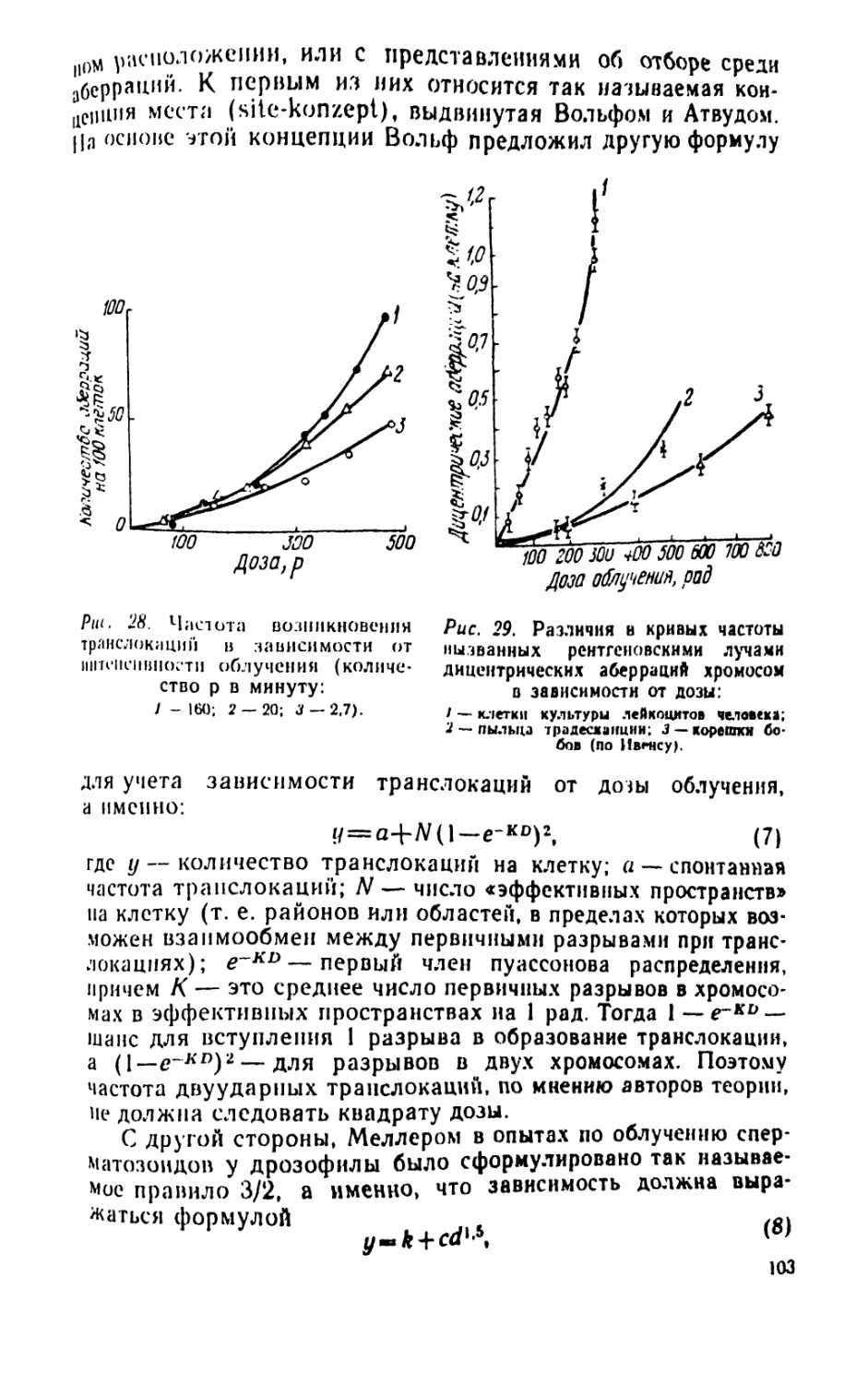
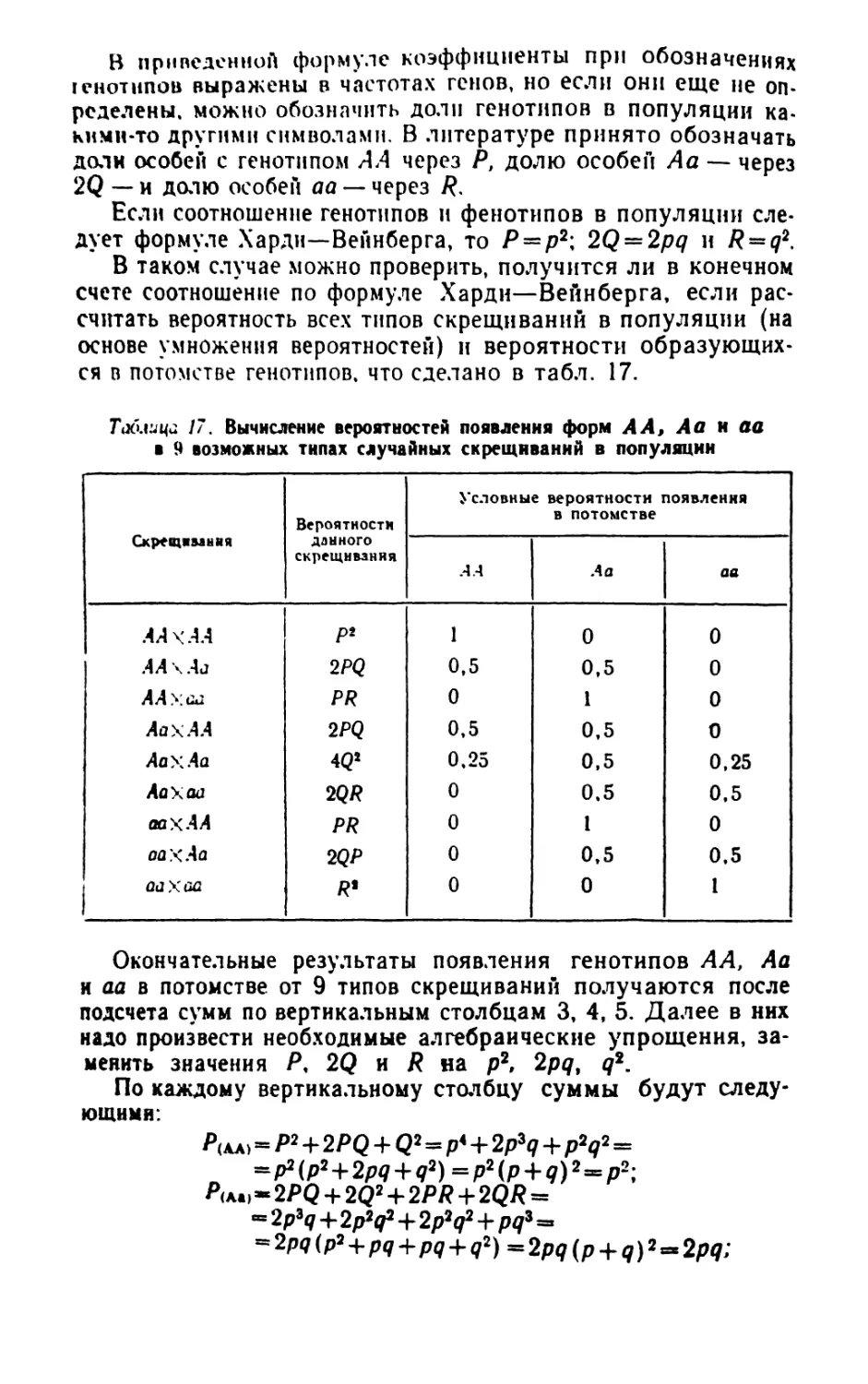
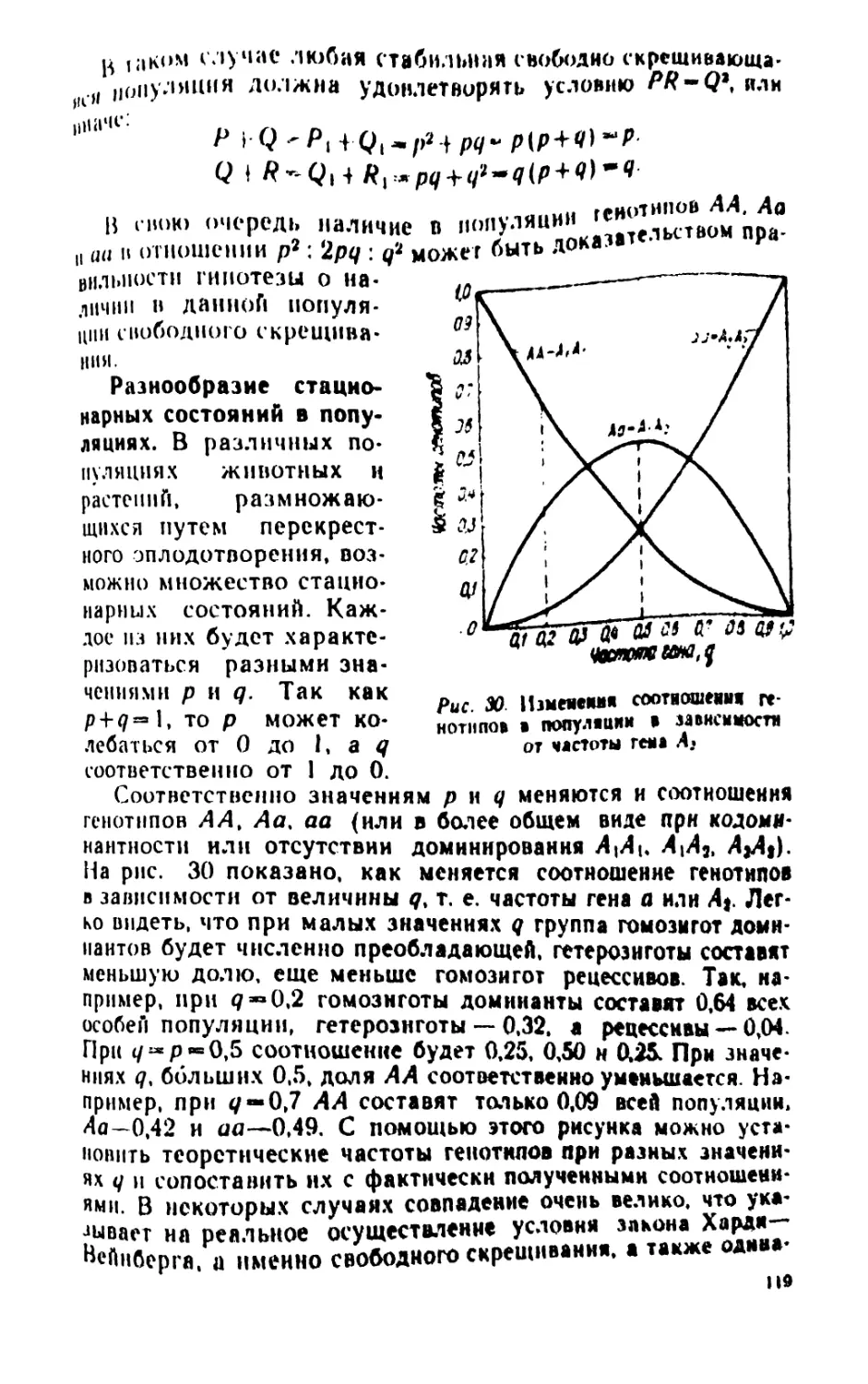
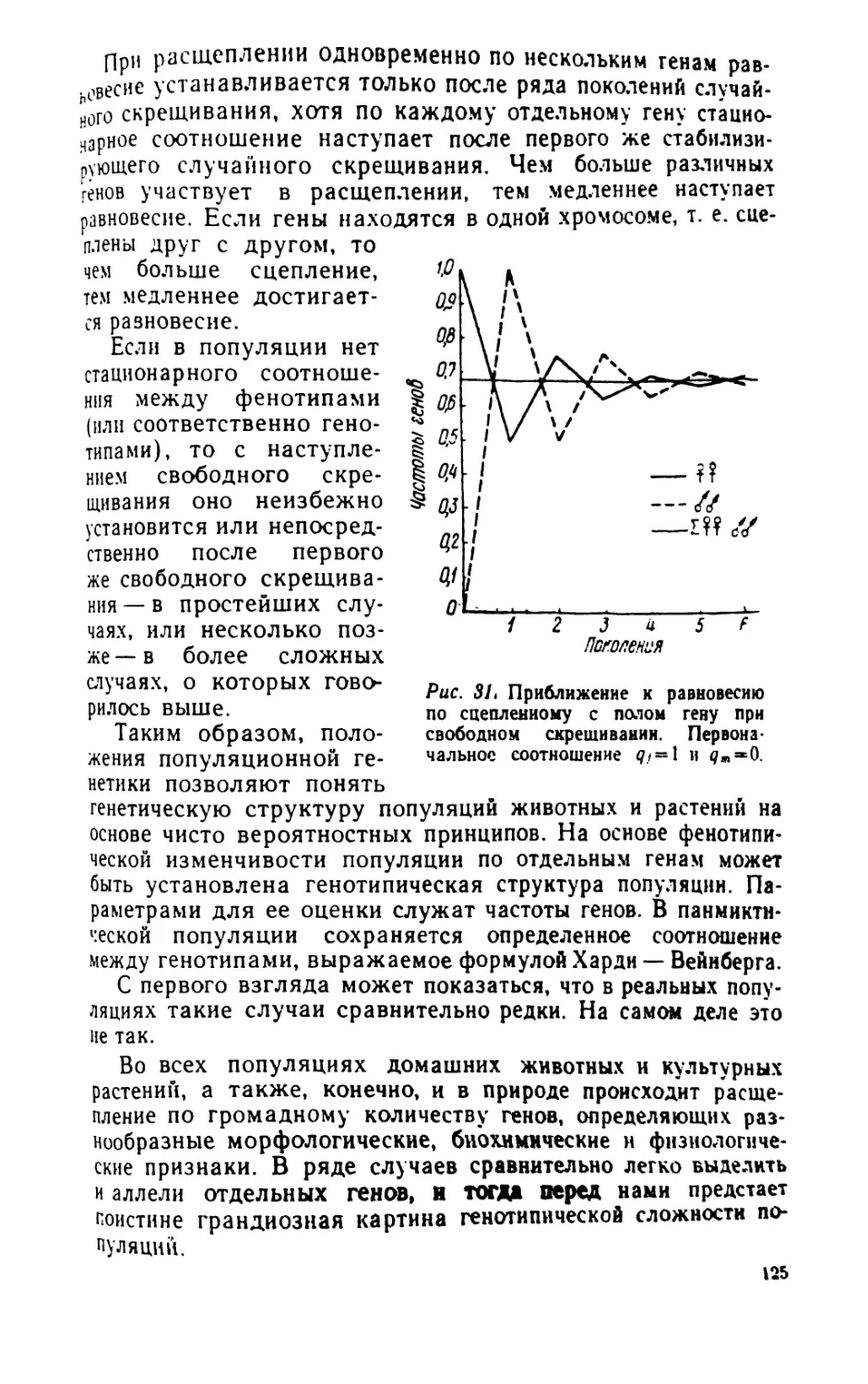
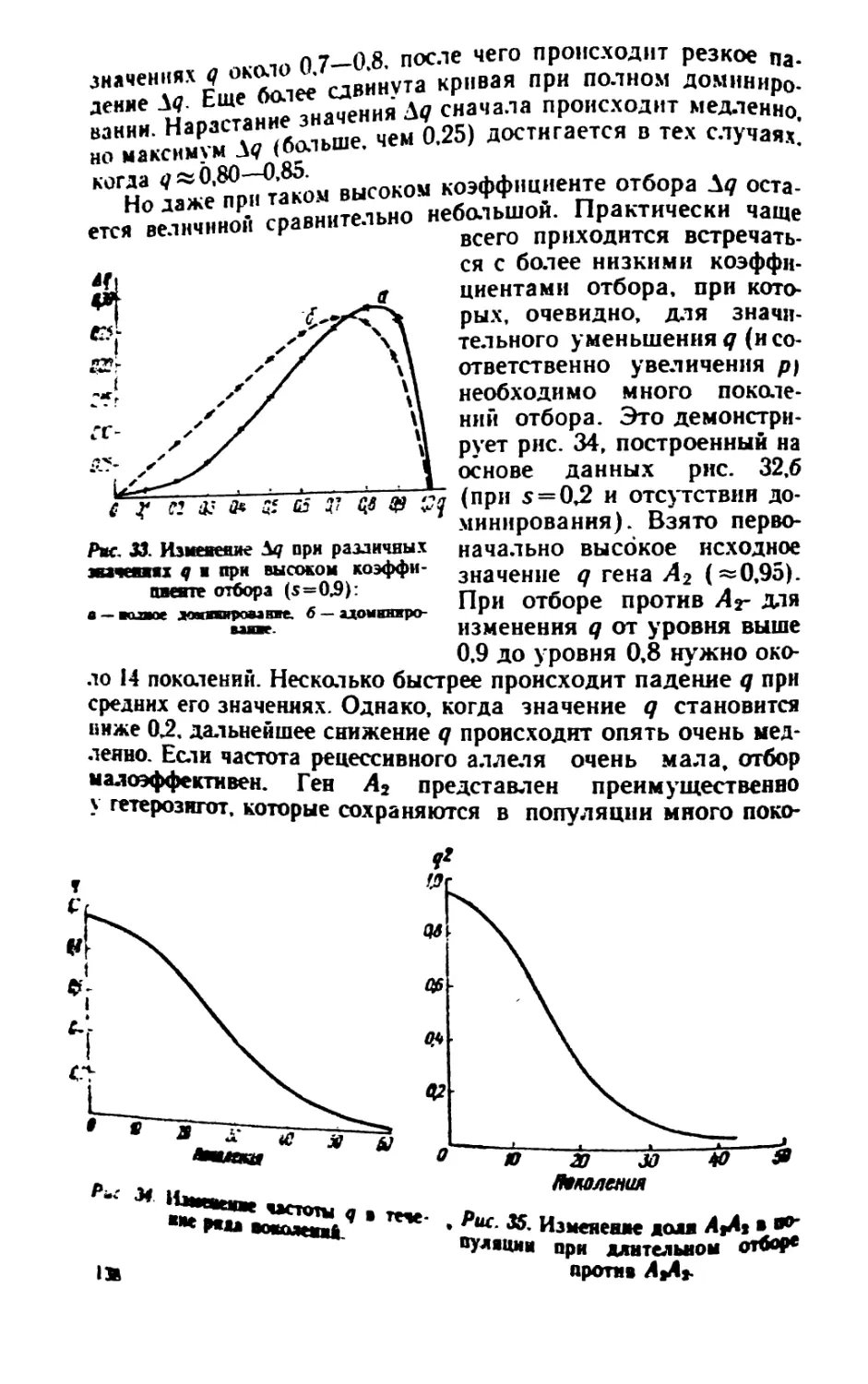
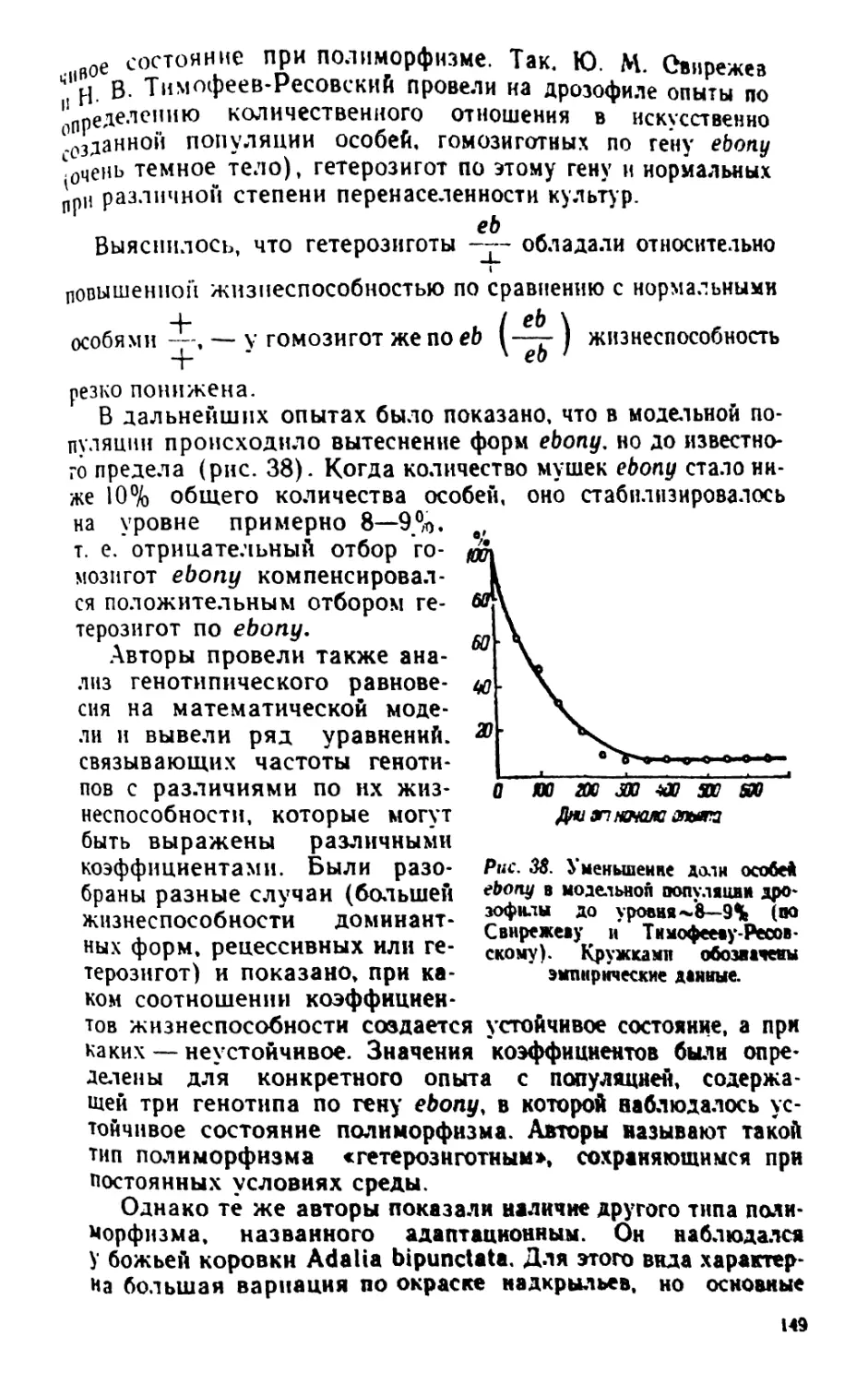
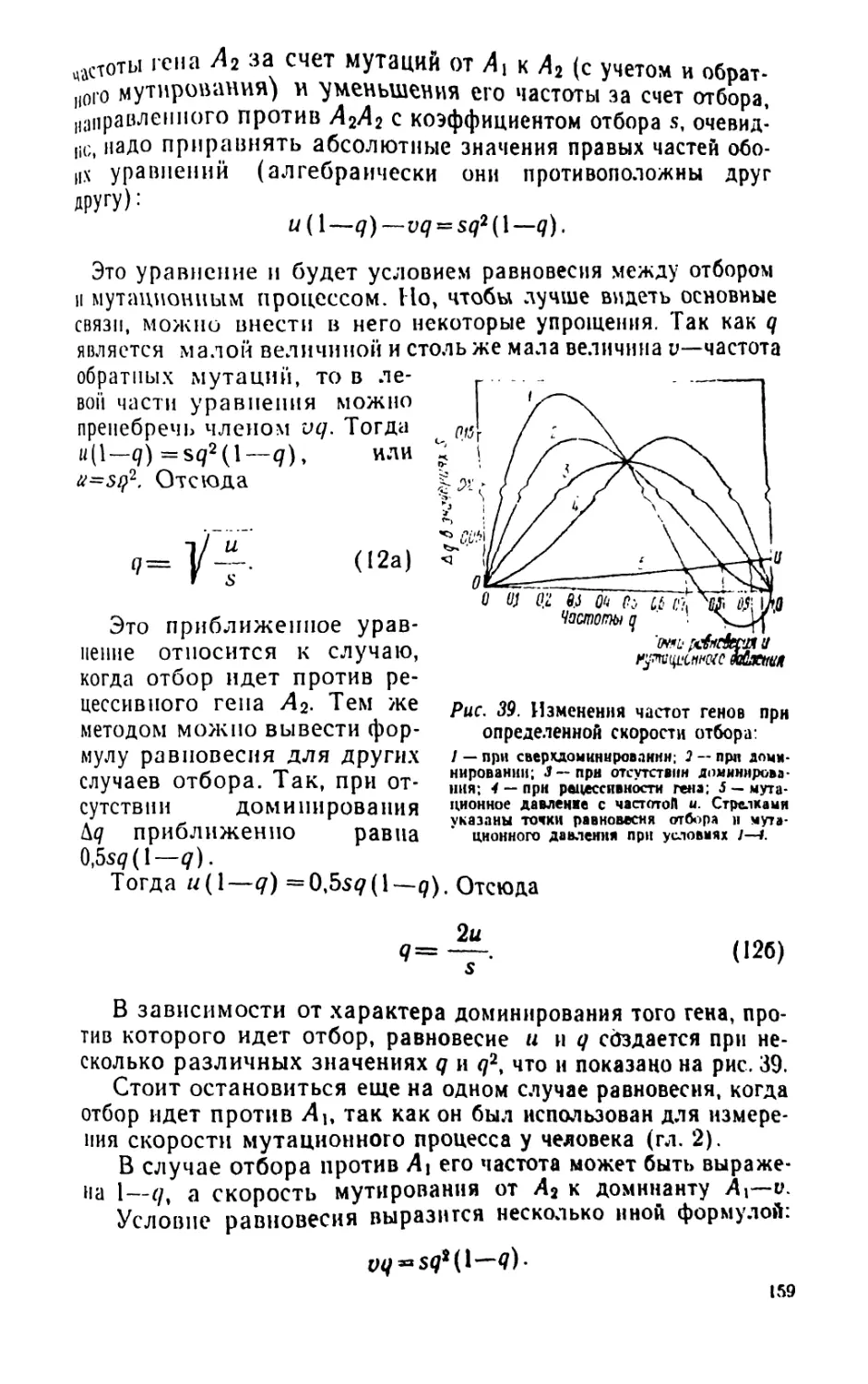
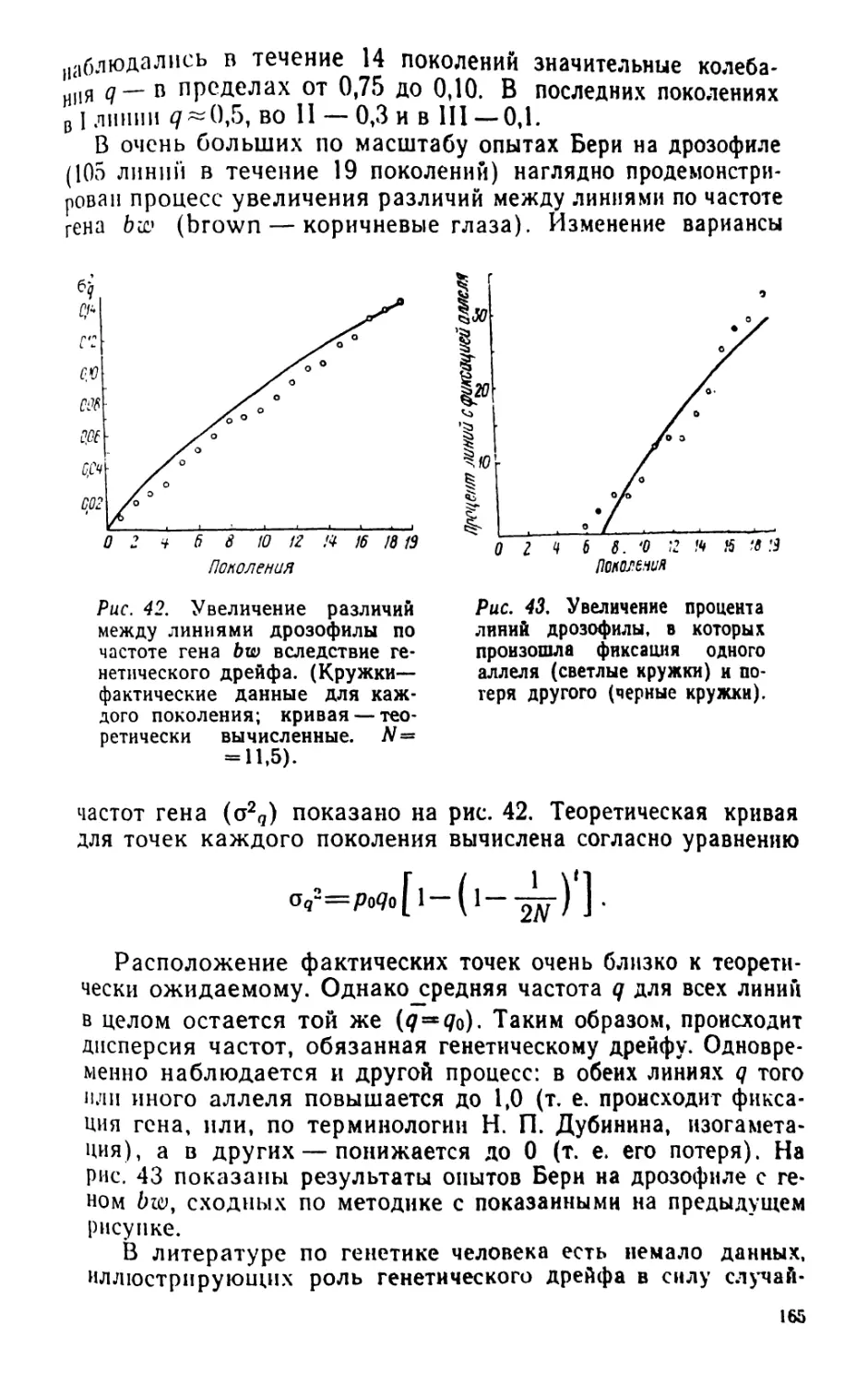
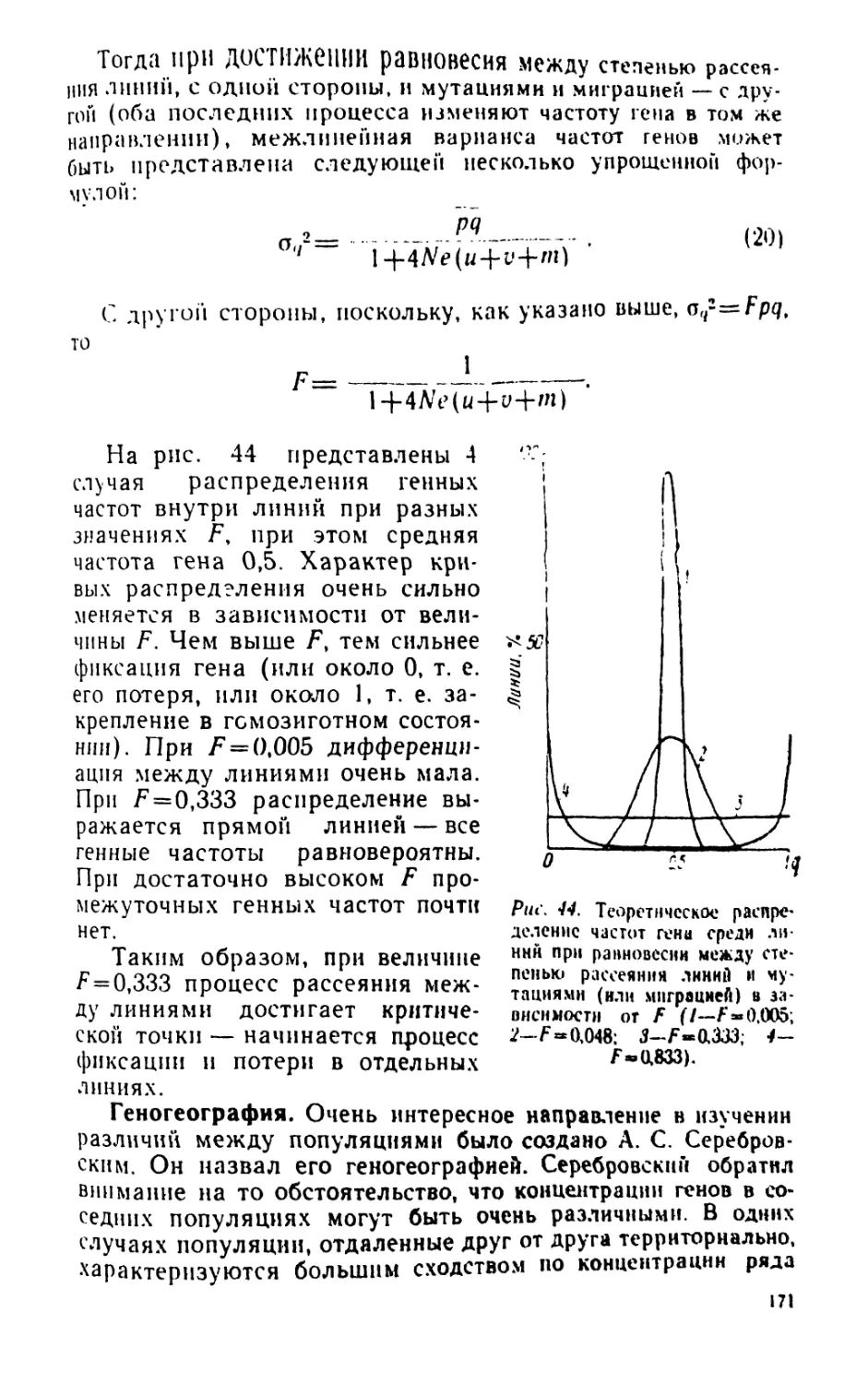
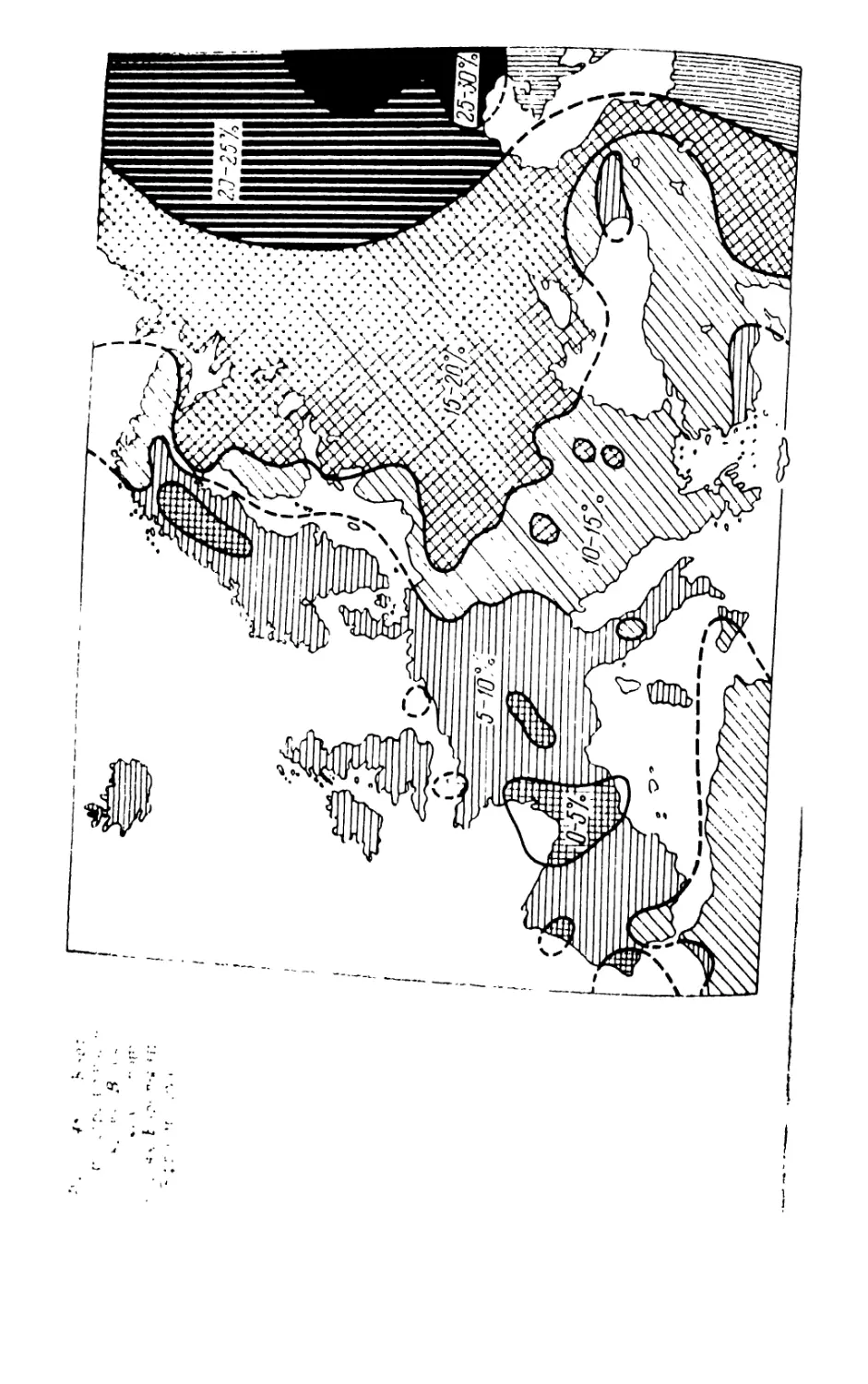
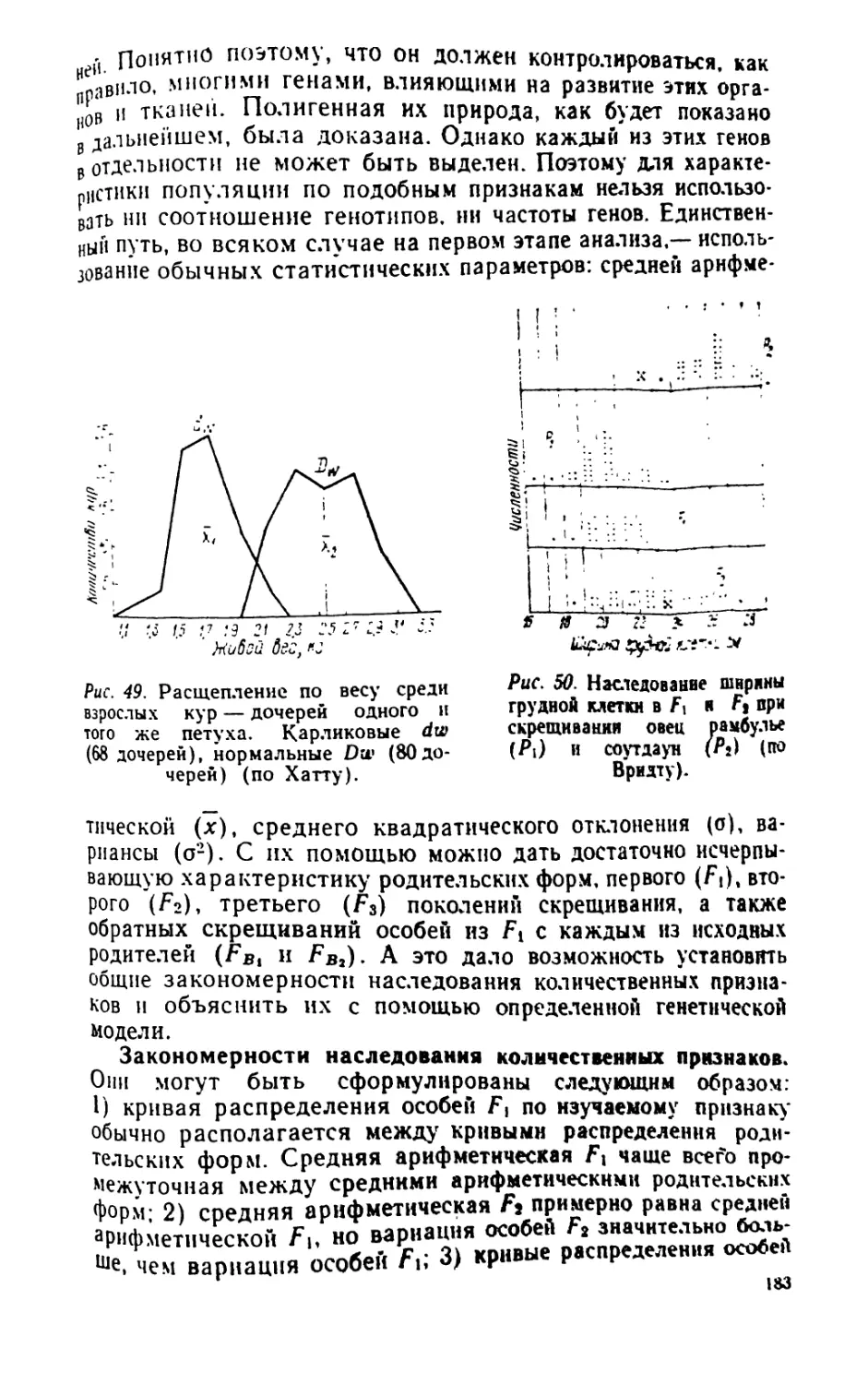
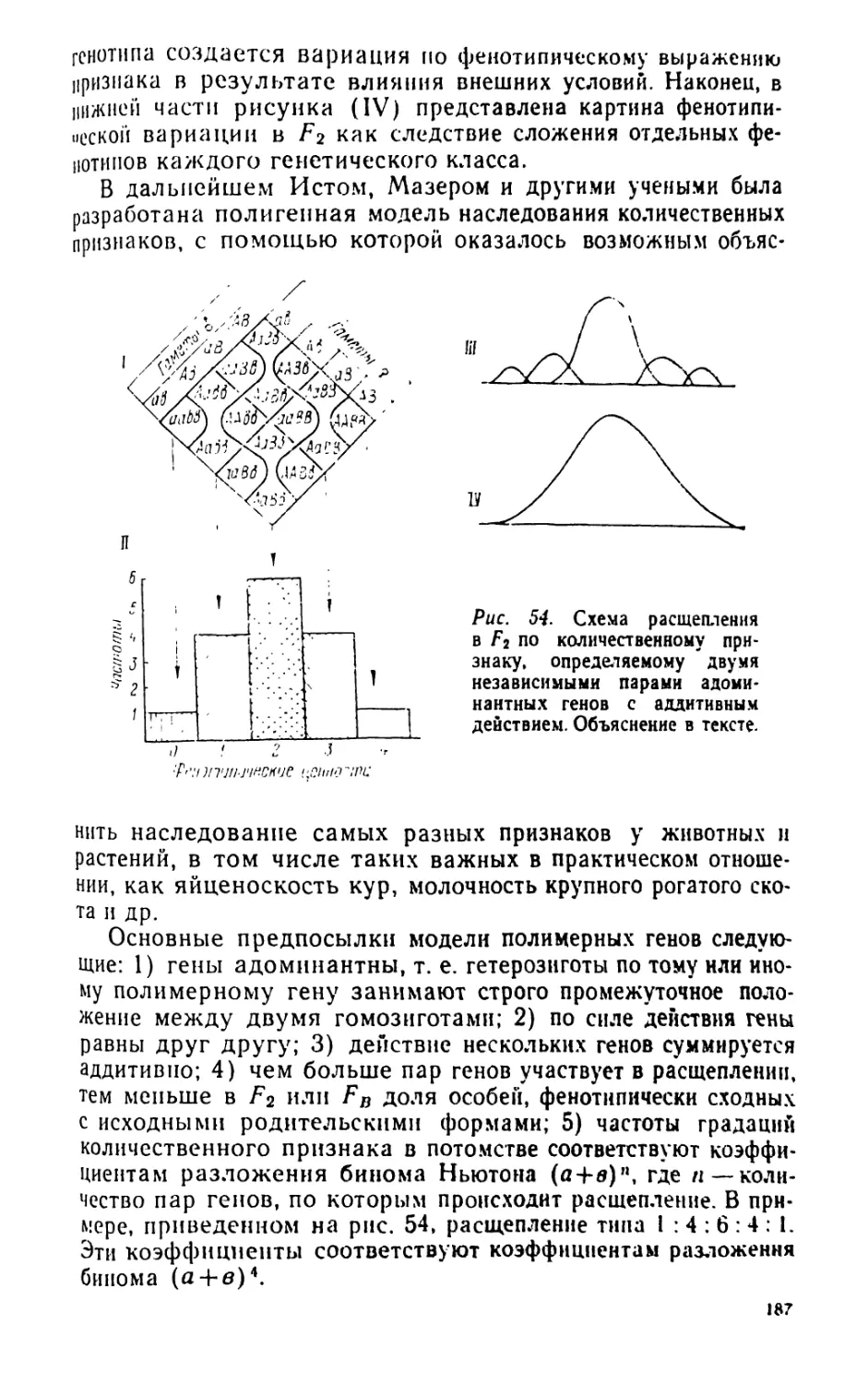
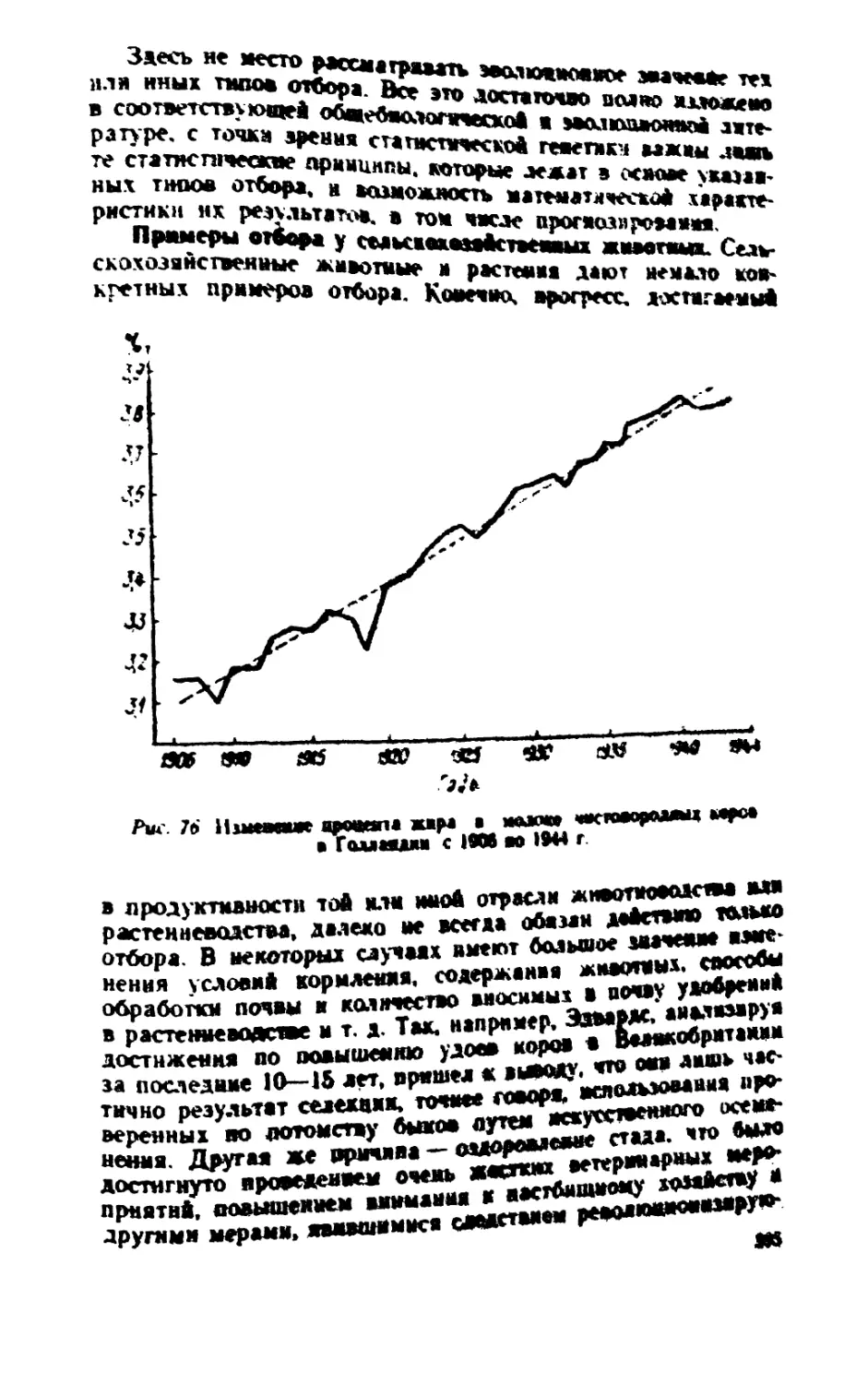
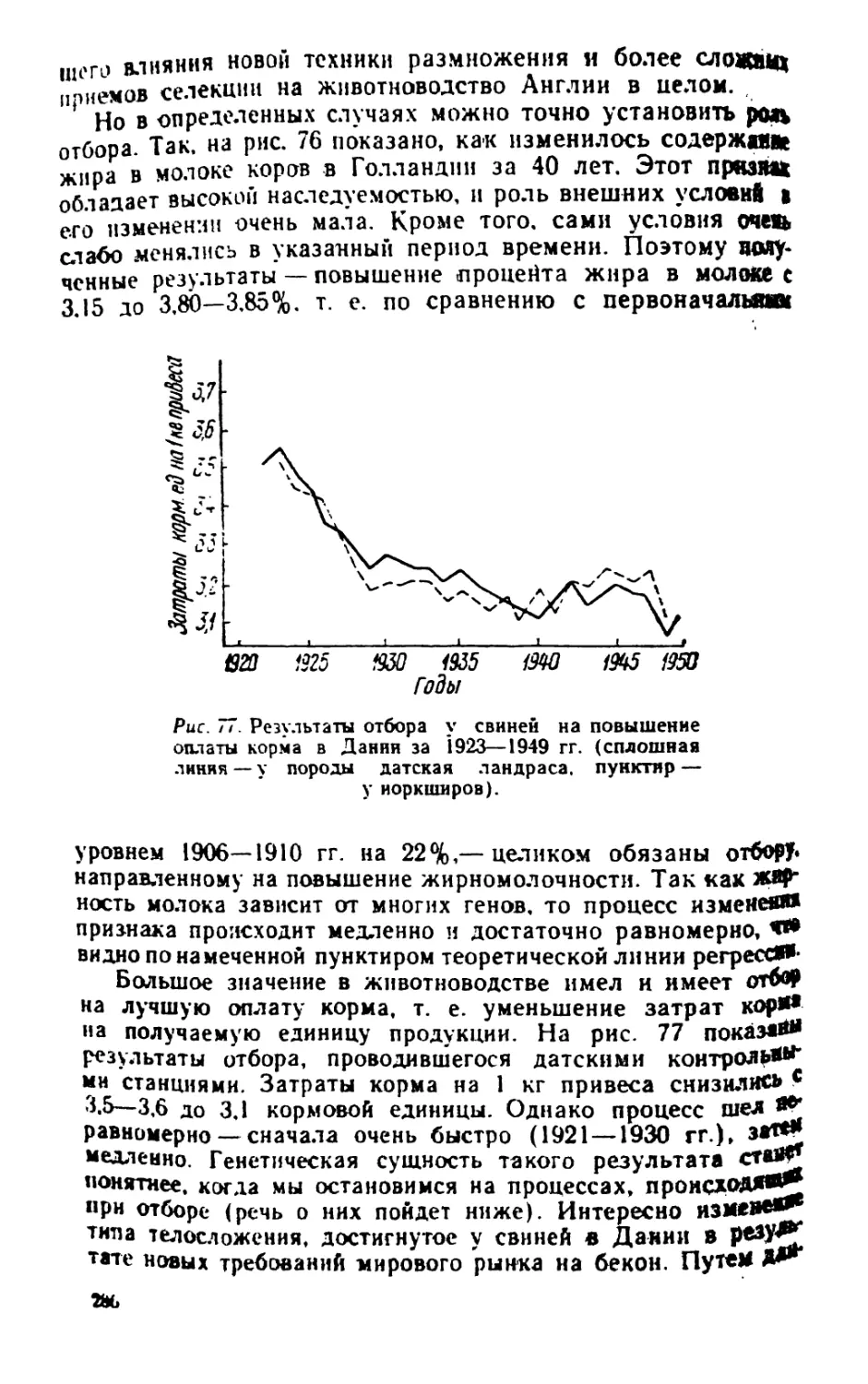
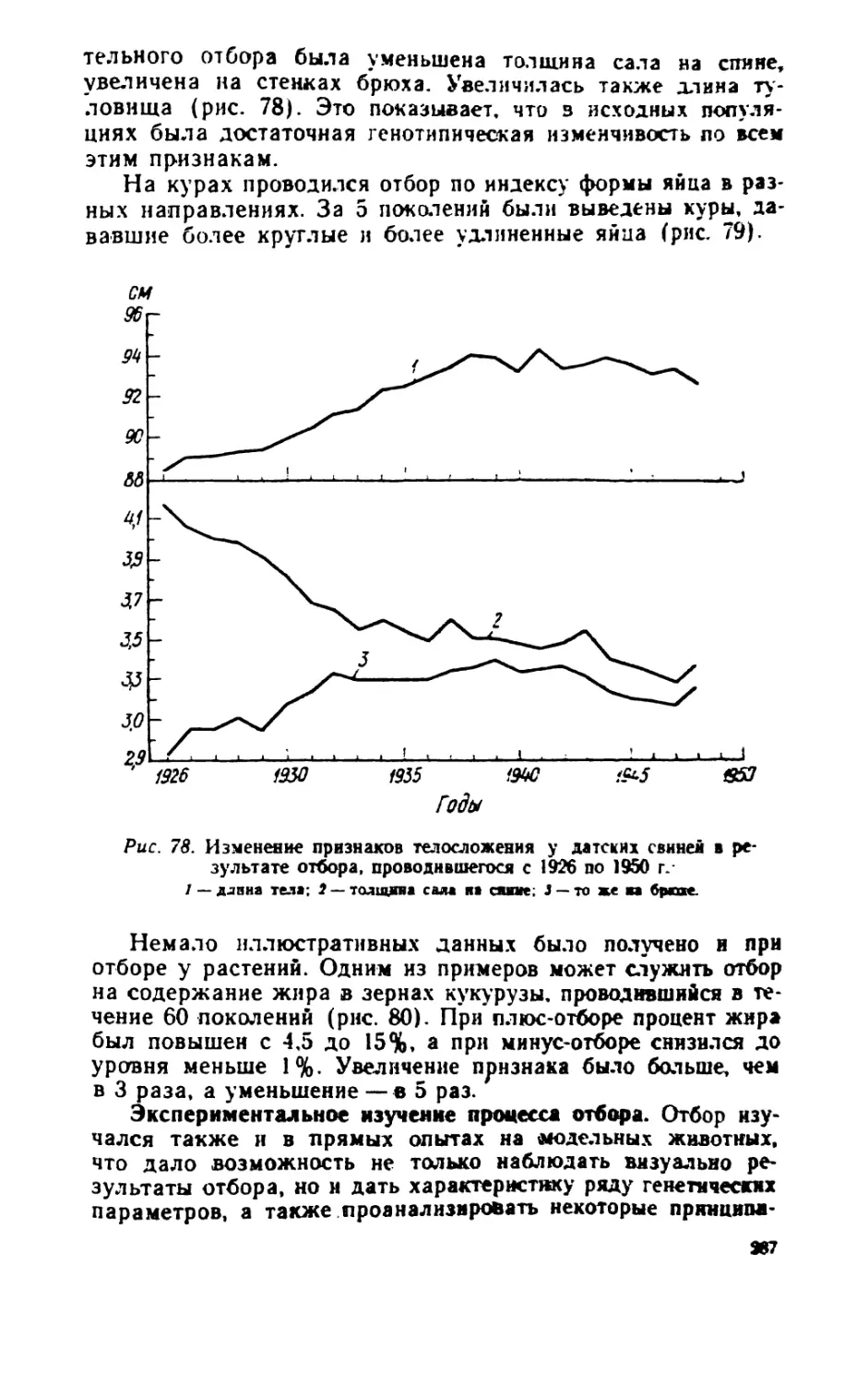
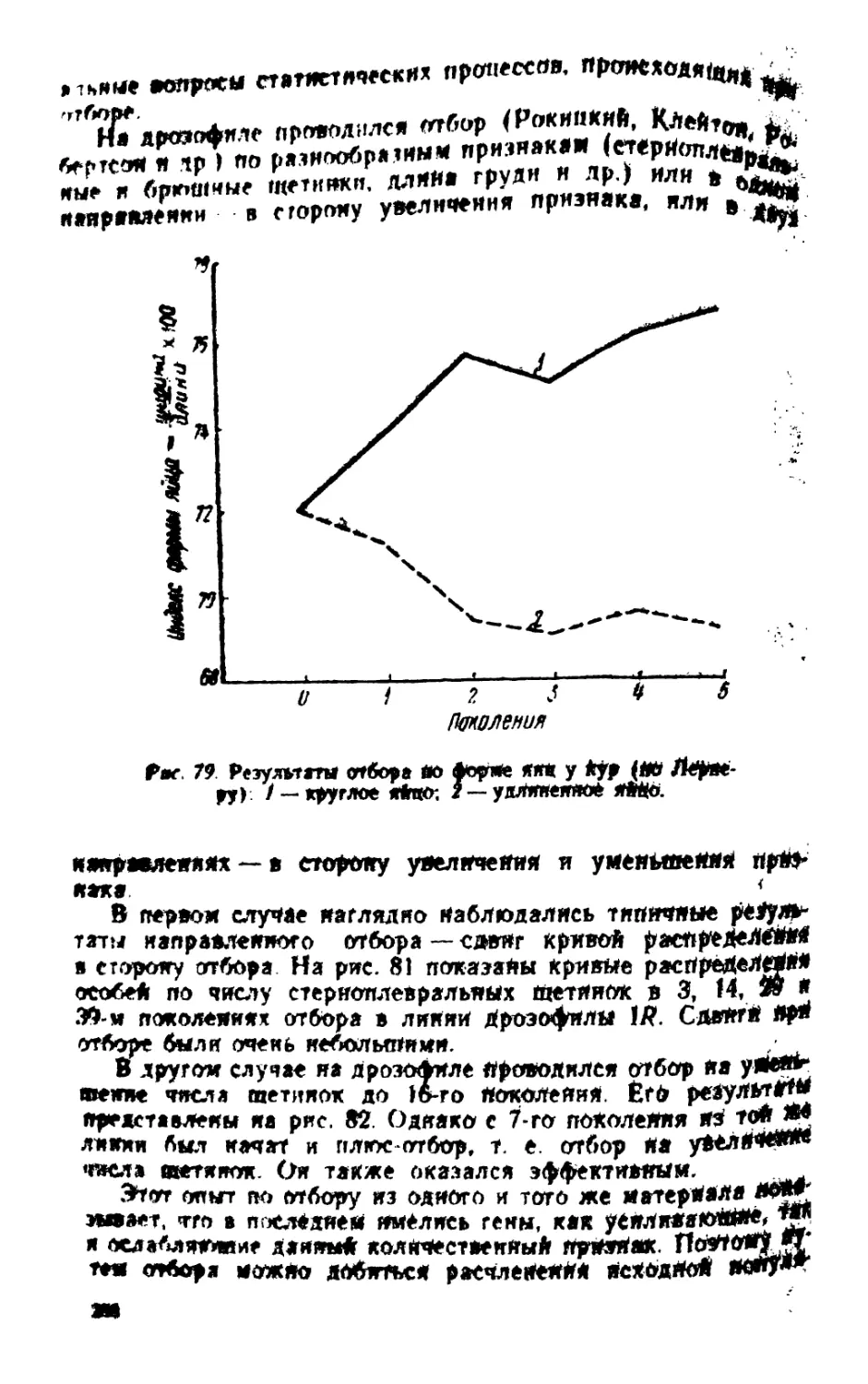
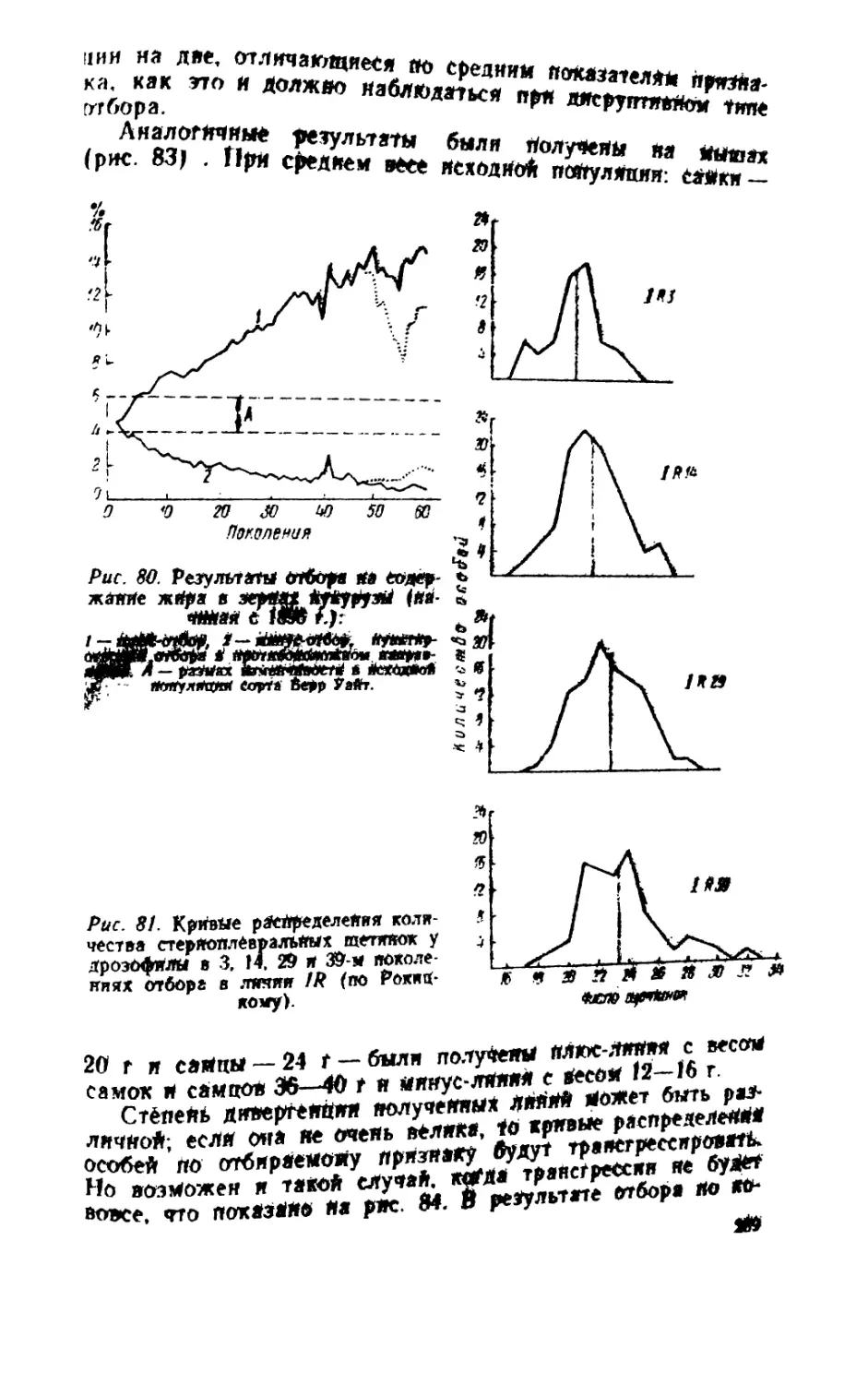
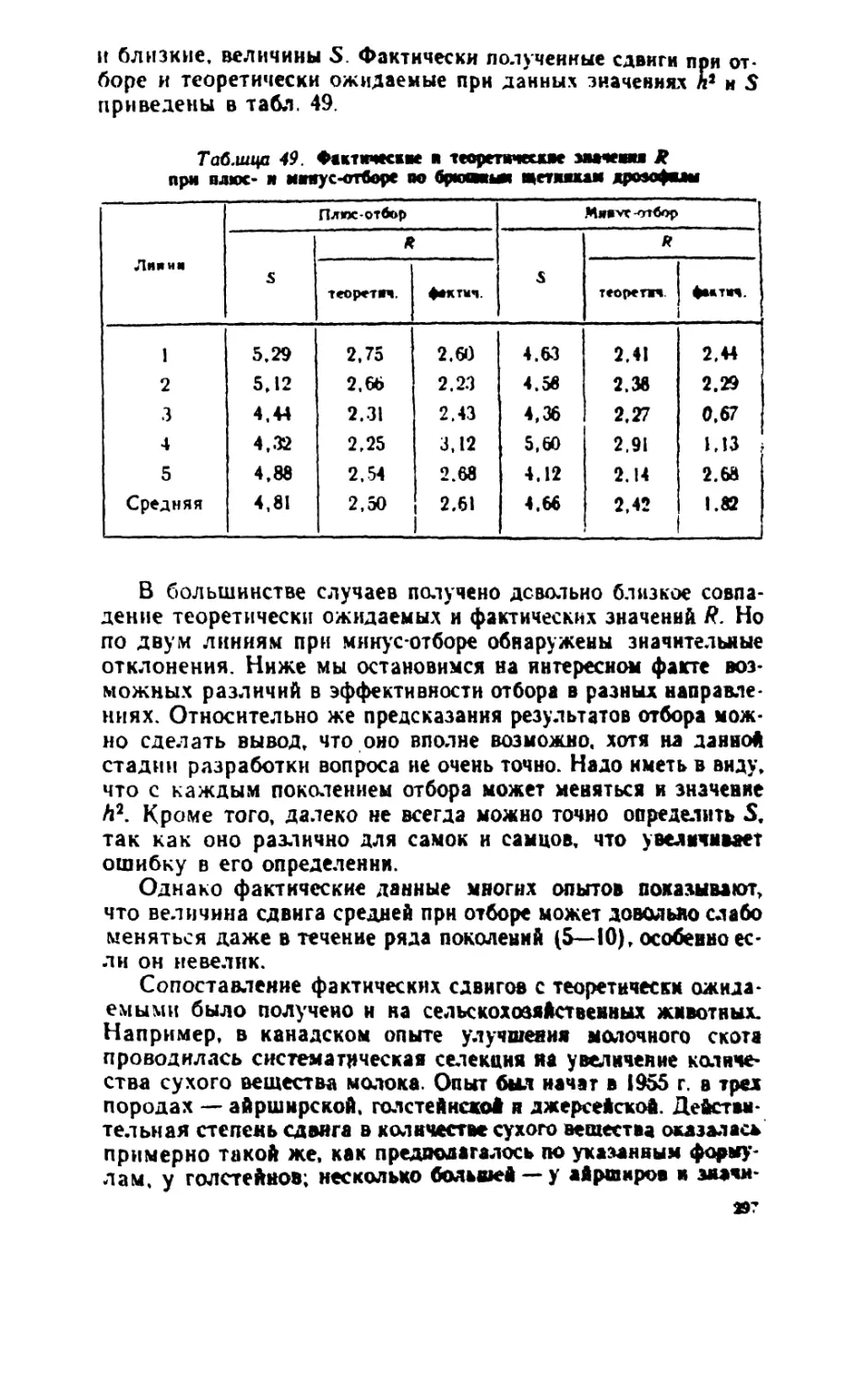
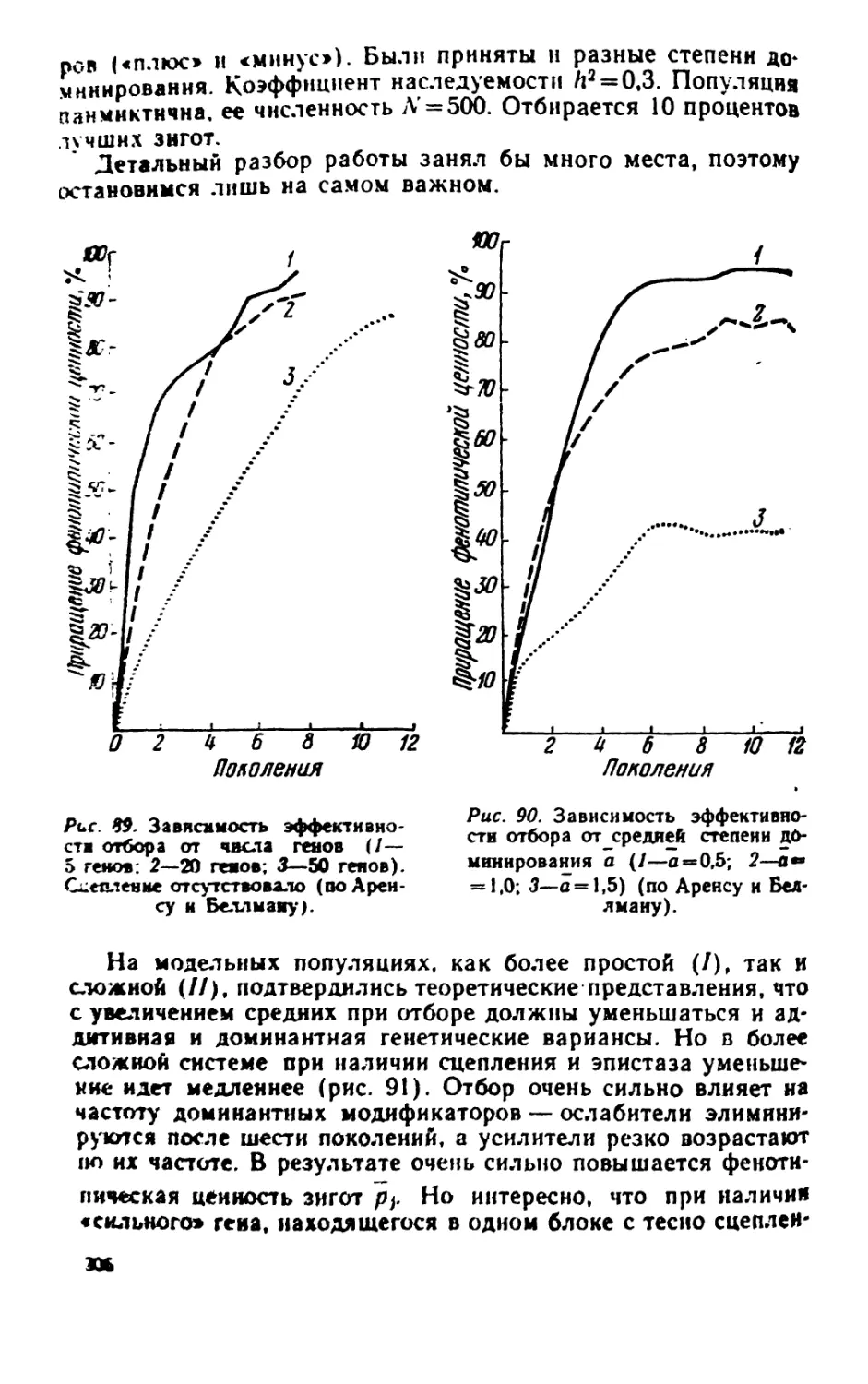
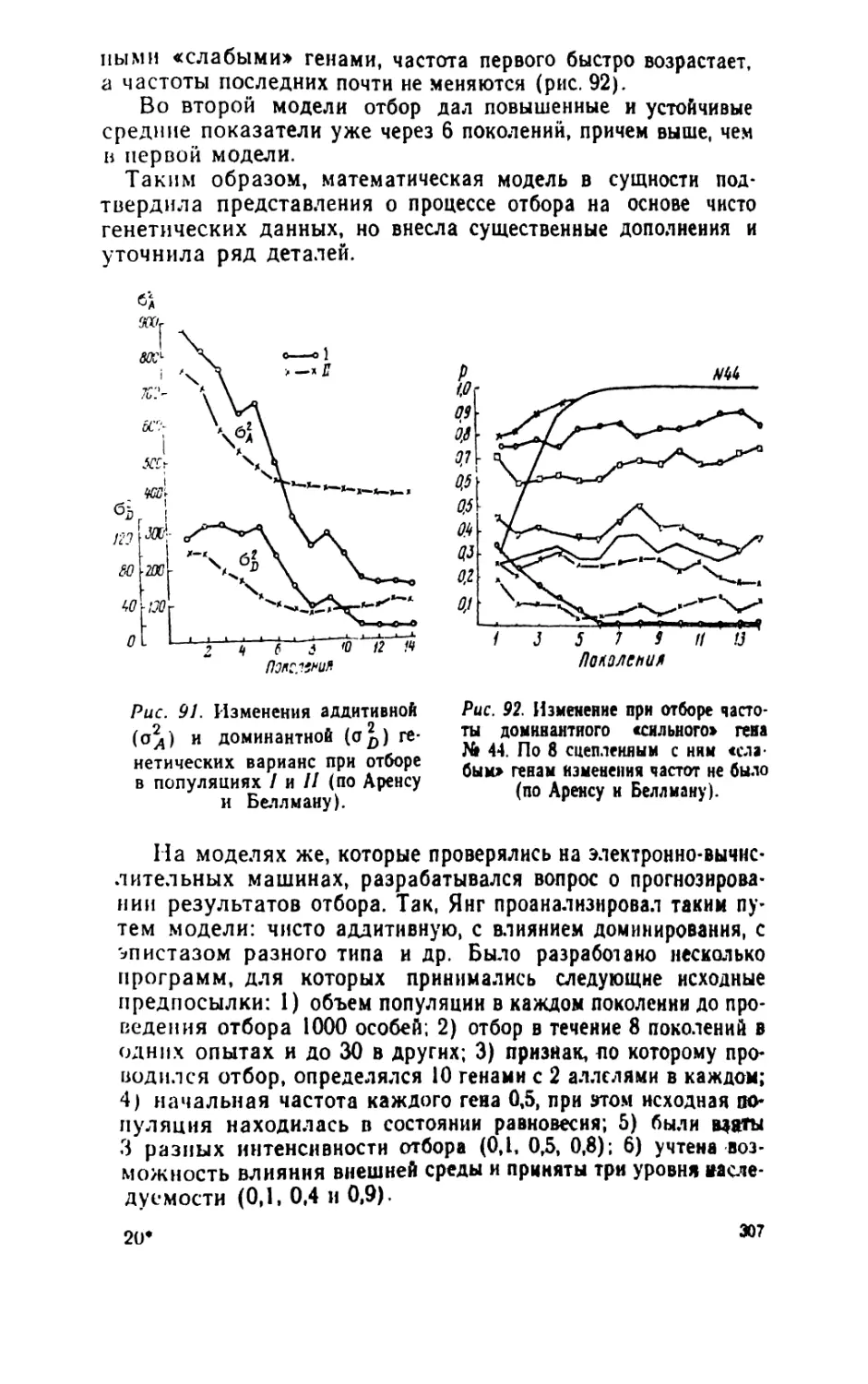
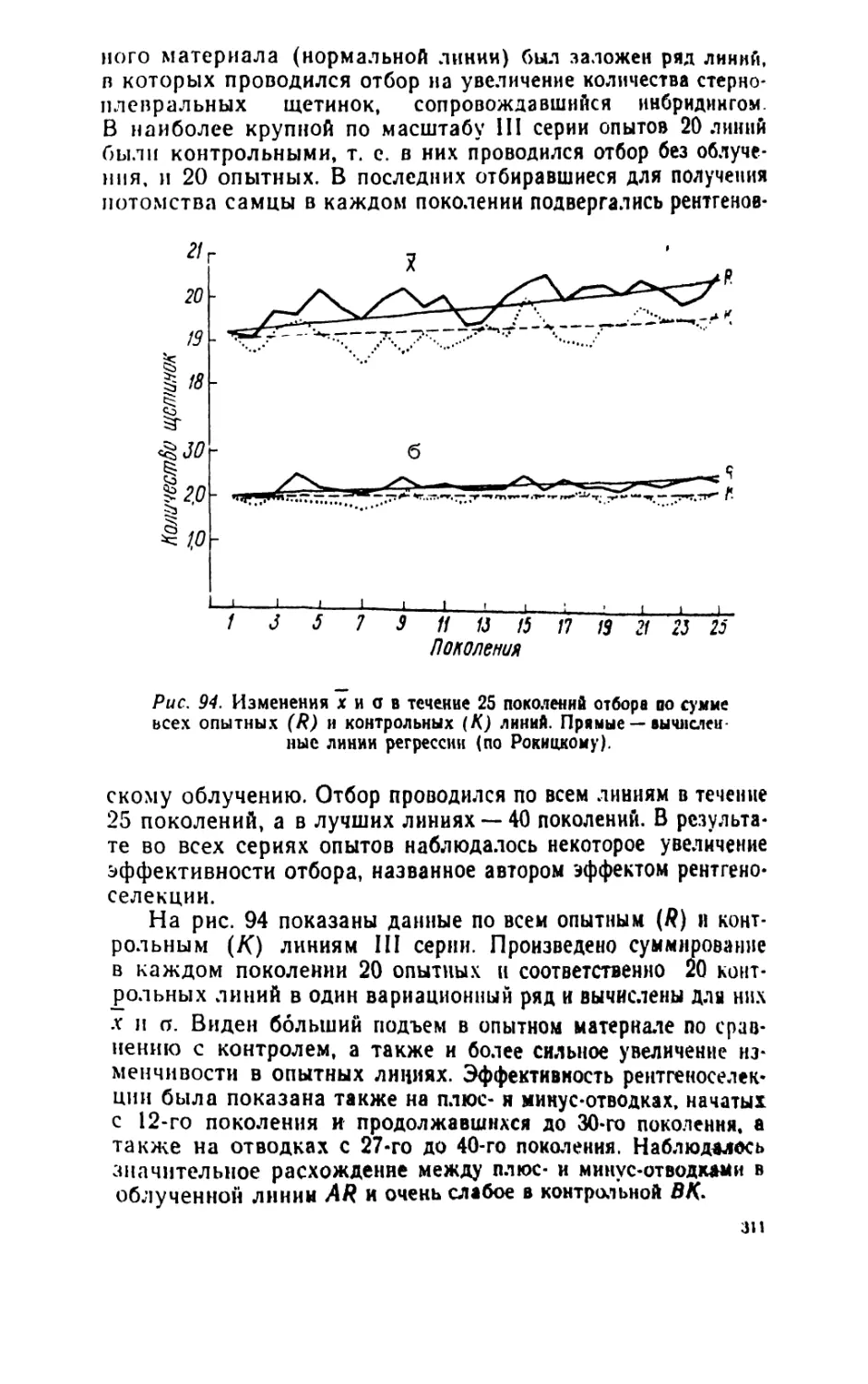
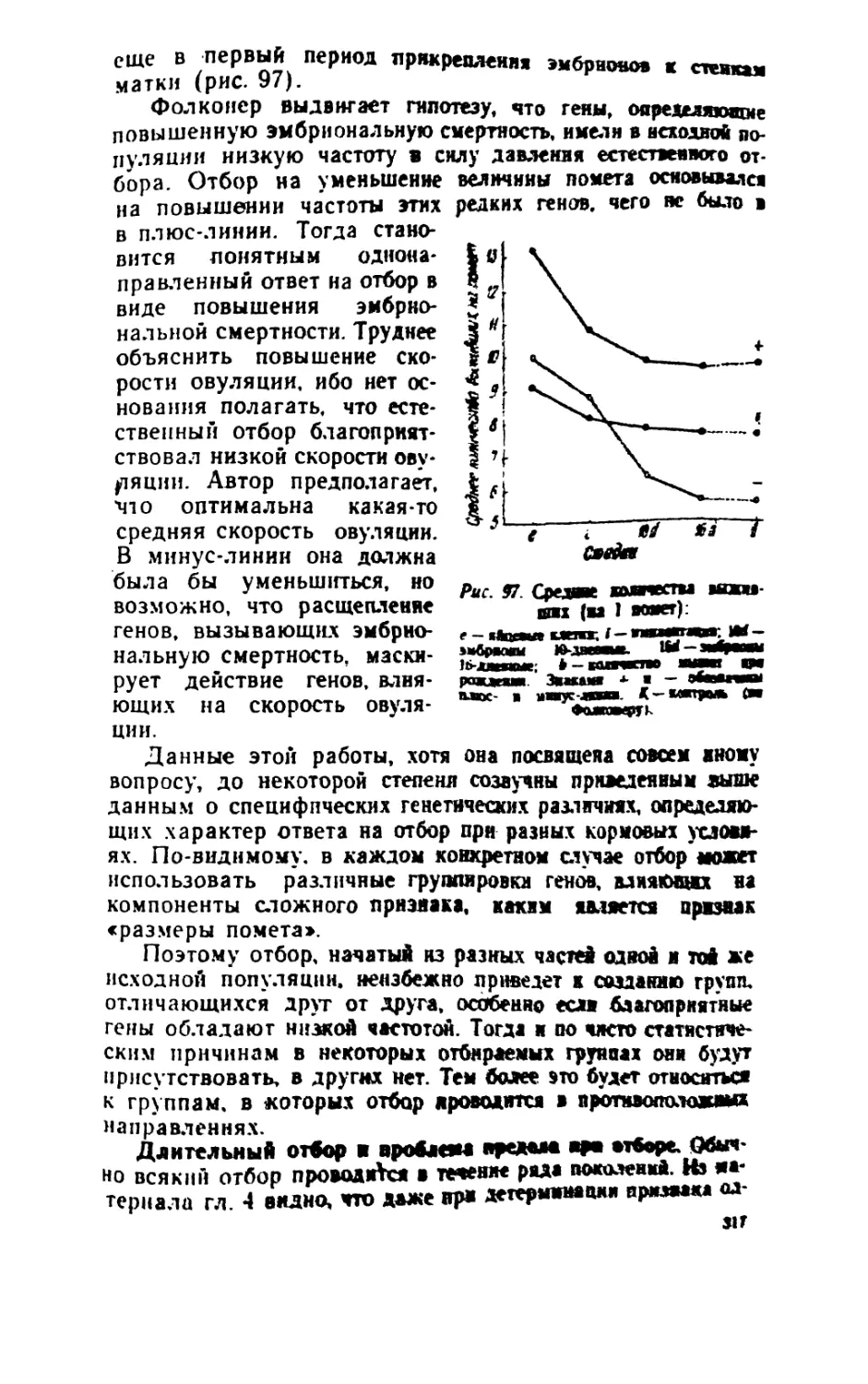
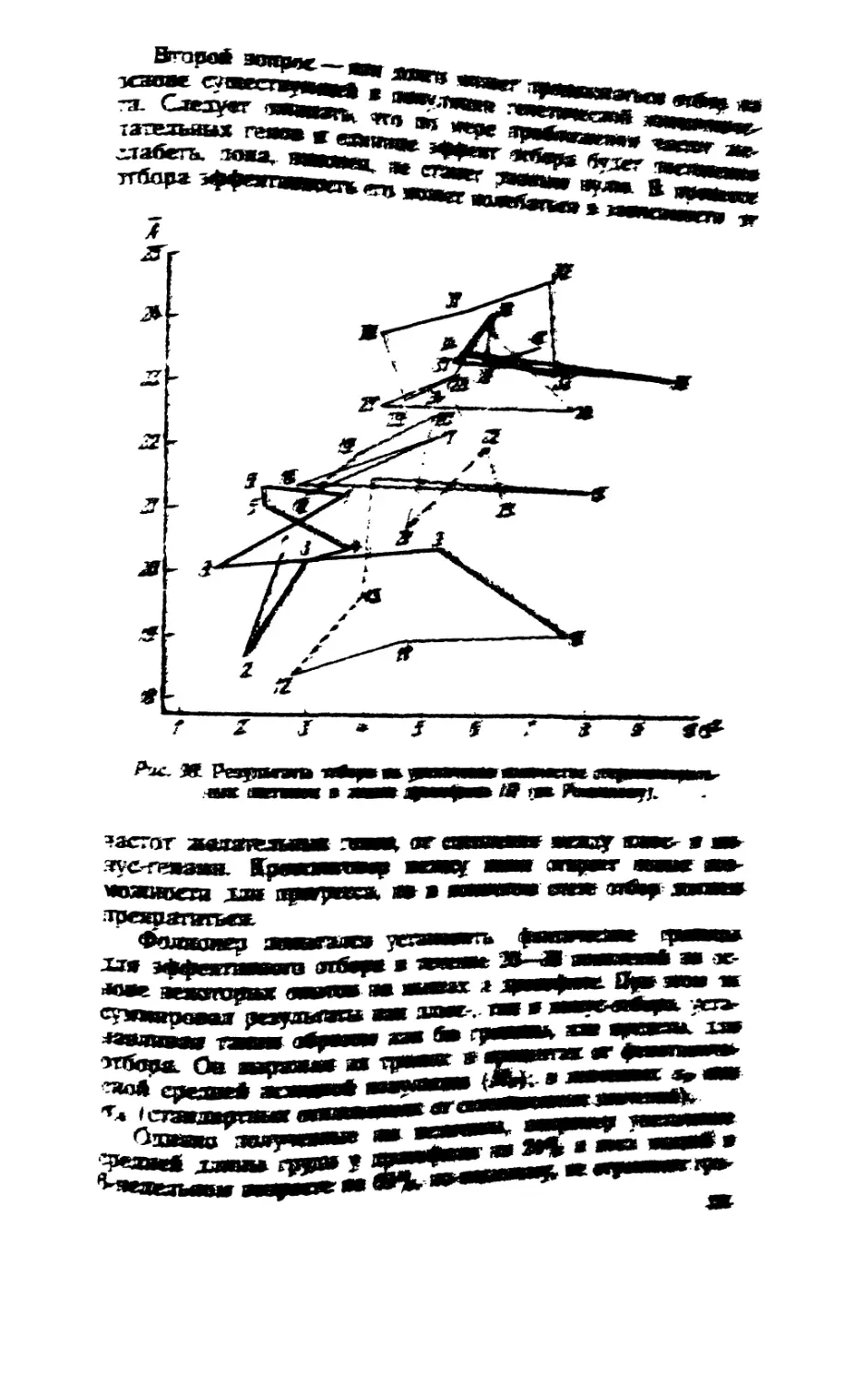
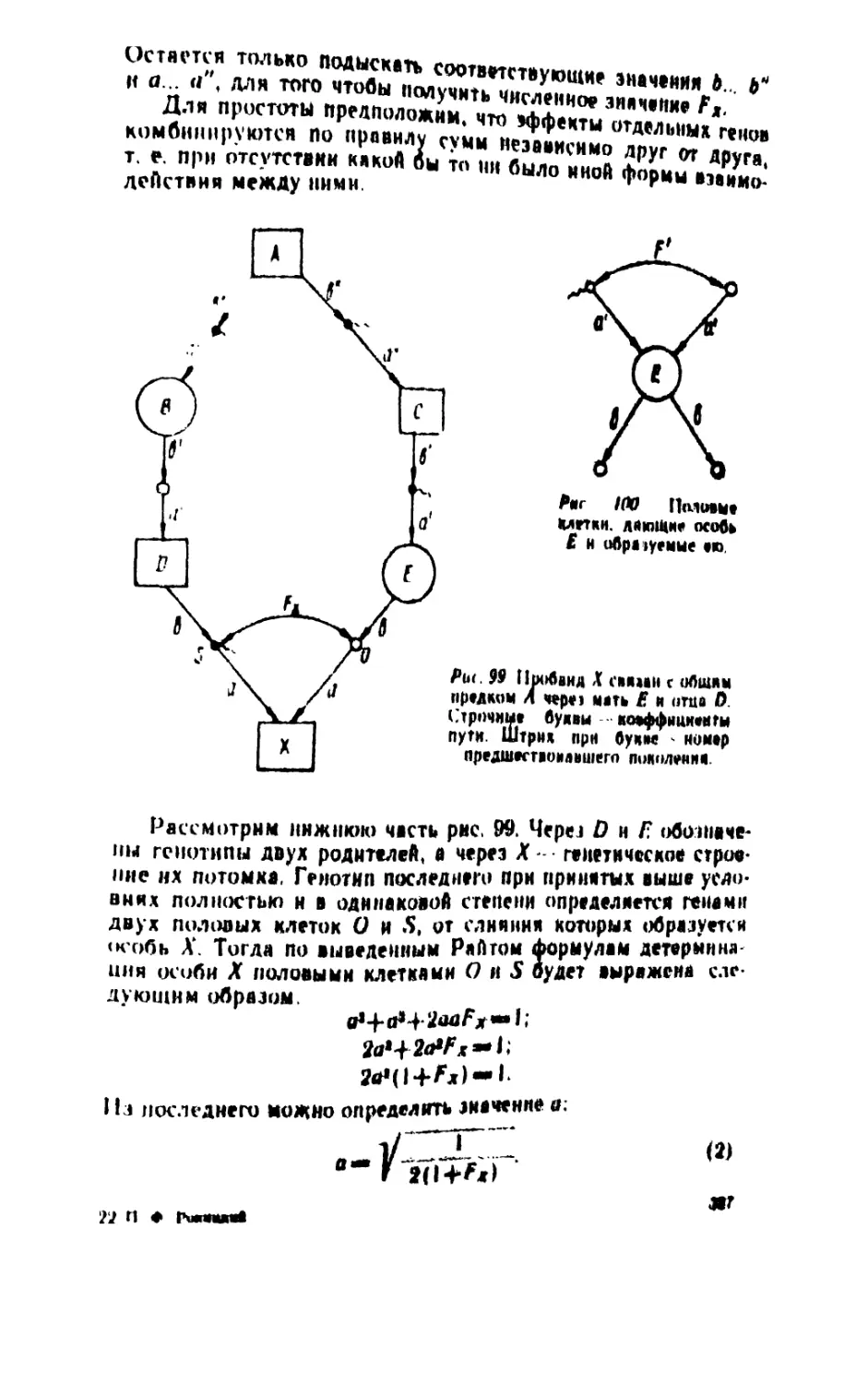
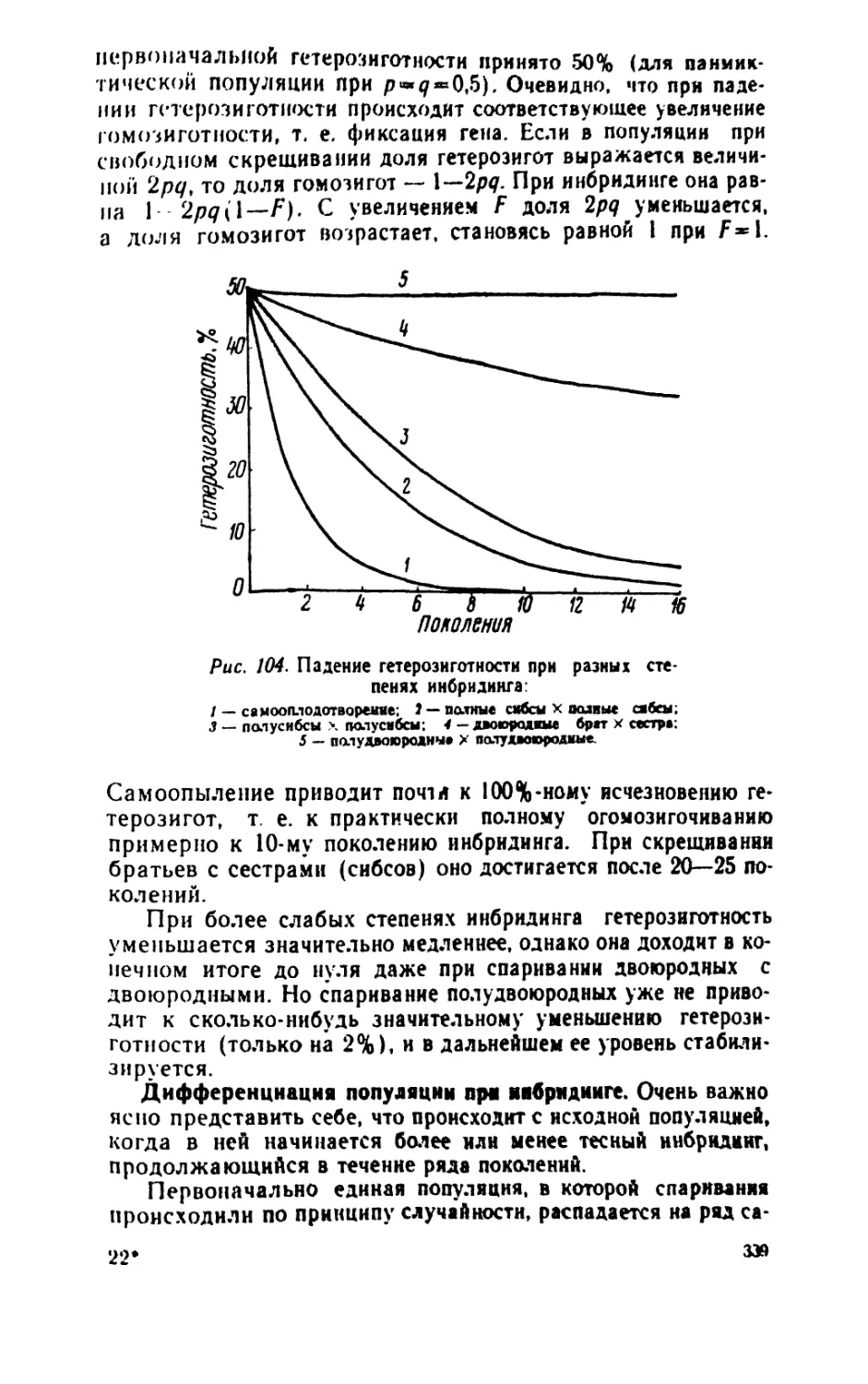
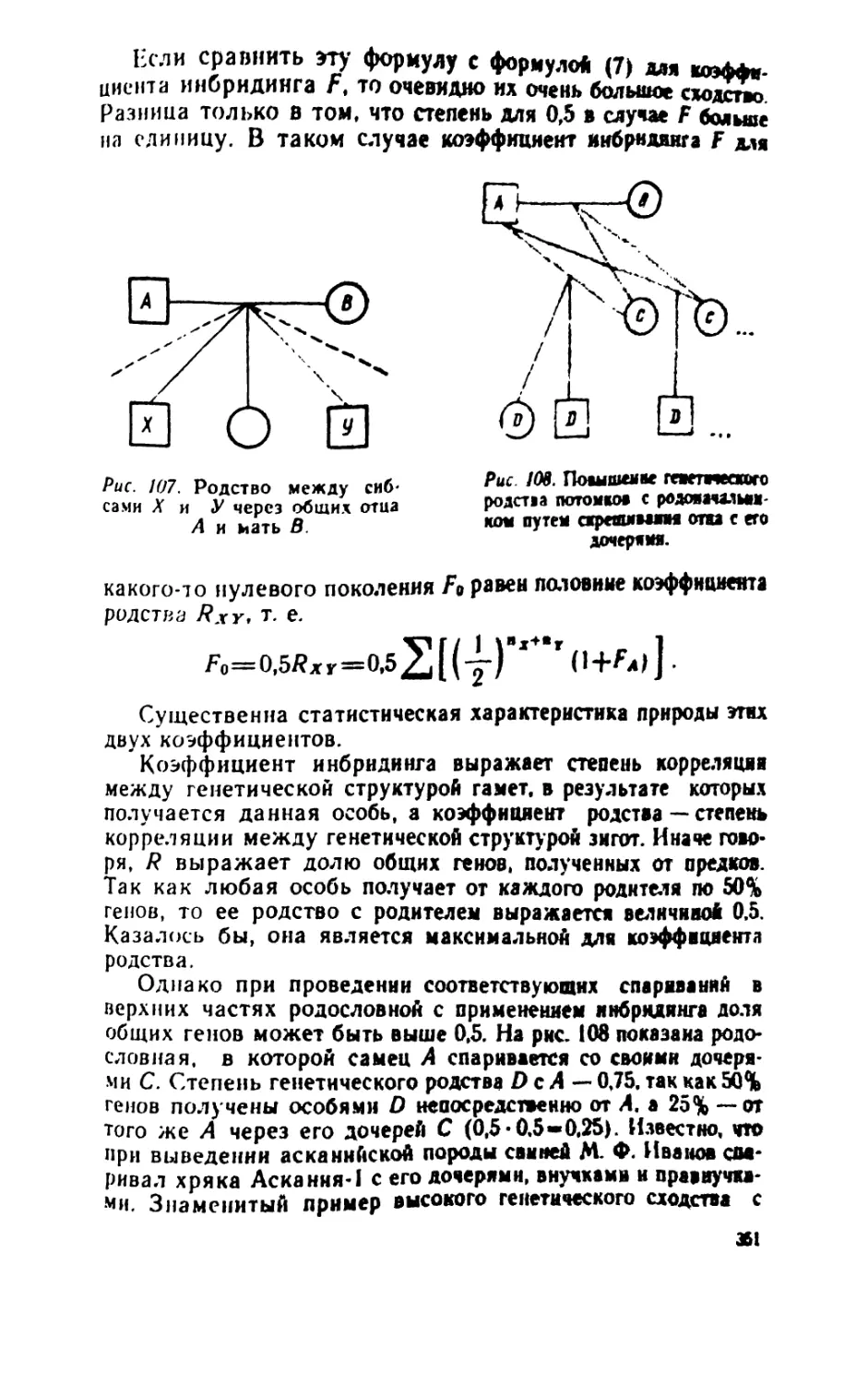
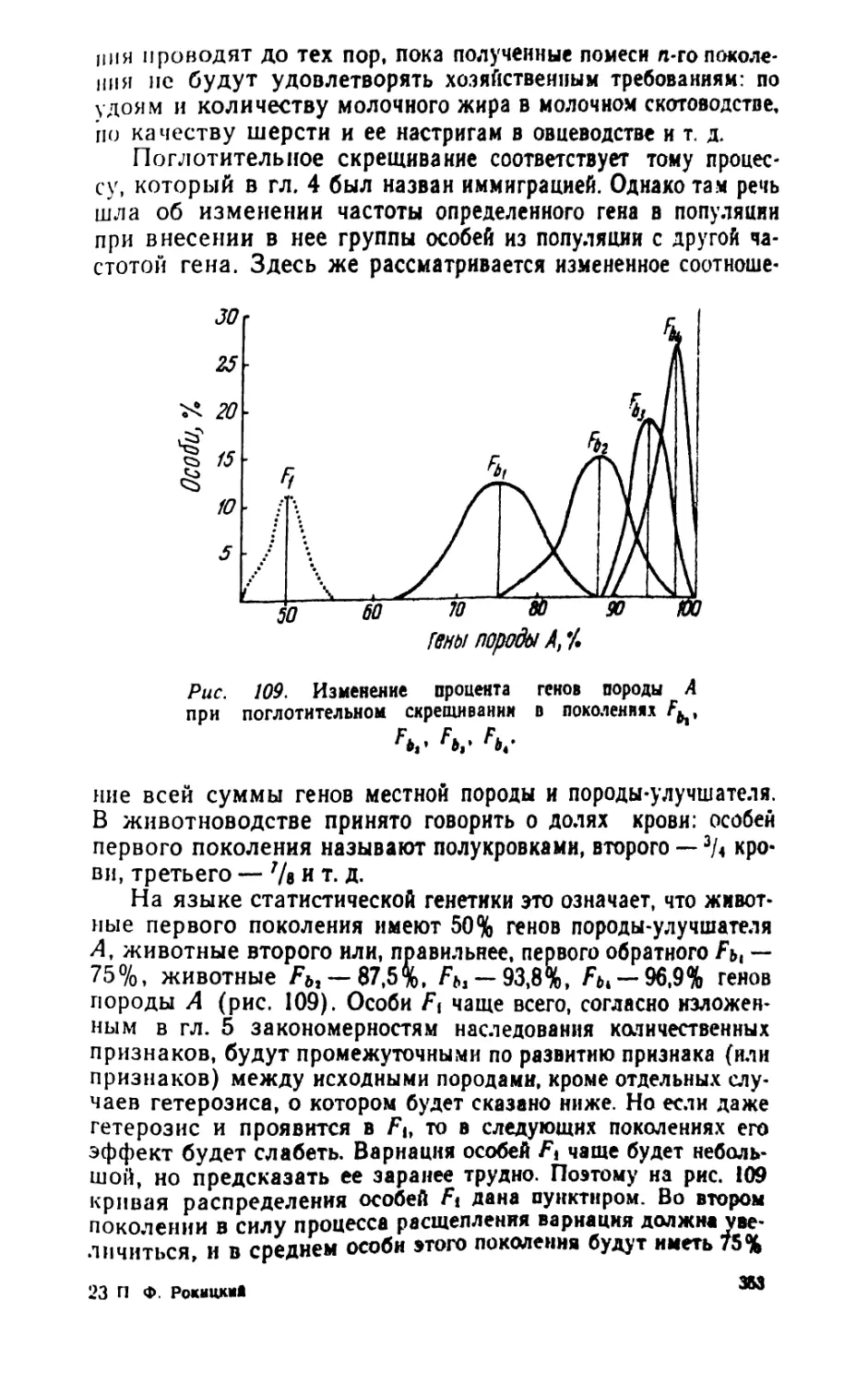
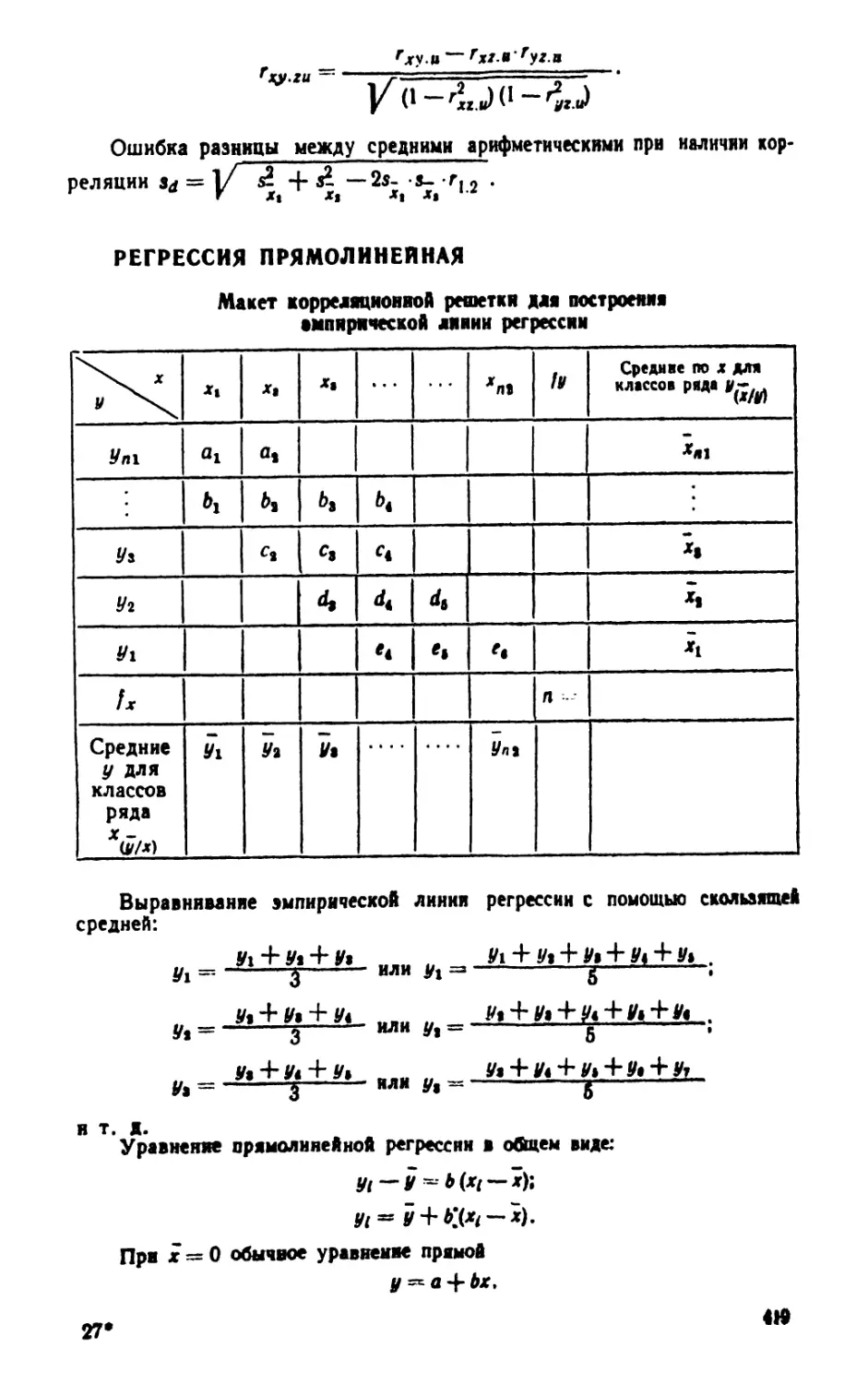
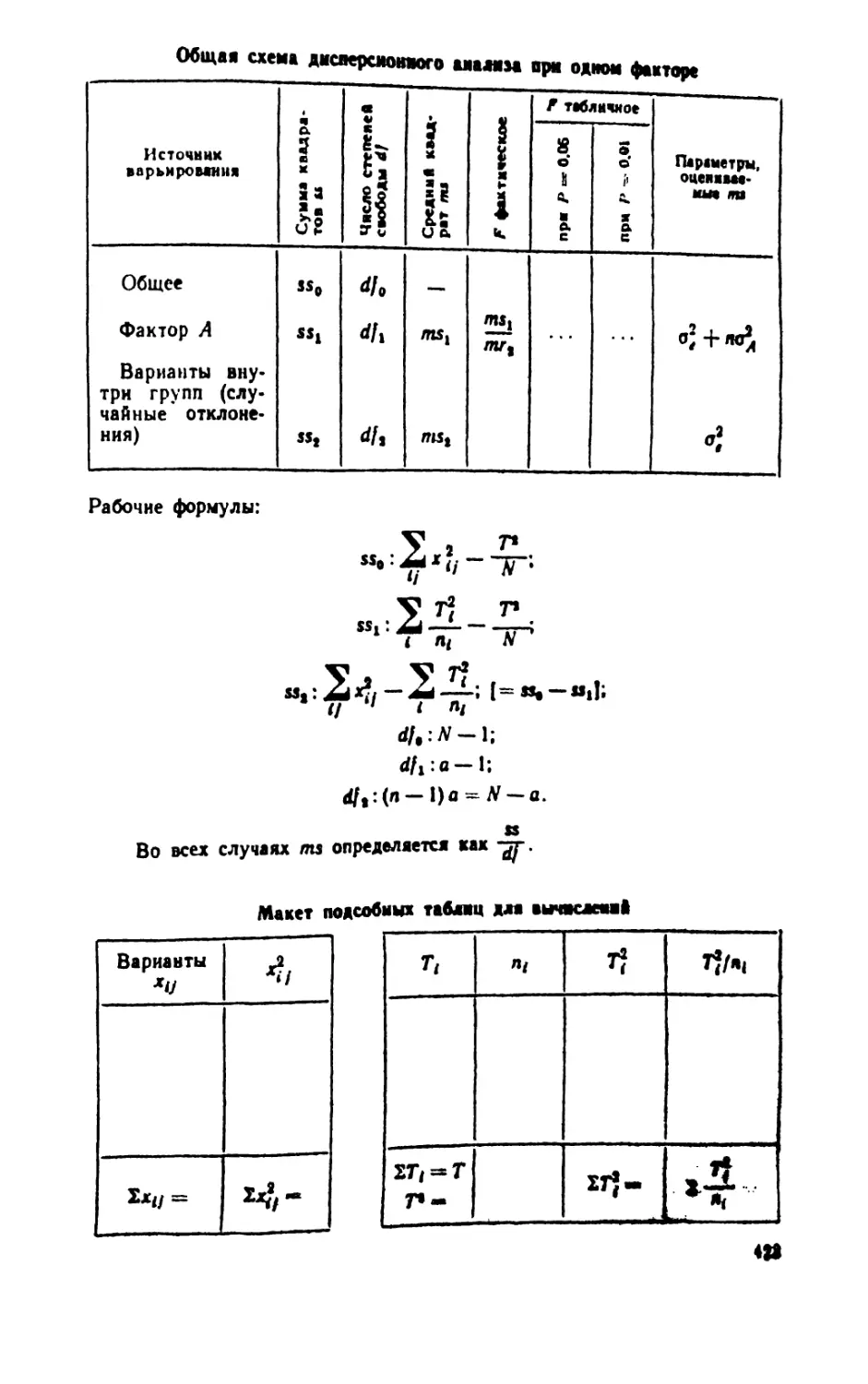
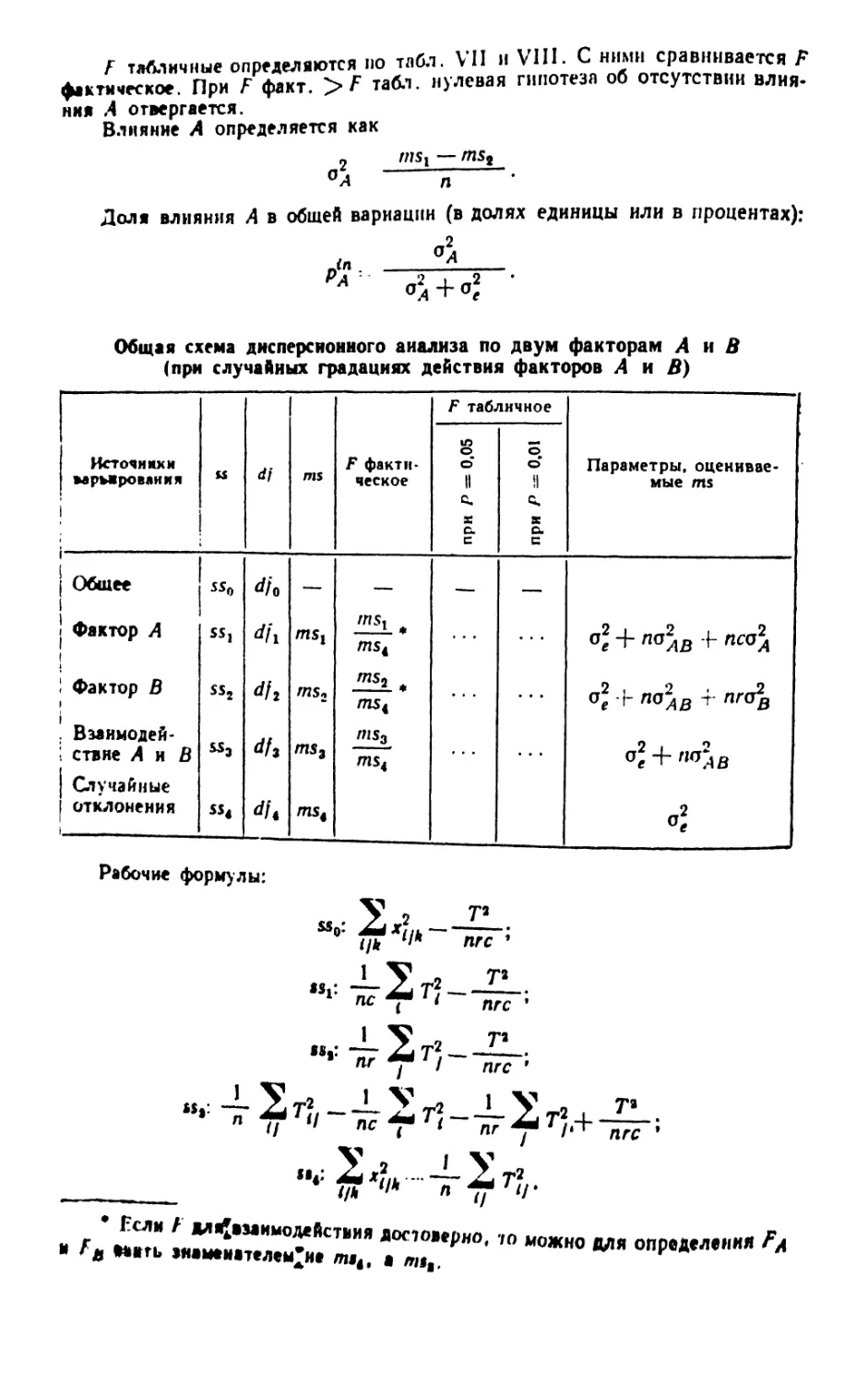
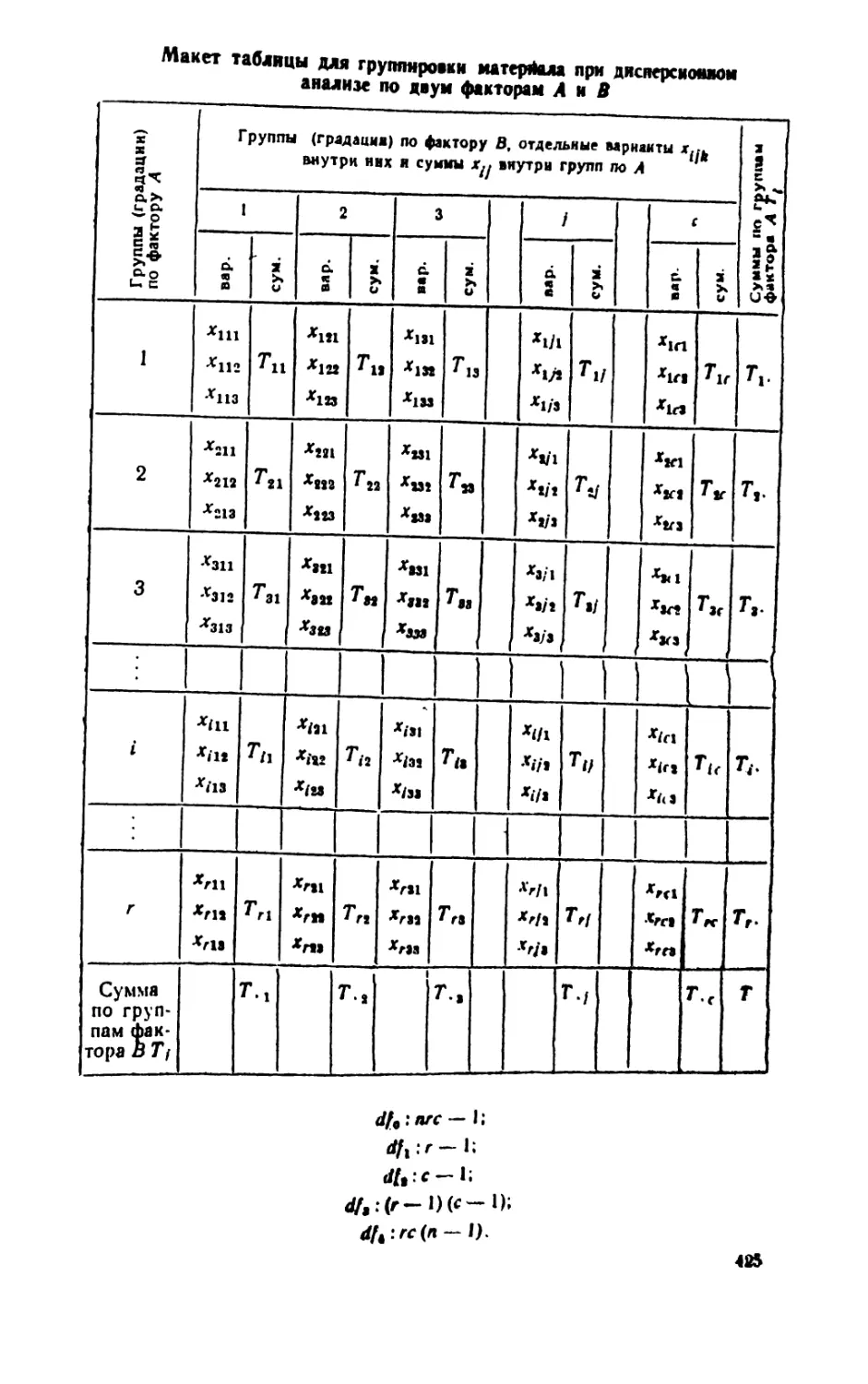
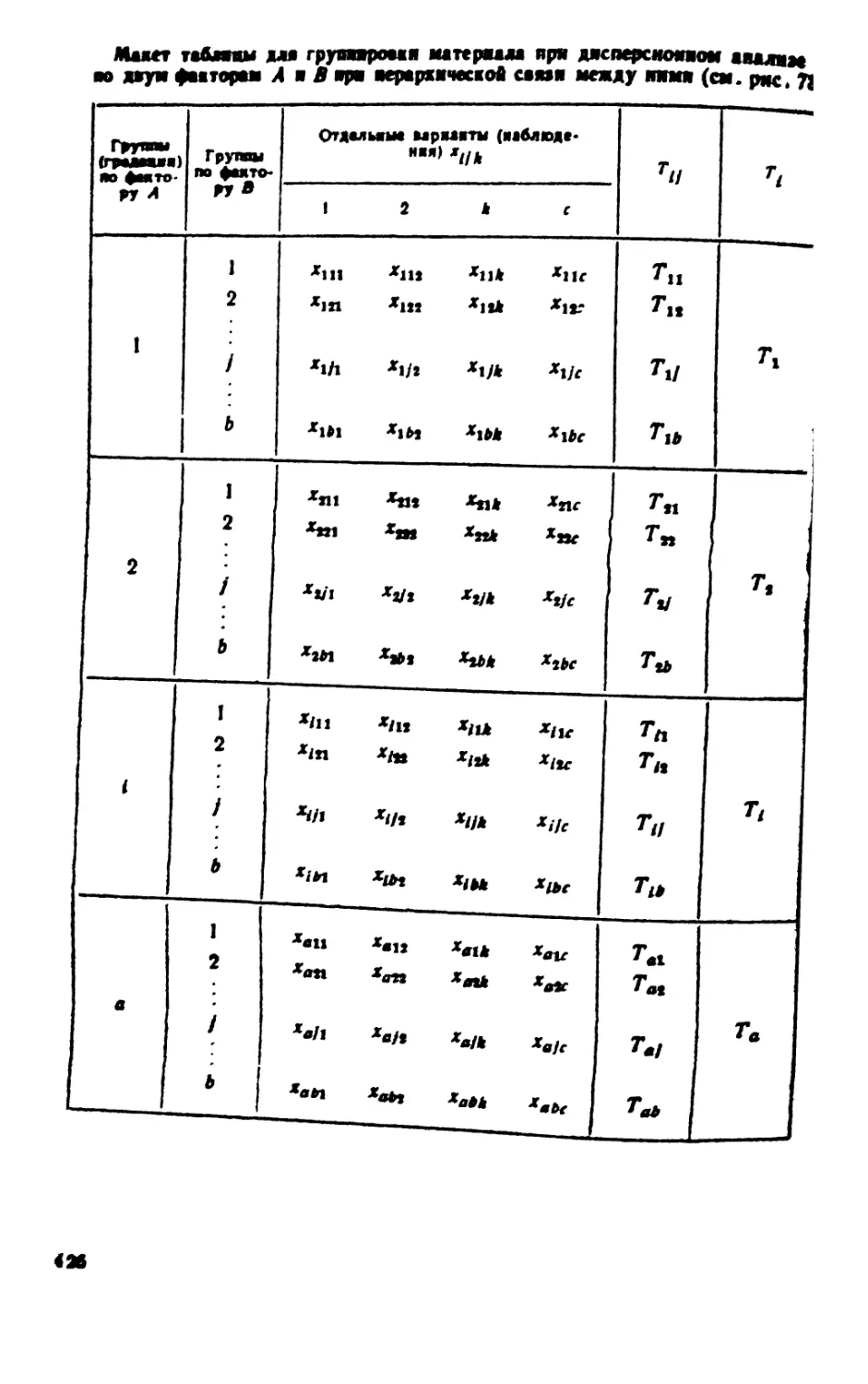
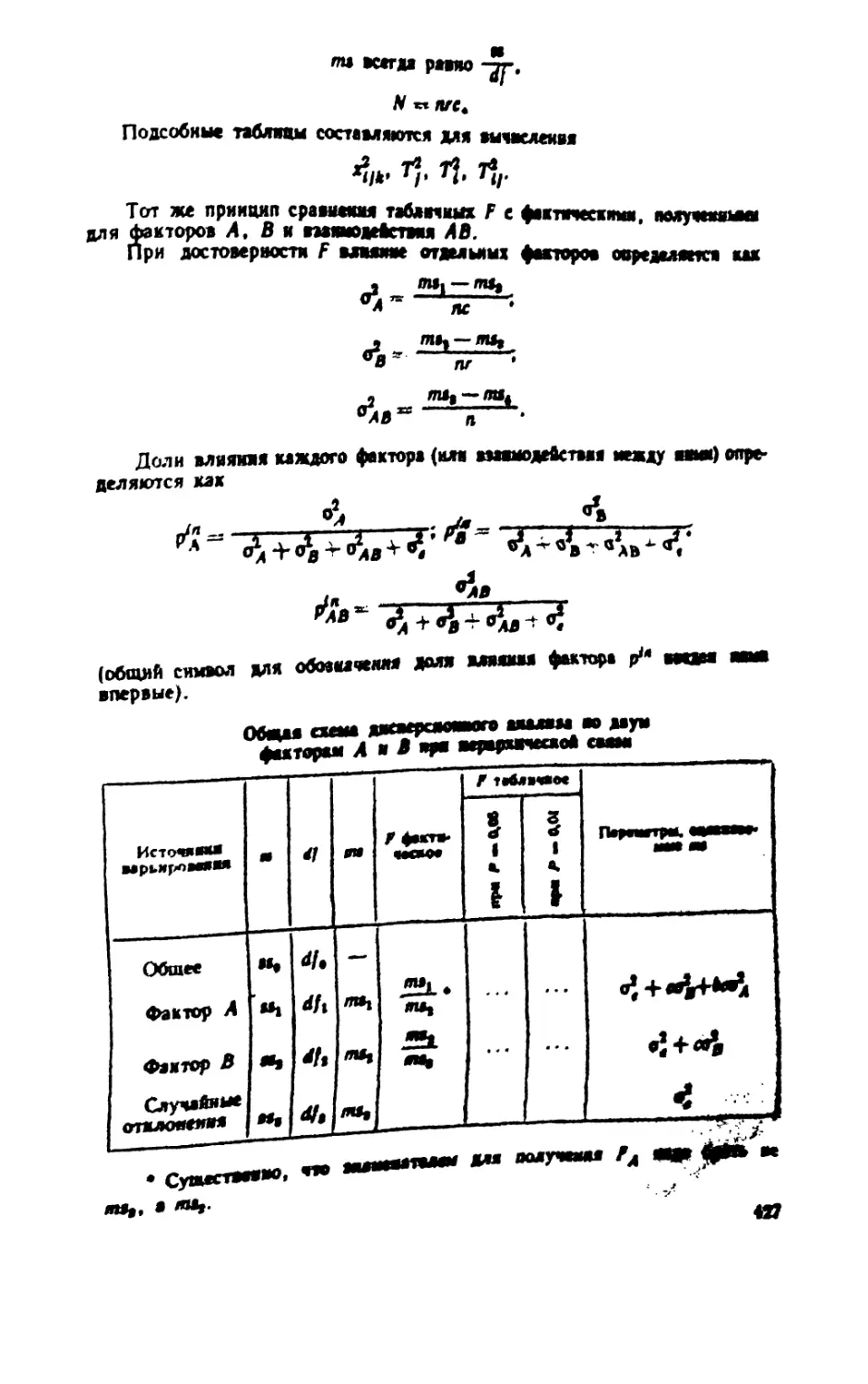
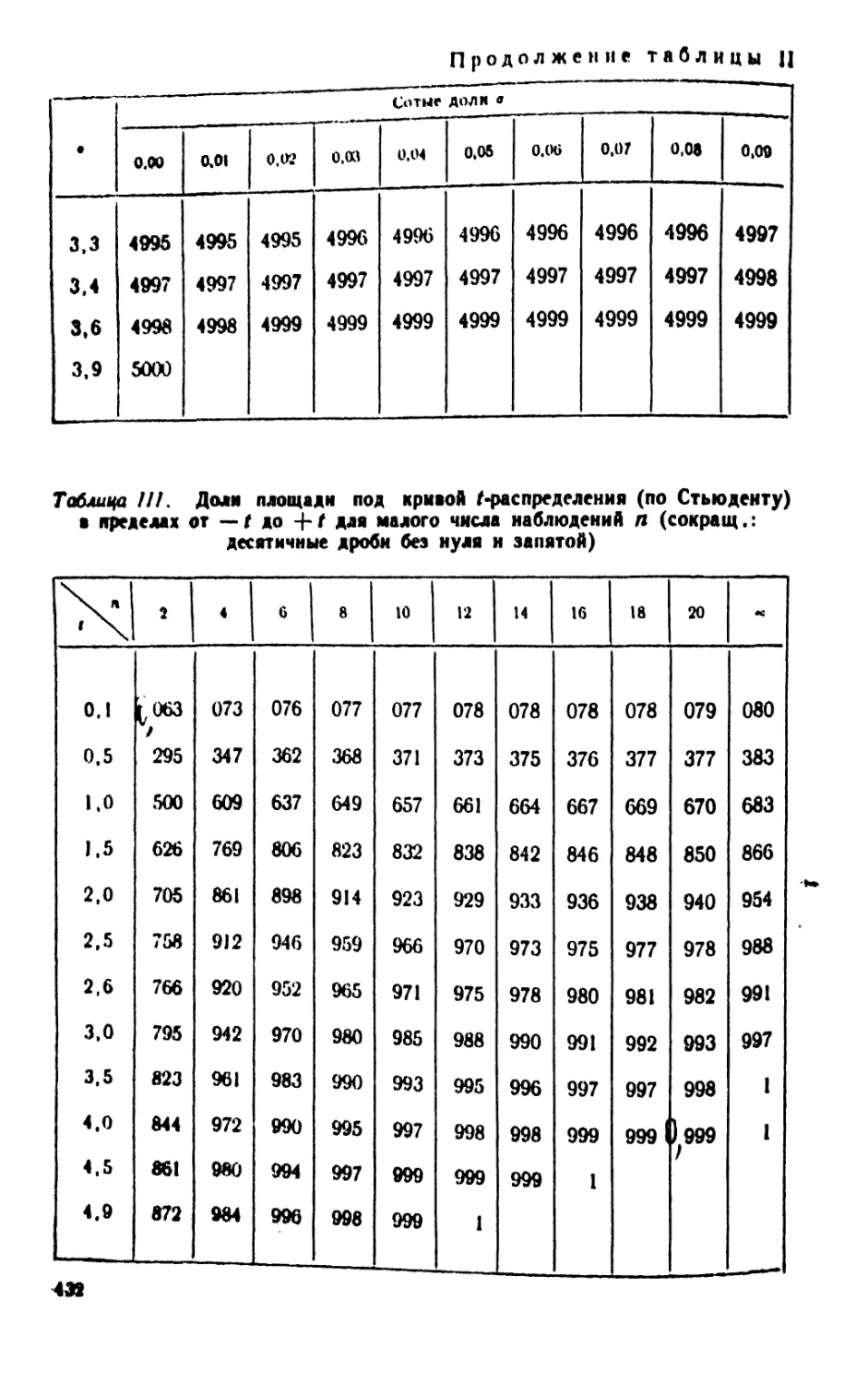
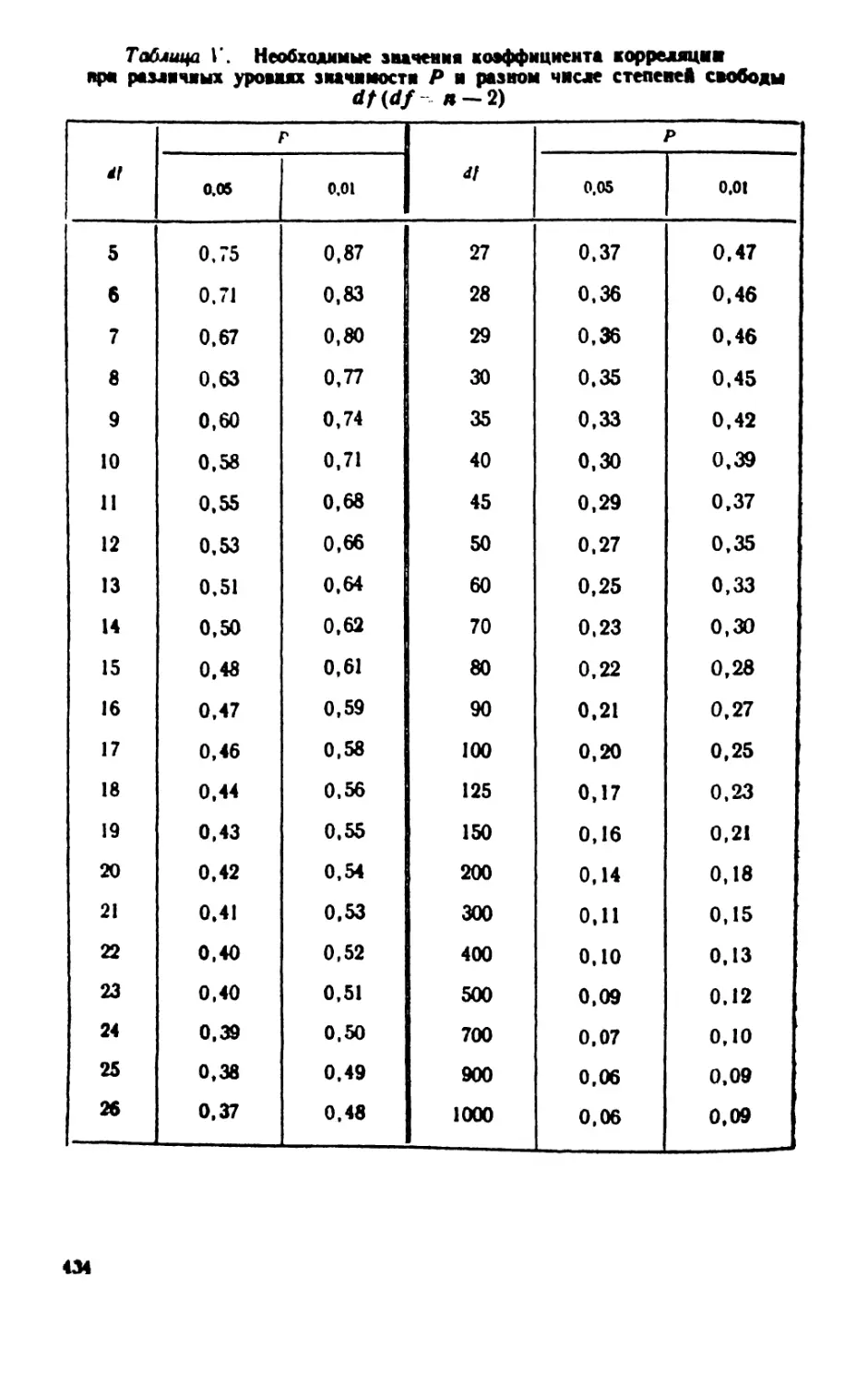
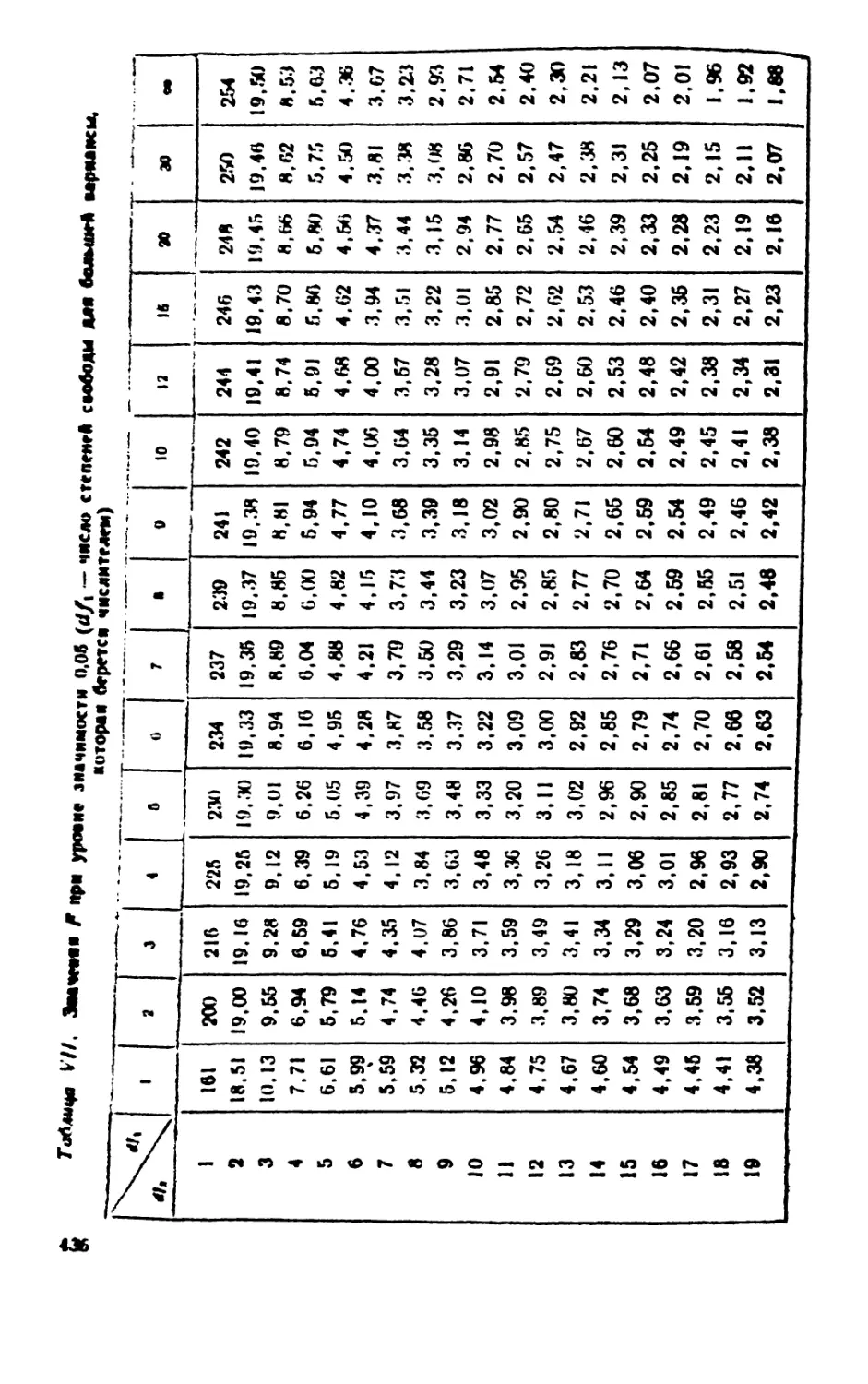
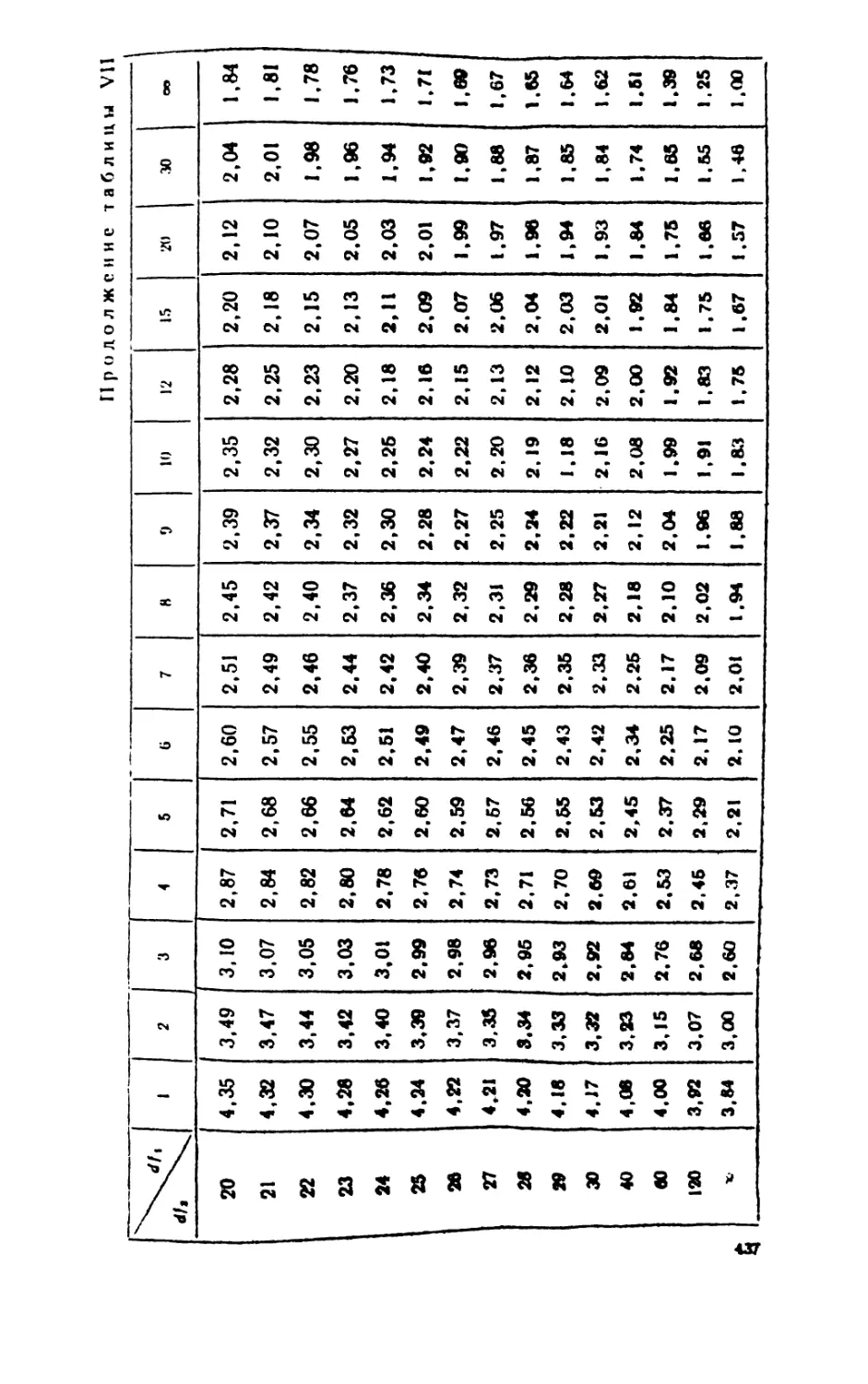
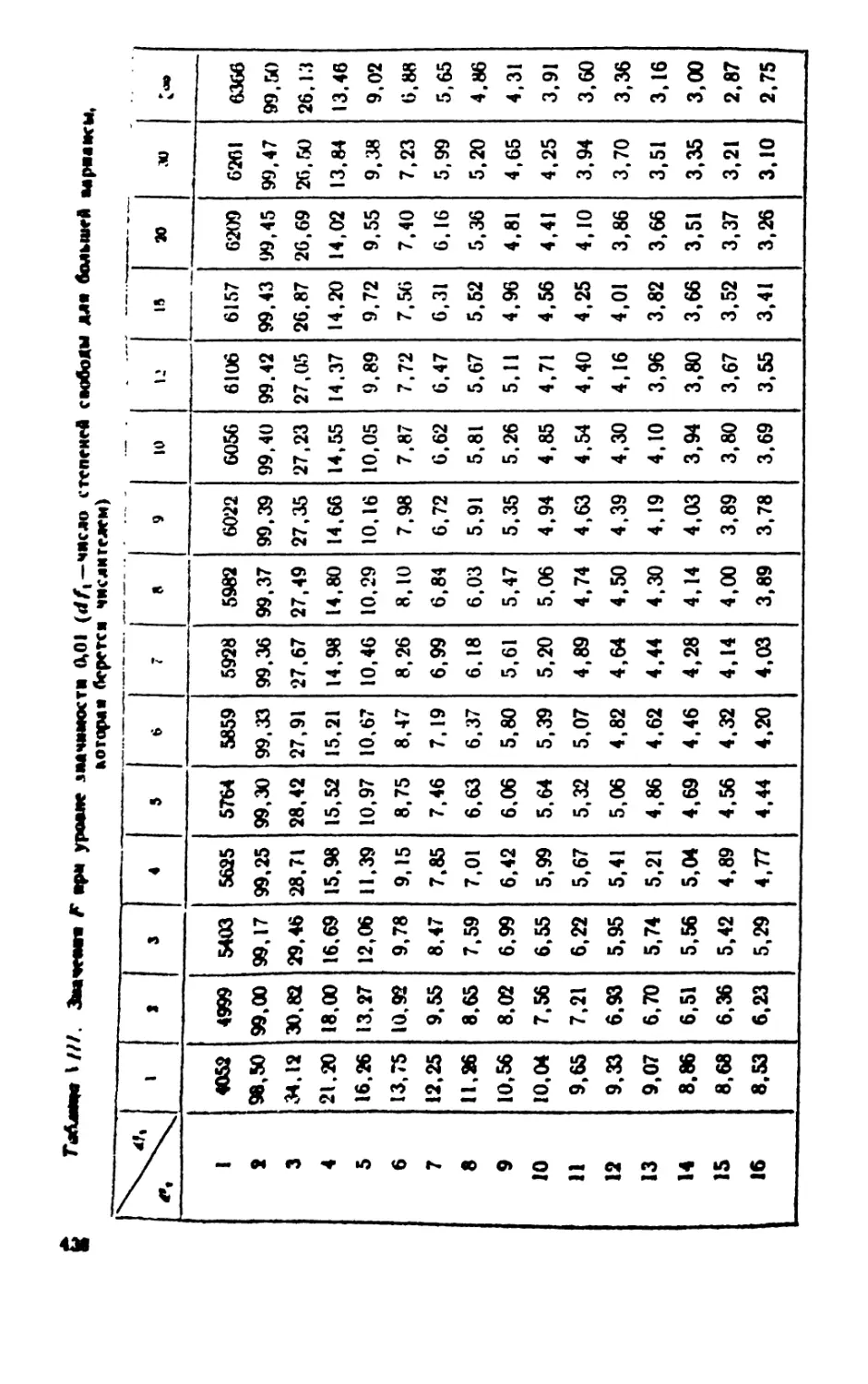
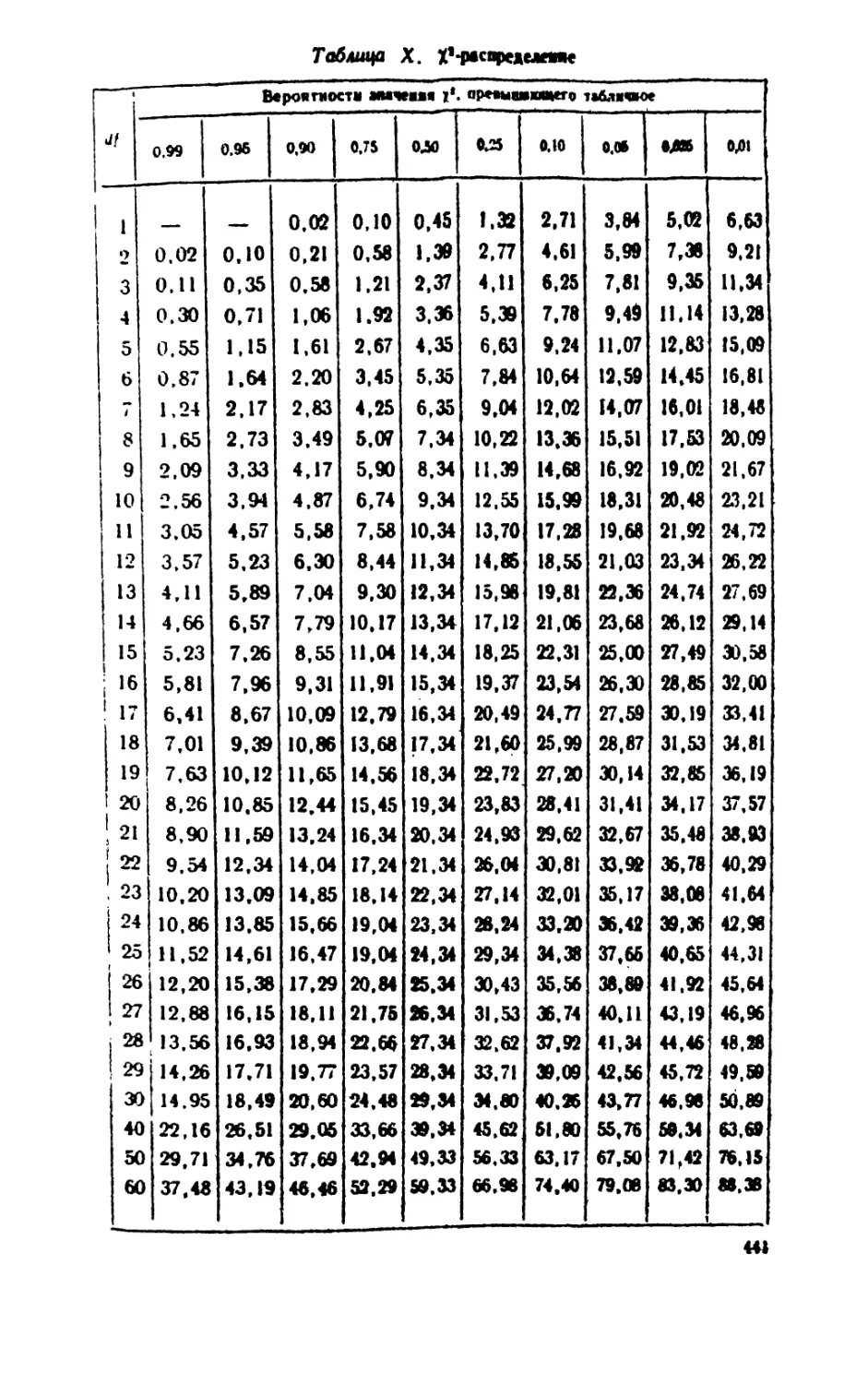
/










































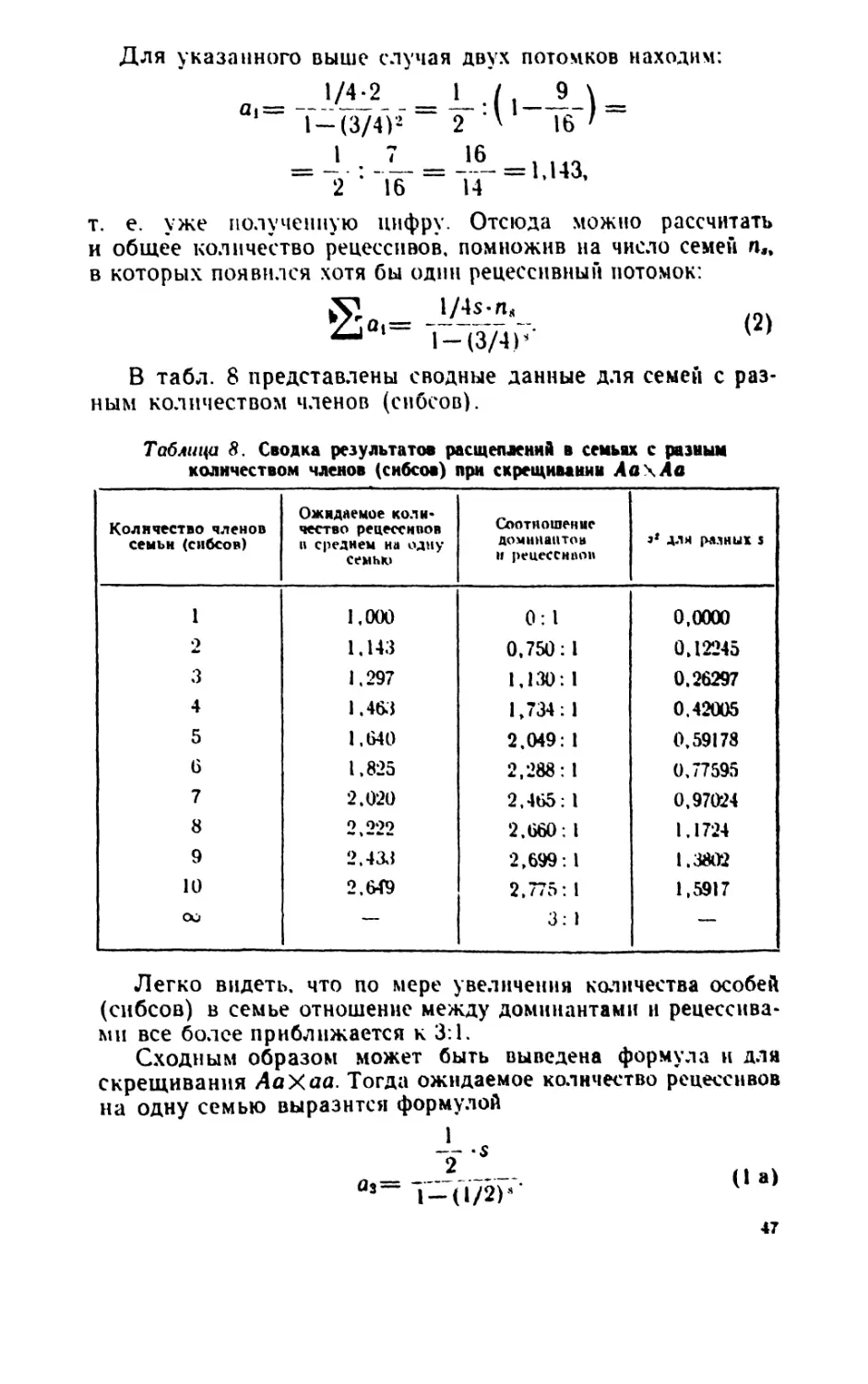


















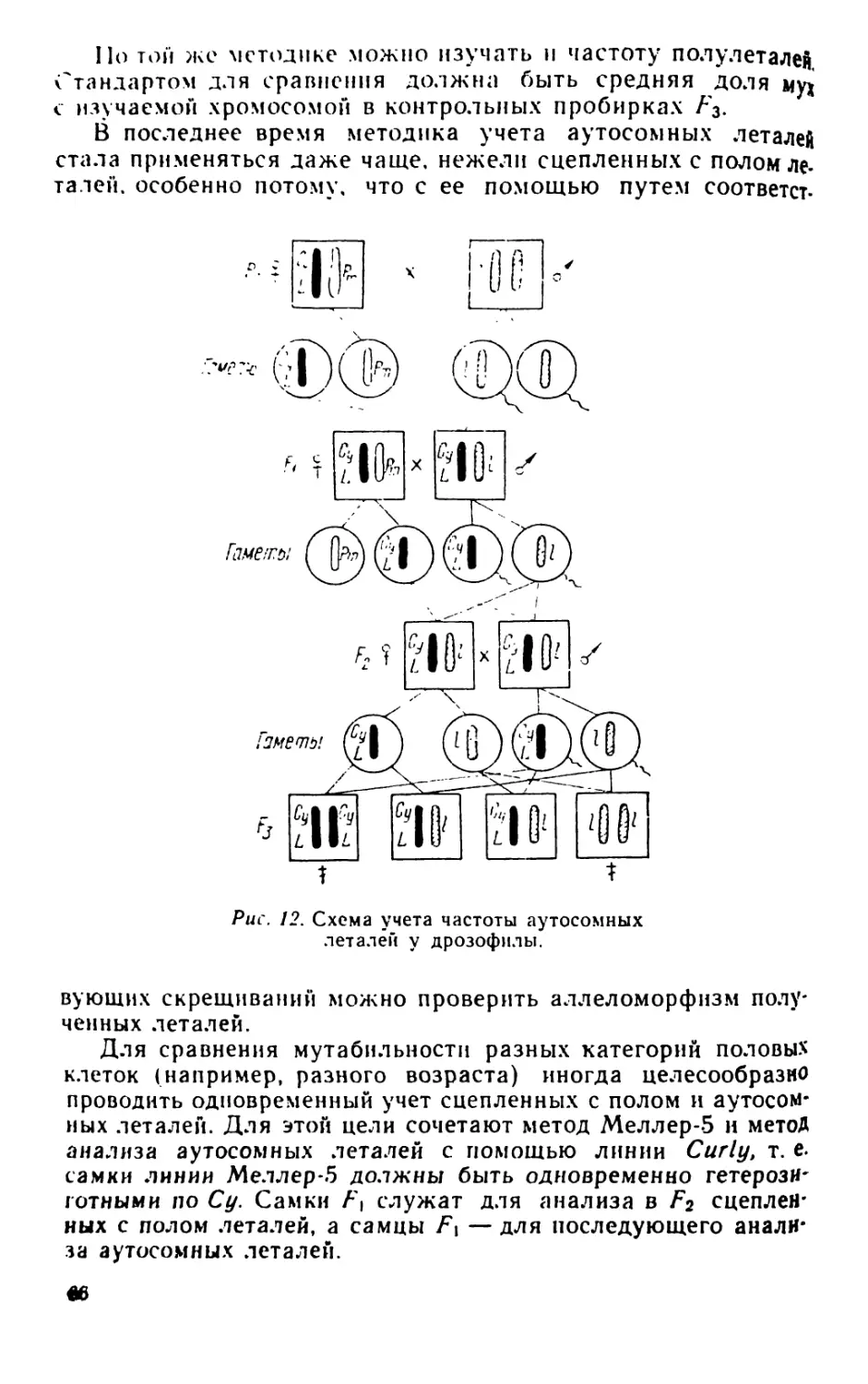





















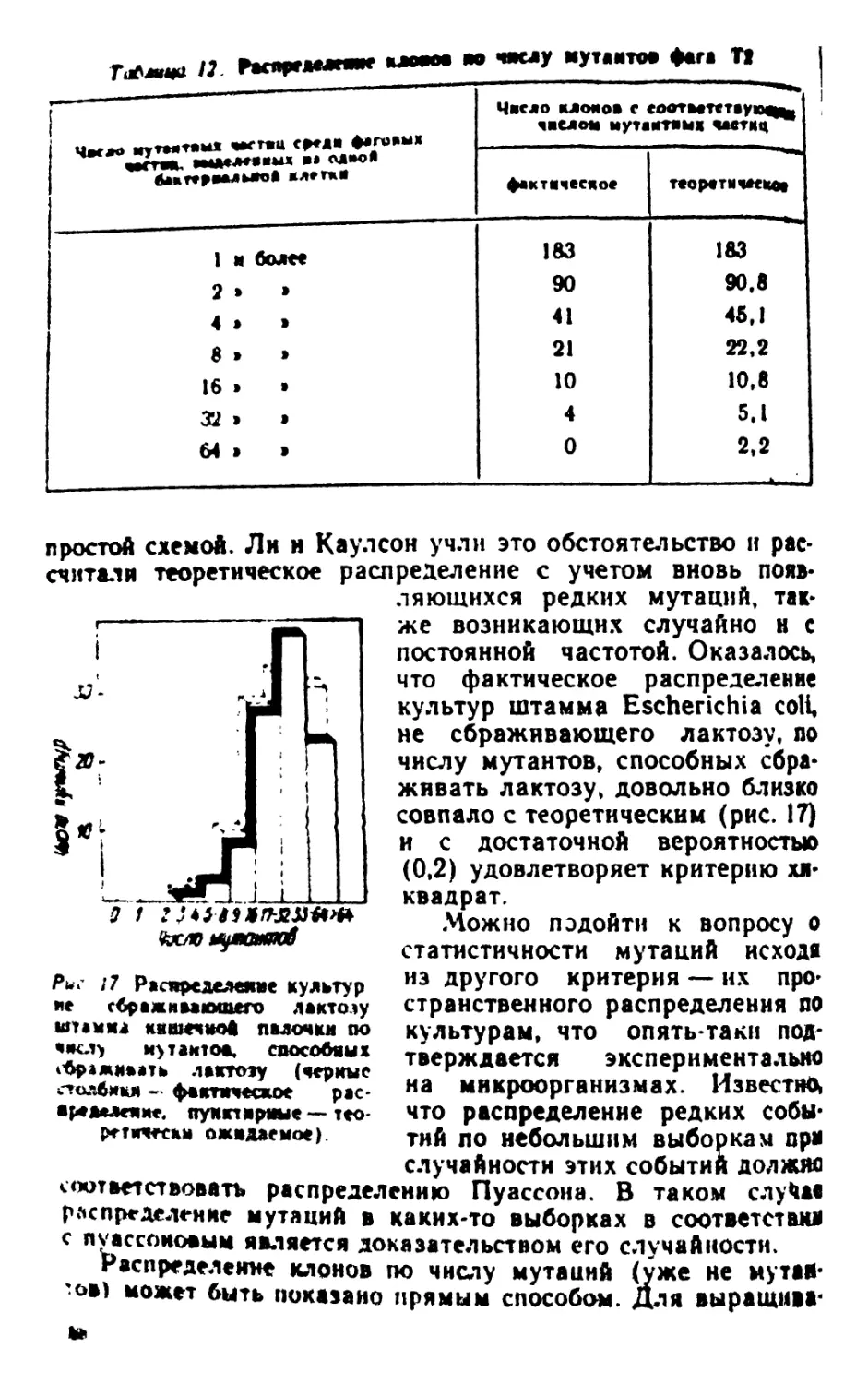

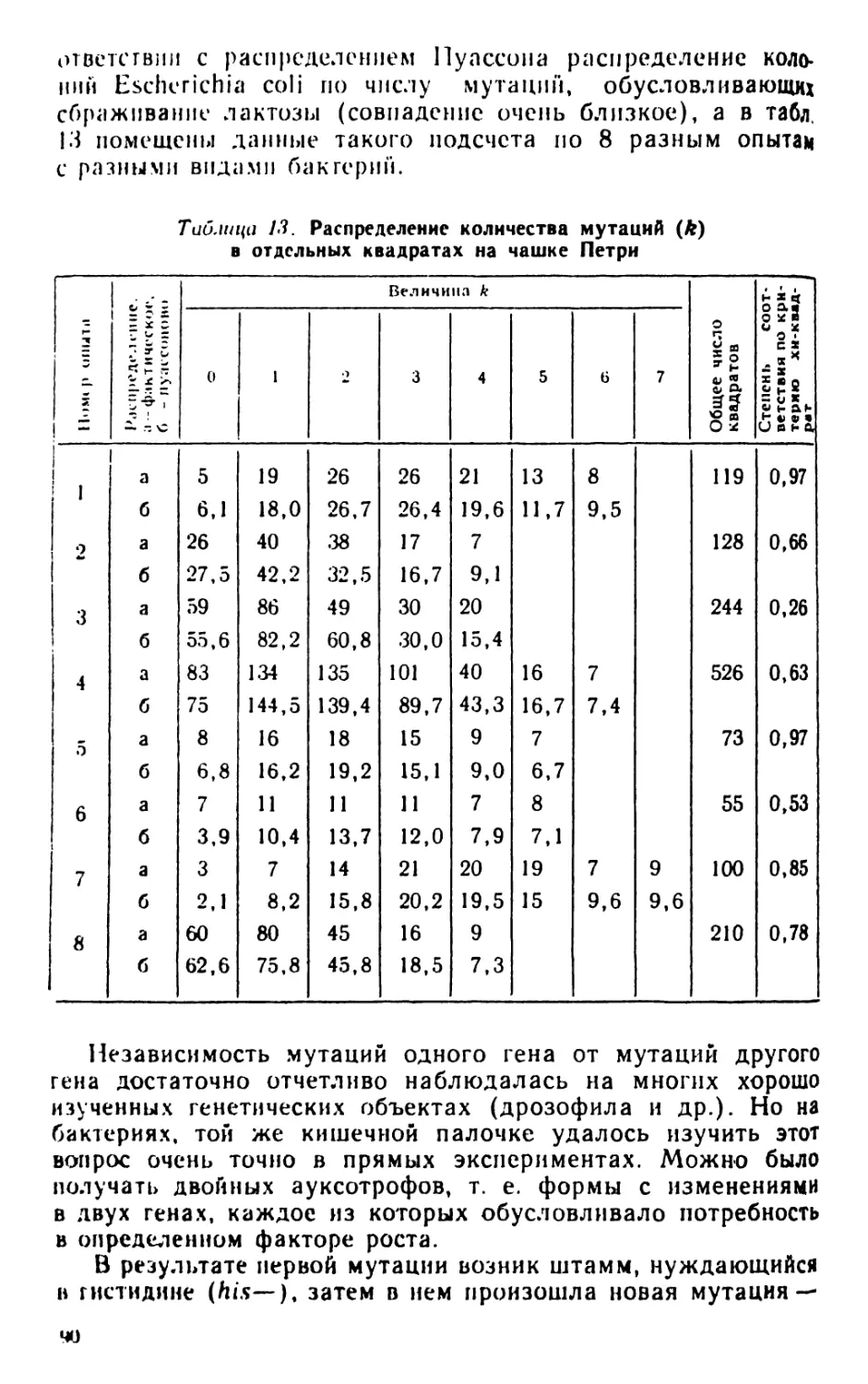














































































































































































































































































































Author: Рокицкий П.Ф.
Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез генетика учебное пособие
Year: 1974




Text
П Ф Рокицкнй
в статистическую
генетику
ИЗДАТЕЛЬСТВО «•ЫШЭЙШАЯ ШКОЛА
57.02
УДК 575(075.8)
Рецензенты: селекции ЛГУ.
- Кафедра генетики и в. Ю. у р б а х
- Проф., докт. биол. н j
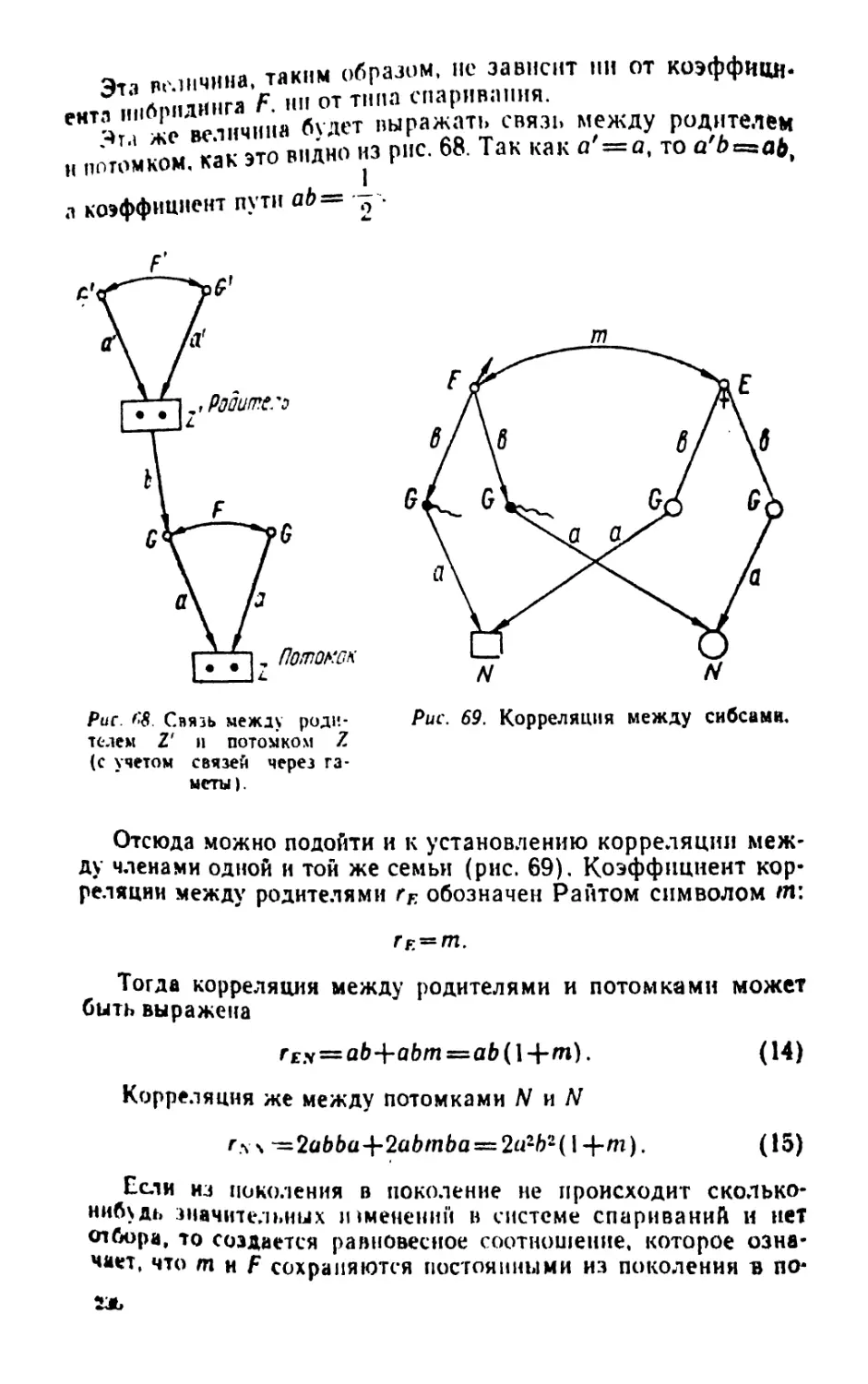
Ропшшй П. Ф. п
Р66 Введение в статистическую генетику. Минск, «ьы-
шэйш. школа», 1974.
448 с с ил.
Книга предназначена в качестве учебного пособия для студентов
биологических cfKtnuAMocrci университетов. Она дает представление о содержании,
методах, теоретическом я практическом значении тех разделов современной
tmrrwut. кторие осиовагеаиттсн яа статистических закономерностях. Особое
акмианве удеяиво анализу явлеияй наследования, мутационному процессу.
геиатичеокоАсгруктуре поиулящгй и ее изменениям под влиянием отбора н
других факторов. статиетическоО характеристике популяций пои непосоыввой
вьрвлци*.
Киигаиож-т был, использована также студентами юотсхвических и евль-
схттияггвеиамх факультетов и вузов. Она может служить руководством для
научных работников к сиециглвстов — генетиков и селекционеров.
? *U04(«)-74 ^73 57.02
*5 Издательство •ВышИкшшя школа». 1074 г.
ПРЕДИСЛОВИЕ
Начиная книгу под названием «Введение в статистическую
генетику», естественно сначала остановиться на вопросе о
том, чем оправдано выделение из генетики такой ее части.
Общеизвестно, что генетика представляет собой
биологическую науку, охватывающую широкий круг явлений
наследственности и изменчивости у всех организмов, начиная от
фагов и вирусов и кончая человеком. Уже сейчас она очень
сильно дифференцирована не только по объектам изучения
(генетика бактерий, генетика человека и т. д.), но и по
применяемым методам, а также по характеру изучаемых
закономерностей. Так, изучение вопросов мутагенеза представляет
крупную и самостоятельную область генетики. Применение
ионизирующих излучений для вызывания мутаций привело к
созданию радиационной генетики. Своеобразие наследственности
и изменчивости в таких сложных системах особей, как семьи
и популяции животных и растений, вызвало необходимость в
создании популяционной генетики. Несколько иная основа для
выделения статистической генетики. Речь идет о тех явлениях
наследственности и изменчивости, которые основываются,
полностью или частично, на статистических закономерностях.
Известно, что проблема статистических, или, иначе,
вероятностных, закономерностей начинает играть все более
значительную роль в современном естествознании.
Напрашивается прежде всего сопоставление с физикой,
внутри которой также создалась особая область
—статистическая физика. Статистическая физика изучает свойства и
поведение систем, состоящих из огромного количества
отдельных частиц — молекул, атомов, электронов. Выяснилось, что
статистические закономерности лежат в основе очень многих
физических явлений, таких, как радиоактивный распад,
термодинамические явления и некоторые другие. Законы
термодинамики были установлены эмпирически, но позднее они
получили более глубокое обоснование и были выведены из
статистических, вероятностных законов.
3
Примерно такое же положение наблюдается сеАчас и в
ряде областей биологии и особенно генетики.
Достойно особого внимания то обстоятельство, что первые
1тапы развития генетики как экспериментальной наукн были
i то же время и началом создания статистической генетики.
Уже Г Менделю было ясно, что наблюдавшиеся им лакано*
мерные численные отношения групп особей в потомстве гнбрн*
дов основываюгся на вероятностных законах* В классической
работе «Опыты над растительными гибридами» (1865) он пи*
сал: «По теории вероятности в среднем каждая форма пыль*
цы А н а соединяется одинаковое число раз с каждой
формой зачатковой клетки А и а; поэтому одна на двух пыльце*
вых клеток А встречается при оплодотворении с зачатковой
клеткой А, другая — с зачатковой клеткой а; таким же обра-
эом одна пыльцевая клетка а соединяется с зачатковой
клеткой Д, другая — с о»,
Поэтому численные отношения в потомстве являются го*
воря современным азыком> статистическими, т. е, отражают»-
мм лежащие в их основе вероятности, чему будет посвящена
специальная глава в книге.
Мендель правильно оценивал необходимость больших
чисел, чтобы добиться приближения фактически получаемых
данных по растеплению к теоретически ожидаемым исходя
из вероятностей образования половых клеток и их
соединения. В отдельных же семьях, т, е, в малых выборках, он
наблюдал значительную вариацию в распределении признаков
среди потомков. Так, он привел данные о десяти растениях, у
которых наблюдались различные отношения, в том числе та»
кие отклонения, как 28:6, т, е, 3,29:0/71. и 19:10, т. е, 2,72:1,31
В настоящее время в генетике очень широко применяют
различные математические, или, точнее говоря, математиков
статистические, приемы, а в ряде случаев идут дальше, соз*
давая определенные математические модели генетических
явлений, которые анализируются в экспериментах. Наконец,
появился и такой круг вопросов, в которых удается провести
чисто математический анализ. Все это привело к созданию
рядя книг и монографий под разнообразными названиями:
«Математическая генетика», «Биометрическая генетика» или*
более просто, «Количественная генетика», В значительной
степени это синонимы, так как фактически в них идет речь об
омом и том же. Мы все же предпочитаем термин
«статистическая генетика»» так как он более отчетливо выражает
основную идею о статистичности ряда генетических явлений как
обзде* закономерности,
Статистическая генетика — это не просто применение ста*
тмстических методов в генетике, а анализ генетических ивл*
ни! со статистической точки зрения, начиная при этом с ***
4
лсиий элементарного расщеплении» или мутаций, и кончав
процессами» протекающими в сложных системах особей —
популяциях или вилах.
Нельзя не отметить» что существует» к сожалению» разрыв
между значительным прогрессом в раэвнтин статистической
генетики, разработкой новых подходов к математическому
анализу генетических явлений и очень большими
трудностями дли большинства генетиков н селекционеров в понимании
работ и статей в этой области.
Причины такого разрыва многообразны, но одна из них.
возможно» заключается в отсутствии переходного звена меж-
ду тем низким уровнем математических и статистических
знаний» которые получают студенты-генетики даже в
университетах (еще более низок он в сельскохозяйственных н меди-
цннских вузах), и большой математической сложностью
некоторых научных работ по статистической генетике, попадаю-
ших в их руки.
В результате очень часто создается ситуация, когда
молодой генетик или селекционер, будучи не в состоянии
разобраться в использованном в статье математическом аппарате,
просто игнорирует ее, а в дальнейшем вообще проходит мимо
статей и исследований, в которых применяют незнакомые ему
приемы и подходы.
Кроме того, нельзя не учесть, что многие генетики, да н
вообще биологи не обладают необходимыми навыками
статистического и математического мышления, выработка которых
совсем не так проста и требует не только соответствующего
образования в вузе, но н длительной тренировки в процессе
работы, привычки к статистическому подходу в понимании
генетических н селекционных процессов.
Эти обстоятельства и явились причиной создания данной
книги как введения в статистическую генетику. Словом
«введение» мы подчеркиваем, что в ней не будет дано со всей
полнотой то, что излагается в существующих больших
зарубежных монографиях» например' Э. Вебср, О. Кемпсорна и
других авторов, но она должна помочь студенту
университета, биологического или сельскохозяйственного вуза, а также
научному работнику — не специалисту по статистической
генетике — приблизиться к статистической интерпретации
генетических явлений и понимать литературу в этой области.
Материал книги был положен в основу читаемого в
течение нескольких лет в Белорусском государственном
университете им. Вч И. Ленина спецкурса под таким же плаванием
для студентов, специализирующихся по генетике и цитолога*.
В настоящее время спецкурс «Статистическая генетика»
читается лишь в немногих университетах Советского Союза. Мы
надеемся, что данная книга явится хорошим стимулом для
S
организации этого курса и послужит учебным пособием дЛя
его слушателей.
Отзывы и пожелания о книге просьба направлять по ал.
рссу: г. Минск, Академическая, 27, Институт генетики и цнто."
логии Академии наук БССР, академику П. Ф. Рокицкому.
Введение
СТАТИСТИЧЕСКИЕ ЗАКОНОМЕРНОСТИ
И ОСНОВНЫЕ ПОНЯТИЯ ТЕОРИИ ВЕРОЯТНОСТЕЙ
Различные формы причинно-следственных связей в природе.
В основе всех явлении природы, органической и
неорганической, лежат причинно-следственные связи и всеобщая
обусловленность» носящая на философском языке название
детерминизма. Детерминизм — общин принцип всякой науки. В
прошлом казалось, что основным характером связи
является простая, элементарная зависимость между причиной и
порождаемым ею следствием и что эта зависимость
однозначна, т. е. одной причине строго соответствует определенное
следствие (так называемый Лапласовский детерминизм).
Сейчас выявляется значение иных, более сложных
причинно-следственных связей и обусловленностей. К их числу относятся
различные формы функциональных зависимостей, которые
могут группироваться в сложные цепи, различные виды
взаимодействий (имеющих разный знак — «плюс» и «минус») и т.п.
Среди них следует также назвать статистические, или
вероятностные, связи. В этом случае конечный результат явления,
обычно обнаруживающегося в больших совокупностях
объектов, создается объединенным действием многих независимых
или почти независимых факторов, каждый из которых
осуществляется случайно, но с определенной вероятностью.
Статистические закономерности — выражение объективной
случайности. Нет сомнения, что статистические
закономерности очень широко представлены в природе. Прежде всего ::ео«".-
ходпмо иметь в виду, что основанные на случайности
статистические процессы детерминнстнчны по своей природе, т. с.
строго обусловлены, но это детерминизм особого рода.
Наряду с необходимыми явлениями существуют и случайные,
т. е. такие, в которых причинно-следственные связи скрыты.
Диалектический материализм признает объективную
случайность. Случайность есть форма проявления необходимости
н в то же время дополнение необходимости. Энгельс писал:
«Случайность — это только один пат юс взаимозависимости,
другой полюс которой называется необходимостью».
7
Лм»лект*к^иатериалистическог представление oo отлк-
*»■•<* сяуцлЬжктл как форм* необходимости дает возмож-
■«crv правили» оценивать многочисленные факты
статистически! лмшоттеряосге* • явления! природы и в том числе в
явлениях наследственности. Однако • реальных явлениях и
ваооессзх происходят очень можно* переплетение закояомер-
мете* когда динамические, кеве^оятиостные закономерности
**кляды»ак>тся на статистические и наоборот. Немало
примеров »того рода дает и статистическая генетика.
Статистические заеааизаизмвостя в биологических явлениях.
Статистические «коиомериоети отнюдь не противоречат
биологическим, как некоторые думают, а входят в них как
органическое ослое. Нередко биологические закономерности
только я осуществляются в форме статистических. Рождение
особи мужского или женского пола биологически обусловлено.
Но к**кр*тяое осу иествлеиве втой биологической
обусловленности возможно лишь в рамках вероятностных законов,
которые воаволяют понять как распределение отдельных семей
во количеству особей женского и мужского пола (в одних
семьях может быть больше мальчиков, в других— девочек),
так в обще* соотношение между особями мужского и
женского яола в сумме всех семей.
Неправильно мнение, что случайность — это отражение не;
еуюеетвеяяых связей. Наоборот, как раз самые существенные
связи могут выражаться в случайности, вероятностной форме,
как гго имеет место, в частности, в явлениях радиоактивного
распада
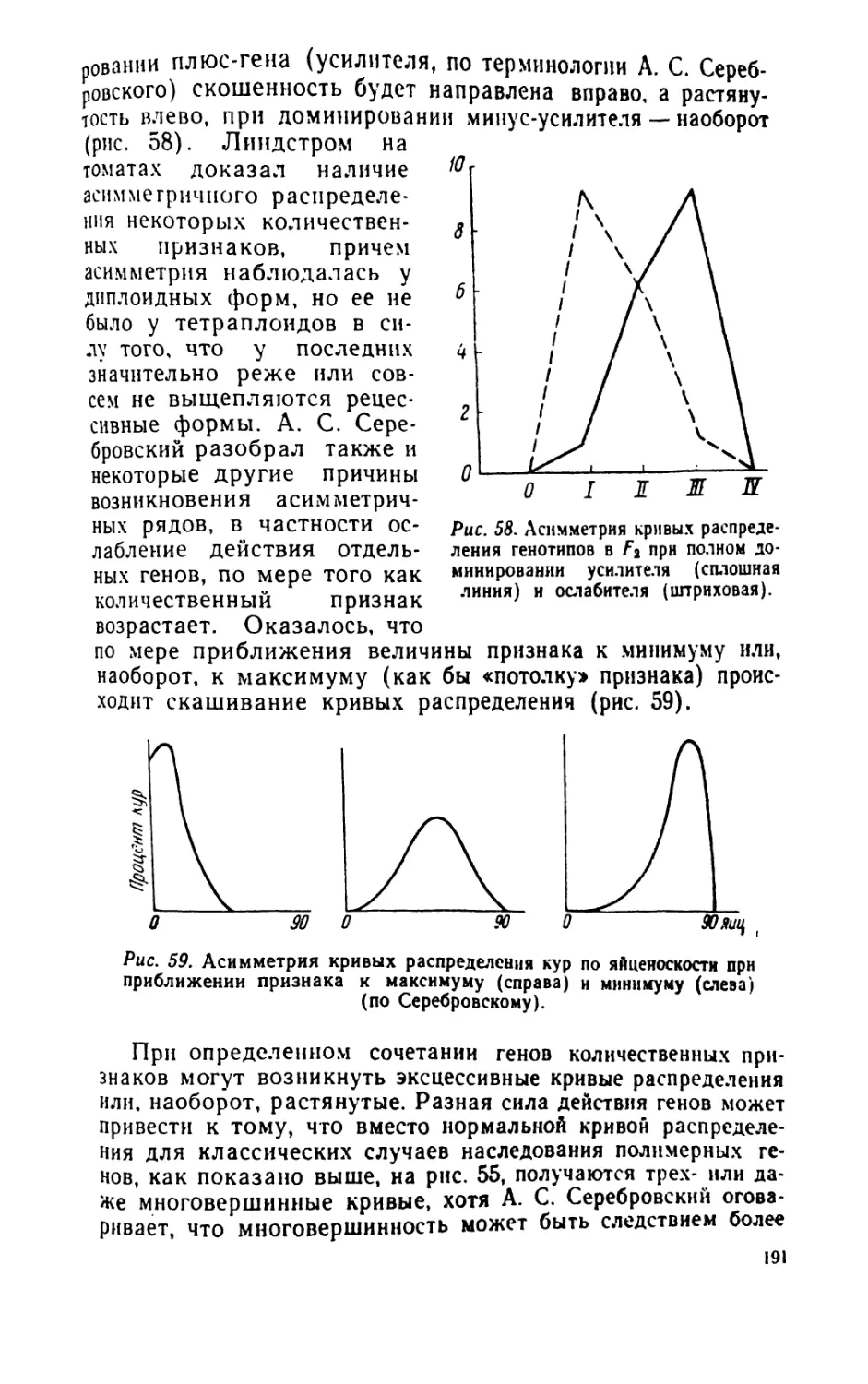
1вав1ввтивс1ь ярсяоязвиия явлений яря статистических
вявикяввмимнктях. Предсказуемость в одинаковой степени
возможна при любых видах закономерностей, как элемеитарио-
явичнийых. твк и статистических, Казалось бы. что
астрономические предсказания, основанные иа известных законах
движения небесных тел, являются наиболее точными. Но это
верно при некоторых идеальных условиях. Оказывается, что
современная астрономия широко использует именно
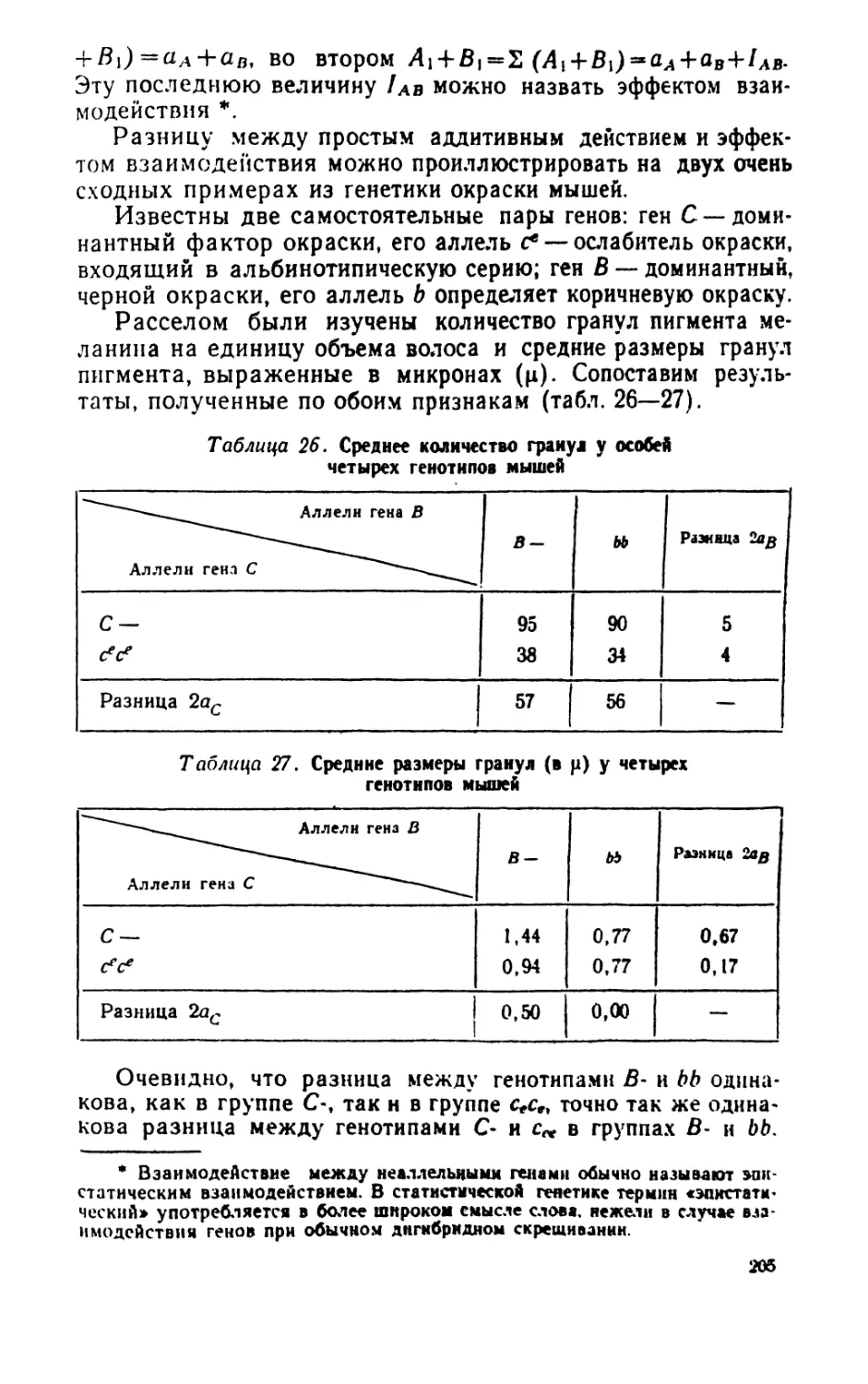
статистические методы, особенно при изучении больших
совокупностей звезд* Еще в начале прошлого века Лаплас в работе
«О кометах» применял методы теории вероятностей.
Конечно, статистические закономерности не дают
возможности предсказать появление отдельных событий. Каждое от-
ильное событие я мест только свою вероятность появления,
бллмвую пая меньшую. В зтом неполнота статистических
законов, впрочем, не большая, чем многих других законов. Но
маяк** мечен** статистических закономерностей
заключаемся в том, что они помогают предвидеть свойства больших
'См М С ШшлвФ€тА. 1и*иш, жамил вазуи. Пая. «-е. М„ 1*46.
совокупностей и предсказать частоту определенных событий
в них.
Изучение вида крота в пределах его ареала во небольшим
выборкам дает возможность судить о вядг в целом, при этом
не только о средних значениях его признаков, но к о граяв-
цах колебаний вокруг средних, о вероятных частотах
отдельных групп кротов.
Дальнейший материал статистической генетики покажет,
что ясное понимание явлений в одной только вероятностно*
форме открывает громадные возможности для предсказания
того, как они будут осуществляться при тех или иных
условиях,
Явления наследственности, как известно, наблюдаются на
разных уровнях, таких, как макромолекулы, хромосомы,
половые и соматические клетки, целостные организмы, семьи,
популяции, виды. На каждом таком уровне проявляются как
вероятностные, так и невероятностные —динамические
—закономерности *, при этом в своеобразной для каждого уровня
форме.
Так как весь материал статистической генетики строится
на принципиальных положениях теории вероятностей и
математической (или биологической) статистики, необходимо
остановиться на некоторых основных понятиях и положениях
теории вероятностей.
Вероятность. Под вероятностью обычно понимается
возможность осуществления определенного события в некотором
количестве случаев из общего количества всех равновозможвых.
Математически она может быть выражена следующим
образом:
Здесь Р(А) — вероятность осуществления события Л; ш —
число благоприятных случаев, т. е. случаев, когда данное
событие А осуществляется; п — общее число всех возможных,
или, правильнее, равновозможных случаев.
Вероятность, как известно из общих курсов статистики,
можно обозначать как большой Р, так и малой буквой
р («пэ»). Но так как значком р в статистической генетике
обозначают частоты генов, удобнее обозначать вероятность
большой буквой Р (кроме тех случаев, когда это специально
оговорено).
* В дальнейшем мы будсы употреблять пот общий термам цпамяче-
скме закономерности» для всеж тех случае», когда ■ яалетшх обмаруаш-
пптсп закономерности, не и|«ШИ^JH»^ Oei могут
быть самыми рш^ичны!^— ф|Ь»оло1^^«>4>С^^ч^с1с»ма и юты*.
й
Прицеленная формула, очевидно, относится к эмпирической
н* роятногти, так кап для определения значения Р нужно было
\АИ- наблюдать осуществление данного события Ач т. е.
должны были реализоваться те условия, которые необходимы для
осуществления события. Кроме того, надо было знать и
относительную частоту явления, установленную эмпирически на
достаточно большом материале. Оцененная таким образом
вероятность называется
апостериорной, т. е. она
является следствием уже
полученных ранее данных. Но
определенная апостериорная
вероятность в свою очередь
дает возможность
предсказать частоту данного
явления или события в
последующих опытах или
наблюдениях при сохранении
первоначальных исходных
условий. Тогда вероятность
становится уже априорной.
При небольшом числе
опытов или наблюдений,
охватывающих малое число случаев, частота события в каждом
из них варьирует и носит в значительной степени случайный
характер. Однако при увеличении количества опытов и их
объема эмпирическая частота события приближается к некоторой
постоянной величине, т. е. к его вероятности. Тогда частоту
события при достаточном числе отдельных наблюдавшихся
случаев можно принять за истинное значение вероятности.
Апостериорная и априорная вероятности могут служить
оценками математической вероятности или математического
ожидания. Чем больше проведено испытаний, тем точнее
оценка последней. Взаимоотношения между ними могут быть
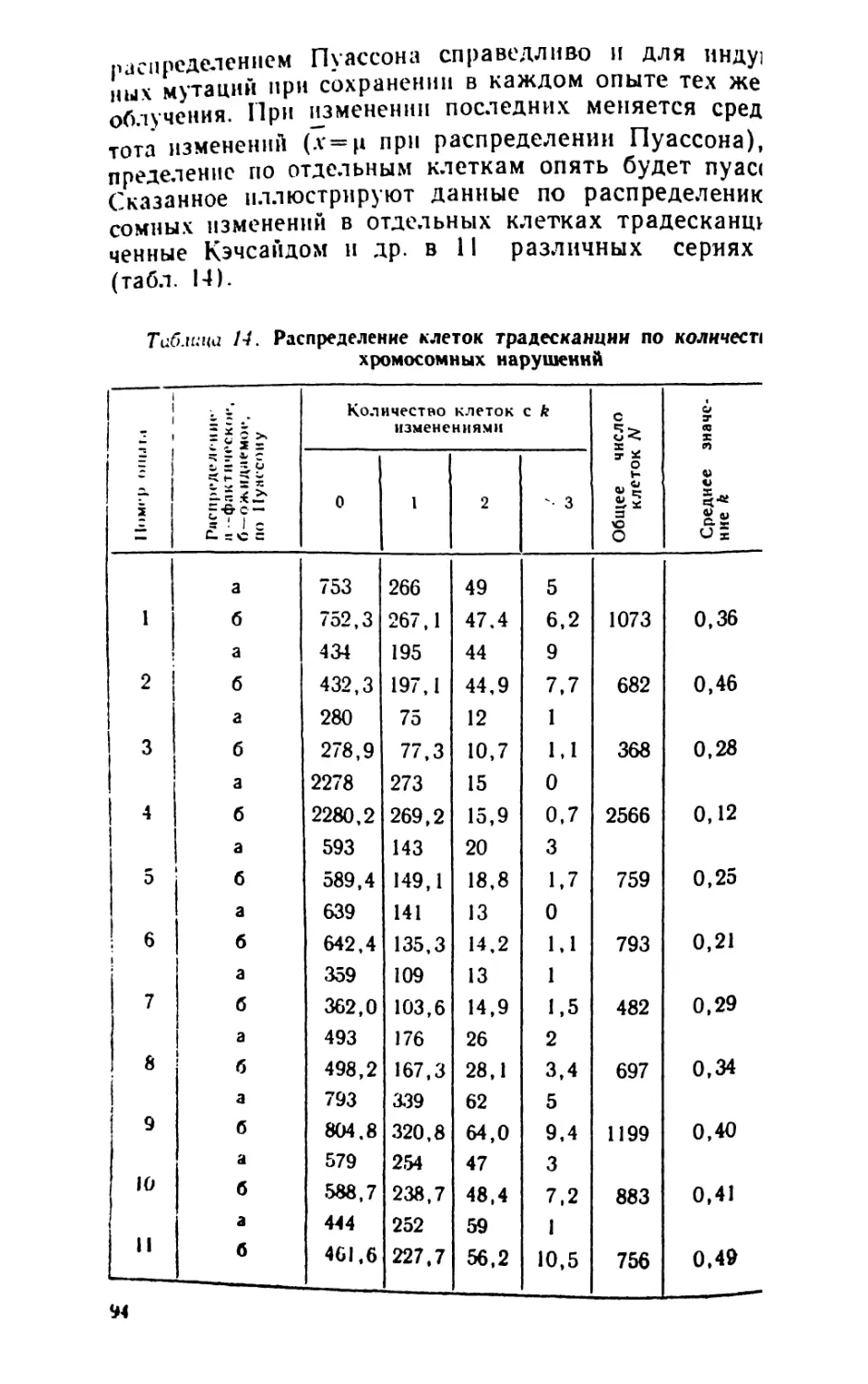
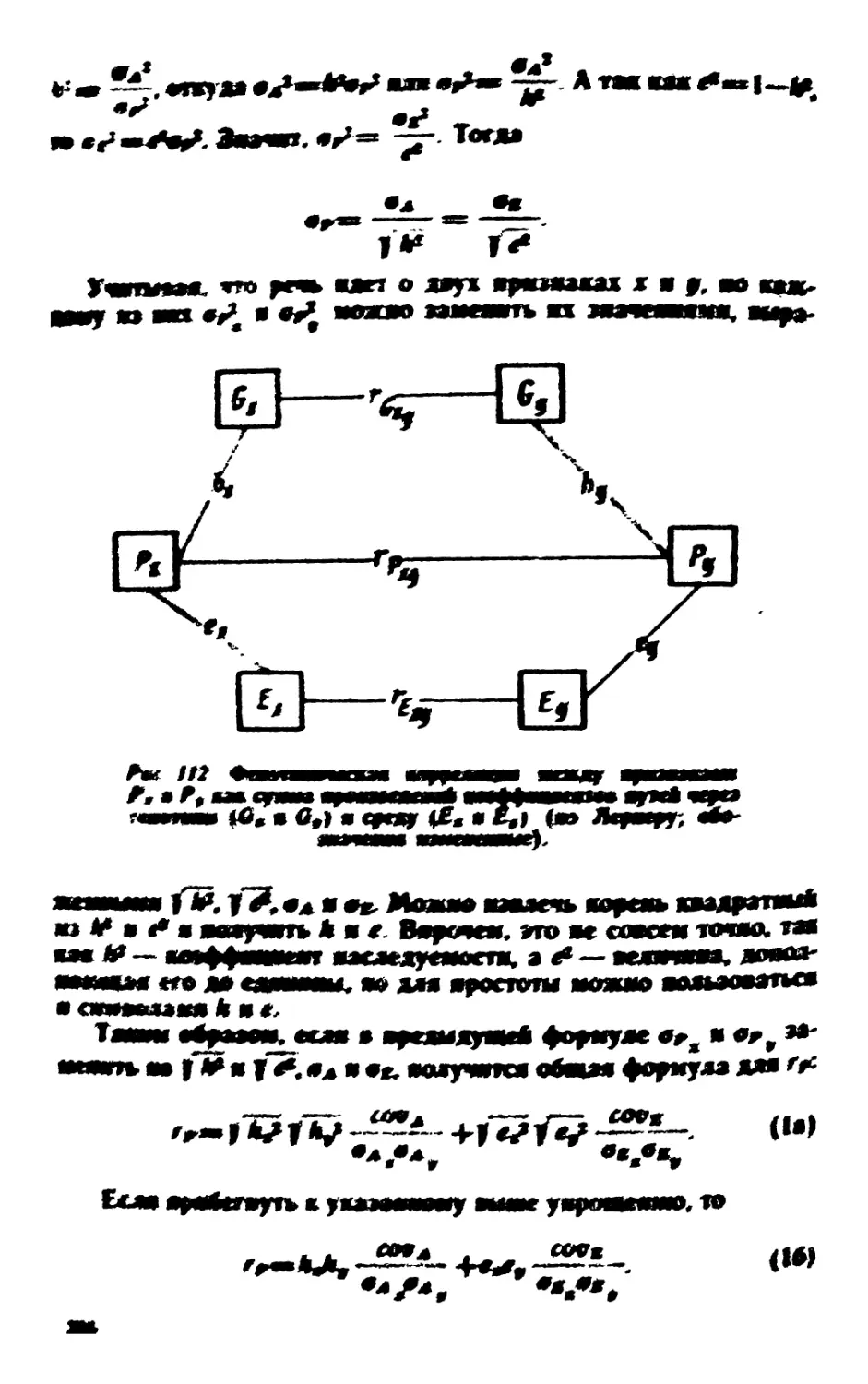
проиллюстрированы рис. 1.
Следует иметь в виду, что для определения вероятности
события далеко не всегда необходимо непосредственное
определение его частоты исходя из опытных данных. Теория
вероятностей дает возможность определять математические
вероятности событий или косвенно, через вероятности других
событий, с ними связанных, или путем построения теоретических
совокупностей всех возможных случаев и вычисления аля них
теоретических вероятностей. Однако и в этом случае для
проверки правильности таких теоретических конструкций надо
обращаться к эксперименту. Многочисленные примеры этого
рола мы увидим в материале статистической генетики.
Вероятность события, как явствует из формулы, обычно
■>£
Эксперимент-
Математическая
вероятность
^
Ч
*feg>-
Рис. I. Сняэь между данными
эксперимента и цероятностями:
апостериорной, априорной и математической.
10
выражается дробью и может колебаться в пределах от ну.?»
(когда событие вообще не осуществляется) до единицы <ссъ
бытие осуществляется во всех случаях), т. е.
O^P^I.
Отсюда возможность оценки событий как менее или более
вероятных. Впрочем, она всегда субъективна, так как между
более вероятными и менее вероятными событиями можно
установить любую границу.
На практике принято пользоваться стандартными уровнями
вероятности (доверительные вероятности): 0.95 и 0.99. Иногда
к ним присоединяют еще 0,999.
Случайная величина и случайное событие. С понятием по-
роятностн тесно связано и другое понятие — понятие
случайной величины или случайного события. Это величина, которая
в результате опыта может принять то или иное значение, но
какое именно, заранее неизвестно.
Допустим, известно, что вес телят при рождении колеблется
в пределах 27—35 кг, но точно предсказать, каким окажется
вес данного конкретного теленка, мы не можем.
Однако наши знания о закономерностях осуществления
данного события значительно увеличатся, если мы сможем
оценить вероятности различных возможных значений
случайной величины, т. е. если сумеем установить, какова
вероятность рождения в помете норок именно коричневой норки, а не
голубой, каковы вероятности рождения телят с живым весом
30 кг, 27 кг или 32 кг. Если полученные вероятности
неодинаковы, то это дает известное основание для конкретной работы
с данным материалом—группой животных или растении
и для предвидения результатов.
Процесс осуществления случайной величины или
случайного события на основе его вероятности называют
вероятностным или стохастическим.
В основе всей статистической теории лежат две теоремы;
сложения вероятностей и умножения вероятностей.
Теорема сложения вероятностей. Вероятность
осуществления двух несовместных событий, т. е. таких, каждое мл
которых осуществляется независимо от другого, равна сумме ihv
роятиостей этих событий:
Приведем пример из генетики. Он будет полезен еще и
полотому, что введет и систему некоторых обозначений,
используемых в статистической генетике. В популяции возможны:
3 генотипа ЛА Аа аа
С частотами
или вероятностичи Р ?Ц R
II
•с
tw»
*,-*,*****« *+*?•
Вашими» • лмиюм случае iwiwuicmr частота*, яла
ллзят T«rj» яра «fmtm идополаг «c«6e* X оккшшм. фар*
ЛА лдея* «am, *А\ ^т A+-SQ4 ш
В «Йцп ми» те***«у скокмям аеусчжчгвв «охаю
опт
Мх**)-^
*л^
«
^—ie««mmciK: 1£— сушо m ootemtiL
Ни ami теорекы шгакат зм шип сждгтжа.
Г Беж «auw uvwm* we ««гогжеслые собмпся «ж Да
«ч Л ^ т* гут** шх wtf&m&cvt& уимгп еднивд. т. е.
E^HN-*,kS»i
pcerreccrv «быта* А. шмяинявня iqm yr mm»*, ч»
В
Мда)-Д***
чЛвхЛ** Всшмнирсяь передаче Kiiifu Лв
JM)a»P ркуезуа А «г *|Х
Учитывая биологический аспект сочетания двух
рецессивных генов, надо иметь в виду возможность меньшей
вероятности одной из гомозигот, например ЬЬ, в силу пониженной
жизнеспособности этих форм \ Рьь = .- вместо Рьь**—)т
В таком случае вероятность
_ JL J — '
РааЬЬ- А '"J -~20"
Полная вероятность. В формуле полной вероятности
используются обе теоремы — сложения и умножения
вероятностей. Смысл ее заключается в том, что если какое-то событие
А может произойти вместе с одним из частных событий Ни
//*, .... Нп (все W,, вместе взятые, составляют полную группу
несовместных событий*), то вероятность события А
вычисляется как сумма произведений вероятности каждого
события Hi на вероятность события А при каждом событии
Ни т. е.
P(A)=i;P(ir,)PH,H).
Эта закономерность существенна в тех случаях, когда надо
рассчитать полные результаты скрещиваний в популяции
между особями разных генотипов. Возьмем приведенный выше
пример, когда популяция состоит из 3 генотипов: АА% Аа и аа
i частотами, т.е. вероятностями, каждого из них
соответственно /\ 2Q. /?. Допустим, что между всеми ими происходят
случайные скрещивания, в том числе между сходными по
генотипу, т. е. ААхАА, АахАа, ааХаа, и несходными — АаУ^аа,
АахАА и т. д.
Тогда можно рассчитать вероятность каждого скрещивания
и условные вероятности появления в этих скрещиваниях форм
АА. Аа и аа. Всего возможны 9 скрещиваний, вероятности
которых можно выразить произведениями вероятностей взятых
для скрещивания генотипов. Например, вероятность для
скрещивания ААхАа равна
P = PaaXPao~Px2Q~2PQ и т. л.
Чтобы получить общую сумму вероятностей всех типов
потомков (эта сумма должна равняться единице), надо
помножить вероятности каждого скрещивании на условные
вероятности появления в потомстве данного типа скрещивания
форм соответствующего генотипа и их просуммировать.
Удобнее это делать для каждого типа потомков: АА% Аа и аа.
# События 11% ■ статистической литературе часто натыюют гипотезами.
14
Расчеты такого типа будут не раз фигурировать ниже при
разборе конкретного материала статистической генетики
Математический аппарат статистической генетики. Теория
вероятностей лежит в основе всех тех статистических
методов, которые широко используются в статистической генетике
и обычно излагаются во всех руководствах по биологической
статистике, а именно: биномиальное, пуассоново и
нормальное распределения и их статистические показатели, критерии
достоверности статистических показателей, корреляция и
регрессия, ковариаитный анализ, ряд разделов дисперсионного
анализа, критерий хи-квадрат и др, В курсе статистической
генетики нет смысла их рассматривать, но, чтобы облегчить
работу генетика и селекционера, в приложении дан
статистический справочник, содержащий макеты таблиц для обычных
вычислений, важнейшие формулы и статистические таблицы.
Более сложные и специфические для статистической
генетики методы излагаются непосредственно в тексте
соответствующих глав книги.
Глава 1
СТАТИСТИЧЕСКИЕ И ДИНАМИЧЕСКИЕ ПРОЦЕССЫ
КАК ОСНОВА ЗАКОНОМЕРНОСТЕЙ
НАСЛЕДОВАНИЯ
Статистические и динамические условия расщепления.
Закономерности наследования (расщепления, независимого или
сцепленного наследования) установлены уже давно, и нет
надобности на них останавливаться. Но существенно, что в
основе этих кажущихся очень элементарными закономерностей
лежат достаточно сложные явления, осуществляющиеся
преимущественно на хромосомном и клеточном уровнях.
Некоторые из них имеют функциональный или динамический
характер, другие же — статистические по своей природе, и их
осуществление может быть оценено только вероятностями.
Классическими считаются следующие отношения: при
скрещивании АахАа— 1:2:1 по генотипу и 3:1 по фенотипу; при
скрещивании АаХаа— 1:1 как по фенотипу, так и по генотк-
пу. Анализ расщеплений проводят, как известно, по
конкретным признакам, изучаемым у отдельных особей потомства
исходных родителей. Однако реализация конкретных
закономерностей расщепления возможна лишь при следующих
условиях:
а) у особей Аа гаметы А и а образуются в равном
количестве, или, иначе говоря, появляются они с одинаковыми
вероятностями (Я = 0,5);
б) соединение гамет разного типа, т. с. Л и А или And
и т. д., происходит при оплодотворении по законам
случайности, т. е. с вероятностью, равной произведению вероятностей
образования данных типов гамет;
в) жизнеспособность особей различной генотппической
конституции, т. е. АА, Аа или аа, одинакова;
г) признаки А или а должны одинаково хорошо
проявляться фенотипически.
Очевидно, что, если хотя бы одно из этих условий не
соблюдено, точность менделевских. отношений будет также
нарушена.
Первое из перечисленных условий может быть расшифрО*
вано на клеточном уровне как одинаковая вероятность попаД*"
16
ния каждой из двух хромосом дайной пары в половую клет-
ку при мейозе. Цитологический анализ показал, что в резуль-
тате двух мейотических делений, происходящих у особи Аа,
из каждой диплоидной половой клетки образуются тетрады,
т. е. четыре клетки, две из которых несут по гену А и две —
по гену а. При этом только в сперматогенезе пег четыре
превращаются в сперматозоиды. В овогенезе же из четырех
клеток сохраняется лишь одна — будущая яйцеклетка, остальные
три — полярные тельца — погибают.
Таким образом при образовании женских половых клеток
осуществляется еще один дополнительный вероятностный
процесс — случайная выборка одной клетки из четырех
возможных.
Второе условие также имеет вероятностную основу на
клеточном уровне. Неоплодотворениая яйцевая клетка любой
структуры по данной паре генов А~ач т. е. имеющая любую
из пары гомологичных хромосом, может быть оплодотворена
сперматозоидом (или опылена пыльцой) опять-таки любой
структуры. Вероятность образования того или иного типа
зигот выражается произведением вероятностей появления двух
типов половых клеток — Я| У Я2, т. е. в данном случае
0,5X0,5-0,25.
Наконец, третье и четвертое условия имеют не
вероятностную, а динамическую основу. Как большая или меньшая
жизнеспособность фенотипов А и а, так и степень проявления
в фенотипе доминантных и рецессивных генов в основном
определяются закономерностями индивидуального развития.
Впрочем, как мы увидим в дальнейшем, и в их осуществлении
участвуют статистические закономерности.
Строгая закономерность менделевских расщеплений.
Казалось бы, что при наличии четырех различных условий для
осуществления расщепления, каждое из которых может быть
выполнено не в полной мере, трудно было бы ожидать строго
закономерной картины расщепления. Тем более удивительно,
что в действительности она наблюдается у громадного
количества видов животных и растений, в том числе и у человека,
к на большом количестве признаков. Некоторые примеры
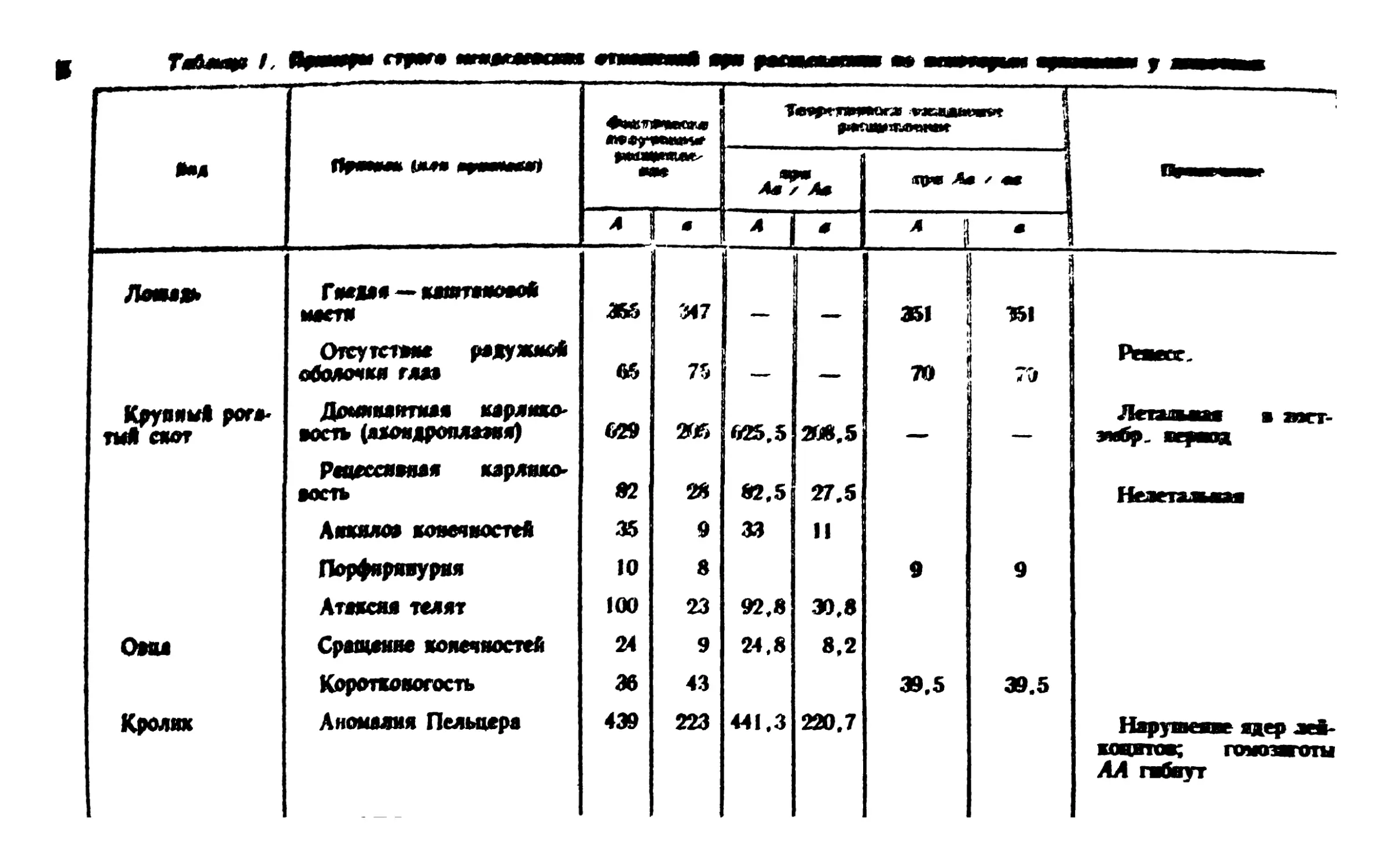
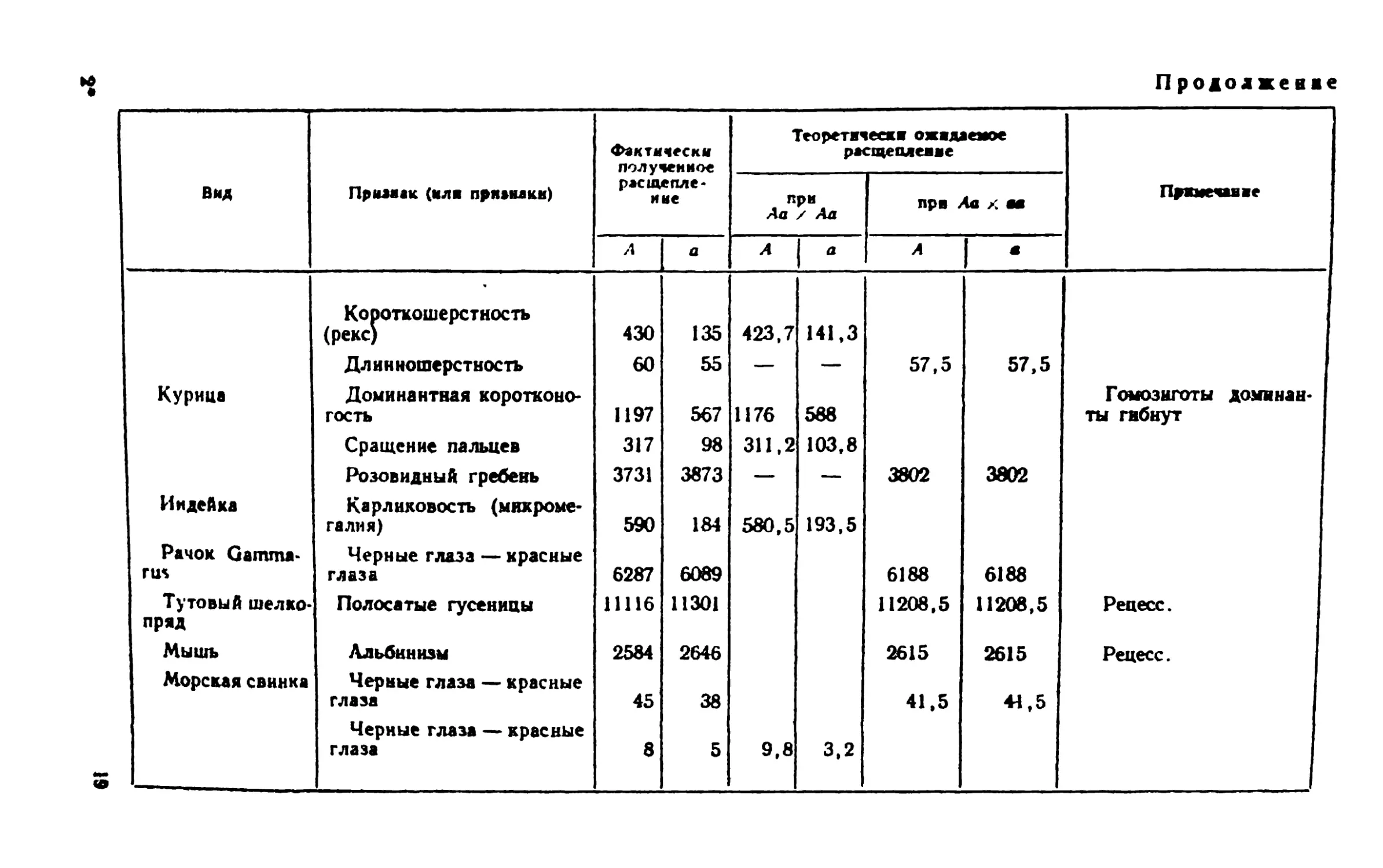
этого рода приведены в табл. 1, но необходимо учитывать, что
они составляют ничтожную долю данных, накопленных в
генетике со времени вторичного открытия законов Менделя. Как
и следовало ожидать, чем больше общее количество особей,
тем ближе фактически получаемое отношение к теоретически
ожидаемому 1:1 или 3:1.
Так, например, у тутового шелкопряда наблюдалось
расщепление по окраске гусениц: полосатых 1! 116 и нормальных
11 301, что составляет соотношение 0,992:1,008; или по
другому признаку: ковровых 5662 и нормальных 5638 (отношение
М
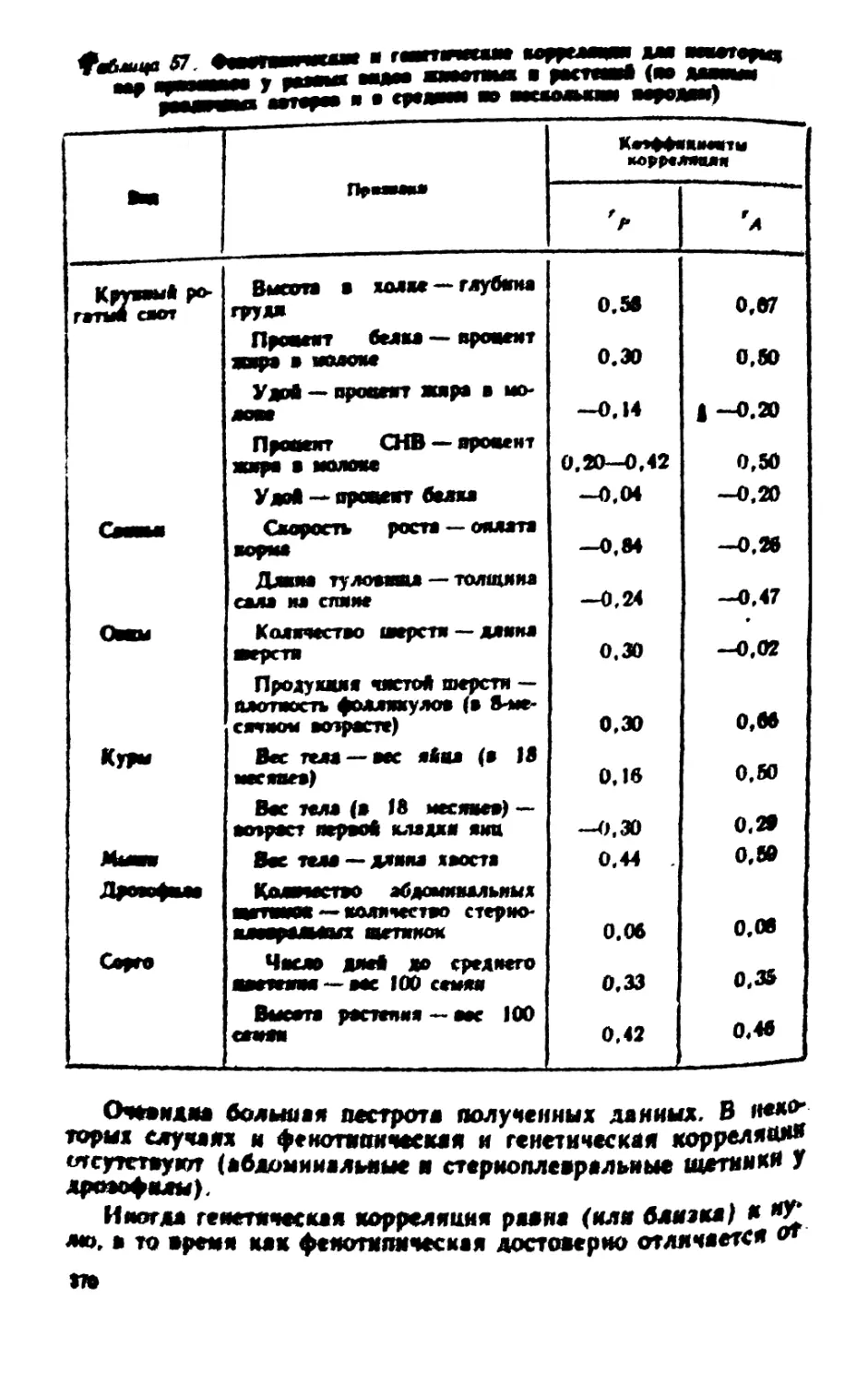
лшваш^щяттшттт* «i i n ** m «
1 *M
JUmm*
КруппыЛ рога*
ты* am
Оме
Кролик
i
Гиеле* — яаттаиооо*
мает*
Огсутетме рмуядей
Домшеитмая карлике-
•есть (алоядом****)
Рецессялиая карляко-
•ость
Аикялое кояеяиостей
Порфирииурия
Атаксия телят
Сращение конечностей
Коротжоиогосп»
Аномалия Пельцера
ФМНПРЯМЗДР
T<aqf* гттосв --«псадоиот*
*** | Л#Т>М |
* I *
i
65
629
82
35
10
100
24
36
439
5Я7
2№
28
9
8
23
9
43
223
ft2S. S
208.5
«.S 27. 5
33 I II
92.»
24.8
441.3
30,8
8,2
220.7
351
70
39,5
"Ю1
7\1
39.5
эмбр..
HapjfUKJUK ядер
коцига;
УМ
Продолжение
Вид
Курица
Индейка
Рачок Gamma-
1 Тутовый
шелкопряд
1 Мышь
1 Морская свинка
Примак (или пряники)
Короткошерстность
(реке)
Дл и н ношерстность
Доминантная коротконо-
гость
Сращение пальцев
Роговидный гребень
Карликовость (микроме-
га ли я)
Черные глаза — красные
глаза
Полосатые гусеницы
Альбинизм
Черные глаза — красные
глаза
1 Черные глаза — красные
глаза
Фактически
полученное
расщепление
А
430
60
1197
317
3731
590
6287
11116
2584
45
8
а
135
55
567
98
3873
184
6089
11301
2646
38
5
Теоретически ожидаемое
рдсщеалеяие
при
Аа / Аа
~А j а
423,7
—
1176
311,2
—
580.5
9,8
141,3
—
588
103,8
—
193,5
3,2
при Аа к ж
А
57,5
3802
6188
11208,5
2615
41.5
| л
57,5
3802
6188
11208,5
2615
44,5
1 Примечание
|
Гомозиготы домннан- 1
ты гибнут
Рецесс. 1
Рецесс. 1
1
1002 0 996) При учете ряда браков людей, имеющих группу
*дои АВ, с лицами других групп было установлено, что
фактор ,4 унаследовало 1570 человек, а фактор В— 1611.
Отношение здесь 0.99:1.01.
Поразительно то, что расщепление очень отчетливо
выражено даже тогда, когда рецессивные формы заведомо
уродливы Если данный геи не затрагивает эмбриональных
стадий, то доминантные и рецессивные особи рождаются в
строго мснделевском соотношении, У свиней есть уродство — гид-
роиефалня (водянка головы). При спаривании животных,
заведомо известных как носителей наследственных задатков
гидроцефалии, т. е. гетерознгот. было получено 136
нормальных и 42 уродливых поросенка. Теоретическое отношение
(при 3:1) должно быть: 133,5 нормальных и 44,5 уродливых.
Степень совпадения очень высокая, что можно легко
проверить и с помощью критерия хн-квадрат.
В связи с тем что одно время высказывалось мнение о
снелшнчкостн» отношения 3:1, полученного Менделем в его
опытах с горохом. В. П. Эфронмсон свел в одной таблице
данные о расщеплении по окраске семядолей (на желтые н на
зеленые), полученные как в опытах Менделя, так и в 14
других опытах, проведенных разными исследователями.
Оказалось, что всего было получено: желтых 152823 и зеленых
50676, т. е. отношение 3,004:0.996. В приведенных отношениях
при расщеплении автор делает ту же ошибку, которая была
допущена и Менделем, а именно: принимает количество ре-
цесснаоа за единицу. Надо же исходить из суммы
доминантой и рецесснвов, взятых за 4. Мы приводим уже
исправленные отношения Само собой разумеется, конечно, что по
отдельным опытам отношения сильно колеблются, т. е.
существует дисперсия отношений в соответствии с
закономерностями случайной вариации.
Таким образом, громадный фактический материал
полностью соответствует предположению, что расщепление
основывается прежде всего на сочетании двух вероятностных
процессов, осуществляющихся на клеточном уровне.
Расщеплете на гаметах. Тетрадный анализ. Изучение
нормальной картины мейоза совершенно точно показало, что
в каждую половую клетку попадает только одна хромосома
на нары гомологичных хромосом. Оставалось неясным,
действительно ли в пределах каждой тетрады также происходит
расщепление по паре генов А и а в отношении 1:1. Однако в
процессе патогенетических исследований были обнаружены
факты, когда можно было наблюдать этн расщепления на
гаметах вполне визуально. Так. у кукурузы была обнаружена
пара генов W и w, определяющих крахмалистый н восковидный
таны эндосперма. Но *тн гены действуют еще к в пыльце.
во
Поэтому, в пыльцевых зернах» несущих ген W9 образуется
крахмал, а в пыльцевых зернах, несущих ген ш, — декстрин.
При обработке иодом пыльца первого типа окрашивается в
синий цвет. В одном из опытов было подсчитано 3437 синих
пыльцевых зерен и 3482 неокрашенных (отношение, очень
близкое к 1 : К а именно 0,994 : 1,006). Подобные же явления
были обнаружены и у других видов растений, например
энотеры, сорго.
Наибольшее значение сыграло открытие так называемого
тетрадного анализа, т. е. возможности обнаружения
расщепления на четырех клетках тетрады после двух делений
созревания. У таких растений, как мхи, дрожжевые грибки,
гаплоидная фаза существует в виде самостоятельной формы. После
прохождения мейоза образуются четыре споры. Их можно
наблюдать самостоятельно, а также можно получать от них
гаплоидные растения (у мхов) или колонии (у дрожжевых
грибков). Расщепление происходит по различным
морфологическим или биохимическим признакам, и тогда наглядно
видно, что расщепление в тетрадах происходит в отношении 1:1.
Нарушения вероятностной картины мейоза. Нерасхожде-
нне хромосом. Наличие закономерности расщепления у
громадного количества видов животных, растений и
микроорганизмов указывает на широкое распространение
вероятностных процессов при образовании половых клеток и при
оплодотворении. В то же время надо иметь в виду, что возможны
отклонения от вероятностных процессов иди их более сложное
сочетание с другими процессами, в результате чего
расщепление может сильно нарушиться или даже исчезнуть вовсе.
Остановимся на некоторых из них и начнем с нарушения
вероятностной картины мейоза.
При нормальном процессе мейоза и образования патовых
клеток каждый ген из данной пары аллелей попадает в
патовую клетку с вероятностью, равной 0,5.
Однако, если мейоз протекает как-то иначе, нарушается
и вероятностная картина. К этой категории относятся факты
так называемого нерасхождення хромосом.
Открытое впервые Брнджесом в 1916 г. явление
нерасхождения патовых хромосом у дрозофилы как причины
нарушения обычной картины наследования сцепленных с палом
признаков было одним из важнейших экспериментальных
доказательств хромосомной теории наследственности н локализации
генов в патовой хромосоме. Но оно интересно и в плане
статистической генетики. Дело в том, что нерасхожденне
патовых хромосом представляет собой событие,
осуществляющееся с очень малой вероятностью. Так, из данных самого Брид
жеса было видно, что случаев первичного нерасхождения
было талько 12 на общее каличество 20 484 изученных особи
II
(обоих полов), что составляет частоту 0,0006 (или 0,06%).
Эта частота, правда, не охватывает всех случаев нерасхожде-
кия. так как яйца без Х-хромосомы (их теоретически должно
быть столько же, сколько и яиц с двумя Х-хромосомами) при
оплодотворении сперматозоидами с У-хромосомон
нежизнеспособны. Очень редко выживают также зиготы, полученные из
инц с двумя Х-хромосомами, оплодотворенных
сперматозоидами с Х-хромосомой. Поэтому указанную частоту
нерасхождения надо увеличить во всяком случае вдвое. Хотя
нерасхождение вызывается какими-то специфическими событиями во
время одного из меиотическнх делений клетки, вероятность
этих событий очень мала.
Тот же процент нерасхождения обнаруживается и при
объединении данных многих исследователей, изучавших не-
расхожденис. Так, на общее количество 170 422 особи было
получено 89 исключительных особей, т. е. 0,0005 (или 0,05%),
величина того же порядка, что и найденная Бриджесом.
Однако в одной линии (eosin-miniature), изученной Сэфиром,
получен значительно больший процент исключительных
особей: 87 на 47 545 особей (0,002 или 0,2%).
Мэйвором было также показано, что облучение
рентгеновскими лучами самок дрозофил увеличивает частоту
первичного нерасхождения — 0,009 от облученных самок по сравнению
с 0,0004 в контроле, т. е. примерно в 20 раз. Наряду с
нерасхождением половых хромосом у дрозофилы хорошо
изучено нерасхождение и аутосом.
Явление нерасхождения известно сейчас у очень многих
видов растений и животных. При участии гамет с одной
лишней хромосомой получаются так называемые трисомики,
т. е. формы, у которых в клетках тела утроена только одна
хромосома (2п-Н), остальные же, как обычно, удвоены.
Расщепление в потомстве трисомиков, очевидно, также будет
нарушенным. Классический пример — трисомия у дурмана
(Datura stramonium): при наличии 12 пар хромосом было
обнаружено 12 типов трисомиков, морфологически
отличающихся друг от друга.
При участии же гамет с отсутствием одной хромосомы
получаются так называемые моносомики (2п—1). Наконец,
при самооплодотворении моносомиков могут появиться нулли-
сомики (2п—2).
Нерасхождение хромосом у человека. Нерасхождение
хромосом встречается и при образовании половых клеток у
людей и лежит в основе так называемых хромосомных
болезнен. Данные но ним получены недавно. Как известно, они
очень важны для медицинской практики. Для
статистической же генетики существенны количественные данные о
частоте нерасхождений.
Я
У мужчин, больных синдромом Клайнфельтера
(характеризуются рядом физиологических и психических дефектов),
оказалось в клетках тела по 47 хромосом вместо
нормальных 46, в том числе две Х-хромосомы и одна У-хромосома.
Частота синдрома Клайнфельтера в популяциях человека
примерно 0,00! (или 0,1%).
Если особь ХХУ получается в результате оплодотворения
яйцезой клетки с двумя Х-хромосомамн сперматозоидом
с У-хромосомой, то, очевидно, такая яйцевая клетка должна
быть результатом нерасхождения половых хромосом в мено-
зе. Но при этом должен возникать не один, а два типа
половых клеток: XX и О. При оплодотворении двух типов
половых клеток двумя типами сперматозоидов — с Х- и с
У-хромосомой следует ожидать образования четырех типов зигот:
XXX; ХХУ; ХО и УО. Последний тип зигот едва лн может
быть жизнеспособным вследствие слишком большого
нарушения хромосомного баланса (полное отсутствие Х-хромо-
сом). Остальные же типы были фактически обнаружены. Так,
особи XXX — Х-трисомики — действительно существуют;
фенотипически они женщины, нередко бесплодные, с
недоразвитыми яичниками и умственно отсталые. Впрочем,
проявление этих аномалий очень изменчиво. В некоторых случаях
женщины с Х-трисомией могут быть и нормальными. По-
видимому, у человека в отличие от дрозофилы прибавка
лишней Х-хромосомы, составляющей только около 5% всего
гаплоидного набора, относительно слабо изменяет генный
баланс.
Наконец, наличие хромосомного набора ХО было
обнаружено у женщин, страдающих синдромом Тернера — Шере-
шевского. Он встречается реже — примерно 1 на 5000
девочек, т. е. 0,0001 на все население (0,01%).
Анализ полового хроматина дал возможность накопить
значительный фактический материал по частоте аномалий
половых хромосом. У особей мужского пола в Европе она
оказалась порядка 0,003, но в других странах света,
по-видимому, ниже. О частоте появления форм ХО и XXX судить
было труднее. Но при осторожной оценке можно принять,
что средняя частота нерасхождений половых хромосом в га-
метогенезе человека, приводящих ко всем видам заболеваний,
около 0,0025, т. е. примерно 0,25%. Эта величина значительно
выше, чем приведенная выше частота нерасхождения у
дрозофилы, но вполне сопоставима с более высокой частотой,
обнаруженной в одной из линий дрозофилы. В дальнейшем
было показано, что у человека существует нерасхождение
и по другим хромосомам, а отсюда, конечно, и появление
трисомиков по ним. Так, дети с болезнью Дауна, которая
диагностируется очень четко (идиотия и целый комплекс
23
конституционных нарушении, в частности головы, лица, глаз),
оказались трисомиками по очень маленькой хромосоме № 21.
По некоторым данным, есть положительная связь частоты
болезни Дауна с возрастом матерей (старение организма
несколько повышает частоту нерасхождения хромосом).
Частоту нерасхождения хромосом № 21 оценивают
величиной примерно 0,003 (0,3%), что очень близко к частоте
нерасхождения Х-хромосом. На количество родов это
составляет 1 : 500-600, т. е. 0,0016 (0,16%).
Была обнаружена трисомня и по хромосоме № 22, также
очень маленькой по размерам, и по хромосомам групп 16—18
и 13—15. Последние хромосомы крупные, но номера
хромосом приходится обозначать условно, так как 16, 17 и 18-я
хромосомы очень похожи друг на друга. То же относится и к
13, 14 и 15-й хромосомам.
Трисомия по хромосомам, входящим в эти группы,
приводит к более сильным нарушениям развития, что можно было
ожидать, учитывая их размеры. Кроме того, по-видимому, во
многих случаях нельзя обнаружить трисомиков, так как они
умирают еще до рождения. Поэтому частота нерасхождения
по этим хромосомам не может быть установлена прямым
путем. Экстраполируя данные о частоте нерасхождения
маленьких хромосом № 21 и крупных Х-хромосом на весь
хромосомный набор человека, можно сделать вывод, что общая
частота нерасхождения должна составлять 0,003X23 = 0,069 —
— 0,07, т. е. около 7% общего числа гамет. По-видимому,
такой величиной можно суммарно оценить нарушение
нормального процесса мейоза у человека, т. е. отклонение от
изложенной выше вероятностной его картины.
Очевидно, что вероятность этого отклонения довольно
высока. Пока нет достаточных данных о факторах,
вызывающих нерасхождение хромосом в обычных условиях жизни.
Несколько в стороне от нашей темы вопрос о
нерасхождении хромосом на том или ином этапе развития эмбрионов,
что приводит к появлению мозаиков, поэтому мы не станем
его рассматривать.
Мейотический дрейф. За последние годы обнаружены
интересные нарушения случайности расхождения хромосом
в гаметах, хотя механизм такого нарушения в некоторых
случаях остается неясным. Эти явления получили название
мейотического дрейфа (со словом «дрейф» мы встретимся
ниже —в главах, посвященных изменениям генетической
структуры популяций). При мейотическом дрейфе
наблюдается постоянное нарушение соотношения при расщеплении,
так как гетерозиготы по определенной паре аллелей
передают эти аллели потомству в отношении, отличающемся
от 1 : 1.
Факты этого рода были обнаружены на дрозофиле,
кукурузе, мышах.
Наиболее отчетливы те из них, когда удается установить
действие определенных генов, ответственных за нарушение
расщепления. Так, у Drosophila melanogaster фактор,
расположенный во 2-й хромосоме, даже был назван нарушителем
расщепления {SD). Он проявляется только у самцов и
только в гетерозиготном состоянии, причем в том случае, когда
хромосома, несущая этот фактор, конъюгнрует с нормальной
гомологичной хромосомой. После нормального мейоза
образуются как будто бы и нормальные сперматозоиды. Однако
те из них, которые несут нормальную хромосому, в
дальнейшем не функционируют. Таким образом из каждого сперма-
тоцита получается 4 сперматозоида, из них 2 —те, в которые
попадают хромосомы с геном SD, будут функционирующими,
а остальные — нефункционирующими.
У нескольких видов Drosophila (D. obscura, D. pseudoob-
scura, D. persimilis) была обнаружена ненормально высокая
частота самок. Оказалось, что в Х-хромосоме имелся фактор,
определявший различную способность к оплодотворению
сперматозоидов с Х- и У-хромосомами. Несмотря на
нормальность процесса мейоза, сперматозоиды с У-хромосомон
оказываются функционально несостоятельными. Поэтому
после оплодотворения образуются зиготы, из большинства
которых получаются самки. Фактор, детерминирующий такое
нарушение соотношения полов, никак не проявляется в
самках, но передается от матерей сыновьям по обычной схеме
сцепленного с полом наследования.
В других случаях мейотический дрейф определялся не
генами, а своеобразной структурой хромосом. Так, у
кукурузы хромосома № 10 имеет две формы: нормальную и
аномальную — с большим гетерохроматиновым вздутием на
конце длинного плеча хромосомы. У особей, гомозиготных
по любой форме хромосом, расщепление но генам 10-й
хромосомы происходит совершенно нормально. У гетерозигот же
менделевские отношения нарушаются: аномальную 10-ю
хромосому получают 70% половых клеток. Все гены, близко
расположенные к участку, где находится вздутие хромосомы,
будут давать ненормальное расщепление. Интересно, что
аномальная 10-я хромосома в гетерозиготном состоянии
влияет на неслучайность расхождения и других хромосом,
характеризующихся наличием вздутии, что в свою очередь
приводит к нарушению отношений при расщеплении по
другим генам. Здесь не место рассматривать цитологические
картины, создающиеся в результате свойств необычной 10-й
хромосомы. Для статистической генетики важно само
наличие фактов, нарушающих статистическую закономерность
25
й»сшг»ы*мия. Существу** и некоторые другие примеры меЛ-
'мического лИФ>. мехвитмм которых очень неясны. Так,
v мышей хромосома с некоторыми аллелями гена бесхвосто-
сти обнаруживаете* а 95% хромосом, У кукурузы известно
неслучайное расхождение по В-хромосоме, но уже при мито-
ти^ских делениях, У Dro*ophila affinis был обнаружен ген,
при котором начинают преобладать сперматозоиды с У-хро-
ШЮИИОЙ И Т. 2L
Нарушения вероятностной картины оплодотворения.
Второй вероятностный процесс, лежащий в основе меиделевсяого
соотношении в потомстве (в простейшем случае при мояо*
гибридном скрещивании),- равная вероятность
оплодотворения яйцеклеток Л или а гаметами такой же возможной
структуры, т. е. Л или а. Многочисленные факты строгих
менделевских соотношений также доказывают бесспорное
его наличие. Однако возникает такой же вопрос, как и в
отношении вероятностного процесса образования гамет
разного типа: насколько часты отклонения от него. Эта
проблема довольно сложна, а изучена еще очень слабо. Прежде
всего бросается в глаза то обстоятельство, что при одно-
временном наличии пылыш или сперматозоидов разных
видов, как это имеет место при опылении растений ветром
или насекомыми, или при оплодотворении икринок рыб в
воде, оплодотворение происходит все же. как правило, пыльцой
или сперматозоид а мн своего вида. Межвидовая
гибридизация происходит только как исключение и при некоторых
специфических условиях. Таким образом, в этом случае
наблюдается некоторая избирательность или селективность в про-
цше соединения гамет. Правда, она ограничивается лишь
принадлежностью половых клеток к одному виду,
Выли получены экспериментальные данные также о
поведении сперматозоидов одной или разных пород, правда,
результаты их довольно противоречивы. Так, В, К* Милова-
моа проводил осеменение крольчих породы шиншилла смесью
спермы от самца шиншилла (брата самок) и самца породы
ал века (черных), В результате большинство потомков по
фенотипу были черные — алиска, В другом опыте осеменили
самок алиска смесью спермы самца окраски аляска (брата
самок) и самца шиншилла, В этом случае большинство
потомков были шиншилл*. Из этих и некоторых других сход*
нмх с ними опытов был сделан вывод, что сперматозоиды
неродственных самцов /обладают преимущественной способ*
поспею оплодотворения по сравнению со сперматозоидами
самцов, родственных самкам. Однако в других опытах, когда
брели смесь спермы от самцов разных пород, но самцы были
не родственны самкам, получалось потомство той же породы,
<ио и самки (сперма смешивалась в одинаковых долях)-
Впрочем, отношения менялись при разных долях
смешивания спермы самцов нескольких пород. Эти факты как
будто бы указывают на наличие некоторой селективности
оплодотворения. Возможно, что оплодотворение осуществляется
скорее всего теми сперматозоидами, которые обладают
наибольшей конкурентоспособностью. Подобно этому у растений
некоторые пыльцевые зерна могут прорастать быстрее, чем
другие, и поэтому скорее достигать яйцеклетки. Однако
сейчас же возникает вопрос о том, существует м корреляция
между этой повышенной конкурентоспособностью и наличием
в сперматозоиде определенного аллеля или аллелей. Так как
расщепляющихся генов очень много (при наличии в геноме
многих тысяч генов), то такая корреляция, если даже она
и существует, может быть только с некоторыми аллелями
(доминантными или рецессивными).
Поэтому по громадному большинству генов будет
осуществляться чисто вероятностный процесс оплодотворения
гаметами генотипически А или а, В или в и г д.
Благоприятствует случайности оплодотворения также и то
обстоятельство, что мужские половые клетки всегда образуются в
количестве во много раз большем, нежели это необходимо для
оплодотворения относительно немногочисленных женских
половых клеток.
Влияние на соотношение при расщеплении равной
жизнеспособности зигот. Нарушение картины расщепления при
моногибридном скрещивании может вызываться не только
отклонениями от вероятностной схемы образования гамет
и оплодотворения, но также различиями в жизнеспособности
ломииантов и рецессивов. Оно улавливается лучше всего
тогда, когда гибнет еще на эмбриональной стадии
определенная геиотипическая группа в целом, например гомозиготы
доминанты АА. Тогда отношение 3: 1 или 1:2; I
превращается в отношение 2:1. Такие гены получили название
доминантных с летальным действием в гомозиготном
состоянии. Старый пример этого рода — скрещивание желтых
мышей, которое давало растепление 2 желтых : I черный.
Сейчас известно много таких фактов. Так, при разведении
серебристо-черных лисиц были обнаружены новые формы, так
называемые платиновые и беломордые. Оказалось, что
скрещивание платиновых (или беломордых) друг с другом всегда
дает расщепление 2 платиновых (или беломордых) к )
серебристо-черной. При этом наблюдается и пониженная
плодовитость платиновых. Если серебристо-черные давали
в среднем около 4,5 щенка на самку, то беломордые — около
3,5 щенка, а платиновые еще меньше.
На классических модельных объектах, в том числе
дрозофиле, изучено значительное количество доминантных генов
*7
с летальным действием в гомозиготном состоянии. Известны
они и у сельскохозяйственных животных, например,
аномалия Пельцера (нарушение ядер лейкоцитов) у кроликов,
доминантная коротконогость у кур (см. табл. 1).
Сложнее решить, что наблюдаемое отклонение отношения
от ожидаемого *— результат разницы в жизнеспособности
в том случае, когда гибель особей происходит не в период
эмбриогенеза, а позднее, на стадии взрослых организмов
и молодняка после рождения.
Интересный пример последнего — наследование серой
окраски шнрази у каракульских овец, изученное Я. Л. Глем-
боцким и Б. Н. Васиным. Впрочем, серая окраска известна
не только у каракульских, но и у других пород овец —
Сокольских и малич, наследуется она так же, как серая
окраска шнрази.
Овцеводов издавна удивлял тот факт, что при разведении
овец ширазн в чистоте, т. е. при скрещивании их друг с
другом, всегда появляются черные ягнята. Все серые бараны,
генетическую структуру которых по данному гену можно
точно определить, так как от каждого получается значительное
потомство, оказывались гетерозиготными. В то же время
обнаруживалось, что черные ягнята рождаются в доле около
25% всего потомства, т. е. соотношение между серыми и
черными при рождении 3 : 1, а не 2 : 1, как у желтых мышей или
платиновых лисиц. Я. Л. Глембоцкий провел очень полный
анализ этого вопроса на значительном материале по
нескольким совхозам и показал следующее.
Расщепление по гену серой окраски (обозначаемому
автором символом We) по двум совхозам («Караул-Базар»
и «Каракуль») было 3081 : 1115, т. е. 2,95: 1,05. Отклонение
от отношения 3 : 1 незначительно. Таким образом, рождаются
как гомозиготы WeWe, так и гетерозиготы Wewe, т. е.
летальное действие в эмбриональном возрасте не
обнаруживается. Это подтверждается и данными о проценте
мертворожденных от разных категорий маток, а также и о
проценте яловости. Не сказывается летальное действие и в первый
месяц жизни ягнят. Однако в дальнейшем, в старших
возрастах, обнаруживается отчетливая разница <в отходе между
ярками ширази и иешнразн, достигающая почти 9%.
Оказалось, что гибель гомозигот по ширази имеет определенное
анатомо-физиологическое основание, а именно недоразвитие
у них рубца. В результате при переходе ягнят-гомозигот на
растительный корм происходит их гибель (чаще всего от
тимпанита).
• Доказать достоверность отклонения от ожидаемого расщепления
можно соответствующими математическими методами (см. ниже).
2В
Поэтому было предложено получать серых каракульских
ягнят в хозяйствах не путем скрещивания серых с серыми
(так как при этом гибнет около lU приплода), а скрещивая
серых с черными. Серые всегда гетерозиготы, поэтому они
дают расщепление в потомстве в отношении 1 : 1 (половина
серых и половина черных ягнят).
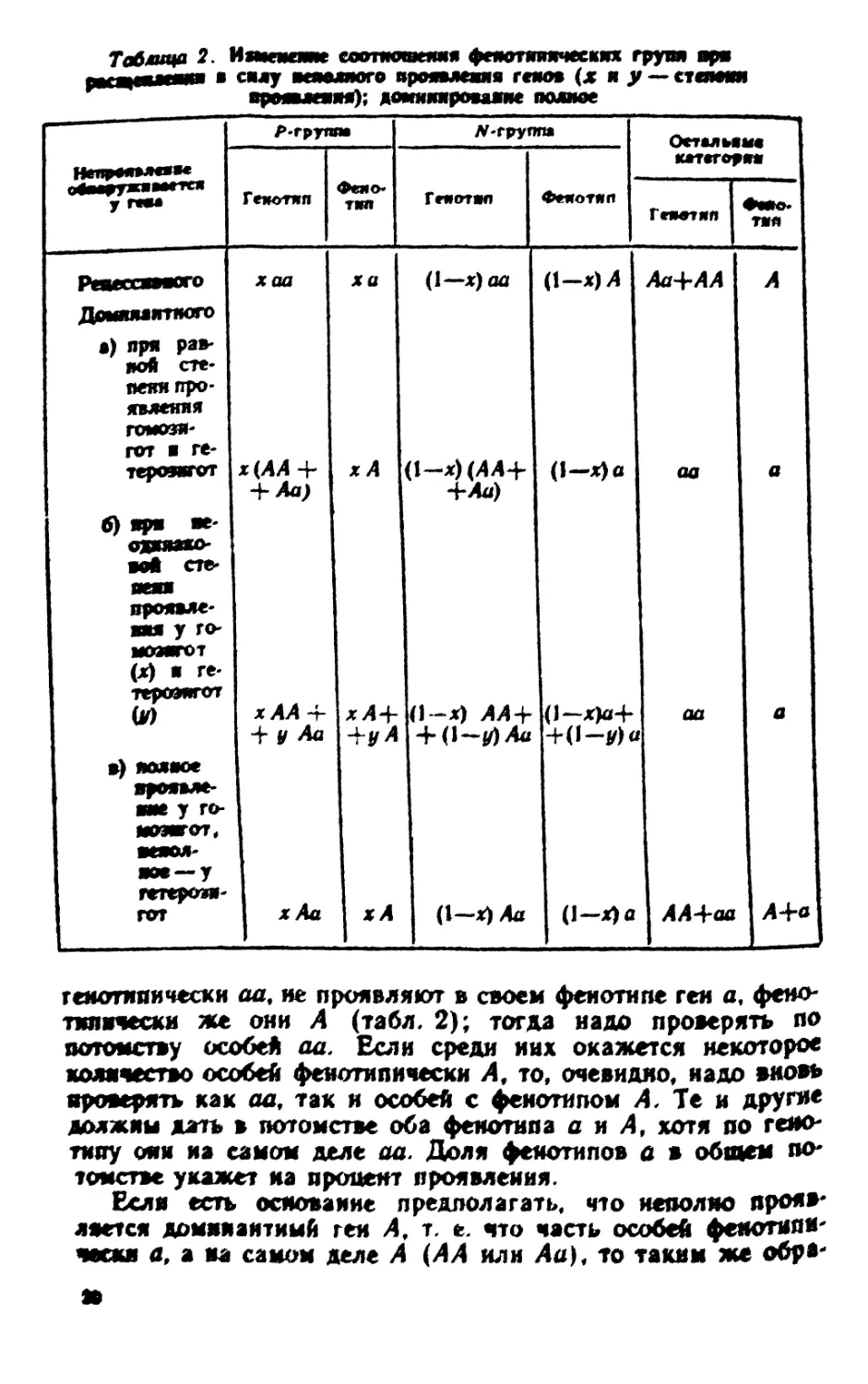
Влияние неполного проявления генов. Наконец, четвертая
группа явлений, могущих повлиять на соотношение
фенотипов в потомстве,— неполное проявление, или, иначе, пене-
трантиость гена. Очевидно, здесь может быть несколько
случаев, отличающихся по результатам. Если неполно
проявляется рецессивный ген а, то среди особей генотипически
аа часть проявляет фенотип а, а часть—фенотип А, тем
самым увеличивается доля фенотипа А. Вместо 3,4 при
моногибридном расщеплении их может быть значительно больше
в зависимости от степени непроявления.
Если неполно проявляется доминантный ген Л, то даже
при скрещивании ААхАА часть потомков окажется по
фенотипу а. При скрещивании ААхаа вместо того, чтобы в F\
(гетерозиготы) все потомство было А по фенотипу, часть
особей будет по фенотипу а. Внешняя картина напоминает
переменное доминирование (гетерозиготы в одних случаях
фенотипически Л, а в других а).
Впервые неполное проявление генов было изучено
Н. В. Тимофеевым-Ресовским и др. на дрозофиле. Но оно
часто наблюдается и у других видов, в том числе и у
человека. Известно, что некоторые наследственные заболевания
у людей, даже гомозиготных по ним, могут не проявляться
вовсе или же проявляются только в пожилом возрасте.
С повышением среднего возраста у людей стало выявляться
значение некоторых доминантных и рецессивных генов, ранее
мало или почти не известных в силу своего неполного
проявления.
Обладают неполным проявлением врожденная миотоння,
множественный неврофиброматоз, отосклероз, шизофрения
и многие другие наследственные заболевания.
У сельскохозяйственных животных также известны
неполно проявляющиеся гены, в том числе и такие, пенетрант-
ность которых меняется с возрастом. Так, у коров,
гомозиготных по рецессивному гену так называемых цистозных
яичников, процент проявления в возрасте 2,5 года только 2—3%,
в возрасте 5,5 лет — около 20% и в возрасте 8,5 лет —44—
57%.
Методы проверки неполной проявляемости гена. Анализ
неполного проявления довольно труден. Если доля
фенотипов А больше, чем ожидается при обычном моногибрндном
проявлении, то можно предполагать, что некоторые особи,
29
Тйблицо 2. Ивмемевле еоотношепя* феяо i иинческяя групп ере
шнвцяиям* ■ сяду неполного проявления генов (л я у — степени
премию**); доминирование полное
у птл 1
|р««а»«го
Дрштшяпюго 1
а) пря
равной
степени
проявлен**
ГОМОЗЯ'
гот ■ ге-
теровикгг
| б) вря не
| в5 ст&
I проявле-
1 вия у го-
1 момгот
« • ге-
1 тероэнгот
1 в) полное
1
проявление у го-
1 М09ИГОТ,
1 венол-
«ж —У
1 гегеро»*-
J гот
Я-груям |
Геяолм 1
хаа 1
х(Л/1 4-
+ Ла)
Феи о* 1
тип
ха
*Л
|
+ у Аа
хАа
I»,
N-tpyntiM 1
Ггяотш? 1
(1—x)aa
(\-x)(AA+\
+Aa)
0 -*) >M +
\ + {\-У)Аи
(\~x)Aa
Фенотип 1
<i-*M
(l-«)a
(J-x)a4-
(1-х) a
Осталыш* 1
icrrtrof** 1
Гсютял I
Ля+ЛЛ
00
1
aa
AA+aa
ТВЛ I
A
a 1
о 1
J J
генотнпически аа9 не проявляют в своем фенотипе ген а, феио-
тилнчески же они Л (табл. 2); тогда надо проверять по
потомству особей aa. Если среди них окажется некоторое
количество особей фенотипическн А, то, очевидно, надо шношь
проверять как aa, так и особей с фенотипом Л, Те и другие
должны дать в потомстве оба фенотипа а н А, хепп по
генотипу они иа самом деле aa. Дол* фенотипов а в общем
потомстве укажет на процент проявления.
Бели есть основание предполагать* что неполно
проявляется доминантный гея Л, т е. что часть особей фенотип*'
часяи а, а на самом деле Л (ЛЛ или Аа)ч то таким же обр*'
ш
юм надо проверить по потомству все категории, как а (ибо
среди них могут оказаться особи геиотипическн АА или
Аа), так и АА (а также, конечно, Аа, если можно отделить
АА от Ли). Иногда может окапаться удобным скрещивание
А А Ума. При полном проявлении надо ожидать, что все
потомство окажется по фенотипу А. Если же среди них часть
особей фенотипически а, то возникает задача проверки таких
особей. Если они на самом деле геиотипическн Аа> то в
потомстве они дадут не только особей А% но и а. Ори этом
будет наблюдаться отклонение от теоретически ожидаемого
отношении 3:1 и превышение доли фенотипов а. Таким
образом, только тщательный аиалнэ по потомству всех особей или
полный анализ родословных в течение нескольких поколений»
если приходится анализировать наследование неполно про-
являющихся генов у человека, позволит оценить степень
проявления,
f//A 4 \ и 1
1 У I I I
Рис 2. Проеме*** шрнтли* и» одной и и-»
обеих ггорои** тел*.
Еше более осложняется картина ирн неодинаковой степе-
ни проявления у гомо- и гетерозигот по доминантному гену
А. Тогда надо проверять по потомству отдельно гомозигот
АА н гетерозигот Аа.
Если обозначить через Р группу особей, проявляющих в
своем фенотипе детерминированный данным геном признак,
а через N ~ группу особей, не проявляющих признака, хотя
и несущих в генотипе этот геи, то можно составить
следующую сводку, показывающую, в какую сторону сдвигаются
соотношения при расщеплении в зависимости от того, какой
признак — рецессивный или доминантный — характеризуется
неполным проявлением (табл. 2).
Анализ испроявлеиия при его несимметричности,
Оригинальный метод определения степени нелроявления или про*
явления rtma был предложен для таких морфологических
особенностей, которые могут проявляться на одной или обеих
сторонах тела. Тогда, очевидно, особи в популяции или семье,
несущие неполно проявляющийся геи (доминантный или
рецессивный), могут быть распределены в четыре группы,
представленные на рис. 2: пропмпютне признак на обеих
сторонах тела (а), проявляющие признак только па правой (а)
или только на левой (с) сторонах; не пропвлпющн* примака,
хотя и несущие данный геи (d). Все вместе они составляют
31
гр\ппу а\ Кроме того, п популяции имеются особи с друщц
аллелем гена, имеющим полное проявление (группа у).
Если имеется в виду неполно проявляющийся доминант,
ный ген (доминирование полное), то х включает генотипы
АЛ и -4а, а у — аа% если рецессивный ген, то наоборот.
Основы л я трудность заключается в установлении
количества особей группы d, так как фенотипически они не
отличаются от особей группы у. Но если предположить, что нет
корреляции между проявлением на правой и на левой
сторонах тела (что действительно было обнаружено во многих
случаях, в том числе на изменениях щетинок и крыльев у
дрозофилы, на пегостях у млекопитающих), то должно быть
верно следующее уравнение: bc*=ad.
Это вытекает 'из анализа корреляции с помощью
четырехпольной решетки
Г^——-—__j_| +
-
—
а
с
—
Ь
d
При отсутствии корреляции произведения по двум
диагоналям должны быть равны друг другу: bc = ad.
Отсюда d= . Тогда можно определить х:
х = а+Ь+с+—.
а
В группе х проявляются особи из групп а, Ь и с. Процент
проявления
р— ^ -100.
х
Непроявление гена как функция онтогенеза. Очевидно, что
проявление или непроявление конкретного гена у отдельно!
особи есть результат ее онтогенеза. Процесс индивидуального
развития у особи с фенотипнческим непроявлением гена как-то
отличается от процесса развития особи, проявляющей
refill данной конкретной ситуации ген может не проявить своего
действия или и силу влияния каких-то факторов, внешних *л*
внутренних, тормозящих или модифицирующих его действ*.
32
пли. что может быть более вероятно, в силу нарушения
синхронности между действием гена на признак и морфогенезом
данного органа. Если, например, у дрозофилы на крыле уже
\спела сформироваться вторая поперечная жилка или
какая-то щетинка на теле, то ген, детерминирующий отсутствие
жилки или щетинки, уже не может проявить своего действия.
Этот пример, конечно, упрощенный, действительные
соотношения между развивающимися в процессе онтогенеза органами
п действием отдельных генов более сложны (см. об этом
в гл. 10).
Вероятностный характер неполного проявления. Хотя само
по себе непроявление—результат нарушения онтогенеза, но
для каждой особи невозможно установить конкретные
причины, почему у нее ген проявился или не проявился, нельзя
п предсказать, проявится ли у нее данный ген или нет.
Поэтому приходится измерять проявление гена (или степень пе-
нетрантностп) по частоте проявления, т. е. и здесь речь идет
о вероятности. При 50% проявления вероятность обнаружения
в данной группе или в популяции особей с данным геном
может быть оценена в 0,5, при 75% проявления —в 0,75,
100%-ное проявление указывает на полную вероятность
(Я= 1) и т. д. Иначе говоря, при неполном проявлении
происходит наложение вероятностного процесса на динамический.
Впрочем, сама проблема проявления (или, как еще его
называют, пенетрантностн) остается в значительной степени
неясной, хотя ее значение бесспорно.
Мы оставляем в стороне близкую к ней проблему
выражения гена, иначе называемую экспрессивностью. В этом
случае также наблюдается широкий диапазон вариации, крайней
степенью которой является нулевое выражение, т. е.
непроявление гена.
Роль вероятностных процессов при наследовании двух или
нескольких генов. Рассмотрим теперь более сложные случаи
наследования с участием двух или нескольких пар генов.
Очевидно, что отношения 1:2:1 по генотипу или 3:1 по фенотипу
должны быть отправными и для них. Указанные выше 4
условия осуществления расщепления действительны также для
днгибридного и пол и гибридного скрещивания. Однако их
дополняют и некоторые другие существенные условия.
Возможность независимого распределения признаков^ потомстве
определяется на клеточном и хромосомном уровнях механизмом
перехода хромосом разных пар, несущих гены А и а ц гены
В и а, в половые клетки. Как известно, при мейозе
гомологичные хромосомы конъюгнруют друг с другом. Однако взаимное
расположение их по отношению друг к другу в пространстве
может быть двояким, как это показано на рис. 3. Оба
положения одинаково вероятны. Прн обоих положениях получаются
33
3 П. ф. РокицкиЛ
дм тмпа га iter. вероятности которых также равны В кооечаси
счете гетерозиготные особи АаВв образуют четыре тип
гамет — АВЩ Авщ аВ и as — в соотношении I : I : I : | ^
с вероятностями их образования Я=0.25).
Таким образом, закономерность независимого наследов*
ива признаков в потомстве основывается еще на одном веро
ятвостном процессе, осуществляющемся на клеточном уровне
а именно: свободном сочетании хромосом разных пар при об
разовавнн валовых клеток, т. е. при переходе в мейозе oi
днолондностн iLieroK к гаплондностн свободное сочетание мо
Рис 3. Схема образовавяя 4 типов гамет (4) в результате разного расоаю-
меппп nap хромосом при конъюгации (2).
жег происходить потому, что кариотип состоит из многих пар
хромосом, между которыми нет физической связи.
Отсутствие этой связи между отдельными парами хромосом
создает в сущности безграничные возможности для возникновения
различных сочетаний генов разных пар хромосом в потомстве
от любых скрещиваний, так как родители всегда отличаются
друг от друга по многим парам аллелей. Как совершенно
правильно указывает В. А. Ратиер *, по мере перехода от одной
генетической единицы к другой в их иерархической системе,
начиная от код он а, рать линейной упорядоченности неуклонно
падает. На уровне генотипа ее нет в соотношении отдельных
сегрегоноя (хромосом). В результате на первый план
выдвигаются чисто статистические принципы регулирования реком-
бинацноиных процессов, как основных элементов попудяцнон-
иого уровня, о чем будет говориться ниже.
Говоря о вероятностной основе дигнбрндных и полнгнбрнд-
mux расщеплений, естественно поставить тот же вопрос, кото-
* См, Вг А, Рйгнлр. Генетические увравляюшие системы. Новое*-.
*tpail«№&.
М
рый был разобрав от »■,„_
«сто* едены »е^«1а^2^^Ш om*,*«^V\uS:
расщеплена*. к «*овда сгатасппесжую жаргш»
разша вар, Одавж аз тааУ «г.^—^ **•*
4?
А ^
Рис. 4. Травсккапия учаспса
длиной хромосомы ва жоротжую (а);
четыре ттша гамет (б).
крепления их к хромосомам других пар. что показам на
рнс. 4. а. На рнс 4, б изображены четыре типа гамет. По
своей генетической структуре они представляются обычны» АВ%
А в, аВ и as. Но в двух из них произошло нарушение генного
баланса: или нехватка генетического материала (гамета 3),
или, наоборот, лпшннй материал (гамета 2) Последствием
этого явится нарушение соотношения 1:1:1:1, которое еледо-
вало бы ожидать при скрешиванин АаВвХааел (рис 5). В
зиготах две хромосомы с транслокацией — часть длинной
хромосомы прикреплена к короткой, однако генный бадане не
нарушен. Поэтому генотипы А В должны быть по фенотипу
нормальными. Такие же фенотнпически нормальные генотипы
аавв. В генотипе аВ — излишек генетического материала за
счет прикрепившегося к короткой хромосоме участка длинной
хромосомы, в генотипе Ае. наоборот, его нехватка Фенопгаи-
ческнй эффект излишка или нехватки будет зависеть от
размеров транслированного участка: при больших Р^ерах
возможна полная летальность, тогда формы А* и шВ —
3е
35
ме появляются, при малых размерах излишка или нехватки
формы аВ и Ав могут появиться, хотя очень велик шанс, что
они будут обладать пониженной жизнеспособностью. Так или
иначе, но основанное на вероятностных законах соотношение
1:1:1:1 будет нарушено.
1 с- fc I Я j<
J J
В
/С13(Г4
Q-c
Ршг. 7. Кольца из 14
хромосом у энотеры (по
Клеланду).
г 2
Л*- * Результат* решшрокво*
траасюкашв:
л -I
*-1
Рш:. 8. Схема соедвяошя половых
меток знотерм, образуемых гетерами
готами гюиш Г
F. Формы О J я KV
Еще: Соане интересны результаты обменных, или реципрок-
а, трааслокац*6~ Простейшая схема обменной транслока-
ни* доказана на рис, 6. Если произошла такая траислокаця*
иеакду хромосомами двух пар гомологичных хромосом, то
ири югыоганин будет наблюдаться характерная крестообраз-
ная фигура расположения хромосом (рис 6. В), приводящая
* ограждению кольца хромосом. На рис, 6, Г в кольце 4 хро-
а^?"!* "° ^f * » ироцесс образования рецнпрокиых транс-
*^V™ ******тс* еще одна яара хромосом, то кольцо н ме*
кЖ^Л^Г Ш"1М»ТЬ У** * хромосом Срис. 6, Д). Подобная
"Гтнна на*аюдается у энотеры На рис 7 представлена фо-
тографня клеток энотеры с кольцами, включающими 14 «^
масон (7 пар,. При разрыве кольца (лева? чат" JinE
количество хромосом можно легко подсчитать ОчевЕ Si
могут выживать только те дочерние um^i^SS^X.
ют полный набор хромосом. Но дальнейшая картина у 5£Д
оказалась еще более сложной из-за гетерозигс^иости SSZ
сом по леталям, У Oenothera lamarckiana образуются тЕдько
лва типа гамет, получившие название Gaudena и Velan*
,«/.мОМ°хИГОТЫ по Gauden* W) и гомозиготы но Velan*
(VV) гибнут в силу наличия в них леталей. попому
выживают только гетероэиготы по этим комплексам VQ4 которые
сохраняются в гетерозиготном состоянии, постоянно отщепляя
от себя гибнущие гомозиготы GG и VV (рис, 8).
У Оел, lamarckiana гомозиготы GG и КК гибнут на стадии
зиготы. У Oen, muricata картина сложнее, так как гомозиготы
вообще не образуются в силу летальности части гамет, Но
результат тот же: сохраняются только гетерозиготы по транс-
локационным комплексам rubena—curvan*. Это явление
получило название эффекта Реннера. Таким образом, включение
леталей предупреждает распад комплекса.
Известны и более простые случаи так называемых
сбалансированных леталей. Например, у дрозофилы линия у X
w Curly Curly л Curly /
X—г^-^—гА так *** гибнут гомозиготы ^ ™ и у-,
II ' Curly /
У высших организмов кольцевые хромосомы являются
аберрациями и, как все хромосомные аберрации, возникают в везна
чнтельном количестве случаев, т, е, обладают малой
вероятностью (об этом более подробно в гл. 2), При
экспериментальном получении они, как правило, элиминируются. Но у вотсрм
и нескольких других видов они сохранились в силу
своеобразия ъволкмии. котла соединились вместе несколько
механизмов, каждый нз которых сан по себе вреден (реиипрокиме
транслокация, накопление деталей, самоопыление), но в сово-
купмостн система приобрела стабильность и гволюиноииую
значимость. —****-*,
ВаанмодеДствме гемм дан фактор нарушим статмспимч
ской картины расщеяаеяи, В общем курсе генетики
излагается немало данных о разлияиыд видах взаимодействия'генов
в развитии. Для статистииеско* генетики важны те нз них.
которые саэдаюг необычную картину Р****™»*™™1^
шую иа той же вероятностной основе, м© осложняемую
взаимодействием геИОВ. s_l±Mutmmmu^
Как известно, классическим оримером «°™^*P%
плидополнительных, геиои™™"^**^£*££
рашеииых иур при сирешииаиии представителе* дауж пор»
17
(*>w% *\р В F7 *? полумаются и цветные и белые в соотио.
шенин 97 Цветная окраска ~ результат взаимодействия дву*
мсчиткмадыА друг от друга доминантных генов А и В. Исход»
нме породы генотнпически ААЬЬ и ааВВ.
Мутднтные гены а \\ Ь блокируют определенные этапы син.
тпа пигментов в перьях кур» поэтому обе эти породы кур бе-
1ые Для нормального синтеза пигмента необходимо наличие
обоих доминантных генов А и В% которые имеются у всех
особей f i и у 9/I6 F* Остальные три категории особей F% aaB.%%
А.ЬЬ и uabb фенотнпически неотличимы, так как все он*
белые.
Куры же дают хороший пример и эпистатического действия
генов, т е. случая, когда при наличии одного доминантного
гена, называемого зпистатическим *, нельзя заметить расщепле*
нкя по генам другой пары. Так, при скрещивании белых
леггорнов и белых виандотов все цыплята в F\ белые, во втором
поколении появляются и окрашенные, но в доле 3/16 всего
потомства. В действительности расщепление днгибридное, но
белая окраска создается в силу действия доминантного гена
/подавителя окраски. Исходные породы ПСС (белая окраска
обязана наличию I) и Нес (с — рецессивный ген белой
окраски). В Fx все ПСс белые, в /*2 формы / ... С..., /... се и Нес
белые, и только ПСс окрашенные. Эпистатический фактор I
не дает возможности проявиться расщеплению по паре генов
С-с.
Случаев взаимодействия генов качественных признаков
(аллельного и неаллельного) очень много, и нет смысла
рассматривать каждый из них в отдельности. Поэтому мы
ограничимся лишь приведением сводной табл. 3, в которой
показаны разные распределения фенотипов при дигибридном
скрещивании в зависимости от характера взаимодействия. Со ста*
гнческой точки зрения существенно лишь следующее. Взаимен
действие обычно приводит к значительному уменьшению
количества разных категорий фенотипов, т. е. к уменьшению
вариации потомства по качественным признакам вплоть до слу*
чая. когда появляются только две категории потомков, т. е.
имитируется моногибридное расщепление, и только
тщательный генетический анализ позволяет в конечном счете доказать
ди1ибрн.гный характер расщепления. В некоторых случаях
также очень релко выражается асимметрия в распределения
потомства по категориям (случаи 15:1; 1Я:3 и др.).
Проявляется та же картина, о которой несколько раз гово*
рмлшгь выше: на статистические, вероятностные закономерно-
# \\рш ««дли* иьгледоядмм* иоличестигихих признаков (i.v 5) *******
дг* t*#r (К'алк.лиш* rnwi, кйрушяк>щ*х аддитиимоеть их действия. *****
»*4ы»4«>т >n*i"tdtU4*c*HU В »тоц о^чае термин опистатич«чкмй» иЫ**
иг» 1*.ч|*« 6rv»rr Ш»(и*м* сыжм
Таблица 3. Разные распределения фенотипов в Fz при диги&ридиом
скрещивании в зависимости от взаимодействия генов (аллельяого
и неаллельного) в расчете на я 16
^\^^ Генотипическне!
ч^ формулы и
^^^^ численности 1
Вилы ^ч.
/ыаимолеиствия ^>^ [
Нет доминантности, нет
взаимодействия
I А домин; нтно, нет
взаимодействия
В доминантно, нет
взаимодействия
А и И доминантны, нет
взаимодействия
А н В доминантны, их
взаимодействие приводит к
появлению нового признака
1 А и В доминантны, ком-
плементарны
1 А и В доминантны,
полимерны
А и В доминантны, А эпи-
1 статично к В/в
А и В доминантны. А
эпистатнчно к В/в, в крнп-
1 томерно к а
1
3
3 '
2
6
3
ее
О 1
•г
1 2
* 1
«2
~ 1
4
6
9
1 9
9
1
1
W [
* 1
« |
-Г J
2 '
3
1 1
2
! 3
3
I
9
1
1
'* 1
з
2
2
2
3
3
3
9 1
1
1
1
! 1
1
1 7
15
12
13
1 А и В домлнантны, а э-ни- |
статично к В/е | 9
1 з
з
1 3
1
1
Г
4
i
I
»
сги накладываются динамические, в данном случае
выражающиеся в сложном взаимодействии продуктов действия генов
и процессе индивидуального развития.
Проверка соответствия фактически получаемого
расщепления теоретически ожидаемому. Существует несколько тестов
для проверки генетических гипотез. Один из них основыва-
J9
ется иа вычислении статистических ошибок, второй — на
применении метола хн-квадрат
Допустим, что надо проверить расщепление во втором
поколении на соответствие его моногибридному 3:1 от
скрещивания дрозофил, нормальных по крыльям ( + ), с особями,
имеющими зачаточные крылья (vg—vestigial). Оно оказалось
следующим: 144 -f и 51 vg. Всего 195 особей.
Можно применить 2 способа: а) исходить из фактических
чисел и сравнить их с теоретически ожидаемыми и б)
рассчитать теоретические отношения и сравнить их с полученными
фактически отношениями. При первом способе нормальных
195-3 195-1
дрозофил и vg следует ожидать —-— = 146,25 и —-— =
Ур(п—р)
где п — общая численность потомства, ар — теоретически
рассчитанная. Ошибки будут одинаковыми для обеих групп:
■*у.
146,25-48,75
195
=*у^
75-146,25
195
=У 36,56= ±6,05.
Можно вычислить ошибки также и для фактических чисел.
Результаты можно свести в следующую таблицу:
Показателям
I Фактические числа
1 Ожидаемые числа н их ошибки
1 Разница (d) и ее ошибка (sd)
Категории особеЛ
нормальные (т)
144
146.25±6,05
1 2.25±6,05
cg
51
43.75±6,05
2,25±6.05
Разница не только не превышает ошибки, но значительно
меньше ее, поэтому можно считать, что полученное отношение
в потомстве вполне совпадает с ожидаемым.
При втором способе определяются доли полученных
величии при гипотезе, что ожидается отношение 3:1 и сумма
их равна 4. Тогда доля нормальных особей от 4 составит
*2#95. и доля vestigial -77^- = 1,05.
195
195
40
Ошибка для них вычисляется по формуле
г 195 г 195
Сводная таблица будет иметь следующий вид:
Гкмшэатяш
Теоретически отношения
Фактически полученные я их ошибки
Разница (d) в ее ошибка (sj)
Категория особей ]
вормыыше (-)
3
2.95^0,13
0,05г0.13
и 1
1
1.05 т 0,13
0,05^0,13
Результаты получены те же, что и при сравнении
фактических и ожидаемых численностей: разница явно не
достоверна. Конечно, если разница превышает ошибку, то надо
определить / (= ). При /^2 разницу следует считать
выходящей за пределы случайности, и тогда, очевидно, полученное
отношение уже достоверно отличается от ожидаемого: при
!ш2 с вероятностью р=0,95, при /=2,58 с вероятностью
р=0,99, а при f>2,58 с вероятностью />, большей 0,99
(р>0,99).
Другой метод — хи-квадрат (х2). широко применяемый
и вошедший во все учебные руководства по генетике. Он дает
возможность: а) оцепить результаты расщепления в целом;
б) установить степень соответствия фактически полученных
данных теоретически ожидаемым по каждой группе особей
и в) оценить результаты отдельных опытов.
Известно, что простейшая формула для хн-квадрата
следующая:
у (О-Е)*
где £ —теоретически ожидаемая численность, а О
—фактически полученная.
С помощью хи-квадрата могут быть проанализированы
случаи расщеплений (моногибридного и дигибридного). а
также, как указано в курсах статистики, любые данные опытов —
степень соответствия полученных данных ожидаемым.
У бобов наблюдалось расщепление по их окраске:
интенсивно окрашенные (АА), слабо окрашенные (Аа) и с
небольшими окрашенными полосами (аа). Всего было изучено 355
41
Лобов Ожидаемые количества всех трех фенотипов при
гипотезе моногнорндного расщепления будут следующими:
£,=£,= 3^ =88.75;
£j==355^=l77,50.
4
Результаты анализа представлены в табл. 4.
Таблица 4. Ашдогз рдсщепжшя у бобов с помощью критерия хмчпадрат
Гсиотаг
АА
Аа
аа
V
Чдсготз
ф*КТМ-
чеекдя О
92
182
81
355
ожидаемая £
88.75
177,50
88,75
355
0-£
3.25
4.50
-7,75
<о-а»
10,56
20.25
60.06
<0-£>»
1 ^
0,1190
0,1141
0,6767
Х*= 0,90981
Число степеней свободы *//=л—1=2. Отсюда по таблице
Е2 устанавливаем, что для отбрасывания нулевой гипотезы нет
основания (вероятность соответствия между 0,75 и 0,50).
Для иллюстрации подобных расчетов при днгибрндном
скрещивании можно использовать пример на дрозофиле по
двум ларам генов: нормальные щетинки — редуцированные
щетннкн н нормальные глаза — уменьшенные глаза. Для
простоты обозначим их обычными символами А— а н В—6. В
обоих случаях доминировали нормальные признаки.
Для расчетов ожидаемых численностен надо общую сумму
особей п разделять на 16 и частное помножить на 9 (группа
АВ) и на 3 (группа АЬ = аВ). Формы ab составляют 1/16 л.
Результаты приведены в табл. 5.
Число степеней свободы в данном случае df=n— 1 =4—I =3.
Вероятность соответствия по таблице х* р ж 0,25.
Оснований для того, чтобы отвергнуть нулевую гипотезу, нет. Однако
бросается в глгз^ высокое значение х* Для последней
строчки — фенотипа аЬ. Фактическое количество особей ab значи-
Т*2£?° ыеньше ожидаемого. По-видимому, двойные рецесенвы
<шЬЪ обладают меньшей жизнеспособностью.
th!M,DT случаи' *°гЛа. несмотря на ожидаемое
расщепление 9:3:3:1 при доказанной генетически независимости ге-
42
Таблица 5. Анализ расщепления у дрозофилы по двум парам
генов (А — а ш В — в)
Фенотип
АВ
аВ
| ив
V
Частота
0
122
41
40
6
| 209
£ !
117.56
.{9.19
39,19
13.06
о - £
4,44
1,81
! 0,81
-7.06
[209 I 0
(О - СГ
<o - £>*
£ 1
\ 1
19,71 j 0,168
3,27 j 0.084
0,66 ! 0,017 j
j 49,84 j 3.817 |
1 X*- 4.086 j
нов (нахождение их в разных группах сцепления), значение
X2 выше уровня, допустимого в силу случайности. Примером
могут быть данные, приведенные в табл. 6.
Таблица б. Анализ расщеплена* у дрозофилы по генам brows
(11 хромосомы) н scarlet (III хромосомы)
Феаотжп
С нормальной
окраской глаз
brown
scarlet
white
1 Х
Частота
О £
1)34
430
465
133
2162
1217
405
405
135
2162
0-£
--83
25
60
_2
<0-£Р
6889
625
3600
4
£
5,7
I.S
8.9
0.03
| j X» 16.13
Табличные значения х2 Для Р-0,05 9,35 и для Р*0,01
11,34. Полученное значение х2 значительно превышает и
последнюю цифру. Нулевая гипотеза должна быть отброшена.
Очевидно, что тать ко биологический анализ, например
установление различии в скорости развития ыутантных и
нормальных форы или их различной жизнеспособности (или. наконец,
какие-то неучтенные ошибки в самой постановке опыта),
может дать ответ на причину такого отклонения в значении х*-
Н а конец, критерий хн-квадрат может быть использован
для установления степени однородности или разнородности
данных разных опытов. Известно, что между семьями иди
43
разными группами животных и растений могут быть
значительные различия по соотношениям в потомстве в силу слу.
чайной их вариации, о чем уже говорилось ранее. Но в
некоторых опытах возможны и различия, выходящие за границы
случайности. Их можно обнаружить с помощью критерия
хи-квадрат. Идея установления разнородности заключается
е том, что определяют х2 двумя способами. По первому из них,
примененному во всех приведенных выше примерах, значение
X* вычислено для каждой группы, а затем путем простого
суммирования определено объединенное значение %2 для всех
групп. По другому способу можно обработать весь материал
в целом, не считаясь с отдельными опытами, получить
соответствующие объединенные эмпирические значения для двух
или нескольких расщепляющихся групп, вычислить
теоретически ожидаемые величины и получить значение х2- Разница
между хи-квадратами, вычисленными двумя способами,
позволяет судить о степени разнородности материала *.
Для иллюстрации приведем данные Мазера по анализу
расщепления в 4 семьях львиного зева (Antirrhinum majusj
(табл. 7).
Таблица 7. Расщепление у львиного зем по окраске цветков:
желтой СУ) и цвета слоновой костя (/)
1 Семья
1
2
3
4
1 Z I
1 Количество растений
У
27
15
24
25
91 1
) ,
2
12
6
5
1
25 1
всего
29
27
30
30 1
116 1
X1
5.07
5,44
0.40
1.11
X* 12.02 J
В последней строчке таблицы приведено значение хи-квад-
рата как суммы хи-квадратов отдельных семей. Но оно
довольно велико, выше табличного даже при Р = 0,01. Иначе
говоря, нулевая гипотеза должна быть отброшена. Но если
вычислить х2 Для расщепления по объединенным данным, то
Х2~0,74, что указывает на полное соответствие гипотезе
растепления 3:1. Таким образом, величина суммарного %2
состоит из 2 компонентов, один из них характеризует степень слу-
* Эти вопросы очень полно и с серьезным теоретическим обоснованием
изложены в кн.: Weber E. Mathematiscne Grundlagea der Genetifc. Iena. 1967.
44
чайного отклонения от ожидаемого отношения, а другой —
гетерогенность, разнородность фактического материала.
Хг суммарная 12,02 <Ц=*
X2 по объединенным данным 0,74 <//=|
X2 за счет гетерогенности 11,28 rf/=>3
Р для гетерогенности 0,01—0,02, в то время как Р для £
объединенного материала 0,30—0,50, Таким образом,
возможно, что соответствие ожидаемому расщеплению по всему
материалу является случайным, а результаты по некоторым
семьям (>fc 1 и Nf 2) не случайно не согласуются с гипотезой.
Степень гетерогенности может быть охарактеризовала
также вариансой. В данном случае, по Мазеру. о*~3,75
Для анализа доли гетерогенности в %* можно применить
специальный метод ортогональных функций (см. Weber).
Метод пробанда Вейнбсрга. Предыдущие методы проверки
расщеплений относились к таким случаям, когда типы
скрещиваний Аа/Аа или АаХаа или какие-либо иные уже
установлены, и поэтому в потомстве могло быть теоретически
ожидаемое расщепление. Очевидно, что наблюдать фактические
расщепления и сопоставлять их с теоретически ожидаемыми
можно только в том случае, если есть уверенность, что
учтены все возможные скрещивания, а так как судить об пом
можно только по потомству, то это требует знания генотипа
родителей, а также значительного количества потомков от
каждого скрещивания. Иное дело, если изучаются животные
с небольшим количеством потомков, к их числу относится
и человек. В этом случае какая-то часть гетерозиготных
родительских пар не будет учтена только потому, что в их
малочисленном потомстве не появилось ни одного рецессивного
потомка.
Для наглядности можно привести такой пример.
Предположим, изучается появление альбиносов у грызунов. Если семья
имеет большое количество детенышей, то вероятность
появления альбиносов 1/4. а нормальных 3/4. Но если семьи
состоят только из двух детенышей (как вто имеет место, например,
у зайцев), то ситуация усложняется. Возможны следующие
случаи: Л .,..,, , ,,,♦
а) оба детеныша альбиносы. Вероятность: 1/4 -1/4—1/16;
б) первый детеныш альбинос, второй нормальный.
Вероятность: 1/4 -3/4—3/16;
в) первый нормальный, второй альбинос. Вероятность:
г) оба детеныша нормальные. Вероятность: 3/4 • 3/4-9/16.
Таким образом, если бы были учтены нее семьи с двум*
детенышами, то следовало бы иметь следующее:
46
1 Тип сече Л
С 2 альбиносами
С 1 альбиносом
С 0 альбиносов
V
Коли честно ccmciI
1
6
9
16
Количество 1
J.'IbOllNOCOU 1
О
6
0
8 1
1
Так как в каждой семье (их 16) 2 потомка, то всего
потомков 32, из них альбиносов 8, т. е. отношение доминантов к ре-
цесенвам 3:1, или в расчете на каждую семью 0,5 рецессива.
Однако в действительности семьи с отсутствием альбиносов
не отличимы от семьи, где вообще нет расщепления
(скрещивания АА\ААУ ААхАа и т. д.). Фактически приведенная
выше таблица превращается в следующую:
1 _ Количество
1 Тип семей 1 семей
С 2 альбиносами
С 1 альбиносом
С 0 альбиносов
V
1
6
?
7
Всего Количество
потомков I альбиносов 1
2
12
?
14 ,
2
6
?
8 1
В таком случае обнаружится 8 альбиносов на 7 семей или
на 14 потомков. В среднем на семью окажется 1,143 потомка
рецессивов вместо ожидаемой теоретически доли 0,50 на
семью из 2 потомков.
Исходя из изложенного, был разработан ряд формул, с
помощью которых можно рассчитать ожидаемое число
рецессивов на одну семью при s-количестве детей (сибсов) в каждой
семье и отсюда ожидаемое соотношение между количеством
доминантных и рецессивных форм при суммировании многих
таких семей.
Ожидаемое рецессивное потомство на одну семью (для
случая скрещивания двух гетерозигот) может быть выражено
формулой
а\= т
l/4s_
1 —(3/4)*
(О
v>
Для указанного выше случая двух потомков находим:
eiae_A/ii2__=L:'(,__L)e
1 —(3/4>* 2 v 16 '
~ 2 • 16 - 14 " ' '
т. е. уже полученную цифру. Отсюда можно рассчитать
и общее количество рецессивов, помножив на число семей па%
в которых появился хотя бы одни рецессивный потомок:
^°'- 1-(3/4)'- {)
В табл. 8 представлены сводные данные для семей с
разным количеством членов (сносов).
Таблица 8. Сводка результатов расщеплений в семьях с разным
количеством членов (сибсоа) при скрещивании Аа\Аа
Количество членов
семьи (сносов)
1
2
3
4
5
6
7
8
9
10
00
Ожидаемое
количество рецессивов
п среднем ни одну
семью
1,000
1,143
1,297
1.463
1.640
1,825
2.020
2,222
2.433
2.649
—
Соотношение
доминантой
и рецессивов
0:1
0.750:1
1,130:1
1,734:1
2,049: 1
2,288:1
2.465: 1
2.660: 1
2,699:1
2.775: 1
3: 1
?' ДЛЯ P4.1HUX 5
0,0000
0,12245
0.26297
0.42005
0,59178
0,77595
0,97024
1.1724
1.3802
1,5917
~~
Легко видеть, что по мере увеличения количества особей
(сибсов) в семье отношение между доминантами и рецессива-
ми все более приближается к 3:1.
Сходным образом может быть выведена формула и для
скрещивания АаХаа. Тогда ожидаемое количество рецессивов
на одну семью выразится формулой
1
fl»eT-o72K l,a)
47
Пользуясь этом формулой, можно рассчитать соответствую,
щие величины для таблицы, аналогичной приведенной выше.
В *том случае, очевидно, предельным отношением явится не
3:1, а 1:1.
Так как p=l—q, то возможна единая формула количества
рецессивов на одну семью для обоих типов скрещиваний,
а именно:
Все вышесказанное относится к случаям изучения
родословных, когда родители изучаемых особей фенотипически
проявляют доминантные (нормальные) признаки, генотип же
их неизвестен.
В качестве наглядного примера можно привести
следующие данные, собранные Хогбеном и др. (Hogben, Worrell a. Zi-
eve) относительно семей, в которых встречались больные аль-
каптонурией. Всего было изучено 37 семей с разным
количеством детей. Фактические и ожидаемые данные приведены
в табл. 9. Ожидаемые количества вычислены по данным табл. 8.
Например, если при 3 сибсах количество рецессивов на одну
Таблица 9. Потомство с алькаптонурией от нормальных родителей
| Количество членов
1 семьи (s)
1
2
3
4
5
6
•
9
11
14
' £
Количество
семей с
данным s (ns)
5
8
5
2
3
3
2
3
1
1
3
1
37
Количество особей
с алькаптонурией
фактическое
5
10
8
4
4
8
5
5
4
2
7
4
66
ожидаемое
5
9,14
6,49
2,93
4,92
5,47
4,04
6,67
2,43
2,65
8,61
3,56
61,91
V2
5
0,00000
0,97960
1,31485
0,84010
1.77534
2,32785
1,94048
3,51720
1,38020
1,59170
5,41590
2,44640
23,52962 1
семью равно 1,297, то для 5 таких семей (3-я строчка
таблицы) количество рецессивов должно быть 1,297X5 = 6,49.
Чтобы определить, достоверно ли отличается фактически
полученное отношение от ожидаемого, надо найти ошибку
фактически полученных величин. Но так как в каждом сибсе
свои о и о2, целесообразно ввести дополнительную графу
в табл. 9, а именно п8оа2, чтобы получить объединенную я* а*2
как результат суммирования частных подвыборок. Так, если
в табл. 9 есть 3 семьи с 5 детьми, то а2 (=0,59178) надо
умножить на 3 (3-0,59178=1,77534). Отсюда а= уТ77534=1,36.
Разница между фактически полученным количеством особей
с алькаптонурией и теоретически ожидаемым равна 0,92,
т. е. она меньше стандартного отклонения о. Если обратиться
к общему итогу по всем семьям (последняя строчка таблицы),
то о2=23,52962, откуда а= УТЗ,52962«4Д Разница же
между фактическим количеством и теоретически ожидаемым
66—61,91=4,09. Она также меньше стандартного отклонения,
т. е. может считаться недостоверной.
Изложенный метод впервые предложен Вейнбергом
(Weinberg) в 1912 г. Он назвал его прямым методом или методом
пробанда. Позднее он был значительно углублен другими
авторами с использованием метода максимального
правдоподобия (maximum likelihood).
Коэффициент рекомбинации и методы его определения. При
скрещивании гетерозиготы АаВЬ с рецессивной формой ааЬЬ
и независимости генов А—а и В—b наблюдается обычное
расщепление 1:1:1:1. Наряду с формами АВ и ab (по фенотипу)
появляются новые сочетания АЬ и аВ. Обычно их называют
рекомбинантами. Доля рекомбинантов во всем получаемом
потомстве может быть названа коэффициентом рекомбинации
тг. Иногда его обозначают также символами г,, в и др.
Коэффициентом рекомбинации может быть названа и доля гамет,
получающаяся в результате перекомбинации хромосом с
генами А и В.
Если обозначить рекомбинанты АЬ и аВ буквами R\ и /?2»
исходные формы АВ и aa — N\ и jV2, to
_ Ri+Ri _ Rt+Rz m
r Ni+N2+Ri+R2~ n • w
Так как R\ и R2 составляют половину всего потомства при
обратном скрещивании и независимом наследовании, то в
данном случае тг=0,5.
При сцепленном наследовании доля рекомбинантов в
потомстве может быть меньше 0,5, а при полном сцеплении их
не будет вовсе, т. е. тг будет равно 0.
Как n.im'TiKi, сцепленное наследование может наблюдаться
при диух рашых типах скрещивании: когда сцепленные
Доминантные гены находится в одной -хромосоме I гетерозиготы
АВ \ I
имеют строение ) пли п разных \ строение гетерози-
аВ \
WT Ah I'
Rami ivimct при коэффициенте рекомбинации тг by&yt
тогда следующими-.
Для особи - --
ab
ЛИ АЬ
2 0-'",) 2 '»,.
Для особи „
об
АВ АЬ
1 1
Гаметы
Их частоты
аВ
1
2 ""'г
ай
yd-mr)
Гаметы
Их частоты
аВ
aft
/г;г
■(1-ю,)
2-О-/"г)
1
•тг
В первом случае рекомбннантами при обратном
скрещивании будут формы Ab и аВ, во втором ЛВ и ab. К ним обоим
может быть применена та же формула (4), только рекомби-
нанты меняются местами.
Остановимся па результатах обратного скрещивания типа
AaBbxaabh. При общем количестве особей п расщепление
при наличии рекомбинации будет следующим:
I Фенотипы
Их доли
ности)
Ожидаемые
ности
Фактические
ности
(вероят-
числен-
числен-
| АВ
1
п
iV»
Ab
1
п
«I
atf
1
/2
*2
a&
-jfO — m,)\
n 1
"2" (1—«r)
;Vf
В этом простейшем случае очевидно, что коэффициент
рекомбинации может быть непосредственно вычислен по доле
фактически полученных рекомбннаитов, так как
ПхЛ-Ri— 0"'иг+ -0 тг = птг.
so
Отсюда
п
Величина тТ будет отражать вероятность появлении реком-
Сикантиых особей.
По можно подойти к решению этой задачи и с помощью
более общего метода — метода максимального правдоподобия,
так как только последний дает возможность устанавливать
коэффициент рекомбинации при более сложных вариантах
скрещиваний.
Сущность метода максимального правдоподобия
заключается в том, что можно рассчитать вероятность различных
сочетаний численностей классов при расщеплении в случае
рекомбинации таким образом, чтобы она была максимальной.
Она может быть получена из разложения многочлена,
состоящего из указанных выше частот, или вероятностей, для
каждого класса, возведенного в степень п:
Г 1 111 1"
I-2-<1-"2''+ у ,Пг+ уш'+ у ('-"МJ •
Исходя из наблюдаемых численностей каждого класса, она
будет иметь вид
/if Г 1 1*./ 1 \н. / 1 \R>
X[T(l-m,)j =-№Жй4*
2d-»Ir>] (-»-,„,) .
Так как метод максимального правдоподобия требует,
чтобы эта вероятность была наибольшей, то используют
величину L—меру правдоподобия.
В данном случае числовые коэффициенты в левой части
выражения не имеют значения и
Максимум правдоподобия достигается при том значении
тг% при котором достигает максимума его логарифм. Поэтому
IogL=(^1+iV2)log-Jr(l-mr) + (/?t+/?2)log^-mr.
4*
51
II далее дифференцируем и приравниваем к нулю: *
А^А =0
dm r
В конечном счете после соответствующих преобразована!
/?"l + /?2
может быть получено значение тг= , соответствующее
уравнению (4).
Так. например, если общее количество особей в четырех
классах равно 2022, а особеи-рекомбинантов 716, то mr=0,35
(или 35%).
Ошибка вычисляется по обычной формуле для доли,
а именно:
„уш
-р)
В данном случае sw = 0,10.
II тогда гНг±5жг=0,35±0Л0.
Как известно, школой Моргана было доказано, что
появляющиеся при сцепленном наследовании рекомбинанты —
результат кроссинговера в участке хромосом между данными
1енамн. Частота кроссинговера, или вероятность разрыва и
обмена участками хромосом, измеряемая коэффициентом
рекомбинации, прямо пропорциональна расстоянию между генами.
Это значит, что вероятности разрывов и обменов участками
хромосом более или менее одинаковы по длине хромосом.
Вычисление коэффициента рекомбинации при более
сложных случаях расщеплений. Коэффициент рекомбинации может
быть вычислен и в тех случаях, когда по каким-то причинам
нельзя получить обратные скрещивания, но есть F2. При этом
надо учитывать две возможности: а) кроссинговер происходит
только у особей одного пола, например только у самок, ка*
это наблюдается у дрозофилы; б) он осуществляется у обонх
полов, одинаково часто или с несколько различной частотой
(млекопитающие, птицы).
В первом случае при скрещивании особей Fu полученных
. АВ ab
от родителей -г— X —г-, самки будут образовывать четыре
типа гамет со следующими теоретически ожидаемыми частей
а самцы —два типа: АВ(1/2) и ab (1/2).
В результате в Ft получится восемь категорий генотипов
четыре - в результате рекомбинации и четыре — без нее.
V?
Распределение их по фенотипу, их доли, ожидаемые и
фактические численности (при общей численности, равной п)
будут следующими:
Фенотипы
Долн (вероятности)
Ожидаемые численности
Фактические
численности
АВ
— (3-/пг)
-J-*3-"1')
<Vi
Ab
1
4 mr
л
~ mr
Rx
cB
1
4 mr
n
~4~ Mr
*,
ab
-J- (1—wr) |
"f (\-*f) 1
A'i J
Применив метод максимального правдоподобия, можно
составить следующее квадратное уравнение (приводим его без
вывода):
nm.2_ (,у,+4Л|+4Л2+3^г)тг+3(«1+/г2)=0.
Подставив в него численности всех четырех групп
(фенотипов), полученных в F1% определяем значение /лг.
Для иллюстрации метода приведем следующие данные,
полученные на дрозофиле. В результате скрещивания
родительских форм — нормальных особей и гомозиготных по генам
black (Ы) и vestigial (vg) — было получено в Fj нормальных
103, vg 5, Ы 9 и Ы—vg 32. Всего 149 особей.
Квадратное уравнение будет иметь вид
149mr2-255mr+42=0.
Отсюда шг=0,18, а его ошибка smr=0,03*.
При наличии кроссинговера у обоих патов в F? получится
уже 8 категорий генотипов, которые сгруппируются в 4
фенотипа:
Фенотипы
Доли
(вероятности)
Ожидаемые
численности
Фактические
численности
АВ
Т~(2-*?)
-г*2*"*
A'i
Ab
-ГО-Ч)
*1
аВ
-го-^
*«
еЬ
I о
~т'г
4 т'
N%
* Мы пользуемся для вычисления ошибки обычном формулой ошибки
доли, но можно вычислять ее по более сложно* форм>ле (об этом см,
специальную литературу).
53
Применение метода максимального правдоподобия
позволит вывести квадратное уравнение дли определения значения
nm^—(Ni—2Ri—2R2—N2)mr—2N2 = 0.
В качестве примера можно использовать расщепление
у кукурузы по генам sh (сморщенный эндосперм) и аи
(желтый хлорофилл). В 1:2 было получено растений: нормальных
2108, нормальных по эндосперму, но с желтым хлорофиллом
311, со сморщенным эндоспермом п зеленых 310 и
проявляющих оба рецессивных признака 492. После подстановки чис-
ленностей каждой группы в указанное выше уравнение, по
которому определяется шг:
3221 Шг2—374тг—984 = 0.
Отсюда
тг=0,613 и smr =0,026.
В задачах этого типа необходимо проверять, какова была
группировка генов у исходных родительских форм: если оба
доминанта были у одного родителя, а оба рецессива — у
другого, то кроссоверными классами в F2 являются формы аВ
и АЬ. Если же родители были аВ и АЬУ то кроссоверными
становятся крайние классы — АВ и ab.
При решении уравнения, записанного в той же форме, как
приведенная выше, получается значение не mr, а 1—mr.
Частота рекомбинации и расстояние между генами.
Изучение различий в значениях коэффициентов рекомбинаций
между разными генами привело к формулировке важнейшей
генетической закономерности, а именно: величина рекомбинации
зависит от расстояния между генами, расположенными в
хромосомах линейно. Значение тпг — это линейная функция
расстояния, поэтому оно может служить мерилом расстояния,
правда, выраженного в генетических величинах, но
отражающего в какой-то степени физическое расстояние. Чем оно
больше, тем больше вероятности разломов между генами и нового
соединения гомологичных хромосом. По предложению Стёр-
тсванта, за единицу измерения генетического расстояния был
принят отрезок хромосомы, в пределах которого происходит
в среднем один кроссинговер на 100 гамет, или 1% кросенн-
говера (в этом случае коэффициент рекомбинации
переводится в проценты); советские же ученые назвали ее
морга пилой.
Следующий шаг в применении положения о зависимости
частоты рекомбинаций от расстояния — установление
взаимного расположения генов. Возникла необходимость учета
растеплений в скрещиваниях с участием трех или нескольких
%4
Экспериментальные данные показали, что выраженное
в тг расстояние между двумя генами Aw В равно с>мме или
разности между каждым из них и третьим геном С, т. е.
>"мв=/и,лг±л»ггя. ("))
Если наблюдается суммирование, то порядок генов
А—С—В, если, наоборот, разность, то А—В—С. Тем самым
были заложены основы для генетического картирования
хромосом.
В последующем выяснилось, что правило простого
арифметического суммирования по формуле (5) верно для
сравнительно коротких расстояний (в пределах 10 морганид). При
больших же расстояниях тгль оказывается меньше
ожидаемого, так как наряду с одиночными кроссинговерами происходят
и двойные, возвращающие гены на старые места. Поэтому,
если отдельно рассматривать интервалы между генами АС и Cfl,
обозначив их соответственно через тг\ и тг^ то в
действительности
mrlW)=m,i+mr2-2 т,г тг*. (6)
Было показано, что между двойными кроссинговерами
происходят довольно сложные взаимоотношения, а именно:
происшедший обмен на одном участке хромосомы тормозит обмен
на другом. Это явление было названо интерференцией.
Степень совпадения двойных кроссипговероп при интерференции
(коиисидемция) обычно обозначают величиной С. поэтому
указанная выше формула (6) должна быть усложнена:
/nr(i+2)==Mrt-f Mrs—2 Cmrrrnr>. (6 а)
На больших расстояниях значение С приближается к
единице, тогда надо считаться только с наличием двойных крос-
енпговеров.
В конечном счете соотношение этих процессов приводит
к тому, что тг для сцепленных генов имеет предел ОД т. с.
гены, находящиеся па большом расстоянии друг от друга
в хромосоме, ведут себя как независимые, В этом случае
вероятность рекомбинации уже не является мерилом расстояния.
Возникает вопрос о том, нельзя ли нее же пользоваться
данными о длинных расстояниях для установления истинных
генетических расстояний (J), вводя какую-то поправку в
данные о первых. Такая поправка была сделана Холлойном,
предложившим формулу
</=:-'- /п(|-2/Я,>. <7)
Но формула Холдейиа ие отражает полностью сложных
отношений между генетическим расстоянием, частого* кроссинге*
яеров. интерференцией и коиисндеяцией, тем 6aiee что ощщ,
по видимому, неодииакоаы у разных видов. Поэтому для онр*
деления длинных расстояний выгоднее пользоваться чисто
эмпирическим методом суммирования частот рекомбинаций на
более коротких промежуточных участках хромосомы.
Этим мы ограничимся при рассмотрении рекомбинаций,
так как онтогенетическая природа внутрихромосомных ре»
комбинаций выходит эа рамки статистической генетики. Для
последней существенно лишь то, что точки обменов
распределены по длине хромосомы в основном случайно н средняя
их частота более или менее постоянна для отдельных
участков хромосом, это является основанием для более сложных
расчетов, применяемых при анализе рекомбинаций. В то же
аремя надо иметь в виду, что статнетичность появления ре-
комбииантов ограничивается по крайней мере двумя
обстоятельствами:
а) Кроссннговеры протекают неравномерно по длине
хромосом; есть участки в хромосомах, где кроссинговер вообще
не может происходить или происходит слабее, чем на других
участках, в силу особенностей их строения; так, в центромер-
иых областях хромосом дрозофилы частота кросеннговера
ниже, чем в днетальных.
б) Кроссинговер может быть вообще заторможен у
одного пола, как это имеет место у дрозофилы в обычных
условиях (в эксперименте все же удается добиться кросеннговера
и у самцов).
Вообще интенсивность процессов рекомбинаций
определяется как структурой хромосом, так и генотипом и даже
конкретными генами. Неизменными являются лишь
относительное положение генов на каоте и сами физические размеры
хромосом и генов.
Таким образом, в процессах обмена участками
гомологичных хромосом наблюдается сложное переплетение
статистических и динамических закономерностей с преобладанием
вторых.
Глава 2
СТОХАСТИКА МУТАЦИОННОГО ПРОЦЕССА
Мутации как ошябм в редунмкамм. Мутации
неразрывно связаны с редупликацией как наиболее важным свойствам
живого. Редупликация наследственных структур обесвечява-
ет при смене поколений сохранение постоянства
морфологических особенностей живых существ, типов обмена,
физиологических и биохимических свойств видов. Выработка
механизма редупликации лежит у истоков происхождения жизни.
Но если бы редупликация была абсолютно жесткой, то
не было бы материала для эволюции, для создания
новообразовании в процессе эволюции, не было бы и самой
эволюции. Таким образом, нарушения редупликации, или, как
иногда говорят, ошибки редупликации, так же необходимы, как
и сама редупликация.
Под мутациями в самом общем смысле слова следует
понимать такое нарушение редупликации, при котором одни
из дочерних реллнкантов отлетается от родительского
элемента. Не имеет принципиального значения в статистическом
смысле слова, когда и как нарушение происходит,
непосредственно ли в процессе редупликации или до него—
неправильный ли это синтез молекулы ДНК или же изменение уже
существующих генов путем замены оснований в триплетах
и т. д. Это уже специальный вопрос, н он детально
рассматривается в литературе по мутагенезу. С точки зрения
статистической генетики существенно лишь то, каков характер
нарушения редупликаций (мутаций), является ли он
статистическим или динамическим, т. е. в какой степени к нему при-
ложнмы законы вероятности.
Как и во многих других рассмотренных выше генетических
явлениях, основное значение и в этом случае приобретают ста*
тистическне, вероятностные закономерности, так как ошибки
редупликации являются в конечном счете случайными
редкими событиями, оцениваемыми вероятностями. Они
реализуются в виде отдельных случаев в громадных по объему
множествах клеток, организмов, фаговых частиц и т. д.
57
Соотношение между частотой нарушений редупликации
и частотой нормального процесса редупликации. Оно должно
оып> таким, чтобы, с одной стороны, обеспечивалась достаточ*
пая стабильность вида, выработавшаяся эволюцпонно (поэ-
Юму-то мутации и являются редкими событиями), а с дру.
гон — оставалась бы возможной дальнейшая эволюция на
основе появления новых изменений.
Сейчас хорошо известно, что уровень мутабильностн,
свойственный различным линиям внутри вида пли отдельным
видам, неодинаков и что ом находится под генетическим
контролем. Очевидно, что этот уровень представляет собой
результат эволюционного процесса, т. е. отбора на необходимый в
данных конкретных условиях уровень мутационной
изменчивости. Результатом отбора является и то, что между
отдельными генами могут быть очень большие различия в мута-
бплыюсти (в крайних случаях па 3 порядка, т. е. в 1000 раз),
о чем речь пойдет ниже.
Но важно основное, что мутации, т. е. ошибки
редупликации, представляют собой редкие события, возникающие
только с известной вероятностью.
Общая характеристика методов учета мутаций. Чтобы
оценить вероятность возникновения мутаций, необходимо
установить методы их учета. С первого взгляда может казаться,
что наиболее простым и универсальным методом служит
подсчет обнаруженных в популяции животных пли растений
мутантов, т. е. форм с новыми признаками, и установление их
доли в общем количестве изученных растений или животных.
Именно по такому пути шел де Фриз, впервые применивший
количественные методы оценки частоты мутаций. Работая с
энотерой (Oenothera lamarckiana), он обнаружил среди
растений с нормальными для этого вида признаками ряд
измененных форм — с широкими листьями, более крупных по
размерам, с красными жилками и т. д. Некоторые из них
появлялись лишь в одном экземпляре на многие тысячи
просмотренных растений, другие повторялись. Это обстоятельство дало
де Фризу основание включить в число формулированных им
положений мутационной теории пункты о том, что мутации
появляются в единичном числе и что они могут иногда
повторяться.
Как выяснилось позднее, так называемые «мутации» у
энотеры в действительности представляют собой результат
сложных хромосомных перестроек ( о чем уже упоминалось
в предыдущей главе) и лишь немногие из наблюдавшихся
до Фризом типов могут быть отнесены к настоящим
мутациям.
В действительности учет мутаций очень редко может быть
слолан прямым путем. Прежде всего он зависит от способов
г. к
размножения объекта изучения. При бесполом и
вегетативном размножении многометочных организмов мутации
обнаруживаются в соматических клетках или побегах, из них
возникающих, или в потомстве одной особи, дающей клоны.
У самооплодотворяющихся форм можно обнаружить мутации
в виде новой формы среди однородных по внешности особей
самооплодотворяющейся линии. Но в этом случае все же нет
абсолютной уверенности, что новая форма появилась в
результате мутации в предыдущем поколении, а не возникла
(пли уже существовала) ранее. Известно, что у растений,
обычно считающихся самоопылителями, иногда происходит
перекрестное опыление, что совершенно меняет ожидаемую
картину.
У перекрестноопыляющихся или раздельнополых
организмов между временем появления рецессивной мутации и
обнаружением мутантных форм может пройти немало
поколений, у которых мутации находятся в гетерозиготном
состоянии до тех пор, пока не наступит случайное скрещивание двух
гетерозигот. В результате в потомстве появляются
гомозиготы по возникшему ранее в порядке мутации рецессивному
гену. Очень велика и вероятность потери возникшей ранее
мутации в силу случайных причин (см. об этом в гл. 4).
Значительно легче обнаружить доминантные мутации, так как они
проявляются в гетерозиготе уже у особи первого поколения.
Поэтому частота видимых доминантных мутаций может быть
определена непосредственно по проценту форм с вновь
возникшими доминантными признаками.
Очень труден вопрос о диагнозе тех признаков,
изменения которых должны быть учтены. Если ярко выраженные
морфологические признаки и изменения в них могут быть
обнаружены визуально, то слабо выраженные легко
пропустить. При учете мутаций по таким признакам приходится
считаться с индивидуальностью исследователя. Чтобы ослабить
этот персональный фактор, прибегают к предварительной
разработке объективных шкал для оценки признаков (например,
окрасок глаз дрозофилы, тонов окраски шерстного покрова
и т. д.). Мутации же физиологических и биохимических
признаков, а также количественных признаков требуют
специальных методов учета.
Чтобы избежать субъективности в оценке, например
хромосомных нарушений при просмотре метафазных плаепшок,
предварительно зашифровывают материал таким образом!
чтобы лицо, наблюдающее нарушения, не знало, с каким
именно материалом, контрольным или опытным, оно имеет
дело. В силу особенностей размножения микроорганизмов
учет мутаций у них связан с известными трудностями и
имеет свою специфику, о чем будет сказано ниже.
59
гаметы
Гаметы
Очевидно, что для точного учета мутации лучше всего
иметь особо разработанные экспериментальные методы длц
каждого конкретного объекта. А чтобы провести количествен-
мое сравнение частот мутаций, определенных для разных
объектов, нужно точно знать, с помощью какого метода они
получены и к чему конкретно относятся (хромосома,
отдельные гены, целый геном, клетка, клеточное поколение и т. д,),
Чаще всего
употребляют термин «частота
мутаций». Но иногда
употребляют и другой —
«скорость мутационного
процесса*. Они очень близки
друг к другу, но
некоторые исследователи
предпочитают второй термин,
как более общую
характеристику регулярно
протекающего у всех
организмов процесса
возникновения мутаций.
Следует иметь в виду
еще одно общее
обстоятельство . Оценка частот
мутаций (например, у
дрозофилы) относится к
популяциям гамет, однако
не всех, а только тех, из
которых образовались
доступные для учета
зиготы. В таком случае
частоты мутаций измеряют
лишь долю жизнеспособных гамет, которые мутировали.
Измерять же частоту мутаций у всех подвергавшихся
воздействию половых клеток становится невозможным.
Однако у микроорганизмов в силу специфических
особенностей их размножения задача не столь безнадежна.
Но если предположить, что примененное воздействие
может убивать в примерно одинаковой степени мутантные и нему-
тантные клетки, определение частоты мутаций на основе только
выживших клеток приобретает достаточный вес.
Характеристика скорости мутационного процесса должна
охватывать сумму мутаций всех категорий. С другой стороны*
могут иметь значение данные о частотах мутаций отдельны*
генов. Этот показатель, который будет рассмотрен специальна
также очень существен для характеристики мутационного
процесса.
Рис. 9. Учет сцепленных с полом
летальных мутаций по методике С1В.
60
Учет частоты сцепленных с полом летальных и видимых
мутаций у дрозофилы. Исторически наиболее ранним (1027)
и достаточно точным был разработанный Мёллером метод учета
сцепленных с полом мутаций, т. е. мутаций, возникающих
в половой хромосоме дрозофилы (Drosophila melanogaster).
Он получил название метода CIB — по структуре половой
хромосомы. Схема его представлена на рис. 9.
Линия С1В характеризуется тем, что в одной из половых
хромосом самок находится доминантный ген Ваг, вызывающий
образование узких, полосковидных глаз (в гомозиготном
состоянии). Гетерозиготы же по гену Ваг имеют глаза формы
фасолин, причем число фасеток в их глазах промежуточное
между числом фасеток (около 800) у нормальных особей
и гомозигот Ваг (число фасеток около 100). Кроме того,
с этой же половой хромосоме, содержащей ген Bar, имеется
инверсия С, препятствующая кроссинговеру и в то же время
обладающая летальным действием в гомозиготном состоянии
(/). Поэтому у самок этой линии, внешне отличимых по глазам
в форме фасолин, при их обычном разведении половина
сыновей., а именно те, которые получают хромосому С/В, гибнет.
Допустим теперь, что в нормальной Х-хромосоме одного из
сперматозоидов отца из поколения Р возникла летальная
мутация. Она обозначена в верхнем правом углу рис. 9 в
хромосоме самца Р буквой /. После оплодотворения яйцевой
клетки она перейдет к одной из самок F\ ( Bar или не-Barj.
Обычно для последующего анализа используются только самки
Ваг. Все скрещивания таких самок должны быть строго
индивидуальными (1 ? X' rf). От каждой самки Ваг из г и которая,
таким образом, имеет одну половую хромосому С/fl, а вторую
с мутантным летальным геном /, получается поколение f2, в
котором не будет ни одной особи мужского пола. Одна половина
самцов гибнет из-за хромосомы С/В, а вторая — из-за
хромосомы с мутантным геном /.
Так как летальная мутация возникла в единичном
сперматозоиде из многих других, то она передается только одной
самке F\ из многих других и эффект ее летального действия
проявится только в одной пробирке F2, полученной от этой
самки, так что особи одной пробирки F2 отображают одну
хромосому сперматозоидов Р. По числу пробирок с отсутствием
самцов можно судить о проценте летальных мутаций,
возникших у Р. Достаточно просмотреть п пробирок f2, полученных
от индивидуальных скрещиваний, и если среди них в т
пробирках не будет самцов, то процент летальных мутаций р
выразится как
/я-100 ,.,
г п
61
Ошибка л.ти р вычисляется по обычной формуле при альтер,
натнпнон изменчивости
' * У п
Разница между р\ опыта и р0 контроля (пли между р\ и /?,
двух опытов) и ее ошибка определяется по обычной формуле
Закономерное, а не случайное отсутствие самцов может
быть доказано только при достаточном количестве особей
в пробирке, а именно: должно быть не менее 8—10 самок при
полном отсутствии самцов.
С помощью метода С1В могут быть учтены и видимые
мутации, возникающие в половой хромосоме дрозофилы. Схема
наследования такая же, как и в случае летальной мутации.
Допустим, что в Х-хромосоме одного из сперматозоидов
возникла видимая мутация а. Тогда у самок Fy она окажется
в гетерозиготном состоянии и проявится в F2 у всех самцов.
Вторая половина самцов F2, как уже сказано, гибнет, так как
она получит хромосому С1В.
Таким образом при методе С1В учитывается процент
мутаций в исходном числе Х-хромосом, бывших в
сперматозоидах * Р. Число пробирок п в F2 соответствует количеству
анализированных хромосом в учтенных самцах Р. Все же метод
С1В чаще всего используется для установления процента
летальных мутаций.
Другой, более простой метод учета сцепленных с полом
видимых мутаций будет изложен ниже.
Учет сцепленных с полом летальных мутаций у дрозофилы
может производиться и по методике Меллер-5, имеющей
несколько модификаций. Схема анализа по одной из них
представлена на рис. 10.
В отличие от линии С1В исходные самки линии Меллер-5
гомозиготны по нескольким генам Х-хромосомы. На рис. 10
отмечены лишь два гена: Ваг (иолосковидпые глаза) и W*
(apricot — глаза желтоватые, напоминающие цвет абрикосов)-
В других модификациях метода Меллер-5 используются и иные
гены, например yellow (желтое тело). Кроме того, обе
Х-хромосомы содержат две инверсии, затрудняющие перекрест между
ними, но не имеющие летального действия. При появлении
летальной мутации / в Х-хромосоме одною из сперматозоидов
исходною самца Р гибнет половина самцов Г2 (самцы пс-Ваг),
И чдесь закономерное, а не случайное отсутствие самцов
не-Лаг может быть доказано только при достаточном ко.чи-
ъч
чествс потомства в пробирке Fb а именно: должно быть не
менее 8—10 самцов Ваг и ни одного самца ис-Вдг. В ггом
случае вероятность появления летальной мутации больше 0,{Ю.
Если же возникает видимая мутация а, то она проявляется
у половины самцов F2 (у самцов не-Вш-).
Таким образом, учет мутаций с помощью обеих методик
цедется на количество Х-хромосом, находившихся в
сперматозоидах самцов Я.
Гамет/
/>?
йвкйИ'Ш»
i
Гаметы
h
<Н1Ш ww
input
или See о
Put. 10. Учет сцепленных с пило* лотымшх
мутаций по мотодико Мсллср-5
Обычно методы С1В и Л\еллер-5 служат для установления
мутагенного действия того или иного физического или
химического фактора. Группа самцов (опытная) подвергается
воздействию и скрещивается с самками тон или иной исходной
линии. Другая группа самцов (контрольная, без воздействия)
также скрещивается с соответствующими самками. Однако
и и контроле могут возникать летальные мутации. Так как ле-
тальная мутация не могла быть в скрытом виде в Х-хромоео-
ме исходного самца, частота летальных мутаций в контроле
является точным показателем спонтанной частоты мутации-
Поскольку процент, выражающий частоту спонтанных
мутации, очень'мал (чаще всего это какая-то доля "Р01**"™/; ™
Для получении достоверных данных количество пробирок г,
63
контрили должно быть, как правило, значительно большим,
нежели количество пробирок /'2 опыта. Численности опытной
и контрольной групп должны быть такими, чтобы обеспечить
достаточную достоиерность различий по проценту мутаций
к'ожду ними.
В силу статистпчмости процесса мутаций не следует
ожидать полною соотнетстпня частот мутаций по отдельным
сериям при пх понторностн. Само же распределение разных частот
-nrb'i *
Pus. //. Учет видимых сцепленных с полом мутаций по методу
сцепленных X-хромосом.
мутации по отдельным опытам как событий, обладающих
малой вероятностью, должно быть пуассоновым.
Линии С1В и Меллер-5 могут быть использованы также
для учета полулетальных мутаций, а именно: в первом случае
вместо полного отсутствия самцов ие-Bar появляется все же
1—2 самца, во втором их окажется меньше ожидаемой
половины. Все такие случаи отклонений по отношению полов
требуют дополнительной проверки в поколениях F3 и F4, и при
повторении наблюдавшихся отклонений появление
полулегальной мутации можно считать доказанным *.
Очень изящный метод учета видимых сцепленных с полон
мутаций стал возможен после получения у дрозофилы линий
так называемых сцепленных Х-хромосом. У самок этой
линии X-хромосомы, содержащие ген yellow (желтое тело),
соединены друг с другом концами — центромерами. Кроме того,
* Впрочем, ааличяе отклонения от ожидаемых отношений среди самад*
■ пробирках Ft или f, может объясняться и некоторыми другими причинам*
например очень редкими случая мл кросеннговера.
W
у этих самок есть У-хромосома. Если в Х-хромосоме одного
из сперматозоидов самца Р возникла видимая мутация а, то
она сразу проявится у одного самца F, (четвертый в нижнем
ряду рис. 11). В силу особенностей линии сцепленных Х-хро-
мосом отклоняющиеся по хромосомному набору формы
(с тремя X или с двумя У) гибнут, и линия разводится в
чистоте (самки у у и самцы + ). Тогда выраженная в процентах
доля самцов с видимыми изменениями в F\ непосредственно
измерит частоту видимых мутаций. Осложняющим бывает то
обстоятельство, что мутантные формы могут обладать
большой длительностью развития и вылупляться из куколок
позднее, нежели нормальные формы. Поэтому целесообразно
просматривать культуры F\ (или F2 при методах С1В и Мел-
лер-5) до тех пор, пока не выведутся мухи из всех имеющихся
куколок.
При появлении видимых мутаций может оказаться, что
некоторые из них затрагивают одни и те же признаки и более
или менее сходны по своему выражению. Тогда возникает
более сложная задача их идентификации (установление алле-
лизма или псевдоаллелизма или, наконец, того, что они не
аллельны друг другу), а это является темой специального
генетического анализа.
Метод учета летальных мутаций в аутосомах дрозофилы.
Для этой цели используются специальные линии, так
называемые сбалансированные. Так, линия CyL содержит в одной из
пары вторых хромосом доминантные гены Curly (загнутые
крылья) и Lobe (маленькие глаза), каждый из которых в
гомозиготном состоянии летален, а в другой хромосоме — также
доминантный ген Plum. В силу наличия инверсий кроссинговер
в этих хромосомах подавлен. На рис. 12 представлена схема
наследования летальной мутации сперматозоида, возникшей
в одной из вторых хромосом. Особенность этой схемы в том,
что из Ft выбираются для скрещивания только определенные
категории особей ^ CyL и Q CyL/Pm, чтобы во втором
поколении скрещивать'друг с другом особей, получивших от
исходного самца Р одну и ту же вторую хромосому. Если в этой
хромосоме возникла летальная мутация, то в Fj не появится вовсе
категория нормальных особей (на рис. 12 в правом нижнем
углу). Если же мутация не возникла, то в FZf кроме особей CyL
(гетерознгот, так как гомозиготы по этим генам погибнут еще
во время эмбриогенеза), будут также и нормальные особи.
Таким образом, по проценту культур F3, в которых отсутствуют
нормальные особи, можно судить о проценте летальных
мутации, возникших в сперматозоидах самца Р. Очевидно, что этот
метод может служить для учета и видимых мутаций во второй
хромосоме — вместо категории нормальных особей появится
категория с каким-то иным мутационным признаком.
5 П. ф. Рокицкий
По тон же методике можно изучать и частоту полулеталей
Стандартом для сравнения должна быть средняя доля мух
с изучаемой хромосомой в контрольных пробирках F$.
В последнее время методика учета аутосомных леталей
стала применяться даже чаще, нежели сцепленных с полом
леталей. особенно потому, что с ее помощью путем соответст-
Л
т
."U? ?'■€
£.?
пщ*т
/
иметы
т шш\т
t
Рис. 12. Схема учета частоты аутосомных
леталей у дрозофилы.
вующих скрещиваний можно проверить аллеломорфизм
полученных леталей.
Для сравнения мутабильности разных категорий половых
клеток (например, разного возраста) иногда целесообразно
проводить одновременный учет сцепленных с полом и
аутосомных леталей. Для этой цели сочетают метод Меллер-5 и метод
анализа аутосомных леталей с помощью линии Curly, т. е.
самки линии Меллер-5 должны быть одновременно
гетерозиготными по Су. Самки F\ служат для анализа в F2
сцепленных с полом леталей, а самцы F\ — для последующего
анализа аутосомных леталей.
т
Изучение частоты летальных мутации обладает тем
преимуществом, что при этом полностью снимается роль
индивидуальности исследователя, чего трудно избегнуть при учете
видимых мутаций. Однако учет видимых мутаций при приме-
пении метода Меллер-5 также облегчает задачу, так как в F2
все самцы будут нести появившееся мутационное изменение,
а при методе сцепленных Х-хромосом — только один самец»
которого легко пропустить.
Метод изучения мутаций определенных генов. Хотя этот
метод имеет более ограниченные цели, он очень точен и
позволяет установить не частоту общей суммы видимых
мутации, а частоты мутаций определенных генов. На дрозофиле
он может быть применен по отношению как к сцепленным
с полом, так и аутосомиым генам. Для этой цели
используются специальные линии со многими генами-маркерами.
Например, ywf (yellow — желтое тело, white—белые глаза
и forked — закрученные щетинки) и т. д. Самки (или самцы)
из таких линии скрещиваются с нормальными по этим генам
особями. В F\ появляются самки, гетерозиготные по
генам-маркерам. Если в нормальной хромосоме (под влиянием
примененного воздействия или спонтанно) произойдет мутация по
одному из генов-маркеров, то она проявится у особи F\.
Таким образом может быть учтена частота мутаций отдельных
генов по доле особей, проявивших этот ген из общего числа
просмотренных гетерозигот.
Этот метод, естественно, требует очень большого объема
экспериментального материала.
Нужно учесть, что наблюдаемые измененные формы могут
Сыть следствием не только генной мутации, но и дефишенси
(выпадения) участка хромосомы с этим геном. Для их
различения может быть применен дополнительный учет
жизнеспособности появляющихся мутантных форм, а также
цитологическое исследование хромосом слюнных желез. Исчезновение
определенных дисков может быть доказательством того, что
в данном случае имеется не аналогичная мутация, а только
выпадение, которое можно проверить при разведении
полученных гетерозигот в следующих поколениях. Метод изучения
хромосом слюнных желез позволяет установить и размеры
выпавшего участка и его точное местоположение.
Подобный же метод изучения мутаций определенных генов
был применен для анализа мутационного процесса у мышей.
Учет доминантных деталей. Обычный метод измерения
частоты доминантных деталей у подвергавшихся воздействию
самцов дрозофилы заключается в сравнении количества нераэ-
вившихся яиц опытной группы с контролем. Это количество
может быть выражено в процентах к общему количеству яиц
в контроле.
5*
d:
Нельзя, однако» не отметить некоторую неопределенность
понятия «доминантные летали». Предусматривается, что это
изменения доминантные, т. е. проявляющиеся в первом
поколении и заключающиеся в смерти особи на какой-то стадии
развития. Хотя генетически границей следовало бы считать
период до оплодотворения, однако на практике чаще всего
границей считают вылупление из куколок дрозофилы. Кроме того,
нет полной уверенности в том, что невылупленпе из яиц не
определяется и другими причинами, например неоплодотворен-
ностью (эта причина бывает наиболее частой в опытах по
облучению незрелых мужских половых клеток). Если же
оплодотворение произошло, то невылупление может
обусловливаться и рядом других причин, не входящих в категорию
доминантных леталей. Наконец, не может быть изучена и
генетическая природа доминантных леталей. Только по косвенным
соображениям можно сделать вывод, что лишь немногие из
них представляют собой генные (точковые) мутации,
большинство же, вероятно, различные хромосомные нарушения.
Несмотря на все эти ограничения, учет доминантных
леталей все же полезен при сравнении результатов действия
различных физических и химических мутагенов.
Учет хромосомных аберраций. Этот учет может быть
произведен с помощью различных сложных линий дрозофилы, но
значительно проще проводить его цитологическими методами,
пользуясь анализом хромосом слюнных желез. Для гигантских
хромосом слюнных желез Бриджесом были составлены точные
карты нормального расположения в них дисков, полос и
светлых промежутков. Позднее в них были отмечены и места
расположения многих изученных генов.
Сравнительно простая методика изготовления препаратов
с хромосомами слюнных желез дает возможность учитывать
хромосомные нарушения всех типов (транслокации, делеции,
инверсии и т. д.) и выразить их количество в процентах к
общему числу изученных хромосом. При этом могут быть учтены
и такие типы хромосомных нарушений (например, дупликации
и делеции очень небольших участков хромосом), которые не
устанавливаются обычными генетическими методами.
Уменьшается и опасность потери изменений вследствие
отбора против нежизнеспособности хромосомных нарушений,
так как анализ проводится по особям F\, хотя некоторые все
же могут быть потеряны как доминантные летали.
Аналогично положение и с культурой тканей (человека,
животных и растений). Визуальные наблюдения метафазных
пластинок отдельных клеток дают возможность обнаружить
различные хромосомные отклонения — разрывы хромосом,
смосты», т. е. соединение хромосом, и т. д. При этом можно
выделять хромосомные изменения одноударного и двуударно-
68
го типов (о чем речь пойдет ниже). При анализе культур
клеток учитывается, таким образом, количество хромосомных
изменений на общее количество изученных клеток.
Мы не будем здесь останавливаться на методах учета
различных хромосомных аберраций у дрозофилы с помощью
специально созданных для чтой цели сложных линий, так как
принцип учета здесь тот же: количество обнаруженных
изменений относится или к общему числу изученных хромосом
(если изменения обнаруживаются лишь в одной хромосоме),
или к общему числу изученных исходных гамет (если
учитываются изменения, захватывающие не одну, а две или больше
хромосом).
Особенности учета мутаций у перепончатокрылых.
Свойственное перепончатокрылым определение пола (диплоидность
самок и гаплоидность самцов) дает ряд ценных
возможностей для определения частоты мутаций, но в то же время
вносит некоторые ограничения. Известно, что наездник Habrob-
гасоп был одним из первых объектов, на котором было
доказано мутагенное действие рентгеновских лучей, а также
быстрых нейтронов. Новым объектом для генетических
исследований стала паразитическая оса Marmoniella. У всех
перепончатокрылых любая мутация сразу же проявляется в F\. Если
мать была гетерозиготна по какой-то рецессивной мутации, то
последняя проявляется у половины самцов первого
поколения.
А. С. Михайлов в 1931 г. обнаружил мутацию белых глаз
у пчел. Применяя технику искусственного осеменения
пчелиных маток, он смог демонстрировать расщепление среди
трутней по окраске глаз на большом материале. Так, только одна
матка № 63 дала 811 белоглазых и 806 черноглазых трутней.
При таком характере определения пола частота видимых
мутаций может быть определена непосредственно по проценту
самок, дающих расщепление среди сыновей в F{. Очень легко
также доказать, что мутация возникла в данном поколении,
а не ранее, так как она была бы обнаружена у самцов —
братьев анализируемой самки.
При гетерозиготности самок перепончатокрылых по
летальному гену число ожидаемых сыновей уменьшается вдвое. Но
практически это означает, что неспособна к развитию
половина неоплодотворенных яиц, а уловить это при способе
размножения перепончатокрылых почти невозможно. Впрочем, у Мог-
moniella с ее 5 хорошо маркированными хромосомами летали,
по которым могут быть гетерозиготны самкн, довольно легко
локализовать.
У ос отсутствуют гигантские хромосомы, как это имеет
место у дрозофил, поэтому к ним не применим прямой
цитологический метод.
69
Тесты для учета доминантных легален у видов с такой
системой определения пола (гаплодиплоидной) вообще
сомнительны.
Методы учета частоты мутаций у млекопитающих. Мышь
пока почти единственный вид млекопитающих, кроме
человека, на котором проводилось более детальное изучение мутаци-
гнного процесса. Подобно дрозофиле, у мышей учитывалось
появление доминантных и рецессивных леталей, видимых му-
1ацнй, дефншенси и транслокаций. Однако надо учесть неко*
торые принципиальные различия между генетикой дрозофилы
и генетикой мыши: 1) у дрозофилы три пары больших хромо-
ir-M и одна маленьких; у мыши 20 пар хромосом небольшого
размера; 2) Х-хромосома дрозофилы несет около 1/5 всех
генов, мыши меньше— 1/20 всех генов. Поэтому у мышей
сцепленные с полом мутации составляют очень маленькую долю
гсех мутаций; 3) у самцов дрозофилы нет кроссинговера,
\ мышей кроссинговер происходит у обоих полов. Все это
значительно осложняет методы учета мутаций у мышей.
Простейший метод определения частоты доминантных
леталей у млекопитающих заключается в сравнении размеров
помета контрольной и подвергавшейся воздействию групп,
однако надо учитывать, что на размеры пометов очень большое
слияние оказывают многочисленные факторы внешней среды.
Коэффициент наследуемости размеров пометов у многоплод-
ных животных (см. гл. 6) 0,10—0,15. Данные, полученные
мною с сотрудниками на кроликах в одной из серий опытов,
таковы: контроль — 7,6 крольчонка на самку; опытная
группа, полученная от осеменения облученной рентгеновскими
лучами спермой,— 1,0 крольчонка на самку. В числе детенышей
в опытной группе были 1 мертворожденный и 1 недоношенный,
что явно указывает на появление доминантных леталей.
Дополнительно применяют также выборочное вскрытие
беременных самок и подсчет живых и мертвых зародышей, а
также сравнение с количеством желтых тел в матке самок.
В силу этих усложнений в методах доминантные летали у
млекопитающих могут быть установлены достаточно точно.
Рецессивные летали у млекопитающих можно обнаружить
только в тех случаях, когда они возникают в
непосредственной близости от генов-маркеров, которые уже известны у
мышей, но так как маркированные хромосомы составляют лишь
небольшую часть общего генома мыши, то данные о
количестве леталей, полученные таким путем, не могут дать точной
характеристики истинной частоты мутаций в целом для вида.
Доминантные видимые мутации могут быть обнаружены
очень легко— в первом же поколении. Таковы некоторые
мутации у норок, например платиновая окраска. Доля мутант-
них форм но отношению к числу разводимых или изученных
?п
порок могла бы характеризовать частоту доминантных
видимых мутаций. Однако такие подсчеты очень приблизительны,
так как норок разводят во многих хозяйствах во всех странах
мира и пет возможности точно учесть, сколько же в
действительности норок изучалось.
Еще труднее обстоит дело с учетом рецессивных видимых
мутаций. Появление рецессивной мутантной формы —
результат скрещивания гетерозигот, сама же мутация могла
возникнуть значительно раньше и оставаться скрытой в течение ряда
прошлых поколений. Только длительный инбридинг может
1арантировать, что мутационное изменение действительно
появилось вновь в какой-то не очень отдаленный период, а это
требует создания специальных инбредных линий и проведения
определенной схемы скрещиваний. Так, чтобы уловить
появление мутации в одном из сперматозоидов облученного
рентгеновскими пли иными лучами самца мыши, взятого из
проверенной линии, надо скрестить его с нормальными самками
и особей первого поколения скрещивать друг с другом. Затем
из F2 следует отобрать по нескольку самок (считается, что их
надо взять не менее 7) и скрещивать их с их отцами. Если
некоторые из самцов F) были гетерозиготны по возникшей
видимой мутации, то в F$ от них выщепятся мутантные формы.
Очевидно, что масштаб таких опытов должен быть очень
большим.
В настоящее время у мышей стал играть большую роль
более простои метод изучения мутаций определенных генов,
описанный выше для дрозофилы, при котором контрольные
или подвергавшиеся воздействию особи (у мышей пол не
имеет значения) скрещиваются с особями, гомозиготными по
многим рецессивным генам, и тогда в F\ будет уловлена любая
мутация в нормальной хромосоме по генам мутантной линии.
Расселом были созданы такие линии, гомозиготные по 7
видимым и хорошо различимым рецессивным генам. Очевидно, что
в этом случае могут быть учтены и любые доминантные
мутации. Но объем таких опытов должен быть достаточно
большим: в одном из опытов Рассела было изучено свыше 80
тысяч особей первого поколения.
Если у дрозофилы вопрос о том, является ли возникшее
в F\ изменение действительно мутацией, аналогичной мутации
гена маркированной хромосомы, пли же это выпадение
участка хромосомы (дефншенси), может быть легко решен
цитологически на хромосомах слюнных желез, то у мышей вывод
можно сделать только после перевода этой мутации в
гомозиготное состояние.
Методы установления частоты мутаций у человека.
Несмотря на громадные и очевидные трудности анализа
мутационного процесса у человека, все же были разработаны методы, по-
71
эволяюшие характеризовать частоту мутаций, правда, далеко
не всех категорий.
Различают прямой и косвенный методы.
Прямой метод заключается в определении отношения
числа новых случаев заболевания или морфологического
изменения, т. е. числа измененных детей, рожденных от здоровых ро-
днтелей. к числу всех генов данного локуса у лиц изученной
популяции. При этом расчет идет на одно поколение (у
человека его длительность принимается равной 30 годам).
Очевидно, что прямой метод может быть применен только для учета
доминантных аутосомных мутаций.
Косвенный метод заключается в определении частоты
мутаций по связи сравнительной жизнеспособности мутационных
форм, их частоты в данной популяции и скорости
мутационного процесса исходя из возможного равновесия в популяции
между отбором и частотой мутаций (об этом более полно
в гл. 4). Применяют при этом формулу Холдейна *:
2v
2Р= —• (2)
s
где v — частота мутаций (от рецессива к доминанту); 5 — се*
лекционное значение мутантных форм; 2Р — доля особей в
популяции с данным доминантным изменением. Величина 5
обычно определяется как относительная разница между пло-
довитостью нормальных особей и лиц с данным доминантным
геном, т. е. 5=1—/, где / — относительная плодовитость
мутантных форм.
Прямой метод был применен Нилом и Фоллсом для
установления частоты появления доминантной мутации ретино*
бластомы. Было обнаружено 49 случаев на 1 054 985 детей,
что дает частоту мутирования данного гена 2,32-10"6.
Косвенный метод с помощью формулы Холдейна может
быть проиллюстрирован на примере доминантной
карликовости (ахондроплазии или хондродистрофии). Мёрком было
установлено, что в Дании доминантная карликовость
встречается с частотой 10,7-1 О**5, а ее селекционное значение 5 =*0,804
(так как /«0,196). Тогда
10'7-|0-'=^|4-
Отсюда
м '^•'"-•"■8М =4,3-Ю-.
ф Принцип установления частоты мутаций исходя из равновесия отбора
и мутационного процесса в популяции был предложен раньше Холдейна
советским ученым В. П. Эфроимсоном.
72
Интересно, что при расчете прямым методом Мерк получил
почти ту же частоту данной мутации. На W075 рождений
в одной из больниц в Копенгагене 8 детей с ахоадроолазией
(Н3 общего числа 10) родилось от нормальных родителей, т.е.
были мутантами, отсюда частота мутации
— *£$*-**■»*■
Оба метода страдают значительной неточностью в силу
ряда причин, достаточно полно освещенных в специальной
литературе по генетике человека, но они все же дают порядок
величин частоты мутаций у человека.
Подобные расчеты делаются и по другим видам мутаций,
и том числе рецессивным, но в этом случае можно
пользоваться только непрямым методом.
Хромосомные мутации у людей обнаруживаются в настоя-
шее время довольно часто в силу того, что многие из них
приводят к определенным морфологическим и
физиологическим нарушениям, получившим в медицине определенные
названия (синдромы Клайнфельтера, Тернера—Шерешевского,
Дауна и т. д.). Учет таких хромосомных болезней в
определенных популяциях дает возможность оценить их частоту
количественно, о чем говорилось ранее, в гл. I, в связи с вопросом
о частоте нерасхожденнй.
Методы учета частоты мутаций у высших растений. Эти
методы довольно разнообразны и специфичны для различных ви-
лов, поэтому остановимся на них очень кратко.
Так как у многих видов растений хромосомы очень круп*
ные, а в хромосомном наборе их немного и одни из них
довольно легко отличимы от других, то открываются широкие
возможности для применения цитологических методов учета
различных хромосомных нарушений в процессах митоза в
тканях и органах (в частности, в корешках или в эндосперме).
Возможно также изучение нарушений в мейозе. Хотя и не все
хромосомные нарушения полностью улавливаются, тем не
менее можно получить достаточно полный фактический
материал для количественной характеристики естественного или
экспериментально вызванного тем или иным воздействием
мутагенеза по частоте хромосомных изменений (всех или
отдельных их типов) на общее число изученных клеток (или,
правильнее, наблюдавшихся митозов).
Генетические методы изучения мутаций очень хорошо
разработаны на кукурузе (школой Стадлера). С их помощью
можно изучать доминантные летали (по количеству погибших
эмбрионов в семенах), дефишенси или мутации специфических
генов (по растениям F\)9 видимые мутации в немаркированных
хромосомах, рецессивные (по расщеплению в потомстве от ге*
:з
L
/ г J
8 6*
Рис, IS. График отбора на папе х — а2 в
линии SK.
тсроииотных по мутации растений F{) и т. д. Ряд
мутационных явлении можно лучше анализировать благодаря тому, что
и семени эмбрион диплоиден, а эндосперм триплоиден. Осо-
6\ю, хорошо учитываемую категорию составляют так
называемые хлорофплльные мутации. Некоторые мутации могут об-
наружнваться непосредственно по пыльце.
У растений-самоопылителей в силу гомозиготностн их
линии мутации можно находить довольно просто, и тогда
частоты мутации
устанавливаются как доли растений
х{ (т. е. выросших из
подвергнутых воздействию
семян), дающих
расщепление в потомстве, в общем
количестве растений.
Учет мутаций генов,
определяющих
количественные признаки. В этом
случае обычные методы
учета мутаций и их
количественной
характеристики непрнложимы.
Попытки учета мутационного возникновения генов, влияющих на
количественные признаки, были сделаны на основе
обнаружения возрастания изменчивости в инбредных и
селекционных линиях. Примером сказанного могут быть работы Ро-
кицкого. а также Клейтона и Робертсона на дрозофиле.
Для одновременного учета изменении средней
арифметической и вариансы Рокицким был применен графический
метод, а именно: на оси абсцисс откладывались значения
вариансы (а2), а на оси ординат — значения средних арифметических
изучаемого признака (х). В данном случае изучалось
количество стерноплевральных щетинок дрозофилы. На рис. 13
показан результат отбора и инбридинга в контрольной
линии SK. После значительной изменчивости в исходной
популяции (0-е поколение) и в 1—2-м поколениях (о2 достигала 7)
линия стабилизировалась на уровне х«16 и о2«2. Это
наиболее простой случаи стабилизации без мутаций. В других
случаях линия сохранялась стабильной много поколений, затеи
подхватывалась отбором мутация данного количественного
признака, резко возрастала изменчивость и отбор приводил
к повышению значения признака. На рис. 14 показано, что
в течение 32 поколений линия оставалась стабильной (х» 19,5;
о^З>, с 33-го поколения произошло увеличение изменчивости
(о- достигла 6), после чего в результате отбора средняя
увеличилась до 22—23. Сходная картина и на рис. 15. Но в этом слу-
A
го\
'<?
—1—2—j—г—j—i—>—J—u*
Рис. 14. Графин отбора иа поле 7— о* дчинл А7/АГ.
-^— Г— о1 а *****
Я:« /5 Граф** отбора *а «** '
П
ц-ле первоначально колебания изменчивости в линии были
значительно больше (особенность данной линии в том, что особи
каждого поколения подвергались рентгеновскому облучению),
Мутация выделилась в 36-м поколении — а2 достигла 8.
Вследствие отбора средняя повысилась до 24, а в 40-м поколении
изменчивость уменьшилась (а2»4), т. е. вернулась к тому же
уровню, который имела линия в 20—35-м поколениях.
Так как в линиях XII/C и V7? проводился длительный
инбридинг и отбор на увеличение числа щетинок, можно с до-
статочной уверенностью предполагать, что в данном случае
произошло не выщепление уже существовавших в исходной
популяции генов, влиявших на количественный признак, а
появились мутации.
Конечно, во многих конкретных случаях диагностировать
возникновение мутации количественного признака нелегко,
\огя еще в классической работе Иоганнсена упоминается о
появлении мутации в чистой линии.
Методы обнаружения мутаций у микроорганизмов и учета
их частоты. Анализ мутаций у микроорганизмов, генетическое
изучение которых стало за последние годы самостоятельной
областью генетики и достигло громадных успехов, связано
с особенностями их размножения. У многих грибов и
водорослей происходит нормальный половой процесс, принципиально
сходный с половым процессом у высших растений. Однако
ситуация для анализа упрощается тем обстоятельством, что
у них имеется гаплоидная фаза. Наличие гаплоидной фазы
дает возможность применить так называемый тетрадный
анализ (о нем говорилось выше, в гл. 1). Всякая рецессивная
мутация, возникшая в одной из хромосом у диплоидной формы,
проявится в гаплоиде таким же образом, как и доминантная.
Поэтому непосредственный подсчет измененных форм
гаплоидов и их доли в общем количестве изученных гаплоидов
дает возможность установить частоту мутаций.
Несколько осложняет картину наличие у некоторых грибов
наряду с нормальным половым процессом так называемого
парасексуального цикла. Дело в том, что гифы мицелия таких
грибов многоядерны, хотя большинство ядер гаплоидно. Если
п ядрах мицелия произошли мутации, то их последующее
выявление осуществляется через стадию, получившую название
гетерокариона. Гетерокарион получается в результате цито-
плазматическнх анастомозов между отдельными гифами с
обменом ядер между ними. Поэтому в нем имеются гаплоидные
ядра разных генотипов. В дальнейшем гетерокарион
распадается на одноядерные конидии, в каждой из которых может
выявиться мутантный генотип.
Простейшая форма размножения бактерий, не образующих
итор.-— многократное повторение делений клеток. В результа*
тс получается громадное количество отдельных бактериальных
клеток, совершенно несоизмеримое с количеством потомства,
которое может быть получено у высших животных и
растений. Поэтому разрешающая способность генетических опытов,
проводимых на микроорганизмах, значительно выше, чем на
любых других организмах. Но в то же время совершенно
очевидно» что при таком громадном количестве отдельных клеток
в культурах микроорганизмов учесть все потомство нет
возможности.
Поэтому были разработаны различные приемы
генетического анализа у бактерий, служащие в то же время и для
учета возникающих мутаций. Одни из них пригодны для учета
морфологических изменений, с помощью других можно
обнаруживать биохимические мутации.
Наиболее общий метод—-метод клонирования.
Клон—-вегетативно размноженная культура из одной клетки (бактерии,
гриба или водоросли) или из одной вирусной частицы. Обычно
в исходной клетке находится одно ядро, или нуклеоид. Все
дочерние клетки клона имеют один и тот же исходный генотип
во всех поколениях. Если же в исходной клетке было не одно,
а два или несколько ядер (или нуклеоидов), то клон
распадается на новые клоны, каждый из которых происходит от
одного первоначального ядра. При различиях по видимым
признакам ( например, форме бактерий или их цвету) можно эти
колонии различать на глаз, высевая их, например, на
искусственную среду. Если клетка имела два генетически
различающихся нуклеоида, то колония будет состоять из двух половин
с разным генотипом. Если в клетке было 4 нуклеоида, но один
из них был мутантным, т. е. измененным, то колония
разделится на две неравных части: 1/4 —с мутантным генотипом,
3/4 — с немутантным.
Известно, что при росте культура бактерий проходит 3 фа
зы: нормальную, лаг-фазу и логарифмическую. Оказалось, что
клетки бактерии кишечной палочки Escherichia coli
различаются по содержанию нуклеоидов: в I фазе — всего I, во II — 2
и в III—4. Так как нуклеоиды гаплоидны, то легко выявить
мутацию нуклеоида по размеру измененного сектора в
колонии (так называемый метод Виткина). При размножении
клеток I фазы клон состоит из однородных клеток, при
размножении клеток II фазы и при возникновении мутации в одном
нуклеоиде клон состоит из двух половинок, в III фазе
возникновение мутации в одном из нуклеоидов обнаруживается по
наличию сектора, занимающего 1/4 часть колонии в чашке
Петри.
Очень интересна и детально разработана методика учета
биохимических мутаций у микроорганизмов. Обычно
микроорганизмы сами способны синтезировать нужные им вещества
77
(аминокислоты, витамины и др.). Такие организмы
называют ирототрофнымп. Прототрофные организмы способны расти
на минимальной среде, т. е. такой, которая содержит только
сахара и соли. Но у них могут появиться мутации,
вызывающие утрату способности к синтезу того или иного вещества.
Очевидно, что они будут способны жить и размножаться
только в том случае, если к питательной среде добавлено
данное необходимое вещество. Микроорганизмы, нуждающиеся
для своего роста в каких-либо компонентах полной среды,
называют ауксотрофнымп. Мутации от прототрофности к ауксо-
трофности можно назвать биохимическими. Это «прямые»
мутации — от базигена к трансгену — по терминологии Серебров-
ского. Наоборот, мутацию от ранее возникшего мутационного
изменения (трансгена) к нормальному типу (базнгену)
можно назвать обратной мутацией.
Чтобы обнаружить мутационные клоны, которые не
способны синтезировать необходимые вещества, анализируемую
смесь бактериальных клеток высевают на средах —
минимальной и полностью питательной. Из клеток с нормальным
генотипом разовьются нормальные колонии на той и на другой
среде, а из клеток с биохимической мутацией колонии
разовьются только на полной питательной среде.
Для ускорения этой довольно кропотливой работы Ледер-
берг предложил метод так называемых отпечатков или реплик.
Применяют особые печатки, соответствующие по размерам
чашкам Петри. Плоскость печатки покрывают бархатом и к
ней прикладывают чашку с анализируемыми колониями,
выросшими на полной питательной среде. На бархате остаются
клетки отдельных колоний. Затем к печатке прикладывают
лве чашки — с полной средой и с минимальной. На них потом
вырастают колонии клеток. Благодаря методу отпечатков
места колоний одинаковы в культурах на обеих чашках. Их
можно просмотреть в проходящем свете и подсчитать процент
неразвнвшихся на минимальной среде колоний, т. е.
возникших биохимических мутаций. В дальнейшем проводится
кропотливая работа по посевам мутантных колоний, выросших
на полной среде, на специальных средах, содержащих только
аминокислоты или только витамины, и т. д. Когда выяснен тип
ьещества, в котором нуждается данный мутант, проверяют
более конкретно, что это за вещество — та или другая
аминокислота или определенный витамин. Есть также ряд
дополнительных методов. В конечном счете можно установить частоту
прямых биохимических мутаций.
Для изучения обратных мутаций применяют метод селек-
тииных сред, т. е. сред без какого-либо определенного
метаболита. Па такой среде отбираются из большого количества
клеток мутантиого штамма лишь единичные, способные расти на
?h
такой среде. Очевидно, и них произошла обратная мутация,
иозвратпншая бактериальным клеткам способность
синтезировать данное вещество.
Методом селективных сред обнаруживают также мутации
устойчивости к различным антибиотикам или ядам. Все
бактериальные клетки нормального генотипа при внесении в
культуру данного вещества должны гибнуть, а те, которые
размножаются, представляют собой мутантов, устойчивых к
данному веществу. Можно подсчитать концентрацию мутантов
п исходной клеточной популяции и таким образом определить
процент мутаций.
Наконец, можно применить способ расчетов, основанных не
на проценте мутантов, а на относительном числе клонов, не
содержащих мутантов. Для этого используется пуассоново
распределение. Выше указывалось, что распределение
мутаций как редких событий по отдельным клонам (или сериям
опытов) должно следовать распределению Пуассона. В
дальнейшем это было доказано экспериментально.
Распределение Пуассона описывают следующей
формулой:
тх
где Я(л-) — вероятность того, что произойдет X событий;
Л'! — факториал (например, 3! = 1.2.3); т — среднее число
событий на какую-то единицу, которая изучается (клетка,
бактерия, клоп); е — основание натуральных логарифмов (2.718).
Формулу можно упростить, приняв X = 0, тогда ш* = Х! = 1
п Pt0)=eL'"'.
Отсюда \gP(0) = —т.
Это значит, что Р«» — количество единиц, в которых не
произошло событие, а если имеют в виду появление мутации, то
это будет то количество культур, в которых не произошло ни
одной мутации. Зная Р(0), можно определить т.
Высевая культуры бактерий на агарнзованиые среды, на
которых выявляются мутанты, легко определить Р(о». Если же
пользоваться селективными средами, не допускающими роста
исходной культуры, то из многих клеток только мутанты
будут образовывать колонии, которые обнаруживаются
визуально. Зная долю колоний, не дающих роста, и принимая ее за
Р(о), можно определить /и, а зная т, а также общее число
прошедших клеточных делений, оценить вероятность (а) мутации
на 1 клетку и на 1 деление по такой формуле:
79
Так как d непосредственно нельзя определить, то
принимают его приблизительно равным числу клеток N в культуре,
имея в виду, что число клеток культуры образуется путем
многократного деления одной клетки. Этот метод может быть
применен не только к бактериям, но и к соматическим клеткам
в культуре тканей, если они образуются путем днхотомическо-
ю деления.
При большом числе клеток популяции d&N. Этот же
способ определения частоты мутаций на 1 клетку и на 1 деление,
очевидно, подходит и к одноклеточным животным (Protozoa)
при условии, что они размножаются только бесполым путем,
т. е. путем деления.
В настоящее время изучено громадное количество мутаций
у микроорганизмов, поставившее генетику микроорганизмов
на первое место по количеству генов с разнообразным
действием.
Частота возникновения спонтанных мутаций как типичный
стохастический процесс. Чтобы получить количественную
характеристику этого процесса, т. е. выразить его в значениях
вероятностей, небесполезно проанализировать некоторые
данные по этому вопросу на разных объектах. Наиболее полно
в этом отношении изучена дрозофила (Drosophila melanogas-
ter), у которой возникновение мутаций наблюдалось в
лабораториях всего мира свыше 50 лет, а также (за последние
20 лет) микроорганизмы.
Естественная частота спонтанных сцепленных с полом
летальных мутации, по данным Меллера (1954), составляет
0.17%, однако в некоторых работах приводились и более
высокие цнфры — до 0,3%. Существенно то обстоятельство, что
она различна для особей, взятых из разных линий. Так, Деме-
рек показал, что в то время как в линии Орегон-Л она равна
только 0,07. в линии Флорида она выше в 15 раз (1,09).
У дрожжей Левиным н Арман показано наличие высокой
частоты спонтанных мутаций от нестимулируемостн к стимули-
руемости (nst-^st) у одной из nsf-линий. Она составила
4,1 • 10~5, в то время как у стабильного штамма — 10-*. Ими же
была обнаружена у одного штамма повышенная частота
обратных мутаций (к прототрофности по аденину) порядка
10~7—1Q-* на клетку на одно деление при частоте спонтанных
реверсий у стабильного ad-мутантного штамма 8,6- Ю"*9.
Было показано, что на частоту' спонтанных мутаций
влияет ие только генотипическая среда в целом, но и отдельные
гены — мутаторы. В то же время интересно, что
морфологически изменения в Х-хромосоме, например включение трансло-
имровамных участков, в общем мало влияли на суммарную
частсгту летальных мутаций в этой хромосоме. Количество воз-
пикающих летальных мутаций во всех хромосомах дрозофилы.
■и
по расчетам ряда авторов, примерно в 6 раз больше
количества мутаций, возникающих в Х-хромосоме, так что для всех
хромосом частоту естественного мутирования (по летальным
мутациям) можно оценить величиной около 1,02% (0,17«6).
Частота видимых мутаций ниже, чем летальных, примерно
в 8 раз (по данным Тимофеева-Ресовского и Дельбрюка), т.е.
составляет около 0,12%, хотя она очень сильно зависит от
частоты мутирования отдельных генов. Кроме того, многие
видимые мутации могут быть не замечены из-за
несовершенства техники обнаружения, так что указываемая суммарная
частота видимых мутаций, вероятно, преуменьшена. Кроме двух
отчетливых категорий мутаций — летальных и видимых,
возможны полулетальные мутации и мутации с очень слабым
видимым действием (а также неполно проявляющиеся). Такие
мутации иногда называют малыми. По-видимому, они
обладают большей частотой, чем указанные выше. Суммарно для
всех хромосом дрозофилы их частоту принимают около 3%,
Наконец, еще менее ясна категория доминантных леталей,
частоту которых оценивают примерно в 1%, а иногда и выше —
до 5%. В таком случае общее количество возможных мутаций
у дрозофилы может быть выражено величиной около 5%. Это
значит, что в каждой из 20 половых клеток дрозофилы может
появиться хотя бы одна генная мутация. Количество
спонтанных хромосомных аберраций труднее поддается учету, но, по
некоторым данным, оно довольно близко к частоте генных
мутаций.
Можно подойти к этому же вопросу и исходя нз частоты
мутирования отдельных генов. По некоторым нз генов Dro-
sophila melanogaster были получены достаточно точные
значения их мутирования. Хотя и есть гены более мутантиые и менее
мутантные, но в среднем, учитывая также возможные различия
б частоте мутаций у самок и у самцов (у последних она выше),
можно считать вероятность мутации на 1 локус равной
2-10-5—3- Ю-5. Если бы мы знали число генов, входящих в
геном, то можно было бы также подсчитать возможную
суммарную частоту возникновения точечных мутаций. Одно время
считали, что у дрозофилы окаю 5000 генов, тогда суммарная
частота генных мутаций должна быть
3.1О-*-5-10»=15-1О-*
т. е. около 15%, значительно больше, чем получаемая путем
фактического подсчета разных типов мутаций. Однако об
истинном числе генов, входящих в геном, приходится судить только
косвенно. В связи с развитием молекулярной генетики и
установлением общей длины цепочек ДНК и средней длины одного
цистрона (гена) были сделаны попытки учесть и общее
количество генов, которое оказывается значительно большим, чем
I, II Ф Р-ъпм***
в>
представлялось ранее. С другой стороны, установление сред,
пси частоты мутирования производится по немногим изучен-
»'ым гонам. Возможно, что они составляют малочисленную вы-
борку генов, обладающих максимальной мутагенностью. По
Гюльшпнств) же генов, обладающих более низкой мутабиль.
постью, просто не удается получить достаточно
исчерпывающий материал, но которому можно было бы судить об их
мутабильностп.
Установление частоты спонтанных мутации у других жи-
потных объектов, кроме дрозофилы, сопряжено с большими
трудностями. Чаще всего здесь можно исходить только из
учета мутации отдельных генов. Гак, в известных опытах
Рассела на мышах, методика которых приведена выше,
учитывалась частота появления мутаций по 7 рецессивным видимым
аутосомным генам. На очень большом материале — свыше 250
тысяч мышей — была установлена частота мутирования
0,7—1,3- Ю-5, т. е. в среднем 1 мутация на 100 тысяч гамет.
Такую же частоту указывают для кукурузы. Картер и другие
исследователи по двум генам мышей наблюдали более
высокую частоту мутирования. Однако и в этом случае приходится
учитывать, что в орбиту исследования попадают
преимущественно более мутабильные гены. Что касается таких категорий
мутаций у мышей, как доминантные летали и хромосомные
аберрации, то для первых обычно называется величина около
1 %, а для вторых — около 2%.
Выше уже указывалось, что у человека частота мутаций
может учитываться как прямым, так и косвенным методом.
Оба они дают приблизительно одинаковые величины —
примерно 2-10~5—3-10~5. По разным генам, определяемым тем или
иным методом (для рецессивных мутаций годится только
косвенный метод), были получены величины, несколько большие
указанных и несколько меньшие. Однако величину 3-Ю"5
можно принять как наиболее характерную для мутационного
процесса у человека. Эта величина примерно того же порядка,
что и для дрозофилы, мышей, а также многих видов растений.
Суммарное количество хромосомных аберраций определить
трудно, но во всяком случае оно не ниже, чем для генных
мутаций.
Поскольку у человека количество генов значительно
больше, чем у дрозофилы и мыши, суммарная частота мутаций всех
генов на каждую гамету становится весьма значительной. По
мнению некоторых исследователей, она может достигать 6-10"'.
Таким образом, у всех рассмотренных видов может быть
определена с большей или меньшей точностью частота
появлении спонтанных мутаций, т. е. их вероятность. Даже при
сравнительно высокой суммарной частоте мутаций общая картина
не меняется: появление мутационного изменения любого гена
82
обладает лишь некоторой вероятностью, хотя она и не
является строго постоянной величиной и может меняться в силу
разных причин — внутренних и внешних.
Некоторые примеры частоты мутаций у разных организмов
приведены в табл. 10.
Таблица 10. Спонтанная мутабильиость отдельных генов
у некоторых организмов (по различным источникам)
Инд
Мутациоииос изменение
Чдстотл
Человек
Культура костного
мо:*га человека
Мышь (Mus musculib)
Дрозофила (Dro^o-
philij melanogaster)
Кукуруза (Zea mays)
Heiipociiopa (Neuro-
spora crassa)
Кишечная палочка
(Escherichia coli)
Бактериофаг Т-2
Вирус мозаики
табака
Альбинизм, А - а
Фенилкетонурия, F — /
Микроцефалия, Мк -* тк
Хоидридистрофия, Ch -* ch
Гемофилия (сценл. с полом)
И >Л
Устойчивость к в-азагуаиоишу,
S — 8 — и -ч—8—я
Светло-серая окраска шерсти,
D-d
Желтая окраска тела (у ),
У — у
Желтая окраска теля {у i ,),
Белые глаза, W -► w
Пурпурный эндосперм, Р -+ р
Потребность в аденине, ade ~
ade л
Потребность в гистидине,' Ль-L
+ -/its-
Чувствительность к
стрептомицину, Str — s ~* sir —d
Круг хозяев, А+ - ЬГ
Аукуба мозаика, Auk - auk
5Г5
8 10~5
2,7.10~5
4,2 Ю'5
3,2-10"*
1 -10—в
3 10"*
1 -10—*
1 1(Г5
4 10~5
1 10~*
4 ИГ8
2 Ю~6
ГКГ1
3-КГ1
1-е
1.6 Ю
Как указывалось выше, методы исчисления частот мутаций
отдельных генов для объектов, указанных в табл. П>. не
одинаковы. Для размножающихся обычным половым путем
животных и растений учитывается частота появления мутантных
гамет (на общее число гамет) в расчете на 1 поколение.
В культуре тканей, а также у кишечной палочки и других
бактерий счет идет на 1 клетку и I деление. У нейроспоры.
размножающейся бесполым путем, учитывается частота появлс-
иия мутантных спор. У бактериофага же учитываются измене- j
ния данного гена на 1 репликацию. I
Мы оставляем в стороне рассмотрение особой категории I
ммацни —так называемых мутабильных генов, особенно хорсь !
шо изученных у кукурузы. Оказалось, что мутабильное состоя- s
ние таких генов индуцируется другими генами, приводящими '
к их генетической нестабильности, причем сама картина
действия генов-индукторов очень сложна и нередко связана со
структурными изменениями хромосом. Точно так же доста-
точно загадочными и. по-видимому, не имеющими ничего
общего с обычными генными мутациями, своеобразными по своей
природе и редкими являются так называемые парамутации —
класс направленных мутации, возникающих с очень большой
частотой у аллелей локуса R (кукуруза), находящегося
в длинном плече 10-й хромосомы. Направленные изменения
в каждом аллеле этого гена возникают в гетерозиготе под
влиянием парамутагенного действия другого аллеля. Эти пока не
поддающиеся объяснению явления описаны в ряде работ
Бринка и др.
Частота прямых и обратных мутаций. Под прямыми
мутациями обычно понимают мутации от дикого типа к
измененному (по терминологии А. С. Серебровского, от базигена
к трансгену), под обратными — от мутантного типа к
нормальному. По-видимому, чаще бывают прямые мутации, но
возможно и равенство частот или даже большая частота
обратных мутаций. Н. В. Тимофеев-Ресовский очень детально,
на громадном фактическом материале, изучал частоту
прямых и обратных мутаций по ряду генов дрозофилы (white —
белые глаза, forked — завернутые щетинки и др.). Правда, он
проводил сравнение частот при воздействии рентгеновских
лучен (доза ^5000 р). Сводные цифровые данные по 5 генам
приведены в табл. 11.
Таблица II. Количество обнаруженных прямых и обратных мутаций
по 5 генам Dr. melanogaster (по Тимофееву-Ресовскому)
Пари влл«лей
U - rf
J 1 '—0
F-I
Р~Р
Прямые мутации
количество
глмет
112000
112000
51500
51500
67 500
число
мутации
44
12
4
12
1
Обратные мутации
количество
гамет
87 500
89 500
48 000
58 500
63 500
число
мутаций
0
3
3
14
10
*4
Если подсчитать на основе конкретных данных частоты
прямых и обратных мутаций по паре аллелей W—«,*, то
обнаруживается следующая разница:
и-ю-*
з,зю-&
Частота прямых мутаций выше частоты обратных примерно
на 1 порядок величин.
Для пары аллелей F—/
2,3-Ю-4
2,4- Ю-4
Частоты прямых и обратных мутаций одинаковы.
Для пары аллелей Р—р
1,5- Ю-5
Pz^=±p.
1,5-Ю-4
В этом случае, наоборот, частота обратных мутаций выше
частоты прямых в 10 раз.
Очень полные данные о частоте прямых н обратных
мутации получены на микроорганизмах. И здесь картина довольно
разнообразная.
Так, у кишечной палочки мутации от устойчивости к
стрептомицину (str—s) к потребности в стрептомицине (str—d\
и обратно обладали следующей частотой:
ью-*
str — s < * str — d.
Ы0~*
Разница на один порядок в пользу обратного мутирования.
Для другого гена кишечной палочки, контролирующего
синтез гнстиднна, соотношения таковы:
2,Ы0~4
his + ^ ' *г. his —.
4.Ы0-1
Вероятность обратной мутации ниже в 50 раз. Впрочем,
проблема не так проста, как может казаться сначала.
Во-первых, нет уверенности в том, что обратная мутация
представляет собой возврат к тому же состоянию, которое было до
прямой мутации, т. е. вновь возникший базнген может быть не
совсем тот, который был ранее, а какой-то новый, фенотипиче-
85
,*. с ним сходный. Во-вторых, могла произойти мутация «
смоги данного гена, а гена супрес сора
Тогда вместо схемы
his- ~* his +
следует писать
Jus -sup — Z- his — swp + -
Наконец если принимать, что ген состоит из многих само-
стмтельио мутирующих единиц, то возможно, что частота
м\тациоииою* изменения /iis-f-Wiis— в действительности
сумма частот изменений многих мутирующих единиц, феноти-
пическое выражение которых одинаково (или почти
одинаково), хот* каждый мутант имеет только одну измененную
пар> муклеондов. а изменение his—*his+ характеризует
изменение только немногих или даже одной мутирующей едя*
ниаы — мутона, при котором восстанавливается функция гена.
Риамс стороны случайности в мутационном процессе
В случайности мутационного процесса можно выделить по
крайней мере 4 различных элемента: а) случайность мутаций
во времени; б) случайность в пространстве; в) независимость
одиой мутации от другой; г) отсутствие закономерного
соотношения между вновь возникшим мутационным изменением
и его адаптивностью к условиям внешней среды.
Все эти особенности в стохастике появления мутаций
фактически наблюдались на большом количестве видов
животных и растениях, но во многих случаях нельзя было точно
доказать их вероятностный характер. Развитие генетики
микроорганизмов дало возможность наблюдать появление
мутаций на клонах, возникающих из одной клетки (так
называемый флюктуационный анализ), что позволило
экспериментально проверить характер распределения мутаций и во
вредив и в пространстве. Так, в работах Ли и Каулсона по
изучению в штаммах кишечной палочки Escherichia coli чис-
'2? "У™11*0*'ггюсобных сбраживать лактозу (штаммы были не*
сбраживающими), и Луриа и Дельбрюка с бактериями, з*
Роенными фагом Т2, было показано, что вероятность
появления мутаций одинакова для всех бактериальных клеток по-
SKI!!?' 10зннкши* из 0АН0* клетки, т. е. для всех клож*
пГГ!™1**10™!1 * ВИДУ ст«ндартныс, нормальные условий,
tllJE^l»!i***uttc* н*кот°Рв* средняя частота спои-
ша^!К?; Т° Же 0Т|10СИТСЯ и к индуцированным мутв-
На?1!?7! HЛРМ определенных дозах воздействия.
ршмыж4tJ*.п^Астам«иа схема возникновения мутаций «
р **«* втавах создания клонов. Для простоты принято, <ио
в каждом клоне только по 4 клетки. Но статистическая
закономерность будет относиться и к клонам, состоящим из
многих клеток. Если мутация возникла в первой (исходной)
клетке клона, все 4 клетки клона окажутся мутантными. При
возникновении мутации в одной из двух клеток, появившихся
после первого деления, получаются только 2 мутанта в клоне,
после второго деления — I мутант и т. д. Таким образом,
хотя в каждом случае происходит лишь единичное событие —
мутаиия, число мутантов в кленах колеблется от 0 до 4
(для схематического случая, изображенного на рис. 16). Если
г о
!>! Ы Й В й й р р is
^ 2 : t i i у с й *мт
Рис. 16. Схема распределения клонов по числу мутантов в эаз^симм;::^ от
возникновения мутации: в одной исходной бактериальной клетке, a uiuo.i
из двух или в одной из четырех.
действительно вероятность возникновения мутации на одну
клетку и на одно поколение одинакова, то чисто клонов с
4 мутантами должно быть вдвое меньше, нежели число
клонов с 2 мутантами, а с 2 — вдвое меньше, чем с 1 мутантом.
В силу редкости мутаций, т. е. малой их вероятности,
останется большая группа клонов, в которых не наблюдалось ни
одного мутанта, счет же, очевидно, надо вести по клонам с
мутантами. В 7 клонах, показанных на рис. 16, имеется не
менее одного мутанта, в 3 — не менее двух, а в I — не менее
четырех. Чем больше численность клонов и популяций в них,
тем это соотношение будет ближе к соотношению 1:2.
И действительно, фактическое распределение клонов по
числу мутантов, проведенное на фаге Т2 (табл. 12), очень
близко к ожидаемому, если допустить одинаковую
вероятность возникновения мутаций для всех периодов роста
культуры и для всех клеток популяции.
Однако надо иметь в виду следующее осложнение. Схема
на рис. 16 предусматривает одноразовое появление мутации.
На самом же деле возможно появление новых мутаций, хотя
тоже статистически очень редких, на каком-то л-м делении
клеток клона. Эти мутации также размножатся, что,
очевидно, увеличит число мутантов по сравнению с принятой выше
* О С С Z С О
• • • : • ее о о се ее се
• ••• ••ее г;фф #ооо cmoz со#с сес#сеое
иоооа во числу мутантов фага Т1
t
1 1 и более
2 » »
1 4 » »
8 » »
16 » •
! за » »
64 » •
Часло ляолош с еоотмтствувца. 1
чаьаом иутаятаыя частиц 1
фактическое
183
90
41
21
10
4
0
Ttop«™«*eK«t I
183
90.8
45.1
22.2
10.8
5.1
2,2
j
простой схемой. Ли и Каулсон учли это обстоятельство и рас-
считали теоретическое распределение с учетом вновь
появляющихся редких мутаций, та*
же возникающих случайно н с
постоянной частотой. Оказалось,
что фактическое распределение
культур штамма Escherichia coli,
не сбраживающего лактозу, по
числу мутантов, способных
сбраживать лактозу, довольно близко
совпало с теоретическим (рис. 17)
и с достаточной вероятностью
(0,2) удовлетворяет критерию хя-
квадрат.
Можно подойти к вопросу о
статистичности мутаций исхода
из другого критерия — нх
пространственного распределения по
культурам, что опять-таки
подтверждается экспериментально
на микроорганизмах. Известно*
что распределение редких
событий ло небольшим выборкам при
случайности этих событий должно
соответствовать распределению Пуассона. В таком случае
распределение мутаций в каких-то выборках в соответствий
с nvaccoHOBUM является доказательством его случайности.
Распределение клонов по числу мутаиий (уже не
мутантов) может быть показано прямым способом. Для выращмвг
£}•
I"'
I
-L
?** П Распределение культур
я* сбражнааюшего лактозу
штамма кишечно* палочки по
чтл) иутантоа, способных
^брджиаать лактозу (черные
столб*** -. фактическое
распределение, пункт арные —
теоретически ожидаемое).
»*
ния культур используют не жидкую, а плотную среду и на
ней подсчитывают мутантные клоны, т. е. клоны, возникшие
из 1 мутантной клетки. Обычно выращивание производится
в чашках Петри с агаром. На поверхности агара находятся
аминокислоты и лактоза. Клетки, образующие колонию,
обладают свойством не усваивать лактозу. Если же в одной из
них возникает мутация, способная усваивать лактозу, то
метки, образующиеся из такой мутантной клетки, размножаются
о I г J о
Рис. 18. Распределена
количества мутантных
колоний бактерий в отдельных
квадратах сетки, наложея-
ной на чашку Петри (в
полных квадратах от 0 до 5
колоний).
Рис. 19. Ркяредедеме
колок** ккшечво! вамп в
квадратах чаша Петра во
чяслу иутаан!. вышваюошх
сбражваапе лактозы
<черные столбшш — фактическое
расврелелевяе. оукктнв-
ные — теоретически
ожидаемое» оо Пуассову).
быстрее, и на поверхности колонии образуются сосочкв.
Чтобы их заметить, включают в среду какое-либо вецество-кра-
ситель, являющееся индикатором. Благодаря индикатору
сосочки краснеют, остальная же поверхность колонии сохрани*
ет белый цвет. Каждый сосочек соответствует одной мутации
независимо от количества в нем мутантных бактерий.
На рис. 18 показана схема подсчета расоредедення
мутаций в виде отдельных окрашенных пятнышек в квадратах се*
ти. наложенной на чашку Петри, в которой разводится
колония бактерий. Количество окрашенных пятнышек, т. е. му-
тантных колоний, каждая из которых соответствует одной
мутации, может быть различным в отдельных квадратах
(учитываются только полные квадраты), В некоторых квадратах
не обнаруживалось ни одной мутации, в других —одна, две
и т. д. На рис. 19 показано наблюдаемое и ожидаемое в со*
а»
ответствии с распределением Пуассона распределение
колоний Escherichia coli по числу мутации, обусловливающих
сбраживание лактозы (совпадение очень близкое), а в табл.
13 помещены данные такого подсчета по 8 разным опытам
с разными видами бактерии.
Таблица /.?. Распределение количества мутаций (k)
в отдельных квадратах на чашке Петри
=
1
2
3
4
5
6
7
8
si ^ 9
- £ Z
Z ° =
S t Р
^ н г:
- £ *Г
5 "5- i
а
б
а
б
а
б
а
б
а
б
а
б
а
б
а
б
| Величина fc
0
5
6,1
26
27.5
59
55,6
83
75
8
6,8
7
3,9
3
2,1
60
62,6
1
19
18,0
40
42,2
86
82,2
1.34
144.5
16
16,2
11
10,4
7
8,2
80
75,8
о
26
26,7
.38
32,5
49
60,8
135
139,4
18
19,2
11
13,7
14
15,8
45
45.8
3
26
26.4
17
16.7
30
30.0
101
89.7
15
15.1
11
12,0
21
20,2
16
18,5
4
21
19,6
7
9.1
20
15,4
40
43.3
9
9,0
7
7.9
20
19,5
9
7.3
5
13
11,7
16
16,7
7
6,7
8
7,1
19
15
б
8
9,5
7
7,4
7
9,6
7
9
9,6
Общее число
квадратов
119
128
244
526
73
55
100
210
Степень соот-
ветствия по
критерию хи-квлд-
0,97
0,66
0,26
0,63
0,97
0,53
0,85
0,78
Независимость мутаций одного гена от мутаций другого
гена достаточно отчетливо наблюдалась на многих хорошо
изученных генетических объектах (дрозофила и др.)- Но на
бактериях, той же кишечной палочке удалось изучить этот
вопрос очень точно в прямых экспериментах. Можно было
получать двойных ауксотрофов, т. е. формы с изменениями
в двух генах, каждое из которых обусловливало потребность
в определенном факторе роста.
В результате первой мутации возник штамм, нуждающийся
в гистидине (his—). затем в нем произошла новая мутация —
чо
потребности в метиониие (met — ). Таким образом был получен
двойной ауксотроф, характеризовавшийся потребностью
одновременно и в гистидине и в метиониие. В дальнейшем в нем
изучались обратные мутации, но все они были только или
мутациями к типу /i/s+, или мутациями к типу met+. Обратной же
мутации одновременно к типу his+ met+ практически
наблюдать не удалось. Это дает полное основание считать, что
каждый из генов мутирует независимо от другого. При вероятности
обратной мутации каждого гена порядка Ю-8 одновременная
мутация обоих генов должна быть равна произведению
вероятностей:
10-*.10-*=10-,e.
При такой низкой вероятности обнаружение двух
мутаций одновременно требует громадного экспериментального
материала. Изучалась также и корреляционная зависимость
между появлением мутантов того или другого типа в
большом числе разных культур. Оказалось, что она отсутствует
(г = —0,06).
Несколько сложнее вопрос об адаптивности мутаций, т. е.
о степени их соответствия конкретным условиям среды.
Эмпирический материал, полученный при изучении мутаций у
животных, уже давно показал, что и в отношении адаптивности
мутации случайны, т. е. они могут быть и вредными, и
нейтральными, и положительными. Однако появление этих трех
категорий мутаций отнюдь не равновероятно. Последняя
очень маловероятна потому, что она происходит в сложных
генетических системах, уже прошедших длительную эволюцию
на адаптивность. Всякое нарушение в них всегда имеет
больше шансов ухудшить систему, нежели улучшить. Поэтому
действие мутации на организменном уровне не является
чисто случайным. Оно определяется специфическим характером
эволюции вида, что обусловливает и характер самих мутаций.
Так как у микроорганизмов трудно разделить
модификации и мутации, то одно время существовало мнение, что у
микроорганизмов возможны чисто адаптивные мутации, т. е.
что под влиянием того или иного фактора, например
химического вещества, могут появляться мутации, требующие
именно данного вещества. Но прямые опыты, проведенные
разными методами, опровергли это мнение. Так,
мутационные формы бактерий, устойчивые к стрептомицину, не
индуцировались непосредственно стрептомицином, а
возникали во время роста популяций бактерий в отсутствие
стрептомицина, что было экспериментально показано Дж. и Э. Ле-
дербергамн. На основе мутаций можно было создать
устойчивые к стрептомицину культуры бактерий, хотя эти культуры
сами никогда не соприкасались со стрептомицином.
$1
Ограничения в статистичности естественного
мутационного процесса. Таким образом, несмотря на вероятностный
характер мутационного процесса в целом, необходимо
учитывать ряд его сторон, которые носят не статистический, а
динамический характер, т. е. определяются историческими, эво*
лкжноннымн и иными причинами. Разнообразие мутаций и
их ненаправленность не означают, что появляющиеся
изменения могут быть любыми. В конечном счете они не могут
выходить за рамки признаков вида или рода, выработавшихся
в процессе эволюции. Впрочем, в силу некоторых условностей
в характеристике систематических различий иногда могут
появляться как мутации и формы с признаками высших
систематических категории, также имеющие генетическую
основу. Однако мутации, приводящие к резким изменениям
фенотипа, редки именно потому, что они сильно нарушают
механизмы, определяющие индивидуальное развитие, и поэтому
чаше всего невыгодны для организма с точки зрения его
выживаемости. Большинство видимых мутаций вредны, так как
существующие в настоящий момент гены, входящие в геном
вида, представляют собой результат прежде происходивших
мутаций, уже прошедших длительный путь естественного
отбора.
Однако некоторые мутации, особенно малые, т. е. слабо
изменяющие фенотип или обладающие некоторым
специфическим действием (например, мутационно возникшая
устойчивость к заболеваниям или химическим веществам), могут
оказаться полезными, особенно при резком изменении внешних
условий. Известно, например, что при обработке деревьев
и кустарников ядохимикатами для борьбы с насекомыми-
вредителями у последних в результате отбора устойчивых к
ядам мутантов насекомых очень быстро вырабатываются
расы, устойчивые к применяемым химическим веществам. То
же наблюдается и при действии антибиотиков на бактерии.
Значит, мутации устойчивости (к антибиотикам или ядам),
бесполезные при отсутствии ядов или антибиотиков,
становятся полезными при новых условиях.
Если некоторые мутации, нарушающие важнейшие
метаболические процессы, всегда губительны для организма, то
есть и такие мутации, которые вредны в одних условиях й
полезны в других.
Наконец, есть мутации, действие которых полезно в
гетерозиготном состоянии, а вредно — в гомозиготном. Таким
образом, хотя возникновение каждой мутации как таковой
представляет собой вероятностный процесс, но категории
мутаций, их типы, соотношение между ними, наконец,
полезность, нейтральность или вредность (вплоть до летальности)
определяются предыдущей историей вида и характером его
92
соотношении с конкретными условиями внешней среды На
потенциальные возможности мутационного процесса
накладывает ограничения исторически сложившийся генотип вида как
интегрированное целое.
Статистичность индуцированных мутаций. Частоту
мутаций увеличивают (в большей или меньшей степени)
различные мутагенные факторы, физические и химические. Иногда
употребляют выражение «искусственное вызывание
мутаций», отсюда может создаться представление, что мутации
уже не появляются по статистическим законам, а каждая из
них вызывается определенным примененным в эксперименте
воздействием. На самом деле статистический принцип
появления мутаций сохраняется и здесь, хотя меняется нх
вероятность. Известно множество примеров из разных областей
науки, техники и обычной жизни, когда удается увеличить или
уменьшить вероятность события. Даже при значительном
повышении общей частоты мутаций по отношению к каждому
отдельному гену сохраняется положение о вероятности
мутирования, хотя и повышаемой под влиянием мутагена. Так как
различные гены обладают различной частотой мутирования,
то можно говорить как бы о спектре генных мутаций,
спонтанном или индуцированном. Спектры мутаций, получаемых
при воздействии ионизирующих излучений (различных
видов), в общем очень близки к спектрам спонтанных мутаций,
что указывает на случайный характер распространения самой
энергии излучения по длине хромосомы (или молекулы
ДНК). Несколько иная картина наблюдается при изучении
влияния некоторых химических веществ, которые могут
давать спектр мутаций, отличающийся от спонтанного или
вызванного радиацией. Очевидно, что в этом случае
наблюдается отклонение от случайности, обязанное способности
данных химических мутагенов взаимодействовать с
определенными химически специфичными частями молекулы ДНК.
Однако пока не известно ни одного случая, когда под
влиянием химического мутагена возникали бы только
определенные немногие мутации. В конечном счете лишь суживаются
границы возможных мутаций. Но в их пределах
возникновение отдельных мутаций остается случайным и может быть
оценено вероятностью. Вероятность возникновения мутаций
может меняться и в зависимости от стадии развития патовых
клеток, на которую влиял примененный мутаген. Чаще всего
они возникают на стадиях, близких к концу гаметогенеза,
т. е. к оплодотворению, причем интересно, что это одинаково
относится к столь далеким видам, как дрозофила и мышь
или мышь и человек.
Указанное выше общее положение о распределении мута-
иионных изменений по отдельным клеткам в соответствии с
93
распределением Пуассона справедливо и для инду]
ных мутаций при сохранении в каждом опыте тех же
облучения. При изменении последних меняется сред
тота изменений (л = ц при распределении Пуассона),
ппеделение по отдельным клеткам опять будет пуас<
Сказанное иллюстрируют данные по распределеник
сомных изменений в отдельных клетках традесканщ
ценные Кэчсайдом и др. в 11 различных сериях
(табл. 14).
Таблица 14. Распределение клеток традесканции по колнчесп
хромосомных нарушений
1
5
1
2
3
| 4
5
! 6
1 7
{ 8
j 9
10
II
Uh
~ — *> с
•" tZ s я
с =с = |
а
б
б
а
б
а
б
а
б
а
б
а
б
1 а
б
а
б
а
б
! а
б
Количество клеток с Л
изменениями
0
753
752,3
434
432,3
280
278,9
2278
2280,2
593
589,4
: 639
642,4
359
362,0
493
498,2
793
804,8
579
588,7
444
401,6
l
266
267,1
195
197,1
75
77,3
273
269,2
143
149,1
141
135,3
109
103,6
176
167,3
339
320,8
254
238,7
252
227,7
2
49
47,4
44
44,9
12
10,7
15
15,9
20
18,8
13
14,2
13
14,9
26
28,1
62
64,0
47
48,4
59
56,2
^ 3
5
6,2
9
7.7
1
1.1
0
0,7
3
1.7
0
1.1
1
1.5
2
3,4
5
9,4
3
7,2
1
10,5
Общее число
клеток N
1073
682
368
2566
759
793
482
697
1199
883
756
ее
X
(О
5-
Ох
0,36
0,46
0,28
0,12
0,25
0,21
0,29
0,34
0,40
0,41
0.49
94
Статистичность индуцированных мутаций выражается и в
других явлениях. Так, сохраняются определенные частоты
летальных мутаций по различным хромосомам дрозофилы
(табл. 15) при облучении, причем они более или менее
пропорциональны длинам хромосом, наиболее точно
измеряемым в гигантских хромосомах слюнных желез. Иначе
говоря, распределение летальных мутаций по хромосомам слу-
чапио% и частота их зависит от размеров хромосом.
Таблица 15. Частоты летальных мутации по различным хромосомам
дрозофилы, отнесенных к лозе 3000 о оеитсенотких луче* ^«о
Тимофееву-Ресовскому и Циммеру), %
\
Шапиро Н. И. и Серебров-
ская Р. И. (1934)
Берг Р. Л. (1937)
Тимофеев-Ресовский Н. В.
(1947)
Отношения частот мутаций
Отношения длин хромосом:
а) в митозе
б) в слюнных железах
1
Хромосомы
• I
8,4
&,4
. 9,0
\
1
1
■ !
21,9
2.3,4
21,6
1 2.5
1,5
2,2
"!,v I
1
1 °<3 1
1
24,2 1
2,6 ( 0,04 |
I
1.5
2,4
0,05 1
0,06 [
5i
г;;
L х
ol
I
4--
11
) |__
X 1
т _о_
-,
1
J^
"it
л
J-£«
'оГ
if
>т
ю
Время, v
Я'*
Рис, 20. Независимость
частоты вызванных
рентгеновским облучением летальных
мутаций от времени (по
данным Тимофеева
-Ресовского, Паттерсона,
Хансена и др.). Горизонтальная
прерывистая линия —
частота мутаций, отнесенных к
дозе 3300р.
Выявилась также независимость индуцированных
генных мутаций от времени (рис. 20). Частота мутации
остается той же независимо от того, в каких интервалах времени
эта доза подана, прерывисто или подряд; малыми дозами,
но длительными или большими» но кратковременными.
Важна лишь суммарная доза облучения.
95
Статистический анализ зависимости между частотой
мутаций и дозой облучения. В ряде работ, начиная с работы
Меллера (1928), изучалась зависимость частоты
экспериментально вызванных мутаций от дозы облучения. Для
тачечных мутаций, как рецессивных видимых, так и летальных,
установлена прямолинейная зависимость. Наиболее полные
данные получены иа дрозофиле Н. В. Тимофеевым-Ресовским
(рис. 21). Лозднсе они были подтверждены многими другн*
/т
'!
$Ю1
**
CIl
Экспериментальные
Теоретически ожидаемые
гооо jooo
то
5№
Рис. 21. Прямолинейная зависимость между частотой
сцепленных с полом летальных мутаций у дрозофилы
с дозой облучения (по Тимофееву-Ресовскому).
ми авторами на самых различных объектах, в том числе и
на низших организмах-вирусах, нейроспоре (рис. 22),
бактериях. Та же прямолинейная зависимость обнаружена и на
частотах мутаций одного гена (рис. 23). Оказалось далее,
что количество возникающих мутаций зависит не от
длительности времени, в течение которого производилось облучение,
а только от суммарной дозы облучения (см. рнс. 20). Все
это привело к выводу, что каждая мутация вызывается
единичной ионизацией наследственных структур половых клеток-
Известно, чго прямолинейная зависимость описываете*
Уравнением регрессии у**а+вх> Применительно к
характеристике мутационного процесса это уравнение лучше заок-
сать таким образом:
у-ft + od, (*)
где у —выход мутаций; * —частота спонтанного
мутационного процесса; ц — коэффициент пропорциональности; &~~
доза облучения.
<*>
Иа рис 21 линии регрессии начинаются с нуля. т. е как
будто бы Л*0. Вообще принимается, что значение к надо вы
читать, и тогда линия зависимости должна проходить через
нулевую точку координат. При учете же всех мутаций она
ion*»8 пересекать ось ординат на расстоянии к от нуля. Но
звестно, что частота спонтанного мутирования очень мала
п0 сравнению с частотой индуцированных мутавдй, поэтому
/ 2 J
Доза, в юооор
Рис. 22. Зависимость между дозой
облучения и частотой видимых
мутации у нейроспоры (по Деме-
реку).
I 4 В
дозсбюоор
Рис. 23. Возрастание частоты
мутаций одного гена (от f
к ш*) с увеличение* дозы
облучаема (оо Тнмофеешу-Ресов»
скопу).
ее нелегко уловить. И вообще установление характера
отрезка прямой (или кривой) для малых доз вызывает большие'
затруднения. Обычно он строится путем экстраполяции от
более высоких доз. К сожалению, непосредственных
экспериментальных данных о сравнительном мутационном действии
малых доз облучения очень немного. Старые опыты
Спенсера и Штерна (1948) показали, что мутагенный эффект дают
очень малые дозы облучения н что количество мутаций
примерно удваивается прн увеличении дозы в 2 раза. Так. при
Дозе облучения 25 р было получено 0,0974% мутаций, а пря
Дозе 50 р — 0.2439% мутаций. "Впрочем, частота мутаций при
Дозе 25 р почти неотличима от частоты споитаиных муташА
Новые данные были получены Я, Л. Глембоцким. Э А Абе-
л*вой и др., при этом был показан кумулятивный и шю»*
2J»j е%„Рзр„ого <*»«— ^Ж*К"ЕЛЕ5
Дрозофилы гамма-лучами доэой i л/ ***
0,220±0,048% рецессивных сцепленных с полом летальных
мутаций, а при четырехкратном облучении половых клеток на
тон же стадии дозами по 5 рад — 0,210±0,048% мутаций.
Демерек смог повысить частоту мутаций у кишечной
палочки Escherichia coli облучением дозой 17 р.
Трудность при сравнении малых и больших доз облучения
связана также с тем, что при больших дозах значительно
возрастает количество грубых нарушений хромосом. Но в то
же время происходит зачатковый отбор, т. е. клетки с
грубыми 'нарушениями хромосом гибнут. Кроме того, по-видимому,
протекают количественно еще плохо оцениваемые процессы
репараций (восстановления). Однако суммирование всех этих
процессов может дать чисто линейную зависимость между
частотой мутаций и дозой облучения.
В наиболее чистом виде анализ может быть проведен
путем изучения частоты обратных мутации при разных дозах
облучения. В опытах Демерека на кишечной палочке была
обнаружена чисто линейная зависимость частоты обратных
мутаций от дозы.
При изучении сравнительного действия различных видов
излучений могут быть получены линии регрессии с
различными углами пересечения оси ординат. Для них соответственно
будут разные уравнения:
#1 = 6 +Mi;
#з = 6 + а3^з и т. д.
Если принять какое-то значение а, например, для
действия рентгеновских лучей за стандарт, то с ним можно сравнить
остальные а. Таким путем приходят к установлению
биологической или генетической эффективности того или другого
вида излучения (ОБЭ):
аз аз
—, и т. д.
ai ai
Так, при сравнении аберраций, вызванных нейтронами
и рентгеновскими лучами (нейтронное действие выражено в
и-единицах, рентгеновское — в рентгенах, причем к» и 1р
выражают примерно одинаковое рассеяние энергии в тканях).
получены значения относительной эффективности нейтронов:
в микроспорах традесканции 2,2—3,6, для лимфомы мышей
2,3—3,5, для клеток кончиков корешков сеянцев разных
растении 2,6. Эти данные указывают на большую эффективность
нейтронов. Однако подобное сравнение может быть сделано
лишь при прямолинейной зависимости. Если же зависимости
98
имеют криволинейную форму, то для сравнения необходимо
учитывать несколько разных параметров уравнений
криволинейной регрессии. Именно эта более сложная картина
наблюдается для различных структурных изменений хромосом. Она
связана с числом разрывов. Поэтому для одних хромосомных
изменений, при которых
происходит одиночный разрыв хромосом,
наблюдается прямолинейная
зависимость, при других — когда
два разрыва — криволинейная с
показателем степени, близким
к 2. Сказанное иллюстрирует
рис. 24 для двух видов
хромосомных разрывов у традесканции, а
также рис. 25, где сопоставлены
хроматидные обмены у
традесканции и транслокации в сперми-
ях дрозофилы, обнаруживающие
криволинейную зависимость.
Если весь механизм
возникновения данного типа аберраций
одноударный, тогда зависимость
их количества от дозы
выражается уравнением y = k + ad (рис. 21).
При полной двуударностн
у=к+$>сР. Если k очень мало, то
им можно пренебречь и тогда
#=pd2. Таковы кривая б рис. 26,
а также кривые для хромосомных
обменов у традесканции и
сперматозоидов дрозофилы рис. 25.
Однако обнаруживаемый в
конкретным опыте суммарный выход аберрации может
включать как одноударные, так и двучдарные аберрации. Тогда
показатель степени не будет равен 2, а окажется
промежуточным между 1 и 2, а уравнение должно быть записано,
согласно Ли и Кэчсайду, следующим образом:
&Шт "4,:f №
Штетовш мйшиА
Рис. 24. Прямолинейная
зависимость между доэоя облучения
нейтронами и количеством
хромосомных транслокаций
(черные кружки) и нзохроматидвых
делецнн (светлые кружки).
Крестиками показано
количество клеток без аберраций (по
Кояджеру н Джайльсу).
J^ft + arf + fc/2,
(5)
где a — коэффициент пропорциональности для одноударного
типа аберраций, р — для двуударного, d—-доза.
Если можно легко расчленить два типа изменений, то,
очевидно, один из них — одноударный — будет давать
прямолинейную зависимость и выражаться величиной си/, а другой —
двуударный, где связь криволинейная, и выражается
полностью через компонент (kfr
7*
99
50 100 150
дозаур
О 1000 2000 JOOO WO 5000
Доза,р
Рис. 25. Зависимость от дозы рентгеновского
облучения частоты возникновения:
а - хроматндных обменов у традесканции; б — транслокацнП
в спермиях дрозофилы.
Таб,шца 16. Частота структурных изменений хромосом дрозофилы,
■озникавишх под действием рентгеновских лучей, и показатели
степени при d. (Из книги Ли; переработано по данным
Тимофеева-Ресовского, Меллера, Хвостовой, Бельговского)
Типы структурных изменении
Хромосомные обмены,
включающие Х-хромосому
Обмены между 11 и III
хромосомами
i Хромосомные обмены,
; затрагивающие IV хромо-
сому
Хромосомные обмены
разного типа
1 Мелкие перестройки в
X -хромосоме
Дозы, р
1500
3000
6000
380
1500
1000
4000
1000
2000
4000
6000
1000
2000
4000
6000
1000
4000
Количество аберраций
на 1 спермин
0,0058 ± 0,002
0.0214 ±0,005
0,0735 ± 0,010
0,0021 ± 0.0008
0,0252 ± 0,003
0,0116 ±0,001
0,0830 ± 0.008
0,0007 ± 0,0002
0.0014 ± 0.0003
0,0036 ± 0,0006
0,0054 ± 0,0010
0,017 ±0,003
0,041 ±0,005
0.105 ±0,008
0,200 ± 0,016
0.00043 ± 0,00007 i
0.00162 ± 0,00022
Показатель
степени при d
1,78
1,80
1,42
1,03
1.04
1.40
0,96
100
не,
Однако ио-лможны случаи, когда степень при d выражается
личиной, не равной 2.
В табл. 16 сведены некоторые данные о частотах
структурных изменений хромосом дрозофилы, возникавших под дейст-
ипем рентгеновских лучей, и показатели степени компонента
уравнения. Только в двух случаях наблюдалась линейная за-
50 100
доза облучения, р
Рис. 26. Зависимость от дозы
рентгеновского облучения частоты
возникновения у традесканции:
а — нзохроматидных делеций (линейная);
6 — хроматидных транслокаций
(экспоненциальная зависимость, показатель степени
примерно 2) (по Саксу и Джайльсу).
О 20 W 60 60 100
Дозо облучения >р
Рис. 27. Зависимость от дозы
рентгеновского облучения
частоты возникновения
хромосомных аберраций в меристеме
корешков бобов:
1 — «хроматидные разрывы (txdW7)'
2 — хроматидные трялслокацян
[piftfy: 3 — нзохроидтидкые делецмн
(erf1»4*); 4 — хроматидные делеини
(rd1»71) (iro Рнаеллу).
висимость, т. е. показатель степени был близок к единице.
Таковы обмены, затрагивающие IV хромосому, и мелкие
перестройки в Х-хромосоме. Во всех остальных случаях показатель
степени был от 1,4 до 1,8.
На рис. 27 показаны зависимости от дозы облучения
рентгеновскими лучами различных хромосомных аберраций у Vicia
faba.
Ахроматидные разрывы (кривая / рис. 27) явно
выражаются прямой линией, т. е. здесь применимо уравнение */ = £ + ad;
хроматидные разрывы (кривая 2) -— целиком функция с
показателем степени, равным 2 (fki2). Что же касается кривых
3 и 4, то показатели степени здесь 1,48—1,71, т. е., возможно»
происходит смешение одноударных и двуударных разрывов»
гДе ad+ pJ2 дают какую-то суммарную величину cdnt где л<2.
101
По ча.шым Пненсн, различные хромосомные изменения по.
».iiih.iii'i-i. п облученных клетках культуры лейкоцитом перифе.
пнчечкоп кроии человека с очень высокой частотой, несмотря
па небольшие до 1Ы облучения (т. е. угн клетки обладают вы-
.окон чувствительностью к оОлученпю). Показатель же
степени Лил очень низким — 1,2 *.
Нсли предполагать, что ccl" - ш/+ \\d2, то возникает задача
выделить ли дна компонента, для чего надо определить а и (J,
И \помянутой работе Ивенса это сделано для двух наиболее
1\щестнеиных категории хромосомных изменений, а именно
для дпцепгрикоп (с соединенными фрагментами) и для колец.
Значении и и \\ но объединенным данным 5 опытов **
оказались следующими;
а Р
Дниептрнкп (2,51 ±0,15) ■ 10 :] (2,22±0,57) • КИ
Кольца (9,19±0,92) • 10 4 (1,31 ±0,37) .|(Н
Днцситрик11 + к()льца (3,42±0,17) • К)-3 (3,51 ±0,68) • 10-в
Фактические точки на корреляционном поле дали
довольно хорошее совпадение (но критерию хи-квадрат) с
теоретическими, вычисленными по уравнению y = /c + ad + fid2. Точно
так же проверка значения р показала, что он вполне
достоверна отличается от нуля.
В то же время очевидно, что удельный вес значения |W2
очень мал в общей величине связи частоты хромосомных
нарушений и дозы облучения, так как р меньше а на 3 порядка.
Впрочем, этого можно было ожидать, учитывая, что
суммарный показатель степени был всего 1,17. По мере увеличения
показателя степени возрастает и доля fid2 в общей связи.
Наконец, если показатель степени становится равным 2, а близко
к нулю и тогда связь может быть выражена уравнением
y**k + pd2. (б)
Выдвигаются также и некоторые другие объяснения,
почему возможны отклонения от квадратичной зависимости (кроме
разобранного выше смешения одиоударпых и двуударных
типов аберраций). Эти объяснения связаны или с
представлениями и более тонких механизмах образования разрывов и
соединения концов хромосом при определенном их пространствеи-
Ппричрм, и oriurux некоторых других шпором на тех же культурах
Kirгок л»'йколнтоц получались и кмлратичные шншеимосгн. Тик, Норма"
лы формулу 1/ —2,7 • 10-ЧУ Но рачбор причин различий и этих опытах но
•полит н нашу чадачу.
и.,, ** °/"0(о6,,х Тг1КОМ) ьыдр.чсини. ь чостнопи, с помощью дисперсной-
• «и-ли1.1 гы п ...поильной сплнпнчеекой литературе.
|1()М расположении, или с представлениями об отборе среди
а6ерраиий. К первым из них относится так начинаемая
концепция мести (site-konzept), выдвинутая Вольфом и Атвудом.
ця основе чтой концепции Вольф предложил другую формулу
%0J
Ь
05
Доза,р
Put. 28. Частот возникновения
трлислокиции и зависимости от
штмк'ииности облучения
(количество р в минуту:
J - 160; 2-20; 3 — 2,7).
v
100 200 SOU +00 Ш ЫЮ Ш 6J0
Доза оФгуч&шя, род
Рис. 29. Различия в кривых частоты
нызванных рентгеновскими лучами
дицентрических аберраций хромосом
в зависимости от дозы:
/ — клетки культури лейкоцитов человека;
2 — пыльца традесканции; J — корешки
бобов (по И вику).
для учета зависимости транслокаций от дочы облучения,
а именно:
y = a+N(\~e-™)\ (7)
где у —количество транслокации на клетку; а —- спонтанная
частота трапелокаций; N — число «эффективных пространств»
на клетку (т. е. районов или областей, в пределах которых
возможен взаимообмен между первичными разрывами при транс-
локациях); e~KD—первый член пуассонова распределения,
причем К — это среднее число первичных разрывов в
хромосомах в эффективных пространствах на 1 рад. Тогда 1 — e~KD —
шанс для вступления 1 разрыва в образование транслокацни,
а (1— e~KD)2 — для разрывов в двух хромосомах. Поэтому
частота двуударных транслокаций, по мнению авторов теории,
не должна следовать квадрату дозы.
С другой стороны, Меллером в опытах по облучению
сперматозоидов у дрозофилы было сформулировано так
называемое правило 3/2, а именно, что зависимость должна
выражаться формулой e Jl4 /g)
у- k + cdx\
103
i iK как сохраняются только такие хромосомные аберрации
(например, симметричные транслокации), которые не влияют
на жизнеспособность клеток, остальные же устраняются
отбором. Наличие такого отбора среди клеток вытекало из
экспериментальных данных Меллера 1940 г.
Наконец, надо иметь в виду, что характер связен и
вытекающих из них кривых может меняться даже для одного и то-
ю же типа хромосомных аберрации в зависимости от
интенсивности облучения (рис. 28) и от специфики вида (рис. 29).
Статистичность и причинность в мутационном процессе.
Как и всякий вероятностный процесс, процесс возникновения
мутаций не является беспричинным. Он происходит в
результате каких-то мпкрособытнй в клетках, хромосомах,
молекулах ДНК или в цитоплазматнческих структурах, эти микросо-
бытня имеют свои микропричины и не могут быть
определенными для каждого конкретного случая мутаций. В процессе
развития генетики высказывались на этот счет различные
точки зрения. Так, еще в 1935 г. Н. В. Тимофеев-Ресовский,
М. Г. Циммер и М. Дельбрюк выдвинули гипотезу, что
причиной спонтанных мутационных изменений наследственных
структур является тепловое движение атомов. В результате
рассеянности этого процесса в отдельных местах клетки и
хромосом возможно случайное повышение уровня энергии,
приводящее к локальной мутации. Однако в дальнейшем
стали указывать на возможную роль естественного (для
спонтанного мутирования) и повышенного (при
экспериментальном воздействии) фона радиации, различных химических
веществ, как поступающих из внешней среды, так и
образующихся в результате метаболических процессов внутри
клетки, и т. д.
В некоторых случаях удалось доказать наличие в тканях
особых мутагенных веществ, как бы аутомутагенов. В этом
отношении особый интерес вызывает работа Н. П. Дубинина
и В. К. Щербакова, показавшая, что в одной из рас
конских бобов повышенная мутабильность обязана веществам,
находящимся в кожуре семян, по-видимому, пигментам.
Все эти и многие другие факторы способны влиять, как
правило, на хромосомы в самых разнообразных фазах
клеточного цикла. Однако установлено наличие и таких веществ,
которые действуют преимущественно в момент репликации
ДНК. В этих случаях в сущности можно говорить о
спонтанных ошибках самого механизма репликации ДНК (или
хромосом).
В характеристику мутационного процесса как
стохастического не вносит изменений и представление о наличии в
организмах естественных аутоантигенных систем, снижающих в
определенных случаях частоту спонтанных мутаций (Щер-
104
баков). Такие системы, очевидно, сложились в процессе
эволюции и не допускают повышения уровня мутационного
процесса сверх оптимальной для каждого вида границы.
Стохастический подход в оценке биологического и
генетического действия ионизирующих облучений. Выше
указывалось на наличие прямой зависимости между дозой облучения-
и частотой мутаций, т. е. на повышение вероятности мутаций
при повышении дозы. Зависимость может быть выражена
графически в виде соответствующей линии, при этом для
определенной категории мутаций — точковых и некоторых
хромосомных аберраций — в виде прямой. Такой характер связи
указывал на одноударный характер воздействия энергии
облучения на наследственные структуры и был обобщен как
теория попадания. Принцип попадания заключается в том,
что при любых радиобиологических реакциях первичными
являются повреждения элементарных биологических структур
клетки. Такие повреждения возникают как следствие
случайных и квантированных взаимодействий вещества ткани с
энергией облучения. Это может быть: ионизация
макромолекулы, ее возбуждение или же непрямое влияние на
макромолекулу химически активными веществами среды,
окружающей молекулу, но возникшими в среде в результате
облучения. Таким образом, в конечном счете речь идет о случайном
попадании одного (в простейшем случае генные мутации
имеют, как правило, одноударный характер) или нескольких
квантов ионизирующих излучений в достаточно малую по
размерам внутриклеточную структуру — «мишень».
Результатом такого попадания является тот или иной эффект —
генетический или радиобиологический (включая, конечно,
прямое разрушение или инактивацию). Теория попадания и
мишени позволяет описывать в математической форме
зависимость частоты поражения различных мишеней от дозы
облучения, т. е. рассчитывать соответствующие кривые
доза—-эффект. Примеры их на мутациях приведены выше. Конечно,
наиболее удобны для анализа с помощью теории попадания
одноударные биологические реакции. Но и в более сложных
случаях, лри многоударностн, основное, как бы первичное
звено процесса — это попадание квантов излучения в
соответствующие мишени.
Однако сейчас все более и более выясняется, что принцип
попадания, ставший классическим после работ Тимофеева-
Ресовского, Краузера, Дессауэра, Циммера и др., является
лишь частным случаем более общего подхода, при котором
учитывается целый ряд стохастических факторов.
В наиболее полной форме этот подход развит в
монографии Хуга и Келлерера «Стохастическая радиобиология*.
Авторы ее исходят из представления о множественности элемен-
105
тарных событий, происходящих в любой живой системе, и
множественности причин, приводящих к ним. Вот почему
описание явлении возможно только с позиций теории
вероятности. Именно случайность в проявлении многих факторов
была взята как общин принцип, общая основа любого жиз-
/енного явления.
При воздействии облучений происходит наложение
нескольких случайностей, или стохастик. Одна из них, которую
авторы считают неотъемлемой характеристикой всех живых
систем, —это стохастика жизненных процессов. На нее же
накладывается случайность взаимодействия частиц или
квантов ионизирующего излучения с веществами пли
компонентами клетки. В силу этого возможны все переходы от
единичных попаданий в отдельные структуры-мишени (в этих
случаях вполне возможен анализ с точки зрения теории попадания)
до самых сложных, когда происходит смена ряда
последовательных состояний. В результате под влиянием облучения
объект переходит из одного состояния в другое. При этом
происходят и процессы репараций. Радиочувствительность
облучаемых объектов также не остается неизменной.
Переходы биологического объекта от одного состояния к
другому могут быть описаны системами дифференциальных
уравнений. Соответствующий математический аппарат для
этой цели и представлен в книге Хуга и Келлерера. Он дает
возможность провести общий анализ дозовых кривых,
учитываемых соответствующими генетическими, физиологическими
и иными методами радиобиологических реакций, и
рассмотреть самые различные ситуации.
Статистический подход позволяет обойти ряд
математических затруднений, встречавшихся при интерпретации
зависимости между дозой и ее эффектом, когда исходили только
из принципа попадания. Таким образом, форма кривой
доза—эффект определяется многими факторами, например
распределением поглощенной энергии в клетке (очевидно, само
по себе пространственно неравномерным), с одной стороны,
а с другой — разнообразными физиологическими процессами,
происходящими в клетке и имеющими стохастическую
природу.
Развитая Хугом и Келлерером стохастическая теория
имеет значение для радиобиологии в целом. Генетические
эффекты облучения являются в этом отношении лишь
частными случаями, и для их анализа теория попадания может
по-прежнему служить неплохой основой.
В общем же виде кривые доза — эффект, если взять весь
интервал воздействий, имеют S-образный характер с
различными относительными значениями крутизны S, которая опре-
106
дсляется как D/o~ (D — средняя доза, а о2 — варианса
распределения экспериментальных точек). На определенных
интервалах они могут быть близки к прямым, что соответствует уже
приведенным выше экспериментальным данным по увеличению
количества мутации с повышением дозы облучения.
Чисто статистический подход отнюдь не снимает
возможности установления минимальных размеров объекта
воздействия. Так, например, на основе математического анализа
распределения поглощенной энергии был сделан вывод, что
минимальный диаметр чувствительной области при
инактивации клеток млекопитающих равен примерно 1 \i. Именно эта
величина была получена в работе по изучению хромосомных
аберраций в лейкоцитах человека в случае образования
хромосомных колец. Для их образования, очевидно, «нужны два
разрыва хромосом и случайное соединение разорванных
колец.
Математическая интерпретация цепных процессов в
мутагенезе. Н. П. Дубининым было развито представление о
цепных процессах в мутагенезе, иначе говоря, о том. что между
начальным действием мутагена и конечным результатом в
виде мутации проходит целый ряд цепных превращений с
образованием активных элементов, которые представляют
собой предмутационные потенциальные изменения.
Вероятность превращения таких потенциальных изменений в
конечный результат — мутации меняется со временем, а общая
сумма возникающих за какой-то период мутаций будет
зависеть от динамики количества активных элементов в
клетках.
Не вдаваясь в обсуждение проблемы предмутацнонных
из1\гененип и теории цепных процессов в мутагенезе, что не
входит в задачу математической генетики, дадим краткую
характеристику предложенной Н. П. Дубининым и Е. П. Ваулнным
математической модели цепного процесса в мутагенезе. Авторы
ее вводят следующие первоначальные обозначения: n(t) —
концентрация активных элементов; Nt* — количество мутант-
ных хромосом; Nt — количество немутантных хромосом; /С«* —
константа скорости появления мутаций в единицу времени на
каждый активный элемент; К* — вероятность столкновения
активного элемента с хромосомой; /(о — скорость размножения
хромосом (принимается как некая постоянная величина);
К— коэффициент размножения активных элементов.
Эти параметры могут быть связаны друг с другом
следующими уравнениями.
Скорость появления мутаций
dNt*
-K-=Kfn(t). (9)
107
С другой стороны, эта же величина должна быть пропор.
шюнальна количеству иемутпровавших хромосом, т. е. Nt
В таком случае
KS = K«Nt. (Ю)
Общее же уравнение для динамики появления мутации
может быть выражено таким образом:
— =KtNtn(t). (И)
Если n(t) не зависит от времени, т. е. оно константно
(No), то
а величина Nt* может быть определена из предыдущего
уравнения.
Отсюда авторы выводят следующий экспоненциальный
закон проявления мутаций во времени:
is
Л^=-^ЛоЛ^о/--1. (12)
До
При этом число мутаций на геном принимается величиной
постоянной.
Однако количество активных элементов в клетке,
вероятно, не остается постоянным во времени. Можно предполагать
две возможности, что оно уменьшается или увеличивается.
Закон изменения величины n(t) со временем может быть
записан так:
dn(t) __<
—: —«п.
at
Если принять, что n(t) представляет собой величину,
выражающую скорость исчезновения этих элементов во времени,
то она выразится как
n(t)=n0e-dt.
Подстановка значения n(t) из уравнения* (9) в
уравнение (11) приводит к следующей формуле, выражающей
скорость проявления мутаций на геном при уменьшении
количества активных элементов:
^=/Oi0e-df. (13)
dtNt
В данном случае скорость будет уменьшаться.
108
Если же л (0 возрастает со временем, то число
проявляющихся мутаций на геном будет возрастать по формуле (3), что
„ объясняется наличием цепных процессов в мутагенезе.
Далее авторы несколько усложняют анализ, предполагая,
что часть активных элементов возникает с какой-то скоростью
(dn), а часть исчезает с иной скоростью (-рп).
Тогда суммарная скорость изменения количества активных
элементов будет равна dn—(V
На дальнейших усложнениях, введенных авторами, можно
не останавливаться. Существен лишь тот общий вывод, что
если активные элементы размножаются быстрее, нежели
распадаются, то наблюдается рост числа мутаций на геном. В
обратном же случае процесс появления мутаций затухает и
число мутаций на геном падает.
Глава 3
ГЕНЕТИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИЙ
Рассмотрение вероятностных законов на уровне особей в
системах популяций. Явления жизни, в том числе и
генетические, предстают на разных уровнях организации живого:
молекулярном, хромосомном, клеточном, организменном. Во
всех случаях, когда в явлениях участвуют большие
совокупности объектов — молекул, хромосом, клеток и т. д.,— на
сцену выступают вероятностные, статистические закономерности
как ведущие. Именно на них основываются элементарные
законы наследственности. В то же время на одни
статистические закономерности накладываются другие
статистические закономерности, как бы второго порядка, а также
динамические, которые осложняют предшествовавшие
закономерности, меняют их форму, частично и даже полностью, как
это было показано в предыдущих главах.
В данной главе мы переходим к новому уровню
организации живого — популяционному и пойдем по тому же пути,
как ранее, а именно: сначала рассмотрим вероятностные
законы, характеризующие основные процессы, происходящие в
популяциях, а затем факторы, осложняющие, изменяющие
эти процессы. Эти факторы в свою очередь могут иметь как
статистическую, так и нестатистическую (динамическую)
природу.
Популяциоиная генетика. Изучение генетических
процессов, которые происходят в больших совокупностях особей,
носящих название популяций, составляет особую область
генетики, получившую название популяционной генетики.
Основы всего последующего развития популяционной генетики,
играющей сейчас громадную роль для понимания процессов
естественной эволюции видов в природе, а также
искусственной эволюции домашних животных и возделываемых
растений под влиянием человека, заложила классическая работа
С. С. Четверикова «О некоторых моментах эволюционного
>чеиня с точки зрения современной генетики»,
опубликовании
ная в 192G г. * Популяционная генетика выросла, с одной сто-
ромы, из генетики, а с другой — из биометрии. Хотя с
первого вчгляда генетический и биометрический подходи кажутся
противоположными друг другу, так как первый имеет дело
с дискретности ми явлений наследственности, а второй с не-
ирсрывностями в явлениях изменчивости, синтез между
ними оказался в высшей степени плодотворным. Развитие по-
пуляционной генетики связано с именами советских ученых —
С. С. Четверикова, Н. П. Дубинина, Н. В.
Тимофеева-Ресовского, Д. Д. Ромашова — и зарубежных — Р. Фишера, Дж.
Холдейна, С. Райта, Дж. Лаша и многих других.
Популяция. Как показывает само название, объект попу-
ляционной генетики не особь, а популяция.
Понятием «популяция» пользуются в различных областях
биологии в несколько разных смыслах. Так, экологи часто
говорят о популяциях планктонных организмов и включают
в такие популяции даже особей разных видов. Ихтиологи
употребляют термин «.популяция» для обозначения
населения рыб определенного водоема. Под человеческой
популяцией понимается совокупность всех людей на земном шаре,
объединяемых одним видовым названием Homo sapiens. Но
можно говорить о популяциях людей в пределах более
ограниченной территории — какой-либо страны или области.
С генетической точки зрения лучше всего рассматривать
популяцию как совокупность особей вида, населяющих
определенную территорию и неодинаковых по своим фенотиличе-
ским и генотипическим свойствам. В качестве исходной для
анализа протекающих в популяции генетических процессов
рассматривают так называемую панмиктическую популяцию,
т. е. популяцию, состоящую из свободно скрещивающихся
особей. Предусматривается, что в панмнктической популяции
все входящие в ее состав особи могут спариваться друг с
другом в любых сочетаниях независимо от их генетической
природы, и вероятность таких спариваний будет целиком
зависеть от вероятности встреч соответствующих категорий
генотипов в популяции. Образуется множество родительских
пар, потомство которых в своей совокупности будет иметь
некоторую генотипическую структуру. Таким образом, в
понятие панмнктической популяции с самого начала вводится
статистический принцип. Очевидно, что такая популяция
является идеализированной моделью, прототипы которой могут
встречаться с той или иной степенью приближения к модели
во многих конкретных популяциях животных и растений или
* В 1965 г. она была переиздана. См.: Бюлл. Моск. о-ва испытателей
пИфоди отд биологии т. 70. выи. 4. 1965; сб. Классики современной гене-
Т,,К11- Л.. 1968.
ill
в популяциях пород животных и сортов растений. Само собой
разумеется, что свободно скрещивающиеся популяции
возможны только у видов, размножающихся половым путем.
Закономерности в популяциях видов, размножающихся
бесполым п>чем, будут иными.
В следующих главах будут показаны те факторы, которые
могут нарушать первоначальное условие свободного или
случайного скрещивания в популяции, и разобраны результаты
их действия.
Частоты генов как параметры популяции. В панмнктиче-
ской популяции, как указывалось выше, все особи имеют
одинаковые вероятности скрещиваться друг с другом и
равную вероятность дать потомство. Но при этом
предусматривается не чисто физическая возможность спаривания любых
самок с любыми самцами, а принципиальная возможность
его осуществления. Отсюда вытекает построение еще одной
модели: всю совокупность половых клеток, образуемых
особями такой популяции, можно рассматривать как единое
целое, как будто все они помещены в один сосуд и
перемешаны друг с другом. Тогда соединение женских и мужских
половых клеток происходит чисто случайно и его результаты
будут зависеть только от частоты (или замеряемой частотой
вероятности) тех иди других типов половых клеток. А
поскольку каждая половая клетка до оплодотворения
содержит, как правило, только один ген из пары или серии
аллелей, то и совокупность генов, находящихся в половых
клетках всех особей популяции, вместе взятых, можно
рассматривать как единое целое, как единый генофонд. Доля всех
отдельных аллельных генов в общем генофонде популяции
называется частотой генов.
В зависимости от частот отдельных генов в популяции
складывается определенное соотношение генотипов и
фенотипов. Зная это соотношение, можно в свою очередь
определить частоты генов.
Определение частоты генов. Для разбора методов
определения частот генов возьмем следующий конкретный пример
В стаде овец было обнаружено 729 длинноухих (АА), 111 ге-
терозигот короткоухих (Аа) и 4 овцы безухих (аа). Если по
количеству образуемых половых клеток все три категории
особей не отличаются друг от друга, то, принимая для простоты
расчета, что каждая особь образует только по 2 половые
клетки, получим следующее количество генов А и а в общем
генофонде:
Ген А: (729 - 2) + 111 - 1569 половых клеток;
Ген и: 111 +м-2). 119 половых клеток.
При переводе в доли получим: р для гена А равно 0,93 и д
длЯ гена а — 0,07. Это и будут частоты генов. Их записывают
следующим образом:
Р(Л)=0,93; <7<о)=0.07.
При этом
В данном примере частоты генов вычислены на основе из-
[.естных численностей или долей генотипическн
отличающихся друг от друга групп особей. Зная же частоты генов, можно
в свою очередь предсказать конкретные соотношения, которые
будут получены в следующем поколении свободно
скрещивающейся популяции. Выгоднее это сделать в общем виде, для
любых значений р и q в генофонде. Как самки, так и самцы
будут образовывать гаметы двух типов — .4 и а — в
соотношении' P(A)'-Q(a)*. Результаты соединения мужских и женских га-
ует легко показать с помощью следующей четырехпольной
таблицы.
^^^-^^^^ Гаметы ?£ ,
! ""~-^-^^^ и их частоты
Гамегы oV "^ ■—^^^ '
II их частоты ^—^^^
Л р
a q
А
Р
А А
1
а
Я
Аа 1
РЯ
Aa I <ы
1 W 1 Я* \
В потомстве образуются 3 генотипа в соотношении,
выражаемом коэффициентами р2, 2pq и q2'-
p2AA + 2pqAa + q*aa. (2)
Формула Харди—Вейнберга. Соотношение p*AA+2pqAa +
-q2aa названо формулой или законом Хардн—Вейнберга.
Дело в том, что эта формула выражает определенную
закономерность соотношения генотипов в популяции при условии
свободного скрещивания в ней.
* В этой главе мы сохраняем обозначения генов одной пары через .4
и а, как это было принято в классической работе С. С. Четверикова. Во
многих монографиях по пепуляционно* генетике принимают более общие
обозначения А{ и А2. В дальнейшем мы будем прибеть именно к этим
°6<лиачсиням.
Ь " Ъ Р «ьимкпП
113
В прнпеденноА формуле коэффициенты при обозначениях
генотипов выражены в частотах генов, но если они еще не
определены, можно обозначить доли генотипов в популяции
какими-то другими символами. В литературе принято обозначать
доли особей с генотипом АА через Р, долю особен Аа — через
2Q — и долю особей аа — через /?,
Если соотношение генотипов и фенотипов в популяции
следует формуле Хардн—Вейнберга, то Я = р2; 2Q = 2pq и R = q2.
В таком случае можно проверить, получится ли в конечном
счете соотношение по формуле Хардн—Вейнберга, если
рассчитать вероятность всех типов скрещивании в популяции (на
основе умножения вероятностей) и вероятности
образующихся в потомстве генотипов, что сделано в табл. 17.
Телица 17. Вычисление вероятностей появления форм А А, Аа н аа
■ 9 возможных типах случайных скрещиваний в популяции
Скрещмвдння
1 ААХАА
1 АА\Аа
AAy.Qxi
АахАА
АаУ.Аа
Аахаа
аахАА
оахАа
аахао
Вероятности
данного
скрещивания
Р2
2PQ
PR
2PQ
4Q*
2QR
PR
2QP
R*
Условные вероятности появления
в потомстве
.4.4
1
0,5
0
0.5
0.25
0
0
0
0
.4 а
0
0.5
1
0.5
0.5
0.5
I
0.5
0
аа
0
0
0
0
0.25
0.5
0
0.5
I
Окончательные результаты появления генотипов АА, Аа
и аа в потомстве от 9 типов скрещиваний получаются после
подсчета сумм по вертикальным столбцам 3, 4, 5. Далее в ннх
надо произвести необходимые алгебраические упрощения,
заменить значения Р, 2Q и R на р2, 2pq, q2.
По каждому вертикальному столбцу суммы будут
следующими:
Л аа) - Р2+2PQ + Q2=р« + 2р3?+р V =
=p*(p* + 2pq + q*) = р2(р + <7)2 = р2;
/\A.,~2PQ + 2Q2 + 2/>tf + 2Q/? =
- 2p*q+2pV+2р V + pq*=
= 2pq(p> + pq+pq + q2)=2pq(p + q)2 = 2pq;
= q2(p2 + 2pq+q>)=q2(p+q)2^
т е. в конечном счете:
p2AA+2pqAa + q*aa.
Таким образом, суммарные результаты, получаемые в
потомстве при случайности всех спариваний исходных форм
в популяции, основываются на чисто вероятностном,
статистическом процессе.
Определение частот генов по доле одного из генотипов. Из
формулы Харди — Вейнберга видно, что частоту генов можно
определить по доле особей /?, проявляющих рецессивный ген а,
т. е. генотипическн аа. Приняв ее за qz, можно определить q как
корень квадратный из R: q= |/?.
4
В примере с овцами доля безухих была -г-г- =0,0048.
Тогда q]a = 0,0048 и д = 0,07.
Отсюда р = 0,93.
Этот способ, очевидно, удобно применять тогда, когда ге-
терозиготы и доминанты гомозиготы не отличимы друг от
друга. Если же они отличаются, то за основу подсчета частот
можно принять любую группу гомозигот — как домннантов,
так и рецессивов.
Приведем следующий пример. У овец было установлено
два типа гемоглобина. Один движется к аноду относительно
быстро Mi), второй относительно медленно {А2)> третий, по-
видимому, смесь первых двух — с промежуточной скоростью,
т. е. АХА2. На основе генетического анализа можно было прийти
к выводу, что различия между овцами определяются двумя
аллелями. У гомозигот А\А\ гемоглобин первого типа, у
гомозигот А2А2 — второго типа; гетерознготы А\А* имеют
смешанный гемоглобин.
В изученной популяции шотландских черноголовых овец
U933 головы) было обнаружено овец А\А\ 60,7%, овец ЛоЛу—
6.1% и овец А\А2 — 33,3%, или в долях единицы, что удобнее
.зля вычисления частот генов: 0,61, 0,06, 0,33.
В таком случае для определения частот генов можно взять
как особей А\Аи так и особей А^А*
По AxAi Р(Л,)= V 0,61 =0,78 и тогда <7Ul)=0,22.
По А2А2 <7(а5)= У 0,06=0,24 и тогда p(Al)=0,76.
В данном случае, при кодоминантности генов А% н Аь
в сущности неважно, что считать за р, а что за q.
Разница между определенными двумя способами частота-
ми генов очень невелика — всего 0,02, но можно думать, что
более точные результаты получены по группе АХАХ% так как
она многочисленнее.
Можно проверить и долю группы А\А2 ( = 2рд). При пер-
вом способе определения (по /Mi):
2/х/- 2- 0,78- 0,22-0,34.
При втором
2р(/= 2-0.76.0,24-0.36.
Фактически получена доля Л Из = 0,33. Первый способ дает
значение 2pq% более близкое к фактически полученному.
Конечно, чем больше объем популяции, в которой изучено
распределение фенотипов, тем точнее могут быть вычислены
частоты генов.
Связь соотношения генотипов и частот генов. Вернемся
опять к формуле Харди— Вейнберга и остановимся более
подробно на связи соотношения генотипов в популяции и частот
генов.
Доли генотипов в популяции мы выражали величинами
Р,\Л, 2()ап, Raa
P+2Q+/?=1. (3)
Если конкретные количества их в популяции jV2, Nu М,
а общее количество iV, то
N ' Ч N ' N
Допустим, что каждая особь образует только 2 гена данной
пары аллелей. Тогда общее число генов в геноАоиде соста-
пит 2Л-'. число генов 4-2jV2+jV, и число генов a—Nt+2No.
В таком случае частоты генов будут следующими:
При образовании гамет происходит случайная выборка
генов, попадающих п гаметы. Вероятность попадания в гамету
гена А равна р, гена а — q. В силу независимости этих
выборок вероятность того, что данная зигота явится результатом
соединений А и А, т. е. будет иметь генотип АА, равна р-р~Рг-
То же относится и к вероятности появления генотипа аа, ко-
не
аЯ будет раина q-q^q2, Вероятность появления генотипа
,]а .произведение pq. Но так как Аа может появиться двумя
^особами, иероятпость надо удвоить: 2pq.
Таким образом, если выполняется исходное условие о
свободном, т. с. случайном, скрещивании и об одинаковой вероят-
пости образования гамет, то потомок может иметь генотипи-
ческую структуру АА% Аа или аа с вероятностями р\ 2pqt q2.
Это верно при любых значениях долей Я, 2Q% R% если
только распределение генотипов в первом поколении
удовлетворяет указанным пыше формулам связи между долями генотипов
и частотами генов:
P + Q = P\ (? + /?-<?. (5)
Если выразить строение популяции в общем виде как
PAA+2QAa + Raa, где Я, 2Q, R —доли в популяции
соответствующих групп гомозиготных и гетерозиготных особей, то
состояние равновесия популяции по формуле Хорда—Вейнбсрга
определится следующим условием:
PR~Q2. (6)
В этом случае, очевидно, Р должно быть равно ра, Q-p</
и /? = </*.
Закон стабилизирующего скрещивания. Легко представить
такой случаи, когда в конкретной популяции по тем или иным
причинам соотношение величин Р% 2Q и R не соответствует
указанному условию, т. с. равенству Я/? — Q2. Оказалось» что
если в такой популяции в дальнейшем будет происходить
свободное скрещивание» то в следующем поколении вновь
установится повое соотношение фенотипов и генотипов,
соответствующее формуле Харли—Вейиберга. В этом заключается закон
стабилизирующего скрещивания Пирсона, который можно счи-
тать как бы вторым законом свободного скрещивания,
Сущность закона можно выразить так. Если в популяции
имеется соотношение генотипов xAA+2yAa+zoa, не
находящееся в состоянии равновесия (т. е. xz+yl)% то а результате
первого же свободного скрещивания в популяции создадутся
новые соотношения гомозигот и гетерозигот хИ>1+2уИа+*|аа.
удовлетворяющие условию XiZ\=*ypt
Новая структура популяции окажется равновесной, т е
соответствующей формуле Харди — Вейиберга.
Проиллюстрируем сказанное следующим примером.
Допустим, что в популяции было 0»\AAt OAAa и 0,5аа. Очевидно,
что в данном случае де^у1. Каким же окажется следующее
поколение, если в данной нестационарной популяции
произойдет свободное скрещивание?
117
Для этого иадо рассчитать соотношение гамет А н а, обрщ.
ауемых самкямм м сами**»- с помощью приведенных выше
формул определим р н q:
pmP + Q-0,\ +0.2-0,3;
,,..<? +Я-0.2+ 0.5-0,7.
Зная частоты генов, легко рассчитать, какое потомство
получится в следующем поколении.
, ^^""^-^^^^ Глисты i V
^^*^*^^^^ И il\ ЧИСТОГЫ
.4 0.3
1 и 0.7
А
0,3
АА
0.09
Ли
0,21
и
0,7
Ли
0.21
0.49
В целом по таблице получается: 0,09/1/1+0.42/10+0.4900,
^=0.49. отсюда </= у 0,49=0.7. а /> = 0,3. Очевидно, что:
1) частоты генов сохранились те же: /?(а)=0,3; </(а)=:0,7 и 2)
полученное соотношение генотипов будет сохраняться и в
следующих поколениях.
Для иллюстрации роли стабилизирующего скрещивания
можно было бы привести много примеров, в том числе и
таких, когда исходные родительские формы состоят только иэ
«иной или двух генотипнческнх групп вместо трех, и во всех
случаях будет наблюдаться та же картина стабилизации
соотношения после первого же свободного скрещивания.
Мехами hi, который стабилизирует соотношение численностей
генотипов в популяции, заложен в самом свободном скрещивании,
саолГУШеНИе его позможно только за счет сил, лежащих вне
oihrnu*1101"0 скРещиоания' К их числу относятся отбор, мутаци*
< **1ан Процесс и искоторые другие факторы, о которых будет
ишмоОп1 Харди"~ВеЙ"беРга можно сформулировать следую-
юшейся f °М ссли в достаточно большой свободно скрещнва-
по двум "аимиктической) популяции происходит расщепление
Рш УстовмИ**есКОЛЬКИМ аллслям одного гена, то при иекото-
лгним ЧШС1ПТ С0*Раняются н* поколения в поколение опреде-
*о*ет бып/1^ пнх аллслей» а соотношение частот генотипов
тт* выражено общей формулой p*AA+2pqAa + q*QQ.
14 uiKoM случае любая стабильная свободно скрещивающа-
k.,i попу-ишня должна удовлетворять условию PR-Q\ ял и
,|Иа1К Р \-Q ' Pi+Qi-pl РЯ~ Р1Р + *)шр'
Q I R~QiiRr»Pq + q'*~q(P + V~q
.,..иы генотипов ДЛ. Аа
\\ спою очередь наличие в популяции геи твом д
„ аа и отношении р2 : 2pq : ^ может быть докажи
вильности гипотеэи о
наличии и данной
популяции свободного
скрещивания.
Разнообразие
стационарных состояний в
популяциях. В различных
популяциях животных и
растений,
размножающихся путем
перекрестного оплодотворения,
возможно множество
стационарных состояний.
Каждое из них будет
характеризоваться разными
значениями р и q. Так как
р + <7=1, то р может
колебаться от 0 до 1, a q
соответственно от 1 до 0.
Соответственно значениям р и q меняются и соотношения
генотипов АА% Аа% аа (или в более общем виде при кодом*-
иаитности или отсутствии доминирования А\Аи Л\Аъ A*At)-
На рис. 30 показано, как меняется соотношение генотипов
в зависимости от величины q% т. е. частоты гена а или А%.
Легко видеть, что при малых значениях q группа гомозигот доми-
иаитов будет численно преобладающей, гетерознготы составят
меньшую долю, еще меньше гомозигот рецессивов. Так,
например, при q «0.2 гомозиготы доминанты составят 0,64 всех
особей популяции, гетерознготы — 0.32. а рецесенвы — 0,04.
При </«р«0,5 соотношение будет 0,25. 0,50 н 0.2& При
значениях ?, больших 0,5, доля АА соответственно уменьшается
Например, при (/ — 0,7 АА составят только 0,09 всей популяции»
Аа—0,42 и аа~0.49. С помощью этого рисунка можно
установить теоретические частоты генотипов при разных
значениях </ и сопоставить их с фактически полученными
соотношениями. В некоторых случаях совпадение очень велико, что
указывает на реальное осуществление условия закона Харда-
Вейиверга. а именно свободного скрещивания, а также олива-
119
Ы02 V (МЕТЯ* в' J5 й$$
Рис. 30. Изменена* соотношеама
генотипов в пооуляаяи • ммеяиостя
01 частоты гена А}
ковой жизнеспособности разных генотипов. В качестве приме-
ра большого соответствия формуле Харди—Вейнберга можно
привести соотношение по группам крови М и N в популяции
из 1279 человек. Генотипы М составляли 0,28, AfiV—0,50
и \—0,22, что соответствует ожидаемому соотношению при
^м«0.53 и (/N-0,47. Впрочем, такое соответствие не всегда
доказывает отсутствие различий по жизнеспособности. Для
установления его надо знать частоты генов и соотношение
генотипов предыдущего поколения.
Установление доли гетерозигот. В большинстве
приведенных выше примеров гетерозиготы были отличимы от гомозигот
доминантов. Однако, как известно, это бывает далеко не
всегда. Популяция может распадаться только на 2 фенотипические
группы: А (среди которых АА и Аа) и а (генетически аа).
Если популяция находится в равновесии или хотя бы
приближается к равновесию, то с помощью формулы Харди—Вейнберга
можно установить, какова доля гетерозигот среди фенотипи-
чески однородных особей, проявляющих доминантный ген,
т. е. сколько скрытых носителей рецессивного гена.
По массовым исследованиям известно, что частота
проявления альбиносов в человеческих популяциях примерно 1 на
20 тысяч, т. е. 0,00005. Если (/-=0,00005, то q = }' 0,00005=
= 0,007. В таком случае частота гетерозигот должна составить
2/><г = 2-0,993-0,007 = 0,014, т. е. 14 гетерозигот на 1000
человек, или, иначе, 1 гетерозиготная особь в среднем на 70
человек.
Подобные расчеты имеют значение в селекции животных,
когда приходится иметь дело с довольно многочисленной
группой наследственных уродств и ненормальностей. Важно
предвидеть, какой процент быков может оказаться носителем
вредного рецессивного гена. Можно привести следующий пример.
У новорожденных телят айрширского скота была обнаружена,
водянка, зависящая от рецессивного гена. Частота появления
ненормальных телят в породе 0,0033. Тогда q= У 0,0033 = 0,057.
Отсюда частота гетерозигот 2^=2-0,947-0,057=0,11. Это
значит, что примерно 1 бык из 10 окажется гетерозиготным по
данному рецессивному гену. Впрочем, расчет этот очень
приблизителен, так как породу нельзя рассматривать как свободно
скрещивающуюся популяцию. Кроме того, часть гомозиготных
особей может умереть еще до рождения. Но в последнем
случае фактически полученное значение q2 окажется заниженным
и действительная доля гетерозигот еще выше. Во всяком
случае приведенный пример показывает, насколько велика в
действительности может быть доля гетерозигот в популяции.
Применение формулы Харди—Вейнберга к более сложным
случаям наследования. До сих пор рассматривалось расщепле-
120
1йе в популяции по одной паре аллельиых генов {А-а иль
j ~~А2)* ПРИ этом не сцепленных с полом, т. е. аутосомных
и0 возможны случаи расщепления по нескольким аллелям од-
(0го локуса; по генам, сцепленным с полом; наконец, по
нескольким генам разных пар хромосом. Наиболее удобный
пример — группы крови системы АВО у человека. Фенотипу О
соответствует генотип 00 (I группа крови), фенотипу А
—генотипы АА и АО (II группа крови), фенотипу В — генотипы
ВВ и ВО (III группа крови) и фенотипу АВ — генотип АВ
/IV группа крови). Универсальные доноры, как известно, люди
I группы крови, так как у них отсутствуют оба антитела
\А и В), универсальные реципиенты — люди IV группы.
Данных по популяционнои генетике групп крови очень много, так
что к ним легко применить формулу Харди — Веинберга.
Частоту аллеля А обозначим через р, частоту аллеля В—q
н частоту аллеля О—г; р + <7-Ьг=1. По каждой паре идет
самостоятельное расщепление. Общая формула структуры попу*
ляции будет следующей: p2AA + 2pqAB + q2BB+2qrBO+
+2рл40 + r200% или для лучшего запоминания: p1AA + qiBB+
-trW0 + 2pqAB + 2prA0+2qrB0.
Допустим, что в определенной популяции (пример
совершенно конкретный, во многих популяциях наблюдается
подобное или близкое к нему соотношение) частоты фенотипов
были следующими: Л—0,36; В—0,23; ЛВ—0,08 и О—0.33.
Учитывая указанную выше структуру по формуле Хардн —
Веинберга, данные о фенотипах и генотипах можно записать
следующим образом:
Фенотипы
Генотипы
Частоты ожидаемые
> фактические
А
АА + АО
р* - 2рг
0.36
В
ВВ-ВО
Я* -г 2qr
0,23
АВ
АВ
Ъ>9
0,08
О
ОО
г«
0.33
Частоты генов можно определить различными способами.
Один из них следующий. По группе О определяем значение г.
г*=0,33; г— ) 0,33=0,5744.
Так как сумма групп В+О дает сумму частот
q2+2qr+r*= (fl+r)^0,56.
то отсюда q + г- У*0|б = 0,7483; <7« 0,7483—0,5744 = 0.1739.
Точно так же сумма Л+О = р2+2р'+г*=1р+п2=0,69. Тог.
Да р+г= >гО*69=0,8307. Откуда р=0,8307-0,5744=0,2563.
Окончательные значения частот будут следующими:
121
Для гена А р = 0,2563
» » В 9=0.1739
» » О г = 0,5744
2=1,0046
То, что сумма не равна 1, объясняется недостаточной
точностью вычисления. Можно проверить степень соответствия по
группе АВ:
2р</ = 2-0,2563-0,1735 = 0,089.
При кодомпнантности изучаемых в популяции аллелей
методы вычисления частот генов и сопоставления фактического
распределения фенотипов с теоретически ожидаемым по
формуле Харди—Вейнберга проще. В качестве примера приведем
данные, полученные в Белорусском научно-исследовательском
институте животноводства по трансферринам крови крупного
рогатого скота на материале 627 голов черно-пестрой породы.
Расщепление идет по 3 аллельным генам, гомозиготы и гете-
розиготы по ним легко отличимы биохимическими методами
(с помощью электрофореза). Поэтому возможны 6 генотипов,
фактическое распределение по которым следующее:
ЛЛ — 130 АЕ — 22
AD — 294 DE — 28
DD— 151 ЕЕ— 2
2 = 627
Частоты аллелей можно вычислить по следующим
формулам:
__ 2nAA+nAD+nAE 2-130+294+22 =
Р{А)~ 2N 2-627
-^_0 459-
~ 1254 -°'45У'
_ 2nDD+nAD+nDE 2-151+294+28 =
РФ)-' 2/7 — 2-627
624 = 0,498;
1254
2nEE+nAE+nDE 4+22+28 _ 54 _nn,q
Р(„= & = 2.627 - 7254" ~°'тб-
На основе определенных частот генов Л, D и Е можно
вычислить и теоретически ожидаемое распределение 6
генотипов. Оно окажется следующим:
122
ЛД-132,1 АЕ-24 7
AD- 286,7 D£-26,9
DP- 155,5 ЕЕ- 1,2
2 = 627,1
Обнаруживается очень большое соответствие фактически
наблюдающихся частот теоретически ожидаемым, очевидное
даже без проверки с помощью критерия хи-квадрат.
Сохранение равновесия в популяции по формуле Харди —
Вейнберга при различиях в частотах генов у самок и самцов.
В приведенных расчетах предусматривалось, что мужская
и женская части популяции обладают одинаковой
генетической структурой, т. е. частоты р и q для самок и самцов одни
и те же. Однако практически возможны и такие случаи, когда
значения р и q в мужской и женской частях популяции
неодинаковы. Особенно часто это может быть у
сельскохозяйственных животных, у которых, как правило, мужская часть
популяции количественно значительно меньше, чем женская. При
применении искусственного осеменения, например у крупного
рогатого скота, количество самок может быть больше
количества самцов в тысячи раз, как это имеет место при высоком
уровне искусственного осеменения в СССР, Новой Зеландии.
США и других странах. Тогда можно ожидать, что значения
/mi q у самок и самцов для различных аллелей по ряду
причин могут оказаться неодинаковыми. Возникает вопрос, в
какой степени при этом соотношение в потомстве будет
соответствовать формуле Харди—Вейнберга.
Допустим, что в мужской части популяции концентрация
гена А\ равна р\ а гена А2 — q\ а в женской соответственно
р" и q". Тогда структура популяции следующего поколения
может быть получена путем перемножения двучленов:
(p'Ai+q'A2)(p"Ai+q"A2)=p'p"AiAi+
+ {p'q"+p"q')AiA2+q'q"A2A2.
Для такой новой популяции можно вычислить и новые
значения р и q (обозначим их p,v и qs), пользуясь указанными
выше формулами, связывающими концентрации генов с
количеством доминантных, рецессивных и гетерозиготных форм.
Заменим значение q через 1 — р> Тогда правая часть
равенства примет следующий вид:
123
Ps = pY+-lp'(]-P")+\p"(\-p')=P'p"+±-p>-.
- \ р'р"+ | р"- | р"р'= т р'+ т р"=
= \(Р'+Р"). (7а)
Таким же образом вычисляем и значение q^:
?.v=y(W). (8)
Новая популяция окажется опять в равновесии с новыми
значениями ps и qs, средними между значениями р и q
родительских групп.
Соотношение в популяции по сцепленным с полом генам.
По гомогаметному полу соотношение между частотой генов
и частотой генотипов такое же, как по аутосомным, но по ге-
терогаметному полу образуются только два генотипа и
каждая особь содержит в своем генотипе только один аллель:
Самки Самцы
Генотипы АА; Аа\ аа А; а
Частоты Я; 2Q; R S; Т
Тогда частота гена А среди самок pf=P-\-Q, а среди
самцов pm=S. Учитывая, что самцы (т) дают вдвое меньше
сцепленных с патом генов, нежели самки (/), общее количество
генов А для всей популяции составит:
Рш= \Р/+ -у Рт= у(2р,+Рт) = y(2P+2Q+S). (9)
Если частоты генов среди самок и самцов неодинаковы, то
гопуляция не будет в равновесии. Хотя общее распределение
1енов в популяции, как целом, не изменяется, но их
распределение между полами колеблется таким образом, что
популяция понемногу приближается к равновесию.
Не вдаваясь в детали, можно указать на следующее.
Разница между частотами сцепленного с полом гена у самок и у
самцов с каждым поколением уменьшается вдвое. Популяция
приблизится к равновесию только через несколько поколений.
На рис. 31 показано приближение к равновесию по
сцепленному с полом гену в том случае, когда в исходном нулевом
поколении ?;=], a qm = Q. Равновесие установится на уровне
</"0,67.
124
При расщеплении одновременно по нескольким генам рав-
ьсвесие устанавливается только после ряда поколений
случайного скрещивания, хотя по каждому отдельному гену
стационарное соотношение наступает после первого же
стабилизирующего случайного скрещивания. Чем больше различных
генов участвует в расщеплении, тем медленнее наступает
равновесие. Если гены находятся в одной хромосоме, т. е.
сцеплены друг с другом, то
чем больше сцепление,
тем медленнее
достигается равновесие.
Если в популяции нет
стационарного
соотношения между фенотипами
(или соответственно
генотипами), то с
наступлением свободного
скрещивания оно неизбежно
установится или
непосредственно после первого
же свободного
скрещивания — в простейших
случаях, или несколько
позже—в более сложных
случаях, о которых
говорилось выше.
Таким образом,
положения популяционной
генетики позволяют понять
генетическую структуру популяций животных и растении на
основе чисто вероятностных принципов. На основе
стенотипической изменчивости популяции по отдельным генам может
быть установлена генотипическая структура популяции.
Параметрами для ее оценки служат частоты генов. В панмиктн-
1:еской популяции сохраняется определенное соотношение
между генотипами, выражаемое формулой Харди — Вейнберга.
С первого взгляда может показаться, что в реальных
популяциях такие случаи сравнительно редки. На самом деле это
не так.
Во всех популяциях домашних животных и культурных
растении, а также, конечно, и в природе происходит
расщепление по громадному количеству генов, определяющих
разнообразные морфологические, биохимические и
физиологические признаки. В ряде случаев сравнительно легко выделить
и аллели отдельных генов, и тогда перед нами предстает
поистине грандиозная картина генотипической сложности
популяции.
125
3 и 5
Поколения
Рис. 8L Приближение к равновесию
по сцепленному с полом гену при
свободном скрещивании.
Первоначальное соотношение q.t — l и <7«~0.
Хотя условием для применения формулы Харди — Вейн-
бсрга является свободное скрещивание в популяции и поэтому
каждая конкретная популяция может быть только некоторым
приближением к такой идеальной модели, осложняемой
многими обстоятельствами, о которых будет говориться ниже, в
действительности есть очень много случаев, когда
соотношение фенотипов и генотипов в популяции по определенной паре
генов (или серии аллелей) очень близко к ожидаемому по
формуле Харди — Вейнберга. Это относится прежде всего к
случаям так называемого сбалансированного полиморфизма,
при котором определенное соотношение между генотипами
сохраняется в течение ряда поколений.
Однако постоянство соотношения между генотипами при
свободном скрещивании отнюдь не означает сохранения тех же
значений Л 2Q и R. И при стационарном распределении
генотипов можно ожидать небольших изменений в частотах
гомозигот и гетерозигот от поколения к поколению, что было
отмечено еще самим Харди. Хотя при наличии случайного
скрещивания нет тенденции к каким-либо систематическим
изменениям соотношений, но так как частоты генов р и q могут
чисто случайно колебаться от одного поколения к другому, то
и генетическая структура популяции может меняться, хотя
и медленно. Существенно то, что, хотя каждое новое
поколение будет отличаться от предыдущего, возврата в старое
состояние уже не будет. Если вместо частоты р в каком-то
поколении случайно появилась частота гена р\ч то дальнейшие
случайные колебания будут происходить уже вокруг этого р\.
Таким образом могут происходить изменения частот генов,
з конечном счете имеющие как бы направление.
Статистическая основа таких процессов —так называемые цепи Маркова,
широко излагаемые в специальной статистической
литературе*.
Отсюда вытекает важный вывод, что популяция — это
система, имеющая определенную структуру и находящаяся в
некотором сбалаисированном состоянии при наличии свободного
скрещивания, но состояние это подвижное, динамическое.
Однако возможны многочисленные процессы (случайные
и неслучайные), которые моту! привести к существенным
нарушениям в структуре популяций и отклонениям соотношений
генотипов от формулы Харди — Вейнберга. Обнаружение
таких отклонений в фактически изучаемых популяциях
естественно приводит к вопросу о их причинах. В последующих
главах они будут подробно разобраны, а сейчас надлежит
Дать лишь краткий их перечень.
Ичл 2*"'н; №?ер Вв*ЛЛ1И* » теорию вероятностей и ее приложеяя*
136
1. Различия в жизнеспособности и плодовитости особей
разных генотипов, в простейшем случае генотипов АгАи АХАЬ
fafa- Они приводят к явлению, носящему общее название
отбора. Своеобразные рэзновидностя отбора — эмиграция и
иммиграция, также нарушающие структуру популяции.
2. Мутационный процесс. Мутации отдельных генов созда*
ют так называемый мутационный пресс, или мутационное
давление, на структуру популяции.
3. Система спариваний. В свободно скрещивающейся
популяции предусматривается равный шанс спаривания друг с
другом любых особей независимо от их генотипа. Но если
будет наблюдаться (или вызываться человеком) особая
тенденция спаривания особей более сходных друг с другом по
доотипу или фенотипу или, наоборот, менее сходных, то она
приведет к отклонению от соотношения по формуле Харди —
Вейнберга.
Влияние типа спаривания довольно легко изучить на
материале домашних животных или возделываемых растений, но
значительно сложнее этот вопрос в природе, где он связан
и с территориальным размещением особей, и с биологией
размножения, и со стадностью или колониализмом и т. д.
Наконец, на структуру популяций влияют колебания их
численности, приводящие к так называемым геяетико-автома*
тическим процессам (по терминологии советских авторов) или
генетическому дрейфу (по терминологии Райта—Фишера).
Эти явления также носят в значительной степени
стохастический характер, если оставить в стороне причины колебаний
численности, которые чаще всего определяются в природе
экологическими факторами.
Глава 4
ИЗМЕНЕНИЯ ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ
ПОПУЛЯЦИЙ
И ОПРЕДЕЛЯЮЩИЕ ИХ ФАКТОРЫ
В предыдущей главе было показано, что в панмиктической
популяции сохраняется стационарное соотношение между
фенотипами и генотипами в соответствии с формулой Харди —
Вейнберга, если только нет никаких внешних сил, внешних по
отношению к популяции факторов, нарушающих это
соотношение. Был дан и перечень таких факторов. Первый из них
и самый существенный — отбор.
Отбор как причина сдвигов в частотах генов и в
соотношении между генотипами. Под отбором в классическом смысле
слова обычно понимается полное или частичное устранение
ьакой-то группы особей от размножения, т. е. от образования
следующего поколения.
В действительности отбор — это суммарный результат ряда
процессов. Какие-то особи или группы особей могут
обладать пониженной по сравнению с другими особями
жизнеспособностью. Они чаще погибают, а в крайних случаях
вообще устраняются из популяции. Поэтому и потомства от них
будет меньше или совсем не будет. Очевидно, не оставят
потомства и те особи или группы особей, которые погибнут по
тем или иным причинам до достижения половой зрелости.
Особи или их группы могут отличаться и по плодовитости.
Более плодовитые дадут больше потомства, менее
плодовитые—меньше. Некоторые вообще не дадут потомства.
Таким образом, несмотря на самые разные причины,
определяющие конечные результаты, эффект отбора сводится к
большей или меньшей вероятности дать потомство.
При отсутствии отбора каждая особь популяции имеет
одинаковые шансы дать потомство. Фактические различия
ь степени размножения отдельных особей, в коэффициенте
размножения существуют и без отбора, но они являются чисто
случайными и, как правило, характеризуются нормальной
кривой распределения. Если же группа особей, генотипически
отличающаяся от других, устраняется от размножения, то на
структуру будущего поколения окажет влис,чие только
оставшаяся часть популяции. Очевидно, что есл! пчяется часть
I2K
гомозигот А2А2, то оставшаяся часть этой группы в общем
размножении особей популяции будет меньше той доли,
которую занимала группа А2А2 до отбора. Отсюда следует, что
процесс отбора должен неизбежно повлиять на частоты генов
в следующем поколении и привести к изменению соотношения
генотипов и фенотипов в популяции. При отборе, как правило,
в популяции очень долго сохраняются те же гены и их
аллели, которые были в ней до отбора, но меняется их частота,
кроме случаев появления мутаций. Мутации могут быть
подхвачены отбором и таким образом войти, как будет показано
ниже, в генетическую структуру популяции.
Изменения, достигнутые в результате отбора, должны
сохраняться и после прекращения отбора. Однако возможны
и исключения, когда после прекращения отбора происходит
возврат к тому положению в популяции, которое
существовало до начала отбора. Они будут разобраны особо.
Отбор по одному гену. Анализ процесса отбора и его
результатов целесообразно провести, начиная с простейшей
модели, когда расщепление в популяции происходит только по
двум аллелям одного гена: А\ и А2*. Допустим, что отбор
идет против гена А2. Это значит, что особи А\АХ и А\А2
обладают каким-то преимуществом в условиях природы или при
разведении их человеком по сравнению с особями А2А2. В
первом случае можно говорить о сравнительной адаптивной
ценности, а при искусственном отборе — о селекционной, или раз-
ьеденческой ценности А\А\ или А\А2 по сравнению с А2А2.
В одной из последующих глав вопрос об адаптивной или
селекционной ценности будет разобран подробнее. Сейчас же
укажем на следующее.
Для каждого генотипа, точнее, для каждой группы особей
определенного генотипа может быть определена адаптивная
или селекционная ценность по сравнению со средней
адаптивной или селекционной ценностью всей популяции.
Эта ценность представляет собой суммарный результат
указанных выи4е многих факторов, включая жизнеспособность
и выживаемость в конкретных условиях среды, плодовитость
и способность дать здоровое потомство и т. д. По всем этим
свойствам варьируют не только особи разных, но и особи
сходных генотипов. Тем самым каждая отдельная особь или
группа особей получает больший или меньший шанс устоять
в условиях борьбы за существование в природе и дать
следующее поколение.
При разведении животных и растении селекционную
ценность определяет селекционер. Он устраняет от размножения
* В этой н ~ • юшнх главах для пары аллелей одного гена будут
применяться о( оозначения Ах и А2 вместо А и а.
У П. Ф. PokuukiiA
129
нежелательные дли него формы и сохраняет только те,
которые считает полезными, лаже если последние обладают мень*
шей выживаемостью или плодовитостью, качествами совер.
шенно необходимыми в условиях естественного отбора в
природе.
Процесс отбора и конечном счете имеет статистическую
природу, хотя и сложную, основанную на нескольких
статистических процессах. Поэтому и результаты отбора могут быть
оценены только статистически, как суммарный итог того
факта, что в силу разной адаптивной пли селекционной ценности
от одних генотипов может остаться в среднем больше
потомков, а от других меньше.
При расщеплении в популяции по одной паре генов А\ н А2
и возникновении трех генотипов А\Аи АХА2 и А2А2
равновесное соотношение между ними будет существовать только при
условии одинаковой селекционной или адаптивной ценности
всех трех генотипов. Если это не так, то неизбежен процесс
отбора. Он может затронуть только одну генотипическую
категорию или две в зависимости от того, какова
относительная селекционная ценность трех генотипов, а также каковы
доминантные отношения между генами А\ и А2.
Простейший случай — полное доминирование А\ над А2.
Тогда особи генотипов А\АХ и А\А2 обладают одинаковой
селекционной ценностью, а особи генотипа А2А2, если отбор идет
против А2А2, меньшей, нежели селекционная ценность А\А\
и А\А2. Возможные различия в селекционной ценности в
зависимости от доминирования могут быть представлены
следующим образом:
Хдраьтер домиииромння
Пал нос
Отсутствие доминирования
Сверхдоминироваиие
Селекционная ценность генотипов
большая
AxAt
A,At
средняя
АхАг
АХАХ
меньшая
А%Аг
А%Аг
При отсутствии доминирования, т. е. когда гетерозигота
А\А% занимает строго промежуточное положение между
гомозиготами А{АХ и А2А2, наименьшая селекционная ценность
характеризует формы А2А2% так как отбор идет против А2%
наибольшая— формы А\А\% 1етерозиготы же занимают по
селекционной ценности среднее положение. Более сложный
случай - сверхдоминнрование, когда наибольшей селекционной
ценностью характеризуются гетерозиготы АХА2.
Коэффициент отбора. Проще всего оценить результат
отбора по степени уменьшения количества гамет, образуемых
той генотнпической группой, против которой направлен отбор,
по сравнению с другими генотипическими группами. Это
уменьшение обозначают символом s и называют
коэффициентом отбора. Если селекционную ценность предпочитаемой при
отборе генотнпической группы считать единицей, то и число
гамет, ею образуемых, можно обозначить единицей. Тогда
селекционная ценность группы, против которой идет отбор,
например А2А2у может быть выражена величиной 1—s.
Конкретно это значит, что если коэффициент отбора s равен единице.
то группа А2А2 полностью устраняется из размножения,
потомства она не даст вовсе, при 5 = 0,5 — на 50%, при s=0,25 —
на одну четверть и т. д.
С первого взгляда может показаться, что оценка
результатов отбора по количеству образуемых гамет представляет
собой нечто искусственное и лучше было бы судить об отборе по
фактической гибели части или доли особей определенного
генотипа. Но конечным результатом такой гибели явится
уменьшение доли потомства от них. Это равнозначно тому, что
особи данного генотипа, против которого идет отбор, дадут
меньше гамет. Таким образом, уменьшение доли гамет
представляет собой итоговый результат, в котором объединяются
воедино все возможные причины действия отбора: физическая
гибель части особей от самых различных причин в природе
или в результате отбраковки при селекции в животноводстве
и растениеводстве, уменьшение плодовитости вплоть до
бесплодия отдельных особей и т. д.
Так как при отсутствии доминирования гетерозиготы /М2
занимают среднее положение по селекционной ценности
между АХАХ и А2А2> а для А2А2 s=l, то для АХА2 s=0,5.
Соотношение между селекционными ценностями трех
генотипов А{А\, А\А2 и А2А2 можно записать в более общем виде
для любой степени доминирования, если обозначить символом
h степень доминирования, отражающуюся в уменьшении
плодовитости гетерознгот. Оно будет следующим:
l:(l-/is):(l-s).
При полном доминировании Л^О, при отсутствии
доминирования /i = 0,5. Чтобы не было недоразумений, надо
оговорить, что иногда эту последнюю форму доминирования
определяют как промежуточное доминирование.
Изменение частоты гена за одно поколение при различных
случаях отбора. Полное доминирование. Для выяснения
результатов отбора надо рассмотреть, как изменится вклад
разных генотипов и общий генофонд для образования следующе-
ю поколения при коэффициенте отбора s и полном
доминировании.
Рассмотрим простейший случай, когда и популяции проис-
холнт пягшенление но одной паре генов Ах и А2 в соответствии
с Формуюй Харли-Вейнберга и ген Ах полностью доминирует
над л/Коэффициент отбора — .v. Вклад разных генотипов
н общий генофонд для образования следующего поколения
можно рассчитать таким образом:
Исходные генотипы ЛХЛХ АХА% А2А2 Сумма
Их доли в популяции р2 2pq q2 I
Селекционная ценность ] 1 1 — s —
Вклад о генофонд для г/фа-
мваин* след>юшего поколения /И 2/н/ Я2 0 —«) 1 — *</*
Так как селекционная ценность форм А2А2 ниже, чем у
остальных групп (на s), то и вклад их в общий генофонд ниже
на величину sq2. Поэтому общая сумма гамет будет не 1,
а только I— sq2. Чтобы установить новые соотношения
генотипов следующего поколения, выраженные в частотах генов р,
и <7i, надо сначала определить их новые доли, имея в виду их
общую сумму 1— sq'2. Тогда
Генотипы
Прежние доли
Вклад в обшнн генофонд
Новые доли
Mi
Рг
Р3
Р2
\~sq*
АХА%
2pq
2pq
2pq
1 - sq2
A%At
Я2
q*(\-~s)
Сумма
!
I — sg*
i
i
Чтобы определить новое значение Ци можно
воспользоваться одной из приведенных в предыдущей главе формул,
связывающих доли генотипов Р, 2Q, R с частотами генов р и q или
pi и qu а именно: P+Q=Pt и Q+R=qit Тогда
Удобнее заменить р на \—q:
(\-q)q+qz-sq* _ q-sq* q(\—sq)
Яг-
\-sqt \—sq* \—sqi
За одно поколение отбора при коэффициенте отбора s
частоте гена Аг в популяции изменилась от q к qt. Это изменение
частоты геиа, обозначаемое через A q, можно определить
следующим образом:
1~*^
(О
Знак минус перед окончательным результатом указывает
то, что частота гена А2 уменьшилась. Далее видно, что
окончательное значение Д</ зависит от коэффициента отбора $
„от первоначальных значений р и q, или, проще, от
первоначального значения q, так как р н q связаны равенством
Таким образом, нами рассмотрен один частный случай
отбора—отбор против А2А2 (при условии полного
доминирования А\ над Лз). Но очевидно, что направления отбора мот
быть различными — против А\АХ или против А2А\ и, далее,
более сложная ситуация, когда отбор идет одновременно против
двух форм с разными коэффициентами отбора $\ и Si и
благоприятствует только одному генотипу. Различные типы
отбора н зависимости от его направления показаны в табл, 18.
Разобранный случай отбора обозначен в таблице под Л* 1а.
Направления отбора № 1 и 3 осуществляются в двух вариантах:
при полном доминировании — 1а и 36 и при отсутствии
доминирования — 16 и За. Следует обратить внимание на различия
в селекционной ценности разных генотипов в связи с
характером доминирования при отборе против А2А2 и А\А\.
Результаты отбора во всех случаях можно рассчитать тем же методом,
Таблица 18. Типы отбора в зависимости от его направления и семщиоши*
(адаптивная) ценность разных генотипов при рамнчмом
характере доминирования. Ковффяциеит отбора $
Номер
°
1 ,й
1 Н)
2
За
36
4
Г,
6
Направление отбора
Отбора нет
Отбор против АгА2
» » Аг
» » АХА2
» » АХАХ
» * Ах
Отбор благоприятствует
А}АХ и идет против АхАг и
А2А2
Отбор благоприятствует
Л,Л, и идет против АХА% и
\А2А2
()тбор благоприятствует
А~А2 и идет против АХАХ и
\АХА2
Генотипы я их
относительна* селекционная
ценность
/Mi
1
1
1
I
J -S
1 — S
1
II-"
/М»
1
l-0,5f
1 -S
1
l-s
1
/М*
1
I-S
J
1
)
1-4
l-h
1
Характер
допенпро-
ваяна
полное
отсутствует
>
>
полное
отсутствует
сверхдоми-
ннровавле
I отсутствует
ш
кочории был применен дли случая 1а. Но на некоторых из них
целесообразно остановиться подробнее, а именно на случае
16, когда отбор идет против Ао при отсутствии доминирования
ото значит, что он захватит не только особей А2А2, но и гсте-
розигот А\А2). и на случае 5, когда отбор идет против обеих
гомозигот в пользу гетерозпгот при сверхдоминировании.
Изменение частоты гена при отборе в отсутствие
доминирования. Если при полном доминировании А\ изменяется
селекционная ценность только одного генотипа А2А2 и колнчест-
lo образуемых им гамет становится равным 1—s вместо 1,
то при отсутствии доминирования формы генотипа А{А2 по
своей селекционной ценности будут находиться точно
посредине между Л|Л, и А2А2. Отбор будет действовать и против А\А2%
но с интенсивностью вдвое меньшей — коэффициент селекции
0.5 s *. Вклад группы АХА2 в генофонд составит 2pq (1—0,5s).
Чтобы установить, каков будет в таком случае суммарный
генофонд следующего поколения, надо сложить вклад в него
трех генотипов, а именно:
p* + 2pq(\— 0,5s)+?2(l— s)=p2 + 2pq—pqs + q2—q2s =
= p2 + 2pq + q2—pqs—q2s.
Так как p2 + 2pq + q2= 1, то это выражение примет более
простой вид:
l—pqs—q2s.
Есть смысл заменить р на \—q, и тогда суммарный
генофонд можно записать в следующем виде:
1 — (1 —q) sq—sq2 = 1 — sq.
Напомним, что в случае 1а табл. 18 он выглядел иначе:
l—sq2.
Новые доли генотипов можно получить с помощью
расчета, аналогичного выполненному выше:
Генотипы
Прежние доли
Селекционная ценность
Вклад в генофонд для
образования следующего
поколения
Новые доли
АгАг
Р2
1
Рг
Р2
1 — sq
А\А2
2pq
1 —0,6s
2pq(\— 0,5s)
2pq(\—0t5s)
1 — sq
f\yj\ 2
Я2
1 —s
<72(l-*)
g2(l-s)
1 —sq
Сумма
1
—
1 — sq
* Этот случай кажется наиболее вероятным. Однако возможно, что и
при отсутствии доминирования коэффициент селекции не будет равен точно
половине s для гомозигот А2А2. Тогда расчеты надо вести по типу отбора,
обозначенному в табл. 18 под № 4.
134
\-sq "♦" \~Sq '
\—sq ^ i_Sfl 4=
= &-^b£qJ+A\-42s-4+№ PJ-0,bpqs+q*-q
l~s4 • - ■ - - -_-^ .
Заменим р на 1— q, откроем скобки и произведем некоторые
сокращения, и тогда
Таким образом, для случая la Aq = — }-, а для
1|Р . 0,bsq(\-q) 1~*4*
сл\чая 16 Л а = .
\—sq
Интересно сопоставить эти два значения Л qy поскольку они
относятся к одному и тому же виду отбора, но различаются
только полным или неполным доминированием Ai no
отношению кА2. Тогда
Подставляя в правую часть разные значения q при одном
и том же значении 5, можно обнаружить, насколько различны
будут кривые изменения Д<7 в зависимости от характера
доминирования (рис.32—33).
Так как значение Д<7, как это будет видно в дальнейшем.
служит важнейшей величиной для характеристики процесса
отбора, есть смысл выразить его более простым,
приближенным образом. Для этого следует обратить внимание на
структуру знаменателя.
В обеих приведенных формулах для Д</ в знаменателях
\—sq или 1—sq2. Так как обычно и s и q меньше единицы, то
их произведение всегда очень малая величина. Тогда для
оценки приближенного значения Д? достаточно
воспользоваться значением числителя, т. е.
при отсутствии доминирования Д(/= ±0,5s?(l— q)\
при полном доминировании Д (/= ±s<72(l— q).
Знаки « + » и «—> относятся к двум противоположным
направлениям отбора: в пользу /Ма ( + ) или против А^А2 (—).
Знак + будет относиться к направлениям отбора,
обозначенным в табл. 18 № За и 36, т. е. если он идет против АХАХ или
Аг и в пользу А2А2.
135
Отбор при сверхдоминировании. Этот случай особенно
интересен и на него в последнее время обращается большое
внимание В табл. 18 он фигурирует под № 5. Селекционным
преимуществом обладают гетерознготы, и отбор идет против
обеих "гомозигот, хотя и с разными коэффициентами s{ и s2.
Все расчеты можно провести так же, как и ранее, хотя они
окажутся несколько сложнее. Опять начнем с исходного
поколения (до отбора):
Исходные генотипы
Их дат и
Селекционная ценность
Вклад в генофонд для
образования следующего
поколения
ЛИ,
р>
l-s.
?s0-si)
А\А%
2pq
1
2pq
2"* 2
Я2
1 —s2
<72(l-s,)
Сумма
1
—
i-pht-<^
Новая сумма включает уже три величины, п в ней
участвуют два коэффициента отбора s\ и s2. Тогда новые доли по
трем исходным генотипам будут следующими:
AiAi А{А2 А2А2
_j4hzsA__ 2p<7 qz{\— s2)
1-/7=51-^2 1—p2Si—q2s2 1 — p2si—q2s2"
Чтобы определить новое значение qir снова воспользуемся
одной из приведенных выше формул, связывающих доли
генотипов с частотами генов: q{=Q+Rm Тогда
*-Q+*« рЛ + W-** =
= ii±_^Z^!i2 (\-q)q+q2-q2s2 q—q2s2
i—p-sx—qis2 \—p2Si—qZS2 i_p2Si_^2S2-
Отсюда мы приходим к значению Д q:
л <?= ...Jz£?i__ _fl== q-q2s2-q+p2q$i+q3sz =
= rrfsi+PlSj+fH _ —^s»+P*gg|+ (1 —P) «fog _
I— p-si—q2sz ~ \—p2st—q2Sz ~~
I —p*st—q42 1 —p2Si—q2s2 '
(3)
jTo6u не усложнять формулу, не будем р преобразовывать
ч »то ^а1Цает вн"мание то обстоятельство, что знак при ЬЯ
m слУчае «плюс», т. е. несмотря на отбор против формы
0,000
^ концентрация гена А2 в популяции повышается, хотя,
как*мы увидим дальше, этот процесс не идет до конца, т. е.
до 100%-ной концентрации гена Л2, а останавливается на ка-
ком-то уровне
Изменение Aq как показатель эффективности отбора.
Значение \qt как видно из разобранных выше случаев отбора,
зависит, с одной стороны,
от исходной величины q A(t\
(или р), а с другой — от °^
коэффициента отбора s,
выражающего его
интенсивность. Чтобы не
усложнять картины, можно
принять для расчетов какое-
то определенное
значение s и установить, как
будет меняться Aq в
зависимости от величины q.
При этом, очевидно,
следует учитывать и
характер доминирования.
На рис. 32 кривая а
показывает изменение Aq
при отборе против А2А2
(доминирование полное,
что соответствует случаю
1а табл. 18) при разных
значениях частоты гена q, а кривая б — подобное же
изменение Aq при отборе против Лг-, т. е. когда доминирование
отсутствует (случай 16 табл. 18). В обоих случаях s=0,2.
Очевидно сходство обеих кривых. При малых и больших
значениях исходного значения q значения Aq очень малы. Они
выше при средних значениях q. Мы не приводим графиков для
случаев отбора в противоположном направлении в пользу
А\А\ или А г (случай За и 36), так как они очень близки
к изображенным на рис. 32. Наблюдается некоторое различие
между кривыми в зависимости от характера доминирования:
при полном доминировании максимальные сдвиги в Aq при
значениях д = 0,6—0,7—0,8, а при отсутствии доминирования —
при <7, близких к 0,5.
При s = 0,2 сдвиги Aq за одно поколение очень малы
\максимум 0,028—0,032, а в большинстве случаев еще ниже).
Однако с повышением коэффициента отбора значения ±q
окажутся более высокими. На рис. 33 показаны изменения Aq
при 5 = 0,9. Aq достигает величины 0,25—0,28, при этом при
отсутствии доминирования уже не наблюдается
куполообразная кривая рис. 33, б, а она сдвинута вправо, максимум Д</прн
О 0J 0,2 0.3 Cfi Ofi Сб С? Цд OS !fil
Рис. 32. Изменение ^ при различных
значениях q:
а — при отборе против /Ы:. Домвняровакке
полное; б — прп отборе против Ах—. Адомн-
кнровакне.
137
п 11ГЛ1П О 7-ОА после чего происходит резкое па-
значениях « okwo . ута крпвая При полном доминиро-
ДеНМ! Няп^тание значения Лд сначала происходит медленно,
п™ю^?Ф™ ЧеМ 0'25) Д0СТИГЭеТСЯ В Тех **»«*.
К°ГНо л^^т^ом высоком коэффициенте отбора Л//
остается веянной сравнительно небольшой. Практически чаще
ется всшчпп г всего приходится
встречаться с более низкими
коэффициентами отбора, при
которых, очевидно, для
значительного уменьшения q (и
соответственно увеличения р)
необходимо много
поколений отбора. Это
демонстрирует рис. 34, построенный на
основе данных рис. 32,6
"п (при 5 = 0.2 и отсутствии
доминирования). Взято
первоначально высокое нсходное
значение q гена Ач (^0,95).
При отборе против А г- для
изменения q от уровня выше
0,9 до уровня 0,8 нужно
около 14 поколений. Несколько быстрее происходит падение q при
средних его значениях. Однако, когда значение q становится
ниже 0.2, дальнейшее снижение q происходит опять очень
медленно. Если частота рецессивного аллеля очень мала, отбор
малоэффективен. Ген А2 представлен преимущественно
> гетерознгот, которые сохраняются в популяции много поко-
Рис 33. Изменение Sq ори различных
звачелгах q ш при высоком
коэффициенте отбора (5=0.9):
б — ядомвнжро-
р« 3* I
I*
Ю 20 30
Пшмления
40
'^1^ш^^Я § "*• . р<* » Иэмеяешк жои» >Mi ■ £
пулядии ПрИ длительном от***
прошв АЙ*
аяй Интересно проследить, как это сказывается на конкрет-
• стрУктУРе П0ПУЛЯ1*ИИ- Иа РИС- 35 представлено изменение
И1,йт0Ты Ф» т- е- доли ген°типов A7A2 (при полном доминиро-
чЗС „ 5 = 0,2). Процесс отбора начался в популяции, состо-
5а"-( преимущественно из гомозигот А2Аг (свыше 90%). Па-
яВ е иХ частоты вначале происходит довольно быстро. Одна-
Дб после того как их доля снижается до 0,16 (что соответ-
-твует значению q, равному 0.4), процесс резко замедляется,
\бор протекает очень медленно и значение ф изменяется в
каждом поколении на ничтожную величину. На сдвиг же от
1^0,95 до q2=0,16 понадобится окаю 30 покалений.
До сих пор результаты отбора оценивались по сдвиг)' в
величине q. т. е. по Aq. Но, очевидно, уменьшение q есть в то же
время увеличение р. Так что можно было бы пользоваться Др.
Селекционный эффект Др может быть определен как
увеличение частоты р за одно покаление.
Выше были разобраны только некоторые случаи отбора
н приведены для них значения Aq. В табл. 19 дана сводка
значений Д<7 для всех типов отбора, указанных в табл. 18.
Средняя приспособленность (fitness) популяции. В силу
происходящего в популяции отбора устраняются менее
приспособленные формы (илн нежелательные при селекции,
проводимой человеком) и создаетсфв каждом поколении новое
соотношение фенотипов и генотипов на основе новых значений
Pi для А\ и q{ для А2. Эти соотношения, вообще говоря, очевь
различны в зависимости от типа отбора и характера межал-
лельного взаимодействия, т. е. типа доминирования Но онн
з каждом случае характеризуют некоторую среднюю
приспособленность (fitness) популяции. Обычно ее обозначают
символом ю. В данном случае, очевидно, речь идет о приложении
этого понятия к анализу изменений в популяции при одной
паре генов.
Если для каждого из генотипов популяции А\А\Щ dИз,.4*4*
обладающих частотами /\ 2Q и /?, принять коэффициенты
приспособленности а, р, у, то средняя приспособленность
популяции
w**aP+$Q+yR. (Hi
Но поскольку, как было показано в предыдущей главе,
пРи наличии панмиксин в популяции Р**р?; 2Q**2pq\ /?^<т*,то
коэффициент средней приспособленности, выраженный в ча-
Стотах аллелей, будет иметь вид
a>«o/>*+20w+w*- (46>
Учитывая, что p+q-\. можно заменить в этой формуле
качение р на 1— q или q на I—л что даст возможность уста-
139
Ttf.mw 19. Частого генов я мх шмгнгпмя при отторг (номгра соответствуют типам отбора, покаюннмм а тобл. \к)
Частоты ггмои (поелг I поколения отоорд)
Номер/
О
1а
At
А»
п
р
I — sq*
P* + pq (1-0,55)
1 — sq
P — spq
1 — 2spq
P — sp*
1 — sp*
p* + 2pq — 2spq
1 — sp* — 2sp?
P — SiP<7
I — 2sxpq — s27a
P — S\P2
1 — s,pa — s2q*
P — S|P* — s&q
1 — stp« — 2s#q
Я~*Я*
1 — sq*
p<7(l-Ot5s) + <7»(l-s)
1 — s</
я — *ря
1 — 2$р?
J — sp*
Я — spq
1 — spa — 2sp?
<7 — *.P? ~ *2<72
1 — 2sxpq — s2</2
У — ЭД»
1 — s,p3 — s^*
9 — *aP9
1 — sxp* — 2sjP7
A*
0
V(l-9)
I —s<7«
0,5*7(1-9)
1 — s?
sp7(2iy— 1)
1 — 2spq
sp*q
1 — sp*
p<7 (ps — s + 2gs)
1 — sp2 — 2sp7
SiPq (Я — p) — s2pq*
1 — 2^р7 — .v/a
p? fop — sa<7)
1 — Slp* — s2q*
*iP29 + 52p9 (? — p)
1 — SiP3 — 2s2pq
Частоты i снов, при которых
достигался рлнновгеие (/> ,
СООТПГТСТИГННО р)
Рп Р
Рп~ I
Рп ^ I
рл-0,5
Рл-0
Р/1-0
я.
1
Рл
Яп
— Рп
Я„
"Яп
— Яп
Рп
So
новить. как меняется значение ш в зависимости от изменений
п пли Я-
Y Для проведения анализа удобнее принять значение
приспособленности одного из генотипов, например А\А\Ч за единицу
(<х=1), как это, в частности, было принято выше при анализе
отбора.
Рис. 35. Зависимость средней приспособленности w от
частоты гена Ах при разных отношениях приспособленности
генотипов АХА2 и А2А2 (по Тимофееву-Ресовскому я Свире-
жеву). Объяснение в тексте.
Н. В. Тимофеевым-Ресовским и Ю. М. Свирежевым были
исследованы изменения w в зависимости от значений р при
трех вариантах соотношений между р и у (во всех случаях
а=1):
1) р>1; p>Y;
2) р<1; p<Y;
3) Y<P<1-
Графики для всех трех случаев показаны на рис. 36. В
случае I имеет место гетерозиготный полиморфизм с частотой
аллеля А\ч равной
P-Y
Р*=
2р—Y—I '
При этом и значение w максимально.
В случае II равновесие в популяции неустойчиво. Величина
Рх сдвигается или в сторону р-*Ь> или в сторону />-*К так как
при значении р*, показанном на рис. 36, значение w
минимально. .
Наконец, в случае III происходит вытеснение аллеля А*
и популяция стремится к состоянию, когда р-* 1. Л\акснм>м
А-1 я w наступает при />= I.
Адаптивная ценность - это та же приспособленность ш для
каждого генотипа, которая равна 1-s. Тогда р может быть
представлена как J-s,. а у как l-s. (случаи № 4 сводной
табл. 18).
Формула для а\ выраженная в значениях частот генов
н с использованием коэффициентов отбора s, примет вид:
ц-=/?2 + 2(1— s,)pq+(l — s2)q2 =
= p2 + 2pq—2slpq + q2—s2q2 =
=p2+2pq+q2—2stpq—s2g2;
ш=1— 2stpq—s2q2. (4в)
Для Л q с учетом средней приспособленности популяции
ав может быть дано следующее уравнение:
Л,-^-.-^. (5)
2ш dq
Эта формула является наиболее общей для всех случаев
отбора организмов — диплоидов. Но ее можно отнести и к
гаплоидам, и к тетраллоидам и т. д. Достаточно только заменить
множитель 2 в знаменателе на 1 (для гаплоидов), на 4 (для
тетраплоидов) и т. д.
При сцепленных с полом генах коэффициент должен
быть 2/3.
Если значение ад различно для самок и самцов, что
встречается довольно часто, общее w устанавливается как среднее
геометрическое из значений w г; и w jj :
;-У
W = f W -Г Г -w J.-?.
Ниже мы вновь вернемся к понятию приспособленности
при характеристике процесса отбора по суммарному генотипу,
т. е. по многим генам, определяющим количественные
признаки.
Экспериментальные доказательства изменения частоты
гена при отборе. Приведенные теоретические расчеты изменений
частот генов при отборе были подтверждены специальными
опытами на модельных объектах (дрозофила, мыши). Так,
Меррслем была взята в качестве исходной популяция Drosop-
hila roelanogaster. которая состояла из особей нормальных,
а также гомо- и гетерознгот по рецессивному гену raspberry
142
..-малиновая окраска глаз). Частота гена ras
обоих
на-
изменениях фактических
патов) и теоретическая
.„5-Md.innvuur. v..r^..„ »*,«,,,. Td^iura гена ras у оС
,о1ов была равна ОД так что первоначальная популяция
Удилась в равновесном сосгоянин. Отбор велся против ген*
ras, что соответствует случаю (а табл. 18, и результат его был
подробно разобран выше.
На рис. 37 нанесены данные оЬ
частот генов (суммарно для обоих
кривая падения частоты
гена ras, т. е. q. Была сделана
проверка жизнеспособности
мутантных и нормальных
особей и их участия в
скрещиваниях. Различия их
жизнеспособности не были
обнаружены, но размножение
самцов ras несколько
слабее, чем самок. Это
обстоятельство было учтено
при расчете теоретической
кривой. Промежуток между
поколениями принимался за
25 дней.
Совершенно очевидно
большое сходство
эмпирической и теоретической
кривых, а также то, что
падение частоты гена ras,
первоначально значительное,
позднее становится все меньше.
Изменение доли гетерозигот при отборе. Приведенные
выше кривые свидетельствуют, что под действием отбора
популяция изменяется быстрее всего при средних значениях
частот генов (р^9^0,5), хотя на это может влиять к степень
доминирования. При значениях же р или </, приближающихся
к 1,0 или 0, сдвиги за одно покаяние очень малы. Чем больше
интенсивность селекции, измеряемая коэффициентом s, тем
быстрее происходит изменение частот р н q.
Для понимания этого процесса существенна разница в
доле гетерозигот. Если сравнить доли гетерозигот при близких
к 0,5 или при очень малых значениях </, то разница между
долями гетерозигот А{А2 и гомозигот A%A2 оказывается очень
значительной. Так, прн <? = 0,6 рецессиаы А*Аъ составляют
0,36, гетерознготы АХА2 — 0,48 всей популяции, последних
немного больше, чем первых. При 9*0,01 доля гомозигот рецес-
снвов только 0,0001, гетерозигот же — 0,0198, т. е. кашчество
гетерозигот больше количества гомозигот рецесснвое почти
в 200 раз.
fkmSufj
Aw Ю0
Рис. 37. Изменение частоты гада ras
у дрозофилы прн отборе протм вето.
В исходной популяции <7»0,5
(ломаная линия — фактические результаты,
плавная — теоретически вычислен-
ные).
143
Ггторсмнготы псе премя сохраняются и большем
количестве Отсюда ясно, как трудно освободить популяцию от
нежелательных рецессипои. Их как бы охраняет доминирование.
Однако в случае полной летальности рецессивного гена А^А2
скорость его потери будет выше, чем для пелеталыюго гена.
Количество поколений, необходимое для определенного
сдвига в значении д. Попытаемся рассчитать число поколений,
необходимое для сдвига в частоте гена q иа какую-то величину.
Для этого можно воспользоваться одной из приведенных выше
формул для </|, в частности для случая полного доминирования
(случай 1а табл. 1<М:
q(\—sq)
(/t= 1-59* '
Примем, что 5 = I. т. с. что группа AqA2 полностью исключается
из размножения, и предварительно преобразуем эту формулу:
Если обозначить частоты генов для ряда последующих
0. 1, 2 t поколении значками qo, q%4 q-i ?«, то
<7о __ _ <h j/o
ч%~ i+qo> 42-l+q% \+2q0 '
qo _ _ qo
l+3^o l+^o
Отсюда можно вычислить / для перехода от q0 к qt\
,«*=* «J L. (б)
qoqt qt Qo
Приведем пример из генетики человека. Допустим, что ген
альбинизма встречается в конкретной человеческой популяции
с частотой (/=0,007. Тогда </2=0,00005, т- е- ' альбинос на
20 тысяч человек. Желательно снизить эту частоту до <7< = 0,005
(4:«0,000025; один альбинос на 40 тысяч лиц).
В таком случае
'= W " W «200-143-67 (поколений).
При длительности человеческого поколения примерно 25
лет это составит около 1400 лет. Конечно, для видов с
меньшей длительностью поколения время соответственно сокра*
шается, но очевидно, что чем меньше величины </0 и qt, тем
144
дольше нрсмспи потребуется для перехода от q*nqt. Процесс
отбора очень длителен, и 'эффективность его по мере
уменьшения значения q становится все меньше.
Условия прекращения действия отбора. Действие отбора
может прекратиться в нескольких случаях. Наиболее
элементарный, когда неличина s становится равной нулю. Тогда
генетическая структура популяции стабилизируется на тех
уровнях рп и £/м. До которых они были доведены происходившим
ранее отбором. С другой стороны, отбор прекратится, если рп
(или соответственно qn) достигнет 1, хотя, как указывалось
выше, достигнуть таких значений нелегко. Эти случаи будут
иметь место при типах отбора, обозначенных в табл. 19 Л 1а
и 16, За и 36 (последняя графа). Однако отбор против гетеро-
зигот Л|Л2 приведет к равновесию, когда рп достигнет 0,5
(=*</п) *. Более сложная картина будет наблюдаться при
типах отбора, отмеченных в табл. 19 № 4—6.
При типе № 4 отбор благоприятствует А\А\Ч но идет против
А\А2 (с коэффициентом st) и против ЛИг U
коэффициентом 52). В этом случае А <7 будет выражаться более сложной
формулой:
S\pq(q-P)-S2pq*
1— <ls\pq—s2ql
Популяция окажется в равновесии, если Л q станет равной
нулю, а это возможно, если числитель ^tp4(9—Р)—5ipq*=0.
Отсюда S\pq(q-—p)~S2pq1-
s{ a
При преобразовании этого равенства получим: ~—■ =
s2 q~p
(точнее, ~ — = —— —). Сходным образом и для типа отбора
А*2 Чп — Рп
№ 6 получим условие равновесия:
Sj Рп—Чп
Si Рп
Заслуживает особого внимания случай сверхдоминнрова-
иия, отмеченный в таблице под № 5. Отбор идет в пользу гете-
розигот Л Из и против форм А{А{ и A2A2 (с разными
коэффициентами отбора $i и s2). Выше уже было выведено
значение Д q\
А P<l(SiP-~s%q)
• Это равновесие нельзя считать стабильным. Случайные отклоним* от
чего могут постепенно накапливаться, пока, маконеи. или р. или <? не
приблизятся к 0.
145
И) П. ф. р.жпикнП
Чтобы \q было равно нулю, надо, чтобы стал равен нулю
числитель. А так как р и q всегда определенные величины, то
н\лю должен быть равен множитель (s{p—s2q):
Sip—s2q = 0 или s2q = sip;
q Si q Si ( qn st \
и далее — = , или -—- = ( точнее, -—— = —I.
p s^ \—q s2 v I—qn s2 '
Так как надо установить, какова связь между значением qn
и соотношением si и s2% то следует произвести дальнейшие
преобразования уравнения, а именно:
qns2 = Si—qnSi; qns2-{-qnSi = Si\
?n=—-. (7)
Si + S2
Это формула равновесия: при таком соотношении Si и s2 qn
становится величиной постоянной. Она может быть выведена
и из указанного выше уравнения для w — средней
приспособленности популяции, ибо состояние равновесия означает или
то, что при отборе достигнуто максимальное значение ад, или,
если оно не максимальное, что нет возможности изменить его.
Анализируя формулу qns2 = pnS\, легко видеть, что если q
превышает то значение, которое необходимо для сохранения
равновесия (однако все же не равно 1), ар соответственно
меньше требующегося (pnS\<qns2), то &q будет иметь
отрицательный знак. Это значит, что q при отборе в пользу гетерози-
гот А]А2 будет увеличиваться, а р — уменьшаться. Точно так
же, если значение q меньше, чем нужно для сохранения
равновесия, то оно будет увеличиваться. Отбор приведет к
некоторому промежуточному значению q (и соответственно р) со-
i л ясно указанному выше уравнению, и оба аллеля окажутся
и популяции в некотором стабильном соотношении. Это же
ьерно в случае расщепления в популяции по нескольким
аллелям одного гена, если только гетерозиготы по каждой паре
аллелей обладают преимуществом по сравнению с гомозиго-
тами. Из формулы (7) видно, что относительная частота q
(т. с. аллеля А2) при стабильном состоянии популяции
зависит п конечном счете от степени различия в селекционной
ценности Si и s2 обеих групп гомозигот (А\А\ и А2А2). Этот случай
отбора особенно интересен тем, что позволяет яснее понять
явления сбалансированного полиморфизма, изученные по многим
признакам в естественных популяциях, а также в популяциях
сельскохозяйственных животных и человека.
Us
Сбалансированный полиморфизм. Различия по группам
ф0ви У человека и^ по биохимическим свойствам крови и мо-
10ка У сельскохозяйственных животных достаточно хорошо из-
вестны. Л\енее известны, но также важны с общебнологиче-
ской и эволюционной точки зрения данные о полиморфизме
п0 морфологическим признакам и признакам окраски у
бабочек, моллюсков и других животных, о которых говорится во
многих сводках и монографиях.
Наиболее отчетливый пример полиморфизма, который
можно оценить и по частоте генов, и по выживаемости разных ге-
нотнпических групп,— серповидно-клеточная анемия у
человека. Ген серповидно-клеточной анемии приводит к
ненормальному типу гемоглобина. У гомозигот эритроциты имеют форму
серпиков. Многие (большинство) из гомозигот умирают, так
как весь гемоглобин у них ненормального типа. Гетерозиготы
не страдают анемией, но оказалось, что в их кровн можно
обнаружить серповидные эритроциты — до 35%.
Таким образом, в отношении синтеза гемоглобина ген
серповидно-клеточной анемии частично доминантен, в отношении
анемии — рецессивен, а в отношении выживаемости можно
предположить, что он сверхдоминантен. Дело в том, что в
различных областях земного шара, преимущественно в Африке,
процент особей, ненормальных по гемоглобину (это
гетерозиготы и немногие выживающие гомозиготы, обладающие
серповидно-клеточной анемией), иногда исключительно высок —до
40%. При сильной нежизнеспособности гомозигот такой
высокий процент гетерозигот нельзя понять без допущения о
каком-то селективном преимуществе гетерозигот перед
нормальными гомозиготами.
Природа этого преимущества оказалась связанной с
устойчивостью к малярии. Гетерозиготы менее чувствительны к
малярии, нежели нормальные особи-гомозиготы.
Можно привести следующие расчеты некоторых
параметров, характеризующих популяцию по гену
серповидно-клеточной анемии для местностей с высоким процентом (до 40)
ненормальных по гемоглобину людей. Процент гомозигот в этом
случае был определен Эллисоном как 2,9,
Тогда частота гомозигот по данному гену в популяции
составит 0,012, а частота гетерозигот — 0,388. Так как
Q-Q + R, то 9 = 0,206. Если данная частота гена
представляет собой величину, более или менее соответствующую
значению для стационарного состояния популяции, которое со-
сдается в результате отбора, благоприятствующего гетерознго-
Si
там, то при наличии свободного скрещивания q= —-—.
Путем сравнения плодовитости и жизнеспособности можно
установить выживаемость гомозигот по гену
серповидно-клеточной анемии по сравнению с гетерозиготами. Она оказалась
равной 0,25. Отсюда s«=QJ5. Зная s2 it </, можно определить st
исходя из расчета 0,206=—ГК=Г- Тогда ^i=0,197. sx пред-
ставляет собой коэффициент отбора гомозигот А\А\ по
сравнению с гетерозиготами А\Аъ Селекционное преимущество по
отношению к нормальным гомозиготам выразится величиной
I ;(1—s,) = l,24. Гетерознготы устойчивее к малярии на 24%
по сравнению с гомозиготными нормальными особями.
Пример с серповидно-клеточной анемией хорошо
иллюстрирует те силы отбора, которые могут удержать в популяции
генетически различные формы в некотором стабильном
соотношении. Но в го же время он недостаточно типичен для
обычных случаев сбалансированного полиморфизма, так как
коэффициент отбора здесь очень велик и один из генотипов
заведомо ненормален. В большинстве же случаев
полиморфизма нормальны все генотипы и нет стать очевидного
преимущества в отношении выживаемости одного генотипа над
другим. Однако очень слабого преимущества гетерозигот
достаточно, чтобы обеспечить сбалансированную систему
полиморфизма с некоторой промежуточной частотой генов.
Заслуживают внимания многочисленные данные по
биохимическому полиморфизму крупного рогатого скота и других
сельскохозяйственных животных. Так, например, известно, что
различия по трасферинам сыворотки крови крупного рогато-
ю скота определяются несколькими аллелями одного и
того же гена. У большинства европейских пород этих аллелей 3,
а у африканского скота их 5. В большинстве случаев
распределение животных различных генотипов в стадах и породах
довольно точно соответствует формуле Харди—Вейнберга
♦ пример приведен в предыдущей главе). Но иногда
наблюдаются и с>щественные отклонения от ожидаемых отношений,
в частности избыток гетерозигот по некоторым сочетаниям
аллелей.
Были попытки объяснить это явление разными причинами,
например, пониженной эмбриональной смертностью
гетерозигот по сравнению с гомозиготами или более высокой опло-
дотворяемостью или даже несовместимостью плода н матери
при определенных типах скрещиваний.
Однако до сих пор не получены достаточно убедительные
доказательства наличия таких факторов, которые
поддерживали бы сбалансированное состояние биохимического
полиморфизма у сельскохозяйственных животных.
Очень ценны экспериментальные работы, проведенные на
модельных объектах, для изучения причин, создающих устой-
14*
ое состояние при полиморфизме. Так. Ю. М. Овирежез
4 Н В. Тимофеев-Ресовский провели на дрозофиле опыты по
Пределен и ю количественного отношения в искусственно
изданной популяции особей, гомозиготных по гену ebony
Lочень темное тело), гетерозигот по этому гену и нормальных
ррИ различной степени перенаселенности культур.
еЬ л
Выяснилось, что гетерозиготы —- обладали относительно
повышенной жизнеспособностью по сравнению с нормальными
*ф
жизнеспособность
особями —, — у гомозигот же по
резко понижена.
В дальнейших опытах было показано, что в модельной
популяции происходило вытеснение форм ebony, но до
известного предела (рис. 38). Когда количество мушек ebony стало
ниже 10% общего количества особей, оно стабилизировалось
на уровне примерно 8—9%,
т. е. отрицательный отбор
гомозигот ebony
компенсировался положительным отбором
гетерозигот по ebony.
Авторы провели также
анализ генотипнческого
равновесия на математической
модели и вывели ряд уравнений,
связывающих частоты
генотипов с различиями по их
жизнеспособности, которые могут
быть выражены различными
коэффициентами. Были
разобраны разные случаи (большей
жизнеспособности
доминантных форм, рецессивных или
гетерозигот) и показано» при
каком соотношении
коэффициентов жизнеспособности создается устойчивое состояние, а при
каких — неустойчивое. Значения коэффициентов были
определены для конкретного опыта с популяцией,
содержащей три генотипа по гену ebony, в которой наблюдалось
устойчивое состояние полиморфизма. Акторы называют такой
тип полиморфизма «гетерозиготным», сохраняющимся при
постоянных условиях среды.
Однако те же авторы показали наличие другого типа
полиморфизма, названного адаптационным. Он наблюдался
У божьей коровки Adalia bipunctata. Для этого вида
характерна большая вариация по окраске надкрыльев, но основные
0 ЮО 200 Ж 400 500 500
Дни эп н&ак опьтъ
Рис. 38. Уменьшение доли особе*
ebony в модельной популяции
дрозофилы до уровня-&—9% (*>
Свнрежеву н
Тимофееву-Ресовскому). Кружками обоэвячевы
эмпирические данные.
149
^^иы r*v чеоная с красными пятнами и красили с черными
п^намЛоминнруст черная окраска. Оказалось, что у Ad. Ь|.
пГсиТа генотипнческий полиморфизм динамичен: » летний
nmu преимуществом обладают черные формы, а в зимний-
ны I1oi-v изменение частот генов в зимний и летний
пенимы приходится описывать разными уравнениями. Наблю-
I2JS? некая фазовая картина, когда система популяции со-
веошает колебания по годам с периодом, равным году. Коле-
"линя в разные годы неодинаковы по размаху, но они происхо-
лвт вокруг некоторого центра, который также медленно
смешается, по-видимому, в результате более глубоких измене-
нин климата от года к году.
Таким образом, проблема сбалансированного
полиморфизма даже создающегося за счет отбора, оказывается более
1-лиАной. чем это кажется с первого взгляда.
В более поздней работе Ю. М. Свирежева и Н. В.
Тимофеева-Ресовского разобран теоретически и экспериментально
епчай рецессивной, сцепленной с полом мутации Саналнзиро-
аалось поведение мутации eversac у Drosophila funebris).
Определены условия как для вытеснения того или иного алле-
ля. так и для создания некоторого состояния полиморфизма,
характеризуемого определенным соотношением коэффициентов
относительной жизнеспособности трех генотипов. При этом
авторы указывают, что сцепление с полом повышает
устойчивость полиморфизма, расширяя его область.
Наряду с анализом конкретной модели, проведенным
Тимофеевым-Ресовским и Свирежевым, есть ряд работ, в которых
рассматривались различные частные случаи сбаланенрованно-
ю полиморфизма: при отборе в гапло- и днплофазе, лрн
различии в коэффициентах отбора у разных полов, при изменении
приспособленности особей разных генотипов в зависимости
от их частот и т. д.
В ряде работ, особенно из школы Ф. Добжанского.
рассматривается еще одна категория полиморфизма —
хромосомный полиморфизм, обеспечивающий приспособление
популяции к меняющимся условиям среды. Выделяют также
инверсионный полиморфизм, т. е. полиморфизм по инверсиям хромо-
юи Обычно такие инверсии находятся в гетерозиготном
состоянии. Однако эту тему как специальную мы рассматривать
не будем
Среди возможных причин полиморфизма следует иметь
* виду и неоднородность условий среды, в которых живет
популяция, в силу чего в одних частях ареала вида или
популяции отбор может благоприятствовать одному ал л ел ю, а в дру-
1нх —другому.
Гетерлиготиы! полиморфизм и моногибридный гетерозис.
'*н iia вопроса тесно связаны друг с другом, так как в обоих
1Учаях им^тся сверхдоминирование определенных генов при-
ио>цее к их сохРаиенню в С^У преимущества гетероэигот
' процессе отбора и гетершисного эффекта. Работами
Ч К Беляева и его сотрудников было доказано наличие моно-
гибрндного гетерозиса у пушных зверей. Накапливаются
данные и по другим объектам.
Таким образом, можно сделать вывод, что рассмотренный
выше случаи отбора в пользу гетерозигот очень существен не
только для теории отбора, но и для важных проблем
современной генетики.
Ниже будет показано, что в создании того или другого
равновесного состояния популяции имеют значение и другие
факторы, изменяющие ее структуру. При этом надо помнить.
что если сбалансированный полиморфизм относится к
категории устойчивого равновесного состояния, то возможно
состояние неустойчивое и частично устойчивое. Во многих работах
рассматривались математически различные условия для
большей или меньшей устойчивости популяции.
Миграция, эмиграция и иммиграция. Под общим термином
«миграция», означающим в точном смысле передвижение,
объединяются два рода явлений: эмиграция и иммиграция.
Эмиграция — естественный уход из данной популяции илн
искусственный вывод из нее какой-то части особен, проводимый
человеком. Иммиграция же —наоборот, вхождение или
внесение в данную популяцию какой-то группы особей из другой
популяции.
Эмиграция и отбор. Если эмиграция происходит случайно
по отношению к отдельным особям, составляющим популяцию.
она не окажет влияния на генетическую структуру
популяции *. Если же эмиграция носит неслучайный характер, т. е.
из популяции эмигрируют особи определенного генотипа, то
оставшаяся часть популяции будет генотипически иной, а
действие такой эмиграции равносильно действию отбора.
Иммиграция — процесс, обратный по сравнению с
эмиграцией: в исходную популяцию включаются особи из других
популяций. В природе такие явления наблюдаются при сильны^
передвижениях особей вида из одной местности в другую.
и которой уже имеются представители данного вида. Иногда
такой перенос в условиях природы производится и человеком.
Например, в Сибири перебрасывали соболей очень темной
окраски — баргузннскнх — для улучшения местных популяций
соболей. Передвижение и включение новых групп в прежние
популяции наблюдается и в человеческих популяциях.
* Однако, если численность популяции ьеледстьие энягрлцяы рез*о
уменьшится, п ней могут проявиться особые процессы, косящие н&эмпк
генетического дрейфа. Они будут рассмотрены няж*
151
Влияние эмиграции и иммиграции на структуру популяций
по большому количеству генов, воздействующих на сложные
признаки, прежде всего количественные, будет рассмотрено
в последующих главах. Здесь же мы ограничимся лишь
рассмотрением влияния иммиграции на простейшую модель
популяции, в которой идет расщепление по одной паре аллелей.
11 в этом случае анализ результатов иммиграции лучше всего
провести путем учета сдвига в частоте гена за одно поколение,
т. е. установления величины Sq.
Допустим, что в основной популяции частота какого-то
гена была до. В группе, вносимой в нее. иммигрируемой,
частота гена qm. Нужно также учесть объем иммигрирующей
популяции и исходной. Если принять суммарный объем за единицу,
а вносимую часть ее за /л, то основная часть популяции (без
иммиграции) составляет 1—ш.
В таком случае частота гена в смешанной популяции
может быть выражена как q\ = tnqm + (1—m)qo=mqm+q0—mq0.
Отсюда Sq = q\~-qo = mqm+q0---niqo--qo = m(qm—qo). (8a)
Этой формуле путем соответствующих преобразований можно
придать и более сложный вид:
Sq = т (рМт—РтЧо) - (86)
Значение \q зависит от трех величин: а) от того, каково
соотношение размеров иммигрируемой группы и размеров
популяции, в которую включается эта группа, т. е. от величины
1:ш; б) от значений ро и <7о в первоначальной популяции;
в) от значений рт и qm в иммигрируемой группе.
На этой основе можно рассчитать, как будут меняться
значения рп и qn в смешанной популяции, т. е. можно
охарактеризовать то, что в литературе называют потоком или течением
генов (gene flow) за ряд поколений.
Очевидно, что чем меньше первоначальная разница между
значениями q в исходной популяции и в иммигрируемой
группе, тем меньше Д</.
Если известны частоты генов в исходной и смешанной
популяциях, то можно определить т —долю иммигрируемой
группы а смешанной популяции, по следующей формуле:
тв10о(|--2=^5.)§ (9)
где </о —частота гена в исходной популяции, qn — в
смешанной. Цт— в иммигрируемой.
Была сделана попытка по этой формуле путем изучения
частот одного из аллелей крови (AgN) установить, какую
долю составляют русские в эскимосском населении Алеутских
1И
оовов. В этом случае qn=0,181 (для смешанной популя-
с2 <7о=0*087 <для ^кимосов) и <jw~0,417 <для русских).
рачение m (в процентах) равно 28,5.
j Впрочем, это только пример для вычислений, так как ис-
ч0дные данные очень неточны.
Наконец, можно связать число поколений, в течение
которых проходила иммиграция, с изменениями частот генов с
подошью формулы
(1-т)*=-^=^, (10)
где п — число поколений.
С каждым поколением разница между </о и qm уменьшается
благодаря «потоку генов».
Гласе сделал попытку охарактеризовать этот процесс путем
сравнения популяции негров и белых в США по одному из
генов резус-фактора. Исходные данные были следующими:
<7n=0,599; qn = 0,433; gm=0,028. Отсюда были получены
ш=3,4°/о и п=10.
Таким образом, при анализе процессов отбора в
популяциях надо учитывать возможные потоки генов между соседними
смешивающимися популяциями, которые могут изменить
первоначальную структуру популяции.
Роль мутаций в изменении структуры популяций. В начале
главы разбирался вопрос об изменении генетической
структуры популяций в результате отбора, при этом принималось
наличие в популяции определенных генов как постоянных.
В действительности же в любой популяции животных и
растений постоянно возникают мутации, которые самим своим
появлением влияют на структуру популяции. Однако роль
мутаций в изменении структуры популяций имеет несколько
сторон: первая из них заключается в самом факте появления
мутаций; вторая — в насыщении популяции мутациями; третья—
во включении мутантных форм в процесс отбора и, наконец.
четвертая — в количественном соотношении между частотой
возникающих мутаций и интенсивностью отбора. Эта
последняя формулируется как проблема равновесия между
мутационным давлением и действием отбора.
Появление мутаций. Насыщение ими популяций. Вопрос
о появлении мутаций специально рассматривался в гл. 2.
Основной вывод заключался в том. что появление мутаций
представляет собой стохастический процесс, который может быть
охарактеризован вероятностями. Эти вероятности измеряются
конкретными частотами разных категорий мутации: отдельных
генов пли генома в целом, а также разных типов мутаций.
Известно, что спонтанные генные мутации могут быть доми-
153
иантнымп. адоминантными и рецессивными. При рецессивно-
сгн они могут очень долго сохраняться в популяции в скрытом
гетерозиготном состоянии, создавая, по терминологии
11. II. Шмальгаузена. так называемый мобилизационный
резерв наследственной изменчивости.
Впервые С. С. Четвериков и его сотрудники (1928)
экспериментально показали, что под покровом внешне однородного
вида скрывается большое количество рецессивных мутации,
находящихся в гетерозиготном состоянии. Несмотря на то что
кажды/i отдельный ген мутирует очень редко (в среднем с
вероятностью порядка 10 5)\ при умножении этой частоты на
общее количество генов в геноме и на число образующихся
половых клеток количество скрытых мутации в любой популяции
должно быть очень большим. Так. Баур показал, что до 15%
особен львиного зева (Antirrhinum) несут какую-либо
мутацию. Природные популяции, а также популяции домашних
животных и культурных растений обладают громадным резервом
наследственной изменчивости, что не может не иметь
значения для естественного и искусственного отбора, если только
отдельные наследственные изменения будут подхвачены
отбором. Однако между появлением отдельных мутации и их
реальным включением в процесс отбора происходит ряд
событий, определяющих дальнейшую судьбу мутантного гена.
Процессы, происходящие с мутантными генами до их
включения в отбор. Если особь — носительница данного
рецессивного гена — не погибнет и даст потомство, то она передаст его
половине своих потомков. Таким образом, судьба каждого
мутантного гена определяется вероятностью для ее носителя
иметь потомство. Очень многое зависит от того, изменяются ли
размеры популяции, т. е. будет ли в следующем поколении
столько же особей, сколько было в предыдущем. При
стабильности популяции по численности (что является в общем
правилом, хотя эта стабильность относительна и между
отдельными популяциями могут быть чисто случайные колебания в
степени стабильности) каждая пара особей даст в среднем двух
потомков. Но в отдельных семьях возможны: ноль потомков,
I потомок, 2 потомка, ..., г потомков. При 0 потомков ген
сразу теряется, при 1 потомке возможность сохранения гена, как
и его утраты, равна 0,5. При 2 потомках вероятность потерн
гена снижается уже до 0,25, что видно из следующего расчета.
Среди 2 потомков возможны или АА + АА — вероятность 0.25,
или АА + Аа — вероятность 0,5, или Аа + Аа — вероятность
0,25. Утеря рецессивного гена наблюдается в первом случае
с вероятностью 0,25. Таким образом, по мере увеличения числа
потомков вероятность потери гена уменьшается от исходной
величины 0,5 при 1 потомке до 0,5 в степени г при г потомках.
Но распределение количества потомков в отдельных семьях
154
целовать распределению Пуассона. На этой основ*
f,y'l-D0M бил сделан расчет средней вероятности исчмиок-
4>" мутации, возникшей у одной особи (табл. 20).
ТчО.шци :>''- Вероятность исчгзмомям мутяцня, ишпмюеА
у одной особи популяция
Поколение
" .
3
7
13
31
63
127
Предел
Кгрочтнооь I
1!рН ОТСУТСТВИИ СгДС*- I
0,3679
0,6259
0,7905
0.8873
0.9411
0,9698
0,9847
1.0000
при ct.ie*™**ftn прг*чу- 1
щест*е rwpouinmi я i% I
0.3642
0.6197
0,7825
0,8783
0,9313
0.9591
0,9729
0.9803
J
В пределе при отсутствии селекционного преимущества rt-
терозигот мутация исчезает. Но при некотором селективном
преимуществе гетерозигот (правый столбец табл. 20)
появляется некоторая, хотя и очень малая, вероятность, что мутации
сохранится. Чем выше селективное преимущество гетерознгог,
тем больше вероятность сохранении мутации. Есть
исследования, преимущественно ътшатического порядка, в которых
проанализированы условия сохранения и популяции вшьъ
возникшего аллсльного гена с селективным преимуществом
гетерозигот и без него.
В связи с этим интересны результаты опытов Расмусом по
поведению летальной системы с определенными генами. Из
одной ящичной популяции дрозофилы было создано 30 линий,
поддерживавшихся в течение 19 поколений. В 4 линиях ле*
талышй геи все же сохранился. В дальнейшем из этих линий
была создана новая ящичная популяция, в которой установи*
лось равновесие на уровне частоты гена —0,14. Это
показывало, что гетерозиготы по летальному гену в исходной
популяции были неодинаковыми по своей приспособленности и сохра*
инлись те, приспособленность которых обеспечила сохранение
Данного гена.
Отношения, создающие возможность сохранения мутанпю-
ю гена в популяции, могут быть очень своеобразны. Так, Ва*
танабе Такао обнаружил и естественных популяциях дршофи-
155
лы и одном из районов Японии особей, гомозиготных по
полностью рецессивному гену гЫ, локализованному во второй
хромосоме. Гомозиготы (и самки, и самцы) были полностью
бесплодны, но частота их в популяциях достигала уровня 3,1 —
7,1% (в различные годы), несмотря на их пониженную
жизнеспособность. Оказалось, что такая высокая концентрация
гомозигот объяснялась сильным гетерозисом по плодовитости
гстерозигот, хотя они и не имели преимуществ по сравнению
с нормальными особями по жизнеспособности и скорости
развития.
Влияние отбора на гетерозпгот является также причиной
эволюции самого доминирования, так как при
благоприятности мутации в процессе естественного отбора будут
сохраняться особи с наиболее сильно выраженным признаком в зиготе,
т. е. эволюция будет идти в сторону повышения
доминантности; при неблагопрнятности мутации — наоборот.
Рассмотрение детален этого вопроса и, в частности, различий во
взглядах на механизм эволюции доминирования Фишера, Ранта,
Холдейна не входит в нашу задачу.
Возможности сохранения появившейся мутации и
увеличения ее концентрации в популяции. Роль мутаций как
основного материала в естественной эволюции, а также в
искусственной эволюции домашних животных и культурных растений
з настоящее время достаточно хорошо доказана. В таком
случае очевидно, что должен быть ряд факторов, противостоящих
общей тенденции к исчезновению появляющихся мутаций.
Первый из них — неполная рецессивность, или, лучше сказать,
некоторая степень доминирования мутантного гена, и второй —
его полезное действие.
Гетерознготы по таким мутантным генам имеют больше
шансов сохраниться, хотя их единичность по-прежнему создает
высокий шанс их элиминации из популяции в силу самой ста-
тистичности попадания их потомству. Но одни и те же
мутации могут многократно повторяться, чем создается некоторое
мутационное давление, которое имеет тенденцию включать
новые аллели в генофонд популяции. Наконец, единичная
мутация, если она нейтральна или даже вредна, может резко
увеличиться в своей численности благодаря особым процессам,
получившим название дрейфа генов или, по советским
авторам, генетико-автоматических процессов. Но так как дрейф
генов может влиять на структуру популяции в целом, он будет
рассмотрен ниже. В связи же с вопросом о мутациях важно
сейчас только то, что в популяции единичная мутация может
размножаться до такого уровня, когда начнут скрещиваться
лругс другом гетерознготы А}А2 и появятся гомозиготы рецес-
гнвы А2А2. Последние попадут в сферу действия отбора.
Учитывая, что большинство мутаций неблагоприятно для жизне-
156
.„особности и выживаемости организмов, гомозиготы по ним
чате всего будут также устраняться отбором. Сохранятся
а1ишь немногие —лучше выживающие, или нейтральные, или,
наконец, те, которые как бы перерабатываются в процессе
отбора и приобретают в сочетании с другими мутациями
полезные свойства. Эта тема особенно хорошо разработана
И. И. Шмальгаузеном.
Все сказанное относится преимущественно к естественной
эволюции. Мутации у разводимых человеком животных и
растении оцениваются по их влиянию на хозяйственно-полезные
признаки. Поэтому возможны случаи, когда искусственным
отбором могут быть закреплены мутации, снижающие общую
жизнеспособность. Современные культурные свиньи — это
результат длительного накопления и сочетания многих мутаций,
обусловивших изменение физиологии и онтогенеза их предков
в сторону способности к быстрому откорму и накоплению
большого количества мяса и сала (с измененными
биохимическими свойствами). Очевидно, что в условиях природы
высококультурные свиньи были бы маложизнеспособиыми. Согласно
определению Д. К. Беляева, стабилизирующий отбор,
сохранявший постоянство видов в природе, заменяется при
одомашнении дестабилизирующим отбором, при котором нарушается
прежде существовавшая слаженность организма. В новых
условиях при разведении человеком закрепляются и мутации,
не имевшие шансов сохраниться в условиях природы. Хорб-
шим примером являются мутации у норок, разводимых на
фермах. Многие из них, выражающиеся в изменении окраски
(серебристо-голубые, паломино и др.) в тон или иной степени
влияют и на физиологические признаки (плодовитость,
устойчивость к заболеваниям и др.). Однако в силу ценности в
хозяйственном отношении их разводят на звероводческих
фермах миллионами голов, хотя при этом подчас приходится
принимать специальные меры для ослабления вредного действия
мутантных генов.
Создание равновесия между отбором и мутационным
давлением. Однако в условиях природы, когда происходит
отрицательный отбор по расщепляющемуся в популяции
рецессивному пли доминантному гену (т. е. против форм А\А\ или /М2).
возможна ситуация, когда отбор становится как бы
неэффективным, так как ему противостоит непрерывно протекающий
мутационный процесс, вновь поставляющий в популяцию тот
аллель, частота которого уменьшается отбором. При
определенном уровне обоих процессов — отбора и мутационного
давления — происходит их уравновешивание.
Допустим, что ген А\ мутирует к Л2 с частотой, равной
о каждом поколении величине и. Это значит, что из всех
имеющихся в генофонде популяции генов Л» доля генов, равная и.
157
мутирует к А2. Ксли частота геиа А\ Сила \\ исходном
поколении />0, то частота вновь появившегося вследствие мутации
гена А2 будет ир0. На эту величину уменьшится частота гена Л,
в следующем поколении. Она будет равна ро—и/?о.
Но так как возможны и прямые и обратные мутации, т. е.
и
v
с разными скоростями мутаций и и с\ то картина несколько
усложнится. Если первоначальная частота гена А2 в
популяции была равна </0. то при скорости обратной мутации к Аи
равной :\ она уменьшится в следующем поколении на
величину vqlu т. с. будет равна q(r—vq0.
В этом случае общее изменение частоты гена (Aq) за одно
поколение Aq = up0—vqo> или Д</ = //(1—q0)—vq0.
Сначала можно поставить вопрос о том, какое должно
быть соотношение между v и и, чтобы не произошло изменения
первоначальных частот, т. е. при условии, что Aq = 0.
Тогда и(\— qQ) —у<7о = 0, или и (l—qo) = vq0.
Преобразуя это равенство, находим:
v 1 — <7о
Отсюда
*--£г- <">
При таком соотношении между частотами прямых и
обратных мутаций qo (и соответственно р0) останется неизменным.
Популяция будет находиться в состоянии равновесия. В
данном случае рассматривается популяция, в которой отбор не
происходит.
Теперь можно осложнить анализ учетом отбора. Как и
ранее, возможен простейший случай — отбор протекает против
форм А2А2% доминирование гена Л, полное. В этом случае
Д q= -—----.
1 —s^-
Если отбросить знаменатель, так как он близок к 1, то
Aq~—sq2(l—q).
С другой стороны, при учете прямых и обратных мутаций
\q~u(\ — q)—vq.
Для установления условия, при котором произойдет
уравновешивание двух противоположных процессов — увеличения
цЛСтоты гена А2 за счет мутации от Ах к А2 (с учетом и
обратно мутирования) и уменьшения его частоты за счет отбора,
^правленного против А2А2 с коэффициентом отбора s, очевид-
liC| надо приравнять абсолютные значения правых частей
обоих уравнений (алгебраически они противоположны друг
другу):
w(l— q) — vq = sq*(\ — q).
Это уравнение и будет условием равновесия между отбором
и мутационным процессом. Но, чтобы лучше видеть основные
связи, можно внести в него некоторые упрощения. Так как q
ямяется малой величиной и столь же мала величина и—частота
обратных мутации, то в
левой части уравнения можно
пренебречь членом vq. Тогда
u(\—q) = sg2(l—g), или
u=sf}2. Отсюда
с w
'- Я
(12а)
Это приближенное
уравнение относится к случаю,
когда отбор идет против
рецессивного гена А2. Тем же
методом можно вывести
формулу равновесия для других
случаев отбора. Так, при
отсутствии доминирования
Д</ приближенно равна
0.5*7(1—?).
Тогда u(l—q) =0y5sq(\—q). Отсюда
ОХ У № Р:> U /??,
Чистоты q
'owl рсёнскрл и
Рис. 39. Изменения частот генов при
определенной скорости отбора:
/ — при сверхдоминировлинн; 2 — при
доминировании; J--при отсутствии
доминирования; 4 — при рецессивности гена; 5 —
мутационное давление с частотой и. Стрелками
укаэаны точки равновесия отбора н
мутационного давления при условиях 1—4.
я=
2и
s
(126)
В зависимости от характера доминирования того гена,
против которого идет отбор, равновесие и и q создается при
несколько различных значениях q и q2% что и показано на рис. 39.
Стоит остановиться еще на одном случае равновесия, когда
отбор идет против Аи так как он был использован для
измерения скорости мутационного процесса у человека (гл. 2).
В случае отбора против А\ его частота может быть выраже-
1—с/, а скорость мутирования от А2 к доминанту Ах—v.
Условие равновесия выразится несколько иной формулой:
на
vq**sq*{l—q)-
159
Отсюда
--=</(1-</), или -~=pq.
s s
Правая часть равенства представляет собой не что иное,
как половину доли гетерозигот в популяции, анализируемой по
данному доминантному гену.
Напомним, что при структуре популяции PA\Ai + 2QAiA2+
+ RA2Ao
2Q = 2pg.
Тогда
— =2Q; v = Qs. (12в)
Это уравнение может быть использовано для установления
частоты мутаций v (от А2 к А\)ч так как если отбор идет
против А 1.4,, то гомозигот в популяции будет очень мало. Гетеро-
зиготы же А{А2 составят львиную долю фенотипов,
проявляющих данный мутантный ген.
В гл. 2 были приведены данные Холдейна об установлении
частоты мутации по гену доминантной карликовости (хондро-
дистрофия) в Дании.
Величина 2Q= 10,7-10"5; s=0,804.
Отсюда
10,7 -10-»-0,804 .. 1П_5
v= =4,3-10-\
Определение скорости мутирования — это как бы
дополнительный результат, вытекающий из основной закономерности-
создания равновесия между мутационным давлением и
отбором при достижении определенного уровня концентрации гена,
против которого идет отбор.
Более общий вывод заключается в том, что скорость
мутационного процесса, которая, как правило, очень низка,
начинает играть роль только при очень малых значениях Д</, т. е.
основным фактором в изменении частот генов в популяции
является отбор. Увеличение частоты мутаций даже в 2—3 раза
очень мало скажется на изменении kq. Однако и в природе,
ив популяциях животных и растений, а также человека
нельзя не считаться с процессом накопления мутаций, который
получил название мутационного груза. Были сделаны попытки
выразить величину мутационного груза математически. Так»
Кимура и Кроу предложили следующую формулу:
Lm = u( l-6± у *?(2+в))
2л(1-2Л) '
160
Здесь и-частота мутаций от А> к Л2; «-коэффициент
отбора; ft — степень доминирования. Знаки «+» и «~» перед
корнем квадратным соответствуют значениям в>0 или в<0.
Случайный дрейф генов. Соотношение генов в популяции
может измениться п результате случайных процессов,
получивших название «генетико-автоматических» (Дубинин,
Ромашов) или «случайного дрейфа генов». Эти процессы
наблюдаются в малых популяциях как следствие самой выборочности
популяции следующего поколения, образующейся из
популяции предыдущего.
Разберем, как происходит образование отдельных, или ча*
стных популяций из основной, или исходной, популяции.
Каждая частная популяция образуется благодаря выборке N
особей из исходной популяции. Так как каждая особь несет два
гена одного локуса, то разделение исходной популяции на
частные равнозначно получению серии случайных выборок,
каждая из которых будет иметь по 2N генов. Каждая такая
выборка будет характеризоваться своими частотами генов —
аллелей: р\ и q{\ р2 и q2\ p3 и ?з и т. д.
Проиллюстрируем сказанное следующим примером.
Допустим, что из исходной популяции, состоящей из особей А\Аи
А\А2 и А2АЪ находящихся в соотношении 1:2:1 (т. е. при
/?о=<7о=0,5), берутся только по две особи для скрещивания.
Результаты всех возможных типов скрещиваний и их
вероятности представлены в табл. 21.
Таблица 21. Результаты возможных скрещиваний любых двух особей
из популяции и их вероятности
Скрещивание
AiAiXAiA* \
AlAtxAlAl J
AxAoXAlA* I
А1А1хАгАг }
AtAtxAlAl |
АхАгхА%Аг )
A9AtxAlAt ]
1 >M,xMi
Их вероятности
1/16
4/16
I 6/I6
4/I6 l
I/» I
Частота генов а гютомстм I
P
l
0,75
0,5
0,25
0
0 I
0
0,25
0.5
1
0.75
1 I
И П. Ф. РокицкиА
Только п 6/16 псех случаев частоты генов остаются
прежними (=0,5). Во всех остальных случаях р и q примут разные
значения между Он 1, при этом в 2/16 случаев один из
аллелей полностью исчезнет, а второй закрепится. В среднем же
по всем скрещиваниям сохраняется равенство ро = ^о = 0,5. Чем
больше количество особей в популяции, тем меньше
вероятность фиксации отдельных генов и, таким образом, утери
генетической вариации вследствие дрейфа генов. В большой
популяции случайность отклонения в частотах генов мала.
Если частоту генов в исходной популяции обозначить ?о,
то в следующем поколении в силу случайных отклонений она
может быть иной, например qu тогда разницу qx—q0 можно
обозначить 6., (малая дельта в отличие от Aq —
статистических сдвигов в значении q вследствие отбора, мутаций или
миграции). Степень вариации отдельных значении б7
характеризуется варнансой
, ро?о ^7о(1 — ^7о) /0
a«=-W- = —2ЛГ~' (13)
Эта формула вытекает из того, что возможные разные
частоты генов соответствуют вероятностям, определенным при
разложении бинома Ньютона (po-\-qo)2N-
Варианса о26 показывает величину возможного разброса
в частотах генов в отдельных частях популяции за одно
поколение в силу чисто статистических процессов распределения
генов по гаметам.
Значение а* обратно пропорционально N, т. е. размерам
популяции, или, точнее, 2N — числу аллельных генов.
Вероятность возможных значений р для популяций разных размеров
(2jV=50 и 2JV=5000) демонстрирует график, представленный
на рис. 40. Он относится к генам, для которых ро и qo в
родительском поколении были равны 0,5. При 2/V = 50 частоты
аллелей данного гена колеблются от 0,30 до 0,70, а при
2/V = 5000 — от 0,48 до 0,52. Количественные значения о2 для
этих двух случаев таковы:
о\ [2Л/ = 50]= M^ilL =0,005;
6<r J 50
о* [2Л/ = 5000]= -;—- =0,00005.
л''1 J 5000
Процесс рассеяния частот генов в отдельных малых
популяциях повторяется в каждом поколении, но он всякий раз
начинается вновь от тех частот, которые создались в дочерней
популяции вследствие рассеяния в предыдущем поколении. Ес-
162
1Й в популяции создалось новое соотношение, например
„,=0,7, а Ц\ =0,3, то дальнейшее рассеяние будет идти уже от
этого соотношения. Так, на основе случайных колебаний частот
генов происходит их изменение, которое и называется дрейфом.
Возможные последствия дрейфа генов в небольших
популяциях представлены в виде схемы, изображенной на рис. 41.
Частоту генов можно уподобить упругому шарику,
который, падая по наклонной доске и ударяясь во вбитые
штифтики, отскакивает от них и занимает различные положения, пока,
№г
рч
%щ
•5
; 1 1
• [
t -У1
Ь
0,30 Q35 O/tO 0fi5 0.50 0$5 0,60 065
Частота гена
Рис. 40. Вероятности разброса
различных частот гена: при 2/V = 50 и
2,V = 5000.
Желоба
Рис. 41. Наглядная модель
генетического дрейфа от первоначальной
частоты 0,6 (шарик /) до частоты,
равной нулю (шарик 2).
наконец, не дойдет до одного из краев доски и не скатится
в желоб. Так же меняется по случайным причинам частота
гена р (или q) в популяции (причем надо помнить, что размах
возможных колебаний этой частоты тем больше, чем
меньше N), пока она не станет равной 1,0 или 0,0. В первом случае
ген А фиксируется в популяции, во втором — исчезает.
Процесс этот необратим. Частота гена А может колебаться вокруг
любой исходной величины, но в конце концов, если при этом
не вмешиваются никакие иные факторы в виде отбора и т. п.,
дойдет до 1 (полная фиксация) или до 0 (утеря гена).
Процессы утраты и фиксации идут со скоростью —т генов
на одно поколение, т.
1
4iV
вероятность утраты и
их вероятность равна
1
2ЛГ
2/V
из них
4N
вероятность фиксации.
Скорость дрейфа в конечном счете зависит от размеров
популяции. Но известно, что она никогда не остается
постоянна
163
нои. В моменты сжатия популяции генетический дрейф
протекает очень интенсивно. Происходит резкая перестройка
генной структуры популяции — потеря большого числа генов,
бывших у тех особен, которые погибли или не дали потомства,
и сохранение немногих генов. Наоборот, в период расширения
популяции немногие сохранившиеся гены передаются уже
многим особям численно возросшей популяции. В свою очередь
распад большой популяции на ряд более мелких нзолятов
приведет к дифференциации в них частот генов, но уже от того
уровня, который был достигнут в предыдущей большой
популяции. Таким образом, для генетического дрейфа большое
значение имеют колебания численности популяции.
Райт предложил следующую формулу для расчета
фиксации и потери генов в малых популяциях:
Фиксация qoSpoqoP
Потеря po—3PoqoP
Ни потери, ни фиксации SpoQoP
Здесь P=(i-^L)'. (и)
Случайный генетический дрейф анализировался Жарнец-
ким на математических моделях с помощью
электронно-вычислительных машин. Предусматривалось, что размеры
популяции небольшие и она замкнутая, первоначальные частоты
генов 0,1 и 0,5. Были взяты также разные соотношения самцов
и самок (в эффективной части популяции): 1 : 25; 5:25 и
25:25. Скорость фиксации аллеля была наибольшей при
низкой частоте гена. Чем больше были эффективные размеры
популяции, тем меньше изменение частот генов, как и следовало
ожидать из самой идеи генетического дрейфа. Самое
существенное в этом математическом опыте — установление
влияния ограниченной численности одного пола на эффективные
размеры популяции и отсюда на скорость фиксации генов.
Известно, что в животноводстве при искусственном
осеменении резко уменьшается количество самцов, что, очевидно,
может привести к значительным последствиям — потере одних
генов и фиксации других.
Наряду с теоретическим анализом вопроса о генетическом
дрейфе, проведенном Р. Фишером, С. Райтом, Н. П.
Дубининым, Д. Д. Ромашовым, были получены и экспериментальные
доказательства его существования на модельных объектах.
Так, Фолконер изучил случайный дрейф в 3 линиях мышей.
Первоначально в линиях было расщепление по паре генов
агути — сплошная окраска. Исходная частота рецессивного
1сна сплошной окраски была равна 0,5. Для получения
следующего поколения брали 6 пар родителей. Во всех линиях
1Ы
„аблюдались в течение 14 поколений значительные
колебания q— в пределах от 0,75 до 0,10. В последних поколениях
в1 линии <7-0Д во 11 — 0,3 и в 111 — 0,1.
В очень больших по масштабу опытах Бери на дрозофиле
(105 линий в течение 19 поколений) наглядно
продемонстрирован процесс увеличения различий между линиями по частоте
гена bw (brown — коричневые глаза). Изменение вариансы
О 2 ч 5 8 10 12 /* 16 18 13
Поколения
Рис. 42. Увеличение различий
между линиями дрозофилы по
частоте гена bw вследствие
генетического дрейфа. (Кружки—
фактические данные для
каждого поколения; кривая —
теоретически вычисленные. N=*
= 11,5).
о г
6 8. '0 1Z Щ № '8 !9
Поколения
Рис. 43, Увеличение процента
линий дрозофилы, в которых
произошла фиксация одного
аллеля (светлые кружки) н
потеря другого (черные кружки).
частот гена (o2q) показано на рис. 42. Теоретическая кривая
для точек каждого поколения вычислена согласно уравнению
a^=Wo[l-(l-^)'].
2N
Расположение фактических точек очень близко к
теоретически ожидаемому. Однако_средняя частота q для всех линии
в целом остается той же {q = qo). Таким образом, происходит
дисперсия частот, обязанная генетическому дрейфу.
Одновременно наблюдается и другой процесс: в обеих линиях q того
или иного аллеля повышается до 1,0 (т. е. происходит
фиксация гена, или, по терминологии Н. П. Дубинина, нзогамета-
Ция), а в других — понижается до 0 (т. е. его потеря). На
рис. 43 показаны результаты опытов Бери на дрозофиле с
геном bw, сходных по методике с показанными на предыдущем
рисунке.
В литературе по генетике человека есть немало данных,
иллюстрирующих роль генетического дрейфа в силу случай-
165
„ых колебаннй частот геиоо D популяции, усиливаемых коле-
л» мня ми п численности.
?"и популяция возникла от небольшого количества осо-
бей то гены, которые были у этих особе ft, могут быть раз-
множены до высокой концентрации (иногда это явление назы-
дамп €э*Фектом родоначальников»). В небольших популяциях
и ИЭ01ЯТМ отделенных теми или иными барьерами, в том
чипе национальными или религиозными, от остального
населения могут распространиться вредные гены. Так, среди буров
Южной Африки и лапландцев Швеции очень высокая частота
генов порфирим. На некоторых островах Аландского
архипелаги частота болезни Внллебрандта больше 0.1.
По данным японских авторов, действие генетического
дрейфа наблюдалось в нзолятах человеческого населения в
горных районах Японии и привело к повышению частоты группы
крови 0 в одном из изолятов до 43,8% и снижению частоты
гр\ппы А и В до 1,8%.
Интересны также результаты изменений численности
популяции островов Тристан-да-К\нья с 1820 по 1961 г. В 1820 г.
было всего несколько человек, к 1850 г. численность возросла
до 100, к 1855 г. резко упала (всего до 20), к 1880 г. опять
увеличилась до 100, а к концу 19 века опять упала до 20. В 20
веке она аала возрастать и сейчас приближается к 300. Было
показано, что вклад генотипов некоторых особей в общий
генофонд популяции островов был очень велик.
Известно, что в некоторых районах США, а также Швеции
очень высока частота альбинизма. Высказывалось мнение, что
первоначально частота мутации альбинизма повышалась
благодаря генетическому дрейфу, а в дальнейшем могло иметь
значение некоторое преимущество гетерозигот по этому гену.
Сложнее обстоит вопрос о роли случайного дрейфа генов
в эволюционном процессе. Некоторые факты, например
различия между колониями улиток Сераеа, ранее трактовавшиеся
как результат генетического дрейфа, в настоящее время
объясняются действием отбора в специфических условиях среды,
где живут колонии Несомненно, что дрейф не имеет значения
в больших популяциях, например при jV>500, но он может
смграть|рать в очень малых популяциях, особенно таких,
которые образуются от небольшого количества особей — сосно-
мтедей», по терминологии Майра. Популяция, образуемая
потомками таких особей, будет содержать небольшое коли-
witbo мутантных генов. Размножение ее может повысить кон-
ii^IU1110 огдельиых генов до такого уровня, когда начнет
готы Ч *ТЬ m6i}V* т е К0ГАа начнут скрещиваться гетерозн-
iJL *>.*** отн<>снтся и к резким колебаниям численности
' генгТ||Ц^*им дрейфом следует считаться и при раэведе*
14*
ими сельскохозяйственных животных. В стадах, малых по
размерам, могут быть нежелательные гены, и тогда генетический
дрейф может сильно поднять частоту этих генов и тем самым
вызвать нежелательные последствия для дальнейшей работы
со стадом.
В термин «генетический дрейф* иногда включают не
только результат случайных изменений в частотах генов в
различных выборках гамет или колебаний численности, приводящих к
потере одних генов и повышению концентрации других, но и
результат других причин: изменений в направлении отбора,
случайной гибридизации, волн миграций, появления
благоприятных мутаций. По-видимому, такое расширение понятия
едва ли оправдано и только запутывает дифференциацию
факторов, изменяющих структуру популяций.
Анализ случайных колебаний в частотах генов в малых
популяциях логически связан с общим вопросом о
результатах расчленения популяции на отдельные группы, благодаря
чему прекращаются свободные скрещивания в популяции и
создаются шансы для спаривания родственных особей, т. е.
инбридинга.
Влияние инбридинга на структуру популяций. Модель
панмнктической популяции предусматривает, что спаривание
между отдельными особями происходит случайно. Среди них
могут быть спаривания как родственных, так и неродственных
особей. Шансы тех и других одинаковы. Однако в малых по
размерам популяциях, где особи связаны между собой через
родителей, дедов и вообще через общих предков, иногда даже
немногих родоначальников, повышается вероятность встречи
одинаковых генов при соединении двух гамет. Мерой
вероятности передачи одинаковых генов особи от предков служит
коэффициент инбридинга F, впервые предложенный Райтом *.
Если коэффициент инбридинга в популяции равен F, это
значит, что вероятность для двух аллелей быть идентичными
при оплодотворении оценивается величиной F, а вероятность
быть иеидентнчными 1 — F. Однако в равновесной популяции,
имеющей структуру p2AiA\+2pqA\A^+q1A%Aii уже есть
вероятность соединения идентичных аллелей: для At oua равна /Я,
а для Аг—q2. Вероятность же соединения разных аллелей
равна 2pq. При наличии инбридинга с коэффициентом F
соответственно снизится вероятность соединения разных аллелей —
на величину 2Fpq — и увеличится вероятность соединения
одинаковых аллелей — на величину Fpq для каждой гомозиготной
группы:
• Инбридинг, его обоснование, результаты я методы намерении будут
изложены более полно а гл. Я. здесь же мы рассмотрим роль ннбрядмнга
как фактора, изменяющего структуру менделеаской ионудтии а простейте*
«с форме, при расщеплении по одно* паре гено*.
167
AiAt p2+Fpq
AiA2 2(\-F)pq
A2A2 q2+Fpq
Группа гетерознгот уменьшится на 2Fpq, а группа
гомозигот возрастет по каждому из генов — А{ и Л2 — на Fpq. Иначе
говоря, в популяции происходит снижение гетерозиготностн
и соответственное увеличение гомозиготности, но соотношение
частот генов р и q в отличие от того, что происходит при отборе»
при этом не изменится.
Долю гетерознгот часто обозначают через Н. При наличии
инбридинга H = 2pq{\—F). Если F = 0, H = 2pq.
Вероятность соединения одинаковых аллелей может
служить характеристикой степени панмнксии в популяции. Ее
называют панмиктическнм индексом и обозначают символом
Р(Р=\— F). Если принять исходную степень гетерозиготностн
при панмнксии Я0 за единицу, то можно рассчитать, на какую
долю будет уменьшаться гетерозиготность при разных F.
Очевидно, что в бесконечно большой панмиктической
популяции F=0. Но чем меньше размеры популяции, тем
больше вероятность соединения одинаковых аллелей, т. е. F>0.
Если обозначить через N количество особей, эффективно
размножающихся в родительском поколении, то они образуют
2Л~ аллелей по каждому аутосомному гену. Вероятность того,
что две случайно соединившиеся гаметы будут содержать
идентичные аллели того же гена, которые окажутся повторением
тех же аллелей в родительском поколении, равна —г-. Во всех
/ 2N-1 \
других случаях ^их доля ———J соединяющиеся гаметы
содержат различные аллели. Но при наличии инбридинга,
измеряемого коэффициентом /\ создается дополнительная
вероятность, что часть случаев из последней группы перейдет
в группу соединяющихся идентичных аллелей. Тогда общая
вероятность для случайно соединяющихся аллелей быть
идентичными составит
2N ^ 2N
F и F — коэффициенты инбридинга двух соседних
поколений, В таком случае общая формула для /-поколения будет
следующей:
F~w+(l-w)-F™ (,5а)
!6в
Выраженный в виде этой общей формулы коэффициент
„нбридинга состоит из двух частей: первоначальная (пра-
рая) — результат предыдущего инбридинга — и
дополнительная—прибавка в величине инбридинга за одно поколение,
которую можно обозначить как A F:
Л f= -sir-
2N
Тогда предыдущее уравнение может быть записано как
Ft = bF+(\-tiF)-Ft-i. (156)
Отсюда можно получить точное значение Д F:
Af=Ilz£zL (16)
Можно также выразить его с помощью Я —уже
упоминавшегося выше панмиктического индекса. Так как P=l—F, то
-£-=l-AF. (17а)
Таким образом панмиктический индекс уменьшается на
постоянную величину в каждом поколении в зависимости
от Л F. Возврат на два поколения приведет к формуле
~i- = (l-AF)«,
а если вернуться к основной популяции, то
Pt=(\-bF)*Po. (176)
Для первоначальной популяции принимается, что
коэффициент инбридинга в ней равен 0, и тогда Р=1.
В таком случае коэффициент инбридинга в каком-то
поколении, отнесенный к основной популяции, может быть
выражен как
F|S=1-(1-AF)«. (18)
Инбридинг требует поправок и в формуле вариансы для
А ^ и q. Так,
-..-ф-*** "9',
169
V = Mo[l-(l--^-) ] =poi/oFt. (196)
Обращаясь к основной указанной выше формуле для Д F,
т. е. Л F= ——, видим, что прибавка в величине инбридинга за
поколение, как и сама величина /\ обратно пропорциональна
размерам популяции. При очень больших ее размерах эффект
инбридинга равен или очень близок к нулю. При iV = 5000
Л F= ———. Если в популяции F уже было равно нулю, то
1000 ' " 1
прибавка Af = 0,001 практически ничего не составляет. Если
первоначально в популяции уже была какая-то величина /\
то она останется неизмененной в большой популяции и
несколько увеличится в малой, при этом тем больше, чем меньше
размеры популяции. Поэтому разбивка большой популяции на
более мелкие, изолированные популяции наряду с уже
указанным случайным дрейфом генов приводит и к закономерному
повышению гомознготности и падению гетерозиготности
вследствие инбридинга.
Говоря о размерах популяции, следует помнить, что имеют
значение не размеры популяции вообще, а только величина той
ее части, которая дает в данном поколении потомство. Если
обозначить ее через Ne, то лучше записать
Впрочем, установление величины Ne возможно только в
строго планируемых опытах, при массовом же размножении
популяций (в лабораториях или в природе) судить о ней
можно лишь приблизительно.
Параметры популяций при учете совокупного действия
мутаций, миграций и инбридинга. Так как в популяции
одновременно действуют разнообразные факторы, влияющие на ее
структуру, целесообразно остановиться на их совокупном
действии, ограничиваясь только некоторыми общими выводами,
так как их математическое обоснование дается в специальных
работах (Райт, Малеко и др.).
Путь рассуждений может быть тот же, что и ранее.
Допустим, что популяция разделена на ряд популяций или
линий с эффективными размерами Ne. В каждой из них долю т
составляют иммигранты, попадавшие в данную линию
случайно ич других линий. В популяции в целом средние частоты
двух аллелей А{ и А2 р и q. Скорости мутаций в обоих
направлениях и и с.
170
Тогда при ЛОСТИЖеИНИ равновесия между степенью
рассеяния линий, с одной стороны, и мутациями и миграцией — с
другой (оба последних процесса изменяют частоту гена в том же
направлении), межлинейная варианса частот генов может
быть представлена следующей несколько упрощенной
формулой:
о _ г п
(20)
С другой стороны, поскольку, как указано выше, at,z=Fpq,
то
/=•= —
1+4№»(ы+и+и»)
*
3
На рис. 44 представлены 4 '",
сличая распределения генных
частот внутри линий при разных
значениях Л при этом средняя
частота гена 0,5. Характер
кривых распред?ления очень сильно
меняется в зависимости от
величины F. Чем выше /\ тем сильнее ^5С
фиксация гена (или около 0, т. е. §
его потеря, или около 1, т. е.
закрепление в гомозиготном
состоянии). При F = 0,005
дифференциация между линиями очень мала.
При У7 = 0,333 распределение
выражается прямой линией — все
генные частоты равновероятны.
При достаточно высоком F
промежуточных генных частот ПОЧТИ р1Н\ 44. Теоретическое распре
Нет. деление частот гена среди ли
Таким образом, при величине ннй ПР" равновесии между сте
f-nw пппиргг пягррянио mpw пеиью Ра«еяния ЛИНИЙ и чу
r-u,666 процесс рассеяния меж- тациямн (нли Mlirpau„eA) в за
ду линиями достигает критиче- онсимостн от F ?/-f «0,005;
ской точки — начинается процесс 2—F-Q.048: 3—^«(ШЗ; 4—
фиксации и потери в отдельных F-M33).
линиях.
Геногеография. Очень интересное направление в изучении
различий между популяциями было создано А. С. Серебров-
ским. Он назвал его геногеографией. Серебровскнн обратил
внимание на то обстоятельство, что концентрации генов в
соседних популяциях могут быть очень различными. В одних
случаях популяции, отдаленные друг от друга территориально,
характеризуются большим сходством по концентрацнн ряда
171
j chop, и л о доказывает, что они составляют одну единую по-
п\линию. В других случаях популяции, расположенные
близко млн даже рядом друг с другом, могут сильно отличаться по
частоте отдельных генов. Материалом для геногеографиче-
1ких исследований Серебровского и его сотрудников (С. Г.
Петров н др.) служили куры различных областей нашей страны
(преимущественно Кавказа). Так как у кур тем же Серебров-
скнм было хорошо изучено наследование ряда генов
качественных признаков (окраски, формы гребня, наличия хохла и
т. д.), то можно было сравнивать структуру двух популяций
по нескольким генам. Простейший способ сравнения
заключался в том, что брались разницы между частотами многих
генов (Л), они возводились в квадрат (Д2) и далее сумма
квадратов разниц делилась на число генов (п). Таким
образом, величина
2Д2 -|/~2Д2~
или I/
п In
могла служить мерилом различий между двумя популяциями
по многим генам. Так как больший вес приобретали гены с
частотами, близкими к 0,5, то А. С. Серебровскии несколько
модернизировал метод сравнения, использовав дробь, в
числителе которой была разница между частотами, а в знаменателе
сумма.
Для сравнения многих популяций кур Серебровскии
применил также графический метод. Для этого он выбирал
несколько наиболее удобных генов, которые легче
диагностировались и частоты которых были не очень малы, и откладывал
значения частот каждого мутантного аллеля в виде векторов.
Векторы начинались из общей начальной точки и шли всегда
в одном направлении, концы же векторов соединялись
линиями, как это показано на рисунках. Получались своеобразные
многоугольники. Формы их отражали специфику структуры
изучаемой популяции. Обычно использовались 8 генов,
векторы которых всегда направлялись в соответствии с
общеизвестной розой ветров (восток, север, северо-восток и т. д.).
Сравнение подобных графиков давало возможность
решать ряд вопросов, касающихся сходства или различия
популяций, их прошлой истории, распада общей популяции на
отдельные изоляты или, наоборот, слияния популяций.
Так, на рис. 45 представлены графики популяций
беспородных кур пяти мест: Черниговской, Полтавской, Московской,
Ярославской и Кировской областей, обнаруживающие
замечательное сходство, явно указывающее на единство громадной
пипуляции кур значительной части Европейской России. На-
«Лорот, графики рис. 46 для пяти селений Дагестана, разде-
лепных рекой Койсу и населенных двумя разными племенами
(аварским и андийским), явно разбиваются на две группы —
аварскую и андийскую.
А. С. Серебровский показал наличие сложных процессов,
приводящих к различному распределению многих генов в
популяциях, т. е. к различиям в генофонде популяций, среди
Рис. 45. Сходные структуры популяций кур (по 8
генам):
/ — Нежин: 2 — Ярославль; 3 — Кировская область; А —
Полтава; 5 —Московская область (по Серебровскому).
Рис. 46. Структура популяций кур в
андийских (I—2) и аварских (3—5)
селениях горного Дагестана.
Рис. 47. Структура популяций кур
с высокой изогаметацией
(пунктирные векторы — высокие частоты
генов) в двух селениях Сисиана
(Армения) (по Серебровскому).
которых можно назвать процессы «генетической диффузии»,
медленного распространения генов во .рее стороны от центра
их возникновения; передвижение самих популяций; миграции
из одной популяции в другую; мутационный процесс,
непрерывно вносящий в генофонд новые гены, и т. д.
В некоторых случаях форма графика позволяет наглядно
наблюдать изогаметацию, т. е. фиксацию частот генов на
высоком или низком уровне, очевидно, как следствие
генетических процессов, шедших в очень малых по размерам
популяциях (рис. 47).
Гсногеография разрабатывалась Серебровским
применительно к сельскохозяйственным животным. В результате был
вскрыт ряд сложных процессов, определяющих различия по
генетической структуре даже очень близко расположенных по-
173
пуляшш. Очень богатый геногеографичеекий материал был
получен позднее, при изучении распространения многих генов у
человека. Большое значение для понимания природы
сбалансированного наследственного полиморфизма имели данные о
распространении генов аномальных гемоглобннов, в частности
серповидно-клеточной анемии, о которой говорилось выше, а
также гена гемоглобина С в Африке, гена Е в Юго-Восточной
Азии и многих других. Существенно, что по многим генам
можно было установить закономерности географического
распространения в рамках не только континентов, но и в
глобальном масштабе, на территории всего земного шара. Так,
выяснился отчетливый градиент падения частоты гена крови б
(обозначается теперь Jn) в Европе с востока, где частота 25%,
па запад—п Испании 5—10%, а в Пиренеях еще ниже,
0-5% (рис. 48).
Для гена же (1Л детерминирующего отрицательный резус
крови, и для гена гаптоглобулпиов Нрх составлены карты,
охватывающие все континенты. Для первого из них
характерна высокая концентрация в Европе и очень низкая в
Австралии, Америке, для второго — максимальное значение частоты
в Центральной Африке.
Популяция как динамическая система. Из всего сказанного
выше следует вывод, что в любой популяции происходят
очень сложные процессы, как стохастические, так и
направленные. Даже в тех случаях, когда в популяции полностью
реализуется закон Хардп — Вейнберга и можно в том или ином
поколении установить строгое соответствие частот генов этому
закону, указывающее на наличие исходных условии, при
которых он может осуществиться, надо отдавать себе ясный отчет
в том, что такая картина отображает лишь один момент в
смене поколений данной популяции. В реальных условиях
наблюдается лишь некоторое приближение к осуществлению этой
формулы. При разбивке большой популяции на субпопуляцин
определенного размера в последних оказываются различные
частоты того же гена п проверить их соответствие ожидаемым
по формуле Хардп — Вейнберга очень трудно. Частоты генов
в смежных поколениях связаны между собой определенной
функцией, отражающей многие стороны жизни популяции, в
том числе явления расщепления, зависящего от стохастики
образования половых клеток и их соединения, мутационного
процесса, давления отбора и др. Связь между частотами
соседних поколении выражается через A q. Если </,Лн|1 не равно
</,". то популяция изменяется из поколения в поколение. При
(//'H~q.fi \q-~tf „ популяция находится в равновесном
состоянии. В статистической генетике разрабатывают системы
Рекуррентных пли дифференциальных уравнений, дающие
возможность выявить движение популяции.
175
Mo и равновесные состояния популяции имеют разную
степень устойчивости. В одних случаях популяция возвращается
н предыдущее состояние при любых отклонениях от него, в
других—не возвращается и переходит в повое состояние
устойчивости.
Наиболее существенным параметром, от которого зависит
в конечном счете движение популяции, является мера ее
средней приспособленности w. В свое время Фишер сформулировал
фундаментальную теорию естественного отбора (имея в виду
панмиктнческую популяцию, в которой происходит постоянный
отбор генотипов по аллелям одного гена): популяция
эволюционирует таким образом, что ее средняя приспособленность
w возрастает, причем степень этого возрастания определяется
аддитивной вариансой по приспособленности отдельных
генотипов популяции. В общем виде это можно записать так:
dw Wa—w 2a«.
^ = 22р,^^р<=-=2=- >0. (21)
w w
Если w достигает максимума, то популяция будет
находиться в состоянии устойчивого равновесия.
Для случая одной пары аллелей Тернер предложил
следующую формулу:
dw Л - - . / 1 dw \
где Wo и W\ — средние приспособленности, a q0 и q\ —
соответствующие им частоты генов. Эту формулу автор связал с
формулой Фишера и показал некоторые случаи, когда последняя
неприменима.
В дальнейшем Кимура и другие авторы распространили
фундаментальную теорему естественного отбора на более
сложные случаи (многие гены, изменения коэффициентов
приспособленности различных генотипов во времени, а также
в связи с колебаниями условий среды). Впрочем, некоторые
авторы выражали сомнения во всеобщности теоремы Фишера
и проанализировали случаи, когда при переходе к равновесию
w не обязательно возрастало •.
Вопрос о характере изменения средней приспособленности
популяции проанализирован Н. В. Тимофеевым-Ресовским
и Ю. М. Свирежевым математически в системе траекторий
• Сводка данных по атому вопросу сделана В. А. Ратпером.
неевклидопого пространства; ими показано, что популяция
всегда выбирает то направление изменения, при котором ее
средняя приспособленность возрастает, причем скорость
движения популяции пропорциональна генетическому
разнообразию популяции а- и обратно пропорциональна средней
приспособленности w в данный момент. Чем популяция ближе
к равновесному устойчивому состоянию, тем меньше ее
генетическое разнообразие, больше средняя приспособленность и тем
меньше скорость отбора, в пределе стремящаяся к нулю.
Интересно также и то, что авторы формулировали эту
задачу в терминах теории игр и пришли к тем же выводам.
Для характеристики популяции существенно понятие
генетического груза (L). Под этим термином понимают отношение
разницы между фактической средней приспособленностью
популяции и наибольшей приспособленностью одного из
генотипов, имеющихся в популяции, к наибольшей
приспособленности:
Иначе говоря, фактическая средняя приспособленность
популяции ниже той, которая была бы, если бы вся популяция
состояла только из наиболее приспособленных генотипов.
Менее приспособленные генотипы составляют как бы груз,
который тянет популяцию вниз. В то же время из поколения в
поколение протекает эволюция популяции в сторону ослабления
влияния генетического груза.
Генетический груз слагается из многих величин. В
популяции постоянно происходит расщепление на генотипы А\Аи
АХА2 и Л2Л2, неодинаковые по своей приспособленности и
поэтому подвергающиеся действию того или иного типа отбора.
При большей приспособленности гетерозигот
(сверхдоминирование) от них постоянно выщепляются гомозиготы с более
низкой приспособленностью. Этот компонент генетического
груза можно назвать сегрегационным грузом (L,).
В результате мутационного процесса в популяции
накапливаются мутантные гены. За счет их также снижается средняя
приспособленность популяции (мутационный груз Lm).
За счет повышения доли гомозигот при инбридинге
создается инбредныи груз (Li), также уменьшающий среднюю
приспособленность популяции, иногда очень резко (инбредная
депрессия).
Порой говорят также об эволюционном грузе {L<), имея
в виду случаи, когда протекает интенсивный, но еще далекий
от завершения отбор в пользу одного аллеля.
177
12 П. Ф. РокицкяА
Для оценки компонентов генетического груза предложены
различные математические формулы. Интересен выдвинутый
Кимура принцип минимальности генетического груза, смысл
которого заключается в том, что в процессе эволюции
происходит изменение всех генетических параметров таким
образом, чтобы суммарный генетический груз был минимальным.
В математические формулы, выражающие этот принцип,
входят частота мутации, степень доминирования, давление
отбора.
Глава 5
СТАТИСТИЧЕСКАЯ ХАРАКТЕРИСТИКА
ПОПУЛЯЦИЙ
ПРИ НЕПРЕРЫВНОЙ ВАРИАЦИИ
Дискретные различия по прерывным признакам и их
наследование. В предыдущих главах говорилось о статистическом
анализе популяций в тех простейших случаях, когда фенотн-
пические различия между особями зависели от различий в 1—
2 генах и когда эти различия не затемнены влиянием среды.
Тогда общая фенотипическая структура популяции легко
разложима па достаточно четкие фенотипическне и генотнпнческие
группы. Характеристика же генотнпической вариации может
быть сделана на основе частот отдельных генов.
Сейчас хорошо известно, что во всех популяциях, как
природных, так и разводимых человеком, наблюдается
изменчивость по большому количеству генов, затрагивающих
разнообразные морфологические признаки. Аллели этих генов, как
правило, могут быть сравнительно легко выделены. За
последние годы выяснилось, что и многие физиологические и
биохимические признаки также находятся под контролем четко мен-
делирующих генов. Таким образом, в популяциях животных и
растении, а также и человека наблюдается расщепление по
огромному количеству генов, определяющих наследственные
различия, при этом не только морфологические, но и
физиологические, биохимические и т. д. Одни из этих различии
устанавливаются относительно легко, и довольно просто
выявляются конкретные определяющие их гены, другие же внешне
плохо выражены, но могут быть вскрыты с помощью тонких
биохимических пли иммунологических методов.
Характеристика популяции по качественным дискретным
признакам или таким количественным признакам, которые
имеют строго определенные градации (например, в ряде
случаев биохимического полиморфизма), может быть сделана по
доле определенных категорий, выделяемых по фенотипу или
генотипу. Эти доли представляют собой параметры,
характеризующие общую изменчивость популяции. Если определенная
фенотипическая группа в популяции в действительности
состоит из двух пли нескольких генотнпических категорий, то
она может быть разложена на эти категории с помощью
формулы Харди — Вейнберга, которая позволяет рассчитать соот-
ношение генотипнческнх категории в популяции, в том числе
и внутри фенотипически однородных групп. Параметрами
структуры популяции во всех таких случаях являются частоты
отдельных генов.
Количественные признаки. Сложнее обстоит дело в тех
случаях, когда приходится оперировать количественными
признаками, вариация которых непрерывна. К их числу относятся
вес животных, длина всего тела к отдельных его частей, вес
различных органов, такие важные в сельскохозяйственном
отношении признаки, как количество молока от коровы за
лактацию, процент жира в молоке, настриг шерсти овец,
яйценоскость кур. количество детенышей в помете и ряд других.
В этом случае различия между особями выражаются какими-то
цифрами, т. е. представляют собой ряд со всеми переходами,
который может быть разбит на любые классы по произволу
исследователя. Поэтому нет возможности выделить отдельные
строго очерченные группы.
Идея разложения количественного признака на отдельные
определяющие его гены в сущности аналогична идее
разложения качественного признака, первоначально реализованной
на примере окраски млекопитающих. Так, обычная «дикая»
окраска кролика оказалась зависящей от самостоятельных
генов: агути (его аллеломорф — сплошная окраска), гена
наличия окраски вообще (его аллеломорф — альбинизм) и далее
ряда генов, определяющих различные этапы пигментации
(образование желтого пигмента, черного и т. д.).
Качественные признаки также можно выражать
количественно. Пегость млекопитающих обычно считается
качественным признаком, однако многие авторы создавали шкалы, в
которых степень развития пегостен можно было выражать
числами или процентами.
Наоборот, такой типично количественный признак, как
высота растений, можно выражать как качественный («высокие»
растения и «карликовые»).
Таким образом, главное заключается в том, насколько
велика разница между особями по изучаемым признакам —
есть ли между ними переходы или резкие границы. Именно
отсутствие границ между ступенями развития признака
затрудняет их анализ и требует иных параметров для
характеристики популяций по количественным признакам и для
вскрытия их генетической природы.
По отношению к количественным признакам приходится
иметь в виду два обстоятельства: а) сильную зависимость их
выражения от условий внешней среды и б) возможную
детерминацию их многими генами. Легко видеть, что даже
простейшее расщепление по одной паре генов может быть полностью
прикрыто вариацией, вызванной средой (табл. 22).
180
Таблица 22. Расщепление по одной паре генов по количественному признаку (значения классов условные4)
' „ I
1 Поколения и их генотипы ,
\ Рг:аа
\ Pt:AA
\ F^.Aa
\ F,: АА 4 2Аа + аа
i
17
1
Классы и их частоты
18
19
20
21
22
23 24
Средовой вариации нет
1
1
1
Сильная вариация, вызванная с
1
1 2
редой
25
1
26 1
1
1 1
1
\
27 1 28 1 29 \
Рх:ла
Р%АА
F,:Ao
Ft:aa
2Ла
АА
XF,
1 l I
1
1 1
1
1
L_
б 1
6
1 6
15 1
15
I
1
15
20 1
1 '
20
2
22
15 1
6
15 1
12
27
6
15
в
.ТО
36
1
1
20
1
40
1
42
Г>
15
12
6
36
15
6
2
15
_ч
20
1
20 1
22 1
1
15
15
1
JLl
6
6 1
_l!
1
1 1
1
I i
Приведенный шпитетнческин пример (нижняя часть
таблицы!, может быть, является крайним. Здесь, говоря словами
\ С. Серебровского, генотипнческин каркас невозможно
вскрыть из-под паралитического, несмотря на его простоту.
В действительности, как мы увидим дальше, при развитии
количественных признаков происходит сочетание сложной гено-
типмческои картины расщепления и средовых влияний, но
первая может быть установлена теми или иными методами.
Случаи моногибридного наследования количественных
признаков. В тех случаях, когда влияние среды не очень сильно,
можно обнаружить и такие, впрочем, редкие случаи, когда
различие в количественном признаке обязано расщеплению по
одному гену. Еще Менделем были описаны результаты
скрещивания двух форм гороха — низкорослой (карликовой) и
нормальной. Гибриды первого поколения были высокими, а в
F2 наблюдалось расщепление на 787 высокорослых и 277
низкорослых, что соответствует простому моногибридному
соотношению 2,96:1,04. Подобные же результаты были получены
Менделем и на фасоли при скрещивании нормальной фасоли
с карликовой формой Phaseolus nanus. Сейчас для нас ясно,
что карликовые формы как гороха, так и фасоли появляются
в результате рецессивных мутаций одного гена, поэтому и
наблюдается доминирование нормальной высоты растения в
первом поколении и моногибрндное расщепление во втором
поколении. Отчетливое расщепление по весу наблюдалось среди
взрослых кур —дочерей петуха, гетерозиготного по гену
карликовости da* (рис. 49). В сходном случае моногибрндного
наследования длины стебелька проростка у томатов
вариация Fi полностью соответствовала вариации родительских
форм с большей длиной стебелька, однако в F2 двувершин-
ность была сильно смазана в силу влияния внешних условий.
Еще менее ясным было расщепление в F2 по ширине грудной
клетки при скрещивании пород овец соутдаун и рамбулье
(рис. 50), интерпретируемое изучавшим это скрещивание Рит-
цманом как моногибрндное. Действительно, особи F\ были
полностью узкогрудыми, в F2 же отделить широкогрудых от
узкогрудых было уже трудно.
Параметры для характеристики непрерывной вариации
количественных признаков. Значительно чаще наблюдаются
количественные признаки, характеризующиеся непрерывной
вариацией, и тогда выделить какие-либо определенные
категории нет возможности. Это приводит к непрерывному ряду
изменчивости у особей любых поколений скрещивания. К
таким относится большинство хозяйственно-полезных признаков
растении и животных. Биологу — животноводу и
растениеводу — ясно, что развитие любого такого признака в конечном
счете зависит от совокупного действия многих органов и тка-
1*2
ией Понятно поэтому, что он должен контролироваться как
Правило, многими генами, влияющими на развитие этих орга-
нов и тканен. Полигенная их природа, как будет показано
в дальнейшем, была доказана. Однако каждый из этих генов
в отдельности не может быть выделен. Поэтому для
характеристики популяции по подобным признакам нельзя
использовать ни соотношение генотипов, ни частоты генов.
Единственный путь, во всяком случае на первом этапе анализа,—
использование обычных статистических параметров: средней арнфме-
?«
/ \ / '
^^ [_ \ i
■:/ rj 15 1i :9 21 25 25 Z<
' Мибоа бдс, и
^j: ^
1>
t
.1 -.:■•.
? п л и * у: :*
Щи*з sy>eJ rst'.'L >
Рис. 49. Расщепление по весу среди
взрослых кур — дочерей одного и
того же петуха. Карликовые dw
(68 дочерей), нормальные Dw (80
дочерей) (по Хатту).
Рис. 50. Наследование ширины
грудной клетки в F\ я Ft ври
скрещивании овей рамбулье
(Pi) и соутдаун (Pi) (по
Вридту).
тической (х), среднего квадратического отклонения (о), ва-
риансы (о2). С их помощью можно дать достаточно
исчерпывающую характеристику родительских форм, первого (F{)%
второго (F2), третьего (F3) поколении скрещивания, а также
обратных^ скрещиваний особей из F{ с каждым нз исходных
родителей (FBl и FBi). А это дало возможность установить
общие закономерности наследования количественных
признаков и объяснить их с помощью определенно»! генетической
модели.
Закономерности наследования количественных признаков.
Они могут быть сформулированы следующим образом:
1) кривая распределения особей F, по изучаемому признаку
обычно располагается между кривыми распределения
родительских форм. Средняя арифметическая Fx чаще всего
промежуточная между средними арифметическими родительских
форм- 2) средняя арифметическая F* примерно равна средней
арифметической F,, но вариация ^обе* F* *»^
ше, чем вариация особей F.; 3) кривые распределения особей
Г 183
из обратных скрещиваний (беккроссов) F} на каждого из
родителей сдвинуты ближе к кривым распределения тех из
родительских форм, с которыми произведено скрещивание;
Г8Г
8 10 12 Ш 16 18 20 22 24 26 28 дО
24
20\
«о
IS 20 22 24
8 10 12 Щ 16 18 20 22 24 26 28 7М
Количестдо щетинок
Рис. 51. Результаты скрещивания линий
дрозофилы, отличавшихся по количеству
стерноплевральных щетинок: Р — плюс-
линия (1+) и минус-линия (Я—); F\,
F% и FB (F\XH—) (по Рокицкому).
4) кривые распределения F2 от особей F2t взятых из разных
частей вариационного ряда F2, располагаются по-разному в
зависимости от того, из какой части ряда взяты особи F2.
Все перечисленные закономерности наглядно
иллюстрируются тремя примерами: на дрозофиле, курах и табаке.
На рис. 51 и в табл. 23 приведены полученные автором
книги данные по наследованию числа стерноплевральных ше-
1М
Таблица 23. Статистические показатели количества стериоплевральиых
щетинок у исходных родительских форм //— и(.-г, Ри Рг и Fb{Fv/H—)
Поколения
Родители: // —
/- лс
г
' 1
''"•;
FbU\:H-)
п
71
37
57
131
64
X • Ь-
~" X
12,44 ; 0,17
25,57 ±0,29
20,33 . 0,19
18,67 0,22
15.78 £0.36
■з 1
1.40
1.76 1
1.42 1
2.55 |
2,88
тинок у Drosophila melanogaster. Средняя арифметическая
для минус-линии (Н—) была 12,44 щетинки, для плюс-линни
(L + ) —25,57 щетинки. Средние квадратические отклонения в
Рзстт-я
20[
П
JZ1
ш=.
га
F^A
42 46
n-гП [Th
50 54 58 62
Ffx8
50 54 58
dec яиц, z
6f
&
ЦО 55 TO SS
oo
Рис. 52. Наследование веса яиц при
скрещивании кур леггорнов из двух
высоко инбридированных линий:
А — с мелкими яйцами. В — с более
крупными (по Робертсону).
Рис. 53. Наследование длины
венчика цветков у табака (по
Исту).
обеих линиях близки — 1,40—1,76. F} занимает
промежуточное положение между исходными линиями (х=20,33), и
вариация в ней невелика (а=1,42). В F2 средняя арифметическая
Слизка к средней арифметической F\4 н вариация резко
возросла (о = 2,55). Средняя арифметическая признака в Fa(f\X
X//-—) сдвинута в сторону средней арифметической минус-
185
.шипи //--. Вариации в FIt тоже большая, что очень рельефно
видно на рис. 51.
В опыте на курах скрещивались две ипбредиые линии,
отличавшиеся по весу яиц: в линии А л#ж46 г, в линии В хжбО г
(рис. 52). В F] средняя арифметическая всех яиц
промежуточная, около 52 г, изменчивость резко возросла, F2 получено не
было, но зато в двух FB кривые распределения сдвинуты или
в сторону линии Л, или в сторону линии В в зависимости от
типа обратного скрещивания.
Наконец, на рис. 53 показано наследование длины венчика
у табака по классической работе Иста. Данные по F\ и F2
соответствуют тому, что было показано выше на дрозофиле. Но
в опыте было получено F3 от особей, взятых от разных групп
F2. Очевидно, что F2 генетически неоднородно, так как в F3
получены отличавшиеся друг от друга кривые распределения,
сдвинутые вправо и влево в соответствии с положением в
вариационной кривой F2 родительских форм, взятых для
получения F3.
Теория полимерных генов. Для объяснения наследования
количественных признаков была предложена теория
полимерных, или множественных, генов. В простейшей форме, при
наличии только двух пар генов, влияющих на признак в
одинаковой степени (их действие суммируется), она была впервые
применена Нильсоном-Эле к окраске зерен и колосковых
чешуи у мягких пшениц. При скрещивании пшениц с красными
и с неокрашенными (белыми) зернами растения первого
поколения имели зерна промежуточной окраски. Во втором же
поколении зерна были окрашены в разной степени — начиная от
темно-красных и кончая белыми. Растения с белыми зернами
составляли примерно Vie всех растений F2\ частоты же
растений с красной окраской зерен по интенсивности ее
распределялись в ряд, примерно соответствовавший отношению 1:4:
: 6: 4. Это дало основание предположить, что в данном случае
происходит расщепление по двум парам полудоминантных
генов с аддитивным действием, т. е. генов, влияние которых
суммируется арифметически.
Схема такого расщепления в F2 и его реализация в виде
генотипов и фенотипов показана на рис. 54. В верхней части
рисунка (I) выписаны генетические формулы особей F2j ниже
(II) — количественное распределение генотипов при
суммарном действии генов и их условная феиотипическая ценность
(принимается, что феиотипическая ценность двойных рецесси-
иов равна нулю, а каждый доминантный геи в единичной дозе
дает 'эффект в виде условной единицы признака). Тогда
особи, у которых в генотипе есть все 4 гена, имеют фепотипиче-
t кую ценность, равную 4. Под номером III дано распределение
фенотипов внутри гепотнпических классов. На основе каждого
\ы>
генотипа создается вариация по феиотипическому выражению
признака в результате влияния внешних условий. Наконец, в
нижней части рисунка (IV) представлена картина фенотипи-
исской вариации в F2 как следствие сложения отдельных
фенотипов каждого генетического класса.
В дальнейшем Истом, Мазером и другими учеными была
разработана полигеиная модель наследования количественных
признаков, с помощью которой оказалось возможным объяс-
Рис. 54. Схема расщепления
в F2 по количественному прн-
знак7, определяемому двумя
независимыми парами адомн-
нантных генов с аддитивным
действием. Объяснение в тексте.
,) ' 2 J
•'Л'.у нтличкекче саше ~ти
нить наследование самых разных признаков у животных и
растений, в том числе таких важных в практическом
отношении, как яйценоскость кур, молочность крупного рогатого
скота и др.
Основные предпосылки модели полимерных генов
следующие: 1) гены адомииантны, т. е. гетерозиготы по тому или
иному полимерному гену занимают строго промежуточное
положение между двумя гомозиготами; 2) по силе действия гены
равны друг другу; 3) действие нескольких генов суммируется
аддитивно; 4) чем больше пар генов участвует в расщеплении,
тем меньше в F2 или FB доля особей, фенотнпически сходных
с исходными родительскими формами; 5) частоты градаций
количественного признака в потомстве соответствуют
коэффициентам разложения бинома Ньютона (а+в)*\ где
«—количество пар генов, по которым происходит растепление. В
примере, приведенном на рис. 54, расщепление типа 1:4:6:4:1.
Эти коэффициенты соответствуют коэффициентам разложения
бинома (а + в)4.
187
Согласно теории полимерных те\\оъ> ^роъенъ разъ\\т\\я
количественного признака в F\ и F2 должен быть
промежуточным между уровнями развития признака исходных
родительских форм. Признаки, которые изучал Нильсон-Эле у овса и
пшеницы, были в сущности качественными (окраска чешуи и
зерен), но были выделены количественные градации по тону
окраски, и тогда обнаруживался промежуточный характер F,
tsao
2000 2500
Вес, г
3Q00
Рис. 55. Наследование веса у кроликов
при скрещивании мелких польских
(Pi) и фландров (Р2) (по Пизу).
9 11 13 15 /7 19
Длина початка, см
Рис. 56. Наследование длины
початка у кукурузы. Исходные
формы: Pi—Самб; Р^—Блэк Мексикен
(по Исту).
и F2. Но есть немало примеров и чисто количественных
признаков, на которых наблюдалась та же закономерность. Так,
в опытах Касла при скрещивании кроликов двух пород,
различавшихся по длине ушей (ПО мм и 220 мм), в F\ были
получены кролики со средней длиной ушей в 164 мм. Такая же
средняя была и в F2. Подобные данные получены в работах
Пиза по скрещиванию кроликов, взятых из пород,
значительно отличавшихся по среднему весу (рис. 55). Однако
осложняющим обстоятельством у животных может быть так
называемый материнский эффект. Оказалось, что если отдельно
рассматривать потомство в Fu полученное от матерей, взятых
из породы кроликов более легкого веса, и от матерей из
породы более тяжелых, то между этими группами потомков
обнаруживается некоторая разница в потомстве в пользу
особей, происходящих от более тяжелых матерей. Но такой раз-
ice
нииы нет между потомством, происходящим от разных отцов.
Впрочем, величина материнского эффекта невелика, и
обнаружить его можно только на значительном материале.
На рис. 56 представлены данные по наследованию длины
початка у кукурузы, полученные Истом и Эмерсоном в их
классических работах по обо-
^ 26 \
ъ A f
§ fit
М
10
8
б
г
о
«п
п
U
снованию теории
полимерных генов.
Модель полимерных
генов позволяет понять и
такие случаи наследования,
когда изменчивость во
втором поколении выходит за
границы изменчивости
скрещивающихся родительских
форм (пород, сортов и т. д.).
На рис. 57 показаны
результаты скрещивания кур
гамбургских и себрайт-бентам
(все веса приведены к весу
петухов). Особи
гамбургской породы весили в
среднем 1100—1300 г (куры
около 1100 г, петухи около
1300 г). Себрайт-бентамы
весили только 600—800 г
(куры около 600 г, петухи
около 750 г). Для
скрещивания в качестве родителя
был взят очень крупный
петух гамбургской породы,
весивший 1350 г. Особи F\ были довольно однородными, со
средним весом примерно 1200 г. Второе же поколение
оказалось очень разнообразным. Наряду с промежуточными по
весу особями были куры и петухи, более крупные, нежели
гамбургские, и более мелкие, чем себрайт-бентамы (минимум
около 650 г и максимум около 1600 г). Для объяснения
такого расщепления в F2 Пеннетт и Бэйли высказали
предположение, что различие в весе между бентамами и гамбургскими
определяется четырьмя парами генов, которые сгруппированы
таким образом, что мелкие бентамы имеют формулу aaeeccDD,
а более крупные гамбургские — AABBCCdd. Кроме того,
учитывая, что помеси первого поколения имеют средний вес,
более близкий к среднему весу гамбургских, можно
предположить, что некоторые из этих генов доминантны или частично
доминантны.
189
юз тюттшъгжшкшшт
Рис. 57. Результаты скрещивания
крупного петуха гамбургской породы
(Pi) и мелких кур бентамок (г2).
Появление во втором поколении особенно тяжелых и
особенно легких кур можно рассматривать как результат
комбинации AABBCCDD и aabbccdd. Последующие скрещивания
подтвердили это предположение. Так, путем скрещивания
самых мелких кур с мелким же петухом из второго поколения
была получена в F3 группа очень мелких кур, достаточно
константных. Некоторые пары особен, промежуточные по весу,
также дали однородное потомство, т. е. была случайно
достигнута гомозиготность по некоторым из генов, влиявших на вес.
Но скрещивание самых тяжелых кур ( Vвесом 1390 г и9?—
1290 г) дало сильное расщепление — от 980 до 1350 г среди 9 2
и от 990 до 1630 г среди .V. Константную группу кур,
которые должны были быть в результате сочетания 4 доминантных
генов, получить не удалось. Возможно, что некоторые из генов
были сверхдоминантными, и тогда максимальный вес был
свойствен гетерознготам.
Подобное явление наблюдал и Пиз на кроликах — от
самых крупных кроликов константные семьи не были получены.
Таким образом, как ни удобна и привлекательна модель
полимерных генов, ряд конкретных фактов вынудил внести в
нее дополнения и усложнения.
Впрочем, надо иметь в виду статистическую природу
явлений, наблюдающихся при наследовании количественных
признаков, вследствие чего внешняя картина наследования в
громадном большинстве случаев (промежуточность первого
поколения и увеличение вариации во втором) может быть как бы
усредненным результатом генетических взаимоотношений при
скрещивании, несмотря на усложнения в ряде деталей.
Усложнения и дополнения к теории полимерных генов.
Вкратце они сводятся к следующему. Отдельные гены могут
обладать разной степенью доминирования, вплоть до еверх-
домннирования. Действия отдельных полимерных генов могут
быть не равны друг другу. Наконец, суммарное действие
нескольких генов может быть не только простым аддитивным, но
и осложняться различными формами межгенного
взаимодействия. В одних случаях одни гены мешают или ослабляют
действие других, в других, наоборот, усиливают. В обоих
случаях суммарное действие двух или нескольких генов
отклоняется от простой арифметической суммы действия отдельных
генов. Все эти усложнения могут отразиться на картине
расщепления в F2, даже если не учитывать влияние внешней
среды.
На модели дигибридного и полигнбридного расщеплений
Л. С. Серебровский показал, как изменяются кривые
распределения генотипов в F2 в зависимости от характера
доминирования, различий в силе действия генов и т. д. Так, при полном
доминировании создается асимметрия кривой, и при домини-
I'JO
ровании плюс-гена (усилителя, по терминологии А. С. Сереб-
ровского) скошенность будет направлена вправо, а
растянутость влево, при доминировании минус-усилителя — наоборот
(рис. 58). Лнндстром на
томатах доказал наличие
асимметричного
распределения некоторых
количественных признаков, причем
асимметрия наблюдалась у
диплоидных форм, но ее не
было у тетраплоидов в
силу того, что у последних
значительно реже или
совсем не выщепляются
рецессивные формы. А. С. Сере-
бровский разобрал также и
некоторые другие причины
возникновения
асимметричных рядов, в частности
ослабление действия
отдельных генов, по мере того как
количественный признак
возрастает. Оказалось, что
по мере приближения величины признака к минимуму или,
наоборот, к максимуму (как бы «потолку» признака)
происходит скашивание кривых распределения (рис. 59).
Рис. 58. Асимметрия кривых
распределения генотипов в F2 при полном
доминировании усилителя (сплошная
линия) и ослабителя (штриховая).
90щ
Рис. 59. Асимметрия кривых распределения кур по яйценоскости при
приближении признака к максимуму (справа) и минимуму (слева)
(по Серебровскому).
При определенном сочетании генов количественных
признаков могут возникнуть эксцессивные кривые распределения
или, наоборот, растянутые. Разная сила действия генов может
привести к тому, что вместо нормальной кривой
распределения для классических случаев наследования полимерных
генов, как показано выше, на рис. 55, получаются трех- или
даже многовершинные кривые, хотя А. С. Серебровскнй
оговаривает, что многовершинность может быть следствием более
191
♦лемснглрнон причины — неправильного шкалирования
признака.
Теория пол и генов Мазера. Мазер выдвинул идею о том, что
количественные признаки детерминируются многими слабо
действующими генами, которые он назвал полигенами. Но так
как группы их могут находиться в одной хромосоме, то
каждая такая группа сцепленных генов будет вести себя, как один
ген. В силу же кроссинговера в хромосоме иногда они могут
отделяться друг от друга и тогда ведут себя как
самостоятельные модификаторы количественного признака.
Не следует путать понятие «полигены» Мазера с понятием
полигенной наследственности в целом *.
Надо помнить о том. что гены, повышающие уровень
развития данного количественного признака (их можно назвать
плюс-генами), могут быть сцеплены и с генами
противоположного действия (минус-генами). Поэтому до тех пор, пока это
сцепление сохраняется, проявляется суммарный эффект всей
хромосомы в целом или блок-участка данной хромосомы.
Однако если при кроссннговере произойдет отрыв плюс-гена
(или плюс-генов) от минус-гена, то появляется возможность
для отбора по плюс-гену и тем самым повышения уровня
развития признака.
Концепция Мазера о полигенах не является общепринятой.
Ряд исследователей считает, что она может быть применима
только к некоторым случаям наследования количественных
признаков. Чаще же всего надо иметь в виду наличие главных
генов, в большой степени влияющих на развитие признака
(олигогенов), затем более слабых, типа полигенов Мазера, и,
наконец, генов-модификаторов, которые, действуя на другие
признаки, одновременно модифицируют действие основных
генов. Теоретические и экспериментальные исследования в
общем подтвердили такое представление, хотя возможны
различные варианты. По-видимому, между главными генами —
олигогенами — и полигенами имеются все переходы, и любые
изменения в генотипической структуре сказываются на
значениях количественных признаков.
Для выяснения вопроса о сравнительной роли генов с
сильным действием и слабым была создана математическая
модель с одной группой сцепления, содержащей 10 генов (8 со
слабым и 2 с сильным действием), и проверены результаты
отбора на приспособленность с помощью вычислительных
машин. Оказалось, что если суммарный эффект 2 олигогенов в 8
и более раз превышал эффект полигенов, то отбирались в
первую очередь олигогены. При меньшей разнице отбирались по-
• Такая путаница имеет место в книге Дж. БрюбеАкера сСельскохоэяА-
ствелмая генетика» (М., 1966),
192
лнгеиы. Подиергалси анализу и вопрос о возможности
создания блока из нескольких генов, который должен вести себя,
как один геи или как бы суперген.
Тернер на математических моделях показал, что
естественный отбор может благоприятствовать созданию таких
блоков. Это соответствует концепции Мазера, однако не во всех
случаях. Расчеты привели Тернера к выводу, что при трех и
более генах отбор благоприятствует не минимальной частоте
рекомбинации, т. е. их более сильному сцеплению» а некоторой
оптимальной частоте. Очевидно, что в этом случае блоки генов
оказываются неполными.
Возможности определения числа генов, влияющих на
количественный признак. Поскольку различия между
используемыми для скрещиваний линиями или породами по тому или
иному количественному признаку определяются, как правило,
многими генами, было бы интересно определить количество
этих генов, но совершенно очевидно, что при этом придется
прибегнуть к известным упрощениям и ограничениям, а
именно: 1) что сила действия изучаемых генов одинакова и они
адоминантны; 2) что скрещиваемые формы гомозиготны по
этим генам; 3) что все гены, усиливающие признак, находятся
у одного родителя, а ослабляющие у другого. Однако
существенно, что невыполнение отдельных ограничивающих условий
влияет на полученный результат, т. е. на определяемое число
генов, только в сторону его увеличения. Таким образом,
применение разбираемых ниже формул дает во всяком случае
минимальное число генов, которыми различаются
скрещиваемые формы по данному количественному признаку.
Впервые такую формулу для определения п генов (не
аллелей, число аллелей тогда 2п) предложили Касл и Райт:
где D — разница между средними родительских пород или
линий; о2 — вариансы F2 и А*.
Для исключения влияния доминирования А. С. Серебров-
ский предложил более общую формулу:
3D»-4DDl+4Dl«
—т^г- <»
В этой формуле есть еще величина Di—разница между
средней величиной признака Ft и признака у рецессивного типа.
• В отношении формулы Касла—Ра1та, • также ■ иных, боле*
сложных, критические замечания высказываясь радом авторов, в том часл«
Ю. А. Фшншчеико, О. Тадмным ■ АР-
193
13 П Ф. Рокнцмй
При адомшшровашш D = 2Duji тогда эта формула
превращается в формулу Касла— Ранта.
При полном* доминировании D = Dit и тогда
3D-
"-щъ-ъу (3)
Вместо F2 можно использовать FB (скрещивание F4 с
родителем, имеющим меньшее развитие признака). В этом случае
доминирование также не будет иметь значения и может быть
использована предыдущая формула.
Несколько более сложная формула была предложена Бер-
тоном (Burton):
= 0,25(0,75-/1+/*2)Д2
4-4, ' [)
Здесь D — разница между средними родительских форм, а
. Xfx—xPx
D '
Эта формула имеет те же ограничения, что и формула Се-
ребровского, и указывает на минимальное число генов,
участвующих в скрещивании.
3. С. Никоро применила формулу Бертона к литературным
данным о наследовании пегости у мышей и получила п = 3,5,
т. е. ~4.
При анализе наследования числа стерноплевральных
щетинок у дрозофилы П. Ф. Рокицкии также применил формулы
Серебровского (неопубл.).
Результаты представлены в табл. 24.
Таблица 24, Данные о числе генов, которыми отличались селектированные
по количеству стерноплевральных щетинок линии дрозофилы
1 Скрещиваемые линии
HlxL\°
uU,.r\4
n:i 'L\ (821)
ъ (823)
H»xL\°
"Z ль1<821)
1 » (823)
D*
П '" 2 2
8 (7Ft - oFi)
6,1
9,10
17,2
3D* — 4DD, -*- 40»
2 2
\b(oF2-oFi)
6,4
9,8
17,8
Те же формулы, но с подстановкой
°Рг+°Рг
вместо oFi величины п—
5,4
4,0
8,0
5,6
4,1
8,2
194
Совершенно очевидно, что в варианте Н\у1? участвовало
меньше генов, чем в остальных скрещиваниях, что
подтверждается и сравнением кривых распределения F\ и F2. Для
скрещивания пл и Li характерна сильная вариация в гь
указывающая на наличие расщепления еще в этом поколении.
Поэтому для включения в формулу вместо on была взята
средняя арифметическая между средними квадратическими
отклонениями родительских форм. Эта замена не повлияла на
примерное значение п для скрещивания п2 и Li, но привела
к уменьшению вдвое значений п для остальных вариантов
скрещиваний.
Несмотря на всю неточность применяемых формул в связи
с ограничивающими их условиями, они дают информацию о
степени генетических различий между скрещивающимися
формами по данному количественному признаку.
Выделение отдельных генов с помощью
хромосом-анализаторов и сигнальных генов. Наиболее точный метод
установления отдельных генов количественного признака — проведение
тщательного генетического анализа с использованием
хромосом, маркированных определенными генами, и сигнальных
генов, находящихся в отдельных хромосомах. Этот метод был
детально разработан А. С. Серебровским, но, к сожалению, он
может применяться только у видов, генетика которых хорошо
разработана (дрозофила, кукуруза). Однако
немногочисленные работы с применением этого метода все же дали довольно
много в познании генетики количественных признаков. Так, в
моей работе (1927) путем скрещивания исходного материала
с линиями-анализаторами, содержащими сигнальные гены
во II и III хромосомах, и анализа реципрокиых скрещиваний
было доказано наличие в половой и III хромосомах в плюс-
линии Li14 генов, влияющих на количество стерноплевральных
щетинок дрозофилы, причем эффект половой хромосомы
измеряется величиной ~ 3,5 щетинки (при х = 21), а III
хромосомы ~ 9 щетинок. Роль II хромосомы была незначительной.
В лаборатории А. С. Серебровского М. Л. Карпом был
проведен очень интересный и поучительный анализ генов III
хромосомы, влияющих на количество стернитальных щетинок.
Для этого были использованы шесть рецессивных генов и
получены многочисленные кроссоверные формы, позволившие
установить роль отдельных участков III хромосомы. Было
показано наличие не менее шести генов, влиявших на количество
Щетинок: двух ослабителей, одного — очень сильного
усилителя и трех-четырех — со слабым действием, причем
расположены они в хромосоме вперемежку.
Но не следует и переоценивать число генов, влияющих на
13* 195
количественный признак. Так, Спикетом и Тудеем был прове-
лен генетический анализ различии между двумя линиями
дрозофилы vg-4 и с£-6, в которых ранее проводился отбор на
увеличение количества стерноплевральных щетинок (подобно
тому, как это было сделано мною в работе 1927 г.), и
установлены конкретные гены. В III хромосоме было обнаружено
только два гена — За и 36, влиявших на щетинки. Суммарный
эффект их был аддитивным. Во II хромосоме — лишь один ген.
Между этим геном и геном За наблюдалось довольно сложное
взаимодействие. 80% генетических различий между
селектированной плюс-линией vgA и инбредной линией Орегон
определялось только этими 3 генами. Кроме того, для одной из
плюс-линий (vg-б) были обнаружены еще два гена,
расположенные в половой хромосоме (в районах около 2-й и около
50-й морганид генетической карты). Таким образом, различие
между линией vgS и Орегон зависит от пяти генов, а между
линией vgA и Орегон от трех. Хотя авторы называют их
полигенами, но предпочтительнее употреблять для них
наименование олигогенов.
Количественные признаки и гены-регуляторы. В свете
современных представлений о существовании генов-регуляторов
и структурных генов интересна попытка перебросить мост от
обычных положений о генах количественных признаков к этим
более сложным представлениям. Уоткин изучал различия
между нормальными формами томатов и мутантными,
которые характеризуются ограниченным ростом центрального
стебля. Оказалось, что в тканях мутантных форм уменьшено
количество пероксидазы. Он объясняет этот факт изменением
действия генов-регуляторов, а не структурных генов, и
считает, что в картине наследования и развития количественных
признаков основную роль играют именно гены-регуляторы.
Метод треугольника. А, С. Серебровский предложил очень
оригинальный метод анализа наследования количественных
признаков, названный им «методом треугольника». Он может
быть применен не только к случаям скрещиваний,
поставленных по определенному плану, когда получают Fx и F2 от
родителей, различающихся по количественному признаку, как
мы видели выше, но и к массовому популяционному материалу
с большим количеством отдельных скрещиваний.
Оказывается, что если разнести данные по сибсам отдельных
скрещиваемых пар на решетку, где на оси абцнес нанесены а2, а на оси
ординат — средние значения признака, то получится очень
интересное и специфическое распределение точек. Но для
этого надо предварительно рассмотреть результаты всех типов
моногибридных скрещиваний, представленных в табл. 25.
Сибсы каждого типа скрещивания характеризуются
разными значениями средних арифметических и варианс — феио-
1%
Таблица 25. Типы мопогибрмдных скрещиваний в популяции, их частоты и биометрические показатели
для потомства каждого типа скрещиваний *
Тип скрещивания
QQXQQ
J авхАа
I аахАА
i АахАа
\ АсхАА
АахАА
Частота скрещивания
ч*(я-ч')
4PV | q22P"
{ 2/>Я Я*
W ( ++
4pV(2pv 2pq)
4. ( 2W p«
рЧр'р')
Состав потомства
ua
аа + Аа
Аа
аа + 2Аа + АА
Аа + АА
АА
1
Л1 сноса потомства
ш
Ма+-у i
Ma + i
Ma + i .
3
Ma + ~y i
Ma + Ж
•t
и
i'*
0
2
■f"
0 I
1
,a.
T
1
4
2
4
1
4
0'
i* + ol
o?
«» + o*
i» + «';
Сохранены все обрзиачения А. С. Серебровского (среднее арифметическое М; омрнансы — гепотшшческаи ог, феко-
типическая а£, паратигжческа* о;; / — действие гена в одной дозе).
(О
типического гснотнпическоЛ и средоной — в зависимости от
генетической структуры. При этом принимается, что
доминантный ген в одной дозе обладает действием иа признак,
(мвным I, а » двух дозах (у гомозиготы) —2 L
Данные таблицы представлены графически на рис. 60. На
нем нанесена решетка М -- о2 •. Следует обратить внимание иа
расположение точек. Первые три точки слева расположены на
линии гч2. В трех скрещиваниях (1, 3 и 6) в потомстве нет
расщепления, полому а,*=адг. Увеличение значения аф2 идет за
счет эффекта гена .4, обозначенного через i\ поэтому градации
Н
Г:
К-'
•VT'
■W
.•>•/.
, I
.*-?. ,
Рис. 60 Теоретическое распределение
точек разных сносов в популяции в
поле Л) -- о* при моногибридном скре-
щяаашм и их частоты (по Серебров-
скому).
Рис. 61. Распределение
эмпирических точек отдельных сибсов
в поле М — о2 (по Серебров-
скому).
1 2
по оси о%г—j- Р и — Р. На первой расположены две точки,
а на второй одна. По оси средней арифметической градации:
Л/а+О (уровень признака у рецессивной гомозиготы аа)\
Affl-f . |. Ма + -1; Afa-f -x- i и Afa-f- --- i (нижняя точка
треугольника). Вершина треугольника (точка справа)
соответствует расщеплению при скрещивании двух гетерознгот, при
2
этом средняя для этого сибса Ma-f «\ а аФ2=ая24- — Л
Поскольку существует вариация между сибсами и но
средним арифметическим, и по фенотипическим вариансам, то
между эмпирическими точками для многих сибсов будет раз-
рос <рис 61), однако они будут группироваться вокруг шести
мгги'чДю* ^4£6J>0,ICK,,ft употреблял старое обозначение для средней ар*Ф'
т
I
11С,пров, соответствующих теоретическим точкам. Сгущение
нокруг правых 3 точек будет больше, нежели "вокруг
остальных.
Различным генотипическим распределениям соответствуют
треугольники Л1 — о2 с разным расположением точек. А. С. Се-
ребровский исследовал, как
влияют на расположение
точек доминирование, сцеп- . . •/•. У
ление с полом, число генов
и их сила, летальность,
сцепление генов и др. (рис. 62).
Метод треугольника очень
интересен, однако основная
трудность для его
использования на практике
заключается в
недостаточности разрешающей
способности, что зависит от
степени вариации в данном
признаке, вызываемой
средой (паратипической). Чем
меньше эта вариация, тем
отчетливее видны центры
сгущения эмпирических
точек. При определенном
соотношении между средовои и
геиотппической вариацией
ряд генотипических групп
или сливаются, или разброс
их становится совершенно
неопределенным и никаких
центров выделить не
удается. Несмотря на эту трудность, метод Серебровского
заслуживает внимания и может быть применен не только для
анализа наследования количественных признаков, но я для
изучения других вопросов, например структуры популяции,
породы, линий, процессов отбора в течение ряда поколений и др.
Разрабатывая метод треугольника, А. С Серебровскнй
очень близко подошел к необходимости выяснить состав фено-
типической вариансы и ее отдельных компонентов — генотипн-
ческон и средовои варианс, но для него основной задачей было
вскрыть отдельные гены, влияющие на количественный
признак, поэтому он не пошел дальше в разработке новых
параметров для характеристики популяции как единого целого,
которые были предложены рядом ученых, в том числе Райтом,
Фишером, Лашсм и др.
199
Ш
Рис. 62. Различия в расположении
точек на поле М — о* при увеличении
числа генов (7), усилении
доминирования (ll)t увеличении разницы
между силами действия генов (III).
Расщепление по двум парам генов (по
Серебровскому).
Разделение общей фенотнпической вариации на отдельные
компоненты — средовую и генотипическую. Выше для
характеристики количественных признаков использовались обычные
биометрические показатели х и а. С их помощью удалось
сделать существенные выводы об общей картине наследования
количественных признаков и лх генетической обусловленности
отдельными генами. Для этого анализировались определенные
скрещивания линий, пород или сортов, отличающихся по
развитию количественного признака. Однако при анализе
популяции, в которой осуществляется большое количество
разнообразных скрещиваний, нет возможности оперировать
отдельными генами и пользоваться такими параметрами, как частоты
генов. Поэтому понадобились новые параметры для
характеристики генетической структуры популяции по количественным
признакам. Подойти к их установлению можно следующим
образом.
Любая варианта в популяции имеет какое-то
стенотипическое значение у. Это фенотипическое значение может быть
выражено как отклонение у от популяционной средней \i и
обозначено условным символом Р:
Так как вариация в популяции определяется в конечном
счете различиями в генетической структуре отдельных особей
и влиянием средовых факторов, то и для каждой отдельной
особи ее фенотипическое значение Р можно рассматривать как
результат суммирования генотипического (6) и средового (Е)
отклонений, т. е.
P=G+E. (5)
Но из этого, конечно, не следует, что в фенотипе каждой
особи можно выделить самостоятельные доли, зависящие от
генотипа яйца, из которого развился данный фенотип, и от
условий среды. Фенотип особей — это единый целостный
организм. О генотипе особей непосредственно, без анализа их
потомства судить невозможно. Выделение же долей вариации,
зависящих от наследственности и от среды, производится
только для популяций.
Чтобы определить вариансы отклонений G и Е для
популяции, надо возвести каждое отклонение в квадрат,
просуммировать все отклонения и разделить суммы на п.
Если мы возводим в квадрат левую половину равенства, то
это же надо сделать и с правой половиной:
Я2=(С+£)2;
P* = G2+E2+2GE.
эд
"алее, разделив каждый член равенства на л, получим:
v/>2 2 G2 I£* 2ZGE
п п п
v рг £ G1
Величина —— представляет собой аР2. может быть
п п
2 Е2
обозначена как ас2, a как <л?. Сложнее вопрос о велп-
п
чине — . Так как G и Е представляют собой фенотипиче-
п
ские и средовые отклонения, то сумма их произведений — кова-
рианса между этими отклонениями (для краткости можно
назвать ее ковариансой между генотипом и средой). Как известно,
коварнанса входит в структуру коэффициента корреляции.
В данном случае удобнее заменить коварнансу значением тса%ч
2 GE 2IGE *
исходя из того, что =rGEj a =2гсЕО0аг.
В таком случае формула примет следующий вид:
Ор2=ос2+Оег+2гСеОсОе. (6)
Если предположить, что корреляция между вариацией генотя-
пических и средовых факторов равна нулю, то формула
упрощается:
ap2=OG2 + 0£2.
Здесь ор2 — общая фенотипияеская варианса; ос2 —доля ва-
риансы, зависящая от различии между отдельными особями
популяции по их наследственности; о** — доля вариансы,
вызванная различиями в условиях внешней среды, при которых
развивались особи популяции.
Соотношения между значениями ос2 и ajs2 могут быть
различными. Так, если растениеводу приходится работать с
чистыми линиями растений-самоопылителей, то внутри чистых
линии величина ос2 будет равна нулю или. с учетом изредка
возникающих мутаций, очень мала. Тогда орг почти полностью
будет сводиться к а*2. В условиях эксперимента с животными
также возможны разные варианты. При максимально
контролируемых условиях внешней среды, например в специальных
камерах, можно достичь большого единообразия условий.
Тогда различия, обнаруживаемые между опытными
животными, будут в большой степени определяться их генотипами.
Наоборот, для изучения влияния различных факторов среды
надо брать животных, максимально однородных генетически,
Для чего используют особей из инбредных линий или однояйце-
20!
вых двоен. В популяциях сельскохозяйственных животных,
состоящих из генетически различных особей, разводимых в
разнообразных условиях кормления и содержания, общая
стенотипическая варианса популяции оР2 представляет собой
сложный результат суммирования различии между входящими в нее
и генотипически отличающимися друг от друга группами или
особями (эти различия измеряются вариансой ас2) и различий,
вызванных условиями внешней среды (измеряются вариансой
внутри каждой группы особей, более или менее сходных
генетически). В этом случае на фенотипической вариансе может
отразиться и наличие корреляции между G и £. т. е. известной
сопряженности в вариации генотипов и характере их
фенотипической реакции на различные условия среды. И тогда надо
учитывать последний член уравнения 2гсео0ое.
Таким образом, получено наиболее общее выражение,
характеризующее фенотипнческую структуру популяции по
количественному признаку. Фенотипическая варианса
распадается на два компонента — генотипическую и средовую ва-
рнансы.
Коэффициент наследуемости как мерило доли генетической
вариации в общей фенотипической вариации популяции.
Какими бы путями ни создавалась фенотипическая
изменчивость в популяции и из каких бы компонентов ни состояла, она
всегда может быть выявлена практически и измерена
вариансой. Самое существенное заключается в выяснении того, какую
долю в общей фенотипической вариансе занимает варианса,
зависящая от генетических различий в популяции, т. е.
уставе2 D
вовление величины —-. В простейшем случае она равна
of2
Эта величина, обозначаемая ft2, получила название
коэффициента наследуемости (heritability, Heritabilitat). Термин
«наследуемость» стал общепринятым, хотя он не совсем удачен,
так как буквальное его понимание может привести к
недоразумениям. Под ним надо понимать только то, что вытекает из
форм\лы Л-= —-. а именно, что кг является мерилом удель-
Орг
кого веса генотнпической изменчивости в общей изменчивости
популяции.
Ввиду того что коэффициент наследуемости принадлежит
к числу важнейших параметров, служащих для
характеристики популяции по количественным признакам, в гл. 6 будет
дано его обоснование, подробное изложение методов
определенна и показано его значение.
702
А сейчас перейдем к вопросу о том, какова генетическая
основа фенотнпической варнансы и ее компонентов. Для этого
сначала определим ряд понятий, таких, как популяционная
средняя, закономерности суммарного действия генов и их
средний эффект, генотипиче-
ская и селекционная
ценности особи и др.
Популяционная
средняя при одной паре
генов. Всякая особь может
быть охарактеризована
по ее фенотнпической
ценности, которая
представляет собой производное
генотипической ценности
и сргдовых влияний. Из
фенотнпической ценности
отдельных особей
складывается фенотипическая
ценность популяции как
целого. Так как
отклонения, вызванные
условиями внешней среды, идут
в разных направлениях,
то, кроме некоторых
специфических случаев,
среднюю средовых
отклонений популяции можно считать равной нулю, и тогда
популяционная средняя отражает одновременно п феноттгнче-
скую и генотипнческую ценность особей популяции. Иначе
говоря, средняя фенотипическая ценность популяции равна
генотипической ценности ее генотипа (или генотипов).
Для простоты можно опять обратиться к модели моногнб-
ридного расщепления в популяции, чтобы в дальнейшем
перебросить мост к системе многих генов. В популяции имеются
генотипы А\А\, А\А% и А*Аг- Если принять, что ген А% в
одиночной дозе вызывает увеличение признака на а, тогда А*А2 имеют
генотипнческую ценность, равную нулю, .4t>li —2а, гетерози-
готы А1-4- при отсутствии доминирования — а. Для расчетов
выгоднее принять за нуль гетерозигот А%Аъ тогда генотмаиче-
ская ценность А%А\ будет выражаться +а, для генотипов же
Л2А2 — величиной — а. Вся генетическая вариация будет
определяться разницей между А*А% и Лу!*. Она равна 2а. Но
ситуация осложняется возможностью разного доминирования,
представленного на рис. 63. Величиной d можно обозначить
разницу между конкретным значением А%Аг (для данного
случая) и нулем, если бы ЛИз находились точно посередине между
Рис. 63, Соотношение между а в d вря
разных стеяевях доамшяроваюи. Объяс-
в тексте.
20J
А\А\ и A<:A*. Тогда при отсутствии доминирования d = 0 (I
случаи), при неполном доминировании йфа, но a>d (II и III
случаи). При полном доминировании d=a (случай IV). Если
i/>ti, наблюдается сверхдомииирование, причем оно может
быть двояким: плюс-сверхдомииироваиием и мииус-сверхдоми-
ннрованием (У и VI случаи). Степень доминирования может
быть выражена отношением —. Генотипическая ценность гете-
а
розиготы А\А2 равна d. Таким образом, зная конкретные
значения признака у гомозигот А\А{ и ЛИ2, а также гетерозигот
/М:> можно определить значения а и d. Так, для веса
карликовых мышей (pygmy — pg) и нормальных были получены сле-
Н~ PS
дующие значения: 14 г, — 6 г. Гетерозиготы имели
+ PS
10 Т 14~6 л -А Ю 14 + 6 О
средний вес 12 г. Тогда а= —-— =4 rt rf= 12 — =2 г.
2
Степень доминирования — • 100=50%.
Так как частоты генов А\ и А2 в популяции равны р и q, то
для получения среднего генотипического и одновременно фено-
типнческого значения всей популяции надо перемножить
частоты каждого генотипа на его ценность:
для A Hi (+a)Xp2=ap2
» AiA2 dX%PQ = 2dpq
» А2А2 (—a)Xq2=—aq2
Z = ffia-q*a+2dpq = a{p*-(p)+2dpq = a(p—q)+2dpq (8)
Это и будет средняя для популяции (хР или Мр). Левая
часть a (p—q) относится к гомозиготам, правая — к гетерози-
готам. При отсутствии доминирования (d=0) MP=a(p—q) =
= a(\—2q). При полном доминировании d=a, и тогда с
учетом обоих членов суммы МР=а{1-— 2q2).
Зная значения and для любой пары генов, а также их
частоты в популяции р и q, можно рассчитать среднюю
генетическую (фенотипнческую) для популяции по гену данного
признака.
Аддитивная и неаддитивная суммация генов. От разбора
ситуации с одной парой генов надо перейти к случаю влияния
многих генов разных аллелей. Возникает вопрос о том, по
какому закону создается совокупный эффект многих генов:
происходит ли простое суммирование действия отдельных
генов. 1. е осуществляется закон аддитивности, или же при этом
суммировании создается дополнительный эффект за счет взаи-
молсйстпии между генами. В первом случае А\ + В\*~Ш (А\ +
ли
+ Й1)=ад+ал, во втором ^i + B,==2 (Ai + Bx) = aA+aB+IAB.
Эту последнюю величину /дв можно назвать эффектом
взаимодействия *.
Разницу между простым аддитивным действием и
эффектом взаимодействия можно проиллюстрировать на двух очень
сходных примерах из генетики окраски мышей.
Известны две самостоятельные пары генов: ген С
—доминантный фактор окраски, его аллель (^ — ослабитель окраски,
входящий в альбинотипическую серию; ген В — доминантный,
черной окраски, его аллель b определяет коричневую окраску.
Расселом были изучены количество гранул пигмента
меланина на единицу объема волоса и средние размеры гранул
пигмента, выраженные в микронах (ц). Сопоставим
результаты, полученные по обоим признакам (табл. 26—27).
Таблица 26. Среднее количество гранул у особей
четырех генотипов мышей
^"~~"^-—^^^ Аллелн гена В
Аллелн генл С "^""*-—^^^
С —
Разница 2ас
1 в-
95
38
57 |
I
ьь
90
34
56
pj
5
4
—
Таблица 27. Средние размеры гранул (в ji) у четырех
генотипов мышей
~~^-^^^^ Аллелн гена В
Аллели гена С *—-^^^^
С —
в-
1,44
0,94
Разница 2аг 1 0,50 |
с 1 !
! •
0,77
0,77
0,00
т 1
Разнице 2ag I
0,67
0,17 J
—
Очевидно, что разница между генотипами 5- и bb
одинакова, как в группе С-, так н в группе с*с», точно так же
одинакова разница между генотипами С- и с« в группах В- и bb.
* Взаимодействие между неаллельными генами обычно называют эок-
статическим взаимодействием. В статистической генетике термин сэпкстатм*
чсскнй» употребляется в более широком смысле слова, нежели в случае
взаимодействия генов при обычном дигмбридном скрещивании.
206
Таким обра «ом, эффекты обоих генов суммируются аддитивно.
Значение признака у особен каждого сложного генотипа
равно простой сумме действия двух генов.
Совсем иная картина наблюдается при изучении действии
тех же генов на средние размеры гранул (табл. 27).
Ршэняиа между С- и Ci* очень значительна при наличии В
и отсутствует при Ык То же относится и к действию гена В—Ь
на фоне генов С—г*. Таким образом, здесь нет простого
аддитивного суммирования действия генов, а налицо
дополнительный эффект взаимодействия генов.
Правда, приведенные примеры относятся к
количественному действию единичных альтернативных генов, но, как мы
увидим дальше, эти же принципы аддитивности и
взаимодействия относятся к суммарному действию многих генов,
определяющих чисто количественную вариацию в популяции.
Принцип аддитивности может проявляться и в суммарном действии
генов одного и того же локуса.
Классический пример такого простого суммирования —
содержание витамина А в эндосперме кукурузы в связи с
наличием гена У (желтый эндосперм):
Генотнп Доза Витамин А, ед/г
УУУ 0 0,05
УУУ 1 2,25
yYY 2 5,00
YYY 3 7,50
Однако есть случаи, когда увеличение количества
доминантных генов в генотипе уже не влияет на усиление признака.
С другой стороны, возможна и такая ситуация, когда
комбинация одних аллелей проявляет полиостью аддитивный
эффект, а комбинация других дает эффект взаимодействия, т. е.
количественное выражение признака оказывается более
высоким, чем это ожидалось бы по аддитивной схеме. Наконец, по
аддитивному признаку могут суммироваться и эффекты сверх-
доминирования. Так, В. И. Поляковским было показано, что
овцематки, гетерозиготные по одному из генов трапсферринов
сыворотки крови, давали больше двоен, чем гомозиготы
(~на 8%), и гетерозиготы по гену гемоглобина НЬ больше,
чем гомозиготы (~на 9%). Автор смог проанализировать
эффект совместного действия обоих генов в гомо- и
гетерозиготном состоянии, который оказался более резко выраженным.
Это видно из табл. 28, охватывающей данные за два года.
Суммарное действие генов выражалось примерно в 18%
прибавки двоен у гетерознгот, т. е. происходило по аддитивной
схеме.
JUfi
ТиПища 28. Количество диоеи в зависимости от эиготиостя
по Т/ и НЬ лом уйм оицшлток
1 иди
1%7
1%8
За 2 года
Зиготнпсть ояцгматок
\ш 7/ н lib локусам
Гомозиготы
Гстерозиготы
Гомозиготы j
Гетерозиготы
Гомозиготы
Гетерозиготы
Вг«го
IMTOK
75
39
78
46 j
153
65
В том числе лиши» д*о#я* j
Ko.m*»ct»o
22
21
22
19
44
40
I Промен т 1
29.3 5,2
53.6 7.9
28.2±4.9 j
41.2:7.2
28,7 3.6
47.1.-5.4
Популяцнонная средняя при многих парах генов. Если
принять для простоты, что действие многих генов суммируется ад-
дитивио, то популяцнонная средняя может быть представлена
как
MP~la(p—q) + 22dpq. (9)
Эта формула аналогична формуле (8) с тон только разницей,
что в нее включены знаки сумм. В правую часть равенства
входит величина d> которая, очевидно, представляет собой
среднюю степень доминирования по всем генам. Если учесть
статистический характер суммирования многих генов с разной
степенью доминирования, то величина d едва ли может быть
значительной. Скорее всего она приближается к нулю. И тогда
популяцнонная средняя МР будет выражаться только
2а (р—q). Она является средним и фенотипнческим и генотн-
ннческим значением. Если бы все гены увеличивали среднюю и
фиксировались бы в популяции, то возможно максимальное
значение средней +2а. Если бы, наоборот, все гены
уменьшали среднюю, то ее значение было бы —£а. Это теоретические
границы возможной вариации в популяции. Выраженная
формулой 2a(p—q) средняя располагается где-то посередине
между ними.
Средний эффект генов и селекционная ценность особей.
Для того чтобы подойти в дальнейшем к пониманию как
структуры популяции по количественным признакам; так н
процессов, в ней происходящих, в частности при отборе, надо
обосновать еще понятия о среднем эффекте генов и о
селекционной (пли разведенческой) ценности генотипов.
От родителей потомству передаются определенные гены
и их сочетания. Поэтому необходимо новое мерило ценности
ЗйГ
особен, выраженное n iепах, а не генотипах, как было выше.
Таким может быть средний эффект гена (или генов) как
среднее отклонение от значения популяционнон средней особей,
получивших этот ген от родителей. Критерием для сравнения
служат особи, которые получили бы этот ген от других
родителей, взятых случайно из популяции. Иначе говоря, это
средний эффект замены одного гена другим. Чтобы уяснить все
сказанное, остановимся на данных табл. 29. Если ранее мы
имели дело с + а и —а как мерилами действия генов
непосредственно данных особей, то теперь вводится новая
величина — средний эффект гена, измеряющий его влияние на
потомство. Он зависит от значений а и d и от частот данных
генов в популяции. Зная эти величины, можно учесть средний
эффект данного гена в следующем поколении.
Таблица 29. Определение среднего эффекта генов аг (для гена Аг)
и ои (для гена А2)
1 2
с
X
Н
Г'
л.
Генотипы, их
оценки и частота
образуемых ими
гамет
р
Я
Р
"?1
I Я
Средние
оценки
получающихся
генотипов
| pa -f- qd
pd + qa
Значение
популяционнон" средмеЛ
a(p — q) + 2dpq
Эффекты генов at и о,
q [a + d (q — p)J = at
— p[a + d(q—p))- a3
Средний эффект для гена вычисляется как разница между
средним значением образуемых генотипов (с участием
данного гена) и вычисленной выше популяционнон средней. Тогда
ai = pa + qd—[a (p—q) + 2dpq].
Пекле раскрытия скобок и преобразований
*\ = q[a + d(q-p)]. (10)
Сходным образом вычисляется и сег:
a2=—p[a+d(q—p)]. (11)
Отсюда может быть получена величина а как средний эффект
замены А\ на А2:
a = a + d(q-p). (12)
Взаимоотношения между ними таковы: a*=ai—0С2;
ai = (/a; a?08— pa.
Таким образом, средний эффект генов родителей
определяет среднюю генетическую ценность потомства. Поэтому
оценка особей, произведенная по среднему значению его
потомков, и представляет собой разведенческую (или
селекционную) ценность особи. Селекционная ценность, таким образом,
может быть измерена конкретным путем. Если данная особь
скрещивается с рядом особей, представляющих случайную
выборку из популяции, то значение селекционной ценности
представляет собой удвоенное среднее отклонение потомства от
популяционной средней. Удвоенное потому, что родитель
передает потомству только половину своих генов, другую же
половину генов потомство получает из случайной выборки
популяции. Селекционная ценность может быть выражена как в
отклонениях от средней популяции, так и в абсолютных
единицах. Как средний эффект является качеством гена и
популяции, так и селекционная ценность представляет собой качество
особи и популяции, с которой особь скрещивается. Нельзя
говорить о селекционной ценности особи без учета специфики
популяции, в которой проводится скрещивание. Если
определять селекционную ценность особи в понятиях средних
эффектов, то она равна сумме средних эффектов генов, причем это
суммирование должно быть сделано по парам аллелей
каждого гена и по всем генам.
Для одного гена с двумя аллелями она может быть
записана следующим образом:
Генотипы Селекционная ценность
АгАг 2ax^=2qa.
АХА2 *! + «!- fa-P)«
А2А2 2а, = — 2ра
Если принимать во внимание все гены, то селекционная
ценность отдельного генотипа представляет собой сумму
средних эффектов имеющихся в нем пар аллелей. При неаддптяв-
ной комбинации генотипических ценностей возникает
некоторое усложнение, приводящее к тому, что оба определения
селекционной ценности, а именно практическое—в измеряемом
значении потомства — и теоретическое — в средних эффектах
генов — становятся неравнозначными.
Если популяция находится в равновесии по формуле Хар-
дн—Вейиберга, среднее значение селекционных ценностей
должно быть равно нулю, если, конечно, выражать их в
отклонениях от популяционной средней. Тогда надо помножить
селекционные ценности каждого генотипа на их частоты:
2p2qa + 2pq(q—p)a—2q*pa. - 2pq<* (p + q—p—q) - 0.
МП Ф РокпикиА 20»
Но селекционные ценности можно выражать в абсолютных
единицах, и тогда средняя селекционная ценность должна
быть равна средней геиотипическоп ценности и средней фено-
тлпическои ценности.
В простейшем случае при простом суммировании средних
эффектов генов по аддитивному принципу селекционную
ценность можно определить как аддитивное значение генотипа
особи или просто как аддитивный генотип, вариацию же в нем
описать в понятиях аддитивных эффектов генов. Для
обозначения селекционной ценности особи можно употребить
символ А как основной компонент для характеристики ее геноти-
пкческой ценности.
Возникает вопрос о том, какие еще элементы надо
учитывать.
Эффекты отклонений, вызванных доминированием и
взаимодействием. В отсутствие доминирования генотипические
и селекционные значения совпадают. Если же имеются
отклонения, вызванные доминированием (т. е. внутриаллельным
взаимодействием) или эпистазом (т. е. межаллельным
взаимодействием), то в общем виде геиотипическое значение для
суммы всех генов выразится формулой
G = A + D + L (13а)
Здесь А — результат аддитивного суммирования действия
всех генов; D — сумма доминантных отклонений н / — сумма
всех отклонений, вызванных различными взаимодействиями
между генами. В дальнейшем мы увидим, что / имеет
сложную природу, причем надо иметь в виду, что и D — тоже
взаимодействие, но между аллелями одного гена. Поэтому
выражение «аддитивное действие» имеет двоякий смысл.
Применительно к аллелям, т. е. генам одного локуса, оно означает
отсутствие доминирования, по отношению к разным генам —
отсутствие эпнетаза. В виде варианс это можно записать так:
ciG2 = <7A2 + aD- + a,2. (136)
А общая фенотнпнческая варианса с учетом средовой равна
ор2 = стл2 + (Тг/Ч0,2 + о*.2. (14)
Рассмотрим теперь сущность отклонений, вызванных
доминированием, в понятиях частот генов и средних эффектов
генов. Сводные данные для модели одной пары генов
представлены в табл. 30.
210
Таблица 30. Генотипические ценности геиотижю ДИи АгАг и А*А7
и их отклонения от популяциониой средней Мр [—а (р — q)-r 2dpq\
вследствие доминирования. Средний эффект замены гена a a-rdiq — p)
Генотипы
АХА2
^2^ 2
Их
частоты
р2
2pq
Яг
Оценка
эффекта
генов
а
d
— а
Отклонения от полулянионяой средлей i
Генотппическое
значение
2q(a-pd)
2q(*-qd)
a(q-p) + d(l-2pq)
(q — p)a + 2pqd
-2p(a + qd)
-2p(a + pd)
Олекцяон-
ное значение
2?a
(q — p)*
— 2p*
' Отклонение j
вследствие 1
доминирования J
2pqd
-2p*d
Здесь генотипические значения превращены в отклонения от
иопуляционной средней в соответствии с тем, что сказано
выше, но при этом они выражены в двоякой форме —или в
терминах а, или в терминах а. В качестве примера возьмем
генотип А\А\. Его генотипическая оценка выражается о. Популя-
ционная же средняя MP=a(p—q)+2dpq. В таком случае-
отклонение генотипической оценки этого генотипа от популя-
ционной средней будет равно
a—[a(p—q) + 2dpq).
После соответствующих преобразований оно примет вид
2q(a—dp).
Если заменить а на его значение в а, исходя из уравнения
a = a—d(q—р), то получается отклонение генотипического
значения в а, равное 2q(a—qd). Если отнять от него значение
селекционной ценности 2qa, то получится величина
отклонения, вызванного доминированием: —2q2d.
Такие же расчеты проделаны для остальных генотипов
АХА2 и Л2Л2. Во всех значениях отклонении, вызванных
доминированием, фигурирует, как и следовало ожидать,
величина d. Если d = 0, отклонение тоже равно нулю, и тогда
селекционные и генотипические значения совпадают.
И наоборот, если генотипические и селекционные значения
равны, это указывает на то, что d=0, и тогда аллельные гены
аддитивны по отношению друг к другу.
Это же может быть верно и но отношению к популяции в
Целом. Если доминантные отклонения данного генотипа
помножить на его частоту в популяции, то —2p2q2d^4p2qid—
14*
«Ml
Все сказанное выше может быть отнесено и к неаллельно-
му взаимодействию. При его наличии будет наблюдаться раз-
f? и и л между генотипическим значением особи с двумя (или
несколькими) генами и селекционным. Эта разница
количественно будет отражать роль взаимодействия.
Так, Фолконер вычислил эти разницы для приведенного
выше примера с неаддитнвно суммирующимися генами,
влияющими на размеры гранул пигментов, и установил степень
влияния взаимодействия между генами В-bb, с одной стороны,
и С-сс* — с другой.
Сущность отдельных варианс, характеризующих
популяцию, и их структура. Остановимся теперь на отдельных
величинах, входящих в формулу (6).
Варнанса ор2 не требует специального разбора. Она
определяется в каждом отдельном случае на основе фенотипиче-
ских значений данного признака у всех особей, составляющих
популяцию, ос2 — варианса генотипических значений особей
популяции. Как уже указывалось, она может состоять из 3
компонентов:
аа2 = ал2 + ая2 + 0/2.
Первая из них — аддитивная варианса
(оа2)—представляет собой важнейший компонент генетической вариансы. В то
же время это варианса селекционных значений, так как она
является главной причиной сходства между родственниками
и именно она по преимуществу определяет генетические
различия в популяции, которые удается наблюдать; она же
определяет ответ популяции на отбор по количественному признаку.
Как мы увидим в дальнейшем, именно аддитивная варианса
легче всего выделяется из фенотипической вариансы *.
Чтобы определить аддитивную вариансу, достаточно взять
выраженные в отклонениях селекционные значения,
указанные в табл. 30, для каждого генотипа, возвести их в квадрат,
умножить на частоты генотипов и разделить на сумму всех
генотипов. В конечном счете она будет следующей:
oA2=aWp2+(q-p)22pq+4p2q2]=
= 2pq a2(2pq+p2+q2-2pq+2pq);
oA2=2pqa2. (15a)
В значениях же р и q
OA2=2pq\a+d(q-p)]\ (156)
• В следующей главе, посвященной наследуемости, будет дан раэбор
Ai>* ее видов: в широком смысле слова I—д~ I и в узком I ""J"") • В п0~
«лгднгм числителем для определения коэффициента наследуемости
являли и о\.
Эта формула для аАг выведена на основе моногибридной
модели. Величина d может быть самой различной. Легко
вывести различные модификации формулы для случаев, когда
d = a (полное доминирование) или d>a
(сверхдоминирование). Но она может быть обобщена: для системы нескольких
аллелей одного гена (локуса), для наличия инбридинга или
отбора и, наконец, для случая, когда учитываются все гены
в популяции. Тогда
о*А =2о>А{ау (15в)
а
Варианса доминантных отклонений (в значениях р, q и d)
oD2 = dn^4qip2 + 8p^+4pAq2)=4p2q2d2(q2+2pq+pi)\
oD*=(2pqd)\ (16)
Так как селекционные, т. е. аддитивные, значения и
доминантные отклонения не коррелируют друг с другом, то геноти-
пическая варианса без учета эпистатического взаимодействия
будет простои суммой:
og2=oa4-od2;
OG2=2pq[a+d(q-p)Y+(2pqd)\ (17)
Более сложной представляется величина а/2. Она состоит
из многих компонентов, так как в конечном счете возможны
взаимодействия между любым количеством генов: двумя
парами, тремя и т. д. Но поскольку взаимодействия между
большим числом генов обычно составляют малую долю общей
варнансы, то с ними можно не считаться. Поэтому чаще всего
ограничиваются двухфакторным или трехфакторным
взаимодействиями. Но даже при двух факторах возможны 3 вида
взаимодействия: между аддитивными эффектами первого
и второго генов, между аддитивным эффектом первого гена
и доминантным отклонением второго, между аддитивным
эффектом второго гена и доминантным отклонением первого
и, наконец, между доминантными отклонениями по обоим
генам.
В упрощенной форме это можно записать следующим
образом:
За пределами правой части уравнения остаются все
остальные, более сложные случаи межаллельных (эпистатнческих)
взаимодействий.
Общий случай для диплоидной популяции с произвольным
числом расщепляющихся генов, любым числом аллелей в ло-
кусе и произвольным эпнстазом был разобран Кемпсорном
213
В *той модели гены обозначены а (а=\, 2, 3, 4 я),
произвольная пара локуса а Аа и Аа . Генотнпическое значение
;а
/1
особпА1 А1 А*гА*2А*3 ... обозначается как JJ АааА*а. Частота
а=1
гена А* —р*-
Генотнпическая структура популяции тогда
П Ра Ра. Аа Аа .
*■-* r xar ja га ja
<х=1
Аддитивная варианса для локуса а
о2 =2£(ав )\
А(в) *^\"Ча' f
где аа — аддитивный эффект гена Аа. .
Доминантная варианса для локуса а
02 =ЕЫа . )2,
где de. . — доминантное отклонение для локуса а.
Произведение аддитивных варианс для двух локусов а и а7
а2 =4E(a*a*'i i ,)2,
АЛ(ол) * а а'' '
где а°oia'iaiat— произведение отклонений, связанных с генами
А* и Ла', в локусах а и а', и т. д.
ia ta J '
Таким образом, компоненты генетической вариансы
согласно модели Кемпсорна в общем виде записываются так:
общая аддитивная варианса популяции
а
общая доминантная варианса популяции
а
общее произведение аддитивных варианс популяции
^л=2^ит.д.
аа'
В конечном счете мы приходим к той же общей формуле
+«-**^>^ллЪ^ы>+*ААА+*ААО+ ... (19)
214
Средовая (иногда ее называют паратипической) варианса
of} может вызываться очень многими причинами, как
внешними (кормление, содержание животных» агротехника
выращивания растений, удобрения), так и внутренними (возраст
матерей, влияние генотипа матерей и среды их тела при развитии
эмбриона, вариация в самом процессе индивидуального
развития и др.)- Ниже, рассматривая конкретные модели и схемы
для анализа компонентов вариансы, на некоторых из этих
причин мы остановимся особо. Сейчас отметим лишь наиболее
общие положения. Одни из причин, влияющих на значение а^2,
являются контролируемыми, т. е. они поддаются
регулированию в полевых условиях или в условиях лабораторного опыта,
другие — неконтролируемыми. Последние представляют собой
многочисленные и подчас не очень сильные по своему действию
внешние условия, которые создают как бы статистический фон
наблюдений или опытов, а также трудно учитываемые
взаимодействия различных генотипов популяции со средой. В
генетических опытах стремятся свести к минимуму изменчивость,
вызываемую средой, во всяком случае в той части среды, которая
поддается контролю.
Обычно считается, что все особи популяции в равной мере
подвергаются влиянию колеблющихся условий внешней среды
На самом деле одни особи (или группа особей) подвергаются
одним влияниям среды, а другие особи — иным.
Поэтому фактически получаемое значение оег может быть
преувеличенным. Чтобы избежать преувеличенной его оценки,
стремятся внести уточнения в виде поправок к исходным
данным, например на возраст животных, на кормовые
рационы и пр.
Наибольшие сомнения всегда вызывал последний
компонент вариансы:
2 COVGE^ZroeOGOE.
Нет сомнения, что такая корреляция существует. Есть
немало ее примеров и из практики животноводства и
растениеводства и на основе лабораторных опытов. Общеизвестно, что
коровы разных генотипов по-разному реагируют на улучшение
кормления *. То же наблюдается и у других видов
сельскохозяйственных животных. Это проводит к корреляции между
фенотипическими значениями и отклонениями в условиях
среды. А так как есть корреляция между фенотипическими и ге-
нотипическими значениями, то существует н корреляция
между генотипом и средой, точнее, между генотипическими и сре-
довымн отклонениями. Но очень часто при конкретном
анализе этот компонент оказывается включенным в генотипическую
вариансу.
* Вопрос о взаимодействии геаотипа ■ среды будет расскагршмтъе*
также в гл< 7.
*!5
Надо n.Mi'ii. в виду несколько разных сторон во
взаимоотношении генотипа и среды, не всегда сводящихся к ковариа-
ции между ними. Поэтому Лерпер считает возможным
включить еще одни член в число компонентов варпаисы и условно
обозначает его символом f(GE).
Разные генотипы могут давать различные значения средо-
Boii вариаисы - - количественные, а иногда и качественные,
т. е. обладать как бы разной чувствительностью к одним и тем
же условиям среды. II наоборот, специфические различия
в среде мог\т дать тот же или обратный эффект у разных
генотипов, т. е. характер отклонения будет определяться
специфическими условиями.
Так. в работе Краутмапа сообщены данные о различиях
разных липни кур породы леггорн, а также их гибридов по
потребностям в питательных веществах. При снижении в
корме содержания белка и метиоиина одни линии или гибриды
снижали интенсивность яйцекладки, другие — вес яиц,
третьи — вес тела. Был один тип, который, несмотря на
снижение содержания белка и метиоиина, сохранял все эти
показатели на прежнем уровне, а другой — снижал одновременно
все три показателя. Таким образом, характер взаимодействия
особей разных генотипов с одним и тем же изменением в
кормлении был различен.
Другой пример приводит Фолконер. Две линии мышей
содержались в условиях хорошего и плохого кормления.
Линия А дала привесы за период с 3 до 6 недель при хорошем
кормлении 17,2, а при плохом— 12,6 г, а линия В— при
хорошем 16,6, а при плохом 13,3 г. Это значит, что линия А
росла лучше, чем линия В при хорошем кормлении, по хуже, чем
линия В при плохом.
Компоненты вариаисы при различных родственных связях
между особями. Изучение сходства между родственниками
дает очень много для понимания природы количественных
признаков и их генетических основ. Оно дает возможность
установить конкретные или фактические доли вариаисы внутри общей
фактической вариаисы и сравнить их с теоретически
ожидаемыми V Эти фактические доли вариаисы могут быть получены
путем сопоставления вариации внутри тех или иных
родственных групп и между ними. Тем самым выясняется разница
между ними. Сходство внутри группы может быть отнесено ко
всей вариансе по общему принципу дисперсионного анализа.
Так, если ои2 *— компонент межгруиповой вариаисы, a aw2 —
• В литературе иногда употребляют двойное обозначение: V — для
общи*, как бы причинных компонентов вариаисы н оа-~ для наблюдаемых
фактически компииентпп. Чтобы не осложнять изложение, мы пользуемся
одним оби значением о*.
внутригрупповой, то можно вычислить так называемый
внутриклассовый коэффициент корреляции / или г,с;
ов2
Межгрупповой компонент, отражающий степень вариации,
общую для членов группы, может рассматриваться как
коварна пса членов группы. Исходя из этого принципа,
используется и сравнение потомства с одним из родителей или со
средней из величин, характеризующих количественный
признак обоих родителей. Как известно, связь между потомками и
родителями выгоднее измерять коэффициентом регрессии, и
тогда
Числитель представляет собой ковариансу между
родителями и потомками, а знаменатель — вариансу родителей.
Таким образом, коварианса между родственными
особями — новый критерий для характеристики генетической
структуры популяции. Необходимо вскрыть ее генетическую
природу для наиболее часто используемых типов родственных
связен, а именно: а) родители и потомство (один или оба); б) по-
лусибсы; в) сибсы.
Выше уже говорилось о том, что генотипическое значение
особи можно рассматривать как отклонение от популяционнон
средней. Тогда коварианса вытекает из сопоставления геноти-
пических значений особей со средней генотипической их
потомства, получаемого путем случайного скрещивания в
популяции. Среднее генотипическое значение потомства
составляет половину селекционной ценности родителей» и коварианса
вычисляется на основе сопоставления гепотнпических
ценностей особен с половиной их селекционной ценности, т. е.
1 „
соиОР= — аА-.
Можно доказать это равенство, пользуясь значениями
Р, q и а.
Воспользовавшись данными табл. 30, составим табл. 31.
характеризующую связь родителей и потомства для разных
генотипов. В последней колонке запишем селекционные
значения, взятые из табл. 30 и разделенные пополам.
Для получения ковариансы нужно перемножить друг с
другом три колонки: частоту, генотипическое значение родителей
2»7
Таб.шца J/. Соотношение между генотипическнми
значениями родителей и потомков
Родители
Ггмотмпы
ЛгЛг
AtAt
А А*
Частоты
Р1
2pq
Генотипнческне значении
2? (a — qd)
(q — p)* - 2pqd
— 2р (а + P<i)
Средние генотмпические
значении потомства
q*
I
-75Г (q — р) а
— Ра
и генотипическое значение потомства — и просуммировать
результаты по всем генотипам:
covOP=pq a2(p2+2pq+q2) +
+2p2q2ad(-q+q-p+p)\
cov0p=pq a2.
Но так как выше было указано, что 2pq а2=ад2, то
1
COV0p= —
(20а)
ол2.
(206)
Это же относится и к суммированию действия многих генов.
И в ьтом случае коварианса между потомством и одним из
родителей равна половине аддитивной вариансы.
То же самое верно и в том случае, если изучается
коварианса потомства со значением не одного из родителей, а со
средней величиной из значений двух родителей, т. е. covop.
Расчеты здесь более сложные, поэтому мы ограничимся
итоговой строчкой:
cov0l> = a2pq—2adpq(p—q)+d2pq(p—q)2=
= pq[a+d(q—p)¥=pq a2;
cou0P = pq a2= -г- о а2 *.
(21)
Ковариация полусибсов (HS), т. е. потомства, у которого
один родитель общий, а вторые — разные, равна
1 г
4
Вывод ее аналогичен описанному выше.
(22)
В следующей главе будет показано, что, несмотря на равенство
тор и сооор, коэффициенты наследуемости, вычисляемые по регрессиям
потомков на одного родителя (F) или на среднего родителя (Я), отлича*
*пса пом «тел** 2.
2ti
Вариаиса средних величин семей полуснбсов равна
— pq a= "зг^л2. Наконец, коварианса между полными сиб-
сами (FS) представляет собой вариансу средних фенотипи-
ческих семей полных сибсов, т, е. потомства, происходящего
от одних и тех же родителей, и имеет следующий вид:
covFB=cov0p+d2p2q2=pq a2+<Pp2q2.
Так как 2pq а2=аА2, a 4<Pp2q2—OD2, то
COvFB=—oA2+—oj>2. (23)
Эта формула относится и к совокупности всех генов.
Возникает вопрос о том, в какой степени включаются в
значения варианс различные виды взаимодействий. Не вдаваясь
в достаточно разработанные математические детали,
ограничимся лишь сводкой для простейшего случая взаимодействий
при двух парах генов (табл. 32), имея в виду, что более
сложные виды взаимодействий обладают незначительным удельным
весом в общей вариансе.
Таблица 32. Ковариансы при разных родственных связях
я вклад в них разных компонентов варнанс
Родственные группы
Потомки — родители (ОР и 6Р)
Пол у сносы (HS)
Полные сносы (FS)
Алгебраическое выражение вклада
Компоненты в§рмансы и
коэффициенты нх вклад*
°~Л
I
2
1
4
1
2
X
2 2
"D 9АЛ
1
4
У
1
4
I
1
4
*« 1
•~AD
1
8
* 1
' 2
I 9DD
1
16
Ул 1
Поскольку о\ и о*дд практически неотличимы, то при
определении ал2 с помощью ковариансы между родителями и
потомками значение а* всегда завышается за счет о* Завы-
А ДА
шается также и значение аг0 в полных енбеах за счет о)ю.
219
В прсдыл.мцнч рассхждепннх предусматривалось, что все
гены, детерминирующие количественный признак, независимы.
Н 1.ШЧН11 сцепления не влияет на ковариансы между
родителями и потомками, по па ковариансах между полусибсами и
полными сибсамп оно должно сказываться (так как влияет
на характер расщепления) в сторону их увеличения. Эта
дополнительная доля ковариансы проявляется в увеличении ком*
понента взаимодействия.
Рассматривая компоненты варнансы у различных
родственных групп, не следует забывать, что во всех случаях должна
быть и средовая варнаиса. Однако сравнение их дает
возможность подразделить ое~ на два компонента, обозначаемые
символами о2у< и о- ,. Первый из них связывают с общей средой,
влияющей па всех особей разных групп. Второй же — с той
средой, которая влияет только в пределах одной группы
it почти не поддается контролю. Примеры такого
подразделения будут даны ниже.
Общие принципы выделения компонентов фенотипической
варнансы. Для выделения отдельных компонентов, входящих
в фенотипическую вариансу, возможны очень многие способы,
основанные на разработке различных математических
моделей и получении необходимых количественных данных путем
постановки опытов. Первый этап заключается в разделении
общей фенотипической вариансы на средовую н геиотипиче-
скую. После такого разделения может быть вычислен
коэффициент наследуемости. Ввиду значения наследуемости как
генетического параметра материал по этому вопросу, в том числе
и различные методы выделения генетического и средового
компонентов, изложен более полно в гл. 6. А сейчас
остановимся только на методах выделения отдельных компонентов
генетической вариации.
Непосредственное определение аддитивной вариансы
достигается лучше всего с помощью коэффициента регрессии
(см. об этом в гл. 6), а по полусибсам — методом
внутриклассовой корреляции.
Статистическая модель опыта для оценки ковариансы
между полусибсами следующая:
Xih~ii + Si + eik,
где Si — вклад отца в общую изменчивость, a e\h — вариация
между полусибсами.
В *том случае соуцз^Ош2, а о.2= ~-Од2.
Таким путем оценивается лишь аддитивная варнаиса.
Применяется модель однофакториого анализа, где фактор
А самцы производители.
220
В дальнейшем возникает необходимость отделить
аддитивную и пеаддитивную часть генотипической вариансы.
Наиболее подходящий для этого метод —оценка коварианс
между полусибсами и сибсами, получаемая в одном и том же
опыте, поставленном по простейшей иерархической схеме. Макеты
необходимых таблиц для вычислений даны в приложении.
Математическая модель следующая:
Xijk^ii+Si+fij+eijk,
где Xijh — фенотипическое значение признака у потомка k от
скрещивания отца * с матерью /; 5,-— вклад отца в
фенотипическое значение признака; [ц — вклад матери; е^к— вариация
внутри сибсов.
Тогда
COVHS = 0$2\ COVpb = Ou7"\'Oj2\
Известно, что
covHS=o*t = ~ о'-А + -^ о'ДА+ ....
а
covFS=ol +°2, = Т°а+ Т02* + Т°" + Т °^ + • • •
Чтобы выключить аддитивную часть вариансы и получить
только неаддитивную, т. е. доминантную плюс взаимодействие,
надо удвоить первую строчку и вычесть ее из второй:
cov,8= y о\ + ~ о», + -i- о\ А + -- о^р
°: +о? -™ =°2, -°\ - т °* + Т ° л.л + т •** •
Таким образом, 4о,3 оценят аддитивную варнансу,
а 4(а/2—о42) —пеаддитивную, включая и взаимодействие.
Знаменателем надо взять aP2=ae2-fo/a+o^2. Надо иметь
в виду, что и а**, измеряющая вариацию среди сибсов, имеет
сложную природу. Кроме того, осложняющими
обстоятельствами могут быть материнский эффект, сказывающийся на
аД и эффект сцепления с полом, который может быть
обнаружен по разнице между самками и самцами внутри каждой
группы. Генетическая структура значений варианс с
указанными усложнениями представлена в табл. 33.
«1
Таблица 33. Генетическая структура варианс при анализе
иерархического дисперсионного комплекса
1
1
Вари-
I »КСЫ
1 *>
о;
1 *'
п/.
Компоненты
А4ДН-
ТИ1-
ныП
1 4
14
,,
доми-
M.IHT-
ими
5Ь
0
1/4
„
взлнмодеЛстннс
аАА
1/16
3/16
3 4
'AD
0
1/8
7/8
«л
'DD
0
1/16
15/16
о
М.4.1
1/64
7/64
7/8
ринский
эффект
о
0
1
0
о
0
0
1
Сцепление
с полом
о
'AS
0
1/2
1/2
9?
"AS
1/2
1/4
1/4
Легко видеть, что прибавка значения ое2 к а*2 и or приводит
к созданию полной структуры ар2, т. е. состава из ас,2 и а^2 по
всем компонентам генетической и средовой вариансы. Но за
счет наличия материнского эффекта и сцепления с полом
возможно некоторое смещение фактического значения а/2. Кроме
того, на ней может сказаться и эффект общей среды (в таблице
он не обозначен, так как его роль спорадическая).
Для учета взаимодействия применяется обычная схема
двухфакторного анализа, причем факторы А и В — это
влияние отцов и влияние матерей # (или у растений опыление
пыльцой семяпочек во всех сочетаниях).
Статистическая модель следующая:
Xijk = [i+Si+fij+(sf)ij+eijk.
Здесь Si — вклад отца, /<j — матери, (sf)o— эффект
взаимодействия родителей, e<j* — вариация среди сибсов от каждой
скрещиваемой пары.
В некоторых модификациях этой модели, например Ком-
стока-Робинсона, учитывают также эффект повторности, ио это
необязательно.
Дналлельные скрещивания далеко не всегда можно
осуществить. Наибольшее распространение они получили при
скрещивании иибредных линий кукурузы, однако этот метод
был использован для анализа вариансы и у животных
(дрозофила, куры. мыши).
Здесь, как и в предыдущей схеме, аА2 = 4оД или ола=4а/*»
ИЛИОд'=т:2(<>,' + 0,2|.
• В приложении даны макеты таблиц и вычислений по этой схема.
222
Величина же 4o\f характеризует взаимодействие. Она
охватывает также ар2, т. е.
Материнское влияние в этом комплексе уже ие
учитывается. При сцеплении с полом закономерности те же, что
и в табл. 33.
Генетическая структура отдельных варианс приведена
в табл. 34.
Таблица 34. Генетическая структура аарнаас пра аналяэе даужфажтормого
дисперсионного комплекса (при дмдллелыюи скрещиташ»)
Ворнаисы
1 °<
*
°1,
°£
2
аА
1/4
1/4
0
1/2
2
0
0
1/4
3/4
2
9АА
1/16
1/16
1/8
3/4
9AD
0
0
J/S
7/в
2
•OD
0
1 0
1/16
15/16
2
»4 4 4
1,64
1,«
3/32
56/64
2
о
0 1
0
1
Наконец, возможно проводить расчленение фенотнпической
вариансы и в тех случаях, когда имеются исходные
родительские формы (Pi и Pa), Fu ffc Fbpi и Рьрз. В простейшей форме *
за основу берется фенотипическая варианса Р3. Принимается,
что она включает в себя следующие компоненты:
аЕг определяется как средняя между о3 исходных
родительских линий (имеется в виду, что они инбредны) и Л. Далее
o2Ft сравнивается с о*, , которая генетически рассматривается
как
Разница между о^, н °f Ааст значение —о^. Подставляя
о2Б и а2А в одну из формул (о*, или сг^ ), можно вычислить
o2D. Тогда о*, получается как сумма в^-Н^-Н*1* *
* Более сложные методы, предложенные Мамром и требующе оесяь
большой вычислительно* техник*, «десь ее рмвнреютс*.
Ж
Для расчленения фенотипнческой варнапсы иа компоненты
мог>т быть применены одновременно разные методы. Так,
Д.псон на Tribolium использовал сраниеине средних и варианс
высокоинбредных линий, регрессии потомков к родителям при
случайном и избирательном скрещивании родителей,
сравнение аутбредиых популяций с нибреднымн и с F\ от
скрещивания инбрелных линий и, наконец, схему дналлельных
скрещивании.
Некоторые примеры фактических соотношений между
компонентами фенотипнческой и генотнпической вариансы.
Выделение отдельных компонентов проводилось во многих опытах.
Очень полные данные были получены рядом авторов на
дрозофиле. Они сведены в табл. 35.
Таблица 35. Компоненты фенотипнческой вариансы (в %) у дрозофилы
по 4 признакам (по данным разных авторов)
Компоненты
BJpttftHCU
Генетическая
аддитивная
Генепсческля
неадднтивная
Срезов* я
Примаки 1
збДОМННЫЬ-
НЫС ЩРТММКИ
52
9
39
длина груди
43
6
51
размеры
янчннкоп
30
40
30
количество яиц,
отложенных
91 3 ДНЯ
18
44
38
По разным признакам удельный вес средовой вариансы
колеблется от 30 до 50%. Изменялось и соотношение между
аддитивной н неадднтивнон частями генетической вариансы.
Для шетинок и длины груди неадднтивная доля была
значительно меньше, чем аддитивная. Для признаков, связанных с
размножением (размеры яичников, количество яиц),
наоборот» неадднтивная доля больше аддитивной.
Но очень многое, по-видимому, зависит от исходного
материала. Так, А. И. Добнна на материале тех же абдоминальных
щетинок дрозофилы, которые изучались и другими авторами,
показала, что при скрещивании различных инбредных линий
получаются разные результаты. В комбинациях линий 50X44,
50у24 наблюдался только аддитивный компонент, в
комбинациях 50x20. 1x10—оба компонента, причем с
преобладанием неаддитивной части. Примером разложения вариансы с
выделением доминантной части могут быть приведенные в
книге Брюбейкера данные Алларда о различии двух сортов
пшеницы оо срокам колошения. Оказалось, что средовой компонент
занимает 12% фенотипнческой вариансы, а 88% приходится
234
nn долю генотнпической варнансы, в том числе 76%
—аддитивный компонент и только 12%—доминирование.
Наконец, о роли взаимодействия. В опытах на курах
Нордским и Кемисорном на основе двухфакторной схемы
дисперсионного анализа был установлен эффект взаимодействия
генотипа исходных линий с условиями содержания кур на
фермах. Наиболее высоким он был для признака смертности
цыплят к 1 мая (31%), по признакам же скорости полового
созревания, яйценоскости, весу яиц —9—10%. Совсем не
сказывалось взаимодействие на качестве яиц.
Глава 6
НАСЛЕДУЕМОСТЬ
Общее определение наследуемости. В предыдущей главе
же было введено понятие о наследуемости как мериле удель-
юго веса генетической изменчивости в общей феиотипической
!Зменчивостн популяции. Наследуемость измеряется коэффи-
шентом, который обычно обозначают символом Л2. В простей-
нем случае
~~~ аР ~~~ og2+Oe2'
Наследуемость ни в коем случае нельзя путать с понятием
наследственности или наследования. Под наследственностью
понимается общебиологнческое явление, заключающееся в
наличии сходных признаков у родителей и потомков вследствие
передачи через половые клетки (эта передача и есть
наследование) определенных материальных структур (хромосомы с
входящими в них молекулами ДНК и дискретными частями
молекул ДНК —генами; определенные цитоплазматические
элементы). Наследственность можно отнести к двум
связанным родством особям (мать—дочь, дед — внук и т. д.).
Наследуемость же — чисто статистическое понятие, всегда
характеризующее группу особей популяции.
Наследуемость в узком и широком смысле слова. Так как
генетическая вариаиса может состоять из трех компонентов,
удельный вес которых в суммарной величине ос2 бывает
разным, то Лаш (Lush) предложил различать два вида
наследуемости— в узком и широком смысле слова. Первая измеряется
долей только аддитивной нариансы в общей феиотипической
оа2
вариансе, т. е. /z2= —-, вторая же соответствует общей фор-
Ор2
а 2
муле коэффициента наследуемости, т. е. Л2= —~. В литера*
аР"
туре применяют общее обозначение h\ но было бы целесооо-
22»i
разно ввести особое обозначение для коэффициентов
наследуемости в узком смысле слова, например hf. Иногда в литера-
л
туре обозначают h2 во втором значении символом №.
Определение Л42, как это будет показано ниже, очень важно
для селекционных целей, так как при отборе используются
главным образом гены с аддитивным действием. Зная Ai2,
можно судить о возможной эффективности отбора. Однако
в конкретных условиях выделение компонентов генетической
вариансы, описанное в предыдущей главе, далеко не всегда
возможно. Поэтому часто приходится пользоваться только
коэффициентом наследуемости в широком смысле слова или
величиной, средней между обоими коэффициентами.
Методы определения коэффициента наследуемости.
Сравнение инбредных и неинбредных линий. Все методы
определения коэффициента наследуемости в конечном счете сводятся к
сравнению степени изменчивости (или сходства) различных
групп животных, связанных между собой или родством, или
общностью внешних условий, при которых они развивались.
Но техника вычислений может сильно варьировать в
зависимости от изучаемого материала и применявшихся методов
разведения животных или растений, а также от степени учета
влияния внешних условий.
Теоретически наиболее простой метод — сопоставление
изменчивости особей из генетически однородных линии,
например инбредных, и особей, получаемых в результате случайных
спариваний в популяции. Так как в нервом случае почти вся
изменчивость зависит от внешних условий, т. е. ар,2 = ая2, а во
втором о2р =0£ +02G, то разность между ог и о2р и дает
величину og2> t .е. долю наследственных различий в общей
изменчивости. Подобный анализ был применен Райтом (Wright) для
изучения роли наследственности и среды в изменчивости пе-
гости у морских свинок. Сопоставлялись данные об
изменчивости пегости в инбредных линиях, полученных путем
скрещивания братХсестра в течение 20—25 поколений (10 лет
разведения), и изменчивости пегости морских свинок,
разводившихся аутбредно (табл. 36).
Общая варианса в результате инбридинга снизилась
и почти сравнялась с величиной вариансы, создающейся в
контроле, т. е. в аутбредной линии, за счет условий внешней среды.
Снижение общей вариансы произошло за счет уменьшения
доли генетической изменчивости. Наследуемость (Л2) в
инбредных линиях -~г- =0,028. В аутбредных же линиях
0,364
0 271
А*=^—-=0,421.
0,643
15* **?
.№Lz
ТаГиица J6\ Относительная poJb наследственности и условий
шкнюей среды ■ изменчивости пегоети морских свинок
!
Иг-г г* ник мрмлцкн
Наследственность
Среда, общая
для ио«та
Среда, не
общая для почета
V
Ми&кгмыс линии
фЛКТИЧОСКОГ
[п >словиnx
единицах)
0.010
0,020
0,334
0,364
в *# к обще Л
нарнднсе
2.8
5,5
91,7
100,0
Контроль (вутбредные
лммии)
фактическое
(в условных
ГДИННГЫХ)
0,271
0,002
0,370
0,643
DapHencc
42,2
0.3
57,5
100,0 1
Сходная методика была применена Робертсоном в
модельных опытах на дрозофиле (Drosophila melanogaster). Изуча-
лась изменчивость длины туловища и количества
откладываемых самками яиц (за определенный период времени). Из
исходной так называемой дикой линии, которая представляла
собой генетически неоднородную популяцию, были заложены
инбредные линии. Затем от скрещивания инбредных линий
получено F\. Результаты представлены в табл. 37.
Таблица 37. Значена* варнавс в генетически однородной н неоднородно!
популяции дрозофил я определение коэффициента наследуемости
| Популяции, их генетическая структура
| а определенпе Ня
Генетически неоднородная популяция
1 (0> Ос + °£>
Генетически однородная популяция (агр ~ v*E)
Разница между ними (о£)
Н*сл<1\(иость h1 I —_ \
Признаки и их варяансы I
длина
туловища
0,366
0,186
0,180
0.49
1
я П цен ос* I
кость 1
43,3
16.6
26,7
0.62
1 . _J
Таким образом, установление удельного веса наследствен*
ной изменчивости в общей фенотипической изменчивости по*
пулицни стало возможным благодаря применению специфичен
тш
ского метода разведения— инбридинга. Но в практике
инбридинг используется редко, поэтому были необходимы другие,
более общие методы определения наследуемости. Первой
попыткой этого рода явились работы Райта. Используемые во
многих исследованиях даже и в настоящее время методы
учета наследуемости основываются в конечном счете на
формулах Райта.
Метод коэффициентов путей. С. Райт разработал
оригинальный метод анализа связей между причинами и
следствиями, получивший название метода коэффициентов путей (path
е
Рис. 64. Схема анализа связи между величина мл
х и у и их причинами А, В, С, D. Односторонние
стрелки — коэффициенты пути, двусторонняя —
коэффициент корреляции.
coefficients). Хотя в дальнейшем он подвергался критике,
однако оказалось, что окончательные выводы, полученные на
основе этого метода или же путем более глубокого
математического анализа, совпадают. В то же время этот метод дал
возможность связать целый ряд явлений* и оказался очень
полезным для анализа причин изменчивости в популяции,
различных связей между родственными особями, разработки
теории наследуемости, а также теории инбридинга. Поэтому
целесообразно дать краткое изложение метода коэффициентов
путей.
В отличие от корреляционного и регрессионного анализа
метод коэффициентов путей учитывает не только коэффициент
корреляции между различными колЕИ^тцеинцмн прИ^иау>ацИ
или пертаенными величинами, но н взяциТОТ^'Ч^""11 ***«£
ЯНМн. Он позволяет определить степень» с которой изменчн-
* Метод коэффициентов путей можно использовать в ряде областей
биологии, а также в экономике.
239
Г ...
вичь данного признака в пределах группы детерминируется
изменчивостью ряда факторов или причин, объединенных в
некую определенную систему.
Возьмем и качестве примера систему, изображенную на
рис. 64.
Эта система достаточно сложна. В ней переменные
величины а* и {/ детерминируются частично независимыми
переменными А и D и частично обшими причинами В и~С, при этом В
и С связаны между собой какими-то более отдаленными
причинами, вызывающими корреляцию между В и С. Это и
показано двусторонней стрелкой гВсу а также пунктирными
линиями к общей причине Е.
Ри:. 65. Влияние на х независимых (слева) и зависимых
(справа) причин.
Очевидно, что в этой сложной системе могут быть
выделены отдельные части, например часть, в которой фигурируют
только независимые причины или же только зависимые
(рис. 65).
Если предположить, что все переменные величины связаны
между собой линейно, а их эффекты комбинируются по
правилу сумм, то можно прийти к установлению понятия
коэффициента пути. Коэффициент пути, измеряющий удельный вес
влияния, например от А до ху представляет собой отношение
среднего квадратического отклонения переменной х,
возникающего только под влиянием А, к общему среднему кзадра-
тическому отклонению того же х, возникающему под влиянием
всех причин, вместе взятых, т. е. А, В, С и т. д.
Сказанное можно записать следующим образом:
р(х+-А)=рхА=-^=а. (1)
Ох
Таким же образом можно рассматривать коэффициенты
пути Ь, с и т. д.
Коэффициент пути отличается от коэффициента
корреляции тем, что он имеет направление, т. е. является векторной
величиной. Далее, он представляет собой некую абсолютную
величину, могущую принимать разные значения. Если
изменить направление пути, то изменится его значение. В общем
Р, л отличается от рЛтХ.
?у>
Райт ввел еще один коэффициент—коэффициент
детерминации d. dx.A — это та часть в определении х, за которую
ответственна причина Л.
Если установлены все причины, то
dx.A + dx.B+dx.c + ...= l. (2)
По Райту, коэффициенты детерминации идентичны
возведенным в квадрат коэффициентам пути:
йх.А=(рх.л)*=Щ-=а*. (3)
Ох"
Легко видеть прямую связь между коэффициентом
детерминации Райта и приведенным в предыдущей главе
разложением фенотипическои вариансы на генетическую и средовую
по формуле
Разделим обе половины равенства на ор2:
Qp2 __ og*_ , 0£?_ ,
Ор2 Ор2 Ор2
—- =dp,Q\ —- =dpmEl dp,chdpmE=l-
Op- Op2
В данном случае можно говорить о полной детерминации
Р через G и Е.
При независимости А и В гАВ=0, и тогда ох2=олг+ов2-
Это показано на рис. 65 (левая часть) стрелками: А-+х;
В-+х. Но никак нельзя представить эту связь тройной,
например А-+х->В.
Если В = const, то о2 =0; а2 л=о2 .
В * х.А А
Можно связать общую формулу коэффициента пути сГ
формулой для коэффициентов детерминации:
___ О х.А О А Ох.В Ов
Ох Ох Ох Ох
rl \А °л2 i °в* I
dx.A+dx.B= — Н г" = ' •
Ox" Ох*
Если х=А+В и гав=0, то х=~А+В. Отсюда
cov(x.A)=oa\ *Т
соо(х.А) ♦
ГхЛ=х
ояоа
233
Использование коэффициентов путей для анализа
различных генетических связей. С помощью метода коэффициентов
nvu-н можно устанавливать многочисленные связи между
родителями и детьми, членами одной и той же семьи, между га*
мигами или между зиготами, а также между зиготами и
гаметами. Остановимся лишь на некоторых примерах, которые
нам понадобятся ниже, в частности в главе, посвященной
инбридингу. При этом принимается простейшая схема аддитив-
п-е
поколение
(пч)-е
поколение
Рис. 66. Связи между зиготой Z
и гаметами G{ и G2 через
коэффициенты пути а и коэффициент
инбридинга F.
Рис. 67. Связи между
зиготой 1, ее гаметами G, и
G2 и гаметами предыдущего
поколения.
ного действия генов: если эффект гена А2 равен 0, то эффект
гена А\ в единичной дозе (у гетерозигот Л Иг) равен 1, а
эффект двух генов А\А\ — 2. Так что для гамет возможны
значения 0 и 1, а для зигот 0, 1 и 2. Разберем простейший случай:
детерминацию зигот двумя гаметами — сперматозоидом и
яйцом (рис. 66). Буквами а, как показано выше, обозначаются
коэффициенты пути от каждой гаметы к зиготе (они
одинаковы, поэтому обозначаются одним символом). Но здесь
введена еще одна связь между гаметами G\ и G2. Она обозначена
через F и выражает возможную корреляцию между
генетической структурой гамет в том случае, если они получили
сходные гены от какого-то общего предка. Райт назвал величину
этой корреляции коэффициентом инбридинга *.
Об ш-тмьтнаиим этого коэффициента см. в гл. 8.
Очевидно, что полная детерминация зиготы, выраженная
в коэффициентах пути, будет следующей:
d zr,+d zrn+d zo^ =&+a*+2a*F=2a*+2c£F= 1.
Отсюда
1 1
а"=-57ГГТГ; a= -===-• (Ю)
20+F) I2(l+F)
Так как F находится в знаменателе, то значение а будет тем
меньше, чем сильнее корреляция между Gt и G2.
Если же G\ и G2 корреляционно не связаны, т. е. F=Qt
то а2= —; а= .
В этом случае а = 0,71.
Если же г %г =1, то а = 0,5. Таким образом, коэффициент
пути а должен иметь значение между 0,50 и 0,71.
Сходным образом можно определить связи между зиготой
и образуемыми ею гаметами, между двумя поколениями,
между членами одной и той же семьи и т. д.
В первом случае (рис. 67) путь от зиготы к гамете G{ равен
b = r.QZ = r Q,z=a'+a'F'=a'(\+F').
Если учесть предыдущую формулу для а', то F—F'
и, значит,
Ь= 1 - .(1" + Р);
V2(l+F)
Весь же путь от G/ до Gi
*-tV-i£-
Так как каждое поколение можно рассматривать как путь
от Gi' до Gi или от G/' до С/, то
"-ЫргУ-
1+F
что после преобразований приводит к выражению общей связи
между любыми двумя гаметами соседних поколений:
2
?зз
a,, n,тчнна таким образом, не зависит пи от коэффиад.
.лм аиига F ни от типа спаривания.
еНТчт". же величина будет выражать связь между родителем
и потомком, как это видно из рис. 68. Так как а =а, то a'b^ab,
л коэффициент пути ab= ^ *
Потомок
Рис. f>8. Связь между
родителем Z' и потомком Z
(с учетом связей через ra-
ыоты).
N N
Рис. 69. Корреляция между сибсама.
Отсюда можно подойти и к установлению корреляции
между членами одной и той же семьи (рис. 69), Коэффициент
корреляции между родителями гЕ обозначен Райтом символом т:
ге — гп.
Тогда корреляция между родителями и потомками может
быть выражена
г£.у = ab+abtn=ab (1+т).
Корреляция же между потомками N н N
r*s~=2abba+2abmba=z2aW(l+m).
(И)
(15)
Если из поколения в поколение не происходит сколько-
нибудь значительных изменений в системе спариваний и нет
отбора, то создается равновесное соотношение, которое
означает, что m и F сохраняются постоянными из поколения в по-
£*,
коление, константны также коэффициенты пути а и Ь. Тогда
Можно выразить ряд значений через ш, а именно:
fc2 L -; (16а)
2—т
F=ft2m== . "*- . (166)
2—т
Точно так же а2= - . В случае равновесия формула
1 lf~\+F'~
. I
сокращается до оа= - -.
При равновесии в популяции формулы корреляций между
членами семей окажутся более простыми:
rK=m = const;
Oe.v= ^-(l+m);
r*„= ~-(|+m). (17)
Мы разобрали лишь некоторые наиболее типичные случаи
приложения метода коэффициентов путей к анализу
генетических связей. Однако возможны различные усложнения:
неодинаковые коэффициенты пути между зиготами и гаметами,
различия в величинах Г. Очень оригинальными оказываются
зависимости при сцепленной с полом наследственности и т. д.
Однако они требуют специального рассмотрения. Для
дальнейшего достаточно ограничиться сводкой основных символов
и их значений (табл. 38), причем в ней предусматривается
полная детерминация наследственностью, без учета
возможных влияний на эти связи внешних условий, которые,
очевидно, могут быть как сводящими, т. е. усиливающими
корреляцию между родственниками, так н разводящими, т. е. ослебля-
• ютим и ее.
23?
7\?r ища №. Важнейшие коэффициенты путей и корреляции
Покадлслн
Коэффициент пути
от гаметы к
jnroTe
от зиготы к
| гамете
от зиготы к
зиготе
Корреляция
между
спариваемыми
генотипами
между соеди-
1 няющимися га-
I метами
между
родителем и потомком
Сим- 1
поли
а
Ь
аЬ
т
F
rNE
1
Значении 1
ирн
инбридинге
1
>2(1+F)
v1-^
JLl/L±£l
2 V l+F
2F
1 + F'
—
ab(\+m)
при
свободном
скрещивании
)/+
YZ
1
2
i °
1
1
2
при равно- 1
весии 1
-tV2-"1
i
УТ^т
1
2
const
Ьгт
Т" (l+m)
Коэффициенты детерминации Райта в приложении к
установлению роли наследственности и среды в изменчивости. Если
пчг — общая фенотипическая варианса популяции *, crv2 —
варианса, определяемая наследственностью, а ал2 — варианса,
определяемая внешней средой, то детерминация
наследственен2 -
ностью выразится как dv= ——, а детерминация средой
Onz
а«^
Таким образом, dy представляет собой современный
коэффициент наследуемости h1.
* В данном случае ми пользуемся обозначениями Райта. В настояшее
ftpt-мн лместо греческих буки пишут латинские.
Райт обращал внимание и на другую величину,
являющуюся дополнением к наследуемости,— долю изменчивости,
зависящую от внешней среды. В работе по анализу пегости морских
свинок он смог разложить ее еще на два компонента:
изменчивость, определяемую средой, общей для помета (на ее долю
в инбредном материале приходится 5% общей вариансы);
и изменчивость, определяемую средой, не общей для помета
(около 92% общей изменчивости). О разложении средовой
компоненты на составные части говорилось и в предыдущей
главе.
Далее Райт ввел так называемый коэффициент корреляции
между генотипом и фенотипом /%4f= ——. Отсюда следует, что
а,
d =г2 =/г2. По смыслу коэффициента ясно, что его нельзя
Матери Доч^и
~1 г(*0,5) 1 renomuno'~]
iPHUIIIUflbl
rtt ГГ*
| Фенотипы] ГмУт,дГч-
Фенотипь
Рис. 70. Корреляция между
фенотипами матерей и дочерей.
применять к особи. Непонимание этого обстоятельства было
основанием многих недоразумений. В действительности же гуч,
как и всякий другой статистический показатель, относится
только к совокупности. Он измеряет степень сопряженности
в распределении особей данной популяции по их фенотнпиче-
ским и генотипическим показателям.
Для анализа связей между генотипами и фенотипами
в пределах одного или нескольких поколении Райт применил
метод коэффициентов путей. Эту связь можно выразить
схематически рис. 70, который представляет собой дальнейшее
развитие схемы, изображенной на рис. 68.
Коэффициент корреляции между генотипами матерей и
генотипами дочерей при свободном скрещивании равен 0,5 (см.
табл. 38). Коэффициент же корреляции между фенотипами
матерей и фенотипами дочерей равен произведению
коэффициентов корреляции по всей цепи, связывающей эти
фенотипы, т. е.
''матери — дочери == Гу$ • 0,5 • Tw.
Отсюда
''матери - дочери === 0vor-^,
ИЛИ
r2w в 2гмат*ри - дочери;
Г2уф —dv = A*.
23*
/lJ== ЗГмаи'рн - дочери- ( 1оЗ)
T.ikhm образом, c/Y( = //-) может быть Определено как уд.
юеннын коэффициент корреляции между фенотипами матерей
н дочерен (или одного iu родителей и потомков).
Основанием для этого расчета, как видно из представленной схемы,
является теоретическая величина генетической корреляции
(корреляции между генотипами) между родителями и потомками.
Но очевидно, что можно воспользоваться коррелятивными
зависимостями между фенотипами любых родственных групп,
вставляя в цепь коэффициентов корреляции коэффициенты
генетической корреляции, которые при различных родственных
связях могут принимать разные значения, а именно:
идентичные двойни — 1;
полные енбеы — 1/2;
один родитель — потомство — 1/2;
средняя между двумя родителями и потомством— 1;
предок — потомки -— (1/2)k (k — степень родства, или
порядковый номер поколения от предка);
полусибсы— 1/4;
дяди — племянники — 1/4;
кузины — 1/8.
Из приведенного разбора коэффициентов путей Райта
видно, что формулы для г между родителем и потомком и между
членами одной и той же семьи (см. рис. 69) совпадают. В
таким случае можно определить Л2 и с помощью коэффициента
корреляции между сибсамн, т. е.
/i2 = 2rFS*.
В животноводстве нередко приходится иметь дело с полу-
сибсами, т. е. с потомками, имеющими одного и того же отца,
но разных матерен (или наоборот). Целесообразно
рассмотреть схему связей между фенотипами полусибсов (рис.71),
чтобы установить, как в данном случае должен быть
определен коэффициент h2.
Коэффициент корреляции между полусибсами и в этом
случае будет равен произведению коэффициентов корреляции по
всему пути, т. е.
О/ s=^ф • 0,5 • 0,5 • rV(p;
rH8=rV 0,25 = ft*. 0,25.
Откуда
/i2 = 4rHs. (186)
* В дальнейшем полных енбеов будем обозначать буквами FS, а полу-
nifccuo - HS.
240
Исходное для вычисления наследуемости значение
коэффициента корреляции между родителями и потомками (или
иными родственными группами) * может, кроме генетического
сходства, отражать также влияние каких-либо общих для обоих
поколений факторов внешней среды (например, недокорма).
На его основе получится преувеличенное представление о
степени наследуемости. Поэтому следует проводить анализ
таким образом, чтобы не так сказывалось влияние средовых
факторов, особенно специфических, на корреляционную
зависимость между родителями и потомками.
Потомки1... п
Отец
".CWCMKUtJi
Генотипы
-г(=о.5)-
Генотип
-П*Ц5)-
Генотипы
'И
U
Фенотип
'И
Фенотипы
lHS~
Фенотипы
Рис. 71. Корреляция между фенотипами полуснбсов.
Ограничения в использовании коэффициентов корреляции
для определения А2. Формула /12=2гмать-дочь. применяемая
для определения наследуемости многими сотнями
исследователей на самых различных объектах (сельскохозяйственных и
лабораторных), в том числе и автором книги, во многих случаях
дает хорошие и четкие результаты. Хотя значение А2 никогда
не может быть абсолютно точным, весьма существенно, каково
это значение — большое, среднее или малое, ибо это позволяет
судить как о генетической структуре популяции, так и о
необходимых методах селекционной работы для достижения
наилучших результатов.
Однако некоторые авторы выражают сомнение в
правильности метода определения наследуемости с помощью
коэффициентов корреляции, так как иногда он приводит как будто бы
к нелепым результатам — к значениям №<0 или А2>1. Такие
результаты чаще всего объясняются весьма простыми
причинами — недостаточностью взятых выборок. Вычисляемые при
этом коэффициенты корреляции имеют довольно широкие до-
* Формулу (18 6) выгоднее использовать не путем
непосредственного определения коэффициентов корреляции между полусибсами, а
применяя дисперсионный анализ.
1(3 II. Ф. РоК1Ш.киЛ
241
верительное границы, как правило, асимметричные. Поэтому
неудивительно, что при слабых связях фактические
коэффициента корреляции могут быть со знаком «lynniyc», а отсюда и
/:*-,0. что в конечном счете указывает на Слизкое к нулю
значение показателя наследуемости. Наоборот, преувеличенные
коэффициенты корреляции при их удвоении дают значения
/г>1. Таким образом, подобные случаи отнюдь не должны
вызывать сомнений в правильности самого метода. Конечно,
надо всегда помнить об условиях применения самой схемы
Райта (свободное скрещивание, т. е. отсутствие отбора и ин-
бридингаТ1ГдддТТТТТвТюсть действия генов). Неполное
выполнение этих условий неизбежно приводит к неточностям в
определении h2 с помощью коэффициентов корреляции между
фенотипами матерен и дочерей.
Возможны и вполне закономерные случаи отрицательных
значений /i2, создающиеся при наличии отрицательных связей
мать — дочь.
Коэффициент отрицательной корреляции между матерями
и дочерями в основном может возникать по двум причинам.
1. Статистическое взаимодействие генотип — среда,
которое проявляется в том, что разные генотипы неодинаково
реагируют на изменение среды вплоть до порядка их
расположения по уровню развития признака. Так, при резком различии
в условиях между поколениями хорошие матери могут дать
дочерей худших, чем дочери от плохих матерей. Впрочем, это
случай наиболее крайний и редкий. Очевидно, что нет смысла
изучать корреляцию между матерями и дочерями или
средними между родителями и потомством, если условия внешней
среды в двух смежных поколениях были очень неодинаковыми.
2. Межаллельное взаимодействие. Этот случай достаточно
подробно изложен в работе 3. С. Нпкоро. Рассмотрим вариант,
когда /i2=2r7n_rf не случайно дает значение больше единицы.
Из схемы Райта вытекает, что /i2= —m~d » где rG — коэф-
rG
фициепт генетической корреляции родитель — потомок,
причем, как указано выше, в равновесной популяции при условии
свободного скрещивания гс = 0,5. Деление коэффициента rw_d
на го равносильно умножению его на 2. Но если два
необходимых условия — равновесная структура или свобода
скрещивания—нарушаются, то rGt естественно, может не быть
равным 0,5- Тогда умножение rm_d на коэффициент 2 может
привести к неожиданным результатам. В эту величину необходимо
внести поправки. Так, например, если равновесие в популяции
нарушено за счет инбридинга, то можно использовать формулу
24'2
где Fm и F(i — коэффициенты инбридинга по материнскому
и дочернему поколениям (о коэффициентах инбридинга F
говорилось выше). Если Fm&Fti, то
/2 О l+F
Л=2Лт-"-ГТзТ-
В тех случаях, когда нет уверенности в осуществлении
условий, предусмотренных моделью Райта, лучше
воздержаться от вычисления коэффициента наследуемости с помощью
коэффициента корреляции между фенотипами родственников.
Однако наличие нулевых или отрицательных коэффициентов
корреляции по каким-либо признакам между смежными
поколениями все же достаточно красноречиво говорит о ничтожной
доле генетической вариации внутри общей фенотипическои
вариации. Отсюда следуют соответствующие выводы о
неблагоприятной ситуации для селекции, о чем речь пойдет ниже.
В конечном счете каждый из методов определения
коэффициента наследуемости: по корреляции между потомством и
одним из родителей (матери — дочери), между потомством
и средним родителем, полными сибсами пли полусибсамп —
имеет свои достоинства и недостатки, зависящие от удельного
веса средовых факторов, влияния наряду с аддитивными
генами также разной степени доминирования, в том числе
сверхдоминирования, материнского эффекта и конкретной ситуации
в анализируемом материале. Так, коэффициент корреляции
между полными сибсами, казалось бы, дает хорошую
возможность определить /i2 по формуле /i2=2rFS. Однако в стадах
крупного рогатого скота нелегко подобрать достаточное
количество семей, в которых было бы не менее двух потомков, при
этом родившихся и воспитывавшихся в достаточно сходные
по условиям годы. Поэтому приходится считать этот способ
не очень надежным в применении к данному виду животных,
хотя он вполне применим к многоплодным животным.
Корреляция же между полусибсамп представляется более
выгодной, но надо иметь в виду, что учет полусибсов по
матерям может показать преувеличенное сходство, так как
потомство одних и тех же матерей, хотя и от разных отцов,
подвергается влиянию общих условий эмбрионального, а в какой-то
части и постэмбрионального развития (кормление молоком
матерей). Кроме того, возможно и влияние генотипа матерей
(так называемый материнский эффект). На полусибсов же по
отцам эти факторы не влияют.
Что же касается использования формул, где фигурирует
корреляция между родственниками, более отдаленными,
нежели полусибсы, то при этом очень сильно возрастает ошибка
выборочностп полученных результатов корреляции, а отсюда
16*
243
и наследуемости, поэтому нет смысла применять их на
практике, кроме каких-либо особых, исключительных случаев.
Поэтому целесообразно наряду с коэффициентами корреляции
применять и другие параметры для определения
наследуемости, в какой-то степени ослабляющие ограничения,
накладываемые на формулы с коэффициентами корреляции.
Коэффициент регрессии как мерило наследуемости.
Используя сходство между родителями для определения
наследуемости, исходят из предположения, что родители
представляют собой случайную выборку из популяции и что
скрещиваются они также случайно. Однако очень часто одно или
оба условия не выдерживаются. Как в условиях опыта, так и
особенно у сельскохозяйственных животных и растений
родители представляют собой отобранную группу, поэтому феноти-
ппческая варианса среди родителей меньше, чем в популяции
в целом, и меньше, чем среди потомков. Спаривание
родителей также производится часто по принципу фенотипического
сходства (так называемое ассортативное спаривание), т. е.
имеется корреляция между фенотипическими значениями
спариваемых особей, что также влияет на соотношение ва-
рианс родителей и потомков.
Эти затруднения преодолеваются другим методом для
определения коэффициента наследуемости — методом регрессии.
Исходная формула в случае регрессии по своему
коэффициенту та же, что при использовании коэффициента корреляции,
а именно: если известна регрессия потомков к матерям, то
№=2Ьом, или /i*=2/?om, (19)
здесь Ь и R — разные обозначения коэффициента регрессии;
О — потомство; Л1 — матери.
Если учет признаков возможен для обоих родителей, то
можно определить коэффициент регрессии потомков (О)
к средней обоих родителей (Я). В этом случае h2=bQ-, т. е.
сама по себе величина коэффициента регрессии дает значение
коэффициента наследуемости. Так, например, изучали у свиней
регрессию числа позвонков потомков по отношению к среднему
числу позвонков v родителей. Было получено /?(й)=0,74.
Тогда Л= = 0.74.
Как указывалось выше, коэффициент регрессии имеет
известные преимущества по сравнению с коэффициентом
корреляции, использование» которого ограничено рядом условий
(родители как случайная выборка из популяции, отсутствие
отбора, одинаковость варианс родительской группы и
популяции и др,). Ь частности, отбор родителей из популяции не
влияет \\л регрессию потомков по родителям — ковариаиса
:м;
уменьшается в той же степени, что и варианса родителей,
поэтому угол наклона линии регрессии, измеряемый
коэффициентом Ь, остается тем же.
Точно так же нарушение случайности скрещивания
родительских пар слабо влияет на величину регрессии потомков к
средней величине признака родителей. На значение
коэффициента регрессии не влияет п степень изменчивости признака.
Регрессия почти целиком определяется наличием генотипиче-
ских различий среди особей родительского поколения и среди
потомков. Впрочем, на нее может повлиять полное или
неполное доминирование. Если доминантные гены передаются
отцами, коэффициент регрессий детей к отцам будет больше, а к
матерям — меньше и наоборот. Поэтому регрессия потомков к
одному из родителей может быть мерилом '/гй2. а регрессия
потомства к средним обоих родителей — Л2.
В методическом отношении следует учесть такие
осложнения. Если численности потомства от разных семей сильно
различаются, то целесообразно ввести поправки в средние
значения каждого потомства согласно их весам. Если вариансы
среди особей двух полов неодинаковы, то использование
формулы /i2=&o- может дать неточные результаты, ибо она
требует равенства варианс обоих полов. Тогда лучше определять
наследуемость отдельно по регрессиям дочерей к матерям
и сыновей к отцам. Если оба значения h2 окажутся более или
менее одинаковыми, можно вычислить и регрессию к средним
значениям родителей без взвешивания средних значений
потомков по количеству самок и самцов среди них.
Интересен случай, когда регрессия потомков к матерям
значительно больше, чем регрессия к отцам, что может быть
доказательством так называемого материнского эффекта, т. е.
влияния фенотипа матерей на фенотип потомства. Факты
материнского эффекта известны для веса и размеров тела у
млекопитающих.
Хорошей иллюстрацией использования регрессии для
определения коэффициента наследуемости могут быть данные Рнва
и Робертсона, полученные на дрозофиле (Drosophila melano-
gaster), графически изображенные на рис. 72. Измеряли
длину крыльев у 37 пар родительских особей н у их потомков.
Точки на корреляционном поле показывают средние значения
длины крыльев двух родителей (шкала на оси х) и средние
значения их потомства (шкала на оси у).
Хотя родители спаривались по принципу «подобные с
подобными», отчетливо видна регрессия потомства по среднему
значению родителей.
Значение коэффициента для теоретической линии регрессии
0,577^0,070. В таком случае и Л2 = 0,577±0,070.
245
Регрессия потомства к матерям внутри группы каждого
отиа. Описанные выше приемы определения наследуемости
п\тем установления регрессии потомства к обоим родителям
не всегда пригодны, особенно к сельскохозяйственным
животным. Так как каждый самец обычно скрещивается со многими
самками, то определение средней между родителями теряет
смысл, а отсюда вытекает, что нельзя применить метод
регрессии потомства к средним по паре родителей. Кроме того,
самцов обычно очень немного. Поэтому более подходящим может
Рис. 72. Регрессия потомков к средней величине крыльев
родительских пар у дрозофилы.
На оси абсцисс средние значения родительских пар, на оси
ординат — потомков каждой пары. Наклонная линия —
теоретическая линия регрессии (по Риву и Робертсону).
оказаться метод определения средней регрессии потомства
к матерям, вычисленной по регрессиям потомства к матерям
внутри группы каждого быка. По этим отдельным значениям
регрессии получают объединенный коэффициент регрессии как
средней взвешенной. Он будет равен половине Л2. Ковариация
>хе потомства и матерей равна половине аддитивной вариансы
популяции как целого, так что регрессия потомства к матерям,
вычисленная по каждому быку отдельно, выражает
аддитивную часть наследуемости, т. е. 6=0,5 —. Успешное исполь-
ар-
з< ванне этого метода зависит от отсутствия материнского
эффекта, повышающего сходство между матерями и дочерями.
Для проверки последнего учитывают значения квадратическях
отклонений для самок и для самцов и в определяемые
коэффициенты регрессии вносят поправку на отношение aV :orf. Этот
метод был применен Клейтоном, Моррисом и Робертсоном при
изучении наследуемости количества брюшных щетинок у дро-
J4tt
зофнлы. Были вычислены коэффициенты регрессии дочерей id)
к матерям (т) и сыновей (s) к материм по каждом) самцу;
bdm = 0,269; Ьт = 0.20С
Так как отношение между сигмами самок и самцоз
равнялось 1,17, то коэффициент регрессии Ь«т был умножен на
1,17 и стал равным 0,241. Были получены следующие
значения ft2:
по bdm h2 -0,54;
по исправленному Ь9т Л2— 0,48;
по объединенным данным Л2«0,51.
Сопоставление результатов определения Л2 с помощью мор-
реляционного и регрессионного анализа. Для изучения
наследуемости обычно стараются использовать различные методы:
определение корреляционных зависимостей между разными
родственными группами (матери — дочери, родители —все
потомство, полные снбсы, полусибсы), а также учет регрессии
тем или иным способом. Сопоставление полученных
коэффициентов наследуемости позволяет судить о степени их
надежности.
Так, в работе Шими Эль Шими, посвященной
наследуемости молочной продуктивности и жирности молока фризского
скота при разведении в местностях с разными почвами,
сравнивались показатели наследуемости, полученные на основе
коэффициентов корреляции и коэффициентов регрессии. В обоих
случаях автор сопоставлял дочерей с матерями, применяя
формулы
A*=2rm.f и h*=2Rdm.
Работа Эль Шими заслуживает особого внимания по
объему изученного материала (около 9000 лактации по матерям
и дочерям от 286 отцов) и по особой тщательности отбора
первичных данных, с тем чтобы освободиться от побочных
осложняющих факторов (различная длина лактационных периодов,
неблагоприятные условия военных лет и др.). Задача же
работы заключалась в изучении влияния на показатели наследу-
емости по двум признакам таких факторов, как возраст коров
(номер лактации от 1 до 3) и различия в почвах лугов и
пастбищ, с которых крупный рогатый скот получал траву и сено.
Оказалось, что эти факторы очень слабо влияли на величину
наследуемости: в пределах сотых долей Л2. Основное же
различие было по признакам: для жирномолочности Л3,
определенный с помощью корреляции, составлял 0.82—0.84, а по
методу регрессии — 0,77—0,85; для обильномодочиости
соответственно 0.37—0,41 и 0,36—0.40.
247
Очень четкие результаты могут дать также опыты на
модельных животных. Так, Клейтон, Моррис н Робертсон
получили очень сходные значения коэффициентов наследуемости
отчества брюшных щетинок у дрозофилы (Drosophila me-
jar.ogaster) тремя методами:
з! по корреляции между сибсамн (полными)
Л2=0,53+0,07;
3) по корреляции между полусибсамн
/z-=0,4S±0,ll;
б) по регрессии потомков к родителям
Л2=0,51 ±0,07.
Однако, как показала Л. А. Васильева, такое совпадение
бызает не всегда. На одном и том же материале крупного
рогатого скота совхоза «Омский» Омской области были
получены оценки Л2, представленные в табл. 39.
Таблица 39. Значения коэффициентов наследуемости для процента
жира и удоя за 300 дней лактации, вычисленные по трем формулам
1 i
1
i
j Поизалки
Процент жнра
Удой за 300 днеЗ лактации
Формулы для Л*
*md
0,416
0,238
2bdm
0,462
0,236
ArHS
0,440
0,124
По первому признаку значения Л2, полученные по всем
трем формулам, очень близки, по второму же оценка по
формуле Л- = 4гн5 оказалась вдвое ниже. Это показывает, что при
анализе по полуенбеам имелись факторы, сильно нарушавшие
исходные предпосылки для применения формулы. Так как
корреляция между полусибсамн учитывается по отцам, то на по-
л>ченное различие мог влиять более интенсивный отбор по
быкам. Вообще при изучении конкретных стад
сельскохозяйственных животных возможны значительные колебания в
значениях h2 как в зависимости от метода его определения, так и
->т конкретных условий содержания стад.
Использование дисперсионного анализа для определенна
коэффициента наследуемости. Наличие данных о родственных
связях у животных и растений и возможность группировки осо-
:■«
беи по характеру родственных связей дают возможность
применить более современные методы изучения наследуемости,
а именно дисперсионный анализ. С его помощью также можно
определять значения вариаис — фенотипической, средовои и
геиотипической — и на их основе — коэффициенты
наследуемости. При этом, как будет показано в дальнейшем,
используются те же сибсы и полусибсы, которых можно
анализировать и корреляционным методом. Поэтому всегда можно
сопоставить результаты, полученные корреляционным и
дисперсионным методами, а также перевести данные дисперсионного
анализа в данные о корреляциях между субсами и полусиб-
сами.
В различных схемах дисперсионного анализа основное
заключается в доказательстве роли того или иного фактора
(или факторов) в общей вариации по данному признаку. Это
доказательство получается с помощью критерия F. Если
фактически полученное значение F превышает табличное (см. в
приложении табл. VII—VIII), влияние данного фактора
считается доказанным, если не превышает —сохраняется
нулевая гипотеза.
Для изучения наследуемости используется и другая
сторона дисперсионного анализа, а именно возможность
установления долей влияния отдельных факторов в общей вариации
(чаще всего речь идет о влиянии отцов и матерей). К
сожалению, в отношении конкретных методов, с помощью которых
можно было бы устанавливать эти доли влияний, была
допущена изрядная путаница, извратившая самую суть
дисперсионного анализа, как известно, хорошо разработанного
математически и вполне обоснованно применяемого в генетике и
селекции. Ниже, после разбора методов дисперсионного
анализа, применяемых для получения значения Л2, остановимся и на
неправильных, на наш взгляд, подходах к оценке
наследуемости.
Структура иерархического комплекса. В обычных
дисперсионных комплексах уровни (градации) одного фактора
сочетаются с уровнями (градациями) любого другого фактора.
Например, изучая одновременно влияние какого-либо
химического вещества и температуры, можно построить план
опытов, при котором будут созданы все сочетания уровней
(градаций) химического вещества и уровней (градаций)
температурного фактора. В результате будут получены группы
вариант, на которые действуют любые сочетания изученных
факторов. Вследствие этого модель дисперсионного комплекса
может быть представлена в отклонениях или в вариансах.
Xijk-v + Ai + Bj + AiBj + e^H.
249
^=o= +o=+°= +o;
*.**
Однако при анализе материала, взятого из природы или
и* хозяйств, обычные факторные схемы часто применить
невозможно, так как градации одного фактора не сочетаются с
градациями другого фактора. В рамках каждого фактора они
различны.
Именно такая ситуация чаще всего имеет место при
изучении наследуемости, и тогда применяется так называемая
иерархическая схема дисперсионного анализа, или просто
иерархический комплекс.
Су?'
*206»U
.\\KV>
."yX^Saii-
Mazxpa | v ^ v< i >-, m /?....£
jbma^l /Л 't\ /7\ /4\ I /f\ '* '* !
f1^ 7J. Слези иерархического дисперсионного комплекса.
Схема иерархического комплекса применительно к
сельскохозяйственным животным и прежде всего к крупному
рогатому скоту представлена на рис. 73. Допустим, что учитываются
следующие факторы: порода, производители, матери, т. е.
самки, покрытые производителями, и, наконец, их потомство. Но
очевидно, что в породе М один производители^ а в породе N
другие. Каждый производитель покрывает определенных
самок (х, у /, — покрытые одним производителем; Л, /,..., /г—
другим и т. д.). Для каждого производителя они различны.
Мы оставляем пока в стороне возможность специальных
опытов, в которых самцы меняются местами: в один сезон группа
самок покрывается самцом I, а во второй — самцом II (ди- и
полналлелыюе скрещивания, о них будет говориться ниже).
Для установления наследуемости с помощью
иерархической схемы достаточно рассмотрения только факторов:
производители и матери. Потомство анализируется по влиянию этих
двух факторов Обычными методами * устанавливается доля
* О** достаточна полно наложены в кн : /7. Ф. Рокицкий. Бйологиче-
«•в гмтвстш, Нэд £« Минск, 1973. Основные формулы и макеты тиблиа
и* а ормдожеяяя.
Ии
1:сли, как указано выше, в £ вхсдит изменчивость,
вызываемая влиянием внешней среды, плюс генетическая
изменчивость, составляющая примерно половину всей генетической
вариации популяции, а в D и в 5 — соответственно по одной
четверти генетической изменчивости популяииидо знаменатель
для вычисления Л2, т. е. общая фенотипическая вариация,
может быть выражен суммой E+S + D. Числитель же может быть
вычислен или только по отцам, тогда значение 5 надо учет-
рерить (45), или по матерям (4D), или, наконец, с учетом
роли и отцов и матерей. В последнем случае он должен быть
2(S + D). Таким образом, возможны три разных формулы для
вычисления А2:
Л2=
4S
Ф
= = 2(5+Р)
D+S+E D+S+E D+S+£
(20)
Выбор той или иной формулы или использование всех трех
для сопоставления результатов зависит от особенностей
изучаемого материала.
Конкретные примеры определения коэффициента
наследуемости методами дисперсионного анализа по иерархической
схеме. Мы разобрали так подробно эту схему анализа потому,
что она очень широко применяется многими исследователями.
Приведем несколько конкретных примеров.
Уитом и Риггсом были вычислены коэффициенты
наследуемости длительности беременности на коровах 5 пород
(табл. 41).
Таблица 41. Дисперсионный анализ длительности бередешюсга
Источник варнаиня
Производители
внутри породы
Матки от отдельных
производителей
Полные сносы
Число
степеней свободы
22
165
165
Средни А
кмдрат
134.3
34.5
31.6
Структур! среднего
къэдрат*
E-r!,37D + 20,985
£ + 1.370
■ Е
Авторы провели также анализ роли пород, но строчка
«пароды» в таблицу не включена. Пропущена также графа
«сумма квадратов» как ненужная для определения наследуемости.
Значения коэффициентов / н * следующие: /«16,4; *« 1,37-
Тогда£«31ДЬ~2.1 и5«*4,&
Значения наследуемости для длительности беременности
коров оказались следующими:
253
по формуле -
40
D+S + 1:
2(S+D)
* * -D+S+E'-0*'
» „ -—- ~ 0,50.
D+S + П
Из данного примера видно, что при использовании различ»
ных методов анализа могут быть получены несколько
отличающиеся значения наследуемости. Значение Л2 по первой фор-
муле обладает более узкими доверительными границами,
поэтому его можно считать более точным. Однако несколько
удивляет, что /?\ вычисленный по самцам, был выше, нежели
вычисленный по самкам. Как будет видно из последующего
изложения, скорее можно было ожидать обратного.
Все же общее количество материала в этом исследовании
было слишком малым, на каждую мать приходилось всего 1,37
дочери, поэтому в определении наследуемости могли быть
неточности. Мы приводим этот пример как достаточно типичный
для метода.
Но есть примеры исследовании, более выдержанных с
методической стороны, в которых было получено очень большое
совпадение результатов по всем трем формулам.
Так. на свиньях породы датская ландрас Фредин и
Джонсон определяли наследуемость ряда признаков. Материал,
взятый из данных многих датских станции, где проводились
испытания по потомству, охватывал 468 самцов, каждый из
которых скрещивался с 2 матками; из потомства учитывалось по
2 самки и 2 самца от каждой матки.
Данные о наследуемости одного из признаков, а именно
длины тела (только по мужскому потомству), приведены в
табл. 42.
Таблица 42. Значения //2 длины тела у датских
свиней, полученные разными методами
Формула
45
D-rS + E
1 D+S+E
| 2 (S -Н D)
D + S + E
к*
0,57
0,48
0,53
254
Результаты определения h2 с помощью дисперсионного
анализа были сопоставлены с данными, полученными путем
установления коэффициентов корреляции между полными сиб-
сами и полуенбеами: Tf8 = 0,263 и r/js = 0.142. В таком случае
1г2 по rFS равен 0,526, а ло rHs — 0,568. Совпадение очень
большое.
Метод внутриклассовой корреляции для оценки
наследуемости. Частным случаем использования иерархического
дисперсионного комплекса для оценки наследуемости является
вычисление коэффициента внутриклассовой корреляции.
В описанном выше комплексе источник вариации 1 порядка —
быки, II порядка — матери-коровы, с ними скрещивавшиеся,
111 — потомки от каждой пары.
В практике иногда трудно подобрать достаточное
количество матерей, имеющих по нескольку лактирующих дочерей от
одного и того же быка. По этой причине может быть
применен упрощенный дисперсионный комплекс, где источник
вариации 1 порядка — быки, II — их потомки. В таком случае
двухфакторный дисперсионный комплекс превращается в од-
иофакторный, в котором весь анализ ведется только по
вариации между быками.
Это и будет случай так называемой внутриклассовой кор-
реляции I Г{С=— 1 . При определении наследуемости
х сга-Н-Ог
коэффициент корреляции умножается на 4:
h2_ 4q,a _ 45
os2+aur op2
Если при этом можно учесть и различия между дочерями,
полученными от отдельных быков, то их можно использовать
для знаменателя (os2+of+Ou>2)> но расчет приводится только
по самцам.
Подобная работа была выполнена Л. А. Васильевой *. Ее
данные по двум совхозам в сокращенном и немного
измененном виде приведены в табл. 43.
Коэффициенты наследуемости по проценту жира в
молоке, вычисленные этим методом, оказались очень сходными для
обоих совхозов. Но интересно отметить, что в этой же работе
Васильевой есть данные о коэффициентах наследуемости
молочной продуктивности. Они оказались резко отличными: по
совхозу «Омский» /t2 = 0,152 и по совхозу «Троицкий» А2« 0,532.
Автор отмечает очень большие колебания в условиях кормле-
* В ней рассматривается также вопрос о коэффициенте повторяемости.
О нем будет идти речь о гл. 10.
255
Тиб.шцч 43. \чоъффнииент наследуемости процента жира в молоке
коров по двум совхозам
Источник iuiptiniuui
Между быками
Между
дочерями от
отдельных быков
Среди дочерей
Между быками
Между
дочерями от
отдельных быков
Среди дочерей
df
щк
Сов.
29
699
2423
1,855
0,259
0,047
Строение «is
\OJ tOMCKUlh
а;, + 4,25о2 +
-|- 100,13a;
4 + 4,250-*
°l
Совхоз «Троицкий*
15
' 262
722
0,920
0,133
0,052
o2w + 4,03a; +
+ 60,33a;
a^ +4,03a;
°l
Значение a'
2
no a,
of- 0,0159
a; = 0,0498
a^--. 0,0471
0,564
o2s --■ 0,013
1 a;-0,020
4~ 0,052
0,612
ния в совхозе «Троицкий», а поэтому полученные по этому
совхозу данные по молочной продуктивности, по его мнению,
не отражают действительного генетического состояния стада.
Роль отдельных компонентов генетической вариансы в
коэффициенте наследуемости. Для определения коэффициента
наследуемости употреблялись три формулы. Одна из них
основывалась только на использовании а*2 (значение о«2
учетверялось), вторая — только на а/2 (значение о/2 тоже учетверялось),
в третьей фигурировали и о.ч2 и о/2. Возникает вопрос о том,
«ака^{ наследуемость (в узком или широком смысле слова) ими
измеряется.
Знаменатель аР2, очевидно, представляет собой сумму о**+
+a!2+otc2 пли. иначе, ол2+спг+аД Компонент различий между
самцами—а,2 —это варианса различий между полусибсовы-
ми семьями п, таким образом, не что иное, как оценка фено-
типической ковариансы между иолусибсами (covHs). Как
показано в предыдущей главе, она представляет собой -ra^
(точнее, ■ олг-\—га'&лл* "о последняя величина настолько
4 16
мала, что ею можно пренебрегать).
Компонент ow2 имеет сложную природу. В него входит как
средовои компонент, так и генетический. Чтобы выяснить его
структуру, можно рассуждать следующим образом. Так как
компонент межгрупповой вариансы равен ковариансе всех
членов всех групп, то внутрнгрупповой компонент должен быть
равен всей вариансе минус коварианса членов группы.
Потомство отдельных самок —это семьи полных сибсов. В таком
случае варианса внутри потомства представляет собой
op2—covFs. Так как covfb— ~ сгЛ2+ ~г°^ (отбрасывая
взаимодействие), то
0vt=(Ja2+od2+o<.2- у ад2- -- ое?= ~ ал2+ Т а/>2+аЛ
В данной случае <jt.2 — это средовая варианса, общая для всех
групп. Выше мы обозначили ее символом oW. Следовательно,
<V2= — Ол2+ — aD2 + G2fU:.
Наконец, вариация между потомством отдельных самок
может быть представлена как
Or2—O*2—Oe'1=C0UFs—C0VHs= — 0А2+ ~- Oir+CT2,,
4 4
(средовои компонент — варианса, создаваемая за счет среды,
не общей для сибсов каждой самки).
Итоги всего представлены в виде табл. 44.
Таблица 44. Компоненты генетической и средовои вариансы,
определяемые при дисперсионном анализе роли производителей и самок
Источники парнацнн н их
обозначения
Производители о2
Самки о?
Потомство a2w
о* + о?+с£
Производители + самкн
1 а? + о?
Копармансы
CWHS
COV^-COV^
02г — со\у s
о
cov^s
Оцсннв.1*мые ими п*рдметрм
Т°'л
•i+el+eJc+eir
т-^д + т-'Ь-Кг
17 П. Ф. РокнцкнЛ
257
Окюлл становится ясным, что о„= н о,- и значения насле-
XM-MiKiH оцениваемые по отцам и матерям, могут быть Ht
•авмы. Коэффициент наследуемости по отцам (при нерархиче»
!-м»м. конечно, комплексе) — это наследуемость в узком смысле
с юна т. с. /i- = - д . Коэффициент наследуемости по матерям,
О,л
как и коэффициент наследуемости по сумме отцов и матерей,
ближе к —;-— коэффициенту наследуемости в широком смыс-
ле слова. Надо иметь также в виду, что в показатель
наследуемости но матерям входит компонент взаимодействия между
генотипами матерей и отцов, который никак не отражен в
таблице и может быть учтен лишь другими, более сложными
методами, в частности системой полиаллельных скрещиваний,
к которым приложим метод обычного двухфакторного анализа
с моделью
Осг=оа2+ов2+о2ав+о€2'
Кроме отклонений за счет доминирования, в показатель а/*
входит средовой компонент, а именно варианса, создающаяся
за счет среды, общей для сибсов каждой самки.
Если между о,2 и о,2 нет существенных различий, то,
очевидно, роль доминирования, взаимодействия и общей среды
невелика, с ними можно не считаться, и оба параметра могут
быть одинаково использованы для определения коэффициента
наследуемости. Однако возможны случаи столь сильного
влияния других факторов, кроме аддитивного компонента, что
использование параметра а/2 становится невозможным. Особенно
это относится к многоплодным животным, у которых может
обнаруживаться сильное влияние средовых факторов, общих
как внутри каждого помета, так и для нескольких пометов
одних и тех же самок. Естественно, что они резко увеличивают
корреляцию между енбеами.
Фолконер приводит в качестве примера неопубликованные
данные Баумана по весу мышей (самок) в 6-недельном
возрасте. Было получено 719 потомков от 74 самцов и 192 самок,
каждая из которых давала один помет. Опыт проводился в
течение 4 поколений *. Не вдаваясь в детали, можно
ограничиться лишь приведением сводной табл. 45.
Очень велики значения сД очевидно, за счет среды, общей
для пометов, и о/2 — за счет среды, не общей для пометов-
Поэтому целесообразно использование для определения А1
только о\2:
"- 5,14 -U,J/-
• Н А Плохинский в книге «Наследуемость» даст нсоерную мнтер1»Рг
танию *того опыта, считая его результаты непонятными.
258
Таблица 4о. Дисперсионный анализ веса мышей в 6-яеде.аыюи
возрасте
Источник парнации
Самцы
Самки
Потомство
df
70
118
527
mv
17,10
10,79
2,19
Структура т%
ol +k'o'j + dk'ol
ol + ko*
1
1 Зиачшае f>
07- 0,48
07 2.47
4 2.19
о2* 5.14
k = 3,48; k' 4,16; d 2.33.
В дальнейшем Фолконер устанавливает соотношение
отдельных компонентов вариации неса мышеи исходя из
указанных выше данных о структуре отдельных показателей
вариации (правда, без учета возможных неаддитивных эффектов).
Если принять ор2 за 100%. то
ал~4а*2=1,92,
что составляет 37%.
агл=о/2-о.2= 1,99 (39%).
Наконец,
a-W=au-2-2oy:==l,23, т. е. 24%.
Очевидно, что варнанса, создаваемая за счет общей среды
(о2сс). больше, нежели варнанса, создаваемая за счет средовых
влияний, обусловливающих различия в потомстве полных сиб-
сов, т. е. пометов.
Из этого примера видно, что применение дисперсионного
анализа по сравнению с другими методами дает в общем более
надежные и достоверные значения коэффициентов
наследуемости, так как при этом можно точнее выделить из общей
вариации роль средовых факторов, что не всегда удается при
пользовании формулами, в которые входят коэффициенты
корреляции. В последнем случае приходится проводить
специальный анализ, чтобы исключить из общего значения
корреляции величину этой средовой корреляции, например, путем
сравнения пар, взятых из одного хозяйства, и пар. взятых из
разных хозяйств. Таким способом П. Р. Лепер вычислил сте^
пень наследуемости молочной продуктивности (по материалам
Балтийской племенной книги). Степень наследуемости
оказалась равной 0,15. Гоуэн (Gowen) в свое время изучил корре-
17*
259
IV утерями и дочерями но дани
-чннм межд> ^т п0ЛУЧИл значении /г
;,%е;ч*йскогоj ско < 0диак0 „рц учете Д
^к1ИВН^Т111мнними имовиямн в общей I
шмн по данным книги рекорда
L> для молочной щ^
доли изменчивости,
- * отними MviW..n - ™~. корреляции между
вызванной втш • ^ iT|| целнчнны пришлось значительно
матерями и *^0о_о;Ь С другой стороны» соотношение меж*
уЖИ^11Ьными Гредовыми факторами и ролью наследстве*
ду Ра~™4' л суммарно, без расчленения на аддитивные, До*
Н^1лнж и эпиотатичеекпе факторы) при конкретном at*
«w становится более сложным.
Роль отдельных средовых факторов при определении в*
едуемости. Хотя общим положением при анализе наследуй
"ости является необходимость максимально уменьшить ром
средовых факторов, особенно плохо учитываемых» однако при*
тически добиться этого нелегко. В приведенной выше работе
Ать Шимн было достигнуто оптимальное положение, поэтому
ему удалось получить очень стабильные значения коэффициент
гов наследуемости, не применяя дисперсионный анализ, н
одновременно показать незначительную роль таких факторов,
как возраст коров и различия в кормах, полученных с
участков, отличавшихся по почвам. Однако литературные даням
на этот счет довольно противоречивы. Так, если по удоям Эль
Шими не отметил роли возраста коров, то по признака*
экстерьера и живому весу крупного рогатого скота Блекмор,
Мак-Гилдарт и Лаш обнаружили явное повышение
коэффициентов наследуемости с возрастом. Например, для глубины
груди телят 6-месячного возраста /Г2 = 0,24, для 12-месячной)
Л-1-0.34 и 24-месячного h2 = 0,79. Иная картина по
наследуемости живого веса: при рождении максимальное /12в»0,8в,.В
б месячном возрасте Л- = 0,14 и в 24-месячном /i2« 0,53..
По-видимому, в послемодочный период резче сказываются средой**
различия в кормлении. К зрелому же возрасту повышакл**
коэффициенты наследуемости и промеров, и живого веса.
Очень важны, но, к сожалению, противоречивы данные 0
коэффициентах наследуемости различных хозянственно-ПйДВ*
ьыч признаков в зависимости от уровня продуктивности.
Для установления роли и удельного веса отдельных фан**
ров, как генетических, так н средовых, в вариации прианвК*»
неоценима роль дисперсионного анализа,
.in!!!Ч^Н1,У Ааа»ые приводит Лаш в своей известной КШИ*
ппми^ Breedin* Plans> no работе Плана (Plum), изучав»**
Гь иапп«^НЧИВОСТИ и™1*™ молочного жнраУ О**
^^SV&Zl НЛД°ЛЮ рамичий МСЖДУ громив n«t*
S^t «^STft ваРиацин- Различия между стада** **
аЕН IJSSIiep> ">*""*** определяют 12% общей М**
До^^^ь^К"рмлении К0Р°" внутри ст?да - в%. •
довадыю большую долю изменчивости (около 30*) *
\далось связать с определенными внешними или внутренними
факторами. Для наследуемости количества молочного жира
(по данным о коровах) //2«0,26. Параллельное изучение
регрессии дочерей к матерям в пределах потомства тех же
производителей дало близкое значение Ла (в пределах от 0,15 до
0,30). Однако надо иметь в виду, что в той части
изменчивости, причины которой остались неустановленными, может быть
некоторая доля генетических причин. Поэтому полученный
коэффициент наследуемости следует считать заниженным.
Неправильные подходы к оценке наследуемости. Все
изложенное выше показывает, что методы определения
коэффициентов наследуемости достаточно хорошо разработаны и
обоснованы теоретически. Но мы уже упоминали, что в нашу
литературу проникли неправильные представления о некоторых
методах. Предложены также новые методы, не имеющие
достаточных оснований. Ввиду того что они получили
распространение среди некоторых генетиков и селекционеров, на них
придется остановиться.
Н. А, Плохинский предложил следующую очень простую
формулу для вычисления Л2, а именно:
Сер
По его мнению, вычисленное таким образом значение Л2
является основным показателем наследуемости, который надо
предпочесть всем остальным возможным методам его
вычисления. Он считает, что тот же принцип может быть применен для
оценки влияния любого фактора х вообще, т. е. цх2= -х*~ где
Су
1}—корреляционное отношение (буквой С обозначена сумма
квадратов отклонении, в литературе обычен символ s$).
Несмотря на то что в статьях В. Ю. Урбаха и Э. X.
Гинзбурга была отчетливо показана математическая
несостоятельность метода Н. А, Плохинского, он по-прежнему
используется, очевидно, из-за простоты, а также потому, что
критика его носила чисто математический характер. Но надо
сказать, что и без специального математического анализа
легко убедиться в ошибочности рассуждений Плохинского,
если разобраться в сущности метода дисперсионного анализа.
Прежде всего относительно величины суммы квадратов
{ss% или С, по терминологии Н. А. Плохинского). Она по
существу своему не может быть мерилом вариации. Если бы это
было так, то незачем определять s* (или а1) как
s =* df n'_i
251
а достаточно было бы пользоваться только числителем. На
самом же деле сумма квадратов отклонений от средней
представляет собой чисто арифметическую величину, зависящую
от объема изучаемой совокупности. Только отнеся ее к числу
степеней свободы, можно получить оценку вариации.
Это же в еще большей степени относится к анализу
дисперсионных комплексов.
Для обоснования своей позиции Н. А. Плохинский
выдвигает положение, что сумма квадратов аддитивна, т. е. всегда
равна сумме вычисленных отдельно по источникам вариации
сумм квадратов. Но в данном случае нет никакого закона
аддитивности, а есть лишь констатация элементарного
арифметического факта, заранее предусматриваемого самим
построением любого дисперсионного комплекса.
Естественно, что суммы квадратов вычисляются для
каждой строчки дисперсионного комплекса отдельно и для каждой
из них есть свои числа степеней свободы. Чтобы сохранить
возможность суммирования результатов, Плохинский делит
суммы квадратов не на каждое соответствующее им число
степеней свободы, а на одну и ту же величину — объем
комплекса минус единица (N— 1). При делении Сх на Си эта
величина сокращается. Таким образом, показатель Н. А. Плохин-
ского Л2=т]2 никак нельзя считать показателем
наследуемости в том общепринятом смысле, который вкладывается в
понятие наследуемости в современной генетике и селекции.
В действительности основной величиной при дисперсионном
анализе служит средний квадрат (ms), получаемый как
частное от деления ss на число степеней свободы df. Именно
отношение средних квадратов, так называемое вариансное
отношение (variance ratio), и есть критерий F, по величине которого
с\дят, можно ли считать влияние того или иного фактора в
опыте или в наблюдении доказанным или нет. И сам
Н. А. Плохинский вынужден пользоваться критерием F для
доказательства достоверности показателя наследуемости,
вычисляя F общепринятым способом, т. е. делением ss на
соответствующее число степеней свободы, а для определения самого
показателя наследуемости прибегает к другому,
противоречащему ему способу и считает нецелесообразным пользоваться
факториальными средними квадратами.
Возникает вопрос, почему же все-таки при пользовании
Сх
формулой hz= — получаются величины, напоминающие зна-
С„
чения h1, получаемые другими способами, а иногда даже
близкие к ним. Объясняется это тем, что ss и ms коррелятивно
связаны между собой. Но формула Н. А. Плохинского, как
правило, дает завышенные величины и может указать на наличие
'М
наследуемости даже тогда, когда на самом деле коэффициент
/i2 равен или близок к нулю.
Этот случай продемонстрирован в статье 3. С. Никоро и
П. Ф. Рокицкого при сопоставлении данных, обработанных по
методу Н. А. Плохннского и путем использования
внутриклассового коэффициента корреляции, роль которого Плохинским
вообще отрицается. В статье показано, что при заведомом
отсутствии влияния показатель Плохннского всегда больше
нуля и имеет значение, определяемое не биологически, а чисто
арифметически. При определенной ситуации он может давать
значение ~ 0,5, хотя критерий F указывает на недоказанность
различии между самцами.
Но даже в тех случаях, когда Н. А. Плохинский пользуется
средними квадратами, он не желает считаться с их сложной
структурой, а поэтому, выделяя компонент аД не указывает,
что его надо умножить на 4.
С
Вот почему показатель Плохннского ft2=Tf= -J дает
небу
верное представление об уровне наследуемости, а его
теоретические и биометрические концепции, относящиеся к другим
методам определения наследуемости, также имеют
существенные недостатки. Этим самым, конечно, не отрицается
необходимость постоянной проверки, в какой степени в реальной
обстановке стада, породы или экспериментальной группы
лабораторных животных сохраняются теоретические предпосылки
применения того или иного метода, о которых говорилось выше.
Возможность использования для определения
наследуемости полиаллельных скрещиваний. При анализе
иерархического комплекса полные сибсы получаются только от матери.
Если же учитывать происхождение по отцу (или отцам), то по
отношению к последнему все они являются только полуснб-
сами, т. е. имеющими разных матерей, но одних и тех же отцов.
Данные о таких полусибсах могут быть проанализированы
методом корреляции. Но в силу самой структуры
иерархического комплекса остается неучтенным взаимодействие между
отцами и матерями. Оно входит как компонент в структуру <тД
\ю не может быть из него выделено.
Значительно более выгоден для анализа обычный двухфак-
торный дисперсионный комплекс, в котором учитывается
влияние факторов А и В раздельно и их совместное действие
(A +J3). Но осуществить его в отношении наследуемости можно
только с помощью так называемого ди- или полиаллельного
скрещивания, при котором самки покрываются попеременно
теми же самцами; у растений же исходные гибридные линии
можно переопылять в любых сочетаниях. На рис. 74 показаны
иерархическая и дналлельная схемы скрещивания. Самка I
скрещивается с самцом А (например, в первый год или оер-
263
вый случной сезон) и с самцом В (во второй год или сезон).
Самка же II покрывается в первый год самцом В, а во
второй—самцом А. Получаются 4 группы полных сибсов: а и Ь
происходят от самки I, но различаются отцами, с и d — от
самки II, но от разных отцов, а и d—от одного отца А} но
разных матерей, b и с — от одного отца б, но разных
матерей I и II. Таким образом, есть полная возможность учета всех
сочетаний отцов и матерей н тем самым определения
коэффициентов наследуемости по модели двухфакторного
дисперсионного анализа.
Полусибсы (hs)
а 6
Рис. 74. Схема скрещиваний:
а — иерархического; б — диаллельиого. Объяснение в тексте.
Этот метод применим только к некоторым видам
сельскохозяйственных животных (куры, свиньи), а также к
лабораторным, хотя далеко не ко всем и не во всех случаях. Кроме
того, он может быть использован при изучении гибридов
самоопыленных линий, например, кукурузы.
На крупном рогатом скоте подобрать группы с полиал-
лельными скрещиваниями трудно. Тем не менее Н. А. Плохин-
ский сообщает о результатах, полученных коллективом
Института цитологии и генетики СО АН СССР. Изучалось 19 групп
в племенном стаде опытного хозяйства СибНИИЖа, каждая
имела трех отцов и несколько матерей, осемененных каждым
из этих отцов. От 142 матерей получено 839 телят. Изучался
признак — живой вес при рождении, в общем
характеризующийся, как показывают литературные данные, довольно
высокой наследуемостью. Правда, количество телят в разных
группах было различным (от 22 до 115). Существенно, что при
среднем показателе наследуемости дс 0,67 (с колебаниями
по группам от 0,39 до 0,87) величина вариансы, выражающей
264
взаимодействие» оказалась довольно значительной -- до 0,27
(с колебаниями от 0,02 до 0,54). Из этого факта следует
вывод, что необходимо считаться с возможностью
взаимодействия между генотипами матерей и отиов. При иерархической
схеме анализа это должно проявиться в том, что /г по
матерям может быть значительно больше Л2 по отцам за счет не
только материнского эффекта или общих средовыч влияний,
но и за счет компонента взаимодействия,
Примером использования полиаллельного метода
являются данные, полученные Л. В. Хотылсвой и Л Л. Тарутиной на
кукурузе. Было использовано шесть инбредных линий
кукурузы. Получено р (р— 1) комбинаций по одному из методов,
разработанных Гриффингом, т. е. 30 комбинаций. Учитывался
ряд признаков, которые будут указаны ниже.
Дисперсионный анализ позволил расчленить общую феио-
типическую варнансу у полученных гибридов на вариансу,
обусловленную генотипическими различиями, и вариансу,
вызванную различными условиями внешней среды. w3a оценку
наследуемости в широком смысле слова можно тогда
принять 9
OG2+0ra
Но дальнейший анализ данных дал возможность
разложить генотипическую вариансу на вариансу обшей
комбинационной способности, т. е. выражающую общий эффект
гетерозиса, и вариансу специфической комбинационной способности,
т. е. выражающую различие между разными сочетаниями
исходных линий. Схема анализа представлена в табл. 46.
Таблица 46. Схема анадиэа мриаис общей и специфической
комбинациемой способности ддя метода, аилючающего
прямые и обратные скрпщгаания
Источник мриацин
| Обща а
комбинационная способность
Специфическая
комбинационная способность
Реципрокние эффекты
Случайные
отклонения
Число rurfi^mefl
свободы d\
2
р(р-\)
Средний
ншлпрлт
пи
mif
rupwrpu. ома-
мети cptдаяи*
клддрагат
oJ + 30,4 1
+ 7ip»2)o2g
p — число родительских линий.
265
Общая комбинационная способность почти нацело зависит
ст аддитивных генов, поэтому Ол':—20к* *.
Влрианса же специфической комбинационной способности
i.'iuhcht от неаддитнвнон части генотипа (доминантные гены
и взаимодействие неаллельных генов): aV-A=cyA
В таком случае о<;-==2а/+аД а вся фенотипическая
вариация аг*=г2а,2+ог+а<?.
Это позволяет дать следующую формулу для Аа в узком
смысле слова.
/.•_ °л2 2a'* - /on
Hi»- Jo*--t-ev-t-cv*
Рабогы названных авторов содержат не только полные
данные о значениях средних квадратов, характеризующих общую
и специфическую комбинационные способности и роль случай-
ны\ отклонений, о соответствующих Л*, но и сопоставление
наследуемости в узком смысле слова (Лг) и в широком (Л2) для
опытов, проведенных в Воронежской области и в Белоруссии.
Ограничимся только последними (табл. 47).
Таблица 47. Значения коэффициентов наследуемости //, и h для
ряда признаков гибридной кукурузы в условиях Белоруссии
Прмшак
Вес первого початка
Длина початка
Ширина початка
Число рядов teptii
Высота растения
Высота прикрепления початка
Значении
"I
0,81
0,55
0.G2
0,53
0,27
0,47
Л»
0,84
0,72
0,64
0,66
0,63
0,58
По большинству признаков значения h- незначительно
превышают значения /i|2. т. е. роль неаддитнвных генетических
факторов невелика. Лишь по признаку высоты растений
разница между А1*»0,27 (самая низкая цифра из всех
полученных величии) и №=0,63 составляет 0,36. Таким образом, по
лому признаку в наибольшей степени сказывается роль
доминантных генов и взаимодействия в специфическом эффекте
• Коэффициент 'i постаплен а сняли с тем. что учитыииются прямые
* "братиме (рецнпрокиые) скрещивании.
ОД
гетерозиса, проявляющемся при скрещивании определенных
липни.
Следует отметить, что чтот метод выделения
специфической комбинационной способности был применен для
отыскания наиболее выгодных сочетании пород свиней при
промышленном скрещивании (работы Белорусского института
животноводства).
Применение формулы Уоррена для определения
коэффициента наследуемости. Эта формула предусматривает
возможность определения коэффициента наследуемости в тех случаях,
когда имеются данные об инбредных линиях В\ и В2, Л и F*
от скрещивания между ними \ По формуле Уоррена
Л2==^=(:^< (22)
G2Ft Op"
НЛП
Ор-
Таким образом, получаемая величина представляет собой
половину наследуемости в узком смысле слова.
о2к2 расшифровывается как характеристика фенотипнческой
вариации. Считается, что она состоит из 0.25о^+0,5ал2+
+0,25охг. Эпнстатическая доля варнансы не учитывается.
Числитель же формулы, т. е. величина ол2» определяется
с помощью сравнения F\ и исходных линий В\ и В2.
Имеется в виду, что o2f( включает компоненты ог2+0,25ол2.
Исходные же родительские линии характеризуются средовой
рариансон оЛ а,.2 для F{ можно считать средней между ое2
исходных линий. В таком случае
2о2*,- (о2в,+о2*а) =2а,2+0,5олг-Ое~о^=0|,5ал2.
Таким образом, по Уоррену:
2оа,|-(о*в.+Л,) 0,5ал2
ft2=
oVj Op2
Формула эта может быть применена к скрещиванию и не-
инбредиых линий, хотя в этом случае она еще менее точна.
Некоторые исследователи, работающие с нерасщепляющн-
мнеи исходными инбредными популяциями и с
расщепляющимися, получаемыми в F2. пользуются более простой формулой
для Аа:
(fin OP*
* Возможно также испалмомние беккроссов с исходными линия мм.
267
Инеется * 1мдч. что гг>, включает в себя генотнпнчесхую
Г/ * я средоачю о,* вариаисы* так что в сумме они
составляет Gp1
Значение же а,г получается как средневзвешенный средний
квадрат из внутригрупповыд средних квадратов для нерасщеп-
лнюанхся популяций — двух родительских (Р% и Р2) и Ff.
Подобные методом Бил н Эткинс вычислили
коэффициенты наследуемости xi* ряда признаков сорго, в том числе для
аеса 100 зерен А* оказался равным 0,6—0,7, для урожайности
по зерну — 0,4—0.8 (различия в зависимости от того, какие
линии скрещивались).
Использование данных по однояйцевым двойням для онре-
доения коэффициентов наследуемое™. Общеизвестно, что
однояйцевые двойни (у человека, у крупного рогатого скота)
являются ярким примером большой роли наследственности в
феиетипическом развитии громадного количества их признаков.
Данных по ним очень мио^о.
Но получаемые при изучении двоен коэффициенты, как
правило, снлыю завышены, поскольку сходство партнеров
однояйцевой пары усиливается общностью среды как до
рождения, так н в период воспитания. Некоторую поправку можно
внести, если параллельно изучать однояйцевых и
неоднояйцевых двоен и определять разницу коэффициентов корреляции
междч партнерами первых и вторых. Данные о двойнях у
людей и у крупного рогатого скота обобщены Фолконером, но
некоторые из них вызывают большие сомнения, в частности,
известно, что у разнояйцевых двоен крупного рогатого скота
нередко встречается соединение кровеносных сосудов, что
повышает сходство между ними. Поэтому Л*, определяемые по
разнице коэффициентов корреляции между парами
однояйцевых и неояиояйцевых двоен, оказываются непомерно низкими.
И* сказанного вытекает, что двойни недостаточно удобны для
изучения наследуемости
Коэффициент наследуемости признаков, не поддающихся
точной величественной оаенке. Такие случаи относительно
редки» но в принципе к ним также может быть применено
установление корреляции между родителями и детьми, только надо
использовать не обычный коэффициент, корреляции, а
коэффициент ранговой корреляции, при котором признаки
родителей я детей выражаются не точными количественными
оценками, а рангами» т, е. порядковыми номерами, оценивающими
их относительное качество. Наиболее часто в биологической
статистике применяется коэффициент ранговой корреляции
Спнрмена (см. приложение). Поскольку он может быть ис*
пользован для установления корреляции между потомками я
обоями родителями, при определении коэффициента
наследуемости нет надобности умножать полученный коэффициент ран-
говой корреляции иа 2 Полученное значение может служить
для ориентировочно* оценки наследуемости
На сходном принципе опенки рантов основана формула
Шварца и Уирдена для показателя наследуемости:
где гл. и г_ — средине рант ддя детей от лучших I -И и худших
(—) родителей; /?+ и /L — средние ранги для лучших и
худших родителей; f — коэффициент, равный 1. если с потомками
сравниваются оба родителя, и равный 2. если с потомками
сравнивается один родитель.
Под лучшими и худшими родителям» понимается половина
популяции (стада), ранжированная от середины ряда в
сторону «плюс» и «мннуо. Этот способ элементарно прост и не
требует разбора.
Следует лишь указать, что. как и все неоараметркческие
методы, он очень неточен.
Мы пока оставляем в стороне еще один способ определения
наследуемости — по сдвигу при отборе, об этом будет
говориться в гл. 7.
Определение достоверности коэффициентов наследуемости.
Так как в различных опытах по определению наследуемости
получают нередко разные значения коэффициентов даже для
одного и того же признака, то возникает настоятельная
необходимость установления достоверности получаемых
выборочных значений Л3, что в конечном счете определяется тем
методом, который был применен для вычисления коэффициента
наследуемости.
При использовании корреляций достаточно определять
достоверность коэффициентов корреляции обычными
статистическими методами (см. приложение)» сравнивая их со средними
ошибками. Эта средняя ошибка является и ошибкой
коэффициента наследуемости.
Если нужно более точно определить достоверные границы,
целесообразно переводить г в г, определять ошибку для *,
установить достоверные границы для г и затем иол ученные
крайние значения г перевести снова в г. Впрочем, можно
ограничиться и простой записью значения А* с его ошибкой.
При вычислении коэффициентов* наследуемости с помощью
коэффициентов регрессии теми же обычными статистическими
методами вычисляются средние ошибки для значений Ь и
записываются как ошибки коэффициентов наследуемости.
Сложнее обстоит дело при использовании метода
дисперсионного анализа, поскольку здесь фигурируют различные
до
источники парнашш. Для доказательства достоверности
влиянии производителен л маток на потомство, а также
взаимодействия между ними (при одной схеме анализа) или только про»
нзводнтелен и маток (при другой схеме) применяется крите»
рий F, т. е. отношение средних квадратов: ms3 к msi, или ms3
к ms2 и т. д. В тех случаях, когда достоверность не доказана,
нет смысла определять и коэффициент наследуемости, хотя
возможны и такие случаи, когда, например, достоверным
оказывается влияние только самцов или только самок. Тогда
наследуемость определяется только по одной формуле (с
помощью as2 или а/2).
Определение ошибок для /i2, вычисленных с помощью
дисперсионного анализа, связано со значительными трудностями,
поэтому значения h2 в этих случаях чаще всего приводятся без
указания на их ошибки. Но в специальной литературе есть
указания на некоторые формулы. Так, например, при
вычислении наследуемости путем анализа внутриклассовой
корреляции ric можно пойти по пути перевода г в г, с тем чтобы
определить ошибку и доверительные границы для z при заданной
вероятности (например, р = 0,95). Однако для случаев, когда
внутриклассовая корреляция вычисляется на выборках
значительного объема, Фишер считает возможным применить
следующую формулу для ошибки 5Г. :
у 1 (к-\)ы
где k — число особей в каждой группе, ал' — число классов.
Фишер все же указывает на известные ограничения в ее
применении. Величина Sric может служить ошибкой и для ft2,
вычисленного, например, по формуле
Op-
Тогда sfci=4srjc.
Хейзелем и Террилом предложена для Л2 следующая
формула ошибки, вычисленной с помощью внутриклассового
коэффициента корреляции:
где sSf и ssa— сумма квадратов для строчек «матери» и
«дочери»; п — число самцов; к — число дочерей от каждого из них.
* Формула нами упрошена: вместо отношений коиарианс представлено
непосредственно значение Нг.
i70
Для обоснования этой и подобных формул следует
обратиться к специальной литературе.
Некоторое представление о величинах ошибок для № может
дать и окольный путь— через коэффициенты корреляции, так
4S
как известно, что hr по формуле-—-^—- равен 4rUs.
a hzs+D=2rFe. Ошибки же для коэффициентов корреляции
определяются легко. Их с некоторой натяжкой можно отнести
и к значениям Л2, вычисленным по той или иной формуле.
Наконец, планируя проведение опытов с целью определения
коэффициентов наследуемости, можно наметить оптимальные
варианты для числа особей на семью, количества семей и т. д.,
которые обеспечивали бы максимальную точность
коэффициентов за счет уменьшения выборочной вариансы.
Так, Фолконер приводит следующую формулу вариансы
(о2) для выборочного внутриклассового коэффициента
корреляции *:
Gr" ~ п(п—1)(Л^ —1) "~~ ""
Здесь N — количество семей, aw — количество особей на
семью. Тогда общее число изучаемых особей составляет nN.
Если размеры ограничиваются этим количеством особей, то
можно показать, что вариаиса для ric будет минимальной,
когда/1» 1//v Таким образом, оптимальные размеры семьи
зависят от величины наследуемости. В случае полных сибсов
/i2=z2rie% а полусибсов Л2=4г1Г. Тогда наиболее эффективными
для получения более точных значений Л* будут размеры семей.
для полных сибсов п— ,
2
4
для полусибсов п=г -- .
Так как априорное представление о величине
наследуемости может быть только приблизительным, то заранее опреде
лить оптимальные размеры семьи невозможно. Однако, по
мнению Фолконера, снижение эффективности будет большим, если
размеры семьи меньше указанного оптимума. Это значит, что
лучше ошибиться в сторону более крупных размеров семей.
Фолконер ссылается на расчеты Робертсона, показавшего, что
если отсутствует предварительное представление о значении
наследуемости, то анализ по полуснбсам должен основываться
на семьях с числом особей примерно 20—30.
* Фолконер обозначает его через / Выше ми давали обозначение для
внутриклассового коэффициента корреляции через г,с.
*71
При наиболее эффективном плане опытов, т. е. когда
п=1/г|г. выборочная варнанса внутриклассового
коэффициента корреляции может быть приблизительно следующей:
Or2. = гг-
■<• nN
Отсюда при оптимальном плане опытов вариансы коэффи-
цяентов наследуемости будут приблизительно следующими:
для семей с полными сибсами
о* =4ог2 с
для семей с полуснбсамн
Это значит, что при прочих равных условиях показатели
наследуемости по семьям с полными сибсами вдвое точнее» чем
по семьям с полуснбсамн.
Сопоставляя эти расчеты с расчетами по определению
наследуемости с помощью коэффициентов регрессии, Фолконер
делает общий вывод, что метод регрессии предпочтительнее,
т. е. точнее, при более высоких значениях коэффициентов
наследуемости, при низких же целесообразнее применять метод
корреляции. Грубой границей при одинаковом объеме
опытного материала является h2j^0,2.
Для опытов, когда семьи с полуснбсамн разбиваются на
подгруппы с полными сибсами (анализ только по отцам или
только по матерям) и нет никаких предварительных данных о
величине наследуемости, лучше всего планировать по 3—4
самки на самца и около 10 потомков на каждую самку.
Значения коэффициентов наследуемости для разных при*
такое. Количество исследований, посвященных изучению
наследуемости у животных и в несколько меньшей степени у
растений, исчисляется в настоящее время многими тысячами.
Самый общий вывод из них заключается в том, что
нельзя,абсолютизировать получаемые конкретные показатели
наследуемости. Так как коэффициент наследуемости — это параметр,
относящийся к популяции, в которой возможны самые
различные сочетания внешних условий, с одной стороны, и разная
степень генетической изменчивости — с другой, то
неудивительна очень большая колеблемость значений коэффициентов
наследуемости применительно к разным породам, стадам, сортам
NT. Д.
Но в то же время, если проводить сравнение в примерно
равных условиях внешней среды и методов разведения, то
16/1-
nN '
326*
nN
*7*
становится очевидной роль самих признаков, для которых
вычисляются коэффициенты наследуемости.
Есть признаки, характеризующиеся очень низким удельным
весом наследственности в изменчивости популяции и
обусловленные в большей мере средовыми факторами. Коэффициенты
наследуемости таких признаков очень невысокие, порядка 0,1 —
0,2. Есть и такие признаки, вариация которых слабо
подвержена влиянию средовых факторов, и коэффициенты
наследуемости их очень высокие — 0,7—ОД а иногда даже 0.9. В табл.
48 приведены значения коэффициентов наследуемости для
некоторых видов животных н растений, преимущественно
сельскохозяйственных *. Они охватывают ничтожную долю
имеющихся данных.
Бросается в глаза, что низкие коэффициенты
наследуемости характеризуют многие признаки, связанные с размножением
(плодовитость овец; количество детенышей в пометал: поросят
у свиней, щенков у лисиц, мышат у мышей; количество
откладываемых яиц у дрозофилы и т. д.). Конечно, это совсем не
значит, что такие признаки не находятся под контролем
наследственности. И по таким признакам можно выделить или
создать отбором наследственно обусловленные линии или
группы. Это только означает, что удельный вес вариации по
ним под влиянием наследственности значительно ниже* чем
под влиянием среды. Он может быть объяснен эволюцнонно,
как результат естественного отбора, возможно,
стабилизирующего, при котором особи, наследственно значительно
отклонявшиеся от оптимального уровня по таким биологически важным
свойствам, как размножение, устранялись отбором. С другой
стороны, было очень важно для сохранения вида, чтобы он мог
легко и быстро отвечать на условия меняющейся среды увели*
челием или уменьшением численности приплода для
сохранения оптимальной и наиболее выгодной численности
популяции.
С другой стороны, по конституционным признакам:
количеству позвонков, длине головы, складчатости кожи у овец,
ширине серебристого кольца у серебристо-черных лисиц, длине
крыльев у дрозофилы — показатели наследуемости высокие.
Это совсем не значит, что по ним особенно велика
наследственная изменчивость. Все дело в том, что удельный вес
последней в общей вариации значительно больше, чем удельный вес
средовых факторов. Сами же признаки относительно
стабильны. Как будет показано в дальнейшем, по признакам с
высокой наследуемостью легче вести отбор, так как условия ввеш-
* Преобладание в сяисЕе видов животных объясяяется сягшшшюсш»
автора.
18 П ♦ Роь:*чм11 273
у вмлиа ■ ркт«м8
1 1
I Крутешй рогат*
| met
Ошы
Сашам
I
1 1
I 1
Курм
"' " ' i i——1——i i i У
1 7
\ *
1 Пегесть
Удой молока ia 1 лдктащяю
I Проктит жара а молоке
Вас ара рождении
Вмети* суточный удой
Способность к оплодотворению
(itpa первой случке)
Содержание балка а шло»
Длята туловище
Скорость молокоотдвчи
Длительность берешяяоетя
Глубина груди
1 Устойчивость
1 Длина голоаы
Живой аас к концу откорма
Длина еэсрети
! Настраг надетой шерсти
Вас ври рождена* '
Саладчатость котя
Плодовмтость i
Вас тала а годичном возрасте
Размерь! помета
Количество noaaowKoa
Толщина шпика на сток?
Суточный привес 1
Оплата корма I
Длина тела 1
Вес к 160 дням 1
Длительность беременности 1
Жи пин нос обнос, ib I
Яйценоскость 1
Вас тала 1
1 Эятим Д»
1 '
о,*>-о.9б
0.2 -0.4
о.е -«,8
0.2 -0,6
0.4 -0.6
0.01-0,06
0.4 -0,8
0.40-0.98
0,38-0,66 1
0.20-0,46
0.30-0,80 1
0.16-0.301
0,60-0.70 1
0.80—0,90 1
0.30-0,70
0,40-0.60 1
0.3 -0.6
0.S -0.8
0.1 —0.2 1
0,10-0.60 I
0,10-0,20 I
0,7 -4,8 1
0.5 —0,7 1
0.4 —0.6 I
0.3 1
0.4 —0,9 1
0.3
0.1
0.1
0.2 -0.3 1
0.2 -0.* 1
I
1
Куры
1
Серебристо-черные
л ненцы
Мыши
1
Крысы
Дрозофила
Кукурум
Сахарив* сае*аа
(полмгибрмдмая)
Прололжеиие
1 1
Вес яАоа
Возраст перюй яйцекладки
Ширям* груди |
Падеж от лейкоза
Жиаиеслособмогп.
Половая зрелость
Форма яйца
Количество шейкой а помете
Ширина серебристого коаыи
Размеры помета
Вес тела в 6-недельном возрасте
Длина хвоста в б-иедмыюи во**
расти
Проявление белых пятен на
голове
Возраст полонов аредостя самой
Ответ яичников на гоивдотрол*
яыи гормон
1 Количество цетинок на сегментах
брюшка
Количество грудных щетиной
Количество откладываемых яиц
Величина иичикиа
/Длина крыльев
Длина грулн
Вес паевого початка
Двина початиа
Высота растении
I Урожая корни!
1 Сахаристость
г I
о.б -М I
0,2 -0.3
0.2 1
0.10-0,20 1
0.1
0.1 -0.4
0.2-0.5
0.1 -0.15 1
0.6
0.1О-О.20
0.30-О.40
0.6
0,4
О.Ю-0.20
0.30-0,40
0.5 -О.б
0.5 -О.б
0.2
0.3
0,6
0.4
0.0
0.6
03*
0.1-
0.4*
Наследуемость ■ У*** с,,мсл
18»
ТП
•ей соеды хотя они всегда в какой-то степени влияют, слабо
масмртат наследственные различия между особями.
Для практики сельского хозяйства очень важно знать, вдт
коза наследуемость хозяйственно-полезных признаков, таких,
как отата корма, маючная продуктивность, жирность
молока содержание белка в молоке, настриг шерсти и т. п. Из
табл 48. в которой сведены очень многие данные различных
исследований, бидно. например, что наследуемость молочной
продуктивности, т. е. общего количества молока за лактацию,
значительно ниже, чем наследуемость процента жира в молоке,
а также некоторых других особенностей молока. В массовых
стадах показатели наследуемости молочной продуктивности
налы — 0.2—0.3. Только в случае большой выравненностн ус*
ловли содержания и кормления они могут быть выше
—порядка 0.3—0.4—0,5. Наследуемость же процента жира в
молоке, как правило, значительно выше. В очень немногих работах
для этого признака приводят Л*, разный 0.3-4),4. Обычно же
к1 равен 0.5—0.6 и даже 0,8.
Для громадного большинства признаков показатели
наследуемости средние. Это показывает, что по ним в реальных
стадах, породах, сортах нет значительной разницы между
вариацией, вызываемой наследственными различиями, и вариацией,
вызываемой средовымн факторами.
Заачеяяе показателя наследуемости. С теоретической точки
зрения главное значение коэффициента наследуемости
заключается в том. что он дает возможность устанавливать степень
надежности суждений о селекционной ценности особей по их
фенотипическон ценности. Фактически можно непосредственно
судить только о фенотнпнческях ценностях особей,
селекционные же ценности определяются по степени их влияния на
следующее поколение. В таком случае селекционер или
экспериментатор проводит отбор родителей по их фенотипнческнм
ценностям Успех же отбора в желательном изменении
популяции может быть предсказан тальк о в том случае, если
известна степень соответствия между фенотипнческнм и и
селекционными значениями Эта степень соответствия измеряется
коэффициентом наследуемости.
Как указывалось выше, наследуемость в узком смысле
стой представляет отношение аддитивной генетической ва-
ряаясы к обшей фенотипической. С другой стороны,
коэффициент наследуемости может быть представлен и как к*—ЬА/рч
т е. как регрессия селекционной ценности к фенотипической
Основания здесь те же, что и при выведения формулы
I t
COUOP ^ -£*AZ-
Ъхлн выделить в фешггвпнческой ценности два компо-
г*
цента — селекционную ценность Л, определяемую суммой
передаваемых потомству аддитивных генов, и некий остаток Р.
включающий все остальные, т. е. определяемые средовыми
факторами, доминантным отклонением и различными неал-
лельнымн взаимодействиями, то можно записать P^A + R.
Так как А н Р не коррелируют друг с другом, то covAp^oA\
и тогда ЬА/р= —- ==л2.
оРг
Так как из общего курса статистики известно, что
то
r=fcx/, .
Ох
В значениях А и Р это равенство может быть записано так:
Глр=Ьа'р- =А,« -г- =Л.
си я
Так как обычно коэффициент наследуемости записывают
как Л2, то глР= у ft*.
Рассматривая же наследуемость как регрессию
селекционного значения (А) к фенотипнческому (Р), в самом общем
виде можно считать, что селекционное значение особи
представляет собой произведение фенотипнческого значения на
коэффициент наследуемости, т, е- А = Рк*.
Естественно, что фенотипические и геиотипические
значения особей не могут оцениваться абсолютно, а только как
отклонения от популяционной средней- Зная ft2 и Р, можно
ожидать с определенной вероятностью какое-то значение .4. На*
следуемость служит для установления степени надежности
суждения о селекционной ценности по фенотнпнческой
ценности или степени соответствия между фенотнпнческой оценкой
и селекционной ценностью. Вот почему коэффициенты
наследуемости включаются почти во все формулы, используемые при
селекции животных и растений, о чем речь пойдет ниже.
Во всяком случае коэффициент наследуемости относится
к числу важнейших параметров при генетическом анализе
количественных признаков в популяции.
Важно иметь в виду, что наследуемость — это не только
характеристика признака, но также популяции и тех средовш
условий, в которых находились особи данной популяции.
Так как коэффициент наследуемости вычисляется на
основе ряда величин, то изменение в любой из них повлияет на его
277
значение. Вес генетические компоненты зависят от частот
геном, а по ним разные популяции могут отличаться в силу своей
прошлой истории, и силу тех процессов, которые в них
происходили. Поэтому неудивительно, что коэффициенты
наследуемости по одному н тому же признаку для разных популяций,
кажущихся даже очень сходными, бывают неодинаковы. Так
как средовой компонент входит в определение коэффициента
наследуемости, то при большей вариации по условиям
содержания, кормления, агротехники коэффициенты наследуемости
будут ниже, а при однородных условиях—выше. Имеют
значение и размеры популяции. Если популяция сравнительно
мала, то более вероятны в ней процессы повышения гомозигот-
ности по ряду генов и отсюда более низкие показатели насле-
д\емостн, чем в больших по размерам популяциях.
Таким образом, обнаруживаемые показатели наследуемое*
м\ всегда относятся к конкретным популяциям, особи
которых находятся в конкретных же внешних условиях. Данные о
гих позволяют наметить наиболее эффективные методы
работы, они позволяют предвидеть, какие результаты могут быть
получены в данной группе животных или растений как при
\лучшенни кормления, содержания, агротехники, так и при
применении различных систем отбора и спаривания.
Глава 7
ОТБОР ПО КОЛИЧЕСТВЕННЫМ ПРИЗНАКАМ,
ЕГО МЕХАНИЗМ И РЕЗУЛЬТАТЫ
Феиотипическая структура популяции и внешние условия.
Феиотипическая структура популяции по количественным
признакам, как и по качественным, создается на основе re-
иотипичеокой структуры. Дополнительный фактор—влияние
условии внешней среды на развитие отдельных особей
популяции. Несмотря на постоянную смену одних поколений
другими, популяция может длительное время сохраняться
стабильной генетически в соответствии с законом Хардп — Венн-
бсрга, если нет внешних сил, нарушающих свободное
скрещивание, и значительных колебаний численности, которые могли
бы привести к генетическому дрейфу. Популяция может
оставаться неизменной и фепотниическн, если более или менее
сохраняются те же условия внешней среды. Однако во ммогил
случаях внешние условия не остаются неизменными.
Например, при улучшении кормления и содержания
сельскохозяйственных животных меняются фенотипнческие свойства
животных и феиотипическая структура популяции.
Возникает вопрос, изменяется ли при этом генетическая
структура популяции.
Вопрос этот очень важен принципиально, так как в
широких кругах работников сельского хозяйства, особенно
зоотехников, до недавнего времени бытовало традиционное и
примитивное представление о том, что улучшение условий
внешней среды одновременно и адекватно меняет и
наследственную структуру животных. Это представление
основывалось на вере в так называемое унаследование приобретенных
признаков.
Рамки н задачи нашей книги не дают возможности
рассмотреть эту проблему подробно*. Необходимо лишь
указать, что громадное количество экспериментальных данных
отвергает представление об унаследовании приобретенных
• См. на эту тему прекрасную книгу Л Я- Бляхерз «Проблема наследо
влния приобретенных'признаков» (At, 1971).
!l|..'«ruHHl. J •'. "б .МСКИЛНЮМ Л«\!<Ч|(»НИН П'НО'ГННИ ОСОбй
iu мм «a изменением ее фенотиинческ-их свойств. Хотя весь
процесс «юлюнни заключился в бесчисленных новообразо*
пиниях, однако появление новых признаков у видов
происходит значительно более сложным путем (прежде всего на
ч'нопс мутаций, рекомбинаций и естественного отбора). Но*
vroMy представление об унаследовании приобретенных
признаков не может быть положено в основу понимания
процессов, приводящих к изменению геноигиичеекой структуры по-
пулиинн. Этим самым, конечно, не отрицается громадная
роль условии внешней среды в мутационном процессе (о чем
говорилось в 1Л. 2), в практике животноводства н растсииё-
нодегнл, в повышении фенотиннческой продуктивности
животных и растений. Кроме того, роль внешней среды
существенна в юм компоненте фенотипичеекой вариации, который
полупи название взаимодействия генотипа и среды.
Так как максимальная продуктивность животных и
растении может быть достигнута лишь при создании для них
необходимых условий (агротехника, удобрения для растений,
кормление и содержание животных и т. д.), то естественно,
что мероприятия по изменению генетической структуры
популяции (породы, сорта и т. д.) должны проводиться
одновременно с мероприятиями по улучшению условии внешней ере*
ды для разводимых животных и растений. Они должны
взаимно дополнять друг друга.
Пути изменения генотипической структуры популяций по
количественным признакам. Они, в сущности, те же, что были
рассмотрены при анализе изменений генотипической
структуры популяций но отдельным генам, а именно: отбор,
мутации, миграции, генетихо-автоматические процессы, с одним
только существенным отличием, что в случае количественных
признаков эти процессы охватывают многие гены, влияющие
на данный признак. Это еще более подчеркивает
статистическую природу явлений, лежащих в основе отбора, хотя и
здесь, как правило, происходит сложное переплетение
статистических и динамических процессов»
Отбор, Как уже указывалось в гл. 4, под отбором в
классическом смысле следует понимать устранение какой-то
группы особей из популяции от размножения. Иногда его
заменяют термином «селекция». Но термин «селекция» понимают
двояко: селекция - только как отбор и селекция как
совокупность всех методов, применяемых для улучшения, породы,
сор?а, вида, культуры и т. д. Именно в последнем значении
говорят о селекции крупного рогатого скота, селекции
овощных культур и г. х Предпочтительнее употреблять термин
«селекция» в широком смысле слова, не заменяя им более
определенного термина «отбор»,
inn
При отсутствии отбора каждая особь имеет одинаковые
возможности для размножения, поэтому фактически
различия между особями по размножению будут случайными. В
больших популяциях эти различия, как правило,
характерною! ся нормальной кривой распределения и взаимно
уравновешиваются. Поэтому они не скажутся на частотах генов в
следующих поколениях. В малых по объему популяциях в
силу'генетического дрейфа чисто случайные отклонения в
скорости размножения влияют на частоту генов в следующих
поколениях. Если часть особей устраняется из популяции
(в природе гибнет, уничтожается хищниками; при
искусственном рлзпедеиии отбраковывается человеком), то
размножается лишь оставшаяся часть- При полном устранении какой-
то генетической группы ее представители могут вообще не
появиться в следующем поколении.
Однако чаще всего устраняется только часть особей
генетической группы, поэтому доля оставшейся части в общем
размножении особей популяции соответственно меньше той
доли, которую занимала эта группа до отбора. Этот процесс
неизбежно скажется на частотах генов в следующем
поколении, как это было показано по отношению к отдельным
генам в гл. 4.
Различие между естественным и искусственным отбором.
Принципиальная основа естественного и искусственного
отбора одна и та же. Но между ними есть существенное
различие. Естественный отбор — это выживание наиболее
приспособленных форм, поэтому от менее жизнеспособных,
частично устраняемых отбором особей автоматически остается
меньше потомков. Искусственный же отбор направлен на
получение более ценных в хозяйственном отношении животных
или растений, размножение особей и оставлеинеот них
потомства регулирует человек исходя из поставленной перед
собой задачи. Повышение жизнеспособности (или
приспособленности), хотя и может учитываться при проведении
искусственного отбора, не является главной задачей.
В практике животноводства и растениеводства при
проведении искусственного отбора необходимо считаться с
возможностью естественного отбора, происходящего вне воли
человека, особенно в условиях внешней среды, далеких от
оптимальных.
При значительной интенсивности естественного отбора и
при противоположности его направления направлению
искусственного отбора хозяйственный эффект последнего мо*
жет оказаться очень ослабленным или даже отсутствовать
вовсе.
Естественный отбор в природе чаще всего основывается
на фенотншгческих свойствах отдельных особей популяции.
281
и» степени их устойчивости в колеблющихся условиях внеш.
ней среды» их выживаемости и способности давать потомства
Хотя отбор может происходить на любых этапах жизни
особи, на любой стадии, н том числе яйца, личинки, куколки,
головастика или эмбриона, но наибольшее значение он имеет,
очевидно, в период до наступления патовой зрелости или в
начале перж)да размножения. Отбор же особей после того>
как от них получено потомство, уже не может сказаться ив.
генотипической структуре следующего поколения. Кроме
отбора по фенотипическим свойствам отдельных особей,
возможны также различные формы группового естественного
отбора, т с отбора по семьям, стадам и т. д.
AAA
J Л Ш Ж I
Рис. 75. Три тяпа отбора:
сг*йш.ъитргжяшм* \1 — Ш), дмеруптнвкмй (IV) н наораялешшА (V).
Наиболее элементарной формой искусственного отбора
также является отбор на основе фенотипа особей. В
практике племенной работы его обычно называют массовым
отбором. Он достаточно эффективен только при некоторых
условиях, в частности, при наличии в популяции большой гено-
тнличеогой варнаиии и если он ведется по признакам с
высокой наслеауем остью.
Поэтому в процессе развития животноводства и
растениеводства и разработки генетнюхелекционной теории были
выработаны и более совершенные методы отбора, в
частности по генотипу, которые сочетаются также с различными
приемами скрещивания особей, чем достигается более высокая
эффективность селекционной и племенной работы.
Характер отбора и его результаты будут различными в
зависимости от того, какие фенотитжческне группы отбирают-
св. а какие элиминируются.
Твяы отбора. Сейчас различают по крайней мере три
основных типа отбора: стабилизирующий, разрывающий
имеруитивиый) и направленный.
Разница между ними показана на рис. 75. Вверху
представлены кривые распределения особей популяции родитель-
ского поколения по какому-либо признаку. Заштрихована
размножающаяся часть популяции, не заштрихована — эли-
минируемая. Внизу — кривые распределения потомства.
При стабилизирующем отборе чаще всего устраняются от
размножения крайние плюб и минус-вар и анты, так как
отбор благоприятствует какому-то среднему, оптимальному
значению признака. Отсюда и название этого типа отбора —
стабилизирующий. Но в зависимости от того, какая часть
особей из кривой распределения родительского поколения
сохраняется, может измениться степень вариации в
последующем поколении. В случае / (рис. 75) получается экснес-
снвная кривая, в случаях // и /// — та же нормальная
кривая распределения- Очевидно, что в / случае отбираются
особи с меньшей генетической изменчивостью, поэтому в
потомстве вариация уже, а кривая распределения более крутая
(эксцесс). В случае // размножающиеся особи средней части
кривой распределения обладают той же вариацией, что и вся
исходная популяция. Тогда они дадут потомство с
распределением, каким характеризовалась родительская
популяция. Но возврат к средней может наблюдаться и тогда, когда
устраняются от размножения особи средней части кривой
распределения родителей, а дают потомство крайние
варианты. Но условием возврата к средней является свободное
скрещивание <нх друг с другом, поэтому они воспроизводят
то же распределемие, которое было у родительского
поколения.
При дисруптивном (разрывающем) отборе
селекционным преимуществом обладают, как и в случае ///
стабилизирующего отбора, крайние плюс- и мннус-вариаиты. а
средние устраняются отбором. Но скрещиваются только
плюс-варианты с плюс-вариантами и минус-варианты с
минус-вариантами. Поэтому каждая группа скрещиваний дает
в' потомстве различные кривые распределения, сдвинутые по
отношению друт к другу. В следующем поколении получается
двухвершинная кривая, являющаяся сложением
варьирующих групп, получающихся от исходных плюс- и
минус-родителей. При дальнейшем продолжении такого отбора новые
группы потомков могут уже не трансгрессировать друг с
другом. Отбор приводит к дивергенции, расчленению на две
группы первоначально единой группы. Отсюда и название —
разрывающий.
Наконец, при направленном отборе потомство даст группа
особей, располагающаяся в правой {V случай, рис. 75) ила
левой части кривой распределения родителей. Потомство
также распреаеляется по нормальной кривой, но его средние
сдвинута в определенном направлении вправо (иди влево)
по сравнению со средней арифметический родителю** груп-
гь: Этот сдвиг будет все более нарастать в последующ**
-^калениях отбора.- Поскольку отбор идет в одном направлю
н га. то и сдвиг кривой будет направленным,
В условиях естественного отбора в природе можно обща,
ружить все три типа отбора, правда, нередко они взаимно
переплетаются.
При искусственном отборе (в животноводстве пли
растениеводстве! наиболее част направленный отбор» так как
обычно селекционер стремится к усилению того или иного
хозяйственно-полезного признака. Д. К- Беляев выдвину*
идею, что при одомашнении животных происходит особый
тип отбора, который он назвал дестабилизирующим. Этот ти
отбора разрушает все те коррелятивные связи, которые
выработались в природе вследствие преимущественно
стабилизирующего отбора. Создается новая система организма, на
фоне которой происходит проводимый селекционером искус*
ственный направленный отбор. В силу дестабилизации
у орган и злое появляются новые свойства (в поведении,
размножении и т. д.), которых не могло быть при наличии
стабилизирующего отбора в природе. Появление этих свойств в
свою очередь облегчает проведение направленного отбора.
Данные о различных типах отбора в природе. Старым,
но достаточно наглядным примером стабилизирующего
отбора являются данные Бампуса, пат ученные на воробьях еще
в конце 19 века. Сравнивались по ряду морфологических
показателей погибшие и выжившие воробьи (после тяжелых
условий зимы 1898 г.). Выжившие воробьи по всем
изученным признакам (длине тела, размаху крыльев, весу тела
и др.) в большей степени приближались к среди ivm
величинам популяции. Во многих других исследованиях было
подтверждено, что отбор часто приводит к элиминации
отклоняющихся особей. Пример направленного отбора —
распространение меланнстических форм бабочек в загрязненных
районах Северной Европы л Северной Америки,
приближающееся к 100%. Так, в районе Манчестера (Англия) за 50 лет
количество темных форм повысилось с 1 до 98%. Однако
в местностях незагрязненных отбор шел в сторону типичных
форм. Были проведены и специальные опыты в природе,
показавшие дифференцированное уничтожение бабочек ърш*
гамн.
Есть несколько интересных фактов микроэволюцми —
у чешуекрылых, улиток, ужей, а также у дрозофилы,
которыми можно было доказать наличие разнонаправленного
отбора — элиминации тех или других генетически
различающихся форм в разных местах их обитания.
Дисруптивиый отбор был хорошо показан в природных
популяциях миметических бабочек.
paw. с«« ^«ssssss isszssi ^
ных тноов отбора, и кадожяоо* вмтмтячесш* хавай*
рнстикн их результат*». • то* WJ* арогвози ровая»
Првмеры отбор* у адмлвммйст^мж жвввтвт. Седь-
скохоэянствеявые животные и растшяя дают немало ков-
кратных npmt^oe отбора. Ковечяо, ввогресс, достагаюый
4?
Jf
l_.
л»
as яо—55 S 5» *« *и
W*
ft* 7«. Иишмше «»«**« «*M • гшют «*тао»м»« «P«
• Гаммам с I9M а» »*« г
в лродуктважкти то* ма тА отрасли *«J252T«lS
растенн»одства. дааеао не ™»«ZZL*SEJ%2
отбора. В векоторь* ^^^^Z^Z^^^
нения условий ^«"""J**?^ ?ЯУ У**»*»"*
обработки п<*вы « «»«ГК~ £^3m!Z ******
в растениеводов* и т. д. Та». J^J^ * К^обвитаим
достижения по ммлим» удоев кори лввв> чК-
г» последние 10-15 *ft. "«^^^^^.в^овава» ваа-
тнчно результат сележаиа. w^J^|rMee«ci»«woro oCt"**
верейных во потомству «и^ «fJ^y^Tw «•*»
нения. Другая же ариадаа - •JgSП»етери*«Р»« ■*•*:
достигнут? вро****** в^/^2^шй«У i***"»*!
ДРУГИМИ 1Мр**«» «ввВШВМаСЯ ,;*Щ* дав
шсго влияния новой техники размножения и более слоодц
шлемов селекции на животноводство Англии в целом.
' Но в определенных случаях можно точно установить po«v
отбора. Так. на рис. 76 показано, как изменилось содержи*
жира в молоке коров в Голландии за 40 лет. Этот прнзюк
обладает высокой наследуемостью, и роль внешних условий в
его изменении очень мала. Кроме того, сами условия очвд
слабо менялись в указанный период времени. Поэтому яаяу*
ченные результаты — повышение лроцейта жира в молоке с
3.15 до 3,80—3,85%, т. е. по сравнению с первоначалыши
I/-
5лП
;9
^ ~л
ъя
\
\**
—1 1 1
1
1
/^
(RS ^ч
L
^•Л
\ 1 »
t
№ 1925
1950 4955
Годы
то № 1950
Рис. 77. Результаты отбора у свиней на повышение
оплаты корма в Дании за 1923—1949 гг. (сплошная
линия — у породы датская ландраса. пунктир —
у йоркширов).
уровнем 1906—1910 гг. на 22%,—целиком обязаны отбору.
направленному на повышение жирномолочности. Так как ЖЯР*
ность молока зависит от многих генов, то процесс измене***
признака происходит медленно и достаточно равномерно, ***
видно по намеченной пунктиром теоретической линии регресс**-
Большое значение в животноводстве имел и имеет отбор
на лучшую оплату корма, т. е. уменьшение затрат кор**
на получаемую единицу продукции. На рис. 77 показ***
результаты отбора, проводившегося датскими контроле**1"
ми станциями. Затраты корма на 1 кг привеса снизились С
3.5—3,6 до 3,1 кормовой единицы. Однако процесс шел в^
равномерно — сначала очень быстро (1921 — 1930 гг.), з***£
медленно. Генетическая сущность такого результата c^JJl
понятнее, когда мы остановимся на процессах, пронсДОА***:
нрн отборе (речь о них пойдет ниже). Интересно изм*й«*^
типа телосложения» достигнутое у свиней в Дании 8 Р^У^
тате новых требований мирового рынка на бекон. Путе* Д***
зд
тельного отбора была уменьшена толщина сала на стане,
увеличена на стенках брюха. Увеличилась также длина
туловища (рис. 78). Это показывает, что з исходных
популяциях была достаточная генотипичеекая изменчивость ло всем
этим признакам.
На курах проводился отбор по индексу формы яйца в
разных направлениях. За 5 поколений были выведены куры,
дававшие более круглые и более удлиненные яйпа (рис. 79).
см
96г-
92\
90
A3
3J9
1926
1930
1935 19W
Годы
№5
«57
Рис. 78. Изменение признаков телосложения у датских свиней в
результате отбора, проводившегося с 1926 по 1950 г.
/ — длина тела; 2 — толщина сала hi сшше; 3 — то же п бршое
Немало иллюстративных данных было получено и при
отборе у растений. Одним из примеров может служить отбор
на содержание жира в зернах кукурузы, проводившийся в
течение 60 поколений (рис. 80). При плюс-отборе процент жира
был повышен с 4,5 до 15%, а при минус-отборе снизился до
уровня меньше 1%. Увеличение признака было больше, чем
в 3 раза, а уменьшение—в 5 раз.
Экспериментальное изучение процесса отбора. Отбор
изучался также и в прямых опытах на модельных животных,
что дало возможность не только наблюдать визуально
результаты отбора, но и дать характеристику ряду генетических
параметров, а также проанализировать некоторые принципа-
*7
^ГГйрЛиы? тгтиякя «им* груди и др.) или * Да"1
;«р;м*С в с%о*у уели<е«н* приэн.к.. или г«**
а.*
п
I
1"
7 —-*-^г "**-*.
// / 2 J * 5
Поколения
Рас. 79 Реэулып-п* опбора во ф<оро«е **Я у typ ДО йф*-
ру> t — *ругж* яйпю; 2 — ywimtrim* *w#.
nmtpmjftnnnx — в етор-ояу уаелячеяяя я уиеньтеяяя rtpJW*
9 перяо* случае тглядко наблюдались тяшютые р**у/№-
таты иаяраял^яяосо отбора — сдаяг крнно* pacfi^JM^**
в сторону отбора На ряс. 81 локазаЯы Крияме paertf^rfflW*
особе* по числу стерноплевралъяых тетянок в 3, 14, да *
3*-м яоиолеяяях отбора в ляяяя Дроэофялм 1/?. Сдяяг* *рт
отборе была очень кеЛольп*и*я. _/^_
В друготя случае nt дрозоАяле Ярсяюдялся отбор я* У****
теяяе числа шетияок до )6-го Лок<*ле*иЯ. £ffr реаулет**»
яредстаалеяы #га рис. 82. Одяако с 7-го пояолеяяя ЯЗ tgw J**
ляяяя был качает и гглюс -отбор, т. е. отбор Я* yftwftw"»
«тела шегяяок. Оя также окзэалс* эффектявяым, ^
Этот ояыт по отбору яз одяого и того же *^еР**;5|**^
эдаает, что в пгзсл#жяе* телясь гены, как У**^^****^ !К
я ослабляя*»** даияы* яолмествеяяы* rtpifcrrffltt. По^О** JT
те* отбор* ложно ж*£я?ься расчлейе*** яс*6д*оА *в*У**
нии на две, отличающие** т> tbenuuu ««*-*.. .
Яиг, Л?. Результаты отбора *а ею-яф-
/-***-<****, /r-i„.. . .
«г**»»-
Яаг. Я/. Кривые рЯгйределе***
количеств* стераоплёвралыгых ямппкж у
яроэефглы в 3, 14. 20 * ЭЭ-я поколе- t , ^ « К > ^ - „ «,
^яхотбор* в ля-пнг /* (по Роки* ' А * * * -tJL?
20 г я с**цМ-24 г-«tan, ^^/Й^иГ*
самок « сам*» Эб-40 f я "«"^TSif S5Jr 6*™ Р**
лячйо*; есл* он* «е мель в*л*к*. *J «рт» a£rpeknpo**iU
особе* V «гбирпыо*,•$ff^%wff£& * W
Но возможен я тавоя «y«J. "f я1-1я;т„Г^бора «в «о*
вовсе. <rro яотса**** M Р* W. В результате к
лнчеству брюшных (стернитальных) щетинок в плюс-линии
г>ыла достигнута средняя — 87 щетинок с размахом нзменчль
вости от 60 до ! 15 и в минус-линии — 11 щетинок с размахо*
изменчивости от 0 до 28 щетинок *.
-| » 1 : : 1 . 1 ! i l I i J—■ *
0 1 2 J - 5 6 7 8 9 Ю 11 12 15 /4 15 16
Поколения
Рис. 82. Результаты отбора на уменьшение количества стерноплевральных
щетинок в линии Я3—. С 7-го поколения был начат и плюс-отбор (по Рокнц-
кому). Пунктир — выровненная линия регрессии.
О 2 Ц б 8 10 12 to 16 18 20 22 £4
Поколения
Рис. 83. Результаты отбора на увеличение и
уменьшение живого веса мышей (по Фолконеру).
Эффективные результаты отбора на увеличение «размеров
белого пятна на голове мыши были получены ГудейлоМ
(рис. 85). Первая мышь, от которой был начат отбор, имела
всего 12 белых волосков на голове, а после 20 поколений
отбора лятно занимало почти всю голову. Отбор начался почти
* Ниже будет специально разобран вопрос о причинах различий при
отборе а плюс- и минус-направлениях.
290
от нуля. Вот почему Гудейл признал невероятной мысль о
том, что в исходном материале \же были все гены, которые
определили развитие белого пятна, покрывшего почти всю
голову. Поэтому он высказал предположение, что отбор
сам создает для себя желательную изменчивость, т. е.
изменчивость в том же направлении, в каком ведется отбор, и
даже создал гипотезу о нескольких состояниях одного и того же
гена.
В дальнейшем, разбирая генетические основы отбора,
придется вернуться к этому вопроЬу и показать, как можно
трактовать эти и им подобные, с первого взгляда непонятные
факты.
-""г
0 5-04 3
Рис. 85. Результаты отбора
на увеличение размеров
белого пятна на голове у
мышей (по Гудейлу).
Статистическая оценка результатов Отбора по
количественным признакам. Чтобы охарактеризовать изменения, которые
происходят при отборе, нужно сравнить признаки двух или
нескольких поколений: если это животные — то oco6eff одного
и того же возраста, а если растения — то экземпляры одной
и той же фазы развития.
Исходной величиной для сравнения служит прежде всего
средняя арифметическая данного признака в популяции или
стаде. При искусственном отборе из родительского поколения
выбирается определенная группа особей, лучших по
фенотипу. Их средняя по данному признаку отличается от средней
популяции. Предполагается, что отобранные особи
отличаются от популяции и генотипически. Очевидно, что если бы
можно было знать их генотипы заранее, задача отбора была бы
очень облегчена. Но мы рассмотрим случай, когда отбор
проводится только по фенотипу. Чтобы не усложнять картины,
принимают, как это было сделано выше при анализе
свободно скрещивающейся популяции, что среди особей селекциони-
0 '0 20 50 W 50 00 ГО 30 90 $? '■'<?
число щетинок
Рис. 84. Результаты отбора в течение
34—35 поколений по количеству стернн-
тальных щетинок у дрозофилы:
О — исходная линяя, «—» — ммнус-лпнпя, « + > —
плюс-линия. Данные только по самкам (по
Клейтону, Моррису н Робертсону)
19*
291
руемон группы нет различий ни по плодовитости, ни по
жизнеспособности потомства. Тогда результаты отбора будут ц*.
ликом следствием генотипических различий между отобран*
ной группой и популяцией.
Результат отбора, или сдвиг при отборе (иногда в
литературе его называют также ответом на отбор), может быть
оценен разницей между средней фенотнпической признака
у полученного потомства и средним значением того же
признака у родительского поколения до отбора, т. е. у исходной
популяции. Обычно его обозначают буквой /?.
Рис. 86. Корреляционное поле с точками средних
значений родителей (ось X) и потомства (ось_У). Крупные
точки — отобранные родители (со_средней Xi). Средняя
их потомства у1ш
Для характеристики же интенсивности отбора вводят так
называемый ^елекщюннын дифференциал, который
представляет разницу "между фёнотипнческой средней отобранной
группы .особей и средней всего родительского поколения до
отбора. По аналогии с коэффициентом отбора, которнй был
рассмотрен выше, его обозначают буквой S.
Для статистической оценки результатов отбора
используют корреляционное поле, на оси абсцисс которого нанесены
значения количественного признака у родителей, а на осе
ординат — потомства. В гл. 6 с помощью такого поля
вычислялись коэффициенты регрессии и наследуемости.
На рис. 72 изображено корреляционное поле для регрессий
потомков к родителям по длине крыльев у дрозофилы. Отбора
не было, поэтому x=t/.
292
Значение Ь0~, равное 0,577±0,07, было мерилом
наследуемости, так как Л*=6 -.
ОР
Корреляционное поле может быть использовано и для
установления взаимоотношений между сдвигом при отборе R и
селекционным дифференциалом S (рис. 86). На нем точками
обозначены средние родительских пар (по оси л:) и их потом-
ci ва (по оси у), а более крупными точками — отобранные особи.
Коэффициент регрессии потомков к средним родительских
пар — Ь -
г ор
Рис 87. Зависимость селекционного дмфферсн
циала 5 от доли пооулацин, остаалаеыой для по
лучвнля потомства, м от степени измени воет и
арнэнака. Объяснение а тексте.
Отобранная для получения следующего поколения группа
особен имеет среднюю арифметическую х{. Популяционмая же
средняя — ж0. Тогда дифференциал селекции S»Xj— *♦.
Отобранная группа дала потомство со средней j/ь Потомство же в
целом имеет среднюю арифметическую у* В таком случае сдвиг
при отборе /? = i/,—//о.
Отношение же R/S представляет собой не что иное, как
коэффициент регрессии Ь -: —
Ь0р R—Sb0~. Так как
boP = hl то
tf=A*S.
(П
Таково общее уравнение связи между сдвигом при отборе и
селекционным дифференциалом с учетом коэффициента
наследуемости. Отклонение потомства отобранной группы от попу-
ляционной средней является в таком случае мерилом
селекционной ценности этой группы по сумме генов. Необходимо
иметь в виду, что использовать это уравнение можно тольк*
при некоторых условиях; 1) если сходство между потомками
и родителями ие определяется какими-либо дополнительными
причинами ненаследственного порядка; 2) если нет естестве
ного отбора, иначе говоря, если плодовитость и жизнеспоса*
ность не связаны коррелятивно с фенотипическим значен»**
количественного признака, по которому проводится отб%
3> если скрещивания в группе особей, отобранных в качеств
родителей следующего поколения» происходят по принцвд
случайности.
Селекционный дифференциал принадлежит к числу ва»
иейшнх параметров для оценки отбора, поэтому важно устами
вить факторы, влияющие на него. Среди них: величина дол*
популяции, включаемой в отбираемую группу, и степень фе**~
типической вариации признака в популяции, которая, как и*
вестна измеряется средним квадратическнм отклонением (о).
Роль этих факторов иллюстрирует рис. 87. В его левой час*
ги [а) показано, что при о=2 единицам и отборе 50% особей
популяции S«h6 единицы. На средней части рисунка (б) им
гс* же значении о и отборе 20% особей S=2t8 единицы, (>
лекционный дифференциал S увеличился более чем на 50V
Наконец» на правой части рисунка (в) среднее квадратическй*
отклонение вдвое меньше, чем в случае б, а да!Я отбираем!*
особей та же. Селекционный дифференциал вдвое меньше. Й*
сопоставления трех едем можно сделать два вывода: при тЛ
ке вариации чем меньше доля отбираемых особей, тем боли
toe S; но ори том же проценте отбираемых особей селекшЯЦ»
шй дифференциал прямо зависит от величины <г. меяьА
в — меньше и 5,
Так как среднее квадратическое отклонение — вели
именованна*» есть смысл выражать сдвиг при отборе и
ииошшй дифференциал не в конкретных значениях отбираемо*
го признака (например, и килограммах молока или в колнчест*
ве снесенных за год анц). а в долях о*у т. е. пользоваться и*
дичинами
R S
к .
Of> Op
Тогда приведенное выше общее уравнение R=h*S шея**
быть представлено езедуншшж образом:
**танд»ртмаоиаянро величину обозначаю* иереи f и cw
н* .:
*шк перше» нпемсиаиосга отбора.
» тшшю случае
ф
В данной формуле, как н в предыдущих, R выражает сдвиг
средней величины признака за одно поколение Но иногда
полезно оценить эффективность отбора за единицу временя,
например за год, тогда надо разделить правую часть равенства
на 7—длительность интервала между поколениями ( в годад):
»-*р. и.
Обычно за J принимают промежуток временя между
рождением родителей я рождением попгомков. Так, для круояого
рогатого скота У*5 лет, для овец J**4 года.
Селекционный дифференциал обычно различен для особей
разных полов. Если самец покрывает много самок, а пря
искусственном осеменении спермой одного производителя
осеменяют сотни я даже тысячи самок, то, очевидно, селекционные
дифференциалы для самцов и самок резко отличаются.
Из последней формулы явствует, что сдвиг при отборе за
год. нлн, иначе, эффект отбора за год, прямо вроаорциояадея
значению i, козффнциенту наследуемости я степени фенотияя
ческой вариации признака, т. е. оР, я обратно пропорционален
длительности поколения. Для оценки же сдвига вря отборе за
поколение делить ва J не нужна
Так как А*= —т 1"»сяедуемосп в узком смысле слова),
то в формуле (3) значение ¥ можно заменять яд — идя яа
¥~£*= —. Тогда
ар %
Ecj» взяп иаоамую общую фодолу Oh те в> ме mmn
быть ояредезеяо звачеше
*»=!- (61
Таким образом. ** ■ведстаывет coftai и шинам сзмга
„р. отборе ■ ажпта*>Щ»**шт»яГ~ *»» *">
грубое оврея*л«*е юэ+фа—мпа «асасдомст», во
, ,,* ■<— амия «ас
дчемости. т. е. наследуемости, которая ответственна за паду,
«генный фактический сдвиг при отборе. Так как обычно сдвиги
при отборе за ряд поколений несколько отличаются друг от
др\га, то на графике они выражаются ломаной линией. При
аырааниваннн ее методом регрессии можно вычистить
значение коэффициента регрессии Ь как среднего значения ~.
г. е. реализованной наследуемости.
Так. для данных по изменению живого веса мышей, пред-
ставленных выше, на рис. 83. Фат кокер вычислил такие
значении коэффициента регрессии:
XI я плюс-отбора 0Л75±:0,016;
для минус-отбора 0.5!8±:<W>23.
Что же касается точности определения h* no R и S, то Белд-
1зном и Грнсом на кукурузе было показано, что при селекции
редкая реализованная наследуемость в каждом поколении
рнмерно равна среднему значению наследуемости,
определенному по полуенбеам.
Связь между сдвигом при отборе R и значениями i, А2 и о#>
лет возможность использовать эту формулу для конкретных
«лекционных расчетов. Так, установив для данного стада или
астн породы коэффициент наследуемости h2 и степень варна-
1ин. т. е. значение о*, можно решить ряд задач, например:
i) каких животных надо отобрать, чтобы получить от них по-
омство с желательным уровнем развития признака, т. е.
определенный сдвиг в следующем поколении *, или б) какова будет
игличина признака у потомства, если известно, какая доля из
юднтельского поколения оставляется для размножения, иначе,
предсказать результаты отбора.
Возможности прогнозирования результатов отбора. Прог-
1оз можно сделать, используя приведенные выше уравнения
П) н (3).
Однако степень точности прогноза зависит от ряда причин.
В специально поставленных модельных опытах (на мышах,
ароэофиле) удавалось достичь довольно большого совпадения
фактических результатов с теоретически ожидаемыми. Так, в
упомянутой уже работе Клейтона, Морриса н Робертсоиа были
вычислены значения R по данным о наследуемости н
селекционном дифференциале в разных линиях плюс- и минус-отбора
по количеству брюшных щетинок. На основе изучения
изменчивости популяции до начала отбора было получено значение
М*0,52 В разных линиях былн несколько различные, хотя я
* Дтда расчете* см. в жявге «Творствчесмае основы седосшю яиюог-
«мх» <*., 1968).
m
и близкие, величины 5. Фактически полученные сдвиги при
отборе и теоретически ожидаемые при данных значениях А* и S
приведены в табл. 49.
Таб.ища 49. Фгктмчкжие и тсореткчесхяе эмчешш J?
при плюс- и имус-отбере по бркимм щешшжлм дроэофим
Линии
1
2
3
4
5
Средняя
Плюс-отбор
5
5.29
5.12
| 4,44
4.32
4.88
4,81
*
теоретвч.
2.75
2,66
2.31
2.25
1 2.54
2.50
ф*ктщ.
2,60
2.23
2.43
3.12
; 2.68
i 2.61
I
Mwmvc-moop j
5
4.63
4.58
4.36
t 5,60
4.12
I 4.66
* i
теоретжч.
2.41
2.38
2.27
2.91
2.14
2.42
фмтмч.
2.44
2.29
0.67 ,
1,13 i
2.68
1.82
В большинстве случаев получено довольно близкое
совпадение теоретически ожидаемых н фактических значений /?. Но
по двум линиям при минус-отборе обнаружены значительные
отклонения. Ниже мы остановимся на интересном факте
возможных различий в эффективности отбора в разных
направлениях. Относительно же предсказания результатов отбора
можно сделать вывод, что оно вполне возможно, хотя на данной
стадии разработки вопроса не очень точно. Надо иметь в виду,
что с каждым поколением отбора может меняться н значение
А2. Кроме того, далеко не всегда можно точно определить 5,
так как оно различно для самок и самцов, что увеличивает
ошибку в его определении.
Однако фактические данные многих опытов показывают,
что величина сдвига средней при отборе может довольно слабо
меняться даже в течение ряда поколений (5—10), особенно
если он невелик.
Сопоставление фактических сдвигов с теоретически
ожидаемыми было получено и на сельскохозяйственных животных.
Например, в канадском опыте улучшения молочного скота
проводилась систематическая селекция на увеличение
количества сухого вещества молока. Опыт был начат в 1955 г. в трех
породах — айршнрекой, голстейнежой и джерсейской.
Действительная степень сдвига в количестве сухого вещества оказалась
примерно такой же, как предполагалось по указанным
формулам, у голстеййов; несколько большей —у айрюнров и *****-
тетьно больше* — у джерсеев. Автор работы Хикмзи обмена
гт такое расхождение у джерсеев использованием в «то* поро-
д^ посторонних быков. А, А. Охкии и Г. А. Стакан провели
прогнозирование результатов отбора по настригу шерсти у
тонкорунных овец ллеммвода «Курьииский* (Алтайский
край) при рваном коэффициенте отбора по маточной части
стада, Очень тщательно рассчитаны значения селекционных
дифференциалов по изученным группам овец.
Для расчета сдвига при селекции была применена
несколько более сложная формула, а именно:
где // — интенсивность селекции по материм; ар — среднее
квадратическое отклонение по всем животным; S,—
селекционный дифференциал по производителям; А* — показатель
наследуемости; /«—число лет, за которое происходит смена
поколений.
Результаты оказались очень обнадеживающими Как по
всему стаду, так и по отдельным генеалогическим группам (ли-
киям) обнаружилось почти полное совпадение фактически по*
лученного ответа на селекцию с теоретически рассчитанным.
Некоторое расхождение оказалось в одной линии при всех
коэффициентах отбора (фактический сдвиг при отборе был иг
сколько выше ожидаемого), особенно при очень высоких.
Подобная работа была проведена Н, Ф. Решетниковой я
3, С. Киселевой на молочном скоте нескольких совхозов
Западной Сибири. Исследовалось содержание жира, белка и
сухих обезжиренных веществ в молоке. В большинстве случаев
было получено удовлетворительное совпадение фактического
ответа и теоретически рассчитанного по общим формулам (I)
и (%), Авторы разобрали также возможные причины
несовпадения прогноза я фактических данных. Отмечается, в
частности, что распределение особей в стаде по тому или иному
признаку иногда не подчиняется закону нормального распределения
и что на степень точности прогноза влияет наличие корреляции
между селекционируемыми признаками и молочной
продуктивностью в целом,
Некоторые данные о том, в какой степени фактические
результаты отбора совладают с прогнозом, были получены иа
герефордах Бриксом, Кларком и Киффером, В опыте,
продолжавшемся 25 лет, более высокие показатели, нежели
ожидалось, наблюдались по признакам животных до отъема (живой
вес при рождении и отъеме, привесы), а значительное
совпадение ожидаемых и фактических результатов отбора — по док**
аателям после отъема молодняка.
iWtQMOBUe!
Orjpa дали ««отдаидсаде JL?.???!!09,,P<*a»»»
т.
/
изменение етру«*уры мжГ л,ш,мх «^ «^«адыев м
<в каждой » 2^т^Гв^ТГ " Д^^*
иие длины беряоаой кости ui^Jl!*1/***<***■*
ложных направлениях ♦ ■*>»**• * —— ■•
штж* mmjm mm
измени
кои шерсти, Насидуеиость
длины 6ерщ,ыА костя О,»
На основе определенного
селекционного Дифференциала
были рассчитаны ожидаемые
иъыенении признаков и оню-
ставлены с фактически до-
лученным сдвигом. На
рис, 88 показано
расхождение (в значениях среднего
квадратн чес кого
отклонения; между плюс- и минус*
лннняни в величине
берцовой кости. Совпадение
фактически полученного
расхождения и теоретически
ожидаемого довольно большое, хотя в есть тенялшяя отставая**
фактических результатов отбора or ожидаемые что можно
объяснить некоторым уменьшением величины А1 яри отборе
по сравнению с первоначальный его значение*
попытки прогноза результатов отбора были сделаны я яа
растениях, В этом отношения очень интересны данные Лента
по анализу результатов отбора на ярвнигние яронента жир*
в зернах кукурузы ten. рис W). Автор ярямеяжч ту же o6*iy»
формулу (3).
Первоначальные результаты «годясь очень °^^Н**Г
вающими: только в 5 случаях из 24 емдзияы* ЯР*^***1
Д.7Н разных отрезков отбора, охватывающих Ю**^* "
блюдалось Достаточно «р^с^ад^^
лаемых результатов с фактически ^У*»™™^- отдельных
конечно^ясинть их истовость» ь ^^^J^^
параметров уравнения. Но^^^^*%2S^^v^^
таты все'же было ™^™££^^
распределении жира но зерну, в °^^^- - до* — пре**у<
в зародыше. Повышенное W^^. /3*
шественио результат У^^^^^^^^п^
жение идет не за с<** нзмеиеяня P**"»f7T.^
снижения процента жира в
• Этот семг ножнв *•***
деде»****»»
При пересчете данных с учетом этих обстоятельств можно
было предсказать повышение содержания жира вплоть до
60 поколений отбора.
Неточность прогнозирования может подчас объясняться и
некоторыми специфическими причинами. Так, Фолконер
показал (в опытах по отбору на величину помета у мышей), что
расхождение между фактическим ответом на селекцию и
ожидаемым на основе оценки наследуемости объясняется
наличием материнского эффекта. Наследуемость величины помета
очень низкая — всего 0,12. При расчете коэффициенты
прямолинейной регрессии дочь— мать были нулевыми, но если
определить частную регрессию дочерей к матерям, то она была
равна 0,13. Таким образом был обнаружен слабый
материнский эффект, что и сказалось на результатах.
Неточность прогноза может вызываться и другими
причинами, в частности изменением внешних условий. Прогноз
действителен только при сохранении одних и тех же условий
среды.
Связь между коэффициентом отбора s и интенсивностью
отбора I. В гл. 4 рассматривались изменения популяции при
отборе, причем мерилом этих изменений была величина \q—
изменение частоты гена за одно поколение. Была установлена
связь между значением q и коэффициентом отбора s. Нет
сомнений, что в основе отбора по количественным признакам
также лежат изменения частот генов, но непосредственно
оперировать этим показателем нет возможности. Для описания
процесса отбора используются другие параметры: изменение
средней арифметической популяции, которая сама по себе
представляет комбинированный эффект всех генов, и
интенсивность селекции i (или селекционный дифференциал S).
Возникает задача перебросить мост между этими разными
параметрами и показать их возможную связь.
Как было показано выше (см. рис. 86), отобранная из
популяции группа отклоняется от фенотнпической средней попу-
ляции на величину S — селекционный дифференциал. В ней
частота гена иная. Для оценки этого сдвига в частоте гена
можно использовать своеобразную величину — регрессию
частоты гена q к фенотипическому значению популяции Р, т. е.
fegp. И тогда новое значение частоты гена q\ в отобранной
группе можно представить с помощью уравнения регрессии:
q\ = q + bqpS.
Можно представить частоты и генотипические значения
трех генотипов в популяции следующим образом •:
• Имеется t bmiv. что нет корреляции между генотипом и срело*
(тоги P-G>
*ю
АьАк р* о а
ЛИ* 2р(/ 0,5 d
АъАг <у* 1 —а
Тогда коварнанса генных частот с фенотипическнми
значениями может быть получена из суммы произведений q и Р
каждое из которых умножается на частоты генотипов в попу'
ляции. Из них можно вычесть произведение средней частоты
гена на фенотипическую среднюю МР. Это приводит к
уравнению
covqp=zpqd—яга—qMPt
В свое время было показано, что
МР=а(р~q)+2dpq.
Подставив это выражение вместо МР, после преобразований
получим:
covqp= -p<A<*+d(q~p\\
Величина, находящаяся ъ квадратных скобках,
представляет собой не что другое, как а, т. е. средний эффект замены
гена (равный разнице между at н сц):
сотряс— pq a. (7)
Отсюда можно определить значение Ь.
к М*
ЬЧР~- _
Здесь ор2 — фенотилическая варианса. Подстамяя значения
b.4i> и S в уравнение для д]% подучим:
бр
Знак минус указывает, что отбор идет против А* частота к
Р°Твдтл. 4 были даны значен» *Я *а*9™»«»1™Г\У$»
с учетом *. Можно ваять формулу ^^1*ы„ииро»я*
относилась к такому же случаю, т. е. полному до»»" к—
[d=a; a=2ga). Тогда
При отсутствии доминирования
ipq - = 0,5*7 (1—9).
О/»
После некоторых упрощений может быть получена общая
формула, связывающая л и г.
2а
5 = /—-. (9)
Ор
Эта формула, естественно, относится к одному гену с двумя
аллелями. Коэффициент отбора прямо пропорционален иитен-
2а „
сивностн отбора, а также величине . В числителе —
разор
нмца между значениями двух гомозигот, выраженная в феноти-
пнческих сигмах.
По остается еще вторая, более трудная задача — перейти
к эффекту многих генов, влияющих на количественный
признак
Полигенная наследственность как основа для отбора.
В предыдущей главе были рассмотрены генетические основы
количественных признаков. Показано, как первоначально
простую модель полимерных генов пришлось в дальнейшем
усложнить, так как гены могут отличаться друг от друга по степени
доминирования, силе действия и характеру суммирования
эффектов отдельных генов — аддитивному и неаддитивному. Все
это дает очень сложную генетическую основу фенотипической
изменчивости в любой популяции, в которой начинается отбор.
В силу этого неудивительно, что результаты отбора в
отдельных линиях, создаваемых из одного и того же исходного
материала, могут быть различными в зависимости от того, какие
конкретно генотипы были у отбираемых особей. Отсюда
вытекает, что при отборе используется далеко не вся генетическая
изменчивость популяции, а лишь какая-то часть генов. Их
можно назвать как бы эффективными генами. Поскольку среди
них могут быть гены и с сильным и со слабым действием,
количество их может быть достаточно большим. Однако были
приведены данные, указывающие на то, что в ряде случаев не
следует и преувеличивать их количество.
Чтобы выявить связь между сдвигом при отборе,
аддитивной вариансой и действием отдельных генов, необходимо
принять некоторые упрощенные предпосылки. В процесс отбора
иоклекаетси п генов, каждый ш них имеет пару аллелей. Раз-
ница между двумя гомозиготами по каждому гену равна 2а.
Отбор прекращается, когда все благоприятные аллели перешли
» гомозиготное состояние. Гены обладают разным действием.
ЗЯ
Тогда суммарное действие всех генов, определяющее общий
сдвиг при отборе /?, можно выразить через R=2na, где~а —
среднее действие гена.
Аддитивная вариаиса по каждому гену (локусу), как
указывалось в предыдущей главе,
Очевидно, что частоты отдельных генов различны, но можно
принять, что в среднем они равны 0,5 (/?=<7=0,5).
В таком случае аддитивная вариаиса для каждого гена
будет
а для всех генов
аА«-у*
0A*:=i-nla*). (Юг)
Если все гены имеют равный эффект, то
ал2=улл*. (106)
Из двух уравнений /?=2яа и оА*= ir™* можно получить
значение пл т. е. общее количество генов, ответственных за
результаты отбора, а именно:
Впрочем, это уравнение дает, как правило, очень высокие
цифры количества генов, но со слабым действием каждого us
них. Вклад каждого гена в общую фенотнпическую сигму
может быть выражен величиной —
С помощью уравнения оАг= — па* можно после ряда
преобразований получить и эначеиие —, а именно:
0> 1П
Очевидно, что при любом значении K°^"*"£a„J£ffi
мости вклад каждого отдельного «на («npoaopwW
«а
чффекг*) в общую вариацию признака обратно
пропорционален числу генов.
Некоторые расчеты Фолкоисра на основе анализа литера-
турнич данных о результатах отбора по различным признакам
у мышей и дрозофилы привели его к значениям количества
генов от 30 до 100, а вклад отдельных генов выражается
величинами ^0,20 фенотипической а.
Однако во всех этих расчетах не учитываются возможные
различия в действии отдельных генов, что может очень сильно
менять картину.
Сели предположить, что есть несколько более сильно
действующих генов (олнгогенов), влияющих на количественный
признак, то следовало бы ожидать возрастания их частоты при
отборе и довольно сильных сдвигов в значении признака в
первых поколениях. При отборе же по слабым генам
(полигенам) эффект отбора должен быть более слабым, а
длительность отбора большей. Осложнения в общей картине могут
вызываться доминированием, взаимодействием между
генами, сильным модифицирующим влиянием внешней среды.
За последние годы все эти вопросы подверглись довольно
основательному математическому анализу с применением
моделирования на электронно-вычислительных устройствах.
Математическое моделирование процессов отбора. В целом
ряде опытов, проведенных на математических моделях,
анализировались самые различные случаи отбора с учетом роли
размеров популяции, количества генов, интенсивности отбора,
влияния на процесс отбора сцепления, доминирования, эпи-
стаза, сверхдоминирования и т. д. Было показано, что картина
отбора в этих случаях более сложна, нежели при анализе
отбора но одной паре генов. Так, при небольшом числе генов
(/1^12) сильное сцепление уменьшает эффект отбора, но оно
не сказывается при большем числе генов. При малом числе
генов и отсутствии сцепления генетическая изменчивость при
отборе уменьшается довольно слабо. При наличии
доминирования наиболее сильный сдвиг в результате отбора происходит
на первом его этапе. В целом между результатами отбора,
сцеплением, размерами популяции существует сложное
соотношение и возможны самые разные варианты ситуации,
складывающейся при отборе, в том числе разные вероятности
фиксации генов, различия в роли гетерознготиости и др.
Очень интересная работа по моделированию процесса
отбора при различных исходных предпосылках проведена
немецкими учеными X. Аренсом и К. Бсллманом *.
Были проверены на вычислительных машинах две модели:
простейшая и сложная. В простейшей модели было принято п
• Опубликована н журнале «Генетика» зи 1969 г.
301
локусов. Зигота, составленная из двух гамет, изображалась
парой двойных чисел (доминантный аллель—1,
рецессивный — 0). Если в гамете пять генов, то в зависимости от
доминантности или рецессивности каждого из них можно было
представить гамету в виде 11001, 01101 и т. д. Зигота же изоб-
01011 __
ражалась двумя рядами цифр: например: imm- Каждый
вертикальный столбец описывает одну пару аллелей.
Принималось некоторое начальное распределение гамет по
соответствующему алгоритму. Селекционная ценность генотипов G
складывалась аддитивно из ценностей отдельных пар аллелей
с учетом доминирования вплоть до сверхдомннирования.
Предусматривалось, что условия внешней среды создают вариацию,
имеющую характер нормального распределения.
Для простоты в этой схеме аддитивное взаимодействие не
принималось во внимание.
Давление отбора измерялось процентом «лучших» зигот,
т. е. имеющих наибольшую фенотипическую ценность.
На простейшей модели изучалась зависимость
эффективности отбора от числа генов, степени сцепления и характера
доминирования. Учитывалось также влияние ограничения
численности популяции.
При отсутствии сцепления эффективность отбора, как
указывалось выше, уменьшалась с увеличением количества генов,
но не очень сильно (рис. 89). При тесном сцеплении
тормозящий эффект количества генов проявлялся слабо, ниже была и
эффективность отбора.
Очень сильное влияние оказывала средняя степень
доминирования (обозначалась символом а). Эффективность отбора
была очень высокой при среднем доминировании а=0,5,
несколько ниже —при полном доминировании (а=1,0) и резко
снижалась при сверхдоминировании (а=1,5), что
иллюстрирует рис. 90. Впрочем, надо сказать, что в некоторых
конкретных условиях очень маловероятен случай, чтобы средняя
сумма доминантного действия многих генов давала эффект
сверхдоминирования. При вариации ряда генов по доминированию
наиболее частой должна быть кривая / рис. 90.
Во второй модели была взята ситуация, очень близкая к
тому, что может быть у какого-либо вида, например кукурузы:
10 пар хромосом с учетом их длины. В них распределены
случайно 99 генов с разными частотами рекомбинаций в каждой
паре хромосом. Принималось, что в 4 хромосомах находятся
тесно сцепленные блоки генов. Из числа 99 генов — 3 ссйль-
ные», т. е. с фенотнпическим действием, в 5 раз превышающим
действие «слабых* генов, 86 сслабых» и 10 генов-модификато-
20 II «I». Ройники*»
305
ров («плюс» и «минус»). Были приняты и разные степени до*
минирования. Коэффициент наследуемости Л2 = 0,3. Популяция
панмнктнчна, ее численность Л'= 500. Отбирается 10 процентов
лучших зигот.
Детальный разбор работы занял бы много места, поэтому
остановимся лишь на самом важном.
г
1»
5 I
1 1
1
\ 1
1/
/
/
/ /
0 2 4 б д Ю 12
Поколения
Рис. 99. Зависимость
эффективности отбора от числа генов (/—
5 генов: 2—20 геяов; 3—50 геяов).
Сиесленке отсутствовало (во Арен-
су и Беллмаву).
2 U 6 8 10 12
Поколения
Рис. 90. Зависимость
эффективности отбора от_средией степени
доминирования а (/—а=0,5; 2—в*»
= 1,0; 3—а = 1,5) (по Аренсу и Бед-
лману).
На модельных популяциях, как более простой (/), так и
сложной (//), подтвердились теоретические представления, что
с увеличением средних при отборе должны уменьшаться и
аддитивная и доминантная генетические вариансы. Но в более
сложной системе при наличии сцепления и эпистаза
уменьшение идет медленнее (рис. 91). Отбор очень сильно влияет на
частоту доминантных модификаторов — ослабители
элиминируются после шести поколений, а усилители резко возрастают
по их частоте. В результате очень сильно повышается феноти-
пическая ценность зигот pj. Но интересно, что при наличия
«сильного» гена, находящегося в одном блоке с тесно сцеплен-
ЭК
пымн «слабыми* генами, частота первого быстро возрастает,
а частоты последних почти не меняются (рис. 92).
Во второй модели отбор дал повышенные и устойчивые
средние показатели уже через 6 поколений, причем выше, чем
в первой модели.
Таким образом, математическая модель в сущности
подтвердила представления о процессе отбора на основе чисто
генетических данных, но внесла существенные дополнения и
уточнила ряд деталей.
Т~ Ц б д f0 U
Попс.цния
J 5 1 S tt U
Поколений
Рис. 91. Изменения аддитивной
(о2А) и доминантной (а^)
генетических варианс при отборе
в популяциях I и II (по Аренсу
и Беллману).
Рис. 92. Изменение при отборе
частоты доминантного «сильного» гена
№ 44. По 8 сцепленным с ним
«слабым» генам изменения частот не было
(по Аренсу и Беллману).
На моделях же, которые проверялись на
электронно-вычислительных машинах, разрабатывался вопрос о
прогнозировании результатов отбора. Так, Янг проанализировал таким
путем модели: чисто аддитивную, с влиянием доминирования, с
*пистазом разного типа и др. Было разработано несколько
программ, для которых принимались следующие исходные
предпосылки: 1) объем популяции в каждом поколении до
проведения отбора 1000 особей; 2) отбор в течение 8 поколений в
одних опытах и до 30 в других; 3) признак, ло которому
проводился отбор, определялся 10 генами с 2 аллелями в каждом;
4) начальная частота каждого гена ОД при этом исходная
популяция находилась о состоянии равновесия; 5) были вдагы
3 разных интенсивности отбора (0,1, ОД 0,8); 6) учтена
возможность влияния внешней среды и приняты три уровня
наследуемости (0,1, 0,4 и 0,9).
20* *°7
Кроме того, была учтена возможность сцепления с 3 коэф*
фициентамн рекомбинации (0,5, 0,2 и 0,05).
Оказалось, что в случае аддитивной модели совпадение
между ожидаемым сдвигом при отборе, предсказываемым в*
(кновё коэффициента наследуемости и селекционного дифф*.
ренциала в предыдущем поколении, и фактическим феиотипн-
ческнм сдвигом было очень большим, хотя прогноз генетиче-
ского сдвига был менее точен при более высоких коэффициент
Рис. 93. Результаты отбора по фенотипу в плюс- и мннуонаправле
ннях:
о — при Л- -1.0; б — при Л3-0.2 (по Лашу).
тах отбора. В модели же с доминированием прогнозирование
вообще было очень неточным.
В моделях с учетом эпистатнческого взаимодействия генов
чмеиьшенне возможности прогнозирования зависело от
вариации по поколениям коэффициентов наследуемости и вариавс
Величина сцепления не оказывала сколько-нибудь
заметного влияния на успех прогнозирования, а также на значения
коэффициентов наследуемости и конечный результат, дости*
гаемый при отборе.
Эффективность отбора и показатели наследуемости при*
мака. Существует бесспорная зависимость между результа*
тами отбора (по фенотипу) и наследуемостью признака.
Отбор лучших по фенотипнческим признакам особей при высокой
их наследуемости дает сильный сдвиг потомства в направлении
отбора. При низкой наследуемости признака происходит по*#
ти полный возврат к средней исходного поколения.
га
Па рис. 93, а показано, что при Л2, близком к 1,0, отбор
очень эффективен. Для большей наглядности представлены
результаты плюс- и минус-отбора, но можно ограничиться
рассмотрением только правых частей диаграмм — при
направленном отборе на повышение данного признака. При Л2=1
средние каждого последующего поколения отбора равны средним
отобранной для размножения группы предыдущего поколения
(заштрихованы части вариационной кривой, оставляемые для
размножения). При значении Л2 = 0,2 (рис. 93,6) средние
последующих поколений незначительно отличаются от средних
предыдущих поколений. Наблюдается очень сильный возврат
к средней родительского поколения.
Фактическим подтверждением приведенной схемы,
принадлежащей американскому генетику и селекционеру Лашу,
являются данные, полученные на курах и серебристо-черных
лисицах. Лернером и др. были показаны большие различия в
результатах отбора по числу яиц, снесенных курами в октябре
второго года жизни, и по среднему их весу в ноябре. Эти
данные получены на значительном материале (600 кур-несушек
леггорн). Наследуемость первого признака около0,25. а
второго приблизительно 0,75. Отбор по второму признаку в отличие
от первого привел к созданию двух групп, которые
различались по весу яиц почти на 8 г. Отсюда следует вывод, что в
простейших случаях отбора наследуемость может быть
определена даже непосредственно по степени сдвига потомства по
сравнению с родительским поколением. С. Г. Давыдовым были
сделаны расчеты степени генотнпнческого сдвига потомства
при отборе особей, отклоняющихся от средней по фенотипу на
1, 2 и 3 о и при разных значениях наследуемости. В
несколько измененном виде они представлены в табл. 50.
Таблица 50. Степень гевогтшческого улушкян потонет щт
отборе по фенотипу ■ эмястюст! от стеаем междуеяоетя
Отбор особе ft,
отклоняющихся от средней
по фенотип)*
На КО о
> 2.0 о
» 3,0 о
Сдваг потомства \1 ооюлеякя) по гаяотяоу (в sq) от
средней восуляов!
А* ~ 0.1
+0.05
-0.09
+0J4
М-0.25
+0J3
+0,25
+0.38
•« - 0.3
+0,25
+0,49 1
+0,74
Таким образом, при малых А1 генотипическое улучшение
потомства невелико. Примером отбора при малом значении
W могут служить данные нашей работы по изучению модо-
309
пнт.чтн серебристо-черных лисиц, проведенной вместе <
11 П. .Чситиион. Диалогичные результаты получены авторо*
при изучении плодовитости свиньи (неопубл.). Было устаноЕ-
.л\<\ что размеры помета серебристо-черных лисиц имеют шц-
к\к> наследуемость. Коэффициент наследуемости плодовитое-
in, т. е. числа детенышей в помете, равен 0,1. При разной
плодовитости матерей-лисиц (от 1 до 8 щенков) их дочери имела
примерно одинаковую плодовитость: 4,43, 4,73, 4,64 щенка я
т. д. В Салтыковском зверосовхозе систематически проводили
работу по увеличению числа щенков в помете. За ряд лет про-
пчошло некоторое повышение средней плодовитости матерей,
от которых получали потомство. За первые четыре года
средняя плодовитость матерей была равна 4,41 щенка, а в
последующие четырехлетия—4,61, 4,79, 4,83. Однако оказалось,
что средняя плодовитость дочерей от изученных матерей
изменилась за все 16 лет очень мало, а именно (по четырехлетним
периодам): 4,68, 4,72, 4,80, 4,79. Если сравнить плодовитость
потомства лисиц, имевших в помете в среднем 6 и больше
летенышей, и лисиц, имевших в потомстве не более 3 щенков,то
гпа оказывается очень близкой. Так, по одной группе (156 пар
матерен — дочерей) соответствующие значения плодовитости
были 4,31 и 4.23 щенка; по другой (103 пары)—4,73 и 4,75; н
только по третьей группе (103 пары) величина плодовитости
матерей и дочерей отличалась более сильно — 4,65 и 3,83. По-
видимому, в последнем случае выделилась какая-то линия с
пониженной плодовитостью. Впрочем, и число пар матерей —
дочерей в этой минус-группе очень мало— всего 17.
Таким образом, эффективность массового отбора, т. е.
отбора лисиц по их непосредственной плодовитости, оказалась
очень малой. При сильном влиянии внешних условий на
изменчивость плодовитости (что и выражается низким
коэффициентом наследуемости) очень велика вероятность того, что
отбираемые лучшие по фенотипу самки генетически не отличаются
или отличаются в незначительной степени от генетической
средней стада. Это вытекает из приведенной выше формулы
(*). При малом h2 мала и величина /?.
Возможности повышения эффективности отбора за счет
увеличения наследственной изменчивости мутагенными
факторами. При анализе результатов отбора имеется в виду, что
отбор основывается на уже имеющейся в исходном материале
наследственной изменчивости и его эффективность зависит от
степени реализации последней. Но возникает вопрос о том,
нельзя ли повысить эффективность отбора, вызывая мутации
и селекционируемом материале каким-либо
экспериментальным воздействием, например ионизирующими облучениями-
Первые опыты в этом направлении были начаты автором эта*
строк еще в \Ш\ г. и проводились до 1940 г. Из одного исход-
З.ч
ного материала (нормальной линии) был заложен ряд линий,
п которых проводился отбор на увеличение количества стерно-
плепральных щетинок, сопровождавшийся инбридингом.
В наиболее крупной по масштабу III серии опытов 20 линий
были контрольными, т. е. в них проводился отбор без
облучения, и 20 опытных. В последних отбиравшиеся для получения
потомства самцы в каждом поколении подвергались рентгенов-
-J 1 1 1 1 1 t . i , |
1 3 5 7 3 И U И 11 13 21 23 25
Поколения
Рис. 94. Изменения х и о в течение 25 поколений отбора по сумме
всех опытных (R) и контрольных (К) линий. Прямые — вычислен
ные линии регрессии (по Рокицкому).
скому облучению. Отбор проводился по всем линиям в течение
25 поколений, а в лучших линиях —40 поколений. В
результате во всех сериях опытов наблюдалось некоторое увеличение
эффективности отбора, названное автором эффектом рентгено-
селекции.
На рис. 94 показаны данные по всем опытным (/?) и
контрольным (К) линиям III серии. Произведено суммирование
в каждом поколении 20 опытных и соответственно 20
контрольных линий в один вариационный ряд и вычислены для них
х и а. Виден больший подъем в опытном материале по
сравнению с контролем, а также и более сильное увеличение
изменчивости в опытных линиях. Эффективность рентгеноселек-
ции была показана также на плюс- н минус-отводках, начатых
с 12-го поколения и продолжавшихся до 30-го поколения, а
также на отводках с 27-го до 40-го поколения. Наблюдалась
значительное расхождение между плюс- и минус-отводками в
облученной линии AR и очень слабое в контрольной ВК.
зи
Ьылн сделаны также расчеты средней эффективности рент*
ли чслекцнн (в виде разницы между рентгеновскими линиями
и контрольными) различными методами — по изменению сред-
iiv\ мл время отбора или по величине коэффициента Ь уравне-
пни регрессии, построенных по всем точкам 25 или 40 поколе*
ини. В большинстве случаев их значения были около 0.028—
0.040. Та же разница между лучшими рентгеновскими линиями
и лучшими контрольными (0,028). Однако в плюс- и мннус-от*
волках обнархжнлась асимметрия в ответе на отбор: разница
уежду St и АЧ в среднем на поколение была 0,089, а между
#- „"Л"— 0.039.
Значения эффективности отбора за счет облучения,
приводимые Клейтоном и Робертсоном (в работе 1955 г.), близки к
нлшич. По данным Лялы. облучение дрозофил видов Drosophila
serrata и Dr. birchii приводило к тому, что в последующих
поколениях в линиях, полученных от облученных популяций,
численность особен была выше, чем в контрольных линиях в
новых условиях (температурных). Автор объясняет это
увеличением генетической изменчивости, вызванным облучением.
Интересные опыты по селекции после воздействия облуче-
ииями (у-лучамн и др.), а также химическими мутагенами
проведены Броком на растениях Arabidopsis. Отбор в Л13- и My
поколениях после воздействия велся по времени зацветания
растений и по весу растении после 22 дней их выращивания.
Эффективность примененных воздействии была неодинакова,
но все они увеличивали изменчивость обоих признаков. Чаще
возникали формы более поздноцветущие и формы с меньшим
весом по сравнению с контролем. Отбор оказался
результативным по времени цветения, особенно в опытах по применению
химического мутагена диэтилметансульфоната. Селекция же
по росту растений была в большинстве случаев
неэффективной, несмотря на увеличение вариансы.
Отбор в различных условиях внешней среды. Этот вопрос
очень важен практически, так как всякий отбор у животных и
у растений проводится в конкретных условиях внешней среды,
л также теоретически, поскольку в структуре фенотипической
вариансы предусматривается компонент взаимодействия
генотипа и среды. Вскрытие этого компонента, как показано в гл. 5.
далеко не всегда возможно. Анализ различий в результатах
отбора, проводимого в неодинаковых условиях внешней среды,
позволяет получить дополнительную информацию о наличии
взаимодействия генотипа и конкретных внешних условий.
Рассуждая логически, можно думать, что в результате
отбора на повышение определенного признака будут получены
животные или растения, которые проявят этот признак именно
в тех условиях, в каких велся отбор. А как быть, если отбор
приходится вести в худших условиях? Будут ли те лучшие
312
fJG
ПО
rc
животные, которые были обнаружены в этих условиях, лучшп-
мн п в других условиях, например в условиях повышенного
\ровня кормления и содержания, и наоборот? Вообще очень
трудно рассчитывать, чтобы отбор всегда велся только при
оптимальных условиях.
Поэтому для практики отбора важно
оценить: а) результативность
отбора при разных условиях
внешней среды и б)
возможность переноса материала,
полученного в результате отбора
в одних условиях, в другие
условия.
Еще в 1928 г. советский
ученый О. В. Гаркави изучил, как
распределяются коровы по
суточному удою в условиях
скудного кормления (в тогдашних
мелких крестьянских
хозяйствах) и как распределяются эти
же коровы при кормлении их
по нормам. Результаты
представлены на рис. 95. В верхней
части рисунка выделены три
группы коров — худшие,
средние и лучшие, а в нижней
показано, что часть худших
остались по-прежнему худшими н в
новых условиях, но у другой
части удои повысились, и они
перешли в категорию средних.
У некоторых из лучших коров
удои повысились очень сильно,
у других — слабо, и они
приблизились к средним по удоям.
Однако полный переход из
группы в группу —из худших
в самые лучшие —не
наблюдался. Коэффициент корреля-
цип между удоями одних и
тех же коров при скудном н хорошем кормлении был высоким
(г = 0.79).
Результаты исследования О. В, Гаркави были
обнадеживающими: животные, отобранные по удоям молока при плохих
условиях, будут чаще всего лучшими н при хороших, хотя в
некотором количестве случаев совпадения не будет. Однако в
последующих работах были получены не столь отчетливые дан-
31Э
Суашаше ydaut фун.7,
Рис. 95. Распределение одних * тех
же коров по суточным удоям при
скудном (а) н хорошем (б)
кормлении (по Гаркави).
,<ыг Иногда, особенно при значительном изменении уоцщ
гюряюк распределения животных по их качеству сильно W
ня-кя Это зависело, с одной стороны, от степени рамнцЛь
\с ювиях, с другой стороны—от наследуемости признаков, lfc%
выше была наследуемость, тем меньше менялась оценил Щ
вотмых в разных условиях.
В ряде работ (на модельных животных- дрозофил*, Чн
шах> изучали результаты отбора в разных условиях, |ц
М. М. Камшилов изучал влияние отбора на степень npoflfe
ния признака. Путем отбора мушек в первые дни вылуллш*
на сиежем корме автор смог повысить процент проявленияЩ
eyeless (безглазые) с 14 до 63%, При отборе же В другИХуОО^
виях сдвиг в проявлении признака был значительно менышш
Наиболее полные опыты были проведены ФолконерОК Ш
мышах. Отбор велся при плохом и хорошем кормлении капа*
укрупнение, так и на умельчение мышей. Кроме того, прНШ
Ая.-шсь методы прямого и косвенного отбора. Последний U
ключа лея в том, что отбор проводился при одних усломиш
например плохих, а затем животные переносились в хорош*
условия и наоборот. Отбор велся на протяжении свыше 20 ЯВ-
колений. Родственные скрещивания не допускались.
Не вдаваясь в детали опыта, ограничимся лишь общДО
выводами, сформулированными самим автором: 1) отбор ops
разных условиях привел к созданию животных, отличавшШ!
друг от друга физиологически, например, в их теле был р&
иый процент жира, воды. Это значит, что отбирались жиаогВД
с таким набором генов, под контролем которых находялю
несколько различные физиологические процессы; 2) отбору
ше всего проводить в таких условиях среды, в которых улу*
шениме животные должны жить; 3) но если животные должш
жить в различных условиях окружающей среды, то отбор ДО
жен проводиться в наихудших из этих условий, тек как Цр
плохом кормлении отбор давал наиболее желательных жна#
шах, т. е, таких, которые оказывались хорошими не только вр*
плохих, но и при хороших условиях.
Фолконер, правда, очень осторожен в перенесении вывоДО
из ею работы на сельскохозяйственных животных, но во вМ"
ком случае результаты опытов очень поучительны.
Из других работ по этому вопросу следует упомянуть с**
дующие.
Результаты, полученные Коркмаиом также м нъШ*
('А поколения отбора), сходны с полученными ФолкошерО*
Они интересны ио признаку, на который велся отбор; ушел***
пие различий между самками и самцами. Лучшие результат*
были получены в линии, в которой отбирались семы* Л««*
самаы и самые тяжелые самки.
Макнутом и Диллом были показаны поразительны* р****'
4М
чм между разными линиями мышей на режим питья Мышам
лапали пить только соленую воду (4%-ный раствор ш*******
соли). В одной линии через неделю смертность достигала 50%
МЫП1И же другой линии не гибли даже после 36 недель такого
питьевого режима. Было показано также доминирование
устойчивости к солевому режиму над чувствительность*,,
fa ким образом, очевидно, что отбор ло *тому признаку, идун^й
и определенных и достаточно специфических условиях, эфф**
тивен именно при >тих условиях.
В целом можно сделать вывод, что к вопросам отбора при
разных условиях следует подходить глубже, с учетом япиыо-
действия генотипа и среды. Последнее зависит как от
признаков, по которым проводится отбор, так и от специфики
внешних условий.
Результаты отбора по одному и тому же признаку в
разных направлениях» В ряде случаев проводили отбор
одновременно в направлении увеличения и уменьшения значения
признака. При достаточном количеств* поколений отбора в
некоторых случаях обнаруживается разница между создаваемыми
отбором плюс- и минус-линиями. Так, у мышей, у которых
велся отбор на увеличение и уменьшение размеров (см. рис. 83ь
фенотипическая изменчивость в плюс-лиини была вдвое
больше, чем в минус-линии, В минус-ликин был выше и
коэффициент наследуемости, больше эффективность отбора.
Аналогичные явления наблюдались и в опытах со свинья-
ми, где пели отбор на большие и меньшие привесы и тем самым
на увеличение и уменьшение живого веса.
Таким образом, можно говорить о своеобразном явления,
получившем название «асимметрии ответа на отбор». Причины
его недостаточно ясны. Возможно, они могут быть
различными. Так, Лериер высказыввл мнение, что асимметрия может
создаваться в тех случаях, когда отбор в одном направлении
основывается на фиксации благоприятных генов, а в другом
идет в пользу гетерозигот по немногим генам; тогда в первом
случае он будет идти быстрее, а во втором медленнее.
Иногда асимметрию можно объяснить материнским
эффектом. Так, на мышах было показано, что асимметрия при отборе
по весу проявляется главным образом в эмбриональный
период, а последний в значительной степени находится под
контролем фенотипа матери. Однако асимметрия в ответе иа отбор
может наблюдаться и по таким признакам, иа которых
заведомо не может проявляться материнский вффеут, например иа
щетинках у дрозофилы,
В качестве одной из причин выдвигается также первой»'
чальная асимметрия в распределении ь исходной популяция
плюс- и минус- генов, влияющих иа селекционируемый признак,
а также их различий в домиивитиом вффесте.
315
Для изучения возможны* причин асимметрии при отбор*
в противоположных направлениях Фолконер провел специалы
ные опыты на мышах по отбору в течение 31 поколения н*
увеличение и уменьшение размеров помета. Результаты
опытов показаны на рис. 96. Надо иметь в виду, что размеры тку
мета, т. е. количество детенышей, обладают очень низкой
наследуемостью, поэтому для получения сдвига лри отборе
понадобилось много поколений, как это вытекает из смысла
формулы (6) о связи А2 с R и 5. При сравнении с конт-
5» J I. , г : 1 1 ■ —
<3- 5 Ю 15 20 25 30
Поколения
Рис. 96. Результаты отбора на увеличение
(плиния) м уменьшение (—линия)
количества мышат в помете (К—контроль) (по Фол-
конеру).
ролем результаты, полученные в плюс- и минус-линиях, был*
одинаковы: наблюдалось увеличение или уменьшение
количества детенышей за весь период отбора на 1,6. Но была
обнаружена значительная разница в величине реализуемой
наследуемости — при плюс-отборе 8% и при минус-отбо-
ре 23%.
В дальнейшем Фолконер установил различия между
плюс- и минус-линиями по тем компонентам, из которых
складывается признак сразмеры помета*, а именно: а) скорость
овуляции; б) доля яиц, оплодотворяющихся и успешно
прикрепляющихся к стенке матки; в) доля имплантированных
'эмбрионов, которые доживают до рождения (иначе говоря,
какова эмбриональная смертность). Изучались путем
вскрытия самки 32-го и 33-го поколений.
Выяснилось, что отбор в противоположных направлениях
приводит к качественно разным ответам. При плюс-отборе
сильно увеличивается скорость овуляции, что не происходит
или. точнее, почти не происходит при минус-отборе.
По-видимому, здесь участвуют разные ттны. В дальнейшем же
резкая разница создается за счет большей смертности эмбрио*
нов (до 34% по сравнению с 11% в контроле) в минус-линн*
.iit>
еще в первый период прикрепления эмбрионов к стопа»
матки (рис. 97).
Фолкоиер выдвигает гипотезу, что гены, определяющие
повышенную эмбриональную смертность, имели в исходной
популяции низкую частоту в силу давления естественного
отбора. Отбор на уменьшение величины помета основывался
на повышении частоты этих редких генов, чего не было в
в плюс-линии. Тогда
становится понятным
однонаправленный ответ на отбор в
виде повышения
эмбриона л ьной смертности. Труднее
объяснить повышение
скорости овуляции, ибо нет
основания полагать, что
естественный отбор
благоприятствовал низкой скорости
овуляции. Автор предполагает,
"что оптимальна какая-то
средняя скорость овуляции.
В минус-линии она должна
была бы уменьшиться, но
возможно, что расщепление
генов, вызывающих
эмбриональную смертность,
маскирует действие генов,
влияющих на скорость
овуляции.
Данные этой работы, хотя она посвящена совсем иному
вопросу, до некоторой степени созвучны приведенным выше
данным о специфических генетических различиях,
определяющих характер ответа на отбор при разных кормовых
условиях. По-видимому, в каждом конкретном случае отбор может
использовать различные группировки генов, влияюадх на
компоненты сложного признака, каким является признак
сразмеры помета».
Поэтому отбор, начатый из разных части одной и то! же
исходной популяции, неизбежно приведет к созданию rpvmu
отличающихся друг от друга, особенно если благоприятные
гены обладают низкой частотой. Тогда и по чисто
статистическим причинам в некоторых отбираемых гртноах они будут
присутствовать, в других нет. Тем более *то будет относиться
к группам, в которых отбор проводите* в противоположна
направлениях.
Длительный ortop ■ проблем проем ■*• orfop*
Обычно всякий отбор проводяУся в течение ряда поколений. №
материала гл. 4 видно, что даже hjmi детерминации ирилам од-
Сведя
Рис. 97. Средине юяиестм
швх (ва 1 влет):
е — яДцсмп Мвшж; 1 — 1
эмбряош Ю-Д1ШПМ lie — :
„<ж марон генов изменение в частоте гена происходит очень
Mriii-нии и для достижения достаточно ощутимого сдвига в
Ч.-Н-71П4» (в сторону уменьшения или увеличения) необходимо
много поколений. Кроме того, на моногибрндной модели
было показано, что при определенных ситуациях отбор может
прекратиться. После достижения определенного соотношения
частот и коэффициентов отбора популяция окажется в рав*
новеспом состоянии. Вес это относится и к случаям отбора
по количественным признакам, определяемым, как правило,
многими генами. Приведенные выше данные, особенно
полученные путем математического моделирования, показывают,
что при отборе по количественным признакам картина очень
усложняется. Однако и здесь процесс отбора имеет ту же
основу - повышение частоты ряда генов и постепенную их
фиксацию, т. е. переход в гомозиготное состояние
определяющих их аллелей. Поэтому сдвиги .при отборе, исходя из
теоретических предпосылок, не могут происходить бесконечно.
Раньше или позже, но надо ожидать, что все благоприятные
для развития данного признака первоначально
расщепляющиеся в популяции гены фиксируются. По мере такой
фиксации должна уменьшаться генетическая варианса и в пределе
дойти до нуля. Останется только средовая варианса. Ответ
же на отбор будет все уменьшаться н уменьшаться и,
наконец, сдвиги при отборе станут равными нулю. Очевидно, что
при достижении такого предела (плато селекции) отбор в
противоположном направлении, т. е. в сторону уменьшения
признака, если проводился плюс-отбор (или наоборот), также
будет неэффективным.
Такова как бы идеализированная картина процессов
отбора. Однако при сравнении фактических результатов с этой
теоретически мыслимой картиной наблюдается ряд
осложнений, на которых следует остановиться. Прежде всего — вопрос
о влиянии отбора на фенотнпнческую вариацию потомства. С
первого взгляда может казаться, что так как для получения
следующею поколения отбирается только часть популяции
с меньшей фенотипнческой и генотнпической вариацией, то н
в следующем поколении должно наблюдаться значительное
уменьшение вариации. Но это произошло бы только в том
случае, если бы были отобраны все гомозиготы АА или аа
и признак зависел бы от одного гена.
В действительности размах фенотипнческой вариации
уменьшится мало. По каждому полимерному гену вновь
произойдет расщепление (возникнет новое соотношение pi8>Mi+
+2pbQ%A%At+qMtAi вместо соотношения p2A\Ai+2pqAtAt+
+<fMHi) Кроме того, сохранится доля изменчивости, визы-
меыа* средой.
Лашем подсчитано, что только при очень высоких ко*
Л»
.ффпииснтах наследуемости в значительной интенсивности
отбора ДОЛЖНО происходить заметное уменьшение феяотюга-
чсской вариации в следующем поколении отбора.
Некоторые из подсчетов Лаша приведены в табл. 51. В расчетах
сравнивалась изменчивость двух соседних поколений отбора.
В действительности отбор проводится в течение ряда
поколений. При этом теоретически следует ожидать постепенную
фиксацию генов, влияющих на развитие количественного
признака, соответствующее уменьшение генетической вариан-
сы и отсюда дальнейшее уменьшение фенотипичеекой
изменчивости.
Таблица 51. Изменчивость потомства (в вдеятах от явшячмостя
родительского поколения) ври развмх юаффицисатах наследуемости
Доля, отбираемая из
родительской популяции
0.5
0.1
Ргзылх нмкучавосп ютоистм
ори М ~ 1
оры V ~ 0.5
63 96
76 J 95
прш fc* » 0J
98
99
Фактические данные многих опытов показывают иное.
В ряде случаев фенотипическая вариация не только не
уменьшается, но увеличивается, что бывает даже тогда, когда,
казалось бы, достигнут предел для отбора- Это видно
непосредственно из рис. 81, 84: кривые распределения становятся более
растянутыми и значения средних квадратнческнх отклонений
увеличиваются. Так как возможна связь между значениями
х и а. то лучше пользоваться коэффициентом вариация.
Но последний также увеличивается, что было показано, в
частности, в опытах по отбору на увеличение веса мышей. В первых
поколениях отбора коэффициент вариации р«6%, в
последующих значение v увеличилось до 12—14%. Такие же факты
наблюдались и у кур при отборе по весу цыплят. При отборе
на уменьшение количества стерноплеврадшм щетниок у
дрозофилы в линии ЛЧ по нашим данным, х уменьшился с Ш
до 14,6. Коэффициент же вариации в I—8-м поколениях
составлял ~8%. а в 13— 16-м повысился до ~16%.
Возможно, что уменьшение генотнпяческой вариация
сопровождается увеличением средовой вариации, как бы «ем»
пененрующей это уменьшение, чтобы сохранить необходимей
уровень фенотипичеекой вариации. Это обстоятельство
можно сопоставить с данными, известными для ряда растив*-
319
тесшт? овбршшягу-
?рашвж гдаюаиптгнасш 9
9 та
та. Свезуег
слабеть, мша,.
мадных потенциальных возможностей отбора. В ряде случ*
ев достигнуты результаты, значительно превысившие лимиты,
указанные*Фолконером для некоторых признаков дрозофил*
м мыши. В этом отношении наиболее разительны результаты
этбора на повышение содержания жира в зернах кукурузы
(см. рис. 80) — достигнуто увеличение процента жира более
чем в 3 раза.
Вопрос о возможном пределе при отборе изучался также
в специальных опытах Робертсона на мышах. В 7 линиях
мышей проводился плюс- и минус-отбор по весу тела, а также
по привесу за период с 3- до 6-недельного возраста на
протяжении 31—52 поколении. Линии былн заложены нз
гетерогенного материала. Результаты оказались неоднородными. В
некоторых линиях был достигнут предел по весу тела через
11—28 поколений (в плюс-линии средние 28—36 г и в минус*
линии 10—14 г). Но в двух плюс-линиях при отборе после
достижения максимума вес тела снизился. Были также
созданы сублинии, в некоторых из ннл был получен большой вес,
но в дальнейшем при изменении направления отбора сдвига
ае было. В тюдлннни же с минус-отбором отбор на повышение
веса все же давал результаты, что указывало на частичное
сохранение аддитивной генетической варнансы, т. е. на не*
полную фиксацию генов, влияющих на вес. В линиях же,
достигших предела, очевидно, такая фиксация произошла.
Проводилось также скрещивание линий, где ранее уже
велся отбор по весу, и в создаваемых после скрещивания
популяциях вновь проводился плюс- и минус-отбор. В результате
был достигнут новый предел, по мнению автора, уже
окончательный. Однако при мннус-отборе результаты были менее
ясными. Во всяком случае очевидно, что скрещиваемые линии
отличались по набору генов, влиявших на вес.
В линиях, в которых отбор не давал результатов, автор
сделал попытку увеличить изменчивость, чтобы вновь
добиться эффективности отбора, двумя способами — локальным
облучением (в области семенников) дозой 600 р и
скрещиванием с мышами неинбредной популяции. В облученных линиях
отбор был по-прежнему неэффективным. Отбор же среди
потомков от аутбредного скрещивания дал возможность
несколько сдвинуть полученный ранее максимум по весу.
Проблеме преодоления потолка при отборе было
посвящено также несколько работ, проведенных на дрозофиле
(Осман, Робертсон и др.). Применялся тот же метод
скрещивания линий, достигших предела при отборе, с другими
линиями, что позволило изменить этот потолок. Робертсон
предположил, что предел при отборе создается в результате тага
что аддитивная генетическая варианса становится близко!
к нулю, однако граница потолка зависит не только сгг инте*
Ж
сивностн отбора, но и от размеров популяции, что нашло
подтверждение и в специальных опытах на дрозофиле Джонса,
Френкхэма и др.
Является ли достижение предела при отборе результатом
перехода в гомозиготное состояние определенных генов?
Положительный ответ был получен в опытах Фразера и Сно-
укрофта по изменению числа окутеллярных щетинок
дрозофилы путем применения метода инвертированных больших
хромосом (половой, II и III), содержавших гены, влиявшие
на количественный признак.
Были точно установлены и проанализированы
генетические системы, влиявшие на увеличение числа щетинок при
отборе. Их переход в гомозиготное состояние приводил к
созданию стабильных линий, далее не изменявшихся при
отборе, однако эти системы были нендентичными для разных
линий.
Обычно о результатах отбора, идущего в одном
направлении, лучше всего судить по регрессии потомков к
родителям. Поэтому изменение средней при длительном отборе
представляется чаще всего прямой, которая только к концу
отбора должна стать более пологой.
Но, как указывает Райт, это верно лишь для сравнительно
небольшого числа поколений. В более широком аспекте
изменения признаков скорее могут быть описаны сигмоидаль-
ной кривой с подъемами и спусками. Высота подъема может
зависеть от самых разнообразных генетических причин.
Отклонения от снгмоидальности могут определяться
различиями в начальных частотах генов, степенью их доминирования,
различием в силе действия. Если имеется один-два основных
гена, сильно влияющих на признак (олигогены), и большое
количество слабых, то на первых этапах отбора будет
наблюдаться быстрое изменение средних значений признака, пока
эти олигогены не будут фиксированы.
Точно так же быстрый прогресс вначале может быть
вызван сочетанием плюс-генов с широким интервалом по
характеру доминирования. Этим же можно объяснить и
вторичный подъем эффективности отбора после периода
некоторого его затухания.
Несмотря на продолжительный и эффективный отбор,
многие линии могут сохранять такой уровень наследственной
изменчивости, который даст возможность изменить
направление отбора в противоположном направлении и достигнуть
при этом достаточно очевидных результатов. Отсюда
вытекает вывод, что признаки, по которым уже достигнут
значительный результат при отборе и которые в то же время
реагируют на отбор в противоположном направлении, находятся
под" контролем таких генетических систем, в которых опта-
наивный эффект при отборе не означает приближения к мак*
chmvmv частоты аллелей определяющих их генов, я основым-
<-н-и на некотором сочетании генов, в том числе и иахЬдя-
щнхея в гетерспнготном состоянии.
Наконец, надо иметь в виду и тькис случаи, когда отбор
постепенно прекращается, несмотря на то чго в
селекционированных линиях сохраняется еще значительная наследствен*
мая изменчивость. Эта несколько парадоксальная ситуация
г01да1ыллеь н результате того, что и процессе отбора
происходило накопление рядя биологически отрицательных явле-
нии снижение жизнеспособности, плодовитости и т. д.
В опытах Лернсра по отбору у белых леггорнов на длин*
шнюьчмь в течение 19 поколений было достигнуто
увеличение средней длины ног (с 9.7 до 11.1 см еще к 10— 12-му по-
каичшю). Кривая вариация признака, как и следовало
ожидать, сдвигалась от поколения к поколению. Появлялись
классы, отсутствовавшие в исходной популяции (в том числе
курица с длиной ног 12.3 см). Но после 12-го поколения
отбор стал неэффективным. Было достигнуто как бы плато для
отбора, однако не потому, что исчерпалась наличная
генетическая изменчивость (А2 длины ног даже увеличился), а в
силу падения воспроизводительной способности. Параллельно
удлинению ног уменьшалось число половозрелых потомков
на одну курицу, поэтому у части материала вообще
пришлось прекратить отбор. В дальнейшем, при разведении без
отбора, длина ног у кур уменьшилась, а плодовитость
увеличилась.
Эти и подобные им факты, полученные на других
объектах, в том числе и на дрозофиле, дали основание Лернеру
сформулировать идею о генетическом гомеостазе. Под ним
следует понимать свойство популяции поддерживать
генетическое равновесие или такое соотношение между генами,
которое наиболее благоприятно для жизни представителей
популяиии в конкретных условиях внешней среды. Всякое
отклонение от него, в том числе в результате отбора, когда
меняются взаимоотношения между генами разных локусов
или соотношение между генотипами в целом и внешней
средой, чаще всего приводит к неблагоприятным последствиям —
понижению жизнеспособности, плодовитости и других жиз*
иенио важных свойств, несмотря на то что конкретные приз*
наки, но которым производится отбор, усиливаются. Насту-
пает своего рода «селекционная депрессия*.
Надо отметить, что среди многочисленных данных есть как
подтверждающие идею о гомеостазе, так и противоречащие
«ft. Кроме того, не все отрицательные последствия,
наблюдаемые при отборе, обязательно объясняются только
нарушением гомсостаза. Так. в Канзао£ком университете (СшА)
J24
проводили отбор а течение 7 поколений а 13 линиях кур на
повышение интенсивности яйцекладки — был получен эяачн-
тсльный сдвиг по Пому признаку. В это же время неуклонно
снижались вес ниц и живой вес кур-несушек в возраст* 32
и 55 недоль, особенно в линиях, где увеличивалась
интенсивность яйцекладки в первые месяцы сезона. Авторы объясняли
это явление отрицательной генетической корреляцией между
селекционируемым признаком, с одной стороны, и живым
весом и несом яиц — с другой. Но можно трактовать его к как
пример нарушения гомеостаза.
В то же время очевидно, что в ряде случаев отбор может
быть эффективным очень долго, несмотря на стремление се*
лекшюнера добиться определенной стабильности создаваемых
им пород животных или сортов растений, и ие приводить ни
к каким вредным результатам. Известно, что у кур леггорнов
за 7 поколений удалось получить стать крайние формы, как
куры с живым весом 1,1 кг и 2.6 кг и со средним весом яиц
45 и 66 г. Подобный же пример успешного отбора на
повышение яйценоскости кур бел всяких нарушений жизненных
функций получен на ферме Калифорнийского университета
(США), об этом же говорят и приведенные выше данные по
отбору на увеличение размеров у мышей,
Поэтому в каждом конкретном случае нет возможности
предвидеть ни тех изменений функции размножения, которые
могут создаться в процессе отбора, ни возможного
предел н о уровне селекционируемого признака. По-видимому,
для признаков с участием в основном аддитивных генов
опасность нарушения гомеостаза невелика. Тогда отбор вполне
результативен без нарушения гомеостаза, пока не
достигается плато селекции, когда исчерпывается имевшаяся
генетическая изменчивость. Но плато селекции в силу различных
генетических механизмов может быть достигнуто н в таких
случаях, когда сохраняется известная степень генетической
изменчивости.
Глава 8
ВЛИЯНИЕ РАЗЛИЧНЫХ СИСТЕМ СПАРИВАНИЯ
НА СТРУКТУРУ ПОПУЛЯЦИИ. ИНБРИДИНГ
И АУТБРИДИНГ
Коэффициент инбридинга. Как >же укалывалось выше.
мидель панмнктнческой популяции предусматривает, что спарн*
ваиия между отдельными особями происходят случайно.
Однако в реальной обстановке в стадах, породах животных, сортах
растений и в природных популяциях, часто разделенных на
большое количество мелких популяций, действительное
положение вещей может быть шачитедьио сложнее. Одним из
таких усложнений является спаривание особен, родственных по
происхождению. Чем более родственны спаривающиеся особи»
тем больше шансов, что они передадут потомству одинаковые
гены и тем выше вероятность появления тмотигот в их
потомстве. Мерой вероятности передачи одинаковых генов особи от
предков служит коэффпписщ инбридинга F Впервые он был
предложен Рантом (1922). Рант обосновал его с помощью paj-
работаниой им методики коэффициентов путей (см. гл iil н
определил как коэффициент корреляции между генетическим
строением неоилодотворенного яйца и генетическим строением
сперматозоида, соединяющихся при образовании тиготы.
Позже Малеко (1948) выра.жл коэффициент инбридинга н
понятиях популициоппой генетики.
(. помощью коэффициента инбридинга можно судии» о том.
в какой степени в данной популяции спаривании отклонялись
от случайных в силу родственности спаривающихся особей и
как меняется при этом соотношение генотипов и фенотипов »
популяции.
Вывод формулы для коэффициента инбридинга с помощью
коэффициентов путей. Допустим, что нам требуется
определить коэффициент инбридинга F для особи Л, имеющий
родословную, изображенную на рис. 99. Иначе говоря, надо
определить коэффициент корреляции между гаметами О и «S. из
которых она обраювалась.
Так как половые клетки егниываст только один сложный
путь, проходящий через общей» предка А, то, согласно теории
Райта, эта корреляция равна
т\
Остается только подыски* т*+^
и а... <Л для tcw^^^^ m4fm»' » •-
Для простоты предположи « X.I£Miwllllf f*
комбинируются no Vw*y ^ r<«™
т. е. при отсутствии какой йы то mt было ииой Е" Друг*'
действия между ними ЫЛо миой *°9*и BiaHMo.
Мгтки. лйюЩи* особь
£ н обриуыше ею
W Пвобыд Л смми с оЛшни
предком Л чер«! мать £ и ото 0.
Строчные бумвм коеффиииеи гн
пути. Штрмк при буж - номер
предшестюидишвгп поколении.
Рассмотрим нижнюю часть рис, 99. Через D н /Г обомаче-
ны генотипы двух родителей, а через Х- генетическое
строение мх потомка, Генотип последнего при принятых выше
условиях полностью и в одинаковой степени определяется генами
двух половых клеток О и J5, от слияния которых образуется
<к обь X Тогда по выведенным РаЛтом формулам детерминя
иия особи X половыми клетками О и S будет выражена
следу ютим образом.
01+04'M*f*<*">•'
JhH+toVx-H;
I! j последнего можно определить значение в:
№
П П + Гшггшгай
МТ
По этой формуле можно найти значение а для любого
поколения, например:
°"= V 2(1+Г") • (3>
Каждая пара патовых клеток, например родителя £,
находятся в точно таком же отношении к его генотипу, как и те две
гголовые клетки, от слияния которых он образовался, т. е.
родителя Е можно рассматривать как синтез собственных
половых клеток, которые образуются им для воспроизводства
следующего покатения. Действительно, если обратиться ж
рис. 100, то станет ясным, что родитель £ образует только
такого качества гаметы, из которых сам образовался *.
Особь Е связана с патовыми клетками, из которых она
подучена, путями а' и а'. С теми же половыми клетками, которые
она сама образует, она связана путями Ь и 6.
Нужно иметь в виду, что при этом не исключена
возможность сцепления между генами и инбредности особей
предшествующего поколения.
Корреляция между £ и гаметой от него равна
коэффициенту пути Ь, так как этот путь является единственным.
Корреляция между £ и яйцеклеткой, из которой она образовалась,
равна o'-l-a'F', так как есть связь между половыми клетками
через £ Но при образовании £ имеются два пути: один путь
между £ и яйцеклеткой, а другой между £ и сперматозоидом.
Л так как эти связи одинаковы, то
b=a'+a'F'=a'( \+Ff)-
°*= Г 2(1+F') *
Следовательно,
Ь=У 2<1+Р) -и+П = У±И+П. (4)
Уравнение (4), так же как и (2), справедливо для любого
звена изучаемой родословной.
Определим теперь, чему равен коэффициент пути между
гаметой, образуемой особью £, и той гаметой, из которой она
образовалась. Величина этого коэффициента равна etb% или Vi-
В самом деле, если выразить с! и Ь через F из формул (2)
и (4) и перемножить их, то
г 2(1+Р) г 2 l T ' 2
* Нддаюш* может бить тоаыш • силу иуташкинюго процессе.
Таким образом, этот коэффициент пути независимо от
степени инбридинга всегда равен 1/2.
В том случае, когда инбридинг отсутствует, каждый из
сомножителей а' и Ь равен корню квадратному из половины,
в чем легко убедиться путем подстановки в формулы значения
F = Q.
Вернемся теперь к родословной, представленной на рис. 99.
Согласно формуле (5), Ьа'=—\ Ъ'а"=—\ Ь"=Уу-
Подставив эти значения в формулу (1), будем иметь
х~ ~2 2*21222 1 2 '
Этот результат получается в том случае, когда предок А не
инбреден и й"= у —.
Если бы предок А был инбреден, то Ъ" по формуле (4) было
бы равно:
*''=Уу(1+Ы
(здесь FA соответствует F" и является коэффициентом
инбридинга предка Л). Тогда
Нетрудно подметить путем вставки дополнительных звеньев
в родословную, что показатель степени 5 образуется по
определенному закону, а именно: он равен л,+л<1+1, где /t,—
число поколений от отца до общего предка (включая и его
самого), а л* — число поколений от матери до А.
Таким образом, общей формулой при инбридинге на одного
общего предка будет
Fx= (у) U+Fa). (6)
Но инбридинг может быть осуществлен на несколько
предков, тогда, применяя правило Райта (корреляция между
гаметами будет равна сумме коэффициентов сложных путей),
будем иметь ^ ^
329
Рассмотрим в качестве примера родословную, изображен-
н\и> на рис. 101. В этом случае гаметы, от слияния которых
образуется особь Л', связаны двумя путями: один путь проходит
П
f-^/i.
<
/J
1 А Ц
/
/ :
п
ч
ч.
Ч^г
\ /
1 х J
Ямг. /0/. Родословная
пробанда А' с общими
предками /4 и В.
Рис. 102. Родословная пробанда X
с отдаленным общим предком А.
через общего предка Ач а другой через В. Следовательно, если
предки А и fl не инбредны, то
Если же предки сами — результат инбридинга, то
Fx=(\)\\+F*)+(\)\\+Fm).
Вывод формулы для коэффициента инбридинга на основе
вероятностей. Возможен и другой способ вывода формулы на
основе не коэффициентов путей, а вероятностей попадания
аллелей от родителей к детям. Разберем его на примере
родословной рис. 102. Родители пробанда X — самка Р и самец <?•
Они имеют общего предка А, отделенного от Р двумя
поколениями, а от Q — одним. Анализ вероятностей попадания
одного и того же аллеля потомку лучше начать с верхней части
родословной, т. е. с самок В и С, имеющих общего отца Д.
Вероятность того, что они получили тот же ген от пред*
ка А, равна '/г. н такова же вероятность, что они полумиле
'Ш
другой аллель. Но если сам предок .4 был уже результатом
инбридинга с коэффициентом FA, то вероятность получения
одинаковых аллелей особями В и С повышается на 4jFA. Таким
образом, общая вероятность становится равной VaO+f.**.
Далее, вероятность того, что В передаст тот же ген от .4 к
D, равна опять V?. от D к Я—'/i и от Р к пробандуХ—Vr
Суммарная вероятность (lk)\ To же относится и к передаче
одинакового аллеля по другой стороне родословной, но здесь
между А и X только два поколения, и отсюда вероятность равна
(7г)2- Окончательная вероятность того, что особь X получит
идентичные аллели от предка А, составит
Показатели степени при величине Vj'• 3 — количество
промежуточных поколений по материнской линии (п\) и 2 —
количество промежуточных поколении по отцовской линии (п>).
Тогда формула для Fx в общем виде может быть
представлена следующим образом:
*,-± (.№, (!Г'
Для практического использования формулы выгоднее первый
(1 \п*+я*
— / :
Формула совершенно аналогична выведенной ранее
формуле (6).
Если ничего неизвестно о родословной предка, можно
предполагать, что в ней не было инбридинга:
^=0; Fx-(±)
Формула для Fx упрощенная.
Если общих предков несколько, коэффициент инбридинга
подсчитывается по каждому из них (с помощью вероятностей),
после чего полученные значения суммируются:
Вероятностным методом получена формула (7).
331
Метод общих предков (coancostry). Наряду с
коэффициентом инбридинга, вычисляемым обычным путем по указанной
нише формуле (7), существует еще одни коэффициент,
идентичный по своему конечному результату коэффициенту
инбридинга потомства /\ но вычисляемый иначе — по количеству
общих предков у двух родителей, т. е, это вероятность того, что
две гаметы, одна из которых получена от одного из родителей,
а другая от другого, содержат идентичные аллели в силу
происхождения. Чтобы отличить его от обычного F, пользуются
символом /.
А ВС D А В А 8
р a Pi
Рис, 103. Две родословные пробандов:
Л'| — без инбридинга; А": — с инбридингом.
Анализ родственных связен через предков позволяет
составить ряд уравнений, с помощью которых довольно легко
вычислить, как изменяется коэффициент инбридинга F в ряду
поколений. Чтобы понять сущность метода, надо разобрать две
родословные, в одной из которых все предки разные, а в
другой — одинаковые (рис. 103). Родители особи Х\—Р и Q, деды
и бабки—А, В, С и D. Происхождение Р полностью
определяется предками А и В, а происхождение Q—С и D. Таким
образом, родство Р и Q представляет собой нечто среднее между
.4, В, С и D. Этот факт может быть выражен в форме
вероятности. Гаметы от Р в половине случаев «есут ген от А и в
половине от В. То же относится и к Q. В таком случае две гаметы:
с дна от Р, а другая от Q — будут нести гены от А и С в !Л
случая, от А и D — в 74, от В и С — в 1/А и от В и D — в У4 случая.
Вероятности того, что в двух гаметах имеются гены А и С
и т. д., соответствуют их происхождению от А и С, что можно
записать как (ас и т. д. В таком случае
Ipq= -г- 1ас+ -~г !ad+ -j- Jbc+ — Ibd,
или
FXx = \PQ~ JL (Uc+UD+fac+fBD)• *8)
4
:m
Это основная формула для всех расчетов предков в ряду
поколений. Из нее могут быть получены формулы для более част-
ных связей, например:
Jpc= -у (fAc+fec);
fpD= — ifAD+fBD)
и т. д.
Обращаясь теперь к правой стороне рнс. 10.3, мы видим, что
у Pi и Яг одинаковые предки А и В, т. е. Р\ и Я2 —полные
сибсы. Их потомок Х2.
Применяя общую формулу (8), можно записать:
/jr2=/piP,= -т- (\aa+Ub+\ab+\ba)\
но так как /вл==/дв, то
4
Величина /ал, как и Jbb, указывает на общих предков особи
как таковой, и, значит, это вероятность, что две гаметы,
полученные от А, будут содержать одинаковые гены. Как было
показано выше, вероятность равна 1/2 (l+FA). Если известны
предки родителей А, то можно установить и значение Яд.
В таком случае
^2=/PlPa=±[i-(l+fA)+l(1+fB)+0] =
Если равны нулю и FA и FB, то получается более простое
выражение
Полученное в данном случае значение Я*„ выраженное
через \PiPn как раз равно значению Fx> вычисляемому по
обычной формуле Райта.
Коэффициент инбридинга и изменение соотношения между
долями гомо- и гетерозигот. Если коэффициент инбридинга
в популяции равен Я, это значит, что вероятность двух аллелей
одного гена быть идентичными при оплодотворении оценивает-
ззз
ся личиной F. а вероятность быть иеидентнчными - ъелшЩ
Н°Й0^ако в популяции, имеющей структуру /AMt+2p?iMi+
^ :^4, >же есТь вероятность соединения идентичных
аллели \л* At она равна р\ для А2 — </2. Вероятность же
соединения неидентнчных аллелей может быть оценена частотой 2р<),
В таком случае, как показано выше, в гл. 4, вероятность
соединения одинаковых аллелей увеличится, а
неодинаковых — уменьшится, т. е. вместо частоты гетерозигот ЛИг,
равной 2м."будет частота 2pq(\-F)=2pq—2pqF.
Величина же 2pqF распределится поровну на генотип ЛИ*
[f+pqF) и генотип A^Az (<?2+р</Л.
Таким образом, при сравнении с панмнксней инбридинг
увеличивает каждую долю гомозигот на pqF н уменьшает дсь
лю гетерозигот на 2pqF. Но общие частоты генов р и q в
отличие от того, что происходит при отборе, ке изменяются.
Долю гетерозигот часто обозначают символом А/.
Значение Н в общем виде может быть выражено как 2pq(\—F)*
При f=0 H = 2pq.
Если принять исходную степень гетерозиготности при паи-
мнксин Н0 за единицу, то можно рассчитать, как будет
уменьшаться гетерозиготность (в далях единицы) при
систематическом инбридинге. Но можно, конечно, вести счет не от //#,
принимаемого за единицу, а от //0= — (при р=^=0,5 в по-
популяции). Хотя цифры будут несколько иными, но характер
изменения гетерозиготности при разных степенях инбридинга
будет тот же.
Однако сначала остановимся на значениях для особей,
получаемых при разных типах спаривания.
Очевидно, что они будут зависеть от степени родственных
связен между родительскими формами. При
самооплодотворении шанс на то. что обе гаметы получают одинаковые
аллели, равен 0,5. Значит, F для потомка в этом случае 0,5. F дли
потомка от спаривания полных сибсов — 0,25 и т. д.
Значения F при спаривании особей, находящихся в разной
степени родства, показаны в табл. 52.
В данном случае речь идет об оценке коэффициентов
инбридинга потомков при однократном спаривании. Значе*
ние F указывает на вероятность для каждой особи данного
типа спаривания получить одинаковые аллели и в то же врем*
на степень инбридинга, характеризующую потомство от
данного спаривания в целом. По отношению к отдельной особи это
лишь вероятность наличия у одной особи одинаковых аллелей
того же гена, но поскольку генов много, то по отношению к су*'
we генов можно говорить о F как о средней мере такой ъёро*
:ш
Таблица У>. Заачетея Mifyw»roi шм»рш«а
потомка при амрааашш особе!, нажоцвдпся ■ р*я
стелем родстъа
Самооплодотворение
Отец \ дочь
Брат \ сестра (FS)
I Пол у братья >; полусестры (HS)
Двоюр. братья X жвоюр. сестры
1
' 1
i
0.50 1
0.25
0.25 1
0.125
0,0625
ятностн для генотипа в целом. По отношению же к популяции
значение F указывает на среднюю степень совпадения
сходных аллелей по многим генам для всех генотипов, входящих в
структуру популяции.
Систематический инбридинг. В предыдущих разделах
рассматривались случаи, когда при уменьшении количества
предков инбридинг происходит автоматически, т. е, спаривание
особей, находящихся в близком родстве, повышает
вероятность соединения гамет с одинаковыми генами.
Но возможно и систематическое спаривание родственных
особей в течение ряда поколений. Тогда будет наблюдаться
постоянное изменение величины коэффициента инбридинга, а
вместе с тем падение гетерознготности и повышение гомозигот-
ностн. Процесс этот будет происходить с разной скоростью в
зависимости от степени родства спариваемых особей.
Существуют различные методы вычисления изменений
коэффициента инбридинга в ряду поколений при применении
систематического инбридинга: на основе коэффициентов
путей (Райт), матричной алгебры (Фишер) и др. Наиболее
простым из них является использование коэффициента / и
уравнений (8), (9), (10), выведенных выше. На рис. 103
представлена родословная особи Jf* полученной путем спаривания сиб-
сов. Допустим, что и предки А и В также были получены
путем спаривания снбсов. т. е. родословная продолжена вверх.
Предпосылки те же: Ua4bb. Fa = F* Если значком /
обозначить поколение, к которому относится X, то
/ab=Fi-i; Faa^Fbb= ~ (l-fft-a).
Подставив эти величины в уравнение (9). подучим уравнение
для любого поколения /: " • »'aoncfW
f,= —(l+f^+2F,M),
335
или
Fi~ ^ (I + 2/Vt+f, »). (11)
Пользуясь этим рекуррентным уравнением, можно вычне*
лить значения F для ряда поколений. Дли нулевого поколения
(!ш*0) ft» О, для первого ft» 0,25 (так как второй и третий
члены уравнения (II), находящиеся в скобках, равны нулю).
Соответственно:
Ft !«0.375; ft-9ш» 0,50; ft^=0,59.
Легко видеть, что изменение F происходит в данном случав
неравномерно. В гл, 4 приведено значение Дг» ■■ т^у "*«
Воспользовавшись значениями F для двух соседних
поколений, можно получить \F для различных промежутков меж*
лу поколениями» например: до спаривания между енбеами н
I м поколением спаривания» между 2-м и 1-м поколениям*
и г. д, В случае спаривания между полными енбеамн
значения \F будут следующими: 0.25, 0,17, 0,20, 0,19 и т. д.
С первого взгляда может показаться, что было бы доста-
точно взять простую разницу между F соседних поколений. На
самом деле это не так.
Уравнение для A F дает более сложную связь между Ft
и /V t Поэтому A F оказываются различными в первых
поколениях инбридинга. Лишь позднее значение A F стабилизирует*
ся окаю величины 0,191.
Рекуррентные уравнения составлены по такому же
принципу для любых систем родственных спариваний, начиная с
самого тесного инбридинга—самоопыления или
самооплодотворения. Они приведены в табл. 53. По этим формулам были
вычислены и коэффициенты инбридинга по поколениям
(табл. 54),
Таблица 53. Реиурремтиме ураавеимя для разных систем иабрадиига
Тип сп*р«*дмм*
1 Сшюоплодотаорепае
Родатма х потомка •
Гкимы* сябсы ч полны* енбеы
Подусмбсы х пмусабсы
*— II.
Урамепк*
1/2 0 + /W
1/8 (1 )■ UF,_{ -\ F,_J
Номер
формум
| j , —|
(13)
(И)
00
• Даа с*яп***иых с иодом генов возрастания инбридинга ияекоа***
136
Таблица 54. Иэмыдеим коэффициентов аибрядяяга по
при рпиых тимк спариваний
Поколении /
0
1
2
3
4
1 5
6
7
8
9
10
П
12
13
14
15
16
17
18
19
20
Эмй.вямя F f ]
С.ИООПЛОДОТ-
RUp«MH<
0
0.500
0.760
0,878
0.938
0.969
0.984
0.992
0,996
0.998
0.999
Полки* tnttit \
ПОЛЯМ* (*бСЫ
0
0.260
0,375
0,500
0.594
0.672
0.734
0.785
0.826
0,859
0.886
0,908
0.996
0.940
0.951
0.961
0,968
0,974
0.979
0.983
0,986
ПолуМкм х оолу.
«■4см
0
0.125
0.219
0,306
0,381
0,449
0,509
0,563
0.611 j
0,654
0.691
0,726
0,755
0,782
0,806
0.827
0.846
0,863
0,878
0.891
0,903
При самооплодотворении уже за 10 поколений значение
Fx практически приближается к единице. При других типах
спариваний возрастание протекает медленнее. В таблице нет
колонки для типа спаривания родителиXпотомки, так как
изменение Fx в этом случае аналогично изменению при
спаривании полных сибсов.
Систематический инбридинг сопровождается столь же
систематическим процессом уменьшения доли гетерознгот и
увеличения доли гомозигот, зависящим от степени родства
спариваемых особей. Проанализировать этот процесс удобнее всего
на модели популяции диплоидных организмов, в которой
проводится самый тесный инбридинг - самооплодотворение <са-
моопыленне у растений).
337
2:1 П. Ф. РоммоимЯ
Гели принять, чго исходная популяция вообще возникла щ
одипй гетерожготной особи А\А2 (очевидно, ее гетерозигот*
ность /Л,- 10()%). то в первом же поколении она даст *U A{A\ +
t ;Л AtAf+Vi А7А2. т. е. процент гетерозиготности будет
ранен 50. Это доля 2pq в общей популяции. Очевидно, что щ
каждом следующем поколении гетерозиготная часть популя*
цин 2pq снова расщепляется на lU A\Ai + 7U A\A2+ XUA2A2, т.е.
к уже имеющимся гомозиготам добавляется по 0,25 доли re*
тероэигот предыдущего поколения. Тем самым доля гетеро*
зигот уменьшается в каждом поколении вдвое: в 1-м
поколении она составляла 7г. во втором — 74. в третьем — 7в» иако*
иен. в л-м поколении — (72)".
«Можно рассчитать возрастание доли каждой категории
гомозигот. Сумма всех генотипов составляет единицу. Ее мож*
2п+i j 2 \
но представить как —^ . Доля гетерозигот -^~ = -t-%
Пели отнять от общей суммы долю гетерозигот и остаток
разделить на 2, получим долю каждой категории гомозигот:
/ 2"" 2 ) .2_
\ Оп+\ Оп + 1 / '
2*+«—2
2"+!-2"~"
2"—1
2п+1
Изменение структуры популяции при самооплодотворении
и процесс уменьшения гетерозиготности представлен в
табл. 55.
Таблица 55. Соотношение генотипов в каждом поколении
при самооплодотворении и падение гетерозиготности
1 Помоле*
i пне
| 1
I 2
3
п
Доля генотипов
Л.*г
1/4
3/8
7/16
2"- 1
2**'
AxAt
2/4
2/8
2/16
1
2я
АгЛг
1/4
3/8
7/16
2"- 1
Доля
гетерозигот п
1/2
1/4
1/8
0/2)"
Подобные расчеты, только более сложные, были проведены
для определения степени падения гетерозиготности при pW
личных системах спариваний (рис. 104). Исходным уровнем
зли
первоначальной гетероаиготности принято 50% (для панмик-
тической популяции при p**q*nQ$). Очевидно, что при
падении гстерозиготности происходит соответствующее увеличение
гомочиготности, т. е. фиксация геиа. Если в популяции при
свободном скрещивании доля гетерозигот выражается
величиной 2pq, то доля гомозигот — \-2pq. При инбридинге она
равна \--2pq\l-F). С увеличением F доля 2pq уменьшается,
а доля гомозигот возрастает, становясь равной 1 при F«l.
Рис. 104. Падение гетерознготности при разных
степенях инбридинга:
/ — самооплодотворение; 1 — полные сябсы х полные сжбеы;
3 — палусибсы х гюлуснбсы; 4 — двоюродные брег х сестре:
5 — палудвоюроднче у палудвоородяые.
Самоопыление приводит почта к 100%-ному исчезновению
гетерозигот, т. е. к практически полному огомозигочиванию
примерно к 10-му поколению инбридинга. При скрещивании
братьев с сестрами (сибсов) оно достигается после 20—25
поколений.
При более слабых степенях инбридинга гетерозиготность
уменьшается значительно медленнее, однако она доходит в
конечном итоге до нуля даже при спаривании двоюродных с
двоюродными. Но спаривание полудвоюродных уже не
приводит к сколько-нибудь значительному уменьшению
гетерознготности (только на 2%), и в дальнейшем ее уровень
стабилизируется.
Дифференциация популяции при инбридинге. Очень важно
ясно представить себе, что происходите исходной популяцией,
когда в ней начинается более или менее тесный инбридинг,
продолжающийся в течение ряда поколений.
Первоначально единая популяция, в которой спаривания
происходили по принципу случайности, распадается на ряд са-
22 • з»
мостоятельных инбредных линий, и каждой из которых сперт
маются родственные особи, например братья и сестры. В
инбредных линиях происходит систематическое падение гетеро-
зиготиости и возрастание гомозиготности но большому коля*
честву генов. Но так как исходные формы, от которых
начались иибредные линии, различны по своей генетической стру*
туре, то в гомозиготное состояние в отдельных линиях пера»
ходят разные гены .Появляются значительные различия меж*
ду линиями по генотипу и фенотипу, что отчетливо наблюдаег*
ся при сравнении различных инбредных групп, созданных н»
единой исходной популяции, у домашних животных и
культурных растений, а также у модельных животных в
экспериментах по изучению результатов инбридинга. Очень показательны
данные, полученные Райтом на морских свинках. Иибредные
линии резко отличались друг от друга по окраске и оттенкам,
по рисункам пегостей, по некоторым морфологическим при*
жакам.
Так как инбридинг приводит к дифференциации исходной
панмиктической популяции на самостоятельные группы, в
каждой из которых возрастает гомозиготность по ряду генов, то
при анализе генетической вариации во всей популяции,
состоящей из инбредных линий, следует отдельно рассматривать:
а) вариацию внутри линий или инбредных групп; б) вариацию
между линиями; в) общую вариацию во всей популяции в
целом.
Если считать, что до или в начале инбридинга общая
генетическая вариаиса была равна ас,2, то вариансы,
характеризующие три указанные вариации, будут иметь следующие
значения:
вариаиса внутри линий (1— F)aG2\
вариаиса между линиями 2Fog2;
общая вариаиса (\+F)o02,
При приближении коэффициента инбридинга к единице, чЮ
наступает, как показано выше, только после длительного
тесного инбридинга, генетическая вариаиса внутри линий должна
стать равной нулю. Каждая линия приближается к полной го*
мозиготиости (чистая линия). Генетическая же вариаиса
между линиями, а также общая вариаиса осей популяции
достигнет аиачеиия 2а<Д т. е. возрастет вдвое по сравнению с гея*
тической вариаисой исходной популяции до начала инбридинга'
Таким образом, роль инбридинга двоякая: в инбредных лЯ'
ниях он приводит к уменьшению генетической вариации (в пр*
деле до иуля), во всей же популяции ~ к возрастанию
генетической вариации за счет дифференциации между линиями.
Анализ результатов инбридинга позволяет подойти к 6&
лее общему его определению, а именно: инбридинг — это
такая система спариваний, при которой особи более родствен**
340
друг другу по сравнению с теин, которые спариваются в
популяции случайно.
Очевидно, что и в панмиктической популяции могут быть
случаи спариваний между родственниками, но они не
нарушают общей картины панмиксии и не приводят к повышению
юмозиготности популяции в целом. Иное дело, есля по каким-
7о причинам естественным (например, изоляция из популяции
численно малых групп, о чем будет говориться ниже) или
искусственным (например, при сознательном проведении
родственных спариваний у сельскохозяйственных животных)
происходит более частое спаривание родственных особей. Если
инбридингом называть спаривание особей, находящихся по
отношению друг к другу в большей степени родства, нежели
среднее родство особей в популяции, то спаривание особей
менее родственных следует называть аутбридиигом •, Но если
критерием инбридинга считать его результаты в виде с ниже*
ния гетерозиготности, то из изложенного выше видно, что
некоторые спаривания родственных особей, даже проводимые
систематически, уже нельзя считать инбридингом. Из рис. 104
видно, что если спаривание организмов, находящихся в
двоюродной степени родства, приводит к полной гомозиготиости,
правда, после многих поколений (их количество не может быть
представлено на оси абсцисс графика), то при спаривании
полудвоюродных снижения гетерозиготности ниже уровня
48,1% не происходит, а соответствующее повышение
гомозиготности достигает лишь 51,9%.
Связь между инбридингом и размерами популяции, В
разобранных выше родословных с инбридингом (рис. 99—102)
проба ид X был результатом спаривания особей, связанных
общим предком А или несколькими общими предками А, В
и т. д. Но возможны и такие случаи, когда общих предков
установить нет возможности, но они существуют в силу
ограниченности размеров популяции.
Приведенная выше формула Д f= ™ относилась к
идеализированной популяции. В действительности же надо иметь
в виду только ту часть популяции, которая в каждом данном
поколении даст потомство. Обозначим ее через N* Тогда
2N,
В ряде случаев участвует в размножении рваное количество
самок (N,) и самцов (Nm).
• Слариммк* иеродстмивых осаб* *«сто шшшшллт сярошммют.
341
1\нмом бы.и» выведена глгдующли формули, iд^ учитц^й
,-цм М>ЛИЧеС1ИО ГЛМОК И ГНМЦОН. Щ'
I I
•l/V, I/V,,, ' <1Л/, '
И Ю1ДН
Л,;-- sNn, ' HN, П*»
\«>гя -на формула иг очсш, точили, по щ нее следует, ц^
чгм меньше /V, или Г00 1МГТГ1ИГ1И1О Nm или N'/, тем больше при*
блика коэффициента инбридинги. Особенное значение им**?
меньший но численности иол, Гели, нлирнмер, и стаде много
гпмок, но и размножении принимает участие только 2 с«мщ,
ю Л/ равна -|/|#,| или (i% увеличения коэффициенте иибр*
дн hi л та одно поколение.
Ноли два ноли не рампы по количеству, ио родительски*
особи равномерно распределены но отдельным семьим (как по
иолу, так и количественно), может быть использована такж*
формула
ЛГ" :,•«... + а», (,в|
Здесь .Vm и Л/, — действительные числа родителей, прнче*
предусматривается, что самок больше, чем самцов.
Установление эффективного количества самок и снмиов и*
всегда возможно (в условиях природы или при массовом pi*
ьедеиии, например, линий дрозогЬилы). Приходится в >Т1П
случаях определять Ne или Nf и Nm приблизительно, на осио*
ве косвенных данных.
Инбредная депрессия. Многочисленные данные,
накопленные в практике животноводства и растениеводства, а также
полученные в прямых опытах по изучению инбридинга, пока*
зали, что во многих случаях в инбредных линиях наблюдаете*
снижение уровня развития ряда признаков, называемое
инбредной депрессией, Однако степень депрессии очень раз*
лична. Сильнее всего она проявляется на признаках, связа*'
ных с размножением или с важными физиологическими осо*
бенностями организма, т. е. определяющих общую приспособ'
лепиость организма (fitness), па признаках же, более или ме*
нее нейтральных по отношению к приспособленности (длине
шерсти у овец, размеры крыльев дрозофилы, вес тела и др-Ь
проявляется слабо или лаже ее пет вовсе. Известно, дале*» "*°
существуют и процветают многие виды растений, нормаль^
размножающиеся путем самоопыления, хотя, с другой СТОр*>*
мы, известно, что в чистых, т. е. полученных путем саиоопЫЛ**
342
мни, линия* 1>№1*Ш'ЩЩ)тн1хтшнм*ь Шлшнмсш
ИИЙр^ЛИИИ Д^НреСГИЯ, В ИНбрЫ/Цщх ЛИНИИ* /l*fr>p*TWHW* **-
НОТНЫХ, ННИ|>ИМ*р МЫМ**Й, 1фЦе, flOOJKWMMI CHHMtHH* ряд* Ш>
каоп*леЙ н парные периоды ннЛрияиши, но ы млин*Ьш*ы
линии гтиби/мнируются ид некотором, бодмг *И4*ом, «** )
иучбркдных жинотиых, урояне гмодояитостн, дя»и#до*>//*г
пи, и их можно бе* труде рязяодить II яиетот* * т*ч*ни* и*/
1их поколений, Тиионы, я честности, иснолмудмм* » медицин*
И 6ИОЛОГИИ ННбреДНЫ* ЛИНИИ ММШДЙ И ДРУГИХ Л*бор»1<>р*М1
животных.
Ношимет вопрос о том, миоям ж* геипичееяяе прннмт
испникионеиии инбредиой ддиреесии?
Для я ого надо снояя обратится н шн* и+ншни* *>**-
ношения генотипов д результате инбрнлмш, нмюртюго *&
гффиииеитом Л
Генотипы Частоты Генотииииесии* Произведение идето?
4нвч^ияя ид их тмтпичлсш
'4НШЧ*НИН
АхАг tyq—ipqP d 'lpqd*~lpqdf
—u{p-q)+2dpijb-Fh
Сумма проюишниЬ средних частот геиоя ш поиудяиян
(онн обозначены р и q, а не р и q) ид их г?нотиннчеою* ****»
иия даст среднюю лолуляшюниую яри инбридинге, Можно ы
обозначить через М?:
M,**QCp-q)+24pq(l-F). (17)
Поиуляииониая же средняя, пан укаэаио ш*ш* (гл. 5, фор»
мула 9), раина
Мгш*аСр-Я)+им
Теперь можно определить, миом рмииид между попул*
иионной средней после пьбршймшгш я до яибридиигя;
M,-MPm*utf-q)+2dM~2<ffir-a(p~h~Mpi—
rm-TdpqF.
мл
Так как величины р, q и F являются вполне значимыми, то,
очевидно, разница между Mf и Мр будет только в том случае!
ко:да J^O, т. е. когда гомозигота но данному локусу
отличается от среднего значения двух гомозигот или, если говорить
о многих генах, среднее значение гетерознгот отличается от
среднего двух гомозигот. Таким образом, возможное
изменение популяционной средней признака при инбридинге зависит
от наличия доминирования (любого), причем знак этого
изменения «минус», т. е. оно идет в сторону рецессива АтАг-
Величина изменения зависит от значений р и q. Она наибольшая
при максимальном значении произведения pqy которое будет
наблюдаться при p = q = 0,5.
Этот вывод можно перенести на случай совместного
действия многих генов на признак. При условии аддитивности
сложения эффектов генов
MF—MP=—2F2dpq. (18)
Для снижения средней в результате инбридинга
необходимо, чтобы гены, усиливающие признак, были преимущественно
доминантны, аллели же их,
ослабляющие признаки,
рецессивны. Как известно, это
условие реально
существует, так. как рецессивные
гены чаще всего действуют в
сторону уменьшения
(ослабления) признака или — в
более обобщенной
формулировке — в сторону снижения
приспособленности. Тогда
становится понятной ий-
бредная депрессия как
следствие сдзига популяционной
средней при инбридинге в
сторону рецессивных
аллелей. Этот сдвиг сильнее при
средних частотах генов, что
чаще всего и наблюдается в
В дальнейшем, когда разни-
О 20 40 60 8QF,%
Рис. 105. Уменьшение количества
мышат в помете при разных степенях
инбридинга (по Фолконеру).
первых поколениях инбридинга
ца между р и q увеличивается, сдвиг становится слабее,
степень инбредной депрессии уменьшается. Это и объясняет
указанный выше факт, что ннбредные линии можно длительно
разводить.
Если действие отдельных генов комбинируется по
аддитивному закону, то изменение средней при инбридинге должно
быть прямо пропорционально изменению коэффициента иибрп-
341
дпнга /-. Если выровнять изменение F, то такой же прямой ли-
ннсп должна выражаться и степень инбредной депрессии
(рис. 105). Когда же изменение F отклоняется от прямой в
сторону большей пологости, что имеет место при высоких
значениях F, линия депрессии должна изменяться таким же
образом.
Эту общую картину могут осложнять многие
дополнительные факторы. Среди них следует назвать несовпадение между
теоретическими и фактическими значениями F в ряду
поколений. Это несовпадение вызывается естественным отбором в
пользу гетерозигот против гомозигот и приводит к замедлению
скорости гомозиготности.
Расчеты показали, что, например, при коэффициенте
отбора против гомозигот, равном 0,2, 4F (в %) при
самооплодотворении будет не 50, а только 44, при скрещивании полны*
сибсов не 19, а 15%.
Поэтому теоретическим кривым падения гетерозиготности,
изображенным на рис. 104, нельзя придавать абсолютного
значения.
В таком случае можно ожидать и несоответствия
фактически определяемых коэффициентов наследуемости теоретически
ожидаемым при принятом теоретически уменьшении степени
гетерозиготности.
Дело в том, что уменьшение коэффициента наследуемости
при инбридинге выражается формулой
ht= 1-VF, ' °9)
Здесь li02 — коэффициент наследуемости признака,
определенный для исходной популяции до отбора; Ft и Л*2 —
коэффициенты инбридинга и наследуемости в какой-то момент
времени в период проведения инбридинга. При F/ = l hr
становится равным нулю. Однако ряд обстоятельств и прежде всего
естественный отбор против гетерозигот могут привести к тому,
что наследуемость уменьшается слабее, чем следовало
ожидать при данном значении F. Это подтверждают и
экспериментальные данные. Так, в опытах Тантави и Рива, проведенных
на дрозофиле, показано, что Л2 выживаемости, плодовитости,
а также средней длины крыльев уменьшался очень мало
вплоть до достижения F«40—50%. Этот эффект авторы
объяснили смягчением влияния инбридинга отбором в пользу
гетерозигот. При увеличении же коэффициента инбридинга
наследуемость уменьшалась очень быстро. Так, Л2 для средней
длины крыльев при F«50—60% составлял 0,27—0,33, а при
F^80—90% только 0Д2—0.17.
Инбридинг и фенотнпическая изменчивость. Приведенные
выше данные указывали на то, что генетический эффект нн-
345
брлдннга сводится: а) к повышению гомознготиости внутри
каждой ипбредной выделенной из популяции группы или
линии; б) к увеличению генетической изменчивости между
отдельными иибредными линиями по сравнению с
изменчивостью исходной популяции. Во втором случае, очевидно,
должна увеличиваться фенотнпическая вариаиса, если
рассматривать сумму инбредных линий как единое целое. В
первом же случае в силу уменьшения доли генетической вариации
в каждой линии и при сохранении вариации, вызываемой
средой, на одном уровне можно было бы ожидать уменьшения и
фепотнпнческой вариации по каждому признаку внутри
н;«бреднон линии.
Однако данные многих авторов (К. М. Лютиков, Н. Н.
Колесник и др.), полученные на сельскохозяйственных
животных, указывали на обратное, а именно на увеличение фенотн-
пической вариации у инбредных животных по сравнению с
а\тбредными.
Более детальные и длительные (по количеству поколений)
опыты были проведены на курах, мышах и дрозофиле. Так,
X. Ф. Кушнер обнаружил повышенную фенотипическую
изменчивость но живому весу и яйценоскости у кур породы белый
леггорн, подвергавшихся длительному инбридингу. Гнльмор
показал, что после 24 поколений инбридинга (F^99%)
фенотнпическая изменчивость по яйценоскости, весу тела была
выше, нежели до инбридинга. Сходные данные были получены
на мышах и крысах. Б часто упоминаемых опытах Кинга по
длительному инбридингу с отбором у крыс коэффициент
изменчивости веса тела увеличился с 3% в первых 7—15
поколениях до 13% к 20—25 поколениям.
Очень тщательные работы были проведены на дрозофиле.
М. Л. Карп изучал яйценоскость в экспериментально
созданных линиях, гомозиготных по большим участкам III
хромосомы, включавшим 8 рецессивных генов-маркеров. В этих
линиях яйценоскость была самой низкой по сравнению с
другими линиями, по коэффициент вариации по этому признаку был
вдвое выше, чем коэффициент вариации гетерозигот.
Подобные данные были получены и в других опытах на дрозофиле
(Гоуэн, Тантавн, Робертсои и Рив). В работе Расмуссон
тесный инбридинг проводился в течение 70 поколений.
Изменчивость возрастала только в пределах первых 10 поколений, а
затем устанавливалась на одном уровне.
Но интересно, что у растений повышение фепотипической
изменчивости при инбридинге наблюдается не у всех видов,
а только у тех, которые в естественных условиях размножают*
си аутбредно, например у ржи, кукурузы, красного клевера-
И самоолмленнмх линиях таких видов вариаиса ряда приэнг
кои, например высоты растений, урожайиости, выше в аут-
.4 И,
бредных, обычно высокогетерозиготных их популяциях. У
видов же растений-самоопылителей такой разницы в
изменчивости нет, т. е. гомозиготные линии изменчивы примерно в
такой же степени, как и гибриды между ними.
Животные, размножающиеся обычным половым путем, в
этом отношении аналогичны перекрестноопыляющимся
растениям.
Из сопоставления этих данных можно прийти к выводу,
что уменьшение наследственной изменчивости в линиях при
инбридинге, снижающее рамки приспособительных
возможностей, компенсируется увеличением средовон компоненты вари-
ансы, вызывающей общее увеличение фенотипической
изменчивости. Инбредиые линии обладают как бы повышенной
чувствительностью к условиям внешней среды, что является
выражением гомеостатической
реакции на инбредную
депрессию, как известно, особенно
сказывающуюся на признаках
приспособленности
(плодовитости, жизнеспособности).
Увеличение фенотипической
изменчивости в инбредных
линиях существенно не только
при практическом
использовании гетерозиса, но важно и
теоретически в связи с
проблемами гомеостаза (по Лернеру)
и канализации развития (по
Уодингтону).
Коэффициент родства R.
С коэффициентом инбридинга
тесно связан коэффициент
родства /?, также предложенный
Рантом. Он характеризует
степень ожидаемой корреляции
между двумя особями,
имеющими общих предков. Нужно
отмстить, что эта корреляция отражает связь между
признаками, которые определяются генотипом. При этом условии в
случайно скрещивающейся популяции корреляция между
родителями и потомками или между сибсами равна 0,5.
Интерпретация этого коэффициента, как н коэффициента
инбридинга, зависит от выбора исходного материала, т. е. той группы.
к которой принадлежали предки. Очень часто фактический
коэффициент будет сильно отличаться от теоретически
ожидаемого, так как на его величину влияет не только
доминирование, но и окружающие условия.
347
Рис. 106. Родство X н .V через
общего предка .4.
В простейших случаях степень родства может быть легко
определена, так сказать, на глаз. Очевидно, что отец и сын (to.
лее родственны друг другу, чем дед и внук. Спаривание
братьев и сестер — это спаривание более близких
родственников, нежели спаривание двоюродных братьев и сестер и т. д.
Но, очевидно, необходимо более точное определение как
того, что надо понимать под родственным спариванием, такн
методов оценки степени родства между особями. Для первой
цели служат разобранные выше коэффициент инбридинга и
коэффициент общих предков. Для второй же существует
несколько критериев. Один из них и получил название
коэффициента родства.
Вывести формулу для него можно следующим образом.
Предположим, что мы желаем определить коэффициент
родства между особями X и Y в родословной, изображенной
на рис. 106, воспользовавшись тем же методом коэффициентов
путей Ранта.
Очевидно, коэффициент корреляции между X и Y будет
равен коэффициенту сложного пути, проходящего через
общего предка Л, т. е.
R„=a.b-a'.b'.a''.b''.bi''-ai"-bt'.ai'-al-bl.
Как мы уже видели выше, 6-а'=6,-а/,=а1'/.6/'с=:
= a1'.ft1'=—; остается определить величины a, ai$ b", b".
Если особи X и Y не инбредны, т. е. FY=Fx=0, то ai=a= -r-
Если же они инбредны, то а= у oi\4-F ) И ai= r 2(14-/7
То же самое нужно сказать и о предке А: если он не инбреД*й»
то b"=bi"=y —; если же он инбреден, то b/f=bi,s^
-у 4-!■+'->•
Подставляя эти величины в предыдущее уравнение, в сЯЧ'
чае отсутствия инбредности А, X и Y будем иметь
1ЖГ
г 2 ' 2 " 2 Г 2 Г 2 *
Х±
■hVr-Ш'
346
При инбредности А, X и Y получаем
Г 2 2 2 Г 2(t+fx, ~
= __П/2)1£ГТ5__
Нетрудно подметить, что показатель степени для — обра-
зуется как сумма пх+пт, где пх- число поколений от X до
общего предка (включая и его самого), а пт- число
поколений от У до Л.
Если бы X и У имели несколько общих предков, то их
соединял бы более чем один путь и корреляция между ними была
бы равна сумме коэффициентов сложных путей. Седова-
тельно, общей формулой является
2[(|Г'о+ы]
Яхг= ======= . (20а)
1i\+Fx)[l+Fr)
Если все F равны 0, то
*"=2(у)ПХ+ПГ- (206)
Если же равны нулю только F* и Fг, то
*«= 2[(уГх+Яг(,+^)]- <**)
Коэффициент родства может высчитываться не только
между X и У, но и между любыми звеньями родословной.
Например, коэффициент родства между д н А
348
Таким образом, коэффициент родстпп R измеряет стеноп
генетического сходства между особями. По нему можно судцц
о том, насколько велико генетическое сходство между даииу.
ми родственными особями, выше оно или ниже, нежели сред.
пни показатель родства между членами свободно скрещиаа-
ющейся популяции.
Между коэффициентом инбридинга и коэффициентом род.
ства есть связь — от одного можно перейти к другому.
Коэффициент родства между родителем и потомком {или
между матерью и дочерью) равен 0,5 (при некоторых
ограничивающих условиях — свободном скрещивании в популяции
и др.). Для установления родства между более отдаленными
друг от друга членами родословной надо 0,5 возвести в степень
л, равную числу поколений между ними. Тогда R между про-
бандом и дедом составит 0,52 = 0,25, пробандом и прадедом —
0,5* = 0,125 и т. д.
Выше разобран случай, когда связь между пробандами X
и Y осуществляется через общего предка А (см. рис. 106).
Тогда при отсутствии инбридинга
*»-2(|р"'
согласно формуле (206).
В более частных случаях, например, для родства между
полными сибсами, а также при отсутствии илбредности
родителей коэффициент родства
#а-у = 2.0,52=0,5.
В данном случае коэффициент 2 введен потому, что полные
сибсы связаны друг с другое двумя цепями — через общего
отца и общую мать (рис. 107).
Для полусибсов же, которые связаны только через одного
родителя, /?д'г=0,52=0,25.
С учетом инбридинга общая формула для Rxy принимает
более сложный вид.
Так, согласно формуле (20а):
YoWxTO'+fV)
согласно формуле (20в):
«»-2[ф""'(|+*.)].
Если сравнить эту формулу с формулой (7) для «***«.
циента инбридинга F, то очевидно их очень большое сходство
Разница только в том, что степень для 0,5 в случае F больше
на елимицу. В таком случае коэффициент инбридинга F для
^Шь
<^ы
-J ...
Рис. 107. Родство между сиб-
сами X и У через общих отца
А и мать В.
Рис 108. Пошшеиве гяншеоютс
родства потомков с родоввшим-
кои путем агрешимлм от с его
дочертим.
какого-то нулевого поколения Fo равен паювине коэффициента
родства Ялг, т. е.
Fo=0,5Rxr=09S%[(jYX**r (\+FAl].
Существенна статистическая характеристика природы этих
двух коэффициентов.
Коэффициент инбридинга выражает степень корреляций
между генетической структурой гамет, в результате которых
получается данная особь, а коэффициент родства — степень
корреляции между генетической структурой зигот. Иначе
говоря, /? выражает долю общих генов, полученных от предков.
Так как любая особь получает от каждого родителя по 50%
генов, то ее родство с родителем выражается величиной 0.5.
Казалось бы, она является максимальной для коэффициента
родства.
Однако при проведении соответствующих спариваний в
верхних частях родословной с применением инбридинга доля
общих генов может быть выше ОД На рис. 108 показана
родословная, в которой самец А спаривается со своими
дочерями С. Степень генетического родства D с А — 0J5. так как 50%
генов получены особями D непосредственно от А. а 25% — от
того же А через его дочерей С (0,5-0,5»0,25). Известно, что
при выведении асканнйской породы саиией М. Ф. Иванов
спаривал хряка Аскания-I с его дочерями, внучками и
правнучками. Знаменитый пример высокого генетического сходства с
36!
шдалгниыми предками -потомки знаменитого быка шорт-
горнской породы Фаворита, жившего в конце 18 века. Было
установлено, что современные шортгорпы Англии имеют
генетическое родство с Фаворитом в среднем 0,55, т. е. большее,
чем родство детей с их отцом.
Очевидно, это стало возможным только потому, что
практиковались различные родственные спаривания в течение
ряда поколений при разведении шортгорнов. Процесс
поддержания генетического сходства с Фаворитом был изучен за
период свыше 100 лет. Оказалось, что средний коэффициент ий-
бридпига был: в 1810 г.—0,17; в 1850 — 0,18; в 1900 — 0,23 и
в 1920 г.—0,26. В эти же годы генетическое сходство с
Фаворитом было: 0,44, 0,50, 0,52, 0,55.
II коэффициент родства R и коэффициент инбридинга/
относят к отдельным особям. Но надо постоянно помнить об
их статистической природе. Так как они являются
коэффициентами корреляции, то относятся к совокупностям. Для
каждой же отдельной особи это только средний шанс, который
надо оценивать как вероятность.
Скрещивания и их разнообразие. Инбридингу как
спариванию родственных особей могут быть противопоставлены
спаривания неродственных особей — аутбредные. По указанному
выше определению, аутбредными следует называть
спаривания особей, менее родственных, нежели средняя степень
родства особей в популяции. Такие спаривания часто называют
одним общим термином «скрещивание». Но под ним
скрывается большое разнообразие, что отразилось в различных
классификациях типов скрещивания. Так, в животноводстве
говорят о межпородных и внутрипородных скрещиваниях,
в растениеводстве — о межсортовых и внутрисортовых. В
одинаковой степени по отношению к животным и растениям раз-
лнчают внутривидовые и межвидовые скрещивания и даже
межродовые.
Рассмотрение всех возможных типов скрещиваний не
входит в нашу задачу. Мы ограничимся лишь теми из них, данные
по которым могут быть выражены в статистических понятиях
или к анализу которых могут быть применены
соответствующие статистические приемы.
Статистическая природа поглотительного скрещивания.
В практике разведения животных для улучшения местного
скота широко применяется поглотительное (или
преобразовательное) скрещивание. Для проведения такого скрещивании
обычно ввозят производителей другой породы того же виде»
но лучшей по своим хозяйственным качествам. Животных пер*
вого поколения, полученных от скрещивания женских особен
местной породы с производителем породы-улучшателя, вновь
покрывают производителем породы-улучшателя. Скр*щя*г
3«tf
пня проводят до тех пор, пока полученные помеси я-го
поколения не будут удовлетворять хозяйственным требованиям: по
удоям и количеству молочного жира в молочном скотоводстве,
по качеству шерсти и ее настригам в овцеводстве и т. д.
Поглотительное скрещивание соответствует тому
процессу, который в гл. 4 был назван иммиграцией. Однако там речь
шла об изменении частоты определенного гена в популяции
при внесении в нее группы особей из популяции с другой
частотой гена. Здесь же рассматривается измененное соотноше-
30
25
X 20
^ 15
10
5
г
-
/у
/
/
—JO Ч 60 70 ~Ю 90 W
Гены породы А,'/.
Рис. 109. Изменение процента генов породы А
при поглотительном скрещивании в поколениях /^,
f»i' V V
ние всей суммы генов местной породы и породы-улучшателя.
В животноводстве принято говорить о долях крови: особей
первого поколения называют полукровками, второго — 3/4
крови, третьего — 7/в и т. д.
На языке статистической генетики это означает, что
животные первого поколения имеют 50% генов породы-улучшателя
А, животные второго или, правильнее, первого обратного Fь, —
75%, животные F&, —87,5%, Z7^ —93,8%, ^6* — 96,9% генов
породы А (рис. 109). Особи F{ чаще всего, согласно
изложенным в гл. 5 закономерностям наследования количественных
признаков, будут промежуточными по развитию признака (или
признаков) между исходными породами, кроме отдельных
случаев гетерозиса, о котором будет сказано ниже. Но если даже
гетерозис и проявится в f|, то в следующих поколениях его
эффект будет слабеть. Вариация особей F% чаще будет
небольшой, но предсказать ее заранее трудно. Поэтому на рис. 109
кривая распределения особей F% дана пунктиром. Во втором
поколении в силу процесса расщепления вариация должна
увеличиться и в среднем особи этого поколения будут иметь 75%
23 П Ф, Рокшцсмй
383
ronon породы-улучшателя; в следующих поколениях вариация
Г)\дс1 уменьшаться. Таким образом, процесс перестройки
генотипа помесей в сторону приближения его к среднему генотипу
породы-улучшателя Л происходит автоматически в силу ряда
обратных скрещиваний, н его можно оценивать статистически.
С увеличением количества поколений вероятность сохранения
генов местной породы становится все меньше. Процесс
поглощения может быть соединен с отбором, направленным или на
ускорение процесса замены генов местной породы генами
породы А, или, наоборот, на сохранение желательных генов
местной породы.
Частным случаем поглотительного скрещивания является
такое, с помощью которого стремятся ввести в местную
породу или один ген, определяющий какой-то качественный
признак (например, комолость), или группу генов, влияющих на
определенный количественный признак, например,
жирномолочность. В этом случае никак нельзя обойтись без отбора. Но
ввести в породу один или несколько генов, не изменяя при
этом остальную часть генотипа, очевидно, очень трудно.
Интрогрессивная гибридизация. Если существуют
популяции одного и того же вида, значительно отличающиеся друг
от друга по многим признакам и даже относящиеся к разным
подвидам, расам, экотипам, но способные давать при
скрещивании плодовитое потомство, то между ними может
происходить обмен генами. Такой процесс обмена генами был назван
Андерсоном интрогрессивной гибридизацией. Он был впервые
изучен на кукурузе. Оказалось, что в так называемом
кукурузном поясе США происходит интрогрессивная гибридизация
между кремнистой кукурузой северных районов и зубовидной
кукурузой Мексики. Существующие сорта кукурузы возникли
от скрещивания сортов белой зубовидной мексиканского
происхождения с сортами северного кремнистого типа, а с
середины 19 века стали проводить преднамеренные скрещивания
между этими сортами, приведшие к новым сочетаниям
признаков. Андерсон доказал наличие интрогрессивной
гибридизации также на видах табака (Nicotiana), на ирисе (Iris) и
пришел к выводу, что она представляет собой важный
механизм не только образования новых сортов и рас, но и
эволюции вообще. В дальнейшем ряд проблем взаимной передачи
генов изучался на популяциях турнепса, редиса, райграса, а
также на хлопчатнике. Выяснилось, что степень
интрогрессивной гибридизации зависит от степени пространственной
изоляции популяций, т. е. от расстояния между ними, плотности
популяции и ряда других причин, но для разных видов роль
этих факторов неодинакова.
Интрогрессивная гибридизация, по-видимому, происходит
у некоторых видов лесных деревьев, а также у животных.
эсл
Скрещивания для использования гетерозис* Общеиз
вестно явление гетерозиса, наблюдаемого в первом поколыши
скрещивания в виде более сильного развития количественного
признака (или признаков) по сравнению с таковым у
исходных форм растений или животных. Гетерозис, или, как ранее
называли, гибридная сила, может проявляться при самых
различных скрещиваниях, в том числе межвидовых
(классический пример — мул; поэтому впервые полученные у растении
гетерозисные формы называли растительными мулами). Ваш*
Цшп
№
т
135
1,Ю
125
120
S
\
\
Ч/
. чх
F,
Рис. ПО. Положительный (а) и
отрицательный (б) гетерозис у томатов.
Точки — средние Ри Р2, F\, Ft.
Горизонтальная линия — средняя между
Pi и Р2 (по Пауэрсу).
Рис. Ш. Сдвиг гетерозиса оа одно
поколение npg материнском
эффекте (шкала на оси ординат
условная):
/ — аффект гетерозиса; 2 — umpimciiA
эффект; 3 — суммарное двйстпе
гетерозиса н материнского эффект*.
вотноводстве, особенно в свиноводстве в птицеводстве,
широко применяется межпородное промышленное скрещивание.
Наибольшее практическое значение имеет гетерозис,
полученный при скрещивании гибридных линий кукурузы. Изучение
гетерозиса у кукурузы помогло сформулировать ряд гипотез
гетерозиса *.
Но в диагностике наличия гетерозиса встречаются
известные трудности и противоречия. Иногда считают гетерозисом
те случаи, когда F} превышает среднюю между родительскими
формами. Но правильнее считать настоящим гетерозисом
только те случаи, когда средняя F{ no данному признаку
выходит за границы средней родительской формы, при этом
возможны два варианта: Л>Рь Р2—положительный гетерозис;
Fi<Pi, Р2— отрицательный гетерозис (рис. ПО). Общее
правило для гетерозиса — его ослабление в Ft н в последующих
поколениях (исключение составляют случаи материнского
* Разбор их» а также вопросов практического использованы* гетерозиса
не входит в задачу книги. Об этом достаточно шстервывающе говорите* s
Ряде монографий (см. список литературы).
365
23*
эффекта, который может наблюдаться у млекопитающих и
передаваться от самок F\ к особям F2 и от F2 к Fz). Поэтому
хотя в F2 эффект собственно гетерозиса снижается, к нему
прибавляется материнский эффект F\. На рис. 111 линией /
показан обычный результат гетерозиса, линией 2—материнский
эффект — передача матерями F] гетерозисного эффекта
особям F2 и матерями F2— особям F3. Величина материнского
эффекта в следующем поколении принята равной гетерозисно-
му эффект)' в предыдущем поколении, но она может быть и
меньшей. Линия 3 дает суммарный результат— сдвиг
эффекта гетерозиса на одно поколение. Максимум гетерозисного
эффекта не в F\, а в F2.
Статистнко-генетические основы гетерозиса. Гетерозис
создается в результате скрещивания особей, принадлежащих к
группам, не родственным друг другу. Это могут быть инбред-
ные линии, породы и даже виды. Для анализа удобнее
рассматривать скрещивание представителей двух популяций,
различающихся такими обычными генетическими параметрами,
как частота генов.
Допустим, что исходные родительские популяции, которые
разводились самостоятельно, как панмиктические,
характеризуются частотами генов: первая — р и q, вторая — р\ и q\.
Различия между ними примем за у=р—Р)=Ч—?»♦ В таком
случае частоты рх и qx можно выразить с помощью у: р—у
nq+y.
Геиотипические значения обозначаем, как и ранее, а,
d.— а. В этом случае учитываются компоненты аддитивный и
доминирования, но не учитывается взаимодействие.
Задача заключается в том, чтобы выразить значения F\
и F2r в которых наблюдается гетерозис, с помощью
отклонения от средних родительских поколений (или от средних
между двумя родителями), которое наглядно представлено на
рис 110, и рассмотреть, какие генетические параметры
ответствен ни *а это отклонение,
В гл. 5 была вывелена формула для популяционной средней:
MPt~a(p—q)+2dpq.
Для второй родительской популяции с частотами р—У
*Я+У
Mpt=aip—y-q—y)+2d[(p—y)(q+y))=
a(pq~~2y)+2d[pq + y(p~y)~tJ2l
Отсюда можно получить среднюю двух родителей:
Особи двух популяций свободно скрещивают» MVr
С ДРУГОМ. r*-w»««/iv* друг
Построим обычную решетку для получения генотипов в Fx.
^^^^^^^^ Гаметы от Рх
Гаметы от Pi ^s*"*>-**^^^
Ах P-V
At Й + У
А%
Р
р{Р~У)
1 Р(ЯЛУ)
*t \
€
Генотипы, их частоты и генотнпическне значения в f, будут
следующими:
Генотипы >М, ЛИ, >М*
Частоты р(р-у) 2рЯ+У{Р-9) W+*>
Генотипические
значения в а —А
В таком случае средняя генотниическая Л
MF% = a(p2~py-q*-qy) +42w+y(p-fl)J-
Разница между Л^< н М- может быть получена путем
вычитания уравнения (21) из уравнения (22).Этой будет мерило
значения гетерозиса Я^,:
Легко видеть, что все части уравнений сокращаются, кроме
одной — йуг\ г
HFi^dy^d(p-pi)\ (23)
Это выражение относится к одной паре генов. Так как р и pi
всегда представляют собой определенные величины, то,
очевидно, значение Нг< целиком зависит от d. Если <f=Q, то я #/,=0,
т. е. не может быть гетерозиса.
Следует обратить внимание иа то, что чем больше d, там
гетерозис выше. Максимальным он будет при й>сц т. е. ори
сверхдом инированин.
Кроме того, степень гетерозиса завесит от квадрата рааа*
иы в частотах р. Еслв иет разницы a **cw«р> м|*^°f J£
жет быть и гетерозиса. Чем башне разница в частота*, тем
больше и степень гетерозиса.
Можно перейти теперь к сумме генов, приняв, что суммиро-
м.чпие происходит аддитивно. Тогда
llr^cltf. (24)
Смысл этой формулы тот же: гетерозис зависит от
доминирования. Однако если бы разные гены обладали разной
степенью доминирования и при этом с разными знаками, то
эффекты их доминирования должны были бы взаимно
погашаться и Id была бы равна нулю. Наличие же гетерозиса
требует, чтобы Id была определенной и вполне значимой
величиной. Значит, доминирование большинства генов должно
идти и одном направлении (плюс — при плюс-гетерозисе или
минус— при минус-гетерозисе). Напрашивается аналогия
с тем, что говорилось выше об инбреднои депрессии, которая
тоже связана с направленным доминированием.
Формула //Fl = 2(i{/2 явилась основанием для гипотезы
о том, что гетерозис зависит от суммирования доминантных
генов. При этом возможно и такое суммирование их действия,
когда Fi не выходит за пределы средних родителей, но
располагается не посередине между Р\ и Рг, а ближе к одному из
них. Такой случай, как мы указывали выше, не следует
называть гетерозисом, хотя принципиальная его основа та же.
Но если при сочетании действия многих генов возникает
компонент взаимодействия, то он прибавляется к значению
Zdy2, хотя это и трудно выразить математически. Однако с
помощью некоторых методов, в частности анализа диаллельных
скрещиваний, о чем будет говориться ниже, можно в
некоторых случаях в эффекте гетерозиса оценить и роль
взаимодействия между генами.
F2 обычно получается путем свободного скрещивания
особен Ft. В этом случае также можно рассчитать MFa.
Не вдаваясь в детальные расчеты, ограничимся
приведением готовой формулы
Л1,, = а (р- 2У-Я-^у) +2d(p-~y)(q+--y) =
^a(p-q-y)+d [2pq+y{p—q) — у I/2 J .
Отсюда
//,,^-Мк,-Му>=:ус/{Г-= \- HFt. (25)
Дли суммы генов
ий--;-2'«•--£ л. ,2б)
Т*
Эли соответствует и фактическим данным, что степень гете-
розиса в г 2 ниже, чем в tu при этом чаще всего вдвое (хотя
из этого правила бывают исключения).
При анализе гетероэисного эффекта были рассмотрены
два варианта — с одной парой генов и со многими. Обычная
интерпретация гетерозиса основывается на втором варианте.
Но за последнее время появилось довольно много
доказательств существования моногибридного гетерозиса. Об этом
упоминалось в гл. 4 в связи с отбором в пользу гетерозигот.
Работами Д. К. Беляева и его сотрудников было доказано
наличие моногибридного гетерозиса у пушных зверей.
Накапливаются данные и по другим объектам — растениям н
животным. Сверхдоминирование могут проявлять не только
рецессивные гены с видимым действием, но даже летальные гены.
Так, М. Д. Голубовский и Г. В. Викторова показали, что
в природных популяциях дрозофил на Украине (близ Умани)
некоторые летали устойчиво сохранялись в разные годы в
разных популяциях и даже возрастала их концентрация (до
1—2%), что указывало на влияние отбора в пользу
гетерозигот по этим летальным генам.
Ли и Редеем на растении Arabidopsis было показано, что
многие гетерозиготы, полученные путем скрещивания ланий,
юмознготных по рецессивным генам (Arabidopsis —
самоопыляющаяся форма), проявляли гетерозис. При этом можно
было доказать биохимические преимущества гетерозигот, что
соответствует концепции гетерозиса, развиваемой В. С. Кир-
пичнпковым.
Общая и специфическая комбинационная способность.
Выше было показано, что в процессе инбридинга происходит
расчленение ранее единой исходной популяции на ряд инбредных
линии. В пределе, т. е. при F=l, каждая линия становится
гомозиготной по определенному подбору генов. Внутри нее нет
генетической вариации. Если скрещиваются такие высоко-
ннбредные линии, то в F\ от каждой пары также нет
генетической вариации. Между отдельными же F{ вследствие геноти-
пических различий скрещиваемых линий должна быть
генетическая вариация. Измеряющая ее варнанса будет отражать,
если, конечно, таких F\ много, генотнпическую вариансу
исходной популяции (до инбридинга). В тех случаях, когда ин-
бредные линии не полностью гомозиготны, в Fх от их
скрещивания сохраняется некоторая генотипическая вариация,
Вариапса же между отдельными F{ будет несколько меньше,
чем при скрещивании полностью гомозиготных линий.
Кроссы, т. е. Fx между полностью гомозиготными линиями»
генетически равнозначны отдельным особям исходной
популяции, при неполной же гомозиготности линий— как бы
отдельным семьям, выделенным из популяции.
359
Влриаиса между Fu полученными от многих скрещиваний
икбредных линий (о1*,), может быть выражена следующий
врачом:
o*rt = FoA*+r-vr,*+nc-AA + FWAD+FWDD+- .. (27)
Здесь Ол1 и oDz — аддитивная и доминантная вариансы
исходной популяции; аглл* o2dd и a2AD — вариансы взаимодействия;
F — коэффициент инбридинга.
Сама формула представляет собой не что другое, как
обычную формулу для aG2 с той только разницей, что в нее включен
коэффициент F. Очевидно, что если /г=1, то o2f%=Og2
(исходно»! популяции).
Но надо иметь в виду известные упрощения, принятые для
этом формулы, а именно: все инбредные линии
характеризуются одинаковыми значениями F, линии не имеют родства
друг с другом и т. д. Поэтому ее надо считать лишь некоторым
приближением к действительности. Однако из ее
рассмотрения можно сделать некоторые существенные выводы. Разные
степени при F показывают, что при увеличении F аддитивный
компонент изменяется прямо пропорционально F, т. е.
зависимость здесь линейная. Компонент же доминантной вариансы,
а также взаимодействия АхА возрастают пропорционально
квадрату F. Все остальные компоненты взаимодействия
пропорциональны третьей степени F. Значит, на низких уровнях
инбридинга (при малых F) роль доминантного компонента и
взаимодействия меньше, чем на высоких (при больших F).
Поэтому если в вариации данного признака первенствующее
значение имеют неаддитивные компоненты, то скрещивания
между линиями с невысокими F будут слабо влиять на
различия между кроссами, а скрещивания между линиями с
высокими F — более сильно.
После этого разбора структуры F\ можно перейти к
выяснению удельного веса отдельных компонентов в вариации
между кроссами. Для этой цели служит разделение
комбинационной способности линий на две части: общую
комбинационную способность и специфическую. Под первой понимается
среднее значение Fi, которое дает данная инбредная линия
со всеми другими, а под специфической — ее отклонение от
общей комбинационной способности, которое она дает при
скрещивании с какой-либо определенной линией. В таком
случае, если выразить среднее значение F\ как отклонение от
общей средней для всех кроссов (Ft), то можно представить
значение каждого кросса как сумму общей и специфической
комбинационной способности этих линий.
Среднее значение кросса между линиями X и Y может быть
выражено следующим образом:
эво
Mxt~GCz+GCy+scxt. (28)
Здесь GCX и GCr — общие комбинационные способносп для
линий X и У, a SCjy —их специфическая комбинационная
способность. Последняя представляет собой не что другое, как
компонент взаимодействия между генотипами двух линий.
Для оценки общей и специфической комбинационной
способности существует ряд методов. Наиболее разработанный
из них — дисперсионный анализ результатов дналлельаых
или, точнее, полналлельных скрещиваний.
Различные методы анализа и соответствующие
математические модели были разработаны Грнффиигом. Так как они
достаточно полно освещены в работах советских авторов
(Н. В. Турбин, Л. В. Хотылева, Л. А. Тарутина и др.), мы
дадим только общие принципы анализа и приведем в качестве
примера одну из моделей.
Допустим, что имеется р инбредных линий и они
скрещиваются друг с другом. Возможны четыре метода. I —
анализируются родительские формы Flf получаемые при прямых и
обратных, т. е. реципрокных, скрещиваниях. В этом случае
возможен р2 комбинаций. II — родительские формы и только
прямые скрещивания для получения F§. Тогда комбинаций
р(р—1)
всего р-^1--^- -. III—прямые и реципрокные скрещивания
на F\. Комбинаций всего р(/>—1). IV —только прямые
скрещивания на Fi. Комбинаций 1/2 р(р— 1).
Выбор той или иной модели определяется конкретным
материалом. Кроме того, надо различать, каким образом
выбирались родительские линии — случайно из некоторой
исходной популяции или же по какому-либо принципу. В последнем
случае их нельзя рассматривать как случайную выборку.
Поэтому внутри каждого метода можно построить две несколько
различающиеся модели.
Во всех случаях проводится двухфакторный дисперсионный
анализ обычного типа с учетом взаимодействия. Если
проводились реципрокные скрещивания, то учитывается и фактор
рецнпрокности, что несколько усложняет картин)*.
Для иллюстрации хода анализа используем простейший
метод IV, в котором не учитываются родители и нет
рецнпрокности (такой анализ был впервые проведен Спрэгом и Тейту-
том, давшими интерпретацию общей и специфической
комбинационной способности в системе диаллельныхскрещнванн»).
Математическая модель для него следующая:
*,,« м+*1+Л+*«+ 7Г -f 2 #|*
ж
Здесь i, j — величины от 1 до р, k — от I до Ь, / от I до с;
и — популяцнонная средняя; g\ и gj — общие комбинационные
эффекты; Sij — специфические комбинационные эффекты
(S{) = sn); eijki — ошибка ijkl-ro наблюдения.
После составления сводной таблицы, в которую вносятся
результаты всех сочетаний линий (№ 1 с № 2, 3, 4, 5, 6 и т. д.;
.V» 2 с № 3, 4, 5 и т. д.), устанавливаются значения общей
специфической комбинационной способности сначала для всего
материала в целом (табл. 56), а затем для каждой отдельной
линии н ее сочетаний с другими.
Таблица 56. Общая схема анализа варианс по IV методу
(по Л. В. Хотылевой)
Источники вариации
Обедая
комбинационная способность
Специфическая
комбинационная
способность
Случайные
отклонения
Степень
свободы
df
р-1
2
m
Сумма
квадратов
S5
ssg
«J
8Se
Средние
квадраты
ms
msg
Оцениваемые
параметры •
0'
• Определение oj: и о^ производится обычным методом.
Первоначально методы выделения общей и специфической
комбинационной способности были разработаны
применительно к кукурузе, у которой выведены многие тысячи инбредных
линий; чтобы планировать наиболее желательные по высоте
гетерозиса скрещивания, нужна была оценка комбинационной
способности отдельных линий. Но эти методы вполне прило-
жимы и к другим объектам, ибо дают значительно большую
информацию о результатах скрещиваний, чем чисто
эмпирический анализ скрещиваний различных рас, линий пли пород.
В, Т. Горин и И. Н. Никитченко провели исследования
различных сочетаний пород свиней для промышленного скрещи-
ьаиия с соответствующей оценкой комбинационной
способности по мясо-сальным качествам, для чего использовали II и
IV методы Гриффинга.
Тсйтум и Спрэг определили генетические компоненты
общей и специфической комбинационной способности следу-
ющим образом:
OCefaA2+/rW>Ui+...f (29)
SC~* F4ii?+F4fiAu+FWim+ ... (30>
:м
Такимобразом, различия между линиями по общей
комбинационной способности сводятся к аддитивной генетической
парна псе, характеризующей исходную популяцию» а рапичия
в специфической комбинационной способности — к нсаддитнв-
пой вариаисе (доминирование-f взаимодействие). В силу этого
при повышении коэффициента инбридинга возрастал роль
именно последней.
Соотношение между общей и специфической
комбинационной способностью может быть различным. Так, в одном из
опытов, проведенном на кукурузе Гриффингом и Линдстро-
мом, варианса общей комбинационной способности по
общему урожаю зерна превышала специфическую почти в.два
раза, а в опытах Гоуэна но яйценоскости они были почти равны
друг другу.
Наряду с диаллельными (полналлельными) скрещиваниями
применяются и другие методы разделения комбинационной
способности на общую и специфическую, например тошроссы,
т. е. скрещивания иибредных линий с одним общим
тестером — своего рода эталоном для сравнения линий. Чаще
всего тестером служат особи из лопуляций *.
* Об *том см. а социальной литератор*
Глава 9
ФЕНОТИПИЧЕСКИЕ И ГЕНЕТИЧЕСКИЕ
КОРРЕЛЯЦИИ
Связи между различными признаками у особей в
популяции и их значение. Во всех предыдущих главах, шла ли речь
о структуре популяций по качественным или количественным
признакам или об их изменении под влиянием различных
причин, обычно рассматривались отдельные, как бы
изолированные признаки. На моделях с такими признаками легче всего
проследить закономерности, лежащие в основе тех или иных
генетических явлений. Однако хорошо известно, что между
разными признаками наблюдаются многочисленные связи,
которые вскрываются с помощью различных статистических
методов — корреляционного, ковариационного и регрессионного.
Так как особи в любой популяции — природной или
разводимой человеком — изучаются по их фенотипу, то, очевидно,
изучение корреляции между двумя признаками х и у (в
простейшем случае корреляционного анализа) позволяет установить
степень фенотипической корреляции, выражаемую простым
коэффициентом корреляции г. Такую фенотипическую
корреляцию можно обозначить символом гР. Вычисление фенотипи-
ческих корреляций очень широко распространено во всех
областях биологии, в том числе в генетике, а также в
животноводстве и растениеводстве. Оно имеет немалое практическое
значение, так как позволяет обнаруживать связи между
различными хозяйственно-полезными признаками и использовать
эти связи для целей селекции, при создании новых пород и
сортов, для ранней оценки продуктивных качеств по
признакам, которые могут быть изучены в более раннем возрасте и
которые коррелятивно связаны с продуктивными качествами
взрослых животных.
Фенотипическая корреляция становится объектом
статистической генетики в том случае, когда могут быть вскрыты ее
причины.
Наряду с коэффициентом корреляции может быть
использована коварианса между фенотипическими признаками х \\ у
J^4
(covp). Как известно, коэффициент корреляции связан с кова-
риансой следующим равенством:
COVp *-
rpS~ 9 ^
ор ор
где ор и ар — средние квадратические отклонения признаков
х и у.
В некоторых случаях надо пользоваться не квадратическя-
ми отклонениями, а вариансамиор\ нор2.
Разложение фенотипической корреляции на генептою!
и средовой компоненты. Подобно другим математико-генетиче-
ским параметрам, фенотипическая корреляция, определяемая
путем измерения двух признаков у одних н тех же особей,
имеет двоякую причину — генетическую и средовую. Если бы
были известны генотипические или селекционные ценности
отдельных особей по двум признакам, то можно было бы
вычислить коэффициент генетической корреляции. Так как
генетические и селекционные ценности могут отличаться друг от
друга за счет таких компонентов, как доминантные отклонения и
отклонения, вызываемые различными видами взаимодействия,
то выгоднее иметь в виду только селекционные значения,
сводящиеся к аддитивным эффектам, и считать генетической
корреляцией (rG) корреляцию между последними (гх). С другой
стороны, фенотипические различия между особями могут
вызываться многочисленными средовыми влияниями,
создающими средовые отклонения. Корреляция между средовыми
отклонениями составляет второй компонент фенотипической
корреляции — средовую корреляцию (гЕ). Практически в этот
компонент входят и те отклонения, которые вызываются
неаддитивными эффектами, так как они не могут быть
непосредственно выделены и не входят в селекционные оценки особей.
То же относится и к коварнансе. Генетическая и средовая
коварнансы соответствуют долям ковариансы, которые можно
выделить как аддитивный генетический компонент, с одной
стороны, и как всю остальную часть ковариансы —с другой.
Последняя включает компонент, зависящий от среды, как
основной, а также неаддитивные генетические влияния:
COVp = COVQ+COV][=COOA+COVg.
Отсюда
covA+covt
ОрОР,
а/*п1 к* можно перейти
С помощью коэффициента нааедуе^^ (м 6)> что
от аР к аЛ и о* Дл" 9Т0ГО з»
Vm
•S
Я»***
, ;= *-£- lot»
«««1-я»
m*mmx*?aM0t?
о дяуж
* ■ я, яо k«»>
чяи iPinrni Я ■ i|| ■ i им тпАчщв»
Ю, и О,) т щпш i£.mt,i (m .tli|iii|j, «*>
fl*. i~&.*A я я*. Можяя i
ю «* я «* я ян ■fwi i» km*. Bmpmtm, tfo me совсем точяо. гш
«я т «аяяяаи, я» дм яяосгош иткяо ямьяояятмя
«мая* я я л
Тяяям illgji вязеля я яяшщяия фоуиуяе я*ж я я*, **'
• я» f k*m f **.я* я •«, «шгучятся ояяая форму** **■ V
*tts «*л
<ЯЯ*
Шлт тртштгшуп *
,те
<••)
о*>
,r**r*n
*-*V**%Vb
т
Эта же+орщла
ев го* ауте* (ряс 1И).
яа*а» х я г ^^VM*^ ^
фежотгалеска* ——— ■ 7
гакгакскв* я
ж к блшзкш к е:
ся rtmeuntcaam
заступает яа эеряое
яяляо кэ яредыхгсип г
шкеты наса*х$€*оетш
хпегл зшАШЯЖйба
Но
<ШКЯ 9
«ств при расезюрсяяв
чер, что ueaur
ложе корреляявя «i
ты здрредяяяя. как
шкет фежггпшчсашА
стон оерде*
действия я
сз/зяящюгя. т. е
кя. Другие, яяоборот.
дорредяшгю
сильно )велв«*ггъ
гляяют *яэ хотя) ту
здгкой дорредяяя* ****У
aa елвж (о6*гпю ©я»
ЧэСКЯХ УСЖУЯЯЯЖ
** яйиеяоекостыр «"С1*
CttrtLibaot «op
*яя врем*
лываеяиж -IT— ^^ ,
3. С Няквро я ^
<*** j
*>»ффяавея?а»
ратяомчесяо*}
лмч$ш во эяяяу »_
**тора* форяу*****^
величины г, от соотношения между знаками и величинами гР
и rit \ ;i именно:
1. При нулевых шачепиих г,, и г<; коэффициент корреляции
г, К1кже равен 0.
1? При нулевых значениях Гр и значениях r(j, отличных от
!1>.1я, коэффициент корреляции rt: имеет знак,
противоположный знаку Гц.
.1 При пулевых значениях гг, и отличных от нуля значениях
г г коэффициент иаратипическоп корреляции имеет знак,
одинаковый со знаком г/>.
4. При равенстве абсолютных значений rv и r(i
коэффициент корреляции ri. имеет знак, совпадающий со знаком гР.
Г). Нел и гР II гс имеют одинаковые знаки, то направление
парашппческоп корреляции непосредственно определить не
удается, за исключением случая, указанного в предыдущем
П\ ПКТе.
6. Нслн гР и Гц имеют разные знаки, то знак гЕ совпадает
со знаком г р.
Из сказанного выше видно, что между значениями этих
трех коэффициентов зависимость довольно сложная. Поэтому
понадобились специальные методы определения прежде всего
значения коэффициента генетической корреляции по
коэффициенту фенотнппческой корреляции, зная же первый и второй,
можно определить третий, т. е. г£.
Методы определения генетической корреляции. Формула
Хейзеля. Подобно тому как с помощью дисперсионного
анализа можно выделить отдельные компоненты вариансы,
зависящие от разных источников вариации по одному признаку,
так и в ковариансе по двум признакам можно выделить
отдельные компоненты. Для этого применяется метод учета
корреляций или ковариаис между особями, например, родителями
и детьми с данными признаками хх и уу и, наоборот, особями
с а' и особями с у.
С использованием ковариаис
1 Л . \w/
У COUxxCOVVy
В данном случае covxx и covvu — ковариансы между
родителями и потомками по каждому признаку, a covxy — ковари-
анса между значениями х у родителей и у у потомков.
Так как у животных, особенно у крупного рогатого скота,
удобнее всего сравнивать матерей и дочерей, то для
вычисления гА (rG) широко применяется формула Хейзеля (Hazel) **:
• Авторы используют только символ rG% а не гА.
" У автора несколько иные обозначения.
36»
В пси г — коэффициент фенотяляческой корреляции между
матерями (М) и дочерями (d) с разным сочетанием признаков
х и у: и числителе — крест-накрест, т. е. у матерей учитываете*
признак xt у дочерей у; наоборот, в знаменателе — это пары
одинаковых признаков.
Если известно значение гл (гс), а также V, ty и />, то по
уравнению (2) может быть вычислено и значение средовой хор.
реляции (г£). В некоторых случаях его можно определить и
непосредственно, например при анализе внбредных линий
и кроссов между ними.
Однако значения коэффициентов генетической корреляции
могут быть не очень точными, так как формулы для их
вычисления предусматривают наличие ряда исходных предпосылок
(свободное скрещивание и отсюда равновесие в популяции,
одинаковые частоты генов у самок и самцов, одинаковые
условия среды в смежных поколениях и др.), которые далеко не
всегда выполняются в конкретных популяциях.
Вопрос о том, в какой степени отразится на величине гА
отсутствие тех или иных предусматриваемых теорией условии
в популяциях, разработан еще не достаточно, хотя эти
формулы широко используются во многих исследованиях.
Ошибку для гА вычисляют по следующей, также
приблизительной формуле:
вг*-^пу*г- (э)
Фактические данные о значениях фенотяпичеша и
генетических коэффициентов корреляция. Фенотилическне
коэффициенты корреляции (гР) всегда вычисляются из
непосредственных фактических данных. Коэффициенты же генетические и
средовые — результат применения тех или иных формул,
поэтому в ряде случаев они вызывают большие сомнения.
Кроме того, они могут значительно отличаться даже для одних и
тех же пар признаков в зависимости от того, на каком
материале вычислены.
Некоторые коэффициенты корреляции, полученные в
опытах различных авторов на разных объектах, представлены а
табл. 57. Поскольку большинство исследователей, особенно
советских, ограничиваются двумя коэффициентами -
фенотияяческой и генетической корреляцией, в таблице нег граф*
ДЛЯ ГЕ. ^
24 П. Ф. РокицкмА
Фшбмща 57
Пм
К«*ффияам«тм
коеееляеян
VLppmti
TwriA cm
к ро-
cmtrt
КУР"
Серго
Высот е жалел — глубине
груда
Провеет белил — процент
У*и) — процент жире в ie>
Процент СНВ — процент
ере и молоие
У щЛ — процент белил
Саярогг* роста — оплата
Длине тулоеиии — толщи не
сале не стшне
Количество шерстн — денне
Продуйте чисто* шерстя —
плотное tv фолликулов (е Ъ-т-
сеннои нагреете)
Вес тела —нес иное (в 16
Вес теле (е 18 несшее)) —
погреет яериоб кладки ниц
Вше теле — длина жеосте
Кавииех >») абдоминальных
— количество стерио-
шгтннок
Число дней до среднего
ее пине — нес 100 седее
Висите растение ~ вес 100
О.ЭО
-0.I4
0.30-0,42
-0.04
—0,64
-«0,24
0,30
0,30
0,16
—0.30
0.44 .
0.06
0,33
0.42
0,67
0.80
1-О.20
0,50
-0.20
-0.26
-0.47
-О.02
0,66
0.М
0.2*
0,50
0.00
0.3S
0,46
ChfeMUM больший нестрог* полученных данных, В ««о-
^****Уад** « фекогшшчмк** и генетическая коррел*****
«ягсутспгукгт (ибдомимдлмдо я стсриоплеврдльиые щ*тын%* У
Ароаое)вш*),
Ияогдд ге*ег«ч«с*ея корреляция ршшпш (наш 6лы**л) * "£
д*>, *• то »ре*я *** фенотипичесмаи достоверно отличаете* &
m
и
лЛЯ (например, количество «рсти-хл,,,, ^
Очевидно, что последняя вызвана н^щ^!!*!" » ««).
„. - I, в работе приведено значение г,^,*Г2!^^
Наличие достоверной отрицатели** гевегнчемл*
иии >называет на противоположное декгвиеrSTl!2!^
На два изучаемых признака, ~^*«аш
Наконец, есть случаи заведомо! йоаожнтеаыю! тмм^
ской корреляции. таци*>
Интересные данные по феяогявяческяя в геяегм*™
корреляциям l^p^^^J^mjn SST»
генетики СО АН СССР, Так, Д. К. Беляев в 3 С Кяееша
изччали фенотипическяе и генетические шютяшмежи
содержанием различных компонентов moaoki (ад. бегаж
обшее сухое веШество, сухое вещество без жяра) * щж
между этими компонеятами малой я до* Существ об-
ший вывод, что величины коэффяяленго» геиелпеске!
корреляции, как правило, выше, чем величины юффкэюгтш
фенглипической корреляции между злим же парами щшу
наков. ^
В потомстве отдельных производителей величин шре-
ляиии сильно колебались, причем оойбы! интерес виэмвш
производители, у которых была боке высош, чем во стаду
в целом, положительная корреляция между шдаржтея жя-
ра и белка.
3. С. Ннкоро л 3. С. Киселева показан нити пик
различия в коэффициентах февотияичеои* a шепчет*
корреляций по тем же парам признаков вежду парадов
Так, между процентом бедка я вронеитеи жяра i ясное
коэффициенты гР и гд был: у а*ршяраа 442 я <Mfi» v черв*
пестрых 0,18 и 0,55; между удоем я вроиеитои жара в иолов*
\ тех же пород соответственно -0,14 в -401 -4в * HDt*
Знаки коэффициентов корреляции чате всего оегавадие» я*
ми же, но колебались их абсолютные значения, Ожввя» Ьж
случаи различий н по знакам. Так, у джерега лдеинкка*
корреляция между удоем я проделом общего сумяо ъет
<тва н молоке была 0,15, у других пород (Чершнкорые. тол-
"ениы) -0.27, —0Л1. Ковффноиевты же г» у асях трех во-
род были со знаком минус ^^
Причины генетической кчрреаяинж H«w» owert a.
*оас-т быть, наиболее часто* ирячяж* ity***»* яарреля-
«"" является плейотропное двктвве <^ * *** *eJ*£
"а ратные признаки. В таком «Г«» JJJ*? JJL^S
<;*идать, что при "«ожятеямк*^^^^^ГК
Синице Данные. лриведестые яJ^zL* треаяцив шее
столь высокие *ю*ффиии«^«*«"^ сл^^ ^ь*
**ляются иенлк^нявми, *** * •***»—
2««
■иНчгя (гплнщия, в силу KoTiipfift 1ГМГП1ЧГГКИС коэффициеи«
ти превышают фсиопшнческис. И при илейозроииом действии
n-I hoi о гена коэффинпеи г I енп пческЫ'! корреляции должен
г»ьпь чаще всего ниже единит.! по ряду причин, Поскольку
речь идет о связи между разными признаками, возможны
различия но иронплению гена па этих признаках: полное
проявление на одном, неполное на другом. Может коле*
Литься и степень выражении гена, что связано с
существованием стхястикн п процессах действии гена па разных, даже
илсшнчиых по своей роли участках тела, например правой
и левой стропе тела, одинаковых сегментах и т. д. (об этом
finei юворшьея в гл. 10). Поэтому неудивительно, что в дей*
сшив одного и того же гена, например, на длину крыла и
.злнну груди у дрозофилы или на образование какого-то хими»
ческою пешее поп в молоке коровы и на процесс роста того же
:м1Возного не будет полного совпадения. Иначе говоря, может
колебаться нлейотроипый эффект.
Генетическая корреляция может определяться и двумя
сцепленными генами. Если они очень близко расположены
друг к другу, то поведение вызываемых ими признаков
практически будет такое же, как и при наличии одного гена. Чем
больше расстояние между генами, тем слабее будет
корреляция между определяемыми ими признаками. Кроме того, она
непостоянна, временна. Такие временные корреляции могут
создаваться в результате предыдущей истории стада
животных или группы растении, а позднее ослабевать и исчезать
вовсе.
Наконец, возможен и такой случай, когда два разных
гена влияют на признак в одном н том же или в разных
направлениях, являясь только их модификаторами, ослабляя их или
усиливая, что также скажется па величине генетической
корреляции. При однонаправленном действии создается
положительная корреляция, при разнонаправленном —
отрицательная.
Поэтому неудивительны приведенные в работах Д. К.
Беляева и 3. С. Киселевой, 3. С. Инкоро и 3. С. Киселевой
факты о различиях в величинах генетической и феиотнинче-
ской корреляции в группах потомков разных производителей
или в ipynnax животных разных пород. Поскольку
количественные признаки детерминируются, как правило, многими
генами, одни из которых могут плейотроипо влиять на оба
изучаемых признака, другие же нет. то при разном сочетании
*гнх геною будет наблюдаться вариация в значении
генетической корреляции.
Возможности косвенного отбора при отборе по одному из
коррелирующих признаков. При отборе по одному из
признаков х можно, очевидно, ожидать сдвига и по второму при*
172
знаку у. коррелятивно связанному с ». к», „.^
сД|,„, при отборе по признаку х равеншщ!п£!2±1* I
,„ачриию отбираемых особей я выражается форм^о!9"*
Изменение же признака у может омть овхяеяо апипаша
pcipcccHH селекционного значения у к селешмжюму ц,^.
лию х, т. е.
СО?д Од,
В таком случае сдвиг признака у-результат яс прямого
отбора по (/, а только его корреляции с х (таю! сдадг цель
сообразно обозначить другим символом — Ctft):
CR,=bu)¥,RM. (7)
Заменив величину J>(a)v* его значением из предыдущей фор.
мулы (6), получим
После сокращения на оА*
Так как ол„=ЛуО/>, то геиотипическую сигму можно заменить
феиотнпической, определяемой фактически, н тогда рабочая
формула для CRy
CR^ihJi/AOpr (9)
С помощью этой формулы можно предсказать аеличияу
коррелированного сдвига признака у, если известен РЯД W*
метров признаков у и ж, а также интенсивность огборл' О»
признаку х).
В то же время очевидно и значение косвенного (непрямого)
отбора. Могут быть ситуации, когда отОор по желательному
признаку затруднителен (по тем или иным причиняй), я тогда
эффективен только непрямой отбор. Не исключен н такой слу*
чан, когда непрямой отбор окажется эффективнее, чем вранов,
что вытекает из анализа следующей формулы:
"ЯГ""- iuKflAM '* *■
В ней через Rx означен c^^np^
самому, а через CRx — ^iKr lw зя
ttflll
оип.аку v. Уравнение ;ыя СЯД соответствует указанному
г (')) Y для W» -- обычное уравнение для сдвига при
^Гг.Грг Непрямой отбор будет эффективнее, если числитель
упитает знаменатель. При /„-<* Ay должен быть больше Их.
При равенстве /i /(У должно быть больше ix.
Сирстад предложил несколько иную формулу для
определении соотношения величины сдвига по признаку у при отборе по
коррелятивно связанному с ним признаку х и сдвига при
прямом отборе по признаку у (с учетом числа потомков от
производи гелей):
л а т/ M4+(ai-T)vj
AG V /i^4+(n-iJM G' '
Здесь Л G, — коррелятивный сдвиг признака у при отборе по
прншлку х\ \G — сдвиг признака у при прямом отборе по
нему •; г с — коэффициент генетической корреляции между
л и I/; /iv2 и Л у2 — коэффициенты наследуемости; п — число
потомков от каждого производителя. Автор проверил
использование этой формулы на примере двух признаков: средней
скорости молокоотдачи х и продолжительности доения у у
красной датской породы. Так как на датских станциях быки
оцениваются по 20 дочерям, то п = 20; hx* = 0,77\ /iy2=0,28; rG = 0,84.
В таком случае
АС, __ уХ77(4+\Т.бЖ 084_, ,7 0 84-0 98
SG -V 0.28(4+19"6,77) -°.84-•-17.0,84-0,98.
Отсюда следует, что сдвиг по у при косвенном отборе по х
и сдвиг при прямом отборе по у были почти одинаковы.
Изучение результатов отбора при наличии генетической
корреляции (на экспериментальных объектах и
математических моделях). На курах породы белый плимутрок Карт и
Сейгель вели отбор в плюс- и мииус-направлениях по весу
и Н-иедельном возрасте с учетом несслекционируемого
признака — угла груди — или, наоборот, по углу груди с учетом
веса, не подвергавшегося в этом случае отбору. Оказалось,
что при отборе только по весу изменения по углу груди были
более сильными, нежели изменения веса при отборе по углу
груди. В то же время коэффициенты наследуемости обоих
признаков были довольно близкими, так что причина
наблюдавшихся различий была иной.
Столь же неожиданными были результаты опытов Старки
по созданию путем отбора линий кур с повышенным и
пониженным кровяным давлением: живой вес птиц в минус-линии
• Обозначении хну обратные по сраниснию с формулой (9).
374
был ниже, чем в плюс-линии, хотя отбор во весу it 0ро^
дИЛСЯ.
Известную ясность в утот вопрос внесли данные,чюлучея-
ные на машинных моделях, в которых анализировались
различные варианты генетических связей я других параметров.
Так, в модели Паркера и др. изучались результаты отбора по
одному из пары признаков, между которыми предусматрш-
лась генетическая корреляция с коэффициентами (Х25, 0,50 в
0,75. Учитывались разные значения фототипической варваясн
при 3 величинах интенсивности отбора. Эффект отбора
измерялся в течение 30 поколений. При слабой интенсивности т-
бора величина генетической корреляции не менялась, но при
сильной она непрерывно уменьшалась. Поэтому опенка ее иа
основе фенотипической ковариансы между признаками
становится ненадежной.
В другие серии опытов были внесены некоторые иные
детали: коэффициент наследуемости в узком смысле слова А
равнялся 0,1, 0,4, 0,7. Считалось, что корреляция между
признаками вызывается только плейотропией. Были созданы два
варианта моделей: аддитивная (фенотнпнческне значения
генотипов 2, 1 и 0) и с доминированием (фенотипическве
значения генотипов соответственно 2, 2 н 0), В аддитивной
модели коррелированные сдвиги по несслекцяоннруемому
признаку у при отборе по х хорошо совпадали с теоретически
ожидаемыми. В модели же с доминированием сдвиги геиотипнче-
ских средних у при отборе по х в противоположных
направлениях вначале были симметричными, а в последующих
поколениях несимметричными. При слабой интенсивности отбора
зависимость была почти линейной, но при более сильной
отклонялась от линейной. Существенно, что сдвиги по у при
отборе по х наблюдались только до 15-го поколения, в
дальнейшем же их не было.
Вопрос об асимметричности результатов косвенного
отбора изучался Бореиом и др. также на математических
моделях с использованием электронных вычислительных машин
Для модели были взяты четыре гена: два *$ них
непосредственно влияли на данный признак, а два других были
модификаторами, причем один с положительным действием, а
второй — с отрицательным. Изучались варианты с
различными исходными частотами генов п с разной интенсивностью
отбора. Длительность отбора —9 поколений. Выяснилось, что
асимметрия в ответе на отбор возникает всегда, когда
имеются гены с положительным н отрицательным вкладом в ков*
риансу между признаками. Кроме того, она зависит от частот
этих генов.
В моделях Синга, Белл мама н др. предусматривалось, что
генетическая корреляция между х и у основывается «а «*
owners
уфлн. Отбор велся по признаку лг, а учытывалось измене-
irte признака у. Были изучены два уровня рекомбинации
(m:=rOf005 и m=r0,05) при вариантах наследуемости или
/:,->/»** или Лдг = А„г. Отбор велся на протяжении 11
поколений При т =0,05 обнаружилось отчетливое уменьшение
корреляции между генетическими значениями хну.
При г,;> равном 0.5—ОД прогресс признака у был очень
невелик (как по фенотнпнческой, так и по генотипической
ценности) При rG<0,3 наблюдались различные сдвиги
средних по обоим признакам.
Определение коэффициента генетической корреляции по
сдвигу при прямом отборе и по коррелятивному сдвигу.
Определить величины сдвигов при отборе как по основному
признак), так и по дополнительному, коррелятивно с ним
связанном), удалось в ряде опытов. Зто дало возможность решить
и обратную задачу — определить по соотношению в
величинах сдвигов коэффициент генетической корреляции гЛу не
пользуясь формулами (3) и (4).
Для выведения формулы достаточно записать
соответствующие равенства, вытекающие из формулы (10), меняя местами
признаки:
Rx = ihxOAx\ Rv=ihyOAyl
CRy = ihxrAOAu\ CRX= ihyrA(jAxl
затем взаимно перемножить два последних и из произведения
получить величину гА2:
г *= CRyCRx _ CRyCRx
ihx<jAyihyGAX RyRx
В неоднократно уже цитированной нами работе Рива и
Робертсона по изучению наследования количественных
признаков длины крыльев и тела у дрозофилы приведены данные
по прямому и косвенному отбору. Оказалось, что все методы
2али очень близкие цифры коэффициента генетической
корреляции (гд равнялся 0,71—0,75).
Значение проблемы фенотипических и генетических
корреляций* Подходы к проблеме фенотнпнческих и генетических
корреляций, разработанные в современной статистической
генетике и кратко изложенные выше, связаны с более обшей
проблемой —■ значением корреляционных зависимостей в ее*
ттвеиной эволюции организмов и в искусственной эволю-
иин -• при создании новых сортов растений и пород
животных, Очевидно, что рассмотрение этой проблемы выходит за
рачки нашей книги Но целесообразно назвать некоторые
более конкретные вопросы - т«ом«
которые связаны с проблем* Д£Е* * *******
а) возможность одновремевиого вт&*>, ^^
признакам, которые суммируете* • bbT£J£ "**»*И1
руемого индекса. При построении ашш^л1^Ьаапп-
тываются связи между призвакамя, в том 4*tLv£!!!0' '*-
ная генетическая корреляция; ^** я ttpvnta*.
б) роль взанмодейстивя геноши и среду в сашм. *-
нотнпических и генетических коррелваяв. Эют юыаи*
вопрос тесно связан с разобранной в гл. 71фобдодГ»в4е£
тивности отбора в различных условиях вводи* сведы/^
Глава 10
НЕКОТОРЫЕ СТАТИСТИЧЕСКИЕ ЯВЛЕНИЯ
В «БИОГЕНЕЗЕ
Запрограммированность процесса онтогенеза я А**™*
генов в рамитин. В настоящее время становится все более
очевидным, что процесс индивидуального Р«*ити* *^
каждого вида запрограммирован в генетической СТРУГГУ!*
оплодотворенного яйца. Генетическая информация,
заключающаяся в основном в ядре оплодотворенного яйца,
реализуется в пространстве и во времени. В результате происходи
сложнейшая цепь формообразовательных реакций наадиая
от ранних периодов развития эмбриона и кончая последиимв
этапами жизни особи. Элементарные звенья этих Реа*ади~
биосинтез различных макромолекул: ДНК, РНК « белков»
который находится под контролем генов, действующих в «е-
которой временной последовательности. По-видимому, !»©•
следняя также запрограммирована в генетической структуре
яйца. Можно предполагать, что большинство генов на каж*
дом этапе развития находится в репрессированном
состоянии, но в соответствии с времеинбй программой раэвятвя
включаются в действие разные гены: одни раньше, ДРУГИ
позже; одни на короткий срок, другие на более длительный.
Таким образом, процесс индивидуального развития должая
регулироваться не только действием отдельных генов, HO if
целой системой или сетью переключателей нх действия.
В процессе наследственного осуществления признаков,
обычно обозначаемого термином «феногенез», участвуют как
динамические, так н статистические процессы, так как дейст*
вне генов может на каждом этапе варьировать в силу очев*
многих внутренних н внешних причин. Поэтому реал и лив*
ге*«т*чес«оА программы не является жесткой. Может
наблюдаться довольно значительная ев статистнчность н лш6ЫАЪ-
ПОСТ*
Рассмотрение вопросов фемогенетаки или, а более Ш*Р*
ком смысле слова, биологии развития, жспгрнмемталИ»
371
а:фаГ,отка которых тол*» Шшаае. ^
По мы сочля возможным включить . «
ск„й генетики несколько частных iftiii"1^ «"лит»*.
к которым можно подойти ео етатн*2£2 *"•««"«№.
Вариации в даяствнн тнт. Еш*Т?!*,твчм »*»•*
0 „сполном проявлении кщ. Шрояал««Г; У90»»****
1аКже пенетрантностыо.) ^pa^cHW^^^**^
v которых проявляете* данный прилива. V^E" "***•
(семья, линия, культура), гомозиготных м rL?^ 0вове|
ШСму этот признак. Для «и» шоп^Д 2**££
полное, т. е. 100%-иое, проявление. Ине-яТи?1^**
наследования таких генов и была выведены "ны* ?Л?
номерности наследования. Однако по кем nnSm^SJST
различных признано» у многих мдоа животных к iS2?
, также у человека было открыто явление неяолногГш*£'
ления. В гл. I показано, как меняется каршараом^Е^
в том случае, когда анализируется «аследо§ажие^*пм!«
проявляющегося гена. ""
И. В. ТнмофеввРесоасюй прок* т»1ым» модов»
н ис неполного проявления гена vU (yenae transvertae
incomplctae) у Dtosophila feoebria. Действие атого гена
выражается в исчезновении яоперечных жалок на крыльях.
Одновременно при изучении этого же гена влбдедиось
и иное явление — у тех особен, у которых армадяется tea
vti, степень его выражения (мспресснмоста) может бить
различна; исчезает лишь одяа из двух поперечных жалок
или становится /'рералмион. Путем длительного отбора было
выведено 30 высокогомозиготных ливни с рамнчноя
степенью проявления — от 100 ю 40—в0%, прячем процент
проявления оказывался а дмьяевщеж довольно стабильны*.
Иными словами, были отобранм различные геаыиолик**»-
торы степени проявления.
Оказалось, что вариация по ировенту проявления зависит
также от внешних условна. Так. проявление гена Ы(
возрастало с понижением температуры Но а |мп*чвом счет* вро-
ивление гена - это вероятмот wo. будет ля обмерки
ваться действие гена у **TtJ??lJ*L "*** **П *"'
Р«итерировано как типичяьЛ "£?"™"*«"* T22L
п.„ «-о объяснения, очевидно, нужны конкретные дмаые
гипотезы по этому поводу 'SLSS^SSS^SSLm.
JX* V22K2 XS&SZSmSi
Hvm?***** ****** гшшгш Н» Щ
r • С* Дж, ***** rZZ&tft***** ****** \шю* » ми»,
А общ бядетм* t It М * f1^
Я»
^даются различия межау генами. В одних случаях имрв-
Кеьие гена (яенетрантиость) очень постоянно^ B^^yj^r
кзиенчиво. Известно, например, что мутантиыйi ген W*u*
(безглазие) у дрозофилы в гомозиготной ^WP{-J2K
ваРы>рует/Изученная Б. Л. Астауровым "У^" ^gj
у Drosophila melanogaster варьировала по своему иырак
Рте
ИЗ. Паля ieftcTBB* ва щепгакя геяов Яо* (в) я I> W
у дрвэофялм.
нйю в двух противоположных направлениях — от почти
ваяно* редукция галтеров до сильно увеличенных размеров!
превращеняя их во вторую пару крыльев, даже с отчетливо
выраженным жилкованием. ■
Степень выражения гена при развитии признака зависит
хаи от генетических причин (влияние других генов, геяон-
модификаторов), так и от внешней среды.
Однако на проявление н выражение влияют разные ген*.
что было показано Тимофеевым-Ресовским путем получение
к>льтур с разным сочетанием степени проявления и
выражения.
В процессе феногенеза обнаруживается специфическое
действие генов на признаки. Чаше всего наблюдается вдмиияе
одного и того же гена на разные признаки, что получило 9М-
зваияе олейотропии. Но даже если геи действует на одни Я to
же признаки, например, на жилки крыла или щетинки у Др°*
здфилы, нш окраску шерстного покрова у млекопитакнШЛ
на окраску крыла бабочек, то действие неравнозначно на р*£
пых участках тела. Это явление было названо нами полем Д***
стаяв гена. На рис, 113 показаны поля действия двух гв"2?|
иляяюшяд на щетяики груди дрозофилы: одни из инх г&
(Pofyduefa) прибавляет количество щетинок — поде *™J£!!'
ствяо довольно узкое; другой D (Dichaeta) — реоио у***^
Ptt£ Ш. Лаж я&стпж ran %p±
Ubpnona:
шает, но поле его очень большое Цифры около пике, обоз**-
чающих отдельные щетинки,—средние количества щетяжж в
данчой точке, приведенные к единице Но возможны к более
сложные случаи, когда одяи в tor же гея увеличивает число
шетинок в одних местах и уменьшает в других Таких
оказался ген sp-b (split-bristles). Его поле изображено иа рис 111
Со статистической точки
зрения существенны два
обстоятельства. Каждое поле имеет
свою структуру — в одних
точках редукция (или
прибавление) шетинок большая, в
других меньшая. Можно заметить
как бы центры с максимумом
действия гена. Однако,
несмотря на кажущуюся целостность
поля, корреляция в эффекте
гена в соседних точках, как
правило, близка к нулю. Так,
между четырьмя точками,
отмеченными на рис ИЗ. а, все
коэффициенты корреляции
недостоверны, как вы численные между
точками на одной стороне тела,
так и между точками иа право! и лево! пороках тела (Об
этом еще ниже). Та же картина и для гена Д кроме охиого
случая положительной корреляции между двумя точками,
расположенными на право! и девой сторонах тела. И только для
гена sp-b в трех случаях получены достоверные коэффициев-
ты корреляции (0.54±0.07; 0,40±0,08 в 0.36+0.09) r в трех
других — маловероятные
Действие геяа иа гомологичных участках тела. Пол
гомологичными участками следует вонимать прежде всего правую
и левую стороны тела у билатеральных животных, а также
одинаковые сегменты у животных с сегиентащюнвой
структурой тела (черви). Сюда же относятся я брюшные сегменты
У насекомых, например у дрозофилы. Б Л. Астауров первым
поставил задачу выяснить закономерности феаотнинческой
изменчивости гомодяиямнчных частей в пределах организма.
На примере обнаруженной им мутации tetraptera у дрозофилы
он показал: а) асимметричное проявление признака
измененных галтеров как наиболее частое; б) отсутспяе корреляции
между проявлением tetraptera на правой и мной сторонах
*ела Этот вывод он подтвердил ряхам литературных данных,
в том числе данными IL Ф. Рокинкого об отсутствии
корреляции между количеством ^^^Г^'и^^Т^^Т^
и левой сторонах тела у дрозофилы. Интересна, что «хп же ам-
вод о самостоятельной изменчивости правой и левой сторон
тела можно сделать на основе данных о развитии пегостей у
млекопитающих, о полидактилии у человека, о лишних сосках
у коровы (работа О. Л. Ивановой).
В то же время есть отдельные случаи, когда корреляция в
проявлении (и выражении) признака на двух сторонах тела
все же существует. Б. Л. Астауров проанализировал эти случаи
и пришел к выводу, что они объясняются или непостоянством
внешних условий, или гетерогенностью изучаемого материала.
Так, если вычислить коэффициенты корреляции между стерно-
плевральными щетинками дрозофилы на разных сторонах
тела в культурах плюс- и минус-отбора, то они равны 0,11 и
0,15. При суммировании же обеих культур г повышается до
0,83. Таким образом, корреляция возникает, если материал
является генетически гетерогенным или если он находится в
разнообразных условиях среды.
Впрочем, равная вероятность проявления гена на правой и
левой сторонах тела может быть сдвинута дополнительными
причинами, действующими в процессе морфогенеза. Это
относится, в частности, к большему развитию праворукости или
леворукости у человека, основанному на морфологической
асимметрии в двигательной и речевой областях центральной
нервной системы. Считают, что есть специфические гены,
определяющие праворукость и леворукость (DD — правша, RR—
левша, DR — промежуточные формы). И. С. Гусева изучала
капиллярный узор на пальцах рук человека и пришла к вы-
Еоду, чтов популяциях чаще всего наблюдается правая
асимметрия (более сложные узоры на пальцах правой руки и др.),
так как аллель D обладает высокой частотой. Кроме того,
накладывается влияние упражнения правой руки. У левшей же
соотношения в асимметрии признаков дерматоглифики
обратные.
Таким образом, отсутствие корреляции в проявлении генов
на двух сторонах тела указывает на автономность развития
признаков в двух половинах тела, которые генетически
одинаковы. Впрочем, в указанное общее правило надо внести
поправку, вытекаюшую из высказанного Стертевантом
положения о существовании двух типов признаков: независимых,
наследственное осуществление которых в группе клеток
определяется только генотипом этих клеток, и зависимых,
проявление которых определяется генотипом не только данных клеток»
но и других, влияющих через какие-то вещества,
передающиеся гуморально. К числу последних следует отнести
киноварную окраску дрозофилы vermilion.
В известных опытах Билля и Эфруссни личинкам мушек
пересаживались имагинальные диски глаз от особей раэны*
генотипов. Во всех случаях развивались глаза с окраской» с*
3*2
ответствующей генотипу пересаженного имагиналыюго диска.
Исключением были имагинальные диски от особей vermilion,
пересаженные особями с нормальной окраской. Из них
развевались глаза с нормальной окраской. Если же личинке
vermilion пересаживались диски от нормальных особей, но не 1, а 5.
то не только имплантированные диски давали глаза с
нормальной окраской, но и глаза хозяина (по генотипу vermilion)
приближались по окраске к нормальным. Отсюда был сделан вы*
вол о существовании гормоноподобных веществ — продуктов
действия генов, способных переходить через ткани. В
дальнейшем такие геногормоны были выделены в изолированном виде
как у дрозофилы, так и у бабочек Ephestia. Отсюда вытекает,
что на признаках, осуществление которых определяется
влиянием подобных геногормонов, можно ожидать корреляцию
между проявлением гена и на разных сторонах тела.
Положение о независимом осуществлении признаков на
разных сторонах тела Б. Л. Астауров перенес и на
изменчивость сходных органов — параподиев некоторых пашет и
ножек у многоножек. И здесь корреляция в развитии признака
на разных сторонах одного и того же сегмента и на соседних
сегментах отсутствовала, что указывало на нх
самостоятельную изменчивость. Астауров рассматривал эти вопросы в
связи с общей проблемой наличия симметрии я асимметрии у
организмов, для нас же достаточен общий вывод в плане аати-
стических закономерностей в феногенезе.
Изучение изменчивости гомодинамичных или метамериых
признаков позволило более дифференцированно подойти и к
выделению компонентов средовой вариации.
Выделение отдельных компонентов в средово! вариаясе,
В одной из ранних своих работ по определению доли
наследственности и среды в изменчивости пегостн у морских свинок,
проведенной на основе сравнения инбредных и неннбредных
липни, Райт разделил средовую долю вариансы на два
компонента: общую и не общую для помета. Очевидно, что вторая
создается за счет различий во внешних условиях развития и
жизни морских свинок, первая же представляет собой
вариацию между отдельными детенышами внутри пометов I
вызывается различиями в условиях развития отдельных детенышей
инутри тела матери, а также спонтанным колебанием самого
процесса индивидуального развития.
Фолконор предложил разделять срздовую вариансу or на
общую oj,g, которая характеризует изменчивость между ос*
бями, и частную о2^, выражающую вариацию между
потомкам}! каждой самки. Величину о^ можно считать находящейся
полностью под влиянием контролируемых факторов amwtei
<реды. Величина же а^ отражает влияние иеконтра*ит*ых
условий среды. Однако такое разделение средовои вариансы
можно сделать далеко не по всем признакам. Так, в
частности, вариация числа детенышей в помете морских свинок
содержит долю и контролируемых средовых факторов, что
было показано Раитом с помощью особых методов анализа.
В этом отношении оказалось выгодным проведение анализа
на метамерных или гомодинамичных признаках. А. И. Добнна
и П. Ф. Рокицкий изучали фенотипическую изменчивость
количества щетинок на брюшных сегментах (стернитах)
дрозофилы. При подсчете числа щетинок на двух соседних сегментах
у особей популяции можно разложить фенотипическую
изменчивость обычными методами дисперсионного анализа на
компоненты вариации «между особями» и «внутри группы особей».
Первый компонент измеряет как генетические, так и средовые
различия между особями, а второй — только такие различия,
которые являются результатом вариации в онтогенезе. Зная
ор2 и og2< можно определить о£. Варианса же разницы между
количеством щетинок на любых двух сегментах может быть
принята за о2 . Тогда о* представляет собой разницу между
°е H °2es- Анализ был проведен на материале, полученном при
скрещивании стандартных нормальных линий и многих инбред-
ных линий дрозофилы. Оказалось, что если о2 в некоторых
случаях может быть незначительной и даже близкой к нулю, то
о1 составляет не менее 50% общей средовои вариансы, а в
некоторых случаях и больше.
Сказанное иллюстрируется двумя таблицами. В табл. 58
представлены данные о доле средовои вариансы ое2 в общей
фенотипической вариансе оРг при скрещивании стандартных
линий D 18 и 39 и ее разложение на о2 и о2 . В табл. 59 при-
Таблица 58. Процент средовои вариансы о2Е в общей фенотипической
вариансе о^> при скрещивании линий D 18 и 39 и доли в нем o\g и oEi
1
' Влрнансы
1
! •»,
J 4
1 °'«
! 4.
Абсолютное значение o2f,
Опыт 1
100
54
0
64
12,41
Опыт 2
J 00
60
25
35
17,77
Опыт 3
100
52
0
52
10.»
384
Таблица 59. Соотношение <j£f в о2^ • полк* сдома
при скрещивают навредит лиши, %
Компоненты
4* 1
Скреидоммыв линля |
50X24
18
82 ,
1
1 60>20
0
100
1 WW
0 |
100 |
1 Ю>1 1^29
19
ai
5
95
1
30
70
».|
ведены соотношения о* и о* (в процентах), полученные пу-
тем анализа результатов скрещиваний между многиин Инбред-
ными линиями. В целях экономки места взята лишь часть
экспериментального материала.
Значение ozEt, таким образом, представляет собой ту часть
средовой вариансы, которая не поддается контролю, так как
она отражает разницу между количеством щетинок иа
сегментах брюшка, создающуюся в процессе феногенеэа. Иначе
говоря, это как бы случайные ошибки в процессе
индивидуального развития.
Из приведенных в табл. 58 и 59 цифр видно, что удельный
вес их в общей изменчивости может быть довольно
значительным.
Нормальная вариационная кривая как выражение
суммарного действия многих внешних факторов иа развитие колишт*
венного признака. В предыдущих главах неоднократно
говорилось о вариации количественных признаков, вызываемых
внешней средой. Так как факторов внешней среды, влияющих на
организмы в процессе индивидуального развития, очень много,
то совершенно естественно выражать эту вариацию иормаль*
ноп вариационной кривой и пользоваться для ее
характеристики обычными параметрами (х, о). Это правильно для
большинства количественных признаков животных н растений. Но
не следует забывать и о возможных отклонениях от
нормального распределения: скошенности кривых или их асимметрии,
'эксцессе и др. (рис. 115). Чаще всего они возникают в силу
специфики вариации изучаемого признака. Поэтому при конкрет*
мом исследовании необходимо проверять, в кавоЛ степени
наблюдаемая вариация соответствует нормальному
распределению.
Повторяемость прививков $о времени, Выше говорилось
о повторяемости признаков на гомологичны* частях тела (ира-
пая и левап стороны, сегменты тела). В силу их автономного
25 U Ф Гомилий **
\
рДОитня корреляция между проявлением и выражением при-
•й'ака. дегермнняроваиного одним или несколькими генами на
этих гомологичных частях тела, может отсутствовать вообще
или создаваться какими-то дополнительными факторами.
Наблюдается несколько иная картина, если изучать
повторяемость признаков во времени. В ряде случаев такая
повторяемость есть результат цикличности в жизнедеятельности
организмов. Например, у коров повторяются лактации, каждая из
-Jfe -25 -£ X 'to *2б *3б
а
Рис. 115. Нормальная вариационная кривая (а);
асимметричные (б—минус, е — плюс); эксцессивные (г-
плюс, д — минус).
которых характеризуется общим удоем, процентом жира в
молоке и т. х Между показателями отдельных лактации
существует вариация — большая или меньшая в зависимости от
признака. В свою очередь между этими показателями может быть
корреляция. Подобно этому, у многоплодных животных
повторяются пометы (окролы, опоросы и т. д.), в каждом из которых
имеется определенное число детенышей. Но возможна
повторяемость и не циклическая; например, у одних и тех же
животных можно изучать, как варьируют во времени те или иные
биохимические или физиологические показатели. В то врем*
как вариация между различными частями организма
определяется преимущественно причинами, создающимися в процесс*
феногекеза, вариация между показателями организма в те**"
иие его жизни зависни главным^ JiOjLajojt, от колеблющихся
условий внешней среды. Но так 1сак зависимость между покз*
зателями во времени можно изучать только на нескольких
ЗВ6
организмах, то в вариацию по мим неизбежно входят а
генетические различия между отдельными особями.
Для изучения повторяемости признаков во временя могут
быть применены различные методы. Простейший яз них —
вычисление коэффициента корреляции. Многие работы
посвящены установлению корреляционных зависимостей между
показателями соседних лактации, процентом жира в молоке,
количеством детенышей в помете н т. д. Так, Касл и Сярл изу-
чали корреляцию по молочному жнру между
последовательными лактацнямн коров; Сякка—корреляцию по удоям. Прм
этом самые высокие коэффициенты корреляция были
обнаружены между соседними лактацнямн, например 2-й я 3-й, 3-й в
4-й. Значения г достигали 0.7—0,8. Но чем дальше отстояли
друг от друга лакташи, тем меньшей аановнлась корреляция
между ними, величина г снижалась до 0,3—0А а иногда для
очень отдаленных, например для 1-й и 10-й, даже становилась
отрицательной.
С другой стороны, зависимость между размерами пометов
у серебристо-черных лисиц, по нашим данным (иеопубл.),
выражалась низкими коэффициентами корреляции: для соседних
пометов г=0,10±0,02 (по 2218 коррелированным парам)
I! между пометами через один (1—3, 2-4, 3—5 в т. д.)
г=0,16:±0,04 (по 513 парам пометов).
Второй метод — использование внутриклассовой
корреляции riCl с помощью которой вычисляется коэффициент
повторяемости rw:
где Ok2 — компонент вариансы между особями, а о,2 —
компонент вариансы изучаемых показателей внутри тех же особей.
Для вычисления rv используется чаше всего однофактор-
ный дисперсионный комплекс. В качестве примера можно
привести данные из работы Унта и Ряггса по изучению
длительности беременности у мясных коров (табл. 60). Авторы примени*
ли также двухфакторный иерархический комплекс, в котором
учитывается роль отцов (табл. 61).
Метод вычисления коэффициента повторяемости с
использованием дисперсионного анализа широко применяется и со-
ветскнми авторами (3. С. Никоро, Л. А. Васильева, А. А.
Ильинский и др.).
Поскольку элементами дисперсионного комплекса являются
не один показатель для каждой особи, а много, то
компонентов (или 0/*+о.г) включает не только генетическую вариацию
между особями (ос*)* но и некоторую долю случайных ередо-
вых влияний, определяющих различия между повторяющимися
Т^.*ш<а 60. Атакриюнмы* шшяи ялипльшктт берсмсшак!*
> ист норм я вычисление коэффициента повторяемости
Корояы
Деятельность Леремен-
ностм олиоя я то* же ко-
рошы по рлмым отелам
Число
степей*А
г»оооды
25в
285
oj - 30,6
о!
'ш
Ср*Д»М
кмдрат
47.8
30,6
Структур» tptgatt*
' о'- 2.08в*
<*
47.8 — 30,6
2.08 " в"2
8.2
" 8.2 + 30.6 -"—
Таблица 61. Дасверсшмяы! аяал-яэ длятелыюстн береметюств
у «ясяых коре* по яерярхяческоя схеме и
1кто«мим мра-Ашм
Прояыюдятеля
Коровы от сяясовшаявя
с хжшлнм яроазяодятедем
Длятедъяость беремвя-
яостя одеон я то* же
*ОроШЫ ПО рвЯМО! <УПЛ*Ы
вояторяемостя
Чшсло
степеней
свободы
20
366
167
of ~ 32,2
о? •*»
.;-
f ф •» •
Средами
КВ1Др«Т
139,9
36.1
32.2
38,1-32.2
—гга—
139,0-36,1
~тя—
2,6 + 4,1
М + 4.6 +
•
Структур*
среднего киДОТ»
o?+l.38tf? +
+ 21,860*
о*+».«<>•/
<*
- 2.»
-4.6
, Л IO)
35^-o.tt ]
показателями особей <*у.л***, ,«,„,„^^^
повторяемости представляет сабо*
Повторяемость выражает юлю вараавш огдеяьянх ш*
торяющихся показателе*, которая «ищется скапввем раза*
чин между особями, как генетическая таи в средовых
Коэффициенты повторяемоств в наследуемого евшим
между собой, что вадио из формулы дав г*. Считают, что *ь>
эффиииент повторяемости указывает как бы ва верхнюю
границ)' наследуемости. Еаа nef левых о наследуемого, то
коэффициент повторяемости аает врвбпзвюшюе представлена*
с величине наследуемости. Мы не останавливаемся баке *-
тально на соотношении между гш в А* Укажем лит, то
следует учитывать различие в методах их вмчведенвя. Коэффявв-
ент повторяемости всегда вычисляете! иа основе ещщчвш
показателей, а коэффициент наследуемости мажет быть ам-
числен на основе и единичных в средвях ноказателе*. Есть
формулы, связывающие гш с иоэффкяеитамв шеледопетн,
вычисляемыми обоими методами.
Но в общем прв низко* иаследуемоста щ шовмрвйшм»
должна быть слабой и наоборот. Этвм объясняются вогучае»
мые в опытах разных авторов различав в юффяажягах
повторяемости содержания вира в мовоке хорош в числа
детенышей в пометах лисив. Как взвество, верам* врдаи
характеризуется значительно более ниеосой наследуемостью, чем
второй.
ЛИТЕРАТУРА
Общая
Актуальные проблемы современной генетики. Под ред. С И. Алши-
яяиа. М". 1966.
Беляев Д. К. О некоторых проблемах корреляционной изменчивости а
ее значении для теории эволюции и селекции животных.— Известия СО АН
СССР. № 10, 1962.
Брюбейкер Дж, Сельскохозяйственная генетика. М.. 1966.
Давыдов С. Г. Селекция сельскохозяйственных животных. М.—Л., 19Э&
Дарвин Ч. Происхождение видов. Под ред. Н. И. Вавилова. М,— Лп
1937.
Дубинин И, П. Эволюция популяций и генетика. М., 1966.
— Глембоцкий Я. Л. Генетика популяций и селекция. М., 1967.
Лепер П. Р., Никоро 3. С Генетнко-математические основы различных
методов оценки племенных качеств животных. Новосибирск, 1966.
Лобашев М Е. Генетика. Л., 1967.
Майр Э. Зоологический вид и эволюция. М, 1968.
Математические методы в биологии. Под ред. В. П. Чтецова. М.г 1969.
Нейман Ю. Вводный курс теории вероятности и математической статн-
стяки. М., 1968.
Никоро 3. С, Стакан Г. А., Харитонова 3. И., Васильева Л. A.t Гимг
збург Э. X., Решетникова Н. Ф. Теоретические основы селекции жиаюткых.
М. 1968.
Рокицкий Л. Ф. Генетика. Общий курс. Изд. 4-е. М., 1937.
— Биологическая статистика. Изд. 3-е. Минск, 1973.
Руководство по разведению животных, т. 11. Генетические основы
продуктивности и селекции. Под ред. X. Ф. Кушнера. M.t 1963.
Серебровский А. С. Генетический анализ. М., 1970.
Смит Дж. Математические идеи в биологии. М-, 1970.
Снедекор Дж. Статистические методы в применении к научным
исследованиям в сельском хозяйстве и биологии, М., 1961.
Сэджер Р., Райн Ф. Цитологические и химические основы наследстве*-
иостн. AL, 1964.
Теоретическая и математическая биология. М, 1968.
Фишер Я. А. Статистические методы для исследователей. М, 1958-
Фриц-Ниггли X. Радиобиология, ее основы и достижения М., 1961.
Холден Дж. Факторы эволюции. М.— Л.. 1935.
Шмальгауэеп И. И. Факторы эволюции. М.—Л., 1946.
— Кибернетические вопросы биологии. Новосибирск, 1968.
Шеффе Г Дисюсяошши анализ. М-, 1963.
Эрлмх П-. Холя Р Процесс эволюция. AL 1966.
AUard *. W. Principles of plant breeding. 1 Wiley a. Sons. N, Y — La*
dor., 1980
Bttfiutbemalks. V. I. Mathematical topics in population Genetics. &*
t> Kohraa К S. Springer Verlag Berlin—N. Y- 1970.
Biometnca! Genetic» Ed. by O. Kemptbonie. Pcrgatnon Pms, LaaiW
Orforo-*. Y.-P»ris. I960.
J9D
Falconer D. 5. Introduction to Quantitative Gcnettci, Oliver and Boyd,
Edinburgh—London, I960.
Fisher R. A. The correlation between relatives on the suppoittlon of Men
delian inheritance. Tranc. roy. Soc. Edtnb.. 52, 1918.
— The Genetical Theory of Natural Selection. 2-nd Ed., New York Publ.,
Inc., 1958.
— The Theory ol Inbreeding, 2-nd Ed., Oliver and Boyi Edinburgh-
London, 1965.
Ford £. B. Ecological Genetics. Methuen a, Co., J. Wiley a. Sons* Ion
don-N. Y., 1964.
Hogben, Lancelot. An introduction to Mathematical Qenttici Norton ami
Co.. N. Y.. 1946.
Huxley /. Evolution. The modern synthesis. G Allen a. llnvtn ltd.,
N. Y.— London, 1963.
Johansson /. Genetic aspects ol dairy cattle breeding, linlv. IIHnoin Pnm,
Urbana, 1961.
Kempthome 0. The correlations between relatives In random mating
populations Cold Spring Harb. Symp. quant. Biol., 20, IBM.
— An Introduction to Genetic Statistics.' F. Wiley and Son» and Ctup-
man. N. Y., 1957.
Lerner L M. Population Genetic» and Animal Improvement. Cambr. Univ.
Press. London, 1950.
— Genetic Homeostasis. Oliver and Boyd, Edinburgh, ИЙ4.
— The Genetic Basis of Selection. J Wiley and Sons, N Y, 1966,
Li С С. Population Genetics. Cambridge linlv. Press, Chicago-London,
1955.
Lush J. L. Animal Breeding Plans. Iowa State College Press. Артеа, IMS
— Breeding structure of populations, I. General consideration. In
«Statistics and mathematics in biology». Ed. b. 0. Kempthoma Iowa State Cnl
lege Press, Ames, 1954.
Malecot G. Les Mathematlquea de I'Htredlte. Masson tt CJe., Paris, 1941.
Mather K. Biometrical Genetics. AMhuen and Co., London. IMfc
— Statistical analysis in biology. Fifth ed., Msthuen, London, JW6.
— Jinks J. L. Biometrkit genetics. Chapman a. Hall, London, 1971
Mather W. B. Principles of quantitative genetic*. Burgees Pub Co, Minn.
USA. 19ГА
Newman W. £,, Freeman F. H„ Hettinger K. I Twins: a study ol here
dity and environment, Univ. of Chicago Press, №37.
RasmuKon At. Genetics on the Population Ые1. Svenske Bokrirllsel,
Stockholm—London. 1961.
Stahl W.> Rasch P. SlUr Ц. Vdchal L Populations GenetUt fur Unu
chter. Landwirtsch. SL-Verlaf. Betlin-Preg, 1900.
Statistic* and Mathematics in Biology, Ed. by Kempttiorne 0. sod otheri
Iowa State College, Ames. Iowa, IMf
Wallace B. Topics in populsW» genetics. W W. Norton a. Company.
N. Y., 1968.
Weber'E. Ma them* ti id* Gmndlsgen dor GeoeUk. f\*Am Vetted lens,
1967.
Wright S Correlation and causation. J Agile, k*. 20, №\
— Systems of Mating. G*ndies,6, 19*1
— Evolution In MendetUn Peftdettooe. <fc"^Je'J*] . ,W-AI
— Inbreeding and outbreeding systepst P«* Nal Acad, to <w*ee^
,9' -^Statistics! genetic. lei reM«» * n*U«< Hemun . G». Ms.
~ Evolution *** tfa* &"*** * Г9рш
Pre**. 1968, 19Ш »i
if Jill I
Г &*;** В И Апа.шэ ****** m*V*m * " ^™ыюго |Лстш Ош*
"TJSS^ "~™"*<* pewnonnaima Л. 1WI.
5СЕ£ К Ж Гевептес!»! ммм • отпошспма ыедлеяяо пдемя***
**7 С*птГпад истштеямсыад гибридами М. 1965,
~ а /С Допамтольстео а^барателыюста омодотаорема у
w m**n«*v-««ponnsauae*, М ^ «W0
Сумм Л. Ш* Т. »** > Цат*<**етпка. Л, 1969.
3nW*ro* В /ГО роли эаспсраиекта а апфр а седкс«>жо*аАстае«па1
1ап^о^Ь*а&МШ^
Btemiee ■ Ntammcxpft генетику. Ил 2-е. Л, 196В,
AVtrff** С & Non-disiunction of the ee*<hromosomee of Draaoputtt
Awn Еж* ZooU Л 191X
Orientf ft. £ The cytogenetics of Oenothera, Advances of genetic*
CaAr 7. L. Sftad««/Dr4 ft. White spotting in the fox Amer. Nat*..
T~ tan
ЛЫ**г ft. The analysts of single factor segregation. Ann, Eugen.. 8» 19W.
Ии> /. M The production of nondisjunction by X—rays. Joura. El».
ZooL 1Ш
Akrfott Г W.. Bridges С В Sturtevnt A fi\ The genetics of Drosqnt*
ia. Bibttagriphu Genetic** 2, 1925l
«Чрашая Н The analysis of tetrad data. Genetics» 37. 1962.
ftnoaccs M M, Dtmifisty E. The effect of abnormal chromosome W en
; rrfmntial Mffapabon and crossingover in Maixe. Genetics, 53. 1966.
Safv S ft Genetic and cytotogtcal examination of the phenomena St
pcmafy nor*-disjunction in Dronophita melanogaster. Genetics, & 1920.
Wtcrdrm В L m 4*r. Mendel's experiments. Centaunts, 12. 196&
T**b€tM Г. Method* vnd Fehlerquellen der Unlersuchung auf Mend*
Khe ZeWen beim Menachcn. Arch I Rass. u. Ges. Biol.. 9, 1912.
- Mathematiscae Grundlegen der Probandmeihode. Z. Indukt Abet, ж,
Vcwbungriebft, Л 1987,
***** 5 The Effects in Combination of the Major Colorfactors in the
future ftg Genetks. 11 1927,
Гли *
Д*еа« А. В , A?man H, П. Поашвеяпаа споатакпая ыугабпяьностк J
apoauaei Seech, camietee*— «Гепетпка». 6.Л9. 197a
ДуВшшт H Я^Мовааударпаа runenuu я действа* падуют* па пава**
niwim ia M., 1963.
^ — ^п»и| Д /J Иване аопросы соаремешоа теорем ыгп
*jELSym^ VSft^,!t—ftMU> (nyweuns. мутация)*. Я, 19»
14 1991* Ппарешщиаа хромосом поюимруюпиша па
*ап» паи еюшмщаа пееяеастштвоа аамеичипости у спаъежохоааит*»*»
Г|И^Йвь €ГвИП|,,|9 • седеаша сааааюхоаяЪстаепкых mnaai*ff»i
па111т.Шг1<'М>->|ИМ1Уа >агУсстпеппого отбора пря астастпаппоп « gjj
qvmwiammo пмапаапяя реатгепоаскпм опд^аавшвм тямтйшшлетш*-** р*****
11ап*лаш*илм Г М . Хршг^с Т, р Шшштмтм nit*
iU .wmmik мутаций у c,i шмтям. CooW 111 Вшп» *ЙЕ2***
; кромка - Бис* журил, т 3. шп\ \пГ^ ********** щ f,
Современные пройдены радлашюишо! геаюкя. м IW
luMofxt* РесоесшнЛ Н >., Wahwv. в. tf<§ JfofwcSii Н Пг»—
принципа попадаииа а оаш6акмюг»а. Я, |*& ' lf?wn«at
Ли/ О.. Кеиерео А Стожастес*** радиюбпоаолм. М 1901
Цимыер М Г. Проблему коджеггаатоа радмбамаолш. 1С щ»
Шгр6а*о* В * Естестаева** vyrwomiiil «ponec Септет**,*
закономерности) ~-Сб «Итоге neyim. ««on mamma (иуташ**, ^у
ции>» М. 1969.
Auerbach Ch. Mutation. Аа MttMlecUon to mwiiili on ranlmumi
Part 1 Methods. Oliver a. Boyd. Edinburgh-Umdom, ДО, —"-*«—«>
BofiniVr G,, Lining К. О Studies on X*at awtatloam te Ц» **fe «j
forked )oci of Drosoprula meUnog aster I. A statistical onalyois of maUitoft
frequences. Hereditas. 3&> 1949,
Ban* Я Л ParamuUUon at the R locus k mte. Cotf Sariuf ft»**
S\mp. Quant Biol, 23, 196ft.
Catchesid* D. G. Uu D. 5,. ffcoday /> *t Type ol ггаомжцщ n^a*
ral change induced by the Irradiation of Tradescaatia «tempore! Joorl oi
Gentt. 47, l9>45-194o.
Clayton G.. /?oo«rf*w A Mutation and ojotntitatfetiwiatta. Amer, Nat,
Conner В V. Ci)« Tbe cytofoneUc «fled of sto neumm Genstks,
35 1950.
£ti2'i£ /f /, Dose respoo» relations from In ribo stod** Hwan rata-
tion cytogenetics. NortbHoUtnd Publ. Сотр.. Amsterdam, 1K7.
Ifaldane /. B. The rate of spontaneous mutation of a faunae до. I 0*.
net.. 31, 3, 1935,
Lea D. E. Catchstde 0. G. The mecfctfdsm of todsdfeo ay ratiatlon of
chromosome aberrations in Tradescantia> J. Genet H 11* 190.
- Co«/5on С Д. The distribution of the Humbert of mrtmts t» Bacterial
population. J. Genet., 4. 1949.
Lederberg /., Uderber* £ Replica plating and tedfaod sake*» of bae-
terial mutants. J. Bad» 3, 1961
Luria S. £.. Mbrick M. Mutations of bacteia horn fin» sensitivity la
virus resistance. Genetics. 99* 1941
MichaUofl A S. Cber die VamtMtf dar WatsxiuBttet bei dor 1Ш+
biene (Apis molH(era). Zetttb. f md. Abi VtHetve. Щ ml
Muller H. J. The problem of genetic modification. L Ы. AbaUoBnoitgs-
u. Vererbungslehre. Supp. 1: Verb. Y Intent Goftgr Vembonfraisa» ДО
— The measurement of ftne mutation rate in DteeapWa, Hi ajfh
nubility and its dependence upon temperatufe Ccnetka» Л Ml
— The manner of production of mutation* by ntftatfa*. Ik OWitboo
biology», 1. Mc Grow НШ, N. Y. 1954,
A eel ! V. The study of Human mutation rates. Ал Nafcrsfe* «i Щ
A ormon A*, Otbrim* R. L. Smrt At. Vaamatf *. С Ito beoueney Ы
dicentrics In human kucocylw IrradiaM in ^ivo and in vUiol RaJtofefy. tl
106. 1964.
RadiaUon biototfy* Ed. by Hofiaander A. Vol. I Mc Grow HUl N. T-
1954
Ы1 S. Я. Evkknce lor a doa^uamdleimin tedoae««^^«w
of real ch^matid disoonttnuWas ««^^ Хча^ ^ аоял W*8"*41
™Ч^**№ a FUeba, V^at.
,fnM,Sif К L G^toHt efkds of ra*thm to «mm* !* UWUtkm
Biologyt, I, N. Y.» ItH. ^
J& К. AnaJy»» oJ X ray induced chromosomal aberrations in Tree*,
^Tu^^ReS^SSs *■ The experimental production of mutaUow.
***"zfaii9??; 6. Biopbysik. Bd. I. Da» Trellenprinzip in der Biologfc *
Hind Vertof. Leipiig. IM7.
Главы З н 4
Леля Б. Н Воэможаость анализа признака в популяция.— Ж. икц
Лшли у 4 ■"-»" I 1928.
^Воробцовс И. £. Гользберг К Л. Влияние индуинроваккых геииы» «у.
гааай в^етероэнготжж состояния на жизнеспособность организма.-«Гене-
Т^ерш*^зо* ' С. М. Генетическое строение природных юшуляциА..-
/•С общ. биологии, т.7,Л1 1946.
Дубинин Н. /7., Ромашов Д. Л- Генетическое строение вида н его эй*,
латая — Бнол. журнал, т. I. вып. S-6, I902.
/>агм*?р В. А. Математические модели в популяцнонной генетике: частот-
ны* дггермяжаротанные модели.— Сб. «Итоги науки. Математические
методы а бнологяи. 1968». R. I960.
Рокмцкий П. Ф Попул-щноиная генетика и ее значение для селекции
мастных,— Сб. сГенетнческне осяоаы селекции животных». М., 1969.
Селрежее Ю М. Нарушение панмиксни в популяциях.— «Генетика»,
т. 4,1* 12. 1968.
— Тимофеев-Ресовский И. В. О равновесии генотипов в модельных т>
•гуляниях.— Сб. «Проблемы кибернетики». St 16. 19G6.
Серебровский А. С. Генетический анализ популяций домашних кур
горцев Дагестана.— ЯС экся. бнол., т. 3, 4. 1926—1927.
— Проблемы н методы геногеографня.— Тр. Всес. съезда по генетике,
селекиян, семе*юводству н племенному животноводству 1929 г., т. 2. Л., 1930.
— Геяогеографяя домашних кур Кабарлы и Балкарин. Успехи зоотех-
явчеосих наук, т. 1. 1935.
— Геиогеографи* кур Армении. Успехи зоотехнических наук, т. 1, 1936*
Тшмафеее-Ресоеашй Н, В.. Свирежее Ю. М. О генетическом
полиморфизме в жтудяняях. Экемеряментально-теоретическое исследование,—
«Генетика», т 3. >■ 10, 1967.
Об адаотаанониом полиморфизме в популяциях Adalia btpuoc-
tata — Сб «Проблемы албернетнкн». Nt 16. 1966.
Поатуляхшонная генетика и оптимальные процессы.— «Генетика».
т. 6. ** 10. 1970.
Четвериков С С. О некоторых моментах эволюционного процесса с
точки зрения совревкимои генетики.- Ж шел. биол.. сер. А, т. 2, 1926 (переил*
* 1ш ^i современной генетики. 1920—1940». Л„ 1968).
ЛШшоп А. С. The tickle<ell and Haemoglobin C in aome African
population*. Ann. Hum, One*, 21, 1926.
Т*/*01? 5?1 «?<*tte-cefl polymorphism. Ann. Hum. Genet., 19, 1954.
tk*. 52 НйбГ terum *r*n*'err,ni; a balanced polymorphism. Оене*
Шл^й^яТ* и^шжУ in ""*" population* of mutant DroaophiU. Bf
rl!lA/ *i ^Л?!"** Ы •■"■PWve polymorphism. Amer. HbU 88. 1954.
mii^/:'-Breejdwg rtnictpr* of populations. |J. Effective populaifr*
P^i\wee *l»i mathematica in biology». Iowa atate ОМЮ
ftfir^Jr^f i'^RP'S^ t>«*>bm% In natural populations Proc 3**
Юга Symp. on Math. StaC Pro*., 4. 1965
The theory o/ genetic lomee Genet, Today. 3, 1966,
— Morton N. E. Measurement of gene it***™*» ^
ons. Evolution, 9. 1966. gW "c**wy Mi • *«*И PPPflUtt-
Gruden D, The compulations of jnfarttrfiae cotfHrfmU bi *t~^ .
tions. J. Heredity. 404 Ш ^ «швеей in t^ed p^*
Dobzhansky Th. Mendelian populations and their evolution, к «g,*.^
in the 20-th centuryj> MacrmUao Co., N. Y. )IM. ' ^nsn
Usher R Л The genetical theory of natural selection 2«d td Dom
N, Y 1958.
Ла/Ш AfoWo and oth. A »tudyof inbreeding in some isolated populations.
Japan J. Human Genet., 12, 4, 1966.
Haldane /. 5. 5.. layakar 5. ft The solution of some eqtutioos occuring
in population genetics. J. Genet., 66,3, 19©.
Hardy G. H. .Mendelian Proportions in a Miied Population. Setae*, 28,
!908.
Kerr W. E, Wright 5. Experimental studies of the distribution ot me
frequences in very small populations of Drosophila mttaoogaster. EtolJion,
8. 1954.
Kimura M. Process leading to quasi fiiation of genes In natural poouli-
lions due to random fluctuation of selection intensities Genetics, 39, 1954,
— Solution of a process of random genetic drift with a continuous
model. Proc. Nat. Acad. Sci. USA, 41, 1955.
— On the change oi population fitness by natural selection. Heredity. 12,
1958.
Li С. С The stability of an equilibrura tnd average fitness of a
populations Amer. Nat.. 89. 1956.
Mac-Arthur R. H. Some generalized theorems of natural selection. Proc.
Nat. Acad. ScL USA, 48, 10$.
Merrel D. I, Selective mating as i cause of gene frequency changes in
laboratory populations of Drosophila meJsnogtster. Evolution, 7, 19(1
Rasmusson M. Variation in persistency d a lethal mutant in
populations oi Drosophila melanogaster. Hereditas, 66,1,1970.
Smith Cedric A. B. A note on testing the HardyWeinberg law. Ann.
Hum. Genet, 33, 4, 1970.
Stern C, Timofeeff-Retsowsky N. W. The HardyWetaberg law. Science,
97, 1943.
Turner /. R, Fundamental theorem of natural selection. Nature (EogL),
215, 5105, 1967.
Wright S. On the roles of direct and random changes in gene frequency
in the genetic population. Evolutioo, 2, 1948.
— The genetical structure of populations. Ann. Eugea*, London. 1& №1.
— The theoretical variance within and among suhfivisioos of a
population that is in a steady state. Genetics, 37, 1962.
— Kerr W. S. Experimental studies of the distribution of мм fmquen-
ties in very small populations of Droiophila mdanogaster. fl. Evolution,
8, 1954.
Zarnecki A. Model losowego dryftu genetycznego w так) populacji
zamkni^tej. Acta agrar. et silvestr. Ser. Zootedm., I, 1967,
Глава 5
Вопросы гснегнкя м селекции. Малсж. 1970.
Добина А. И. Рааложе*** гешт/ьаей мршасы is юуаяаяиты, »а*
сящис от аддитивного м иеаддигимого дебетам г*яо*,~С& *Вошш
математической гелегимл». Маис*. 1969 г
— Некоторые результаты мсосряиаетадмтго иучвям июоаинтгт
геиетическиЛ «ершила — Сб. «Вопроси геяетиал и оглашай*. Маю. 1970.
Игнатьев М В. Колщвтаешш* шшллшл деАстеяа аасаедетаещяот и
среды.— Биаг журнал, т. 2, шыя, 4—& |Ш
m
Яолчковекий В. И. Наследственный полиморфизм крупного рогатого
с«*Га и овец БССР по некоторым белкам крови. Автореф. канд. длсс
Минск, 1971.
Рокицкий /7. Ф Генетический анализ числа грудных щетинок у Огою.
phi/a melanogaster.— Ж эксп. биол.. сер. А, т. 3, вып. 3—4, 1927.
Тарутина Л. А. Генетический анализ свойств инбредных линий кукурузы
в системе дмаллельных скрещиваний. Автореф. канд. днес. Минск, 1969.
Турбин Н. В., Хотьиева Л. В., Тарутина Л. А. Генетический анализ
некоторых количественных признаков у кукурузы.—Сб. сВопросы математ».
ческой генетики» Минск, 1969.
Анализ доминирования в системе диаллельных скрещиваний.
Там же.
Филиппенко Ю. А. Изменчивость количественных признаков у мягких
пшениц — Изв. бюро генетики, J& 4, 1926.
— Частная генетика. Ч. I. Растения. Л., 1927.
— Частная генетика. Ч. II. Животные. Л., 1928.
Хотъигва Л. В., Тарутина JI. А. Оценка наследуемости количественных
признаков в популяциях кукурузы на основе диаллельных скрещиваний,—
Сб. «Вопросы математической генетики». Минск, 1969.
Anderson V. Z.t Kempthorne О. A model for the study of quantitative
inheritance. Genetics, 39. 1954.
Baker M. L., Наге1 L X., Reinmiller G. F. The relative importance of
heredity and environment in the growth of pigs at different ages. J. Anim. Scl,
2, 3, 1943.
Becker W. A. Manuals of procedure in quantitative genetics. Washington*
1964.
Beil G. AT, Atkins R. E. Inheritance of quantitative characters in gain
sorghum. Iowa State Journ. of ScL 39. 3, 1965.
Binnet F. E. On total additive genetic variance in the case certain
mating systems. J. Genet., 58. 2, 1963.
— Morris J. A. On the total hereditary variance in the case of certain
mating systems. J. Genet., 58. I, 1962. _
Castle V. E. Heredity in Rabbits and guinea pigs. Bibl. Genetics, 1T1926.
— A Further Study of Size Inheritance in Rabbits with Special Refereo-
ce to the Existence of Genes for Size Characters. Journ. Exp. Zool, 5. 1929.
Cockerham С. С Partitions of hereditary variance for various genetic
models. Genetics. 44. 6, 1959.
Comstock R. £.. Robinson H, F. The components of genetic variance in
populations of biparental progenies and their use in estimating the average
degree of dominance. Biometrics. 14, 1948.
Cooke P., Jones R. M.. Mather K. et al. Estimating the components of
continuous variation. I. Statistical. Heredity. 17, 1, 1962.
— Mather K. Estimating the components of continuous variation. If. Ge-
r.etical. Heredity, 17. 2. 1962.
Dawson P. S. Estimating of components of phenotypic variance for
developmental rate in Tribolium. Heredity. 20, 3, 19&.
East E. M. A men deli an interpretation of variation that is apparently
continuous. Amer. Nat., 44, 1910.
— Studies on Size Inheritance on Nicotiana. Genetics, 1, 1916.
Fisher R. A The correlation between relatives on the supposition of men*
dehtn inheritance. Trans. Roy. Soc Edinburgh. 52. II, 1918.
— tmmer F. R., Tedin C. On genvtical interpretation of statistics ot the
third degree in study of quantitative inheritance. Genetics, 17, 1932.
Fraser A. S., Hansche P. E. Simulation of genetic systems. Major ana
mwor loci. Genet. Today. 3. 1965.
Haiei L. N.. Lamoreaux W. F. lleretabilities, maternal effects end nlcklfl*
?«ffUe*?1 lo **1ив1 maturity and body weight in white leggoms. Poultry
Sci- 26. 6. 1947.
i954W«^e« B '• т** theory tnd analy* of «m c^ j. o*^ a,
-The separation of epistasia from .<ШКе and dttnmanee variation Hi
Klпегаtion means. I. Heredity, 12, 1968.
— The separation of epistaait from additive and dommtnet variation in
generations means. II. Genet ica, 31. I960.
Jinks J. L. The analysis of continuous variation m a dUtWIaf Ntofoaa
rustica varieties. Genetics, Э9, 1054.
— Perkins I. M., Breese E. L. A genera! method of detection additive,
dominance and epistatic variation for metrical traits. Heredity, 24, 1962.
Kan /.. Kruger W. F., QuUenbtrry I. H. Nooeddttive gene effect! of si»
broiler traits as studied from a seriet of diaUtl mating*. Poottry SeL 38,
1959.
Kearsey M. J. Biometrical analysis of a random mtmtjmAilWL
a comparison of a five experimental designs. Heredity, 20, 21, 1965.
— Jinks J. L. A general method of detecting additive. <кнпшаяее aid
epistatic variation for metrical traits. I. Theory. Heredity, Д 3, 1961
Kempthorne 0. The theoretical values of correlations behm relatives
in random mating populations. Genetics, 40, 2, 1955.
— The correlation between relatives in random mating populations. Cold
Spr. Harb. Symp. quant. Biol., 20, 1956.
Kidvell /. F„ kempthorne О. Ал experimental teat of quantitative
genetic theory. Zuchter, 36, 4. 1966.
King S. C, Henderson C. R. Variance components aaelvsji in heriUbiaty
studies. Poultry Sci.. 3-3, 1954.
Kopec St. Studies on the inheritance of the weight of newborn rabbits.
journ. of Genet., 14, 2, 1924.
Lindstrom E. W. First Chromosome Genes in the Tomato. Genetics. 17,
1932.
Mac Arthur I. W., Butler L Silt inheritance in the toaatft. Genetics,
23, 1938.
Mather K. The genetical theory of continuous variation. Hmtttas. SuppL
V, 1949.
— The genetical units of continuous variation. Proc 94b lnttmt Congr.
Genetics. Caryologia. 6, suppl. I, I9S4.
Miller R. #.. Legates /. £., Coekerham С С. Estimation of «maddttifi
hereditary variance in traits of mice. Genetics, 48, 2, 1963.
N elder /. A. The estimation of variance components in certain types of
experiment on quantitative genetics. In: cBtomeirieil Genetics». Pergamen
Press. London—N. Y.f 1960.
Nilsson—Ehle #. Krcuzungsontersuchimgtn an Hafer iaad Wettet. f.
Lunds Univ. Arsskr. Adt, 5, 2, 1909.
— Kreuzungsuntersuchungen an Hafer und Weuen II, Liteds Univ.
Arsskr. Adt., 7. 6\ 1911.
Nordskog A. W.t Kemthorm O. Gewtype-environment interacts» i* rao-
dom sample poultry test. In: cBtotnetrical genetics», Pergaaaon Press,
London — N. Y., 1960.
Pease Af. Experiments on the inheritance of «efght in rabbits. Journ. of
Genet., 20. 2, 192a
Prabhu 5. S, Norain P., Stofk H P.. /Свое/ 5 Analysis of 6X6 diafkt
cross for hatchability in Drosophila meUnagister. J. Gee*t, 80, 1.1968
Punnett /?.. Baihy *. On the inheritance of weight Joan», of GwL
'Redman С S.t Shoffner R. ff. Estimates ^ е«Ф"№у pwmetws ***
zing a poryallel crossing system. Podtrv «jJ^JEL,^ ,« .w шш
Ritzman E. О The Inheritance of ste аЫ ceafcrmitton to aheeiv New
Memo. St. Techn. ML 35, 1823. " шпЛш^т «j -^^^ Dtflw^
Robertson A. Experimental design h» the evaMtoon « genet*
parameters. Biometrics, 15, t Ив* ж
Rofiton h\ V. Studies in quantitative inheritance. XI. Genetic and tn-
\iiwin:<.nta! correlation between body size and egg production in Drosophila
OKtanogasUT. J. Genetics. 55. 1957.
Ssisfert IF. Untersuchungen uber die Vcrerbung quantitativer Charactere
an \rabidopsis thaliana. Zeitsch. f. Pfl-zuchtungt 42. 1960.
Sheridan A. A'.. Hrankham /?.. Jones L. P., Rathie /(. A., Barker I. S, F.
Partitioning of variance and estimation of genetic parameters for various
bristle number characters of Drosophila melanogaster. Theoret. and Appl.
Genetics. 38. 5. 1968.
Spickett S. G.. Thoday A Af. Regular responce to selection. 3. Interaction
between located polygenes. Genet. Res., 7, 1, 1966.
Taneja G. C. Ravat S.. Kumar S. Further estimates of covariances
between full sibs and half sibs for bristle number and wing length samples of
Drosophila bred to varying levels of inbreeding. J. Genet., 59, 2, 1965.
Turner J. R.'G. On supergenes. 1. The evolution of supergenes. Агпег,
Nat.. 101. 919. 1967.
Vat kin W. Quantitative inheritance and the implications of gene action.
Gene e cult., 19, 1. 1967.
Vessety E. Ch. Ein Beitrag zur Schatzung der nicht additiven genotypi-
schen Varianz bei Grosstieren. Abh. Dtsch. Acad. Wiss. Berlin, KI. Math, Phys.
und Tcchnik. 4. 1967—1968.
Vright S. Analysis of variance and correlations between relatives with
respect to deviations from an optimum. J. Genet, 30, 1935.
— The genetics of quantitative variability. Quantitative inheritance.
Papers read at a colloquium at Edinburgh (1950). Ed. by Reeve E.C.R. and
Waddington С H.t London. 1952.
Глава 6
Беляев Д. К. Об интенсивности серебристости серебристо-черных
лиенц,— Тр. ЦНИЛ Главпушнины, Л? 3, 1940.
— Киселева 3. С. Генетические параметры основных компонентов
молока у крупного рогатого скст а.—с Генетика», т. 2, № 10, 1966.
Васшъе'ва Л. А. Использование коэффициентов внутриклассовой
корреляции для оценки наследуемости и повторяемости.— Сб. «Вопросы
математической генетики». Минск, 1969.
Гинзбург Э. X. Сравнение показателей силы влияния.— «Генетика»,
т. 5. Л* 4, 1969.
Игнатьев Af. В. Количественный анализ действия наследственности
и среды — Бнол. журнал, т. 2, вып. 4—5, 1933.
— Определение генотипической н паратипической обусловленности
количественных признаков при помощи близнецового метода.— Тр. меди*
ко-бяол. ин-та, т. 3. М.. 1934.
Занертяев Б. П. Наследуемость оплодотворяемости у телок.— «Сель-
скохоэ биология», т. 6. Л? 3. 1971.
Лепер П. Р. Значение корреляции между продуктивностью родственных
особей в селекционно-племенной работе по крупному рогатому скоту.
«Сб. иауч.-исслед. работ ЛКВСХШ им. Кирова», 1936.
пикоро 3. С. О некоторых случаях отрицательной генетической
коррекции между родителем и потомком у крупного рогатого скота.— Сб.
«Генетические основы селекции животных». Новосибирск, 1965.
— Рокицкий П. Ф. Применение н способы определения коэффициента
наследуемости.-- «Генетика», т. 8, >Ь 2, 1972.
ki с f^fi14*1**^ Н А- Показатели силы влияния.—«Генетика», т. 2,
** 5. 1966,
— Наследуемость. Новосибирск, 1964.
мши ^икИшслелуемосгъ и повторяемость.— Сб «Генетические основы селе**
*** жиютиых и растений». М., 1969.
М
Рокицкий /7. Ф. Селекция сельсжожоа^шч^
мость признакоя.— Сб. «Вопросы геят»7*1^1 *****mx v вас*^
Стакан Г. А., Соскин™Тв£^ф%С^^^ Я?4*
некоторых признаков у тонкорумяых oeeiL-Бюлд. 3tfwniUeiye?0CTl
35, 2. 1963. ^wnn, отх йюл.
Горбин Я. В.. Совчгяко В. К., Бормото* В. £. Няеледуеиость вошга
.чей продуктивности полигибридной сахарное саеклы.— Сб. «ВопвоаГкате-
мэтической генетики». Минск, 1969,
Урба* В. Ю. О показателях силы влияния я хкоерсяоюои аяалхэе,-
«Генетика», т. 4, № 8. 1968.
Хотылева Л. В., Тарутина Л. Л. Оценка наследуемости яодотествепш*
признаков в популяции кукурузы па основе лнаягелышя скрешяаяявй.—
Сб. сВопросы математическое генетики». Мина. /9Ю.
Abplanalp И. Linear heritability estimates. Gene!. Res. 2, 3.1961.
Beardleu I. P., Bratton Я. W.t Salisbury G, V. The carvilmearitT о<
heritability of "butter-tat production. J. Dairy Sd, 33, 1950.
Вей G. M., Atkins R. E. lnheritince о/ quantitative characters in grab
sorghum. Iowa State Journal of Science, 39, 3, 1965.
Bohren В. В., Mc Keen H. £., lamada lukio. Relative effkiences of
heritability estimates based on regression of offsprings on parent. Biometrics, 17,
3. 1961.
Brumby P. /.. Hancock /. A preliminary report of growth and milk
production in identical and fraternal twins of dairy cattfe. *V. Z. I Sd. lech.
Agric, 38, 1956.
Clayton G. A., Morris J. A>, Robertson A. An experimental check on qn-
antitative genetical theory I. Short-term responses to selection. J, Genet. 55,
1957.
Corter R. C., Kincaid C. M. Estimates of genetic ami phenotypic
parameters in beef cattlq. II. Heritability estimates from parent-offspring and half-
sib resemblance. J. Anim. Sci., 18, 1, 1959.
Dempster E. R., Lerner I. Af. HeritabiKtv of threshold character.
Celtics, 35. 1950.
Fisher R. The correlation between relatives on the supposition of Meode-
lian inheritance. Trans. Roy. Soc Edinburgh, 52, 1918.
Freeden H. 7\, Jonsson P. Genie variance and covariance in Danish
Landrace Swine as evaluated under a svstem of individual feeding of progeny
test groups. Z. Tierz. Zuchtbiolog.. 70. 1957.
Gowen J. W. The influence of inheritance and environment on the milk
production and butterfat percentage of Jersey cattle J Agric. ReM9,5» 1934.
Graybill F. A., Robertson W. H. Calculating confidence intervals for
genetic heritability. Poultry Sci.. 36. % 1957.
Graven H. 0. Uniersuchungen fiber die HeritabHiW der BetteffettW-
stung. Z. Tierz. u. Zuchtungbiologie. 71, 2, 1958.
Graybill F. A.. Frank M., Godfrey G. Confidence interval for finance
ratios specifying genetic heritability. Biometrics, 12, 2, 1966.
Griffing B. Concept of general and specific combining ability in relation
to diallel crossing systems. Ausfr. J. Biol. Sci., 9. 1965.
И ay man B. I, The theory and analysis of diaHel crosses, II. Genetics,
43. 1958.
Hazel L. N.. ТеггШ С £. Herilabiiltv of weaning weight and staple length
»n range Rambouillet Lambs. J. Anim. Sci. 4, 1945.
Jerome F. N.. Henderson С ft. King & С HeritabOities. gene
interactions and correlations associated with certain traits in domestic fowL Post*?
Sci.. 35, 5. 1956.
Jinks J. L, Broadhurst P. L Diallel analysis ol litter sue and body
weight in rats. Heredity, 18, 1963.
Johansson Л The heritability Ы mtfk and butter fat yttUL Anim. Breeding
Abstr., 18. 1950.
399
ftkansuw I The manifestation and heritability of quantitative characters
in 4tfr\ cattle under different environmental conditions. Acta genet., 4. 195Э,
—'Cntersuchungen uber die Variation in der Euter und Strlchform det
Kuhe Z Ticrzucht u Zuchtungsbiologie. 70, 3, 1957.
Kempthome О'.. Tandan 0. The estimation of heritability by regression
of offspring on parent Biometrics, 9, 1953.
King 5. C. Heritability studies of egg production in the domestic fowl.
Poultry ScL 33. 1954.
— Henderson С R. Variance components analysis in heritability studies.
Poultry ScL 33, 1954.
Knapp В.. Nordskog A. W. Heritability of growth and efficiency of beef
cattle J. Anim. ScL 5, 1946.
— Clark R. T. Revised estimates of heritability of economic characters
in beef cattle. J. Anim. Sci., 9, 1950.
Ltmer I. M., Cruden D. The heritability of egg weight: the advantages of
mass selection and of early measurements. Poultry ScL 30, 1951.
Le Roy H. L., Lortsher H. Die wichtigsten Methodcn der Heritabilitfitbes-
timmung. Z. Tierzucht u. Zuchtungbiologie, 66, 1956.
Li С. C. The concept of path coefficients and its impact on population
genetics. Biometrics. 12, 1956.
Lush J. L. .Method of measuring the heritability of individual differences
among farm animals. Proc 7-th Intern. Congr. of Genetics, 1939.
— Heritability of quantitative characters in farm animals. Proc. 8-th Int.
Congress Genetics, Hereditas, suppl. I. 356, 1949.
— Hetzer H. 0.. Culbertson с. С Factors affecting birth weigts in swine.
Genetics, 19. 1934.
Mahadeuan P. The effect of environment and heredity on lactation. J, Ag-
ric. Sci.. 41, 1951.
Osborne R., Paterson W. S. On the sampling variance of heritability eati»
mates derived from variance analysis. Proc. Roy. Soc. Edinb., 64, 1952.
Reeve E. С R., Robertson F.' W. Studies in quantitative inheritance. II.
Analysts of a strain of Drosophila melanogaster for long wings. J, Genet.,
Rendel J. M. The use of regression to increase heritability. Aust. J. biol.
Sci. 7, 1954.
— Hobertson A.. Asner A., Khlshin S., Rogab M. The inheritance of milk
production characteristics. J. Dairy ScL 48, 4, 1954.
Robertson A., Lerner /. M. The heritability of all-or-none traits: viability
of poultry. Genetics, 34. 1949.
Robinson H. F., Comstock R. £.. Harvey P. H. Estimates of heritability
and the degree of dominance in corn. Agron. J., 41, 1949.
Sastri A. V. K. Estimation of components of variance and of heritabilitv
with their standard errors. J. Indian Soc. Agric. Statist., 8. J— 2, I95&
Schwartz L., VPeardon S. A distribution — free asymptotic method of
estimating, testing and setting confidence limits of heritability. Biometrics, 15.
*• 195У.
Shelby С £., Clark R. Т., Quesenberry /. /?., Woodward R. R. Heritablli-
t/ of some economic characteristics in record of performance bulls. J. Animal
Sci, 1. 2. I960,
Shimy Abdel Fattah El Shimy. The heritability of milk yield and fat per-
c«Kage in the Frie^ian cattle in the province of Friesland. Z. TierzGcht und
Zfidiiungsbiologie. 69, 4, 1956.
*"**¥ J w. Causation, regression and path analysis» In: «Statistics and
mathematics in biology» ed. by O. Kempthome. Iowa State College Press,
Warner S Л', A method for estimating heritability. Agron. J.. 44. 1983.
Шй л w*bfr f- Dk stalistischen und genetischen Grundlagen von Korpfrmel-
•ungen am kind Z TiertQcht и ZOchtungablologie, 69, 3, J9M.
400
Wheat 7, D., Rlggs /. Я. Heritabiitty end п»«иы»и ^
n bcef cattle. J. Animal Sci., 17. I, «il "^^Ш •«•*» fa*»
1 Уг/дЛ! 5. The relative importance of tertdHy «^ ^vUm-,,.
mining the piebald pattern of guinea pp. РтоюС the Н*1ЕГ*ыЪ
1920. -viaa. ^
__. Correlation and causation. J. Afric, R», 20. №\
— The theory path coefficients. Gaelics, & WO,
— The method of the pith coefficients. Ann. Math. Statist, J, №i
— The analysis of variance and the corretatk» bet**» reaUna wiik
reSpect to deviations from an optimum. J. Genet. 30, tft&
Глава 7
Альтшулер В. £., Никитина Л. Л. Сисюинмваа о(ш xiserm вв
разных уровнях продуктивности стада.— сГшикаэ, S, М в. 1967.
Арене X.t Беллман К. ЮнЗерветшчесяяе методы в айая«гввш| гяввта-
ке __ «Генетика», 5. JA 1ft 1969.
Гаркави О. В. Основы селекция а молоти скотоводстве,- Сб.
«Пламенное дело в крестьянском хозяйстве». М.. 192В.
Кимшилов М. М. Отбор в различима условии проявлял арпвдо.-
Бнол. журнал, т. 4, М б, 1935.
Кушнер X. Ф. Среда я сслеашювии овевал аииогш1.-Тр. Ия-та re-
нетики АН СССР. М 15. 194&
Решетникова Н. Ф., Киселева 3. С. Использован* ковффшаеяп алела-
дуемости для прогнозирования признаков молочко* ародутнмюст* вря л-
данном коэффициенте отбора.—Сб. «Воовош матемапчваво!
Минск. 1969.
Рокицкий П. Ф. Экспериментальный дяаляэ проблемы реитга
I. Сравнение рентгенизированных и версаттеляфовавамх Авш! Ycam
зоотехнических наук. т. 2, 1936.
— Изучение процесса искусственного отбора вря естгвшый в агав-
рнментально вызванной рентгеноваош сблучеияем вдевтавостя--Бвш-
МОИП, отд. биол.. т. 64, J* 4. 1969.
Соскин А. А., Стакан Г. А. О орогяоэяроваянв реяудьтаюв отбора во
количественным признакам у овео,~Сб, «Вопросы млтемлтячашя гавегя-
кн». Минск. 1969.
Стакан Г. А. Значение взаямодейстям геяотяяа со средой в племя**
работе с животными.—Сб. «Генетическая оаювм селекция яшвопяя*».
М., 1969.
Фолконер Д. С. Отбор в различных условиях окрунииоюей средл-
Ж. обш. биологии, т. 23, М б, I9&
Ayala F. /. Evolution of fitness. 1.Improvement to tat arododtvity sod
нге of irradiated populations of Drosophita serrata and Dr. ЫкМ. GtHttic*.
53. 5. 1966.
Barnes B. VP. Stabilising selection in Droeophila metenogtsfer. HtitdHy,
••*, 3, 1968.
Brock R. D. Induced quantiUlfo variation to Arabdafsls tbattana Ab-
handl Dtsch. Acad. Wis* Bertio. 1 19(7.
Clayton G. A.. Morris I. A. Roberto* Д. Ae experimental свес* on
quantitative genetical theory. 1—11. J Genet., 65,1187,
- Robertson A. Mulatto* and H>ann4alfct wtttoo, Аикг. Natur. H.
84G, |955
Edwards /. Scientific trends to *** pwdeeUoi to Omt Bert** VaaK
"Falconer D. S. Selection for large and smalt aiaa to ml*. 1 QeaU 51.
,962~ P.Uern of r^oo» «n **£•< **«~* ^ "** ** **•
Harbor Syrop. QuenHt. BK»t. » I"*
20 n * pd*«om>
•9 The genetics of litter *ut in mice J. Cell end Сотр. Phyalol., 58,
aflflp) 1. I960.
\^~ Qualitatively different response* of selection in oppoaltf directions, tn:
«Statistical Genetics and PUnt breeding», Washington, 19bU
-- Maternal effects and selection response. Genet. Today, 3, 1966,
Frankkam R,t Jones L. P.. Barker /. 5. F. The effect of population sits
and selection intensity in selection for a quantitative character in Drosophila,
1, II Genet. Res., 12/3, 1968.
GoodaU h D Can artificial selection produce unlimited change. Amer,
Natur.. 71, 736, 1937.
Kotkman \\ Selection with regard to the sex difference of body weight
in mice. Hereditas, 43, 1957.
—■ Selection siie in mice in different nutritional environments. Hereditas,
47. 1961.
Krautman B. A. Genetics nutrient interactions in laying hens. Fed. Proc,
30. I. 1971.
Leng L R. Predicted and actual responses during long-term selection (or
chemical composition in maize. Euphytica, 10, 1961.
Lush /.. I. Genetic aspects of the Danish system of progeny testing
swine Iowa agr. exp. Sta Res. Bull. 204. 1931.
Ли AW// JT. Ш/ #. 2\ Strain differences in mice on a high sail diet Ge-
nr)>c and ph\$>o)og>ca) comparisons between two straws. J. He/rd, 64, 6,
1963
Morris /. .4. Continuous selection for egg production using short-term
records. Austr. I Agric. Res,, 14. 1963.
Roberts R. C. The limits to artificial selection for bodv weight in the mouse.
Ml. Genet. Res., 8, 3. 1966
— The limits to artificial selection for bodv weight in the mouse. I If—
IV. Genet Res.. 9, 1, 1967.
Robertson A. A theory of limits in artificial selection. Proc Roy. Soc„
159. I960.
— Artificial selection in plants and animals. Proc. Roy. Soc.. 164, 995»
1966.
Wood worth C. M, Leng E. R.. Jugenheimer /?. W. Fifty generations oJ
selection for protein and oil in corn Agronomy J., 44. 1952.
Wright S. The theoretical course of directional selection. Amer. Natur.,
10J, 934, 1969
Joung S S Computer simulation of directional selection In large
population I. The programme, the additive and the dominance models. Genetics,
S3, ). 1966.
— Computer simulation of directional selection in large population. II. The
iddiriveXadditive and mixed models. Genetics, 56, 1, 1967.
Глам 8
Белле в Д. /С, Евсиков В. И. О влиянии гетерозиготносги материнского
организма иа ж к неспособность потомства. Докл. АН СССР. т. 146, № 6,
Шумный В. /С. Генстико селекционные аспекты проблемы моно*
гибридного гетеромса — «Генетика», т 4. hk 12, 1968.
Ворошило* И. В Изменчивость при инбридинге. - «Генетика», т. &
л» IQ. 19ва
Генетика ггтеромса. Под ред. Н. В Турбина. Минск, 1964.
Гетерозис. Под ред. Н В. Турбина Минск. 1961.
Гетерозис; теории и практика. Л . 1968.
ГолуЬо*с*ий М, Д.. Вилгороеа Г. В. Концентрация и аллелшш
легальны* мутации а соседних природных популяциях Dr. melanoirMSier и ралные
годы ~ «Геиетика», т. 4, М & 1968.
402
Горин В. M.. Hukui4*H*o ft ft Оцоша конбтммошнй
Iia,.in«iuux пород синие* no мясочшишм мче«тм.Л*Г«На
'иЫ рццшгия животноводства в БССРц шып. I. Минск, 1970
Сравнительна* оценки I! и 111 методов Гедффиага «q iieajum
Ki»m6iiM'»imoiuiuA споеибностя сямоопыденимх лани! кукурузы врвиекиод
и(, к i нниоводству.— Сб. «Научные основы развития жяеотиоюдгиэ
„ ЬССГ**, вып. I. Минск. 1970.
Ршмцкий П. Ф.. Хотыл**а Л. в. Математические вопросы гетерозиса
Со *1\и'розис. теория м практика». Л., 196И.
Математические вопросы гетерозиса-:аВестиик «Acmutoij^
(гиенипй науки», М 3. 1967.
Карп М Л. Инцухт и гетерозис, I. Влияние ввутрвхавмосокяото к веж
хромосомного взаимодействии на имаухмевресеяю я гетерозис — Им, АН
ХССР, сер. биол., М 2. 219. 1940.
' Кушнер X. Ф., Зубар€Фа Л. А. Некоторые результаты прняемевия ивбе*
ДШП.-1 н птицеводстве.—Тр. Иита генетики An сССР, 31. 1964.
Кушнер X. Ф. Генетические основы использования гетерозиса в ясяаог-
ноиидстце.-- Сб. «Генетические основы селекции животных»* М*. 1969.
Лютиков К. М, Об инбридинге яри разведении селаодоэяятеяицх
жншм них,—«Успехи современной биологии», выи 3, 1946.
Турбин //. В., Тарутина Л А.ш Хотылеаа Л. В. Результат затри пая
тплышн пропрркп математических моделей олределеявя коибшшюииов
способности.— Изв. АН БССР, сер. биол., М J, 1965.
Сравнительная оценка методов анадии иоиОииадяшпо! cjb>
собиости у растений.— «Генетика», т. 2, М 8. 1966.
Хогылева Л. В., Тарутина Л. А. Изучение комбинационно! ценности са<
моопылеиных линий кукурузы в диадледьяых схрещиваияяг— Сб, кГеяетя-
ка гетерозиса». Минск, 1964.
Bartlett M. S., Haidant /, в. 5. The theory о! Inbreeding with lotted
heterozygosis. J. Genet.. 31, 1935.
Carpenter J. R. Gruntberg H, Rumll E. S Genetkil dfflerentatfen
involving morphological characters In an inbred strain of mice. J. Мопйц 100»
1957.
CluuRh Af.. Cock A. Q. Variability of inbred and incfos* rtjidtara.
Nature, 179. 1030. 1957.
Cruden D. The computation of Inbreeding coefficients In doacd popvlatl*
ons. J. Heredity, 40, 1949,
Etnik L. О.. ТшШ С. В Systematic procedure* for calculating inbrtedtof
coefficients. J. Hered.. 40, 1949,
lisher R. Л. The theory of inbreeding. Oliver a. Boyd, EdtobMrgh^Len
don, 1965.
Grilling B. Concept of general and specific combfotag ibrtlty in reWlon
to dinllel crossing systems. Austr.Journ. Biol Sdn 9, 1 Itffc
— A generanied treatment of diatle) crosses hi quantitative Inheritance.
Heredity, 12, I960.
— Hayman B. I. The theory and analysis of diatteJ стенав Genetic».
.49. 1954.
— Heterosis and quantitative tnhefitanct. Heredity, II I960.
— Mather К The progress of Inbreeding when Wwtygote» are at a
disadvantage. Heredity. 7, 1953.
Jmks I L, Broadhunt P. L ШЫ anaryiia of Utter sue and body
weight in rats. Heredity. 18, 1963.
- Heterosis in dlatlel cross**. Heredity, vol fc IStt
Kina ft D. Studies an Inbreeding. J. Eiper ZooL 26-27. 1919.
U S. L.. R*d*i 0. P, Direct evidenct for models of beterosfs ргетнМ by
mutants of Arabldopsis blocked In the thiamine paitr*ey Thtoret and
applied. Genet.. 39, % 19». ......
Luth /, L Empirical teat of tt» approximate method of ceicttUtmg coef
2ГИ «
ficients of inbreeding and relationship from livestock pedigrees. J. Agr. Re^
4* 1932.
Mc Phee //. C, Wright S. Mendelian analysis of the pure breeds ol
livestock. Ill-IV. Journ. of Hered., IG, 17, 1925—1926.
Mather K. The genetical basis of heterosis. Proc. Roy. Soc, 144. 1965.
Rojas B. A.. Sprague G. F. A comparison of variance components in corn
vield trials. III. General and specific combining ability. Agron. J., 44, 1962.
Rasmuson M. Variation in inbrecd lines of Drosophila melanogaster. He-
reditas. 37. 1951.
Robertson A. The effect of inbreeding on the variation due to recessive
genes. Genetics, 37. 1952.
Robertson /'. №.. Reeve E. С R. Heterozygosity, environmental variation
and heterosis. Nature {Lond.), 170, 1952.
Robinson //. F., Comstock R. £., Khalil A., Harwcy P. H. Dominance ver»
sus over-dominance in heterosis. Amer. Nat., 90, 1956.
Sprafiue G. F., Tatum L. A. General vs. specific combining ability In
single crosses of corn. J. of Amer. Soc. Agr., 34. 1942.
Stubbe H. Uber mono- und digenbedingte Heterosis bei Antirrhinum
majus. Z. ind. Abst. u. Vcrerbungslehre. 85, 1953.
Tantawy A. 0. Genetic variance of random inbred lines of Drosophila
melanogaster in relation to coefficients of inbreeding. Genetics, 42. 1952.
— Hcterisis and genetic variance in hybrids between inbred lines of
Drosophila melanogaster in relation to the level of hornozygotv. Genetlcf,
^2, 1957.
— Reeve £. C. R. Studies in quantitative inheritance. IX. The effects of
inbreeding at different rates in Drosophila melanogaster. Z. Indukt. Abst tt.
Vcr-lehre. 87, 1956.
Wrigth S. Coefficients of inbreeding and relationship. Amer. Nat,
56. J 922
— Inbreeding and homozygosis. Proc. Nat. Acad. Sci. (Wash.), 19, №33.
— Mc Phee H. С An approximate method of calculating coefficients of
inbreeding and relationship from livestock pedigrees. J. Agric. Res., 31 ,192&
Глава 9
Беляев Д. К. О некоторых проблемах коррелятивной изменчивости и их
значение для теории эволюции и селекции животных. — Изв. Сиб. от*
АН СССР. Nt 10, 1962.
— Киселева 3. С. Некоторые генетические параметры основных кОШЮ»
ментов молока у крупного рогатого скота.— сГенетика», т. 2. Л& 9. 1966.
Никоро 3. С, Киселева 3. С. Соотношение генетических и фенотипичае-
ких корреляций. — Сб. «Вопросы математической генетики». Минск. 1969.
Abplanalp //. Genetic and environmental correlations among productSof
traits of poultry. Poultry Sci., 36, 2, 1957.,
Beil G. M., Atkins R. E. Inheritance of quantitative characters in grafr
sorghum. Iowa State. Journal of Sci., 39, 3, 1965.
Bohren В. В.. Hill W. G., Robertson A. Some observations on asyrranet*
ncal correlated responses to selection. Genet. Res., 7, 1, 1966.
Clayton G. A., Knight G. /?., Morris J. A.. Robertson A. An experiment*!
check on quantitative genetical theory. III. Correlated responses. J. Genet»
56, 1957. _
Craig I. V., Biswas D. K.. Saadeh H. K. Genetic variation and ЛГ*
related responses in chickens selected for partyear rate of egg product^**
Poultry Sci.. 48. 4, 1969. ,
Falconer D. S. Validity of the theory of genetic correlation. An expert*
mental test with mice. J Hered., 45, 1954.
Haul L. N, The genetic basis for constructing selection Indexes. GenetM*
26, 1943.
404
Morley F. H. W. Selection for economic character* in Australian Merino
«ocep. V. Further estimates of phenoiypk and genetic pararfteters. Aiistr. J.
\gric. Res , 6, 1955.
Parker R. /., Mc Gilliard L. D„ GUI I. L Genetic correlation and юрок*
tr. selection in simulated populatkma. U. Model of complete dominance, theor.
and Appl. Genet., 40, 3, 1970.
Genetic correlation and response to selection in simulated
populations 111. Correlated response to selection. Theor. and Appl Genetics. 40.
4. 1970.
Reeve E. C. R. The variance of the genetic correlation eoeffidem.
Biometrics. П. 1955.
— Robertson F. W. Studies in quantitative inheritance. II. Analysis o1
. strain of Drosophila melanogaster selected for long wmgs. J. Genet,
'A. 1953.
Robertson A. The sampling variance of the genetic correlation coefficient.
Biometrics, 15, 1959.
Robertson F. W. Studies in quantitative inheritance. XL Genetic and
environmental correlation between body and egg production in Drosophila
melanogaster. J. Genet.. 55, 1957.
Singh R. K.. Bellman K. A., Ahrens H. Die Abhangfjrkeh der komherten
•\ntwort auf die Selektion von Kopplung und Htretabuitit. (Erne Sfenalati-
«.nsstudie). Biometr. Z., 9, 4. 1967.
Sturkie P. D. Seven generations of selection for high and low blood
pressure in chickens. Poultry Sci., 49, 4, 1970.
Syrstad Ola. Estimating direct and correlated response to selection. A note
for clarification. Acta agr. Scand^ 20, 3, 1970.
Taneja G., Negi S„ Kumar 5. Phenotiplc genetic and environmental
correlations between chaeta number and wing length in Drosophila mdanogaf-
ler. .1. Genet., 59, 1, 1964.
Глава 10
Астаиров Б. Л. Исследования наследственного изменении галтеро»
у Drosophila.— Ж. эксп. бмол., сер. А, т. 3, sun 1—2, 1927.
— Фенотипкческая изменчивость гомодннамичных частей а пределах
организма.—Тр. Всес съезда по генетике, селекции, сеаяюмдетау в
племенному животноводству, т. % 1930.
Балкашина Е. И. Случай наследственного гоиеозисэ (гаювариашк)
«Aristopedia» у Dr. melanogaster.— Ж эксе. биол* сер. А. N 4, 1926
Васильева JI. А. Использование «оаффкинентоа втгтрихяассово!
корреляция для оценки наследуемости и повторяемости,— Сб. «Вопросы
математической генетики». Минск, 1969.
Гусева И. С. Наследственная основа морфологическое и функднояаль-
мой асимметрии рук человека.—Сб. сВопросы мшшпкиов генетик* и гене-
тнки человека». Минск, 1971.
Добина Л. И.. Рокицкий П. Ф. Рол» контролируемых я
«контролируемых факторов среды в фенотнпическоА изменчивости на примере одного из
количественных признаков Drosophila melanogaster.— Сб. «Проблемы weene-
риментальноА генетики». Минск, 1971.
Иванов В. И. Некоторые вопросы феногеметнт— Сб. «Актуальны*
вопросы современной генетики». М., 1966.
Иванова О. А. О наследования многовосновоггн у крупного рогатого
скота. —Ж. эксп биол., т. 2» 1926.
Ильин Н. А. Распределение и наследование белых нитей у морской
саамки.— Тр. лабор. эксп. биол. Моем, зоопарка, т. 4, 192&
Ильинский А. А. Повторяемость удоев и содержания жира в моложе за
300 дней лактации коров племзавода «Кдр#авнво».— «Геяияижлок т. 6.
М I 1970.
Концов Н. К. Роль генов п физиологии развития.—Биол. журнал
т. 4.1? о
Рчкицкий П. Ф. Области действия генов. — Тр. Весе, съезда геиет., т. 2
Л. П'<0
- Поле действия генов.— Ж. эксп. биол., сер. Л, т. 5, вып. 3—4, 1929.
— Ген и признак.—Ж. эксп. биол., т. 7, 1931.
Тимофеев-Ресовский Н. В. О феиотмпическом проявлении генотипа.—Ж.
жоп. биол., сер. А, т. 1, вып 3—4. 1925.
Astauroff В. L. Analyse der erblichen Storungsfiillc der bilateralen
Symmetric in Zusammenhang mit der selbstiindigen Variabitat ahnlicher Struktu-
len. Zeitschr. f. ind. Abst.- u. Vererbgl. 55. 3, 1929.
Castle 0. M. Searle S. R. Repeatability of dairy butterfat records in New
Zealand. J. Dairy Sci.. 40, 10, 1957.
Guthrie /. The asymmetry of the small—eyed condition in eyeless
Drosophila. Journ. of Exp. Zool.. 42, 1925.
Jonson L Д., Cortey £. A. Heritability and repeatability of first, second,
third and forth records of varying duration in Brown Swiss Cattle. J. dairy
Sci. 4-1, 1961.
Plunkett Ch. K. The interaction of genetic and environmental factors in
development. Journ. Exp. Zool., 46. 2, 1926.
Reeve £. C. R., Robertson F. W. Studies in quantitative inheritance. VI.
Sternite chaeta number in Drosophila a matameric quantitative character.
Zeitschr. f. indukt. Abstam. und Vererb., 86, 3, 1954.
Rukuky P. Th. Dber die differentielle Wirkung des Gens auf verschiedene
Korpergtgenden. Zeitschr. f, ind. Abst.- u. Vererblehre, 57, 1, 1930.
Timofeeff—Resson'skij N. W. Studies on the phenotypic manifestation of
hereditary factors. I. Genetics, 12, 1927.
Wheat J. D.t Riggs /. K. Heritabilitv and repeatability of gestation length
in boef cattle. J. Animal Sci., 17, 1, 1958.
УКАЗАТЕЛЬ СИМВОЛОВ
t\ — a\ Ах— А2— аллельные гены.
А —селекционная ценность (=(?)•.
А —отклонение, вызванное аддитивными генам.
а — генетическая ценность гомозиготы АХАХ как отклоневне
от средней ценности двух гомозигот.
а — среднее действие гена.
а — среднее доминирование d, выраженное в значениях о.
а, Ьу с... —коэффициенты пути (по Райту).
а — средний эффект замены гена Ах на ,42, средний эффект
генной структуры.
ai. «о — средние эффекты аллелей Ах и Ла.
а, р, у— коэффициенты приспособленности разных генотипов.
Ь0Р — регрессия потомков к одному из родителей.
bQ-p — регрессия потомства к средней обоих родителей.
bQM — регрессия потомства к матерям.
bdM — регрессия дочерей к матерям.
covGE — коварнанса между генотипом и средой.
cov0p — коварианса между потомками и родителями.
covFS — коварианса между сибсами.
covHS — коварианса между полусибсамн.
CRy — коррелированный ответ признака у на отбор по
признаку х.
D — доминантное отклонение.
D —разница между средними родительских линий, пород.
D — сокращенное обозначение о*.
d — геиотипическое значение гетерозиготы АХА% ***
отклонение от средней ценности двух гомозигот (— л).
dxA -— коэффициент детерминации признака А фактором х.
А — сдвиг в величине какого-то показателя за поколение.
Д<7 — изменение в частоте гена.
AF — изменение в величине инбридинга. ^.
6(7 — сдвиг в значении q за поколение при генетическом №*$*•
Е — отклонение от популяцноиной средней, вызванное
неким-то фактором среды или средой в целом.
£ — сокращенное обозначение of.
£0 — отклонение, вызванное общесредовыми факторами.
£w —отклонение, вызванное специфическими средовыми W**'
В скобках (-...) символы, используемые другимГавторами.
407
/", Fx, FA — коэффициент инбридинга (по Райту).
FY — первое поколение от скрещивания двух линий, рас,
пород или популяции.
F, —второе поколение от свободного скрещивания особей Fx,
fe — обратное скрещивание особей Ft с особями одной нэ ро.
днгельскнх групп (Pt или Pj).
FS — парные сибсы.
/, fp p —коэффициент инбридинга на основе общ их. предков.
С — генотипнческая ценность (= Ge), генотипическое
отклонение.
GC —общая комбинационная способность.
Н — частота гетерозиготного генотипа АхАг,
HF —мера гетерозиса, т. е. отклонение средней Fx от средней
максимального родителя.
Я5 — полуснбсы.
л — степень доминирования,
л* — коэффициент наследуемости.
А* — коэффициент наследуемости в узком смысле слова
/ — отклонение в результате взаимодействия неаллельных
генов (эпистатическое).
/ — индекс отбора.
1 —действие гена в одной дозе (по А. С. Серебровскому).
i — интенсивность отбора в единицах фенотипического квад-
ратического отклонения (= Г).
У — длительность интервала между поколениями.
L — генетический груз.
^ — эволюционный груз.
£/ — инбредный груз.
Lm — мутационный груз.
Ls — сегрегационный груз.
MF — популяционная средняя при инбридинге.
Мр — популяционная средняя, ц, хр.
Мх — среднее значение кросса между линиями,
/л — общая частота мутаций,
/л—-доля иммигрирующей популяции.
тг — коэффициент рекомбинации.
N — общий объем популяции
Ые — эффективная величина популяции,
п — количество особей в выборке, иногда среднее количество
потомков на семью. г
О — потомство любой особи или пары родителей.
РА — частота гена А.
Р — родитель.
Р — среднее значение признака двух родителей.
Я—-частота в популяции генотипа АХАХ.
Я — паимиктический индекс, равен \—F.
Р — фенотипическое значение особи.
р — частота гена Ах(^и).
р — доля особей, отбираемых в качестве родителей из попу-
ляции с нормальным распределением ( ~ V).
Рл ' Pjf —доля влияния фактора А% В и т. д. в общей
изменчивости признака.
Ю*
Q — половика частоты в потгм...
-М, <- Н). И°"ул««" г««Р«»»тго **„„.
q — частота геиа А^(- ъ)
пР~~ Ч!^а В ««У*"» генотип, ЛЛ. Г- Л
Я vy, - коэффициент родства ытпГхшУ
R — сдвиг при отборе за одно повали (~ а/л
rie — внутриклассовый коэМнвтт «»!!: 1'
повторяющимися измерениями то же осоЫ "^
гл -аддитивная генетическая корреляция. ^^
г0Е- корреляция между генетическими и средриыми отж*^
ниями. **оие-
/•£ — средовая корреляция.
гр — фенотипическая корреляция (= г).
rG — генетическая корреляция.
S — селекционный дифференциал, выражешши в фантаста
единицах измерения (= i).
S — сокращенное обозначение oj.
SC — специфическая комбинационная способность,
s — коэффициент отбора против определенного генотипа,
уменьшение доли его гамет.
Oq —общая варнавса (-of), сумма всех компонентов,
о, — варнанса между производителями.
о* — варианса между самками внутри производителей.
Gu7» о^ —варнанса внутри группы или семей.
о2еш — средовая варнанса, общая внутри групп,
о^ — средовая варнанса, не общая внутри грулв.
c2pt о* —фенотипическая варианса (= Vp).
o2Qt с1 — генотилическая варианса (= Ко).
с\ — аддитивная геиетнческая варианса (^ Ул).
о^ — варианса, зависящая от доминирования (=■- VD).
а) - варианса, зависящая от взаимодействия меаиу иеалдеяь,
ними генами (= V'/).
варианса за вычетом аддитивной.
о^ — средовая варнанса (= Ve).
о* — паратипнческая варнанса.
, — варианса изменчивости между
о
и с — pay папка iw«wn>«"^ —
о^.—варванса потомства отдельной особ»
ol-utp* рассеяли частот гявморв J^^^^
suss tssss ^*S£* ^ч
ш _ приепособлеяноел- ^^
»_ средняя приелособ*«я«сл-
СТАТИСТИЧЕСКИЙ СПРАВОЧНИК
СОВОКУПНОСТЬ И ЕЕ СТАТИСТИЧЕСКИЕ ПОКАЗАТЕЛИ
Совокупность — всякое множество отдельных отличающихся друг от
др\га и в то же время сходных в некоторых существенных отношениях
объектов.
Частичная или выборочная совокупность (или просто выборка) —
конкретная группа изучаемых объектов (объем п).
Генеральная совокупность —группа объектов, обычно очень большая
(объем Л'-*сс), из которой взята оЛна или несколько выборок.
Стохастическая совокупность — это совокупность, искусственно
создаваемая in всех теоретически возможных случаев осуществления данного
явления (объем оо).
Вариация может быть качественная (различия между вариантами
выражаются путем качественных определений) и количественная (различия
выражаются числами). Количественная вариация может быть прерывной,
или дискретной (варианты характеризуются только целыми числами), и
непрерывной (количественная характеристика вариант выражается любыми
числами).
Вариационный ряд — распределение вариант дп, х2, ..., хп по классам
(классовый промежуток i) или в определенном порядке. Графическое
выражение вариационного ряда (кривая распределения): при прерывной
вариации—полигон, при непрерывной —гистограмма или столбчатая диаграмма.
Число классов в зависимости от количества вариант можно наметить
по'следующей таблице:
Количество вариант Число классов
25-40 5—6
40-60 6—8
60-100 7—10
100-200 8—12
более 200 10—15
Статистические показатели:
а) для характеристики центральной тенденции в совокупности (или в
вариационном ряде) — мода (Мо), медиана (Me), средняя арифметическая
(*). средняя геометрическая (хи);
б) для характеристики степени вариации в совокупности или
вариационном ряде — вариационный размах (11т), среднее отклонение, среднее клад-
ратическос отклонение (о или я), варианса (а2 или *2), коэффициент
вариации (v), коэффициент асимметрии (g\), коэффициент эксцесса (gt).
Формулы для вычисления основных показателей (параметров) для
совокупности, не сгруппированной в вариационный ряд;
410
для машинных вычислений удобны также:
Степени свободы df(- п — 1),
Сумма квадратов (отклонений от средней jr)«[ - 2(х, — i)*i
Макет таблицы для составления вариационного ряда по данным
обладающим дискретной вариацией
Дискретное значение класса X
Частота /
S/(=«)«
/*
2/Х
/*•
Ш*-
Макет таблицы для составления вариационного ряда по данным,
обладающим непрерывной вариацией
Классы*
1
!ш
А = ...
Частота /
...
л
Условные
отклонения в
0
/*
0
2/а-
/л'
0
Х/а*-
х A + i п ,о , у ^-__^—j J-7T=T-
В «вешенная средняя арифметическая
пх + "• + • • • + п*
(к — количество частных совокупи остей).
* Каждый последующий класс долям начинаться с***** 6*******
•юм последняя цифра предыдущего класса (ив I или ^ "^J^ |НИК)срсд
•• Эта графа нтЛпыг^ьнш. Ъ^т^^^^^ Щ9ЛМ^.
чш'нни как среднее
значение
одного и* классов с наибольшей
4II
Средняя геометрическая
x0 1 xrxrx9...xn у TT*7;
logxc -L (log jt, + log xt f log x, + ... + log xn).
Коэффициент асимметрии
пЦх(-х)*
gx (n-\)(n-2)V!o*ir
или
Коэффициент эксцесса
Z(xi-x)* -
Коэффициент вариации
■ —£■».<*).
БИНОМИАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
Относится к дискретным признакам, по формуле бинома Ньютона
(Р + ЧУ.
гае п — чвсло независимых случайных событий; р и q — вероятности
благоприятных н неблагоприятных событий.
Прв р — q = 0,5 распределение симметрично.
ТРЕУГОЛЬНИК ПАСКАЛЯ
Коэффициенты бинома (Сц.х)
п
0 (P + q? I
1 (Р + Я)1 II
2 (P + q)* 1 2 1
3 {P + q? 1 3 3 1
4 (P + q? I4641
в (/> + ?)* 1 5 10 Ю 5 I
6 IP+qf I в 15 20 15 в 1
с»*~ jrf <л — ж> I •
м.»_ г 51 12 345
Нгаряивр. С,., - -щг—щ- . -,,j,jjj ». 10.
412
Вкроит мости каждого члена ряда
Параметры биномиального рвдрадежв, пывишл,^.
" *«" кколвлпп «поп
i
i
для относительных частот:
I» --•• гг.
Л
РАСПРЕДЕЛЕНИЕ ПУАССОНА
Относится к дискретним признак»!, применяется ope
и большом л. вместо биномиального распределена.
Р —. где X — положительна! константа.
п
При возрастании л р становится очень калов велячяио*.
X — яр =t p;
(X \Я х
1 ~~ I - * t
то
Хг _х X*
Тогда теоретические частоты можно вмраэятъ следую»» ряяР*
JL. -IL-. Л— -JL_ v т. Д.
^ ■ / ■ V ТзГ
(яу лею!)
член)
0*»ЛР9.
Так как <; = I — р, то в»-Х[1— —I.
v ' _* -« .щчи вуассоиова
При большом п о» а X. Это равенство укаамм" вя ■■■—
распределения.
НОРМАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
Применяется ■ тех случая*. *■**»
Л - -х.
Уравнение яряяой нормального
Графическое выражение на рис. По.
УЬртфоымное от*ло\гс*ше — отклонение любой варианты от средне
арифметической, выраженное в величине среднего кьадратического отклони
м (или /)
*!•
Приняв а 1 и заменив
\*i —х) величиной /. получили у ^ -,-^е
о }2п
Вероятность любого отклонения от средней как функция нормирован-
кого отклонения в стандартизированном виде представлена в табл. I (в ней
лишь десятичные знаки без нуля и запятой).
В пределах ± 1с — 68.34 вариант
1 г 2о—95.5% »
» г: Зо —99.7% >
Доверительные
вероятности
0.95
0,99
0.999
Уровень
значимости
0.05
0.01
0.001
Доверительные интервалы
— 1.96о ...+1,96о
— 2.58о ... +2,58а
— З.ОЗо ...+3,03о
Средняя ошибка для х : s-
Vn
Критерий / по Стьюденту — Фишеру:
/ =
л-ц
Доли площади под кривой /-распределения по Стьюденту в табл. Ш
(даны лишь десятичные знаки без нуля и запятой).
Отсюда минимальные значения /. обязательно требующиеся для данжит
вероятности при разных л (степенях свободы), в табл. IV.
Доверительный интервал для ц:
x-t.
Уп
Уп
Необходимый объем для выборочной совокупности
А»
-, где A«/s-.
Ошибки разниаы между средними арифметическими:
ПРИ бОЛЬШЛХ П %4 \ £ +
>*.•
ПР* МАЛЫХ Я 14 У Д^-^У + Д^-^в)1 4|+"l
Критерия достоверности рази яды
JL— *i d
414
СЛЛТИСНЧЕСКНЕ **Ш№*Ш „ади.
Доля />( *) ■•= —J-.
Среднее киадратнческое отклонение^ ^ у^.
Средняя ошибка для доли $р •-• 1/51.
Средняя ошибка для р% выраженного в процента! s^=r l/pQw-^
Средняя ошибка для абсолютных чясленностей групп
с , 1/ЗЕ5С
Доверительные границы для доля
p-t%p<P%<P -ttp.
Необходимая численность выборки
" '*[ p('aTp) ]■ гдеА" 'V'
>(i-p)
Достоверность разницы между марочными долтпс
d = px-py; ч~У *ps~% '• '-"•Г"
При малых выборках
/ 71 ГТ P,*x+Py*f „ *»+*«
*r + **
КОРРЕЛЯЦИЯ
Макет таблицы да» •^^"■^^^Г1'1 ЕвРРетМ '
лра*ыи сяосава* (Ц
Рабочая формула г
Макет таблицы для вычисления коэффициента корреляции г
прямым способом (II)
ft» О
1
2
i *
л
i
*i
Z=-
X --
У(
4* --
5 =
V . .
2 --=
*(У1
2 -■:
Рабочие формулы;
2лг,у,- — пху
Zxii/r
y'H-^-jH-^i
Макет корреляционной решетки
X. X
j У*1
*i
1 а*
1 | *»
1 *f
l
| Ух
1 /г
*г
1 "*
*» |
с* J
*i
*,
<*
1 dt
'1
4 111
| /А |
LJ^'J
i I I
i i i
1 *•
<•«
d*
хп%
1
Cb |
*
d.
1 1 '
_лл
/у
п =
11 ^ 1
j z = 1
а,
/yfly /y°J
Z» 1 Z» 1
1 i
1 -1
Г
416
рабочая формула для , „рн ^^ „^^^ ^^
Средние ошибки для г:
при больших выборках и не очень высоком значении г
s, »■
при различных размерах выборок, в том числе малых, н разнообразии
значениях г
г
Отсюда / ~- —.
Для определения достоверности г без определения ошибки см. табл. V.
Перевод г в г и обратно — табл. VI.
Ошибка для г
1
5, -
Определение достоверности разницы между г.
г -*
Отсюда / =
2\ — *%
**г
Доверительные границы для р:
(р-козчМ>иииент корреляции генеральной совокупности).
г-**.
г;<р<^-
Коварнанса covx9 — я —I
: ^.тель . «** формуле к<»фф««*"т< «W"«"
Коварианса как чяслятель в дай *~
4IT
27 П. Ф. Ромики*
Макет корреляционной решетки
при альтернативной вариации
"^-^ X
0
1
2
0
а
с
а + с
I
Ь
d
b + d
2
a + b 1
c + d 1
\n = a + b + c + d\
ad — be
У (a + b)(c + d)(a + c)(b + d) '
Коэффициент корреляции с поправкой Йетса на размеры выборки
|ad-6c|—J-
V (a + b)(c + dHa + c)(b-fZT'
Макет таблицы для определения коэффициента ранговой
корреляции Спирмэиа (rs)
Номер
пары
1 1
2
п
п
X
У
Оценка
равгов
*i
«t
(х{ - У() = d
<*, — У()г - Л*
2 1
_ 62 (л,-</<)« 62<Я
rs~i— n(n* — 1) ~1— л(п« —1) '
где xi и I// —ранги^по первому и второму признакам» а не значения оря*
>наков, как при обычной корреляции.
Ошибка г,— по табл. V.
Коэффициенты частной корреляции:
- гху — rxzryi
rxy.x - ——ч »
, г**~ Г'У г>* _ riy—rxxryx
ГХХ.У j ж . 'jJ,.X ~ = г- J
418
гху.ги —*
гхуи~~гх1.л'гуг.п
V^-^J^-^zJ
Ошибка разницы между средними арифметическими при наличии
корреляции Sd = у S? + & — 2S- •*- Г| 2 .
РЕГРЕССИЯ ПРЯМОЛИНЕЙНАЯ
Макет корреляционной решетки дли построения
•ипнрической линии регрессии
^S. X
У ^S^
Упх
Уг
Уг
Ух
fx
Средние
У для
классов
ряда
1 (У/*)
*i
fli
*i
Ух
*i
<Ч
*»
^2
Уг
1 *§
*з
С»
Ь
~Уг
...
1 *4
<4
4
U
...
4
и
хт
*%
Уп%
fy
п
Среднее no x дли
классов ряда а;-
**1
•
**
ц
*i
Выравнивание эмпирической линии регрессии с помощью скользящей
средней:
ц- *+$ + * или yt~ Л + Ц + J^ + fi + fc.
„ ^1 + У> + У| UJIU „ ft + ft + ft + ft+ft .
Уа = з Уэ ^ б •
Уа + Уа + Уа вяи и Уа + Уа + Уа + Уа + Ут
№ = 7 илн л e J
и т. д.
Уравнение прямолинейной регрессии в общем виде:
У1 — У~Ь(х( — *);
И* ? + *:(*< — *).
При х — 0 обычное уравнение прямой
у^а + Ьх.
М
Система уравнений для определения значений а и Ь:
1) па + (2х,)6=2й;
2) (Zxi)a + pxf)b = lxlyt.
Макет таблицы необходимых величин для решения уравнений
*t
2х{ =
л —
х =
У1
2У1 ----
п =
У =
*?
*? =
*1У1
%*№ =
Коэффициент регрессии R(— b)t выраженный через г:
R -т^-
Kxiv - г ау •
R -г °у
Рабочие формулы при наличии г:
*,„=,-./-*&=.'.>•.
v
уу/х
2 (у,-у)*
г-»/ Z(yi-yF
V 2(х,-*)»
Рабочие формулы для /?(= Ь), если г неизвестно:
Д(«|-*)(У<~У)
ЬУ/х =
2(J:L~X)2 _
S (xf — х) (у,- — у)
"» ~ Z (у, _ у)'-
Макет таблицы для определения Ьх/у и &^/л. по рабочим формулам
*1
1 2х,=
1 п —
1 х :
Vi
| Л --
1 ~у--
х1~* '
(х.-х)*
2 =
1
*,-»
1
(У; - У)%
2 =
(х,— х)<»/,— у)
2 =
4Й)
Ошибка коэффвадента
* s <*t — jt)* Г
Агрессии b(R)
2 (V. — V)« —
jSjXf - s) (yi - y)j*
(п-Я)^-*»1
Если известны г, 0х, <jv, то
ч,.ж-?>. 1/IE21
ох Г n —2
Сравнение коэффициентов регрессии
Ошибка разницы
при малых п
t л/ $\ 5 .
t =,
*1-У
Связь между регрессией и корреляцией
криволинейные зависимости
Уравнение параболы 2-го порядка
у --=а + 6* + «*.
Система уравнений для определения параметров а, 6 и с;
1) /ш + <2*<)* + №фс~2*/;
2) (Гд/) а + <I*f) * + (**?)с ~ 2**М
3) (X*?) а + (ЗЦф * + (**}) с - I*fу,.
Макет таблицы для вычисления необходимых величии
рабочих урввнемнй параболы
*/
У,
2У1 =
А
£■-*
А
2-
xi
2.
*М
Х-
**№
1 -
т
Уравнение экспоненциальной кривой
Г - АВХ.
Логарифмическое преобразование (приведение к уравнению прямой
у а -г Ь х):
\ogW - log Л + (log Я) .г.
ДИСПЕРСИОННЫЙ (ВАРИАНСНЫИ) АНАЛИЗ
При учете влияния одного фактора А отклонение варианты х от средней
ж —ц - Л+ *;
при учете влияния двух факторов А и В
x — \i = A + B + AB + e\
при учете влияния трех факторов А, В. С
х-ц А + В^С+АВ + ВС + АС + АВС + е и т. д.
То же в значениях варманс:
co=:a5i + 0e + a*a + 0* и т- Д-
Макет таблицы для группировки материала при дисперсионном
анализе по одному фактору А
J Группы
1 (градиша)
I во
фактору л
I
2
f
I а
Отдельные варианты
(наблюдена) JT^
1
2
*11
3
*11
*••
/
*1/
л
х\п
хт
*in
хпп
1
Суммы по
группам Г
2^ = 74
2х, = Т,
2*,-Г,
Гха- Г„
bi/-r
1 Средние по
группам Xj
^1
*и J
jr I
_ ., iem
Количество всех вариант N *■- an.
Змачеиие л можат быть не для всех строчек одинаково, для квихди**
строчла ояредаляетса свое л,. Тогда N -. ап[. я****09** *"» ~~-
(Ища» оси, дисероюотог, амм» „р. оиом ф,,^
Источник
варьирования
Общее
Фактор А
Варианты
внутри групп
(случайные
отклонения)
1 !
*
ы
sst
•
г"
*и
1
*
msx
1
я
»•
1
ms,
J
| *• табличное
1 8
о
11
a,
a
e.
с
_J
s
о
a
a
с
...
Параметры,
оценввае-
ммв т
0» + **
Рабочие формулы:
2 Г? Р
- i 1Г-АГ
dU:N-Y.
dft:{n-l)a = N-a.
а
Во всех случаях ms определяется как -щ-.
Макет подсобных табмц дм мгакаеш!
Т,
1 2Г| = Г'
LiirJ
1 я/
Tf
it?- j
1 L II *i
г?/«|
■44
4tt
Варианты
**U = .
*?,
4-
F табличные определяются по табл. VII и VIII. С ними сравнивается F
ф.ктичесТое При /факт. >F табл. нулевая гипотеза об отсутствии влия-
имя .4 отвергается.
Влияние А определяется как
? пщ —rns2
Доля влияния А в обшей вариации (в долях единицы или в процентах):
Ра
°2а
о\ + о]
Общая схема дисперсионного анализа по двум факторам А и В
(при случайных градациях действия факторов А и В)
t Источинхи
1 мръвроваимя
Обшее
1
Фактор А
Фактор В
. Вэанмодей-
. ствне А и В
Случайные
отклонения
и
550
55,
552
«Э
«4
di
dh
dj,
dU
dh
dl.
ms
—
msl
ms2
msz
ms4
F
фактическое
—
L *
III
F табличное
= 0.05
с.
при
—
о
о
!1
С
при
—
1 Л
Параметры,
оцениваемые ms I
°] + псЛВ + псаА
°1 I- паАВ + пгаВ
2 1 2 1
К + ™АВ
о] J
Рабочие формулы:
"о- 2dx\
J2
ijk
Ijk'
nrc
1 nc ~4 nrc »
1 nr +fl / nrc .
"•;*'?»-42ir?,.
7*a
лгс •
ilk ilk"" n f ' ij>
r'.LZ 'JZSSSTZ TZT*,0 "0""°""onp""""""'"
Макет таблицы для ттирошм материала при ляакрсшошт
анализе по дауи факторам Л и В
ни)
гппы (градац
фактору А
"§
1
2
3
1 '
г
Сумма
по
группам
фактора В Tf
/ ■ ■ —
Группы (градации по фактору В, отдельные варианты х
внутри них я суммы хц виутри групп по А Ц
I
(0
*111
*112
ХШ
*211
*212
*213
*311
Г312
-*313
*m
*/13
xn\
xr\\
1
1
Г *
1 °
Г11
kl
l'«
_
Г/1
y'rl
r.J
2
*m
*122
*123
*22l
xm
xtta
^ati
*aai
xsta
xm
Xi%2
xli*
xrti
*rn\
хпш
1
ж
>»
1 °
Iй
'22
I9"
tJ
tJ
r.f|
1 3
1 ft
•
0
*18l
xm
xm
xu\
*13l
*ззз
•^331
1 JTau
*лда
*/»! 1
*Й1
*/ЭЗ
•^ЛЗ!
*rs2 I
^/-за
1
>»
1 °
\ти
1 I
Г"
D
<
Ч
г"
г.,|
U
г
1 у 1
1 s
*i7i
и*
Х1/3
1 xiii
1 xi/j
1 *i/i
xm
1 >»
1 ° 1
Ы
H
г./
XU\
xUt
AV/i
1
1
T"
H
Г.Л
1 f
1 4
1 e
xlfl
xiri
1 *lr3
*lfl
*trf
hti
X3f?
I 1
xic\
x«l
1 ХЛ| 1
хм\
1
«ГГ | J
хю\
1 1
2
1 °
r1'
к
>*
1
r"
J
гЦ
1 1
3
с
- e*-
- *■ •
i ex
k.
r"
r,.
Ъ.
r,.
г
J
dftinrc — I;
4f,:r-l;
4f,:«-l:
«</4./r(/t-l>.
485
Мажет та&нпдо ждя груяяярояяя матерядда яря дясперсяояяом шпшлшы
но дяум фяяторям Л я В яря яердржячесаоЛ сяяая между нямя (см. ряс, 71
Гаушш
|(гры*а»я
1 яо фанто-
1
2
1
|
t
_ 1
я
1
L
L
А Группы
по фаято*
1
2
/
i •
1
1 2
' /
_и
I
2
У
ь
1
2
/
; 1
_и
1 Отдалыгкм мрядатм
(наблюдена*) Ж|/А
1
1
*in
Х1Я
*1/1
Х1М
1 ХЯ1
1 Xf21
Xjj/,
*аи
*tfi
Х«11
xati
*oh
*ab\
2 к
ХШ хик
Xltl х\1к
хМг xMk
Х\Ъ\ х\Ьк
*m *я«
х» хпк
xZ/l Хуь
Х»9 JCtbk
xllt Xilk
xtn xtlk
ХЧ* *ljk
xun xibk
x«« xa\k
xan Xgfr
x*i% хЫк
x**t xakk
с
xllc
x\v
xUc
x\bc
• T"
Тгг
Тгг
Тг1
Тгь
xnc 1 Tn
Т'
Тг
*nc f Tn |
xt/c
xtbc
xlu
xltc
xUc
\тЛ
Тгь
Т,г
Тц
xlbe Tlb
xa\c 1
***
XQlc I
Xatx I
тл I
Таг
Т"
Tab \
г, ]
т,
Та
J
4Ж
т всегда равно -jr.
N ъгПГС*
Подсобные таблицы составляются для вуяяелеивя
*и» г*,. П. П,.
Тот же принцип сравнения табдяянл* F с фактическими пояутшшшл
для факторов А, В и вэаямодевствия ЛВ.
При достоверности F влияние отдвяышя факторов ооведадвекя как
• Bt$t — ff&i
Л ПС
*лв
m$t —-flail
л
«ДУ
Доли влияния каждого фактора (или ввашюдеветаия
деляются как
«U ^ ^
(обЩЙЙ С1ШКИ ДЛЯ OfeMMMH ЯМ «Ш" *«"И *»" ««
впервые).
вявлнаа во двум
фаитора* Л ш В шрш
Исто*****
Обшее
Фактор А
Фактор £
Случай***
отнло*****
m
\*h
|«i
\m*
*н
*f
\dl.
\dh
\dh
*Ь
9* {
i "~
ПШ\
\mH\
msA
F t«блдеос
I
S
i
«W|
Работ фориулы:
V, JL.
sb'jbiXijk abc •
df9:abc— U
dfi'.a — U
dft:a(b-D\
dU:ab(c—l).
ss
ms всегда равно -щ-.
N=-abc = a-brCi
(Ь^средвее число групп В ва каждое а. ^-среднее число вариаит иа
***** 6)* J* т<2 т2
Подсобвш таблшш составляют для ХуА, Т(9 Тц.
Прюшип сраввения F фактических с F табличными тот же.
При достоверности F влияние отдельных факторов определяется над
- msi — m*t
М- be
А~
/njg— ЛЩ
Доля влияния в общей вариации (в долях единицы или в процентах):
лщ А ,„ А
* в А+А+°Г * А + А+°Г
Достоверность разницы между средними арифметическими отдельных
гр>пп в дисперсионном комплексе определяется путем деления d на %л.
л/Я
В длиной случае принимается %d-t-** I/ — , где п — число вариант
> каждой группе или средине число вариант на группу.
Для установления граничных значений для разницы, обеспечивают**
Р--»0.06, ушюшжкп ошибку на коэффициент Q из табл. IX. гдв учтен*
исм1в<мн1во групп и число степеней свободы.
4ЯЙ
КРИТЕРИИ СООТВЕТСТВИЯ (ХИ-КВАДРАТ)
Общая формула
*-,J4E
Хн-квадрат распределение при разных вероятностях приведено в табл. X.
Для отбрасывания нулевой гипотезы нужно, чтобы фажтичесяое
значение хи-квадрата (при данном значении df) превышало табличное при
вероятности 0,05 н ниже. При большей вероятности считается, что соответствие
имеет место.
Число степеней свободы:
а) при 2 классах равно 1;
б) в решетке с 4 и более полями — (г — \)(с— l)t гме r—чясло
горизонтальных рядов, а с — вертикальных столбцов;
сравниваемых классов).
Упрощенная формула хи-квадрата для анализа расохешкшй
Х ~ (о + Ь) •
где а и Ъ — фактические числа в каждом классе, а г — теоретическое
отношение.
Для четырехпольной таблицы
lad — bcp-n
%%ss (a + b)(c + d)(a + c)(b + d) •
с поправкой на непрерывность
Х* = (а+ *)(<; + <0 (а+ <*<*+*>"•
При сравнении двух эмпирических распределений
.-.дело
где /, „ ,,—cnm. ««со. пер^го я .торого P«c % - %'
особей в каждом из них. 429
Таблица I. Доли площади под нормальной кривой в пределах
от — t до +t
/
0,0
0,1
0,2
0.3
! 0.4
0.5
0.6
0,7
0,8
0.9
1.0
1.1
1,2
0
0000
0797
1585
2358
3108
3829
4515
5161
5763
6319
6827
7287
7699
I 1,3 j 8064
1 М
1 ,f5
1 1>6 1
8385
8904
/ 1,7 | 9109
Сотые доли /
1
0080
0876
1663
2434
3182
3899
4581
5223
5821
6372
6875
7330
7737
8098
8415
8690
8926
9127
I 1.8 | 9281 9297
1 1.9 j 9426 9439
' 2%0 \ 9545 9556
2.1 \ 9643 j 9651 |
2.2 1
2,3 1 i
2.4 <
9722
Г?86
№36
2.5 [ 9876 I
2.6 ! 9907 J
2,7 j 9
2.в ( 9
2.9 9
з.<
Ф 9
«1
949 1
963
973
9729
9791
шии 1
9879
<ЛЛ/У 1
9933
9960
W64
9981
г
0160
0955
1741
2510
3255
3969
4647
5285
5878
6424
6923
7373
7775
8132
8444
8715
> 8948
9146
9312
9451
9566
9660
9736
9797 |
9845
9883
9912
9935
99S2
9965
9986
3
0239
1034
1819
2586
3328
4039
4713
5346
5935
6476
6970
7415
7813
8165
8473
8740
8969
9164
9327
9464
9576
9668
9743
9802
9849
9886
9915
9937
9953
QCftfiA
9990
' 4
0319
1113
1897
2661
3401
4108
4778
5407
5991
6528
7017
7457
7850
8198
8501
8764
8990
9181
9342
9476
9586
' 9676
9749
9807
9863
9889
9917
99Э9
9965
9967
9993
5
0399
1192
1974
2737
3473
4177
4843
5467
6047
6579
7063
7499
7887
8230
8529
8789
9011
9199
9357
9488
9596
9684
9756
9812
9857
9892
9920
iW4U
9956
9968
9995
6 1
0478
1271
2051
2812
3545
4245
4907
5527
6102
6629
7109
7540
7923
8262
8557
8812
9031
9216
9371
9500
9606
9692
9762
9817
9861
9895
9922
9942
9958
9969
9997
7
0558
1350
2128
2886
3616
4313
4971
5587
6157
6680
7154
7580
7959
8293
8584
8836
9051
9233
9385
9512
9616
9700
9768
9822
9665
9898
9924
9944
9959
9970
оооя
8
0638
1428
2205
2961
3688
4381
5035
5646
6211
6729
7199
7620
7995
8324
8611
8859
9070
9249
9399
9523
9625
9707
9774
9827
9869
9901
9926
«Лт4©
9960
9971
9999
~
*
0717
1507
2282
3035
3759
4448
5098
5705
6265
6778
7243
7660
8029
8355
8638
8882
9090
9265
9412
9534
9634
9715
9780
9832
9872
9904
9929
9947
9961
9972
9999
430
Таблица //. Частоты (мюпмиш*) мримыюго ргсяредмот, п №000
э
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
ьо
1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
2,0
2,1
2,2
2,3
2,4
2,6
2,6
2,7
2,8
2.9
3,0
3,1
3,2
СВТЫ* ДОЛ! f
0.00 ;
0000
0398
0793
1179
1551
1915
2257
2580
2881
3159
3413
3643
3849
4032
4192
4332
4452
4554
4641
4713
4772
4821
4861
4893
4918
4938
4953
4965
4974
4981
4987
4990
4993
0.01
0040
0438
0832
1217
1591
1950
2291
2611
2910
3186
3438
3665
3869
4049
4207
4345
4463
4564
4649
4719
4778
4826
4865
4896
4920
4940
4955
4966
4976
4982
4987
4991
4993
0,03
0080
0478
0871
1255
1628
1985
2324
2642
2939
3212
1 3461
3686
3888
4066
4222
4357
4474
4573
4656
4726
4783
4630
4868
4698
4922
4941
4956
4967
4976
4962
4987
4991
4904
0,03
0120
0517
0910
1293
1664
2019
2357
2673
2967
3238
3485
1 3708
3907
4082
4236
4370
4484
4582
| 4664
4732
! 4788
4834
4871
4901
4925
4943
| 4967
4968
4977
4983
4ЙАА
4991
4994
0,04
0160
0557
0948
1331
1700
2054
2389
2704
2995
3264
3508
1 3729
3925
4099
4251
4382
4495
4591
4671
4738
4793
4838
4875
4904
4927
4945
4959
4960
4977
4964
4988
4992
4994
0,06
0199
0596
(0987
1368
1736
2088
2422
2734
3023
3289
3531
3749
3944
4115
4265
4394
4505
4599
4676
4744
4798
4842
4676
4906
4929
4946
4960
4970
4976
4984
4989
4992
4994
0.06
0239
0636
1026
1406
1772
2123
2454
! 2764
3061
3315
3554
3770
3962
4131
4279
4406
4515
4608
4686
i 4750
4803
4846
4881
4909
4931
4948
1 4961
4971
4979
4986
4989
4992
4994
0,07
0279
0675
1064
1443
1608
2157
2486
2794
3078
3340
3577
3790
3980
4147
4292
4418
4525
4616
4693
4756
4808
4850
4884
4911
4932
4949
4962
4972
4979
4986
4980
4908
4900
<М* 1
0319
0714
1103
1480
1844
2190
2517
2823
3106
3365
3599
3810
Э997
4162
4306
4429
4536
4625
4699
4761
4812
4864
4887
4913
4034
4951
4003
4073
4080
4008
14000
4000
4006
0,0»
0359
0753
1141
1517
1679
2224
2549
2852
3133
3389
3621
3830
4015
4177
4319
4441
4545
4633
4706
4767
4817
4867
4890
4016
4096
4002
4064 1
4074
,4081
■ЛЯзд 1
4MQ
4«И
LJ
431
Продолжение таблицы Л
•
3.3
3.4
3,6
3,9
Сотые долн о
0.00
4995
4997
| 4998
5000
0.01
4995
4997
4998
0,02
4995
4997
4999
0.03
4996
4997
4999
0.04
4996
4997
4999
0,06
4996
4997
4999
0.00
49%
4997
4999
о.ог
4996
4997
4999
о.оа
4996
4997
4999
0,00
4997
4998
4999
Таблица ///. Доли площади под криаой (-распределения (по Стьюденту)
■ пределах от —f до -f ' для малого числа наблюдений п (сокращ.:
десятичные дроби без нуля н запятой)
Г1
0.!
0.5
1,0
1.5
2.0
2.5
2,6
3.0
3.5
4.0
4.5
4.9
2
fc063
295
500
626
705
758
766
795
823
844
861
872
4 1
073
347
609
769
861
9J2
920
942
961
972
980
984
о
076
362
637
806
898
946
952
970
983
990
994
996
8
077
368
649
823
914
959
%5
980
990
995
997
996
ю
077
371
657
832
923
966
971
985
993
997
999
999
12
078
373
661
838
929
970
975
988
995
998
999
1
14
078
375
664
842
933
973
978
990
9%
| 998
999
16
078
376
667
846
936
975
980
991
997
999
1
18
078
377
669
848
938
977
981
992
997
999!
20
079
377
670
850
940
978
982
993
998
\т
«с
080
383
683
866
954
988
991
997
1
1
_J
438
Таблица /V.
11 не ли
CT«4H'HCft
споболи
3««ч««ни t цм рилшзди
I
2
3
4
5
6
7
8
9
10
II
12
13
14
15
1G
17
18
19
20
21
22
23
24
25
20
27
28
29
30
JO
0.1
6.31
2,92
2,35
2,13
2.02
1,94
1,90
1,86
1,83
1.81
1,80
1.78
1.77
1.76
1,75
1.75
1.74
1.73
1.73
1.73
1.72
1.72
1.71
1.71
1.71
1.71
1.70
1.70
1.70
1.70
1.64
УрО»*Я|, «пищал* p
0.06
12,7
4,30
3,18
2,78
2.57
2,45
2,37
2,31
2.26
2.23
2,20
2.18
2.16
2.15
2.13
2.12
2.П
2.10
2,09
2.09
2,08
2.07
2,07
2.06
2.06
2,06
2.05
2.05
2.05
2,04
1.96
от
0,01
31.82
6.97
4,54
3,75
3.37
3.14
3,00
2.90
2,82
2,76
2.72
2,68
2,65
2.62
2,60
2.58
2.57
2,55
2,54
2,53
2.52
2.51
2.50
2.49
2.49
2.48
2.47
2.47
2.46
2.46
2,33
63,06
9.93
6.84
4,60
4,03
3.71
3,50
3,36
1 3.25
3.17
3.11
3.06
3,01
2.98
2,96
2.92
2,90
2.88
2,86
2,85
2.83
2.82
2,81
2.80
2,79
2.78
2,77
2.76
2.76
2.75
2.58
3,tf
з.<*
3.66
3.20 1
ДО
38 П. Ф Рокицки*
Таблица Г. Необходимые значения коэффициента корреляция
ври различных уровнях значимости Р и разном числе степеней свободы
df{df л-2)
it
5
6
7
8
1 9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
Г j
0.05
0,75
0.71
0,67
0.63
0,60
0,58
0,55
0,53
0,51
0,50
0,48
0.47
0,46
0.44
0,43
0,42
0.41
0,40
0,40
0,39
0,38
0,37
0,01
0,87
0,83
0,80
0,77
0,74
0.71
0,68
0,66
0.64
0.62
0,61
0,59
0,58
0.56
0.55
0,54
0,53
0,52
0,51
0,50
0.49
0.48
df
27
28
29
30
35
40
45
50
60
70
80
90
100
125
150
200
300
400
500
700
900
1000
- . 1
0,05
0,37
0.36
0.36
0,35
0,33
0,30
0,29
0,27
0,25
0,23
0,22
0,21
0,20
0,17
0,16
0,14
0,11
0,10
0,09
0,07
0,06
0,06
0.01
0,47
0,46
0,46
0,45
0.42
0,39
0,37
0,35
0,33
0,30 1
0,28
0,27
0,25 Г
0.23
0,21 1
0,18 I
0,15 |
0.13
0.12
0,10 [
0,09
0,09 I
Таблица VI. Змчеяаа г ара pun* аелгапи * (от 0 до t»).
Для араткоста вол веред доаффащката, «ччвдаа £Ц£
поэтому 06 идо чататъ ш ОДИ
*
0.0
0.1
0.2
0.3
0,4
0,5
0.6
0.7
0,8
0,9
1.0
1(1
1.2
1.3 1
1.4
1.5
1.6
1.7
1,8
1.9
2,0
2.1
2.2
2.3
2.4
2.5
2,6
2.7
2.8
2.9
0,00
00
10
20
29
38
46
56
60
66
72
76
80
83
86
89
90
92
94
95
96
96
97
98
98
98
99
99
99
99
99
0.01
01
11
1 21
30
39
47
54
61
67
72
77
80
84
86
89
91
92
94
95 |
96
96
97
98
98
98
99
99
99
99
99
0.02
02
12
22
31
40
48
55
62
68
! 73
77
81
84
87
89
91
92
94
95
96
97
97
98
98
98
99
99
99
99
99
0.03
03
13
23
32
41
49
56
62
68
73
77
81
84
87
89
91
93
94
95
96
97
97
98
98
98
99
99
99
99
99
Сотые
0.04
04
14
24
33
41
49
56
63
69
74
78
! 81
85
87
89
91
93
94
95
96
97 j
97
98
98
98
99
99
99
99
99
ДОЛ! *
0.06
05
15
25
34
42
50
57
64
69
74
78
82
85
87 1
90
91
93
94
95
96
97
98
98
99
99
99
99
99
99
0,06
1
06
16
25
35
43
51
58
64
70
74
79
82 1
85
88
90
92
93
94
95
96
97
97
98
98
99
99
99
99
99
л1
0.07
07
17
26
35
44
52
59
65
70
75
79
82
85
88 1
90
92
93
94
95
96
97
97
98
98
99
99
99
99
99
99
0.06
' 08
18
27
36
45
52
59
65
71
75
79
83
86
88
90
92
93
94
95
96
97
97
98
98
99
99
99
99
99
99
0.09
I 09
1 I9
1 28
1 37
45
53
60
66
71
76
80
83
86
88
90
92
93
95
96
96
97
98
98
98
99 j
99
99
99
99
i r
g
§
«с
19.50
1С
"М»
*
см
$
8
8
S
'Г
S
3.07
ас
СО
15
С)
ГО
СО
5
СМ
*
ГО
2
2.71
8
см
г з s
см см см
о г** г**
N ift f
CM CM CM
h* О S
1 2.21
5
CM
2,13
Fo
CM
CO
j 2,07
2.0!
8 2
CM CM
я a
* a s
« = 6
« N «
C? Ol 40
ем — •»
см с* ас о ч* «^
сосмсмсмсмсмсмсмсмсмсмем
* \ *
^COOCCM^f — CM — *ЛСМСМГО;0©»0 — Г-*0
«о^^аеосЪиосмосе^оо^'-^сосоемем
NOeDCtPJrtW^NNMNNNWNNN
!
S2
i?"
; — ^ — acot^oor^ — ооосоаосмс©*» —
_. !^^г^са«го«осмоо>г^оФю,ч,,^сосоео
' смсаао*очг^сосососмсмсмсмемсмсмсмсмсм
I
I
Ш
• »
см^г^ог^оосо — слас-г-фф10'«э»'^,<чгсо
см с* so" о" V V со* со* со" cm" cm* cm cm" cm* cm cm" cm" cm" cm
ac — ^г^оарооосмоо —• о q> f Ф о N
— со эс о r^ — О со — oooor^Oibub^r^r^'
см ©" ac* to v* V со* со* со" со* см* см* см* см* см* см* см* см см*
Г»»1СОСМ1СГО^^СОГ^Ю1^Г^. О ^f С^ VO — 00
см с* х* в" V V со* со* со* со* см* см* см* см см* см* см* см с*
t^ox/oScMt^Sc*--oc»a6r^t*eSoS"o
см оГ ас о V V со* со* со* го* со* см* см* см см* см см* см* см
см с* ас о* V V со" го* со* со" со* со* см* см* см* см см* см* см
см с" еГ «о* to V со* го* со* со* со* со* со* см* см" см" см* см см
«сем — со — ю — ас о чг со см — — о о ел
см о* с* аГ %о <*•" V со* со* со* со* со* со* со* со" со* см" см см
S 8
смслслвюмг'в-^со'сососо'сососоооеососо
i n
[
/ * 1
/ ^1
©*о^р»^^о^©аосло^оосоа>юсм
CMoTcft^?tcTto^'4p-4r^,co*rocococococococo 1
— Со — t*T S §* «oB-S3n4>83t**8 I
- вб С N Ю Й Л Ю* Ю* V У V t" * * V V Г * 1
-.cif)*iovcseco>o-wrj*o«osia» I
436
8
S
23
сч
X
t^.
1 о
1
*
1
' чг
л '
сч
-
1 *"" /
"о /
\ / ^
2
S
СМ
см
см
о
см
см
со
см
см
СО
см
со
см
см
1С
см
с
со
см
rt
см
00
см
о
СО
СП
СО
8
8
00
о
см
о
см
00
см
а
см
см
СО
см
55
см
см
см
Cft
см
см
S
см
г
см
о
СО
СО*
8
см
00
S
о
см
ю
см
3
см
8
см
Я
см
о
**•
см
СО
см
8
см
8
см
см
00
см
8
со
со
8
8
8
8
8
см
СО
см
8
см
£
см
8
см
со
см
5
см
S
см
S
см
S
см
3
со
9
со
S
а
й
3
S
см
тт
СМ
00
см
см
см
3
см
8
см
см
см
3
см
S
См
15
см
о
со
9
со
8
S
к
S
о
см
8
см
<0
см
а
см
3
см
35
см
3
см
01
см
8
см
»
см
г
см
8
3
8
*
S
8
&
см
ю
см
а
см
г»
см
см
8
см
Й
см
Г*
см
г
СМ
см
8
см
со
8
а
г*
CD
S
&
8
м
00
см
8
еч
см
см
со
см
&
см
(О
см
г*
см
со
г*
см
$
см
те"
•1
к
т
&
S
5
см
см
см
о>
см
S
см
8!
см
8
см
ю
см
8
см
£
см
8
««
*
to
а
8
3
3
3
г
см
о
см
со
—
а
см
8
см
$
см
со
см
S
см
g
см
S
см
3
со
«0
к
8
00
СО
CD
о
см
г
ем
о
см
см
см
£
см
я
см
см
см
2
см
г
см
8
см
и
СО*
«
S
S
3
8
—
8
ем
S
ем
см
см
оо
mm
ем
*
ем
3
см
см
3
см
S
см
8
СО*
8
3
з
8
е
г
—
8
—
$
—
S
см
о
см
к
см
я
сч
{о
см
з
см
«О
см
СО
S
8
8 8 1
3 5 1
S & 1
Й В
« —
г к
- - 1
Ф об
~ ""
8 S 1
— —
2 3 1
см* — 1
8 о 1
см" ем"
£ о I
см" см 1
8 5 1
см см" 1
<* СО J
см см* |
S S |
w см" |
&. 8 1
П СО* I
8 3
<о со 1
8 »
чзт
'
i числителем)
берете■
ш
1
о
1
9
4 ,/
Я
!
i «
"i
! 2
i *
!
\
1
1 »^
! *
10
«•
•* 1
** 1
-
1 <5 Я ~ *° £2 Ж
о Д — -* о об
I 8 с? со* го" о>" ;о
| ft d —
6261
99,47
26,50
13.84
9.38
7,23
6209
99,45
26.69
14,02
9,55
7.40
6157
99.43
26.87
14.20
9.72
7.50
1 «о с* »л n- ft cs
| 5 * d n oo s
со ft г- -*• с* n-
) ft C4 -
со о го «л ьга ь-
t5 -^ см 1.0 о ос
S Л* N* <<• О* N*
О N - -
6022
99.39
27.35
14.66
10.16
7.98
5982
99.37
27.49
14.80
10.29
8.10
5928
99.36
27.67
14.98
10.46
8.26
ft Q — — Г- N.
lg ГО О» СЧ СО -Г
3 ft S «ft О* АО
ft CI — —
5764
99.30
28.42
15.52
10.97
8.75
5625
99.25
28.71
15.98
11.39
9.15
5403
99,17
29.46
16.69
12.06
9.78
*r ft о oo со о
Я82881!
— 04 *0 «*■ Ю CO
5,65
5,99
6,16
6,31
6.47
6.62
6.72
6.84
6.99
7.19
7.46
7.85
8.47
9.55
12,25
N.
4.86
5.20
5.36
5.52
5.67
5.81
5,91
6.03
6.18
6,37
6.63
7.01
7.59
8.65
Я
«0
4,31
4,65
4.81
4.96
5.11
5.26
5.35
5.47
5,61
5,80
90*9
6,42
6.99
8.02
10,56
ft
3.91
4.25
4.41
4,56
4.71
4,85
S
5.06
5,20
5,39
5.64
5,99
6,55
7.56
10,04
о
3.60
3.94
4,10
4,25
4,40
4,54
s
4,74
4,89
5,07
5.32
5.67
6.22
7,21
9,65
-
3,36
3.70
3,86
4,01
4,16
4,30
со
4,50
4,64
S
5,06
5,41
5,95 ■
6.93
9.33
a
3.16
3,51
3,66
3,82
3,96
4,10
ft
4,30
4.44
s
4,86
5,21
5.74
6,70
9,07
CO
3,00
3,35
3,51
3,66
3,80
3,94
S
4,14
4,28
CO
4,69
5,04
5,56
6,51
8.86
■*
n. m
00 N.
СЧ СЧ*
3,21
3,10
3.37
3,26
3,52
3,41
3,67
3.55
3.80
3.69
3.89
3,78
4,00
3,89
4,14
4,03
3 8 1
4,56
4.44
4,89
4,77
5,42
5,29
6.36
6,23
8.68
8.53
Ю 40
£
СЧ
8
СО
1Л
СЧ
SJ
СЧ
Ч|»
сч
3
сч
сч
сч
00
сч
8
сч
р
сч
со
сч
г*.
(О
сч
8
сч
3
сч
сч
сч
8
сч
ь-
сч
3
сч
2
сч
8
сч
о
сч
«г
сч
8
сч
ч*
чг
сч
8 о 8 2 S б
сч сч -« - - -
588382
сч сч сч сч — -
2 8
ео" со*
8 а
со" сч*
S 8 R
сч" сч* сч
е s з
сч* сч* сч"
S ю So
сч* сч" сч"
К s зs.
сч" сч* сч" —
— СО
со сч
£ 8
CO* СО
8 8 8
со еч" еч
г а
оо г-
сч сч сч сч
Ю ео р
К Г*. Р
сч" сч* сч"
8 8 2 8
сч* сч* сч" сч*
ч* СО
8 «
сч
со со"
о 6S ел ел 6» <ю «Б 25 35 со —
со* со" со* со* сч* сч* сч* сч* сч* сч* сч* сч" сч* сч*
N N N
— •— о
О) —
со г*»
•*• СО
со со со ео
со 8 сч
со со со
2 8 8
со со со
8 8 8
со" со" сч*
8 3 S Я
сч* сч* с* сч*
г г
СЧ <0
Ю ч*
со со со со
4j« со со
со со со
8
сч со ю
со со со
2 8 S
со со со
ел сч (Л —
оо к Л 'Г
сч" сч" сч* сч"
3 8
со* со*
— ю —
Ю ч* ч*
8
со со со со
8 8 8
со со со
a a t
со со со
$ Э $ ю
сч" сч" сч* сч"
со чг
0> 00
с*- О
3 ю ю
8
со со сососоеососо
«О сч ел
<* <г со
со со со
со со ео
со со ео
сч -л сп -<
— 55 f^ О
со* сч" сч" сч*
S о
СЛ 00
со" со"
-. <о —
00 N N
со со со со
г s s
со со со
а 8 ч
со со со
8 S 8 S
ео" со" сч" сч
сч
С- О
а 8 а
ч* со" со*
8
8 8 R
СО* со" со"
со со со
"о «О »- О
со со со со
S 8 5fe «US J:feSg
8 8 9 8
со со со со
2 8
ю* ю"
о а^
ю" Ч|Г
о) S ё ? 3 S SloSS
S л £ «
СО •- 9) t-
ч? чг CO* СО*
"1 Я.
<о* «о
8 8
1Л* ift*
SS8
tft «А Ю
•=38
1Л ift Ift
$98
IA Ю U)
s s й s
»о V V ч?
9 а
«О во
2 2gS$Uf!$
00 О
оо* оо*
СО N N
S 8 S
tw h" l>
со о об <о
г*Г isT <o" «о"
bS2as88888Ua8?8§8
439
ftm»
я»*
6
Т
*
9
to
It
12
13
М
IS
н
IT
1
19
30
м
Эв ■
*
t
э.$
|з.»
1.3
а.»
3.1 j
3.1
3.1
3.1
з.о
.*••
з.#
3,0
3.0
з.» !
э.о
*•*
*.9
**•
в } *.*
»
1
U7
4.2
4.0
4.0
3.9
3.*
з,в
3.7
3.7
3.7
3.7
3,6
3.6
з.«
3.*
3.5
3.S {
1
3.4
3.4
1^
} 4
4.9
4.7
4.5
4.4
4.3
4.»
4.2
4.2
4.1
4.1
4.1
4.0
4.0
4.0
4.0
3.9
з.а
з.в;
3.7
3.7
t
5.3
5.1
4.9
4,8
4.7
4.6
4.5
4.5
4.4
4.4
4.3
4.3
4.3
4.3
4.1
4.»
4.1
4.0 ]
4.0
3.9
с
5.6
5.4
5.»
5.0
4.9
4.0
4.8
4.7
4.6
4.6
4.6
4.5
4.5
4.5
4.5
4.4
4.3
4.»
4.2
4.1
т
5.9
6.6
5.4
5.2
5.1
5.0
5.0
4-9
4.8
4.8 j
4.7
4.7
4.7
4.6
4.6
4.5
4.5
4.4
4.3
4.2
•
6.1
5.8
5.6
5.4
5.3
5.2
5.«
5.1
5,0
4.9
4.9
4.9
4.8
4.8
4.8
4.7
4.6
4.5
4.4
4.4
в
6.3
6.0
5.8
5.6
5.5
5.4
5.3
5.2
5.1
5,1
5.0
5.0
5.0
4.9
4.9
4.8
4.7
4.6
4.6
4.5
10
6.5
6,2
5.9
5.7
5,6
5.5
5.4
5.3
5.3
5.2
5.2
5.!
5.1
5.0
5.0
4.9
4.8
4.7
4.7
4.6
11
6.7
6.3
6.1
5.9
5.7
5.6
5.5
5.4
5.4
5.3
5.3
5.2
5.2
5,1 1
5.1
5.0
4.9
4.8
4.7
4.6
11
м
м
6.2
6.0
5.8
5.7
5,6
5.5
5.5
5.4
5.4
5.3
5.3
5.2
5.2
5.1
5,0
4.9
4.8
4.Т
-J
Таблица X. Х'-ркцкдслемк
2
3
4
5
6
t
8
1 9
10
11 '
1 12
I 13
14
1 15
1 ,6
1 17
1 18
19
1 20
1
' 21
I22
! 23
i24
1 25
( 26
! 27
Бврояшости ашпт* i*. аДОышшшщего тблмчанх 1
0.99
0,02
0.U
0.30
0.55
0.87
1,24
1.65
2.09
2,56
3.05
3.57
4,11
4,66
5,23
5,81
6,41
1 7,01
7,63
I 8,26
8,90
1 9.54
10.20
10,86
JH.52
I12,20
12.88
j 28'13.56
! 29
30
40
50
60
[14,26
(14.95
122,16
29.71
37,48
0.96
о.ю
0.35
0,71
U5
1.64
2.17
2.73
3.33
3.94
4,57
5,23
5,89
6,57
7.26
7,96
8,67
9,39
10,12
10.85
11.59
12.34
13.09
13.85
14,61
15,38
16,15
(16.93
17.71
18,49
26.51
34,76
43.19
0.90
0.02
0,21
0.58
1,06
1.61
2.20
2,83
3.49
4,17
4.87
5,58
6.30
7.04
7,79
8,55
9,31
110,09
110.86
11,65
12.44
13.24
14,04
14,85
15,66
16.47
17,29
18,11
18,94
19.77
20,60
129.05
37,69
[46,46
0.75
0,10
0,58
1.21
1.92
2.67
3,45
4,25
5.W
5,90
6,74
7,58
I 8,44
9,30
110.17
11.04
11,91
12.79
13,68
14.56
15,45
16.34
17,24
18.14
19,04
19,04
20.84
21.75
ЯД
23,57
24.48
33,66
42.94
152,29
0J0
0,45
1.»
2.37
3.36
4.35
5,35
6,35
7.34
8.34
9.34
10,34
11,34
12,34
13.34
14.34
15,34
16,34
17.34
18,34
19,34
20.34
21.34
22,34
23,34
24,34
125.34
26,34
27,34
28,34
29,34
39.34
49,33
59.33
0*25
1.32
2,77
4,11
5,39
6.63
7,84
9,04
10.22
11.39
12.55
13,70
М.86
15,96
17,12
18,25
19.37
20,49
21,60
22.72
23,83
24,93
26,04
27,14
28,24
29,34
30,43
31,53
32,62
33.71
34.80
45,62
56,33
j 66,98
0.10
2.71
4,61
6,25
7.78
1 9,24
10.64
12,02
13.36
14,68
15.99
17.28
18,55
19,81
21,06
22.31
23,54
24.77
25.99
27,20
28,41
29,62
30,81
32,01
33,20
34,38
35,56
36.74
37,92
39,09
40.36
61.80
63,17
74.40
0.0» ftjtt
3,84
5,99
7.81
9,49
11.07
12,59
14,07
15,51
16.92
18.31
19.68
21,03
22,36
23,68
25,00
26.30
27.59
28,87
30,14
31,41
32,67
33.92
35,17
36,42
37,66
38,89
40.11
41,34
42,56
43,77
55,76
67.50
79.0В
I
5,02
7,38
9,35
11.14
12,83
14.45
16,01
17.53
19,02
20,48
21.92
23,34
24,74
26,12
27,49
28,85
30,19
31,53
32.85
34,17
35.48
36,78
38,08
39,36
40.65
41,92
43,19
44,46
45,72
46.98
59,34
71,42
83.30
0,01 1
6.63
9,21
| U.34
13,28
15.09
16,81
18,48
20.09
21.67
23.21
24,72
26,22
27,69
29,14
30,58
32,00
33,41
34.81
36,19
37,57
38,93
40,29
41,64
' 42,98
44,31
45,64
46,96
48,28
49,59
50.89
63,69
76,15
88.38
441
Продолжение таблицы X
Л/
70
80
90
100
Вероятности значения £а, и ре пытающего табличное
0.09
45.44
53,54
61.75
70.06
0,05
51.74
60.39
69,13
77.93
0,90
55,33
64,28
73,29
82,36
0,75
61,70
71,14
80,62
90,13
0.50
69,33
79,33
89,33
99,33
0,25
77,58
88.13
98.64
109.14
0,10
85.53
96.58
107,56
118,50
0,05
90,53
101,88
113,14
124,34
0,026
95.02
106.63
118.14
129.56
0.01
100.42
112,33
124.12
135.81
Таблица XL Значения ординат (в долях единицы)
для разных /*
/
0,0
0.1
0.2
0,3
0.4
0.5
0.6
0.7
0.8
0,9
1.0
1.1
1.2
1.3
1.4
1.5
Ординаты
0.3989
0,3970
0.3910
0.3814
0.3683
0.3521
0,3332
0.3123
0,2897
0.2661
0.2420
0.2179
0.1942
0.1714
0.1497
0.1295
/
1.6
1.7
1.8
1.9
2.0
2,1
2.2
2.3
2,4
2.5
2,6
2.7
2,8
2,9
з.о
3.9
4.0
Ординаты
0.1109
0.0940
0.0790
0.0656
0,0540
0,0440
0.0355
0,0283
0,0224
0.0175
0.0136
0.0104
0.0079
0,0060
0,0044
0,0002
0,0001
- - 1
• Частоты (вакоплениые) нормального распределения (п •= 10000) пред-
стащены • табл. П.
442
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аддитивность
действия генов 204—206
суммирование 206
Вариапса
аддитивная 212, 217, 218, 220,
256, 276. 303
взаимодействия 212, 223
геиотипическая 201
доминантная 213—214
и инбридинг 340
компоненты 212,216, 220—224,
25G
между кроссами 360
методы вычисления 212—216
неаддитивная 212, 221
средовая 201, 215
структуры разных варианс
212—213, 257
фенотипическая 201
Вариация
качественная 179
количественная 180—182
непрерывная 180—182
прерывистая 179
в развитии 385
Вероятность 9—15
апостериорная 10
априорная 10
математическая 10
полная 14
теоремы 11—13
условная 13
эмпирическая 10
Взаимодействие геном
внутриаллельиое 213
межаллелыюе 37, 38
зпистатическое 38
Генетика статистическая
обоснование 3—5
Генетико-автоматическне
процессы, см. дреАф генов
Генетическая корреляция
методы вычисления 368
причины 371—372
Генетический эффект
и доза облучения 96
сравнение мутагенов 98
Геногеографпя 171—175
Геногормоны 383
Гены
вариация п действии 379
модификаторы 192, 306
неполное проявление 29—32,
379
поле действия 380—381
средний эффект 207—209
Гетерозиготы
изменение доли при отборе
143
при инбридинге 167—168
селекционные преимущества
147-149
установление доли в
популяции 120
Гетерозис
комбинационная способность
359-363
я материнский эффект 336
моногибридиый 389
определение 356
отрицательный 355
положительный 355
статистические основы 356—
358
Гибридизация иитрограсенвяая 354
Гомеостаз
генетический 324
пояуляцяоииый 324
443
1>>*
генетически! \и
иибреднын 177
мутационный 177
сотрет ациовны А 177
гголюоиоииыа 177
Действие генов
дернацня в 279
ел ммацня
аддитивная 204, 344
пеаддитнвиая 206
Доминирование
взаимоотношение с а 203
м гетерозис 357
и нибредная депрессия 358—
359
и отбор 305
н приспособленность 139
разные степени 130
в селекквоиная ценность 130
уволюдня 156
Дрейф генов, случайный 161
и размеры популяции 162—163
Дрейф менотическиа 24—25
Мибредные линии
возрастание гоиозаготностн 340
енотишпеская
фсиотишг
345—346
средовая изменчивость 347
Инсридинг
Депресеки 342—345
дифференциация вовулядни
варнавсы 340
гомо- в гетерози-
готности 333—334
кошффкдиип FK 326—331
/>Л 332--333
и коэффициент наследуемости
345
и аоэффииленты пути 327
метод общих предков 333—333
размеры поаулиднн 167—170,
при разных степенях родства
Ж»
систематический 326—335
и февотипическаи
вость 345-046
формулы для F.
С BOtMMHJbJO КФЗффнЦВСВТОН
иути 319
И в**0** «poHiHucidl 331
Инмигранши 151
Коварнанса
генетическая 221—223
потомков-родителей 217, 218
между полусибсами 218
сибсамн 219
средовая 215—216
фенотнпическая 220—224
Количественные признаки
метод треугольника 196
наследование 183—185
полнгены по Мазеру 192
сигнальные гены 195
теория полимерных генов
186—189, 302—303
число генов 193—195. 303
Комбинационная способность
генетические основы 359—360
методы анализа 265» 361
специфическая 266, 362
Коррелирующие признаки 364
Корреляция
генетическая 365
методы определения 368,379
причины 371
ошибка 369
между родственниками 240
соотношение между разными
видами К. 367
средовая 365
Коэффициенты
нпбрндинга 326—333
отбора 131, 300-302
родства 349—350
корреляции, разной 368
пути. см. путевые
коэффициенты
Кроссы
вариация внутри кроссов 359—
360
междинеиные 359
Материнское влияние 245
и гетерозис 355—356
при наследовании
количественных признаков 188—189
в структуре варнанс 221—222
Метод пробаяда ВеДнберга 45—47
Миграции 151
и отбор 151—153
Мутации
баланс с отбором 157—160
и инбридинг 170—171
индуцированные 93
статнетичноеть 80. 95—*95
связь с дозой, крнвпаниич1
нан 99—103
вримвлвненяаи 96—98
методы учета 58—60
высшие растения 73
доминантные летая 67—68
дрозофила 61—66
количественные признаки
74-75
микроорганизмы 76—60
млекопитающие 70—71
перепончатокрылые 69
хромосомные аберрации 66
человек 71—73
принцип попадания 105
слччайяость 86
взаимная 86. 90
во времени 86—87
ограничения 92
в пространстве 87
сохранение в популяции 154—
156
спонтанные 80
стохастические подходы 105—
106
цепные процессы 107—109
частота
общая 80
отдельных генов 81—86
Мутационное давление 127, 157—
160
Мутационный груз 177
Наследуемость
в внутриклассовый
коэффициент корреляции 217. 255
н диаллелъное (полналдедь-
ное) скрещивание 263—265
дисперсионный анализ; метод
24S-254
достоверность 269—272
для качественных признаков
268-269
как коэффициент корреляции
240-241
между полусябсамн 240
между потомками и
родителями 239
между сносами 240
как коэффициент регрессии
244—247. 293-295
и компоненты генетическом
варнансы 256—258
методы определения 206, 227,
239—240. 244э 253
неправильные пол годы 361—
263
однояйцевые двоим 266
разных признаки* 2П—Ж
как сдвиг ври отборе 296.
308—309
сравнение имбиоших и агг-
бредимх линии 227—221
в > жом смысле 226. 2»
формулы, огракячеявя в яри
иеиеиия 241—243
Формула Уоррена 267
в пафосом смысле 22& 265
Неаддитивность
влияние на варяавеу 213
в штоииаиии генов 291—266
Нерасхождение хромосом, частота
21-22 «~~—
У человека 22—24
Олигогеды 192. 304. 306
Оплодотворение
веройтиоинад картина 26
отклонения от случайности
26-27
Отбор
асимметрия ответа 315—316
во адоминаитиому гену 134—
135
дестабилизирующая 157, 284
диср>птяваыя 282—283
so доминантному гену 131—
133
естественный 281
искусственный 281
во комбинационным
способностям 362—363
косвенный 372—375
коэффициент 131
в малых вооуляджях
свое 384
ыкяернмеяталвяое 267—291,
310-^312
направленный 283
и наследуемость 28, 294, 295.
306-309
Н1Ш1ПЩИИ1 изменчивости ври 0
318-ЗГ9
пределы 317—325
вримеры
в природе 281
у седьскоюзийг i и i тан ■ жи-
вотиых и растеинй
и (трисоособлениость 981
как ирнчниа сдвигов в юоу-
дяннях 128, 292-293
прогнозирование резнаьтаяив
296-298
в разных вжлрааленашх 315—
317
среды
«46
зю-зм
равновесие с мутационным
давлением 157—159
размеры 317
по сверхдоминантному гену
136
селекционный дифференциал
293
стабилизирующий 283
типы в зависимости от
доминирования 130, 133
фиксация и потеря генов 163
фундаментальная теория
естественного отбора 176
число поколений 144
эффективность 137—138, 308
Отклонение от популяционной
средней 210—212
доминантное 211
средовое 215—216
эйнстатическое 213
Ланмнксяя 111, 168, 169
Плейотропия 372, 380
Повторяемость 385—389
Полигены 192
Полимерные гены
предпосылки теории 186—189
усложнения 190—191
Полиморфизм
адаптационный 149
и гетерозис 150—151
инверсионный 150
сбалансированный 147
хромосомный 150
Популяции
н внешние условия 279—280
геиотипическая структура 113
как динамическая система 170,
175
изменения при отборе 129—
136, 144
и иибридииг 167—169
малые 170
насыщение мутациями 153—
155
оаимяктическая 111
параметры 112—113
размеры 166
равновесие 145
неустойчивое 141
при различиях в q у 99 я oV
123
по сцепленным с полом генам
124-125
устойчиво* 141. 145
структура ИЗ
стационарные состояния 119
ъффеятимын объем 170
Понудяояоияа* средняя 203, 207
Поток генов 152
Предки общие 347
Приспособленность
коэффициент 139
средняя популяционная по
пр. 139
и Ад 142
Проявление генов
неполное 30
неп роя влей не 31
и онтогенез 33, 379
и характер расщепления 29,
30
Путевые коэффициенты (К. пути)
между гаметами 235
зиготами 234
зиготами и гаметами
234
н инбридинг 235—237
как коэффициенты регрессии
328
методы определения 229—232
множественные 233
между родителями и потом*
ками 236
Расщепление
и нежизнеспособность зигот
27
и неполное проявление 29—33
проверка достоверности 39—
условия
динамические 17
статистические 16, 17, 33
Рекомбинация 49—56
Рентгеноселекция 310—312
Родства коэффициент 349
связь с коэффициентом
инбридинга 351
Сверхдомниирование
и гетерозис 367
и приспособленность 136, 350
при отборе 136
Селекционные значения 130—131,
207, 277
Селекционный дифференциал
292—294
и интенсивность отбора 300—
302
Селекция, см, отбор
Скрещивание
для использования rerepot**
са 355
поглотительное 363
446
типы 352
Средний эффект гена 207—208
Стабилизирующего скрещивания
закон 117—118
Статистические закономерности
в биологии 3, 7, 8
возможность предсказания 8
Статистические модели
при диаллельном
скрещивании 265, 361
при двухфакторном
дисперсионном анализе 222, 263, 361
при иерархической схеме 249,
261, 263
при однофакторном
дисперсионном анализе 250
для оценки коварианс 257
Тетрадный анализ 21
Транслокация
их влияние на расщепление 35
обменные 36
Феногенез
запрограммированность 378
случайные ошибки 385
21тиГнчеС|сие иРоа*«и при
379—382
Харди—Вейиберга аакои 113, 120.
121 #
отклонения 126—127
условия осуществления 118—
119
Ценность
адаптивная 129
генотнпнческая 211
селекционная 207—209
фенотипическая 129, 203
Частота гена
изменение при отборе 131—
137
определение в популяции 115
связь с частотой фенотипов
и генотипов 116— II/
соотношение с коэффициентом
отбора 142
Эмиграция 151
Энотера 36—37
ОГЛАВЛЕНИЕ
Предисловие» 9
Введение. Статистические закономерности и основимс понятия теории
вероятностей 7
Глааа I. Статистические и динамические процессы как основа яако*
момертячеи масле доивтм 16
Глааа 1 Стохлстнкл мутационного процесс* .*••♦.,.. 67
Глааа 3. Генетическая структура ПОЛулЯщА ,Г#: '. . ПО
Глааа 4 Изменении генстическе* СТрГЯЯ^ШЩтцАъ определяю-
клине их факторы , . . «** -*.*■** v .#. J 126
Глааа 5. Статистическая x^paJCT^piWtl^ Лй^Д*ЙЯЛ Яри иелрерыа
иоЛ вариация .*.*,,.....,.>>.. 170
Главе б. Наследуемость ....**,.. 226
Глааа 7. Отбор по колкистеемШН прививкам. *го механизм и
результаты . , , 279
Глааа $ Влияние рваличяы* систем спаривании ия структуру
популяций. Инбридинг и вутбридииг . , . , 326
Глава 9. Фемотипическис и теистические корреляции 364
Глава 10, Некоторые статистические явления » феиогеиез* Я7Н
Литература • * » . • W)
Указатель символов , , , ......».>> 407
Статистический справочник 4)0
Предметами улетел»* , 444
Роницкий Петр Фомин
ВВЕДЕНИЕ В СТАТИСТИЧЕСКУЮ ГЕНЕТИКУ
%1&$т i*AM0ttip 0. H. ba/i**frxu4
K/jppmh<rp Ь, И Гашсгнва
AT НЧаи Сйлш> * m&jp A/Vf-ltf» г, Пашис»*'; ш «гчми W/JJ 1W4 f, йумега «н/ЯР/и
те»*». * I. tU* л » У**»* л *,1 И«* * 7? 4В Tea яла. В0в- Ти|Ы* WW etf*.
II**» I trrO 21 *'<п
ИмДОММРВЛ Hh**»4l0*# ЮШ,*Ш» Т(#)ГЛМр*1Ш*Нф*1* */««НИ* »/i 4ИИ*М ИЦ«)МШ«,
*м*г#*ф## * мим** ?</^<»«* **»*#»*»« лю9рчурц ш, *,i*rr****M*ir, ПШЬ,
Мм**, t* *мрт*. « <ф«*м« 1wn+'tt* %p§tm*<9 Ьымштм **т*евфн* т**г«1ьт*
1Ш К/1 B**ojrH«»», М«М, /tMBMMiaa «|N*aeftf. ft