. Головная капсула (рис. 71) с дорсаль¬
ной стороны разделена теменным и лобными швами или только
лобными швами. Последние ограничивают срединный участок
головной капсулы — лоб. Кпереди от лобной пластинки располо¬
жена поперечная пластинка — наличник, наиболее четко обособ¬
ленная у личинок Pleciidae, Hesperinidae и некоторых других.
В тех случаях, когда клипеус не обособлен, срединная пластинка
головы двукрылых рассматривается как единый фронто-клипе-
альный склерит (рис. 71). Примыкаюш,ие к лобной пластинке ча¬
сти капсулы, называемые боковыми пластинками, массивные, на
вентральной стороне широко расставлены или слиты. Затылочное
отверстие крупное. Тенториум (внутренний скелет головы) обыч-
205
Рис. 67. Общий вид личинок двукрылых сверху
1 — Lonchoptera sp. (Lonchopteridae); 2 — Galomyia amoena
Melg. (piatypezidae); 3 — Fannia sp. (Fannlldae)
НО хорошо развит, расположен в пределах головной капсулы. Ме¬
ста прикрепления передних и задних рук тенториума определяют¬
ся по положению тенториальных ямок. Передние находятся на
головной капсуле сразу за первым мандибулярным сочленением,
задние — около затылочного отверстия с вентральной стороны.
Хорошо развитая головная капсула характерна для личинок
большинства длинноусых двукрылых, за исключением предста¬
вителей некоторых семейств.
2. Группа Hemicephala. Степень и характер редукции головной
капсулы личинок двукрылых очень разнообразны. Наблюдаются
все переходы от частично редуцированной головной капсулы
(преимущественно в заднем отделе) до почти полностью редуциро¬
ванной, с сохранением лишь небольшой дорсальной крышки. Го¬
лова гемицефалического типа характерна для личинок ряда се¬
мейств длинноусых двукрылых и почти всех короткоусых прямо-
шовных двукрылых. Так, например, относительно хорошо разви¬
тая, массивная головная капсула с несклеротизованным задним
отделом, частично втягивающаяся в грудные сегменты, характерна
для личинок комаров-долгоножек (Tipulidae), многих болотниц
(Limoniidae), аксимиид (Axymyiidae) и др. (рис. 72). У личинок
206
Рис. 68. Общий вид личинок двукрылых сбоку
1 — Hyperoscelis eximia Bohem. (Hyperoscelididae); 2 — Paraclusia tigrina Fll. (Clusi
dae); 3 — Temnostoma vespiforme L. (Syrphidae)
некоторых семейств, например галлиц (Cecidomyiidae), она сильно
уменьшена в размерах. Сильно редуцированная головная кап¬
сула (рис. 73, 1, 2, 4—6) характерна для личинок отдельных родов^
болотниц (Limoniidae), бекасниц (Rhagionidae), ктырей (Asili-
dae), зеленушек (Dolichopodidae) и др.
Частичная редукция головной капсулы приводит к возникно¬
вению у большинства групп дополнительных опорных образова¬
ний — стержней, отходящих от задних отделов головной капсулы.
Возникновение стержней у разных групп личинок происходило
различными путями, хотя функционально они эквивалентны и во
всех случаях служат опорой для мускулатуры головного отдела.
Так, у личинок галлиц (Cecidomyiidae) стержни (рис. 73, 3) воз¬
никли в результате усиления тенториального скелета и одновре¬
менной редукции задних отделов головной капсулы (экзоцефаль-
ные стержни; Мамаев, Кривошеина, 1965). У многих личинок ко¬
роткоусых прямошовных двукрылых (Asilidae, Xylophagidae,
Dolichopodidae, Empididae) стержни (рис. 73, 5, 6) представляют
собой видоизмененный задний отдел головной капсулы (метаце-
фальные стержни).
3. Группа Acephala, Головная капсула полностью редуциро¬
вана. Головной сегмент личинок этой группы носит название лож¬
ноголовы (псевдоцефалон, рис. 73, 7), она не отличается по строе¬
нию от покровов остальных сегментов тела, но несет усики и раз¬
личные сенсорные папиллы. Полная редукция головной капсулы
207
Рис. 69. Общий вид личинок двукрылых сбоку
I — Aulaoigaster leuoopeia Melg. (AuUclgastridae); й — Megaeella ер. (Phorldae)} « — Chy
liza sp« (Psilidae)
Рис. 70. Общий вид личинок двукрылых сбоку
1 — Phyllomyza sp. (Mllichlidae); s—Strongylophthalmyia ustulata Zett, (Strongylophthal-
mylidae); з — Phaonia sp. (Muscidae)
наблюдается у личинок отдельных семейств длинноусых двукры¬
лых (Hyperoscelididae, Leptoconopidae) и почти всех круглошов¬
ных двукрылых (за исключениехм Lonchopteridae). У последних
редукция головной капсулы ведет к образованию внутреннего
Рис. 71. Голова личинки двукрылых Pergratospes holoptica Kriv. et Мат.
(Cramptonomyiidae) сверху (7) и снизу (2)
ani — усики; frcl — фронтоклипеальная пластинка; Ibr — верхняя губа; md — мандибу¬
ла; тх — максилла; ос — глазки; t — тенториальные стержни
209
Рис. 72. Голова личинки двукрылых Libnotes undulata \fats. (Limoniidae)
сверху (1) и снизу (2)
irci — фронтоклипеальная пластинка; Лр. р1 — гипостомальная пластинка; №г — верх¬
няя губа; I. р1 — боковая пластинка головной капсулы
опорного скелета мышц (фарингеальных склеритов), входящего
в состав сложного ротоглоточного аппарата (рис. 74).
Ротоглоточный аппарат (рис. 74) у взрослых личинок состоит
из системы склеритов, соединенных различными группами связок*
Располагается ротоглоточный аппарат в головном отделе и двух
первых сегментах груди. Ротовые крючки (важнейшая часть
ротоглоточного аппарата) серповидно изогнуты, как правило, за¬
острены в дистальной части и расширены у основания. По вен¬
тральному краю крючки могут быть снабжены зубовидным или
угловидным выступом. Вентральнее крючков расположены два
мелких так называемых зубных склерита, имеющих неправильна
треугольную или трапециевидную форму. Развита пара обычно
маленьких лабиальных склеритов, достигающих сильного развития
у личинок некоторых высших двукрылых. Задние концы ротовых
крючков подвижно сочленяются с передними концами следующей
пары склеритов, носящих название гипостомальных. Задний от¬
дел гипостомальных склеритов прямой или угловидно скошенный.
Как правило, он подвижно сочленяется со следующей наиболее
массивной частью ротоглоточного аппарата — фарингеальными
склеритами. В ряде случаев (Sarcophagidae, Calliphoridae и др.)
задний отдел гипостомального склерита входит в развилок, обра¬
зованный тонким парастомальным склеритом и дистальным кон¬
цом фарингеального склерита. У личинок некоторых семейств^
210
Рис. 73. Голова и головные сегменты личинок двукрылых
1 — Gnophomyia tristis AI. (Limoniidae); 2 — Chrysopilus sp. (Rhagionidae); 3 — Arthro-
cnodax sp. <Cecidomyiidae); 4, 5 — Laphria flava L. (Asilidae) с вентральной и дорсаль¬
ной сторон; 6 — Xylophagus compeditus Wied. (Xylophagidae); 7 — головной сегмент ли¬
чинки круглошовных двукрылых (схема) (7 — по Зимину, 1948)
crd — кардиальный валик, «подушечка»; тс — м^тацефальные стержни
Рис. 74. Схема строения рото¬
глоточного аппарата кругло-
пювпых двукрылых (по Зими-
пу, 1948)
ак — добавочные (акцессорные) скле¬
риты;
d — дорсальный отросток фаринге¬
ального склерита;
d<—зубные склериты;
/ip — гипостомальный склерит;
/ — фарингеальный склерит;
7Tid—ротовые крючки;
рг — передние отростки фаринге¬
ального склерита;
ps — парастомальный склерит;
V — вентральный отросток фарин¬
геального склерита
иапример настоящих мух (Muscidae), парастомальный склерит
отсутствует (рис. 87, 7).
Фарингеальные склериты — парное образование, имеющее
небольшие передние отростки (к которым примыкает гипостомаль-
ный склерит) и более массивные дорсальные и вентральные зад¬
ние. Передние отростки фарингеального склерита снизу соединены
вентральной перемычкой, задний край которой служит местом
прикрепления фарингеальной мембраны. В некоторых случаях
фарингеальные склериты очень слабо склеротизованы, почти не¬
заметны, как, например, у личинок пятнокрылок (Clusiidae).
Выделенные три типа головы не охватывают всего многообра¬
зия форм головы, встречающегося среди личинок двукрылых. Так,
например, изолированное положение занимают личинки лжекты-
рей (Therevidae), у которых наряду с хорошо развитой головной
капсулой имеется мощный метацефальный стержень; головная
капсула личинок Leptoconopidae редуцирована, но развита це¬
лая система своеобразных внутренних опорных стержней и т. д.
212
Тем не мепее, рассмотренные три типа головной капсулы явля¬
ются наиболее характерными для личинок двукрылых.
Ротовой аппарат личинок двукрылых очень разнообразен.
Наблюдаются все переходы от хорошо развитого ротового аппа¬
рата грызущего типа до сильно измененного колюще-сосущего и,
наконец, до полностью редуцированного (у некоторых групп с внеки-
течным пищеварением). Элементы ротового аппарата представлены
верхней губой, мандибулами, максиллами и нижней губой.
Верхняя губа у личинок длинноусых двукрылых в виде не¬
большой широкой хорошо выраженной пластинки (рис. 75, 1).
Она неподвижно соединена с фронто-клипеусом, но ясно отделена
от него швом (рис. 71, 1\ 72, 1), Боковые стороны верхней губы
могут нести пигментированные склериты, называемые тормой.
1^ептральная сторона губы с развитой в разной степени мягкой сре¬
динной лопастью — энифаринксом (рис. 75, 1). На вентраль¬
ной стороне у личинок некоторых видов имеются массивные
зазубренные склериты-нремандибулы (рис. 75, 1). У личинок
короткоусых прямошовных двукрылых верхняя губа имеет вид
клювообразного выроста передней части головной капсулы
(рис. 75, 2),
Мандибулы личинок двукрылых очень разнообразны. В преде¬
лах отряда наблюдаются все переходы от одночлениковых массив¬
ных мандибул грызущего типа (гомологичных жвалам личинок
жуков) до двучлениковых удлиненных мандибул и одночлениковых
ротовых крючков (рис. 75, 3—6), Мандибулы грызущего тина со¬
стоят из одного крупного склерита и несут на вершине один или
несколько зубцов (рис. 75, 5). В отдельных случаях, например
у личинок ряда мицетофилоидных двукрылых, мандибулы свое¬
образной серповидной формы с рядом зубцов по краю. У личи¬
нок мокрецов (Ceratopogonidae) и комаров-звонцов (ChirojioDidae)
одночлениковые мандибулы сильно удлинены, а у личинок гниль-
ниц (Scatopsidae) могут иметь обособленный, но не отчлененный
вершинный отдел (рис. 75, 4). Для ряда групп длинноусых дву¬
крылых (Psychodidae, Anisopodidae, Trichoceridae, некоторые
Limoniidae и т. д.) и всех короткоусых прямошовных двукрылых
(Xylophagidae, Rhagionidae, Tabanidae и др.) характерны дву-
члениковые мандибулы (рис. 75, 5, 6), представленные базальным
и дистальным склеритами (Hennig, 1948Ь, 1952; Schremmer, 1951;
Кривошеина, 1964, 1969).
Максиллы (рис. 76, 1, 3, 5) состоят из небольшого склерита —
кардо, стипеса с одночлениковым щупиком и лацинии (вместе
с галеа). Для личинок длинноусых двукрылых характерны разно¬
образные формы максилл. У личикок комаров-долгоножек
(Tipulidae) и близких групп семейств, у полосатых комариков (Di-
tomyiidae), аксимиид (Axymyiidae) и др. максиллы в виде неболь-
ншх лопастевидных перепончатых пластинок с сильно редуциро¬
ванным кардо (рис. 76, 5, 5). Для небольшой группы семейств
бибионоидного комплекса характерны максиллы с хорошо обособ-
213
Рис. 75. Элементы ротового аппарата (верхняя губа, мандибулы, максиллы)
личинок двукрылых
1 — верхняя губа и клипеус Scatopse fuscipes Meig. снизу; 2,3 — верхняя губа и манди¬
бул ярно-максиллярный комплекс Rhagio scolopaceus L. (Rhagionidae); 4—6 — манди¬
була Scatopse fuscipes Meig. (Scatopsidae), Pachyneura fasciata Meig. (Pachyneuridae),
Mycetobia sp. (Mycetobiidae)
b. sc — базальный склерит мандибул; crd — кардиальный валик, «подушечка»; gal — га-
леа; md — дистальный склерит мандибул; р. тх — щупики максилл; pmd — преманди-
-була
ленными всеми 3 элементами: стипес, лациния, кардо (рис. 76, 1),
Все элементы максилл склеротизованы, но степень склеротизации
различна. Так, у личинок Pachyneura (Pachyneuridae) лациния
с перепончатым передним краем, а у личинок Hesperinus (Hespe-
rinidae), наоборот, сильно склеротизована, с зубчатым гребнем по
переднему краю. У личинок мицетофилоидного комплекса (Мусе-
tophilidae, Sciaridae и др.) лациния, как и мандибула, преобра¬
зована в серповидную склеротизованную пластинку. У личинок
короткоусых прямошовных двукрылых максилла в виде единого
лопастевидного образования с хорошо развитым максиллярным
214
Рис. 76. Максиллы п нижняя губа личинок двукрылых
1,2 — максилла и нижняя губа Plecia nigra Lund. (Pleciidae); 3,5 — максилла и нижняя
губа Symmerus annulatus Meig. (Ditomyiidae) и Protaxymyia melanoptera Mam. et Kriv.
(Axymyiidae); 4 — нижняя губа Mycetobia sp. (Mycetobiidae)
crd — нардо; hyp — гипофариннс; lac — лациния; Ib — нижняя губа; р. mx — щупик
мансилл; si — стипес
щупиком (рис. 75, 3), У личинок некоторых семейств (Rhagioni-
dae, Tabanidae) имеется специфическое образование — «подушеч¬
ка», расположенное под усиками около основания верхней губы
и усаженное твердыми слабоизогнутыми шипами (рис. 73, 2), Па
21S
мнению некоторых авторов (Bischoff, 1925), это — видоизме-
ненЕшй кардо.
Строение частей нижней губы у личинок двукрылых сильно
варьирует. Нет еще единой точки зрения, что же представляют со¬
бой склеротизованные образования вентральной стороны головы —
нижнюю губу или лишь функционально заменяющие ее склериты.
При рассмотрении различных структур на вентральной стороне
головы за нижнюю губу мы принимаем самостоятельные, обособ¬
ленные от головной капсулы склериты. В простейшем случае ниж¬
няя губа представлена прементумом, ментумом и субментумом
(Ptychopteridae). Однако в большинстве случаев субментум или
прементум редуцирован и нижняя губа настолько видоизменена по
сравнению с тем типом, который характерен для других насеко¬
мых (например, личинок жуков), что гомологизация отдельных
ее частей сильно затрудняется. Наиболее развита нижняя губа
у личинок длинноусых двукрылых. У личинок многих предста¬
вителей бибионоидного комплекса нижняя губа представлена чет¬
ко очерченной и сильносклеротизованной пластинкой (рис. 76, 2).
У личинок большой группы семейств, таких, как Anisopodidae,
Axymyiidae, Ditomyiidae и др., нижняя губа в виде небольшой
лопастевидпой пластинки (рис. 76, 3—5). У таких групп, как Ti-
pulidae, Limoniidae, обособленная пластинка нижней губы силь¬
но редуцирована и смещена внутрь капсулы. На ее месте располо¬
жена сильносклеротизованная зазубренная пластинка, слитая
с капсулой. Эта пластинка, носящая название гипостомальной,
функционально заменяет нижнюю губу (рис. 72, 2), У личинок
короткоусых прямошовных двукрылых нижняя губа, как прави¬
ло, редуцирована, но при этом в ряде семейств (Xylophagidae,
Asilidae, Therevidae, Tabanidae) на вентральной стороне головы
имеется специфическое образование в виде обособленной пластин¬
ки, которую мы называем вентральной пластинкой головы
(рис. 73, 4).
Усики (рис. 78, 10—12), как правило, небольшие, одно-трех-
члениковые, расположены в переднем отделе головы. Они представ¬
лены или хорошо развитыми удлиненными члениками (Scatopsi-
dae, Anisopodidae, Psychodidae, Rhagionidae и др.), или группой
сенсорных папилл, расположенных на плоском основном членике
(Trichoceridae, многие Psychodidae и др.).
Небольшие простые глазки, развитые не у всех личинок, рас¬
положены обычно снаружи от усиков. У личинок отдельных
семейств, например мицетофилоидных двукрылых, глазки, как
правило, хорошо развиты, крупные.
Грудь представлена 3 сегментами, которые у большинства
видов по строению не отличаются от брюшных. Только в пределах
отдельных семейств, например у личинок настоящих комаров
(Culicidae), грудные сегменты значительно расширены по срав¬
нению с брюшными и, сливаясь, образуют единый комплекс.
Брюшко у большинства личинок длинноусых двукрылых
216
(Bibionidae, Mycetophilidae и близкие семейства, Chironornidae,
Ceratopogonidae и т. д.) образовано 9 хорошо развитыми сегмен¬
тами. У некоторых личинок, например гнильииц (Scatopsidae),
многих галлиц (Cecidomyiidae), 9-й брюшной сегмент небольшой
и сдвинут па вентральную сторону тела. Очень большая группа
личинок, например Trichoceridae, Tipulidae, Limoniidae, Cylin-
drotomidae и все личинки короткоусых двукрылых, имеют лишь
8 брюшных сегментов. В некоторых случаях число видимых сег¬
ментов тела сокращается до 7 (Axymyiidae, рис. 63, 2).
В ряде случаев число видимых сегментов тела более 12 (рис. 61,
1—3\ 62, i, 2, 4), что объясняется вторичной сегментацией боль¬
шинства (Therevidae, Anisopodidae, Mycetobiidae) или отдельных
сегментов тела (Pleciidae и др.)*
Сегменты тела, как правило, четко отграничены друг от друга,
но иногда границы между сегментами неясны, и трудность опреде¬
ления границ сегментов усугубляется вследствие наличия много¬
численных складок на теле (например, у личинок Axymyiidae,
Tipulidae). У личинок некоторых групп двукрылых, например ко-
маров-толстоножок (Bibionidae) и бабочниц (Psychodidae), грани¬
цы между сегментами четко выражены лишь с дорсальной стороны,
а иногда, например у личинок журчалок (Syrphidae), определяют¬
ся лишь по положению щетинок на теле.
Личинки большинства групп двукрылых обладают слабосклс-
ротизованными покровами, что в значительной степени объяс¬
няется скрытым образом жизни. Покровы тела могут быть упру¬
гими, эластичными, например, у личинок лжектырей (Therevidae),
бекасниц (Rhagionidae), ктырей (Asilidae), зеленушек (Dolichopo-
didae) и др. Нежные и тонкие покровы наблюдаются у многих обита¬
телей сильно увлажненных субстратов (например, влажной дре¬
весины): личинок галлиц — Kronomyia (Cecidomyiidae), комаров-
долгоножек (Tipulidae), болотниц (Limoniidae), мохнатокрылок —
Trichomyia (Trichomyiidae). У личинок некоторых семейств наблю¬
дается склеротизация отдельных участков покровов. Например,
мощные коричневатые пластинки имеются на дорсальной и отчасти
вентральной сторонах грудных сегментов личинок стволоедок —
Xylophagidae (рис. 64, 1). Хорошо развитые склеротизованные пла¬
стинки покрывают дорсальную поверхность тела большинства личи¬
нок бабочниц Psychoda (Psychodidae, рис.66, 2). У многих личинок
склеротизация покровов наблюдается лишь на заднем конце тела
в области стигмального поля. Так, у личинок ктырей Laphria,
Choerades на конце тела имеется склеротизованная пластинка.
Частичная склеротизация стигмального поля характерна для
личинок отдельных видов комаров-долгоножек (Tipulidae), болот¬
ниц (Limoniidae; рис. 77, 1, 2). В некоторых случаях поверхность
тела значительно уплотняется вследствие отложения извести в ку¬
тикуле и на ее поверхности (личинки бабочниц и львинок — Psycho¬
didae и Stratiomyidae). Иногда наслоения извести бывают на¬
столько значительными, что оседают на поверхности тела и
217
^f — 5
Рис. 77. Конец тела личинок двукрылых
1 — Dictenidia bimaculata L. (Tipulidae); 2 — Gnophomyia tristis Ai. (Limoniidae); з —
Sarcophaga liaemorrhoidalis Meig. (Sarcophagidae); 4 — Chrysopilus sp. (Rhagionidae);
5 — Solva marginata Meig. (Xylomyidae); 6 — Thereva sp. (Therevidae)
щетинках в виде хлопьев, придавая телу причудливую форму, как,
например, у некоторых Pericoma (Psychodidae).
Поверхность тела личинок гладкая или несет различное число
самых разнообразных выростов и кутикулярных образований,
которые по своей форме и размерам могут быть разделены на не¬
сколько категорий.
У многих личинок тело несет различные крупные лопасти и вы¬
росты. Удлиненные разветвленные листовидные выросты придают
личинкам Liogma (Cylindrotomidae) причудливый вид, что в со¬
вокупности с общей зеленоватой окраской тела маскирует личинок
среди окружающего мха. Различные лопастевидные выросты раз¬
виты на боковых сторонах сегментов многих дорсовентрально
уплощенных личинок, как бы еще более расширяя их тело. В этом
отношении характерны, например, Lonchoptera (Lonchopteridae),
Calomyia amoena Meig. (Platypezidae), Fannia (Fanniidae) и др.
(рис. 67). Очень разнообразные выросты расположены на послед¬
нем сегменте тела. Парные выросты окружают стигмальное поле
личинок Tipula, Phoroctenia и других комаров-долгоножек (Ti-
218
pulidae), личинок бекасниц (Rhagionidae), зеленушек (Dolichopo-
didae) и др. (рис. 77). Выросты различной длины характерны для
последнего сегмента тела личинок болотниц (Limoniidae). Разно¬
образные по форме и размерам выросты расположены на последнем
сегменте тела личинок галлиц (Cecidomyiidae). Два цилиндри¬
ческих выроста, направленные вентральпо и выполняющие роль
подталкивателей, имеются у лр:чинок лжектырей — There va (The-
revidae; рис. 77, 6). Мягкие выросты различной формы и раз¬
меров (рис. 65, 3) имеются у многих личинок журчалок из родов
Brachyopa, Hammerschmidtia (Syrphidae).
Тело большинства личинок с определенным количеством сим¬
метрично расположенных щетинок. По своему положению щетин¬
ки делятся на дорсальные, латеральные и вентральные. Дорсаль¬
ные образуют, как правило, один поперечный ряд в средней части
сегмента. Латеральные представлены в большинстве случаев 1—3
щетинками с каждой стороны. Расположение вентральных щетинок
на грудных и брюшных сегментах различно. На грудных они обра¬
зуют по 2 симметричные группы, каждая из которых образована
2—3 щетинками. Эти группы связаны с имагинальными дисками
конечностей и развиты у большинства личинок длинноусых дву¬
крылых. На вентральной стороне брюшных сегментов щетинки
расположены обычно в 2 ряда. Как правило, хорошо развиты раз¬
розненные или собранные в пучки терминальные щетинки послед¬
него сегмента.
Длина щетинок обычно в 2—3 раза уступает длине сегмента
тела, но в отдельных случаях они могут достигать больших раз¬
меров. Так, у личинок некоторых видов мокрецов рода Forcipo-
myia (Ceratopogonidae) дорсальные щетинки по размерам дости¬
гают длины тела. Длинные щетинки (рис. 65, 2) характерны для
личинок галлиц рода Parepidosis (Cecidomyiidae). В ряде случаев
щетинки, особенно терминальн1>1е, преобразуются в различные по
форме папиллы, выросты (рис. 78, 2—5, 7, 13), что характерно для
личинок галлиц (Cecidomyiidae), некоторых родов мокрецов (Ce¬
ratopogonidae), некоторых журчалок (Syrphidae) и др.
Поверхность сегментов часто несет различные мелкие кутику-
лярные образования. Поверхность тела многих личинок густо
покрыта мелкими шипиками. Различные, часто 2—3-зубчатые
структуры имеются на теле личинок Psychoda (Psychodidae),
Bibio (Bibioiiidae), Scatopse (Scalopsidae) и др. (рис. 78, 6', (S).
Звездообразные кутикулярные структуры покрывают тело многих
личинок Fannia (Fanniidae; рис. 78, 9). Своеобразные по форме об¬
разования — кистевидные, ланцетовидные, чешуевидные покры¬
вают тело личинок многих галлиц (Cecidomyiidae) и мокрецов —
Forcipomyia (Ceratopogonidae).
Покровы тела личинок, как правило, не oKpameiuj. Цвет тела
(желтоватый или беловатый) чаще всего зависит от окраски жи¬
рового тела и поглощенной пищи, иногда он определяется окра¬
ской гемолимфы. У многих светлоокрашенных личинок просве-
219
Дорсальные папиллы: 1 — Winnertzia rubricola Мат., 2 — Holoneurus fulvus Kieff., 3 —
Peromyia perpusilla Kieff., 4 — Dicroneurus montanus Mam. (Cesidomyiidae), 5 — Forcipo-
myia kaltenbachi Winn. (Ceratopogonidae); кутикулярные структуры: 6 — Psychoda se¬
verint Tonn. (Psychodidae), 7 — L3jota ruficornis Zett. (Syrphidae), « — Scatopse sp.
(Scatopsidae), 9 — Fannia sp. (Fannlidae); усики; lo — Trichocera annulata Meig. (Tricho-
ceridae), JJ — Scatopse fuscipes Meig. (Scatopsidae), — Forcipomyia paradoxa Kriv.
(Geratopogonidae); 13 — терминальные папиллы Tricholaba trifolii Rubs. (Cecidomyiidae)
(6 — no Satchell, 1947)
чивает темный кишечник, например, у личинок детритниц (Sci-
aridae), грибных комариков (Mycetophilidae) и др. Характерную
расцветку имеют белые личинки туманниц (Hyperoscelididae)
с просвечивающим черно-фиолетовым кишечником. Пожалуй, на¬
ибольшего разнообразия достигает окраска тела личинок Chiro-
nomidae — от желтоватой до голубоватой, фиолетовой и красной.
Живущие на поверхности мха личинки Cylindrotomidae, как пра¬
вило, зеленоватые. У поверхностно живущей личинки мокреца
Forcipomyia monilis Goetgh. (Geratopogonidae) тело изумрудно¬
зеленого оттенка.
Окраска тела часто зависит от характера и густоты покрова из
различных кутикулярных образований. Сероватый оттенок телу
почвообитающих личинок комаров-долгоножек Tipula придают
густые прилегающие волоски, в то же время личинки-ксилофаги
этого семейства (Ctenophora, Tanyptera) беловатые, полупроз¬
рачные; покровы их тела лишены таких волосков. Густые волосо¬
видные кутикулярные образования придают золотистый оттенок
220
телу многих личинок болотниц (Limoniidae), напри1мер Elephan-
tomyia.
В целом личинки, живущие в глубине субстратов, например
в толще древесины и т. д., окрашены монотонно — беловатые или
желтоватые. Разнообразие окрасок наблюдается у личинок, оби¬
тающих под корой, особенно у личинок, развивающихся на по¬
верхности мха, коры и т. д.
У личинок двукрылых, лишенных ног, грудные конечности
иногда функционально заменяются различными двигательными
валиками, группами щетинок и ложными ножками. Наиболее
обычны ползательные валики, расположенрпле на вентральной сто¬
роне преимущественно брюшных сегментов. Валики расположены
на границах сегментов или в их передних отделах. Поверхность
валиков густо покрыта шипиками или бугорками, собранными в
поперечные ряды (рис. 69 и др.). Такие валики характернгл для
личинок большинства короткоусых и многих длинноусых двукры¬
лых. У личинок некоторых галлиц, например Holoneurus (Ceci-
domyiidae), на вентральной стороне сегментов имеются крупные
серповидно изогнутые, склеротизованньте крючки (рис. 79, 6*).
Они могут быть расположены как на грудных, так и на брюшных
сегментах тела. При их наличии поля шипиков на ползательных
валиках отсутствуют. Иногда, например у личинок ктырей (Laph-
riinae), слепней (Tabanidae), на сегментах имеются ползательные
бугорки, способные частично втягиваться внутрь тела (рис. ()4, 3).
В некоторых случаях, у личинок комаров-звонцов (Chironomidae),
многих мокрецов (Ceratopogonidae) и др., на 1-м грудном и послед¬
нем брюшном сегментах тела имеются хорошо выраженные лож¬
ные ножки, выполняющие роль подталкивателей (рис. 79, 2—4),
У личинок отдельных видов, обитающих в сильно увлаж[[енных
средах (натеках сока, пропитанной водой трухе дупел), ложные
ножки могут быть развиты на нескольких брюшных сегментах.
Они представлены или тупым бугорком с венцом темных крючков
на вершине — журчалки Myiatropa, Eristalis (Syrphidae), некото¬
рые виды из рода Phaonia (Muscidae) и др., или длинными конусо¬
видными бугорками с пучком волосков на вершине (по 2 на груд¬
ных и по 3 на брюшных сегментах) — галлицы рода Lestodiplosis
(Cecidomyiidae; рис. 79, 7).
Роль своеобразного подталкивателя выполняет специфическое
образование — «лопаточка», характерное для личинок галлиц
(Cecidomyiidae). Оно представляет собой склеротизованную, обыч¬
но удлиненную пластинку, расположенную на вентральной сто¬
роне 1-го грудного сегмента.
Для трахейной системы личинок двукрылых в целом характер¬
ны массивные дорсальные продольные стволы, соединенные дор¬
сальными анастомозами, и латеральные, соединенные с дорсальны¬
ми стволами поперечными комиссурами (VVhitteii, 1955). От ла¬
теральных ветвей в грудных и брюшных сегментах отходят ветви
покровам, несущие на конце дыхальца. Наиболее характерные
221
особенности строения трахейной системы различных групп дву¬
крылых рассмотрены в работе Н. П. Кривошеиной (1969).
У личинок двукрылых в зависимости от числа и характера рас¬
положения дыхалец на сегментах намечается несколько типов
трахейной системы (рис. 80).
1. Голопнейстический тип характеризуется наличием дыха¬
лец на 1-м и 3-м грудных и первых 8 брюшных сегментах тела. Та¬
кой тип трахейной системы присущ очень немногим группам дву¬
крылых (Hesperinidae, Pachyneuridae). В некоторых случаях у
более специализированных групп наблюдается перемещение пос¬
ледней пары дыхалец с 8-го на 9-й брюшной сегмент (Bibionidae,
Pleciidae).
2. Перипнейстический тип характеризуется наличием дыха¬
лец на 1-м грудном и первых 8 брюшных сегментах. Таким обра¬
зом расположены дыхальца у личинок многих длинноусых
двукрылых (Ditomyiidae, Cecidomyiidae, Scatopsidae, Нурего-
scelididae и др.). У личинок некоторых семейств происходит редук¬
ция последней пары дыхалец и сохраняются, помимо грудных,
лишь первые семь пар брюшных дыхалец (Mycetophilidae, Sciari-
dae).
3. Амфипнейстический тип характеризуется наличием лишь
двух пар дыхалец, расположенных на первом и последнем сег¬
ментах тела. Такое положение дыхалец характерно для личинок
длинноусых (Anisopodidae, Trichoceridae, Mycetobiidae) и боль¬
шей части короткоусых двукрылых.
4. Метапнейстический тип характеризуется наличием лишь
одной пары дыхалец, расположенных на конце тела. Трахейная
система такого типа характерна для большинства обитающих
в воде личинок (Tipulidae, Limoniidae, Culicidae и др.).
5. Апнейстический тип характеризуется полным отсутствием
дыхалец на теле. Он присущ из свободноживущих личинок оби¬
тателям влажных сред (Chironomidae, Ceratopogonidae, Масго-
ceridae, Ceroplatidae и др.).
Таким образом, наибольшее число дыхалец, характерное для
личинок двукрылых — 10 пар. Дыхальца, как правило, распо¬
ложены на боковой стороне сегментов. В отдельных случаях, как,
например, у личинок Agromyzidae, наблюдается перемещение
дыхалец 1-го грудного сегмента (передние дыхальца) на дорсаль¬
ную сторону сегмента. В наибольшей степени варьирует поло¬
жение последней нары дыхалец (задних дыхалец). Так, у личинок
с голопнейстическим типом трахейной системы (Bibionidae, Ple¬
ciidae и др.) задние дыхальца смещены на дорсальную сторону
сегментов. У многих личинок с амфи- или метапнейстическим ти¬
пом трахейной системы (Tipulidae, Anisopodidae, Mycetobiidae,
Hhagionidae, Tabanidae, Calliphoridae, Muscidae и др.) задние ды¬
хальца расположены на конце тела.
В простейшем случае дыхальца расположены непосредственно
на поверхности тела, что характерно для личинок большинства
222
Рис. 79. Анальные папиллы н локомоторные выступы — «ложные ножки»
личинок двукрылых
1,2 — конец тела Georthocladius luteicornis Goetgh. и Metriocnemus sp. (Chironomidae);
3, 4 — передняя ложная ножка Forcipomyia sp. (Ceratopogonidae) и Eiiphaenocbdius aqua¬
tilis Goetgh. (Chironomidae); 5 — анальные папиллы Protaxymyia mclanoptera Mam, et
Kriv. (Axymyiidae); 6 — серповидные вентральные крючки Holoneurus fulvus Kieff.
(Cecidomyiidae); 7 — локомоторные выступы брюшных сегментов Lestodiplosis polypori
Loew (Cecidomyiidae) (J, 4 — no Hennig, 1950)
an. p — анальные папиллы
Рис. 80. Тины Tpaxoiinoii системы личинок двукрылых
1 — голопнейстический; 2 — перипнейстический; З — амфипнейстический; 4 — ме-
тапнейстический; 5 — пропнейстический; 6 — апнейстический
S — дыхальце
длинноусых и короткоусых прямошовных двукрылых, в отдель¬
ных случаях, как, например, у личинок гнильниц (Scatopsidae),
ксиломиид (Xylomyidae), все дыхальца расположены на неболь¬
ших склеротизованных бугорках. Наиболее специфично голо-
жение задних дыхалец. Так, у личинок болотниц (Limoniidae)^
комаров-долгоножек (Tipulidae), бабочниц (Psychodidae) и др.
задние дыхальца расположены на особом стигмальном поле, ок¬
руженном широкими лопастями или пучками волосков (рис. 77,
7, 2). У личинок целого ряда семейств, например, львинок (Stra-
tiomyidae), ксиломиид (Xylomyidae), слепней (Tabanidae), сар-
кофагид (Sarcophagidae) и др., задние дыхальца размещены на дне
глубокой полости (рис. 77, 3).
У личинок сравнительно большой группы двукрылых — оби¬
тателей преимуш,ественно сильно увлажненных субстратов —
задние дыхальца расположены на конце парных выростов или уд¬
линенной дыхальцевой трубки, образованных предпоследним или
последним сегментами тела. Так, парные выросты, несущие ды¬
хальца, расположены на предпоследнем сегменте тела личинок
гнильниц (Scatopsidae; рис. 65, 1), некоторых галлиц (Cecidomy-
iidae) и др. Непарная трубка, являющаяся продолжением послед¬
него сегмента тела, характерна для личинок многих семейств.
Иногда трубка короткая (личинки многих бабочниц —- Psycho¬
didae, журчалок — Syrphidae и др.), а в некоторых случаях,
наоборот, длинная, эластичная (рис. 63, 2), значительно превы¬
шающая размеры тела личинки (Axymyiidae, многие Syrphidae).
Последние часто встречаются в толще сильно увлажненной древе¬
сины или различных субстратов на дне микроводоемов, а трубка
своим свободным концом, несущим дыхальца, выдвигается при
этом к поверхности субстрата.
Строение дыхалец у личинок двукрылых очень разнообразно,
но все они могут рассматриваться в пределах двух больших
групп — симметричных и асимметричных (Кривошеина, 1969).
Симметричные дыхальца (рис. 81, 3) представлены овальной
или округлой пластинкой, состоящей из нескольких отделов.
Центральную часть занимает стигмальная пластинка, несущая
обычно извилистый линочный шов. К периферии от него следует
участок, несущий многочисленные, в виде узких или овальных
щелей, дыхальцевые отверстия. Стигмальную пластинку ограни¬
чивает более или менее сильно склеротизованная полоска — пе-
ритрема. Такого типа дыхальца характерны для большинства ли¬
чинок с голо- или перипнейстическим типом трахейной системы
(Hesperinidae, Pachyneuridae, Ditomyiidae и др.). У личинок
Bibionidae задние дыхальца усложненные (рис. 81, 4).
Симметричные дыхальца характерны также для некоторых
групп с амфи- и метапнейстическим типом трахейной системы
(Axymyiidae, некоторые Psychodidae, Trichoceridae), хотя в
строении передних дыхалец, например Trichoceridae, наблюдает¬
ся некоторая асимметрия. Близки по строению к рассматривае-
224
Рис. 81. Строение дыхалец личинок двукрылых
Переднее дыхальце: 1 — Pseudowallacea hungarica Kert. (Stratiomyidae), 2 — Trichosia
sp. (Sciaridae); заднее дыхальце: 3 — Pachyneura fasciata Meig. (Pachyneuridae), 4 —
Bibio pomonae F. (Bibionidae), 5 — Fannia leucosticta Meig. (Fannlidne), 6 — Texara com¬
pressa Walk. (Megamerinidae), 7 — Temnostoma vespiforme L. (Syrphidae) {5 — no
Зимину, 1948)
1
мому типу дыхальца личинок комаров-долгоножек (Tipulidae),
подробно описанные некоторыми авторами (Brown, 1910; Keilin,
1944, и др.).
Асимметричные дыхальца имеют в большинстве случаев не¬
большое число дыхальцевых отверстий, смещенных к одному
краю пластинки, на другом конце которой расположен линочный
шов. Усложняется строение полости атриума. Такие дыхальца
характерны для личинок многих длинноусых двукрылых, напри¬
мер, детритниц Trichosia (Sciaridae), некоторых Phronia (Мусе-
lr)[)liilidae). Асимметричность в строении передних дыхалец
четко выражена у личинок с трахейной системой амфипнейсти-
ческого типа (рис. 81, 1), У личинок некоторых групп коротко¬
усых прямошовных двукрылых проявляется тенденция к пере¬
стройке также и задних дыхалец (зеленушки — Dolichopodidae
и лжектыри — Therevidae).
Дальнейшая специализация дыхалец наблюдается у личинок
круглошовных двукрылых. Наиболее простые передние дыхальца
личинок журчалок (Syrphidae) — овальные дыхальцевые отвер¬
стия и удлиненная полость атриума. Для личинок многих групп
характерны вееровидные, древовидные дыхальца (Odiniidae,
Drosophilidae, Lonchaeidae, Anthomyiidae и др.) (рис. 83, 5;
85, 6 и др.).
Задние дыхальца личинок круглошовных двукрылых, как пра¬
вило, лишь с 3 дыхальцевыми отверстиями. Отверстия могут быть
8 Б. м. Мамаев и др.
225
овальными, расположенными па общей пластигше, как, например,
у личинок Tephritidae, Otitidae, Lonchaeidae, Pallopteridae и мн.
др. (рис. 86, 2, 6), или изолированными, расположенными обо¬
собленно на выростах, как у личинок Fannia (Fanniidae; рис. 81,
5). У очень многих групп дыхальцевые отверстия в виде длинных
извилистых щелей, как, например, у личинок настоящих мух
(Muscidae), многих журчалок (Syrphidae). Во всех случаях каждое
дыхальцевое отверстие с многочисленными внутренними склеро-
тизованными перегородками, разделяющими его на большое число
мелких камер. Стигмальный диск, несущий линочный шов, сдви¬
нут к внутреннему краю дыхальца.
В некоторых случаях число дыхальцевых отверстий значи¬
тельно превосходит 3, как, например, у личинок Temnostoma
(Syrphidae). Иногда обычное расположение дыхальцевых отвер¬
стий нарушается и они покрывают почти всю поверхность ды-
хальцевой пластинки, как, например, у личинок Megamerinidae
(рис. 81, 6, 7).
У личинок двукрылых, обитающих в различных увлажненных
средах, особенно у апнейстических форм, развиты специфические
образования, которым многие авторы придают большое значение
в процессах дыхания. Это анальные напиллы и в некоторых слу¬
чаях особые вентральные выросты (рис. 79, i, 2, 5).
Наиболее хорошо развиты анальные папиллы у амфибиоти-
ческих и водных форм. Папиллы обычно окружают анальное от¬
верстие и представляют собой нежные тонкостенные образования,
в спокойном состоянии в большинстве случаев втянутые внутрь
тела. Они характерны для личинок многих длинноусых двукры¬
лых. Так, у личинок комаров-звонцов (Chironomidae) имеются
удлиненные парные, иногда четковидные, разделенные перетяж¬
ками анальные папиллы. У личинок мокрецов Forcipomyia (Сега-
topogonidae) они имеют вид широких, обычно закругленных на
конце лопастей. У личинок Tipulidae и Limoniidae — это меш¬
ковидные парные образования в виде бугорков или удлиненных
выростов. У всех видов этих семейств, живущих в слабо увлаж¬
ненной разлагающейся древесине, папиллы редуцированы до ок
руглых бугорков (Tipula flavolineata Meig., Tanyptera, Ctenopho-
га). Очень своеобразны папиллы у личинок Axymyiidae (рис. 79,
5). Несомненно, что такие образования, как анальные папиллы,
носят вторичный характер. Они сильно варьируют в зависимости
от условий обитания и возникают как в пределах определенных
систематических групп, так и у личинок филогенетически далеких
друг от друга групп, но обитающих в сходных условиях.
Таблица для определения семейств отряда
Diptera — Двукрылые
1 (72) Фарингеальные склериты отсутствуют. Голова, как пра¬
вило, имеется, но может быть втянута в грудные сегменты
и снаружи плохо заметна. Если голова сильно редуциро-
226
вапа или вообще отсутствует и заменена светлым головным
сегментом, то брюшные сегменты с хорошо развитыми
боковыми дыхальцами.
2 (55) Голова хорошо развита, массивная. Верхняя губа попереч¬
ная, в виде пластинки. Мандибулы и максиллы обособ¬
лены друг от друга. Метацефальные стержни отсутствуют.
Если головная капсула очень небольшая (Cecidomyiidae;
рис. 73, 5), иногда с тонкими стержнями, отходящими от ее
заднего края, или полностью редуцирована и головной
сегмент светлый (Hyperoscelididae), то имеется не менее
12 сегментов тела (3 грудных и 9 брюшных).
3 (12) Личинки голопнейстические (дыхальца расположены на
1-м, 3-м грудных сегментах и 8 брюшных сегментах тела).
Нижняя губа в виде прямоугольной удлиненной пла¬
стинки, соединенной с широкой перепончатой лопастью
(рис. 76, 2). Максиллы сильно склеротизованы, пред¬
ставлены широкой прямоугольной лацинией, удлинен¬
ным стипесом и, как правило, склеротизованным
стержневидным кардо (рис. 76, 1).
4 (7) Дыхальца, помимо 2 грудных сегментов, расположены
на первых 7 и 9-м брюшных сегментах. Сегменты тела,
по крайней мере последние, с мягкими, хорошо разви¬
тыми конусовидными выростами (рис. 61, 7, 3).
5 (6) 1-й грудной сегмент без склеротизованной пластинки
(рис. 61, 3). Задние дыхальца с 2—3 линочными шва¬
ми, окруженными дыхальцевыми отверстиями. Клипеус
слит с лобной пластинкой, расположен в одной с ней
плоскости. В сильно разложившейся рыхлой древесине
стволов и пней, в подстилке, некоторые виды в почве
в ризосфере корней травянистых растений. Ксило-
фильные личинки — сапроксилофаги, обитатели ризо¬
сферы — факультативные фитофаги
Bibionidae — Толстоножки
6 (5) На 1-м грудном сегменте с дорсальной стороны имеется
склеротизованная пластинка, ограниченная выростами
(рис. 61, 1), Задние дыхальца с 1 линочпым швом, окру¬
женным дыхальцевыми отверстиями. Клипеус четко от¬
делен от лобной пластинки швом, оба расположены под
углом друг к другу. В сильно разложившейся рыхлой
древесине. Сапроксилофаги
Pleciidae — Чернокрылки
7 (4) Дыхальца, помимо 2 грудных сегментов, расположены
на первых 8 брюшных сегментах, или последняя пара на
границе 8-го и 9-го сегментов. Сегменты тела гладкие,
без мягких конусовидных выростов, иногда с длинными
щетинками (рис. 61, 2).
8 (9) 1-й грудной сегмент с дорсальной стороны со склероти¬
зованной пластинкой. Задние дыхальца расположены на
£♦ 227
границе 8-го и 9-го брюшных сегментов. Дыхальца
3-го грудного сегмента очень маленькие, слабо заметны.
Развиваются в заболони лиственных деревьев. Ксило¬
фаги . . . Cramptonomyiidae — Комарики-древесницы
9 (8) 1-й грудной сегмент без склеротизованной пластинки.
Задние дыхальца расположены па 8-м брюшном сег¬
менте. Дыхальца 3-го грудного сегмента относительно
хорошо заметны.
10 (11) Тело с короткими микроскопическими щетинками. Кли-
пеус четко отделен от лобной пластинки швом. Кардо
в виде удлиненного склерита. Лациния по переднему
краю с рядом склеротизованных зубцов. Анальное поле
не ограничено темной ли11ией. В разлагающейся, но
довольно плотной светлой и черноватой древесине пней,
ветвей и стволов. 1^силофаги
Hesperinidae — Медлянки
11 (10) Тело с ДЛИНН1ЛМИ щетинками, иногда достигающими тре¬
ти длины сегмента. Клипеус сливается с лобной пластин¬
кой, их граница нечеткая. Кардо в значительной сте¬
пени редуцирован, короткий. Лациния по переднему
краю не склеротнзована и без зубцов. Анальное поле
четко ограничено темной линией. В светлой и темной
древесине разлагающихся стволов и пней. Ксилофаги
Pachyneuridae — Стройнокрыл ки
12 (3) Личинки периннейстические — дглхальца расположе¬
ны на 1-м грудном и первых 8 брюшных сегментах,
или дыхалец еще меньше. Нижняя губа в виде очень
небольшой округлой или овальной пластинки. Максил-
лы или склеротизованы, тогда серповидные, зазубрен¬
ные по краю, или в виде широкой, поперечной слабо-
склеротизованной пластинки.
13 (20) Хорошо развиты 8 пар брюшных дыхалец. Максиллы
слабо склеротизованы, в виде мягкой лопасти (рис. 7G, 3).
14 (15) Тело на концах тупое (рис. 63, 3). Голова массивная,
широкая, ее ширина почти не уступает ширине тела,
мандибулы одночлениковые, сильно склеротизованы,
максиллы представлены 2 удлиненными лопастями, из
которых наружная, несущая щупик, значительно
длиннее внутренней (рис. 76, 5). В размягченной пре¬
имущественно светлой древесине или в грибах. Ксило-
и мицетофаги ... . Ditomyiidae — Комарики полосатые
15 (14) Тело кпереди несколько сужено (рис. 62, 5; 68, 1). Го¬
лова или небольшая, ее ширина в несколько раз мень¬
ше ширины тела, или сильно редуцирована. Мандибулы
если имеются, то или массивные удлиненные с обособ¬
ленной вершинной частью, или игловидные одночлени¬
ковые.
16 (17) Между головой и 1-м грудным сегментом имеется допол-
228
пительный шейный сегмент (рис. 62, 3\ ()5, 2), На вен¬
тральной стороне 1-го грудного сегмента, как правило,
хорошо развита склеротизованная «лопаточка». Голова
небольшая, ротовые части колюще-сосуш,его типа, ман¬
дибулы игловидные. Под корой и в толш,е разлагаюш,ейся
древесины, в почве, подстилке, растениях. Мицетофаги
и фитофаги, немногие ксилофильные формы (Lestodip-
losis Kieff.)—хиш,ники . . . Cecidomyiidae — Галлицы
17 (КЗ) Дополнительный шейн1>ш сегмент, а также лопаточка
на вентральной стороне 1-го грудного сегмента не вы¬
ражены.
18 (19) Головная капсула имеется, темная, склеротизованная
(рис. 65, 1). Мандибулы массивные, удлиненные, с обо¬
собленной верши1нюй частью, несущей зубцы (рис. 75,
4). Тело слегка уплош,е1ю, сзади с бугорковидными
выступами, без крючков, покровы скульптурированные.
Под корой, в трухе дупел, в различн]|1х разлагающихся
остатках растительного происхождения. Сапрофаги.
Scalopsidue — Гнильницы
19 (18) Головная капсула и ])отовой аппарат полностью реду¬
цированы (рис. 68, 7), имеется лишь небольшой нескле-
ротизованный головной сегмент, несущий короткие уси¬
ки. Задний конец тела с двумя темными склеротизо-
ваннымн крючками, покровы тела гладкие. В сильно
разложившейся светлой или бурой древесине хвойных
и лиственных пород. Сапрохмицетофаги
Hyperoscelididae — Тумапницы
20 (13) Имеется не более 7 пар брюшных дыхалец.
21 (34) Максиллы с серповидной зазубренной по краю лаци-
нной. Пи>1чняя губа редуцирована до очень небольшой
поперечной пластинки. Покровы тела гладкие, нежные.
22 (23) Усики хорошо развиты, трехчлениковые. Голова корот¬
кая и широкая, ее ширина в 1,5—2 раза превосходит
длину. В плодовых телах грибов. Мицетофаги ....
Bolilophilidae — Грибнянки
23 (22) Усики рудиментарные, представлены широкой оваль¬
ной перепончатой пластинкой, несущей небольшие
точечные сенсиллы. Голова удлиненная, ее длина, по
крайней мере, равна ширине.
24 (31) Дыхальца развиты на 1-м грудном и первых семи брюш¬
ных сегментах или же только на 1-м грудном сегменте.
25 (26) Боковые пластинки головы соприкасаются на вентраль¬
ной стороне в 2 точках. Их передние отделы сближены
на очень незначительном протяжении. В разлагаю¬
щейся древесине, под корой, в подстилке, богатой гу¬
мусом почве. Сапрофаги, обитатели древесины — сап-
роксилофаги Sciaridae — Детритпицы
26 (25) Боковые пластинки головы на вентральной стороне
229.
широко расставлены или сближены, соприкасаясь в од¬
ной точке или на значительном протяжении. Если бо¬
ковые пластинки соприкасаются в двух точках, то,
кроме того, их передние отделы сближены на значитель¬
ном протяжении.
27 (28) Боковые пластинки головы соприкасаются на вент¬
ральной стороне в одной точке. Личинки, как правило,
с ползательными валиками на вентральной стороне тела.
В плодовых телах грибов или на поверхности остатков
разлагающейся древесины под щитком. Мицетофаги
Mycelophilidae — Грибные комарики
28 (27) Боковые пластинки головы или широко расставлены,
или соприкасаются на значительном протяжении в пе¬
реднем отделе головы. Ползательные валики не разви¬
ты. На поверхности субстрата (древесины, коры, дре¬
весных грибов, подстилки), свободно или в трубочках.
29 (30) Личинки перипнейстические, помимо грудных, имеется
еще 7 пар брюшных сегментов. Боковые пластинки го¬
ловы на вентральной стороне или широко расставлены,
или сближены па значительном протяжении. Максиллы
часто с длинным сенсорным придатком. Под корой или
в древесине, пронизанной мицелием. Взрослые личинки
пекрофаги Sciophilidae — Тенекрылки
30 (29) Личинки пропнейстические. Боковые пластинки головы
широко расставлены. Максиллы с короткими сенсорными
придатками. Личинки в темной древесине гниющих
опавших веток, где обитают в шелковистых трубочках.
Пищевые связи точно не установлены
Diadocidiidae — Тонконожки
31 (24) Дыхальца отсутствуют.
32 (33) Задний конец тела с 2 парами выростов, из которых
2 направлены в стороны, а 2 — назад. Личинки слиз¬
невидные, живут на поверхности плодовых тел грибов-
трутовиков и в древесине па плесени. Пекрофаги, фа¬
культативные хищники . . Ceroplatidae — Плоскоуски
33 (32) Задний конец тела с 2 парами направленных назад вы¬
ростов. Под корой и в толще рыхлой древесины. Неко¬
торые виды — некрофаги и факультативные хищники
Macroceridae
34 (21) Максиллы лопастевидные, на вершине перепончатые.
Нижняя губа, как правило, хорошо развита, лопасте¬
видная, склеротизована в различной степени. Покровы
тела личинок прочные, эластичные, часто с различными
кутикулярными образованиями.
35 (46) Трахейная система амфипнейстического тнпа (дыхаль¬
ца расположены на первом и последнем сегментах тела).
36 (43) Тело вторично сегментировано или с многочисленными
поперечными складками. Верхняя губа на вентральной
230
стороне с хорошо развитыми премандибулами (как на
рис. 75, 1). Мандибулы (рис. 75, 6) удлиненные, дву-
члениковые. Конец тела с 4—5 небольшими лопастями,
окружаюш,ими дыхальца. Последние расположены не¬
посредственно на поверхности тела или на конце корот¬
кой дыхательной трубки, длина которой в несколько раз
меньше длины тела.
37 (40) Тело личинок удлиненное, змеевидное (рис. 62, 7, 2),
На последних сегментах имеется характерное анальное
поле, окружаюш,ее анальное отверстие. Брюшные сег¬
менты состоят из короткого переднего кольца и следую¬
щего более крупного, длина которого равна или несколь¬
ко превышает ширину, границы между кольцами очень
четкие на всем протяжении.
38 (39) Последний сегмент тела сзади обрублен, с четкими хо¬
рошо развитыми лопастями по краю, разделен на 5 ко¬
лец (рис. 62, 1). Передние дыхальца с 3 дыхальцевыми
отверстиями. Нижняя губа представлена 2 несклероти-
зованными лопастями, несущими по краю волоски.
Под корой на участках со скоплениями бродйщего сока,
в сильно разложившихся плодовых телах древесных
грибов и других жидких разлагающихся растительных
остатках. Сапромицетофаги, с элементами некрофа-
гии Anisopodidae — Разноножки
39 (38) Последний сегмент тела закруглен, без четких лопастей
вокруг дыхалец, разделен на 4 кольца. Передние ды¬
хальца с многочисленными дыхальцевыми отверстиями.
Нижняя губа в виде единой склеротизованной по краю
пластинки (рис. 76, 4). Личинки в очень влажной, силь¬
но разложившейся древесине, а также в микроводо¬
емах в дуплах. Сапромицетофаги с элементами некро-
фагии Mycetobiidae — Сокоедки
40 (37) Тело личинок короткое, слегка уплощено дорсовентраль-
но. Анальное поле отсутствует. Тело с многочисленными
поперечными складками, брюшные сегменты, как прави¬
ло, разделены на 3 одинаковые нечетко обособленные
кольца.
41 (42) Боковые пластинки головы на вентральной стороне
широко расставлены и разделены слабосклеротизован-
ным участком, покрытым вдоль переднего края волоска¬
ми. Внутренний скелет головы хорошо развит, в виде
темных стержней. Задние дыхальца расположены на
конце тела и окружены 4 стигмальными выростами. Сег¬
менты тела без склеротизованных пластинок. Личинки
крупные, длина тела не менее 8—9 мм. Развиваются
под корой, в разлагающихся плодовых телах древесных
грибов. Сапромицетофаги
Trichoceridae — Зимние комары
231
42 (41) Боковые пластинки головы на вентральной стороне со¬
прикасаются, передний край капсулы с вентральной сто¬
роны голый, без волосков. Внутренн^ш скелет головы
не выражен. Задние дыхальца расположены на конце
короткой терминальной дыхательной трубки и окруже¬
ны стигмальными выростами или непосредственно на
дорсальной стороне последнего сегмента, в этом случае
стигмальные выросты отсутствуют. Каждый сегмент
с дорсальной стороны несет по одной или несколько
склеротизованных пластинок, которые у отдельных ви¬
дов могут быть частично редуцированы (рис. 66, 2).
Длина тола не более О—7, обычно 4—5 мм. Во влажной
разлагающейся древесине, подстилке, грибах, во влаж¬
ных моховых подушках по берегам ручьев. Сапромнцо-
тофаги Psychodidae — Бабочпицы
43 (36) Тело без вторичной сегментации, иногда имеются лишь
вторичные (неполные) ложные складки. Верхняя губа
на вентральной стороне без премандибул. Мандибулы
массивные, нерасчлененные. Конец тела закругленный,
без лопастей или с длинной дыхательной трубкой, длина
кото])ой превьииает длину тела (рис. 63, 2).
44 (45) Тело удлиненное, без ложных складок (рис. 63, 1). Зад¬
ние дыхальца расположены на боковых сторонах послед¬
него сегмента. Головная капсула не замкнута, боковые
пластинки головы на вентральной стороне расставлены,
сильно склеротизованы по краю. Во влажной сильно раз-
ложивн1ейся древесине лиственных по])од. Сапроксило-
фаги Trichomyiidae — Мохнатокрылки
45 (44) Тело вальковатое, короткое, с короткими ложными
складками (рис. 63, 2). Задние дыхальца расположены
на конце дыхательной трубки, длина которой превышает
длину тела. Головная капсула замкнута, боковые пла¬
стинки головы на вентральной стороне слиты. Во влаж¬
ной разлагающейся древесине лиственных пород. Ксило-
мнцетофаги Axymyiidae — Древпекрылки
46 (35) Личинки мета- или апнейстические (дыхальца имеются
только на последнем сегменте тела или вообще отсут¬
ствуют).
47 (52) Личинки метапнейстические. Голова (рис. 72) массивная,
частично втянута в грудные сегменты, вдоль заднего
края с перепончатыми участками, иногда занимающими
значительные пространства'головы. В этих случаях скле-
ротизованными остаются лишь небольшие продольные
стержни (рис. 73, 1). Личинки крупные, длина их тела не
менее 10—12 мм.
48 (49) Сегменты тела с листовидными или нитевидными выро¬
стами, расположенными, по меньшей мере, на боковых
и спинной сторонах. Голова полностью втянута в груд¬
232
ные сегменты. Последний сегмент тела с 2 длинными
склеротизованными сзади выростами, действующими
как подталкиватели. Личинки зеленовато-желтые, по внеш¬
нему виду напоминают веточки мха. Среди мха на ство¬
лах деревьев и нней, а также в небольншх временных
водоемах. Фитофаги
Cylindrotomidae — Цилинд])отомиды
49 (48) Сегменты тела без выростов. Если имеются небольшие
листовидные выросты по бокам тела, то последний
сегмент тела с 6 выростами вокруг стигмального поля.
50 (51) Стигмальное ноле (рис. 77, /) окружено 6 выростами, ко¬
торые могут быть сильно укорочены, но всегда хорошо
заметны. Головная капсула массивная, на большем про¬
тяжении склеротизована. Лож'тле ножки, ползательные
валики и бугорки не развиты. Иод корой, в толще разла¬
гающейся Д1)евесины, в трухе в дуплах, в моховых по¬
душках на стволах, в подстилке, почве и в н])нбрежных
илистых отложениях. Санрофаги, ксилофильные фор¬
мы — сапроксило- и сапромицетофаги
Tipulidae — Долгоножки
51 (50) Стигмальное поле (рис. 77, 2) oKpy>jxeiio не более чем
5 выростами, иногда выросты полностью редуцированы.
Головная капсула у большинства представителей сильно
удлинена, на значительном протяжении перепончатая:
склеротизованные участки иногда сохраняются лишь
в виде удлиненных топких стержней (рис. 73, 1). У боль¬
шинства форм хорошо развиты ползательные валики,
иногда имеются бугорки, расположенные не только па
вентральной, но и на боковых сторонах тела, или
ложные ножки. Иод корой, в толще разлагающейся дре¬
весины, в подстилке, грибах, в почве, в П1)ибрежных
илистых отложениях временных водоемов и других
сильно увлажненных субстратах. Сапромицето- и сап-
роксилофаги с элементами некрофагии, некоторые вод¬
ные формы — факультативные хищники
Limoniidae — Болотницы
52 (47) Личинки аннейстические. Голова хорошо развита, не
втянута в грудные сегменты, без перепончатого участка
вдоль заднего края. Личинки мелкие, их длина, как пра¬
вило, не превышает 10 мм.
53 (54) Боковые пластинки головы на вентральной стороне, сбли¬
жены, но не слиты, разделены швом. В переднем отделе
вентральной стороны — сильносклеротизованная тре¬
угольная и зазубренная гипостомальная пластинка.
Сегменты тела на всем протяжении голые, иногда на 2 по¬
следних сегментах тела имеются пучки щетинок. На
вентральной стороне первого и последнего сегментов
тела обычно имеются хорошо развитые ложные ножки
233
(рис. 79, 2^ 4), Под корой, в рыхлой влажной древе¬
сине, в дуплах, в подстилке и в прибрежной зоне различ¬
ных водоемов. Сапро- и мицетофаги, некоторые водные
формы — хищники . . Chironemidae — Комары-звонцы
54 (53) Боковые пластинки головы на вентральной стороне
слиты, шов между ними не выражен. Гипостомальная
пластинка, если имеется, небольшая, прямоугольная
или закругленная, без зубцов по краю. Сегменты тела
на всем протяжении голые, иногда с терминальным
пучком ш,етинок. Если имеются передняя и задняя лож¬
ные ноички, то тело с разнообразными кутикулярными
структурами на всех сегментах. Под корой, в толще
разлагающейся древесины, особенно в старых ходах
ксилофильных насекомых, в трухе в дуплах, в бродящем
соке, в подстилке, а также наносах ила в прибрежной
зоне различных водоемов. Сапро- и мицетофаги, с эле¬
ментами некрофагии . . Ceratopogonidae — Мокрецы
55 (2) Голова небольшая (рис. 73, 2, 5, 6*). Верхняя губа
клювообразная, сжата с боков. Мандибулы и максиллы
слиты друг с другом по меньшей мере в основании и
образуют единый комплекс (рис. 75, о). Имеются или
метацефальные стержни, или обособленная вентральная
пластинка головы (рис. 73, 4). Тело состоит из И сегмен¬
тов (3 грудных и 8 брюшных) или многократно вторично
сегментировано и образовано 19 однотипными сегментами
(Therevidae).
56 (61) Задние дыхальца расположены непосредственно на по¬
верхности боковой или дорсальной стороны последних
сегментов (рис. 62, 4), Голова (рис. 62, 4; 64, i), не
втянута в грудные сегменты, сильно склеротизована,
темная; от заднего края головы внутрь сегментов отходят
один или два удлиненных метацефальных стержня (73, 6),
57 (58) Голова (рис. 73, 6) конусовидная, заостренная, сзади
с 2 метацефальными стержнями. Дорсальная сторона
грудных сегментов и дорсальная сторона последнего
сегмента с темно-коричневыми скле])отизованными пла¬
стинками (рис. 64, 1). Конец тела с 2 склеротизованными
темными крючками. Тело без вторичной сегментации.
Под корой и в толще разлагающейся древесины. Обли¬
гатные хищники
Xylophagidae — Стволоедки (стр. 249)
58 (57) Голова (рис. 73, 5) широкая, овальная, сзади с 1 непар¬
ным метацефальным стержнем. Сегменты тела светлые,
без склеротизованных пластинок (иногда имеется лшиь
небольшая терминальная пластинка).
59 (60) Тело толстое, вальковатое, вторично не сегментировано,
дополнительная складка имеется лишь на дорсальной
стороне последнего сегмента. Конец тела закруглен, иног¬
234
да с бугорками, или с небольшим склеротизоваыным
шипом. Метацефалышй стержень массивный, прямой,
иногда на конце расш,еплен. Задние дыхальца расположе¬
ны на дорсальной стороне последнего сегмента. У кси-
лофильных форм сегменты тела, как правило, окольцо¬
ваны бугорками. Под корой и в толще древесины входах
различных ксилофильных насекомых, в корнях расте¬
ний, в почве. Облигатные хищники
■ Asilidae — Ктыри (стр. 292)
60 (59) Тело (рис. 62, 4) удлиненное, змеевидное, вторично сег¬
ментировано. Конец тела с 2 светлыми цилиндрическими
придатками — подталкивателями (рис. 77, 6), Метаце-
фальный стержень лопатообразно расширен на конце.
Задние дыхальца расположены на боковых сторонах
тела. В трухе в дуплах, в рыхлой сухой древесине, в поч¬
ве. Облигатные хищники
Therevidae — Лжектыри
61 (56) Задние дыхальца расположены на конце последнего сег¬
мента на его поверхности или в особом углублении, если
смещены на дорсальную сторону сегмента, то находятся
на дне особой стигмальной камеры. Голова полностью
или частично втянута в грудные сегменты и на этих
участках соответственно слабо склеротизована, мета-
цефальные стержни отсутствуют. Если метацефальные
стержни имеются (просвечивают через покровы), то го¬
ловная капсула редуцирована до небольшой дорсальной
пластинки (рис. 64, 2),
62 (69) Головная капсула удлиненно-овальная, полностью или
частично втянута в грудные сегменты, метацефальные
стержни отсутствуют (рис. 73, 2).
63 (66) Тело удлиненное, цилиндрическое. Голова более чем
наполовину втянута в грудные сегменты, слабо склеро¬
тизована. Под усиками, по бокам от верхней губы распо¬
ложен валик, густо усаженный щетинками (рис. 73, 2).
Брюшные сегменты с хорошо развитыми ползательными
валиками или окольцованы бугорками. Покровы тела
гладкие, эластичные. Задние дыхальца располо^/кены на
конце последнего сегмента, на его поверхности или на дне
глубокой вертикальной щели.
64 (65) Тело сужено на переднем конце. Задние дыхальца рас¬
положены на конце последнего сегмента тела, окру¬
жены лопастевидными или крючковидными выступами
(рис. 77, 4). На вентральной стороне сегментов имеются
хорошо развитые ползательные валики. Развиваются
в сильно разложившейся древесине, где сравнительно
редки, в подстилке, почве. Облигатные хищники . .
Rhagionidae — Бекасницы
65 (64) Тело веретеновидное, сужено на обоих концах (рис. 64, 3).
235
Задние дыхальца расположены на дне стигмалыюй
вертикальной щели, открывающейся на конце послед¬
него сегмента. Сегменты тела с поперечными поясками
ползательных бугорков. В толще рыхлой древесины,
где сравнительно редки; обычны в подстилке, почве.
Облигатные хищники Tabanidae — Слепни
66 (63) Тело дорсовентрально уплощено (рис. 66, /, 3). Голова
на одну треть втянута в грудные сегменты, хорошо
склеротизована. Под усиками, по бокам от верхней
губы щетинистый валик отсутствует. Ползательные
валики также отсутствуют. Покровы тела с многочис¬
ленными пластинчатыми кутикулярными структурами,
жесткие. Задние дыхальца расположены на конце послед¬
него сегмента тела на дне глубокой горизонтальной
щели.
67 (68) Перед анальной щелью самое большее расположен ряд
из небольших округлых бугорков. Стигмальная камера
смещена на дорсальную сторону последнего сегмента.
Под корой, иногда в трухе дупел и в ходах ксилофиль-
ных насекомых в толще древесины. Сапроксилофаги
с элементами некрофагии
Stratiomyidae — Львинки (стр. 269)
68 (67) Перед анальной щелью расположен ])яд из крупных
конусовидных зубцов. Стигмальная камера расположена
непос])едственно на конце последнего сегмента тела
(рис. 77, 5). Под корой, сапроксилофаги с элементами не¬
крофагии Xylomyidae — Осовидки (стр. 2()2)
69 (62) Головная капсула не втянута, в виде небольнюй плоской
дорсальной пластинки, от которой в глубь сегментов
отходят длинные парные метацефальные и тенториальные
стержни. На вентральной стороне головы имеется V-об-
разный нпжнегубной склерит.
70 (71) Метацефальные стержни расширены на конце. Склериты
головной капсулы черные или черно-коричневые. Послед¬
ний сегмент тела различной формы. Задние дыхальца
расположены или непосредственно на поверхности округ¬
лого последнего сегмента тела, или в небольшом углуб¬
лении и окружены широкими лопастями. В почве,
прибрежных наносах ила, ксилофильные личинки —
в ходах короедов, в натеках сока и в трухе дупел. Свя¬
занные с короедами виды — облигатные хищники
Dolichopodidae — Зеленушки (стр. 309)
71 (70) Метацефальные стержни на конце не раснпфены. Скле¬
риты головной капсулы коричневые или светло-ко1)ич-
невые. Последний сегмент тела округлый, с продольны¬
ми бороздками. Задние дыхальца расположены на тер¬
минальном конце ок])углого последнего сегмента тела.
В почве, в прибрежных наносах ила, под корой и в толще
236
древесины. Педо- и ксилобионты преимущественно хищ¬
ники. . . Empididae — Толкунчики
72 (1) Голова отсутствует. Тело состоит из 3 грудных, 7—8
брюшных сегментов и светлого головного сегмента, не
отличающегося по окраске от остальных сегментов тела
(рис. 69, 70). Имеется группа темных фарингеальных
склеритов, расположенных внутри головного и первых
грудных сегментов тела (рис. 74, 86, 5, 9, 11). Трахей¬
ная система амфи-, редко метапнейстического типа
(рис. 80).
73 (74) Тело личинок мокрицеобразной формы (рис. 67, i),
уплощено дорсовентрально, с четкими темными пла¬
стинками на дорсальной стороне. Два первых и послед¬
ний сегменты тела с длинными тонкими парными при¬
датками. На поверхности головного сегмента имеется
остаток головной капсулы в виде небольшой темной
пластинки. В поверхностных слоях коры на стволах,
в подстилке. Сапрофаги
Lonchopteridae — Острокрылки
74 (73) Тело удлиненное, округлое или овальное в сечении, если
уплощено, то различной формы выросты имеются, по
меньнгей мере, на всех или нескольких брюшных сегмен¬
тах тела. Головная капсула полностью редуцирована.
75 (76) Передние дыхальца сильно смещены на дорсальную
сторону тела, находятся друг от друга на расстоянии,
не превышающем двойной диаметр дыхальца (рис. 82,
i, 3). Ротовые крючки гребневидной формы, с несколь¬
кими крупными зубцами (рис. 82, 2). Задние дыхальца на
}[сбольших бугорках (рис. 82, 4). В тканях живых расте¬
ний, фитофаги . . . Agromyzidae — Минирующие мун1ки
76 (75) Передние дыхальца, если и смещены на дорсальную сто¬
рону тела, находятся друг от друга на расстоянии, зна¬
чительно превышающем двойной диаметр дыхальца.
77 (80) Задние ; ыхальца расположены на конце неразветвлен-
ной трубки различной длины. Если трубка очень корот¬
кая, по п])евышающая диаметр дыхальцевых пласти¬
нок (])ис. 68, 5), то тело бочонкообразное, в переднем
отделе с массивными склеротизованными гребнями (Тет-
nosloma).
78 (70) К})ун11ые (свыше 5 мм), массивные; тело с многочислен¬
ными складками, вальковатое или уплощено дорсовент¬
рально, с тупым широко закругленным передним концом
(рпс. 68, 5; 65, 3), На теле обычны различные по форме
кутнкулярные структуры, склеротизованные крючки
(на переднем конце) и перистые выросты (на боковых
сто]фнах особенно последних сегментов). Задние ды¬
хальца расположены непосредственно на вершине ды-
хальцевой трубки. Анальная пластинка удлиненно¬
237
Рис. 82. Обь>;яй вид и детали строения личинок Agromyzidae (1—4) и Aula-
cigastridae (Aulacigaster leucopeza Meig.— 5, 6)]
1 — общий вид сбоку; 2,5 — ротоглоточный аппарат; 3 — передние дыхальца; 4, б — ко¬
нец тела снизу
овальная, не заходит на боковые стороны сегмента.
В древесине, натеках сока, в различных богатых разлагаю¬
щимися растительными остатками субстратах. Санро- и
ксиломицетофаги Syrphidae — Журчалки
79 (78) Мелкие (до 5 мм), тело без складок, удлиненное, с заост¬
ренным передним концом, без четких кутикулярных
структур или выростов. Задние дыхальца расположены
на вершине трубки на расходящихся в стороны лонасте-
видных выступах (рис. 69, 1). Анальная пластинка
сильно вытянута в поперечном направлении (рис. 82, б),
далеко заходит на боковые стороны сегмента. Ротогло¬
точный аппарат с небольшими ротовыми крючками и
гиностомальным склеритом. В натеках сока на стволах.
Сапрофаги Aulaeigastridae
80 (77) Задние дыхальца расположены на поверхности тела или
на парных четко обособленных друг от друга, иногда
только сближенных в основании, выступах или бугор¬
ках.
238
Рис. 83. Детали строения личинок Megaselia sp. (Phoridae, 1—3) и Odinia
sp. (Odiniidac‘,1 4—10)
1 — передний конец тела; 2,8,9 — анальная пластинка; 3, 7 — ротоглоточный аппарат;
4у 6 — передний и задний конец тела сбоку; 5 — переднее дыхальце; i 10 — шипики
ползательных валиков
ап, р1 — анальная пластинка
81 (86) Анальная щель поперечная (рис. 83, 2). Если кажется
продольной, то ограничена 2 сильно выступающими
над поверхностью тела анальными долями (рис. 83, 6, 8).
82 (83) Ползательные валики с очень крупными, хорошо замет¬
ными темными шипиками (рис. 83, 4, 6, 10), Тело без
папилл. Задние дыхальца расположены на конце тела,
на вершине широких конусовидных выступов. Анальное
отверстие в виде короткой продольной щели, ограничен¬
ной 2 сильно выступающими анальными долями. Обычно
отверстие и доли погружены в глубокую поперечную
складку (рис. 83, Р). Ротоглоточный аппарат массивный,
239
хорошо развит (рис. 83, 7). В ходах ксилофильных на¬
секомых под Kopoii и в толще древесины. Хищники,
паразиты Odiniidae
83 (82) Ползательные валики без 1нипиков (рис. 69, 2), поверх¬
ность тела голая или с многочисленными разнообразны¬
ми кутикулярными структурами. Тело, как правило,
с x0])0Jii0 развитыми паниллами (рис. 83, 1,2).
84 (85) Тело без вторичной сегментации. Ротовые крючки
с К])упными зубчиками по нижнему краю, гребневид¬
ные. Тело удлиненно-овальное, в разной степени уплоще¬
но дорсовеитрально и равиоме])но широко закруглено
спереди и сзади, с четкими выростами и кутикулярными
структурами на всех сегментах, или с характернымр!
2 сбли>1чеиными терминальными напиллами. В плодовых
телах и мицелии древесных грибов. Мицетофаги ....
Platypezidae — Грибные мушки
85 (84) Тело вторично сегментировано, грудные сегменты со¬
стоят из 2 колец, брюншые — из 3. Ротовые крючки
простые (рис. 83, о) или с пильчатым нижним краем.
Тело удлиненное, слегка уплощено дорсовеитрально,
несколько сужено на переднем конце и 1нироко закруг¬
лено сзади; без выростов и кутикулярных структур,
терминальные паничлы, если имеются, не сближены.
В пронизанной мицелием древесине, в почости тела насе¬
комых. Ксиломицетофаги, некрофаги, паразиты ....
Phoridae — Горба гки
86 (81) Анальная щель продольная.
87 (88) Тело короткое, унло1цено дорсовеитрально, с длинными,
обычно перистыми боковыми выростами на всех сегмен¬
тах (рис. 67, 3). Задние дыхальца расположены на боко¬
вых или дорсальной стороне переднего отдела последнего
сегмента. Дыхальн,евые отверстия разобщены, открыва¬
ются на обособленных бугорках (рис. 81, 5). Во влажной
сильно разложившейся древесине преимущественно
лиственных деревьев, в различных разлагающихся ве¬
ществах, реже в живых тканях растений и животных.
Сапро-, некрофаги Fanniidae
88 (87) Тело удлиненное, не уплощено, слегка сужено на перед¬
нем конце и несколько расширено на заднем. Задние ды¬
хальца расположены на конце тела.
89 (90) Задние дыхальца расположены на конце тела на уров¬
не верхней границь! последнего сегмента, являясь как
бы его продолжением. К дыхальцам прилегают (являясь
продолжением дыхальцевых пластинок) различные по
форме и степени склеротизации выступы. Анальная
пластинка поперечно-овальная, четко очерчена, ограни¬
чена рядами шипиков. Под корой и в рыхлой древесине,
в разлагающихся фруктах, травянистой растительности.
240
Сап])о-, некрофаги, факультативные хищники ....
Lonchaeidae — Копьехвостки (стр. 328)
90 (89) Задние дыхальца расположены на конце тела заметно
ниже уровня верхнего края носледнего сегмента. Ды¬
хальца без выступов, расположены или непосредственно
на поверхности сегмента тела или на различных по
форме возвышениях. Если небольшие выступы или
бугорки имеются, то они но являются продолжением
дыхальцевой пластинки, а ])асположены у основания
дыхалец.
91 (96) Последний сегмент тела с темными склеротизованными
крючковидными выступами ])азной длины (рис. 69, 3).
92 (93) Последний сегмент тела с короткими заостренными вы¬
ступами, длина которых значительно менее диаметра
дыхалец (рис. 84, 2, 4). Дыхальцевые отверстия в ви¬
де 3 извилистых щелей покрывают всю поверхность
дыхальцевой пчастинки. Переднее дыхальце (рис. 84, 1,3)
крупное, поперечное, с многочисленными дыхальцевыми
отверстиями. 1^товые крючки и гипостомальные скле¬
риты сильно удлиненные (рис. 84, 5). Под корой лист¬
венных деревьев. Сапрофаги
Calobatidae — Ходуленожки
93 (92) Последний сегмент тела с длинными крючковидными
темными выступами, длина которых заметно превосхо¬
дит диаметр дыхалец (рис. 84, 7, 11).
94 (95) Задние дыхальца расположены на поверхности тела
у основания крючковидных выступов с внутренней
стороны (рис. 84, 11). Последний сегмент тела без па-
пилл (рис. 84, 14). Передние дыхальца вееровидные
(рис. 84, 13). Ротоглоточный аппарат слабо склеротизо-
ван, светло-серый, практически не заметен. В толще
влажной, сильно разложившейся древесин]л. Сапро-
ксилофаги Clusiidae — Пятнокрыл ки
95 (94) Задние дыхальца расположены неносредстве11но на
крючковидных выступах вблизи их вершины (рис. 84, 7).
Последний сегмент тела с 2 рядами четких папилл
(])ис. 84, fV, !)). Передние дыхальца конусовидные, уд¬
линенные (рис. 84, 6). Ротоглоточный аппарат темный
(рис. 84, /0). Под корой. Санрофаги Psilidae
96 (91) Последний сегмент тела гладкий или с небольшими,
различными по форме, но не крючковидными бугорками,
несущими дыхальца.
97 (106) Задние дыхальца расположены на вершине цилиндриче¬
ских, конусовидных или округлых бугорков (рис. 85, 1).
Если бугорки отсутствуют и дыхальца расположены на
поверхности тела, то последний сегмент сильно скошен
и уплощен в верхней части, а дыхальца расположены
ниже средней линии тела (рис. 85, ,9).
241
Рис. 84. Детали строения личинок Rainieria latifrons Loew (Calobatidae,
i—6), Chyliza sp. (Psilidae, 6—10) и Hendelia sp. (Clusiidae, 11—14)
1,2 — передний и задний концы тела сбоку; 3, б, 13 — переднее дыхальце; 4, 7, 12 —
задние дыхальца; 5, ю — ротоглоточный аппарат; 8,9 — последний [сегмент тела сверху
и снизу; 11, 14 — конец тела снизу и сбоку
jJQS ^99)2Последний сегмент тела с четким рядом бугорковидных
папилл, окружающих стигмальные бугорки (рис. 85,
1,5). Бугорки, несущие задние дыхальца, сближены
в основании, если параллельные, то расстояние между
ними не превышает диаметра дыхальцевой пластинки.
Передние дыхальца обычно в виде цилиндрических бу-
Рис. 85. Общий вид и детали строения личинок Drosophila phalerata Meig.
{1—5), Stegana sp. (6, 7) (Drosophilidae) и Phyllomyza sp. (8—10) (Milichiidae)
1 — общий вид сбоку; г, б — переднее дыхальце; 3 — заднее дыхальце; 4, в — ротогло¬
точный аппарат; 5,7 — конец тела сверху; 9, Ю — конец тела сверху и снизу
243
горков, или уплощеииые конусовидные (рис. 85,2,6).
Под корой, в натеках сока, гниющих фруктах, грибах.
Сапрофагп, мицетофаги
Drosophilidae — Плодовые мушки
99 (98) Последний сегмент тела без четких панилл. Бугорки, не¬
сущие задние дыхальца, нгироко расставлены, расстоя¬
ние между дыхальцами не менее чел1 в 2 раза превышает
диаметр дыхальцевой пластинки.
100 (101) Анальная пластинка смещена вперед, находится на
границе 7-го и 8-го брюшных сегментов тела, с расчле¬
ненными долями характерной формы (рис. 85, 10). Зад-
irae дыхальца раснолонхоны или на косо с])озанной поверх¬
ности последнего сегмента (ррю. 70, 1\ 85, .9), или на
внутренней стороне ншроких закругленных буго]жов.
Ротовые крючки массивные (рис. 85, 8). Под корой
лиственных деревьев. Сапрофаги .... Milichiidae
101 (100) Анальная пластинка расположена в среднем отделе по¬
следнего сегмента (рис. 70, 2), с 2 простыми полуок-
ругльтми долями. Задние дыхальца расположены на вер-
н1ине трубчатых или конусовидных бугорков (рис. 87, <2).
102 (103) Передние дыхальца лопастевидные, двойные (рис. 70, 2),
Дорсальные отростки фарингеального склерита с обыч¬
ным заостренным передним выступом (рис. 87, 1). Зад¬
ние дыхальца с овальн1>1мн отве1)стиями (рис. 87, 3). Под
корой лиственных деревьев. Сапрофаги
Strongylophlhalmyiidae
103 (102) Передние дыхальца вееровидные (рис. 86, У).
104 (105) Верхний отдел последнего сегмента почти прямоуголь¬
ный (вид сбоку), участок над задними дыхальцами
как бы вздут или несет по одному небольшому, изолиро¬
ванному от дыхальцевого бугорка, лопастевидному вы¬
ступу. Дорсальные отростки фарингеального скле])ита
с обычным заостренным передним выступом. Анальная
пластинка четко очерчена, несколько вытянута в попе¬
речном направлении. Под корой, в т])авянистых расте¬
ниях. Ксилобионты — мицето-, некрофаги и хищники .
Pallopteridae — Бледнокрылки (стр. 353)
105 (104) Верхний отдел последнего сегмента тела закруглен, не
вздут и не несет лопастсвидные выступ1>1 над дыхальцами.
Дорсальные отростки фарингеального склерита с мас¬
сивным закругленным передним выступом (рис. 87, 4).
Анальная пластинка четко очерчена, овальная (рис. 87,
5). Под корой, в отмирающих и живых тканях травя¬
нистых растений. Сапро- и фитофаги
Chloropidae — Злаковые мухи
106 (97) Задние дыхальца расположены непосредственно на
поверхности сегмента (рис. 86, 4, 8). Иногда во впадине.
Последний сегмент закруглен, не скошен.
244
Рис. 86. Детали строения личинок Acanthoneura pteropleuralis Hend.
(Tephritidae, i—5), Myennis tricolor Hond. (Otitidae, 6—10) и Physiphora sp.
(Ulidiidae, 11-13)
1, 10, 13 — переднее дыхальце; 2, б, 12 — задние дыхальца, з, 4,7,8 — последний
сегмент тела снизу и сверху; 5, 9, 11 — ротоглоточный аппарат
107 (108) Конец тела заметно уплощен дорсовентрально. Задние
дыхальца вытянуты в поперечном направлении (рис. 81,
6), сильно затемнены, расположены на общей темной пла¬
стинке. Узкая анальная пластинка ограничена рядами
крупных шипиков. Под корой лиственных деревьев.
Некрофаги Megamerinidae — Шипоножки
245-
Тис. 87. Детали строения личннок Strongylophthalmyia ustulata Zett.
(Strongylophthalmyiidac, .1—3), Gaurax sp. (Chloropidae — 5), Tachinidae
(6) и Phaonia sp. (Muscidae — 7, S)
1, 4, 6, 7 — ротоглоточный аппарат; 2,5 — конец тела снизу |(5 — с пупария); 3,8 —
-заднее дыхальце
108 (107) Тело не уплощено дорсовентрально. Задние дыхальца
овальные или округлые, общая для обоих дыхалец
темная пластинка отсутствует.
109 (110) Анальная пластинка массивная, трапециевидная (рис. 8G,
5), ее длина составляет не менее 2/3 длины последнего
сегмента (длина анальной щели не менее 1/3 длины
сегмента). Дыхальцевые отверстия задних дыхалец рас¬
положены горизонтально, параллельно друг другу
(рис. 86, 2). Под корой лиственных деревьев, в плодах,
соцветиях, в галлах на стеблях и корнях травянистых
растений. Сапро- и фитофаги
Tephritidae — Пестрокрылки.
110 (109) Анальная пластинка овальная, с короткими или удли¬
ненными боковыми складками (рис. 86, 7), ее длина
значительно менее половины длины последнего сег¬
мента тела. Дыхальцевые отверстия задних дыхалец
радиально расходящиеся, а если параллельные, то рас¬
положены но в горизонтальной плоскости.
'246
111 (114) Дыхальцевые отверстия задних дыхалец ориентированы
вверх и внутрь. Срединные оси, проходящие чере»
срединное дыхальцевое отверстие и стигмальный диск,
пересекаются над дыхальцами. Анальная пластинка
удлиненно-овальная, четко очерчена, без дополнитель¬
ных анальных лопастей. От заднебоковых углов плас¬
тинки отходят поперечные ряды очень мелких птипп-
ков, расположенных в форме треугольника. Анальные
бугорки перед пластинкой не развиты. Тело голое, без
каких-либо папилл или бугорков (кроме шипиков полза-
тельных валиков).
112 (ИЗ) Дыхальцевые пластинки задних дыхалец четко очер¬
чены темной склеротизованной полоской с внутренней
стороны (рис. 86, 6). Дыхальцевые отверстия парал¬
лельны друг другу. Передние дыхальца (рис. 86, 10}
вееровидные, длина их дыхальцевой пластинки прибли¬
зительно в 2,5 раза уступает ширине. Под корой лист¬
венных деревьев, в различных богатых растительными
остатками субстр^гтах. Сапро-, ксилофаги
Otitidae — Лентокрылки
113 (112) Дыхальцевые пластинки задних дыхалец четко очер¬
чены темной полоской с верхней стороны (рис. 86, 12).
Внутренняя сторона дыхальцевой пластинки не очер¬
чена. Дыхальцевые отверстия не параллельные. Перед¬
ние дыхальца сильно вытянуты в поперечном направле¬
нии (рис. 86, 13). Длина их дыхальцевой пластинки не
менее чем в 3 раза уступает ширине. Под корой листвен¬
ных деревьев, в различных богатых растительными остат¬
ками субстратах. Сапро-, ксилофаги
Ulidiidae — А1ухи-блестянки
114 (111) Дыхальцевые отверстия задних дыхалец ориентированы
вниз и внутрь; срединные оси, проходящие через средин¬
ное дыхал ьцевое отверстие и стигмальный диск, рас¬
положены на одной горизонталыгой прямой или пере¬
секаются под дыхальцем. Анальлая пластинка или не
выражена, или с удлиненными боковыми анальными
лопастями. Если небольшая и овальная, то без каких-
либо рядов шипиков в задне-боковых углах. Перед пла¬
стинкой, как правило, имеются хорошо развитые аналь¬
ные бугорки.
115 (116) Задние дыхальца расположены на конце последнего
сегмента на дне глубокой поперечно-овальной впадины.
Дорсальный отросток фарингеального склерита рас¬
щеплен. Некрофаги, хищники, паразиты, обитающие
в самых разнообразных средах. Ксилобионты разви¬
ваются в различных беспозвоночных, преимуществен¬
но личинках насекомых
Sarcophagidae — Серые] мясные мухи
24Т
116 (115) Задние дыхальца расположены непосредственно на по¬
верхности тела, иногда на слегка вдавленной плоской
площадке.
117 (120) Задние дыхальца расположены на небольшой плоской
или слегка вдавленной поперечно-овальной площадке,
окруженной валиком с четкими конусовидными или
тупыми слабо выступающими бугорками. Парастомаль-
ные склериты хорошо развиты, длинные, стержневид¬
ные.
118 (119) Анальная пластинка отсутствует, узкая анальная щель
ограничена несколькими рядами разбросанных мелких
шиников. Хищники, паразиты, некрофаги, в самых
разнообразных субстратах животного происхождения.
Ксилобионты — преимущественно паразиты личинок
различных насекомых и дождевых червей
Calliphoridae — Сине-зеленые мясные мухи
119 (118) Анальная пластинка хорошо развита, с длинными
боковыми лопастями. В различных разлагающихся ве¬
ществах растительного и >1*швотного происхождения.
Санрофаги, хищники, паразиты
Anthomyildae — Цветочницы
120 (117) Задние дыхальца расположены на несколько выпуклой,
голой без бугорков поверхности последнего сегмента
тела. Если бугорки вокруг дыхалец имеются, то очень
небольшие, слабо заметные, и парастомальные склериты
не развиты.
121 (122) Ротовые кргочки с дополнительными акцессорными
склеритами (рис. 87, 7). Перитрел1а задних дыхалец
массивная, сильно склеротизована, черная (рис. 87, 8).
Дыхальцевые отверстия короткие и прямые. Передний
конец тела сильно заострен, задний — широко закруг¬
лен (рис. 70, 3). У видов, развивающихся в дуплах,
имеются хорошо развитые парные ложные но>1чКи,
конел, тела конусовидно вытянут. В различных разла¬
гающихся веществах растительного и животного проис¬
хождения, под корой, в трухе дупел. Хищники ....
Muscidae (Phaoiiiinae) — Настоящие мухи
122 (121) Ротовые крючки без дополнительных акцессорных скле¬
ритов. Леритрема задних дыхалец, как правило, слабо
развита, коричневатая. Если имеются рудименты акцес¬
сорных склеритов или перитрема массивная, то или
дыхальцевые отверстия щелевидные, извилистые, или
дорсальный отросток фарингеальных склеритов массив¬
ный, больше вентрального.
123 (124) Дорсальные отростки фарингеального склерита не более
вентральных. Гипостомальные склериты обособлены.
Дыхальцевые отверстия задних дыхалец извилистые.
В различных разлагающихся веществах растительного
248
и животного происхождения. Сапро-, нокрофаги . . .
Muscidae (Muscinae) — Настоящие мухи
124 (123) Дорсальные отростки фарингеального склерита значи¬
тельно массивнее вентральных (рис. 87, 6). Гипосто-
мальные склериты слиты с фарингеальными, если обо¬
соблены, то ротовые крючки короткие и тупые. Дыха-
льцевые отверстия задних дыхалец разнообразной формы:
от щелевидных извилистых до небольших округлых
пор, в большом количестве покрывающих поверхность
сгигмальной пластинки дыхалец. Паразиты. Ксилобион-
ты развиваются в личинках различных насекомых . . .
Tachinidae — Тахины
СЕМЕЙСТВО XYLOPHAGIDAE - СТВОЛОЕДКИ
Род Xylophag-us iXfeig. в течение длительного времени рассмат¬
ривался в составе семейства Rhagionidae (Lindner, 1925; Leonard,
1930; Brindle, 1961а, и др.). В последние годы род выделяется
в самостоятельное семейство Xylophagidae, однако объем семейст¬
ва большинством исследователей трактуется достаточно нгироко.
Во многих работах в семейство, помимо рода Xylophagus Meig.,
включается также род Coenomyia Latr. (Hennig, 1952; Trojan^
1956; Кривошеина, 1964; Иарчук, 1969а), а нередко и целый ряд
групп двукрылых, обладающих некоторым внешним сходством.
Так, например, в семейство включались роды Arthropeas Loew,
Artliroceras Will., GluLops Biirg. (Curran, 1956; James, 1965a).
Работы последних лет показали, что палеарктические роды Coeno¬
myia Latr., Arthropeas Loew, Aiiacanthaspis Rod., Glutops
Burg. и другие существенно отличаются от Xylophagus Meig. и
должны рассматриваться в качестве представителей самостоятель¬
ных семейств (Nagatomi, Saigiisa, 1970; Кривошеина, 1971в). В на¬
стоящее время в семейство с достоверностью может быть включен
лишь род Xylophagus JMeig., имеющий голарктическое распро¬
странение. В составе этого рода насчитывается 10 неарктических
(James, 1965а) и 12 палеарктических видов (Nagatomi, Saigusa,
1969; Кривошеина, Мамаев, 1972а). Такой объем рода установился
лишь в последнее время. В сводке Линднера (Lindner, 1925) для
Палеарктики указывались всего 2 вида: X. ater JMeig. и X. cin¬
ctus Deg., а X. compeditus Wied. рассматривался как синоним
X. ater J\feig. Сцилади (Szilady, 1932) принимает X. compeditus
Wiecl. в качестве самостоятельного вида и описывает четвертый
европейский вид — X. junki Szil. Результаты работы Сцилади
были признаны правильными А. А. Штакельбергом (1954) и Троя¬
ном (Trojan, 1956), которые сохранили в роде 4 вида, хотя Генниг
(Hennig, 1952) вернулся к объему рода, принятому Линднером
(Lindner, 1925).
Изучение биологии группы показало, что виды X. ater Meig. и
X. compeditus Wied. являются, несомненно, самостоятельными и
249
достаточно хорошо различаются не только по имагинальным, но и
по личиночным признакам (Кривошеина, Мамаев, 1966, 1972а).
Позднее в Тульских засеках (Тульская область) был зарегистри¬
рован пятый встречающийся в Европе вид — X. matsumurae Miy-
at. Этими 5 видами пока исчерпывается список европейских предста¬
вителей рода. Из них один — X. matsumurae Miyat., ранее извест¬
ный из Японии под названием X. maculatus (Matsumura, 1916),
видимо, встречается по всей Палеарктике.
Ф. Плеске (Pleske, 1925), ревизуя палеарктические виды рода,
описал из Уссурийского края два вида — X. bungei Р1. и X. sacha-
linensis Pl. В дальнейшем Ф. Плеске пришел к поспешному выводу
об идентичности X. ater Meig. и X. bungei Pl., в результате чего
появились ошибочные указания на встречаемость X. ater INIeig.
в Уссурийском крае (Pleske, 1930). В последние годы Миятаке
(Miyatake, 1965) описал 2 новых вида из Японии. Из них X. albo-
pilosus Miyat. оказался валидным, а X. omogensis Miyat. был позд¬
нее сведен в синоним (Nagatomi, Saigusa, 1969). В последней рабо¬
те было описано еще 2 новых вида, причем один из них — X. nuda¬
tus Nagat. et Saig. действительно новый, второй же — X. merus
Nagat. et Saig. оказался синонимом X. sachalinensis Pl. (Криво-
шеина. Мамаев, 1972а). Нами по материалам из Приморского
края было дополнительно описано 2 новых вида: X. admirandus
Kriv. et Мат., X. signifer Kriv. et Mam. Таким образом, в настоя¬
щее время в Палеарктике известны 4 европейских, 1 кавказский,
1 восточносибирский, 5 дальневосточных и 1 транспалеарктиче-
ский вид.
Личинки европейских видов, за исключением X. junki Szil.,
описаны несколькими авторами (Hennig, 1952; Brindle, 1961а;
Krivosheina, Mamajev, 1966; Кривошеина, Мамаев, 1967, 1972а),
личинки транспалеарктического и дальневосточных видов опи¬
саны Н. П. Кривошеиной и Б. М. Мамаевым (1972а).
Экология. Представители рода Xylophagus Meig. являются
истинно ксилофильными формами, развитие всех видов проис¬
ходит только в древесине. Личинки встречаются под корой (где
наиболее обычны), в толще разлагающейся древесины, в трухе
дупел, иногда проникают в различные пограничные с древесиной
субстраты, например, в толщу плодовых тел грибов-трутовиков.
Они являются облигатными хищниками, активно уничтожающими
самых разнообразных ксилофильных насекомых: разные фазы
развития короедов, усачей, златок, рогачей, хрущей и других
жуков, а также личинок самых разнообразных двукрылых (Tipuli-
dae, Sciaridae, Bibionidae, Pachyneuridae и др.). Личинки активны
на всех этапах своего развития, при этом даже в младших возра¬
стах сравнительно легко расправляются с крупными жертвами.
Этому способствуют особенности строения специализированной
(длинной заостренной и сильносклеротизованной) головы, дейст¬
вующей как бурав. Личинки очень подвижны, легко перемещают¬
ся по участкам ствола, используя различные полости и ходы или
250
прокладывая их самостоятельно. Ушющепность и склеротиза-
Л.ИЯ переднего конца тела облегчают передвижение. fi
В целом, личинки каждого вида способны питаться самыми
разнообразными насекомыми, но круг объектов ограничивается
в природе определенной экологической специализацией боль-
(пинства видов, выражающейся в приуроченности отдельных ви¬
дов к определенным породам и зонам ствола.
Выявляется четкая закономерность в распределении видов по
лиственным и хвойным породам. Так, личинки X. albopilosiis
Miyat., X. admirandus Kriv. et Mam. и X. signifer Kriv. et Mam.,
так же как и личинки X. ater Meig., развиваются только в лист¬
венных породах. Личинки X. sachalinensis Р1. и X. cinctiis
Deg. развиваются в хвойных породах. Наконец, личинки X. mat-
sumurae Miyat. и X. compeditus Wied. развиваются в разлагающей¬
ся древесине как лиственных, так и хвойных пород, по предпочи¬
тают лиственные.
Для большинства видов характерна специализация к развитию
в разных зонах ствола. Личинки таких видов, как X. sachalinensis
Р1., X. admirandus Kriv. et Мат., X. compeditus Wied., X. cinc¬
tus Deg., развиваются под корой. Личинки X. matsumurae Miyat.,
X. ater Meig., наоборот, развиваются в толще сильно разложив¬
шейся древесины (светлые и бурые гнили). В толще плотной
древесины, преимущественно в бурых гнилях, развиваются ли¬
чинки X. signifer Kriv. et Mam. и X. albopilosus Miyat. Таким об¬
разом, личинки всех известных в настоящее время видов могут
быть подразделены на 2 экологические группы: обитателей под-
коровой зоны и обитателей толщи древесины.
Личинки каждой группы обладают определенной морфологи¬
ческой спецификой. Для подкорных личинок X. admirandus
Kriv. et Мат., X. sachalinensis Pl., X. compeditus Wied., X. cinc¬
tus Deg. характерны заостренные склеротизованные выросты
последнего сегмента тела и склеротизация всех трех грудных
сегментов. Для личинок, развивающихся в толще древесины —
X. signifer Kriv. et Мат., X. matsumurae Miyat. и X. ater Meig.
характерны тупые выросты последнего сегмента тела и склероти¬
зация лишь двух грудных сегментов тела. X. albopilosus Miyat.
по морфологическим признакам занимает промежуточное положе¬
ние, хотя ближе к последней группе.
Морфология. В связи с тем, что в Палеарктике семейство пред¬
ставлено одним родом Xylophagus Meig., ниже приводится ха¬
рактеристика рода и входящих в его состав видов.
Род Xylophagus Meig.
Личинки с узкой заостренной и сильносклеротизованной голов¬
ной капсулой, уплощенными и сильносклеротизованными, по мень¬
шей мере, 2 грудными сегментами.
251
Головная ];апсула (рис. 73, 6) залткпутая, колусовидная, ее
длина не менеэ чем в 3 раза превышает ширину у основания. Зад¬
ний край головной капсулы лишь слегка втянут в грудные сегмен¬
ты. От него в глубь сегментов отходят 2 длинных равномерно
утолщенных метацефальных стержня.
Все три или 2 первых грудных сегмента с дорсальной и 1-й
грудной с вентральной стороны склеротизованы.
Последний сегмент тела с дорсальной стороны с темной плас¬
тинкой, цо длине не превосходящей половину длины сегмента.
Конец пластинки слегка вытянут и несет 2 коротких выроста. На
дорсальной стороне последнего сегмента тела 2 группы мелких
склеротпзованных точек расположены у переднего края сегмента
и два крупных овальных пятна — перед пластинкой.
Трахейная система амфиппсйстического типа. Передние ды¬
хальца грушевидной формы, расположены на боковых сторонах
1-го грудного сегмента; задние дыхальца удлиненно-овальные,
располо>1ач1Ы на склеротизованной пластинке последнего сег¬
мента тела.
Таблица для опр^'делении видов рода Xylophagus Meig.
1 (8) С дорсальной стороны склеротизованы только 2 грудных
сегмента тела. Выросты последнего сегмента тела тупые,
закругленные или с небольшим коротким зубчиком
(рис. 89, о, 5).
2 (3) Срединная склеротизованная пластинка 2-го грудного сег¬
мента тола с дорсальной стороны прямоугольная, плотно
прилегает к боковым пластинкам (рис. 89, 1). На вен¬
тральной стороне 1-го грудного сегмента между склероти-
зованными пластинками точечные склериты отсутствуют,
имеются лишь небольшие узкие полоски между передними
выступами пластинок (рис. 89, 2). Склеротизованные вы¬
росты последнего сегмента удлиненные, на вершине не
утолщены (рис. 89, 3) X. albopilosus Miyal.
3 (2) Срединная склеротизованная пластинка 2-го грудного
сегмента тела небольшая, треугольная или неправильной
формы, не примыкает к боковым, пространство между ними
светлое или с многочисленными точечными склеритами.
На вентральной стороне 1-го грудного сегмента простран¬
ство между склеротизованными пластинками полностью
или на ^/з заполнено удлиненными узкими и точечными
мелкими склеритами (рис. 88, 4\ 91,2). Склеротизованные
выросты последнего сегмента тела на вершине утолщены.
4 (5) На вентральной стороне 1-го грудного сегмента ме>1^ду
склеротизованными полосами расположен светлый нескле-
ротизованный участок прямоугольной формы (рис. 88, 4).
Склеротизованные . выросты последнего сегмента тела
обычно разделены относительно неширокой выемкой и на¬
252
ходятся друг от друга па расстоянии, не нревыигающем
двойную ширину выростов (рис. 88, 6)
X. signifer Kriv. et Мат.
5 (4) На вентральной стороне 1-го грудного сегмента все прост¬
ранство между склеротизованными полосами заполнено
небольшими точечными склеритами (рис. 91, 2). Склероти-
зованные выросты последнего сегмента тела широко расстав¬
лены, обычно находятся друг от друга на расстоянии,
превышак)ш,ем двойную ширину выростов.
6 (7) Передний край склеротизованной пластинки последнего сег¬
мента тела закругленный, без четкого выступа (рис. 89, J).
Дыхальца, расположенные на склеротизованной пла¬
стинке последнего сегмента тела, небольшие, овальные,
удалены от края пластннки на расстояние, но менее чем
в 2 раза превышающее ншрину дыхальца
X. ater-AIeig.
7 (fi) Передний край склеротизованной пластинки последнего
сегмента тела с четким выступом (рис. 91, 3). Дыхальца,
расположенные на склеротизованной пластинке послед¬
него сегмента тела, крупные, удалены от переднего края
пластинки на расстояние не более чем в 1,5 ])аза превы-
шаюш;ее ширину дыхальца ... X. matsumurae AHyat.
8 (1) С дорсальной стороны склеротизованы все 3 грудных
сегмента тела. Выросты последнего сегмента тела крючко¬
видно загнуты вверх, с зубчиком на вершине (рис. 90, 5,
9J, 7).
9 (12) Передний край склеротизованной пластинки последнего
сегмента тела с крупным хорошо обособленным выступом
(рис. 90, J). Задние дыхальца крупные, удалены от перед¬
него края пластинки на расстояние, не превышаюш,ее
ширину дыхалец. Склеротизованная полоска, располо¬
женная у переднего края пластинки последнего сегмента,
как правило, не сливается с пятнами, расположенными
перед пластинкой. Последние узкие, удлиненные.
10 (И) На вентральной стороне 1-го г])удного сегмента точечные
склериты расположены только между передними выступа¬
ми склеротизованных пластинок (рис. 88, 2). Светлый
несклеротизованный участок между пластинками непра¬
вильной формы. Выступ склеротизованной пластинки по¬
следнего сегмента широкий и тупой (рис. 88, 3). Располо¬
женная перед пластинкой полоска иногда сливается с пят¬
нами (рис. 88, 5) X. admirandus Kriv. et Мат.
И (10) На вентральной стороне 1-го грудного сегмента между бо¬
ковыми склеротизованными пластинками расположены
многочисленные точечные склериты. Светлый несклероти¬
зованный участок между пластинками прямоугольной
формы, его длина равна или несколько более длины склеро-
тизованного участка, расположенного перед ним (рис. 90, 4),
253
Рис. 88. Детали строения личинок Xylophagus admirandus Kriv. et Мат.
(i—5) и X. signifer Kriv. et Mam. (4—6} (Xylophagidae)
j — три грудных сегмента тела сверху; г, 4 — 1-й грудной сегмент^ела^снизу; з, в — по¬
следний сегмент тела сверху; б — 2-й грудной сегмент тела сверху
Рис. 89. Детали строения личинок Xylophagus albopilosus Miyat. (/—5)'и
X. ater Meig. 5) (Xylophagidae)
1 — два первых грудных сегмента сверху; г — 1-й грудной сегмент .‘снизу; з, б — послед¬
ний сегмент тела сверху; 4 — 2-й грудной сегмент сверху
Выступ склеротизованпой пластипки последнего сег¬
мента тела заострен. Расположенная перед пластинкой
склеротизовапная полоска изолирована от пятен
(рис. 90, 5) X. compeditus Wied.
12 (9) Передний край склеротизованпой пластипки последнего
сегмента тела с небольшим нечетко обособленным высту¬
пом (рис. 90, 3). Задние дыхальца удалены от не])еднего
края пластинки на расстояние, превышающее ширину
дыхальца. Склеротизовапная полоска, расположенная у
переднего края пластинки последнего сегмента, массивная,
у взрослых личинок сливается с пятнами, располо>1чОнными
перед пластинкой. Пятна массивные, овальные.
13 (14) Ме>1чду склеротизованными пластинками i-го грудного сег¬
мента с вентральной стороны расноложсшл узкие удлинен¬
ные склеротизованныо тяжи и пятна, заполняющие при¬
близительно ^/5 ДЛНН1.1 пластинки (рис. 91, б)
X. sachalineiisis Р1.
14 (13) Между склеротизованными пластинками 1-го грудного
сегмента с вентральной стороны располоихены узкие корот¬
кие склеротизованные тяжи и пятна, занимающие лишь
небольшое пространство между передними выступами
пластинок (рис. 90, 2) X. cinclus Deg.
X. admirandus Kriv. et Мат. Склеротизованные пластинки
имеются на всех 3 грудных сегментах тела, при этом 2 первых
грудных сегмента тела с дорсальной стороны склеротизованы
почти полностью (рис. 88, 1).
Личинки развиваются под корой лиственных, преимущественно
специфических для Приморского края пород: под корой чозении
крупночешуйчатой (в заповеднике «Кедровая Падь»), под корой
ореха маньчжурского, реже под корой черемухи Маака, маакии
амурской, бархата амурского, ильма долинного, липы амурской
(Комарово-Заповедное). Несмотря на то, что вид в целом не специа¬
лизирован в отношении какого-то узкого круга жертв, на боль¬
шинстве из перечисленных пород он связан с конкретными груп¬
пами насекомых. Под корой маакии амурской личинки X. admiraii-
(liis питаются личинками своеобразных реликтов1.1х двукрылых —
PergraLospes holoptica Kriv. et Marn. (CramptoHomyiidae),
проделывающими ходы в плотных слоях заболони. В черемухе
Маака личинки развиваются в заболони в ходах личинок комаров-
долгоножек — Танур1ега nigricornis Meig. и Т. parva Portsch.
(Tipulidae). Под корой бархата амурского и ореха маньчжурского
личинки обычны в поселениях различных видов двукрылых
(Stratiomyidae, Xylomyidae, Megamerinidae). В отдельных случа¬
ях личинки способны проникать непосредственно в толщу плот¬
ной древесины по ходам других насекомых. Так, во влажных,
лежащих на земле колодах липы амурской личинки были обнару¬
жены в ходах личинок слоников — Sipalinus gigas Fabr. и Crasso-
tarsus sp. (Curculionidae), в массе заселенных также личинками
256
львинок — XylopachygasLer inamaevi Kriv. (Slral iomyidae) и
в меньшей степени личинками журчалок — Brachyopa sp. (Syr-
phidae).
X. albopilosus Miyat. Склеротизованные пластинки имеются
только па 2 первых грудных сегментах тела (рис. 89, 7). Склероти-
зованная пластинка последнего сегмента тела (рис. 89, 3) с не¬
большим, но хорошо выраженным выступом. Задние дыхальца
небольшие, овальные, удалены от переднего края пластинки на
расстояние, слегка превышаюш,ее ширину дыхальца.
Личинки развиваются в эндемичных для Приморского края
породах — черемухе Маака и маакии амурской (Комарово-Запо-
ведное), а также под корой чозении крупночешуйчатой (заповед¬
ник «Кедровая падь»). Личинки наиболее характерны для поверх¬
ностных участков заболони черемухи Маака, где они развиваются
в ходах личинок комаров-долгоножек — Tanyplera parva Portsch.
и Т. iiigriconiis Meig. (Tipiilidae), и маакии амурской, где разви¬
ваются в ходах Pergratospes holoptica Kriv. el Мат. (Cramp-
tonomyiidae). Обычно встречаются совместно с личинками Ху1о-
phagus admirandus Kriv. et Мат.
Личинки, по всем признакам близкие расссматриваемому
виду, собраны под корой лиственных деревьев (ивы, ольхи и топо¬
ля лавролистного) на территории Тувинской АССР (Иштии-Хем).
В тополе лавролистном личинки зарегистрированы в поселениях
личинок галлиц (Cecidomyiidae).
X. ater Meig. С дорсальной стороны склеротизованы лишь
2 первых грудных сегмента тела, срединная склеротизованная
пластинка 2-го сегмента изолирована от боковых (рис. 89, 4),
Личинки развиваются в толш,е древесины различных листвен¬
ных деревьев (березы, ольхи, бука европейского, осины, липы
мелколистной, клена, вяза обыкновенного), заселяя как светлые,
так и бурые гнили. В буке европейском встречаются совместно
с личинками двукрылых: комаров-долгоножек — Tanyplera atrata
L. (Tipulidae), пятнокрылок — Paraclusia tigrina Fll., Clusiodes
sp. (Clusiidae) и галлиц (Cecidomyiidae). В отдельных случаях
зарегистрированы в рыхлой сильно увлажненной древесине сов¬
местно с личинками Fannia sp. (Fanniidae) и Temnostoma (Syr-
phidae). Серии личинок собраны в лесных районах Московской,
Тульской и Воронежской областей, в предгорных и горных райо¬
нах Краснодарского края, в горных и равнинных районах За¬
карпатской области.
X. cinctus Deg. Все 3 грудных сегмента тела с дорсальной
стороны склеротизованы почти полностью (рис. 90, 1).
Личинки развиваются под корой хвойных деревьев (сосна
обыкновенная, кедр сибирский, лиственница сибирская, ель
европейская, пихта кавказская). Заселяют стволы вслед за корое¬
дами, встречаясь совместно с личинками короедниц (Medetera
Fisch., Dolichopodidae), ктырей рода Laphria Meig. (Asilidae) и
львинок — Zabrachia minutissima Zett. (Stratiomyidae). В этот
9 Б м. Мамаев и др. 257
Рис. 90. Детали строения личинок Xylophagus cinctus Deg. (1—S) и X. com¬
peditus Wied. (4—6) (Xylophagidae)
1 — 2-й и 3-й грудные сегменты сверху; 2,4 — 1-й грудцой сегмент тела снизу; 8,5 —
последний, сегмент тела сверху; 6 — 3-й грудной сегмент тела сверху
период под корой иногда встречаются личинки детритниц (Sciari-
dae), мокрецов — Forcipomyia Meig. (Ceratopogonidae), галлиц
(Cecidomyiidae) и других ксилофильных насекомых. Встречаются
также в поселениях дровосеков и златок, часто совместно с ли¬
чинками ктырей Choerades Walk. (Asilidae). Серии личинок соб¬
раны в лесных районах Архангельской, Вологодской, Ярославской,
Московской и Воронежской областей (ель европейская, сосна
обыкновенная), в горных районах Краснодарского края (пихта
258
кавказская), Закарпатской области (Квасы), на высоте свыше
1000 м (ель европейская) и в горах Тувинской АССР (Иштии-Хем)
на высоте свыше 1500 м (ель, лиственница сибирская и кедр си¬
бирский).
X. compeditus Wied. Склеротизованные пластинки имеются на
всех 3 грудных сегментах. 3-й грудной сегмент (рис. 90, 6) склеро-
тизован на большем протяжении, у взрослых личинок все 3 пла¬
стинки этого сегмента соприкасаются вдоль боковых сторон.
Личинки развиваются преимуш,ественно под корой разлагаю-
ш,ихся деревьев, лишь в отдельных случаях проникая в толщу
сильно разложившейся рыхлой древесины. Довольно обычны
под корой ольхи, березы, липы мелколистной, дуба черешчатого,
бука европейского и восточного, а также ели европейской и
пихты европейской и кавказской. В отдельных случаях зарегист¬
рированы под корой осины, ивы, клена, ясеня, вяза обыкновен¬
ного и шершавого, граба обыкновенного, караганы. Личинки по¬
являются под корой в момент интенсивного заселения короедами,
когда под кору проникают различные сопутствующие им виды, на¬
пример, личинки короедниц (Medetera Fisch., Dolichopodidae),
различные детритницы (Sciaridae), мокрецы рода Forcipomyia
iVIeig. (Ceralopogonidae) и, наконец, львинки из подсемейства
Pachygasterinae (Straliomyidae). Среди последних для хвойных
характерен вид Zabrachia minutissima Zett., для лиственных
(особенно осины) — Neopachygaster meromelaena Duf. В ряде
случаев личинки X. compeditus Wied. зарегистрированы непосред¬
ственно в ходах ксилофильных жуков: короедов-древесинников
(Scolylidae), сверлильщиков (Lymexylonidae). Личинки — поли¬
фаги, питаются личинками самых разнообразных насекомых, но
предпочитают более крупные жертвы. Довольно обычны явления
каннибализма, причем, как правило, нападает более мелкая ли¬
чинка. Серии личинок собраны в лесных районах Ярославской,
Московской, Тульской и Воронежской областей, в предгорных и
горных лесах Северного Кавказа (1^раснодарский край), в равнин¬
ных и горных лесах Закарпатской области. Во всех точках обна¬
ружены в лиственных породах, кроме того, в Московской обла¬
сти, а также в Закарпатье и на Северном Кавказе — в хвойных
породах (соответственно ели европейской и пихте кавказской).
X. matsumurae Miyat. С дорсальной стороны склеротизованы
лишь 2 первых грудных сегмента тела (рис. 91, /). Склеротизо-
ванная пластинка последнего сегмента тела (рис. 91, 3) с четким
обособленным выступом. Дыхальца крупные, удлиненно-оваль-
ные, удалены от переднего края пластинки на расстояние, не более
чем в 1,5 раза превышающее ширину дыхальца.
Личинки развиваются как в лиственных (береза, липа амур¬
ская, чозения крупночешуйчатая, маакия амурская, орех маньч¬
журский, ильм долинный, осина, тополь лавролистный), так и
хвойных (кедр корейский) деревьях. Взрослые личинки весной
перед окукливанием встречаются непосредственно под корой.
9* 2Г)9
Рис. 91. Детали строения личинок Xylophagus matsumurae Miyat. (1—4)
йс. 91. Детали строения личинок лу1орПа|
X. sachalinensis PL (5—7) (Xylophagiaae)
1 — два первых сегмента тела сверху; г, б — 1-й сегмент тела снизу; з, 7 — последний
сегмент тела сверху; 4 — конец тела снизу; 5 — 2-й и 3-й грудные сегменты сверху |
Личинки предпочитают светлую увлажненную древесину лист-
яеиных пород: липы амурской и ильма долинного в Приморском
крае и березы и тополя лавролистного в Тувинской АССР.
В ильме долинном и липе амурской встречаются совместно с ли¬
чинками комаров-долгоножек — Ctenophora Meig. (Tipulidae),
журчалок — Temnostoma St. Farg. et Serv. (Syrphidae), a также ли¬
чинок жуков-древоедов (Eucnemidae). В березе личинки встре¬
чаются в достаточно увлажненных как светлых, так и бурых гни¬
лях древесины, заселенной личинками двукрылых рода Synneuron
Liind. (Hyperoscelididae), галлиц (Cecidomyiidae), болотниц —
Elephantomyia sp. (Limoniidae) и журчалок — Temnostoma St.
Farg. et Serv. (Syrphidae). Личинки были зарегистрированы также
в более сухой светлой древесине, заселенной личинками жуков-
дровосеков (Cerambycidae), чернотелок — Upis ceramboides L.
(Tenebrionidae), тенелюбов (Melandryidae). В древесине тополя
лавролистного личинки встречаются в достаточно плотной дре¬
весине, заселенной преимущественно личинками двукрылых —
Epiphragma Ost.-Sack. (Limoniidae) и Pachyneura Zett. (Pachy-
neiiridae), и в более рыхлой древесине, источенной личинками
жуков (Curculionidae, Eucnemidae, Oedemeridae). В специфиче¬
ских бурых гнилях кедра корейского личинки питаются личин¬
ками жуков Nematopliis semenovi Nik. (Nematoplidae). Личинки
рассматриваемого вида довольно обычны в Приморском крае, где
зарегистрированы на территории Комарово-Заповедное и запо¬
ведника «Кедровая падь», а также в Тувинской АССР (Иштии-Хем),
зарегистрированы также в Тульской области (Тульские засеки).
X. sachalinensis Р1. Вид настолько близок к X. cinctus Deg.,
что его самостоятельность может быть подвергнута сомнению,
тем более что у некоторых самок X. sachalinensis Р1. отмечено
побурение стернитов брюшка (у самок X. cinctus Deg. этот приз¬
нак более резко выражен). Отличия личинок, также очень незна¬
чительные, приводятся в определительной таблице.
Все 3 грудных сегмента тела личинок с дорсальной стороны
склеротизованы почти полностью. Срединные пластинки массив¬
ные, прямоугольные, плотно примыкают к боковым (рис. 91, 5).
Личинки довольно обычны в Приморском крае, где развивают¬
ся под корой хвойных деревьев (кедра корейского и пихты цель¬
нолистной). Для личинок установлены связи со стволовыми вреди¬
телями. Так, они заселяют кору на церамбицидной стадии ее раз¬
ложения и питаются личинками дровосеков. В отдельных случаях
зарегистрированы на заключительных этапах сколитидной стадии
разложения коры, встречаясь в лубе совместно с личинками
короедниц — Medetera Fisch. (Dolichopodidae), являюш,имися спе¬
цифическими хищниками короедов. Взрослые личинки обычны
в стволах, уже отработанных короедами, когда под корой скапли¬
вается значительное количество темной трухи. В целом личинки
предпочитают достаточно увлажненные участки стволов, заселен¬
ные личинками болотниц рода Libnotes Westw. (Limoniidae),
261
различными видами комаров-долгоножек рода Tipula L. (Tipuli-
dae), личинками детритниц (Sciaridae).
X. signifer Kriv. et Мат. С дорсальной стороны склеротизова-
ны лишь 2 первых грудных сегмента: 1-й склеротизован почти
полностью, на 2-м сегменте срединная пластинка очень небольшая,
изолированная от боковых (рис. 88, 5). Точечные склериты распо¬
ложены также вдоль заднего края сегмента. Склеротизованная
пласт'нка последнего сегмента тела (рис. 88, 6) вытянута впе¬
ред, но без четко обособленного выступа. Дыхальца небольшие,
овальные, удалены от переднего края пластинки на расстояние,
не менее чем в 2 раза превышающее ширину дыхальца.
Личинки в Приморском крае развиваются в толще преимущест¬
венно черненых гнилей древесины различных лиственных деревь¬
ев (ильм долинный, ясень маньчжурский, береза). Личинки про¬
никают в толщу довольно плотной древесины по ходам различных
ксилофильных личинок насекомых, например Hesperinus roh-
dendorfi Kriv. et Mam. (Hesperinidae). В более рыхлой древесине
питаются личинками комаров-долгоножек Ctenophora Meig-
(Tipulidae), перемещаясь по ходам последних. Иногда встречают¬
ся в сильно разложившейся древесине около скоплений личинок
толстоножек — Bibio sp. (Bibionidae).
СЕМКЙСТВО XYLOMYIDAE -ОСОВИДКИ
Семейство ксиломиид — сравнительно небольшое по объему
(насчитывает до 70 видов на всем земном шаре). Представители
семейства в настоящее время известны из различных зоогеогра-
фических областей. Есть данные о наличии монотипных родов,
например в Бразилии, но статус большинства из них требует
уточнения (Arthropeina Lind., Subulonia End., Prista End.).
Так, например, род Arthropeina Lind. сведен сравнительно недав¬
но в синоним Xylomya Rond. (Lindner, 19G9). Палеарктические
представители семейства входят в состав 3 родов: Xylomya
Rond., Solva Walk. и Nematoceropsis Pl. Но до сих пор сущест¬
вуют самые разнообразные взгляды на систематическое положе¬
ние семейства в целом и входящих в его состав родов. Лишь
сравнительно недавно эта своеобразная группа двукрылых, вклю¬
чаемая ранее в состав семейства львинок (Stratiomyidae), получи¬
ла право на самостоятельное существование (Seguy, 1951; Штакель-
берг, 19Г)4; Родендорф, 19G4; James, 1965Ь, и др.). Самостоятель¬
ность входящих в семейство родов также оспаривается. Одни
авторы признают роды Xylomya Rond. и Solva Walk. (Steyskal,
1947; James, 1965b; Кривошеина, 1972а). Другие рассматривают
все палеарктические виды семейства в пределах одного рода Solva
Walk. (Lindner, 1938; Nagatomi, Tanaka, 1971; Rozkosny, 1973).
При этом название Xylomya Rond. заменяется па Macroceromys
Big. (Nagatomi, Tanaka, 1971). Такая точка зрения возникла
262
Ii связи с неправильными определениями типового вида рода
Xylomya Hoiul. Так, в ряде работ (James 1965Ь; Nagatomi, Та¬
нака, 1971) в качестве типового вида указывается X. varia Meig.,
по ряду признаков тяготеющий больше к Solva Walk., чем к ос¬
новной группе видов Xylomya Rond. Но дело в том, что типовым
видом Xylomya Rond. следует считать не varia (Rondani, 1856),
а maculata (Westwood, 1840), намеченный значительно ранее.
Следует также отметить, что различия между родами Xylomya
Rond. и Solva Walk. настолько существенны, что их самостоя¬
тельность не вызывает сомнений.
Личинки семейства изучены еще далеко не достаточно. Сведе¬
ния по морфологии личинок отдельных видов Xylomya Rond. и
Solva Walk. имеются в работах ряда авторов (Brindle, 1981Ь;
Uu^ek et Rozkosny, 1963; Кривошеина, 1965; Rozko8n>% 1973).
Детальные описания с определительными таблицами большинства
видов приведены в работе Н. П. Кривошеиной (1972а). Личинки
Nematoceropsis Р1. неизвестны.
Экология. Личинки изученных видов развиваются в разлагаю¬
щихся растительных остатках, преимущественно под корой лист¬
венных деревьев, а также в разлагающихся корнях травяни¬
стых растений. Встречаются личинки обычно в больших коли¬
чествах, целыми поселениями, предпочитают сильно увлажненные
лубяные волокна и развиваются преимущественно в упавших
и лежащих на земле стволах. Молодые личинки сосредоточиваются
на пограничных со свежей корой участках, обильно смачивае¬
мых соком. Взрослые личинки встречаются в самых разнообраз¬
ных местах, часто на отработанных другими насекомыми участ¬
ках коры.
Личинки обычно развиваются совместно с целым комплексом
двукрылых насекомых, таких как болотницы (Limoniidae), львин-
ки (Stratiomyidae), лентокрылки (Otitidae) и другие, но не
характерны для поселений короедов, усачей и тем более златок.
Личинки-сапронекрофаги. Молодые личинки, скорее всего, пита¬
ются древесным соком, взрослые — некрофаги. Возможно, способ¬
ность питаться погибшими или поврежденными личинками дала
основание некоторым авторам (Hennig, 1952) считать их хищ¬
никами.
Морфология. Тело личинок дорсовентрально уплощено, с удли¬
ненно-овальной головой. Задний отдел капсулы, втянутый в груд¬
ные сегменты, слабо склеротизован, светлый. Фронто-клипеаль-
ный отдел головной капсулы четко отделен швами от боковых
пластинок головы. На вентральной стороне головы расположена
массивная пластинка. Ротовой аппарат представлен удлиненной
клиновидной верхней губой и массивными склеритами мандибу-
лярно-максиллярного комплекса. Тело состоит из И сегментов.
Поверхность сегментов с округлыми и продолговатыми кутику-
лярными пластинками, сплошь покрывающими тело. Только на
1-м и 2-м грудных сегментах с дорсальной стороны и на 1-м
263
грудном с вентральной расположены крупные гладкие поля
(рис. 66, 3).
Хетотаксия тела хорошо выражена. На 1-м и 2-м грудных сег¬
ментах с дорсальной стороны соответственно по 2 и 4 щетинки, на
остальных — по 6 щетинок. Такое же число щетинок на вентраль¬
ной стороне брюшных сегментов. Щетинки обозначаются как
краевые, промежуточные и срединные. Последний сегмент тела
короткий, тупо обрублен на конце, где расположены 2 овальные
гладкие стигмальные пластинки, ограничивающие щель в стигмаль-
ную полость. На вентральной стороне сегмента расположена
анальная щель, окруженная рядами крупных конусовидных зуб¬
цов, расположенных в виде треугольника. Массивные тупые
зубцы имеются также в заднебоковых углах сегмента.
Хорошо развитые дыхальца расположены на боковых сторо¬
нах 1-го грудного сегмента и в стигмальной полости на конце
сегмента. Передние дыхальца представлены стигмальной пластин¬
кой неправильной формы, несущей 2 продолговато-овальных ды¬
хал ьцевых отверстия. Рудименты дыхалец имеются также на бо¬
ковых сторонах брюшных сегментов.
Таблица для определения родов Xyloinyidae
1 (2) Последний сегмент тела с тупыми заднебоковыми углами
(рис. 92, 1). Стигмальные пластинки последнего сегмента
слабовыступающие. Дорсальная стигмальная пластинка на¬
правлена непосредственно вниз (рис. 92, 5). Крупные ли¬
чинки, длина тела не менее 13, в среднем 15 мм
Xylomya Rond. (стр. 267).
2 (1) Последний сегмент тела с четкими прямыми заднебоковыми
углами (рис. 92, 2). Стигмальные пластинки последнего сег¬
мента крупные, сильновыступающие. Дорсальная стигмаль¬
ная пластинка направлена назад и вниз (рис. 92, 6). Средних
размеров личинки, длина тела до 12 мм
Solva Walk. (стр. 264).
Род Solva Walk.
Основные характерные черты личинок приведены в определи¬
тельной таблице. Род представлен в фауне СССР 4—5 ви¬
дами.
Таблица для определения видов рода Solva Walk.
1 (2) Вентральная пластинка головы вытянутая, ее длина не¬
сколько превосходит ширину в основании. Дорсальная
стигмальная пластинка без бугорков (рис. 92, 9). Дыхальце-
вые щели передних дыхалец расположены в центре дыхальца,
окружающая их спереди перитрема хорошо развита, массив¬
ная (рис. 92, 7). Все 6 дорсальных щетинок сегментов тела
хорошо развиты. Длина самых коротких краевых щетинок
264
Рис. 92. Детали строения личинок Xylomya czekanovskii Pl. (i,5), X. yasu-
matsui Nag. etTan. (3), X. moiwana Mats. (4), Solva marginata Meig. (2, 6, 8,
10, 13), S. semota Kriv. (7, 9), S. harmandi Set^uy (11, 14), S. schnitnikovi Pl.
(12) (Xylomyidae)
1, 2, 5, 6 — конец тела сверху и сбоку; 5, 4, 11, 12 — заднебоковые углы конца тела
сверху; 7,8 — дыхальца 1-го грудного сегмента; 9, ю — дорсальная стигмальная пла¬
стинка сзади; 13, 14 — дорсальная пластинка 1-го грудного сегмента
ПОЧТИ достигает половины длины сегмента, а самых длинных
промежуточных — несколько превосходит его длину. По¬
кровы последнего сегмента тела сверху с однотипными плас¬
тинками S. semola Kriv.
2 (1) Вентральная пластинка головы поперечная, ее длина в 1,5—
2 раза менее ширины в основании. Дорсальная стигмальная
пластинка с 2 бугорками (рис. 92, 10), Дыхальцевые щели
передних дыхалец смещены вперед, перитрема, окружаю¬
щая их спереди, узкая, слабо выражена (рис. 92, S). Развиты
и заметны лишь 2 промежуточные дорсальные щетинки тела,
достигающие половины длины сегмента, остальные —
в 5—6 раз короче. Структура покровов последнего сегмента
различна.
265
3 (4) Вентральная пластинка головы разделена четкой продоль¬
ной бороздой на 2 отдела. Дорсальная пластинка 1-го
грудного сегмента гладкая, выпуклая, с очень небольшими
точечными вдавлениями на поверхности (рис. 92, 13). По¬
следний сегмент тела с дорсальной стороны с крупными
гладкими и выпуклыми пластинками
S. marginata Meig.
4 (3) Вентральная пластинка головы выпуклая, без срединной
продольной борозды. Дорсальная пластинка 1-го грудного
сегмента с поперечной бороздой, идущей параллельно перед¬
нему краю и прерванной в центре (рис. 92, 14). Длина
отрезка борозды составляет не менее V3 ширины плас¬
тинки.
5 (6) Передние отделы 2—0 брюшных сегментов с четким попереч¬
ным рядом суженных к вершине и приподнятых над поверх¬
ностью тела пластинок. Дорсальная сторона последнего сег¬
мента тела с несколькими гладкими крупными пластинками,
разбросанными по поверхности сегмента и не образующими
четких рядов. Заднебоковые углы последнего сегмента
с одним конусовидным и несколькими тупыми закруглен¬
ными бугорками (рис. 92, 11). Бугорки на дорсальной
стигмальной пластинке крупные, массивные
S. harmandi Seguy
6 (5) Передние отделы 2—6 брюшных сегментов с поперечным
рядом округлых, слегка приподнятых над поверхностью сег¬
ментов пластинок. Дорсальная поверхность последнего сег¬
мента тела без выделяющихся крупных пластинок, имеются
только мелкие темные пластинки, хорошо заметные на свет¬
лом фоне сегмента. Такие же пластинки расположены на
боковых сторонах сегментов с дорсальной стороны. Задне¬
боковые углы последнего сегмента тела с одним массивным
широко закругленным бугорком (рис. 92, 12). Бугорки на
дорсальной стигмальной пластинке небольшие, в виде ва¬
ликов S. schnitnikovi Р1.
S. harmandi Seguy. Этот вид зарегистрирован на Куриль¬
ских островах (о. Кунашир) и в Японии (Nagatomi, Тапака,
1971). Вид под названием S. umbrosa Kriv. был описан из
Приморского края (Кривошеина, 1972а).
Передние отделы 2—6-го брюшных сегментов с дорсальной
стороны с поперечным рядом из 12—14 хорошо выделяющихся
овальных пластинок. Пластинки светлее и крупнее окружающих
их кутикулярных структур, с овальным темным зубцом на заднем
крае. По такому же ряду пластинок имеется в передних отделах
брюшных сегментов с вентральной стороны. Каждый ряд образо¬
ван 18—19 пластинками, несущими более вытянутый и светлый
(почти белый) зубец.
По нашим данным, в Приморском крае (Комарово-Заповедное)
личинки развиваются под корой маакии амурской, ильма долин-
266
iioi о, чозении крупночешуйчатой, бархата амурского, на Куриль¬
ских островах (о. Кунашир) под корой бархата сахалинского.
S. marginata Meig. Довольно обычный в Европе вид, есть
данные о нахождении вида в Северо-Восточном Китае (Lindner,
И)Г)Г)) и на Алтае (Нарчук, 1972). Личинка описана несколькими
авторами (Brindle, 1961Ь; DuJek, Rozkosn>% 1963; Кривошеина,
190Г); Кривошеина, Мамаев, 1967).
На 2—7-м брюшных сегментах с дорсальной и вентральной
сторон ряд крупных овальных пластинок, несущих четкий зубчик.
На конце последнего сегмента на его заднебоковых углах располо¬
жено по 1 крупному тупому зубцу.
По литературным данным, личинки развиваются под корой
различных видов тополей, конского каштана (Brindle, 1961D;
Du^ek, Rozkosny, 1963; Rozkosny, 1973). На территории нашей
страны личинки обитают преимущественно под корой осины
(в Краснодарском крае), тополя лавролистного (Тувинская
АССР), но встречаются также под корой клена, ивы, граба обык¬
новенного, акации желтой (Краснодарский край. Воронежская,
Закарпатская области). В единичных случаях личинки зареги¬
стрированы в толще древесины, в ходах короедов-древесинников.
5. semota Kriv. описан по материалам из Приморского края.
Личинка изучена Н. П. Кривошеиной (1972а).
В переднем отделе брюшных сегментов тела с дорсальной и
вентральной сторон имеется ряд из 6—8 овальных пластинок, отно¬
сительно более крупных и более выпуклых, чем окружающие
пластинки, густо покрывающие сегменты тела. Пластинки с вент¬
ральной стороны выражены более отчетливо. Личинки развивают¬
ся под корой ясеня маньчжурского и чозении крупночешуйчатой.
S. schnitnikovi PL известен из горных районов Средней Азии
(Памир, Тянь-Шань). Вид очень близок S. flavoscutellaris Mats.,
зарегистрированному в Японии (включая о. Хоккайдо) (Nagato-
mi,Tanaka, 1971). Основные отличительные признаки личинок при¬
ведены в определительной таблице. Личинки развиваются в разла¬
гающихся корнях ферулы, тип питания точно не установлен.
Род Xylomya Rond.
Основные отличительные черты личинок приведены в определи¬
тельной таблице. Род представлен в фауне СССР 3—4 видами.
Таблица для определения видов рода Xylomya Hond.
1 (4) На дорсальной стороне брюшных сегментов имеется лишь по
2 хорошо выраженные промежуточные щетинки, но и они
достигают не более чем V5—V4 длины сегмента. На дорсаль¬
ной стороне последнего сегмента имеется несколько пласти¬
нок, выделяющихся более крупными размерами.
2 (3) Заднебоковые углы последнего сегмента тела с одним не¬
большим слабо заметным бугорком около щетинки. Попереч¬
267
ные ряды укрупненных пластинок вблизи переднего края
2—7-го брюшных сегментов слабо заметны, особенно дор¬
сальные. Пластинки поперечно-овальные, окрашены как и
окружающие покровы, но с темным ободком по заднему
краю, слабо приподняты над поверхностью сегментов . . .
X. czekanovskii Р1.
3 (2) Заднебоковые углы последнего сегмента тела с 2 конусовид¬
ными бугорками около щетинки (рис. 92, 3). Поперечные
ряды укрупненных пластинок вблизи переднего края сегмен¬
тов относительно хорошо выражены. Пластинки не очень
большие, овальные, но заметно приподняты над поверхностью
сегмента и значительно светлее окружающих покровов. За
первым рядом на средних сегментах заметен второй ряд
таких же светлых, но не возвышающихся над поверхностью
тела пластинок X. yasiimatsui Nag. et Тан.
4(1) На дорсальной стороне брюшных сегментов имеется по
() хорошо выраженных щетинок, из которых наиболее длин¬
ные промежуточные составляют не менее половины длины
сегмента. Дорсальная сторона последнего сегмента покрыта
однотипными, приблизительно равными по размерам пла¬
стинками. В передних отделах сегментов имеются поперечные
ряды относительно слабо заметных овальных слегка припод¬
нятых над поверхностью тела светловатых пластинок. Они
наиболее хороню выражены на 2—4-м брюшных сегментах
с дорсальной стороны и на 1 —5-м—с вентральной. Заднебоко¬
вые углы последнего сегмента с 2 далеко отстоящими друг от
друга короткими конусовидными бугорками (рис. 92, 4) . .
X. moiwana Mats.
X. czekanovskii PL ( interrupta Pl.) — обычный транспалеарк-
тнческий вид. Личинка описана Н. П. Кривошеиной (1965).
Личинки развиваются под корой лиственных деревьев, прей
муществонно осины и тополя. Под корой осины обычны в средней
полосе Европейской части СССР, под корой тополя лавролистного
зарегистрированы в Тувинской АССР, ильма долинного, чозении
крупночешуйчатой и в единичных случаях под корой ореха маньч¬
журского — в Приморском крае.
X. moiirana Mats. ( ussuriensis PL). 1^ид известен с о. Куна-
шир (Курильские острова) и о. Хоккайдо (Япония) (Nagatomi,
Танака, 1971). Под названием X. ussuriensis Pl. описан по мате¬
риалам из Приморского края и Северо-Восточного Китая. Личин¬
ка ;)того вида описана в работе П. П. Кривошеиной (1972а).
Личинка внешне очень похожа на личинку'Х. czekanovskii Pl.,
отличия приведены в определительной таблице. Личинки разви¬
ваются среди разлагающихся лубяных волокон маакии амурской.
X. yasuwatsui Nag. et Топ. описан с о. Хоккайдо. Имаго этого
вида выведены из личинок, собранных на о. Кунашир под корой
бархата сахалинского. Основные морфологические черты личинок
приведены в определительной таблице.
268
СЕМЕЙСТВО STHATIOMYIDAE - ЛЬВИНКИ
Львипки — обширное семейство прямошовных короткоусых
двукрылых, характеризуюш,ееся большим морфологическим и эко¬
логическим разнообразием входяш,их в его состав видов. Семейство
насчитывает более 1500 видов, распространенных главным образом
в тропической и субтропической зонах земного шара, на террито¬
рии нашей страны известно около 200 видов. Представители семей¬
ства образуют несколько четко обособленных не только морфоло¬
гически, но и, как правило, экологически, групп, составляющих
основу таких наиболее обширных подсемейств, как Beridinae, Sira-
tiomyinae, Sarginae, Clitellariinae и Pachygasterinae. Следует от¬
метить, что положение отдельных родов в системе семейства оконча¬
тельно еще не определено. Так, род Nemotelus Geoffr. одними
авторами включается в состав подсемейства Clitellariinae (James,
1965с; Rozkosny, 1973), а другими (McFadden, 1907) выделяется
в самостоятельное подсемейство Nemotelinae; род Hermetia Latr.,
выделяемый одними в самостоятельное подсемейство (James,
1965с), другими специалистами (McFadden, 1967) включается
в состав подсемейства Clitellariinae, и т. д.
Нами подробно характеризуется лишь ксилофильное подсемей¬
ство Pachygasterinae. В общих разделах, где в той или иной мере
освещается объем различных подсемейств, мы придерживаемся
системы, приведенной Роскошным (Rozkosny, 1973) для европей¬
ских видов львинок.
Подсемейство Beridinae представлено в фауне СССР родами
Allognosta Ost.-Sack., Actina Meig., Beris Latr., Chorisops Rond.,
Exodontha Rond., известными no единичным видам. Наиболее
обширный из названных родов — Beris Latr. включает около
10 видов. Подсемейство Sarginae представлено родами Sargus
Fabr., Microchrysa Loew, Chloromyia Dunc., Filiptschenkia Pl.
и Chrysochroma \\ill., также включающими небольшое число
видов. Среди представителей подсемейства Stratiomyinae следует
в первую очередь упомянуть роды Stratiomys Geoffr., Odonto-
myia Meig., включающие по 15—20 видов. Такой же объем имеют
роды подсемейства Clitellariinae — Adoxomyia Bezzi, Охусега
Meig., Nemotelus (ieoffr. (Зстальные роды этого подсемейства —
Lasiopa Brulle, Taurocera Lind., Clitellaria Meig. представлены
небольшим числом видов.
Подсемейство Pachygasterinae до самого последнего времени
было представлено в Палеарктике небольшим числом монотипных
или небольших по объему родов: Pachygaster Meig. и Pseudowai-
lacea Kert. из Европы, Kolomania Pl. и Wallacea Dol. с Дальнего
Востока и Raphanocera Pl. из Средней Азии. В последние годы
на территории нашей страны были дополнительно зарегистриро¬
ваны или описаны представители родов Craspedometopon Kert.,
Maackiana Kriv., Ornopyramis Kriv., Cechorismenus Kert.,
Xylopachygaster Kriv. (Кривошеина, 1973а). Кроме того, в рабо-
2()9
тах последнего времени род Pachygaster подразделен на 4 рода:
Zabrachia Coq., Pachygaster Meig., Neopachygaster Aust. и Prao-
myia Kert. (Kraft, Cook, 1961; Кривошеина, 1973а; Rozkosny,
1973). В последнее время на территории СССР зарегистрированы
представители еще 2 родов: Eupachygaster Kert. и Aspidacantha
Kert. (Кривошеина, 1975).
Большинство родов львинок распространено в лесной зоне,
где их личинки приурочены к почве, разлагающейся древесине,
различным водоемам. В составе семейства имеются также харак¬
терные обитатели открытых пространств, их личинки развиваются
в прибрежных участках различных часто солонцеватых водоемов
или в скоплениях разлагающихся органических веществ, часто на
пастбищах или вблизи животноводческих помещений. Отдельные
виды, например Micro^rysa polita L., стали синантропами, а неко¬
торые львинки — обитателями пустынных и полупустынных
ландшафтов, где развиваются в разлагающихся корнях пустын¬
ных растений (Adoxomyia Bezzi) или в кубышках саранчовых
(Chrysochroma Will.).
Биология львинок изучена довольно неравномерно, более из¬
вестны обитатели различных водных и полуводных сред. Ксило-
фильные личинки изучены слабее. Краткие сведения о личинках
неарктических видов имеются в работах ряда авторов (Malloch,
1917; Соок, 1953; Kraft, Соок, 1961, и др.)* Данные о палеаркти-
ческих группах изложены в работах Бриндля (Brindle, 1962а),
Н. П. Кривошеиной (1965, 1975, 1976а) и Н. П. Кривошеиной и
Б. М. Мамаева (1967).
Экология. По образу жизни личинки львинок делятся на 2
крупные экологические группы: водных и полуводных, с одной
стороны, и наземных, заселяющих самые разнообразные субст¬
раты, с другой. К первой группе относятся личинки подсемейств
Stratiomyinae и Clitellariinae (Neinotelus Geoffr. и Oxycera Meig.).
В состав второй группы входят представители остальных подсе¬
мейств, а также виды рода Clitellaria Meig. из Clitellariinae.
Для большинства львинок связи с древесиной или случайны,
или недостаточно изучены. Так, для представителей подсемейства
Beridinae характерно развитие в различных разлагающихся суб¬
стратах растительного происхождения, в том числе во влажных
опавших листьях, подушках мха (Meijere, 1916; Johannsen, 1922;
Lenz, 1923; Bischoff, 1925). В то же время личинки Allognosta
Ost.-Sack. и Exodontha Rond. зарегистрированы в разлагающейся
древесине (Malloch, 1917; McFadden, 1967). Личинки Actina Meig.
в Австралии развиваются в больших количествах в почве среди
разлагающихся корней и стеблей травянистых растений (Pu¬
ller, 1934). Личинки Beris Latr. обнаружены под корой деревьев,
лежащих на болотистых участках (Lenz, 1923). Нами личинки
Beris Latr. неоднократно собирались в сильно увлажненных субст¬
ратах, в первую очередь в сильно разложившихся, представляю¬
щих рыхлую массу плодовых телах различных древесных
270
грибов. Личинки представителей подсемейства Sarginae (Micro-
chrysa Loew, Sargus F., Chloromyia Dunc.) развиваются в скоп¬
лениях гниющего органического материала, преимущественно
в навозе, в почве (Вгаиег, 1883; Lundbeck, 1907; Peterson, 1951;
Ilennig, 1952; McFadden, 1967; Rozkosny, 1973, и др.). Для личи¬
нок Clitellariinae характерно развитие в разнообразных органи¬
ческих материалах как растительного, так и животного происхо¬
ждения, в том числе экскрементах, трупах и т. д. Но в то же вре¬
мя, например, личинки Clitellaria Meig. встречаются в корнях
грецкого ореха (Rozkosn>% 1973), личинки Cyphomyia Wied. и
Adoxomyia Bezzi развиваются в разлагающихся и поврежденных
кактусах, а также под корой папайи (Brunetti, 1923; McFadden,
1967). Нами личинки последнего рода обнаружены в больнтх
количествах в разлагающихся корнях ферулы (Ferula sp.).
Истинно ксилофильными формами являются только предста¬
вители подсемейства Pachygasterinae. Все изученные в настоящее
время виды этого подсемейства развиваются в разлагающихся
стволах деревьев. Они встречаются как в хвойных, так и листвен¬
ных породах, предпочитая последние. Собственно, преимуществен¬
но к хвойным породам приурочены лишь личинки рода Zabrachia
Coq. Личинки отдельных родов, например Pareidalimus Lind. и
Eidalimus Kert., зарегистрированы как под корой хвойных,
так и лиственных пород, но основная масса всех изученных в на¬
стоящее время родов и видов подсемейства зарегистрирована в лист¬
венных породах. При этом у некоторых групп львинок наблю¬
дается определенная избирательность в отношении пород. Так,
например, Pseudowallacea hungarica Kert. развивается под ко¬
рой различных видов тополей. Предпочитает эти же породы Neopa-
chygaster meromelaena Duf., но в целом личинки подсемейства оби¬
тают в самых разнообразных породах.
Личинки заселяют различные участки ствола, встречаясь обыч¬
но в больших количествах, до 100 и более экземпляров в одном
поселении. Наиболее обычны они под корой, где встречается боль¬
шинство зарегистрированных в настоящее время на территории
СССР видов, относящихся к родам Aspidacantha Kert., Cechoris-
menus Kert., Craspedometopon Kert., Maackiana Kriv., Neopachy-
gaster Aust., Ornopyramis Kriv., Wallacea Dol., Pseudowallacea
Kert. Под корой развиваются представители родов и видов, из¬
вестных из Северной Америки (Eidalimus Kert., Pareidalimus
Lind. и др.; Kraft, Соок, 1961). Обитатели лиственных пород
(Neopachygaster Aust., Pseudowallacea Kert. и др.) предпочитают
увлажненные лубяные волокна преимущественно лежащих
на земле стволов. Личинки рода Eupachygaster Kert. обитают
в заполненных древесной трухой дуплах, а личинки отдельных
видов Neopachygaster Aust. обычны в натеках сока на коре и под
корой. На участках с натеками сока предпочитают концентриро¬
ваться личинки Craspedometopon Kert., Meackiana Kriv. и др.
В отдельных случаях личинки встречаются в толще плотной дре¬
271
весины, куда проникают по ходам ксилофильных насекомых (на¬
пример, виды родов Neopachygaster Aust., Zabrachia Coq.). В толще
рыхлой сильно разложившейся древесины личинки львинок под¬
семейства Pachygasterinae не зарегистрированы. Личинки отдель¬
ных видов, в первую очередь Zabrachia minutissima Zett., реже
Neopachygaster meromelaena Duf. и Pseudowallacea hungarica
Kert., заселяют подкоровую зону в местах поселения короедов.
Начальное заселение стволов совпадает с периодом активного их
заселения короедами и продолжается после появления целого ряда
вторичных групп ксилофильных насекомых, в том числе различных
двукрылых (Xylomyidae, Scatopsidae, Lonchaeidae и др.).
Пищевые связи личинок изучены очень слабо. Предполагается,
что большинство из них сапрофаги в широком смысле, питающие¬
ся различными разлагающимися веществами растительного про¬
исхождения. Некоторые группы, например представители подсе¬
мейства Chiromyzinae (Raff, 1931), известны в качестве фитофагов,
вредящих травянистой растительности. В роли потенциальных
вредителей рассматриваются личинки Clitellariinae — Hermetia
palmi vora James, развивающиеся на пальме Elaeis guineensis
(James, 1972). Большая группа личинок питается остатками не
только растительного, но и животного происхождения, даже
тканями живых организмов. Так, Hermetia illucens L. не только
является некрофагом, но способна вызывать миазы у человека
(Dunn, 1916; James, 1947).
Хищничество для личинок львинок не характерно. По нашим
данным, хищничество не свойственно также и ксилофильным фор¬
мам. На определенных, особенно заключительных этапах разви¬
тия, личинки могут питаться погибшими или поврежденными ли¬
чинками ксилофильных насекомых. Такие наблюдения имеются
по личинкам Zabrachia minutissima Zett., Neopachygaster Aust.,
Wallacea nigrotibialis Pl., Pseudowallacea hungarica Kert. и др.
Особенно активны обитатели хвойных пород. Так, личинки Z. mi¬
nutissima Zett. способны питаться поврежденными насекомыми
на всех этапах своего развития. Личинки проникают через по¬
врежденные покровы внутрь тела жертвы, где могут концентриро¬
ваться в больших количествах (до нескольких десятков экземпля¬
ров). Возможно, что им’енно такое поведение личинок Z. minu¬
tissima Zett. давало основание отдельным авторам делать заклю¬
чение о хищничестве ксилофильных личинок львинок (Malloch,
1917; Hennig, 1952; Харитонова, 1972).
По нашим данным, многие обитатели лиственных пород на
начальных этапах своего развития концентрируются на участках
со скоплениями древесного сока. Это дает основание предполагать
возможность питания молодых личинок растительной пищей и пере-
^«лючение на питание животной пищей на заключительных этапах
развития. При этом некоторые виды, например Xylopachygaster
mamaevi Kriv., по-видимому, способны питаться только соком и
различными микроорганизмами.
272
Таким образом, на основании всего сказанного тип питания
ксилофильных личинок львинок может быть охарактеризован
как сапронекрофагия, с преобладанием некрофагии у отдельных
видов.
Морфология, Тело личинок (рис. 66, 1) уплощено дорсовент-
рально, веретеновидное или эллиптическое, более вытянутое у вод¬
ных и амфибиотических форм и укороченное у почвообитающих
и ксилофильных. Окраска тела — от сероватой до коричневой,
часто с различным рисунком, образованным более светлыми или
темными пятнами и полосами. Покровы тела личинок пропитаны
известью, что придает им жесткость и хрупкость.
Голова личинок массивная, удлиненно-овальная, вследствие
значительного уплотнения покровов способна лишь незначительно
втягиваться в грудные сегменты. Задний отдел головы, втянутый
в 1-й грудной сегмент, значительно светлее, более слабо склеро-
тизован по сравнению с передним. Головная капсула представлена
удлиненной срединной (фронтоклипеус), 2 боковыми пластинками
и сильно укороченной почти квадратной вентральной пластинкой.
Внутренний скелет головы представлен 2 тенториальными стерж¬
нями. В передней части головы расположены глазки и небольшие
чаще всего двучлениковые усики.
В качестве верхней губы рассматривается непарный срединный
выступ, являющийся продолжением фронто-клипеальной пластин¬
ки. У личинок львинок имеется лишь одна пара челюстей (манди-
булярно-максиллярный комплекс), состоящая из двух члеников.
Широкий трапециевидный базальный членик по дорсальному
(наружному) краю с сильными направленными назад щетинками.
Дистальный членик удлиненный или квадратный, на вершине
обычно с несколькими рядами и одним пучком изогнутых длин¬
ных щетинок. В целом эти образования служат для соскребания
пищи с поверхности. Щупик максилл расположен на наружной
стороне дистального членика. За нижнюю губу большинство
авторов принимает небольшой выступ, усаженный короткими, на¬
правленными вперед волосками.
Тело образовано 11 сегментами, иногда имеется небольшой 12-й
сегмент (Nemotelus Geoffr.). Поверхность сегментов тела с различ¬
ными многоугольными, квадратными и округлыми пластинками
различных размеров и окраски. В ряде случаев дополнительно
имеются поперечные ряды выступающих пластинок или бугорков.
Хетотаксия тела однородна. Сегменты тела с дорсальной и вент¬
ральной сторон несут обычно по 6 щетинок, ориентированных в 1
или 2 поперечных ряда (на 1-м грудном с дорсальной и всех груд¬
ных сегментах с вентральной стороны щетинок обычно больше);
имеется также по 3—4 латеральных щетинки. На последнем сег¬
менте тела хорошо развиты 2 дорсальные, 6—8 терминальных и
несколько пар вентральных щетинок (рис. 93, 7, 5). Относитель¬
ная длина щетинок может быть различной. У личинок Beridinae
щетинки окружены пучками волосков.
273
Трахейная система голопнейстического типа, одпако функцио¬
нально это амфипнейстическая (для наземных) и метапнейстиче-
ская (для водных) система, так как хорошо развиты и функциони¬
руют лишь передние и задние или только задние дыхальца, а
остальные сильно редуцированы, сохранились лишь в виде неболь¬
ших бугорков. Передние дыхальца грушевидной формы, находятся
непосредственно на поверхности сегмента, задние — овальные^
расположены в стигмальной камере, открывающейся поперечной
ш,елью. Камера находится на дорсальной стороне последнего
сегмента (рис. 93, 5) (у большинства ксилофильных и почвооби-
таюш,их форм) или на конце сегмента (у водных и амфибиотических
форм). У некоторых водных форм последний сегмент, несуш,ий
венчик волосков, вытянут в длинную трубку. Анальное отверстие
удлиненное, ш^елевидное, расположено в среднем отделе послед¬
него сегмента с вентральной стороны (рис. 93, 1). Для львинок
характерно развитие куколки внутри шкурки личинки последнего
возраста. При вылете имаго шкурка личинки раскрывается
Т-образной ш,елью.
В приводимую ниже определительную таблицу не включены
роды Pachygaster Meig. и Praomyia Keri., личинки европейских
видов которых (Pachygaster atra Pz. и Praomyia leachii Curt.)
изучены недостаточно и в наших материалах не представлены.
Таблица для определения родов семейства Stratiomyidae
1 (2) Щетинки сегментов тела окружены волосками, которые об¬
разуют более или менее четкий пучок. Последний сегмент
тела закруглен, по краю с тонким опушением, образуемым
многочисленными волосками. В разложившихся пропи¬
танных влагой плодовых телах различных древесных гри¬
бов, иногда под корой деревьев, разлагаюш,ихся вблизи
воды Beris Latr.
2 (1) Щетинки сегментов тела не окружены волосками. Послед¬
ний сегмент тела без краевого опушения (рис. 93, 7).
3 (6) Стигмальная ш,ель, хотя и находится на дорсальной сто¬
роне последнего сегмента, смещена к его концу (рис. 94,
7; 95, i). Анальная щель ограничена массивными плас¬
тинками; сходные структуры расположены на дуговидной
складке, ограничивающей анальную щель спереди
(рис. 94, 5). Средние отделы 3—6-го брюшных сегментов с
12—16 темными овальными пластинками или вообн^е без
них.
4 (5) Тело покрыто однотипными по строению, размерам п
ориентации неправильными пластинками, не собранными
в поперечные ряды. Их размеры лишь слегка уменьшаются
к краям сегментов. В средней части дорсальной стороны
3—6-го брюшных сегментов четко выделяется поперечный
ряд из 12—16 темных овальных пластинок (рис. 94, У). На
274
последнем сегменте тела хорошо заметны лишь 2 пары тер¬
минальных ш,етинок (рис. 94, 7)
Craspedometopon КеП. (стр. 281)
5 (4) Тело покрыто двумя группами кугикулярных структур,
резко различаюш,ихся по строению, размерам и ориента¬
ции друг относительно друга (рис. 95, 4). В средней части
дорсальной стороны 3—6-го брюшных сегментов в беспо¬
рядке расположены крупные многоугольные, равномерно
окрашенные пластинки, их окружают в несколько раз мень¬
шие, преимущественно прямоугольные пластинки, обра¬
зующие в задней трети сегментов четкие поперечные ря¬
ды. Поперечный ряд темных пластинок отсутствует.
На последнем сегменте тела хорошо заметны, по меньшей
мере, 3 пары терминальных щетинок, 4-я (срединная)
пара — очень короткая (рис. 95, 1)
Maackiana Kriv. (стр. 283).
С (3) Стигмальпая щель заметно удалена от заднего края сег¬
мента, не смещена к его концу (рис. 93, 5, 7). Анальная
щель по краю с небольшими пластинками или длинными
зубцевидными выступами. Структуры складки, ограни¬
чивающей спереди анальную щель, не отличаются от
структур основной поверхности сегмента. Средние от¬
делы 3—6-го брюшных сегментов, самое большее, с 8 ши¬
роко расставленными темными пластинками.
7 (10) Светлое поле 6-го брюшного сегмента находится перед по¬
перечным рядом вентральных щетинок (рис. 98, 4), Аналь¬
ная щель по краю с длинными зубцевидными выступами,
длина которых более чем в 2 раза превышает их ширину
(рис. 98, 2).
8 (9) Краевые дорсальные щетинки расположены вблизи круп¬
ной овальной темной пластинки боковой группы (отделе¬
ны 1—2 рядами кутикулярных структур; рис. 98, 6).
На 3-м грудном сегменте основание краевых и промежу¬
точных щетинок окружено лишь одним концентрическим
рядом мелких пластинок (рис. 98, 5). Следующие ряды
пластинок крупные. 2 дорсальные срединные темные плас¬
тинки 2-го межсегментного ряда отделены от ближайших
темных пластинок ряда не более чем 3 рядами кутикуляр¬
ных структур (рис. 98, 6) . . .Wallacea Dol. (стр. 290)
Я (8) Краевые дорсальные щетинки удалены от крупной оваль¬
ной темной пластинки боковой группы (отделены 4—5 ря¬
дами кутикулярных структур; рис. 98, 8). На 3-м грудном
сегменте между основаниями краевых и промежуточных
дорсальных щетинок имеется поле мелких пластинок
(рис. 98, 9). 2 дорсальные срединные темные пластинки
2-го межсегментного ряда отделены от ближайших тем¬
ных пластинок ряда не менее чем 5 рядами кутикулярных
структур Pseudowallacea Kert. (стр. 288)
275
10 (7) Светлое поле Г)-го брюшного сегмента расположено в сред¬
нем отделе сегмента, на уровне поперечного ряда вентраль¬
ных щетинок (рис. 93, 2, 6). Анальная щель по краю с не¬
большими квадратными или прямоугольными пластинка¬
ми, длина которых не более чем в 2 раза превышает их
ширину (рис. 93, 1).
И (16) Две последние пары терминальных щетинок (^з и ^4) длин¬
ные, их длина, по меньшей мере, достигает ^/3 длины по¬
следнего сегмента.
12 (13) Стигмальная щель с короткой стигмальной складкой, не
доходящей до края сегмента (рис. 95, 5). Краевые дор¬
сальные щетинки брюшных сегментов очень короткие, в
несколько раз короче остальных. Вентральные щетинки
6-го брюшного сегмента расположены на уровне заднего
края светлого поля (рис. 95, 6)
.... Eupachy^j aster Kerl. (стр. 281).
13 (12) Стигмальная щель с длинной стигмальной складкой, до¬
ходящей до края сегмента (рис. 93, 7; 96, !)). Все 6 дор¬
сальных щетинок брюшных сегментов длинные. Вентраль¬
ные щетинки 6-го брюшного сегмента находятся на уровне
последней трети светлого поля (рис. 96, 7).
14 (15) Из 4 пар терминальных щетинок наиболее длинные 2 по¬
следние пары, достигающие ^/3 длины сегмента (рис. 9(), -7).
1-й грудной сегмент с 10 дорсальными щетинками, рас¬
положенными следующим образом: 2 пары щетинок —
в первом и 3 пары щетинок — во втором ряду (как на
рис. 93,5) Xylopachytaster Kriv. (стр. 291).
15 (14) Все 4 пары терминальных щетинок почти равных разме¬
ров, очень длинные, превосходят длину сегмента (рис. 93,
7). 1-й грудной сегмент с 10 дорсальными щетинками, рас¬
положенными следующим образом: 3 пары щетинок рас¬
положены в первом, 2 пары щетинок — во втором ряду
(рис. 93, ^) Zabrachia Coq. (стр. 291)
16 (И) Из двух последних пар терминальных щетинок (/3 и /4)
одна пара в несколько раз короче другой, а длина наи¬
более длинных не превышает половин1.1 длины последнего
сегмента (рис. 93, 1).
17 (18) Последний сегмент тела тупо обрублен и широко закруг¬
лен на конце (рис. 93, /, 5). Три из 4 пар терминальных
щетинок смещены к заднему краю сегмента (рис. 93, .5).
Длина анальной щели составляет не более V3 длины сег¬
мента (рис. 93, 7) Aspidacaniha Kert. (стр. 277)
18 (17) Последний сегмент тела постепенно сужается к концу
(рис. 94, 77, 12). Терминальные щетинки не смещены к зад¬
нему краю сегмента. Длина анальной н;ели составляет
не менее ^^2 длины сегмента.
19 (20) Срединные терминальные щетинки (/4) заметно длиннее
соседних (/3) (рис. 94, 77). Поперечные ряды темных плас-
276
ТИ1ЮК в межсегментных складках на дорсальной стороне
4—6-го брюшных сегментов отсутствуют
Ornopyramis Kriv. (стр. 288).
20 (19) Срединные терминальные щетинки {t^) заметно короче
соседних (^з) (рис. 94, 1). Поперечные ряды темных плас¬
тинок в межсегментных складках всех сегментов хорошо
развиты.
21 (22) На дорсальной стороне брюшных сегментов среди кути-
кулярных структур выделяются, крупные ромбовидные
и многоугольные пластинки, образуюц;ие 2 четких сбли¬
женных поперечных ряда (рис. 94, 5)
Cechorismenus Kert. (стр. 279).
22 (21) Кутикулярные структуры дорсальной стороны брюшных
сегментов тела однородные, могут лишь несколько разли¬
чаться относительными размерами
Neopachygaster Aust. (стр. 283)
Род Aspidacantha Kert.
Анальная щель по краю с небольшими прямоугольными плас¬
тинками. Из 4 пар терминальных щетинок 3 пары длинные, их
размеры возрастают от 3-й к 1-й паре, срединные — короткие.
Светлое поле на вентральной стороне 6-го сегмента (рис. 93, 2}
расположено в среднем отделе сегмента, удлиненно-овальное,
с перетяжкой в задней трети, на уровне которой расположены
срединные вентральные щетинки. Покровы тела преимуществен¬
но с однородными многоугольными пластинками (рис. 93, 4).
В настоящее время мы располагаем данными лишь по морфологии
и экологии личинок А. atra Kerl. (Кривошеина, 1975), который
является типовым видом рода и единственным его палеарктиче-
ским представителем.
А. atra Kert. (рис. 93, 1—5). Тело достигает в длину 6 мм, ко¬
ричневатое, однородно окрашено. Второй ряд темных пластинок,
расположенных в межсегментных складках на дорсальной сторо¬
не, выгнут дугообразно вблизи средней линии тела. Анальная
щель по краю с 13—14 прямоугольными пластинками.
Личинки этого вида собраны в тугаях по Амударье, в окрест¬
ностях г. Дейнау. Личинки развиваются под корой разлагающих¬
ся деревьев туранги, где встречаются совместно с личинками ленто-
крылок Myeunis tricolor Hend. (Otitidae). Взрослые личинки
были многочисленны в течение апреля, в этот период отмечено
питание поврежденными личинками лентокрылок.
277
Рис. 93. Детали строения личинок Aspidacantha atra Kert. (l—б) и Zabra-
chia minutissima Zett. (6—10) (Stratiomyidae)
1 — конец тела снизу; 2,6 — светлое поле на вентральной стороне 6-го брюшного сег¬
мента; 3, 8 — 1-й грудной сегмент сверху; 4 — 4-й брюшной сегмент сверху; 5,7 — по¬
следний сегмент тела сверху; 9, ю — кутикулярные структуры вблизи средней линии
тела на дорсальной стороне 3-го грудного и 4-го брюшного сегментов
а — срединная темная пластинка второго межсегментного ряда; ап — анальная щель;
с — межсегментные ряды темных пластинок; di— d, — краевая, промежуточная и средин¬
ная дорсальные щетинки; I — боковые группы темных пластинок; 8t. р1 — стигмальная
пластинка; U — i* — пары терминальных щетинок iU — краевые, tt — срединные); г^_5 —
вентральные щетинки посчеднего сегмента тела1(г1 — первая, г» — последние пары)
.278
Род Cechorismenus Kert.
Последний сегмент тела (рис. 94, /, 2) с четкими угловидными
выступами около 2-й и 3-й пар терминальных щетинок, между ко¬
торыми расположена стигмальная щель; стигмальная складка
небольшая, оканчивается на уровне 3-й парЫ щетинок. Длина
анальной щели составляет почти длины сегмента, по ее краям
расположены прямоугольные пластинки, размеры которых слег¬
ка превышают размеры окружающих кутикулярных структур.
Из 4 пар терминальных щетинок 3 пары длинные, их размеры
возрастают от 3-й к 1-й паре, срединные слабо заметны.
Светлое поле на вентральной стороне О-го брюшного сегмента
(рис. 94, 4) расположено в средней части сегмента, удлиненно¬
овальное, с перетяжкой в задней трети, вблизи которой располо¬
жены срединные вентральные щетинки. Тело покрыто прямоуголь¬
ными или многоугольными кутикулярными структурами, образую¬
щими характерный рисунок (рис. 94, 3). На дорсальной и
вентральной сторонах грудных сегментов преобладают многоуголь¬
ные пластинки, ориентированные однородно (за исключением об¬
ласти линочного шва). Пластинки в средних отделах сегментов
крупные и значительно, в 2—3 раза, уменьшаются в размерах к
периферии. На дорсальной стороне средних брюшных сегментов
в их задней ^/4 2 поперечных ряда своеобразных пластинок. Первый
из этих рядов образован ромбовидными, второй — крупными
многоугольными пластинками. На вентральной стороне, в задней
четверти сегментов пластинки прямоугольные и образуют четкие
поперечные ряды.
С. flavicomis Kert. Единственный известный вид этого рода,
описан по самцу с о. Тайвань (Kertesz, 1916). Наши материалы
из Приморского края (Комарово-Заповедное), включающие сам¬
цов и самок, мы относим, хотя и с некоторой долей сомнения, к
С. flavicomis Kert. Самцы полностью идентичны с С. flavicortiis
Kert. (описанному по самцам), лишь 3-й членик усика несколько
крупнее. Самки несколько сближаются с представителями рода
Diargemus Kert. (описанного по самкам). Личинка (рис. 94, i —4)-
достигает в длину 5—6 мм, тело светлое, слегка коричневатое.
Многоугольные крупные пластинки расположены вблизи средней
линии тела. Поперечный ряд из светлых ромбовидных и темных
многоугольных пластинок относительно короткий, не достигает
боковых краев сегментов. Ромбовидные пластинки с четким вы¬
ступом сзади. Щетинки тела широкие, слегка уплощенные, па
вершине закругленные (рис. 94, 5), вентральные щетинки не¬
сколько уже дорсальных.
Личинки довольно обычны под корой бархата амурского и
ильма долинного, предпочитают влажные лубяные волокна ле>1ча-
щих на земле стволов, где встречаются совместно с целым ком¬
плексом двукрылых — типичных обитателей зоны коры. Тип пи¬
тания не установлен.
279
Рис. 94. Детали строения личинок Cechorismenus flavicomis Kert. {1—4),
Craspedometopon ussuricum Kriv. (5—9) и Ornopyramis tener Kriv. (10—13)
(Stratiomyidae)
1, 2, 7, 8, 11, 12 — конец тела сверху и снизу; 3, 9, 13 — 4-й брюшной сегмент тела свер¬
ху; 4, б, 10 — светлое поле на вентральной стороне 6-го брюшного сегмента; 5 — аналь-
лая щель с окружающими ее кутикулярными структурами
•<1 — и — пары терминальных щетинок
Род Craspedometopon Kert.
Последний сегмент тела (рис. 94, 7, 8) обрублен и широко
закруглен, короткий, его длина заметно меньше ширины. Свет¬
лое поле на вентральной стороне 6-го брюшного сегмента (рис. 94,
6) овальное, без перетяжки, занимает среднюю четверть сегмента.
Срединные вентральные ш,етинки находятся почти на уровне его
среднего отдела. Кутикулярные структуры тела (рис. 94, 9) в ви¬
де многоугольных и прямоугольных неправильной формы плас¬
тинок.
Род представлен в Палеарктике 2 видами: С. ussuricum Kriv.
и С. fronlale Kert. Первый вид описан из Приморского края
(Кривошеина, 1973а), второй — с о. Тайвань (Kertesz, 1909),
известен из Индии (Brunelti, 1920), Японии (Nagatomi, 1975).
Биология его представителей не изучена. Ниже приводятся крат¬
кие сведения по виду С. ussuricum Kriv.
С. ussuricum Kriv. Личинка (рис. 94, 5—9) достигает длины
9—10 см, светло-коричневая с темно-коричневым рисунком на
дорсальной и вентральной сторонах. На дорсальной стороне на
обш,ем темном фоне выделяются светлые округлые пятна, рас¬
положенные в основании щетинок, и светлые участки вдоль перед¬
него и заднего края сегментов. Отсутствуют какие-либо группы
пластинок, резко отличающиеся по размерам от остальных. Ще¬
тинки тела заметно утолщены в средней части.
Личинки в Приморском крае развиваются под корой листвен¬
ных деревьев, первоначально заселяя участки по границе со
свежим лубом, например, расщепы пней дуба. Встречаются под
корой бархата амурского и ильма долинного. Взрослые личинки,
вероятнее всего, являются некрофагами.
Род Eupachygaster Kert.
Последний сегмент тела закругленный, полусферический
(рис. 95, 5). Анальная щель по периферии без особых кутикуляр-
ных структур. Все 4 пары терминальных щетинок длинные, при¬
чем 2 пары промежуточных (^2 и t^) щетинок длиннее сегмента.
Светлое поле на вентральной стороне 6-го брюшного сегмента уд¬
линенное (рис. 95, 6), расположено в средней части сегмента,
его задний край находится на уровне поперечного ряда вентраль
ных щетинок. Все тело личинок покрыто многоугольными, плотно
примыкающими друг к другу и одинаково ориентированными
пластинками.
Е. tarsalis Zett. является типовым видом рода и единственным
его налеарктическим представителем. Вид недавно зарегистриро¬
ван на территории пашей страны (Кривошеина, 1975).
JFhmhhkh (рис. 95, 5—7) достигают длины 9—10 мм, темно-
ко^ ..чиевые, с 2 продольными полосами как с дорсальной, так
и вентральной сторон. Щетинки тела закруглены на вершине,
281
eoe о о о Q
Рис. 95. Детали строения личинок Maackiana laminiformis Kriv. (/—4)
и Eupachygaster tarsalis Zett. (5—7) (Stratiomyidae)
1,5 — последний сегмент тела сверху; 2 — то же, снизу; 3,6 — светлое поле на вент¬
ральной стороне 6-го брюшного сегмента; 4 — 4-й брюшной сегмент тела сверху; 7 — ще¬
тинки 1-го грудного сегмента
на всем протяжении с редким опушением. Подробные материалы
по морфологии личинки приведены в работе Н. П. Кривошеиной
(1975).
Сведения по биологии личинок в литературе очень скудны.
Указывается на развитие вида под корой различных лиственных
пород — тополя, дуба, ильма, яблони и др. (Brindle, 1962а; Roz-
kosny, 1973), а также на возможность развития в разлагающейся
древесине (Rozkosny, 1973). В Копет-Даге (урочище Ипайкала)
личинки Е. tarsalis Zett. обнаружены во влажной трухе дупел гре¬
цкого ореха (Juglans regia L.). Процесс образования этих дупел
связан с санитарными рубками — спиливанием усыхающих вет¬
вей. На незащищенных спилах нередко поселяются личинки дрово¬
секов, проделывающие ходы в толщу древесины и способствующие
282
проникновению внутрь ствола различных микроорганизмов. Вслед
за ними разрушающиеся участки древесины заселяют личинки
>1хуков-узконадкрылок (Oedemeridae), которым сопутствуют ли¬
чинки Е. tarsalis Zett. Личинки были многочисленны в течение
мая и июня. Тип питания не установлен.
Род Maackiana Kriv. ( = Takagimyia Nag., syn. nov.)
Последний сегмент тела (рис. 95, 1) на конце тупо обрублен
и закруглен, его ширина заметно превосходит длину. Светлое поле
на вентральной стороне 6-го брюшного сегмента (рис. 95, 3) рас¬
положено в средней трети сегмента, удлиненное, с перетяжкой в
передней трети, вблизи которой расположены срединные вентраль¬
ные щетинки. Тело личинок покрыто очень разнородными
по форме и размерам кутикулярными структурами (рис. 95, 4).
Род с единственным представителем был описан из Комарово-
Заповедное Приморского края (Кривошеина, 1973а). Описанный
несколько позднее из Японии род Takagimyia Nag. (Nagatomi,
1975), несомненно, является синонимом Maackiana Kriv. Вопрос
о близости видов М. laminiformis Kriv. и Т. circularis Nag. может
быть решен после детального сравнения типовых материалов.
М. laminiformis Kriv. Личинка (рис. 95, i—^) достигает длины
8 мм, тело коричневатое, кажется пестрым благодаря наличию
затемненных крупных пластинок в средних отделах сегментов. Все
щетинки тела утолщены в основной половине и сужены и за¬
острены у вершины. Анальная щель с каждой стороны ограниче¬
на 7—8 прямоугольно-овальными пластинками.
Личинки обычны в маакии амурской, где развиваются в толще
лубяных волокон совместно с личинками других видов двукры¬
лых, в том числе львинок. Сапронекрофаги.
Род Neopachygaster Aust.
Последний сегмент тела (рис. 96, 1, 2) в задней половине тра¬
пециевидной формы с нечетко выраженными, но заметными угла¬
ми в местах прикрепления терминальных щетинок. Анальная
щель по краю с прямоугольными пластинками. Из 4 пар терминаль¬
ных щетинок последнего сегмента срединные щетинки очень
короткие. Светлое поле па вентральной стороне 6-го брюшного
сегмента удлиненное, с небольшой перетяжкой в задней трети,
расположенной на уровне вентральных щетинок (рис. 96, 3).
Все тело личинок покрыто многоугольными, плотно примыкаю¬
щими друг к другу и несколько варьирующими в размерах свет¬
лыми пластинками и выделяющимися на их фоне темными оваль¬
ными пластинками (рис. 97, 4, 5). Детально личинки рода опи¬
саны в работе Н. П. Кривошеиной (1976а).
В Палеарктике известно 4 вида этого рода. Род широко рас¬
пространен в Северной Америке, известен в Эфиопской области.
283
Рис. 96. Детали строения личинок Neopachygaster intermedia Kriv.
л Xylopachygaster mamaevi Kriv. (5—<‘?) (Stratiomyidae)
1^2, 5, 6 — последний сегмент тела сверху и снизу; з, 7 — светлое поле на вентральнФЙ
стороне 6-го брюшного сегмента тела; 4, в — 4-й брюшной сегмент тела сверху
ti — и — пары терминальных щетинок
Таблица для определения видов рода Neopachygaster Aust.
1 (2) Ряды темных пластинок в межсегментных складках между
1-м и 2-м брюшными сегментами с дорсальной стороны разде¬
лены 5, а между 2-м и 3-м брюшными сегментами — 7 ря¬
дами кутикулярных структур. Срединные дорсальные тем¬
ные пластинки 2-го межсегментного ряда средних брюшных
сегментов сильно смещены назад (рис. 96, 4), отделены от
соседних пластинок не менее чем 3 рядами кутикулярных
структур. На вентральной стороне средних сегментов тела
темные пластинки 2-го межсегментного ряда образуют четкий
выступ вблизи средней линии .... N. intermedia Kriv.
2 (1) Ряды темных пластинок в межсегментных складках между
1-м и 2-м брюшными сегментами с дорсальной стороны разде¬
лены 4, а на границе следующих сегментов — 5 рядами ку¬
тикулярных структур. Срединные дорсальные темные плас¬
тинки 2-го межсегментного ряда средних брюшных сегмен¬
тов отделены от соседних пластинок не более чем 2 рядами
кутикулярных структур (рис. 97, 4). На вентральной сто¬
роне средних сегментов тела темные пластинки 2-го межсег¬
ментного ряда не образуют выступ вблизи средней линии тела
(рис. 97, 6).
3 (4) Ряды темных пластинок на вентральной стороне в межсег¬
ментных складках между 2-м и 3-м грудными сегментами
разделены 2 рядами кутикулярных структур. Срединные
дорсальные темные пластинки 2-го межсегментного ряда сред¬
них брюшных сегментов отделены от соседних пластинок
1 рядом кутикулярных структур (рис. 97, 2)
N. stackelbergi Kriv.
4 (3) Ряды темных пластинок на вентральной стороне в межсег¬
ментных складках между 2-м и 3-м грудными сегментами
разделены 1 рядом кутикулярных структур. Срединные дор¬
сальные темные пластинки 2-го межсегментного ряда сред¬
них брюшных сегментов отделены от соседних пластинок
2 рядами кутикулярных структур (рис. 97, 4).
5 (6) На дорсальной стороне 3-го грудного сегмента между крае¬
выми и промежуточными дорсальными щетинками располо¬
жена группа очень мелких кутикулярных структур, более
мелких, чем окружающие пластинки (рис. 97, 3). Поверх¬
ность сегментов с дорсальной стороны с относительно круп¬
ными кутикулярными структурами, в ряде случаев почти
достигающими размеров темных пластинок
N. meromelaena Duf.
6 (5) На дорсальной стороне 3-го грудного сегмента между крае¬
выми и промежуточными щетинками кутикулярные струк¬
туры такого же размера, как и на других участках сегмен¬
тов. Поверхность сегментов с дорсальной стороны с относи¬
тельно мелкими кутикулярными структурами, которые в
285
несколько раз меньше темных пластинок
х\. secernibilis Kriv.
N, intermedia Kriv. описан из горных районов Средней Азии
(Зеравшанский хребет) (Кривошеина, 19()5). В настоящее время
найден также в горах Киргизии (заповедник Сары-Челек),
Таджикистана (ущелье Кондара) и в Туркмении под г. Дейнау.
Вид, очевидно, широко распространен в горных районах Средней
Азии.
Тело личинок достигает длины 8 мм. Все сегменты тела покрыты
многоугольными, равномерно окрашенными коричневатыми плас¬
тинками (рис. 96, 2).
Личинки развиваются под влажной корой разлагающихся
деревьев грецкого ореха, ивы, туранги. И Туркмении под г. Дей¬
нау личинки обычны во влажной трухе в дуплах н в натеках сока
на стволах туранги и только в единичных случаях встречаются под
ее корой. Видимо, в условиях засушливого климата личинки раз¬
виваются в не свойственных в целом для рода средах, какими яв¬
ляются натеки сока, сохраняющие высокую влажность в течение
летнего периода.
N. meromelaena Duf. встречается в северной и средней полосе
Западной Европы, на территории Европейской части СССР за¬
регистрирован от Ленинградской области до предгорий Северного
Кавказа.
Тело личинок, достигающее длины 7 мм, темно-коричневое.
Срединная часть сегментов выделяется более крупными разме¬
рами пластинок (рис. 97, 3).
Имеющиеся в литературе сведения об образе жизни личинок
очень отрывочны. Указывается на развитие N. meromelaena Dh1‘.
под корой падуба (Verrall, 1909; Brindle, 1962а), а также тополя
(Кривошеина, 1965; Rozkosny, 1973). На территории нашей
страны личинки зарегистрированы нами под корой различных
лиственных деревьев. Наиболее обычны личинки под корой то¬
поля и осины, в единичных случаях отмечены в иве, клене, вя¬
зе шершавом, дубе черешчатом, грабе обыкновенном, буке восточ¬
ном. Личинки, как правило, концентрируются под корой, но в
отдельных случаях проникают в толщу древесины, например по
ходам короедов-древесинников. Под корой личинки развиваются
в сообществе различных насекомых, в том числе львинок, пред¬
почитают влажные участки коры. Сапронекрофаги, в лаборатор¬
ных условиях взрослые личинки питались погибшими личин¬
ками насекомых.
N. secernibilis Kriv. Вид описан из Приморского края (Криво¬
шеина, 1973а), в настоящее время найден также на Курильских
островах (о. Кунашир) и в Тувинской АССР.
Тело личинок, достигающее длины 6 мм, коричневатое, с одно¬
родными равных размеров пластинками (рис. 97, 5), лишь слегка
увеличенными на грудных сегментах, вблизи линочного шва.
Личинки развиваются под корой тополя лавролистного (Ту¬
286
Рис. 97. Кутикулярные структуры на поверхности сегментов личинок
Neopachygaster stackelbergi Kriv. (i, 2), N. meromelaena Duf. (3, 4) и N.
«ecernibilis Kriv. (5, 6) (Stratiomyidae)
1,2, 4, 5 — область темных боковых пластинок (О и средней линии тела (с — л) на дор¬
сальной стороне 4-го брюшного сегмента тела; J — то же, на дорсальной стороне 3-го
грудного сегмента; б — то же, на вентральной стороне 4-го брюшного сегмента тела
dj, d, — краевая и промежуточная дорсальные щетинки
винская АССР, Иштии-Хем), под корой тополя Максимовича и
ильма долинного (Приморский край, Комарово-Заповедное), под
корой бархата сахалинского (Курильские о-ва, о. Кунашир).
Личинки предпочитают влажные лубяные волокна лежащих на
земле стволов. Сапронекрофаги, взрослые личинки питаются по¬
гибшими личинками различных ксилофильных насекомых.
N, stackelbergi Kriv, Вид описан из Приморского края (Криво-
иеина, 1973а), зарегистрирован на о. Кунашир.
Тело личинки, достигающее длины 5 мм, с дорсальной стороны
окрашено неравномерно: на светлом желтоватом фоне сегмента
выделяются более темные участки в переднем и среднем отделах
сегмента. Пластинки, расположенные на дорсальной стороне
сегментов вблизи средней линии, укрупнены (рис. 97, 2, 2),
Личинки развиваются преимущественно под корой ильма до¬
линного, встречаются под корой бархата амурского и тополя
Максимовича. Предпочитают участки, обильно смачиваемые
287
соком, откуда перед окукливанием переходят в темную труху в тре¬
щинах коры. Обычно встречаются в сообществе с личинками
ксиломиид (Xylomyidae) и различных львинок (подсем. Pachy-
gasterinae). Сапронекрофаги. Личинки близкого северо-американ-
ского вида — N. maculicornis Hine обнаружены под корой то¬
поля канадского — Populus delLoides March. (Kraft, Соок, 1961).
Род Ornopyramis Kriv.
Последний сегмент тела (рис. 94, 11, 12) конусовидный, на
вершине закруглен. Светлое поле (рис. 94, 10) расположено в
среднем отделе вентральной стороны б-го брюшного сегмента,
удлиненное, с перетяжкой. Срединные вентральные щетинки на¬
ходятся вблизи поля, за его серединой. Все тело личинок покрыто
однородными мелкими многоугольно-овальными кутикулярными
структурами, расположенными в шахматном порядке и не об¬
разующими четких рядов (рис. 94, 13).
Род с единственным видом О. tener Kriv. описан по материалам
из Приморского края (Кривошеина, 1973а).
О. tener Kriv. (рис. 94, 10—13). Тело личинок достигает длины
5—6 мм, светлое, коричневатое. Для вида характерны своеобраз¬
ные утолщенные дорсальные и латеральные щетинки.
Личинки в Приморском крае (Комарово-Заповедное) разви¬
ваются в лубе ильма долинного. Обычно встречаются совместно
с личинками других видов львинок, преимущественно рода Neo-
pachygasLer Aust. Для личинки этого вида характерна сильно вы¬
тянутая и несколько заостренная голова, что может указывать
на питание животной пищей, но прямые наблюдения о характере
питания отсутствуют.
Род Pseudowallacea Kert.
Последний сегмент тела широко закруглен, на конце без чет¬
ких выступов. Стигмальная щель расположена между 2-й и 3-й
парами терминальных щетинок. По краям анальной щели распо¬
ложены зубцевидные, тупые удлиненные выступы, направленные к
средней линии тела. С каждой стороны от анальной щели 16—
18 таких выступов. Светлое поле на вентральной стороне 6-го
брюшного сегмента овальное, смещено к переднему краю сегмен¬
та и находится перед поперечным рядом вентральных щетинок на
расстоянии, не уступающем длине самого поля. Поверхность сег¬
ментов тела равномерно покрыта относительно мелкими неправиль¬
ной формы пластинками, не образующими четких рядов. На об¬
щем фоне кутикулярных структур сегментов выделяются относи¬
тельно более крупные и более темные пластинки (рис. 98, 8),
2 срединные дорсальные темные пластинки 2-го межсегментного ря¬
да на средних сегментах тела заметно смещены назад (рис. 98, 8),
Щетинки тела расширены в нижней половине и сужены и заост¬
рены к вершине.
288
Гис. 98. Детали строения личинок Wallacea nigrotibialis Pl. (i—6) и Pseudo-
wallacea hungarica Kert. {J—9) (Stratiomyidae)
{,2 — последний сегмент тела сверху и снизу; з — 4-й брюшной сегмент тела сверху;
4 — светлое поле на вентральной стороне 6-го брюшного сегмента; кутикулярные струк¬
туры: 5, 9 — на дорсальной стороне 3-го грудного сегмента, б, 5 — на дорсальной сторо¬
не 4-го брюшного сегмента; 7 — ряды темных пластинок в межсегментных складках дор¬
сальной стороны брюшных сегментов
Обозначения см. на рис. 93.
В роде Pseudowallacea известен лишь 1 вид — Р. hungarica
Kert., зарегистрированный в настоящее время в Румынии, Швеции,
Финляндии (Frey, 1941; Hanson, 1942; Rozkosny, 1973), а также
в Европейской части СССР (Кривошеина, 1965; Нарчук, 19696).
Роскошный (Rozkosny, 1973) считает вид синонимом северо-
10 Б. м. Мамаев и др.
289
aiiiepHKaHCKoro вида Berkshiria albistyliim Johns. В данной работе
вид рассматривается в качестве самостоятельного, впредь до де¬
тального сравнительного изучения типовых материалов.
P.Mmgarica Kert. Личинка достигает в длину 5—6 мм, равно¬
мерно коричневатая. Тело покрыто однородными и однообразно
ориентированными пластинками. На дорсальной стороне брюш¬
ных сегментов между поперечными рядами овальных темных плас¬
тинок межсегментных складок расположено 2—3 ряда значитель¬
но более мелких кутикулярных структур (рис. 98, 7).
Личинки развиваются преимущественно под корой различных
видов тополей. Собраны нами в Вологодской, Московской и в За¬
карпатской областях, Краснодарском крае, в Тувинской АССР.
Предпочитают вла>1чные лубяные волокна лежащих на земле ство¬
лов, где развиваются в сообществе с комплексом насекомых, ти¬
пичным для Wallacea nigrotibialis Pl. и Neopachygaster merome-
laena Duf. Личинки — некрофаги, но, очевидно, с элементами
сапрофагии. Личинки Berkshiria albi stylum Johns. развиваются
под корой тополя канадского (Populus deltoides March.) и вяза
приземистого (Ulmus pumila L.) (Kraft, Соок, 1961).
Род Wallacea Dol.
В роде Wallacea Dol. в настоящее время известно около 10 ви¬
дов, распространенных в различных районах Юго-Восточной Азии.
В Палеарктике известен 1 вид — W. nigrotibialis Р1. Признаки
рода во многом сходны с признаками рода Pseudowallacea Kert.,
основные выявленные отличия указаны в таблице.
W, nigrotibialis Pl. (рис. 98, 1—6), Личинка достигает в дли¬
ну () мм, темно-коричневая, с небольшими светловатыми пятнами
в основании дорсальных щетинок и светлым желтоватым послед¬
ним сегментом тела. Тело покрыто однородными, однообразно
ориентированными многоугольными пластинками, так что поверх¬
ность тела кажется мелкоячеистой (рис. 98, 3). Вокруг дорсаль¬
ных щетннок пластинки несколько укрупнены. На дорсальной
стороне брюшных сегментов между поперечными рядами оваль¬
ных темных пластинок межсегментных складок расположено 2 ря¬
да почти таких же крупных, но более светлых кутикулярных
структур.
Вид описан по материалам из района Тигровой (Pleske, 1930)
и обычен в Комарово-Заповедное (Приморский край), а также на
о. Кунашир (Курильские о-ва). Личинки W. nigrotibialis Pl.
развиваются под корой бархата амурского (Комарово-Заповед-
пое) и бархата сахалинского (о. Кунашир). В отдельных случаях
встречаются под корой ильма долинного, маакии амурской и оре¬
ха маньчжурского. Личинки предпочитают влажные лубяные
волокна лежащих на земле стволов, где они обычно встречаются
« сообществе с личинками копьехвосток (Lonchaea F11.), мегаме-
ринид (Megamerinidae), болотниц (Limoniidae) и других дву¬
^0
крылых. Личинки — сапроиекрофаги. В сходных условиях иод
корой различных лиственных деревьев, в том число в папайе
(Brunetti, 1920), развиваются личинки близких видов этого рода.
Род Xylopachygaster Kriv. ( = Yamalomyia Nat>., syn. nov.)
Последний сегмент тела (рис. 96, 5, 6) слегка обрублен и ши¬
роко закруглен. Светлое поле (рис. 9Г>, 7) на вентральной стороне
Г)-го брюипюго сегмента крупное, удлиненно-овальное, без пере¬
тяжки. Его задний край расположен за поперечным рядом вен¬
тральных щетинок. Все тело покрыто однородными многоуголь¬
ными кутикулярными структурами, размеры которых слегка
уменьшаются к периферии сегментов.
Род с единственным видом был описан по материалам из Ко-
марово-Заповедное Приморского края (Кривошеила, 1973а). Не¬
сомненно, что синонимом этого рода является род Yamatomyia
Nag. (Nagatonii, 1975), описанный из Японии. Вопрос о близости
X. mamaevi Kriv. и Y. japonica (Miyat.) может быть решен лишь
при сравнении типовых материалов. Биология японского вида не
изучена.
X, mamaevi Kriv. (рис. 96, 5—8). Тело личинок длиной до
10 мм, темное, коричневато-серое. Между поперечными рядами
крупных овальных темных пластинок межсегментных складок
(с) расположены значительно более мелкие кутикулярные струк¬
туры.
Личинки в Приморском крае развиваются в плотной светлой
древесине лежаш^их на земле стволов липы, проникая в толш,у
древесины по ходам амброзийных насекомых — личинок жуков
Crassotarsus sp. (Platypodidae) и Sipalinus gigas Fabr. (Curculio-
nidae). Личинки, видимо, питаются продуктами распада древеси¬
ны, мицелием амброзийных грибов, развивающихся в ходах.
Род Zabrachia Coq.
Последний сегмент тела (рис. 93, 7) закругленный, постепенно
сужается от основания к вершине. Светлое поле на вентральной сто¬
роне 6-го брюшного сегмента (рис. 93, 6) удлиненно-овальное,
срединные вентральные щетинки находятся на уровне среднего
отдела поля и приближены к нему. Все тело личинок покрыто
мелкими, неправильной формы кутикулярными структурами,
ориентированными однотипно, но не образующими четких рядов
(рис. 93, 9, 10).
Известны личинки североамерианских видов: Z. polita Ccq.
и Z. plicata Kraft et Соок (Kraft, Соок, 1961). Краткие не совсем
точные сведения о личинке палеарктического вида Z. minutissima
Zett. имеются в работах ряда авторов (Seguy, 1926; Heiinig,
1952; Rozkosny, 1973). Детально личинка этого вида описана
Бриндлем (Brindle, 1962а) и Н. П. Кривошеиной (1965 и др.).
10* 291
Z. minutissima Zett, (рис. 93, в—10), Тело личинок достигает
длины 9 мм, коричневатое, однородно окрашено. Темные пластин¬
ки Л1ежсегментных складок понеречно-овальные, расположенные
между ними кутикулярные структуры в несколько раз мельче.
Две срединные пластинки 2-го межсегментного ряда на дорсаль¬
ной стороне тела сильно смещены назад и находятся от него на
расстоянии, в 1,5 раза превышающем расстояние между пластин¬
ками.
Вид описан из ТПвеции, широко распространен в лесной зоне
Палеарктики. На территории нашей страны зарегистрирован от
Закарпатья до Курильских островов и от Архангельской области
до Северного Кавказа. Личинки в массе развиваются под корой
хвойных деревьев (ель, пихта, лиственница, сосна). В отдельных
случаях личинки обнаружены нами в лиственных деревьях, на¬
пример, под корой и в ходах короедов-древесинников в березе
(Тувинская АССР). Личинки сопутствуют различным видам коро¬
едов. Типичные некрофаги, питающиеся погибшими или повреж¬
денными личинками ксилофильных насекомых, но, видимо, ни¬
когда не переходящие к активному хищничеству.
СЕМЕЙСТВО ASILIDAE -КТЫРИ
Семейство ктырей представляет собой большую и хорошо обо¬
собленную группу двукрылых насекомых, включающую в настоя¬
щее время более 4700 видов (Hull, 1962), распространенных на
всех континентах, кроме Антарктиды. Представленные в фауне
нанюй страны виды входят в состав 4 подсемейств: Asilinae, Dasypo-
goninae, Laphriinae и Leptogastrinae. Подсемейство Leptogastri-
nae некоторые авторы (например, Martin, 1968; Oldroyd, 1969)
рассматривают в качестве самостоятельного семейства. Представи¬
тели подсемейств Asilinae и Dasypogoninae не рассматриваются
в данной работе, так как их личинки не связаны с древесиной.
Подсемейство Laphriinae представлено в фауне СССР родами,
относящимися к 4 трибам (Рихтер, 1968): Maira Schin., Laphria
Meig., Choerades Walk., Niisa Walk. (Laphriini), Andrenosoma
Rond., Pogonosoma Rond. (Andrenosomini), Ctenota Loew (Cte-
notini) и Loewinella Herm. (Atomosiini). Подсемейство Lepto¬
gastrinae, распространенное в основном в тропиках, включает
7 родов, известных в Палеарктике, из которых 4 зарегистриро¬
ваны на территории нашей страны (Лер, 1961): Euscelidia Westw.,
Psilonyx Aldr., Leptogaster Meig., Mesoleptogaster Frey.
Экология. Ктыри встречаются во всех ландшафтных зонах,
но наиболее характерны для открытых пространств, в первую
очередь степей и полупустынь. Существует определенная при¬
уроченность различных систематических групп ктырей к разным
ландшафтным зонам, что в значительной мере определяется обра¬
зом жизни личинок, которые делятся на две большие группы: оби¬
татели почвы (педобионты) и обитатели древесины (ксилобионты).
292
в почве развиваются широко распространенные в различных
:зонах земного шара представители подсемейств Asilinae, Dasypo-
^oninae и отдельные группы Leptogastrinae. Из последних извест¬
на биология лишь 2 родов. Личинки Leptogaster Meig. развива¬
ются в почве (Melin, 1923; Brindle, 1962b; Кривошеина, Мамаев,
1967), личинки Mesoleptogaster eous Lehr — в толщ,е разлагаю-
ш,ейся древесины (Кривошеина, Мамаев, 1967). Из подсемейства
Laphriinae в древесине зарегистрированы личинки всех известных
в настоящ,ее время видов из родов Laphria, Choerades, Andreno-
soma и Pogonosoma (Кривошеина, Мамаев, 1967, 1973, 197Г)а,
б). Личинки Ctenota molitrix Loew и С. riificornis v. d. Wulp,
распространенные в Средней Азии, развиваются в корнях одно¬
летних солянок Salsola и Suaeda (Кривошеина, 1974 г). Таким об¬
разом, с разлагающейся древесиной связаны личинки Laphriinae
и некоторые представители Leptogastrinae. Именно этим фактом
объясняется приуроченность Laphriinae в основном к лесам (как
умеренного, так и тропического поясов), а в степях и полупусты¬
нях — к участкам с древесно-кустарниковой растительностью.
Личинки ксилофильных ктырей развиваются как в лиственных,
так и хвойных породах. Типичными обитателями хвойных пород
являются отдельные представители родов Andrenosoma, Choera¬
des и Laphria, в то же время все изученные виды Mesoleptogaster
и Pogonosoma развиваются только в лиственных деревьях.
Личинки встречаются как в стоящих сухостойных, так и ле¬
жащих на земле стволах, обычны в пнях, особенно хвойных де¬
ревьев, где могут встречаться в очень больших количествах. В це¬
лом предпочитают умеренно увлажненную древесину. Личинки
заселяют слабо разложившиеся стволы с плотными корой и дре¬
весиной. Развиваются как в толще древесины, так и коры, куда
проникают по ходам различных ксилофильных насекомых, пре¬
имущественно дровосеков (Cerambycidae), златок (Buprestidae)
и слоников (Curculionidae). В то же время опредолоипая группа
видов способна развиваться под корой как хвойных, так и лист¬
венных деревьев, при массовом развитии в ней подкорных ксило¬
фильных насекомых. В таких условиях, например, под корой ду¬
ба в Краснодарском крае развивались личинки Pogonosoma и под
корой лиственницы сибирской в Тувинской АССР — виды Choera¬
des. Для личинок Mesoleptogaster характерно обитание в толще
плотной, но уже достаточно сильно разложившейся древесины.
В подобных условиях развиваются личинки отдельных видов
Laphria, например L. rufa Rod.
Личинки ксилофильных ктырей Laphriinae очень подвижны,
легко перемещаются по ходам различного диаметра благодаря
наличию на теле специфических бугорков, но способны проде¬
лывать ходы также самостоятельно. Личинки Leptogastrinae
(Mesoleptogaster) менее подвижны, бугорки на теле развиты сла¬
бее, личинки перемещаются в древесине лишь по ходам других
ксилофильных насекомых.
293
Имеющиеся в литературе сведения о типе питания личинок
ктырей очень противоречивы. Вплоть до самого последнего вре¬
мени преобладало мнение о сапро- и фитофагии большинства ли¬
чинок этих двукрылых, основанное прежде всего на исследова¬
ниях Мелина (Melin, 1923) и вошедшее в целый ряд сводок (Se-
giiy, 1927; Hennig, 1952, и др.)* Отрицалась возможность хиш,ни-
чества даи;е для личинок Laphria и Leptogaster.
Несмотря на отсутствие, за небольшим исключением, специаль¬
ных работ, освеш,аюш,их пищевые связи личинок ктырей, постепен¬
но накапливались данные, свидетельствующие о хищном образе
жизни личинок (Clausen, 1940; Seguy, 1950; Шуровенков, 1961,
19G2, 1973; Daniels, 1966; Knutson, 1972, и др.). Установлено, что
хищниками являются личинки Asilinae — Machimus Loew (Шуро¬
венков, 1962) и Promachus Loew (Bromlei, 1930), личинки Dasy-
pogoninae — Stenopogon sp. (Creathead, 1963), личинки Lepto-
c^astrinae — Leptogaster cylindrica Deg. (Черепанов, 1957; Шуро¬
венков, 1962) и Mesoleptogaster eous Lehr (Кривошеина, Мамаев,
1967). Личинки Mesoleptogaster eous Lehr питаются личинками
разнообразных двукрылых и жуков, развивающихся в древесине
на луканидной стадии разрушения. Облигатными хищниками
являются все известные в настоящее время представители под¬
семейства Laphriinae — виды Laphria, Choerades, Andrenosoma,
Pogonosoma и Ctenota (Кривошеина, Мамаев, 1967, 1973, 1975а, б;
Кривошеина, 1974г). Личинки этих ктырей питаются личинками
различных ксилофильных насекомых, в первую очередь дро¬
восеков (Cerambycidae), златок (Buprestidae), слоников (Cur-
culionidae), а на поздних стадиях разрушения древесины — ли¬
чинками рогачей (Lucanidae). Распространенные в Африке виды
рода Hyperechia Schin. питаются личинками пчелы Xylocopa
(Poulton, 1924; Thorpe, 1927; Tsacas et al., 1970).
Таким образом, в настоящее время доказано хищничество ли¬
чинок отдельных представителей всех 4 подсемейств ктырей. Ка¬
жется вполне вероятным предположение, что личинки большин¬
ства видов семейства — хищники, на что в последнее время ука¬
зывали некоторые исследователи (Лер, 1964).
Морфология, Тело личинок цилиндрическое, слегка сужено к
переднему концу, с небольшой темной головой. Головная капсула
короткая, представлена маленькой лобной пластинкой и более
широкими боковыми, образующими вместе дорсолатеральную
пластинку. Длина пластинки приблизительно равна или несколь¬
ко меньше ее ширины. На вентральной стороне головы распо¬
ложена небольшая, поперечная, обособленная вентральная плас¬
тинка, в некоторых случаях редуцированная (Pogonosoma, An¬
drenosoma). От заднего края дорсолатеральной пластинки отходит
массивный метацефальный стержень, длина которого в несколько
раз превышает ширину. У некоторых личинок задний край стерж¬
ня расщеплен. Верхняя губа клювообразная, заостренная. По бо¬
кам от нее расположены двучлениковые мандибулы, каждая из
294
1^оторых представлена почти прямоугольным базальным склеритом
II узким стилетообразным дистальным. Максиллы обычно массив¬
ные, по переднему краю одно-трехзубчатые, к их внутренней сто¬
роне плотно примыкает дистальный конец мандибулы. Щупики
максилл 2-члениковые, как правило, расположены в вырезке
максилл. Усики хорошо выражены, но небольшие, одночленико-
Бые, расположены у основания базального членика мандибул.
Тело личинок состоит из 11 четких сегментов. Грудные сег¬
менты короткие, их длина меньше ширины. Они обычно голые,
иногда с полями мелких бугорков или шипиков, окольцовываю¬
щих их передние отделы. Брюшные сегменты более удлиненные,
как правило, с хорошо развитыми валиками и бугорками. Полза-
тельные валики в виде удлиненных вздутых участков расположе¬
ны на дорсальной и боковых сторонах передних отделов сегмен¬
тов. На вентральной, а в ряде случаев окольцовывая сегменты,
расположены парные ползательные бугорки. Подобные образова¬
ния отсутствуют на 8-м и, как правило, 7-м брюшных сегментах.
Ползательные бугорки, облегчаюш,ие передвижение по ходам
разного диаметра, особенно сильно развиты у ксилофильных форм.
Последний сегмент тела удлиненный, с дорсальной стороны раз¬
делен складкой на 2 отдела. Передний отдел несет овальные ды¬
хальца, задний — с 4 парами длинных ш,етинок. Из них 2 пары,
расположенные терминально, в дальнейшем обозначаются как
1-я и 2-я пары терминальных щетинок. Конец последнего сегмента
закругленный, гладкий, иногда с небольшими светлыми или тем¬
ными склеротизованными бугорками, зубчиками или валиком.
Личинки имеют 2 пары функционирующих дыхалец: на боковых
сторонах 1-го грудного сегмента и на дорсальной стороне послед¬
него сегмента. Кроме того, рудименты дыхалец расположены на
боковых сторонах 3-го грудного и 1—7-го брюшных сегментов те¬
ла. Передние дыхальца удлиненно-овальные, задние дыхальца —
округлые.
Первые подробные описания личинок 5 видов Laphria и Choe-
rades (L. ephippium F., L. flava L., L. gibbosa L., Ch. gilvus L.
и Ch. marginatus L.) опубликованы Мелином (Melin, 1923). Позд¬
нее Бриндль (Brindle, 1962b) использовал эти материалы и, исклю¬
чив L. ephippium F., опубликовал определительную таблицу ли¬
чинок 4 видов, широко распространенных в Европе и встречаю¬
щихся в Англии. В дальнейшем эта определительная таблица бы¬
ла дополнена новыми признаками (Кривошеипа, Мамаев, 1967),
но список известных личинок остался прежним. В последнее вре¬
мя были получены новые данные по морфологии личинок не толь¬
ко Laphria и Choerades, а также Andrenosoma и Pogonosoma
(Кривошеина, Мамаев, 1973, 1975а, б). На основании всех этих
материалов составлены приводимые ниже определительные таб¬
лицы ксилофильных личинок ктырей.
295
Таблица для определения родов семейства Asilidae
1 (2) Брюшные сегменты тела с хорошо развитыми ползательными
бугорками только на вентральной стороне (рис. 99, 1).
Мандибулы с редуцированным дистальным члеником. Мак-
силлы широко расставл*ены, не соприкасаются по средней
линии (рис. 99, 2), Последний сегмент с почковидным выступом
на конце (рис. 99, 5) Mesoleptogaster Frey (стр. 307).
2(1) Брюшные сегменты тела с хорошо развитыми ползательными
бугорками на дорсальной, вентральной и боковых сторо¬
нах (рис. 64, 4), Мандибулы хорошо развиты. Макснллы со¬
прикасаются по средней линии (рис. 73,5). 11оследн1Й сег¬
мент тела с небольшой склеротизованной пластинкой, не¬
сущей зубчик, или закруглен, без почковидного выступа.
3 (6) Последний сегмент тела закруглен, без кутикулярных струк¬
тур, несет 4 пары терминальных щетинок (рис. 99, 8), Бо¬
ковые участки головного сегмента, примыкающие к максил-
лам, не склеротизованы (рис. 99, 4,5 ^ 5, 10). Максиллы с зам¬
кнутой овальной щупиковой впадиной. Склеротизованная
вентральная пластинка головы отсутствует.
4 (5) Максиллы по переднему краю с 3 зубцами — длинным внут¬
ренним и 2 более короткими наружными (рис. 99, 9), Наруж¬
ный край максилл лишь слегка выступает за край базаль¬
ного склерита мандибул. Щупиковая впадина расположена
в заднем отделе максиллы на уровне выемки, разделяющей
2 наружных зубца. Метацефальный стержень с широкой тем¬
ной срединной полосой, доходящей до конца стержня; длнна
переднего затемненного участка значительно меньше ши¬
рины стержня (рис. 99, 6) . . Andrenosoma Rond. (стр. 297)
5 (4) Максиллы по переднему краю с 2 зубцами: длинным внут¬
ренним и более коротким наружным (рис. 99, 4, 5, 10).
Наружный край максилл заметно выступает за край базаль¬
ного склерита мандибул. Щупиковая впадина расположена
в заднем отделе максиллы на уровне вершины наружного
зубца. Метацефальный стержень с широкой темной средин¬
ной полосой в переднем отделе и узкой — в заднем; длина
переднего затемненного участка значительно больше ши¬
рины стержня (рис. 99, 7) . . Pogonosoma Rond. (стр. 308)
Г) (3) Последний сегмент тела, помимо терминальных щетинок,
песет склеротизованную терминальную пластинку со сре¬
динным зубцом и краевыми бугорками, расположенными в
верхнебоковых углах пластинки. Боковые участки головного
сегмента, примыкающие к максиллам, склеротизованы
(рис. 73, 5). Максиллы по бокам с глубокой незамкнутой
щупиковой впадиной. Склеротизованная вентральная пластин¬
ка головы имеется (рис. 73, 4).
7 (8) Боковые стороны 5—6-го брюшных сегментов с небольши¬
ми, но хорошо развитыми двигательными бугорками, т. е.
296
каждый сегмент несет по 8 бугорков. Терминальная плас¬
тинка последнего сегмента тела хорошо развита, четко очер¬
чена. Длина взрослых личинок обычно не превышает 18—
22 мм Ghoerades Walk. (стр. 297).
8 (7) Боковые стороны 5—6-го брюшных сегментов без двигатель¬
ных бугорков или лишь с рудиментами бугорков, выражен¬
ных в виде концентрических складок, т. е. каждый сегмент
несет по 6 бугорков. Терминальная пластинка последнего
сегмента в большинстве случаев склеротизована слабо;
склеротизация хорошо гыражена лишь в переднем отделе
пластинки, перед срединным зубцом. Длина взрослых ли¬
чинок обычно не менее 25 мм . . . Laphria Meig. (стр. 302)
Род Andrenosoma Rond.
Род распространен во всех зоогеографических областях земно¬
го шара, в Палеарктике представлен 10 видами (Рихтер, 1968).
Основные морфологические признаки рода приведены в опре¬
делительной таблице. В Европейской части СССР в настоящее
время зарегистрировано 2 вида: А. albibarbc Meig. и А. atrum L.,
но известна личинка лишь последнего вида (Кривошеина, Ма¬
маев, 1973).
А. atrum L. (рис. 99, б, Р). Личинка светлая, беловатая. Го¬
ловная капсула небольшая, короткая, поперечная. Максиллярный
щупик расположен в несклеротизованной щупиковой впадине.
На 1—6-м брюшных сегментах по 6 (2 дорсальных и 4 вентраль¬
ных) бугорков.
Вид широко распространен в лесных районах Европейской
части СССР и Кавказа. Личинки были обнаружены в древесине
сухостойной пихты кавказской (Красная Поляна, Краснодар¬
ский край) в июне 1967 г. Сходные по строению личинки, которых
не удалось воспитать до имагинальпой фазы, были обнаружены в
древесине сосновых пней (Левинская, Московская обл.). Личинки
являются характерными обитателями древесины на церамбицид-
ной стадии ее разложения. Питаются личинками дровосеков, в
частности, домового дровосека (Hylotrupes bajulus L.).
Род Ghoerades Walk.
Виды рода распространены в Палеарктической, где известно
более 10 видов, и Индо-Малайской областях. По признакам има¬
го и личинок род очень близок Laphria. Личинки целого ряда ви¬
дов детально описаны в работе Н. П. Кривошеиной и Б. М. Ма¬
маева (1975а).
Для личинок характерны хорошо развитые склеротизованные
^центральная пластинка головы и терминальная пластинка тела,
297
Рис. 99. Общий вид и детали строения Mesoleptogaster eous Lehr (1—3)^
Pogonosoma maroccanum F. (4), P. minus Loew (5), P. lugens Loew (7, S, 10)
и Andrenosoma atrum L. (6‘, 9) (Asilidae)
1 — общий вид сбоку; 2 — головная капсула и метацефальный стержень; з, 8 — конец
тела сверху; 4, 5, 9, Ю — левая половина головной капсулы сверху; 6,7 — метацефаль-
ные стержни
несущая срединный зубец. Головная капсула с массивным, лишь
слегка суженным в среднем отделе метацефальным стержнем.
Вентральная пластинка головы поперечная, прямоугольная или
с закругленными углами. Дистальный членик мандибул хорошо
развит. Максиллы соприкасаются друг с другом, по боковому краю
ограничены склеротизованными пластинками, являющимися час¬
тями расширенной дорсальной пластинки головы. На 1—6-м
брюшных сегментах тела по 8 бугорков (2 дорсальных, 2 латераль¬
ных и 4 вентральных бугорка), размеры которых увеличиваются
к концу тела. Последний сегмент тела разделен с дорсальной
стороны складкой на 2 неравных отдела, из которых меньший
передний несет дыхальца. Концевой отдел сегмента с небольшой
склеротизованной терминальной пластинкой, несущей крупный
срединный зубец или валик и 2 краевых заостренных или тупых
бугорка. Срединный зубец, а иногда и бугорки сильно склероти-
зованы, темно-коричневые или черные.
Таблица для определения видов рода Chooradeft Walk.
j (10) Срединный зубец и краевые бугорки терминальной плас¬
тинки хорошо выражены, с четкой вершиной (рис. 100, 7).
2 (3) Терминальтгая пластинка прямоугольная, короткая и ши¬
рокая, ее ширина в 2,5 раза больше длины. Краевые бу¬
горки массивные, широкие, с уплощенной вершиной, их
ширина значительно превосходит ширину срединного зуб¬
ца (рис. 100, 7, 22) Ch. igneus Meig.
3 (2) Терминальная пластинка овальная, ее ширина лишь в
1,5 раза больше длины. Краевые бугорки конусовидные,
с заостренной или закругленной вершиной, их ширина не
превышает пли лишь слегка превышает ширину срединного
зубца (рис. 100, 2),
4 (5) Срединный зубец терминальной пластинки удлиненный и
изогнутый, его длина более чем в 2 раза превышает ши¬
рину в основании (рис. 100, 2). Терминальная пластинка
наиболее широкая в переднем отделе, на уровне краевых
бугорков, с небольшой выемкой по переднему краю
(рис. 100, 7). Краевые бугорки широкие, крупные и закруг¬
лены на вершине, их окраска по интенсивности почти не от¬
личается от окраски окружающей пластинки .....
Ch, amurensis Herm.
5 (4) Срединный зубец терминальной пластинки массивный, его
длина менее чем в 2 раза превышает ширину в основании
(рис. 100, 4). Терминальная пластинка иная.
6 (7) Терминальная пластинка с четким расширением в перед¬
нем отделе за краевыми бугорками. Краевые бугорки силь¬
но заострены, их вершины черные (рис. 100, 6, 70) . . .
Ch. gilvus L.
7 (6) Терминальная пластинка с четким расширением в среднем
отделе. Краевые бугорки тупые, закругленные, если слегка
заострены, то умеренно склеротизованы, светло-бурые.
8 (9) Краевые бугорки широкие, закругленные, с четко очерчен¬
ной темной вершиной (рис. 100, 3, 4), Срединный зубец
изогнут в вершинной трети. Терминальная пластинка с
хорошо выраженной склеротизацией в задней половине . .
Ch. fimbriatus Meig.
^ (8) Краевые бугорки конусовидные, лишь со слегка затемнен¬
ным вершинным отделом (рис. 100, 5, 9), Срединный зу¬
бец изогнут в среднем отделе, дуговидный. Терминальная
пластинка со слабой склеротизацией в задней полови¬
не Ch, fulvus Meig.
10 (1) Срединный зубец терминальной пластинки не выражен.
На месте зубца расположен удлиненный склеротизованный
валик, вместо краевых бугорков — широкие овальные воз¬
вышения (рис. 100, 8, 12) Choerades sp.
Ch. amurensis Herm, Вентральная пластинка головной капсу¬
лы закруглена с боков и сзади, с ровным передним краем, несу-
299
щим небольшой срединный выступ. Ее ширина несколько менее
чем в 2 раза превосходит длину. Терминальная пластинка (рис. 100^
7, 2) с косо срезанными боковыми сторонами. Окрашена пластинка
сравнительно слабо (желтая), наиболее темная вокруг срединного
зубца. Задний край пластинки светлее переднего.
Вид распространен в Сибири и Приморском крае. Личинки в
Приморском крае развиваются в поверхностных слоях древесины
черемухи Маака, проникая в толщу древесины по ходам личинок
долгоножек — Tanyptera parva Portsch. и Т. nigricornis Meig.
(Tipulidae), которыми, по всей вероятности, и питаются.
Ch, fimbriatus Meig, Вентральная пластинка головной кап¬
сулы почти прямоугольная, с небольшим широким выступом спе¬
реди и сзади. Длина пластинки почти в 3 раза менее ее ширины.
Терминальная пластинка (рис. 100, 3, 4) поперечно-овальная, наи¬
более широкая в среднем отделе. Ее боковые края заметно высту¬
пают за краевые бугорки. Пластинка склеротизована сравнитель¬
но равномерно. Наиболее темная полоса между краевыми бугор¬
ками и срединным зубцом. Светлые пятна вокруг терминальных
щетинок сравнительно небольшие.
Вид нередок в Европейской части СССР и на Кавказе. Личинки
развиваются в толще древесины вяза обыкновенного (Теллерманов-
ское лесничество, Воронеллская область) и светлой древесине бу¬
ка восточного (Гузерипль, Краснодарский край), проникая в нее
по ходам Sinodendron cylindricum L. (Lucanidae).
Ch, fulvus Meig, Вентральная пластинка головной капсулы поч¬
ти прямоугольная, со слегка волнистым задним краем, ее ширина
немногим более чем в 2 раза превосходит длину. Терминальная
пластинка (рис. 100, 5, 9) поперечно-овальная, наиболее широ¬
кая в среднем отделе. Ее боковые края лишь слегка выступают
за краевые бугорки. Центральная часть пластинки окрашена рав¬
номерно, задний край пластинки более светлый. Переход от тем¬
ного к светлому участку постепенный, задний край пластинки без
четких границ. Светлые пятна вокруг терминальных щетинок
сравнительно большие.
Вид распространен в южных областях Европейской части
СССР, в Крыму и на Кавказе. Личинки развиваются в толще
древесины лиственных пород, зарегистрированы в светлых гнилях
древесины граба обыкновенного, бука восточного и в бурых гни¬
лях древесины дуба черешчатого (Ново-Прохладное, Убинская,.
Гузерипль, Краснодарский край). Отмечено питание личинками
жуков Sinodendron cylindricum L. и Dorciis parallelopipedus L.
(Lucanidae).
Ch, gilvus L, Вентральная пластинка головной капсулы поч¬
ти прямоугольная, со слегка волнистым передним краем, превос¬
ходит длину. Терминальная пластинка (рис. 100, 6, 10) попереч¬
но-овальная, наиболее широкая в переднем отделе. Срединная
часть пластинки затемнена, ее задний край — более светлый^
между обоими участками — довольно четкая граница.
300
Рис. 100. Конец тела личинок Choerades Walk. сзади (i, 5, 5—8) и сбоку
{2, 4, 9—12) (Asilidae)
i, 2 — Ch. amurensis Herm.; 3, 4 — Ch. fimbriatus Meig.; 5, 9 — Ch. fulvus Meig.; 6, 10 —
Ch. gilvus L., 7, 11 — Ch. igneus Meig.; 8^ 12 — Choerades sp.
a.t — передний отдел терминальной пластинки; l. t — краевые бугорки; р. t — задний
отдел терминальной пластинки; tu U— 1-я и 2-я пары терминальных щетинок
Вид широко распространен по всей Палеарктике. По литера¬
турным данным (Perris, 1870; Beling, 1882; Melin, 1923; Brindle,
19G2b), личинки развиваются преимущественно в трухе под корой
или в толще коры, предпочитая хвойные породы. По нашим дан¬
ным, обычны в пнях сосны обыкновенной и ели европейской (Ар¬
хангельская обл.). Питаются личинками деревенского дровосека
(Ci iocephalus rusticus L.) и других видов дровосеков. В хвойных
лесах Сибири (Богданова, Коломиец, 1974) развиваются на де¬
ревьях, пораженных дровосеками (Monochamus urussovi Fisch.
и Rhagium inquisitor L.) и смолевками (Pissodes gyllenhali Gyll.).
В отдельных случаях уничтожают до 15% личинок дровосеков.
Ch. igneus Meig. Вентральная пластинка головной капсулы
прямоугольная, с ровным передним краем и четким срединным
выступом. Ее ширина в 2,5 раза превышает длину. Терминальная
пластинка (рис. 100, 7, 11) поперечная, почти прямоугольная,
интенсивно окрашена, задний край пластинки более светлый,
хорошо отграничен от темного участка. 2-я пара терминальных ще¬
тинок — за пределами темного участка пластинки. Вокруг всех
терминальных щетинок очень слабые, небольшие светлые пятна.
Личинка близка описанной Мелином личинке Ch. marginatus L.,
отличаясь более сильной склеротизацией и формой терминальной
пластинки, а также несколько более массивным срединным зубцом.
Вид встречается в Европейской части СССР, включая северо-
западные районы, а также на Кавказе. Личинки развиваются в
древесине сосновых пней, проникая в толщу древесины по хо¬
дам личинок дровосека деревенского (Criocephalus rusticus L.)
и других видов.
Choerades sp. Вентральная пластинка в форме многоугольника
с прямым задним краем, ее ширина в 2 раза превосходит длину.
Терминальная пластинка (рис. 100, S, 12) поперечная, наиболее
широкая в передней половине. Передний край пластинки с чет¬
кой выемкой, задний почти прямой. Пластинка на большем про¬
тяжении темно-коричневая, равномерно окрашенная. Вокруг ще¬
тинок имеются небольшие овальные светлые пятна. На терминаль¬
ной пластинке вместо обычного срединного зубца имеется удли¬
ненный сильно склеротизованный валик, а вместо краевых бу¬
горков — широкие плоские возвышения.
Вид зарегистрирован в Приморском крае (Комарово-Запо-
водное), где его личинки развиваются в древесине пней хвойных
пород: кедра корейского и пихты цельнолистной.
Род Laphria Meig.
Род объединяет значительное количество видов (более 250),
распространенных во всех зоогеографических областях земного
шара. В Палеарктике насчитывается более 50 видов этого рода.
Род по признакам имаго и личинок близок роду Choerades Walk.
Отличительные морфологические признаки приведены в опреде¬
302
лительной таблице. Детальное описание личинок ряда видов при¬
ведено в работе Н. П. Кривошеиной и Б. М. Мамаева (19756).
Таблица для определения видов рода Laphria Meig.
1 (2) Ширина вентральной пластинки головной капсулы не
менее чем втрое превосходит ее длину. Терминальная плас¬
тинка (рис. 101,5, i2) очень слабо выраячена, маленькая,
ограничена срединным зубцом и краевыми бугорками.
Срединный зубец терминальной пластинки неправильной
формы, короткий и тупой, его длина не превышает ши¬
рину L. gibbosa J..
2 (1) Ширина вентральной пластинки головной капсулы по
более чем вдвое превосходит длину. Терминальная плас¬
тинка четкая, поперечная, доходяш,ая, по крайней мере,
до 1-й пары терминальных ш,етинок. Срединный зубец
терминальной пластинки заострен, постепенно сужается
от основания к вершине, его длина значительно превышает
ширину.
3 (10) Вершины краевых бугорков терминальной пластинки ту¬
пые, закругленные и лишь немного темнее окружаюш^их
участков. Темный срединный участок терминальной плас¬
тинки в заднем отделе расширен и заходит за уровень 1-й
пары терминальных щетинок (рис. 101, 6).
4 (5) Передний край терминальной пластинки тела выпуклый,
заметно выступает за уровень краевых бугорков, с широкой
выемкой в средней части (рис. 101, 6'), склеротизован так
же, как и остальная поверхность пластинки. Краевые бу¬
горки удлиненные, их длина заметно превышает ширину
(рис. 101, 7) L. rufa Rod.
5 (4) Передний край терминальной пластинки тела прямой или
волнистый, оканчивается на уровне краевых бугорков,
если выпуклый, то склеротизован заметно слабее осталь¬
ной поверхности пластинки. Краевые буго]жи у^меренной
длины, не превышающей их ширину.
6 (7) Вершины краевых бугорков направлены назад, бугорки
продольно вытянутые (рис. 101, i, 2).Терминальная плас¬
тинка сравнительно узкая, почти квадратная
L. ephippium F.
7 (6) Вершины краевых бугорков направлены в стороны, бу¬
горки поперечно вытянутые (рис. 101, 9, 13). Терминаль¬
ная пластинка расширена, прямоугольная или поперечно¬
овальная.
8 (9) Срединный темный участок терминальной пластинки за
срединным зубцом четко очерчен, в средней части с не¬
большой выемкой (рис. 101, 9). Вокруг 1-й пары терми¬
нальных щетинок крупные, неправильной формы светлые
пятна. Срединный зубец узкий и заостренный (рис. 101,
13) L. vulpina Meig.
эоз
9 (8) Срединный темный участок терминальной пластинки за
срединным зубцом без четкой границы и без выемки в
средней части (рис. 101, 10). Вокруг 1-й пары терминаль¬
ных щетинок небольшие овальные светлые пятна. Сре¬
динный зубец широкий и тупой (рис. 101, 14)
.Laphria sp.
10 (3) Вершины краевых бугорков терминальной пластинки
заостренные, темные. Темный срединный участок терми¬
нальной пластинки в заднем отделе не расширен и не за¬
ходит за уровень 1-й пары терминальных ш,етинок (рис. 101,
S, 11),
И (12) Вершинная половина краевых бугорков сильно склероти-
зована, темная. Срединный зубец терминальной пластинки
направлен назад (рис. 101, 8, 11) . . . L. varia Loew
12 (11) На краевых бугорках сильно склеротизован лишь неболь¬
шой вершинный отдел. Срединный зубец терминальной
пластинки направлен вверх (рис. 101, 3,4)
L. flava L.
L, ephippium F, Вентральная пластинка головы с закруглен¬
ным задним краем, несущим небольшой выступ в средней части.
Терминальная пластинка (рис. 101, i, 2) хорошо развита; слабо
склеротизова}! лишь задний отдел пластинки, где расположена
2-я пара терминальных щетинок. Поверхность переднего отдела
пластинки окрашена равномерно, краевые бугорки тупые, за¬
кругленные, их вершины немного затемнены.
Вид распространен в центральных и южных районах Европей¬
ской части СССР, зарегистрирован в Западной Европе. Личинки
идентифицированы по литературным данным (Melin, 1923), они
развиваются в толще древесины хвойных пород (сосна) (Melin^
1923), в ]1ашем распоряжении имелись личинки, собранные в Тел-
лермановском лесничестве Воронежской области в сухостойном
вязе шершавом в сентябре 1960 г. Личинки проникают в толщу
древесины по ходам личинок дровосеков, златок и горбаток, ко¬
торыми питаются.
L. flava L, Вентральная пластинка головы поперечная, с за¬
кругленным задним краем. Терминальная пластинка последнего
сегмента тела (рис. 101, 5, 4) четко очерчена лишь в переднем
отделе и вокруг срединного зубца. Задний отдел пластинки, ог¬
раничивающий 2-ю пару терминальных щетинок, окрашен, как
правило, слабо, желтоватый.
Вид широко распространен в Палеарктике. Личинки разви¬
ваются в толще древесины как хвойных (сосна, ель), так и лист¬
венных (береза) пород (Melin, 1923; Brindle, 1962b). Нами ли¬
чинки зарегистрированы в толще древесины стволов и пней сос¬
ны обыкновенной и ели европейской (Левинская, Московская
обл.; Волково, Ярославская область; Убинская, Краснодарский
край). Питаются личинками усачей и златок.
L. gibbosa L. Вентральная пластинка головы почти прямоуголь-
304
Рис. 101. Конец тела личинок Laphria Meig. сзади (1, 5, 5, 6, 8—10) и сбоку
<2, 4, 7, 11-14) (Asilidae)
1( £ — L. ephippium F.; 5, 4 ~ L. flava L.; J, 12 — L. gibbosa L.; в, 7 — L. rufa R6d.;
St 11 — L, varia Loew; o, 75 — L. vulpina Melg.; lo, J4 — Laphria sp.
пая, сзади прямая. Термилальная пластинка окрашена равно¬
мерно, выступы пластинки тупые, закругленные, лишь слегка за¬
темнены.
Вид широко распространен в Палеарктике. Личинки предпо¬
читают древесину хвойных пород (Melin, 1923; Кривошеина,
Мамаев, 19756). На территории нашей страны личинки собраны
в толще древесины пней сосны обыкновенной (Левинская, Москов¬
ская обл.). К этому же виду предположительно отнесены личинки,
развивающиеся в бурой древесине ясеня маньчжурского (Кома-
рово-Заповедное, Приморский край). В сосновых пнях личинки
встречаются под корой и реже в прочной древесине, куда они про¬
никают главным образом по ходам личинок дровосека деревен¬
ского (Criocephalus rusticus L.).
L. rufa Rod, Вентральная пластинка головы массивная, с круп¬
ным каудальным выступом. Терминальная пластинка последнего
сегмента тела (рис. 101, 6, 7) хорошо выражена.
Вид распространен на востоке Палеарктики. Личинки в При¬
морском крае (Комарово-Заповедное) развиваются в светлых и бу¬
рых гнилях умеренно плотной древесины различных лиственных
деревьев (ильма долинного, чозении крупночешуйчатой, ясеня
маньчжурского). Питаются личинками жуков-дровосеков (Сегат-
bycidae), комаров-долгоножек (Tipulidae) и других развивающих¬
ся в древесине насекомых.
L. varia Loew. Вентральная пластинка головы умеренной ши¬
рины, закруглена сзади. Терминальная пластинка (рис. 101,
8, 11) небольшая.
Согласно данным Энгеля (Engel, 1930), внд распространен па
всей Палеарктике, однако не исключено, что в Приморском крае
обитает морфологически уклоняющаяся популяция (Рихтер, 19(38).
Личинки в Приморском крае развивались в древесине лиственных
пород (ильм долинный, осина, тополь Максимовича, ольха), где их
пищей служили главным образом личинки различных дровосеков.
L, vulpina Meig. Вентральная пластинка головы многоугольная,
с выступающим задним краем. Терминальная пластинка тела
поперечная, с коричневатым срединным отделом и крупными свет¬
лыми участками вдоль боковых и заднего края пластинки (рис. 101,
.9, 13), Вершины краевых бугорков тупые, закругленные, лишь
слегка темнее самой пластинки, иногда с темным точечным пятном
на вершине.
Вид известен из средних и южных районов Западной Европы,
зарегистрирован в Талыше (Азербайджанская ССР). Личинки со¬
браны в сухобочине ольхи в ходах ольховой златки (Dicerca alni
Fisch.) (Красная Поляна, Краснодарский край).
Laphria sp. Вентральная пластинка головы с овальным зад¬
ним краем. Терминальная пластинка (рис. 101, 10, 14) с темным,,
коричневым срединным отделом и светлым задним краем. Вер¬
шины краевых бугорков тупые, закругленные, несколько темнее
поверхности пластинки.
306
Вид зарегистрирован в высокогорном Кавказе в урочище «Мед¬
вежьи Ворота» (высота около 2000 м) и в окрестностях пос. Гузе-
рипль Краснодарского края. Личинки развиваются в древесине
пихты кавказской, где питаются личинками араратской златки
(Ancylocheira araratica Mars.). По имагинальным признака^м оп¬
ределяется как L. flava L., однако существенные морфологические
отличия личинок заставляют предполагать, что это иной вид.
Род Mesoleplogasler Frey
Род включает виды, известные с о. Мадагаскар и Филиппин¬
ских островов (Frey, 1937), Китая (llsia, 1948) и Дальнего Востока
(Лер, 1961). Биология рода детально не изучена. Личинка извест¬
ного с территории нашей страны М. eous Lehr подробно описана
Н. П. Кривошеиной и Б. М. Мамаевым (19П7) под названием
Leptogaster sp.
М. eous Lehr (рис. 99, 1—3), Личинка желтоватая, относитель¬
но короткая, с хорошо развитыми ползательными бугорками
лишь на вентральной стороне брюшных сегментов, последний сег¬
мент тела с закругленным выступом. Все 3 грудные и 1-й брюш¬
ной сегменты без ползательных бугорков и валиков, боковые сто¬
роны грудных сегментов несут по 1 длинной щетинке. На
6 брюшных сегментах, начиная со 2-го, ползательные валики рас¬
положены на дорсальной и боковых сторонах сегментов, а полза¬
тельные бугорки — па их вентральной стороне. Все локомоторные
выступы смещены к переднему краю сегментов. Последний сег¬
мент разделен с дорсальной стороны складкой на 2 равных отдела
(у Leptogaster Meig. передний отдел значительно меньше заднего).
Задний отдел несет 4 пары щетинок, из них 2 пары на своеобраз¬
ном закругленном почковидном выступе (рис. 99, 3),
Личинки в Приморском крае развиваются в плотной древесине
(бурые или черные гнили) различных как хвойных, так и лист¬
венных деревьев (пихты цельнолистной, ореха маньчжурского,
ясеня маньчжурского, ильма долинного, маакии амурской). Ли¬
чинки относительно малоподвижны, в толще древесины переме¬
щаются по ходам различных ксилофильных насекомых или про¬
делывают ходы самостоятельно в более рыхлых полостях, запол¬
ненных трухой. Обнаружены на участках со скоплениями личинок
двукрылых из семейств болотниц (Limoniidae), детритниц (Sciari-
dae) и туманниц (Hyperoscelididae). Неоднократно наблюда¬
лось питание личинками двукрылых и личинками и особенно
куколками различных жуков, в первую очередь жуков-древо-
едов (Eucnemidae). Личинки встречались в древесине в самых
разнообразных участках леса, но во всех случаях в колодах бы¬
ли единичными.
307
Род Pogonosoma Rond.
Преимущественно индо-малайский род, в Палеарктике пред-
(тавленный 4, в Европейской части СССР — 3 видами. Некоторые
авторы рассматривают род в качестве подрода Andrenosoma Rond.
(Martin, 1966), что кажется вполне вероятным. Личинки морфоло¬
гически очень близки личинкам Andrenosoma Rond. Голова без.
развитой склеротизованной вентральной пластинки. Щупиковая
впадина максилл замкнута. Последний сегмент тела закруглен,,
светлый, без склеротизованной пластинки на конце.
Таблица для определения видов рода Pogonosoma Rond.
1 (2) Склериты ротового аппарата удлиненные, их длина более
чем в 2 раза превосходит длину дорсальной пластинки головы
(рис. 99, 4), Зубцы максилл удлиненные. Краевой зубец
лишь слегка шире внутреннего, удлиненно-овальный, его
длина в 1,5—2 раза превосходит ширину. Выемка между
зубцами остроугольная. Щупиковая впадина занимает зад¬
нюю половину краевого зубца. Краевой зубец с четкой вер¬
шиной, расположенной асимметрично и приближенной к
внутреннему зубцу Р. maroccanum F.
2 (1) Склериты ротового аппарата умеренной длины, их длина
не более чем в 2 раза превосходит длину дорсальной плас¬
тинки головы. Зубцы максилл короткие, широкие и тупые.
Краевой зубец заметно шире внутреннего, овальный, его
длина лишь слегка превосходит ширину. Выемка между
зубцами прямо- или тупоугольная. Щупиковая впадина за¬
нимает задние ^/3 краевого зубца.
3 (4) Краевой зубец неправильной формы, асимметричен, его вер¬
шина несколько смещена к внутреннему краю. На наружной
стороне зубца расположен небольшой тупой выступ (рис. 99,
10) Р. lugens Loew.
4 (3) Краевой зубец правильной формы, овальный, без выступа
(рис. 99, 5) Р. minus Loew.
Р, lugens Loew. Вид распространен в Средней Азии и сравни¬
тельно недавно был зарегистрирован также на Северном Кавказе
(Рихтер, Мамаев, 1971). Под корой и в древесине усыхающего
лоха (Марыйская область. Туркменская ССР) их пищей были ли¬
чинки пестрого лохового дровосека Xylotrechus grumi Sem.
В пнях туранги в тугаях по Амударье под г. Дейнау питались ли¬
чинками радужнойтурангиловой златки (Eurythyrea oxiana Sem.).
В тонких усохших ветвях лоха нападали на личинок дровосека —
лохового клита (Chlorophorus elaeagni Plav.).
Р. maroccanum F. Вид распространен в центральных и южных
областях Европейской части СССР. Личинки развиваются под ко¬
рой разлагающихся стволов дуба черешчатого (Убинская, Крас¬
нодарский край), лежащих на открытых, прогреваемых солнцем
участках. Концентрируются в подкоровой зоне, питаясь личин¬
ками дровосеков, в частности дубового красного — Pyrrhidium
sanguineum L. (Cerambycidae) и ямчатокрылых златок Chrysobo-^
thris affinis F. (Buprestidae).
P. minus Loew. Вид зарегистрирован в Крыму и на Кавказе.
Лнчинки развиваются под корой дуба черешчатого (Убинская,
Краснодарский край), часто совместно с личинками предыдущего
вида при аналогичном наборе жертв.
СЕМЕЙСТВО DOLICHOPQDIDAE — ЗЕЛЕНУШКИ
Большое по объему семейство зеленушек, насчитывающ,ее толь¬
ко в фауне СССР более 50 родов, включает своеобразную и хорошо
обособленную группу двукрылых с разнообразной биологией.
Личинки зеленушек — хищники, обитают в самых разнообразных
субстратах, ярко выраженные гигрофилы. Большую группу со¬
ставляют обитатели водной среды, различных придонных субстра¬
тов (Diaphorinae, Hydrophorinae, Aphrosylinae и др.). Предста¬
вители родов — Aphrosylus Walk., Thinophilus Wahlb., Machae¬
rium Hal. приурочены к прибрежной зоне морей и различных соло-
юватых водоемов (Wheeler, 1897; Негробов, Штакельберг, 1969).
Личинки многих видов (Dolichopus Latr., Neurogona Rond.,
Porphyrops Meig.) (Hennig, 1952 и др.) обитают во влажной почве
по берегам водоемов, в лесной подстилке и т. д. Некоторые виды,
как, например, представители рода Thrypticus Gerst., приспосо¬
бились к обитанию в тканях различных околоводных растений —
камыша, тростника и других (Liibben, 1908; Johannsen, Crosby,
1933). Личинки Campsicnemus fumipennis Far. на Гавайских ос¬
тровах зарегистрированы в стеблях банана (Williams, 1938).
Непосредственно связь с разлагающейся древесиной зарегистри¬
рована лишь у представителей трех голарктических родов семей¬
ства: Medetera Fisch., Systenus I^oew и Sciopus Zeli.
Биология личинок семейства, особенно ксилофильных групп,-
изучена слабо. В литературе имеются краткие описания личинок
отдельных видов (Tragardh, 1914; Hubault, 1925; Beaver, 1966а;
Schmid, 1971). Описания и определительная таблица 16палеаркти-
ческих видов рода Medetera приведены в работе Н. П. Кривошеи-
ной (1974д). По роду Systenus имеются общие сведения о личинках
(Dyte, 1967) и краткие описания и определительная таблица личи¬
нок 3 палеарктических видов (Кривошеина, 19736).
Экология. Личинки короедниц (Medetera) — наиболее обычные
представители ксилофильных зеленушек. Они обитают под корой
самых разнообразных как лиственных, так и хвойных деревьев.
Так, из 16 рассматриваемых нами видов 10 развиваются под ко¬
рой хвойных пород, а 6 зарегистрированы под корой лиственных
деревьев. Личинки рода являются типичными обитателями коро-
едных поселений, встречаясь в ходах короедов на самых различ-
309
ных этапах развития короедного сообщества. Но в отдельных слу¬
чаях личинки встречаются вне поселений последних (Beaver,
1966а). Так, есть указания на нахождение личинок М. impigra
СоП. в плодовых телах грибов Daedalea sp. (Dyte, 1959). В сту¬
денистых плодовых телах грибов на клене обнаружены личинки
М. pallipes Zeli, в Краснодарском крае (Гузерипль). Среди рас¬
сматриваемых нами видов связь с короедами не установлена у
М. feminina Neg. и М. krivosheinae Neg.
Род Systenus Loew включает виды, приуроченные в своем раз¬
витии преимущественно к специфическим жидким средам — на¬
текам древесного сока. Личинки развиваются непосредственно
в соке, заполняющем разнообразные полости и впадины в коре
стволов деревьев (Engel, 1916; Keilin, 1921; Cole, James, Streams,
1970; Steyskal, 1970; Кривошеина, 19736). Кроме того, личинки
встречаются во влажной трухе дупел, что отмечено как для не¬
которых палеарктических (Кривошеина, 19736), так и неаркти¬
ческих (Wirth, 1952) видов. Личинки обитают в рассматриваемых
субстратах в сообществе с целым комплексом разнообразных на¬
секомых, в первую очередь различных двукрылых (Geratopogo-
nidae, Psychodidae, Limoniidae, Stratiomyidae, Drosophilidae и
другими). Под корой разлагающейся древесины, очевидно, раз¬
виваются некоторые виды рода Sciopus. Так, личинки Sciopus sp.
были зарегистрированы под корой ясеня обыкновенного (Гузе¬
рипль, Краснодарский край). Вполне вероятно, что именно таким
образом жизни личинок объясняется приуроченность отдельных
видов рода, например S. platypterus F., к лиственным лесам (Не-
гробов, Штакельберг, 1969).
Личинки представителей рода Medetera широко известны как
хищники, уничтожающие короедов (Perris, 1870; Штакельберг,
1933; De Leon, 1935; Nuorteva, 1956, 1957, 1959, 1963, 1971; Зи¬
новьев, 1957; Johnsey, Nagel, Rudinsky, 1965; Кривошеина,
1971а, б, 1974д; Негробов, 1971; Коломиец, Богданова, 1973;
Арефин, 1974а, и др.), способные истреблять последних во всех
фазах их развития, включая молодых неокрепших жуков. В от¬
дельных случаях личинки уничтожают до 30% популяции коро¬
едов (Богданова, Коломиец, 1974). Но в целом при оценке воз¬
можной роли личинок короедниц решающее значение имеет син¬
хронность сроков лета вредителя и хищника. Личинки мух пре¬
следуют короедов, проникая по стволу на высоту до 10—13 м,
;а иногда до самой вершины (Зиновьев, 1957; Пряхина, Огибин,
1970). Но плотность личинок на разной высоте сильно колеблется
(Schmid, 1971).
Накопленные в литературе сведения говорят о том, что личин¬
ки короедниц не связаны с определенными видами короедов. Пер-
востепенное значение имеет специализация, определяемая породой
дерева, размером ствола и ветвей, типом заселения (Зиновьев, 1957;
Nuorteva, 1959; Beaver, 1966а). Личинки короедниц могут питаться
не только короедами, они нападают на мелких личинок различных
;ио
насекомых, развивающихся в древесине, папример детритпиц
(Sciaridae), смолевок (Pissodes), а также златок и дровосеков
(Nuorteva, 1959; Kline, Rudinsky, 1964; Beaver, 1966а). Имеются
ссылки, к сожалению без указания видовой принадлежности, на
питание трупами личинок насекомых (Kishi, 1969). Тип питания
личинок Systenus точно не установлен. Вполне возможно, что
личинки младших возрастов питаются жидкими органическими
веществами растительного происхождения. Но можно предполо¬
жить, что они также являются хищниками, по меньшей мере, на
заключительных этапах своего развития.
Морфология. Тело личинок (рис. 64, 2\ 107, 1) удлиненное, без
заметных выростов или бугорков, сужено на переднем конце и
пшроко закруглено на заднем. Последний сегмент несет неболь¬
шие, как правило, парные выступы, окружающие задние дыхаль¬
ца.
Головная капсула (рис. 102, 7, 5, 6) сильно редуцирована и
представлена лишь небольшими дорсальными пластинками, пере¬
ходящими спереди в склерит верхней губы. Сзади к ним причле-
няются передние концы метацефальных стержней, уходящих в
глубь грудных сегментов. Задние концы стержней заметно рас¬
ширены. Верхняя губа (рис. 102, 5) клиновидная, мандибулы
двучлениковые, дистальный членик мандибул серповидный, опи¬
рается на массивный базальный членик. Максиллы представлены
несколькими небольшими склеритами. В вентральном отделе го¬
ловного сегмента расположен характерный V-образный склерит
нижней губы (по Bischoff, 1925 — гипофаринкс), опираюшийся на
стержни тенториума. По бокам головы на уровне вершинных от¬
делов дорсальных пластинок расположены 1сГо.:ы1гие одночле-
никовые усики.
Сегменты тела с вентральной стороны с обычными но.,натель¬
ными валиками, несущими различные по форме кутикулярные
структуры. У личинок-амфибионтов могут быть развиты особые
локомоторные выступы, напоминающие ложные ножки. Кути¬
кулярные структуры валиков представлены одним поперечным
рядом крупных бугорков, окаймляющим ряды более мелких ши-
пиков. Ряды кутикулярных структур в большинстве случаев
разобщены вдоль средней линии.
Последний сегмент тела с различным числом небольших вы¬
ступов, окаймляющих задние дыхальца. В типичных случаях
дыхальца окружены 4 симметрично расположенными выступами:
2 дорсальными и 2 вентральными, из которых последние всегда
несколько крупнее. Иногда между дорсальными и вентральными,
а также между дорсальными выступами имеются небольшие до¬
полнительные зубчики (Johannsen, 1935). Для личинок отдель¬
ных групп характерны лишь небольшие дополнительные зубчики
между дорсальными выступами (Argyra Macq., Porphyrops Meig.,
некоторые виды Dolichopus Latr.; Hennig, 1952). Для личинок
Systenus LoewH Medetera Fisch., так же как и для представителей
311
Рис. 102. Передний конец тела и склеротизованпые пластинки головного и
1-го грудного сегментов Medetera Fisch. сверху (Dolichopodidae)
г — M.lblspinosa Neg.; 2 — Ж. dlchrocera Kow.; 5 — М. fasciata'Frey; [4 — M. penicillata
Neg.; J — M. pinlcola Kow.; б —. M. ravida Neg.; 7 — M. signaticomis Loew; — M. stria¬
ta Par.; 9 — Medetera sp.
Ibr — верхняя губа; d. pl — дорсальная пластинка^головы; [тс — метацефальные стерж¬
ни; wd — мандибула; pZ, — скл^)0ти80ванная пластинка головного сегмента; р1^— склеро-
тизованная пластинка 1-го’грудного сегмента
некоторых других родов (Dolichopus Latr., Sciopus Zeli., Asyn¬
detus Loew, Hydrophorus FU.), характерны 4 каудальных высту¬
па, развитых в различной степени.
Таблица для определения родов семейства Dolichopodidae
1 (2) 1-й брюшной сегмент с 2 удлиненными локомоторными вы¬
ступами, несущими на конце и вдоль переднего отдела длин¬
ные заостренные крючки (рис. 107,1). 1-й ряд кутикуляр-
ных структур на ползательных валиках 2—7-го брюшных
сегментов образован простыми шипиками (рис. 107, 5—7).
Боковые отделы дорсальной пластинки головы сильно удли¬
нены, почти достигают основания усиков
Systenus Loew (стр. 325)
"2 (1) Брюшные сегменты тела без локомоторных выступов. 1-й
ряд кутикулярных структур на ползательных валиках 2—
-312
7-го брюшных сегментов образован сдвоенными шипиками.
Боковые отделы дорсальной пластинки головы короткие и
массивные, оканчиваются на значительном расстоянии от
основания усиков Medetera Fisch. (стр. 313)
Род Medetera Fisch.
Личинки беловато-желтые, иногда совсем белые, удлиненные,
с темными, коричневыми или черными элементами ротового аппа¬
рата. Верхняя губа и дорсальная пластинка головы сильно скле-
ротизованы, черные (рис. 102, 7, 5, 6). Последний сегмент тела с
хорошо развитыми вентральными и очень короткими, часто в виде
небольших вздутий дорсальными лопастями (рис. ()4, 2).
На дорсальной стороне головного и 1-го грудного сегментов у
многих видов имеются различной формы склеротизованные плас¬
тинки (рис. 102, 2, 4). Грудные сегменты гладкие, первые 7 брюш¬
ных сегментов с ползательными валиками, покрытыми шипиками.
Шипики на валиках собраны в ряды, разделенные вдоль средней
линии тела. Шипики ползательного валика образуют 2 разные
группы. 1-я группа шипиков образует ряд, окаймляюш^нй валик
спереди и отличающийся по строению от следуюш^их рядов
(рис. 103, 2). В среднем отделе ряда (рис. 103, 2, а) ]1ебольшой
срединный участок покровов голый или покрыт мельчайшими за¬
метными лишь при больших увеличениях шипиками. Далее с
каждой стороны участка расположены группы из 1—5 одиночных
шипиков (рис. 103, 2, Ь). За ними следуют группы сдвоенных
шипиков (рис. 103, 2, с) (2 шипика, расположенные друг за дру¬
гом в продольном направлении и размепценные на общей пластин¬
ке). Передний шипик этих групп, как правило, меньше заднего
или редуцирован.
Вторая группа шипиков (рис. 103, 2, е) образует непосредст¬
венно шипиковое поле, в котором все шипики одиночные, т. е.
четко изолированы друг от друга. Шипики поля собраны в ряды,
разобщенные вдоль средней линии, где расположена ]1ебольшая
срединная пластинка (рис. 103, 2, d), несущая 2—0 зубчиков.
Только у личинок некоторых видов отдельные ряды правой и ле¬
вой сторон тела сближены. С каждой стороны от средней линии
тела имеется по 6—9 рядов, число которых может колебаться как
на правой и левой сторонах сегментов, так и на разных сегментах,
но в целом на средних сегментах число рядов наибольшее. Непо¬
средственно к средней линии тела па средних сегментах прибли¬
жено 3—4, иногда 5 рядов. Личинки перечисленных ниже видов
детально описаны в работе Н. 11. Кривошеиной (1974д).
Таблица для определения видов рода Medelera Fisch.
1 (2) Шипиковое поле ползательпых валиков вблизи средней
линии тела с 5—6 рядами шипиков (рис. 103, 4). Голов¬
ной сегмент с четко очерченной, но обычно слабо-
313:
Рис. 103. Ряды шипиков на ползательпых валиках брюшных сегментов
Medelera Fisch. (Dolichopodidae)
j — М. bispinosa Neg.; 2 — M. dichrocera Kow.; 3 — M. excellens Frey; 4 — M. fasciata
Frey
a — срединный участок покровов с мельчайшдми шипикалш; Ь — одиночные шипики
примыкающие к срединному участку покровов; с — сдвоенные шипики ряда, окаймляю¬
щего шипиковое поле; d — срединная пластинка, разделяющая ряды шипиков шипико-
вого поля; е — ряды шипиков шипикового поля; f,g,h — сдвоенные шипики 2-го, 4-го и
5-го брюшных сегментов;.с — л — средняя линия тела
склеротизованной светло-желтой дорсальной пластинкой
(рис. 102, 3), а 1-й грудной сегмент без пластинки ....
М. fasciata Frey
2 (1) Шипиковое поле ползательпых валиков вблизи средней
линии т£ла с 3—4 рядами шипиков. Если имеется 5 рядов
314
шипиков, то головной и 1-й грудной сегменты с четкими
склеротизованными дорсальными пластинками.
3 (16) Головной и 1-й грудной сегменты тела с дорсальной сто¬
роны с коричневато-желтыми пластинками: массивной
овальной в заднем отделе головного сегмента и узкой, э ви¬
де полоски, на переднем крае 1-го грудного сегмента
(рис. 102, 5).
4(11) иЬтиковые поля средних брюшных сегментов вблизи
средней линии с 4—5 рядами шипиков (рис. 103, 2),
Склоротизованная дорсальная пластинка 1-го грудного
сегмента хорошо выражена, шире пластинки головного
сегмента (ее боковые края выступают за боковые края
лежаш,ей перед пей пластинки; рис. 102, 2, 4),
5 (6) Шипиковые поля на средних брюшных сегментах тела вбли¬
зи средней линии с 5 рядами шипиков. Склеротизованная
пластинка 1-го грудного сегмента тела тонкая, ее длина
вдоль продольной оси тела, самое меньшее в 4 раза, усту¬
пает длине склеротизованной пластинки головного сег¬
мента (рис. 102, 7) М. signaticornis Loew
6 (5) Шипиковые поля на средних брюшных сегментах тела вбли¬
зи средней линии с 4 рядами шипиков. Склеротизованная
пластинка 1-го грудного сегмента тела относительна
массивная, ее длина вдоль продольной оси тела состав¬
ляет не менее длины склеротизованной пластинки голов¬
ного сегмента (рис. 102, 2, 4),
7 (8) Ширина склеротизованной пластинки головного сегмен¬
та в 2 или немного более чем в 2 раза превосходит длину.
Ряд шипиков, ограничивающий шипиковое поле, образо¬
ван группами из 2 шипиков (рис. 103, 2), Вдоль средней
линии тела ряд разделен группами микроскопических
шипиков, выделяющихся в виде серых пятен
М. dichrocera Kow.
8 (7) Ширина склеротизованной пластинки головного сегмента,.
по меньшей мере в 2,5 раза, превосходит длину. Ряд ши¬
пиков, ограничивающий шипиковое поле, образован груп¬
пами из 2—3 шипиков (105, 1). Микроскопические шипика
вблизи средней линии тела в ряду выражены слабо.
9 (10) Склеротизованная пластинка 1-го грудного сегмента мас¬
сивная, ее длина вдоль средней линии тела составляет
почти V2 длины пластинки головного сегмента (рис. 102, 4).
Ряды шипиков ползательных валиков не соприкасаются
вдоль средней линии (рис. 105, 1). Помимо 4 рядов вблизи,
средней линии, в полях шипиков средних брюшных сег¬
ментов имеется еще 4—5 рядов . . М. penicillafa Neg.
10 (9) Склеротизованная пластинка 1-го грудного сегмента зна¬
чительно меньше, ее длина вдоль средней линии тела со¬
ставляет около ^/3 длины пластинки головного сегмента
(рис. 102, 8). 3-й илй 4-й ряд шипиков ползательных ва¬
315г
ликов па части сегментов не разделен вдоль средней ли¬
нии (рис. 106, 2), Помимо 4 рядов вблизи средней линии,
в полях шипиков средних брюшных сегментов имеется
еще 3—4 ряда М. striata Раг.
11 (4) Шипиковые поля средних брюшных сегментов вблизи
средней линии тела только с 3 рядами шипиков (рис. 105,
2). Склеротизованная дорсальная пластинка 1-го грудного
сегмента сравнительно небольшая, не шире, обычно за¬
метно уже пластинки головного сегмента (ее боковые
края но В1>1ступают за края лежащей перед ней пластин¬
ки).
12 (13) Склеротизованная пластинка головного сегмента значи^
тельно шире пластинки 1-го грудного сегмента (рис. 102,
5) М. pinicola Kow.
13 (12) Склеротизованная пластинка головного сегмента равна
по ширине склеротизованной пластинке 1-го грудного
сегмента, с небольшой, но хорошо выраженной выемкой
в середине заднего края.
14 (15) Склеротизованная пластинка 1-го грудного сегмента с не¬
сколькими небольшими вглступами вдоль заднего края
(рис. 102, 6). Группы сдвоенных шипиков, ограничиваю¬
щие спереди шипиковое поле, хорошо выражены, удли¬
ненные, передние лишь слегка короче задних (рис. 105, 5).
Шипики поля темно-коричневые, хорошо выделяются на
светлом фоне сегментов М. ravida Neg.
15 (14) Склеротизованная пластинка 1-го груд?того сегмента с
ромбовидным расширением в средней части (рис. 102, 9).
Группы сдвоенных шипиков, ограничивающие спереди ши¬
пиковое поле, развиты неравномерно, передние очень сла¬
бые, небольшие, тупые или же вообще отсутствуют
(рис. 106, 4). Шипики поля светло-желтые, сравнительно
слабо заметны на светлом фоне сегментов
Medetera sp.
16 (3) Головной и 1-й грудной сегменты тела с до])сальной сто¬
роны без склеротизованных пластинок; иногда на голов¬
ном сегменте имеется лишь очень светлая желтоватая плас¬
тинка, почти не отличающаяся по окраске от окружающих
покровов тела.
17 (20) Верхняя губа массивная, утолщена в вершинной полови¬
не, шире не только дистальных, но и базальных склеритов
мандибул (рис. 102, 1). Личинки крупные, до 10 мм.
18 (19) Ряд шипиков, ограничивающий спереди шипиковое поле,
образован группами, в которых ближе к боковой стороне
тела по 3 шипика (рис. 105, 1) . . М. stackelbergiana Neg.
19 (18) Ряд шипиков, ограничивающий спереди шипиковое поле,
образован группами, в каждой из которых только по
2 шипика (рис. 103, 1) М. bispinosa Neg.
20 (17) Верхняя туба обычных размеров, заостренная, тонкая, ее
316
ширина ne превышает ширины дистальных склеритов
мандибул. Длина тела личинок не более 7 мм.
21 (22) На шипиковом поле средних брюшных сегментов вблизи
средней линии тела по 4 ряда шиников (рис. 104, 7) . . .
М. feminina Neg.
22 (21) На шипиковом поле всех брюшных сегментов вблизи сред¬
ней линии тела по 3 ряда шипиков.
23 (24) Ряд шипиков, окаймляюш;ий шипиковое поле, разделен
по средней линии тела участком, густо покрытым мель¬
чайшими шипиками. Сдвоенные шиники на всех сегмен¬
тах, кроме 1-го брюшного, почти равных размеров, перед¬
ние иногда лишь несколько уже задних (рис. 104, 4) .
М. krivosheinae Neg.
24 (23) Ряд шипиков, окаймляющий шипиковое поле, разделен
по средней линии тела участком, лишенным мельчайших
шипиков. Сдвоенные шипики разных размеров, передние,
по крайней мере на 2-м и 3-м брюшных сегментах, меньше
задних.
25 (26) В ряду сдвоенных шипиков, окаймляющих шипиковое но¬
ле, передрше очень небольшие, короткие и тупые. Их дли¬
на на 2—5-м брюшных сегментах менее ширины. Вблизи
средней линии тела сдвоенным шипикам предшествуют
4—5 слабосклеротизованных шиников, последние мельче
шиников шиппкового поля (рис. 103, 3)
М. excellens Frey
26 (25) В ряду сдвоенных шипиков, окаймляющих шипиковое по¬
ле, передние удлиненные, конусовидные. Их длина на
2—5-м брюшных сегментах равна ширине. Вблизи сред¬
ней линии тела сдвоенным шипикам предшествуют 1—4
простых шипика, последние по размерам равны шипикам
шипикового поля.
27 (30) Около средней линии тела группам сдвоенных шипиков
предшествуют 1—2 простых шипика. Передние из сдвоен¬
ных шипиков широкие и тупые, их длина не превышает
ширину в основании. Шипиковое поле образовано конусо¬
видными, слегка соприкасающимися в основании шипи¬
ками.
28 (29) Головной сегмент с небольшой прямоугольной слабо-
склеротизованной пластинкой с дорсальной стороны. Ря¬
ды шипикового поля средних сегментов тела удлиненные.
1-й ряд образован 6—9 шипиками, 2-й — 12—14, 3-й
включает до 20 шипиков на сред[1их сегментах тела . . .
М. thunebergi Neg.
29 (28) Головной сегмент без слабосклеротизованной пластинки с
дорсальной стороны. Ряды шипикового поля средних
сегментов тела более короткие (рис. 104, 2), 1-й ряд обра¬
зован 6—7 шипиками, 2-й — 10, а 3-й включает до 13 ши¬
ников М. impigra СоЛ.
317
30 (27) Около средней линии тела группам сдвоенных шипиков
предшествуют 3—4 простых шипика (рис. 104, 3). Перед¬
ние из сдвоенных шипиков удлиненные, их длина зтгачи-
тельно превышает ширину в основании. Шипиковое поле
образовано удлиненными, плотно примыкающими друг к
другу шипиками М. inspissata СоП.
М. bispinosa Neg. Длина тела личинок достигает 9—10 мм.
Головной сегмент (рис. 102, 1) со слабосклеротизованной светло-
желтой овальной пластинкой. Ползательные валики с рядами
светло-коричневых шипиков (рис. 103, 1), Шипиковое поле с канч-
дой стороны от средней линии с 7 (на 1-м брюшном) и 8 (на осталь¬
ных) рядами шипиков. На средних брюшных сегментах вблизи
средней линии 4 ряда шипиков. г
Вид описан из Воронежской области. По нашим данным, ли¬
чинки развиваются под корой вяза обыкновенного (Теллерманов-
ское лесничество, Воронежская область) и бука восточного (пос.
Гузерипль Краснодарского края). На буке обнаружены в посе¬
лениях двуцветного короеда (Taphrorychus bicolor Hbst.), реже
в ходах заболонника (Scolytus scolytus F.) и древесинников (Тгу-
podendron domesticum L. и Т. signatum F.).
М, dichrocera Kow. Личинка светло-желтая, крупная, длина
тела до 8 мм. Головной и 1-й грудной сегменты с хорошо развиты¬
ми пластинками (рис. 102, 2). Ползательные валики с хороша
развитыми коричневыми шипиками (рис. 103, 2), Шипиковое
поле с коричневатыми конусовидными шипиками, образующими
на средних сегментах по 8—9 симметричных рядов, из них вблизи
средней линии лишь 4 разобщенных симметричных ряда.
Вид характерен для лесных районов Европы. В Закарпатской
области (Квасы) личинки развиваются под корой лежащих на
земле стволов ели европейской в ходах короедов Ips typographus
L., Dryocoetes autographus Ratz. и Dryocoetes hectographus Rtt.
В литературе есть указание на связь этого вида с лубоедом Hylur-
gops palliatus Gyll. (Nuorteva, 1959).
M. excellens Frey. Личинка мелкая, до 5 мм, беловатая. Го¬
ловной и 1-й грудной сегменты с дорсальной стороны без склеро-
тизованных пластинок. Ползательные валики с рядами светло-
коричневых шипиков (рис. 103, 3). Шипиковое поле с каждой
стороны от средней линии образовано 8—9 рядами, причем вбли¬
зи средней линии лишь по 3 ряда шипиков.
Вид известен из северо-западных районов Европы. Личинки в
Приморском крае (Комарово-Заповедное) развиваются под корой
хвойных, в частности кедра корейского, в ходах лубоедов Hylur-
gops imitator Rtt., Н. interstitialis Chap. В Западной Сибири
личинки на хвойных породах встречаются в поселениях короеда
Orthotomicus proximus Eichh. (Коломиец, Богданова, 1973).
В Польше личинки этого вида зарегистрированы в ходах короеда
Ips cembrae Heer. (Schimitschek, 1930).
М. fasciata Frey, Личинка беловатая, длина тела до 7 мм.
318
Рис. 104. Ряды шипиков на ползательных валиках брюшных сегментов
Medetera Fisch. (Dolichopodidae)
i — м. feminina Neg.; В — U. impigra Coli.; з — M. inspissata Coli.; 4 — M. krivosheinae
Neg.
Обозначения см. на рис. 103
Головной сегмент с желтой, иногда коричневатой склеротизован-
ной пластинкой в форме полуовала (рис. 102, 3). Ползательные
валики со своеобразно расположенными светло-коричневыми
шипиками, относительно хорошо заметными на общем светлом
фоне тела (рис. 103, 4). Шипиковое поле образовано 9 симметрич¬
ными рядами шипиков, вблизи средней линии расположено 5 ря¬
дов, из них 5-й не разобщен. Иногда очень близко к средней
линии тела подходит 6-й ряд.
Вид известен из северных районов Европы. По нашим
данным, личинки в Краснодарском крае (Убинская) развиваются
под корой сосны обыкновенной в ходах короедов — Ips sexden-
Я19
tatus Boem., I. acumiiiatus GylL, Blaslophagus piiiiperda L.
В Западной Сибири вид встречается в поселениях Ips acuminatus
Gyll., Pityogenes bidentatus Hrbst. , Hylurgops palliatus Gyll.
(Коломиец, Богданова, 1973).
М, feminina Neg, Личинка молочно-белая, длина тела дости¬
гает 6 мм. Головной и 1-й грудной сегменты с дорсальной стороны
без склеротизованных пластинок. Ползательные валики с корич¬
неватыми шипиками (рис. 104, 7). Шипиковое поле образовано
11 — 12 симметричными рядами шипиков, причем вблизи средней
линии тела на средних брюшных сегментах по 4 ряда, на осталь¬
ных — по 3 ряда шипиков.
Вид известен из центральных и юго-западных районов Евро¬
пейской части СССР. Связь с короедами точно не установлена.
Личинки в Краснодарском крае (Убинская) собраны под корой
вяза шершавого в колониях личинок галлиц (Cecidomyiidae),
М. impigra СоИ. Личинки беловатые, длина тела достигает
5 мм. На дорсальной стороне головного и 1-го грудного сегментов
склеротизованные пластинки отсутствуют. Ползательные валики
со светло-коричневыми шипиками (рис. 104, 2). Шипиковое поле
образовано 8—9 симметричными рядами шипиков, причем вблизи
средней линии имеется лишь 3 ряда.
Вид распространен в средней полосе Европы, в Крыму, а так¬
же в Сибири. Личинки в Закарпатской области (Хуст) развиваются
под корой бука европейского и в Краснодарском крае (Убинская)
под корой граба обыкновенного в ходах короеда Taphrorychus
villifrons Duf. Вид зарегистрирован также на вязе в ходах за-
болонника — Scolytus scolytus F. (Beaver, 1966а), встречается
на лиственнице (Collin, 1941; Beaver, 1966а).
М. inspissata Coli. Личинка беловатая, мелкая, достигает в
длину лишь 4 мм. На дорсальной стороне головного и 1-го груд¬
ного сегментов склеротизованные пластинки отсутствуют. Пол¬
зательные валики с рядами коричневатых умеренно склеротизо¬
ванных шипиков (рис. 104, 3). Шипиковое поле образовано 8—9
симметричными рядами шипиков, причем вблизи средней линии
тела по 3 ряда шипиков с каждой стороны.
Вид описан из Англии. Личинки в Краснодарском крае (Убин¬
ская) развиваются под корой осины в ходах короедов Trypophloeus
granulatus Ratz.
М. krivosheinae Neg. Личинка желтоватая, длиной до 5 мм.
Головной и 1-й грудной сегменты без склеротизованных пласти¬
нок. Ротовой аппарат склоротизован умеренно, коричневый. Пол¬
зательные валики с хорошо развитыми темно-коричневыми ши¬
пиками (рис. 104, 4). Шипиковое поле образовано 8—9 симмет¬
ричными рядами шипиков, из которых вблизи средней линии
тела расположено лишь 3 ряда.
Личинки развиваются под корой осиновых колод (Волково,
Ярославская область), связь с короедами не установлена.
М. penicillata Neg. Личинки молочно-белого цвета, длина те-
320
Рис. 105. Ряды шипиков на ползательных валиках брюшных сегментов
Medetera Fisch. (Dolichopodidae)
1 — М. penicillata Neg.; 2 — М. pinicola Kow.; 3 — M. ravida Neg.; 4 — M. signatlcomls
Loew
Обозначения см. на рис. 103
ла до 7 мм. Склеротизованные пластинки на дорсальной стороне
головного и 1-го грудного сегментов имеются (рис. 102, 4). Пол-
зательные валики с рядами довольно крупных, массивных за¬
остренных темных шипиков (рис. 105, 1). Шипиковое поле обра¬
зовано 7—9 симметричными рядами простых шипиков, причем
вблизи средней линии тела расположено 4 ряда шипиков. Шипики
удлиненные, сближены друг с другом, первый ряд образован 13—
1 \ Б. М. Мамаев и др.
321
14 шипиками. Срединная пластинка шипикового поля с 3 зуб¬
чиками.
Вид описан из Приморского края (Комарово-Заповедное).
Личинки развиваются под корой хвойных деревьев (например,
кедра корейского) в ходах короеда (Ips sexdentatiis Boern.) и
лубоедов (Ilylurgops interslitialis Chap. и Н. imitator Rtt.).
М. pinicola Kow. Личинка светло-желтая, длина тела достигает
7—8 мм. Головной и 1-й грудной сегменты с хорошо развитыми
темными склеротизованными пластинками (рис. 102, 5). Ползатель-
ные валики с темно-коричневыми шипиками. Сдвоенные шипики
расположены на темных, четко очерченных плош,адках. Шипиковое
поле образовано 7—8 симметричными рядами шипиков, вблизи
средней ЛИНИР1 лишь 3 ряда шипиков (рис. 105, 2).
Вид обычен в лесной зоне. Личинки в Московской области и
в предгорных районах Краснодарского края развиваются под
корой сосны обыкновенной, в ходах короедов Orthotomicus sutu-
ralis Gyll. и Ips sexdentatus Boern., малого елового лубоеда
Hylurgops palliatus Gyll. и большого лесного садовника Blas-
tophagus piniperda L. Ha нахождение личинок в ходах двух
последних видов указывается в литературе (Nuorteva, 1959, 1964).
В сходных условиях вид развивается в Западной Сибири, где
встречается в ходах В. piniperda L. и Orthotomicus proximus
Eichh. (Коломиец, Богданова, 1973).
М. ravida Neg. Личинка очень близка по строению к личинке
Ы. signaticornis Loew. На дорсальной стороне головного и 1-го
грудного сегментов расположены темные склеротизованные рав¬
ные по ширине пластинки (рис. 102, 6).
На ползательных валиках шипики темно-коричневые, хо¬
рошо выделяются на светлом фоне сегментов (рис. 105, 3). Шипи¬
ки в рядах шипикового поля удлиненные, в большинстве слу¬
чаев соприкасаются в основании. Вблизи средней линии по 3 ря¬
да шипиков. Срединная пластинка шипикового поля с 2 зубцами.
Вид описан по материалам из Приморского края (Комарово-
Заповедное). Личинки развиваются под корой хвойных деревьев
(пихты цельнолистной, кедра корейского) в ходах короедов Ips
sexdentatus Boern. Обычно встречаются совместно с М. penicil-
lata Neg. и М. signaticornis Loew.
М, signaticornis Loew. Личинка беловатая, длина тела до 7 мм.
Головной и 1-й грудной сегменты с хорошо развитыми склероти¬
зованными пластинками (рис. 102, 7). Пластинка головного сег¬
мента поперечная, несколько расширена в заднем отделе, с ров¬
ным задним краем. Пластинка 1-го грудного сегмента сужена к
концам, несколько шире пластинки головного сегмента. Элементы
ротового аппарата коричневые, с узкой, тупо закругленной на
вершине верхней губой. Ползательные валики с коричневатыми
шипиками (рис. 105, 4), Группы сдвоенных шипиков хорошо раз¬
виты, тупые, передние шипики в большинстве случаев редуциро¬
ваны. Срединйый участок покровов без четких микроскопических
322
111ИПИК0В, ограничен группами из 3, чаще 2 простых шипиков.
Щипиковое поле с 8 симметричными рядами шипиков на средних
брюшных сегментах тела и с 6—7 рядами на остальных. Вблизи
средней линии тела 5 рядов шипиков, из них 4-й ряд не разобщен
по средней линии. Срединная пластинка шипикового поля с 2
зубчиками. Шипики 1-го ряда заметно крупнее остальных.
Вид обычен для лесной зоны. В Приморском крае личинки
развиваются под корой хвойных деревьев (пихты цельнолистной,
кедра корейского), в ходах короеда Ips sexdentatus Boern. и
лубоеда Hylurgops interstilialis Chap. В Архангельской области
предпочитают ходы Ips typograplius Т.. (Пряхина, Огибин, 1970).
В Тувинской АССР вид поселяется под корой лиственницы сибир¬
ской в ходах большого лиственничного короеда Ips subeloiigatus
Molsch. (Яновский, 19746). Данные по личинке М. signaticornis
Loew (Triigarclh, 1914; Ilubault, 1925), возможно, относятся к
личинке М. striata Parent — относительно близкому М. sigiiati-
coruis Loew и широко распространенному в Европе виду, описан¬
ному сравнительно недавно.
М. stackelbergiana Neg. Личинки желтоватые, крупные, длина
тела достигает 10 мм. Головной сегмент со слабосклеротизован-
ной, плохо заметной пластинкой, на 1-м грудном сегменте плас¬
тинка отсутствует. Ползательные валики со светло-коричневатыми
или желтоватыми шипиками (рис. 106, 1). Шипиковое поле об¬
разовано 8 симметричными рядами шипиков. Вблизи средней ли¬
нии тела на средних брюшных сегментах по 4 ряда, а на 1-м и
7-м по 2—3 ряда шипиков. Личинки этого вида очень близки
личинкам М. bispinosa Neg., отличаясь лишь строением перед¬
него ряда шипиков, окаймляющего шипиковое поле.
Вид описан из Воронежской области, зарегистрирован в При¬
морском крае (Комарово-Заповедное). Личинки развиваются под
корой вяза обыкновенного и ильма долинного в ходах заболон-
ников Scolytiis sulcifrons Rey. и S. jacobsoni Spess.
M. striata liarent. Личинка беловатая, достигает длины 0,5—
7 мм. Головной и 1-й грудной сегменты с хорошо развитыми скле-
ротизованными пластинками (рис. 102, 8), Ползательные валики
с темно-коричневыми шипиками (рис. 106, 2). Сдвоенные шипики
расположены на хорошо развитых удлиненных пластинках. Ши¬
пиковое поле образовано 6—8 симметричными рядами простых
шипиков. Вблизи средней линии тела имеется 3—4 ряда шипиков,
причем 3-й или 4-й ряд не разобщен вдоль средней линии.
Вид широко распространен во всех районах Европейской час¬
ти СССР, на Кавказе, в Западной Сибири. Личинки в Красно¬
дарском крае (Гузерипль, Убинская) развиваются под корой хвой^
ных деревьев (сосны обыкновенной, пихты кавказской) в ходах
лубоедов — Blastophagus piniperda L. , Cryphalus orientalis Eggi,
короедов — Pityokteines cnrvidens Germ. и Orthotomicus siitu-
ralis Cyll. Личинка, описанная из ходов Ips typographus L. (Gab-
ler, 1953), скорее всего относится к данному виду.
!!♦ 323
Рис. 106. Ряды пшпиков на ползательных валиках брюшных сегментов
Medetera Fisch. (Dolichopodidae)
J — м. stackelbergriana Neg.; г — M. striata Par.; 3 — M. thunebergi Neg.; \4 — Mede-
teralsp.
Обозначения см. на рис. 103
м. thunebergi Neg. Личинка желтоватая, длина тела до 8 мм.
1’оловной сегмент с небольшой сероватой прямоугольной плас¬
тинкой, оканчивающейся на одном уровне с боковыми выступами
дорсальной пластинки головы. Ее длина в 2 раза уступает ши¬
рине. На 1-м грудном сегменте иногда имеется очень небольшая,
почти незаметная пластинка. Ползательные валики с коричне¬
выми шипиками (рис. 106, 3). Шипиковое поле состоит из 8—9
симметричных рядов шипиков, вблизи средней линии лишь 3 ря¬
да шипиков. Шипики в рядах разобш,ены или слегка соприкаса¬
ются, все приблизительно равных размеров, относительно круп¬
ные.
Вид описан из Ленинградской области, распространен в лес¬
ных районах Европейской части СССР, в Сибири и на Дальнем
Востоке. Личинки в Закарпатской области (Квасы) развиваются
под корой упавших стволов ели европейской в ходах короедов
Ips typographus L., Dryocoetes autographus Ratz. и D. hecto-
graphus Rtt. В Западной Сибири личинки на хвойных встреча¬
ются в ходах короедов — Ips acuminatus Gyll. и Orthotomicus
proximus Eichh. (Коломиец, Богданова, 1973).
Medetera sp. Личинка светло-желтая, длина тела достигает
С) мм. Передний конец тела с хорошо развитыми склеротизован-
ными пластинками равной ширины (рис. 102, 9). Ползательные
валики со светло-коричневыми, но сравнительно хорошо замет¬
ными на светлом фоне покровов шипиками (рис. 106, 4). Шипи¬
ковое поле образовано 7, иногда 8 симметричными рядами, вбли¬
зи средней линии тела 3 ряда шипиков.
Личинки в Краснодарском крае (Убинская) развиваются под
корой сосны обыкновенной совместно с личинками М. pinicola Kow.
Род Sysfenus Loew
Небольшой по объему род, включающий несколько палеар-
ктических и неарктических видов. Для личинок рода характерен
целый ряд общих признаков, довольно хорошо отличающий их
от личинок других родов зеленушек. Личинки беловато-желтые,
удлиненные, с коричневой головной капсулой. Характерно срав¬
нительно слабое развитие срединного отдела дорсальной пластин¬
ки головы. Боковые отделы дорсальной пластинки (рис. 107, 2)
головы сильно удлинены по сравнению, например, с таковыми
личинок Medetera Fisch. Последний сегмент тела с хорошо раз¬
витыми 4 лопастями, но дорсальные лопасти взрослых личинок
значительно короче вентральных, широкие и тупые (рис. 107, 4).
1-й брюшной сегмент, как указывается и в литературе (Dyte,
1967), с 2 удлиненными локомоторными выступами —«ложными
ложками», несущими на вершине и вдоль переднего отдела длин¬
ные загнутые на конце крючки (рис. 107, 3). Кроме того, у осно¬
вания локомоторных выступов, по их переднему краю располо¬
жено 5 рядов мелких шипиков. На 2—7-м брюшных сегментах
325
Рис. 107. Общий вид и детали строения личинок Systenus pallipes Ros
(1—4^ 6), S. tener Loew (o) и S. scnoltzii Loew (7) (Dolichopodidae)
1 — общий вид тела сбоку; 2 — передний конец тела с дорсальной стороны; 3 — локомо¬
торный выступ; 4 — конец тела сбоку; 5—7 — шипики ползательных валиков средних
брюшных сегментов тела
(рис. 107, 1) хорошо развиты ползательные валики, несущие
очень слабо выраженные, почти не склеротизованные светлые
шипики, образующие разобщенные вдоль средней линии тела
ряды (рис. 107, 5—7). Шипики двух размеров. Основная масса
мелких шипиков собрана в 8—9 следующих друг за другом ря¬
дов, образующих шипиковое поле. Последнее спереди ограничено
одним рядом более крупных шипиков, также разобщенных вдоль
средней линии. Шипики этого ряда простые, не сдвоены, как это
наблюдается, например, у личинок Medetera Fisch. Характерная
особенность личинок Systenus Loew — наличие хорошо развитых
локомоторных выступов на 1-м брюшном сегменте.
326
Таблица для определения видов рода Systenus Loew
1 (2) Длинные загнутые на вершине крючки, собранные в 2 груп¬
пы, расположены не только на локомоторных выступах
1-го брюшного сегмента, но также и на ползательных ва¬
ликах 2—6-го брюшных сегментов . . S. scholtzii Loew
2 (1) Длинные загнутые на вершине крючки расположены только
на локомоторных выступах 1-го брюшного сегмента. На
следующих сегментах, если и имеются, то очень короткие,
в 2—3 раза короче крючков, расположенных на локомотор¬
ных выступах.
3 (4) Шипики, образующие ряды шипикового поля, разных раз¬
меров, по периферии поля крупнее. Шипики лишь слегка
соприкасаются у основания, конусовидные. Их длина в
большинстве случаев равна или в 1,5 раза превосходит
ширину в основании. В первом ряду шипикового поля 8—9,
во втором — 13 шипиков с каждой стороны от средней ли¬
нии (рис. 107, 6). Ряд крупных шипиков, ограничивающих
спереди шипиковое поле, хорошо выражен, включает до
15 шипиков с каждой стороны от средней линии ....
S. pallipes Ros.
4 (3) Шипики, образующие ряды шипикового поля, мелкие, рав¬
ных размеров, лишь в первом ряду несколько крупнее
остальных. Все шипики сильно сближены друг с другом,
на большем протяжении соприкасаются, параллельносто¬
ронние (рис. 107, 5). Их длина, по меньшей мере, в 2 раза
превышает ширину в основании. В первом ряду шипикового
поля И, во втором до 20 шипиков с каждой стороны от
средней линии. Ряд крупных шипиков, ограничивающих
спереди шипиковое поле, выражен нечетко, хорошо заметны
лишь 4—5 шипиков с каждой стороны от сродней линии
тела S. tener Loew
S. pallipes Ros. Широко распространен в лесных районах
СССР. Личинки достаточно хорошо изучены и описаны в ряде
работ (Engel, 1916; Dyte, 1967; Кривошеина, 19736). У личинок
на 3—4-м брюшных сегментах на ползательных валиках перед
шипиковым полем имеется по две группы крючков, длина кото¬
рых не более половины длины крючков локомоторных выступов
1-го брюшного сегмента.
Личинки являются типичными обитателями натеков сока на
деревьях (Engel, 1916; Keilin, 1921), обычны в специфических
полужидких натеках на стволах ильма долинного (Комарово-
Заповедное, Приморский край), а также в бродящем соке на иве
(урочище Ипайкала, Копет-Даг). В отдельных случаях встречают¬
ся в трухе дупел, например бука восточного (Гузерипль, Красно¬
дарский край). В натеках сока на иве личинки довольно обычны и
встречаются совместно с личинками длинноусых двукрылых
(Ceratopogonidae и Anisopodidae). В натеках сока на ильме до-
327
ЛИННОМ личинки встречаются в небольших количествах, как пра¬
вило, сопутствуя личинкам мокрецов (Culicoides fagineus Edw.
и Dasyhelea paludicola Kieff.). В период наступления замороз¬
ков личинки мигрируют в более глубокие слои, уходя в толщу
коры по пропитанным соком трещинам.
S. tener Loew. Вид известен из Англии, на территории нашей
страны зарегистрирован в Копет-Даге. Для личинок характерно
развитие загнутых крючков лишь на 1-м брюшном сегменте.
Личинки развиваются в трухе в дуплах грецкого ореха (Кри-
вошеина, 19736) совместно с личинками бабочниц (Psychodidae),
мокрецов рода Forcipomyia Meig. (Ceratopogonidae), львинок Eupa-
chygaster tarsalis Zett., личинок жуков-узконадкрылок (Oedeme-
ridae). Вследствие того, что по периферии дупел, в значительной
степени в толще живой древесины, проделывают ходы личинки
дровосеков, обеспечивается постоянный приток сока в дупла.
S. scholtzii Loew. Вид сравнительно широко распространен в
Европе. На территории нашей страны зарегистрирован в Копет-
Даге (Кривошеина, 19736). У личинок шипики 1-го ряда шипи-
кового поля крупные, равных размеров, 2-го — лишь слегка ме¬
нее первого, а остальных — значительно мельче (рис. 107, 7).
В первом ряду шипикового поля 8, во втором — 13 шипиков с
каждой стороны от средней линии. Ряд крупных шипиков, огра¬
ничивающих спереди шипиковое поле, хорошо выражен, с каждой
стороны включает до 13 шипиков.
В Копет-Даге (урочище Ипайкала) личинки развиваются в
трухе в дуплах грецкого ореха совместно с S. tener Loew (Кри¬
вошеина, 19736). Личинки зарегистрированы также в натеках
сока (Keilin, 1921).
СЕМЕЙСТВО LONCHAEIDAE - КОПЬЕХВОСТКИ
Копьехвостки — небольшое по объему семейство, включаю¬
щее в настоящее время 9 родов и около 240 видов, из них в Пале-
арктике известно около 80 видов. Систематика группы разраба¬
тывалась многими авторами (Вескег, 1895; Czerny, 1934; Hennig,
1948а), но наиболее интенсивное ее изучение началось в последнее
десятилетие (Morge, 1961, 1963, 1967а и др.; iMcAlpine, 1956а, Ь,
1958 и др.; Ковалев, 1973, 1974), что дало возможность внести
ряд исправлений и дополнений в сведения по биологии копье-
хвосток. Так, например, исследования последних лет (McAlpine,
Morge, 1970) показали, что вид L. zetlersledli Веек, в действитель¬
ности представляет собой группу близких видов. На основании
проведенной ревизии выяснено, что данные Геннига (Hennig,
1948а) относятся не к L. zetterstedti Веек., а к L. bruggeri Morge
(Morge, 1967а). Сведения о личинках копьехвосток, приводимые
Г. А. Зиновьевым (1957), помимо L. zetterstedti Веек, и L. scutel-
laris Rond., следует отнести к L. collini Наск. и L. fugax Веек.
328-
Данные Морге (Morge, 1963) по L. zelterstedii Веек, относятся к
двум видам: L. zettersledti Веек, и L. seitneri Ilend. и т. д.
Экология. Иеемотря на уенешное изучение взрослых форм,
биология группы в целом изучена очень слабо. Это в значитель¬
ной степени определяется как разнообразием пищевых связей
личинок коньехвосток, так и слабой изученностью имагинальной
систематики в недалеком прошлом. К настоящему времени в ли¬
тературе накопился материал по образу жизни личинок отдель¬
ных видов, но он в значительной мере не систематизирован и
не подвергся критическому пересмотру, мало данных о характере
питания личинок, практически не изучены комплексы ксило¬
фил ьных насекомых, в составе которых развиваются копье-
хвостки.
Личинки коньехвосток в целохМ заселяют разнообразные раз¬
лагающиеся вещества растительного происхождения. Так, в
различных фруктах, особенно инжире (Silvestri, 1917; Bezzi,
1919; McAlpine, 195Gb, 1960), развиваются личинки Silba Macq.,
преимущественно в лимонах и апельсинах — личинки Lampro-
lonchaea Bezzi (Bezzi, 1919). В ягодах и абрикосах обычны неко¬
торые виды Dasiops Boiid. (McAlpine, 1961; Moffitt, Varuss, 1961).
В различных однолетних растениях развиваются личинки Dasiops
(в подсолнечнике; McAlpine, 1961) и Setisquamolonchaea Morge
(в кормовой свекле, спарже, петруише, дурмане; Hering, 1954;
Smith, 1957).
С древесной растительностью связана сравнительно небольшая
группа родов. Нанример, под корой ели зарегистрированы ли¬
чинки Silvestrodasiops Morge (Morge, 1963), некоторых видов
Dasiops Bond. (Morge, 1963). Отдельные виды этого рода разви¬
ваются в грецких орехах (McAlpine, 1961), в опунции (McAlpine,
1961; Moffitt, Varnss, 1961), в некоторых случаях способны обра¬
зовывать галлы (Bezzi, 1919). В шишках хвойных пород разви¬
ваются личинки Earomyia Zett. (Bezzi, 1919; Seitiier, 1925; Czerny,
1934; Seguy, 1934; McAlpine, 1956a; Morge, 1963; Голутвина, 1973).
Типично ксилофильными формами являются личинки рода
Lonchaea. Только для личинок отдельных видов, например L. cho¬
rea F., хотя и имеются данные о связях с древесиной (Tiiomikoski,
1930; Gabler, 1953, и др.), в качестве сред обитания более ха¬
рактерны разнообразные разлагающиеся, богатые органическими
веществами субстраты, например, коровий навоз (Hennig, 1953;
Лобанов, 1962, 1972), гниющие свекла, луковицы (Hennig, 1948а,
1953). В сходных субстратах в Индии развивается L. chalybeata
Wied. (Bezzi, 1919; Blanchard, 1948).
Виды, развивающиеся в древесине, встречаются как на хвой¬
ных, так и на лиственных породах. С хвойными породами связан
небольшой, но довольно четко очерчиваемый в систематическом
плане комплекс видов. Это — виды группы corticis (L. zetterstedti
Веек., L. seitneri Hend., L. defecta McAlp.), a также L. brug-
geri Morge и L. collini Hack. Как в хвойных, так и в листвен¬
329
ных породах развиваются личинки L. sylvatica Bel. Группа ви¬
дов, связанная с лиственными породами, более многочисленна и
разнообразна. В литературе имеются некоторые данные о связях
нескольких видов копьехвосток с лиственными породами, преи¬
мущественно с осиной и тополем. Так, с осиной связывается
развитие L. stackelbergi Czerny (Tuomikoski, 1930; L. tarsala),
L. sylvatica Bel. (Hackman, 1956; L. lucidiveiiLris Beck,). Име¬
ются данные о развитии под корой осины (Hackman, 1956; Nuor-
teva, 1967) и тополя (Smith, 1957) L. fugax Beck., в этих же по¬
родах развиваются Tj. palposa Zett. (Bezzi, 1920; Losowski, 1922,
ЦИТ. no Родендорфу, 1950; Czerny, 1934; Smith, 1957) и L. pe¬
regrina Beck. (Tuomikoski, 1930; Nuorteva, 1967; Gollin, 1953).
G тополем связано развитие группы scutellaris в целом (McAIpine,
Munroe, 1968).
В литературе имеются также единичные указания еще на 2—3
породы, с которыми связано развитие копьехвосток: L. peregrina
Beck. и L. scutellaris Rond. развиваются в ясене (Czerny, 1934),
L. palposa Zett.— в березе и боярышнике (Collin, 1953).
Наши материалы позволили значительно расширить перечень
лиственных пород, в которых встречаются личинки копьехвосток.
В настоящее время мы располагаем данными о разнообразном
видовом составе копьехвосток, развивающихся, например, на
ильме долинном и буке. Так, для ильма долинного характерны
виды: L. stackelbergi Czerny, L. freyi Czerny, L. mcalpineiana
Kov., L. palpata Czerny, L. atronitens Kov. Ряд видов, разви¬
вающихся на осине, оказался характерным для бука европейско¬
го и восточного (L. peregrina Beck., L. fugax Beck., L. sylvati¬
ca Bel.). В настоящее время можно указать около 20 пород лист¬
венных деревьев, в том числе маакия амурская, бархат амурский
и сахалинский, чозения крупночешуйчатая и др., в которых
развивается около 20 видов копьехвосток.
До последнего времени ксилофильные личинки копьехвосток
рассматривались лишь в качестве обитателей зоны коры. Наши
наблюдения показали, что личинки рода Lonchaea F11. обычны так¬
же в толще разлагающейся, довольно рыхлой древесины. Так,
личинки L. carpathica Kov. развиваются в толще рыхлой и влаж¬
ной древесины бука европейского, а личинки L. caucasica Kov.—
бука восточного. В своеобразных условиях, в сильно разложив¬
шейся влажной древесине лиственницы сибирской развиваются
личинки Lonchaea sp., близкие L. laticornis Meig.
Значение личинок копьехвосток в ксилофильных сообществах
в значительной степени определяется тем, какие породы (хвойные
или лиственные) они заселяют, на каких этапах разложения коры
и древесины развиваются личинки и с какими видами ксило-
фильных насекомых возможны в этих условиях контакты.
Можно указать лишь на небольшое число видов копьехвосток,
для которых установлена связь с короедными сообществами. Это
преимущественно виды, развивающиеся под корой хвойных де¬
330
ревьев. Мы располагаем данными о связях с поселениями коро¬
едов видов, для которых подобный образ жизни уже освещен в
литературе. Так, в поселениях короедов развиваются личинки
J.. zetlerstedti Веек. (Зиновьев, 1957; Novak, 1960), L. seilueri Hend.
(Ликитюк, 1957; Morge, 1967а), L. defecta McAlp. (McAlpine, 1964;
Xuorleva, 1967) и североамериканский вид L. corticis Taylor
(Taylor, 1929, 1930). Все перечисленные виды относятся к группе
«corticis». В ходах короедов развиваются личинки L. collini
Паск. (Hackman, 1956; Зиновьев, 1958; Nuorteva, 1967; Гаприн-
дашвили и др., 1967; Берозашвили, 1968) и L. bruggeri Morge
(Morge, 1967а). Нами установлена связь с короедными поселе¬
ниями при развитии как на хвойных, так и лиственных породах у
личинок L. sylvatica Bel. В сообществе с различными видами
заболонников (Scolytidae) в ильме долинном развиваются личинки
L. mcalpineiana Kov. и L. palpata Czerny. Вблизи поселений ко¬
роедов на осине зарегистрированы личинки L. fugax Веек. В
литературе имеются сведения о связях с короедными поселениями
личинок L. scutellaris F. (Родендорф, 1950; Kishi, 1972).
Следует отметить, что в лиственных породах, даже заселенных
короедами, личинки копьехвосток сосредоточиваются преимущес¬
твенно на сильно увлажненных участках, часто на нижней сто¬
роне лежащих па земле стволов деревьев и, как правило, не кон¬
тактируют с короедными поселениями. Наконец, целый ряд ви¬
дов развивается в коре на более поздних этапах разложения,
в сообществе с рядом других ксилофильных насекомых, в том
числе златками, дровосеками, различными двукрылыми. Так,
например, личинки L. peregrina Веек, обычно встречаются под
корой совместно с личинками Megamerina dolium F., личинки
L. subneatosa Kov. на сильно увлажненных, например сокото-
чивых, участках — с личинками львинок: Pseudowallacea hun-
garica Kert., Neopachygaster secernibilis Kriv. (Stratiomyidae)
и Texara compressa Walk. (Megamerinidae). Личинки L. krivo-
sheinae Kov. развиваются совместно с целым комплексом различ¬
ных видов львинок (Stratiomyidae), болотниц (Limoniidae), кси-
ломиид (Xylomyidae) и др.
В вопросах пищевых связей личинок копьехвосток до насто¬
ящего времени нет достаточной ясности. Ряд авторов относит
всех личинок к хищникам (Mokrzecki, 1933; Leon, 1934, и др.),
некоторые полностью отрицают возможность хищничества (Bezzi,
1919). В. Ю. Фридолин (1936) указывает на питание мицелием
грибов. Многие авторы (Vimmer, 1926; Taylor, 1929, 1930; Не-
смерчук, 1948; Никитюк, 1957; Негробов, 1971) называют личи¬
нок копьехвосток паразитами, что не соответствует ни их образу
жизни, ни типу питания. Личинки копьехвосток, нападая на
жертву, разрывают ее покровы и высасывают содержимое.
Г. А. Зиновьев (1957), указавший на ошибочность оценки дея¬
тельности личинок копьехвосток как паразитов, подробно описал
их способ питания,
331
в настоящее время в качестве хищников короедов с полной
достоверностью зарегистрированы обитающие на хвойных поро¬
дах личинки L. zellerstedti Веек., L. seitneri Ilend., L. collini
Наск. и L. bniggeri Morge (Morge, 19G1, 1963, 1967a, b; Гаприн-
дашвили и др., 1967; Берозашвили, 1968). Следует отметить, что
в группе «corticis», куда входят первые два вида, личинки неарк¬
тических видов также хищники (McAlpine, 1964; McAlpine, Morge,
1970). Так, личинки L. corticis Taylor, тесно связанные со смо¬
левкой Pissodes strobi Реек., уничтожают до 5,5% личинок и
куколок (Taylor, 1929, 1930). По сообщениям некоторых авторов
(Morge, 1967а, Ь), личинки L. seitneri Ilend. активно истребляют
короедов и могут регулировать их численность.
Имеющиеся в литературе сведения о хищничестве личинок не¬
которых других связанных с хвойными видов ошибочны или
требуют уточнения. Так, например, указание на хищничество
личинок L. fugax Веек, в поселениях короедов Jps typographus L.
(Seitner, 1924) следует отнести к L. brnggeri Morge. Сообщение о
том, что данные по этому же виду (L. fugax Веек.; Seitner, 1925)
следует отнести к L. seitneri Hend. (Hendel, 1928), также ошибоч¬
но (Morge, 1967а). Требуют уточнения сведения о хищничестве
личинок L. palposa Zett. (Losowski, 1922), так как видовое оп¬
ределение материала вызывает сомнение (Mokrzecki, 1933), а также
о хищничестве личинок копьехвосток, питающихся личинками
Hylemyia (Seitner, 1929; Kangas, Leskinen, 1943).
Для личинок, развивающихся на лиственных породах, в ли¬
тературе имеются достаточно противоречивые сведения. Так,
для личинок L. sylvatica Bel. и L. peregrina Веек, указываются
способность как к сапрофагии, так и хищничеству. В качестве
хищников многими авторами рассматриваются личинки L. scu-
tellaris Rond. Подробно эти сведения приводятся в характеристи¬
ках видов. Среди обитателей лиственных пород значительную
группу составляют виды, личинки которых относятся нами к не-
крофагам или сапронекрофагам. Личинки этой группы питаются
поврежденными или погибшими личинками и куколками различ¬
ных ксилофильных насекомых. Пожалуй, впервые серьезное вни¬
мание этой специфике питания личинок уделил Г. А. Зиновьев
(1957). По нашим данным, к некрофагам следует отнести личинок
L. sylvatica Bel., L. caucasica Kov., L. scutellaris Rond., к сапро¬
некрофагам — L. peregrina Beck. и L. krivosheinae Kov.
Анализ имеющихся в пашем распоряжении хотя и небольших
материалов говорит о достаточном разнообразии характера пи¬
тания личинок копьехвосток рода Lonchaea: от сапропекрофагии
до хищничества. Для обитателей хвойных пород хищничество
является твердо установленным фактом. Между тем у обитателей
лиственных хищничество не зарегистрировано. Значительное ме¬
сто среди них занимают личинки-некрофаги. Значение личинок-
некрофагов как в короедных, так и в любых других ксилофиль¬
ных сообществах не может оцениваться в том же плане, как ли-
332
чипок-хищ^иков lia хвойных. Личипки-некрофаги, нападая на
поврежденных или погибших личинок, куколок, а в ряде слу¬
чаев и имапз насекомых, могут выполнять роль санитаров, т. е.
благотворно влиять в определенные периоды на состояние сооб¬
ществ ксилофильных насекомых, например, весной, когда после
зимнего периода в поселениях значительный процент составляют
погибшие личинки.
Значение в целом различных видов личинок копьехвосток в
ксилофильных сообществах в значительной степени определяет¬
ся тем, на каких этапах разложения коры и древесины посе¬
ляются личинки и с какими видами ксилофильных насекомых
возможны в этих условиях контакты. В литературе, особенно при¬
кладного характера, проявляется тенденция к преувеличению ро¬
ли личинок копьехвосток в сообществах ксилофильных насеко¬
мых. Так, данные, известные о типах питания личинок отдельных
видов или о связях с теми или иными поселениями ксилофиль¬
ных насекомых (например, короедов), переносятся на весь ком¬
плекс копьехвосток. В результате личинки всех обнаруживае¬
мых под корой видов обычно рассматриваются, по меньшей мере,
как потенциальные хищники короедов. Неверна и другая край¬
няя точка зрения, что большинство личинок копьехвосток не име¬
ет практического значения (Morge, 1963). Экологические связи
личинок копьехвосток весьма сложны и следует подчеркнуть,
что их роль в сообществах ксилофильных насекомых очень разно¬
образна и не может сводиться лишь к их деятельности в качест¬
ве энтомофагов.
Морфология. Личинки удлиненные, с заостренным передним и
тупо обрубленным задним концом тела. Ротоглоточный аппарат
(рис. 108, 3, 4) хороню развит, сильно склеротизован. Ротовые
крючки в большинстве случаев серповидные, массивные в осно¬
вании, с коротким или удлиненным вершинным отделом. Гино-
стомальные склериты удлиненные, массивные, парастомальные —
тонкие, но хорошо выраженные. Фарингеальные склериты разно¬
образного строения.
Сегменты тела четкие, обособленные, на значительном протя¬
жении голые, имеются лишь небольшие поля шипиков или бугор¬
ков на вентральной стороне сегментов. Поля расположены в пе¬
редних отделах сегментов н образованы несколькими рядами
шипиков (рис. 108,5). Последний сегмент тела, тупо обрублен и
закруглен, на вентральной стороне с хорошо развитой анальной
пластинкой, как правило, окруженной рядами шипиков. Перед¬
ние дыхальца вееровидные, могут иметь до И выростов (Hennig,
1952), несущих дыхальцевые отверстия. Задние дыхальца рас¬
положены на коротких выростах, иногда снабженных различной
формы небольшими лопастями. Вершинная часть выроста и ло¬
пасти светлые или сильносклеротизованные, темные. Дыхальца
округлые или слегка удлиненные, с 3 овальными дыхальцевыми
отверстиями, расположенными радиально (рис. 108, 8).
333
Рис. 108. Общий вид и схема строения личинок копьехвосток (Lonchaeidae)
Общий вид сбоку: J — L. ragnari Наск.; 2 — L. fugax Веек.; 3,4— ротоглоточный аппарат
L. sylvatica Bel. сбоку и снизу; схемы: 5 — шипикового поля на ползательных валиках,
6,7 — расположения мыиняных окончаний на среднем сегменте тела сбоку и Гдвух по¬
следних сегментах сверху, 8 — заднего дыхальца и окаймляющего его выступа. 9 —
переднего дыхальца
а — простой ряд шипиков; Ь — сложный ряд шипиков; с — вторичные косые ряды ши-
пиков; d — дорсальный отросток фарингеального склерита; d. Ь — дорсальная перемычка;
d. t — зубные склериты; / — ф фингеальный склерит; h — высота стигмального выступа;
hp — гипостомальный склерит; I — ползательные валики; 1Ь — лабиальный] склерит;
vi, п, р — рисунок мышечных окончаний (т — боковые пятна, п — срединная двойная
линия пятен, р — дорсальные пятна); md — ротовые крючки; рг — перитрема; ps — па-
растомальный склерит; st. d — стлгмальный диск; st.f — стигмальное отверстие; st. р1 —
стигмальная пластинка; ss — продольная ось, проходящая через отверстия; v — вент¬
ральный отросток фарингеальной мембраны
Морфология личинок семейства изучена очень слабо. Имеются
описания личинок отдельных видов, относящихся преимущест¬
венно к родУ Lonchaea. Сводные данные по личинкам рода при¬
ведены в рабо^гах Морге (Morge, 1963) и Н. П. Кривошеипой (19766).
Описаны личинки отдельных видов рода Dasiops Rond. (McAlpi-
ne, 1961; Мог^, 1963), кратко описана личинка Spermatolonchaea
flavidipennis Zett. (Hering, 1954). В целом имеющиеся сведения
очень отрывочны и трудно сравнимы ввиду разнородности опи¬
саний.
Ниже рассматривается лишь 1 род копьехвосток Lonchaea
F11. Личинки родов Earomyia Zett., Silvestrodasiops Morge и
Dasiops Rond., частично связанные с древесиной, изучены недо¬
статочно.
Род Lonchaea FI1.
Личинки молочно-белые или слегка желтоватые, по форме те¬
ла подразделяются на две обособленные группы. Первую группу
образуют личинки, обладающие вальковатым, утолщенным, иног¬
да дорсовентрально уплощенным телом, тупым на переднем кон¬
це и закругленным или немного уплощенным на заднем (рис.
108, i). Анальная пластинка у этих видов плоская, слегка высту¬
пающая над поверхностью тела. Длина тела относится к его по¬
перечнику, как 6 : 1. К этой группе относятся личинки L. bukow-
skii Czerny, L. ragnari Hack., L. palpata Czerny, L. peregrina Beck.,
L. mcalpineiana Kov., L. subneatosa Kov., L. atronitens Kov.,
L. freyi Czerny, L. stackelbergi Czerny. Вторую группу образуют
личинки, обладающие более тонким, заостренным спереди и ту¬
по обрубленным на конце телом. Анальная пластинка, как пра¬
вило, сильно выступает над поверхностью тела (рис. 108, 2).
Длина тела относится к его высоте, как 10 : 1. К этой группе
относятся личинки L. carpathica Kov., L. caucasica Kov., L. fugax
Beck., L. sylvatica Bel., L. scutellaris Rond., L. palposa Zett.,
L. collini Hack., L. zetterstedti Beck.
Ротоглоточный аппарат личинок копьехвосток обладает ти¬
пичным для круглошовных двукрылых планом строения. Имеют¬
ся хорошо развитые ротовые крючки и зубные склериты, а такнче
небольшие по размерам лабиальные склериты, добавочные скле¬
риты отсутствуют. Гипостомальные и парастомальные склериты
хорошо развиты, имеется небольшой прементум. Дорсальные от¬
ростки фарингеального склерита несколько короче вентральных.
Форма ротовых крючков очень разнообразна (Hennig, 1948а;
Smith, 1957; Morge, 1963; Кривошеина, 19766).
Поверхность тела личинок копьехвосток с характерными вда-
влениями, образующими определенный рисунок. Это — места
прикрепления мышц. 3 пятна имеются на боковых (рис. 108, 6—
7, т) и несколько пятен на дорсальной стороне средних сегмен¬
тов тела (рис. 108, 6—7, р). На дорсальной стороне последнего
335
сегмента тела имеется четкий ряд (одинарный или двойной) вдоль
средней линии (рис. 108, 7, п) и короткие ряды пятен вокруг стиг¬
ма л ьных выростов. /
На вентральной стороне брюшных сегментов -уела имеются
хорошо выраженные ползательные валики (108, 5)1 несущие по
нескольку рядов светлых или темных шипиков различных раз¬
меров. Ряды шипиков простые {а) или сложные (6), образованные
вторичными более короткими рядами (с). Шипики окольцовывают
также анальную пластинку, образуя вокруг нее целые поля или
четко ориентированные ряды. Довольно часто шипики образуют
также выступ вблизи задпебокового угла пластинки (рис. 109,
9).
Последний сегмент тела с небольшими выростами, несущими
задние дыхальца и различные выступы (рис. 108, 8), валики или
крючки. При описаниях личинок используется такой признак,
как высота выступа, представляющая собой расстояние от на¬
ружного края перитремы дыхальца до вершины выступа
(рис. 108, 8).
Личинки копьехБосток обладают трахейной системой амфип-
нейстического типа. Передние дыхальца с расширенным и удли¬
ненным атриумом, несущим преимущественно 4—8 пальцеоб¬
разных выступов (рис. 108, 9). Задние дыхальца массивные,
овальные или округлые (рис. 108, 8), Дыхальцевые отверстия
расходятся радиально, причем таким образом, что 2 прилежащих
к диску отверстия обычно расположены друг против друга. Ось,
проходящая через эти отверстия, может быть ориентирована по-
разному (рис. 108, 5, 55), что связано с положением стигмального
диска. Личинки рода детально описаны в работе Н. П. Криво-
шеиной (1976 б).
Таблица Д1я оиределения видов копьехвосток рода
Lonchaea F11,
1 (16) Тело личинок толстое, массивное, с тупым передним кон¬
цом и закругленным или уплощенным дорсовентрально
задним (рис. 108, 1). Длина тела относится к его высоте,
как 6:1. Анальная пластинка плоская, слабо выступаю¬
щая. Как передний, так и задний отделы ползательных ва¬
ликов с вторичными рядами шипиков (рис. ИЗ, iO). Места
прикрепления мышц, как правило, образуют в переднем
отделе дорсальной стороны последнего сегмента тела двой¬
ную линию (рис. 108, 7; 111, 7).
2 (15) Места прикрепления мышц образуют в переднем отделе дор¬
сальной стороны последнего сегмента тела четкую двой¬
ную линию. Как передний, так и задний отделы полза¬
тельных валиков с многочисленными вторичными рядами
шипиков. Стигма л ьные выросты черные или светлые,
такой же окраски, как остальная поверхность тела.
336
\
3 (6) Последний сегмент тела с длинными крючковидными вы¬
ростами (рис. ИЗ, 2,11), дыхальца расположены на внут-
ренй^ей стороне выростов, обращенной к средней линии
тела.1
4 (5) Крю^овидные выросты на конце тела стройные, удли¬
ненные, их ширина на уровне дыхалец меньше их диа¬
метра, а длина в 1,5—2 раза превосходит диаметр дыхалец
(рис. ИЗ, 1, 2). Выемка между стигмальными выро¬
стами широкая, ее ширина не менее глубины и, как пра¬
вило, равна диаметру дыхалец . . . . L. palpata Czerny
5 (4) Крючковидные выросты на конце тела массивные, широ¬
кие в основании, их ширина на уровне дыхалец не меньше
их диаметра, а длина не более чем в 1,3 раза превосходит
диаметр дыхалец (рис. ИЗ, 3). Выемка между стигмаль¬
ными выростами относительно узкая, ее ширина значи¬
тельно менее глубины и, как правило, менее диаметра ды¬
хальца L. peregrina Веек.
6 (3) Последний сегмент тела с небольшими округлыми или за¬
остренными выступами над дыхальцами; дыхальца рас¬
положены на задней поверхности стигмальных выростов
(рис. 109, 11] 112, 6).
7 (10) Анальная пластинка окружена многочисленными шипи-
ками (рис. 109, 4), Имеются массивные удлиненные вы¬
ступы, образованные шипиками, вблизи заднебоковых уг¬
лов анальной пластинки. Выступы над дыхальцами за¬
острены на вершине, у взрослых личинок и у пупариев
черные (рис. 109, 1J),
8 (9) Выступы последнего сегмента тела на вершине загнуты
вперед (рис. 109, 2). Длина выступа (от периферии ды¬
хальца до вершины выступа) заметно больше диаметра
дыхальца. Выемка между стигмальными выростами от¬
носительно узкая, ее глубина почти в 2 раза превосхо¬
дит ширину (рис. 109, 3) L. bukowskii Czerny
9 (8) Выступы последнего сегмента тела прямые, направлены
вверх (рис. 109, 6, 7). Длина выступа (от периферии
дыхальца до вершины выступа) меньше диаметра дыхаль¬
ца. Выемка между стигмальными выростами относительно
широкая, ее глубина не превосходит ширину (рис. 109, 11)
L. ragnari Наск.
10 (7) Анальная пластинка окружена 1 рядом шипиков, иногда
имеются немногочисленные группы шипиков вблизи ее
заднебоковых углов. Выступы над дыхальцами тупые,
закругленные, светлые.
И (14) Выступы над дыхальцами широкие, уплощенные на вер¬
шине (рис. 112, 6, 7; 115, 6,8), Стигмальный диск распо¬
ложен в верхнебоковом углу дыхальцевой пластинки.
Продольные оси, проходящие через прилежащие к нему
верхние дыхальцевые отверстия двух симметричных ды¬
337
халец, образуют при пересечении тупой угол./Анальная
пластинка с небольшими, но четкими выступаД1И, образо¬
ванными шипиками в заднебоковых углах (рис. 112, S;
115, 9).
12 (13) Выросты, несущие дыхальца, короткие, неб(/льшие, лишь
слегка приподняты над поверхностью сегме^нта. Над ды-
хальцевой пластинкой в верхнебоковом углу имеется чет¬
кий конусовидный выступ \^ис. 112, 6)
L. mcalpineiana Kov.
13 (12) Выросты, несуш,ие дыхальца, заметно вытянуты вверх и
сильно уплош,ены (вид сбоку). Дыхальцевая пластинка
сверху и сбоку окружена ровным без конусовидного вы¬
ступа валиком (рис. 115, 6*) L. subneatosa Kov.
14 (И) Выступы над дыхальцами закругленные, слегка конусо¬
видные (рис. 111, 8, 11). Стигмальный диск расположен
на внутренней стороне дыхальцевой пластинки. Продоль¬
ные оси, проходяш,ие через прилежаш^ие к нему верхние
дыхальцевые отверстия двух симметричных дыхалец, па¬
раллельны. Анальная пластинка без выступов, образо¬
ванных шипиками вблизи заднебоковых углов пластин¬
ки (рис. 111, 5). Ползательные валики и анальная пла¬
стинка окружены относительно крупными и хорошо раз¬
витыми шипиками L. freyi Czerny
15 (2) Места прикрепления мышц образуют в переднем отделе
дорсальной стороны последнего сегмента одинарную сре¬
динную линию (рис. 115, 4). Ползательные валики с хо¬
рошо выраженными вторичными рядами шипиков в зад¬
ней половине валиков, в передней представлены лишь
2—3 короткими рядами в периферических отделах полей
(рис. 115, 2), Стигмальные выросты и поверхность тела
вокруг них слегка затемнены, коричневатые
L. staekelbergi Czerny
16 (1) Тело личинок вытянутое, спереди заостренное, сзади не-
уплощенное (рис. 108, 2). Длина тела относится к его вы¬
соте не менее как 10 : 1. Анальная пластинка вздутая^
выступающая над поверхностью сегмента. Вторичные ря¬
ды коротких шипиков имеются только в задних отделах
ползательных валиков (рис. 110, 4). Места прикрепле¬
ния мышц образуют в переднем отделе дорсальной сторо¬
ны последнего сегмента тела простую линию (рис. 110, 5).
17 (20) Дыхальцевая пластинка угловатая, с широко расстав¬
ленными дыхальцевыми отверстиями, с трех сторон окру¬
жена хорошо обособленным валиком (рис. 110, i, 2).
18 (19) Дыхальце почти квадратное; окружающий его валик зна¬
чительно шире сверху и с наружной стороны, чем с внут¬
ренней. На ползательных валиках особенно выделяется
лишь один ряд шипиков в задней половине. Выступы^
образованные шипиками вблизи заднебоковых углов
338
анальной пластинки, относительно хорошо выражены
(рис. 110, 5) L. carpathica Kov.
19 (18) Дыхальце вытянуто в длину, по форме приближается
к прямоугольнику, окружающий его валик лишь слегка
расширен с наружной стороны по сравнению с внутрен¬
ней (рис. 110, 7). На ползательных валиках выделяются
3 ряда шипиков. Выступы, образованные шипиками
вблизи заднебоковых углов анальной пластинки, выра¬
жены слабо (рис. 110,9) L. caucasica Kov.
20 (17) Дыхальцевая пластинка удлиненно- или округло-оваль¬
ная, без четких углов, окружена выступом или валиком
самое большее сверху и снаружи (рис. 112, 2, 5).
21 (26) Анальная пластинка окружена одним рядом шипиков,
иногда имеется второй ряд точечных шипиков только
перед пластинкой (рис. 112, 5). Выступы, образуемые ши¬
пиками вблизи заднебоковых углов анальной пластинки,
не выражены.
22 (23) На четырех последних сегментах тела в задней полови¬
не полей шипиков вторичные ряды дуговидные, образо¬
ванные темными шипиками, хорошо выделяюш,имися на
светлом фоне покровов. Перед анальной пластинкой имеет¬
ся два ряда темных шипиков, резко различающихся по
размерам, за пластинкой шипики образуют один ряд
(рис. \\2,3) L. fugax Веек.
23 (22) На всех сегментах тела, включая четыре последних, вто¬
ричные ряды шипиков обычные, прямые (рис. 116, 5).
Анальная пластинка окружена лишь одним рядом шипи¬
ков (рис. 116, 4).
24 (25) Задние дыхальца ограничены сверху тупыми, конусовид¬
ными выступами (рис. 116, 2, 2). Продольные оси, прохо¬
дящие через прилежащие к стигмальному диску дыхаль-
цевые отверстия, пересекаются под острым углом на зна¬
чительном удалении от дыхалец (рис. 116, 1)
L. sylvatica Bel.
25 (24) Задние дыхальца ограничены сверху небольшим валиком
(рис. 114, 6,7). Продольные оси, проходящие через при¬
лежащие к стигмальному диску дыхальцевые отверстия,
пересекаются под тупым углом вблизи дыхалец (рис. 114, 6)
L. scutellaris Rond.
26 (21) Анальная пластинка окружена несколькими рядами ши¬
пиков. Выступы, образуемые шипиками вблизи заднебо¬
ковых углов пластинок, хорошо выражены.
27 (28) Стигмальный диск расположен на внутренней стороне
дыхалец так, что оси, проходящие через прилежащие к дис¬
ку дыхальцевые отверстия, пересекаются на значительном
удалении от дыхалец под острым углом. Дыхальца сверху
ограничены тупыми конусовидными выступами, ширина
которых в основании менее диаметра дыхалец (рис. 114,
339
7, 2). Личинки развиваются под корой листзенных де-
ревьев L. palposa Zett.
28 (27) Стигмальный диск расположен в верхнебоковом углу пла¬
стинки так, что оси, проходящие через прилежащие к дис¬
ку дыхальцевые отверстия, пересекаются вблизи дыхалец
под прямым или тупым углом. Личинки развиваются под
корой хвойных деревьев.
29 (30) Задние дыхальца сверху и с наружной стороны ограниче¬
ны валиком, несколько расширенным в верхнебоковом
углу (рис. 111, i, 2). Места прикрепления мышц на дор¬
сальной стороне последнего сегмента в виде поперечных
точек хорошо заметны (рис. 111, 6), Дыхальца удлинен¬
но-овальные (рис. 111, 1) L. collini Наск.
30 (29) Задние дыхальца сверху ограничены длинным конусо¬
видным выступом (рис. 116, 6, 7). Места прикрепления
мышц на дорсальной стороне последнего сегмента тела
в виде продольных точек выражены слабо. Дыхальца ок¬
руглые (рис. 116, 6) L. zetterstedti Веек.
L. bukowskii Czerny. Анальная пластинка (рис. 109, 4) слаба
вытянута в поперечном направлении, округло-овальная, с удли¬
ненной щелью. Шиники образуют вокруг анальной пластинки
широкое поле, в котором намечается 4—5 рядов. Ползательные
валики (рис. 109, 5) с небольшим числом шипиков в переднем от¬
деле и более многочисленными шипиками в заднем отделе.
Вид сравнительно редок, зарегистрирован в северо-западных
и южных районах нашей страны, известен из Финляндии (Шта-
кельберг, 1958). В Приморском крае личинки развиваются под
корой чозении крупночешуйчатой (Комарово-Заповедное).
L. carpathica Kov, Анальная пластинка (рис. 110, 3) попереч¬
но-овальная, выступающая, окружена сравнительно светлыми
слабо заметными шипиками. На ползательных валиках поля не¬
большие, шипики светлые и плохо заметны (рис. 110, 4),
Вид описан по материалам из Закарпатской области (Квасы),
где его личинки развиваются в толще светлой, влажной и рых¬
лой древесины бука европейского совместно с личинками Рага-
clusia tigrina Fll. (Clusiidae). Возможно, они питались личинка¬
ми Р. tigrina Fll. или их остатками.
L. caucasica Kov, У личинок младших возрастов валик замет¬
но уже, чем у взрослых (рис. 110, 6, 7). Анальная пластинка
(рис. 110, 9) поперечно-овальная, выступающая, окружена од¬
ним рядом светлых и слабо заметных крючков. Ползательные
валики с относительно небольшими полями очень светлых шипи¬
ков (рис. 110, 8).
Вид описан из Краснодарского края (Гузеринль), где личинки
развивались совместно с личинками Clusiidae в буке восточном
в толще рыхлой светлой древесины на границе с плодовым телом
гриба-трутовика и в измельченной трухе в основании плодового
тела. В литературе (Ковалев, 1974) для данного вида неверно
340
Рис. 109. Детали строения личинок Lonchaea bukowskii Czerny (1—5) ir
L. ragnari Hack. (6—11) (Lonchaeidae)
1, 2, 6, 7 — конец тела сзади и сбоку; 3, 11 — стигмальные выросты снизу; 4,9 — аналь-
пая пластинка; 5, Ю — ряды шипиков на ползательных валиках; 8 — последние сегмен¬
ты тела сверху
указано место обитания личинок (на поверхности растущего на
буке трутовика). Личинки, собранные в первой декаде июля^
продолжали интенсивно питаться в лабораторных условиях пов¬
режденными личинками усачей, куколками слоников и другими
насекомыми. Через 1—2 суток 8—10 личинок полностью уничто-
341
Рис. 110. Детали строения Lonchaea carpathica Kov. (1—6) и L. caucasica
Kov. (6—9) (Lonchaeidae)
1, 2, 6, 7 — конец тела сзади и сбоку; 3, 9 — анальная пластинка; 4,8 — ряды шипиков
ползательных валиков; 5 — последний сегмент тела сверху
жали жертву, оставляя лишь пустую шкурку. Следует отметить,
что личинки данного вида развиваются на заключительных эта¬
пах разложения древесины и не могут играть какую-либо роль
в сообш,ествах различных стволовых вредителей, например корое¬
дов, усачей, златок.
L, collini Наск, Анальная пластинка (рис. 111, 5, 9) попереч¬
ная, выступающая. Выступы вблизи задне-боковых углов плас¬
тинки небольшие, короткие и образованы мелкими шипиками.
Ползательные валики с небольшими полями шипиков (рис. 111, 4),
Имеется всего 2 (1 простой и 1 сложный) ряда крупных шипиков.
Вид зарегистрирован в Центральной и Северной Европе, Перм¬
ской, Куйбышевской областях, в Грузии и на Дальнем Востоке.
Личинки развиваются на различных хвойных в Сибири (Коломиец,
Богданова, 1973; Богданова, Коломиец, 1974), под корой сосны
М2
Рис. 111. Детали строения Lonchaea collini Hack. (7—6, 9) и L. freyi
Czerny (5, 7, <9, 10, 11) (Lonchaeidae)
J, 2, 8, 11 — конец тела сзади и сбоку; 3, 5 анальная пластинка; 4, ю — ряды шипи-
ков на ползательных валиках; 6,7 — последние сегменты тела сверху; 9 — то же, сбоку
на территории Финляндии (Hackman, 1956; Nuorteva, 1967); до¬
вольно тесно связаны с короедами — Blastophagus piniperda L.
и смолевками — Pissodes pini F. (Hackman, 1956; Nuorteva, 1967),
с Ips typographus L. (Зиновьев, 1958; Богданова, Коломиец,
1974), Dendroctonus micans Kug. (Гаприндашвили и др., 1967;
Берозашвили, 1968). Нами в Европейской части СССР личинки
обнаружены под корой ели европейской, пихты кавказской, сос¬
ны обыкновенной, в Приморском крае (Комарово-Заповедное) —
под корой кедра корейского. В последнем развиваются в поселе¬
ниях Ips sexdentatus Boern. совместно с личинками двукрылых —
львинок (Zabrachia minutissima Zett.) и зеленушек (Medetera
ravida Neg.). Личинки — хищники, активно уничтожают корое¬
дов, например большого елового лубоеда (Dendroctonus micans
Kug.) (Гаприндашвили и др., 1967; Берозашвили, 1968), а также,
по нашим данным, личинок и куколок Ips sexdentatus Boern.
Вид является одним из важнейших энтомофагов стволовых вреди¬
телей.
L. freyi Czerny. Ползательные валики с массивными полями
шипиков, образованными многочисленными вторичными рядами
(рис. 111, 10).
Вид сравнительно редок, известен из северо-западных районов
СССР и из Финляндии. Личинки, по нашим данным, развиваются
под корой ели европейской (Рахов, Закарпатская обл.), ильма
долинного, ясеня маньчжурского (Комарово-Заповедное, Примор¬
ский край). В трухе среди сильно разложившихся лубяных воло¬
кон коры ясеня личинки встречались в сообществе с личинками
двукрылых Empididae, а также комаров-толстопожек — Dilop-
hus tibialis Loew (Bibionidae). Материалы из Приморского края,
возможно, относятся к новому, близкому L. freyi Czerny виду.
L. fugax Веек. Места прикрепления мышц выражены слабо
(рис. 112, 4). Ползательные валики с небольшими, но своеобраз¬
ными полями шипиков (рис. 112, 2).
Вид известен преимущественно из северных и центральных
районов Европы. В литературе имеются данные о нахождении ли¬
чинок под корой осины (Hackman, 1956; Nuorteva, 1967) и тополя
(Smith, 1957). По нашим данным, развиваются под корой различ¬
ных лиственных пород, в том числе бука восточного и европей¬
ского, граба, дуба (Московская обл., Закарпатье, Краснодарский
край), но предпочитают осину и тополь. В осине личинки обнару¬
жены в сильно увлажненных волокнах коры вблизи поселений
короедов — Trypophloeus granulatus Ratz. Из-за неточного опре¬
деления имагинальных материалов В. Г. Ковалевым, сведения
об обитании личинок L. fugax Веек, под корой маакии амурской
(Кривошеина, 1972д) следует отнести к другому виду. Точные
данные о питании личинок в литературе отсутствуют. Имею¬
щиеся сведения (Seitner, 1924) о хищничестве личинок этого вида
в поселениях Ips typographus L. следует отнести к L. bruggeri
Morge. Сообщение о том, что данные по L. fugax Веек. (Seitner,
344
Рис. 112. Детали строения Lonchaea fugax Веек. (^—5) и L. mcalpineiana
Kov. (6—10) (Lonchaeidae)
1, 5, 6,7 — конец тела сзади и сбоку; 2, 9 — ряды шипиков ползательных валиков;
3,8 — анальная пластинка; 4, ю — последние сегменты тела сверху
1925) следует отнести к L. seitneri Hend. (Hendel, 1928), некото¬
рыми авторами отрицается (Morge, 1967а).
L, mcalpineiana Kov. Места прикрепления мышц у взрослых
личинок на дорсальной стороне последнего сегмента хорошо за¬
метны (рис. 112, 10). Ползательные валики с большими полями,
вторичные косые ряды расположены как в передней, так и задней
половине полей (рис. 112, 9).
345
Личинки развиваются под корой ильма долинного (Комарово-
Заповедное, Приморский край), во влажной волокнистой коре
лежащих на земле колод в сообществе с личинками ксиломиид —
Xylomya czekanovskii Pl. (Xylomyidae), ходуленожек — Rainieria
latifrons Loew (Calobatidae) и мегамеринид — Техага compressa
Walk. (Megamerinidae). Более сухие участки коры были заселены
заболонниками — Scolytus sulcifrons Rey., S. jacobsoni Spess.
(Scolytidae) и личинками зеленушек — Medetera stackelbergiana
Neg. (Dolichopodidae).
L. palpata Czerny. Ha последнем сегменте тела сбоку от дыхалец
расположены длинные, направленные назад и дуговидно загнутые
вверх крючковидные выросты (рис. ИЗ, i, 2, 8), Поверхность вы¬
ростов, несущих дыхальца (рис. ИЗ, 4), а также небольшой уча¬
сток поверхности сегмента вокруг выростов сильно склеротизо-
ваны, черные. Выросты в основании не уплощены, расположены
сбоку от дыхалец и не охватывают своим основанием дыхальца
спереди (рис. ИЗ, 4). Места прикрепления мышц хорошо замет¬
ны (рис. ИЗ, 2). Анальная пластинка (рис. ИЗ, 7) поперечно¬
овальная. Ползательные валики с многочисленными рядами ши-
пиков (рис. ИЗ, б).
Вид, описанный по материалам из Приморского края (Czerny,
1934), был сведен в синоним к L. peregrina Веек. (Morge, 1963),
а позднее вновь восстановлен в качестве самостоятельного (Ко¬
валев, 1973). Выведен из личинок, собранных под корой ильма
долинного, липы и бархата амурского (Комарово-Заповедное,
Приморский край). Личинки встречались в толще лубяных воло¬
кон коры лежащих на земле стволов бархата амурского, одновре¬
менно заселенных также личинками различных видов львинок,
в том числе VVallaceanigrotibialis Р1. (Stratiomyidae), а также бо-
лотниц — Gnophomyia tristis AI. (Limoniidae). Под корой ильма
долинного встречались совместно с личинками L. mcalpineiana
Kov. Как и предыдущий вид, в ильме долинном встречаются в со¬
обществе с заболонниками (Scolytus sulcifrons Rey, S. jacobsoni
Spess.), личинками зеленушек — Medetera stackelbergiana Neg. и
личинками других перечисленных выше двукрылых.
L. palposa Zett, Точки, соответствующие местам прикрепления
мышц, образуют одну срединную полосу (рис. 114, 4), Анальная
пластинка (рис. 114, 5) поперечно-овальная, выступающая. Пол¬
зательные валики с относительно небольшими полями шипиков
(рис. 114, 5).
Вид обычен в Европе. В литературе есть указание на нахожде¬
ние личинок под корой березы и боярышника (Collin, 1953), оси¬
ны (Czerny, 1934), а также тополя (Bezzi, 1920; Smith, 1957).
Нами личинки зарегистрированы совместно с личинками L. scu-
tellaris Rond. под корой ивы белой среди скоплений личинок гал-
лиц — Winnertzia sp. (Убинская, Краснодарский край). Близкие
личинки обнаружены под корой чозении круппочешуйчатой (Кед¬
ровая Падь, Приморский край). Точные данные о питании личи-
346
Рис. из. Детали строения Lonchaea palpata Czerny (i, 2, 4, 6—8) и L. pere¬
grina Beck. (3, 5, 9—11) (Lonchaeidae)
1,5 — крючковидные выросты последнего сегмента тела снизу; 3,4 — крючковидный
вырост с внутренней стороны; 2, п — последний сегмент тела сверху; 6, 10 — ряды шипи¬
ков ползательных валиков; 7,9 — анальная пластинка; '8 — крючковидные выросты
сбоку
Рис. 114. Детали строения Lonchaea palposa Zett. (1—5) и L. scutellaris
Rond. (6—10) (Loncnaeidae)
1,2, 6,7 — конец тела сзади и сбоку; 3,9 — ряды шипиков ползательных валиков;
4, 10 — конец тела сверху; 5,8 — анальная пластинка
нок отсутствуют. Личинки вида относятся одними авторами
(Morge, 1963) к сапрофагам, другими — к хищникам, уничтожаю¬
щим короедов (Losowski, 1922), но в последнем случае правиль¬
ность определения вида вызывает сомнение (Mokrzecki, 1933).
L, peregrina Бес/с. На последнем сегменте тела в переднебоко¬
вых углах вблизи дыхайец расположены длинные направленные
назад и слегка загнутые на вершине крючковидные выросты
(рис. ИЗ, 5, 5). Места прикрепления мышц хорошо заметны
(рис. ИЗ, 11). Анальная пластинка (рис. ИЗ, 9) окружена одним
рядом шипиков спереди и двумя — сзади. Ползательные валики,
как и у L. palpata, с 2 группами многочисленных рядов шипиков
{рис. ИЗ, 20). Каких-либо четких различий в строении полей у ли¬
чинок L. palpata Czemy и L. peregrina Веек, найти не удалось.
Следует отметить, что материалы, определенные по имаго
В. Г. Ковалевым как L. peregrina (Ковалев, 1974), по признакам
личинок и пупариев распадаются на 2 резко различные между
348
собой серии. В настоящей работе под названием L. peregrina опи¬
саны личинки, которые в литературе уже фигурируют под этим
названием (Ковалев, 1974). У личинок второй серии крючковидные
выступы направлены в стороны и вверх, относительно короткие
Именно к этой серии, судя по рисункам, близки пупарии L. pere¬
grina, описанные Морге (Morge, 1963). Вполне возможно, что рас¬
сматриваемая нами личинка относится не к L. peregrina, а к ново¬
му виду.
Вид широко распространен в Европе, где зарегистрирован под
корой осины (Nuorteva, 1967), тополя (Collin, 1953) и ясеня (Czer-
пу, 1934). На территории СССР личинки развиваются под корой
осины, тополя, встречаются под корой бука европейского и вос¬
точного и граба обыкновенного (Московская, Закарпатская об¬
ласти, Краснодарский край). Личинки обычны в сильно увлаж¬
ненных волокнах коры, где сосредоточиваются на нижней стороне
лежащих на земле стволов и в комлевой части пней. В тополе встре¬
чаются совместно с личинками мегамеринид — Megamerina dolium
F. (Megamerinidae). В буке обнаружены на участках, пронизан¬
ных грибным мицелием. Точные сведения о питании личинок
отсутствуют. По данным некоторых авторов (Czerny, 1934; Роден-
дорф, 1950), личинки L. peregrina Веек.— хищники. Личинок это¬
го вида скорее всего следует рассматривать в группе сапронекро-
фагов. В отдельных случаях наблюдалось питание взрослых
личинок в лабораторных условиях трупами личинок различных
насекомых, хищничество не зарегистрировано.
L. ragnari Наск, Места прикрепления мышц довольно хоро¬
шо заметны (рис. 109, 8), схема их расположения близка таковой
L. bukowskii. Анальная пластинка (рис. 109, 9) слегка вытянута
в поперечном направлении. Ползательные валики с относитель¬
но небольшим числом шипиков в переднем отделе и более много¬
численными шипиками в заднем (рис. 109, 10).
Вид описан из Финляндии, где места обитания личинок не
известны. Нами личинки собраны под разлагающейся корой кле¬
на (Красная Поляна, Краснодарский край).
L, scutellaris Rond. Задние дыхальца с очень короткими ва¬
ликообразными возвышениями (рис. 114, 6, 7). Места прикрепле¬
ния мышц в виде небольших точек, образующих на дорсальной
поверхности последнего сегмента одну срединную продольную
полосу (рис. 114, 10). Анальная пластинка (рис. 114, 8) слегка вы¬
ступающая. Ползательные валики с небольшими полями шипи¬
ков (рис. 114, 9).
Вид зарегистрирован в Ленинградской обл., на Кавказе, в
Средней Азии, известен из многих точек Западной Европы. В ли¬
тературе имеются данные о развитии вида под корой ясеня (Czer¬
ny, 1934). Нами личинки зарегистрированы под корой ивы белой
совместно с личинками L. palposa Zett., а также под корой вяза
шершавого и дуба черешчатого (Убинская, Краснодарский
край). Из-за неточного определения имагинальных мате-
349
риалов В. Г. Ковалевым, сведения об обитании личинок L. scutel-
laris Rond. под корой маакии амурской (Кривошеина, 1972д)
следует относить к другому виду. В ряде работ личинки указывают¬
ся в качестве хищников короедов (Родендорф, 1950; Зиновьев,
1957; Morge, 1963; Кривошеина, 1971а; Харитонова, 1972). По
нашим наблюдениям последних лет, личинки являются типич¬
ными некрофагами. Взрослые личинки питаются трупами личи¬
нок узкотелых златок, а также галлиц — Winiiertzia sp., в сооб¬
ществе с которыми развиваются. О питании личинок этого вида
не живыми, а погибшими насекомыми свидетельствуют спе¬
циально проведенные эксперименты (Kishi, 1972). Указание на
питание личинок трупами короедов имеется также у Г. А. Зи¬
новьева (1957).
L. stackelbergi Czerny. Задние дыхальца со своеобразными ко¬
нусовидными выступами (рис. 115, 7, 5), Анальная пластинка
(рис. 115, 3) поперечно-овальная, не выступающая. Ползатель-
ные валики с массивными полями шипиков, имеющими вторичные
косые ряды как в передней, так и задней половинах (рис. 115, 2).
Вид довольно обычен в Сибири и в Приморском крае, откуда
описан (Czerny, 1934), известен из Ленинградской области. Ли¬
чинки в Приморском крае (Комарово-Заповедное) развиваются
под корой лиственных: ильма долинного, тополя Максимовича^
липы амурской, предпочитают влажные волокна луба, концент¬
рируясь вблизи комлевой части сухостойных деревьев. Данные
о питании личинок отсутствуют.
L, subneatosa Кои. Места прикрепления мышц на дорсальной
стороне последнего сегмента образуют рисунок из двух линий
в передней половине сегмента и одной — в задней (рис. 115, 7).
Анальная пластинка (рис. 115, 9) поперечно-овальная, относитель¬
но небольшая. Поля шипиков на ползательных валиках относи¬
тельно большие, короткие косые вторичные ряды шипиков рас¬
положены как в переднем, так и заднем отделах ползательных
валиков (рис. 115,70).
Вид описан по материалам из Московской области. Краснодар¬
ского края, Грузии (Ковалев, 1974). По нашим данным, личинки
развиваются под корой осины, тополя, липы (Московская область.
Краснодарский край). В осине личинки встречаются в сообщест¬
ве с личинками львинок Pseudowallacea hungarica Kert. (Stratio-
myidae) и других двукрылых, типичных обитателей коры. Точ¬
ные данные о питании личинок отсутствуют. При содержании ли¬
чинок в лаборатории хищничество не зарегистрировано.
L. sylvatica Bel. Места прикрепления мышц слабо заметные
(рис. 116, 5). Анальная пластинка (рис. 116, 4) поперечно-оваль¬
ная, выступающая. Ползательные валики с хорошо выраженными
полями шипиков (рис. 116, 5).
Вид довольно обычен в Палеарктике, в том числе на террито¬
рии Европейской части СССР. Личинки развиваются под корой
хвойных: ели европейской, сосны обыкновенной, пихты кавказ-
350
Рис. 115. Детали строения Lonchaea stackelbergi Czerny (1—5) и L. sub-
neatosa Kov. (6—10) (Lonchaeidae)
1, 5, 6, 8 — конец тела сзади и сбоку; 2, 1 о — ряды шипиков ползательных валиков;
3,9 — анальная пластинка; 4,7 — последние сегменты тела сверху
ской (Рахов, Закарпатская область; Убинская, Краснодарский
край) и различных лиственных пород: ольхи, бука, граба, клена,
дуба (Квасы, Рахов, Закарпатская область; Убинская, Гузерипль,
Краснодарский край; Московская область). В литературе есть
указания на связь вида с осиной (Насктап, 1956; L. lucidiventris
Веек.). Личинки предпочитают увлажненные субстраты, встре¬
чаясь преимущественно на нижней стороне лежащих на земле ко¬
351
Рис. 116. Детали строения Lonchaea sylvatica Bel. (1—6) и L. zetterstedti
Веек. (6—10) (Lonchaeidae)
1, 2, 6, 7 — конец тела сзади и сбоку; 3,9 — ряды шипиков ползательных валиков;
4, 10 — анальная пластинка; 5,8 — последние сегменты тела сверху
лод. Установлена четкая связь с поселениями различных видов
ксилофильных насекомых. В буке личинки встречались на уча¬
стках, заселенных короедом Taphrorychus bicolor Hbst., заболонни-
ком Scolytus scolytus F. (Scolytidae), личинками галлиц (Cecidomyii-
dae), a также личинками усачей Morimus verecundus Fald. (Ceramby-
cidae). В сосне личинки зарегистрированы в поселениях короеда
Ips acuminatus Gyll. (Scolytidae). В колодах бука, пронизанных
грибным мицелием, встречаются совместно с Lonchaea peregrina
Веек. Морге (Morge, 1963) относит личинок к сапрофагам, однако
возможность развития личинок как сапрофагов требует уточне¬
ния. Личинки, по меньшей мере, некрофаги. Неоднократно отме¬
чалось питание личинок, собранных как под корой хвойных, так
352
и лиственных пород, поврежденными или недавно погибшими ли¬
чинками различных видов дровосеков, златок, хрущей. Принимая
во внимание широкое распространение вида, развитие под корой
самых разнообразных пород деревьев, преимущественное заселе¬
ние коры на сколитидной стадии разложения, а также тип пита¬
ния, следует отметить возможное существенное значение его ли¬
чинок в сообществах ксилофильных насекомых в качестве потре¬
бителей погибших личинок насекомых, в первую очередь, коро¬
едов и дровосеков.
L. zetterstedti Веек, Места прикрепления мышц на дорсальной
стороне последнего сегмента тела в виде одной срединной линии
(рис. 116, <^). Анальная пластинка (рис. 116, 10) поперечная, вы¬
ступающая. Ползательные валики с хорошо развитыми полями
светлых шипиков, вторичные косые ряды имеются лишь в зад¬
ней половине полей (рис. 116, 9),
Вид характерен для северных районов нашей страны (Шта-
кельберг, 1958), широко распространен в обоих полушариях
(McAlpine, Morge, 1970), развивается на самых разнообразных хвой¬
ных породах. Нами в Архангельской обл. (Обозерская) личинки
зарегистрированы под корой сосны обыкновенной. На хвойных
встречается в поселениях Trypodendron lineatum 01. (Novak,
1960) и различных других видов короедов, а также дровосеков
(Morge, 1967а, Ь). Данные Гепнига (Hennig, 1948а) о связях вида
с Ips typographus L. относятся к другому виду (L. bruggeri Morge).
Вид, несомненно, играет важную роль в регуляции численности
короедов и, возможно, других стволовых вредителей.
СЕМЕЙСТВО PALLOPTERIDAE - БЛЕДНОКРЫЛКИ
Небольшое семейство бледнокрылок представлено в Пале-
арктике одним родом — Palloplera, включающим около 20 па-
леарктических видов.
Экология. Образ жизни личинок практически не изучен.
В литературе имеются краткие данные о регистрации личинок от¬
дельных видов в древесной и травянистой растительности. Под
корой деревьев развиваются такие виды, как Р. usta Meig., Р. ustu¬
lata Fll., Р. muliebris Harr. (Perris, 1870; Seitner, 1924; Morge,
1956, и др.). Личинки таких видов, как Р. umbellatarum F.,
Р. arcuata Fll., Р. parallela Loew, зарегистр|ировацы в соцветиях
сложноцветных (Balachowsky, Mesnil, 1935; Collin, 1951; Hennig,
1952). Виды Р. trimaculaMeig. и Р. sajltuum L. выведены из стеблей
травянистых растений (Meijere, 1944; Collin, 1951). Интересно,
что для ксилофильного вида Р. usta Meig. имеются также данные
о .развитии личинок в соцветиях сложноцветных (Asteraceae)
(Czeijny, 1934).
Кс.илобионтные личинки встречаются как под корой хвойных,
так ,и лиственных деревьев. По нашим наблюдениям, личинки
Б. М. Мамаев и др. 353
р. usta Meig. предпочитают хвойные породы, а Р. ustulata F11.-—
лиственные. Но в литературе есть указание на обнаружение ли¬
чинок Р. ustulata F11. также под корой хвойных пород, точнее, ели
европейской (Morge, 1956). Личинки Р. usta Meig. и Р. ustulata
F11. обычны в стволах, пораженных короедами, кроме того, личин¬
ки Р. ustulata Fll. обнаружены вблизи поселений личинок галлиц
(Seitner, 1924; Morge, 1956), часто встречаются вблизи участков
с разросшимся мицелием грибов. С поселениями короедов связа¬
ны также личинки Р. muliebris Нагг. (Perris, 1870).
Ксилофильные личинки рода рассматриваются в качестве воз¬
можных хищников короедов. Личинки некоторых видов, напри¬
мер Р. usta Meig., зарегистрированы в отдельных случаях в ка¬
честве активных хищников, значительно снижающих численность
короедов (Morge, 1961, 1967а, Ь). Но в целом наличие сравнитель¬
но небольнюго количества данных о хищничестве личинок может
указывать на возможность смешанного питания — от некрофагии
до хипщпчества.
Род Palloptera Fll.
Тело личинок удлиненное, сужено на переднем конце и ши¬
роко закруглено на заднем (рис. 117, 1,2), Для личинок характер¬
но положение задних дыхалец на небольших трубковидных
выступах. Ротоглоточный аппарат (рис. 117, 5) с сильносклеротизо-
ванными ротовыми крючками и передним отделом гипостомаль-
ного склерита. Ротовые крючки слегка серповидно загнуты на
верпшпе, относительно короткие. Лежащие под крючками зубные
склериты маленькие, но хорошо развиты. Гиностомальные и
нарастомальные склериты относительно длинные, хорошо раз¬
виты. Фарингеальные склериты светлые, склеротизованы лин1ь
но переднему краю и на границе дорсальных и вентральных от¬
ростков. Дорсальные отростки короче и уже вентральных. Пос¬
ледние с большим, но светлым, слабо заметным выступом. Тело
личинок на большем протяжении голое. Ряды шипиков имеются
только на нолзательных валиках, которые хорошо развиты на
границе брюшных сегментов (между грудными и 1-м брюшным от¬
сутствуют). Валики относительно небольшие. Поля шипиков со¬
стоят из 4—5 рядов, из которых один образован более крупными,
собранными в группы по 3—5, шипиками. Остальные ряды пред¬
ставлены точечными равномерно расположенными шипиками. По¬
следний сегмент тела характерной формы, тупо обрублен и закруг¬
лен на конце, так что верхний угол сегмента (вид сбоку) но
форме приближается к прямому. Конец сегмента слегка раздвоен
(рис. 117,5). На дорсальной стороне имеется продольное углубление,
соответствующее местам прикрепления мышц. Анальная пластин¬
ка поперечно-овальная, слегка выступающая, четко очерчена, ог¬
раничена отдельными группами мелких слабо заметных шипи¬
ков (рис. 117, 10), Передние дыхальца (рис. 117, 6) вееровидные,
354
Рис. 117. Детали строения личинок Palloptera ustulata Fll. (i—7'), Р. ori-
entana Kov. (<5) и P. usta Meig. 10) (Palloptoridao)
1^2 — передний и задний концы тела сбоку; 3,4 — последний сегмент тела сверху и они»
зу; 5 — ротоглоточный аппарат; б — переднее дыхальце; 7—9 — последний сегмент тела
сбоку; 10 — последний сегмент тела снизу
С 5—8 короткими округлыми выступами, несущими дыхальцевые
отверстия. Задние дыхальца расположены на очень коротких труб¬
ковидных склеротизованных бугорках. Их длина менее ширины
дыхальцевой пластинки. Над дыхальцем в отдельных случаях рас¬
положены небольшие лопастевидные выступы. В ртличие от личи¬
нок копьехвосток (Lonchaeidae), эти выступы расположены не
в одной плоскости с дыхальцевыми пластинками, а смещены к ос-
12*
355
новацию стигмального бугорка. Дыхальца округлые, с прямыми
удлипеппо-овальпыми отверстиями. Последние расноло>]чены под
прямым углом друг к другу. Линочиый шов находится па внут¬
ренней стороне дыхальцевой пластинки.
В целом представители семейств изучеп1>1 очень слабо. В ли¬
тературе имеются детальные описания личинок Palloptera nsla
IMeig. и Р. ustulata Fll. (Morge, 195()), краткое описание пупария
Р. orientana Kov. (Ковалев, 1972). К сожалению, описания доволь¬
но общие, и дифференциальные диагнозы отсутствуют. Поэтому
предложенная ниже таблица, составленная на основе имеющих¬
ся в нашем распоряжении небольших материалов, носит предва¬
рительный характер.
Таблица для опргдолс’нии видов рода Ра11ор1(та FI1.
1 (()) Последний сегмент тела с лопастевидным выступом над
дыхальцем (рис. 117, 9).
2 (3) Лопастевидный выступ над задним Д1.1хальцем тупой и не¬
большой, четко проявляющийся ли]нь на пупариях
(рис. 117, 8). Переднее дыхальце с 6 отверстиями ....
Р. orientana Kov.
3 (2) Лопастевидный выступ над задним дыхальцем удлинен¬
ный, хорошо заметный. Переднее дыхальце с 5 или
с 7 дыхальцевыми отверстиями.
4 (5) Переднее дыхальце с 5 дыхальцевыми отверстиями
Р. usta Meig.
5 (4) Переднее дыхальце с 7 дыхальцевыми отверстиями . . .
Palloptera sp.
6 (1) Последний сегмент тела без лопастевидного выступа над
задним дыхальцем (рис. 117, 2), в виде небольшого бу¬
горка заметен лишь на нунарин (рис. 117, 7). Переднее
дыхальце с О дыхальцевыми отверстиями (рис. 117, 6)
Р. ustulata Fll.
orientana Коо. Вид описан из Приморского края. Личиики
зарегистри|)ованы под корой ореха мапьч/1чурского на участках'
с грибн1.1М м,ице.11ием среди’ поселопий крупн1.гх личинок галлиц
Wyallella iissiirieiisis Маш. (Cecidomyiidae), погибших пЬсле зи¬
мовки, а также вблизи носелемий личинок лентокрШюк — Pseu-
doseioplera in^n-ica Slack. (O(ilidae). Связи с поселениями корое¬
дов не выявлены. KouKpeTHFiie данные о типе питания отсутствуют.
Обнаружение личинок вблизи участков с грибным мицелием и по¬
селением личинок галлиц могут указывать на возможность мице-
тофагии, некрофагин или хищничества. Сообщение В. Г. Кова¬
лева (1972) о питании личинок бледнЬкрь^лки личинками галлиц
неточно и должно быть дополнено указанием на то, что галлицы
представляли собой колонию личинок, погибшую послё перезимов¬
ки, причем разлагающиеся личинки бйли погружены'в отмершие’
пленки мицелия.
356
р. usta Ме1ц. Вид характерен для северной и средней полосы
Европы. J[h4hhkh зарегистрированы под корой хвойных пород:
ели европейской, сосны обыкновенной, лиственницы европейской.
Обычны в стволах, заселенных короедами. В ряде случаев прояв¬
ляют себя как активные хищники (Morge, 195()).
ustulata FU, Довол1,но обычный вид, встречающийся по всей
территории Европейской части СССР. Личинки зарегистрирова¬
ны под корой разнообразных лиственных деревьев: березы боро¬
давчатой, граба обыкновенного, дуба черешчатого, тополей, кле¬
нов, ивы и т. д. в Закарпатской и Л1осковской областях. Под корой
граба обыкновенного личинки концентрировались в больших коли¬
чествах на участках, не заселенных личинкахми каких-либо других
насекомых. В тополе личинки обнаружены под корой вблизи поселе¬
ний личинок детритниц (Sciaridae). По данным литературы (Mor¬
ge, 195()), личинки встречаются в поселениях короедов — Ips
typographus L. и Pityogenes chalcographus L. (Scolytidae); обна¬
ружены как под корой хвойных, так и лиственных деревьев.
Palloptera sp. Личинки с признаками, приводимыми в лите¬
ратуре для Р. saltuum fj., т. е. с лопастевидными выступами над
задними дыхальцами и 7 дыхальцевыми отверстиями на передних
дыхальцах. Собраны под корой пихты цельнолистной в Примор¬
ском крае. Но для Р. saltuum L. указано развитие в травянистых
растениях (Meijere, 194^i).
ЛИТЕРАТУРА
Арефин В, С, Энтомофаги короедов хвойных пород на юге Приморского края. —
В кн.: Фауна и экология насекомых Сибири. Новосибирск, «Паука»,
1974а, с. 166-173.
Арефин В, С. О биологии Thanasimus siibstriatus Gebl. (Colcoptera, Cleri-
dac) — массового хищника короедов в Приморском крае. —В кн.: Насеко¬
мые — разрушители древесины в лесных биоценозах Южного Приморья.
М., «Наука», 19746, с. 105—110.
Берозашвили Г. И. Материалы по изучению местного естественного врага
большого елового лубоеда. — «Труды Ин-та защиты раст. Груз. ССР»,
1968, 20, с. 141-145.
Богданова Д, А, Значение двукрылых в регуляции численности насекомых-
ксилофагов.— В кн.: Вопросы энтомологии Сибири. Новосибирск,
«Наука», 1974, с. 78—79.
Богданова Д. А., Коломиец Н, Г, Паразиты п хищники стволовых вредите¬
лей хвойных пород в предгорьях Енисейского кряжа.— В кн.: Фауна
и экология насекомых Сибири. Новосибирск, «Наука», 1974, с. 154—166.
Буковский В. И, Некоторые данные о врагах и сожителях короедов в Крыму.—
«Труды Крымского гос. заповедника», 1940, 2, с. 170—188.
Бызова Ю, Б., Келейникова С. И, Семейство Tenebrionidae — чернотелки.—
В кн.: Определитель обитающих в почве личинок насекомых. М.,
«Наука», 1964, с. 463—496.
Воронцов А. И, Использование хищных жуков для борьбы с короедами.—
В кн.: Результаты работ ВНИИЛХ за 1941 — 1945 гг. М.— Л., Изд-во
ВНИИЛХ, 1949, 27, с. 49-52.
Воронцов А. //., Харитонова II, 3. К биологии мурапьежука.— «Научные
труды Моск. лесотехн. ин-та», 1971, 38, с. 27—36.
Гаприндашвили II, /Г., Гумберидзе О, Д., Харазошвили /Г. В, Материалы
к изучению видового состава естественных врагов большого елового
лубоеда в Грузии.— «Сообщ. ЛП Груз. ССР», 1907, 47, 1, с. 107—172.
Гиляров М, С. CoMeiicTBO Cuciijidae — нлоскотелки.- В кн.: Определитель
обитаюпи1х в почве личинок насекомых. М., «Паука», 1904а, с. 414—
415.
Гиляров Л/. С, CeMeiicTBO Nitidulidae — блестянки.— В кн.: Определитель
oбитaroи^иx в ночпе личинок насекомых. М., «Наука», 19046, с. 420—421.
Гиляров М. С. Семейство Pythidae.— В кн.: Определитель обитающих в поч¬
ве личинок насекомых. М., «Наука», 1964в, с. 453—454.
Гириц Л. А. Основы биологическо!'! борьбы с короедом-тииографом (Ips
typographiis I.., Coleoptera, Jpidae). Львов, «Вшца школа» при Львов¬
ском гос. ун-те, 1975.
Голутвина Л, С, Вредители шишек и семян хвойных пород в Иркутской
области. Автореф. канд. дисс. Л., 1973, с. 1 — 19.
Гурьева Е, Л. Сем. Elateridae — щелкуны,— В кн.: Определитель насекомых
Европейской части СССР, т. 2. М.— Л., «Наука», 1965, с. 266—280.
Гусев В. И. Полезные насекомые, встречающиеся на деревьях, заселенных
короедами.— «Изв. Лепингр. лесн. ин-та», 1928, 36, с. 133—153.
358
Долин В, Г. Личинки жуков-щелкунов (проволочники) Европейской части
СССР. Киев, «Урожай», 1964а.
Долин В. Г. Семейство Elateridae — щелкуны.— В кн.: Определитель обита¬
ющих в почве личинок насекомых. М., «Наука», 19646, с. 376—405.
Земкова Р. И. Стволовые вредители темнохвойных лесов Западного Саяна.
Красноярск, «Наука», 1965.
Зиновьев Г. А. Материалы к н;^учению роли биотических факторов в регуля¬
ции численностн скрытностволовых вредителе11.— «Энтомол. обозр.»
1957, 36, 2, с. 322—354.
Зиновьев Г. Л. О структуре, динамике и типологии очагов размножения коро¬
едов.— «Зоол. журн.», 1958, 37, 3, с. 379—393.
Зиновьев Г. А, О значении комплекса онтомофагов в ограничении размноже¬
ния короедов хвойного леса.— В кн.: Чтения памяти Н. А. Холодков-
ского. М.— Л., Изд-во ЛН СССР, 1959, с. 62—86.
Зимин Л- С, Определитель лнчинок синантронных мух Таджикистана. Опре¬
делители но фауне СССР, т. 28. Л., Изд-во АН СССР, 1948, с. 1 — 114.
Исаев Л. С., Яновский В, М. Сложение комплексов энтомофагов и микроор¬
ганизмов на деревьях лиственницы, заселенных стволовыми вредителя¬
ми.— В кн.: Биологические методы борьбы с вредителями сельского
хоз-ва, т. 1. Тан1кент, «ФАН», 1966, с. 110—112.
Келейпикова С, Я., Мамаев U. М. Биология дальневосточных представителей
чернотелок трибы Misolampini (Coleoptera).- «Научп. докл. высш. шко¬
лы., биол. науки», 1972, 3, с. 27—31.
Ниршенблат Я. Д. Новый вид рода Meloponcus Кг. (Со1., Staph.) из ходов
дальневосточных короедов.— «Энтомол. обозр.», 1948, 30, 1—2, с. 48—
49.
Киртенблат Я. Д. Семейство Stapliylinidae — стафилины, или коротко-
падкрылыо жуки.— В кн.: Определитель насекомых Европейской части
СССР, т. 2. М., «Наука», 1965, с. 116—156.
Кобахидзе Д. II.у Супаташвили 1И, М, Некоторые данные по изучению
Rhizophagiis grandis Gyll. в Г1>узин.— «Сообщ. АН Груз. ССР», 1967,
48, 2, с. 443—448.
Ковалев В. 1\ Новый вид рода Palloplera F11. (Diptera, Pallopteridae) из Юж¬
ного Приморья.— «Зоол. журн.», 1972, 51, 5, с. 755—757.
Ковалев В, Г. Описание новых и малоизвестных видов мух рода Lonchaea
F11. (Diptera, Lonchaeidae) из Южного Приморья.— «Энтомол. обозр.»,
1973, 52, 1, с. 201-214.
Ковалев В. Г. Ревизия налеарктических мух-коньехвосток группы Lonchaea
peregrina (Diptera, l.onchaeidac).— В кн.: Насекомые — разрушители
древесины в лесных биоценозах Южного Приморья. М., «Наука», 1974,
с. 86-95.
Козак В, Т, Короеды хвойных пород Волынского Полесья, их пнтомофаги
и меры борьбы с ними. Автореф. канд. дисс. Киев, 1972.
Козак В, Т. Хищники короедов хвойных пород в условиях Волынского
Полесья.— «Лесоводство и агролесомелиор.», 1974, 37, с. 99—104.
Козаржевская Э, Ф., Мамаев Б, Л/. Сукцессия насекомых и других беспозво¬
ночных в древесине ели и их роль в разрушении валежа и порубочных
остатков.— «Изв. АН СССР, сер. биол.», 1962, 3, с. 449—454.
Коломиец IL 1\, Богданова Д, А, Паразиты и хищники стволовых вредителей
хвойных пород в Западной Сибири.— «Труды Биол. ин-та». Новосибирск,
«Наука», 1973, 14, с. 69—82.
Криволуцкая Г, О. Скрытностволовые вредители в темнохвойных лесах За¬
падной Сибири. М.— Л., «Наука», 1965.
Кривошеина II, Я. Отряд Diptera — двукрылые.— В кн.: Определитель оби¬
тающих в почве личинок насекомых. М., «Наука», 1964, с. 605—808.
Кривошеина II. Я. Новые данные по систематике дендрофильных львинок
(Diptera, Stratiomyiidae) и их личинок.— «Энтомол. обозр.», 1965, 44,
3, с. 652—664.
359
Кривошеина Н. П. О биологии и морфологии малоизученных мокрецов рода
Forcipomyia Mcig. (Diptcra, Crratopogonidae).— «Зоол. жури.», 1968,
47, 4, с. 578-590.
Кривошеина Н. П. Онтогенез и эволюция двукрылых насекомых. М., «Наука»,
1969.
Кривошеина Н, П. Значение плодоядных личинок двукрылых в ксилофиль-
ных сообществах.— В кн.: Защита леса от вредных насекомых и болез¬
ней, т. 3. М., МЛТИ, 1971а, с. 62-65.
Кривошеина Н. Я. Хищные личинки двукрылых в coo6niecTBax насекомых-
ксилобионтов.— В кн.: Проблемы защиты таежных лесов. К'расноярск,
19716, с. 84-85.
Кривошеина Н. Я. Семейство Glutopidae fam. n. и его положение в системе
отряда двукрылых (Diptera, Bracliycera Orthorrhapha).— «Энтомол. обозр.»,
1971в, 50, 3, с. 681-694.
Кривошеина Н. Я. Новые данные по систематике и биологии двукрылых се¬
мейства Xylomyidae Советского Союза. — «Зоол. журн.», 1972а, 51,
1, с. 69-78.
Кривошеина Я. Я. К биологии видов рода Tipulodina Alex. (Diptera, Tipii-
lidae), нового для фауны ССС1\— «Докл. АН СССР, сер. биол.», 19726,
202, 1, с. 241-243.
Кривошеина Я. 7/. Насекомые — ризобиопты аридных облаете!*!.— В кн.:
Проблемы почвенной зоологии. Материалы 4-го Всес. совещания. Баку,
1972в, с. 82-83.
Кривошеина Я. Я. Экологические связи двукрылых насекомых, развиваю¬
щихся под корой и в древесине ореха маньчжурского.— «Труды БПИ
ДВНЦ». Владивосток, 1972 г, 7(110), с. 87—97.
Кривошеина II. II, Комплексы двукрылых насекомых, развиваюпи^хся в ма-
акии амурской.— «Труды БПИ ДВНЦ». Владивосток, 1972д, 7(110),
с. 121 — 128.
Кривошеина Я. Я. К экологии ксилобионтных личинок типулид (Diptera,
Tipulidae).— «Экология», 1972е, 3, с. 45—52.
Кривошеина II. Я. Премагинальные фазы комаров-долгоножек (Diptera,
Tipulidae) Дальнего Востока.— «Зоол. журн.», 1972ж, 51, 4, с. 534—
546.
Кривошеина II. II. Новые данные о львинках подсемейства Pachygasterinae
(Diptera, Stratiomyidae) Советского Союза.— «Энтомол. обозр.», 1973а,
52, 1, с. 178-193.
Кр1.вошеина II. Я. К биологии зелепупюк рода Systenus Lw. (Diptera, Doli-
chopcdidae).— «Зоол. журн.», 19736, 52, 7, с. 1095—1096.
Кривошеина Я. Я. К‘омары-долгоножки рода Tipula L. (Diptera, Tipulidae),
развиваюп1иеся в разлагающейся древесине в Южном Приморье.—
«Бюлл. МОИП, отд. биол.», 1973в, 77, 3, с. 45—55.
Кривошеина II. Я. Бибионоидные двукрылые заповедника Кедровая Падь.—
В кн.: «Энтомологические исследования на Дальнем Востоке», т. 2. Вла¬
дивосток, «Наука», 1973г, с. 10—19.
Кривошеина II. II. Двукрылые насекомые — обитатели коры и древесины
тополя Максимовича и родственных пород.— В кн.: Насекомые — раз-
руи)ители древесины в лесных биоценозах Южного Приморья. М.,
«Наука», 1974а, с. 31—40. '
Кривошеина II. II. Ксилофильные нас:комые, развиваюии^еся в ольхе воло¬
систой в Южном Приморье.— В кн.: Насекомые — разрушители древе¬
сины в лесных биоценозах Южного Приморья. М., «Наука», 19/46,
с. 56—64. I . i :
Кривошеина Я. Я. Особенности заселения двукрылыми насцкомыми разла¬
гающихся стволов бархата аму1^ского и других энде1ццчных для Даль¬
него Востока древесных пород.— В кн.: -Нас^комь^е — разрушители дре¬
весины в лесных биоценозах Ю|жйогЬ,Йриморья. М., <Шаука», 1974в,
с. 96-104. ' ' ■ ' '
Криео^шеина Я. Я. Личинки ктырей (Asilidae, |Ctei^otjni) — ризо^ион^ы
360
пустынпых ])астений.—«Научн. докл. высш. школы. Биол. наукш>, 1974 г.,
4, с. 16-21.
Кривошеина Н. П. Морфология личинок зеленушек рода Medetcra Fisch.
(Diptera, Dolichopodidae).— «Энтомол. обозр.», 1974д, 53, 2, с. 309—
323.
Кривошеина Н, П. Новые данные о двукрылых насекомых (Stratiomyidae,
Pachygasterinae), развивающихся в тугайной растительности Средней
Азии.— «Научн. докл. высш. школы, Биол. науки», 1975, 10, с. И—17.
Кривошеина Н. Л. Ксилофильные львинки рода Neopachygaster (Diptera,
Stratiomyidae), их родственные связи и биология. — «Зоол. журн.»,
1976а, 55, 4, с. 559—565.
Кривошеина Н. П. Сравнительно-морфологическая характеристика личинок
палеарктических видов мух-копьехвосток рода Lonchaea F11. (Diptera,
Lonchaeidae).— В кн.: Эволюционная морфология личинок насекомых.
М., «Наука», 19766, с. 22—55.
Кривошеина Н. П., Мамаев Б. М. Личинки европейских сирфид рода Temno-
stoma (Syrphidae, Diptera). — «Энтомол. обозр.», 1962, 41, 4, с. 921—
930.
Кривошеина Н. Я., Мамаев Б. М. Определитель личинок двукрылых насеко¬
мых — обитателе!! древесины. М., «Наука», 1967.
Кривошеина Н. Я., Мамаев Б. М. Новое для фауны СССР семейство двукры¬
лых Cramptonomyiidae (Diptera, Nematocera), его морфология, экология
и филогенетические связи. — «Энтомол. обозр.», 1970, 49, 4, с. 923—935.
Кривошеина Я. Я., Мамаев Б. М. Обзор палеарктических видов двукрылых
рода Xylophagus Moig. (Diptera, Xylophagidae). — «Энтомол. обозр.»,
1972а, 51, 2, с. 430-445.
Кривошеина Я. Я., Мамаев Б. М. Новые данные о реликтовых двукрылых
семейства Pachyneuridae (Diptera).— «Научн. докл. высш. школы.
Биол. науки», 19726, И, с. 13—18.
Кривошеина Я. Я., Мамаев Б. М. Личинки KTbipeii трибы Andrenosomini
(Diptera, Asilidae) как ксилофильные формы.— «Научн. докл. высш. шко¬
лы. Биол. науки», 1973, 9, с. 20—23.
Кривошеина Я. Я., Мамаев Б. М. Ревизия материалов по морфологии и
экологии ктырей трибы Laphriini (Diptera, Asilidae). Сооб. I. Род Choera-
des Walk.— «Вестник зоол.», 1975а, 2, с. 66—72.
Кривошеина Я. Я., Мамаев Б. М. Ревизия материалов по морфологии и эко¬
логии ксилофильных личинок ктырей трибы Laphriini (Diptera, Asi¬
lidae). Сообщ. II. Род Laphria Mg.— «Вестник зоол.», 19756, 6, с. 46—
50.
Кривошеина Я. Я., Мамаев Б. М., Ягдыев Л. Насекомые-ксилофаги, разви¬
вающиеся на эллении малолистной Aellenia subaphylla (С. А. Меу)
Aellen.— В кн.: Насекомые как компоненты биогеоценоза саксаулового
леса. М., «Наука», 1975, с. 207—211.
Кривошеина Я. Я., Ремм X. Я. Новые виды ксилофильных мокрецов (Dipte¬
ra, Ceratopogonidae) из Приморского края.— В кн.: Насекомые — разру¬
шители древесины в лесных биоценозах Южного Приморья. М., «Наука»,
1974, с. 116—124.
Крыжановский О. Л, Семейство Histeridae — карапузики.— В кн.: Опреде¬
литель насекомых Европейской части СССР, т. 2. М.— Л., «Наука»,
1965а, с. 95—104.
Крыжановский О, Л, Семейство Ostomatidae (Trogossitidae)—щитовидки.—
В кн.: Определитель насекомых Европейской части СССР, т. 2, М,— Л.,
«Наука», 19656, с. 239—240.
Крыжановский О. Л, Сем. Cucujidae — плоскотелки.— В кн.: Определитель
насекомых Европейской части СССР, т. 2. М.— Л., «Наука» 1965в,
с. 309-313.
Крыжановский О, Л,, Тихомирова А, Л,, Филатова Л. Д, Стафилиниды (Coleop-
tera, Staphylinidae) Южного Приморья. — В кн.: Экология почвенных
беспозвоночных. М., «Наука», 1973, с. 144—173.
13 в. В. Мамаев и др. 361
Куренцов А. И, Короеды верховий реки Супутинки.— «Труды горнотаежной
станции Д. Ф. АН СССР», 1. Хабаровск, Дальгиз, 1936, с. 185—205.
Куренцов А, И. Короеды Дальнего Востока СССР. М.— Л., Изд-во АН СССР,
1941.
Лер П, А. Ктыри подсемейства Leptogastrinae (Diptera, Asilidae) фауны
СССР.— «Энтомол. обозр.», 1961, 40, 3, с. 686—703.
Лер П. А, О питании и значении ктырей.— «Труды н.-и. ин-та защиты расте¬
ний». Алма-Ата, Изд-во Сельхозлит., 1964, 8, с. 213—244.
Лобанов А. М. Места зимовок преимагинальных стадий синантропных мух.—
«Научн. докл. высш. пшолы, Биол. науки», 1962, 2, с. 36—40.
Лобанов А. М, О морфологии взрослой личинки Tetanops sintenisi (Diptera,
Otitidae).— «Зоол. журн.», 1972, 51, 1, с. 146—149.
Малахова В, П. Некоторые данные о видовом составе энтомофагов стенографа
в Приморье.— «Сообщ. ДВФ. АН СССР». Владивосток, 1962, 15, с. 89—
94.
Мамаев Б, М. Личинки рогачей (Coleoptera, Lucanidae) как разрушители
гниющей древесины в дубравах Европейской части СССР.— «Зоол.
журн.», 1960а, 39, 6, с. 873—881.
Мамаев Б. М. Зоологическая оценка стадий естественного разрушения дре¬
весины — «Изв. АН СССР, сер. биол.», 19606, 4, с. 610—617.
Мамаев Б. М, Приспособительные особенности беспозвоночных-ксилобион-
тов и их роль в естественном разрушении древесины. Автореф. канд. дисс.
М., 1960в.
Мамаев Б. М. Деятельность крупных беспозвоночных — один из основных
факторов естественного разрушения древесины.— «Pedobiologia», 1961,
1, 1, с. 38-52.
Мамаев Б. М, Явления конвергенции у насекомых как результат одинакового
использования среды обитания (на примере ксилофагов).—«Журн.
общ. биол.», 1966, 27, 4, с. 457—462.
Мамаев Б. М, Использование насекомыми смолы хвойных деревьев как среды
обитания (с обзором насекомых-резиниколов).— «Журн. общ. биол.»,
1971а, 32, 4, с. 501-507.
Мамаев Б. М. Основные закономерности эволюции сообществ насекомых —
разрушителей древесины.— В кн.: Защита леса от вредных насекомых.
(Всес. научн.-техн. конф. «Применение новых химич. и биол. методов
борьбы с вредит, и болезн. леса».— Тез. докл.), 19716, 2, с. 113—117.
Мамаев Б. М. Экологические связи насекомых-ксилобионтов, развивающихся
на японской ольхе.— «Экология», 1971 в, 2, с. 50—53.
Мамаев Б. М. Японская щитовка — враг короедов.— «Защита раст.»,
1971г, 9, с. 50.
Мамаев Б. М. Зоогеографическая характеристика комплексов насекомых,
формируюшихся в древесине клена маньчжурского в Южном Приморье.—
«Изв. АН СССР, сер. биол.», 1972а, 2, с. 265-268.
Мамаев Б. М. Насекомые — разрушители древесины пихты и сопутствую¬
щие им энтомофаги в Южном Приморье.— «Лесоведение», 19726, 4,
с. 67—72.
Мамаев Б. М. Определитель насекомых по личинкам. М., «Просвещение»,
1972в.
Мамаев Б. М. Общие закономерности формирования сообществ беспозвоноч¬
ных — разрушителей древесины в Южном Приморье.— «Труды БПИ
ДВНЦ АН СССР», 1972г, 7(110), с. 5—15.
Мамаев Б. М. Насекомые — разрушители древесины кедра корейского и
сопутствующие им виды.— «Труды БПИ ДВНЦ АН СССР», 1972д,
7(110), с. 44—56.
Мамаев Б. М, Видовой состав и экологические связи насекомых — разруши¬
телей древесины ильма долинного.— «Труды БПИ ДВНЦ АН СССР»,
1972е, 7(110), с. 106-120.
Мамаев Б, М. Энтомофаг против стволовых вредителей.— «Защита раст.»,
1973а, 1, с. 42.
362
Мамаев Б. М. Материалы по экологии реликтового таракана.— «Экология»,
19736, 6, с. 76—79.
Мамаев Б. М, Морфология личинки жука Nematoplus Lee. и филогенетиче¬
ские связи некоторых семейств Heteromera (Coleoptera, Cucujioidea).—
«Энтомол. обозр.», 1973в, 52, 3, с. 586—598.
Мамаев Б. М. Новые находки реликтовых насекомых в заповеднике «Кедро¬
вая падь».-«Труды БПИ ДВНЦ АН СССР», 1973, 5, с. 20-23.
Мамаев Б. М. Зоогеография ксилофильных сообществ Южного Приморья.—
В кн.: Насекомые — разрушители древесины в лесных биоценозах
Южного Приморья. М., «Наука», 1974а, с. 5—30.
Мамаев Б. М. Новые виды пятнокрылок (Diptera, Clusiidae) из Южного
Приморья.— В кн.: Насекомые — разрушители древесины в лесных
биоценозах Южного Приморья. М., «Наука», 19746, с. 111 — 115.
Мамаев Б. М. Основы биологических связей насекомых с разрушающейся
древесиной.—«Журн. общ. биол.», 1974в, 35, 1, с. 51—57.
Мамаев Б. М. Беспозвоночные как индикаторы стадий естественного разру¬
шения древесины.— В кн.: Вопросы экологической физиологии беспо¬
звоночных. М., «Наука», 1974г, с. 198—212.
Мамаев Б. М. Преимагинальные фазы развития жука Syntelia histeroides
Lewis (Synteliidae) в сравнении с некоторыми Histeridae (Coleoptera).—
«Энтомол. обозр.», 1974д, 53, 4, с. 866—891.
Мамаев Б. М, Адаптация личинок слоников трибы Sipalini и близких групп
к жизни в древесине.— «Научн. докл. высш. школы, биол. науки»,
1974е, 6, с. 11-13.
Мамаев Б. М. Географические закономерности эволюции ксилофильных
сообществ насекомых. — «Матер, седьмого съезда Всес. энтомол. общ.»,
1, 1974ж, с. 79—80.
Мамаев Б. М. Комплексы насекомых на завершающих стадиях разрушения
древесины в Палеарктике и Южной Азии.— В кн.: Проблемы почвенной
зоологии. Вильнюс, 1975, с. 221—222.
Мамаев Б, М., Антонова Е. Б, Экологическая специализация ксилофильных
детритниц Палеарктики.— «Экология», 1974, 3, с. 88—90.
Мамаев Б. Л/., Данилевский М. Л, Новые данные по системе подсемейства
Aseminae (Coleoptera, Cerambycidae) в связи с особенностями строения
личинок.— «Зоол. журн.», 1973, 52, 8, с. 1257—1261.
Мамаев Б. Л/., Данилевский М. Л. Личинки жуков — дровосеков. М.,
«Наука», 1975.
Мамаев Б. Л/., Кравченко М. А, Личинки рогохвостов (Hymenoptera, Siri-
cidae).— В кн.: Насекомые — разрушители древесины в лесных биоцено¬
зах Южного Приморья. М., «Наука», 1974, с. 74—85.
Мамаев Б, М., Кривошеина Н. П. Личинки галлиц (Diptera, Cecidomyiidae).
Сравнительная морфология, биология, определительные таблицы. М.,
«Наука», 1965.
Мамаев Б. М., К ривошеина Я. П, Новые данные по систематике и биологии
длинноусых двукрылых сем. Axymyiidae.— «Энтомол. обозр.», 1966,
45, 1, с. 168-180.
Мамаев Б. М., К ривошеина Н. Я. Новые данные по морфологии и экологии
длинноусых двукрылых сем. Hyperoscelididae (Diptera, Nematocera).—
«Энтомол. обозр.», 1969, 48, 4, с. 933—942.
Мамаев Б, М.у Никитский Я. Б. Особенности географического распростра¬
нения хищных жуков, уничтожающих короедов.— «Проблемы защиты
таежных лесов». Красноярск, 1971, с. 95—97.
Мамаев Б. М., Соколов Д, Ф, Участие беспозвоночных животных в естествен¬
ном разрушении древесины дуба.— «Почвоведение», 1960, 4, с. 24—30.
Машнина Т, Я. Естественные враги вредителей стволов сосны в лесах Бело¬
русского Полесья.— В кн.: Научн. раб. Белор. НИИЛХ. Минск, Изв.
Академии с.-х. наук БССР, 1960, с. 205—209.
Медведев Л, Я. Сем. Pythidae — трухляки. — В кн.: Определитель насеко¬
мых Европейской части СССР, т. 2. М.— Л., «Наука», 1965, с. 341—
342.
13* 363
Царчук Э. П, Сем. Xylophagidae (Coenomyiidae, Erinnidac).— В кн.: Опреде¬
литель насекомых Европейской части СССР, т. 5, 1. М.— Л., «Наука»,
1969а, с. 453-454.
Царчук Э. П. Сем. Stratiomyidae — львинки.— В кн.: Определитель насеко¬
мых Европейской части СССР, т. 5, 1. М.— Л., «Наука», 19696, с. 454—
481.
Царчук Э. П. Находка Solva marginata Meigen (Diptera, Xylomyidae) в Мон¬
гольской Народной Республике.— В кн.: Насекомые Монголии. Л.,
«Наука», 1972, с. 1—784.
Негробов О. П. Виды рода Medetera (Dolichopodidae, Diptera) как энтомофаги
короедов.— В кн.: Защита леса от вредных насекомых и болезней, т. 3.
М., МЛТИ, 1971, с. 89—90.
ЦегробовО. Я., Штакельберг А, А, Семейство Dolichopodidae — зеленушки.—
В кн.: Определитель насекомых Европейской части СССР, т. 5, 1.
М.— Л., «Наука», 1969, с. 671—751.
Цесмерчук С. М. Основные энтомовредители тяньшанской ели и меры борьбы
с ними.— «Труды Алма-Атинск. гос. зап.», 1948, 7, с. 5—54.
Цикитский Н. Б. Видовой состав и некоторые особенности экологии насе¬
комых, селящихся в ходах двухцветного короеда (Taphrorychus bicolor
Hbst.).— «Научн. труды МЛТИ», 1971, 38, с. 37—46.
Цикитский И. Б. Морфология и экология Aulonium ruficome 01. (Coleopte-
га, Colydiidae) — энтомофага, уничтожающего сосновых короедов.—
«Научи, докл. высщ. школы. Биол. пауки», 1972, 4, с. 20—26.
Цикитский Н. Б. Морфология личинок и образ жизни Nemosoma Latr.
(Colcoptera, Trogossitidae) — хищников короедов Северо-Западного
Кавказа.— «Зоол. журн.», 1974, 53, 4, с. 563—569.
Цикитский //. Б. Морфология личинок и образ жизни чернотелок рода
Hypophloeus F. (Coleoptera, Tenebrionidae).— «Зоол. журн.», 1976,
55, 1, с. 41-51.
Цикитюк А, И. Хищные и паразитические насекомые как регуляторы вредо¬
носной деятельности и распространения короедов хвойного леса.
Сообщ. I.— «Бюлл. МОИП, отд. биол.», 1951, 56, 5, с. 58—63.
Цикитюк А. И. Хищные и паразитические насекомые как регуляторы вредо¬
носной деятельности и распространения короедов хвойного леса. Сообщ.
III.— «Бюлл. МОИП, отд. биол.», 1957, 62, 2, с. 51—55.
Огибин Б. Ц, О регуляции плотности популяции Ips typographus (Coleoptera,
Ipidae) на преимагинальных стадиях развития.— «Зоол. жури.», 1974,
53, 1, с. 40-49.
Паньшин И, В. Естественное регулирование численности массового соснового
лубоеда и обоснование методов борьбы с вредителями. Автореф. канд.
дисс. Воронеж, 1964.
Плешаков А, С. Хищные насекомые — истребители большого лиственнич¬
ного короеда в Прибайкалье.— «Изв. СО АН СССР», 1966, 3, 12, с. 124—
126.
Погориляк И. М. Жуки, обитающие в короедных гнездах Украинских Кар¬
пат.— «Тез. докл. и сообщ. XVII научн. конф., сер. биол.», Ужгород,
1963, с. 77-80.
Померанцев Д, К. Биологические заметки о жуках, полезных в лесоводстве,
живущих под корою деревьев. I.— «Русск. энтомол. обозр.», 1902а, 2,
2, с. 87-91.
Померанцев Д. К. Биологические заметки о жуках, полезных в лесоводстве,
живущих под корою деревьев. П.— «Русск. энтомол. обозр.», 19026,
2, 3, с. 151-156.
Померанцев Д. К. Биологические заметки о жуках, полезных в лесоводстве,
живущих под корою деревьев. III.— «Русск. энтомол. обозр.», 1902в,
2, 6, с. 328-332.
Померанцев Д. Я. Биологические заметки о жуках, полезных в лесоводстве,
живущих под корою деревьев. IV.— «Русск. энтомол. обозр.», 1903а,
3, 1, с. 22-23.
Померанцев Д, К. Биологические заметки о жуках, полезных в лесоводстве,
364
живущих под корою деревьев. V.— «Русск. энтомол. обозр.», 19036,
3, 2f с. 77—78.
Померанцев Д. К, Биологические заметки о жуках, полезных в лесоводстве,
живущих под корою деревьев. VI.— «Русск. энтомол. обозр.», 1903в
3, 3-4, с. 200-202. ^
Померанцев Д. К. Биологические заметки о жуках, полезных в лесоводстве,
живущих под корою деревьев. VII.— «Русск. энтомол. обозр.», 1904
4, 2-3, с. 85-89.
Потоцкая В, А. Определитель личинок коротконадкрылых жуков Staphy-
linidae Европейской части СССР. М., «Наука», 1967.
Потоцкая В, А, Некоторые данные об экологии стафилинид, развивающихся
в древесине. — «Экология», 1973а, 2, с. 61—65.
Потоцкая В, А. Морфология личинок некоторых ксилобионтных стафилинид
(Coleoptera, Stapnylinidae).— «Зоол. журн.», 19736, 52, 1, с. 54—63.
Потоцкая В, А. Сравнительная характеристика ксилобионтных личинок
стафилинид в связи с особенностями пищевой специализации.— В кн.:
Насекомые — разрушители древесины в лесных биоценозах Южного
Приморья. М., «Наука», 1974, с. 65—73.
Потоцкая В. А, Личинки жуков-стафилинид (Coleoptera, Staphylinidae),
развивающихся в древесине.— В кн.: Эволюционная морфология личи¬
нок насекомых. М., «Наука», 1976, с. 156—174.
Пряхина Е. Ф.у Огибин Б. //. К фауне и экологии двукрылых насекомых
(Diptera), обитающих под корой ели в Архангельской области.— «Тез.
докл. к отчетной сессии лаборатории лесоведения и лесоводства за 1969 г.».
Архангельск, 1970, с. 58—63.
Рейхардт А. Я. Фауна СССР. Жесткокрылые Sphaeritidae и Histeridae,
т. 5, 3. М.— Л., Изд-во АН СССР, 1941, с. 1—420.
Рихтер В. А, Жуки-пестряки (Coleoptera, Cleridae) фауны СССР.— «Труды
Всес. энтомол. общ-ва», 1961, 48, с. 63—123.
Рихтер В, А, Хищные мухи-ктыри (Diptera, Asilidae) Кавказа. Л., «Наука»,
1968.
Рихтер В. А., Мамаев Б, М. Новые ктыри подсемейства Laphriinae (Diptera,
Asilidae) в фауне Кавказа.— «Зоол. журн.», 1971, 50, 10, с. 1589—1591.
Родендорф Б, Б. О новом виде Lonchaea (Diptera, Lonchaeidae) из ходов ко¬
роеда Scolytus scolytus. — «Энтомол. обозр.», 1950, 31, 1—2, с. 80—81.
Родендорф Б. Б. Историческое развитие двукрылых насекомых. М., «Наука»,
1964.
Родендорф Б. Б., Калугина Н. С, Находка своеобразных двукрылых нимфо-
мийид (Diptera, Nymphomyiidae) в Приморье.— «Энтомол. обозр.», 1974,
53, 3, с. 686—694.
Скопин Нш 1\ Морфологические особенности личинок и положение в системе
некоторых надродовых групп Ulomini (Coleoptera, Tenebrionidae).—
«Труды XIII Междунар. энтомол. конгр.», 1971, 1, с. 302—303.
Старк В. Н. Влияние жука-карапузика Platysoma oblongum F. на развитие
на сосновых пнях Blastophagus piniperda. L.— «Защита раст.», 1926,
3, 4-5, с. 339-342.
Стриганова Б. Р. Морфо-функциональные особенности личинки Prionocy-
phon serricornis Miill. (Coleoptera, Helodidae) в связи с обитанием в воде.—
«Энтомол. обозр.», 1961, 40, 3, с. 557—583.
Тарасова Д. А, Паразиты и хищники стволовых вредителей в сосново-лист¬
венных лесах Салаира.— В кн.: Исследования по биол. методу борьбы
с вредителями сельского и лесного хозяйства. Новосибирск, 1965,
с. 120—128.
Тарасова Д. А. Стволовые вредители и их энтомофаги на вырубках Средне¬
обской котловины. Автореф. канд. дисс. Новосибирск, 1968.
Филиппенкова В, В, Некоторые биоценотические взаимоотношения шести¬
зубого короеда в насаждениях лесостепного Заволжья.— «Уч. зап.
Ку11бышевского пед. ин-та им. В. В. Куйбышева. Вопросы зоологии»,
1970, 83, с. 48-54.
Фридолин В, /О. Животно-растительное сообщество горной страны Хибин:
365
биоценологические исследования 1930—1935 гг.— «Труды Кольск.
базы АН СССР», 1936, 3, с. 1-195.
Харитонова Н, 3. Энтомофаги короедов хвойных пород. М., «Лесная промыш¬
ленность», 1972.
Черепанов А, И, Жуки-щелкуны Западной Сибири. Новосибирск, Книжное
изд-во, 1957.
Черепанов А, И, Проволочники Западной Сибири. (Определитель). М.,
«Наука», 1965.
Шарова И, X. Личинки жуков-жужелиц, полезных и вредных в сельском
хозяйстве.— «Уч. зап. МГПИ им. В. И. Ленина», 1958, 124, 7, с. 4—165.
Шиперович В, Я. Энтомофауна лесорубочных остатков и условия ее исполь¬
зования в биологическом методе борьбы. — «Тез. докл. 3-й эколог, конф.»,
т. I. Киев, 1954, с. 299—300.
Шовен Р, Физиология насекомых. М., ИЛ, 1953.
Штакельберг А, А. Определитель мух Европейской части СССР. Л., Изд-во
АН СССР, 1933.
Штакельберг А, А, Материалы по фауне двукрылых Ленинградской области.
II. Díptera, Brachycera. — «Труды Зоол. ин-та АН СССР», 1954, 15,
с. 199-228.
Штакельберг А, А, Материалы по фауне двукрылых Ленинградской области.
III. Díptera, Acalyptrata, ч. I.—«Труды Зоол. ин-та АН СССР», 1958,
24, с. 159-163.
Шуровенков Б. Г. Энтомофаги личинок хлебных жуков и проволочников.—
«Защита раст.», 1961, 12, с. 45—46.
Шуровенков Б, Г. Полевые хищные энтомофаги (Coleóptera, Carabidae и
Díptera, Asilídae) и факторы, определяющие их эффективность.—
«Энтомол. обозр.», 1962, 16, 4, с. 763—780.
Шуровенков Б. Г, Энтомофагия мух-ктырей и их личинок в связи с биоло¬
гическим обоснованием мер борьбы с почвообитающими вредителями.—
«Научн. труды Воронеж, с.-х. ин-та.», 1973, 8, 1, с. 43—52.
Юрченко Г. И. Об энтомофагах стволовых вредителей в усыхающих ельниках
Хабаровского края.—В кн.: Итоги изучения лесов Дальнего Востока.
Владивосток, 1967, 281—283.
Яновский В, М, Некоторые особенности формирования комплексов энто-
мофагов стволовых вредителей.— В кн.: Исследования в лесах Сибири,
т. 2. Красноярск, 1968, 225—230.
Яновский В, М. О воздействии энтомофагов на динамику численности ство¬
ловых вредителей.— В кн.: Проблемы защиты таежных лесов. Красно¬
ярск, 1971, с. 131—133.
Яновский В, М. Изменение численности энтомофагов стволовых вредителей
в различных ландшафтно-экологических условиях горных лесов Тувы
и Монголии.— В кн.: Вопросы энтомологии Сибири. Новосибирск,
1974а, с. 103-105.
Яновский В, М, Роль энтомофагов в динамике численности большого лист¬
венничного короеда.— В кн.: Экология популяций лесных животных
Сибири. Новосибирск, «Наука», 19746, с. 189—205.
Areekul S. The comparative internal larval anatomy of several genera of Sca-
rabaeidae (Coleoptera).— «Ann. Entomol. Soc. America», 1957, 50, 6,
562—677.
Arnett R. H. The beetles of the United States. Michigan, 1968.
Aspock Я., Aspöck £/., Rausch H, Bestimmungsschlüssel der Larven der Raphî-
diopteren Mitteleuropas (Insecta, Neuropteroidea).— «Z. angew. Zool.»,
1974a, 61, 1, 46-62.
Aspöck Я., Rausch Я., Aspöck U, Untersuchungen über die Ökologie der Raphi-
diopteren Mitteleuropas (Insecta, Neuropteroidea).— «Z. angew. Entomol.
1974b, 76, 1, 1—30.
Bagnall R. Epuraea angustula Er. and Acrulia inflata Gyll. coleopterous parasi¬
tes on species of the Stephensian genus Trypodendron.— «Trans. Natur.
Hist. Soc., Northumberland», n. s., 1906, 1, 416.
366
Balachowsky A., Mesnil L. Les insectes nuisibles aux plantes cultivées. Paris,
1935.
Dalazy iS., Michalski J, Materialy do znajomosci chrsaszczy (Coleoptera) wyste-
pujachych w zerowiskach kornikow (Scolytidae).— «Polskie Pismo Ento¬
mol.», 1960, 30, 9, 133—144.
Beaver R, A, The biology and immature stages of two species of Medetera
(Diptera, Dolichopodidae) associated with the bark beetle Scolytus scoly-
tus (F.).—«Proc. Roy. Entomol. Soc. London», A, 1966a, 41,10—12,145—154.
Beaver R, A, Notes on the fauna associated with the elm bark beetles in Wyt-
ham Wood, Berks. I. Coleoptera.— «Entomol. Monthly Mag.», 1966
(1967) b, 102, 1226-1228, 163-170.
Beaver R. A, The regulation of population density in the bark beetle Scolytus
scolytus.— «J. Animal Ecol.», 1967, 36, 2, 435—451.
Becker G. Grundzüge der Insekten-Sukzession in Pinus-Arten der Gebirge von
Guatemala.— «Z. angew. Entomol.», 1955, 37, 1—28.
Becker T. Dipterologische Studien. III. Lonchaeidae.— «Berlin. Entomol. Z.»,
1895, 40, 2, 313-344.
Beling T. Beitrag zur Metamorphose der zweiflügeliger Insekten aus den Famili¬
en Tabanidae, Leptidae, Asilidae, Empididae, Dolichopodidae und Syr-
phidae.— «Arch. Naturgesch.», 1882, 44, 187—240.
Bezzi M. Two new Ethiopian Lonchaeidae with notes on other species (Dipt.).—
«Bull. Entomol. Res.», 1919, 9, 3, 241—254.
Bezzi M. Further notes on the Lonchaeidae (Dipt.), with description of new
species from Asia and Africa.— «Bull. Entomol. Res.», 1920, 11, 1, 2, 199—
210.
Bischoff W, Über die Kopfbildung der Dipterenlarven. III. Teil. Die Köpfe der
Orthorrhaphen — Brachycera — Larven.— «Arch. Naturgesch.», Abt. A,
1925, 90, 8, 1—105.
Blackman M. W., Stage H. H, Notes on insect bred from the bark and wood
of the American larch.— «N. Y. State Coll. Forestry. Techn. Publ.»,
1918, 10, 11-115.
Blackman M. W,, Stage H. H, On the succession of insects living in the hart
and wood of dying, dead and decaying hickory.— «N. Y. State Coll. Fo¬
restry. Techn. Publ.», 1924, 17, 3—268.
Blanchard E. E, Synopsis de los Loncheidos Argentinos. — «Rev. Invest. Agrie.»,
1948, 2, 4, 157-178.
Böving A, G.j Champlain A, B, Larvae of North American beetles of the family
Cleridae.—«Proc. U.S. Nat. Mus.», 1920, 57, 575—649.
Böving A., Craighead F. Illustrated synopsis of principal larval forms of order
Coleoptera.— «Entomol. Americana», 1931, 11, 1—351.
Böving A., Rozen J, Anatomical and systematic study of the mature larvae of
the Nitidulidae (Coleoptera).— «Entomol. medd.», 1962, 31, 4, 265—299.
Brauer F. Die Zweiflügler der keiserlichen Museums zu Wien. III. Systemati¬
sche Studien auf Grundlage der Dipteren-Larven nebst einer Zusammenstel¬
lung von Beispielen aus der Literatur über dieselben und Beschreibung neuer
Formen. — «Denkschr. k. Akad. Wiss.», Math.-Naturwiss. CL, 1883, 47,
1-100.
Brindle A. Taxonomie notes on the larvae of British Diptera. 2. The genus Xylop-
hagus Meigen (Erinna Meigen) (Rhagionidae).— «Entomologist», 1961a,
94, 144-148.
Brindle A, Taxonomic notes on the larvae of British Diptera. 3. The genus Solva
Walker (Xylomyia Rondani : Subula Meigen) (Stratiomyidae).—«Entomo¬
logist», 1961b, 94, 202-205.
Brindle A. Taxonomic notes on the larvae of British Diptera. 7. The genus
Pachygaster Meigen (Stratiomyidae).— «Entomologist», 1962a, 95, 77—82.
Brindle A. Taxonomic notes on the larvae of British Diptera. 10. The Asili¬
dae.— «Entomologist», 1962b, 95, 1193, 242—247.
Bromlei S. W, Bee-killing robber-flies.—«J. New York Entomol. Soc.», 1930,
38, 159-173.
367
Brown J. M. Some points in the anatomy of the larva of Tipula maxima.—
«Trans. Linnean Soc. London», ser. 2, 1910, 11, 7, 125—135.
Brues C, T. Insect dietary. Cambridge, 1946.
Brunetti E, Uiptera Brachycera. I. The fauna of British India incl. Ceylon
and Burma, 1920, 1—96.
Brunetti E. Second revision of the oriental Stratiomyidae.— «Rec. Indian Mus.»,
1923, 25, 45-179.
Burakowski B. Obserwacje biologiczno-morfologiczne nad Pytho kolwensis
C. Sahib. (Coleoptera, Pythidae) w Polsce.—«Fragmenta faunistica», 1962,
10, 2, 173-204.
Clausen P, Entomophagous insects. New York and London, 1940.
Cole E., James Jr., Streams Fr. Insects emerging from brown slime fluxes
in southern New England.— «Cañad. Entolologist», 1970, 102, 3, 321 —
333.
Collin J. E, The British species of the dolichopodid genus Medetera Fisch.
(Dipt.).— «Entomol. Monthly Mag.», 1941, 77, 141 — 153.
Collin J. E. The British species of the genus Palloptera Fallen (Diptera).—
«Entomol. Rec.», 1951, 63, 1—6.
Collin J, E, A revision of the British (and notes on the other) species of Lon¬
chaeidae (Diptera).—«Trans. Soc. Brit. Entomol.», 1953, 11, 181—207.
Cook E. F. On the early stages of Neopachygaster maculicornis (Hino) and
Bcrkshiria aldrichi (Malloch): Diptera, Stratiomyidae.—«Ann. Entomol.
Soc. America», 1953, 46, 2, 293—299.
Creathead D. A review of the insect enemies of Acridoidea (Orthoptera).—
«Trans. Roy. Entomol. Soc. London», 1963, 114, 14, 437—517.
Crowson R. A. The natural classification of the families of Coleoptera. London,
1955.
Crowson R, A, A. review of the classification of Cleroidea (Coleoptera), with'
description of two new genera of Peltidae and of several new larval types.—
«Trans. Roy. Entomol. Soc. London», 1964, 116, 12, 275—327.
Crowson R. A. Further observations on Cleroidea (Coleoptera). — «Proc. Roy.
Entomol. Soc. London», 1970, 39, 1—2, 1—20.
Curran C, H, The families and genera of North American Diptera, 1965,
1-146.
Czerny L. Lonchaeidae. In: E. Lindner, Die Fliegen der palaearktischen Region,
Bd. 43, Stuttgart, 1934, S. 1—40.
Dajoz R. Révision des Colydiidae anophtalmes de la faune paléarctique (Col.).
l\\ Etudes sur les genres Anommatus et Langelandia.—«Ann. Soc.
entomol. France», n. s., 1968, 4, 3, 975—988.
Daniels N. E. The association of a rangeland grub, Phyllophaga koehleriana
(Coleoptera : Scarabaeidae), with asilid larvae and with mites.— «Ann.
Entomol. Soc. America», 1966, 59, 5, 1021.
Derksen W. Die Sukzession der pterygoten Insekten im abgestorbenen Buchen¬
holz.-«Z. MorphoL, Okol. Tiere», 1941, 37, 683-734.
Deschamps P, Note sur la cellulase chez les larves des insectes xylophages de la
famille des Cerambycides.—«Bull. Soc. zool. France», 1949, 74, 2, 103—
108.
Deschamps P. Contribution à l’étude de la xylophagie. La nutrition des larves
des Cerambycides.— «Ann. sei. natur.», 1953, 15, 449—553.
Downes J. A. Notes on the liie-cycle of Xylophagus ater Meig. (Dipt., Rhagio
nidae).— «Entomol. Monthly Mag.», 1953, 89, 136—137.
Dunn L. H. Hermetia illucens breeding in a human cadaver.— «EntomoL
News», 1916, 27, 59—61.
Dusek Rozkosny R. Revision mitteleuropäischer Arten der Familie Stratio¬
myidae (Diptera) mit besonderer Berücksichtigung der Fauna der CSSR.
I. — «Cas. Cs. Spol. entomol.», 1963, 60, 3, 202—221.
Dyte C. E. Some interesting habitats of larval Dolichopodidae (Diptera).—
«Entomol. Monthly Mag.», 1959, 95, 139-143.
Dyte C. E. Some distinctions between the larvae and pupae of the Empididae
368
and Dolichopodidae (Díptera).— «Proc. Roy. Entomol. Soc. London»
1967, 42, 7—9, 119—128.
Eidmann E. Sukzessionen westafrikanischer Holzinsekten.— «Mitt. Akad
Dtsch. Forstwissensch.», 1943, 2, 240—271.
Elton C, S. The pattern of animal communities. New York — London, 1966
Emden F. , van. Larvae of British beetles. 4. Various small families.— «Ento
mol. Monthly Mag.», 1943, 79, 209—223, 259—270.
Emden F. I. van. Larvae of British beetles. 5. Elateridae.— «Entomol. Month
ly Mag.», 1945, 81, 13-37.
Emden F. I. van. Larvae of British beetles. 6. Tenebríonidae.— «Entomol
Monthly Mag.», 1947, 83, 154—171.
Emden F. /. van. On the larvae oí Palorus (Coleoptera, Tenebríonidae). A supp
lement to «Larvae of British Beetles». 6.— «Entomol. Monthly Mag.», 1948
84, 10.
Emden //. F. Morphology and identification of the British larvae of the genus
Elater (Coleoptera, Elateridae).— «Entomol. Monthly Mag.», 1956, 92
167-187.
Engel E, O. Beiträge zur Kenntnis einiger Dipterenlarven.— «Mitt. Münchn
Entomol. Ges.», 1916, 7, 68—76.
Engel E, O. Asilidae. In: Lindner E. Die Fliegen der paläearktischen Region
IV. Stuttgart, 1930 1—491.
Escherich K. Die Forstinsekten Mitteleuropas, Bd. 2. Berlin, 1923.
Frey R. Über orientalische Leptogaster-Arten (Dipt., Asilidae).— «Notulae
Entomol.», 1937, 17, 38—52.
Frey R. Diptera-Brachycera (excl. Muscidae, Tachinidae).— In: Enumeratio
Insectorum Fenniae — VI. Diptera. Helsingfors., 1941.
Fukuda /1. In: Kawada A. Illustrated insect larvae of Japan. Tokyo, 1959.
Fuller M. The early stages of Actina incisuralis.— «Proc. Linnean Soc. New
South Wales», 1934, 59, 190—196.
Gäbler H. Dipterenlarven als Parasiten und Synöken des Buchdruckers, Ips
typographus L.— «Z. angew. Entomol.», 1953, 35, 1, 55—62.
Gardner J. C. M. Immature stages of Indian Coleoptera (9).— «Indian Forest
Rec.», 1931, 16, 4, 1-21.
Gardner J. C. M. Immature stages of Indian Coleoptera (21). Cleridae.— «In¬
dian Forest Rec.», n. s., 1937, 3, 2, 31—46.
Gardner J. C. M. A list of described immature stages of Indian Coleoptera.—
«Indian Forest Rec.», n. s., 1948, 7, 5, 163—191.
Hackman W. The Lonchaeidae (Diptera) of Eastern Fennoscandia.— «Notulae
Entomol.», 1956, 36, 89—115.
Hanson B. H. Das bisher unbekannte Männchen einer für Schweden neuen FJie-
genart (Dipt., Strat.).— «Entomol. tidskr.», 1942, 63, 63—64.
Hayashi N. On the larvae of some species of small families of Cucujoidea in
Japan (Coleoptera).— «Insecta Matsumurana», suppL, 1969, 7, 1—9.
Hayashi N. On tne larvae of some species of Colydiidae, Tetratomidae and
Aderidae occurring in Japan (Coleoptera; Cucujoidea).— «Kontyú», 1972,
40, 2, 100-111.
Hendel F. Eine neue europäische Lonchaea-Art, deren Larve Borkenkäfer-
Räuber ist.— «Konowia», 1928, 7, 37—39.
Hennig W. Beiträge zur Kenntnis des Kopulationsapparates und der Syste¬
matik der Acalyptrata. Lonchaeidae und Lauxaniidae.— «Acta zool. Lil-
loana», 1948a, 6, 333—429.
Hennig W. Die Larvenformen der Dipteren. I. Teil. Berlin, 1948b.
Hennig W. Die Larvenformen der Dipteren. 2. Teil. Berlin, 1950.
Hennig W. Die Larvenformen der Dipteren. 3. Teil. Berlin, 1952.
Hennig W. Diptera, Zweiflügler. In: Sorauer. Handbuch der Pflanzenkrankhei-
ten, Berlin, 1953, 5, 5, 108-110.
Hering E. M. Lebensweise und Larven morphologic von Lonchaea flavidipennis
Zett. (Diptera).—«Dtsch. Entomol. Z.», 1954, 1, 86—89.
Hickin N. E. The insect factor in wood decay. London, 1963.
Hinton H. E. On the structure, function and distribution of the Prolegs of the
369
Panorpoidea, with a criticism of the Berlese — Imms theory.— «Trans.
Roy. Entomol. Soc. London», 1955, 106, 455—545.
Houlbert C., Bétis L. Fam. Clerides.— «Trav. sclent. Univ. Rennes», 1905, 4,
suppl. 52, 119-139.
Hsia K. L, Studies on Chinese Asilidae. L Leptogastrinae.—«Sinensia», 1948,
19, 1—6, 23—56.
Hubault E, Contribution á la biologie du genre Medeterus Fischer.— «Ann.
Sei. natur. Zool.», 1925, 8, 3—6, 133—141.
Hull F, M. Robberflies of the world. — «Bull. Smithson. Inst. U. S. Nat. Mus.»,
1962, 224, 1, 1-430.
James M. T. The flies that cause myiasis in man.—«U. S. Dept. Agrie. Misc.
Publ.», 1947, 631, 1-175.
James M. T. Family Xylophagidae (Coenomyiidae, Erinnidae).— In: Stone et
al. A catalog of the Diptera of America north of Mexico. Washington,
1965a, 296-298.
James M. T, Family Xylomyidae. In: Stone et al. A catalog of the Diptera of
America north of Mexico. Washington, 1965b, 298—299.
James M. T. Family Stratiomyidae In: Stone et al. A catalog of the Diptera
of America north of Mexico. Washington, 1965c, 299—317.
James M. T. Contribution to our knowledge of the Nearctic Pachygasterinae
(Diptera, Stratiomyidae).—«Ann. Entomol. Soc. America», 1965d, 58, 6,
902—908.
James M. T. A new Hermetia of potentional economic importance (Diptera,
Stratiomyidae).— «Pacif. Insects», 1972, 14, 1, 73—75.
Johannsen 0. A. Stratiomyid larvae and pupae of the Northeastern States.—
«J. New York Entomol. Soc.», 1922, 30, 141—153.
Johannsen O.A. Aquatic Diptera. II. Orthorrhapha.—«Mem. Cornell Univ.
Agrie. Exper. Stat.», 1935, 177, 1—62.
Johannsen 0. A., Crosby C. R. The life history of Thryptieus muhlenbergiae sp.
nov. (Diptera) — «Psyche», 1913, 20, 164—166.
Johnsey R. L., Nagel W. P., Rudinsky J. A. The diptera Medetera aldrichii
Wheeler (Dolichopodidae) and Lonchaea furnissi Me Alpine (Lonchaeidae)
associated with the Douglas-fir beetle in Western Oregon and Washington.—
«Cañad. Entomologist», 1965, 97, 5, 521—527.
Kangas E., Leskinen K. Pegohylemyia anthracina Czerny (Dipt., Muscidae) als
Zapfenschädling an der Fichte.— «Ann. Entomol. fennica», 1943, 9, 3,
125-212.
Kasule F. K. The subfamilies of the larvae of Staphylinidae (Coleoptera) with
keys to the larvae of British genera of Steninae and Proteininae.— «Trans.
Roy. Entomol. Soc. London», 1966, 118, 8, 261—283.
Kasule F, K. The larval characters of some subfamilies of British Staphylinidae
(Coleoptera) with keys to the known genera.— «Trans. Roy. Entomol. Soc.
London», 1968, 120, 4, 115—138.
Kawada A. 1959. Illustrated insect larvae of Japan. Tokyo.
Keilin D. On the life-history of Dasyhelea obscura Winnertz (Diptera, Nema-
tocera, Ceratopogonidae), with some remarks on the parasites and heredita¬
ry bacterian simbiont of this midge.— «Ann. and Mag. Natur. Hist.», ser.
9, 1921,8, 47, 576-590.
Keilin D. Respiratory systems and respiratory adaptations in larvae and pupae
of Diptera.— Parasitology, 1944, 36, 1—65.
Kemner A. Vara Clerider deras lefnadssätt och larver.— «Entomol. tidskr.»,
1913, 34, 191-210.
Kertész K. Vorarbeiten zu einer Monographie der Notacanthen XII—XXIL —
«Ann. Mus. Natur. Hung.», 1909, 7, 369—397.
Kertész K. Vorarbeiten zu einer Monographie der Notacanthen. XXXVI—
XXXVIII.— «Ann. Mus. Natur. Hung», 1916, 14, 123—218.
Kishi Y. A study on the ability of Medetera sp. (Diptera: Dolichopodidae) to
prey upon the bark and wood boring Coleoptera.— «Appl. Entomol.
and Zool.», 1969, 4, 4, 177-184.
370
Kishi Y, Ability of Lonchaea scutellaris Rondani (Diptera, Lonchaeidae) to
prey upon bark weevils and bark beetles.— «Kontyú», 1972, 40, 1, 1—6.
Kleine R. Die europäischen Borkenkäfer und ihre Feinde aus den Ordnungen
Coleopterenund Hymenopteren.—«Entomol. Blätter», 1908, 4, 11,205—208.
Kleine R. Die europäischen Borkenkäfer und ihre Feinde aus den Ordnungen
Coleopteren und Hymenopteren.— «Entomol. Blätter», 1909a, 5, 3, 41—50.
Kleine R. Die europäischen Borkenkäfer und ihre Feinde aus den Ordnungen
Coleopteren und Hymenopteren.— «Entomol. Blätter», 1909b, 5, 4, 76—69.
Kleine R. Die europäischen Borkenkäfer und die bei ihnen lebenden Räuber,
Parasiten und Kommensalen.— «Entomol. Blätter», 1944a, 40, 3/4, 68—83.
Kleine R. Die europäischen Borkenkäfer und die bei ihnen lebenden Räuber, Pa¬
rasiten und Kommensalen.— «Entomol. Blätter», 1944b, 40, 5/6, 125—133.
Kline L, TV., Rudinsky J. A, Predators and parasites of the Douglas-fir beetle:
description and identification of the immature stages.— «Oregon St. Univ.
Agrie. Exper. Stat. Techn. Bull.», 1964, 79, 1—52.
Knutson L, F. Pupa of Neomochtherus angustipennis (Hine), with notes on
feeding habits of robber flies and a review of publications on morphology
of immature stages (Diptera, Asilidae).— «Proc. Biol. Soc. Washington»,
1972, 85, 13, 163-178.
Korschefsky R. Tenebrioniden und Alleculiden Larven.— «Arb. physiol, angew.
Entomol.», 1943, 10, 1, 58—68.
Kraft K. /., Cook E. F, A revision of the Pachygasterinae (Diptera, Stratiomyi¬
dae) of America north of Mexico.— «Misc. Publ. Entomol. Soc. America»,
1961, 3, 1, 2—24.
Krivoscheina N, P., Mamajeu B. M. [Кривошеина H. П., Мамаев Б. М.] Die
Larven der europäischen Arten der Gattung Xylophagus Meigen (Diptera,
Xylophagidae).— Beitr. Entomol., 1966, 16, 3/4, 275—283.
Krogerus R, Beobachtungen über die Sukzession einiger Insektenbiocoenosen
in Fichtenstümpfen.— «Notulae Entomol.», 1927, 7, 121 — 126.
Larsson S, G. Larverne. Malacodermata atc.— «Denmarks Fauna», 1938, 10, 5,
193-310.
Lenz F. Metamorphose der Stratiomyiden.— «Arch. Naturgesch.», 1923, 89A,
2, 39-62.
Leon de. An annotated list of parasites predators, and other associated fauna
of the mountain pine beetle in western white pine and lodgepole pine.—
«Cañad. Entomologist», 1934, 66, 3, 51—61.
Leon D. de. A study of Medetera aldrichii Wh. (Diptera: Dolichopodidae) a
predator of the mountain pine beetle (Dendroctonus monticolae Hopk.,
Coleo.— Scolytidae).— «Entomol. americana», 1935, 15, 59—90.
Leonard M, D, A revision of the dipterous family Rhagionidae (Leptidae) in
the United States and Canada. — «Amer. Entomol. Soc. Mem.», 1930, 7,
1-181.
Lindner E. Rhagionidae.— In: Lindner E. Die Fliegen der palaearktischen
Region, Bd. 4, 20. Stuttgart, 1925, 3—5.
Lindner E. Stratiomyidae.— In: Lindner E. Die Fliegen der palaearktischen
Region, Bd 4, 18. Stuttgart, 1938, 1—218.
Lindner E. Stratiomyidae. In: Ruwenzori expedition 1934—1935. British Muse¬
um (Natur. Hist.), 1939, 2, 1—2, 1 — 11.
Lindner E. Zur Kenntnis der ostasiatischen Stratiomyiiden. (Dipt.).— «Bon¬
ner zool. Beitr.», 1955, 3—4, 6, 220—222.
Lindner E. Fünfter Beitrag zur Kenntnis der südamerikanischer Stratiomyiden-
fauna (Dipt.).— «Stuttg. Beitr. Naturkunde», 1969, 203, 1 — 15.
Lindner W. Ökologie und Larvenbiologie einheimischer Histeriden.— «Z. Morp¬
hoL, Okol. Tiere», 1967, 59, 341-380.
Lohse G. A. Die Käfer Mitteleuropas, Bd 4, Staphylinidae. I. Krefeld, 1964.
Lübben H. Thrypticus smaragdinus Gerst. und seine Lebensgeschichte.—
«Zool. Jahrb. Syst.», 1908, 26, 319—332.
Lundbeck W, Diptera Danica, genera, and species of flies hitherto found in
Denmark, I. Stratiomyidae, Xylophagidae, Coenomyidae, Tabanidae, Lep¬
tidae, Acroceridae, 1907, 1 — 114.
371
Lundblad О. Nágra skalbaggsfynd.— Entomol. tidskr., 1943, 64, 177—184.
Malloch J. R. A preliminary classification of Diptera, exclusive of Pupipara,
based upon larval and pupal characters, with keys to imagines in certain
families.- Part I.— «Bull. Illinois State Lab. Naturi Hist.», 1917, 12 (3),
161—409.
Martin Ch, Я. The status of Andrenosoma Rondani, Pilica Curran, and Pogo-
nosoma Rondani, with two new species (Diptera: Asilidae).— «Pan-Pacif.
Entomol.», 1966, 42, 4, 323—328.
Martin Ch. H. The new family Leptogastridae (the Grass-Flies) compared
with the Asilidae (Robber Flies) (Diptera).— «J. Kansas Entomol. Soc.»,
1968, 42, 1, 70-100.
Matsumura S. Thousand insects of Japan. Additamenta 2. Tokyo, 1916.
Me Alpine J. F. Cone infesting lonchaeids of the genus Earomyia Zett., with
description of five new species from Western North America (Diptera : Lon¬
chaeidae).— «Cañad. Entomologist», 1956a, 88, 4, 178—196.
McAlpine J. F. Old World lonchaeids of the genus Silba Macquart (—Carpo-
lonchaea Bezzi) with descriptions of six new species (Diptera : Lonchae-
idae).- «Cañad. Entomologist», 1956b, 88, 9, 521—544.
McAlpine J. F. Identities of lonchaeid flies described by Zetterstedt, with
notes on related species (Diptera).— «Cañad. Entomologist», 1958, 90,
7, 402—418.
McAlpine J. F. Chapter XVI. Diptera (Brachycera) Lonchaeidae. In: Hanst-
röm В., Brinck P., Rudebeck G. «South African animal life», v. 7, Uppsala,
1960, 327—376.
McAlpine J. F. A new species of Dasiops (Diptera, Lonchaeidae) injurious to
apricots.— «Cañad. Entomologist», 1961, 93, 7, 539—544.
McAlpine J. F. Descriptions of new Lonchaeidae (Diptera). L— «Cañad. En¬
tomologist», 1964a, 96, 4, 661—700.
McAlpine J. F. Descriptions of new Lonchaeidae (Diptera). IL—«Cañad.
Entomologist», 1964b, 96, 5, 701—757.
McAlpine J. F., Morge G. The identity, distribution and biology of Lonchaea
zetterstedti with notes on related species (Diptera, Lonchaeidae).—«Cañad.
Entomologist», 1970, 102, 12, 1559—1566.
McAlpine J. F.j M unroe D.D. Swarming of lonchaeid flies and other insects,
with descriptions of four new species of Lonchaeidae (Diptera).— «Cañad.
Entomologist», 1968, 100, 11, 1154—1178.
McFadden M. W. Soldier fly larvae in America north of Mexico.— «Proc. Nat.
Mus.», 1967, 121, 3569, 1—72.
Meijere J, C. II. Beiträge zur Kenntnis der Dipterelarven und Puppen.
«Zool. Jahrb.», 1916, 40, 177-322.
Meijere J. C. H. Over de metamorphose van Metopia leucocephala Rossi,
Cacoxenus indagator Löw, Palloptera saltuum L., Paranthomyza nitida
Mg. en Hydrellia nigripes Zett. (Dipt.) — Tijdschr. v. Entomol. 1944
(1943), 86, 57—61.
Melin D. Contributions to the knowledge of the biology, metamorphosis and
distribution of the Swedish Asilids in relation to the whole family of Asi-
lids.— Zool. Bidrag, 1923, 8, 1—317.
Metzger R. Die Kamelhalsfliegen (Neuroptera, Raphidiidae).— Wittenberg
Lutherstadt, Neue Brehm-Bücherei. 1960.
Miyatake M. Six new species and one new name of some Dipterous families from
Japan.— «Trans. Shikoku Entomol. Soc.», 1965, 8, 105—114.
Moffitt H. R., Varuss F. R. Dasiops alveofrons a new pest of apricots in Cali¬
fornia.— «J. Econ. Entomol.», 1961, 54, 3, 504—505.
Mokrzecki L. Raub- und Schmarotzer-Insekten des Buchdruckers Ips typograp¬
hus L. in Polen.— «Polskie pismo entomol.», 1933, 12, 1—4, 275—289.
Morge G. Über Morphologie und Lebensweise der bisher unbekannten Larven
von Palloptera usta Meigen, P. ustulata Fallén und Stegana coleoptrata
Scopoli.— «Beitr. Entomol.», 1956, 6, 124—137.
Morge G. Die Bedeutung der Diptera im Kampf gegen die Borkenkäfer.—
«Arch. Forstwesen», 1961, 10, 505—511.
372
Morge G. Dio Lonchaeidae und Pallopteridae Österreichs und angrenzenden
Gebiete. L TejL Die Lonchaeidae.— «NaturkundL Jahrb. Stadt Linz»,
1963, 9, 123-312.
Morge G. Eine Beobachtung zur Grundfrage der Abhängigkeit von Wirkungsg¬
rad und Wert natürlicher Feinde gegenüber Schädlingen.—«Beitr. EntomoL»,
1967a, 17, 1—2, 225—233.
Morge G. Die Pallopteridae. Linz, 1967b.
Mulsant £*., Rey C. Histoire naturelle des Coléoptères de France. Paris, 1863.
Nagatomi A, The Sarginae and Pachygasterinae of Japan (Diptera, Stratio¬
myidae).— «Trans. Roy. Entomol. Soc. London», 1975, 126, 3, 305—421.
Nagatomi Л., Saigusa Г. The Xylophagidae of Japan (Diptera).— «Mem. Fac.
Agrie. Kagoshima Univ.», 1969, 7, 1, 177—188.
Nagatomi Л., Saigusa T. The Coenomyiidae of Japan (Diptera).— «Mem. Fae.
Agrie. Kagoshima Univ.», 1970, 7, 2, 257—292.
Nagatomi Л., Tanaka A. The Solvidae of Japan (Diptera).- «Mushi», 1971,
45, 6, 101-146.
Novak V. Pfirozeni nepfátelé a nemoei dfevokaza cárkového Trypodendron line-
atum Oliv.— «Zool. Listy», 1960, 9 (23), 309—322.
Nuorteva M. Über den Fiehtenstamm-Bastkäfer, Hylurgops palliatus Gyll.,
und seine Insektenfeinde.—«Acta Entomol. fenniea», 1956, 13, 1, 1 — 118.
Nuorteva M. Über die Voraussetzungen der biologischen Bekämpfung der Bor¬
kenkäfer mit Hilfe ihrer Insektenfeinde in der finnischen Waldpflege.—
Verhandl. IV. internat. Pflanzensehutz-Kongr. Hamburg, 1957, 1, 1025—
26.
Nuorteva M. Untersuchungen über einige in den Frassbildern der Borkenkäfer
lebende Medetera-Arten (Dipt., Dolichopodidae).— «Ann. Entomol.
fenniea», 1959, 25, 192—210.
Nuorteva M. Die Borkenkäfer (Col., Scolytidae) von Virolahti, Südostfinnland
und deren natürliche Insektenfeinde.— «Ann. Entomol. fenniea», 1963, 29,
281—288.
Nuorteva M. Über den Einfluss der Menge des Brutmaterials auf die Vermehrlich-
keit und die natürlichen Feinde des grossen Waldgärtners, Blastophagus
piniperda L. (Col., Scolytidae).-«Ann. Entomol. fenniea», 1964, 30, 1—17.
Nuorteva M. On the habitats of some Lonchaea species in Fennoscandia (Dip¬
tera, Lonchaeidae).- «Ann. Entomol. fenniea», 1967, 33, 118—121.
Nuorteva M. Die Borkenkäfer (Col., Scolytidae) und deren Insektenfeinde im
Kiehspiel Kuusamo, Nordfinnland.—«Ann. Entomol. fenniea», 1971, 31,
65-74.
Oldroyd H. The family Leptogastridae (Diptera). — «Proe. Roy. Entomol. Soe.
London», 1969, 38, 1-2, 27—31.
Palm Th. Über die Entwicklung und Lebensweise einiger wenig bekannten
Käferarten im Urwaldgebiet am Fluss Dalälven. 3.— «Opuse. Entomol.»,
1940, 5-7, 21-26.
Palm Th. Die Holz- und Rinden-Käfer der nordsehwedisehen Laubbäume.
Stockholm, 1951,
Palm Th. Die Holz- und Rinden-Käfer der süd- und mittelsehwedisehen Laub¬
bäume.— «Opuse. Entomol.», 1959, suppl. 16.
Parkin E. A. The digestive enzymes of some woodboring beetle larvae.—
J. Exper. Biol., 1940, 17, 4, 364-377.
Paulian R. Les premiers états des Staphylinoidoa.— Mém. Mus. nat. hist,
natur., 1941, 15, 1—361.
Perris E. Histoire des insectes du pin maritime.— «Ann. Soe. entomol. France»,
ser. 3, 1854, 2, 593—646.
Perris E. Histoire des métamorphoses de divers insectes.- «Mém. Soe. scient.
Liège», 1855, 10, 238.
Perris E. Les insectes du pin maritime. Paris, 1863.
Perris E. Histoire des insectes du pin maritime: Diptères. — «Ann. Soc. entomol.
France», sér. 4, 1870, 10, 133—366.
Perris E. Larves de Coléoptères. Paris, 1877.
373
Peterson A. Larvae of insects. An introduction to Nearctic sp^ies, v. 2. Ohio,
1951, 1-416. /
Pleske T. Révision des espèces paléarctiques des familM Erinnidae et Coe¬
nomyiidae.— Encyclop. entomoL, sér. B. 2. Dipt. 1925, 161 — 184.
Pleske T, Résultats scientifique des Expéditions eMomologiques du Musée
zoologique dans la région de l’Oussouri. II. Dip^tera. Les Stratiomyiidae,
Erinnidae, Coenomyiidae et Oestridae.— Ежегодник Зоомузея АН
СССР, 1930, 181-206.
Poulton E. В. The relation between the larvae of the asilid genus Hyperechia
(Laphriinae) and those of xylocopid bees.— «Trans. Entomol. Soc. London»,
1924, 1, 11, 121-123.
Raff J. W. Notes on Chiromyza australis Macq.— «Victorian Naturalist», 1931,
47, 213—214.
Reitter E, Fauna Germanica, Bd 2. Stuttgart, 1909.
Rondani C, Dipterologiae Italicae prodromus, 1, 1856, p. 172.
Rozko^ÿ R. Tne Stratiomyioidea (Diptera) of Fennoscandia and Denmark.—
«Fauna entomol. scand.», 1973, 1, 117—131.
Saalas U. Fichtenkäfer Finnlands. I.— «Ann. Acad. sei. fennici», ser. A, 1917,
8, 1-298.
Saalas U. Fichtenkäfer Finnlands. П. — «Ann. Acad. sei. fennici», ser. A, 1923,
22, 1-746.
Satchell С, H. 1947. The larvae of the British species of Psych oda (Diptera,
Psychodidae).— «Parasitology», 38, 51—69.
Savely H. R. Ecological relations of certain animals in dead pine and oak logs.—
«Ecol. Monogr.», 1939, 9, 321—385.
Schedl K. E. Breeding habits of arboricole insects in Central Africa.— Proc.
10th Internat. Congr. Entomol. (1956), 1958, 1, 183—197.
Scheerpeliz O. In: Blunck H. Syllabus der Insektenbiologie. Coleoptera, v. 1,
1925, 58-112.
Schimitschek E. Tetropium gabrieli Weise und Tetropium fuscum F. Ein Beit¬
rag zu ihrer Lebensgeschichte und J.ebensgemeinschaft.— «Z. angew.
Entomol.», 1929, 15, 2, 229-334.
Schimitschek E. Die achtzähnige Lärchenborkenkäfer Ips cembrae Heer. Zur
Kenntnis seiner Biologie und Ökologie sowie seines Lebensvereines.— «Z.
angew. Entomol.», 1931 (1930), 17, 2, 259—355.
Schimitschek E. Forstentomologische Studien in Urwald Rotwald.— «Z. angew.
Entomol.», 1952-1953, 34, 178-215, 514-542; 35, 1-54.
Schlottke E. Über die Verdauungsfermente in holzfressender Käferlarven.—
«Zool. Jahrb.», Abt. allg. Zool. Physiol. Tiere, 1948, 61, H. 1—3, 88—140.
Schmid J. M. Medetera aldrichii (Diptera: Dolichopodidae) in the Black Hills.
II. Biology and densities of the immature stages.— «Cañad. Entomolo¬
gist», 1971, 103, 6, 848-853.
Schremmer F. Die Mundteile der Brachyceren-Larven und dor Kopfbau der Lar¬
ven von Stratiomys chamaeleon L.— üstcrr. zool. Z.», 1951, 3, 3/4, 326—
397.
Séguy E. Diptères Brachycères (Stratiomyidae). Faune de France. Paris, 1926,
13, 1—305.
Séguy E. Diptères Brachycères (Asilidae). Faune de France. Paris, 1927, 17,
1-184.
Séguy E. Diptères (Brachycères) (Lonchaeidae). Faune de France. Paris, 1934,
28, 171-186.
Séguy E. La biologie des Diptères.— Encyclop. entomol., 1950, A 26, p. 1—609.
Séguy E. Ordre des Diptères (Diptera, Linné, 1758).— In: Traité de zoologie,
anatomie, systématique, biologie, v. 10, Grassé P. P. (Ed.). Paris, 1951,
1, 449—744.
Seitner M. Beobachtungen und Erfahrungen aus dem Auftreten des achtzäh-
nigen Fichtenborkenkäfers Ips typographus L. in Oberösterreich und Steier¬
mark in demj Jahren 1921 bis einschl. 1923.— «Zbl. ges. Forst- und Holz¬
wirtschaft», 1924, 50, 1, 2—23.
374
Seitner M, Lonchaea viridana Meig. ein Tannenzapfen- und Samenschädling.—
«Zbl. ges. Forst'wesen»» 1925, 51, 5/6, 159—161.
Seitner M. ChortophMa laricicola Karl — die Lärchenzapfen-und Samenflie¬
ge, und ihre Feinde; Parasiten und Räuber.— «Zbl. ges. Forstwesen», 1929,
55, 5/6, 153-167.
Silvestri F, Sulla Lonchaea aristella Beck.— «Boll. Lab. zool. gen. tgr. Portia»,
1917, 12, 123-146.
Smetana A. Drabcikoviti-Staphylinidae.— «Fauna CSR», 1958, 12, Praha.
Smith K. G. V. Notes on the immature stages of four British species of Lonchaea
FIL (Diptera. Lonchaeidae).— «Entomol. Monthly Mag.», 1957, 92, 402—
405.
Steel W. O. The larvae of the genera Omaliinae (Coleoptera, Staphylinidae)
with particular reference to the British fauna.— «Trans. Roy. Entomol.
Soc. London», 1970, 122, 1, 1—47.
Steyskal G. C, A revision of the nearctic species of Xylomyia and Solva (Diptera,
Erinnidae).— «Michigan Acad. Sei., Arts and Letters, Papers», 1947, 31,
181 — 190.
SteyskalG. C. R evi si onary notes on the genus Systenus Loew with the description
of a new species (Diptera, Dolichopodidae). —«Proc. Entomol. Soc. Washing¬
ton», 1970, 72, 1, 107-111.
Szilady Z. Holzfliegen, Erinnidae (Xylophagidae).— In: «Die Tierwelt Deutsch¬
lands», Bd 2, 1932, 2-7.
Taylor R, L, The biology of the white pine weevil (Pissodes strobi Peek.) ami
a study of the insect parasites from economic standpoint. — «Entomol.
americana», 1929, 9, 167—246.
Taylor R, L. The biology of the white pine weevil (Pissodes strobi Peek.) und
a study of the insect parasites from economic standpoint.— «Entomol.
americana», 1930, 10, 1—86.
Thorpe W, H, The larvae and pupae of the genus Hyperechia.— «Trans. Ento¬
mol. Soe. London», 1927, 75, 177—185.
Thuneberg E. A revision of the Palaearetie species of the genus Medetera Fi¬
scher (Dipt., Dolichopodidae).— «Ann. Entomol. fennica», 1955, 21, 3,
130—157.
Trägärdh /. Skogsentomologiska bidrag. 1—5.— «Entomol. tidskr.», 1914,
35, 188—209.
Trojan P. Klueze do oznaezania owadow Polski, 28.— Muehowki — Diptera,
Erinnidae. Warszawa, 1956, 19, 1—21.
Tsacas L., Desmier de Chenon /?., CoutinR, Observationssur le parasitisme lar-
vaire d’Hyperechia bomboides (Dipt., Asilidae).— «Ann. Soe. entomol.
France», 1970, 6, 2, 493—512.
Tuomikoski R. Havaintoja puunrungoilta tavatuista kaksisüpisistä.— «Notulae
Entomol.», 1930, 10, 126-127.
Verhoeff K, W. Studien über die Organisation und Biologie der Staphylinoidea.
IV. Zur Kenntnis der Staphyliniden-Larven. V. Zur Kenntnis der Oxyteli-
den-Larven.— «Arch. Natur. Berlin», 1919, 85, Abt. A, 6, 1 — 111.
Verrall G, H. Stratiomyidae. «Brit. Flies», 1909, 5, 75—76.
Vimmer A. Muse parast kurovce Ips typographus nalezen Cechäch.— Lesn.
praee, 1926, 5, 451—555.
Vite J. P. Die holzzerstörenden Insekten Mitteleuropas. Göttingen, 1952.
Wallace H. R, The ecology of the insect fauna in pine stumps.— J. Animal Ecol.,
1953, 22, 154-168.
Wensel R. L. On the classification of the histerid beetles.— «Field Mus. Publ.
Zool.», Chicago, 1944, 28, 41—101.
Westwood J, O. Introduction to the modern classification of insects. Synopsis
of the genera of British insects. London, 1840, 1 — 158.
Wheeler W, M, A genus of maritime Dolichopodidae new of America.— «Proc.
California Acad. Sei. (3) Zool.», 1897, 1, 145—152.
Whitten J, M. A comparative morphological study of the tracheal system of
larval Diptera. Part L— «Quart. J. Microseop. Sei.», 1955, 96, 2, 257—278.
375
Wi(}ckowski S. Entomofauna pniakow sosnowich w zalezno^i od wicku i roz-
miaru pniaka.— «Ekol. Polska», 1957, 5, 13—140. /
Wichtman H, E. Untersuchungen an Ips typographus iJ und seiner Umwelt.
Die Kamelhalsfliegen.— «Z. angew. Entomol.», ^57, 40, 3, 433—440.
Wiedemann W. M. Die Zellulose-Verdauung bei Lani^llicornienlarven (Scara¬
baeidae).— Z. Morphol., Okol. Tiere, 1930, 19,/228—258.
Williams F, X. Biological studies in Hawaiian water-living Insects. Part.
III. Diptera or Flies.— «Proc. Hawaii Entomol. Soc.», 1938, 10, 85—129.
Winkler J. R. Die Buntkäfer (Cleridae).— Neue Brehm — Bücherei, 1961,
281, 3-108.
Wirth W. W, Three new nearctic species of Systenus with description of the
immature stages from tree cavities (Diptera, Dolichopodidae).— «Proc.
Entomol. Soc. Washington», 1952, 54, 236—244.
Wyniger R. Beobachtungen an Raphidia maior Burm.— «Mitt, entomol. Ges.
Basel», N. F., 1959, 9, 1--13.
\ АЛФАВИТНЫЙ УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ НАСЕКОМЫХ
abieticola J. Sahlb., Pytho 183, 184,
185
abietis L., Hylobius 85, 164
abietis Wank., Laemophloeus 167,
174, 175
Abracinae 87
Acanthocinus 16, 34, 40, 50, 93, 140
Acanthoneura 38, 41, 47, 245
Acephala 204, 207
Acicnemis 14
Acritus 86, 87, 88, 89, 92
Acrulia 98, 103
Actina 269, 270
acuminata Deg., Melanophila 16
acuminatus Gyll., Ips 93, 95, 114,
166, 320, 325, 352
Adelocera 34, 56, 81, 140, 141, 142,
144, 145, 14Г), 146
Adephaga 58, 63
Aderidae 76
Adoxomyia 269, 270, 271
admirandus Kriv. et Mam., Xylo-
phagus 37, 45, 250, 251, 253, 255,
256, 257
aedilis L., Acanthocinus 50, 93
aeneus Richt., Rhizophagus 164, 165,
165, 166
Aeolesthes 47
aequata Er., Atheta 110
aethiops Lac., Ampedus 149, 149
Aesalus 24, 45
affinis F., Chrysobothris 16, 18, 133,
309
affinis Pk., Atrocus 105, 110
Aglaophis 129
Agnatus 185, 186, 187
Agrilus 16, 40, 47
Agriotini 141
Agromyzidae 222, 237, 238
Agrypnini 141
alabeta Begny, Graptomyza 38
Alaus Ш, 142, 145, 146
albibarbe Meig., Andrenosoma 297
albistylum Johns., Berkshiria 290
albopilosus Miyat., Xylophagus 37,
45, 250, 251, 252, 255, 257
Aleochara 97, 101
Aleocharinae 97, 99
Alleculidae 59, 63, 64, 72, 76, 77
Allognosta 269, 270
alni Fisch., Dicerca 306
altaica Cand., Adelocera 144, 145, 145
alternans Er., Laemophloeus 167,
174, 174, 175
Amartus 157
amicula Steph., Atheta 105, 110
amitinus Eichh., Ips 93
amoena Meig., Calomyia 206, 218
Ampedus 24,50,140,141,143, Ш,147,
147, 148, 149, 149, 150,151, 152, 153
amurensis Herm., Choerades 38, 299,
301
amurensis Heyd., Phellopsis 73
amurensis Portsch., Acanthoneura 38,
47
amurensis Rtt., Hololepta 90, 95
Anacanthaspis 249
Anaspis 77
Ancylocheira 22, 307
Andrenosoma 34, 38, 44, 292, 293,
294, 295, 296, 297, 298, 308
Andrenosomini 292
angusta Aube., Leptusa 107, 112
angustatum Hoffm., Cylister 92, 93
angustula Sturm., Epuraea 161, 162,
162, 163
Anisandrus 19
Anisopodidae 202, 203, 213, 216, 217,
222, 231, 327
Anobiidae 47, 64, 67, 68, 118
Anobium 68, 132
Anommatus 177
Anomognathus 105, 107, 109
anophthalma Aube., Langelandia 176,
177
Anthicidae 77, 186
Anthomyiidae 225, 248
Anthribidae 37, 67
annulata Meig., Trichocera 220
annulatus Meig., Symmerus 202, 215
aperta Lund., Bolitophila 43
Aphrosylinae 309
377
Aphrosylus 309
aquatilis Goetgh., Euphaenocladius
223
araratica Mars., Ancylocheira 22, 307
Archostemata 57
arcuata Pz., Strangalia 22, 129
arcuata Fll., Palloptera 353
arcuatus L., Plagionotus 56
Argyra 311
Arthroceras 249
Arthrocnodax 211
Arthropeas 249
Arthropeina 262
Asemum 22, 34
asiaticus Reiche., Trigonurus 102
Asilidae 38, 39, 41, 45, 203, 207, 211,
216, 217, 235, 257, 258, 292, 296,
298, 301, 305
Asilinae 292, 293, 294
Aspidacantha 37, 270, 271, 276, 277,
278
Asyndetus 312
ater Meig., Xylophagus 27, 37, 45,
203, 249, 250, 251, 253, 255, 257
ater Pk., Hylastes 164
aterrimus L., Ectinus 145
Atheta 104, 105, 110
Athouini 141
Athous 143, 152, 153
Athomosiini 292
atra Kert., Aspidacantha 277, 278
atra Pz., Pachygaster 274
atrata L., Tanyptera 257
Atrecus 105, 109, 110, 111
atronitens Kov., Lonchaea 330, 335
atrum L., Andrenosoma 34, 297, 298
Aulacigaster 208, 238
Aulacigastridae 208, 238, 238
Aulonium 14, 41, 177, 179, 180, 181,
182
aurata L., Cetonia 68
autographus Ratz., Dryocoetes 92,
93, 110, 113, 318, 325
Axymyiidae 47, 202, 206, 213, 215,
216, 217, 223, 224, 226, 232
bajulus L., Hylotrupes 22, 34, 128,
297
balteatus L., Ampedus 147, 149,
149, 151
barbatus Schall., Serropalpus 22, 44
bartelsi Fald., Gnorimus 24
bellicosa Lew., Phloetrya 22
Beridinae 269
Beris 269, 270, 274
Berkshiria 290
Bibio 44, 219, 225, 262
Bibionidae 44, 48, 201, 217, 219, 222,
225, 227, 250, 262, 344
bicolor Fll., Brachyopa 204
bicolor Hbst., Tii^phrorychus 116,
138, 318, 353
bidentatus F., /Silvanus 30
bidentatus Hbst., Pityogenes 320
biguttata Gyll., Tomoxia 74
biguttatus Pili., Drapetes 81
bimaculata L., Dictenidia 218
bipustulatus F., Rhizophagus 159,
165, 165
bipustulatus F., Tachinus 102
bipustulatus L., Calambus 152, 153
bispinosa Neg., Mcdetera 37, 312,
314, 316, 318, 323
Bitoma 31, 177, 178, 179
Bittacus 54
Bius 78, 188, 192
Blastophagus 93, 95, 96, 97, 113,
114, 163, 164, 166, 320, 322, 323,
344
Bledius 97
Bolitochara 102, 107
Bolitophagus 190
Bolitophila 43
Bolitophilidae 43, 229
Boreus 54
Boridae 39, 75
Boros 19, 20, 39, 75
Bostrychidae 64, 67, 68, 87, 118
Bostrychoidea 58
Brachycera — Cyclorrhapha 201
Brachycera — Orthorrhapha 201
Brachyopa 41, 43, 47, 204, 219, 257
Brachypterolus 157
Brachypterus 157
braueri Alb., Inocellia 199, 200
Brontes 41, 79, 79, 167, 170
bruggeri Morge, Lonchaea 37, 52,
328, 329, 331, 332, 344, 353
brunneus Steph., Lyctus 68
Bryocharis 104
bukowskii Czerny, Lonchaea 335, 337,
340, 341, 349
bungei Pl., Xylophagus 250
Buprestidae 44, 47, 62, 63, 85, 86,
293, 294, 309
buprestoides L., Spondylus 22, 93
Buprestis 22, 85
Byrrhidae 61, 67, 68
Byrrhus 68
cadaverinus Hoffm., Hister 88, 94
Caenosciara 26
caesus Hbst., Plegaderus 36
Calambus 140, 143, 152, 153
Calitys 70, 71
Calliphoridae 210, 222, 248
Callipogon 22
Calomyia 206, 218
Calobatidae 41, 241, 242, 346
Calopus 22, 69
378
Camponotus 26
Camptomyia 41, 42
Campsicnemus 309
Cantharidae 56, 60, 61, 65, 78, 83,
84
Cantharis 84
Carabidae 56, 60, 61, 62, 63, 66, 66
Cardiophorinae 58, 72, 141, 142
Cardiophorus 81, 144, 153
cardoni Lew., Melambia 137
carinatus Lew., Opilo 126, 128
carpathica Kov., Lonchaea 330, 335,
339, 340, 342
Carpelimus 97
Carpophilus 79, 157, 158, 160, 161
castaneum Hbst., Colydium 177
caucasica Kov., Lonchaea 330, 332,
335, 339, 340, 342
caucasicum Men., Nemosoma 138
caucasicus Motsch., Cucujus 56, 171,
172
Cechorismenus 269, 271, 277, 279,
280
Cecidomyiidae 41, 42, 43, 44, 202,
202, 204, 204, 207, 211, 217, 219,
220, 221, 222, 223, 224, 227, 229,
257, 258, 261, 320, 352
cembrae Heer., Ips 318
cembrae Motsch., Pissodes 113
Cephaloidae 80, 81
Cephalon 81
ceramboides L., Upis 188, 189, 261
Cerambycidae 36, 44, 47, 48, 51,
56, 62, 63, 65, 66, 70, 87, 261, 293,
294, 306, 309, 352
Cerambyx 47
Ceratopogonidae 41, 201, 203, 213,
217, 219, 220, 220, 221, 222, 223,
226, 234, 258, 259, 310, 327, 328
Cercyon 63, 86
Ceroplatidae 222, 230
Ceruchus 11, 12, 24, 45
cervus L., Lucanus 68
Cerylon 59, 84
Cerylonidae 59, 83, 84, 177
Cetonia 68
chalcographus L., Pityogenes 93, 95,
112, 113, 175, 185, 186, 357
chalybeata Wied., Lonchaea 329
Chiromyzinae 272
Chironomidae 201, 213, 217, 220, 221,
222, 223, 226, 234
Chloromyia 269, 271
Chlorophorus 308
Chloropidae 36, 244, 246
Choerades 16, 24, 34, 38, 44, 45, 203,
217, 258, 292, 293, 294, 295, 297,
299, 300, 301, 302
chorea F. Lonchaea 329
Chorisops 269
christophi Rtt., Librodor 29
Chrysobothris 16, 18, 40, 133, 309
Chrysochroma 269, 270
chrysomelinus Hochw., Ceruchus 11,
12, 24
Chrysopilus 50, 211, 218
Chyliza 208, 242
cinctus Deg., Xylophagus 27, 37,
249, 251, 256, 257, 258, 261
cinctus Pk., Hypoganus 152, 154,
155
cinereus Hbst., Crypturgus 97
cinnabarinus Esch., Ampedus 147,
149, 150
cinnabarinus Scop., Cucujus 171, 172,
172
circularis Nag., Takagimyia 283
Gisidae 87
clavipes F., Cucujus 171
Cleridae 36, 38, 41, 49, 58, 59, 60,
62, 63, 72, 80, 82, 83, 117, 120,
121, 122, 122
Cleroidea 58, 64
Clerus 16, 36, 118, 121, 123, 124, 125,
130
Clitellaria 269, 270, 271
Clitellariinae 269, 270, 271, 272
Clusia 26
Clusiidae 44, 207, 212, 241, 242, 257
Clusiodes 26, 257
Clytus 128
coccinea L., Pyrochroa 19, 75
Coenomyia 249
coerulea L., Trogossita 135, 140
coeruleus Deg., Korynetes 125, 126
coeruleus L., Helops 24, 128, 188,
189, 191
cognata Ramb., Raphidia 197, 198,
199
Coleoptera 53, 57, 65
Colobicus 176, 181
colobotheoides Bat., Aglaophis 129
collini Hack., Lonchaea 14, 37, 52,
328, 329, 331, 332, 335, 340, 342,
343
Colydiidae 41, 59, 70, 73, 73, 75,
76, 176, 179, 180
Colydium 43, 59, 73, 75, 177, 180,
181, 182
comes Rtt., Rhysodes 66
compeditus Wied., Xylophagus 27,
39, 211, 249, 251, 256, 258, 259
complanata Er., Placusa 69, 108,
114
compressa Walk., Texara 225, 331,
346
compressum Hbst., Platysoma 96
conica Pz., Brachyopa 43, 47
Cononotidae 186
379
conspersa Gyll., Adclocera 34, 56,
81, 144, 145, 145
coriarius L., Prionus 39
cornutum Sturm., Ncmosoma 136,
138
cornutus F-W., Cryphaeus 189
corticis Taylor, Lonchaea 37, 329,
331, 332
Corymbitini 141
Cossus 54
cossus L., Cossus 54
Cramptonomyiidae 38, 43, 48, 201,
201, 209, 228, 256, 257
Cratomerus 133
Craspedometopon 37, 269, 271, 275,
280, 281
crassicollis Er., Melanotus 152, 156
crassicornis Schum., Inocellia 199,
200
Crassotarsus 11, 19, 43, 256, 291
Crematogaster 26
crcnata F., Bitoma 31, 177, 178
crenatus Pk., Dendrophagus 167
Criocephalus 22, 34, 302, 306
Cryphaeus 189, 192
Cryphalus 109, 323
Cryptarcha 157, 158, 160, 161
Cryptarchips 157, 158, 160
Cryptocercus 12, 24, 45
Cryptohypninae 141
Cryptorrhynchus 47
Crypturgus 92, 95, 97, 112, 166
Ctenophora 220, 226, 261, 262
Ctenophorinae 44, 48
Ctenota 292, 293, 294
Ctenotini 292
Cucujidae 19, 40, 41, 42, 45, 51, 56,
59, 64, 65, 79, 79, 166, 168, 170,
172, 174, 176
Cucujus 19, 50, 56, 59, 79, 167, 169,
170, 171, 172, 172, 173
Culicidae 36, 216, 222
Culicoides 41, 328
culinaris F., Uloma 78, 188, 189
Cupedidae 57, 63, 65, 66
Cupes 66
Curculionidae 43, 62, 64, 85, 86, 87,
256, 261, 291, 293, 294
cursor L., Toxotus 22
curvidens Germ., Pityokteincs 323
cuspidata Er., Anomognathus 105,
107, 109
Cyclorrhapha 45, 201
cylindrica Deg., Leptogaster 294
cylindricum L., Sinodendron 11, 24,
44, 300
Cylindrotomidae 217, 218, 220, 233
cylindrus Hbst., Platypus 19
Cylistcr 14, 27, 29, 30, 31, 32, 33,
87, 88, 89, 90, 92, 93, 94, 95
Cyllodcs 157
Cyphaea 106
Cyphomyia 271
czekanovskii Pl., Xylomya 17, 265,
268, 346
Dasiops 329, 335
Dasyhelea 41, 328
Dasypogoninae 292, 293, 294
dealbatus Kr., Clerus 125
Dechomus 73, 177, 180, 181
decoratus Germ., Agnatus 185, 186,
187
defecta McAlp., Lonchaea 329, 331
Dendroctonus 113, 158, 166, 344
Dendrophagus 79, 167, 170
Dendrophilinae 87
Dendrophilopsis 86
Dendrophilus 86
Denops 118, 120, 121, 124, J25, 132
Denticollis 16, 49, 143, 152, 153,
154, 154
deplanatum Gyll., Platysoma 29, 96
depressa Miirkl., Placusa 98
depressus F., Rhizophagus 165, 165,
166
depressus Hbst., Pediacus 168, 175,
176
depressus L., Pytho 20, 31, 33, 34,
75, 183, 184, 185, 185, 186
depsarium L., Tragosoma 22
Dermestidae 72, 74
dermestoides F., Pediacus 167, 175,
176, 176
dermestoides L., Elateroides 19, 21,
74
Diadocidiidae 230
Diaphorinae 309
Diargemus 279
Dicerca 22, 306
dichrocera Kow., Medetera 37, 312,
314, 315, 318
Dicroneurus 220
Dictenidia 26, 218
dilatatus Rtt., Xylogenes 125
Dilophiis 44, 201, 344
Diptera 55, 200, 226
Dircaea 22
discicollis Lee., Lecontia 183
dispar Pk., Rhizophagus 32, 164,
165, 166
Ditomyiidae 202, 202, 213, 215, 216,
222, 224, 228
Ditylus 22
Dolichopodidae 37, 41, 45, 202, 203,
207, 217, 219, 225, 236, 257, 259,
261, 309, 312, 312, 314, 319, 321,
324, 326, 346
Dolichopus 309, 311, 312
dolium F., Megamerina 331, 349
380
domesticum L., Trypodondron 98,
138, 318
domesticus Sturm., Opilo 128
Dorcus 11, 24, 44, 128, 301
Drapetes 80, 81
Dromius 56
Drosophila 243
Drosophilidae 225, 243, 244, 310
Dryocoetos 14, 92, 93, 110, 113, 163,
186, 318, 325
dubia Schall., Melandrya 71
dubius F., Thanasimus 130
duodecimstriatus Muli., Anommatus
177
duplicatus Sahlb., Ips 93, 164
Earomyia 329, 335
Ectaetia 44
Ectinus 143, 145
Eidalimus 271
Elacatidae 80
elaeagni Plav., Chlorophorus 308
elaeagni Richt., Cratomerus 133
Elateridae 45, 49, 56, 58, 59, 60, 61,
64, 72, 78, 80, 81, 140, 142, 145,
147, 149, 151, 152, 154
Elateroides 11, 19, 21, 74
elegantulus Schonh., Ampedus 149,
150
Elephantomyia 44, 221, 261
elongatum F., Colydium 75, \11,180,
182
elongatum L., Nemosoma 135, 138
elongatus L., Tillus 59, 80, 120, 122,
126, 131
emarginatus Latr., Colobicus 176
Empididae 207, 237, 344
Enchodes 22
Endomychidae 83
Endophloeus 176, 181
Enneadesmus 125
eous Lehr, Mesoleptogaster 50, 293,
294, 298, 307
ephippium F., Laphria 39, 295, 303,
304, 305
Epidapus 26
Epiphragma 26, 201, 261
Epuraea 14, 41, 59, 79, 157, 158, 161,
162, 162, 163, 164
Ergates 22
Eristalis 221
erythrogonus Miill., Ampedus 147,
147, 149
Eucephala 204, 205
Eucnemidae 58, 59, 85, 86, 261, 307
Euphaenocladius 223
Eupachygaster 270, 271, 276, 281,
282, 282, 283, 328
Eurythyrca 22, 23, 308
Eurytracholus 11, 44
Euscelidia 292
excellens Frey, Modetera 314, 317, 318
eximia Bohem., Hyperoscelis 207
Exodontha 269, 270
expectatus Smet., Gabrius 107, 111
faber L., Ergates 22
fagineus Edw., Culicoides 328
Fannia 206, 218, 219, 220, 225, 226,
257
Fanniidae 206, 218, 219, 220, 225, 226,
240, 257
fasciata Frey, Medetera 37, 203, 312,
314, 314, 318
fasciata L., Adelocera 144, 145, 146
fasciata Meig., Pachyneura 214, 214,
225
fasciculare 01., Nosodendron 84
febrilis Loew, Dilophus 201
feminina Neg., Medetera 310, 317,
319, 320
fenestralis Scop., Sylvicola 202
ferrugineus L., Ludius 149, 155
ferrugineus L., Pityophagus 160, 164
ferrugineus Steph., Laemophloeus
167, 174
filiforme F., Colydium 177
Filiptschenkia 269
fimbriatus Meig., Choerades 38, 299,
300, 301
flabellicornis Schn., Elateroides 19
flava L., Laphria 39, 211, 295, 304,
305, 307
flavicomis Hbst., Paromalus 91, 95
flavicomis Kert., Cechorismenus 279,
280
flavidipennis Zett., Spermatolon-
chaea 335
flavipes Stein., Raphidia 195
flavolineata Meig., Tipula 226
flavoscutellaris Mats., Solva 267
Forcipomyia 41, 219, 220, 220, 223,
226, 258, 259, 328
formicarius L., Thanasimus 27, 32,
33, 82, 130, 131
fraxini Kug., Hypophloeus 27, 29,
32, 191, 193, 194
freyi Czerny, Lonchaea 330, 335, 338,
343, 344
frontale Kert., Craspedometopon 281
fugax Beck., Lonchaea 330, 331, 332,
334, 335, 339, 344, 345
fulvus Kieff., Holoneurus 220, 223
fulvus Meig., Choerades 38, 299, 300,
301
fumipennis Par., Campsicnemus 309
fusca L., Cantharis 84
fuscicornis F., Tremex 137
fuscipes Meig., Scatopse 214, 220
fuscus Er., Pediacus 167, 176, 176
381
Gabrius 98, 107, 109, 111
Gaurax 246
Georthocladius 223
gibbosa L., Laphria 38, 295, 303, 304,
305
gigas F., Sipalinus 11, 19, 21^ 43,
256, 291
gigas L., Urocerus 54
gilvus L., Choerades 34, 38, 295, 299,
300, 301
glabratus Zett., Hylurgops 110, 113,
166, 186
Glischrochilus 157, 158, 159, 160
Glutops 249
Gnathoncus 86, 89
Gnophomyia 211, 218, 346
Gnorimus 24
grandis Gyll., Rhizophagus 157, 164,
165, 166
granulatus Ratz., Trypophloeus 320,
344
Graptomyza 38, 41
grotnii Hagel, Cryphalus 109
grumi Sem., Xylotrechus 34, 139,
308
Grynocharis 56, 80, 82, 133
gyllenhali Gyll., Pissodes 302
Habrocerinae 99
haematodes Er., Cucujus 19, 79, 167,
171, 172, 172, 173
haemorrhoidalis Meig., Sarcophaga
218
Hammerschmidtia 41, 47, 219
Hapalaraea 103, 106, 112
harmandi Seguy, Solva 265, 266
hectographus Rtt., Dryocoetes 163,
186, 318, 325
Helodidae 62, 72, 73
Helopini 44
Helops 24, 128, 188, 189, 191, 192
Hemicephala 204, 206
hemipterus L., Carpophilus 79, 160
Hendelia 242
Hermetia 269, 272
Hesperinidae 36, 44, 48, 205, 214,
222, 224, 228, 262
Hesperinus 26, 50, 214, 262
Hetaeriinae 87
Heteromera 58
himalayensis Stebb., Thanasimus 130
Hister 87, 88, 89, 90, 94
Histeridae 36, 49, 60, 61, 62, 69, 70,
86, 88, 89. 90, 91
Histerini 86
histeroides Lew., Syntelia 68
hispidulus Thoms., Crypturgus 166
Hololepta 36, 69, 87, 89, 90, 94, 95
Holometabola 57
Holoneurus 220, 223
holoptica Kriv. et Mam., Pergratospes
38, 43, 201, 209, 256, 257
hungarica Kert., Pseudowallacea 225,
271, 272, 289, 289, 290, 331, 350
Hydrophilidae 63, 86
Hydrophorinae 309
Hydrophorus 312
Hylastes 14, 92, 97, 164, 185
Hylemyia 332
Hylobius 14, 37, 40, 85, 164
Hylotrupes 22, 34, 128, 297
Hylurgops 14, 29, 92, 95, 110, 113,
164, 166, 186, 318, 320, 322, 323
Hymenoptera 54, 55
Hyperechia 294
Hyperoscelididae 45, 207, 209, 220,
222, 227, 229, 261, 307
Hyperoscelis 45, 207
Hypoganus 140, 143, 152, 154, 155
Hypophloeus 27, 29, 30, 31, 32, 33,
43, 76, 78, 188, 190, 191, 191,
193, 194, 194, 195
Hypulus 22, 43
igneus Meig., Choerades 38, 203^
299, 301, 302
illucens L., Hermetia 272
inflata Gyll., Acrulia 98
imitator Rtt., Hylurgops 318, 322
impigra Coli., Medetera 39, 310, 317,
319, 320
Inocellia 199, 200
Inocellidae 196, 200
ingrica Stack., Pseudoseioptera 356
inquisitor L., Rhagium 18, 34, 93,
302
inspissata Coli., Medetera 318, 319^
320
intermedia Kriv., Neopachygaster
205, 284, 285, 286
interrupta Pl., Xylomya 268
interstitialis Chap., Hylurgops 318,
322, 323
intricatus Ratz., Scolytus 96
ioptera Steph., Hapalaraea 106, 112
Ipidia 157, 158, 160
Ips 14, 30, 32, 33, 92, 93, 94, 95,
96, 97, 109, 112, 113, 114, 116,
125, 140, 164, 166, 175, 183, 186,
318, 319, 320, 322, 323, 325, 332,
344, 352, 353, 357
Istrisia 24, 45
jacobsoni Spess., Scolytus 125, 323,
346
japonica Miyat., Yamatomyia 291
japonica Rtt., Trogossita 14, 82,
140
jelskii Wank., Lado 177, 183
382
jezoensis Кбпо, Pytho 183
juglandis Helw., Synchita 177
junki Szil., Xylophagus 249, 250
kaltenbachi Winn., Forcipomyia 220
kariyana Alex., Libnotes 41, 43
karybenthinus Szil., Tabanus 203
kerteszi Duda, Mesaxymyia 202
Kolomania 269
koltzei Heyd., Sachalinobia 22
kolwensis C. Sahlb., Pytho 183, 185,
185, 186
Korynetes 123, 125, 126
Korynetinae 118
krivosheinae Kov., Lonchaea 331,
332
krivosheinae Neg., Medetera 310, 317,
319, 320
Kronomyia 217
Lado 177, 180, 182, 183
ladogensis Lacksch., Libnotes 43
Laemophloeus 42, 167, 168, 169, 170,
173, 174, 174, 175
laevis F., Ditylus 22
laeviuscula Gyll., Epuraea 162, 162,
163
Lagriidae 186
laminiformis Kriv., Maackiana 38,
282, 283
Lampra 16, 40
Lampyridae 61, 78, 83
Lamprolonchaea 329
Langelandia 176, 177, 181
lapathi L., Cryptorrhynchus 47
Laphria 24, 38, 39, 44, 211, 217, 257,
292, 293, 294, 295, 297, 302, 303,
304, 305, 306, 307
Laphriinae 38, 39, 44, 50, 221, 292,
293, 294
laricis F., Orthotomicus 92, 93, 95,
109, 113, 114, 164, 166
Lasiopa 269
Lasiotrichius 24
Lasius 26
laticeps Ramb., Medon 107, 113
laticornis Meig., Lonchaea 330
latifrons Loew, Rainieria 242, 346
leachii Curt., Praomyia 274
Lecontia 183
Lejota 220
lentus Grav., Nudobius 32, 56, 98,
108, 113
Lepidoptera 54, 54
lepidoptera Pz., Adelocera 144, 146
Lepidopteryx 59, 135, 135, 136, 136
Leptochirus 98
Leptoconopidae 209
Leptogaster 292, 293, 294, 307
Leptogastrinae 292, 293, 294
Leptura 22
Leptusa 106, 107, 112
Lestodiplosis 221, 223, 229
leucopeza Meig., Aulacigaster 208,
238
leucosticta Meig., Fannia 225
lewisi Jacobs., Thanasimus 130
Libnotes 19, 21, 41, 43, 47, 210, 261
Librodor 29, 157, 158, 160, 160
limbatus F., Thymalus 71
Limoniidae 38, 41, 43, 44, 47, 201,
201, 206, 207, 210, 211, 213, 216,
217, 218, 219, 221, 222, 224, 226,
233, 261, 263, 290, 307, 310, 331,
346
lineare Er., Cylister 29, 88, 92, 93
linearis F., Hypophloeus 32, 78, 188,
193, 194, 194
linearis L., Denticollis 152, 154,
154
lineatum 01., Trypodendron 93, 163,
166, 353
Liogma 218
Lissodema 128
livida Sahlb., Xylita 71
Loewinella 292
Lonchaea 14, 17, 32, 33, 37, 38, 39,
41, 51, 52, 290, 328, 329, 330, 332,
334, 335, 336, 341, 342, 343, 345,
347, 34S, 351, 352, 352
Lonchaeidae 37, 38, 39, 41, 45, 225,
226, 241, 272, 328, 334, 341, 342,
343, 345, 347, 348, 351, 352, 355
Lonchoptera 206, 218
Lonchopteridae 204, 206, 209, 218,
237
longicollis F.-W., Denoip3l20,121, 125
longicollis Gyll., Ortnotomicus 95,
96
longula Er., Epuraea 162, 163
longulus Gyll., Hypophloeus 188, 193,
194
Lophocateridae 56, 80, 82, 133
Lucanidae 45, 48, 67, 68, 294, 300
Lucanus 68
lucidiventris Beck., Lonchaea 330,
351
Ludius 140, 141, 143, 149, 155
lugens Loew, Pogonosoma 34, 298,
308
luteicornis Goetgh., Georthocladius
223
Lycidae 59, 61, 76
Lyctidae 67, 68, 87, 118
Lyctocoris 29
Lyctus 68, 125
Lymexylon 118, 129
Lymexylonidae 43, 47 , 72, 74 , 259
Maackiana 38, 269, 271, 275, 282, 283
383
Machaerium 309
Machimus 294
Macroceridae 222, 230
Macroceromys 262
maculata Meig., Xylomya 263
maculatus Mats., Xylophagus 250
maculicornis Hine, Neopachygaster
288
Magdalis 14
Maira 292
maior Burm., Raphidia 196, 197, 198,
199
Malthodes 56
mamaevi Kov., Lonchaca 38
mamaevi Kriv., Xylopachygaster 19,
43, 47, 257, 272, 284, 291
mandibularis F., Prostomis 166
marginata Meig., Solva 205j 218,
265, 266, 267
marginatus L., Choerades 295
mariana L., Buprestis 22, 85
maroccanum F., Pogonosoma 34, 298,
308
matsumurae Miyat., Xylophagus 39,
250, 251, 253, 259, 260
mauritanicus L., Tenebroides 133,
135, 139
mcalpineiana Kov., Lonchaea 330,
331, 335, 338, 345, 345, 346
Mecoptera 54
Medetera 14, 28, 32, 33, 37, 39, 41,
50, 203, 257, 259, 261, 309, 310,
311, 312, 313, 314, 314, 315, 316,
317, 318, 319, 320, 321, 322, 323,
324, 325, 326, 344, 346
Medon 107, 108, 113
Megamerina 349
Megamerinidae 41, 225, 226, 245, 256,
290, 331, 346, 349
Megapenthes 143
Megasemum 22
Megaselia 208, 239
Megatoma 74
megerlei Lac., Ampedus 146, 147,
149
Megopis 22
Melambia 133, 135, 136, 136, 137
Melandrya 22, 71, 128, 137
Melandryidae 43, 44, 63, 64, 70, 71,
72, 128, 261
Melanophila 16
melanoptera Mam. et Kriv., Prota-
xymyia 215, 223
Melanotus 140, 141, 143, 151, 152,
155, 156
Melasinae 58
Meligethes 157
Melyridae 59, 82
mendax AI., Metalimnobia 38
meromelaena Duf., Neopachygaster
259, 271, 272, 285, 286, 287, 290
merus Nagat. et Saig., Xylophagus
250
Mesaxymyia 47, 202
Mesoleptogaster 45, 50, 292, 293, 294,
296, 298, 307
Mesosa 16, 66, 137
Metalimnobia 38
Metriocnemus 223
micans Kug., Dendroctonus 113, 158,
166, 344
Microchrysa 269, 270, 271
Milichiidae 208, 24'i, 244
minor Hart., Blastophagus 93, 113,
114, 164, 166
minus Loew, Pogonosoma 298, 308,
309
minutissima Zett., Zabrachia 14, 31,
37, 51, 257, 259, 272, 278, 291, 292,
344
minutus Hbst., Acri tus 88, 92
mirabilis Motsch., Toxotus 22
Misolampidius 12, 24, 188, 189, 192
Misolampini 12, 44
mniszechii Grouv., Cucujus 171
modesta Will., Trichopteromyia 202
moiwana Mats., Xylomya 38, 265,
268
molitor L., Tenebrio 189
molitrix Loew, Ctenota 293
mollis L., Opilo 126, 128
monilicornis Gyll., Xylostiba 115,
116
monilis F., Laemophloeus 167, 173,
174
monilis Goetgh., Forcipomyia 220
Monochamus 16, 34, 113, 302
monographus F., Xyleborus 92
montanus Mam., Dicroneurus 220
mordax Rtt., Prostomis 24, 168
Mordellidae 43, 72, 74
Morimus 16, 352
muliebris Harr., Palloptera 353, 354,
multistriatus Marsch., Scolytus 15
Muscidae 204, 208, 212, 221, 222. 246,
248, 249
Muscinae 249
muticus F., Laemophloeus 167, 168,
174, 174
mutilatus Rosenh., Athous 153
mutillarius F., Clerus 16, 118, 121 ^
125
Mycetobia 202, 214, 215
Mycetobiidae 202, 203, 214, 215,
217, 222, 231
Mycetophagidae 77, 78
Mycetophagus 78
Mycetophilidae 214, 217, 220, 222,
225, 230
Myennis 245, 277
384
Myiairopa 221
myops Dalm., Mesosa 66^ 137
Mynnica 26
namanganensis Sem., Xylotrechiis
133, 137
navale L., Lymexylon 118, 129
Neatiis 188
Necrobia 49, 117
Nematocera 200
Nematoceropsis 262
Nematoplidae 45, 83, 84, 261
Nematoplus 24, 45, 83, 84, 261
Nemosoma 133, 135, 136, 136, 137,
138
Nemotelinae 269
Nemotelus 269, 270, 273
Neopachygaster 37, 205, 259, 270,
271, 272, 277, 283, 284, 285, 286,
287, 287, 288, 290, 331
Neurogona 309
Neuroptera 53
niger Kby., Pytho 183, 185
nigerrimus Lac., Ampedus 146, 151
nigra Lund., Plccia 44, 201, 215
nigricollis Alb., Raphidia 196, 197,
198
nigricornis Meig., Tanyptera 256, 257,
300
nigrinus Hbst., Ampedus 147, 148,
151
nigroflavus Goeze., Ampedus 151,
152, 153
nigrotibialis Pl., Wallacea 272, 289,
290, 346
Nitidulidae 41, 42, 51, 59, 79, 80,
157, 159
nitidulus F., Rhizophagus 165, 165,
166
nivalis Lew., Pytho 183, 186
Nosodendridae 83, 84
Nosodendron 84
notata F., Raphidia 197, 198, 199
nudatus Nagat. et Saig., Xylophagus
250
Nudobius 32, 56, 98, 108, 109, 110,
113, 114
Nusa 292
Nymphomyiidae 200
Nyphades 14
obliqua Er., Bolitochara 102
oblonga L., Grynocharis 56, 82
oblongum F., Cylister 27, 32, 33, 88,
92, 93, 94
ocellaris L., Epiphragma 201
ochrinulus Rtt., Ampedus 152
ochropterus Germ., Ampedus 147,
150
Odinia 43, 239
Odiniidae 43, 225, 239, 240
Odontomyia 269
Oedemeridae 44, 63, 69, 70, 261, 283,
328
Olisthaerus 98, 102, 104
Omalium 97
omogensis Miyat., Xylophagus 250
Omosita 157
opacus Er., Hylastes 92, 97
opacus Lew., Cucujus 171, 172
ophiopsis L., Raphidia 198, 199
Opilo 80, 123, 126, 126, 128
Orchesia 22
orientalis Egg., Cryphalus 323
orientalis Nik., Enchodes 22
orientana Kov., Palloptera 355, 356
Ornopyramis 269, 271, 277, 280, 288
Orthopleura 58, 118, 120, 121, 123,
126, 128, 129
Orthotomicus 14, 92, 93, 95, 96,
109, 113, 114, 163, 164, 166, 318,
322, 323, 325
Osmylus 53
Otho 85
Otitidao 36, 41, 226, 245, 247, 263, 356
oxiana Som., Eurythyrca 23, 308
Oxycera 269, 270
Oxytclinae 99
Oxytelus 97
Pachygaster 269, 270, 274
Pachygasterinae 41, 259, 269, 271,
272, 288
Pachyneura 50, 214, 214, 225, 261
Pachyneuridae 36, 44, 48, 201, 214,
214, 222, 224, 225, 228, 250, 261
Paederinae 97, 99
palliatus Gyll., Hylurgops 29, 92,
95, 113, 164, 166, 318, 320, 322
pallida Pasc., Acicnemis 14
pallipes Ros., Systenus 326, 327
pallipes Zett., IMedetera 310
Pallodcs 157
Palloptera 353, 354, 355, 356, 357
Pallopteridae 41, 226, 244, 353, 355
palmivora James, Hermetia 272
Palorus 189, 192
palpata Czerny, Lonchaea 330, 331,
335, 337, 346, 347, 348
palposa Zett., Lonchaea 33, 330,
332, 335, 340, 346, 348, 349
paludicola Kieff., Dasyhelea 328
Panesthiinae 12
Paraclusia 26, 207, 257
paradoxa Kriv., Forcipomyia 220
parallela Loew, Palloptera 353
parallelepipedus Hbst., Paromalus
29, 30, 91, 95
parallelopipedus L., Dorcus 11, 24,
44, 128, 300
385
Pareidalimus 271
Parepidosis 204, 219
Paromalus 14, 29, 30, 87, 89, 91,
95
parreysii Stev., Alaus 145, 146
parva Portsch., Tanyptera 256, 257,
300
Paururus 24
pectinicomis F., Ptilinus 132
pectinicornis L., Schizotus 19
Pediacus 167, 168, 170, 175, 176,
176
Pcdilidae 76, 185, 186, 187
Pedilus 185, 186, 187
Pcltidae 64, 70, 71, 71, 133
penicillata Neg., Medetera 312, 315,
320, 321, 322
peregrina Beck., Lonchaea 330, 331,
332, 335, 337, 346, 347, 348, 349,
352
Pergratospcs 38, 43, 201, 209, 256,
257
Pericoma 218
Peromyia 41, 220
perroudi Muls., Uloma 188
persuasoria L., Rhyssa 54
perpusilla Kieff., Peromyia 220
pfeili Ratz., Xyleborus 187
phalaenoides L., Psychoda 205
phalerata Meig., Drosophila 243
Phaonia 208, 221, 246
Phaoniinae 248
Phellopsis 73
Phloeonomus 98, 102, 103, 116
Phloeopora 107
Phloetrya 22
Pholioxenus 86
Phoridae 208, 239, 240
Phoroctenia 25, 26, 218
Phronia 225
Phyllomyza 208, 243
Physiphora 245
piceae 111., Pissodes 14
picipes Hbst., Neatus 188
picipes Mnnh., Quedius 115
pilicornis Pk., Atrecus 105, 110, 111
pini L., Pissodes 15, 93, 344
pini Pz., Hypophloeus 33
pinicola Kow., Medetera 32, 312,
316, 321, 322
piniperda L., Blastophagus 93, 95,
96, 97, 113, 114, 163, 164, 166, 320,
322, 323, 344
piniphilus Hbst., Pissodes 93, 113
Pissodes 14, 15, 37, 40, 93, 113, 302,
311, 332, 344
Pityogenes 93, 95, 112, 113, 175,
185, 186, 320, 357
Pityokteines 323
Pityophagus 157, 158, 160, 161, 164
Placusa 27, 29, 32, 33, 69, 98, 106,
108, 114
plagiatus Mnnh., Quedius 114, 115,
115, 116
Plagionotus 16, 40, 56
plana Sulz., Hololepta 69, 90, 94, 95
planatus L., Brontes 7P, 167
planulata Er., Epuraea 162, 163
Platypezidae 204, 206, 218, 240
Platypodidae 43, 47, 86, 291
platypterus F., Sciopus 310
Pia typus 19
Platysoma 29, 87, 89, 91, 95, 96
Platysomini 87
Platystethus 97
Plecia 44, 'JOl, 215
Pleciidae 44, 48, 201, 201, 205, 215,
217, 222, 227
Plegaderus 14, 36, 69, 87, 89, 91,
92, 96, 97
plicata Kraft et Cook, Zabrachia 291
Pogonocerus 56
Pogonosoma 16, 34, 38, 41, 292, 293,
294, 295, 296, 298, 308, 309
polita L., Microchrysa 270
polita Coq., Zabrachia 291
Polygraphiis L., 14, 32, 95, 110, 113,
164, 166, 175, 183, 185
polygraphiis L., Polygraphus 113,
164, 175, 183
Polyphaga 58
polypori Loew, Lestodiplosis 223
pomonae F., Bibio 225
pomonae Steph., Ampedus 147, 148
pomorum Hbst., Ampedus 147, 151,
152
Porphyrops 309, 311
praeusthus F., Ampedus 148
Praomyia 270, 274
Prionocyphon 73
Prionus 39
Prismognathus 11, 24, 25, 44
Prista 262
Procraerus 143
prolongata Geoffr., Xiphydria 43
Promachus 294
Prostomidae 167
Prostomis 24, 166, 168, 169, 170
Protaxymyia 47, 215, 223
proximus Blandf., Polygraphus 95
proximus Eichh., Orthotomicus 93,
113, 318, 322, 325
Pseudopyrochroa 19
Pseudoseioptera 356
Pseudowallacea 225, 269, 271, 272,
275, 288, 289, 289, 290, 331, 350
Psilidae 208, 241, 242
Psilonyx 292
Psychoda 205, 217, 219, 220
Psychodidae 204, 205, 213, 216
386
217, 218, 219, 220, 224, 232, 310,
328
pteroplouralis Hend., Acanthoneura
47, 245
Ptilinus 132
Ptychopteridae 216
punctata Hbst., Adelocera 144, 145,
146
puncta tum Deg., Anobium 68, 132
puncticollis C. Sahlb., Rhizophagus
33
punctifrons Thoms., Polygraphus 110
pusiHa 111., Epuraea 161, 162, 163
pusillus Grav., Phloeonomus 102
pusillus Gyll., Crypturgus 92, 95, 97,
112
pygmaea Pk., Hapalaraea 112
Pyrochroa 19, 75
Pyrochroidae 36, 40, 56, 64, 75, 76
Pyrrhidium 16, 23, 34, 125, 139, 309
Pythidae 36, 40, 51, 59, 74, 75, 183,
185
Pytho 20, 31, 33, 34, 36, 50, 59,
75, 183, 184, 185, 185, 186
quadricostulatum Kr., Megasemum 22
quadrifasciata L., Strangalia 22
quadriguttatus F., Librodor 160
quadrimaculata Rtt., Epuraea 162,
163
quadripunctatus L., Glischrochilus
157, 159
Quedius 98, 109, 114, 115, 115, 116
quercea Hbst., Adelocera 144, 145,
146
quercinus Guens., Hypulus 43
ragnari Hack., Lonchaea 334, 335,
337, 341, 349
Rainieria 242, 346
Raphanocera 269
Raphidia 54, 195, 196, 197, 198, 199
Raphidiidae 196
Raphidioptera 53, 54, 195, 196
ratzeburgi Brauer, Raphidia 197, 198,
199
ravida Neg., Medetera 312, 316, 321,
322, 344
relictus B.-Bien., Cryptocercus 12, 24,
45
relictus Sem., Callipogon 22
repandus Horn., Thanasimus 130
Rnagio 50, 214
Rhagionidae 50, 202, 207, 211, 213,
214, 215, 216, 217, 218, 219, 222,
235 249
Rhagium 16, 18, 34, 40, 93, 113, 302
Rhizophagidae 157
Rhizophagus 14, 32, 33, 41, 42, 157,
158, i5P, 159, 164, 165, 166
Rhysodes 66
Rhysodidae 58, 66, 66
Rhyssa 54
rohdendorfi Kriv. et Mam., Hespe-
rinus 26, 262
rubens Pili., Denticollis 152, 154,
154
rubricola Mam., Winnertzia 220
rubrofemoratus Woll., Eurytrachelus
11, 44
rubronotata Rtt., Epuraea 162, 163
rufa Rod., Laphria 293, 303, 305,
306
ruficollis L., Cardiophorus 153
ruficorne 01., Aulonium 181, 182
ruficornis V. d. Wulp, Ctenota 293
ruficornis Zett., Lejota 220
rufipes Brahm., Thanasimus 33, 130,
131
rufipes Deg., Necrobia 117
rufipes Hbst., Melanotus 151, 155,
156
rufiventris Esch., Athous 152, 153
rufobrunnea Lew., Istrisia 24, 45
rufus Deg., Stenagostus 156
rusticus L., Criocephalus 22, 34, 302,
306
sachalinensis Pl., Xylophagus 37,
250, 251, 256, 260, 261
Sachalinobia 22
Salpingidae 45, 76, 128, 183
saltuum L., Palloptera 353, 357
sanguineum L., Pyrrhidium 23, 34,
125, 139, 309
sanguineus L., Ampedus 147, 150,
151
sanguinicollis L., Orthopleura 58, 118,
120, 121, 126, 129
sanguinolentus Schrk., Ampedus 147,
148, 151
Saperda 40
Saprinus 89
Sarcophaga 218
Sarcophagidae 210, 218, 224, 247
Sarcinae 269, 271
Sargus 269, 271
Sarpinini 86
satrapa Kiesw., Ampedus 147, 150,
151
saucius Er., Plegaderus 36, 91, 96
saxeseni Ratz., Xyleborus 138
scabra Thunb., Calitys 71
scabricornis Scop., Megopis 22
Scaphidiidae 70
Scarabaeidae 48, 51, 62, 64, 67, 68
Scatopse 41, 204, 214, 219, 220
Scatopsidae 41, 44, 203, 204, 213, 214,
216, 217, 219, 220, 222, 224, 229,
272
387
Scenopinidae 50, 203
Schizotus 19
schncideri Pz., Boros 19, 20, 39, 75
schnitnikovi Pl., Solva 265^ 266, 267
scholtzii Loew, Systenus 326, 327,
328
Sciaridae 42, 44, 45, 141, 201, 214,
220, 222, 225, 229, 250, 258, 259,
262, 307, 311
Sciophilidae 230
Sciopus 309, 310, 312
scitus Grav., Quedius 114, 115, 116
scolopaceus L., Rhagio 214
Scoloposcelis 30
Scolytidae 36, 43, 47, 86, 259, 331,
346, 352, 357
Scolytus 14, 15, 33, 125, 318, 320,
323, 346, 352
scolytus L., Scolytus 33, 318, 320,
352
scopini Furs., Enneadesmus 125
scripticollis Hochh., Zeteotomus 115,
110
scutellaris Rond., Lonchaea 328, 330,
331, 332, 335, 339, 346, 348, 349,
350
Scythropochroa 26
secernibilis Kriv., Neopachygaster
286, 287, 331
seitneri Hend., Lonchaca 37, 329,
331, 332
semenovi Nik., Nematoplus 24, 45,
84, 261
semota Kriv., Solva 265, 265, 267
separanda Rtt., Synchita 73
serraticornis L., Calopus 22, 69
serricornis Miill., Prionocyphon 73
Serropalpus 22, 44
Setenis 12
Setisquamalonchaea 329
severint Tonn., Psychoda 220
sexdentatus Boern., Ips 32, 93, 94,
95. 97, 113, 114, 125, 140, 166,
318, 322, 32.3, 344
Siagonium 98, 102, 103
signaticornis Loew, Medetera 32, 33,
37, 312, 315, 321, 322, 323
signatum F., Trypodendron 166, 318
signifer Kriv. et Mam., Xylophagus
37, 250, 251, 253, 255, 262
Silba 329
Silphidae 69
Silvanidae 84, 166
Silvanus 30, 41, 167
Silvestrodasiops 329, 335
Sinodendron 11, 24, 44, 301
Sipalinus 11, 19, 21, 43, 256, 291
Sirex 24
Siricidae 44, 48
Siricinae 48
Solva 205, 218, 262, 263, 264, 265,
265, 266, 267
Soronia 157
spectabilis Kr., Tiichodas 120, 121,
132, 133
Spermatolonchaea 335
spinosulus Latr., Endophloeus 176
splendidulus Grav., Gabrius 107, 111
Spondylis 22, 93
spondyloides Germ., Otho 85
squamulosa Gebl., Lepidopteryx
59, 135, 136, 136
stackelbergi Czerny, Lonchaea 330,
335, 338, 350, 351
stackelbergi Kriv., Neopachygaster
285, 287, 287
stackelbergiana Neg., Medetera 37,
316, 323, 324, 346
Staphylinidae 49, 56, 59, 61, 65, 69,
97, 100, 101, 102, 105, 106, 107,
108, 115
Staphylininae 97, 99
Stegana 243
Stelidota 157
Stenagostus 16, 34, 39, 49, 140, 141,
143, 151, 154, 156
Stenophanes 12, 192
Stenopogon 294
stenostyla Sav., Tipula 44
Stenotrachelidac 59, 72, 77
stepanovi Denis., Hypoganus 155
Stephanopachys 68
Strangalia 22, 129
Stratiomyidae 37, 38, 41, 43, 47, 51,
205, 217, 224, 225, 236, 256, 257,
259, 262, 263, 269, 278, 280, 282,
284, 287, 289, 310, 331, 346, 350
Stratiomyinae 269, 270
Stratiomys 269
striata Par., Medetera 37, 312, 316,
323, 324
strobi Peck., Pissodes 332
Strongylophthalmyia 208, 246
Strongylophthalmyiidae 41, 208, 244,
246
subaeneus Motsch., Prismognathus 11,
24, 25, 44
subdepressus WolL, Palorus 189
subelongatus Motsch., Ips 30, 33,
93, 94, 96, 97, 323
subneatosa Kov., Lonchaea 331, 335,
338, 350, 351
subnitens Verr., Scatopse 204
subopacus Thoms., Polygraphus 32,
166, 183
substriatus Gyll., Olisthaerus 102
substriatus Gebl., Thanasimus 32,
122, 130, 131
substriatus Pk., Stephanopachys 68
Subulonia 262
388
sulcatum 01., Aulonium 177
sulcicollis Germ., Dechomus 177, 180
sulcifrons Rey, Scolytus 323, 346
suturalis Gyll., Orthotomicus 95, 96,
109, 113, 114, 163, 322, 323
sylvatica Bel., Lonchaea 39, 52, 330,
331, 332, 334, 335, 339, 350
Sylvicola 202
Symmerus 202 у 215
Synchita 73, 177, 179
Synneuron 261
Syntelia 67, 68
Synteliidae 49, 64, 68
Syrphidae 38, 41, 43, 47, 204, 207,
217, 219, 220, 221, 224, 225, 225,
226, 238, 257, 261
Systenus 50, 309, 310, 311, 312, 325,
326, 326, 327, 328
Tabanidae 202, 203, 213, 215, 216,
221, 222, 224, 236
Tabanus 203
Tachinidae 246, 249
Tachinus 101, 102, 104
Tachyporinae 99
tachyporoides Waltl., Placusa 33, 98
Takagimyia 283
Tanyptera 26, 220, 226, 256, 257, 300
Taphrorychus 14, 116, 138, 318, 320,
352
tarsalis Zett., Eupachygastcr 281,
282, 282, 283, 328
tarsata Fll., Lonchaea 330
Tarsostenus 120, 124, 129
Tauroccra 269
tekkensis Коп., Melambia 135, 136,
137
Temnostoma 47, 207, 225, 226, 257,
261
Tenebrio 188, 189, 190
Tenebrionidae 43, 44, 5J, 63, 64,
72, 76, 77, 78, 188, 189, 190, 191,
194, 261
tenebrosus Er., Melanotus 155, 156
Tenebroides 133, 134, 135, 139
tener Kriv., Ornopyramis 280, 288
tener Loew, Systenus ^26, 327, 328
Tenerus 38, 72, 118, 120, 122, 123,
129
tentyrioidcs Sols., Misolampidius 24,
188, 189
Tephritidae 36, 38, 41, 47, 226, 245,
246
Teretrinae 87
Teretrius 89
testaceus F., Laemophloeus 167,
174
Tetropium 16, 40, 113, 140
Texara 225, 331, 346
Thanasimus 14, 27, 30, 31, 32, 33,
36, 41, 58, 82, 118, 122, 123, 130,
131
Thereva 202, 218, 219
Therovidae 50, 202, 203, 212, 216,
217, 218, 219, 225, 234, 235
Thinophilus 309
thoracica Cr., Strangalia 22
thoracica Tourn., Epuraea 162, 164
thoracicus F., Bius 78, 188
thoracicus Fisch., Pogonocerus 56
Thrypticus 309
thunebergi Neg., Medetera 317, 324,
325
Thymalus 70, 71
tibialis Loew, Dilophus 344
tigrina Fll., Paraclusia 207, 257
Tillinae 118
Tilloidea 118, 124, 132
Tillus 59, 80, 118, 120, 122, 124, 126,
131, 132
Tipula 44, 218, 220, 226, 262
Tipulidae 44, 206, 213, 216, 217,
218, 218, 222, 224, 225, 226,
233, 250, 256, 257, 261, 262, 300,
306
Tomoxia 74
Toxotus 22
Trachodes 14, 40, 138
Tragosoma 22
Trechus 66
Tremex 24, 137
Trichius 24, 44
Trichocera 220
Trichoceridae 41, 213, 216, 217, 220,
999 99Л 941
Trichodes 118, 120, 121, 122, 123,
132, 133
Tricholaba 220
Trichomyia 44, 202, 217
Trichomyiidae 44, 202, 217; 232
Trichopteromyia 202
Trichosia 26, 225, 225
tricolor Hcnd., Myennis 245, 277
trifolii Riibs., Tricholaba 220
Trigonurus 70, 98, 101, 102
trimacula Meig., Palloptera 353
tristis AI., Gnophomyia 211, 218,
346
trisulcum Geoffr., Aulonium 14, 41.
180, 181, 182
Trixagidae 59, 76, 81
Trogossita 14, 82, 133, 135, 136, 139.
140
Trogossitidae 49, 59, 60, 61, 62, 63,
71, 72, 82, 83, 133, 134, 135,136
trossulus Nordm., Gabrius 107, 111
Trypodendron 19, 93, 98, 138, 163,
166, 318, 353
Trypophloeus 320, 344
Turanium 133
389
turkestanicus Ball., Tenebroides 139
typographus L., Ips 33, 92, 93, 94,
95, 96, 97, 109, 112, 113, 114,
116, 164, 175, 183, 186, 318, 323,
325, 332, 344, 353, 357
ulanowskyi Gglb., Aesalus 24
Ulidiidae 245, 247
ulmicorticis Mam., Parepidosis 204
Uloma 76, 78, 188, 189, 192
ulrikae Asp., Raphidia 198, 199
umbellatarum F., Palloptera 353
umbratus Motsch., Nudobius 113, 114
umbrosa Kriv., Solva 266
undata L., Megatoma 74
undulata Mats., Libnotes 43, 210
undulatus Deg., Stenagostus 34, 39,
151, 154, 156
unicolor 01., Epuraea 79, 162, 163,
164
unicolor Pili., Hypophloeus 193, 194,
195
unidentatus F., Silvanus 167
unifasciata F., Tilloidea 132
univittatus Rossi, Tarsostenus 120,
129
Upis 188, 189, 190, 261
Urocerus 24, 54
urussovi Fisch., Monochamus 34, 302
ussuricum Kriv., Craspedometopon
280, 281
ussuriense Kov., Lonchaea 38
ussuriensis Mam., Wyattella 356
ussuriensis Pl., Xylomya 268
usta Meig., Palloptera 353, 354, 355,
356, 357
ustulata Fll., Palloptera 353, 354,
355, 356, 357
ustulata Zett., Strongylophthalmyia
208, 246
varia Loew, Laphria 38, 304, 305,
306
varia Meig., Xylomya 263
variabilis Motsch., Cephalon 81
varians Germ., Denticollis 154
variegatus Roel., Nyphades 14
variicornis Dalm., Leptura 22
verecundus Fald., Morimus 352
vespiforme L., Temnostoma 207, 225
villifrons Duf., Taphrorychus 320
villosus Geoffr., Stenagostus 151,
154, 156
vittata Meig., Phoroctenia 25
vittatum Fauv., Siagonium 102
vulneratus Pz., Plegaderus 36, 69,
91, 96, 97
vulpina Meig., Laphria 303, 305,
Wallacea 269, 271, 272, 275, 289,
290, 346
Winnertzia 41, 42, 220, 346, 350
Wyattella 41, 356
xanthopus Er., Quedius 114, 115,
116
xanthostigma Schum., Raphidia 197,
198, 199
Xiphydria 24, 43
Xyleborus 19, 92, 138, 187
Xylita 44, 70, 71
Xylocopa 294
Xylogenes 125
Xylomya 17, 38, 262, 263, 264, 265,
267, 268
Xylomyidae 36, 38, 51, 205, 218,
224, 236, 256, 262, 264, 265, 272,
288, 331, 346
Xylopachygaster 11, 19, 43, 47, 257,
269, 272, 276, 284, 291
Xylophagidae 37, 39, 41, 45, 49, 202,
203, 211, 213, 216, 217, 234, 249,
255, 258, 260
Xylophagus 16, 17, 27, 37, 39, 45,
49, 50, 203, 211, 249, 250, 251,
252, 253, 255, 256, 257, 258, 259,
260, 261, 262
Xylostiba 103, 115, 116
Xylotrechus 16, 34, 133, 137, 139,
308
Yamatomyia 291
yasumatsui Nag. et Tan., Xylomya
265, 268
Zabrachia 14, 31, 37, 51, 257, 259,
270, 271, 272, 276, 278, 291, 292, 344
Zeteotomus 98, 109, 115, 116
zetterstedti Beck., Lonchaea 37, 52,
328, 329, 331, 332, 335, 340, 352, 353
Zilora 70
Zopheridae 59, 64, 72, 73, 77
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ
Г л а в а 1
ОСНОВЫ ФОРМИРОВАНИЯ СООБЩЕСТВ КСИЛОФИЛЬ-
НЫХ НАСЕКОМЫХ
Общая характеристика этапов сукцессии ксилофильных на¬
секомых 8
Насекомые как индикаторы стадий разрушения коры и дре¬
весины 12
Хищные насекомые как компоненты ксилофильных сообществ 26
Глава 2
ЗАКОНОМЕРНОСТИ ЭКОЛОГИЧЕСКОЙ СПЕЦИАЛИЗА¬
ЦИИ КСИЛОФИЛЬНЫХ НАСЕКОМЫХ
Основные направления специализации ксилофильных насеко¬
мых 35
Характеристика пищевых режимов ксилофильных насекомых 45
Глава 3
СИСТЕМАТИЧЕСКИЙ ОБЗОР ЛИЧИНОК
Группы ксилобионтных насекомых 53
Введение в морфологию личинок насекомых 55
Отряд Coleoptera — Жесткокрылые, или жуки 57
Сравнительная морфология личинок жесткокрылых 58
Таблица для определения семейств отряда Coleoptera — Жест¬
кокрылые 65
Семейство Histeridae — Карапузики 86
Семейство Staphylinidae — Стафилиниды 97
Семейство Cleridae— Пестряки 117
Семейство Trogossitidae — Темнотелки 133
Семейство Elateridae — Щелкуны 140
Семейство Nitidulidae — Блестянки 157
Семейство Cucujidae — Плоскотелки 166
Семейство Colydiidae — Узкотелки 176
Семейство Pythidae — Трухляки 183
Семейство Pedilidae — Лжебыстрянки 186
Семейство Tenebrionidae — Чернотелки 188
Отряд Raphidioptera — Верблюдки 195
391
Таблица для определения семейств отряда Raphidioptera—Вер-
б ЛЮДКИ 196
Семейство Raphidiidae — Верблюдки настоящие 196
Семейство Inocellidae — Малоглазки 200
Отряд Diptera — Двукрылые, или комары и мухи 200
Сравнительная морфология личинок двукрылых 201
Таблица для определения семейств отряда Diptera — Двукры¬
лые 226
Семейство Xylophagidae — Стволоедки 249
Семейство Xylomyidae — Осовидки 262
Семейство Stratiomyidae — Львинки 269
Семейство Asilidae — Ктыри 292
Семейство Dolichopodidae — Зеленушки 309
Семейство Lonchaeidae — Копьехвостки 328
Семейство Palloptcridae — Бледнокрылки 353
ЛИТЕРАТУРА 358
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
НАСЕКОМЫХ 377
Борис Михайлович Мамаев,
Нина Павловна Кривошеина,
Варвара Александровна Потоцкая
ОПРЕДЕЛИТЕЛЬ ЛИЧИНОК ХИЩНЫХ НАСЕКОМЫХ-
ЭНТОМОФАГОВ СТВОЛОВЫХ ВРЕДИТЕЛЕЙ
Утверждено к печати Институтом эволюционной морфологии
и экологии животных им. А, Н. Северцова Академии наук СССР
Редактор издательства Э. А, Фролова. ^Судожник А. Г. Кобрин
Художественный редактор Н. Н. Власик. Технический редактор Р. Г. Грузинова
Корректоры М. С. Бочарова, А. А. Смогилева
Сдано в набор 20/ХII 1976 г. Подписано к печати 6/IV 1977 г.
Формат бОхРоЧхе. Бумага типогр.Л 1. Уел. печ. л. 24,5. Уч.-изд. л. 26,9.
Тираж 1360. Т-08506. Тип. аак. 1674. Цена 3 р. С5 к.
Издательство «Наука*. 103717 ГСП, Москва^ К-62, Подсосенский пер., 21
а-я типография издательства «Наука*. 121099, Москва, Г-99, Шубинскцй пер., 10