/










































































































































































































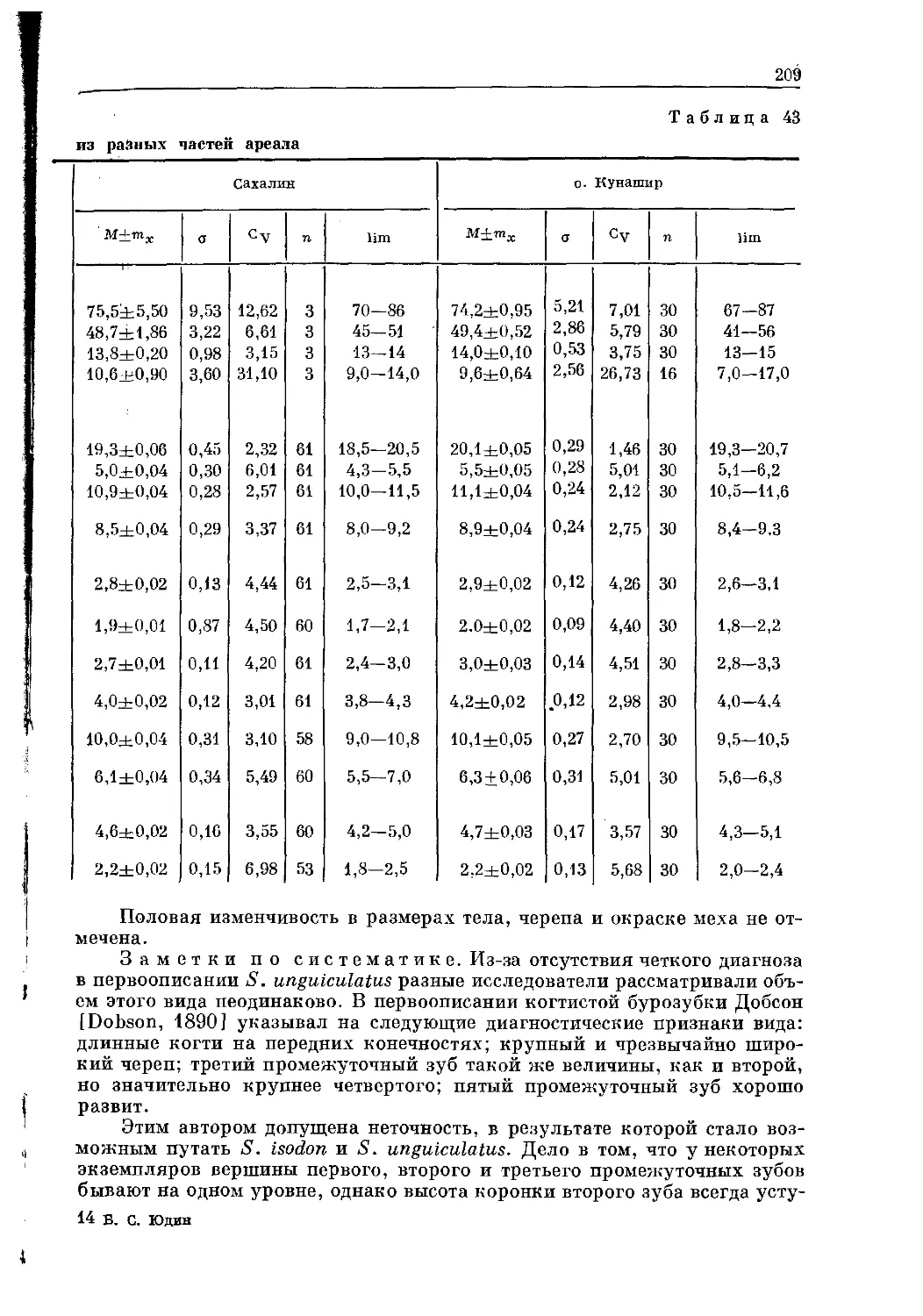





























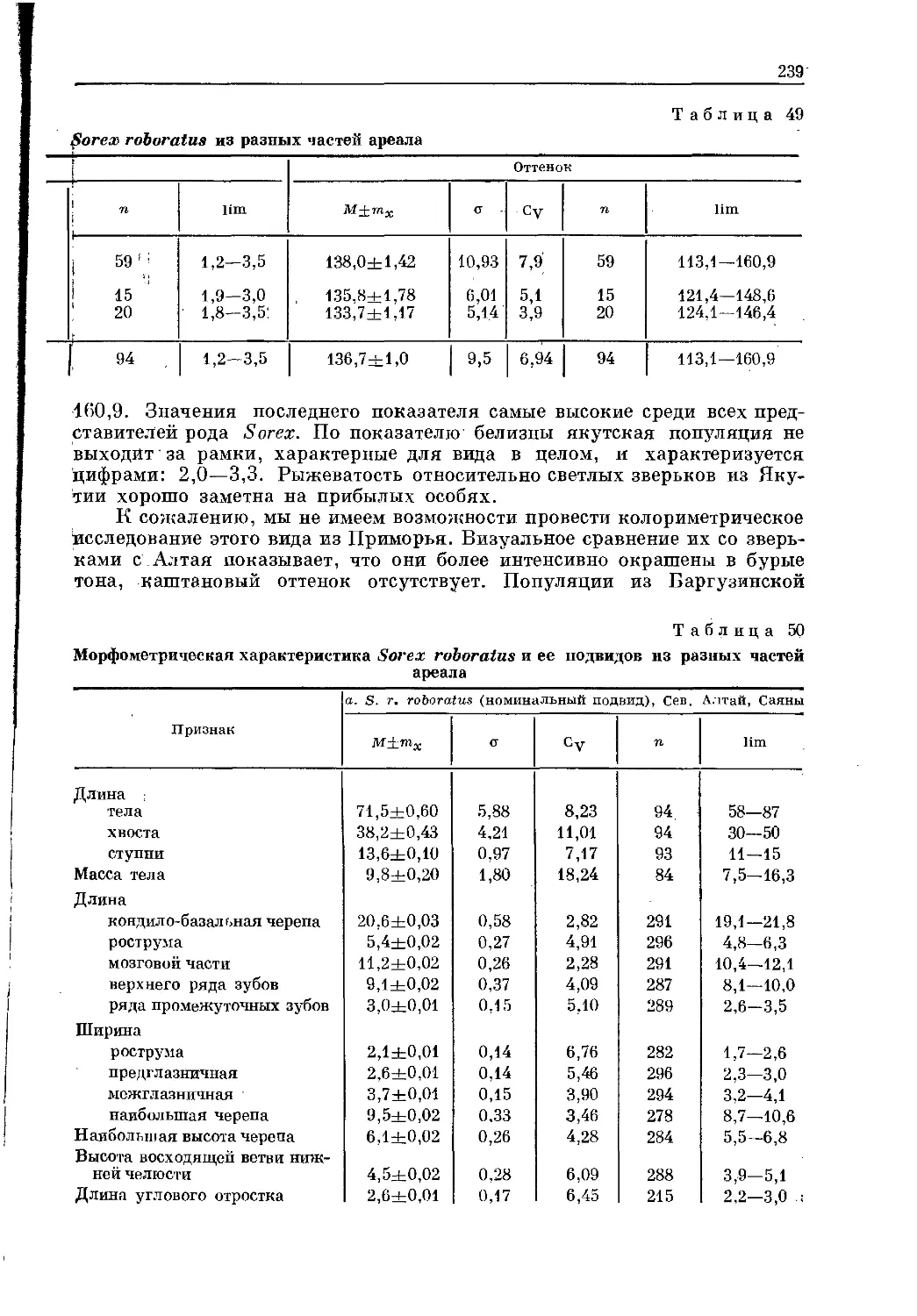
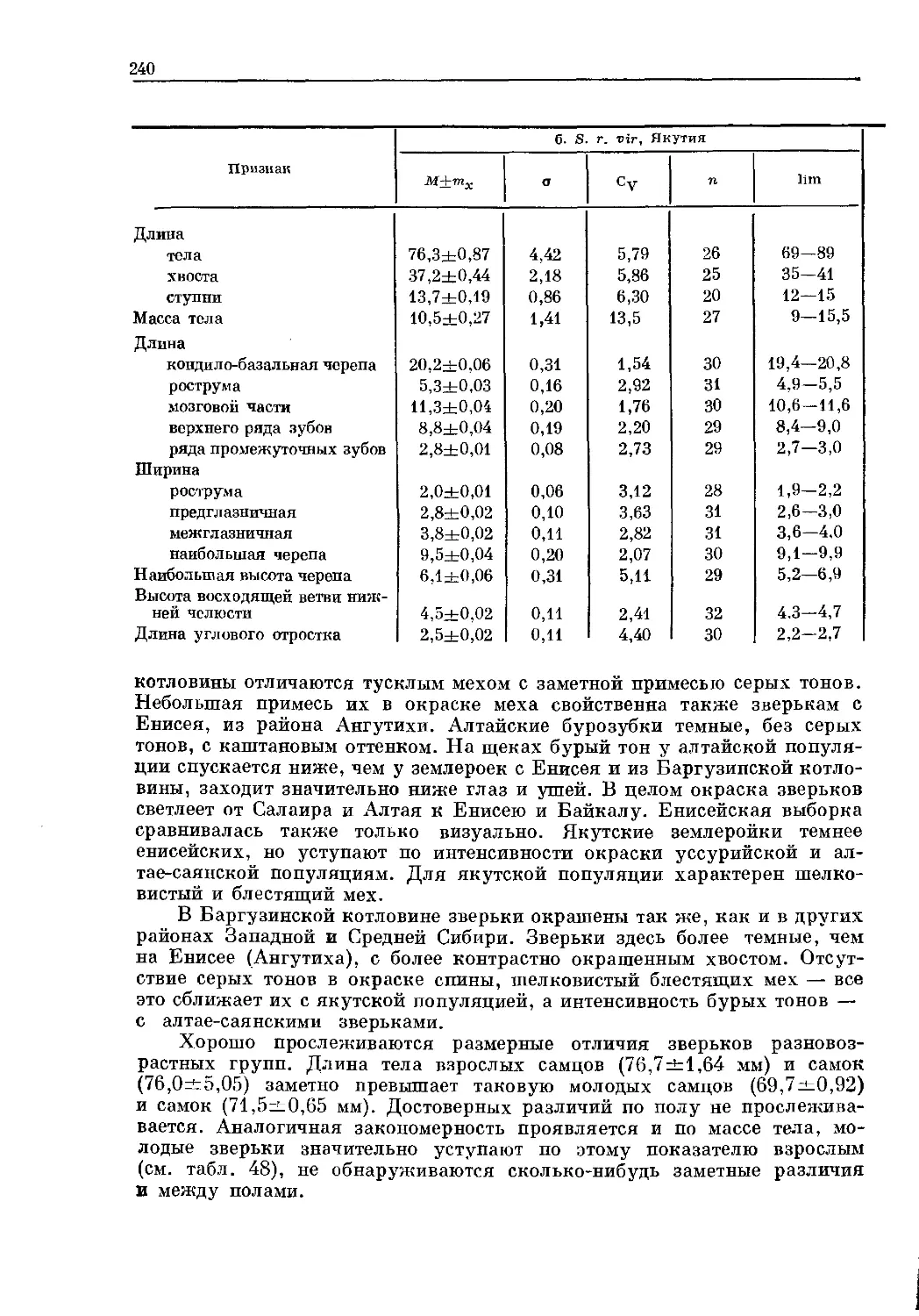





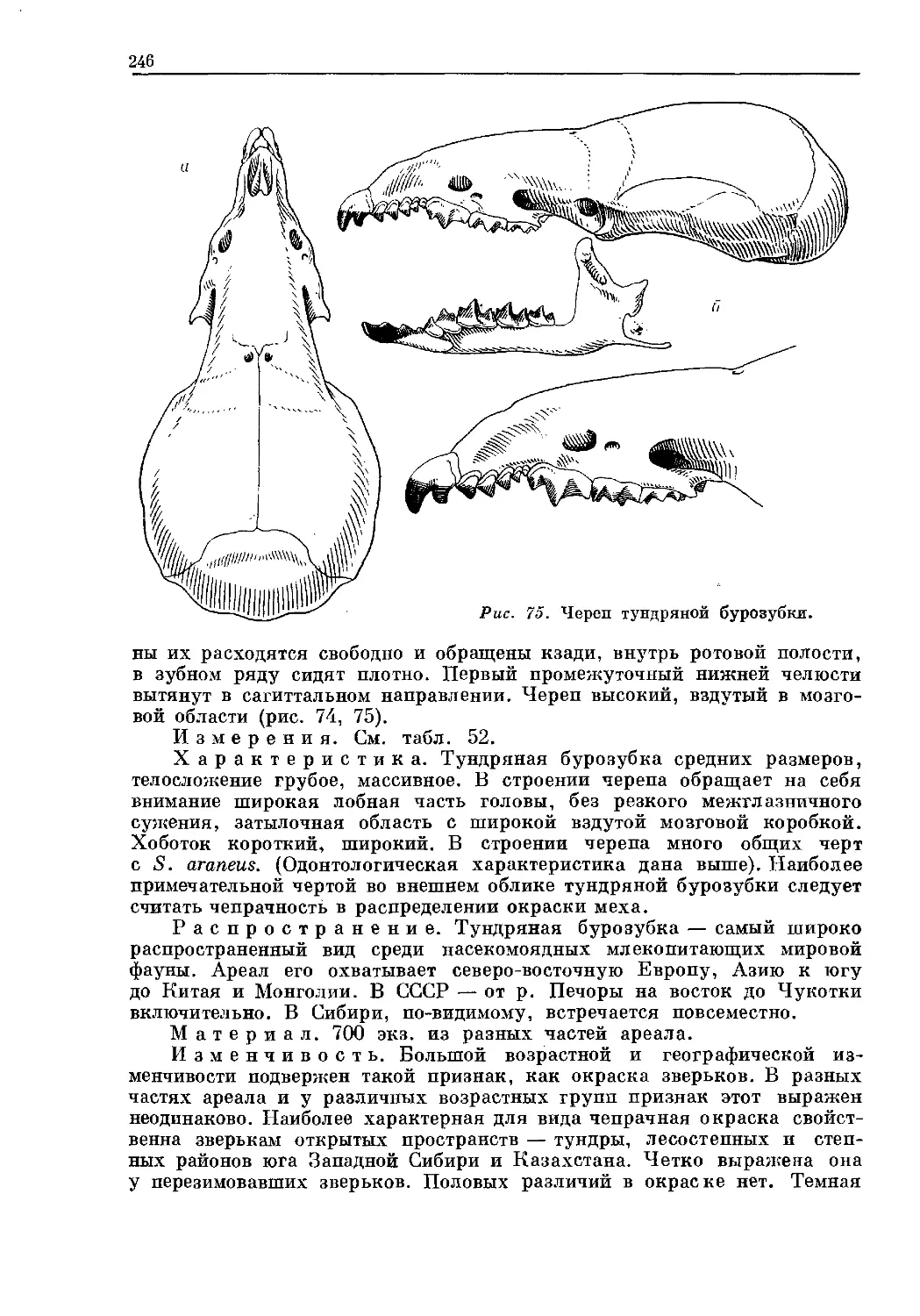



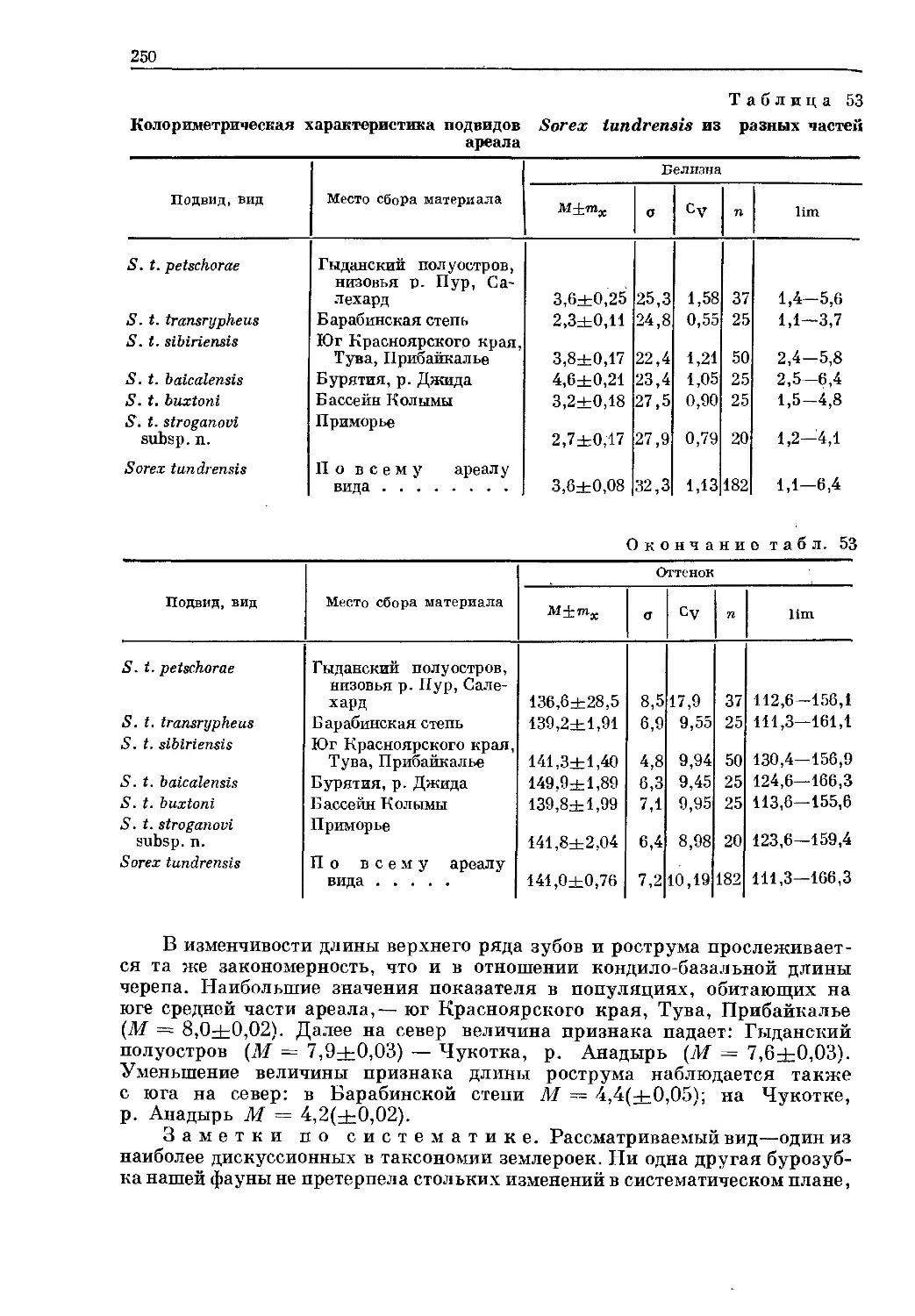























































































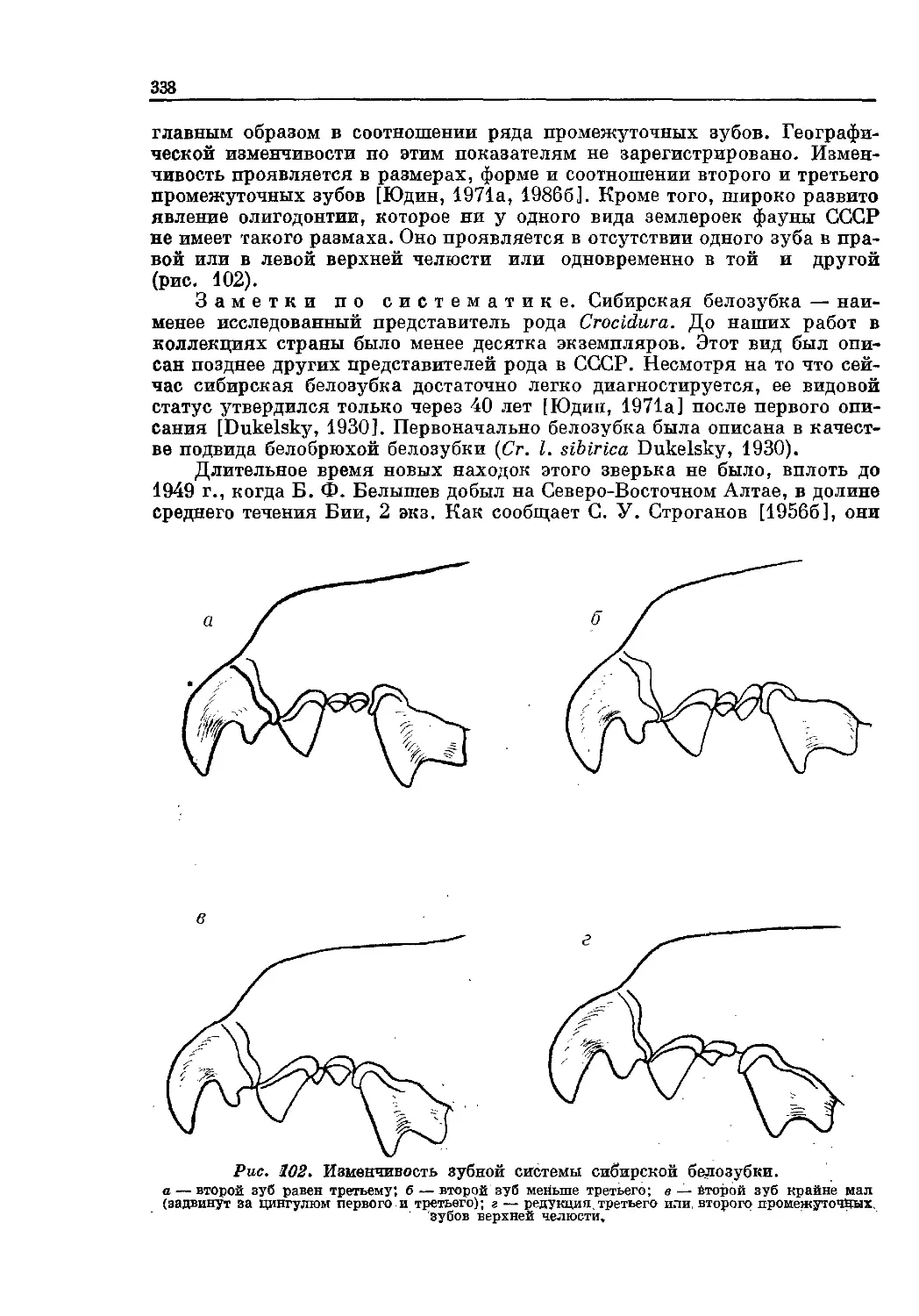


























Text
Б. С. Юдин
НАСЕКОМОЯДНЫЕ
МЛЕКОПИТАЮЩИЕ
СИБИРИ
'*--АУКЛ>
СИБИРСКОЕ ОТДЕЛЕНИЕ
Б. С. Юдин
НАСЕКОМОЯДНЫЕ
МЛЕКОПИТАЮЩИЕ
СИБИРИ
Борис Степанович Юдин был крупным
специалистом по систематике и экологии
насекомоядных млекопитающих. Его рабо-
ты широко известны в нашей стране и за ру-
бежом. Наряду с разработкой вопросов сис-
тематики, фауны и экологии мелких млеко-
питающих Б. С. Юдин занимался изучением
ресурсов диких копытных Сибири и разра-
боткойпутей их рационального использова-
ния. Большой вклад внес Борис Степанович
в расширение научных исследований в Си-
бири. Он был организатором и председате-
лем Сибирского отделения Всесоюзного те-
риологического общества, активным пропа-
гандистом научных достижений, участником
всесоюзных и международных конферен-
ций, конгрессов, симпозиумов, публиковал
популярные книги, брошюры, статьи, вы-
ступал по телевидению и в периодиче-
ской печати. Много сил и энергии отдал
Б. С. Юдин делу охраны природы сибир-
ского региона. Он инициатор работ по
акклиматизации выхухоли и реакклима-
тизации кабана на территории Новоси-
бирской области и прилежащих районов.
За успешную научную и общественную
деятельность Б. С. Юдин награжден ме-
далью «За трудовую доблесть».
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
БИОЛОГИЧЕСКИЙ ИНСТИТУТ
Б. С. ЮДИН
НАСЕКОМОЯДНЫЕ
МЛЕКОПИТАЮЩИЕ
СИБИРИ
Ответственный редактор
академик В. Н. Большаков
о
НОВОСИБИРСК
«НАУКА»
СИБИРСКОЕ ОТДЕЛЕНИЕ
1989
УДК 599.33
Насекомоядные млекопитающие Сибири/Юдин Б. С.— Но-
восибирск: Наука. Сиб. отд-ние, 1989.— 360 с.
ISBN 5-02—028930—2.
Монография представляет собой итог 30-летнего изучения автором
отряда Insectivora. В ней обобщены сведения по систематике, географиче-
скому распространению насекомоядных. Приводятся оригинальные данные
по морфологии, экологии этой своеобразной группы плацентарных млеко-
питающих. Наиболее подробно характеризуются землеройки рода Sorex.
Дано описание новых подвидов, вида Sorex beringianus и подрода Stroga-
novia Yudin, subsp. n. Значительное внимание уделено изучению внутри-
видовой изменчивости морфометрических показателей. Благодаря большо-
му количеству иллюстраций, определительным таблицам семейств, родов,
видов мoнoгpaфияJ может использоваться в качестве практического
пособия.
Книга рассчитана на зоологов, преподавателей и студентов биоло-
гических факультетов вузов, специалистов в области биоценологии, охото-
ведения и охраны природы.
Табл. 71. Ил. 102. Библиогр.: 345 назв.
Рецензенты
доктора биологических наук И. П. Лаптев, Ю. С. Равкин
Утверждено к печати
Биологическим институтом СО АН СССР
£63*0
ЛИчги , •
^1907000000—751
055(02)—89
599—89, кп. 2
©Издательство «Наука», 1989
ISBN 5—02—028930—2
Б. С. Юдин (1928—1986)
ОТ РЕДАКТОРА
Насекомоядные млекопитающие — наиболее древний отряд пла-
центарных животных — долгое время оставались слабо изученной груп-
пой, особенно в систематическом плане. Во многом этому способствовало
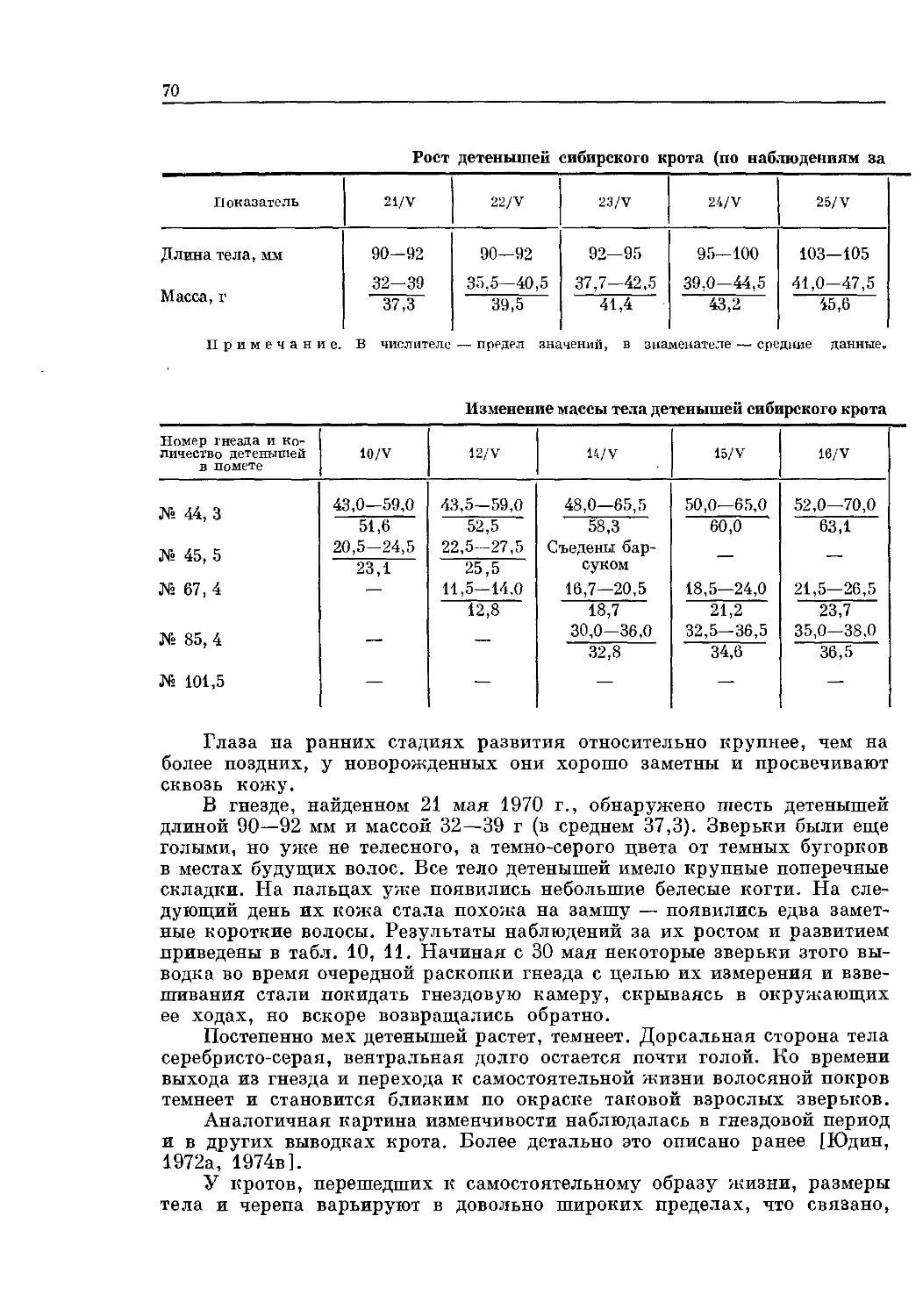
значительное морфологическое сходство близких видов. Выход в свет фун-
даментальной монографии С. У. Строганова [1957] позволил вести даль-
нейшие углубленные исследования экологии, морфологии, таксономии
этой группы животных. За последние 30 лет после опубликования указан-
ной сводки накоплен значительный новый материал, требующий обобще-
ния. Данная книга представляет собой итог научной деятельности извест-
ного зоолога Б. С. Юдина, ушедшего из жизни в расцвете творческих сил.
Он автор более 100 публикаций. В его работах «Экология бурозубок За-
падной Сибири», «Мелкие млекопитающие севера Дальнего Востока»,
«Млекопитающие Алтае-Саяцской горной страны» детально описана те-
риофауна этих регионов, дан обстоятельный анализ ее генезиса и связи с
фаунами сопредельных территорий, приводятся обширные оригинальные
сведения по экологии млекопитающих. Им опубликован новый определи-
тель насекомоядных млекопитающих Сибири — группы, наиболее труд-
ной для таксономических разработок.
Б. С. Юдин с 1951 г. работал в Биологическом институте
СО АН СССР. Закончил аспирантуру под руководством С. У. Строганова,
который определил его дальнейший научный путь, и защитил кандидат-
скую диссертацию на тему «Экология бурозубок (род Sorex) Западной Си-
бири и их практическое значение». С 1961 г. Б. С. Юдин — ученый секре-
тарь института, а с 1965 г. в течение 7 лет — заместитель директора по
научной работе. С 1964 г. он заведовал зоологическим музеем Биологи-
ческого института, с 1970 г. был бессменным руководителем лаборатории
(вначале териологии с зоологическим музеем, затем — экологических ос-
нов охраны генофонда млекопитающих).
Особенно велик вклад Б. С. Юдина в изучение систематики насекомо-
ядных млекопитающих Сибири. В результате многолетней и кропотливой
работы на основе больших серийных коллекций стало возможным пере-
смотреть систематический статус отдельных видов насекомоядных, уточ-
нить внутривидовую таксономию, что нашло отражение в данной моно-
графии. В частности, на основе исследования новых материалов с Сахали-
на и из Приморья показана видовая самостоятельность дальневосточной
бурозубки, которая считалась подвидом малой бурозубки. Анализ об-
ширного материала, выполненный непосредственно автором, позволил
также выделить особый подрод (Stroganovia) рода Sorex, описать новый
вид — берингийская бурозубка — с Курильских островов.
Подтверждено обитание в Палеарктике представителей подрода
Otisorex, считавшегося эндемиком Неарктики. Б. С. Юдиным описан так-
же ряд подвидов, таких как S. cinereus kamtschatica, S. minutissimus cauda-
6
ta, S. m. stroganovi. Наряду co сведениями по таксономии и внутривидо-
вой изменчивости насекомоядных Сибири, их географическому распро-
странению, а также определительными таблицами семейств, родов и видов
книга содержит данные по экологии отдельных видов, приведена их мор-
фологическая характеристика.
Фактической основой для изучения насекомоядных млекопитающих
послужила коллекция зоологического музея БИ СО АН СССР (свыше
30 тыс. экз.), основанная С. У. Строгановым и дополненная автором сов-
местно с другими сотрудниками института: Л. И. Галкиной, О. В. Гри-
горьевым, Н. П. Григорьевой, А. А. Максимовым, А. Б. Маркиной,
А. С. Николаевым, В. В. Николаевым, Т. В. Поймуратовой, А. Ф. Потап-
киной, В. Е. Сергеевым, А. К. Тарховой, В. И. Телегиным, С. В. Трегу-
бенко, Ю. Г. Швецовым, К. Т. Юрловым и др. В ней представлены живот-
ные Сибири и Дальнего Востока от Урала до Чукотки, Камчатки, При-
морья и Курильских островов включительно. Кроме того, исследован кол-
лекционный материал зоологических музеев МГУ и ЗИНа АН СССР.
Часть коллекций получена из ряда учреждений Сибири как результат
комплексных работ. Использованы также сборы Ф. Ф. Буйдалиной,
Л. В. Викторова, Г. А. Воронова, Н. Ф. Реймерса, Ю. Ф. Марина,
М. В. Охотиной и др.
К сожалению, смерть прервала плодотворную деятельность Б. С. Юди-
на, не дав ему в полной мере осуществить задуманное. Он не увидел окон-
чательный вариант своей рукописи, ее техническую подготовку осущест-
вляла жена и товарищ по работе С. А. Юдина при участии И. О. Григорье-
вой, Т. М. Карташовой, В. И. Унжаковой, М. Г. Чераневой. Научно-ме-
тодическая помощь при оформлении рукописи оказана коллегами:
Л. И. Галкиной, В. И. Фалеевым, А. Ю. Харитоновым, Ю. Г. Швецовым.
Все, содействовавшие выходу в свет монографии, стремились к максималь-
ному сохранению авторского текста. В том случае, когда в общем вариан-
те рукописи имелись лишь черновые записи по каким-либо разделам, для
более полного и логичного построения книги они были скомпонованы в
соответствии с авторским планом. С. А. Юдиной на основе опубликованных
статей автора и его черновиков составлены следующие разделы: харак-
теристика отряда Насекомоядные; семейства: Ежи, Выхухоли, Кроты;
виды: тундряная бурозубка, кутора обыкновенная; подвиды бурой буро-
зубки и малой белозубки; характеристика и определительные таблицы
подрода Sorex. Материал по общей характеристике таксонов взят глав-
ным образом из ранних работ Б. С. Юдина, в частности «Насекомоядные
млекопитающие Сибири. Определитель» [1971].
Подавляющее большинство рисунков оригинальны, они выполнены
художником В. Н. Ляховым по карандашным эскизам автора. Кроме то-
го, использованы иллюстрации Н. Н. Кондакова и Г. В. Чайкина, а так-
же покойного С. У. Строганова, переданные его женой А. Ф. Потапкиной.
Все фотографии принадлежат автору.
Б. С. Юдин не только полно представил систематику насекомоядных
Сибири с позиций современного состояния науки, но и существенно до-
полнил имеющиеся сведения по экологии и морфологии этой своеобразной
группы животных. Большой объем материала, представленный в книге,
позволяет надеяться, что она послужит хорошим подспорьем в дальней-
ших исследованиях в области систематики, морфологии и изменчивости
мелких млекопитающих.
В. Н. Большаков
7
ВВЕДЕНИЕ
Среди плацентарных млекопитающих насекомоядные представляют
собой наиболее древнюю и примитивную группу. Наиболее ранние находки
относятся к среднемеловым отложениям мезозоя [Строганов, 1957]. Не-
которые исследователи считают, что они дали начало позднейшим отрядам
плацентарных [Барбаш-Никифоров, Формозов, 1963]. Примитивные ежи
известны с нижнего, а кроты и землеройки — с позднего эоцена [Строга-
нов, 1957]. В ходе длительной эволюции насекомоядные сильно диверги-
ровали, современные представители далеко отошли друг от друга, вслед-
ствие чего таксономические подразделения в ранге семейства имеют кар-
динальные отличия. С одной стороны, насекомоядные сохранили некоторые
древние признаки, с другой — обладают высокой специализацией. Бла-
годаря узкой специализации они заняли ниши, в которых не имеют
конкурентов и дошли до наших дней, сохранив некоторые примитивные
черты организации. В фауне СССР насекомоядные представлены ежами
(Erinaceidae), выхухолью (Desmana), кротами (Talpidae), землеройками
(Soricidae). В Сибири зарегистрировано 25 видов насекомоядных, при-
надлежащих к 9 родам и 4 семействам.
Насекомоядные распространены по всему земному шару, кроме
Австралии и большей части Южной Америки. В Сибири встречаются
повсеместно.
При описании приняты следующие измерения: длина тела
с головой — расстояние от конца носа до анального отверстия. Для
измерения зверька кладут на спину, выпрямляя при этом его позвоноч-
ник; длина хвоста — расстояние от анального [отверстия до кон-
ца стержня хвоста, без концевых волос; длина ступни — рас-
стояние от задней стороны пятки до самого конца длинного пальца, без
когтей; кондило-базальная длиначерепа — расстояние
от наиболее выступающей вперед части межчелюстной или предчелюстной
кости до задней стороны затылочных мыщелков (рис. 1,2); длина
рострума — расстояние от наиболее выступающей вперед части меж-
челюстной или предчелюстной кости до for. anteorintalia; щирица
рострума — расстояние между внешними сторонами рострума в об-
ласти клыков у ежей и кротов, первого резца у выхухолей и первого проме-
жуточного у землероек; длина мозговой части чере-
па — расстояние от лобных отверстий до задней стороны затылочных мы-
щелков; скуловая ширина — наибольшее расстояние•. между
внешними сторонами скуловых дуг; наибольшая ши р ин а
черепа — расстояние между наиболее выдающимися в бока сторона-
ми мозговой коробки; межглазничная ширина —наимень-
шее расстояние между внутренними стенками глазниц (измеряется свер-
ху); выс, ота черепа — наибольшая высота в области слуховых
костей; длина верхнего ряда з у б о>в — расстояние от резцов
до задней стороны последнего коренного зуба; длина ряда про-
межуточных зубов — расстояние между передней стороной пер-
вого одновершинного зуба и задней стороной последнего одновершинного
зуба верхней челюсти (только для землероек); высота коронар-
ного отростка — расстояние от вершины proc, angularis дб его
8
Рис. 2. Схема промеров черепа (на примере
землеройки).
а — сверху, б — сбоку. 1 — кондило-базальная дли-
на; 2 — ширина рострума; 3 — длина рострума; 4 —• пред-
глазничная ширина; 5 — межглазничная ширина в —
длина мозговой части; 7 — наибольшая ширина; 8 — наи-
большая высота; 9 — длина верхнего ряда зубов; 10 —
длина промежуточных зубов; 11 — высота proc, coronoi-
deus; 12 — длина proc, angularis. На всех рисунках с изо-
бражением черепа а — сверху, б — сбоку, в — снизу.
основания; длина углового отростка — расстояние от вер-
шины proc, angularis до его основания; предглазничная шири-
на — расстояние между предглазничными отверстиями (foramen anteor-
bitalia); высота восходящей ветви нижней челюс-
т и — расстояние между наружным краем восходящей ветви и основанием
нижней челюсти; индексы частей туловища высчитаны по
отношению к длине тела, %; индексы черепа рассчитаны по от-
ношению к кондило-базальной длине черепа, %.
Рис. 1. Череп ежа (ориг.).
а — сверху: 1 — межчелюстная кость — os intermaxillare(-praemaxillare); 2 — переднее небное
отверстие — foramen palatinum anterius; з — носовая кость — os nasale; 4 — предглазничное от-
верстие — foramen infraorbitale; 5 — шов между верхнечелюстной и лобной костями — sutura ma-
xillo-frontalis; 6 — скуловая кость — os zygomaticum(-jugale); 7 — скуловой отросток чешуйчатой
кости — processus zygomaticusossiss squamosi; в — межтеменная кость — os interparietale; 9 —
верхнезатылочный гребень — crista supraoccipitalis; 10 — теменная кость — os parietale; 11 —
венечный шов — sutura coronalis; 12 — лобная кость — os frontale; 13 — сагиттальный шов — su-
tura sagittalis; 14 — шов между носовыми костями —• sutura internasalis; 15 — верхнечелюстная
кость — os maxsillare; 16 — шов между верхнечелюстной и межчелюстной костями — sutura maxil-
lo-intermax il laris;
6 — снизу: 1 — межчелюстная кость — os intermaxillare(=praemaxsillare); 2 — верхнече-
люстная кость — os maxillare; з — шов между небной и верхнечелюстной костями — sutura pajato-
maxillaris; 4 — небная кость — os palatinum; 5— скуловая кость — os zygomatic( = jigale);
в — сошник — vomer; 7 — отросток крыловидной кости — proc, hamularis; в — сочленовная ям-
ка — fossa glenoidea; 9 — послесочленовный отросток — proc, postglenoideus; 10 — слуховой бара-
бан — bulla tympani; 11 — сосцевидный отросток — proc, mastoideus; 12 — предзатылочный от-
росток — proc, paroccipitalts; 13 — верхнезатылочная кость — os supraoccipitale; 14 — затылочный
мыщелок — condylus occipitalis: 15 — затылочное отверстие — foramen magnum; 16 — основная
затылочная кость — os basioccipitale; 17— основная клиновидная кость — os basisphenoideum’,
18 — передняя клиновидная кость — os praesphenoideum; 19 — крыловидная ямка — fossa ptery-
goidea; 20 — хоанальный вырез; 21 — небный шов — sutura palatlna; 22 — небное отверстие —
foramen palatinum; 23 — переднее небное отверстие — foramen palatinum anterius;
в — сбоку: 1 — межчелюстная кость — os itermaxillare (= praemaxillare); 2 — предглазнич-
ное отверстие — foramen infraorbitale; 3 — верхнечелюстная кость — os maxillare; 4 — лобная
кость — os frontale; 5 — теменная кость — os parietale; в — межтеменная кость — os interparietale;
7 — верхнезатылочный гребень — crista supraoccipitalis; з — верхняя затылочная кость — os
supraoccipitalis; 9 — затылочный мыщелок — condylus occipitalis; 10 — боковой затылочный отро-
сток — proc, paroccipitalis; и — слуховой барабан — bulla tympan; 12 — слуховой проход —
meatus auditorius extemus; 13 — заднесочленовный отросток — proc, postglenoideus; 14 — сочленов-
ная ямка — fossa glenoidea; 15 — крыловидная кость — os pterygoideum; 16 — скуловая кость —
оз jugale(=zygomaticus); 17 — венечный отросток—proc, coronoideus; 18 — сочленовный отрос-
ток — proc, articilaris; 19 — угловой отросток — proc, angularis.
9
ОТРЯД INSECTIVORA BOWDICH, 1821 —
НАСЕКОМОЯДНЫЕ
К отряду насекомоядных относятся разнообразные по размерам,
внешнему виду и анатомическому строению звери, приспособившиеся к
наземному, подземно-роющему, земноводному и древесному образу жизни
(рис. 3). Обитание насекомоядных в различных экологических условиях
наложило отпечаток на их общий облик. Землеройки, ведущие наземный
и полуроющий образ жизни, маленькие, мышеобразной внешности.. Кро-
ты, проводящие жизнь в подземных ходах, плотные, вальковатого тело-
сложения, с массивными расширенными передними конечностями. Вы-
хухоль, ведущая земноводный образ жизни, имеет клинообразную форму
тела, расширенные задние конечности с плавательными перепонками и
сжатый с боков хвост, что способствует нырянию и плаванию в воде. Не-
уклюжие, малоподвижные ежи защищены от врагов надежным иглистым
панцирем.
Насекомоядные характеризуются следующими признаками. Зверьки
небольших размеров, самый крупный вид в фауне СССР афганский еж
(Erinaceus megalotis Blyth) достигает размеров 265—300 мм, самый мел-
кий — карликовая белозубка (Suncus etruscus Savi) — не более 45 мм.
Конечности относительно короткие, стопоходящие, редко полустопохо-
дящие, в большинстве случаев пятипалые. Пальцы вооружены когтями,
большой палец не противопоставляется другим.
Морда, за небольшим исключением, вытянута в подвижный хоботок.
Ушные раковины рудиментарны у кротов, слабо развиты у землероек и
хорошо — у ежей. Волосяной покров насекомоядных варьирует от барха-
тистого меха кротов до иглистого панциря ежей. Окраска меха неяркая,
бурая, коричневая, буро-серая. Ключицы обычно хорошо развиты. Лице-
вой отдел длинный, составляет 1/2—2/3 черепа, мозговая коробка отно-
сительно небольшая. Череп клиновидной формы, сужающийся в лицевом
отделе. Барабанная кость чаще всего кольцевидной формы. Скуловые дуги
или слабо развиты, или вовсе отсутствуют. В строении нижней челюсти
характерно сильное развитие углового отростка, высокого венечного и
высоко поднятого суставного отростков.
Зубная система дифиодонтная 1 (две смены зубов: молочные и по-
стоянные), гетеродонтная (зубы разных категорий). Число зубов варьи-
рует от 44 до 26. Резцы часто уменьшены в числе. Передняя пара резцов
крупная, клыки мелкие, по форме и величине зачастую похожи на резцы
или предкоренные. Передние коренные простые, конусовидные, в разной
степени моляризованы. Задние коренные с острыми вершинами, которые,
соединяясь гребнями, образуют секториальный (режущий) тип зуба,
имеющий в плане V- или W-образный вид на жевательной стороне коронки.
1 И. И. Стогов [1969] приходит к выводу о наличии лишь одной генерации зубов.
и,
Рис. 3. Биологические типы насекомоядных (по С. У. Строганову [1957]). Дре-
весный: 1 — тупайя (Tupaia]; наземный: 2 — пустынный — прыгунчик [Macros-
се tides)', 3 — лесной — землеройка (Sorex)', 4 — пустынно-степной — еж (Не-
miechinus); подземно-роющий: 5 — крот (Talpa)-, 6 — златокрот (Chrysochlorts)-,
земноводный: 7 — водоплавка (Neomys)-, 8 — выдровая землеройка (Potomogale)-,
9 — выхухоль (Desmana).
12
Полушария головного мозга развиты слабо, с гладкой поверхностью,
не прикрывают мозжечка. Матка насекомоядных двурогая. Кость копу-
лятивного органа (os penis) отсутствует. Плацента дискоидальная. Число
сосков от трех до шести пар. У многих видов развиты боковые, анальные и
хвостовые железы, выделяющие секрет, издающий резкий мускусный
запах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1(2). Верхняя сторона тела покрыта короткими иглами (колючка-
ми) ............................................Erinaceidae — Ежи
2(1). Все тело покрыто мягкими волосами.....................3
3(4). Между пальцами передних и задних конечностей хорошо выра-
жена плавательная перепонка. Хвост сжат с боков, покрыт чешуй-
ками ......................................Desmanidae — Выхухоли
4(3). Между пальцами плавательных перепонок нет. Скуловые дуги
или отсутствуют, или очень слабые, тонкие. Задненебных отверстий
нет ............................................................5
5(6). Кисти передних конечностей широкие, лопатообразные. Когти
уплощенные, не сжатые с боков. Ушных раковин нет. Хвост короткий,
круглый в сечении, покрыт волосками. Скуловые дуги есть. Передние верх-
ние резцы прямые, без добавочных зубцов, посажены вертикально; ниж-
ние резцы малы и слабо наклонены вперед .... .Talpidae — Кроты
6(5). Кисти не широкие. Когти передних конечностей узкие, сжатые
с боков. Ушные раковины есть. Скуловых дуг нет. Передние верхние рез-
цы серповидно изогнуты, с добавочной лопастью позади главной вершины,
посажены с наклоном вперед; нижние резцы велики, посажены более или
менее горизонтально . ................... . Soricidae — Землеройки
СЕМЕЙСТВО
ERINACEIDAE FISCHER VON WALDHEIM, 1817 —
[ЕЖИ
ПОДСЕМЕЙСТВО
[ERINACEINAE FISCHER VON WALDHEIM, 1817
Диагноз. Среди насекомоядных ежи резко выделяются своеоб-
разной внешностью: сравнительно крупными размерами, наличием иг-
листого кожноГо покрова.
Зубная формула однотипна для всех видов:
i4; с 4-; Pm 4; м-|- = зб.
di Z d
Передние верхние резцы не примыкают друг к другу, выполняют
функцию Клыков (см. рис. 1).
Х а р акт е р и с т и к а. Телосложение плотное, внешне ежи не-
уклюжи. Голова с конусообразно вытянутой мордой; ушные раковины
покрыты волосами и выступают из-под меха. Ноги широко расставленные,
короткие, стопоходящие. Хвост очень короткий. Окраска меха и игл варь-
ирует от светло-серого до желто-коричневого, в зависимости от вида и
возраста, .Есть ежи хромисты, меланисты, альбиносы и пегие.
13
Рис. 4. Форма головы, ушной раковины и иглистого панциря ежей.
а — еж обыкновенный — Erinaceus europaeus L.; б — ушастый -*• Hemiechinus auritus Gmelin*
Лицевой отдел черепа массивный, лицевые швы хорошо заметны;
скуловые дуги достаточно развиты; слуховые пузыри неполные, строение
слуховых косточек (область среднего уха) служит одним из видовых при-
знаков.
Обе половины таза срощены в лобковом симфизе. Строение гениталий
самцов специфично для каждого вида. На брюхе у самок находится пять
пар сосков.
Распространение. Европа, Азия, Африка.
Заметки по систематике. Систематика ежей фауны СССР
до сих пор — один из сложных вопросов, требующих своего решения.
Нет общепринятого мнения о числе видов ежей в нашей стране и их ро-
довой принадлежности. С. И. Огнев [1928] приводил 13 видов ежей,
относящихся к четырем родам. Более поздними исследователями неод-
нократно пересматривалась внутривидовая таксономия в объеме видов, их
статус в ранге рода. Достаточно указать, что для территории СССР
Н. А. Бобринский, Б. А. Кузнецов и А. П. Кузякин [1944] приводят
четыре вида ежей, относящихся к одному роду: Ё. europaeus, Е. dauricus,
Е. auritus, E.hypomelas. В «Определителе...» этих авторов более позднего
издания [Те же, 1965] Е. dauricus переведен в ранг подвида обыкновенно-
го ежа и вводится новый вид — афганский еж (Е. megalotis Blyth, 1845).
А. А. Гуреев [1963] свел всех ежей в четыре вида, объединив их в
два рода. Из этого далеко не полного перечня можно видеть, насколько
разноречивы взгляды исследователей на объем видов и родов ежей. Сле-
дует добавить, что все эти перестройки малоубедительны из-за отсутствия
у авторов новых фактических материалов.
Большинство зоологов считают, что в современной мировой фауне
семейство Ежи насчитывает шесть родов, около 36 видов. Из них в преде-
лах Сибири встречается три вида, принадлежащих к двум родам.
14
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. ERINACEIDAE
1(2). Иглистый панцирь на голове иногда разделен на две части (см.
рис. 4, а). Поверхность Колючек гладкая, без зернистых бугорков, про-
дольных бороздок и валиков. Послесочленовный отросток (proc, postgle-
noideus) меньше и короче сосцевидного (proc, mastoideus), внутри не име-
ет полости; glans penis на дорсальной стороне несет валики, покрытые ро-
говыми бугорками................Ertnaceus L.— Ежи обыкновенные
2(1). Иглистый панцирь на голове не расчленен пробором на две части
(см. рис. 4, б). На поверхности колючек имеются продольные бороздки и
15
между ними валики,
на гребнях которых
рассеяны зернистые
бугорки. Послесочле-
новный отросток ра-
вен сосцевидному и
лишь немного уже
его, внутри имеет хо-
рошо обозначенную
полость; на дорсаль-
ной стороне имеются
два валика с хорошо
выраженными остры-
ми шипами ....
. . . . Hemiechinus
Fitz.— Ежи ушастые
Род' Erinaceus
Linnaeus, 1758 —
Ежи обыкновенные
Тип рода.
Erinaceus europaeus
L„ 1758.
Диагноз. См. таблицу для определения родов ежей.
Характеристика. Окраска зверьков от светло-серой и бурой
до черно-бурой, встречаются альбиносы. Мех, покрывающий голову,
бока и низ тела, длинный грубый.
Череп относительно низкий с широкой и массивной ростральной
частью, ширина рострума над клыками примерно в 1,5 раза меньше шири-
ны между внутренними стенками предглазничных отверстий (for. infraor-
bitale); высота черепа в области клыков лишь немного меньше половины
его высоты в области слуховых барабанов. Отростки основной клиновид-
ной кости (os basisphenoideum) в виде пластинок, охватывают слуховую
полость только с внутренней стороны. Послесочленовный отросток (proc,
postglenoides) массивный, без вдавленно-овальной полости внутри; он
меньше и короче сосцевидного (proc, mastoides), последний обычно короче
направленного назад предзатылочного отростка (proc, paraoccipitalis).
Слуховые барабаны небольшие, уплощенные. Верхние клыки в большин-
стве случаев с одним корнем.
Распространение. Европа и Азия, от Испании до Китая
включительно. В фауне Сибири род представлен одним видом.
Erinaceus europaeus Linnaeus, 1758 — Обыкновенный еж
Голотип. Описан по экземпляру ,из окрестностей Упсалы,
Швеция.
Диагноз. Признаки вида совпадают с таковыми рода, указаны в
характеристике последнего.
Измерения (мм). Длина тела с головой 237—272, п = 10, М =
= 251; длина хвоста 19,0—30,0, 0, п = 10, М = 23,8; длина ступни 34—
47,0, п = 7, М = 23,0; высота уха 24,0—34,0, п = 10, М = 27. Масса
16
Рис. 6. Череп обыкновенного ежа.
240—350 г. Кондило-базальная длина черепа 41,8—58,7, п — 25, М =
= 54,7; длина рострума 15,2—17,5, п = 24, М = 16,1; длина верхнего
ряда зубов 26,7—30,3, п = 24,0, М = 28,3; ширина рострума 10,3—
13,7, п = 24,0, W = 12,1; предглазничная ширина 12,6—16,5, п = 24,0,
М — 18,4; наибольшая ширина 24,5—30,0, п = 24,0, М = 28,3; скуло-
вая ширина 29,2—38,6, п — 24, М = 34,3; наибольшая высота черепа
17,7—22,1, п = 22, М = 18,4; высота proc, coronoideus 16,7—19,7, п =
= 23, W = 19,0.
Характеристика. Настоящие, или обыкновенные, ежи (Eri-
ч naceus) известны с олигоцена [Строганов, 1957]. Это наиболее крупные
представители сем. Erinaceidae. В окраске меха зверьков преобладают
17
Рис. 7. Строение glans penis обыкновенного ежа.
А —- Erinaceus europaeus’, Б — Е. е. amurensis; а — латеральная; б •— дорсальная;
в — вентральная сторона.
бурые и серовато-охристые тона разной интенсивности, иногда на боках —
с желтовато-ржавым оттенком. Волосы щетинистые, жесткие. Окраска
иглистого панциря буроватая с беловатыми штрихами (рис. 4, 5). С воз-
растом окраска светлеет и приобретает характерную желтизну с палевым
оттенком. Среди окрашенных игл часто встречаются непигментированные,
особенно отмечаются они у ежей, обитающих на Дальнем Востоке. В ре-
зультате общая светло-серая окраска пестрит белыми вкраплениями не-
окрашенных игл. Длина игл колеблется от 25 до 35 мм. Уши короткие,
менее 35 мм. Череп массивный, носовая часть его широкая, без сужений.
В лобной части череп вытянутый (рис. 6). Послесочленовный отросток с
с едва заметной выемкой или без нее. Glans penis короткая и толстая (дли-
на 14,0, толщина 7,4 мм), на ее дорсальной стороне имеются два роговых
валика без шипов (рис. 7, а).
Распространение. В Западной Сибири заселяет пространст-
во между Уралом и Иртышом. Из этого района известен из окрестностей
Кургана, Туринска, Тюмени, Ялуторовска и Тары, из долины Концы,
верховьев Васюгана. Далее на восток ареал европейского ежа прерывает-
ся до Дальнего Востока. Там он заселяет Амуро-Уссурийский край (до-
лины среднего Амура, Уссури, Имана, район оз. Ханка, до Тихоокеан-
ского побережья).
Материал. Исследовано 194 экз. из разных частей ареала.
Заметки по систематике. Количество подвидов для Си-
бири остается не выясненным, далее приводятся материалы по одному
из них.
Erinaceus europaeus amurensis Schrenks, 1858 —
Дальневосточный, или амурский, еж
Голотип. Описан по экземпляру с Айгуна (Амур).
Диагноз. Зверьки выделяются крупными размерами, более свет-
лой окраской, строением glans penis.
2 Б. С. Ю'ТУ .. j . . пи—
БИБЛИОТЕКА
(пшииическлгв иястжтут*
СО АН СССР
18
Измерения (мм.) Длина тела с головой 245—272, п = 3, М —
= 261,7; длина хвоста 19—28,0, п — 2, М = 22,4. Кондило-базальная
.длина черепа 51,0—58,0, п = 11, М — 56,0; длина рострума 15,0—17,5,
п = 11, М — 16,2; длина верхнего ряда зубов 27,0—30,3, п = 11, М =
= 28,6; ширина рострума 10,3—13,7, п = 11, М = 12,1; предглазничная
ширина 14,4—19,9, п = 11, М = 16,1; межглазничная ширина 17,0—
20,7, п = 11, М = 18,4; наибольшая ширина 26,4—30,0, п — 11, М =
— 28,3; скуловая ширина 32,8—38,6, п = 9, М = 34,8; наибольшая вы-
сота черепа 19,0—20,9, п = 8, М = 19,7; высота proc, coronoideus 18,0—
19,7, п = 9, М = 18,9; длина proc, angularis 5,0—6,8, п = 9, М = 5,9.
Характеристика. Иглистый покров светлой окраски, боль-
шое число игл без пигмента, что создает общий светлый тон с буроватым
оттенком. Волосяной покров крайне жесткий, щетинистый, интенсивно
бурого цвета. Особенно жесткая щетина на лбу, едва обозначен небольшой
пробор между колючками, однако голое пятно отсутствует. У некоторых
экземпляров окраска передней нижней части тела светлее, чем задней.
Длина волос 3,5—4,5 см. Существенных вариаций в окраске иглистого
панциря и волос зверьков из Приморья не обнаружено.
На валиках крупного glans penis (длина 19,5, толщина 8,9 мм) очень
мелкие шипики, несколько сот штук (рис. 7, б).
Распространение. Обитает на юге Приморья, в заповедни-
ке «Кедровая Падь», добыт в окрестностях Владивостока, в районе бухты
Анна.
Материал. 13 экз.
Род Hemiechinus Fitzinger, 1866 —
Ушастые ежи
Тип рода. Erinaceus platyotis Sundevall (1841).
Диагноз. Иглистый панцирь меньшего размера, чем у ежей рода
Erinaceus, и не заходит на бока. На поверхности игл имеются продольные
бороздки, а между ними — валики, на гребнях которых рассеяны зер-
нистые бугорки. Уши обычно длиннее половины головы, исключение со-
ставляет даурский еж. Диагностические признаки в строении черепа:
послечленовый отросток по величине равен сосцевидному и лишь немного
уже его, имеет внутри хорошо обозначенную полость. Glans penis на дор-
сальной стороне имеет два валика с хорошо выраженными острыми
шипами.
Характеристика. Иглистый панцирь окрашен неравномерно,
на каждой игле несколько темных колец — от 1 до 3.
Обращает на себя внимание узкая ростральная часть черепа, ширина
рострума под клыками почти в 2 раза уже ширины между внутренними
краями for. infraorbitale. Высота черепа в области клыков равна прибли-
зительно одной трети высоты в области слуховых барабанов. Последние
сравнительно большие и изнутри значительно вздутые. Верхние клыки в
большинстве случаев с двумя корнями.
Распространение. Юго-восточная часть Европы, Казахстан,
Средняя Азия, на юг до Северной Индии и Египта.
19
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА HEMIECHINUS
1(3). На поверхности игл хорошо выражены продольные валики с бу-
горками и борозды............................................... 2
2(3). Уши короткие. Отогнутое вперед ухо не заходит за глаза. Воло-
сяной покров грубый, бурого тона. Головка копулятивного органа с длин-
ными тонкими шипами, несущими на конце бородки......................
..................................Н. dauricus Sundv.— Даурский еж
3(2). Уши длинные. Отогнутое вперед ухо заходит за глаза .... 4
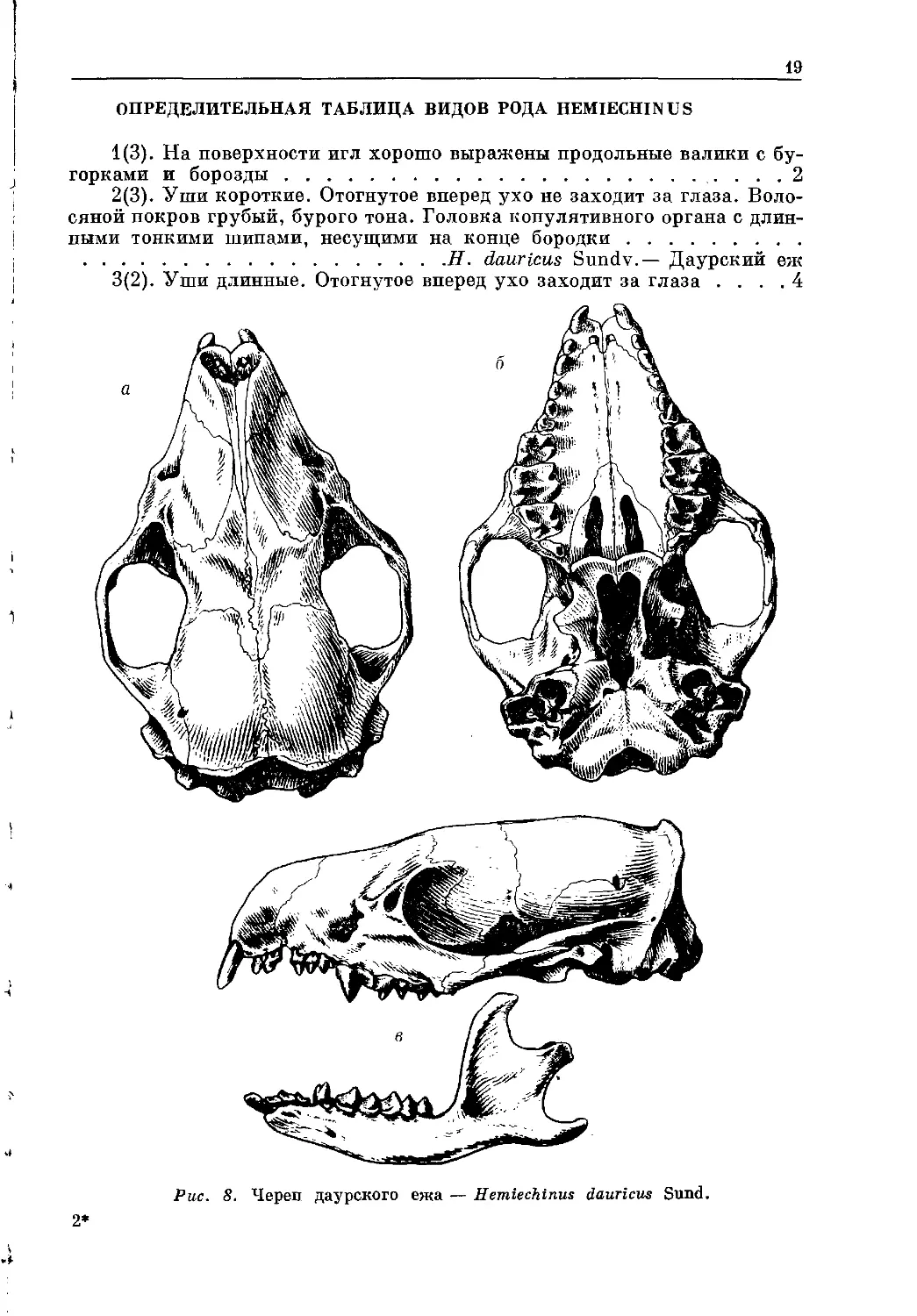
Рис. 8. Череп даурского ежа — Hemiechinus dauricus Sund.
2»
20
4(2). На поверхности игл чаще один или два темных пояска. Мех на
нижней стороне тела мягкий, светлых тонов. Головка копулятивного ор-
гана несет толстые короткие шипы. Средние шипы выше крайних . . . .
.................................Н. auritus Gmelin — Ушастый еж
Hemiechinus dauricus Sundvall, 1841 —
Даурский еж
Г о л о т и п. Описан по экземпляру из Даурии.
Диагноз. Уши короткие, отогнутые вперед, не доходят до глаз,
размеры крупные. Иглы с продольными бороздками и валиками, относи-
тельно короткие.
Измерения (мм). Длина тела с головой 233—270, п = 17, М =
= 250; длина хвоста 24,0—38,0, п = 14, М — 32; длина ступни 33,0—
45,0, п — 15, М = 41,1; высота уха 29,0—34,0 п = 9, М — 31,6. Масса
600—800 г. Кондило-базальная длина черепа 50,0—57,5, п = 29, М =
= 53,1; длина рострума 13,7—15,7, п — 21, М — 14,5; длина верхнего
ряда зубов 26,2—29,5, п = 30, М = 27,3; ширина рострума 7,0—8,6, п =
= 30, М = 7,8; предглазничная ширина 12,8—14,6, п = 30, М — 13,0;
межглазничная ширина 12,4—18,3, п = 30, М = 18,3; наибольшая ши-
рина 25,0—30,0, п = 30, М = 26,5; скуловая ширина 31,7—34,5, п =
— 21, М = 32,0; наибольшая высота черепа 16,5—20,0, п = 30, М =
— 18,3; высота proc, coronoideus 15,3—20,5, п = 29, М'= 18,8.
Характеристика. Зверьки по внешнему виду похожи на
обыкновенного ежа. Иглистый панцирь окрашен в серо-бурый цвет, не
колючий, так как иглы в отличие от игл европейского ежа как бы при-
глажены, направлены назад. Окраска меха темная, насыщенного серо-бу-
рого тона. Волосы жесткие, но несколько мягче, чем у Е. е. amurensis,
и более короткие. Лишь отдельные волосы достигают 3—3,5 см. Нижняя
часть тела всегда окрашена одноцветно, тогда как у европейского ежа
передняя часть нижней стороны тела светлее, а задняя — темнее. Вокруг
глаз и на конце морды мех серо-коричневый. Череп крупный, массивный,
•с широко расставленными скуловыми дугами (рис. 8). Ростральная часть
черепа резко сужается к пе-
реду. Между носовыми и че-
люстными костями тонкие
отростки края лобных кос-
тей, носовые кости очень уз-
кие. Передний конец верх-
ней челюсти сильно опущен
вниз. Передние резцы далеко
выдаются вперед. Glans pe-
nis характерна для предста-
вителей рода Hemiechinus.
Для вида специфично нали-
чие темных валиков на дор-
сальной стороне penis и стро-
ение шипиков, у даурского
Рис. 9. Строение glans penis да-
урского ежа.
а — дорсальная, б — латеральная сто-
рона.
21
ежа они длинные, тонкие, некоторые из них на конце с бородками (рис. 9).
Распространение. Даурский еж заселяет Южное и Цент-
ральное Забайкалье и сопредельные районы Монголии. С запада ареал
ежа ограничен оз. Торей, р. Джида, д. Покровкой в долине р. Про. Север-
ная граница ареала проходит по побережью Гусиного озера, через устье
р. Удинка и западному побережью Убукуна. Восточная граница проходит
по линии: устье Оронгоя, Тунгуя через села Усть-Киран, Шарагол, далее
на восток от Кепчайского и Яблонового хребта. Южная граница уходит в
пределы Монголии. Даурский еж — обычный вид для Ангинской степи
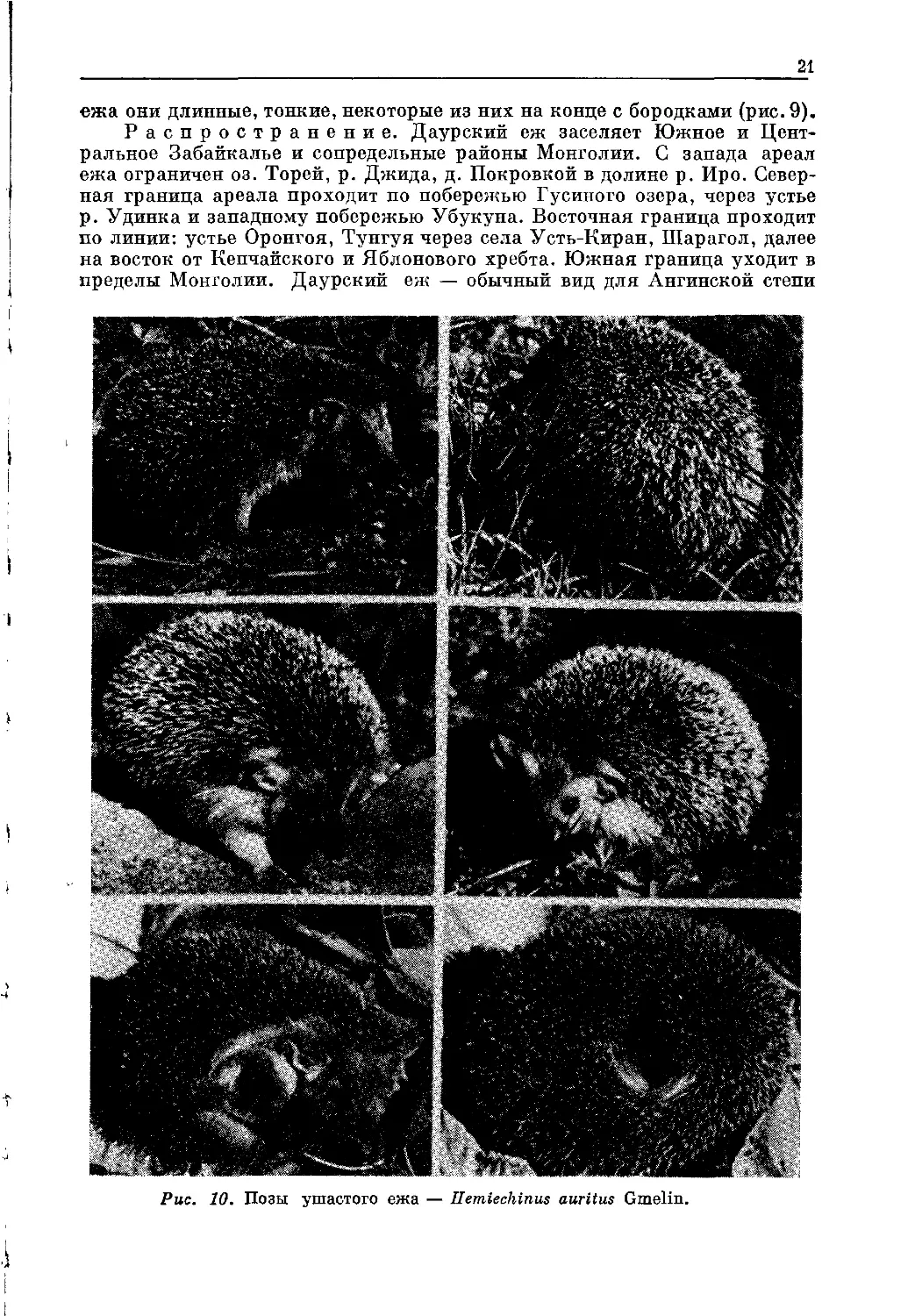
Рис. 10. Позы ушастого ежа — Hemiechinus auritus Gmelin.
22
Рис. 11. Череп ушастого ежа.
долины рек Опок, Борзя. В пределах Сибири даурский еж представлен
номинальным видом. -Л Л 4
Материал. 23 экз. преимущественно из Забайкалья (Борзинско-
го района) и Китая.
23
Заметки по систематике. Многие авторы по разному
трактовали систематическое положение даурского ежа [Строганов, 1957].
Hemiechinus auritus Gmelin, 1770 —
Ушастый еж
Голотип. Описан по экземпляру с низовьев Волги.
Диагноз. Уши длинные, отогнутые вперед, они заходят за глаза.
Колючки с продольными бороздками и валиками, вдоль которых рассеяны
бугорки. На колючках обычно два или один темный поясок. Мех на брюш-
ке серовато-бурый или молочно-белый (рис. 10).
Измерения (мм). Длина тела с головой 127—183, п = 5, М =
= 166; длина хвоста 17,0—23,0, п = 5, М — 20; длина ступни 31,0—37,0,
п = 5, М = 33; высота уха 37,0—45,0, п = 4, М — 39,0. Кондило-ба-
зальная длина черепа 41,3—46,4, п = 22, М = 44,0; длина рострума
10,7—12,0, п — 22, М = 11,4; длина верхнего ряда зубов 21,0—23,0,
п — 23, М = 22,2; ширина рострума 6,9—8,6, п = 22, М = 7,8; пред-
глазничная ширина 9,7—11,3, п = 25, М = 10,4; межглазничная ширина
12,7—14,7, п = 23, М = 14,0; наибольшая ширина 20,7—23,5, п = 23,
М — 21,9; скуловая ширина 24,0—27,4, п = 21, М — 25,9; наибольшая
высота черепа 16,2—17,5, п = 22, М = 16,9; высота proc, coronoideus
13,4—23,0, п = 23, М = 14,2.
Характеристика. Ушастый еж — самый мелкий представи-
тель ежей, обитающих в Сибири. Внешность его настолько характерна,
что видовую принадлежность можно установить с первого взгляда. Обра-
щают на себя внимание большие уши, иглистый покров головы не разде-
лен пробором, иглы короче, чем у обыкновенного ежа, и торчат в разные
стороны, образуя плотный густой панцирь. Волосяной покров тонкий,
пушистый, волосы относительно короткие.
Череп умеренной длины, с довольно широкими скуловыми дугами.
Носовые кости в средней части сужены. Ростральный отдел сужается к
концу (рис. 11). Glans penis более вытянута, чем у европейского ежа,
длиной 22,5, толщиной 8 мм. На ее дорсальной стороне два параллельных
валика, вооруженных рядами массивных шипиков, с загнутыми вершина-
ми в направлении к основанию копулятивного органа (рис. 12).
Р аспространение.
Ареал вида простирается по
Юго-Восточной Европе, Ка-
захстану, Передней и Цент-
ральной Азии на юг до Егип-
та й Индии. В Сибири засе-
ляет сопредельные с Казах-
станом районы от Южного
Урала на север до Омска,
Кулундинскую степь до Ка-
расука, с. Локоть и Змеино-
горска. В Туве найден в Уб-
сунурской котловине и в
окрестностях Кызыла.
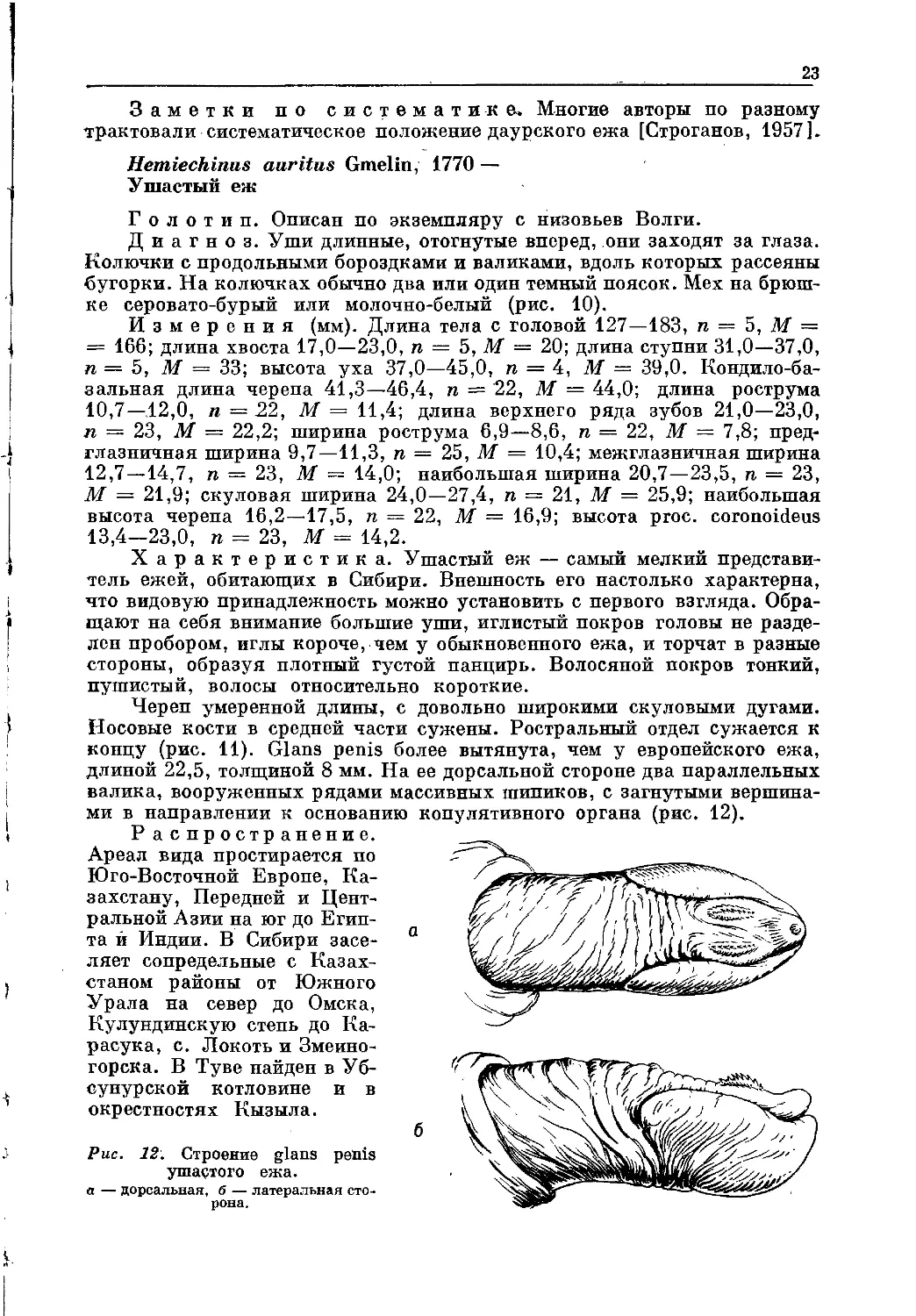
Рис. 12. Строение glans penis
ушастого ежа.
а — дорсальная, б — латеральная сто-
рона.
24
Заметки по систематике. В Сибири встречаются два
подвида ушастого ежа: Н. a. auritus Gmelin, 1770, южные районы Запад-
ной Сибири и Н. albulus Stoliczka, 1872, Тува.
Hemiechinus auritus auritus Gmelin, 1770 —
Северный ушастый еж
Голотип. См. в описании вида.
Диагноз. Отличается от других подвидов размерами, темной ок-
раской волосяного и иглистого покровов.
Измерения. Имеющиеся данные автором не систематизированы.
Характеристика. Окраска головы от серовато-ржаво-пале-
вой до темно-бурой, с более светлыми ушами. Окраска брюшка светлая
или совсем белая, иногда со светло-бурыми тонами. Иголки опоясаны дву-
мя черными поясами. По кондило-базальной длине черепа, длине ростру-
ма, скуловой ширине этот подвид хорошо отличим от Н. a. albulus.
Распространение. Южные районы Западной Сибири.
Материал. 7 экз.
Hemiechinus auritus albulus Stoliczka, 1872 —
Беловатый ушастый еж
Голотип. Описан по экземпляру из Яркенда.
Диагноз. Зверьки крупных размеров, по всем краниометрическим
показателям превосходят номинальный подвид.
Измерения. Нет данных (прим. ред.— В. Б.).
Характеристика. Иглистый покров отличается бледностью
и тусклостью, мех пышный и шелковистый, уши окрашены более светло,
чем у предыдущего подвида. Брюшко чисто белого цвета, бока со слабо-
ржавым налетом.
Распространение. За пределами РСФСР широко распростра-
нен в Туркменистане, Монголии.
СЕМЕЙСТВО DESMANIDAE GRAY, 1821 —
ВЫХУХОЛИ]
|ПОДСЕМЕЙСТВО DESMANINAE THOMAS, 1912
Диагноз. В облике зверьков ярко выражено приспособление к
водному образу жизни (рис. 13). Туловище клиновидной формы массивное,
покрыто густым мягким мехом, плохо намокающим в воде. Передние ко-
нечности значительно меньше задних. Между пальцами — хорошо выра-
женная кожистая плавательная перепонка (рис. 14). Голова конусовид-
ная, морда вытянута в длинный подвижный хоботок, глаза очень малень-
кие (см. рис. 14, а).
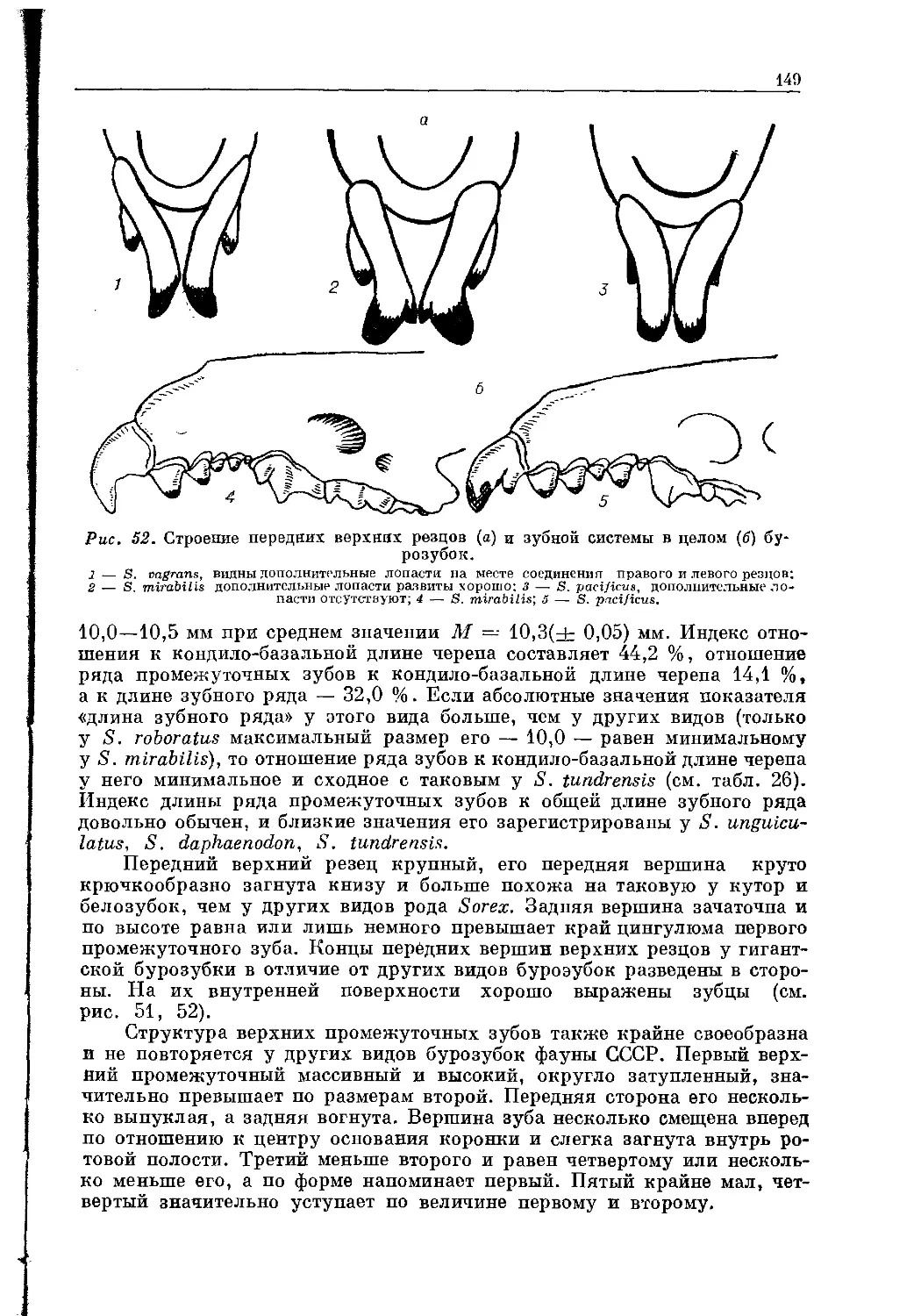
Зубная формула: I-|-; С-^-; Pm-|-; М = 44.
Передние резцы в верхней челюсти очень большие, трехгранные.
Клыки и передние предкоренные слабо дифференцированы, коренные
широкие, многовершинные (рис. 15).
Характеристика. Длина тела зверьков достигает 12—22 см,
масса до 500 г. Строение конечностей обусловлено водным образом жизни:
наличие плавательных перепонок, кисти конечностей обрамлены щеткой
из жестких и щетинистых волос; задние конечности массивные, широкие
25
Рис. 13. Русская выхухоль — Desmana moschata L.
и далеко отодвинуты назад, поставлены «по-гагарьи». Хвост почти равен
длине туловища, покрыт плоскими чешуйками, между которыми видны ред-
кие короткие волосы. Шея короткая, без перехвата, переходящая в конусо-
видную голову. Зрение в значительной степени редуцировано, ушных ра-
ковин нет, но слух у зверьков хорошо развит.
Череп вытянут в длину (до 50—58 мм), монолитный, с хорошо выра-
женными гребнями, широкий в мозговой части и узкий — в скуловой.
Распространение. Европейская часть СССР (бассейны
Дона, Волги и Урала); акклиматизирована в Западной Сибири в пойме
Оби; юго-западная Франция, Испания, Португалия.
Заметки по систематике. В систематическом отношении
сем. Desmanidae близко к сем. Talpidae. Их генеалогическая связь просле-
живается через самые примитивные формы подсем. Urotrichinae, с кото-
рыми имеют сходство в строении черепа и плечевого аппарата [Строганов,
1948]. Обнаруживаются некоторые сходные черты их строения с сем.
Soricidae. Связь с примитивными кротовыми указывает на древность про-
исхождения выхухолей, резко выраженные признаки адаптации к полу-
водному образу жизни — недалеко зашедшую специализацию этой группы
насекомоядных млекопитающих.
Пюиссюгюр [Puissegur, 1935] считает, что миоцен был эпохой про-
цветания выхухоли. Появление собственно Desmana Симпсон [Simpson,
1945] относит к нижнему плиоцену. Из позднеплейстоценовых отложений
известны Myogale moschata (юг СССР) и D. tegelensis из западных пред-
26
Рис.
14. Голова (а), кисть
и ступня (б) русской выхухоли.
горий Внутренних Карпат [Пидопличко, 1954]. В плейстоцене известны
D.magna, D. tegelensis, D. thermalis, D. moschata, представленные несколь-
кими подвидами [Пидопличко, 1951,1954; Heller, 1954; Верещагин, 1959].
Из голоценовых отложений Венгрии описан Desmana crassidens [Kretzoi,
1953].
В составе семейства два современных монотипических рода — Des-
mana Guld. и Galemys Каир.
27
Рис. 15. Череп русской выхухоли.
28
Род Desmana Giildestaedt, 1777 —
Выхухоль
Тип рода. Castor moschatus L., 1758 ( = Desmana moschata L.).
Диагноз. Основные признаки — характерные для семейства.
Характеристика. Относительно крупные зверьки, длина те-
ла до 22 см. Хвост сжат с боков, длинный, у самого основания с перетяж-
кой, а его проксимальная часть грушевидно расширена. Здесь находятся
мускусные железы, из которых через многочисленные мелкие отверстия
выделяется резко пахнущий маслянистый секрет. По гребню хвоста тя-
нется киль из упругих волос. Хвост выполняет роль локомоторного орга-
на и руля при движении выхухоли в воде.
От представителей рода Galemys Капр животные рода Desmana отли-
чаются более крупными размерами, сжатым с боков массивным хвостом,
более уплощенным хоботком и окраской меха. Четкие различия просле-
живаются в конфигурации черепа, строении промежуточных зубов и осо-
бенно коренных нижней челюсти. Зубная формула общая для всего се-
мейства.
Распространение. В настоящее время выхухоль локально
распространена в бассейнах Дона, Волги, Урала и акклиматизирована в
Западной Сибири — в среднем течении Оби.
В составе рода один современный вид.
Desmana moschata Linnaeus, 1758 — Русская выхухоль
При описании вида Линней в качестве «terra typica» отметил: «Habi-
tat in Russial aquosis» (c. 59). Эту цитату привел С. И. Огнев [1928].
Голотип. Не установлен.
Диагноз. Основные признаки такие же, как и у рода (см. харак-
теристику рода и семейства).
Измерения (мм). Длина тела с головой 194—221 (М = 208),
длина хвоста 181—200 (М — 192), длина ступни 53—58 (М — 56,4). Масса
339—471 (М = 408) г. Кондило-базальная длина черепа 51,5—55,5 (М =
= 54,1), длина верхнего ряда зубов 28,0—30,8 (М = 29,7), наибольшая ши-
рина черепа 28,5—31,6 (М=30,2), межглазничная ширина 8,9—10,4 (М =
= 9,4), ширина скуловых дуг 21,0—22,2 {М = 21,6), наибольшая высота
18,8-20, 0 (М = 19,2).
Характеристика. Выхухоль — зверек плотного телосло-
жения, приспособленный к жизни в воде. Форма тела валькообразная,
несколько сплюснутая в спинно-брюшном направлении. Шея короткая,
голова конической формы и оканчивается длинным коническим хоботком.
Рис. 16. Строение glans pe-
nis русской выхухоли.
29
Хоботок сжат в горизонтальной плоскости, оголен и несколько расширен
к концу. Находящиеся в его передней части ноздри снабжены закрываю-
щейся кожистой перепонкой, что является приспособлением для передви-
жения в воде. Хоботок служит для поиска пищи в мягком илистом дне
водоемов.
Ноги выхухоли также носят отпечаток приспособления к обитанию в
водной среде — они снабжены оторочкой из жестких волос, а пальцы со-
единены плавательной перепонкой и выполняют роль весел. На земле выху-
холь передвигается крайне медленно и ее без труда может догнать чело-
век, зато в воде она плавает стремительно и легко маневрирует среди вод-
ной растительности. Передние лапы зверька значительно меньше задних
и вместе с хоботком выполняют роль «руля глубины».
Хвост длинный, мощный, сжат с боков и покрыт чешуистой, темно-
бурой кожей, напоминает таковой у ондатры. У основания хвоста на ниж-
ней поверхности имеется мускусная железа, выделяющая жирообразный,
сильно пахнущий мускус. При помощи задних лап выхухоль смазывает
свою шерсть. Хвост служит для поворотов в стороны и движения вперед.
Быстро плывущая выхухоль активно работает хвостом, это хорошо видна
при наблюдении за зверьком.
Тело выхухоли покрыто густым шелковистым и блестящим мехом.
Спина окрашена в темный серо-каштановый цвет, а брюшная сторона свет-
лая, беловато-серая с серебристым оттенком. Мех гуще на брюшке и реже
на спине. Молодые зверьки серые сверху и грязно-белые снизу. Подпушь-
по всему телу матово-серая. Шерстный покров выхухоли хорошо сохраня-
ет зверька от низких температур, так как способен удерживать большое
количество воздуха и не намокать. Зрение у выхухоли развито слабо.
Хрусталик глаза устроен примитивно, выхухоль близорука. Из органон
чувств хорошо развито осязание. Главным органом осязания служит хо-
боток, которым выхухоль обследует встречающиеся на пути предметы,
и в меньшей мере — вибрисы. Обоняние развито хорошо, но выхухоль,
особенно при опасности, больше полагается на слух и быстро реагирует
на звуки.
Сосков 4 пары. Glans penis с многочисленными шипами (рис. 16).
Распространение. Как показывают находки ископаемых
остатков, в древние времена выхухоль была распространена значительно
шире, чем сейчас, и, видимо, заселяла всю Европу. В настоящее время рус-
ская выхухоль обитает лишь на незначительной территории в пределах
СССР. Современный естественный ареал выхухоли занимает бассейн Дона,
Волги, Урала.
Для восстановления и увеличения численности выхухоли ведутся ра-
боты по ее расселению в границах естественного ареала и за его предела-
ми. В результате численность зверьков во многих местах их прежнего оби-
тания возросла. В Западной Сибири, где выхухоль раньше не обитала, пи
нашим рекомендациям она была выпущена в водоемы Томской (1958 г.)
и Новосибирской областей (1968 г.). Зверьки быстро освоились в новых
условиях, размножились и широко расселились. Сейчас район их обита-
ния только на Оби имеет протяженность около 400 км. В 1968 г. партия
зверьков из 114 особей, привезенных из Владимирской области, выпущена
на р. Тартас в Новосибирской области [Юдин, 1962а—в — 1964а—в].
Учеты, проведенные нами в 1980 г., показали, что здесь зверьки также
хорошо расселились по долине реки, освоив и ее притоки, но численность
их незначительна в связи с отсутствием природоохранных мероприятий.
30
По данным обследования местности, в Алтайском крае имеется боль-
шое количество водоемов, пригодных для обитания выхухоли в этом райо-
не. На III Всесоюзном совещании по выхухоли создание очагов в Сибири
было оценено как большое достижение в деле спасения ценного вида,
занесенного в Красную книгу СССР и Международную красную книгу.
Итоги работы за 20 лет по акклиматизации выхухоли в Сибири позволяют
сделать вывод о возможности существования этого зверька в Сибири при
условии соблюдения ряда природоохранных мероприятий [Юдин, 1988].
Заметки по систематике. См. сводку А. А. Гуреева
«Фауна СССР. Млекопитающие» [1979].
СЕМЕЙСТВО
TALPIDAE FISCHER VON WALDHEIM, 1817 —
КРОТЫ
Диагноз. Зверьки средних и мелких размеров. Большинство име-
ет типичную «кротовую» внешность, реже — мышеобразную. Длина тела
60—230 мм. Глаза мелкие, едва заметны. Ушные раковины чаще руди-
ментарны, в виде кожного валика вокруг слухового отверстия, реже раз-
виты и выступают из волосяного покрова. У большинства форм ладони
лопатообразно расширены, приспособлены к рытью. Тело покрыто гус-
тым, бархатистым волосяным покровом.
Череп с удлиненным лицевым отделом. Скулы тонкие, расстояние
между ними уже мозговой части черепа. Резцы долотообразные,- заднеко-
ренные с тремя корнями и с W-образной жевательной поверхностью.
Таз длинный, узкий, без вентрального симфиза.
Зубная формула:
'fer- ртЙЬ мА-44 - 34.
Характеристика. В семейство объединены роющие, редко —
бегающие формы мелких и средних насекомоядных млекопитающих, веду-
щих преимущественно подземно-роющий образ жизни.
Представители семейства имеют сходный внешний облик — валько-
видное тело, на коротких ногах, с чрезвычайно расширенными лопато-
образными передними лапами, приспособленными для рытья, с повернуты-
ми наружу ладонями. Голова коническая, широкая в основании, в рост-
ральной части вытянутая в хоботок, что отражает общий для семейства
адаптивный тип. Шейный отдел не выражен. Глаза скрыты в волосяном
покрове и едва заметны. Иногда они затянуты кожистой перепонкой. Мех
короткий, ровный, бархатистый и лишен ворса. Окраска одноцветная,
чаще всего черная или черно-бурая. Передние конечности у специализи-
рованных видов короткие, мощные, вооружены уплощенными сильными
когтями, задние значительно слабее и уступают им в размерах. По строе-
нию они сходны с конечностями землероек. У бегающих форм обе пары
ног напоминают таковые землероек. Хвост широко варьирует по длине и
форме: у роющих {Talpa, Asioscalops) он короткий, покрыт равномерно
щетинистыми волосами, у представителей рода Eoscalops — рудиментарен,
а у Urotrichus и Uropsilus — длинный, почти равный длине туловища
(рис. 17).
Череп конический, удлиненный в ростральной части, со слабо выра-
женными гребнями, сходный по общей конфигурации у большинства пред-
31
ставителей семейства. Кости лицевого отдела плотно срастаются, и швы
между ними незаметны. Хоботок поддерживается предносовой косточкой
(os praenasale).
Нижняя челюсть длинная, чаще всего с невысокой восходящей ветвью
и небольшим поперечным мыщелком. Вертикальная ось коронарного от-
ростка у разных представителей семейства имеет определенный угол на-
клона к горизонтальной оси нижней челюсти.
Резцы, клыки и предкоренные весьма изменчивы по числу, форме и
расположению. Резцы чаще всего уплощены, долотообразной закруглен-
ной формы. У представителей Talpinae клыки верхней челюсти хорошо
развиты, но значительно варьируют по размерам и форме. Pm1-3 — про-
стые, конические, сходные между собой по форме; Рш4 чаще крупный, ха-
рактерный по конфигурации для насекомоядных; С1 и Рш1-3 с одним или
двумя корнями. Заднекоренные крупные, константные по числу и форме,
туберкуло-секторального типа, W-образные. Коронки их низкие и с линг-
вальной стороны уже, чем с наружной. Пара- и метаконус расположены
около середины коронки. Талонид не развит или в виде небольшого высту-
па протокона. Рш4 и моляры с тремя корнями.
Нижние резцы меньше верхних, направлены вперед, округлой, доло-
тообразной или конической формы, уплощающиеся спереди назад. Клык
рудиментарен, сходен по размерам и форме с резцами и прижат к ним.
Рш1 крупный и выполняет функцию клыка, Рт4 крупнее и более сложна
устроен, чем предыдущие зубы. Моляры крупные, жевательная поверх-
ность их W-образной формы.
В строении посткраниального скелета разных представителей семейст-
ва много общих черт, что связано со специализацией подавляющего числа
видов к подземно-роющему образу жизни.
Позвоночник состоит из 7 укороченных шейных позвонков, 13—
14 грудных, 5—6 поясничных, четырех крестцовых и 8—20 хвостовых. Груд-
ная клетка сужена кпереди и расширена кзади, что связано с мощным раз-
витием пояса передних конечностей. Рукоятка грудины заметно вытянута
вперед. Степень ее развития и гребней на ее поверхности находится в пря-
мой зависимости от специализации: роющей деятельности или бегающего
образа жизни. Среди насекомоядных у кротов лопатка самая вытянутая
в длину, с высоким гребнем в задней части, расположена параллельно оси
тела. У роющих форм ключица своеобразной формы, короткая и толстая.
Нередко ее ширина превышает длину, а у бегающих — длинная и тонкая,
сходная с таковой у землероек. Она непосредственно сочленуется с пле-
чевой костью, что является уникальным явлением среди млекопитающих.
Локтевая кость у роющих форм укорочена, с мощным гребнем и хороша
развитым локтевым отростком. У высокоспециализированных, видов кис-
ти крупные, широкие, с большими фалангами пальцев и широко варьиру-
ют в деталях строения. Таз длинный и узкий, непосредственно срастается
с позвоночником. Два крестцовых и три каудальных позвонка срослись
между собой и подвздошной костью в единое образование. Сросшиеся
остистые отростки этих позвонков образуют мощный гребень, придаю-
щий крестцовому отделу дополнительную жесткость. Лобковые кос-
ти не образуют вентрального симфиза, что также специфично для
семейства.
Бегающие формы, по внешнему виду похожи на землероек, но по
структуре черепа, зубной системе и многим элементам скелета они сходны
с роющими формами.
32
Рис. 17. Представители сем. Talpidae (по С. У. Строганову [1948]).
1 — Talpa europaea L J 2 — Eoscalops longirostris M.-Edw.; 3 — Scalopus aquaticus L«> 4 Con
dylura cristata Ll 5 — Uropsilus soricipes M.-Edw.J 6 —Urotrichus-Talpoides Temmt
33
Рис. 18. Типы строения тазовых костей кротов, обитающих в Сибири
(по С. У, Строганову [1957]).
1 — европеоидный — Talpa europaea L.; 2 — цекоидный — Asioscalops altaica Nik; 3 — моге-
роидный — Mogera robusta Nehring.
3 Б. С. Юдин
34
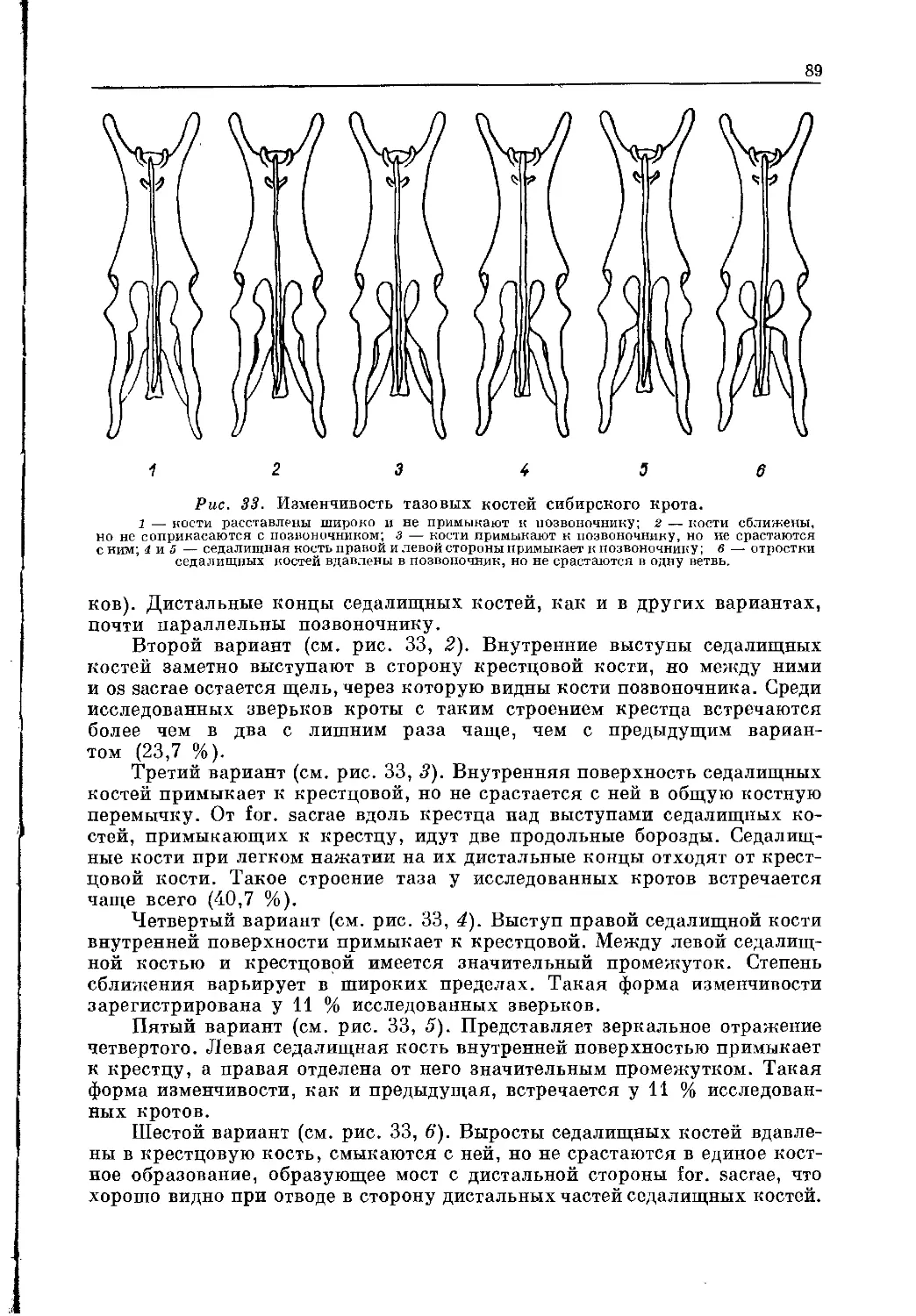
Грулих [Grulich, 1971], исследовав большую серию тазовых костей
разных видов кротов и проанализировав имеющуюся литературу, пришел
к выводу о большой таксономической ценности этого признака. Им выде-
лено три типа строения таза кротов: цекоидный, европеоидный и могеро-
идный (рис. 18). К цекоидному типу он относит такие виды, как Т. соеса
Savi, Т. caucasica Satunin, Т. minuta Blainv. (Г. levantis), Т. romana
Thomas, As. altaica Nik., Parascaptor grandidens Strog. и некоторых пред-
ставителей подсем. Urotrichinae (Dymecodon pilirostris True, Urotrichus tal-
poides Temm.). Костный мост у них неразвит. К европеоидному типу отнес
Talpa europaea L., Parascaptor leucura Blyth, Eoscalops longirostris M.-Edw.
и Eoscalops micrura Hodgson. Кроты, объединенные этим типом строения
таза, имеют хорошо выраженный костный мост, ограничивающий позади
for. sacrae. К могероидному типу относятся формы подсем. Scalopinae
(Scalopus aquaticus L.) и Scapanus orarius True, а в подсем. Talpinae —
Parascaptor grandidens Strog., Mogera robusta Nehring, M. wogura Temm. и
Euroscaptor mizura Gunther.
У кротов с цекоидным типом строения таза мускулатура задних ко-
нечностей развита гораздо слабее, чем у видов с могероидным [Там же].
Цекоидный и европеоидный типы таза развиты лишь у тех форм в подсем.
Talpinae, у которых в процессе филогенеза не произошло редукции числа
зубов, и зубной аппарат остается примитивным, в то время как у форм с
могероидным типом таза произошло значительное уменьшение числа зу-
бов, число которых у видов рода Scaptochirus сократилось до 40, а челюсти
стали значительно укороченными, большая часть восточно-палеарктиче-
ских форм подсем. Talpinae имеет могероидный тип таза, а у западно-
палеарктических видов таз европеоидного или цекоидного типов. Виды с
могероидным строением таза имеют другое строение слуховых косточек и
другой тип строения penis, чем западно-палеарктические виды рода Talpa.
Распространение. Представители сем. Кроты широко рас-
пространены в Евразии и Северной Америке. В ископаемом состоянии их
остатки известны с эоцена. Широкое распространение кроты получили в
олигоцене [Строганов, 1948]. Судя по палеонтологическим данным, уже в
миоцене они специализировались в двух направлениях: роющая деятель-
ность и бег. В это время известны представители родов Talpa L., Scapto-
chirus M.-Edw., Domninoides Green, Scapanus Pomel, Scalopus Geoff. Со-
временная фауна Talpidae сложилась в плейстоцене [Строганов, 1948].
В Европе она представлена видами рода Talpa (4—5 видов), в Азии—родов
Rhinchonax Thomas (1 вид), Nasillus Thomas (2 вида), Uropsilus M.-Edw.
(1 вид), Urotrichus Temm. (1 вид), Talpa L. (7 видов), Parascaptor Gill.
(1 вид), Scaptochirus M.-Edw. (1 вид), Mogera Pomel (3 вида), Scapanulus
Thomas (1 вид). В Северной Америке обитают представители сем. Talpi-
dae: роды Neurotrichus Giint. (1 вид), Scapanus Pomel (3 вида), Parascalops
True (1 вид), Scalopus Geoff. (3 вида), Condylura liliger (1 вид) [Строганов,
1948].
Подсемейство Uropsilinae формировалось в Азии, Talpinae — общее
для Европы, Азии и Северной Америки. Роды Scaptonyx, Urotrichus и Sca-
panulus представлены только в Азии, род Neurotrichus — эндемик северо-
американского континента. Представители рода Talpa заселяют Европу и
Азию, а родов Mogera, Parascaptor и Scaptochirus — только Азию [Allen,
1938; Строганов, 1948, 1957; Hall, Kelson, 1959; Walker et al., 1964; Гуре-
ев, 1979; и др.].
35
Образ жизни. Наиболее характерная черта сем. Кроты —
адаптация к подземно-роющему образу жизни. Эволюция наиболее спе-
циализированных видов шла в направлении развития приспособлений к
рытью и питанию почвообитающими животными. Поэтому решающее зна-
чение в их распространении, исключая непреодолимые преграды, играл
характер почвенного покрова. Жизнь в толще почвы отложила отпечаток
на всю организацию кротов: они в значительной мере утратили зрение;
сформировался мощный пояс из передних конечностей и таза, приспособ-
ленный для рытья почвы; волосяной покров без ворса (адаптация к пе-
редвижению в узких подземных галереях) и ряд других приспособлений.
В связи с этим тело приобрело вальковидную форму с короткими конеч-
ностями. Кроты приспособлены к жизни в ландшафтах с рыхлыми почвами,
преимущественно в лесных и кустарниковых зарослях и по прилежа-
щим к ним опушкам. Эта связь обусловлена наличием здесь влажных мест,
изобилующих почвообитающими беспозвоночными, составляющими осно-
ву рациона кротов.
Основным кормом для кротов в разных частях ареала служат мало-
щетинковые черви, запасы которых в биотопах суши составляют самую
большую по массе долю среди всех животных. Вероятно, приспособление
к жизни в почве, где они практически не имеют конкурентов, к питанию
самыми распространенными и многочисленными червями обеспечило кро-
там возможность широкого распространения по суше и освоения различ-
ных биотопов. Стабильность экологической обстановки и наличие одно-
образного корма, с одной стороны, обеспечило их процветание, с другой —
позволило сохранить ряд примитивных черт организации и выработать
сходные конвергентные признаки. Это обстоятельство в значительной сте-
пени усложняет разработку системы кротовых. Учитывая это, при по-
строении системы следует по-иному подходить к оценке признаков, иначе,
чем у групп, ведущих наземный образ жизни.
Наличие или отсутствие червей — решающий критерий в освоении
кротами территории и фактор, определяющий плотность ее заселения.
Для добывания пищи кроты устраивают сложную систему подземных
«охотничьих» ходов. Их протяженность на индивидуальном участке прямо
пропорциональна обилию червей, последние обусловливают и их числен-
ность на единицу площади [Юдин, 19706, 1972е].
Зверьки почти всю жизнь проводят под землей и крайне редко выходят
на дневную поверхность. Более чем за 30-летнюю практику автора в отло-
ве мелких млекопитающих цилиндрами и конусами в разных районах
страны кроты в них попадали крайне редко, даже при высокой числен-
ности. Отлавливались главным образом звери в таежных биотопах, где
хорошо развиты подстилка или моховый покров, преимущественно в дож-
дливую погоду. Вероятно, это были расселяющиеся прибылые зверьки.
Как приспособление к подземному образу жизни у обыкновенного
крота следует рассматривать относительно высокое развитие легких,
которые составляют 1,28—1,95 % к массе тела. Так, у куторы (N. j adi-
eus), ведущей полуводный образ жизни, этот показатель заметно ниже
(1,06 %).
У крота, добывание пищи которого связано со значительными локо-
моторными нагрузками в среде с низким содержанием кислорода, уровень
развития сердца выше, чем у полуроющих землероек и наземных ежей.
Эта же закономерность прослеживается в развитии почек и в длине ки-
шечника.
3*
36
Как уже было сказано, протяженность охотничьих ходов зверьков
зависит от обилия почвообитающих беспозвоночных, главным образом от
дождевых червей, и составляет сотни метров. Кроме охотничьих ходов
на индивидуальных участках между участками отдельных зверьков
имеются магистральные ходы [Юдин, 1972е], соединяющие их в одно по-
селение. Последние служат для передвижения самцов, посещающих не-
сколько участков самок во время гона, а позднее — путями расселения
молодняка. Самки ведут более оседлый образ жизни, они сильнее привя-
заны к своему гнезду и прилежащему индивидуальному охотничьему
участку. Есть основание полагать, что и самцы не совершают значитель-
ных кочевок. Помеченный нами в гнезде недавно родившийся самец был
пойман через год уже взрослым самцом в состоянии гона вблизи гнезда,
где он был помечен [Юдин, 1972д]. С другой стороны, помеченных одно-
временно с ним собратьев, а также кротов, меченых в других гнездах, на
следующий год повторно отловить не удалось.
После распада выводка прибылые кроты расселяются по магистраль-
ным ходам в поиске свободной подходящей территории, где.и оседают, со-
вершенствуя старую или выстраивая новую систему ходов и гнездо. Гнез-
до крота располагается чаще всего в сложном кротовом выбросе — кро-
товине, в заброшенном муравейнике земляных муравьев, реже — под
корнями деревьев. Жилье крот всегда делает на положительных элемен-
тах рельефа, что спасает его от затопления вешними или ливневыми вода-
ми. Гнездовая камера находится в возвышении на уровне почвы или не-
сколько выше ее, реже — ниже уровня почвы. Вокруг гнездовой камеры
вырывается сложная система горизонтальных кольцевых ходов, окру-
жающих ее в разных плоскостях и соединяющихся между собой радиаль-
ными ходами. Такая система обеспечивает хорошую вентиляцию камеры
и сухость окружающей ее почвы. Зимой в этих ходах, как правило, хра-
нятся запасы дождевых червей [Юдин, 1972в, г].
Внутри гнездовая камера выстлана растительной ветошью, чаще все-
го (у сибирского крота) листьями березы и лисохвоста. Исследование вы-
стилки почти из 300 гнезд крота в окрестностях Новосибирска показало,
что, несмотря на большое видовое разнообразие травянистых растений в
местах устройства гнезд, на выстилку использовались почти исключи-
тельно эти два вида, обладающие гигроскопичностью и антисептическими
свойствами [Юдин, 19706].
Размножаются кроты один-два раза в году. Детеныши родятся голы-
ми, беспомощными, выкармливаются в гнезде молоком матери почти до
размеров взрослых. Зверьки гнездового периода, достигшие уже почти
размеров матери, еще отказываются есть дождевых червей. Вероятно,
как только они переходят на питание червями, выводок распадается. У си-
бирского крота известен феномен латентного периода в развитии опло-
дотворенного яйца. Это единственный случай среди насекомоядных млеко-
питающих мировой фауны. В конце мая — начале июня перезимовавшие
и уже родившие самки кроются вторично. Рост и развитие оплодотворен-
ного яйца тормозится на стадии бластулы. Зимой определить беременность
у сибирского крота невооруженным глазом не представляется возможным.
Активный рост и развитие яйца начинается только в марте следующего
года [Бородулина, 1951; Юдин, 1972а, 1974в, 1980а]. Прибылые самки
кроются в возрасте несколько более месяца, вскоре после выхода из
гнезда.
37
Рождение приурочено к апрелю — маю следующего года, т. е. через
8—10 мес после оплодотворения яйца. Такая особенность размножения,
на наш взгляд, адаптация к обитанию в Сибири в годы с суровыми
малоснежными зимами, вызывающими глубокое промерзание почвы, де-
лающей невозможным или затруднительным добычу корма и ведущую к
гибели кротов на значительных территориях. В этом случае больше всего
шансов выжить имеют самые мелкие прибылые самки, для поддержания
жизни которых требуется меньшее абсолютное количество пищи, чем дру-
гим сочленам популяции. Способность уходить в зиму беременными сам-
кам всех возрастных групп обеспечивает больший «запас прочности» вида
в годы суровых зимовок [Юдин, 1972г, 1974в, 1980а].
У других видов кротов этой особенности в репродуктивном цикле нет.
Гон у них проходит ранней весной (в разных частях ареала неодновремен-
но), самки кроются и вскоре приносят приплод. Некоторые самки успе-
вают дать два помета за сезон размножения.
Заметки по систематике. Кроты, обитающие в Си-
бири, относятся к подсем. Talpinae. Несмотря на довольно много-
численные публикации по частным вопросам морфологии и таксономии,
анализу фаун разных регионов, систематика сем. Talpidae до сих пор
оставалась неразработанной. Это объясняется, с одной стороны, обитанием
разных групп кротов в довольно близких экологических условиях, что
наложило отпечаток на их организацию и привело к вторичному появле-
нию сходных признаков, с другой — вследствие особенностей экологии
они сохранили ряд примитивных черт организации при общем расцвете
группы. В оценке признаков при построении системы сем. Talpidae на эти
обстоятельства должно обращаться больше внимания, чем при разработке
систематики чисто наземных форм. Существенным тормозом в разработке
систематики является недостаточность коллекционных материалов, по-
зволяющих строить не умозрительные классификации, а базирующиеся
на новой, солидной фактической основе. Недостаточны и палеонтологи-
ческие материалы для разработки филогенетических схем.
Выдвинут ряд в разной степени аргументированных классификации
сем. Talpidae, но убедительной, общепризнанной системы пока не пред-
ложено. Все они могут рассматриваться в лучшем случае на уровне рабо-
чих гипотез.
Первую, наиболее исчерпывающую сводку по систематике кротовых
в объеме семейства предложил Добсон [Dobson, 1883]. Она сыграла поло-
жительную роль в разработке классификации этой сложной в таксономи-
ческом отношении группы, но, естественно, за 100 лет существенно уста-
рела. Благодаря обстоятельным сводкам True [1897], Jackson [1915],
а также монографии Hall, Kelson [1959], посвященной американским
кротам, и Аллена [Allen, 1938] по кротам Монголии и Китая систематика
этой группы существенно продвинулась вперед. В объеме мировой фау-
ны систематика кротов разработана С. У. Строгановым [1948] и приведе-
на в обзорных монографиях Эллермана и Моррисон-Скотта [Ellerman,
Morrison-Scott, 1951], В. Е. Соколова [1973], А. А. Гуреева [1979].
В большинстве случаев это компилятивные работы, не базирующиеся на
новых, оригинальных, фактических материалах, специально подобранных
для этих исследований.
Начало изучения систематики кротов уходит своими корнями к осно-
воположнику систематики Карлу Линнею [Linneus, 1758], описавшему
Talpa europaea, Scalopus aquaticus и Condylura cristata. Geoffroy [1803]
38
описал род Scalopus, liliger [1811] выделил род Condylura, Fischer
[1814] — автор первоописания трибы Talpini, Murray [1866] обосновал
выделение подсем. Talpinae, Thomas [1912] внес заметный вклад в сис-
тематику семейства, выделив и описав подсемейства Uropsilinae, Scalopi-
nae, Condylurinae. С. У. Строганов [1948] на основании изучения ори-
гинальных материалов провел ревизию систематики кротовых в объеме
мировой фауны, выделил подсем. Urotrichinae и описал род Asioscalops
[Строганов, 1941, 1957]. Reed [1961] описал подсем. Proscalopinae.
На основе этих и более частных исследований современных кротов и
палеонтологического материала разработан ряд систем семейства. Прежде
всего следует указать на схемы, предложенные Добсоном [Dobson, 1883],
Симпсоном [Simpson, 1945], С. У. Строгановым [1948], Штейном [Stein,
1960], Ван Валеном [Van Valen, 1967] и А. А. Гуреевым [1979]. Поскольку
мы не ставили задачу анализа классификации сем. Talpidae в объеме миро-
вой фауны, взгляды названных авторов здесь не обсуждаются. Читатель для
знакомства с ними может обратиться к первоисточникам или к моногра-
фии А. А. Гуреева [1979], где довольно детально представлены названные
систематические построения. Уместно лишь обратить внимание на су-
щественные различия в представлении систематики Talpidae с позиций
разных авторов. Они касаются всех таксономических уровней — от вида
до подсемейства включительно.
Отсутствие серийных материалов из разных частей ареала по подав-
ляющему числу видов семейства, а следовательно, незнание диапазона
их изменчивости обусловило произвольную трактовку объема видов, ро-
дов, подсемейств. Иллюстрацией сказанному может служить сопоставле-
ние результатов монографических исследований сем. Talpidae, увидев-
ших свет одновременно: англичанина Шварца [Schwarz, 1948] и
С. У. Строганова [1948]. Шварц, сторонник широкой концепции вида,
свел всех кротов подсем. Talpinae в один род с тремя видами (Т. еигораеа,
Т. соеса, Т. micrura), из которых два заселяют Европу, а третий живет в
Азии. Близких взглядов на систему кротовых придерживаются Эллерман
и Моррисон-Скотт [Ellerman, Morrison-Scott, 1951]. Все виды известных
кротов Азии (Т. micrura Handson, Т. klossi Thomas, Euroscaptor grandis
Miller, T. longirostrisM.-Edv., T. parvidens Miller, T. leucura Blytch, Scap-
tochirus moschatus M.-Edw., T. wogura, T. insularis Swinhoe, Mogera lato-
uchei Thomas, Mogera hainana Temm. и т. д.), которых ранее относили к
пяти родам — Talpa, Euroscaptor, Parascaptor, Scaptochlrus, Mogera,
оказались объединены в один вид Т. micrura.
С. У. Строганов [1948] в составе подсемейства рассматривает шесть
родов и 19 видов, из которых представители только одного рода живут в
Европе, а остальные — в Азии. Позднее он выделил сибирского крота в
самостоятельный вид. Штейн [Schtein, 1960] в состав подсемейства вклю-
чает четыре рода с восемью видами. Уже из этой краткой справки видно,
насколько разные позиции в оценке системы подсемейства у названных ав-
торов. К чему приводят произвольные «укрупнения» видов, можно про-
иллюстрировать одним примером. Ряд авторов, в том числе А. П. Кузякин
[Бобринский и др., 1965], вслед за Штейном [Stein, 1960] объединили в
один вид Т. minima Deparma и Т. hercegovinensis Bolkay, затем включи-
ли в его состав Т. mizura Gunther. Другие авторы североиспанских кар-
ликовых кротов, греческого крота Т. olympica Chaworth-Musters и мел-
ких кротов из Югославии [Niethammer, 1964а, б; Felten, Scorch, 1965,
1966; Todorovic, 1965] объединили в гигантский гетерогенный «вид»,
39
отдельные популяции которого оторваны друг от друга на тысячи кило-
метров. Подобной внутривидовой дизъюнкции у млекопитающих ранее
не было известна. Однако с накоплением новых материалов японский зо-
олог Имаицуми [Imaizumi, 1955] убедительно доказал видовую само-
стоятельность японского горного крота (Т. mizura), а Корбет [Corbet,
1967 ] указал на наличие существенных различий между Т. mizura и
Т. olympica. Chaworth-Musters (последний, по мнению Грулиха [1970],
экоморфа Т. соеса). В более поздней работе Имаицума и Ешиуки [Imai-
zumi, Yoshiuki, 1961] снова отнесли японского крота к роду Euroscaptor,
в составе которого он и был описан Гюнтером [1880]. Реальность такой
позиции разделяет Грулих [Grulich, 1971], изучивший на большом мате-
риале особенности строения и изменчивости тазовых костей у разных пред-
ставителей подсем. Talpidae. Зиглер [Ziegler, 1971], изучавший молочную
систему зубов различных представителей сем. Talpidae, пришел к выводу
о неправомерности выводов Шварца [Schwarz, 1948].
Примеров подобной путаницы в систематике кротов, когда авторы
произвольно, без каких-либо веских аргументов, подтвержденных факти-
ческим материалом, «укрупняют» или «дробят» виды, можно привести
много. Этот пример иллюстрирует беспочвенность и невозможность серь-
езной ревизии семейства без накопления новых фактических материалов
по изменчивости видов. Основой для такой ревизии могут служить лишь
скрупулезные исследования на новой фактической основе.
Нет однозначного толкования видового статуса и родовой дифферен-
циации кротов фауны СССР, в том числе не однозначны представления и о
кротах Сибири [Огнев, 1928; Строганов, 1948, 1957; Бобринский и др.,
1944, 1965; Гуреев, 1963, 1979; и др.]. В монографии С. И. Огнева [1928]
в составе семейства рассматриваются два подсемейства (Talpinae, Desma-
ninae) с двумя родами кротов и могер (Talpa, Mogera) в подсемействе на-
стоящих кротов. В состав рода Talpa включены четыре вида (Т. еигораеа,
Т. altaica, Т. caucasica, Т. соеса), а в состав рода Mogera — один вид —
Mogera robusta. В «Определителе млекопитающих СССР» [Бобринский и
др., 1944 ] в состав семейства также включено подсемейство выхухолей
(Desmaninae), собственно кротовые представлены двумя родами: Talpa
как монотипический род с одним сборным видом Т. еигораеа и род Moge-
ra, также с одним видом (М. robusta). Европейский крот в понимании
Н. А. Бобринского и А. П. Кузякина [Там же] представлен как конгло-
мерат, состоящий из разных по сущности и объему систематических еди-
ниц, и не регламентирован реальным систематическим статусом. В этот
«вид» включены все кроты фауны СССР, кроме могер. По С. У. Строгано-
ву [1948, 1957 ], в СССР обитает пять видов кротов рода Talpa, один вид
рода Asioscalops и один — рода Mogera.
Во втором издании «Определителя млекопитающих СССР» один из
авторов — А. П. Кузякин [Бобринский и др., 1965] — отказался от ра-
нее принятой концепции, в результате чего выхухолевые в полном соот-
ветствии с действительным положением выделены в самостоятельное
сем. Выхухолевые — Desmanidae, а в составе подсемейства кротовых при-
знано уже четыре рода с 13—15 видами. В род Talpa вошли обитающие в
СССР три вида (Т. hercegovinensis, Т. еигораеа, Т. altaica) вместо одного
Т. еигораеа в первом издании и род Mogera — с двумя видами (М. robusta,
М. wogura). Впервые для фауны СССР им отмечена средняя могера
(М. wogura).
40
Таким образом, в фауне СССР оставались не установленными ни чис-
ло родов, ни число видов. С. И. Огнев [1928] принимал два рода с пятью
видами, Н. А. Бобринский, Б. А. Кузнецов и А. П. Кузякин [1944] —
два рода и два вида, С. У. Строганов [1948, 1957] в роде Tapia рассматри-
вает пять видов, а в роде Mogera — один. Причем объем вида этим иссле-
дователем принимается иначе, чем С. И. Огневым, Н. А. Бобринским и
А. П. Кузякиным. Сибирский крот выделен С. У. Строгановым ]1941]
сначала в подрод, а позднее [Строганов, 1957] — в самостоятельный род.
А. А. Гуреев [1963, 1979] в составе семейства рассматривает пять видов
кротов (Т. europaea, Т. altaica, Т. caucasica, Т. romana, Т. соеса) рода
Talpa и два вида (М. robusta, М. wogura) рода Mogera. Из представителей
рода Talpa только два — общие с видами, приведенными в определителе
[Бобринский и др., 1965].
Из краткого перечня представлений о систематике кротов видно, что
таксономия этой группы далека от совершенства, и выдвинутые в назван-
ных работах варианты не нашли общего признания. Больше того, приме-
нение хромосомного анализа для систематики позволило все виды кротов
Кавказа (Г. orientalis, Т. romana, Т. minuta, Т. hercegovinensis, Т. соеса)
объединить в два (Т. caucasica и Т. levantis), как это показано в работах
Р. И. Дзуева [1972, 1976, 1977, 1980], Дзуева и соавторов [Dsuev et al.,
1978], А. И. Козловского [1972].
Мы рассмотрели разработку систематики на видовом и родовом уров-
нях; что же касается подвидов кротов, то здесь дело обстоит еще сложнее.
Отсутствие серийных материалов по большинству видов не позволяло ис-
следователям дать сколько-нибудь удовлетворительный анализ внутриви-
довых градаций, поскольку неясны и границы видов, что привело к выде-
лению ряда нереальных подвидов. Эти разногласия обусловлены почти
полной неизученностью внутривидового полиморфизма, что затрудняет
определение таксономической значимости признаков и способствует произ-
вольной оценке ранга отдельных систематических категорий.
В систематике сибирских представителей подсем. Talpinae наиболее
спорным следует считать правомерность выделения в самостоятельный род
(Asioscalops) сибирского крота, внутривидовую систематику Т. europaea
и As. altaica. Для решения этих вопросов нами изучена индивидуальная
и популяционная изменчивость зубной системы сибирского крота [Юдин,
1977], строение тазовых костей кротов и могер Сибири [Юдин, 1975], на
обширном материале методом колориметрирования изучена вариабель-
ность окраски сибирского и европейского кротов, изменчивость зубной
системы европейского крота. На больших сериях прослежена изменчи-
вость меристических признаков сибирского и европейского кротов в пре-
делах Сибири. Результаты этих исследований приводятся в соответст-
вующих очерках.
ПОДСЕМЕЙСТВО
TALPINAE FISCHER VON WALDHEIM, 1817 —
НАСТОЯЩИЕ КРОТЫ
Тело вальковидное, на коротких ногах. Ладони чрезвычайно расши-
рены, лопатообразные, в несколько раз шире ступни. Ушных раковин
нет. Звездообразного кожистого выроста на конце морды нет. Хвост ко-
роткий, короче головы, покрыт густыми, жесткими волосами. Зубов
40—44. Верхние резцы мелкие, незначительно убывают по размерам от
41
первого к третьему, сходны по форме. Верхние клыки большие кинжало-
образные, резко выделяются по размерам и конфигурации среди других
категорий зубов. В нижней челюсти клыки мелкие, прижаты к резцам
и почти не отличаются от них, Рт15 несущий функцию клыка, крупный,
выделяется среди других зубов по размерам и форме, Pm4 и Рш4 крупные,
своеобразного строения.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ПОДСЕМ. TALPINAE
1(4). Всего зубов 44, в нижней челюсти 22; нижние клыки есть —
примыкают к резцам и похожи на них по размерам и форме.....2
2(3). Зубы относительно размеров черепа крупные, заднекоренные
широкие. Вертикальная ось восходящей ветви нижней челюсти сильно
наклонена вперед по отношению к продольной оси нижней челюсти
(рис. 19).........................Talpa L.—Кроты обыкновенные
3(2). Зубы относительно размеров черепа мелкие, заднекоренные
узкие (рис. 20). Вертикальная ось восходящей ветви нижней челюсти
перпендикулярна продольной оси нижней челюсти................
............................Asioscalops Strog.— Кроты сибирские
4(1'). Всего зубов 42, в нижней челюсти нет клыка........
............................................Mogera L.— Могеры
Род Talpa Linnaeus, 1758 —
Кроты обыкновенные
Тип рода. Talpa еигораеа L., 1758.
Диагноз. Зубная формула:
14; с -I; PmA; м-1 = 44.
У кротов рода Talpa сохранилась зубная формула, типичная для
плацентарных млекопитающих. Зубы крупные. Восходящая ветвь нижней
челюсти относительно узкая, наклонена вперед.
Характеристика. Кроты средней величины. В облике об-
ращает на себя внимание удлиненная ростральная часть головы, вытя-
нутая в виде хоботка, и сравнительно длинный хвост. Глаза видны снару-
жи и либо имеют подвижное веко, либо закрыты кожистой перепонкой.
Череп средних размеров, у некоторых форм кротов Кавказа он мелкий.
По сравнению с сибирским кротом череп крота обыкновенного менее вы-
тянут, сужен в ростральной части и менее угловат, со сглаженными шва-
ми и вздутой мозговой коробкой. Если череп рассматривать сверху, то
хорошо видны выступающие в стороны нижние края foramina onteor-
bitalia.
Характерной чертой обыкновенных кротов является крупнозубость
по отношению к размерам черепа. Длина верхнего ряда зубов по сред-
ним показателям колеблется по отношению к кондило-базальной длине
черепа (%): в Центральной и Восточной Европе — 42,5, Польше — 42,2,
Башкирии — 41,2, в районах Центрального Урала — 41,5, Тюменской
области — 40,8. Верхние клыки саблевидные, без значительного расши-
рения в основании по сравнению с кротами рода Asioscalops. Нижние
клыки малы, почти не отличаются по размерам от резцов, плотно к ним
прижаты и выполняют их функцию. Ргщ чрезвычайно велик по сравнению
с двумя другими зубами этой категории и несет функцию клыка. Между
клыком и Рпц всегда имеется более или менее значительный промежуток.
42
Ри,с. 19. Череп обыкновенного крота — Talpa еигораеа.
Премоляры верхней челюсти пропорциональны размерам коренных; сре-
ди них более развит Pm4. Верхние заднекоренные крупные и высокие.
Восходящая ветвь нижней челюсти (proc, coronoideus) узкая, ее верти-
кальная ось имеет сильный наклон вперед относительно продольной ниж-
ней челюсти.
Распространение. Европа, Малая Азия, Кавказ и Запад-
ная Сибирь к востоку до бассейна Иртыша и нижнего течения Оби.
Заметки по систематике. Число видов в роде точно не
установлено. В мировой фауне известно около 12 современных видов.
В СССР, по данным разных авторов,— от 1 до 5. В Сибири род представ-
лен одним видом.
Talpa еигораеа Linnaeus, 1758 — Крот обыкновенный
Голотип. Первоописание сделано по экземпляру из Швеции,
окрестности Упсалы. Типовой экземпляр утрачен.
Диагноз. Признаки вида указаны в характеристике рода.
Измерения. См. табл. 1.
Характеристика. По общему габитусу похож на сибирского
крота, но заметно уступает ему в размерах. Длина тела 128—162 мм.
Зверьки имеют значительно вытянутую ростральную часть головы. Мех
бархатистый, мягкий, как и у других кротов лишен ворса. На ногах и
хвосте волосяной покров более грубый. Длина волос летнего меха 6—7,
43
зимнего 11 —12 мм. Окраска меха у сибирских представителей вида изу-
чена недостаточно из-за малочисленности коллекционных материалов.
По имеющимся данным можно сказать, что она более вариабельна,, чем у
Ля. altaica', возможно, это связано с небольшим объемом исследованной
выборки. Она колеблется от черной до серо-черной. Для вида известны
индивидуальные цветовые аберрации: альбинизм, хромизм, дымчато-
серые и пятнистые с разной интенсивностью и соотношением цветовых
гамм. Однако это редкие случаи, встречающиеся единично. Наиболее
частое отклонение в окраске — наличие рыжевато-желтого пятна на вент-
ральной части тела разной величины, формы и интенсивности цвета.
Окраска нижней стороны тела сходна с таковой спины, но более светлого
тона. Взрослые зверьки в свежем невыношенном меху блестяще-черные.
Потертый мех становится матовым, приобретает серый оттенок. У моло-
дых зверьков волосяной покров тусклый.
Нами установлены популяционные вариации в окраске обыкновенно-
го крота. Зверьки, добытые на правом берегу Иртыша (Уватский р-н,
окр. пос. Миссея), на ростральной части головы имели рыжевато-охристую
окраску меха, простирающуюся до глаз и ушей. На вентральной стороне
рыжее пятно захватывает нижнюю губу или идет до самой груди. Кроты,
обитающие в бассейне рек Конца, Сосьва, Демьянка и в Тобольском райо-
не, охристо-рыжего цвета в окраске меха головы не имели.
Колориметрическое исследование волосяного покрова обыкновенного
крота дало возможность установить, что зверьки из Калининской области
(71. е. europaea) более темные по сравнению со зверьками из Тюменской
области. По показателю белизны они достоверно отличаются (табл. 2)
на уровне подвидов. При этом следует иметь в виду, что сравниваемые
выборки малы (12 и 14 экз.). Эти популяции различаются также досто-
верно по оттенку (см. табл. 2).
44
Таблица 1
Морфометрическая характеристика Talpa еигораеа
Возрастная и половая группа М±тх СТ cv п lim
1 * 3 4 . 5 6
Взрослые самцы Длина тела 145,2±2,42 с головой 5,13 3,54 13 137,0—157,0
самки 128,0 — —* 1 —-
В ц е л о м по виду 133,5±3,89 23,64 17,71 37 128,oJ162,O
Взрослые самцы Длина х 40,2± 1,98 воста 7Д5 17,81 13 30,0—49,0
самки 41,0 —• — 1 -—
В целом по виду .... 34,0± 1,13 6,90 20,27 37 25,5—49,0
Взрослые самцы Длина а 20,2±0,17 пупни 0,60 2,96 13 19,0—21,0
самки 18,0 — —- 1 —
В целом по виду .... 19,6±0,17 1,04 5,28 37 18,0—21,0
Взрослые самцы Масса 98,4±2,32 тела 7,71 7,83 11 84,5—109,5
самки 67,0 — — 1 —
В целом по виду .... 94,6+3,33 12,0 12,69 13 67,0—109,5
Кон Самцы прибылые дило-бавальна. 35,4+0,36 я длина че 0,62 репа 1,76 3 34,5—36,1
взрослые 35,7+0,21 0,97 2,72 22 33,0—37,5
Самки прибылые 34,0±0,18 0,44 1,30 6 33,5—34,7 '
взрослые 33,3 — — 1 —
В целом по виду .... 35,0+0,14 1,21 3,45 72 33,0—37,5
Д Самцы прибылые лина верхнег( 14,3+0,17 > ряда зуб 0,38 ов 2,61 5 13,9—14,9
взрослые 14,4+0,11 0,53 3,69 23 12,9—15,0
Самки прибылые 14,0±0,06 0,14 0,98 6 13,8—14,2
взрослые 14,0 — — 1 —
В ц е л о м по виду 14,3+0,05 0,45 3,13 76 12,9—15,3
Длина верхнего ряда заднекоренных зубов
Самцы прибылые 6,4+0,10 0,21 3,32 5 6,1—6,6
взрослые 6,4+0,03 0,15 2,31 22 6,2—6,6
Самки прибылые 6,2+0,04 0,10 1,59 6 6,0—6,3
взрослые 6,2 — — 1 —
В ц е л о м по виду 6,3+0,03 0,23 3,57 54 5,8—6,7
Ширина рострума
Самцы прибылые 4,4±0,06 0,14 3,21 5 4,2—4,5
взрослые ' 5,1+0,10 0,47 9,23 23 3,9—5,6
Самки прибылые 4,3+0,21 0,52 12,09 6 4,1—5,4
взрослые 5,0 — 1
В ц е л о м по виду ' 4,8±0,О5 0,44 9,14 76 3,9—5,6
45
Окончание табл. 1
1 2 | 3 | 4 | 5 | в
Иредглазничная ширина
Самцы прибылые 6,9+0,08 0,17 2,51 5 6,8—7,2
взрослые 6,9±0,06 0,29 4,13 23 6,2-7,6
Сайки прибылые 6,5+0,05 0,12 1,79 6 6,3—6,6
взрослые 6,3 — — 1 —
В ц е л о м по виду 6,6±0,04 0,34 5,10 76 6,0—7,6
Скуловая ширина
Самцы прибылые 12,1+0,12 0,20 1.65 3 11,9—12,3
взрослые 12,5+0,10 0,45 3,58 20 11,9—13,7
Самки прибылые 11,6+0,18 0,45 3,83 6 11,0-12,0
взрослые 11,5 — — 1 —
В Ц е л о м по виду 12,0+0,07 0,55 4,56 60 11,0-13,7
Наибольшая ширина черепа
Самцы прибылые 16,8±0,07 0,12 0,69 3 16,7-16,9
взрослые 17,2+0,07 0,34 1,56 23 16,3-17,7
Самки прибылые 16,2+0,09 0,19 1,19 5 16,0-16,5
взрослые 16,2 — — 1 —
В ц е л о м по виду 16,7+0,08 0,66 3,95 71 15,0-17,7
Наибольшая высота черепа
Самцы прибылые 10,9+0,10 0,23 2,10 5 10,6—11,2
взрослые 10,6+0,08 0,36 3,36 23 10,0—11,3
Самки прибылые 10,8+0,11 0,26 2,44 6 10,3—11,0
взрослые 10,0 — — 1 —
В ц е л о м по виду 10,7+0,05 0,43 4,08 74 9,7-11,3
Высота восходящей ветви нижней челюсти
Самцы прибылые 7,0±0,07 0,15 2,11 5 6,8—7,2
взрослые 7,2+0,06 0,30 4,12 23 6,3—7,6
Самки Прибылые 6,5+0,07 0,18 2,75 6 6,3—6,8
взрослые 6,6 — — 1 —
В ц е л о м по Виду 6,9+0,05 0,43 6,25 75 5,8-7,6
Примечание. Здесь и далее в аналогичных таблицах линейные измерения —
масса тела — г. Взрослые зверьки — все особи рождения Предыдущих лет, в возрасте от года (один
раз перезимовавшие) и более, прибылые — все особи рождения текущего года.
Таблица 2
Сравнительная характеристика окраски обыкновенного и сибирского кротов
Окраск а Калининская обл., Т, еигораеа, (1) Тюменская обл. Г. еигораеа (2) Зап., Ср. Сибирь, As. altaica (3) Достовер- ность различий средних
cv п Су п М±тпж Су X е* 1 пз bi СО е* 43 b
Белизна 1,63+0,13 27,8 12 2,12+0,18 32,3 14 1,69+0,06 51,42 137 2,2 2,1
От тенек 125,0+2,37 6,6 12 116,3+1,88 6,1 14 125,7+0,87 8,10 137 2,9 2,2
46
Белизна
Рис. 21. Сравнительная характеристи-
ка окраски обыкновенного и сибирского
кротов.
1 — Т. europaea (Тюменская обл.); г — Т-
europaea (Калининская обл.); 3 — As. altaica
(в целом по виду).
Окраска обыкновенного кро-
та, обитающего в Сибири, по срав-
нению с As. altaica более светлая.
По белизне Td = 2,1, а по оттен-
ку Td — 2,2 (см. табл. 2). Размах
изменчивости окраски меха у
обыкновенного крота значительно
ниже, чем у сибирского (рис. 21).
Возможно, это вызвано количест-
венным различием сравниваемых
выборок.
При сравнении внешнего
строения европейского и сибирско-
го кротов обращают на себя вни-
мание следующие особенности.
Как уже отмечалось, европейский крот по размерам заметно уступает си-
бирскому, эти показатели отличаются с большой достоверностью (табл. 3).
Значительно превосходит сибирский крот европейского и по массе (Td=
= 2,7). Европейский заметно более длиннохвостый {Td = 6,9) и имеет
относительно большую ступню {Td = 2,3).
Коренные качественные и количественные различия прослеживаются
в строении черепа и соотношении его отдельных элементов. Череп евро-
пейского крота составляет 25,9 % от длины тела, а сибирского — 23,6 %.
В то же время абсолютные показатели по этому признаку больше у сибир-
ского крота (соответственно 35,8 и 34,6 мм). Таким образом, европейский
крот относительно более крупноголовый.
Самобытно строение зубной системы у зверьков этого вида. Зубы
европейского крота относительно размеров черепа крупные, с высокими
коронками. Так, длина верхнего ряда зубов в абсолютных показателях
больше, чем у сибирского крота (14,2 и 13,9 мм). По отношению к разме-
рам черепа она составляет 41 %, а у сибирского — 38,8 %. Особенно ве-
лик ряд трех заднекоренных зубов, превосходящий этот показатель у
сибирского крота как по абсолютной величине (6,3 и 5,5 мм), так и по
отношению к кондило-базальной длине черепа (18,2 и 15,4 %). Различия
выборок по этому признаку высоко достоверны {Td = 3,7). В конфигура-
ции черепа обращает на себя внимание узость лицевой части — ширина
рострума над клыками. По этому показателю виды достоверно отличают-
ся {Td = 2,3). Предглазничная, скуловая и наибольшая ширина про-
порциональны размеру черепа и имеют одинаковые относительные показа-
тели у обоих видов. То же самое можно сказать и о величине восходящей
ветви нижней челюсти. Однако по форме последняя коренным образом
отличается от таковой у сибирского крота: у Т. europaea она имеет за-
метный наклон вперед и ее вертикальная ось по отношению к продольной
оси нижней челюсти сильно наклонена вперед.
У сибирского крота она широкая, иной конфигурации, а ее верти-
кальная ось перпендикулярна продольной оси нижней челюсти (рис. 22).
47
Таблица 3
Сравнительная характеристика пропорций тела и черепа европейского и сибир-
ского кротов (на примере прибылых зверьков по материалам из Сибири)
Признак Т. europaea As. altaica Достоверность различий (но Стьюденту)
Значение, мм Индекс, % Значение, мм Индекс, %
Длина тела с головой 133,5+3,886 25,9 * 151,8+1,679 23,6 * 4,3
хвоста 34,1+1,134 25,5 24,6+0,771 16,2 6,9
ступни 19,6+0,170 14,7 20,2+0,20 13,3 2,3
Масса тела, г 94,6+3,324 70,9 107,4+3,470 70,8 2,7
Длина кондил о-базал ь- ная черепа 34,6+0,304 — 35,8+0,145 — 3,6
верхнего ряда зубов 14,2+0,323 ' 41,0 13,9+0,047 38,8 0,9
верхнего ряда заднекоренных зубов 6,3+0,217 18,2 5,5+0,025 15,4 3,7
Ширина рострума 4,4±0,396 12,7 5,3+0,035 14,8 2,3
предглазничная 6,9+0,233 19,9 7,0+0,032 19,6 0,4
скуловая 11,8±0,434 34,1 12,3+0,056 34,4 1,1
наибольшая чере- па 16,5+0,383 47,7 17,0+0,059 47,7 1,3
Наибольшая высота черепа 10,9+0,237 31,5 11,1+0,039 31,0 0,8
Высота восходящей ветви нижней че- люсти 6,8+0,429 19,7 7,0+0,42 19,6 0,5
Примечание. Индексы промеров черепа берутся относительно его кондило-ба-
зальной длины, длина хвоста, размер ступни — относительно длины тела. Звездочкой поме-
чены данные отношения кондило-базальной длины к длине тела.
Строение таза у обыкновенного крота специфично. Характерная его
черта — наличие сплошного широкого костного моста (см. рис. 18),
ограничивающего for. sacrae с дистальной стороны. Костный мост обра-
зован в результате срастания в одно костное образование os sacrae и
os ischii. Признак этот стойкий, его изменчивость у сибирских предста-
вителей вида нами не отмечена. Грулих [Grulich, 1971] исследовал
Рис. 22. Строение венечного отростка некоторых видов кротов родов Talpa и ylsiosca-
lops.
1 — Т« minuta; 2 — Т. соеса; 3 — Т. caucasica; 4 — Т. europaea', 5 — As. altaica.
48
624 экз. европейского крота из Франции, Австрии, Югославии, Чехосло-
вакии, Болгарии и Румынии. Из них только у 1 экз. костный мост от-
сутствовал. Судя по рисунку таза, приведенному И. Грулихом [Там же],
этот экземпляр скорее относится к Т. соеса, чем к Т. еигораеа.
Из других особенностей строения таза европейского крота обращает
на себя внимание прочность конструкции, относительно большая ширина
и массивность. Наибольшей ширины таз достигает в области вертлужных
впадин. Дистальные концы os ischium сужены в направлении к позвоноч-
нику, а не отведены в стороны. Строение таза европейского крота — на-
дежный признак, позволяющий безошибочно идентифицировать вид. Его
изменчивость выражается лишь в колебании размеров при сохранении
особенностей строения, что не снижает таксономической значимости этого
признака.
В составе рода Talpa имеются виды (Т. caucasica, Т. levantis, Т. соеса,
Т. romana), у которых седалищные кости не образуют сплошной пере-
мычки позади for. sacrae. Поэтому наличие моста между os sacrae и os
ischii — признак видового ранга, свойственный только Т. еигораеа.
Общим для всех видов рода Talpa является относительная крупнозубость
и строение коронарного отростка [Юдин, 1971а, 1975в].
Таз сибирского крота имеет иное строение: седалищные кости у не-
го не срастаются с позвоночником и не образуют костного моста, ограни-
чивающего for. sacrae дистально. Дистальные концы их чаще всего парал-
лельны позвоночнику. Нами изучено строение таза у 148 экз. As. altaica
из разных пунктов его ареала и ни у одного экземпляра не зарегистриро-
вано указанной костной перемычки. Детальное описание строения таза и
его изменчивости приведено в соответствующем видовом очерке.
В идо специфично строение glans penis обыкновенного крота. Голов-
ка копулятивного органа слабо коническая, постепенно сужающаяся от
основания к свободному концу и несколько сжатая с боков по направле-
нию от округлого основания к дистальному концу. По всей ее цоверх-
ности густо разбросаны шипики, чаще всего сидящие парами (рис. 23).
От сибирского крота Т. еигораеа существенно отличается и по этому при-
знаку. У обыкновенного крота glans penis тонкая и более короткая, чем
у Ах. altaica (11,3—15,5 мм; М = 13,3 мм при толщине основания 1,9 мм),
покрывающие его шипы мелкие и иной формы. Мочеполовое отверстие
открывается на самом конце головки. Утонченный участок ее свободного
конца шипиками не покрыт. Нет их и у основания головки.
Обыкновенный крот хорошо отличается от других видов и по хромо-
сомному набору. В диплоидном наборе у него 34 хромосомы при NF = 68.
Сравнительная характеристика кариотипа некоторых видов кротов при-
ведена в табл. 4.
Распространение. Европа и Западная Сибирь. Распростра-
нение этого вида в Западной Сибири выяснено недостаточно. Ареал его
простирается между Уралом и Иртышом. На севере граница распростра-
нения обыкновенного крота пересекает Урал приблизительно у 64° с. ш.
и тянется к Березову, откуда спускается к Ирбиту и Тюмени. По-видимо-
му, в районе Тобольска, Тюмени и Ишима ареал обыкновенного крота
перекрывается с ареалом крота сибирского.
Изменчивость. Изменчивость наблюдается в широких пре-
делах. Недостаточно репрезентативный материал по кротам из Сибири не
позволяет провести детальную характеристику возрастной и половой
изменчивости размеров тела. Однако по имеющимся сведениям очевидно,
49
Рис. 23. Строение glans penis обыкновенного (Л) и сибирского кротов (5).
Усл. обозн. см. рис. 7.
что взрослые самцы — самые крупные особи популяции, менее круп-
ные — прибылые самцы и взрослые самки и самые мелкие — прибылые
самки (см. табл. 2). В этом отношении изменчивость аналогична таковой
у сибирского крота. Та же закономерность прослеживается ив распреде-
лении весовых групп. Масса самцов (прибылых и взрослых) колеблется от
84,5 до 109,5 М = 98,4 Д=2,3 г, а у самок она значительно меньше
4 Б. С. Юдин
50
Таблица 4
Сравнительная характеристика кариотипов некоторых видов кротов
Вид, подвид 2n NF Автор
Talpa europaea L. 34 68 Borgaonkar [1965], Mey-
Talpa levantis Thomas 34 66 Jan [1966] Дзуев [1972a, 1981]
T. romana stankovici V. et 34 6 Todorovic [1965]
E. Martino T. hercegovinensis Bolkay 36 68 Todorovic [1969]
T. соеса Savi 36 70 Meylan [1966]
T. caucasica Satunin 38 66 Дзуев [19726]
(в табл. 2 включены не все имеющиеся в нашем распоряжении материа-
лы). По данным К. Я. Фатеева [1966], в Костромской области масса сам-
цов колеблется по годам: от 80 до 95 г (11m 47—109), а самок — 71—86
(lim 39-109).
Окраска варьирует в широких пределах. В коллекции зоологическо-
го музея Биологического института СО АН СССР хранится уникальная
серия шкурок, демонстрирующая цветовые вариации обыкновенного кро-
та. Есть хромисты, альбиносы, зверьки «голубые», пестрые на фоне золо-
тистого или белого цвета с пятнами разной конфигурации, разных разме-
ров и интенсивности окраски — от голубовато-серого до черного. Однако
все они относятся к зверькам, добытым в европейской части ареала,
и отобраны из больших серий типично окрашенных зверьков. В природе
такие отклонения крайне редки. Из-за небольших размеров ареала обык-
новенного крота в Сибири, а главное, скудных коллекционных сборов
цветовые вариации неизвестны.
Обычная окраска обыкновенного крота близка к таковой сибирского.
Белизна составляет 2,12 (±0,18), а оттенок 116,3 (±1,88) (по материалам
из Тюменской обл.). При сравнении окраски кротов (см. табл. 2) из Тю-
менской и Калининской областей достоверность различий составляет
{Td = 2,2). Кроты из Сибири светлее, чем из западных районов. Что ка-
сается показателя оттенка, то у кротов из Калининской области он со-
ставляет 125,0 (±2,37), а из Сибири 116,3 (±1,88), т. е. насыщенность
оттенка выше в западных районах ареала. Достоверность разности сред-
них значений по этому показателю равна Td = 2,9. Однако при этом сле-
дует иметь в виду небольшие объемы сравниваемых выборок и значитель-
ный полиморфизм в окраске. Учитывая это обстоятельство, сделанные
выводы можно считать предварительными.
При сравнении окраски обыкновенного крота из Тюменской об-
ласти с таковой сибирского крота выявлено, что первый светлее (белизна
2,12±0,18), чем второй (1,69±0,06). Достоверность различий средних
значений довольно высокая {Td = 2,1). Сибирский крот отличается и по
насыщенности оттенка (125,7±0,87) по сравнению с европейским (116,3±
±1,88). Достоверность разности средних значений значительна {Td =
= 2,2). Таким образом, обыкновенный крот из Западной Сибири досто-
верно отличается по окраске от особей из более западных районов ареала.
Эти отличия достигают подвидового ранга. Хорошо отличаются европей-
ские кроты по окраске и от сибирского. Для более детальной характе-
ристики полиморфизма окраски обыкновенного крота в Сибири необхо-
дим сбор новых серийных материалов.
51
Полиморфизм размеров черепа обыкновенного крота также рассмат-
ривается преимущественно на сибирском материале. Кондило-базальная
длина колеблется в пределах 33,0—37,5 (М = 35,1 =±=0,1) мм. По обра-
ботанным материалам из Костромской области (сборы Ф. А. Соколова,
п = 146 экз.) этот показатель колеблется от 32,0 до 36,8 (М = 32,2=±=
=t0,08), т. е. кроты из Сибири достоверно отличаются по этому признаку
от кротов из европейской части ареала в пределах СССР. Кондило-базаль-
ная длина черепа обыкновенного крота из Польши (Беловежская Пуща)
колеблется в пределах 31,7—36,1 (М = 33,7=*=0,1; при п — 51). Досто-
верность различий средних — Td = 2,8, т. е. уровень различий высокий.
По величине признака одновозрастные и взрослые прибылые самцы и
самки между собой практически не отличаются (см. Табл. 1). Половые от-
личия по этому признаку значительны (Td — 8,6), самцы имеют заметно
более крупный череп по сравнению с самками. Взрослые звери (самцы и
самки) также обладают более крупными черепами, чем прибылые
(Td = 2,5).
Длина зубного ряда подвержена меньшему половому и возрастному
полиморфизму. Достоверных различий не прослеживается. Одним из
стабильных признаков у обыкновенного крота является длина заднекорен-
ных зубов верхней челюсти. Достоверных различий по нему нет в разных
возрастных и половых группах (lim 5,8—6,7; М = 6,3=±=0,031 мм).
Ширина рострума колеблется значительно больше. Этот показатель
у взрослых зверьков достоверно выше (Td = 4,5), чем у прибылых. Раз-
личий по полу у одновозрастных зверьков нет.
Весьма стабильна и предглазничная ширина черепа, рано достигаю-
щая размеров взрослых. Скуловая ширина больше (Td — 4,7) у самцов,
чем у самок, но достоверен этот показатель только для взрослых. Мало
подвержен изменчивости и такой признак, как высота черепа. Высота вос-
ходящей четви черепа у самок меньше, чем у самцов. Ширина черепа ме-
няется с возрастом и неодинакова у зверьков разного пола. У взрослых
зверьков она достоверно больше (Td — 4,1), чем у прибылых, а у самцов
значительно превосходит таковую у самок (Td = 4,2).
Различия метрических признаков черепа между сеголетками и зверь-
ками в возрасте года достоверны. В возрасте 1 года значения их стабили-
зируются и в дальнейшем мало подвержены изменениям.
Как видно из приведенных примеров, размах полиморфизма по боль-
шинству признаков значителен, в то же время ряд признаков довольно
стабилен и мало подвержен возрастной и половой изменчивости.
Строение таза европейского крота типично для вида в целом и не
подвержено сколько-нибудь заметным качественным изменениям. Поли-
морфизм сводится к изменению размеров: длина колеблется от 23,5 до
31,0 (М = 26,9) при ширине 6,0—10,0 (М = 8,8) мм (табл. 5).
Изменчивость зубной системы обыкновенного крота не изучена. Что
касается сибирского крота, то она подробно рассмотрена на обширном
фактическом материале (2402 экз.) из разных частей ареала вида с исклю-
чительно широким диапазоном проявления [Юдин, 1977]. Детальное
описание этого явления приведено в очерке о сибирском кроте.
У обыкновенного крота аберрации зубной системы невелики. Нами
изучена серия зверьков из 826 экз. из разных частей ареала. Среди 51 экз.
обыкновенного крота из Польши (Беловежская Пуща) ни у одного не
оказалось отклонений в строении зубов. Из табл. 6 видно, что с продви-
жением на восток от центра ареала вида к его периферии изменчивость
4*
52
Таблица 5
Характеристика тазовых коетей кротов
Призйак Т. leva-ntiSi п=10 Т, caueasica, п=20 Т. еигораеа^ n=G8 Ав. altaica, п=35
Кавказский заповедник Урал, Зап. Си- бирь Иркутская обл., Бурятия
Длина таза Ширина таза 17,8—21,5 19,5 4,6—6,5 5,5 21,5—25,0 23,3 6,4-9,0 7,6 23,5—31,0 26,9 6,0—10,0 8,8 28,0—34,0 30,8 8,8—11,2 9,6
зубов возрастает. В Костромской области она отмечена у 1,8 % исследо-
ванных зверьков, в Пермской — у 12,5 в Свердловской — у 8,2, в бассей-
не рек Конда и Сосьва — у 24, в Тюменской области — 33,3 %. Всего
аберрации зубов оказались в 62 случаях, что составляет 1,6 %, в то вре-
мя как у сибирского крота изменчивость была зарегистрирована у 32,9 %
[Юдин, 1977]. Отклонение от нормы в числе зубов в сторону олигодонтии,
полиДонтии и двувершинности составляло среди резцов (Is)— три случая
(4,8 % от числа всех отклонений) и премоляров (Pm1-4, Pih1j4)— 59 слу-
чаев (95,2 %). Изменчивость заднекоренных у обыкновенного крота не
зарегистрирована.
Наиболее значительное отличие было у одного зверька из Тюменской
области: в верхней челюсти четвертый премоляр был двухвершинный,
а в нижней оказался дополнительный третий зуб этой категории. У Asios-
calaps altaica, кроме того, зарегистрирована изменчивость заднекоренных
(дополнительная 4-я пара), частые нарушения в положении зубов на че-
люсти и сложные отклонения одновременно в левой и правой нижней и
верхней челюстях, охватывающие одновременно до восьми зубов разных
категорий [Юдин, 1977]. Обращает на себя внимание не только увеличе-
ние числа случаев изменчивости в направлении с запада на восток к пе-
риферии ареала, но и подверженность этому явлению зубов разных кате-
горий (см. табл. 6). Полиодонтия отмечена в 7 случаях (11,3 %) от числа
всех отклонений, олигодонтия — в 48 (77,4 %) и двухвершинность —
в 11 случаях (17,7 %)2 *.
Рис. 24. Резцы и Клыки обык-
новенного крота.
а — прибылого (вверху) и
старого (внизу); б — четвер-
тый верхний предкоренной
(Pm4); 1 — Т. е. uralensis Og-
nev; 2 — Т. е. transutalensis
Strog. (По Строганову [19571).
2 Отклонения в строении какого-либо зуба одновременно в двух челюстях рас-
сматриваются как Два случая.
53
Таблица 6
Меристическая изменчивость зубной системы Talpa europa-
ea из разных частей ареала
Показатель Костром- ская обл., п = 215 Пермская обл.. п = 48 Свердлов- ская обл., п-476 Заповед- ник «Ма- лая Сось- ва», п = 27 Тюмен- ская обл., Тобол, г — 9
+ Р1Г 0 0 1 1 0
+РЛ 0 0 0 1 0
—РтЧТ 1 0 0 0 0
—Рт’Л 1 0 1 0 0
—РгщП 1 0 1 0 0
—PmjJI 1 0 0 0 0
—Рт2П 0 0 1 0 0
—РпРЛ 0 0 4 0 0
—Рт2ПЛ 0 1 0 0 0
Н-РшгЛ 0 0 1 0 0
—Рт2П 1 2 11 0 0
—Рт^Л 0 3 12 0 0
—Рт»Л 0 0 1 0 0
—Рт3П 0 0 2 0 0
—Рт3П 0 0 1 0 0
—Рт4Л 0 0 1 1 0
+Рт,П 0 0 2 0 0
—Pm4nW 0 0 0 3 0
PirATIW 0 0 0 2 0
PmfllflW 0 0 0 1 2
Pm4nW Рт3П 0 0 0 0 1
Количество
отклоне- ний, % 1,8 12,5 8,2 24,0 33,3
Примечание. П — правый, Л — левый, ПЛ — одновре-
менно правый if левый зубы, Ч---наличие дополнительного, — отсут-
ствие зуба соответствующей категории; W — ДвухвершиНность одно*
вершинного в «норме» зуба. В показателе, изображенном в виде дроби,
в числителе — изменения в верхней челюсти, в знаменателе — в ниж-
ней.
Возрастная изменчивость зубов сводится к снашиванию их свобод-
ных концов у резцов, клыков и премоляров и стиранию и сглаживанию
жевательной поверхности коренных. Резцы, имеющие лопатовидное рас-
ширение от основания к режущей поверхности, у молодых зверьков обра-
зуют плотный, почти сплошной ряд. С изнашиванием широкой части рез-
цов их основания в виде пеньков оказываются отделенными друг от друга
небольшими диастемами (рис. 24).
Популяционная изменчивость, кроме того, выражается в наличии у
Т. е. transuralensis, заселяющего самую восточную часть ареала, талона и
зачаточного гипоконуса у четвертого верхнего предкоренного (Рш4),
не встречадмцихся у других подвидов.
54
Таблица 7
Морфометрическая характеристика подвидов Talpa еигораеа
Признак М тх п а cv lim
1 2 3 5 6 7
а. Т. е. еигораеа (по'. [Grulich, 1969]), Центр, и Вост. Европа
Длина тела с головой хвоста ступни 126,4 36,8 18,2 0,26 0,16 0,08 909 907 903 7,89 4,87 2,26 6,24 13,2 12,43 —
Масса тела Длина 69,1 0,59 904 15,47 22,39 —
кондило-базальная черепа 33,6 0,055 789 1,42 4,23 ——
верхнего ряда зубов верхнего ряда заднекорен- 14,2 0,02 824 0,55 3,88 —-
ных зубов Ширина 4,5 0,01 821 о,з0 6,7 —
скуловая 11,6 0,02 819 0,58 5,02 —
наибольшая черепа 16,0 0,02 808 0,63 3.93 —
Наибольшая высота черепа 10,1 0,02 817 0,44 4,37 —
б. Т. е. еигораеа, Польша (Беловежская Пуща)
Длина кондило-базальная черепа верхнего ряда зубов Ширина 33,7 14,2 0,09 0,03 51 51 0,676 0,229 2,00 1,61 31,7—36,1 13,0—15,0
рострума 4,7 0,03 51 0.195 4,16 4,4-5,3
предглазничная 6,5 0,12 51 0,106 1,63 6,1—7,0
скуловая 11,9 0,04 51 0,360 3.2 11,1—13,6
наибольшая черепа 16,2 0,04 51 0,282 1,73 15,0—17,3
Наибольшая высота черепа 10,5 0,01 51 0,103 0,98 9,0—11,3
Высота коронарного отростка 6,7 0,03 51 0,193 2.89 5,9—7,4
е. Т- е. еигораеа, Костромская обл.
Длина
тела с головой 100,9 0,95 51 0,95 9,4 82—117
кондило-базальная черепа 32,2 0,08 146 0,93 2,7 32,0—36,6
верхнего ряда зубов 14,1 0,04 146 0,43 3,0 13,0—15,0-
верхнего ряда заднекорен- ных зубов 5,9 0,02 145 1,19 3,1 5,1-6,3
Ширина
рострума 4,8 0,02 146 0,26 5,4 4,1—5,5
предглазничная 6,5 0,04 146 0,28 4,3 5,7—6,7
скуловая 11,7 0,05 118 0,39 3,3 10,8-12,5-
наибольшая черепа 16,2 0,04 141 0,46 2,8 15,0—17,1
Наибольшая высота черепа 10,3 0,02 146 0,61 6,0 9,3—10,6
Высота коронарного отростка 6,6 0,04 130 0,29 4,4 6,1—7,1
г. Т. е. uralensis, Юж. Урал, Башкирия
Длина
кондило-базальная черепа 35,0 0,08 334 1,40 3,99 31,6—37,6-
верхнего ряда зубов 14,4 0,02 334 0,41 2,82 13,1—15,4
Ширина
рострума наибольшая черепа 4,8 0,02 334 0,27 5,71 4,1—5,4
16,4 0,03 334 0,53 3,21 15,3—17,8
Наибольшая высота черепа 10,7 0,02 334 0,41 3,78 9,6—11,6
55
Окончание табл. 7
1 I 2 | 3 | 4 | 5 | 6 | 7
д. Т. е. uralensis, Центр. Урал
Длина
тела с головой 133,7 2,76 19 12,04 9,0 111,0—162,0
хвоста 30,6 0,74 19 3,23 10,6 26.0-40,0
ступни 19,4 0,28 19 1,16 6,0 18,0-21,0
Длина
кондило-базальная черепа 34,9 0,3 20 1,34 3,9 33,0—37,5
верхнего ряда зубов 14,5 0,1 20 0,45 3,1 13,8—15,3
Ширина
рострума 4,7 0,08 20 0,33 7,1 4,0-5,2
предглазничная 6,5 0,09 20 0,38 5,8 6,0-7,5
скуловая 11,8 0,14 14 0,51 4,3 11,2—13,0
наибольшая черепа 16,3 0,18 19 0,77 4,8 15,0—18,0
Наибольшая высота черепа 10,7 0,12 19 0,29 4,9 9,7—11,6
Высота коронарного отростка 6,7 0,09 19 0,43 6.5 5,8-7,3
е. Т. е. ob crisis, Сея . Приобъе
Длина тела с головой 130,6 7,6 5 17,14 13.1 114,0—157,0
хвоста 28,5 1,14 5 2.55 8,9 25,5—32,0
ступни 19,6 0,49 5 1,05 5,6 18,1—21,0
Длина кондило-базальная черепа 35,2 0,49 8 1,39 4,0 33,4—37,5 '
верхнего ряда зубов 14,4 0,13 8 0,36 2,5 14,0—15,0
верхнего ряда заднекорен- ных зубов 6,4 0,1 8 0,29 4,6 6,0-6,7
Ширина рострума 4,7 0,1 8 0,29 6,4 4.3-5,1
предглазничная 6,5 0,06 8 0,17 2,7 6,2—6.7
скуловая 11,8 0,19 7 0,52 4,4 11,3—12,4
наибольшая черепа 16,5 0,17 8 0;49 3,0 15,9—17,2
Наибольшая высота черепа 10,3 0,18 8 0,47 4,6 9,7—11,0
Высота коронарного отростка 7,0 0,19 8 0,55 8,0 6,3—7,8
ж. Т. е. transuralensis, Тюменская обл.
Длина
тела с головой 143,8 1,7 12 5,99 4,2 128,0—150,0
хвоста 42,2 1,53 12 5,28 12,5 33,0—49,0
ступни 19,9 0,23 12 0,79 4,0 18,0—21,0
Масса тела Длина 95,0 3,60 12 12,45 13,1 67,0—109,5
кондило-базальная черепа 35,0 0,17 44 1,13 3,2 32,7—36.8
верхнего ряда зубов 14,3 0,07 48 0,44 3,1 12,9—15,0
верхнего ряда заднекорен- ных зубов 6,3 0,03 46 0,2 3,3 5,8—6,6
Ширина
рострума 4,9 0,07 48 0,49 10,1 3,9—5,6
предглазничная 6,7 0,05 48 0,32 4,8 6,1—7,6
скуловая 12,1 0,09 39 0,55 4,6 11,0—13,7
наибольшая черепа 16,9 0,09 44 0,56 3,4 15,5—17,7
Н аиболыпая высота черепа 10,8 0,05 47 0,36 3,4 9,8—11,3
Высота коронарного отростка 7,0 0,06 48 0,38 5,5 6,2—7,6
56
Таблица 7а
Достоверность различий (по Стьюденту) между подвидами Talpa europaea
Признак В —Г в-д в—е в—ж Д-е д—ж е—ж
Длина тела с головой 11,2 3,9 21,7 0,4 3,1 1,7
хвоста — — — — 1,5 6,8 7,2
ступни — — — — 0,4 1,4 0,6
Длина кондило-базальная черепа 3.6 0,9 1,2 2,1 0,5 0,3 0,4
верхнего ряда зубов 6,6 3,7 2,2 2,6 0,6 1,7 0,7
верхнего ряда заднекорен- ных зубов — — 4.7 11,1 — — 0,9
Ширина рострума 0 1,6 0,4 1,4 0 1,9 1,6
предглазничная — 0 0 3,2 0 2,1 2,6
скуловая —• 0,7 0,5 4,2 0 1,8 1,4
наибольшая черепа 4,1 0,6 1,7 7,5 0,8 2,0 2,1
Наибольшая высота черепа 14,1 3,3 0 9,3 1,9 0,8 2,8
Высота коронарного отростка — 0,9 1,9 5,6 1,4 2,7 0
Примечание.
Измерения даны в миллиметрах
и граммах (масса тела).
Кроме того, хорошо прослеживается географическая изменчивость
в изменении величины тела и размеров черепа от более мелких форм на
западе к самым крупным на востоке ареала. Длина тела обыкновенно-
го крота из Центральной и Восточной Европы [Grulich, 1969а] составляет
126,4 (±0,262) мм (табл. 7, 7а) с Урала — 133,7 (±2,762), а из южных
районов Тюменской области — 143,7 (±1,729) мм. Эти данные разли-
чаются с большой достоверностью (Td1_2 = 2,6; Tdx_3 = 10,0; Td2_3 =
= 3,1). К сожалению, выборки по размерам тела кротов с Урала (19 экз).
и из Тюменской области (12 экз.) недостаточны для статистической обра-
ботки. Тем не менее указанная закономерность реально существует,
что подтверждается аналогичной направленностью изменчивости кондило-
базальной длины черепа. В Центральной и Восточной Европе [Grulich,
1969] этот показатель равен 33,6±0,051 мм (при п = 789 экз.), в Польше
(наши данные)— 33,7±0,095 (п — 51), в Костромской области 32,2±
±0,08 (п = 146), Башкирии — 35,0±0,076 (п — 334), в районах Цент-
рального Урала — 34,9±0,301 (п = 20), юг Тюменской области — 35,0±
±0,492 (и = 44) и на севере Приобья — 35,2±0,492 мм (п = 8). Досто-
верность различий крайних вариантов велика (Td = 7,9). Аналогичная
направленность изменчивости прослеживается и по другим признакам.
Заметки по систематике. Видовая самостоятельность
обыкновенного крота не вызывает сомнений. Это один из более четко диф-
ференцированных и примитивных видов рода Talpa. Глаза у него откры-
ты, резцы устроены более просто, в архитектуре черепа выражены юве-
нильные черты [Строганов, 1957]. Что касается объема вида, то он при-
нимается разными авторами весьма неоднозначно. Чаще всего в состав
его произвольно без достаточной аргументации, подкрепленной факти-
ческим материалом, включали не только другие самостоятельные виды,
но и объединяли с видами, которые относятся, по мнению ряда авторов,
к другому роду [Бобринский и др., 1944, 1965].
57
Исследования большого коллекционного материала показали, что
обыкновенный крот обладает хорошо выраженными самобытными черта-
ми. Поскольку в нашу задачу входит исследование систематики видов
кротов, обитающих только на территории Сибири, то мы будем сравни-
вать специфичные структуры только обыкновенного и сибирского кротов,
объединяемых многими авторами в один род Talpa, а иногда и в один
вид Т. еигораеа. Из европейских кротов он ближе всего к Т. соеса. Иссле-
дованный материал из Италии позволяет говорить о близости в строении
их зубной системы и значительном сходстве в строении коронарного
отростка и таза (см. рис. 22, 18).
Н. А. Бобринский и А. П. Кузякин [Бобринский и др., 1944] всех
кротов рода Talpa и сибирского крота (Asioscalops) объединили в один
гетерогенный, сборный вид — Т. еигораеа. Позднее А. П. Кузякин [1966]
частично отказался от этой произвольной трактовки. Во втором издании
«Определителя млекопитающих СССР» в состав рода Talpa им были
включены в качестве видов Т. еигораеа, Т. hercegovinensis, Т. altaica.
В Т. еигораеа в качестве подвидов вошли Т. caucasica, Т. ledantis. Автор
отрицает подвидовую самостоятельность Т. е. uralensis и Т. е. trans-
uralensis.
Подчеркивая специфику сибирского крота, С. У. Строганов [1941]
выделил его сначала в подрод, а позднее [Строганов, 1957] — в отдель-
ный род Asioscalops.
Отличия обыкновенного и сибирского кротов лучше всего воспри-
нимаются при сравнении их отличительных признаков, как в свое время
это сделал С. У. Строганов [1948]. Приводим сравнение диагностических
признаков обыкновенного и сибирского кротов:
Признак
Длина тела, мм
Кондило-базальная
длина черепа, мм
Зубы и отношение (%)
к кондило-базаль-
ной длине черепа
Клык
Рш1
Первый верхний ко-
ренной (М1)
Верхние molaris
Нижние края foramen
infraorbitale
Т. еигораеа
110—160 (М = 133)
32—37,5 (М = 34,6)
Крупные, 41
Не имеет заметного рас-
ширения основания ко-
ронки; продольный
диаметр клыка у осно-
вания не превышает
2 мм.
На его задней стороне
нет полулунной выем-
ки. Зуб саблевидной
формы
Между клыком и Pm1
всегда есть более менее
значительный промежу-
ток
Широкий с хорошо раз-
витым протоконусом
Высокие и крупные: ши-
рина М1 1,7—1,8 мм;
ширина М2 2,1—2,3 мм
Конусы очень высокие с
глубокими впадинами
между ними
Отодвинуты в стороны и
хорошо видны при рас-
смотрении черепа сверху
As. altaica
125—195 (М = 155,7)
30,2—41,4 (М = 36,7)
Мелкие, 38,8
С чрезвычайно расширенным
основанием коронки: про-
дольный диаметр клыка у
основания равен 2,5—3 мм.
На его задней стороне хоро-
шо выраженная полулун-
ной формы выемка. Зуб
крючкообразной формы
Тесно прижат к верхнему
клыку: между ними пет
свободного промежутка
Узкий с очень слабо разви-
тым конусом
Низкие и мелкие: ширина
М1 не превышает 1,2 мм;
ширина М2 до 1,9 мм. Ко-
нусы их сравнительно не-
высокие с неглубокими
впадинами между ними
Не выступают в стороны и
почти не видны при рас-
смотрении черепа сверху
58
Признак
Восходящая ветвь
нижней челюсти
Седалищные кости (is-
chium) в области
вертлюжных впадин
Glans penis
Т. еигораеа
Узкая сильно накло-
нена вперед по отноше-
нию к продольной оси
нижней челюсти
Соединены широкой кост-
ной перемычкой с поз-
воночником: дисталь-
ные концы этих костей
заметно сведены
внутрь, к позвоночнику
Тонкая, слабо кониче-
ская. Ее длина 11,3—
15,5 (М = 13,3) мм.
Покрывающие ее поверх-
ность шипики сидят по
одному или группами
по два
As. altaica
Широкая, ее вертикальная
ось перпендикулярна про-
дольной оси нижней че-
люсти
Не соединены костной пере-
мычкой с позвоночником:
дистальные концы этих
костей параллельны про-
дольной оси позвоночника
Толстая, более коническая.
Ее длина 17—17,5 (М —
— 17,1) мм. Покрывающие
ее поверхность шипики
сидят группами по 2—
4 шт.
Перечисленные признаки со всей убедительностью доказывают ви-
довую самостоятельность обыкновенного и сибирского кротов.
С. И. Огнев [1928] рассматривал Т. еигораеа в объеме, принимаемом
специалистами в настоящее время. Сибирского крота он рассматривал
как самостоятельный вид рода Talpa. Этот автор в составе вида в фауне
СССР различал три подвида: Т. е. еигораеа, Т. е. brauneri, Т. е. uralensls.
Еще Паллас [Pallas, 1811] обратил внимание на то, что уже в Прикамье
обыкновенные кроты крупнее, чем в западной части ареала. На самобыт-
ность крота Зауралья обращал внимание Н. Ф. Кащенко [1905], но не-
правильно трактовал таксономический статус, считал его промежуточной
формой между Т. еигораеа и Т. altaica. Еще в 1925 г. С. И. Огнев кротов,
обитающих в восточных районах европейской части страны, выделил
в подвид уральского обыкновенного крота — Т. е. uralensls, охарактери-
зовав его как более крупную форму, отличающуюся от Т. е. brauneri
по структуре зубов, а от сибирского крота — крупнозубостью и меньши-
ми размерами тела. Хотя кротов из районов Южного и Среднего Зауралья
он не имел, по, ссылаясь на личное сообщение Н. Ф. Кащенко, исследо-
вавшего материалы из этого района, он прозорливо отметил их отличие
от уральского подвида.
В. Н. Скалой и В. В. Раевский [1940] описали подвид обского обык-
новенного крота Т. е. obensis из бассейна р. Малая Сосьва. В его диагнозе
указано, что новый подвид отличается от Т. е. uralensls мелкими размера-
ми и особенностями строения верхних резцов. Исследование материалов
из terra typica показало, что по размерам тела он мало уступает ураль-
скому подвиду, а по кондило-базальной длине черепа — это самая круп-
ная популяция. Строение резцов молодых зверьков не отличается от
таковых других подвидов. Вероятно, ошибка в диагноз вкралась вслед-
ствие исследования старых зверьков со стертыми зубами и незнания воз-
растной изменчивости этого признака (см. рис. 24). В результате стира-
ния широкой части резцов оставшиеся от них «пеньки» оказываются почти
равными по ширине.
Таким образом, подвид уральского обыкновенного крота следует
считать нереальным.
Учитывая, что кроты, обитающие в южных районах Тюменской об-
ласти в бассейнах рек Конда, Тавда, Ишим, Тобол и Демьянка и на вос-
точных склонах Южного и Среднего Урала, относятся к подвиду Т. е. trans-
59
uralensis, то крота, описанного В. Н. Скалоном и В. В. Раевским, вероят-
но, следует отнести к подвиду Т. е. uralensis.
Таким образом, в пределах Сибири обитает один подвид обыкно-
венного крота.
Talpa еигораеа transuralensis Stroganov, 1956 —
Зауральский обыкновенный крот
Голотип. № 6694 (кол. 151), , ad., 27/VIII 1950 г. Коллектор
В. И. Телегин, Тюменская обл., Уватский р-н, д. Соровая (среднее те-
чение Демьянки). Шкурка с черепом. Коллекция зоологического музея
Биологического института СО АН СССР.
II а р а т и п ы. № 6690 (кол. 2304), 27/VIII 1950.; № 6259 (кол.
157), Тюменская обл., д. Кирюхино, 7/VIII 1955, sex?; № 6160 (кол. 170),
там же, 8/VIII 1955, sex? Коллектор В. И. Телегин. Три шкур-
ки с черепом.
Диагноз. Отличается от прочих подвидов обыкновенного крота
наличием талона и зачаточного гипоконуса у четвертого верхнего пред-
коренного (Pm4) (см. рис. 24).
Измерения (мм). Длина тела с головой 128,0—150,0 (М —
= 143,8=*=1,729); хвоста 33—39 (М — 42,2=*=1,527); ступни 18—21 (М —
= 19,9)3. Масса 67—109,5 (7И = 95=t3,595) г. Кондило-базальная длина
черепа 32,7—36,8 (М — 35,0=1=0,171); длина зубного ряда 12,9—15,0
(М = 14,3=t0,065); заднекоренных 5,8—6,6 (М = 6,3=1=0,031); ширина
рострума над клыками 3,9—5,6 (IV = 4,9=+=0,071); предглазничная ши-
рина 6,1—7,6 (IP = 6,7=t0,047); скуловая ширина 11,0—13,7 (М = 12,1);
наибольшая ширина черепа 15,5—17,7 (М = 16,9=±=0,085); высота коро-
нарного отростка 6,2—7,6 (М = 7,0=t0,565).
Распространение. Восточные склоны Южного и Среднего
Урала, бассейн рек Конца, Тавда, низовья Тобола, Ишима, Иртыша
и Демьянки на север до Березовского района.
Материал. 73 экз., бассейн Демьянки, долина нижнего тече-
ния Иртыша, реки Конца, Тавда, Тобол, (Тобольский, Уватский р-ны
Тюменской обл.).
Заметки по систематике. Впервые на самобытные чер-
ты организации этого подвида — наличие на Pm4 талона и зачаточного
гипоконуса у двух исследованных зверьков — обратил внимание
С. У. Строганов [1948], но счел их за индивидуальное отклонение и кро-
тов юга Тюменской области отнес к подвиду Т. е. uralensis. Как уже от-
мечалось выше, С. И. Огнев [1928] обратил внимание на крупные разме-
ры этого крота, но не придал ему подвидового таксономического статуса.
С поступлением новых материалов С. У. Строганов [1956] установил стой-
кость отмеченного им признака, свойственного всей исследованной серии
и описал на этом основании новый подвид зауральского обыкновенного
крота. С. У. Строганов [1957] отмечает, что подобная структура Pm*
характерна для южно-азиатских и дальневосточных кротов родов Mogera,
Parascaptor, Eoscalops и не отмечалась ранее у кротов родов Talpa и Asi-
oscalops.
Поскольку, по указанию С. У. Строганова [1957], кроты, обитающие
севернее ареала зауральского обыкновенного крота и описанные
3 Размеры заимствованы с этикеток коллекторов. Вероятно, при измерении
допущена ошибка и данные этого промера завышены.
60
В. Н. Скалоном и В. В. Раевским [1940] как обской обыкновенный крот,
не обладают названными особенностями строения Pm4, а их подвидовая
принадлежность, установленная названными авторами, признана не-
реальной, их следует рассматривать в составе подвида зауральского
обыкновенного крота.
Род Asioscalops Stroganov, 1941 — Кроты сибирские
Тип рода. Talpa altaica Nik., 1883.
Диагноз. Зубная формула: I С j-; Pm М = 44. Зубы мел-
кие. Коронарный отросток относительно широкий, его вертикальная
ось перпендикулярна продольной оси нижней челюсти.
Характеристика. По габитуальным признакам похож на
представителей рода Talpa, отличается от них большими размерами тела,
укороченной мордой и меньшей длиной хвоста. Глаза видны снаружи
и снабжены подвижным веком.
Череп крупный, кондило-базальная длина его составляет 30,2—41,4
(М = 36,7^±=0,07) мм. В конфигурации черепа обращает на себя внимание
угловатость в затылочной области, значительная вытянутость при до-
вольно уплощенной мозговой коробке и широкий носовой отдел. В скульп-
туре черепа характернр сильное сжатие maxilla снизу, под foramen ан-
teorbitalia и расширение сверху над ним настолько, что нижние края
предглазничных отверстий не видны при рассмотрении черепа сверху.
Коренным образом отличается форма proc, coronoideus. В отличие от
видов рода Talpa он массивный, широкий и его вертикальная ось пер-
пендикулярна продольной оси нижней челюсти (см. рис. 22).
Относительно размеров тела череп сибирского крота несколько мень-
ше (23,6 %), чем у европейского (25,9 %), хотя по абсолютным показа-
телям он значительно крупнее. Достоверно отличается этот вид от евро-
пейского крота и шириной рострума (Td = 2,3), а в предглазничной
области он относительно уже. По наибольшей ширине черепа пропорции
этих видов различимы. По отношению наибольшей высоты черепа к его
кондило-базальной длине видно, что у сибирского крота череп более
плоский.
Зубная формула та же, что и у рода Talpa. Несмотря на большие
размеры тела и черепа зубы сибирского крота мелкие, незначительная
высота их стоит в резкой диспропорции с крупными размерами черепа.
В целом зубная система крайне своеобразна. Редукция molares достигла
таких размеров, что они в подсем. Talpidae не только относительно, но
и абсолютно мельче, чем мелких кротов подсемейств Uropsilinae и Uro-
trichinae [Строганов, 1957]. При средних значениях кондило-базальной
длины черепа сеголеток у Т. europaea — 34,6(^0,304) — длина ряда
коренных зубов равна 6,3(^=0,217) мм, что составляет 18,2 % (см. табл. 3).
В то же время у сибирского крота при средней кондило-базальной длине
черепа 35,8(±0,145) длина ряда коренных зубов равна всего
5,5(^0,025) мм, что составляет только 15,4 %. По результатам обобщен-
ной выборки всех исследованных сибирских кротов (п = 546) при сред-
нем показателе кондило-базальной длины черепа 36,7(±0,07) мм ряд
коренных равен 5,6(=t0,01) мм или 15,1 %.
Верхние резцы относительно короткие и широкие. Верхние клыки
крупные с чрезвычайно расширенным основанием (до 3 мм) и с полулун-
61
ной выемкой на задней стороне основной трети зуба. Верхние предкорен-
ные относительно мельче заднекоренных, довольно крупные и широкие,
хотя и уступают по размерам таковым у обыкновенного крота. Рш1 всегда
плотно прижат к основанию клыка. Заднекоренные зубы мелкие: низкие
и узкие, с невысокими конусами и неглубокими впадинами между ними.
Foramen sacrae дистально не ограничена костным мостом из срос-
шихся os ischium и os sacrae. Дистальные концы os ischium параллельны
продольной оси позвоночника.
Распространение. Западная и Средняя Сибирь, Юго-За-
падное Забайкалье.
Заметки по систематике. Подчеркивая самобытность
сибирского крота, С. У. Строганов [1941] вначале выделил его в особый
подрод Talpa, а позднее [Строганов, 1957] возвел в ранг рода Asioscalops.
Основанием для такого выделения послужил комплекс охарактеризо-
ванных выше признаков (своеобразная структура зубов, строение черепа
и таза). Важное значение С. У. Строганов придавал и такой существен-
ной биологической особенности сибирского крота, как латентная фаза
в развитии оплодотворенного яйца — феномен, свойственный среди пред-
ставителей мировой фауны насекомоядных млекопитающих только си-
бирскому кроту [Бородулина, 1951, 1953; Депарма, 1951, 1953; Баевский,
1967, 1970; Юдин, 1972д, 19746, 1980а.]. Нами отмечается как весьма
существенная структурная особенность — строение восходящей ветви
нижней челюсти [Юдин, 1971а]. У всех видов рода Talpa proc, coronoi-
deus узкий и сильно наклоненный вперед по отношению к продольной
оси нижней ветви челюсти. У сибирского крота он массивный, широкий,
а его вертикальная ось перпендикулярна горизонтальной оси нижней
челюсти (см. рис. 22). Признак этот весьма стойкий и в сочетании с мел-
кими зубами указывает на коренные отличия в структуре всего жеватель-
ного аппарата сибирского крота. Мы придаем им диагностическое зна-
чение родового уровня.
Подтверждением реальности выделения сибирского крота в ранг
рода служит обстоятельство, что специфичные черты, свойственные всем
видам рода Talpa (конфигурация черепа, крупнозубость, строение proc,
coronoideus), не свойственны роду Asioscalops. Это положение представ-
ляется весьма существенным, учитывая, что специфика сибирского крота
касается столь жизненно важных структур, как зубная система, строение
нижней челюсти и таза. Наконец, такая экологическая особенность, как
наличие диапаузы в развитии оплодотворенного яйца, ставит сибирского
крота в совершенно особое положение среди не только представителей
сем. Talpidae, но и всех насекомоядных, подчеркивая самобытный путь
их эволюции.
Нам представляется выделение сибирского крота в отдельный род
вполне обоснованным. В этом качестве он занимает свое естественное
место в системе. Род представлен одним видом.
Asioscalops altaica Nikolsky, 1883 — Сибирский крот
Г о л о т и п. Вид описан по экземпляру из окрестностей д. Тоу-
раки, Алтай.
Диагноз. Диагностические признаки вида те же, что и рода.
Измерения. Морфометрическая характеристика сибирского кро-
та приведена в табл. 3, 5, 8. Как можно видеть из данных таблиц,
62
Таблица 8
Морфометрическая характеристика Asioscdlops altaica
Возрастная и половая группа ст Су п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые взрослые 155,0+2,36 162,4+1,30 13,75 11,68 8,9 7,2 34 81 135—195 143—190
Самки прибылые 147,5+2,73 10,57 7,2 15 132—173
взрослые 152,6+2,07 12,24 8,0 35 136—171
В ц е л о м по виду 155,7+0,97 Длина хвоста 15,29 9,8 251 125—195
Самцы прибылые 24,7±0,84 4,87 19,7 34 19—39
взрослые 25,4+0,58 5,16 20,3 80 16—47
Самки прибылые 24,5+1,79 6,94 28,4 15 17—39
взрослые 25,6+0,80 4,79 18,7 36 20—38
В ц е л о м по виду 25,7+0,37 Длина ступни 5,85 22,8 251 16—47
Самцы прибылые 20,7+0,28 1,58 7,6 33 18—25
взрослые 21,7+0,19 1,72 7,9 81 18—26
Самки прибылые 19,6+0,30 1,15 5,9 15 17—21
взрослые 20,7+0,15 0,89 4,3 36 19—23
В ц е л о м по виду 21,3+0,42 Масса тела 1,66 7,8 250 17—26
Самцы прибылые 116,0+4,82 25,96 22,4 29 60—176
взрослые 148,9+3,55 30,55 20,5 74 83—230
Самки прибылые 93,8+4,44 14,71 15,7 И 70—115
взрослые 107,6+3,13 17,99 16,7 33 72—152
В ц е л о м по виду 125,2+2,27 32,95 23,6 210 60—230
Кондило-базальная длина черепа
Самцы прибылые 36,2+0,20 1,46 4,0 56 33,4—39,0
взрослые 37,8+0,12 1,39 3,7 144 33,5—41,4
Самки прибылые 35,2+0,21 1,45 4,1 46 30,2—38,0
взрослые 36,5+0,15 1,30 3,6 75 34,0—40,5
В ц е л о м по виду 36,7+0,07 1,66 4,5 546 30,2—41,4
Наиболыиая длина черепа
Самцы прибылые 37,0+0,18 1,36 3,7 56 33,8—39,3
взрослые 38,2+0,10 1,28 3,4 144 34,3—41,9
Самки прибылые 35,8+0,22 1,47 4,1 46 31,2—38,6
взрослые 37,0+0,14 1,19 3,2 76 34,8—40,5
В ц е л о м по виду 37,3+0,06 1,52 4,1 538 31,2—41,9
Длина верхнего ряда зубов
Самцы прибылые 14,0+0,06 0,45 3,2 56 13,0—14,9
взрослые 14,6+0,03 0,48 3,3 148 13,3—15,8
Самки прибылые 13,7+0,08 0,53 3,9 46 12,2—14,6
взрослые 14,1+0,05 0,43 3,1 80 12,9—15,1
В ц е л о м по виду 14,1+0,02 0,55 3,9 550 12,2—15,8
63
Окончание табл. 8
1 | 2 | 3 | 4 | 5 | 6
Длина верхнего ряда заднекоренных зубов
Самцы прибылые 5,6+0,04 0,29 5,2 57 5,1—6,5
взрослые 5,7+0,02 0,27 4,7 148 5,1—6,5
Самки прибылые 5,4±0,04 0,24 4,4 46 4,9-5,9
взрослые 5,5±0,03 0,29 5,2 82 4,9-6,2
В ц е л о м по виду 5,6+0,01 0,28 5,1 553 4,8-6,8
Ширина рострума над клыками
Самцы прибылые 5,3+0,05 0,37 7,0 54 4,5—6,2
взрослые 5,8+0,03 0,34 5,9 149 4,5-6,6
Самки прибылые 5,2±0,06 0,37 7,1 43 4,3-6,1
взрослые 5,4±0,04 0,37 6,7 80 4,1-6,6
В ц е л о м по виду 5,5+0,02 0,41 7,4 544 4,1—6,6
Предглазничная ширина
Самцы прибылые 7,1+0,05 0,35 5,0 56 6,0—8,0
взрослые 7,3+0,03 0,33 4,5 149 6,2-8,6
Самки прибылые 6,9+0,05 0,31 4,5 46 6,4—7,9
взрослые 7,0+0,04 0,33 4,7 82 5,8—7,8
В ц е л о м по виду 7,1+0,01 0,37 5,2 561 5,8—8,6
Наибольшая ширина черепа
Самцы прибылые 17,3+0,09 0,63 3,6 55 15,8—18,5
взрослые 17,8±0,05 0,59 3,3 145 16,0-19,2
Самки прибылые 16,8+0,08 0,50 3,0 45 15,6—17,7
взрослые 17,2+0,06 0,52 3,0 77 16,0—19,0
В ц е л о м по виду 17,4+0,03 0,69 4,0 545 15,6—19,2
Наибольшая высота черепа
Самцы прибылые 12,6+0,08 0,50 4,0 43 11,6—13,4
взрослые 13,0+0,06 0,64 5,0 144 8,2—14,2
Самки прибылые 12,2+0,09 0,50 4,1 34 10,8—13,0
взрослые 12,5+0,05 0,40 3,2 63 11,6—13,4
В ц е л о м по виду 12,6+0,03 0,63 5,0 439 8,2—14,2
Высота восходящей ветви нижней челюсти
Самцы прибылые 7,1+0,06 0,45 6,3 56 6,3—8,0
взрослые 7,5+0,04 0,45 6,0 148 5,7-8,6
Самки прибылые 6,7+0,06 0,37 5,5 45 5,8—7,6
взрослые 7,0±0,05 0,43 6,2 81 5,6-8,4
В ц е л о м по виду 7,2+0,02 0,52 7,2 547 5,6-8,6
отличается
от обыкновенного
сибирский крот с большой достоверностью
по размерам тела, черепа и отдельных его структур.
Характеристика. Как уже говорилось при описании рода,
во внешнем облике сибирского крота обращают на себя внимание крупные
размеры тела, крепкое телосложение и укороченная, толстая ростральная
часть головы. Самым характерным признаком, по которому он отличается
64
от всех других представителей сем. Talpidae, является качественное
различие, проявляющееся в своеобразной форме и мелких размерах зубов
по отношению к крупному черепу. В отличие от видов рода Talpa сибир-
ский крот имеет самобытное строение восходящей ветви нижней челюсти.
Кроме признаков, указанных в характеристике рода, он отличается и
по ряду других метрических показателей (см. табл. 3). Отличия сибир-
ского и обыкновенного кротов подробно рассмотрены в видовом очерке
последнего.
Головка копулятивного органа своеобразного строения. В плане она
коническая, массивная, толстая у основания и постепенно сужающаяся
к концу. В последней трети конец glans penis тонкий округлый. В целом
тело копулятивного органа слегка сжато в латеральном направлении.
В отличие от glans penis обыкновенного крота шипы покрывают всю ее
поверхность, кроме тонкого свободного конца и продольного валика
вдоль вентральной стороны, распределяются они реже, размеры имеют
более крупные, образуя группы по 2—4 в ряд (см. рис. 23, б). Длина
головки значительно больше, чем у обыкновенного крота, и составляет
17—17,5 (Л7 = 17,1) мм, при толщине у основания 3 мм.
Распространение. Ареал сибирского крота простирается
на восток от Иртыша, занимая обширную территорию Западной и Средней
Сибири и Южного Забайкалья. Западная граница ареала точно не уста-
новлена. В бассейне Оби крот заселяет территории, прилегающие к рекам
Васюган, Нюролька, Чижапка, Чузик, Парабель, Чая, Шегарка, Тым,
Кеть, Чулым, Томь [Строганов, 1957; Лаптев, 1958].
В Западной Сибири северная граница идет южнее впадения Иртыша
в Обь. Восточнее поднимается до верховьев Таза, на Енисее — до Ени-
сейской губы, что значительно выше Северного полярного круга. На
востоке граница идет по Нижней Тунгуске до Вилюя и верховьев Лены,
спускаясь до Байкала, где крот заселяет районы Прибайкалья и юго-
западного Забайкалья до р. Селенги.
Южная граница точно не установлена. В северной части Барабинской
и Кулундинской степей крота нет. От Шегарки по р. Бакса крот спускает-
ся до широты с. Колывань (севернее Новосибирска). Здесь он переходит
на правый берег Оби, распространяясь по приобским сосновым борам
до Барнаула [Юдин, 1969а]. На Алтае крот широко расселен как в лес-
ной зоне, так и в южных безлесных районах. Здесь граница его распрост-
ранения уходит в пределы Монголии. Крот широко распространен в горах
Салаира, Кузнецкого Алатау, в Саянах.
Материал. 2600 экз. Из таежной и лесостепной зон Западной
Сибири, из Салаира, Кузнецкого Алатау, Горной Шории, Алтая, Запад-
ного и Восточного Саяна, Прибайкалья и Забайкалья, а также ряда райо-
нов Средней Сибири и Якутии.
Изменчивость. Мех сибирского крота сравнительно длинный
и пушистый. Зимний мех гуще и длиннее летнего, длина его 11—12,5 мм
по сравнению с более коротким летним (5,5—6,5 мм). Окраске волосяного
покрова свойствен широкий диапазон индивидуальной и популяцион-
ной изменчивости. Индивидуальная изменчивость весьма значительна и
сводится к появлению цветовых аберраций, проявляющихся в виде час-
тичного или полного альбинизма, хромизма и пятнистости. Наиболее
часто она выражается в отклонениях интенсивности окраски. Среди цве-
товых отклонений чаще всего встречаются зверьки с желтовато-охристы-
ми пятнами, как правило, локализованными в области вентральной части
65
шеи, груди, живота. Тем не менее такие отклонения встречаются единично
на сотни исследованных зверьков.
Характеризуя географическую изменчивость окраски меха сибир-
ского крота, С. У. Строганов [1957] указывает, что она имеет закономер-
но выраженный характер: «Серии исследованных сибирских кротов по
окраске представляют два хорошо различимых типа, причем каждый
из них в свою очередь распадается на темноокрашенные и светлоокрашен-
ные формы, локализированные географически» (с. 66). Цитируемый ав-
тор прав лишь отчасти. Действительно, на большом серийном материале
(нами просмотрено около 1 тыс. экз.) более темные формы преобладают
в районах Салаира, Кузнецкого Алатау, Саян, т. е. в горных районах
юга Сибири. Однако среди них обнаруживается вся гамма переходов
как резко выделяющихся отдельных особей серого цвета (аберраций),
так и постепенные переходы от темных к более светлым формам с менее
насыщенными бурыми тонами. Вопреки утверждению С. У. Строганова,
среди сравнительно светлоокрашенных дымчато-бурых саянских кротов
(As. a. suschkini) иногда попадаются темноокрашенные особи, весьма
близкие к алтайскому кроту (As. a. altaica), или дымчато-серые экземпля-
ры, не отличающиеся от салаирского крота (As. a. salairica).
На Алтае, Салаире и в Саянах преобладают темноокрашенные особи,
но по степени насыщенности тонов не отличаются друг от друга. Самые
темные из них добыты на Салаире и в окрестностях Новосибирска. Особи
дымчато-серые встречаются в популяциях салаирских кротов, но они
не являются преобладающими. Причем в разное время года и по годам
эти соотношения меняются, так как молодые зверьки окрашены в более
светлый и тусклый наряд по сравнению со взрослыми разновозрастны-
ми зверями.
Можно с некоторой условностью выделить лишь два преобладающих
типа окраски: темный с коричнево-бурыми тонами, охватывающий кротов
из горных районов юга Сибири, и более светлый с сероватым оттенком,
приуроченный преимущественно к равнинным районам Западной и Сред-
ней Сибири, до бассейна Лены включительно. Однако абсолютизировать
их нельзя. Прав был С. У. Строганов [1957], который указывал, что из-
менчивость окраски меха сибирского крота весьма значительна и нередко
в популяциях одного подвида проявляется тип окраски, свойственный
представителям другого подвида. Подведя итог сказанному, можно за-
ключить, что проведенное ранее выделение подвидовых категорий у си-
бирского крота по различиям в окраске при изучении новых обширных
материалов не подтвердилось. Окраска варьирует в широких пределах
(рис. 25).
Колориметрический анализ окраски меха кротов из значительно
удаленных участков ареала вида, относящихся, по мнению С. У. Стро-
ганова, к разным подвидам, показал, что достоверных различий в их
окраске нет (см. рис. 25, б), их показатели в значительной мере перекры-
ваются. Достаточно указать, что на больших сериях кротов, собранных
в окрестностях Новосибирска в 1971—1972 гг. и относящихся к подвиду
As. a. altaica, показатели окраски практически перекрывали таковые
других «подвидов». По размаху изменчивости показателя белизны и от-
тенка кроты имеют самый широкий спектр — от 0,8 до 4,1. Только 2 экз.
из всех исследованных зверьков (что составляет лишь 1,5 %) из разных
районов не укладывались в пределы полученных характеристик. Это
демонстрирует довольно широкий диапазон изменчивости окраски у си-
5 Б. С. Юдин
66
белизна
Рис. 25. Колориметрическая характеристика сибирского крота.
а: 1 — в целом для вида; 2 — As. alt. altaica (Алтай); 3 — As. alt. salairica (Присалаирье); 4 —
As. alt. gusevi (Хамар-Дабан); 5 — As. alt. salairica (равнинные районы юга Зап. Сибири);
б: 1 — As. alt. altaica (Телецкое озеро); 2 — As. alt. altaica (Томская обл.); 3 — As. alt. altaica
(Кожевникове); 4 — As. alt. salairica (окр. Новосибирска); 5 — As. alt. gusevi (Забайкалье).
бирского крота и отсутствие четкой географической приуроченности
этой изменчивости.
Методом колориметрирования не исследованы зверьки, относящиеся
к подвиду Xs. a. tymensis, и выделены на основании светлой окраски меха
и мелких размеров. Имеющийся в нашем распоряжении материал, в том
числе и типовая коллекция, послужившая Н. Ф. Егорину [1936] основа-
нием для описания подвида, из-за длительного хранения выцвел и для
изучения окраски не пригоден. Что касается размеров зверьков, то эти
кроты по некоторым средним показателям значительно отличаются от
других подвидов (табл. 9).
В то же время, анализируя колориметрические характеристики,
нельзя не отметить существенное отличие отдельных популяций по диа-
пазону изменчивости. Так, серия с юга Томской области (Кожевников-
ский р-н) имеет довольно монотипную окраску. Показатель белизны у
зверьков этой серии колеблется между 1 и 1,6, а оттенка — между 120
и 138,9. Вторая серия — из Томского района и бассейна р. Кеть (более
северного) — имела значительно больший разброс показателей (0,8—3,0
и 113,2—165,8), причем определенно намечается сдвиг в сторону увели-
чения белизны. Крайние варианты отклонения чаще всего характерны
для отдельных оербей.
В целом для вида колориметрические значения показателей близки
к таковым обыкновенного крота (см. рис. 21). При этом меньший размах
изменчивости окраски у обыкновенного крота можно объяснить неболь-
шими по территории районами и малыми размерами выборки (в 5 раз
меньшей, чем по сибирскому кроту).
В довольно широких пределах варьируют размеры тела и черепа.
Молодые кроты, рожденные в апреле — мае, уже в июне переходят к
67
Рис. 26. Детеныши одного выводка сибирского крота.
самостоятельной жизни [Юдин, 1972а, 1974в], имея массу около 60—
100 г. К осени они достигают размеров 100—150 г. В возрасте одного года
зверьки достигают максимальных размеров, и в последующие годы
их размеры остаются стабильными. Масса зверьков в возрастной
выборке 1—5 лет колеблется у самок — от 72 до 152, у самцов —
от 83 до 230 г.
Возрастная изменчивость крота выражается в изменении размеров
тела, окраски, конфигурации и размеров черепа, а также его отдельных
элементов.
Детеныши у сибирского крота рождаются в апреле — мае голыми,
слепыми и не способными к самостоятельной жизни без длительного вы-
кармливания материнским молоком [Юдин, 1972а]. Максимальная длина
тела эмбрионов, зафиксированная нами, равна 40,5 мм, а максимальные
размеры найденных в гнезде детенышей составляют 42,5 мм (рис. 26).
Следовательно, размер новорожденных находится в интервале между
этими двумя величинами. Эмбриональное и постэмбриональное развитие
и рост сибирского крота описаны нами ранее [Юдин, 1972а, 1974в]. Мак-
симальная масса эмбриона (без оболочки) составляет 3,8 г, а минимальная
масса детеныша из гнезда — 4,3 г. С полной уверенностью можно ска-
зать, что сибирский крот при рождении в длину несколько более 40 мм
и весит около 4 г. Тело новорожденных светлого телесного цвета. Кожа
сухая, в многочисленных поперечных складках, полностью лишенных
волосяного покрова. У новорожденных детенышей нет когтей и зубов,
но их внешность уже типично «кротовая». Все пропорции тела и формы
близки к таковым взрослых зверей, отличается лишь укороченная лице-
вая часть головы. Еще в утробе матери передние конечности уже «вы-
вернуты» в стороны и с расширенными кистями и укороченным плечевым
отделом, характерным для роющих форм; задние — типичные конечности
бегательного типа.
Таблица 9
Морфометрическая характеристика «подвидов» Asioscalops altaica по смешанным выборкам зверьков (разного пола и возраста)
Признак As. a. altaica As. a. salairica Аз. а. suschkini
М±т п а Cv lim М±т п О Су lim М±т п О Су lim
Длина тела с головой 155,6+1,92 52 13,82 8,86 120195 157,5+1,21 167 15,70 9,96 99-182 153,6+5,81 7 15,37 10,0 126-178
хвоста 23,3+0,67 52 4,86 20,85 16-39 26,8+0,47 167 6,0 22,6 17-47 29,6+1,98 7 5,23 17,67 22-39
ступни 20,3+0,25 52 1,78 8,76 17-25 22,1+0,62 166 8,02 36,5 19—26 20,7+0,49 7 1,19 6,22 19—22
Масса тела 116,9±5,53 28 29,25 25,02 80-196 128,9+2,71 144 32,50 25,21 60-230 124±14,64 6 35,87 28,77 99-180
Длина кондило-базаль- ная черепа 36,2+0,15 115 1,58 4,36 33,2-40,7 37,0±О,О8 390 1,58 4,25 32-41,4 36,0+0,29 27 1,50 4,16 34,1-39,1
наибольшая че- репа 36,9+0,13 107 1,39 3,76 34,4-40,9 37,5+0,07 390 1,67 4,45 31,2-41,9 36,7+0,28 28 1,50 4,09 34,7—39,7
верхнего ряда зубов 14,0+0,05 108 0,48 3,72 13,0-15,5 14,2+0,03 401 0,53 3,73 12,2-15,8 14,0+0,10 28 0,52 3,71 13,3-15,2
верхнего ряда заднекоренных зубов 5,5+0,03 108 0,26 4,72 5,0-6,0 5,7+0,01 404 0,29 5,09 4,9-6,8 5,4+0,05 28 0,29 5,4 5,1-6,1
Ширина рострума 5,4±0,04 109 0,37 6,85 4,5-6,4 5,5±0,02 395 0,41 7,45 4,1-6,6 5,3+0,08 28 0,42 7,92 4,7-6,1
предглазничная 6,9+0,04 116 0,40 5,79 6,2-8,5 7,1+0,02 404 0,35 5,07 5,8-8,1 7,0+0,09 26 0,51 7,29 6,4-8,6
скуловая 12,5+0,07 85 0,65 5,20 11,6-14,3 12,7+0,03 324 0,62 4,88 8,2-14,2 11,2+0,09 28 0,50 4,46 11,6-13,6
наибольшая че- репа 17,2+0,09 75 0,68 3,95 15,2-19,2 17,5+0,03 355 0,64 3,66 15,6-19,2 17,3+0,19 14 0,71 4,10 16,0-18,2
Наибольшая высота черепа Высота восходящей 11,2±0,04 112 0,43 3,83 10,3-12,0
ветви нижней че- люсти 7,1+0,05 75 0,54 7,60 5,7-8,6
11,0±0,02 399 0,41 3,72 9,0-12,2 11,2±0,09 28 0,50 4,46 10,3-11,8
7,2+0,03 399 0,50 6,9 6,0-8,7 7,1+0,15 13 0,54 7,60 6,3-8,3
Окончание табл. 9
As. a. tymensis Ав. a. gusevi
Признак М±т 71 а Cv lim М±т п а Су lim
Длина
тела с головой 144,0±2,00 25 10,0 6,94 117—160 153,3+2,32 16 9,27 6,01 140-165
хвоста 22,0+0,42 25 1,83 8,32 19—26 31,6±1,57 16 6,26 19,8 19-41
ступни 19,5+0,223 25 1,12 5,75 17-22 20,7+0,31 16 1,25 6,03 19-23
Масса тела Длина — — — — — 116,7±9,59 16 38,35 32,86 83-200
кондило-базальная черепа 35,9+0,23 65 1,83 5,10 31,8-40,0 36,1+0,31 22 1,45 4,01 34,1-38,8
наибольшая черепа 36,4+0,21 65 1,66 4,56 32,5-40,0 36,60+0,28 22 1,32 3,60 34,9-39,1
верхнего ряда зубов 13,8+0,07 65 0,58 4,20 12,8—15,8 13,9±0,11 22 0,54 3,88 12,7-14,8
верхнего ряда заднекоренных зу- бов 5,5±0,03 65 0,26 4,72 4,8-6,0 5,4+0,03 22 0,14 2,59 5,1-5,6
Ширина
рострума 5,3+0,05 65 0,42 7,92 4,4-6,1 5,5+0,08 22 0,37 6,73 5,0-6,4
предглазничная 7,0+0,39 65 0,31 4,42 6,3—7,8 7,1+0,08 22 0,35 4,93 6,2-7,7
скуловая 12,4+0,08 56 0,62 5,00 11,0-13,5 12,8+0,12 17 0,49 3,83 12,0-13,6
наибольшая черепа 17,1±0,09 61 0,72 4,21 15,0-18,7 17,1±0,15 22 0,68 3,98 16,2-18,7
Наибольшая высота черепа 10,9±0,04 64 0,35 3,21 10,0—11,7 11,2±0,10 22 0,45 4,02 10,0—12,0
Высота восходящей ветви нижней че- люсти 7,0±0,07 60 0,54 7,71 5,5—8,0 7,2±0,12 22 0,57 7,92 6,5-8,6
70
Рост детенышей сибирского крота (по наблюдениям за
Показатель 21/V 22/V 23/V 24/V 25/V
Длина тела, мм 90—92 90—92 92—95 95—100 103—105
Масса, г 32—39 37,3 35,5—40,5 39,5 37,7—42,5 41,4 39,0—44,5 43,2 41,0—47,5 45,6
Примечание. В числителе — предел значений, в знаменателе — средние данные,
Изменение массы тела детенышей сибирского крота
Номер гнезда и ко- личество детенышей в помете 10/V 12/V 14/V 15/V 16/V
№ 44, 3 43,0—59,0 51,6 43,5—59,0 52,5 48,0—65,5 58,3 50,0—65,0 60,0 52,0—70,0 63,1
№ 45, 5 20,5—24,5 23,1 22,5—27,5 25,5 Съедены бар- суком — —
№ 67, 4 — 11,5—14.0 16,7—20,5 18,5—24,0 21,5—26,5
12,8 18,7 21,2 23,7
№ 85, 4 — — 30,0—36,0 32,8 32,5—36,5 34,6 35,0—38,0 36,5
№ 101,5 — — — — —
Глаза на ранних стадиях развития относительно крупнее, чем на
более поздних, у новорожденных они хорошо заметны и просвечивают
сквозь кожу.
В гнезде, найденном 21 мая 1970 г., обнаружено шесть детенышей
длиной 90—92 мм и массой 32—39 г (в среднем 37,3). Зверьки были еще
голыми, но уже не телесного, а темно-серого цвета от темных бугорков
в местах будущих волос. Все тело детенышей имело крупные поперечные
складки. На пальцах уже появились небольшие белесые когти. На сле-
дующий день их кожа стала похожа на замшу — появились едва замет-
ные короткие волосы. Результаты наблюдений за их ростом и развитием
приведены в табл. 10, 11. Начиная с 30 мая некоторые зверьки этого вы-
водка во время очередной раскопки гнезда с целью их измерения и взве-
шивания стали покидать гнездовую камеру, скрываясь в окружающих
ее ходах, но вскоре возвращались обратно.
Постепенно мех детенышей растет, темнеет. Дорсальная сторона тела
серебристо-серая, вентральная долго остается почти голой. Ко времени
выхода из гнезда и перехода к самостоятельной жизни волосяной покров
темнеет и становится близким по окраске таковой взрослых зверьков.
Аналогичная картина изменчивости наблюдалась в гнездовой период
и в других выводках крота. Более детально это описано ранее [Юдин,
1972а, 1974в].
У кротов, перешедших к самостоятельному образу жизни, размеры
тела и черепа варьируют в довольно широких пределах, что связано,
71
Таблица 10
выводком из шести зверьков с 21 мая по 4 июня 1970 г.)
26/V 28/V 30/V 31/V 2/VI 4/VI
103—105 105—110 110—120 110—125 130 125
42,5—48,0 46,6 48,5—53,5 52,0 50,5—65,5 54,1 52,0—58,0 56,2 60,5 59,5
Таблица 11
(по наблюдениям за пятью выводками в мае 1971 г.)
21/V 23/V 26/V 30/V
Перешли к самостоя- тельной жизни — —
38,5—41,0 43,5—46,0 47,5—56,5 73,0—74,0
39,7 44,7 50,2 73,5
Съедены землеройками — — —
— — 14,5—20,2 17,6 Съедены землеройками
с одной стороны, с возрастными и половыми отличиями, с другой — с
особенностями отбора в популяции, особенно в малоснежные и суровые
зимы. Самостоятельно живущие зверьки разного возраста различаются
по длине тела (см. табл. 8), хотя сеголетки к концу года могут достигать
размеров взрослых. Наименьшие размеры взрослых зверьков бывают
меньше наибольших размеров сеголеток.
Особенно отчетливо возрастная изменчивость проявляется в массе
тела. Средняя масса прибылых зверьков (самцы и самки вместе) состав-
ляет 107,3(±3,47) г, перезимовавших одну или несколько зим —
135,9(±2,98) г. Еще больше возрастные различия при сравнении крайних
показателей. У прибылых зверьков (самцы и самки вместе) масса колеб-
лется от 60 до 176 г, а у зверьков в возрасте 1—5 лет — от 72 до 230 г.
В последнем случае минимальная масса относится к самкам в возрасте
1 года, добытым в начале апреля второго сезона их жизни. С развитием
эмбрионов масса самок быстро возрастает.
Если размеры и масса тела позволяют фиксировать значительные
возрастные изменения, то длина ступни у прибылых (20,7±0,28; 19,6 ±
±0,30) и у взрослых (21,7±0,19; 20,7±0,15), а также хвоста (24,7±0,84;
24,5±1,79) и (25,4±0,58; 25,6±0,80) у этих групп животных самцов и
самок соответственно достоверно не различается.
К концу лета у родившихся весной зверьков, особенно прибылых
самцов, длина тела, размер хвоста и ступни достигают размеров, близких
к таковым взрослых, значительно уступают они им лишь в массе. Один
12
из помеченных в гнезде детенышей был вторично пойман в возрасте одного
года, длина его тела равнялась 161,2 мм, масса — 148,3 г; кондило-ба-
зальная длина черепа составляла 36,5 мм.
Такие показатели, как длина ряда коренных зубов, ширина ростру-
ма, предглазничная ширина черепа, скуловая ширина, наибольшая вы-
сота черепа у зверьков разных возрастных групп практически не разли-
чаются (см. табл. 8). Лишь незначительно кондило-базальная и наиболь-
шая длина черепа у перезимовавших самцов превышает этот показатель
у прибылых зверьков: соответственно 37,8.1.0,12 —36,5110,20—36,5±0,20
и 38,2±0,10-37,01-0,18.
Череп формируется в основном в первые месяцы жизни зверьков,
в дальнейшем лишь увеличивается в длину. В возрасте одного года он
достигает максимальных размеров. У молодых зверьков несколько короче
длина зубного ряда.
Зубы у молодых кротов появляются в последние дни пребывания
в гнезде, в возрасте немногим более 1 мес. В период лактационного вы-
кармливания на челюстях зверьков видны лишь прозрачные хрящевые
утолщения, даже отдаленно не напоминающие зубы. Первыми прореза-
ются конусы коренных зубов и Pm4. Несколько позднее появляются
клык, резцы и остальные предкоренные. В первый год жизни зубы кротов
острые, с хорошо выраженными вершинами коренных. У зверька в воз-
расте 1 года зубы были острыми, без заметного притупления вершин.
У двухлетних кротов вершины резцов передне- и заднекоренных несут
явные следы стирания. На клыке выемка с задней стороны заметно «под-
работана». У трехлетних кротов хорошо заметны следы стирания на рез-
цах, они становятся разомкнутыми за счет стирания расширенных частей
свободного конца коронки. Клык заметно утончается за счет стирания
в области задней выемки. Коронки предкоренных сношены примерно
на 1/3 их первоначальной высоты. Конусы на коренных зубах значитель-
но сглажены. В возрасте 4 лет продолжает увеличиваться стирание рез-
цов, они.становятся совершенно разомкнутыми и свободно сидят в зубном
ряду. Клык становится тупее и короче. Вследствие компенсаторного
роста он как бы утолщается, делается массивнее. Переднекоренные стер-
ты почти до половины высоты коронок. На заднекоренных конусы и греб-
ни сильно сглажены.
У кротов в возрасте 5 лет от резцов остаются лишь короткие разомк-
нутые «пеньки». Клык верхней челюсти становится коротким и широким,
напоминающим по очертаниям Рп^. Предкоренные сильно стерты, но
их остатки еще сохраняются за счет компенсаторного роста, вследствие
чего совершенно изменяют первоначальную форму (рис. 27).
Половая изменчивость выражается в меньших размерах тела самок
по сравнению с самцами.
Половые и возрастные различия длины тела у сибирского крота:
Группа п М~т Td
Прибылые самцы 34 155,0 2,358 2,1
самки 15 147,5 2,729
Взрослые самцы 81 162,4 1,297 4,0
самки 35 152,6 2,069
Самцы прибылые 34 155,0 2,358 2,7
взрослые 81 162,4 1,297
Самки прибылые 15 147,5 2,729 1.5
взрослые 35 152,5 2,069
73
Рис. 27. Возрастные изменения в строении зубов сибирского крота (Asioscalops altaica
Nik.).
1—2— до 1,г> мес (гнездовой период); 3—2 мес (переход к самостоятельной жизни); 4—6—8 мес;
5—1 год; 6—2 года; 7—3 года; 8—4 года.
Прибылые самцы крупнее самок, взрослые самцы по этому показателю
достоверно (Td =- 4,0) отличаются от взрослых самок.
Кондило-базальная длина черепа весьма изменчива и колеблется
от 30,2 до 41,4 (М = 36,7 + 0,07) мм (см. табл. 9). В работах предыду-
щих исследователей [Егорин, 1936; Строганов, 1957; и др.] выдвинут
тезис о географической изменчивости длины черепа. По С. У. Строганову
[1957], наиболее крупный он у As. a. suschkini, близок к нему номиналь-
ный подвид, значительно мельче он у As. a. salairica и самый мелкий
у As. a. tymensis и As. a. gusevi, последним в этом ряду можно назвать
4s. a. subsp.— забайкальского крота. В результате исследования боль-
шой серии кротов установлено, что столь четко выраженного проявле-
ния географической изменчивости по этому признаку, как это указано
у С. У. Строганова [1957], нет. Сказанное нами иллюстрируют данные
табл. 9 (приведены средние показатели по всей выборке без подразделе-
ния на возраст и пол) и 15 (размеры только взрослых самцов — от года
до пяти лет).
У As. a. salairica значения кондило-базальной длины черепа наимень-
шие — 32,0 мм, что лишь на 0,2 мм больше, чем у As. a. tymensis, а
наибольшее — 41,4 — максимальная для всех подвидов. В то же время
средние показатели примеров у As. a. altaica, As. a. salairica, As. а.
74
suschkini достоверных отличий не имеют. Только As. a. tymensis досто-
верно отличается от других кротов меньшими размерами.
Подвиды кротов различаются главным образом по длине тела, массе,
длине черепа и зубного ряда. Как показано выше, различия эти не досто-
верны и в данном случае мы имеем дело не с подвидами, а с фенотипами.
Вариабельность признаков у кротов наряду с возрастными и половы-
ми особенностями в значительной мере определяется количественным
соотношением разновозрастных особей, переживших годы с разными
погодными условиями. В неблагоприятные для кротов холодные и мало-
снежные зимы гибнут в первую очередь наиболее крупные особи и попу-
ляция «мельчает». Впоследствии, если погодные условия в течение ряда
лет будут благоприятными, размеры особей популяции восстанавлива-
ются. Поэтому, анализируя какую-либо выборку, нужно помнить, что
мы имеем дело с «временным срезом», отражающим количественное соот-
ношение различных фенотипов и характер погодных условий предыду-
щих лет. При этом средняя величина размерных показателей значительно
колеблется в пределах крайних вариантов, свойственных виду. Она
зависит от количественного соотношения в исследуемой выборке тех
или иных фенотипов. Преобладание мелких особей в популяциях на
периферии ареала мы объясняем большей вероятностью возникновения
там неблагоприятной обстановки для жизни кротов.
Индивидуальная изменчивость зубной системы крота крайне велика.
Нам неизвестно ни одного вида млекопитающего, имеющего столь широ-
кий диапазон изменчивости зубов [Юдин, 1977]. Здесь мы сталкиваемся
с явлением совершенно феноменальным. Поэтому этот аспект изменчи-
вости нами рассмотрен детально.
С целью изучения индивидуальной и популяционной изменчивости
зубов нами исследовано 2402 экз. кротов. Отклонения, проявляющиеся
в изменении числа зубов и их формы, зарегистрированы в 792 случаях
у 478 особей (19,9 % от числа исследованных зверьков), т. е. каждый
пятый крот имел отклонение от «нормального» положения зубов в челюс-
ти. Кроме того, довольно часто встречались отклонения в положении
зубов в челюсти, но они нами не учитывались, поскольку имели посте-
пенные переходы от явных, резких отклонений к едва заметным и норме.
Эта форма изменчивости количественно не охарактеризована. За норму
нами принималась формула зубной системы сибирского крота (см. диаг-
ноз). Что касается изменения формы зубов, то во внимание принимались
только резкие отклонения — двухвершинность у переднекоренных, а
за нормальное положение зубов в челюсти принималось их расположение
у подавляющего большинства исследованных особей.
Исключительно большая изменчивость зубов у сибирского крота —
явление не случайное — проявляется во всем ареале вида, в разных
популяциях, постоянно, из года в год. Причем отдельные популяции
характеризуются качественной и количественной неравнозначностью
изменчивости. Мы рассматриваем это явление как одну из форм прояв-
ления эпигенетического полиморфизма. Если некоторые отрывочные
сведения по изменчивости зубов европейского крота опубликованы [Stein,
1963], то по сибирскому кроту таких сведений нет.
Для изучения изменчивости зубной системы сибирского крота авто-
ром была специально подобрана и исследована обширная коллекция
зверьков этого вида из разных частей его ареала (равнинная часть За-
падной Сибири, Алтай, Салаир, Кузнецкий Алатау, Саяны, Забайкалье,
75
Рис. 28. Зубная система си-
бирского крота.
I1, I2, I3— резцы верхней и 115 Т2*
13— нижней челюсти; С — клыкй;
Pm1, Pm2, Pm3, Pm4— передне-
коренные верхней и Pmt, Pm-,, Pm3,
Pm4 •— нижней челюсти: M*, М2,
М3—коренные зубы верхней и Мо
М.,, М3 — нижней челюсти.
бассейн Енисея, юг Яку-
тии)4, а также изучены со-
ответствующие собрания
в зоологических музеях
Московского и Томского
университетов. Исследо-
ван материал, собранный
в период с 1930 по 1973 г.
Учитывая отсутствие в ли-
тературе детального опи-
сания зубов сибирского
крота, целесообразно дать
краткую характеристику
категорий и отдельных зу-
бов. С. У. Строганов
[1948, 1957] в обстоятель-
ных монографиях по насе-
комоядным не приводит
Л. П. Кузякин [Бобринский и др., 1965] указывает лишь па колебания
числа зубов от 40 до 44.
Характеризуя зубную систему крота, автор оперирует общеприня-
тым обозначением зубов (см. рис. 28). Для удобства введены некоторые
символы. Их значение оговорено в описании изменчивости зубной си-
стемы обыкновенного крота.
Весь исследованный материал из 21 пункта Сибири сгруппирован
в восемь более крупных районов (табл. 12). Поскольку анализ изменчи-
вости зубов у зверьков разного пола не дал достоверных различий, все
зверьки из одного места рассматриваются в целом, без дифференциации
по полу. Материалы, приведенные в табл. 13. под рубрикой «Юг Западной
Сибири», получены из старых сборов ВНИО (Новосибирск), более точной
географической привязанности не имеют (наиболее вероятно, они собраны
на Северо-Восточном Алтае).
Изменчивость зубов верхней челюсти. Наиболее вариабельны пред-
коренные зубы верхней челюсти. Ни разу не отмечена редукция резцов
(I1, I2, I3), редукция клыков (С) и редукция коренных (М1, М2, М3). Это
наиболее константные категории зубов. Только у одного крота отмечена
полифиодонтность клыка и у восьми — полиодонтия коренных (добавоч-
ные М4).
Резцы (I1, I2, I3). В верхней челюсти три пары резцов долотообразной
формы, слегка выпуклых спереди и скошенных к середине (рис. 28). Пло-
ская, расширенная часть свободного конца зуба, постепенно сужаясь,
4 Коллекция хранится в зоологическом музее Биологического института
СО АН СССР (Новосибирск),
“76
Число отклонений отдельных зубов (I) и их отношение к общему числу
Символ зуба Томская обл. Окрестности Новосибир- ска, п=233 Салаир, Куз- нецкий Ала- тау, та=496
северные районы, п=133 южные райо- ны, п=254
I | II 1 1 и 1 1 11 1 1 п
Верхняя
V* Рш1 26 * 48,2 57 36,3 17 25,8 78 43,1
Pm2 9 16,7 2 1,3 — — 4 2,2
Pm3 1 1,9 2 1,3 10 15,2 5 2,8
Pm4 2 3,6 2 1,3 2 3,0 4 2,2
М4 — — 4 2,5 2 3,0 — —
Нижняя
КМз 2 3,6 — — — — — —
Рш2 6 11,1 53 33,8 29 43,9 48 26,5
Рт3 3 5,6 16 10,2 5 7,6 18 9,9
Рт4 5 9,3 21 13,3 1 1,5 24 13,3
Всего 54 100 15'/ 100 66 100 181 100
Отклонения от числа ис-
следованных зверь-
ков, % 40,6 61,1 28,2 38,5
* Указывается не число отклонений левого и правого зубов, а процент по отношению к об
** Левая челюсть сломана, возможен дополнительный М4 с той и другой стороны.
*** В коллекции из этого района часть нижних челюстей утрачена, показатель может быть
приобретает к основанию цилиндрическую форму. Расширенные части
зуба сомкнуты и образуют плотный ряд из мономорфных элементов. Меж-
ду резцами (I3) и клыком (С1) небольшой промежуток, «образованный раз-
нонаправленным изгибом этих зубов. Основания зубов в челюсти сидят
свободно, не касаются друг друга, так как они значительно уже соприка-
сающихся между собой вершин. Вследствие этого в процессе стирания
широких концов зубы у старых особей в возрасте 4—5 лет становятся
разомкнутыми, существенно изменяются по форме, представляя прозрач-
ные, округлые в поперечном сечении «пеньки», свободно сидящие в че-
люсти. В этом отношении изменчивость верхних и нижних резцов сходна.
Первый резец (I1) лишь немного шире и выше двух последующих.
Второй резец (I2) и третий (I3) по ширине почти равны между собой.
Верхние резцы шире соответствующих зубов нижней челюсти. Как уже
отмечалось выше, зубы этой категории малоизменчивы. Ни одного от-
клонения в числе зубов у 2402 экз. исследованных кротов не зарегистри-
ровано. Их изменчивость проявляется в крайне незначительных колеба-
ниях ширины отдельных резцов и их постановки в челюсти.
Клыки (С1). Высокие, кинжаловидной формы, с характерной полу-
лунной выемкой на задней стороне и чрезвычайно расширенным осно-
ванием коронки. Они в 2—3 раза выше зубов,других категорий с чрез-
77
Таблица 12
отклонений (II, %) У сибирского крота из разных частей ареала
Юг Зап. Сибири, п=^692 Алтай, Тува, п—332 Юг Ср. Сибири, Якутия, п=238 Забайкалье, и=24 Итого по районам, «=2402
I 1 п I 1 н I 1 II I 1 И I 1 П
челюсть
—- — 1 1,3 — — — — 1 0,1
75 71,4 40 50,0 20 19,6 16 34,0 329 41,5
1 1,0 1 1,2 4 3,9 1 2,1 22 2,8
6 5,7 9 11,3 2 2,0 2 4,3 37 4,7
3 2,9 3 3,8 3 2,9 — — 19 2,4
1 0,9 1 ** 1,2 — — — — 8 1,0
челюсть
— — — — 2 2,0 3 6,4 7 0,9
7 *** 6,7 10 12,5 33 32,4 8 17,0 194 24,5
2 1,9 6 7,5 16 15,7 13 27,7 79 10,0
10 9,5 9 11,2 22 21,5 4 8,5 96 12,1
105 100 80 | 100 102 | 100 47 100 792 100
15,1 24 42,8 195,8 32,9
щему числу отклонений.
занижен.
вычайно расширенным основанием коронки, по ширине составляющим
около 2/3 его высоты. Вдоль внутренней средней части клык имеет утол-
щение в виде валика, ограниченного спереди глубокой бороздой, по
которой во время жевательного движения происходит соприкосновение
с ним противостоящего клыка нижней челюсти, а сзади постепенно утон-
чается к внутренней режущей поверхности, по которой скользит Pmr.
Клык и верхние челюсти по числу и форме наиболее стабильные из всех
категорий зубов. Только у одного крота оказались два клыка, параллель-
но расположенные в челюсти (полифиодонтность).
Верхние переднекоренные зубы (Pm1, Pm2, Pm3, Pm4). Переднеко-
ренные верхней челюсти — наиболее вариабельная категория зубов,
они подвержены широкой изменчивости как по числу, так и по форме
и постановке в челюсти. Отмечены следующие варианты изменчивости
переднекоренных зубов (рис. 29):
Олигодонтия:
— Pm1 левый или правый
— Pm1 левый и правый одновременно
— Pm1 и Pm2 правые
— Pm2 левый или правый
— Pm2 левый и правый одновременно
— Pm3 левый или правый
78
— Pm3 левый и правый одновременно
— Pm4 левый или правый
— Pm4 левый и правый одновременно
— Pm1 и Pm2 левой и правой челюсти одновременно
Д вухвершинность:
Pm2 и Pm3 правой стороны
Pm2 левый и Pm3 левый и правый
Pm3 и Pm4 правые
Полиодонтия:
4- Pm2 левый
+ Pm3 левый или правый
+ Pm3 левый и правый одновременно
+ Pm4 левый или правый
+ Pm4 левый и правый одновременно
- j- Pm4 правый и Pm4 правый двухвершинный
Изменчивость формы.
Предкоренные зубы крайне вариабельны не только по числу зубов,
но и по форме. Первые три предкоренных довольно сходны по высоте и
конфигурации (см. рис. 28). Они одновершинные, копьевидной формы,
с едва заметным изгибом вершины внутрь ротовой полости. Первый из
них чаще более строен и несколько напоминает клык в миниатюре. Вто-
рой чаще всего уступает по высоте и ширине первому и третьему, но
нередко равен им. Третий мало чем отличается от двух предыдущих,
но, как правило, выше второго. Отмечены следующие вариации:
Рис. 29. Положение зубов на челюсти при полиодонтии и олигодонтии (а) и варианты
полиодонтии переднекоренных верхней челюсти (б).
1 — характерное положение переднекоренного верхней челюсти при полиодонтии; 2 —• Pm, пред-
ставлен двумя зубами, в результате Pm, вытеснен из ряда; з — за счет редукции Pm* второй зуб
Риг2 смещен вперед; 4 — четвертый; 5 — третий переднекоренные.
79
1. Нормальная форма зубов, присущая большинству исследованных
особей (см. рис. 28).
2. Недоразвитость одного из зубов, едва видного над челюстью.
3. Вытянутость зубов в высоту в виде стройных, острых конусов.
4. Смещение вершин к передней трети зуба.
5. Двухвершинность варьирует от едва заметной дополнительной
вершины до полного разделения, имеющих лишь общее основание. Про-
является только у Pm3 и Pm4.
6. Изменчивость положения зубов в челюсти. Наряду с указанными
выше отклонениями отмечается изменчивость в положении зубов. С одной
стороны, это явление часто связано с редукцией или полиодонтией, с
другой — оно проявляется и при нормальном числе зубов.
При редукции одного или нескольких зубов наблюдается смещение
оставшихся. Так, при редукции первого предкоренного второй и третий,
как правило, посажены более свободно и как бы сдвинуты вперед
(см. рис. 29). Аналогичная картина наблюдается и при редукции других
зубов. Однако такое смещение происходит не всегда. Надо полагать,
что это обусловлено временем начала торможения роста редуцировав-
шегося зуба на ранних стадиях эмбрионального развития, в этом случае
другие зубы располагаются в челюсти более свободно. При полиодонтии,
напротив, зубы посажены плотно, что нередко вызывает их отклонение
от нормального положения. При нормальном числе зубов в верхней
челюсти у подавляющего большинства кротов продольная ось передне-
коренных зубов (особенно Pm1, Pm2, Pm3) почти параллельна продоль-
ной оси челюсти, с едва заметным отклонением у Pm3 и довольно замет-
ным у Pm4. Задние концы последних расставлены шире, чем передние
(см. рис. 29,6). При полиодонтии часто наблюдаются изменения положе-
ния зубов в челюсти. Из-за появления «лишнего» элемента в зубном ряду
становится тесно и продольная ось отклоняется, располагаясь под углом
к челюсти. Такие изменения варьируют в широких пределах, и угол
отклонения зависит от сместившихся зубов — одного, двух, трех в од-
ной челюсти или в двух одновременно.
Эта категория зубов варьирует в широких пределах как по числу
случаев, так и по форме проявления. На их долю приходится 98 % абер-
раций. Ниже приводится характеристика изменчивости отдельных зубов.
Первый верхний предкоренной Pm1. Это самый вариабельный эле-
мент зубной системы крота. Из 792 исследованных случаев отклонений
от нормы (олиго-, полиодонтия, двухвершинность; положение в челюсти
не учитывалось) на долю этого зуба приходится 329 (41,5 %). Чаще всего
это — редукция зубов, зарегистрированная в 310 случаях, в том числе
в 71 одновременно в нижней и верхней челюстях. Наряду с этим имели
место случаи смещения продольной оси зуба по отношению — челюсти
а также отклонения в форме зуба. Случаев двухвершинности у Pm1 не
отмечено.
Второй верхний предкоренной (Pm2). Как правило, этот зуб несколь-
ко ниже Pm1 и Pm3, реже все три зуба равны по высоте. Из 2402 исследо-
ванных зверьков изменчивость этого зуба зарегистрирована лишь у
14 экз. в 22 случаях из 792 (2,8 %), из них олигодонтия в 2 раза пре-
обладает над полиодонтией.
Третий верхний предкоренной (Pm3). Этот зуб несколько выше,
и, как правило, шире второго. Существенным отличием в характере
изменчивости представляется наличие двухвершинности этого зуба.
80
Кроты с изменчивостью Pm3 зарегистрированы в 37 случаях (4,7 %),
в том числе у шести зверьков олигодонтия одного зуба, полиодонтия у
23, трех из них одновременно в левой и правой челюстях. Двухвершин-
ность отмечена пять раз.
Четвертый верхний предкоренной (Рш4). Самый крупный предко-
ренной зуб с мощной вершиной, находящейся на одном уровне с про-
токонусами коренных зубов. Коронка расширена в продольном направ-
лении пропорционально величине зуба. Талон отсутствует, гипоконус
зачаточный. По форме почти не отличается от предыдущего зуба. Pm4 ма-
ло подвержен изменчивости. Полиодонтия отмечена у восьми зверьков,
в том числе два раза в той и другой челюсти одновременно. Двухвер-
шинность отмечена у девяти зверьков. Из 792 случаев отклонений на
долю этого зуба приходится 19 (2,4 %).
Подводя итог сказанному, следует подчеркнуть, что редукция зубов
челюсти идет главным образом за счет Рш1, а полиодонтия появляется
преимущественно у Рш3.
Коренные зубы М1, М2, М3. Коренные зубы сибирского крота тубер-
куло-секторального типа, относительно мелкие и низкие. По размерам
самый крупный — первый моляр, второй уступает ему в длине, но зна-
чительно шире. Протоконус развит слабо. Пара- и метаконусы более вы-
сокие, чем на последних коренных. W-образное строение менее выражено.
Второй коренной с хорошо развитым талоном и мощным протоконусом
почти одинаковы в длину и ширину. Жевательная поверхность его более
четко выраженной W-образной формы по сравнению с первым коренным.
Как и у предыдущего зуба, гребни, соединяющие конусы, хорошо разви-
ты, но конусы М2 ниже, чем у первого коренного. Третий коренной
рудиментарен, он в 2 раза короче М1, имеет низкие конусы и неглубокие
впадины между ними.
Эта категория зубов весьма константна по числу. Среди исследо-
ванной нами серии редукция зубов не зарегистрирована. Только у одного
крота с Алтая, из района Телецкого озера, обнаружено нарушение,
являющееся, вероятно, следствием травмы, полученной в раннем возрасте.
Повреждена скуловая и лобная кости. Кость на месте М2 и М3 гладкая,
без следов корней. Зубы выросли в противоположную сторону в пере-
вернутом на 180° положении, в глазницу. В восьми случаях (1,0 %)
отмечена полиодонтия в ряду моляров, связанная с появлением допол-
нительного зуба позади М3, из них в трех случаях дополнительно зубы
были одновременно в левой и правой челюстях у зверька с четырьмя
коренными в одной челюсти, вторая челюсть была сломана и установить,
был ли в ней дополнительный коренной зуб не удалось. В другом случае
дополнительный моляр был с одной стороны (справа). Дополнительный
коренной зуб рудиментарен как по размерам (0,6—0,8 мм), так и по форме
(рис. 30). Он имеет только один корень, слабо развитый талон с рудимен-
тарным протоконусом. Наиболее четко выражен протоконус, метаконус
значительно уступает ему в размерах и сглажен, а иногда совсем
зачаточен.
Появление М4 мы расцениваем как проявление атавизма, выражаю-
щееся в частичной реализации более многочисленных закладок эмбрио-
нальных зубов. Это положение согласуется с мнением Aruback Chris-
tie-Linde [1912], указывающего, что у эмбрионов землероек закладыва-
ется число зубов больше, чем их имеется у взрослых зверьков. В ходе
эмбрионального развития часть из них редуцируется. Уместно отметить,
81
ветви нижней челюсти.
что эта точка зрения не общепризнанная, она встречает серьезные возра-
жения [Kindal, 1959].
Кроме указанных случаев изменчивость коренных зубов проявля-
ется в крайне незначительных отклонениях положения и высоты кону-
сов, конфигурации соединяющих их гребней. Возрастная изменчивость
весьма значительна. В возрасте до года следов стирания зубов почти не
видно (см. рис. 27), у старых особей в возрасте 4—5 лет коренные зубы
настолько изнашиваются, что иногда коронка их полностью стирается
и от них остаются лишь «пеньки» корней, выдающиеся из челюсти в ре-
зультате компенсаторного роста или резорбции костной ткани челюсти.
Судя по числу случаев изменчивости зубов, эти процессы в верхней
челюсти идут более интенсивно (52,5 %), чем в нижней (45,5 %). Однако
как уже отмечалось в примечании к табл. 14, коллекционные материалы
из одного района были получены автором с частично утраченными ниж-
ними челюстями, что могло в какой-то мере повлиять на общую оценку
интенсивности процессов изменчивости зубов нижней в верхней челюсти.
Изменчивость зубов нижней челюсти. Число зубов нижней челюсти
равно таковому верхней и представлено зубами тех же категорий и их
одинаковым соотношением (см. рис. 28). Наиболее примечательной осо-
бенностью является смена функций у клыков и первым предкоренным.
Клык нижней челюсти выполняет функцию резца и имеет сходную с
ним форму и размеры. Ргп, выполняет функцию клыка и резко выделя-
ется по размерам среди зубов этой категории.
Резцы нижней челюсти 1Х, 12, 13. Эта категория зубов похожа на
таковую верхней челюсти, но они длинные и посажены более наклонно,
образуют полукруг из плотно прижатых друг к другу долотообразных
вершин. Соприкасаясь с первым резцом, второй заходит за него внутрь,
внешняя сторона последнего накрывает внутреннюю сторону резца. В то
же время второй резец находится в таком же отношении к третьему.
6 Б. С. Юдин
82
Клык мало чем отличается от резцов. На его внешней стороне заметен
гребень, придающий зубу трехгранную форму в поперечном сечении.
В отличие от клыка резцы имеют долотообразную форму и не несут на
внешней поверхности гребень. Три пары резцов и пара клыков, сходных
по форме, образуют плотную, мономорфную группу, обрамляющую пе-
реднюю часть нижней челюсти (см. рис. 30). По форме резцы довольно
однотипны. Их вершины долотообразно расширены и асимметрично
скошены кнаружи. Они постепенно сужаются к корню, приобретая
цилиндрическую форму. Если рассматривать зубы левой челюсти спере-
ди, то левая (внутренняя) более утолщенная сторона каждого зуба по-
степенно утончается кнаружи. Спереди они имеют выпуклую поверх-
ность и слегка вогнуты с лингвальной стороны. По ширине резцы после-
довательно убывают от первого к третьему.
Зубы этой категории малоподвержены изменчивости. Из 2402 иссле-
дованных кротов отмечено лишь семь случаев (0,9 %) отклонения в ко-
личестве резцов. В сборах из Томской области (окр. с. Парабель) у самки
в левой нижней челюсти оказалось не три, а четыре резца (см. рис. 30, в).
Второй резец был парным. Одновременно отмечена редукция Рт2. В этом
же районе пойман самец с парным первым резцом в левой челюсти и ре-
дукцией Рш4 в правой. В первом случае дополнительный резец распола-
гался в одном ряду с остальными зубами, а во втором — параллельно
своему двойнику во втором ряду (полифиодонтность). В Забайкалье
добыт крот с редукцией I2 и два крота с редукцией I3, в Ермаковском
районе Красноярского края — один крот с дополнительным Г2 с левой
стороны.
Изменчивость, выражающаяся в постановке резцов в челюсти, крайне
незначительна и ею можно пренебречь. Возрастная изменчивость сводится
к постепенному стиранию верхней, расширенной части зубов. В резуль-
тате у кротов в возрасте 4—5 лет резцы не смыкаются в сплошной ряд,
а представляют разрозненные «пеньки» корней, имеющих цилиндрическую
форму. Изношенные зубы становятся стеклянно-прозрачными.
Клыки (С4). В отличие от верхней челюсти клык по форме едва отли-
чим от резцов и выполняет их функцию. Он менее уплощен, трехгранный
в поперечном сечении, что обусловлено гребнем, развитым на внешней
поверхности зуба, и несколько вогнут с лингвальной стороны. Иногда
вершина его слегка отогнута кзади или в бок от ряда резцов, но, как
правило, клык плотно прилегает и прикрыт спереди третьим резцом.
Ширина клыка варьирует в незначительных пределах. Между клыком
и Рпц имеется заметный разрыв (см. рис. 30, б).
Клык всегда представлен одной парой, по одному зубу в челюсти,
изменений количества зубов не отмечено.
Переднекоренные нижней челюсти Pmx, Pm2, Pm3, Рш4. Эти зубы
довольно сходны по форме, но резко различаются по величине. Самый
крупный из них Ршц затем идет самый мелкий Рш2, следующий Рш3
несколько крупнее предыдущего, но значительно уступает по размерам
хорошо развитому, крупному Рш4. Это категория зубов за исключением
первого переднекоренного наиболее вариабельна в нижней челюсти как
по числу зубов, так и по их форме. Положение в челюсти предкоренных
зубов, особенно Рш2 и Рш3, несколько отличное от коренных. Их про-
дольная ось находится под углом к оси челюсти. Передний край направ-
лен к лингвальной, а задний — к наружной стороне челюсти. Кроме
того, передний край Рш4 заходит за задний край Рш3, в свою очередь
83
передний край последнего заходит соответственно за Рш2, а у Рт2 —
за Рт4. Нередко коронка одного зуба располагается в челюсти парал-
лельно ее оси.
Первый нижний переднекоренной (Рт4). Pm4 выделяется среди
других зубов этой категории крупными размерами. Он в 2,5 раза шире
Рш2 и Рл13 и даже несколько шире Рш4. Копьевидная форма зуба доволь-
но постоянна. Это один из менее вариабельных зубов. В челюсти всегда
представлен одной парой. При исследовании 2402 экз. кротов отклонений
в числе не обнаружено.
Второй нижний предкоренной (Рш2). По размерам значительно
меньше Рт4 и почти равен Рш3. По форме три первых переднекоренных
имеют большое сходство. Рш2 — наиболее вариабельный зуб нижней
челюсти. Из 792 случаев отклонений в зубной системе кротов 194 (24,5 %)
падает на этот зуб. Редукция зубов чаще всего осуществляется за счет
Рш2. Отмечена редукция левого, правого, левого и правого зубов одно-
временно (10 случаев) и в сочетании с редукцией других зубов. Полио-
донтия за счет появления двух дополнительных Рш2 зарегистрирована
всего один раз, и два раза — появление дополнительного зуба в одной
челюсти. Из 2402 исследованных кротов изменчивость Рт2 отмечена
у 160 экз. (6,6 %), главным образом это редукция зубов. Форма зуба
довольно устойчива. Двухвершинность отмечена только у 3 экз.
Третий нижний предкоренной (Рт3). По размеру, форме и харак-
теру изменчивости близок к предыдущему. Среди исследованных измен-
чивость этого зуба зарегистрирована в 79 случаях (10 %), т. е. более
чем в 2 раза реже, чем у предыдущего. Чаще всего это редукция одного
из зубов, но нередко отсутствуют зубы одновременно в обеих челюстях
или в сочетании с изменчивостью других зубов. Иногда зуб недоразвит
и едва заметен над поверхностью челюсти. Четыре раза встречались
двухвершинные зубы и 8 случаев полиодонтии, в том числе один раз
одновременно в обеих челюстях (рис. 31, 32).
Четвертый нижний предкоренной (Рш4). Среди предкоренных зубов
Рш4 наиболее сложно устроен и значительно крупнее двух предыдущих
зубов. У исследованных кротов изменчивость отмечена в 96 случаях
(12,1 %). Наиболее характерная форма изменчивости этого зуба — полио-
донтия. Дополнительные зубы отмечены в 44 случаях, один раз одновре-
менно в двух челюстях было по два Рш4. Почти столь же часто отмечена
двухвершинность (39 случаев) с разной степенью выраженности от пол-
ного разделения зуба на две вершины, соединенные лишь в области цин-
гулюма, до едва заметной добавочной вершины со всеми переходами
между этими крайними вариантами (см. рис. 31, 32). В трех случаях
двухвершинность Рш4 отмечена одновременно в правой и левой челюстях.
Значительно реже этот зуб подвергнут редукции (13 случаев, в том числе
одновременно в левой и правой челюстях в 4 случаях). Из сказанного
видно, что формы изменчивости Рш4 более разнообразны. В отличие
от предыдущих зубов, редукционный процесс уступает полиодонтии.
В нижней челюсти олигодонтия идет преимущественно за счет второ-
го премоляра, функционально соответствующего Pm1 верхней челюсти.
Полиодонтия свойственна главным образом Рш4, подтверждением чему
служит высокая частота двухвершинности этого зуба, представляющая
завершающую стадию срастания одновершинных зубов.
Изменчивость зубов одновременно верхней и нижней челюстей. Сте-
пень отклонения в зубной системе крота проявляется по-разному и коли-
6*
84
а б
Рис. 31. Отклонения в количестве зубов и их положении (а) и в числе и форме пе-
реднекоренных нижней челюсти (б).
1 — одновременная редукция Рш1, Рш2 и Рш3, Рш,; 2 — одновременная редукция Рш, в левой
и правой ветви нижней челюсти; 3 — редукция Рта; 4 — смещение Pm,; 5 — в правой ветви предко-
ренные представлены пятью зубами, в левой Pmf— двухвершинный; в и 7 — Pmt и Pm,— представ-
лены двумя зубами.
Рис. 32. Отклонения в количестве и форме переднекоренных зубов нижней челюсти.
1 — Pm4 представлен двумя полностью изолированными зубами; 2 — Рш, разделен почти
до цингулюма на две вершины; 3—5 — разные варианты двухвершинности у Рш4; 6 — Pm. и Рщ»
срослись в один двухвершинный зуб, на Рпц— видна зачаточная дополнительная вершина.
чественно неоднозначна. Чаще всего изменчивость связана с каким-либо
одним зубом. Это — частичная или полная редукция, появление допол-
нительного зуба, отклонения в форме или положении в челюсти. Нередко
такие изменения происходят одновременно в верхней и нижней челюстях.
Довольно часто изменению одного зуба сопутствует отклонение соседнего
зуба или зуба другой категории и даже одновременно в верхней и нижней
челюстях (табл. 13). Ниже приводятся примеры наиболее сложных от-
клонений в числе зубов. В числителе отражена изменчивость зубов верх-
ней челюсти, в знаменателе — нижней, а множитель указывает на то,
сколько раз такая форма изменчивости встречалась среди 2402 иссле-
дованных кротов. Положение зубов в челюсти и характер изменчивости
описаны в очерке по обыкновенному кроту.
85
Таблица 13
Коррелятивная зависимость частоты изменчивости первого и вто-
рого переднекоренных зубов верхней и нижней челюстей сибирского
крота, %
Район сбора материала, число обследо- ванных зверьков Изменчивость
Рт1 Рт2 общая Рт1 и Рт
Томская обл., северные районы; п = 133 48,2 11,1 59,3
южные районы; п = 254 36,3 33,8 70,1
Новосибирская обл.; п = 233 25,8 43,9 69,7
С ал аир, Кузнецкий Алатау; п — 496 43,1 26,5 69,6
Юг Западной Сибири; п = 692 71,4 6,7 78,1
Алтай, Тува; п = 332 50,5 12,5 62,5
Юг Средней Сибири, Якутия; п — 238 19,6 32,4 52,0
Забайкалье; п = 24 34,0 17,0 51,0
По всем районам 41,0 24,5 66,0
Нами зарегистрированы следующие отклонения в зубной системе
верхней и нижней челюстей:
—Pm1 Л 9. —Pm1 Л —Pm1 Л —Pm1 Л —РпРЛП
—РтаЛ ’ —Рт2П’ —Рт2ЛП’ —Рт3П’ —Рт2ЛП ’
—Рт’П е. —Рт’ЛП — Рт’ЛП „ —РпРЛП - —Рт1ЛП
---------о; --.------------3: -------о; ----- -2;
—Рт4Рт3Л +11Л ’ —Рт2Л —Рт2П —Рт2ЛП
—РпРЛП 9 —РпРЛП —РтС! —РпРЛП —РпРЛП —Рт2ЛП
—Рт3П ’ —Рт3ЛП’ —Рт2ЛП’ —Рт2ЛП’ +Рт3Л ’ —Рт2П ’
—Рт2—Рт4Л +Рт*Л Рт4Л\У PnrTIW +М4ЛП +М4П
—Рт2—Рт3—Рт4ЛП ’ —Рт2Л11’ —Рт4ЛГР +Рт4Л’ —Рт2Л’ —Рт3Л’
—Рт1—Рт2—Рт3Л
—Рт2П
С одной стороны, зубная система сибирского крота по числу зубов
и соотношению их категорий близка к наиболее примитивным формам
млекопитающих. В ней представлены все элементы исходной для многих
млекопитающих формулы, состоящей из 44 зубов. С другой — это высоко
дифференцированная зубная система. В постнатальный период развития
у кротов известна лишь одна генерация зубов — постоянная. Зубной ряд
полный, без диастемы.
На основании исследования изменчивости зубной системы сибирского
крота установлено большое число случаев отклонений от нормы, прояв-
ляющееся в колебании количества зубов в широких предедах (от 34 до
47), непостоянство их формы и положения в челюсти, выяснено своеобра-
зие изменчивости зубов разных категорий.
Такие жизненно важные категории зубов, как резцы, клыки и корен-
ные, устойчивы по числу и форме, их стабильность генетически закрепле-
на. Верхние резцы, клыки нижней челюсти и первый переднекоренной
нижней челюсти, несущий функцию клыка, по числу зубов не меняются.
Только у 1 экз. отмечена полифиодонтность клыка верхней челюсти и
у 7 экз. полиодонтность резцов нижней. Восемь зверьков имели дополни-
тельные коренные зубы в верхней челюсти (М4). Изменчивость зубов
86
названных категорий незначительна и составляет всего 1 %. Наиболее
вариабельными оказались переднекоренные зубы. Число экземпляров
с изменчивостью зубов этой категории составляет 98 % среди особей,
имевших отклонения в числе зубов. У кротов Сибири наиболее подвержен
изменчивости первый переднекоренной зуб верхней челюсти (Рш1) —
41,5 %, затем идет второй предкоренной нижней — Рт2, функционально
связанный с Рш1. На долю этих зубов приходится 66 % случаев отклоне-
ния от нормы. У других зубов частота отклонений идет в убывающем
порядке (%): Рш4 (12,1), Рт3 (10), Рт3 (4,7), Рт2 (2,8) Рш4 (2,4), М3 (1,0),
11-э (0,9).
В популяциях из разных частей ареала зубы разных категорий меня-
ются не с одинаковой частотой. Так, Рт1 при среднем показателе измен-
чивости 41,5 % в разных частях ареала изменяется от 19,6 до 71,4 %,
с наименьшим показателем на юге Средней Сибири и наибольшей частотой
отклонений в южных районах Западной Сибири.
Изменчивость наиболее вариабельных Рш1 и Рш2 коррелятивно
связана между собой, о чем можно судить по данным табл. 14. Увеличе-
ние показателя изменчивости одного зуба влечет за собой снижение пока-
зателя изменчивости второго, и наоборот. Суммы показателей изменчиво-
сти по районам довольно устойчивы и колеблются незначительно. Только
в двух случаях изменчивость Рш1 меньше, чем Рш2 (Новосибирская обл.
и юг Ср. Сибири).
Следовательно, подавляющее число случаев отклонений в числе зубов
связано с изменчивостью Рт1 и Рт,. На долю этих зубов приходится
66 % изменчивости зубов всех категорий.
Весьма характерно, что изменчивость зубов имеет определенную
направленность. Редукция зубов происходит главным образом за счет
Рш1, Рш2, а полиодонтия и двухвершинность проявляются преимущест-
венно у третьего и четвертого ложнокоренных. В верхней челюсти редук-
ция Рш1 [зарегистрирована в 310 случаях из 329 всех отклонений этого
зуба (93,9 %). В то же время у Рт3 из 37 случаев отклонений редукция
отмечена только в шести (16,2 %). Совершенно очевидно, что тенденция
к сокращению числа зубов идет преимущественно за счет редукции неспе-
циализированных, одновершинных зубов. Есть все основания рассмат-
ривать преобладание редукционных процессов над полиодонтией как
прогрессивную тенденцию эволюции зубной системы сибирского крота, ве-
дущую к сокращению числа ее элементов за счет конических, одновершин-
ных зубов и генетического закрепления специализированных зубов раз-
ных категорий. Полиодонтия проявляется значительно реже и отражает
ход филогенетического преобразования в процессе эволюции этого вида.
Функциональная нагрузка на зубы разной категории у крота неоди-
накова. Крайне важны в процессе поедания пищи передние резцы. При
поедании дождевых червей, а сибирский крот питается преимущественно
ими [Юдин, 1972е], он, как правило, удерживает червя резцами, выдав-
ливая мозолистыми выростами на внутренней стороне кистей содержимое
кишечника жертвы, и с помощью клыков верхней челюсти и Ргщ нижней
откусывает куски тела. Учитывая, что в желудке крота остатки червей
представлены фрагментами, составляющими 1/3 — 1/4 часть червя, то,
надо полагать, дальнейшей значительной обработки пищи зубами не
происходит. Правда, иногда встречаются кусочки в 1—2 см и меньше.
Отсюда можно сделать вывод об исключительной важности зубов этой
категории для первоначального расчленения червя.
87
Как показали наши исследования, резцы менее всего подвергнуты
изменчивости. Предкоренные (кроме Рш1) и коренные, вероятно, служат
больше всего для дробления и перемалывания хитина жуков. В отличие
от европейского крота у сибирского последние не играют существенной
роли в рационе и поедаются весьма редко.
Функциональная зависимость в изменчивости зубов сибирского крота
хорошо иллюстрируется количественными изменениями этого процесса
при смене функций отдельных зубов. Как уже указывалось выше, в верх-
ней челюсти наиболее стабилен клык, а наиболее изменчив Pm1. Дубли-
рующий его в нижней челюсти Ртх несет функцию клыка. В результате
исследования 2402 экз. установлено, что это один из самых стабильных
зубов. Зато Рш2, функционально заменяющий Ртх,— наиболее изменчи-
вый зуб нижней челюсти.
Одновременное существование разнонаправленной изменчивости зуб-
ной системы (олиго- и полиодонтия), вероятно, можно объяснить действи-
ем ведущего и стабилизирующего отбора [Шмальгаузен, 1939, 1946].
Изменчивость уменьшается лишь при усилении стабилизирующего отбора
[Шмальгаузен, 1939]. Следовательно, изменчивость зубной системы кро-
тов зависит не только от действия отбора, но и от его направленности.
Учитывая исключительно высокую изменчивость зубной системы крота
и преобладание редукционных процессов, можно считать, что проявляется
преимущественное действие ведущего отбора.
Последовательность редукции зубов у сибирского крота можно ис-
пользовать для расшифровки гомологии зубов у более прогрессивных
форм плацентарных млекопитающих и познания путей эволюции зубной
системы. При заселении новых территорий изменчивость проявляется
в широких масштабах, в разных формах, в каждом конкретном случае
реализуется по-своему. Своеобразие ее направления в разных районах —
суть популяционная изменчивость, эволюционирующая по-своему в каж-
дом участке ареала. В центральных частях ареала (Алтае-Саянская гор-
ная система) изменчивость мало выражена. Если учесть, что большая
часть ареала сибирского крота связана с относительно молодым ландшаф-
том Западной Сибири (в плейстоцене равнинная территория Западной Си-
бири еще была покрыта морем), правомерно считать популяцию вида,
занимающую теперь эту территорию, более молодой по сравнению с ис-
ходной, с еще активно идущими процессами формообразования. Широкая
изменчивость, в частности, зубной системы сибирского крота подтвержда-
ет этот тезис. Любопытной чертой изменчивости, связанной, вероятно,
со временем заселения кротом той или иной территории, является показа-
тель ее частоты в разных частях ареала.
По нашим представлениям, род Asioscalops формировался в районах
Алтае-Саянской горной страны. Отсюда он проник на юг Средней и Во-
сточной Сибири, на сопредельные территории Монголии и в равнинную
часть Западной Сибири. Генетическая связь флоры и фауны этого региона
с Алтае-Саянской горной страной подтверждена многими исследователями
[Огнев, 1933; Крыштафович, 1945; Крылов и др., 1964; Куминова, 1960;
Юдин, 1964в; и др.].
Ареалогический анализ изменчивости зубов сибирского крота указы-
вает на различное проявление ее в разных частях ареала. В горных райо-
нах юга Сибири (Алтай, Саяны) изменчивость составляет 15,1—25,0 %
случаев от числа исследованных зверьков и достигает максимума на пери-
ферии ареала вида в равнинных районах Западной Сибири, граничавших
88
с Васюганскими болотами. Здесь число случаев изменчивости отмечено от
40,6—61,4 % (Томская обл.) до 100 % (в Забайкалье) от числа исследо-
ванных зверьков. Промежуточное положение занимает Присалаирье (Но-
восибирск), Салаир и Кузнецкий Алатау (38,3 %). Редкие отклонения от
нормы характерны для зверьков равнинных районов Западной Сибири
и Забайкалья (см. табл. 12).
Уместно отметить, что среди землероек изменчивость зубов, прояв-
ляющаяся в полной редукции или рудиментарности промежуточных,
редка, чаще всего зарегистрирована у сибирской белозубки (Crocidura
sibirica), ареал которой также простирается в пределах Алтая, Саян,
Салаира, Кузнецкого Алатау и равнинной части Западной Сибири, приле-
гающей к Алтаю и Салаиру, т. е. налагается на ареал сибирского крота
[Юдин, 1971а]. Изучение изменчивости близких видов белозубок из евро-
пейской части страны, с Кавказа, из Средней Азии, с Дальнего Востока
показало, что в этих регионах, для которых характерны более «старые»
ландшафты, эта форма изменчивости проявляется крайне редко.
В заключение следует отметить, что у кротов довольно часто встре-
чаются различные механические повреждения зубов, особенно верхних
и нижних премоляров (вероятно, облом о твердые частицы, попадаемые
в ротовую полость при поедании пищи). Они хорошо отличаются от реду-
цированных зубов по остаткам корней и наличию альвеол в челюсти.
Такие отклонения нами во внимание не принимал'ись. Кариоз зубов край-
не редок.
При исследовании большой серии кротов разного возраста из разных
районов (в том числе в гнездовой период) визуально обнаружить молоч-
ные зубы у этого вида не удалось. Вероятно, молочная система редуциру-
ется в эмбриональный период на ранних стадиях ее развития.
При исследовании молочной системы у Scapanus latimanus [Ziegler,
1971, 1972] выяснено, что эти кроты покидают гнездо с полным набором
молочных зубов и теряют их, за исключением первого премоляра, в тече-
ние первых дней самостоятельной жизни. Последовательность смены
молочных зубов происходит в следующем порядке: третий резец, второй и
третий премоляры выпадают первыми, затем клык и четвертый премоляр
и, наконец, последними заменялись первый и второй резцы.
Изменчивость тазовых костей. При выделении рода Asioscalops
С. У. Строганов [1957] одним из существенных признаков считал своеоб-
разное строение таза, отличающегося от такового представителей рода
Talpa и Mogera. С целью оценки таксономической ценности этого призна-
ка нами изучен диапазон его изменчивости у сибирского крота из разных
частей ареала.
В выборках из разных районов Сибири соотношение вариантов измен-
чивости тазовых костей оказалось неодинаковым, но из-за небольшого
объема материала мы не проводим анализа географической изменчивости.
В результате изучения 236 экз. кротов, собранных в разных пунк-
тах ареала вида, установлено шесть вариантов взаимоположения os
sacrae и os ischii (рис. 33), оказавшимися более изменчивыми структу-
рами таза.
Первый вариант (см. рис. 33, 1). Между верхней частью седалищных
костей и крестцом, позади for. sacrae — широкая щель, через которую
хорошо видны подвздошные кости. Седалищные кости за os sacrae лишь
незначительно вытянуты в сторону крестцовой кости, но не соприкасаются
с ней. Такой вариант встречается не часто (у 10,6 % исследованных зверь-
89
Рис. 33. Изменчивость тазовых костей сибирского крота.
1 — кости расставлены широко и не примыкают к позвоночнику; 2 — кости сближены,
но не соприкасаются с позвоночником; 3 — кости примыкают к позвоночнику, но не срастаются
с ним; 4 и 5 — седалищная кость правой и левой стороны примыкает к позвоночнику; 6 — отростки
седалищных костей вдавлены в позвоночник, но не срастаются в одну ветвь.
ков). Дистальные концы седалищных костей, как и в других вариантах,
почти параллельны позвоночнику.
Второй вариант (см. рис. 33, 2). Внутренние выступы седалищных
костей заметно выступают в сторону крестцовой кости, но между ними
и os sacrae остается щель, через которую видны кости позвоночника. Среди
исследованных зверьков кроты с таким строением крестца встречаются
более чем в два с лишним раза чаще, чем с предыдущим вариан-
том (23,7 %).
Третий вариант (см. рис. 33, 3). Внутренняя поверхность седалищных
костей примыкает к крестцовой, но не срастается с ней в общую костную
перемычку. От for. sacrae вдоль крестца над выступами седалищных ко-
стей, примыкающих к крестцу, идут две продольные борозды. Седалищ-
ные кости при легком нажатии на их дистальные концы отходят от крест-
цовой кости. Такое строение таза у исследованных кротов встречается
чаще всего (40,7 %).
Четвертый вариант (см. рис. 33, 4). Выступ правой седалищной кости
внутренней поверхности примыкает к крестцовой. Между левой седалищ-
ной костью и крестцовой имеется значительный промежуток. Степень
сближения варьирует в широких пределах. Такая форма изменчивости
зарегистрирована у 11 % исследованных зверьков.
Пятый вариант (см. рис. 33, 5). Представляет зеркальное отражение
четвертого. Левая седалищная кость внутренней поверхностью примыкает
к крестцу, а правая отделена от него значительным промежутком. Такая
форма изменчивости, как и предыдущая, встречается у 11 % исследован-
ных кротов.
Шестой вариант (см. рис. 33, 6). Выросты седалищных костей вдавле-
ны в крестцовую кость, смыкаются с ней, но не срастаются в единое кост-
ное образование, образующее мост с дистальной стороны for. sacrae, что
хорошо видно при отводе в сторону дистальных частей седалищных костей.
90
Это самый редкий вариант изменчивости (3,4 %). Только у одного крота
с Хамар-Дабана (юго-восточная периферия ареала вида) внутренние по-
верхности выступов седалищных костей плотно срослись с позвоночником,
но при этом for. sacrae осталось открытым с дорсальной стороны. Образо-
вавшаяся перемычка, как бы подстилая его, лежит ниже, а от отверстий
кзади идут две продольные борозды вдоль гребня остистых отростков.
Тип изменчивости тазовых костей обусловлен не только индивидуаль-
ными отклонениями, географической изменчивостью, пока еще недоста-
точно ясной, но, вероятно, меняется и во времени. Исследованные нами
ископаемые остатки костей таза 38 экз. сибирского крота из сборов
Н. Д. Оводова в пещерах Дивногорская-11 и из Каменного Мешка, что
в устье Бирюсы при ее впадении в Енисей, показали, что все они относят-
ся к первому варианту, довольно редко встречающемуся у рецентных осо-
бей сибирского крота. У кротов, добытых в сопредельных районах, из
исследованных 52 экз. такой вариант изменчивости тазовых костей отме-
чен только у одного (1,9 %).
Широкая вариабельность в строении таза не перекрывает специфиче-
ской особенности, свойственной виду,— седалищные кости не образуют
с крестцовой общего костного моста в виде единой костной перемычки,
как у европейского крота или могер. Специфичное строение таза позволя-
ет надежно отличать его от других представителей сем. Talpidae, оби-
тающих в Сибири.
Кроме вариабельности в архитектонике таза сибирского крота ему
присуща широкая изменчивость размеров. Несмотря на это пропорции
отдельных элементов всегда укладываются в общую схему — таз слабой,
ажурной конструкции, узкий, с параллельно направленными седалищ-
ными костями. Кроме того, на дорсальной стороне проксимальной части
тазовых костей у Т. еигораеа видна лишь одна, передняя, пара попереч-
ных отростков, а у сибирского крота их всегда две пары (см. рис. 18).
Заметки по систематике. Положение вида в системе
описано выше. Рассмотрим вопросы внутривидовой таксономии. Впервые,
как уже говорилось, вид сибирского крота был описан А. М. Никольским
[1883] с Алтая. Позднее из разных частей ареала в его составе был выде-
лен ряд форм подвидового ранга. В 1905 г. Н. Ф. Кащенко описал подвид
саянского крота Talpa altaica var. suschkini из окрестностей с. Ермаков-
ского (Западный Саян). В 1936 г. Н. Ф. Егорин описал салаирского крота
(Т. a. salairica), а в 1937 г.— нарымского (Т. a. tymensis) и сибирского
(Т. a. sibirica). В 1956 г. А. С. Фетисов описал из Забайкалья забайкаль-
ского крота (As. a. gusevi).
Выделение подвидового таксономического ранга проводилось на осно-
вании анализа размеров тела, черепа и окраски меха. Уместно отметить,
что окраска определялась глазомерно, без приборного измерения, а дан-
ные размеров кротов или отдельных структур не обрабатывались стати-
стически.
Ниже приводится оценка разработанной ранее внутривидовой таксо-
номии сибирского крота, базирующаяся на изучении новых фактических
материалов, в частности, на колориметрическом изучении изменчивости
окраски зверьков из разных частей ареала.
Номинальный подвид (As. a. altaica) отличается от прочих подвидов,
по мнению С. У. Строганова [1957], более темной окраской летнего меха
с ярко выраженным коричневым оттенком. Зверьки сравнительно круп-
ные. Диагноз подвида довольно расплывчат. При нашей оценке этого
91
Таблица 14
Колориметрическая характеристика подвидов Asioscalops altaica из разных частей
ареала
Место сбора материала Белизна Оттенок
М±т Су а п М±т Су а п
1. As. a. altaica
Алтай, Телецкое озеро Томская обл.: окр. Томска и Кол- 1,63+0,12 2. As. 36,0 a. sal 0,5 airica 25 122,7+2,29 9,3 10,3 25
пашево 1,95+0,13 33,6 0,7 25 129,4+2,29 8,9 11,5 25
Кожевниковский р-н Окрестности Новосибир- 1,32+0,04 14,2 0,2 20 131,8+1,12 3,8 4,9 20
ска По подвиду в це- 1,69+0,12 47,6 0,8 47 122,0+1,43 8,0 9,7 20
лом 1,68+0,11 59,2 1,1 92 126,1+1,08 8,2 10,4 92
3. As. a. gusevi
Забайкалье, Хамар-Да- бан 1,83+0,10 24,4 0,4 20 127,4+1,54 5,4 6,8 20
По в и д у в целом 1,69+0,06 51,4 0,7 137 125,7+0,87 8,1 10,1 137
Достоверность различий выборок 1—2 1,9 1—3 1,3 2—3 1,0 1-2 1-3 1,9 1,7 2-3 0,2
подвида мы опирались на обширный фактический материал из разных
районов Алтая, в том числе и из terra typica. Окраска зверьков из района
Телецкого озера достоверно не отличается от таковой особей As. a. salai-
rica и As. a. gusevi [Там же] (табл. 14, см. рис. 25, б). Несколько больший
размах изменчивости окраски у кротов, относимых к As. a. sdlairica,
объясняется, видимо, большей выборкой номинального подвида. Кроме
того, материал собран в районах с различными ландшафтными условиями
(горы и предгорья Салаира, равнинные районы юга Западной Сибири).
Средние показатели этих выборок весьма близки (см. табл. 14, рис. 25).
Так что говорить о более темной окраске номинального подвида фактиче-
ски нет оснований. По морфометрической характеристике он сходен
с As. a. altaica (см. табл. 10).
По окраске номинальный подвид не отличается и от забайкальско-
го — показатели его полностью укладываются в таковые As. a. altaica.
Одновозрастные зверьки одного пола алтайского и салаирского кротов
не отличимы по размерам тела и черепа. Таким образом, нет оснований
выделять салаирского крота (As. a. salairicd) в самостоятельный подвид.
Саянский крот (As. a. suschkini) был выделен в подвид на основании
более светлой окраски меха и «в среднем более крупных размеров тела
и черепа» [Строганов, 1957]. В нашем распоряжении не было достаточно
репрезентативного материала для объективной оценки окраски саянского
крота, как не было его и у С. У. Строганова [Там же]. Небольшая серия
зверьков визуально не отличалась по окраске от номинального подвида.
По размерам (см. табл. 10) кроты с Алтая, Салаира, Кузнецкого Алатау
и равнинных районов юга Западной Сибири не отличаются. По минималь-
92
ным размерам кондило-базальной длины черепа, длине и массе тела зверь-
ки подвида уступают алтайскому и салаирскому. Следовательно, выделе-
ние этого подвида в самостоятельную таксономическую единицу не право-
мерно. Следует сказать, что материала из Саян недостаточно.
На специфику забайкальского крота обратил внимание еще
С. У. Строганов [1957], а А. С. Фетисов [1956] описал его в качестве
подвида (As. a. gusevi).
В диагнозе подвида салаирского крота (As. a. salairica) в качестве
диагностических признаков, по которым он отличается от номинального
и саянского кротов, приводятся сравнительно небольшой рост и отсутст-
вие в окраске коричневых оттенков. В диагнозе автора первоописания
[Егорин, 1936] указывается, что размеры крупные (туловище и ступня),
череп крупный, менее массивный и уплощенный. Не ясно, что автор имеет
в виду, указывая на крупный, но «менее массивный» череп, вероятно, по
сравнению с описанным выше номинальным подвидом. В то же время
сравнение одновозрастных самцов не подтверждает этого положения,
а по объединенной выборке разновозрастных и разнополых зверьков
кондило-базальная длина черепа салаирского крота несколько больше,
чем у номинального подвида. С. У. Строганов [1957] указывает на отсут-
ствие в окраске коричневых тонов.
По средним показателям (см. табл. 9) это самый крупный «подвид»
(кондило-базальная длина черепа составляет 37,0(±0,08), наибольший
показатель этого промера 41,4 мм; длина тела 157,5(±=1,21), максималь-
ный показатель 182 мм; масса — 128,9(±2,70), максимальная — 230 г.
Бурые тона в окраске зверьков с территории, на которой, по мнению
С. У. Строганова [Там же], обитает салаирский крот, имеют значитель-
ный разброс, а у некоторых экземпляров близки к показателям для вида
в целом. Средняя величина оттенка самая высокая среди всех подвидов —
127,7±1,61. В отдельных выборках со значительно удаленных друг от
друга территорий этот показатель довольно сильно варьирует: от 122
до 131,8.
Таким образом, диагностические признаки, на основании которых
крот, заселяющий Салаир, Кузнецкое Алатау, Кузбасс, бассейн р. Томь
до Кети включительно и районы равнинной части юга Западной Сибири
на восток до Вилюя и Тунгуски, не подтверждаются, и эта группировка
не может характеризоваться как самостоятельный подвид, достоверно
отличающийся от номинального. Все указанные в диагнозе признаки
варьируют в широких пределах.
Выделение нарымского крота (As. a. tymensis) в качестве подвида
основано на «более светлой окраске меха и в среднем более мелких разме-
рах». Средние показатели размера черепа его, действительно, самые мел-
кие для кротов Сибири — кондило-базальная длина черепа 35,9(=t0,22),
длина тела 144,0(=t2,00) мм. В описании этого подвида [Егорин, 1937]
обращает на себя внимание следующее обстоятельство. Вся типовая серия
состоит из одних самок, а они, как можно видеть из данных приведенного
выше вывода, уступают по размерам самцам. Вероятно, это прибылые
зверьки, т. е. самые мелкие особи в популяции. Отсюда наряду с мелкими
размерами становится очевидным и второй диагностический признак —
светлая, серовато-аспидная (светло-стальная) окраска меха, свойственная
прибылым зверькам. Кроме того, в распоряжении автора кроме типовой
серии было 62 зверька с р. Тым; при описании размеров не указано число
исследованных экземпляров.
93
Шкурки этого подвида, имеющиеся в коллекции, выцвели от длитель-
ного хранения и оказались непригодными для колориметрического иссле-
дования.
Нам представляется, что в окраске кротов, обитающих в довольно
однородных, стабильных условиях почвенного субстрата, трудно ожидать
четких географических закономерностей ее изменчивости. Широкий диа-
пазон разброса показателей белизны и оттенка в разных популяциях
можно считать как биотопическую изменчивость. Это подтверждается
колориметрическими исследованиями. Верхняя граница значений оттенка
у пяти исследованных выборок довольно близка. Весьма плотно лежит
на графике минимальный показатель белизны. При общем разбросе значе-
ний этого показателя от 0,8 до 4,1 границы всех пяти популяций проходят
между 0,8 и 1,4 (см. табл. 14, рис. 25, б). Около половины исследованных
зверьков из всех популяций приходится на этот интервал значений. Наи-
более широкий диапазон у показателей оттенка и максимальной белизны.
Зверьки с меньшим показателем оттенка больше всего находятся в выбор-
ках из популяций As. a. altaica и As. a. salairica.
Для реальной оценки окраски с целью надежного ее использования
при таксономических построениях нужно иметь достаточно репрезента-
тивные выборки из разных популяций прибылых и перезимовавших
зверьков.
Необоснованное выделение подвидов сибирского крота стало возмож-
ным, вероятно, вследствие недостаточного объема фактического материа-
ла, имевшегося в распоряжении исследователей. Значительные половые
и возрастные колебания в размерах и окраске Зверьков не учитывались
при первоописаниях. Кроме того, для оценки вариабельности окраски
меха крота, имевшей существенное значение при выделении подвидов,
нельзя использовать давно хранящиеся в музеях сборы, так как со време-
нем они меняют оттенок (светлеют). Непригоден для разграничения слабо
выраженных цветовых вариаций визуальный метод оценки, ранее приме-
нявшийся всеми исследователями, так как в этом случае результат в
большой степени зависит от индивидуального восприятия цвета. Приме-
ненный нами колориметрический метод оценки свободен от субъективно-
сти и дает реальные количественные значения. Используя этот метод,
разграничить ранее описанные подвиды оказалось невозможным. Можно
лишь говорить о некоторых тенденциях в изменчивости окраски отдель-
ных популяций (широта разброса, посветление, насыщенность оттенка).
Анализ морфометрических показателей также не подтверждает право-
мерности выделения подвидов. По этим параметрам отличаются популя-
ции периферии ареала. Форма As. a. tymensis, занимающая территорию
на северо-западном и северном пределе распространения вида в Западной
Сибири, и As. a. gusevi — на юго-восточной его периферии отличаются
более мелкими размерами, чем кроты других территорий. Здесь же силь-
но выражены и другие формы изменчивости [Юдин, 1975в]. Мы рассмат-
риваем это явление как реакцию вида на неблагоприятные условия обита-
ния. Так как материал из этих двух районов недостаточен, окончательно
судить о правомочности выделения подвидов трудно, так же, как и отри-
цать реальность их существования.
Следует сказать, что имеющиеся в нашем распоряжении материалы
не подтверждают реальность существования подвидов, описанных с тер-
ритории Алтае-Саянской горной страны и сопредельных. Кроты As. a. al-
taica, As. a. salairica, As. a. suschkini, As. a. sibirica, по H. Ф. Егорину
94
[1936] и С. У. Строганову [1957], мы относим к номинальному подвиду
As. a. altaica. Что касается кротов Васюганья, Тыма, Якутии и Забай-
калья, их подвидовой статус не ясен. В этом отношении мы стоим на по-
зиции А. П. Кузякина [Бобринский и др., 1965], объединяющего кротов
Алтая и Саян в номинальный подвид. As. a. salairica — скорее переходная
форма (особенно по окраске), по размерам не отличающаяся от номиналь-
ного подвида. Подтверждением может служить такой стойкий показатель,
как средняя кондило-базальная длина черепа, которая колеблется незна-
чительно: от 35,9(±0,23) до 37,0(^0,08). У одновозрастных и однополых
зверей этот показатель колеблется еще меньше.
При небольших выборках возможна ошибка в оценке популяции того
или иного региона. Особенно значительные погрешности могут быть допу-
щены, если выборки взяты асинхронно или условия зимовки в этих райо-
нах в момент сбора материала существенно отличались. Весной, когда
самки еще кормят детенышей и привязаны к гнезду, а самцы уже в со-
стоянии гона, в ловушки, поставленные в магистральные ходы, ловятся
почти исключительно перезимовавшие (одну или несколько зим) самцы,
т. е. самые крупные представители популяции. Сравнение такой выборки
с выборкой из этого же места, но полученной, скажем, в июне, будет
характеризоваться широким разбросом морфометрических показателей
и заниженными их средними величинами. В августе — сентябре, когда
в уловах преобладают прибылые особи, сравнение дает статистически
достоверно отличающиеся результаты.
Дело в том, что после распада выводков [Юдин, 1972, 1974а], когда
взрослые самки покрыты, наблюдается некоторый спад активности самцов
(до момента покрытия прибылых самок) и интенсивное расселение молод-
няка. Когда старые и молодые самки покрыты, взрослые самки и самцы
«оседают» на своих индивидуальных участках, и в уловах оказываются
почти исключительно кроты рождения этого года, т. е. самая мелкая
часть популяции. Сравнение этих показателей дает наглядное подтверж-
дение сказанному. Весенние и осенние выборки зверьков из одного
места по морфометрическим показателям будут достоверно различаться
больше, чем кроты из значительно удаленных участков ареала.
Общеизвестно, что большинство подвидов животных — географи-
ческие изоляты. Сибирский крот — выходец из Алтае-Саянской горной
страны. Все его популяции более или менее постоянно связаны (или
имели связь в недалеком прошлом), что обеспечивало свободный обмен
генами на протяжении длительного времени. Выделенные подвиды сибир-
ского крота — не географические изоляты, а результат адаптивной реак-
ции на местные условия. Разграничить такие группировки и дать номенк-
латурную оценку трудно и вряд ли целесообразно с позиций систематики.
Это, скорее, задача экологическая, чем таксономическая, из области вы-
яснения степени воздействия климатических и ландшафтных условий
на вид.
Наглядным подтверждением сказанному может служить следующий
пример. Н. Ф. Егорин [1936] описал подвид Talpa (Asioscalops) a. sibirica
из окрестностей ст. Тяжин, основываясь на очень крупных размерах и
более темной окраске по сравнению с As. a. salairica (табл. 15).
Автор не приводит длины тела этого подвида, а лишь указывает на
то, что они такие же, как у As. a. altaica, поэтому мы и заимствовали этот
показатель из описания номинального подвида, приведенного в той же
работе. Различия действительно значительные (см. табл. 15). Но, если
95
Таблица 15
Сравнительная морфометрическая характеристика двух подвидов кротов,
по Н. Ф. Егорину [1936], мм
Промер Т. a. salairica Т. a. sibirica
м lim м lim
Череп общая длина 38,8 36,4—40,7 40,3 39,8-41,6
наибольшая ширина 17,9 17,0—18,6 18,5 17,8—18,9
Общая длина тела 165,0 158-183 215 191—232
обратиться к характеристике материала, на основании которого
Н. Ф. Егорин [1936] описал новый подвид, все становится на свои места.
В распоряжении автора была небольшая серия из взрослых самцов, в то
время как описание As. a. salairica делалось по особям разного пола и
разных возрастных групп. Это обстоятельство и привело Н. Ф. Егорина
к ошибке — возрастные и половые различия им были оценены как так-
сономические. Что касается размеров тела сибирского крота, то они фан-
тастически велики и, несомненно, являются результатом ошибки, допу-
щенной при измерении.
По нашим сведениям (п = 546), эти показатели для всех возрастов
самцов и самок (включая материалы по As. a. sibirica) иные (мм):
общая длина черепа — 32,0—41,4 (М = 37,0); наибольшая ширина
черепа — 15,2—19,2 (М = 17,4); длина тела с головой — 99—195
(М = 156,6).
Таким образом, анализ обширного материала свидетельствует о ши-
роком размахе изменчивости признаков у крота этой группировки
(см. табл. 9). Если сравнить наши данные только по перезимовавшим
самцам из всех пунктов ареала в объеме, принимаемом С. У. Строгановым
[1957] для подвидов, то они практически не различаются (см. табл. 8)
и в то же время значительно выше общих показателей (по всем возрастам
для самцов и самок) для подвида (см. табл. 9).
Средний показатель наибольшей длины черепа взрослых самцов весь-
ма стабилен (см. табл. 8). Три выборки, взятые из разных пунктов ареа-
ла подвида, практически неразличимы по этому показателю (38,3; 38,4;
38,8) и заметно превышают средний показатель в целом по виду (37,3).
Размах вариации признака во всех трех выборках весьма близок (наимень-
шие показатели 35,5; 35,8; 36,6; наибольшие — 40,8; 40,3; 40,4). Раз-
брос составляет 4,9 мм, в то время как для подвидов в целом он равен
10,7 мм. Материал собран в разные годы в период с 1949 по 1972 г. Отсю-
да можно сделать вывод, что средний показатель для кротов этой катего-
рии весьма устойчив и значительно превышает таковой для подвидов.
Следовательно, от соотношения в выборке зверьков разного возраста и
пола в значительной мере зависят средние значения признака.
Аналогичная картина получена и при анализе размеров наибольшей
величины черепа, признака еще более стабильного ( М= 17,9; 17,9; 17,9).
Что касается размеров тела, то этот показатель более вариабелен и при
измерении зверьков возможны ошибки. У самцов из Салаира средний
размер тела оказался ниже среднего по подвиду (п = 8), а в остальном он
отражает приведенную выше закономерность при характеристике измен-
чивости черепа.
96
Размеры тела сибирского крота, приведенные Н. Ф. Егориным [1936],
явно завышены. По нашим данным, они значительно меньше: М —
= 155,5, max — 195 мм.
Как уже указывалось в литературе [Stein, 1963; Юдин, 1974, 1980],
в неблагоприятные зимы в первую очередь элиминируются наиболее
крупные особи популяции, а выживают самые мелкие, для поддержания
жизни которых требуется меньше пищи. Исходя из этого морфометри-
ческие показатели популяции меняются во времени. После благоприят-
ных зим, особенно нескольких подряд, популяция «укрупняется», а после
неблагоприятных — «мельчает». Следовательно, для выявления степени
сходства или различия популяций необходимо сравнивать единообраз-
ные выборки по возрасту, полу, сезону. Вероятно, показатели груп-
повой выборки, взятые одновременно в один сезон, будут отражать спе-
цифику изучаемой группировки, но наиболее достоверные показатели для
целей систематики может дать сравнение прибылых зверьков одного пола.
Во избежание ошибок при оценке подвидовых различий следует со-
поставлять показатели только взрослых самцов. Размеры взрослых сам-
цов, а также кондило-базальная длина и наибольшая ширина черепа из
разных районов Салаира, Предсалаирья и равнинных районов юга Сиби-
ри достоверно не отличаются.
Указание Н. Ф. Егорина [1936] на закономерность увеличения раз-
меров кротов в направлении с запада на восток не подтверждаются. Во-
первых, нельзя сравнивать европейского крота и сибирского, как это
делает названный автор, во-вторых, из-за неполноты имеющихся в то
время материалов он не знал, что кроты Якутии и Забайкалья, заселяю-
щие восточную периферию ареала вида, значительно мельче, чем живущие
в Западной Сибири. Анализ обширного материала показал, что популя-
ции кротов северной, северо-восточной и восточной периферии ареала
мельче, чем юга Западной Сибири (см. табл. 9). Вероятно, именно здесь,
на пределе существования вида, более жестко проявляется действие эли-
минирующих факторов среды, когда кротам зимой трудно добывать пищу
и идет отбор наиболее крупных особей.
Подводя итог анализу изменчивости сибирского крота, можно ска-
зать, что для выделения таких подвидов, как As. a. salairica, As. a. susch-
kini, As. a. sibirica, нет достаточных оснований. Можно отметить лишь
тенденцию убывания размеров зверьков в направлении на север и на
северо-восток.
Род Mogera Pomel, 1848 — Могеры
Тип рода. Talpa wogura Temm., 1833, Япония.
Диагноз. Зубная формула:
1'4; сф; P»i М| = 42.
Зубы значительно крупнее, чем у представителей рода Talpa и
Asioscalops, Pm2 меньше Pm1 и Pm3, Pm1 крупный. Клыки в нижней че-
люсти отсутствуют, их функцию выполняет Рт2. Череп массивный с отно-
сительно широким рострумом. Головка копулятивного органа в отличие
от таковой представителей рода Talpa и Asioscalops короткая, грушевид-
ной формы и покрыта редко сидящими крупными шипами.
97
Характеристика. Род включает относительно крупных и
мелких зверей кротообразной внешности. Длина хвоста менее длины го-
ловы. Глаза закрыты кожистой перепонкой. Окраска меха буровато-ко-
ричневая разных оттенков. В нижней челюсти отсутствуют клыки. Pm*
имеет небольшой талон и гипоконус.
Таз могероидного типа — os ischium полностью срастаются с позво-
ночником, образуя две монолитные широкие костные перемычки (см.
рис. 18). Наряду с указанными особенностями в строении таза характер-
на его относительная узость. По структурным особенностям тазовых кос-
тей могеры ближе всего стоят к американским кротам родов Scapanus
и Scalopus.
Распространение. Ареал рода охватывает Японию, Тай-
вань, Китай, Корею. В СССР встречается в Приморье, на севере до р. Би-
ра (47°20 с. ш.) [Строганов, 1957].
Заметки по систематике. В систематическом отношении
род Mogera ближе всего стоит к южно-азиатским кротам Eoscalops, с ко-
торыми у него одинаковые слуховые косточки и сходное строение Pm1.
Однако существенно отличается по структуре таза и зубной формуле.
По этому признаку они близки с американскими кротами родов Scapanus
и Scalopus. От представителей родов Talpa и Asioscalops могеры
отличаются отсутствием клыков в нижней челюсти, особенностями
строения слухового аппарата и совершенно иной конструкцией таза (см.
рис, 18).
В современной фауне род представлен пятью видами: М. wogura,
Temm., М. robusta Nehring, М. latouchei Thomas, M. tokudae Kuroda,
M. kobeae Thomas. Род могер выделен в составе подсем. Talpinae в 1848 г.
Помелем [Pomel, 1848]. Позднее взгляды исследователей на правомер-
ность такого выделения могер неоднократно менялись. Одни систематики
[Огнев, 1928; Allen, 1938; Бобринский и др., 1944, 1965; Simpson, 1945;
Строганов, 1948, 1957; Stein, 1960; Охотина, 1966; Юдин, 1971а] безо-
говорочно признали реальность этой систематической категории, другие
[Schwarz, 1948; Ellerman, Morriison-Scott, 1951] рассматривали могер в
составе рода Talpa, а Имаизуми [Imaizumi, 1955] свел роды Euroscaptor,
Mogera и Parascaptor в ранг подродов рода Talpa.
Род могер хорошо морфологически очерчен. Все виды имеют специ-
фическую окраску меха, волосы у них растут пучками, а не поодиночке,
как у представителей рода Talpa, имеют специфическую зубную форму-
лу. В отличие от представителей родов Talpa и Asioscalops у них 42 зуба
(в нижней челюсти нет клыков). Косточка среднего уха имеет самобытное
строение [Строганов, 1948]. Отличаются они и по хромосомному набору:
у Т. еигораеа число хромосом 34 (NF=68), у Т. caucasica—38 у Т. levantis
—34 (2VF=66), у Т. romana — 34 (NF=68), у Т. hercegovinensis — 36, у
Т. соеса — 36, а у Mogera wogura insularis —32 [Matthey, 1958].
Большинство авторов признает в составе рода три вида (М. wogura,
М. robusta, М. latouchei). Томас [Thomas, 1912] описал с полуострова
Хайнань М. hainana, которую С. У. Строганов [1948] рассматривает в
качестве подвида большой могеры (М. wogura hainana). В сводке по на-
секомоядным Японии Хисасе Абе [Abe, 1971] приводит еще два вида —
М. tokudae Kuroda и М. kobeae Thomas. В СССР род представлен двумя
видами: средней (М. wogura Temm.) и большой (М. robusta Nehring)
могер ам и.
7 Б. С. Юдин
98
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА MOGERA
1(2). Размеры (мм) средние: длина тела 122—152, длина ступни 17—
19, кондило-базальная длина черепа 31,5—37,6......................
.............................Mogera wogura Temm.— Средняя могера
2(1). Размеры (мм) крупные: длина тела 138—206, длина ступни 21 —
28, кондило-базальная длина черепа 38,6—45,3 .............' ... »
..........................Mogera robusta Nehring — Большая могера
Mogera wogura Temminck, 1833 — Средняя могера
Голотип. Не установлен. Вид описан по экземпляру из Японии
(окрестности Нагасаки, Кюсю).
Диагноз. Звери средних размеров. Значительно уступают по ве-
личине большой могере. Длина тела 120—156, длина ступни 17—19, кон-
дило-базальная длина черепа 31,5—37,7 мм (рис. 34). Различия размеров
тела, черепа и отдельных его элементов достоверны (см. табл. 16).
Характеристика. Габитуально похожи на других предста-
вителей рода. Значительно уступают по размерам большой могере, но
крупнее М. latouchei. Количественные показатели таких признаков, как
кондило-базальная длина черепа, длина ступни, даже не перекрываются
соответствующими показателями большой могеры. По большинству при-
знаков это перекрывание незначительное. Оба вида с высокой степенью
достоверности отличаются по всем морфометрическим показателям (кроме
длины хвоста). Различия двух видов проявляются не только в абсолют-
ных количественных показателях признаков, но и в неодинаковых, сво-
Морфометрические различия-
Признак М. wogura
М+тх п , о cv lim
Длина тела с головой 136,0+ 5,468 5 12,23 9,0 122—152
хвоста 22,0±3,54 5 7,91 35,9 14—32
ступни 18,4+0,400 5 0,89 4,9 17—19
Масса тела 89,0+4,000 2 5,66 6,4 85—93
Длина кондило-базальная черепа 33,8±0,805 8 2,28 6,7 31,5—37,6
верхнего ряда зубов 14,4+0,374 8 1,06 7,3 13,4—15,9
верхнего ряда заднекорен- ных зубов 6,4+0,141 8 0,40 6,3 5,8—6,9
Ширина рострума 5,0+0,247 8 0,70 13,9 4,2—6,0
предглазничная 6,8+0,219 8 0,62 9,1 5,9—7,6
скуловая 13,0±0,464 7 1,23 9,4 11,3—14,8
наибольшая черепа 17,1+0,461 8 1,31 7,6 15,4—19,0
Наибольшая высота черепа 11,0+0,222 8 0,63 5,7 10,1—11,8
Высота восходящей челюсти 7,5+0,214 8 0,61 8,0 6,7—8,4
99
еобразных для каждого вида пропорциях. Так, если отношение длины
черепа к длине тела довольно близко, то относительная длина как верх-
него ряда зубов, так и заднекоренных к кондило-базальной длине черепа
у средней могеры значительно больше, чем у большой. Для конфигурации
черепа средней могеры характерна относительная узость рострума при
относительно большом значении показателей предглазничной, скуловой и
наибольшей высоты и ширины черепа к кондило-базальной длине. Отно-
сительная высота восходящей ветви нижней челюсти лишь незначительно
превышает ее у большой могеры (табл. 17).
Тазовые кости в общем плане сходны с таковыми большой могеры,
но они меньше по размерам: короче и уже; отличаются деталями строения.
В частности, их относительная ширина больше, чем у М. robusta.
Мех средней могеры мягкий и шелковистый, с большим количеством
пуховых волос в пучке. Цвет меха у средней могеры темно-серо-коричне-
вый, с преобладанием насыщенных темных тонов. На вентральной сторо-
не тела ряд ярко окрашенных охристо-золотистых пятен, тянущихся в
виде небольшой полосы от передних лап и груди к задней части живота.
На горле мех рыжевато-коричневый. У старых самцов эта расцветка
спускается до груди, а у молодых — охватывает губу.
Специфическим для средней могеры является отсутствие прианаль-
ных желез. Поэтому самцы даже в состоянии гона в отличие от самцов
большой могеры не обладают специфическим «чесночным» запахом.
Основные отличия этих двух видов приведены в очерке по сис-
тематике.
Головка копулятивного органа короткая, грушевидной формы, рас-
ширена к свободному концу. Большая часть ее покрыта крупными шипа-
ми, посаженными расширенными основаниями в овальные лунки. Верши-
Таблица 16
видов могер
М. robusta Достоверновть различий сред- них (Td)
п а Су lim
170,5±1,630 71 13,73 8,0 138—206 6,0
25,7±0,547 70 4,58 17,8 17—37 1,0
23,4±0,183 71 1,54 6,6 21—28 12,4
180,3±5,570 90 47,26 26,2 90—299 13,3
41,9±0,174 90 1,65 3,9 38,6—45,3 9,8
17,3±0,061 86 0,56 3,7 15,0—18,4 6,8
7,2±0,026 86 0,24 3,3 6,6-7,9 5,6
6,5±0,039 90 0,37 5,7 5,8—7,5 5,9
7,8±0,032 90 0,30 3,9 5,8—8,3 6,4
15,9±0,074 64 0,60 3,7 14,6—17,0 6,2
20,0±0,087 90 0,83 4,1 18,2—21,7 6,2
12,4±0,038 90 0,36 2,9 11,4—13,3 6,3
9,2±0,060 88 0,57 6,2 8,1—10,2 7,8
7*
100
Рис. 34. Череп средней могеры — Mogera wogura.
ны шипов направлены к основанию копулятивного органа и, видимо, не-
сут роль фиксатора во время спаривания. Дорсальная часть glans penis
расчленена рядом продольных складок, не доходящих до основания.
У основания головки, на дорсальной стороне (рис. 35), имеется полулун-
ное углубление с несколькими овальными бугорками на дне 5.
Распространение. Японские острова, Восточный Китай,
Корейский полуостров. СССР: крайний юг Приморья, оз. Хасан [Юдин,
1971а].
Материал. 3 экз., окрестности оз. Хасан, коллекторы Д. И. Би-
биков и Б. С. Юдин; 1 экз., Корея, коллектор Н. Дарская; 5 экз., Япо-
ния, коллекторы Хисасе-Абе и К. Таката.
Заметки по систематике. Средняя могера — тип рода
Mogera. Впервые вид описан из окрестностей г. Нагасаки Темминком
? Описание копулятивного органа и рисунок сделаны по спиртовому экземпляру.
101
Таблица 17
Морфометрическая характеристика обыкновенного, сибирского кротов,
средней и большой могер
Признак Т. еигораеа As. altai- ca М. wogura М. robusta
Длина тела с головой 25,8* 23,6 24,9 24,6
хвоста 25,5 16,2 16,2 15,1
ступни 14,7 13,3 13,5 13,7
Масса тела 70,9 70,8 65,4 105,7
Длина верхнего ряда зубов 41,0 38,8 42,6 41,3
верхнего ряда заднеко- ренных зубов 18,2 15,4 18,9 17,2
Ширина рострума 12,7 14,8 14,8 15.5
предглазничная 19,9 19,6 20,1 18.6
скуловая 34,1 34,4 38,5 37,9
наибольшая черепа 47.7 47,7 50,6 47,7
Наибольшая высота черепа 31,5 31,0 32,5 29,6
Высота восходящей ветви нижней челюсти 19,7 19,6 22,2 22,0
Примечание. Показатели взяты по отношению к кондило-базальной
длине черепа; размер хвоста, ступни и масса — по отношению к длине тела, %.
Звездочкой помечены данные — отношение кондило-базальной длины черепа к дли-
не тела.
[Temminck, 1833]. Маак [59] и Радде [Radde, 1862] отнесли к этому виду
большую могеру из Приморья. После описания ее как самостоятельного
вида Нерингом [Nehring, 1891 ] она была выделена из состава М. wogura,
хотя позднее еще неоднократно делались попытки их объединения в один
вид [Kuroda, 1939]. Имаизуми [Imaizumi, 1955] не только объединил
их в один искусственный композиционный вид, но и отнес к роду Talpa
{Talpa wogura robusta). Добсон [Dobson, 1883], С. У. Строганов [1948],
Бобринский [1965], М. В. Охотина [1966], Б. С. Юдин [1971а],
А. А. Гуреев [1979] и др. рассматривают их как самостоятельные виды.
С. У. Строганов [1948] установил видоспецифичное строение слуховых
косточек большой и средней могер, М. В. Охотина [1966] и Б. С. Юдин
[1971а] показали различия в строении glans penis, Б. С. Юдин [1975в] —
различия в строении тазовых костей. Кроме того, как сказано в характе-
ристике вида, средняя и большая могеры отличаются с большой достовер-
ностью не только по абсолютным морфометрическим показателям, но и
своеобразием пропорций отдельных частей тела и черепа. Для удобства
сравнения мы приводим некоторые видоспецифичные показатели:
Сравнительная характеристика видов
Признак М. robusta М. wogura
Длина тела с головой,.мм У самцов 182,5, у самок 164,2 У самцов 174,7, у самок 161,0
Масса тела, г У самцов 256,5, у самок 146,3 У самцов 179,6, у самок 142,0
Кондило-базальная длина черепа, мм Длина верхнего ряда зу- бов, мм Преобладающий цвет ок- раски меха У самцов 44,6, у самок 41,8 16,7 Коричневый У самцов 40,9, у самок 39,3 15,9 Темно-серый с примесью коричневого
102
Признак
Collum mallei processus
manubrium
Apophysis orbicularis
Прианальные железы
Подкожная мускулатура
Головка копулятивного
органа
М. robusta
Образуют прямой угол
Отсутствует
Есть
Развита хорошо
Массивная, приостренная
на конце. На дорсаль-
ной стороне у ее осно-
вания нет углубления
М. wogura
Образуют тупой угол
Имеется
Нет
Развита слабо
Меньше, притупленная
на конце. На дорсаль-
ной стороне у ее осно-
вания есть углубление
Все это позволяет рассматривать М. wogura как самостоятельный
вид. От М. latouchei отличается большими размерами тела и черепа, строе-
нием Pm1 [Строганов, 1948]. Впервые на обитание средней могеры в
СССР указал А. П. Кузякин [Бобринский и др., 1965]. В СССР, вероят-
но, обитает средняя могера, относящаяся к подвиду М. w. согеапа
[Thomas, 1907].
Mogera robusta Nehring, 1891 — Большая могера
Голотип. Вид описан по экземпляру из окрестностей Влади-
востока.
Диагноз. Самый крупный представитель сем. Talpidae: длина
тела 151—206, длина ступни 20—28 мм, масса 122—299 г. Pm1 выше
Pm2, последний уступает по высоте Pm3. Прианальные железы от-
сутствуют.
Измерения. См. табл. 18.
Характеристика. Размеры крупные (см. табл. 18), тело
вальковидное, в задней части несколько сдавленное в дорсовентральном
направлении. В целом внешность типично кротовая. Зверек более тя-
желого телосложения, чем обыкновенный и сибирский крот. Отношение
массы тела к его длине составляет 105,7 %, в то время как у рассмотрен-
ных выше видов оно равно 70,9 и 70,8 %. Хвост очень короткий — 15,1 %
от длины тела (у обыкновенного крота — 25,5, у сибирского крота —
Л
Рис. 35. Строение glans penis могер.
А — средняя могера; Б — большая могера; а — дорсальная, б — вентральная, в — латераль-
ная сторона.
103
Т а’б л и ц а 18
Морфометрическая характеристика Mogera robusta
Возрастная и половая группа М±тж CF CV п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 175,3+2,41 10,77 6,14 20 156,0—200,0
взрослые 178,2+3,44 15,38 8,63 20 140,0—206,0
Самки прибылые 162,4+1,73 8,83 5,44 26 138,0—178,0
взрослые 169,8+6,81 11,79 6,94 3 157,5-181,0
В целом по виду ... 170,5±1,63 13,73 8,10 71 138,0—206,0
Длина хвоста
Самцы прибылые 27,9+1,00 4,49 16,10 20 21,0—35,0
взрослые 27,8+0,98 4,29 15,43 19 21,0—37,0
Самки прибылые 23,1+0,59 3,0 12,99 26 16,5-28,7
взрослые 19,0+1,50 2,60 13,67 3 17,5—22,0
В целом по виду ... 25,7+0,55 4,58 17,84 70 16,5—37,0
Длина ступни
Самцы прибылые 24,0+0,22 0,98 4,07 20 21,0—25,6
взрослые 24,5±0,33 1,46 5,96 20 21,0-28,0
Самки прибылые 22,1±0,11 0,57 2,57 26 20,5—23,5
взрослые 23,5+1,80 3,12 13,29 3 21,0-27,0
В целом по виду ... 23,4+0,18 1,54 6,60 71 20,5-28,0
Масса тела
Самцы прибылые 186,6+6,50 29,08 15,58 20 90,0—219,0
взрослые 226,3+10,41 47,72 21,08 21 110,0—299,0
Самки прибылые 144,2±3,56 18,15 12,59 26 107,0—175,0
взрослые 171,5+13,35 23,11 13,48 3 155,5—198,0
В целом по виду ... 180,3+5,57 47,27 26,22 72 90,0-299,0
Кондило-базальная длина черепа
Самцы прибылые 42,4+0,17 0,76 1,80 20 40,3—43,5
взрослые 43,5+0,27 1,25 2,86 22 40,5—45,0
Самки прибылые 40,3+0,11 0,59 1,47 28" 38,7—41,5
взрослые 42,5+1,42 2,46 5,79 3 40,9—45,3
В целом по виду ... 41,9+0,17 1,66 3,95 90 38,6—45,3
Наиболыиая длина черепа
Самцы прибылые взрослые 43,1+0,15 44,3+0,24 0,66 1,14 1,53 2,57 20 22 41,4-44,1 41,6—45,7
Самки прибылые 41,1+0,13 0,70 1,70 27 39,2—42,5
взрослые 43,3+1,43 2,49 5,73 3 41,8—46,2
В целом по виду ... 42,8+0,17 1,60 3,73 89 39,2—46,2
Длина верхнего ряда зубов
Самцы прибылые 17,5+0,07 0,32 1,84 19 16,7—18,1
взрослые 17,7+0,10 0,45 2,53 22 16,7-18,4
Самки прибылые 16,8+0,09 0,46 ' 2,71 27 15,0-17,5
взрослые 17,3+0,59 1,03 5,94 3 16,4—18,4
В ц е л о м по виду ... 17,3+0,06 0,57 3,26 86 15,0—18,4
104
Окончание табл. 18
1 2 3 4 5 6
Длина верхнего ряда заднекоренных зубов
Самцы прибылые 7,2+0,04 0,15 2,10 19 6,9—7,4
взрослые 7,4±0,04 0,20 2,77 22 7,1—7,9
Самки прибылые 7,1+0,06 0,27 3,83 27 6,6—7,5
взрослые 7,2+0,17 0,29 4,03 3 7,0—7,5
В целом по виду ... 7,2±0,03 0,42 3,35 86 6,6—7,9
Ширина рострума
Самцы прибылые 6,5+0,05 0,21 3,16 20 6,0—6,8
взрослые 6,8±0,06 0,29 4,20 22 6,2—7,5
Самки прибылые 6,1+0,03 0,17 2,75 28 5,8—6,5
взрослые 6,8+0,27 0,46 6,83 3 6,5—7,3
В целом по виду ... 6,5+0,04 0,37 5,69 90 5,8—7,5
Предглазничная ширина
Самцы прибылые 7,8+0,09 0,41 5,19 20 6,6—8,3
взрослые 7,8+0,06 0,28 3,55 22 7,0—8,2
Самки прибылые 7,7+0,05 0,25 3,30 28 7,1—8,1
взрослые 7,7+0,15 0,27 3,44 3 7,5—8,0
В целом по виду ... 7,8+0,03 Скуловс 0,30 1Я ширина 3,87 90 6,6—8,3
Самцы прибылые 15,8+0,09 0,34 2,17 15 15,2—16,2
взрослые 16,3±0,14 0,63 3,87 19 14,6—17,0
Самки прибылые 15,4+0,12 0,48 3,08 15 14,8-16,6
взрослые 17,0 —• — 1 —
В целом по виду ... 15,9+0,08 Наибольшая 0,60 ширина ч 3,75 ерепа 64 14,6—17,0
Самцы прибылые 20,2+0,09 0,39 1,92 20 19,3—20,7
взрослые 20,8+0,13 0,58 2,80 22 19,2—21,6
Самки прибылые 19,2+0,08 0,44 2,27 28 18,2—20,0
взрослые 20,4+0,67 0,16 5,67 3 19,7—21,7
В целом по виду ... 20,0+0,09 Наибольшая 0,83 высота 41 4,12 ’репа 90 18,2—21,7
Самцы прибылые 12,5+0,06 0,28 2,27 20 12,0—13,0
взрослые 12,5+0,08 0,37 2,98 22 11,4—13,0
Самки прибылые 12,2+0,05 0,28 2,32 28 11,8—12,7
взрослые 12,3+0,51 0,89 7,23 3 11,6—13,3
В целом по виду ... Высот 12,4+0,04 и восходящей 0,36 ветви ним 2,89 ‘ней челюс 90 ти 11,4—13,3
Самцы прибылые 9,4±0,08 0,36 3,83 20 8,6—10,0
взрослые 9,8±0,06 0,28 2,80 22 9,2—10,2
Самки прибылые 8,7+0,05 0,26 2,98 28 8,2—9,2
взрослые 9,3+0,48 0,83 8,98 3 8,6—10,2
В целом по виду ... 9,2+0,06 0,57 6,17 88 8,2—10,2
105
16,2 %). Лапы мощные, бледно-желтого цвета. Во внешнем облике боль-
шой могеры обращает на себя внимание окраска меха, отличающаяся от
таковой у представителей рода Talpa и Astbscalops буровато-коричневым
цветом. Дорсальная сторона тела имеет более насыщенную окраску, чем
вентральная, с металлическим отливом невыношенного меха.- Вентраль-
ная сторона с охристо-серебристым оттенком. На подбородке, горле и
груди яркое ржаво-охристое пятно. У взрослых зверьков на черепе
яркое охристое пятно разных размеров и конфигурации, у самцов оно
крупнее.
Выношенный мех теряет металлический блеск и становится тусклым,
а вновь выросший после линьки волосяной покров буровато-коричневый,
блестящий. Ювенильный мех прибылых зверьков, ведущих самостоятель-
ную жизнь, со значительной примесью серых тонов на шее, груди, вокруг
передних ног, белесовато-серый. На вентральной стороне тела ярких пя-
тен нет, кроме небольшого участка на задней части живота. Через
1,5 мес после выхода из гнезда мех у зверьков становится таким, как
у взрослых.
Волосяной покров состоит из волос трех категорий: направляющих,
остевых и пуховых [Охотина, 1966]. Волосы двух последних категорий
располагаются пучками — остевой волос окружен несколькими пуховы-
ми, а между пучками растут одиночные направляющие волосы. Остевой
волос в летнем меху почти вдвое короче, чём в зимнем. Длина летних во-
лос 4,3—8,2, зимних •— 6—14 мм. Самые длинные остевые волосы распо-
ложены на боках, загривке, хребте и огузке, а самые короткие — на лбу
и вокруг передних ног [Охотина, 1966].
Череп крупный, массивный, значительно превосходящий по разме-
рам таковой у других кротов. Ростральная часть черепа значительно
расширена (15,5 %) по отношению к кондило-базальной длине, в то вре-
мя как у обыкновенного крота это соотношение составляет 12,7, а у си-
бирского — 14,8 %. По относительной длине зубного ряда (41,3 %)
большая могера близка к обыкновенному кроту (41,0 %) и заметно пре-
восходит сибирского (38,8 %), а по показателю отношения длины ряда
коренных зубов превосходит оба этих вида (см. табл. 18). Предглазничный
отдел черепа стройный и его отношение к длине черепа уступает (18,6 %)
таковому обыкновенного (19,2 %) и сибирского (19,6 %) кротов. В то же
время в скуловой области он более широк, а по наибольшей ширине имеет
одинаковые пропорции, как и у этих двух видов, при заметно более низ-
кой относительной высоте (29,6 % — у большой могеры, 31,5 — у обык-
новенного крота и 31,0 % — у сибирского). Обращает на себя внимание
чрезвычайно мощный и высокий коронарный отросток нижней челюсти
(22 % — у могеры, 19,7 — у обыкновенного и 19,6 % — у сибирского
кротов). Его вертикальная ось перпендикулярна продольной оси ниж-
ней челюсти.
Зубы крупные. Средние верхние резцы сравнительно высокие, не-
сколько шире боковых. Верхние клыки высокие и крупные. Рш1 с двумя
корнями; Рш1 и Рш3 крупнее Рш3; Рш4 крупный и широкий. Нижние
резцы сравнительно длинные, косо направлены вперед. Первый нижний
предкоренной (Pmj) чрезвычайно крупный и выполняет функцию отсутст-
вующего в нижней челюсти клыка. Proc, manubrium и collum mallei
образуют прямой угол; apophysis orbicularis отсутствует (рис. 36).
Glans penis короткая, грушевидная, сжатая в дорсовентральном направ-
лении. Тело ее покрыто крупными, редко посаженными шипами, кроме
106
Рис. 36. Череп большой могеры — Mogera robusta.
передней трети дорсальной стороны и самого конца копулятивного органа
с вентральной стороны (см. рис. 35).
Распространение. Ареал вида простирается в пределах
Юго-Восточной Азии, охватывая лесные территории Северо-Восточного
Китая, Корейского полуострова и юга Дальнего Востока СССР. К запа-
ду граница распространения идет примерно до 105° в. д., к востоку до Ти-
хоокеанского побережья. На юге Дальнего Востока проходит северная
граница распространения вида. Он известен из бассейна рек Бекин, Уссу-
ри, оз. Ханка, долины Одарки. Могера заселяет южные районы Приморья
и западные склоны Сихотэ-Алиня, к северу до 47°20' с. ш. (долина
р. Бира).
Детали распространения вида охарактеризованы в работе М. В. Охо-
тиной [1966] и ряда других исследователей [Маак, 1859; Radde, 1862;
Sowerby, 1923; Огнев, 1928; Плятер-Плохоцкий, 1936; Вендланд, 1938;
Abe, 1971; Вон Хон Гу, 1955; Строганов, 1948, 1957; Колосов, 1954; Ку-
ренцов, 1959; и др.].
107
Таблица 19
Морфометрическая характеристика Mogera robusta с юга Дальнего Востока (по
М. В. Охотиной [1966], п = 255)
Признак Иол Возраст
Взрослые (1—4 года) Прибылые (1—11 мес)
Длина
тела <? 165-206 (М = 182) 151—200 (М = 178)
$ 155—195 (М = 173,8) 138—181 (М = 164,2)
хвоста <? 26—37 (М = 30) 24-38 (М = 28,9)
? 15,5—31,0 (М = 23,8) 16,5—33,0 (М = 24,31)
ступни а 24,5—28,0 (М = 25) 20—27 (М = 23,9)
? 18—26 (М = 22,2) 20,5—27,0 (М = 22,4)
Масса тела <? 166—299 (М = 256,5) 122—287 (М = 205)
? 140—280 (М = 178,5) 102—185 (М = 146,3)
Кондило-базальная дли- <? 43,4—46,0 (М = 44,6) 38,9—45,7 (М = 43,1)
на черепа $ 40,2—44,8 (М = 41,8) 38,8—45,2 (М = 40,5)
Наибольшая ширина че- а 20,5—22,0 (М = 21,5) 18,3—21,7 (М = 20,6)
репа 2 19,1—21,0 (М = 20,0) 18,3—21,2 (М= 19,3)
Наибольшая высота че- <? 12,1—13,4 (М = 12,7) 11,3-13,4 (М = 12,6)
репа 2 11,2—13,2 (М = 12,1) 11,5—13,2 (М= 12,2)
Ширина рострума <? 6,5—7,5 (М = 7,1) 6,0—7,5 (М = 6,6)
2 6,1—7,6 (М = 6,9) 5,8-7,3 (М = 6,3)
Длина верхнего ряда зу- <? 16,3—17,6 (М = 16,7) 14,8—17,3 (М = 16,3)
бов 2 15,4—16,6 (М = 15,9) 14,4—16,6 (М = 15,5)
Примечание. Измерения даны в миллиметрах и граммах (масса тела).
Зверьки по территории распределены спорадично, заселяя преиму-
щественно лесные местообитания и периферии прилежащих к ним участ-
ков других ландшафтов. Предпочитают долинные широколиственные леса
и подножия гор с аллювиальными почвами с мощным верхним горизонтом.
Это обстоятельство и определяет характер распространения и распределе-
ния вида по территории.
Материал. 216 экз. из разных районов Приморья и Маньчжурии.
Изменчивость. Половая и возрастная изменчивость большой
могеры во многом сходна с таковой обыкновенного и сибирского кротов.
Размеры тела взрослых самцов достоверно крупнее, чем самок и при-
былых самцов (табл. 19); прибылые самцы крупнее прибылых самок.
Длина хвоста не подвержена большой изменчивости — различия в вели-
чине этого признака у зверьков разного возраста и пола незначительны.
Практически не различается длина ступни. Широко варьирует масса
тела. Самые высокие значения массы у взрослых самцов, затем по убы-
вающей идут прибылые самцы, взрослые самки и прибылые самки (см.
табл. 19).
Аномальная окраска у большой могеры встречается крайне редко.
М. В. Охотина [1966] из 495 исследованных шкурок обнаружила частич-
ный альбинизм только на одной (бело-розовое пятно, простирающееся
на грудь, шею, вокруг одной передней ноги и на загривок).
Изменчивость окраски меха выражается в смене ювенального серо-
вато-бурого меха на блестящий буро-коричневый мех. В ювенальном
108
Рис. 37. Возрастная изменчивость зубов у большой могеры.
мехе на вентральной стороне нет ржаво-оранжевых пятен. У прибылых
зверьков в окраске меха в большой степени проявляются серые тона, осо-
бенно на шее, груди и вокруг передних ног. Выношенный мех взрослых
могер серый, тусклый. После линьки он становится блестящим с преобла-
данием коричневато-бурых тонов.
Возрастная изменчивость прослеживается в степени стирания зубов
и образования гребней.
Кондило-базальная длина черепа у взрослых самцов больше по срав-
нению с другими сочленами популяции. Половая и возрастная изменчи-
вость значения этого признака проявляется в той же последовательности,
что и размеры тела (см. табл. 19). Значения этого признака уменьшаются
от взрослых самцов к прибылым самцам, старым и прибылым самкам.
Возрастная изменчивость значительна и выражается в степени износа
зубов у зверьков разных возрастных групп (рис. 37). У прибылых зверь-
ков, перешедших к самостоятельной жизни, зубы до зимы остаются
островершинными.
У могер в возрасте одного года на всех зубах видна сглаженность
конусов и незначительная притупленность всех вершин, внутренняя сто-
рона клыка несет следы стирания. В возрасте двух лет все вершины ко-
ренных зубов заметно притуплены. Следы стирания хорошо видны и на
премолярах. Заметен компенсаторный рост клыка верхней челюсти. Вер-
шины Рт1 — Рт4 значительно притуплены, а резцы еще мало изменены.
Вершина Рт, заметно сношена. У трехлетних могер вершины всех зубов
сильно сглажены, кроме клыка, вершина которого, хотя и носит следы
изнашивания, но еще довольно острая. Его высота близка к его ширине.
Все коренные сильно сглажены. За счет компенсаторного роста зубы зна-
чительно выступают из альвеол.
В возрасте четырех лет все зубы верхней челюсти, исключая клык
и Рш4, стерты до основания. Вершины коренных сношены до общего
уровня. Расширенная часть резцов полностью сносилась, и они пред-
ставляют несомкнутый ряд пеньков. В нижней челюсти почти все зубы
выровнены. Клык верхней челюсти настолько сношен, что его ширина
у основания превышает длину. Желобок, идущий по наружной боковой
поверхности середины зуба, разделяет его как бы на две части. Все зубы
еще больше выдвинуты из альвеол и стерты почти до десен. Это предель-
ный возраст для большой могеры [Охотина, 1966].
Изменчивость зубной системы, связанная с полиодонтией, олиго-
донтией и многовершинностью, проявляется в значительно меньшей
109
степени, чем у сибирского крота. У 192 экз. большой могеры из Приморья
и Маньчжурии нами зарегистрированы только 11 случаев (5,7 %) откло-
нений. Из них из Маньчжурии у 71,4 %, из Сихотэ-Алиня у 40 %, из
Судзухинского заповедника у 10 и из заповедника Кедровая Падь у
0,6 %. Обнаружены следующие аномалии (усл. обозн. те же, что и для
обыкновенного крота, см. раздел «Изменчивость» в очерке по обыкновен-
ному кроту): — Рт2Л (два случая); — Рт2П (пять); — Рт2ЛП (три);
PrnllW (один). Кроме того, отмечены значительные колебания размеров
Рш4. Примечательно, что все аномалии зубов у большой могеры отно-
сятся только к верхней челюсти, что, вероятно, связано с редукцией ниж-
него клыка, характерной для рода Mogera.
Pm2 и Pm3 имеют одну или две дополнительные вершины. Отмечено
неоднократное отклонение в постановке в челюсти верхних предкорен-
ных: оси их коронок находятся под углом к продольной оси челюсти.
В нижней челюсти такая постановка зубов является нормой. При от-
сутствии одного зуба (Рт2) ряд оставшихся зубов всегда прямой. При
редукции предкоренного зуба одновременно в левой и правой частях че-
люсти зубы расположены параллельно оси челюсти рядами.
Кости тазового пояса, как уже говорилось выше, образуют прочную
монолитную конструкцию с двумя поперечными мостами. Изменчивость
их выражается в вариации размеров, в связи со значительной изменчи-
востью размеров тела у зверьков разного возраста и пола, при сохранении
общего строения и пропорций, характерных для вида.
Заметки по систематике. Впервые описавшие могеру с
юга Дальнего Востока Маак [1861] и Радде [Radde, 1862] ошибочно от-
несли к М. wogura, описанной Темминком [Temminck, 1833]. Видовая
самостоятельность большой могеры была установлена Нерингом [Neh-
ring, 1891 ] по экземпляру из окрестностей Владивостока. Основой для
выделения вида послужили крупные размеры тела, особенности строе-
ния черепа и скелета. Большая могера отличается от средней по ряду
признаков. (Основные различия приведены при описании средней моге-
ры.) Верхние резцы у средней могеры образуют между клыками более
дугообразно вытянутый ряд, чем у большой могеры.
Взгляды исследователей на систематическое положение вида неодно-
значны. Большинство исследователей [Nehring, 1891; Огнев, 1913, 1928;
Sowerby, 1923; Howell, 1929; Kishida, 1930; Строганов, 1948, 1957; Stein,
1960; Охотина, 1966; Abe, 1971; Юдин, 1971а; Grulich, 1971; и.др-] под-
держивает правомерность выделения в составе рода М. robusta. Курода
[Kuroda, 1939] объединил большую могеру со средней и рассматривал
ее в качестве подвида последней (М. w. robusta). Эту позицию принял
Вон Хон Гу [1955] и Имаизуми [Imaizumi, 1955], но последний еще усу-
губил ошибку предыдущего исследователя, отнеся среднюю могеру к ро-
ду Talpa (Т. w. robusta), рассматривая большую могеру в качестве под-
вида средней.
Шварц [Schwarz, 1948] оба вида могер, как и ряд других южно-
азиатских видов подсем. Talpinae, рассматривал как подвиды Eoscalops
micrura [Hodgson, 1840], причем последний им отнесен к роду Talpa.
Убедительная критика несостоятельности этих построений имеется в мо-
нографии С. У. Строганова [1957], который считает неправомерным и
произвольным сведение 12 видов кротов разных родов (Mogera, Parascaptor,
Euroscaptor, Eoscalops и Scaptochirus) в один вид рода Talpa. Поэтому здесь
нет необходимости проводить все контраргументы. Сейчас несостоятельность
110
этой позиции совершенно очевидна. Фактически Шварц [Там же] вер-
нулся к давно отвергнутой систематиками доктрине О. Клейншмидта,
признававшей не реальные биологические виды, а «круг форм» («formen
kreis») и в свое время широко бытовавшей особенно среди немецких систе-
матиков. Работами С. У. Строганова [1948, 1957], Грулиха [Grulich,
1971 ] убедительно доказана самостоятельность рода Mogera. С одной
стороны, представители перечисленных выше родов не являются викари-
рующими формами [Строганов, 1948, 1957], а с другой — они достаточ-
но хорошо очерчены морфологически [Grulich, 1971 ].
В работе М. В. Охотиной [1966] и Б. С. Юдина [1971а] приведены
аргументы в пользу самостоятельности этого вида.
СЕМЕЙСТВО
SORICIDAE FISCHER VON WALDHEIM, 1817 —
ЗЕМЛЕРОЙКИ
Диагноз. Зверьки чаще мелкие, реже средних размеров, мыше-
образной внешности. Ростральная часть головы вытянута в хоботок.
Длина тела 34—180 мм. Глаза маленькие, едва заметные среди волося-
ного покрова. Мех густой, бархатистый. Череп с вытянутым лицевым
отделом. Скуловые дуги отсутствуют. Первые верхние резцы длинные,
с зубцом у основания. Верхние коренные зубы в плане четырехугольные,
с W-образными гребнями на жевательной поверхности, имеют четы-
ре корня.
Зубная формула:
I 3~ °; С 1 —- ? • Pm --~~1 • М 3~2 =44 — 24
3 — 0 ’ 1 — 0 ’ 4 — 1 ’ v 3 — 2 4
Барабанная кость кольцеобразная, расположена горизонтально и
не образует барабанной камеры.
Характеристика. Внешний вид землероек, относящихся
к разным родам, довольно сходен. Размеры землероек варьируют в ши-
роких пределах. Самый мелкий представитель семейства — беЛозубка
малютка (S uncus etruscus) имеет длину тела 34—44 мм, вес 1,2—1,7 г,
самая крупная землеройка из современных — белозубка-голиаф (Ргае-
sorex goliaph) в длину 180 мм, длина хвоста 110 мм, размером с крысу.
Они ведут наземный, полуроющий и полуводный образ жизни [Стро-
ганов, 1957].
Тело землероек умеренно удлиненное. Ушные раковины по размерам
варьируют от рудиментарных, скрытых в меху (Blarina Gray, Апои-
rosorex M.-Edw., Sorex L., Neomys Каир), до хорошо развитых и значи-
тельно выступающих из меха (Crocidura Wagl., Suncus Ehrenb., Diplo-
mesodon Brandt и др ). Конечности у большинства представителей
бегательные, а у приспособленных к водному образу жизни имеют плава-
тельные перепонки (Nectogale M.-Edw.) или щетинистые оторочки вокруг
лап (Neomys Каир.). Длина хвоста варьирует в широких пределах от
очень короткого, не превышающего длину задней ступни (Anourosorex
M.-Edw.), до умеренно длинного, не превышающего длину туловища
(Sorex L., Neomys Каир, Crocidura Wagler и др.), и очень длинного
(рис. 38), превышающего длину тела (Soriculus Blyth, Chodsigoa Kastsch).
Ill
Рис. 38. Хвост и одновершинные зубы у землероек разных родов.
а — Sorex L.; б — Neomys Каир; в — Crocidura Wagler.
Чаще всего стержень хвоста равномерно покрыт короткими упругими
волосами (Sorex L. и др.). У некоторых среди коротких волос хвоста
выступают длинные жесткие щетинки (Crocidura Wagler, Podihik Dera-
niyag.) или на его вентральной стороне развит киль из длинных волос
(Neomys Каир).
Тело зверьков покрыто мягким бархатистым мехом, чаще жестким
и длинным. На губах, лапах и хвосте волосяной покров более жесткий.
Вибриссы развиты хорошо, нередко достигают длины головы. Чаще всего
окраска меха однотонная, бывает чепрачная (Sorex L., Neomys Каир)
или пятнистая (Diplomesodon. Brandt). У большинства видов нижняя
часть тела окрашена светлее верхней. На подошвах развиты мозоли раз-
ной величины и формы. Половое и анальное отверстия окружены общим
кожным кольцом, образующим иногда клоаку. Строение копулятивного
органа отличается большим разнообразием по размерам и конфигурации.
Особенно варьирует форма glans penis. У некоторых она покрыта шипами.
На боках тела или у основания хвоста расположены железы, издаю-
щие специфический мускусный запах.
Череп конический, относительно узкий и длинный: лицевой отдел
в разной степени вытянут у представителей разных родов, умеренно ши-
рокий и сравнительно низкий в затылочной области. Кости черепа еще
в раннем возрасте плотно срастаются и многие швы становятся незамет-
ными. Скуловая дуга редуцирована, имеются лишь скуловые отростки
верхнечелюстной кости.
Своеобразно строение барабанной кости (os tympanicum), она коль-
цеобразная, расположена в отношении основания черепа горизонтально,
не слита с ним и не образует барабанной камеры. Дорсальная стенка
полости среднего уха представлена перепонкой между крыльями клино-
видной (os alisphenoides) и каменной (os petrosum) костями. Основная
клиновидная кость (os basisphenoideum) не имеет слухового отростка.
В отличие от представителей сем. Talpidae в области крыловидной кости
(os pterygoideum) нет вздутия. Предзатылочного отверстия (proc, рагос-
cipitalis) нет.
Нижняя челюсть с полным двойным сочленением. Сочленовный от-
росток (proc, articularis) с двумя разобщенными суставными фасетками —
верхняя сочленяется с чешуйчатой костью, нижняя — с заднесочленов-
ным отростком (proc, postglanoideus). Угловой отросток (proc, angularis)
вытянутый, палочковидный. Венечный отросток развит сильно. Зубы
112
или окрашены в красно-бурый цвет, или белые, или смешанной окраски:
резцы, клыки, предкоренные окрашены, а коренные нет. Редукция идет
преимущественно за счет промежуточных-зубов. Число зубов варьирует
от 44 до 24. Передние резцы верхней челюсти крупные, сильно наклонены
и вытянуты вперед; они серповидно изогнуты с двумя вершинами в виде
лопастей или зубцов. Боковые резцы, клыки и передние предкоренные
одновершинные, конические, широко варьируют по числу зубов, высоте,
соотношению и форме коронок. Они объединены под условным названием
«промежуточные» зубы. У рецентных видов число промежуточных в верх-
ней челюсти 5—2, а в нижней — 3—2. Последний премоляр значительно
моляризован. Коронки верхних коренных брахиодонтные, с семью ко-
нусовидными бугорками на жевательной поверхности, соединены между
собой острыми гребнями. Пара- и метаконус занимают почти срединное
положение. Зубы в плане четырехугольные с РР-образным узором на
вершинах и соединяющих их гребнях. М3— трехвершинный с тремя или
двумя корнями, вытянут в поперечном направлении.
Нижние резцы очень крупные, сильно удлиненные и наклонены
вперед по направлению к оси челюсти; их внутренняя поверхность глад7
кая или с 1—3 зубцами. Рпц— с одной, реже с двумя вершинами.
Рш2— короткий и высокий, с одной или двумя вершинами.
Mj— Ms с ТУ-образной жевательной поверхностью и двумя корнями.
Рукоятка грудины (manubrium sterni) с хорошо развитыми боковы-
ми лопастями на проксимальном конце. Ключица (clavicola) тонкая,
длинная и слегка изогнута. Она причленяется к внутреннему отростку
вилочки акромиального отростка и к рукоятке грудины. Лопатка (sca-
pula) очень длинная, узкая, с хорошо развитым гребнем на ее средней
части и каракоидшям отростком (proc, caracoides). Плечевая кость (hu-
merus) умеренной длины с довольно крупными буграми в проксимальной
и дистальной частях. Бедренные кости с хороню развитыми вертлугами.
Соотношение длины бедра и голени варьирует в зависимости от харак-
тера аллюров и особенностей локомоторных функций у разных предста-
вителей семейства. Шейные позвонки с узкими дугами без остистых от-
ростков. У атланта хорошо развиты боковые лопасти, .сочленяющиеся
с затылочной костью. На пятом позвонке есть боковые отростки, направ-
ленные назад. В отличие от них грудные позвонки несут остистые отрост-
ки, сильно наклоненные кзади, и 14 пар ребер. Крестец образован
четырьмя сросшимися позвонками. Количество и форма хвостовых
позвонков широко изменчивы. Своеобразно строение позвоночника у Scu-
tisorex Thomas: тела позвонков, их дуги и отростки разрастаются, образуя
сплошную ажурную конструкцию- Таз относительно широкий: ширина
его составляет не менее трети длины. Лонное сращение (simphisis pubis)
отсутствует (исключение составляет Anourosorex squamipes M.-Edw.) Боль-
шая и малая берцовые кости в нижней части сращены.
Os penis отсутствует.
Обращают на себя внимание крупные размеры печени, почек, что
обусловлено высоким уровнем обмена веществ у зверьков.
Распространение. Землеройки заселяют обширную тер-
риторию суши, охватывающую Европу, Азию, Африку, Северную и Цент-
ральную Америку и крайний северо-запад Южной Америки. Они за-
селяют все природные зоны от тундр до пустынь включительно, в горах
поднимаются до гольцового пояса. Тем не менее распространение боль-
шинства видов темнейшим образом связано с лесной растительностью-
из
Образ жизни. Исключительно высокий уровень обмена, ха-
рактерный для землероек, обеспечивает им высокую .активность, а.малый
рост — возможность довольствоваться мелкими жертвами и небольшим
количеством пищи при частом употреблении. Мелкие размеры, особен-
ности строения конечностей дают землеройкам возможность проникать
в самые укромные уголки почвенного слоя, завалы растительной ветоши,
пространство между корнями и в щели среди камней, «минировать» снег,
подстилку и рыхлые слои почвы в поисках пищи и укрытий. Особенно-
сти строения зубного аппарата наряду с вытянутой ростральной частью
головы, стремительностью движений обеспечивают возможность быстро
схватывать, удерживать и умерщвлять жертву, расчленять животных
с мягкими тканями и перетирать твердую пищу. Своеобразно устроенные
передние резцы служат универсальным средством для захватывания
жертвы, извлечения содержимого коконов или жирового тела и крупных
насекомых, доставать их из щелей, трещин. Массивный Pm4 с хорошо
развитым прото-, метаконусом и парастилем выполняет функцию «хищ-
ного зуба» и приспособлен для дробления твердых частиц пищи, а остро-
бугорчатые широкие коренные зубы — для ее перетирания. Поэтому,
несмотря на примитивные черты строения зубной системы, в целом это
универсальный аппарат, обеспечивающий потребление зверьками разно-
образной пищи животного и растительного происхождения.
Исключительно важное значение имеет способность зверьков пере-
ключаться с поедания одних кормов на другие. Так, тундряная буро-
зубка в степных и лесостепных районах Западной Сибири предпочитает
поедать различных насекомых и избегает дождевых червей. Этот же вид
на Таймыре, где выбор кормов крайне ограничен, питается исключитель-
но дождевыми червями, которые занимают значительный удельный вес
в питании и других видов землероек в этом регионе. Установлена способ-
ность видов рода Sorex переключаться на более массовые в местах обита-
ния и в определенное время кормовые объекты и в других частях обшир-
ного ареала зверьков — [Юдин, 1955, 1956а, 1957в, 1962в; и др.]. Эти
адаптации в общих чертах сформировались на заре становления семей-
ства, а частные модификации в морфологии отдельных групп возникли
в связи с особенностями конкретных мест обитания. Как адаптацию к
жизни в слое подстилки следует рассматривать и редукцию скуловых
костей, что способствовало возможности проникновения землероек в
различные узкие пространства.
Таким образом, сохранение примитивных черт организации не только
не препятствовало их расцвету, но и способствовало освоению разных
экологических ниш в биогеоценозах. Основные типы строения зубо-че-
люстного аппарата у землероек сложились уже к концу миоцена [Гу-
реев, 1979]. Для Soricini свойствен менее мощный зубо-челюстной аппа-
рат с тонкими ветвями челюстей, приспособленных для удержания и
расчленения мелких, с мягкими тканями животных, а для Heterosoricini,
Blarinini и Anourosoricini характерна мощная нижняя челюсть, приспо-
собленная для удержания большой добычи. Представители подсем. Сго-
cidurinae занимают как бы промежуточное положение между ними, но
ближе стоят к Soricinae. По сравнению с бурозубками у них более укоро-
ченная лицевая часть черепа, челюсти и зубы более мощные, зубной ряд
короткий.
Это достигается разными путями: за счет сокращения числа проме-
жуточных зубов или резкого сокращения размеров коронок отдельных
8 Б. С. Юдин
114
зубов и одновременного сочетания того и другого. Для бурозубок рода
Sorex характерно наличие пяти промежуточных зубов (варианты соотно-
шения размеров различные — размер зубов последовательно убывает от
первого к пятому, они образуют парные группы, отдельные зубы выше
или ниже других); у представителей Blarina, Microsorex промежуточных
зубов также пять, но третий и пятый сильно редуцированы; у Neomys
и Suncus промежуточных четыре; для Crocidura, Notiosorex, Chimarrogale
и других характерно наличие трех промежуточных, первый очень круп-
ный и два мелких; у Diplomesod.on, Anourosorex — два промежуточных
зуба.
Такое разнообразие строения зубного аппарата — результат пище-
вой специализации видов. В то же время, представители рода Sorex, про-
межуточные зубы и резцы которых весьма полиморфны у разных видов,
распространены циркумполярно, заселяют разные природные зоны и
ландшафты, в разных районах питаются разнообразно, что свидетель-
ствует об универсальности их зубного аппарата, позволяющего охотиться
за животной добычей и поедать растительный корм. Однако обитание их
в сходных условиях делает сходной их кормовую базу. Универсальность
зубо-челюстного аппарата, мелкие размеры и высокая активность играют
решающую' роль в их жизни.
Современные представители подсем. Soricinae расширили свой ареал
за счет проникновения в более северные районы, а белозубые землеройки
(подсем. Crocidurinae) сократили его по сравнению с плиоценом и плей-
стоценом, исчезнув в его северных частях [Гуреев, 1971].
В настоящее время наиболее процветающими являются бурозубые
землеройки, а среди них представители рода Sorex, и белозубые — род
Crocidura. Первые заселяют преимущественно умеренную зону Пале-
арктики и Неарктики, а вторые главным образом — тропики и субтропи-
ки Старого Света.
Филогенетические связи разных групп бурозубок достаточно под-
робно рассмотрены А. А. Гуреевым [1979].
Заметки по систематике. В работах, посвященных
систематике землероек Старого и Нового Света, как частных, так и об-
зорных, недостатка нет. Однако сложность таксономии группы настоль-
ко велика, что представления о системе землероек, ранжировке отдельных
таксонов (от вида и до подсемейства включительно) у разных исследова-
телей нередко противоречивы.
Изучение систематики землероек началось с работ К. Линнея [Lin-
naeus, 1758], описавшего обыкновенную бурозубку (Sorex araneus L.),
и продолжается по настоящее время. Кауп [Каир, 1829] описал род Neo-
mys, а на протяжении следующей половины второго столетия были опи-
саны еще 12 родов, два подсемейства: Soricinae Murray, 1866, Crocidurinae
M.-Edw., 1868—1874 и одно надсемейство Soricoidea Gill, 1872. В первой
половине нашего столетия были описаны еще семь родов рецентных видов
землероек и подсем. Scutisoricinae Allen, 1917. В 1971 г. А. А. Гуреев
описал подсемейство вымерших землероек — Saturniniinae (верхний
эоцен, Европа).
Ряд авторов разработали и предложили несколько классификаций
землероек. Так, Симпсон [Simpson, 1945] в составе семейства выделял
три подсемейства: Soricinae Murray, Crocidurinae M.-Edw. и Scutisoricinae
J. Allen. Репеннинг [Repenning, 1967] и Ван Вален [Van Valen, 1967]
все рецентные виды объединяли в два подсемейства — Soricinae и Сто-
115'
Таблица 20
Структура фауны землероек СССР по представлениям разных
исследователей
Автор Число видов в роде Всего
Sorex N eomys Suncus Crocidura Diplome- sodon
С. И. Огнев [1928, 1931, 1935] 21 3 1 10 1 36
С. И. Огнев [1951] 17 2 1 8 1 29
Н. А. Бобринский и др. [1944] 6 2 1 4 1 14
Бобринский и др. [1965] 9 2 1 4 1 17
Эллерман и Моррисон- Скотт [Ellerman, Mor- rison-Scott, 1951] 8 2 1 4 1 16
А. А. Гуреев [1971] 18 3 1 9 1 32
А. А. Гуреев [1979] 15 3 1 11 1 31
А. А. Гуреев [1981 ] 16 3 1 10 1 31
Б. С. Юдин [1965а, б, 19706] 17 3 1 — 1 —
cidurinae. А. А. Гуреев [1979] всех современных землероек свел в одно
подсем. Soricinae Fischer, выделив в его составе ряд триб (Soricini Fi-
scher, Crocidurini Stirton, Blarinini Stirton, Anourosoricini Stirton).
Исследователи, предложившие эти классификации, опирались на
разные признаки и имели противоречивые взгляды на филогенетические
взаимоотношения и объем таксонов. В нашей работе нет необходимости
обсуждать достоинства и недостатки предложенных построений. Они
сопоставляются в монографии А- А. Гуреева [1979], посвященной насе-
комоядным мира. Здесь лишь уместно еще раз подчеркнуть сложность
таксономии этого семейства, что хорошо иллюстрируется анализом взгля-
дов разных исследователей на его систематику.
Нет единого,взгляда и на систематику землероек СССР. С. И. Огнев
[1928], которого с полным правом можно считать основоположником
систематики насекомоядных отечественной фауны, внес выдающийся вклад
в разработку этого вопроса. Им приводится (включая дополнение ко II
и III томам) 20 видов в роде Sorex L., три — в роде Neomys Капр, 10 —
в роде Crocidura Wagler, один — в роде Pachyura (Suncus) Ehrenb. и
один — в роде Diplomesodon Brandt. В последней работе взгляды С. И. Ог-
нева [1951] на систематику землероек значительно изменились: число
видов во всех родах сократилось (табл. 20).
Как видно из данных табл. 20, общности взглядов на видовой состав
трех родов из пяти, представленных в фауне СССР, нет не только у раз-
ных авторов, но и у одних и тех же исследователей. Так, С. И. Огнев
[1928, 1931, 1935] в роде Sorex принимает 21 вид, Neomys — 3, Crocidu-
ra — 10, а в 1951 г. [Огнев, 1951] в этом роде число видов сократилось
до 17, Neomys — до 2, Crocidura — до 8. У Н. А. Бобринского и А. IL Ку-
зякина [Бобринский и др., 1944, 1965], стоящих на позиции «укрупне-
ния» видов, число их значительно меньше, чем у других исследователей,
причем в первом издании «Определителя...» [Бобринский и др., 1944]
8*
116
было всего 14 видов землероек (по сравнению с 36 и 29, по С. И. Огневу
[1928—1951]), а во втором — уже 17. В трех из четырех публикаций
А. А. Гуреева приводится разный состав бурозубок (15, 18, 16 видов) и
белозубок (9, 11, 10 видов).
Все это еще раз указывает на сложность таксономии землероек и
необходимость продолжения исследований в этом направлении. Расхож-
дение в видовом составе во многом объясняется еще и тем, что нередко
авторы строят свои представления, не опираясь на новый фактический
материал, а умозрительно, без должных оснований, произвольно пере-
тасовывая виды землероек. Причина такого положения в значительной
мере кроется в отсутствии необходимых коллекционных материалов в
хранилищах страны, отражающих размах изменчивости видов.
В изучении систематики сибирских бурозубок большой вклад внес
ученик и последователь С. И. Огнева профессор С. У. Строганов, опуб-
ликовавший серию частных исследований и обобщающую монографию
[Строганов, 1957], в которых на новой фактической основе была сделана
попытка ревизии таксономии сибирских насекомоядных. Эта работа,
ценная сама по себе, послужила основой для дальнейшей разработки
систематики насекомоядных Сибири, в том числе и землероек.
Взгляды разных исследователей на объем видов землероек, обитаю-
щих в Сибири, здесь не обсуждаются. Они детально рассмотрены ниже,
в соответствующих видовых очерках.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
ПОДСЕМЕЙСТВ И РОДОВ СЕМ. SORTCIDAE
1(4). Вершины зубов окрашены в красно-бурый цвет. В заглазничной
области крыши черепа имеются два сближенных отверстия. Хвост равно-
мерно покрыт короткими, равными по длине волосами; киля или высту-
пающих длинных волос на хвосте нет.............................
............................Soricinae — Бурозубые землеройки.
2(3). На нижней стороне хвоста волосы не удлинены и не образуют
«киля» (см. рис. 38, а). Ступня необрамлена плавательной оторочкой из
жестких волос. Промежуточных зубов в верхней челюсти по пять с каж-
дой стороны (см. рис. 38, а). Всего зубов 32...................
........................................Sorex L.— Бурозубки
3(2). На нижней стороне хвоста волосы удлинены и образуют «киль»
(см. рис. 38, б). Ступня обрамлена плавательной оторочкой из жестких
волос (рис. 39). Промежуточных зубов в верхней челюсти четыре с каж-
дой стороны (см. рис. 38, б). Всего зубов 30...................
.......................................Neomys Капр — Куторы
4(1). Вершины зубов не окрашены в красно-бурый цвет, белые. В за-
глазничной области крыши черепа отверстий нет. В волосяном покрове,
равномерно покрывающем хвост, среди коротких волос редко рассеяны
длинные, торчащие в стороны жесткие волосы (см. рис. 38, в) ... .
.............................Crocidurinae — Белозубые землеройки
5(3). Промежуточных зубов в верхней челюсти по три с каждой сто-
роны (см. рис. 38, в). Всего зубов 28..........................
...................................Crocidura Wagler — Белозубки
117
ПОДСЕМЕЙСТВО
SORICINAE FISCHER VON WALDHEIM, 1817 —
БУРОЗУБЫЕ ЗЕМЛЕРОЙКИ
Род Sorex Linnaeus, 1758 —
Бурозубки
Тип рода. Sorex araneus L., 1758.
Диагноз. Промежуточных зубов по пять с каждой стороны верх-
ней челюсти. Вершины зубов окрашены в красно-бурый цвет.
Зубная формула:
I -Ь Din -Ь Pm М = 32.
Ввиду отсутствия у исследователей единого мнения о порядке ре-
дукции резцов, клыков и премоляров (промежуточных зубов) мы вслед
118
119
Рис. 40. Строение glans penis бурозубок.
2 — S. unguiculatus Dobson; 2 — S. isodon Turov; 3 — S. caecutiens Laxm.', 4 — S. roboratus
HoH.; 5 — S. tundrensis Mer.; 6 — S. araneus L.; 7 — 8. gracillimus Thomas; л — S. minutus L.; 9 —
8. cinereus Kerr; 10 — S. beringianus Yudin; 11 — 8. minutissimus Zimm. ,12 — 8. daphaenodon Thomas.
за А. П. Кузякиным [Бобринский и др., 1965] объединяем их (кроме
Рт4) в группу dentes intermediens (Din).
Характеристика. Бурозубые землеройки преимущественно
мелкие или средние зверьки. В составе рода самый мелкий представи-
тель — крошечная бурозубка (S. minutissimus Zimmer.). Длина тела
30—53, кондило-базальная длина черепа 12,0—14,3 мм, масса тела 1,4—
4,5 г*. Самый крупный — гигантская бурозубка (5- mirabilis Ognev).
Длина ее тела достигает 72—91, кондило-базальная длина черепа 22,7—
24,2 мм, масса тела 11—14 г. По габитуальным признакам все сочлены
рода очень схожи между собой, их внешние видовые отличия невелики.
Для них характерна типичная «землеройковая» внешность: вытянутый
в хоботок конец морды, почти полностью скрытые в меху ушные ракови-
ны, в виде невысокого кожистого валика, лапы без плавательных пере-
понок или щетинистых оторочек (кроме S- cinereus Kerr). Хвост равномер-
но покрыт жесткими волосами с длинной кисточкой на конце
(см. рис. 38, а). Длина хвоста у разных видов и даже подвидов колеблет-
ся в широких пределах — от 1/2 до 2/3 длины тела. Мех, покрывающий
тело, мягкий, бархатистый. Окраска волосяного покрова верхней стороны
тела коричнево-бурая разной интенсивности и оттенков. Низ тела чаще
всего окрашен светлее, чем дорсальная сторона, иногда почти чисто белый.
Сосков три пары. Половое и анальное отверстия не образуют клоаки.
Penis относительно размеров тела большой, S-образно изогнутый. Строе-
ние glans penis разнообразно и специфично для каждого вида (рис. 40),
что является надежным критерием для установления видового статуса
перезимовавших самцов [Юдин, 19656; Долгов, Лукьянова, 1966].
В строении черепа бурозубок обращает на себя внимание сильно
вытянутая, резко сужающаяся тонкая и длинная лицевая часть. Мозго-
вая капсула резко расширена, овальная, у разных видов значительно
варьирует по высоте и ширине (табл. 21). На лобных костях два неболь-
ших сближенных отверстия. Краниометрические признаки видов под-
вержены значительной географической и возрастной изменчивости [Deh-
nel, 1949; Строганов, 1957; Межжерин, 1964, 1965а; Долгов, 1966; и др.].
Вершины всех зубов красновато-коричневые, с большим размахом
видовой, популяционной и индивидуальной изменчивости интенсивности
окраски. Промежуточных зубов по пять с каждой стороны верхней че-
люсти (см. рис. 38, а). Конфигурация, размеры, соотношение и форма
зубов в пределах рода весьма изменчивы, но устойчивы для каждого вида.
Изменчивость проявляется в отклонениях размеров отдельных элементов
и изменчивости окраски и в исключительно редких случаях в виде олиго-
и полиодонтии. Первый верхний резец крупный, с двумя вершинами.
Соотношение их размеров у разных видов неодинаково. Нижний резец
сильно вытянут в длину и наклонен вперед по отношению к продольной
оси нижней челюсти. На его верхней, режущей, поверхности не менее
трех ясно выраженных лопастей. Рт4 крупный, сильно моляризован,
6 Это максимальная масса, которая характерна для перезимовавших беременных
самок.
120
Таблица 21
Сравнительная характеристика соотношения отделов черепа разных видов бу-
розубок
Вид Кондило-ба- зальная дли- на, мм Наибольшая ширина, мм Отношение наибольшей ширины к кондило-ба- зальной дли- .не, % Межглавнпч- ная ширина, мм Отношение межглазнич- ной ширины к кондило- базальной длине, %
У. mirabilis 23,3+0,10 10,6+0,06 45,4 4,8+0,04 20,6
S. roboratus 20,3+0,03 9,5+0,02 46,7 3,7+0,01 18,2
S. unguiculatus 19,9+0,05 10,2+0,03 51,3 4,2+0,01 20,2
S. araneus 19,7+0,03 9,7±0,02 49,2 3,8±0,02 19,2
S. isodon 19,5±0,11 10,0+0,20 51,0 3,9±0,01 20,0
S. daphaenodon 17,9±0,12 9,1+0,02 50,8 3,7+0,01 20,6
S. tundrensis 17,7+0,02 8,8+0,01 49,7 3,4+0,01 19,2
S. caecutiens 17,4±0,03 8,7+0,01 50,0 3,3+0,01 18,9
S. cinereus 16,2±0,03 7,9+0,02 48,7 2,9+0,02 17,9
S. gracillimus 15,7±0,05 7,5±0,03 47,7 2,9+0,02 18,4
S .beringianus 15,5+0,14 7,6+0,13 49,0 2,8+0,07 18,1
S. minutus 15,1±0,03 7,3+0,02 48,3 2,7+0,01 17,8
S. minutissimus 13,3+0,03 6,3+0,02 47,3 2,8±0,01 21,0
Окончание табл. 21
Вид Предглазничная ширина, мм Отношение пред- глазничной ши- рины к кондило- бавальной длине, % Ширина ростру- ма, мм Отношение шири- ны рострума к 'к ондило-базаль- ной длине, %
5. mirabilis 3,6+0,04 15,4 2,6+0,03 11,1
S. roboratus 2,6+0,01 12,8 2,0±0,02 9,8
S. unguiculatus 2,8+0,08 14,0 2,0+0,01 10,0
S. araneus 2,7±0,02 13,7 1,9±0,02 9,6
S. isodon 2,7±0,01 13,8 1,9±0,01 9.7
S. daphaenodon 2,4±0,01 13,4 1,7+0,01 9,4
S. tundrensis 2,4+0,01 13,5 1,7+0,01 9,6
S. caecutiens 2,3+0,01 13,2 1,6+0,01 9,1
S. cinereus 2,2+0,01 13,5 1,4+0,01 8,6
S. gracillimus 2,0+0,01 12,7 1,4+0,01 8,9
S. beringianus 2,1+0,03 13,5 1,3+0,04 8,4
S. minutus 2,0±0,01 13,2 1,4±0,01 9,2
S. minutissimus 1,9+0,01 14,2 1,3+0,01 9,7
на задней стороне коронок Pm4, М1, М2 широкая и глубокая борозда,
проходящая по всему талону, начиная от основания протоконуса между
мезостилем и гипоконусом. Вследствие этого талон скошен сверху вниз.
Если зубы рассматривать сверху, то борозда имеет вид полулунной вы-
резки, через которую видна верхнечелюстная кость. Величина ее у раз-
ных видов неодинакова, наиболее сильно она выражена у S. roboratus
и менее всего у S- daphaenodon. На жевательной стороне коронок между
121
Рис, 41. Строение зубов верхней челюсти бурозубок,
а — S* daphaenodon; б — S. araneus.
мезостилем и протоконусом расположена глубо-
кая пирамидальная впадина или воронка с ром-
бическим основанием.
У S. daphaenodon в отличие от других ви-
дов бурозубок гипоконус последнего верхнего
предкоренного (Pm4) всегда хорошо развит и
густо пигментирован. Метаконус этого зуба и ме-
таконусы М1 и М2 массивные, с тупыми краями.
Полулунная выемка (рис. 41) на задней стороне
коронок многовершинных зубов верхней челю-
сти слабо выражена. В отличие от всех других
видов пирамидальная впадина на жевательной
стороне между мезостилем и протоконусом отсут-
ствует на первом и втором коренных, а на третьем
она зачаточна. Все конусы на верхних коренных
низкие и массивные по сравнению с таковыми
других видов. Зубной ряд широкий, зубы в нем
плотно сомкнуты- Не менее характерно строение
нижних коренных. Петли на жевательной сто-
роне первого и второго коренного не равнове-
лики: задние петли значительно крупнее перед-
них (приблизительно на 1/3 своей величины).
Задняя петля третьего нижнего коренного лишь
немного уступает по величине передней. Пигментация эмали зубов
верхней и нижней челюстей интенсивная, в подавляющем большинст-
ве случаев кроваво-темного цвета [Строганов, 19571. Структура
зубного аппарата крупнозубой бурозубки настолько самобытна, непов-
торима, в отличие от других видов рода, что мы нашли необходимым
выделить ее в ранг особого подрода, назвав в честь известного советского
териолога, большого знатока насекомоядных млекопитающих С. У. Стро-
ганова — Stroganovia Yudin, subgen. n.
У всех бурозубок воронки заднего нижнего коренного с пятью ко-
ническими бугорками. Первый нижний предкоренной зуб специфичен,
У разных видов соотношение высоты и длины его коронки, а также ко-
личество дополнительных вершин и степени их развития не одинаковы.
Представители рода известны в ископаемом состоянии с нижнего
миоцена в Европе и с верхнего плиоцена в Азии и Северной Америке
[Огнев, 1933а, б; Гуреев, 1963, 1979]. В составе рода объединены наименее
специализированные виды насекомоядных. Они эволюционировали в
направлении приспособления к жизни в лесных биогеоценозах, питанию
мелкими беспозвоночными и охоте на них в слое подстилки, мохового
покрова, в естественных нишах, норах и добыче их на поверхности почвы.
Мелкие размеры бурозубок — результат адаптации к жизни в слое под-
стилки и снижению абсолютного количества потребления пищи [Меж-
жерин, 1962; Юдин, 1962].
Распростр анение. Область распространения представи-
телей рода охватывает обширную территорию Европы, Азии до Северо-
122
Западного Памира и Тибета включительно, Северную Америку й север-
ную оконечность Южной Америки.
Заметки по систематике. Точное число видов не уста-
новлено. В фауне СССР, по нашим представлениям, обитает 18 видов,
из них 13 — в Сибири.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА SOREX
1(2). В нижней челюсти постмандибулярного отверстия нет
(рис. 42, а)...................................Otisorex De Kay
2(1). В нижней челюсти постмандибулярное отверстие есть . . . 3
3(4). На внутренней стороне переднего верхнего резца есть хорошо
выраженный зубец, направленный вперед и в сторону такого же зубца
на соседнем зубе...........................Ognevia Dolg. et Heptn.
4(3). На внутренней стороне переднего резца хорошо выраженного
зубца нет.................................................... 5
5(6). Второй промежуточный зуб верхней челюсти ниже первого и
третьего или равен третьему. Размеры тела мелкие, кондило-базальная
длина черепа не превышает 18,3 мм...............Eurosorex Strog.
6(7). Соотношение промежуточных зубов иное. Второй промежуточ-
ный зуб выше первого и третьего, вершины зубов овально притупленные,
округлые. Коренные зубы массивные, ряд их широкий. Метаконусы Рт1,
М1, М2 хорошо развиты, с тупыми краями, пирамидальной впадины на
жевательной стороне коронок, М1 и М2 между мезостилем и протоконусом
нет........................................... Stroganovia Yudin
7(2). На жевательной стороне коренных зубов хорошо выражена пира-
мидальная впадина. Размеры крупные и средние .... Sorex L.
В составе рода в свое время было описано три подрода. Однако в
практике таксономических работ отечественных систематиков они, за
небольшим исключением [Юдин, 1971а], не использовались, в то время
как американские териологи применяют эту категорию для выделения
внутри рода группировок генетически близких видов со сходными при-
знаками.
К подроду Otisorex De Kay относится 20 видов бурозубок [Hall, Kel-
son, 1959], 19 из которых эндемичны для североамериканского Конти-
нента, один обитает в Палеарктике и Неарктике (5. cinereus Kerr), и один
пока известен только из Палеарктики (8. beringianus Yudin). Подрод
выделен на основании отсутствия в нижней челюсти постмандибуляр-
ного канала.
В подрод Ognevia Dolg. et Hept, пока входит один резко уклоняю-
щийся по ряду габитуальных признаков и строению зубного аппарата
вид — гигантская бурозубка (S. mirabilis Ognev), заселяющая Вьетнам,
Корею и юг советского Дальнего Востока. Основанием для выделения
подрода послужило наличие дополнительного зубца на внутренней сто-
роне переднего верхнего резца. В американской фауне есть вид (8. vag-
rans Baird), имеющий сходное строение переднего резца [Юдин, 19696].
В подроде Eurosorex Strog. объединены мелкие виды бурозубок (5. mi-
nutissimus Zimm., S. minutusY,., S. buchariensis Ognev, 8. kozlovi Strog.),
у которых второй промежуточный зуб верхней челюсти меньше первого
и третьего. Кроме того, у них много общих черт в строении glans penis
123
и мелкие размеры тела. Такое соотношение зубов и у когтистой буро-
зубки (S. unguiculatus Dobson), но она значительно крупнее их по разме-
рам и характеризуется совершенно иным типом строения glans penis.
Подрод Stroganovia Yudin, subsp. n. включает только один, резко
уклоняющийся вид — бурозубку крупнозубую (5. daphaenodon Thomas),
характеризующуюся совершенно иным строением зубной системы, чем
все остальные виды рода Sorex Палеарктики и Неарктики.
Подрод Sorex объединяет зверьков крупных и средних размеров, об-
щим для всех является наличие на нижней челюсти постмандибулярного
отверстия, на жевательной стороне коренных зубов хорошо выражена
пирамидальная впадина. Подрод объединяет семь видов бурозубок, оби-
тающих в Сибири.
Подрод Otisorex De Kay, 1842
Представители подрода характеризуются отсутствием постманди-
булярного отверстия в нижней челюсти (рис. 42, а). В составе подрода
19 видов, широко распространенных в Неарктике, один из них заходит
в Палеарктику. В Палеарктике два вида.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА OTISOREX
1(2). Первый промежуточный зуб крупнее второго, третий крупнее
четвертого. Кондило-базальная длина черепа 14,6—16,8 (М = 16,2) мм.
Первый верхний резец крупный, его задняя вершина значительно меньше
первой и отогнута вбок (наружу), сверху оторочена бороздкой
(см. рис. 44). Череп широкий и высокий в области мозговой капсулы . .
...............Sorex cinereus Kerr — Трансарктическая бурозубка
2(1). Соотношение промежуточных зубов иное. Первый промежуточ-
ный зуб меньше или равен второму, третий меньше двух предыдущих
и четвертого. Кондило-базальная длина черепа 15,0—15,8 (М = 15,5) мм.
Окраска меха трехцветная, чепрачная..............................
............Sorex beringianus Yudin — Берингийская бурозубка
Рис. 42. Нижняя челюсть бурозубок.
а — Otisorex (постмандибулярного канала нет): б — Sorex (постмандибулярный канал есть) .
124
Sorex cinereus Kerr, 1792 —
Трансарктическая бурозубка
Голотип. Вид описан по экземпляру из форта Северная Канада.
Диагноз. Относительно мелкая бурозубка рода Sorex. По разме-
рам она немного уступает таким видам, как S. daphaenodon Thomas, 1907;
S. tundrensis Merr., 1900; S. caecutiens Laxm., 1788 и лишь немногим
больше S. gracillimus Thomas, 1907; S. beringianus Yudin, 1967 и S. mi-
nutus L., 1766.
Измерения. Длина тела с головой 45—66 (М = 58,4+0,25),
длина хвоста 26—54 (М = 44,4+ 0,36), кондило-базальная длина черепа
14,6—16,8 (М = 15,8) мм. См. также табл. 22.
Характеристика. Зверек стройного телосложения (рис. 43,
44), о чем можно судить уже по тому, что при почти равных со средней
бурозубкой размерах она значительно уступает последней по массе. Масса
средних бурозубок из бассейна Колымы, добытых в одних биотопах и
в одно время с S. cinereus, достигает 10 г, редко бывает менее 5 г, в то
время как подавляющее большинство трансарктических бурозубок весит
4—4,5 г и только отдельные зверьки достигают массы 5,5 г. Отношение
массы к длине тела у прибылых зверьков составляет 7,4 %, для S- cae-
cutiens этот показатель равен 8,0 %. По внешнему виду она больше всего
напоминает берингийскую бурозубку (S. beringianus Yudin).
Окраска меха дорсальной стороны тела землисто-бурая с примесью
пепельного цвета, распространяющаяся на более светлые бока коричнево-
палевых тонов. Окраска брюшной стороны светло-серая. Колориметри-
ческая характеристика меха дорсальной части тела указывает на высокие
значения показателя белизны — 3,8—6,1, М = 4,95(±0,12) и незначи-
тельную насыщенность рыжих тонов — 114,3—138,8, М — 125,4(± 1,20).
Рис. 43. Трансарктическая бурозубка — У. cinereus Кегг.
125
Таблица 22
Морфометрическая характеристика Sorex cinereus
Возрастная и половая группа а cv п lim
1 2 3 5 6
& шна тела с головой
Самцы прибылые 58,3+0,32 1,85 3,2 34 46—62
взрослые 59,1±1,41 4,22 7,1 9 45—66
Самки прибылые 58,5+0,36 1,92 3,3 29 56—63
взрослые 56,5+1,05 2,10 3,7 4 45—59
В целом по виду ... 58,4+0,25 2,32 4,0 85 45-66
Длина, хвоста
Самцы прибылые 45,3+0,64 3,79 8,4 35 33-54
взрослые 44,0+0,80 2,52 5,7 10 26—48
Самки прибылые 44,9±0,59 3,18 7,1 29 39—53
взрослые 43,0+1,06 2,13 4,9 4 41—45
В целом по виду ... 44,6+0,36 3,31 7,4 87 26—54
Длина ступни
Самцы прибылые 13,2+0,08 0,47 3,5 35 12—14
взрослые 13,0+0,12 0,39 3,0 10 13-14
Самки прибылые 13,1+0,09 0,51 3,9 29 12—15
взрослые 13,1+0,10 0,20 1,5 4 13—14
В целом по виду ... 13,1+0,05 0,47 3,6 87 12-15
Масса, тела
Самцы прибылые 4,3+0,05 0,30 7,1 34 4-5
взрослые 4,8+0,29 0,87 18,2 9 3,7—6,0
Самки прибылые 4,4+0,07 0,35 8,0 28 4—5
взрослые 4,1+0,13 0,25 6,1 4 4-4,5
В целом по виду ... 4,4+0,05 0,45 10,2 83 3,7—6,0
Кондило-базальная длина черепа
Самцы прибылые 16,1+0,05 0,31 1,9 46 15,3—16,7
взрослые 16,1+0,11 0,29 1,8 7 14,6—16,5
Самки прибылые 16,2+0,05 0,25 1,6 32 15,7—16,8
взрослые 16,1+0,03 0,06 0,4 3 16—16,1
В целом по виду ... 16,2+0,03 0,28 1,7 96 14,6—16,8
Длина рострума
Самцы прибылые 4,1+0,03 0,23 5,5 46 3,7—4,7
взрослые 4,1+0,06 0,18 4,3 9 3,9-4,4
Самки прибылые 4,2±0,04 0,21 4,9 32 3,7—4,6
взрослые 4,3+0,03 0,06 1,4 4 4,2—4,3
В целом по виду ... 4,2±0,02 0,21 5,0 99 3,7—4,7
Длина мозговой части черепа
Самцы прибылые 9,3+0,05 0,29 3,1 39 8,8—10,2
взрослые 9,0±0,09 0,25 2,8 8 8,5—9,2
Самки прибылые 9,3±0,04 0,20 2,1 27 9,0—9,6
взрослые 8,9±0,07 0,12 1,3 3 8,8—9,0
В целом по виду ... 9,3+0,03 0,28 3,0 84 8,5—10,2
126
Продолжение табл. 22
1 I 2 | 3 | 4 | 5 | 6
Длина верхнего ряда зубов
Самцы прибылые 7,0+0,03 0,17 2,5 46 6,6—7,5
взрослые 6,6+0,11 0,34 5,2 9 6,6—7
Самки прибылые 7,0+0,03 0,17 2,4 32 6,7—7,5
взрослые 6,9+0,05 0,10 1,4 4 6,8—7,0
В целом по виду ... 6,9±0,02 0,23 3,2 99 6,0—7,5
Длина ряда промежуточных зубов
Самцы прибылые 2,2+0,02 0,12 5,5 46 2—2,5
взрослые 2,2±0,04 0,13 5,9 9 2—2,4
Самки прибылые 2,2±0,02 0,11 4,8 32 2—2,4
взрослые 2,3+0,06 0,13 5,5 4 2,1-2,4
В целом по виду ... 2,2+0,01 0,12 5,2 99 2—2,5
Ширина рострума
Самцы прибылые взрослые 1,4+0,02 1,4±0,04 0,16 0,11 11,1 7,6 46 9 1,2—1,8 2—2,4
Самки прибылые 1,4±0,02 0,12 8,9 32 1,2-1,8
взрослые 1,4+0,03 0,05 3,6 4 1,3—1,4
В целом по виду ... Пре 1,4+0,01 •дглазничная 0,14 миринс 9,6 99 1,2—1,8
Самцы прибылые 2,1+0,02 0,13 6,1 46 1,9—2,5
взрослые 2,1±0,05 0,15 7,0 9 2,0—2,4
Самки прибйлые 2,2+0,02 0,13 6,1 32 2,0—2,5
взрослые 2,3+0,03 0,66 2,6 4 2,2—2,3
В целом по виду ... 2,2+0,01 0,13 6,0 99 1,9—2,5
Межглазничная ширина
Самцы прибылые 2,9+0,02 0,17 5,7 46 2,6—3,4
взрослые 2,9±0,05 0,15 5,0 9 2,7—3,1
Самки прибылые 2,9+0,03 0,15 5,1 32 2,7—3,4
взрослые 2,9±0,05 0,10 3,3 4 2,8—3,0
В целом по виду ... 2,9±0,02 0,15 5,2 99 2,6—3,4
Наибольшая ширина черепа
Самцы прибылые 7,9+0,03 0,19 2,4 46 7,5-8,4
взрослые 7,7+0,08 0,22 2,8 8 7,4—8,0
Самки прибылые 7,9+0,04 0,21 2,5 32 7.5—8,3
взрослые 7,6+0,03 0,06 0,8 4 7,5—7,6
В целом по виду ... 7,9±0,02 0,21 2,6 98 7,4—8,4
Наибольшая высота черепа
Самцы прибылые 5,3±0,03 0,17 3,2 46 5,0—5,7
взрослые 4,6+0,08 0,23 4,9 9 4,2—4,8
Самки прибылые 5,3+0,04 0,20 3,8 32 4,7—5,6
взрослые 4,4±0,10 0,19 4,3 4 4,3—4,7
В целом по виду ... 5,2±0,03 0,32 6,1 99 4,2—5,7
127
Окончание табл. 22
2 3 4 5 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 3,2±0,02 0,12 3,7 46 3,0—3,5
взрослые 3,2±0,03 0,10 3,1 10 3,0-3,4
Самки прибылые 3,3±0,02 0,10 2,9 32 3,1—3,5
взрослые 3,3±0,03 0,05 1,5 4 3,2-3,3
В целом по виду ... 3,2±0,01 0,11 3,3 100 3,0-3,5
Длина углового отростка
Самцы прибылые 1,9±0,02 0,12 6,0 38 1,7—2,2
взрослые 1,9±0,04 0,10 4,9 7 1,8—2,0
Самки прибылые 2,0±0,02 0,09 4,5 27 1,7—2,1
взрослые 1,9±0,03 0,06 3,0 3 1,9—2,0
В цел ом по виду ... 1,9±0,01 0,10 5,3 83 1,7—2,2
По сравнению со средней бурозубкой из этого же района (рис. 45)
зверьки достоверно отличаются как по оттенку, так и по белизне —
у 5. caecutiens 2,7-5,0, М = 4,08(±0,11) и 127,0-150,0, М =
*= 136,9 (±1,19) — признаки эти изменчивы. Диапазон изменчивости
окраски S. cinereus по этим двум признакам почти одинаков. При этом
нужно иметь в виду, что колориметрическое исследование окраски S. ci-
nereus проведено только из одного пункта и у одного подвида (S. с. camts-
chatica). При исследовании материала из разных частей ареала, возмож-
но, широта изменчивости окажется большей.
Рис. 44. Череп трансарктической бурозубки.
128
Рис. 45. Сравнительная характеристи-
ка окраски средней и камчатской транс-
арктической бурозубок из бассейнов
рек Колымы и Омолон.
1 — S. caecutiens; 2 — S. с. camtschaiica
Yudin*
Хвост по длине весьма измен-
чив. У зверьков из тундры он до-
стоверно короче, чем у бурозубок
таежных популяций из более юж-
ных районов (табл. 23). Стержень
хвоста покрыт редкими, не торча-
щими в стороны волосами, сквозь
которые видны кожные покровы.
Окраска хвоста двухцветная:
верхняя сторона его темно-бурая,
нижняя — белесая. Этот вид по
максимальным показателям отно-
сится к наиболее длиннохвостым бурозубкам (табл. 24): по отношению
длины хвоста к длине тела он уступает (М. 76,3 %) только гигантской
бурозубке (81,6 %), дальневосточной (77,4%) и берингийской (77,4%). При-
чем различия в длине хвоста с двумя последними видами минимальные.
Череп относительно размеров зверька короткий (27,7 %), меньше
он только у S. unguiculatus (табл. 25), широкий и высокий в мозговой
части (48,7 %). Мозговая капсула округлая, ее длина лишь немногим
превышает ширину. В межглазничпой области череп чрезвычайно узкий
(17,9 %) и уступает по этому показателю только черепу S. minutus. Ростг
рум короткий (25,9 %). По отношению ширины рострума к кондило-
базальной длине этот вид относится к числу зверьков, обладающих че-
репом с наиболее узкой ростральной частью (8,6 %). Более узкий рост-
рум имеет только S. beringianus (см. табл. 21).
Обращает на себя внимание относительно короткий зубной ряд.
Индекс этого показателя к кондило-базальной длине черепа — один из
самых небольших (42,5 %) и только едва превышает таковой у S. minutus
(42,4 %) и равен этому показателю у S. beringianus. Отношение длины
промежуточных зубов к кондило-базальной длине черепа несколько мень-
ше, чем у S. cinereus (13,5 %), только у S. beringianus (12,9) и 5. minutis-
simus (12,8), также как и длина промежуточных зубов к длине зубного
ряда (у S. cinereus 31,8 %) (табл. 26).
Первый верхний резец трансарктической бурозубки по длине состав-
ляет столько же, сколько три следующих промежуточных зуба вместе
взятые (см. рис. 44). Характерный признак этого вида — желобок на
наружной боковой поверхности верхнего резца, особенно хорошо выра-
женный у S. с. porterikol между его передней и задней вершинами. Задняя
вершина несколько отогнута наружу (рис. 46, а), а желобок охватывает
ее спереди и сверху. С внутренней стороны на передних вершинах рез-
цов в месте их соприкосновения имеются хорошо развитые дополнитель-
ные лопасти (см., рис. 46, б). Первый двухвершинный резец наиболее
интенсивно пигментирован, четыре промежуточных зуба окрашены почти
одинаково, пятый — белый.
Промежуточные зубы по сравнению с мелким черепом относительно
крупные, плотно посажены в зубном ряду, с высокими коронками и дву-
129
Таблица 23
Морфометрическая характеристика подвидов Sorex cinereusns разных частей ареала
S. с. camtschatica
Признак S. cinereus, Аляс- ка, юго-восточная Канада, США S- с. portenkoi, СССР, Чукотка, бас- сейн р. Анадырь СССР, северо-вос- ток Сибири, бас- сейн р. Омолон СССР, Камчатка, бухта Камбальная и оз. Ажабачье
Крайние варианты п X Крайние варианты п X Крайние варианты п X Крайние варианты п X
1 2 3 4 5 6 7 8 9 10 11 12 13
Длина тела 44—49 15 46 45—50 12 48 55—66 64 58 54-62 21 57
хвоста 35—42 15 40 26-34 12 28,9 39—47 65 43 42-54 22 48
ступни 10—11 15 11 10—11 12 10 12—14 65 13 11—14 22 14
Масса тела — — — — 1 3,7 3,7—5,5 64 4,3 4,0—6,0 20 5,0
Длина кондило-ба- зальная черепа 15,4-16,6 14 15,9 14,6-15.8 12 15,3 15,6—16,7 63 16,1 15,3-16,8 34 16,0
рострума 3,5—4,6 15 4,1 3,7—4,0 12 3,8 3,8—4,7 64 4,2 3,7—4,5 35 4,0
мозгов ой части 8,7—9,6 14 9,0 8,4-9,5 12 8,9 8,5-9,5 63 9,0 8,9—10,2 33 9,4
Ширина рострума 1,2—1,4 И 1,3 1,3—1,6 12 1,4 1,2-1,5 64 1,4 1,2—1,8 35 1,3
предглазнич- ная 2,0—2,3 И 2,0 2,0—2,2 12 2,0 2,0—2,4 64 2,2 2,0—2,5 35 2,1
межглазнич- ная 2,7—3,2 15 2,9 2,7—3,1 12 2,9 2,6—3,1 64 2,9 2,7—3,4 35 3,0
наибольшая черепа 7,6—8,2 14 7,8 7,3—8,2 12 7,9 7,4—8,3 63 7,8 7,7—8,4 34 8,0
Наибольшая вы- сота черепа 4,1-5,3 14 4,5 4,7—5,2 10 5,1 4,2-5,7 52 5,3 4,7-5,7 35 5,2
Длина верхнего ря- да зубов 6,3—7,2 11 6,9 6,5—6,9 12 6,7 6,3*—7,5 64 6,9 6,7—7,2 35 6,8
промежуточ- ных зубов 2,1—2,4 И 2,2 2,0—2,2 12 2,0 2,0—2,5 64 2,1 2,0-2,4 35 2,2
Высота венечного отростка 3,2—3,6 15 3,3 3,0—3,2 к 12 3,0 3,0—3,4 65 3,2 3,0—3,5 37 3,2
Длина углового отростка 1,7—2,0 11 1,8 1,6—2,0 12 1,8 1,7-2,0 63 1,9 1,7—2,2 20 1,8
* Этот показатель значительно ниже, чем у других зверьков (6,7—6,9), измерен старый зверек с
сильно стертыми зубами.
парной группировкой первых четырех (см. рис. 44). Несмотря на то
что первый промежуточный несколько крупнее второго, вершина послед-
него несколько выше, что обусловлено более высоким его положением
на челюсти. Этот признак весьма характерен для диагностики вида и
представляется надежным отличием от S. caecutiens и S. minutus. Много-
вершинные зубы относительно размеров черепа крупные, в отличие от
промежуточных расставлены редко, с хорошо выраженной полулунной
выемкой на задней стороне, через которую видны небные кости. Гипо-
конусы многовершинных развиты слабо и не пигментированы.
Рт, вытянут в сагиттальном направлении: длина основания его ко-
ронки в 2 раза больше высоты. Высота восходящей ветви нижней челюсти
не более 3,5 мм.
Видоспецифично для трансарктической бурозубки строение полово-
го аппарата самцов и самок. У перезимовавших самцов glans penis корот-
9 Б. С. Юдин
130
Таблица 24
Сравнительная характеристика некоторых мсрфсметркческих признаков
разных видов бурозубок
Вид Длина тела с голо- вой, мм Длина хвоста, мм Отношение длины хвоста к длине тела, %
S. mirabilis 83,6+ 1.56 68,3+0,75 81,6
S. roboratus 72,0±0,45 37,7+0,28 52,4
S. unguiculatus 74,4+0,76 48,3+0,48 64,9
S. araneus 68,9+0,45 42,5+0,22 65,6
S. isodon 69,9+0,45 44,1+0,23 63,0
S. daphaenodon 60,1±0,53 32,3±0,28 53,7
S. tundrensis 63,4±0,26 33,9+0,23 53,4
S. caecutiens 60,5±0,28 36,1±0,22 59,6
S. cinereus 58,4+0,25 44,6+0,36 76,3
S. gracillimus 54,9+0,94 43,5+0.42 79,2
S. beringianus 52,3+2,25 40,5+0,29 77,4
S. minutus 52,2+0,59 36,9+0,32 70,6
S. minutissimus 44,8+0,35 24,2+0,27 54,0
кая, цилиндрическая, без обособленной коронки. Мочеполовой канал
открывается в центре тупо обрезанного переднего конца головки, на со-
сочке выступающем из небольшого углубления. По конфигурации ко-
пулятивного органа этот вид ближе всего к S. beringianus, относящегося
к одному с ним подроду (Otisorex). Clans penis этих двух видов резко отли-
чается по форме от других представителей рода фауны СССР. Строение
копулятивных органов североамериканских видов, за небольшим исклю-
чением [Юдин, 1967а, 19696, 19726], не изучено.
Таблица 25
Сравнительная характеристика некоторых морфометрических признаков раз-
ных видов бурозубок
Вид Длина тела с голо- вой, мм Кондило-базальная длина черепа, мм Отношение кондило- базальной длины к длине тела, %
S. mirabilis 83,6±1,56 23,3+0,01 27,8
S. roboratus 72,0+0,45 20,3+0,03 28,1
S. unguiculatus 74,4+0,76 19,9+0,05 26,7
S. araneus 68,9+0,45 19,7+0,03 28,5
S. isodon 69,9±0,45 19,5±0,11 27,8
S. daphaenodon 60,1+0,53 17,9+0,12 29,7
S. tundrensis 63,4+0,26 17,7+0,02 27,9
S. caecutiens 60,5+0,28 17,4+0,03 28,7
S. cinereus 58,4+0,25 16,2+0,03 27,7
S. gracillimus 54,9+0,94 15,7+0,05 28,5
S. beringianus 52,3+2,25 15,5±0,14 29,6
S. minutus 52,2+0,59 15,1+0,03 28,9
S. minutissimus 44,8+0,35 13,3+0,03 29,6
131
Таблица 26
Сравнительная характеристика зубного ряда разных видов бурозубок
Вид Длина, мм Отношение, %
кондило-ба- зальиая верхнего ря- да зубов ряда проме- жуточных зу- бов зубного ряда к кондило- Зазальной длине ряда про- межуточ- ных зубов к конди- ло-базаль- ной длине ряда про- межуточ- ных зубов к длине зубного ряда
5. mirabilis 23,3+0,01 10,3+0,05 3,3+0,04 44,2 14,1 32,0
S. roboratus 20,3+0,03 9,0±0,02 3,0+0,01 44,3 14,8 33,3
S, unguiculatus 19,9+0,05 8,8±0,35 3,0±0,01 44,2 14,6 32,9
S. araneus 19,7+0,03 9,0±0,02 3,1+0,01 45,7 15,7 34,4
S. isodon 19,5±0,ll 8,8±0,02 3,0±0,01 45,1 15,3 34,0
S. daphaenodon 17,9±0,12 8,0±0,02 2,6+0,01 44,7 14,5 32,5
S. tundrensis 17,7+0,02 7,8±0,01 2,5±0,01 44,0 14,1 32,0
S. caecutiens 17,4+0,03 7,7+0,02 2,7+0,01 44,2 15,5 35,1
S. cinereus 16,2+0,03 6,9±0,02 2,2+0,01 42,5 13,5 31,8
S. gracillimus 15,7+0,05 6,8±0,02 2,5±0,01 43,9 15,9 36,8
S. beringianus 15,5+0,14 6,6+0,05 2,0+0,02 42,6 12,9 30,3
S. minutus 15,1+0,03 6,4±0,01 2,2+0,01 42,4 14,5 34,3
.S', munutissimus 13,3+0,03 5,7+0,03 1,7+0,01 42,8 12,8 29,8
Своеобразна форма семенников взрослых самцов трансарктической
бурозубки. Они в отличие от таковых других видов почти шаровидные,
с незначительной разницей продольного и поперечного диаметров.
В строении матки обращают на себя внимание непропорционально
длинные, тонкие рога, при весьма коротком влагалище, что соответст-
вует короткой головке копулятивного органа (см. рис. 40).
Изменчивость. Несмотря на большой ареал этой бурозубки
на территории СССР, изменчивость ее морфологических признаков весьма
широкая. Возрастная изменчивость проявляется так же, как и у других
видов: стертый волосяной покров на хвосте, кистях, ступнях не восста-
навливается, поэтому степень его износа пропорциональна возрасту
зверьков.
Распространение и характер соотношений темной и светлой окраски
у зверьков разного возраста следующие: у прибылых зверьков окраска
меха по белизне близка к таковой у перезимовавших в летнем наряде
(4,8 и 4,7), по несколько уступает окраске старых зверьков в зимнем на-
ряде (5,2). Показатель оттенка также близок у прибылых зверьков и
перезимовавших в летнем наряде (126,1 и 125,1), а зимний наряд послед-
них значительно темнее (115,1).
Рис. 46. Строение передних
верхних резцов S. cinereus раг-
tenkoi Strog.
а — дополнительные лопасти хоро-
mo развиты; б — то же, на месте
соединения правого и левого рез-
цов.
9*
132
По размерам тела прибылые самцы несколько уступают перезимо-
вавшим, прибылые самки, напротив, немного крупнее. Прибылые разно-
полые зверьки между собой по длине тела не различаются. Взрослые сам-
цы намного больше одновозрастных самок. Длина хвоста у землероек
этого вида изменчива в широких пределах: у прибылых самцов и самок
достоверно больше, чем у перезимовавших (см. табл. 22).
Возрастные различия по массе тела более заметны у самцов. При-
былые самцы и самки по этому признаку не различаются, а перезимо-
вавшие самцы крупнее самок.
Возрастная специфика в стирании зубов заключается в том, что быст-
рее других снашивается самая высокая вершина второго промежуточного
зуба. В результате этого у старых особей вершины первых четырех про-
межуточных выравниваются, зубной ряд становится похож на таковой
средней бурозубки, отличаясь лишь более плотной постановкой зубов в
челюсти.
Наряду с возрастной, половой и сезонной изменчивостью у этого вида
четко выражена и географическая. К сожалению, серии шкурок 5. cine-
reus имеются только из бассейна Колымы, поэтому мы не можем говорить
о географической изменчивости окраски.
Хорошо выражена географическая изменчивость длины тела зверь-
ков из разных популяций. Более мелкие зверьки обитают на северо-
американском континенте (см. табл. 23). В фауне СССР самые мелкие
землеройки заселяют тундры крайнего северо-востока Сибири. По ма-
териалам, собранным автором в бассейне р. Анадырь, длина тела ко-
леблется от 45 до 50 мм, а кондило-базальная длина черепа — 14,6—
15,8 мм. Значительно крупнее землеройки из таежной зоны северо-восто-
ка Сибири. Серия бурозубок, собранных нами в бассейне Колымы
(р. Омолон, долина р. Кегали), отличается большей длиной тела (55—66)
и кондило-базальной длиной черепа (15,6—16,7 мм). Близкие значения
этих признаков имеют зверьки с Камчатки.
Географическая изменчивость лучше всего прослеживается в длине
хвоста (см. табл. 23). Самые крупные зверьки по длине тела, ступни,
кондило-базальной длине черепа обитают в бассейне Колымы, а самые
мелкие — в тундре Чукотского полуострова. Наиболее длиннохвостые
бурозубки заселяют Камчатку. Аналогичная закономерность постепен-
ного перехода короткохвостых зверьков к длиннохвостым от тундры к
югу прослеживается и на североамериканском континенте.
Распространение. Трансарктическая бурозубка — пред-
ставитель неарктической фауны на азиатском континенте — относится к
подроду Otisorex, представители которого широко распространены на
территории американского континента от Аляски до 35° с. ш. на юг и на
прилежащих островах. В СССР она известна с крайнего северо-востока
Сибири и с Камчатки, где была собрана в бассейне р. Палеваам, в районе
поселков Уэлеп и Эгвекинот, на побережье зал. Креста [Долгов, Криво-
шеев, 1973]; в бассейне верхнего течения р. Омолон (в долине ее правого
притока — р. Кегали); в бассейне р. Анадырь, в районе Марково, р. Снеж-
ная, Танюрер, на побережье Анадырского залива, близ пос. Анадырь.
На Камчатке зверьков этого вида ловили в районе оз. Ажебачье и бухты
Камбальная [Юдин, 1969, 1973].
Заметки по систематике. Систематическое положение
трансарктической бурозубки (5. cinereus Kerr) до последнего времени бы-
ло предметом широкой дискуссии и оставалось неопределенным. В ряде
133
работ отрицалась ее видовая самостоятельность. Бринк [Brink, 1952,
1953] без достаточных оснований произвольно объединил такие гетеро-
генные элементы, как S. caecutiens Laxm., S. cinereus Kerr, S. lyelli Mere.,
S. preblei Jackson, в искусственный композиционный вид — S. caecutiens,
в состав которого кроме палеарктических видов вошли и неарктические.
Сведения о S. cinereus в фауне нашей страны появились в печати не-
давно. С. У. Строганов [1956а, 1957] по материалам Л. А. Портенко и
нашим сборам впервые установил обитание S. cinereus на северо-востоке
Сибири и описал новый подвид S. cinereus portenkoi Strog. Отсутствие в
его распоряжении взрослых самцов лишило описание S. cinereus убеди-
тельности, что послужило причиной для разногласий. Строение glans
penis этою вида стало изучаться лишь недавно [Юдин, 19696, 1972д].
Позднее систематическое положение 5. cinereus рассматривалось с раз-
ных позиций и неоднозначно [Гуреев, 1963; Бобринский и др., 1965;
Долгов, 1966; Юдин, 1965а, 1969а — 1973в]. Вследствие отсутствия но-
вых фактических материалов, а также из-за тенденциозного подхода к
систематике бурозубок большинство авторов не признавало самостоя-
тельности этого вида и вслед за Бринком [Brink, 1952, 1953] считало
его синонимом S. caecutiens и даже — S. minutus [Бобринский и др.,
1965].
А. А. Гуреев [1963] не упоминает об обитании S. cinereus в СССР,
хотя работы С. У. Строганова [19566, 1957] к этому периоду уже уви-
дели свет. С. С. Шварц [1963] выразил сомнение в реальности ее обита-
ния в Палеарктике, а А. П. Кузякин [Бобринский и др., 1965] произ-
вольно отнес землероек из долины р. Анадырь (5. с. portenkoi) в качестве
подвида S. minutus. Вслед за ним В. А. Долгов [1966] поставил под сом-
нение реальность обитания в СССР S. cinereus, считая ее спорным видом
нашей фауны, но в более поздних работах [Долгов, Кривошеев, 1973;
Долгов, Реймерс, 1979] ее присутствие в Сибири уже не вызывало
сомнений.
В настоящее время собран достаточно большой новый фактический
материал, позволивший разобраться не только в систематическом поло-
жении вида, но и во внутривидовой таксономии. Автором исследованы
155 экз. S. cinereus из Северной Америки (Канада, США), северо-восточ-
ной Сибири и с Камчатки. В результате установлена ее видовая самостоя-
тельность и несостоятельность априорных выводов Бринка [Brink, 1952,
1953 ] о ее идентичности S. caecutiens. Специфичные для каждого вида га-
битуальные, краниологические и одонтологические признаки, а также
кардинальные отличия в строении половых органов самцов и самок де-
лают видовую самостоятельность S. cinereus, S. caecutiens и S. minutus
очевидной [Юдин, 1969, 1971—1973]. В отличие от других представите-
лей рода Sorex, обитающих в СССР (кроме S. beringianus), трансаркти-
ческая бурозубка относится к подроду Otisorex, a S. caecutiens — подроду
Sorex. Американские зоологи Хоффманн и Петерсон [Hoffmann, Peter-
son, 1967], ознакомившись с бурозубкой из долины р. Анадырь, также
отнесли их к S. cinereus.
Трансарктическая бурозубка отличается от S. caecutiens мелкими раз-
мерами (длина тела колеблется от 45 до 66 (М = 58,4±0,25), в то время
как длина тела средней бурозубки составляет 51—76 (М = 60,5±
±0,28) мм. Кондило-базальная длина черепа S. cinereus, по нашим дан-
ным колеблется от 14,6 до 16,8 (М = 15,8) мм, а по Холлу и Кельсону
134
[Hall, Kelson, 1959] — 14,6—16,9 мм. У средней бурозубки кондило-
базальная длина черепа составляет 16,1—19,1 (М = 17,4±0,02) мм.
(табл. 27).
Обнаруживаются отличия этих видов, выражающиеся в конфигура-
ции черепа и строении зубов. Череп средней бурозубки более массивный
и широкий. Ширина черепа S. caecutiens 7,8—10,0 (М — 8,7±0,01),
а у S. cinereus — 7,4—8,4 (М = 7,9—0,02) мм.
Приводим основные различия между трансарктической и средней
бурозубками:
Признак
В нижней челюсти
постмандибуляр-
ный канал
Передняя верши-
на первого верх-
него резца
Передние вершины
левого и правого
резцов
г <
Промежуточные зу-
бы
S.cinereus
Нет (см. рис. 42, а)
Крупнее и выше задней, по-
следняя отогнута наружу.
Желобок на наружной бо-
ковой поверхности этого
зуба, между передней и
задней вершинами, выра-
жен хорошо и, огибая зад-
нюю вершину, заходит до
края зуба (см. рис. 44)
Расставлены широко, между
ними видны хорошо раз-
витые дополнительные ло-
пасти (см. рис. 46)
Относительно крупные, с вы-
сокими коническими ко-
ронками. Вершина второго
промежуточного зуба выше
таковой первого и треть-
его. В зубном ряду они си-
дят плотно (см. рис. 44)
Glans penis (см.
рис. 40)
4,4—5,3 мм, поперечный диа-
метр 1 мм. Матка инфан-
тильных самок с длинными
рогами (8,2—10,8 мм) и
коротким влагалищем
(3,2—4,0 мм). Стержень
копулятивного органа без
заметного разграничения
постепенно переходит в
округлую головку
S. caecutiens
Есть (см. рис. 42,6)
Незначительно превышает
по размерам и высоте зад-
нюю. Последняя не отог-
нута наружу. Желобок на
наружной боковой поверх-
ности этого зуба не выра-
жен (см. рис. 81)
Плотно прижаты между со-
бой, дополнительные ло-
пасти на их внутренней
поверхности зачаточны
Относительно мелкие, низ-
кие, с сильно расширен-
ными в сагиттальном на-
правлении основаниями.
Вершина второго проме-
жуточного зуба ниже пер-
вой или на одном уровне
с ним. В зубном ряду они
сидят свободно (см.
рис. 81)
5,0—8,5 (М = 6,4) мм, тол-
щина основания 0,8—1,1
(М = 0,9) мм. Матка ин-
фантильных самок с уме-
ренной длины рогами
(6,0—6,6 мм) и удлинен-
ным влагалищем (5,0—
6,2 мм). Головка цилинд-
рическая, с хорошо выра-
женной, сферически закруг-
ленной коронкой на сво-
бодном конце. С дорсаль-
ной стороны свободного
конца колонки хорошо вы-
ражен крючкообразный
вырост
Приведенные характеристики убедительно показывают на коренные
отличия в организации S. cinereus и S. caecutiens.
А. П. Кузякин [Бобринский и др., 1965] рассматривает S. cinereus
в качестве подвида S. minutus. Позиция этого исследователя базируется
на некотором внешнем сходстве этих двух видов.
Прежде всего эти два таксона относятся к двум разным подродам
(Sorex и Otisorex), что само по себе является решающим аргументом в
135
пользу их различия на видовом уровне. Постмандибулярный канал у
5. cinereus отсутствует, у S. minutus он есть. Однако они отличаются не
только габитуально. По размерам тела S. cinereus крупнее S. minutus
(у первого длина тела с головой М = 58,4±0,25, кондило-базальная
длина черепа М = 16,2±0,03), у второго соответственно (52,9=1=0,59 и
15,14=0,03 мм). Лишь землеройки из зоны тундры близки по размерам к
малой бурозубке, а подвид, заселяющий таежную зону, отличается зна-
чительно больше, чем по приведенным выше средним показателям вида.
Длина хвоста у S. cinereus, особенно у S. с. camtschatica, значительно
больше, чем у S. minutus. Имеются убедительные отличия в строении по-
ловых органов самца и самки.
Описанные выше отличия не оставляют сомнений в видовой самостоя-
тельности этих двух таксонов. Кроме того, прослеживаются со всей
определенностью отличия и в структуре зубной системы: у S. cinereus
вершина второго зуба выше таковой первого и третьего, у S. minutus
высота второго зуба всегда меньше высоты первого и третьего.
Копулятивный орган взрослых особей этих видов отличается как по
размерам, так и по форме. У 5. minutus, уступающей по размерам тела
S. cinereus, копулятивный орган непропорционально крупный (его длина
8,3—11,4 (М = 9,3) мм, а толщина у основания 1,0—1,1 мм). Кроме того,
они отличаются и по форме (см. рис. 40). Все это еще раз указывает на
отсутствие оснований для их объединения в один вид.
Видовая самостоятельность S. cinereus подтверждается и кариологи-
ческими исследованиями. Эта бурозубка входит в группу видов с высо-
ким диплоидным числом (2п — 58—66) хромосом, к которой кроме S. ci-
nereus относится североамериканский вид S. fumeus Miller и западный
палеаркт S. alpinus Schinz. В то время как S. caecutiens и S. minutus,
с которым некоторые авторы объединяют рассматриваемую землеройку,
относятся к группе с 2п = 42.
Хромосомный набор S. cinereus, исследованный у зверьков из Север-
ной Америки [Meylan, 1968] и с территории СССР [Воронцов, Крал,
1986], одинаков. У этого вида 2п = 62 (NF = 70), в то время как у
S. caecutiens 2п = 42, NF — 68, а у S. minutus 2п = 42, NF{ — 56.
Обзор подвидов
В Северной Америке описаны 11 подвидов S. cinereus [Hall, Kelson,
1959]. В СССР — три: S. с. portenkoi Strog., S. с. camtschatica Yudin и
S. c. cecutinoides Strog. С. У. Строганов [1956a] описал подвид транс-
арктической бурозубки S. с. portenkoi Strog. с Чукотского полуострова
(по материалам из бассейна р. Анадырь) и S. с. cecutinoides Strog. [Стро-
ганов, 1957] из Якутии. В результате сравнительного изучения новых ма-
териалов из районов северо-востока Сибири, послуживших автору для
выделения S. с. cecutinoides, и дополнительных материалов из места пер-
воописания мы пришли к выводу о том, что все исследованные зверьки
относятся не к S. cinereus, а к S. caecutiens. В нижней челюсти у них
имеется постмандибулярный капал. Сомнение в реальности этого подви-
да, вероятно, возникло у автора еще при его первоописании, о чем сви-
детельствует само название «cecutinoides», т. е. похожая на S. caecutiens.
Поэтому S. с. cecutinoides как подвид S. cinereus нами не обсуждается.
Трансарктические бурозубки с Камчатки значительно отличаются от
тундровых популяций. У них большие размеры тела, черепа и особенно
136
Морфометрическая характеристика
Признак S. сгпетеиз
М±тх а Су п lim
Длина
тела 58,4±0,25 2,32 4,0 85 45—66
хвоста 44,6±0,36 3,31 7,4 87 26—54
ступни 13,1±0,05 0,47 3,6 87 12—15
Масса тела Длина 4,4±0,05 0,45 10,2 83 3,7—6,0
кондило-базальная черепа 16,2±0,03 0,28 1,7 96 14,6—16,8
рострума 4,2±0,02 0,21 5,0 99 3,7—4,7
мозговой части Ширина 9,3±0,03 0,28 3,0 84 8,5-10,2
рострума 1,4+0,01 0,14 2,6 99 1,2—1,8
предглазничная 2,2+0,01 0,13 6,0 99 1,9—2,5
Межглазничная 2,9—0,02 0,15 5,2 99 2,6—3,4
наибольшая черепа Длина 7,9+0,02 0,21 2,6 98 7,4—8,4
верхнего ряда зубов 6,9±0,02 0,23 3,2 99 6,0—7,5
ряда промежуточных зубов 2,2+0,01 0,12 5,2 99 2,0—2,5
1 Гаиболыпая высота черепа Длина восходящей ветви ниж- 5,2±0,03 0,32 6,1 99 4,2—5,7
ней челюсти 3,2+0,01 0,11 3,3 100 3,0—3,5
Длина углового отростка 1,9+0,01 0,10 5,3 83 1,7—2,2
длина хвоста. По этим признакам они выделены в самостоятельный под-
вид— S. с. camtschatica Yudin [Юдин, 1972д]. Позднее [Юдин, 1973а]
установлена принадлежность к нему бурозубок из районов таежной зоны
северо-востока Сибири (бассейн р. Омолон).
Sorex cinereus portenkoi Stroganov, 1956 —
Восточносибирская трансарктическая бурозубка
Голотип. Описан по экземпляру, добытому Б. С. Юдиным
12/VIII 1954 г. на побережье Анадырского залива, близ пос. Анадырь.
№ 2328, коллекция С. У. Строганова. Голотип хранится в Зоологическом
институте АН СССР, № 45059 (Ленинград) (рис. 47).
Диагноз. Размеры мелкие. Длина тела не превышает 50 мм, дли-
на хвоста не более 35 мм, что составляет в среднем 59 % от длины тела.
Кондило-базальная длина черепа 14,6—15,8 (М = 15,3) мм.
Измерения. См. табл. 23.
Характеристика. Зверьки отличаются стройным телосложе-
нием, длинным хвостом, отороченным хорошо выраженной щеткой волос,
большой ступней. Длина верхнего ряда зубов 6,5—6,9 (М — 6,7), длина
ряда промежуточных зубов 2—2,2 (М = 2,0) мм.
Распространение. Западная часть Чукотского полуостро-
ва, р. Палеваам близ впадения в Чаунскую губу; восточная часть полу-
острова — долина р. Анадырь от с. Марково до г. Анадырь.
137
Таблица 27
Sorex cinereus и Sorex caecutiens
s. caecutiens Достоверность различий по Стьюденту (Td)
О CV п lim
60,5±0,28 6,53 10,79 556 45—78 5,6
36,1±0,22 5,15 14,27 555 26—60 20,1
11,7±0,05 1,05 9,01 527 10—14 19,8
5,3±0,07 1,49 27,97 521 3,0—11 10,5
17,4±0,03 0,87 4,97 812 16,1—19,4 28,3
4,6±0,61 0,34 7,39 834 3,9—5,6 17,9
9,8±0,02 0,48 4,84 812 8,8—11,4 13,9
1,6±0,01 0,13 8,10 834 1.1—1,9 14,1
2,3±0,01 0,30 12,93 834 2,0—2,8 7,1
3,3±0,01 0,18 5,64 834 2,7—3,7 17,9
8,7±0,01 0,42 4,48 809 7,8—10,0 35,8
7,7±0,02 0,46 5,98 831 6,7—8,8 28,3
2,7±0,01 0,23 8,36 832 2,2—3,2 35,4
5,6±0,01 0,35 6,31 808 4,8—6,3 12,7
4,0±0,01 0,29 7,22 823 3,5—4,6 56,6
2,1±0,01 0,13 6,19 702 1,7—2,4 14,1
Материал. 1 экз., Чукотка, р. Палеваам, 12 экз., р. Анадырь.
Заметки по систематике. С. У. Строганов 11956а], вы-
деляя трансарктическую бурозубку с Чукотского полуострова в само-
стоятельный подвид, отличал ее от неарктических бурозубок этого вида
по малым размерам тела и черепа. По размерам тела восточносибирская
бурозубка слабо отличается от своих североамериканских сородичей:
длина ее тела 45—50 мм, а североамериканских землероек 46—61 мм;
длина хвоста соответственно 26—34 и 35—42 мм, длина ступни одного
размера 10—11 мм IHall, Kelson, 1959]. Данные по североамериканским
землеройкам приводятся для вида в целом (см. табл. 23). Если взять
крайние значения, то наименьшие мало чем отличаются от таковых чу-
котских бурозубок. По этим признакам они близки между собой. Отсутст-
вие серийных материалов из Северной Америки не позволяет сделать
определенный вывод о их взаимоотношении с S. с. portenkoi.
Sorex cinereus camtschatica Yudin, 1972 —
Камчатская трансарктическая бурозубка
Г о л о т и п. Описан по экземпляру из сборов В. Г. Воронова. № 481
(кол.), д', 18/VIII 1967 г., Камчатка (бухта Камбальная). Коллекция
зоологического музея Биологического института СО АН СССР, инв.
№ 2342 (Новосибирск).
138
Рис. 47. Череп восточносибирской трансарктической бурозубки.
Диагноз. Отличается относительно крупными размерами тела,
длинным хвостом, длинной ступней. Кондило-базальная длина черепа
колеблется от 15,3 до 16,8 (ЛГ = 16,1) мм.
Измерения. См. табл. 23.
Характеристика. Основанием для выделения камчатской
трансарктической бурозубки в таксон подвидового ранга послужили
крупные размеры зверька, особенно длинный хвост, составляющий 79 %
от длины тела. У восточносибирской бурозубки длина хвоста составляет
59 % длины тела. По этому признаку землеройки хорошо различаются
между собой. Для камчатских землероек характерна сильно удлиненная
задняя ступня и большая, чем у землероек с Чукотского полуострова,
кондило-базальная длина черепа.
Наиболее примечательны для камчатского подвида длинный хвост —
на Камчатке 42—54 (М = 48), в долине Омолона 39—47 (М = 43) мм и
большая ступня — 11—14 (М = 13,5) мм с хорошо выраженной щеткой
упругих щетинистых волос, обрамляющих ее, как у куторы. Тот и дру-
гой признаки характерны для островных форм землероек.
Распространение. Известна из бассейна р. Омолон (долина
р. Кегали), южной части Камчатки (зал. Камбальный) и восточного по-
бережья Камчатки (оз. Ажабачье), окрестности Мильково.
Материал. 75 экз., бассейн р. Омолон. Камчатка: 7 экз., зал.
Камбальный и 29 экз., оз. Ажабачье.
Заметки по систематике. S', с. camtschatica обнаружи-
вает сходство с близким американским видом S. hydrodromus Dobson с
о. Уналашка (Алеутские острова). Последнюю Хелле и Кельсон [Hall,
Kelson, 1959] считают идентичной S. arcticus Kerr [Hoffmann, Peterson,
1967] и рассматривают ее как синоним S. pribilofensis Mere., а А. А. Гу-
139
Рис. 48. Строение верхней
челюстей Sorex hydrodromus
лашка).
(а) и^нижней (б)
Dobson (о. Уна-
реев [1971] полагает, что она должна быть отнесена в качестве подвида
S. cinereus. Независимо от исхода решения таксономической стороны
вопроса эта землеройка по ряду внешних признаков близка к камчатско-
му подвиду трансарктической бурозубки и американским видам, засе-
ляющим острова, расположенные в непосредственной близости от северо-
восточной окраины нашей страны и северо-западного побережья Аляски.
Исследованный нами экземпляр S. hydrodromus (рис. 48) с о. Уналаш-
ка имел сходные с S. с. camtschatica габитуальные данные [Юдин, 19696].
У североамериканских бурозубок наблюдается та же тенденция широт-
ной изменчивости некоторых внешних признаков, что и у бурозубок се-
веро-востока азиатского континента. У S. cinereus с Аляски длина хвоста
значительно меньше, чем у форм, распространенных в более южных
районах. Аналогичная закономерность наблюдается и у трансарктической
бурозубки Северо-Востока СССР [Юдин, 1972д]. В Анадырском крае оби-
140
Рис. 49. Череп берингийской бурозубки.
тает мелкая короткохвостая форма, а южнее, в верховье Омолона и на
Камчатке,— более крупная длиннохвостая бурозубка. Создается впечат-
ление, что эти две формы проникли в азиатскую часть ареала вида разны-
ми путями и формировались автономно — S. с. portenkoi внедрилась на
континент через некогда существовавший Берингийский мост между
Аляской и Азией, a S. с. camtschatica — через гряду на месте современ-
ной системы Алеутских, Командорских островов и п-ова Камчатки, види-
мо, служивших в свое время вторым мостом для взаимопроникновения
азиатских и американских форм растений и животных [Юдин, 1973а].
Sorex beringianus Yudin, 1967 —
Берингийская бурозубка
Голотип. № 152 (кол.), <?, ad., шкурка, череп и спиртовый
препарат копулятивного органа. Коллектор А. Г. Велижанин. Коллек-
ция зоологического музея Биологического института СО АН СССР,
№ 2341 (Новосибирск). Курильская гряда, о. Парамушир, устье р. Ту-
харка, 5 августа 1966 г.
Паратипы. 4 зкз.
Диагноз. Мелкая длиннохвостая бурозубка: длина тела 46—56,
хвоста 40—41 мм (78 % от длины тела). Кондило-базальная длина черепа
15,0—15,8 мм. Первый и второй промежуточные зубы крупные, почти
равны между собой; третий меньше двух предыдущих и четвертого. Чет-
вертый хорошо развит и значительно крупнее крайнего мелкого пятого
(рис. 49).
Измерения. Длиннохвостая бурозубка по размерам несколько
крупнее только крошечной и малой. Ввиду незначительного количества
исследованного материала ниже приводится таблица с результатами изме-
141
Таблица 28
Морфометрическая характеристика Sorexberingianus со. Парамушир (по Б. С. Юдину
[1967])
Признак р. Шум- ная, К« 650 Р, Тухарка М lim
К" 103 № 104 № 152 | Кв 233
9 9 О’ С I а
Длина тела 52 56 55 46 52,3 46—56
хвоста — 41 40 40 40,5 40,4 40—41
Ступни — 11 11,2 11 11,2 11,1 11—11.2
Масса тела — 3,6 — 4,5 2 4,5 3,6-4,5
Длина кондил о-базальная черепа 15,0 15,8 15,6 15,6 15,3 15,6 15,0—15,8
рострума 4,0 4,0 4,1 4,0 3,9 4,0 3,9-4,1
мозговой части 8,6 8,8 9,0 8,7 8,8 8,8 8,6—9,0
верхнего ряда зубов 6,5 6,7 6,7 6,5 6,7 6,6 6,5-6,7
промежуточных зубов 2,0 2,0 2,0 2,0 2,1 2,0 2,0—2,1
Ширина рострума 1,2 1,4 1,3 1,3 1,2 1,3 1,2—1,4
предглазничная 2,1 2,1 2,1 2,2 2,0 2,1 2,0-2,2
межглазничная 2,9 2,7 3,0 2,9 2,6 2,8 2.6—3,0
наибольшая черепа 7,2 7,7 7,8 7,4 7,9 7,6 7,2-7,9
Наибольшая высота че- репа 4,4 4,8 4,8 4,5 5,0 4,7 4,4-5,0
Высота восхоцящей вет- ви нижней челюсти 3,0 3,1 3,1 3,1 3,0 3,1 3,0—3,1
Длина углового отростка 2,0 2,0 1,9 1,9 2,0 2,0 1,9—2,0
рений всех зверьков этого вида, имеющихся в нашем распоряжении
(табл. 28).
Характеристика. По внешнему виду берингийская бурозуб-
ка напоминает дальневосточную (5. gracillimus), но в отличие от нее имеет
более широкую и укороченную ростральную часть головы. Дальневосточ-
ная бурозубка стройнее, изящнее, с узкой, вытянутой в длинный хобо-
ток мордой и с более длинным хвостом. У этих двух видов разные строе-
ние зубной системы, форма коронок, соотношение промежуточных зубов
и их постановка в челюсти. Копулятивный орган имеет существенные раз-
личия в строении [Юдин, 1967]. По длине хвоста (составляющего 78 %
от длины тела) берингийская бурозубка уступает только S. mirabilis и
S. gracillimus (см. табл. 24) и близка к S. cinereus. Голова относительно
крупная: отношение кондило-базальной длины черепа к длине тела та-
кое же, как у самых крупноголовых видов, таких как S. daphaenodon и
S. minutissimus (см. табл. 25).
Окраска меха чепрачная, трехцветная: на спине мех бурых тонов,
с примесью палево-серых оттенков, значительно темнее, чем на боках и
брюшной стороне тела. Темный спинной чепрак резко отграничен от более
светлых палево-серых боков. Вентральная сторона тела серая, с незначи-
тельной палевой примесью. Колориметрический анализ двух зверьков
свидетельствует о их очень светлой окраске (см. табл. 30). Хвост покрыт
короткими волосами с хорошо выраженной кисточкой на конце. Ступни
и кисти ног светлые, палево-серые.
142
Таблица 29
Сравнительная характеристика размеров отделов черепа и нижней челюсти у разных
видов бурозубок
Вид Кондило-ба- зальная дли- на черепа, мм Наибольшая вы- сота черепа, мм Отношение наи- большей высоты черепа к конди- ло-базальной длине, % Высота восходя- щей ветви ниж- ней челюсти, мм Отношение вы- соты восходя- щей ветви к кондило-базаль- ной длине, % Длина углового отростка, мм Отношение дли- ны углового от- ростка к конди- ло-базальной длине, %
5. mirabilis 23,3+0,10 5,9 25,3 5,1+0,04 21,9 2,8+0,07 12,0
S. roboratus 20,3+0,03 6,1 30,0 4,5±0,01 22,2 2,6+0,01 12,8
S. unguiculatus 19,9+0,05 6,2 31,2 4,7+0,02 23,6 2,2+0,01 11,1
S. araneus 19,7+0,03 6,1 31,0 5,0+0,02 25,4 2,1+0,01 10,7
S. isodon 19,5±0,11 6,3 32,3 4,8±0,01 24,6 2,1+0,01 10,8
S. daphaenodon 17,9+0,12 6,0 33,5 4,8+0,02 26,8 2,3+0,01 12,8
S. tundrensis 17,7+0,02 5,7 32,2 4,2+0,02 23,7 2,1+0,01 11,9
S. caecutiens 17,4+0,03 5,6 32,2 4,0+0,01 23,0 2,1+0,01 12,1
S. cinereus 16,2+0,03 5,2 32,1 3,2+0,01 19,8 1,9+0,01 11,7
S. gracillimus 15,7+0,05 4,8 30,6 2,9+0,02 18,5 1,9+0,02 12,1
S. beringianus 15,5+0,14 4,7 30,3 3,1+0,02 20,0 2,0+0,02 12,9
8. minutus 15,1+0,03 4,7 31,1 3,0+0,01 19,9 1,8+0,01 11,9
S. minutissimus 13,3+0,03 3,8 28,6 2,8+0,01 21,1 1,6±0,01 12,0
Кондило-базальная длина черепа 15,0—15,8 (М = 15,5) мм. Только
два вида среди палеарктических землероек (S. minutissimus и S. minu-
tus) имеют меньшие значения этого признака. По пропорциям отдельных
показателей череп берингийской бурозубки близок к таковому у S. ci-
nereus и лишь незначительно шире в мозговой и межглазничной областях,
ниже в мозговой части, с относительно более высокой восходящей ветвью
нижней челюсти и длинным угловым отростком (табл. 29).
Зубной ряд относительно короткий, только у S. minutus он несколь-
ко меньше, а по длине ряда промежуточных зубов уступает лишь S. mi-
nutissimus (см. табл. 26). При этом нужно иметь в виду малый объем иссле-
дованного материала (5 экз.) и возможность отклонений (уплощение
черепа, укорочение ряда промежуточных зубов) за счет наличия среди
них двух перезимовавших зверьков. Крайне самобытно соотношение зу-
бов у берингийской бурозубки: третий промежуточный зуб уступает по
размерам второму и четвертому. Среди палеарктических бурозубок та-
кое соотношение специфично только для этого вида и свойственно ряду
видов неарктической фауны. В частности, берингийская бурозубка обна-
руживает большое сходство со строением промежуточных зубов и конфи-
гурации черепа с такими неарктическими видами, как S. lyelli Мег.,
1902 и S. longirostris Bachman, 1837. Отсутствие в хранилищах нашей
страны серийных материалов неарктических видов землероек, а в литера-
туре — описания строения копулятивного органа североамериканских
бурозубок не позволяет высказать более определенного мнения о взаимо-
отношениях этих видов с берингийской бурозубкой.
От других видов фауны СССР берингийская бурозубка надежно отли-
чается соотношением промежуточных зубов и строением копулятивного
143
органа (см. рис. 40). Он короткий (3,8—3,9 мм) и толстый (1,1—1,4 мм),
несколько загнут книзу и увенчан на свободном конце массивной, сфери-
чески закругленной коронкой.
Распространение. Известна только из мест первоописания
вида — о. Парамушир.
Материал. 4 экз. типовой серии, среди них два перезимовав-
ших самца.
Изменчивость. Не изучена из-за малочисленности материала.
Окраска взрослых зверьков чепрачная, трехцветная.
Заметки по систематике. Вид описан позднее всех дру-
гих представителей бурозубок [Юдин, 19676]. Основанием для описания
землероек, собранных А. Г. Велижаниным на о. Парамушир, послужили
особенности строения зубной системы и копулятивного органа. Как уже
говорилось выше, подобного соотношения промежуточных зубов и строе-
ния glans penis не имеет ни один вид палеарктических бурозубок. Что ка-
сается взаимоотношений с неарктическими представителями рода, то
вполне вероятно, что берингийская бурозубка проникает на северо-
американский континент. Возможно, ранее была описана как вид, но
отсутствие необходимых сравнительных материалов не позволяло сделать
какого-либо определенного заключения по этому поводу.
М. В. Охотина [1977] считает берингийскую бурозубку нереальным
видом, относя ее к S. cinereus. Мы не исследовали материал, имевшийся
в распоряжении М. В. Охотиной с о. Парамушир. Не исключено, что там
наряду с берингийской бурозубкой обитает и трансарктическая. Дейст-
вительно, берингийская бурозубка близка к трансарктической и в отли-
чие от всех других видов бурозубок Палеарктики относится вместе с ней
к подроду Otisorex. Наряду с этим имеются и существенные отличия.
Структура ряда промежуточных зубов у берингийской бурозубки иная,
чем у трансарктической. Трудно представить, чтобы столь жестко фикси-
рованная структура, как промежуточные зубы, была подвержена у земле-
роек о. Парамушир такой внутривидовой изменчивости, если встать на
позицию названного автора и признать берингийскую бурозубку как
представителя вида S. cinereus.
Наряду с этим имеются кардинальные отличия и в строении копуля-
тивного органа. Правда, исследованные нами половые органы транс-
арктической бурозубки, вероятно, значительно трансформированы
вследствие фиксации. Однако размер glans penis у нее значительно боль-
ше, чем у берингийской, учитывая даже то, что от фиксации она могла
уменьшиться в размерах. Судя по описанию копулятивного органа транс-
арктической бурозубки с Камчатки [Долгов, Реймерс, 1979], по форме
он действительно во всех деталях похож на таковой берингийской, но
имеет большие размеры. Авторы считают, что по строению гениталий
трансарктические бурозубки с Камчатки сходны с описанной нами бе-
рингийской бурозубкой, и предполагают их видовое тождество и подви-
довой статус описанной нами землеройки.
Специфичное строение промежуточных зубов у землероек с о. Пара-
мушир не позволяет согласиться с этим предположением. Имеется ряд
других признаков, по которым эти два вида различаются: окраска меха
берингийской бурозубки чепрачная, самая светлая среди всех бурозу-
бок (табл. 30), размеры тела меньше, а относительная длина хвоста боль-
ше, отношение длины черепа к длине тела больше, а зубной ряд и ряд
промежуточных зубов меньше как по абсолютным, так и по относитель-
144
Таблица 30 Колориметрическая характеристика бурозубок
Вид Белиана Оттенок
M±mx cv a n M±mx cv a n
5. daphaenodon 2,9+0,18 23,4 0,7 14 138,7±2,29 6,2 8,4 14
S. mirabilis 4,4±0,56 28,8 1,2 5 121,9+3,09 5,7 6,8 5
S. araneus 2,8+0,08 33,3 0,9 150 134,8+0,81 7,3 9,9 150
S. tundrensis 3,5±0,08 32,3 1,1 182 141,0+0,76 7,2 10,2 182
S. unguiculatus 3,3+0,13 24,2 0,8 40 136,3+0,99 4,6 6,3 40
S. isodon 2,2+0,05 26,4 0,6 130 131,4+0,91 7,9 10,4 130
S. roboratus 2,4+0,05 21,7 0,5 94 136,7±0,98 6,9 9,5 94
S. caecutiens 3,4+0,05 20,7 0,7 204 136,6+0,56 5,8 8,1 204
S. gracillimus 4,l±0,13 16,1 0,7 25 132,4+1,12 4,2 5,6 25
S. minutissimus 5,l±0,19 29,3 1,5 60 122,8±0,97 6,1 7,5 , 60
S. minutus 4,9+0,14 22,6 1,1 67 133,5+0,87 5,1 6,9 67
S. cinereus 5,0±0,12 11,7 0,6 25 125,4+1,20 4,8 6,1 25
5. beringianus 6,9±1,23 25,3 1,7 2 129,6+4,48 4,9 6,3 2
ным показателям (см. табл. 26). В мозговой и межглазничной областях
череп у берингийской бурозубки более широк и низок в затылочной об-
ласти. Существенно отличается большими размерами углового отростка.
Имеющийся в нашем распоряжении материал пока не позволяет го-
ворить о полной тождественности этих видов, а лишь о их несомненной
близости, что вполне понятно, так как оба вида относятся к одному под-
роду и только у них одних из палеарктических бурозубок в нижней че-
люсти нет постмандибулярного канала.
В составе подрода один вид.
Подрод Ognevia Dolgov et Heptner, 1967
Sorex mirabilis Ognev, 1937 —
Гигантская бурозубка
Г о л о т и п. Из сборов С. И. Огнева. № 247 (кол.), б1, 31/VII 1936 г.
Коллекция Зоологического музея МГУ. Добыт С. А. Надецким в долине
Кишинки, Приморский край.
Диагноз. Самая крупная и длиннохвостая бурозубка среди пале-
арктических видов рода Sorex (рис. 50, 51). Длина тела от 72 до 90 (М =
= 83,6+1,56), хвоста 64—74 (М — 68,3+0,75), ступни 16—18 (М =
= 17,0+0,18) мм. На внутренней стороне передней вершины первого
верхнего резца имеется зубец. Задняя вершина этого зуба зачаточна.
Измерения. См. табл. 31.
Характеристика. Длина тела зверьков по средним значе-
ниям значительно превышает таковую других видов бурозубок. Несмот-
ря на то что минимальные значения этого признака перекрываются с наи-
большей величиной у других видов бурозубок, по массивности и особен-
ности габитуса гигантская бурозубка четко обособлена. В первую оче-
редь она отличается чрезвычайно длинным для землероек хвостом. Длина
хвоста составляет 81,7 % длины тела, в то время как у других представи-
145
Рис. 50. Гигантская бурозубка — Sorex mirabilis Ognev.
телей рода Sorex Сибири это отношение колеблется в пределах 52—79,2 %.
Гигантская бурозубка надежно диагностируется по этим признакам без
привлечения других.
Мех тусклый, землисто-буро-серого однотонного цвета на всем теле
(см. рис. 50). Такой же интенсивности или едва более светлая окраска
на боках и вентральной стороне тела. Чаще всего она однотонная по всему
10 в. С. Юдин
146
Таблица 31
Морфометрическая характеристика Sorex mirabilis
Возрастная и половая группа М±тпж О cv п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 82,7+3,05 7,47 9,04 6 72-90
взрослые — — —— — —
Самки прибылые 87,5-1-1,19 2,38 2,72 4 86—91
взрослые 83,3+4,41 7,64 9,17 3 75-90
В целом по виду ... 83,6+1,56 6,24 7,47 16 72—91
Длина хвоста
Самцы прибылые 70,0+0,93 2,28 3,26 6 68—74
взрослые — —• — —— —
Самки прибылые 65,8+0,75 1,50 2,28 4 64—67
взрослые 71,0+1,00 1,73 2,44 3 69—72
В целом по виду ... 68,3+0,75 3,00 4,40 16 64—74
Длина ступни
Самцы прибылые 17,3+0,21 0,52 3,0 6 17—18
взрослые — — —— —
Самки прибылые 16,8+0,48 0,96 5,72 4 16—18
взрослые 17,3+0,33 0,58 3,33 3 17—18
В целом по виду ... 17,0+0,18 0,73 4,30 16 16—18
Масса тела
Самцы прибылые 11,9+0,62 1,25 10,46 4 10,7—14,2
взрослые — —- — — —
Самки прибылые 15,0+1,47 2,94 19,54 4 11,9—18,0
взрослые 13,3+0,95 1,34 10,14 2 12,3—14,2
В целом по виду ... 13,2+0,66 2,28 17,26 12 10,7—18,0
Кондило-базальная длина черепа
Самцы прибылые 23,3+0,14 0,33 1,42 6 22,7—23,6
взрослые — — — — —
Самки прибылые 23,2+0,07 0,12 0,50 3 23,1—23,3
взрослые 23,7+0,32 0,56 2,35 3 23,1—24,2
В целом по виду ... 23,3+0,09 0,40 1,73 18 22,7—24,2
Длина рострума,
Самцы прибылые 6,8+0,06 0,15 2,17 6 6,6—7,0
взрослые —• — — — —
Самки прибылые 7,1+0,27 0,47 6,69 3 6,7—7,6
взрослые 7,0+0,09 0,15 2,17 3 6,9-7,2
В целом по виду ... 6,8+0,07 0,31 4,62 18 6,6—7,6
Длина мозговой части черепа
Самцы прибылые 12,4+0,08 0,19 1,51 5 12,2—12,7
взрослые —— — — — —
Самки прибылые 12,4+0,06 0,10 0,81 3 12,3—12,5
взрослые 12,2+0,26 0,45 3,69 3 11,8—12,7
В целом по виду ... 12,3+0,07 0,27 2,18 17 11,8—12,7
147
Продолжение табл. 31
1 2 3 4 5 6
Длина верхнего ряда зубов
Самцы прибылые 10,4+0,07 0,18 1,70 6 10,1—10,5
взрослые — — — — —
Самки прибылые 10,4+0,13 0,23 2,23 3 10,1—10,5
взрослые 10,4+0,10 0,14 1,36 2 10,3—10,5
В целом по виду ... 10,3+0,05 0,18 1,73 14 10,0—10,5
Длина ряда промежуточных зубов
Самцы прибылые 3,4+0,08 0,19 5,59 6 3,1—3,6
взрослые — — — — —
Самки прибылые 3,3±0,06 0,10 3,03 3 3,2-3,4
взрослые 3,4+0,06 0,10 2,94 3 3,3-3,5
В целом по виду ... 3,3+0,04 0,17 5,30 17 3,0—3,6
Ширина рострума
Самцы прибылые 2,6±0,04 0,10 3,81 6 2,4—2,7
взрослые — — — —— —
Самки прибылые 2,6±0,03 0,06 2,19 3 2,6-2,7
взрослые 2,7 — -— 1 —
В целом по виду ... 2,6+0,03 0,12 4,62 12 2,3—2,7
Предглазничная ширина
Самцы прибылые 3,7+0,07 0,17 4,68 6 3,5-4,0
взрослые —- — — — —
Самки прибылые 3,7±0,03 0,06 1,58 3 3,6—3,7
взрослые 3,6±0,07 0,12 3,18 3 3,5—3,7
В целом по виду ... 3,6±0,04 0,14 3,86 15 3,5-4,0
Межглазничная ширина
Самцы прибылые 4,8+0,07 0,18 3,67 6 4,5—5,0
взрослые — — — — —
Самки прибылые 4,8+0,06 0,10 2,08 3 4,7—4,9
взрослые 4,9+0,06 0,10 2,04 3 4,8-5,0
В целом по виду ... 4,8+0,04 0,15 3,07 18 4,5—5,0
Наибольшая ширина черепа
Самцы прибылые 10,6+0,09 0,20 1,85 5 10,4—10,9
взрослые — — — — —
Самки прибылые 10,5+0,17 0,30 2,86 3 10,2—10,8
взрослые 11,0+0,12 0,20 1,82 3 10,8—11,2
В целом по виду ... 10,6+0,06 0,26 2,45 17 10,2—11,2
Наибольшая высота черепа
Самцы прибылые взрослые 6,2+0,15 0,34 5,38 5 5,8—6,6
Самки прибылые 6,1+0,09 0,15 2,52 3 5,9—6,2
взрослые 5,7+0,23 0,40 7,02 3 5,3—6,1
В целом по виду ... 5,9+0,09 0,38 6,36 17 5,3—6,6
Высота восходящей ветви нижней челюсти
Самцы прибылые 5,2+0,04 0,11 2,04 6 5,0-5,3
взрослые — — — — ——
Самки прибылые 5,0±0,06 0,13 2,50 4,9—5,2
взрослые 5,3+0,07 0,12 2,17 3 5,2—5,4
В целом по виду ... 5,1+0,04 0,17 3,32 19 4,8—5,4
10*
148
Окончание табл. 31
1 I 2 I 3 I 4 I 5 | 6
Длина, углового отростка
Самцы прибылые 2,8+0,10 0,25 8,84 6 2,4—3,1
взрослые — — — — —
Самки прибылые 2,8±0,19 0,32 11,35 3 2,6—3,2
взрослые 2,5+0,07 0,11 4,68 3 2,4—2,6
В целом по виду ... 2,8+0,07 0,29 10,23 18 2,4-3,2
телу. Рострум окрашен светлее. Светлые и длинные, достигающие 30 мм
вибрисы четко выделяются на фоне общей темной окраски меха. Хвост
покрыт редкими, очень короткими щетинистыми волосами. Кисточки из
удлиненных волос на конце хвоста нет. Сквозь редкие волосы видны че-
шуйки кожи хвоста. Окраска его темная, одноцветная сверху и снизу.
Череп массивный, длинный, грубой архитектуры. Кондило-базаль-
ная длина его составляет 22,7—24,2 (М = 23,3) мм. Лицевой отдел мас-
сивный, длинный, с очень широким рострумом. По отношению к кондило-
базальной длине черепа значения этого показателя самые высокие среди
сибирских видов бурозубок (табл. 32). В предглазничной области череп
очень широкий. Череп обращает на себя внимание своей относительной
узостью и уплощенностыо. Нижняя челюсть относительно крупного че-
репа кажется слабой. Высота ее восходящей ветви в среднем составляет
М = 5,1(± 0,04) мм. Proc, coronoideus наклонен вперед. Отношение вы-
соты восходящей ветви нижней челюсти к кондило-базальной длине черепа
составляет 21,8 %, в то время как у S. isodon, обитающей вместе с гигант-
ской бурозубкой, этот показатель равен 24,6 % (см. табл. 29).
По одонтологическим признакам этот вид также надежно диагности-
руется. Длина зубного ряда гигантской бурозубки колеблется в пределах
Таблица 32
Сравнительная характеристика отдельных элементов черепа бурозубок
Вид Нондило-ба- зальная дли- на черепа, мм Длина рост- рума, мм Отношение дли- ны рострума к кондило-базаль- ной длине, % Длина мозго- вой части, мм Отношение моз- говой части к кондило-базаль- ной, %
5. mirabilis 23,3+0,01 6,8+0,07 29,1 12,3+0,07 52,7
S. roboratus 20,3+0,03 5,3±0,02 26,1 11,2+0,01 55,1
S. unguiculatus 19,9±0,05 5,3+0,03 26,6 11,2+0,03 56,2
S. araneus 19,7+0,03 5,3+0,02 26,9 10,8+0,03 54,0
S. isodon 19,5±0,11 5,4±0,13 27,6 10,9+0,02 55,8
S. daphaenodon 17,9+0,12 4,5+0,02 25,1 10,5+0,02 58,6
S. tundrensis 17,7+0,02 4,4+0,01 24,8 10,1+0,01 57,0
S. caecutiens 17,4±0,03 4,6+0,01 26,4 9,8±0,02 56,3
S. cinereus 16,2+0,03 4,2+0,02 25,9 9,3+0,03 57,4
S. gracillimus 15,7+0,05 4,2+0,04 26,8 8,7+0,04 55,4
S. beringianus 15,5+0,14 4,0+0,03 25,8 8,8+0,07 56,8
S. minutus 15,1+0,03 3,9+0,02 25,8 8,6+0,02 56,9
S. minutissimus 13,3+0,03 3,3+0,03 24,8 7,7+0,02 57,8
149
Рис. 52. Строение передних верхних резцов (а) и зубной системы в целом (б) бу-
розубок.
1 — S. vagrans, видны дополнительные лопасти на месте соединения правого и левого резцов;
2 — S. mirabilis дополнительные лопасти развиты хорошо: 3 — S. pacificus, дополнительные ло-
пасти отсутствуют; 4 — S. mirabilis; 5 — S. pacificus.
10,0—10,5 мм при среднем значении М — 10,3(± 0,05) мм. Индекс отно-
шения к кондило-базальной длине черепа составляет 44,2 %, отношение
ряда промежуточных зубов к кондило-базальной длине черепа 14,1 %,
а к длине зубного ряда — 32,0 %. Если абсолютные значения показателя
«длина зубного ряда» у этого вида больше, чем у других видов (только
у S. roboratus максимальный размер его — 10,0 — равен минимальному
у S. mirabilis), то отношение ряда зубов к кондило-базальной длине черепа
у него минимальное и сходное с таковым у S. tundrensis (см. табл. 26).
Индекс длины ряда промежуточных зубов к общей длине зубного ряда
довольно обычен, и близкие значения его зарегистрированы у S. unguicu-
latus, S. daphaenodon, S. tundrensis.
Передний верхний резец крупный, его передняя вершина круто
крючкообразно загнута книзу и больше похожа на таковую у кутор и
белозубок, чем у других видов рода Sorex. Задняя вершина зачаточна и
по высоте равна или лишь немного превышает край цингулюма первого
промежуточного зуба. Концы передних вершин верхних резцов у гигант-
ской бурозубки в отличие от других видов бурозубок разведены в сторо-
ны. На их внутренней поверхности хорошо выражены зубцы (см.
рис. 51, 52).
Структура верхних промежуточных зубов также крайне своеобразна
и не повторяется у других видов бурозубок фауны СССР. Первый верх-
ний промежуточный массивный и высокий, округло затупленный, зна-
чительно превышает по размерам второй. Передняя сторона его несколь-
ко выпуклая, а задняя вогнута. Вершина зуба несколько смещена вперед
по отношению к центру основания коронки и слегка загнута внутрь ро-
товой полости. Третий меньше второго и равен четвертому или несколь-
ко меньше его, а по форме напоминает первый. Пятый крайне мал, чет-
вертый значительно уступает по величине первому и второму.
150
Распространение. Гигантская бурозубка относится к чис-
лу наименее изученных землероек. В настоящее время известна из При-
морья и Северной Кореи. Впервые добыта С. А. Надецким в долине
Кишинки (окр. оз. Ханка), позднее ее собирали в разные годы в заповед-
нике «Кедровая Падь» и Лазовском. В Северной Корее добыта в 15 км
к югу от Иотоху, в окрестностях Игена и Бьектана.
Материал. 19 экз., Приморье, Корея.
Изменчивость. Гигантская бурозубка относится к числу наи-
более редких видов мировой фауны. В коллекциях СССР нам известно
всего 20 экз., поэтому сведения о изменчивости вида крайне скудны.
Размеры зверьков колеблются в значительных пределах. Так, длина
тела изменяется от 72 до 91 (М = 83,6) мм, масса — 10,7—18,0 (М =
= 13,2) г. Взрослые зверьки по средним значениям показателя «размер
тела» не отличаются от молодых. Длина хвоста у взрослых больше (71 —
=1=1,0), чем у молодых (М = 65,8±0,75). Несколько больше ширина че-
репа у взрослых зверьков. По остальным показателям существенных
различий пет. Создается впечатление, что зверьки быстро достигают
размеров взрослых. Кроме того, в отличие от других видов индивидуаль-
ная жизнь гигантской бурозубки, вероятно, длится дольше, чем можно
объяснить ее малочисленность. Эфемерные виды зверьков для воспол-
нения высокой гибели вынуждены размножаться более интенсивно. Нуж-
но сказать, что предположение о большем долголетии мы делаем на осно-
вании характера стирания зубов, однако требуется материал для под-
тверждения этого вывода, в частности данные гистологических иссле-
дований.
Наблюдается некоторый полиморфизм окраски. Так, у типового
экземпляра окраска на боках тела заметно светлее, чем на дорсальной
стороне. У большинства добытых нами зверьков окраска была однотон-
ной. Выношенный мех старых зверьков становится серым, с большей бе-
лизной, чем у молодых зверьков. По данным колориметрического анализа
обнаруживается довольно широкий диапазон изменчивости белизны (от
2,4 до 5,5) при среднем показателе 4,4(_0,56). Верхний предел значений
показателя оттенка самый низкий среди всех бурозубок: он укладывается
в пределах между 116 и 132 (при средней величине 121,9+3,09), что,
вероятно, является показателем стенотопности вида. У такого полигон-
ного вида, как S. tundrensis, значение этих показателей варьирует в
пределах от 1,3 до 6,5 (М = 3,5=t0,08) по белизне и от 112 до 165 (М =
= 141=t0,12) по оттенку (см. табл. 30).
В структуре ряда промежуточных зубов прослеживается изменчи-
вость в соотношении величины третьего и четвертого зубов. Чаще эти зу-
бы равны, но у некоторых экземпляров, в том числе и у типового, третий
зуб меньше четвертого. Из 19 исследованных зверьков третий зуб был
меньше четвертого только у 2 экз. В. А. Долгов [1968] отмечает случай
редукции левого и правого пятых промежуточных зубов верхней челюсти
у одного зверька.
Заметки по систематике. Гигантская бурозубка на-
столько самобытна, что объем вида и его естественные границы, каза-
лось бы, не должны вызывать сомнений. Даже по внешнему виду ее не-
возможно спутать с другими представителями рода. Тем не менее едино-
го мнения о ее видовом статусе и объеме вида нет. Существуют две край-
ние точки зрения. Одна из них рассматривает S. mirabilis как самостоя-
тельный вид в роде Sorex, имеющий ограниченный ареал в юго-восточ-
151
ной части Палеарктики, т. е. в том объеме, как его представлял автор
первоописания [Огнев, 1937] и рассматривают большинство специалис-
тов. В. Г. Гептнер и В. А. Долгов [19G7 ] в полном соответствии с дейст-
вительным положением в системе выделили гигантскую бурозубку в
отдельный подрод (Ognevia). О необходимости выделить гигантскую буро-
зубку в подрод в личпой беседе неоднократно говорил и С. У. Строганов,
но не успевший этого сделать сам.
Вторая точка зрения принадлежит Н. А. Бобринскому и А. П. Кузя-
кину [Бобринский и др., 1944], без достаточных на то оснований объеди-
нивших ее с американским видом S. pacificus Coues, 1877, который встре-
чается в северо-западной Калифорнии и на юго-западе Орегоны, и та-
ким образом расширивших ареал созданного ими композиционного вида
до Неарктики и Палеарктики.
Гигантская бурозубка была описана С. И. Огневым [1937] по экземп-
ляру из Приморья, из долины Кишинки. Несмотря на единственный
экземпляр, бывший в распоряжении автора первоописания, четкая обо-
собленность вида была настолько очевидной, что не вызывала ни у кого
возражений. Н. А. Бобринский и А. П. Кузякин [Бобринский и др.,
1944] произвольно объединили ее с американским видом S. pacificus,
не имея в руках сравнительного материала и базируясь только на описа-
нии из монографии Джексона [Jackson, 1928].
Убедительная критика несостоятельной этой позиции дана
С. У. Строгановым [1957]. Точку зрения последнего автора отстаивают и
В. Г. Гептнер и В. А. Долгов [1967], а также Б. С. Юдин [19696, 1971а].
Нами исследованы новейшие литературные данные, а также экземпляр
S. pacificus, хранящийся в Зоологическом музее АН СССР (Ленинград).
Findley [1955], Hall, Kelson [1959] S. pacificus включили в качест-
ве подвида S. vagrans Baird, не имея на то убедительных оснований. Зем-
леройки S. vagrans, исследованные нами по материалам из Канады и
США (присланные д-ром В. Прюитом и д-ром Р. Гоффманном),— мелкие,
кондило-базальная длина черепа их колеблется от 16,7 до 17,4 (наши изме-
рения) и от 16,1 до 19,1 мм — по Джексону [Jackson, 1928]. Конфигура-
ция черепа у них иная, они существенно отличаются и по деталям строе-
ния зубов. Кондило-базальная длина черепа 5. pacificus колеблется от
20 до 23 мм. Главным отличием является отсутствие дополнительного
зубца на внутренней стороне первой вершины переднего верхнего резца
(см. рис. 52, 3), в то время как у S. vagrans он есть. Первый промежуточ-
ный зуб у S. pacificus несколько меньше второго или равен ему. Второй
зуб массивный, третий составляет только половину четвертого, почти
равного по высоте первому и второму, пятый крайне мал (см. рис. 52, 3).
Кроме того, гигантская бурозубка крупнее тихоокеанской. Кондило-
базальная длина черепа у нее равна 22,7—24,2 (М = 23,3±0,10) мм,
т. е. даже средние значения этого показателя у гигантской бурозубки вы-
ше наибольших тихоокеанской.
Основные отличия этих двух видов следующие:
Признак S. pacificus S. mirab Us
Дополнительный зубец
на внутренней стороне
первой вершины перво-
го верхнего резца (см.
рис. 52,3) Нет Есть
152
Признак
Вторая (задняя) вершина
первого верхнего резца
Первый промежуточный
зуб
Четвертый промежуточ-
ный зуб
S. pad ficus
Развита хорошо и дости-
гает более половины
высоты правого проме-
жуточного зуба (см.
рис. 52, 5).
Меньше второго или ра-
вен ему
Почти равен первому (см.
рис. 52, 5)
8. mirabilis
Зачаточна, по высоте рав-
на или лишь немного
превышает верхний
край цингулюма пер-
вого промежуточного
зуба (см. рис. 52, 4).
Массивный, значительно
крупнее второго
Менее половины размера
первого (см. рис. 52, 4)
Перечисленные признаки настолько существенны и самобытны для
каждого вида, что можно с полной уверенностью считать вопрос о их
таксономической самостоятельности решенным. Совершенно прав
С. У. Строганов [1957], указывая, что Н. А. Бобринский и А. П. Кузякин
[Бобринский и др., 1944] обратили внимание лишь на сходство размеров
и оставили без внимания признаки, качественно характеризующие эти
виды: особенности строения зубной системы. А. П. Кузякин [1965] про-
должает считать гигантскую бурозубку с юга Дальнего Востока конспеци-
фичной 5. pactficus и рассматривает ее в качестве подвида тихооке-
анской — 5. pactficus mirabilis.
Аналогичную позицию занимает и Бринк [Van Den Brink, 1953,
1967]. Известный американский зоолог Хоффманн [Hoffmann, 1971]
доказал умозрительность подобных выводов, не базирующихся на факти-
ческом материале [Бобринский и др., 1944, 1965; Ellerman, Morrison-
Scott, 1951; Van Den Brink, 1953, 1967] и объединяющих неарктический
вид 5. pactficus с палеарктическим — 5. mirabilis. В то же время, по его
мнению [Hoffmann, 1971], к 5. mirabilis более близка другая американ-
ская землеройка 5. bendirii Мег., относящаяся к числу самых крупных
представителей рода Sorex. Как и гигантская бурозубка, она имеет одно-
тонную и даже еще более темную окраску. Оба вида обитают в остаточ-
ных лесах древнего Берингового мезофильного леса, связь между участ-
ками которого прекратилась в позднем миоцене [Wolfe, Leopold, 1967].
У S. bendirii, как и у S. mirabilis, довольно хорошо развит дополни-
тельный зубец на внутренней стороне первой вершины верхнего передне-
го резца. Однако соотношение промежуточных зубов, их форма, строение
переднего резца (его задней вершины), конфигурация черепа специфичны
для каждого вида. Особенно обращает на себя внимание очень массивная
нижняя челюсть S. bendirii и ее высокая восходящая ветвь. Существен-
ным отличием этой землеройки является отсутствие постмандибулярного
канала, признака, имеющего подродовой уровень (подрод Otisorex). Мы
склонны считать, поддерживая мнение Хоффманна, что внешнее сходство
(размеры и окраска) — результат конвергенции, адаптации двух само-
стоятельных видов к обитанию в экологически сходных условиях. В этой
же работе [Hoffmann, 1971] автор поддерживает наши взгляды [Юдин,
19696] на взаимоотношения видов 5. mirabilis и S. pactficus.
В Приморье обитает номинальный подвид S. т. mirabilis. С. У. Стро-
ганов [1956] описал подвид корейской гигантской бурозубки — S. mi-
rabilis kutscheruki Strog., отличающийся несколько меньшими размерами.
Этот подвид заселяет Северную Корею. Малочисленность материала
не позволяет достоверно охарактеризовать размеры полиморфизма ги-
гантской бурозубки, поэтому трудно судить о реальности описанного
подвида.
153
Подрод Eurosorex Stroganov, 1952
Представители подрода характеризуются мелкими размерами, общи-
ми чертами в строении glans penis. В составе подрода четыре вида бурозу-
бок, на территории Сибири встречаются два.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА EUROSOREX
1(2). Размеры мелкие: длина тела с головой 30—53 (М=44,8±0,35),
кондило-базальная длина черепа 12,0—14,5 (М== 13,3+0,03), длина хвоста
18—37 (М^ 24,2+ 0,27) мм. Промежуточные зубы с округлыми вершинами
в зубном ряду сидят плотно, вертикальная ось их коронок наклонена
внутрь ротовой полости (см. рис. 54). Первый зуб значительно крупнее
второго и третьего ....................................... .
.................Sorex minutissimus Zimm.— Крошечная бурозубка
2(1). Размеры несколько крупнее: длина тела с головой 40—62 (М=
= 52,2±0,59), кондило-базальная длина черепа 13,9—15,8 (М=15,1±
+0,03) мм. Промежуточные зубы в зубном ряду сидят свободно. Верти-
кальная ось их коронок перпендикулярна продольной оси челюсти
(см. рис. 55). Первый зуб почти одинаковых размеров с третьим и значи-
тельно крупнее второго, второй намного меньше третьего.......
..............................Sorex minutus L.— Малая бурозубка
Sorex minutissimus Zimmermann, 178(Г—
Крошечная бурозубка
Голотип. Типовой экземпляр неизвестен. Вид описан по материа-
лу из окрестностей с. Кийского, р. Кия (ныне г. Мариинск, Кемеров-
ская обл.).
Диагноз. Самая мелкая бурозубка. Длина тела 30—53 (М=44,8±
+ 0,35), длина хвоста 18—37 (М=24,2+0,27) мм, что составляет 54 %.
Череп широкий в ростральном, пред- и межглазничном отделах, узкий
и низкий в мозговой части.
Измерения. См. табл. 33.
Характеристика. Крошечная бурозубка самая мелкая, с хо-
рошо очерченными признаками. От других представителей рода отличает-
ся рядом габитуальных черт: наряду с мелкими размерами относительно
широкая и крупная голова с коротким хоботком, короткий или средних
размеров хвост. Размеры тела, хвоста и окраска меха изменчивы в широ-
ких пределах в зависимости от географической локализации той или иной
популяции вида. У разных подвидов отношение длины хвоста к длине те-
ла колеблется от 43,6 до 68,6 %. Окраска меха двухцветная: спина бу-
рая, буро-серая, кофейно-бурая разной интенсивности (рис. 53, табл. 34),
иной характер локализации темной окраски. У одних подвидов темная
окраска спины постепенно переходит в более светлые бока и белесую
вентральную часть тела, у других при таком же соотношении вентраль-
ной части окрашена в относительно темные тона, у третьих темная окрас-
ка образует чепрак, резко отграниченный от более светлых боков.
Череп описываемого вида самый мелкий среди всех бурозубок: его
кондило-базальная длина колеблется от 12,0 до 14,5 (М=13,3±0,03) мм
(см. табл. 33). Относительно размеров тела череп довольно крупный —
154
Таблица 33
Морфометрическая характеристика Sorex minutissimus
Возрастная и половая группа М±тх <3 Cv п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 45,4±0,49 3,45 7,59 50 37—52
взрослые 45,5+0,93 4,34 9,53 22 37—53
Самки прибылые 45,1+1,08 4,83 10,72 20 30—50
взрослые 48,0+1,19 3,95 8,23 11 42—53
В целом по виду ... 44,8+0,35 4,38 9,77 155 30—53
Длина хвоста
Самцы прибылые 24,3+0,46 3,54 14,61 59 18—37
взрослые 23,6+0,02 4,88 20,64 23 18—37
Самки прибылые 23,8+0,55 2,76 11,61 25 19—30
взрослые 23,5+0,93 3,21 13,64 12 20—32
В целом по виду ... 24,2+0,27 3,60 14,86 182 18—37
Длина ступни
Самцы прибылые 8,2±0,08 0,62 7,55 60 7,0—10,0
взрослые 8,0±0,14 0,68 8,44 23 7,0—9,0
Самки прибылые 8,3+0,15 0,77 9,22 25 7,0—10,0
взрослые 8,3+0,22 0,75 9,14 12 7,0—9,0
В целом по виду ... 8,2±0,05 0,69 8,44 181 7,0—10,0
Масса тела
Самцы прибылые 2,1+0,05 0,28 13,37 31 1,5—2,5
взрослые 2,6+0,15 0,60 23,19 16 1,7—3,8
Самки прибылые 2,2+0,07 0,25 11,78 13 1,8—2,6
взрослые 3,1+0,27 0,82 25,99 9 1,8—4,5
В целом по виду ... 2,3+0,05 0,49 21,87 97 1,5—4,5
Кондило-базальная длина черепа
Самцы прибылые 13,4+0,06 0,47 3,49 64 12,2—14,5
взрослые 13,1+0,10 0,58 4,42 32 12,0—14,1
Самки прибылые 13,5+0,10 0,52 3,87 29 12,2—14,5
взрослые 13,4+0,16 0,62 4,61 14 12,3—14,2
В целом по виду ... 13,3+0,03 0,50 3,76 231 12,0—14,5
Длина рострума
Самцы прибылые 3,3+0,02 0,19 5,77 67 2,8—3,8
взрослые 3,2+0,06 0,34 10,67 33 2,2—3,7
Самки прибылые 3,4+0,08 0,42 12,60 29 3,0—5,8
взрослые 3,2+0,08 0,30 9,46 15 2,7—3,8
В целом по виду ... 3,3+0,02 0,29 8,78 245 2,2—5,3
Длина мозговой части черепа
Самцы прибылые 7,7+0,05 0,28 3,67 39 7,0—8,5
взрослые 7,6±0,06 0,29 3,81 27 7,0-8,1
Самки прибылые 7,8±0,09 0,38 4,90 18 6,7—8,4
взрослые 7,8+0,10 0,33 4,25 12 7,2—8,2
В целом по виду ... 7,7±0,02 0,32 4,10 175 6,7—8,5
155
Продолжение табл. 33
1 2 3 4 5 6
Длина верхнего ряда зубов
Самцы прибылые 5,8+0,03 0,24 4,12 64 5,1—6,2
взрослые 5,5±0,06 0,32 5,80 30 4.7—6,0
Самки прибылые 5,8+0,04 0,21 3,62 28 5,4—6,1
взрослые 5,5+0,10 0,36 6,42 12 5,0-6,0
В целом по виду ... 5,7+0,03 0,42 7,34 231 4,7-6,2
Длина ряда промежуточных зубов
Самцы прибылые 1,7+0,03 0,19 11,03 40 1,0—2,0
взрослые 1,7±0,04 0,19 11,53 27 1,2-2,0
Самки прибылые 1,8+0,03 0,14 8,10 17 1,5—2,0
взрослые 1,6+0,04 0,16 9,79 13 1,3—1,9
В целом по виду ... 1,7±0,01 0,16 9,13 177 1,0—2,0
Ширина рострума
Самцы прибылые 1,3+0,01 0,09 6,56 38 1,2—1,5
взрослые 1,3+0,02 0,11 8,47 26 1,0-1,5
Самки прибылые 1,3+0,02 0,07 5,56 18 1,2-1,5
взрослые 1,4+0,04 0,13 9,14 10 1,3—1,6
В целом по виду ... 1,3+0,01 ОДО 7,29 171 1,0—1,6
Предглазничная ширина
Самцы прибылые 1,9+0,02 0,11 5,53 40 1,7—2,1
взрослые 1,9±0,03 0,14 7,44 26 1,5—2,1
Самки прибылые 1,9+0,03 0,10 5,36 18 1,7—2,1
взрослые 1,9+0,03 0,11 5,54 10 1,7—2,1
В целом по виду ... 1,9+0,01 0,12 6,31 175 1,5—2,1
Межглазничная ширина
Самцы прибылые 2,8+0,02 0,19 6,83 68 2,4—3,1
взрослые 2,7+0,03 0,18 6,62 33 2,2—3,1
Самки прибылые 2,8+0,04 0,19 6,78 29 2,5-3,1
взрослые 2,8+0,04 0,14 5,12 15 2,6—3,0
В целом по виду ... 2,8+0,01 0,18 6,36 245 2,2-3,1
Наибольшая ширина черепа
Самцы прибылые 6,5±0,04 0,29 4,40 64 5,6—7,0
взрослые 6,1+0,06 0,33 5,40 33 5,2—6,7
Самки прибылые 6,5±0,04 0,20 3,13 29 6,2—6,8
взрослые 6,3+0,10 0,37 5,90 14 5,5-6,8
В целом по виду ... 6,3+0,02 0,32 5,04 233 5,2—7,0
Наибольшая высота черепа
Самцы прибылые 4,0±0,03 0,23 5,75 63 3,4-4,5
взрослые 3,7±0,04 0,23 6,32 31 3,2-4,1
Самки прибылые 3,9±0,06 0,29 7,59 28 3,2-4,4
взрослые 3,8+0,08 0,28 7,56 14 3,4—4,5
В целом по виду ... 3,8+0,02 0,34 8,87 218 2,9-4,7
156
Окончание табл. 33
1 1 2 | 3 | 4 | 5 | 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 2,7+0,02 0,10 3,50 41 2,4—2,9
взрослые 2,7±0,02 0,11 4,21 27 2,5—3,0
Самки прибылые 2,7±0,03 0,11 3,97 18 2,5—2,9
взрослые 2,8±0,06 0,21 7,55 13 2,5—3,4
В целом по виду ... 2,7+0,01 0,13 4,61 179 2,4-3,4
Длина углового отростка
1,6±0,02
1,5±0.04
1,6±0,03
1,5+0,03
1,6±0,01
Самцы прибылые
взрослые
Самки прибылые
взрослые
В целом по виду ...
0,13 8,27 30 1,3-1,9
0,18 11,42 20 1,3—2,0
0,10 6.52 12 1,5—1,8
0,08 5,52 8 1,4—1,6
0,14 8,71 131 1,3—2,0
отношение кондило-базальной длины черепа к длине тела составляет
29,6 %. По этому показателю S. minutissimus едва уступает только S. da-
phaenodon (29,7 %) и очень близка у S. beringianus (см. табл. 25). Размер
черепа у зверьков из разных частей ареала колеблется. Лицевой отдел
черепа широкий, рострум короткий. Отношение ширины рострума к кон-
дило-базальной длине характеризуется высокими значениями (9,7 %),
а длина рострума — низкими (24,8 %). Аналогичное соотношение этих
показателей только у 5. tundrensis (см. табл. 32). Мозговой отдел черепа,
напротив, довольно удлинен (57,8 %). В предглазничной области череп
Рис. 53. Крошечная бурозубка — 5. minutissimus Zimm.
157
Таблица 34
Колориметрическая характеристика подвидов Sorex minutissimus из разных частей
ареала
Район сбора материала Белизна Оттенок
Су п 11m Су п lim
S. т. minutissimus, таежные и подта- ежные леса Запад- ной Сибири 3,4+0,26 29,4 15 2,0—5,8 122,9±2,36 7,7 15 104,2-135,4
S. т. barabensis, Ба- рабинская степь 4,6+0,52 19,6 3 3,9—5,6 121,4+3,40 4,9 116,0—127,7
S. т. caudata, Севе- ро-Восточный Ал- тай, Телецкое озеро 5.7±0,23 8,0 4 5,2—6,3 130,4+3,40 5,2 4 123,8—137,0
S. т. stroganovi, Юго- Восточный Алтай, Чуйская степь 6,6+0,28 8,4 4 5,9—7,1 128,5±4,36 6,8 4 119,0—140,0
S. т. tschuktschorum, Чукотка, Якутия 6,2+0,18 14,8 26 4,7—8,2 122,2+1,17 4,9 26 109,1—133,2
5. т. tscherskii, Приамурье, бас- сейн р. Зоя 3,1±0,06 6,8 11 2,7—3,4 129,9±1,54 3,8 И 122,8-135,8
S. т. tscherskii, При- морье 4,7±0,22 11,7 6 3,8—5,4 121,3+1,75 3,5 6 115,2—126,7
также весьма широк (14,2 %), по этому признаку крошечная буро-
зубка уступает лишь S. mirabilis (см. табл. 21), а по межглазничной ши-
рине не имеет себе равных — 21,0 %. Череп довольно узок (см. табл. 29)
и сильно уплощен (28,6 %) в мозговой части (самый низкий абсолютный
показатель), и только у 5. mirabilis относительная высота черепа меньше
(25,3 %).
Верхний резец небольших размеров. Первый верхний промежуточ-
ный зуб крупный, заметно больше второго и третьего; второй меньше
третьего, хотя нередко его вершина не уступает по высоте первому и
второму промежуточным зубам. Четвертый и пятый зубы последователь-
но уменьшены. Соотношение промежуточных зубов близко к таковому
у 5. minutus. Отличие заключается в более крупном первом промежуточ-
ном зубе, форме коронок, более плотном положении зубов в зубном ряду
и конфигурации коронок: они более узкие в основании, вытянуты, их
вершины несколько затуплены и наклонены кзади (внутрь ротовой поло-
сти) по отношению к продольной оси челюсти (рис. 54).
У крошечной бурозубки не только самый меныпий абсолютный пока-
затель длины ряда зубов и длины ряда промежуточных зубов, но и отно-
сительно короткий зубной ряд и ряд промежуточных зубов (29,8 %).
Строение glans penis резко отличается от таковой всех других видов пале-
арктических бурозубок (см. рис. 40). В основной части головка копуля-
тивного органа цилиндрическая, слабо коническая в направлении к сво-
бодному концу. В дистальной трети несколько расширена за счет лате-
ральных валиков, слегка сжата и косо срезана в дорсовентральном на-
правлении. Ее свободный конец копьевидной формы. Размер головки ма-
158
Рис. 54. Череп крошечной бурозубки.
ленький: длина 3,8—6,1 (Л/=4,8)мм,— толщина у основания 0,6—1,1
(М= 0,8) мм.
Распространение. Можно без преувеличения сказать, что
5. minutissimus — один их видов бурозубок с широким ареалом. Она засе-
ляет обширную территорию от Скандинавского полуострова до Чукотки
и Камчатки. На севере граница ареала проходит далеко за Северным по-
лярным кругом (на Таймыре до 71° с. ш.), а на юге — по северу Монго-
лии, на юго-востоке — в Приморье, на Сахалине и в Японии.
В Сибири крошечная бурозубка обитает на большей части террито-
рии. Она встречается во всех природных зонах от северной лесотундры
до гор юга Сибири. Зверьки добывались в Зауралье в бассейне рек Конда,
Сосьва, Тобол, Ишим, в низовьях Оби у Салехарда, на Тазовском и Гы-
данском полуостровах, в окрестностях пос. Самбург на р. Пур. Южнее
этих пунктов она известна из многих районов бассейна Оби и ее прито-
ков — Иртыша, Васюгана, Тыма, Кети, Чулыма, Томи, Шегарки, Баксы,
Кинды, Оми, Тартаса, Уени, Ини, Верди и др. В степных и лесостепных
районах зарегистрирована в бассейне Ишима, окрестностях озер Чаны,
Сартлан.
В Казахстане крошечная бурозубка отмечена близ оз. Кургальджино.
В Средней Сибири этот вид встречается на Таймыре, далеко за Север-
ным полярным кругом (р. Черная, приток Пясины), на границе северной
лесотундры и тундры. Это самый северный ныне известный пункт обита-
ния вида [Юдин, 1980]. Она добыта в ряде мест бассейна Енисея, в том
числе на Нижней Тунгуске, в верхнем течении Енисея в пределах Тувы,
в бассейне Байкала. В Восточной Сибири распространение крошечной
бурозубки изучено менее полно. В бассейне Лены она добыта от широты
Якутска до ее верхнего течения в Прибайкалье. На Яне добывалась в
159
верхнем течении реки, а на Колыме — по всему бассейну на север до
Северного полярного круга.
В горах юга Сибири крошечная бурозубка распространена от запад-
ных предгорий Алтая, в Салаире, Кузнецком Алатау, Западном и Восточ-
ном Саяне, на Хамар-Дабане, в горах Забайкалья, Витимском плоско-
горье и далее на восток до Тихоокеанского побережья [Юдин и др., 1970].
На крайнем севере Дальнего Востока зверьки этого вида заселяют бас-
сейн р. Анадырь и побережье Охотского моря (близ Магадана) [Юдин,
и др., 1976]. На Камчатке добыта в ряде пунктов от северной части полу-
острова до мыса Лопатка. На юге Дальнего Востока встречается всюду
от Приамурья до границы с Кореей.
Материал. 315 экз. из разных районов Сибири и Дальнего Во-
стока.
Изменчивость. Крошечная бурозубка — один из самых поли-
морфных палеарктических видов рода Sorex. Широкой изменчивости под-
вержена окраска. Она проявляется, с одной стороны, в соотношении
интенсивности оттенка и белизны, с другой — в характере распределения
окраски по телу (наличие или отсутствие чепрака) и закономерно прояв-
ляется в разных частях ареала и у разных возрастных групп и по сезо-
нам. В таежной зоне Западной Сибири, от Урала до Енисея, чепрачность
в окраске меха не выражена: темная окраска спины постепенно переходит
в более светлую на боках и вентральной стороне. У перезимовавших буро-
зубок контраст между темной спиной и боками виден довольно хорошо,
хотя граница между ними нечеткая, расплывчатая, а у прибылых она
практически неуловима.
Чепрачная окраска характерна для бурозубок степной и лесостеп-
ной зон Западной Сибири, Казахстана, Северо-Восточного Алтая и северо-
востока Сибири, а также Сахалина.
В лесостепной и степной зонах Западной Сибири и в Казахстане тип
окраски иной. В распределении окраски перезимовавших зверьков четко
выражена чепрачность: темная окраска спины с ярким шоколадным от-
тенком четко отграничена от светлых с палевым оттенком боков. Вент-
ральная часть тела белесая. У прибылых замлероек сохраняется тот же
тип распределения окраски, но чепрачность выражена менее контрастно.
Обитающая в Казахстане (о. Кургальджино) крошечная бурозубка окра-
шена еще более контрастно.
Весьма вариабельна окраска у крошечной бурозубки из разных
частей гор юга Сибири. На Западном и Северо-Западном Алтае (до лево-
бережья Катуни включительно) окраска у землероек чепрачная. В Цент-
ральном Алтае (Усть-Коксинский р-н) она ничем не отличается от таковой
у зверьков из равнинных районов лесостепи и степи Западной Сибири
и Казахстана: у взрослых чепрак яркий, хорошо очерчен, бока и вентраль-
ная сторона тела светлые. Далее в направлении на северо-восток (Шаба-
линский р-н) чепрачность сохраняется, но из-за значительного потемне-
ния окраски боков и вентральной стороны тела констрастность менее
заметна. У перезимовавших бурозубок ниже темного чепрака на боках
хорошо заметна переходная полоса палевого цвета. У отдельных особей
прибылых зверьков чепрачность едва улавливается, у других четко прояв-
ляется. У зверьков на правобережье Катуни (район Каракольских озер,
Телецкое озеро) граница чепрака едва заметна даже у перезимовавших
зверьков за счет более широкой палевой полосы на боку и буровато-серых
тонов в окраске боков и вентральной части.
160
На Юго-Восточном Алтае (Чуйская степь, долины рек Бугузун, Юс-
тыд, Чулышман, близ оз. Джулу-Куль) крошечная бурозубка окрашена
по другому типу. Чепрачность в распределении окраски по телу отсутст-
вует, мех светлый, серовато-бурый на спине и светлый, серебристо-серо-
вато-белесый без палевого оттенка на боках и брюшке. Окраска хвоста
резко двухцветная.
На Салаире, в Кузнецком Алатау, Саянах, в Туве и на восток до
Иркутской области включительно окраска близка к таковой у крошечной
бурозубки из таежных районов равнинной части Западной Сибири. Пере-
ход темных тонов спины к бокам у большинства зверьков постепенный,
без четкой границы. Бока и низ тела окрашены светлее, но со значитель-
ной примесью бурых тонов. Хвост слабо двухцветный. Отдельные особи
даже среди прибылых имеют хорошо выраженную чепрачность окраски.
Это зверьки из степных ландшафтов предгорных остепненных районов
или межгорных котловин. В зимнем наряде чепрак отграничен от боков
более четко, палевая полоса на боках неширокая, мех длинный, шелко-
вистый. Светлая окраска меха на боках со слабым палевым оттенком
и поднимается выше, чем у зверьков в летнем наряде.
На Мал. Хамар-Дабане (р. Самхат) переход темной окраски спины
на латеральную и вентральную части тела постепенный. У перезимовав-
ших бурозубок из других районов Бурятии (Темник, Улзар, Байкаль-
ский заповедник) двухцветность окраски хорошо заметна. У бурозубки
с о. Ольхон, добытой в марте (зимний мех), чепрачность была выражена
довольно четко, но бока имели грязно-серую окраску, хотя и более свет-
лую, чем в летнем меху. Палевая переходная полоса на боках очень уз-
кая. У прибылых бурозубок из Забайкалья (Сохондинский заповедник)
темная окраска спины постепенно переходит в чуть более светлую на бо-
ках и вентральной стороне.
У крошечной бурозубки, обитающей в высоких широтах Таймыра,
Якутии, Чукотки окраска спины светлая со значительной примесью се-
рых тонов. Чепрак есть, но граница между окраской спины и боков раз-
мытая. В зимнем меху (сентябрь) у землероек с Таймыра (Хантайское
озеро, бассейн р. Пясина) и Чукотки (долина р. Омолон) чепрачность
четкая, бока белесые с серым оттенком, а вентральная часть светлее, чем
в летнем меху. Хвост резко двухцветный. У перезимовавших зверьков
летний мех намного светлее, как бы «выцветший». Чепрак четко очерчен
в зимнем и заметен в летнем меху (см. рис. 53).
Совершенно обособлена по характеру окраски крошечная бурозубка
из Приморья: мех темный, переход от спины к бокам и вентральной части
тела постепенный. Вентральная часть бурая, того же цвета, что и спина,
лишь светлее по тону. Зимний мех светлее летнего, с палевым оттенком
и длинным волосом.
Иная окраска у крошечных бурозубок Сахалина. У них окраска
меха спины менее насыщена, чем у зверьков из Приморья, с хорошо выра-
женной чепрачностью. Переход окраски спины к более светлым бокам
имеет четкую границу как у прибылых, так и у перезимовавших особей.
Вентральная часть летом светлая, белесая, зимой серебристо-белая. Хвост
резко двухцветный. Переходной полосы между темной спиной и светлыми
боками в зимнем и летнем меху перезимовавших зверьков нет, а у прибы-
лых она очень узкая.
В пределах обширного ареала вида показатели оттенка и белизны
очень изменчивы: значения первого — 2,0—8,2, второго — 104,2—140,0
161
(см. табл. 34). Для детального обсуждения изменчивости окраски мы не
располагаем достаточным материалом, так как крошечная бурозубка
относится к наиболее редким видам насекомоядных и до недавнего време-
ни в коллекциях насчитывались единичные особи. По имеющимся данным,
можно говорить о тенденции увеличения значений показателя белизны
в направлении с запада (равнинные районы Западной Сибири) на северо-
восток (Чукотка, Камчатка), а также на юг (горы Северо-Восточного
и Юго-Восточного Алтая). Окраска зверьков из Приморья близка к тако-
вой западно-сибирских представителей, но из-за малочисленности мате-
риала для окончательного суждения о ней нет оснований.
Что касается размаха изменчивости показателя оттенка, то он близок
по крайним и средним значениям как в Западной Сибири (104,2—135,4;
М= 122,9=2,36), так и на севере Дальнего Востока (109,1—133,2;
М= 122,2=1,17). Меньший размах значений этих показателей изменчи-
вости в других районах скорее всего обусловлен не диапазоном полимор-
физма, а малым объемом исследованных выборок (см. табл. 34). У земле-
роек таежной зоны окраска меха варьирует от матовой, серовато-бурой
до насыщенной кофейно-бурой, что обусловлено возрастной и сезонной
изменчивостью. Летний мех, особенно у прибылых зверьков, более туск-
лый, менее пышный и короткий, а зимой блестящий и более длинный.
Значение белизны колеблется от 2,0 до 5,8(37 = 3,4=0,26), оттенок от
104,2 до 135,4 (М =122,9=2,36), т. е. показатели эти изменчивы в до-
вольно широких пределах. Здесь, а также в Приморье, локализуются
популяции наиболее темноокрашенных зверьков (см. рис. 56).
Географическая изменчивость окраски проявляется и в изменении
ее интенсивности. Так, зверьки севера Дальнего Востока и Камчатки
окрашены в более светлые тона, чем таежных и подтаежных районов За-
падной Сибири или юга Дальнего Востока. Они достоверно отличаются
по белизне (Td = 7,2), но близки по оттенку (Td — 0,27). Общее посвет-
ление окраски меха дорсальной стороны тела прослеживается и в направ-
лении к югу. Зверьки из Северного Казахстана и Чуйской степи имеют
светлую «пустынную» окраску.
Возрастная изменчивость окраски меха сводится к смене более туск-
лого и редкого волосяного покрова прибылых зверьков на интенсивную
взрослых. В популяциях, где для взрослых характерна четкая чепрач-
ность в распределении окраски, у молодых она выражена слабо, особен-
но в первый гнездовой период. Кроме того, с возрастом у зверьков воло-
сяной покров на ушах, лапах и хвосте снашивается и не восстанавливает-
ся. Поэтому у старых зверьков ушные раковины и лапки почти голые.
В первую очередь снашиваются торчащие в сторону волосы на хвосте,
кончик хвоста у очень старых зверьков совершенно голый. Зимний мех
у видов всех популяций с блеском, с длинным волосом, пушистый.
Весьма значительна вариабельность размеров тела и соотношения
размеров отдельных его частей. Наиболее крупные зверьки заселяют
Юго-Восточный Алтай: длина их тела составляет 48—53 (М = 49,2=
=0,04) мм. Далее, по убывающей, следуют зверьки из Северо-Восточно-
го Алтая, равнинных таежных районов Западной Сибири, Приморья,
Чукотки, лесостепных и степных зон Западной Сибири. Казахстана и
Западного Алтая (для последнего характерны самые мелкие формы).
Географическая изменчивость прослеживается и в размерах хвоста.
Самые длиннохвостые землеройки населяют Центральный и Северо-Во-
сточный Алтай (М = 30,8=0,39; lim 28—37 мм), затем идут по убываю-
И Б. С. Юдин
162
Морфометрическая характеристика подвидов
Признак a. S. т. minutissimus, таежная зона Зап. Сибири
М±тх О Су п lim
Длина
тела 46,1±0,56 3,86 8,4 47 33—53
хвоста 23,8±0,30 2,26 9,5 56 21—34
ступни 8,4±0,07 0,56 6,6 55 8—10
Масса тела Длина 2,2±0,05 0,30 13,4 37 1,5—3,0
кондило-базальная черепа 13,5±0,04 0,33 2,4 71 12,7—14,0
рострума 3,2±0,02 0,21 6,5 77 2,6—3,5
мозговой части черепа 7,8±0,06 0,28 3,6 27 7,1—8,3
верхнего ряда зубов 5,9±0,02 0,20 3,4 75 5,3—6,1
промежуточных зубов Ширина 1,7±0,02 0,13 7,7 27 1,3-1,9
рострума 1,4±0,03 0,02 8,6 26 1,2-1,5
предглазничная 1,9±0,03 0,13 6,8 26 1,7—2,2
межгл азничная 2,9±0,02 0,16 5,6 76 2,4—3,1
наибольшая черепа 6,5±0,03 0,25 3,8 71 6,0—7,0
Наибольшая высота черепа Высота восходящей ветви ниж- 3,9±0,04 0,33 8,2 68 3,0-4,5
ней челюсти 2,8±0,03 0,15 5,1 24 2,5—3,1
Длина углового отростка 1,5±0,04 0,16 12,7 18 1,1—1,8
щей (табл. 35) землеройки Приморья и Хабаровского края (71/ = 28,0±
±0,53; lim 22—32), Юго-Восточного Алтая (М = 27,5±0,67; lim 25—
30), таежной зоны Западной Сибири, Зауралья, заповедника «Малая
Сосьва» (71/ = 25,3±0,85; lim 20—29), Приобья (71/ = 23,8±0,30;
lim 21—34), Тувы (М = 24,4; lim 20—29), Таймыра {М = 24,5; lim 23—
25), Прибайкалья (М = 23,4±0,78; lim 19—31), лесостепи Западной Си-
бири (71/ = 22,3±0,34; lim 19—26), Западного Алтая (М = 22,2±0,47;
lim 49—28) и самые короткохвостые землеройки живут на севере Дальне-
го Востока (71/ = 22,0±0,28; lim 18—25) и в Читинской области, в Со-
хондинском заповеднике (71/ — 21,1 ±0,78; lim 19—31 мм).
Весьма значительная изменчивость формы и соотношения размеров
промежуточных зубов у этого вида. Первый верхний промежуточный зуб
всегда крупнее остальных зубов этой категории, но коронка его может
быть более или менее вытянутой по высоте. Второй промежуточный чаще
всего меньше первого и третьего, реже он равен третьему (не по уровню
вершин, апо абсолютным размерам). Изменчива форма коронок: промежу-
точные зубы всегда вытянуты, но количественные значения этого призна-
ка весьма изменчивы. В зубном ряду они сидят всегда плотно, однако в
разных популяциях этот признак проявляется по-разному. У подвидов
с более коротким рядом промежуточных зубов коронки их выше, а зубы
сидят плотнее и наоборот.
Значительна изменчивость размеров черепа у крошечной бурозубки
из разных районов обширного ареала вида (см. табл. 35). Наибольшие
163
Sorex minutissimus из разных частей ареала
Таблица 35
б. S. т . barabensis, Сев. Бараба в. S. т. abnormis, Юго-Западный и Центр. Алтай
М±тх СТ cv п lim СТ CV п lim
42,1+0,80 3,83 9,1 23 30—47 44,1±0,63 3,85 8,7 38 33—53
22,3+0,34 1,85 8,3 30 19—26 22,2±0,47 2,88 12,9 38 19—28
7,7±0,11 0,59 7,7 32 7,0—9,0 7,4±0,23 1,39 12,7 38 7-8
2,5±0,23 0,87 34,0 14 1,4-3,8 2,0±0,02 0,15 7,5 27 1,5-3,7
12,7±0,07 0,37 2,9 30 12—13,6 12,8±0,07 0,33 2,6 21 12,2—13,8
3,0±0,05 0,27 8,8 31 2,2—3,4 2,8±0,02 0,13 3,9 22 2,7—3,2
7,5±0,04 0,21 2.7 22 7,2—8,0 7,2±0,03 0,14 1,9 21 7,0—7,6
5,5±0,06 0,30 5,5 29 4,7—6,0 5,9±0,03 0,13 2,1 22 5,7-6,4
1,5+0,04 0,17 11,4 23 1,0-1,7 1,3+0,02 0,08 6,4 22 1,1—1,4
1,3±0,02 0,09 6,7 22 1,0-1,4 1,2±0,01 0,06 4,7 22 1,0—1,2
1,9±0,03 0,15 7,6 22 1,5-2,1 1,7±0,02 0,08 4,6 22 1,6-1,9
2,7±0,04 0,20 7,4 31 2,2—3,1 2,5±0,03 0,12 4,8 22 2,2—2,8
6,1±0,07 0,37 6,1 31 5,2—6,9 5,6±0,04 0,17 3,1 22 5,1—5,7
3,9±0,05 0,28 7,3 29 3,2—4,7 3,5±0,03 0,14 3,9 22 3,1-3,6
2,8±0,03 0,16 5,5 23 2,7—3,4 2,7±0,02 0,08 3,0 20 2,6—2,9
1,4±0,02 0,06 4,4 15 1,4—1,6 1,3±0,02 0,08 6,1 17 1,1—1,4
размеры кондило-базальной длины черепа зарегистрированы на южной
периферии ареала, в Юго-Восточном Алтае (М = 14,2±0,04; lim 14,0—
14,3 мм), а самые мелкие — в лесостепи Западной Сибири (М = 12,7 —
0,07; lim 12,0—13,6) и в Западном Алтае (М = 12,8; lim 12,2—13,8 мм).
По величине этого показателя близка к ним крошечная бурозубка с Чу-
котского полуострова (М = 13,1 ±0,05; lim 12,2—14,0 мм). Довольно
крупным оказался череп у землероек этого вида с Таймыра, самой север-
ной периферии ареала (Л/ = 13,7; lim 13,6—13,8 мм).
Географическая изменчивость ряда признаков крошечной бурозубки
проявляется наиболее четко, чем у других видов. Иногда зверьки разных
подвидов заселяют сопредельные территории, разделенные всего несколь-
кими километрами или десятками километров. Так, границы номинально-
го подвида и S. т. barabensis на юге Западной Сибири, вероятно, соприка-
саются, проходя в районе р. Омь. Зверьки, обитающие севернее этой ре-
ки, относятся к номинальному подвиду, а южнее — к барабинскому.
Расстояние между крайними находками этих двух подвидов составляет
менее 30 км. Отличия подвидов не вызывают сомнений: у номинального
чепрак не выражен, у барабинской крошечной бурозубки он очерчен
хорошо; у первого хвост довольно длинный, у второго — короткий.
По долине р. Чулышман по остепненным ландшафтам с монгольскими
элементами растительного покрова до устья р. Башкаус проникает и
S. т. stroganovi, в то время как в окружающей Телецкое озеро черневой
тайге и по водоразделу рек Чулашман и Башкаус обитает S. т. caudata.
11*
164
г. S. т. caudata, Алтай, Саяны
2И^тя ст cv п Нш
Признак
Длина тела 45,2+0,53 3,28 7,23 38 40-53
хвоста 30,8+0,39 2,47 8,03 40 28—37
ступни 8,9±0,75 0,47 5,33 40 8-10
Масса тела 2,2±0,22 0,75 33,50 12 1,5—3,8
Длина
кондило-базальная черепа 13,8+0,04 0,34 2,43 59 12,0—14,2
рострума 3,4+0,02 0,18 5,35 59 2,8—3,8
мозговой части черепа 7,9+0,03 0,24 3,02 51 7,1—8,3
верхнего ряда зубов 5,8+0,02 0,18 3,08 59 5,0-6,1
промежуточных зубов 1,8+0,02 0,14 7,79 51 1,5—1,9
Ширина
рострума 1,4+0,01 0,10 7,66 51 1,1—1,6
предглазничная 2,0+0,01 0,09 4,46 51 1,7—2,2
межгл азничная 2,8+0,01 0,08 2,83 59 2,6—3,0
наибольшая черепа 6,4+0,03 0,23 3,60 59 5,8—6,8
Наибольшая высота черепа 3,7+0,03 0,24 6,29 59 3,1—4,2
Высота восходящей ветви ниж-
ней челюсти; 2,8+0,01 0,09 3,21 49 2,7—3,0
Длина углового отростка 1,5+0,02 0,12 7,58 39 1,5-3,8
Последняя встречается вплоть до правого берега Катуни, а на левобере-
жии обитает уже другой подвид — S. т. abnormis. Они надежно диффе-
ренцируются по ряду признаков. Так, длина хвоста бурозубок этих под-
видов характеризуется не перекрывающимися значениями (у S. т. cau-
data — 29—35, у S. т. abnormus — 19—28).
Наряду с популяциями номинального подвида, заселяющего черне-
вую тайгу близ Мариинска и горы Кузнецкого Алатау, встречается бара-
бинская крошечная бурозубка (добыта серия близ оз. Учум, в 120 км
южнее Мариинска).
Таким образом, можно с уверенностью говорить о четкой приурочен-
ности подвидов к определенным ландшафтам, хорошим индикатором
которых они являются.
Возрастная изменчивость, кроме упомянутого выше полиморфизма
в окраске волосяного покрова, прослеживается в размерах тела и черепа.
По массе тела перезимовавшие самцы и самки крупнее прибылых. Конди-
ло-базальная длина черепа с возрастом становится меньше, различия эти
достоверны (Td — 2,6). С большой достоверностью различается длина
зубного ряда у перезимовавших самцов (меньше таковой у прибылых),
а череп значительно уже и ниже (см. табл. 33). В зубной системе за счет
стирания в первую очередь снашиваются вершины первого верхнего рез-
ца, первого и третьего промежуточных зубов. В нижней челюсти также
в первую очередь снашиваются выступающие из ряда вершины зубов.
Конусы и гребни коренных зубов сглаживаются. У самых старых зверь-
ков все коронки зубов верхней челюсти снашиваются почти до одинако-
165
Продолжение табл. 35
д. S- т. stroganovi, Юго-Восточный Алтай е. S. т. ischuktschorum, Колыма, Чукотка
СТ cv п 1 im СТ cv п lim
49,2±0,97 2,17 4,41 5 48—53 43,8±0,70 4,55 10,4 42 30—52
27,5±0,67 1,64 5,48 6 25—30 22,0±0,28 1,91 8,68 48 18-25
9,2±0,31 0,75 8,21 6 8—10 8,1±0,08 0,57 7,03 46 6—9
2,3±0,55 1,23 53,18 5 1,5-4,5 2,1±0,09 0,44 20,38 23 1,6—3,5
14,2±0,04 0,10 0,70 7 14,0—14,3 13,1±0,05 0,39 3,04 54 12,2—14,0
3,6±0,03 0,09 2,49 7 3,5—3,8 3,3±0,04 0,34 10,20 60 2,5—3,5
8,1±0,03 0,09 1,11 7 6,2—6,7 7,7±0,04 0,28 3,71 50 6,7—8,3
6,0±0,05 0,13 2,10 7 5,7-6,1 5,5±0,02 0,16 2,89 55 5,1—5,8
1,9±0,03 0,08 4,24 7 1,8—2,0 1,7±0,01 0,08 5,06 51 1,5—1,8
1,4±0,02 0,05 3,94 7 1,3—1,4 1,3±0,02 0,09 7,06 51 1,1—1,5
2,0±0,04 0,12 5,77 7 1,8—2,1 1,9±0,02 0,11 5,86 55 1,5—2,1
2,5±0,03 0,08 2,67 7 2,7—2,9 2,6±0,02 0,13 5,08 61 2,4-3,1
6,5±0,07 0,18 2,77 7 6,2—6,7 6,2±0,03 0,26 4,28 56 5,5—7,0
3,9±0,09 0,24 6,28 7 3,5-4,2 3,6±0,05 0,34 9,55 52 2,9-4,2
2,7±0,02 0,06 2,14 7 2,6—2,8 2,7±0,01 0,10 3,74 56 2,4-2,9
1,6±0,03 0,07 4,42 5 1,5—1,7 1,6±0,02 0,14 8,59 43 1,3—2,0
вой высоты, их первоначальное соотношение угадывается только по вели-
чине оснований коронок.
Glans penis неполовозрелых зверьков мелких размеров полупро-
зрачная и еще не имеет видоспецифичного строения, наблюдающегося
у перезимовавших зверьков к весне.
Заметки по систематике. Впервые крошечная бурозуб-
ка была добыта знаменитым исследователем нашей фауны Палласом
[Pallas, 1773] на р. Кия у с. Кийского (ныне г. Мариинск) и на Енисее.
Хотя П. С. Паллас и обратил внимание на ее крайне мелкие размеры,
уступающие уже известной в то время S. minutus, он не дал ей номенкла-
турного обозначения. На основании опубликованных им диагностиче-
ских признаков в ранг вида ее выделил Циммерманн [Zimmermann,
1780] и дал название S. minutissimus Zimm.
Таксономически этот вид настолько хорошо очерчен, что его само-
стоятельность в течение 200 лет у исследователей не вызывала сомнений.
Иоган-Фридрих Гмелин в 1789 г. в 13-м издании «Systema naturae» опи-
сал землеройку под названием S. exilis, а в 1793 г. во французском изда-
нии этой книги описал S. minimus. Судя по диагнозам, это один и тот же
вид — крошечная бурозубка, и по приоритету за ней должно было остать-
ся название, предложенное Циммерманном — S. minutissimus. Долгое
время эти две формы (S. exilis, S. minimus') неверно относили в число си-
нонимов малой бурозубки [Miller, 1912; Огнев, 1928; Ellerman, Morrison-
Scott, 1951; и др.]. С. И. Огнев описал крошечную бурозубку под назва-
нием S. tscherskii Ognev, 1913 и потом — S. ussuriensis Ognev, 1921. Пер-
166
Окончание табл. 35
Признак мс. S. тп. ussuriensis, Приморье
СТ Су п lim
Длина
тела 45,4+0,93 4,69 10,32 23 38—53
хвоста 28.0ч-0т53 2,77 9,80 27 22—32
ступни 8,5±0,13 0,72 8,45 27 7—10
Масса тела Длина 2,2±0,12 0,37 16,91 10 1,9—3,1
кондило-базальная черепа 13,5±0,07 0,50 3,66 42 12,6—14.5
рострума 3,4±0,04 0,26 7,60 43 2,7—3,8
мозговой части черепа 7,7±0,06 0,37 4,74 42 7,0-8,5
верхнего ряда зубов 5,7±0,04 0,26 4,48 38 5,1—6,2
промежуточных зубов Ширина 1,8±0,02 0,15 8,26 42 1,5-2,0
рострума 1,4±0,01 0,08 5,54 38 1,2—1,6
предгл азничная 1,9±0,02 0,11 5,49 38 1,7—2,1
межглазничная 2,7±0,02 0,14 5,22 43 2,4-3,0
наибольшая черепа 6,2±0,05 0,30 4,83 41 5,6-6,7
Наибольшая высота черепа Высота восходящей ветви ниж- 3,6±0,04 0,24 6,73 35 3,0—4,1
ней челюсти 2,7+0,02 0,10 3,75 43 2,4—2,8
Длина углового отростка' 1,6±0,02 0,12 7,41 29 1,3—1,8
вое название многие годы фигурировало в отечественной зоологической
литературе.
Согласно приоритету название вида — 8. minutissimus Zimm., 1780 —
восстановил С. У. Строганов [1957]. Он считал, что на основании упло-
щенного черепа эта бурозубка систематически ближе всего к S. vir Allen
(= S. roboratus Hollister, 1913) и будто бы у них сходное строение glans
penis. Мы не разделяем этого мнения. Уплощенность черепа — единствен-
ный признак сближающий эти два вида. На наш взгляд, С. У. Строганов
[1957] придает ему необоснованно большое таксономическое значение.
Достаточно напомнить, что даже внутри вида, но у разных подвидов кро-
шечной бурозубки, он колеблется в значительных пределах. Кроме того,
еще более уплощенный череп, чем эти два вида, имеет S. mirabilis Ognev,
но она находится настолько в обособленном положении, что по ряду струк-
тур была совершенно правильно выделена в отдельный подрод [Гептнер,
Долгов, 1967] и наделена рядом признаков, коренным образом отличаю-
щих ее от этих двух видов.
Крошечная и бурая бурозубки существенно различаются не только
по размерам (одна из них самая мелкая, а вторая одна из самых крупных
землероек), но и по конфигурации черепа, структуре зубной системы,
строению гениталий и другим показателям. У крошечной бурозубки
ростральная часть черепа относительно короткая, а у 8. roboratus длин-
ная; зубной ряд у крошечной бурозубки относительно кондило-базальной
длины черепа короткий (42,8 %), а у 8. roboratus — длинный (44,3 %).
Зубы у этих видов имеют совершенно не сходную конфигурацию коронок
и разное соотношение их в зубном ряду: зубы крошечной бурозубки сидят
167
очень плотно, а у S. roboratus растянуты сильнее, чем у любого другого
вида бурозубок (см. рис. 70).
Габитуально крошечная бурозубка похожа на S. minutus, но их сход-
ство ограничивается главным образом малыми размерами. Даже по внеш-
нему виду эти два вида различаются безошибочно. У малой бурозубки
ростральная часть головы и череп вытянуты, узкие, с хорошо выражен-
ным перехватом в области глаз. Отношение межглазничной ширины
к кондило-базальной длине черепа составляет у малой бурозубки 17,8,
у крошечной — 21,0 % (см. табл. 21). Череп крошечной бурозубки более
уплощен в мозговой области (28,6 %) по сравнению с черепом малой буро-
зубки (31,1 %). У крошечной бурозубки хоботок в отличие от S. minutus
короткий и широкий, ростральная часть головы шире и короче. Длина
хвоста у S. minutus составляет 70,6, у S. minutissimus — 54,0 %
(см. табл. 24). Первый промежуточный зуб у S. minutus равен третьему,
у S. minutissimus он превосходит его по величине. Свободно сидящие
в зубном ряду коронки промежуточных зубов у малой бурозубки широкие
в основании, с острыми вершинами, а их вертикальная ось перпендику-
лярна продольной оси челюсти. У крошечной бурозубки строение зубной
системы иное: промежуточные зубы вытянуты, узкие, с притупленными
вершинами, направленными внутрь ротовой полости, в зубном ряду сидят
очень плотно. Ряд промежуточных зубов относительно короткий (29,8 %)
по сравнению с этой категорией зубов малой бурозубки (34,3 %).
Головка копулятивного органа этих видов различного строения:
у крошечной бурозубки она относительно малых размеров и ее коронка
имеет «копьевидную» форму, у малой бурозубки головка непропорцио-
нально больших размеров, в виде длинного, слабо сужающегося к свобод-
ному концу стержня, увенчанного на конце небольшим кольцевым вали-
ком (см. рис. 40).
В пределах ареала вида описано девять подвидов крошечной буро-
зубки, из них семь обитает в Сибири.
Обзор подвидов
Sorex minutissimus minutissimus Zimmermann, 1780 —
Обыкновенная крошечная бурозубка
Голотип. Западная Сибирь, бассейн р. Кия, окрестности с. Кий-
ское (ныне г. Мариинск, Кемеровская обл.).
Диагноз. Окраска дорсальной части тела характеризуется мень-
шими значениями белизны, чем у большинства других подвидов
(см. рис. 56), летний мех на спине довольно тусклый, чепрачность не
выражена: темный, серовато-бурый оттенок спины, постепенно светлея
на боках, без резкой границы переходит в серовато-белую окраску вент-
ральной части тела. Размеры средние.
Измерения. См. табл. 35.
Распространение. Ареал обыкновенной крошечной буро-
зубки охватывает обширную территорию центральных и восточных райо-
нов европейской части СССР и таежную зону Сибири к востоку до бассей-
на Енисея и Прибайкалья. На севере граница ареала полярного круга
доходит до Северного (66° с. ш., Самбург на р. Тур). В Западной Сибири
зверьки этого подвида заселяют леса до широты верховьев р. Тара
(р. Бол. Ича, окр. д. Украинка), сел Базой и Кожевникове (на Оби),
Томска, Мариинска. Восточная граница подвида не ясна.
168
Таблица 36
Достоверность различий (по Стьюденту) между подвидами Sorex minutissi-
mus
Признак а — б а — в а — г а — д а — е а — ж
Длина тела 4,1 2,4 1,2 2,8 2,6 0,6
хвоста 3,3 2,9 14,2 5,0 4,4 6,9
ступни 5,4 4,2 0,7 2,5 2,8 0,7
Масса тела 1,3 3,7 0 0,2 0,9 0
Длина кондило-базальная черепа 9,9 8,7 5,3 12,4 6,3 0
рострума 3,7 14,1 7,1 11,1 2,2 4,5
мозговой части че- репа 4,2 8,9 1,5 4,5 1,4 1,2
верхнего ряда зубов 6,3 0 3,5 1,9 14,1 4,5
промежуточных зу- бов 4,5 14,1 3,5 5,5 0 3,5
Ширина рострума 2,8 6,3 0 0 2,8 0
пред гл азничная 0 5,6 3,2 2,0 0 0
межгл азничная 4,5 11,1 4,5 2,8 10,6 7,1
наибольшая черепа 5,3 18,0 2,4 0 7,1 5,2
Наибольшая высота че- репа 0 8,0 4,0 0 4,7 5,3
Высота восходящей вет- ви нижней челюсти 0 2,8 0 2,8 2,2 2,2
Длина углового отростка 2,2 4,5 0 2,0 — 2,2
Примечание.. Названия подвидов соответственно буквенным обозначениям
см. в табл, 35,
Материал. 77 экз., Тюменская, Томская, Новосибирская, Ир-
кутская области и Красноярский край.
Заметки по систематике. Крошечные бурозубки номи-
нального подвида хорошо отличаются от распространенных южнее
S. т. barabensis и S. т. abnormis большими размерами тела, хвоста и отсутст-
вием четко выраженной чепрачности в окраске. От барабинской крошечной
бурозубки номинальный подвид достоверно отличается большими разме-
рами тела (Td = 4,1), хвоста (Td = 3,3), ступни (Td = 5,4), кондило-ба-
зальной длиной черепа (Td = 9,9), а также рядом других показателей
(табл. 36). От S. т. abnormis отличается также большим телом и более
длинным хвостом, отсутствием четко выраженной чепрачности и ярких
палевых тонов в окраске меха спины. От длиннохвостой бурозубки
(S. т. caudata) при сходных размерах тела отличается более коротким
хвостом (Td = 14,2) и меньшими размерами кондило-базальной длины
черепа (Td = 5,3). У этих подвидов сходный тип окраски — темный цвет
спины без резкой границы переходит в более светлый на боках, а затем в
белесый на брюшке, но у S. т. caudata последнее относительно более тем-
ного серовато-палевого тона.
От крошечной бурозубки Строганова (S. т. stroganovi) обыкновенная
крошечная бурозубка отграничена с юга популяциями 5. т. abnormis и
S. т. caudata. Она достоверно меньше, чем зверьки Юго-Восточного
Алтая, по размерам тела (Td = 2,8), хвоста, ступни и кондило-ба-
169
зальной длине черепа (Td=12,4) и его отдельных элементов, а также по
более темной окраске меха. От граничащего с номинальным подвидом на
востоке S. т. tschuktschorum отличается более светлой окраской (Td =
— 8,9) меха (см. табл. 34, рис. 56). Эти два подвида имеют почти не пере-
крывающиеся значения белизны при довольно сходных значениях от-
тенка. Наряду с этим чукотская крошечная бурозубка имеет хорошо вы-
раженную чепрачность в окраске. Кроме того, у нее достоверно меньшие
размеры тела (Td = 2,6), хвоста (Td — 4,4), ступни (Td— 2,8) и кондило-
базальная длина черепа (Td = 6,3).
С крошечной бурозубкой Черского (S. т. ussuriensis) обыкновенная
крошечная бурозубка сходна по характеру окраски спины. Однако у
S. т. ussuriensis переход от темной спины к бокам и низу тела еще более
постепенный, а вентральная сторона окрашена темнее, чем у всех осталь-
ных подвидов, со значительной примесью бурых тонов. Хвост слабо двух-
цветный. Колориметрические исследования проведены на небольшой вы-
борке, тем не менее можно говорить, что по окраске спины это наиболее
темные подвиды в Сибири. Кроме того, этот подвид достоверно отличается
от номинального более длинным хвостом (Td = 6,9).
Sorex minutissimus barabensis Stroganov, 1956 —
Барабинская крошечная бурозубка
Голотип. $ , ad., 20/VIII 1952 г. Сборы А. А. Максимова. Озе-
ро Сартлан, Барабинский р-н, Новосибирская обл. Коллекция зоологи-
ческого музея Биологического института СО АН СССР.
Диагноз. Самый мелкий зверек среди подвидов крошечной буро-
зубки: длина тела 30—47 (М = 42,1 ± 0,80) мм. Кондило-базальная дли-
на черепа 12—13,6 (М — 12,7 =t 0,07) мм. Летний мех на спине — ко-
фейно-бурого цвета, чепрачность хорошо выражена. Окраска боков посте-
пенно светлеет и переходит в белесо-серую с палевым отливом окраску
вентральной части тела. Хвост слабо двухцветный, соответственно в
тон спины и нижней части тела.
Измерения. См. табл. 35.
Распространение. Ареал подвида очерчен нечетко. Бара-
бинская бурозубка — обитатель лесостепных ландшафтов. Она известна
из ряда пунктов Ишимской и Барабинской степей, Макушинского района
Курганской области. В Барабинской степи добыта в среднем течении
р. Омь, у д. Чумаково, близ южной границы номинального подвида (у
поселков Горелая Грива, Зоново), близ оз. Чаны (у пос. Квашнино),
оз. Сартлан (пос. Эгербеш). В долине Оби она добыта близ пос. Черный
Мыс (севернее Новосибирска). Восточным районом ее обитания являются
степи между Салтонским и Батеневским горными кряжами. Здесь крошеч-
ная бурозубка добыта в окрестностях оз. Учум (Ужурский р-н Краснояр-
ского края). Таким образом, по имеющимся в настоящее время данным,
барабинская крошечная бурозубка заселяет полосу между 55 и 56° с. ш.
в лесостепной зоне Западной Сибири от Тобола на западе и до Енисея
на востоке. Северная граница подвида проходит примерно по 56° с. ш.,
ареалы этого и номинального подвида соприкасаются. Южная граница
выяснена.
Материал. 32 экз., лесостепные и степные районы юга Западной
Сибири.
Заметки по систематике. Один из четко очерченных под-
видов S. minutissimus. Прежде всего обращают на себя внимание мелкие
170
размеры и чепрачная окраска спины. От номинального подвида достовер-
но отличается длиной тела, хвоста и размером черепа. Наибольшее сходст-
во прослеживается с S. т. abnormis. Оба эти подвида характеризуются
мелкими размерами тела, хвоста и черепа, а также чепрачностью окраски.
С. У. Строганов [1957] одной из характерных черт подвида считает нали-
чие ювенильных черт в строении черепа во всех возрастных группах. На-
ши материалы не подтверждают этого тезиса: наибольшие значения высо-
ты черепа барабинской крошечной бурозубки не уступают таковым номи-
нального подвида, но по отношению к кондило-базальной длине черепа
у первой он составляет 30,7, а у номинального подвида — 28,9 %. Юве-
нильные черты в строении черепа С. У. Строганов [Там же] рассматрива-
ет как проявление примитивности. По характеру окраски и длине хвоста
она близка к S. т. tschuktschorum, уступая ей в размерах. С другими под-
видами по размерам она различается еще более (см. табл. 35).
Sorex minutissimus abnormis Stroganov, 1949 —
Казахстанская крошечная бурозубка
Голотип, с?, лето 1932 г., окрестности Челкара, Казахстан. Сбо-
ры Е. П. Лутовой.
Диагноз. По размерам близка к барабинскому подвиду. Средние
размеры тела (М = 44,1 =t 0,63), относительно короткий хвост (М =
«= 22,2 ± 0,47), небольшая ступня (М = 7,4 0,28) мм, хорошо выра-
жена чепрачность.
Измерения. См. табл. 35.
Распространение. Ареал обширен: от юго-западных райо-
нов (окр. Челкара, Актюбинская обл.), Центрального Казахстана (окр.
оз. Кургальджин) до Юго-Западного и Центрального Алтая (окр. Усть-
Коксы, Абая). Отмечено обитание данного подвида в Шебалинском районе
Горно-Алтайской автономной области (окр. сел Черга, Камай, Актел,
Мухор-Черга)7. Возможно проникновение подвида в северо-западную
часть Монголии.
Материал. 1 экз., Центральный Казахстан, коллекция Зооло-
гического музея МГУ. Алтай: 3 экз., окр. пос. Усть-Кокса, 46 экз., Ше-
балинский район, зоологический музей БИ СО АН СССР (Новосибирск).
Заметки по систематике. Отличительные признаки под-
робно приведены при рассмотрении номинального и барабинского подви-
дов. Однако следует отметить, что это самый «яркий» подвид, с хорошо
выраженной чепрачностью. Светлый серебристо-серый тон боков и брюха
резко отграничен от яркой окраски спины, хвост сверху светлый, пале-
вый, а снизу серебристо-палевый. Голова окрашена светлее спины. Из
трех подвидов крошечной бурозубки, обитающих на Алтае (S. т. caudata,
S. т. stroganovi, S. т. abnormis), данная форма наиболее мелкая (см.
табл. 36). По длине хвоста близка к чукотскому, а по величине зубов —
к номинальному подвиду [Строганов, 1949] (см. табл. 36).
Sorex minutissimus caudata Yudin, 1964 —
Длиннохвостая крошечная бурозубка |
Голотип. Описан по материалу из Северо-Восточного Алтая,
(р. Бия, окр. с. Кебезень). № 501 (кол.), сборы И. В Лукьяновой. Типо-
’ В прежние годы крошечная бурозубка в данном районе не обнаруживалась
[Юдин и др., 1979].’
171
вой экземпляр (шкурка и череп) хранится в зоологическом музее АН СССР
(Ленинград).
Паратипы. 3 шкурки с черепами, № 7553 (кол. 804), № 7554
(кол. 844), № 7552 (кол. 2751). Зоологический музей БИ СО АН СССР
(Новосибирск).
Диагноз. Характеризуется длинным хвостом — (28—37) {М =
= 30,8 zt 0,39) мм и чепрачной окраской меха дорсальной части тела
перезимовавших зверьков. Длина хвоста по отношению к длине тела сос-
тавляет 68 %.
Измерения. См. табл. 35.
Характеристика. По размерам тела близка к номинальному
подвиду, отличается от него значительно более длинным хвостом и не-
сколько более крупным черепом (см. очерк S. т. minutissimus).
Распространение. В настоящее время известна из Северо-
Восточного Алтая, из районов, прилегающих к северной части Телецкого
озера, и верховий р. Бия (окр. сел Артыбаш, Кебезень, Турочак), из от-
рогов Салаирского хребта (верховья р. Бердь), восточных склонов Куз-
нецкого Алатау (р. Аскыз) и Западного Саяна (р. Табат).
Материал. 68 экз., Алтай, Салаир, Кузнецкий Алатау, Запад-
ный Саян.
Заметки по систематике. Ранее с Алтая крошечная бу-
розубка была известна по единственной находке Л. М. Шульпина из до-
лины р. Чулышман, но она относится к другому подвиду — S. т. stroga-
novi. Зверьки из Северо-Восточного Алтая (район Телецкого озера,
пос. Артыбаш) и верховьев р. Бия (пос. Кебезень, с. Турочак) отнесены к
подвиду S. т. caudata [Юдин, 19646]. Наиболее примечательной особен-
ностью длиннохвостой крошечной бурозубки, как явствует из ее русского
названия, является длинный хвост, составляющий в среднем 68 % от дли-
ны тела.
Всех крошечных бурозубок с Алтая С. У. Строганов [1957] относил к
номинальному подвиду, хотя в его распоряжении из этого региона был
всего 1 экз., судя по месту находки, относящийся к подвиду S. т. stro-
ganovi. Нами установлено, что на Алтае обитают три подвида крошечной
бурозубки: в Юго-Западном и Центральном Алтае — S. т. abnormis,
в Юго-Восточном — S. т. stroganovi и в Северо-Восточном Алтае —
S. т. caudata. Как уже указывалось выше, в очерке, посвященном номи-
нальному подвиду, различия между ними статистически достоверны по
многим морфометрическим показателям и, кроме того, подчеркиваются
разным типом окраски меха дорсальной стороны тела.
Что касается отличий от S. т. stroganovi, заселяющей сопредельные
районы Юго-Восточного Алтая, то они выражаются в мелких размерах
тела длиннохвостой крошечной бурозубки {Td = 3,6), большой длине
хвоста {Td = 4,3) и меньших размерах черепа {Td = 6,1). Наряду с этим
она характеризуется наличием темноокрашенного чепрака на спине. У
крошечной бурозубки Строганова спина окрашена в «пустынный», туск-
лый, серовато-бурый цвет. В отличие от S. т. tschuktschorum она надежно
диагностируется по ряду признаков: значительно более длинному хвосту
{Td = 1'8,1), большим размерам кондило-базальной длины черепа {Td —
= 12,4) и ряду других показателей (см. табл. 35). От S. т. ussuriensis
длиннохвостая крошечная бурозубка достоверно отличается более длин-
ным хвостом {Td — 3,0), большим черепом {Td — 3,7) и характером окрас-
ки волосяного покрова дорсальной части тела. У уссурийской крошечной
172
бурозубки распределение окраски на спине не чепрачное: темные тона
постепенно, без заметных резких границ, переходят в более светлую ок-
раску боков и вентральной стороны тела, которые значительно более тем-
ные, чем у других подвидов.
Sorex minutissimus stroganovi Yudin, 1964 —
Крошечная бурозубка Строганова
Голотип. Юго-Восточный Алтай, верховья р. Юстыд, 9/VIII
1962 г. Типовой экземпляр (шкурка, череп) хранится в Зоологическом
музее АН СССР (Ленинград), № 140 (кол.). Сборы Б. С. Юдина.
Паратипы. 3 экз. (шкурки с черепами), № 7540 (кол. 522),
№ 7541 (кол. 627), № 7542 (кол. 777). Зоологический музей БИ СО АН
СССР (Новосибирск).
Диагноз. Характеризуется крупными размерами тела, черепа,
тусклой и светлой окраской меха. Длина тела 48—53 (М = 49,2 =t
=ь 0,97), длинный хвост 25—30 (М = 27,5 ± 0,67) и большая ступня
8—10 (М = 9,2 ± 0,31) мм. Кондило-базальная длина черепа 14,0—
14,3 (М — 14,2 ± 0,04) мм — самая большая среди всех известных под-
видов крошечной бурозубки (см. табл. 35).
Измерения. См. табл. 35.
Характеристика. Найденные в 1962 г. в Юго-Восточном
Алтае бурозубки характеризуются самобытными чертами, ранее не из-
вестными для крошечной бурозубки. Они отличаются от всех описанных
ранее подвидов крупными размерами тела, хвоста, ступни и черепа, «пус-
тынной» окраской меха, со значительной примесью серых тонов. Отличия
этого подвида от обыкновенной, барабинской и длиннохвостой крошечных
бурозубок приведены в соответствующих очерках. От S. т. tschuktscho-
rum она отличается большими размерами тела, хвоста, ступни и значе-
ниями показателей черепа (см. табл. 35). Своеобразна окраска зверьков,
чепрак отсутствует как у перезимовавших, так и прибылых зверьков,,
а в окраске верхней части спины отчетливо заметна значительная при-
месь светлых, серых тонов.
Распространение. В настоящее время известна из восточной
части Чуйской степи и прилежащих районов. Добыта в верховьях р. Юс-
тыд и в пограничном с Тувой районе по р. Бугузун, против устья ее пра-
вого притока р. Байлюгем (в районе горы Талдуаир). Вероятно, зверьки
этого подвида заселяют всю юго-восточную часть Алтая и прилежащие к
ним районы Тувы. По р. Чулышман она спускается в сторону Телецкого
озера и известна из района правого притока р. Чульча (сборы Л. И. Бар-
совой) и близ устья р. Башкаус — левого притока р. Чулышман (сборы
Б. С. Юдина).
Материал. 7 экз., реки Юстыд, Бугузун и Чулышман.
Заметки по систематике. До наших исследований одна
землеройка этого подвида была найдена Л..М. Шульпиным в долине р. Чу-
лышман. С. У. Строганов [1957] отнес ее к номинальному подвиду, как и
всех прочих крошечных бурозубок, заселяющих Алтай, Саяны и Туву»
Sorex minutissimus tschukschorum Stroganov,. 1949 —
Чукотская крошечная бурозубка
Голотип. <5*, ad., 9/Г 1932 г. Сборы Л. А. Портенко. Чукотский
полуостров, долина р. Анадырь близ с. Марково.
173
Диагноз. Размеры относительно мелкие, хвост короткий. Ок-
раска светлая, резко двухцветная. Чепрак от серо- до шоколадно-бурого.
Во взрослом наряде граница между темным верхом спины и серовато-
белыми боками четкая.
Измерения. См. табл. 35.
Характеристика. Размеры хвоста (Td = 10,5), ступни
(Td = 2,6) и кондило-базальной длины черепа (Td = 4,7) значительно
больше, чем у чукотской бурозубки (см. табл. 35). С. У. Строганов [1957],
характеризуя чукотскую крошечную бурозубку как короткохвостую фор-
му, указывает, что «короткохвостость»— свойство арктических и субарк-
тических землероек. Анализ морфометрических показателей разных под-
видов крошечной бурозубки не подтверждает этого тезиса. Самые ко-
роткохвостые формы заселяют не только северные районы, но и южные
(Ишимская и Барабинская степи, Казахстан, Западный Алтай) (см.
табл. 35). Причем граница между ареалами короткохвостой S. т. Ьага-
bensis и длиннохвостой S. т. minutissimus подвидами практически сопри-
касается, при этом у обитающей севернее землеройки номинального под-
вида хвост длиннее, чем у заселяющей более южные районы барабинской
крошечной бурозубки. Этот признак, вероятно, связан с характером ланд-
шафтов, заселяемых зверьками, а не только их широтным положением.
Аналогичные «несоответствия» имеются и у подвидов других видов зем-
лероек.
Распространение. Ареал этого подвида охватывает обширную
территорию Восточной Сибири, севера Дальнего Востока и Камчатки.
Материал. 78 экз., районы Восточной Сибири, Чукотки и Кам-
чатки.
Заметки по систематике. Впервые на самобытность кро-
шечной бурозубки северо-востока Сибири обратил внимание С. И. Огнев
[1926, 1928, 1941]. Таксономический ранг ей был присвоен С. У. Строга-
новым [1949, 1957], выделившим ее в качестве подвида.
Нами собрана большая серия зверьков в бассейне р. Омолон, позво-
лившая более детально охарактеризовать окраску и габитуальные осо-
бенности этой бурозубки. Ее отличия от S. т. minutissimus, S. т. Ъага-
bensis, S. т. abnormis, S. т. caudata, S. т. stroganovi описаны в соответст-
вующих подвидовых очерках. От S. т. ussuriensis, заселяющей юг Даль-
него Востока, она четко отличается по характеру окраски: у крошечной
бурозубки Черского окраска значительно темнее (см. рис. 56), нет чепра-
ка, а вентральная часть тела значительно более темная.
Sorex minutissimus ussuriensis Ognev, 1921 —
Уссурийская крошечная бурозубка
Голотип. Долина р. Бекин, среднее течение р. Уссури. Коллек-
ция Зоологического музея МГУ (Москва).
Диагноз. Окраска меха темная, тусклая. Серовато-бурая окрас-
ка на дорсальной стороне тела постепенно переходит в относительно тем-
ную окраску вентральной стороны. Хвост слабо двухцветный, относитель-
но длинный.
Измерения. См. табл. 35.
Характеристика.- Звери средних размеров с относительно
длинным хвостом.
Распространение. Этот подвид заселяет Приморье и Саха-
лин. Точно границы очертить нельзя из-за малочисленности материала.
В Сунгаро-Ханкайской провинции обитает бурозубка Черского.
174
Материал. 43 экз., Приморье, Сахалин.
Заметки по систематике. Подвид хорошо отличается по
ряду признаков от других подвидов. Отличия приведены выше. Колори-
метрические показатели интенсивности белизны и оттенка близки к тако-
вым у номинального подвида (см. рис. 56, табл. 35).
Впервые на специфические особенности этой землеройки обратил
внимание С. И. Огнев [1921], описав ее в качестве вида. Из диагностиче-
ских признаков, приведенных для этой землеройки, заслуживают внима-
ния относительная длина рострума (по сравнению с номинальным подви-
дом) (Td = 4,5) и окраска спины — черновато-серая, с сильным палево-
бурым оттенком и буро-серой темной нижней стороной. Бока и брюшко
темные. Хвост слабо двухцветный. Эти признаки полностью укладывают-
ся в параметры, полученные при изучении собранных нами крошечных
бурозубок из Приморья. Размеры тела и черепа, приведенные С. И. Ог-
невым [1928], характеризуют крайние мелкие варианты из наших сборов,
что связано с малочисленностью имевшегося в его распоряжении материа-
ла (всего три зверька).
С. У. Строганов [1957] землероек с Дальнего Востока относил к под-
виду S. т. tscherskii, описанному С. И. Огневым [1913] по экземпляру из
долины р. Одарка (бассейн оз. Ханка). Сравнивая диагнозы и характе-
ристики, приведенные для этого подвида С. И. Огневым [1928] и
С. У. Строгановым [1957], нетрудно уловить в них существенные разли-
чия. По С. И. Огневу [1928], крошечная бурозубка Черского (S. т. tscher-
skii) значительно меньше крошечной темноцветной бурозубки (S. ussu-
riensis), как назвал ее С. И. Огнев [1928], она имеет меньшие размеры че-
репа и короткий лицевой отдел. Судя по описанию, окраска тела светлая
и чепрачная: «Окраска летней шерсти на спине довольно светлая, серо-
буроватая с легким палево-оливковым оттенком. Бока тела и щеки сереб-
ристо-серобелесые, одинакового цвета с брюшной областью» (с. 255).
С. У. Строганов [1957] приводит совершенно иное описание окраски, под-
черкивая ее темные тона, постепенный переход от спины к бокам и темно-
окрашенной вентральной части тела, т. е. окраску, которую описал
С. И. Огнев [1928] для S. ussuriensis и которую мы обнаружили на серии
зверьков из Приморья. Показатели, приведенные для крошечной бурозуб-
ки Черского, полностью укладываются в таковые Чукотской крошечной
бурозубки. Поэтому к подвиду S. т. ussuriensis мы относим только зверь-
ков широколиственных лесов Приморья и Сахалина.
Sorex minutus Linnaeus," 1766
Малая бурозубка
ИЖ Голотип. Типового экземпляра нет. Голотип описан К. Линнеем
на основании письменного сообщения, сделанного Лаксманном, который
поймал малую бурозубку близ Б рнаула 8.J
8 А. А. Гуреев [1971 ] неверно ук1зывает, что тип вида выделен на основании опи-
сания Лаксманна [Laxmann, 1769], опубликованного в его знаменитых «Письмах из
Сибири» (Sibirische Briefe). На самом деле Лаксманн сообщил описание вида К. Линнею
письмом,ина этом основании в 1766 г., т. е. за три года до выхода в свет указанной
А. А. Гуреевым работы [Laxmann, 1769], был описан новый вид. Лаксманн в своих
«Письмах из Сибири» дал описание этого вида S. pygmaeus. Согласно приоритету за
видом сохранилось название, данное Линнеем — S. minutus L.
175
Диагноз. Мелкая, сравнительно длиннохвостая бурозубка. Дли-
на тела с головой 40—62 (М — 52,2 ztz 0,59), длина хвоста 30—42 (М =я
— 36,9 zfc 0,32) мм. Кондило-базальная длина черепа 19,9—15,8 (М =
= 15,1 ± 0,03) мм. Промежуточныеfзубы в зубном ряду расположены
свободно. Второй промежуточный зуб верхней челюсти меньше первого
и третьего, четвертый меньше предыдущего, пятый мал (рис. 55).
Измерения. См. табл. 37.
Характеристика. Телосложение стройное: отношение массы
тела к длине у прибылых зверьков составляет 6,5. Даже у такого строй-
ного зверька, как S. gracillimus, значения этого показателя несколько
больше — 6,8. Малая бурозубка хорошо диагностируется по размерам
тела. По величине ей уступает только крошечная бурозубка S. minutissi-
mus Zimm. Во внешнем облике наряду с мелкими размерами обращает на
себя внимание довольно длинный хвост (71 % от длины тела с головой),
покрытый торчащими в стороны волосами с хорошо обозначенной кисточ-
кой на конце. Хоботок длинный, его длина подчеркивается наличием ха-
рактерного для малой бурозубки сужения в области глаз. Мозговая часть
головы выпуклая, округлая.
Окраска меха двухцветная, незначительно варьирующая по характе-
ру распределения на теле темных и светлых тонов. Интенсивность окраски
меха дорсальной части тела довольно вариабельна (рис. 57). Колори-
метрическая характеристика окраски приведена в табл. 38. Буро-серая
с рыжеватым оттенком окраска спины постепенно переходит в более свет-
лую на боках, а затем в серовато-белесую или грязно-серую на вентраль-
ной стороне -тела.
176
Таблица 37
Морфометрическая характеристика Sorex minutus
Возрастная и половая группа М±тх О Су п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 51,1+0,51 2,52 4,93 24 45—57
взрослые 57,2+0,91 2,23 3,90 6 54—60
Самки прибылые 51,0+0,89 3,22 6,31 13 44—56
взрослые 57,5+2,50 3,54 6,15 2 55—60
В целом по виду ... 52,2+0,59 4,51 8,63 58 40—60
Длина хвоста
Самцы прибылые 37,1+0,53 2,60 7,02 24 33-42
взрослые 37,3+1,15 2,81 7,51 6 32—40
Самки прибылые 36,9+0,38 1,38 3,74 13 35—39
взрослые 39,0+0,0 0,0 0,0 2 —
В целом по виду ... 36,9+0,32 2,41 6,52 58 30-42
Длина ступни
Самцы прибылые 10,7+0,09 0,46 4,32 24 10,0—11,0
взрослые 10,6+0,20 0,49 4,65 6 10,0—11,0
Самки прибылые 10,6+0,15 0,56 5,25 13 9,8—11,5
взрослые Ю,5±0,50 0,71 6,73 2 10,0—11,0
В целом по виду ... 10,6+0,08 0,57 5,36 57 9,8—11,5
Масса тела
Самцы прибылые 3,2+0,08 0,36 11,27 19 2,5—4,0
взрослые 4,4+0,21 0,46 10,42 5 4,0—5,0
Самки прибылые 3,2+0,16 0,51 16,16 10 2,5—4,0
взрослые 3,4+0,10 0,14 4,16 2 3,3-3,5
В целом по виду ... 3,4±0,09 0,60 17,6 47 2,5—5,0
Кондило-базальная длина черепа
Самцы прибылые 15,2+0,06 0,30 1,99 27 14,3—15,8
взрослые 15,2+0,06 0,15 0,96 7 15,0—15,4
Самки прибылые 15,3+0,09 0,36 2,37 17 14,3—15,8
взрослые 15,2+0,15 0,21 1,40 2 15,0—15,3
В целом по виду ... 15,1+0,03 0,33 2,22 185 13,9—15,8
Длина рострума
Самцы прибылые взрослые 4,0+0,03 3,9±0,11 0,15 0,30 3,65 7,73 27 8 3,5—4,2 3,4-4,2
Самки прибылые 4,0±0,02 0,08 2,07 17 3,8—4,1
взрослые 4,1±0,15 0,21 5,24 2 3,9—4,2
В целом по виду ... 3,9±0,02 0,20 5,18 192 3,5—4,2
Длина мозговой части черепа
Самцы прибылые 8,7±0,04 0,21 2,41 27 8,1—9,1
взрослые 8,7+0,06 0,15 1,73 7 8.5—8,9
Самки прибылые 8,7+0,06 0,23 2,69 17 8,3—9,2
взрослые 8,4±0,25 0,35 4,23 2 8,1—8,6
В целом по виду ... 8,6+0,02 0,23 2,7 185 8,1—9,2
177
Продолжение табл. 37
1 2 з * 5 6
Длина верхнего ряда зубов
Самцы прибылые 6,5+0,02 0,10 1,52 27 6,2—6,7
взрослые 6,3+0,03 0,09 1,45 8 6,2—6,5
Самки прибылые 6,5±0,03 0,14 2,15 17 6,2—6,8
взрослые 6,4+0,0 0,0 0,0 2 —
В целом по виду ... 6,4+0,01 0,14 2,20 191 6,2-6,8
Длина ряда промежуточных зубов
Самцы прибылые 2,3±0,02 0,09 3,76 27 2,1—2,4
взрослые 2,2±0,04 0,10 4,76 8 2,1—2,4
Самки прибылые 2,2±0,02 0,09 3,86 17 2,1—2,4
взрослые 2,2+0,0 0,0 0,0 2 —
В целом по виду ... 2,2+0,01 , 0,11 4,90 192 2,1—2,4
Ширина рострума
Самцы прибылые 1,4+0,01 0,07 4,93 27 1,3-1,5
взрослые 1,4±0,05 0,12 8,45 6 1,3—1,6
Самки прибылые 1,4+0,02 0,10 6,82 17 1,3—1,6
взрослые 1,4±0,05 0,07 5,24 2 1,3-1,4
В целом по виду ... 1,4+0,01 0,08 5,54 190 1,3-1,6
Предглазничная ширина черепа
Самцы прибылые 2,0+0,02 0,10 5,05 27 1,7—2,2
взрослые 2,1+0,03 0,09 4,44 8 2,0—2,2
Самки прибылые 2,0+0,02 0.07 3,45 17 1,9-2,2
взрослые 2,1+0,0 0,0 0,0 2 —
В целом по виду ... 2,0+0,01 0,10 5,01 192 1,7-2,2
Межглазничная ширина черепа
Самцы прибылые 2,8+0,03 0,14 4,96 27 2,4-3,0
взрослые 2,8+0,05 0,13 4,68 8 2,6—3,0
Самки прибылые 2,8+0,03 0,12 4,45 17 2,6—3,0
взрослые 2,8+0,05 0,07 2,58 2 2,7—2,8
В целом по виду ... 2,7+0,01 0,11 4,15 192 2,4-3,0
Н аиболыиая ширина черепа
Самцы прибылые 7,4+0,05 0,23 3,13 27 7,0—7,8
взрослые 7,4+0,07 0,19 2,52 7 7,0-7,6
Самки прибылые 7,5+0,05 0,19 2,54 17 7,0—7,8
взрослые 7,4±0,15 0,21 2,89 2 7,2—7,5
В целом по виду ... 7,3+0,02 0,21 2,82 183 7,0-7,8
Высота черепа
Самцы прибылые 4,8+0,05 0,23 4,79 26 4,2-5,2
взрослые 4,7+0,07 0,19 3,96 7 4,4-5,0
Самки прибылые 4,7±0,05 0,21 4,48 17 4,2-5,0
взрослые 4,5±0,0 0,0 0,0 2 —
В целом по виду ... 12 Б. с. Юдин 4,7+0,02 0,23 4,84 183 4,2—5,2
178
Окончание табл. 37
1 2 3 4 5 6
Высота восходящей ветви нижней челюсти
Самцы прибылые Ц 3,0+0,02 0,09 3,01 25 2,9—3,2
взрослые 3,1+0,03 0,07 2,22 7 3,0—3,2
Самки прибылые 3,0+0,02 0,07 2,26 17 2,9—3,2
взрослые 3,1±0,0 0,0 0,0 2 —
В целом по виду ... 3,0±0,01 0,09 3,06 188 2,7—3,2
Длина углового отростка
Самцы прибылые
взрослые
Самки прибылые
взрослые
В целом по виду ...
1,8±0,02
1,7±0,06
1,8±0,03
1,7±0,10
1,8±0,01
0,08 4,66 14 1,6—1,9
0,10 5,88 3 1,6-1,8
0,08 4,20 8 1,7—1,9
0,14 8,32 2 1,6—1,8
0,09 5,09 120 1,6-1,9
Утверждение В. А. Долгова [1972] о более резком переходе между
темной и светлой окраской меха у малой бурозубки по сравнению
с крошечной не отвечает действительности. У малой бурозубки четко
выраженной чепрачности в окраске нет — переход от темной спины к
светлой нижней части тела постепенный. Для крошечной бурозубки ха-
рактерна широкая вариабельность распространения окраски по телу:
у одних подвидов переход постепенный, у других ясно прослеживается
наличие чепрачности и резкой границы между темной спиной и светлым
брюшком. Кроме того, характер распределения темной и светлой окраски
неодинаков у разных возрастных групп (см. очерк о крошечной буро-
зубке).
По колориметрической характеристике малая бурозубка занимает
среднее положение среди представителей рода. Значения оттенка —
133,5+0,87, белизны — 4,9+044 (рис. 57). Темная окраска головы спу-
скается далеко ниже глаз и уха, а со спины заходит на плечи и бедра. Зим-
ний мех длинный (6,5 мм), шелковистый и более светлый. Значения бе-
лизны меха спины перезимовавших зверьков составляют 3,8±6,0 (М =
= 4,9), а у прибылых зверьков — 5,1—10,2 (М — 7,2). Окраска волося-
ного покрова хвоста менее контрастна, чем летом, за счет бурых тонов.
Череп мелкий с расширенной и вздутой мозговой капсулой. Кон-
дило-базальная длина черепа меньше только у крошечной бурозубки. Ин-
декс высоты черепа к кондило-базальной длине составляет 31,4 %, в то
время как у крошечной бурозуб’
ки он равен 28,4 %. Лицевой от-
дел черепа длинный и узкий: у ма-
лой бурозубки индекс его показа-
телей к кондило-базальной длине
Рис. 56. Сравнительная характеристи-
ка окраски подвидов крошечной буро-
зубки.
2 — S, т. minutissimus', 2 — S. т. baraben-
sis; 3 — S. т. caudata\ 4 — S. т. stroganovi'
5 — S. т. tschukrshorum’, 6 — S. т. tscher-
Ьелизна sfcii.
179
Таблица 38
Колориметрическая характеристика Sorex minutus
Район сбора материала Белизна Оттенок
Су п Ит М±тх Су п lim
Западная Сибирь, Томская, Новоси- бирская обл. 4,1±0,22 21,1 16 2,7—4,4 136,8±2,21 6,5 16 117,4—153,6
Прибайкалье, Хамар- Дабан 4,6±0,17 18,6 25 2,9—6,6 132,2±0,82 3,1 2,5 126,5—143,5
Алтай, Телецкое озеро 5,7±0,19 16,7 26 4,1—7,7 132,7±1,4 5,4 26 116,6-148.3
черепа составляет 26,5 и 10,0 %, а у крошечной соответственно — 24,8 и
10,0 %.
По конфигурации череп малой бурозубки похож на череп средней
(S. caecutiens), но отличается меньшими размерами.
Структура зубов крайне специфична и может быть надежным при-
знаком, позволяющим безошибочно определять видовую принадлежность
зверьков. Промежуточные зубы мелкие, стройные. Вертикальная ось ко-
ронок первого, второго и третьего промежуточных перпендикулярна про-
дольной оси челюсти; четвертый несколько наклонен назад, а ось пятого
перпендикулярна таковой трех первых или несколько наклонена вперед.
Наиболее характерно для структуры зубной системы этого вида соотноше-
ние трех первых промежуточных. Первый и третий зуб по величине равны
между собой, второй заметно уступает им по высоте (см. рис. 55). Перед-
ний резец умеренных размеров. Его вторая (внутренняя) вершина значи-
тельно меньше и тоньше первой. По высоте он равен или несколько усту-
пает первому промежуточному зубу. Вершины промежуточных зубов
острые, с ровными передними и задними сторонами коронок, имеющими
довольно правильную конусовидную форму. Вершины промежуточных
зубов относительно оси коронки лишь незначительно сдвинуты кпереди.
Четвертый промежуточный зуб почти вдвое уступает по размеру
третьему, а пятый находится в таком же соотношении к четвертому. Ги-
поконусы многовершинных
зубов развиты слабо, не пиг-
ментированы. Первый пред-
коренной нижней челюсти
(Рпц) вытянут в сагитталь-
ном направлении.
Весьма своеобразно
строение копулятивного орга-
на взрослых особей. Голов-
ка его почти цилиндрическая,
лишь слегка утончающаяся к
Рис. 57. Колориметрическая ха-
рактеристика окраски малой буро-
зубки.
1 — равнинные районы Западной
Сибири (Томская, Новосибирская об-
ласти); 2 — Прибайкалье (Хамар-Да-
бан); 3 — Алтай (Телецкое озеро).
12*
180
свободному концу, увенчаному коронкой в виде узкого кольцевого валика
по переднему краю резко притупленного свободного конца члена. Моче-
половое отверстие открывается сосочком в центре переднего среза (см.
рис. 40). Glans penis крупная относительно мелких размеров тела зверь-
ка. По отношению к длине тела этот орган у малой бурозубки составляет
17,8 %, в то время как у крошечной — 11, а у средней — только 10,5 %.
Длина головки 8,3—11,4 (М = 9,3) мм, толщина у основания 1,0—1,1 мм.
Распространение. Малая бурозубка — типичный западный
палеаркт. Она заселяет Европу, Западную и Среднюю Сибирь, Казах-
стан, Тянь-Шань, Заилийский Алатау, Алтай, Саяны, Прибайкалье.
Указание А. П. Кузякина [Бобринский и др., 1965] на распространение
малой бурозубки в Центральной Якутии, на Чукотском полуострове,
в бассейне р. Анадырь, на Шантарских островах, Амуре, в Приморье, на
Сахалине и Курильских островах обязано путанице в таксономии вида.
Автор объединял, как уже было сказано выше, малую бурозубку в один
композиционный «вид» с S. gracillimus и S. cinereus. Включение
С. У. Строгановым [1957] в ареал малой бурозубки районов Дальнего
Востока также объясняется объединением этого вида с S. gracillimus.
Проведенные исследования последних лет показали, что на восток от Бай-
кала малой бурозубки нет.
Она населяет обширную территорию на восток от Урала, ареал сужа-
ется в направлении на юго-восток к Байкалу. На Урале S. minutus
заселяет разные ландшафты от Полярного Урала, Ямала до южных отро-
гов хребта между 50° и 70° с. ш., область распространения заходит в рав-
нины Казахстана. Самый северный пункт этой части ареала, откуда из-
вестна малая бурозубка, находится на полуостровах Ямал и Таймырском,
за Северным полярным кругом. Она добыта в ряде мест Гыданского и Та-
зовского полуостровов, в бассейне рек Ныда, Таз, на широте Северного
полярного круга. Из более южных районов имеются сборы из низовий j
р. Пур (близ Самбурга). На Оби она добыта близ Ниж. Киевата, в Ямало- ]
Ненецком национальном округе, южнее, у Ларь-Егана, в бассейне рек !
Таз, Кеть, в районе Колпашево, на Чулыме, Васюгане и поднимается по j
долинам Оби и Иртыша до Алтая включительно. Граница распространения
вида на севере между Тазовским и Таймырским полуостровом не ясна.
Самая северная точка нахождения вида на Таймыре — Хантайское водо-
хранилище, где малая бурозубка добыта близ горы Турмакит, у пос. Снежно-
горска. Далее восточная граница по Енисею спускается к Ангаре, Чуе (пра-
вому притоку Лены). Самый восточный пункт нахождения малой буро-
зубки — восточное побережье Байкала и р. Селенга. В наших обширных
многолетних сборах из Северного Забайкалья и Витимского плоскогорья
ее нет.
Южная граница проходит в Казахстане: Кургальджинский заповед-
ник, г. Каракалинск, о. Зайсан (окр. Баян-Аула), Заилийский и Терскей-
Алатау, Саяны и Хамар-Дабан.
В пределах очерченного ареала в Сибири она распространена спора-
дически, предпочитает заселять лесные ландшафты, а в них — долины рек
и ручьев. Численность всюду не велика.
Материал. 192 зкз., Западная Сибирь, Прибайкалье, Алтай.
Изменчивость. Возрастная и сезонная изменчивость малой
бурозубки такого же характера; как и у других представителей рода.
Наиболее специфична изменчивость волосяного покрова хвоста. У пере-
зимовавших и даже совсем старых зверьков на хвосте сохраняются не
181
совсем сношенные волоски. В меньшей степени по сравнению с другими
видами они снашиваются и на тыльной стороне лап, и на ушах [Маркова,
Долгов, 1972]. Возрастная изменчивость проявляется в увеличении раз-
меров и массы тела у перезимовавших зверьков по сравнению с прибылы-
ми. Например, длина тела прибылых самцов составляет 51,1 zt 0,51 , а пе-
резимовавших — 57,2 0,91 мм. Половых различий по этому показате-
лю не наблюдается (см. табл. 37).
Аналогичная картина прослеживается и при анализе половой и
возрастной изменчивости массы тела, однако, масса перезимовавших сам-
цов (М = 4,4 0,21 г) несколько превышает таковую перезимовавших
самок (М = 3,4 =±= 0,10 г), разнополые прибылые зверьки по массе не раз-
личаются. Достоверных возрастных и половых различий по краниологи-
ческим признакам нет (см. табл. 37). Только зубной ряд за счет стирания
передних резцов с возрастом укорачивается. Кондило-базальная длина
черепа зверьков в выборках разных лет неодинакова: значения этого по-
казателя могут быть большими или меньшими то у прибылых, то у пере-
зимовавших землероек. Такая изменчивость, как справедливо замечает
В. А. Долгов [1972], отражает степень благоприятности того или иного
года для процветания вида. В оптимальные годы молодые зверьки имеют
размеры, превышающие таковые у их родителей, и наоборот.
Возрастная изменчивость в стирании промежуточных зубов несколь-
ко иная, чем у других видов, и во многом сходна с таковой у S. unguicu-
latus, для которого характерен общий с S. minutus тип соотношения трех
первых промежуточных зубов. Первый и третий промежуточные зубы как
более высокие в зубном ряду снашиваются быстрее, чем второй, четвертый
и пятый. В результате этого с возрастом различия в высоте коронок этих
зубов исчезают и они становятся почти одной высоты или равномерно убы-
вают от первого к пятому. Однако первоначальное соотношение улавлива-
ется по ширине коронок. Свойственная для вида последовательность из-
менения их величины — 1 > 2 < 3 >4 >5 — для опытного глаза за-
метна даже у очень старых зверьков с сильно стертыми зубами.
Географическая изменчивость окраски зверьков этого вида весьма не-
значительна и не имеет четкой направленности. Малая бурозубка из раз-
ных частей ареала характеризуется довольно стабильным распределением
темной и светлой окраски. Интенсивность ее меняется в более широких
пределах. Наиболее темноокрашенные зверьки из сибирских выборок об-
наружены в бассейнах рек Конда и Сосьва. Серия зверьков с Енисея
(Ангутиха) отличается общим посветлением меха в сторону развития се-
рых тонов с шоколадным (теплым или рыжеватым) оттенком. Вопреки мне-
нию С. У. Строганова [1957], утверждавшего, что окраска меха подверже-
на незначительной индивидуальной, сезонной и географической измен-
чивости, необходимо отметить ее относительную стабильность по сравне-
нию с таковой многих других видов землероек. Характер распределения
темного и светлого тонов стабилен. В то же время нельзя не указать на
довольно широкую изменчивость интенсивности окраски зверьков из раз-
ных частей ареала. Значение показателя оттенка колеблется от 116,6 до
153,6 (М = 133,9), белизны — от 2,7 до 7,7 (М = 4,8). Крайние показа-
тели выборок из разных частей ареала в значительной мере перекрывают-
ся. Зверьки из равнинной тайги юга Западной Сибири по количественным
значениям показателя оттенка неотличимы от таковых с Алтая (Телецкое
озеро). Можно говорить лишь о некоторой тенденции усиления интенсив-
ности этого признака. По белизне различия более существенны.
182
Равнинная популяция из Приобья и Присалаирья более темная (4,1
± 0,22 и 136,8 2,21), чем с Северо-Восточного Алтая (5,7 ± 0,19 и
132,7 ± 1,41). Выборка из Забайкалья (хр. Хамар-Дабан) характеризует-
ся сходными средними значениями этого показателя (4,6 ± 0,17 и 132,2 zt
±0,82). Вариабельность признака насыщенности у зверьков из равнинной
части Западной Сибири и с Алтая близка. У зверьков с Хамар-Дабана
диапазон изменчивости этого признака меньше. Поэтому говорить в насто-
ящее время о направленной географической изменчивости окраски нет
оснований. Утверждение С. У. Строганова [1957] о широкой вариабель-
ности окраски малой бурозубки объясняется более широким пониманием
объема вида — в его состав он включал в качестве подвида S. gracillimus,
рассматриваемый нами [Юдин, 1965а, 1971а] как самостоятельный вид.
Четкой географической изменчивости размеров тела и черепа также
не прослеживается. Это обстоятельство отмечено и в работе В. А. Долгова
[1972], в которой указаны самые низкие значения кондило-базальной дли-
ны черепа для зверьков из Карелии. Наши материалы подтверждают дан-
ные В. А. Долгова [1972] об однородности популяций средней полосы лес-
ной зоны европейской части СССР, Западной Сибири, Северного Казах-
стана и Алтая. Изменчивость длины ряда зубов проявляется еще меньше,
чем кондило-базальная длина черепа.
Незначительна индивидуальная и географическая изменчивость зуб-
ной системы малой бурозубки по сравнению с другими видами. Она вы-
ражается в некоторых отклонениях в размере отдельных промежуточных
зубов, в пределах типичной для вида их категории.
В целом в ареале вида изменчивость кондило-базальной длины черепа
и одонтологических признаков не равнонаправлена, подвержена различ-
ным флюктуациям и носит гетероклинальный характер [Долгов, 19666,
19686].
Заметки по систематике. Малая бурозубка относится
к числу немногих видов представителей рода Sorex, видовой статус кото-
рой не вызывал сомнения у исследователей. Что касается его объема и ес-
тественных границ, то разные авторы понимали его не равнозначно.
Лаксманн, собравший серию зверьков из окрестностей Барнаула,
дал ей первую характеристику и назвал S. pygmaeus. Линней в 1766 г.
описал ее как S. minutus. Позднее в работах Лаксманна [Laxmann, 1769],,
Палласа [Pallas, 1811], Э. Эверсмана [1850] она фигурировала под
видовым названием S. pygmaeus Laxm. После публикации каталога
млекопитающих Миллера [Miller, 1912] за малой бурозубкой прочно
укрепилось ее первоначальное название — S. minutus L., сохранившееся
до наших дней.
С. И. Огнев [1928], Н. А. Бобринский, А. П. Кузякин [Бобринский,
и др., 1944], С. У. Строганов [1957], А. А. Гуреев [1963] принимали объ-
ем вида более широко, чем он рассматривается сейчас [Юдин, 1965, 1971а].
В его состав они включали в качестве подвида дальневосточную бурозубку
(5. т. gracillimus Thomas), выделенную нами [Юдин, 1965а] в самостоя-
тельный вид (S. gracillimus Thomas). Дальневосточная бурозубка S. gra-
cillimus хорошо морфологически выделяется как самобытный таксон ви-
дового ранга. Она крупнее малой бурозубки (длина тела с головой 49—
64 (М = 54,9 ± 0,94 мм), кондило-базальная длина ее черепа 15—16,5
(М = 15,7 0,05) мм, более длиннохвостая (79 % от длины тела).
Структура промежуточных зубов и конфигурация черепа иные. Зу-
бы этой категории сидят в зубном ряду еще более свободно, чем у малой.
183
бурозубки. Основания их коронок чрезвычайно вытянуты в сагитталь-
ном направлении, а вершины расставлены широко и значительно смеще-
ны кпереди по отношению к вертикальной оси зуба. Передняя сторона
конуса зуба короче задней (внутренней). Длина ряда промежуточных зу-
бов по отношению к кондило-базальной длине черепа больше у дальне-
восточной бурозубки: 36,8 % от длины зубного ряда или 15,9 % от конди-
ло-базальной длины, у малой соответственно 34,3 и 14,5 %. Промежуточ-
ные зубы последовательно убывают от самого крупного первого к пятому,
последний развит хорошо (см. рис. 67). Коренные отличия видов кроются
в строении головки копулятивного органа (см. диагноз и характеристику).
Выделение дальневосточной бурозубки в самостоятельный вид позво-
лило рассматривать малую в объеме естественных границ вида. Наряду с
таксономическим значением это обстоятельство существенно изменило
представление о распространении вида. Ранее восточная граница этого
западного палеаркта проводилась по Курильским островам и Японии, что
позволяло считать его транспалеарктом. В действительности вид рас-
пространен на восток только до Байкала, что имеет принципиальное зна-
чение для оценки путей формирования фауны Сибири и Дальнего Востока.
А. П. Кузякин [Бобринский и др., 1965], для взглядов которого на
таксономию насекомоядных млекопитающих характерна крайняя позиция
«укрупнителя» объема видов, под видом S. minutus понимал не только
собственно малую бурозубку, но и S. gracillimus и даже S. cinereus, относя-
щуюся не только к самостоятельному виду неарктического происхождения,
но и к другому подроду (Otisorex). Столь произвольная трактовка вида не
соответствует его естественному объему и расширяет ареал малой бурозуб-
ки не только до Чукотского полуострова включительно, но предполагает
включение в него северной части Северной Америки.
Нами [Юдин, 1965а, б, 19696—1973а] показано, что трансарктиче-
ская бурозубка (S. cinereus Kerr)— вид с хорошо очерченными признака-
ми, по которым она безошибочно отличается от видов подрода Sorex.
А. П. Кузякин [Бобринский и др., 1965] в характеристике несуществую-
щего подвида S. т. portenkoi Strog. (подвид относится к S. cinereus) сам
указывает на один из признаков, исключающих возможность объедине-
ния ее с малой бурозубкой: «третий верхний промежуточный зуб меньше
второго». У малой бурозубки такого соотношения зубов не бывает — тре-
тий всегда больше второго. Кроме того, существенное отличие этих двух
видов заключается в отсутствии в нижней челюсти S. cinereus постманди-
булярного канала, признака, свойственного всем представителям подрода
Sorex и отсутствующего у видов подрода Otisorex. У S. minutus такой. канал
есть. Совершенно различное строение имеет копулятивный орган у этих
видов (см. рис. 40). Отличаются они и по хромосомному набору: у S. mi-
nutus 2п = 42, a NF = 56; у S. cinereus — 2п = 66, a NF — 70 [Меу-
1ап, 1965, 1968].
Правомерность выделения дальневосточной бурозубки в самостоя-
тельный вид подтверждена исследованиями В. А. Долгова [19666, 19686,
1972], ранее считавшего эту форму подвидом малой бурозубки. В коллек-
ции зоологического музея Биологического института есть экземпляры
. S. gracillimus (сборы Н. Ф. Реймерса с Сахалина), определенные В. А. Дол-
говым как S. minutus.
По размерам тела в фауне Сибири к малой бурозубке близка крошеч-
:ная. По внешнему облику она отличается от последней более вытянутым
_ хоботком, резким сужением в предглазничной области, округлой мозго-
184
вой частью головы, длинным волосистым хвостом и более стройным тело-
сложением. Размеры тела у них трансгрессируют, но хорошо различимы
по средним величинам: малая бурозубка имеет длину тела 40—62 (М—52,2
0,59), крошечная — 30—52 (М — 44,8 z±z 0,35) мм, т. е. близкие к низ-
шему показателю у малой бурозубки. Хорошо различаются средние зна-
чения длины хвоста у этих видов, имеющие незначительную трансгрессию
меньших (для малой) и больших (для крошечной) величины. У малой
бурозубки длина хвоста колеблется в пределах 32—42 (М — 36,9) мм, у кро-
шечной — 18—37 (М = 24,2 ±. 0,27) мм.
Довольно хорошо они различаются и по размерам черепа: у малой
бурозубки 14,3—15,8 (М = 15,1 0,03), у крошечной — 12,0—14,5
(М = 13,3 ± 0,03) мм. У некоторых подвидов крошечной бурозубки
(S. т. barabensis, S. т. abnormis) они не перекрываются с таковыми S. mi-
nutus. В среднем для малой бурозубки отношение длины хвоста к длине
тела составляет 71, у крошечной — 54 %. У некоторых короткохвостых
подвидов этот показатель еще меньше. В целом для S. minutus характерна
минимальная длина хвоста — 30 мм, в то время как у S. т. barabensis его
размеры колеблются от 19 до 26, у S. т. abnormis — 18—22, у S. т. stro-
ganovi 25—30, у <S. т. tschuktschorum 18—27 мм.
Весьма существенны различия этих видов и по длине зубного ряда:
у малой бурозубки он равен 6,2—6,8 (М = 6,4 ± 0,01), у крошечной —
4,7—6,2 (М = 5,7—0,03) мм. Аналогичны различия и по другим призна-
кам, а степень их трансгрессии крайне незначительна. Весьма существен-
но, что зубы этих видов имеют несходную структуру.
Наиболее близки по размерам тела к малой бурозубке S. cinereus,
S. beringianus и S. gracillimus. От двух первых она отличается наличием
постмандибулярного канала, а самобытная организация дальневосточной
бурозубки и их различия описаны выше.
От средней бурозубки S. minutus отличается меньшими размерами и
более стройным телосложением. Кондило-базальная длина черепа этих
видов не трансгрессирует (у средней бурозубки значения этого признака
колеблются в пределах — 16,1—19,4 (М = 17,4± 0,03) мм). От тундря-
ной бурозубки (S. tundrensis) она отличается более узким черепом (у ма-
лой — 6,7—7,8 (М = 7,3 zt 0,02); у тундряной — 8,0—10,0 (М — 8,8
0,01), мм структурой зубов и glans penis.
От остальных представителей рода S. minutus отличается значитель-
но меньшими размерами и другими особенностями строения.
Описанный С. И. Огневым [1921] подвид малой бурозубки S. т. vol-
nuchini отличается от S. minutus по ряду морфологических признаков и
кариотипу. У номинальной формы S. т. minutus 2п = 42, NF = 56. Из
аутосом — 6 пар мета-, одна пара субмета- и 13 пар акроцентрических.
Гетерохромосомы акроцентрические, Х-хромосома не превышает 5 % от
длины гаплоидного набора. В диплоидном наборе у S. т. volnuchini
2п — 40, NF = 60. Среди аутосом 8 пар мета-, одна пара субмета- и 10 пар
акроцентрических; Х-хромосома метацентрическая, от длины гаплоидного
набора составляет 40,4 %; Y-хромосома акроцентрическая [Козловский,
1973а, б].
Хромосомный набор у малой бурозубки из Швейцарии [Meylan,
1965], в СССР из Московской [Орлов, Аленин, 1968] и Кемеровской об-
ластей [Козловский, 1968; Орлов, Козловский, 1971] оказался идентич-
ным. Таким образом, S. volnuchini выделена из состава малой бурозубки
как самостоятельный вид.
185
В 1905 г. Н. Ф. Кащенко по экземпляру из сборов Н. М. Пржеваль-
ского из Цайдама описал подвид малой бурозубки S. т. thibetanus. Ис-
следованиями В. А. Долгова, Р. С. Хоффмана [1977] убедительно доказа-
на видовая самостоятельность этой формы. Авторы объединили в один вид
ряд форм видового и подвидового рангов из высокогорий Центральной
Азии (S. т. thibetanus Kastsch., S. planiceps Miller, S. buchariensis Ognev,
S. kozlovt Strog., по приоритету назвав его S. thibetanus Kastsch.). He вда-
ваясь в детали правомерности такого объединения, факт признания ви-
довой самостоятельности формы, столь далеко географически локализо-
ванной и ранее признаваемой Н. Ф. Кащенко как подвид малой бурозуб-
ки, представляется весьма реальным. А. А. Гуреев [1979] не принимает
предложенный этими исследователями объем вида, считая, что S. т. thi-
betanus — типичный S. minutus, ссылаясь на типовой экземпляр, хра-
нящийся в ЗИН АН СССР (№ 6557). Экземпляр, послуживший Н. Ф. Ка-
щенко типом для описания S. т. thibetanus, мы не видели.
Подводя итог анализу таксономии малой бурозубки, можно считать,
что в настоящее время она очерчена в реальных, естественных границах
вида, что подтверждается последними исследованиями хромосомного на-
бора малой бурозубки, стабильного на всем протяжении ее ареала [Меу-
1оп, 1965; Орлов, Аленин, 1968; Орлов, Козловский, 1971]. Формы, вклю-
чавшиеся ранее разными авторами в состав вида малой бурозубки (S.
gracillimus, S. cinereus, S. volnuchini, S. thibetanus), квалифицируются в
настоящее время как самостоятельные видовые таксоны.
В Сибири вид представлен номинальным подвидом — S. т. minutus L.
Подрод Stroganovia Yudin, subsp. n.
Sorex daphaenodon Thomas, 1907 —
Крупнозубая бурозубка
Голотип. Вид описан по экземпляру из окрестностей близ Корса-
кова (Южный Сахалин). Самка хранится в Британском музее.
Диагноз. Размеры средние. Длина тела 47—75 (М = 60,1),
хвост короткий —26—39 (М = 32,3) мм, что составляет 53,7 % от длины
тела. Кондило-базальная длина черепа 16,8—18,8 (М = 17,9 dt 0,03) мм.
Окраска спинной стороны тела темно-коричневая, в летнем меху прибылых
зверьков чепрачность не выражена. Ростральная часть относительно дли-
ны черепа короткая (25,1 %), мозговая — самая длинная среди бурозубок
(58,6 %) и самая высокая (33,5 %). Восходящая ветвь нижней челюсти
выше, чем у других видов (26,8 %), угловой отросток относительно длин-
ный (12,8 %). Промежуточные зубы массивные, слабо сужаются к закруг-
ленной вершине и плотно прижаты друг к другу. Вершина второго проме-
жуточного зуба выше вершины первого и третьего. Коренные — массив-
ные, широкие, пирамидальная воронка на их жевательной стороне от-
сутствует. Рт1 низкий с вытянутым основанием коронки.
Измерения. См. табл. 39.
Характеристика. Во внешнем облике крупнозубой бурозуб-
ки обращает на себя внимание относительно крупная голова, с коротким
и тупым хоботком, темная спина и относительно других видов темная ниж-
няя часть тела, темные лапы и относительно короткий хвост (рис. 58, 59).
По размерам тела этот вид ближе всего к средней бурозубке, а габи-
туально — к тундряной. По средним значениям размера тела он несколько
186
Рис. 58. Крупнозубая бурозубка — 5. daphaenodon Thomas.
уступает тундряной, а от средней отличается более коротким хвостом и тя-
желым телосложением. Голова по очертаниям сходна с таковой крошеч-
ной и тундряной бурозубок. Сужение в области глаз не выражено. Отно-
шение кондило-базальной длины черепа к длине тела у этого вида самое
большое (29,7 %) среди бурозубок.
Окраска спины от коричнево-буро-серой до еще более темной землисто-
бурой с кофейно-коричневым оттенком. Окраска вентральной стороны
уступает по насыщенности окраске только равнозубой, длиннокоготной и
гигантской бурозубок. В окраске боков, на границе между темной спи-
ной и более светлыми боками, хорошо видна примесь ржавых тонов, осо-
бенно хорошо заметная у перезимовавших зверьков. У прибылых зверь-
ков переход от темной спины к бокам постепенный, а у перезимовавших
эта граница более четкая.
Колориметрические исследования проведены на небольшом материа-
ле из разных пунктов ареала, тем не менее, сопоставляя их с результата-
ми визуальных исследований, можно сказать, что диапазон изменчивости
окраски у этого вида невелик (рис. 60). Значения оттенка колеблются от
130 до 160 (М=138,7 =t= 2,29), а белизна — от 1,5 до 3,6 (7И=2,9 =ь
0,18). Зимний мех значительно темнее, длиннее (до 12 мм) и пышнее
летнего, с более заметным присутствием рыжих тонов на боках и пале-
вых — на вентральной стороне тела. Хвост темноокрашенный, покрыт
длинными волосами, с более развитой, чем у других видов, кисточкой.
Мех на верхней стороне лап такой же темный, как на спине.
Череп своеобразной конфигурации. Он монолитен, с выпуклой моз-
говой капсулой (самая большая относительная высота среди бурозубок —
187
Таблица 39
Морфометрическая характеристика Sorex daphaenodon
Возрастная и половая группа M+m-j. О cv п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 57,4+0,72 4,50 7,85 39 51,0-68,0
взрослые 66,5+1,79 5,62 8,46 10 59—74
Самки прибылые 58,9+0,99 5,26 8,93 28 47-70
взрослые 68,5±5,60 11,21 16,3 4 52—75
В целом по виду ... 60,1+0,53 5,81 9,66 119 47—75
Длина хвоста
Самцы прибылые 32,0±0,48 2,93 9,16 38 26—37
взрослые 30,8±1,0 3,16 10,24 10 26-34
Самки прибылые 33,7+0,55 2,87 8,51 27 30-39
взрослые 33,3+1,8 3,59 10,81 4 30—38
В целом по виду ... 32,3+0,28 3,05 9,44 116 26-39
Длин 1 ступни
Самцы прибылые 11,4+0,17 0,91 8,04 30 9-13
взрослые 11,7+0,17 0,5 4,29 9 11—12
Самки прибылые 11,7+0,19 0,98 8,39 27 9-14
взрослые 12,0±0,0 0,0 0,00 3 —
В целом по виду ... 11,6+0,09 0,84 7,24 106 9-14
Мас са тела
Самцы прибылые 5,5+0,16 0,71 12,89 21 4,0-7,0
взрослые 8,0+0,35 0,93 11,67 7 7,0—9,5
Самки прибылые 5,9+0,35 1,20 20,29 12 4,5—8,0
взрослые 8,7+0,15 0,21 2,45 2 8,5-8,8
В целом по виду ... 6,1+0,20 1,59 25,97 64 4,0-9,5
1 (ондило-базал ъная длин 1 черепа
Самцы прибылые 17,9+0,05 0,35 1,94 42 16,8—18,7
взрослые 17,8+0,14 0,51 2,89 13 17,1—18,5
Самки прибылые 17,9+0,07 0,38 2,14 35 17,1—18,7
взрослые 17,7+0,13 0,41 2,35 10 17,1—18,5
В целом по виду ... 17,9+0,03 0,40 2,23 206 16,8—18,7
Длина рострума
Самцы прибылые 4,5±0,04 0,25 5,6 44 3,7-5,1
взрослые 4,5±0,08 0,3 6,6 14 4,2—5,0
Самки прибылые 4,5±0,04 0,23 5,03 40 4,5-5,0
взрослые 4,3+0,09 0,28 6,45 10 4,0—5,0
В целом по виду ... 4,5+0,02 0,26 5,71 224 3,7-5,1
188
Продолжение табл. 39
1 ' 2 3 * 5 6
Длина мозговой части черепа
Самцы прибылые 10,5±0,04 0,28 2,7 42 9,8—11
взрослые 10,3±0,09 0,33 3,22 13 9,5—10,8
Самки прибылые 10,4±0,05 0,27 2,64 36 9,9—10,9
взрослые 10,2±0,10 0,30 2,99 10 9,7—10,6
В целом по виду ... 10,5±0,02 0,30 2,88 206 9,5—11
Длина верх чего ряда зубов
Самцы прибылые 8,0±0,04 0,23 2,87 42 7,5—8,6
взрослые 7,7±0,01 0,38 4,95 14 7,0—8,3
Самки прибылые 8,1±0,04 0,21 2,64 38 7,7—8,5
взрослые 7,8±0,09 0,27 3,49 9 7,5—8,4
В целом по виду ... 8,0±0,02 0,27 3,32 217 7,0—8,6
Длина ряда промежуток 1ЫХ зубов
Самцы прибылые 2,6±0,02 0,15 6,04 44 2,3—2,8
взрослые 2,5±0,04 0,15 5,81 14 2,3—2,7
Самки прибылые 2,6±0,02 0,13 5,02 39 2,2—2,8
взрослые 2,6±0,03 0,10 3,81 10 2,4—2,7
В целом по виду ... 2,6±0,01 0,14 5,32 223 2,2—2,8
Ширина рострума
Самцы прибылые 1,7±0,02 0,08 4,66 25 1,5-1,8
взрослые 1,7±0,03 0,08 4,86 И 1,6—1,8
Самки прибылые 1,7±0,02 0,12 7,1 38 1,5—2,0
взрослые 1,7+0,03 0,09 5,09 9 1,6—1,8
В целом по виду ... 1,7±0,01 0,10 5,75 198 1,5—2,0
Предглазничная ширина
Самцы прибылые 2,4±0,02 0,13 5,43 44 2,2—2,8
взрослые 2,5±0,05 0,17 6,93 14 2,2—2,7
Самки прибылые 2,4+0,02 0,13 5,37 40 2,2—2,7
взрослые 2,4+0,04 0,13 5,28 10 2,3—2,7
В целом по виду ... 2,4+0,01 0,14 5,97 224 2,2—2,8
Межглазничная ширина
Самцы прибылые 3,7±0,03 0,19 5,23 44 3,1—4,0
взрослые 3,7±0,05 0,19 5,25 14 3,5—4,1
Самки прибылые 3,7±0,02 0,12 3,37 40 3,3—3,9
взрослые 3,7±0,05 0,16 4,33 10 3,4-3,9
В целом по виду ... 3,7±0,01 0,17 4,54 224 3,1—4,1
Наибольшая ширина черепа
Самцы прибылые 9,1 + 0,05 0,31 3,38 39 8,5—9,9
взрослые 9,2±0,08 0,28 3,07 13 8,6—9,6
Самки прибылые 9,0±0,05 0,29 3,24 37 8,5—9,6
взрослые 9,1±0,12 0,38 4,18 10 8,6-9,8
В целом по виду ... 9,1±0,02 0,30 3,30 205 8,5-9,9
189
Окончание табл. 39
1 2 1 3 1 4 5 6
Наибольшая высота черепа
Самцы прибылые 6,1+0,05 0,28 4,67 35 5,3-6,7
взрослые 5,7+0,07 0,26 4,48 12 5,3—6,1
Самки прибылые 6,0±0,06 0,35 5,76 35 5,3-6,6
взрослые 5,8+0,09 0,25 4,36 9 5,5—6,2
В целом по виду ... 6,0±0,02 0,29 4,87 197 5,3-6,7
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,7±0,05 0,30 6,32 42 3,9-5,4
взрослые 4,7+0,09 0,31 6,63 13 4,1-5,2
Самки прибылые 4,7+0,05 0,32 6,65 40 4,0—5,4
взрослые 4,7±0,09 0,28 5,85 10 4,3-5,2
В целом по виду ... 4,8±0,02 0,32 6,73 215 3,5-5,4
Длина углового отростка
Самцы прибылые 2,3+0,02 0,13 5,76 33 2,0—2,6
взрослые 2,2+0,05 0,16 7,27 11 1,8-2,4
Самки прибылые 2,3+0,03 0,15 6,76 33 1,9—2,5
взрослые 2,3±0,06 0,16 6,97 8 2,0-2,5
В целом по виду ... 2,3±0,01 0,15 6,78 163 1,8-2,6
33,5 %), с высокой лицевой частью, клювовидно скошенной кпереди.
В области соединения лобных и носовых костей череп несколько вздут.
Ламбоидальный и сагиттальный гребни развиваются рано и хорошо выра-
жены даже у неперезимовавших зверьков.
Показатель кондило-базальной длины черепа несколько выше, чем
у S. tundrensis (17,7 ± 0,02 мм) и S. caecutiens (17,4 0,03 мм), хотя по
размерам тела S. daphaenodon уступает тому (63,4 0,26 мм) и другому
(60,5 0,28 мм). Восходящая ветвь нижней челюсти самая высокая
среди бурозубок (см. табл. 29), а по длине углового отростка она не усту-
пает (12,8 %) и S. beringianus (12,9 %).
Совершенно своеобразна структура зубной системы. Самобытные
черты организации проявляются как в структуре промежуточных зубов,
так и многовершинных. Для всех категорий зубов этого вида характерна
притупленность их структурных элементов, сглаженность вершин, кону-
сов, гребней и интенсивная окраска. Ряд многовершинных зубов чрезвы-
чайно широк, все зубы крупные, квадратные (см. рис. 59).
Первый верхний резец развит относительно слабо. Его второй зубец
лишь немного уступает в размерах первому и по высоте равен первому
промежуточному зубу. Промежуточные зубы крайне своеобразны по кон-
фигурации: они массивны, слабо сужаются от основания коронок к округ-
лым тупым вершинам и тесно поставлены в зубном ряду. Вершины их
сближены и имеют слабую тенденцию к нацлону вертикальной оси в сторо-
ну многовершинных зубов. Вершины располагаются почти над серединой
основания коронок. Первые три зуба по размерам близки, но вследствие
разного их положения на челюсти вершина второго зуба всегда несколько
190
Рис. 59. Череп крупнозубой бурозубки.
Усл. обозн. см. рис. 1.
выше первой и третьей; третий зуб по размерам едва уступает второму,
четвертый в такой же мере меньше предыдущего зуба, пятый зачаточ-
ный — он в несколько раз меньше четвертого. Задняя часть его коронки
задвинута за передний выступ парастиля соседнего с ним многовершин-
ного зуба. Длина ряда промежуточных зубов колеблется от 2,2 до 2,9
(М= 2,6+0,01) мм, составляя в среднем для вида 32,5 % от длины зуб-
ного ряда и 14,5 % от кондило-базальной длины черепа (см. табл. 26). По
этому показателю крупнозубая бурозубка ближе всего к тундряной.
В структуре многовершинных зубов обращает на себя внимание
следующее:
1) чрезвычайно большая
Белизна
ширина ряда многовершинных зубов;
2) низкие, сильно притупленные ко-
нусы и соединяющие их гребни;
3) отсутствие пирамидальной ворон-
ки на жевательной стороне между мезо-
стилем и протоконусом на первом и вто-
ром многовершинных;
4) интенсивная пигментация эмали,
даже на жевательной поверхности зу-
бов;
5) хорошо развитый и интенсивно
пигментированный гипоконус первого
многовершинного зуба;
Рис. 60. Колориметрическая характеристика
крупнозубой бурозубки.
191
6) слабая выраженность серповидной выемки на задней поверхности
многовершинных зубов, вследствие чего ряд зубов плотно сомкнут, а
зубы в плане имеют почти квадратную форму.
Названные структурные особенности настолько самобытны, что
позволяют безошибочно отличить этот вид от других. Pm1 вытянут в
длину: высота его коронки почти в 2 раза уступает длине ее основания.
Головка копулятивного органа чрезвычайно длинная и тонкая —
нитевидно вытянутая, почти цилиндрическая. Glans penis крупнозубой
бурозубки самая длинная и тонкая среди бурозубок фауны СССР: длина
11—13 (М=11,7), толщина у основания всего 0,5—0,7 (М=0,6) мм
(см. рис. 40).
Распространение. По характеру распространения крупно-
зубая бурозубка — типичный восточный палеаркт. Ее ареал простира-
ется от Сахалина, Камчатки, севера Дальнего Бостока, Охотского по-
бережья и Приморья до Урала на западе, включая тундровую, лесотунд-
ровую, таежную и лесостепную зоны.
Наиболее западной точкой распространения этого вида следует
считать районы Среднего Урала. Здесь она добыта в ряде пунктов Сверд-
ловской, а также Челябинской областей (р-н оз. Шугуняк). На запад
от Урала сведений о крупнозубой бурозубке нет. Зверьки широко рас-
пространены от Южного Ямала до Полярного и Центрального Урала
(Свердловская и Челябинская области). Восточнее этих пунктов она
известна из ряда сопредельных территорий низовий^Оби (пос. Лабытнан-
ги, Шурышкарский р-н, пос. Мужи), южнее — из ряда пунктов бассейна
р. Конда и Сосьва, Тюменской области (Абатский и Вагайский р-ны,
окр. г. Ишим). В бассейне Тобола она обычна близ Тобольска, южнее —
в Упоровском районе и в лесостепных районах бассейна р. Ишим юга
Тюменской области.
Указание С. У. Строганова [1957] о ее распространении в Куста-
найской области ошибочно, основано на неточном материале. Далее от
Тюмени южная граница распространения вида идет на широте оз. Чаны,
Новосибирска, по Приобским борам спускается выступом на юг до Бар-
наула, но обходит Алтай, Салаир, Горную Шорию и Кузнецкий|Алатау
с севера. Известна она из района г. Анжеро-Судженска и из Хакасии.
В горах Алтае-Саянской горной страны ее нет [Юдин и др., 1979]. В этом
районе наиболее южные пункты находок вида известны из северных
отрогов Кузнецкого Алатау близ г. Анжеро-Судженска и из сопредель-
ных районов Хакасии. В обширных сборах с северных склонов Западного
Саяна (пос. Майна), из ряда районов Тувы и Восточного Саяна ее нет.
Далее на восток она отмечена в ряде пунктов Прибайкалья, Бурятии,
Забайкалья, сопредельных территорий Монголии, Маньчжурии, по
всему Приамурью и Приморью до Сахалина включительно [Юдин,
1971а, г, 19736, 1975а]. На восток от Полярного Урала и нижнего При-
обья северная граница спускается южнее, так как в обширных сборах
с Гыданского полуострова (Бол. Хета), в бассейнах рек Полуй, Пур
(Самбург) крупнозубая бурозубка не обнаружена. Она известна с Тазов-
ского полуострова и верховьев р. Таз (устье р. Мольтки). В таежных
районах Западной Сибири, вероятно, распространена спорадически по
всей территории. В бассейне Васюгана она малочисленна и известна
из окрестностей пос. Чижапка, а в низовьях коллектировалась близ
пос. Ларино. На правобережье добывалась на р. Кеть и ее притоках.
Обычна в прилежащих к Оби районах близ г. Колпашево.
192
На севере Новосибирской области обычна в бассейне р. Тара (при-
ток Бол. Ича) и р. Майзас в среднем течении р. Омь, в урочище Горелая
Грива, восточнее — в бассейне р. Каргат (с. Ровенское), в Куйбышевском
районе близ оз. Мохнатое, на северном берегу оз. Чаны (пос. Квашнино),
на оз. Сартлан (пос. Эгербаш). Далее на юг в Кулундинскую спепь она
не идет: в сборах из разных районов за последние 30 лет, насчитывающих
несколько тысяч землероек, крупнозубая бурозубка ни разу не встрече-
на. Отсюда граница идет по широте оз. Мензелинское, севернее Новоси-
бирска, здесь пересекает Обь и, как уже говорилось, спускается по сос-
новым борам до Бийска [Юдин, 1969а].
В Средней Сибири распространение этого вида изучено слабо. В мно-
голетних обширных сборах из разных пунктов тундровой, лесотундро-
вой и таежной зон Таймыра [Юдин, 1980в] она не обнаружена. Несомнен-
но, что в этом районе северная граница ее распространения проходит
южнее плато Путорана. В Якутии распространена широко, доходит
до арктического побережья. Она добывалась в бассейне р. Яна. По Ин-
дигирке граница спускается до дельты [Михель, 1938; Млекопитающие
Якутии, 1971 ], на Колыме встречается всюду и коллектировалась многи-
ми исследователями. В Центральной и Южной Якутии это обычный
вид в районе Станового хребта (р. Токо). Известна с Вилюя. Добывалась
на р. Алазея, между Сватой и Ардахтах близ Верхоянска. Известна из
многих пунктов с о. Омолон, из бассейна р. Анадырь (Марково, Ваеги,
Танюрер).
На Охотском побережье близ Охотска и г. Олы (к северу от Магада-
на). На Аянском побережье Охотского моря не обнаружена [Алина,
Реймерс, 1975]. На Камчатке она, видимо, распространена повсеместно;
коллективировалась в западной части полуострова (р. Утхолок), на
юго-западе (бухта Камбальная), в центральной (пос. Мильково) и восточ-
ной частях у оз. Ажабачье. По трассе БАМ отлавливалась сотрудниками
нашей лаборатории в Северном Прибайкалье и Забайкалье, на Витимском
плоскогорье, в ряде пунктов в долине р. Амалат, в Амурской области
на междуречье Алдана и Зеи, на р. Нюкже, в отрогах хр. Тукурингра,
на р. Уркан (бассейн Зеи), в ряде пунктов по Зее. В нижнем Приамурье,
в низовьях Амгуни. Кроме того, она известна из района Хабаровска
и Комсомольска (пос. Пивень, Комсомольский заповедник), близ д. Ки-
сел евки, ст. Лазо, из ряда пунктов бассейна Уссури, р. Кия, из Прихан-
кайской низменности, близ оз. Ханка. Зверьки заселяют острова
Бол. Шантар и Сахалин. На Курильских островах ее нет. На Сахалине
распространена повсеместно в северной, центральной и южной частях
острова.
Известна из Монголии, Маньчжурии, Кореи.
Материал. 184 экз. из разных частей ареала.
Изменчивость. Географическая и возрастная изменчивость
весьма значительна, индивидуальная проявляется слабо.
Возрастная изменчивость крупнозубой бурозубки хорошо выражена
в размерах тела: перезимовавшие зверьки крупнее молодых (см. табл. 39).
Различия эти статистически достоверны. Среди перезимовавших бурозу-
бок самки несколько крупнее (М = 68,8 мм) самцов (66,5 мм). Аналогич-
ная закономерность прослеживается и среди прибылых зверьков; самки
крупнее (58,9 мм), чем самцы (57,4 мм), различия заключаются лишь
в величине разрыва этих показателей (см. табл. 39). Хорошо выражена
географическая изменчивость в размерах бурозубок из разных частей
193
ареала. Наиболее крупные землеройки заселяют Западную Сибирь:
по этому признаку они с большой достоверностью (Td = 3,9) отличаются
от зверьков из Якутии, северо-востока Сибири, Приамурья и Приморья.
Достоверные различия установлены и по большей величине хвоста, ступни
и массы тела.
Длина хвоста колеблется в значительных пределах — от 26 до 39 мм.
Наблюдается хорошо выраженная географическая изменчивость этого
признака: наиболее короткий хвост (в среднем 31 мм) имеют зверьки,
обитающие в Забайкалье, Якутии, бассейне рек Колыма и Анадырь.
У номинального подвида (Сахалин, Приморье) хвост заметно длиннее
(М — 34,3 мм) и близок к таковому бурозубок, населяющих Западную
Сибирь (34,4 мм). Отличия эти статистически достоверны (Td — 8,8 и
5,9). Отношение длины хвоста к длине тела западно-сибирских бурозу-
бок составляет 65 %, у зверьков из Приморья и Сахалина — 52, а в
Приамурье и северо-восточных районах Сибири — 50 %. Размеры хвоста
и ступни у зверьков разного пола и возраста достоверно не различаются.
Изменчивость массы зверьков разного пола и возраста та же, что и раз-
меров тела (см. табл. 39).
Возрастная и половая изменчивость размеров черепа невелика.
Достоверных различий по этому признаку между молодыми и взрослыми
(самцы и самки) нет, так же как не различаются эти показатели у зверьков
разного пола как у взрослых, так и молодых (см. табл. 39). Различия
статистически недостоверны. Аналогичная закономерность выявлена и
Таблица 40
Морфометрическая характеристика Sorex daphaenodon и ее подвидов из разных
частей ареала
Признак S. daphaenodon
М ± тх а Cv п lim
Длина
тела 60,1+0,53 5,81 9,66 119 47—70
хвоста 32,3+0,28 3,05 9,44 116 26—39
ступни 11,6+0,09 0,84 7,24 106 9—14
Масса тела Длина 6,1+0,20 1,59 25,97 64 4-12,5
кондило-базальная черепа 17,9+0,03 0,40 2,23 206 16,8—18,8
рострума 4,5±0,02 0,26 5,71 224 3,7—5,5
мозговой части 10,5±0,02 0,30 2,88 206 9,0—11,5
верхнего ряда зубов 8,0+0,02 0,27 3,32 217 7,0—8,8
ряда промежуточных зубов Ширина 2,6+0,01 0,14 5,32 223 2,2—2,9
рострума 1,7+0,01 0,10 5,75 198 1,5-2,0
предглазничная 3,7+0,01 0,17 4,54 224 3,1—4,6
межглазничная 2,4±0,01 0,14 5,97 224 2,2-2,9
наибольшая черепа 9,1±0,02 0,30 3,30 205 8,3—9,9
Наибольшая высота черепа Высота восходящей ветви ниж- 6,0+0,02 0,29 4,87 197 5,3—6,9
ней челюсти 4,8+0,02 0,32 6,73 215 4,1—5,6
Длина углового отростка 2,3+0,01 0,15 6,78 163 1,8-2,7
13 Б. С. Юдин
194
a. S. d. daphaenodon, Сахалин, Приморье
М ± 7ПХ О Су п 11m
Признак
Длина тела 64,8+0,95 3,01 4,65 10 61—70
хвоста 34,3+0,90 2,83 8.25 10 23—38
ступни 11,9±0,31 0,99 8,36 10 10—13
Масса тела 7,1±0,40 1,13 15,80 8 5—8
Длина
кондило-базальная черепа 17,8+0,09 0,36 2,02 18 17,3—18,6
рострума 4,6±0,05 0,21 4,62 22 4,1—4,9
мозговой части 10,4+0,06 0,24 2,28 18 9,9—10,8
верхнего ряда зубов 8,1±0,01 0,33 4,06 21 7,0—8,4
ряда промежуточных зубов 2,7±0,04 0,17 6,20 21 2,2—2,8
Ширина
рострума 1,7±0,03 0,13 7,66 22 1,5—2,0
предглазничная 2,4±0,02 0,10 4,13 22 2,3—2,6
межглазничная 3,7+0,03 0,12 3,36 22 3,4—3,8
наибольшая черепа 9,0±0,08 0,37 4,08 19 8,6-9,9
Наибольшая высота черепа 5,9±0,07 0,32 5,46 19 5,3-6,3
Высота восходящей ветви ниж-
ней челюсти 4,6±0,07 0,33 7,03 22 4,1—5,4
Длина углового отростка 2,2±0,05 0,21 9,33 18 1,8—2,7
при анализе морфометрических показателей черепа. Только длина зубного
ряда старых самцов (7,7 =±=0,1) и самок (7,8 =t 0,09 мм) значительно ус-
тупает таковой у молодых (соответственно 8,0 =*= 0,04 и 8,1 =t 0,04). Высота
черепа разная у зверьков разного возраста и одинаковая у зверьков
одной возрастной группы (см. табл. 39). Наиболее крупный череп у
бурозубок этого вида из Западной Сибири — кондило-базальная длина
черепа равна 18,2 ±= 0,05 мм. Номинальная форма и зверьки из северо-
восточных районов Сибири по величине этого показателя не различаются
между собой (соответственно 17,8 ±= 0,09 и 17,8=1=0,03 мм). Их различия
с западно-сибирской формой крупнозубой бурозубки статистически
достоверны (соответственно Td = 3,9 и 6,9) (табл. 40).
На серийном материале хорошо видна географическая изменчивость
в окраске меха: наиболее темные зверьки населяют Западную Сибирь,
несколько более светлые бурозубки обитают в Якутии и на севере Дальне-
го Востока. Бурозубки из Приморья и Сахалина еще светлее. Самые
светлые крупнозубые бурозубки зарегистрированы в Забайкалье — в
окраске их меха прослеживается значительная примесь шоколадных
и палевых тонов.
Сезонная изменчивость в окраске связана с появлением более длин-
ного, пушистого, и темного зимнего меха. Темный оттенок меха у земле-
роек в зимнем наряде спускается со спины на бока ниже, чем у зверьков,
добытых летом, без ясной границы переходит на вентральную сторону,)
также более темную, чем летом.
195
Окончание табл. 40
б. S. d. sanguinidens, северо-восток Сибири в. S. d- scaloni, лесотундровая и таеж- ная зоны Зап. Сибири Достоверность различий по Стьюденту (Т<Г)
М ± тх а cv п lim М ± тх а Су п lim \ а—б 1 а—в б—в
61,7±0,60 5,34 8,67 79 51-75 64,7±0,48 1,97 3,10 57 50—67 2,8 0,1 3,9
31,2±0,32 2,76 8,82 76 26—39 34,4±0,44 2,40 6,97 30 30—39 3,3 0,1 5,9
11,4±0,10 0,83 7,30 66 9—13 11,8±0,13 0,57 4,80 49 11—14 1,5 0,3 2,5
6,0±0,22 1,61 26,8 54 4—12,5 5,7±0,12 0,51 8,9 19 4,2—6,2 2,4 3,3 1,2
17,8±0,03 0,39 2,16 127 16,8—18,8 18,2±0,05 0,35 1,94 56 17,4—18,8 0 3,9 6,9
4,4±0,02 0,24 5,55 136 3,7—5,5 4,6±0,03 0,25 5,33 61 4,0—5,2 3,7 0 5,5
10,4±0,03 0,30 2,86 128 9,0—11,5 10,6±0,04 0,30 2,86 55 9,8—11 0 2,8 4
8,0±0,02 0,26 4,23 132 7,3—8,8 8,2±0,03 0,22 2,70 59 7,7—8,6 4,5 3,2 5,5
2,6±0,01 0,15 5,60 136 2,3—2,9 2,6±0,01 0,12 4,40 61 2,2—2,9 2,4 2,4 0
1,7±0,01 0,09 5,10 110 1,5—1,9 1,7±0,01 0,10 5,61 61 1,5—1,9 0 0 0
2,4±0,01 0,14 5,69 136 2,2—2,8 2,5±0,02 0,17 6,93 61 2,2—2,9 0 3,5 4.5
3,8±0,01 0,15 3,98 136 3,3—4,1 3,7±0,02 0,18 4,78 61 3,4—4,6 3,2 0 4,7
9,1±0,03 0,29 3,22 127 8,3—9,8 9,3±0,03 0,24 2,56 54 8,6—9,8 1,2 3,5 4,5
6,0±0,02 0,25 4,23 123 5,3-6,5 6,2±0,04 0,30 4,78 51 5,5—6,9 1,4 3,7 14,1
4,7±0,02 0,23 4,89 129 4,3-5,4 5,1±0,02 0,19 3,63 59 4,9—5,5 1,4 6,9 4,5
2,3±0,01 0,13 5,51 100 2,0—2,6 2,4+0,02 0,10 4,17 40 2,2—2,6 1,9 3,7 0
Летом хвост двухцветный, сверху окрашен в тон дорсальной стороны
тела, снизу — вентральной. Зимой темнеет, граница между темной и
светлой окраской смазывается, а у зверьков из некоторых районов
полностью исчезает. У прибылых зверьков кисточка хвоста из темных
волос более длинная, чем у других видов бурозубок. С возрастом она
постепенно снашивается, а у старых зверьков стирается почти пол-
ностью.
Заметки по систематике. Крупнозубая бурозубка по ряду
охарактеризованных выше признаков настолько четко очерчена и само-
бытна среди видов рода Sorex, что не вызывает сомнения не только в
реальности статуса вида, но возникла необходимость выделения его
в отдельный подрод, чтобы подчеркнуть особое положение этой бурозуб-
ки в системе. Все признаки позволяют надежно диагностировать крупно-
зубую бурозубку. Уместно напомнить, что только для нее характерны
описанные выше крайне уклоняющиеся от других видов структурные
особенности зубной системы: форма промежуточных зубов, широкие
квадратные в плане и интенсивно окрашенные коренные, отсутствие
ромбовидной воронки и полулунной выемки на зубах. Самый крупный
(относительно размеров тела) череп, относительно самый высокий череп,,
с самой длинной мозговой частью и с самой высокой восходящей ветвью
нижней челюсти и ряд других особенностей. Все это ставит крупнозубую
бурозубку особняком от других сочленов рода как сильно уклоняющийся,
высокоспециализированный вид. На этом основании нами она выделяется
13*
196
в отдельный подрод (Stroganovia), названный в память о большом знатоке
насекомоядных млекопитающих С. У. Строганове.
Наиболее близким видом по внешнему складу, конфигурации че-
репа, соотношению размеров промежуточных зубов следует считать
тундряную бурозубку. Во многом сходен и характер ее распростра-
нения. Однако по форме промежуточных зубов, и особенно по структур-
ным особенностям многовершинных зубов, эту бурозубку нельзя сбли-
зить ни с одним палеарктическим видом.
Впервые крупнозубая бурозубка была описана Томасом [Thomas,
1907] с Сахалина. Вид сразу же был принят как «надежный» и не вызы-
вал возражений [Сатунин, 1914]. Тем не менее более поздние исследо-
ватели внесли ряд неточностей в его трактовку, а некоторые вообще
отрицали его реальность [Бобринский и др., 1944]. Описанная Алленом
[Allen, 1914] Sorex sanguinidens с Колымы, несомненно, синоним круп-
нозубой бурозубки и рассматривается нами как ее подвид. Ее диагно-
стические признаки со всей определенностью подтверждают этот тезис:
строение промежуточных и многовершинных зубов, их пигментация,
соотношение и размеры, морфометрические данные и описание внешности
зверька идентичны таковым номинальной формы.
С. И. Огнев [1928] поставил в системе эти два вида рядом, считая
их близкими, но диагностировать как один вид не смог. Больше того,
описанная им форма 5. d. sibiriensis [Огнев, 1928], которую ранее [Огнев,
1921 ] он рассматривал в ранге видового таксона, в полном соответствии
с его действительным положением (= 5. arcticus) в самом деле — один
из подвидов арктической бурозубки [Строганов, 1957]. Все эти недора-
зумения возникли вследствие ничтожно малого фактического материала,
имевшегося в его руках и не позволившего изучить пределы полимор-
физма рассматриваемых видов. Так, при описании 5. sanguinidens он
располагал лишь тремя черепами взрослых зверьков. Отсюда происхо-
дят и полученные им различия: несколько меньшие размеры черепа
и его отдельных структур, что вполне укладывается в пределы возрастной
изменчивости. Этот вопрос со всей убедительностью освещен в моногра-
фии С. У. Строганова [1957]. Наши исследования обширных материалов
из разных частей ареала подтвердили правильность этих суждений.
Знакомство с экземпляром 5. sibiriensis из типовой серии, хранящейся
в Зоологическом институте АН СССР, подтвердило его принадлежность
к S. arcticus, = 5. tundrensis. Н. А. Бобринский и А. П. Кузякин [Боб-
ринский и др., 1944] не только не признают видовой самостоятельности
крупнозубой бурозубки, но не выделяют ее даже на уровне подвида,
включая в искусственно созданный ими композиционный вид — Sorex
araneus, в котором произвольно объединили всех бурозубок, не относя-
щихся к крошечной, малой, бухарской и средней. Такой огульный под-
ход к систематике землероек уже нашел соответствующую оценку [Стро-
ганов, 1957; Юдин, 1971а; Долгов, Юдин, 1975; и др.]. Во втором изда-
нии «Определителя млекопитающих СССР» [Бобринский и др., 1965]
А. П. Кузякин принимает объем вида и внутривидовых категорий, при-
нятых в монографии С. У. Строганова [1957].
А. А. Гуреев [1963, 1971, 1979], признавая видовую самостоятель-
ность крупнозубой бурозубки, без всякой аргументации относит S. d. ьап-
guinidens к 5. arcticus. Поскольку в литературе [Строганов, 1957] было
высказано иное мнение о систематическом статусе этой формы, столь
произвольное перенесение подвида из состава одного вида в другой, без
197
фактического подкрепления этого действия, неубедительно. Окраска
меха, не говоря уже о полной тождественности структуры зубной систе-
мы у S. d. sanguinidens и крупнозубой бурозубки, убедительно свиде-
тельствует о их полном тождестве и не имеет отношения к тундряной
бурозубке. У последней из районов Колымы, откуда описан S. d. sangui-
nidens, в окраске меха четко выражена чепрачность, особенно у пере-
зимовавших особей. В диагнозе S. sanguinidens этот признак не отмечен,
хотя были исследованы зимние шкурки [Огнев, 1928], на которых чеп-
прачность видна более четко.
В настоящее время выделяется три подвида крупнозубой бурозубки.
Обзор подвидов
Sorex daphaenodon daphaenodon Thomas, 1907 —
Дальневосточная крупнозубая бурозубка
Голотип. См. в описании голотипа вида.
Диагноз. Размеры относительно других подвидов мелкие. Длина
тела 61—70 (Л/ = 64,8j_0,95), кондило-базальная длина черепа 17,3—
18,6 (М = 17,8±0,09), хвоста — 23—28 (М = 34,3^0,90) мм. Окраска
спины более светлая, серовато-бурая, со значительным коричневым
оттенком, вентральная сторона грязно-коричнево-серая с примесью
палевого цвета.
Измерения. См. табл. 40.
Характеристика. По размеру тела зверьки относятся к
наиболее мелким формам. Причем бурозубки из Приамурья и с Витим-
ского плоскогорья еще более мелкие, чем из Приморья, и имеют меньшую
длину хвоста. По этим показателям они достоверно отличаются от дальне-
восточных землероек из Приморья. Их следует выделить в самостоятель-
ный подвид.
Визуальное изучение окраски 72 зверьков из Приморья дало сле-
дующие результаты. Окраска серовато-бурая с хорошо выраженным
коричневым оттенком на дорсальной части тела. Бока окрашены в тот же
цвет, но со значительным преобладанием светлых тонов. У некоторых
особей довольно явственно улавливается граница между темной спиной
и более светлыми боками, чаще переход этот почти не заметен. Зимний
мех более длинный (М = 6,5 мм). Волосяной покров более темный,
особенно заметно меняется в сторону потемнения окраска нижней
части тела. Темная окраска спины спускается на бока и вентральную
часть тела.
Лапы и ступни сверху темные. Хвост у зверьков в летнем наряде
двухцветный, в тон верха и низа тела, а зимой почти одноцветный, тем-
ный, чуть светлее на нижней стороне. Подбородок и губы окрашены
в светлые серовато-палевые тона.
Забайкальские землеройки окрашены светлее, с большой примесью
палевых и шоколадных тонов.
Распространение. Дальний Восток: Сахалин, Приморье,
Забайкалье.
Материал. 72 экз.
Заметки по систематике. От 5. d. sanguinidens отличается
меньшими размерами, более светлой окраской, меньшей пышностью,
густотой и длиной зимнего меха и большей длиной хвоста. По размерам
198
черепа они не различимы, только длина рострума номинального подвида
достоверно больше, чем у 5. d. sanguinidens.
От 5. d. scaloni дальневосточная крупнозубая бурозубка отличается
меньшими размерами и более светлой окраской. По длине хвоста между
ними обнаруживается большое сходство.
С. У. Строганов [1957 ] в составе этого подвида рассматривает
S. d. orri, описанный в 1933 г. Куродой [Kuroda, 1933] как самостоя-
тельный вид 5. orri. Насколько мне известно, С. У. Строганов фактиче-
ский материал по этому «виду» не исследовал, а сделал заключение по
описанию [Там же]. Исследование материалов из Японии, а также обоб-
щение новейших данных японских систематиков [Abe, 1971] привело
нас [Юдин, 1975а] к выводу, что эта форма относится к S. caecutiens,
а не к 5. daphaenodon. Обитание крупнозубой бурозубки на Курильских
островах, откуда описан этот вид, фактическим материалом не под-
тверждено.
Sorex daphaenodon sanguinidens Allen, 1914 —
Восточносибирская крупнозубая бурозубка
Голотип.?, ad., 11/XII 1911 г., Нижнеколымск, близ устья
Колымы. Хранится в Музее сравнительной зоологии в Кембридже
(США).
Диагноз. Характеризуется коротким хвостом, отношение длины
хвоста к длине тела составляет 50 %. Более темной окраской, чем у но-
минального подвида и более резким разграничением темной окраски
спины и светлых боков, длинным и пушистым зимним мехом.
Измерения. См. табл. 40.
Характеристика. Наиболее характерной чертой этого под-
вида является короткий хвост — 26—32 (М = 31,2=±=0,32) мм. При
сравнении серийных материалов видно, что окраска спины заметно тем-
нее, чем у номинального подвида. У перезимовавших особей в летнем
меху довольно четко обозначена чепрачность, а переход от боков к вен-
тральной части постепенный. На границе темного чепрака и боков узкая
полоса ржавого оттенка, благодаря которой сглаживается переход от
спины к бокам. В области лопаток и огузка темная окраска спускается
до лап. Хвост слабо двухцветный.
Летний мех не длиннее такового номинального подвида, а зимний —
длиннее и пушистее (до 12 мм). Указание С. У. Строганова [1957] на более
светлый пушистый и длинный мех, свойственный этому подвиду, спра-
ведливо только в отношении длины и пушистости зимнего меха. Ошибка
в оценке окраски меха вкралась вследствие исследования зимних, выцвет-
ших шкурок, до набивки долгое время хранившихся в спирте.
Из 26 зверьков из окрестностей Якутска четыре по насыщенности
окраски не отличались от таковых из Приморья.
Распространение. Ареал подвида охватывает обширную
территорию северо-восточной части Сибири, от среднего течения Вилюя,
Якутска и долины Яны до Колымы и побережья Охотского моря.
Материал. 136 экз.
Заметки по систематике. Наиболее характерная черта
этого подвида — короткий хвост, статистически с большой достовер-
ностью отличающийся от хвостов других подвидов. Зимний мех чрез-
вычайно длинный и пушистый, на спине достигает 10—12 мм. .
199
Sorex daphaenodon scaloni Ognev, 1933 —
Западносибирская крупнозубая бурозубка
Голотип. № 3539 (кол. С. И. Огнева), шкурка, череп; устье
р. Мольтка, бассейн р. Таз, 28/IX 1929 г. Коллектор В. Н. Скалой. Кол-
лекция Зоологического музея МГУ, № 13428.
Диагноз. Крупнее двух предыдущих подвидов. Длина тела
50—67 (М = 64,7^0,48) мм, кондило-базальная длина черепа 17,4—18,8
(М = 18,2zt0,05) мм. Череп более массивный и удлиненный. Окраска
меха спины интенсивно темная, черно-бурая.
Измерения. См. табл. 40.
Характеристика. По окраске меха дорсальной части тела
самый темноокрашенный подвид крупнозубой бурозубки. Мех интенсив-
но бурый, в зимнем наряде черно-бурый. В летней окраске меха заметен
постепенный переход от темноокрашенной спины к бокам, в зимней обо-
значена чепрачность. Подбородок и губы мышино-серые с палевым оттен-
ком. Серый мех от горла заходит почти до глаз и ушей, образуя вокруг
глаз серое обрамление. В зимнем наряде хвост резко двухцветный. Бурые
тона со спины, в области плеч и от огузка спускаются до лап. Вентраль-
ная сторона дымчато-белесая с примесью палевого цвета. Длина волос
в зимнем меху достигает 9,5 мм.
Размеры тела и хвоста достоверно больше, чем у S. d. sanguinidens.
Череп статистически достоверно больше, чем у других подвидов.
Восходящая ветвь нижней челюсти по отношению к кондило-базальной
длине черепа у 5. d. scaloni составляет — 28,0 %, у S. d. daphae-
nodon — 26,4, у S. d. sanguinidens — 25,8 %.
Распространение. Урал, Зауралье, лесотундровая и таежу
ная зоны Западной Сибири, лесостепь, на юг до Бийска, Анжеро-Суд-
женска, Хакасии и далее на восток до Енисея.
Материал. 61 экз.
Заметки по систематике. Этот подвид очерчен более
четко в системе внутривидовых таксонов крупнозубой бурозубки.
С. У. Строганов [1957] упоминает об экземпляре крупнозубой бурозубки
из окрестностей Аксуат, Кустанайской области, хранящемся в зоологи-
ческом музее Биологического института СО АН СССР. Для него харак-
терна совершенно необычная окраска шкурки со светлым, ярким окрасом
спины, совершенно четко очерченным чепраком и контрастно отличаюу
щимися светлыми боками. Мы исследовали этот экземпляр и пришли
к выводу, что к типичной шкурке 5. tundrensis transuralensis ошибочно
привязан череп 5. daphaenodon неизвестного происхождения, в природе
зверьков такой окраски, относящихся к крупнозубой бурозубке, не
существует.
Подрод Sorex Linnaeus, 1758
Представители подрода характеризуются крупными и средними
размерами, наличием на нижней челюсти постмандибулярного отверстия,
на жевательной стороне коренных зубов хорошо выражена пирамидаль-
ная впадина. На территории Сибири встречается семь видов этого
подрода.
200
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА SOREX
1(3). Второй промежуточный зуб верхней челюсти меньше первого
и третьего ................................................... 2
2(6). Размеры крупные, длина тела с головой 65—91 (Л/ = 74,4+
^=0,76) мм; масса 6—18,5 (М = 10,1+0,37) г; телосложение тяжелое, на
передних конечностях когти длиной до 5 мм. Череп массивный, грубый,
угловатой конфигурации, кондило-базальная длина его 18,5—21,2 (М =
= 19,9+0,05) мм. Второй промежуточный зуб верхней челюсти меньше
первого и третьего. Окраска меха темно-бурая, glans penis самая массив-
ная среди таковых всех видов рода Sorex..........................
.................Sorex unguiculatus Dobson — Когтистая бурозубка
3(1). Соотношение промежуточных зубов иное.................4
4(6). Промежуточные зубы мелкие, последовательно и равномерно
убывающие в размере от первого к третьему......................5
5(4) Размеры крупные, длина тела с головой 51—88 (М — 69,4 +
+ 0,45); кондило-базальная длина черепа 17,4—21,0 (М = 19,5—0,11) мм,
череп широкий, угловатый. Промежуточные зубы относительно мелкие,
последовательно и равномерно убывающие в размере от первого к пятому,
в зубном ряду сидят свободно. Окраска меха темная, однотонная . . .
.......................Sorex isodon Turov — Равнозубая бурозубка
6(5). Размеры мелкие, длина тела с головой 49—64 (М = 51,9 +
—0,94); хвост длинный 39—47 (М = 43,5+0,42); кондило-базальная
длина черепа 15,0—16,5 (М = 15,7±0,05) мм. Ряд промежуточных
зубов сильно вытянут в сагиттальном направлении. Зубы низкие, с рас-
ширенными основаниями коронок и широко расставленными вершинами,
последовательно уменьшаются от первого к пятому .................
...........Sorex gracillimus Thomas — Дальневосточная бурозубка
7(4). Соотношение промежуточных зубов иное.............. .8
8(10). Второй промежуточный зуб выше первого, третий и четвертый
равны между собой..............................................9
9(8). Размеры крупные, длина тела с головой 58—89 (М = 72,0+
+ 0,45); кондило-базальная длина черепа 18,8—21,8 (М = 20,3+0,03) мм;
телосложение тяжелое, хоботок широкий, вытянутый. Череп относи-
тельно узкий, вытянут в длину. Зубы относительно черепа мелкие,
в зубном ряду сидят свободно, второй промежуточный зуб выше первого,
третий и четвертый равны между собой, окраска меха трехцветная . .
..........................Sorex roboratus Holl.— Бурая бурозубка
10(8). Соотношение зубов иное.............................11
11(13). Зубы по величине пропорциональны размерам черепа. Два
первых промежуточных равны между собой, зубы высокие, в зубном
ряду сидят плотно ........................................... 12
12(11). Размеры средние, длина тела с головой 46—82 (М = 63,4+
+ 0,26); длина хвоста 24—47 (М = 33,9+0,23) мм; масса тела 3—11
(М = 6,1+0,07) г; кондило-базальная длина черепа 15,7—19,4 (М =
17,6+0,02) мм. Голова широкая, массивная, череп вздутый в мозговой
области, сужение в области глаз выражено слабо; хоботок короткий
и широкий. Зубы по величине пропорциональны размеру черепа. Два
первых равны между собой, промежуточные высокие, в ряду сидят плотно.
Окраска меха чепрачная ..........................................
.......................Sorex tundrensis Мегг.— Тундряная бурозубка
13(11). Соотношение промежуточных зубов иное..............14
201
14(16). Зубной ряд вытянут, зубы мелкие, в зубном ряду сидят сво-
бодно, первые четыре промежуточные зуба образуют первые группи-
ровки....................................................... 15
15(14). Размеры средние, длина тела с головой 45—78 (М = 60,5±
±0,28); кондило-базальная длина черепа 16,1—19,4 (М = 17,4±0,03) мм.
Зубной ряд вытянут в сагиттальном направлении. Коронки промежуточ-
ных зубов мелкие, свободно сидят в зубном ряду. Первые четыре про-
межуточных зуба образуют парные группировки. Окраска меха трех-
цветная. . . ................................................
........................Sorex caecutiens Laxm.— Средняя бурозубка
16(14). Соотношение промежуточных зубов иное............,17
17(16). Зубной ряд самый длинный среди всех представителей рода
и составляет 45,7 % от кондило-базальной длины черепа. Зубы в зубном
ряду сидят свободно, наблюдается диспропорция в размерах двух пер-
вых крупных промежуточных зубов, пятый очень мал..........18
18(17). Размеры крупные, длина тела с головой 51—86 (ЛГ = 68,9±
±0,45); длина хвоста 34—49 (М — 42,5± 0,22) мм; масса тела 5,5—15,5
(М = 8,8±0,15) г; кондило-базальная длина черепа 18,7—20,8 (ЛГ =
9,8±0,06) мм. Череп умеренно вздутый в мозговой части, вытянут в ли-
цевой части. Зубной ряд длинный, зубы сидят свободно, в строении двух
первых промежуточных зубов наблюдается диспропорция. Окраска
трехцветная..................................................
........................Sorex araneus L.— Обыкновенная бурозубка
Sorex unguiculatus Dobson, 1890 —
Когтистая бурозубка
Голотип. Типовой экземпляр не выделен. Вид описан по серии
из коллекции, собранной Л. И. Шренком на о. Сахалин. Типовая серия
хранится в Зоологическом институте АН СССР (Ленинград).
Диагноз. Одна из самых крупных бурозубок фауны Сибири.
По размерам тела лишь немного уступает гигантской бурозубке —
S. mirabilis. Длина тела с головой 65—91 (М = 74,4±0,76) мм, масса
тела 6—18 (М = 10,1) г, тяжелого телосложения, с длинными когтями
на передних конечностях (до 5 мм). Череп массивный, грубый, угловатой
конфигурации (кондило-базальная длина 18,5—21,2 (М = 19,9±0,05),
наибольшая ширина 9—11 (М = 10,2±0,03) мм). Второй промежуточный
зуб верхней челюсти меньше первого и третьего (см. рис. 62). Темно-
бурая окраска спины постепенно переходит в бурую с примесью ржаво-
коричневых тонов на боках и грязновато-бурую на вентральной стороне
тела.
Измерения. См. табл. 41.
Характеристика. Во внешнем облике бурозубки обращают
на себя внимание крупные размеры и тяжелое плотное телосложение
(рис. 61, 62). Длина хвоста составляет значительно больше половины
длины тела — 64,9 %. Кисти ног крупные, широкие. У взрослых зверь-
ков на передних конечностях длинные когти, иногда до 5 мм. Окраска
хвоста двухцветная: сверху темная, в тон спины, а снизу светлая с па-
левым оттенком.
По интенсивности окраски спины (рис. 63) когтистая бурозубка
мало отличается от S. araneus и S. isodon. Колориметрическая характе-
ристика окраски полностью укладывается в таковую этих двух видов.
202
Таблица 41
Морфометрическая характеристика Sorex unguiculalus
Возрастная и половая группа М±тх О Су п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 72,1±0,89 4,34 6,03 24 65—79
взрослые 75,0± 5,57 9,64 12,86 3 68—86
Самки прибылые 72,0± 1,05 4,57 6,36 19 65—87
взрослые 80,9+1,30 5,03 6,21 15 73—90
В целом по виду . . . 74,4+0,76 6,23 8,38 67 65—90
Длина хвоста
Самцы прибылые 48,9±0,70 3,44 7,03 24 43—56
взрослые 44,0±2,08 3,61 8,19 3 41—48
Самки прибылые 47,0+1,05 4,58 9,75 19 40—53
взрослые 48,5±0,52 2,90 4,11 15 46—52
В целом по виду . . . 48,3+0,48 3,91 8,11 67 40—56
Длина ступни
Самцы прибылые 14,0+0,17 0,83 5,96 24 12-15
взрослые 13,7±0,33 0,58 4,23 3 13—14
Самки прибылые 14,1±0,13 0,58 4,15 19 13—15
взрослые 13,9+0,17 0,64 4,62 15 13—15
В целом по виду . . . 13,9±0,08 0,69 4,92 67 12—15
Масса тела
Самцы прибылые 9,29±0,26 1,14 12,26 20 6,8—11,0
взрослые 15,7±1,86 3,22 20,52 3 12,0—18,0
Самки прибылые 8,7±0,55 1,91 21,98 12 7,0—14,0
взрослые 11,5±0,67 2,21 19,25 11 9,0—15,5
В целом по виду . . . 10,1+0,37 2,68 26,51 52 6,8—18,0
Кондило-базальная длина черепа
Самцы прибылые 20,1+0,08 0,43 2,13 29 19,2—21,0
взрослые 20,4+0,15 0,21 1,04 2 20,2—20,5
Самки прибылые 20,0+0,09 0,42 2,09 21 19,1—20,7
взрослые 20,0+0,10 0,43 2,14 16 18,9—20,5
В целом по виду . . . 19,9+0,05 0,61 2,08 151 18,5—21,0
Длина рострума
Самцы прибылые взрослые 5,4±0,05 5,5+0,25 0,29 0,44 5,37 7,93 30 3 5,0—6,2 5,2—6,0
Самки прибылые 5,4±0,06 0,29 5,39 22 4,8—6,0
взрослые 5,4±0,04 0,18 3,27 16 5,1-5,7
В целом по виду . . . 5,3+0,03 0,38 7,21 155 4,8—6,2
Длина мозговой части черепа
Самцы прибылые 11,3+0,05 0,28 2,50 29 10,5—11,8
взрослые 11,5+0,30 0,42 3,69 2 11,2—11,8
Самки прибылые 11,1+0,05 0,23 2,04 21 10,8—11,7
взрослые 11,0+0,06 0,25 2,30 16 10,4—11,4
В целом по виду . . . 11,2+0,03 0,31 2,83 151 10,4—11,8
203
П родолжение табл. 41
‘ 2 ' 3 4 5 6
Длина верхнего ряда зубов
Самцы прибылые 8,9+0,06 0,33 3,67 29 8,1—9,5
взрослые 8,7+0,09 0,15 1,76 3 8,5—8,8
Самки прибылые 8,8±0,06 0,28 3,17 20 8,2-9,3
взрослые 8,7±0,05 0,21 2,49 16 8,3—9,0
В целом по виду . . . 8,8±0,03 0,35 3,99 152 8,1—9,5
Длина промежуточного ряда зубов
Самцы прибылые 3,0+0,03 0,14 4,66 30 2,7—3,2
взрослые 3,0±0,06 0,10 3,33 3 2,9-3,1
Самки прибылые 2,9+0,03 0,13 4,47 20 2,7—3,2
взрослые 2,9+0,03 0,14 4,78 16 2,6—3,1
В . д е л о м по виду . . . 2,9±0,01 0,15 5,12 153 2,6—3,2
Ширина рострума
Самцы прибылые 2,0+0,02 0,10 4,83 30 1,8—2,2
<= взрослые 2,1±0,09 0,15 7,39 3 1,9—2,2
Самки прибылые 2,0+0,02 0,11 5,31 . 22 1,8—2,2
взрослые 2,0±0,03 0,12 5,64 16 1,8—2,2
В целом по виду . . . 2,0±0,01 0,11 5,38 154 1,8—2,2
Предглазничная ширина
Самцы прибылые 2,9±0,03 0,17 5,89 30 2,6—3,3
взрослые 2,9±0,12 0,21 7,26 3 2,7—3,1
Самки прибылые 2,9±0,04 0,19 6,72 22 2,4—3,1
взрослые 3,0+0,04 0,14 4,80 16 2,7—3,2
В целом по виду . . . 2,8±0,01 0,17 6,10 155 2,4—3,3
Межглазничная ширина
Самцы прибылые 4,2+0,03 0,17 4,01 30 3,9—4,5
взрослые 4,3+0,06 0,10 2,33 3 4,2—4,4
Самки прибылые 4,1+0,03 0,13 3,03 22 3,9—4,4
взрослые 4,3+0,04 0,15 3,54 16 4,0—4,5
В целом по виду . . ". 4,2+0,01 0,16 3,93 155 3,9—4,5
Наибольшая ширина черепа
Самцы прибылые 10,4+0,04 0,23 2,20 29 9,6—10,4
взрослые 10,4 — — 1 —
Самки прибылые 10,0+0,06 0,28 2,76 20 9,5—10,3
взрослые 10,2+0,05 0,19 1,86 13 9,8—10,5
В целом по виду . . . 10,2+0,03 0,32 3,16 143 9,5—10,5
Наибольшая высота черепа
Самцы прибылые 6,4+0,05 0,27 4,16 28 5,7-6,8
взрослые 6,2 — — 1 —
Самки прибылые 6,3+0,06 0,26 4,18 21 5,8-6,7
взрослые 6,0+0,05 0,18 2,99 14 5,6—6,4
В целом по виду . . . 6,2+0,03 0,33 5,22 145 5,6—6,8
204
Окончание табл. 41
1 | 2 I 3 J 4J 5 | 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,8±0,03 0,17 3,53 30 4,3—5,1
взрослые 4,9±0,12 0,21 4,28 3 4,7—5,1
Самки прибылые 4,7±0,03 0,14 3,08 22 4,5—5,0
взрослые 4,7±0,03 0,15 3,14 16 4,3—4,9
В целом по виду . . . 4,7±0,02 0,19 4,11 154 4,3-5Д
Длина углового отростка
Самцы прибылые 2,2±0,03 0,14 6,27 30 2,0—2,5
взрослые 2,2±0,10 0,17 7,87 3 2,1—2,4
Самки прибылые 2,3±0,02 0,14 4,59 32 2,1—2,4
взрослые 2,2±0,02 0,09 4,28 16 2,0—2,3
В целом по виду . . . 2,2±0,01 0,14 6,44 145 2,0—2,5
Мы сравниваем данные виды потому, что многие исследователи рассмат-
ривали их в качестве одного. Что касается характера распределения
окраски по телу, то она близка к таковой у S. isodon, отличается лишь
наличием ржаво-коричневых тонов в окраске меха боков. Как у и пос-
ледней переход от темно-бурой спины к более светлому брюшку посте-
пенный, без резкой смены. Вентральная сторона относительно темно-
окрашенная. В отличие от S. araneus, для которой характерно наличие
чепрака, особенно хорошо выраженного у перезимовавших зверьков
и прибылых в зимнем меху, когтистой бурозубке чепрачность не свойст-
венна. По средним значениям белизны она светлее, чем обыкновенная
и равнозубая бурозубка, а по интенсивности оттенка более темная
(см. табл. 30, рис. 63). По величине показателя оттенка она уступает
только S. tundrensis, S. daphaenodon и близка 5. roboratus и S. caecutiens.
Рис. 61. Когтистая бурозубка — Sorex unguiculatus Dobson.
205
Рис, 62. Череп когтистой бурозубки.
Усл. обозн, см. рис. 1.
В особенностях черепа наиболее достопримечательна массивность
и угловатость при относительно короткой длине. Отношение кондило-
базальной длины черепа к длине тела составляет 26,7 %, это самый низ-
кий показатель среди всех сибирских представителей рода Sorex (см.
табл. 25). При средней кондило-базальной длине черепа (19,9 мм) ширина
мозговой части значительна — 10,2 мм, что составляет 51,3 %, в то вре-
мя как у всех остальных видов бурозубок этот показатель значительно
ниже. Череп также широкий (к кондило-базальной длине, %) в меж-
глазнпчной области — 20,2, ростральной части — 10, мозговой (взду-
тый)— 31,1. Соотношение длины мозговой и лицевой частей черепа,
а также длины зубного ряда и кондило-базальной длины довольно про-
порциональное, по этим показателям она занимает промежуточное по-
ложение среди других видов бурозубок. Восходящая ветвь нижней че-
люсти низкая, составляет 23,6 % от кондило-базальной длины черепа,
в то время как у S. isodon — 24,6, у S. araneus — 25,4, у 5. daphaeno-
don— 26,8, рг. angularis самая низкая — 11,1 %—среди всех видов
бурозубок (см. табл. 29).
Самобытные черты организации свойственны и зубной системе этого
вида. Передний резец верхней челюсти хорошо развит. Его второй (внут-
ренний) зубец лишь немного меньше первого и незначительно уступает
по величине первому промежуточному зубу. Соотношение промежуточ-
ных зубов стойкое и специфично для вида. Только по одному этому приз-
наку, а также по размеру зверька когтистая бурозубка надежно диаг-
ностируется.
206
S. isodon различия в их
Рис. 63. Колориметрическая характе-
ристика бурозубок.
1 — S. isodon; 2 — S. araneus; 3 — S. ungu-
iculatus.
Первый одновершинный зуб
крупнее остальных. Второй замет-
но меньше первого и третьего.
Постановка зуба такова, что это
соотношение хорошо улавливает-
ся даже у старых особей, с силь-
но стертыми коронками зубов,
его основание на челюсти всегда
выше оснований первого и третье-
го зубов (см. рис. 62). Если при
стирании зубов у 5. araneus и
в значительной мере нивелируют-
ся, то у старых когтистых бурозубок специфика этих зубов остается
заметной. Подобное соотношение промежуточных зубов свойственно
и S. minutus, но она легко отличается по небольшим размерам, морфо-
метрические признаки этих двух видов не перекрываются. По размерам
промежуточные зубы пропорциональны размерам черепа, что отличает
их от S.isodon, для которой характерны мелкие зубы, при большом че-
репе. Первые три промежуточных зуба обычно со слегка вогнутой перед-
ней и задней сторонами, а у четвертого и пятого они не вогнуты. Верши-
ны лишь слегка сдвинуты вперед по отношению к вертикальной оси
коронок. Вертикальные оси первых трех промежуточных параллельны
между собой, у четвертого она наклонена кзади (внутрь ротовой полости),
а у пятого — кпереди. Первый предкоренной зуб нижней челюсти (Рпц)
вытянут в сагиттальном направлении.
Отношение верхнего ряда зубов к кондило-базальной длине черепа
составляет 44,2, а промежуточных зубов к длине зубного ряда — 14,6 %
(см. табл. 26). Надежно отличается от других видов когтистая бурозубка
по размерам и строению glans penis, которая самая массивная среди
таковых всех видов бурозубок. Свободная часть ее толстая, почти ци-
линдрическая, несколько загнутая вниз в дистальной части, с резко
притупленным свободным концом. В поперечном разрезе у основания
головка яйцевидной формы несколько сжата с боков. При рассмотрении
ее сверху видно слабое утончение с боков к дистальному концу. Спереди,
в центре тупого свободного конца головки, находится углубление с вы-
ходом мочеполового канала. Длина glans penis 9,3—12,5 (М = 11,1),
толщина у основания 1,9—2,4 (М = 2,0) мм (см. рис. 40).
Распространение. Когтистая бурозубка — типичный во-
сточный палеаркт. Ареал этого вида в настоящее время очерчен весьма
схематично. Известно его обитание в Приамурье, Приморье, на Шан-
тарских островах, Сахалине, Курильских островах, в Маньчжурии,
Корее и на Хоккайдо [Юдин, 1971а]. Наиболее западная точка распро-
странения — район Мал. Хингана (д. Сотниково), несколько северо-
восточнее известна из верховьев р. Бурея. Южнее коллективировалась
в Хабаровском, Комсомольском (Пивань), Лазовском районах. В нижнем
течении Амура добыта близ с. Софийского, на западных, восточных,
северных и южных склонах Сихоте-Алиня по рекам Илюно, Гоба, Иод-
207
вуха, Муравейка, в верховьях Комаровки, на р. Одарке, близ оз. Ханка,
в Сучанском и Шкотовском районах, в заповедниках Уссурийском,
«Кедровая Падь», близ г. Находки.
На Сахалине она, по-видимому, распространена повсеместно, зверь-
ки добывались на Северном, Среднем и Южном Сахалине. Здесь они
зарегистрированы в ряде пунктов: у Невелика, Чехова, Ново-Александ-
ровска, Южно-Курильска и др.
Материал. 155 экз. Приморье, Сахалин, Кунашир.
Изменчивость. Когтистая бурозубка монотипический вид,
что уже само по себе говорит о незначительном размахе изменчивости
по сравнению с политипическими видами. Изменчивость окраски выраже-
на слабо. С возрастом мех на хвосте, ступнях и кистях стирается. К вес-
не зимний мех у зверьков вынашивается и становится более светлым.
Колориметрические исследования показали хорошо выраженный рыжий
оттенок. Значения оттенка у зверьков этого вида колеблются от 120,6
до 148,1. В географической изменчивости проявляется некоторая тен-
денция увеличения насыщенности рыжих тонов у зверьков, обитающих
в Приморье, по сравнению с таковыми с о. Кунашир (табл. 42), эти от-
личия достоверны (Td — 3,5). Достоверны отличия и по интенсивности
оттенка (Td = 3,2).
По морфологическим показателям популяции из Приморья (Уссу-
рийский заповедник, Шкотовский и Хасанский р-ны), островов Саха-
лина и Кунашира достоверно не различаются. Наибольшей длиной те-
ла обладают зверьки с Сахалина (М = 75,5=ь5,5) мм при наименьшей
длине черепа (М — 19,3=t0,06) мм. На основании кондило-базальной
длины черепа можно говорить о намечающейся тенденции сокращения
размеров черепа бурозубок, обитающих на островах, по сравнению с
таковыми на материке, но различия эти крайне незначительны. Длина
хвоста возрастает у островных форм (табл. 43).
Таким образом, географическая изменчивость в окраске меха прояв-
ляется довольно хорошо, различия островных и материковых популяций
достоверны. В размерах зверьков различия менее выражены, несмотря на
их изоляцию, что свидетельствует о недавней их разобщенности. Острова
Кунашир, Малая Курильская гряда, Хоккайдо, Сахалин в недалеком:
прошлом представляли полуостров Приамурья [Велижанин, 1970].
Возрастная изменчивость в размерах зверьков выражается в следую-
щем: средние значения кондило-базальной длины черепа у молодых и ста-
рых зверьков, самцов и самок, практически одинаковы. Аналогичная кар-
тина возрастной изменчивости наблюдается и по большинству других па-
раметров (см. табл. 41). Исключение составляют длина верхнего ряда зу-
бов и высота черепа: у перезимовавших зверьков они несколько меньше,
чем у прибылых. Наблюдаются существенные отличия в размерах тела:
Таблица 42
Колориметрическая характеристика Sorex unguiculatus
Место взятия выбор- ки Белизна Оттенок
М+тпх а Су п а Су п
Приморье 3,2+0,18 0,68 21,1 14 140,4± 1,38 5,18 3,7 14
О. Кунашир 3,4±0,17 0,89 26,1 26 134,1+1,13 5,76 4,3 26
208
Морфометрическая характеристика Sorex unguiculatus
Признак Приморье, уссурийский заповедник Приморье, Шкотовский и Хасанский р-ны г
М±тх Су п lim М±тх Су п lim
Длина тела 74,2+2,06 7,42 9,99 13 65—90 74,6+1,48 6,80 9,12 21 65—91
хвоста 47,5±1,22 4,39 9,23 13 41—57 47,0+1,02 4,69 9,98 21 40—57
ступни 13,9±0,18 0,64 4,6 13 13—15 13,9+0,20 0,94 6,79 21 12—15
Масса тела 10,3+0,89 3,22 31,4 13 6,8—18,0 10,4+0,56 2,52 24,31 20 8—18,5
Длина кондило-ба- зальная че- репа 20,3±0,08 0,37 1,81 19 19,6—21,0 20,4+0,05 0,31 1,54 41 19,8—21,2
рострума 5,6±0,07 0,32 5,75 20 5,3-6,7 5,5+0,03 0,23 4,18 44 5,1—6,0
мозговой части 11,4±0,06 0,27 2,35 19 11,0—11,8 11,3+0,03 0,22 1,93 41 10,8—11,8
верхнего ряда зубов 9,0±0,05 0,24 2,67 20 8,6—9,3 8,9±0,04 0,27 3,02 41 8,4-9,6
ряда проме- жуточных зу- бов 3,1±0,03 0,11 3,71 20 2,8—3,2 3,0+0,02 0,12 4,03 42 2,7—3,2
Ширина рострума 2,0+0,02 0,11 5,13 20 1,9—2,2 2,0+0,01 0,10 4,97 44 1,8-2,3
предглазнич- ная 3,0+0,03 0,11 3,73 20 2,8—3,1 2,9+0,02 0,12 4,01 44 2,7—3,1
межглазнич- ная 4,3+0,04 0,18 4,14 20 3,9-4,5 4,3+0,02 0,13 3,05 44 4,0—4,5
наибольшая черепа 10,3±0,05 0,21 2,00 16 9,9—10,0 10,4+0,04 0,26 2,51 39 9,9—11,0
Наибольшая высо- та черепа 6,2+0,06 0,25 4,01 16 5,7—6,6 6,4±0,05 0,29 4,47 39 5,9—6,9
Высота восходя- щей ветви ниж- ней челюсти 4,8+0,05 0,20 4,20 20 4,5—5,1 4,8+0,02 0,16 3,27 44 4,5—5,1
Длина углового от- ростка 2,2+0,02 0,09 3,90 20 2,0—2,4 2,3—0,02 0,15 6,52 42 2,1—2,8
перезимовавшие самцы и самки заметно крупнее (75,0 + 5,57 и 80,9 =t
+ 1,30 мм), чем прибылые (72,1 0,89 и 72,0 + 1,05 мм соответственно).
Особенно заметны различия в массе зверьков разных возрастных групп:
прибылые — 9,29 ztz 0,26 и 8,7 zt 0,55, перезимовавшие — 15,7 =t 1,86
и 11,5 + 0,67 г (самцы и самки соответственно).
По данным Ф. Р. Штильмарка и В. А. Долгова [1974], в районе Ком-
сомольска (пос. Пивань) 5. unguiculatus имела следующие размеры: кон-
дило-базальная длина черепа 19,6—21,1 (М = 20,0 zt 0,07) мм у прибылых
зверьков (п = 22) и 19,8—20,6 (М = 20,1 zt 0,12) мм — у перезимовав-
ших (п = 22); длина зубного ряда 8,4—9,1 (М = 8,7 ztz 0,03) и 8,3—8,9
(М = 8,5 zt 0,07 )мм. Как характерную черту вида авторы указывают
малую изменчивость промежуточных зубов. Эти данные полностью со-
гласуются с нашими, свидетельствующими о стабильности соотношения
и формы промежуточных зубов у этого вида.
209
Т а блица 43
из разных частей ареала
Сахалин о. Кунашир
М+тх а cv п lim М±таж а CV п lim
75,5+5,50 9,53 12,62 3 70—86 74,2+0,95 5,21 7,01 30 67—87
48,7± 1,86 3,22 6,61 3 45—51 49,4+0,52 2,86 5,79 30 41—56
13,8+0,20 0,98 3,15 3 13—14 14,0±0,10 0,53 3,75 30 13—15
10,64=0,90 3,60 31,10 3 9,0-14,0 9,6±0,64 2,56 26,73 16 7,0—17,0
19,3+0,06 0,45 2,32 61 18,5—20,5 20,1±0,05 0,29 1,46 30 19,3—20,7
5,0+0,04 0,30 6,01 61 4,3—5,5 5,5+0,05 0,28 5,01 30 5,1-6,2
10,9+0,04 0,28 2,57 61 10,0—11,5 11,1+0,04 0,24 2,12 30 10,5—11,6
8,5+0,04 0,29 3,37 61 8,0—9,2 8,9±0,04 0,24 2,75 30 8,4-9,3
2,8+0,02 0,13 4,44 61 2,5-3,1 2,9+0,02 0,12 4,26 30 2,6—3,1
1,9+0,01 0,87 4,50 60 1,7—2,1 2.0+0,02 0,09 4,40 30 1,8—2,2
2,7+0,01 0,11 4,20 61 2,4-3,0 3,0+0,03 0,14 4,51 30 2,8—3,3
J 4.0ч~0,02 0,12 3,01 61 3,8—4,3 4,2+0,02 0,12 2,98 30 4,0—4,4
10,0+0,04 0,31 3,10 58 9,0—10,8 10,1+0,05 0,27 2,70 30 9,5—10,5
6,1+0,04 0,34 5,49 60 5,5—7,0 6,3 + 0,06 0,31 5,01 30 5,6—6,8
4,6+0,02 0,16 3,55 60 4,2—5,0 4,7+0,03 0,17 3,57 30 4,3—5,1
2,2+0,02 0,15 6,98 53 1,8-2,5 2,2+0,02 0,13 5,68 30 2,0-2,4
Половая изменчивость в размерах тела, черепа и окраске меха не от-
мечена.
। Заметки по систематике. Из-за отсутствия четкого диагноза
в первоописании S. unguiculatus разные исследователи рассматривали объ-
’ ем этого вида неодинаково. В первоописании когтистой бурозубки Добсон
[Dobson, 1890] указывал на следующие диагностические признаки вида:
длинные когти на передних конечностях; крупный и чрезвычайно широ-
кий череп; третий промежуточный зуб такой же величины, как и второй,
но значительно крупнее четвертого; пятый промежуточный зуб хорошо
I развит.
Этим автором допущена неточность, в результате которой стало воз-
(| можным путать S. isodon и 5. unguiculatus. Дело в том, что у некоторых
экземпляров вершины первого, второго и третьего промежуточных зубов
бывают на одном уровне, однако высота коронки второго зуба всегда усту-
14 Б. С. Юдин
i
210
пает таковой первого и третьего: один уровень их вершин достигается бо-
лее высоким положением на челюсти меньшего по размерам второго про-
межуточного зуба. Допустимо и другое предположение — серия, имев-
шаяся в распоряжении Добсона, состояла из экземпляров, относящихся
к двум видам: S. unguiculatus и S. isodon. Если описание сделано не по ти-
повому экземпляру, а по серии, то вследствие этого в диагнозе могла быть
допущена ошибка. Равнозубая и когтистая бурозубка габитуально и по
окраске схожи, поэтому при том уровне знаний таксономии такая ошибка
не исключается.
Так, Томас [Thomas, 1907], который исследовал серию зверьков, со-
стоявшую явно из двух видов, объединил в один «сборный вид» под назва-
нием S. unguiculatus. Позднее эту же ошибку повторил Скарен [Skaren,
1964], безосновательно объединивший S. isodon и S. unguiculatus в один
вид, обозначив его ареал от Финляндии до Тихоокеанского побережья.
Сиивонен [Siivonen, 1965] детально изучил серию исследованных в свое
время Томасом и Скареном землероек, привлек новые сравнительные ма-
териалы и со всей убедительностью доказал несостоятельность системати-
ческих построений этих исследователей. На основании таких признаков,
как окраска меха, архитектура черепа, строение зубного аппарата, а так-
же морфометрии Сиивонен пришел к выводу о наличии в изучаемой серии
двух самостоятельных видов: 5. unguiculatus и S. isodon. Утверждение
Бринка [Brink, 1957, 1967] и Скарена [Skaren, 1964] о конспецифичности
S. isodon и S. unguiculatus и североамериканского вида S. obscurus Merr.
неверно. Это положение убедительно доказано Хоффманном [Hoffmann,
1971].
С. И. Огнев [1928] признал видовую самостоятельность 5. unguicy-
latus, не указывая в диагнозе на ее основное отличие от S. isodon,— соот-
ношение трех первых промежуточных зубов. Судя по рисунку (с. 205), он
не различал S. isodon (в то время известную как подвид обыкновенной)
и S. unguiculatus. Соотношение зубов, изображенное на рис. 74 [Там же],
характерно для равнозубой бурозубки, а не для когтистой, т. е., признавая
видовую самостоятельность S. unguiculatus, С. И. Огнев рассматривал в ее
составе и имевшиеся в его распоряжении особи равнозубой бурозубки,
в то время еще не выделенной в самостоятельный вид.
Н. А. Бобринский и А. П. Кузякин [Бобринский и др., 1944] без вся-
ких оснований и не утруждая себя аргументацией, объединили S. unguicu-
latus в один вид с обыкновенной бурозубкой (S. araneus), понимая объем
последней весьма широко и включая в искусственно созданный ими компо-
зиционный вид кроме собственно S. araneus еще и 5. isodon, S. unguicula-
tus, S. daphaenodon, S. tundrensis, S. raddei, S. caucasicus. Во втором изда-
нии пособия «Определитель млекопитающих СССР» [Бобринский и др.,
1965] эта ошибка отчасти была исправлена, так как в полном соответствии
с фактическим положением S. unguiculatus была выведена из состава ком-
позиционного вида и принималась в качестве самостоятельного вида.
С. У. Строганов [1957], признавая видовую самостоятельность 5. un-
guiculatus, принимал его в очень широком объеме, включая в него и S. iso-
don. Он ошибочно описал равнозубую бурозубку (смотри систематику
5. isodon) в подвид обыкновенной (S. a. isodon), в результате чего послед-
нюю на Дальнем Востоке, где нет обыкновенной бурозубки, рассматривал
как 5. unguiculatus. В диагнозе когтистой бурозубки у С. У. Строганова
[1957] нет указания па специфичное строение и соотношение промежуточ-
ных зубов этого вида. В его характеристике в качестве существенной
211
черты вида приводится указание на относительно мелкие зубы, что более
свойственно S. isodon, чем S. unguiculatus. Этот факт подтверждает нераз-
работанность систематики двух видов в работе С. У. Строганова [1957].
В коллекции Зоологического музея АН СССР хранится серия равно-
зубых бурозубок, собранных Ю. В. Авериным на Камчатке, а также ма-
териалы из Приморья, определенные С. У. Строгановым как S. unguicu-
latus. Аналогичные ошибочные определения есть и в коллекции С. У. Стро-
ганова, хранящейся в зоологическом музее Биологического института СО
АН СССР (Новосибирск).
Таким образом, под S. unguiculatus, в понимании С. У. Строганова
[1957], имеется в виду систематически не регламентированная группиров-
ка из двух видов (5. unguiculatus и S. isodon), не столь широкая, как это
представляют Н. А. Бобринский и А. П. Кузякин [Бобринский и др.,
1944], но все же обладающая чертами обыкновенной, равнозубой и ког-
тистой бурозубок. В монографии С. У. Строганова [1957] на рис. 64
(с. 161), изображающем череп и зубы S. unguiculatus, дано характер-
ное для S. isodon соотношение промежуточных зубов, не имеющих ничего
общего с таковыми у S. unguiculatus.
Неточен диагноз и описание этого вида и в работах А. А. Гуреева
[1963, 1971], в которых сказано, что «второй промежуточный зуб равен
третьему, или немного меньше его» (с. 97). В диагнозе данного вида во вто-
рой работе [Гуреев, 1971] как один из признаков приводится «...первый
верхний промежуточный в 1,5 раза больше второго» (с. 98), а в описании
сказано: «...второй промежуточный зуб равен третьему или немного мень-
ше его». На рисунках когтистой бурозубки (в той и другой работах) изоб-
ражены зубы, не существующего в природе вида, по соотношению их раз-
меров они были бы более близки к S. isodon, если бы не их форма и малый
пятый промежуточный зуб. С соотношением и формой зубов у S. unguicu-
latus они не имеют ничего общего.
Как уже сказано выше, А. П. Кузякин [Бобринский и др., 1965] дает
тот Hie диагноз — «...второй верхний промежуточный зуб значительно
(раза в полтора) меньше первого и равен или меньше третьего». Если бы
не это «равен», диагноз был бы точен. Такая неточность, допускаемая ря-
дом исследователей, на наш взгляд, может происходить по ряду причин.
Во-первых, авторы путают габитуально похожих равнозубую и когтистую
бурозубок; во вторых, они неправильно интерпретируют положение, ког-
да вершина второго зуба находится на одной высоте с вершиной первого
и третьего, или только третьего, как «равные зубы», не учитывая, что ос-
нование второго зуба всегда находится заметно выше основания первого
и третьего, а следовательно, при равном положении вершин абсолютные
размеры второго зуба всегда остаются меньше таковых соседних; в-третьих
при изучении зубов старых зверьков когтистой бурозубки не учитывается
охарактеризованная выше специфика снашивания зубов (более высокий
первый и третий стираются быстрее, что приводит к их уравниванию по
высоте с другими).
Впервые диагноз этого вида с учетом указанной выше специфики был
сформулирован в нашей работе [Юдин, 1965а]. При исследовании серии
зверьков (более 450 экз.) из разных районов Приамурья, Приморья, Са-
халина, Курильских островов и из Японии во всех случаях было уста-
новлено стойкое соотношение промежуточных зубов: второй промежуточ-
ный зуб уступает по размеру первому и третьему. Аналогичное соотноше-
ние отмечено Сиивоненом [Siivonen, 1965] у всех исследованных им зверь-
14*
212
ков этого вида. О видовой самобытности S. unguiculatus со всей определен-
ностью свидетельствует специфичное строение glans penis.
В настояще время по комплексу признаков этот вид четко очерчен,
его таксономическое положение и границы не вызывают сомнений. Ана-
логичные взгляды на объем вида изложены в работах Сиивонена [Siivo-
nen, 1965], В. А, Долгова [19686, 1972], Ф. Р. Штильмарка, В. А. Дол-
гова [1974].
Кариологическими исследованиями японских генетиков Такаги и Фу-
имаки [Takagi, Fujimaki, 1966] установлен кариотип S. unguiculatus.
По их данным, у S. unguiculatus и S. shinto saevus Thomas 2п = 42, a NF =
= 62. Однако Fredga [1968], исследовав по фотографиям кариотипы сред-
ней и когтистой бурозубок, нашел число плеч у них равным 68. Названные
японские кариологи пришли к выводу, что кариотипы S. shinto saevus Tho-
mas и S. unguiculatus Dobson из Японии идентичны по всем показателям.
Нам представляется, что в их исследование вкралась ошибка, связанная
с неточностью таксономического определения исходного материала.
S. q. shinto из Японии и S. s. saevus с Сахалина, описанный Томасом в 1905
и 1907 гг., оказался нереальным видом, впрочем, как и подавляющее боль-
шинство форм, диагностированных этим автором. Выделенная им форма
относится к S. caecutiens и имеет подвидовой уровень. Следовательно, ка-
риотип S. s. shinto должен быть аналогичным таковому S. caecutiens, что
и было доказано Фредга [Fredga, 1968]. Им также обнаружено неквалифи-
цированное обсуждение материала Такаги и Фуимаки, указывающих на
наличие у того и другого видов NF = 62, а на приведенных в работе фото-
графиях Фредга обнаружил их равными 68, что соответствует данному
числу у S. caecutiens и согласуется с теоретическим предположением.
К тому же S. unguiculatus — настолько четко охарактеризованный
вид, что трудно предположить наличие у него хромосомного набора, пол-
ностью идентичного с таковым у средней бурозубки. Нам представляется,
что исследователи располагали неправильно определенным материалом,
относящимся к S. shinto (-S. caecutiens). Для уточнения кариотипа S. un-
guiculatus необходимо провести исследование хромосомного набора этого
вида и равнозубой бурозубки из мест их совместного обитания (Приморье,
Сахалин). У последней также 2п = 22 и NF = 68, но от средней бурозуб-
ки она отличается иным соотношением мета- и субметацентрических хро-
мосом среди первых 9 пар крупных аутосом.
Sorex isodon Turov, 1924 —
Равнозубая бурозубка
Голотип. № 13486, шкурка, череп. Вид описан по экземпляру
с оз. Байкал, Северо-Восточное Забайкалье, баргузинская тайга,
14 VIII 1923 г. С. С. Туров (из коллекции С. И. Огнева, № 1461).
Диагноз. Размеры крупные. Длина тела с головой 51—88 (М =
= 69,4 + 0,45) мм. Окраска меха темная, однотонная на дорсальной и
вентральной стороне тела. Четкой границы между окраской спины, боков
и вентральной части тела нет. Кондило-базальная длина черепа 17,4—21,0
(М = 19,5 + 0,11) мм. Промежуточные зубы по отношению к крупному
черепу мелкие, пирамидальной формы, с острыми вершинами, последо-
вательно и равномерно убывающие по размерам от самого крупного пер-
вого к пятому. Коронки в зубном ряду сидят свободно (см. рис. 65).
Измерения. См. табл. 44.
213
Таблица 44
Морфометрическая характеристика Sorex isodon
Возрастная и половая группа М±тх а Су п lim
1 2 3 5 6
Длина тела с головой
Самцы прибылые 67,8+0,70 6,19 9,14 79 51—85
взрослые 76,4+0,99 4,72 6,18 23 65—86
Самки прибылые 68,1+0,69 4,98 7,32 52 55—77
взрослые 79,0+1,76 6,33 8,02 13 64—88
В целом по виду . . . 69,9±0,45 6,59 9,43 213 51—88
Длина хвоста
Самцы прибылые 43,9+0,36 3,15 7,18 79 36-51
взрослые 43,2+0,86 4,01 9,29 22 36—53
Самки прибылые 43,6+0,47 3,36 7,69 52 35—51
взрослые 44,6±0,66 2,39 5,36 13 40—48
В целом по виду . . . 44,1±0,23 3,28 7,43 212 35—53
Длина ступни
Самцы прибылые 13,5+0,10 0,92 6,83 80 11—15
взрослые 13,5+0,19 0,91 6,75 23 11—15
Самки прибылые 13,6±0,12 0,92 6,77 55 11—15
взрослые 14,0±0,43 1,54 11,01 13 12—16
В целом по виду . . . 13,5±0,07 0,96 7,07 217 11,0—16
Масса тела
Самцы прибылые 8,3±0,24 2,05 24,86 72 6,5—14,5
взрослые 12,1+0,39 1,79 14,76 21 8,0—23,5
Самки прибылые 8,0±0,13 0,95 11,85 53 5,5—10,0
взрослые 11,5+0,71 2,36 20,53 11 6,2—15,0
В целом по виду . . . 8,7+0,16 2,16 24,74 193 5,5—23,5
Кондило-базальная длина черепа
Самцы прибылые взрослые 19,5±0,05 19,8±0,10 0,53 0,52 2,71 2,65 105 28 18,5—21,0 18,7—20,5
Самки прибылые 19,6±0,06 0,55 2,82 85 18,4—20,8
взрослые 19,6+0,09 0,37 1,87 17 19,0—20,2
В целом по виду . . . 19,5+0,03 0,56 2,86 429 18,4—21,0
Длина рострума
Самцы прибылые взрослые 5,4±0,03 5,5±0,05 0,30 0,28 5,53 5,12 109 28 4,6—6,0 4,4—5,8
Самки прибылые 5,4+0,03 0,27 4,89 87 5,0—6,1
взрослые 5,4+0,05 0,20 3,68 17 5,0—5,8
В целом по виду . . . 5,4+0,13 0,28 5,27 441 4,4—6,1
Длина мозговой части черепа
Самцы прибылые 10,9+0,03 0,32 2,93 105 10,0—11,7
взрослые 10,9+0,07 0,38 3,48 28 9,7—11,5
Самки прибылые 10,8±0,04 0,33 3,01 85 9,9—12,0
взрослые 10,8+0,06 0,24 2,17 17 10,5—11,3
В целом по виду . . . 10,9+0,02 0,31 2,85 430 9,7—12,0
214
Продолжение табл. 44
1 2 3 4 5 6
Длина верхнего ряда зубов
Самцы прибылые 8,8+0,03 0,30 3,42 104 8,1—9,7
взрослые 8,6±0,06 0,32 3,65. 27 8,0—9,1
Самки прибылые 8,9±0,04 0,33 3,73 85 8,2—9,8
взрослые 8,7+0,06 0,25 2,86 17 8,3—9,1
В целом по виду . . . 8,8±0,02 0,31 3,56 427 8,0—9,8
Длина ряда промежуточных зубов
Самцы прибылые 3,1+0,02 0,17 5,54 107 2,7—3,7
взрослые 3,0+0,03 0,18 6,02 28 2,7—3,3
Самки прибылые 3,1+0,02 0,17 5,69 86 2,7—3,6
взрослые 3,1+0,03 0,13 4,31 17 2,8—3,3
В целом по виду . . . 3,0+0,01 0,16 5,26 432 2,7—3,6
Ширина рострума
Самцы прибылые 1,9+0,01 0,15 7,77 103 1,5—2,5
взрослые 1,9+0,01 0,10 4,87 26 1,7—2,1
Самки прибылые 1,9+0,01 0,12 6,35 82 1,6—2,2
взрослые 1,9±0,09 0,10 5,17 17 1,7—2,0
В целом по виду . . . 1,9+0,01 0,13 7,20 427 1,5-2,5-
Предглазничная ширина
Самцы прибылые 2,7+0,01 0,14 4,29 108 2,5—3,1
взрослые 2,9+0,03 0,15 5,01 28 2,7—3,2
Самки прибылые 2,8±0,01 0,13 4,86 87 2,4—3,0
взрослые 2,8+0,04 0,14 5,13 17 2,5—3,0
В целом по виду . . . 2,7+0,01 0,16 5,85 440 2,4—3,2
Межглазничная ширина
Самцы прибылые 3,9+0,01 0,14 3,50 108 3,7—4,2
взрослые 4,1+0,03 0,13 3,30 28 3,7—4,3
Самки прибылые 3,9+0,01 0,13 3,27 87 3,7—4,2
взрослые 3,9+0,03 0,11 2,69 17 3,7—4,1
В целом по виду . . . 3,9±0,01 0,14 3,57 439 3,7—4,3
Наибольшая ширина черепа
Самцы прибылые 9,8±0,03 0,28 2,83 105 9.0—10,5
взрослые 10,0+0,08 0,43 4,26 28 8,7—10,6
Самки прибылые 9,9+0,03 0,33 3,29 133 8,7—10,6
взрослые 9,9+0,05 0,19 1,88 17 9,5—10,2
В целом по виду . . . 10,0+0,20 0,40 4,09 426 8,7—10,6
Наиболъшая высота черепа
Самцы прибылые 6,3+0,03 0,32 5,04 103 5,1—7,0
взрослые 6,2+0,04 0,23 3,66 28 6,0—6,9
Самки прибылые 6,3+0,03 0,30 4,74 85 5,3—6,9
взрослые 6,2+0,06 0,26 4,26 17 5,7—6,6
В целом по виду . . . 6,3+0,01 0,29 4,64 426 5,1—7,0
215
Окончание табл. 44
1 1 2 1 3 1 4 1 5 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,8±0,03 0,26 5,39 107 4,3-5,7
взрослые 5,0±0,04 0,23 4,55 28 4,6—5,5
Самки прибылые 4,8±0,02 0,22 4,60 87 4,3—5,3
взрослые 4,9±0,03 0,14 2,79 17 4,6—5,1
В целом по виду . . . 4,8±0,01 0,24 4,92 433 4,3-5,7
Длина углового отростка
Самцы прибылые взрослые 2,1±0,01 2,1±0,03 0,14 0,13 6,79 6,51 98 26 1,4-2,5 1,7—2,3
Самки прибылые 2,1±0,02 0,14 6,73 83 1,6-2,4
взрослые 2,0±0,02 0,07 3,42 13 1,9—2,1
В целом по виду . . . 2,1+0,01 0,14 6,74 371 1,4-2,5
Характеристика. Во внешнем облике равнозубой бурозубки
обращает на себя внимание характер окраски —мех зверьков очень темный
и почти одноцветный на всем теле. Переход между насыщенной темно-бу-
рой окраской меха спины, боков и вентральной части тела неуловим.
Брюшная сторона имеет тот же окрас, но иногда несколько меньшей сте-
пени насыщенности. Встречаются особи с одинаковой интенсивностью
окраски верхней и нижней части тела. Мех живых или только что погиб-
ших зверьков с едва уловимым малиновым или сиреневым оттенком. Зим-
ний мех гуще, значительно длиннее летнего, шелковистый и блестящий.
На вентральной стороне увеличивается примесь серых тонов. Как видно
из графика колориметрической характеристики окраски меха спины рав-
нозубой бурозубки (см. рис. 63), она относится к наиболее темноок ращен-
ным видам бурозубок. О характере ее окраски можно судить также по дан-
ным табл. 45.
Мех взрослых зверьков темнее, чем у молодых, а различия в окраске
спины и нижней части тела еще менее заметны. Если сравнивать окраску
меха равнозубой и обыкновенной бурозубок, то пределы изменчивости
этого признака у первой значительно шире (см. рис. 63), что объясняется,
вероятно, большим набором ландшафтов в пределах обширного ареала
вида, занимающего территорию с разными природными условиями.
При этом обращает на себя внимание, что равнозубая бурозубка, оби-
тающая в пределах ареала обыкновенной бурозубки, темнее и менее из-
менчива (см. рис. 63). По сравнению с S. unguiculatus у нее большие зна-
чения показателя оттенка. В целом значения ее показателей полностью
укладываются в таковые равнозубой бурозубки. Нижний предел измен-
чивости оттенка у когтистой бурозубки лежит значительно выше, значе-
ния его не менее 120,6, в то время как у равнозубой бурозубки он состав-
ляет 109,4. Средний показатель оттенка (137,2) также значительно выше,
чем у обыкновенной и равнозубой бурозубок.
Хвост средних размеров (63 % от длины тела, по пропорции), близок
тс таковому обыкновенной бурозубки (65,6 %), двухцветный — сверху
‘окрашен в тон спины, а снизу с преобладанием более светлых тонов, с хо-
рошо заметным рыжевато-розовым оттенком.
216
Таблица 45
Колориметрическая характеристика подвидов Sorex isodon из разных частей ареала
Место сбора мате- риала Белизна Оттенок
Су lim п Су lim п
Приморье, запо- ведник «Кедро- вая Падь» 1,93±0,09 23,3 1,1—2,5 25 134,2± 1,49 5,6 118,1—146,5 25
Салаир (Тогучин- ский р-н) 1,96+0,12 29,9 0,9—2,6 25 129,0+2,84 11,0 115,8—155,3 25
Приобье (юг Том- ской обл., с. Ко- жевникове) 2,06+0,09 20,6 1,2—3,1 20 136,1+2,04 6,7 122,2—153,5 20
Алтай, Телецкое озеро 2,18+0,09 19,6 1,4—2,9 25 125,9+2,11 8,4 109,4—139,7 25
Камчатка, о. Па- рамушир 2,52+0,14 19,6 1,9—3,4 12 134,9+2,89 7,4 119,7—150,9 12
Приамурье, бас- сейн р. Зея 2,70+0,07 12,7 2,1-3,3 25 130,5+1,36 5,2 118,6—144,6 25
Бурятия, хр. Ха- мар-Дабан 2,95+0,08 13,1 1,7—3,5 23 130,7+1,12 4,1 119,2—140,0 23
Приамурье, низо- вье р. Амур (Хабаровский край) В ц е л о м по си- бирской части ареала .... 4,5 2,60 2,8—6,4 0,9—6,4 23 178 330,7 131,5 110,5—148,3 109,4—155,3 23 178
Для внешнего облика равнозубой бурозубки характерны приземис-
тость (по сравнению с обыкновенной, средней, тундряной), валькова-
тость, широкая голова, без заметного сужения в области глаз, относитель-
но короткий и тупой хоботок и хорошо развитая широкая грудь
(см. рис. 64). Все это в совокупности со спецификой окраски позволяет
правильно определить равнозубую бурозубку по внешнему облику, не
привлекая другие признаки.
Череп S. isodon по размерам близок к таковому обыкновенной буро-
зубки, средних размеров (27,8 % от длины тела), но несколько массивнее
и угловатей (см. табл. 32). В скульптуре черепа обращает на себя внима-
ние компактность, монолитность (рис. 64, 65).
Самобытно и весьма характерно для вида строение промежуточных
зубов. При диагностике вида заслуживают внимания следующие моменты:
1) первый верхний промежуточный зуб всегда значительно выше и массив-
нее последующих; 2) коронки промежуточных зубов по высоте и массив-
ности равномерно и последовательно уменьшаются от первого к пятому;
3) пятый зуб хорошо развит и по высоте лишь немного уступает четвер-
тому; 4) промежуточные зубы по отношению к размеру черепа мелкие;
5) коронки промежуточных зубов имеют вид равнобоких, островершин-
ных пирамид, с ровными, не выгнутыми краями; 6) промежуточные зубы
в ряду стоят свободно; 7) первый предкоренной нижней челюсти (Ргщ)
вытянут в сагиттальном направлении, иногда со слабо выраженной второй
217
Рис. 64. Равнозубая бурозубка — Sorex isodon Turov.
вершиной; 8) гипоконусы коренных верхней челюсти, как правило, не
пигментированы.
Длина зубного ряда в среднем для зверьков с территории Сибири co-
составляет 8,8±0,02 мм, длина ряда промежуточных зубов 3,0+0,01 мм.
Зубной ряд и ряд промежуточных зубов довольно длинные. Отношения:
длина зубного ряда — кондило-базальная длина черепа и то же к длине
ряда промежуточных зубов — составляют 45,1 и 15,3 % соответственно.
У обыкновенной бурозубки эти показатели таковы: 45,7 и 15,7 %, т. е.
близки (см. табл. 26).
Восходящая ветвь нижней челюсти у S. isodon хорошо развита, но бо-
лее низкая, чем у обыкновенной бурозубки, ее отношение к кондило-ба-
зальной длине черепа составляет 24,6, а у последней — 25,4 %.
В. А. Долгов [1964] указывает на специфическую структуру безымян-
ной кости равнозубой бурозубки.
Весьма видоспецифично строение головки копулятивного органа, ко-
ренным образом отличающейся от таковой других видов. По массивности
она уступает только glans penis когтистой бурозубки. Массивная головка,
почти цилиндрическая в основной трети, саблевидно изогнута в дисталь-
ной части, заканчивается опущенным вниз тупым, округлым концом, с не-
большим углублением в месте выхода мочеполового канала. При рассмот-
рении сбоку видно, что она почти одинакового диаметра по всей длине,
постепенно слабо загибается вниз и незначительно сужается лишь в по-
следней трети. При рассмотрении сверху она несколько сжата с боков, по-
степенно утончается к свободному концу. В поперечном разрезе головка
в основном овальная с более длинным вертикальным диаметром и несколь-
ко сжата с боков. Ее свободный конец не увенчан коронкой или утолще-
218
нием в виде валика. Длина свободной части копулятивного органа взрос-
лых самцов 9,7—12 (М = 10,8), толщина у корня 1,3—1,7 (М = 1,4) мм.
Равнозубая бурозубка отличается от обыкновенной по хромосомному
набору, имеющему 2п = 42 и NF = 68 [Орлов, Козловский, 1971; Орлов,
Булатова, 1983]. В хромосомном наборе 9 пар метацентрических аутосом,
4 пары субметацентрических и 7 пар субтело- и телоцентрических аутосом.
Х-хромосома телоцентрическая, Y-хромосома субметацентрическая.
У обыкновенной бурозубки из Сибири число хромосом у самцов 25,
27, у самок 24, 27 при NF = 40 [Козловский, 1969, 1970]. От средней
бурозубки (2н = 42 и NF = 68) отличается иным соотношением мета-
и субметацентрических хромосом среди первых 9 пар крупных аутосом.
Перечисленные выше признаки позволяют квалифицировать равно-
зубую бурозубку как самостоятельный вид.
Распространение. Равнозубая бурозубка — широко рас-
пространенный вид, ее ареал охватывает обширную территорию северо-
восточной Евразии, вклиниваясь на северо-западе до Финляндии вклю-
чительно, Чукотский и Камчатский полуострова, северные Курильские
острова (Шумшу, Парамушир), Охотское побережье, Приморье, Шантар-
ские острова, Сахалин. Она заселяет всю Восточную и Среднюю Сибирь,
горы юга Сибири, таежную и подтаежную зоны Западной Сибири, Урал,
Зауралье.
В Сибири равнозубая бурозубка известна из ряда районов Северного
Урала, с территории, прилегающей к низовьям Оби, из бассейна
рек Ныда, Таз, Пур (до Самбурга на север). На севере Средней Сибири
сведения о ее распространении крайне скудны. Нами она добыта на Тай-
мыре, близ оз. Турочедо, на широте пос. Потапово — г. Норильск. В Яку-
тии коллектировалась на широте Якутска, на реках Кенкеме, Лене,
в устье р. Леписке, в 50 км севернее устья Вилюя, в западных отрогах
219
Верхоянского хребта. Известна она из бассейна Олекмы, на юге Яку-
тии из бассейна рек Чуйка и Пилька. В бассейне Колымы, по данным
обширных сборов В. Г. Кривошеева, собранных в течение пяти лет, ее
не оказалось. Нами найдена в бассейне р. Омолон, в долине р. Кегали
и у пос. Сеймчан, близ Магадана. В бассейне р. Анадырь коллектирова-
лась от пос. Новый Еропол до устья этой реки, по берегам Красного
озера, в долинах рек Ламутская и Великая, против устья р. Таню-
рер, в окрестностях поселков Марково, Усть-Белая и Ваеги [Юдин,
1971а; Охотина, 1973; Кривошеев и др., 1978]. Известна с полуострова
Тайганос, а на Камчатке это — один из фоновых видов. Широко расп-
ространена в Приморье и Хабаровском крае, известна из многих пунк-
тов Приамурья и Читинской области (на юг до Сохондинского заповед-
ника включительно).
Южная граница распространения вида в Западной Сибири в деталях
не ясна. Равнозубая бурозубка добыта во многих местах в бывшем Кондо-
Сосьвинском заповеднике (заповедник «Малая Сосьва») в бассейне р. Та-
ра, по Иртышу она обычна в районах севернее Тобольска. При детальном
изучении мелких млекопитающих территорий, прилежащих к р. Омь, этот
вид отмечен лишь на правобережье. По-видимому, южная граница на се-
вере Барабинской низменности идет по междуречью рек Омь и Тара.
Южнее 55 ° с. ш- равнозубая бурозубка не встречается. Обь южная гра-
ница распространения вида пересекает по 55 ° с. ш., затем резко спускает-
ся на юг по Присалаирью, Салаиру и Кузнецкому Алатау до Алтая, вклю-
чая горы Сибири (Алтай, Саяны, Хамар-Дабан, Витимское плоскогорье)
и Приамурья. В горах Алтае-Саянской страны встречается почти повсе-
местно. В западных предгорных степях и остепненных горных районах
ее нет. Юго-западная граница распространения проходит в пределах ка-
захстанской части Алтая и Калбинских гор [Юдин и др., 1979].
Материал. 440 экз. из разных частей ареала вида.
Изменчивость. Возрастная изменчивость равнозубой буро-
зубки, как и у других видов, проявляется в стирании зубов, уплощении
черепа, стирании волосяного покрова на хвосте и лапках. Существенные
различия обнаруживаются в размерах и массе тела. Длина тела взрослых
зверьков достоверно больше (см. табл. 44), чем молодых. Эта же законо-
мерность четко прослеживается и по массе разных возрастных групп сам-
цов и самок. По другим параметрам достоверных различий нет. Возраст-
ная изменчивость проявляется в окраске меха зверьков: значительное по-
темнение и более равномерная окраска на дорсальной и вентральной сто-
роне тела у перезимовавших землероек.
Колориметрические данные большинства зверьков исследованных
выборок из разных пунктов ареала довольно плотно укладываются на гра-
фике в три категории (рис. 66). Крайние варианты в окраске характерны,
с одной стороны, для выборок из Приморья и Западной Сибири (наиболее
темные) и, с другой — из низовий Амура (наиболее светлые). Промежу-
точное положение занимают зверьки из Бурятии, верховьев Амура (р. Зея),
с Камчатки и о. Парамушир (см. рис. 66). Отличия эти весьма значитель-
ны. Так, крайние значения показатели белизны зверьков западно-сибир-
ских и нижнеамурских выборок не перекрываются. Весьма заметны раз-
личия и средних значений.
Прослеживается тенденция посветления меха зверьков в направлении
с запада на восток. Исключение составляют бурозубки из Приморья (см.
табл. 45). более темноокрашенные среди всех исследованных выборок
220
Рис. 66. Колориметрическая характе-
ристика равнозубой бурозубки из раз-
ных частей ареала.
] — Западная Сибирь; 2 — Приморье; 3 —
Бурятия, Верхнее Приамурье (р. Зея), Кам-
чатка, о. Парамушир; 4 — Нижнее При-
амурье (Хабаровский край).
и практически по этому признаку
не отличающиеся от западно-си-
бирских (см. рис. 66). Наиболее
темную окраску имеют зверьки и
из равнинных и горных районов
юга Западной Сибири, особенно с
Салаира, прилежащих районов
Приобья и Северо-Восточного Ал-
тая. Вероятно, характер окраски
обусловливается не просто прост-
ранственной изоляцией, а природными условиями конкретного района
обитания. Замечено, что на Салаире и Северо-Восточном Алтае многие
виды землероек окрашены темнее, это же отмечается и для землероек
из Приморья,
Экологическая обстановка для землероек в Приморье: таежные ланд-
шафты, жаркое лето, высокая увлажненность мест обитания — гораздо
более близка таковой на Салаире и Алтае, чем в Средней и Восточной Си-
бири, где распространена осветленная лиственничная тайга. Довольно
однородную группировку по интенсивности окраски меха составляют
зверьки этого вида из Бурятии, бассейна Зеи, с Камчатки и о. Парамушир,
окрашенных значительно светлее, чем западно-сибирские. Совершенно
обособленное положение занимают бурозубки из низовий Амура (Хаба-
ровский край), даже крайние варианты их окраски не перекрываются с та-
ковыми западно-сибирских выборок (см. рис. 66). Значения показателя
белизны для зверьков, обитающих в Сибири, колеблются от 0,9 (Салаир)
до 6,4 (низовья Амура), а оттенка — от 109,4 (Алтай, Телецкое озеро)
до 155,3 (Салаир). Наиболее темные особи с насыщенными бурыми тонами
встречаются на Салаире, самые светлые — в низовьях Амура, причем ве-
личины белизны и оттенка здесь колеблются в широких пределах. Наблю-
дается прямая зависимость: с расширением ареала вида увеличивается
диапазон изменчивости окраски.
Средние значения признака оттенка в выборках из разных частей
ареала лежат в близких пределах — 130,3—134,2, а значения белизны
колеблются от 1,9 до 4,5 (см. табл. 45). Верхние и нижние границы измен-
чивости оттенка достаточно близки в разных выборках (см. рис. 66).
Индивидуальная изменчивость окраски сводится главным образом
к наличию редких аномалий: встречаются зверьки с белой кисточкой на
хвосте.
Стойких показателей, имеющих географическую направленность из-
менчивости, у вида обнаружить не удалось. Статистический анализ много-
численных выборок из разных районов Западной, Средней Сибири, Яку-
тии, гор юга Сибири, с Чукотки, Камчатки, Охотского побережья и из
Приморья показал, что отклонения от средних значений отдельных выбо-
рок из разных частей ареала и от общей видовой характеристики вида ста-
221
тистически недостоверны, что свидетельствует о его мономорфизме. Эти
данные согласуются и с материалами других исследователей.
По материалам Ф. Р. Штильмарка и В. А. Долгова [1974], изучавших
биологию равнозубой бурозубки (у этих авторов S. centralis) в районе
г. Комсомольска (окр. пос. Пивень), она характеризуется следующими
морфометрическими показателями: кондило-базальная длина черепа
у прибылых зверьков колеблется в пределах 18,8—19,5 (М = 19,2±0,07),
у перезимовавших зверьков — 18,6—20,2 (М = 19,5+0,16) мм; длина
зубного ряда соответственно 8,3—8,6 (М = 8,4 ± 0,03) и 7,8—8,6 (М =
— 8,3±0,08) мм. Эти данные близки к нашим материалам (см. табл. 44).
В зубной системе крайне редко проявляются отклонения в положе-
нии зубов, олиго- и полиодонтия. Так, у зверьков из заповедника «Кед-
ровая Падь» (№ 1174, кол. ЗМБИ) пятый промежуточный зуб оказался
задвинутым за первый многовершинный зуб, а соответствующий зуб с ле-
вой стороны отсутствовал. У зверька с Камчатки (№ 213, кол. ЗМБИ)
в правой верхней челюсти между вторым и третьим промежуточными зу-
бами оказался дополнительный (шестой по счету), по размерам заметно
ниже и уже соседних. Такие или подобные отклонения крайне редки и
встречаются единично в выборках из нескольких тысяч зверьков.
Заметки по систематике. Равнозубая бурозубка отно-
сится к числу сравнительно недавно описанных видов [Туров, 1924], но
за столь незначительный промежуток времени вокруг нее разгорались
серьезные споры и высказывались взаимоисключающие мнения. Вид пере-
жил «взлеты» и «падения» во взглядах систематиков. Достаточно убедитель-
но это положение можно иллюстрировать следующими примерами. За пе-
риод в восемь лет исследователи относили его к пяти разным видам:
С. У. Строганов [1957] — к S. araneus; Скарен [Skaren, 1963,1964], Халк-
ка, Скарен [Halkka, Skaren, 1964] — к 5. unguiculatus; В. А. Долгов
[1964] — к 5. centralis; Б. С. Юдин [1965а, 1971а], Сиивонен [Siivonen,
1965, 1969] и др.— к S. isodon. Больше того, по Скарену [Skaren, 1963,
1964], равнозубая бурозубка не только один вид с 5. unguiculatus,
но и синоним североамериканского вида 5. obscurus. Все это привело к пу-
танице в оценке таксономического положения и объема рассматриваемого
таксона.
Впервые на таксономическую обособленность равнозубой бурозубки
обратил внимание С. С. Туров [1924], выделив, ее главным образом опи-
раясь на структурные особенности зубной системы, в качестве natio с си-
бирской обыкновенной бурозубкой (S. araneus tomensis isodon Turov).
Материалы, послужившие для описания новой формы, были собраны в бар-
гузинской тайге. С. С. Туров [1924] точно подметил и описал характер-
ные черты, свойственные этой бурозубке, сравнивая ее с обыкновенной,
но квалифицировал их не как видовые. В этом же качестве принимал ее
и С. И. Огнев [1928], ссылаясь на описание С. С. Турова [1924], и вероят-
но, не имея в руках оригинальных материалов. В 1933 г. Гудвин [Good-
win, 1933] собрал на Сахалине серию зверьков, среди которых оказались
особи, относящиеся к неизвестной ему форме. Он описал новый вид (5. gra~
vest), по морфологическим признакам тела, конфигурации зубов и харак-
теру окраски полностью идентичный с равнозубой бурозубкой и являю-
щийся ее синонимом.
Автор первоописания в более поздней работе [Туров, 1936] рассмат-
ривал равнозубую бурозубку уже в качестве подвида (S. isodon isodon).
Одновременно С. У. Строганов [1936], изучавший фауну мелких млеко-
222
питающих Валдайской возвышенности, собрал серию зверьков равнозу-
бой бурозубки и, сравнив их с другими таксонами рода, пришел к выводу
о ее видовой самостоятельности (S. isodon rutenus Strog.). Позднее в ра-
ботах сторонников укрупнения видов равнозубая бурозубка как вид вновь
«исчезла» и рассматривалась в составе обыкновенной бурозубки — 5. ara-
neus [Бобринский и др., 1944,1965; Ellerman, Morrison-Scott, 1951; и др.].
Эти авторы не признали равнозубую бурозубку даже в качестве подвида
и совершенно немотивированно отнесли ее (из Забайкалья) к 5. araneus
roboratus. С. У. Строганов [1957], впервые выделивший равнозубую буро-
зубку в самостоятельный вид, в монографии по сибирским насекомоядным
отказался от первоначальных взглядов и рассматривал 5. isodon уже как
подвид обыкновенной бурозубки (S. a. isodon Turov), хотя им первона-
чально были правильно интерпретированы ее видовые отличия от 5. ara-
neus.
Как уже говорилось выше, Скарен [Skaren, 1963, 1964], Халкка
и Скарен [Halkka, Skaren, 1964] без всяких оснований, умозрительно,
объединили равнозубую бурозубку с 5. unguiculatus. Последняя имеет ряд
четких признаков, не позволяющий спутать ее с другими видами. Прежде
всего это — строение зубной системы (второй промежуточный зуб меньше
первого и третьего, у равнозубой промежуточные зубы последовательно
убывают от первого к пятому), специфическое строение копулятивного
органа (см. рис. 40), наличие длинных когтей на передних лапках и др.
Объединение этих четко очерченных видов, отличающихся рядом дискрет-
ных признаков, не имеет под собой фактической основы.
Впервые на необходимость восстановления видовой самостоятельности
равнозубой бурозубки указал И. И. Стогов [1961]. В. А. Долгов [1964],
признавая видовую самостоятельность равнозубой бурозубки по сущест-
ву, отнес ее к 5. centralis Thomas. Под этим названием она фигурирует
во многих публикациях этого исследователя и ряда последовавших за ним
авторов. Ошибка была допущена из-за нечеткого диагноза 5. centralis,
описанного Томасом [Thomas, 1971]. «Вид» этот оказался нереальным,
а описание относится к крупным популяциям средней бурозубки ($. cae-
cutiens), обитающим на Саянах и в окрестностях Байкала. Как сообщил
мне В. А. Долгов (в личной переписке), ознакомившись с типовым экзем-
пляром, послужившим для описания S. centralis, хранившимся в Лон-
доне в коллекции Британского музея, он убедился в неральности этого
таксона (синоним 5. caecutiens), ничего общего не имеющего с равнозубой
бурозубкой.
В наших работах [Юдин, 19656, 1971а] впервые был опубликован ри-
сунок glans penis равнозубой бурозубки и дана оценка видовых призна-
ков. Сиивонен [Siivonen, 1965, 1969] в обстоятельных исследованиях, по-
священных равнозубой бурозубке, убедительно показал несостоятель-
ность высказываний, отрицающих ее видовой статус и объединение
с S. araneus, S. unguiculatus, S. centralis. Детальное исследование систе-
матики, проведенное в нашей работе по насекомоядным Сибири [Юдин,
1971а], еще раз подтвердило правомочность этих выводов.
Прослеживаются совершенно четкие кариологические различия в
хромосомном наборе равнозубой бурозубки, обыкновенной и когтистой.
Число хромосом у 5. isodon значительно больше, чем у 5. araneus. У рав-
нозубой бурозубки число хромосом у самок 40 и у самцов 41 [Halkka,
Skaren, 1964], а по В. Н. Орлову и А. И. Козловскому [1971] число хро-
мосом у обыкновенной бурозубки колеблется от 20 до 31 [Воронцов, 1958;
223
Tobias, 1957; Matthey, 1958; Ford et al., 1957; Ford, Hamerton, 1958; Bor-
gaoncar, 1965; Halkka, Skaren, 1964; Meylan, 1964, 1965; Козловский,
1969, 1970, 1972]. У 5. unguiculatus число хромосом 42 [Takagi, Fujimaki,
1966]. Скарен [Skaren, 1963, 1964] же установил существенные различия
мускусного запаха, издаваемого обыкновенной и равнозубой бурозубками.
В настоящее время видовой статус равнозубой бурозубки у специалис-
тов не вызывает сомнений. Вид имеет самобытные габитуальные черты,
окраску, структуры и соотношения зубов, форму копулятивного органа
и хромосомный набор.
Внутривидовая систематика равнозубой бурозубки не разработана.
Описаны следующие подвиды: номинальная форма из баргузинской тай-
ги — 5. i. Isodon Turov, 1936; с Северо-Восточного Алтая — 5. г. prin-
ceps Scalon et Rajevski, 1940; из европейской части СССР (Валдайская
возвышенность) — S. i. rutenus Strog., 1936.
Колориметрические исследования, проведенные нами, позволяют бо-
лее реалистично оценить внутривидовую изменчивость. Совершенно оче-
видно, что зверьки из Бурятии, Читинской области, Камчатки и о. Пара-
мушир значительно светлее и могут быть выделены в самостоятельный
подвид. Для окончательного решения этого вопроса необходимо прове-
дение дополнительных колориметрических исследований равнозубой бу-
розубки из типичного местообитания (баргузинская тайга). Совершенно
очевидна подвидовая самостоятельность светлоокрашенных землероек
из низовий Амура и темных зверьков с юга Приморья, изолированных
от таковых в Западной Сибири более светлоокрашенными популяциями
Средней и Восточной Сибири.
Sorex gracillimus Thomas, 1907 —
Дальневосточная бурозубка
Голотип. Томас [Thomas, 1907] по черепу и задней лапке зверь-
ка, добытого в окрестностях Дарине, близ Корсакова (Сахалин), описал
подвид Sorex minutus gracillimus. Голотип хранится в Британском музее.
Б. С. Юдин [1965а] исследовал серию собранных Н. Ф. Реймерсом
в разных районах Сахалина зверьков и описал вид 5. gracillimus.
Паратипы. Хранятся в Зоологическом музее АН СССР (Ленин-
град) и в зоологическом музее Биологического института Сибирского от-
деления АН СССР (Новосибирск).
Диагноз. Мелкая, длина тела 49—64 (М = 54,9±0,94) мм, чуть
крупнее 5. minutus, длиннохвостая — 39—47 (М = 43,5±0,42) мм — зем-
леройка. Кондило-базальная длина черепа 15,6—16,5 (М — 15,7±
±0,05) мм. Ряд промежуточных зубов сильно растянут (36,8 % от длины
зубного ряда, или 15,9 % от кондило-базальной длины черепа), зубы низ-
кие, с чрезвычайно расширенными в сагиттальном направлении основа-
ниями коронок и широко расставленными вершинами, последовательно
уменьшаются от первого к пятому (см. рис. 68).
Измерения. См. в табл. 46, 47.
Характеристика. Зверек стройного телосложения с сильно
вытянутым, тонким и подвижным хоботком. При внешнем осмотре обра-
щает на себя внимание длинный хвост, составляющий 79,2 % длины
тела (39—47; М = 43,5+0,42 мм). По этому показателю уступает лишь
S. mirabilis. Хвост с хорошо развитой кисточкой на конце, покрыт
224
Таблица 46
Морфометрическая характеристика Sorex gracillimus
Возрастная и половая группа СТ Cv п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 52,5+0,76 2,14 4,07 8 50—57
взрослые 59,2 + 1.64 4.02 6,80 6 53—64
Самки прибылые 52,1±0,83 2,19 4.21 7 49—55
взрослые 61,5+2,50 3,54 5,75 2 59—64
В целом по виду . . . 54,9±0,94 4,52 8,23 23 49—64
Длина хвоста
Самцы прибылые 44,0+0,70 2,00 4,55 8 41—47
взрослые 42,0+0,91 2,23 5,29 6 39—45
Самки прибылые 43,6+0,61 1,62 3,71 7 41—45
взрослые 45,5+0,50 0,71 1,55 2 45—46
В целом по виду . . . 43,5± 0,04 2,02 4,64 23 39—47
Длина ступни
Самцы прибылые 11,1+0,24 0.68 6,13 8 10—12
взрослые 10,8+0.17 0,41 3,77 6 10—11
Самки прибылые 11,4+0,30 0,79 6,88 7 10—12
взрослые 12,0+0,0 0,0 0,0 2 —
В целом по виду . . . 11,2+0,14 0,69 6,13 23 10—12
Масса тела
Самцы прибылые 3,5±0,13 0,32 9,04 6 3,0—4,0
взрослые 4,7±0,11 0,26 5,53 6 4,5—5,0
Самки прибылые 3,7±0,12 0,27 7,40 5 3,5—4,0
взрослые 6,5+0,0 0,0 0,0 1 —.
В целом по виду . . . 4,1+0,20 0,83 20,25 18 3,0—6,5
Кондило-базальная длина черепа
Самцы прибылые 15,9+0,08 0,24 1,50 8 15,6—16,3
взрослые 15,9+0,08 0,21 1,30 6 15,6—16,2
Самки прибылые 16,0+0,10 0,26 1,63 7 15,7—16,5
взрослые 15,8+0.20 0,28 1,79 2 15,6—16,0
В целом по виду . . . 15,7+0,04 0,32 1,91 45 15,6—16,5
Длина рострума
Самцы прибылые взрослые 4,4+0,04 4,3+0,01 0,11 0,04 2,55 0,95 8 6 4,2—4,6 4,3-4,4
Самки прибылые 4,4±0,03 0,09 2,04 7 4,3—4,5
взрослые 4,5±0,10 0,14 3,14 2 4,4—4,6
В целом по виду . . . 4,2+0,04 0,27 6,46 45 4,2—4,6
Длина мозговой части черепа
Самцы прибылые 8,8+0,07 0,21 2,38 8 8,5—9,1
взрослые 8,7+0,05 0,13 1,53 6 8,6—8,9
Самки прибылые 8,9±0,08 0,21 2,37 7 8,5—9,1
взрослые 8,8±0,05 0,07 0,80 2 8,7—8,8
В целом по виду . . . 8,7±0,04 0,24 2,75 45 8,5—9,1
Длина верхнего ряда зубов
Самцы прибылые 6,7+0,05 0,13 1,91 8 6,6—7,0
взрослые 6,7±0,03 0,08 1,12 6 6,6—6,8
Самки прибылые 6,8+0,05 0,12 1,80 5 6,6—6,9
взрослые 6,6±0,10 0,14 2,14 2 6,5—6,7
В целом по виду . . . 6,8±0,02 0,13 1,87 43 6,5—7.0
225
Окончание табл. 46
1 2 з 4 5 6
Длина ряда промежуточных зубов
Самцы прибылые 2,5±0,03 0,07 2,80 8 2,4—2,6
взрослые 2,5±0,04 0,09 3,59 6 2,4—2,6
Самки прибылые 2,6±0,02 0,04 1,60 6 2,5—2,6
взрослые 2,5+0,0 0,0 0,0 2 —
В целом по виду . . . 2,5±0,01 0,08 3,30 44 2,4—2,6
Ширина рострума
Самцы прибылые 1,3+0,02 0,05 3,49 8 1,3—1,4
взрослые 1,4±0,02 0,04 2,88 6 1,4—1,5
Самки прибылые 1,3±0,01 0,05 3,67 7 1,3—1,4
взрослые 1,4±0,0 0,0 0,0 2 —
В целом по виду . . . 1,4±0,01 0,07 4,86 45 1,3—1,5
Предглазничная ширина
Самцы прибылые 2,0±0,02 0,05 2,67 8 1,9—2,1
взрослые 2,0±0,0 0,0 0,0 6 —
Самки прибылые 2,0±0,0 0,0 0,0 7 —
взрослые 2,1+0,0 0,0 0,0 2 —
В целом по виду . . . 2,0±,001 0,06 2,84 45 1,9—2,1
Межглазничная ширина
Самцы прибылые 2,9±0,03 0,09 3,25 8 2,7—3,0
взрослые 2,9±0,03 0,06 2,18 6 2,8—3,0
Самки прибылые 2,9±0,03 0,09 3,11 7 2,8—3,0
взрослые 2,9±0,0 0,0 0,0 2 —
В целом по виду . . . 2,9+0,02 0,10 3,50 45 2,7—3,0
Наибольшая ширина черепа
Самцы прибылые 7,5±0,06 0,16 2,12 8 7,2—7,7
взрослые 7,6±0,09 0.21 2,75 6 7,3—7,8
Самки прибылые 7,6+0,8 0,21 2,78 7 7,4—8,0
взрослые 7,5±0,05 0,07 0,95 2 7,4—7,5
Б целом по виду . . . 7,5±0,03 0,21 2,73 45 7,2—7,8
Наибольшая высота черепа
Самцы прибылые 4,9±0,06 0,18 3,64 8 4,5—5,0
взрослые 4,6±0,08 0,19 4,20 6 4,3—4,8
Самки прибылые 4,9±0,06 0,15 3,06 7 4,7—5,1
взрослые 4,6+0,15 0,21 4,66 2 4,4—4,7
В целом по виду . . . 4,8±0,03 0,21 4,42 45 4,3-5,1
Высота восходящей ветви нижней челюсти
Самцы прибылые 2,9±0,03 0,09 3,14 8 2,8—3,0
взрослые 2,9±0,04 0,09 3,04 5 2,8—3,0
Самки прибылые 3,0±0,03 0,07 2,3 7 2,9-3,1
взрослые 2,9+0,0 0,0 0,0 1 —
В целом по виду . . . 2,9±0,02 0,10 3,29 43 2,8-3,1
Длина углового отростка
Самцы прибылые 1,9±0,03 0,08 4,36 8 1,8—2.0
взрослые 1,8±0,04 0,09 4,86 5 1,7—1,9
Самки прибылые 1,9±0,03 0,08 4,05 7 1,8—2,0
взрослые 1,8±0,0 0,0 0,0 1 —
В целом по виду . . . 1,9+0,02 0,10 5,22 38 1,7—2,0
15 Б. С. Юдин
226
Морфометрическая характеристика
Признак Хабаровский край, Приморье Сахалин
М^7ПХ а Су . п lim Xf±mx,, а
Длина тела 47,0±2,65 4,58 9,74 3 43-52
хвоста 37,1±0,67 1,15 3,11 3 36—38 — —
ступни 10,4±0,3 0,51 4,90 4 10—11 —
Масса тела —. —. — —
Длина кондило-базальная черепа 15,7±0,06 0,24 1,53 18 15,4—16,2 15,5±0,06 0,25
рострума 4,3±0,08 0,33 7,70 18 3,9—5,4 3,9±0,03 0,11
мозговой части 8,7±0,06 0,26 2,99 18 8,4—9,1 8,5±0,05 0,20
верхнего ряда зубов 6,7±0,06 0,25 3,73 18 5,9—6,9 6,8±0,03 0,14
ряда промежуточ- ных зубов 2,7+0,02 0,07 2,59 18 2,6—2,8 2,5±0,02 0,10
Ширина рострума 1,4±0,03 0,11 7,86 18 1,2—1,6 1,4±0,01 0,08
предглазничная 1,9±0,02 0,09 4,74 18 1,8—2,1 1,9±0,01 0,05
межглазничная 2,8±0,03 0,14 5,0 18 2,4—3,0 2,9±0,03 0,12
наибольшая черепа 7,3±0,06 0,26 3,56 18 6,7—7,8 7,5±0,05 0,24
Наибольшая высота че- репа 4,7+0,09 0,37 7,84 18 4,0—5,2 4,8±0,04 0,17
Высота восходящей вет- ви нижней челюсти 2,9±0,02 0,09 3,10 16 2,7—3,0 2,9+0,02 0,09
Длина углового отростка 1,8±0,04 0,13 7,22 10 1,7—2,0 2,0±0,03 0,11
торчащими в стороны удлиненными волосами, слабо двухцвет-
ный (рис. 67, 68).
Окраска меха на дорсальной стороне тела темная, буровато-оливко-
во-серая. У перезимовавших зверьков довольно явственно виден чепрак.
Зимний мех ярче — кофейно-бурый. Бурые тона простираются на плечи
и бедра. Значения показателей белизны и оттенка дорсальной стороны
М ~ 4,1 ±0,13 и М = 132,4±1,12. Вентральная сторона покрыта более
светлым серовато-белесым мехом с бурым оттенком. Окраска хвоста
сверху в тон спины темная, на нижней стороне светлая. По колоримет-
рической характеристике ее параметры целиком укладываются в таковые
малой бурозубки (рис. 69).
Череп по величине незначительно крупнее (М = 15,7±=0,05), чем
у малой бурозубки (М = 15,1 ±0,03), вытянутый, узкий в ростральной
части, со вздутой мозговой капсулой (см. табл. 32). Все эти показатели
перекрываются, но хорошо улавливаются на серийном материале. Отно-
шение длины рострума к кондило-базальной длине черепа у S. gracillimus
26,8, у S. minutus 25,8 %.
Передний верхний резец относительно крупный. Его второй (внут-
ренний) зубец хорошо развит. По высоте первый резец, первый и второй
промежуточные зубы почти равны между собой. Коронки промежуточных
конусовидные, низкие, сильно вытянуты в сагиттальном направлении
227
Таблица 47
Sorex gracillimus из разных частей ареала
Сахалин о. Кунашир В целом по виду
Су п lim М±тх а Су п lim 1И±тх а Су п Пт
54,9±0,94 4,52 8,23 23 49—64 54,9±0,94 4,52 8,23 23 49—64
, — — — 43,5±0,42 2,02 4,64 23 39—47 43,5±0,42 2,02 4,64 23 39—47
— — — И,2±0,14 0,69 6,13 23 10—12 11,2±0,14 0,69 6,13 23 10—12
— — — 4,1±0,20 0,83 20,25 18 3—6,5 4,1±0,20 0,83 20,24 18 3,0-6,5
1,64 20 15,0—15,9 15,9±0,05 0,24 1,50 25 15,6—16,5 15;7±0,05 0,30 1,91 45 15—16,5
2,88 20 3,8—4,2 4,4±0,02 0,10 2,23 25 4,2—4,6 4,4±0,04 0,27 6,46 45 3,8-4,6
2,35 20 8,1—8,8 8,8±0,04 0,19 2,18 25 8,5-9,1 8,7±0,04 0,24 2,75 45 8,1—9,1
2,03 20 6,5—7,0 6,7±0,02 0,12 1,70 23 6,5—7,0 6,8±0,02 0,12 1,87 43 6,5—7,0
3,91 20 2,4—2,7 2,5±0,01 0,07 2,77 24 2,4—2,6 2,5±0,01 0,08 3,30 44 2,4—2,7
5,60 20 1,2—1,5 1,4±0,01 0,06 4,30 25 1,3—1,5 1,4±0,01 0,07 4,86 45 1,2-1,5
2,63 20 1,9—2,0 2,0±0,01 0,04 2,19 25 1,9—2,1 2,0±0,01 0,06 2,83 45 1,9-2,1
4,34 20 2,7—3,0 2,9±0,01 0,08 2,71 25 2,7—3,0 2,9±0,02 0,10 3,50 45 2,7—3,0
3,14 20 7,1—7,8 7,5±0,04 0,18 2,39 25 7,2—8,0 7,5 + 0,03 0,20 2,73 45 7,1—8,0
3,60 20 4,4—5,0 4,8±0,05 0,24 5,05 25 4,3—5,2 4,8±0,03 0,21 4,42 45 4,3—5,2
3,05 20 2,7—3,0 3,0±0,02 0,09 2,87 23 2,8—3,1 2,9±0,02 0,10 3,29 43 2,7—3,1
5,66 15 1,8—2,2 1,9±0,02 0,06 4,49 23 1,7—2,0 1,9±0,02 0,10 5,21 38 1,7—2,2
и сидят свободно в относительно длинном зубном ряду. Индекс ряда
промежуточных зубов к кондило-базальной длине черепа у дальневосточ-
ной бурозубки самый большой среди сибирских представителей рода
и составляет 36,8 % (см. табл. 26). Вершины коронок промежуточных
зубов острые, широко расставлены и несколько сдвинуты вперед по
отношению к оси зуба. Вертикальные оси трех первых промежуточных
зубов почти параллельны между собой и направлены перпендикулярно
к оси челюсти, в то время как четвертый зуб имеет наклон кзади (внутрь
ротовой полости), а пятый — вперед.
Строение копулятивного органа дальневосточной бурозубки совер-
шенно иное, чем у малой. При почти равных размерах тела зверьков
этих видов, даже при несколько больших у дальневосточной бурозубки,
чем у малой, размеры копулятивного органа последней непропорцио-
нальны: у 5. gracillimus он в 2 раза короче, чем у 5. minutus, и при этом
в 1,5 раза толще у основания. Отношение длины glans penis к длине
тела составляет у дальневосточной бурозубки 8,8, у малой — 17,8%.
Если у малой бурозубки член вытянутый, цилиндрический, с тупо за-
канчивающимся свободным концом, увенчанным обрамляющим его
валиком, то у дальневосточной он конический сжатый с боков и заострен-
ный на свободном конце.
15*
228
Рис. 67. Дальневосточная бурозубка'— Sorex gracillimus Thomas.
Дальневосточная бурозубка также стройна и изящна, как малая.
Ее масса колеблется от 3—6,5 (М = 4,1) г, у малой — 2,5—5 (М = 3,4) г.
Отношение массы тела к длине прибылых зверьков выражается пока-
зателем 6,8, а у малой — 6,5.
Распространение. Ареал дальневосточной бурозубки охва-
тывает крайний юго-восток азиатской части СССР: Приамурье, Охотское
Рис. 68. Череп дальневосточной бурозубки.
229
Рис. 69. Сравнительная характеристи-
ка окраски дальневосточной и малой
бурозубок.
1 — S. gracillimus', 2 — S. minutus.
делизна
низовьях Амура близ пос. Пивень
побережье, Приморье, Шантарс-
кие острова, Сахалин, Южные Ку-
рилы, а также Хоккайдо и Се-
верную Корею.
В очерченных пределах она
известна из ряда пунктов. В Ха-
баровском крае зарегистрирована
в районе ст. Лазо и Киселевка, з
(коллектор Р. Ф. Штильмарк). Зверьки добыты в верховьях р. Кия
(В. Н. Добросельский), в бассейне Уссури, у г. Советская Гавань
(Аникеев). В Северном Приамурье коллектировалась сотрудниками на-
шей лаборатории Л. И. Галкиной и В. В. Николаевым в отрогах
хр. Тукурингра по рекам Уркан, Гилюй, Эракингра (близ Зейского
заповедника) и в Нижнем Приамурье на р. Амгунь. На западном по-
бережье Охотского моря этот вид известен из окрестностей Аяна [Рей-
мерс, Алина, 1967].
Самая северная точка нахождения вида — долина р. Кухтуй, выше
Охотска, близ с. Резиденция (около 59° с. ш.) [Юдин и др., 1976]. В бас-
сейне Амура ее собирала в ряде пунктов С. С. Водовоз (Ульчский, Ха-
баровский, Комсомольский, им. Лазо, Верхнебуреинский р-ны). В При-
морье добыта близ г. Ванино (Р. Ф. Штильмарк), в заповедниках —
быв. Супутинском (Уссурийском) и «Кедровая Падь» (А. П. Новик).
На побережье Татарского залива ее добывал Н. П. Крылов. В верховьях
Имана, близ оз. Кизи, найдена Н. А. Формозовым. На о. Бол. Шантар
бурозубка собрана В. В. Кучеруком и А. Г. Кузнецовым. На Сахалине
обнаружена рядом исследователей (Н. Ф. Реймерс, Г. А. Воронов) в райо-
не Даринэ, в центральной части Сахалина, на р. Кеури, у поселков Но-
во-Александровск, Куликово, Оха, и г. Южно-Сахалинска.
На о. Кунашир в ряде пунктов колллектировалась нами, а на о. Ши-
котан — А. И. Гизенко.
Материал. 235 экз., Сахалин; 25 экз., о. Кунашир; 18 экз.,
Приморье, Хабаровский край.
Изменчивость. Возрастная изменчивость проявляется, как
И У других видов бурозубок, в стирании волосяного покрова на лапках,
хвосте и в снашивании зубов. С возрастом меняется окраска зверьков:
у перезимовавших особей темный мех спины довольно четко отделен
от светлых боков. У прибылых землероек эта граница более расплывча-
та. Зимний мех длиннее и более насыщен бурыми тонами. Хвост, ступни
и кисти с вытертым волосяным покровом. Хорошо прослеживается воз-
растная изменчивость интенсивности окраски. Так, у перезимовавших
бурозубок значения показателя белизны колеблются в пределах 2,7—
4,3 (М = 3,5), а оттенок 128—151 (М — 135,8), в то время как для
прибылых они составляют соответственно 3,8—5,2 (М = 4,4) и 125—
140 (М = 131), т. е. перезимовавшие особи более темно-бурые, нежели
прибылые зверьки. Длина тела перезимовавших особей достоверно боль-
ше, чем у прибылых (см. табл. 46).
230
Половых различий у одновозрастных групп зверьков нет или они
незначительны: прибылые самцы в длину (М = 52,5^=0,76) такие же,
как и самки (М = 52,1=1=0,83). Можно считать, что и перезимовавшие
самцы (М = 59,2=1=1,64) немного отличаются по этому признаку от са-
мок того же возраста (М = 61,5=1=2,5). Что касается размеров черепа,
то достоверных различий между особями разного возраста и пола не
обнаружено почти по всем изученным параметрам (см. табл. 46). Следует
обратить внимание только на хорошо заметные различия в высоте чере-
па: у перезимовавших зверьков череп достоверно ниже по сравнению
с молодыми.
Четко проявляется возрастная изменчивость в стертости зубов.
Уже осенью у зверьков первого выводка заметна стертость вершин про-
межуточных зубов. От осени к весне этот процесс идет довольно равно-
мерно, но сохраняются различия, особенно в начале зимы, между зверь-
ками разных генераций — у бурозубок более ранних пометов изношен-
ность зубов заметнее. Весной эти различия практически нивелируются.
В результате стирания промежуточных зубов без того вытянутые в са-
гиттальном направлении коронки у старых особей кажутся крайне низ-
кими и еще более вытянутыми.
Индивидуальная и географическая изменчивость промежуточных
зубов незначительна и проявляется у небольшого числа особей. Полио-
донтия и олигодонтия не отмечены. Первый зуб несколько массивнее
последующих, второй выше третьего, нередко они равны между собой,
а четвертый уступает предыдущему и почти равен пятому, который всег-
да хорошо развит и не задвинут за парастиль первого многовершинного.
Второй и третий зубы наиболее вариабельны по размерам. Заметной
географической изменчивости (материк, острова Сахалин, Кунашир,
Хоккайдо) не обнаружено (см. табл. 47).
Заметки по систематике. Первым на самобытность даль-
невосточной бурозубки обратил внимание Томас [Thomas, 1907]. Имея
в своем распоряжении только череп и ступню одного зверька, он сумел
даже на столь скудном материале уловить самобытные черты организа-
ции бурозубки и на этом основании выделил ее в подвид, отнеся к малой
бурозубке — S. minutus gracillimus. В этом качестве она принималась
всеми последующими исследователями [Огнев, 1928, 1935; Строганов,
1957; Гуреев, 1963; Бобринский и др., 1965; и др.], что было обусловлено
главным образом малочисленностью материалов, имевшихся в распоря-
жении исследователей, и общей слабой разработанности систематики
землероек. Мелкие землеройки, заселявшие Приамурье, Приморье,
Сахалин, Шантарские, Курильские острова и Хоккайдо, рассматривались
как подвид малой бурозубки, якобы имеющей дизъюнктивный ареал
в районе между Байкалом и Приамурьем.
Исследование большой серии землероек с Сахалина (235 экз.), лю-
безно переданной Н. Ф. Реймерсом, позволило нам детально охаракте-
ризовать эту форму и выделить в самостоятельный, таксономически
хорошо очерченный вид, названный по приоритету 5. gracillimus Thomas
[Юдин, 1965а]. Основой для его диагностирования как самостоятельной
формы видового ранга послужили описанные выше самобытные черты
строения glans penis, особенности строения черепа, форма, размеры и
своеобразное соотношение промежуточных зубов.
Головка копулятивного органа совершенно не схожа с таковой у
малой бурозубки. Она коренным образом отличается по форме и разме-
231
рам. Этот орган у описанного нами вида значительно короче, толще,
имеет абсолютно несхожие форму и детали строения (см. рис. 40). По
форме и размерам glans penis дальневосточная бурозубка ближе
к 5. raddei, S. roboratus, S. asper Thomas, чем к S. minutus.
Различия с малой бурозубкой, к которой ранее ее относили, в ос-
новном следующие:
Признак S. minutus
Отношение длины хвоста 71
к длине тела, %
Второй промежуток- Ниже третьего
ный зуб верхней че-
люсти
Зубцой ряд Относительно короткий
Индекс длины ряда 14,6, а к длине зубного ря-
прОмежуточных зу- да — 34,3
бов к кондило-ба-
зальной длине чере-
па, %
Восходящая ветвь Относительно высокая. Ее
нижней челюсти отношение к кондило-ба-
зальной длине черепа со-
ставляет 19,9 %
Копулятивный орган Цилиндрический, длинный,
8,3—11,4 (М = 9,3) мм,
относительно тонкий (1,0—
1,1) мм с тупым, увенчан-
ным валиком, свободным
концом
S. gracillimus
79,2
Выше или равен третьему
Длинный, зубы в челюсти
сидят свободно
15,9, а к длине зубного ря-
да — 36,8
Относительно низкая. Ее от-
ношение к кондило-базаль-
ной длине черепа состав-
ляет 18,5 %
Конический, короткий, 4,0—
5,5 (М = мм, относи-
тельно толстый у основа-
ния (1,4—1,6 мм) с заост-
ренным свободным кон-
цом, без валика
Более поздние исследования новых материалов подтвердили реаль-
ность существования нового вида [Долгов, Лукьянова, 1966; Гуреев,
1971, 1979; Юдин, 1971а], ставшего общепризнанным.
Sorex roboratus Hollister, 1913 —
Бурая бурозубка
Голотип. Вид описан по одному экземпляру с дефектным чере-
пом, добытым на Алтае (пос. Тапуча, близ Телецкого озера), сГ, ad.,
7/VIII 1912 г. Коллекция Национального музея США, № 175436.
Д И а г н о з. Крупная бурозубка, уступающая в размерах только
5. mirabilis. Длина тела 58—89 (М = 72,0=t0,45) мм, кондило-базаль-
ная длина черепа 19,0—21,8 (М = 20,3±0,03) мм. Телосложение тяже-
лое, голова длинная, с постепенным переходом от мозговой области в
чрезвычайно вытянутый, широкий хоботок. Окраска меха двухцветная,
переход постепенный от темно-бурой спины к более светлому, серовато-
буро-белому брюшку. Череп вытянут в длину, относительно узкий в
мозговой части (индекс к кондило-базальной длине черепа составляет
46,5, у 5. araneus — 49,6 %) и широкий в ростральной. Первый верхний
резец крупнее этого зуба других бурозубок, вытянут вперед. Вторая
(задняя) вершина его ниже первой и равна высоте первого промежуточ-
ного зуба. Зубы относительно черепа мелкие, в зубном ряду сидят сво-
бодно. Основания коронок промежуточных зубов вытянуты в сагиттальном
направлении, вследствие чего вершины широко расставлены. Второй про-
межуточный зуб выше первого, его вершина по высоте находится на одном
232
Таблица 48
Морфометрическая характеристика Sorex roboratus
Возрастная и половая группа М±тх а CV п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 69,7± 0,92 6,02 8,64 43 58—82
взрослые 76,7± 1,64 6,55 8,54 16 62—87
Самки прибылые 71,5+0,63 5,06 7,08 61 60—82
взрослые 76,0± 5,05 10,1 13,29 4 68—89
В целом по виду . . . 72,0±0,45 6,05 8,41 181 58—89
Длина хвоста
Самцы прибылые 38,3+0,66 4,30 11,24 43 31—50
взрослые 35,4+0,75 2,99 8,47 16 30-41
Самки прибылые 37,2+0,47 3,69 9,92 61 30—48
взрослые 37,5+1,66 3,32 8,84 4 33—41
В целом по виду . . . 37,7+0,28 3,79 10,05 180 30—50
Длина ступни
Самцы прибылые взрослые 13,2+0,18 13,4+0,21 1,14 1,82 8,61 6,08 40 15 11—15 12—15
Самки прибылые 13,2+0,12 0,86 6,54 52 11—15
взрослые 13,9±0,43 0,85 6,15 4 13-15
В целом по виду . . . 13,4+0,07 Масса 0,95 тела 7,10 167 11—15
Самцы прибылые 9,2+0,25 1,54 16,64 39 5,1—13,8
взрослые 11,11+0,62 2,30 20,73 14 7,0—15,5
Самки прибылые 9,1+0,19 1,39 15,30 53 7,0—12,7
взрослые 12,0+1,65 3,31 27,59 4 8,8—16,2
В целом по виду . . . 9,6+0,15 1,88 19,56 163 5,2—16,2
Кондило-базальная длина черепа
Самцы прибылые 20,3+0,07 0,65 3,22 73 19,0—21,6
взрослые 20,2+0,12 0,57 2,82 29 19,0—21,3
Самки прибылые 20,1+0,07 0,65 3,21 83 19,0—21,8
взрослые 20,0+0,16 0,48 2,40 9 19,1—20,8
В целом по виду . . . 20,3+0,03 0,66 3,22 428 19,0—21,8
Длина рострума
Самцы прибылые 5,3+0,03 0,27 5,03 73 4,5-6,0
взрослые 5,2+0,05 0,25 4,71 29 4,6—5,8
Самки прибылые 5,3+0,03 0,26 4,99 86 4,5—6,0
взрослые 5,2+0,07 0,21 3,92 9 4,9—5,5
В целом по виду . . . 5,3+0,02 0,30 5,72 434 4,5-6,0
Длина мозговой части черепа
Самцы прибылые 11,2+0,03 0,26 2,33 73 10,6—11,7
взрослые 11,1+0,04 0,24 2,14 29 10,6—11,6
Самки прибылые 11,1+0,03 0,30 2,68 83 10,5—12,1
взрослые 11,0±0,1 0,30 2,72 9 10,4—11,3
В целом по виду . . . 11,2+0,01 0,27 2,45 428 10,4—12,1
233
Продолжение табл. 48
1 | 2 | з 1 4 1 5 1 6
Наибольшая ширина черепа
Самцы прибылые 9,4±0,04 0,31 3,32 68 8,7—10,0
взрослые 9,6±0,06 0,31 3,21 28 9,0—10,4
Самки прибылые 9,3+0,03 0,30 3,25 79 8,7—10,0
взрослые 9,6±0,09 0,27 2,82 9 9,1—9,8
В целом по виду . . . 9,5±0,02 0,33 3,48 404 8,7-10,4
Межглазничная ширина
Самцы прибылые 3,7±0,02 0,14 3,82 71 3,4-4,0
взрослые 3,8±0,03 0,14 3,62 29 3,5—4,1
Самки прибылые 3,7±0,02 0,16 4,21 86 3,4—4,0
взрослые 3,7±0,05 0,15 3,92 9 3,5-4,0
В целом по виду . . . 3,7±0,01 0,14 3,91 432 3.4—4,1
Предглазничная ширина
Самцы прибылые 2,6±0,02 0,14 5,65 73 2,2—2,9
взрослые 2,7±0,02 0,13 4,75 29 2,4—2,9
Самки прибылые 2,6+0,02 0,14 5,63 86 2,2—2,9
взрослые 2,6±0,05 0,16 6,0 9 2,4—2,9
В целом по виду . . . 2,6±0,01 0,15 5,82 434 2,2—2,9
Ширина рострума
Самцы прибылые 2,0±0,02 0,12 6,14 67 1,7—2,2
взрослые 2,1+0,03 0,13 6,03 26 1,9—2,3
Самки прибылые 2,0±0,02 0,14 6,93 80 1,8—2,6
взрослые 2,1±0,06 0,16 7,62 8 1,8—2,3
В целом по виду . . . 2,0+0,02 0,13 6,53 416 1,7-2,6
Длина верхнего ряда зубов
Самцы прибылые 9,0±0,05 0,39 4,29 71 8,1—10,0
взрослые 8,6+0,07 0,35 4,11 29 7,8—9,2
Самки прибылые 9,0±0,04 0,35 3,86 79 8,2—10,0
взрослые 8,7±0,1 0,30 3,45 9 8,2—9,0
В целом по виду . . . 9,0±0,02 0,42 4,64 420 7,8—10,0
Длина ряда промежуточных зубов
Самцы прибылые 3,0±0,02 0,21 6,96 72 2,6—3,5
взрослые 2,9±0,03 0,16 5,42 29 2,6—3,2
Самки прибылые 3,0±0,02 0,20 6,54 78 2,5—3,4
взрослые 2,9+0,09 0,26 8,90 9 2,5—3,2
В целом по виду . . . 3,0+0,01 0,17 5,73 422 2,5-3,5
Наибольшая высота черепа
Самцы прибылые 6,1+0,03 0,26 4,21 71 5,6-6,7
взрослые 5,7±0,04 0,23 4,03 29 4,9-6,0
Самки прибылые 6,0±0,03 0,28 4,53 78 5,0—6,5
взрослые 5,7±0,06 0,19 3,34 9 5,3—5,9
В целом по виду . . • 6,1+0,01 0,29 4,76 417 4,9—6,7
234
Окончание табл. 48
1 | 2 | 3 | 4 | 5 | 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,4±0,03 0,24 5,43 70 4,0—4,9
взрослые 4,5±0,05 0,27 5,98 29 4,0—5,0
Самки прибылые 4,4±0,03 0,25 5,78 85 3,9—5,1
взрослые 4,5±0,07 0,20 4,52 9 4,2—4,7
В целом по виду . . . 4,5±0,01 0,26 5,82 425 3,9—5-1
Длина иглового отростка
Самцы прибылые 2,6±0,03 0,21 8,03 60 2,0—3,0
взрослые 2,6±0,03 0,17 6,48 25 2,4—3,0
Самки прибылые 2,5±0,02 0,18 7,19 80 2,2—3 0
взрослые 2,6±0,05 0,15 5,23 8 2,4—2,8
В ц ел ом ‘ по виду . . . ’ 2,6±0,01 0,18 7,08 327 2,0—3,0
уровне с первой вершиной переднего резца. Третий и четвертый почти
равны между собой и значительно уступают по высоте и массивности
первому и второму. Пятый Значительно меньше четвертого.
Измерения. См. табл. 48.
Характеристика. Во внешнем облике бурой бурозубки в
первую очередь обращают на себя внимание крупные размеры, темно-
бурая окраска, постепенно светлеющая на боках и переходящая в серо-
вато-буро-белесую на вентральной стороне тела, длинная голова без
заметного сужения в области глаз и длинный, расширенный в области
промежуточных зубов хоботок (рис. 70, 71). Только вышедшие из гнезда
зверьки по размерам превышают взрослых зверьков многих видов. Те-
лосложение массивное, тяжелое, Хвост умеренной длины — 30—50
(7И = 37,7=t0,28) мм, немногим более половины длины тела. Длина
хвоста подвержена значительной изменчивости. У зверьков из северо-
восточной части ареала (Якутия) короткий хвост, на юге ареала —
длинный (30—50 мм). У обыкновенной бурозубки изменчивость этого
признака несколько меньшая (34—49 мм). Ступни крупные (М — 13,4=ь
—0,07). По общему облику эта бурозубка похожа на обыкновенную, но
отличается от последней большими размерами, тяжелым телосложением,
более развитой грудной частью, формой головы и рострума и более ко-
ротким хвостом (у 5. araneus 34—49 (М = 42,5z±?0,22) мм, по отношению
к длине тела — 61,6 %). ;
В отличие от обыкновенной у бурой бурозубки светлая окраска
вентральной стороны тела почти совсем не заходит на бока, но прости-
рается до кистей и ступней, задняя треть спины окрашена более интен-
сивно, чем голова и плечи. На щеках бурый тон спускается ниже глаз
и ушей. По насыщенности окраски дорсальной части тела, средним зна-
чениям колориметрических данных 5. roboratus близка к 5. isodon, S. ara-
neus, S. unguiculatus и 5. daphaenodon (см. табл. 30). Наиболее темные
особи ее близки к таковым S. isodon, S. araneus и 5. tundrensis, а наибо-
лее светлые — к 5. isodon н S. daphaenodon. Диапазон изменчивости
белизны этого вида колеблется в пределах 12 и 3,5, а оттепка —
113,1 — 160. ; : : . .
235
Рис. 70. Бурая бурозубка — S. robaratus Holl.
У S. araneus при почти совпадающей величине оттенка белизна бо-
лее изменчива в сторону посветления — ее максимальный показатель
равен 5,3, а у 3. tundrensis изменчивость того и другого признаков варьи-
рует в больших пределах — оттенок от 112 до 166, а белизна от 1,3 до
236
6,5. Можно сказать, что изменчивость окраски 5. roboratus, учитывая
ее обширный ареал, охватывающий различные природные зоны Сибири,
довольно незначительна. У вида сохраняется специфика окраски в раз-
ных природных условиях.
Характерным для вида является строение черепа. Его размеры
перекрываются с таковыми обыкновенной, когтистой и равнозубой бу-
розубок, однако в конфигурации имеется ряд специфичных черт. Череп
удлиненный, мозговая коробка узкая, постепенно переходящая в доволь-
но широкую ростральную часть. В затылочном отделе череп уплощен
(отношение высоты черепа к кондило-базальной длине составляет 30,3,
а у S. araneus — 31,2 %). Нижняя челюсть легкая, передний резец вы-
тянут в длину. Восходящая ветвь нижней челюсти относительно низкая
(индекс высоты к кондило-базальной длине черепа 22,0 %, а у 5. arane-
us — 25,4 %). Этот показатель весьма характерен для S. roboratus. Proc,
angularis тонкий, почти прямой и длинный (12,7 % от кондило-базаль-
ной длины черепа, а у S. araneus — 10,8 %).
Специфические особенности в структуре зубов кардинально отли-
чают эту землеройку от других видов. По длине зубного ряда и разме-
рам черепа эта землеройка занимает второе место среди представите-
лей рода в пределах Сибири. Длина зубного ряда S. roboratus 7,8—10,0
(М = 9,0±0,02) мм. Точно такой же длины зубной ряд у обыкновенной
бурозубки (8,2—9,9 мм; М = 9,0±0,02), а ряд промежуточных зубов
у бурой бурозубки несколько уступает (2,5—3,5 мм; М = 3,0±0,01) та-
ковому обыкновенной (2,8—3,6 мм; М = 3,1 ±0,01). Зубы довольно
мелкие.
Специфика зубной системы заключена в форме зубов и их соотноше-
нии. Первый верхний резец чрезвычайно вытянут вперед. При рассмот-
рении черепа сверху резцы далеко выступают вперед. Между их внут-
ренней стороной и челюстной костью виден большой просвет. Передняя
вершина этого зуба значительно выше и шире второй (задней). Послед-
няя, в свою очередь, равна по высоте первому промежуточному зубу.
Промежуточные расположены в ряду свободно, едва касаясь основания
предыдущего зуба. Основания коронок зубов сильно вытянуты в сагит-
тальном направлении. Вершины зубов заметно сдвинуты вперед по отно-
шению к середине основания коронки.
Размеры, форма и соотношение весьма своеобразны, особенно первых
двух промежуточных. Первый по высоте равен второй (задней) вершине
переднего резца и уступает по высоте второму, равному по высоте или
чуть ниже первой вершины переднего резца. Если зубной ряд рассматри-
вать в профиль, повернув череп зубами вверх, то создается впечатление
«провала» между первой вершиной резца и вторым промежуточным зубом
за счет более низкой второй вершины и первого промежуточного зуба.
Третий и четвертый промежуточные зубы почти вдвое уступают первому
и второму по массивности и высоте, почти равны между собой, пятый
мал. Коронки первых четвертых зубов вытянуты в высоту, сосцевидной
формы, с округло-затупленными вершинами.
Направление вертикальных осей весьма изменчиво. Два первых
зуба имеют заметный наклон внутрь ротовой полости, два последующих
чаще всего имеют вертикальную ось, перпендикулярную продольной
оси черепа. На задней стороне первых двух промежуточных зубов сильно
развиты вырезки, за счет которых усиливается впечатление их наклона
по направлению внутрь ротовой полости. Многовершинные зубы по срав-
237
нению с таковыми других видов расставлены свободно, между задней
стороной предыдущего многовершинного зуба и передней последующего
имеются значительные полулунные промежутки, через которые видны
небные кости. Гипоконус на последнем предкоренном и коренных зача-
точный, не пигментированный. Пирамидальная воронка хорошо развита.
Талом этих зубов развит сильнее, чем у других видов бурозубок.
Pm! с удлиненной низкой коронкой и заметной второй вершиной.
Головка копулятивного органа устроена специфично. В общем ко-
ническая опа сравнительно короткая и толстая, округлая в поперечном
сечении (см. рис. 40), в дистальной части утончающаяся, со слегка заг-
нутым вниз притупленным концом. В сравнении с другими видами земле-
роек бурая бурозубка имеет непропорционально небольшой копулятив-
ный орган. Длина glans penis 5,4—6,8 (М = 6,2), толщина у основания
1,2—1,7 (М = 1,5) мм. По отношению к размерам тела длина копулятив-
ного органа составляет всего 8,6 %, в то время как у S. caecutiens —
10,5, S. araneus — 13,9, 5. isodon — 15,4, S. unguiculatus — 14,9, у
S. minutus — 17,8 %.
Впервые 5. roboratus описал Холлистер [Hollister, 1913]. Позднее
Аллеп [Allen, 1914] квалифицировал ее как S. rir, а С. И. Огнев [1928]—
как подвид S. araneus, назвав его S. a. roboratus. С. У. Строганов [1957]
считает S. roboratus подвидом S. araneus. Нами при сопоставлении морфо-
метрических характеристик установлено, что экземпляр, имевшийся
в распоряжении Холлистера и послуживший ему для первоописания,
относился не к S. araneus, а к самостоятельному виду — S. roboratus
[Юдин, 1965а, 1971а и др.). Самостоятельность этого вида была под-
тверждена и кариологическими исследованиями [Орлов, Козловский,
1971 ].
Распространение. Бурая бурозубка — восточный пале-
аркт, близкий по характеру распространения к крупнозубой бурозубке.
Ареал ее простирается на восток от Оби до Чукотки и Приморья.
В нижпем течении Оби бурая бурозубка добыта в Шурышкарском
районе [Николаев, 1966]. Северная граница распространения — Ямало-
Ненецкий автономный округ, Гыданский полуостров; зверьки обна-
ружены в окрестностях с. Ларино Томской области, что на р. Ларь-
Еган, в окрестностях Колпашево, Томска, Новосибирска. С. У. Строганов
[1957] приводит окрестности Среднеколымска и Нижнеколымска как
пункты находок этого вида. В. Г. Кривошеев коллектировал ее в среднем
течении Колымы от пос. Зырянка до Среднеколымска Б. С. Юдин [1975а]
собрал серию зверьков в долине рек Омолон и Кегали, а В. Г. Беляев —
у Сеймчана. В приобских сосновых борах между Новосибирском и Бар-
наулом 5. roboratus не встречена. Эта бурозубка заселяет лесотундровую
и лесную зоны, проникает в горы юга Сибири и сопредельные районы
Монголии.
Материал. 498 экз., Забайкалье, Алтай, Салаир, Саяны, Якутия.
Изменчивость. Волосяной покров землеройки варьирует от
относительно грубого до шелковистого. Окраска меха весьма стабильная
и не подвержена значительной изменчивости, но имеет свои особенности
в разных популяциях (рис. 72). Окраска зверьков разных популяций
(Алтай, Салаир, Забайкалье, Якутия, Чукотка) весьма слабо различается
и имеет незначительную изменчивость по белизне и оттенку. Наиболее
темные особи встречены на Салаире и его отрогах. Серия зверьков из
окрестностей Новосибирска и пос. Мирный (Тогучинский р-н Новоси-
238
Колориметрическая характеристика подвидов
Подвид Место сбора материала Белизна а Су
S. г. roboratus Алтай, Салаир, Саяны, Яку- 2,2+0,07 0,54 24,0
S. г. vir тия Бассейн Колымы 2.4+0,08 0,32 12,8
S. г. thomasi Забайкалье 2,06±0,10 0,44 17,2
П о в и д у в целом . . . 2,4±0,05 0,51 21,7
бирской обл.) наиболее темная. Значения белизны у S. roboratus пере-
крываются таковыми бурой бурозубки других популяций менее чем на
50 %. По вариабельности оттенка она отличается только от якутских
бурозубок этого вида.
Исследование шести выборок, относящихся к трем подвидам, пока-
зало их чрезвычайную близость (табл. 49). Исключая якутскую и салаир-
скую популяции номинального подвида, остальные выборки совпадают
почти полностью. Можно лишь говорить, что крайне темные варианты
у всех (кроме салаирских) выборок идентичны. Что касается светлых
особей, то наблюдается следующая тенденция. Самые темные по такому
признаку, как белизна, с очень ограниченным диапазоном изменчивости
салаирские особи, значения этого показателя составляют 1,2—2,2
(рис. 72). Алтайские и саянские зверьки несколько светлее и имеют
больший размах изменчивости: 1,7—3,5, хотя значения других показа-
телей в значительной мере трансгрессируют. Совершенно четко уклады-
вается в эти рамки характеристика окраски как по белизне, так и по
Белизна
оттенку у [зверьков из бассейна
Омолона (западная Чукотка):
1,9-3,0.
Намечается незначительная
склонность в сторону посветления
у забайкальских землероек с
хр. Хамар-Дабан: 1,8—3,5, но эти
значения также укладываются в
таковые у бурозубок этого вида
с Алтая и Саян. По интенсивности
оттенка несколько выделяются
землеройки Якутии (Горный р-н).
Оттенок этой популяции харак-
теризуется величинами 135,2 и
Рис. 72. Сравнительная характеристика
окраски подвидов бурой бурозубки из
разных частей ареала.
1 — S. г. vir (Чукотка, бассейн р. Омолон);
2 — S. г. thomasi (Забайкалье); л—в —
S. г. roboratus (3 — Сал аир; 4 — Якутия;
5 — Тува, Саяны; 6 — Юго-Восточный Алтай,
р. Бугузун, оз. Джулукуль)
239
Таблица 49
Sorex, roboratus из разных частей ареала
1 Оттенок
' п lim М±тх а Су п lim
j 59 1 ’ 1 15 ' 20 1,2—3,5 1,9—3,0 1,8—3,5: 138,0+1,42 , 135,8+1,78 133,7+1,17 10,93 6,01 5,14 7,9 5,1 3,9 59 15 20 113,1—160,9 121,4—148,6 124,1—146,4
94 1,2—3,5 136,7+1,0 9,5 6,94 94 113,1—160,9
160,9. Значения последнего показателя самые высокие среди всех пред-
ставителей рода Sorex. По показателю белизны якутская популяция не
выходит за рамки, характерные для вида в целом, и характеризуется
цифрами: 2,0—3,3. Рыжеватость относительно светлых зверьков из Яку-
тии хорошо заметна на прибылых особях.
К сожалению, мы не имеем возможности провести колориметрическое
исследование этого вида из Приморья. Визуальное сравнение их со зверь-
ками с Алтая показывает, что они более интенсивно окрашены в бурые
тона, каштановый оттенок отсутствует. Популяции из Баргузинской
Таблица 50
Морфометрическая характеристика Sorex roboratus и ее подвидов из разных частей
ареала
Признак a. S. г, roboratus (номинальный подвид), Сев. Алтай, Саяны
М+тх а Су п lim
Длина ;
тела 71,5+0,60 5,88 8,23 94 58—87
хвоста 38,2+0,43 4,21 11,01 94 30—50
ступни 13,6+0,10 0,97 7,17 93 11—15
Масса тела 9,8+0,20 1,80 18,24 84 7,5-16,3
Длина
кондило-базальная черепа 20,6+0,03 0,58 2,82 291 19,1—21,8
рострума 5,4+0,02 0,27 4,91 296 4,8—6,3
мозговой части 11,2+0,02 0,26 2,28 291 10,4—12,1
верхнего ряда зубов 9,1+0,02 0,37 4,09 287 8,1—10,0
ряда промежуточных зубов 3,0+0,01 0,15 5,10 289 2,6-3,5
Ширина
рострума 2,1+0,01 0,14 6,76 282 1,7—2,6
предглазничная 2,6+0,01 0,14 5,46 296 2,3—3,0
межглазничная 3,7+0,01 0,15 3,90 294 3,2—4,1
наибольшая черепа 9,5+0,02 0,33 3,46 278 8,7—10,6
Наибольшая высота черепа 6,1+0,02 0,26 4,28 284 5,5—6,8
Высота восходящей ветви ниж-
ней челюсти 4,5+0,02 0,28 6,09 288 3,9—5,1
Длина углового отростка 2,6+0,01 0,17 6,45 215 2,2—3,0
240
Признак б. S. г. vir, Якутия
М+тпх а CV п lim
Длина тела 76,3±0,87 4,42 5,79 26 69—89
хвоста 37,2±0,44 2,18 5,86 25 35-41
ступни 13,7±0,19 0,86 6,30 20 12-15
Масса тела 10,5±0,27 1,41 13,5 27 9—15,5
Длина кондило-базальная черепа 20,2+0,06 0,31 1,54 30 19,4—20,8
рострума 5,3±0,03 0,16 2,92 31 4,9—5,5
мозговой части 11,3+0,04 0,20 1,76 30 10,6—11,6
верхнего ряда зубов 8,8±0,04 0,19 2,20 29 8,4—9,0
ряда промежуточных зубов 2,8±0,01 0,08 2,73 29 2,7—3,0
Ширина рострума 2,0±0,01 0,06 3,12 28 1,9—2,2
предглазничная 2,8±0,02 0,10 3,63 31 2,6—3,0
межглазничная 3,8±0,02 0,11 2,82 31 3,6—4.0
наибольшая черепа 9,5+0,04 0,20 2,07 30 9,1—9,9
Наибольшая высота черепа 6,1±0,06 0,31 5,11 29 5,2—6,9
Высота восходящей ветви ниж- ней челюсти 4,5+0,02 0,11 2,41 32 4,3-4,7
Длина углового отростка 2,5±0,02 0,11 4,40 30 2,2—2,7
котловины отличаются тусклым мехом с заметной примесью серых тонов.
Небольшая примесь их в окраске меха свойственна также зверькам с
Енисея, из района Ангутихи. Алтайские бурозубки темные, без серых
тонов, с каштановым оттенком. На щеках бурый тон у алтайской популя-
ции спускается ниже, чем у землероек с Енисея и из Баргузинской котло-
вины, заходит значительно ниже глаз и ушей. В целом окраска зверьков
светлеет от Салаира и Алтая к Енисею и Байкалу. Енисейская выборка
сравнивалась также только визуально. Якутские землеройки темнее
енисейских, но уступают по интенсивности окраски уссурийской и ал-
тае-саянской популяциям. Для якутской популяции характерен шелко-
вистый и блестящий мех.
В Баргузинской котловине зверьки окрашены так же, как и в других
районах Западной и Средней Сибири. Зверьки здесь более темные, чем
на Енисее (Ангутиха), с более контрастно окрашенным хвостом. Отсут-
ствие серых тонов в окраске спины, шелковистый блестящих мех — все
это сближает их с якутской популяцией, а интенсивность бурых тонов —
с алтае-саянскими зверьками.
Хорошо прослеживаются размерные отличия зверьков разновоз-
растных групп. Длина тела взрослых самцов (7б,7±1,64 мм) и самок
(76,0=t5,05) заметно превышает таковую молодых самцов (69.7^0,92)
и самок (71,5±0,65 мм). Достоверных различий по полу не прослежива-
вается. Аналогичная закономерность проявляется и по массе тела, мо-
лодые зверьки значительно уступают по этому показателю взрослым
(см. табл. 48), не обнаруживаются сколько-нибудь заметные различия
и между полами.
241
Продолжение табл. 50
в. S. г. thomasi. Забайкалье г. S. г . platicranius, Приморье
М±тх О Су п lim М+тпх О Су п lim
66,9±1,27 5,22 7,81 17 58—79 72,8± 1,20 2,68 3,69 5 70—76
33,7±0,34 1,40 4,14 17 30—36 36,1+0,87 1,95 5,36 5 35-39
12,7±0,18 0,70 5,57 16 12—14 — — — — —
7,9±0,27 1,12 14,14 17 6,8—10,9 7,8±0,33 0,74 9,51 5 7,0—8,9
19,4+0,08 0,42 2,18 29 18,8—20,4 20,1±0,09 0,54 2,66 35 19,2-21,0
4,8±0,05 0,25 5,14 29 4,4-5,5 5,2±0,04 0,24 4,61 35 4,3—5,5
11,0±0,06 0,30 2,69 29 9,9—11,4 11,3±0,05 0,30 2,67 35 10,6-11,9
8,4±0,05 0,25 9,01 29 7,8—8,8 8,8±0,04 0,23 2,61 32 8,5-9,4
2,8±0,02 0,10 3,44 29 2,6—3,0 3,0±0,03 0,15 5,21 32 2,7—3,3
1,9±0,02 0,12 6,26 28 1,8—2,3 2,0±0,01 0,06 3,09 35 1,8—2,1
2,5±0,03 0,17 6,53 29 2,2—2,8 2,7±0,02 0,10 3,86 35 2,4-2,9
3,6±0,03 0,15 4,27 29 3,3-4,0 3,7±0,03 0,14 3,67 35 3,5—4,0
9,2±0,06 0,29 3,13 28 8,6-9,8 9,4±0,06 0,32 3,45 31 8,6—9,9
6,0±0,05 0,27 4,55 29 5,4-6,6 6,1±0,06 0,34 5,56 33 4,9—6,6
4,2+0,03 0,15 3,51 29 4,0—4,5 4,4±0,02 0,14 3,13 34 4,2—4,7
11 2,4±0,04 0,18 7,57 22 2,1—2,8 2,5±0,03 0,15 6,13 27 2,2—2,8
Высота черепа с возрастом уменьшается 'табл 50). По остальным
параметрам возрастных а половых различий не обнаруживается.
Географическая изменчивость размеров весьма своеобразна. G одной
1 стороны, выборки из популяций, находящихся в одном районе, по некото-
рым показателям достоверно различаются, с другой стороны, далеко
отстоящие друг от друга популяции по многим параметрам оказываются
неразличимыми. Прослеживается изменчивость кондило-базальной дли-
; ны черепа у S. roboratus из разных частей ареала (см. табл. 50, рис. 73).
По средним значениям длины тела достоверные различия между выбор-
ками из разных участков распространения вида существуют только у 5. ro-
boratus (Td = 3,3) и S. roboratus vir (Td — 4,5) (см. табл. 51). Более мел-
। кие зверьки живут в Забайкалье — 66,9 (±12,7) мм при среднем пока-
। зателе для вида по всему ареалу — 72,0 (±0,45) мм. Длина хвоста колеб-
лется в значительных пределах. Наиболее короткий хвост у землероек
из Забайкалья (33,7-}-0,34) мм, они достоверно отличаются по этому
’ признаку от зверьков из всех выборок, кроме землероек из Приморья,
также имеющих относительно короткий хвост. Зверьки с Хамар-Дабана
достоверно отличаются меньшими размерами ступни, массой тела, кон-
дило-базальной длиной черепа, длиной рострума, верхнего и промежу-
точного рядов зубов и рядом других показателей (см. табл. 51).
5 Заметки по систематике. Бурая бурозубка относится
к числу видов, наиболее четко очерченных и легко распознаваемых среди
( других представителей рода. Объединение ее с другими видами: 5. ага-
1 neus, S. asper, S. isodon, S. arcticus, S. vir(=S. roboratus) [Бобринский
и др., 1944, 1965] — не обосновано.
16 Б. С. Юдин
242
Окончание табл. 50
Признак д. S. г. subsp., Бугузун
О Су п lim
Длина
тела 72,4+0,99 6,19' 8,54 39 60—87
хвоста 38,7+0,52 3,28' 8,46 39 33—46
ступни 12,9+0,13 О',77 5,95 38 11—14
Масса тела 9,5+0,40 2,19 22,98 30 5,2—16,2
Длина
кондило-базальная черепа 19,6+0,05 0,35 1,78 43 19,0—20,5
рострума 5,1+0,03 0,19 3,76 43 4,5—5,4
мозговой части черепа 10,9+0,03 0,17 1,59 43 10,6—11,4
верхнего ряда зубов 8,6+0,04 0,26 2,99 43 8,1—9,0
ряда промежуточных зубов 2,8+0,02 0,13 4,64 43 2,5—3,0
Ширина
рострума 2,0+0,02 0,12 5,77 43 1,9—2,3
предглазничная 2,5+0,02 0,11 4,32 43 2,2—2,7
межглазничная 3,7+0,02 0,14 3,76 43 3,4—4,0
наибольшая черепа 9,3+0,04 0,27 2,90 37 8,8—9,8
Наибольшая высота черепа 5,8+0,04 0,27 4,64 42 5,0—6,3
Высота восходящей ветви ниж-'
ней челюсти 4,3+0,03 0,21 4,92 42 4,0—4,7
Длина углового отростка 2,5+0,03 . 0,16 6,34 33 2,0—2,7
В пределах Сибири выделены четыре подвида S. roboratus, есть пред-
положение о существовании еще одного подвида — S. г. subsp., обитаю-
щего на границе с Западной Тувой в бассейне Бугузуна 9.
Обзор подвидов
Sorex roboratus roboratus Hollister, 1913 —
Сибирская горная бурозубка
Диагноз. Зверьки относительно крупные, по окраске темнее
других подвидов, показатель оттенка (М = 138,0+1,42) самый высокий,
а показатель белизны (М = 2,4+0,07) самый низкий, мех грубый.
Измерения. Длина тела — М = 71,5—0,6, длина хвоста —
М = 38,2—0,43, длина ступни — М = 13,6+0,1 мм. См. также табл. 50.
Характеристика. Зверьки, населяющие Салаир и его отро-
ги, наиболее темные, алтайские популяции несколько светлее и отлича-
ются большим размахом изменчивости по белизне (1,9—3,5). Эта популяция
отличается от енисейской и баргузинской более насыщенным бурым тоном.
На щеках бурый тон спускается значительно ниже, чем у названных
выше зверьков. В окраске наблюдается постепенное посветление популя-
ций от Алтая к Енисею и далее на восток.
Распространение. От Салаира через Горную Шорию си-
бирская горная бурозубка проникает на Алтай. Характерная черта рас-
пространения подвида — приуроченность на большей части ареала к 8
8 Подробное описание подвида автором не дано.
243
Рис. 73. Изменчивость кондило-базальной
длины черепа бурой бурозубки из разных
частей ареала.
1 — S.r. р. (Алтай, оз. Джулукуль); 2 — S. г. vir
(Якутия); 3 — S. г. platicranius (Приморье); 4 — S.r.
roboratus (Алтай, Телеикое озеро); 5 — S. г. thomasi
(Забайкалье, Сохондо).
горным ландшафтам. В Алтае-Саянской
горной стране этот подвид встречается
во всех крупных горных системах: на
Алтае, в Горной Шории, на Салаире,
Кузнецком Алатау, Западном и Во-
сточном Саянах, в ряде районов Тувы.
Западная граница распространения —
Тигирецкий и Колыванский хребты.
Восточнее зверьки добыты в бассейне
Катуни, в районе с. Майма. В Цент-
ральном Алтае этот подвид распростра-
нен от Усть-Коксы по хребтам Терек-
тинскому, Кумирскому, Тонгош, всюду
встречается в Северо-Восточном Алтае,
по хребтам, окружающим Телецкое
озеро. В Туве заселяет все районы с
хвойными лесами.
Кондило-базальная длина черепа, «ч
Материал. 296 экз., различные части ареала этого подвида.
Sorex roboratus vir Allen, 1914 —
Якутская бурая бурозубка
Диагноз. Размеры довольно крупные, длина тела М — 76,3±
±0,87 мм. По этому показателю подвид достоверно отличается от других
представителей вида.
Измерения. См. табл. 50.
Характеристика. Зверьки темные, по насыщенности окраски
уступают только S. г. roboratus, показатель оттенка — М = 135,8±
±1,78, а у S. г. roboratus М = 138,0±1,42, мех шелковистый в отличие
от 5. г. roboratus. Ржавых тонов в окраске спины нет. Бока окрашены
в светло-бурые тона, слабеющие по насыщенности от спины к брюшку,
окрашенному в серовато-палевый тон. По морфометрическим показа-
телям достоверно отличается по ряду признаков от других подвидов
(табл. 51).
Распространение. Встречается по всей Колыме, от пос. Зы-
рянка до Нижнеколымска, а также в бассейне Омолона. В Анадырском
крае зверьки добыты в низовьях Анадыря (сборы Н. П. Сокольникова),
близ. пос. Еропол [Кривошеев и др., 1978].
Материал. 30 экз., Якутия.
Sorex roboratus thomasi Ognev, 1921 —
Забайкальская бурая бурозубка
Диагноз. Зверьки наиболее мелкие среди всех подвидов S. ro-
boratus. Длина тела — М = 66,9±12,7, длина хвоста — М — 33,7±
±0,34, кондило-базальная длина черепа — М = 19,4±0,08 мм.
Измерения. См. также табл. 50.
16*
244
Таблица 51
Достоверность различий (по Стьюденту) между подвидами Sorex roboratus
Признак а—б а—в а—г а—д б—д б-г б—д
Длина
тела 4,5 3.3 0,9 0,8 6,1 2,4 2,9
хвоста 1,6 8,2 1,9 0,7 6,3 0,8 2.2
ступни 0.5 4,4 — 4,3 3,8 — 3.5
Масса тела Длина 2,1 5,7 5,2 0,7 6,8 6,3 2,1
кондило-базальная черепа 6,0 14,2 2.1 17,2 8,0 0,9 7,7
рострума 2,8 11,1 4,7 8,3 8,6 2,0 4,7
мозговой части 2,2 3,2 1,9 8,3 4,2 — 8.0
верхнего ряда зубов 6,7 12,9 6,7 11,2 6,3 0 3,5
ряда промежуточных зубов Ширина рострума 14,1 8,9 0 8,9 0 6,3 0
7,1 8,9 7,1 4,5 4,5 0 0
пре дгл азнична я 8,9 3,2 4,7 4,7 8,3 3.5 10,6
межгл азничная 4,5 3,2 0 0 5,5 2,8 3,5
наибольшая черепа 0 4,7 1,6 4,5 4,2 1,4 3,5
Высота черепа Высота восходящей ветви ниж- 0 1,9 0 6,7 1,3 0 4,2
ней челюсти 0 8,3 3,5 5,6 8,3 3,5 5,5
Длина углового отростка 4,7 4,9 3,2 3,2 2,2 0 0
Примечание. Буквенные обозначения подвидов см. табл. 50.
Характеристика. Указание С. У. Строганова, что зверьки
имеют монотонную тусклую окраску, подтверждается нашими материала-
ми. Окраска зверьков из Баргузинской котловины не отличается от
таковых из других частей ареала. В Западной и Средней Сибири зверьки
скорее более темные с более контрастно окрашенным хвостом. Отсутствие
серых тонов в окраске спины, шелковистый блестящий мех ставят их
ближе всего к якутской бурозубке (S. г. vir).
Распространение. Подвид широко распространен в лесных
районах байкальского бассейна. Бурозубки отлавливались на хребтах
Байкальском, Приморском, Баргузинском, Улан-Бургасы, Морском,
Северо-Муйском, Бол. и Мал. Хамар-Дабане и др. Очень редка в Север-
ном Прибайкалье, где добыта в долине Будурмана (приток Сосновки)
3. Ф. Сватош (по С. У. Строганову [1957]). В Баргузинской котловине
живет преимущественно в долинных биотопах [Лямкин, 1977]. На Запад-
ном побережье Байкала подвид не обнаружен [Литвинов, 1972; Швецов,
1977]. Относительно много этого зверька на восточном побережье Байка-
ла [Лямкин, 1978], южнее, на территории МНР, обнаружена недавно
в западных предгорьях Хэнтэя [Shotolchu, Stuble, 1971]. Эта бурозубка
поймана в небольшом количестве в Прихубсугулье [Литвинов, 1972].
Материал. 29 экз. Баргузинская котловина.
Sorex roboratus platicranius Ognev, 1921 —
Уссурийская бурая бурозубка
Диагноз. Длина тела — М = 72,8±1,2, относительно короткий
хвост — М = 36,4zt0,87.
Измерения. См. табл. 50.
Характеристика. Самая темная форма из всех S. roboratus.
Окрас спины темно-бурый, матовый, без примеси каких-либо тонов,
245
зверьки близки по окрасу к S. г. roboratus (см. рис. 72). По морфометри-
ческим показателям занимает среднее положение между S. г. thomasi
и S. г. roboratus (см. табл. 50). По большинству показателей достоверно
отличается от других подвидов (см. табл. 51).
Распространение. Зверьки занимают территорию по восточной
границе ареала вида, известны из бассейна Уссури (район г. Дальнере-
ченска, г. Уссурийска). Сотрудниками Биологического института
СО АН СССР в 1966 г. отловлены зверьки в окрестностях с. Раздольное
(Надеждинский р-н), с. Ново-Хатуничи (Шкотовский р-н), добыты зверь-
ки и из Амурской области (г. Зея, бассейн Уссури, р. Кия).
Материал. 22 экз., Приморский край, 5 экз., Амурская область,
8 экз., бассейн Уссури.
Sorex tundrensis Merriam, 1900 —
Тундряная бурозубка
Г о л о т и н. Вид описан с о. Св. Михаила (Аляска).
Диагноз. Размеры средние. Длина тела с головой 46—82 (М =
=63,4±0,26), длина хвоста 24—47 (М = 33,9±0,23), длина ступни 8—14
(М = 11,6±0,5) мм, масса 3—11 (М = 6,1 ±0,07) г. Кондило-базальная
длина черепа 15,7—19,4 (М = 17,6± 0,02) мм. Голова широкая, массив-
ная. Сужение в области глаз выражено слабо. Хоботок короткий и
широкий. В окраске у большинства подвидов более или менее выражена
чепрачность. Зубы по величине пропорциональны размерам черепа.
Два первых по величине равны между собой. Вершина первого верхнего
резца находится на одном уровне со второй вершиной этого же резца.
Из-за особенности положения зубов в челюсти вершина первого всегда
ниже вершины второго, третий зуб ниже второго, четвертый уступает
по величине третьему, пятый мал. Промежуточные зубы высокие, верши-
Рис. 74. Тундряная бурозубка — Sorex tundrensis Мегг.
246
ны их расходятся свободно и обращены кзади, внутрь ротовой полости,
в зубном ряду сидят плотно. Первый промежуточный нижней челюсти
вытянут в сагиттальном направлении. Череп высокий, вздутый в мозго-
вой области (рис. 74, 75).
Измерения. См. табл. 52.
Характеристика. Тундряная бурозубка средних размеров,
телосложение грубое, массивное. В строении черепа обращает на себя
внимание широкая лобная часть головы, без резкого межглазничного
сужения, затылочная область с широкой вздутой мозговой коробкой.
Хоботок короткий, широкий. В строении черепа много общих черт
с S. araneus. (Одонтологическая характеристика дана выше). Наиболее
примечательной чертой во внешнем облике тундряной бурозубки следует
считать чепрачность в распределении окраски меха.
Распространение. Тундряная бурозубка — самый широко
распространенный вид среди насекомоядных млекопитающих мировой
фауны. Ареал его охватывает северо-восточную Европу, Азию к югу
до Китая и Монголии. В СССР — от р. Печоры на восток до Чукотки
включительно. В Сибири, по-видимому, встречается повсеместно.
Материал. 700 экз. из разных частей ареала.
Изменчивость. Большой возрастной и географической из-
менчивости подвержен такой признак, как окраска зверьков. В разных
частях ареала и у различных возрастных групп признак этот выражен
неодинаково. Наиболее характерная для вида чепрачная окраска свойст-
венна зверькам открытых пространств — тундры, лесостепных и степ-
ных районов юга Западной Сибири и Казахстана. Четко выражена она
у перезимовавших зверьков. Половых различий в окраске нет. Темная
247
Таблица 52
Морфометрическая характеристика Sorex tundrensis
Возрастная и половая группа М±тж (Г Су п lim
1 3 4 5 6
Длина тела с головой
Самцы прибылые 61,9+0,37 4,20 6,79 126 46,0-71,0
взрослые 67,9+0,71 6,00 8,84 72 53,0—79,0
Самки прибылые 62,9+0,47 5,33 8,47 129 48,0—77,0
взрослые 72,7+1,22 6,59 9,07 29 61,0—83,0
В целом по виду . . . 63,4+0,26 6,16 9,72 574 46,0-83,0
Длина хвоста
Самцы прибылые 34,2+0,41 4,60 13,45 126 22,0-45,0
взрослые 33,5±0,59 4,94 14,73 71 20,0—44,0
Самки прибылые 34,2+0,46 5,25 15,36 129 22,0—47,0
взрослые 33,0+0,59 3,13 9,51 29 25,0—41,0
В ц е л о м по виду . . . 33,9+0,23 5,42 15,99 576 20,0—47,0
Длина ступни
Самцы прибылые 11,6+0,1 1,06 9,1 118 8,0-13,5
взрослые 12,0+0,11 0,95 7,88 60 10,0—14,5
Самки прибылые Н,7±0,09 1,04 8,92 125 8,0—14,5
взрослые 11,6+0,25 1,28 11,11 27 9,0—14,0
В целом по виду 11,6+0,05 1,09 9,38 562 8,0—14,5
Масса тела
Самцы прибылые взрослые 5,5+0,07 7,7+0,17 0,76 1,35 13,67 17,55 113 63 4,0-8,8 4,0—10,0
Самки прибылые 6,0±0,12 1,22 20,45 105 3,8—10,0
взрослые 8,7+0,52 2,54 29,11 24 4,0—16,0
В целом по виду . . . 6,1+0,07 1,61 26,27 481 3,8—16,0
Кондило-базальная длина черепа
Самцы прибылые 17,8+0,05 17,7±0,07 0,58 0,61 3,25 3,47 150 72 16,4—19,2 16,0—19,0
взрослые
Самки прибылые 17,7±О,05 0,57 3,20 135 16,4—19,4
взрослые 17,6+0,10 ' 0,51 2,89 24 16,6—18,8
В ц е л о м по виду . . . 17,7+0,02 0,59 3,33 656 16,0—19,4
Длина рострума
Самцы прибылые взрослые 4,5±0,02 4,4±0,03 0,58 0,01 3,25 3,47 150 72 4,0—5,0 3,7—4,9
Самки прибылые 4,5+0,02 0,57 3,20 135 3,6—5,4
взрослые 4,3±0,06 0,51 2,89 24 3,6-5,0
В целом по виду . . . 4,4±0,01 0,59 3,33 656 3,6—5,4
Длина мозговой части черепа
Самцы прибылые 10,2+0,03 0,30 2,97 151 9,4—11,0
взрослые 10,1+0,04 0,35 3,50 73 8,9-10,0
Самки прибылые 10,1+0,03 0,33 3,24 132 8,8—11,0
взрослые 10,0±0,05 0,26 2,63 24 9,5-10,0
В целом по виду . . , 380 8,8—11,0
248
Продолжение табл. 52
1 | 2 1 3 1 4 1 5 | в
Наибольшая ширина черепа
Самцы прибылые 8,9±0,03 0,31 3,52 149 8,1—9,9
взрослые 9,0±0,04 0,33 3,70 72 8,0—9,7
Самки прибылые 8,9±0,03 0,33 3,67 133 8,2—10,Р
взрослые 8,8±0,08 0,39 4,47 23 8,0—9,4
В целом по виду . . . 8,8±0,01 0,32 3,62 644 8,0—10,0
Межглазничная ширина
Самцы прибылые 3,4±0,01 0,16 4,68 154 3,0—4,0
взрослые 3,5±0,01 0,13 3,57 76 3,3—3,9
Самки прибылые 3,5±0,01 0,15 4,32 146 3,2—2,8
взрослые 3,5±0,03 0,14 3,97 28 3,2—3,8
В целом по виду . . . 3,4±0,01 0,15 4,30 695 3,0—4,0
Предглазничная ширина
Самцы прибылые 2,4±0,01 0,14 6,02 152 2,1—2,8
взрослые 2,5±0,02 0,19 5,41 74 2,2—2,9
Самки прибылые 2,4±0,01 0,16 6,57 140 2,0—2,8
взрослые 2,5+0,01 0,13 5,42 25 2,2—2,8
В целом по виду . . . 2,4±0,01 0,15 6,37 672 2,0—2,9
Ширина рострума
Самцы прибылые 1,7±0,01 0,10 5,79 152 1,4—2,0
взрослые 1,7±0,01 0,07 3,95 74 1,6—1,9
Самки прибылые 1,7+0,01 0,09 5,17 138 1,5—1,9
взрослые 1,7±0,01 0,06 3,52 24 1,6—1,8
В целом по виду . . . 1,7+0,01 0,10 5,72 669 1,4—2,0
Длина верхнего ряда зубов
Самцы прибылые 7,9±0,02 0,29 3,70 152 7,2—8,5
взрослые 7,6±0,04 0,36 4,78 74 6,7—8,4
Самки прибылые 7,9±0,03 0,34 4,24 139 7,0—9,1
взрослые 7,6+0,06 0,28 3,65 24 7,1—8,1
В целом по виду . . . 7,8±0,01 0,34 4,37 669 6,7—9,1
Длина ряда промежуточных зубов
Самцы прибылые 2,5+0,01 0,15 5,90 154 2,2—2,8
взрослые 2,5+0,02 0,19 7,49 77 2,1—3,0
Самки прибылые 2,5±0,02 0,18 7,16 145 2,1—3,3
взрослые 2,5+0,03 0,14 5,50 28 2,2—2,9
В целом по виду . . . 2,5+0,01 0,18 7,15 695 2,1-3,3
Наибольшая высота черепа
Самцы прибылые 5,8+0,02 0,30 5,08 147 4,2-6,6
взрослые 5,5±0,02 0,21 3,76 72 4,9—5,9
Самки прибылые 5,8±0,02 0,26 4,50 130 4,9—6,4
взрослые 5,5±0,04 0,18 3,36 24 5,0—5,8
В ц е л о м по виду . . . 5,7±0,01 0,31 ' 5,46 650 4,2—6,6
249
Окончание табл. 52
1 Г 2 з | 4 | 5 | 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,2±0,02 0,23 5,48 153 3,8-4,7
взрослые 4,3±0,03 0,23 5,29 75 3,9—4,7
Самки прибылые 4,2±0,02 0,24 5,61 143 3,8—4,8
взрослые 4,2±0,04 0,22 5,16 28 3,9-4,6
В целом по виду . . . 4,2±0,01 0,24 5,64 682 3,8—4,8
Длина углового отростка
Самцы прибылые 2,1±0,01 0,14 6,72 130 1,7—2,5
взрослые 2,1±0,01 0,12 5,80 66 1,7—2,4
Самки прибылые 2,1±0,01 0,12 5,85 121 1,7—2,4
взрослые 2,1±0,02 0,08 3,94 21 2,0—2,3
В целом по виду . . . 2,1±0,01 0,13 6,17 567 1,7-2,5
окраска спины в виде узкого чепрака четко по прямой линии грани-
чит со светлой окраской боков, перехода в окраску брюшной стороны
нет. В области передних конечностей темная окраска спускается на
плечи.
Молодые зверьки окрашены, как и перезимовавшие, но из-за менее
четкой окраски брюшка и боков контрастность слабо выражена, однако
чепрак заметен. Тундряная бурозубка лесной зоны окрашена темнее
и менее контрастно. Наличие чепрака едва улавливается даже у взрослых
зверьков вследствие темной окраски боков и вентральной стороны тела.
Наиболее темноокрашенные бурозубки этого вида заселяют леса Салаира
и прилежащих районов лесной зоны. В целом окраска спины весьма из-
менчива и значения белизны колеблются от 1,1 до 6,4 (М = 3,5±0,08),
а оттенка — 111,3—166,3 (М = 141,0±0,76) (табл. 53). По средним зна-
чениям показателя оттенка тундряная бурозубка имеет самую насыщен-
ную бурыми тонами окраску (см. табл. 30, рис. 76).
Возрастная изменчивость хорошо прослеживается в длине тела взрос-
лых самцов (М = 67,9±0,71) и самок (М = 72,7±1,22), для молодых —
М = 61,9(+0,37) и М = 62.9(^0,47) соответственно. Аналогичная зако-
номерность наблюдается и в отношении массы тела (см. табл. 52). По дру-
гим показателям сколько-нибудь заметных возрастных различий не про-
слеживается.
Тундряная бурозубка по сравнению с другими видами рода характе-
ризуется ярко выраженной географической изменчивостью по морфомет-
рическим признакам (табл. 54). Наименьшая длина тела присуща зверь-
кам популяций южной части ареала (М = 57,0±1,59), далее на север
(бассейн Колымы, Гыданский полуостров) значения этого показателя уве-
личиваются: М=66,1(±0,63), 66,9(±0,7). По средним показателям конди-
ло-базальной длины тела наиболее крупные зверьки обитают в южных час-
тях ареала. На Юго-Восточном Алтае, в Туве, Прибайкалье этот показатель
больше — М =18,1(4=0,12), 18,0(±0,03), чем на западе — М = 16,9(±0,09)
в Барабинской степи. Уменьшается кондило-базальная длина черепа и по
направлению на север: М = 17,4(+0,03) на Чукотке, р. Анадырь и М =
= 16,9(+0,06) в Бурятии.
250
Таблица 53
Колориметрическая характеристика подвидов Sorex tundrensis из разных частей
ареала
Подвид, вид Место сбора материала Белизна
М±тх О Cv п lim
S. t. petschorae Гыданский полуостров, низовья р. Пур, Са- лехард 3,6±0,25 25,3 1,58 37 1,4—5,6
S. t. transrypheus Барабинская степь 2,3+0,11 24,8 0,55 25 1,1—3,7
S. t. sibiriensis Юг Красноярского края, Тува, Прибайкалье 3,8+0,17 22,4 1,21 50 2,4-5,8
S. t. baicalensis Бурятия, р. Джида 4,6+0,21 23,4 1,05 25 2,5—6,4
S. t. buxtoni Бассейн Колымы 3,2+0,18 27,5 0,90 25 1,5-4,8
S. t. stroganovi subsp. n. Приморье 2,7+0,17 27,9 0,79 20 1,2—4,1
Sorex tundrensis По всему ареалу вида 3,6+0,08 32,3 1,13 182 1,1—6,4
Окончание табл. 53
Подвид, вид Место сбора материала Оттенок
М±тх а Су п lim
S. t. petschorae Гыданский полуостров, низовья р. Пур, Сале- хард 136,6±28,5 8,5 17,9 37 112,6—156,1
S. t. transrypheus Барабинская степь 139,2+1,91 6,9 9,55 25 111,3—161,1
S. t. sibiriensis Юг Красноярского края, Тува, Прибайкалье 141,3+1,40 4,8 9,94 50 130,4—156,9
S. t. baicalensis Бурятия, р. Джида 149,9± 1,89 6,3 9,45 25 124,6—166,3
S. t. buxtoni Бассейн Колымы 139,8+1,99 7,1 9,95 25 113,6—155,6
S. t. stroganovi subsp. n. Приморье 141,8±2,04 6,4 8,98 20 123,6—159,4
Sorex tundrensis По всему ареалу вида 141,0+0,76 7,2 10,19 182 111,3—166,3
В изменчивости длины верхнего ряда зубов и рострума прослеживает-
ся та же закономерность, что и в отношении кондило-базальной длины
черепа. Наибольшие значения показателя в популяциях, обитающих на
юге средней части ареала, — юг Красноярского края, Тува, Прибайкалье
(М = 8,0±0,02). Далее на север величина признака падает: Гыданский
полуостров (М = 7,9±0,03) — Чукотка, р. Анадырь (М = 7,6±0,03).
Уменьшение величины признака длины рострума наблюдается также
с юга на север: в Барабинской степи М = 4,4(410,05); на Чукотке,
р. Анадырь М = 4,2(±0,02).
Заметки по систематике. Рассматриваемый вид—один из
наиболее дискуссионных в таксономии землероек. Ни одна другая бурозуб-
ка нашей фауны не претерпела стольких изменений в систематическом плане,
251
Рис. 76. Сравнительная характеристика окраски тундряной бурозубки из разных
частей ареала.
1 — S.t. transrypheus (Барабинская степь); 2—S. t. stroganovi (Приморье); 3 — S. t. petschorae (Гы-
данский полуостров); 4 — St 1. sibiriensis (Тува, Красноярский край); 5 — S. I. baicalensis (Бурятия);
6 — S. t. buxtoni (бассейн Колымы).
сколько выпало на долю 5. tundrensis. Долгое время тундряная бурозубка
в фауне СССР была представлена как 5. arcticus. Арктическая бурозубка
впервые была описана Керром (Kerr, 1792] из Канады. Позднее с территории
Аляски был описан как самостоятельный вид тундряная бурозубка (5. tu-
ndrensis Merriam, 1900). С. У. Строганов в 1936 г. отнес к этому виду буро-
зубку с низовий Колымы (S. araneus ultimus Gl. Allen, 1914). Позднее ряд
авторов [Строганов, 19566; Bee, Hall, 1956; Hall, Kelsen, 1959; и др.] объеди-
нили тундряную бурозубку с арктической (S. arcticus Kerr, 1792). Под этим
названием она существовала до 1980 г., пока В. Е. Соколов и В. Н. Ор-
лов не восстановили видовой статус тундряной бурозубки для территории
Палеарктики и Аляски, считая тундряную и арктическую бурозубок са-
мостоятельными видами. Однако названные исследователи не привели
достаточных обоснований такого разделения.
Кариологические исследования [Иваницкая, Козловский, 1983; Охо-
тина, 1983] подтвердили точку зрения об отсутствии в фауне Палеарктики
арктической бурозубки, распространение которой ограничено Канадой.
Для палеарктических бурозубок, неоправдано отнесенных к 5. arcticus
Kerr, восстановлено их прежнее название — тундряная бурозубка
(S. tundrensis Метт., 1900). В подтверждение этому нами проведено сравне-
ние ряда морфометрических признаков тундряной бурозубки из Пале-
арктики (Аляска, Сибирь, Дальний Восток) и арктической бурозубки из
Неарктики (штат Мичиган). По большинству признаков виды хорошо
различаются (табл. 55).
252
Морфометрическая характеристика
Признак S. tundrensis
(J Су п lim
Длина тела 63,4+0,26 6,16 9,72 574 46—82
хвоста 33,9+0,23 5,42 15,99 576 24-47
ступни 11,6+0,05 1,09 9,38 562 • 8—14
Масса тела 6,1+0,07 1,61 26,27 481 3—12
Длина кондило-базальная черепа 17,6+0,02 0,59 3,33 656 15,7—19,4
рострума 4,4+0,01 0,59 3,33 655 3,5—5,4
мозговой части 10,1+0,01 0,33 3,31 655 8,8—11,0
верхнего ряда зубов 7,8±0,01 0,34 4,37 669 6,3-9,1
ряда промежуточных зубов 2,5+0,01 0,18 7,15 695 1,7—3,3
Ширина рострума 1,7+0,01 0,10 5,72 669 1,4—2,0
предглазничная 2,4±0,01 0,15 6,37 672 2,0—2,9
межглазничная 3,3+0,01 0,15 4,30 695 3,0—4,0
наибольшая черепа 8,8±0,01 0,32 3,62 644 8,0—10,0
Наибольшая высота черепа 5,7±0,01 0,31 5,46 650 4,2—6,6
Высота восходящей ветви ниж- ней челюсти 4,2+0,01 0,24 5,64 682 3,8—4,8
Длина углового отростка 2,1=1=0,01 0,13 6,17 567 1,7—2,5
Признак в. S. t. sibiriensis, юг Красноярского края, Алтай, Ту- ва, Прибайкалье
° Су п lim
Длина тела 62,7+0,40 5,16 8,22 167 50—82
хвоста 37,5±0,25 3,20 8,55 171 29—46
ступни 12,1+0,10 1,28 10,57 159 9—14
Масса тела 6,5±0,12 1,42 22,02 135 4—11
Длина кондило-базальная черепа 18,0±0,03 0,48 2,67 238 16,8—19,2
рострума 4,6±0,01 0,22 4,79 249 4,1—5,1
мозговой части 10,2+0,02 0,32 3,15 239 9,0—11,0
верхнего ряда зубов 8,0±0,02 0,28 3,46 248 7,0—8,6
ряда промежуточных зубов 2,6±0,01 0,01 6,57 249 2,1—2,9
Ширина рострума 1,7±0,01 0,09 5,45 249 1,5—2,0
предглазничная 2,5+0,01 0,15 6,29 249 2,0—2,9
межглазничная 3,5±0,01 0,14 3,93 249 3,1—4,0
наибольшая черепа 9,0±0,02 0,28 3,14 236 8,0—9,9
Наибольшая высота черепа 5,8+0,02 0,33 5,60 240 4,9—6,6
Высота восходящей ветви ниж- ней челюсти 4,4±0,01 0,22 5,10 244 3,9—4,8
Длина углового отростка 2,1±00,1 0,12 5,74 210 1,8—2,4
253
Таблица 54
Sorex tundrensis и ее подвидов
a. S. t. borealis, Чукотка, р. Анадырь б- S. t. buxtoni, бассейн р. Колымы
М±тх а Су п lim т-±Мх а Су п lim
64,5+0,59 5,56 8,61 89 51—81 66,1+0,63 6,61 9,99 111 48—80
30,6+0,23 2,19 7,17 88 25-39 32,1+0,25 2,62 8,16 но 24-37
11,3+0,06 0,51 4,55 83 10—13 11,7+0,06 0,62 5,37 111 10—13
5,7+0,14 1,29 22,63 81 4—11 6,6+0,19 1,97 29,80 109 4-10
17,4+0,03 0,30 1,71 95 16,7—18,0 17,6+0,03 0,34 1,93 126 16,6—18,4
4,2+0,02 0,20 4,69 92 3,8—4,7 4,5±0,02 0,21 4,65 126 3,4—5,0
ю,о±о,оз 0,24 2,40 91 9,1—10,6 10,0+0,02 0,23 2,26 126 9,5—10,7
7,6+0,03 0,25 3,34 91 6,7—8,0 7,7±0,02 0,23 2,99 126 7,1—8,2
2,5+0,01 0,13 5,15 91 2,2—2,7 2,5+0,01 0,10 4,16 126 2,1—2,8
1,7+0,01 0,08 4,70 91 1,5—1,8 1,7+0,01 0,10 5,76 126 1,4—2,0
2,3+0,01 0,13 5,75 91 2,0—2,7 2,4±0,01 0,14 5,93 126 2,0—2,7
3,4±0,01 0,11 3,33 91 3,2—3,7 3,4+0,01 0,14 4,03 126 3,0—3,8
8,7+0,02 0,24 2,69 89 8,0—9,4 8,8+0,02 0,24 2,74 125 8,1—9,4
5,7±0,03 0,26 4,55 91 5,0—6,3 5,7+0,02 0,25 4,39 125 4,7—6,2
4,0+0,01 0,10 2,45 91 3,8—4,2 4,1+0,01 0,15 3,65 123 3,8-4,8
2,0+0,02 0,13 6,32 70 1,7—2,4 2,1±0,01 0,10 5,16 113 1,8-2,3
Продолжение табл. 54
г. S. t. transrypheus, Барабинская степь д. S. t. petschorae, Гыданский полуостров
М+тх <т Су п lim М+тпж а Су п lim
57,0+1,59 7,44 13,05 22 46—72 66,9+0,70 4,29 6,41 38 60-82
33,1+0,61 2,82 8,50 21 27—37 39,1+0,60 3,70 9,47 38 34-46
10,6+0,19 0,90 8,48 22 9—12 11,8±0,09 0,58 4,88 38 11—13
6,5+0,73 1,26 19,46 3 5,0—8,0 6,6+0,25 1,52 22,98 36 4-11
16,9+0,09 0,44 2,57 26 16,0—17,5 17,9+0,04 0,28 1,54 57 17,3-18,5
4,4+0,05 0,34 8,11 39 3,5—4,7 4,7+0,02 0,14 2,91 59 4,3-5,0
9,7+0,07 0,37 3,84 26 8,8—10,2 10,1+0,03 0,24 2,33 58 9,3—10,5
7,4+0,05 0,27 3,58 27 6,9—8,2 7,9±0,03 0,23 2,95 59 7,4—8,3
2,3+0,02 0,13 5,36 39 2,0—2,6 2,6+0,01 0,08 2,95 59 2,4—2,8
1,7+0,02 0,10 6,12 27 1,5—1,9 1,7+0,01 0,07 4,21 59 1,5-1,8
2,4+0,03 0,16 6,76 28 2,1—2,8 2,4±0,01 0,11 4,40 59 2,1—2,6
3,4+0,02 0,12 3,61 39 3,2—3,7 3,4±0,02 0,12 3,70 59 3,1-3,6
8,7+0,06 0,27 3,10 23 8,1-9,2 8,8+0,03 0,22 2,54 57 8,2—9,4
5,5+0,05 0,24 4,37 23 5,0-5,9 5,7±0,04 0,29 5,10 57 5,2—6,3
4,1+0,02 0,13 3,20 36 3,8—4,4 4,1+0,02 0,12 3,10 58 3,8—4,3
2,0+0,03 0,16 8,10 22 1,7—2,4 2,0+0,01 0,10 4,83 47 1,8-2,3
254
Признак ж. 8. t. schnitnikovi, Центр, и Юж. Алтай
(J cv п lim
Длина тела 64,1+0,76 5,00 7,78 43 52—74
хвоста 40,5±0,50 3,36 8,30 45 32—47
ступни 12,0±0,15 0,96 8,02 44 10—14
Масса тела 5,7+0,18 0,83 14,60 22 5—7
Длина кондило-базальная черепа 18,1±0,12 0,63 3,50 30 16,7—19,4
рострума 4,4+0,06 0,42 9,50 42 3,6-5,4
мозговой части 10,3+0,05 0,29 2,85 30 9,5—11,0
верхнего ряда зубов 8,1±0,06 0,32 3,93 30 7,5-9,1
ряда промежуточных зубов 2,6±0,04 0,28 10,69 43 2,4-3,0
Ширина рострума 1,8±0,03 0,14 7,96 28 1,5—2,0
предглазничная 2,4±0,04 0,19 7,79 30 2,2—2,8
межгл азничная 3,5±0,02 0,14 4,08 42 3,2—3,8
наибольшая черепа 9,1 ±0,06 0,34 3,75 30 8,3—10,0
Наибольшая высота черепа 5,7±0,06 0,33 5,77 29 4,2—6,2
Высота восходящей ветви ниж- ней челюсти 4,5±0,03 0,17 3,73 41 4,2—5,0
Длина углового отростка 2,1±0,03 0,15 6,91 25 1,8—2,5
Таблица 55
Сравнительная характеристика арктической
и тундряной бурозубок
Признак S. tundrensis, Сибирь, Дальний Вос- ток S. tund- rensis, Аляска, Xs 40771 S. arcticus lariocum, штат Мичиган
Xs 551720 X» 551722
Длина кондило-базальная черепа 17,6 17,6 19.2 19,0
рострума 4,4 4,6 5,0 5,1
мозговой части 10,1 10,0 10,9 10,8
верхнего ряда зубов 7,8 6,4 8,3 8,3
ряда промежуточ- ных зубов 2,5 2,5 2,7 2,8
Ширина рострума 1,7 1,5 1,7 1,7
пред гл азничная 2,4 2,2 2,5 2,5
межгл азничная 3,4 4,3 3,5 3,4
наибольшая черепа 8,8 9,0 9,5 9,4
Наибольшая высота че- репа 5,7 6,0 6,3 6,2
Высота восходящей вет- ви нижней челюсти 4,2 4,0 4,4 4,4
Длина углового отростка 2,1 2,2 2,4 2,3
255
Окончание табл. 54
3. S. t. baicalensis, Бурятия, р. Джида е. S. t. stroganovi subsp. n., Приморье
М+тх а Су п lim M±mx а cv n lim
60,3+0,70 6,33 10,49 82 48—80 60,8±0,79 3,94 6,48 25 55-72
25,8+0,35 3,15 12,20 81 20—33 35,7+0,51 2,56 7,16 25 31—40
10,6±0,13 1,14 10,71 82 8—12 12,0±0,13 0,65 5,38 25 11—13
5,4±0,17 1,44 26,79 76 3—10 5,4±0,28 1,35 25,04 23 4-8
16,9+0,06 0,47 2,80 67 15,7—17,6 17,4+0,07 0,34 1,96 21 17—18
4,2±0,03 0,26 6,12 67 3,5—4,6 4,4+0,03 0,14 3,13 25 4,2-4,7
9,7±0,04 0,34 3,52 67 8,0—10,9 10,0+0,04 0,20 2,01 21 9,7—10,4
7,4+0,04 0,30 4,09 67 6,3—8,0 7,7+0,05 0,24 3,10 24 7,3—8,1
2,3+0,02 0,19 8,44 67 1,7—2,6 2,4+0,02 0,11 4,52 24 2,1—2,5
1,7+0,01 0,09 5,31 67 1,5—2,0 1,7+0,02 0,08 4,49 25 1,5-1,8
2,3±0,02 0,13 5,74 67 2,0—2,6 2,4+0,02 0,11 4,76 25 2,1—2,5
3,4±0,02 0,13 4,07 67 3,1—3,7 3,4+0,03 0,13 3,89 25 3,1—3,7
8,6±0,03 0,27 3,10 66 8,0—9,0 8,6+0,04 0,17 1,98 21 8,3-9,0
5,5±0,04 0,34 6,17 67 6,3—8,0 5,7+0,04 0,19 3,32 21 5,3—6,1
4,1±0,02 0,16 3,83 59 3,8—4,6 4,2+0,03 0,16 3,82 25 3,8—4,4
2,0+0,01 0,10 5,20 59 1,7—2,2 2,0+0,02 0,11 5,84 24 1,7—2,2
Рис. 77. Череп арктической бурозубки (S. arcticus lariocum Кегг), штат Мичиган
(США).
256
Различия в строении черепа сводятся к следующему:
Признак S. arcticus
(штат Мичиган)
Череп Крупный, широкий в мозговой
части, узкий в ростральной, с
овальной мягких очертаний
мозговой капсулой, высокий
(рис. 77)
Постмандибу- Нет
лярное отвер-
стие
Передние резцы Средних размеров, их вторая
вершина лишь немного усту-
пает по величине первой, на
внутренней стороне передней
лопасти хорошо развиты до-
полнительные зубцы. Канав-
ки, разделяющей лопасти,
нет
Промежуточ- Крупные, постепенно уменыпа-
ные зубы ются по высоте, лишь пятый
в 3 раза меньше четвертого,
хотя и хорошо развит. Пиг-
ментация уменьшается от пер-
вого зуба к третьему, четвер-
тый и пятый белые
8. tundrensis
(Аляска)
Такой же, как и у бурозубок с
территории СССР, т. е. мень-
ших размеров, чем у S. arc-
ticus, широкий, массивный,
сужение в ростральной части
выражено слабо (рис. 78).
Есть
Немного меньше такового у
У. arcticus, с хорошо развитой
передней лопастью и разделя-
ющей канавкой
Риц
Умеренно вытянут в длину,
довольно высокий и массив-
ный, без дополнительной
вершины (см. рис. 77)
Менее массивные и с острыми
вершинами. Первые два зуба
почти равны по высоте. Вер-
шина внутренней лопасти пе-
реднего резца ниже вершины
первого промежуточного зуба,
который ниже вершины вто-
рого; третий промежуточный
меньше второго (его внутрен-
няя сторона выгнута, благо-
даря чему он выглядит изящ-
нее двух предыдущих), пя-
тый хорошо развит и направ-
лен в противоположную сторо-
ну от предыдущих промежу-
точных зубов. Пигментация
постепенно убывает от перво-
го к пятому
Вытянут в длину, с небольшой
дополнительной вершиной
(см. рис. 78)
По окраске меха и строению glans penis тундряная бурозубка близка
к S. araneus. Размеры черепа близки таковым средней бурозубки, отличия
заключаются в его конфигурации и в строении зубов. У 5. tundrensis он
толстокостный, массивный, с высокой мозговой и лицевой областями.
Ростральная часть более короткая и широкая, чем у средней бурозубки.
Верхние промежуточные зубы в челюсти сидят плотно (у средней бурозуб-
ки они редко расставлены). Их вершины направлены внутрь ротовой по-
лости (у средней вертикальная ось коронок перпендикулярна оси челю-
сти). У последней зубы в виде изящных островершинных пирамид,
у тундряной они более грубой конфигурации, с менее острыми загнутыми
вершинами. В ряду верхних промежуточных зубов выше других поднима-
ется вершина второго зуба. Вершина первого промежуточного примерно
на одном уровне с задней вершиной первого резца и вершиной третьего
промежуточного зуба. Четвертый меньше третьего; пятый уступает по
размерам и высоте четвертому. Различия в высоте четвертого и пятого зу-
257
Рис. 78. Череп тундряной бурозубки (5. tundrensis Метт.) с Аляски,
ба меньше, чем у S. caecutiens. Рш1 значительно вытянут в сагиттальном
направлении.
С территории Сибири и Дальнего Востока описано восемь подвидов
тундряной бурозубки.
Таблица 56
Достоверность различий (по Стьюденту) между подвидами Sorex tundrensis
Признак а—б а—в а—г а-д а—ж
Длина
тела 1,9 2,5 4,2 2,6 0,4
хвоста 4,4 20,3 3,8 13,2 17,9
ступни 4,7 6,9 3,5 4,6 4,3
Масса тела 3,8 4,3 1,1 3,1 0
Длина
кондило-базальная черепа 4,7 14,1 5,3 10,0 5,7
рострума 10,6 17,9 3,7 17,8 3,2
мозговой части 0 5,5 3,9 2,4 5,2
верхнего ряда зубов 2,8 11,1 3,4 7,1 7,5
ряда промежуточных зубов 0 7,1 8,9 7,1 2,4
Ширина
, рострума 0 0 0 0 3,1
предглазничная 7,1 14,1 3,2 7,1 2,4
межглазничная 0 7,1 0 0 4,5
наибольшая черепа 3,5 10,6 0 2,8 6,3
Наибольшая высота черепа 0 2,8 3,4 0 0
Высота восходящей ветви нижней
челюсти 7,1 28,3 4,5 4,5 15,8
Длина углового отростка 4,5 4,5 0 0 2,8
17 Б. С. Юдин
258
Признак а—з а—е б—в б—г б-д б—ж
Длина
тела 4,6 3,8 4,6 5,7 0,8 2,1
хвоста 11,5 9,1 15,3 1,5 10,8 15,0
ступни 4,1 4,9 3,4 5,5 0,9 1,9
Масса тела Длина кондило-базальная черепа 1,4 0,9 0,5 0,1 0 3,4
7,5 0 9,4 7,4 6,0 4,1
рострума 0 5,5 4,5 7,4 7,1 1,6
мозговой части 6,7 0 7,1 0,8 2,8 5,6
верхнего ряда зубов 4,0 1,7 10,6 5,6 5,5 6,3
ряда промежуточных зубов Лирина 8,9 4,5 7,1 8,9 7,1 2,4
рострума 0 0 0 0 0 3,2
предглазничная 0 4,5 7,1 0 0 0
межглазничная 0 0 7,1 0 0 4,5
наибольшая черепа 2,8 2,2 7,1 1,6 0 4,7
Наибольшая высота черепа Высота восходящей ветви ниж- 4,0 0 3,5 3,7 0 0
ней челюсти . 4,5 6,3 21,2 0 0 12,6
Длина углового отростка 0 0 0 0 7,1 0
Обзор подвидов
Sorex tundrensis borealis Kastshenko, 1905 —
Северная тундряная бурозубка
Голотип. Описан по экземплярам из коллекции А. Ф. Мидден-
дорфа, находящейся в ЗИНе АН СССР.
Диагноз. Зверьки средней величины, хвост относительно корот-
кий, окраска чепрачная, с подпалинами.
Измерения. См. табл. 54.
Характеристика. Среди других форм 5. tundrensis бурозуб-
ка хорошо обособлена. Ее характерной чертой является наличие резко
очерченного чепрака. Темный тон верхней части тела почти не спускается
на бока, проходит на уровне глаз и ушей, бока светлые, переходов нет.
Окраска спины и боков устойчивая. На плечах расположено по бурова-
то-коричневому пятну, которые не сливаются с темной спиной. Бурые
пятна на боках связаны с темной окраской задней части спины. Окраска
старых зверьков более яркая и контрастная. Длина зимнего меха — 8,2,
летнего — 3,2 мм. Окраска зимнего меха тусклая. По морфометрическим
признакам подвид хорошо обособлен от других (табл. 56).
Распространение. Ареал северной тундряной бурозубки ох-
ватывает зону тундр от Енисейского залива и низовий Енисея до Чукотки
и Камчатки включительно.
Материал. 140 экз., Якутия, Таймыр, Чукотка.
Sorex tundrensis buxtoni Allen, 1903 —
Восточносибирская тундряная бурозубка
Голотип. $, ad., 27/VII 1901 г., из сборов Н. Г. Бекстона
(N. G. Buxton). Окрестности Гижиги. Американский музей естественной
истории, № 18655.
Диагноз. По размерам близка к предыдущему подвиду.
259
Продолжение табл. 56
б—3 б—е В—Г в-д в—Ж В—3 в—е Г-д Г —Ж Г—3 г—е Д—ж
6,2 5,2 3,5 5,2 1,6 2,9 2,1 5,7 4,0 1,9 2,1 2,7
15,1 6,3 6,7 2,5 5,4 27,2 3,2 7,0 0,4 10,4 3,3 1,8
4,8 2,1 6,9 2,2 0,6 9,1 0,6 5,7 5,8 0 6,1 1,1
4,7 3,5 0 0,4 3,7 5,3 3,6 0,1 1,0 1,5 1,4 2,9
10,4 2,6 11,6 2,0 0,8 16,4 7,9 10,2 8 0 4,4 1,6
8,3 2,8 9,8 4,5 3,3 12,6 6,3 11,1 3,8 1,7 5,2 4,7
8,3 0 6,9 2,8 1,9 11,2 4,5 5,3 6,9 0 3,7 3,4
6,7 0 3,2 2,8 1,6 13,4 5,6 8,6 8,9 0 4,2 2,9
8,9 3,5 11,1 0 0 13,4 8,9 9,5 6,7 0 3,5 0
0 0 0 0 3,2 0 0 0 2,8 0 0 3.2
4,5 0 3,2 7,1 2,4 8,9 4,5 0 0 0,5 0 0
0 0 4,5 4,5 0 4,5 3,2 0 3,5 0 0 3,5
5,5 4,5 4,7 5.5 1,6 11,1 8,9 1,5 4,7 1,5 1,4 4,5
4,5 0 5,6 2,2 1,6 6,7 2,2 3,1 2,6 0 3,1 0
0 3,1 13,4 13,4 3,2 13,4 6,3 0 11,1 0 2,8 11.1
7,1 4,5 3,2 7,1 0 7,1 4,5 0 2,4 0 0 3,2
Окончание табл. 56
Признак д—3 д-е Ж—3 ж—е з—е
Длина 0,5
тела 6,7 5,8 3,7 3,0
хвоста 19,1 4,8 24,1 6,7 16,0
ступни 7,6 1,3 7,1 0 7,7
Масса тела Длина кондило-базальная черепа 3,9 3,2 1,2 0,9 0
13,9 6,2 8,9 5,1 5,4
рострума 13,9 ' 8,3 2,9 0 4.7
мозговой части 8 2,0 9,4 4,7 5,3
верхнего ряда зубов 10,0 3,4 9,7 5.1 4,7
ряда промежуточных зубов Ширина 13,4 8,9 6,7 4,5 3,5
рострума 0 0 3,2 2,8 0
предглазничная 4,5 0 2,2 0 3,5
межгл азничная 0 0 3,5 2,8 0
наибольшая черепа 4,7 4,0 7,5 6,9 0
Наибольшая высота черепа Высота восходящей ветви нижней 3,5 0 2,8 0 3,5
челюсти 0 2,8 11,1 7,1 28
Длина углового отростка 0 0 3,2 2,8 0
Примечание. Буквенные обозначения подвидов см. табл. 54.
Измерения. См. табл. 54.
Характеристика. Темная окраска спины в области плеч
переходит на лапы в виде размытого бурого пятна различной интенсивно-
сти, у молодых зверьков зто либо совсем не выражено, либо едва заметно.
Граница между окраской спины и боков не резкая. У старых зверьков
окрас контрастный, на границе спины и боков — лимонно-палевые под-
палины, брюшко грязно-серого цвета со слабой примесью палевых тонов.
17*
260
Бока у молодых зверьков буровато-серые. По морфометрическим показа-
телям подвид близок к номинальному (см. табл. 54).
Распространение. Лесная полоса Сибири, от бассейна Вилюя
и нижнего течения Витима на восток до побережья Охотского моря на
север до криволесья, а на юг — до лесостепного Забайкалья и Амурской
области.
Материал. 75 зкз., Чукотка; 36 зкз., Якутия.
Sorex tundrensis sibiriensis Ognev, 1921 —
Западносибирская тундряная бурозубка
Голотип. $, 17/VIII 1899 г., с. Кольчугино Кузнецкого уезда
Томской губернии (ныне ст. Кольчугино, в 4 км к северо-востоку от г. Ле-
нинск-Кузнецкий, Кемеровская обл.). Коллекция ЗИН АН СССР,
№ 6527.
Диагноз. Для зверьков характерны темная окраска меха и длин-
ный хвост.
Измерения. См. табл. 54.
Характеристика. Окраска меха старых зверьков чепрачная.
Спина темно-бурая без рыжих оттенков, отсутствует резкая граница ме-
жду спиной и боками. Темная окраска спины заходит по плечам на бедра.
Бока темные буровато-палево-серые с грязным оттенком. Хвост контраст-
но окрашен в тон, соответствующий спине и брюшку. Лапы буровато-
серые. В популяции доминируют особи без чепрака. Окраска подвида
стабильна в разных частях ареала (рис. 79). По морфометрическим пока-
зателям этот подвид отличается от других крупными размерами, длиной
хвоста (см. табл. 54), ближе всех по ряду признаков он стоит к семире-
ченской тундряной бурозубке.
Распространение. Таежная зона Западной и Средней Сиби-
ри, Кузнецкий Алатау, Северо-Восточный Алтай, Саяны, Тува; на во-
сток — до Байкала, Нижней Тунгуски и нижнего течения Енисея.
Мат е р и а л. 26 экз., Тува; 6 зкз., Алтай; 22 экз., Тюменская об-
ласть; 274 экз., Иркутская область.
Sorex tundrensis transrypheus Stroganov, 1956 —
Зауральская тундряная бурозубка
Голотип, d1, ad., 24/IV 1952 г. Окрестности г. Кустаная. Из кол-
лекции С. У. Строганова, № 1608.
Диагноз. Отличается от других подвидов небольшими размерами
и длинным хвостом. У большинства
особей расцветка чепрачная, с желто-
вато-охристыми подпалинами.
Измерения. См. табл. 54.
Характеристика. Чеп-
рачность хорошо выражена, спина
Рис. 79. Сравнительная характеристика
окраски западносибирской и забайкаль-
ской тундряных бурозубок.
1 — S. t. sibiriensis (Тува, юг Красноярского
края); 2 — S.t. sibiriensis (Малый Хамар-Дабан,
Самхат, Забайкалье); 3 — S. t. baicalensis (Буря-
тия, р. Джида, с. Дарестуй).
261
темно-бурая с блестящим шелковистым мехом, на боках он светлый, с
едва уловимым палевым оттенком. Хвост резко двухцветный, уши бурые.
Окраска меха вокруг ушей светлая. На плечах различимы бурые пятна,
у молодых они сливаются с чепрачной окраской меха. Подвид характери-
зуется самыми низкими значениями белизны (М = 2,3±0,11). Заураль-
ская тундряная бурозубка — самый мелкий, хорошо очерченный подвид,
достоверно отличается от других подвидов по ряду морфометрических
признаков (см. табл. 56).
Распространение. Широко распространен в лесостепной зо-
не Западной Сибири и Северного Казахстана от Кустаная на восток до
Алтая, а на севере — от 56° с. ш. к югу до оз. Кургальджин (Акмолин-
ская обл.) и южнее до Карагандинской области.
Материал. 38 экз., Барабинская степь.
Sorex tundrensis petschorae Ognev, 1921 —
Печорская тундряная бурозубка
Голотип. $ , ad., 1906 г., А. В. Журавский. Низовье Печоры.
Коллекция ЗИН АН СССР, № 8459.
Диагноз. По внешнему виду и окраске близка к S. t. borealis,
по более длиннохвостая и темнее окрашена.
Измерения. См. табл. 54.
Характеристика. В окраске меха резко выделяется чепрач-
ность. Светлые тона распространяются от щек, между ушей и глаз на
лоб. В области плеч едва заметны бурые пятна, не соединяющиеся с чеп-
раком. Бока и брюшко серебристо-белые. Средние значения оттенка
(М = 136,6+28,5)— самые минимальные из всех подвидов. По совокуп-
ности внешних признаков этот подвид близок к S. t. schnitnikovi. Различия
недостоверны по длине тела, хвоста, кондило-базальной длине черепа
и одонтологическим признакам (см. табл. 54).
Распространение. Ареал подвида охватывает тундровую зо-
ну европейского Севера и Западной Сибири, на восток идет до Енисея.
Материал. 20 экз., Гыданский полуостров; 40 экз., Ямал,
Самбург.
Sorex tundrensis schnitnikovi Ognev, 1921 —
Горная, или семиреченская, тундряная бурозубка
Голотип. <5*, 1910 г., из сборов В. Н. Шнитникова. Окрестности
г. Копала, Семиречья. Коллекция ЗИН АН СССР, № 8641.
Диагноз. Из немногих крупных подвидов самый длиннохвостый
зверек. Чепрак отсутствует.
Измерения. См. табл. 54.
Характеристика. Летняя окраска меха спины и боков одно-
тонная, брюшко дымчато-серое. Семиреченская тундряная бурозубка
горная форма, характеризуется крупными телом (М = 64,1 ±0,76), чере-
пом (М — 18,1±0,12) и чрезвычайно длинным хвостом (М = 40,5±0,50).
По морфометрическим признакам наиболее близка к S. t. sibiriensis. Не-
смотря на это по ряду показателей достоверно отличается от него длиной
хвоста (Td = 5,4), длиной (Td = 3,3) и шириной рострума (Td = 3,2).
По большинству показателей различия недостоверны (см. табл. 56).
Распространение. Населяет центральные и южные районы
Алтая, долину Чулышмана; окрестности оз. Джулу-Куль; добывалась на
18 Б. С. Юдин
262
территории Алтайского заповедника, отлавливалась на хребтах Тарба-
гатай, Саур и Джунгарский Алатау.
Материал. 24 экз., Джунгарский хребет, Колбинский р-н;
20 экз., Алтайский заповедник.
Sorex tundrensis baicalensis Ognev, 1913 —
Забайкальская тундряная бурозубка
Голотип. $ , 1891 г., Ю. Н. Вагнер. Забайкалье, Горный Зерен-
туй. Коллекция Зоологического музея МГУ, № 2710.
Диагноз. Зверьки мелкие, короткохвостые. Окраска меха очень
светлая.
Измерения. См. табл. 54.
Характеристика. Забайкальская тундряная бурозубка от-
личается от других подвидов мелким черепом, отсутствием или слабо вы-
раженной чепрачностью в окраске меха, коротким хвостом. По большин-
ству морфометрических признаков близка к зауральской тундряной буро-
зубке, достоверно отличается от этого подвида лишь длиной хвоста
(М ~ 25,8+0,35; Td — 10,4), по другим показателям достоверных отли-
чий нет (см. табл. 56). Для подвида характерны самые высокие значения
показателей белизны (М = 4,6±0,21) и оттенка (М — 149,9±1,89)
(см. рис. 79). Бока и брюшко палево-серые. Хвост двухцветный. Летний
мех короткий (3,2 мм), блестящий, зимний на спине длинный (6,8 мм).
Среди светлоокрашенных особей встречаются полностью или частично
выраженные меланисты (добыты Ю. Г. Швецовым в районе Селенги близ
Наушки).
Распространение. Южные степные районы Забайкалья, от
долины Селенги к востоку до Бурей; Северная Монголия.
Материал. 62 экз., долина Джиды; 2 экз., Наушки, р. Селенга;
4 экз., Монголия, Южный Хангай, Уган-Тур.
Sorex tundrensis stroganovi subsp. n.—
Тундряная бурозубка Строганова 10
Голотип. № 11426, коллекция ЗИН АН СССР, А. И. Черский.
Окрестности оз. Ханка.
Диагноз. Длина тела с головой 55—72 (М = 60,8+0,79) мм.
От восточносибирской и южнозабайкальской форм отличается длиной
хвоста (М ~ 35,7±0,51) мм. Длина ступни 11—13 (М = 12,0+0,13),
кондило-базальная длина черепа 17—18 (М — 17,4+0,7) мм; масса 4—8
(М = 5,4±0,28) г. Характерна сильная пигментация эмали коронок
зубов.
Измерения. См. табл. 54.
Характеристика. Окраска чепрачная, коричневого тона, пе-
редняя часть спины светлее задней, окраска меха до ушей и за ушами
светлая. Бока палево-серые, брюшко такого же цвета, но светлых тонов.
Хвост почти однотонный, бурый, чуть светлее с вентральной стороны.
Значения показателя белизны низкие (М = 2,7±0,17), оттенка — М =
= 141,8±0,04. По последнему признаку уступает лишь 5. t. baicalensis —
М = 149,9+1,89 (см. табл. 53).
10 С. У. Строгановым [1957] сделано описание подвида по одному экземпляру.
Диагноз нами уточнен и дополнен на основании изучения 28 экз. Дано название.
263
Распространение. Приморье, с. Троицкое близ оз. Ханка;
заповедник «Кедровая Падь»; Надеждинский р-н, с. Раздольное.
Материал. 4 экз., Кедровая Падь; 19 экз., окрестности оз. Хан-
ка; 5 экз., с. Раздольное.
Sorex caecutiens Laxmann, 1788 —
Средняя бурозубка
Голотип. Описан по экземпляру с юго-западного побережья
Байкала. Типовой экземпляр был передан Лаксманном в Академию наук,
но был утрачен.
Диагноз. Бурозубка средних размеров. Длина тела с головой
45—47 (М = 60,5^-0,28) мм. Голова широкая в затылочной части с тон-
ким лицевым отделом. Ряд промежуточных зубов сильно вытянут в длину
(35,1 % от кондило-базальной длины). Коронки промежуточных зубов
мелкие, чрезвычайно вытянуты в сагиттальном направлении, свободно
сидящие в зубном ряду (см. рис. 81). Первые четыре прмежуточных зуба
образуют парные группировки: два первых, а также третий и четвертый
почти равны между собой. Пятый крайне мал и резко уступает в размерах
четырем предыдущим, как бы создающим единую группу.
Измерения. См. табл. 57.
Характеристика. Во внешнем облике зверька обращает на
себя внимание стройное телосложение, хорошо заметное сужение головы
в области глаз, вздутость мозгового отдела черепа и хорошо развитый хо-
боток (рис. 80, 81).
Окраска тела трехцветная: спина темная с преобладанием бурого
и серого цветов, на боках к ним примешиваются рыжие тона, особенно
Рис. 80. Средняя бурозубка — S. caecutiens Laxm.
18*
264
Таблица 57
Морфометрическая характеристика Sorex caecutiens
Возрастная и половая группа М±тх а Су п lim
Самцы прибылые Длина тела 58,3+0,43 с головой 4,36 7,47 105 45—69
взрослые 67,1±0,49 3,92 5,84 63 52—74
Самки прибылые 57,3±0,40 4,25 7,41 110 46—70
взрослые 68,3+0,81 5,18 7,59 41 55—78
В ц е л о м по виду . . . 60,5+0,28 6,53 10,79 556 45—78
Длина хвоста
Самцы прибылые 38,9+0,13 0,99 8,16 105 26—60
взрослые 35,7+0,45 3,59 10,04 63 30—47
Самки прибылые 36,9+0,46 4,84 13,12 109 30—58
взрослые 36,1+0,84 5,41 15,0 42 27—57
В целом по виду , . . 36,1+0,22 5,15 14,27 555 26—60
Длина ступни
Самцы прибылые 12,1+0,10 0,71 15,23 104 10—14
взрослые 11,9+0,08 0,61 5,15 63 11—14
Самки прибылые 11,9+0,09 0,87 7,34 99 10—14
взрослые 11,8+0,13 0,84 7,10 42 10—14
В целом по виду . . • 11,7+0,05 1,05 9,01 527 10—14
Масса тела
Самцы прибылые 4,7+0,07 0,71 15,23 96 3,5—7,0
взрослые 7,2+0,12 0,98 13,60 61 4,0—9,0
Самки прибылые 4,5+0,08 0,83 18,26 99 3,0—7,0
взрослые 7,0±0,24 1,41 20,35 35 4,0—11,0
В целом по виду . . . 5,3+0,07 1,49 27,97 521 3,0—11,0
Кондило-базальная длина черепа
Самцы прибылые 17,6±0,06 0,75 4,23 134 16,4—19,3
взрослые 17,5+0,07 0,57 3,26 62 16,2—18,5
Самки прибылые 17,6+0,06 0,69 3,93 131 16,1—19,4
взрослые 17,4+0,10 0,65 3,74 45 16,2—18,9
В целом по виду . . . 17,4+0,03 0,87 4,97 812 16,1—19,4
Длина рострума
Самцы прибылые взрослые 4,7+0,03 4,6±0,03 0,34 0,27 7,11 5,86 137 66 3,9—5,5 4,1—5,6
Самки прибылые 4,7+0,03 0,33 7,07 135 4,0—5,5
взрослые 4,6±0,05 0,31 6,70 43 3,9—5,4
В целом по виду . . . 4,6+0,01 0,34 7,39 834 3,9—5,6
Длина мозговой части черепа
Самцы прибылые 9,9±0,04 0,41 4,16 134 8,9—10,8
взрослые 9,9+0,03 0,30 3,03 62 9,3—10,6
Самки прибылые 9,9±0,04 0,41 4,11 131 8,8—11,4
взрослые 9,8+0,05 0,34 3,44 41 9,0-10,5
В целом по виду . . . 9,8+0,02 0,48 4,84 812 8,8—11,4
265
Продолжение табл. 57
1 3 3 * 5 6
Наибольшая ширина черепа
Самцы прибылые 8,8+0,03 0,31 3,51 134 7,8-9,5
взрослые 8,8+0,03 0,26 2,97 62 8,2—9,5
Самки прибылые 8,7+0,03 0,32 3,66 131 8,0-10,0
взрослые 8,7+0,05 0,34 3,94 41 8,0—9,8
В целом по виду . . . 8,7+0,01 0,42 4,48 809 7,8—10,0
Межглазничная ширина
Самцы прибылые 3,3+0,01 0,16 4,71 137 2,7—3,7
взрослые 3,4±0,01 0,12 3,58 66 3,1—3,6
Самки прибылые 3,3+0,01 0,17 5,08 135 2,7—3,7
взрослые 3,3±0,02 0,14 4,18 43 3,0—3,7
В целом по виду . . . 3,3+0,01 0,18 5,64 834 2,7—3,7
Предглазничная ширина
Самцы прибылые 2,3+0,01 0,16 6,72 137 2,0—2,8
взрослые 2,4±0,01 0,12 5,17 66 2,1—2,7
Самки прибылые 2,3+0,05 0,64 7,19 135 2,0—2,7
взрослые 2,3+0,02 0,14 5,99 43 2,0—2,7
В целом по виду . . . 2,3+0,01 0,30 12,93 834 2,0—2,8
Ширина рострума
Самцы прибылые 1,5±0,01 0,10 6,28 137 1,3—1,7
взрослые 1,6±0,01 0,09 5,64 66 1,4—1,9
Самки прибылые 1,5+0,01 0,11 7,0 135 1,1—1,8
взрослые 1,6+0,01 0,06 3,74 43 1,5—1,7
В целом по виду . . . 1,6+0,01 0,13 8,10 834 1,1-1,9
Длина~верхнего ряда зубов
Самцы прибылые 7,9±0,03 0,38 4,78 135 6,9—8,6
взрослые 7,6+0,04 0,35 4,61 66 6,7—8,3
Самки прибылые 7,9±0,03 0,40 5,04 134 7,1—8,8
взрослые 7,6±0,06 0,40 5,21 43 7,0—8,4
В целом по виду . . . 7,7±0,02 0,46 5,98 831 6,7—8,8
Длина ряда промежуточных зубов
Самцы прибылые 2,7±0,01 0,18 6,75 135 2,2—3,1
взрослые 2,7±0,02 0,18 6,57 66 2,3-3,0
Самки прибылые 2,8±0,02 0,22 7,76 135 2,3—3,2
взрослые 2,7+0,03 0,20 7,46 43 2,4-3,2
В целом по виду . . . 2,7±0,01 0,23 8,36 832 2,2—3,2
Наибольшая высота черепа
Самцы прибылые 5,7±0,02 0,28 4,85 133 4,8—6,3
взрослые 5,4±0,02 0,18 3,40 61 5,0—6,0
Самки прибылые 5,6±0,03 0,30 5,04 130 4,9—6,5
взрослые 5,3+0,04 0,23 4,35 41 4,8—5,9
В целом по виду . . . 5,6±0,01 0,35 6,31 808 4,8—6,3
266
Окончание табл. 57
1 2 3 4 5 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,1±0,02 0,27 6,54 136 3,5-4,6
взрослые 4,1±0,03 0,26 6,28 66 3,7—4,6
Самки прибылые 4,1±0,02 0,26 6,47 134 3,5—4,6
взрослые 4,1±0,03 0,22 5,33 42 3,8-4,5
В целом по виду . . . 4,0±0,01 0,29 7,22 823 3,5—4,6
Длина углового отростка
Самцы прибылые 2,1±0,01 0,11 5,20 112 1,7-2,3
взрослые 2,1±0,01 0,09 4,42 58 1,9-2,3
Самки прибылые 2,1±0,01 0,10 4,88 107 1,8—2,4
взрослые 2,1±0,02 0,10 4,86 36 1,8—2,2
В целом по виду . . . 2,1±0,01 0,13 6,19 702 1,7—2,4
хорошо заметные у взрослых зверьков. Насыщенная бурая окраска спины
постепенно, без резкой границы, переходит в более светлую на боках, но
с примесью рыжих тонов, и в светлую, серовато-белесую на вентральной
стороне тела. В пределах этой характеристики распределение окраски на
теле зверьков, интенсивность ее на дорсальной стороне меняются в широ-
ких пределах в разных частях ареала. Белизна колеблется от 1,5 до 5,2
(М = 3,4^0,05), оттенок — от 117,9 до 166,4 (М = 136,6±=0,56).
По размерам тела S. caecutiens занимает среднее положение среди
13 видов бурозубок, населяющих Сибирь. По длине тела с головой —
45—78 (М — 60,5) мм — она уступает шести видам, с одним видом (S. da-
phaenodon) имеет равные размеры и крупнее пяти. По длине хвоста —
25—60 (М — 36,1) мм — она также занимает среднее положение среди
сибирских представителей рода Sorex, по средним значениям этого показа-
теля даже несколько ближе к короткохвостым формам. В структуре чере-
па прежде всего обращает на себя внимание его относительная ширина
в мозговой части, вздутость, резкое сужение в межглазничном отделе
и в ростральной части.
Череп умеренных размеров, кондило-базальная длина его колеблется
от 16,1 до 19,4 (М = 17,4^:0,03) мм. По этому признаку бурозубка зани-
мает восьмое место. Относительно размеров тела череп крупнее (28,7 %)
и уступает только четырем видам. Необходимо отметить, что большей
относительной длиной черепа обладают преимущественно мелкие формы
бурозубок. Самый высокий индекс отношения кондило-базальной длины
черепа к размерам тела имеет S. daphaenodon, а затем по убывающей 5. mi-
nutissimus и 5. beringianus, имеющие равные индексы, и S. minutus. Са-
мый низкий индекс у 5. unguiculatus (см. табл. 25).
Ростральная часть черепа средней бурозубки умеренно вытянута в
длину (26,4 % к кондило-базальной длине черепа), уступает по этому
признаку пяти видам, а по размерам мозгового отдела (56,3 %) — шести.
В какой-то мере о пропорциональном соотношении размеров лицевого
и мозгового отделов черепа свидетельствует средний индекс зубного ряда
(44,2 %). У четырех видов сибирских бурозубок этот показатель выше,
два имеют такую же величину индекса, как средняя бурозубка, пять
уступают ей по величине этого признака. Что касается длины ряда проме-
267
жуточных зубов, то по этому показателю средняя бурозубка занимает
одно из первых мест (35,1 %) как по отношению к кондило-базальной дли-
не черепа (15,5 %), так и к длине ряда зубов (35,1 %). По последнему
показателю она уступает лишь 5. gracillimus (36,8 %). Для промежуточ-
ных зубов этих двух видов характерен не только отнсительно длинный
ряд, но и свободная постановка коронок в ряду, довольно близкое соот-
ношение, размеры и форма коронок (см. табл. 26).
Для вида характерна наибольшая ширина черепа (50 %), по этому
признаку он уступает только трем видам бурозубок (см. табл. 21). По
отношению межглазничной ширины к кондило-базальной длине (18,9 %)
занимает седьмое место, по индексу предглазничной ширины (13,2 %)
делит 10—11-е место, а по ширине рострума (9,1 %) стоит на 10-м из 13.
Таким образом, череп средней бурозубки отличается пропорциональным
отношением лицевого и мозгового отделов, относительной шириной и высо-
той в мозговой области и резким сужением в ростральной части
(см. табл. 21).
Для этой бурозубки характерны мелкие размеры коронок промежу-
точных зубов, свободное положение их в зубном ряду, вытянутость осно-
ваний в сагиттальном направлении и их парная группировка. Вертикаль-
ные оси этих зубов параллельны друг другу и перпендикулярны продоль-
ной оси челюсти. В соотношении зубов наблюдается изменчивость, но
видовая специфичность их при этом остается неизменной. Соотношение
величины отдельных элементов промежуточных зубов у этого вида де-
тально описано В. А. Долговым [1968а, б].
Передний верхний резец относительно массивный. Его первая верши-
на шире задней (внутренний зубец). По высоте они почти равны между
собой и с первым промежуточным зубом. Вследствие этого при рассмотре-
268
нии зубного ряда в профиль вершины резца и слабо убывающие по высоте
вершины первых четырех промежуточных как бы образуют ровный ряд,
резко обрывающийся между относительно высоким четвертым промежу-
точным и зачаточным пятым. По этому признаку средняя бурозубка хоро-
шо отличается от других видов. Парность группировки зубов условная,
но тем не менее чаще всего первый зуб выше второго, третий выше чет-
вертого. Однако высота коронки не всегда соответствует положению вер-
шины, так как в этом большую роль играет положение зуба на челюсти.
Видоспецифично строение копулятивного органа размножающихся
зверьков. Головка его цилиндрическая, одинакового диаметра на всем
протяжении, увенчанная на свободном конце массивной, сферически за-
кругленной коронкой. Стержень glans penis более всего схож с таковым
малой бурозубки, но отличается меньшими размерами, несмотря на боль-
шую длину зверька. Длина его составляет 5—8,5 (М = 6,4) мм, при тол-
щине у основания 0,8—1,1 (М = 0,9) мм, у малой бурозубки — 8,3—
11,4 (М - 9,3) мм и 1,0 мм. Отношение длины glans penis к длине тела
с головой у средней бурозубки составляет 10,5 а у малой — 17,8 %. Кро-
ме того, у нее коронка glans penis шире, чем у малой бурозубки, резко
скошена в вентральном направлении и с большим крючкообразным выро-
стом снизу, отсутствующим у малой бурозубки. Охарактеризованные чер-
ты копулятивного органа настолько видоспецифичны, а из-за малой из-
менчивости абсолютно надежны, что позволяют хорошо диагностировать
взрослых самцов (см. рис. 40).
Распространение. Ареал средней бурозубки охватывает се-
вер СССР от западных границ до Чукотки, Курильских островов, Сахали-
на и Приморья включительно. К югу граница ареала вида идет до Украи-
ны, Тамбовской области, южной границы Башкирии. В Барабинской сте-
пи до широты оз. Чаны, Новосибирска, по приобским борам и Салаиру
спускается до Алтая включительно. Встречается всюду в горных райо-
нах Сибири.
Материал. 676 экз., Читинская, Амурская, Магаданская обла-
сти, Якутия, Хабаровский край, Курильские острова, Камчатка,
Сахалин.
Изменчивость. Для средней бурозубки характерна возраст-
ная, сезонная и географическая изменчивость в пропорциях тела.
Возрастная изменчивость проявляется в стирании меха на хвосте,
ступнях, кистях и стирании зубов, а также в изменении размеров тела.
Характер стирания меха аналогичен таковому у других видов бурозубок.
Взрослые самцы и самки достоверно больше по длине тела с головой
(см. табл. 57) по сравнению с прибылыми. Длина хвоста несколько боль-
ше у прибылых (самцы: М — 38,9—^0,13), чем у перезимовавших бурозу-
бок (М = 35,7+0,45). Особенно заметны возрастные изменения в массе
зверьков. Средний вес прибылых бурозубок этого вида почти вполовину
меньше такового перезимовавших (как самцов, так и самок). В размерах
черепа и соотношении его элементов возрастная изменчивость не просле-
живается, кроме некоторого изменения длины зубного ряда: у прибылых
зверьков она несколько больше — М = 7,8, у перезимовавших — М —
= 7,5 мм (и у самцов, и у самок).
Половая изменчивость проявляется меньше. Длина тела прибылых
самцов лишь немного больше (М = 58,3 мм) по сравнению с самками
(М = 57,3 мм). Среди перезимовавших бурозубок самки заметно крупнее
(М = 68,3 г), чем самцы (М = 67,1 г), а масса больше у самцов
269
(см. табл. 57). По длине хвоста, ступни половые различия недостоверны.
Географическая изменчивость размеров тела выражается в увеличе-
нии значений этого показателя по направлению с запада на восток и севе-
ро-восток. Наименьший средний показатель характерен для зверьков из
равнинной части Западной Сибири (М = 58,9±0,49 мм). У подвида
5. с. caecutiens, заселяющего горы юга Сибири и Среднюю Сибирь, он
несколько больше (М = 60,1 ±0,48 мм), у тундрового S. с. koreni не отли-
чается от такового предыдущего подвида (М = 60,1 ±0,81 мм). Восточно-
сибирская средняя бурозубка (5. с. macropygmaeus) заметно крупнее
(61,3±0,54 мм), самые крупные зверьки вида заселяют восточную пери-
ферию ареала — на о. Кунашир сахалинская средняя бурозубка
(У. с. shinto) достоверно крупнее всех остальных подвидов (62,5±0,13 мм).
Еще более проявляется изменчивость длины хвоста. Географическая
изменчивость по этому признаку имеет несколько иную направленность.
Самые короткохвостые землеройки занимают северные пределы ареала
и характерны для S. с. koreni (М — 32,2±0,47 мм), несколько больше
этот показатель у восточносибирской средней бурозубки (М — 34,5±
±0,25 мм). Подвиды средней бурозубки, заселяющие Западную и Сред-
нюю Сибирь, по этому признаку практически не различимы (М = 37,2
(±0,37) и 37,0(±0,34)). Хвост S. с. shinto исключительно длинный, в сред-
нем 37,9 мм, что составляет 60,7 % от длины тела.
Географическая направленность изменчивости кондило-базальной
длины черепа средней бурозубки также своеобразна. Минимальные значе-
ния этого признака характерны для бурозубок равнинной части Запад-
ной (М = 16,8±0,04 мм) и Восточной Сибири (М = 16,9±0,07 мм). Зем-
леройки гор юга Сибири имеют достоверно большие размеры кондило-
базальной длины черепа (W = 18,0±0,03 мм). По этому показателю среди
последних выделяются зверьки с о. Кунашир (М = 18,5±0,03 мм)
(рис. 82, табл. 58).
В широких пределах изменяется и окраска зверьков. Возрастная
изменчивость сводится главным образом к появлению более яркой рыже-
ватой примеси в окраске меха боков
вых отличий по этому признаку
не обнаруживается. Возрастная
изменчивость характеризуется за-
метным уменьшением белизны и
некоторым увеличением интенсив-
ности оттенка, т. е. общим по-
темнением окраски меха дорсаль-
ной части тела у взрослых зверь-
ков.
В целом по виду показатель
белизны колеблется в значитель-
Рис. 82. Изменчивость кандило-ба-
зальной длины черепа средней буро-
зубки из разных частей ареала.
1 — S. с. caecutiens (горы юга Сибири, Ср. Си-
бирь); 2 — S. с. pleskei (равнинная часть
Зап. Сибири); 3 — S. с. koreni (бассейны рек
Омолон, Колыма); 4 — S. с. macropygmaeus
(Приморье); 5 — S. с. shinto (о. Кунашир).
у перезимовавших зверьков. Поло-
КонОило-базальная Олина черепа, мм
270
Морфометрическая характеристика Sorex caecutiens
S. caecutiens
Признак а Су п lim
Длина
тела 60,5±0,28 6,53 10,79 556 45—78
хвоста 36,1±0,22 5,15 14,27 555 25—60
стухши 11,7±0,05 1,05 9,01 527 10—14
Масса тела 5,3±0,07 1,49 27,97 521 2,8—11,0
Длина
кондило-базальная черепа 17,4±0,03 0,87 4,97 812 16,1—19,4
рострума 4,6±0,01 0,34 7,39 834 3,9—5,6
мозговой части 9,8±0,02 0,48 4,84 812 8,8—11,4
верхнего ряда зубов 7,7±0,2 0,46 5,98 831 6,7—8,8
ряда промежуточных зубов 2,7±0,01 0,23 8,36 832 2,2—3,2
Ширина
рострума 1,6±0,01. 0,13 8,10 834 1,1—1,9
предглазничная 2,3±0,01 0,30 12,93 834 2,0—2,8
межглазничная 3,3±0,01 .0,18 5,64 834 . 2,7—3,7
наибольшая черепа 8,7±0,01 0,42 4,48 809 7,8—10,0
Наибольшая высота черепа 5,6±0,01 , 0,35 6,31 808 4,3—6,6
Высота восходящей ветви ниж-
ней челюсти 4,0±0,01 . 0,29 7,22 823 3,4-4,7
Длина углового отростка 2,1±0,01 0,13 6,19 702 1,7—2,4
вых пределах—от 1,5 до 5,2 (Л/=Э,4± 0,05), также как и оттенка —117,9—
166,4 (М = 136,6±0,56). Наиболее темноокрашенные бурозубки обитают
в восточной части ареала в широколиственных лесах Приморья и относят-
ся к подвиду S. с. macropygmaeus. Так, зверьки, добытые на территории
заповедника «Кедровая Падь», имеют самые низкие значения показателя
белизны — 1,5—3,5 (М — 2,7±0,09) и самые высокие насыщенности от-
тенка — 126,0—166,4 (М = 144,4^1,96). Затем идут зверьки из влажных
лесов таежной части Салаира и Присалаирья и тайги Среднего Приобья.
Значения белизны колеблются од 1,8 до 4,3 (М = 2,9±0,09), а насыщен-
ности оттенка — от 122,1 до 155,8 (М = 136,6±1,24). По возрастанию
значений белизны следуют зверьки о. Кунашир, в окраске которых за-
метны примесь серых тонов на дорсальной стороне тела, палевый оттенок
на вентральной части, общее просветление и тусклость меха. Белизна
в окраске меха зверьков с о. Кунашир колеблется от 2,2 до 4,1
(М = 3,3±0,13), насыщенность оттенка — 127,8 до 150,0 (М = 137,3±
±1,95). ‘
Близкие средние значения показателя белизны и оттенка у землероек
гор юга Сибири: 2,4—5,2 (М = 3,4±0,06) и 117,9—139,8 (М = 133,4±
±0,94). Самые светлоокрашенные зверьки заселяют северные и северо-
восточные части ареала вида в Сибири. Средние значения показателей
окраски разных подвидов разнятся между собой меньше, чем крайние
271
Таблица 58
и подвидов из разных частей ареала
a. S. с. caecutiens, горы юга Сибири, Ср. Сибирь б. S. с. pleskei, равнинная часть Зап. Сибири
М±тж а Су п lim М±7ПХ а cv п lim
60,1±0,48 6,36 10,58 178 46—78 59,9±0,49 4,51 7,7 86 50—69
37,0±0,34 4,55 12,30 175 25-47 37,2+0,37 3,48 9,4 88 30—46
12,0±0,09 1,15 9,53 160 10—14 11,6+0,06 0,57 4,9 84 10—13
5,5±0,10 1,31 23,80 177 2,8—10 5,1±0,19 1,60 31,6 74 3—8,8
18,0±0,03 0,47 2,58 249 16,6—19,1 17,4±0,04 0,43 2,5 151 16,2—18,5
4,9±0,01 0,22 4,44 252 4,2-5,6 4,7±0,02 0,23 4,8 157 4,1—5,4
10,1±0,02 0,32 3,15 249 8,8—11,4 9,9±0,02 0,26 2,6 151 9,2—10,5
8,0±0,02 0,27 3,36 252 7,6—8,8 7,6±0,02 0,28 3,6 157 7,0-8,5
2,9±0,01 0,13 4,38 252 2,6—3,2 2,7±0,01 0,15 5,5 157 2,4-3,1
1,6±0,01 0,11 6,70 252 1,3-1,9 1,5±0,01 0,10 6,8 157 1,1—1,9
2,4±0,01 0,13 5,52 252 2,0-2,8 2,3±0,05 0,59 25,4 157 2,0-2,5
3,3±0,01 0,15 4,55 258 2,2—3,7 3,2+0,01 0,13 4,0 157 3,0-3,5
8,9±0,02 0,29 3,27 248 7,8—10 8,7±0,02 0,22 2,6 148 8,0-9,2
5,6±0,02 0,28 5,0 248 4,8—6,5 5,7±0,02 0,29 5,0 150 4,8-6,2
4,3±0,01 0,23 5,50 249 3,5—4,7 3,9±0,01 0,18 4,6 156 3,5-4,4
2,1±0,01 0,10 4,97 208 1,7—2,3 2,1±0,01 0,10 4,9 123 1,8—2,3
у отдельных подвидов, например у номинального (рис. 83, 84)
(табл. 59, 60).
Хвост резко двухцветный: верхняя часть окрашена в тон спины в
темные тона, нижняя — в светлые. У молодых зверьков окраска более
контрастна за счет того, что нижняя часть серовато-белая, у перезимовав-
ших зверьков она палевая, и окраска менее контрастная. Кисточка на
хвосте у прибылых зверьков хорошо выражена, образована темными
длинными волосами, а стержень покрыт относительно короткими, приле-
гающими волосами. У перезимовавших зверьков кисточка отсутствует,
а стержень хвоста как бы «побрит» или совсем голый. У наиболее длин-
нохвостых землероек (S. с. shinto) отношение длины хвоста к длине тела
составляет 60,7, у более короткохвостых (S. с. koreni) — 53,5 %.
Кондило-базальная длина черепа у землероек разного возраста и пола
достоверно не отличается (см. табл. 57). Индивидуальная и географиче-
ская изменчивость этого признака колеблется от 16,1 до 19,4 (М =
— 17,4) мм. Эти признаки, как и длина тела, у средней бурозубки пере-
крываются с таковыми тундряной, крупнозубой и в меньшей мере —
обыкновенной, равнозубой и другими бурозубками. Наибольшая величи-
на этого показателя наблюдается на крайней восточной периферии ареа-
ла у землероек S. с. shinto 17,7—19,4 (М = 18,5) мм. У этого подвида
самые высокие значения среднего показателя среди всех остальных под-
272
Признак в. S. с. koreni, бассейны рек Омолон, Колымы
а CV п lim
Длина
тела 60,1±0,81 6,26 10,4 60 47—72
хвоста 32,2±0,47 3,61 11,2 60 25—38
ступни 11,3+0,13 0,97 8,6 60 10—13
Масса тела 4,6±0,16 1,10 23,8 50 3,5—8,0
Длина
кондило-базальная черепа 16,8±0,04 0,37 2,2 77 16,1—17,7
рострума 4,3±0,02 0,19 4,4 77 3,9—4,7
мозговой части 9,7±0,03 0,29 3,0 77 8,8—10,3
верхнего ряда зубов 7,3±0,02 0,18 2,5 75 6,9—7,8
ряда промежуточных зубов Ширина 2,5+0,01 0,12 4,5 75 2,2—2,7
рострума 1,6±0,01 0,12 7,4 77 1,4—1,9
предглазничная 2,2±0,01 0,11 5,2 77 2,0—2,5
межглазничная 3,2±0,02 0,13 4,1 77 3,0—3,6
наибольшая черепа 8,4±0,03 0,27 3,2 77 7,9-9,0
Наибольшая высота черепа 5,5±0,04 0,30 5,5 76 5,0-6,6
Высота восходящей ветви ниж-
ней челюсти 3,8+0,02 0,15 3,9 74 3,4-4,7
Длина углового отростка 2,1+0,01 0,09 4,5 57 1,8—2,3
видов. Минимальные значения показателя длины тела у средних бурозу-
бок, заселяющих север и северо-восток сибирской части ареала (М ==
= 16,8±0,04) и относящихся к подвидам S. с. koreni и S. с. macropyg-
maeus (М = 16,9±0,07). В нарастающем порядке изменяется этот пока-
затель в направлении к равнинным районам Западной Сибири (S. с. ple-
skei — М = 17,4:^0,04) и к горам юга Сибири (S. с. caecutiens — М —
= 18,0=t0,03) мм. Таким образом, определенной географической направ-
ленности в изменении этого признака не прослеживается. Мелкие зверьки
заселяют северо-восток и Приморье, а рядом, несколько восточнее, на
островах обитают самые крупные землеройки. В направлении на запад
этот показатель сначала заметно возрастает у номинального подвида,
но снова снижается в пределах Западной Сибири, хотя и остается более
высоким, чем у зверьков с северо-востока Сибири и из Приморья
(см. табл. 58).
Эти материалы демонстрируют видовую специфичность направлен-
ности изменчивости размеров тела. Они опровергают заключение некото-
рых исследователей [Niethammer, 1956; Межжерин, 1964, 19656] о якобы
существующей закономерности уменьшения массы тела бурозубок одного
вида в направлении с юга на север и с запада на восток. Нельзя согласить-
ся и с В. А. Долговым [1972], что популяция этого вида самых крупных
бурозубок обитает на Алтае. Как показали исследования обширных мате-
риалов, наиболее крупные зверьки заселяют южные острова Курильской
гряды, т. е. юго-восточную периферию ареала, в то же время на сопре-
273
Продолжение табл. 58
г. S. р. macropygmaeus, Приморье Д. S. с. shinto, о. Кунашир
М±тж а cv п lim М±тх а CV п Нт
61,3+0,54 7,53 12,27 197 45-78 62,5+0,13 5,42 8,68 20 58-78
34,5+0,25 3,44 9,99 197 28-43 37,9+0,49 1,95 5.14 20 51-60
11,4+0,07 1,00 8,78 188 10—13 11,9+0,13 0,50 4,21 20 13—14
5,3+0,01 1,60 30,1 195 3,2—8,8 5,2+0,12 0,43 8,24 13 48—7
16,9+0,07 1,10 6,50 253 16,4—17,9 18,5±0,03 0,21 1,2 19 17,7—19,4
4,4+0,02 0,33 7,43 262 4,1-4,9 4,8+0,03 0,14 2,9 22 4,5—5,0
9,6±0,04 0,65 6,77 253 9,1—10,2 10,0+0,03 0,13 1,3 19 9,7—10,3
7,4±0,03 0,49 8,22 252 6,7—8,0 7,6+0,03 0,15 1,93 22 7,5-8,1
2,6+0,01 0,20 7,92 262 2,3—2,8 2,8±0,02 0,09 3,39 22 2,9-2,5
1,5±0,01 0,15 9,76 262 1,3—1,8 1,8±0,02 0,08 4,9 22 1,4—1,6
2,2+0,01 0,18 8,04 262 2,0—2,6 2,4+0,02 0,08 3,13 22 2,2—2,5
3,2+0,01 0,24 7,46 262 2,9—4,3 3,3+0,02 0,09 2,75 22 3,2—3,4
8,6+0,04 0,58 6,76 254 8,0—9,6 8,6+0,05 0,20 2,33 19 8,5—8,8
5,5+0,03 0,45 8,22 252 4,3—6,6 5,7+0,06 0,26 4,52 19 5,3-5,9
3,9+0,01 0,28 7,18 259 3,6—4,6 4,1+0,02 0,09 2,08 21 4,1—4,3
2,1+0,01 0,16 7,98 230 1,8—2,4 2,1±0,02 0,07 3,44 22 2,0—2,2
дельных территориях в Приморье и Приамурье живут землеройки, зна
чительно уступающие по морфометрическим показателям.
Как уже говорилось выше, для промежуточных зубов характерна
парная группировка, свободное положение в ряду и значительная вытя-
нутость основания коронок в сагиттальном направлении. Соотношение
размеров отдельных промежуточных зубов у этого вида детально описано
В. А. Долговым [19686]. Им выделены три типа промежуточных зубов.
Тип Г. Первый промежуточный равен второму, третий заметно мень-
ше второго и равен четвертому. Пятый очень мал или заточен. Нередко
третий зуб несколько меньше четвертого.
Тип II. Первый промежуточный по высоте и ширине крупнее трех
последующих зубов, которые в целом равны между собой и образуют
единую группу. Третий довольно часто меньше второго и четвертого.
Пятый очень мал.
Тип III. Первые четыре промежуточных зуба образуют единую груп-
пу и по величине незначительно убывают от первого к четвертому. Иногда
третий меньше второго и четвертого. Пятый развит слабо.
Географической приуроченности зверьков с описанными типами зубов
не обнаруживается. Наиболее часто во всех исследованных выборках
встречаются (до 93 %) бурозубки с первым типом.
Заметки по систематике. Средняя бурозубка —морфоло-
гически четко очерченный вид и при современном уровне изученности из-
менчивости признаков и таксономии бурозубок надежно диагностируется.
274
Окончание табл. 58
Признак Достоверность разности средних по верны при СД > Майру (различия досто- 1,28)
а—б | а—в | а—г | а—д I б—в I б—г | б-Д | в—г 1 в—д I г-д
Длрна тела 0,11 0,01 0,09 0,19 0,12 0,20 0,55 0,09 0,19 0,08
хвоста 0,03 0,58 0,31 1,38 0,70 0,39 0,13 0,32 1,06 0,63
ступни 0,23 0,36 0,28 0,06 0,25 0,14 0,29 0,08 0,41 0,33
Масса тела 0,14 0,35 0,06 0,17 0,16 0,07 0,05 0,25 0,39 0,049 -
Длина кондило-базальная че- репа 0,60 1,31 0,70 0,41 0,69 0,37 0,47 0,01 1,55 0,61 ,
рострума 0,46 1,40 0,86 0,28 0,88 0,47 0,27 0,20 1,52 0,85 '
мозговой части 0,34 0,65 0,44 0,22 0,37 0,26 0,26 0,04 0,71 0,51
верхнего ряда зубов 0,06 0,21 0,18 0,95 0,26 0,22 — 0,01 0,91 0,31
ряда промежуточных зубов 0,52 1,56 0,87 0,45 0,90 0,49 0,42 0,06 1,9 0,69 ,
Ширина рострума 0,33 0,09 0,29 — 0,22 0,02 1,67 0,20 1,0 1,3
предглазничная 0,07 0,73 0,46 — 0,18 0,12 0,15 0,12 1,05 0,77
межглазничная 0,29 0,40 0,24 — 0,12 0,04 0,45 0,05 0,45 0,30
наибольшая черепа 0,35 0,77 0,35 0,41 0,51 0,15 0,23 0,15 0,43 — t
Наибольшая высота черепа 0,78 1,65 1,15 0,21 0,70 0,49 — 0,07 0,4 0,31 ;
Высота восходящей ветви нижней челюсти 0,75 1,19 0,73 0,61 0,44 0,14 0,09 0,19 1,25 0,54
Длина углового отростка 0,08 0,02 0,03 — 0,10 0,03 — 0,04 — —
Впервые ее видовая самостоятельность была установлена Лаксман-
ном [Laxmann, 1788] по материалам из Прибайкалья. Однако длительное
время она была известна под названием S. macropygmaeus Miller, 1901.
Приоритет был восстановлен Чейворт-Мастерсом [Chaworth-Musters],
и ей было присвоено название, данное первым автором, описавшим вид
[Ellermann, Morrison-Scott, 1951]. В кратком описании Э. Л. Лаксманн
дал сжатую характеристику, рисунок зверька, указал размер. Еще со
времени П. С. Палласа [Pallas, 1811] реальность этого вида вызывала
сомнения главным образом из-за утраты голотипа. П. С. Паллас отнес
ее к S. pygmaeus. Как показали исследования С. У. Строганова [1957],
под этим названием он принимал произвольный, композиционный вид,
включающий «S', minutus, S. minutissimus и S. caecutiens в современном
понимании объема этих видов.
Доводы С. У. Строганова [Там же] о том, что если бы Паллас не игно-
рировал данные Лаксманна о размерах средней бурозубки и подобной
путаницы могло не быть, вполне убедительны. Первоописание и приводи-
мый в нем рисунок весьма информативны и не оставляют сомнений, что
Лаксманном был описан новый вид. Выдвинутое П. С. Палласом положе-
ние было без должной ревизии принято другими исследователями [Fischer,
1814; Brandt, 1853; Кащенко, 1905; Огнев, 1913; и др.]. В отечественной
литературе впервые и убедительно аргументированное название S. caecu-
tiens ввел С. У. Строганов [Там же], проделав кропотливое исследование,
275
ОттёнОк
Белизна
Рис. 83 Сравнительная характеристика
окраски подвидов средней бурозубки.
1 — S caecutiens', 2 — S. с. pleskei’, 3 —
St с. koreni’, 4 — S. с. maeropygmaeus; 5 —
St c, shinto.
Белизна
Рис. 84. Колориметрическая характе-
ристика горносибирской средней бу-
розубки (5. с. caecutiens) из разных ча-
стей ареала.
1 — Северо-Восточный Алтай (Телецкое озеро);
2 — Тува (Вост. Танну-Ола); з — Бурятия
(реки Джида, Улаар); 4 — Прибайкалье (Бар-
гу зинский заповедник); 5 — бассейн р. Зеи
(Зейский заповедник).
подтвердившее приоритет данного Лаксманном названия. Дополнитель-
ные данные, полученные в последние годы, убеждают в правильности трак-
товки систематической природы S. caecutiens, а восстановление приорите-
та не противоречит систематической номенклатуре. Сейчас всеми исследо-
вателями этот вид принимается в такой трактовке.
С. И. Огнев [1928] принимал среднюю бурозубку в объеме, близком
к современному пониманию. Однако он не дает общего описания вида,
а принимает его как «круг форм» подвидового ранга (S. т. maeropygmaeus,
S. т. rozanovi, S. т. koreni, S. т. pleskei, S. т. altaicus, S. т. aranaides).
Подвид S. с. shinto С. И. Огнев [1928] считал самостоятельным видом
S. shinto, приводя его описание по Томасу [Thomas, 1907], поскольку сам
не располагал материалом. Такой вывод не является неожиданностью,
учитывая резкое уклонение подвида и недостаточную изученность в то
время полиморфизма вида. Тем не менее от внимания этого выдающегося
систематика не ускользнуло большое сходство описанной Томасом земле-
ройки с S. caecutiens.
Н. А. Бобринский и А. П. Кузякин [Бобринский и др., 1944] основ-
ным отличительным признаком средней бурозубки (S. maeropygmaeus,—
S. arcticus) считали размер тела, отвергая такой надежный признак, как
видоспецифическая структура промежуточных зубов. В результате этого
под названием S. maeropygmaeus в понимании названных авторов прини-
мался искусственный «композиционный вид», включающий собственно
средних бурозубок и зверьков средних размеров (S. arcticus), а одну из
форм средней бурозубки (5. т. aranoides) они отнесли к группе S. araneus.
Во втором издании «Определителя млекопитающих СССР» [Бобринский
и др., 1965] А. П. Кузякин формально принял видовое название средней
бурозубки (S. caecutiens), но по существу не отказался от своих необосно-
ванных позиций. Больше того, в разборе представлений С. У. Строганова
[1957 ] об объеме этого вида и его внутривидовой таксономии А. П. Кузя-
276
Таблица 59
Колориметрическая характеристика Sorex caecutiens из разных частей ареала
Подвид Место сбора материала Белизна
М±тх О cv п lim
S. с. caecutiens Телецкое озеро 3,1±0,1 0,5 16,2 25 2,4-4,3
Тува 4,0±0,11 0,55 13,8 25 3,0-5,2
Хамар-Дабан 3,7=4=0,11 0,55 14,7 25 2,8—4,8
Баргузинский хребет 3,5±0,09 0,45 13,9 25 2,4-4,7
По подвиду . . . 3,4±0,06 0,55 16,4 100 2,4—5,2
S. с. pleskei Предгорья Салаира Томская обл. (южная и 2,9=4=0,11 0,42 14,4 15 1,9-3,5
средняя тайга) 2,9±0,12 0,6 20,7 25 1,8—4,3
По подвиду . . . S. с. koreni Бассейны рек Колымы 2,9±0,09 0,53 18,4 40 1,8—4,3
S. с. macropyg- и Омолона Приморье, Кедровая 4,08±0.11 0,55 13,7 25 2,7—5,0
таиез Падь 2,7±0,09 0,45 17,4 25 1,5—3,5
S. с. shinto О. Кунашир 3,3±0,13 0,48 15,3 14 2,2—4,1
П о виду ... 3,4±0,05 0,69 20,72 204 1,5-5,2
Окончание табл. 59
Подвид Место сбора материала Оттенок
М±тж а Су п lim
S. с. caecutiens Телецкое озеро 131,7±1,47 7,35 5,6 25 119,7—146,8
Тува 136,5=4=1,06 5,3 3,9 25 125,5—146,7
Хамар-Дабан 134,1±1,13 5,65 4,2 25 120,9—144,3
Баргузинский хребет 131,1±1,17 5,85 4,5 25 117,9—139,8
По подвиду . . . 133,4±0,94 9,4 4,5 100 117,9—139,8
S. с. pleskei Предгорья Салаира Томская область (южная 139,8±1,85 7,03 5,1 15 125,6—149,4
и средняя тайга) 139,5=1=1,67 8,35 6,0 25 122,1—155,8
По подвиду . . . S. с. koreni Бассейны рек Колымы 136,6±1,24 7,94 5,6 40 122,1—155,8
S. с. macropyg- и Омолона Приморье, Кедровая 136,9=1=1,19 5.95 4,4 25 127,0—150,0
maeus Падь 144,4± 1,96 9,8 6,8 25 126,0—166,4
S. с. shinto О. Кунашир 137,3±1,95 7,41 5,3 14 127,8—150,0
Повиду ... 136,6±0,56 8,06 5,89 204 117,9—166,4
кин продемонстрировал полное непонимание вопроса, объединив вместе
виды S. caecutiens, S. arcticus, S. minutus, S. araneus. Наша оценка взгля-
дов этого автора на систематику бурозубок нашла отражение в специаль-
ной публикации [Долгов, Юдин, 1973].
Средняя и тундряная бурозубки, несмотря на сходство в размерах
тела, четко различаются между собой по ряду других признаков. Даже
внешний облик их различен, и опытный специалист безошибочно различа-
ет их. Средняя бурозубка стройного, легкого телосложения с широкой
выпуклой в мозговой части головой, сильно вытянутым хоботком и резким
277
Таблица 60
Достоверность различий (по Стьюденту) между подвидами
5. caecutiens
Подвид а б В г д
а б в г Д тенс S. с. caecutiens S. с. pleskei S. с. koreni S. с. macropygmaues S. с. shinto Примечание. В верхней к. 0,46 0,94 0,7 0,1 0,19 1,76 0,2 0,39 0,16 0,02 1,38 0,75 0,57 0,44 0,5 0,65 0,23 0,45 0,02 0,41 части — белизна, в нижней — от-
сужением в области глаз. Темная окраска спины постепенно переходит
в более светлую с примесью рыжего цвета на боках и белесо-серую на
[ брюшке. Характер окраски устойчив на всем протяжении обширного
। ареала вида. Изменчивость проявляется только в степени насыщенности
оттенка и белизны.
У тундряной бурозубки тяжелое телосложение, короткий, широкий
хоботок. Резкого сужения в области глаз нет. В окраске меха в большей
или меньшей степени выражена чепрачность. О несостоятельности сужде-
ния о видовом статусе средней бурозубки только по размерам тела можно
I судить на основании того, что один из ее подвидов (5. с. shinto) по боль-
шинству морфометрических признаков отличается от остальных больше,
чем многие виды между собой, и в то же время по ряду других признаков
имеется полное сходство.
Существенное различие этих двух видов прослеживается в структуре
j промежуточных зубов и строении черепа. Череп тундряной бурозубки
| более толстокостный, компактный и широкий в ростральной области.
$ Особенно четкие различия в строении промежуточных зубов: у средней
бурозубки вершины первого и трех последующих по высоте почти равны,
пятый крайне мал. Зубы относительно мелкие и образуют парные группи-
ровки, описанные выше. Зубы в зубном ряду сидят свободно, основания
их сильно вытянуты в сагиттальном направлении. Вертикальные оси
коронок перпендикулярны продольной оси челюсти и параллельны между
। собой. Длина промежуточных зубов по отношению к длине зубного ряда
I составляет 35,1 %. У тундряной бурозубки вершина второго промежу-
; точного зуба всегда выше первого. Третий, четвертый и пятый последова-
5 тельно убывает по высоте. Зубы средней величины в зубном ряду сидят
,i плотно. Отношение длины ряда промежуточных к длине зубного ряда
। составляет 32 %. Конфигурация коронок промежуточных зубов у тунд-
ряной бурозубки иная, а вертикальные оси расходятся веером.
1 Копулятивный орган отличается коренным образом (см. рис. 40).
По этим признакам средняя и тундряная бурозубки надежно диагно-
стируются как самостоятельные виды. Еще более четкие различия суще-
ствуют между средней и обыкновенной бурозубками. Специфичность и са-
мостоятельность этих видов убедительно доказана С. У. Строгановым
Г [1957], полемизирующим с Н. А. Бобринским и А. П. Кузякиным [Боб-
| ринский и др., 1944].
j Существенные трудности в понимании таксономии вида объясняются
тем, что Brink [1952, 1953] объединил с S. caecutiens такие виды, как
S. cinereus, S. lyelli Merr., 5. preblei Jackson, создав искусственный ком-
19 Б. С. Юдин
278
позиционный вид с циркумполярным распространением. Включенные в не-
го гетерогенные элементы относятся не только к разным видам, но и раз-
ным подродам {Otisorex и Sorex). Эта позиция Бринка получила отрица-
тельную оценку в работах Хоффманна, Петерсена [Hoffmann, Peterson,
1967], Б. С. Юдина [1969а, б] и Хоффманна [Hoffmann, 1971]. Несостоя-
тельность подобного объединения очевидна. Подробный анализ взглядов
Бринка дан названными исследователями. Причина неправомерного объе-
динения резко дифференцированных видов кроется в формальном под-
ходе к таксономической оценке только с позиции размерных признаков,
внесенных в систематику землероек Н. А. Бобринским и А. П. Кузякиным
[Бобринский и др., 1944]. На это мы уже указывали ранее [Юдин,
19726, д]. Эллерман и Моррисон-Скотт [Ellerman, Morrison-Scott, 1951],
ссылаясь на работу Н. А. Бобринского и А. П. Кузякина [Там же], пол-
ностью приняли объем вида в их понимании. Под названием S. caecutiens
они сконструировали несуществующий в природе сборный вид, включаю-
щий кроме собственно средней бурозубки S. arcticus и S. araneus.
К средней бурозубке относится S. centralis, описанный Томасом
[Thomas, 1911] с Саян. В настоящее время мы относим эту форму к но-
минальному подвиду. Сам О. Томас указывал на близость описанного им
«вида» к средней бурозубке. С. И. Огнев [1928] рассматривал ее в качест-
ве самостоятельного вида. Ссылаясь на описание О. Томаса и не имея
фактического материала, он все же отметил ее сходство с крупными фор-
мами средней бурозубки {S. т. aranoides). Принять окончательное реше-
ние ему мешало указание О. Томаса о том, что пятый промежуточный
зуб лишь немного уступает четвертому, что не свойственно землеройкам
этого вида. С. У. Строганов [1957] объясняет это индивидуальной измен-
чивостью. Нам представляется, что в распоряжении О. Томаса оказались
старые особи со стертыми зубами, у которых различия в высоте четвертого
и пятого зубов нивелировались вследствие более сильного стирания пер-
вых четырех промежуточных. По остальным признаком S. centralis пол-
ностью аналогичен S. с. caecutiens(см. табл. 58). По Томасу [Thomas, 1911 ],
S. centralis имеет длину тела 63—68, хвоста — 39—41, задней ступни —
12 мм. Кондило-базальная длина черепа 18,6, наибольшая ширина 9,2,
высота 5,3, длина верхнего ряда зубов 8,4 мм.
Реально представлял объем и внутривидовую таксономию этого вида
С. У. Строганов [1957]. Он впервые четко установил границы вида, дал
его исчерпывающую характеристику и анализ взглядов на этот вид его
предшественников.
Вероятно, тот же признак, который помешал С. И. Огневу быть до
конца последовательным в оценке S. centralis (относительно хорошо раз-
витый пятый промежуточный зуб), ввел в заблуждение В. А. Долгова
[1964], необоснованно присвоившего равнозубой бурозубке S. isodon
название S. centralis. Правильно понимая объем и границы вида, В. А. Дол-
гов ошибочно дал ему название, относящееся к одной из форм средней бу-
розубки. Ознакомившись с типовым экземпляром, послужившим О. То-
масу для первоописания S. centralis и хранящимся в Британском музее
естественной истории, В. А. Долгов (личн. сообщ.) убедился в ее принад-
лежности к средней бурозубке.
Географическая изменчивость средней бурозубки проявляется в ши-
роких пределах. С. У. Строганов [1957] указывал на небольшую амплиту-
ду колебаний размеров у разных подвидов: длина хвоста от 5 до 8 мм,
кондило-базальная длина черепа от 0,7 до 1,5 мм. По его данным, самая
279
крупная форма средней бурозубки населяет горы юга Сибири. По нашим
данным, самый крупный подвид S. с. shinto заселяет острова Сахалин,
Шантарские, Кунашир и Японские. Для него характерны самые широкие
колебания метрических показателей как внутри подвида, так и между
ними. Амплитуда колебаний длины тела составляет 45—78 мм, при этом
минимальные значения для разных подвидов разнятся на 11, а максималь-
ные на 9 мм. Показатель длины хвоста колеблется от 25 до 60 мм, соответ-
ственно минимальные и максимальные показатели отличаются на 26
и 22 мм. Кондило-базальная длина черепа изменчива в целом для вида
(16,1—19,4 мм), а диапазон минимальных и максимальных значений со-
ставляет 1,6—1,7 мм.
Географическая изменчивость проявляется в изменении общих раз-
меров тела и черепа, соразмерности отдельных частей его и в окраске.
Разные авторы принимают число подвидов средней бурозубки и их
объем неодинаково. С. И. Огнев [1928] в составе вида выделял шесть под-
видов: S. т. macropygmaeus Miller, S. т. koreni Allen, S. m. pleskei Ognev,
S. m. altaicus Ognev, S. m. aranoides Ognev, S. m. rozanovi Ognev. Кроме
того, две формы, относящиеся к этому виду (S. centralis Thomas и S. shin-
to Thomas), он рассматривал как самостоятельные виды. С. У. Строгано-
вым [1957] три последних подвида и S. centralis были отнесены к одному
подвиду S. с. caecutiens, a S, shinto рассматривалась им в качестве хорошо
обособленного подвида 5. с. shinto. Н. А. Бобринский и А. П. Кузякин
[Бобринский и др., 1944], не имевшие реального представления не только
о внутривидовой таксономии, но даже и об объеме вида, включали в его
состав ряд форм, относящихся к средней и «арктической» бурозубкам, не
давая какого-либо убедительного объяснения. Во втором издании этой
книги А. П. Кузякин [Бобринский и др., 1965] в составе вида рассматри-
вал три подвида: S. с. caecutiens Laxmann, S. с. buxtoni Allen, =S. arcti-
cus, и S. c. shinto Thomas. С. У. Строганов [1957] выделял пять подвидов
средней бурозубки: S. с. caecutiens, S. с. pleskei, S. с. koreni, S. с. macro-
pygmaeus, S. с. shinto.
Проведенная нами ревизия внутривидовой таксономии этого вида
показала, что средние бурозубки, относимые G. У. Строгановым [Там же]
к подвиду S. с. pleskei, ни по одному показателю достоверно не отличают-
ся от номинального подвида. Коэффициент достоверности (СД — 1,28,
по Майеру) по всем метрическим показателям размеров тела и черепа
не превышает 0,78, т. е. только по одному признаку (длина ряда промежу-
точных зубов) можно различить менее 80 % выборок сравниваемых. Еще
менее достоверны различия в окраске. Коэффициент достоверности раз-
личий по белизне у этих подвидов 0,46, а по оттенку — 0,19 (см. табл. 60).
Таким образом, относившиеся к этому подвиду землеройки (S. с. ples-
kei) мы рассматриваем в составе номинального подвида. Различия между
ними по диагнозам, приводимым С. У. Строгановым [1957], сводились
к указанию, что номинальный подвид крупнее предыдущего и «... в общем
бледнее окрашен». Все остальные признаки, приводимые в диагнозах,
равнозначны для того и другого подвидов. Статистически обработан ма-
териал из 157 экз. S. с. pleskei (по С. У. Строганову [1957]) и 252 экз.
S. с. caecutiens из разных пунктов их ареалов. Показано отсутствие досто-
верных различий по всем параметрам (см. табл. 60, рис. 85).
Колориметрическое исследование окраски S. с. pleskei из предгорий
Салаира, таежных районов Томской области и S. с. caecutiens с Алтая,
из Тувы, Прибайкалья не дало достоверных различий. Можно говорить
19*
280
Рис. 85. Колориметрическая характери-
стика западной средней бурозубки (S. с.
pleskei). Равнинная южная тайга (Том-
ская, Новосибирская области).
1 — в целом по виду (8. caecutiens Laxm.);
2 — изменчивость окраски S. с. pleskei.
лишь о тенденции к потемнению
окраски у равнинных форм и по-
светлению у горных, но различия
эти недостоверны, также как и не-
которое уменьшение размеров те-
ла, черепа и соотношения их от-
дельных элементов (см. табл. 58).
В пределах ареала S. с. caecu-
tiens, в понимании G. У. Строгано-
ва [1957], амплитуда колебаний средних значений выборок из разных
пунктов больше (0,9), чем между средними показателями рассматриваемых
подвидов (0,5). Графическая характеристика окраски, полученная путем
обработки колориметрических данных (см. рис. 83, 84), подтверждает
справедливость этого заключения.
Таким образом, отличия этих подвидов по величине тела и окраске
действительно существуют, но они недостоверны и не могут быть положены
в основу выделения подвидов. Базируясь на этом заключении, мы вклю-
чаем в ареал номинального подвида территорию от Урала до Енисея, к се-
веру до бассейна Нижней Тунгуски и районы Прибайкалья. Исследова-
ние средних бурозубок этого подвида из европейской части ареала нами
не проводилось и вопрос о его реальности в целом остается открытым.
Для Сибири описано четыре подвида.
Обзор подвидов
Sorex caecutiens caecutiens Laxmann, 1788 —
Горносибирская средняя бурозубка
Голотип. Вид описан по экземпляру из юго-западного побе-
режья Байкала.
Диагноз. По морфометрическим показателям эти зверьки ближе
всего к номинальному подвиду. Отличий в окраске меха от других пред-
ставителей не обнаружено. Достоверно отличается (СД = 1,3) этот подвид
от S. с. koreni по кондило-базальной длине черепа и длине рострума
(СД = 1,4), с 5. с. shinto достоверны различия по длине хвоста. Показа-
тель оттенка самый низкий их всех подвидов (М = 133,4±0,94).
Измерения. См. табл. 58.
Характеристика. В пределах ареала подвида (см. рис. 84)
изменяется окраска дорсальной стороны тела по белизне. Колориметри-
ческому анализу были подвергнуты зверьки из разных частей ареала:
Северо-Восточного Алтая, Тувы, Бурятии, Прибайкалья, Зейского за-
поведника.
Распространение. Ареал подвида охватывает Северо-Во-
сточный Алтай, Туву, Бурятию, Прибайкалье, бассейн Зеи.
281
Материал. 255 экз. из разных частей ареала.
Заметки по систематике. Горносибирская средняя бу-
розубка по ряду метрических показателей (длина тела, ступни, хвоста)
близка с 5. с. pleskei, S. с. koreni, S. с. macropygmaeus, достоверных отли-
чий не обнаружено (см. табл. 58). Однако существуют различия с S. с. ko-
reni по кондило-базальной длине черепа (СД = 1,3) и длине рострума
(СД = 1,4), различаются эти подвиды также по длине ряда промежуточ-
ных зубов (СД = 1,56) и наибольшей высоте черепа (СД = 1,65). Значи-
тельно уступает этот подвид по названным показателям 5. с. shinto — са-
мому крупному подвиду, населяющему Курильские острова (о. Кунашир).
Sorex caecutiens koreni Gl. Allen, 1914 —
Северосибирская средняя бурозубка 11
Голотип. $, ad., 19/X 1911 г., Нижне-Колымск, близ устья
Колымы. Коллекция Кембриджского музея сравнительной зоологии.
Диагноз. Отличается от других подвидов несколько меньшими
размерами тела, хвоста, массы и кондило-базальной длиной черепа, дли-
ной зубного ряда и ряда промежуточных зубов (см. табл. 58).
Измерения. См. табл. 58.
Характеристика. Мех самый светлый среди всех подвидов
(белизна 4,08). Достоверно отличается от 5. с. macropygmaeus по белизне
(см. табл. 60), от 5. с. caecutiens меньшей кондило-базальной длиной че-
репа, коротким рострумом, длиной ряда зубов верхней челюсти и проме-
жуточных зубов (см. табл. 58).
Распространение. Ареал подвида охватывает тундровую
зону от Печоры на западе до р. Анадырь на востоке и лесотундру Восточ-
ной Сибири. Более точные границы подвида, по имеющимся материалам,
установить пока трудно.
Материал. 91 экз., Якутия, р. Анадырь.
Заметки по систематике. По размерам ближе всего
к 5. с. macropygmaeus, но достоверно отличается от нее более светлой
окраской (значения белизны у 5. с. koreni 4,08, у 5. с. macropyg-
maeus — 2,7). Указания Аллена [Allen, 1914], С. И. Огнева [1928] и
С. У. Строганова [1957] на короткохвостость этого подвида справедливы,
но статистически отличия недостоверны.
Sorex caecutiens macropygmaeus Miller, 1901 —
Восточносибирская средняя бурозубка
Голотип. сГ, ad., 23/IX 1897 г., Петропавловск-Камчатский.
Коллекция Национального музея США (Вашингтон), № 84012.
Диагноз. Отличается от других подвидов средней бурозубки
более темной окраской меха. По этому показателю различия с североси-
бирской средней бурозубкой достоверны (СД = 1,38).
Измерения. См. табл. 58.
Распространение. Лесотундра и тайга на восток от Лены
до Камчатки включительно, Приамурье и Приморье.
Материал. 228 экз., о. Парамушир, низовья Амура, Охотское
побережье, заповедник «Кедровая Падь», Камчатка, окрестности Магадана.
11 У С. И. Огнева [1928] русское название этого подвида — короткохвостая
бурозубка Корона. Нам представляется более приемлемым название, употребляемое
С. У. Строгановым, в котором отражена территориальная локализация подвида.
282
Характеристика. Восточносибирская средняя бурозубка от-
носится к числу наиболее мелких подвидов. Однако различия эти стати-
стически достоверны только по отношению к S. с. shinto (см. табл. 58).
От северосибирской средней бурозубки по метрическим показателям тела
и черепа, а также его пропорций не отличается. У восточносибирского
подвида несколько большая длина хвоста (М = 34,5) по сравнению с се-
веросибирской (М = 32,2), но значения этих показателей широко пере-
крываются и достоверно не различаются (СД = 0,08). Ошибка в оценке
таксономической значимости этого признака, считавшегося основным при
выделении подвида, очевидно, происходит от скудности материала, имев-
шегося в руках исследователей. Неточность в описании цветовых вариа-
ций этих двух подвидов связана с тем, что С. У. Строганов [1957] исследо-
вал выцветшие от долгого хранения шкурки.
Заметки по систематике. Колориметрические исследо-
вания, проведенные нами на свежем, специально собранном материале
из ряда районов Приморья и Чукотки (см. табл. 60), показали четкие цве-
товые различия, позволяющие с достоверностью различить эти два подви-
да. Нельзя не подчеркнуть их близость, на которую указывал и С. У. Стро-
ганов [1957]. Упоминание С. У. Строгановым [Там же] о длиннохвостых
землеройках этого подвида с Сахалина (с. 215) совершенно непонятно,
так как они относятся к другому подвиду — 5. с. shinto, о чем он сам пи-
шет на следующей странице названного сочинения.
Что касается 5. с. buxtoni, включенного А. П. Кузякиным [Бобрин-
ский и др., 1965] в качестве подвида caecutiens, он равнозначен S. с. ko-
reni. Номенклатурно такое название не правомерно, так как под S. buxtoni
Аллен [Allen, 1903] описал арктическую бурозубку, которая в настоящее
время и рассматривается как подвид 5. tundrensis.
Sorex caecutiens shinto Thomas, 1905 —
Сахалинская средняя бурозубка
Голотип. Описание сделано по экземпляру, добытому на Саха-
лине, близ г. Корсакова и хранящемуся в Британском музее.
Диагноз. Резко выделяется от других представителей вида круп-
ными размерами и очень длинным хвостом (60,7 % от длины тела).
Измерения. См. табл. 58.
Характеристика. Сахалинская средняя бурозубка, имея
близкий ареал с восточносибирской средней бурозубкой, достоверно отли-
чается по ряду метрических показателей. Окраска зверьков тусклая,
буровато-серая, довольно светлая, переход от спины к бокам грязно-серо-
го цвета, брюшко палевых тонов. По данным колориметрического исследо-
вания, средние значения окраски «S’, с. shinto и других подвидов близки
(см. рис. 83). Достоверных различий по окраске нет.
Распространение. Острова Курильской гряды (о. Куна-
шир), Сахалин.
Материал. 22 экз., Курильские острова (о. Кунашир), Сахалин.
Sorex araneus Linnaeus, 1758 —
Обыкновенная бурозубка
Голотип. Вид описан по экземпляру из Швеции (окр. г. Упсалы).
Д и а г н о з. Крупная бурозубка: длина тела с головой 51—86 (М =
а= 68,9±0,45) мм, масса тела 5,5—15,5 (М = 8,8±0,15) г. Хвост умерен-
283
Рис. 86. Обыкновенная бурозубка — S. araneus L.
ной длины, составляет более 65,6 % длины тела. Голова широкая в мозго-
вой части с хорошо выраженным перехватом в области глаз и умеренно
длинным хоботком. Окраска чепрачная: у перезимовавших зверьков чеп-
рак темно-бурый, четко отделен от более светлых боков.
Измерения. См. табл. 61.
Характеристика. Хвост оканчивается у прибылых зверьков
хорошо выраженной кисточкой. Во внешнем облике бурозубки обращает
на себя внимание наряду с крупными размерами тела специфическая окра-
ска меха: спина окрашена значительно темнее боков и образует резко очер-
ченный чепрак темно-бурого цвета (рис. 86).
На боках: мех окрашен светлее. У перезимовавших бурозубок в лет-
нем наряде хорошо видна узкая переходная полоса ржаво-бурого цвета
с примесью рыжих тонов или палево-бурого цвета, разделяющая темный
чепрак и светлую окраску боков и нижней части тела.
Колориметрическое изучение окраски меха дорсальной стороны тела
обыкновенной бурозубки из разных частей сибирской части ареала позво-
ляет сделать заключение о широкой изменчивости показателя белизны и
оттенка (см. рис. 63). Несмотря на то что ареал обыкновенной бурозубки
в Сибири значительно уступает таковому /?. isodon, размах изменчивости
окраски у них остается близким, что, вероятно, связано с большим разно-
образием мест обитаний, осваиваемых S. araneus. В то же время у S. un-
guiculatus, занимающей ареал, по площади и разнообразию природных
условий значительно уступающий этим двум видам, довольно более широ-
кий размах изменчивости, чем можно было ожидать. Величина показателя
белизны обыкновенной бурозубки колеблется в пределах 1,3—5,5 (М =
— 2,8±=0,08), а оттенка — 122,3—155,5 (М = 134,8±0,81). Возрастная
и географическая изменчивость окраски рассмотрена ниже.
Зимний мех длиннее, ярче и значительно темнее летнего, чепрачность
выражена более четко, а палевые оттенки на боках чаще всего отсутствуют
или едва уловимы. У молодых зверьков мех тусклый, светлый, а палевая
284
Таблица 61
Морфометрическая характеристика Sorex araneus
Возрастная и половая группа М+тх О Су п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 68,3±0,69 5,49 8,04 64 55,0—83,0
взрослые 74,1 ±1,06 5,59 7,55 28 63,0—83,0
Самки прибылые 67,2±0,52 4,17 6,20 65 57,0—75,0
взрослые 77,5+1,71 6,16 7,95 13 64,0—86,0
В целом по виду . . . 68,9+0,45 6,41 9,30 204 55,0—86,0
Длина хвоста
Самцы прибылые 43,5±0,37 3,03 6,98 66 36,0—48,0
взрослые 40,9±0,64 3,41 8,34 28 34,0—49,0
Самки прибылые 42,6+0,39 3,14 3,38 65 35,0—49,0
взрослые 42,1+0,84 3,04 7,23 13 39,0—47,0
В целом по виду . . . 42,5±0,22 3,20 7,54 206 34,0—49,0
Длина ступни
Самцы прибылые 13,2+0,10 0,81 6,15 63 12,0—14,0
взрослые 12,8±0,12 0,64 5,01 27 12,0—14,0
Самки прибылые 13,0+0,10 0,77 5,94 64 12,0—14,0
взрослые 13,4±0,16 0,56 4,15 12 12,5—14,0
В целом по виду . . . 13,0±0,06 0,77 5,93 200 12,0—14,0
Масса тела
Самцы прибылые 8,2+0,18 1,26 15,44 52 5,5—11,7
взрослые 10,4+0,37 1,63 15,65 20 7,3—14,0
Самки прибылые 8,0+0,19 1,18 14,76 40 6,0—12,5
взрослые 11,9±0,46 1,61 13,49 12 9,0—15,5
В целом по виду . . . 8,8±0,15 1,83 20,79 147 5,5—15,5
Кондило-базальная длина черепа
Самцы прибылые 19,8+0,06 0,52 2,64 80 18,7—20,8
взрослые 19,8±0,07 0,40 2,02 36 18,9—20,4
Самки прибылые 19,7±0,06 0,53 2,70 76 18,8—21,3
взрослые 19,8±0,10 0,45 2,28 21 19,1-20,5
В целом по виду . . . 19,7+0,03 0,49 2,50 341 18,7—21,3
Длина рострума
Самцы прибылые взрослые 5,4±0,04 5,4+0,05 0,32 0,32 5,87 5,99 81 37 4,6—6,0 4,6—5,8
Самки прибылые 5,3+0,05 0,41 7,79 76 4,2—6,1
взрослые 5,4+0,05 0,22 4,09 21 5,1-5,9
В целом по виду . . . 5,3±0,02 0,31 5,80 243 4,2-6,1
Длина мозговой части черепа
Самцы прибылые 10,9±0,04 0,36 3,34 80 9,8—11,7
взрослые 10,8±0,05 0,30 2,75 36 10,3—11,4
Самки прибылые 10,8±0,04 0,33 3,01 76 10,1—11,6
взрослые 10,8±0,06 0,29 2,72 21 10,2-11,2
В целом по виду . . . 10,8±0,02 0,32 2,96 341 9,8—11,7
2S5
Продолжение табл. 61
1 2 3 * 5 6
Длина верхнего ряда зубов
Самцы прибылые 9,1±0,03 0,28 3,05 76 8,4-9,7
взрослые 8,8+0,05 0,26 2,91 30 8,3—9,4
Самки прибылые 9,1±0,05 0,37 4,08 62 8,2—9,9
взрослые 8,9+0,04 0,18 1,97 21 8,6—9,3
В целом по виду . . . 9,0±0,02 0,29 3,26 317 8,2-9,9
Длина ряда промежуточных зубов
Самцы прибылые 3,2+0,02 0,16 4,87 81 2,8—3,6
взрослые 3,1+0,03 0,15 4,78 37 2,8—3,5
Самки прибылые 3,2+0,02 0,17 5,15 76 2,8—3,5
взрослые 3,1+0,03 0,14 4,38 21 2,8-3,4
В целом по виду . . . 3,1+0,01 0,15 4,72 343 2,8-3,6
Ширина рострума
Самцы прибылые 1,9+0,01 0,10 5,59 80 1,7-2,1
взрослые 1,9+0,01 0,08 4,06 31 1,7—2,0
Самки прибылые 1,9+0,01 0,10 5,40 63 1,7—2,0
взрослые 1,9±0,02 0,08 3,84 21 1,8—2,1
В целом по виду . . . 1,9+0,01 0,10 5,17 323 1,7—2,1
Предглазничная ширина
Самцы прибылые 2,8+0,02 0,14 4,88 80 2,4-3,1
взрослые 2,8+0,02 0,12 4,18 31 2,5-3,1
Самки прибылые 2,8+0,02 0,13 4,79 63 2,5—3,1
взрослые 2,9+0,03 0,12 4,14 21 2,7—3,1
В целом по виду . . . 2,8+0,01 0,14 5,04 323 2,4-3,1
Межглазничная ширина
Самцы прибылые 3,8+0,02 0,14 3,80 81 3,5-4,1
взрослые 3,9±0,02 0,14 3,66 37 3,5—4,1
Самки прибылые 3,8+0,02 0,14 3,52 76 3,5—4,1
взрослые 3,9+0,03 0,12 3,09 21 3,7—4,1
В целом по виду . . . 3,8+0,01 0,14 3,65 343 3,5-4,1
Наибольшая ширина черепа
Самцы прибылые 9,8+0,04 0,33 3,33 80 9,1—10,7
взрослые 9,9±0,04 0,21 2,16 36 9,4—10,4
Самки прибылые 9,8±0,03 0,29 2,96 75 9,0—10,5
взрослые 9,9+0,08 0,35 3,56 21 9,2-10,5
В целом по виду . . . 9,8+0,02 0,32 3,32 337 9,0—10,7
Наибольшая высота черепа
Самцы прибылые 6,2+0,03 0,25 4,03 80 5,6-6,6
взрослые 6,0+0,04 0,22 3,73 36 5,2-6,5
Самки прибылые 6,2+0,03 0,23 3,76 73 5,7—6,7
взрослые 5,9+0,06 0,29 4,87 20 5,3—6,5
В целом по виду . . . 6,1+0,02 0,27 4,37 335 5,2-6,7
286
Окончани е табл. 61
1 2 3 4 5 6
Высота восходящей ветви нижней челюсти
Самцы прибылые 5,0±0,03 0,26 5,11 81 4,3—5,6
взрослые 5,1+0,04 0,22 4,29 37 4,6-5,4
Самки прибылые 5,0±0,05 0,45 9,08 76 4,0—6,1
взрослые 5,1±0,05 0,23 4,58 21 4,7—5,4
В целом по виду . . . 5,0±0,02 0,29 5,84 341 4,0—6,1
Длина углового отростка
Самцы прибылые 2,2±0,02 0,14 6,39 79 1,9—2,5
взрослые 2,1±0,02 0,12 5,53 30 1,9—2,3
Самки прибылые 2,2±0,02 0,12 5,51 60 1,8—2,3
взрослые 2,1+0,03 0,12 5,54 21 1,8—2,2
В целом по виду . . . 2,1±0,01 0,12 5,77 305 1,8—2,3
полоса у только что перешедших к самостоятельной жизни землероек
почти не видна, но более явственно проявляется после первой летней
линьки.
Хвост двухцветный, окрашен соответственно в тон дорсальной и вен-
тральной частей тела.
Череп крупный: кондило-базальная длина 18,7—21,3 (М = 19,7±
±0,03) мм, умеренно вздутый в мозговой части и вытянут в лицевой (см.
табл. 32). Очертания его довольно мягкие и сглаженные, умеренная ши-
рина в межглазничной области (19,2 %). Восходящая ветвь нижней че-
люсти мощная и довольно высокая (25,4 %), уступает по высоте только
S. daphaenodon. Угловой отросток относительно короткий и изогнутый
(рис. 87), имеет наименьшую относительную длину среди всех видов (см.
табл. 29). По длине зубного ряда обыкновенная бурозубка не уступает
ни одному виду рода Sorex (см. табл. 26). Зубной ряд длинный, зубы в нем
сидят свободно (45,7 %). Ряд промежуточных зубов также самый длин-
ный среди бурозубок (15,7 %), а по отношению длины ряда промежуточных
зубов к длине зубного ряда она уступает только S. gracillimus и S. caecu-
tiens.
Коронки зубов по отношению к размерам черепа умеренной величины.
Обращает на себя внимание прежде всего диспропорция в размерах двух
крупных первых промежуточных зубов и малые размеры пятого, а также
их грубая конфигурация (см. рис. 87). Весьма характерно наличие двух
парных группировок: первые две заметно крупнее двух последующих и их
вертикальная ось как бы наклонена назад, внутрь ротовой полости. Два
последующих значительно уступают по размерам предыдущим и образуют
вторую пару. Соотношение их размеров варьирует в широких пределах,
но чаще всего третий зуб несколько крупнее четвертого. Вертикальная
ось их коронок перпендикулярна продольной оси верхней челюсти или
даже слегка наклонена вперед. Вследствие этого при рассмотрении чере-
па в профиль создается впечатление излома зубного ряда между вторым
и третьим промежуточными зубами. Этот признак характерен только для
обыкновенной бурозубки и не повторяется у других видов.
287
Многовершинные зубы имеют хорошо выраженные острые конусы,
а пирамидальные воронки между мезостилем и протоконусом хорошо вы-
ражены. Предкоренной зуб нижней челюсти конический, не вытянут в ос-
новании и никогда не имеет дополнительной вершины. Его высота лишь
немного меньше продольной оси основания. Glans penis в виде утончаю-
щегося к дистальному концу конуса. Его головка в поперечном сечении
у основания округлая, постепенно сужающаяся к свободному концу и слег-
ка сжатая с боков (см. рис. 40). Копулятивный орган перезимовавших
зверьков 8,2—10,8 (М = 9,6) мм.
Распространение. Обыкновенная бурозубка по характеру
распространения — типичный западный палеаркт. В Сибири находится
лишь восточный участок ее обширного ареала, простирающегося через
всю Европу (кроме Ирландии и юга Пиренейского полуострова), лесотунд-
ровую, таежную, лесостепную и степную зоны и горы Западной Сибири,
Северного Казахстана, юг Средней Сибири до Прибайкалья включитель-
но и сопредельных районов северно-западной части Монголии. Указания
на распространение обыкновенной бурозубки в Приморье основаны на не-
правильной интерпретации систематического статуса S. isodon, ошибочно
принимавшегося за подвид обыкновенной бурозубки [Бобринский и др.,
1944; 1965; Строганов, 1957; и др.].
Ареал вида по конфигурации характерен для западных палеарктов,
проникающих в Сибирь: он имеет форму клина с широким основанием
в Европе и узкой вершиной в направлении на юго-восток, в то время как
для восточных палеарктов широкое основание клина находится на восто-
ке, а узкая часть — на северо-западе [Юдин, 1974а].
288
От Ямала на восток, в районах, примыкающих к нижнему течению
Оби, она обычна близ пос. Лабытнанги, в Шурышкарском районе близ по-
селков Мужи и Питлер, коллектировалась в районе Вапзеват, Ниязмы,
Поливат, далее на восток граница идет по нижнему течению р. Пур (у
пос. Самбург она крайне редка). На Гыданском полуострове в долине
р. Бол. Хета этой бурозубки уже нет, граница ее распространения про-
ходит южнее, спускаясь к Енисею. На правобережье Енисея она встре-
чается по Подкаменой Тунгуске и на восточном побережье Байкала. В се-
верном Прибайкалье и на Витимском плоскогорье обыкновенной бурозуб-
ки нет. Далее на юг она заселяет западные склоны хр. Улан-Бургасы (реки
Хырянка, Карпушка, Турку), здесь находится восточный предел распро-
странения вида. На хребтах Бол. и Мал. Хамар-Дабан она обычна.
На Урале обыкновенная бурозубка распространена повсеместно от Ямала
и Полярного Урала до южных районов Челябинской области, бассейна
Урала.
В Северном Казахстане она обитает в бассейне Иртыша и известна
из Курганской, Кустанайской, Кокчетавской, Павлодарской и Семипа-
латинской областей. Обыкновенная бурозубка заселяет Калбинские горы
и весь Казахстанский Алтай. Южная граница идет по отрогам Калбинско-
го и Курчумского хребтов. В Зайсанской котловине, а также в горах к за-
паду и югу от нее этого вида нет. В Алтае-Саянской горной стране она рас-
пространена повсеместно, кроме пограничных с Монголией районов Юго-
Восточного Алтая, Юго-Восточной Тувы, ряда районов Восточного Саяна
и высокогорий.
В пределах очерченной территории она широко распространена по
разным природным зонам и ландшафтам, доминируя по численности в по-
давляющем большинстве районов.
Материал. 464 экз., Пермская область, Башкирия, Южный
Урал, Западная Сибирь, Алтай, Салаир, Красноярский край.
Изменчивость. Структурная изменчивость [Яблоков, 1966]
обыкновенной бурозубки проявляется в виде сезонной, возрастной, поло-
вой, географической и травматической.
Изменчивость размеров тела в разные сезоны и в разных популяциях
изучена у 204 бурозубок. Длина тела взрослых зверьков (перезимовавших)
достоверно больше таковой прибылых (в возрасте от 1 до 6 мес) и состав-
ляет: М = 74,1+1,06 (д'), М = 77,5+1,71 ($) и М = 68,3+0,69 ( д'),
М = 67,2+0,52 ($) соответственно. По длине хвоста эта закономерность
выражена несколько слабее и имеет обратную направленность: длина
хвоста прибылых зверьков незначительно больше таковой перезимовав-
ших (см. табл. 61). Достоверных различий в размерах задней ступни
у зверьков разного возраста не обнаруживается. Различия по массе тела
весьма существенны. У перезимовавших зверьков она колеблется от 7,3
до 15,5 г. Максимальная масса у перезимовавших беременных самок:
М = 11,9+0,46 (?), М = 10,4+0,37 (д'), а у прибылых М = 8,0+0,19
($), М = 8,2+0,18 г (д').
Череп у прибылых зверьков достигает размеров черепа взрослых еще
в гнездовой период или вскоре после выхода из гнезда, поэтому сколько-
нибудь заметных возрастных различий по этому признаку не обнаружи-
вается. Кондило-базальная длина черепа перезимовавших особей колеб-
лется в пределах 18,9—20,5, М = 19,8+0,07 (д') и М = 19,8+0.10 ($),
а прибылых — 18,4+21,3, М = 19,8+0,06 (д') и М = 19,7+0,06 ($).
Максимальные размеры черепа наблюдаются у зверьков до зимовки. Зи-
289
мой череп становится более монолитным, его части плотно срастаются, на
месте швов образуются гребни, общие размеры его уменьшаются. Не об-
наруживается достоверных отличий и по большинству других показателей
в структуре черепа.
Наибольшие различия видны в длине зубного ряда (см. табл. 61),
наименьшие — в длине ряда промежуточных зубов: у перезимовавших
2,8—3,5 (М = 3,1), у прибылых 2,8—3,6 (М = 3,2) мм. Отмечается воз-
растная изменчивость и высоты черепа (см. табл. 61), однако размах из-
менчивости может быть более широк не между перезимовавшими и при-
былыми, а внутри группы прибылых зверьков, состоящей из особей разных
генераций.
Размах изменчивости (Cv* S. Cv) длины и массы тела обыкновенной
бурозубки из разных районов:
Группа Масса, г Длина, мм
Самцы (п = 72)
прибылые 8,2*0,18 68,3*0,69
взрослые Самки (п = 52) 10,4*0,37 74,1*1,06
прибылые 8,0*0,12 67,2*0,52
взрослые 11,9*0,46 77,5*1,71
Изменчивость массы тела у молодых в 3—5 раз больше по сравнению
с изменчивостью длины, и значительно выше у самок (молодых и перези-
мовавших), что связано с беременностью. Это подтверждает известное по-
ложение о большем размахе.изменчивости массы по сравнению с линейной.
Значительный размах имеет и возрастная изменчивость окраски. Мо-
лодые зверьки при выходе из гнезда и до первой осенней линьки тусклые,
бледные. Полоса на боку с примесью рыжих или палевых тонов едва за-
метна, а у некоторых зверьков не улавливается вовсе. Чепрачность нечет-
ко выражена и переход от темноокрашенной спины к более светлой вен-
тральной части тела на боках не столь резко очерчен. Зимняя окраска бо-
лее темная, мех длинный, до 6,5 мм, в то время как летом — 3—4 мм
(рис. 88). Чепрачность хорошо заметна. К осени прибылые зверьки при-
обретают более темную окраску, чепрак четко очерчивается, а палевая
полоса на боках хорошо выражена. Окраска хвоста у молодых зверьков
резко двухцветная, соответственно в тон спины и вентральной части тела,
а у перезимовавших он темноокрашен, почти лишен волосяного покрова
и без кисточки на конце, хорошо выраженной у прибылых зверьков.
Процесс изменения окраски и опушенности хвоста происходит постепенно
в течение зимы и весны.
Нами прослежено различие в окраске меха землероек разного воз-
раста. Оно значительно перекрывает географическую изменчивость. Раз-
личия по белизне и оттенку обыкновенной бурозубки разных возрастов
значительно перекрывают таковые у разных подвидов территориально
значительно удаленных популяций. Поэтому при оценке географических
различий необходимо сравнивать между собой только одновозрастные вы-
борки из разных мест.
Половая изменчивость обыкновенной бурозубки выражена менее чет-
ко. Длина тела прибылых самок (67,2*0,52) и прибылых самцов (68,3*
*0,69) разнится значительно меньше, чем зверьков разных возрастных
групп (см. табл. 61). Длина тела взрослых самок 77,5 (*1,75), самцов —
74,1 (*1,06) мм. Размах изменчивости этого показателя шире, более чет-
290
Рис. 88. Сезонная изменчивость окраски
обыкновенной бурозубки на Алтае
(Телецкое озеро).
1 — в целом по виду (для сибирской части
ареала); г — зимний мех; 3—летний мех.
Рис. 89. Колориметрическая характе-
ристика обыкновенной бурозубки из
разных частей ареала.
1 — Алтай (Телецкое озеро); 2 — Алтай (Те-
лецкое озеро, зима); з — Новосибирская об-
ласть (Тогучин); 4 — Прибайкалье (Хамар-
Дабан); .5 — окрестности Оренбурга; 6 —
Томская область (с, Поздняково),
ко выражен, что подтверждается и сравнением лимитов: у самок 64—86»
у самцов 63—83, достоверность различий также велика. Это мы связываем
с довольно быстрой элиминацией самцов, ведущих более активную жизнь,
вынужденных больше передвигаться в поисках самок и, следовательно,
имеющих больший шанс стать жертвой хищников, по сравнению с самка-
ми, предпочитающими относительно оседлый образ жизни и сильнее при-
вязанными к гнездовому участку.
Длина хвоста у прибылых самцов несколько больше (43,5=^0,37), чем
у прибылых самок (42,6±0,39), а у перезимовавших зверьков наблюдается
обратная закономерность: хвост у взрослых самок длиннее (42,1 zt0,84),
чем у самцов соответствующей возрастной группы (40,9±0,64). Возможно,
это связано с незначительным объемом выборки, так как по лимитам эти
показатели у взрослых самцов перекрывают изменчивость самок как в сто-
рону меньших, так и в сторону больших размеров (см. табл. 61). Половых
различий в длине ступни у прибылых зверьков нет, а у взрослых значения
этого показателя несколько выше у самок.
Кондило-базальная длина черепа прибылых самок достоверно мень-
ше (17,7^=0,06), чем самцов (19,8=t=0,06), а у взрослых половых различий
по этому показателю не прослеживается (соответственно 19,8±0,07
и 19,8±0,10). По другим краниологическим показателям половые разли-
чия также не обнаруживаются.
Географическая изменчивость окраски выражается в постепенном
увеличении белизны в направлении с запада на восток. Более темноокра-
шенные зверьки заселяют Урал и прилежащие районы Западной Сибири
(рис. 89). Бурозубки из равнинной части Западной Сибири несколько
светлее и самые светлые на восточном пределе распространения вида
29t
Таблица 62
Колориметрическая характеристика Sorex araneus из разных частей ареала
Белизна
Оттенок
Место сбора
материала
М±тх a Cv
lim
lim
Урал, окр.
Оренбурга
Томское При-
обье, пос.
Поздняково
Салаир, Тогу-
чинский р-н
Северо-В осточ-
точный Ал-
тай,
Телецкое озе-
ро (зимний
мех)
Там же (летний
мех)
Хамар-Дабан,
окр. пос. Тан-
хой
2,4+0,110,7
2,83+0,15 0,7
2,9+0,14 0,7
23,7 25
25,7 25
23,2 25
1,3—3,8 138,9+1,12
1,3-4,5 138,5±1,68
1,4—3,9 134,9+1,44
129,5-150,0
127,8—155,5
120,0—152,8
1,9+0,08 0,4
2,9+0,18 0,9
3,7+0,20 0,7
20,2 25
30,4 25
27,9 25
1,1-2,6 119,3+1,38
1,3-4,3 138,6+1,44
2,3—5,5 138,5+1,33
6,9
7,2
6,6
5,8 25 107,2-135,0
5,2 25 118,2-147,5
4,8 25 129,5-155,3
ЛГ±тж
в Прибайкалье (хр. Хамар-Дабан). В Западной Сибири темноокрашенные
бурозубки заселяют Верхнее Приобье, на Салаире и Алтае бурозубки
лишь немного светлее, чем в Томском Приобье (табл. 62).
Случаи альбинизма крайне редки [Долгов, 1968а]. Нами частичные
альбиносы отлавливались в разных районах Сибири. Чаще всего зто были
зверьки с белым кончиком хвоста, реже — с белым пятном на морде.
В Томской области в один цилиндр одновременно были пойманы две обык-
новенные бурозубки, имевшие ровный пепельно-серый светлый окрас
всего тела. Такие случаи также крайне редки.
Для обыкновенной бурозубки характерен широкий диапазон измен-
чивости промежуточных зубов. Эти вопросы детально освещены
В. А. Долговым [19686]. Первый резец верхней челюсти всегда хо-
рошо развит. Его второй зубец весьма изменчив: иногда он равен первому
зубцу и первому промежуточному зубу, иногда уступает им по высоте.
Передняя и задняя стороны коронок промежуточных зубов обычно слег-
ка вогнуты, вершины притупленные и едва сдвинуты вперед по отношению
к середине коронки. Вертикальная ось двух первых промежуточных не-
сколько наклонена в сторону многовершинных зубов, а оси третьего и чет-
вертого перпендикулярны челюсти или слегка наклонены вперед: в зави-
симости от степени проявления этих признаков между первой и второй-
парой зубов в той или иной мере выражен как бы передом зубного ряда.
Пятый крайне мал, как правило, в разной степени задвинут за парастиль
первого многовершинного зуба, так что при рассмотрении зубного ряда
сбоку он или едва виден, или не виден совсем.
Значительной изменчивости подвержена высота промежуточных зу-
бов, но все же они всегда довольно высокие по сравнению с зубами рав-
нозубой или когтистой бурозубок. Гипоконусы многовершинных зубов
292
всегда хорошо развиты, вершины их пигментированы, а степень окраски
и цвет подвержены широкой изменчивости: от почти коричневых, темных
до светлых желтоватых.
Как уже указывалось выше, наиболеечасто встречающийся тип соот-
ношения промежуточных зубов у обыкновенной бурозубки — попарная
группировка: два первых равны смежду собой и заметно крупнее двух по-
следующих, тоже равных между собой. Пятый зуб всегда развит крайне
слабо. Причем различия в величине этих пар могут быть неодинаковыми.
Второй тип изменчивости характеризуется теми же показателями, но при
одном исключении — во второй паре четвертый зуб меньше третьего. Тре-
тий тип отличается от первого несколько более слабым развитием второго
промежуточного зуба. Иногда встречается соотношение зубов, когда каж-
дый второй зуб пары меньше предыдущего. Наконец, существует вариант,
когда парная группировка практически не улавливается и зубы постепен-
но уменьшаются от первого к четвертому. Однако пятый всегда значитель-
но меньше четвертого.
Крайне редки случаи полиодонтии и олигодонтии как у обыкновенной
бурозубки, так и у других видов. Из нескольких тысяч просмотренных
землероек обнаружена редукция одного из промежуточных зубов
(третьего, четвертого зубов, пятого в одной челюсти, левой или правой,
или в обеих одновременно). При редукции одного из зубов оставшиеся
расположены в челюсти более свободно. Также редки случаи появления
шестого промежуточного зуба. Дополнительный зуб не достигает размеров
соседних и вытеснен из зубного ряда внутрь ротовой полости. При этом,
как указывает В. А. Долгов [1968а], изменяется положение на челюсти
и форма коронок соседних с дополнительным зубом.
Случаев редукции или появления дополнительных зубов в нижней
челюсти нам неизвестно.
Травматологическая изменчивость редка и чаще всего связана с час-
тичной потерей фаланг пальцев на передних или задних лапах, одного или
одновременно нескольких пальцев, потеря части хвоста. Иногда попада-
ются зверьки с обрубком хвоста, напоминающим таковой у леммингов,
или с надломленным, как бы «горбатым» хвостом. Во всех случаях трав-
мы старые. В. А. Долгов [1968в, г] указывает на случаи травм скелета
и даже копулятивного органа бурозубок.
Обыкновенная бурозубка отличается от других видов рода по хромо-
сомному набору. Для нее характерно неодинаковое число хромосом у зверь-
ков разного пола и в разных частях ареала. В Европе у самцов 2п = 21 —
32, у самок 2п = 20—31 при NF — 40; в Сибири у самцов 2п = 25—27,
у самок 2п — 24—27 при NF = 40 [Meylan, 1964, 1965; Козловский,
1969, 1970]. Число аутосом в европейских популяциях этого вида варь-
ирует от 18 до 29 [Meylan, 1964, 1965]. В Сибири популяция из Кемеров-
ской области мономорфная, у самок в диплоидном наборе 24, у самцов —
25 хромосом. Число аутосом у S. araneus под Иркутском варьирует от 22
до 25 [Орлов, Козловский, 1969; Козловский, 1969, 1970].
Полиморфизм по центрическим соединениям (робертсоновский) у мле-
копитающих наиболее полно изучен у обыкновенной бурозубки. К настоя-
щему времени кариологически исследованы несколько десятков популя-
ций этого вида на большей части ареала вида [Орлов, 1974]. Порядок и по-
следовательность слияния одноплечих хромосом в двуплечие происходит
по одному плану, и по этой причине структура кариотипа полиморфных
популяций едина. Meylan [1965] описал две хромосомные расы, кариоти-
293
пические различия которых состояли в различном порядке слияния одно-
плечих хромосом в двуплечие. Их видовая самостоятельность была под-
тверждена морфологическим анализом [Ott, 1968; Plert, 1973]. Впоследст-
вии кариотипические отличия от расы Б были обнаружены в одной скан-
динавской [Fredga, 1973] и нескольких сибирских популяциях [Козлов-
ский, 1970; Fedyk, Ivanitskaya, 1972] этих бурозубок.
Кариотипическая структура обыкновенной бурозубки сибирских по-
пуляций описана В. Г. Волобуевым и В. М. Анискиным [1981]. Б. Крал
и С. И. Раджабли [Kral, Badjabli, 1974] обнаружили различия в морфоло-
гии четырех пар аутосом. В результате исследований авторами установле-
но, что сибирская популяция обыкновенной бурозубки характеризуется
двумя уровнями хромосомной изменчивости — внутри- и межпопуляцион-
ной. Первая обусловливается полиморфизмом робертсоновского типа.
Размах вариаций в каждой из популяций при неизменном числе плеч, рав-
ном 40, зависит от количества пар хромосом, вовлеченных в процесс цент-
рических слияний. В отдельной популяции число таких пар изменяется
от 0 до 5, что ведет к соответствующей вариации числа аутосом от 18 до
24. Установлены различия и в поперечной исчерченности хромосом (диф-
ференциальное окрашивание: 1, 2, 9-я пары). Морфология оставшихся
пяти пар аутосом различна в разных популяциях. По структуре кариоти-
па все исследованные популяции распадаются на три пространственно
отграниченные друг от друга формы: новосибирскую, томскую, ермаков-
скую. Новосибирская имеет восточную границу по Томи (4 точки);
томская — от Томска до Телецкого озера на юге (5 точек), западная гра-
ница идет по Томи, восточная, видимо, до Енисея; ермаковская (1 точ-
ка) — на правобережье Енисея, в 90 км от томской формы на левобережье
Енисея (пос. Майма).
По мнению В. Г. Волобуева и В. М. Анискина [1981], описанные хро-
мосомные формы обыкновенной бурозубки либо уже достигли репродук-
тивной изоляции, либо ее становление близко к завершению, что и под-
нимает вопрос о таксономическом статусе этих форм.
Заметки по систематике. Видовая самостоятельность
обыкновенной бурозубки с момента ее описания до наших дней не вызыва-
ла сомнений у исследователей. Тем не менее долгое время она была одним
из сложных видов, объем которого оставался неясным. Лишь благодаря
исследованиям последних 15 лет, в том числе с привлечением кариологи-
ческих методов, достигнут значительный прогресс в «очищении» вида
от чуждых ему элементов. После описания вида К. Линнеем [1758] в то
или иное время в его состав включали все виды «крупных бурозубок», но
постепенно исследователи выделяли искусственно внесенные в его виды,
и в настоящее время они ближе всего к представлениям автора его перво-
описания.
С. И. Огнев [1928, 1933, 1935] объем 5. araneus принимал в весьма ши-
роких границах, существенно отличающихся от современных представ-
лений. Недостатком его концепции было восприятие немецкой системы
«форменкрайс», не признающей четко очерченных видов, а заменяющих
его «кругом форм» подвидового ранга. В монографии С. И. Огнева [1928]
нет характеристики вида, а лишь приведен ряд подвидовых очерков.
Кроме того, в состав вида Обыкновенная бурозубка он включал равнозу-
бую бурозубку — 5. isodon (=S. a. tomensis isodon), частично арктическую
(5. arcticus schnitnikovi Ognev) и кавказскую 5. caucasicus (S. araneus sa-
tunini Ognev). Впервые на обособленность равнозубой бурозубки обратил
294
внимание С. С. Туров [1924], выделивший ее по материалам из Баргузин-
ского заповедника в качестве natio — 5. araneus tomensis isodon, т. e. вна-
чале рассматривал ее в составе обыкновенной бурозубки. Позднее он воз-
вел равнозубую бурозубку в ранг вида [Туров, 1936]. Одновременно ее
видовая самостоятельность была подтверждена С. У. Строгановым [1936]
по материалам с Валдайской возвышенности. В. Н. Орлов и А. И. Козлов-
ский [1971 ] установили отличие ее кариотипа на видовом уровне (42 хро-
мосомы) от такового обыкновенной (самец — 25—27, самка — 24—27),
и таким образом этот вид был исключен из состава 5. araneus.
А. П. Кузякин [Бобринский и др., 1944, 1965] рассматривал в составе
вида обыкновенной бурозубки такие хорошо консолидированные видо-
вые формы, как S. isodon, S. ungulatus, S. raddei, S. asper, S. roboratus,
S. caecutiens, S. daphaenodon, частично — 5. arcticus (S. a. schnitnikovi).
Основой для объединения столь разных видов в одну систематически не
регламентированную группу «5. araneus» всех «относительно крупных зем-
лероек без деления ее на виды и подвиды» [Бобринский и др., 1944, с. 52]
послужила имеющаяся трансгрессия в размерах тела и черепа. Однако,
как совершенно справедливо отметил С. У. Строганов [1957], размеры эти
сами по себе для видовой дифференциации в данном случае самостоятель-
ного значения не имеют. Виды, объединенные в эту искусственную компо-
зиционную группу, обладают дискретными качественными отличиями,
заключающимися в структурных одонтологических и краниологических
особенностях, строении органов половой системы, окраске, пропорциях
отдельных элементов тела и других. Несмотря на хорошо аргументиро-
ванную критику, данную в монографии С. У. Строганова [Там же], в ко-
торой автор на основе анализа большинства доступного в то время мате-
риала приводит убедительную ревизию систематики этой группы, во вто-
ром выпуске «Определителя...» эти ошибки повторяются почти в том же
объеме.
Все это заставляет нас провести детальный разбор взглядов А. П. Ку-
зякина [Бобринский и др., 1944, 1965] на систематику обыкновенной буро-
зубки. Этого можно было бы и не делать, так как «Определитель...» не яв-
ляется научной работой, а лишь учебное пособие для студентов. Однако
взгляды этих исследователей были широко и не критически восприняты
Эллерманом и Моррисон-Скоттом [Ellerman, Morrison-Scott, 1951], а так-
же рядом отечественных зоологов, преимущественно неспециалистов по
систематике насекомоядных, что внесло существенную дезинформацию.
В частности, тем самым была вызвана путаница в оценке места фауны на-
секомоядных млекопитающих Палеарктики в фауне мира и ее связей с дру-
гими регионами, были обесценены экологические работы по ряду видов,
так как полученные сведения оказались обезличенными.
Прежде всего следует указать на основную слабую сторону систе-
матики, предлагаемой А. П. Кузякиным [Бобринский и др., 1965]. Она
строится на чисто количественных оценках размеров тела и черепа, в то
время как виды представляют качественно различные категории. При
одних и тех же метрических показателях они могут коренным образом от-
личаться по ряду других более существенных параметров, в том числе
кариологических, что и было доказано на примере обыкновенной бурозуб-
ки [Meylan, 1964, 1965; Козловский, 1969, 1970; и др. ].
А. П. Кузякин объединяет в «виде» обыкновенной бурозубки наряду
с собственно 5. araneus еще восемь перечисленных выше видов и частично
S. caecutiens.
295
Рассмотрим основные отличия обыкновенной бурозубки от других
видов, включенных этим исследователем [Бобринский и др., 1965] в ка-
честве «подвидов». Размеры тела и черепа, на основе которых построена
система Soricidae, в данном случае не имеют диагностического и систе-
матического значения. Для обыкновенной бурозубки, как и для других
искусственно включенных в ее состав видов, наряду с крупными размера-
ми тела свойственны и самобытные, присущие только им черты. Первое,
на что обращает внимание исследователь, это специфическая окраска меха
зверьков. Для обыкновенной бурозубки характерно наличие чепрачности
в распределении окраски меха по телу. Спина, окрашенная в темно-бурые
тона, резко отграничена от светлой серой или свинцово-серой вентраль-
ной стороны. В этом отношении она близка к тундряной, но отличается
от нее наличием на боках буровато-палевой полосы, меняющейся по ин-
тенсивности от едва заметной у прибылых зверьков в первом наряде до
более яркой к осени. Максимальных размеров интенсивности окраски
полоса достигает у перезимовавших землероек в летнем наряде. В зимнем
наряде она почти незаметна. Ни один из видов, включенных А. П. Кузя-
киным [Там же] в состав 5. araneus, кроме собственно обыкновенной бу-
розубки, такой окраски меха не имеет. Признак этот весьма устойчив,
и его было бы вполне достаточно для отрицательного решения вопроса
о правомочности объединения в один вид этих гетерогенных форм.
Надежным признаком, позволяющим выделить 5. araneus среди дру-
гих видов бурозубок, является своеобразная форма Рпц. Он высокий, вы-
тянут в вертикальном направлении, примерно равен продольной своей
оси. Дополнительной вершины на его внутренней стороне нет. У всех ви-
дов, произвольно включенных А. П. Кузякиным [Там же] в состав вида
обыкновенной бурозубки, этот зуб слабее, коронка его вытянута в сагит-
тальном направлении и имеет в той или иной мере развитую дополнитель-
ную вершину (см. очерки соответствующих видов).
Наконец, существенное значение в классификации бурозубок имеет
строение половых органов самцов, а в некоторых случаях и самок. Копу-
лятивный орган обыкновенной бурозубки отличается по строению от та-
кового других видов (см. рис. 40). Все эти признаки дискретны, и каж-
дого из них достаточно, чтобы обосновать видовую самостоятельность
обыкновенной бурозубки. Кроме того, уместно отметить, что включение
в качестве подвидов бурозубку Радде, кавказскую, равнозубую, крупно-
зубую, бурую нарушает принцип викарности ареалов подвидов. Все они
обитают в одних биотопах с обыкновенной бурозубкой.
В диагностике вида имеет значение не только абсолютная величина
признака, но и относительная. Так, обыкновенная бурозубка имеет самый
малый относительный размер углового отростка (10,7 %), самый большой
относительный показатель длины зубного ряда (45,7 %) и ряда проме-
жуточных зубов (15,7 %) к кондило-базальной длине черепа. Все эти при-
знаки, взятые безотносительно, ни о чем не говорят (см. табл. 26).
Весьма характерно для обыкновенной бурозубки и соотношение про-
межуточных зубов верхней челюсти. По этому признаку вид надежно диаг-
ностируется. Зубы имеют грубую конфигурацию, два первых, как прави-
ло, равны между собой и значительно крупнее двух последующих, также
равных между собой, или чаще всего третий несколько крупнее четвер-
того. Пятый крайне мал. Вертикальные оси первых двух несколько накло-
нены внутрь ротовой полости, а двух последующих — перпендикулярно
продольной оси верхней челюсти. Вследствие этого зубной ряд как бы
296
Рис. 90. Структура промежуточных зубов бурозубок.
а — Sorex raddei; б — S. alpinus; в — S. araneus; г — S. asper.
«переломлен» между вторым и третьим одновершинными зубами. Совер-
шенно иная структура промежуточных зубов у тянь-шаньской бурозубки
[Строганов, 1957]. Зубы ее, особенно три первых, значительно массив-
нее, а соотношение их иное (рис. 90).
Поскольку мы не даем очерка о тянь-шаньской бурозубке (S. asper
Thomas), эндемика Тянь-Шаня, то следует более детально охарактеризо-
вать ее основные черты и отличия от обыкновенной бурозубки.
Промежуточные зубы непропорционально высокие и широкие и резко
контрастируют с размерами черепа и многобугорчатыми зубами. Как и у
всех видов бурозубок, у нее пропорционально размерам черепа сильно
развиты три первых зуба.
Тянь-шаньская бурозубка распространена только в горах Тянь-Шаня.
Она характеризуется весьма самобытной организацией и легко дифферен-
цируется среди представителей других видов рода Sorex. Наиболее харак-
терной чертой вида является ее исключительная крупнозубость. Ни один
вид не имеет столь мощных, крупных одновершинных зубов, как тянь-
шаньская бурозубка. Обращает на себя внимание диспропорция размеров
одновершинных (промежуточных) и коренных зубов. Как правило, зубы
интенсивно пигментированы. Соотношение размеров зубов между собой
у этой бурозубки такое же, как у тундряной, но отличаются они по раз-
мерам и форме. Рш! в отличие от такового 5. araneus вытянут в сагитталь-
ном направлении. Описанные структуры одновершинных зубов настолько
отличны от всех известных вариаций зубной системы обыкновенной буро-
зубки, что говорить о их систематической близости не приходится.
Копулятивный орган тянь-шаньской бурозубки имеет своеобразное
строение и несхож с таковым обыкновенной бурозубки (рис. 91). Свое-
образна и окраска зверьков. В отличие от обыкновенной бурозубки, имею-
щей чепрачный характер распределения темной окраски на спине, пале-
вые бока и серовато-белесое брюшко разной интенсивности, но всегда свет-
лое, тянь-шаньская бурозубка окрашена иначе. Мех дорсальной части
тела буровато-серой, тусклой окраски, постепенно переходящей на боках
в более светлую по тону окраску того же цвета, а затем в серую с палево-
буроватой примесью относительно темную окраску вентральной стороны.
Зверьков обыкновенной бурозубки ни в одной части ее обширного ареала
такой окраски меха не встречается.
297
Рис. 91. Строение glans penis
бурозубок.
А — Sorex raddei; Б — S. araneus',
В — S. alpinus; Г — S. asper. Ос-
тальные усл. обозн. см. рис. 7-
Таким образом, по
структуре зубов, выража-
ющейся в крупных раз-
мерах промежуточных, их
своеобразной форме, соот-
ношению и диспропорции
с многобугорчатыми зуба-
ми тянь-шаньскую буро-
зубку нельзя сблизить ни
с одним известным видом.
Строение надежно отличает
ее от обыкновенной. Несхожи они и по строению головки копулятивного
органа. Все это дает право считать объединение 5. asper с обыкновенной
бурозубкой лишенным фактического основания. Уместно отметить,
что С. И. Огнев [1928, 1940] не был последователен в оценке этого
таксона. Сначала он считал ее самостоятельным видом, а позднее [Огнев,
1940] без должной аргументации свел в подвид обыкновенной бурозубки.
Следует более подробно остановиться на систематическом отношении
S. araneus и S. raddei. поскольку очерка о бурозубке Радде, эндемика
Кавказа, нет. Систематике 5. raddei, ее взаимоотношению с альпийской
(5. alpinus), кавказской (S. caucasicus) и обыкновенной бурозубками нами
посвящена специальная статья [Юдин, 19726]. Этот самобытный вид на-
столько четко охарактеризован систематически, что спутать его с каким-
либо другим представителем рода невозможно. Нами была собрана обшир-
ная коллекция из разных районов Кавказа, а также изучены сборы, хра-
нящиеся в музеях страны.
Основные отличия бурозубок Радде и обыкновенной сводятся к сле-
дующему. Габитуально первая хорошо отличается от второй несколько
меньшими размерами тела и значительно большей длиной хвоста: длина
тела 58—80 (М = 66), хвоста 39—59 (М = 48) мм, что составляет 72 %
от длины тела (для обыкновенной этот показатель составляет 65,6 %).
Указанные отличия вполне достоверны и достаточны для диагностирова
ния вида. Кроме того, во внешнем облике землеройки Радде обращает
на себя внимание своеобразие окраски меха, также подчеркивающее само-
бытность вида. В отличие от большинства видов рода Sorex мех этой буро-
зубки на всем теле одноцветный, матовый, не блестящий, лишь слегка от-
личается более светлым тоном на вентральной стороне. Окраска меха
темная, буровато-серая с шоколадным отливом. Тыльные стороны ступ-
ней и кистей темные, покрыты бурым волосом. Хвост резко четырехгран-
ный в поперечном сечении, покрыт короткими волосками. Ни у одного
из многих тысяч исследованных зверьков обыкновенных бурозубок этих
признаков нет.
Во внешнем строении черепа бурозубки Радде наглядны некоторая
угловатость и широкая ростральная часть,более короткая, чем у обыкно-
венной бурозубки. Специфичным отличительным признаком служит
структура зубов. При сравнении с другими видами бурозубок обращает
20 Б. С. Юдин
298
на себя внимание относительно короткий передний резец верхней челюсти.
Его вторая внутренняя вершина значительно меньше первой и ниже пер-
вого промежуточного зуба. В целом соотношение промежуточных зубов
ближе к таковому у тундряной бурозубки (S. tundrensis Merr.) и коренным
образом отличается от их структуры у обыкновенной. Весьма характерна
массивность второго зуба. В зубном ряду его вершина, как правило, выше
других зубов, хотя по высоте он иногда почти равен первому, что дости-
гается благодаря более высокому его положению в челюсти. Нижняя че-
люсть массивна, с высоким коронарным отростком, с коротким, толстым
и загнутым вверх на свободном конце угловым отростком. У обыкновенной
бурозубки массивности нижней челюсти не наблюдается, Рш± значительно
вытянут в длину, с едва заметной дополнительной вершиной. Этот признак
К. А. Сатунин [Satunin, 1895] считал весьма существенным, выделяя бу-
розубку Радде в самостоятельный вид. У обыкновенной бурозубки, как
сказано выше, Ршх имеет иную конфигурацию.
Головка копулятивного органа по размерам и форме существенно от-
личается от таковой обыкновенной бурозубки: она почти вдвое уступает
по размерам и имеет иное строение (см. рис. 91). Отличается S. raddei
и по кариотипу, у нее 36 хромосом [Козловский, 1973а].
Живые и недавно пойманные зверьки издают сильный, довольно
приятный мускусный запах, не свойственный другим видам. Только взрос-
лые самцы средней бурозубки (S. caecutiens) имеют почти такой же силь-
ный, но иной запах.
О самобытности бурозубки Радде говорит уже тот факт, что первона-
чальное ее описание сделано К. А. Сатуниным [Satunin, 1895] по единст-
венному сильно выцветшему от давности хранения в спирте экземпляру.
При этом автором первоописания точно подмечены наиболее характерные
диагностические признаки — специфичность строения зубов и габитуаль-
ный склад. К. А. Сатунин не имел возможности опереться на особенности
окраски меха, весьма характерный признак при диагностике этого вида,
и строения копулятивного органа, использованных более поздними иссле-
дователями. Как наиболее специфичные признаки он выделял строение
Ртх и длинный хвост. С. И. Огнев [1926, 1928] считал наиболее стойким
отличием бурозубки Радде от обыкновенной конфигурацию черепа, специ-
фическое строение одновершинных зубов и окраску меха. Однако, объеди-
няя бурозубку Радде с кавказской (S. caucasicus), а последнюю с 5. a. sa-
tunini, С. И. Огнев [Там же] допускал ошибку. В описании 5. caucasicus
К. А. Сатунин [1913, 1915] дает для нее иную конфигурацию промежуточ-
ных зубов и отмечает светлую окраску меха нижней части шеи, хвоста.
Позднее [Долгов, Лукьянова, 1966] установлено отличие в окраске
меха и строении копулятивного органа этого вида. 5. caucasicus отличается
и по кариотипу (25 хромосом, 33 аутосомы, А'Е = 46). В целом интер-
претация С. И. Огневым бурозубки Радде как самостоятельного вида вер-
на, но им внесены неточности при описании якобы существующего вариан-
та окраски со светлой вентральной частью и палевыми боками. В дейст-
вительности на больших сериях зверьков из разных мест обитания нами
установлена устойчивость одноцветной окраски меха зверьков этого вида.
Наблюдаются лишь небольшие тональные отличия верхней и нижней час-
тей тела при очень темной общей окраске. Примеси рыжего и палевого цве-
тов в окраске меха бурозубки Радде нами не отмечено ни у одного экземп-
ляра и нет зверьков с белесым брюшком. Все это говорит о несостоятель-
ности объединения бурозубки Радде в один вид с обыкновенной бурозубкой.
299
Сначала А. П. Кузякин [Бобринский и др., 1944] свел бурозубку
Радде в подвид обыкновенной, объединив ее в нерегламентированную сис-
тематическим рангом группу, включающую «особи почти одноцветной ок-
раски»— S. isodon, S. unguiculatus, S. raddei, представляющую четко оха-
рактеризованные самостоятельные виды. Во втором издании этой книги
А. П. Кузякин выделил из этой группы в полном соответствии с действи-
тельно существующим положением как самостоятельный вид 5. unguicu-
latus, но объединил 5. raddei с 5. caucasicus (= S. a. satunini), рассматри-
вая эту сборную группу в качестве подвида обыкновенной бурозубки —
5. a. raddei Satunini, чем внес дополнительную путаницу не только в так-
сономию бурозубки Радде, но и в систематику бурозубок в целом. Как уже
указывалось выше, Эллерман и Моррисон-Скотт [Ellerman, Morrison-
Scott, 1951], во всей полноте воспринявшие систематические построения
А. П. Кузякина [Бобринский и др., 1944], также рассматривали ее в ка-
честве подвида обыкновенной бурозубки. В этом плане весьма любопытно
отметить, что в своих ранних работах С. К. Даль [1940, 1948] признавал
видовую самостоятельность бурозубки Радде, а позднее [Даль, 1954],-
видимо, под влиянием работы А. П. Кузякина [Бобринский и др., 1944]
объединил ее с обыкновенной бурозубкой, чем обесценил приводимые ори-
гинальные материалы по их образу жизни. Его указание о наличии в Ар-
мянской ССР обыкновенной бурозубки с однотонной окраской спины и вен-
тральной части тела и с темной спиной и серебристо-белым брюшком сви-
детельствует о путанице, так как первый вариант окраски несомненно
относится к зверькам бурозубки Радде.
В более поздних исследованиях видовая самостоятельность бурозуб-
ки Радде в полном соответствии с фактическим положением дела была
восстановлена [Spitzenberger, Steiner, 1962; Гуреев, 1963; Лаврова, За-
жигин, 1965; Юдин, 19656; 19726; Долгов, Лукьянова, 1966; Долгов,
19686; Spitzenberger, 1968; и др.].
К. А. Сатунин [1905], характеризуя строение верхнего переднего рез-
ца, указывает на сходство бурозубки Радде с альпийской (S. alpinus),
считая ее «заменителем последней на Кавказе». Однако он отметил отличия
в соотношении одновершинных зубов, в структуре Рпц и в строении бу-
горков на кистях передних ног. Кроме того, уместно отметить, что для аль-
пийской бурозубки характерна еще большая длина хвоста, охристо-бело-
ватый цвет кистей и ступней. Как показали наши исследования альпий-
ских бурозубок, любезно присланных из Венского музея естественной
истории природы доктором Ф. Шпиценбергером, головка копулятивного
органа зверьков этого вида коренным образом отличается по размерам
и строению от таковой бурозубки Радде и обыкновенной бурозубки (см.
рис. 91). Подводя итог сказанному, можно с полной уверенностью считать
установленной видовую самостоятельность бурозубки Радде [Юдин, 19726].
Не выдерживает критики и утверждение А. П. Кузякина [Бобрин-
ский и др., 1965] об идентичности 5. arcticus schnitnikovi Ognev и обыкно-
венной бурозубки. Это, действительно, существенно уклонившаяся, наи-
более крупная форма арктической бурозубки, никакого отношения к
обыкновенной не имеющая. Характеристика этого вида приведена ниже в
соответствующем очерке.
Бурая бурозубка (S. roboratus, = S. vir Alien), также включенная
в состав 5. araneus, настолько самобытна, что безошибочно отличается даже
по габитуальным признакам (см. соответствующий очерк), не прибегая к
не менее своеобразным одонтологическим и краниологическим показателям.
20*
300
Холлистер [Hollister, 1913] описал 5. roboratus по одному экземпляру
из окрестностей Топуча с Алтая. С. И. Огнев [1928] отнес ее к 5. araneus
в качестве подвида 5. a. roboratus. Эта форма характеризуется большими
размерами черепа (кондило-базальная длина 22 мм) и мелкими зубами.
Размеры кондило-базальной длины не укладываются в лимиты этого при-
знака для обыкновенной бурозубки из разных районов ее ареала в Сиби-
ри. Максимальный известный показатель не превышает 21,3 (М =
— 19,7) мм. Из этого совершенно очевидно, что экземпляр, имевшийся
в распоряжении Холлистера, относится не к 5. araneus, а к самостоятель-
ному виду — 5. roboratus [Юдин, 19656, 1971а], у которого большие раз-
меры кондило-базальной длины черепа (18,8—21,8, М — 20,3 мм) и более
мелкие, чем у обыкновенной бурозубки, зубы.
У 5. roboratus 42 хромосомы [Орлов, Козловский, 1971] в отличие
от 5. araneus, имеющей кариотип из 24—27 хромосом.
Одонтологические черты бурой бурозубки настолько самобытны, что
объединение ее с обыкновенной бурозубкой по этому признаку совершен-
но нереально. Передний резец этого вида отличается от такового всех дру-
гих видов бурозубок. Первый промежуточный зуб значительно меньше
второго. Все коронки промежуточных зубов имеют совершенно иную кон-
фигурацию по сравнению с обыкновенной бурозубкой. Они сильно вытя-
нуты в основании в сагиттальном направлении и имеют сосцевидные вер-
шины. Коренные зубы расставлены редко. На Рт2 имеется четко выра-
женная дополнительная вершина, а основание его очень сильно вытянуто
в длину. Череп длинный, относительно узкий, плоский в мозговой части
и широкий в ростральной (см. табл. 29).
Несостоятельность объединения 5. araneus с 5. unguiculatus была
настолько очевидной, что во втором издании «Определителя...» [Бобрин-
ский и др., 1965] А. П. Кузякин рассматривал их уже как самостоятель-
ные виды. Уместно напомнить, что у S. unguiculatus 42 хромосомы. Все это
говорит о несостоятельности взглядов А. П. Кузякина [Там же] на си-
стематику бурозубок, в частности, на объем вида обыкновенной бурозубки.
Нужно отметить непоследовательность в оценке объема вида и
С. У. Строгановым [1957], необосновано включившем в его состав равно-
зубую бурозубку, видовой статус которой признавался им ранее [Строга-
нов, 1936]. Эта ничем не обоснованная непоследовательность совершенно
правильно была отмечена в рецензии на его монографию П. Б. Юргенсо-
ном [1959], также как и то, что своей публикацией С. У. Строганов внес
весомый вклад в разработку систематики данной группы.
Характеристика 5. isodon приведена в очерке вида. Здесь упомянем
лишь наиболее характерные черты, по которым она хорошо отличается от
обыкновенной бурозубки. В окраске меха отсутствуют чепрачность и па-
левая полоса между темной спиной и светлым брюшком. Переход темной
окраски спины к более светлой, но того же бурого тона осуществляется
постепенно от спины к вентральной части тела. Зубы имеют иные конфи-
гурацию, соотношение и размеры. Первый промежуточный всегда больше
последующих, равномерно убывающих по размерам от первого к пятому.
Пятый хорошо развит и немногим уступает четвертому. Зубы «пильчатые»,
в виде равнобедренных треугольников. В зубном ряду промежуточные си-
дят свободно. Pmj сильно вытянут в сагиттальном направлении и имеет
дополнительную вершину.
Головка копулятивного органа коренным образом отличается от тако-
вой обыкновенной бурозубки по длине, массивности и форме (см. рис, 40).
301
В таежных равнинных и горных районах и в лесотундре виды не викари-
руют, заселяют одни и те же биотопы. Переходных особей по любому из
перечисленных признаков не зарегистрировано.
В настоящее время самостоятельность этого вида признана большин-
ством исследователей. Работами кариологов [Орлов, Козловский, 1969;
Козловский, 1970] доказана специфичность кариотипа обыкновенной буро-
зубки из Кемеровской и Иркутской областей, отличающая ее от обыкно-
венной бурозубки из европейской части страны. В. И. Орлов [1974] на этом
основании рассматривает ее в качестве самостоятельного вида — S. gemalus
Ott., хотя далее указывает, что видовая самостоятельность ее должным
образом не обоснована.
Обзор подвидов
В сводке С. У. Строганова [1957] для Сибири приводится четыре под-
вида обыкновенной бурозубки (S. a. araneus, S. a. tomensis, S. a. roboratus
и S. a. isodon). Внутривидовая систематика базируется преимущественно
на размерах и специфике окраски.
Рассмотрим диагностические признаки этих подвидов.
Номинальный подвид, по С. У. Строганову [1957], имеет следующий
диагноз: «Характеризуется относительно мелкими размерами тела и чере-
па (см. измерения). Окраска летнего меха на спинной стороне темная,
серовато-бурая, на боках светлее, с примесью палевого цвета. Зимний
мех на спине темно-бурый, на боках несколько рыжеватый» (с. 122).
Далее автор приводит размеры тела и черепа зверьков. Длина тела с
головой 60,0—80,0 (М = 70,0), длина хвоста 35—48 (М = 40,0), длина
задней ступни 12,0—13,0 (М = 12,3) мм. По нашим материалам с Урала
(окр. Бирска и Миаса), длина тела с головой 57—72 (М = 64,2); длина
хвоста 37—47 (М = 42,5); длина ступни 12—13 (М = 12,6) мм. По дан-
ным В. А. Межжерина [1965], на Украине длина хвоста обыкновенной
бурозубки в среднем равна 33—50 (М = 42,7), задней ступни 11,5—15
(М = 13,3) мм. Кондило-базальная длина черепа номинального подвида,
по С. У. Строганову [1957], в среднем равна 18,4 (18,0—19,2) мм. По дан-
ным В. А. Межжерина [1965], на Украине — 19,3 (18,6—20,8) мм.
В. А. Долгов [1963] для подвида в целом приводит значения этого пока-
зателя у бурозубок Карпат 19,5, Карелии — 18,4 мм, т. е. величины весь-
ма близкие. По нашим материалам с Урала, этот показатель выражается
М = 19,4 (18,9—20) мм, что значительно больше по сравнению с данными
С. У. Строганова [1957], приводимыми для номинального подвида из
районов Тавды, Туры, Исети и Тобола.
Анализ метрических показателей размеров тела, а также окраски
зверьков из популяций, относящихся, по С. У. Строганову [1957], к раз-
ным подвидам, позволяет сделать иные выводы.
Достоверных критериев для выделения в ранг подвидов зверьков но-
минального подвида, по С. У. Строганову [Там же], и западносибирской
обыкновенной бурозубки нет. Разберем этот тезис на конкретном материа-
ле. Метрические характеристики номинального подвида из европейской
части СССР приведены в ряде исследований. По литературным сведени-
ям, средние значения размеров тела и черепа не дают оснований для их
разделения на два подвида. Кондило-базальная длина черепа обыкновен-
ной бурозубки с Украины, по В. А. Межжерину [1965а],— 19,3; для
бурозубок всей европейской части страны, по В. А. Долгову [1963],—
19,5; для западных районов Сибири, по С. У. Строганову [1957],—
302
18,4 мм. По нашим данным, для зверьков с Урала этот показатель состав-
ляет 19,4, а для западносибирского подвида (наши данные) — 19,5 мм.
Таким образом, по этому признаку нет оснований для выделения двух
подвидов.
Аналогично положение и при анализе других показателей. Длина те-
ла для номинального подвида, по С. У. Строганову [1957], составляет
М = 70 мм. По нашим данным, для западносибирской бурозубки из
разных районов подвидового ареала М = 67,8 мм. Длина хвоста у
номинального подвида на Украине равна 42,7 [Межжерин, 1965], на
Урале (по нашим материалам) — 42,3, по С. У. Строганову [1957],—
40,0, а у западносибирского подвида (по нашим сведениям) — 41,8 мм.
Таким образом, достоверных различий по этим двум признакам найти не
удается, как и в случае длины ступни (табл. 63).
Колориметрические данные обыкновенных бурозубок из разных
районов Сибири также не дают оснований для разделения по признаку
окраски обыкновенных бурозубок европейской части ареала, Западной
Сибири и зверьков, заселяющих восточную часть ареала, кроме горных
районов, где обитают землеройки, относящиеся к другому подвиду — си-
бирской горной бурозубке. Сравнение обыкновенных бурозубок из райо-
нов, где, по мнению С. И. Огнева [1928] и С. У. Строганова [1957], оби-
тают разные подвиды, надежных оснований для разделения не дало.
На нереальность существования подвида обыкновенной бурозубки
со Среднего Урала, описанного С. И. Огневым [1933а] как S. a. uralen-
sis, указывал еще С. У. Строганов [1957]. Мы полностью разделяем это
мнение. Оказалась нереальным как подвид и бурозубка из Катон-Карагая7!
описанная С. И. Огневым [1933а, 1935] как S. a. iohanseni Ognev, 1933.
При исследовании типовой серии, послужившей для ее описания, нами
установлено, что она неоднородна и состоит из бурозубок двух видов —
обыкновенной и тундряной. Эта путаница в исходном материале повлекла
утверждение о якобы меньших размерах черепа у вновь описанного под-
вида «обыкновенной бурозубки» из этого района и его уплощенности (за
счет возрастной изменчивости). Определенный С. И. Огневым [Там же]
экземпляр как S. a. iohanseni под № 49476, хранящийся в коллекции МГУ,
принадлежит не обыкновенной, а тундряной бурозубке. Таким образом,
нереальность этого подвида обыкновенной бурозубки вполне очевидна.
Среди типовой серии часть относится к S. araneus, а другая — к S. tund-
rensis. К такому же выводу пришел С. У. Строганов [1957], исследовав-
ший серию зверьков, собранных здесь позднее и хранящихся в коллекции
Зоологического музея АН СССР (Ленинград). Это — обыкновенная буро-
зубка из Катон-Карагая относится к S. a. tomensis.
От первого издания «Определителя...» [Бобринский и др., 1944,
1965] ко второму представления А. П. Кузякина менялись в направлении
приближения к реальному положению вещей, но тем не менее они сущест-
венно отстают от развития взглядов систематиков в последние годы. Цити-
руемый автор не мог освободиться от своих ошибок и во внутривидовой
систематике обыкновенной бурозубки, как, впрочем, и других видов зем-
лероек и во втором издании. Отчасти мы разделяем представление
А. П. Кузякина [Там же] о расширении границ номинального подвида
обыкновенной бурозубки. Как показано нами выше, западносибирская
бурозубка и номинальный подвид достоверно не отличаются ни по одному
признаку. Поэтому, следуя правилу приоритета, западносибирских
обыкновенных бурозубок следовало бы отнести к номинальному подвиду.
303
Однако во второй части этого тезиса А. П. Кузякин допускает ряд оши-
бок. В частности, относя в подвид обыкновенной бурозубки S. arcticus
schnitnikovi Ognev из Семиречья. Таким образом, он объединяет ее с тунд-
ряной бурозубкой.
Убедительное обоснование этого положения дает С. У. Строганов
[1957], правильность взглядов которого подтверждается новыми обшир-
ными фактическими материалами, исследованными нами из Терскей Ала-
тау. А. П. Кузякин [1965] правильно оценивает подвидовой статус S. а.
roboratus Hollister, 1913, но допускает ошибку, объединяя ее с S. a. to-
mensis. Последняя, вероятно, вообще реально не существует. Отдельные
популяции равнинной части Западной Сибири, ранее объединявшиеся
С. И. Огневым [1933а, 1935] и С. У. Строгановым [1957] в этот подвид,
относятся к номинальному подвиду, а другие — к горной обыкновенной
бурозубке.
Таким образом, некритическим объединением ряда форм подвидового
и видового ранга в один композиционный вид обыкновенной бурозубки
А. П. Кузякин [Бобринский и др., 1965] внес серьезную путаницу не только
в разработку его внутривидовой таксономии, но и систематики рода в целом.
По данным колориметрического анализа обыкновенной бурозубки из
разных частей ареала вида в Сибири, значения показателя оттенка зверь-
ков из горных районов, относящихся к подвиду S. a. roboratus Holl., 1913,
следующие: для Алтая М = 138,55 (±1,44), для Хамар-Дабана М =
= 138,50 (±1,53); у западносибирского подвида S. a. araneus L. из При-
обья М — 138,52 (±1,68), с Урала (окр. Оренбурга) М — 138,89 (±1,12),
т. е. достоверных отличий нет. По показателю белизны бурозубки с Урала
Таблица 63
Морфометрическая характеристика Sorex araneus и ее подвидов_
Признак S. araneus
М±тх а cv п lim
Длина 51-86
тела 68,9±0,45 6,45 9,30 204
хвоста 42,5±0,22 3,20 7,54 206 34—49
ступни 13,0±0,06 0,77 5,93 200 11—15
Масса тела Длина 8,8±0,15 1,83 20,79 147 5,5—15,5
кондило-базальная черепа 19,7±0,03 0,49 2,50 341 18,4—21,3
рострума 5,3±0,02 0,31 5,80 243 4,2—6,1
мозговой части 10,8±0,02 0,32 2,96 341 9,8—11,7
верхнего ряда зубов 9,0±0,02 0,29 3,26 317 8,2—9,9
ряда промежуточных зубов Ширина 3,1±0,01 0,15 4,72 343 2,8—3,6
рострума 1,9±0,01 0,10 5,17 323 1,7—2,2
предглазничная 2,8±0,01 0,14 5,04 323 2,4-3,1
межглазничная 3,8±0,01 0,14 3,65 343 3,3-4,2
наибольшая черепа 9,8±0,02 0,32 3,32 337 8,8—10,7
Наибольшая высота черепа Высота восходящей ветви ниж- 6,1±0,02 0,27 4,37 335 5,0—6,8
ней челюсти 5,0±0,02 0,29 5,84 341 4,0—5,6
Длина углового отростка 2,1±0,01 0,12 5,47 305 1,8—2,5
304
Признак a. A. a. araneus, лесные и лесостепные районы Зап. и Ср. Сибири
О Су 71 lim
Длина тела 64,2±1,61 5,11 7,95 10 57—72
хвоста 42,5±0,98 3,08 7,25 10 37—47
ступни 12,6±0,10 0,32 2,52 10 12—13
Масса тела 7,0 — — 1 —
Длина кондило-базальная черепа 19,4±0,09 0,36 1,85 15 18,9—20,0
рострума 5,3+0,05 0,19 3,52 15 4,8-5,5
мозговой части 10,8+0,11 0,44 4,05 15 10,3—11,6
верхнего ряда зубов 8,7±0,06 0,23 2,62 15 8,2-9,1
ряда промежуточных зубов 3,1±0,02 0,06 2,07 15 3,0-3,2
Ширина рострума 1,8+0,02 0,09 4,96 15 1,7—2,0
предглазничная 2,8±0,02 0,07 2,67 15 2,7—2,9
межгл азничная 3,8±0,03 0,03 0,10 15 3,7—4,0
наибольшая черепа 9,8±0,06 0,22 2,20 15 9,4—10,2
Наибольшая высота черепа 6,1±0,08 0,31 5,13 15 5,6—6,5
Высота восходящей ветви ниж- ней челюсти 4,7±0,03 0,12 2,50 15 4,6—5,0
Длина углового отростка 2,0+0,03 0,09 4,59 14 1,8—2,2
несколько темнее {М = 2,38 ± 0,11), чем из других частей ареала, в част-
ности Западной Сибири (М = 2,83 0,15) (см. табл. 62).
Анализ этих материалов приводит нас к убеждению, что на террито-
рии восточнее Уральских гор, в равнинной части обыкновенная бурозуб-
ка представлена одним подвидом, вероятно, номинальным, что было ранее
отмечено А. П. Кузякиным [Бобринский и др., 1965]. В Алтае-Саянской
горной стране до Хамар-Дабана включительно обитает S. a. roboratus.
Таксономическая самостоятельность последнего не вызывает сомнений
и подтверждается метрическими показателями размеров тела, черепа и
окраски, но номенклатурный вопрос остается нерешенным. Дело в том,
что подвидовое название оказалось преаккупировано. Как нами показано
ранее [Юдин, 1971а], под этим названием Холлистер [Hollister, 1913]
описал не обыкновенную бурозубку, а бурую, позднее описанную Алле-
ном [Gl. Allen, 1914] с Колымы в качестве вида — S. vir Allen, 1914. Так
как зверьки оказались идентичными описанным с Алтая, то они должны
называться S. roboratus Holl., 1913. Реальный подвид обыкновенной бу-
розубки из горных районов Сибири оказался без названия. Поэтому мы
предлагаем называть его горной обыкновенной бурозубкой — S. а. гу-
pheus subsp. п. Систематическая самостоятельность этого подвида хорошо
демонстрируется данными табл. 63 и рис. 89. Это самый крупный подвид
обыкновенной бурозубки (длина тела с головой М = 70,47 + 0,640), наи-
более длиннохвостый (М = 43,13 ± 0,326) и имеет наибольшие размеры
ступни (М — 13,32 ± 0,084) и массу тела (М = 9,17 + 0,191). Череп ха-
рактеризуется самым большим показателем кондило-базальной длины
305
Продолжение табл. 63
б. S- a. tomensis, Томская обл. в. S. a. rypheus subsp. п., горные районы Сибири
М±тк <3 Cv п lim М±тж О CV п lim
67,8±0,63 6,29 9,26 99 51—83 70,5±0,64 6,24 8,84 95 67—86
41,8±0,31 3,14 7,50 101 34—49 43,1±0,33 3,18 7,36 95 35—49
12,7±0,06 0,62 4,89 95 11—14 13,3±0,08 0,82 6,18 95 12—15
8,4±0,23 1,94 22,17 71 5,5—15,5 9,2±0,19 1,66 18,06 75 8,5—14,0
19,5±0,03 0,42 2,14 185 18,5—20,6 20,0±0,39 0,39 1,92 141 19,2—21,3
5,3±0,02 0,25 4,71 187 4,2—5,9 5,4±0,03 0,36 6,60 141 4,4-6,1
10,8±0,02 0,33 3,01 185 9,0—11,6 11,0±0,02 0,27 2,42 141 9,8—11,7
8,9±0,02 0,25 2,77 177 8,2-9,4 9,2±0,02 0,26 2,82 125 8,6—9,9
3,1±0,01 0,12 3,89 187 2,8—3,4 3,2±0,01 0,13 4,04 141 3,0-3,6
1,9±0,01 0,10 5,27 183 1,7—2,2 1,9±0,01 0,08 4,39 125 1,7-2,1
2,7±0,01 0,13 4,67 183 2,4-3,1 2,8±0,01 0,13 4,77 125 2,4—3,1
3,8±0,01 0,14 3,63 187 3,3—4,2 3,9±0,01 0,12 3,20 141 3,5—4,1
9,6±0,02 0,28 2,94 182 8,8—10,2 9,9+0,03 0,30 3,05 140 9,2-10,7
6,1±0,02 0,27 4,45 181 5,3—6,8 6,2±0,02 0,02 4,14 139 5,0-6,7
4,9±0,02 0,21 4,36 156 4,0—5,4 5,2+0,03 0,32 6,10 141 4,8—5,6
2,1±0,01 0,12 5,81 171 1,8—2,5 2,2±0,01 0,12 5,40 120 1,9—2,4
Окончание табл. 63
Признак Достоверность различий (по Стьюденту)
а-б а—в б—в
Длина
тела 2,0 3,6 3,0
хвоста 0,7 0,8 2,9
ступни 0,9 5,5 6,0
Масса тела — — 2,7
Длина
кондило-базальная черепа 1,1 1,5 1,3
рострума 0 1,7 0,3
мозговой части 0 1,8 7,1
верхнего ряда зубов 3,2 7,9 7,1
ряда промежуточных зубов 0 <4,7 3,5
Ширина
рострума 4,5 4,5 10,6
предглазничная 4,5 0 7,1
межглазничная 0 3,2 7,7
наибольшая черепа 3,2 1,5 8,3
Наибольшая высота черепа 0 1,2 8,3
Высота восходящей ветви нижней челюсти 5,6 11,8 7,1
Длина углового отростка 3,2 6,3 3,4
306
(М = 20,0 it 0,030) при лимите 19,1—20,8 мм. Это наиболее четко очер-
ченный подвид обыкновенной бурозубки.
Утверждение С. У. Строганова [1957] о более светлой окраске сибир-
ской горной бурозубки основано на изучении серий землероек из Алтай-
ского заповедника, хранящейся в Зоологическом музее МГУ. Из-за дли-
тельного хранения зверьки выцвели, утратили первоначальную окраску.
Собранный нами материал (несколько тысяч зверьков) на территории
заповедника, прилежащих к нему участках и из других районов Сибири
не подтверждает этого заключения. Колориметрическое исследование
100 экз. с Алтая, Салаира и Хамар-Дабана показало, что по окраске меха
зверьки этого подвида не отличаются от других. При этом следует иметь
в виду широкий диапазон колебаний коэффициента вариации белизны и
существенные отличия в окраске зверьков в разные сезоны. Отличия зим-
ней и летней окраски горных обыкновенных бурозубок из одного и того
же места (в частности, с побережья Телецкого озера) значительно боль-
шие, чем между выборками разных подвидов, собранных в один сезон
(см. табл. 62, рис. 88, 89). Среднеарифметические величины оттенка раз-
ных подвидов у прибылых землероек очень близки, и совершенно обособ-
ленно они находятся у зверьков в зимнем меху (см. рис. 89).
В монографии С. У. Строганова [1957 ] приводится описание еще од-
ного подвида обыкновенной бурозубки — S. a. isodon Turov. Как показа-
но ранее, S. isodon — самостоятельный вид, поэтому во внутривидовой
таксономии S. araneus этот подвид не обсуждается.
Sorex araneus araneus Linnaeus, 1758 —
Европейская обыкновенная бурозубка
Голотип. См. голотип вида.
Ди а г но з. Характеризуется относительно мелкими размерами
тела и черепа.
Измерения. См. табл. 63.
Р а с п р о с т ранение. Европейская часть СССР, Северный Ка-
захстан, Урал, лесные и лесостепные равнинные районы Западной и
Средней Сибири.
Материал. 530 экз., Южный, Северный Урал, Западная и Сред-
няя Сибирь.
Заметки по с и с тематике. Как уже сказано выше, под-
вид отличается главным образом меньшими размерами тела и черепа.
До наших исследовайий в его состав включались зверьки, заселяющие
европейскую часть ареала, Урал, включая районы бассейнов Тавды,
Туры, Исети и Тобола. Как показали исследования С. У. Строганова
[1957], подтверждающиеся нашими данными, бурозубки с Северного Ура-
ла, выделенные С. И. Огневым [1933] в особый подвид — S. a. uralensis,
не отличаются от номинального подвида. Наши данные не позволяют вы-
делить в самостоятельный подвид и землероек, описанных С. И. Огневым
[Там же], в качестве подвидов S. a. tomensiswS. a. iohanseni. Обоснова-
ние их объединения сделано выше. S. isodon princeps [Скалой, Раевский,
1934], S. i. р. montanus [Скалой, Раевский, 1940] и S. i. rutenus [Строганов,
1936К включаемые С. У. .Строгановым [1957] в состав обыкновенной
бурозубки, относятся к равнозубой бурозубке — S. isodon Turov.
На основании исследований В. М. Анискина и В. Т. Волобуева [1981 ]
обыкновенные бурозубки, заселяющие территорию на запад от р. Томь,
307
выделены как «новосибирская» хромосомная форма, характеризующаяся
отличиями в морфологии аутосом. Таким образом, установлены кариоло-
гические отличия этого подвида от горной обыкновенной бурозубки.
Sorex araneus rypheus subsp. n.—
Обыкновенная горная бурозубка
Голотип. Тип не выделен, подвид описан по экземплярам, добы-
тым на Алтае (Прителецкая тайга).
Диагноз. Размеры крупнее, чем у предыдущего подвида. Длина
тела 57—86 (М = 70,5 0,64), кондило-базальная длина черепа 19,2—
21,3 (М = 20,0 ± 0,39) мм.
Измерения. См. табл. 63.
Р а с пространение. Заселяет восточную часть ареала вида, свя-
зан с горами юга Сибири: Салаир, Кузнецкий Алатау, Алтай, Западный
и Восточный Саян, Хамар-Дабан, вероятно, хребты Прибайкалья.
Материал. 141 экз., горные области Западной и Средней Сибири,
Алтай, Саяны, Прибайкалье.
3 а'м етки по систематике. В систематике подвида весьма
существенная путаница. Тщательное изучение первоописания S. robora-
tus, сделанного Холлистером [Hollister, 1913], убедило нас, что под этим
названием автор описал самостоятельный вид бурой бурозубки [Юдин,
1971а] — S. roboratus, а не зверьков, ныне относимых к обыкновенной
бурозубке в качестве подвида. Позднее Аллен описал [Gl. Allen, 1914]
S. roboratus с Колымы под названием S. vir. По праву приоритета, нами
восстановлено название S. roboratus Holl. В то же время на Алтае и в дру-
гих горных системах юга Сибири реально существует особая форма обык-
новенной бурозубки, выделенная С. И. Огневым [1928] в качестве подви-
да. Поскольку ее подвидовое название «roboratus» преаккупировано и от-
носится к другому реальному виду — S. roboratus, этот подвид обыкновен-
ной бурозубки остался номенклатурно не обозначенным. Восполняя этот
пробел и учитывая характер распространения подвида, связанного пре-
имущественно с горными районами, мы предлагаем оставить неизменным
ее русское название — обыкновенная горная бурозубка и присвоить но-
вое латинское название — S. a. rypheus subsp. n.
В. Т. Волобуев и В. М. Анискин [1981], исследовавшие структуру
кариотипа обыкновенной бурозубки из ряда пунктов юга Западной Си-
бири, выделили хромосомную форму «томскую», распространенную от
Томска до Телецкого озера, с западной границей ареала по р. Томь и на
восток, видимо, до Енисея. Таким образом, можно говорить, что обыкно-
венная горная бурозубка (5. a. rypheus subsp. п.) характеризуется кроме
крупных размеров тела и черепа и своеобразной структурой кариотипа.
Род Neomys Каир, 1829 — Куторы, или водоплавки
Тип рода. Sorex fodiens Pennant, 1771.
Диагноз. Череп крупный массивный, с широкой сферической
мозговой капсулой. На лобных костях имеются два небольших отверстия.
Зубная формула:
1-1; с4; Рт4; мА = 30.
1 ’ U ' Z 1 о ,
308
Рис. 92. Водяная кутора — Neomys fodiens Pennant.
Вершины большинства зубов буро-красные, передняя вершина пер-
вого верхнего резца приблизительно в 4—5 раз крупнее задней. Верхняя
сторона нижнего резца без зазубрин, гладкая или слегка волнистая,
у основания иногда есть небольшой округлый выступ. Промежуточных
зубов в верхних челюстях по 4 с каждой стороны.
X арактеристика. Крупные землеройки (размер тела 60—
100 мм), ведущие полуводный образ жизни, что отложило отпечаток на
всю их организацию (рис. 92). В общем габитусе зверьков бросается в гла-
за]уплощенность задней части тела, «гагарий» тип постановки задних
конечностей, наличие оторочек из жестких волос, окаймляющих края ступ-
ней и пальцев. На нижней стороне хвоста хорошо развит киль из удлинен-
ных жестких волос, хвост служит рулем и выполняет локомоторные функ-
ции. Ушные раковины малы и почти полностью скрыты в густом барха-
тистом водонепроницаемом меху. Все зти особенности носят адаптивный
характер к плаванию и нырянию. Окраска меха зверьков двухцветная:
верхняя сторона тела черная, нижняя — белая, иногда со слабой приме-
сью оранжевых или светло-коричневых тонов.
Половое и анальное отверстия у обоих полов окружены общим кож-
ным кольцом. Penis прямой со сложно устроенной, расширенной голов-
кой. Сосков 5—6 пар. Самобытность и систематическая обособленность не
вызывают сомнений в правильности выделения кутор в качестве само-
стоятельного таксона в ранге рода.
Распространение. Европа, Малая Азия, Кавказ и Закав-
казье, Казахстан, Сибирь, Дальний Восток.
Заметки по систематике. Если выделение кутор в ка-
честве самостоятельного рода не вызывает сомнения, то в отношении
видовой таксономии до сих пор нет единого мнения. Большинство авторов
рассматривает в составе рода два вида: N. fodiens, N. anomalus. Другие
исследователи приводят для рода три вида, признавая видовую само-
стоятельность N. schelkovnikovi. Затруднения в таксономии кутор вызваны
тем, что обитание в сходных условиях наложило отпечаток на строение
зверьков и в значительной степени нивелировало видовые габитуальные
309
отличия. Кутора Шелковникова (N. schelkovnikovi) впервые была описана
К. А. Сатуниным в 1913 г. С. И. Огнев [1928] рассматривает в составе рода
Neomys три вида. Эллерман и Моррисон-Скотт [Ellerman, Morrison-Scott,
1951] относят кутору Шелковникова к одной из форм N. fodiens. Это мне-
ние разделяют С. У. Строганов [1957], Н. А. Бобринский, Б. А. Куз-
нецов, А. П. Кузякин [1965] и др. В сводке о фауне млекопитающих
СССР N. schelkovnikovi [Млекопитающие..., 1963] рассматривается
как самостоятельный вид. В качестве основных диагностических
признаков приводится характер волосяного покрова, степень развития
рулевых волос на хвосте, опушенность кистей и ступней.
Нами проведена ревизия рода Neomys. Наряду с метрическими пока-
зателями и признаками, ранее использованными другими исследователя-
ми, в основу классификации положено строение glans penis [Юдин, 1965б,:
19706]. В результате установлено, что род Neomys включает три современ-
ных вида, из которых один широко распространен в Сибири.
Neomys fodiens Pennant, 1771 —
Обыкновенная кутора
Г о л о т и п. Описан по экземпляру из окрестностей Берлина.
Диагноз. Сравнительно крупная землеройка, хорошо развит греб-
невидный киль на нижней стороне хвоста, занимающий обычно не менее
двух третей дистальной части хвоста (см. рис. 38). Пальцы и края лап
окаймлены хорошо развитой плавательной оторочкой из упругих щети-
нистых волос (см. рис. 39).
Измерения. См. табл. 64.
Таблица 64
Морфометрическая характеристика Neomys fodiens
Признак М+тПд. <3 Су п lim
Длина
тела 80,5± 1,01 0,09 0,11 73 65,6—103
хвоста 63,6+0,68 5,3 8,33 61 44—72
ступни 19,1+0,12 0,93 4,87 60 17—20,5
Масса тела 14,4+0,65 4,46 30,97 41 8,5—25,0
Длина
кондило-базальная черепа 21,4±0,07 0,62 2,89 80 19,8—22,2
рострума 6,6±0,03 0,29 4,39 90 6,0—7,1
мозговой части 11,3+0,04 0,30 3,19 80 10,3—12
верхнего ряда зубов 10,5±0,04 0,36 3,43 83 10—11
промежуточных зубов 3,6+0,02 0,19 5,28 88 3,2—4,0
Ширина
рострума 2,2±0,02 0,18 8,18 84 2,0—2,6
предглазничная 3,3+0,02 0,18 5,45 84 3-4
межглазничная 4,5+0,04 0,37 8,22 85 4-4,8
наибольшая черепа 10,9+0,05 0,44 4,04 76 10,2—11,6
Наибольшая высота черепа 6,7±0,04 0,34 5,07 74 6,2—7,2
Высота восходящей ветви ниж-
ней челюсти 4,7+0,02 0,18 3,83 85 4,2—5,0
Длина углового отростка 2,01+0,01 0,08 3,98 66 1,9—2,2
310
Характеристика. Кутора обыкновенная хорошо отличается
от прочих наших землероек характерным внешним обликом, особенности
эти связаны с наземно-водным образом жизни зверька. Мех густой барха-
тистый, с ярко выраженной контрастной окраской, чепрачностью. Позади
глаз небольшое светлое пятно. Окраска верхней и нижней части хвоста
соответствует таковой спинной и брюшной сторон тела. Задняя ступня ку-
торы значительно удлинена и вывернута слегка кнаружи, по краям об-
рамлена волосяной оторочкой. Плавательные оторочки, киль на хвосте,
наличие клапанов, закрывающих слуховое отверстие при нырянии, сви-
детельствуют о специализации куторы к жизни в воде.
Череп сильно вздут и расширен, длина мозговой капсулы (от лобных
отверстий) равна ее ширине (рис. 93). Первый верхний резец крупный,
длинный. Его передняя вершина в 4—5 раз крупнее задней. Верхние про-
межуточные зубы с чрезвычайно расширенными в сагиттальном направ-
лении основаниями коронок и резко скошенными вершинами далеко от-
стоят друг от друга и последовательно уменьшаются от первого к четверто-
му. Удлиненный верхний резец с сильно развитой передней лопастью и
удлиненный ланцетовидный нижний резец свидетельствуют о приспособле-
нии их к выполнению хватательной функции и удержанию добычи.
Glans penis своеобразно и сложно устроена. Центральный стержень
на свободном конце увенчан сложным образованием (orifieium urethae
ext) с двумя ветвистыми как с дорсальной, так и с латеральной сторон
придатками [Юдин, 19656, 19706]. Латеральные складки плотно облега-
ют центральный стержень, срастаясь с ним и соединяясь между собой на
вентральной стороне, у основания glans penis (рис. 94). Их внутренние
края расчленены на множество длинных зубцов. На латеральной стороне
копулятивного органа, в его основной части, видны хорошо развитые пост-
латеральные ушеобразные выросты. По этому признаку N. fodlens легко
отличается от двух других кутор. Шипы на вентральной и латеральной
311
Рис. 94. Строение glans pe-
nis водяной куторы.
а — дорсальная, б — вентраль-
ная сторона.
сторонах мелкие. На
дорсальной стороне ши-
пы лишь на стержне
копулятивного органа
у основания головки.
Распростра-
нение. Обыкновен-
ная кутора — транспа-
леарктический вид, за-
селяющий Европу, За-
падную, Среднюю и Во-
сточную Сибирь, на во-
стоке — Сахалин вклю-
чительно. Ранее счита-
лось, что самая север-
ная граница распространения вида идет по р. Пур в Западной Сиби-
ри и близ Магадана на севере Дальнего Востока [Юдин, 1971а; Юдин и
др., 1976]. В результате исследований, проведенных сотрудниками Био-
логического института в течение пяти лет на территории п-ова Таймыр,
установлены северные точки обитания куторы. Она была отловлена в под-
зоне северной тайги в районе Норильска у пос. Валек и на ключевом уча-
стке «Турмакит» на берегу Хантайского водохранилища в отрогах плато
Путорана у горы Турмакит [Юдин, 1980в; Юдин и др., 1986]. На севере
Дальнего Востока, представляющем северо-восточную периферию ареала
вида, она добыта в лесной зоне Охотского побережья (Магаданская обл.)
до истоков рек Туя, Олы, Яма [Васьковский, 1966]. На обширной терри-
тории юга Восточной Сибири, от Байкала до Амура, кутора зарегистриро-
вана лишь на юге Читинской области, в Сохондинском заповеднике (сборы
и коллекция Л. И. Галкиной). На Дальнем Востоке она известна из ни-
зовий Амура и окрестностей Владивостока, обитает на юге Сахалина.
Южная граница в лесостепной и степной зонах Западной Сибири про-
ходит по верховью Ишима, районе Караганды, по Иртышу до государст-
венной границы.
Материал. 104 экз., Алтай, Кемеровская, Новосибирская обла-
сти, п-ов Таймыр, север Дальнего Востока, Приморский край.
Заметки по систематике. В Сибири зарегистрировано
два подвида: N. f. fodiens Pennant, 1771 и N. f. orientis Thomas, 1914.
Обзор подвидов
Neomys fodiens fodiens Pennant, 1771 —
Западная кутора
Г о л о т и п. См. в описании вида. >
Диагноз. Характеризуется сравнительно мелкими размерами.
Измерения. См. табл. 65.
312
Морфометрическая характеристика
Признак Д’. fodiens
М±тх а Су п lim
Длина
тела 80,5±0,01 0,09 0,11 73 65,6—103
хвоста 63,6±0,68 5,31 8,33 61 44—72
ступни 19,7±0,12 0,93 4,87 60 17,0—20,5
Масса тела Длина 14,4±0,65 4,46 30,97 47 8,5—25,0
кондило-базальная черепа 21,4+0,07 0,62 2,89 80 19,8—22,2
рострума 6,6±0,03 0,29 4,39 90 6,0—7,1
мозговой части 11,3±0,04 0,3 3,19 80 10,3—12
верхнего ряда зубов 10,5±0,04 0,36 3,43 83 10,0—11,0
промежуточных зубов Ширина 3,6±0,02 0,19 5,28 88 3,3—4,0
рострума 2,2±0,02 0,18 8,18 84 2,0—2,6
предглазничная 3,3+0,02 0,18 5,45 84 3,0—4,0
межглазничная 4,5+0,04 0,37 8,2? 85 4,0—4,8
наибольшая черепа 10,9+0,05 0,44 4,04 76 10,2—11,6
Наибольшая высота черепа Высота восходящей ветви ниж- 6,7±0,84 0,31 5,07 74 6,2—7,2
ней челюсти 4,7±0,02 0,18 3,83 85 4,2—5,0
Длина углового отростка 2,01±0,04 0,18 3,98 66 1,9—2,2
Характеристика. От восточной куторы отличается по ряду
морфометрических показателей (см. табл. 65).
Распространение. Западная кутора заселяет предуральские
таежные лесостепные районы, Барабинскую степь, распространена к
востоку до Оби и приалтайских степей.
Материал. 13 экз. из Кондо-Сосьвинского заповедника.
Neomys fodiens orientis Thomas, 1914 —
Восточная кутора
Голотип. Описан по экземпляру из района р. Каменная, близ
Джаркента.
Диагноз. От номинального подвида отличается большими раз-
мерами и рядом других морфометрических признаков (см. табл. 65).
Измерения. См. табл. 65.
Характеристика. Реальность признаков, на основании кото-
рых восточная кутора может рассматриваться в качестве обособленного
подвида, показана на большом материале. По некоторым морфометри-
ческим показателям различия между подвидами недостоверны (см.
табл. 65).
313
Таблица 65
Neomys fodiens и ее подвидов
a. N. /. fodiens, номинальный подвид, Кондо-Сосьвинский заповедник б. N. f. orientis. Кемеровская обл., Алтай Достовер- ность раз- личий (Td) по Стью- денту
M±mx а Су п lim М±тх С Су п lim а-б
90,0± 1,86 5,3 5,89 8 82,0—97,0 85±2,6 8,9 10,47 12 70,5—109,0 1,7
61,0±1,56 3,6 5,9 8 52,0-66,0 66,2=1=1,1 3,81 5,76 12 60,0—72,0 2,7
19,2+0,27 0,76 3,96 8 18,0—20,0 19,4=1=0,21 0,69 3,56 11 18,0—20,5 6,6
— — — — — 19,6+1,9 4,66 24,02 6 14,3—24,1 —
21,6+0,1 0,04 0,17 13 20,2—22,0 21,9=1=0,16 0,56 2,56 12 20,8—22,8 1,6
6,7+0,07 0,25 3,73 13 6,2—7,0 6,9±0,07 0,53 7,68 13 6,5—7,2 2
11,4=1=0,12 0,43 3,77 13 10,2—11,6 11,5+0,09 0,31 2,69 12 11,0—11,9 0,7
10,3+0,06 0,22 2,14 13 10,1—11,0 10,8+0,13 0,43 3,98 И 10,0—11,3 3,5
3,7+0,03 0,11 7,3 13 3,5—3,9 3,7+0,36 1,3 35,14 13 3,4-3,9 0
2,3+0,04 0,14 6,9 13 2,0—2,3 2,2+0,23 0,79 35,91 12 2,0—2,3 0,4
3,4+0,04 0,14 4,11 13 3,1-3,6 3,4+0,34 1,18 34,71 12 3,2—3,5 0
4,6+0,04 0,14 3,04 13 4,0—4,7 4,6+0,06 0,21 4,57 12 4,2—4,8 0
10,9+0,05 0,18 1,65 13 10,8—11,8 11,1=1=0,09 0,29 2,61 11 10,8—11,6 1,9
6,7+0,09 0,32 4,72 12 6,3—7,2 6,6+0,1 0,3 4,55 9 6,2—7,0 0,7
4,7+0,03 0,11 2,34 13 4,6-5,0 4,8±0,38 1,31 27,29 12 4,6—5,0 0,3
2,0+0,02 0,06 3,0 8 2,0—2,1 2,1+0,04 0,10 4,76 6 2,0—2,2 2,2
Распространение. К востоку от Оби и предалтайских сте-
пей до побережья Охотского моря, Приморья и Сахалина включительно.
Материал. 90 экз. из различных частей ареала.
ПОДСЕМЕЙСТВО
CROCIDURINAE MILNE-EDWARDS, 1868—1874]—
БЕЛОЗУБЫЕ ЗЕМЛЕРОЙКИ
Род Crocidura Wagler, 1832 — Белозубки
Тип рода. Sorex leucodon Hermann, 1780.
Диагноз. Землеройки средних, реже крупных размеров, веду-
щие наземный образ жизни. Самая мелкая белозубка в Сибири Crocidura
suaveolens Pallas, 1811, длина ее тела — М = 59,2+1,56, кондило-ба-
зальная длина черепа — М = 16,3 =t 0,18 мм, а наиболее крупная
Cr. lasiura Dobson, 1890 — длина М = 87,6±2,24, кондило-базальная
длина черепа — М = 21,6+0,21 мм.
Череп массивный, угловатый, уплощен, с широкой и короткой рост-
ральной частью. Коронки зубов не пигментированы, чисто белые.
21 в. С. Юдин
314
Зубная формула:
l|; С±; Pm-1; м|-28.
Отличительная черта рода — наличие в верхней челюсти трех про-
межуточных зубов с каждой стороны.
Характеристика. По телосложению белозубки стройнее и
изящнее бурозубок. Хорошо развиты ушные раковины, которые заметно
выступают из меха. Хвост покрыт равномерным волосяным покровом,
с жесткими, торчащими во все стороны щетинками. Киль отсутствует,
длина хвоста значительно варьирует: от М = 30,6±=0,48 у малой до
М = 34,4^0,31 у сибирской белозубки, но не превышает 2/3 тела зверь-
ка. На лапах плавательных оторочек нет. Сосков три пары. Половое и
анальное отверстия открываются в неглубокую клоаку. Glans penis срав-
нительно короткая, сложного строения, покрыта шипами. Особенности
строения головки копулятивного органа, форма и характер расположе-
ния шипов — систематические признаки видов белозубок [Виноградов,
1958; Юдин и др., 1978].
Характерные для рода черты заключены и в строении зубов. Это вы-
ражается в резкой диспропорции по высоте передней и задней вершин
первого резца верхней челюсти. Передняя вершина круто загнута вниз и
во много раз превышает заднюю. Нижний первый резец длинный, без
дополнительных лопастей. Еще одной примечательной чертой рода являет-
ся наличие в верхней челюсти трех промежуточных зубов с каждой сторо-
ны (а не пяти, как у бурозубок — род Sorex, и не четырех, как у кутор —
род Neomys). Первый резец очень крупный с хорошо развитым передним
зубцом, второй резец намного уступает первому, иногда несет на себе
дополнительный бугорок. Первый промежуточный, коренные верхней
челюсти тоже очень крупные. Структура промежуточных зубов под-
вержена значительной изменчивости, часто наблюдается редукция одно-
го из двух последних промежуточных зубов.
Распространение. Представители рода широко распростра-
нены в тропических и субтропических зонах, несколько видов встречает-
ся в умеренных областях Европы и Азии. Южная Сибирь и Приморье
представляют северную периферию ареала рода. Ареал охватывает Афри-
ку и Евразию от Атлантического побережья до Японии включительно
[Строганов, 1957].
Заметки по систематике. Белозубки — одна из самых
трудных и наименее разработанных в таксономическом отношении групп
насекомоядных млекопитающих [Долгов, Юдин, 1975]. Взгляды иссле-
дователей на объем и число видов белозубок фауны СССР коренным
образом расходятся: по С. И. Огневу [1928, 1951] их 10 и 8; по Н. А. Боб-
ринскому, Б. А. Кузнецову и А. П. Кузякину [1944, 1965] — 4;
по Б. С. Виноградову [1958] — 10, по А. А. Гурееву [1963, 1971]
— 9 и 12.
В последнее время с целью уточнения таксономии белозубок про-
ведены исследования, позволяющие на новой фактической основе очер-
тить объем ряда видов [Richter, 1970; Долгов, 1971, 1974; Vlasak, 1972;
Юдин и др., 1978; и др.]. Определенный вклад в систематику белозубок
внесен кариологическими исследованиями [Bovey, 1949а, б; Воронцов,
1958; Matthey, 1958; Tobias, 1962; Borgaonkar, 1965; Meylan, 1966; Ива-
ницкая и др., 1977], с помощью которых стало возможным уточнить
315
реальность существования таксонов, выделенных с помощью класси-
ческих методов изучения систематиков.
В пределах Сибири, по последним данным, обитает три вида бело-
зубок.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА CROCIDURA
1(2). Размеры мелкие, зверьки стройного телосложения. Кондило-
базальная длина черепа не более 17, ширина мозговой капсулы меньше
8,5 мм. Glans penis не разделена на проксимальный и дорсальный отделы.
«Окно» на дорсальной стороне копулятивного органа сильно смещено к
свободному концу (рис. 95, а)....................................
. . . ...............Crocidura suaveolens Pallas — Малая белозубка
2(1). Размеры крупные, телосложение массивное. Окраска меха одно-
тонная, темно-бурая, почти черная; хвост толстый у основания, кони-
чески утончающийся к концу. Кондило-базальная длина черепа не менее
19, длина зубного ряда более 9, длина промежуточных зубов более 2,4 мм
.............................................................. 3
3(4). Glans penis разделена на проксимальный и дистальный отделы.
«Окно» округлой формы, широкое (см. рис. 95, б) . ...............
.....................Crocidura lasiura Dobson — Большая белозубка
4(3). Размеры средние. Окраска меха дорсальной стороны темно-
бурая, вентральной — беловато-серая. Кондило-базальная длина черепа
больше 17, но менее 19 мм. Glans penis не разделена на переднюю и зад-
нюю части. На вентральной стороне головки нет глубоких поперечных
складок. Шипы, покрывающие всю поверхность головки копулятивного
органа, заходят на стержень лишь с боков, не образуя позади головки
замкнутого кольца (см. рис. 95, в) .................
..................Crocidura sibirica Dukelsky — Сибирская белозубка
Crocidura suaveolens Pallas, 1811 —
Малая белозубка
Голотип. Вид описан по экземпляру, добытому в Крыму, близ
Херсонеса.
Диагноз. Коричневато-бурая или палево-серая окраска спины
постепенно переходит в темную на боках и в белесую на брюшке. Хвост
короткий, длина его не превышает половины длины тела и составляет
не более 36 мм, тонкий, без перетяжки у основания. Зверьки мелких раз-
меров, длина тела с головой от 47 до 67 (М = 59,2±1,56), кондило-ба-
зальная длина черепа (М = 16,3±0,18) мм, ширина черепа не превы-
шает 8,5 (ЛГ = 7,9±0,07) мм. Масса тела 3—6 (М = 4,6±0,62) г.
Измерения. См. табл. 66.
Характеристика. Самая мелкая белозубка фауны Сибири
(рис. 96). Хвост тонкий, равномерной толщины, короткий и составляет
не более половины длины тела с головой. От других видов белозубок Си-
бири отличается стройным телосложением и вытянутым хоботком.
О стройности телосложения зверьков можно судить по отношению массы
к длине тела. У малой белозубки этот показатель самый низкий —
7,8 %. Звери покрыты мягким, блестящим мехом, окрашенным от блед-
но-палево-серого и светло-песчано-палевого до коричнево-сероватого.
21*
316
л
317
Рис. 96. Малая белозубка — Crocidura suaveolens Pallas.
Среди сибирских белозубок это самые светлые зверьки. По результатам
колориметрических исследований пяти зверьков, показатель белизны
укладывается в пределы 4,8—6,8 (М = 5,45—0,36), а оттенок — между
120 и 133 (М = 126,7±1,92). Средние значения показателя оттенка близ-
ки к таковым других белозубок Сибири, а по белизне они представляют
самые светлые формы (рис. 98). Зимний мех темнее, длиннее и мягче
летнего.
Череп тонкостенный, стройный, узкий в мозговой и лицевой части
и не превышает 8,5 мм (рис. 97). Кондило-базальная длина колеблется от
14,8 до 17,6 мм. Максимальные значения этого промера лишь едва пере-
крываются с минимальными других видов белозубок. Отношение наи-
большей ширины черепа к кондило-базальной длине в среднем состав-
ляет 48,5 % (табл. 67). Признак этот трансгрессирует с таковым других
белозубок Сибири и служит надежным критерием при диагностировании
вида. Ростральная часть черепа узкая — 1,8—2,2 (М = 2,0±0,03) мм,
ее отношение к наибольшей ширине черепа составляет в среднем 25,3 %,
что меньше, чем у других сибирских видов белозубок (см. табл. 67). Вы-
сота восходящей ветви нижней челюсти не превышает 4,6 (М = 4,0±
±0,08) мм, в' то время как у других видов величина этого показате-
ля чаще всего больше 4,6 мм. Отношение высоты челюсти в области вос-
ходящей ветви к кондило-базальной длине меньше, чем у других бело-
зубок, и составляет 24,5 %.
Многовершинные зубы весьма стабильны по строению и расположе-
нию в челюсти, но они не имеют диагностического значения. Передняя
лопасть первого верхнего резца стройная, несколько загнута внутрь.
Вторая вершина его равна второму промежуточному зубу или несколько
меньше его. Ряд промежуточных зубов относительно длинный (М = 2,1).
Индекс отношения его к верхнему ряду зубов составляет 28,0 %. Струк-
тура промежуточных зубов подвержена значительной изменчивости. Вто-
Рис. 95. Строение glans penis белозубок.
А — Ст, suaveolens Pallas; Б — Cr. lasiura Dobson; В.— Cr. sibirica Dukelsky; а — дорсальная,
б — вентральная, в — латеральная сторона.,
318
Таблица 66
Морфометрическая характеристика Crocidura suaveolens
Возрастная и половая группа М±тпд. а Су п lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 66,0 — — 1 —
взрослые 58,0±1,0 1,41 2,44 2 57,0—59,0
Самки прибылые 57,0±2,35 6,66 11,68 8 47,0—67,0
В целом по виду . . . 59,2±1,56 5,85 9,90 14 47,0—67,0
Длина хвоста
Самцы прибылые 36,0 — — —
взрослые 31,0±1,00 1,41 4,56 2 30,0—23,0
Самки прибылые 30,2±0,10 2,77 9,18 8 24,0—33,0
В целом по виду . . . 30,6±0,98 3,53 11,53 13 24,0—36,0
Длина ступни
Самцы прибылые 11,0 — — 1 —
взрослые 11,0±0,0 0,0 0,0 2 —
Самки прибылые 10,9±0,28 0,79 7,24 8 9,0—11,5
В целом по виду . . . 10,8±0,17 0,64 5,89 14 9,0—11,5
Масса тела
Самцы прибылые 3,0 — — 1 —
взрослые — — — — —
Самки прибылые 3,5 — — 1 —
В целом по виду . . . 4,6±0,62 1,39 30,16 5 3,0—4,5
Кондило-базальная длина черепа
Самцы прибылые 17,4 — — 1 —
взрослые 16,3+0,66 1,14 6,97 3 15,7—17,6
Самки прибылые 16,4±0,37 1,06 6,46 8 14,8—17,5
В целом по виду . . . 16,3±0,18 0,85 5,22 23 14,8—17,6
Длина рострума
Самцы прибылые 4,8 — — 1 —
взрослые 4,9±0,12 0,21 4,28 3 4,7—5,1
Самки прибылые 4,8±0,15 0,42 8,90 8 4,1—5,3
В целом по виду . . . 4,7±0,07 0,34 7,30 23 4,1—5,3
Наибольшая ширина черепа
Самцы прибылые 8,1+ — — 1 —
взрослые 7,8±0,32 0,55 7,09 3 7,4—8,4
Самки прибылые 7,9+0,13 0,36 4,62 8 7,4—8,5
В целом по виду . . . 7,9±0,07 0,34 4,32 29 7,4—8,5
Межглазничная ширина
Самцы прибылые 3,8 — — 1 —
взрослые 3,7±0,15 0,27 7,15 3 3,5-4,0
Самки прибылые 3,8±0,10 0,30 7,79 8 3,4—4,2
В целом по виду . . . 3,8±0,05 0,22 5,73 23 3,4—4,2
319
Окончание табл. 66
1 2 з 4 5 в
Предглазничная ширина
Самцы прибылые 3,2 — — 1
взрослые 3,0+0,03 0,06 1,90 3 3,0—3,1
Самки прибылые 3,0±0,04 0,12 3,92 8 2,7—3,1
В целом по виду . . . 3,1±0,03 0,14 4,57 23 2,7—3,2
Ширина рострума
Самцы прибылые 2,2 — — 1 —
взрослые 2,1±0,03 0,06 2,79 3 2,0—2,1
Самки прибылые 2,0+0,04 0,9 5,41 8 1,8—2,1
В целом по виду . . . 2,0+0,03 0,12 6,04 23 1,8—2,2
Длина верхнего ряда зубов
Самцы прибылые 7,6 — — 1 —
взрослые 7,4±0,34 0,59 7,88 3 7,0-8,1
Самки прибылые 7,6+0,24 0,68 8,97 8 6,7—8,4
В целом по виду . . . 7,5+0,11 0,54 7,18 23 6,7—8,4
Длина ряда промежуточных зубов
Самцы прибылые 2,4 — — 1 —
взрослые 2,1+0,06 0,10 4,76 3 2,0—2,2
Самки прибылые 2,2+0,07 0,19 8,96 8 1,9—2,4
В целом по виду . . . 2,1±0,04 0,18 8,28 23 1,9—2,4
Наибольшая высота черепа
Самцы прибылые 5,0 — 1
взрослые 4,4±0,27 0,47 10,82 3 4,0—4,9
Самки прибылые 4,6±0,12 0,33 7,27 8 4,1-5,0
В цел ом по виду 4,6±0,07 0,35 7,61 23 4,0—5,0
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,3 — —— 1 —
взрослые 3,9±0,30 0,52 13,32 3 3,6-4,5
Самки прибылые 3,9±0,14 0,41 10,44 8 3,4—4,6
В ц е л о м по виду . . . 4,0±0,08 0,36 9,06 22 3,4—4,6
Длина углового отростка
Самцы прибылые 1,7 — —— 1 —
взрослые 2,0±0,03 0,06 2,94 3 1,9—2,0
Самки прибылые 1,9±0,04 0,12 6,08 7 1,8—2,1
В целом по виду . . . 1,9±0,03 0,14 7,56 20 1,7—2,2
Примечание. Данные по взрослым самкам отсутствуют.
рой и третий промежуточные относительно крупные. Соотношение их
между собой варьирует. С. И. Огнев [1928] приводит семь типов различ-
ных соотношений высоты промежуточных зубов для вида из разных
частей ареала.
Головка копулятивного органа около 2 мм в длину и 1 мм в ширину.
Поверхность головки взрослых самцов, кроме вентрально-проксимального
320
отдела и «окна», на дорсальной стороне покрыта шипами, имеющими треу-
гольную форму и черепицеобразно налегающими друг на друга. На вент-
ральной стороне шипы иногда заходят на corpus penis. С дорсальной сто-
роны головка четко отграничена от corpus penis рядом шипов и превы-
Сравнительная характеристика некоторых
Отношение
Cr. suaveolens
Длины хвоста к длине тела
Наибольшей ширины черепа к конди-
ло-базальной длине
Ширины рострума к кондило-базаль-
ной длине черепа
Ширины рострума к наибольшей ши-
рине черепа
Длины верхнего зубного ряда к кон-
дило-базальной длине
Высоты восходящей ветви нижней че-
люсти к кондило-базальной длине
24—36(М = 30,6±0,98)
47—67(М = 59,2±1,56) ~ 50,0
7,4—8,5(М = 7,9±0,07)
14,8—17,6(Af = 16,3±0,18) ~ 48’5
1,7—2,2(М = 2,0±0,03)
14,8—17,6(М = 16,3±0,18) - 12,2
1,7—2,2(М = 2,0±0,03)
7,4—8,5(М = 7,9±0,07) - 25,3
6.7—8,4(М = 7,5±0,11)
14,8—17,6(М = 16,3±0,18) - 46,0
3,4—4,6(М = 4,0±0,08)
14,8—17,6(ЛГ = 16,3±0,18) “ 24,5
321
Рис. 98. Колориметрическая характе-
ристика белозубок Сибири.
1 — Cr. sibirica; 2 — Cr. Zasiura; 3 — Cr. su-
aDeolens; 4 — Cr. sibirica; 5 — Cr. lasiura.
Для 4 и 5 — усредненный показатель окраски
зимнего меха.
шает его по ширине. На вентраль-
ной стороне четкой границы меж-
ду головкой и стержневой частью
нет. Покрытый шипами поясок по-
зади головки corpus penis отсут-
ствует [Долгов, 1971]. По форме
головка члена вытянутая, остро
притупленная или округлая, в за-
висимости от состояния органа
несколько сжатая в дорсовентральном направлении и не разделена по-
перечной складкой на передний и задний отделы. На вентральной и ла-
теральной стороне шипики заходят к основанию члена дальше пределов
головки.
По сообщению В. А. Долгова [1971], при фиксации на головке иног-
да образуются складки. В вентро-проксимальном отделе головка косо
срезана (см. рис. 95, а), здесь расположен выход мочеполового канала,
обрамленного снизу двумя крупными овальными сосочками, а сверху —
рядом складчатых структур. Весь «срез» окружен валиком. Обрамлен-
ные им детали органа не несут шипиков или чешуек. G дорсальной сто-
роны головка открыта в виде вытянутого в продольном направлении
«окна», из которого выступают мягкие ткани органа, не покрытые шипи-
ками. Кромка «окна» шипов не имеет, форма его варьирует. По данным
В. А. Долгова [1971], у половозрелых самцов glans penis меньше, его
длина около 1,5 мм. Валик, окружающий структуры, расположен вокруг
выхода мочеполового канала, развит слабо и сдвинут к переднему концу.
Сосочки, обрамляющие мочеполовой канал, часто сращены с венчиком,
а складки под выходом мочеполового канала не развиты. На головке ши-
Таблица 67
метрических показателей белозубок
Cr. sibirica Cr. leucodon
30—39(47 = 34,7+0,31) 24—36(47 = 30,6+0,98)
53—80(47 = 66,9+0,76) “ и1’8 8,6—9,3(47 = 8,9+0,02) 68—136(47 = 87,6+2,27) ‘1о’° 8,7—12,0(47 = 10,1+0,09)
17,2—18,8(47 = 18,2+0,04) ~ 2,1—2,7(47 = 2,3+0,01) 19,5—26,3(47 = 21,6+0,21) 2.0—3,1(47 = 2,6+0,03)
17,2—18,8(47 = 18,2+0,04) ~ 12,6 2,1—2,7(47 = 2,3+0,01) 19,5—26,3(47 = 21,6+0,21) “ 1“’° 2,0—3,1(47 = 2,6+0,03)
8,6—9,3(47 = 8,9+0,02) — 25,8 7,8-8,8(47= 8,3+0,03) 8,7—12,0(47 = 10,1+0,09) i'5,b 9,4—11,0(47= 10,0+0,05)
17,2—18,8(47 = 18,2+0,04) ~ 45’6 4,6—5.2(47 = 4,9+0,02) 19,5-26,3(47= 21,6+0,21) 5,0—7,3(47 = 6,6+0,05)
17,2—18,8(47 = 18,2+0,04) 26,9 19,5—26,3(47 = 21,6+0,21) “
Колориметрическая характеристика белозубок
се
ЕГ
К
Ч
V0
д
Н
Ст. lasiura lim 2,1-3,7 125-141
Т-1
Су СО о о? ьГ т-1
с> 0,5 9,2
к g -н СО 00 о см -Н +1 см S3
I Ст. sibirica 1 lim 2,9-6,5 106-149
Й О о см см
Су 22,0 10,4
с> 1,0 12,9
к g -Н ем ** ем о о см +1 +1 LQ СО СМ
I Cr. suaveolens I lim 4,8-6,8 120-133
LQ Ю
Су СО ** со тн
о см о
к g -Н 5,5±0,36 126,7±1,92
Признак Белизна Оттенок
нов нет, а развита зернистость. Головка неполо-
возрелых самцов длиной около 1,2 мм, покрыта
мало заметными бугорками, из которых затем
развивается зернистость, переходящая с возра-
стом в шипы. Мочеполовой канал не смещен
вентрально, а открывается на конце головки (см.
рис. 95), как у взрослых. Сосочки не развиты,
валик едва намечается, «окно» на дорсальной сто-
роне округлой формы. По строению головка ко-
пулятивного органа малой белозубки наиболее
близка к таковой сибирской белозубки, чем бе-
лобрюхой (см. рис. 95).
Распространение. Малая белозуб-
ка занимает обширный ареал, охватывающий Се-
верную Африку, Европу и большую часть Азии.
В СССР она заселяет Кавказ, южные районы ев-
ропейской части страны, север Эстонской ССР,
Подмосковье, Средний и Южный Урал (Свердлов-
ская и Челябинская области), Казахстан, Забай-
калье, Приморье. В горных районах Северо-За-
падного Алтая, на территории между Обью и
Байкалом этой белозубки нет. Вероятно, связь
с восточным участком ареала вида осуществляет-
ся через Китай и Монголию. В юго-западной Мон-
голии, в Забайкалье она добыта в бассейне Джи-
ды, в Южном Приморье в долине Сучана и Ту-
мангана, Уссурийском (бывш. Супутинском) за-
поведнике, окрестностях Находки, заповеднике
«Кедровая Падь», в окрестностях ст. Хасан.
Материал. Нами исследовано 36 экз.
разных подвидов малой белозубки из разных ча-
стей ареала.
Изменчивость. Как уже говорилось
выше, в коллекциях имеются весьма скудные сбо-
ры по этому виду из Сибири, поэтому изменчи-
вость окраски исследовалась на весьма ограни-
ченном материале. По нашим данным, это наи-
более светлоокрашенные землеройки среди бело-
зубой Сибири. По средним значениям насыщенно-
сти оттенка все три вида существенных отличий
не имеют и различаются лишь широтой измен-
чивости (табл. 68). В пределах обширного ареа-
ла окраска малой белозубки варьирует очень
широко. Зверьки из Забайкалья значительно свет-
лее, чем особи из Приморья. С накоплением ма-
териалов из разных частей ареала, возможно,
границы изменчивости окраски будут уточнены.
Изменчивость размеров малой белозубки в
Сибири невелика. Возможно, это также связано
со сравнительно небольшим материалом, доступ-
ным для исследования. Длина тела зверьков варь-
ирует от 47 до 67 (М = 59,2) мм, существен-
ных половых и возрастных различий в длине
тела не зафиксировано.
323
Заметки по систематике. По морфологическим показа-
телям отдельные подвиды малой белозубки трансгрессируют с другими
видами. Вследствие этого возникают сложности при диагностике вида из
некоторых частей его ареала. Чаще всего значения показателей наиболее
крупных экземпляров малой белозубки перекрывают наименьшие бело-
брюхой (Cr. leucodon Hermann, 1780). Однако эти две формы хорошо
диагностируются по ряду признаков, в том числе по строению копулятив-
ного органа. У белобрюхой белозубки на копулятивном органе позади
головки хорошо выражен кольцевой валик, покрытый шипами. У малой
белозубки его нет. В Сибири малая белозубка диагностируется легко.
С большой белозубкой они четко различаются по величине тела, черепа,
внешнему облику, окраске и другим признакам. Наиболее близки они с
сибирской белозубкой, но отличаются структурой промежуточных зубов,
шириной черепа. Соотношения между ними, как справедливо отмечал
С. У. Строганов [1957], еще недостаточно выяснены.
В Сибири описаны три подвида малой белозубки, слабо различаю-
щихся между собой.
Обзор подвидов
Crocidura suaveolens suaveolens Pallas, 1811 —
Малая южнорусская белозубка
Голотип. Сведения приведены в описании вида.
Диагноз. Характеризуется относительно темной окраской меха,
укороченным хвостом и малыми размерами черепа. Летний мех на спине
шелковистый, с блеском, коричнево-сероватый, на брюшной стороне —
светло-серый, белесый.
Характеристика. При сопоставлении измерений, приводи-
мых С. У. Строгановым [1957], с нашими данными обнаружено, что мак-
симальная длина тела у этой белозубки, по С. У. Строганову, несколько
больше полученной нами для вида (в Сибири) в целом, а средние значения
длины несколько уступают. По кондило-базальной длине черепа они ста-
тистически не различимы (М = 16,2 и М = 16,3). Череп у них более
узкий в мозговой части (М = 7,5), но шире в области for. anteorbitalla
(М = 3,4). Длина верхнего ряда зубов у этого подвида больше, чем по
нашим данным — М = 7,7 и М = 7,5.
С. У. Строганов [Там же] указывает, что этот подвид близок к семи-
реченской белозубке, отличается от нее лишь незначительным потемне-
нием окраски и несколько меньшими размерами.
Распространение. Подвид заселяет европейскую часть
СССР и Кавказ. В Сибири известен только с восточных склонов Южного
и Среднего Урала.
Crocidura suaveolens ilensis Miller, 1901 —
Малая семиреченская белозубка
Голотип. Известен из Куктурука на р. Или, хранится в Бри-
танском музее.
Диагноз. Окраска меха спинной стороны бледная от дымчато-
серой до серо-буроватой, брюшной — серовато-белесая.
Характеристика. Размеры несколько крупнее, чем у номи-
нального подвида. По С. У. Строганову [19571, длина тела с головой 55—
324
62 (М — 59) мм, что соответствует нашим данным по виду в целом, хвост
несколько длиннее (М = 33). Масса тела М = 6,5 г.
Кондило-базальная длина черепа у этого подвида больше (16,7—
17,8 (М — 17,2) мм), чем у номинального подвида. От номинального под-
вида отличается «пустынным» типом окраски — бледной и тусклой.
Распространение. В Сибири подвид занимает юг Западной
Сибири, от Челябинской области на восток по Северному Кузбассу до
Балхаша.
Crocidura suaveolens shantungensis Miller, 1901 —
Дальневосточная малая белозубка
Голотип. Описан из окрестностей Шаньдуна, Северный Китай,
№ 86151, сборы П. Д. Бергена, хранится в коллекции Национального
музея США.
Диагноз. Характеризуется меньшими размерами, чем предыду-
щий подвид, от которого отличается также более бледной окраской меха.
Спина серовато-бурая, тусклая, брюшко серо-белесое.
Измерения. Длина тела с головой 53—62 (М = 56,0); длина
хвоста 25—36 (М = 30,0); длина задней ступни 10,6—11,6 (М = 11,0);
кондило-базальная длина черепа 15,8—17,3 (М — 16,5); наибольшая ши-
рина 7,4—8,2 (М = 7,8); межглазничная ширина 3,3—3,5 (М — 3,4);
длина верхнего ряда зубов 6,9—7,8 (М = 7,2) мм.
Заметки по систематике. С. У. Строганов [1957] считает
малую белозубку из Приморья и Кореи идентичными, рассматривает виды
Cr. corea Thomas и Cr. longicaudata Mori как синонимы и по приоритету
относит малых белозубок из Приморья Cr. s. orientis Ognev к подвиду
Cr. s. shantungensis Miller, широко распространенному в Приморье, Корее
и Восточном Китае. А. П. Кузякин [Бобринский и др., 1965], признавая
реальность подвидов Cr. s. suaveolens Pall, и Cr. s. ilensis Miller, рассмат-
ривает их в ином объеме. В частности, белозубок с Дальнего Востока от-
носит к номинальному подвиду. В нашем распоряжении недостаточно
материала, чтобы провести ревизию внутривидовой систематики с примене-
нием статистических методов. Окончательное решение этого вопроса мы
оставляем до накопления новых материалов.
Распространение. Ареал подвида охватывает Приморье,
юго-восточную Монголию, Китай [Строганов, 1957].
Crocidura lasiura Dobson, 1980 —
Большая белозубка
Голотип. Описан по экземпляру из Приморья (долина Уссури).
Голотип хранится в коллекции Зоологического музея АН СССР (Ле-
нинград).
Диагноз. Размеры крупные: длина тела с головой (М = 87,6=±
±=2,27 мм). Хвост толстый, относительно короткий (М = 37,7±=0,93),
составляет меньше половины длины тела (43,0 %), постепенно утончает-
ся к концу. Длина его не превышает 58 мм.
Окраска меха темная, почти черная, однотонная на всем теле, penis
массивный, с широким «окном» на дорсальной стороне головки. Головка
копулятивного органа половозрелых самцов плотно покрыта налегаю-
щими друг на друга шипами, направленными вершинами к основа-
нию penis.
325
Таблица 69
Морфологическая характеристика Crocidura lasiura
Возрастная и половая группа М^тпх а Cv 71 lim
1 2 3 4 5 6
Длина тела с головой
Самцы прибылые 81,1±1,77 6,38 7,87 13 73,0—94,0
взрослые 98,0±3,73 9,14 9,33 6 81,0—106,0
Самки прибылые 88,0±11,36 19,67 22,36 3 70,0—109,0
взрослые 95,0±7,98 21,12 22,23 7 68,0—136,0
В целом по виду . . . 87,6±2,27 13,80 15,75 37 68,0—136,0
Длина хвоста
Самцы прибылыо 35,1±1,0 3,59 10,24 13 28,0—40,0
взрослые 42,5±1,38 3,39 7,98 6 39,0—47,0
Самки прибылые 35,0+0,0 0,0 0,0 4 —
взрослые 43,1±3,09 8,17 18,95 7 31,0—58,0
В целом по виду . . . 37,7+0,93 5,82 15,44 39 28,0—58,0
Длина ступни
Самцы прибылые 14,0+0,36 1,29 9,22 13 11,0—16,0
взрослые 16,0+0,45 1,10 6,85 6 15,0—18,0
Самки прибылые 15,0+0,0 0,0 0,0 2 —
взрослые 15,7+1,17 2,28 18,35 6 12,0—20,0
В целом по виду . . . 14,7+0,29 1,72 11,74 36 11,0—20,0
Масса тела
Самцы прибылые 13,2+0,78 2,19 16,62 8 9,2—16,0
взрослые 21,1+3,32 6,64 31,42 4 12,0—27,5
Самки прибылые 14,0+5,02 10,03 71,67 4 8,0—29,0
взрослые 24,6+3,86 7,72 31,53 4 17,0—35,0
В целом по виду . . . 17,2+1,69 7,58 44,04 20 8,0—35,0
Кондило-базальная длина черепа
Самцы прибылые 21,3+0,36 1,25 5,90 12 19,5—23,2
взрослые 23,1+0,45 1,36 7,89 9 21,1—24,9
Самки прибылые 20,7+0,17 0,48 2,33 8 19,8—21,2
взрослые 22,2+0,64 1,92 8,67 9 19,7—26,3
В ц е л о м по виду . . . 21,6+0,21 1,49 6,92 49 19,5-26,3
Длина рострума
Самцы прибылые взрослые 6,1+0,07 6,4+0,12 0,30 0,38 4,97 5,98 18 И 5,5—6,9 6,0—7,1
Самки прибылые 5,8+0,09 0,26 4,54 8 5,5-6,2
взрослые 6,2+0,20 0,62 10,09 10 5,3-7,4
В ц е л о м по виду . . . 6,1+0,05 0,41 6,75 62 5,3-7,4
Длина мозговой части черепа
Самцы прибылые 11,5+0,33 0,73 6,36 5 10,5—12,2
взрослые 12,4+0,38 0,76 6,14 4 11,3-13,0
Самки прибылые 10,6+0,20 0,44 4,13 5 10,2—11,2
взрослые 11,7+0,28 0,56 4,74 4 11,2—12,3
В целом по виду . . . 11,5+0,20 0,87 7,51 18 10,2—13,0
326
Продолжение табл. 69
1 2 3 4 5 6
Наибольшая ширина черепа
Самцы прибылые 9,8±0,10 0,36 3,64 14 8,8—10,3
взрослые 10,5±0,18 0,56 5,32 10 9,9—11,4
Самки прибылые 9,7±0,08 0,23 2,34 8 9,4—10,1
взрослые 10,6±0,27 0,77 7,32 8 9,5—12,0
В целом по виду . . . 10,1±0,09 0,60 5,95 50 8,8—12,0
Межглазничная ширина
Самцы прибылые 4,9±0,04 0,17 3,56 18 4,5—5,2
взрослые 5,0±0,05 0,18 3,64 11 4,7-5,3
Самки прибылые 4,8±0,05 0,15 3,12 8 4,5—5,0
взрослые 5,0±0,10 0,30 6,05 10 4,5-5,5
[В целом по виду . . . 4,9±0,03 0,21 4,39 61 4,5—5,5
Предглазничная ширина
Самцы прибылые 3,8±0,04 0,18 4,65 18 3,5-4,1
взрослые 3,9±0,04 0,13 3,34 11 3,6—4,0
Самки прибылые 3,7±0,07 0,18 4,99 8 3,3—3,8
взрослые 4,0±0,08 0,27 6,68 10 3,5—4,5
В целом по виду . . . 3,8±0,03 0,19 5,04 61 3,3-4,5
Ширина рострума
Самцы прибылые 2,5±0,05 0,20 7,98 18 2,0—2,9
взрослые 2,7±0,07 0,22 8,16 И 2,3—3,1
Самки прибылые 2,4±0,05 0,14 5,85 8 2,2—2,6
взрослые 2,6±0,07 0,23 8,88 10 2,3-3,1
В целом по виду . . . 2,6±0,03 0,23 8,98 62 2,0—3,1
Длина верхнего ряда зубов
Самцы прибылые 10,0±0,06 0,26 2,62 18 9,5—10,5
взрослые Ю,2±0,13 0,44 4,26 И 9,5—11,0
Самки прибылые 9,8±0,09 0,26 2,67 8 9,4—10,1
взрослые 10,1±0,14 0,45 4,49 10 9,5—11,0
В целом по виду . . . 10,0±0,05 0,35 3,50 62 9,4—11,0
Длина ряда промежуточных зубов
Самцы прибылые 2,9±0,04 0,17 5,95 18 2,6—3,2
взрослые 3,0±0,04 0,12 4,14 11 2,8—3,2
Самки прибылые 2,8±0,05 0,14 4,92 8 2,6—3,0
взрослые 2,9±0,05 0,16 5,35 10 2,7—3,2
В целом по виду . . . 2,9±0,02 0,18 6,14 62 2,6—3,2
Наибольшая высота черепа
Самцы прибылые 6,0±0,09 0,33 5,42 14 5,6—6,6
взрослые 6,3±0,13 0,38 6,03 9 5,8—7,0
Самки прибылые 5,9±0,06 0,17 2,84 8 5,8—6,2
взрослые 6,1±0,15 0,46 7,51 9 5,7—7,2
В целом по виду . . . 6,1±0,05 0,33 5,50 51 5,6—7,2
327
Окончание табл. 69
1 2 3 4 5 6
Высота в эсходящей ветви нижней челюсти
Самцы прибылые 5,6±0,08 0,33 5,81 18 5,0-6,2
взрослые 5,8±0,15 0,47 8,12 10 5,0-6,7
Самки прибылые 5,5±0,08 0,21 3,92 8 5,1-5,8
взрослые 6,0±0,19 0,60 10,11 10 5,3—7,3
В целом по виду . . . 5,6±0,05 0,42 7,42 60 5,0—7,3
Длина углового отростка
Самцы прибылые 2,6±0,08 0,30 11,81 16 2,0-3,0
взрослые 2,5±0,08 0,24 9,72 9 2,0—2,7
Самки прибылые 2,2+0,07 0,21 9,60 8 2,0—2,5
взрослые 2,6±0,14 0,39 15,32 8 1,9-3,3
В целом по виду . . . 2,5±0,04 0,30 12,29 53 1,9-3,3
Измерения. См. табл. 69.
Характеристика. Большая белозубки — самый крупный
представитель рода в фауне СССР (рис. 99). Для нее характерно тяжелое
телосложение, мощная грудь, отношение массы к длине тела составляет
19,6 %, в то время как у малой и сибирской белозубок он равен 7,8 и
10,3 %. Мех густой, длинный. По характеру волосяного покрова она
больше похожа на полевку, чем на землеройку. Окраска меха бурая или
черно-бурая, на нижней стороне с серым оттенком. По окраске этот вид
отличается от малой и сибирской.
Волосяной покров неоднороден. Наиболее длинные волосы окрашены
в черно-бурый цвет с интенсивным коричневым оттенком. Основа их
светлее, значительна примесь аспидно-серого тона. Среди них есть более
короткие и тонкие волосы рыжевато-бурого цвета. Сочетание оттенков
темно-бурого с темно-коричневым и ржаво-бурым придает волосам общий
темный коричневый тон. Волосы на спине длиннее, чем на боках и
брюшке.
По С. И. Огневу [1928], зимний мех длиннее летнего, его отдельные
волосы на спине достигают 11—12 мм. Общий тон спины темнее, чем у
летних особей, брюшко отличается серо-коричневато-свинцовым тоном.
Кисти и ступни черно-бурые со светлыми когтями (см. рис. 99). Хвост
одноцветный темный, довольно густо покрыт волосами, среди которых
разбросаны различной длины (до 9 мм) черные волосы.
Череп крупный стройный, узкий в мозговой части (рис. 100). По
конфигурации он хорошо отличается от черепа малой и сибирской бело-
зубок. Отношение наибольшей ширины к кондило-базальной длине чере-
па составляет 46,8 %, в то время как у малой и сибирской белозубок
этот индекс соответственно равен 48,5, 48,9 %, что свидетельствует о его
относительной узости. По этому признаку зверьки хорошо отличаются от
других белозубок. Череп вытянутый и весьма массивен.
Рострум относительно короткий — он составляет 28,2 %, у малой
белозубки 28,8 и у сибирской — 28,9 %.
По кондило-базальной длине черепа большая белозубка надежно
диагностируется от других видов, так как показатели не перекрываются.
Для черепа характерно значительное развитие сагиттального и ламбдо-
328
Рис. 99. Большая белозубка — Crocidura lasiura Dobson.
Рис. 100. Череп большой белозубки.
329
видного гребней. Зубной ряд несколько более вытянутый. Отношение его
длины к кондило-базальной длине черепа 46,3 %, а ряда промежуточ-
ных зубов к этому промеру — 13,4 %, что больше, чем у двух других ви-
дов белозубок. Ряд промежуточных зубов длиннее (29 %), чем у других
белозубок (см. табл. 67).
Передний верхний резец по конфигурации сходен с таковым других
белозубок. Специфичным является наличие на внутренней стороне вто-
рого зубца и хорошо выраженного дополнительного бугорка [Долгов,
1971]. Вторая вершина переднего верхнего резца значительно уступает
первому промежуточному зубу и примерно равна, или лишь незначитель-
но больше второго и третьего. Первый промежуточный зуб по высоте в
2—2,5 раза выше второго и третьего. Чаще всего второй и третий проме-
жуточные равны между собой, иногда второй больше третьего, и в редких
случаях третий зуб больше второго. В целом для промежуточных зубов
наиболее характерен меньший разрыв в размерах между первыми и двумя
последующими промежуточными зубами.
Головка копулятивного органа равномерно покрыта довольно круп-
ными шипами, крупная — ширина 1,2—1,6, длина 1,9—2,4 мм. Длина
penis 3,2—3,7 мм (см. рис. 95, Б). Шипы плотно покрывают почти всю по-
верхность головки, налегая друг на друга. «Окно» на дорсальной стороне
головки круглое спереди и прямоусеченное сзади. Ограничивающая его
узкая полоска лишена шипиков. Выступающая через «окно» часть копуля-
тивного органа, как и у других белозубок, не имеет на поверхности шипи-
ков или чешуек. Головка резко сплющена в дорсовентральном направле-
нии и четко разделена поперечной складкой на передний и задний отделы.
В. А. Долгов [1971] считает, что складки эти появились вследствие фик-
сации, а их наличие обусловливается состоянием члена. Во всяком случае
у просмотренных нами самцов головка имела строение, изображенное на
рис. 95, б. Проксимальная часть головки нечетко отграничена рядом круп-
ных шипиков от стержня члена, лишенного шипиков или чешуек. У ис-
следованных нами экземпляров не обнаружено участка на дорсальной
стороне проксимальной части головки, лишенного шипиков. Здесь имеет-
ся лишь незначительная выемка. Вероятно, это одна из форм изменчивости
в строении glans penis.
Мочеполовое отверстие расположено на кососрезанном переднем кон-
це и обрамлено двумя сосочками и рядом складок. Вся эта часть лишена
шипиков и состоит из мягкой складчатой ткани, обрамленной вздутым
валиком, на внутренней стороне которого шипиков нет, а на наружной
есть мелкие шипики, постепенно увеличивающиеся к проксимальной час-
ти. Головка покрыта шипами довольно равномерно на дорсальной, вент-
ральной и латеральной сторонах. На стержень члена шипы не заходят.
Структуры, ограниченные «окном» на дорсальной стороне головки и вали-
ком на кососрезанной передней части головки, лишены шипиков или че-
шуек.
Распространение. Ареал большой белозубки охватывает
районы южной части Приморья и Приамурье, Корею, Северо-Восточный
Китай, Японию и другие районы Юго-Восточной Азии.
В СССР наиболее западный пункт распространения вида — г. Ха-
баровск (15 км по Чернореченскому шоссе), р. Самар-Орловка у с. Благо-
датное Хабаровского края. Наиболее широко распространена и чаще до-
бывалась белозубка в районах Приморья. Есть предположение, что на
Дальнем Востоке под названием Cr. lasiura два вида: собственно Cr. lasiu-
22 Б. С. Юдин
330
га и Cr. thomasi, отличающаяся от первой меньшими размерами. В одном
месте обитают крупные особи с ювенильными чертами {Cr. lasiura) и мел-
кие — со стертыми зубами (Cr. thomasi). Недостаточное количество мате-
риала не позволяет нам это утверждать. Большая белозубка добывалась
в Хорольском районе близ Краскино в долине Уссури, Туман-Гана, Су-
кутинки — близ оз. Ханка, на побережье р. Одарки, из ряда пунктов
Хасанского района, в заповеднике «Кедровая падь», окрестности оз. Ха-
сан, в районе залива Посьета, у мыса Басаргин.
Материал. 66 экз., преимущественно из Приморского края.
Изменчивость. Поскольку большая белозубка занимает в
СССР небольшую территорию на юго-востоке Сибири, а коллекционных
материалов из Корей и Китая недостаточно, проследить закономерности
географической изменчивости этого вида в настоящее время не представ-
ляется возможным.
Окраска большой белозубки довольно стабильная, темно-бурая.
Сибирские зверьки этого вида более темные. Показатель белизны колеб-
лется от 2,0 до 3,7 (М = 2,7 0,16). Признак этот не перекрывается с та-
ковым у малой белозубки, и лишь крайние, более светлые, варианты пере-
крываются со значениями показателя белизны сибирской белозубки
(см. табл. 68). Величины показателя оттенка колеблются от 125 до 141
(М = 131,4 — 2,77), что лишь незначительно выше таковых у других ви-
дов белозубок Сибири (см. табл. 68). Однако в марте 1962 г. на р. Самар-
Орловка (Хабаровский край) добыта беременная самка, имеющая серую
окраску по всему телу, ростральная часть головы и передние лапы были
темно-серыми.
Наибольшие значения оттенка имеют зверьки в зимнем наряде,
т. е. их волосяной покров зимой светлее и более насыщен бурыми тонами.
Показатель белизны у летнего меха равен 2,0—3,3, у зимнего — 3,1—3,7.
Оттенок у летних зверьков колеблется в пределах 125—135, а у зимних —
140-141.
Молодые зверьки по сравнению со старыми наиболее темные. Так,
молодые особи из района оз. Ханка имели почти черный цвет. Колори-
метрическим методом исследовалась небольшая серия из 11 зверьков.
Визуальное изучение окраски хранящихся в музеях зверьков этого вида
подтверждает правильность заключения о стабильности окраски меха
Cr. lasiura (см. рис. 98, 2).
Возрастная изменчивость вида весьма значительна. В год рождения
зверьки не достигают размеров взрослых перезимовавших особей. Разме-
ры белозубок этого вида колеблются в широких пределах (см. табл. 69):
так, длина тела варьирует от 68 до 136 мм. В. Н. Долгов [1971 ] это связы-
вает со временем полового созревания: если зверек созревает в год рожде-
ния, то он прекращает рост и остается «мелким» всю жизнь. Особи, у кото-
рых половое созревание наступает на следующий год, имеют большее вре-
мя для роста. Автор указывает, что среди особей с нестертыми зубами
крупных экземпляров не встречено. Однако среди перезимовавших зверь-
ков встречаются особи с зубами разной степени стертости и значительно
варьирующие по размерам тела. Мы полагаем, что у белозубок больше
продолжительность жизни по сравнению с бурозубками, вероятно, они зи-
муют не один раз. Если это так, то становится понятной широкая измен-
чивость размеров тела.
Масса тела изменчива в еще больших пределах — 8—35 (М = 17,2
± 1,69) г. Значительна изменчивость морфометрических признаков при-
331
былых и взрослых (перезимовавших) зверьков. Так, средний показатель
длины тела молодых зверьков равен (М = 83,1 zt 2,37), а взрослых (М =
= 96,4 + 4,47) мм, длина хвоста соответственно равна (М = 35,6 ±:
=tz 0,9) и (М = 42,8 ztz 1,72) мм. Изменчивость этих показателей в целом
для вида колеблется в широких пределах: длина тела — 68—136, длина
хвоста — 28—58 мм, масса тела — 8—35 г. Так же широка изменчивость
и по краниологическим признакам.
Кондило-базальная длина черепа варьирует от 19,5 до 26,3 (М =
= 21,6 ztz 0,21) мм. У прибылых зверьков этот показатель соответствует
(Л/ = 21,0 zt 0,2), а у взрослых (М = 22,6 zt 0,4) мм. Череп с возрастом
становится не только массивнее, но в отличие от бурозубок и выше (см.
табл. 69).
Наиболее стабильным признаком является длина верхнего ряда зу-
бов. Изменчивость этого признака укладывается в пределах от 9,4 до
11 мм. Возрастные и половые различия по этому признаку не значитель-
ны — средние показатели разных половых и возрастных групп колеблются
в небольших пределах — 9,8—10,2. Весьма стабильна длина ряда проме-
жуточных зубов — средний показатель для разных возрастных и половых
групп колеблется от 2,8 до 3,0 мм.
Возрастная изменчивость зубной системы в общем сходна с таковой
для всех землероек. С возрастом зубы постепенно стираются, причем на-
иболее интенсивно изнашиваются вершины верхнего резца, первый про-
межуточный и конусы первого многовершинного зуба. В результате с
возрастом диспропорция в размерах зубов нивелируется — передний верх-
ний резец и первый промежуточный становятся значительно мельче,
а второй и третий, на которые, вероятно, приходится меньшая функцио-
нальная нагрузка, стираются незначительно. Возрастная изменчивость
зубов большой белозубки описана В. Н. Долговым [1971 ]. По данным это-
го автора, у молодых особей передний резец плотно прилегает к верхне-
челюстной кости, корни не видны. Стертые зубы с возрастом выдвигаются
вперед за счет роста корней, чем компенсируется стирание резца. В ре-
зультате корни зуба обнажаются, но постепенно зарастают за счет роста
верхнечелюстной кости. Подвержено изменчивости и соотношезие разме-
ров промежуточных зубов [Долгов, 1971].
Заметки по систематике. Большая белозубка — самый
крупный представитель рода Crocidura фауны СССР. Ее видовые
признаки настолько четки, что это один из немногих видов, таксоно-
мическое положение которого не вызывает сомнений у разных ис-
следователей.
Описанные из Кореи и Китая виды Cr. thomasi Sower by, 1917; Cr.
lizenkani Kischida, 1931; Cr. yamashinai Kuroda, 1934; Cr. campus-lincol-
nensis Sowordy, 1945, судя по диагнозам, несомненно относятся к Cr. la-
siura. Внутривидовых форм не описано. Недостаток в наших коллекциях
материалов не позволяет однозначно решить вопрос о монотипичности ви-
да. Н. А. Бобринский, А. П. Кузякин [1944] объединяют большую бело-
зубку с серой (Cr. lasia Thomas), базируясь только на некотором сходстве
в размерах. Однако, как справедливо отмечает С. У. Строганов [1957],
такое объединение недостаточно аргументировано. Большая белозубка
отличается иным типом окраски и расцветки, строением хвоста,
столь характерного для большой белозубки. Позднее А. П. Кузя-
кин [1965] пришел к выводу о видовой самостоятельности большой
белозубки.
22*
332
Crocidura sibirica Dukelsky, 1930 —
Сибирская белозубка
Голотип, с? , ad., 16/VII 1928 г. Описан по экземпляру из окрест-
ностей с. Означенное на Енисее, в 96 км к югу от Минусинска. Сборы
Н. М. Дукельской. Коллекция Зоологического музея МГУ, № 14176 (из
собрания С. И. Огнева, № 3343).
Диагноз. Размеры средние. Длина тела 53—80 (66,9 0,76) мм.
Окраска спины буро-серая или черновато-серая с вкраплением волос
с блестящими окончаниями («искры»), постепенно переходящая в серовато-
белесую на вентральной стороне тела. Кондило-базальная длина че-
репа 17,2—18,8 (18,2 ± 0,04) мм. Череп массивный, относительно широ-
кий. При пропорциональной длине зубного ряда верхней челюсти ряд
промежуточных зубов короткий, составляет 9,3—11,7 (10,9 %) от конди-
ло-базальной длины черепа и 20,5—25,0 (24 %) от длины верхнего зубно-
го ряда. Второй и третий промежуточные зубы крайне малы, в 4—5 раз
уступают по высоте первому (рис. 101). Восходящая ветвь нижней челю-
сти относительно высокая: 26,7—27,7 (26,9 %) от кондило-базальной дли-
ны черепа (табл. 70). Головка копулятивного органа грушевидная, не раз-
делена на проксимальный и дорсальный отделы, покрыта шипами. Ее
длина 3 мм (см. рис. 95, в).
Измерения. См. табл. 71.
Характеристика. По размерам тела близка к малой белозуб-
ке, отличается более тяжелым сложением. Отношение к длине тела состав-
ляет 12 %, у малой 7,8 %. Голова грубая с расширенной, укороченной
а
Рис. 101. Череп сибирской белозубки.
333
Таблица 70
Сравнительные морфометрические показатели сибирской и ма-
___________________лой белозубок_________________
Признак Cr. sibirica Сгя tuaveolent
Длина кондило-базальная черепа 17,2—18,8 18,2 14,8—17,6 16,3
верхнего ряда зубов 7,8—8,8 8,3 6,7—8,4 7,5
ряда промежуточных зубов 1,6—2,2 2,0 1,8-2,5 2,1
Высота восходящей ветви нижней че- 4,6-5,2 3,4—4,6
люсти 4,9 4,0
Отношение к кондило-базальной дли- 45,3-46,8 45,2—47,7
не черепа длины ряда зубов 45,6 46,0
длины ряда промежуточных зу- бов 9,3—11,7 10,9 12,1—14,0 12,8
высоты восходящей ветви нижней 26,7—27,7 23,0—26,1
челюсти 26,9 24,5
Отношение длины ряда промежуточ- ных зубов к длине зубного ряда 20,5—25,0 24,1 26,9—29,8 28,0
Примечание. Измерения — в мм, индексы — в %; в числите-
ле лимит, в знаменателе — средний показатель.
мордой. Хвост 30—39 (34,7 st 0,31) мм, в среднем 51,8 % от длины тела
темно-бурый, интенсивность окраски на верхней и нижней стороне почти
одинакова. Четкой границы между темной дорсальной и светлой вентраль-
ной сторонами тела нет. Значения оттенка и белизны колеблются в доволь-
но значительных пределах (см. рис. 98): от 106 до 149 (М = 126,4 st 2,94)
и от 2,9 до 6,5 (М = 4,5 st 0,22). Несмотря на столь широкий диапазон
изменчивости окраски средний показатель оттенка такой же, как у малой
белозубки (М = 126,7 st 1,92) и незначительно уступает таковому у
большой (131,7 st 2,78). Все особи окрашены светлее большой белозубки
и несколько темнее малой (см. рис. 98, 4). В окраске нижней части тела
иногда улавливаются охристые оттенки. Нельзя согласиться с утвержде-
нием А. П. Кузякина [Бобринский и др., 1965], что переход в окраске от
спины к брюшку имеет отчетливую границу. Нами просмотрено более
120 зверьков этого вида из разных районов, добытых во все сезоны года,
и ни у одного экземпляра четкой границы не обнаружено. Этот признак
свойствен белобрюхой, а не сибирской белозубке.
Череп. От большой белозубки сибирская отличается меньшими раз-
мерами черепа. При рассмотрении черепа сбоку обращает на себя внимание
прямая линия профиля лицевой части черепа и значительная высота рост-
рума. Примечательно строение нижней челюсти: высота ее восходящей
ветви у сибирской белозубки выше, чем у других видов. Отношение этого
показателя к кондило-базальной длине черепа составляет у малой бело-
зубки — 24,5, у большой — 25,9, у сибирской — 26,9 %.
Видоспецифична структура зубной системы сибирской белозубки.
Первый промежуточный крупный, второй и третий мелкие. Чаще, чем у
334
Таблица 71
Морфометрическая характеристика Crocidura sibirica
Возрастная и половая группа а Су п 11m
Длина тела с головой
Самцы прибылые 67,0±1,05 4,68 6,98 20 58,0—73,0
взрослые 64,0±1,45 5,23 8,17 13 53,0—71,0
Самки прибылые 64,4±2,32 6,57 10,20 8 54,0—74,0
взрослые 69,4±2,27 5,08 7,32 5 63,0—76,0
В целом по виду . . . 66,9±0,76 5,67 8,47 56 53,0—76,0
Длина хвоста
Самцы прибылые 34,3±0,49 2,20 6,42 20 31,0—38,0
взрослые 36,1±0,43 1,61 4,46 14 32,0—38,0
Самки прибылые 33,9±0,92 2,59 7,64 8 30,0—39,0
взрослые 33,2± 1,08 2,64 7,96 6 30,0—37,0
В целом по виду . . . 34,7±0,31 2,36 6,79 59 30,0—39,0
Длина ступни
Самцы прибылые 11,9±0,20 0,91 7,66 20 11,0—14,0
взрослые 11,9±0,14 0,54 4,51 14 11,0—13,0
Самки прибылые 11,5±0,19 0,54 4,65 8 11,0—12,0
взрослые 11,8±0,31 0,75 6,36 6 11,0—13,0
В целом по виду . . . 11,9±0,09 0,71 5,95 59 11,0—14,0
Масса тела
Самцы прибылые 6,6+0,21 0,82 12,45 16 5,3—8,2
взрослые 6,5±0,54 1,43 21,93 7 5,0—9,5
Самки прибылые 7,8±0,65 1,31 16,78 4 6,5—9,6
взрослые 6,0 — — 1 —
В целом по виду . . . 6,8±0,18 1,07 15,85 37 5,0—9,6
Кондило-базальная длина черепа
Самцы прибылые 18,2±0,06 0,30 1,64 24 17,4—18,8
взрослые 18,3±0,08 0,29 1,60 15 17,7—18,7
Самки прибылые 18,1±0,15 0,30 1,65 4 17,8—18,5
взрослые 18,1±0,08 0,26 1,45 12 17,7—18,6
В целом по виду • . . 18,2±0,04 0,33 1,84 76 17,4-18,8
Длина рострума
Самцы прибылые 5,2±0,06 0,30 5,80 24 4,6—5,6
взрослые 5,3±0,09 0,37 6,89 16 4,7—6,0
Самки прибылые 5,2±0,16 0,35 6,69 5 4,9—5,8
взрослые 5,1±0,06 0,23 4,50 13 4,8—5,5
В целом по виду . . . 5,2±0,03 0,30 5,87 82 4,6—6,0
Наибольшая ширина черепа
Самцы прибылые 8,9±0,03 0,17 1,87 24 8,6—9,2
взрослые 8,9±0,05 0,18 2,02 15 8,6—9,3
Самки прибылые 8,9±0,08 0,17 1,87 5 8,8—9,2
взрослые 8,9±0,05 0,16 1,82 12 8,6—9,1
В целом по виду . . . 8,9±0,02 0,16 1,84 76 8,6—9,3
335
Окончание табл. 71
1 2 3 4 5 6
Межглазничная ширина
Самцы прибылые 4,3±0,02 0,11 2,63 24 4,0—4,5
взрослые 4,3±0,04 0,15 3,46 16 4,1-4,6
Самки прибылые 4,2±0,08 0,18 4,28 5 4,0—4,5
взрослые 4,3±0,03 0,11 2,56 13 4,1—4,5
В целом по виду . . . 4,3±0,02 0,14 3,21 82 4,0—4,6
Предглазничная ширина
Самцы прибылые 3,5±0,03 0,13 3,77 24 3,2—3,7
взрослые 3,6±0,02 0,06 1,78 16 3,4—3,6
Самки прибылые 3,5±0,03 0,06 1,55 5 3,5—3,6
взрослые 3,6±0,03 0,11 3,11 13 3,4—3,7
В целом по виду . . . 3,5±0,02 0,14 3,94 82 3,2—3,9
Ширина рострума
Самцы прибылые 2,3±0,03 0,12 5,17 24 2,1—2,7
взрослые 2,3±0,02 0,10 4,10 16 2,2—2,5
Самки прибылые 2,2±0,06 0,13 5,99 5 2,1—2,4
взрослые 2,3±0,03 0,12 5,10 13 2,2—2,6
В целом по виду . . . 2,3±0,01 0,12 5,10 82 2,1—2,7
Д глм«я верхнего ряда зубов
Самцы прибылые 8,4±0,05 0,22 2,65 24 7,8—8,6
взрослые 8,4±0,06 0,23 2,75 15 8,0—8,7
Самки прибылые 8,2—0,15 0,34 4,18 5 7,8—8,7
взрослые 8,3±0,06 0,23 2,81 13 7,8—8,6
В целом по виду . . . 8,3±0,03 0,23 2,81 81 7,8—8,8
Длина ряда промежуточных зубов
Самцы прибылые 2,0±0,03 0,14 7,16 24 1,6—2,2
взрослые 2,0±0,03 0,14 6,66 16 1,8—2,2
Самки прибылые 1,9±0,10 0,22 11,29 5 1,6—2,2
взрослые 2,0±0,03 0,12 6,0 13 1,8—2,2
В целом по виду . . . 2,0±0,02 0,13 6,68 82 1,6—2,2
Высота черепа
Самцы прибылые 5,0±0,03 0,14 2,78 22 4,8—5,3
взрослые 5,2±0,04 0,17 3,26 15 5,0—5,5
Самки прибылые 5,1±0,10 0,21 4,02 4 4,9-5,3
взрослые 5,1 ±0,06 0,19 3,79 12 4,8-5,5
В целом по виду . . . 5,1±0,02 0,19 3,64 74 4,6-5,5
Высота восходящей ветви нижней челюсти
Самцы прибылые 4,9±0,03 0,12 2,49 24 4,7—5,2
взрослые 4,9±0,03 0,11 2,17 16 4,7—5,1
Самки прибылые 4,8±0,10 0,23 4,76 5 4,6-5,2
взрослые 4,8±0,05 0,19 4,01 13 4,6—5,1
В целом по виду . . . 4,9±0,02 0,14 2,94 82 4,6-5,2
Длина углового отростка 2,1—2,8
Самцы прибылые 2,5±0,05 0,24 9,93 21
взрослые 2,4±0,05 0,17 7,42 14 2,0—2,6
Самки прибылые 2,4±0,08 0,18 7,70 5 2,1—2,6
взрослые 2,3±0,05 0,16 6,91 11 2,1—2,5
В целом по виду . . . 2,4±0,02 0,21 8,77 75 2,0—2,8
336
других представителей семейства, у этого вида наблюдается редукция од-
ного из промежуточных зубов (иногда одновременно в обеих челюстях).
Другой особенностью зубной системы сибирской белозубки следует
считать относительно короткий ряд промежуточных зубов. Если отноше-
ние его длины к кондило-базальной длине черепа (45,6 %) уступает тако-
вому у малой (46 %) и большой (46,3 %) белозубок незначительно, то от-
ношение ряда промежуточных зубов отличается значительно больше,
соответственно составляя: 11,0; 12,8; 13,4 %, а по отношению к длине
зубного ряда — 24,1; 28,0 и 29,0 %. Укорочение ряда промежуточных
зубов связано прежде всего с мелкими размерами второго и третьего зу-
бов. Тенденция к олигодонтии, или уменьшению размеров промежуточ-
ных зубов,— характерная черта этого вида. Чаще всего поперечный диа-
метр третьего зуба больше, чем второго. Сумма продольных диаметров
второго и третьего зубов равна или меньше, чем первого.
Строение головки копулятивного органа весьма характерно. С дор-
сальной стороны она грушевидной формы, а с латеральной как бы не-
сколько сжата в дорсовентральном направлении. Передний ее конец тупо
приострен и скошен в дорсовентральном направлении. Головка не расчле-
нена на передний и задний отделы, чем наряду с размерами отличается от
таковой у большой белозубки. От стержня она четко отграничена только с
дорсальной стороны. Ряд крупных шипов, покрывающих всю головку с
дорсальной стороны, кроме участка, отграниченного «окном», образует
четкую границу между головкой и corpus penis. На переднем срезе между
двумя крупными сосочками располагается выход мочеполового канала.
Мелкие структуры этой части головки не покрыты шипами и обрамлены
слабо выраженным валиком, несущим с наружной стороны шипы. На
вентральной стороне члена головка переходит в стержень без заметной
границы. При этом по бокам стержня шипы заметно заходят за линию
головки.
Б. С. Виноградов [1958], впервые описавший строение копулятивно-
го органа сибирской белозубки по добытому нами близ Бийска экземпля-
ру, допустил неточность. При фиксации головка перекрутилась, в резуль-
тате чего форма ее изменилась. Это послужило основанием для утвержде-
ния о якобы существующем продольном разделении задней части головки
на обособленные доли. Знакомство с экземпляром, послужившим Б. С. Ви-
ноградову [1958] для описания белозубки Огнева (Cr. ognevt Strog.,
1956), хранящегося в спиртовой коллекции Зоологического музея АН
СССР в Ленинграде, позволяет убедиться в ошибочности первоописания.
Описанный экземпляр относится к сибирской белозубке и имеет типич-
ное для вида строение копулятивного органа. Размеры головки члена:
длина 3, ширина 2 мм.
Распространение. Ареал сибирской белозубки охватывает
территорию юго-восточной части Западной Сибири между Иртышом на за-
паде и Енисеем на востоке. Северная граница распространения идет по
широте г. Томска. На юг простирается до дельты Черного Иртыша. На
территории Казахстана известна в правобережье Северо-Казахстанской
области близ с. Убинского и из Ивановского хребта, в окрестностях Ле-
ниногорска, Зыряновска, Болыпенарымского. Распространена в Колы-
ванском хребте, близ Горной Колывани и в отрогах Тигирецкого хребта.
В Северо-Западном Алтае собиралась в бассейне Чарыша; в Центральном
Алтае по левому притоку Катуни р. Черге, в отрогах Куминского хребта
по р. Чемал и в бывшем Старо-Бардинском районе; в Уймонской котло-
337
вине; в северной части Алтая в районе пос. Майма, г. Горно-Алтайска;
в Северо-Восточном Алтае в ряде пунктов на р. Бие у впадения в нее
р. Ненинка близ пос. Кубия, Кунаган; в среднем течении Бии; в верхо-
вьях у пос. Кебезень, Иогач, Артыбаш, в ряде пунктов на Телецком озере
(Яйлю, урочище Турочак, гора Колюшту). В Юго-Восточном Алтае не об-
наружена.
В Горной Шории заселяет бассейн Кондомы у Темиртау, окрестности
сел Подкатунь, Таштагол. В Приобье граница идет по левобережью, от
с. Кожевникове (Томская обл.) к пос. Черный Мыс, от с. Колывань до
Новосибирска и далее по приобским борам до Барнаула, Бийска и Горно-
Алтайска. Эта линия образует западную границу распространения вида.
От Кожевникове северная граница проходит на Томск, в направлении на
юго-восток, к Салаиру и Кузнецкому Алатау. В Салаире зверьки добы-
ты близ пос. Мирного в верховьях Ини, по западным отрогам, у Новоси-
бирска и в верховьях Верди у пос. Серебряниково; в Кузнецком бассей-
не — в районе р. Кара-Чумыш, на правом притоке Томи р. Тайден и р. Ас-
кыз, в Новокузнецком районе близ д. Узково и в прилегающих районах
на Малой Речке [Юдин и др., 1979]. Вид распространен спорадично, на-
селяет территорию крайне неравномерно и всюду малочислен. Сведения
о характере распределения по ландшафтам и обилии вида разных биото-
пах охарактеризованы нами ранее [Там же].
Указания С. У. Строганова [1957], А. П. Кузякина [Бобринский и др.,
1965] о ее обитании в Забайкалье неверны, там живет малая белозубка.
Материал. Более 150 экз. из разных пунктов ареала.
Изменчивость. Колориметрическая изменчивость окраски си-
бирской белозубки проявляется в более широких пределах, чем у других
видов белозубки, населяющих Сибирь. Однако при оценке сравнительной
изменчивости окраски нужно иметь в виду, что материал по малой и боль-
шой белозубкам незначителен. С накоплением новых материалов, возмож-
но, изменится и представление о диапазоне изменчивости их окраски.
У сибирской белозубки величина показателя оттенка колеблется от
106 до 149 (М = 126,4 ± 2,94), а белизна - от 2,9 до 6,5 (М = 4,5
±= 0,22). Зимний мех длиннее, пышнее и шелковистее, а по окраске более
темный, с насыщенным бурым оттенком, характеризующийся крайними
вариантами колориметрических показателей окраски. Белизна меха
зверьков, добытых в зимние месяцы, характеризуется показателем 2,9—
3,3, а оттенок 123—148. У зверьков, добытых в середине октября 1981 г.,
мех темный, шелковистый, но еще не достиг длины, характерной для бо-
лее поздних месяцев. Только на огузке выступает длинная ость. Хорошо
заметна «искристость» меха.
Возрастная и половая изменчивость выражена крайне слабо (см.
табл. 71). Длина тела взрослых самцов мало отличима от таковой прибы-
лых. Взрослые самки крупнее (69,4 =t 2,27), чем молодые (64,4 =ь 2,32).
Молодые самцы по величине этого показателя несколько больше (67,0
=±г 1,05) взрослых (64,0 ± 1,45). У взрослых самцов хвост длиннее, чем
у молодых. Половых и возрастных различий в размере ступни не просле-
живается. Изменчивость массы тела зверьков разного возраста нечеткая.
Самки несколько тяжелее самцов.
Метрическая изменчивость краниологических показателей у зверь-
ков разных возрастных и половых групп недостоверна.
Строение зубов — характерное для рода в целом, но имеет видоспе-
цифичные черты, охарактеризованные выше. Изменчивость проявляется
333
главным образом в соотношении ряда промежуточных зубов. Географи-
ческой изменчивости по этим показателям не зарегистрировано. Измен-
чивость проявляется в размерах, форме и соотношении второго и третьего
промежуточных зубов [Юдин, 1971а, 19866]. Кроме того, широко развито
явление олигодонтии, которое ни у одного вида землероек фауны СССР
не имеет такого размаха. Оно проявляется в отсутствии одного зуба в пра-
вой или в левой верхней челюсти или одновременно в той и другой
(рис. 102).
Заметки по систематике. Сибирская белозубка — наи-
менее исследованный представитель рода Crocidura. До наших работ в
коллекциях страны было менее десятка экземпляров. Этот вид был опи-
сан позднее других представителей рода в СССР. Несмотря на то что сей-
час сибирская белозубка достаточно легко диагностируется, ее видовой
статус утвердился только через 40 лет [Юдин, 1971а] после первого опи-
сания [Dukelsky, 1930]. Первоначально белозубка была описана в качест-
ве подвида белобрюхой белозубки (Cr. I. sibirica Dukelsky, 1930).
Длительное время новых находок этого зверька не было, вплоть до
1949 г., когда Б. Ф. Белышев добыл на Северо-Восточном Алтае, в долине
среднего течения Бии, 2 экз. Как сообщает С. У. Строганов [19566], они
Рис. 102. Изменчивость зубной системы сибирской белозубки.
а — второй зуб равен третьему; б — второй зуб меньше третьего; в — Второй зуб крайне мал
(задвинут за цингулюм первого и третьего); г — редукция, третьего или второго промежуточных.
зубов верхней челюсти.
339
предназначались С. И. Огневу для новоописания. Однако из-за кончины
С. И. Огнева описание не состоялось. Три зверька, добытые нами в 1953 г.
в Верхнеобском бору, между Бийском и Барнаулом, и в 1956 г. в Томской
области (с. Кожевниково), а также сборы из окрестностей Томска, а так-
же из Кемеровской, Новосибирской областей послужили для выделения
С. У. Строгановым [19566] белозубки в особый подвид — Cr. I. ognevi
Strog., 1956. В монографии С. У. Строганова [1957] приводятся оба под-
вида Cr. I. sibirica Dukelsky, 1930 и Cr. I. ognevi Strog., 1956 как таксоны
подвидового ранга одного вида. В качестве подвида белобрюхой белозуб-
ки рассматривает ее и Эллерман и Моррисон-Скотт [Ellerman, Morrison-
Scott, 1951].
Б. С. Виноградов [1958], исследовавший экземпляр самца, добытого
нами в Верхнеобском бору, выделил эту белозубку в качестве самостоя-
тельного вида — Cr. ognevi. О несостоятельности этого выделения гово-
рилось выше. А. А. Гуреев [1963] отнес зверьков с Енисея к Cr. I. sibiri-
ca, а из другой части ареала — к Cr. ognevi, т. е. к двум разным видам,
повторив эту ошибку и в более поздней работе [Гуреев, 1971]. А. П. Ку-
зякин [Бобринский и др., 1965], являющийся крайним представителем
«укрупнителей» видов, тем не менее правильно рассматривает Cr. I. si-
birica как синоним Cr. ognevi, но относит ее к виду белобрюхой белозубки.
Позиция эта несостоятельна, так как сибирская белозубка отличает-
ся от белобрюхой характером распределения окраски на дорсальной и
вентральной частях тела (у нее нет четкой границы между темной спиной
и светлым брюшком), особенностями зубной системы, краниометрическими
показателями и различным строением головки копулятивного органа.
У белобрюхой белозубки позади покрытой шипами головки на дорсаль-
ной стороне corpus penis имеется отделенный от нее полоской, свободной
от шипов, поясок, охватывающий член и загибающийся в вентро-прокси-
мальном направлении. На вентральной стороне копулятивного органа он
разомкнут. Поверхность пояска покрыта крупными, как и на головке,
шипами. Вентропроксимальный отдел головки также покрыт шипами.
Размеры головки крупнее (длина 4, ширина 2 мм), чем у сибирской бело-
зубки. Коренное отличие заключается в отсутствии у нее позади головки
поперечного пояска, вооруженного шипами. Однако А. П. Кузякин не
мог не заметить ряда специфических черт, характеризующих этих зверь-
ков, и в полном соответствии с фактическим положением отметил: «Си-
бирская форма белобрюхой белозубки отличается от других форм этого
вида, как и от других близких видов лучше, чем последние различаются
между собой... Накопляющийся материал, быть может, даст основание
для выделения этой формы в качестве особого вида» [Бобринский и др.,
1965, с.. 77]. Он оказался прав — собранный в последние годы новый
фактический материал позволил выделить сибирскую белозубку в са-
мостоятельный вид. Зверьки из новых мест находок оказались идентичны-
ми с белозубкой, описанной Н. М. Дукельской [Dukelsky, 1930] с Енисея,
и объединены нами [Юдин, 1971а] в самостоятельный видовой таксон, на-
званный по приоритету — Cr. sibirica Dukelsky, 1930 — сибирской бело-
зубкой. Описанные С. У. Строгановым [19566, 1957] подвиды этой бело-
зубки выделены по деталям окраски. Однако признаки, на которые опи-
рается названный автор при их выделении, укладываются в пределы по-
лиморфизма окраски зверьков из одной точки ареала, поэтому их нельзя
принять как реальные. Имеющиеся в настоящее время материалы позво-
ляют считать, что вид на протяжении всего ареала представлен номи-
нальной формой.
340
СПИСОК ЛИТЕРАТУРЫ
Алина А. В., Реймерс Н. Ф. Наземные млекопитающие (Mammalia) Айнского побе-
режья Охотского моря // Систематика, фауна, зоогеография млекопитающих и
их паразитов.— Новосибирск: Наука. Сиб. отд-ние, 1975.— С. 127—140.
Анискин В. М., Волобуев В. Т. Хромосомный полиморфизм в сибирских популяциях
бурозубок araneus — arcticus комплекса (Insectivora, Soricidae). Сообщ. III.
Три хромосомные формы обыкновенной бурозубки Sorex araneus L. // Генети-
ка,— 1981.— Т. 17, № 10.— С. 1784—1791.
Баевский Ю. Б. Цито- и кариометрические исследования бластоцист (Talpa altaica)
периода диапаузы // Докл. АН СССР.— 1967.— Т. 176, № 4.— С. 5—6.
Баевский Ю. Б. Эмбриональная диапауза млекопитающих: Автореф. дис. ... канд.
биол. наук.— Новосибирск, 1970.— 31 с.
Банников А. Г. Млекопитающие Монгольской Народной Республики.— М.: Изд-во
АН СССР, 1954.— 669 с.
Барабаш-Никифоров И. И., Формозов А. Н. Териология: Учеб, пособие для ун-
тов СССР.— М.: Высш, шк., 1963.— 396 с.
Бобринский Н. А., Кузнецов Б. А., Кузякин А. П. Определитель млекопитающих
СССР. Насекомоядные/Под ред. проф. Н. А. Бобринского.— М.: Изд-во
АН СССР, 1944.— 440 с.
Бобринский Н. А., Кузнецов Б. А., Кузякин А. П. Определитель млекопитающих
СССР. Насекомоядные/Под ред. А. П. Кузякина.— М.: Просвещение, 1965.—
384 с.
Бородулина Т. Л. О латентном периоде в развитии эмбриона алтайского крота // Докл.
АН СССР.—1951.—Т. 80, № 4,—С. 689—692.
Бородулина Т. Л. К биологии алтайского крота // Тр. ин-та/Ин-т эволюционной мор-
фологии животных им. А. Н. Северцова.— 1953.— Вып. 9.— С. 250—280.
Велижанин А. Г. Пути становления фауны Курильских островов // Бюл. МОИП.
Отд. биол.— 1970.— Т. 75, вып. 4.— С. 5—16.
ВендландО. В. Некоторые материалы об уссурийском кроте (Mogera robusta Nehring) 11
Вести. Дальневост. фил. АН СССР.— 1938.— Т. 31, вып. 4.
Верещагин И. К. Млекопитающие Кавказа // История формирования фауны.— М.;
Л.: Изд-во АН СССР, 1959.— 704 с.
Виноградов Б. С. Млекопитающие Минусинского края и Уренхоя // Ежегодник му-
зея им. Н. М. Мартьянова.— Минусинск, 1927.— Т. 5, вып. 1.— С. 33—50.
Виноградов Б. С. О строении наружных гениталий у землероек-белозубок (род Croci-
dura, Insectivora, Mammalia) как диагностическом признаке // Зоол. журн,—
1958,— Т, 37, вып. 8,— С. 1236—1248.
Виолович Н. А., Юдин Б. С., Поляков Е. Ф. К познанию блох (Aphoniptera) сибирско-
го крота (Asiocalops altaica Nikolski, 1883) юга Западной Сибири // Системати-
ка, фауна, зоогеография млекопитающих и их паразитов.— Новосибирск: Нау-
ка. Сиб. отд-ние, 1975.— С. 225—229.
Вольцерт Я. Л., Юдин Б, С, Пространственные изменения фаунистических комплек-
сов мелких млекопитающих (Micromammalia) Якутии //‘ Охотничъе-промысло-
вые ресурсы Сибири.— Новосибирск: Наука. Сиб. отд-ние, 1986.— С. 198—201.
Вон Хон Гу. Млекопитающие Кореи.— Пхеньян: Изд-во АН КНДР, 1955.
Воронцов Н. Н. Значение изучения хромосомных наборов для систематики млеко-
питающих // Бюл. МОИП. Отд. биол,— 1958.— Т. 63, выц. 1,— С. 5—36.
Воронцов Н. Н., Крал Б. Кариологическая дифференциация и систематика рода So-
rex И Четвертый съезд Всесоюз. териол. о-ва.— М., 1986.— Т. 1.— С. 48—50.
Галкина Л. И., Потапкина А. Ф., Юдин Б. С. Эколого-фаунистический очерк мелких
млекопитающих (Micromammalia) Юго-Восточной Тувы // Фауна и системати-
ка позвоночных Сибири.— Новосибирск: Наука. Сиб. отд-ние,. 1977.—
С. 60—80.
341
Гептнер В. Г., Долгов В. А. О систематическом положении Sorex mirabilis Ognev,
1937 (Mammalia, Soricidae) // Зоол. журн.— 1967.— T. 46, вып 9.— С. 1419—
1422.
Гуреев A. A. Insectivora. Насекомоядные // Млекопитающие фауны СССР.— М.;
Л.: Изд-во АН СССР, 1963.— Ч. 1.— С. 52—122.
Гуреев А. А. Землеройки (Soricidae) фауны мира.— Л.: Наука. Ленингр. отд-ние,
1971,— 254 с.
Гуреев А. А. Насекомоядные (Mammalia, Insectivora). Ежи, кроты и землеройки (Eri-
naceidae, Talpidae, Soricidae) // Фауна СССР. Млекопитающие.— Л.: Наука.
! Ленингр. отд-ние, 1979.— Т. 4, вып. 2.— 501 с.
1 Даль С. К. Насекомоядные и рукокрылые Армянской ССР и Нахичеванской АССР.
(Данные по распространению, систематике и биологии) И Зоологический сбор-
ник.— Ереван, 1940.— Вып. 2.— С. 5—26.
Даль С. К. Позвоночные животные Тамбакского хребта // Там же, 1948.— Т. 5.
Даль С. К. Насекомоядные // Животный мир Армянской ССР. Т. 1: Позвоночные жи-
вотные.— Ереван, 1954.— С. 137—141.
Депарма Н. К. Крот.— М.: Заготиздат, 1951.— 47 с.
Депарма Н. К. Особенности строения полового аппарата и биология размножения
европейских и сибирских кротов // Тр. ин-та/Всесоюз. НИИ охот, промысла.—
1953,— Вып. 131.— С. 98—115.
Дзуев Р. И. Описание кариотипов двух форм кавказских кротов И Фауна, экология и
охрана животных Северного Кавказа.— Нальчик, 1972,— Вып. 1.— С. 44—50.
Дзуев Р. И. О распространении кариотипических форм кротов Кавказа // Там же.—
Вып. 3. — С. 147—153.
Дзуев Р. И. Распространение кариотипических форм кротов Кавказа // Экология,
методы изучения и организация охраны млекопитающих горных областей.—
Свердловск, 1977.— С. 71—72.
Дзуев Р. И. К изучению популяционной изменчивости рода Talpa Кавказа // Фауна,
экология и охрана животных Северного Кавказа.— Нальчик, 1980.— С. 3—16.
Дзуев Р. И., Темботов А. К., Иванов В. Г. Кариологические исследования кротов Се-
верного Кавказа И Бюл. МОИП.— 1972а.— Т. 77, вып. 1.— С. 33—36.
Дзуев Р. И., Темботов А. К., Иванов В. Г. Новые данные по кариологии кротов Кав-
каза // Зоол. журн.— 19726.— Т. 51, вып. 12.— С. 188—189.
Долгов В. А. Об изменчивости бурозубок поймы реки Оки (Mammalia, Soricidae) //
Бюл. МОИП. Отд. биол.— 1963.— Т. 64, вып. 4.— С. 135—140.
Долгов В. А. Равнозубая бурозубка — Sorex centralis Thomas, 1911 (Mammalia, Insec-
tivora) в фауне Советского Союза // Зоол. журн,— 1964,— Т. 43, вып. 6.—
С. 898—903.
Долгов В. А. О некоторых закономерностях географической изменчивости млекопитаю-
щих И Докл. АН СССР,— 1966а,— Т. 171, вып. 5,— С. 1230—1237.
Долгов В. А. Виды палеарктических землероек рода Sorex и их индивидуальная и по-
пуляционная изменчивость: Автореф. дис. ... канд. биол. наук.— М., 19666.—
35 с.
Долгов В. А. Распространение и численность палеарктических бурозубок (Insectivo-
ra, Soricidae) // Зоол. журн.— 1967.— Т. 46, вып. 11.— С. 1701—1712.
Долгов В. А. Аномалии зубной системы бурозубок (Mammalia, Sorex) И Тр. Зоол.
музея Моск, ун-та.— 1968а.— Т. 10.— С. 222—224.
Долгов В. А. Особенности и изменчивость одонтологических признаков палеаркти-
ческих бурозубок (Mammalia, Sorex) // Там же,— 19686.— С. 179—199.
Долгов В. А. Травмы скелета обыкновенной бурозубки // Acta Thcriol.— 1968в.—
Vol. 13, N 33.— Р. 355—356.
Долгов В. А. Травмы glans penis бурозубок (Sorex) // Там же.— 1968г.— С. 554—555.
Долгов В. А. Случаи альбинизма у обыкновенной бурозубки // Там же.— 1968д.
Долгов В. А. Изменчивость большой белозубки Crocidura lasiura (Mammalia, Insecti-
vora) И Зоол. журн.— 1971.— Т. 50, вып. 5,— С. 783—785.
Долгов В. А. Краниометрия и закономерности географической изменчивости кранио-
метрических признаков палеарктических бурозубок (Mammalia, Sorex) // Тр.
Зоол. музея Моск, ун-та.— 1972.— Т. 13.— С. 150—186.
Долгов В. А. Диагностика малой (С rocidura suaveolens) и белобрюхой (С. leucodon)
белозубок (Insectivora, Soricidae) // Зоол. журн.— 1974.— Т. 53, вып. 6.—
С. 912—918.
Долгов В. А. Белобрюхая белозубка (Crocidura leucodon) в Копет-Даге (Insectivora,
Mammalia) И Тр. Зоол. музея МГУ.— 1977.— Вып. 19.
Долгов В. А., Киселев В. Е. Изменчивость крота (Talpa europaea L., 1758) в Башки-
рии И Науч. докл. высш. шк. Биол. науки.— 1971.— № 1.— С. 29—37.
342
Долгов В. А., Кривошеев В. Г. К фауне землероек-бурозубок Чукотки (Mammalia,
Sorex) И Биологические проблемы Севера.— Магадан, 1973.— Вып. 2.—
С. 74—76.
Долгов В. А., Лукьянова И. В. О строении гениталий палеарктических бурозубок {So-
rex, Insectivora) как о систематическом признаке // Зоол. журн.— 1966.— Т. 45,
вып. 12.— С. 1852—1861.
Долгов В. А., Хоффман Р. С. Тибетская бурозубка — Sorex thibetanus Kaschenko,
1905 (Soricidae, Mammalia) // Зоол. журн.— 1977.— T. 56, вып. 11.— С. 1687—
1692.
Долгов В. А., Юдин Б. С. Рецензия // Зоол. журн.— 1972.— Т. 51, вып. 11.—
С. 1754—1757.— Рец. на кн.: Гуреев А. А. Землеройки (Soricidae) фауны ми-
ра.— Л.: Наука. Ленингр. отд-ние, 1971.— 254 с.
Долгов В. А., Юдин Б. С. Состояние и задачи исследований насекомоядных млекопи-
тающих (Mammalia, Insectivora) СССР // Систематика, фауна, зоогеография мле-
копитающих и их паразитов.— Новосибирск: Наука. Сиб. отд-ние, 1975.—
С. 5—40.— (Фауна Сибири; Вып. 23).
Дулькейт Г. Д. Охотничья фауна, вопросы и методы оценки производительности охот-
ничьих угодий алтайско-саянской тайги.— Красноярск, 1964.— 351 с.
Егорин Н. Ф. Крот Салаирского кряжа и промысел на него И Тр. ин-та/Биол. н.-и.
ии-т при Том. уи-те.— 1936.— Т. 3.— С. 136—158.
Золотарев Н. Г. Млекопитающие бассейна реки Имана.— М.; Л.: Изд-во АН СССР,
1936,— С. 55—57.
Иваницкая Е. Ю., Исаков С. И., Коробицкая К. В. Хромосомные наборы двух видов
землероек из Таджикистана Sorex buchariensis Ognev, 1921 и Crocidura suaveolens
Pallas, 1811 (Soricidae, Insectivora) // Зоол. журн.— 1977.— T. 56, вып. 12.—
С. 1896—1899.
Иваницкая Е. Ю., Козловский А. И. Кариологические доказательства отсутствия в
Палеарктике арктической бурозубки {Sorex arcticus) II Там же.— 1983.—
Т. 62, вып. 3.— С. 399—408.
Исаков С. И., Юдин Б. С. Систематика и географическое распространение ушастого
ежа {Hemiechinus auritus Gmelin, 1770) в Таджикистане // Изв. АН ТаджССР.
Отд. биол.— 1971.— № 2.— С. 61—67.
Кащенко И. Ф. Обзор млекопитающих Западной Сибири и Туркестана И Изв. Том.
ун-та.— 1905,— Т. 27, вып. 1.— С. 4—102.
Кириллова Г. А., Ильин В. А., Юдин Б. С. О новых природно-очаговых инфекциях на
Таймыре // Материалы IV Междунар. симпозиума по приполярной медици-
не.— Новосибирск: Наука. Сиб. отд-ние, 1977.
Козловский А. И. Хромосомные наборы некоторых видов землероек Sorex.— М.:
Изд-во МГУ, 1968.— С. 136—138.— (Научная конференция молодых ученых
Московского университета).
Козловский А. И. Хромосомный полиморфизм иркутской популяции обыкновенной
бурозубки — Sorex araneus L. // Материалы Второго всесоюзного совещания по
млекопитающим.— Новосибирск: Наука. Сиб. отд-ние, 1969.— С. 10—11.
Козловский А. И. Хромосомный полиморфизм в восточно-сибирских популяциях
обыкновенной бурозубки И Цитология.— 1970.— Т. 12, вып. 11.— С. 1459—
1464.
Козловский А. И. Хромосомный анализ полиморфной популяции обыкновенной буро-
зубки И Цитология.— 1972,— Т. 14, вып. 6.— С. 761—768.
Козловский А. И. Соматические хромосомы двух видов землероек-бурозубок Кавка-
за // Зоол. журн.— 1973а.— Т. 52, вып. 4.— С. 571—576.
Козловский А. И. Результаты кариологического обследования аллопатрических форм
малой бурозубки {Sorex minutus) И Там же.— 19736.— Т. 52, вып. 3.—
С. 390—398.
Колосов А. М. Биология крота-могеры // Тр. ин-та/Моск. пушно-меховой ин-т.—
1954,— № 5,— С. 16—24.
Кривошеев В. Г., Чернявский Ф. Б., Железнов Н. К. и др. Новые данные по фауне
млекопитающих Анадырского края И Фауна и зоогеография млекопитающих
северо-востока Сибири.— Владивосток: Дальневост. кн. изд-во, 1978.—
С. 66—94.
Крылов Г. В., Градобоев Н. Д., Юдин Б. С. Новая монография о природе Западной
Сибири И Изв. Сиб. отд-ния АН СССР. Сер. биол.-мед. наук.— 1964.— № 8,
вып. 2.— С. 150—153.— Рец. на кн.: Западная Сибирь/Отв. ред. Г. Д. Рихтер.—
М.: Изд-во АН СССР, 1963.— 488 с.
Крыштафович А. Н. Палеоботаника.— М., 1945.— 650 с.
343
Кузнецов Б. А. Млекопитающие Казахстана // Материалы к познанию фауны и флоры
СССР.— М.: МОИП, 1948,— Вып. 13.— 236 с.
Кумипова А. В. Растительный покров Алтая.— Новосибирск: РИО СО АН СССР.—
I960,— 450 с.
Куренцов А. И. Вопросы зоогеографии южных частей Дальнего Востока // Зоол.
журн.— 1959.— Т. 28, вып. 2,— С. 153—166.
Куренцов А. И. Животный мир Приамурья и Приморья.— Хабаровск, 1959.— 262 с.
Лаврова М. Я., Зажигин В. С. О систематике и биологии землероек Краснодарского
края с оценкой их роли в лептоспирозных очагах И Зоол. журн.— 1965.—
Т. 14, вып. 1.— С. 101—-109.
Лаптев И. П. Млекопитающие Александровского района Томской области // Заметки
по фауне и флоре Сибири.— Томск: Изд-во Том. ун-та, 1953.—Вып. 17.—
С. 3—13.
Лаптев И. П. Млекопитающие таежной зоны Западной Сибири.— Томск: Изд-во Том.
ун-та, 1958.— С. 19—20.
Летов Г. С., Летова Г. И. Материалы к фауне насекомоядных Тувы и их эктопарази-
тов И Докл. ин-та/Иркут. н.-и. цротивочум. ин-т.— 1962.— Вып. 4.—
С. 164—166.
Литвинов Н. И. Заметки о фауне позвоночных островов Байкала // Хозяйственное
использование охотничьей фауны.— Иркутск, 1972.— С. 57—87.
Лукьянова И. В. Ландшафтное распространение бурозубок в Северо-Восточном
Алтае // Вопросы зоологии,— Томск: Изд-во Том. ун-та, 1966.— С. 213—
214.
Лямкин В. Ф. Зоогеография млекопитающих и птиц Баргузинской котловины // Ре-
гиональные биогеографические исследования в Сибири.— Иркутск, 1977.—
С. 111—117.
Лямкин В. Ф. Мелкие млекопитающие восточного побережья озера Байкал // Второй
съезд Всесоюзного териологического общества.— М.: Наука, 1978.—
С. 103—104.
Маак Р. Путешествие на Амур.— СПб., 1859.
Маак Р. Путешествие по долине реки Уссури.— Спб., 1861.
Матурова Р. Т. Мелкие млекопитающие хребта Улан-Бургасы (Восточное Прибай-
калье).— Новосибирск: Наука. Сиб. отд-ние, 1982.
Межжерин В. А. Роль пищевого фактора в эволюции землероек-бурозубок // Вопр.
экологии.— 1962.— Т. 6.— С. 98—99.
Межжерин В. А. Явление Денеля и его возможное объяснение // Acta theriol.—
1964,— Vol. 8, N 6.
Межжерин В. А. Очерк четвертичной истории и происхождения современной фауны
землероек-бурозубок (Sorex, Insectivora, Mammalia) // Материалы по четвертич-
ному периоду Украины (к VII конгрессу ИНКВА в США).— Киев: Наук, дум-
ка, 1965а.— С. 164—173.
Межжерин В. А. О смене адаптаций в процессе эволюции (на примере рода Sorex,
Insectivora, Mammalia) // Материалы зоологического совещания по проблеме
«Биологические основы реконструкции, рационального использования и охраны
фауны южной зоны европейской части СССР».— Киев: Наук, думка, 19656.—
С. 76—81.
Михель Н. М. Промысловые звери Северо-Восточной Якутии.— Л.: Изд-во Главсев-
морпути, 1938.— 175 с.
Млекопитающие фауны СССР/Сост.: И. М. Громов, А. А. Гуреев, Г. А. Новиков.—
М.; Л.: Изд-во АН СССР, 1963.— Ч. 1.— 639 с.
Млекопитающие Якутии/В. А. Тавровский, О. В. Егоров, В. Г. Кривошеев, М. В. По-
пов, Ю. В. Лабутин.— М.: Наука, 1971.— 660 с.
Наумов Н. П. Млекопитающие Тунгусского округа И Тр. Полярной комиссии
АН СССР,—Л., 1934,—Вып. 4.
Николаев А. С. Мышевидные и насекомоядные в предгорьях Салаира // Вопр. зооло-
гии.— Томск: Изд-во Том. ун-та, 1966.— С. 243—245.
Никольский А. М. Путешествие в Алтайские горы летом 1882 // Тр. СПб. о-ва естест-
воиспытателей.— 1883.— Т. 14, вып. 1.— С. 165—170.
Огнев С. И. Заметки по фауне летучих мышей (Chiroptera) и насекомоядных (Insecti-
vora) Уссурийского края П Ежегодник Зоол. музея Академии наук.— М.,
1913.— Т. 18.— С. 410—419.
'Огнев С. И. Опыт описания фауны Московской губернии //Млекопитающие москов-
ской губернии.— М., 1913.— Т. 1, ч. 1.— С. 97—148.
гОгнев С. И. Материалы для систематики насекомоядных России // Ежегодник Зоол.
музея Академии наук.— М:, 1921.—. Т. 22, № 4.—С. 31,1—350.
344
Огнев С. И. Материалы к познанию млекопитающих Кавказа И Учен. зап. Северо-
Кавказ. ин-та краеведения.— 1926.— Т. 1.— С. 45.
Огнев С. И. Млекопитающие северо-восточной Сибири. Насекомоядные.— Влади-
восток, 1926.— С. 17—27.
Огнев С. И. Звери СССР и прилежащих стран (звери Восточной Европы и Северной
Азии).— М.; Л.: Изд-во АН СССР, 1928.— Т. 1.— 631 с.
Огнев С. И. Географическое распространение насекомоядных млекопитающих в свя-
зи с их палеонтологией И Изв. Ассоц. н,- и. ин-та Моск, ун-та.— 1929.— Т. 2,
вып. 1.— С. 107—133.
Огнев С. И. Звери Восточной Европы и Северной Азии. Т. 2: Хищные млекопитаю-
щие.— М.; Л.: Госиздат, 1931.— 776 с.
Огнев С. И. Материалы по систематике палеарктических землероек // Бюл. н.-и. ин-
та зоологии Моск. гос. ун-та им. М. Н. Покровского.— М.; Л.: Медгиз, 1933а.—
С. 57—63.
Огнев С. И. Материалы по систематике, морфологии и географическому распростране-
нию землероек // Зоол. журн.— 19336.— Т. 12, вып. 4.— С. 8—16.
Огнев С. И. Звери СССР и прилежащих стран (приложение).— М.: Изд-во АН СССР,
1935,— Т. 3,— С. 601—641.
Огнев С. И. Новый замечательный вид землеройки (Sorex mirabilis sp.) // Бюл. МОИП.
Отд. биол,— 1937.— Т. 16, вып. 5.— С. 268—271.
Огнев С. И. Млекопитающие центрального Тянь-Шаня (Заилийского и Кунгей-Ала-
тау): Материалы к познанию фауны и флоры СССР.— М.: МОИП, 1940.—
250 с.
Огнев С. И. Заметки по систематике Анадырских млекопитающих, собранных
Л. А. Портенко И Портенко Л. А. Фауна Анадырского края.— М.; Л., 1941.—
Ч. 3: Млекопитающие.— С. 94—105.— (Тр. ин-та/ Н.-и. ин-т полярного зем-
леделия, животноводства и промыслового хоз-ва. Сер. промысл, хоз-во;
Вып. 14).
Огнев С. И. Экология млекопитающих: Материалы к познанию фауны и флоры
СССР,— М.: МОИП, 1951,— 253 с.
Орлов В. Н. Кариосистематика млекопитающих И Цитогенетические методы в систе-
матике млекопитающих.— М.: Наука, 1974.— 207 с.
Орлов В. Н., Аленин В. П. Кариотипы некоторых видов землероек рода Insectivora,
Soricidae // Зоол. журн.— 1968.— Т. 47, вып. 7.— С. 1071—1074.
Орлов В. Н., Булатова Н. Ш. Сравнительная цитогенетика и кариосистематика.— М.:
Наука, 1983.— 405 с.
Орлов В. Н., Козловский А. И. Хромосомные наборы двух географически удаленных
популяций и их место в общей системе хромосомного полиморфизма обыкновен-
ной бурозубки И Цитология.— 1969.— Т. 2, вып. 9.— С. 1129—1136.
Орлов В. Н., Козловский А. И. Обзор хромосомных наборов землероек рода Sorex //
Вести. МГУ. Отд. биол.-почв.— 1971.— № 2.— С. 12—16.
Орлов С. И. Северные границы распространения некоторых видов млекопитающих //
Изв. Сиб. краевед, ст. защиты растений от вредителей.— Новосибирск, 1930.—
Вып. 1: Зоол., № 4(7).— С. 90—94.
Охотина М. В. Дальневосточный крот и его промысел.— М.: Наука, 1966.— 136 с.
Охотина М. В. Бурозубки (Insectivora, Sores) Анадырской низменности И Вести,
зоологии.— 1973.— № 5.— С. 30—34.
Охотина М. В. Землеройки (Insectivora, Soricidae) острова Сахалин И Зоол. журн.—
1977.— Т. 56, вып. 2,— С. 243—249.
Очиров Ю. Д., Башанов К. А. Млекопитающие Тувы.— Кызыл: Тувин. кн. изд-во,
1975.— 138 с.
Пидопличко И. Г. О ледниковом периоде.— Киев: Изд-во АН УССР, 1951.— Вып. 2.—
264 с.; 1954,— Вып. 3.— 280 с.
Плятер-Плохоцкий К. А. Вредные и полезные млекопитающие в сельском хозяйстве
ДВК.— Хабаровск, 1936.— С. 26—28.
Попов М. В. Фауна насекомоядных Якутии И Вопр. зоологии.—Томск: Изд-во Том.
ун-та, 1966.— С. 283—240.
Разоренова А. П. Кротовый промысел в Западной Сибири // Зоол. журн.— 1932.—
Т. 11, вып. 2.
Реймерс Н. Ф. Птицы и млекопитающие южной тайги Средней Сибири.— М.; Л.:
Наука, 1966.— 418 с.
Реймерс Н. Ф., Алина А. В. Млекопитающие Аяиского побережья Охотского моря И
Бюл. МОИП. Отд. биол.— 1967.— Т. 72, вып. 5.— С. 154—155.
Сатунин К. А. Обзор млекопитающих в Закаспийской области // Зап. Импер. отдела
Импер. Рус. геогр. о-ва.— Тифлис, 1905.— Т. 25, вып. 3.— С. 1—56.
345
Сатунин К. А. Фауна Черноморского побережья Кавказского края.— Тифлис, 1913.—
Т. 2,— С. 24—25.
Сатунин К. А. Определитель млекопитающих Российской империи. Рукокрылые, на-
секомоядные и хищные.— Тифлис, 1914.— Вып. 1.— С. 45—82.
Сатунин К. А. Млекопитающие Кавказского края (Chiroptera, Insectivora, Carnivo-
ra) И Зап. Кавказ, музея. Сер. А.— Тифлис, 1915.— С. 63—113.
Скалой В. Н. Материалы к познанию фауны южных границ Сибири // Изв. н.-и. про-
тивочум. ин-та Сибири и Дальнего Востока.— 1936.— Т. 3.— С. 106—118.
Скалой В. Н., Раевский В. В. Sorex idoson princeps montanus И Науч.-метод, зап.
Гл. упр. по зоопаркам, зоосадам и заповедникам.— М., 1934.— Вып. 7.—
С. 198—199.
Скалой В. Н., Раевский В. В. Новые формы млекопитающих из Кондо-Сосьвинского
заповедника // Науч.-метод, зап. Гл. упр. заповедников.— М., 1940.— Вып. 7.—
С. 193—200.
Соколов В. Е. Систематика млекопитающих.— М-: Высш, шк., 1973.— 430 с.
Соколов Г. А., Швепева В. Я., Балагура Н. Н. Опыт учета абсолютной численности
мелких млекопитающих в лесах Западного Саяна И Экология популяций лес-
ных животных Сибири.— Новосибирск: Наука. Сиб. отд-ние, 1974.— С. 77—86.
Стогов И. И. Систематика и географическое распространение бурозубок Казахста-
на // Тезисы докладов Первого Всесоюзного совещания по млекопитающим.—
М.: Изд-во МГУ, 1961,—Т. 1,— С. 145—147.
Строганов С. У. Фауна млекопитающих Валдайской возвышенности И Зоол. журн.—
1936.— Т. 15, вып. 1,— С. 128—142.
Строганов С. У. Насекомоядные млекопитающие фауны СССР // Докл. АН СССР.—
1941,— Т. 33, № 3,— С. 270—272.
Строганов С. У. Систематика кротовых (Talpidae) И Тр. ин-та/Зоол. ин-т АН СССР.—
1948.— Т. 8, вып. 2,— С. 286—405.
Строганов С. У. Обзор подвидов и географического распространения крошечной буро-
зубки (Sorex tscherskii Ognev) // Учен. зап. Том. ун-та.— 1949.— Т. 12.—
С. 183—192.
Строганов С. У. Новый для фауны Сибири вид землеройки И Тр. ин-та/Биол. ин-т
Зап.-Сиб. фил. АН СССР,— 1956а,— Вып. 1.— С. 11—14.
Строганов С. У. Материалы по систематике сибирских млекопитающих И Там же.—
19566,— С. 3—10.
Строганов С. У. Звери Сибири. Насекомоядные.— М.: Изд-во АН СССР, 1957.—
267 с.
Тихомиров А. А., Корчагин А. Н. Список и описание ноллекпий млекопитающих му-
зея // Изв. о-ва Петербургских любителей естествознания,— 1889.— Т. 2.
Туров С. С. О фаунё позвоночных животных северо-восточного побережья оз. Бай-
кал // Докл. Рос. Акад. наук.— 1924.— С. 109.
Туров С. С. Материалы по млекопитающим северо-восточного побережья Байкала и
Баргузинского хребта // Тр. зоол. музея Моск, ун-та.— 1936.— Вып. 3.—
С. 25—40.
Фатеев К. Я. Особенности внутривидовой изменчивости крота Talpa еигораеа И Внут-
ривидовая изменчивость наземных позвоночных животных и микроэволюции.—
Свердловск, 1966.— (Тр. Всесоюз. совсщ.).
Федоров К. П., Юдин Б. С. Гельминты сибирского крота (Asioscalops altaica Nikolski,
1883) И Систематика, фауна, зоогеография млекопитающих и их паразитов.—
Новосибирск: Наука. Сиб. отд-ние, 1975.— С. 181—192,— (Тр. Биол. ин-та
СО АН СССР; Вып. 23).
Фетисов А. С. Современное зоогеографическое районирование Селенгинской Даурии
на основании териологических данных // Зоол. журн.— 1956,— Т. 25, вып. 10.—
С. 1535—1540.
Черепанов А. И., Лаптев И. П., Юдин Б. С. Состояние и задачи зоологических иссле-
дований в Сибири И Вопросы зоологии.— Томск: Изд-во Том. ун-та, 1966.—
С. 5—10.
Шварц С. С. Пути приспособления наземных животных к условиям существования в
Субарктике // Тр. ин-та/Ин-т биологии УФАН СССР.— 1963.— Вып. 33.—
132 с.
Швецов Ю. Г. Мелкие млекопитающие Байкальской котловины.— Новосибирск: Нау-
ка. Сиб. отд-ние, 1977.— 148 с.
Швецов Ю. Г., Галкина Л. И., Юдин Б. С. и др. Население наземных позвоночных
средней части Хамар-Дабана // Фауна и экология позвоночных Сибири.— Но-
восибирск: Наука. Сиб. отд-ние, 1980.— С. 98—110,— (Тр. Биол. ин-та;
Вып. 44).
23 б. С. Юдин
346
Шмальгаузен И. И. Основы сравнительной анатомии.— М.: Изд-во АН СССР, 1938.—
253 с.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса.— М.; Л.: Изд-
во АН СССР, 1939.— 231 с.
Шмальгаузен И. И. Факторы эволюции.— М., Л.; Изд-во АН СССР, 1946.— 396 с.
Штильмарк Ф. Р. Материалы о фауне и размещении по биотопам грызунов верхне-
ленской тайги // Защита лесов от насекомых-вредителей.— М.: Наука, 1963.—
С. 208—215.
Штильмарк Ф. Р., Долгов В. А. Землеройки (Mammalia, Soricidae) Нижнего При-
амурья.— Владивосток, 1974.— С. 58—64.— (Тр. ин-та/Биол.-почв. ин-т. Нов.
сер.; Т. 17).
Эверсман Э. Естественная история Оренбургского края И Естественная история мле-
копитающих животных Оренбургского края.— Казань, 1850.— Ч. 2.— С. 86.
Юдин Б. С. Материалы по экологии некоторых видов бурозубок лесостепной полосы
Западной Сибири // Тр. Том. ун-та.— 1955.— Т. 131.— С. 262—268.
Юдин Б. С. Материалы по питанию бурозубок (Sorex L.) Западной Сибири // Там же.—
1956а,— Т. 142,— С. 295—302.
Юдин Б. С. Териофауна южной части Томской области // Тезисы докладов на X отчет-
ной научной сессии Западно-Сибирского филиала АН СССР.— Новосибирск,
19566.— С. 1—3.
Юдин Б. С. К экологии крошечной бурозубки (S. minutissimus Limmerman) в Запад-
ной Сибири И Тезисы докладов на IX научной конференции.— Новосибирск:
Новосиб. с.-х. ин-т, 1957а.— С. 49—51.
Юдин Б. С. Роль бурозубок в лесных ценозах И Тезисы докладов на VII научной кон-
ференции, посвященной 40-летию Октября.— Томск: Изд-во Том. ун-та,
19576,— С. 55—56.
Юдин Б. С. Значение бурозубок в истреблении насекомых-вредителей сельского хо-
зяйства И Тезисы докладов на совещании зоологов Сибири.— Новосибирск:
Кн. изд-во, 1957в.— С. 82—83.
Юдин Б. С. Акклиматизация выхухоли в пойменных водоемах верхней Оби и перспек-
тивы ее дальнейшего расселения // Проблемы биологических исследований в Си-
бири.— Горно-Алтайск: Горно-Алт. кн. изд-во, 1962а.— С. 266—269.
Юдин Б. С. Значение внутривидовых отношений в расселении вида (на примере обык-
новенной бурозубки — Sorex araneus L.) // Проблемы внутривидовых отноше-
ний организмов.— Томск: Изд-во Том. ун-та, 19626.— С. 164—167.
Юдин Б. С. Экология бурозубок (р. Sorex) Западной Сибири // Тр. ин-та/Биол. ин-т
СО АН СССР,— 1962в,— Вып. 8,— С. 33—134.
Юдин Б. С. Результаты выпуска выхухоли в Томской области и возможности искус-
ственного расширения ее ареала в Западной Сибири И Акклиматизация живот-
ных в СССР.— Алма-Ата: Изд-во АН КазССР, 1963.— С. 205—207.
Юдин Б. С. Выхухоль в Западной Сибири И Охота и охотничье хоз-во.— 1964а,—
№ 3,— С. 21—22.
Юдин Б. С. Географическое распределение и внутривидовая таксономия крошечной
бурозубки в Западной Сибири // Acta Theriol.— 19646.— Vol. 8, N 10.—
P. 167—179.
Юдин Б. С. Итоги исследований насекомоядных фауны СССР и задачи их дальнейше-
го изучения // Тезисы докладов зоологического совещания.— Томск: Изд-во
Том. ун-та, 1964в.— С. 178—180.
Юдин Б. С. Новый для фауны СССР вид землеройки (Insectivora, Soricidae) // Новые
малоизвестные виды фауны Сибири.— Новосибирск: РИО СО АН СССР,
1965а,— С. 44—47.
Юдин Б. С. Структура гениталий как основной признак в классификации землероек И
Изв. СО АН СССР. Сер. биол.-мед,— 19656,— Вып. 3, № 12,— С. 61—75.
Юдин Б. С. Выхухоль И Книга охотника.— 2-е изд.— Новосибирск: Новосиб. кн.
изд-во, 1966.— С. 142—444.
Юдин Б. С. Выхухоль поселилась в Сибири // Знай свой край.— Новосибирск: Зап.-
Сиб. кн. изд-во, 1967а.— С. 43.
Юдин Б. С. Новый для Палеарктики вид бурозубки (Mammalia, Sorex) Курильских
островов И Изв. СО АН СССР. Сер. биол.-мед.— 19676.— Вып. 1, № 5.—
С. 155—157.
Юдин Б. С. Комплексы насекомоядных млекопитающих (Mammalia, Insectivora)
в ландшафтах Новосибирской области // Биологическое районирование Ново-
сибирской области.— Новосибирск: Наука. Сиб. отд-ние, 1969а.— С. 131—143.
Юдин Б. С. Новые данные по систематике некоторых видов землероек (Soricidae, Insec-
tivora) Палеарктики и Неарктики //Acta theriol.—19696.—Vol.14, N3.— Р. 21—34.
347
Юдин Б. С. Архитектура гнезда сибирского крота // Изв. СО АН СССР. Сер. биол.—
1970.— Т. 15, вып. 5.— С. 98—109.
Юдин Б. С. Зоологический музей Биологического института Сибирского отделения
АН СССР // Фауна Сибири.— Новосибирск: Наука. Сиб. отд-ние, 1970а.—
С. 7—9.
Юдин Б. С. Обзор видов рода Neomys Каир (Soricidae, Insectivora) // Там же.—
19706,— С. 247—251.
Юдин Б. С. Насекомоядные млекопитающие Сибири: Определитель.— Новосибирск:
Наука. Сиб. отд-ние, 1971а.— 170 с.
Юдин Б. С. Результаты акклиматизации выхухоли в Западной Сибири и ее охрана //
Рациональное использование и охрана живой природы Сибири.— Томск: Изд-
во Том. ун-та, 19716.— С. 170—172.
Юдин Б. С. Фауна землероек (Mammalia, Soricidae) севера Западной Сибири И Био-
логические проблемы Севера.— Магадан, 1971в.— С. 48—53.— (Тр. ин-та/
Сев.-Вост, компл. ин-т ДВНЦ АН СССР; Вып. 42).
Юдин Б. С. Экология бурозубок (род Sorex) Западной Сибири // Тр. ин-та/Биол. ин-т
СО АН СССР,— 1971г,— Вып. 8,— С. 33—134.
Юдин Б. С. Биологические особенности сибирского крота (Asioscalops altaica Nikols-
ку, 1883), связанные с размножением И Изв. СО АН СССР. Сер. биол.— 1972а.—
№ 10, вып. 2.— С. 64—74.
Юдин Б. С. Бурозубка Радде — Sorex raddei Satunin, 1895 (Mammalia, Soricidae) в
фауне Советского Союза // Aquilo. Ser. Zool.— 19726.— N 13.— P. 69—76.
Юдин Б. С. Запасание сибирским кротом дождевых червей как одна из адаптаций к
жизни в условиях климата Сибири // Изв. СО АН СССР. Сер. биол.— 1972в.—
№ 15, вып. 3.— С. 133—136.
Юдин Б. С. К методике изучения зимней биологии крота (Asioscalops altaica Nikolsky,
1883) И Териология.— Новосибирск: Наука. Сиб. отд-ние, 1972г.— Т. 1.—
С. 354—355.
Юдин Б. С. К систематике трансарктической бурозубки (Sorex cinereus Kerr, 1792)
фауны СССР // Там же, 1972д.— С. 45—50.
Юдин Б. С. Фаунистические связи насекомоядных млекопитающих Камчатки, Ку-
рильских островов, Сахалина, Приморья и Японии // Зоологические проблемы
Сибири.— Новосибирск: Наука. Сиб. отд-ние, 1972е.— С. 506—507.
Юдин Б. С. К характеристике трансарктической бурозубки (Sorex cinereus Kerr,
1792) северо-востока Сибири и Камчатки // Фауна Сибири.— Новосибирск:
Наука. Сиб. отд-ние, 1973а.— С. 269—279.— (Тр. ин-та/Биол. ин-т СО АН
СССР; Вып. 16, ч. 2).
Юдин Б. С. Фауна насекомоядных млекопитающих (Mammalia, Insectivora) Пред-
байкалья и Забайкалья // Там же, 19736.— С. 280—296.
Юдин Б. С. Экологические исследования в Биологическом институте Сибирского
отделения АН СССР за 25 лет (1944—1969) // Проблемы экологии.— Томск:
Изд-во Том. ун-та, 1973в.— Т. 3.— С. 257—270.
Юдин Б. С. Ареологическая структура фауны насекомоядных млекопитающих (Mam-
malia, Insectivora) Сибири и взаимосвязь слагающих ее элементов // Symposium
Theriologicum 11 in Brno, 1971.— Praha, 1974a.— P. 109—116.
Юдин Б. С. Изменчивость зубной системы сибирского крота (Asioscalops altaica Ni-
kolsky, 1883) // Aquilo. Ser. Zool.— 19746.— N 2.— P. 59—63.
Юдин Б, С. Особенности размножения сибирского крота (Asioscalops altaica Nikols-
ку, 1883) // Acta Tneriol.— 1974b.— Vol. 19, N 24.— P. 255—366.
Юдин Б. С. Состояние запасов выхухоли в Западной Сибири // Материалы III Все-
союзного совещания по охране и восстановлению выхухоли.— Воронеж, 1974 г.
Юдин Б. С. Эколого-фаунистические комплексы (Insectivora) Дальнего Востока //
Первый международный конгресс по млекопитающим.— М.: Наука, 1974д.—
С. 355—366.
Юдин Б. С. Комплексы насекомоядных млекопитающих (Mammalia, Insectivora)
Дальнего Востока // Систематика, фауна, зоогеография млекопитающих и их
паразитов.— Новосибирск: Наука. Сиб. отд-ние, 1975а,— С. 41—69.— (Тр.
ин-та/Биол. ин-т СО АН СССР; Вып. 23).
Юдин Б. С. Перспективы акклиматизации выхухоли на Алтае И Охрана, рациональ-
ное использование и воспроизводство природных ресурсов Алтайского края.—
Барнаул: Алт. кн. изд-во, 19756.— С. 283—285.
Юдин Б. С. Строение тазовых костей кротов и могер (Mammalia, Talpa, Asioscalops,
Mogera) и их таксономическое значение // Систематика, фауна, зоогеография
млекопитающих и их паразитов.— Новосибирск: Наука. Сиб. отд-ние, 1975в.—
С. 41—69.— (Тр. ин-та/Биол. ин-т СО АН СССР; Вып. 23).
348
Юдин Б. С. Индивидуальная и популяционная изменчивость зубной системы сибир-
ского крота в связи с проблемой происхождения многовершинных зубов И Фау-
на и систематика позвоночных Сибири.— Новосибирск: Наука. Сиб. отд-ние,
1977,— С. 178—199,— (Тр. ин-та/Биол. ин-т СО АН СССР; Вып. 31).
Юдин Б. С. Адаптация сибирского крота (Asioscalops altaica Nikolsky, 1883) к зим-
ним условиям И Материалы советско-финского симпозиума «Адаптация живот-
ных к зимним условиям».— М.: Наука, 1980а.— С. 146—151.
Юдин Б. С. Географическая изменчивость видового состава млекопитающих гор Юга
Алтая И Проблемы зоогеографии и истории фауны.— Новосибирск: Наука.
Сиб. отд-ние, 19806.— С. 190—204.
Юдин Б. С. Зональные и ландшафтные группировки мелких млекопитающих (Micro-
mammalia) Таймыра // Фауна и экология позвоночных Сибири.— Новосибирск:
Наука. Сиб. отд-ние, 1980в.— С. 5—31.— (Тр. ин-та/Биол. ин-т СО АН СССР;
Вып. 44).
Юдин Б. С. Зональные и ландшафтные особенности структуры населения насекомо-
ядных млекопитающих в Западной Сибири // Материалы совещания «Вид и его
продуктивность в ареале».— Свердловск, 1984.— Ч. 1.— С. 86—87.
Юдин Б. С. Зональные особенности фауны и населения насекомоядных млекопитаю-
щих Средней Сибири И Четвертый съезд Всесоюзного териологического общест-
ва,— М.: Наука, 1986а.— Т. 1.— С. 148—149.
Юдин Б. С. Сибирская белозубка (Crocidura sibirica Dukelsky) в фауне Советского Сою-
за // Фауна, таксономия, экология млекопитающих и птиц.— Новосибирск:
Наука. Сиб. отд-ние, 1987.— С. 43—54.
Юдин Б. С. Состояние сибирской популяции выхухоли и перспективы дальнейших
работ по спасению вида // Редкие наземные позвоночные Сибири.— Новоси-
бирск: Наука. Сиб. отд-ние, 1988.— С. 287—292.
Юдин Б. С., Барсова Л. И. Землеройки кедровых лесов Прителецкого очага клещевого
энцефалита // Природа очагов клещевого энцефалита на Алтае,— Новосибирск:
Наука. Сиб. отд-ние, 1967.— С. 51—59.
• Юдин Б. С., Галкина Л. И., Потапкина А. Ф. Млекопитающие Алтае-Саянской гор-
ной страны.— Новосибирск: Наука. Сиб. отд-ние, 1979.— 293 с.
Юдин Б. С., Золотаренко Г. С. Типовые коллекции животных, хранящихся в зооло-
гическом музее Биологического института // Фауна Сибири.— Новосибирск:
Наука. Сиб. отд-ние, 1970.— С. 10—17.
Юдин Б. С., Исаков С. И. Систематика и фауна землероек (Soricidae, Mammalia) Тад-
жикистана. Сообщ. 1. Род Suncus и Sorex // Изв. АН ТаджССР. Отд. биол.—
1975,— № 4,— С. 47—53.
Юдин Б. С., Исаков С. И., Долгов В. А. Систематика и фауна землероек (Mammalia,
Soricidae) Таджикистана // Фауна и экология животных Таджикистана.— Ду-
шанбе: Дониш, 1978.— С. 92—116.
Юдин Б. С., Кривошеев В. Г., Беляев В. Г. Мелкие млекопитающие Севера Дальнего
Востока.— Новосибирск: Наука. Сиб. отд-ние, 1976.— 269 с.
Юдин Б. С., Литвинов Ю. Н., Юдина С. А. Фауна мелких млекопитающих (Micromam-
malia) таежной зоны Таймыра и ее связь с населением сопредельных террито-
рий И Охотничье-промысловые ресурсы Сибири.— Новосибирск: Наука. Сиб.
отд-ние, 1986.— С. 192—202.
Юдин Б. С., Москвитина Н. С., Власов В. В. Опыт абсолютного учета численности
мелких млекопитающих на изолированной площади // Экология.— 1972.—
№ 2,— С. 95—97.
Юдин Б. С., Николаев В. В. Сообщества мелких млекопитающих (Micromam-
malia) вертикальных поясов Центральной части Восточного Саяна (Тофа-
лария) И Фауна и систематика позвоночных Сибири.— Новосибирск: Наука.
Сиб. отд-ние, 1977а.— С. 81—93.— (Тр. ин-та/Биол. ин-т СО АН СССР;
Вып. 31).
Юдин Б. С., Потапкина А. Ф. Территориальные группировки мелких млекопитающих
(Micromammalia) в Кузнецком Алатау и Западном Саяне И Там же.— С. 32—59.
Юдин Б. С., Потапкина А. Ф., Галкина Л. И. Эколого-фаунистический анализ населе-
ния мелких млекопитающих (Micromammalia) Центрального Алтая//Там же.—
С. 5-31.
Юргенсон П. Б. Материалы к познанию млекопитающих Прителецкого участка Алтай-
ского госзаповедника // Тр. Алт. заповедника.— М., 1938.— Вып. 1.—С. 92 —
169.
Юргенсон П. Б. Рецензия И Бюл. МОИП. Отд. биол.— 1959.— Т. 64, вып. 1.—
С. 145—147.— Рец. на кн.: Строганов С. У. Звери Сибири. Насекомоядные.—
М.: Изд-во АН СССР, 1957,— 267 с.
349
Юрлов К. Т., Юдин В. С., Потапкина А. Ф. и др. К характеристике фауны мелких
млекопитающих северной лесостепи Барабинской низменности // Животный
мир Барабы.— Новосибирск: РИО СО АН СССР, 1965.— С. 184—207.
Яблоков А. В. Изменчивость млекопитающих.— М.: Наука, 1966.— 363 с.
Яковлев Е. К границам распространения промысловых зверей и птиц в Туруханском
крае И Тр. Зоол. секции Средне-Сиб. отд. Рус. геогр. о-ва.— Красноярск,
1930.— Вып. 1.
Янушевич А. И. Материалы по позвоночным Тувинской области // Изв. Зап.-Сиб.
фил. АН СССР.— Новосибирск, 1948.— Т. 2, вып. 2.— С. 3—27.
Янушевич А. И. Фауна позвоночных Тувинской области.— Новосибирск: Зап.-Сиб.
кн. изд-во, 1952.— 142 с.
Янушевич А. И., Юрлов К. Т. Вертикальное распространение млекопитающих и птиц
в Западном Саяне // Изв. Зап.-Сиб. фил. АН СССР, зоол.— Новосибирск,
1950,—Т. 3, вып. 2,—С. 3—33.
Abe Н. Classification and biology of Japanese Insectivora (Mammalia). I. Studies on
variation and classification // J. Fac. Agric. Hokkaido imp. Univ.— Sapporo,
1967.— Vol. 55, N 3,— P. 191—265.
Abe H. Classification and biology of Japanese Insectivora (Mammalia). II. Biological
aspects // Ibid, 1968,— Vol. 55, N 4.— P. 429—458.
Abe H. Small Mammals of Central Nepal // Ibid, 1971.— Vol. 56, N 4.— P. 367—423.
Allen Gl. M. Notes of the birds and mammals of the Arctic coast of East Siberia Mam-
mals // Proc. New Engl. Zool. Club.— 1914.— Vol. 5.— P. 49—66.
Allen Gl. M. The mammals of China and Mongolia.— N. Y., 1938.— Vol. 11, pt. 1.—
620 p.
Allen Gl. M. Checklist of African mammals // Bull. Mus. Comp. Zool.— Cambridge,
1939.— Vol. 82,— P. 5—53.
Allen J. A. Report of the Mammals collected in Northeastern Siberia by Jesup Pacific
expedition, with field, potes by N. g. Buxton // Bull. Amer. Mus. Nat. Hist.—
1903,— Vol. 19.— P. 101—184.
Allen J. A. The skeletal characters of Scutisorex Thomas // Ibid.— 1917.— Vol. 37.—
P. 769—784.
Bee J. W., Hall E. R. Mammals of Northern Alaska on the Arctic slope // Univ. Kans.
Pubis. Natur. Hist.— 1956.— Vol. 8.— 356 p.
Bovey R. La formule chromosomique de quelgues Insectivores indigeues // Experientia.—
1949a.— Vol. 5, N 1.— P. 72—73.
Bovey R. Les chromosomes des Chiropteres et des Insectivores // Rev. Suis. Zool.—
1949b.— Vol. 56,— P. 371—460.
Borgaonkar D. S. A List of Chromosome Numbers in the Order Insectivora // J. Heredi-
ty.— 1965,— Vol. 56, N 6,— P. 266.
Brandt J. Bemerkungen fiber d. Gatt. Sorex. // Melanges biologiques de L’Acad.— St.-
Petersb., 1853.
Brink F. H. Une nouvelle musaraigne dens les Pays-Bas//Proc. Konink. Nederl. Akad.
Wetten Schappen-Amsterdam. Ser. C.— 1952.— Vol. 55, N 4.— P. 370—374.
Brink F. H. La Musaraigue uspece circum-boreale // Mammalia.— 1953.— Vol. 17,
N 2,— P. 96—125.
Brink F. H. Saugetiere Europas.— Hamburg; Berlin, 1957.— 350 S.
Brink F. H. Ginide des Mammiferes sauvages de 1’Europe occidentale.— Neuchatel:
Delachaux Niestle, 1967.
Corbet G. B. The pygmy moles of Europe and Japan // J. Zool.— 1967.— Vol. 153,
N 4,— P. 567—568.
Dehnel A. Badania nad rodzajem Sorex L. // Ann. Univ. Mariae Curie-Sklodowska.
Cect. C.— 1949,— Vol. 4, N 2.— P. 17—102.
DobsonG. E, A monograph of the Insectivora, systematics and anatomical.— L., 1883.—
Pt. 2,— P. 126—172.
Dobson G. E. A synopsis of the genera of the family Soricidae // Proc. Zool. Soc. Lond.—
1890,— Vol. 5,— P. 48—51.
Dolhnan G, On the african shrews belonging to the genus Crocidura 11 Ann. Mag. Natur.
Hist. Ser. 8.— 1915, 1916,— Vol. 15—17.— P. 188—209.
““Dukelsky N. Zur Kennthis der Saugetierfauna Westsibiriens // Zool. Anzeiger.— 1930.—
Bd 88, H. 1/4.— S. 75—76.
Dzuev R. T., Tembotov A. K., Grulich I. Structure of ranges of caucasien moles //
II Congre.su; theriologicus.— Brno, 1978.— P. 216.
Ellerman J. R., Morrison-Scott T. C. Checklist of Palaeartic and Indian Mammals 1758
to 194.6, Berlin Museum.— L., 1951.— 810 p.
350
Fedyk S., Ivanitskaya E. Y. Chromosomes of Siberian shrews // Acta Theriol.— 1972.—
vol. 17, N 36.— P. 475—492.
Felten H., Storch G. Insectifresser und Nagetiere aus N. Griechenland und Jugoslavien
(Mammalia: Insectivora und Rodentia) // Seneckenbergiana biologica.— 1965.—
Vol. 46, N 5,— P. 341—367.
Felten H., Storch G. Kleinsauger com Pelister-Massiv in Slid-Jugoslavien // Natur und
Museum.— 1966.— Vol. 96, N 4.— P. 129—135.
Findley J. S. Speciation of the Wandering shrew // Univ. Kane. Publ. Mus. Natur.
Hist.— 1955.— Vol. 9, N 1.— P. 1—68.
Fischer F. Zoognosie.— 1814.— Vol. 3.
Ford С. E., Hamerton J. L. A system of chromosomal polymorphism in the common
shrew (Sorex araneus L.) // XV Int. Congr. Zool.— 1968.— Sect. 2.
Ford С. E., Hamerton J. L., Sharman G. B. Chromosome polymorphism in the common
shrew // Nature (bond.).— 1957.— Vol. 180, N 4582,— P. 392—393.
Fredga K. Chromosomes of the Masked Shrew (Sorex caecutiens Laxm.) // Hereditas.—
1968,— Bd 60, H. 1/2,— P. 269—271.
Fredga K. A new chiomosome rase of the common shrew (Sorex araneus) in Sweden //
J Hereditas.— 1973.— Vol. 73, N 1,— P. 153—156.
Geoffroy S. H. Catalogue Mammals // Amer. Mu.,. Natur. Hist.— 1803.— 77 p.
Goodwin G. G. A new species of shrew from Eastern Siberia // Amer. Mus. Novit.—
1933.— N 657.
Grulich I. Zu den allometrischen Beziehungen einiger systematisch-taxonomischer Merk-
male der Vertreter dec Gattung Talpa (Mammalia, Insectivora) // Zool. Listy.—
1969.— Vol. 18, N 2.— P. 163—184.
Grulich I. Die Standortanspriiche von Talpa ca.eci Savi (Talpidae, Insectivora)// Ibid.—
1970,— Vol. 19, N 2,— P. 199—219.
Grulich I. Zum Bau des Beckens (pelvis), eines systematisch-taxonomischen Merk-
males, bei der Unterfamilie Talpinae // Ibid.— 1971.— Vol. 20, N 1.— P.
15—28.
Halkka O., Skaren U. Evolution chromosomique chez genre Sorex: nouvelle informa-
tion // Experientia.— 1964.— Vol. 20, N 6.— P. 314—315.
Hall E. R., Kelson K. R. The Mammals of North America.— N. Y.: Ronald Press,
1959.— Vol. 1/2,— 1083 p.
Heller F. Neue Fundstellen altdiluvialer Desmana-Reste in Siidwestdeutschland Neues
Jahr II Geol. Palaeontol. Monatsch.— 1954.— N 10.
Hoffman R. S. Relationships of centrain Holarctic shrews genus Sorex // Saugetier.—
1971,— Bd 36, H. 4,— S. 193—200.
Hoffman R. S., Peterson R. S. Systematics and zoogeography of Sorex in the Bering
Strait area // Syst. Zool.— 1967,— Vol. 16, N 2.— P. 127—136.
Hollister N. Two new mammals from the Siberian Altai // Smith. Misc. Coll.— Was-
hington, 1913.— Vol. 60, N 2.
Howell. Mammals from China // Pioc. Natl. Mus. USA.— 1929.— Vol. 75.— P. 7.
liliger C. Prodromus systematicus Mammalium et Avium.— 1811.— P. 125—127.
Imaizumi Y. Taxonomic studies on the Japanese mountain (Talpa mizura), with descrip-
tions of two new subspecies // Bull. Natl. Mus. (Tokyo). Sect. Zool.— 1955.—
Vol. 1,— P. 26—38.
Imaizumi Y., Yoshiuki M. A comparative osteology of the pelvic girdle of the Japanese
Talpidae. Pt. 1 // J. Mammal. Soc. Japan.— 1961.— Vol. 2, N 1.— P. 10—14.
Jackson H. H. A review of the American moles // North. Amer. Fauna.— Washington,
1915,— Vol. 38,— P. 3—98.
Jackson H. H. A taxonomic review of the American longtailed shrews // Ibid.— 1928.—
Vol. 51,— 238 p.
Junge J. A., Hoffmann R. S., Debry R. W. Relationships within the Holarctic Sorex
arcticus—Sorex tundrensis species complex // Acta Theriol.— 1983.— Rc. 28,
fc. 21/31.— P. 339—350.
Каир J. Ent-Gesch. Naturl. System. Europa // Thierwelt.— 1928.— Vol. 1.— P. 117.
Kral B., Radjabli S. Banding patterns and Robertsonian fusion in the Western Siberian
population of Sorex araneus (Insectivora, Soricidae) // Zool. Listy.— 1974.—
Vol. 23, N 3.— P. 217—227.
Kretzoi M. Jelentes a Kislangi Kalabrial (villafrankai) fauna feltarasarol // Madyar alla-
mi foldt. int. evi jelentese.— 1953,— Vol. 1.
Kretzoi M. Drepanosorex — neu definiert // Vertebrata Hungarica.— 1965.— T. 7,
fc. 1/2,— P. 117—129.
Kindal M. Some aspects of the tooth development in Soricidae // Acta Odontol.
(Seand).— 1959.— Vol. 17, pt. 4,— P. 203—237.
351
Kishida К. The mammals fauna of Northern Japan II Dobutsugaku Zaschi.— Tokyo,
1930.— Vol. 42, N 15.
Kuroda N. Mammals of the Northern Kurile Islands H Bull. Biogeogr. Soc. Japan.—
1933.— Vol. 4, N 1 — P. 47—48.
Kuroda N. Mogera Wogura robusta 11 Ibid.— 1939.— Vol. 9, N 1.— P. 39.
Laxmann E. Sibirische Briefe, herausgeg H A. L. Schlozer. Cottingen und Gotha.—
1769.
Laxmann E. Sorex caecutiens. P. 1. Natur. Hist, of Central Asia, Nova acta // Acad. Sci.
Petropol.— 1788.— Vol. 3.— P. 54—83.
Linnaeus C. Systema Naturae. 10-ed.— 1758.— Vol. 1.— 824 p.
Matthey R. Les chromosomes des mammiferes Eutheriens. Liste critique et assai zur
1’evolution chromosomique // Arch. d. Jul. Klaus Stift Vererb.— 1958.—
Vol. 33.— P. 253—297.
Meylan A. Le polymorphysme chromosomique de Sorex araneus L. (Mammalia, Insecti-
vora) H Rev. Suis. Zool.— 1964,— Vol. 71, N 4,— P. 903—983.
Meylan A. Repartition geographique des races chromosomiques de Sorex araneus L. en
Europe (Mammalia, Insectivora) // Ibid.— 1965.— Vol. 72, N 4.— P. 634—646.
Meylan A. Donnees nouvelles sur les chromosomes des Insectivores europeens (Mamma-
lia) // Ibid.— 1966,— Vol. 73.— P. 548—558.
Meylan A. Donnes neuvelles sur les chromosomes des Insectivores europeens (Mamma-
lia) // Ibid— 1968— Vol. 75, N 3— P. 691—696.
Miller G. S. List of North American land mammals in the United States // Bull. Natl.
Mus. USA— 1912— Vol. 79.
Murray A. The geographic distribution of Mammals.— L., 1866.— Vol. 16.— 420 p.
Nehring A. U ber Mogera robusta n. sp. und fiber Meles sp. n. Wladivostok. Litzb. //
Ges. Naturforsch. Freunde—Berlin, 1891.— S. 95—103.
Niethammer J. Insectenfresser und Nager Spaniens // Bonn. Zool. Beitrag.— 1956.—
Bd 7— S. 249—295.
Niethammer J. Ein Beitrag zur Kenntnis der Kleinsauger Nordspaniens // Z. Sauge-
tier— 1964a— Bd 29— S. 193—220.
Niethammer J. Der Zwergmaulwurt (Talpa mizura) ein neues Relikt aus Spanien // Na-
turwis.— 1964b.— Jg. 51/74.— S. 148—149.
Olert J. Schadelmessugen an rheinischen Wald und Schabrackenspitzmausen // Bonn.
Zool. Beitrag.— 1973.— Bd 24, H. 4.
Ott J. Nachweis natiirlicher reproduktiver Isolation zwischen Sorex gemellus sp. n. und
Sorex araneus Linnaeus, 1758 in der Schweiz (Mammalia, Insectivora) // Rev.
Suis. Zool.— 1968.— Vol. 75, N 1.— P. 53—75.
Pallas P. S. Reisen dutch verschiedene Provinzen des Russischen reichs.— 1773.— Bd 2.
Pallas P. S. Zoographia Rosso-Asiatica // Betropoll.— 1811.— T. 1.
Pomel A. Etudes sur les cornassiers insectivores // Bull. Univ. Arch. Sci. Sect. Phys,
et Natur— 1948— T. 9— P. 247.
Puissegur C. Recherches sur le Desman des Pyrenees.— Toilouse, 1935.
/Tladde G. Reiseni im Stiden von Ost-Sibirien in Jahren 1855—1859.— St.-Petersb.,
I 1862— S. 115—116.
TFeed К. M. The Proscalopinae new subfamily of Talpid insectivores // Bull. Mus. Comp.
Zool— 1961— Vol. 125, N 14— P. 1—11.
Repenning Ch. A. Palearctic — Nearctic mammalian dispersal in the late Cenozoic //
Bering Land Bridge.— Stanford, 1967.— P. 288—311.
Richter H. Systematisch taxonomische untersuchungen an palearctischen Weibzahn-
spitzmausen der gattung Crocidura (III). Zur Taxonomie und Verbreitung der pa-
learctischen Crociduren (Mammalia, Insectivora, Soricidae) // Zool. Alh. Staate.
Mus. Tierk— Dresden, 1970— Bd 31, H. 17— S. 293—304.
Satunin K. A. Sorex raddei Satunin n. sp. und Melies taxus aranarius Satunin n. subsp.
Zwai neue Siiugethierarten ous dem Kaukasus und aus dem nuteren Wolggebiete //
Arch. Naturgesch.— 1865.— Bd 1, H. 1.
Schwarz E. Revision of the Old-Word of the genus Talpa Linnaeus // Proc. Zool. Soc.
Lond.— 1948.— Vol. 118, pt. 1.
Shotolchu N., Stuble M. Zur Saugetierfauna der Mongolei // Erstnachweise von zwei
Sorex-Arten // Mitt. Zool. Mus. Berlin.— 1971.— Bd 47, H. 1.— S. 43—45.
Siivonen L. Sorex isodon Turov (1924) and S. unguiculatus Dobson (1890) as independent
shrew species // Aquilo. Ser. Zool.— 1965.— T. 4.— P. 3—49.
Siivonen L. Sorex isodon Turov is not synonymous with S. centralis Thomas // Ibid.—
1969— T. 7— P. 42—49.
Simpsom G. G. The principles of classification and a classification of Mammals // Bull.
Amer. Mus. Natur. Hist— 1945— Vol. 85— 350 p.
352
Skaren U. Variation in two shrews, S. unguiculatus Dobson and S. a. araneus L. // Ann.
Zool. Fenn.— 1964.— Vol. 1.— P. 94—124.
Sowerby A. de C. The Naturalist in Manchuria // Corea and China.— 1923.— P. 21.
*—-Spitzenberger F. Zur Verbreitung und Systematik tiirkischer Soricinae (Insectivora, Mam-
malia) // Ann. Naturch. Mus. Wien.— 1968.— Bd 72.— S. 273—289.
— Spitzenberger F., Steiner H. Uber Insektenfresser (Insectivora) und Wiihlmause (Micro-
tinae) der nordosttiirkischen Feuchtwalder // Bonn. Zool. Beitrag.— 1962.—
Bd 13,— P. 284—310.
Stein G. H. W. Schadelallometrien und Systematik bei altwetlichen Maulwiirfen (Talpi-
dae) H Mitt. Zool. Mus.— Berlin, I960,— Bd 36, H 1 — P. 1—49.
Stein G. H. W. Anomalien der Zahnzahl und ihre geographische Variabilitat die Insecti-
voren. I. Maulwurf, Talpa еигораеа L. // Ibid.— 1963.— Bd 39, N 1.—
S. 223—240.
Stirton R. A. A new genus of Soricidae from the Barstow Miocene of California II Bull.
Univ. Calif. Publ. Dept. Geol. Sci.— 1930,— Vol. 19, N 8,— P. 217—228.
Takagi N., Fujimaki Y. Chromosomes of Sorex shinto saevus Thomas and Sorex unguicu-
latus Dobson // Jap. J. Gen.— 1966.— Vol. 41.— P. 109—113.
Temminck. Talpa wogura // Fauna Japonica.— 1833.— Vol. 19, pt. 4.
Tobias Ph. V. Chromosomes sex-cells and evolution in a mammals, based mainly on stu-
dies of the reproductive glands of the gebrids and a new list of mammals.— L.,
1956— 420 p.
Tobias Ph. V. Chromosomes sex-cells and evolution in a mammalia // Биология.—
1957— N 17— P. 725—781.
Todorovic M. Boundaries of two populations of the species Talpa еигораеа еигораеа L.
and T. mizura hercegouinensis Bolkay in Herzegovina // Arh. Biol.— 1965.—
Rc. 17, fc. 4— P. 29—30.
Thomas O. Mammals from the Island of Sachalien and Hokkaido // Proc. Zool. Soc.—
L., 1907.
Thomas O. A new Chinese mole of the genus Scaptochirus // Ann. Mag. Natur. Hist.—
1910— Vol. 8, N 5— P. 350—351.
Thomas O. New mammals from Central and Western Asia, mostli collected by Mr. Doug-
las Carrthers // Ibid.— 1911.— Ser. 8.— Vol. 7.
Thomas O. On a collection of small mammals from the Tsin-Ling Mountains, Central
China // Ibid— 1912— Vol. 8, N 10— P. 395—398.
True F. A revision of the American moles // Proc. Natur. Mus. USA.— 1897.— Vol. 19.—
112 p.
Van Valen L. New Paleocene insectivores and insectivore classification // Bull. Amer.
Mus. Natur. Hist— 1967— Vol. 135, art. 5— P. 214—284.
Vlasak P. The biology of reproduction and postnatal development of Crocidura suaveolens
Pallas, 1811 under laboratory conditions // Acta Univ. Carolinae. Ser. Biol. —
1972— Vol. 3— P. 207—292.
Walker E. P., Wanick F., Lange R. et al. Mammals of the World. // Baltimore1 Acad.
Press, 1964— Vol. 1.— 664 p.
Wolfe J. A., Leopold E. B. Neogene and Early Quaternary vegetation of northwestern
North America and northeastern Asia // The Bering Land Bridge.— Stanford,
1967— P. 193—206.
Ziegler A. C. Dental homologies and possible relationships of recent Talpidae // J. Mam-
mal— 1971—Vol. 52, N 1— P. 50—58.
Ziegler A. C. Processes of mammalian tooth development as illustrated by dental onto-
geny in the mole Scapanus latimanus (Talpidae, Insectivora) // Arch. Oral. Biol.—
1972— Vol. 0—P. 61—76.
Zimmermann E. A. Geographische des Menschen und der viersiissigen Tiere.— Leipzig,
1780.
353
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
Белозубка 88, 314, 331, 339
— белобрюхая 323, 333, 338, 339
— большая 315, 324—331, 333, 336, 337
— голиаф 110
Огнева 336
— малая 315, 317, 320, 322—324, 327,
333, 336, 337
----южнорусская 323
----семиреченская 323
----дальневосточная 324
— сибирская 88, 314, 315, 323, 327,
330, 332, 333, 336, 339
Белозубки 116, 117, 119
Бурозубка 5, 116, 117, 119, 120, 121,
122, 123, 130,131,132,144,147,186,
219, 229, 230, 265, 266, 283, 300
— альпийская 266, 283, 297, 300
— арктическая 196, 251, 279, 293, 299
— берингийская 5, 123, 128, 140—144
— бурая 166, 200, 231, 235—238, 241,
243, 295, 299, 300, 300, 307
---- сибирская горная 242
---- забайкальская 243
---- уссурийская 244
----якутская 243, 244
— бухарская 196
— гигантская 119, 122, 128, 144—146,
149, 150—152
---- корейская 152
— дальневосточная 128, 141, 183, 223,
227, 228, 231
— кавказская 295, 298
— когтистая 123, 200, 201, 205—206,
910 911 915 917
— крошечная 119, 153, 157, 158—163,
165—168, 171, 173,175,178,180,183,
184, 196
----барабинская 163, 164, 168, 169,
170, 172
----длиннохвостая 170—172
----казахстанская 170
----обыкновенная 167, 169, 172, 178,
196
---- Строганова 172
---- уссурийская 171, 173
----Черского 174
----чукотская 169, 170, 172, 174
— крупнозубная 121, 123, 185, 186,
190, 191, 192, 194—197 199,271,295
----восточносибирская 198
----дальневосточная 182, 183, 197,
200, 223, 227
---- западносибирская 199
— малая 153, 167, 174, 175, 178—
185, 196, 227, 228,229,230,231,268
— обыкновенная 114, 201, 204, 210, 211,
215, 216, 217, 218, 221, 222, 223, 234,
236, 271, 282, 283, 286, 287, 288—301,
306, 307
---- горная 303, 307
---- европейская 306
— Радде 295, 297—299
— равнозубая 200, 204, 210, 211, 212,
215—223, 271, 293, 294, 295, 300
— средняя 124, 128, 133, 134, 180,
184, 185, 186,196,201,216,256,263,
266, 267—280, 298
----восточносибирская 269, 281, 282
----горносибирская 280, 281, 275
---- сахалинская 269, 282
----северосибирская (тундровая) 281,
282
— тихоокеанская 151
— трансарктическая 123, 124, 127 —
133, 134, 135, 137, 139, 143
----восточносибирская 136, 137, 138
----камчатская 137, 138, 139
— тундряная 113, 184—186, 196, 197,
200, 216,245, 246,249,251,256,257,
271, 276, 277, 295, 296, 298
----восточносибирская 260, 258
----горная (семиреченская) 261
---- забайкальская 260, 262
---- западносибирская 260
---- зауральская 260
---- печерская 261
---- северная 257, 258
— — Строганова 262
— Тянь-шанская 296
Водоплавка 11, 307
Выхухоли 12, 24, 25, 39
Выхухоль 6, 7, 11, 25, 27, 28, 29, 30
— русская 25, 26, 27, 29, 28
Еж 6, 7, 9, 10, 11, 12, 13
— амурский 17
— афганский 13
— дальневосточный 17
— даурский 19, 20, 21, 22, 23
— обыкновенный 14, 15, 16, 17, 42
— ушастый 13, 15, 18, 20, 21, 22, 23
----беловатый 24
----северный 24
1 Полужирным выделены страницы, на которых приводится описание или раз-
вернутая характеристика таксона.
354
Землеройки 7, 10, 11, 12, 110—116,
133, 143
— белозубые 313
— бурозубые 116, 117, 119
— выдровые 11
Златокрот 11
Крот 11, 58, 87, 88
— обыкновенный 34, 35, 41, 42, 43,
102, 105, 107
---- обской 58
----зауральский 59—60
— европейский 39, 40, 41, 46, 47 , 48,
96, 51
— сибирский 38, 40, 42, 45, 47, 48,
50, 51, 56, 60, 61—96, 102, 105
----алтайский 90, 92, 65
---- забайкальский 73, 91, 92
----нарымский 90, 92
----салаирский 65, 90, 91, 92
----саянский 65, 91
— японский горный 39
Кроты 7, 12, 31—41
— настоящие 40, 41
— обыкновенные 41, 42—43
— сибирские 41, 60—61
Кутора 116, 307, 314
— водяная 117
— обыкновенная 308, 309—311
---- западная 311—312
----восточная 312—313
Куторы 116, 307—309
Могера 96—97, 105, 108, 109
— большая 97, 98, 99, 101, 102—НО
— средняя 97, 98—102, 109
Могеры 41, 96, 97
Насекомоядные 5, 6, 7, 10, 61, 116,
121, 122
Прыгунчик 11
Тупайя И
355
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Anourosorex НО, 114
— squamipes 112
Anourosoricini ИЗ, 114, 115
Asioscalops 30, 37, 38, 39, 40, 41, 42, 47,
57, 59, 60, 61, 87,88, 94,96,97,105
.— altaica 33, 34, 40, 41, 42 , 43, 44, 45,
46, 47, 52, 57, 58,61,62,73, 91,101
----altaica 65, 66, 68, 73, 90, 93, 94
----qusevi 66, 69, 73, 90, 91, 92, 93
----tymensis 66, 67, 73, 74, 92, 93
----salairica 65, 66, 68, 73, 90, 91, 92,
93, 94, 95, 96
----sibirica 93, 94, 95, 96
---- subsp. 73
----suschkini 65, 68, 73, 74, 91, 93, 96
Blarina 110, 114
Blarinini 113, 115
Chimarrogale 114
Chodsigoa 110, 111
Chrysochloris 11
Condylura 34, 37
— cristata 32, 37
Condylurinae 38
Crocidura 110, 111, 114, 115, 116, 313,
315, 331, 338
— campus-lincolnensis 331
— corea 324
— lasia 331
— lasiura 313, 315, 317, 321, 322, 324,
325, 328, 329, 330, 331
— leucodon 321, 323
---- ognevi 339
----sibirica 338, 339
— lisenkani 33
— longicaudata 324
— ognevi 336, 339
— sibirica 88, 315, 317, 321, 322, 332,
333, 334
— suaveolens 313, 315, 317, 318, 320,
322, 323, 333,
----ilensis 323, 324
----suaveolens 323, 324
----shantungensis 324
----orientis 324
— thomasi 330
— yamaschinae 331
Crocidurinae 113, 114, 116, 313
Crocidurini 115
Desmana 7, 11, 25, 26, 28
— crassidens 26
— magna 26
— moschata 25, 26, 28
— tegelensis 25, 26
— thermalis 26
Desmanidae 12, 24, 39
Desmaninae 24, 39
Diplomesodon 110, 111, 114, 115
Dominoides 34
Eoscalops 30, 32, 59, 97, 109
— longirotris 32, 34
— micrura 34, 109
Erinaceidea 7, 12, 16, 24, 25
Erinaceinae 12
Erinaceus 14, 15, 16, 18
— auritus 13
— dauricus 13, 19
— europaeus 13, 15, 17
----amurensis 17, 20
— hupoaelas 13
— megalotis 10, 13
— platyotis 18
Euroscaptor 38, 39, 97, 109
— mizura 34
— grandis 38
Eurosorex 122, 153
Galemys 26, 28
golliaph, Praesorex 110
Hemiechinus 11, 15, 18, 19, 20
— alhulus 24
— auritus 13, 20, 21, 23
albulus 24
auritus 24
— dauricus 19, 20
Heterosoricini 113
Insectivora 10
latimanus, Scapanus 88
Macroscelides 11
Microsorex 114
Mogera 34, 38, 39, 40, 41, 59, 88, 96, 97,
98, 100, 109, 110
— hainana 38, 97
— insularis 97
. kobeae 97
— latouchei 38, 97, 98, 102
— robusta 33, 34, 39, 40, 97, 98, 99,
101, 102, 103, 106, 107, 109‘ 110
—tokudea 97
— wogura 34, 39, 40, 97, 98, 100, 101,
102, 109
---- coreana 102
------hainana 97
----robusta 109
moschata, Myogale 25
356
moschatus, Castor 28
Nasillus 34
Nectogale 110
Neomys 11, 110, 111, 114, 115, 116,
307, 309 314
— anomalus 308
— fodiens 35, 117, 308, 309, 310, 312,
313
----fodiens 311, 313
----orientis 311, 312, 313
— schelkovnikovi 308, 309
Neurotrichus 34
Notiosorex 114
Ognevia 122, 144, 151
Otisorex 122, 123, 130, 132, 133, 134,
143, 183, 278
Pachyura (Suncus) 115
Parascalops 34
Parascaptor 34, 38, 59, 97, 109
— grandidens 34
— leucura 34
pilirostris, Dymecodon 34
Podihik 111
Potomogale 11
Proscalopinae 38
Rinchonax 34
Saturniniinae 114
Scalopinae 34, 38
Scalopus 34, 37, 97
— agitations 32, 34, 37
Scapanulus 34
Scapanus 34, 97
— latimanus 88
— orarius 34
Scaptochirus 34, 38, 109
— moschatus 38
Scaptonyx 34
Scutisorex 112
Scutisoricinae 114
Sorex 5, 11, 110, 111, 113, 114, 115,
116, 117, 122,123,124,133,134, 144,
145, 149, 150,152,159,182,183, 195,
199, 200, 205, 239, 266, 278, 279, 286,
296, 297, 314
— alpinus 135, 296, 297, 299
— araneus 114, 117, 119, 120, 121, 130,
131, 142, 144,148,201,204,205,206,
210, 221, 222, 231,234,235,236,237,
241, 246, 256, 275,276,278,282,283,
284, 291, 292, 293, 294, 295, 296, 297,
299, 300, 303, 306
---- araneus 301, 303, 304, 306
— — raddei 299
— — roboratus 222, 237, 300, 301, 303,
304
---- rypheus subsp. n. 304, 305, 307
---- satunini 293, 298, 299
----isodon 210, 222, 301, 306
— — iohanseni 302, 306
----tomensis 293, 294, 301, 302, 303,
305, 306
--------isodon 221, 293, 294, 306
----ultimus 251
----uralensis 302, 306
— arcticus 138, 196, 241, 251, 255,
256, 275, 276, 278, 279, 294
----lariocum 254, 255, 275
----schnitnicovi 293, 294, 299, 303
— asper 231, 241, 294, 296, 297
— bendirii 152
— beringianus 119, 120, 122, 123, 124,
128, 131, 133, 140,141 ,’142,144,148,
156, 157, 184, 189, 266,
— buchariensis 122, 185
— buxtoni 282
— caecutiens 199, 120, 124, 127, 128,
129,130, 131, 133,134,137,142,144,
146,148, 179, 189, 198,201,204,212,
222,237, 256, 263,264,266,270,274,
275,276, 277, 278,279,280,282,286,
294, 298
----buxtoni 279, 282
----caecutiens 269, 271, 272, 275, 276,
277, 278, 279, 280, 281
----koreni 269, 271, 272, 275, 276,
277, 279, 280, 281, 282
----maeropygmaeus 269, 270, 272, 273,
275, 276, 277, 279, 281
----pleckei 269, 271, 272, 275, 276,
277, 279, 280, 281
----shinto 269, 271, 273, 275, 276,
277, 279, 280, 281, 282
— caucasicus 210, 293, 297, 298, 299
— centralis 221, 222, 278, 279
— cinereus 119, 120, 128, 129, 130, 131,
132, 133, 135, 136,137,139,141,142,
143, 144, 148, 180,183,184,185,277
----camtschatica 127, 128, 129, 135,
136, 137, 138, 139, 140
----cecutinoides 135
----partenkoi 128, 129, 131, 133, 135,
136, 140
— daphaenodon 119, 120, 121, 122, 123,
124, 127, 130, 131,141,142144,146,
147, 148, 149,156,185,186,187,189,
193, 198, 199,204, 205, 210,234,266,
286, 294
---- daphaenodon 194, 197, 199
----sanguinidens 195, 196, 197, 198,
199
----scaloni 195, 198, 199
----sihiriensis 196
----orri 198
— exilis 165
— fodiens 307
— fumeus 135
— gemalus 301
— gracillimus 119, 120, 124, 130, 131,
141, 142, 144, 148,175,180,182,183,
184, 185, 200, 223, 224, 226, 227, 228,
229, 230, 231, 266, 267, 286
— gravesi 221
— hydrodromus 138, 139
— isodon 119, 120, 130, 131, 142, 144,
146, 148, 200,201,204,205,206,209,
210, 211, 212,213,216,217,221,222,
357
234, 237, 241,278,283,287,293, 294,
299, 300, 306
-----isodon 221, 223
-----princeps 223, 306
--------montanus 306
----- rutenus 222, 223, 306
— kozlovi 122, 185
— leucodon 313
— longirostris 142
— lyelli 133, 142, 227
— macropygmaeus 274, 275
-----aranoides 275, 278, 279
— — altaicus 275, 279
----- koreni 275, 279
-----macropygmaeus 275, 279
-----pleskei 275, 279
----- rozanovi 275, 279
— mirabilis 119, 120, 122, 130, 131,
141, 147, 148, 150,152, 166,201,231
-----kutscheruki 152
-----mirabilis 152
— minimus 165
— minutissimus 119, 120, 121, 122, 128,
130, 131, 141, 142, 144, 148,153,154,
156, 157, 158,165, 166,167,168,169,
175, 266, 274
-----abnormis 163, 164, 168, 170, 171,
173, 184
-----barabensis 157, 163, 168, 169, 173,
178, 184
-----caudata 157, 163, 164, 168, 170,
171, 173, 178
-----minutissimus 157, 162, 167, 171,
173, 178
-----stroganovi 157, 165, 168, 170, 171,
172, 173, 178, 184
-----tscherskii 157, 174, 178
— — tshuktschorum 157, 165, 169, 170,
171, 172, 178, 184
-----ussuriensis 169, 171, 173, 174
— minutus 119, 120, 122, 124, 128,
129, 130, 131,133,134,135,142,144,
148, 153, 157,165,167,174,176,179,
180, 181, 182,183,184,185,206,223,
226, 227, 229,231,237,266,274,276
— — gracillimus 182, 223, 230
— — minutus 184, 185
-----portenkoi 183
-----thibetanus 185
— mirabilis 130, 131, 144, 145, 146,
149, 150, 152, 157, 201, 223, 231
-----kutscheruki 152
— obscurus 210; 221
-----volnuchini 184
— pacificus 149, 151, 152, 301
-----mirabilis 152
— planiceps 185
— prcblei 133, 277
— pygmaeus 174, 182, 274
— pribilo fensis 138
— raddei 210, 231, 294, 296, 297, 298, 299
— roboratus 119, 120, 130, 131, 142,
144, 148, 149, 166,167,200,204,231,
232, 234, 235, 236,237,238,239,241,
242, 244, 245, 294, 299, 300, 307
-----platicranius 241, 243, 244
----- roboratus 238, 239, 242, 243, 245
-----subsp. 242, 243
-----thomasi 238, 241, 243, 245
----- vir 238, 240, 241, 243, 244
— sanguinidens 196, 197
— shinto 212, 275, 279
-----shinto 212
----- saevus 212
— sibiriensis 196
— thibetanus 185
— tsherskii 165
— tundrensis 119, 120, 124, 130, 131,
142, 144, 148,149,150,184,189,196,
200, 204, 210,234,235,241,245,247,
250, 251, 252,253,254,256,257,258,
282, 298, 302
-----baicalensis 250, 251, 255, 260,
262
-----borealis 253, 257, 261
-----buxoni 250, 251, 253, 258
— — petschorae 250, 251, 253, 261
-----schnitnikovi 254, 261
-----sibiriensis 250, 251, 252, 260, 261
-----stroganovi subsp. n. 251, 255, 262
-----transrypheus 251, 253, 260
-----transuralensis 199
— unguiculatus 119,120, 123, 128, 130,
131, 142, 144,148,149,181,200, 201,
202, 204, 206,207,208,209,210, 211,
212, 215, 221,222,223,234,237,266,
283, 294, 299, 300
— ussuriensis 165, 174
— vagrans 122, 149, 151, 152
— vir 166, 237, 241, 299, 304, 307
— volnuchini 184, 185
Soricidae 7, 12, 25, 110, 116, 295
Soricinae 113, 114, 115, 116, 117
Soricini 113, 114, 115
Soricoidea 114
Soriculus 110
Stroganovia 121, 122, 123, 185, 196
Suncus 110, 114, 115
— etruscus 10, 110
Talpa 11, 30, 34, 38, 39, 40, 41, 42, 43,
47, 48, 56, 57, 58, 59, 61, 64, 88, 94,
96,97,101, 105, 109
— altaica 34, 39, 40, 57, 58, 60, 90
— — salairica 90, 95
-----sibirica 90, 94, 95
-----suschkini, var. 90
-----tymensis 90
— caucasica 34, 39, 40, 47, 48, 50, 52,
57, 97
— coeca 34, 38, 39, 40, 47, 48, 50, 57,97
— еигораеа 32, 33, 38, 39, 40, 41, 42,
43, 45, 47, 50, 52, 53, 54, 56, 57, 58,
60, 90, 97, 101
-----brauneri 58
----- еигораеа 43, 54, 58, 60
-----obensis 55, 58
-----transuralensis 52, 53, 55, 57, 58,
59
— — uralensis 52, 54, 55, 57, 58, 59
358
— hercegovinensis 38, 40, 50, 57, 97
— ingularis 38
— klossi 38
— leucura 38
— levantis 34, 40, 48, 50, 57, 97
— longirostris 38
— micrura 38
— minima 38, 47
— minuta 34, 40, 47
— mizura 38, 39
— olympica 38, 39
— orientalis 40
— parvidens 38
— romana 34, 40, 48, 50, 97
---stancovici 50
— wogura 38, 96
-----robusta 101, 109
Talpidae 7, 12, 25, 30, 32, 34, 37, 39, 60,
61, 64, 90, 102, 109, 111
Talpinae 31, 34, 37, 39, 40, 41, 97, 109
Talpini 38
Tupaia 11
Uropsilinae 34, 38, 60
Uropsilus 30, 34
— doricipes 32
Urotrichinae 25, 34, 38, 60
Urotrichus 30, 34
— talpoides 32, 34
359
ОГЛАВЛЕНИЕ
ОТ РЕДАКТОРА............................................................ 5
ВВЕДЕНИЕ................................................................ 7
ОТРЯД INSECTIVORA BOWDICH, 1821 — НАСЕКОМОЯДНЫЕ ....................... 10
Определительная таблица семейств.................................... 12
Семейство Erinaceidae Fischer von Waldheim, 1817 — Ежи .... —
Подсемейство Erinaceinae Fischer von Waldheim, 1817.................. —
Определительная таблица родов сем. Erinaceidae...................... 14
Род Erinaceus Linnaeus, 1758 — Ежи обыкновенные................ 15
Erinaceus europaeus Linnaeus, 1758 — Обыкновенный еж......... —
Род Hemiechinus Fitzinger, 1866 — Ушастые ежи.................. 18
Определительная таблица видов рода Hemischinus.................... 19
Hemiechinus dauricus Sundvall, 1841 — Даурский еж........... 20
Hemiechinus auritus Gmelin, 1770 — Ушастый еж............... 23
Семейство Desmanidae Gray, 1821 — Выхухоли........................ 24
Подсемейство Desmaninae Thomas, 1912............................... —
Род Desmana Giildestaedt, 1777 — Выхухоль...................... 28
Desmana moschata Linnaeus, 1758 — Русская выхухоль........... —
Семейство Talpidae Fischer von Waldheim, 1817 — Кроты .... 30
Подсемейство Talpinae Fischer von Waldheim, 1817 — Настоящие кроты 41
Определительная таблица родов подсем. Talpinae.................
Род Talpa Linnaeus, 1758 — Кроты обыкновенные.................. 42
Talpa europaea Linnaeus, 1758 — Крот обыкновенный........... 43
Род Asioscalops Stroganov, 1941 — Кроты сибирские ............. 60
Asioscalops altaica Nikolsky 1883 — Сибирский крот.......... 61
Род Mogera Pomel, 1848 — Могеры................................ 96
Определительная таблица видов рода Mogera......................... 98
Mogera wogura Temminck, 1833 — Средняя могера —
Mogera robusta Nehring, 1891 — Большая могера.............. 102
Семейство Soricidae Fischer von Waldheim, 1817—Землеройки . . . 110
Определительная таблица подсемейств и родов сем. Soricidae....... 116
Подсемейство Soricinae Fischer von Waldheim, 1817—Буроэубые землеройки 117
Род Sorex Linnaeus, 1758 — Бурозубки............................ —
Определительная таблица подродов рода Sorex...................... 122
Подрод Otisorex De Kay, 1842 ................................. 123
Определительная таблица видов подрода Otisorex..................... —
Sorex cinereus Kerr, 1792 — Трансарктическая бурозубка .... 124
Sorex beringianus Yudin, 1967 — Берингийская бурозубка .... 140
Подрод Ognevia Dolgov et Heptner, 1967 ....................... 144
Sorex mirabilis Ognev, 1937 — Гигантская бурозубка........... —
Подрод Eurosorex Stroganov, 1952 ............................. 153
360
Определительная таблица видов подрода Eurosorex . ................... 153
Sorex minutissimus Zimmermann, 1780 — Крошечная бурозубка ... —
Sorex minutus Linnaeus, 1766 — Малая бурозубка................. 174
Подрод Stroganovia Yudin, subsp. n................................ 185
Sorex daphaenodon Thomas, 1907 — Крупнозубая бурозубка ... —
Подрод Sorex Linnaeus, 1758 ...................................... 199
Определительная таблица видов подрода Sorex.......................... 200
Sorex unguiculatus Dobson, 1890 — Когтистая бурозубка . . . 201
Sorex isodon Turov, 1924 — Равнозубая бурозубка............... 212
Sorex gracillimus Thomas, 1907 — Дальневосточная бурозубка . . 223
Sorex roboratus Hollister, 1913 — Бурая бурозубка............. 231
Sorex tundrensis Merriam, 1900 — Тундряная бурозубка......... 245
Sorex caecutiens Laxmann, 1788 — Средняя бурозубка............ 263
Sorex araneus Linnaeus, 1758 — Обыкновенная бурозубка .... 282
Род Neomys Каир, 1829 — Куторы, или водоплавки.................... 307
Neomys fodiens Pennant, 1771 — Обыкновенная кутора............. 309
Подсемейство Crocidurinae Milne-Edwards, 1868—1874 —Белозубые зем-
леройки ............................................................. 313
Род Crocidura Wagler, 1832 — Белозубки.............................. —
Определительная таблица видов рода Crocidura........................ 315
Crocidura suaveolens Pallas, 1811 — Малая белозубка.............. —
Crocidura lasiura Dobson, 1980 — Большая белозубка............. 324
Crocidura sibirica Dukelsky, 1930 — Сибирская белозубка .... 332
СПИСОК ЛИТЕРАТУРЫ......................................................... 340
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ . . 353
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ.............................................. 355
Научное издание
Юдин Борис Степанович
НАСЕКОМОЯДНЫЕ
МЛЕКОПИТАЮЩИЕ
СИБИРИ
Редактор издательства В. В. Болковинцер
Художественный редактор В. В. Седунов
Художник Н. А. Пискун
Технический редактор Т. Н. Драгун
Корректоры Л. Л. Михайлова, Е. Н. Зимина
ИБ № 34370
Сдано в набор 02.08.88. Подписано в печать 28.03.89.МН-01426.
Формат 70х100'/1». Бумага книжно-журнальная. Обыкновенная
гарнитура. Высокая печать. Усл. печ. п. 29,34-вкл. на мел.
бумаге. Усл. кр.-отт.29,4. Уч.-изд. л. 29,7. Тираж 1250 экз.
Заказ ЗМ» 288. Цена 6р. 30 к.
Ордена Трудового Красного Знамени издательство «Наука»,
Сибирское отделение. 630099 Новосибирск, ул. Советская, 18.
4-я типография издательства «Наука». 630077 Новосибирск,
ул. Станиславского, 25.