














































































































































































































































































Author: Фурса Т.Б. Филов А.И.
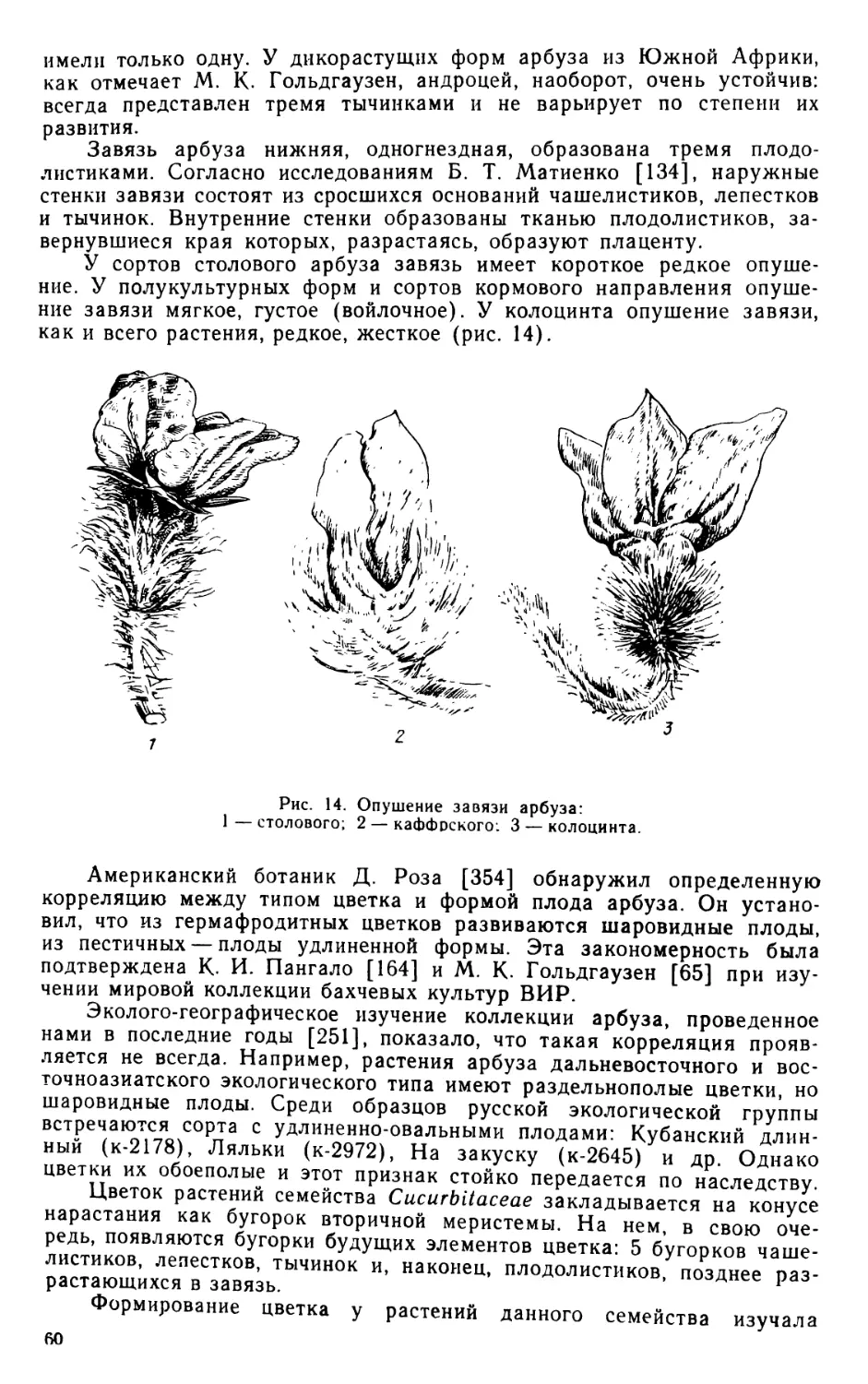
Tags: растения со съедобными семенами и плодами бобовые растения ботаника флора растениеводство культурные растения флора ссср
Year: 1982




FLORA OF CULTIVATED PLANTS
Published under the general editorship of
D. D. Brezhnev, Academician of VASKHNIL,
Hero of Socialist Labour of The USSR
T. B. FURSA, A. I. FILOV
XXI
CUCURBITACEAE
(Cifrullus, Cucurbita)
Editors of the volume
r ... O’ N. Korovina,
Candidate of biological science
r *' “• Fursa,
Candidate of biological science
moskow
KOLOS» 19^2
КУЛЬТУРНАЯ ФЛОРА СССР
Под общим руководством академика
ВАСХНИЛ, Героя Социалистического
Труда Д. Д. Брежнева
Т. Б. ФУРСА, А. И. ФИЛОВ
XXI
ТЫКВЕННЫЕ
( арбуз. 1ыква)
Редакторы тома
кандидаты биологических наук
О. Н. Коровина, Т. Б. Фурса
МОСКВА «КОЛОС» 1982
УДК 635.615/618 + 635.62
Фурса Т. Б., Филов А. И. Культурная флора СССР. Т. 21. Тыквенные. — М.: Ко-
лос, 1982, 279 с.
Настоящий том посвящен двум родам семейства тыквенных: арбузу (Citrullus)
и тыкве (Cucurbita).
В разделе «Арбуз» дано ботаническое описание рода Citrullus и его видов, при-
ведены предложенные автором ботанические классификации столового, кормового ар-
буза и колоцинта, используемого в медицине. Указаны биологические свойства арбуза,
освещены история его культуры, достижения селекции.
В разделе «Тыква» приведена новая ботаническая классификация возделывае-
мых видов, изложены история их изучения и распространения, биологические особен-
ности.
Наряду со сводкой литературных данных в книге представлены оригинальные
исследования авторов по биологии названных культур, являющиеся результатом мно-
голетнего изучения коллекций в полевых условиях.
Книга рассчитана на селекционеров, научных работников, преподавателей и уча-
щихся высших и средних учебных заведений сельскохозяйственного и биологического
профиля.
Таблиц — 35, иллюстраций — 62, библиография — 596 названий.
Одобрена и рекомендована для издания бюро Отделения растениеводства и се-
лекции ВАСХНИЛ.
3803080300—288
038(01 )-82 83-82
© Издательство «Колоо, 1982
ПРЕДИСЛОВИЕ
многолаетТнеЯг^И1лямИ?УЛЬТУРНОЙ флоры СССР является продолжением
многолетнего издания, инициатором которого был академик Н. И. Ва-
вил предвоенные годы под его руководством вышло 7 томов.
После длительного перерыва выпуск Культурной флоры СССР был
возобновлен под общим руководством академика ВАСХНИЛ П. М. Жу-
ковского, а затем академика ВАСХНИЛ [д, Д, Брежнева") В последние
годы среди других были выпущены два тома по овощным культурам:
«Корнеплодные растения» (1971) и «Лук» (1978). Третий том по на-
званной группе культур — «Тыквенные» — посвящен арбузу и тыкве,
имеющим важное народнохозяйственное значение. Относящиеся к тому
же семейству огурец и дыня, также широко распространенные в СССР,
будут рассмотрены в другом томе.
Раздел «Арбуз» подготовлен кандидатом биологических наук
Т. Б. Фурса, раздел «Тыква» — доктором биологических наук А. И. Фи-
ловым. Оба автора — сотрудники Всесоюзного научно-исследователь-
ского института растениеводства им. Н. И. Вавилова, длительное время
изучавшие указанные культуры. Раздел «Кариотип и методы его изуче-
ния» написан на основе материала кандидата биологических наук
Г. А. Шелепиной.
Ценность книги заключается в том, что такое полное монографи-
ческое описание названных выше культур публикуется впервые.
Ни в нашей стране, ни за рубежом не издавались подобные моногра-
фии. Наряду с полной сводкой литературных данных в книге представ-
лены оригинальные исследования авторов по систематике, филогении,
биологии цветения, биологии развития, анатомии, физиологии и другим
аспектам.
Особо следует отметить новые ботанические классификации арбуза
и тыквы, разработанные в соответствии с международными правилами
ботанической номенклатуры на основании многолетнего изучения кол-
лекций в полевых условиях и критического пересмотра ранее предло-
женных классификаций.
Представлена также эколого-географическая классификация ар-
буза, которая позволила вскрыть его популяционную структуру, про-
следить пути распространения по странам земного шара и установить
определенные закономерности экологической изменчивости.
5
систему, предложенную Нодэном, но классификации их базировались
в основном на морфологических критериях. Следует отметить, что си-
стема Пакса и Мюллера, основанная преимущественно на строении
андроцея, и в настоящее время признается многими ботаниками. По-
следователем этого направления является Д. Хатчинсон [312].
Современный период в развитии систематики тыквенных знаме-
нуется появлением классификации Ч. Джеффри [315, 316], принципи-
ально отличающейся от прежних построений. Помимо морфологических
критериев, Джеффри использовал новейшие данные, полученные в об-
ласти анатомии, эмбриологии, цитологии и палинологии тыквенных рас-
тений.
В создании его системы большую роль сыграли исследования
А. Циммермана [378] и X. Чакраварти [286] по эволюционной анато-
мии и морфологии цветка, В. Пури [349] по изучению тонких структур
андроцея и генецея. Богатый материал дали сравнительно-анатомиче-
ское изучение строения семенной кожуры у разных родов тыквенных
Б. Сингхом [362], В. Н. Сойфером [202], генетические исследования
Т. Вайтекера [375], сравнительно-эмбриологическое изучение семей-
ства В. А. Гуляевым [73], биохимическое изучение плодов и семян
Cucurbitaceae С. Рэмом [352], П. Даннели и Л. Фоуденом [295] и,
наконец, палинологические исследования, проведенные в последние
годы Ч. Мартикорена [333], Л. А. Алешиной [6].
Проанализировав данные названных авторов и дополнив их био-
логическим изучением живых коллекций, Джеффри счел возможным
критически пересмотреть существующие системы и предложил новую
классификацию, более полно отражающую естественные филогенетиче-
ские связи внутри обширного полиморфного семейства Cucurbitaceae.
Согласно классификации Ч. Джеффри [315], Cucurbitaceae подраз-
деляется на 2 подсемейства: Cucurbitoideae, включающее 8 триб,
18 подтриб и 85 родов, и Zanonioideae, включающее 18 родов, объеди-
ненных в одну трибу.
Роды Citrullus Schrad, Cucurbita L, Lagenaria Ser, Luffa Adans,
виды которых возделываются в СССР, первоначально были отнесены
им к трибе Cucurbiteae, так же как в системе Пакса и Мюллера, а род
Cucumis L. к трибе Melothrieae. После тщательного анализа новых
палинологических данных, которым Джеффри придает решающее зна-
чение в таксономии, в предложенную систему были внесены некоторые
уточнения. Подтрибы Benincasinae и Cucurbitinae возведены в ранг
триб, отдельные подтрибы, например Acanthosicyoinae. упразднены
В последней классификации Джеффри [316] роды Citrullus. Lagenaria
и Luffai отнесены к трибе Benincaseae Ser, Cucurbita — к трибе Сисиг-
biteae Ser, Cucumis — к трибе Melothrieae Endl.
АРБУЗ—CITRULLUS SCHRAD.
Schrader 1836, in Ecklon et Zeyh., Enum Pl. Afr. Austr. 2:279;
id 1838, Linnaeae, 12:412; id,, 1953, Taxon 2, 4:99, nom. conserv.—
Anguria Mill. 1754, Gard. Diet. Abridg. ed. 4. — Citrullus Necker, 1790,
Elem. 1 : 240. — Colocynthis Mill. 1754, Gard. Diet. Abridg. ed. 4.
Typus: Citrullus vulgaris Schard. ( = C. lanatus (Thunb.) Matsum.
et Nakai).
ОБЗОР РОДА
Род Citrullus относится к трибе Benincaseae Ser., подтрибе Be-
nincasinae (Ser.) C. Jeffr. В эту подтрибу включены еще 11 родов:
Benincasa Savi, Bryonia L., Ecballium A. Rich., Acanthosicyos Hook, f.,
Praecitrullus (Stocks) Pang., Diplocyclos (Endl.) Post et Kuntze, La-
genaria Ser., Coccinia Wight et Arn., Cogniauxia Bail!., Eureiandra
Hook, f., Raphidiocystis Hook. f.
Название Citrullus впервые употребил Форскал в 1775 г., однако
он не дал диагноза рода. Это сделал позже X. Шрадер, ставший един-
ственно законным автором рода Citrullus. Название Citrullus Schard.
было узаконено Восьмым международным ботаническим конгрессом в
1954 г. и включено в список сохраняемых названий — nomina conser-
vanda (III приложение к Международному кодексу ботанической но-
менклатуры, 1959).
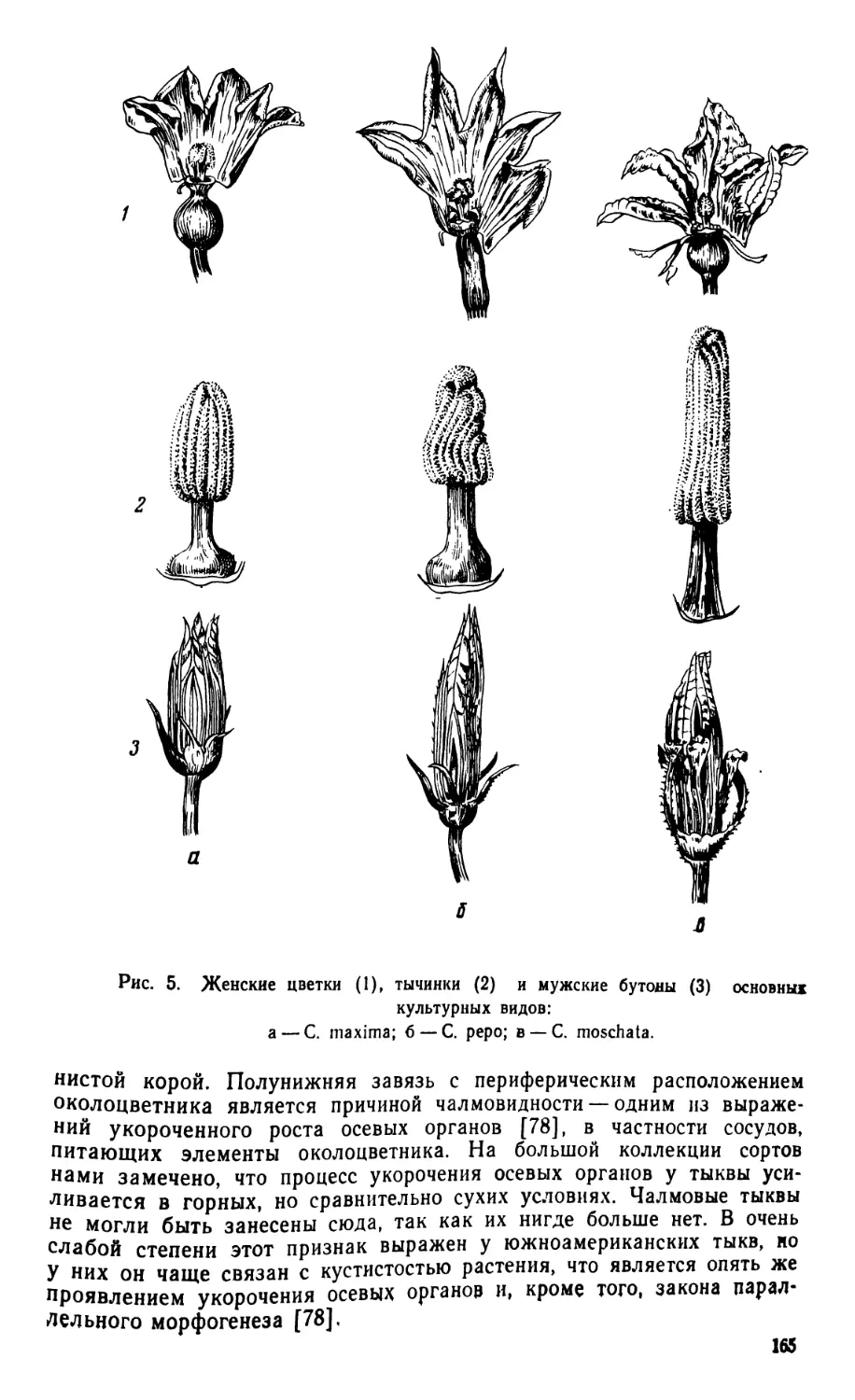
Растения однолетние или многолетние, стелющиеся. Стебель раз-
ветвленный, с мягким или жестким опушением, 2—5-раздельными уси-
ками. Листья обратнояйцевидной формы, трехнервные, рассеченные,
очень редко цельнокрайные. Цветки одиночные, раздельнополые или
обоеполые. Чашелистики «и лепестки срастаются у основания. Венчик
желтый, 5-лепестный, тычинок 5, из них 4 сросшиеся попарно, одна
свободная. Рыльце пестика 3-лопастное, завязь нижняя, 3-гнездная.
Плод — тыквина, многосемянный, сочный, не отпадает от плодоножки.
Семена удлиненные, с твердой кожурой. Число хромосом 2п = 22.
Происходит из Африки. Распространен в тропических и субтропи-
ческих областях земного шара. Представлен дикорастущими, полукуль-
турными и возделываемыми формами.
ЭВОЛЮЦИЯ, РАСПРОСТРАНЕНИЕ И ЭКОЛОГИЯ
Эволюция рода. При установлении филогенетических связей и сте-
пени родства различных форм очень важно правильно выбрать кри-
терий оценки.
В современной систематике наряду с морфологическими призна-
ками большое таксономическое значение придается строению пыльцы.
Использование палинологических данных позволило монографу Сисиг-
bitaceae Ч. Джеффри [315, 316] внести в классификацию тыквенных
9
растений существенные изменения, более точно отражающие филогене-
тические связи между различными группами семейства.
Для выяснения филогенетических связей Citrullus с другими ро-
дами Cucurbitaceae мы обратились прежде всего к исследованиям
Л. А. Алешиной по морфологии пыльцы тыквенных [6, 7, 8], дополнив
их данными по эмбриологии [72, 73], анатомии плода [135, 137] и се-
мян [202], строению нектарников цветка [150] различных представи-
телей Cucurbitaceae, Сопоставление результатов, полученных разными
исследователями, дало возможность сделать некоторые выводы отно-
сительно характера эволюции и связи Citrullus с другими родами се-
мейства.
Трибу Benincaseae Ser., к которой относится арбуз, можно считать
достаточно древней, так как она объединяет роды, встречающиеся в
Новом и Старом Свете. Большинство родов подтрибы Benincasinae
сосредоточено в Африке и на Мадагаскаре, некоторые встречаются
в Азии. Среди представителей этой подтрибы можно выделить роды,
обнаруживающие большее пли меньшее сходство с Citrullus. По строе-
нию пыльцы наиболее близки к нему роды Acanthosicyos Hook. f. и
Eureiandra Hook. f. из Западной Африки [8]. Сходство в строении
пыльцы послужило Джеффри основанием отнести один из видов Cit-
rullus— С. naudinianus (Sond.) Hook. f. к роду Acanthosicyos [315].
Палинологические данные позволяют сближать Citrullus и с родом
Benincasa Savi, распространенным на Мадагаскаре и в Восточной Азии.
Л. А. Алешина [8] отмечает, что Citrullus является как бы свя-
зующим звеном между подгруппами Bryonia и Benincasa палингруппы
Bryonia, Сходны роды Citrullus и Benincasa также по типу нектарников
[150], гистологическому строению плода [135, 137], анатомии кожуры
семян [202]. У того и другого рода встречаются семена с рубчиком и
без рубчика, причем семена бенинказы, не имеющие рубчика, об-
наруживают большое морфологическое сходство с семенами С. naudi-
nianus— наиболее примитивного вида Citrullus. Анатомическое строе-
ние кожуры семян без рубчика отличается более слабой дифференциа-
цией слоев, что свидетельствует о большей древности вида. Сходство
анатомии и морфологии семян Benincasa и Citrullus заставляет пред-
полагать какую-то общность в эволюции этих родов.
По анатомии плода и семян, а также в какой-то мере по строению
пыльцы обнаруживается близость Citrullus и с родом Lagenaria, про-
исходящим из Африки, но широко расселившимся в далекие времена
по всему земному шару. Таким образом, сходство родов Citrullus,
Acanthosicyos, Eureiandra, Benincasa и Lagenaria по ряду важных так-
сономических признаков дает основание говорить об их филогенетиче-
ской близости и предполагать происхождение от общего корня.
Нельзя оставить без внимания входящий в ту же подтрибу род
Praecitrullus, поскольку само название говорит о его родстве с Citrul-
lus. Мысль о том, что этот индийский эндем является предком Citrullus,
высказал К. И. Пангало [165]. Однако дальнейшие исследования ‘[8,
72, 361] убедительно доказали ошибочность такого мнения.
Praecitrullus — очень древний, возможно, даже верхнемеловой ре-
ликт, эволюция которого шла, однако, совсем иными путями, чем эво-
люция Citrullus. В. А. Гуляев [72] отмечает, что зародышевый мешок
его развивается не по типу Polygonum, характерному для Cucur-
bitaceae, а по АШит-типу. Эндоспермальные гаустории по форме и раз-
мерам сильно отличаются от таковых у Citrullus. Пыльца его имеет
шиповатую экзину, свойственную примитивному роду Sicyos, распро-
страненному в Австралии, Полинезии и Америке, а также эволюционно
продвинутому американскому роду Cucurbita.
Ю
Можно предполагать, что дифференциация Praecitrullus началась
еще в те далекие геологические времена, когда Азия и Америка имели
тесную связь между собой, т. е. в конце мелового периода. Л. А. Але-
шина [8] отмечает, что палингруппа Praecitrullus — промежуточное
звено в развитии палингруппы Cucurbita. Возможно, Praecitrullus за-
нимает промежуточное положение и в более широком эволюционном
аспекте — между Cucurbitaceae Старого и Нового Света. Заблуждение
относительно его родства с Citrullus возникло, вероятно, на основе
большого морфологического сходства семян этих родов.
О древности подтрибы Benincasinae свидетельствует также род
Diplocyclos, распространенный в Африке, Юго-Восточной Азии и Ав-
стралии. По данным Л. А. Алешиной 1[8], он занимает промежуточное
положение между палингруппами с шиповатой и сетчатой экзиной
пыльцы.
Таким образом, сопоставление данных палинологии, эмбриологии
и анатомии дало возможность выявить в подтрибе Benincasinae роды
Acanthosicyos, Eureiandra, Benincasa, наиболее близкие к Citrullus,
окончательно исключить Praecitrullus из числа его предков и, наконец,
показать древность этой подтрибы. Последнее обстоятельство наводит
на мысль, что предковые формы Citrullus следует искать среди более
примитивных групп Cucurbitaceae, а именно в трибе Fevilleae [315].
По строению пыльцы последняя неоднородна [7]: часть входящих в
нее родов имеет пыльцевые зерна с сетчатой экзиной, другие — с поло-
сатой. У некоторых представителей этой трибы, например подрода
Microlagenaria С. Jeffr. рода Thlandiantha Bunge и в какой-то мере
рода Gerrardanthus Harv., пыльца сходна с пыльцой Citrullus.
Плод Gerrardanthus по анатомическому строению отнесен Б. Т. Ма-
тиенко [136] к гистологическому типу Citrullus. Это один из немногих
представителей Cucurbitaceae, имеющий высыхающие плоды типа ко-
робочки, более древние в эволюционном отношении, чем сочные плоды
типа тыквины. Б. Т. Матиенко считает, что склереидный слой, свой-
ственный плодам Citrullus, можно рассматривать как остатки механи-
ческой ткани, хорошо развитой в плодах типа коробочки. Следует от-
метить, что гистологическое строение плода Citrullus отличается слабой
специализацией, что свидетельствует о его малой эволюционной про-
двинутостк.
Таким образом, данные по морфологии пыльцы и анатомии плода
Citrullus дают возможность считать этот род довольно древним и эво-
люционно мало продвинутым. Есть основания предполагать, что Cit-
rullus филогенетически связан с такими примитивными представителями
Cucurbitaceae, как Thladiantha, особенно Т. africana С. Jeff, и, воз-
можно, Gerrardanthus. Следует отметить, что некоторые ботаники пы-
таются сблизить Citrullus с Cucumis L. и найти связующее звено между
ними: К. И. Пангало [166] называет такой формой Praecitrullus,
А. Д. Меузе [334]— Citrullus naudinianus. По строению пыльцы эти
роды не обнаруживают филогенетической близости: Cucumis можно
считать эволюционно более продвинутым и молодым родом, чем Cit-
rullus.
Экология и распространение. Предки арбуза, так же как и других
тыквенных растений, были мезофитами и имели форму лиан [44, 100].
Черты лиан проявляются и у современных представителей Cucur-
bitaceae. Им свойственны тонкие эластичные стебли с полостью, круп-
ные листья, развитые усики, слабое развитие механической ткани и
хорошее развитие лубяных волокон. Проводящие сосуды стебля круп-
ные, приспособленные для подачи воды на большую высоту.
11
О мезофильном характере предков арбуза свидетельствуют и встре-
чающиеся среди культурных сортов цельнолистные формы, обнаружен-
ные нами также у полукультурных форм арбуза из Судана. Этот ре-
ликтовый. архаический признак в последние годы широко применяется
в селекции гетерозисных гибридов арбуза. Генетическая устойчивость
и рецессивный характер наследования позволяют использовать нерас-
сеченный лист в качестве сигнального признака для выявления в Fi
негибридных форм.
Климат на земле в конце мелового и начале третичного периода
был влажным и умеренно теплым. Эоцен и начало олигоцена ознаме-
новались значительным потеплением и уменьшением влажности в сред-
них широтах [114]. Здесь развивается субтропическая растительность,
местами принимающая ксерофитный характер. Появляются травянистые
равнины и саванны. На смену мезофильным лианам тропического леса
приходят ксерофиты, заселяющие саванны и пустыни. Видимо, в это
время и начинает формироваться современный облик Citrullus.
Ареалы и эволюция видов. Род Citrullus насчитывает 4 вида:
С. lanatus (Thunb.) Matsum. et Nakai, C. colocynthis (L.) Schard.,
C. ecirrhosus Cogn., C. naudinianus (Sond.) Hook. f. Современный ареал
наиболее полиморфного из них С. lanatus (шерстистого арбуза) про-
стирается в границах Судано-Анголезской флористической области,
охватывающей почти всю Африку, южную часть Аравии, Иран, Афга-
нистан, западный Пакистан; С. ecirrhosus (безусиковый арбуз) сосре-
доточен в пустыне Намиб, там же преимущественно встречается и
С. naudinianus (арбуз Нодэна). Вид С. colocynthis (колоцинт) рас-
пространен в основном в Сахаро-Гобийской флористической области
лишь местами внедряясь в пределы Судано-Анголезской области
(рис. 1).
Рис. 1. Распространение дикорастущих видов Citrullus Schrad:
1—Сахаро-Гобийская флористическая область; 2 — Судано-Анголезская;
3 — область Намиба-Карру; а — С. laffatus; б — С. colocynthis; в — С. ecirrho-
sus; г — С. naudinianus.
12
Таким образом, род Citrullus в диком состоянии сосредоточен пре-
имущественно в Африке, лишь частично заходя в Азию. Правда, один
из видов — С. colocynthis встречается и в Австралии. Но попал он
туда, вероятно, сравнительно недавно. Легкие сухие плоды колоцинта
могли быть занесены из Индии волнами океана. Таким же путем, воз-
можно, распространилась по земному шару и лагенария, происходящая
из Африки.
Можно предполагать, что древний ареал Citrullus был также со-
средоточен в Африке и именно там происходила основная дифферен-
циация этого рода. Видимо, Африку можно считать родиной многих
тыквенных, так как здесь зарегистрировано большое число эндемичных
видов данного семейства. Среди видов Citrullus эндемами Африки яв-
ляются С. naudinianus и С. ecirrhosus. Первый из них встречается
в Анголе, Намибии, Ботсване, Мозамбике, Южно-Африканской Респуб-
лике. Ареал второго ограничен районом пустыни Намиб.
С. naudinianus — ярко выраженный ксерофит, приспособившийся
к существованию в пустыне. Он имеет многолетний клубневой корень,
сильно рассеченные листья, усики, редуцированные до шипов, мелкие
бугорчатые плоды. Характерная особенность С. naudinianus — двудом-
ность, несвойственная другим видам Citrullus. Кожура семени пред-
ставлена у него однородной тканью, состоящей из каменистых клеток.
Морфологические, анатомические и другие особенности этого вида сви-
детельствуют о его примитивности и слабой эволюционной продвину-
тости. Видимо, С. naudinianus обособился в самом начале дифферен-
циации Citrullus и дальнейшая его эволюция шла по пути специализа-
ции, приспособления к пустынным условиям. О ранней обособленности
С. naudinianus свидетельствует и его слабая перекрестная совмести-
мость с другими видами Citrullus [361]. Своеобразие и обособленность
дают возможность считать его наиболее древним видом среди Citrullus.
С. ecirrhosus также обладает рядом своеобразных черт. Он при-
способился максимально использовать очень короткий в пустыне Намиб
период дождей благодаря интенсивному росту вегетативной массы и
одновременному раскрытию большого количества женских цветков на
растении. Своим названием вид обязан отсутствию усиков. Однако наш
опыт выращивания этого растения в более влажном климате (на юге
Украины) показал, что С. ecirrhosus образует нормально развитые
усики. Таким образом, отсутствие последних имеет не генетический ха-
рактер, а объясняется неблагоприятными для развития этого признака
экологическими условиями.
С. ecirrhosus, так же как С. naudinianus, плохо скрещивается
с другими видами Citrullus. У гибридов обнаруживаются нарушения
в мейозе, следствием чего является низкая фертильность пыльцы [361].
М. Шимотзума на основании цитологического изучения гибридов, по-
лученных при скрещивании разных видов Citrullus, высказал предпо-
ложение, что в дифференциации этого рода наряду с географической
изоляцией значительную роль играли, видимо, хромосомные пере-
стройки.
Третий дикорастущий вид—С. colocynthis — наиболее эволюционно
молодой и процветающий. Ареал его огромен и простирается от пустынь
Северной Африки через Аравию, Иран, Афганистан в Индию и Австра-
лию. Известен колоцинт и в СССР. Заросли его встречаются в южной
Туркмении по берегам реки Теджен. На месте значительной части
современного ареала колоцинта в палеогене простиралось море, от-
ступившее только в неогене. Можно предполагать, что дифференциация
этого вида Citrullus происходила позже, в конце третичного периода,
когда на месте отступившего моря образовалась пустыня.
13
Об эволюционной продвинутое™ С. colocynthis свидетельствуют
отчетливо выраженная дифференциация слоев кожуры семени и дру-
гие анатомические признаки. Колоцинт легко скрещивается со столовым
аобузом и дикорастущими формами С. lanatus, давая фертильные гиб-
риды. Вид очень полиморфен, особенно его африканские формы, обна-
руживающие большое разнообразие по признакам листа и плода. Азиат-
ские формы колоцинта более однотипны.
По своей экологии колоцинт — типичный ксерофит, хорошо приспо-
собленный к суровым условиям пустыни: растения компактные, с мел-
кими, сильно рассеченными листьями и крепкими усиками, цепляю-
щимися за выступы почвы и соседние кусты. Многочисленные высыхаю-
щие плоды колоцинта легко перекатываются на большие расстояния,
обеспечивая широкое расселение вида.
С. lanatus наиболее полиморфен среди Citrullus. Он включает ди-
корастущие и возделываемые формы арбуза. Благодаря последним
этот вид вышел далеко за пределы Африканского континента и рас-
пространился по всему земному шару. В отличие от дикорастущих
возделываемые формы арбуза известны не только в Старом, но и в
Новом Свете.
Современный ареал дикорастущих форм С. lanatus невелик и к
тому же разорван (см. рис. 1), что свидетельствует о большой древ-
ности этого вида. Со времени путешествия Д. Ливингстона заросли
диких форм арбуза известны в Калахари (ныне Ботсвана), небольшие
очаги встречаются в Танзании, Кении, Судане, а также в Нигерии,
Гане, Сенегале. Дикорастущие формы С. lanatus распространены в
основном в Южной и Восточной Африке. В западной части континента
они встречаются значительно реже.
Западноафриканские представители этого вида весьма своеобразны
по морфологии и анатомии семян, которые по форме и размеру напо-
минают семена тыквы; в плоде они заключены в толстые слизистые
капсулы. Защитный слой семенной кожуры представлен только одним
рядом каменистых клеток в отличие от других форм данного вида,
у которых кожура семени состоит из разнородных тканей. Своеобразие
этих форм С. lanatus дало нам основание выделить их в самостоятель-
ный подвид subsp. mucosospermus — слизистосемянный [252]. Растения
С. lanatus имеют более мезофильный облик: они, как правило, мощные,
с длинными густоопушенными плетями, крупными листьями и плодами,
достигающими иногда 20 кг. В дикорастущем состоянии известны горь-
кие и негорькие (съедобные) формы этого вида. Генетически они
близки между собой и по плодам не всегда различаются.
Предками столового арбуза были, вероятно, съедобные формы
типа tsamma, сохранившиеся до сих пор в Калахари. Но не отсюда
произошли культурные формы его, так как на тысячи километров
вокруг Калахари не было и признаков земледелия. В этих районах
издавна обитали кочевые племена. Ареал свободного произрастания
съедобных форм арбуза был на заре земледелия, несомненно, намного
шире и простирался вдоль берегов Восточной Африки до Судана и
Египта, а возможно, и за пределы континента до Индии [365]. Предки
столового арбуза, или формы «эволюционного перепутья» (по К- И. Пан-
гало), сохранились здесь только потому, что впадина Калахари оказа-
лась как бы естественным заповедником, которого в течение многих
тысячелетий не касались никакие геологические бури. Со времени оли-
гоцена климат Калахари также не претерпел резких изменений, по-
этому и дошли до нас эти реликты третичного периода.
Таким образом, путь эволюции рода Citrullus от примитивных
предковых форм до дифференциации видов и начала эволюции куль-
14
турного арбуза в общих чертах представляется нам в следующем виде
(табл. 1).
Таблица 1. Примерная схема эволюции рода
Эра Период Эпоха Продолжитель- ность Появление родов и видов
Кайнозой- ская Четвертичный Современная Плейстоцен 1 млн. лет Культурные формы Citrullus lanatus Современные виды Citrullus
Третичный Плиоцен Неоген 25—30 млн. лет Citrullus colocynthis
Миоцен I 1 Citrullus ecirrhosus i ж Citrullus lanatus J Citrullus naudinianus 1
Олигоцен Палеоген 30—35 млн. лет Citrullus. Acanthosicyos t
Эоцен _J J 1 1 1 : 1 Thladiantha airicana, Г* 1 j | Eureiandra ( j
Палеоцен i L- Thladiantha
Наиболее примитивным и своеобразным является С. naudinianus.
возникший, вероятно, в самом начале дифференциации рода, когда
признаки видов только начинали формироваться. Эволюция С. naudi-
nianus пошла по пути резкой специализации и не дала новых форм.
Наименее специализирован среди Citrullus С. lanatus.
Можно предполагать, что именно этот вид оказался исходным для
других. В процессе эволюции довольно рано обособился и С. ecirrhosus.
Однако он все же сохранил определенное сходство с С. lanatus по
морфологии плода и семян. Позднее других, в конце третичного пе-
риода, сформировался С. colocynthis.
В третичный период и более позднее время С. lanatus отличался,
вероятно, большим многообразием форм и обширным ареалом. Бла-
годаря высокому полиморфизму он оказался исходным не только для
других видов, но и для культурного арбуза. Термин ancestralis, перво-
начальный, предложенный М. К. Гольдгаузен для форм арбуза из Ка-
лахари, по своей сути очень верно отражает исходное положение
С. lanatus.
КЛАССИФИКАЦИЯ РОДА CITRULLUS SCHRAD.
ИСТОРИЯ ИЗУЧЕНИЯ
Первой попыткой дифференциации Citrullus была классификация
X. Шрадера [355], разделившего этот род на 4 вида: С. vulgaris
15
Schrad., C. caff er Schrad., C. atnarus Schrad., C. colocynthis L. (Schrad.).
Два нз них (C. vulgaris и C. colocynthis) вошли в известную моно-
графию А. Коньо и X. Хармса [290]\ в которой указываются еще два
вида: С. ecirrhosus Cogn. и С. naudinianus (Sond.) Hook. f.
Стокс описал так называемый пустотелый (фистулезный) арбуз —
С. fistulosus Stocks, эндемичный для Индии. Это растение заинтересо-
вало своим своеобразием К- И. Пангало, изучавшего его в посеве.
Оказалось, что по целому ряду признаков фистулезный арбуз выходит
за рамки рода Citrullus. К. И. Пангало [165] выделил его в самостоя-
тельный род Praecitrullus с одним видом Р. fistulosus (Stocks) Pang.,
считая прапращуром современных видов арбуза и в то же время свя-
зующим звеном между Citrullus и Cucumis. По мнению Пангало,
Praecitrullus — реликт той отдаленной эпохи, когда названные роды
еще не обособились. Исследованиями А. Г. Алиевой [9], Т. Кошо [320],
В. А. Гуляева [72, 73], М. Шимотзума [361], В. Н. Сойфера [202],
Ч. Джеффри [316] убедительно доказано, что Praecitrullus по числу
хромосом, их структуре, морфологии пыльцы, строению зародышевого
мешка, кожуры семени и другим ботаническим признакам не имеет
ничего общего с Citrullus и не может рассматриваться как его пред-
ковая форма.
Ч. Джеффри [316] на основании новых палинологических данных
считает Praecitrullus самостоятельным родом, близким к Diplocyclos.
Японский генетик М. Шимотзума придерживается такого же мнения.
Одним из первых ботаников, попытавшимся создать внутривидо-
вую классификацию столового арбуза (С. vulgaris), был Ф. Алефельд
[280]. Однако его классификация не нашла применения. По мнению
К. И. Пангало она представляла просто краткое описание нескольких
сортов, никак между собой не связанных.
Ф. Рэн [350] дал довольно четкую классификацию североамери-
канских сортов арбуза по форме и окраске плода. Выделенные им
группы в современном понимании следует рассматривать как сорто-
типы. Однако эта классификация не могла претендовать на признание,
так как была построена на очень ограниченном материале. Л. Бейли
[282] дифференцировал С. vulgaris на две разновидности: var. lanatus
и var. citroides, но не выделил столовый арбуз, поэтому его классифи-
кация неполная.
К. И. Пангало четко разграничил столовый и кормовой арбуз,
разделив С. vulgaris на два вида: С. aedulis Pang, (затем С. edulis
Pang.) и С. colocynthoides Pang. Однако выделение новых видов было
сделано в нарушение правил Международного кодекса ботанической
номенклатуры. Вид С. edulis Pang, оказался поздним гомонимом, опуб-
ликованным ранее Е. Шпахом (1838). Г. Швайнфурт [357] выделил
разновидность var. colocynthoides среди форм С. colocynthis, возде-
лываемых в одном из оазисов Египта.
Немецкий ботаник Р. Мансфельд вместо С. vulgaris Schrad. ввел
в 1959 г. новый биноминал —С. lanatus (Thunb.) Mansf. Основанием
для такой комбинации послужило более раннее название Momordica
lanata Thunb., данное Тунбергом в 1794 г. Позднее Хара [308] уста-
новил, что японские ботаники Матзумура и Накаи ввели биноминал
С. lanatus раньше Р. Мансфельда и предложил узаконить их авторство.
Р. Мансфельд предложил внутривидовую классификацию С. lana-
tus, согласно которой этот вид подразделяется на три разновидности:
var. lanatus, var. caffer и var. citroides. Первая из них включает дико-
растущие формы арбуза Южной Африки, вторая — культурный сладкий
съедобный арбуз каффрский, третья — кормовые и цукатные формы.
Мы считаем неправильным объединять в одну разновидность арбуз
16
столовый и каффрский. Столовый арбуз имеет иной ареал, отличает
большим разнообразием форм, яркоокрашенной сладкой мякотью
другими морфологическими особенностями. Он, несомненно, заслуж
вает выделения в самостоятельный таксон. Классификация Маг
фельда, по нашему мнению, не отражает всего разнообразия форм д
корастущего и культурного арбуза и потому не может быть приня
полностью.
НОВАЯ КЛАССИФИКАЦИЯ
Мы предлагаем новую классификацию рода Citrullus, разработа
ную на основе многолетнего изучения мировой коллекции арбуза с уч
том систем, предложенных ранее. Коллекция арбуза, имеющаяся ]
Всесоюзном научно-исследовательском институте растениеводсп
им. Н. И. Вавилова (ВИР), собрана более чем из 60 стран. Она н
считывает в настоящее время свыше 2400 образцов возделываемых
дикорастущих форм и охватывает все разнообразие видов данного род
Система рода Citrullus Schrad. представляется нам в следующе
виде:
Citrullus lanatus (Thunb.) Matsum. et Nakai
subsp. lanatus
var. lanatus
var. capensis (Alef.) Fursa
var. citroides (Bailey) Mansf.
subsp. vulgaris (Schrad.) Fursa
var. vulgaris
var. cordophanus (Тег-Avan.) Fursa
subsp. mucosospermus Fursa
var. mucosospermus
var. senegalicus Fursa
Citrullus colocynthis (L.) Schrad.
subsp. colocynthis
subsp. insipidus (Pang.) Fursa
Citrullus ecirrhosus Cogn.
Citrullus naudinianus (Sond.) Hook. f.
Ключ для определения видов
1. Усики развитые, 2—4-раздельные; плоды гладкие......................1
+ Усики короткие, одиночные, шиповидные; плоды бугорчатые............
...................................IV. С. naudinianus (Sond.) Hook. f.
2. Листья негофрированные ............................................Г
+ Листья широкие, гофрированные; плоды угловатые.....................
............................................III. С. ecirrhosus Cogn.
3. Опушение редкое или густое, мягкое; плоды обычно без горечи.......
................................I. С. lanatus (Thunb.) Matsum et Nakai
4- Опушение чаще жесткое; плоды горькие или безвкусные...............
.......................................II. С. colocynthis (L.) Schrad.
ОПИСАНИЕ ВИДОВ И ВНУТРИВИДОВЫХ ТАКСОНОВ
I. Арбуз обыкновенный (шерстистый) — Citrullus lanatus (Thunb.).
Matsum. et Nakai, 1920, Cat. Sem. Spor. Hort. Bot. Univ. Imp. To-
kyo : 38; Mansf. 1959, Kulturpfl. 2 : 421. — Momordica lanata Thunb.
1794, Prodr. pl. cap. : 13. — Citrullus battich Forsck. 1775, Fl. aeg-
arab. : 167, sine descr. generica. — C. pasteca Sager. 1826, Ann. Sci. Nat.
(Paris.) ser. 1, 8:312. — C. chodospermus Fal. et Dunal., 1836, Bull.
Soc. agric. Herault: 264.— C. citrullus Kars. 1883, Pharm. med.
2 Зак. 2323
17
Bol : 889. — Anguria citrullus Mill. 1768. Gard. Did. Abridg. ed. 8, jn
corr - I. indica sen batteca Rumph. 1747, Herb. amb. 5 : 400, — co/o.
„unrhis ainarissitna Schrad. 1833, Index Sem. Golting. ; 2; id.,
I nmaea. 10 : 70. — C. citrullus Kuntzc, 1891. Revis, 1 : 259. — Cticuiius
Jolocunlhis Thunb. 1794, 1. c. : 13, — C. giganlea Salisb. 1796, Prodr. : 157
— C. amarissimus Schrad. 1827, Index Sem. Golting. : 2. — C. dissectus
Decnc. 1834. Nouv. Ann. Mus. Paris. 3:449. — Cocurbita citrullus L.
1753, Sp. Pl. : 1010.— C. anguria Duch in Lam. 1786, Encycl. 2: 158^
nomen illegit.— C. pinnatifida Schrank, 1824, Syll. Pl. Nov. Ratisbon.
1 : 190. — C. caffra Schrad. 1836. Eckl. et Zey., Enum. : 279.
Typus: «Cucurbita citrullus: foliis multipartitis. Habitat in Apulia,
Calabria, Sicilia» (L.).
Полиморфный вид, включающий дикорастущие и возделываемые
формы пищевого и кормового назначения.
По нашей классификации С. lanatus включает три подвида, раз-
личающихся по ареалу, биологическим особенностям и морфологиче-
ским признакам: subsp. lanatus, subsp. vulgaris и subsp. mucososper-
mus. Каждый подвид дифференцирован на разновидности.
Рис. 2. Листья видов Citrullus:
верхний ряд: С. lanatus — subsp. lanatus, subsp, vulgaris, subsp. mucoso-
spermus; нижний ряд: C colocynthis, C. ecirrhosus, C. naudinianus.
Первый из них объединяет дикорастущие формы из Южной Аф-
рики (рис. 2. 3, 4), кормовые и цукатные формы. Среди этого подвида
мы выделяем var. lanatus ( = caff er)— арбуз каффрский, описанный
Д. Ливингстоном [121] в его путешествии по Южной Африке как «са-
мое удивительное растение пустыни». Второй подвид включает столовый
сладкий и полукулътурный суданский арбуз. Все имеющиеся сорта сто-
лового арбуза с ботанической точки зрения однотипны, поэтому отне-
сены нами к одной разновидности.
Третий подвид выделен нами впервые. Он объединяет не описан-
ные в ботанической литературе формы арбуза Западной Африки, от-
личающиеся необычными семенами, похожими на семена тыквы.
В плоде они заключены в толстые слизистые капсулы, благодаря чему
подвил получил название слизистосемянный (niucosospertnus).
1к
Рис. 3. Плоды видов Citrullus:
ряд: С. lanatus — subsp. lanatus, subsp. mucosospermus, subsp. vulgaris;
нижний ряд: C. colocynthis (2 плода), C. ecirrhosus, С. naudinianus.
верхний
Эмм
Рис. 4. Семена видов Citrullus и Praecitrullus:
верхний ряд: 1, 2, 3—С. lanatus, subsp. lanatus—var. lanatus, capensis, citroides;
4, 5 —subsp. vulgaris — var. cordophanus, vulgaris; 6, 7 — subsp. mucosospermus var
mucosospermus, var. senegalicus; нижний ряд: 1, 2 —C. colocynthis subsp. colocynthis,
subsp. insipidus; 3 — C. ecirrhosus; 4 — C. naudinianus; 5 — Praecitrullus fistulosus.
oc 0
2*
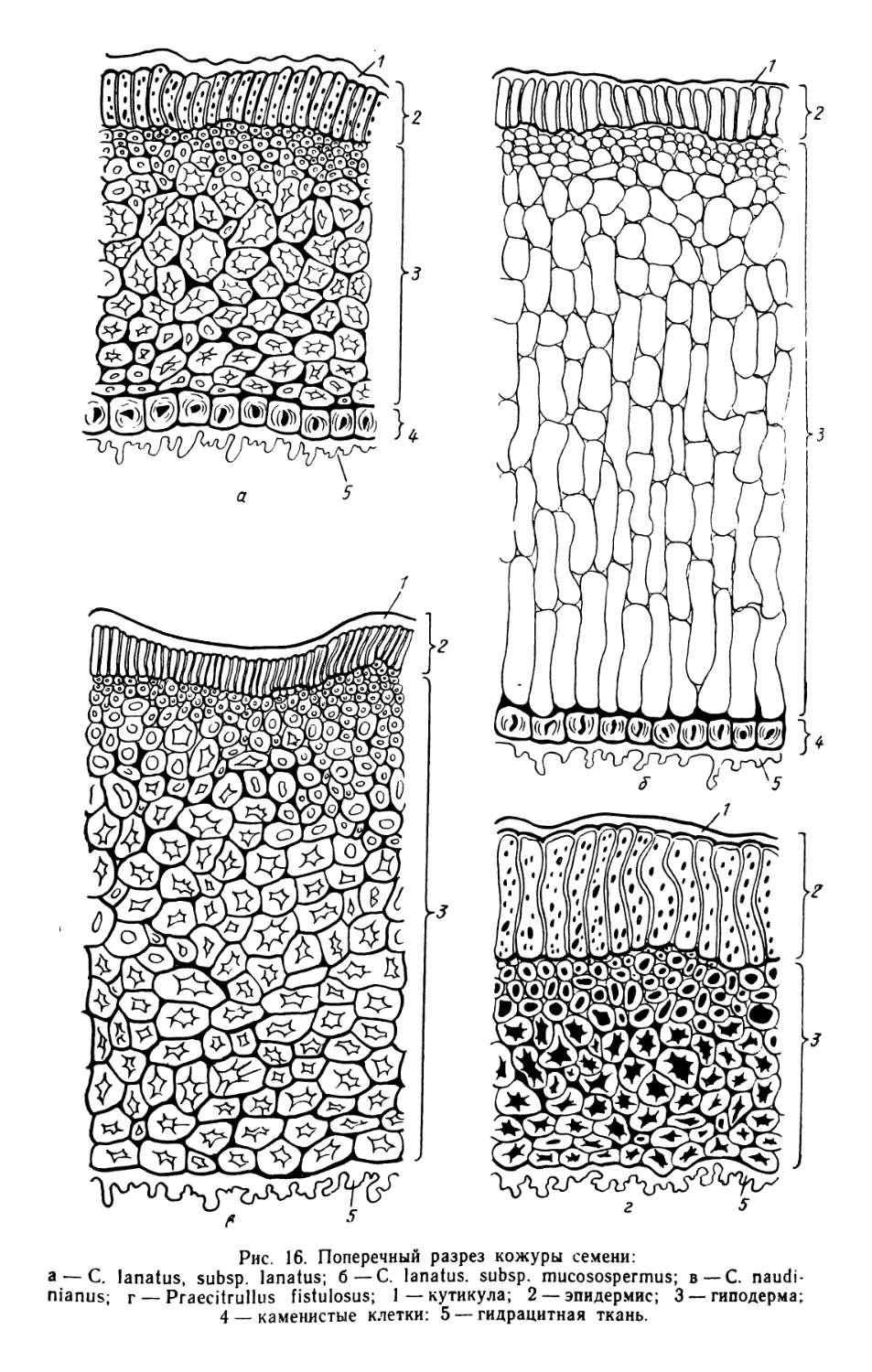
Анатомическая структура кожуры семян у этого подвида очень
своеобразна. Непосредственно под тонкостенным эпидермисом, не имею-
щим кутикулы, расположена многослойная ткань, состоящая из тонко-
стенных, легко ослизняющихся клеток. Эта ткань вместе с эпидермисом
и образует слизистую капсулу. Свойственная семенам других форм
арбуза многослойная склеренхима имеется только в рубчике семени.
Защитный слой кожуры представлен лишь одним рядом каменистых
клеток.
В Нигерии и Гане описанные формы слизистосемянного арбуза
возделывают для получения семян, богатых маслом и белком. Неко-
торые сорта содержат до 60% масла.
Изучение структуры хромосом subsp. mucosospermus выявило на-
столько резкие отличия его от других представителей Citrullus [230],
что возник вопрос о выделении этого подвида в самостоятельный вид.
Ключ для определения подвидов арбуза шерстистого —
Citrullus lanatus (Thunb.) Matsum. et Nakai
1. Опушение густое, войлочное; листья широкие, с округлыми долями; венчик ярко-
желтый; семена без рубчика; плоды крупные, не меньше 20 см в диаметре;
мякоть плотная, пресная или горькая, иногда сладковатая .............
.................................................. I. subsp. lanatus.
4- Опушение редкое; листья более узкие; венчик светло-желтый; семена с рубчиком 2.
2. Плоды средние и крупные (15—40 см в диаметре); мякоть сочная, сладкая,
иногда безвкусная; кожура семени толстая, твердая...................
..................................II. subsp. vulgaris (Schrad.) Fursa.
4- Плоды мелкие, до 15 см, или средние, до 20 см в диаметре; мякоть горькая,
безвкусная, реже сладковатая; кожура семени тонкая; семена заключены
в студенистую оболочку..................... III. subsp. mucosospermus Fursa.
I. Подвид шерстистый — subsp. lanatus.— C. vulgaris
var. lanatus Bailey, 1930, Gent. Herb. 2: 186. — C. lanatus var. lanatus,
Mansf. 1959, Kulturpfl. 2 : 421.
Характерный признак подвида — густое мягкое опушение растений,
особенно молодых побегов и завязи. Отсюда название lanatus — шер-
стистый.
Растения длинноплетистые, с мягкими крупными, слабо рассечен-
ными листьями. Цветки ярко-желтые, с заостренными лепестками. Мя-
коть плода плотная, белая или желтоватая, иногда горькая. Семена
выпуклые, без рубчика.
Объединяет дикорастущие формы Южной и Юго-Западной Аф-
рики, а также возделываемые за пределами этого континента кормо-
вые и цукатные формы арбуза. Включает три разновидности: var. lana-
tus (арбуз каффрский), var. capensis (арбуз капский) и var. citroides
(арбуз цитронный).
Ключ для определения разновидностей арбуза шерстистого — subsp. lanatus
1. Листья крупные, длиной более 15 см; плоды крупные, 20—40 см в диаметре,
съедобные, иногда горькие; семена крупные, длиной 1,5—2 см...........2.
4- Листья мелкие, 12—15 см; плоды мелкие, 10—15 см, горькие; семена средние,
1,0—],5 см..................................var. capensis (Alef.) Fursa.
2. Дикорастущий; мякоть сочная...........................var. lanatus.
4- Возделываемый; мякоть плотная, менее сочная......................
.......................................var. citroides (Bailey) Mansf.
Разновидность каффрская — var. lanatus Mansf.
1959, Kulturpfl. 2:421. — C. caffer Schrad. 1834, Index Sem. Gotting.
1836, Linnaea, 10: 109. — C. caffrorum Schrad. 1838, Linnaea, 12:413.—
C. lanatus var. caffer (Schrad). Mansf. 1959, 1. c. : 421.
Растения мощные, с густым войлочным опушением. Стебель тол-
стый, отчетливо граненый. Листья крупные до 25 см, слаборассеченные,
с округлыми долями и неприятным специфическим запахом. Цветки
20
крупные, ярко-желтые, чаще обоеполые. Плоды массой до 20 кг обычно
неправильной формы, с нечетким рисунком, в виде разорванных полос
или пятен. Мякоть сочная, сладковатая, содержит до 4—5% раство-
римого сухого вещества. Семена крупные, красные или коричневые.
Дикорастущие съедобные арбузы Калахари и прилежащих к ней
областей. Местное название tsamma.
Разновидность капская — var. capensis (Alef) Fursa,
comb. nov. — Citrullus colocynthis (L.) Schrad. var. capensis Alef. 1866,
Landwirtschaftl. Fl. : 211. — C. amarus Schrad. 1834, 1. c. : 413.
Растения длинноплетистые, с более мелкими, чем у каффрского
арбуза, листьями и плодами. Цветки раздельнополые, плоды шаровид-
ные, с четким рисунком из мозаичных полос на розовато-желтом фоне.
Мякоть твердая, белая, обычно горькая. Семена оливковые или ко-
ричневые.
Разновидность особенно широко распространена в бывшей Кап-
ской области, откуда и получила свое название. Часто встречается как
сорняк на полях.
Разновидность цитр о иная — var. citroides (Bailey)
Mansf. 1959, Kulturpfl. 2:421. — C. vulgaris var. citroides Bailey 1930,
Gent. Herb, 2 : 186. — C. colocynthoides auct. non Schweinf. : Pang. 1930,
Tp. по прикл. бот. ген. сел. 23, 3 : 66, 81.
По морфологическим признакам разновидность близка к каффр-
ской, но более выравнена. Цветки всегда раздельнополые. Плоды ци-
линдрические или шаровидные, с пятнистым или полосатым рисунком.
Мякоть белая или желтоватая, плотная, с высоким содержанием пек-
тина. Семена узкие, оливковые, реже красные.
К этой разновидности относятся кормовые и цукатные формы ар-
буза, возделываемые в СССР и США. Они очень неприхотливы, растут
в засушливой степи и полупустыне. Плоды могут храниться в течение
нескольких месяцев.
II. Подвид обыкновенный — subsp. vulgaris (Schrad.) Fursa,
comb. nov. — C. vulgaris Schrad. 1836 in Eckl. et Zey., Enum. : 279;
id., 1838, Linnaea, 12:412; Naud. 1859, Ann. Sci. Nat. Paris 4, 12:10,
Pax in. Engl. 1894, Pflanzenfam. 4, 5:27; Cogn. et Harms, in Engl. 1924,
Pflanzenr. 88 :103; Васильченко, 1957, Фл. СССР, 24:111. —C. edulis
Spach., 1838, Hist. nat. veg. Phan. 6:214.— C. edulis Pang. 1930. Tp.
прикл. бот. ген. сел., 23, 3:44, 81; Вавилов, 1937, Теорет. осн. сел.
3:144; Pang. 1945, Бюлл. Моск. общ. исп. прир. отдел, биол. 50,5 —
6:80.
Стебель средней длины, округло-граненый, слабоопушенный. Ли-
стья сизо-зеленые, без запаха, средне- и сильнорассеченные. Цветки
обоеполые или раздельнополые. Венчик светло-желтый, с округлыми
лепестками. Плоды разнообразны по форме и окраске. Мякоть соч-
ная, сладкая, иногда безвкусная. Семена с рубчиком.
Объединяет культурные и полукультурные возделываемые формы
арбуза. Первые широко распространены в субтропическом поясе, вто-
рые сосредоточены в Северо-Восточной Африке.
Ключ для определения разновидностей арбуза обыкновенного —
subsp. vulgarus (Schrad.) Fursa
1. Возделываемый; мякоть розовых и красных тонов, иногда желтая, сладкая;
десертный ............................................ var vulgaris.
Полукультурный; мякоть бледно-желтая, несладкая; источник влаги.
.................................var. cordophanus (Ter-Avan.) Fursa.
Разновидность столовая — var. vulgaris. Стебель
округло-граненый, длиной до 2 м, слабоопушенный. Листья рассечен-
21
ные, иногда цельнокрайние. Цветки обоеполые, реже раздельнополые.
Мякоть зернистая, сочная, разных оттенков (от лимонной до малино-
вой), содержит до 10—11% сахара. Семена разнообразны по цвету и
размеру.
Столовый десертный арбуз, широко возделывается в странах суб-
тропического пояса от 25° до 48° с. ш. Особенно распространен в СССР
и США, где занимает большие площади.
Разновидность кордофанская — var. cordop hanus
(Тег-Avan.) Fursa, comb. nov.— C. lanatus subsp. cordophanus Ter-Avan.,
1966. Бот. журн. 51, 3:423. — C. vulgaris var. sudanicus Filov et Fursa,
1962, Бот. журн. 47 : 1039.
Ввиду того что нами не обнаружен гербарий типа subsp. cordopha-
nus Ter-Avan., мы выделяем неотип: «Происхождение — Судан, репро-
дукция— Геническая опытная станция, 4/VIII 1966, герб, лист № 5252,
№ по каталогу ВИР 4102, собр. Т. Б. Фурса» (ВИР).
Растения длинноплетистые, с прижатыми к земле грубыми рас-
сеченными листьями. Цветки обоеполые. Плоды шаровидные, с поло-
сатым рисунком или однотонные. Мякоть желтоватая, содержит 4—6%
сахара и пектиновые вещества. Плоды могут храниться несколько ме-
сяцев, при этом мякоть мацерируется.
Представлена полукультурными формами, встречающимися в Су-
дане, Верхнем Египте, Кении. В засушливой саванне плоды этого рас-
тения служат источником воды.
111. Подвид с л и з и ст о с е м я н н ы й —subsp. mucoso-
s р е г m u s Fursa, subsp. nov.—Caulis tenuis, 1,5 m long internodiis longis.
Flores diclini, sublutei. Fructus globosi, 12—14 cm diam., pupla dura alba
amara vel insipida. Semina plana, magna cucurbitoidea, in capsulis
mucosis, pelliculae structura propria. Formae spontaneae et subspontaneae
Africae occidentalis (Gana, Gvinea, Senegal).
Typus: Forma macrosperma. Origo — Gana. Reproductio— SAS
WIR, 5/VIII 1957, specimen herbaceim, N 3833, catalogi numerus WIR
k-3742, 1g. N. P. Oltarshevskyi (WIR).
Paratypus: Forma microsperma. Origo — Gvinea. Reproductio — Sta-
tic experimentalis Genizeskaja, 25/VII 1970, introductionis numerus
289367, 1g. T. B. Fursa (WIR).
Стебель тонкий, длиной 1,5 м, с длинными междоузлиями. Цветки
раздельнополые, светло-желтые. Плоды мелкие, шаровидные, 12—14 см
в диаметре или средние, с твердой горькой или безвкусной мякотью.
Семена плоские, крупные, как у тыквы, заключены в слизистые кап-
сулы. Имеют своеобразное строение кожуры.
Тип: крупносемянная форма. Происхождение — Гана. Репродук-
ция— САС ВИР, 5/V1II 1957, герб, лист № 3833, № по каталогу ВИР
3742, собр. Н. П. Олтаржевский (ВИР).
Паратип: мелкосемянная форма. Происхождение — Гвинея. Репро-
дукция— Геническая опытная станция, 25/VII 1970, интродукционный
№ 289367, собр. Т. Б. Фурса (ВИР).
Дикорастущие и полукультурные формы Западной Африки (Гана,
Нигерия, Гвинея, Сенегал).
Ключ для определения разновидностей арбуза слизистосемяиного —
subsp. mucosospermus Fursa
1. Мякоть горькая, плотная, белая; семена крупные, длиной 1,5—2 см, редко мел-
кие, заключены целиком в студенистую оболочку....var. mucosospermus.
+ Мякоть без горечи, бледно-розовая; семена средние, 1,0—1,2 см; оболочка
в виде двух студенистых подушечек на плоских сторонах семени.....
.......................................... var. senegallcus Fursa.
22
Разновидность слизистосемянная — var. mucoso-
spermus. Стебель отчетливо граненый, тонкий. Листья сильнорас-
сеченные, темно-зеленые, прижатые к земле. Плоды однотонные или
с полосатым рисунком, 12—14 см в диаметре. Мякоть очень твердая,
чаще горькая. Семена крупные, плоские, в слизистых капсулах.
Представлена дикорастущими и полукультурными формами. Рас-
пространена в Гане и Нигерии. В пищу используют семена, богатые
маслом и белком.
Разновидность сенегальская — var. senegalicus
Fursa, var. nov. — C. vulgaris var. senegalicus Fursa, 1962, Бот. журн.
47:1039, sine typo — Folia erecta dissecta. Flores parfecti. Fructus
18—20 cm diam., saepe striati. Pulpa alba vel helvola, edulis sed sub-
dulcis. Semina parva et plana. Formae subspontanea.
Typus: Origo — Senegal. Reproductio— Astrachanskij opornyi punkt
WIR, 5/VIII 1976, catalog! numerus WIR k-3837, 1g. T. B. Fursa (WIR).
Тип: Происхождение — Сенегал. Репродукция — Астраханский опор-
ный пункт ВИР, 5/VIII 1976, № по каталогу ВИР 3837, собр. Т. Б. Фур-
са (ВИР).
Черты ксерофильной организации в общем облике растения выра-
жены слабее, чем у предыдущей разновидности. Листья прямостоячие,
рассеченные. Цветки обоеполые. Плоды до 20 см в диаметре, чаще
с полосатым рисунком. Мякоть бледно-розовая, съедобная, но мало-
сладкая. Семена более мелкие, чем у предыдущей разновидности, дли-
ной 0,7 см.
II. Арбуз колоцинт — С. colocynthis (L.) Schrad.— 1838, Linnaea,
12:414. — Cucumis colocynthis L. 1753, Sp. PL: 1010. — C. pseudocolo-
cynthis Wender. 1834, Sei. sem. hort. Marb.: 2; Naud., 1859, Ann. Sci.
Nat. (Paris) 4 ser. 12:99; Pax, 1894, Pflanzenfam. 4, 5 :27; Cogn. et
Harms, in Engl. 1924, Pflanzenr. 88 : 109; Пангало, 1930, Tp. прикл.
бот. ген. сел. 23, 3:70; Никитин, 1954, Фл. Туркм. 6:366; Васильченко,
1957, Фл. СССР, 24:113. — Colocynthis vulgaris Schrad. 1833, Index
Sem. Hort. Cotting.: 2. — C. officinalis Schrad. 1838, Linnaea, 12:421.—
C. officinarum Cogn. in DC. 1881, Mon. Phan. 3:511. —Mansf. 1959,
Kulturpflanz. 2:421.
Typus: «Cucumis colocynthis: foliis multifidis, pomis globosis» (K).
Встречается в пустынях Северной Африки, Аравии. Ирана, Афга-
нистана, Передней Индии, Австралии. Обнаружен Н. Н. Карягдыевым
[102] на территории СССР в Туркмении, в долине реки Теджен.
Колоцинт известен преимущественно в дикорастущем состоянии.
Плоды его ядовиты, используются в медицине как желудочное сред-
ство.
Вид полиморфен. Еще Ф. Алефельд [280] дифференцировал его
на несколько разновидностей. К. И. Пангало [160], изучая формы ко-
лоцинта разного происхождения, выделил четыре разновидности: var.
insipidus, var. stenotomus, var. pachyrhizus и var. vulgaris. Мы счи-
таем целесообразным оставить две из них: var. insipidus и var. steno-
tomus, но рассматриваем их в ранге подвидов. Две другие разновид-
ности не имеют таксономических отличий от var. stenotomus и при-
соединены к ней. Var. stenotomus включает тип вида, поэтому новый
подвид автоматически получает название subsp. colocynthis.
Subsp. insipidus отличается от типичного колоцинта. Г. Швайнфурт
[357] встречал эту форму в Египте в качестве возделываемой. Внеш-
ний облик растения имеет много общего со столовым арбузом, что
дало основание К. И. Пангало [160] считать эту разновидность группой
23
cultus вида C. colocynthis, а А. И. Филову [240]—рассматривать ее
как дикорастущую форму столового арбуза.
Подвид полиморфен, имеет формы, приближающиеся к колоцинту
и к столовому арбузу, легко скрещивается с ними, образуя растения
гибридного характера. Вероятно, этот подвид гибридного происхожде-
ния, возник в результате одичания культурных сортов вследствие пере-
опыления их с колоцинтами. Пангало упоминает только о формах
с пресной мякотью, отсюда и название insipidus, но среди этого под-
вида встречаются и образцы с горькой мякотью.
Ключ для определения подвидов арбуза колоцинта —
Citrullus colocynthis (L.) Schrad.
1. Растения компактные; листья очень мелкие, до 10 см, сизо-зеленые, жесткоопу-
шенные; плоды мелкие, 5—12 см в диаметре, горькие; семена очень мелкие —
длиной 0,5—0,7 см.................................I. subsp. colocynthis.
+ Растения плетистые; листья средние—13—18 см, зеленые, слабоопушенные;
плоды средние, до 18 см, безвкусные, реже горькие; семена средние, 0,9—2 см,
иногда с рубчиком.....................II. subsp. insipidus (Pang.) Fursa.
I. Подвид узколистный — subsp. colocynthis. — C. colocynthis var.
stenotomus Pang., 1930, var. pachirhizus Pang., var. vulgaris Pang.
1930, Tp. прикл. бот. ген. сел. 23, 3: 72.
Однолетние или многолетние растения, иногда с толстым деревя-
нистым корнем. Стебель короткий, сильно разветвленный. Листья мел-
кие, до 10 см, сизо-зеленые, жесткоопушенные, сильнорассеченные.
Усики двураздельные, хорошо развиты, обладают свойством гаптотро-
пизма. Цветки раздельнополые, мелкие, до 2,5 см, с округлыми бледно-
желтыми лепестками. Плоды мелкие, 5—12 см в диаметре, высыхаю-
щие. Окраска зрелого плода тускло-лимонная. Мякоть плотная, белая,
сухая, горькая, ядовитая. Семена мелкие, 0,5—0,7 см, без рубчика,
коричневые.
Распространен от Северной Африки до Индии.
II. Подвид безвкусный — subsp. insipidus (Pang.) Fursa, comb. nov.
— C. colocynthis var. insipidus Pang. 1930, Tp. прикл. бот. ген. сел. 23,
3: 73.
Стебель тонкий, более длинный, чем у типичного колоцинта. Ли-
стья прямостоячие, мягкоопушенные. Цветки раздельнополые. Плоды
средние, до 18 см в диаметре, часто неправильной угловатой формы,
розово-охристые, с полосатым мозаичным рисунком. Мякоть белая или
розовая, безвкусная, реже горькая. Семена крупнее, чем у типичного
колоцинта, иногда с рубчиком.
Распространен по берегам Средиземного моря: в Алжире, Тунисе,
Египте, Иордании.
III. Арбуз безусиковый — С. ecirrhosus Cogn. Verh. Bot. Ver. P.
Brandenb. 30 :151; id, in Engler 1883, Bot. Jahrb, 10:270; Dinter,
1919, Feddes Repert. 15 :432; Cogn. et Harms, in Engl. 1924, Pflanzenr.
88 : 115; Meeuse, 1962, Bothalia, 8:58. — Colocynthis ecirrhosus (Cogn.)
Chakrav. 1949, Sci. St. Cult. 15 : 32.
Typus: «near Swakopmund, Marloth 1192». Юго-Западная Аф-
рика (В).
Многолетнее растение с деревянистым корнем. Стебель длинный,
около 3 м, округло-граненый, толстый, с редким, но жестким опуше-
нием. Междоузлия длинные, 13—15 см, листья мелкие, 6—8 см, силь-
норассеченные, с округлыми долями, гофрированные, обладают непри-
ятным запахом, полуприжаты к земле. Усики двураздельные, вполне
развитые. Цветки раздельнополые, крупные, 3,5—4 см, с желтыми
округлыми лепестками. Плоды средней величины, 15—17 см в диа-
24
метре, угловатые, серо-зеленые, с тусклыми полосами. Мякоть белаяг
плотная, горькая. Семена мелкие, широкие, темно-коричневые.
Вид известен только в дикорастущем состоянии, зарегистрирован
в пустыне Намиб (Юго-Западная Африка). Эндем. Ареал его почти
ограничен районом пустыни.
Изучая этот вид в культуре, К. И. Пангало [166] пришел к вы-
воду, что он отличается от колоцинта лишь более слабым развитием
усиков и его следует считать разновидностью последнего.
Более детальное изучение заставляет пересмотреть мнение
К. И. Пангало. С. ecirrhosus отличается от колоцинта рядом морфоло-
гических признаков, имеет узкий замкнутый ареал, с трудом скрещи-
вается с колоцинтом. Это, несомненно, самостоятельный вид, однако
неправильно называть его безусиковым арбузом: при выращивании в
культуре он образует вполне развитые двураздельные усики. Отсут-
ствие последних в естественных местах его обитания связано с эколо-
гическими условиями.
У другого обитающего здесь вида арбуза — С. naudinianus усики
тоже редуцированы почти до колючек, а у Acanthosicyos horrida пре-
вратились в острые шипы.
IV. Арбуз Нодэна — С. naudinianus (Sond.). Hook. f. on Oliv. 1871,
Fl. Trop. Afr. 2:549; Cogn. in DC 1881, Mon Phan. 3:511; id., 1880,
Verh. Bot. Ver. P. Brandenb. 30 : 150; Cogn. et Harms, in Engl. 1924,
Pflanzenr. 88:114; Meeuse, 1962, Bothalia, 8:55. — Cucumis dissecti-
folius Naud. 1859. Ann. Sci. Nat. (Paris), 4 ser. 11:23. — C. naudinianus
Sond. in Harv. et Sond. 1862, Fl. cap. 2:496. — Colocynthis naudinianus
(Sond). Kuntze, 1891, Rev. Gen. Bot. (Paris) 1 : 256.
Lectotypus: «Cucumis dissectifolius Naud: Transvall — Potehef-
stroom: Mooi River, Burke 488» (K).
Двудомное многолетнее растение с клубневидным корнем. Сте-
бель тонкий, очень длинный, 3—4 м, почти голый. Листья мелкие,
длиной 6—8 см, грубые, рассеченные почти до основания, с узкими
удлиненными долями. Усики простые, редуцированные до колючек.
Цветки раздельнополые, со светло-желтыми лепестками. Завязь и плод
покрыты короткими коническими шипами (бугорками). Плоды эллип-
тические, мелкие, длиной 6—12 см, шириной 4—8 см. Мякоть сочная,
кисловатая, съедобная. Семена выпуклые, гладкие, белые, длиной
0,8 см. Прорастание спорадическое.
Вид зарегистрирован в Анголе, Замбии, Зимбабве, Намибии, Мо-
замбике, провинции Трансвааль (ЮАР). Известен только в дикорас-
тущем состоянии.
Арбуз Нодэна своеобразен по биологическим особенностям и мор-
фологии. В отличие от других видов Citrullus — это двудомное расте-
ние. Плоды его продолговатые, мелкие, покрыты толстыми кониче-
скими бугорками. Кора плода снимается, как у апельсина; мякоть
съедобная, кисло-сладкая, с острым привкусом. Семена твердые, дере-
вянистые.
М. Шимотзума [361] отмечает, что обычным способом семена
этого вида прорастить не удается. Для их проращивания был исполь-
зован новый метод скарификации твердых семян, разработанный нами
совместно с отделом семеноведения ВИР. Нам удалось получить
всходы, вырастить в культуре мужские и женские экземпляры этого
вида и собрать гербарий. Характерная особенность растений — быстрое
укоренение стеблей в местах их соприкосновения с землей. Вид не по-
ражался болезнями, которые были отмечены у столового арбуза [259].
Очень своеобразным оказалось строение кожуры семян этого вида.
25
Таблица 2. Таксономические признаки
Признак Так
С. lanatus (Thunb.) Matsum.
subsp. lanatus subsp. vulgaris |
var. lanatus var. capensis var. citro- ides var. cordop- hanus var. vulgaris
Распространение Страны Южной Африки Страны Южной Африки США, СССР, Венгрия Судан, Кения, Эфиопия Страны субтропи- ческого пояса
Образ жизни Однолетний Однолетний Однолетний Однолетний Однолетний
Использование Дикорасту- щий Дикорасту- щий Кормовой и цукат- ный Источник биологиче- ской воды Десертный
Форма стебля Толстый граненый Тонкий граненый Толстый округло- граненый Тонкий округло- граненый Тонкий округло- граненый
Длина » Очень длинный Длинный Очень длинный Длинный Длинный и средний
Опушение Сильное войлочное Среднее войлочное Среднее войлочное Слабое жесткое Редкое мягкое
Размер листа Крупный Средний и мелкий Крупный и средний Средний От мелкого до круп- ного
Рассеченность Слабая и средняя Слабая и средняя Слабая Средняя От сильно- рассечен- ного до цельного
Доли листа Широкие округлые Широкие округлые Широкие округлые Вытянутые Вытянутые
Усики 2—4-раз- дельные 2-раздель- ные 2—3-раз- дельные 2-раздель- ные 2-раздельные
Половой тип Андромо- нойкист и монойкист Монойкист Монойкист Андромо- нойкист и монойкист Андромоной- кист и мо- нойкист
Форма лепестков Широкие округлые Округлые и заост- ренные Длинные заострен- ные Округлые Округлые
Окраска венчика Ярко- желтый Ярко- желтый Ярко- желтый Сернисто- желтый Сернисто- желтый
Опушение завязи Густое войлочное Войлочное Г устое войлочное Редкое войлочное Редкое торчащее
Размер плода Крупный (до 20 кг) Средний и мелкий (0,5—2 кг) Крупный (до 15 кг) Средний (2—4 кг) От мелких до крупных
Форма » Неправиль- но удли- ненная Шаровид- ная Удлинен- ная и ша- ровидная Шаровид- ная и овальная Шаровидная и удлиненная
Характер мякоти Плотная, но сочная Плотная Плотная Плотная, но сочная Сочная зернистая
Окраска > Желтова- тая Белая Белая и желтова- тая Белая и желтова- тая Яркоокра- шенная
Вкус Слабослад- кий, съе- добный Горький, реже без- вкусный Пресный Слабо- сладкий Сладкий
26
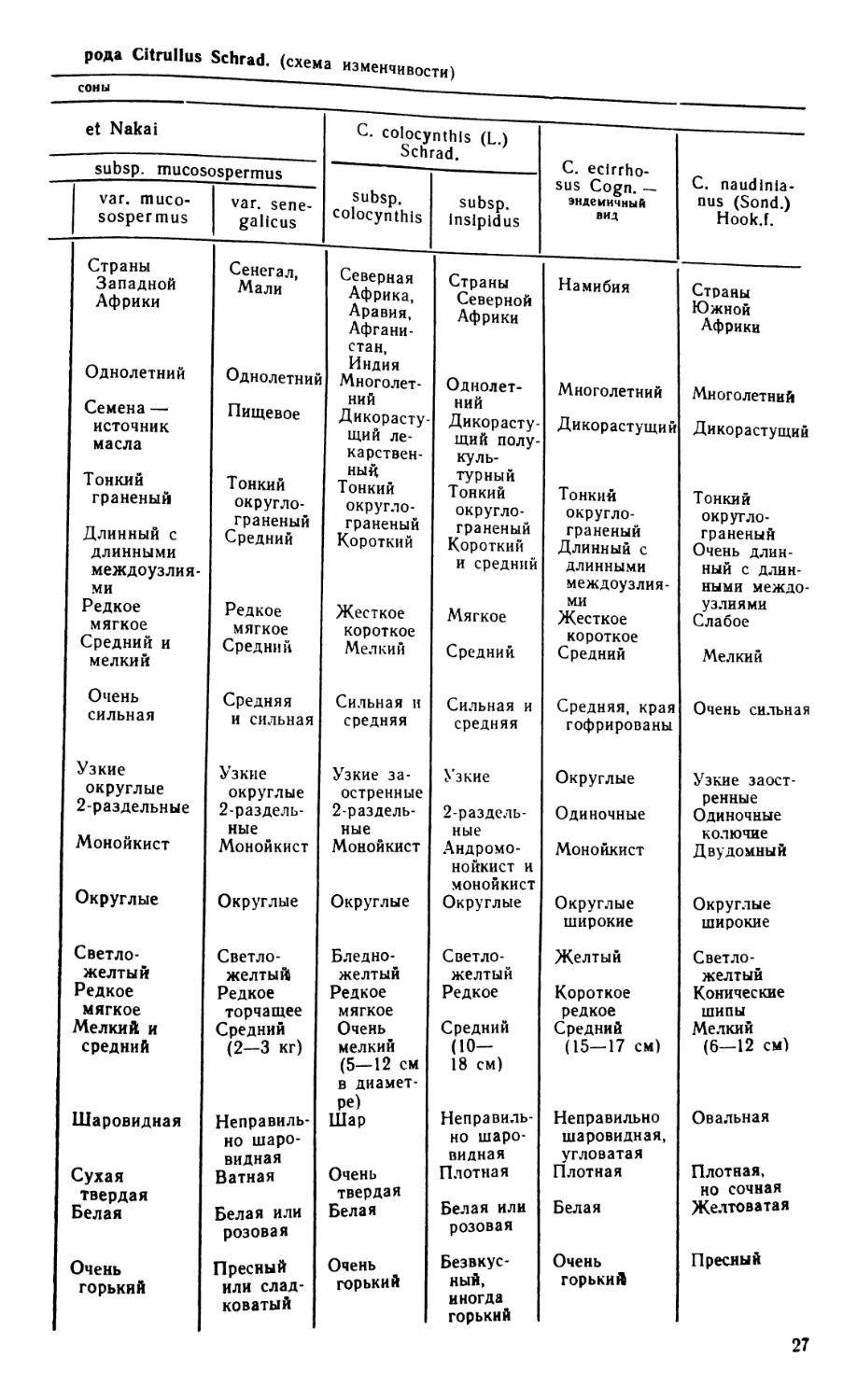
рода Citrullus Schrad. (схема изменчивости)
соны
et Nakai С. colocynthis (L.) Schrad. С. ecirrho- sus Cogn. — эндемичный вид —
subsp. mucosospermus С. naudinia-
var. muco- sospermus var. sene- galicus subsp. colocynthis subsp. insipidus nus (Sond.) Hook.f.
Страны Западной Африки Сенегал, Мали Северная Африка, Аравия, Афгани- стан, Индия Многолет- ний Дикорасту- щий ле- карствен- ный Тонкий округло- граненый Короткий Жесткое короткое Мелкий Страны Северной Африки Намибия Страны Южной Африки
Однолетний Семена — источник масла Тонкий граненый Длинный с длинными междоузлия- ми Редкое мягкое Средний и мелкий Однолетний Пищевое Тонкий округло- граненый Средний Редкое мягкое Средний Однолет- ний Дикорасту- щий полу- куль- турный Тонкий округло- граненый Короткий и средний Мягкое Средний Многолетний Дикорастущий Тонкий округло- граненый Длинный с длинными междоузлия- ми Жесткое короткое Средний Многолетний Дикорастущий Тонкий округло- граненый Очень длин- ный с длин- ными междо- узлиями Слабое Мелкий
Очень сильная Средняя и сильная Сильная и средняя Сильная и средняя Средняя, края гофрированы Очень сильная
Узкие округлые 2-раздельные Монойкист Округлые Узкие округлые 2-раздель- ные Монойкист Округлые Узкие за- остренные 2-раздель- ные Монойкист Округлые Узкие 2-раздель- ные Андромо- нойкист и монойкист Округлые Округлые Одиночные Монойкист Округлые широкие Узкие заост- ренные Одиночные колючие Двудомный Округлые широкие
Светло- желтый Редкое мягкое Мелкий и средний Светло- желтый Редкое торчащее Средний (2—3 кг) Бледно- желтый Редкое мягкое Очень мелкий (5—12 см в диамет- Светло- желтый Редкое Средний (10— 18 см) Желтый Короткое редкое Средний (15—17 см) Светло-^ желтый Конические шипы Мелкий (6—12 см)
Шаровидная Сухая твердая Белая Очень горький Неправиль- но шаро- видная Ватная Белая или розовая Пресный или слад- коватый ре) Шар Очень твердая Белая Очень горький Неправиль- но шаро- видная Плотная Белая или розовая Безвкус- ный, иногда горький Неправильно шаровидная, угловатая Плотная Белая Очень горькиА Овальная Плотная, но сочная Желтоватая Пресный
27
Признак Так
С. lanatus (Thunb.) Matsum.
subsp. lanatus subsp. vulgaris |
var. lanatus var. сареп- sis var. citro- ides var. cordop- hanus var. vulgaris
Размер семян Крупные Средние Крупные Средние От мелких до крупных
Форма » Широкие, без руб- чика Без руб- чика Узкие, без рубчика Узкие, с рубчиком С рубчиком
Окраска » Красные или корич- невые Оливковые и корич- невые Оливковые и красные Кремовые с темным пятном Разнообразны по окраске
Непосредственно под эпидермисом, состоящим из коротких клеток с
толстой кутикулой, расположена многослойная ткань из каменистых
клеток. Дифференциация субэпидермальной ткани, свойственная дру-
гим видам Citrullus, у С. naudinianus отсутствует.
По структуре кожуры семени вид близок к роду Acanthosicyos.
Отсутствие дифференциации слоев семенной кожуры свидетельствует
о древности этих форм.
Положение С. naudinianus в системе Citrullus еще окончательно
не определено. Монограф Cucurbitaceae Южной Африки А. Д. Меузе
[334] считает его связующей формой между Citrullus и Cucumis и
выделяет в особый подрод Pseudocitrullus. Основанием для этого слу-
жат простые шиповидные усики, бугорчатые плоды и химический со-
став горького вещества.
Ч. Джеффри [315], основываясь на палинологических данных, вы-
водит С. naudinianus за пределы рода Citrullus и относит к роду
Acanthosicyos. Он считает, что нельзя рассматривать этот вид как пе-
реходную форму между Cucumis и Citrullus, так как пыльца его не
имеет ничего общего с пыльцой Cucumis.
М. Шимотзума [361] рассматривает С. naudinianus в системе рода
Citrullus, так как он скрещивается, хотя и с трудом, со всеми видами
этого рода. Он отметил довольно хорошее завязывание плодов (50—
80%) и высокую всхожесть гибридных семян (65—83%). Однако фер-
тильность пыльцы Fi межвидовых гибридов во всех случаях оказалась
пониженной и в большинстве комбинаций составляла 20—30%. Автор
объясняет это транслокациями хромосом, особенно характерными для
гибридов между дикими видами, и высказывает мысль, что видообразо-
вание в роде Citrullus могло быть связано с хромосомными перестрой-
ками. Им же выяснено, что С. naudinianus относительно легко скрещи-
вается со столовым арбузом (завязывание составляет 60%, всхожесть
гибридных семян — 70%, фертильность пыльцы гибридов Ft— 47,6%),
благодаря чему открываются реальные перспективы использования
данного вида в селекции.
Знаменательно, что ботаники стремятся найти связующее звено
между двумя родами, центры происхождения которых находятся в Аф-
рике и ареалы тесно соприкасаются. К. И. Пангало указывал на
Praecitrullus fistulosus, А. Меузе — на Citrullus naudinianus.
Своеобразие этих видов, естественно, вызывает мысль об их осо-
бом положении. По своей экологии они диаметрально противоположны.
28
Продолжение таблицы 2
соны
et Nakai С. colocynthis (L.) Schrad. С. ecirrhosus Cogn. — эндемичный вид С. naudinianus (Sond.) Hook. f.
subsp. mocosospermus subsp. colocynthis subsp. insipidus
var. muco- sospermus var. sene- galicus
Очень крупные Плоские, с выпуклым рубчиком Кремовые в студенистой оболочке Средние и мелкие Плоские, с рубчиком Кремовые Очень мелкие Узкие, без рубчика Коричневые Средние и мелкие Без руб- чика или с рубчи- ком Коричневые и черные Мелкие Широкие, без рубчика Темно-корич- невые Средние Выпуклые гладкие Белые
Если Praecitrullus — мезофит, приспособившийся к муссонному климату
Индии, то С. naudinianus — типичный ксерофит. Многолетний клубне-
видный корень и твердые каменистые семена обеспечивают выживание
его в пустыне Намиб и Калахари. С. naudinianus — форма, несомненно,
древняя. Возможно, он является реликтовым видом, сохранившимся
с тех далеких времен, когда род Citrullus еще не обособился.
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ВИДОВ
Чтобы наглядно представить различия видов и внутривидовых так-
сонов арбуза, была составлена схема изменчивости основных диагно-
стических признаков, представленная в таблице 2. Для сравнительной
характеристики таксонов использованы 22 преимущественно морфоло-
гических признака.
Из таблицы видно, что виды четко различаются между собой по
ареалу, габитусу растений, характеру опушения, типу листа и цветка,
размеру и форме плода, характеру мякоти, морфологии семян.
Подвиды имеют различия по ареалу, опушению стебля и завязи,
форме и рассеченности листа, окраске и форме лепестков венчика, на-
личию рубчика у семян.
Разновидности можно определить по морфологии плода, типу мя-
коти, размеру и окраске семян, а некоторые также по ареалу и харак-
теру использования плодов.
Представленная схема может быть использована как определитель
таксонов арбуза в предложенной нами ботанической классификации
этого рода в дополнение к приведенным в тексте ключам.
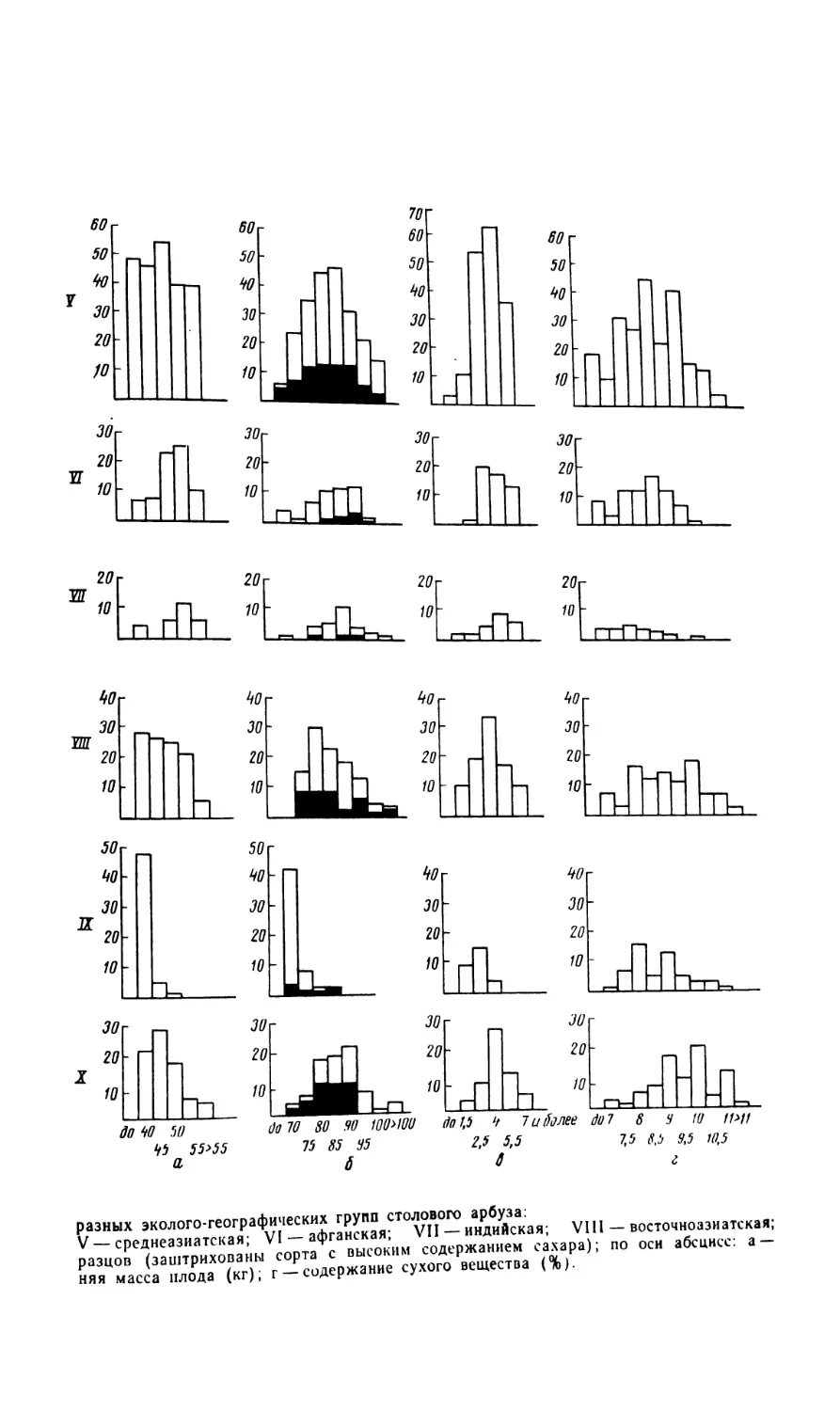
ЭКОЛОГО-ГЕОГРАФИЧЕСКИЕ ГРУППЫ СТОЛОВОГО АРБУЗА
И ЭКОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ
Все разнообразие форм столового арбуза охватывается одним бо-
таническим таксоном var. vulgaris. Для более детальной систематиза-
ции мы использовали эколого-географический принцип. Все известные
на земном шаре сорта столового арбуза были разбиты на 10 эколого-
географических групп: русская, малоазиатская, закавказская, средне-
азиатская, афганская, индийская, восточноазиатская, дальневосточная,
американская, западноевропейская. Между группами не всегда суще-
ствуют четкие морфологические различия, хотя они и не исключаются.
29
Группы различаются по экологии, географии, физиологии и представ-
ляют как бы естественные сложные эколого-географические популяции,
возделываемые в том или ином районе земного шара.
Изложенная ранее ботаническая классификация на внутривидовом
уровне дополняется, таким образом, эколого-географической, в основу
которой положено учение Е. Н. Синской [192] об экотипах.
Развивая положения Е. Н. Синской, Н. И. Вавилов неоднократно
обращал внимание на то, что применительно к культурным растениям
правильнее употребить термин «агроэкотип», так как в его формирова-
нии играют роль не только климатические условия, но и определенная
направленность искусственного отбора. Мы использовали для арбуза
термин «эколого-географическая группа», а не более узкое понятие
«экотип», хотя разница между ними невелика. Оба термина мы пони-
маем в толковании, предложенном Е. Н. Синской и Н. И. Вавиловым.
Районы возделывания арбуза резко различаются между собой по
климатическим условиям и прежде всего по увлажнению. Если для
Японии и Индии характерен муссонный климат с чередованием зимнего
сухого периода и летнего периода дождей, то в южных степях СССР,
в Турции, Иране, Афганистане лето чрезвычайно засушливо, а для юго-
восточных районов США характерно равномерное увлажнение в течение
всего года. Так, в Индии сформировался тип арбузов с ярко выражен-
ным мезофильным строением органов, в СССР (Поволжье) и Турции —
с более ксерофильным.
Экологические группы арбуза были выделены нами на основании
изучения образцов различного происхождения в течение 1956—1961 гг.
в Херсонской области.
При выделении эколого-географических групп использованы сле-
дующие критерии: 1) происхождение и география образца; 2) преоб-
ладание ксеро- и мезоморфных черт (размер и рассеченность листа,
число устьиц на единицу поверхности и др.); 3) физиологические по-
казатели (сосущая сила, водоудерживающая способность листа);
4) генетические показатели (половой тип растений).
Морфологические и хозяйственно-ценные признаки при выделении
групп имели второстепенное значение, но также учитывались в ряде
случаев, особенно консистенция и окраска мякоти, содержание сахара
в плоде и другие, свидетельствующие о степени окультуренности сорта.
ЭКОЛОГО-ГЕОГРАФИЧЕСКИЕ ГРУППЫ
Русская. Включает сорта арбуза Поволжья, Северного Кавказа,
Украины, а также группы скороспелок из средней полосы и Сибири.
Район ее распространения простирается на юге до 45°, на севере до
50° с. ш. Севернее в небольшом количестве выращивают только наибо-
лее скороспелые формы.
Сорта арбуза русской группы растут в основном в степях По-
волжья и Украины, для которых характерно жаркое засушливое лето
с сильными восточными ветрами типа суховеев. Годовое количество
осадков в Нижнем Поволжье не превышает 250 мм, на Украине и
Северном Кавказе — 300—350 мм. Среднесуточная температура июля
22° С. В условиях жаркого и засушливого климата при интенсивной
селекции на повышение сахара и улучшение вкуса плодов и сформи-
ровался тип отечественных сортов арбуза, заслуженно считающихся не-
превзойденными среди мирового сортимента.
Для большинства сортов арбуза этой группы характерны высокая
окультуренность и выравненность. Преобладают сорта с шаровидными
плодами, с розовой или малиновой мякотью. Чаще встречаются сред-
30
неспелые и скороспелые формы, вызревающие на 70—80-й день с мо-
мента всходов. Вегетативные органы отличаются тонким, нежным ти-
пом строения (gracilior, по определению К. И. Пангало). Листья, как
правило, среднего размера, сильно или даже очень сильно изрезанные,
ажурные. Плети тонкие, средней длины (не более 2 м). По половому
типу андромонойкисты (на одном растении формируются мужские и
обоеполые цветки).
Общему облику растений арбуза русской группы присущи черты
ксерофильной структуры, что проявляется в анатомическом строении
органов и в сравнительно высокой сосущей силе листа (15,4 атм*).
Среди них имеются образцы с повышенной устойчивостью к засухе,
которые могут служить исходным материалом для создания засухо-
устойчивых сортов арбуза.
Малоазиатская. Формы арбуза из Малой Азии в общем сходны
с русской группой, но менее окультурены. По-видимому, в южнорусские
степи они проникли через греческие колонии на Черном море в древ-
ности, нашли здесь благоприятные условия и дали начало новому,
более культурному типу. От малоазиатских форм, вероятно, произошли
также сорта арбуза Армении, близкие к русской группе.
Климат той части Турции, где возделывают арбуз, сходен с кли-
матом наших южных степей, но характеризуется еще более жарким
и засушливым летом. Температура июля от 23,6° до 28° С, годовая
сумма осадков 300—400 мм, но выпадают они преимущественно в зим-
ние месяцы. В отдельные годы почти 5 месяцев не бывает дождей.
Бахчеводство в Турции долгое время находилось на низким уровне,
поэтому чистых выравненных образцов, а тем более селекционных сор-
тов здесь почти нет. Сортовой состав представлен более или менее
пестрыми популяциями и довольно однообразен. Плоды преимущест-
венно шаровидные, с полосатым рисунком, часто переходящим в мо-
заику. Мякоть обычно розовая, очень редко красная. В плодах ко вре-
мени созревания семян часто образуются пустоты и мякоть становится
малосъедобной, иногда плаценты расплываются. Но среди этих в об-
щем малокультурных форм встречаются иногда сорта с нежной сладкой
мякотью, содержащие 10—11% сахара. В последние годы местные
сорта почти вытеснены инорайонными.
Общий облик растений малоазиатской группы арбуза, как и рус-
ской, носит черты gracilior —тонкого нежного строения. По типу пола
это андромонойкисты.
Ксерофильность организации проявляется сильнее, чем у русской
группы: сосущая сила листа составляет 18,4 атм. Среди малоазиат-
ской группы арбуза много образцов, отличающихся устойчивостью к за-
сухе, а некоторые из них еще и высоким содержанием сахара и хоро-
шими вкусовыми качествами. Такие формы, несомненно, перспективны
для селекции на засухоустойчивость.
Закавказская. По типу близка к среднеазиатской. Объединяет
сорта арбуза Азербайджана, Грузии, Армении. Они сформировались
в условиях высокой влажности и потому имеют четко выраженный ме-
зофильный облик. Листья крупные, слабо- или среднерассеченные,
плоды обычно крупные, толстокорые, чаще овальные или цилиндриче-
ские. Мякоть яркая, но грубоватая и малосладкая. Содержание сахара
не превышает 8—9%.
Мезофильный характер растений арбуза закавказской группы очень
четко проявляется в анатомическом строении листа: клетки эпидермиса
♦ Согласно Международной системе единиц (СИ, SI) 1 атм = 101,325 кПа
(1,01 325 • 108 Па).
31
крупные, устьиц сравнительно мало, губчатая ткань развита лучше
столбчатой, сосущая сила листа самая низкая среди всех изученных
сортов (9,6 атм). В формировании сортов этой группы участвовали
среднеазиатские, иранские и, возможно, малоазиатские формы.
Среднеазиатская. Включает сорта арбуза республик Средней Азии
и частично Казахстана.
Лето здесь сухое и очень жаркое. В районе Ташкента годовая
сумма осадков 348 м. Среднесуточная температура июля очень высокая
(27—28°С). Арбузы выращивают преимущественно в условиях ороше-
ния, что проявляется во внешнем облике и биологических особенностях
сортов.
Среди образцов арбуза Средней Азии много хороших выравненных
селекционных сортов, но встречаются и пестрые популяции, не имею-
щие сортовых названий. Арбузы среднеазиатской группы отличаются
грубым типом строения (rigidus, по определению К. И. Пангало),
что отчетливо заметно во внешнем облике растений, проявляется в осо-
бенностях плодов, характере мякоти. Плети, как правило, длинные, тол-
стые, листья крупные и средние, реже сильнорассеченные, клетки тка-
ней листа и проводящие сосуды крупные. Плоды шаровидные или не-
сколько вытянутые, обычно крупные, реже среднего размера, с толстой
или средней по толщине корой; мякоть грубая, волокнистая, розовая
или карминная, часто с желтым оттенком. Содержание сахара невысо-
кое. Вкусовые качества посредственные, но некоторые селекционные
сорта, особенно новые, имеют хороший вкус, высокое содержание са-
хара и яркую красивую мякоть. Арбузы Средней Азии более поздне-
спелые и созревают в условиях Херсонской области на 80—95-й день
с момента всходов. Поэтому, вероятно, среди них много лежких сортов,
выдерживающих хранение в течение нескольких месяцев без потери
вкусовых качеств.
По половому типу растения этой группы, как правило, андромоной-
кисты, но встречаются сорта, особенно западнокитайские, — монойкисты
(на одном растении формируются мужские и женские цветки).
Афганская. В целом близка к среднеазиатской, но менее окульту-
рена. По-видимому, большинство среднеазиатских форм арбуза про-
исходит из Афганистана и Ирана. Возможно, афганские и иранские
сорта арбуза проникли также в республики Закавказья и южнорусские
степи. Иранские формы арбуза отличаются от афганских гораздо боль-
шим разнообразием сортового состава, лучшими вкусовыми качествами,
более нежной мякотью, большей сладостью. Но так как их общий
облик сходен, мы считаем возможным объединить эти формы в одну
экологическую группу.
Возделывают арбузы преимущественно в северных районах Ирана
и Афганистана (35—37° с. ш.), близких по климату к нашим южным
степям, но отличающихся очень жарким летом. Среднесуточная тем-
пература июля здесь не ниже 25° С. Годовое количество осадков около
250 мм. Арбузы выращивают преимущественно при орошении, но встре-
чается и богарная культура.
Экотип в целом грубый» мало окультуренный [38]. Плоды средней
величины или крупные, обычно не выравнены по форме, чаще встре-
чаются овальные. В окраске преобладает сетчатый рисунок на светло-
зеленом или зеленом фоне. Мякоть грубая, волокнистая, малосладкая
(особенно у афганских сортов, сахаристость плодов которых не пре-
вышает 5—6%). Содержание сахара в плодах иранских форм арбуза
7—9%. Сорта с красной мякотью встречаются редко (чаще среди иран-
ских форм). Обычно мякоть белая, желтоватая, бледно-розовая. Иногда
32
при созревании около семян образуются пустоты. Среди афганских
форм арбуза встречаются образцы с совершенно необычной ярко-оран-
жевой окраской плодов, по внешнему виду похожих на тыкву. По ха-
рактеру мякоти и семян они не отличаются от других форм арбуза.
Интересной физиологической особенностью этих образцов является ран-
нее пожелтение листьев и плетей, которые к моменту созревания ста-
новятся совершенно желтыми. Плоды оранжевой окраски выглядят
среди них очень эффектно.
Грубая организация заметна во внешнем облике растений и в
анатомическом строении органов: плети обычно длинные, листья круп-
ные, рассеченность их средняя, клетки эпидермиса листа также круп-
ные. Цветки, как правило, гермафродитные (половой тип — андромо-
нойкисты), но встречаются цветки и без тычинок. По продолжитель-
ности периода вегетации арбузы афганской группы следует отнести к
позднеспелым.
Формирование растений в условиях жаркого и сухого лета способ-
ствовало появлению форм с высоким осмотическим давлением клеточ-
ного сока и большой сосущей силой. По нашим данным, сосущая сила
листа у афганских форм арбуза составляет 19,6, у иранских—19,1 атм.
Среди мирового сортимента столовых сортов арбуза они отличаются
наиболее высокими показателями.
Дальневосточная. Район ее распространния—Приморский, в мень-
шей степени Хабаровский край. Эта зона характеризуется муссонным
климатом с сухим зимним и влажным летним периодами. Годовая
сумма осадков в Приморье 500—600 мм, 70% их выпадает летом. Сред-
несуточная температура июля 18—20° С.
В условиях умеренно влажного климата Приморья сформировался
своеобразный экологический тип арбуза, сильно отличающийся от эко-
типов сухих районов Азии и Европы.
Дальневосточная. Район ее распространения—Приморский, в мень-
рового сортимента. Период вегетации 60—65 дней. Растения характери-
зуются очень слаборазвитой вегетативной массой. Плети короткие
(1,0—1,5 м); листья мелкие, рассеченность пластинок средняя; цветки
мелкие; масса плода около 1 кг; форма шаровидная. Мякоть малень-
кая, водянистая и малосладкая (7—8% сахара), невкусная. Но отдель-
ные отселектированные образцы имеют приятную сладкую мякоть.
Плоды заполнены большим количеством крупных семян, что делает их
малопригодными для употребления. Характерная особенность дальне-
восточных образцов арбуза — лимонно-желтая окраска мякоти. Поло-
вой тип — монойкисты. В данном случае наблюдается исключение из
общей закономерности, так как шаровидные плоды обычно коррели-
руют с наличием тычинок в женских цветках. Следует отметить, что
очень скороспелые образцы с шаровидными плодами иного происхож-
дения, например из США, также являются монойкистами.
Группа представлена в основном невыравненными сортами-популя-
циями, не имеющими сортовых названий. Но отдельные скороспелые
образцы, селектируемые Дальневосточной опытной станцией ВИР, по
выравненное™, вкусовым качествам и сахаристости не уступают скоро-
спелым сортам русской группы. Благодаря исключительной скороспе-
лости образцы этой экологической группы представляют большой ин-
терес для селекции.
Восточноазиатская. Включает сорта арбуза, возделываемые в Япо-
нии, восточном Китае и на полуострове Корея. Осадки выпадают здесь
преимущественно в летний период, но из-за очень высокой температуры
(27—28° С) арбузы выращивают при орошении.
3
Зак. 2323
33
Арбуз в этих районах возделывают с давних пор, и, судя по на-
званию си-гуа (западный плод), он проник из стран древней земле-
дельческой культуры Азии. Преобладают выравненные, хорошо отсе-
лектированные сорта, плоды которых имеют хорошие вкусовые ка-
чества.
Плоды, как правило, среднего размера или крупные, чаще оваль-
ные или тупо-эллиптические. Мякоть нежная, сахаристая, но содержит
мало кислоты и кажется пресной. Очень часто встречаются сорта с не-
обычной желтой и оранжевой мякотью, реже с красной; сортов с розо-
вой мякотью почти нет. Содержание сахара в плодах 8—9%, с оран-
жевой мякотью до 10%. Кожура семян многих китайских сортов по-
крыта трещинками. Вероятно, вследствие интенсивности культуры и
определенного направления селекции большинство сортов скороспелые
(продолжительность вегетации в среднем 75—80 дней).
Внешний облик растений носит черты мезофильной организации:
листья крупные, среднерассеченные, пластинка тонкая; клетки эпидер-
миса листьев крупные, с небольшим количеством устьиц. Цветки круп-
ные, плети довольно толстые. Сосущая сила листьев сравнительно не-
велика (11,9 атм). Тычинки в женских цветках, как правило, отсут-
ствуют. Половой тип — монойкисты.
Индийская. В Индии арбузы выращивают преимущественно в се-
веро-западных штатах, меньше — в центральных и южных. Климат
здесь тропический, муссонный, характеризуется чередованием периодов
засухи и ливней. Дождливый период продолжается с июня по сентябрь,
за это время выпадает более 75% годового количества осадков, кото-
рое в среднем по стране составляет 1134 мм. В северо-западных райо-
нах Индии влияние муссонов сказывается слабее: годовое количество
осадков здесь меньше (700—900 мм) и период дождей короче. При
запаздывании муссонов посевам угрожает гибель от засухи, так как
период вегетации характеризуется высокими температурами (28—
30°С). Арбуз и другие культуры выращивают частично при орошении.
Строение органов растений имеет резко выраженный мезофильный
характер: плети толстые, длинные (более 2 м); листья очень крупные,
со слаборассеченной пластинкой; цветки и плоды крупные. Половой
тип, как правило, андромонойкисты. Мезофильность типа проявляется
и в анатомическом строении органов, и в физиологических показателях:
сосущая сила листьев невелика и составляет 11,3 атм.
Арбузы индийской группы не отселектированы. Популяции обычно
не выравнены по форме и окраске плодов, не имеют сортовых назва-
ний. Плоды крупные (10—12 кг), чаще шаровидные или сплюснуто-ша-
ровидные, преимущественно с однотонной темно- или светло-зеленой
окраской. Кора обычно очень толстая, эластичная; мякоть грубоватая,
несочная, карминного или шарлахового оттенка. Содержание сахара не-
высокое (7—8%), вкусовые качества посредственные.
По продолжительности периода вегетации индийские арбузы можно
отнести к среднеспелым. Отсутствие позднеспелых форм, по-видимому,
объясняется тем, что муссонный период в северных районах Индии
сравнительно короткий, не обеспечивает их вызревания.
В условиях нашей страны арбузы индийской группы могут исполь-
зоваться как сочный калорийный корм для скота. Благодаря толстой
коре и плотной мякоти плоды их сохраняются в течение нескольких
месяцев.
Американская. Включает в основном одинаковый сортимент арбу-
зов США и Канады. В Америку арбуз завезен из Европы и Азии,
34
и здесь в течение сравнительно короткого периода (менее 100 лет) был
создан высококультурный, селекционно отработанный тип американ-
ских сортов. Арбузы выращивают на всей территории США, от 30 до
40° с. ш., но основной район их распространения — юго-восточные штаты
(Флорида, Джорджия, Алабама, Техас) и в меньшей степени Кали-
форния. Эти районы характеризуются большим количеством осадков
(1000—1200 мм), выпадающих равномерно в течение года. Темпера-
тура летних месяцев высокая (26—28°С). Климат зоны субтропический.
Американские сорта арбуза отличаются большим содержанием са-
хара (10—12%), хорошими вкусовыми качествами, яркой окраской и
нежной мелкозернистой консистенцией мякоти.
Высокая окультуренность сортов сближает американскую группу
с русской, но они имеют и свои специфические особенности, например,
удлиненную, часто цилиндрическую форму плодов, редко встречаю-
щуюся у сортов иного происхождения. С удлиненными плодами кор-
релирует отсутствие тычинок в женских цветках; половой тип — мо-
нойкисты.
Внешний облик растений в общем имеет мезофильный характер,
но выраженный слабее, чем, например, у индийской группы. Плети
обычно средней длины, листья средние или крупные, чаще слаборассе-
ченные, сосущая сила листа 13 атм.
Сорта арбуза американской группы с успехом выращивают в на-
шей стране.
Западноевропейская. Объединяет сорта Болгарии, Венгрии, Румы-
нии, Греции, Италии, Югославии. В отличие от других групп сортимент
арбузов Западной Европы не имеет специфических черт, видимо, по-
тому, что в его создании участвовали русские, американские и мало-
азиатские формы. Но все же климатические условия накладывают от-
печаток на характер возделываемых здесь сортов. В целом они имеют
мезофильный облик, что проявляется не только в морфологии расте-
ний, но и в анатомической структуре листа.
Венгерские сорта, как правило, ранне- и среднеспелые. Плоды их
имеют тонкую хрупкую кору, красную мякоть и высокое содержание
сахара. Сорта Италии, Югославии, Болгарии более позднеспелые.
Плоды их обычно с розовой мякотью и менее сахаристые. В Венгрии
распространены почти исключительно селекционные сорта, в Болгарии
и Югославии преобладают местные популяции. Болгарские цельнолист-
ные раздельнополые сорта арбуза используют в последнее время для
получения гетерозисных гибридов.
ЭКОЛОГИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ ПРИЗНАКОВ
Разделение всего существующего разнообразия возделываемых
форм столового арбуза по эколого-географическим группам оказалось
удобным для изучения коллекции. Однако необходимо было выяснить,
насколько эти группы соответствуют естественным сложным популя-
циям арбуза, являющимся формой существования вида, а также опре-
делить характерные для каждой группы особенности и установить гра-
ницы между ними.
С этой целью была сделана попытка применить математические
методы исследования. Статистическому анализу были подвергнуты мно-
голетние данные изучения коллекции арбуза, насчитывающей более
1500 образцов. Использование математических методов позволило вы-
явить определенные закономерности изменчивости, которые найдут при-
менение не только в ботанических исследованиях, но и в практической
селекционной работе.
3*
35
цветки (половой тип —
Рис. 5. Распределение эколого-географических
групп столового арбуза в зависимости от ча-
стоты встречаемости образцов с определенным
половым типом:
1 — 1 — восточноазиатская, 2 — американская.
3 — дальневосточная; 11—4 — среднеазиатская,
5 — афганская, 6 — индийская; III—7 —мало-
азиатская, 8 — русская.
Для статистического анализа были взяты морфологические при-
знаки, характеризующие эколого-географическую группу: половой тип
растений, форма плода, цвет мякоти, окраска рыльца.
Все разнообразие форм столового арбуза укладывается в два по-
ловых типа: монойкисты (растения имеют мужские и женские цветки)
и андромонойкисты (растения формируют мужские и обоеполые
цветки).
По форме плода разнообразие арбуза невелико. Преобладают ша-
роплодные сорта, но встречаются арбузы с цилиндрическими, оваль-
ными, грушевидными, очень редко со сплюснутыми плодами. Цвет мя-
коти имеет целую гамму оттенков: от бледно-розового до малинового,
от бледно-лимонного до оранжевого. Часто встречается мякоть шарла-
ховой и карминной окраски.
Для удобства обработки признаки были сгруппированы следую-
щим образом:
I — половой тип: 1 — андромонойкист, 2 — монойкист, 3 — смешан-
ный (т. е. имеются растения 1-го и 2-го типа);
II — форма плода: 1—шаровидная, 2 — удлиненная;
III — окраска мякоти: 1 — розовая или малиновая, 2 — все оттенки
желто-красной окраски (шарлаховая, карминная, желтая, оранжевая).
Математическими методами (с помощью коэффициента сопряжен-
ности Пирсона, отражающего превышение фактической частоты встре-
чаемости над теоретической) было доказано, что у русской и мало-
азиатской экологических групп чаще встречаются гермафродитные
"у афганской, индийской и
среднеазиатской — с одина-
ковой вероятностью оба типа
цветков. У американской,
восточноазиатской и дальне-
восточной групп преобла-
дают пестичные цветки (по-
ловой тип — монойкист). Ко-
эффициент Пирсона, отра-
жающий связь данного при-
знака с экологией, равен 0,53.
На рисунке 5 изученная
по этому признаку коллек-
ция арбуза разбивается на
группы в системе координат.
По оси абсцисс откладывают
частоту встречаемости в эко-
логической группе образцов
с половым типом андромо-
нойкист, а по оси ординат —
частоту встречаемости об-
разцов арбуза с половым ти-
пом монойкист. Координа-
тами точек являются разно-
сти между теоретическими
и фактическими частотами
встречаемости образцов с
тем или иным половым ти-
пом.
Если проанализировать
изученную коллекцию арбуза по вероятностям встречаемости двух по-
ловых типов в каждой экологической группе, то вся она разобьется на
36
три типа: ксерофильный, мезофильный и ксеромезофильный. У первого
чаще встречаются монойкисты, у второго—андромонойкисты, у тре-
тьего— оба половых типа.
Четкое географическое распределение групп на рисунке 5 дает воз-
можность сделать определенные выводы. Половой тип монойкист свой-
ствен сортам американской, восточной и дальневосточной групп, куль-
тивируемым в районах с достаточным увлажнением и имеющим мезо-
фильный характер. Половой тип андромонойкист характеризует сорта
русской и малоазиатской групп, возделываемые в сухих районах и
имеющие ксерофильный облик. Промежуточное положение афганской,
индийской и близкой к ним среднеазиатской групп, неоднородных по
типу пола, свидетельствует о слабой селекционной отработанности и
подтверждает их исходное положение (особенно первых двух) в эво-
люции культурного арбуза.
Преобладающая форма плода у арбуза шаровидная, но встречаются
и другие. Важно было выяснить, существует ли определенная законо-
мерность в географическом распределении арбуза с различными фор-
мами плодов.
Определенную форму плода имеют сорта лишь тех групп, в кото-
рых ведется соответствующая селекция. Так, для образцов русской
группы характерны шаровидные плоды, среди американских сортов
чаще встречаются длинноплодные формы.
Коэффициент сопряженности этих двух качественных признаков
равен 0,44, что указывает на существенную, хотя и не очень тесную
связь. Шаровидной форме плода чаще предшествует тип гермафродит-
ного цветка, эллипсовидной — пестичного. При смешанном половом
типе встречаются разные формы плода. Для русской и малоазиатской
групп характерны гермафродитный тип цветка и шаровидная форма
плода.
Следует отметить, что по всем изученным морфологическим
признакам русская и малоазиатская группы близки. Это подтверждает
предположение, что в формировании русской экологической группы
большую роль играли формы из Малой Азии, проникавшие в южнорус-
ские степи через греческие колонии на Черном море.
Многообразие оттенков окраски мякоти затрудняет установление
наиболее распространенной из них в данной экологической группе. Гео-
графически окраска мякоти распределяется следующим образом: азиат-
ские формы имеют желто-красные оттенки, причем к востоку усили-
вается желтый цвет, европейские — розовые и малиновые, американ-
ские—красные оттенки.
При изучении экологической изменчивости рассматривались также
количественные признаки арбуза: продолжительность периода вегета-
ции и составляющих его фаз, средняя масса плода и содержание са-
хара в плоде. Статистические характеристики этих признаков (средние
их значения) в целом для экологических групп сведены в таблицу 3.
Группы расположены в порядке убывания продолжительности периода
вегетации.
Из таблицы видно, что и средняя масса плода коррелирует с дан-
ным признаком. Хорошо заметна тенденция повышения содержания са-
хара у среднеспелых групп сортов и снижения его как у позднеспелых,
так и у скороспелых.
Для оценки степени разнообразия каждой экологической группы
по рассматриваемым признакам были рассчитаны коэффициенты ва-
риации (V) (табл. 4). Независимо от группы наиболее изменчивым
оказалась средняя масса плода. Наибольшая изменчивость этого при-
37
kOr
30-
20-
10 -
a
30-
20-
10-
до1,5 9 7и$олее Oo7 8 9 W 11>11
2>5 8,5 7,5 8,5 9,5 10,5
0 г
Рис. 6. Средние характеристики количественных признаков
I — русская, II — американская; III — малоазиатская; IV — закавказская;
IX — дальневосточная; X — западноевропейская. По осп ординат: число об
число дней от всходов до цветения; б — число дней до созревания; в — сред-
разных эколого-географических групп столового арбуза:
V — среднеазиатская; VI — афганская; VII — индийская; VIII — восточноазиатская;
разцов (заштрихованы сорта с высоким содержанием сахара); по оси абсцисс: а—'
няя масса плода (кг); г —содержание сухого вещества (%).
знака наблюдалась в восточноазиатской и индийской группах. Менее
изменчивые признаки у арбуза — продолжительность периода вегета-
ции и содержание сахара.
Таблица 3. Средние значения количественных признаков арбуза различных
эколого-географических групп
Эколого-географическая группа Продолжительность периода, дней Средняя масса плода, кг Содержание сахара, % Процент высокоса- харистых форм
всходы — цветение цветение- созревание всходы— созревание
Афганская 50 ±0,7 38 г 0,7 88± 1,0 4,9 ±0,20 8,4-10,12 11
Индийская 50± 1,2 36 ± 1,7 86± 1,7 4.7 ±0,50 8.0 1- 0,30 10
Среднеазиатская 47 ±0,6 39 ±0.6 86 ±0,6 4,5 ±0,10 8,9 ±0,09 33
Американская 46 ±0,6 41+0,7 86 ±0,7 4.2 ±0,14 9.4 ±0.80 42
Русская 464 0,3 40 ±0,3 86 ±0,4 3,5 ±0,06 — 33
Восточноазиатская 45 ±0,6 38 ±0,6 83 ±0,7 3.6 + 0,06 9,0 ±0,11 34
Западноевропейская 44 ±0,7 40 ±0,8 84 ±0,9 3,6 + 0,20 9,4 + 0,12 47
Закавказская 44 ±0,9 38 ± 1,0 82 ± 1,1 4.4 ±0,20 8,9 + 0,12 27
Малоазиатская 42 ±0,5 37 ±0,5 79±0,6 4,1 +0.20 8,4 7 0.09 14
Дальневосточная 34 ±0.7 33 ±0,7 66 ±0,8 1,9±0,13 8,5 ± 0,13 13
Таблица 4. Коэффициент вариации (V) количественных признаков арбуза
Эколого-географическая группа Продолжительность периода Средняя масса плода Содержание сахара
всходы—цветение |всходы- созревание
Афганская 13 10 36 13
Индийская 13 11 50 17
Среднеазиатская 17 10 35 19
Американская 19 13 43 12
Русская 15 11 36 15
Восточноазиатская 15 9 49 13
Западноевропейская 15 10 43 11
Закавказская 18 12 38 12
Малоазиатская 14 10 36 13
Дальневосточная 16 9 38 12
Более заметно выступают различия между группами на гистограм-
мах распределения сортов в экологической группе по градациям при-
знака (рис. 6). Так, период всходы—цветение в среднем для культуры
длится 40—60 дней, но у большинства сортов индийской и афганской
групп продолжительность его составляет 50—55 дней, у дальневосточ-
ной— до 40, у остальных — 40—45 дней. Несколько выделяется сред-
неазиатская группа арбуза. Гистограмма указывает на очень большую
неоднородность этой группы по данному признаку. Такая же законо-
мерность проявляется по продолжительности периода всходы—созре-
вание.
Дальневосточные и восточноазиатские формы арбуза созревают на
60—70-й день, большинство сортов американской, западноевропейской,
малоазиатской и закавказской групп — на 80—90-й, а индийской и аф-
ганской на 85—95-й день. Русские и среднеазиатские сорта неодно-
родны по скороспелости.
Очень важным показателем является коэффициент корреляции (г)
между двумя основными фазами развития: всходы—цветение и всходы___
созревание, что дает возможность определять скороспелость сорта по
началу цветения. Для культуры в целом он равен 0,6. Наибольшая
корреляция (табл. 5) отмечена у закавказской, восточноазиатской,
дальневосточной, американской эколого-географических групп (г =
= 0,73—0,77). Для них время зацветания может служить показателем
скороспелости. Менее тесная связь отмечена у сортов русской и спел-
неазиатской групп (г = 0,68) и слабая (г = 0,42-0,56)-у остальных
40
групп. Выявляется интересная закономерность. Все сорта из групп с
высоким коэффициентом корреляции относятся к мезофильному типу
и возделываются в районах, достаточно обеспеченных влагой. В этом
случае происходит естественное завершение вегетации арбуза, т. е. вто-
рая половина периода развития не сокращается искусственно за счет
недостатка влаги и повышенных температур. Поэтому наблюдается тес-
ная корреляция между продолжительностью первой половины вегета-
ции и всего периода. В засушливых условиях эта связь ослабевает, так
как происходит искусственное сокращение второй половины вегетации
арбуза, что приводит к нивелированию различий между сортами по
скороспелости.
Таблица 5. Коэффициент корреляции (г) между признаками арбуза
Эколого-географическая группа Продолжительность периодов всходы— цветение—всходы — созревание Продолжительность периода всходы— созревание—средняя масса плода Средняя масса плода —содержание сахара
Афганская 0,57 ±0,09 0,16 ± 0,14 —0,09±0,12
Индийская 0,48±0,15 0,20 + 0,20 0,20 ±0,18
Среднеазиатская 0,68 ±0,03 0,36 ±0,07 -0,01 ±0,06
Американская 0,73+0,03 0,06 ±0,08 -0,01 -0,07
Русская 0,68 + 0,04 0,09 । 0,07 0,01-0,07
Восточноазиатская 0,76 + 0,04 0,08 ±0,10 —0,17 + 0,09
Западноевропейская 0,42 10,09 -0,20 ±0,11 - 0,02 2:0,13
Закавказская 0,78 + 0,04 0,25-0,11 —0,21 ±0,14
Малоазиатская 0,56 ±0,06 -0,25 ±0,09 0,02-0,08
Дальневосточная 0,74 ±0,06 0,39 ±0,11 0,20 ±0,13
Велики различия мсжлу группами по средней массе плода. Высо-
кий коэффициент вариации указывает на значительную изменчивость
этого признака также в пределах группы (см. табл. 4). Это хорошо
видно и на рисунке 6: наиболее крупноплодные — афганские, средне-
азиатские и индийские сорта, самые мелкоплодные — дальневосточные.
Корреляция между величиной плода и продолжительностью периода
вегетации очень слабая (г = 0,06—0,39), иногда выражена отрицатель-
ной величиной, что свидетельствует о независимости этих признаков
(см. табл. 5).
На рисунке бив таблице 3 хорошо видны различия эколого-гео-
графических групп по содержанию сахара в плоде. Сорта с высоким
его содержанием (более 9,5%) встречаются чаще всего в западноев-
ропейской и американской группах. Довольно высок процент таких
форм среди русских, среднеазиатских и восточноазиатских сортов. Са-
мая низкая сахаристость отмечена в индийской и афганской группах,
сорта которых меньше подвергались селекции. Они, по-видимому, яв-
ляются исходными и для всего разнообразия сортов столового арбуза.
В каждой группе можно проследить приуроченность сахаристых форм
к определенным географическим районам. Так, среди русской группы
они встречаются чаще всего в Нижнем Поволжье, на Северном Кав-
казе, несколько реже на Украине. Из сортов арбуза Средней Азин
самые сахаристые выращивают в Туркмении и Таджикистане. В За-
падной Европе самые сладкие арбузы возделывают в Болгарии, Юго-
славии, Венгрии. Среди американской группы выделяются сорта южных
и западных штатов США. Высокосахаристые формы восточноазиатской
группы встречаются почти исключительно в Японии.
Прямая корреляция между содержанием сахара в плоде и про-
должительностью периода вегетации фактически отсутствует (см.
табл. 3). Максимум высокосахаристых форм отмечен у среднеспелых
сортов (см. рис. 6). Эта закономерность четко прослеживается во всех
41
эколого-географических группах, хотя в каждой из них проявляется
своя специфика. Так, у наиболее сахаристых сортов русской группы
период от всходов до созревания составляет 85—95 дней, американской
и западноевропейской — 80—90 дней. У сортов восточноазиатской
группы высокая сахаристость сдвигается в сторону среднеранних форм
(период от всходов до созревания 75—85 дней).
Установленная закономерность имеет очень важное значение в се-
лекции арбуза. Она позволяет в значительной степени исключить эле-
мент случайности при отборе высокосахаристых форм и сортов-улуч-
шателей, используемых в селекции.
ИСТОРИЯ ВВЕДЕНИЯ АРБУЗА В КУЛЬТУРУ
Исходным для возделываемого арбуза оказался вид С. lanatus, и
поныне встречающийся в дикорастущем состоянии. Естественные за-
росли этого арбуза были впервые обнаружены Д. Ливингстоном [121]
в безводной пустыне Калахари. Позднее разными исследователями
[293, 332, 358] были найдены другие очаги дикого арбуза не только
в районах, прилегающих к Калахари, но также в Восточной Африке
вплоть до Судана. Местное население называло эти арбузы tsamma
(цамка), а также etanga, kerne, kengwa. Иногда они встречались как
сорняки на полях, иногда возделывались, но по качеству не отличались
от дикорастущих. Д. Ливингстон и другие исследователи отмечали, что
дикие арбузы встречаются в виде двух форм: горькой и лишенной
горечи, съедобной, однако внешне не отличающихся. Установить при-
годность плода в пищу можно было, только пробуя его на вкус.
Генетические исследования таких форм [361] показали, что они
легко скрещиваются между собой, обнаруживая большую близость.
По-видимому, мы имеем дело с естественной гетерогенной популяцией,
разные биотипы которой постоянно переопыляются, образуя формы с
различной степенью горечи в плоде.
Современный ареал естественного произрастания С. lanatus не-
велик и к тому же разорван, что свидетельствует о большой древности
этого вида. В отдаленные геологические эпохи он, несомненно, был
значительно шире и, возможно, как полагает Ф. Штульман [365], про-
стирался за пределы Африки вплоть до Индии. Мнение Ф. Штульмана
достаточно обосновано, и с ним вполне можно согласиться.
В конце неогена произошли грандиозные тектонические разломы
земной коры, в результате которых образовался ряд впадин — грабенов,
в том числе грабен Красного моря [208]. В результате нарушилась
тесная связь Африки с Азией, сообщающихся теперь только через Суэц-
кий перешеек. На севере Индии поднялись Гималайские горы, надежно
изолировавшие Индостанский полуостров с суши. Единый ранее ареал
арбуза оказался разорванным, и дальнейшая эволюция его африканских
и индийских форм шла уже разными путями. В условиях муссонного
климата Индии сформировался иной экологический тип арбуза, чем
в Африке, возможно, легче поддающийся окультуриванию.
Проследить путь культурной эволюции арбуза нелегко, и только
по косвенным данным можно восстановить отдельные его этапы.
Наиболее достоверные материалы дают археологические находки.
Большую роль играют также данные лингвистики, помогающие просле-
дить пути распространения культуры по разным странам.
Одним из первых очагов возделывания арбуза был, вероятно, Еги-
пет— страна, давшая миру многие культурные растения. Е. Н. Син-
ская [195] сообщает, что при раскопках здесь были найдены зерна
пшеницы, возраст которых определяется в 10 тыс. лет, следовательно,
42
первые шаги земледелия в дельте Нила восходят к глубокой древности.
Археологические раскопки на территории Египта помогли установить
преемственную связь его земледельческой культуры с еще более древ-
ними цивилизациями — тазианской и бадарийской, развивавшимися юж-
нее Египта на территории нынешнего Судана. Во флоре этих стран
арбуз и ныне известен в дикорастущем состоянии. Можно предполагать,
что в далекие времена он был здесь типичным растением, известным
местному населению.
Г. Швейнфурт [358] обнаружил в гробнице египетского жреца хо-
рошо сохранившиеся листья арбуза, оказавшегося идентичным тому,
который и теперь возделывается в Верхнем Египте для получения мас-
личных семян. Автор отмечает также, что найденная форма близка
к дикорастущему арбузу Центральной Африки. Эти находки относятся
ко времени XX династии, т. е. примерно к XXI в. до н. э. Следова-
тельно, арбуз был известен в Египте уже 4000 лет назад, но еще мало
отличался от форм, встречающихся в окружающей дикой флоре. К ска-
занному можно добавить, что листья и семена арбуза обнаружены и в
других староегипетских курганах того периода.
Древний Египет имел обширные связи не только с соседними го-
сударствами, но и со странами Междуречья — Ассирией и Вавилоном,
а также с царством Хеттов, занимавшим территорию современной Тур-
ции. Вполне вероятно, что арбуз попал в эти страны из Египта уже
как возделываемое растение. Во всяком случае египетское battich
встречается в несколько измененном виде (bartich, belikt, batteke) в
ряде стран арабского Востока. Название это, в свою очередь, происхо-
дит от древнееврейского abbatitchim. Ниже приводится перечень на-
званий арбуза на разных языках мира, помогающий проследить пути
его распространения:
древнееврейский — abbatitchim, abbatichim, battichim; коптский — betucke; совре-
менный арабский — battich, batteca, batrich; берберский — tadellaat, della; языки на-
родов Северной Африки, близкие к берберскому, — dellaa, hadadja; туарегский —
tiled jest; семитский — samoka.
индийские языки: древнеиндийский санскрит—chaya-pula, tarambya, kutuwombi;
современные индийские — tarmuj, turbus, tarmus, tarbus; бенгальский — tarbuj;
иранские языки: персидский — hindevane, hinduahen, tarbuzah; таджикский —
тарбуз;
греческий язык — carpusia;
славянские языки: русский — арбуз; украинский — кавун; болгарский — диня, лю-
беница; польский — arbuz, kavon jadalny; словацкий—dina; чешский — lubenice, dyne,
meloun vodni; сербский — lubenice;
романские языки: латинский — Citrullus; французский — melon d’eau (старое —
pasteque); итальянский — melone, 1, acqua, anguria, cocomero (старое—pastecca);
португальский — melancia, belancia; испанский — melon de aqua, sandia, cindia; сар-
динский — sindria; румынский — bubenita, pepene verde;
германские языки: английский — watermelon; немецкий — Wassermelon (старое —
pastequegurke); голландский — watermelon; датский — vandmelon; норвежский —
vannmelon; шведский — vattenmelon;
угорские языки: венгерский — gorogdinnue;
тюркские языки: турецкий — карпуз; туркменский — тарбуза, гарпыз; казах-
ский— карбыз; киргизский — тарбаз, дарбыз; узбекский — тарвуз, тарбуз; татарский—
карбуз; азербайджанский — гармыз; монгольский — тербас;
китайский — си-гуа; японский — suikwa; корейский — subak;
языки народов Южной Африки — kengoue, kerne, etanga, tsamma.
На территории Малой Азии арбуз известен с давних пор. При
раскопках армянской крепости Кармир-Блур, существовавшей в VIII—
VII вв. до и. э. в период расцвета государства Урарту, среди других
растительных остатков были найдены в окаменевшем желудке коровы
семена арбуза, близкие к современным [173]. Таким образом, в
II—I тысячелетиях до н. э. арбуз был известен не только в Египте,
но и в ближайших к нему азиатских странах.
43
На территории Африки, несомненно, существовали и другие очаги
возделывания арбуза. Об этом свидетельствуют самостоятельные на-
звания его у разных племен, населявших северную и восточную Аф-
рику. Однако небольшие локальные очаги не имели и не могли иметь
решающего значения в широком распространении культуры арбуза.
Вторым крупным центром формирования и расселения арбуза была
Индия, также страна древнейшей цивилизации. Известны индийские
археологические находки, относящиеся к III—II тысячелетиям до н. э.
Трудно предположить, что культурный арбуз попал в Индию непо-
средственно из Египта: слишком велико расстояние между этими стра-
нами; к тому же индийское название арбуза не имеет ничего общего
с арабским. Видимо, надо согласиться с мнением Ф. Штульмана, что
древний ареал естественного произрастания съедобного арбуза охва-
тывал не только африканские страны, но и простирался в Азию вплоть
до Индии.
В древнеиндийском языке — санскрите, возникшем 4000 лет назад,
наряду с другими названиями арбуза употреблялось слово tarambuya,
созвучное современным индийским названиям turbus, tarbus, tarmus.
Следовательно, арбуз был известен здесь уже во II тысячелетии
до н. э., почти так же давно, как в Египте. Из Индии культурный
арбуз расселился по многим странам Азии. Возможно, индийский очаг
сыграл даже большую роль в широком распространении этой культуры,
чем арабский.
В Персию арбуз попал, несомненно, из Индии. Тесные торговые
связи между этими государствами существовали в течение многих ве-
ков, к тому же территория нынешнего Западного Пакистана, где арбуз
и сейчас широко возделывается, в VI в. до н. э. была частью Персид-
ского царства. Персидское название арбуза hindevane означает «при-
шедший из Индии». Оно встречается здесь наряду со словами turbuz
и tarbuzah, также заимствованными из индийского языка.
В Китай арбуз проник, вероятно, из Индии или из Персии, о чем
свидетельствует его название си-гуа — западный плод. Из Персии же
арбуз распространился по многим среднеазиатским странам, а позднее
попал в приволжские степи. Известно, что в VI—V вв. до н. э. Персия
была могущественным государством, поддерживавшим торговые связи
не только с азиатскими, но и с европейскими народами. В более позднее
время, в VIII—X вв. н. э., Персия и Средняя Азия вели оживленную
торговлю с Киевской Русью. Торговые пути проходили через столицу
Хазарского царства Итиль (ныне Астрахань), и весьма вероятно, что
арбуз попал на Волгу именно таким путем вместе с другими замор-
скими товарами. Названия арбуза на русском, узбекском, туркменском,
таджикском, казахском языках с небольшими изменениями повторяют
персидское слово tarbus.
Предположение о том, что арбуз был занесен в русские степи та-
тарами, маловероятно, так как татары были кочевым народом и зем-
леделием не занимались. Слово арбуз, возможно, было заимствовано
ими у народов Средней Азии.
В Причерноморье арбуз попал, вероятно, не только с берегов
Волги, но и другим путем — через греческие колонии на Черном море.
При раскопках древнего Херсонеса среди остатков различных возделы-
ваемых растений были обнаружены семена арбуза, возраст которых
восходит к I—II вв. н. э. Семена эти сохраняются в Херсонесском ар-
хеологическом музее в Крыму. При раскопках в районе Цимлянского
моря были найдены семена арбуза, относящиеся к VIII в. н. э., близкие
современным формам.
В страны Южной Европы: Испанию, Грецию, Италию, Францию
44
арбуз проник, вероятно, еще до нашей эры из Северной Африки, о
чем свидетельствуют названия его в этих странах (с. 43). Французское
pasteque созвучно староитальянскому pastecca, а оно, в свою очередь,
арабскому batteke. Испанские названия арбуза sandria и sandia, со-
звучные чилийскому sandia, близки к tadellaat, встречающемуся у на-
родов Северной Африки. Древние греки называли арбуз pepones.
Ф. Штульман [365] считает это слово вольным переводом арабского
названия арбуза. Позднее из pepones образовалось слово реро, которым
стали называть тыкву.
К. Линней считал родиной арбуза Италию, так как здесь это рас-
тение обычно. Арбуз был известен уже в Древнем Риме. Он фигури-
рует в указе Диоклетиана о ценах (301 г.).
В Западную и Северную Европу арбуз проник значительно позже,
чем дыня. Об этом красноречиво говорят его названия, переводимые
как водяная дыня: английское — watermelon, французское — melon
d’eau (наряду с pasteque), немецкое — Wassermelone и т. д. (с. 43).
Эти названия свидетельствуют также, что арбуз по качеству был, не-
сомненно, хуже дыни, высоко ценимой в европейских странах.
В Соединенные Штаты Америки арбуз попал, вероятно, совсем не-
давно вместе с переселенцами из других стран уже как вполне куль-
турное растение и нашел здесь весьма благоприятные условия. Благо-
даря интенсивной и целенаправленной селекции в США были созданы
за короткое время разнообразные сорта арбуза, получившие признание
не только у себя на родине, но и за ее пределами.
В России арбуз долгое время был локализован в небольших оча-
гах и получил известность лишь к началу XVII в. К этому времени он
широко распространился по берегам Волги, Дона и Кубани. Русские
арбузы по своим качествам были, несомненно, лучше европейских, о
чем не раз упоминали путешественники, посетившие Россию в XVII—
XVIII вв. [157, 303], и русские историки, в частности В. Н. Татищев
[2Ю].
Хотя на юго-востоке страны арбуз в начале XVIII в. был широко
распространен, в столице он все еще считался диковинкой. Петр I,
побывавший в Камышине в начале своего царствования, был поражен,
узнав, что на русской земле растут такие чудесные плоды, велел дать
в честь арбуза салют и на шпиле городского магистрата водрузить мед-
ный шар — символ арбузной державы [209].
Долгое время крестьяне выращивали арбузы только для собствен-
ных нужд, позднее стали использовать полученную продукцию для
продажи. Постепенно с приусадебных участков и огородов арбуз пере-
шел на поля и приобрел значение промышленной культуры.
В середине XIX в. товарное бахчеводство в России получило зна-
чительное развитие благодаря появлению железных дорог, связавших
зоны выращивания бахчевых с крупными промышленными городами
страны.
Основным районом товарного бахчеводства в конце прошлого века
было Нижнее Поволжье. Здесь засевали бахчевыми культурами более
6% всей пахотной земли. К 1916 г. эта площадь возросла вдвое. Волж-
ские арбузы прочно завоевали славу не только в России, но и за ру-
бежом.
В годы Советской власти бахчеводство в нашей стране получило
дальнейшее развитие.
Современное распространение культурного и дикого арбуза по райо-
нам мира представлено на рисунке 7.
45
Рис. 7. Современный ареал возделываемых и дикооастущих форм Citrullus
lanatus:
1 — столовый арбуз; 2 — дикорастущий.
В процессе многовекового отбора столовый арбуз настолько изме-
нился по сравнению со своим дикорастущим предком, что черты род-
ства можно уловить лишь с трудом. У кормового арбуза, также про-
исходящего от диких африканских форм, это сходство проступает до-
вольно отчетливо (рис. 8).
Кормовой арбуз как культурное растение появился не более 40—
50 лет назад, но его исходная форма была известна крестьянам давно.
Рис. 8. Плод кормового арбуза С. lanatus var. citroides, сорт Зеленокорый (к-3282).
46
В посевах столовых сортов иногда спонтанно появлялись отдельные
растения с длинными крупными плодами и белой невкусной мякотью.
Появление таких сорняков, по-видимому, можно рассматривать как не-
кий атавизм, свидетельствующий о большой гетерогенности столовых
сортов и подтверждающий происхождение их от форм, известных и
поныне в дикой флоре Южной Африки.
В нашей стране созданы сорта кормового арбуза.
В США плоды дикорастущих арбузов из Африки, отличающиеся
плотной белой мякотью и хорошей лежкостью, используются в конди-
терском производстве для приготовления цукатов, поэтому они полу-
чили название цукатных или цитронных арбузов. В отличие от кор-
мовых сортов цукатные арбузы имеют, как правило, шаровидную форму
плода. От аналогичных дикорастущих форм капской разновидности они
отличаются большей выравненностью и отсутствием горечи. Надо по-
лагать, они были введены в культуру совсем недавно.
Дикорастущие арбузы из Южной Африки в последние десятилетия
все шире применяются в селекции столового арбуза для создания сор-
тов, устойчивых к наиболее вредоносным заболеваниям. Почти все но-
вые сорта арбуза, выведенные в США и широко распространенные во
многих странах мира, созданы с использованием дикорастущих форм
из Южной и Центральной Африки.
НАРОДНОХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Арбуз получил широкую известность благодаря сочным сладким
плодам неповторимого вкуса, которые используют обычно на десерт.
Среди растворимых сухих веществ в плодах арбуза преобладают
сахара (6—11 %): сахароза, глюкоза, фруктоза. Наиболее сладкая из
них фруктоза, составляющая 50—60% общей суммы сахаров. На сред-
ний плод массой 3—4 кг приходится 200—300 г сахара.
В плодах арбуза имеются и необходимые организму кислоты:
аскорбиновая, никотиновая, фолиевая. Последняя играет большую
роль в регулировании жизненно важных функций: участвует в синтезе
белка, необходима для кроветворения, обладает антисклеротическим
действием, способствует регулированию жирового обмена. Фолиевая
кислота, содержащаяся, например, в шпинате, зеленом горошке, цвет-
ной капусте, разрушается при варке, поэтому арбуз считают одним из
наиболее ценных источников ее.
Арбуз богат также солями железа и щелочными веществами, ней-
трализующими избыток кислот, вносимых с основными продуктами пи-
тания (мясо, рыба, хлеб, яйца). Легкая усвояемость позволяет реко-
мендовать его людям любого возраста. Норма потребления арбуза,
разработанная Институтом питания, составляет 10—12 кг в год.
Кроме пищевой и диетической ценности, арбуз имеет большое ле-
карственное значение. Он является хорошим мочегонным средством,
способствующим выведению из организма многих вредных веществ, по-
этому особенно рекомендуется при почечнокаменной болезни. Известно
целебное действие арбуза при заболеваниях печени, атеросклерозе, ги-
пертонии. В последнее время установлено, что семена арбуза, так же
как семена тыквы, обладают глистогонными свойствами.
В настоящее время арбуз используется в нашей стране почти ис-
ключительно в свежем виде, но может служить и сырьем для перера-
ботки. В конце прошлого века и в начале нынешнего из сока арбуза
нередко изготовляли мед —нардек. Были попытки получать столовое
вино из мякоти арбуза. Из корок варили цукаты, а мелкие недозрелые
47
плоды солили, как огурцы. Сейчас эти способы переработки сохрани-
лись лишь у отдельных любителей.
В последнее время делаются попытки наладить производство кон-
сервированного арбузного сока как диетического напитка, употребляе-
мого при заболеваниях почек, печени и других недугах. Но его вы-
пускают пока в очень ограниченном количестве. В США развито про-
изводство цукатов из арбуза.
В последние годы и в СССР начато производство цукатов из пло-
дов арбуза. В качестве сырья используется, как правило, кормовой
арбуз, однако делаются попытки заменить его специальными сортами.
Внедрением цукатного арбуза в производство занимается ВИР.
В народном хозяйстве СССР арбуз применяется и как сочный корм
для скота. Сорта кормового арбуза имеют крупные плоды с плотной
несладкой мякотью, которые могут сохраняться до весны. По химиче-
скому составу плодов кормовой арбуз приближается к тыкве. Он ис-
пользуется не только в свежем виде, но и как компонент силосной
массы, повышая усвоение грубых кормов. Кормовой арбуз отличается
сильным молокогонным действием, поэтому особенно ценен в рационе
крупного рогатого скота. Благодаря высокой засухоустойчивости и не-
прихотливости он может возделываться на бедных песчаных почвах
полупустыни, где другие культуры погибают. За свою исключительную
выносливость кормовой арбуз получил название «верблюда раститель-
ного царства».
Кроме пищевого и кормового использования, арбуз играет опре-
деленную роль в агротехнике полевых культур. Он считается одним
из лучших предшественников для зерновых и прежде всего для пше-
ницы, так как, будучи пропашной культурой, оставляет поле чистым от
сорняков и выносит при этом мало питательных веществ из почвы.
В народном хозяйстве нашей страны находят применение и дико-
растущие виды арбуза, например колоцинт, очаги которого встречаются
на юге Туркмении, в долине реки Теджен. Плоды его ядовиты, вызы-
вают очень тяжелые отравления, но в малых дозах успешно исполь-
зуются в медицине как эффективное средство при желудочно-кишечных
заболеваниях. Лечебные свойства колоцинта известны давно. В неко-
торых странах Европы и в Индии его даже специально культивируют
как лекарственное растение. Исследования последних лет показали, что
горькие вещества из плодов колоцинта препятствуют росту опухолевых
образований. Таким образом, область применения его в медицине имеет
перспективы расширения.
В настоящее время принимаются меры для сохранения очагов
естественного произрастания колоцинта в нашей стране.
РАСПРОСТРАНЕНИЕ АРБУЗА
Площадь, занятая бахчевыми культурами, на всех континентах
нашей планеты составляет около 2 млн. га. Привести более точную
цифру не представляется возможным, так как в мировой статистике
отсутствуют данные по многим странам. В настоящее время самые
крупные производители продукции бахчеводства — СССР, США, Япо-
ния, Турция, Иран, Афганистан. Значительные площади бахчевые зани-
мают в Индии, Китае, Австралии, Ираке, Сирии, Египте, Югославии,
Италии, Греции, Болгарии.
СССР занимает первое место в мире по площади (около
400 тыс. га) и валовому сбору столового арбуза. Основные площади
его посевов сосредоточены в европейской части. Они прилегают к бас-
сейнам крупных рек: Дона, Волги, Днепра. Значительные массивы бах-
48
чевых расположены по берегам Азовского и Черного морей. Районы
наиболее интенсивного бахчеводства сосредоточены в Астраханской,
Волгоградской, Ростовской, Херсонской, Запорожской областях, а также
в Краснодарском и Ставропольском краях (рис. 9). Здесь же нахо-
дятся основные научно-исследовательские и селекционные учреждения,
ведущие работу с бахчевыми
культурами. Это зона засушли- \ ,
вых степей, где годовое количе- \
ство осадков не превышает 300— д.
400 мм, а местами составляет,
100—150 мм. Сумма эффектив-
ных температур достигает 3dUU— \
3500°С, средняя температура в 7
летние месяцы (июль и август)
обычно выше 30° С. \
Дальнейшие перспективы раз-
вития бахчеводства в нашей стра- г===\ i—i X
не связаны именно с этими райо- &&Й&1t ь_. .Iz EZZZh J у
нами. Отсюда поступает в города \ г
и промышленные центры основная
часть продукции бахчевых. Только
одна Астраханская область дает рис. 9. Основные районы возделывания
более 3/4 арбузов ПО РСФСР. Од- столового арбуза ^^европейской части
НИМ ИЗ главных поставщиков ИХ | — наиболее интенсивное бахчеводство; 2—
В Ленинград, Москву И другие развитое бахчеводство; 3 — встречается
крупные города является Лиман- местами,
ский район этой области, где дан-
ная культура признана ведущей.
В Средней Азии, где выращивают преимущественно дыню, в по-
следние годы несколько увеличились площади, занятые арбузом, так
как он меньше поражается распространенным здесь фузариозным увя-
данием, от которого дыня сильно страдает.
Среди Закавказских республик первое место по производству бах-
чевых принадлежит Азербайджану. Здесь выращивают как арбуз, так
и дыню. Особенно славятся бахчевыми культурами Шамхорский и Са-
бирабадский районы, а также Нахичеванская АССР. Азербайджан дает
около 2% общего сбора бахчевой продукции. В Армении и Грузии
бахчеводство развито меньше. Здесь преобладает дыня, но в Эчмиад-
зинском районе Армянской ССР имеются крупные массивы арбуза.
Посевы кормового арбуза размещены в засушливых степях Север-
ного Кавказа, Заволжья и Казахстана.
В СССР выращивают почти исключительно селекционные сорта
арбуза, выведенные научно-исследовательскими институтами и опыт-
ными станциями. Местные сорта-популяции встречаются очень редко,
и районы их распространения строго локализованы. Значительная часть
сортов (около ’/з) создана при использовании мировой коллекции
ВИР.
Среди капиталистических стран первое место по производству арбуза принадле-
жит США. Площадь, занимаемая им, составляет здесь более 100 тыс. га. Арбуз
выращивают преимущественно в юго-восточных штатах, расположенных вдоль побе-
режья Мексиканского залива. Природные условия здесь наиболее благоприятны для
возделывания этой культуры [103]. Основными поставщиками продукции являются
Флорида, Джорджия, Южная Каролина, Алабама, Техас. Они дают 70% валового
сбора арбуза. Половина ранней продукции выращивается во Флориде и Техасе.
Климат основной бахчевой зоны США довольно влажный. Количество осадков
на юго-востоке достигает 1000 мм, что является причиной широкого распространения
многих болезней арбуза, в том числе наиболее опасных — антракноза и фузариоза.
49
Здесь ведется большая работа по созданию сортов, устойчивых к болезням. Число
районированных сортов невелико, но они характеризуются также морфологической вы-
равненностью, высоким содержанием сахара в плодах и их прекрасным вкусом.
Один из главных селекционных центров находится в г. Чарлстон (штат Южная Ка-
ролина).
Американские сорта арбуза представляют большую ценность как сорта-улуч-
шатели. Они широко используются в селекции, а во многих странах выращиваются
наряду с местными сортами или взамен их.
Сорта арбуза, выведенные в США, широко распространены на Кубе, в Мексике,
Бразилии, Аргентине, странах Северной Африки, Турции, Ираке, Иране, Индии, Ав-
стралии.
На второе место по производству арбуза среди капиталистических стран следует
поставить Японию, хотя площадь под этой культурой здесь сравнительно невелика
(40 тыс. га). Среди возделываемых в Японии 46 овощных культур арбуз по значи-
мости и посевной площади занимает одно из первых мест, конкурируя с такими
распространенными в этой стране овощами, как редька и капуста.
Посевы арбуза размещены в основном на острове Хонсю в умеренной зоне, где
климат наиболее благоприятен для его возделывания. Культура ведется как при оро-
шении (на тяжелых почвах), так и без него (на легких почвах). Эта особенность
агротехники учитывается при выведении новых сортов. Селекцией арбуза занимаются
в основном частные фирмы, из которых самая крупная фирма «Takii».
В последнее время сортовые посевы все более заменяются гибридными, поэтому
основным направлением в селекции становится создание высокопродуктивных гиб-
ридов F]. Япония занимает первое место в мире по использованию гибридов овощных,
культур, в том числе арбуза, однако способы их получения пока несовершенны.
Основным методом до сих пор остается искусственное скрещивание сортов. Крестьяне,
занимающиеся выращиванием гибридов, предъявляют к ним очень высокие требова-
ния, допуская не более 1% примеси, поэтому гибриды арбуза в Японии очень вырав-
нены. Наиболее популярны в настоящее время мелкоплодные формы массой 2—3 кг
с тонкой корой и нежной мякотью плодов. Большим спросом пользуются арбузы
с желтой корой и желтой мякотью.
Триплоидный бессемянный арбуз, впервые полученный японским генетиком Ки-
хара, широкого распространения не получил, несмотря на высокое качество плодов.
Возделывание его трудоемко и сопряжено с большими затратами, поэтому выращи-
ванием бессемянных арбузов занимаются лишь отдельные любители.
Значительную роль играет арбуз в сельском хозяйстве Турции. Площади бахчи,
по данным мировой статистики, достигают здесь 200 тыс. га. Арбузы возделывают
повсеместно, но преимущественно в азиатской части страны. Сухой континентальный
климат с жарким продолжительным летом благоприятен для бахчевых культур. Для
современной Турции характерен ограниченный сортимент арбуза. По всей стране воз-
делывают лишь 2—3 сорта. Широко распространен американский сорт Вашингтония,
отличающийся превосходным вкусом плодов. Местные сорта возделывают в меньшем
объеме, они также довольно однотипны. Ограниченность сортового состава арбуза
в Турции была отмечена еще П. М. Жуковским [89] во время его экспедиции в эту
страну.
Хорошо развито бахчеводство и в соседних с Турцией странах Сирии и Ираке.
Площадь, занятая бахчевыми культурами, достигает здесь нескольких десятков ты-
сяч гектаров. В Ираке посевы бахчевых расположены преимущественно в северной
части страны. Выращивают в основном американские сорта.
С древнейших времен развито бахчеводство в Иране. Сухой и жаркий конти-
нентальный климат позволяет выращивать здесь отличные арбузы, в основном мест-
ные сорта-популяции. К. И. Пангало [160] неоднократно отмечал богатство местного
сортимента бахчевых культур, представляющего ценный исходный материал для се-
лекции. Однако в последнее время все большие площади занимают привозные сорта,
преимущественно американские. В западные районы проникают также сорта из Тур-
ции.
Арбуз выращивают в Иране почти повсеместно и на довольно большой площа-
ди, достигающей 100 тыс. га, но лучшая продукция поступает из северных районов:
Тегерана, Мешхеда, Нишапура, Горгана. Одним из крупных поставщиков арбуза
является район Ахваза (центральный Иран). В целом следует отметить, что развитие
бахчеводства в этой стране в значительной степени лимитировано недостатком воды,
так как бахчевые культуры нормально развиваются только при орошении.
По природным условиям к Ирану близок соседний Афганистан, также являющий-
ся страной древней земледельческой культуры. Климат Афганистана еще более за-
сушливый. Значительные площади заняты здесь безводными пустынями, поэтому
богарное земледелие возможно лишь в отдельных районах. Сельскохозяйственные
культуры возделывают в основном при орошении. Бахчеводство известно в поливной
и богарной культуре, последняя особенно распространена на северных склонах Гин-
дукуша. Как и в Иране, здесь выращивают в основном местные сорта-популяции,
часто очень пестрые. Лучшие арбузы поступают из северной части страны (Акча..
50
Шибирган, Мазари-Шариф, Ханабад, Файзабад, Меймене). Плоды афганских арбузов
обычно крупные, толстокорые, с грубоватой мякотью. Разнообразие форм огромно,
так же как и в Иране.
Индия и Пакистан являются основным очагом введения арбуза в культуру.
Выходец из Африки, он нашел здесь свою вторую родину как культурное растение
и отсюда распространился по многим странам мира, сохранив свое древнее санскрит-
ское название.
Индия — страна муссонного климата, отличающаяся разнообразием природных
условий. Арбузы выращивают здесь преимущественно в северных и северо-западных
штатах: Уттар-Прадеш, Раджастхан, Пенджаб, Джамму. Для этой зоны характерны
летний теплый дождливый сезон и зимний холодный сухой. Арбузы выращивают в
первую половину летнего сезона, с апреля по июнь, когда осадков выпадает несколь-
ко меньше, чем во вторую, а температура достигает 35—40° С [106]. В производстве
доминируют местные сорта-популяцин с характерными признаками, свойственными
только этой эколого-географической группе. В последние годы начинают распростра-
няться (особенно в Пакистане) американские сорта, постепенно вытесняя местные
самобытные формы. Площадь, занятая бахчевыми культурами, в Индии и Пакистане
составляет более 100 тыс. га.
Китай — огромная страна с разнообразными почвенно-климатическими зонами
и богатой растительностью. Среди сельскохозяйственных культур арбузу принадлежит
здесь довольно скромное место. Можно отметить два основных района его возделы-
вания: северо-восточные провинции, особенно по нижнему течению рек Хуанхэ и ча-
стично Янцзы, а также Синьцзян-Уйгурский автономный район в Западном Китае.
В восточных районах производство арбуза сосредоточено вокруг крупных горо-
дов (Аньшань, Пекин, Тайюань и др.). Здесь встречаются сорта, близкие по типу
к японским и американским. Нередки формы с желтой и оранжевой мякотью плодов.
Очень своеобразен синьцзянский очаг бахчеводства. Это самая засушливая зона
страны, большая часть которой занята пустынями. Климат здесь резко континенталь-
ный, осадков выпадает очень мало (60—100 мм), почвы засоленные. Земледелие воз-
можно только при орошении. Набор культур весьма ограничен, и среди них одно из
первых мест принадлежит арбузу, который выращивают почти повсеместно. Наиболее
значительные массивы сосредоточены в районе Кашгара, Кулъджи, Урумчи. Арбузы
Синьцзяна относятся к другому типу, чем восточнокитайская группа, они ближе
к среднеазиатской группе. Селекционных сортов здесь нет, повсюду выращивают мест-
ные весьма пестрые популяции. Плоды их, как правило, очень крупные (достигают
20 кг), отличаются хорошей лежкостью и транспортабельностью, поэтому представля-
ют ценность как исходный материал для селекции.
Небольшие площади арбуза имеются и в других азиатских странах: Монголии,
Вьетнаме, Индонезии, на Филиппинах, Корейском полуострове, но они не играют
существенной роли в сельскохозяйственном производстве. То же можно сказать о
странах центральной и южной Америки: Мексике, Бразилии, Аргентине, Кубе. Арбузы
здесь выращивают, но на сравнительно небольшой площади, при этом используют
почти исключительно сорта из США.
Довольно хорошо налажено производство арбузов в Австралии. Бахчеводство
сосредоточено в юго-восточных районах страны. Основным поставщиком продукции
является штат Новый Южный Уэльс, преимущественно прибрежная его часть, и в
меньшей мере штат Виктория. Распространены здесь главным образом американские
сорта арбуза, но встречаются и местные селекционные, созданные с привлечением
американского сортимента.
Арбузы выращивают и в южных странах Западной Европы. Наиболее значи-
тельные площади, достигающие нескольких десятков тысяч гектаров, имеются в Бол-
гарии, Греции, Италии, Югославии, Румынии. Однако характерный для данного райо-
на земного шара умеренно теплый климат в сочетании с высокой влажностью воз-
духа не особенно благоприятен для бахчевых культур. Плоды арбуза здесь обычно
имеют невысокое содержание сахара и отличаются посредственным вкусом. Исклю-
чение составляют тщательно отселектированные сорта Венгрии, созданные, вероятно,
с привлечением лучших иностранных образцов. Так же как в Японии, здесь все более
популярными становятся мелкоплодные формы. В последние годы несколько улучшил-
ся сортимент Болгарии, созданный с привлечением зарубежных (из СССР и США)
сортов.
Арбуз возделывают также в странах северной Африки: Египте, Алжире, Тунисе,
Марокко. Наиболее значительные площади его в Египте. Бахчеводство в этих странах
исключительно поливное. В посевах преобладают американские сорта, но встречают-
ся и неплохие местные формы, например сорт Гиза, возделываемый не только в Егип-
те, но и в соседних странах.
Довольно хорошо развито производство арбуза в ЮАР — на исконной родине
этого растения. Площади под ним достигают здесь 35 тыс. га.
4*
51
БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА АРБУЗА
ТРЕБОВАНИЯ К УСЛОВИЯМ СРЕДЫ
Как уже было сказано, родина арбуза — пустынные районы Юж-
ной Африки, поэтому он весьма теплолюбив и требует для своего
развития длительного периода с температурой выше 15° С. Арбуз
можно успешно возделывать лишь в тех районах, где сумма эффек-
тивных температур достигает 3200° С и выше. В нашей стране наибо-
лее благоприятные условия для выращивания арбуза имеются на юге
Украины, Северном Кавказе и в Нижнем Поволжье, где и сосредото-
чены основные площади этой культуры.
Арбуз — светолюбивое растение, и потому посевы его размещают
всегда на открытых местах. Фотопериодическая реакция его исследо-
вана слабо, однако имеются данные, свидетельствующие о положитель-
ном влиянии укороченного 12-часового дня в первые фазы роста, до
появления 4—5-го настоящего листа [95].
Арбуз растет лучше всего на легких, хорошо прогреваемых и
аэрируемых почвах, достаточно богатых питательными веществами.
При этом он очень чувствителен к реакции почвенной среды и хорошо
развивается лишь на нейтральной и слабощелочной почве.
В условиях экстенсивного крестьянского хозяйства культура арбуза
издавна велась на целине или многолетней естественной залежи. Ста-
ропахотные земли считались для него непригодными [158]. В настоя-
щее время, когда земледелие приобрело интенсивный характер и не-
распаханной земли почти не осталось, арбуз, естественно, вводится в
севооборот. Однако там, где есть возможность (например, в Лиманском
районе Астраханской области), посевы его размещают по многолетней
залежи. Несмотря на высокую требовательность арбуза к плодородию
почвы, для него в то же время совершенно непригоден свежий навоз,
на внесение которого он реагирует более поздним созреванием и ухуд-
шением качества плодов.
Арбуз — засухоустойчивое растение и обычно выращивается на су-
ходольных (неполивных) землях. К избытку влаги он относится отри-
цательно. Во влажные годы созревание начинается позже и качество
плодов ухудшается. Поэтому во влажных субтропиках, характеризую-
щихся высокой суммой эффективных температур и большим количе-
ством осадков, арбуз возделывают лишь на незначительных площадях.
Тем не менее освоение новых земель в засушливых районах заставляет
все чаще прибегать к искусственному орошению бахчевых культур. Ар-
буз при этом образует мощную вегетативную массу, дает хороший уро-
жай крупных плодов, которые, однако, оказываются менее сладкими,
чем при возделывании на суходоле [23]. Чтобы качество плодов при
поливе не снижалось, необходимо вносить повышенные дозы минераль-
ных удобрений, особенно фосфорных. Как показывают многие исследо-
вания [110, 276 и др.], правильное сочетание минеральных удобрений
с орошением не только не снижает сахаристости плодов, но, наоборот,
повышает ее. При внесении минеральных удобрений лучше разви-
ваются корневая система и надземная часть растений, увеличивается
площадь ассимиляционного аппарата, а следовательно, повышается
урожай. Минеральные удобрения наиболее эффективны при глубокой
заделке под основную вспашку, а также при внесении в рядки одно-
временно с посевом. Наибольшую потребность арбуз испытывает в
азоте и фосфоре, причем особенно благоприятным оказывается сочета-
ние компонентов с некоторым превышением фосфора, примерно 1 : 1,3.
Без полива минеральные удобрения дают несколько меньший эффект,
52
но тоже способствуют повышению урожайности и улучшению качества
плодов.
Многолетней практикой бахчеводства и прямыми опытами дока-
зано, что арбуз довольно устойчив к почвенному засолению, особенно
сульфатному [47, 97]. Некоторые авторы считают, что устойчивость
обусловлена сосущей силой корней и листьев, обеспечивающей нормаль-
ную водообеспеченность в условиях острого водного дефицита, вызы-
ваемого засолением. По данным В. Д. Знаменского [97], небольшие
количества сернокислого и хлористого натрия оказывают даже стиму-
лирующее влияние на рост растений и ускоряют наступление основных
фаз развития. Более того, они способствуют накоплению сахаров и осо-
бенно фруктозы, повышая тем самым сладость и улучшая вкус плодов.
Практикам-бахчеводам также хорошо известно, что самые сладкие и
вкусные арбузы получаются на слабозасоленных землях.
По данным Астраханской опытной станции ВИР, содержание хло-
ридов не более 0,02% при плотном остатке 0,15—0,20% не вызывает
угнетения растений. Засоление же 0,1—0,2% при плотном остатке бо-
лее 1% часто приводит к полной гибели растений арбуза.
Основные районы бахчеводства в нашей стране расположены в
зоне засушливых степей. В связи с этим такие признаки, как засухо-
устойчивость и жаростойкость, имеют для арбуза немаловажное зна-
чение. П. А. Генкель [56] отмечал, что засухоустойчивость — это не
признак растения, а процесс его активного приспособления к засухе.
Она определяется целым рядом анатомических, морфологических и фи-
зиологических свойств: развитием корневой системы, величиной сосу-
щей силы листа и корня, интенсивностью транспирации, водоудержи-
вающей способностью листа, толщиной кутикулы и развитием опушения,
наличием эфирных масел, содержанием осмотически активных веществ,
соотношением свободной и связанной воды и т. д.
Засухоустойчивость изучалась нами на образцах арбуза коллекции
ВИР [248]. Из перечисленных выше показателей мы избрали прежде
всего сосущую силу листа как показатель, характеризующий результат
взаимодействия всех осмотических, коллоидно-химических и биохими-
ческих процессов, влияющих на активность воды в клетке [74]. Сосу-
щую силу определяли методом компенсации по Н. А. Максимову и
Н. С. Петинову, позволяющим проводить исследования в полевых усло-
виях. Всего было изучено 36 сортов арбуза разного происхождения;
основные данные получены в 1960 г., на отдельных сортах определения
повторяли в 1961, 1963, 1964 гг.
Несмотря на то что все образцы были выращены в совершенно
одинаковых условиях, экологические особенности районов их происхож-
дения очень четко проявлялись в величине сосущей силы листа. У изу-
ченных образцов она колебалась в значительных пределах (от 9,6 до
22,7 атм). Образцы из влажных мест земного шара (Индии, Китая,
Дальнего Востока, Закавказья) имели более низкие показатели, чем
сорта из районов с сухим климатом: Поволжья, Малой Азии, Ирана,
Афганистана (рис. 10). Наиболее высокую сосущую силу, достигаю-
щую 22 атм, имели листья арбуза из Судана — страны жаркого и су-
хого климата.
Изучение сосущей силы листа по фазам развития растений пока-
зало, что она постепенно возрастает, достигая максимума в период
цветения и снижаясь ко времени созревания плодов.
Как было отмечено выше, среди показателей, определяющих засу-
хоустойчивость растений, большую роль играет соотношение свободной
и связанной воды. Уменьшение количества свободной воды и увеличе-
53
ние связанной ведет к нарушению хода физиологических процессов,
а иногда и к гибели растений. Способность растений удерживать во
время засухи свободную воду является основным условием их выжи-
вания.
Рис. 10. Сосущая сила листа (в атм) у сортов арбуза разных эколого-географических
групп:
1—среднеазиатская; II — американская; III—дальневосточная; IV — индийская; V —
восточноазиатская; VI — русская; VII — малоазиатская; VIII — афганская; IX — су-
данская.
Нами исследована водоудерживающая способность 15 сортов ар-
буза разных эколого-географических групп [248]. Для определения
использован метод А. А. Ничипоровича, основанный на учете потери
воды срезанными листьями. При изучении водоудерживающей способ-
ности листа четко определились различия между сортами разных эко-
лого-географических групп. Листья растений из районов с влажным
климатом (более мезофильные формы) теряли больше воды, чем из
районов с сухим климатом. Самую низкую водоудерживающую способ-
ность имели листья сортов дальневосточной эколого-географической
группы. За 6 ч завядания они потеряли до 50% воды, в то время как
листья сортов русской эколого-географической группы только 22—23%
полного насыщения. Особенно интенсивная потеря воды листьями на-
блюдалась в первые 30 мин завядания. У растений арбуза дальневос-
15%, русской — всего 4% (рис. 11).
Из других признаков, характеризую-
щих водный >ежим и засухоустойчивость
растений арбуза, была исследована сосу-
щая сила кпрней [27]. Выяснено, что
наиболее засрхоустойчивые сорта арбуза
в условиях богарной (неорошаемой)
культуры в Средней Азии имеют мощную
корневую систему и отличаются высокой
интенсивностью выделения пасоки кор-
нями, так называемого «плача». Установ-
лена положительная тесная корреляция
между этими двумя признаками. Тем же
автором проводились исследования по
жаростойкости. Этот показатель опреде-
точнои группы она достигала
Продолжительность забивания
Рис. 11. Потеря воды срезанными
листьями арбуза у сортов различ-
ного происхождения:
1—Мелитопольский 143 (русская
группа); 2 — к-2011 (дальнево-
сточная группа).
54
ляли методом Ф. Ф. Майкова по температуре свертывания водораство-
римых белков листьев. Установлено, что более жаростойкие сорта ар-
буза отличаются высокой температурой свертывания белка (69—71°С).
Среди них одно из первых мест занимает сорт Кузыбай богарный, пер-
спективный для возделывания на богаре.
К аналогичным выводам о связи жаростойкости арбуза с темпера-
турой свертывания белка пришла и М. И. Малинина [132], изучавшая
коллекцию арбуза в засушливой степи Приаралья. Автор отмечает, что
наряду с высокой температурой свертывания белка (65—67° С) жаро-
стойкие сорта арбуза характеризуются высоким уровнем транспирации,
предохраняющей растения от перегрева в жаркие часы дня.
Отмечена также тесная корреляция между жаростойкостью и за-
сухоустойчивостью арбуза.
БИОЛОГИЯ РАЗВИТИЯ
Рост и развитие растений арбуза в первую очередь зависят от ка-
чества семян. Благодаря твердой оболочке, обладающей слабой водо-
проницаемостью, семена арбуза сохраняют жизнеспособность длитель-
ное время. Проведенное нами изучение долговечности семян разных
сортов показало, что они могут сохраняться без потери всхожести
7—8 лет, но энергия прорастания снижается уже через 4—5 лет хра-
нения [256].
На долговечность семян большое влияние оказывают условия вы-
ращивания и уборки, а также степень их зрелости. Семена, выращен-
ные во влажный год, быстрее теряют всхожесть, чем семена, получен-
ные в сухой год. То же можно сказать о семенах, плохо высушенных
после уборки и недостаточно вызревших. На долговечность семян ар-
буза оказывают влияние и сортовые особенности. Согласно нашим дан-
ным, более отработанные сорта дольше сохраняют высокие посевные
качества, чем местные сорта-популяции.
Прорастание. Для прорастания семян арбуза требуется довольно
высокая температура. Нижний ее предел равен 12—14° С, но быстрее
всего семена прорастают при 25—30° С. Такая температура при лабо-
раторной проверке всхожести семян обеспечивает прорастание их в ус-
ловиях оптимальной влажности уже на 3—5-й день. При посеве семян
арбуза в открытый грунт всходы появляются на 10—12-й день, но при
неблагоприятных условиях прорастание может затянуться до 20 дней
и более. Посев проводят чаще всего в первой декаде мая (редко в
конце апреля), когда почва достаточно прогреется, чтобы обеспечить
быстрое появление всходов. Резкие понижения температуры отрица-
тельно сказываются на проростках, вызывая различные физиологиче-
ские нарушения, а иногда и полную их гибель.
В области корневой шейки проростка образуется утолщение стеб-
левого происхождения, которое мы условно называем пяточкой (в бо-
танической литературе нет специального термина для этого выроста).
Пяточка играет большую роль в раскрытии створок семенной кожуры,
помогая выходу зародышевого корешка. Вероятно, она выполняет так-
же всасывающую функцию, пока корешок проростка развит слабо. Об
этом свидетельствует густое опушение, развивающееся по краю пяточки
в условиях высокой влажности.
Форма ее и степень развития могут быть различными. Отмечено,
что у сильных проростков пяточка более развита, чем у слабых [250].
Возможно, существует определенная зависимость между развитием пя-
точки и силой роста проростков, что имеет существенное значение для
раннего прогнозирования гетерозиса гибридов.
55
Фаза семядолей. Изучение ранних этапов онтогенеза растения
представляет интерес для разработки теоретических вопросов в области
систематики и для решения практических задач селекции.
Ценный материал для исследования дает семядольная фаза про-
ростков. В этот период развития у растений могут проявляться черты
древней организации, характеризующие внешний и внутренний облик
далеких предков, поэтому сравнение морфологических и анатомических
признаков проростков позволяет судить о филогенетической близости
видов и форм.
Своеобразие проростков арбуза может проявляться во внешнем
виде семядольных листьев, в различной форме выроста на корневой
шейке, в характере опушения и т. д. Эти признаки могут быть исполь-
зованы и в селекционной работе, например при отдаленной гибридиза-
ции для раннего выявления характера наследования.
Изучение семядольной фазы проростков очень важно с точки зре-
ния биологии развития. Семядольные листья играют большую роль в
формировании корневой системы проростка, оказывая, таким образом,
влияние на последующее развитие молодого растения. Если семядоли
повреждены или слабо развиты, корни развиваются плохо, растение от-
стает в росте и развитии, что ведет к снижению его продуктивности.
Существенные различия отмечены по величине семядольных ли-
стьев. Наиболее крупные семядоли характерны для тех форм Citrullus,
которые имеют хорошо развитые корни (subsp. lanatus var. lanatus,
subsp. vulgaris var. cordophanus, subsp. mucosospermus), что полно-
стью согласуется с имеющимися в литературе указаниями о тесной
взаимосвязи между развитием семядолей и корневой системы про-
ростка [43].
Рис. 12. Размер и форма семядо-
лей:
1—С. ecirrhosus; 2 —С. lanatus.
Очень своеобразную форму семядолей, не встречающуюся у Citrul-
lus и близких ему родов, имеет С. ecirrhosus: узкая пластинка семя-
дольного листа ложкообразной формы постепенно переходит у него в
длинную, лишенную пигмента черешковую часть, семядоли при этом
всегда приподняты (рис. 12). Форма семядолей этого вида служит хо-
рошим диагностическим признаком, по-
зволяющим определять его типичность
уже в период всходов.
Среди видов арбуза наиболее раз-
витые корни имеют проростки С. lana-
tus, особенно арбуза слизистосемян-
ного (subsp. mucosospermus) и дико-
растущих форм арбуза шерстистого
(subsp. lanatus). Следует подчеркнуть,
что развитые растения этих подвидов
отличаются мощной вегетативной мас-
сой, плети их достигают 4—5 м. У про-
ростков С. colocynthis, С. ecirrhosus,
первичный корень развит относительно
С. naudinianus и Praecitrullus
слабо. Некоторые различия между видами отмечены по длине гипоко-
тиля: наиболее длинный гипокотиль имеют проростки арбуза шерсти-
стого, самый короткий — С. naudinianus. ? и
пкАп^м3ЛЬа,1С!1еМчД0ЛЬНЫХ листьев тыквенных растений продолжается
шимся и чтлми патем они °™ирают, передавая свои функции появив-
различия пазных₽епМеНИ НДСТО”ЩИМ листьям. Нами отмечены некоторые
вания семядолей РямВ ^lrU us по продолжительности функциониро-
С ecirrhosus Этот ДРУГИХ начинается отмирание семядолей у
с. ecirrhosus. Этот вид обитает в пустыне Юго-Западной Африки. Ве-
56
гетация его приурочена к очень короткому периоду дождей, и поэтому
весь цикл развития растение проходит ускоренно. Очень быстро
растет стебель, появляются новые листья, одновременно и в большом
количестве раскрываются мужские и женские цветки, обеспечивая за-
вязывание сразу многих плодов. Видимо, раннее отмирание семядолей,
связанное с быстрым появлением настоящих листьев, является в ка-
кой-то мере отражением ускоренного темпа развития растений этого
вида. Позже других отмирают семядоли у столового арбуза, что, ви-
димо, связано с благоприятными условиями питания.
Рост вегетативной массы. Цветение. Первый настоящий лист появ-
ляется через 7—10 дней после всходов, вслед за ним, примерно через
3—5 дней, второй и последующие листья. С появлением 5—6 листьев
наступает фаза шатрика, после чего начинается образование плетей.
Семядоли к этому времени обычно отмирают. В первое время рост над-
земной части идет медленно, но зато усиленно развивается корневая
система, главным образом за счет образования многочисленных боко-
вых корешков. С началом ветвления и особенно появлением первых
цветков вегетативная масса интенсивно нарастает. От главной плети
отходят плети I порядка, которые, в свою очередь, тоже ветвятся, об-
разуя плети II и 111 порядка. В течение вегетации растение образует
до 40 м стеблей и до 900 листьев. Поверхность ассимиляционного ап-
парата развитого растения может достигать 3 м2 [2, 64, 199].
Фаза плетеобразования начинается через 17—37 дней после появ-
ления всходов. Время наступления ее зависит от района возделывания,
погодных условий, сортовых особенностей. Так, в Астрахани у боль-
шинства изучаемых образцов в 1970 г. число дней от всходов до пле-
теобразования составило 17—23, в Геническе (Херсонская область) в
тот же год — 23—40, а в Самарканде — 29—30 [27].
Через 7—19 дней после начала плетеобразования на растении на-
чинают появляться мужские цветки (в пазухе 4—12-го листа главной
плети), а вслед за ними женские и обоеполые (в пазухе 5—30-го листа
главной плети и на плетях I порядка). Продолжительность периода
всходы — раскрытие первого женского цветка колеблется от 36 до
65 дней и, так же как начало плетеобразования, зависит от сорта,
почвенно-климатических и погодных условий. Если в Астрахани скоро-
спелые формы арбуза зацветают уже в середине июня, а среднеспелые
сорта — в конце июня — начале июля, то в Геническе эта фаза насту-
пает несколько позднее: скороспелые формы начинают зацветать в на-
чале— середине июля, а средне- и позднеспелые — в конце июля — на-
чале августа. Жаркая сухая погода ускоряет появление цветков, сырая
и холодная задерживает. Цветение продолжается обычно до конца
вегетации, но интенсивность его после завязывания первых плодов зна-
чительно снижается.
Формирование плода. Период завязывание — созревание плода про-
должается 32—46 дней, в зависимости от скороспелости сорта, но чаще
всего длится 32—35 дней. Рост плодов происходит особенно интенсивно
в первые 3 недели, затем заметно приостанавливается и начинается их
созревание. Мякоть становится сочной, рыхлой, приобретает окраску,
свойственную сорту. Одновременно идет и созревание семян. Как пра-
вило, семена приобретают полную спелость несколько позже потреби-
тельской готовности мякоти, поэтому плоды, предназначенные для се-
менных целей, выдерживают в поле некоторое время после уборки пли
снимают позже. На растении обычно созревают 1—2. редко 3—4 плода.
Внешние признаки зрелости арбуза не так отчетливы, как у дыни,
у которой спелость определяют чаще всего по изменению окраски плода.
57
У зрелого арбуза более четко проявляется рисунок коры, несколько
усыхают плодоножка и ближайший к ней усик, но главным признаком
спелости является изменение звука при постукивании: глухой звук
означает, что плод созрел и готов к употреблению.
Продолжительность периода вегетации. Период от всходов до нача-
чала созревания колеблется в широких пределах (60—120 дней). На
продолжительность его влияют те же факторы, которые определяют
наступление других фаз. Большую роль играют почвенно-климатические
условия района возделывания. Так, сорт Муравлевский (к-3563) в
1972 г. начал созревать в Астрахани через 69 дней после появления
всходов, в Харькове через 76 дней, а в Тирасполе через 108 дней, об-
разец к-3622 соответственно на 67, 71, 97-й день. Приведенные цифры
свидетельствуют о том, что условия Молдавии менее благоприятны
для возделывания арбуза, чем Нижнее Поволжье и Украина.
Период от всходов до созревания значительно меняется и в зави-
симости от погоды. Например, на Кубанской опытной станции ВИР
созревание сорта Огонек было отмечено в сухом и жарком 1972 г. на
68-й день, а в холодном и дождливом 1973 г. на 89-й, а созревание
сорта Мелитопольский 142 началось соответственно на 83-й и 102-й
день.
Но основным фактором, определяющим созревание, является сорт.
Средне- и позднеспелые сорта в любых условиях созревают позже, чем
раннеспелые, так как цикл развития определяется биологическими осо-
бенностями растений.
Разница в созревании скороспелых и позднеспелых сортов, выра-
щенных в одинаковых условиях, может достигать 30 и более дней. Так,
на Кубанской опытной станции ВИР в 1972 г. созревание самого ран-
него сорта арбуза (вр. к-422) было отмечено на 61-й день, а самого
позднего (к-4291)—на 87-й день. На Астраханском опорном пункте
ВИР в том же году плоды наиболее раннего сорта (к-3781) начали
созревать через 67 дней после появления всходов, а наиболее позднего
(вр. к-391) — через 98 дней. На Генической опытной станции ВНИИК
в 1968 г. созревание самого раннего сорта (к-3471) отмечено через
74 дня, а наиболее позднего (к-348) —через 101 день. В условиях Алма-
Аты период от всходов до созревания у скороспелых сортов был равен
79—83 дням, а у позднеспелых—114—116 дням [1].
Развитие корней. Корневая система растений развивается парал-
лельно с надземной частью. Стержневой корень арбуза сравнительно
короткий и углубляется в почву не более чем на 1 м, но чаще всего
достигает глубины лишь 30—40 см. Однако боковые корни могут сильно
разрастаться вширь или вглубь в зависимости от типа почвы и ее во-
дообеспеченности. Без полива (на богаре) корневая система более раз-
вита, чем при орошении. Корни иногда достигают глубины 2 м и про-
стираются в стороны на 8—10 м [27]. Для богарных условий При-
аралья, по данным М. И. Малининой [132], характерно распростране-
ние корней преимущественно в верхнем горизонте (от 10 до 30 см).
Автор делает вывод, что такое расположение корней позволяет им бо-
лее эффективно использовать влагу редких летних осадков.
При орошении, особенно на полях с близким залеганием грунтовых
вод, что характерно для пойменных земель Астраханской области, кор-
невая система арбуза обычно развивается в самых верхних горизонтах,
на глубине 8—12 см, а главный корень не проникает глубже 40 см [49].
БИОЛОГИЯ ЦВЕТЕНИЯ И ОПЫЛЕНИЯ
Цветок и его развитие. У растений рода Citrullus Schrad. встре-
чаются три типа цветка: мужской, женский и гермафродитный.
58
Рис. 13. Цветки арбуза:
1 — мужской; 2 — женский, 3 — обоеполый.
У большинства сортов арбуза на одном растении образуются муж-
ские и гермафродитные (обоеполые) цветки. Такой половой тип назы-
вается мужской однодомностью (андромонойкист). Растения, у которых
формируются мужские и женские цветки, классифицируются как
истинно однодомные, или монойкисты.
Цветок арбуза (рис. 13) имеет 5-лепестный широковоронковидный
венчик с лепестками, сросшимися у основания, и прицветниками лодоч-
кообразной формы. У столового арбуза венчик имеет сернисто-желтые
широкие округлые лепестки
3—4 см в диаметре. У полу-
культурных форм и сортов
кормового направления ле-
пестки венчика более узкие,
заостренные, ярко-желтые,
4—5 см в диаметре. Муж-
ской цветок арбуза всегда
несколько крупнее женского,
имеет 5 тычинок: 4 срос-
шиеся попарно, одна сво-
бодная. Пыльники хромово-
желтые, петлеобразно изогнутые, растрескивающиеся с внешней сто-
роны. Рыльце 4-угольное или округлое, покрытое сосочками, зеленое,
желтое или оранжевое.
В наших исследованиях обнаружена некоторая зависимость окраски
рыльца цветка от экологических условий. Так, у арбуза русской эколо-
гической группы преобладают сорта с зеленым рыльцем, а индийской
группы — с желтым и оранжевым.
Была установлена также определенная связь окраски рыльца
цветка и мякоти плода у столовых сортов арбуза [251]. Оказалось, что
сорта с зеленым или зеленоватым рыльцем имеют обычно розовую или
малиновую мякоть плода, это, вероятно, обусловлено преобладанием
в них пигмента ликопина. Сортам с желтым и оранжевым рыльцем в
большей мере свойственна желтая, шарлаховая и карминная мякоть,
что связано, возможно, с повышенным содержанием каротина.
При изучении степени корреляции этих признаков было установ-
лено, что у сортов с зеленым рыльцем соотношение форм с розовой
и малиновой мякотью и форм с мякотью других тонов равно 6:1.
У сортов с желтым рыльцем это соотношение оказалось равным 2:1,
т. е. изменилось в сторону увеличения числа сортов с карминной и шар-
лаховой мякотью.
Установленная нами корреляция может быть использована в се-
лекционной работе, поскольку она позволяет проводить предваритель-
ный отбор по окраске плода уже в период цветения.
Исследования К. И. Пангало [162] и М. К. Гольдгаузен [65] по-
казали, что в цветках арбуза всегда имеются элементы противополож-
ного пола. Андроцей в женских цветках присутствует обычно в виде
тычиночных бугорков — стаминодиев, гинецей в мужских цветках — в
виде бугорков плодолистиков — пистиллодиев. У сортов с гермафродит-
ными цветками можно обнаружить все ступени перехода от стаминодиев
к нормально развитым тычинкам с фертильной пыльцой. На развитие
андроцея большое влияние оказывают погодные условия. При холодной
погоде тычинки развиваются слабее. Так, в Херсонской области в 1969 г.
средняя температура июля была на 2° С ниже средней многолетней,
и многие коллекционные образцы арбуза, которым свойственны герма-
фродитные цветки, в этом году совсем не образовали тычинок или
59
имели только одну. У дикорастущих форм арбуза из Южной Африки,
как отмечает М. К. Гольдгаузен, андроцей, наоборот, очень устойчив:
всегда представлен тремя тычинками и не варьирует по степени их
развития.
Завязь арбуза нижняя, одногнездная, образована тремя плодо-
листиками. Согласно исследованиям Б. Т. Матиенко [134], наружные
стенки завязи состоят из сросшихся оснований чашелистиков, лепестков
и тычинок. Внутренние стенки образованы тканью плодолистиков, за-
вернувшиеся края которых, разрастаясь, образуют плаценту.
У сортов столового арбуза завязь имеет короткое редкое опуше-
ние. У полукультурных форм и сортов кормового направления опуше-
ние завязи мягкое, густое (войлочное). У колоцинта опушение завязи,
как и всего растения, редкое, жесткое (рис. 14).
Рис. 14. Опушение завязи арбуза:
1—столового; 2 — каффрского: 3 — колоцинта.
Американский ботаник Д. Роза [354] обнаружил определенную
корреляцию между типом цветка и формой плода арбуза. Он устано-
вил, что из гермафродитных цветков развиваются шаровидные плоды,
из пестичных — плоды удлиненной формы. Эта закономерность была
подтверждена К. И. Пангало [164] и М. К. Гольдгаузен [65] при изу-
чении мировой коллекции бахчевых культур ВИР.
Эколого-географическое изучение коллекции арбуза, проведенное
нами в последние годы [251], показало, что такая корреляция прояв-
ляется не всегда. Например, растения арбуза дальневосточного и вос-
точноазиатского экологического типа имеют раздельнополые цветки, но
шаровидные плоды. Среди образцов русской экологической группы
встречаются сорта с удлиненно-овальными плодами: Кубанский длин-
ный (к-2178), Ляльки (к-2972), На закуску (к-2645) и др. Однако
цветки их обоеполые и этот признак стойко передается по наследству.
Цветок растений семейства Cucurbitaceae закладывается на конусе
нарастания как бугорок вторичной меристемы. На нем, в свою оче-
редь, появляются бугорки будущих элементов цветка: 5 бугорков чаше-
листиков, лепестков, тычинок и, наконец, плодолистиков позднее раз-
растающихся в завязь.
Формирование цветка у растений данного семейства изучала
60
И. Н. Львова [129]. Она установила, что в эмбриональном развитии
цветка можно различить следующие фазы.
I. Образование участков меристемы, дающих начало цветочным бу-
горкам, при появлении первого настоящего листа.
II. Образование цветочных бугорков.
III. Дальнейшее развитие конуса нарастания, в процессе которого
он принимает бокаловидную форму и начинает дифференцироваться на
чашечку, венчик, тычинки и плодолистики.
Зачаточный цветок потенциально обоеполый. Из него может сфор-
мироваться мужской, женский и гермафродитный цветок. Развитие по-
следнего наиболее продолжительно.
На формирование того или иного типа цветка большое влияние
оказывают окружающие условия: температура, влажность и особенно
свет. И. Н. Львова [129] отмечает, что на V этапе морфогенеза на цве-
ток существенно влияет спектральный состав света, изменением кото-
рого можно регулировать пол цветка. Так, при освещении зеленым све-
том увеличивается продолжительность всех фаз развития и пестичные
цветки в таком случае могут совсем не образоваться. Преобладание
красного света создает предпосылки для преимущественного развития
тычиночных цветков, а при синем наблюдается ускоренное развитие
пестичных.
Среди ботаников нет единого мнения об эволюционно более древ-
нем цветке — раздельнополом или гермафродитном. Имеются доводы в
пользу того и другого положения. Видимо, этот вопрос нельзя решать
односторонне, поскольку гермафродитизм может быть первичным и вто-
ричным, эволюционно молодым. Раздельнополость же возникла не
только в процессе эволюции, но в значительной степени благодаря ин-
тенсивному искусственному отбору.
Особенности цветения. Цветки арбуза располагаются одиночно
в пазухе листа. Первые женские и гермафродитные цветки заклады-
ваются не на боковых, а на главной
плети и равномерно распределяются
по всему растению (рис. 15).
На высоту заложения генератив-
ных органов большое влияние оказы-
вают погодные и климатические усло-
вия. В годы с холодной весной первые
цветки раскрываются в узлах, дальше
отстоящих от семядольных листьев,
чем в годы с теплой сухой весной.
Сорта разных экологических групп
различаются по высоте заложения
мужских и женских цветков на стебле.
Так, у сортов дальневосточной и вос-
точноазиатской групп первый женский
цветок часто раскрывается уже в па-
зухе 4—6-го листа. Растения арбуза
индийской экологической группы отли-
чаются, наоборот, поздней закладкой
генеративных органов. Первые жен-
ские и гермафродитные цветки раскры-
ваются у них в пазухе 17—20-го листа,
а у образцов из Судана даже на 20—30-м узле. Пестичные цветки, как
правило, раскрываются ближе к семядольным листьям, чем обоеполые.
Период от всходов до начала цветения женских цветков служит
у арбуза показателем скороспелости сорта. По нашим данным, этот
Рис. 15. Расположение обоеполых
цветков на растении арбуза.
61
период в условиях степи Украины составляет 36—65 дней, а у боль-
шинства коллекционных образцов —40—50. Во влажных районах про-
должительность периода всходы — цветение увеличивается, а в засуш-
ливых сокращается. Период от начала цветения до созревания продол-
жается 32—46 дней и в меньшей степени определяется сортовыми осо-
бенностями, чем продолжительность периода всходы — цветение.
Мужской цветок арбуза остается открытым только один день.
К вечеру он закрывается, засыхает и отваливается. Женский цветок
обычно тоже остается открытым 12—15 ч. Если же он оказался неопы-
ленным, то на следующий день раскрывается снова, но рыльце тем-
неет, подсыхает и его жизнеспособность снижается. При опылении
цветков на второй день раскрытия завязывается всего 1—8%
плодов.
Как показали наши наблюдения, наиболее интенсивное цветение
мужских цветков раннеспелых сортов арбуза бывает в первые две не-
дели, затем число раскрывающихся цветков уменьшается. У средне- и
позднеспелых сортов интенсивное цветение мужских цветков продол-
жается значительно дольше.
Женские цветки у раннеспелых сортов арбуза начинают раскры-
ваться через 4—6 дней после появления мужских, а у средне- и поздне-
спелых сортов — через 10—12 дней. Завязывание плодов лучше всего
происходит в первые 10—12 дней после начала цветения, но если рас-
тения развиты слабо, то первые завязи обычно опадают.
Важные факторы, влияющие на эффективность опыления, — влаж-
ность и температура воздуха. Оптимальной является температура 18—
20°С при относительной влажности воздуха не ниже 35%. Слишком
сухая и жаркая погода, так же как холодная и сырая, вызывает опа-
дение завязей.
Завязывание плодов арбуза при естественном опылении может до-
стигать 50% и более, при искусственном составляет 10—20% и лишь
в отдельных благоприятных случаях — 40—50% числа опыленных
цветков.
Мужских цветков на растении арбуза образуется намного больше,
чем женских, и располагаются они по всему растению. Одно растение
арбуза может нести 100—400 мужских цветков и всего 10—30 женских.
Соотношение тех и других значительно меняется под влиянием условий
выращивания, что создает широкие возможности для направленного
сдвига пола в сторону увеличения числа женских цветков. Разные ис-
следователи приводят следующие соотношения мужских и женских
цветков у арбуза, свидетельствующие об изменчивости этого показа-
теля: Р. Хенц и X. Моор — 6:1, Р. Хоутори и Р. Поллард — 7:1,
А. И. Филов [238]— 15 : 1, А. В. Снегирева [199]— 10—12 : 1, Ф. А. Тка-
ченко [221] —20:1. У дикорастущих форм арбуза из Калахари (var.
lanatus) женских цветков на одно растение приходится значительно
меньше: на 450—600 мужских цветков 15—20 женских. Соотношение
тех и других составляет у них примерно 30: 1, у полукультурных форм
арбуза из Судана — 26: 1.
Изучение большого коллекционного материала, проводимое нами в
течение многих лет, показало, что соотношение мужских и женских
цветков на растении зависит от многих причин: происхождения, скоро-
спелости сорта, погодных условий и т. д. Если у скороспелых сортов
арбуза оно равно 6—10: 1, то у позднеспелых — 20—25: 1. По высокой
насыщенности растений женскими цветками выделяются образцы из
Японии: отношение мужских цветков к женским составляет у них
6—7: 1.
62
Регулирование соотношения цветков. Существенное влияние на пол
растений семейства Cucurbitaceae оказывают фотопериод и темпера-
тура. Установлено, что длинный день (до 16 ч) и высокая температура
(около 30° С) вызывают, например, у огурца интенсивный рост и обиль-
ное образование мужских цветков, короткий (8-часовой) день и отно-
сительно низкая температура (16°С), наоборот, способствуют увели-
чению числа женских цветков и более раннему их появлению.
О. В. Юрина [279] использовала эти факторы для получения скоро-
спелой формы арбуза, формирующей женские цветки в 3—5 узлах глав-
ной плети.
Определенное влияние на образование цветков того или иного пола
оказывает также влажность воздуха и почвы. По нашим наблюдениям,
в засушливые годы число мужских цветков на растениях арбуза умень-
шается и возрастает соответственно насыщенность женскими цветками.
Так, у скороспелого сорта New Hampshire midget во влажном 1966 г.
отношение мужских цветков к женским было 7:1, в засушливом
1968 г. —4,6: 1.
Многочисленными работами доказано, что изменить пол растений
можно и путем воздействия физиологически активными веществами
типа ингибиторов и ауксинов. Действие этих веществ на растения ар-
буза изучали С. Рэм, Р. Хенц, X. Моор и К. Е. Дютин.
В опытах Рэма еженедельное опрыскивание всходов арбуза раство-
ром 2,4-Д вызывало торможение роста растений и частичную стериль-
ность пыльцы мужских цветков. При опрыскивании всходов дикорасту-
щего арбуза раствором ТИБ резко повысилось число женских цветков:
соотношение мужских и женских цветков изменилось с 10:1 в конт-
рольном варианте до 0,9:1. Использование гидразида малеиновой кис-
лоты привело к таким же результатам, но действие его оказалось
слабее.
Р. Хенц и X. Моор применяли раствор препарата FW-450 в кон-
центрации 0,5% для раскрытия максимального числа женских цветков.
К. Е. Дютин пытался получить формы арбуза со стерильными муж-
скими цветками воздействием физиологически активными веществами.
Однако он пришел к выводу о недостаточной эффективности этого спо-
соба, так как стерильные мужские цветки возникали лишь на короткий
период после обработки.
В отличие от таких культур, как огурец и кабачок, у которых
используются молодые плоды — зеленцы, у арбуза нет прямой связи
между насыщенностью растений женскими цветками и количеством за-
вязавшихся плодов.
Исследованиями А. Г. Авакяна [2], И. Ф. Лященко [130], А. И. Фи-
лова [238] было установлено, что растущие плоды у арбуза, дыни и
тыквы не только тормозят раскрытие женских цветков и завязывание
новых плодов, но и задерживают рост боковых побегов. В период роста
плода пластические вещества растения расходуются в основном на то,
чтобы обеспечить его нормальное созревание. А. Хиббард пришел к
выводу, что на растении целесообразно оставлять по 2 плода, иначе
они получаются уродливыми и зачастую как следует не вызревают.
А. Г. Авакян [2] отмечает, что при наличии трех завязей на одной
плети арбуза вторая и третья не развиваются, пока не будет снят пер-
вый созревший плод.
Подытоживая эти исследования, можно сказать, что урожай арбуза
зависит не столько от количества женских цветков на растении, сколько
от успешного их оплодотворения и количества плодов, которое расте-
ние может обеспечить необходимыми питательными веществами.
63
Развитие пыльцевого зерна. Пыльца арбуза сравнительно крупная.
Пыльцевое зерно эллиптической формы с полярной осью, 49—52 мкм,
в экваториальном диаметре 52—57 мкм, трехборозднопоровое. Экзина
сетчатая. Пыльцевые зерна столового, кормового и дикорастущего ар-
буза отличаются по рисунку экзины [6]. Пыльца содержит белки, ами-
нокислоты, жиры и сульфгидрильные группы. Крахмал и сахара в ней
отсутствуют.
Микроспорогенез у арбуза в основных чертах проходит так же,
как у других двудольных растений [70, 71, 145]. В тычиночных бугор-
ках обозначаются лопасти тычинок, дифференцируются эпидермальный
и субэпидермальный слои. В последнем выделяются клетки первичного
археспория. Затем обособляются гнезда пыльников по два в каждой
лопасти и слой выстилающих клеток с секреторной функцией — тапе-
тум. Далее образуется вторичный археспорий, дающий начало материн-
ским клеткам микроспор. Они претерпевают два мейотических деления,
в результате которых образуются тетрады — четырехъядерные микро-
споры. Оболочка тетрады исчезает, и она распадается на 4 микроспоры,
дающие начало одноядерной пыльце. Тапетум к этому времени начинает
разрушаться.
В результате деления ядра микроспоры образуется двухъядерная
пыльца, состоящая из двух клеток: генеративной (спермиогенной) и ве-
гетативной. Генеративная клетка значительно меньше и располагается
у оболочки пыльцевого зерна.
Весь цикл развития пыльцевого зерна, начиная от заложения пер-
вичного археспория до образования двуклеточной пыльцы, длится
6—7 дней. Перед растрескиванием пыльников спермиогенная клетка
делится с образованием двух спермиев, которые представляют собой
веретеновидные клетки с овальными ядрами. Попадая на рыльце пе-
стика, пыльца соприкасается с его липкими сосочками, набухает и на-
чинает прорастать. По данным С. К. Цыганкова [266], на рыльце
цветка арбуза попадает до 120 000 пыльцевых зерен, т. е. количество
во много раз большее, чем необходимо для нормального завязывания
семян, однако этот избыток пыльцы нельзя рассматривать как ненуж-
ное излишество.
Прорастание пыльцы на рыльце столового арбуза начинается уже
через 15 мин после опыления. По данным А. Г. Алиевой [10], через 2 ч
пыльцевые трубки достигают столбика и здесь их рост несколько за-
медляется. До первых семяпочек они дорастают через 9 ч. По данным
Д. Р. Портера [347], пыльцевые трубки достигают семяпочек за более
короткий срок — 3—4 ч, по данным В. А. Гуляева [71], — не более
чем за 7—8 ч.
Развитие семяпочки. Зародышевый мешок у растений семейства
Cucurbitaceae развивается из халазальной макроспоры по типу Poly-
gonum [73]. В субэпидермальном слое обособляются нуцеллус и архе-
спориальные клетки, которые становятся материнскими клетками макро-
спор — макроспороцитом. В результате двух мейотических делений об-
разуется тетрада с линейным расположением спор. Три верхние споры
дегенерируют, а из халазальной макроспоры путем ряда последователь-
ных делений образуется восьмиядерный зародышевый мешок. В нем
дифференцируются клетки яйцевого аппарата, центральная клетка за-
родышевого мешка и антиподы. Длина готового к оплодотворению за-
родышевого мешка равна у столового арбуза 65—70 мкм, кормового
70—80, колоцинта 66—90 мкм. Существенных различий в строении за-
родышевого мешка разных видов Citrullus не обнаружено.
Яйцевой аппарат довольно крупный при сравнительно небольших
размерах зародышевого мешка и находится на микропилярном его
64
конце. Он состоит из двух синергид и яйцеклетки, которая распола-
гается между ними и несколько ниже.
На халазальном конце зародышевого мешка формируются 3 анти-
поды. В центре, непосредственно под яйцевым аппаратом, распола-
гаются полярные ядра с очень крупными ядрышками. Слияние их про-
исходит уже после внедрения пыльцевых трубок.
Развитие семяпочки идет параллельно развитию зародышевого
мешка. Семяпочки закладываются уже в завязи диаметром 2 мм. Ну-
целлус принимает грушевидную форму. Верхний конец его, суженный
в носик, входит в микропиле. Внутренний интегумент утолщается выше
носика нуцеллуса. Сильно развивается внешний интегумент. Созревшая
семяпочка у всех представителей трибы Cucurbiteae крассинуцелляр-
ная, двухпокровная. В микропиле семяпочки может входить несколько
пыльцевых трубок, но зародышевого мешка в большинстве случаев до-
стигает только одна. Диаметр ее составляет примерно 7з диаметра за-
родышевого мешка. Проникая внутрь, пыльцевая трубка повреждает
одну из синергид, внедряясь в нее своим кончиком. Содержание ее из-
ливается или внутрь синергиды, или в щель между яйцеклеткой и си-
нергидами.
Собственно процесс оплодотворения у представителей семейства
Cucurbitaceae исследователям пока не удалось наблюдать. Но, по кос-
венным данным, можно заключить, что он проходит быстро и относится
к премитотическому типу [73]. Полярные ядра сливаются, вероятно,
после слияния ядра спермия с одним из них. Пыльцевые трубки у
Citrullus после выполнения своей основной функции не сплющиваются,
как у других тыквенных, а продолжают существовать в тканях ну-
целлуса.
Зародыш у всех тыквенных формируется по типу Onagraceae. Ну-
целлус сохраняется длительное время и начинает дегенерировать только
под влиянием развивающегося эндосперма. Кожура семени образуется
в результате дифференциации внешнего интегумента семяпочки.
Жизнеспособность пыльцы. Биологические свойства пыльцы арбуза
отражены в работах Т. Е. Пащенко [171], А. Г. Алиевой [52], И. М. Са-
фарян [189, 190], Т. Б. Фурса [251].
Жизнеспособность пыльцы и ее фертильность изучали проращива-
нием на искусственных питательных средах, а также путем хранения
ее в различных условиях. А. Г. Алиева проращивала пыльцу во влаж-
ной камере методом висячей капли. По ее данным, оптимальной пита-
тельной средой для проращивания пыльцы арбуза является 10%-ный
раствор сахара с добавлением 2%-ной желатины при температуре
18—20° С. Т. Е. Пащенко применила более простой способ. Питатель-
ную смесь наливали тонким слоем на предметные стекла, которые по-
мещали в чашки Петри, выложенные влажной фильтровальной бумагой.
Лучшей средой для проращивания оказался 20%-ный раствор сахара
с добавлением 2%-ного агара. Через 1 ч в опытах Т. Е. Пащенко про-
растало 59% пыльцевых зерен столового арбуза, 38% кормового и 24%
колоцинта. Пыльца последнего прорастала значительно медленнее. Та-
кой же способ проращивания использовали И. М. Сафарян и
Т. Б. Фурса. В наших опытах оптимальной средой оказался 15%-ный
раствор сахара с добавлением 0,01%-ного раствора борной кислоты.
Проращивание проводили при температуре 26—28° С. Прорастание
пыльцы начиналось через 6—8 мин после посева: через 1 ч проросло
44—65% пыльцевых зерен.
В опытах И. М. Сафарян пыльца сохранялась в цветках в обычных
комнатных условиях при температуре 22—26° С и влажности 60—65%.
5 Зак. 2323
65
При опылении цветков пыльцой, хранившейся 10 ч, процент завязыва-
ния резко падал, а после 30 ч хранения пыльца полностью теряла свою
жизнеспособность.
В исследованиях Т. Е. Пащенко пыльца при хранении в эксикаторе
сохраняла способность к прорастанию на искусственной питательной
среде в течение 6—8 недель, но для опыления оказалась пригодной
только в течение 18 дней.
Многих исследователей [36, 53, 119, 127 и др.] интересовал воп-
рос, равноценна ли пыльца из гермафродитных цветков арбуза по
оплодотворяющей способности пыльце из мужских цветков. Одни счи-
тали пыльцу из гермафродитных цветков не способной к оплодотворе-
нию, другие полностью признавали такую возможность. И. М. Сафарян
[190] и Т. Б. Фурса [249] провели сравнительное изучение пыльцы из
мужских и гермафродитных цветков арбуза. Фертильность пыльцы опре-
деляли путем самоопыления гермафродитных цветков и проращиванием
ее на искусственной питательной среде.
В наших опытах и в работе И. М. Сафарян пыльца из гермафро-
дитных цветков оказалась вполне фертильной. При проращивании на
искусственной питательной среде всхожесть пыльцы из мужских и гер-
мафродитных цветков почти не различалась, но в наших опытах про-
явилось различие в энергии прорастания.
Пыльца из гермафродитных цветков прорастала медленнее, осо-
бенно в первые 30 мин после посева. Через 1 ч длина пыльцевых тру-
бок из гермафродитных цветков была 150—200 мкм, из мужских цвет-
ков— 200—500 мкм, т. е. почти в 2 раза больше.
При опылении гермафродитных цветков своей пыльцой (автога-
мия) процент завязывания в опытах И. М. Сафарян был несколько
ниже, чем при опылении пыльцой из мужских цветков. В наших опы-
тах разницы не отмечено, завязывание плодов при автогамии состав-
ляло 40—60%. Такой высокий результат был связан, возможно, со спо-
собом нанесения пыльцы на рыльце. Опыление цветка проводили опу-
шенным кончиком плети, который в данном случае уподоблялся мохна-
тому брюшку пчелы. Пыльца при этом способе нанесения легко стря-
хивалась на рыльце, не травмируя его.
Опыление. Арбуз — энтомофильное растение, о чем свидетельству-
ют открытое цветение, крупный яркий венчик, липкая тяжелая пыльца,
а также хорошо развитые нектарники в мужских и женских цветках.
Нектарники в мужских цветках представлены выростом пистилло-
дия, сросшимся основанием с боковыми стенками и околоцветником.
Поверхность их имеет небольшую воронкообразную выемку. В жен-
ских цветках нектарники представлены валиком, плотно окружающим
столбик, сросшимся с околоцветником и основаниями стаминодиев.
По данным С. К. Цыганкова [266], цветки арбуза обладают огром-
ными запасами нектара (до 100 кг/га). При этом нектар их содержит
до 22% сахара. Т. И. Казиев и С. С. Сеидова отмечают, что содержа-
ние сахара в нектаре цветков арбуза может достигать даже 50%,
причем мужские цветки отличаются большей нектаропродуктивностью,
чем женские. Выделение нектара начинается в 8—9 ч утра, максималь-
ное его количество выделяется в 11 —14 ч, с наступлением сумерек
секреция прекращается. Женские цветки выделяют нектар только до
опыления. Установлено, что цветки бахчевых культур посещают около
150 видов насекомых, но основную группу (30—40%) опылителей со-
ставляют пчелы, причем из 17 видов пчел наибольшее значение как
опылитель имеет медоносная пчела (Andrena mellifera L.). Второе
место занимают степные муравьи (Lasius niger L.). Некоторую роль
66
играют также трипсы, мухи и клопы, но они чаще находятся в преде-
лах одного цветка, способствуя самоопылению.
Пчелы посещают цветки тыквенных ради нектара и пыльцы, ко-
торую они собирают в особые корзиночки третьей пары ног. Одна пчела
может нести на себе до 70 тыс. пыльцевых зерен, в то время как мура-
вей — лишь 360, а трипс — всего 6.
Деятельность пчел начинается около 8 ч утра, наиболее активны
они в период с 9 до 14 ч. Как показали опыты многих исследователей,
в том числе наши, наиболее высокая завязываемость плодов прихо-
дится на период с 8 до 13 ч (время наилучшего физиологического со-
стояния цветка и наибольшего посещения его пчелами). Каждая пчела
за день совершает 4—5 вылетов из улья и при этом многократно по-
сещает цветки. Опытами А. Н. Невкрыта [149] установлено, что для
получения полноценного урожая плодов арбуза кратность посещений
женского цветка пчелами должна быть примерно равна 36, а мужского
цветка — 20.
Большинство исследователей отмечают, что деятельность пчел в
поле не беспорядочна, а, наоборот, планомерна. В разгар цветения
пчела, перелетая с цветка на цветок, двигается вдоль рядка. Хаотиче-
ское движение пчел бывает только тогда, когда цветущих растений
в поле мало. Закончив облет одного поля, пчела всегда счищает пыльцу
в корзиночки и на другое поле летит уже чистая. На активность пчел
большое влияние оказывают погодные условия. При холодной дождли-
вой или ветреной погоде количество пчел резко сокращается, тогда,
видимо, возрастает роль муравьев, трипсов, клопов.
Деятельность муравьев начинается раньше, чем пчел. Они появ-
ляются одновременно с раскрытием цветков, в 6—7 ч утра. Трипсов
можно обнаружить не только в раскрывшихся цветках, но и в бутонах
накануне раскрытия [221].
У растений семейства Cucurbitaceae возможны 3 способа опыле-
ния: автогамия — самоопыление гермафродитных цветков, гейтенога-
мия — опыление пыльцой с других цветков того же растения и ксено-
гамия— опыление пыльцой с других растений. Первые два способа
можно рассматривать как самоопыление, третий следует отнести к пе-
рекрестному опылению.
Вопрос о способах опыления бахчевых растений интересовал мно-
гих исследователей. Н. И. Вавилов [36] и Р. К. Лангельд [119] счи-
тали арбуз типичным перекрестником, так как его цветки и пыльца
привлекательны для насекомых. Большинство исследователей [61, 87,
127, 164] полагают, что арбуз скорее можно отнести к факультативным
перекрестникам, так как многие его сорта имеют гермафродитные
цветки, что не исключает возможности самоопыления. Вероятно, пра-
вильнее признать возможность разных способов опыления. Оба отли-
чаются известной лабильностью, поэтому трудно сказать, какой способ
преобладает. При достаточном количестве пчел цветки арбуза опы-
ляются преимущественно перекрестно. При малом числе насекомых
сорта с гермафродитными цветками опыляются в основном автогамно,
иногда, возможно, с помощью ветра. В этом, видимо, и следует ви-
деть эволюционное назначение гермафродитизма.
Известно, что перекрестное опыление резко повышает гетерозигот-
ность материала, самоопыление и особенно автогамия, наоборот, ведут
к его гомозиготности и могут фиксировать те благоприятные изменения,
которые иногда возникают путем естественного переопыления различ-
ных биотипов популяции. Сочетание обоих видов опыления поддержи-
вает популяцию в равновесии, обеспечивая ее жизнеспособность.
5*
67
Нами были изучены два способа опыления арбуза (сорт Мелито-
польский 60): гейтеногамия и ксеногамия. Опыт проводили в течение
двух лет (1966 и 1968). В обоих случаях процент завязывания при
гейтеногамии был несколько выше: в первый год при опылении растений
своей пыльцой он был равен 20, при опылении пыльцой с других рас-
тений — 10, во второй год соответственно 60 и 40.
Как видно из полученных данных, гейтеногамия оказалась не-
сколько более результативным способом опыления, чем ксеногамия.
Следует отметить, что к такому же выводу пришла и А. С. Щукина
[277], изучавшая способы опыления среднеазиатских сортов дыни. Од-
нако эти данные нельзя рассматривать как окончательные. Необходимо
дальнейшее углубленное изучение способов опыления арбуза на боль-
шом коллекционном материале.
Наряду с гейтено- и ксеногамией определенную роль в процессе
воспроизведения семян играет автогамное опыление. В наших опытах
растения, выращенные из семян от автогамного опыления, развивались
вполне нормально, а по некоторым показателям даже превосходили
растения контрольного варианта, полученные из семян от свободного
опыления (табл. 6).
Таблица 6. Влияние автогамного опыления на развитие и продуктивность потомства
Сорт Вариант опыта Число растений Длина главной плети, см Число плетей на растении Число плодов на делянке Продук- тивность одного растения, кг
Стокса Контроль 5 142 3,6 11 1,29
« Опыт 5 172 3,6 25 1,66
Любимец хутора Контроль 5 174 3,8 9 1,90
Пятигорска
То же Опыт 5 276 4,4 12 2,47
Из таблицы 6 видно, что растения опытных вариантов у обоих сор-
тов превосходили контрольные по длине плетей, числу плодов и про-
дуктивности. Возможно, автогамия не всегда дает такие высокие ре-
зультаты, но во всяком случае этот способ опыления следует признать
полноценным, существующим наравне с другими. В неблагоприятные
годы с малым числом насекомых автогамия обеспечивает, вероятно,
нормальное воспроизведение потомства. В селекционной работе авто-
гамное опыление вполне можно использовать для получения выравнен-
ных линий от сортов, обладающих хозяйственно-ценными признаками
и свойствами.
Изучение способов опыления у арбуза тесно связано с решением
многих важных вопросов селекционной и семеноводческой работы и
прежде всего спорного вопроса о нормах пространственной изоляции
в семеноводческих посевах. Несколько лет назад в семеноводстве бах-
чевых культур была принята изоляция посевов 2000 м на открытой
местности и 1000 м на защищенной. В настоящее время нормы изо-
ляции сокращены до 1000 м для открытой местности и 500 м для
защищенной. Многие исследователи и селекционеры [69, 112, 124, 204,
262 и др.] считают, что и эти нормы сильно завышены и могут быть
сокращены еще более: до 500 м на открытой местности и 200—300 м
на защищенной.
Л. Е. Кревченко [112] на основе многолетнего опыта селекцион-
ной и семеноводческой работы с бахчевыми культурами полагает, что
даже небольшая изоляция, равная 50 м, надежно гарантирует чистоту
сорта. Основанием для мнения о возможности сокращения норм про-
бе
странственной изоляции в селекционных и семеноводческих посевах
служат многочисленные исследования, проводимые в нашей стране и
за рубежом. Опытами М. К. Гольдгаузен установлено, что перекрестное
опыление между сортами практически отсутствует уже на расстоянии
5—7 м. Исключение составляют лишь длинноплодные сорта арбуза с
раздельнополыми цветками, которые дают довольно высокий процент
гибридов при совместном посеве их с другими сортами.
Изучение биологии цветения арбуза связано также с проблемой
гетерозиса и разработкой новых, более совершенных способов получе-
ния гибридных семян.
АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ ОРГАНОВ АРБУЗА
Значимость анатомических признаков определяется возможностью
использования их для целей систематики и классификации, а также
для выявления форм, ценных для селекции.
При изучении внутренней структуры отдельных органов серьезное
внимание обращено на строение кожуры семени арбуза, имеющей важ-
ное таксономическое значение. Полученные данные использованы при
разработке новой внутривидовой классификации С. lanatus.
Было проведено анатомическое изучение проростков арбуза, кото-
рое помогло проследить развитие проводящей системы стебля и корня,
а также выявить различия в строении этих органов у разных видов,
подвидов и разновидностей.
Изучение анатомии листа у сортов арбуза разных эколого-геогра-
фических групп позволило установить четкие количественные различия
между ними.
При изучении анатомии плода основное внимание было обращено
на структуру механической ткани, определяющей прочность коры плода,
и на строение мякоти плода у сортов арбуза различного происхождения.
Результаты наших исследований дополнены литературными дан-
ными по анатомии разных органов арбуза.
КОЖУРА СЕМЕНИ
Многие растения семейства Cucurbitaceae, в том числе арбуз, —
выходцы из засушливых степей и пустынь. Семена их имеют, как пра-
вило, толстую твердую кожуру (спермодерму), надежно защищающую
зародыш от воздействия неблагоприятных факторов. Мощный слой по-
ристой одревесневшей ткани играет роль термоизолятора, обеспечивая
сохранение жизнеспособности семян в период знойного безводья, когда
температура на поверхности почвы достигает 60—70° С, и при резких
понижениях температуры до заморозков. Хорошо развитая проводящая
система способствует быстрому набуханию и прорастанию семян при
появлении хотя бы небольшого количества влаги. По данным Ф. А. Тка-
ченко [224], для прорастания семян арбуза достаточно 85—90% влаги
от массы сухого семени, в то время как для гороха требуется 116—
141%, а для томата —325—364% влаги. Кожура семян арбуза играет,
таким образом, важную биологическую роль, обеспечивая их длитель-
ное сохранение и быстрое прорастание при наступлении благоприятных
условий.
Строение кожуры семени арбуза описано в работах Д. Фикеля
[300], Ч. Харца [309], Ф. Нетолицкого [341], С. В. Рутковской [183],
В. Н. Сойфера [202], Н. М. Ткаченко [225]. Все названные исследова-
тели, кроме С. В. Рутковской, изучали только столовый арбуз. Опуб-
ликованные ими материалы дают представление о характере семенной
69
кожуры арбуза, но, к сожалению, не отражают разнообразия типов ее
строения у разных форм Citrullus.
На основании литературных данных и наших исследований общий
план строения спермодермы арбуза можно представить в следующем
виде. Кожура семени состоит из нескольких гистологических зон. Сна-
ружи находится палисадный эпидермис, в отдельных случаях замещаю-
щийся изодиаметрическим, иногда отпадающий. Клетки его имеют пиг-
мент, от распределения и концентрации которого зависит окраска се-
мени; у темноокрашенных семян он сконцентрирован в наружной части
клеток. Основные пигменты ликопин, каротин, хлорофилл.
Тангентальные стенки эпидермальных клеток покрыты обычно
слоем кутикулы, толщина которого может быть различной. Трещинки
на поверхности семян, встречающиеся часто у западнокитайских форм
арбуза, обусловлены разрывами слаборазвитой кутикулы и расхожде-
нием эпидермальных клеток. У столового арбуза клетки эпидермиса
длиннее, чем у дикорастущих форм.
Ниже расположена склеренхимная ткань — гиподерма, по строению
которой виды Citrullus заметно различаются. Гиподерма состоит из
6—18 рядов клеток с одревесневшими стенками, пронизанными много-
численными порами и имеющими воздухоносные полости. Как исклю-
чение, встречаются формы, у которых гиподерма состоит из тонкостен-
ных клеток с целлюлозными оболочками. Строение гиподермы указы-
вает на ее защитную и теплоизолирующую функции. В какой-то мере
эта ткань способствует также набуханию семян при попадании их во
влажную среду.
Под гиподермой расположен ряд сильно одревесневших каменистых
клеток с очень толстыми стенками и маленькой сдавленной полостью.
Благодаря тесному сочленению клеток, хорошо заметному на танген-
тальном срезе, этот слой представляет собой очень прочное образова-
ние и служит основной опорной тканью семенной кожуры. От гипо-
дермы он отделяется узкой, сильно преломляющей свет полосой.
Еще глубже расположены тонкостенные гидрацитные трахеиды —
ткань, выполняющая проводящую и всасывающую функции. Этот слой
однотипен у всех видов арбуза.
Анатомическое строение семенных покровов растений сравнительно
редко используется в систематике как таксономический признак.
Имеется небольшое число работ, авторы которых на основании струк-
туры семени уточняют систематическое положение и филогенетические
связи некоторых семейств, подсемейств и родов: В. Ваюган, Д. Вайтхаус
[367], А. П. Меликян [141, 143], А. Л. Тахтаджян и А. П. Меликян
[214], В. Н. Сойфер [202]. Для диагностики видовых различий этот
признак применяли лишь немногие исследователи: С. В. Рутковская
[183], А. П. Меликян [142], 3. В. Редькина [180], Н. С. Морозова [147].
Наибольшую ценность для нас представляет работа С. В. Рутковской,
изучавшей строение семенных покровов разных видов Citrullus на ма-
териале коллекции ВИР. Полученные ею данные должны были под-
твердить правильность существующей классификации арбуза, но это
не всегда становилось возможным. Так, виды С. colocynthoides Pang,
и С. ancestralis Gold, оказались идентичными по строению семенной
кожуры, а С. colocynthis, наоборот, отличался разнообразием ее струк-
туры. По мнению С. В. Рутковской, среди этой гетерогенной группы
в дальнейшем можно ожидать выявления новых видов. Последний вы-
вод нашел подтверждение и в наших исследованиях.
Необходимо отметить, что в коллекции арбуза отсутствовали в то
время многие африканские формы, имеющие большое значение для
70
уточнения классификации рода. Не были представлены виды С. naudi-
nianus и С. ecirrhosus, отсутствовали образцы из Судана, Сенегала,
Ганы, Гвинеи, Мали, привлеченные в коллекцию позднее. Поэтому ра-
бота С. В. Рутковской, несмотря на отдельные важные выводы, пред-
ставляется все же незаконченной. Наши исследования кожуры семени
арбуза можно считать логическим продолжением этой работы. Тип
строения семенной кожуры был использован нами наряду с другими
признаками при разработке новой классификации арбуза. Именно этот
показатель оказался решающим при выделении нового, ранее не опи-
санного в ботанической литературе подвида арбуза слизистосемян-
ного — ssp. mucosospermus Fursa [252].
В настоящей работе приводится анатомическая характеристика се-
менной кожуры всех форм Citrullus, имеющихся в коллекции, и пред-
ставителей некоторых близких родов.
Кожура семени арбуза очень твердая, режется с трудом и для ана-
томических исследований требует специальной подготовки. В течение
месяца семена выдерживали в смеси равных частей глицерина, воды и
спирта. После такой обработки кожура несколько размягчалась и ста-
новилась пригодной для изготовления срезов. Срезы делали от руки
бритвой и сразу же просматривали под микроскопом марки МБД-1 при
увеличении 9X7, 9X10, 9X15. Лигнификацию тканей выявляли путем
окрашивания смесью флороглюцина с соляной кислотой. Для окраши-
вания кутикулы применяли судан-Ш, в отдельных случаях метиленовый
синий. Промеры проводили с помощью окуляр-микрометра. Для зари-
совок использовали рисовальный аппарат РА-6.
Характеристика анатомических признаков семян изученных образ-
цов представлена в таблице 7, где приведены количественные показа-
Таблица 7. Анатомические показатели семян Citrullus и близких родов
Изученный таксон Толщина
кожуры, мкм эпидермиса, мкм эпидермиса к толщине кожуры, % кутикулы, мкм гиподермы, мкм гиподермы к толщине кожуры, % каменистых клеток, мкм
Столовый арбуз с окра- 640-705 300-375 51 19 255 36 60
шенными семенами
Столовый арбуз с бе- 515 75—150 или 20 Нет 350 68 60
лыми семенами отсутствует
Суданский 525-605 195 35 30 245 44 60
Каффрский 530-620 90—150 21 28 360 63 60
Кормовой 510 90 17 30 330 64 60
Капский 470 75 16 15 330 70 50
Слизистосемянный 735-1880 75-150 7 Нет 525—1700 89 60
Сенегальский 1035 75 7 900 90 60
Колоцинт горький 443 45 10 38 315 70 45
Колоцинт пресный 555 150 29 15 345 62 45
Безусиковый 495 60 12 30 345 70 60
Арбуз Нодэна 970 30-70 7 34 850—935 90 Нет
Фистулезный 450 245 40 28 240 53
Бенинказа восковая 750 Нет — Нет 650 86 80
Бенинказа мохнатая 400 150 37 170 43 80
Нара 495 120 24 - 375 76 Нет
Кукумис фиголистный 375 150 40 » 225 60 Не от- делялся
тели, позволяющие судить о толщине семенной кожуры и развитии со-
ставляющих ее слоев. Темносемянные и белосемянные сорта столового
арбуза рассматриваются отдельно, так как имеют некоторые различия
71
в строении семенной кожуры. Из данных таблицы 7 видно, что первая
группа сортов отличается более толстой кожурой благодаря лучшему
развитию эпидермиса. У белосемянных сортов эпидермис обычно пред-
ставлен нежными тонкостенными клетками, не имеющими кутикулы.
При высыхании семени эпидермис часто отпадает или разрушаются
только наружные тангетальные стенки клеток. У некоторых белосемян-
ных сортов палисадный эпидермис заменяется изодиаметрическим. Сла-
бое развитие эпидермиса у белосемянных форм арбуза в какой-то мере
компенсируется более развитой гиподермой.
По строению кожуры семени к столовому арбузу близки полукуль-
турные формы из Судана, занимающие промежуточное положение
между сортами с темноокрашенными и светлыми семенами. Суданские
образцы арбуза имеют много общего со столовым и по другим призна-
кам, поэтому отнесены нами к тому же подвиду subsp. vulgaris.
По строению кожуры близки между собой все три разновидности
арбуза шерстистого (subsp. lanatus, рис. 16,а). Гиподерма кожуры се-
мени у них развита очень хорошо и отличается сильным одревеснением
клеточных стенок. Эпидермис тоньше, чем у столового арбуза, кутикула
толще. Строение семенной кожуры свидетельствует о ксерофильности
этих форм, что подтверждается и образом их жизни. Арбузы каффрский
и капский встречаются в виде естественных зарослей в саваннах и
пустынях Южной Африки, кормовой арбуз (var. citroides) возделы-
вается в засушливых степях нашей страны и считается одним из самых
засухоустойчивых культурных растений.
Совершенно своеобразны внешний вид и анатомическая структура
семян арбуза слизистосемянного (subsp. mucosospermus, см. рис. 16,6).
Эта форма была вначале зарегистрирована как образец тыквы. У дан-
ного подвида встречаются крупно- и мелкосемянные формы. Те и другие
имеют одну характерную особенность: свежие семена заключены в тол-
стую студенистую оболочку, которая при высыхании семени остается
в виде тонкой сухой пленки, легко набухающей в воде. У крупносемян-
ных форм из Ганы оболочка покрывает все семя, у мелкосемянных
сенегальских образцов представлена в виде двух студенистых подуше-
чек на плоских сторонах семени; носик и рубчик при этом остаются
свободными.
Общая структура семени у арбуза слизистосемянного такая же, как
у других форм, но гиподерма состоит из слегка вытянутых тонкостен-
ных клеток без каких-либо признаков одревеснения. Толщина ее до-
стигает 1000 мкм и более, в 3—4 раза превышая толщину этого слоя
у столового арбуза. Вместе с эпидермисом, развитым довольно слабо,
она образует студенистую капсулу, обволакивающую семя. У формы
из Ганы, изображенной на рисунке 16,6, эпидермис палисадный, у се-
негальских форм арбуза на плоских сторонах семени он заменяется
изодиаметрическим. Защитный слой кожуры семени представлен у ар-
буза слизистосемянного только одним рядом клеток, поэтому семена
его легко режутся ножом. В рубчике семени строение кожуры ме-
няется. Гиподерма здесь состоит из клеток с одревесневшими стенками,
слой каменистых клеток увеличивается до 2—3 рядов, иногда больше.
У других форм арбуза утолщения каменистого слоя в зоне рубчика
не отмечено.
Своеобразие семян, обособленность ареала и некоторые морфоло-
гические признаки растений послужили основанием для выделения этой
формы в самостоятельный подвид ssp. mucosospermus [252].
Среди колоцинтов известны две группы форм: собственно колоцинт
с горькими плодами и пресный колоцинт. Они различаются по многим
72
Рис. 16. Поперечный разрез кожуры семени:
а —С lanatus subsp lanatus; б—С. lanatus. subsp. mucosospermus; в —C. naudi-
nianus; г —Praecitrullus fistulosus; 1—кутикула; 2 —эпидермис; 3 —гиподерма;
4 —каменистые клетки: 5 — гидрацитная ткань.
признакам (с. 73), в том числе по строению кожуры семени. Формы
с плодами без горечи по структуре спермодермы занимают как бы
промежуточное положение между горьким колоцинтом и столовым ар-
бузом. Природа их пока остается неясной, но скорее всего они яв-
ляются гибридами.
У типичного колоцинта, обитающего в пустынях Северной Африки,
Аравии и Афганистана, в структуре кожуры семени отчетливо выра-
жены черты ксерофильной организации: тонкий эпидермис, толстая ку-
тикула, мощная гиподерма, состоящая из сильно одревесневших клеток.
По характеру семенной кожуры колоцинт близок к другому ксеро-
фиту— арбузу безусиковому (С. ecirrhosus), произрастающему в южно-
африканской пустыне Намиб.
Интересен во многих отношениях, в том числе по морфологии и
анатомии семян, еще один дикорастущий вид, произрастающий в пу-
стынных районах Южной Африки, — арбуз Нодэна (С. naudinianus).
Семена его белые, выпуклые, гладкие, блестящие, отличаются от семян
всех других видов арбуза и напоминают семена близкого к Citrullus
рода Benincasa. В коллекции ВИР вид представлен двумя формами:
крупносемянной и мелкосемянной, различающимися по степени одревес-
нения кожуры. На рисунке 16, в изображен срез кожуры крупносемян-
ной формы С. naudinianus. Кожура семени очень толстая (достигает
почти 1000 мкм) и фактически состоит из однородной склеренхимы.
Клетки ее настолько сильно одревеснели, что почти не имеют полостей.
Эпидермис составляет лишь 7% толщины кожуры, но покрыт хорошо
развитой кутикулой. Характерная особенность семян этого вида — от-
сутствие слоя каменистых клеток, имеющегося у всех других видов
арбуза.
Мощная, слабо дифференцированная кожура семени С. naudinia-
nus свидетельствует о примитивности и малой эволюционной продвину-
тости этого вида, являющегося, несомненно, наиболее древним среди
Citrullus.
По структуре кожуры семени арбуз Нодэна близок к монотипному
роду Acanthosicyos (нара), также произрастающему в пустынях Юж-
ной Африки. Семена этого растения тоже характеризуются слабой гис-
тологической дифференциацией кожуры, которая состоит из однородной
склеренхимной ткани. Слой каменистых клеток в кожуре отсутствует,
эпидермис не имеет кутикулы и обычно отпадает. Некоторые ботаники
считают более правильным отнести арбуз Нодэна к роду Acanthosicyos,
так как у них много общего. Мы полагаем, что для такого перемеще-
ния нет достаточных оснований.
Из других родов необходимо остановиться прежде всего на
Praecitrullus — эндемичном для Индии растении, которое называют ар-
бузом пустотелым (фистулезным) и считают предком арбуза обыкно-
венного [165]. Семена его по форме и размеру напоминают семена ар-
буза обыкновенного, отличаясь лишь рельефным рубчиком, несвойст-
венным последнему.
Внешнее сходство семян является, пожалуй, единственным общим
признаком этих родов. Расположение семян в плоде уже свидетельст-
вует о своеобразии арбуза фистулезного. Каждое семя заключено в
плотную кожистую капсулу, которая не является, однако, частью се-
менной кожуры, как у арбуза слизистосемянного.
Структура семенной кожуры арбуза фистулезного представлена на
рисунке 16,г. Здесь видно, что эпидермис развит хорошо и составляет
почти половину толщины спермодермы. Клетки его более широкие, чем
у столового арбуза, кутикула довольно толстая. Гиподерма состоит из
74
6—8 рядов клеток и без окрашивания выглядит очень однородной. При
воздействии флороглюцином с соляной кислотой четко выявляются
гистологические различия в химическом составе клеточных оболочек:
верхние 2—3 ряда клеток окрашиваются в оранжево-желтые тона, ниж-
ние слои — в кирпичный цвет. У арбуза обыкновенного гиподерма окра-
шивается равномерно в малиновый цвет. Полости клеток гиподермы у
арбуза фистулезного заполнены темным пигментом, что делает струк-
туру его кожуры еще более своеобразной. Слой каменистых клеток
отсутствует, что свидетельствует о древности этой формы.
Таким образом, род Praecitrullus как бы сочетает черты примитив-
ности, с одной стороны, и глубокой специализации, с другой. Эволюция
этого растения шла, несомненно, совсем иными путями, чем эволюция
арбуза, и нет никаких оснований считать арбуз фистулезный предком
арбуза обыкновенного. Внешнее сходство семян этих родов можно рас-
сматривать лишь как конвергентное. Относительно слабое одревеснение
клеточных стенок и хорошо развитый эпидермис свидетельствуют о ме-
зофильном характере растений арбуза фистулезного, что подтверж-
дается и образом его жизни. Этот вид возделывают как овощ в пе-
риод дождей, преимущественно на северо-западе Индии и в Паки-
стане.
По ряду важных таксономических признаков (морфология пыльцы,
анатомия плода) к Citrullus близок род Benincasa, виды которого вы-
ращивают во влажных субтропиках как овощные растения. Изученные
нами два вида бенинказы различаются по строению семян.
В. cerifera (восковая) имеет гладкие белые выпуклые семена. Для
вида В. hispida (мохнатая) характерны плоские семена с хорошо вы-
раженным рубчиком, напоминающие семена столового арбуза. Однако
сходство семян обоих родов ограничивается только внешними призна-
ками. По структуре кожуры семени оба вида бенинказы заметно отли-
чаются от арбуза. Прежде всего отмечается мезофильность семян. Они
сравнительно мягкие и легко режутся ножом. Гиподерма кожуры се-
мени, особенно у вида В. cerifera, почти полностью сложена из тонко-
стенных клеток. Небольшое одревеснение заметно только в 2—3 ниж-
них рядах.
Все клетки гиподермы В. hispida имеют одревесневшие, но очень
тонкие оболочки. Эпидермис у бенинказы восковой обычно отсутствует.
У бенинказы мохнатой эпидермис развит хорошо, но наружная стенка
его отпадает и он остается в виде длинных тонких волосков, благодаря
чему семена выглядят опушенными. Слой каменистых клеток у обоих
видов представлен в отличие от арбуза 2—3 рядами.
Некоторые ботаники сближают род Citrullus с родом Cucumis, к
которому относятся огурец и дыня. Однако строение кожуры семени
трех дикорастущих видов Cucumis, которые были нами исследованы,
не дает основания говорить о близости их к Citrullus. Чтобы сде-
лать окончательные выводы, необходимо изучить этот род более
полно.
Таким образом, проведенное нами сравнительно-анатомическое изу-
чение семян разных видов Citrullus и представителей близких родов
показывает, что характер семенной кожуры, степень ее гистологической
дифференциации, развитие отдельных слоев могут служить хорошими
таксономическими признаками, позволяющими судить о филогенетиче-
ской близости, эволюционной продвинутости или, наоборот, примитив-
ности изученных форм. В сочетании с другими признаками строение
семенной кожуры целесообразно использовать в систематике при уточ-
нении и разработке ботанических классификаций.
75
ПРОРОСТКИ
Нами проведено сравнительное изучение анатомии проростков че-
тырех видов Citrullus-. С. lanatus, С. colocynthis, С. ecirrhosus, С. naudi-
nianus, а также некоторых близких ему родов: Benincasa, Praecitrul-
lus, Cucumis. Вид С. lanatus, включающий возделываемые и дикорасту-
щие формы арбуза, был исследован более детально по подвидам и раз-
новидностям. Всего изучено 12 образцов различных представителей
Cucurbitaceae. Исследовали 6—8-дневные проростки, выращенные в
растильнях с прокаленным песком при естественном освещении.
Анатомическое изучение проростков проводили с помощью микро-
скопа МБИ-1 при увеличении 9X7 и 40x7; для измерений пользовались
окуляр-микрометром, приведенным в соответствие с объект-микромет-
ром, при указанных выше увеличениях. Окрашивали срезы флороглю-
цином с концентрированной НС1. Были приняты во внимание следую-
щие признаки: толщина листа и составляющих его тканей — столбча-
той и губчатой, число устьиц на нижнем эпидермисе листа, форма во-
лосков. При изучении проводящей системы проростков учитывали число
и характер проводящих пучков на различных ярусах гипокотиля, в се-
мядольном узле, корневой шейке и корне.
В процессе изучения было выяснено, что проростки разных видов
и родов существенно различаются между собой по ряду признаков, в
пределах же вида различия носят в основном количественный характер.
Анатомическая структура семядольного листа видов Citrullus до-
вольно проста. С верхней и нижней стороны он одет однослойным эпи-
дермисом, состоящим из довольно крупных клеток, покрытых тонким
слоем кутикулы. Под верхним эпидермисом расположены 2—3 ряда
столбчатой хлорофиллоносной ткани, состоящей из вытянутых, плотно
прилегающих друг к другу клеток. Далее следует более рыхлая губча-
тая паренхима, насчитывающая 10—12 рядов клеток. Клетки ее сверху
вниз постепенно уменьшаются и, наконец, становятся мелкими. На ниж-
нем эпидермисе листа (в меньшей степени и на верхнем) развиваются
волоски, которые у разных видов различаются по форме.
Столбчатая или палисадная ткань по отношению к толщине семя-
доли составляет у Citrullus 20—25%, губчатая ткань — 70—75%; у
Benincasa и Cucumis sauclexii палисадная ткань более развита и со-
ставляет 28—30% толщины листа.
Толщина семядолей обусловлена в основном развитием губчатой
паренхимы. У форм с толстыми семядолями слой губчатой ткани равен
600 мкм и более, у тонколистных образцов соответственно 400—500 мкм
(табл. 8).
Столбчатая ткань наиболее развита у ксерофильных форм Citrul-
lus — С. naudinianus, С. colocynthis, С. ecirrhosus, что особенно за-
метно по отношению к толщине семядольного листа. Следует отметить,
что у настоящих листьев этих видов губчатая паренхима или вообще
отсутствует, или развита очень слабо. У колоцинта, например, листовая
паренхима состоит только из столбчатой ткани, клетки которой лишь
несколько укорачиваются в нижних рядах, но также плотно прилегают
друг к другу. Такое строение листа колоцинта как форма адаптации
к суровым условиям пустыни отчетливо свидетельствует о высокой сте-
пени его ксерофильности. Наличие губчатой паренхимы в семядолях
колоцинта можно рассматривать как своеобразный атавизм, отражаю-
щий более мезофильную структуру листа далеких предков арбуза,
имевших облик лиан тропического леса.
Опушение семядолей 8-дневных проростков видов Citrullus только
начинает формироваться. Волоски обычно трехклеточные, причем верх-
76
Таблица O я.п««и«« ппооостков Citrullus и близких родов
*_______. Анатомическая структура семядолей _
Таксон Номер no каталогу ВИР толщина семядолей, мкм толщина палисад- ной ткани, мкм отношение толщины палисад- ной ткани к толщине листа, % число слоев палисад- ной ткани толщина губчатой ткани, мкм форма клеток палисадной ткани число устьиц на нижнем эпидермисе волоски
Citrullus lanatus subsp. vulgaris var. vulgaris C. lanatus subsp. vulgaris var. cordophanus 557 3846 756 756 162 180 21 23 2 2-3 576 594 Вытянутая 270 Неоднород- ные по форме
C. lanatus subsp. lanatus var. lanatus 2724 900 180 20 — 666 Сильно вытянутая 270 —
C. lanatus subsp. lanatus var. 3890 720 180 25 2—3 540 Узкая, вытянутая 500 —
capensis
C. lanatus subsp. mucosospermus 1298 864 180 20 2—3 649 Сильно вытянутая —
C. colocynthis 4339 594 144 24 3 414 — 500 —
C. ecirrhosus 4251 756 180 23 3 468 Укороченная — —
C. naudinianus Bp. 250 684 162 24 2 468 Вытянутая — Узкие, длинные
Praecitrullus fistulosus 1954 612 144 23 2 468 Широкая 300 Слабо развитые
шипикм
Cucumis sauclexii 5 590 170 28 2 450 — 500 —
Benincasa cerifera 1 450 126 29 2 324 — — Узкие, острые, длинные
няя кпетка сначала округлая, затем заостряется По форме ее виды
арбуза различаются между собой (рис. 17). Своеобразно^опушение Се.
мядолей С. naudinianus, имеющее вид густо сидящих ков.
Проводящая система про-
ростков слагается из листовых
следов семядолей. Имеется два
узла дифференциации проводя-
щих пучков: в основании семядо-
лей и в области корневой шейки.
Подсемядольный узел, согласно
исследованиям В. К. Василевской
[43], развивается первым, и от
Рис. 18. Проводящий пучок гипокотиля 8-днсв-
ного проростка Citrullus:
1—склеренхима; 2 — флоэма; 3 — камбий: 4—
ксилема; 5 — паренхима, окружающая пучок.
Рис. 17. Форма волосков на верх-
нем эпидермисе семядолей:
1—С. lanatus; 2—С. naudinianus;
3 — Benincasa cerifera.
него сосуды направляются вниз и в семядоли. В центральном цилиндре
первичного корня, отграниченном от кормовой паренхимы перициклом,
проходит один проводящий пучок. У тыквенных растений первичная
ксилема имеет 4-лучевое (тетрархное) строение. Лучи ксилемы в пучке
чередуются с зонами флоэмы. Полости в корне нет, вся центральная
часть его заполнена сосудами и механической тканью. В области кор-
невой шейки картина размещения элементов проводящей системы
совсем иная. Многочисленные, близко расположенные проводящие пучки
почти сливаются в сплошное кольцо. Центральная часть заполнена не-
одревесневшими паренхимными клетками. Полости еще нет. В отличие
от корня в переходной зоне ксилема и флоэма в пучке имеют радиаль-
ное расположение. Пучок, таким образом, приобретает коллатеральную
структуру.
В гипокотиле проводящий пучок постепенно усложняется. В ниж-
ней его части внутренняя флоэма развита еще очень слабо, так что
пучок фактически коллатеральный. В верхней части гипокотиля пучок
становится отчетливо биколлатеральным. Кроме наружной флоэмы, в
нем появляется внутренняя, хотя она развита несколько слабее. Вся
верхняя часть пучка заполнена склеренхимной тканью. Сформировав-
шийся биколлатеральный пучок 8-дневного проростка Citrullus изобра-
жен на рисунке 18.
В центре гипокотиля проходит полость, которая появляется только
у 4 5-дневных проростков. Гипокотиль С. naudinianus в этом возрасте
полости еще не имеет.
В подсемядольном узле пучки, как и в корневой шейке, становятся
78
многочисленными и почти сливаются в сплошное кольцо. В отличие
от нижнего узла они располагаются здесь в два круга, причем наруж-
ный состоит из крупных пучков, внутренний — из мелких. Следует от-
метить, что расположение проводящих пучков в два круга характерно
для развитого стебля тыквенных растений, но там мелкие пучки рас-
полагаются в наружном круге, а крупные — во внутреннем [81].
Описанный тип проводящей системы характерен для всех изучен-
ных нами видов и форм Cucurbitaceae. Некоторые различия отмечены
по числу проводящих пучков гипокотиля. В нижней его части все ис-
следованные формы имеют 4 пучка, в средней зоне у С. naudinianus
и В. cerifera число их остается таким же, но пучки сложные, образо-
вавшиеся от слияния четырех крупных с четырьмя мелкими, у С. ecirrho-
sus и С. lanatus 6 пучков, а у подвида mucosospermus даже 10. Своеоб-
разие проводящей системы этой формы также подтверждает правиль-
ность выделения ее в самостоятельный таксон.
Изученные виды отличались по месту слияния мелких пучков семя-
дольных следов в более крупные. У С. naudinianus и Benincasa идущие
вниз пучки сливаются уже в верхней зоне гипокотиля, у других видов
слияние происходит ниже. В семядольном узле число пучков у всех
форм возрастает, у С. ecirrhosus даже до 20, причем 10 мелких пучков
внутреннего круга образуют почти сплошное кольцо.
Анатомическое строение семядольного листа позволяет судить о сте-
пени ксерофильности той или иной формы, что, несомненно, имеет
значение в селекции такой культуры, как арбуз, возделываемой в за-
сушливых зонах страны.
ЛИСТ
Анатомия листа арбуза изучалась нами в связи с водным режи-
мом растений и засухоустойчивостью [247, 248].
Лист арбуза с обеих сторон покрыт эпидермисом. На нижней сто-
роне клетки его мельче и более извилистые. Эпидермис несет тонкие
многоклеточные волоски, особенно густые на нижней стороне листа.
У дикорастущих и полукультурных форм арбуза они приобретают ха-
рактер шипов. Устьица сосредоточены преимущественно на нижней сто-
роне листа.
Наши исследования, проведенные на 27 сортах арбуза 8 разных
эколого-географических групп, показали, что размеры клеток эпидер-
миса и число устьиц на единицу поверхности в значительной степени
определяются условиями влажности в период их заложения. Во влаж-
ные годы число устьиц уменьшается, в сухие увеличивается. Например,
у сорта Монастырский (к-529, русская эколого-географическая группа)
на нижнем эпидермисе листа во влажном 1960 г. (опыт проводили на
Генической опытной станции ВНИИК) было 300 устьиц на 1 мм2, в
засушливом 1963 г.— 527, в умеренно сухом 1964 г. — 410. Аналогичные
цифры получены и по другим сортам. Так, у сорта Любимец Флориды
(к-72, американская эколого-географическая группа) число устьиц в те
же годы было соответственно 200, 420 и 363, у сорта Сяо-хун-цзер
(к-3507, восточноазиатская эколого-географическая группа)— 233-
288, 360.
Лист арбуза имеет толщину 250—350 мкм и состоит из двух типов
тканей: палисадной и губчатой. Палисадная (или столбчатая) ткань
представлена 1—2 рядами радиально вытянутых и плотно прилегаю-
щих друг к другу клеток, насыщенных хлорофильными зернами. Тол-
щина ее колеблется от 70 до 200 мкм. Губчатая ткань более рыхлая,
состоит из клеток округлой формы с большими межклетниками. Тол-
79
щина ее равна 100—150 мкм. Содержание хлорофилла в губчатой
ткани значительно ниже, чем в палисадной.
Ксерофильные формы имеют, как правило, более развитую пали-
садную ткань, чем мезофильные. Соотношение двух типов тканей листа
служит довольно постоянным для сорта анатомическим показателем и
может быть использовано при выявлении засухоустойчивых форм [41].
Наши исследования показали, что у сортов арбуза русской эколо-
го-географической группы палисадная ткань листа развита несколько
лучше губчатой. Соотношение двух типов тканей в среднем для не-
скольких сортов этой группы равно 1,5: 1,4. У сортов восточноазиат-
ской и дальневосточной групп губчатая ткань листа, наоборот, не-
сколько толще, чем палисадная. Соотношение тканей в среднем для
нескольких сортов у этих групп составляет соответственно 1,2: 1,4 и
1,2: 1,5. У дикорастущих видов арбуза (С. colocynthis и С. ecirrhosus)
лист состоит фактически только из палисадной ткани. На поперечном
срезе листа этих видов видно 5—6 рядов радиально вытянутых клеток,
длина которых постепенно уменьшается от верхнего эпидермиса к ниж-
нему. У листа арбуза ssp. mucosospermus вида С. lanatus палисадная
ткань состоит всего из двух рядов клеток, но по толщине слоя значи-
тельно превосходит губчатую ткань (соответственно 195 и 120 мкм).
Интересно, что у гибридов с дикорастущими формами палисадная
ткань листа развита лучше, чем у столовых сортов. Здесь как бы со-
храняются черты ксерофильной организации, свойственные диким фор-
мам арбуза. Примером может служить американский сорт Конго
(к-3848), полученный от скрещивания столового сорта с дикорастущим
арбузом из Африки. У сорта Конго толщина палисадной ткани листа
равна 165 мкм, а губчатой— 105 мкм. У столового сорта из США Клек-
лей (к-137) палисадная и губчатая ткани листа развиты одинаково.
ПЛОД
Плод тыквенных растений многосемянный, ягодообразный, пара-
карпный, развивается из нижней завязи. Для обозначения такого плода
у ботаников принят термин «тыквина». От плода типа ягода он отли-
чается кожистой или твердой периферической частью и полостью вну-
три плода у многих видов. Плод образован тремя плодолистиками,
сросшимися краями, но фактически он одногнездный [134, 137].
В плоде различают три типа тканей: экзокарпий — наружный эпи-
дермис; эндокарпий — внутренний, имеющий характер тонкой пленки;
мезокарпий — мякоть плода, составляющая его основную массу. Внеш-
нюю плотную часть мезокарпия вместе с эпидермисом называют корой.
Снаружи плод покрыт кутикулой, играющей защитную роль в ре-
гуляции водо- и газообмена. Кутикула покрывает не только верхние
стенки клеток эпидермиса, но часто распространяется и на боковые.
Эпидермальные клетки обычно квадратные или слегка неправильной
формы. Эпидермис плода имеет меньше устьиц, чем лист. На 1 мм2
здесь приходится не более 40 устьиц, причем на темных плодах их
меньше, чем на светлых. Непосредственно под эпидермисом располо-
жены 2—3 ряда клеток колленхимы с утолщенными оболочками,
ниже—10—12 рядов хлорофиллоносной паренхимы и под нею — скле-
реидный слой.
В темноокрашенных плодах арбуза масса хлоропластов, содержа-
щих пигмент, находится ближе к поверхности плода, в светлоокрашен-
ных — в более глубоких слоях паренхимы. Интенсивность окраски плода
зависит от количества пластид. В полосатых плодах хлоропласты лока-
лизованы преимущественно в зоне полосы, где проходят также прово-
дящие пучки.
80
Склереидный слой в коре плода свойствен не всем тыквенным рас-
тениям, Среди возделываемых видов его можно обнаружить у арбуза,
тыквы, лагенарии, люффы, бенинказы, арбуза фистулезного. У дыни и
огурца этот слой отсутствует [138]. Слой механической ткани в коре
плода арбуза выполняет защитную и опорную функции [177]. Он обра-
зуется из живых паренхимных клеток в результате одревеснения их
стенок и последующего исчезновения содержимого. Оболочки клеток
при этом утолщаются и пропитываются лигнином. Отдельные одревес-
невшие клетки можно обнаружить уже в завязи диаметром 2x2,5 см
через 2—3 дня после оплодотворения [244]. Интересно, что процесс
лигнификации клеточных оболочек обратим. В зрелых плодах, храня-
щихся некоторое время, оболочки клеток становятся тоньше и весь
слой механической ткани уменьшается, происходит как бы частичное
раздревеснение [206, 244]. Слой механической ткани состоит из отдель-
ных более или менее крупных групп склереид, между которыми имеются
промежутки неодревесневшей ткани.
От развития склереидного слоя и компактности расположения от-
дельных групп зависит прочность коры плода, а следовательно, при-
годность сорта для транспортировки и в какой-то мере для хранения
[29, 169, 177, 179, 206]. Таким образом, этот признак можно рассмат-
ривать как хозяйственно-важный, представляющий определенный ин-
терес при селекции сортов, пригодных для длительной транспортировки.
Нами были изучены характер развития и строение механической
ткани плода у 130 сортов арбуза различного происхождения. Выяснено,
что мощность склереидного слоя колеблется от 75 до 250 мкм. В зави-
симости от величины групп склереид и компактности их расположения
выделено три типа склереидного слоя: прерывистый, сплошной, проме-
жуточный.
Прерывистый слой на тангентальном срезе представлен простыми,
реже сложными группами склереид, состоящими из 2—3 простых
(рис. 19,а). Границы между группами четкие, заполнены живыми клет-
ками. Такой тип механической ткани был обнаружен у 45 образцов
из изученных 130.
Сплошной слой состоит из крупных групп склереид, включающих
до 30 простых. Границы между сложными группами нечеткие, запол-
нены частично одревесневшей тканью (см. рис. 19,в). Этот тип был
обнаружен у 25 образцов арбуза. Он более характерен для кормового
(С. lanatus subsp. lanatus var. citroides). У столовых сортов чаще всего
встречается промежуточный тип, представленный сложными, но более
мелкими группами склереид (см. рис. 19,6). Он был обнаружен у
55 образцов.
У сортов арбуза с прерывистым склереидным слоем мощность его
больше, чем у сортов со сплошным панцирем, что как бы компенсирует
недостаточную компактность одревесневших участков.
Выявлена корреляция между формой плода арбуза и типом скле-
реидной ткани. Установлено, что у длинноплодных форм чаще встре-
чается прерывистый слой и почти не встречается сплошной. Выяснено,
что характер механической ткани плода в какой-то мере связан с про-
исхождением сорта. У форм арбуза из влажных районов земного шара
слой механической ткани в плоде, как правило, менее развит, чем у
сортов из местностей с сухим климатом. Та же закономерность про-
является в отношении погодных условий: в засушливые годы механи-
ческая ткань развивается лучше, чем в дождливые [170].
Среди отечественных форм арбуза с хорошо развитым склереид-
ным слоем, а следовательно, прочной, упругой корой плода выделены
81
сорта, заслуживающие внимания: Волжский 7
(к-3614). Гибрид 23 (к-3566), Муравлевскии (к-3563), Пионер пустыни
(к-2866), Мелитопольский 143 (к-2840) и др. паГ1РНУИМНму uno
Мякоть плода арбуза состоит из крупных паренхимных клеток
с тонкими стенками, большими вакуолями и
ними воздухом Клетки содержат пигменты, среди которых преобладает
ликопщ? НО часто встречается и каротин. В зрелом плоде пигменты
представлены преимущественно игольчатыми кристаллами, заполняю-
щими вакуоли клеток. В плодах с бледной мякотью и недозрелых пиг-
Рис. 19. Типы склереидного слоя коры плода столового
а — прерывистый; б —
мент находится в аморфном состоянии в пластидах. Интенсивность
окраски мякоти зависит от количества пигмента, его локализации и в
какой-то мере от формы кристаллов [137].
Консистенция мякоти может быть плотной или рыхлой, грубой или
нежной. Чем мельче клетки и чем они плотнее расположены, тем неж-
нее мякоть. Крупные клетки с большими межклетниками, наоборот,
свойственны грубой мякоти. На характер мякоти влияет также сеть
проводящих сосудов, пронизывающих плод. Волокнистая, грубая мякоть
имеет сильно разветвленную проводящую систему, для нежной, таю-
щей мякоти характерно сравнительно слабое развитие проводящих
пучков, почти лишенных механических волокон. От структуры мякоти
плодов в значительной степени зависит их лежкость и транспортабель-
ность. Сорта, плоды которых имеют крупноклетную мякоть с боль-
шими межклетниками, более пригодны для хранения и перевозок, чем
сорта с нежной мелкоклетной мякотью плодов.
ХИМИЧЕСКИЙ СОСТАВ ПЛОДОВ И СЕМЯН АРБУЗА
БИОХИМИЧЕСКИЕ ОСОБЕННОСТИ ПЛОДА
Арбуз возделывают преимущественно ради сочных сахаристых пло-
дов, поэтому биохимия плода столовых сортов изучена более детально.
Плод содержит 87—97% воды, на долю сухого вещества приходится
3—13%. У столовых сортов сухое вещество мякоти представлено почти
исключительно растворимыми углеводами, главным образом сахарами,
составляющими, по данным А. И. Ермакова [85], 86—88%. Сочная
мякоть плода и высокое содержание растворимых сахаров дают воз-
можность проводить оценку качества плода при массовых анализах с
помощью полевого рефрактометра. Между содержанием сухого веще-
ства, определяемого рефрактометром и высушиванием, существует тес-
ная корреляционная связь: коэффициент корреляции (г) 0.87—0,95
арбуза на радиальном (1) и тангентальном (2) срезах:
промежуточный; в — сплошной.
83
[52] . Такая же связь обнаружена между растворимой частью сухого
вещества и суммой сахаров: г = 0,81—0,91. Как указывают авторы, со-
держание сахаров, определяемое по Бертрану, ниже показаний рефрак-
тометра лишь на 1,0—1,5%.
Наиболее полные исследования сахаров в плодах арбуза проведены
С. Н. Лутохиным [128], проанализировавшим 89 сортов разного про-
исхождения. Согласно его определениям, общее количество сахаров в
плоде арбуза колеблется от 5,5 до 9,5% (по другим данным, от 6 до
10%), в зависимости от сорта. В пределах сорта также могут быть
значительные колебания (от 0,25 до 2,5%).
Средний плод арбуза содержит около 200 г сахаров с колебаниями
от 50 до 400 г. Сахара распределяются следующим образом (в %):
фруктоза — 2,92—4,90, глюкоза—1,10—3,38, сахароза — 0,24—2,63. Пре-
обладает фруктоза, составляющая около половины общей суммы саха-
ров. По определениям М. А. Кудрявцевой [116], количество сахарозы
в зрелом плоде арбуза может достигать 4,63%, по данным Н. Н. Ива-
нова [99], —5,47%.
Разноречивость этих данных объясняется тем, что накопление са-
харозы больше, чем других сахаров, зависит от условий созревания
плода, в первую очередь от температуры. Н. Н. Ивановым и
Р. С. Александровой [99] введен термин «созревание сахаров» и от-
мечено, что низкое содержание сахарозы характеризует недозрелые
плоды, внешне имеющие все признаки спелости (красная мякоть, зре-
лые семена).
На спелость арбуза, по данным названных авторов, указывает
также определенная кислотность клеточного сока. Недозрелым плодам
свойственна более высокая активная кислотность (pH 4,6—4,8), при
созревании она снижается (pH 5,2—5,4, иногда достигает 6). Из орга-
нических кислот в плодах арбуза имеется только яблочная.
Наиболее сладкий сахар — фруктоза. А. А. Рихтер ввел единый
показатель сладости, согласно которому фруктоза имеет 220 единиц
сладости, сахароза—145, глюкоза—100. Умножая процентное содер-
жание отдельных сахаров на соответствующий коэффициент, получают
суммарный показатель сладости, который для очень сладких плодов вы-
ражается цифрой, близкой к 2000.
Сахара в плоде распределяются неравномерно [111]. Наиболее са-
харистая центральная часть плода вокруг семян, ближе к коре саха-
ристость падает. Цветочный конец плода содержит также больше са-
харов, чем часть, прилегающая к плодоножке. Сторона плода, обращен-
ная к солнцу, накапливает больше сахаров, чем соприкасающаяся с
землей. Особенности распределения сахаров учитывают при взятии
средней пробы для анализа.
Содержание сахаров определяет качество плода арбуза как де-
сертного продукта, но не исчерпывает его ценности. Арбуз содержит
довольно много витаминов группы В, в частности фолиевую кислоту,
участвующую в процессе кроветворения, нужные для организма мине-
ральные соли, каротиноиды, среди которых преобладает ликопин. Они
играют важную роль в качестве исходных веществ, из которых обра-
зуются витамины группы А, а также «зрительный пурпур», участвую-
щий в зрительном акте.
Содержание витаминов в плодах арбуза (в мг/100 г) следующее:
витамина С — 7, каротина—1, витамина Bi — 0,05, витамина В2 — 0,07,
фолиевой кислоты — 0,15. Мякоть плода содержит (в %): азотистых
веществ — 0,76, липидов — 0,6, клетчатки — 0,4, золы — 0,36, в которой
преобладают К2О (61%), F2O3 (10,3%) и MgO (6,8%) [17, 172, 265].
84
ХИМИЧЕСКИЙ СОСТАВ СЕМЯН
Ценным пищевым продуктом являются и семена арбуза, которые
содержат высококачественное масло и легкоусвояемые белки. По дан-
ным Р. Хирохата (у В. В. Арасимович) [15], белки семян арбуза со-
стоят в основном из глобулинов, содержащих аминокислоты аргинин,
цистин, гистидин, лизин, тирозин, триптофан и др. Среди них преобла-
дает аргинин, содержание которого достигает 15,4%. Сходные данные
по химическому составу белков семян арбуза получены Н. А. Бывших
[35]. Опытами ряда исследователей подтверждена тождественность бел-
ков арбуза, дыни, тыквы.
По данным Н. А. Бывших, семена арбуза содержат 25% масла
от массы всего семени и 50—52% от массы сухого ядра. Близкие
цифры (50—55 и 51,4%) приводят И. Ф. Леонтьев [120] и Д. Бхата
[284]. Промышленный выход масла составляет 20% массы сырья.
Наиболее полное и детальное изучение химического состава масла
из семян арбуза проведено индийскими исследователями [120]. Со-
гласно их данным, в состав арбузного масла входят насыщенные и не-
насыщенные жирные кислоты с преобладанием последних. Содержание
жирных кислот в масле семян арбуза (в %) следующее (в 1-й графе
приведены цифры, полученные Д. Бхата, во 2-й — С. Дхингру):
пальмитиновая 15,5 7,3
стеариновая 5,8 0,8
олеиновая 16,8 43,2
линолевая 62,1 45,2
Данные существенно различаются, но в обоих случаях ясно видно
преобладание ненасыщенных жирных кислот с одной (олеиновая) и
двумя (линолевая) двойными связями.
йодное число арбузного масла составляет 117—123, т. е. сравни-
тельно невысокое [265]. Известно, что чем выше йодное число, тем
более жидкий жир и тем более он пригоден для приготовления лаков,
красок, олифы, но тем ниже его пищевая ценность.
Масло из семян арбуза относится к полувысыхающим. По хими-
ческим и физическим свойствам оно сходно с подсолнечным, в меди-
цине может заменять миндальное масло [120].
Йодное число масла в семенах кормового арбуза несколько выше
и составляет в среднем 133 [15]. Оно относится к числу высыхающих
масел, и пищевая ценность его ниже.
Исследованиями Н. А. Бывших установлено, что биологическая
спелость семян арбуза наступает примерно через 1—1,5 мес. после
технической спелости плодов. Именно в это время биохимические по-
казатели семян становятся характерными для данного сорта.
В некоторых странах Западной Африки (Камеруне, Нигерии, Гане)
арбуз культивируют ради семян, богатых белком и маслом [299, 314,
342]. Мякоть плодов возделываемых здесь форм арбуза горькая, в пищу
не используется. Семена же размалывают в муку и употребляют для
заправки супа (egusi— суп). Здесь возделывают арбуз слизистосемян-
ный, выделенный нами в самостоятельный подвид (subsp. mucososper-
mus Fursa). Семена его более крупные, чем у обычного арбуза, плоские
и заключены в слизистую капсулу.
Анализ семян арбуза из Нигерии показал, что они содержат около
32% белка и 60% масла на сухое вещество ядра. Масло используют
как заменитель пальмового. Калорийность таких семян очень высокая
и составляет 566 калорий [342]. По содержанию фосфора семена ар-
буза занимают первое место среди всех возделываемых здесь овощей.
85
Около половины его находится в виде фитиновой кислоты, содержание
которой достигает 60 мг/100 г. Известно, что фитин широко исполь-
зуют в медицине в качестве лечебного препарата.
ВЛИЯНИЕ ГЕОГРАФИЧЕСКИХ И ЭКОЛОГИЧЕСКИХ ФАКТОРОВ
НА химический состав плодов
Уже в первые годы существования ВИР лабораторией биохимии,
возглавляемой профессором Н. Н. Ивановым, было обращено серьез-
ное внимание на биохимическое изучение бахчевых культур, в том
числе арбуза. Тогда же были сформулированы основные задачи иссле-
дований.
1. Изучение влияния географических факторов на химический со-
став плодов.
2. Исследование динамики накопления сахаров в процессе созре-
вания и превращения веществ во время хранения.
3. Определение химического состава плодов ближайших сородичей
столового арбуза.
Намеченная лабораторией обширная программа исследований была
осуществлена лишь частично, но и те данные, которыми мы распола-
гаем, представляют большую ценность для характеристики сортов и
форм арбуза, имеющихся в коллекции. Исследованиями было установ-
лено, что одни и те же сорта арбуза, выращенные в разных районах,
накапливают различное количество сахара. В таблице 9, составленной
Таблица 9. Влияние географического фактора на сахаристость плодов арбуза,
% сахаров на сырое вещество
Сорт Сумма сахаров Глюкоза Фруктоза Сахароза
Мурашка 8,26/7,06 1,76/1,32 3,79/2,49 2,99/2,67
Белосемячко 8,50'6,02 2,15/1,56 3,48/3,86 2,72/0,54
Желтокорый 6,75/5,19 1,37/1,06 2,98/2,74 2,39/1,95
Малоазиатский 7,80/6,39 1,94/1,49 3,44/3,56 2,31/1,27
по данным Н. Н. Иванова и Р. С. Александровой, приведены резуль-
таты химического анализа сортов арбуза, выращенных в Астрахани и
Харькове. Содержание общего сахара и отдельных сахаров у всех сор-
тов было выше в Астрахани (первые цифры). В Харькове наблюдалось
как бы неполное дозревание плодов, что особенно сказывалось на со-
держании сахарозы.
Закономерность, обнаруженная в 20-е годы Н. Н. Ивановым и
Р. С. Александровой, подтвердилась и в наших исследованиях, про-
веденных в 1973—1974 гг. При сравнении сахаристости арбузов на
Астраханском опорном пункте и Кубанской опытной станции ВИР было
установлено, что по сумме сахаров различия невелики, но на Кубани,
как правило, накапливается меньше (до 2%) сахарозы, лишь у от-
дельных сортов содержание ее здесь достигало 2,3—2,5%.
Сахаристость арбуза меняется не только в разных зонах выращи-
вания, но и в одном пункте в зависимости от метеорологических усло-
вий. В 1972—1973 гг. при биохимическом изучении 30 сортов арбуза
на Кубанской опытной станции ВИР получены следующие данные.
В благоприятном 1972 г. содержание фруктозы в плодах всех сортов
преобладало над содержанием глюкозы: соответственно 1,9—5,98 и
0,26— 2,64%. В менее благоприятном 1973 г. наблюдалась обратная
картина: содержание фруктозы составляло 1,6—2,4%, глюкозы —
86
3,2—6,6%. В 1972 г. плоды были слаще за счет большего накопления
фруктозы, однако по сумме сахаров различия отсутствовали: в 1972 г.—
5,14—8,86%, в 1973 г. —5,4—8,9%.
Полученные результаты свидетельствуют о том, что нельзя судить
о сладости плодов того или иного сорта только по сумме сахаров, не-
обходимо учитывать содержание отдельных фракций.
Детальное изучение сахаристости арбуза в Нижнем Поволжье, где
издавна сосредоточено основное товарное бахчеводство страны, пока-
зало, что в накоплении сахаров большую роль играет не только сумма
активных температур, но и тип бахчи [29]. В Астраханской области
встречается бугровая поливная, бугровая неполивная и ильменная
бахча; первая из них преобладает. В таблице 10 показана сахаристость
плодов арбуза на бахче разных типов.
Таблица 10. Содержание сахаров в плодах сорта арбуза Мурашка
в зависимости от типа бахчи, % на сырое вещество
(по Н. Н. Иванову)
Бахча Сумма сахаров Глюкоза Фруктоза Сахароза
Бугровая поливная 8,04 2,40 3,27 2,25
Бугровая неполивная 8,44 1,34 3,72 3,22
Ильменная 6,59 2,51 4,06 1,92
Степная 8,53 1,31 2,81 3,39
Как видно из таблицы 10, более высокой общей сахаристостью от-
личались арбузы с бугровой неполивной и степной бахчи, самой низ-
кой— с ильменной бахчи. Такая же закономерность наблюдалась и по
содержанию сахарозы. Полученные результаты дают возможность го-
ворить о более быстром накоплении сахаров на суходольной бахче и
замедленном на поливной.
Почти такие же данные были получены В. В. Арасимович [17]
в Молдавии: полив, незначительно снижая общую сахаристость, за-
метно сказывался на содержании сахарозы. К. Беляков [26], изучав-
ший влияние различных режимов орошения на сахаристость арбуза,
отмечает некоторое снижение суммы сахаров при частых поливах, но
не приводит цифрового материала, позволяющего делать определенные
выводы.
Вопрос о влиянии поливов на сахаристость арбуза приобретает
особую значимость в связи с расширением орошаемых площадей бах-
чевых культур, но, как видно из приведенных данных, пока недоста-
точно изучен. К этой проблеме в Нижнем Поволжье тесно примыкает
другая — засоление земель в связи с высоким стоянием сильно мине-
рализованных грунтовых вод. Интенсивное орошение часто приводит
к вторичному засолению полей, вследствие чего снижается урожай,
иногда довольно значительно. В связи с этим очень важно изучать от-
рицательное действие засоления на качество и сахаристость арбуза —
одной из ведущих культур в этой зоне.
В. Д. Знаменский [97], изучавший влияние засоления на качество
арбуза путем вегетационных опытов, пришел к выводу, что небольшой
избыток солей NaCl и Na2SO4, повышающий осмотическое давление
почвенного раствора до 2—3 атм, оказывает даже стимулирующее влия-
ние на рост и развитие арбуза и при этом несколько повышает саха-
ристость плодов за счет большего накопления фруктозы. Дальнейшее
повышение концентрации солевого раствора оказывало отрицательное
87
действие, которое проявлялось особенно заметно при хлористом засо-
лении.
А. С. Морозов [146] подвергает сомнению данные предыдущего
автора, считая их недостаточно обоснованными, и приводит результаты
своих исследований, также проведенных в вегетационных сосудах, но
с применением повторностей. В его опытах сахаристость арбузов сни-
жалась уже при концентрации солей, равной 0,1 М. Концентрация
0,2 М оказывала отрицательное влияние на накопление сахарозы, сни-
жая при этом общую сахаристость на 2,5—3,0% (в расчете на сырое
вещество).
Недостаточно полные и трудно сравнимые данные названных авто-
ров можно рассматривать лишь как предварительные, имеющие ре-
когносцировочный характер и требующие проведения более широких
исследований с привлечением сортового разнообразия арбуза из миро-
вой коллекции ВИР.
ДИНАМИКА СОДЕРЖАНИЯ САХАРОВ ПРИ СОЗРЕВАНИИ
И ХРАНЕНИИ ПЛОДОВ
Лабораторией биохимии ВИР было изучено накопление сахаров
в процессе созревания арбуза, а также превращение веществ в период
хранения и транспортировки готовой продукции (табл. 11).
Таблица 11. Динамика содержания сахаров в плодах сорта арбуза
Белосемячко (в Ташкенте), % на сырое вещество (по М. А. Кудрявцевой)
Возраст плода, дней Сумма сахаров Г люкоза Фруктоза Сахароза pH
10 2,88 1,92 0,96 0 4,9
17 4,24 2,12 2,28 0 4,9
24 6,98 2,64 3,50 0,69 5,1
31 7,79 2,77 3,40 1,40 5,2
40 6,68 1,57 2,89 2,10 4,8
До 30-дневного возраста в плоде идет непрерывное накопление са-
харов, после чего сахаристость начинает снижаться. Если в 10-дневной
завязи преобладает глюкоза, то в 17-дневной глюкоза и фруктоза от-
мечены в равных количествах. Сахароза появляется лишь на 24-й день
в небольшом количестве, и содержание ее продолжает увеличиваться
не только до созревания плода (31-дневный плод), но и в первое время
после его снятия (40-дневный плод) за счет уменьшения моносахаров.
Кислотность клеточного сока снижается по мере созревания плода.
Анализ динамики накопления и превращения сахаров в созреваю-
щих плодах арбуза позволил обрисовать их ход следующим образом.
В молодые завязи из листьев поступает глюкоза, за счет которой позже
образуется фруктоза. Затем из глюкозы и фруктозы синтезируется са-
хароза, накопление которой продолжается и в первое время после сня-
тия зрелого плода. Низкое содержание сахарозы в более северных
районах возделывания и в годы с недостаточным количеством тепла
можно объяснить замедленным превращением сахаров.
Дозревание и перегруппировка сахаров происходят и после снятия
зрелых плодов, особенно в первые 2—3 недели. При этом заметно уве-
личивается количество сахарозы; сумма сахаров или остается на том
же уровне, или даже несколько повышается. Так, по данным В. В. Ара-
симович [17], сахаристость плодов арбуза сорта Пионер пустыни в
Молдавии повысилась в течение первого месяца после уборки с 6,5 до
7,3%, а сорта Мраморный —с 8,8 до 9,4%. Автор отмечает, что в на-
88
чале хранения в плодах происходит усиленный гидролиз полисахари-
дов, в частности пектиновых веществ, которые играют, по-видимому,
большую роль в лежкости сорта.
Перегруппировка веществ, изменение их содержания после уборки
свидетельствуют о важности этого периода в лежкости плодов. Нема-
лую роль в сохранении плодов без ухудшения качества играет, видимо,
пониженная активность окислительных ферментов, свойственная, как
отмечает В. В. Арасимович, в большей мере кормовому арбузу, но
характерная и для столового.
После дозревания и перегруппировки сахаров в зрелом плоде про-
исходит снижение общей сахаристости и особенно сахарозы [116]. При
изучении динамики содержания сахаров в плодах арбуза в процессе
хранения на Астраханском опорном пункте ВИР в 1974 г. была отме-
чена большая роль сорта. Из трех испытанных сортов лучшим ока-
зался Крупноплодный 174-175 (Краснодарской селекционной овоще-кар-
тофельной опытной станции НИИОХ), у которого сумма сахаров через
месяц хранения (с 16 сентября по 14 октября) оказалась на том же
уровне.
П. В. Сокол и В. И. Васина [203] изучали превращение сахаров
арбуза в процессе хранения при разной температуре. Опыт был про-
веден на трех сортах, плоды которых хранились 3 мес. Через месяц
хранения при 6—14° С в плодах всех сортов при небольшом уменьше-
нии общей сахаристости увеличилось содержание сахарозы и резко
снизилось количество глюкозы. Эти данные вполне согласуются с при-
веденными выше и свидетельствуют о замедленном превращении са-
харов в первое время хранения. Температура ниже 6° С подавляла син-
тез сахарозы, способствуя ее гидролизу. Плоды при таких условиях
удалось сохранить только в течение месяца. Оптимальной для хранения
оказалась температура 6—8° С, при которой в течение 3 мес. оставался
удовлетворительный баланс сахаров.
Аналогичные данные приводит Ч. Холл [306], отмечая темпера-
туру 6,2° С как наиболее благоприятную для хранения плодов арбуза.
ХИМИЧЕСКИЙ СОСТАВ ПЛОДОВ РАЗНЫХ ВИДОВ АРБУЗА
Лабораторией биохимии ВИР уже в первые годы работы было на-
мечено изучение химического состава плодов ближайших сородичей
столового арбуза — кормового арбуза и колоцинта (табл. 12).
Таблица 12. Содержание сахаров (% на сырое вещество) и полисахаридов
(% на сухое вещество) в плодах разных видов арбуза (по М. А. Кудрявцевой и
В. В. Арасимович)
Таксон Сумма сахаров Глюкоза Фрук- тоза Саха- роза Пек- тины 1 Гемицел- люлоза 1 I Целлю- лоза pH
Столовый арбуз (С. lanatus subsp. vulgaris) 8,59 1,24 1,84 4,63 0,7 0,8 0,6 5,5
Кормовой арбуз (С. lanatus subsp. lanatus) 1,88 0,99 0,80 0,19 18,0 9,0 11,0 4,8
Колоцинт (С. colocynthis) 1,01 0,64 0,29 0,0 10,0 3,35 11.0 4,4
Таблица 12 наглядно иллюстрирует различия в химическом составе
плодов столового, кормового и дикорастущего арбуза. Сумма сахаров
у первого намного больше, чем у двух других. Если у столового арбуза
89
фруктоза преобладает над глюкозой, то у кормового и колоцинта на-
блюдается обратная картина. Сахароза в плодах колоцинта отсутствует
полностью, а у кормового имеется в очень незначительном количестве.
В отличие от столового арбуза плоды этих видов богаты полисахари-
дами, причем у кормового преобладают пектиновые вещества, а у ко-
лоцинта — клетчатка. Последний отличается и наиболее высокой кис-
лотностью клеточного сока. Аналогичные данные приводят 3. Н. По-
пова и Е. Ю. Сосенская [176].
При химическом анализе полукультурного арбуза из Судана, про-
веденном в 1970 г., были получены цифры, близкие к тем, которые
характеризуют кормовой арбуз. Суданский образец арбуза имел не-
сколько большую сахаристость, чем кормовой (2,9%), но такое же
низкое содержание сахарозы (0,19%). Биохимические данные наряду
с другими подтверждают переходный характер арбуза из Судана в эво-
люции культурных форм.
Высокое содержание пектиновых веществ в плодах кормового ар-
буза обусловливает его засухоустойчивость и способность к длитель-
ному хранению. Пектины обладают высокой гидрофильностью. В со-
четании с водоудерживающими белковыми веществами они прочно
связывают огромные количества воды и придают мякоти плода боль-
шую плотность. Кормовой арбуз содержит 96—97% воды, но благодаря
тому, что она находится не в свободном, а в связанном состоянии, рас-
ход ее на испарение очень незначителен. Плоды кормового арбуза
могут сохраняться в течение 2—3 лет при комнатной температуре без
потери тургора. Колоцинт содержит меньше пектинов и больше клет-
чатки. Плоды его при созревании быстро высыхают и таким образом
сохраняются.
Различия между видами арбуза очень четко проявляются и при
анализе превращения веществ в
л 1—1 I I I I I 1 r~r.^Hu I .
и 10 15 20 25 30 35 40 45 50 55 60
Дни
Рис. 20. Динамика накопления
сахаров в плодах столового (1),
кормового (2) арбуза и колоцин-
та (3).
1
процессе созревания плодов (рис. 20).
На рисунке видно, что у столового
арбуза по мере созревания плода
наблюдается накопление сахаров, у
кормового арбуза и колоцинта, на-
оборот, отмечается уменьшение их
количества. К моменту созревания
плоды двух последних видов содер-
жат столько же сахара, сколько
10-дневная завязь столового арбуза.
Это происходит потому, что глю-
коза, притекающая из листьев, сра-
зу же потребляется у них на синтез
полисахаридов, не успевая образо-
вать фруктозу и сахарозу.
Характерной особенностью химического состава плодов кормового
арбуза является большое количество пектиновых веществ, о чем было
сказано выше. Благодаря высокой желирующей способности пектины
находят широкое применение в пищевой промышленности. Природных
источников пектина очень немного — это отходы от переработки яблок,
свекловичный жом и обмолоченные корзинки подсолнечника. Однако
потребность пищевой промышленности в желирующих веществах столь
велика, что не удовлетворяется существующим сырьем. В поисках но-
вых источников пектина было обращено внимание на кормовой арбуз,
который оказался ценным сырьем.
По желирующим свойствам пектин арбуза превосходит свеклович-
ный и подсолнечный, кроме того, он не вносит в готовую продукцию
90
постороннего вкуса и аромата, что очень важно при производстве кон-
дитерских изделий. Детальное изучение пектиновых веществ различных
сортов кормового арбуза, проведенное в Молдавском НИИ орошаемого
земледелия и овощеводства, позволило выявить формы с высоким со-
держанием пектиновых веществ (до 16,5% на сухое вещество) [20].
На основе одной из них был создан сорт Пектинный [168].
Лабораторией технологической оценки этого института отработан
способ получения арбузного пектина в заводских условиях, достигнуты
высокий выход и хорошее качество продукта. Перспективность исполь-
зования кормового арбуза как сырья для промышленного получения
пектина ставит перед селекционерами задачу углубленного изучения по-
лукультурных и дикорастущих форм арбуза с целью выведения новых
сортов с высоким содержанием пектиновых веществ.
СОДЕРЖАНИЕ ГОРЬКИХ ВЕЩЕСТВ В РАСТЕНИИ АРБУЗА
Почти у всех видов тыквенных растений листья и корни содержат горькие ве-
щества, объединяемые общим названием кукурбитацины. большинство из них ядови-
ты, вызывают тяжелые желудочно-кишечные расстройства, но в малых дозах явля-
ются эффективными лечебными препаратами. Горькие вещества тыквенных изучены
еще недостаточно, хотя актуальность таких исследований несомненна.
Наиболее полно изучены кукурбитацины у 45 видов тыквенных растений С. Рэ-
мом с сотрудниками [352]. Они обнаружили большое разнообразие горьких веществ
в листьях, корнях, плодах, установили, что содержание их может значительно ме-
няться с возрастом растений.
Выявлены три основных типа кукурбнтацинов (В, С, Е), которые, вероятно,
являются исходными для всего разнообразия этих веществ. В корнях молодых сеян-
цев обнаружены только вещества В и Е. Кукурбитацин В обнаружен почти у всех
исследованных видов, чаще всего он встречается в группе с типами D, G, Н (его про-
изводные), а кукурбитацин Е — вместе с I, J, К. С. Рэм выявил определенные зако-
номерности в содержании и распределении горьких веществ у разных видов, связав
их таким образом с таксономией. Им было установлено, что Citrullus содержит в ос-
новном кукурбитацин типа Е, Cucumis — типа В. Исключение составляет огурец
С. sativus, для которого характерен тип С, отсутствующий у других видов рода
Cucumis. Средн видов Citrullus наибольшее разнообразие горьких веществ было об-
наружено у С. naudinianus и С. ecirrhosus, больше всего их оказалось в корнях.
В проростках всех видов арбуза были найдены горькие вещества типа Е и В. У воз-
делываемых сортов горечь позднее исчезала, а у некоторых дикорастущих форм со-
хранялась до конца вегетации.
ГЕНЕТИКА И ЦИТОЛОГИЯ
КАРИОТИП И МЕТОДЫ ЕГО ИЗУЧЕНИЯ*
Кариотип представляет собой характерный для соматической
клетки организма набор хромосом, определяемый их числом, формой,
величиной. Сведения о кариотипе данного индивидуума помогают опре-
делить его взаимоотношения с близкими формами того же вида и по-
ложение в таких систематических группах, как род, семейство. Виды
одного рода и даже представители разных родов могут иметь одинако-
вое число хромосом, но иногда в пределах рода этот признак проявляет
заметную изменчивость. Поэтому наряду с числом хромосом очень
важно учитывать их морфологические особенности (размеры, форма,
положение центромеры и т. п.). Морфология хромосом каждого вида
отличается высокой устойчивостью, а потому является надежным си-
стематическим признаком [213].
Идентификация отдельных хромосом в кариотипе, необходимость
их индивидуализации всегда были основной проблемой при кариологи-
ческих исследованиях. Хромосомы легко идентифицировать, если они
* Раздел подготовлен на основании материала, представленного кандидатом
биологических наук Г. Л. Шелепиной.
91
различаются по величине, первичному и вторичному членению, по на-
личию спутников и т. п. Однако у многих видов растений хромосомы
по этим показателям не идентифицируются.
В отечественной, а в основном в зарубежной литературе имеются
работы, посвященные кариологии отдельных представителей сем. Си-
curbitaceae Juss. [108, 286, 294, 366 и др.]. Однако сведения о цито-
генетике тыквенных культур носят редкий и фрагментарный характер.
Большая часть работ посвящена лишь подсчету чисел хромосом или
простому описанию хромосомных наборов.
Сравнительные цитогенетические исследования по всему видовому
разнообразию рода Citrullus ранее не проводились. Этот род, а также
род Praecitrullus кариологически изучены слабо, что объясняется из-
вестными методическими трудностями. К их числу, как справедливо
указывают М. Шимотзума [359], Р. Н. Триведи и Р. П. Рой [366],
а также подтверждают наши данные, в первую очередь надо отнести
мелкие размеры хромосом, их плохое окрашивание красителями. Иден-
тификация хромосом затруднена, так как они имеют большое сходство
по морфологии и величине. В связи с этим в задачу нашего исследо-
вания входила также разработка методики, обеспечивающей получение
метафазных пластинок с четко выраженной структурной морфологией
хромосом.
Для всех изучавшихся образцов применяли единую методику ци-
тологического исследования. Семена проращивали при 28° С в чашках
Петри на увлажненной фильтровальной бумаге. При длине корешков
проростков 60—80 мм, соответствующей высокой митотической актив-
ности клеток в зоне меристемы, материал помещали в насыщенный рас-
твор а-бромнафталина на 4 ч при 5° С. После этого фиксировали спир-
тоуксусной смесью [1:1] в течение 24—48 ч. Перед окрашиванием
корни мацерировали в 3%-ном растворе NHC1 в течение 30 мин при
комнатной температуре с последующим отмыванием от НС1 45%-ной
уксусной кислотой при той же температуре. Окрашивали 5%-ным ук-
сусным кармином с добавлением хлорного железа 0,02 %-ной концентра-
ции. При этом способе хромосомы окрашивались по длине неравно-
мерно. Метафазные хромосомы изучали в первичной меристеме кореш-
ков проростков на временных давленых препаратах.
Микроскопический анализ проводили на микроскопе NFPK-2
фирмы «Цейсс» (фазовоконтрастный объектив 90Х, окуляр 12,5Х).
Метафазные пластинки фотографировали с помощью микрофотонасадки
«Цейсс».
По каждому образцу просмотрено более 80 метафазных пласти-
нок. Для анализа отобрано по 10 метафаз, в которых хромосомы ле-
жали в одной плоскости и имели сравнительно четкий рисунок диффе-
ренцированного окрашивания. При измерениях длину спутничной нити
и спутника не включали в общую длину хромосомы. При сравнитель-
ном морфологическом анализе учитывали следующие параметры хро-
мосом:
La — абсолютная длина хромосом, мкм;
Е La— длина диплоидного набора хромосом, мкм;
La
L —относительная длина хромосом, 100%;
к о длина короткого плеча
1 —центромерный индекс,--------------------- 100%;
»ch „ л „ длина темноокрашенного района
1 —хроматический индекс, -------------[а------------100%.
92
Таблица 13. Средние значения относительных длин хромосом, их центромерных
и хроматических индексов в кариотипе разных подвидов С. lanatus
Пара Относительная длина хромосом % (Lr±a) Центромерный индекс, > i (Iе ± а) Хроматический индекс, % i (lCh±o)
хромосом III I 1 11 1 111 I 1__+ I 41
1-Я 11,0±0,10 10,8±0,17 10,5±0,13 48 ±1,0 41.5 + 1,5 40,1 ±1.6 47 ±0,9 41 ± 1,8 48± 1,9
2-я 10,6 ±0,09 10.3 + 0,14 10,2±0,16 32 ± 1,2 41,1 ±1,0 38,4 ±1.5 38 ±0,9 57 ±0,9 38 + 0,9
3-я 10,6 ±0,08 10,0 ±0,16 9,8 ±0,27 46± 1,2 43,3 ±1,2 37,4 ±1,8 39± 1,1 50+1,1 60± 1,2
4-я 10,6±0,11 8,6±0,15 9.3 + 0.15 9,5 ±0,19 42± 1,6 39,5 ±1,3 39,8 ±1,5 49± 1,4 40± 1,1 53± 1,8
5-я 9,3±0,16 9,2±0,11 47± 1.2 43.7 ±1,6 38,5 ±1,2 69 ±1,6 45± 1.6 62 ± 1,8
6-я 8,6±0,12 9,0 ± 0,12 9,0 ±0,09 47± 1,4 41,8± 1,4 39.9 ±1,7 51 ± 1,1 56± 1,3 31 ±1,3
7-я 8,6±0,14 8.5 ±0,09 8.8 ±0.07 46± 1,1 42,1 ±1,5 40,3 ±1.3 50± 1,3 56± 1.8 40± 1,4
8-я 8,6 ±0,07 8,210.13 8.7 ±0.09 47 ± 1.3 37,6 ±1,7 40,4 ±1,7 41 ±0,8 57±1,9 100 + 0,0
9-я 8,4±0,10 8.2±0,13 8,4 ±0,20 43±1,2 41,7± 1,1 37.0 ±0,6 61 ±1,2 60±1,1 70±1,9
10-я 7,5 ±0,08 8,0± 0,10 7,9±0,10 30 ±1,2 39,0 ±1,7 40,0 ±1,7 52 ±1,8 30+1,4 70±1,9
11-я 6,4±0,14 7,5 ±0.08 7,2 + 0.16 31 ± 1,5 42,4 ±1,2 41,8± 1,6 48±1,4 50±1,7 , 45±1,6
Примечание I — subsp. lanatus; II — subsp. vulgaris; HI — subsb. mucosospermus.
Таблица 14. Средние значения относительных длин хромосом, их центромерных
и хроматических индексов в кариотипе С. colocynthis, С. ecirrhosus, С. naudinianus
Пара хромосом Относительная длина хромосом, % (Lr ± о) Центромерный индекс, % (1е + а) Хроматический индекс, % (1сЛ ±
1 " 1 1 1 1 11 1 1 1 » / 111
1-Я 11,2±0,10 10,7±0,08 10,5 + 0,03 36.8+1,6 36,8 ± 1,2 40,1 ±0,8 60 ±2,0 57±1,7 I 32±0,9
2-я 10.5 + 0,15 10,4±0.12 10,3 + 0.10 37,4+1.9 41,5± 1,8 39,7 ±1,2 56 +1,1 54±1,2 1 60±1,6
3-я 9.8 i 0,11 9,8 + 0.10 9,8±0,06 40,0+1,5 39,0 ±0.7 41,2 ± 1,3 51+0,9 45± 1,6 1 35+1,3
4-я 9,6±0,14 9,3 ±0,09 9,2±0,07 34.8±2,0 38,6 ±1,0 42,5 ±0,9 39+1,8 65±0,9 55 + 0,9
5-я 9,1 ±0,16 9,0±0,18 8,9±0,14 35,8±1,8 39,8 ±0,9 41,6± 1,2 56 +1,0 65 + 1,0 64+1,3 ГЛ . 1 7
6-я 8,9 ±0,08 8,7 + 0,07 8,9±0.07 39.4± 1,1 39,0 ±1.5 42,4 ±1.0 76 ± 1,3 80 + 0,9 50 ± 1,7 59 ±1,5 35+1,8 54+1,1 60±1,4 30±1,2
7-я 8,8±0,09 8,6 + 0.10 8,9 + 0,08 38,1 + 1.4 42,4 ±1.3 42,9 ±1,3 37±1,2 100±0.0
8-я 8,3 ±0,10 8,5 + 0,14 8,8+1,03 41.6+1.3 45,9 ±0,8 41,8± 1,3 56 ± 2,3 49 ± 2,0 68+1.5 35± 1,5 80+1.3
9-я 8,3 ±0,15 8,5 + 0,08 8,4 + 0,06 40,2+1,6 40.1 ±1,3 41,5± 1.1 66+1,0
10-я 11-я 7,9+0,12 7,4 ±0,14 8,2 + 0.10 7.6 + 0,06 8.2 + 0,08 7.8 + 0,07 42,1 + 1,2 41,4+1,2 40,8 ±1,2 39,7 ±1,0 41,8± 1,2 42,3 ±0.09 84 + 0.8 70+1,6
Примечание: I—С. colocynthis; II — С. ecirrhosus; III — С. naudinianus.
За основу гомологизации хромосом при анализе структуры карио-
типов брали рисунок дифференцированного окрашивания: учитывали
число, величину, форму и расположение различающихся по окрашива-
нию сегментов.
Хромосомы исследованных образцов родов Citrullus и Praecitrul-
lus дифференцированы по длине на хроматические (темноокрашенные)
и ахроматические (светлоокрашенные) участки. Линейная дифференциа-
ция окрашивания позволяет использовать эту качественную характе-
ристику наряду с морфометрическими параметрами как маркер при
идентификации хромосом.
Результаты измерений представлены в таблицах 13, 14 и 15 как
средние значения относительных длин пар гомологов, их центромерных
и хроматических индексов.
Таблица 15. Средние значения относительных длин хромосом, их центромерных
и хроматических индексов в кариотипе Praecitrullus
Пара хромосом Относительная длина хромосом, % (Lr±j) Центромерный индекс, % (1с±4) Хроматический индекс, % (lch±s)
1-я 11,0 + 0,19 45,6+0,7 80 ± 1,7
2-я 10,1 ±0,08 41,4± 1,4 —
3-я 9,5 + 0,14 40,7 ±1,8 83 ±0,9
4-я 9,3 + 0,12 28,4 ±1,2 —
5-я 8,8 + 0,08 40,6 ±2,0 74± 1,4
6-я 8,2 + 0,12 38,5 ±1,8 —
7-я 7,6± 0,10 43,8 ±1,1 71 1,5
8-я 7.5 ±0,07 43,0 ±1,7 56± 1,1
9-я 7,0 + 0,10 41.3 ±1,6 —
10-я 6,9±0,12 43,6 ±0,9 —
11-я 6,7±0,15 41,5 ±1,7 —
12-я 6,1 ±0,10 41,8+1,0 —
Средняя длина хромосомного набора каждого изучаемого ооразца
вычислена путем определения длин хромосомных наборов каждой ме-
тафазной пластинки. Общие длины кариотипов изображены в виде
диаграммы (рис. 21), иллюстрирующей вариабельность этого показа-
теля у изучаемых образцов (цифры вы-
Рнс. 21. Сравнение длин хро-
мосомных наборов видов Cit-
rullus и Praecitrullus:
1 — С. lanatus subsp. mucoso-
spermus; 2 — С. naudinianus;
3 — С. ecirrhosus; 4 — С. colo-
cynthis; 5—С. lanatus subsp'
vulgaris.
ражают процент по отношению к длине
хромосомного набора subsp. mucososper-
mus, принятой за 100%).
Для определения предела ошибки
приводимых средних величин относитель-
ной длины хромосом, центромерного и
хроматического индексов, длины карио-
типов всех образцов были вычислены
средние квадратические ошибки (±о).
При изучении хромосомных наборов
использовали классификацию хромосом
по Левану [327]: метацентрические хро-
мосомы— Iе = 38,5—50%, субметацентри-
ческие хромосомы — Iе = 10,0—38,5%.
Предварительная обработка мате-
риала а-бромнафталином в сочетании с
пониженной температурой дает возмож-
ность получать в каждом отдельном пре-
парате достаточное количество четких
метафазных пластинок, в которых хромо-
94
гомы оказываются дифференцированно окрашенными. Способностью к
такому окрашиванию обладают все хромосомы набора, но в разной
степени. Сходство рисунка окрашивания гомологичных хромосом из раз-
ных метафазных пластинок позволяет заключить, что структурная не-
однородность хромосом является выражением реально существующей
линейной дифференциации. Хромосомы считались гомологичными и по-
лучали номер соответствующей пары, если при последовательном
сравнении с остальными хромосомами набора они оказывались сход-
ными по рисунку дифференцированного окрашивания, величине, место-
положению центромеры.
МОРФОЛОГИЯ ХРОМОСОМ РАЗНЫХ ТАКСОНОВ
Арбуз шерстистый — subsp lanatus. Вид включает три подвида:
арбуз шерстистый (subsp. lanatus), арбуз обыкновенный (subsp. vul-
garis), арбуз слизистосемянный (subsp. mucosospermus). Соматическое
число хромосом subsp. lanatus равно 22. Размер их 1,5—2,8 мкм, длина
диплоидного набора 48,5 ± 0,6 мкм. Суммарная длина темноокрашен-
ных участков составляет 48,8% общей длины хромосомного набора. По
местоположению центромеры можно выделить два типа хромосом:
метацентрический — М (1, 3—9-я пары) и субметацентрический — Sm
(2, 10, 11-я пары).
На основе сопоставления морфометрических параметров относи-
тельной длины, центромерного и хроматического индексов проведена
идентификация каждой пары гомологов всех подвидов (см. табл. 13).
Арбуз обыкновенный — С. lanatus subsp. vulgaris. Соматическое
число хромосом равно 22 [272, 321, 329]. На рис. 22,/ представлена
метафазная пластинка соматических хромосом этого подвида.
Размер хромосом 1,8—2,7 мкм, средняя длина диплоидного набора
49,8 ± 0,9 мкм. По местоположению центромеры в кариотипе можно
выделить два типа хромосом: метацентрический (1—7, 9—11-я пары)
и субметацентрический (8-я пара).
Арбуз слизистосемянный — С. lanatus subsp. mucosospermus. Сома-
тическое число хромосом равно 22. Число хромосом и кариотип этого
подвида приводятся впервые. Размер хромосом 1,3—1,8 мкм, средняя
длина кариотипа 34,3 + 0,4 мкм. По местоположению центромеры в ка-
риотипе можно выделить два типа хромосом: метацентрический (1, 4,
6—8, 10, 11-я пары) и субметацентрический (2, 3, 5, 9-я пары). Пары
3-я и 9-я имеют макроспутники на коротких плечах (рис. 22,2).
Изучение кариотипа ssp. mucosospermus показывает, что он от-
личается от других форм арбуза по величине и морфологии хромосом,
длине диплоидного набора (см. табл. 13), наличию двух пар спутнич-
ных хромосом.
Арбуз колоцинт — С. colocynthis. Широко распространен в пусты-
нях Северной Африки, Аравии, Ирана, Афганистана, Индии, встре-
чается даже в Австралии. Известен и на территории нашей страны в
южной Туркмении.
Соматическое число хромосом равно 22 [109, 160, 374]. Размер
хромосом 1,8—2,7 мкм, средняя длина диплоидного набора 48,4 ±
± 0,3 мкм. По местоположению центромеры в кариотипе можно выде-
лить два типа хромосом: метацентрический (3, 6, 8—11-я пары) и суб-
метацентрический (1, 2, 4, 5, 7-я) (см. табл. 14).
Арбуз безусиковый — С. ecirrhosus. Соматическое число хромосом
равно 22 [359]. Размер хромосом 1,8—2,5 мкм, средняя длина диплоид-
ного набора 46,5±0,6 мкм (см. табл. 14). По местоположению центро-
96
меры в кариотипе можно выделить два типа хромосом: метацентриче-
ский (2—11-я пары) и субметацентрический (1-я пара).
Арбуз Нодэна — С. naudinianus. Соматическое число хромосом
равно 22 [359]. Размер 1,7—2,3 мкм, средняя длина кариотипа 44,6 ±
±0,4 мкм (см. табл. 14). По местоположению центромеры кариотип
характеризуется одним типом метацентрических хромосом.
Рис. 22. Метафазная пластинка соматических хромосом:
1—С. lanatus subsp. vulgaris; 2 — С. lanatus subsp. mucosospermus.
Арбуз фистулезный — Praecitrullus fistulosus. Соматическое число
хромосом равно 24, суммарная длина диплоидного набора 61,9±0,7 мкм,
их размер 1,9—3,4 мкм, больше, чем у Citrullus. По местоположению
центромеры в кариотипе можно выделить два типа хромосом: мета-
96
центрический (1—3, 5, 7—12-я пары) и субметацентрический (4-я и
6-я пары).
Хромосомы Praecitrullus имеют своеобразный рисунок дифферен-
цированного окрашивания: темноокрашенные сегменты у них распреде-
лены по всей длине хромосомы, чем напоминают рисунок окрашивания
хромосом Cucumis sativus L. и отличаются от хромосом Citrullus,
имеющих околоцентромерное расположение окрашенных зон.
По числу, размерам, форме, морфологии, специфическому рисунку
окрашивания хромосомы Praecitrullus отличаются от кариотипов иссле-
дованных видов Citrullus. Отдаленность родства между этими родами
подкрепляется и гибридологическими экспериментами Т. Кошо и С. Вий
[321], которым не удалось получить гибриды, несмотря на большое
число скрещиваний. Мейотические исследования [361] также подтверж-
дают обособленность Praecitrullus.
Сравнительная характеристика кариотипов всех изученных таксо-
нов представлена в таблице 16.
Таблица 16. Морфометрическая характеристика кариотипов видов
Citrullus и Praecitrullus
№ по каталогу ВИР Таксон 2n Абсолютная длина хромо* сом, мкм Длина ди- плоидного набора, мкм Число хромосом
м Sm
2220 С. lanatus subsp. vulgaris 22 1,8-2,7 49,8 ±0,9 10 1
1298 С. lanatus subsp. mucosospermus 22 1,3—1,8 34,3 ±0,4 7 4
4339 C. colocynthis 22 1,8-2,7 48,4 ±0,3 6 5
4251 C. ecirrhosus 22 1,8-2,5 46,5 ±0,6 10 1
4578 C. naudinianus 22 1,7-2,3 44,6 ±0,4 11 0
3877 Praecitrullus 24 1,9—3,4 61,9±0,7 10 2
Примечание. М-метацентрический тип; SГЛ—субметацентрический тип.
Таким образом, комплексный подход к изучению структуры карио-
типа (использование рисунка линейной дифференциации хромосом с
одновременным учетом их морфометрических параметров) позволил
идентифицировать хромосомы каждого изученного образца и дать
сравнительную характеристику исследованных таксонов, причем впер-
вые дано описание кариотипов С. lanatus subsp. mucosospermus,
С. ecirrhosus и С. naudinianus.
Для всех видов рода Citrullus характерна повторяемость типов
хромосом: метацентрического и субметацентрического. У видов С. la-
natus и С. ecirrhosus отмечено одинаковое соотношение метацентриче-
ских и субметацентрических хромосом (10:1). Наибольшее число суб-
метацентрических хромосом обнаружено у С. lanatus subsp. mucosos-
permus и С. colocynthis.
Было установлено, что различия между видами по размеру хро-
мосом также невелики и варьируют в пределах 1,3—2,7 мкм. Самые
короткие хромосомы отмечены у С. lanatus subsp. mucosospermus. Этот
подвид имеет также две спутничные хромосомы, в то время как другие
изученные таксоны лишь одну.
Кариотипические особенности subsp. mucosospermus подкреп-
ляются ранее отмеченным анатомо-морфологическим и эмбриологиче-
ским своеобразием этого таксона и наводят на мысль о возможности
выделения его в самостоятельный вид. Кариотипы двух других подви-
дов С. lanatus (subsp. lanatus и subsp. vulgaris), наоборот, очень
близки между собой по всем показателям, что подтверждает правиль-
ность объединения их в один вид.
97
Для всех представителей рода Citrullus характерно околоцентро-
мерное расположение темноокрашенных районов. Наименьшее количе-
ство хроматина отмечено у С. naudinianus, наибольшее — у С. ecir-
rhosus.
Число, форма, размер хромосом Praecitrullus, особенности поведе-
ния их в мейозе свидетельствуют об отсутствии его родства с Citrullus.
Хромосомы Praecitrullus по морфологии напоминают хромосомы Cucu-
mis sativus, что отмечено 3. А. Кожуховым [1081 и М. Шимотзума
[361].
полиплоидия
Естественная полиплоидия в семействе Cucurbitaceae не зарегистри-
рована. Все известные полиплоиды возделываемых форм получены
искусственным путем. Среди них наибольшей известностью пользуется
триплоидный арбуз, впервые синтезированный японским генетиком
X. Кихара [322] в начале 40-х годов нашего столетия путем опыления
тетраплоидной формы пыльцой диплоидного сорта.
Триплоидный арбуз наряду с бессемянностью обладает и другими
достоинствами. Для него характерны гибридная мощность, высокая уро-
жайность, устойчивость к болезням. Плоды триплоидного арбуза содер-
жат больше сахара, чем диплоидные, и способны длительное время
сохраняться, не теряя своих вкусовых качеств.
Ценные свойства триплоидного арбуза вызвали большой интерес
исследователей. Работы по полиплоидии арбуза широко развернулись
в Японии [324] и были начаты в других странах: СССР [76, 77, 78, 231],
США [281, 369], Индии [288], Румынии [12, 351], Болгарии [122, 125].
Методика получения тетраплоидов арбуза, применяемая разными
исследователями, имеет много общего и отличается лишь в деталях.
Она сводится к обработке проростков в фазе семядолей 0,2—0,4%-ным
водным раствором колхицина. Иногда применяют спиртовой раствор.
Проростки обрабатывают в течение нескольких дней, нанося на точку
роста несколько капель раствора или накладывая ватный тампон, смо-
ченный тем же раствором. Чтобы избежать повышения концентрации,
тщательно контролируют влажность среды, а также оберегают про-
ростки от попадания прямых солнечных лучей.
Существует и другой способ обработки — намачивание семян в рас-
творе колхицина в течение 12—24 ч, последующая промывка их и про-
ращивание в термостате. Иногда применяют намачивание наклюнув-
шихся семян в растворе колхицина в течение 48 ч с последующим про-
ращиванием обычным способом.
Некоторые исследователи [231] считают, что обработка семян дает
более высокий процент тетраплоидов, а также исключает появление
растений-химер.
Тетраплоиды имеют ряд морфологических особенностей, по кото-
рым их легко отличить от диплоидов. Стебли у них короче и толще,
с хорошо выраженными гранями; листья крупные, мясистые, широкие
(индекс листа близок к единице); цветки также крупнее диплоидных.
Плоды по величине почти не отличаются, но имеют несколько сплюс-
нутую форму. Цитологическое изучение показало, что у тетраплоидов
увеличены также размеры клеток и ядер.
Пыльца тетраплоидов заметно крупнее пыльцы диплоидов. По на-
шим данным, диаметр гаплоидного пыльцевого зерна составляет 50—
60 мкм, а диплоидного — 70—75 мкм. Такие же цифры приводит
Н. Кондо, указывая, что размер диплоидной пыльцы примерно на 38%
98
больше, чем гаплоидной. Очень характерны семена тетраплоидов: они
крупнее, чем у диплоидных форм, часто имеют продольные трещины
на кожуре.
Тетраплоидные растения развиваются медленно, особенно вначале,
поэтому выращивать их можно только рассадным способом. В первое
время высаженную рассаду необходимо притенять и поддерживать опти-
мальную влажность почвы.
Многие исследователи отмечают плохую завязываемость плодов и
низкую семенную продуктивность тетраплоидов, сдерживающую их ши-
рокое распространение. По данным Н. Кондо, тетраплоиды содержат
лишь 15—16% семян от числа семян диплоидов. Их малосемянность
обусловлена пониженной фертильностью пыльцы, связанной с наруше-
ниями в мейозе [12], а также ее медленным прорастанием.
Сравнивая прорастание пыльцы диплоидов и тетраплоидов арбуза
на искусственной питательной среде (15%-ный раствор сахарозы при
pH 6), мы обнаружили, что последняя прорастает значительно слабее.
Первые признаки прорастания появлялись лишь через 30 мин после
посева, в то время как гаплоидная пыльца начинала прорастать уже
через 10 мин. Через 1,5—2 ч процент проросшей пыльцы тетраплоидов
был таким же, как у диплоидов через 1 ч после посева. При этом
среди диплоидной пыльцы наблюдалось много деформированных пыль-
цевых трубок, прекративших рост. Длина пыльцевых трубок диплоид-
ной пыльцы через 1,5 ч составляла 300—400 мкм. Такой же длины до-
стигали пыльцевые трубки гаплоидной пыльцы через 1 ч после начала
роста. Таким образом, пыльца тетраплоидов прорастала почти в 2 раза
медленнее, чем пыльца диплоидов.
Из-за слабого роста пыльца тетраплоидов в основной массе до-
стигает семяпочки тогда, когда готовность последней к оплодотворению
значительно снижается. Это, видимо, и является основной причиной
плохого завязывания семян. Поддерживать тетраплоиды особенно
трудно в первых поколениях, но постепенно фертильность их повы-
шается.
Как показали наши опыты, один из путей повышения завязывае-
мости плодов тетраплоидов —скрещивание разных форм и опыление
смесью пыльцы с разных растений. Процент завязывания плодов в этих
вариантах был равен 50—60, в то время как при использовании пыльцы
с того же растения наблюдалась самая низкая завязываемость, не
превышавшая 20%. Высокая самостерильность тетраплоидов отмеча-
лась многими исследователями. Н. Кондо [324] замечает, что иногда
вообще не удается получить плоды тетраплоидов от искусственного
опыления и приходится использовать плоды от свободного опыления.
Как показал опыт японских исследователей, опыление оказывается
более эффективным в начале цветения, а через 2—3 недели почти не
дает результатов. Для ускорения роста тетраплоидов и повышения
фертильности в первых поколениях П. Лозанов [122] предлагает при-
вивать их на другие тыквенные растения, отличающиеся быстрым рос-
том. В качестве подвоев можно использовать крупноплодную тыкву,
лагенарию, бенинказу. Однако опыт болгарского исследователя пока
не нашел применения в работе с полиплоидами.
В повышении продуктивности тетраплоидов большую роль играет
сорт. П. Лозанов [124] среди изученных нм тетраплоидов выделяет
форму Тетра американского происхождения как наиболее перспектив-
ную для получения триплоидных форм. К. Андрус [281] отмечает, что
линии этого сорта (Тетра 1, Тетра 2, Тетра 3) могут быть использо-
ваны не только для получения триплоидов, но и как самостоятельные
7*
99
сорта, обладающие рядом положительных качеств (твердая кора, плот-
ная мякоть, высокое содержание сахара, способность к длительному
хранению и перевозкам). Малое количество семян с потребительской
точки зрения тоже явление положительное, хотя и усложняет семено-
водство сорта.
При скрещивании тетра- и диплоидов плоды завязываются хо-
рошо, но семян бывает меньше, чем у диплоидов (хотя больше, чем
у тетраплоидов). Однако искусственное опыление, применяемое на
опытных посевах, непригодно для массового получения триплоидных
семян, так как требует больших затрат квалифицированного ручного
труда.
Д. Волл [369] предлагает использовать для получения триплоидов
арбуза генетический маркер — темные полосы на плодах отцовского
сорта, которые наследуются доминантно и проявляются, следовательно,
только у гибридов. Используя этот метод, он установил, что 80% по-
лученных при свободном опылении растений были триплоидными. Пред-
ложенный способ позволяет одновременно получать гибридные три-
плоидные формы и поддерживать тетраплоиды.
Очень важный момент при получении триплоидного арбуза — со-
отношение растений отцовской и материнской формы. М. Шимотзума
[360] на основании специально поставленных опытов пришел к заклю-
чению, что самые лучшие результаты дает посев диплоидных и тетра-
плоидных семян в отношении 1 :3. Высевают их по определенной схеме
с таким расчетом, чтобы каждое диплоидное растение было окружено
тетраплоидными. Триплоидные семена используют для выращивания
бессемянных арбузов, реализуемых как продукция высокого качества.
Причины стерильности триплоидного арбуза, дающего бессемян-
ные плоды, детально изучены А. К. Дзелвалтовским [76, 77, 78]. Он уста-
новил, что процессы микро- и макроспорогенеза триплоидов характе-
ризуются резкими нарушениями I и II делений мейоза. Основные из
них: образование унивалентов, неравномерное расхождение хромосом
к полюсам, формирование крупных или многочисленных мелких ядер.
В результате второго деления мейоза часто образуются не тетрады, а
полиады. Следствием этих нарушений является дегенерация элементов
зародышевого мешка и пыльцы, которые у триплоидов оказываются
полностью стерильными.
Плоды триплоидных арбузов — это разросшиеся завязи. Семена
их представлены рудиментами семенной кожуры. Однако в некоторых
случаях кожура достигает почти полного развития, становится твердой,
что снижает товарные качества плода. Это явление считается нежела-
тельным, и селекционеры пытаются его преодолеть путем подбора сор-
тов-опылителей.
Плоды бессемянного арбуза могут развиваться только из опылен-
ного цветка. Оплодотворения семяпочки при этом не происходит, но
пыльца стимулирует разрастание завязи. В качестве опылителей ис-
пользуют обычные диплоидные сорта. От подбора сортов-опылителей
в значительной мере зависят урожайность и товарные качества бес-
семянного арбуза. По данным М. Шимотзума, среди японских сортов
хорошим опылителем является сорт Кахо.
Выращивание триплоидов сопряжено с определенными трудностями.
Семена их имеют очень твердую кожуру, поэтому обязательна скари-
фикация, которую проводят обычно путем расщелкивания носика на-
бухшего семени. Проращивают семена в термостате при 30° С. Наклю-
нувшиеся семена высаживают в стаканчики, поддерживая при этом вы-
сокую температуру окружающей среды. При появлении 3—4 настоящих
100
листьев рассаду высаживают в грунт. Вначале она развивается очень
медленно, так как для нее необходима высокая температура, но затем
догоняет диплоидные растения, и к моменту завязывания плодов раз-
витие триплоидов становится нормальным.
При правильном подборе опылителей триплоиды характеризуются
гибридной мощностью и превышают по урожайности диплоидные сорта.
Важный фактор повышения урожайности триплоидов — размещение
их в посеве. В Японии наиболее распространена следующая схема: че-
редование двух рядков триплоидных растений с одним рядком опыли-
теля. При достаточном количестве семян можно высевать по 3—4 рядка
триплоидов. Сорта-опылители сеют прямо в грунт, оставляя пустые
рядки для высадки триплоидной рассады.
Таким образом, бессемянный арбуз хотя и обладает целым рядом
преимуществ, но семеноводство его сопряжено со значительными труд-
ностями. Главные из них —низкая фертильность тетраплоидов и от-
сутствие простых способов получения триплоидных семян. Дальнейшие
исследования в области использования полиплоидии у арбуза необхо-
димо проводить именно в этих направлениях.
По мнению большинства исследователей, в ближайшее время бес-
семянный арбуз не получит широкого распространения и будет возде-
лываться лишь в ограниченных масштабах.
НАСЛЕДОВАНИЕ ПРИЗНАКОВ
Знание закономерностей наследования признаков имеет существен-
ное значение при создании новых сортов и гетерозисных гибридов. Све-
дения по частной генетике арбуза разбросаны по отдельным источни-
кам и являются подчас противоречивыми, поэтому особый интерес пред-
ставляет обзорная статья Р. Робинсона и др. «Гены тыквенных» [353].
В ней не только проанализированы основные работы зарубежных ав-
торов по генетике тыквенных культур, но также даны таблицы генов,
установленных в настоящее время для огурца, дыни, арбуза, тыквы.
В таблице 17 приведены данные по генам арбуза, взятые из работы
Робинсона.
Ниже рассматриваются характер наследования и генетический кон-
троль некоторых признаков, имеющих практическое значение.
Рассеченность листа. Подавляющее число сортов арбуза характе-
ризуются рассеченными листьями, но изредка встречаются цельнолист-
ные формы. Этот признак благодаря рецессивному характеру наследо-
вания оказался хорошим маркером проростков при производстве гиб-
ридных семян. Установлено, что нерассеченный лист контролируется
единственным геном nl—nonlobed leaves [335]. Рецессивный характер
наследования этого признака подтверждается и нашими опытами. Мы
проводили скрещивание сортов Дынный лист и Желтокорый, обладаю-
щих сигнальными признаками на предварительно инцухтированном ма-
териале. В F] все растения имели рассеченные листья и темные плоды,
в F2 наблюдалось расщепление по типу листа и окраске плода, харак-
терное для дигибридных скрещиваний. Выделены 4 фенотипические
группы, в том числе форма, сочетающая два рецессивных признака:
1) рассеченный лист, темный плод; 2) рассеченный лист, желтый плод;
3) цельный лист, темный плод; 4) цельный лист, желтый плод. В циф-
ровом выражении расщепление укладывалось в схему 12:3:4:1, близ-
кую к классической для дигибридных скрещиваний п отражающую,
по-видимому, эпистатическое действие доминантного гена рассеченно-
сти листа.
101
Таблица 17. Гены, контролирующие признаки арбуза
Символ | Синоним Признак Характер действия гена
а Аг С d dw-1 dw-2 е f g gs go gms L m nl 0 P pm г s su t W Wf Y (B)GC (t) (D) (d3) (c) (msg) (suBi) (b‘) (W) (0 Андромонойкист (andromonoecious) Устойчивость к антракнозу (antracnose resistance) Канареечно-желтая мякоть (Canary yellow flesh) Крапчатая семенная кожура (dotted seed coat) Карликовость (dwarf-1) Карликовость (dwarf-2) Растрескивающаяся кора (explosive rind) Сегментированная поверх- ность плода (furrowed fruit surfase) Светло-зеленая кора (light green skin) Зеленые полосы на коре (striped green skin) Золотистый плод (golden) Неопушенный стебель, расте- ния с мужской стериль- ностью (glabrous male sterile) Длинные семена (long seed) Мозаичная кора (mottled skin) Нерассеченный лист (nonlobed leaves) Овальный плод (oval fruit) Нитчатые полосы (pencilled lines on skin) Восприимчивость к мучнистой росе (powdery mildew subceptibility) Красная кожура семени (red seed coat) Короткие семена (short seeds) Отсутствие горечи (suppressor of bitterness) Бурые семена (tan seed coat) Белые семена (white seed coat) Белая мякоть (White flesh) Желтая мякоть (yellow flesh) Рецессивен по отношению к мо- нойкисту Устойчив к Glomerella cingulata var. orbiculare Доминантна по отношению к ро- зовой мякоти Черная крапчатость семян доми- нантна по отношению к генам г, t и W Короткие междоузлия обуслов- лены более короткими клетка- ми Короткие междоузлия обуслов- лены меньшим числом клеток Слабая, лопающаяся при разре- зе кора Рецессивна по отношению к гладкой Светло-зеленая окраска плода рецессивна по отношению к темно-зеленой Рецессивны к темной, доминант- ны к светлой окраске плода Желтый цвет старых листьев и зрелых плодов Листья слабоопушенные, расте- ния с мужской стерильностью Удлиненные семена рецессивны по отношению к семенам сред- ней длины, взаимодействует с геном s Зеленая мозаика на коре плода Листья слаболопастные, домини- рование неполное Неполное доминирование по от- ношению к шаровидному плоду Незаметные полосы на коре пло- да, рецессивны по отношению к сетке Восприимчивость к Sphaerotheca fuliginea Взаимодействует с генами W и t Эпистатичен по отношению к гену L Негорькие плоды Взаимодействует с генами г и w Взаимодействует с генами г и t Доминантна по отношению к красной мякоти Рецессивна по отношению к красной мякоти
102
Форма плода. Исследованиями Л. М. Витмана [372] установлено,
что овальная форма плода контролируется единственным, не полностью
доминантным геном O-oval fruit [344].
В наших опытах при скрещивании сортов с шаровидной и удли-
ненной формой плода наблюдался промежуточный характер наследо-
вания. Однако в межвидовых скрещиваниях столового арбуза с коло-
цинтом, которые проводила М. К. Гольдгаузен [63], всегда доминиро-
вала шаровидная форма плода, свойственная колоцинту. Видимо, на-
следование этого признака при внутри- и межвидовых скрещиваниях
контролируется разными генами.
Окраска и рисунок плода. Установлено, что зеленая окраска арбу-
за доминирует над светлой и контролируется единственным геном G —
green [343, 346, 372].
Плоды с доминантным аллелем гена имеют темно-зеленую окраску,
с рецессивным — светло-зеленую. Тот или иной оттенок плода опре-
деляется генами-модификаторами. Что же касается рисунка плода в
виде полос, наиболее распространенного среди сортов арбуза, то он
доминантен по отношению к светлоокрашенным плодам и рецессивен
по отношению к темным. Л. М. Витман объясняет эти варианты до-
минирования тем, что фактор полосатости является одним из множе-
ства аллелей локуса g, определяющего окраску коры плода, или на-
ходится в другом локусе, тесно связанном с первым. Для обозначения
гена полосатости Ч. Пул предложил символ gs — striped green skin.
Он же подчеркнул, что узкие нитчатые полосы на коре плода арбуза
контролируются геном р — pencilled.
Окраска мякоти плода. Среди различных окрасок мякоти арбуза
доминирует белая. Для обозначения гена, контролирующего белую мя-
коть, предложен символ Wf — White flesh [353]. Доминирование белой
мякоти по отношению к красной четко проявляется в межвидовых скре-
щиваниях, где особенно заметно эпистатическое действие данного гена
[361]. Такой же характер наследования окраски мякоти у межвидовых
гибридов отмечает М. К. Гольдгаузен [63].
Красная мякоть, будучи рецессивной по отношению к белой, в то
же время доминирует над желтой, о чем сообщает Д. Р. Портер [346],
предложивший для гена желтой мякоти символ у — yellow. Ч. Пул
[343] отмечает, что существует другой оттенок желтой мякоти арбуза,
доминирующий над розовой, который он обозначил символом С — Ca-
nary yellow.
Горькие вещества. Плоды дикорастущих видов и форм арбуза, как
правило, горькие и стойко сохраняют этот признак при скрещивании
со столовыми сортами, что является серьезным препятствием для ис-
пользования таких форм в селекции. Генетики [301, 361], изучавшие
наследование горечи у колоцинта, сообщают о доминантном характере
этого признака, контролируемого одним геном.
В то же время П. Ензлин и С. Рэм [352], а также М. Шимотзума
обнаружили, что отсутствие горечи в плодах столового арбуза подав-
ляется единственным геном, обозначенным О. Чамлисом и др. [287]
символом su — suppressor.
Длина и число стеблей. М. К. Гольдгаузен [63] проанализировала
наследование таких количественных признаков, как длина и число стеб-
лей в межвидовых скрещиваниях арбуза. Установлено, что длинный сте-
бель доминирует над коротким, а большое число плетей — над малым.
При этом многочисленные плети удлиняются до 10—12 м за счет вы-
тягивания междоузлий. Длинные междоузлия с точки зрения селекции
являются отрицательным признаком. Желательно иметь сорта арбуза
103
с компактным кустом, облегчающим механизированную обработку и
уборку. При межвидовой гибридизации такой куст можно получить
повторными беккроссами.
Устойчивость к болезням. Имеющиеся в литературе данные о на-
следовании устойчивости арбуза к болезням немногочисленны и ка-
саются в основном антракноза, особенно распространенного в США
[305, 326, 353, 376]. Отмечают, что наследование устойчивости к ан-
тракнозу носит у арбуза доминантный характер и контролируется одним
геном Аг — antracnose resistance. Н. Винстед подчеркивает, что этот
ген определяет устойчивость к расам антракноза 1 и 3, но не к расе 2.
Сложнее обстоит дело с фузариозным увяданием, наиболее рас-
пространенным в нашей стране. Закономерности наследования устойчи-
вости к этому заболеванию до сих пор неясны, хотя некоторые авторы
предполагают, что оно носит полигенный характер. Н. М. Сазанова
[185], изучавшая устойчивость сортов и гибридов арбуза к этому за-
болеванию, отмечает, что при гибридизации иммунные свойства лучше
передает материнская форма, а М. Д. Дзензелевская [79] указывает
на высокую способность отечественных сортов арбуза Байрачский и
Пионер пустыни передавать признак устойчивости.
КОМБИНАЦИОННАЯ СПОСОБНОСТЬ СОРТОВ АРБУЗА
Гетерозис находит все более широкое применение как один из пу-
тей повышения урожайности сельскохозяйственных культур. У бахче-
вых гетерозис проявляется довольно отчетливо: прибавка урожая может
достигать 50% и более. Многие научно-исследовательские учреждения
нашей страны, работающие с бахчевыми культурами, в том числе и
ВИР, в последние годы широко развернули исследования по этой проб-
леме. Среди нерешенных вопросов на первое место выступает подбор
исходных форм, обеспечивающих повышение продуктивности гибридов,
а также совершенствование способов получения гибридных семян.
Подбор пар тесно связан с изучением комбинационной способности
сортов, участвующих в скрещиваниях, т. е. их способности передавать
свои качества потомству.
Понятие комбинационной способности возникло в 40-е годы при
работе по созданию гибридов кукурузы. Тогда же Г. Спрейг и 3. Татум
предложили различать общую и специфическую комбинационную спо-
собность сортов: первую как выражение средней величины гетерозиса
по всем гибридным комбинациям, вторую как отклонение отдельных
комбинаций от этой величины.
В настоящее время существует только один надежный путь выяв-
ления комбинационной способности сортов — скрещивание с последую-
щим испытанием гибридов. Среди разных типов скрещиваний наиболь-
шую популярность завоевал метод топкроссов, позволяющий при
сравнительно небольшом объеме работ определить комбинационную
способность многих сортов. При этом способе сорт, используемый в ка-
честве тестера (анализатора), скрещивают со многими сортами или ли-
ниями, у которых хотят определить комбинационную способность. Же-
лательно, чтобы сорт-тестер имел широкую генетическую основу, т. е.
был представлен популяцией, включающей несколько биотипов. Видо-
измененный метод топкроссов — свободное переопыление сортов, вы-
сеянных в одном питомнике, которое наиболее целесообразно исполь-
зовать при работе с зерновыми культурами. При небольшом объеме
работы применяют метод диаллельных (парных) скрещиваний в пря-
мых и обратных комбинациях. В этом случае каждый сорт является:
тестером по отношению к другому.
104
В отечественной литературе почти нет данных по изучению ком-
бинационной способности сортов арбуза. Отдельные упоминания о сор-
тах, отличающихся комбинационной ценностью, встречаются в работах
И. С. Вернидубова [48] и Н. М. Сазановой [187].
Пары для скрещиваний подбирают обычно эмпирически. Способ
получения гибридных семян должен быть достаточно простым и эф-
фективным. В последнее время используют метод «сигнального при-
знака». У арбуза им служит нерассеченная пластинка листа. Цельно-
листные (материнские) и рассеченнолистные (отцовские) формы высе-
вают чередующимися рядками, что дает возможность получать гибриды
при свободном переопылении сортов. Благодаря рецессивному харак-
теру наследования цельнолистности все негибридные формы выбрако-
вывают в первом поколении задолго до цветения, оставляя только гиб-
риды.
Этот метод был предложен X. Мором и X. Блекхарстом, затем по-
лучил развитие в работах Ф. А. Ткаченко, А. И. Кононенко, К- П. Синчи,
П. Лозанова, С. В. Шейнфельда. По данным исследователей, гибрид-
ность потомства, полученного таким способом, не превышает 48%.
Чтобы увеличить ее, необходимо проводить дополнительное пчелоопы-
ление и ручное доопыление цветков, что значительно удорожает произ-
водство гибридных семян. Совершенствование существующего способа
является, таким образом, одной из актуальных задач селекции арбуза
на гетерозис.
Целесообразно использовать в качестве материнских компонентов
сорта, сочетающие цельнолистность с раздельнополостью. Такие формы,
будучи облигатными перекрестниками, дают более высокий процент
гибридов при свободном опылении. Затруднение заключается в том,
что большинство отечественных сортов арбуза имеют мужские и гер-
мафродитные цветки, что снижает вероятность скрещивания.
Работа по изучению комбинационной ценности сортов арбуза и су-
ществующих способов получения гибридных семян была начата в ВИР
А. А. Дюкарем под руководством Э. Т. Мещерова. Гибриды получали
на основе районированных сортов. Испытание их проводили на Гени-
ческом ГСУ в течение двух лет (1968 и 1970). Эту работу позднее
продолжили на Астраханском опорном пункте ВИР в несколько дру-
гом плане и на другом материале.
На Геническом ГСУ было проведено испытание двух гибридов ар-
буза: Пионер Харьковский и Проминь, полученных в Украинском НИИ
овощеводства и бахчеводства путем свободного переопыления сорта
Дынный лист с рассеченнолистными сортами. Первый из названных
гибридов был тогда районирован в некоторых областях УССР.
Двухлетнее испытание, проводимое в трехкратной повторности, по-
казало, что число гибридов среди взошедших растений составляло 33—
42%- Несмотря на то что урожай гибридных растений был на 20—30%
выше, чем у исходных форм, образец в целом не имел преимуществ.
Этот опыт убедил, что сорт Дынный лист нецелесообразно использовать
для массового получения гибридных семян.
Испытание ряда других гибридов, но полученных путем искусствен-
ного опыления, обнаружило, что гетерозис по урожайности проявляют
все комбинации, в которых участвовал сорт Стокса киевский. Среди них
особенно выделялись гибриды Стокса киевский X Мелитопольский 60
и Стокса киевский X Любимец хутора Пятигорска: они в течение двух
лет (1968 и 1970) превышали по урожайности стандарт на 69—158%-
Гетерозис гибридов проявлялся ярче в прямой комбинации, но сохра-
нялся и в обратной. Полученные результаты дают возможность гово-
ю&
рпть о высокой комбинационной ценности сорта Стокса киевский и ре-
комендовать его как исходную форму для получения гетерозисных гиб-
ридов арбуза, отличающихся скороспелостью. По качеству плодов и
скороспелости выделялись гибриды с сортом Любимец хутора Пяти-
горска, особенно если он выступал в качестве материнской формы.
Этот сорт целесообразно использовать в скрещиваниях как улучшатель.
В дальнейшей работе по изучению комбинационной способности
сортов арбуза была широко использована мировая коллекция ВИР.
В качестве материнских компонентов подбирали в основном формы
с сигнальными признаками, в том числе образцы, сочетающие цельно-
листность с раздельнополостью: Цера, Цера 21 (Болгария), Цельно-
листный 215 (Быковская бахчевая селекционная опытная станция),
Грибовский цельнолистный (быв. Грибовская овощная селекционно-
опытная станция). Были использованы также цельнолистные аналоги
районированных сортов Мелитопольский 142 и Степной 64—Г-42,
Г-146 и другие, полученные на Быковской бахчевой селекционной опыт-
ной станции.
В качестве отцовских компонентов использовали сорта коллекции,
выделенные по комплексу хозяйственно-ценных признаков и свойств:
New Hampshire midget (Хампширский карлик, США), отличающийся
ультраскороспелостью и многоплодностью (рис. 23); Crimson Sweet,
Рис. 23. Сорт арбуза New Hampshire Midget (США).
Sugar Baby и Shipper WR (США) — среднеранние урожайные сорта с
высоким качеством плодов; Роза Юго-Востока (Быковская бахчевая
селекционная опытная станция) — среднеранний сорт с высоким каче-
ством плодов; Мраморный — высокоурожайный сахаристый сорт селек-
ции ВИР и др. Первоначально скрещивание проводили по схеме топ-
кросса: материнскую форму использовали в качестве тестера для опре-
деления комбинационной способности нескольких отцовских форм. При
таком способе оценивалась общая комбинационная способность сортов.
Позднее схема была несколько видоизменена, введены также парные
106
скрещивания в прямых и обратных сочетаниях, что позволило оценить
сорта не только по общей, но и по специфической комбинационной спо-
собности, а также выявить ряд перспективных гибридов. Полученные
гибриды высевали на делянках площадью 40 м2 и сравнивали по хо-
зяйственно-ценным признакам и свойствам с родительскими формами
и с районированными сортами, взятыми в качестве стандартов (Мели-
топольский 142 для среднеспелых и среднепоздних форм, Победитель 395
для ранних и среднеранних).
В течение четырех лет изучено 39 комбинаций, выделены сорта
с высокой комбинационной способностью, перспективные для использо-
вания в качестве исходных форм при получении гетерозисных гибри-
дов. Хорошей общей комбинационной способностью обладают отечест-
венные сорта арбуза Мраморный (к-2159), Мелитопольский 143
(к-2840), Раздельнополый (к-4616), Грибовский цельнолистный (к-4333),
а также Цера 21 (к-4375) из Болгарии и Sugar Baby (к-3854) из США.
Следует подчеркнуть, что наиболее ценный из них сорт Цера 21. Все
комбинации с этим сортом показали высокий гетерозис. Высокая спе-
цифическая комбинационная способность была отмечена у цельнолист-
ных сортов, выведенных в последние годы на Быковской бахчевой се-
лекционной опытной станции: Г-146 (к-4301), Г-72 (к-4617), Цельно-
листный 215 (к-4307). Ниже приводятся результаты конкурсного
испытания лучших гибридных комбинаций, неоднократно проявивших
гетерозис по основным хозяйственно-ценным признакам (табл. 18).
Таблица 18. Перспективные гибриды арбуза (1975 г.)
Гибрид, сорт Повторности
1-я 2-я 3-я
А1 Б В А Б В А 1 Б1 В
Цера 21хМраморный 83 6,4 610 84 6,3 575 84 6,6 560
Цера 21 X Роза Юго-Востока 78 5,9 635 79 6,1 565 82 5,4 640
r-146XKlondike Striped 96 5,1 560 97 6,4 770 95 6,8 715
Цера 21X Мелитополь- ский 143 88 5,6 505 87 5,5 470 87 5,8 520
Мелитопольский 142 (стандарт) 90 5,1 408 90 5,5 495 89 5,4 460
Продолжение таблицы 18
Гибрид, сорт Повторности Среднее Прибавка урожай- ности к стандарту, %
4-я
А Б 1 в А Б В
Цера 21хМраморный 83 6,0 600 84 6,6 586,2 33,8
Цера 21X Роза Юго-Востока 87 5,8 645 82 5,8 621,3 41,5
T-146XKlondike Striped 96 6,4 640 96 6,2 671,3 53,2
Цера 21X Мелитополь- ский 143 87 5,4 615 87 5,6 502,5 14,6
Мелитопольский 142 (стандарт) 90 4,9 390 90 5,2 438,3 —
Обозначения: А - число дней от всходов до созревания; Б — средняя
масса плода, кг; В — урожайность, ц/га.
Как видно из таблицы 18, все приведенные гибриды превышали
стандарт по урожайности. Наибольший гетерозисный эффект был от-
мечен у гибрида Г-146 X Klondike Striped.
Способность сорта передавать свои качества потомству следует
рассматривать дифференцированно. Есть сорта бахчевых культур, ко-
107
торые передают гибридам комплекс свойств, но большинство из них
заслуживает внимания по одному-двум ценным признакам или свой-
ствам. Поэтому при изучении комбинационной способности существен-
ную роль играет знание закономерностей наследования того или иного
признака или свойства, помогающее в какой-то мере прогнозировать
гетерозис гибридов. Имеющиеся в литературе данные позволяют про-
следить некоторые особенности наследования продуктивности, скороспе-
лости, устойчивости к болезням и других ценных свойств.
Многие авторы [27, 55, 234, 269] считают, что урожайность у ар-
буза наследуется промежуточно. Гетерозис по этому признаку может
проявляться в тех случаях, когда сорта различаются по морфологии,
биологии, экологии. Наибольший эффект дает скрещивание местных
форм с экологически отдаленными. Вероятность проявления гетерозиса
выше, если в скрещиваниях участвуют высокоурожайные сорта. Наи-
более крупноплодные гибриды получаются при скрещивании шароплод-
ных форм с длинноплодными. Имеются сведения, что хорошие резуль-
таты получаются при сочетании многоплодности одного родительского
компонента с крупноплодностью другого, что подтверждается и нашими
данными [260]. Эти факторы всегда необходимо учитывать при под-
боре пар для скрещивания.
При создании скороспелых гибридов желательно подбирать сорта,
как бы дополняющие друг друга по кратковременности основных фаз
развития. Многие исследователи считают такой способ подбора пар
весьма эффективным. Небезразлично, в качестве какого компонента
взят сорт. И. С. Вернидубов [48], В. П. Тощев [220], С. В. Шейнфельд
[274] отмечают, что скороспелость лучше наследуется по отцовской ли-
нии. И. М. Сафарян [188], Н. М. Сазанова [187] считают, что в пере-
даче этого признака преимущество имеет материнская форма. Наши
данные позволяют сделать такое же заключение. Это, вероятно, объ-
ясняется возможностью более полной реализации материнского гено-
типа у гибридов. Мы считаем, что для получения раннеспелых гибридов
в качестве материнской формы целесообразно использовать скороспе-
лый сорт.
Один из важных сортовых показателей арбуза — содержание в
плоде не менее 8—9% сахара. Для получения высокосахаристых гиб-
ридов необходимо знать закономерности наследования этого признака.
По данным В. В. Арасимович [14], содержание сахара у гибридов ар-
буза наследуется промежуточно. В отдельных случаях наблюдается ге-
терозисное увеличение количества сахаров, что можно объяснить не-
зависимостью наследования содержания глюкозы, фруктозы и саха-
розы. При удачном сочетании отдельных форм количество сахара в
плоде резко возрастает. По этой причине сочетание средне- и низко-
сахаристого сортов иногда может дать больший эффект, чем скрещи-
вание высокосахаристых сортов. Но все-таки многие ученые оценивают
последний вариант как более перспективный.
Известно, что существуют сорта-улучшатели, хорошо передающие
высокую сахаристость гибридам, среди них Десертный 83 селекции Би-
рючекутской овощной селекционной опытной станции. Т. С. Вернидубов
считает, что он особенно эффективен в качестве отцовской формы.
И. М. Сафарян выделяет сорт Мелитопольский 142, Н. М. Сазанова —
Дынный лист, Донской 39, Любимец хутора Пятигорска, Муравлевский.
В наших опытах гетерозисное увеличение сахаристости наблюда-
лось почти у всех выделенных гибридов, что свидетельствует о высокой
комбинационной ценности исходных форм. В качестве сортов-улучша-
телей заслуживают рекомендации Мелитопольский 143, Мраморный,
108
Грибовский цельнолистный, Роза Юго-Востока, Любимец хутора Пя-
тигорска, Sugar Baby.
Содержание сахара и вкусовые качества плода тесно связаны со
структурой мякоти: высокосахаристые сорта имеют, как правило, зер-
нистую рыхлую мякоть, малосахаристые — более плотную и грубую,
обычно доминирующую в потомстве; эти особенности необходимо учи-
тывать при подборе родительских пар для скрещивания.
В наших исследованиях для определения характера наследования
некоторых хозяйственно-ценных признаков арбуза были проанализиро-
ваны трехлетние данные по 25 гибридным комбинациям. Установлена
степень доминантности таких количественных признаков, как средняя
масса плода, урожай с растения (продуктивность), содержание сахара
в плоде.
Для определения степени доминантности была использована сле-
дующая формула:
где hp — степень доминантности; Fi — средний показатель первого по-
коления гибридов; МР— средний показатель двух исходных форм;
HP — средний показатель лучшей исходной формы.
Учитывались основные значения степени доминантности: hp = 0 —
доминирование отсутствует; hp>0 < 1—промежуточное наследование
или частичное доминирование; hp>l—сверхдоминирование, гете-
розис.
По продуктивности гетерозис проявился у 17 комбинаций из 25
(hp> 1), частичное доминирование отмечено у 4 комбинаций (hp>0),
промежуточное наследование также у четырех.
По средней массе плода гетерозис наблюдался у 7 гибридов, 9 ком-
бинаций показали частичное или полное доминирование и 9 — проме-
жуточное наследование.
По сумме сахаров гетерозис был отмечен у 15 гибридов, в 3 ком-
бинациях наблюдалось частичное доминирование, в 7 — промежуточное
наследование.
Продуктивность растения арбуза определяется числом плодов и
массой одного плода. Анализ полученных данных показал, что в ком-
бинациях, в которых отмечалось гетерозисное увеличение массы плода,
продуктивность растений была выше (в абсолютном выражении), чем
в тех случаях, когда увеличивалось только число плодов. Наиболее
высокий гетерозис наблюдался при увеличении числа плодов на рас-
тении и массы каждого плода.
ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ АРБУЗА
Изменчивость не однозначное понятие, ее можно рассматривать
в разных аспектах.
Мы изучали изменчивость признаков арбуза в следующих формах:
1) экологическая изменчивость —разнообразие признаков и свойств у
сортов разных эколого-географических групп (см. выше); 2) сортовая
изменчивость — разнообразие сортовых признаков арбуза; 3) геогра-
фическая изменчивость — изменение количественных показателей при
выращивании одних и тех же сортов в различных географических
районах.
109
СХЕМА СОРТОВОЙ ИЗМЕНЧИВОСТИ СТОЛОВОГО АРБУЗА
(С. LANATUS SUBSP. VULGARIS VAR. VULGARIS)
Признаки растения: мощность — мелкое, среднее, крупное; число
плетей — малое (3), среднее (4—5), большое (5).
Признаки стебля: длина главной плети — очень короткая (<1 м),
короткая (1 —1,4 м), средняя (1,5—2 м), длинная (2—2,5 м), очень
длинная (>3 м); форма — округлая, округло-граненая, граненая; тол-
щина— очень тонкий, тонкий, средний, толстый; междоузлия — корот-
кие (<5 см), средние (6—10 см), длинные (>10 см); опушение —
отсутствует, редкое, густое; волоски — мягкие, грубые; усики — одиноч-
ные, двойные, множественные.
Признаки листа: длина черешка — короткий (<5 см), средний
(5—8 см), длинный (>8 см); положение черешка — вертикальное, на-
клонное, горизонтальное; тип листа — нерассеченный, рассеченный;
окраска — желтая, с белыми жилками, зеленая, темно-зеленая, сизо-
зеленая, пестрая; размер (по продольному диаметру)—очень мелкий
(<10 см), мелкий (10—12 см), средний (13—18 см), крупный (18—
20 см), очень крупный (>20 см); рассеченность (рис. 24)—отсутст-
вует, очень слабая, слабая, средняя, сильная, очень сильная; доли лис-
та— узкие, средние, широкие.
Признаки цветка: размер венчика — мелкий (<2 см), средний
(2—3 см), крупный (>3 см); окраска — светло-желтая, желтая, ярко-
желтая; цветоножка — короткая (<1,5 см), средняя (1,6—3,0 см),
длинная (>3 см); окраска рыльца — зеленая, желто-зеленая, желтая,
оранжевая; половой тип — монойкист, андромонойкист.
Признаки завязи: форма — шаровидная, овальная, цилиндрическая;
опушение — отсутствует, очень редкое, редкое, густое.
Признаки плода: форма — сплюснутая, шаровидная, неправильно-
шаровидная, тупо-эллиптическая, овальная, грушевидная, цилиндриче-
Рис. 24. Лист столового арбуза-
1 ' Не^ееЧе5- сильно 3 Г слабоРассеченный;
, _ ери,... ™
ческая; размер — мел-
кий (< 15 см), средний
(15—25 см), крупный
(>25 см); поверхность —
гладкая, сегментирован-
ная, с венообразными вы-
пуклостями, бугристая;
цвет фона — белый, свет-
ло-зеленый, лимонно-жел-
тый, оранжевый, зеленый,
темно-зеленый, черно-зе-
леный; рисунок — отсут-
ствует, сетка, сетчатые
полосы, нитчатые полосы,
мраморность, шиповатые
полосы (узкие или ши-
рокие), размытые по-
лосы, ситцевость (точеч-
ность в сочетании с раз-
мытыми полосами), мо-
заика (мелкая или круп-
ная); окраска рисунка —
светло-зеленый, зеленый,
темно-зеленый, черно-зе-
леный; толщина коры —
по
тонкая (<1 см), средняя (1—1,5 см), толстая (1,5—2 см), очень толстая
(>•2 см); характер коры —хрупкая, гибкая, плотная, деревянистая;
окраска мякоти — лимонно-желтая, бледно-розовая, розовая, апельси-
новая, шарлаховая, карминная, малиновая; консистенция мякоти — зер-
нистая, нежная, ослизняющаяся, грубая, плотная, ватная; сочность мя-
коти— малосочная, сочная, очень сочная; вкус — горький, пресный, ма-
лосладкий, сладкий, очень сладкий.
Признаки семян: размер —очень мелкие (<5 мм), мелкие (5—
10 мм), средние (10—15 мм), крупные (15—20 мм), очень крупные
(>20 мм); форма — узкие, широкие, очень широкие; окраска — белая,
кремовая, серая, табачная, светло-коричневая, красная, коричневая,
черная; поверхность — гладкая, шероховатая, с трещинами; рисунок —
отсутствует, со светлым носиком и ободком, с темным носиком и обод-
ком, с темным пятном, крапчатый.
ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ СОРТОВ
Выращивание одних и тех же сортов арбуза в разных климатиче-
ских зонах позволяет проследить географическую изменчивость при-
знаков и выявить районы, более благоприятные для возделывания этой
культуры.
Мы изучали изменчивость основных хозяйственно-ценных призна-
ков (скороспелости, урожайности, содержания сахара в плоде) у ряда
сортов арбуза при посеве их в Астраханской, Харьковской, Херсонской
областях, на Северном Кавказе и в Молдавии. В таблице 19 приведены
результаты изучения 9 сортов арбуза в условиях Астрахани, Тирасполя
и Харькова. Самый короткий период от всходов до созревания отмечен
почти у всех сортов в Астрахани, наиболее продолжительный — в Мол-
давии. У некоторых сортов разница достигала 40 дней. В Астрахани
были зарегистрированы также более высокая продуктивность и содер-
жание сухого вещества в плоде.
В таблице 20 представлены данные изучения 9 районированных
сортов арбуза в основных зонах товарного бахчеводства: в Нижнем По-
волжье (Астраханская опытная станция ВИР), на Северном Кавказе
(Кубанская опытная станция ВИР) и на юге Украины (Херсонский
опорный пункт ВИР), различных по климатическим и почвенным усло-
виям. Наиболее влажный климат этих районов на Северном Кавказе.
В Нижнем Поволжье зарегистрирована самая высокая сумма активных
температур. Для районов бахчеводства Херсонской области характерны
легкие почвы, хорошо прогреваемые солнцем.
Как видно из приведенных данных, у большинства сортов наиболее
раннее созревание отмечено на Астраханском опорном пункте, здесь
же зарегистрирована самая высокая продуктивность растений. В усло-
виях Херсона почти все сорта оказались более позднеспелыми и менее
урожайными, но в то же время накапливали большое количество са-
хара в плоде. Кубанская опытная станция по всем приведенным по-
казателям заняла промежуточное положение. Наибольшую изменчи-
вость проявили сорта Сухумис и Любимец Краснодара, пластичнее дру-
гих оказался сорт Багаевский мурашка 747/749, имевший высокую про-
дуктивность на всех трех пунктах. Новые сорта Астраханский и Табо-
линский отличались высоким содержанием сахара независимо от места
выращивания. Полученные данные позволяют сделать вывод, что наи-
более благоприятны для возделывания арбуза условия Астрахани, в
Молдавии же целесообразно выращивать лишь скороспелые сорта. Луч-
шие условия для накопления сахара в плоде создаются в Херсонской
области, чему способствуют, видимо, легкие почвы и отсутствие оро-
111
Таблица 19. Географическая изменчивость признаков у различных сортов арбуза (1972 г.)
М по ката- логу ВИР Сорт Происхождение Период от всходов до созревания, дней Урожай с расте- ния, кг Содержание сухого вещества в плоде, % по рефрактометру
Астра- хань Тира- споль Харь- ков Астра- хань Тирасполь Астрахань Тирасполь
4342 Гибрид А-48 Дальний Восток 64 90 76 5,5 3,5 | 8,9 —
3748 New Hampshire midget США 56 73 70 1,8 1,5 9,6 —
2208 Грачевский полосатый Поволжье 67 108 76 9,8 1.4 8,3 7,0
3563 Муравлевский 140 Северный Кавказ 69 108 76 5,6 3,0 10,5 10,0
4247 Klondike WR-65 США 79 94 74 1,8 2,7 11,8 —
3527 Быковский 22 Поволжье 77 108 80 5,1 1,2 9,8 7,5
4104 Король кубанский Северный Кавказ 67 108 76 3,0 1,2 10,5 6,0
3622 Местный Азербайджан I 67 97 71 4,8 1 4,3 9,6 I Ю,0
4303 Волгоградский 61 Поволжье 88 108 78 4,4 2,3 10,5 1 ю,о
Таблица 20. Географическая изменчивость признаков у районированных и находившихся
в государственном сортоиспытании сортов арбуза в основных зонах товарного бахчеводства (1977 г.)
№ по ката- логу ВИР Сорт Происхождение Период от всходов до созревания, дней Урожай с од- ного растения, кг Содержание сухого вещества в плоде, % по рефрактометру
Астра- хань Кубань Херсон Астра- хань Ку- бань Хер- сон Астра- хань Кубань Херсон
3367 Багаевский мурашка Северный Кавказ 74 96 102 13,0 9,2 8,2 9,6 9,8 8,0
3764 Сухумис Абхазия 65 91 102 9,5 11,1 5,2 7,6 8,2 11,0
3992 Любимец Краснодара Северный Кавказ 76 105 97 8,3 6,8 1,6 10,6 10,0 13,0 13,0
4179 Вахшский темнокорый Таджикистан 76 112 97 11,2 3,5 2,2 10,3 10,4
4337 Черносемянный Украина 69 90 97 7,3 4,6 3,5 11,0 9,0 13,0 11,0
4543 Ранний Казахстан 77 105 102 5,2 5,5 4,3 9,0 9,0
4579 Подарок Холодова Поволжье 76 105 108 7,2 5,7 4,2 11,4 10,0 9,0
4619 4633 Астраханский Таболинский > 65 78 105 111 108 114 10,4 8,9 4,2 ) 4,8 6,7 8,8 10,0 11,0 9,6 10,8 10,0 12,0
шения. Однако наряду с почвенно-климатическими условиями районов
возделывания арбуза следует учитывать также биологические особен-
ности сортов.
СОРТОТИПЫ СТОЛОВОГО АРБУЗА
В основу дробной дифференциации коллекции был положен сорто-
тип — единица, принятая для многих культурных растений.
Типизация образцов коллекции столового арбуза, проводимая в те-
чение многих лет, позволила выявить в каждой эколого-географической
группе определенные морфологические типы сортов. Для выделения
сортотипов были использованы только наиболее яркие, так называемые
апробационные признаки, позволяющие без труда отличить один сорт
от другого: форма, окраска и рисунок плода.
При этом мы столкнулись с определенными трудностями, вызван-
ными тем, что сорта арбуза представлены, как правило, популяциями,
иногда довольно пестрыми. В таких случаях мы определяли сортотип
по основному типу окраски плода данной популяции, не принимая во
внимание примеси. Если же популяция представляла смесь разных
форм, что иногда наблюдается у дальневосточных, западнокитайских и
афганских форм, ее просто не типизировали.
Следует также подчеркнуть, что сходство сортов арбуза по окраске
и форме плода, как правило, не отражает их генетического родства,
хотя в некоторых случаях оно несомненно. К таким внешне похожим
и родственным сортам относятся: в русской эколого-географической
группе Мелитопольский 142, Мелитопольский 60, Степняк, Астрахан-
ский (сортотип 25); в среднеазиатской Кузыбай 30, Тез-Пишар (сорто-
тип 29); в американской группе Candy Red, Charleston gray, Sweet
Princess (сортотип 6) и др. У большинства же сортов сходство плодов
фенотипическое и потому выделение сортотипов имеет лишь приклад-
ной характер: помогает устанавливать типичность сорта и выявлять
примеси,
Сортотипы были выделены нами в каждой эколого-географической
группе, однако сопоставление показало, что многие из них повто-
ряются. Наблюдается определенный параллелизм изменчивости окраски
плода арбуза в разных эколого-географических группах. Суммируя все
разнообразие признаков столового арбуза, мы установили 31 сортотип.
Сортотип Характерные признаки плодов
1 Шаровидные, белые, без рисунка или с элементами сетки
2 Шаровидные, золотисто-желтые, без рисунка или с неясными полосами
3 Шаровидные, оранжево-желтые, без рисунка, сегментированные
4 Шаровидные, светло-зеленые или салатные, без рисунка, сегментированные
5 Шаровидные, светло-зеленые, с сеткой или сетчатыми полосами
6 Цилиндрические, светло-зеленые (реже белые), с сеткой или сетчатыми полосами
7 Шаровидные, светло-зеленые, с узкими сетчатыми (нитчатыми) полосами
8 Овальные, реже шаровидные, серо-зеленые, с сеткой нли сетчатыми полосами
9 Шаровидные, светло- или серо-зеленые, с темно-зелеными мазками — пятнами у
плодоножки (мраморный рисунок)
10 Шаровидные или овальные, серо-зеленые, с темно-зелеными нитчатыми полосами
11 Шаровидные, белесые или светло-зеленые, с белесыми шиповатыми полосами
12 Шаровидные, белесые или светло-зеленые, с узкими темно- или черно-зелеными
шиповатыми полосами
13 Овальные или цилиндрические, белесые, с узкими темно- или черно-зелеными ши-
поватыми полосами
14 Шаровидные, светло-зеленые, с широкими шиповатыми зелеными или темно-зелены-
ми полосами, часто смыкающимися отростками (мозаичный рисунок)
15 Цилиндрические, светло-зеленые, с широкими зелеными шиповатыми полосами
16 Шаровидные, светло-зеленые, с зелеными размытыми полосами
17 Шаровидные, светло-зеленые, с точечностью и темно-зелеными размытыми полосами
(ситцевый рисунок)
113
Сортотнп Характерные признаки плодов
18 Шаровидные, светло-зеленые, с широкими темно-зелеными размытыми полосами,
почти закрывающими фон
19 Цилиндрические, светло-зеленые, с зелеными широкими размытыми полосами
20 Шаровидные или овальные, грушевидные, зеленые, с сеткой или сетчатыми поло-
сами
21 Цилиндрические, зеленые, с сеткой или сетчатыми полосами
22 Шаровидные, темно-зеленые, с неясным рисунком
23 Цилиндрические или овальные, зеленые, с неясным рисунком
24 Шаровидные, зеленые, с черно-зелеными узкими шиповатыми полосами
25 Шаровидные, зеленые, с темно- или черно-зелеными широкими шиповатыми поло-
сами, иногда сливающимися отростками
26 Цилиндрические (реже овальные), зеленые, с широкими темпо-зелеными шипова-
тыми полосами
27 Шаровидные, зеленые, с тускло-зелеными размытыми полосами
28 Шаровидные, зеленые, с темно-зелеными размытыми полосами
29 Шаровидные, темно-зеленые, с черно-зелеными шиповатыми полосами
30 Шаровидные, черно-зеленые, со скрытым рисунком или без рисунка
31 Цилиндрические, реже овальные, черно-зеленые, со скрытым рисунком или без ри-
сунка
Наибольшее разнообразие окрасок и форм плода встречается в
американской, среднеазиатской и русской группах, полнее других пред-
ставленных в коллекции. Слабый полиморфизм наблюдается в индий-
ской, восточноазиатской и дальневосточной группах.
Самые распространенные (по числу групп, где они отмечены, и по
количеству образцов в каждой группе) сортотипы 5, 12, 16, 24, 30
(рис. 25).
Наиболее распространенным оказался сортотип 12. Встречается во
всех 10 группах, причем в 9 из них очень часто. Сортотип 24 отличается
от предыдущего более темной окраской фона плода, что дает основа-
ние считать самым распространенным типом рисунка плода столового
арбуза узкие шиповатые полосы. Обнаружен в 7 группах и во всех
отмечен часто. Сортотип 16 встречается в 9 группах и в 6 из них часто
Рис. 25. Наиболее распространенные сортотипы столового арбуза:
]—сортотип 24; 2 — сортотип 5; 3 — сортотип 12; 4 — сортотип 16; 5 — сор-
тотип 30.
114
Сортотип 5 установлен в 8 группах и в 6 из них распространен
широко. В этом сортотипе мы объединили формы с сеткой и с сетча-
тыми полосами, так как оба рисунка легко переходят один в другой и
бывают трудно различимы, особенно если сорт не выравнен. Сорто-
тип 30 отмечен в 7 группах, в 5 из них встречается часто.
Названные сортотипы наиболее широко представлены в коллекции,
но каждая из групп имеет и свойственные только ей типы окраски и
формы плода. Так, в русской эколого-географической группе, кроме
указанных, часто встречаются белоплодные сорта (сортотип 1).
Для малоазиатской группы арбуза характерны плоды с темными
нитчатыми полосами (сортотип 10) и с мозаичным рисунком (сорто-
тип 14), для западноевропейской — зеленые или темно-зеленые (сор-
тотипы 20 и 22), для афганской — сегментированные (сортотипы 2 и 4),
для американской — цилиндрические светло-зеленые (сортотип 6).
Среди дальневосточной группы арбуза преобладают зеленоплодные
формы с нитчатыми или размытыми полосами (сортотипы 10 и 27).
Для всех установленных сортотипов в каждой эколого-географиче-
ской группе выделены типичные сорта, являющиеся эталонами опре-
деленного сортотипа. В таблице 21 даны названия типичных сортов
(эталоны) и номера по каталогу ВИР для всех эколого-географических
групп столового арбуза.
МЕТОДЫ И ДОСТИЖЕНИЯ СЕЛЕКЦИИ АРБУЗА
ЗАДАЧИ СЕЛЕКЦИИ
Селекция арбуза началась в России с тех пор, как зародилось бах-
чеводство. Вначале она осуществлялась бессистемно, путем отбора луч-
ших плодов, позднее для создания новых сортов стали использовать
гибридизацию. Методы селекционной работы постепенно совершенство-
вались, обогащались данными генетики, физиологии, анатомии и дру-
гих дисциплин. Из искусства отдельных любителей селекция бахчевых
культур постепенно превращалась в науку. Первые селекционеры-опыт-
ники пытались лишь повысить качество и урожайность существующих
сортов-популяций. Позднее определились различные направления селек-
ции, учитывающие развитие бахчеводства и специфику создаваемых сор-
тов. Так, в связи с увеличением вывоза продукции в другие районы
появилась необходимость создания транспортабельных сортов, плоды
которых должны были иметь прочную кору, более плотную мякоть и
предпочтительно темную окраску. Возникла задача создания сортов ар-
буза, пригодных для длительного хранения: позднеспелых, устойчивых
к грибным болезням, имеющих определенную структуру мякоти плода.
Распространение болезней, особенно таких вредоносных, как ан-
тракноз и фузариоз, вызвало необходимость начать работу по созданию
устойчивых сортов. В качестве источников иммунитета сначала исполь-
зовали ранее выведенные устойчивые американские сорта, а позднее и
дикорастущие формы. Создание сортов, устойчивых к болезням, одно
из основных направлений в селекции этой культуры.
В последнее время все острее ставится вопрос о механизации тру-
доемких процессов в бахчеводстве, особенно уборки продукции. В связи
с этим встает задача выведения сортов арбуза, пригодных для машин-
ной уборки, к которым предъявляется целый ряд специфических тре-
бований: они должны дружно созревать, иметь шаровидные плоды сред-
него размера с прочной корой. Крупноплодные сорта непригодны для
машинной уборки, так как получается высокий процент брака.
8*
115
о
Таблица 21. Эталоны сортотипов эколого-географических групп столового арбуза
№ сортотипa
Русская Мало- азиат- ская Западноевропейская Закавказская
Aft по ката- логу ВИР сорт Aft по ката- логу ВИР А& по ката- логу ВИР сорт Aft по ката- логу ВИР сорт
Среднеазиатская Ин- дий- ская Афганская Американская Восточно- азиатская
Aft по ката- логу ВИР сорт АЛ по ката- логу ВИР № по ката- логу ВИР сорт № по ката- логу ВИР сорт АЛ по ката- логу ВИР сорт
Дальневос-
точная
сорт
1 348 Мурашка 1076 4299 Цера 3749 Самарканд- 3939 Bush Desert 3509 Сан-
ский белый King Сянь
2 398 Желто- 4080 Golden 4173 The sun
4187 корый midget
3 2578 1422
4 Красавчик 3667 Ербентский 3734 413 Хиндоне 246 Zitkiva
черносе- мянный
5 3490 Сквире кий 4267 Eni Dunia 4442 Таузский 2612 Беш-тухум 4501 414 2822 Early 3736 Сяо-
скороспе- лый белый Canada Цзы
6 3904 Белый 4195 4128 Charleston
1083 длинный gray
7 4495
8 3781 New Hamp- shire midget 4149 Kaxo
9 2159 Мраморный 1006 3001 Дини Мра- 2866 Пионер
1368 морни пустыни
10 3767
11 2838 Белосемяч- 1062 2815 Ак-Чинны 4490 242 Winter
ко 187 melon
12 2481 Король 1020 4219 Anguria di 386 Борчалин- 452 Узбекский 4505 4480 4628 Lousiana 2114 Kanro
Кубы 92 Bagnoco- vallo ский Queen Klondike Striped
13 3547 Длинный полосатый 2712
14 2840 Мелито- 1353 3396 Джафар- 3049 Мозаичный 4201
польский 143 ханский
3631
3691
3688
15
16 3523 Крымский 1520 2005 Лубеница 4344 Сабирабад- ский 3668
17 1013
18 19 4253 Роза Юго- Востока
20 21 22 23 24 25 26 27 28 3614 2692 2220 3567 2652 Быков- ский 199 Любимец хутора Пятигор- ска Мелито- польский 142 Донской 39 Стокса 1358 1375 3674 3862 2577 3580 Дини Чен- джелий- ски Арадский Неаполи- танский Szentesi 3517 3764 3548 282 3615 3149 Грузинский длинный серый Сухумис Широко- овальный полосатый Кахетин- ский Скороспел- ка 675 2816 3895 2878
29 30 31 3588 Огонек 1071 1053 3581 Marsowszky 3518 3519 Кахетин- ский круг- лый черный Кахетин- ский длин- ный черный 3543 2890
Победитель пустыни 4208 72 4297 166 Florida Favorite Crimson Sweet Phiney Early 254 Сигув 3497 Ско- ро- спел- ка
4242 Tender- sweet
Кембаев- ский 1154 2574 216 Stone mountain Tom Watson 3875 Гиб- рид А-44
137 Klekley 3455
Хиндване- Мекке 429 Ниша- пур- ский 19 Chilian Black 3448
Мокки Ак-урук 4212 3848 Congo 2633
Чарджуй- ский зимний Кузыбай 30 Хаит-кара 4509 3785 4191 3854 Sugar Baby 2044 Китай- ский 163
4202 4371 Shipper 2046 Китай- ский 165
Намечается тенденция к расширению площадей защищенного
грунта не только в овощеводстве, но и в бахчеводстве. Сейчас в нашей
стране арбуз только начинают выращивать под временными пленоч-
ными укрытиями и в пленочных теплицах [25, 30, 182, 237]. В ближай-
шее время площади под арбузом в защищенном грунте, несомненно,
расширятся, так как это дает возможность получать раннюю продук-
цию, обеспечивающую высокую прибыль хозяйствам. Выращивание ар-
буза под пленкой вызывает необходимость подбора и выведения соот-
ветствующих сортов: они должны быть раннеспелыми, иметь короткие
плети и небольшие плоды. Сейчас в защищенном грунте используют
преимущественно сорт Огонек Украинского НИИ овощеводства и бах-
чеводства. Небольшой опыт, проведенный нами, показал, что для за-
щищенного грунта перспективен также сорт Сибиряк Западно-Сибир-
ской овоще-картофельной опытной станции.
Не потеряла своего значения и селекция раннеспелых сортов ар-
буза для открытого грунта. Это направление особенно развито в север-
ной зоне бахчеводства, главным образом в Молдавии, Харьковской об-
ласти и других районах с относительно низкими суммами активных
температур, где вызревают лишь скороспелые сорта арбуза.
МЕТОДЫ СЕЛЕКЦИИ
Основными методами селекционной работы с бахчевыми, так же
как с другими культурами, являются гибридизация и отбор, сочетанием
которых создано все разнообразие существующих сортов. В последнее
время наряду с межсортовой гибридизацией все шире начинают исполь-
зовать отдаленные скрещивания с дикорастущими формами арбуза из
Африки. Ценный исходный материал в этом отношении представляет
коллекция арбуза, сосредоточенная в ВИР.
В процессе индивидуального отбора селекционеры чаще всего ис-
пользуют «способ половинок», при котором часть семян из плода вы-
севают, а часть оставляют в качестве контроля для сравнения с полу-
ченной репродукцией и для последующего размножения.
Для создания на основе сортов-популяций выравненных линий при-
бегают к инбридингу, который проводят в течение 3—4 поколений. Бо-
лее длительное близкородственное размножение приводит к снижению
продуктивности растений. Методы селекции, применяемые при созда-
нии гетерозисных гибридов и бессемянных форм арбуза, освещены в
соответствующих разделах.
ДОСТИЖЕНИЯ СЕЛЕКЦИИ
Планомерная селекционная работа с бахчевыми культурами была
начата в 30-е годы на Быковской бахчевой селекционной опытной стан-
ции в Нижнем Поволжье и Бирючекутской овощной селекционной
опытной станции в Ростовской области. Организатором и первым ди-
ректором Быковской бахчевой селекционной опытной станции, создан-
ной в 1931 г., был известный селекционер С. И. Лутохин, сделавший
очень много для умножения славы советского бахчеводства. Выведен-
ные им сорта арбуза Мелитопольский 142 и Мелитопольский 143 и по-
ныне занимают значительные площади в нашей стране. Селекционерами
этой же станции Д. Г. Холодовым, К. П. Синча и другими создано
много сортов арбуза, получивших заслуженное признание: Быков-
ский 22, Роза Юго-Востока, Волжский 7, Подарок Холодова и др.
Бирючекутская овощная селекционная опытная станция, организо-
ванная в 1923 г. профессором А. И. Носатовским, стала ведущим се-
118
лекционным учреждением по бахчевым культурам на Северном Кав-
казе. Селекционерами станции проведена большая работа по восстанов-
лению местных сортов арбуза и дыни, созданы сорта: Десертный 83,
считающийся одним из лучших по вкусовым качествам, Бирючекут-
ский 775, Донской 39 и др. Здесь начал свою трудовую деятельность
старейший селекционер-бахчевод Л. Е. Кревченко. Сорта бахчевых куль-
тур, выведенные им на Бирючекутской, а позднее на Краснодарской
селекционной овоще-картофельной опытной станции, районированы
сейчас во всех областях и краях Северного Кавказа. Среди созданных
им в последнее время сортов арбуза заслуживают внимания Ранний,
Любимец Краснодара, Комсомольский 2.
На Украине селекционную работу с бахчевыми культурами ведет
Днепропетровская овоще-бахчевая опытная станция, организованная в
1937 г. Селекционером А. Т. Галка создан здесь ряд сортов арбуза,
дыни, тыквы, занимающих значительные площади в южных областях
республики, среди них сорта арбуза Мелитопольский 60, Туман днепро-
петровский и др. В настоящее время станция ведет работу по выве-
дению сортов, устойчивых к фузариозному увяданию, и созданию гиб-
ридов арбуза.
В послевоенное время сеть научно-исследовательских учреждений,
работающих с бахчевыми культурами, значительно расширилась. На
базе некоторых опытных станций были организованы институты, соз-
дан Всесоюзный научно-исследовательский институт орошаемого ово-
щеводства и бахчеводства (ВНИИООБ), призванный стать центром
научной координации всех исследований по бахчевым культурам. Ин-
ститут, расположенный недалеко от Астрахани, в зоне товарного бахче-
водства, успешно ведет селекционную работу по созданию сортов бах-
чевых культур, устойчивых к болезням. Исследования проводятся в тес-
Рис. 26. Сорт Мраморный.
119
ном сотрудничестве со Всесоюзным научно-исследовательским институ-
том растениеводства им. Н. И. Вавилова. Богатейший генетический
фонд мировой коллекции ВИР используется в качестве исходного ма-
териала для выведения новых сортов. Изучение устойчивости сортов к
болезням проводится также на Херсонской селекционной опытной стан-
ции бахчеводства ВНИИССОК. В последнее время здесь начаты ис-
следования по механизации трудоемких процессов в бахчеводстве и
прежде всего по созданию машин для уборки арбузов.
Недалеко от Харькова также на базе опытной станции еще раньше
был организован Украинский научно-исследовательский институт овоще-
водства и бахчеводства, занимающийся селекцией и семеноводством
бахчевых культур для северных областей Украины. Здесь выведены
широко распространенные сорта арбуза Скороспелка харьковская, Ого-
нек, Харьковский 54, Черносемянный.
В Средней Азии еще в начале 20-х годов большую научно-иссле-
довательскую работу с бахчевыми культурами стали проводить на Сред-
неазиатской опытной станции ВИР недалеко от Ташкента. Профессор
К. И. Пангало вывел здесь на базе мировой коллекции много сортов
арбуза. Некоторые из них до сих пор районированы и занимают зна-
чительные площади в Узбекистане: Мраморный (рис. 26), Хаит-кара,
Узбекский 452, Белый длинный.
Также создан на базе опытной станции Узбекский научно-исследо-
вательский институт овоще-бахчевых культур и картофеля, работаю-
щий в тесном сотрудничестве со Среднеазиатской опытной станцией
ВИР. С использованием образцов мировой коллекции здесь созданы
сорта арбуза Белый длинный 107, Кузыбай 30 и др.
Вскоре после Великой Отечественной войны был организован Мол-
давский научно-исследовательский институт орошаемого земледелия и
овощеводства, где селекцию бахчевых культур вел профессор К. И. Пан-
гало. Им выведен здесь ряд сортов столового и кормового арбуза.
Среди последних надо отметить сорт Пектинный, используемый в рес-
публике не только для кормовых целей, но и как сырье для промыш-
ленного получения желирующего вещества пектина. В настоящее время
институт успешно продолжает селекционную работу, создавая скоро-
спелые сорта арбуза и дыни для Молдавии.
Научно-исследовательская и селекционная работа с бахчевыми
культурами проводится и в других учреждениях страны.
БОЛЕЗНИ АРБУЗА И ИСТОЧНИКИ УСТОЙЧИВОСТИ К НИМ
Известно более ста болезней, поражающих бахчевые культуры. Из
них в нашей стране наиболее распространены следующие: увядание,
антракноз (розовая пятнистость), мучнистая роса, милдью (ложная
мучнистая роса), макроспориоз, черная гниль, бактериоз и вирусная
мозаика. Болезни причиняют огромный ущерб урожаю бахчевых куль-
тур.
ГРИБНЫЕ, БАКТЕРИАЛЬНЫЕ И ВИРУСНЫЕ БОЛЕЗНИ
Наиболее вредоносные заболевания арбуза — увядание и антрак-
ноз.
Болезнь увядание появилась в СССР сравнительно недавно,
в книге С. Ю. Шембеля [275] она не упоминается. На Быковской бах-
чевой селекционной опытной станции болезнь впервые была обнаружена
в 1931 г. В Средней Азии в послевоенные годы она распространилась
широко.
120
Возбудителем увядания является гриб Fusarium oxysporum
Schlecht. f. niveum. Л. M. Мосиевская [148] установила, что в неко-
торых районах Казахстана (северное Прибалхашье) причиной заболе-
вания может быть гриб Verticillium dahliae Kleb. Фузариум более теп-
лолюбив. Вероятно, поэтому он так быстро распространился в Средней
Азии, куда был занесен из других мест. Вертициллиум устойчив к по-
ниженной температуре, в связи с чем встречается в более северных
районах.
Большинство исследователей считают оба вида гриба широко спе-
циализированными паразитами, поражающими не только тыквенные
растения, но также томат, баклажан, картофель. Однако в этом воп-
росе нет единого мнения. Так, Л. Т. Лукина [126] в Молдавии выде-
лила штаммы Fusarium oxysporum, специфичные только для дыни и
только для арбуза. Узкую специализацию этого патогена подтверждает
и В. И. Тимченко [218]. Д. Кролл [292] выявил для условий Флориды
две расы патогена. По его данным, раса 1 вызывает 100%-ное увядание
восприимчивых сортов арбуза (Florida giant) и очаговое поражение
устойчивых сортов (Charleston gray, Summit). Раса 2 также полностью
поражает восприимчивые сорта, но безвредна для устойчивых.
Увядание может поражать растения арбуза в разном возрасте: в
фазе семядолей и 3—4 настоящих листьев, во время плетеобразования,
в период роста и созревания плодов. У взрослых растений увядают лишь
отдельные плети. Наиболее опасная форма болезни — внезапное увяда-
ние. Листья при этом не теряют зеленой окраски, внешние признаки по-
ражения отсутствуют. Если растение не погибнет сразу, то рост его
замедляется, листья становятся карликовыми.
Другое опасное заболевание арбуза в нашей стране — антрак-
ноз, или розовая пятнистость, вызываемая грибом Colletotrichum lage-
narium (Pass.) Ell. et Halst. Впервые болезнь была обнаружена в
1912—1913 гг. в Поволжье, Крыму, Закавказье. Сейчас распространена
повсюду, за исключением Средней Азии и Армении, где препятствием
для ее развития служит, видимо, высокая сухость воздуха. Частые по-
ливы (особенно дождеванием) и обильные росы способствуют развитию
антракноза. Болезнь поражает стебли, листья, плоды. Наибольший вред
приносит поражение плодов, на которых болезнь проявляется в виде
пятен — язвочек, покрытых розовым налетом. Плоды становятся при
этом несъедобными. Заболевание может проявиться не только в поле,
но при транспортировке и особенно при хранении продукции. В годы
вспышек антракноза погибает до 30—40% урожая. Источником забо-
левания и распространения инфекции могут быть семена пораженных
плодов [178].
Из других болезней следует отметить специфическую для арбуза
черную гниль плодов, возбудителем которой является Fusarium
wolgense Rodigin [181]. Она была обнаружена впервые на Быковской
бахчевой селекционной опытной станции в 1931 г. [46]. Возбудитель
болезни узкоспециализирован и поражает только длинноплодные аме-
риканские сорта. По этой причине один из них — Charleston gray был
забракован Государственной комиссией по сортоиспытанию сельскохо-
зяйственных культур, несмотря на высокую урожайность и хорошие ка-
чества плода. Заболевание проявляется в усыхании и почернении (му-
мификации) цветочного конца плода. Рост плодов при этом замед-
ляется. Высокая влажность воздуха способствует развитию болезни.
Отечественные сорта, преимущественно шаровидные, пммунны к чер-
ной гнили.
121
Из других грибных болезней арбуза известен макроспориоз
(возбудитель Macrosporium cucumerinum Ell. et Ev.). Болезнь пора-
жает листья и плоды, на которых образуется вдавленное темное пятно
с бархатистым налетом. Они становятся несъедобными.
Мучнистая роса в большей степени поражает дыню, но иногда
причиняет существенный ущерб и посевам арбуза. Болезнь широко
распространена по всему земному шару. В нашей стране зарегистри-
рована в 1901 г. Возбудителями являются Erysiphe cichoracearum DC.
и Sphaerotheca fuliginea Poll. Первый в большей степени поражает огу-
рец и арбуз, второй — дыню и тыкву, но в ряде случаев их обнаружи-
вали на одном и том же растении. Болезнь проявляется в виде белого
или рыжеватого налета на листьях, которые при этом теряют зеленый
цвет, увядают и отмирают.
Из бактериальных заболеваний арбуза очень опасен для человека
токсический бактериоз [59, 271]. Впервые болезнь в нашей
стране обнаружена в 1941 г. Употребление пораженных плодов вызы-
вает тяжелое отравление, иногда с летальным исходом. Бактериоз про-
является на плодах в виде резко очерченных пятен-бородавок с темной
точкой в центре. Мякоть таких плодов быстро разлагается, превращаясь
в розово-молочную жидкость.
До последнего времени возбудителем болезни считали бактерии из
группы Proteus [154]. Н. А. Сизовой [191] уточнено систематическое
положение возбудителя. Им оказалась бактерия Erwinia carotovora
(Jones) f. citrullus.
Наряду с токсическим бактериозом известны и другие бактериаль-
ные заболевания арбуза: угловатая пятнистость листьев,
бактериальное увядание, бактериальный некроз
коры. Возбудителями этих болезней являются бактерии из родов
Erwinia и Pseudomonas, а также Bacillus mesentericus. В нашей стране
бактериозы распространены довольно широко, но изучены очень слабо.
Между тем на Северном Кавказе болезнь иногда принимает характер
эпифитотий, развитию которых способствуют пониженная температура
и высокая влажность воздуха.
По данным К- В. Никитиной [152], арбуз чаще всего поражается
бактериальной пятнистостью, возбудитель которой Вас. me-
sentericus хорошо развивается лишь в сухую и жаркую погоду. Холод-
ная погода при высокой влажности способствует развитию бактерий из
рода Pseudomonas. Инфекция может сохраняться в семенах.
Из вирусных болезней арбуз поражают огуречная мозаика
и мозаика арбуза [11]. К характерным признакам их относятся
карликовость растений, пятнистость и деформация листьев. Плоды на
таких растениях обычно не завязываются или бывают мелкими, урод-
ливыми. В отдельных случаях заболевание принимает характер эпиде-
мии, как было отмечено, например, в Новой Зеландии [51]. Источни-
ками заражения являются сорняки: осот, мелотрия и др. Переносчиком
заболевания служит тля.
Среди болезней и вредителей бахчевых культур несколько особня-
ком стоит цветковый паразит — заразиха (Orobanche aegyptiaca
Pers.). В Астраханской области и в Средней Азии она наносит боль-
шой вред бахчевым. Это растение с фиолетово-голубыми цветками и
чешуйками вместо листьев не имеет корней. Своими луковицеобразными
присосками оно присасывается к корням растения-хозяина и таким об-
разом паразитирует на нем. Семена заразихи мелкие, как пыль, сохра-
няют всхожесть до 13 лет. Эффективные меры борьбы пока не найдены.
122
ИСХОДНЫЙ МАТЕРИАЛ ДЛЯ СЕЛЕКЦИИ НА УСТОЙЧИВОСТЬ К БОЛЕЗНЯМ
Наиболее надежное мероприятие в борьбе с болезнями бахчевых
культур — внедрение устойчивых сортов, при создании которых основ-
ную роль играет подбор исходного материала. В настоящее время об-
щепризнанной является теория, согласно которой устойчивые формы
вероятнее всего могут быть обнаружены на родине паразита и хо-
зяина— в центрах происхождения культурных растений [93]. Основной
генный фонд рода Citrullus сосредоточен на Африканском континенте,
в Южной и Западной Африке. Именно здесь следует искать иммунные
и устойчивые к заболеваниям формы арбуза.
Первыми начали использовать генные богатства этого континента
американские селекционеры. В США — стране с теплым и влажным
климатом болезни сельскохозяйственных культур являются настоящим
бедствием, уничтожающим иногда весь урожай. Увядание и антракноз
арбуза были известны здесь значительно раньше, чем у нас. По данным
Д. Портера [345], только в штате Айова площадь бахчевых культур
в 1926 г. сократилась из-за распространения увядания почти на 90%
по сравнению с 1920 г.
Работу по созданию сортов арбуза, устойчивых к фузариозу, начал
здесь в начале века В. Ортон (у Т. Вайтекера) [375]. Среди дикорас-
тущих форм арбуза Южной Африки он обнаружил два образца, им-
мунных к данному заболеванию. От скрещивания одного из них со
столовым сортом Eden был получен сорт Conqueror, широко используе-
мый в дальнейшем в качестве источника устойчивости. Позднее эту ра-
боту продолжили Д. Портер и Д. Мелус [346]. Было выяснено, что
Fusarium требует для нормального развития относительно высокой тем-
пературы (27—30° С) и что растения, зараженные грибом, внешне могут
быть совершенно здоровыми. Гриб, проникая через кончики корней
или раневую поверхность, распространяется с ксилемным током по
сосудам, зачастую только в них и локализуется. Отдельные иммунные
растения были выявлены среди обычных коммерческих сортов Клеклей,
Клондайк, Том Ватсон. На основе их созданы одноименные устойчивые
линии (WR). В качестве источника устойчивости при выведении новых
сортов широко использовали также австралийский сорт Hawkesbury.
В настоящее время в США создан целый ряд сортов (Summit,
Calhoun grey и др.) арбуза, отличающихся повышенной устойчивостью
к фузариозному увяданию.
В. Хендерсон и др. [310] считают сорт Summit особенно перспек-
тивным для использования в дальнейшей селекции. Устойчивость его
наследуется доминантно, что может быть использовано и для получе-
ния гибридов Fi, не поражающихся фузариозом.
Селекция сортов, устойчивых к антракнозу, была начата
в США сравнительно недавно, в 30-е годы. Иммунные к антракнозу
формы были обнаружены также среди дикорастущих африканских. Об-
разец, названный Африка-8, послужил исходной формой для сорта
Congo — одного из лучших современных источников устойчивости к ан-
тракнозу.
Дальнейшие исследования позволили выявить три физиологические
расы возбудителя, из которых раса 2 оказалась наиболее вирулентной
[296, 304, 376]. Затем были идентифицированы еще 4 расы патогена
[319], но наиболее распространенными остаются расы 1 и 2, которые
чаще других вызывают патологические изменения растений. Четких фи-
зиологических различий между расами антракноза обнаружить не уда-
лось. Их можно выявить лишь с помощью сортов-дифференциаторов по
степени устойчивости последних [307].
123
Можно определенно сказать, что современными источниками устой-
чивости арбуза к фузариозу и антракнозу служат почти исключительно
новые американские сорта. Они имеются и в коллекции ВИР.
В последние годы селекционеры всех стран обращают большое вни-
мание на оценку устойчивости сортов бахчевых культур к заболеваниям.
М. Витанов и П. Лозанов [50] в Болгарии испытали по устойчи-
вости к антракнозу более 150 сортов арбуза различного происхожде-
ния. Оценку проводили при искусственном заражении. Все изученные
сорта Болгарии и СССР оказались восприимчивыми к болезни. Среди
сортов США выделены устойчивые Charleston gray, Fairfax, Congo.
При испытании американского сортимента в условиях нашей страны
на Северном Кавказе и в Астрахани эти же сорта были отмечены в
числе лучших. Charleston gray и Fairfax использованы К. Е. Дютиным
[84] при выведении отечественных сортов арбуза, устойчивых к ан-
тракнозу. Один из них — Астраханский успешно прошел государствен-
ное сортоиспытание и с 1977 г. районирован в Астраханской, а позднее
в других областях (рис. 27).
В Австралии в течение трех лет испытывали новые американские
сорта арбуза на устойчивость к фузариозному увяданию [313]. Среди
прочих выделился сорт Calhoun gray, у которого поражение не пре-
вышало 15%. Он отличался также устойчивостью к антракнозу и муч-
нистой росе. Хорошо проявили себя сорта Summit, Shipper, Verona.
Все они по устойчивости превышали стандарт — сорт Candy Red.
В нашей стране поиски надежных источников устойчивости к фу-
зариозному увяданию ведутся все более активно и настойчиво. В Мол-
давском НИИ орошаемого земледелия и овощеводства проведена оценка
150 сортов арбуза [153]. Все американские сорта оказались поражен-
ными. Среди отечественного сортимента меньшей восприимчивостью вы-
делялись Быковский 22, Победитель 395, Красавчик 55. Образец из
Южной Африки (к-2725), имеющийся в коллекции ВИР, также за-
служивает внимания как источник устойчивости к фузариозному увя-
данию.
Американские сорта арбуза, испытываемые на Бирючекутской овощ-
ной селекционной опытной станции в условиях жесткого инфекционного
фона, также оказались восприимчивыми. Из отечественных сортов вы-
делялся Муравлевский [185]. На Днепропетровской овоще-бахчевой
опытной станции А. И. Унковская [235] испытала при искусственном
заражении 40 сортов арбуза раз-
ного происхождения. Иммунных
форм обнаружено не было, но
Рис. 27. Сорт Астраханский.
некоторые американские сорта
(Congo, Klondike, Hawkesbury
Purdue, Takii gem, Harris ear-
list) поражались в слабой сте-
пени. Оценку районированных
сортов арбуза отечественной се-
лекции проводил В. И. Тимченко
[219], иммунных сортов он не об-
наружил. Изучение устойчивости
сортов арбуза к фузариозному
увяданию на искусственном ин'
фекционном фоне начато на КУ"
байской опытной станции ВИ
[257]. Выделен ряд сортов 113
США, у которых поражение не
124
превышало 20%: Congo, Leesburg, Klondike, Klondike Blue Ribbon
striped.
Как показывают приведенные данные, совершенно иммунных к фу-
зариозу сортов арбуза пока не обнаружено. Сорта, считающиеся в
США устойчивыми к болезни, в условиях СССР оказываются воспри-
имчивыми. Видимо, в нашей стране преобладают другие, более виру-
лентные расы возбудителя. Дальнейшие исследования должны быть на-
правлены на их выявление и дифференцированную оценку устойчивости
сортов к определенным расам фузариума.
В настоящее время селекция на иммунитет предусматривает созда-
ние сортов бахчевых культур с комплексной устойчивостью
к нескольким заболеваниям. В этой связи большое значение приобре-
тает выявление источников устойчивости не только к антракнозу и фу-
зариозу, но и к мучнистой росе, бактериозу, вирусной мозаике.
Трехлетнее изучение японских сортов арбуза из коллекции ВИР
в условиях Молдавии показало, что все они устойчивы к мучнистой
росе, сильно здесь распространенной [184]. Невосприимчивость япон-
ских сортов к этой болезни отмечена и на Астраханском опорном пункте
ВИР в 1972—1973 гг. Среди них выделены образцы, заслуживающие
внимания для использования в селекции: к-4153 (Ямато желтый),
к-4157, к-4370 (Кодама суйка), к-4371 (Асахи Мияко №2). Устойчиво-
стью к мучнистой росе отличались образцы иного происхождения:
к-3738 (Китай), к-4434 (Перу), к-4450 (Венгрия), к-4474 (Афганистан),
к-4509 (Индия). Они также могут быть рекомендованы как исходный
материал для селекции.
Источники устойчивости к различным заболеваниям обнаружены
не только среди столовых сортов, но также среди полукультурных и ди-
корастущих форм арбуза, представленных в коллекции. Так, полукуль-
турные образцы из Судана (к-4102 и др.) совершенно не поражаются
мучнистой росой и очень слабо фузариозом. Высокой устойчивостью
к фузариозному увяданию отличаются образцы дикорастущего арбуза
из Южной Африки к-2725 и 2814 (var. lanatus), к-1682 (var. capensis),
образцы кормового арбуза к-54, к-3378, к-3514 (var. citroides). Арбузы
первых двух разновидностей сочетают устойчивость к фузариозу с им-
мунностью к мучнистой росе.
Иммунитетом к антракнозу и мучнистой росе отличаются образцы
из Западной Африки к-1298, к-3837, вр. к-643, 646 (subsp. mucososper-
mus). Последние два оказались устойчивыми и к бактериальному увя-
данию. Образец к-1298 сочетает устойчивость к болезням с устойчиво-
стью к тле — переносчику вирусных болезней [84]. Данные относительно
иммунных свойств образцов арбуза из Западной Африки дополнительно
подтверждают правильность выделения их в самостоятельный таксон.
Приведенный материал показывает, что сосредоточенная в ВИР
коллекция арбуза, ежегодно пополняемая новыми образцами из раз-
личных стран мира, представляет ценный генофонд, содержащий источ-
ники устойчивости ко всем основным заболеваниям арбуза.
Работа в этом направлении начата во Всесоюзном НИИ овощевод-
ства и бахчеводства, на Быковской бахчевой селекционной и Днепро-
петровской овоще-бахчевой опытных станциях.
Ниже приводится описание имеющихся в коллекции ВИР сортов
и форм арбуза, устойчивых к заболеваниям.
Сорта из США. Charleston gray 133 (к-4128)—один из лучших сортов,
выведенных в США в последнее время. Получен путем отбора из сорта Charleston
gray. Устойчив к антракнозу и фузариозному увяданию. Плоды крупные (5—6 кг),
цилиндрические, светло-зеленые, с сетчатым рисунком (сортотип 6). Кора средней
толщины, мякоть малиновая, сочная, прекрасного вкуса, содержит 11—12% сахара.
125
о
L
10
JL.
1
Рис. 28. Сорт Charleston gray
Рис. 29. Сорт Congo.
Семена средние, коричневые, крапчатые. Среднеспелый (от всходов до созревания
80—84 дня) (рис. 28).
Gray belle (к-4298) получен от скрещивания Sugar Baby и Charleston gray, отно-
сится к тому же морфологическому типу. Устойчив к антракнозу и фузариозу.
Calhoun gray (к-4340) получен от скрещивания Calhoun sweet и Charleston gray.
Устойчив к антракнозу и фузариозу.
Congo (к-3848) — один из наиболее устойчивых к антракнозу сортов. Выведен
в США путем скрещивания формы Африка-8 с сортами Jowa Belle и Garrison. Пре-
красно хранится, пригоден для длительной транспортировки. Плоды крупные (5—
6 кг), цилиндрические, темно-зеленые, с широкими расплывчатыми полосами темнее
фона (сортотип 26). Кора очень твердая, мякоть малиновая, прекрасного вкуса, содер-
жит 11 —12% сахара. Семена светло-серые, с желтым ободком. Среднеспелый (от
всходов до созревания 78—80 дней) (рис. 29).
126
b ’ ib ’ ~20
Рис. 30. Сорт Slades majorda.
Garrisonian (к-4328) выведен с участием сорта Congo. Отличается устойчивостью
к антракнозу и фузариозному увяданию. Плоды крупные (7—8 кг), цилиндрические,
светло-зеленые, с широкими темными полосами (сортотип 15). Кора твердая, мякоть
малиновая, содержит 10—11% сахара. Семена средней величины, белые. Среднепозд-
ний (от всходов до созревания 95—100 дней), принадлежит к тому же морфологи-
ческому типу.
Сорта Fairfax (к-4244) и Crimson sweet (к-4297) получены от сложного скрещи-
вания (MilesXPeacockXCharleston gray). Отличаются хорошей транспортабельностью,
а также устойчивостью к антракнозу и фузариозному увяданию. Близки им морфо-
логически сорта Tendersweet (к-4242), устойчивый к антракнозу, Klondike Blue Ribbon
striped (к-4246) и Jubilee (к-4329), устойчивые к фузариозному увяданию.
Hawkesbury (к-4054) выведен в Австралии, завезен в США. Устойчив к фуза-
риозному увяданию. Плоды крупные (5—6 кг), цилиндрические, светло-зеленые, с сет-
кой (сортотип 6). Кора тонкая, мякоть малиновая, содержит 9—11% сахара. Семена
средней величины, коричневые, крапчатые. Среднеспелый (от всходов до созревания
80—85 дней). С участием этого сорта выведены близкие ему сорта Hawkesbury Purdue
F. W. R. (к-4282) и Candy Red W. R. (к-4145).
Klekley’s N6 resistant (к-3851)—линия старого американского сорта Клеклей,
устойчивая к фузариозному увяданию. Плоды среднего размера (3—5 кг), цилиндри-
ческие, темно-зеленые (сортотип 23). Мякоть красная, содержит 9—11% сахара. Се-
мена средней величины, светло-серые. Среднеспелый (от всходов до созревания
85—90 дней).
Klondike R-1 (к-4056), Klondike WR-3 (к-4246), Klondike WR-7 (к-4249), Klondike
R-57 (к-4278), Klondike WR-65 (к-4247) — линии американского сорта Клондайк,
устойчивые к фузариозному увяданию. Морфологически однотипны. Плоды крупные
(5—8 кг), короткоцилиндрические или овальные, черно-зеленые, со скрытой сеткой
сортотип 31). Кора тонкая, мякоть малиновая, содержит 11—12% сахара. Семена
мелкие, коричневые, крапчатые. Среднеспелый (от всходов до созревания 85—87 дней).
К сорту Клондайк по морфологическим признакам близок сорт Blacklee WR (к-4142),
также устойчивый к фузариозному увяданию.
Black diamond Shipper WR (к-4279) устойчив к фузариозному увяданию. Плоды
короткоовальные, массой 4—5 кг, черно-зеленые, без рисунка (сортотип 31). Мякоть
интенсивно розовая, содержит 9—10% сахара. Семена средней величины, светло-корич-
невые. Среднепоздний (от всходов до созревания 90—95 дней).
Verona (к-4352) получен от скрещивания сортов Shipper Blackstone и Charleston
gray, устойчив к антракнозу и фузариозному увяданию. Относится к тому же морфо-
логическому типу, что и предыдущий сорт. Близок к ним сорт Calhoun sweet (к-4341),
также устойчивый к обоим заболеваниям.
127
Рис. 31. Арбуз кордофанский.
Мякоть
Calhoun gray (к-4340) получен путем скрещивания сортов Calhoun sweet и
Charleston gray, устойчив к фузариозному увяданию и антракнозу. Плоды цилиндри-
ческие, светло-зеленые, с зеленой сет-
кой, массой 5—6 кг (сортотип 6).
Мякоть розовая, сочная, содержит
10% сахара. Семена средней величи-
ны, коричневые, крапчатые. Средне-
поздний (от всходов до созревания
95—100 дней).
Дикорастущие и полу-
культурные формы. Slades
majorda (к-2725) —var. lanatus (из
Южной Африки) (рис. 30). Растения
мощные, длинноплетистые, стебли
толстые, отчетливо граненые. Листья
крупные, широкие, с округлыми до-
лями. Цветки ярко-желтые. Плоды
крупные (до 10 кг и более), цилин-
дрические, расширяющиеся к цветоч-
ному концу, светло-зеленые, с зеле-
ным мраморным рисунком,
светло-желтая, менее плотная, чем у
кормового арбуза, сочная, без горечи,
съедобная. Семена крупные, красные,
без ободка. Устойчив к фузариозно-
му увяданию.
Wild Bitter Tsamma (к-2814) —
var. lanatus (из Южной Африки).
Растения того же типа . Плоды
крупные, неправильно шаровидные,
белесые, со светло-зелеными тусклы-
ми полосами. Мякоть желтоватая,
горькая. Устойчив к фузариозу.
К-1298 — var. mucosospermus.
Растения длинноплетистые, стебли
тонкие, округло-граненые. Пластинка
листа вытянутая, темно-зеленая,
сильнорассеченная. Благодаря на-
клонным черешкам положение листь-
ев горизонтальное. Плоды шаровидные, тускло-зеленые, полосатые. Мякоть белая,
плотная, горькая. Семена крупные, плоские, заключены в студенистую оболочку.
Устойчив к антракнозу и тле.
К-1682 (из Египта)—var. capensis. Растения длинноплетистые. Листья средней
величины, с округлыми долями, слаборассеченные. Плоды шаровидные, кремово-жел-
тые, с тусклыми широкими полосами. Мякоть белая, плотная, горькая. Устойчив
к фузариозному увяданию.
Арбуз кордофанский (к-4102) (из Судана)—var. cordophanus. Растения длинно-
плетистые, с длинными междоузлиями. Листья с наклонными черешками сизоватого
оттенка. Плоды короткоовальные, желтовато-светло-зеленые, с зелеными полосами.
Мякоть белая, плотная, в процессе хранения мацерируется, и плод заполняется соком.
Семена кремовые, с коричневым пятном посередине. Устойчив к мучнистой росе и
фузариозу (рис. 31).
СПИСОК ЛИТЕРАТУРЫ
1. Абилов Д. Особенности роста и развития некоторых сортов арбуза и их уро-
жайность в условиях предгорья Алма-Аты. Автореф. дис. на соиск. учен, степени
канд. с.-х. наук. Алма-Ата, 1972.
2. А в а к я н А. Г. Биологические особенности роста и развития важнейших овощ-
ных и бахчевых культур. Автореф. дис. на соиск. учен, степени д-ра биол. наук.
Л., 1965.
3. А кс юк И. Целебный напиток. — Сельская жизнь, 1973, 10 окт.
4. Алексеев А. М. Итоги и перспективы работ по изучению водного режима ра-
стений, проводимых в Биологическом институте Казанского филиала АН СССР. —
В кн.: Изв. Казанского филиала АН СССР, 1958.
5. Алексеева М. В. О глобулинах семян видов арбуза. — Труды по химии при-
родных соединений, 1966, вып. 6.
6. Алешина Л. А. О пыльце тыквенных.— Бот. журн., 1964, т. 49, № 12.
128
7. Алешина Л. А. Палинологические данные к систематике трибы Fevilleae Pax.
сем. Cucurbitaceae Juss. — Бот. журн., 1966, т. 51, № 2.
8. Алешина Л. А. Палинологические данные к систематике и филогении сем.
Cucurbitaceae Juss. — В кн.: Морфология пыльцы. Л., 1971.
9. Алиева А. Г. Цитологическое исследование арбузов. — Соц. наука и техника,
1939, № 9.
10. Алиева А. Г. Наблюдения над ростом пыльцевых трубок у арбузов. — Сборник
науч. тр. по бахчевым культурам. Ташкент, 1952.
11. Алманиязов Э. А. Вирусные болезни бахчевых культур в условиях Казах-
ской ССР. Автореф. дис. на соиск. учен, степени канд. биол. наук. Л., 1970.
12. Ангел И. Сравнительные исследования форм 2п, Зп и 4п Citrullus vulgaris.—
В кн.: Генетика, 1969, т. 5, № 8.
13. Анюховская И. В. Селекция столового арбуза на скороспелость в условиях
Молдавии. Автореф. дис. на соиск. учен, степени канд. с.-х. наук. Кишинев, 1973.
14. Арасимович В. В. Закономерности в наследовании химических признаков у
бахчевых культур в связи с селекцией на химический состав. — Изв. АН СССР.
Сер. биол., 1937, № 6.
15. Арасимович В. В. Биохимия арбуза.— В кн.: Биохимия культурных расте-
ний, т. 4. М.—Л, 1938.
16. Арасимович В. В. Пектиновые вещества кормового арбуза. —Тр. Молд. НИИ
орош. земледелия и овощеводства, 1960, т. 2.
17. А р а с и м о в и ч В. В., Савченко А. П. Биохимия арбуза С. edulis Pang.—
В кн.: Биохимия культурных растений Молдавии. Биохимия бахчевых. Кишинев,
1966, вып. 4.
18. Архангельский С. И. Указ Диоклетиана о таксах 301 г. — Изв. Нижегород-
ского гос. ун-та, 1928, вып. 2.
19. Базилевская Н. А. Растениеводственные ресурсы Южной Африки. — Тр. по
прикл. бот., ген. и сел., 1929, т. 22, № 4.
20. Балтага С. В. Биохимия кормового арбуза. — В кн.: Биохимия культурных
растений Молдавии. Биохимия бахчевых. Кишинев, 1966, вып. 4.
21. Барановский П. М., Семенова Д. Н. Оценка гибридных комбинаций ку-
курузы на гетерозис методом определения силы начального роста растений. —
Вести, с.-х. науки. Алма-Ата, 1963, № 12.
22. Бахчеводство. Под ред. А. И. Филова. М., 1959.
23. Белик В. Ф., П о д м о г а е в а М. И., Сазанова Н. М. Влияние орошения на
продуктивность растений и качество плодов бахчевых культур. — Науч. тр.
НИИОХ. Бахчевые культуры, 1965, т. 3.
24. Белик В. Ф., Соломина И. П. Влияние условий выращивания на некоторые
физиологические особенности растений бахчевых культур. — Науч. тр. НИИОХ.
Бахчевые культуры, 1965, т. 3.
25. Белик В. Ф. Приемы выращивания ранних арбузов и дынь. — Картофель и ово-
щи, 1974, № 5.
26. Б е л я к о в К. Режим орошения и химический состав арбузов. — Степные про-
сторы, 1972, № 6.
27. Бессонова Е. И. Биологические особенности арбузов и дынь иа богаре Узбе-
кистана и их селекция. Автореф. дис. на соиск. учен, степени д-ра с.-х. наук.
Ташкент, 1971.
28. Бессонова Е. И. Характер наследования некоторых признаков столового ар-
буза.— В кн.: Богарное бахчеводство в Узбекистане. Ташкент, 1971.
29. Бойко И. К вопросу об анатомическом строении плодов арбузов. — Науч. тр.
Днепропетр. ун-та, 1940, т. 21, вып. 1.
30. Борисова Р. Л. Арбузы и дыни под пленкой в Крыму. — Картофель и овощи,
1968, № 4.
31. Брежнев Д. Д. Овощеводство в США. М., 1961.
32. Брежнев Д. Д., Шмараев Г. Е. Растениеводство Австралии. М., 1974
33. Б р и т и к о в Е. А. К физиолого-биохимическому анализу прорастания пыльцы и
роста пыльцевых трубок в тканях пестика. — Тр. ин-та физиологии растений
им. К. А. Тимирязева, 1954, т. 8, вып. 2.
34. Бубенцов С., Мосиевская Л. М. Увядание бахчевых культур в Казахста-
не.— Науч. тр. Узб. НИИ овоще-бахчевых культур и картофеля, 1972, вып. 9.
35. Бывших Н. А. Биохимические изменения в созревающих семенах арбуза и роль
плода в послеуборочном их дозревании. Автореф. дис. на соиск. учен, степени
канд. с.-х. наук. Мичуринск, 1954.
36. Вавилов Н. И. Полевые культуры Юго-Востока. М.—Л., 1922.
37. В а в и л о в Н. И. О междуродовых гибридах дынь, арбузов и тыкв. — Тр. по
прикл. бот., ген. и сел., 1925, т. 14, вып. 1.
38. Вавилов Н. И. Земледельческий Афганистан. М.~Л., 1929.
39. Вавилов Н. И. Проблема происхождения культурных растений в современном
понимании. — В кн.: Достижения и перспективы в области прикладной ботаники,
генетики и селекции. Л., 1929.
129
40 Вавилов Н. И. Ботанико-географические основы селекции. М.—Л., 1935.
41. Василевская В. К. О значении анатомических коэффициентов как признака
засухоустойчивости растений. — Бот. журн. СССР, 1938, т. 23, № 4.
42. Василевская В. К. Изучение онтогенеза как один из методов экологической
анатомии. — Проблемы ботаники, 1950, вып. 1.
43. Василевская В. К. Анатомическое строение зародыша и проростка некото-
рых травянистых растений. — Вести. ЛГУ. Сер. биол., 1959, № 3, вып. 1.
44. Васильченко И. Т. Арбуз как «болотное» растение. — Природа, 1939, № 3.
45. Васильченко И. Т. Тыквенные — Cucurbitaceae. — В кн.: Флора СССР. М.—
Л., 1957, т. 24.
46. В е р н е р А. Р., Клинг Е. Г. Причины устойчивости плодов арбуза к черной
гнили. — В кн.: Отчет комиссии по ирригации Нижневолжской эксп. АН СССР
(Тр., вып. 3), 1934.
47. Вернер А. Р., Бугаев Н. Е. К вопросу о солеустойчивости и сосущей силе
бахчевых. — Тр. Быковской зональной опыт, ст., 1935, вып. 2.
48. В е р н и д у б о в И. С. Гетерозис столового арбуза в условиях Волгоградского
Заволжья. Автореф. дис. на соиск. учен, степени канд. с.-х. наук. Волгоград, 1972.
49. Веселовский А. А. Обоснование площадей питания, схем посева и способов
механизированной уборки арбуза для условий дельты Волги. Автореф. дис. на
соиск. учен, степени канд. с.-х. наук. М., 1972.
50. В и т а н о в М., Лозанов П. Изучение восприимчивости различных сортов ар-
буза к антракнозу (Colletotrichum lagenarium Pass.). — Градинарска и лозарска
наука, 1966, т. 3, вып. 6. На болг. яз.
51. Вирусная мозаика арбуза на тыквенных в Новой Зеландии. — Реф. журн.
Овощные и бахчевые культуры, 1972, № 4.
52. Г а б ч е н к о А., Сизов В. Содержание сухих веществ и сахаров в арбузах. —
Картофель и овощи, 1972, № 9.
53. Галка А. Т. Биология цветения и оплодотворения у бахчевых культур. — В кн.:
Итоги работ Днепропетр. овоще-бахчевого отд. ВНИИЗХ за 1937—1939 гг., 194 К
ч. 4.
54. Галка А. Т. Селекция бахчевых культур на длину вегетационного периода. —
Науч. тр. Укр. НИИ овощеводства, 1950, т. 2.
55. Галка А. Т. Селекция арбуза и дыни на Днепропетровской овоще-бахчевой се-
лекционной станции. — Науч. тр. УкрНИИ овощеводства, 1954, № 3.
56. Г е н к е л ь П. А. Развитие учения о засухоустойчивости растений в Советском
Союзе. — С.-х. биология, 1967, № 5.
57. Герасимова-Навашина Е. Н. К цитолого-эмбриологическому пониманию
процесса опыления. — Тр. Бот. ин-та АН СССР, 1952, сер. 7, вып. 3.
58. Герасимова-Навашина Е. Н. Еще об одной возможной биологической
функции нектарников. — Бот. журн., 1966, т. 51, К» 12.
59. Гну те н ко М. П. Токсический бактериоз арбузов на Юго-Востоке европейской
части РСФСР и перспективы борьбы с ним. — В кн.: Всесоюз. симпозиум по бак-
териальным заболеваниям растений. (Тезисы докл.). Киев, 1966.
60. Г о л ь д г а у з е н М. К. Американские сорта столовых арбузов и их значение для
бахчеводства СССР.— Докл. ВАСХНИЛ, 1941, вып. 8.
61. Гольдгаузен М. К. О пространственной изоляции у бахчевых культур.—Сад
и огород, 1949, № 4.
62. Гольдгаузен М. К. Вегетационный период арбузов и дынь. — Изв. Молд. фи-
лиала АН СССР, 1952, № 4—5.
63. Гольдгаузен М. К. Межвидовая гибридизация арбузов — Тр. Молд. НИИ
орош. земледелия и овощеводства, 1959, т. 1.
64. Гольдгаузен М. К. Стеблевая система арбузов и урожай. — Тр. объед. научн.
сессии. Кишинев, 1959, т. 1.
65. Гольдгаузен М. К. Об изменчивости андроцея в обоеполых цветках арбуза.
— Тр. Молд. НИИ орош. земледелия и овощеводства, 1960, № 2.
66. Гольдгаузен М. К- История возделывания арбуза в СССР. — Тр. Молд. НИИ
орош. земледелия и овощеводства, 1960, т. 2.
67. Гольдгаузен М. К. О межвидовых гибридах Praecitrullus с Citrullus. —
Тр. Молд. НИИ орош. земледелия и овощеводства, 1961, т. 3.
68. Гольдгаузен М. К. Из истории арбуза как культурного растения. — Тр.
Молд. НИИ орош. земледелия и овощеводства, 1962, т. 4, вып. 1.
69. Г о л ь д г а у з е и М. К., Ершов Е. М. О выработке норм пространственной
изоляции у арбузов. — В кн.: Орошаемое земледелие и овощеводство. Кишинев,
1968.
70. Гуляев В. А. Применение сравнительно-эмбриологического метода к система-
тике рода Citrullus Schrad. — Тр. Плодоовощного ин-та им. И. В. Мичурина,
1955, т. 8.
71. Гуляев В. А. К описанию процесса оплодотворения у Citrullus vulgaris Schrad.
Бюл. Всесоюз. НИИ растениеводства, 1961, № 9.
130
72. Гуляев В. А. О типе зародышевого мешка Praecitrullus fistulosus (Stocks) Pang.
— ДАН СССР, 1962, т. 146, № 5.
73. Гуляев В. А. Сравнительная эмбриология Cucurbitaceae и ее значение для си-
стематики семейства. — Бот. журн., 1963, т. 48, вып. 1.
74. Гусев Н. А. Современное состояние работ по изучению водного режима расте-
ний и дальнейшее их развитие. — В кн.: Биологические основы орошаемого зем-
леделия. М., 1957.
75. Декандоль А. Местопроисхождение возделываемых растений. Пер. с франц.
СПб., 1885.
76. Дзелвалтовский А. К. Редукционное деление при макроспорогенезе у
триплоидной формы арбуза Асахи Ямато. — Цитология и генетика, 1968, т. 2, № 5.
77. Дзелвалтовский А. К. Особенности развития женского гаметофита у три-
плоидной формы арбуза Асахи Ямато. — Цитология и генетика, 1969, т. 3, № 1.
78. Дзелвалтовский А. К. Особенности формирования пыльцы у эксперимен-
тально-полиплоидного арбуза. — Цитология и генетика, 1970, т. 4, № 5.
79. Дзензелевская М. Д., Лукина Л. Т. Наследование ряда признаков у ги-
бридов столового арбуза Fi и F2. — В кн.: Селекция и семеноводство. Кишинев,
1973.
80. Дорофеев П. И. О плиоценовой флоре нагавских глин на Дону.— ДАН СССР,
1957, т. 117, вып. 1.
81. Дюгаева Т. М. О строении листа, междоузлия и узла у некоторых видов сем.
Cucurbitaceae. — Учен. зап. Ленингр. пед. ин-та, 1966, т. 310.
82. Дютин К. Е. Гетерозис и получение гибридных семян арбузов и дынь. Автореф.
дис. на соиск. учен, степени канд. с.-х. наук. М., 1969.
83. Д ю т и н К. Е. Селекция арбуза на устойчивость к антракнозу (обзор иностр,
лит.). — Новости с.-х. науки и практики, 1971, № 11.
84. Дютин К. Е., Богоявленская С. М., Калчанов В. А. Предварительные
итоги селекции арбуза на устойчивость к антракнозу. — Сборник науч. тр.
ВНИИОБ, 1974, вып. 2.
85. Ермаков А. И., Луковникова Г. А. Химический состав плодов бахчевых
культур и простейшие методы его определения. — Науч. тр. НИИОХ. Бахчевые
культуры, 1965, т. 3.
86. Е р ш о в Е. М. Кормовой арбуз — ценный корм для животных. — Земледелие и
животноводство Молдавии, 1960, № 4.
87. Ершов Е. М. О пространственной изоляции семеноводческих посевов арбузов.—
Картофель и овощи, 1968, № 7.
88. Ершов Е. М. Об автогамии у столового арбуза в условиях Молдавии. — Тр.
Молд. НИИ орош. земледелия и овощеводства, 1969, т. 10, вып. 1.
89. Жуковский П. М. Земледельческая Турция. М.—Л., 1933.
90. Жуковский П. М. Триплоидный арбуз.—Бюл. Всесоюзн. НИИ растениеводства,
1958, № 4.
91. Жуковский П. М. Природа и объем вида у культурных растений. — Бот.
журн., 1967, т. 52, № 10.
92. Жуковский П. М. Новые очаги происхождения и генцентры культурных ра-
стений и узкоэндемичные микроцентры родственных видов. — Бот. журн., 1968,
т. 53, № 4.
93. Жуковский П. М. Культурные растения и их сородичи. Л., 1971.
94. Захаров А. Ф., Барановская Л. И., Деминцева В. С., Ибраимов
А. И. Дифференциация хромосом человека по длине и проблема их идентифика-
ции. 1. Общая картина дифференциации, выявляемая с помощью 5-бромдезоксиу-
ридина. — Цитология, 1973, т. 15, № 5.
95. 3 а у р а л о в О. А. Особенности стадийного развития арбузов и дынь. — Агро-
биология, 1956, № 5.
96. Знаменский В. Д. О сахаристости донских арбузов. — Тр. с.-х. опыт, учреж-
дений Сев. Кавказа, 1927, т. 10.
97. Знаменский В. Д. Влияние солей на развитие и сахаристость арбуза.— Тр.
по прикл. бот., ген. и сел., 1930, т. 23, вып. 3.
98. Знаменский В. Д. Кормовой арбуз. Ростов-на-Дону, 1940.
99. Иванов Н. Н., Александрова Р. С. Проблема сахаристости бахчевых куль-
тур.— Изв. Гос. ин-та опытн. агрономии, 1928, т. 6, № 3—4.
100. Камилова Ф. Г. Особенности строения и формирования стебля бахчевых
тыквенных. — Тр. Ин-та бот. АН УзССР, 1955, вып. 3.
101. Камилова Ф. Г. Строение пятидневных проростков некоторых видов плети-
стой и кустовой форм бахчевых. — Узб. бот. журн., 1966, № 4.
102. К ар ягдыев Н. Н. Новый вид для флоры СССР Citrullus colocynthis (L.)
Schrad. — Бот. материалы герб. Бот. ин-та АН СССР, 1950, т. 12.
103. Кич у нов Н. И. Культура арбузов в Америке. — Вести, садоводства, плодовод-
ства и огородничества. СПб, 1908, № 5.
104. Келлер Б. А. Об анатомическом строении листьев, устойчивых к жаре и засу-
хе. — Сов. ботаника, 1938, № 2.
131
105. Ковалевский Г. В. Земледельческая Индия. — Тр. по прикл. бот., ген. и сел.,
1929, т. 21, вып. 5.
106. Ковалевский Г. В. К истории культуры арбуза и дыни в европейской части
СССР. — Тр. по прикл. бот., ген. и сел., 1930, т. 23, вып. 3.
107. Кожухов 3. А. Кариотипические особенности культурных тыквенных. — Тр. по
прикл. бот., ген. и сел., 1925, т. 14, вып. 2.
108. Кожухов 3. А. Кариологическое исследование культурных и диких видов и
разновидностей рода Cucumis. — Тр. по прикл. бот., ген. и сел., 1930, т. 23, вып. 3.
109. Кожухов 3. А. К кариосистематике семейства тыквенных. — Журн. Ин-та бот.
ВУАН, 1934, т. 1, вып. 9.
ПО. Константинов Б. С. Особенности минерального питания арбуза при ороше-
нии на буграх Бэра в Астраханской области. Автореф. дис. на соиск. учен, сте-
пени канд. с.-х. наук. Астрахань, 1972.
111. Корейша 3. И. Биохимическая характеристика бахчевых культур. — В кн.:
Сборник научн. работ по бахчевым культурам. Ташкент, 1953.
112. Кревченко Л. Е. Сортопрочистки, примеси, пространственная изоляция.—
Картофель и овощи, 1967, № 4.
113. Кревченко Л. Е. Бахчеводство Северного Кавказа. Краснодар, 1972.
114. Криштофович А. Н. Палеоботаника. Л., 1957.
115. Крылов Т. В., Положи й А. В. Об объеме и структуре вида у растений.—
Изв. Сиб. отд. АН СССР. Сер. биол.-мед. наук, 1967, вып. 3.
116. Кудрявцева М. А. Опыт изучения сахаристости мирового сортимента арбу-
зов и дынь. — Тр. по прикл. бот., ген. и сел., 1930, т. 23, вып. 3.
117. Кузнецов А. В., Сагалович Е. Н. Овощеводство в Китае. М., 1959.
118. Курс ботаники. Под ред. Л. И. Курганова и М. И. Голенкина, 1940, т. 1.
119. Лангельд Р. К. К биологии цветения и оплодотворения у тыквенных.—Тр. Бы-
ковской зональной ст. бахчеводства, 1935, вып. 2.
120. Леонтьев И. Ф. Химия масла из семян арбуза. — Природа, 1948, № 5.
121. Ливингстон Д. Путешествие по Южной Африке с 1840 по 1856 г. М., 1947.
122. Лозанов П. Бессемянные арбузы. — Градинарство, 1965, т. 7, кн. 5. На болг. яз.
123. Лозанов П. Использование сортов с генетическим маркером для производства
гибридных семян арбуза. — Градинарство, 1967, кн. 6. На болг. яз.
124. Лозанов П. Пространственная изоляция при культуре тыквенных (Cucurbita-
сеае). — Градинарство, 1968, кн. 5. На болг. яз.
125. Лозанов П., Георгиев X., Даскалов С. Использование полиплоидов в
селекции арбуза. — Генетика и селекция, София, 1968, т. 1, № 4.
126. Лукина Л. Т. Фузариозное увядание дынь и арбузов в Молдавии и меры
борьбы с ним. Автореф. дис. на соиск. учен, степени канд. с.-х. наук. Кишинев,
1969.
127. Лутохин С. Н. Об автогамии у арбуза. — Тр. агрохимической лаб. Гос. поли-
техи. музея, 1927, вып. 1.
128. Лутохин С. Н. Исследования по качественному содержанию сахаров в раз-
личных сортах арбуза и дыни. — Тр. агрохимической лаб. Гос. политехи, музея,
1928, вып. 1.
129. Львова И. Н. Органогенез бахчевых культур. — Науч. тр. НИИОХ, 1965, т. 3.
130. Лященко И. Ф. Влияние удаления цветков на цветение и плодоношение у бах-
чевых растений. — Тр. Рост. обл. биол. общ-ва. Ростов-на-Дону, 1938.
131. Малви хи л У. Пески Калахари. Л., 1969.
132. Малинина М. И. Особенности роста и развития арбуза на богаре в полупу-
стынной зоне Западного Казахстана. Автореф. дис. на соиск. учен, степени канд.
с.-х. наук. Л., 1960.
133. Маршак М. С. Лечебные свойства тыквенных овощей. — Овощевод, 1959, № 8.
134. М атиен к о Б. Т. Об анатомо-морфологической природе цветка и плода тыквен-
ных.— Тр. Бот. ин-та им. Комарова, 1957, сер. 7, вып. 4.
135. М атиен к о Б. Т. Анатомия плода Benincasa Savi и ее сходство с анатомией
плода тыквы и арбуза. — Изв. АН Молд. ССР, 1963, т. 10.
136. Матиенко Б. Т. Сравнительная анатомия и ультраструктура плодов тыквен-
ных. Автореф. дис. на соиск. учен, степени д-ра биол. наук. Киев, 1967.
137. Матиенко Б. Т. Сравнительная анатомия и улътраструктура плодов тыквен-
ных. Кишинев, 1969.
138. Матиенко Б. Т. Анатомическая характеристика плодов культурных и дикора-
стущих тыквенных. Кишинев, 1972.
139. Международный кодекс ботанической номенклатуры. Л., 1959, 1974.
140. М е ж д у н а р о д н ы й кодекс номенклатуры культурных растений. Л., 1969.
141. Меликян А. П. О возможности применения признака строения спермодермы
для систематики нимфейных. — Изв. АН Арм. ССР. Биол. науки. 11964, т. 17, № 1.
142. Меликян А. П. Значение признаков анатомического строения семенной кожу-
ры для видовой таксономии (на примере рода Alisma L.).—Учен. зап. Ерев.
ун-та, 1968, вып. 2.
132
143. Меликян А. П. Типы семенной кожуры Hamamelidaceae и близких семейств
в связи с их систематическими взаимоотношениями. — Бот. журн., 1973, т. 58, № 3.
144. Мировой агроклиматический справочник. Под редакцией Г. Т. Селянинова.
М.—Л, 1937.
145. М н а ц а к а н я н О. А. Исследование микроспорогенеза и мужского гаметофита
у культурного арбуза. — В кн.: Тезисы докладов конференции молодых науч, со-
трудников и аспирантов НИИ земледелия Арм. ССР. Эчмиадзин, 1967.
146. Морозов А. С. Влияние солей NaCl и Na2SO4 на сахаристость арбузов и дынь.
— ДАН СССР, 1938, т. 21, № 6.
147. Морозова Н. С. Сравнительно-анатомическая характеристика семенной кожу-
ры некоторых видов Caltha. — Бот. журн., 1973, т. 58, № 4.
148. М ос невская Л. М. Увядание арбузов и дынь в северном Прибалхашье. Ав-
тореф. дис. на соиск. учен, степени канд. биол.наук. Л., 1968.
149. Невкрыта А. Н. Насекомые, опыляющие бахчевые культуры. Киев, 1953.
150. Немирович-Данченко Е. Н. Морфолого-систематическое изучение нектар-
ников семейства тыквенных. — Изв. Сиб. отд. АН СССР. Сер. биол.-мед. наук,
1965, вып. 2, № 8.
151. Немирович-Данченко Е. Н. К вопросу о происхождении нектарников тык-
венных Cucurbitaceae Juss.— Бот. журн. 1966, т. 51, № 3.
152. Никитина К. В., Бычков В. П. Изучение бактериальных болезней бахчевых
культур в условиях Кубани.— Бюл. ВИР, 1977, № 74.
153. Никифорова Н. А. Фитопатологическая характеристика сортов арбузов в Мол-
давии.— Тр. Молд. НИИ орош. земледелия и овощеводства, 1968, т. 8, вып. 2.
154. Никулина Н. К., Шворнева А. М. Токсический бактериоз арбузов. — Защи-
та растений от вредителей и болезней, 1957, № 5.
155. Определитель болезней растений. Под ред. М. К. Хохрякова. Л., 1966.
156. Павлов И. П. Селекция и семеноводство овощных культур. М, 1963.
157. П алл ас П. С. Путешествие по разным провинциям Российского государства.
СПб, 1788, ч. 3.
158. Пангало К. И. Арбузы. Л, 1927.
159. Пангало К. И. Бахчевые культуры Анатолии. — В кн.: Земледельческая Турция.
Под ред. П. М. Жуковского. М, 1933.
160. Пангало К. И. Арбузы северного полушария. — Тр. по прикл. бот, ген. и сел,
1930, т. 23, вып. 3.
161. Пангало К. И. Бахчеводство СССР. М.—Л, 1934.
162. Пангало К. И. О многообразии проявления пола у растений на примере
Cucurbitaceae. — ДАН СССР, 1936, т. 3, № 2.
163. Пангало К. И. Ныне живущие предки возделываемых арбузов. — ДАН СССР.
Нов. сер, 1938, т. 20, № 7—8.
164. Пангало К. И. Пол и цветение у возделываемых Cucurbitaceae. — Бот. журн.
СССР, 1943, т. 28.
165. Пангало К. И. Новый род сем. Cucurbitaceae Praecitrullus — прапращур со-
временных арбузов (Citrullus Forskal). — Бот. журн. СССР, 1944, т. 29, № 5.
166. Пангало К. И. Состав и эволюция рода Citrullus Forsk. Бюл. МОИП. Отд.
биол, 1945, т. 50, вып. 5—6.
167. Пангало К. И. Новые принципы внутривидовой систематики культурных ра-
стений.— Бот. журн, 1948, т. 33, № 1.
168. Пангало К. И. Пектин из кормового арбуза. — Картофель и овощи, 1960, № 7.
169. Пантелеева Н. И. Анатомические структуры коры арбуза, определяющие ее
прочность (к проблеме транспортабельности). — Тр. Молд. НИИ орош. земледелия
и овощеводства, 1959, т. 1.
170. Пантелеева Н. И. Влияние внешних условий на прочность коры арбуза.—
Тр. Молд. НИИ орош. земледелия и овощеводства, 1960, т. 2.
171. Пащенко Т. Е. Морфобиологические особенности пыльцы дынь, арбузов и тыкв.
— Научн. тр. Ташк. ун-та, 1966, вып. 301.
172 Петровский К. Арбузы, дыни, помидоры, баклажаны. — Наука и жизнь, 1964,
№ 8.
173. П и о т р о в с к и й Б. Б. Кармир-Блур. Ереван, 1950.
174. Подмогаева М. И, Сазанова Н. М. Изменить нормы пространственной
изоляции. — Картофель и овощи, 1965, № 2.
175. Попов В. В. Географическое распространение и кормовые связи рода Cteno-
plectra F. Sm, (Hymenoptera, Melittidae). — ДАН СССР, 1956, т. 108, вып. 5.
176. Попова 3. Н, Сосенская Е. Ю. Химическая характеристика видов арбузов
и дынь. — Тр. по прикл. бот, ген. и сел, 1936, т. 5, сер. 3, № 15.
177. Попова 3. Н. Сравнительное анатомическое исследование строения коры и мя-
коти различных сортов арбузов. — Учен. зап. Сарат. ун-та, 1937, т. 1 (14), вып. 1.
178. Прокудина Ф. В, Смирина Л, Чулкова Т. О распространении антрак-
ноза арбузов семенами. — Картофель и овощи, 1972, № 1.
179. Рахимбаев X. Развитие механического слоя в стенке плода двух видов арбу-
за.— Сборник студенческих работ Средиеаз. ун-та, 1955, вып. 2.
133
180. Редькина 3. В. Морфолого-анатомическая характеристика семян вик. — Тр. по
прикл. бот., ген. и сел., 1971, т. 44, вып. 3.
181. Родигин М. Н. Обзор исследований по иммунитету бахчевых культур. — Научн.
тр. НИИОХ, 1965, № 3.
182. Рудь П. Арбузы выращиваем под пленкой. — Картофель и овощи, 1971, № 4.
183. Р у т к о в с к а я С. В. Об анатомическом строении семенной кожуры рода Cit-
rullus. Сборник н.-и. работ Азово-Черноморского СХИ, 1948, вып. 12.
184. Савчук С. А. Японские арбузы в Молдавии. — Тр. Молд. НИИ орош. земледе-
лия и овощеводства, 1970, т. 11, вып. 2.
185. Сазанова Н. М. Селекция арбуза на устойчивость к фузариозному увяданию.
— Науч. тр. Бирючекутской овощной сел. опыт, ст., 1972, вып. 2.
186. Сазанова Н. М. Селекция арбуза на лежкость и транспортабельность. — Кар-
тофель и овощи, 1973, № 2.
187. Сазанова Н. М. Селекция скороспелых сортов арбуза и дыни в условиях
Ростовской области. Автореф. дис. на соиск. учен, степени канд. с.-х. наук. М., 1974.
188. Сафарян И. М. Гетерозисная селекция арбузов в условиях Араратской равни-
ны Армянской ССР. Автореф. дис. на соиск. учен, степени канд. биол. наук.
Ереван, 1966.
189. Сафарян И. М. Влияние возраста рыльца и пыльцы на оплодотворение арбу-
за.— Сборник науч. тр. НИИ земледелия Арм. ССР, Эчмиадзин, 1966.
190. Сафарян И. М. Самооплодотворение и фертильность пыльцы гермафродитного
цветка у арбуза. — Биол. жури. Арм., 1966, т. 19, № 1.
191. Сизова Н. А. О природе возбудителя токсического бактериоза арбузов.—
Науч. докл. высшей школы. Биол. науки, 1965, № 3.
192. С и иска я Е. Н. Учение об экотипах в свете филогенеза высших растений.—
В кн.: Успехи современной биологии, 1938, т. 9, вып. 1 (4).
193. Синская Е. Н. Принципы классификации культурных растений в связи с зада-
чами внутривидовой систематики. — Бот. журн., 1948, т. 33, № 1.
194. Синская Е. Н. Учение о виде и таксонах (конспект лекций). Л., 1961.
195. Синская Е. Н. Историческая география культурной флоры (на заре земледе-
лия). Л., 1969.
196. Синча К. П. Использование сортов бахчевых культур с сигнальными признака-
ми для получения гибридных семян.—Сборник науч, информ, по овощеводству. М.,
1966.
197. Синча К- П. Выведение и внедрение новых сортов арбуза в Нижнем Поволжье.
Докл. по совокупности опубликованных работ на соиск. учен, степени канд. с.-х.
наук. Волгоград, 1967.
198. Синча К. П. Некоторые биологические особенности цветения и приемы гибри-
дизации в условиях Заволжья. — Науч. тр. Быковской бахчевой опыт, ст., 1969,
вып. 5.
199. Снегирева А. В. Биологические особенности сортов столового арбуза.—
Учен. зап. Тираспольского пед. ин-та, 1958, вып. 4.
200. С к в ор ц о в А. К. Сущность таксона и проблемы внутривидовой систематики
растений. — Бюл. Моск, общ-ва испытателей природы, 1971, т. 76, вып. 5.
201. Словарь полезных растений на двадцати европейских языках. Под ред.
Л. Л. Балашева. М., 1970.
202. Сойфер В. Н. Анатомия семян Cucurbitaceae Juss. как систематический при-
знак.— Бюл. Моск, общ-ва испытателей природы. Отд. биол., 1964, т. 69, вып. 1.
203. Сокол П. В., Васина В. И. Изменение содержания сахаров в мякоти арбузов
в зависимости от различных температурных условий. Докл. ВАСХНИЛ, 1973, № 7.
204. Соколова В. К. Пространственную изоляцию можно сократить. — Картофель и
овощи, 1967, № 6.
205. Сотченко В. С. Сравнительная оценка методов изучения комбинационной спо-
собности линий кукурузы. Автореф. дис. на соиск. учен, степени канд. с.-х. наук.
Л., 1970.
206. Старчевая К. П. К анатомической характеристике арбузов. — Науч. зап.
Дяепропетр. ун-та (сб. раб. биол. фак.), 1940, вып. 3.
207. С т и в е н с о н Ф., Г. Джонс. Некоторые источники устойчивости культурных
растений к болезням. — В кн.: Болезни растений. М., 1956.
208. Страхов Н. М. Историческая геология. М., 1938.
209. Суханов Я. Салют арбузу. — Сельская жизнь, № 252, 1967, 3 окт.
210. Татищев В. Н. Лексикон Российской исторической, географической, политиче-
ской и гражданской. СПб., 1793, ч. 1.
211. Тахтаджян А. Л. Система и филогения цветковых растений. М.—Л., 1966.
212. Тахтаджян А. Л. Биосистематика: прошлое, настоящее, будущее. — Бот. журн.,
1970, т. 55, вып. 3.
213. Тахтаджян А. Л. Происхождение и расселение цветковых растений. Л., 1970.
214. Тахтаджян А. Л., Меликян А. П. Сравнительное изучение семенной обо-
лочки Leontice, Gumnospermum, Caulophyllum и близких родов в связи с их си-
стематикой. — Бот. журн., 1972, т. 58, вып. 10.
134
215. Тер-Аванесян Д. В. Сельское хозяйство Индии. М., 1961.
216. Тер-Аванесян Д. В. Арбуз кордофанский Citrullus lanatus (Thunb.) Mansf.
ssp. cordophanus Ter-Avan. Бот. жури., 1966, т. 51, № 3.
217. T ex а н о в и ч Г. А., Фурса Т. Б. Оценка коллекции арбуза на устойчивость
к фузариозному увяданию. — Картофель и овощи, 1975, № 11.
218. Тимченко В. И., Бейдер А. М., Кононенко А. Г. Фитопатологическая
оценка сортов и гибридов арбуза на устойчивость против фузариозного увядания.
— Овощеводство и бахчеводство, 1972. Киев, вып. 14. На укр. яз.
219. Тимченко В. И. Фузариозное увядание арбузов на Украине. — Науч. тр.
Укр. НИИ овоще-бахчевых культур и картофеля, 1972, вып. 9.
220. Т о щ е в В. П. Проявление гетерозиса у дынь и тыкв в зависимости от система-
тических, экологических и морфологических различий родительских пар. — Авто-
реф. дис. на соиск. учен, степени канд. биол. наук. Л., 1967.
221. Ткаченко Ф. А. Биологические особенности цветения, опыления и плодообра-
зования арбуза в лесостепной зоне УССР. — Науч. тр. Укр. НИИ овощеводства,
1954, т. 3.
222. Ткаченко Ф. А. Использование гетерозиса в овощеводстве и бахчеводстве. —
В кн.: Генетика — сельскому хозяйству. М., 1963.
223. Ткаченко Ф. А., Кононенко А. И. Новый способ получения гибридных се-
мян арбузов. — В кн.: Использование гетерозиса в овощеводстве. Краснодар, 1963.
224. Ткаченко Ф. А. Семеноводство и семеноведение овощных и бахчевых куль-
тур в Украинской ССР. Автореф. дис. на соиск. учен, степени д-ра с.-х. наук.
Л., 1967.
225. Ткаченко Н. М. Морфолого-анатомическое строение и некоторые биологические
особенности семян овощных, бахчевых и лекарственных растений. Автореф. дис.
на соиск. учен, степени д-ра с.-х. наук. Харьков, 1971.
226. Турбин Н. В., Хоты л ев а А. В. Гетерозис. — Цитология и генетика, 1967,
т. 1, № 6.
227. Турбин Н. В. Генетика гетерозиса и методы селекции на комбинационную цен-
ность. — Вести, с.-х. науки, 1967, № 3.
228. Турков В. Д., Шелепина Г. А. Кариологические исследования огурцов
С. sativus L. — Докл. ВАСХНИЛ, 1972, вып. 3.
229. Турков В. Д., Шелепина Г. А. К методике идентификации хромосом
С. sativus L. по качественным и количественным характеристикам. — Докл.
ВАСХНИЛ, 1973, № 5.
230. Турков В. Д., Шелепина Г. А. Цитогенетическое исследование рода Citrul-
lus Schrad. — Генетика и селекция, София, 1975, т. 8, № 1.
231. Тюрин Т. А. Некоторые данные получения тетраплоидных форм арбузов,—
Учен. зап. Оренбургского пед. ин-та, 1970, вып. 29.
232. У н к о в с к а я А. И. Изучение влияния условий воспитания и места репродукции
у гибридов огурцов.—В кн.: Тезисы докладов иа научно-технической конференции
молодых ученых. Днепропетровск, 1967.
233. Унковская А. И. Абсолютный вес семян и гетерозис арбузов. — Картофель и
овощи, 1969, № 7.
234. Унковская А. М. Некоторые вопросы гетерозиса и селекции арбузов в север-
ной степи УССР. Автореф. дис. на соиск. учен, степени канд. с.-х. наук. Харьков,
1971.
235. Унковская А. И., Артамонова В. Оценка сортов арбуза на устойчивость
к увяданию. — Картофель и овощи, 1974, № 9.
236. Фефер И. И., Миндлин М. 3., Прокопович И. И. О противоглистных
свойствах семян арбуза. — Фармакология и токсикология, 1954. № 5.
237. Филиппова Н. П. Арбузы и дыни в пленочной теплице. — Картофель и овощи,
1972, № 3.
238. Филов А. И. О биологии цветения и оплодотворения у бахчевых. — Соц. расте-
ниеводство, 1935, № 10.
239. Филов А. И. Отношение гермафродитных сортов бахчевых культур к самоопы-
лению.— Докл. ВАСХНИЛ, 1939, № 12.
240. Филов А. И. Новые данные о происхождении и классификации арбузов (Citrul-
lus Forsk.). — Бот. журн., 1962, т. 47, вып. 7.
241. Филов А. И. О происхождении и классификации арбузов. — Тр. респ. опыт,
ст. картофельного и овощного хоз-ва. Алма-Ата, 1969.
242. Филов А. И., Виленская Г. М. Культурные тыквенные на различных языках
мира. — Тр. по прикл. бот., ген. и сел., 1972, т. 47, вып. 2.
243. Фирсова М. К. Методы определения качества семян. М., 1959.
244. Фурса Т. Б. Транспортабельность арбузов в зависимости от анатомического
строения коры плода. — Тр. по прикл. бот., ген. и сел., 1962, т. 35, вып. 1
245. Фурса Т. Б. Экотипы арбузов.— Тр. по прикл. бот., ген. и сел., 1965, т. 37,
вып. 2.
246. Фурса Т. Б. Раздел «Бахчевые». — В кн.: Гетерозис в овощеводстве. Л.. 1966.
135
247. Фурса Т. Б. Анатомо-физиологические различия экотипов арбузов. — Докл. сов.
ученых XVII Международному конгрессу по садоводству. М., 1966.
248. Фурса Т. Б. Анатомо-физиологические особенности листа у арбузов различного
происхождения. — Тр. по прикл. бот., ген. и сел., 1968, т. 40, вып. 1.
249. Фурса Т. Б. Об опылении арбузов. — Картофель и овощи, 1969, № 8.
250. Фурса Т. Б., Гвоздева 3. В. Повышение всхожести семян некоторых видов
семейства Cucurbitaceae. — Тр. по прикл. бот., ген. и сел., 1971, т. 44, вып. 3.
251. Фурса Т. Б. Биология цветения и опыления арбуза. — Тр. по прикл. бот., ген.
и сел., 1971, т. 45, вып. 1.
252. Фурса Т. Б. К систематике рода Citrullus Schrad. — Бот. журн., 1972, т. 57,
№ 1.
253. Фурса Т. Б. К эволюции рода Citrullus Schrad. — Бот. журн., 1972, т. 57, №11.
254. Фурса Т. Б. К истории введения арбуза в культуру. — Тр. по прикл. бот., ген. и
сел., 1973, т. 49, вып. 2.
255. Фурса Т. Б. Сравнительная морфология и анатомия проростков Citrullus
Schrad. — Тр. по прикл. бот., ген. и сел., 1973, т. 49, вып. 2.
256. Фурса Т. Б. Долговечность семян арбуза разного происхождения при откры-
том хранении. — Тр. по прикл. бот., ген. и сел., 1974, т. 52, вып. 2.
257. Фурса Т. Б., Малинина М. И, Артюгина 3. Д., Теханович Г. А. Изу-
чение коллекции бахчевых культур. — Тр. по прикл. бот., ген. и сел., 1974, т. 52,
вып. 3.
258. Фурса Т. Б. Кожура семени арбуза как таксономический признак. — Тр. по
прикл. бот., ген. и сел.. 1974, т. 51, вып. 3.
259. Фурса Т. Б., Щеглов С. В. Арбуз Нодэна под Астраханью. — Тр. по прикл.
бот., ген. и сел., 1975, т. 54, вып. 1.
260. Фурса Т. Б., Щеглов С. В. Оценка комбинационной способности сортов ар-
буза.— Тр. по прикл. бот., ген. и сел., 1975, т. 55, вып. 2.
261. Фурса Т. Б. Классификация столового арбуза.— Тр. по прикл. бот., ген. и сел.,
1977, т. 61, вып. 1.
262. Холодов Д. Т. О пространственной изоляции на семеноводческих посевах бах-
чевых культур. — Науч. тр. НИИОХ, 1965, т. 3.
263. Хохряков М. К. и др. Определитель болезней растений. Под ред. М. К- Хох-
рякова. Л., 1966.
264. Хромосомные числа цветковых растений. Л.. 1969.
265. Церевитинов Ф. В. Химия и товароведение свежих плодов и овощей. Т. 2,
М„ 1949.
266. Цыганков С. К. Опылители бахчево-огородных культур. — Сов. агрономия,
1947, № 4.
267. Черноглазое Л. А., Кичу нов Н. И. Огурцы, дыни, арбузы, тыквы. СПб.,
1887.
268. Ч опа нов П., Оразмухомедов. Новые местообитания дикого арбуза ко-
лоцинта Citrullus colocynthis (L.) Schrad. в Туркмении. — Изв. АН Туркм. ССР,
1959, № 6.
269. Ч и ж к о в Л. В. Характеристика гибридов столового арбуза и их родительских
форм на богаре Узбекистана. Автореф. дис. на соиск. учен, степени канд. с.-х.
наук. Ташкент, 1970.
270. Шаханов Е. Гетерозисный эффект гибридов от различных комбинаций скре-
щивания сортов дыни. Автореф. дис. на соиск. учен, степени каид. биол. наук.
Алма-Ата, 1972.
271. Ш вор нева А. М. Токсический бактериоз арбуза. — Науч. тр. Быковской опыт,
ст., 1969, вып. 5.
272. Шелепина Г. А., Фурса Т. Б. Изучение хромосомных наборов арбузов. —
Докл. ВАСХНИЛ, 1971, № 12.
273. Шелепина Г. А. Кариологическое исследование семейства Cucurbitaceae Juss.
(Cucumis L., Citrullus Schrad, Praecitrullus Pang.). Автореф. дне. на соиск. учен,
степени канд. биол. наук. Л., 1973.
274. Шейяфельд С. В. Использование эффекта гетерозиса у столового арбуза на
основе выведенных цельнолистных форм в условиях Молдавии. Автореф. дис. на
соиск. учен, степени каид. с.-х. наук. Кишинев, 1973.
275. Шембель С. Ю. Грибные болезни арбузов, дынь, огурцов и тыкв и меры борь-
бы с ними. Астрахань, 1915.
276. Щербаков В. И. Влияние минеральных удобрений на урожай и качество сто-
лового арбуза в орошаемых и неорошаемых условиях южной степи Украины.
Автореф. дис. на соиск. учен, степени канд. с.-х. наук. Симферополь, 1966.
277. Щукина А. С. О перекрестном опылении у среднеазиатских дынь. — В кн.: Ми-
ровые растительные ресурсы в Средней Азии. Ташкент, 1968.
278. Юзепчук С. В. Тезисы по проблемным вопросам о виде у культурных расте-
ний и о принципах в систематике. — Бот. журн., 1948, т. 33, № 1.
279. Юрина О. В. Селекция и семеноводство тыквенных культур. М., 1966.
280. Alef eld F. Landwirtschaftliche Flora. Berlin, 1886.
136
281. Andrus C. F. Spevial types of watermelon. USD. A. Crops Research Division.—
Agr. Research Service, 1964.
282. В ai ley L. H. Three discussions in Cucurbitaceae. — Cent. Herb., 1930, v. 2, fasc. 4.
283. Beattie F. H., Doolittle S. P. Watermelons. — Depart, of agricult. Farmers'
bulletin, 1953, N 1394.
284. Bhatia J. S., Grewal S. S., Sukhija P. S. Polar lipids of watermelon (Cit-
rullus vulgaris) and squash-melon (Citrullus vulgaris, var. fistulosus) seeds. —
J. Sci. Food Agr., 1971, v. 22.
285. Cable С. C. Arisona producers provide variaty of melons. — Progr. Agr. in Ari-
sona, 1973, v. 25, N 2.
286. Chakravarty H. L. Monograph of Indian Cucurbitaceae (Taxonomy and distri-
bution). — Rec. Bot Sue, India, 1959, v. 17, N 1,
287. Chambliss 0. Z., Erickson H. T., Jones С. M. Genetic control of bitter-
ness in watermelon fruits. — Proc. Amer. Soc. Hort, Sci., 1968, v. 93.
288. Chopra V. Z., Swaminathan F. A. Induction of polyploidy in watermelon. —
Proc. India Acad. sci. 1960, v. 51, sect B, N 2.
289. Cogniaux A. Cucurbitacees. — De Candolle Monographiae Phanerogamarum,
1881, t. 3.
290. Cogniaux A., Harms H. — Pflanzenreich, 1924, v. 4, H. 88.
291. Ch r al 1 J. M. History and present status of watermelon improvement by breeding.
— Proc. Soil. Sci. Fla., 1953, v. 13.
292. C h г a 1 1 J. M. Physiologia specialization in Fusarium oxysporum v. niveum. -
Phytopathology, 1963, N. 1.
293. Din ter K. Deutsch Sudwest Africa. Leipzig, 1909.
294. Dumas R. de Vaulx. Mise au point d’une me’thode de denom brement d
chromosomes chez le melon. (Cucumis melo L.). — Ann. Amelior Plant.. 1970, v. 2<
N 3.
295. Dunnill P., Fowden L. The aminoacids of seeds of the Cucurbitaceae. —
Phytochemistry, 1965, v. 4.
296. D u 11 a S. K-, Hale С. V., H a j ne E. G. Observations on the physiological Races
of Colletotrichum lagenarium. — Botan. Gaz., 1960, v. 21, N 3.
297. E 1 m s t г о m G. W., Hopkins D. Z, Variable suseptibility to bacterial rind ne-
crosis in watermelon. — Hort. Sci., 1973, v. 8, N 1.
298. Emonson K. Watermelon production and marketing in Victoria. —Veget. Growers
Dig., 1971, N 30.
299. Epenhuijsen C. W. Growung nafive vegetables in Nigeria. — Roma. FAO, 1974.
300. F i с к e 1 J. F. Ueber die Anatomic una Entwicklungsgeschichte der Samenschale
einiger Cucurbitaceaen. — Botan. Zeitung, 1876, v. 34, N 47, 48, 49.
301. Fosberg F. R. Proposal to conserve the generic name Citrullus Schrader. Pro-
posal no 8 submitted to the Paris congress. — Taxon, 1953, v. 2. N. 4.
302. Furusato K., Miyazawa A. Crossing exsperiments with Citrullus vulgaris
Schrad and Citrullus colocynthis Schrad. — Natl. Inst. Genetics (Japan) Ahn. Rpt.,
1957, v. 7.
303. Georgi I. G. Geographisch—physikalische und Naturhistorische Beschreibung des
Russischen Reichs zur Uebersicht bischeriger Kenntnisse von demselben. Konigsberg,
1800, t. 3, Bd. 5.
304. Goode M. G. Physiological specialisation in Colletotrichum lagenarium. — Phyto-
pathology, 1958, v. 48, N 2.
305. Hall C. J., Dutt a S. K. Inheritance of resistance to the fungus Colletotrichum
lagenarium (Pass.) Ell. and Halst. in watermelons. — Proc. Amer. Soc. Hort. Sci.,
1960, v. 75.
306. Hall C. Producing watermelons in Kansas. — Kansas state Univ, agric., 1970, Bull.
532.
307. H a d w i g e г L. A., H a 11 С. V. A biochemical study of the hostparasite relationship
between Colletotrichum lagenarium and cucurbit hosts. — Proc. Amer. Soc. Hort.
Sci., 1963, v. 82.
308. Нага H. The correct autor’s name of Citrullus lanatis (Cucurbitaceae). — Taxon,
1969, v. 18, N 3.
309. Harz C. D. Landwirtsvhaftliche Samenkunge. Berlin, 1885.
310, H e n d e r s о n W. R., J e n к i n s S. F., R a w 1 i n g s I. O. The inheritance of Fu-
’ sarium wilt resistance in watermelon, Citrullus lanatus (Thunb.) Mansf. — J. Amer
Soc. Hort. Sci., 1970, v. 95.
311. Hooker J. D. Cucurbitaceae. — In G. Benthama et J. Hooker. Genera plantarum.
London, 1867, v. 1.
312. Hutchinson J. The genera of flowering plants. Oxford. 1967.
313. Inch A. et al. Wilt resistant watermelons. — Queensland, agr. J.. 1972, v 98.
N 7.
314. Ito P., Atubra O. Watermelon culture in Ghana. — Chana Farmer, 1972. \ 16
N 2.
137
315. Jeffrey C. Notes on Cucurbitaceae including a proposed new classification of the
Family.—Kew Bull., 1962, v. 15, N 3.
316. Jeffrey C. A note on pollen morphology in Cucurbitaceae.—Kew. Bull., 1964,
v. 17, N 3.
317. Jeffrey C. On the Classification of the Cucurbitaceae.—Kew. Bull., 1966, v. 20,
N 3.
318. Jeffrey C. Systematic categories for cultivate plants. — Taxon, 1968, v. 17.
319. Jenkins S. F., Winstead N. N., McCombs C. L. Pathogenic compari-
son of three new and four previously described races of Glomerella angulata, var.
orbiculare. — Plant Dis. Reptr., 1964, v. 48.
320. Khoshoo T. H. Cytotaxonomy of Indian species of Citrullus. — Current. Sci.,
1955, v. 24.
321. Khoshoo T. N., V i j S. P. Biosystematics of Citrullus vulgaris var. fistulosus,—
Caryologia, 1963, v. 16, N 2.
322. Kihara H. Triploid watermelons. — Proc. Amer. Soc. Hort. Sci., 1951, v. 58.
323. Kirkwood J. C. The comparative embryology of the Cucurbitaceae. — Bull. New
York. Bot Gard., 1905, N 3.
324. Kondo N. Studies on the Triploid watermelon. — Inst. Breed Res., Tokyo, Agric.
Univ., 1961, N 1.
325. Kuroiwa T., Tanaka N. DNA replication pattern in somatic chromosomes of
Crepis capillaris. — Cytologia, Tokyo, 1970, v. 35.
326. Layton D. V. The parasitism of Colletotrichum lagenarium (Pass.) Ell and
Kalst. — Res. Bull. Ja. Agric. Exp. Sta., 1937, v. 233.
327. Levan A., Fredga K., Sandberg A. Nomenclature for centromeric position
on chromosomes. — Hereditas, 1964, v. 52, N 1.
328. Linneus C. Species plantarum. Stockholm, 1753.
329. McKay J. W. Chromosome numbers in the Cucurbitaceae. — Bot. Gaz., 1930,
v. 89, N 4.
330. McKay J. W. Chromosome studies in the Cucurbitaceae. — Univ. Calif. Publ. Bot.,
1931, v. 16, N 9.
331. Mansfeld R. Vorlaufiges verzeichnis landwirtschaftlich und gartnerisch kultivier-
ter. pflanzenarten. — Die Kulturpflanze, Berlin, 1959, Beih. 2.
332. M a r 1 о t h R. Das Kapland. Pretoria, 1908, Bd. 2, T. 3.
333. Marficorena C. Material para una monografia de la morfologia del polen de
Cucurbitaceae.—Grana Palynologica, 1963, v. 4, N 1.
334. Me euse A. D. Cucurbitaceae of the Southern Africa.—Bothalia, 1962, N 8, part. 1.
335. M о h r H. C. A mutant leaf form in watermelon. — Proc. Assn. So. Agr. Workers,
1953, v. 50.
336. Mohr H. C., Blackhurst H. T., Jensen E. R. Fi hybrid Watermelons from
openpollinatet seed by use of a genetic market.—Proc. Amer. Soc. Hort. Sci., 1955,
v. 65.
337. Muller E., Pax F. Cucurbitaceae. — Jn A. Engler und K. Prantl. Die naturli-
chen Pflanzenfamilien, Leipzig, 1894, Bd. 4, Arb. 5.
338. N a i t h a n i S. P., Das P. Somatic chromosome numbers in some cultivated Cucur-
bits. — Current. Set, 1947, v. 16, N 6.
339. Nath P., Dutt а О. P. Heterosis in watermelon. — Jndian J. Hortic., 1970, v. 27,
N 3/4.
340. N a u d i n Ch. Revue des cucurbitaceaes cultivees an museum. — Ann. Sci. Nat.
Bot., 1859, t. 12.
341. Netolitzky F. R. Anatomie der Angiospermen—Samen.—Handbuch der Pflan-
zenanatomie, Berlin, 1926, v. 10.
342. О к e O. Z. Nutritive value of Nigerian watermelon. — Nature, 1965, v. 207, N 4993.
343. Poole C. F. Genetics of cultivated cucurbits. — J. Hered., 1944, v. 35.
344. Poole C. F., C i r i m b a 11 P. C. Interaction of sex, shape and weight genes in
watermelon. — J. Agr. Res., 1945, v. 71.
345. Porter D. R. Inheritance of certain fruit and seed characters in watermelons.—
Higardia, 1937, v. 10.
346 Porter D. R. Breeding high—quality wilt resistant watermelons. — Bull. Calif.
’ Agric. Exp. Sta., 1937, N 614.
347. Porter D. R. Today’s watermelon varieties how they were developed. — Am.
Veget. Grower, 1969, v. 17, N 6.
348. P ro v vi den ti R., Schroeder T. Epiphytotic of watermelon mosaic among
Cucurbitaceae in central New York in 1969. — Plant Dis. Report., 1970, v. 54, N 9.
349. Puri V. Studies in floral anatomy. VII. On placentation in the Cucurbitaceae.—
Phytomorphology, 1954, v. 4, N 3.
350. Rahe F. Growing watermelons in the North states and classification of waterme-
lons.— New Hampsh. Coll. Agr. Exp. Sta. Bull., 1901, v. 86.
351. Raicu P., Anghel J. Doua linii noi autotetraploide de pepene verde (Citrullus
vulgare). — Gradina via livada, 1966, An. 15, N 10.
138
qco R e h m S., E n s 1 i n P. К., M e e u s e A. D. J., W e s s e 1 n J. H, Bitter principles
° ’ of the Cucurbitaceae. — J. Sci., Food Agrig., 1957, v. 8, N 12.
353. Robinson R. W., Munger H. M., Whitaker T. W., Bohn G. W. Genes
of the Cucurbitaceae. — Hort Sci., 1976, v. 11, N 6.
354. Rosa J. T. The inheritance of flower types in Cucumis and citrullus. — Hilgardia,
1928, v. 3, N 9.
355. Schrader H. A. Cucurbitaceae. Linnaea, 1938, Bd. 12.
356 Schenck N. C. Resistance of commercial watermelon varieties to Fusarium wilt.
’ — Proc. Fla State Hort. Soc., 1961, v. 74.
357. Schweinfurt G. Monatschrift des Vereines zur Beforderung des Gartenbauen,
Leipzig, 1876.
358. Schweinfurth G. Ueber Pflanzenreste aus altaegyptischen Grabern. — Ber.
Deutsch. Bot. Gesellsch, 1884, Bd. 2, Heft 7.
359. Shimotsuma M. Chromosome numbers of Citrullus species. — Chrom. Into
Serv., 1961, N 2.
360. Shimotsuma M. Studies on Triploid seed production in Watermelons. — Report
Kihara Inst. Biol. Res. Jokohama, 1961, v. 14.
361, Shimotsuma M. Cytogenetic and Evolutionary Studies in the genus Citrullus.
— Report Kihara Inst. Biol. Res., 1963, v. 15.
362. Singh B. Studies on the structure and development of seeds of Cucurbitaceae. —
Phytomorphology, 1953, v. 3, N 3.
363. S i 11 e r 1 у W. Breeding for disease resistance in cucurbits. — Ann. Rev. Phyto-
pathol.., 1972, v. 10.
364. Sparague G. F., Tatum Z. A. General vs. specific combining ability in single
crosses of corn.— J. Amer. Soc. Agr., 1942, v. 34, N 10.
365. Stuhlman F. Beitrage zur Kulturgeschichte von Ost-Africa. — Deutsch Ost-
Africa, Berlin, 1909, Bd. 10.
366. T r i v e d i R. N., Roy R. P. Cytological Studies in Cucumis and Citrullus. —
Cytologia, 1970, v. 35, N 4.
367. Vaughan V. C., Whitehouse J. M. Seed structure and the taxonomy of the
Cruciferas. — Bot. J. Linn Soc. 1971, v. 64, N 4.
368. V о s a C. G., M a r c h i P. Quenacrine fluorescence and Giemsa statining in
plants. — Nature (Lond.), New Biol., 1972, N 237.
369. Wall J. R. Use of marker gene in producing triploidi watermelons. — Proc.
Amer. Soc. Hort Sci., 1960, v. 76.
370 Watermelons for the garden. Washington, U. S., Dep. Agr. Leaflet, 1966,
’ N 528.
371. Watts V. M. A market male sterile mutant in Watermelon. — Proc. Am. Soc.
Hort. Sci., 1962, v. 81.
372. Weetman L. M. Inheritance and correlation of Shape size, and color in the
watermelom Citrullus vulgaris Schrad.—Agr. Sta. Res. Bui., Jowa, 1937, N 228.
373. Whitaker T. W. Chromosome numbers in the cultivated Cucurbits. — Amer.
Jour. Bot., 1930, v. 18.
374. Whitaker T. W. Cytological and phylogenetic studies in the Cucurbitaceae. —
Bot. Gaz. 1933, v. 94, N 4.
375. Whitaker T. W., Davis G. N. Cucurbits. Botany. Cultivation and Utilization.
New York, 1962.
376. Winstead N. N., G oo d e M. G., Barham W. S. Resistance in watermelon to
Colletotrichum lagenarium. Rases 1, 2 and 3. —Plant. Dis. Reptr., 1959, v. 43.
377. Woe nig F. Die Pflanzen im alten Aeqypten. Leipzig, 1886.
378. Zimmerman A. Die Cucurbitaceae Beitrage zu Anatomie, Phisiologie, Morpholo-
gic, Biologie, Pathologie und Systematic Jena, 1922, H. 2.
Характеристика районированных сортов арбуза
Столовый арбуз
ПРИЛОЖЕНИЕ
Сорт Сортотнп Морфологические признаки Хозяйственно-биологические признаки и свойства
плода мякоти семян период вегетации устойчивость к болезням лежкость и транспорта- бельность другие признаки и свойства
масса, кг форма окраска окраска консистенция размер и окраска
Астрахан- 25 Крупный Тупоэллип- Зеленый, с Ярко- Нежная Средние, Средне- Устойчив Транспор- Отзывчив
ский (5—6) тическая темно-зеле- ными шипо- ватыми по- лосами красная сладкая коричневые, крапчатые спелый к антрак- нозу табельный на оро- шение
Багаевский мурашка 747/749 5 Средний (3-4) Шаровид- ная Светло-зеле- ный, с сет- кой Ярко- розовая Нежная Средние, кремовые с черным ободком > — Лежкий, транспор- табельный —
Белый длинный (Амери- канский белый) 6 Очень круп- ный (7—10' Цилиндри- ческая Зеленовато- белый Малино- вая Плотная Средние, белые Поздне- спелый Транспор- табельный Засухо- устойчив
Бирючекут- ский 775 12 Средний (4-5) Шаровид- ная Светло-зеле- ный, с узки- ми темно-зе- леными по- лосами Густо- розовая Нежная Мелкие, черные Средне- поздний Лежкий, транспор- табельный
Быков- ский 22 12 То же > То же Малино- вая Зернистая Средние, светло-ко- ричневые » То же
Быков- ский 199 20 Крупный (5-6) > Зеленый, с сеткой Красная Мелкозер- нистая Крупные, красные Средне- спелый — Транспор- табельный —
Вахшский темноко- рый Волжский 7 30 Средний (3-4) > Черно-зеленый без рисунка Малино- вая Зернистая Средние, темно-ко- ричневые Средне- ранний —
1 Крупный (5-6) > Белый, без рисунка Розовая Волокни- стая Средние, черные Средне- поздний Лежкий, транспор- табельный
Десерт- ный 83 Зимний 344 30 5 Средний (3—5) Крупный (5-6) Тупоэллип- тическая Черно-зеле- ный, со скрытым рисунком Светло-зеле- ный, с сет- кой Малино- вая Розовая
Комсомоль- ский 2 28 Средний (3-4) Сплюснуто- шаровид- ная Зеленый, с темно-зеле- ными раз- мытыми по- лосами »
Король Кубы 92 12 Средний (3-4) Шаровид- ная Светло-зеле- ный, с тем- но-зелеными шиповатыми полосами »
Кузыбай 30 29 Очень круп- ный (8-10) Темно-зеле- ный, с чер- но-зелеными размытыми полосами
Любимец Красно- дара 18 Средний (2—4) > Светло-зеле- ный, с тем- но-зелеными широкими полосами »
Любимец хутора Пятигор- ска 286 24 Средник! (3-5) Темно-зеле- ный, с чер- но-зелеными шиповатыми полосами »
Мелитополь- ский 60 25 Крупный (5-7) Тупоэллип тическая Зеленый, с черно-зеле- ными шипо- ватыми по- лосами Малино- вая
Зернистая, очень сладкая Крупные, коричневые, крапчатые Средне- спелый — Для мест- ного упот- ребления —
Слабово- локнистая Средние, светло-ко- ричневые с темными пятнами Поздне- спелый — Очень лежкий, транспор- табель- ный —
Зернистая Крупные, кре- мовые с темным ободком и носиком Средне- ранний
Нежная Мелкие, черные, с трещинами Средне- спелый
Волокни- Крупные, бе- Поздне- Относи- Лежкий, Отзывчив
стая лые с тем- ным носи- ком спелый тельно устойчив к мучни- стой росе и фуза- риозу транспор- табельный на оро- шение
Зернистая Средние, светло-ко- ричневые Средне- ранний
Средние, кре- мовые с темным ободком и носиком Средне- спелый Относи- тельно устойчив к антрак- нозу — —
» Средние, красные » Лежкий, транспор- табельный Засухо- устойчив
Сорт Сортотип Морфологические признаки
плода мякоти
масса, кг форма окраска окраска консистенция
Мелитополь- ский 142 25 Крупный (5-7) Тупоэллип- тическая Зеленый, с черно-зеле- ными шипо- ватыми по- лосами Малино- вая Зернистая
Мозаичный местный 14 Средний (4-5) Шаровид- ная Светло-зеле- ный, с зеле- ным мо- заичным рисунком Розовая Мелкозер- нистая
Мраморный 9 То же » Светло-зеле- ный, с тем- но-зелеными мазками Малино- вая Нежная
Муравлев- ский 28 Средний (3-4) Зеленый, с черно-зеле- ными размытыми полосами Розовая Волокни- стая
Мухианский местный 16 То же » Светло-зеле- ный с тем- но-зелеными размытыми полосами Бледно- розовая Зернистая
Огонек 30 Мелкий (2-3) > Черно-зеле- ный, со скрытым рисунком Ярко- крас- ная Нежная
Победитель 395 29 То же > Темно-зеле- ный, с чер- но-зелеными полосами Малино- вая Зернистая
Продолжение
Хозяйственно-биологические признаки и свойства
семян размер и окраска период вегетации устойчивость к болезням лежкость и транспорта- бельность другие признаки и свойства
Средние, красные Средние, черные Мелкие, красные Крупные, коричневые, крапчатые Средние, светло-жел- тые с ко- ричневым ободком Мелкие, светло-ко- ричневые Мелкие, желтовато- серые Средне- спелый Скороспе- лый Средне- поздний Средне- спелый » Средне- ранний Скоро- спелый Относи- Транспор- — тельно 1 табельный устойчив к фуза- риозу — 1 Для мест- Засухо- 1 ногоупот- устойчив 1 ребления — 1 Лежкий 1 — Относи- 1 Транспор- — тельно 1 табельный устойчив 1 1 к фуза- 1 1 риозу 1 — 1 * 1 — _ Дружное созрева- ние
Подарок 28 Средний Тупоэллип- Зеленый, с Красная Слабо-
Холодова (4-5) тическая темно-зеле- ными раз- мытыми полосами волок- нистая
Ранний 29 Средний (3-4) Шаровид- ная Темно-зеленый с черно-зе- леными по- лосами > Зернистая
Роза Юго- Востока 18 Крупный (4-6) » Светло-зеле- ный, с темно-зеле- ными широ- кими поло- сами Ярко- крас- ная >
Самарканд- ский белый 1 Крупный (6-7) > Светло-зеле- ный Розовая Плотная
Спутник 29 То же Тупоэллип- тическая Темно-зеле- ный, с узки- ми шипова- тыми поло- сами Густо- розо- вая Волокни- стая
Стокса 647/649 28 Мелкий (1-2) Шаровид- ная Темно-зеле- ный^ с ши- рокими размытыми полосами Шарла- ховая Нежная
Стокса Киевский 27 Средний (2-4) » Темно-зеле- ный, с туск- ло-зелеными полосами >
Сухумис 22 Крупный (6-7) Тупоэллип- тическая Темно-зеле- ный, без рисунка Красная Мелкозер- нистая
Узбекский 452 ft 12 Крупный (4-7) Шаровид- ная Светло-зеле- ный, с зеле- ными шипова- тыми полосами Розовая Зернистая
Мелкие,
коричневые
Поздне-
спелый
Лежкий,
транспор-
табельный
Средние,
кремовые
Средне-
спелый
Крупные,
светло-ко-
ричневые
Скоро-
спелый
Транспор-
табельный
Мелкие Крупные, кремовые Средне- спелый Средне- поздний — Лежкий Для бога- ры Засухо- устойчив
Мелкие, желто-ко- ричневые Скоро- спелый — Для мест- ного упот- ребления —
То же > Относи- тельно устойчив к антрак- То же —
Мелкие, Средне- нозу Относи- — Отзывчив
светло-ко- ричневые спелый тельно устойчив к фуза- риозу Транспор- на оро- шение
Средние, кре- мовые с темным носиком Средне- поздний табельный
Продолжение
£ с Морфологические признаки Хозяйственно-биологические признаки и свойства
1 плодя мякоти семян период вегетации устойчивость к болезням лежкость и другие признаки и свойства
Сорт о О масса, кг форма окраска окраска консистенция размер и окраска транспорта- бельность
Хант-кара Цельнолист- ный Черносемян- ный Юбилей- ный 72 30 16 29 12 Очень крупный (6—10) Средний (3-4) Мелкий (2-3) Крупный (5-6) Шаровид- ная > » Черно-зеле- ный, со скрытым рисунком Светло-зеле- ный, с зеле- ными раз- мытыми по- лосами Темно-зеле- ный, с раз- мытыми полосами Светло-зеле- ный, с тем- но-зелеными шиповатыми полосами Розовая Густо- розо- вая Розовая Ко Волокни- стая Мелкозер- нистая Крупнозер- нистая Зернистая р м о в о й а Крупные, белые Мелкие, темно-ко- ричневые, крапчатые Средние, черные Средние, бе- лые с тем- ным носи- ком । р б у 3 Поздне- спелый Скоро- спелый Средне- спелый Средне- поздний Относи- тельно устойчив к фуза- риозу Очень лежкий и тран- спорта- бельный Отзывчив на оро- шение
Морфологические признаки
Сорт плода мякоти семян Засухоустойчивость Использование
масса, кг | форма | окраска
Богарный 112 Бродский 37-42 Дисхим Пектинный Крупный (7-8) Средний (5-6) Крупный (8-10) Крупный (6-7) Цилиндрическая Шар Цилиндрическая Белесый, со скрытым рисунком Светло-зеленый, с шиповатыми полосами Белесый, со скры- тым рисунком Светло-зеленый, со скрытым рисунком Бледно-желтая, плотная Белая, плотная Желтоватая, плотная Белая, плотная Средние оливковые Средние, ярко-зеле- ные Средние, оливковые Средние, зеленые Засухоустойчив » » > Для возделыва- ния на богаре, на корм На корм На корм, в цу- катном про- изводстве Сырье для полу- чения пектина
ТЫКВА— CUCURBITA L.
Cucurbita L., 1753, Sp. Pl.: 1010; idem, 1754, Gen. Pl. ed. 5:441,—
Pepo Adans., 1763, Fam. 2:138. — Ozodykus Ratin., 1832, Atlant.
Journ. : 145.— Sphenantha Schrad., 1838, Linnaea, 12:416.— Pyleocaeygs
Casp., 1847, Rend. Acad. Sci. Napoli, 6:409. — Tristemon Scheele, 1848,
Linnaea, 21 : 586.
Typus: Cucurbita pepo L.
ОБЗОР РОДА
Род тыквы относится к семейству Cucurbitaceae, трибе Cucurbiteae,
субтрибе Cucurbitinae-, последняя отличается от родственной Сисите-
rinae колокольчатым венчиком и происхождением.
Описание рода. Травянистые длинноплетистые (лиановые) однолет-
ние или многолетние (в тропиках) растения. Стебель и боковые побеги
стелющиеся или лазящие, сравнительно тонкие, длинные, чаще ребрис-
тые, отчетливо или неясно 5-гранные. Междоузлия большей частью
удлиненные. Листья очередные, цельные, крупные, чаще с надрезной
пластинкой, от округлой до 5-угольной и 3-угольной формы, зеленые,
иногда с белой пятнистостью в углах жилкования. Усики пазушные,
многораздельные, редко они отсутствуют. Все надземные органы сильно
опушены. Корневая система простратная, боковые корни несколько
длиннее побегов стебля, некоторые (тропические) дикие виды со зна-
чительными корневыми утолщениями, обеспечивающими им многолет-
ний образ жизни.
Цветки однополые (иногда с недоразвитым андроцеем или гине-
цеем), одиночные, реже (тычиночные) в пучках, пазушные, крупные,
желтые; чашечка колокольчатая, чашелистиков 5 (4—7). Венчик коло-
кольчатый, 5(4—7)-лопастный до раздельного, лепестки на верхушке
отогнутые наружу, чаще с бахромчатыми краями, реже прямые. Тычи-
нок 5, они сросшиеся. Пыльники, спирально закрученные и сросшиеся
в центральную колонку. У пестичных цветков столбик желобчатый,
рыльце 3—5-раздельное (по числу семенных камер плода), ярко-оран-
жевое. Стаминодии треугольные, маленькие, в количестве трех, завязь
3—5-гнездная, нижняя, от сплюснутой до удлиненной.
Плод мясистый, обычно крупная тыквина различной формы и
окраски, с пятнисто-полосатым рисунком или без него. Кора деревяни-
стая или мягкая. Мякоть белая, кремовая или желтая. У большинства
форм плод имеет полость, в которой заключены плаценты с семенами.
Семена многочисленные, овально-сплюснутые, чаще с боковым рубчи-
ком, белые, серые, желтые или черные (редко).
Отличительный признак рода — очень крупные плоды у культурных
форм (доходящие до самых крупных размеров в растительном мире)
и твердая деревянистая кора шаровидных или яйцевидных мелких пло-
дов у всех диких видов.
145
Географическое распространение. Род тыквы представляют как.
культурные формы, так и дикие. Культурные тыквы распространены
всюду, где развито земледелие, за исключением северных районов, ле-
жащих за пределами 62—63° с. ш. Дикие виды и формы тыкв распро-
странены в Центральной и Южной Америке, на различных плато ска-
листых гор и на территории современных штатов Калифорнии, Техаса
и Флориды (США). Ряд декоративных мелкоплодных форм гибрид-
ного происхождения разводится преимущественно в ботанических садах
всех континентов.
В нашей стране в культуре тыквы наиболее широко распространены
на Украине, Северном Кавказе, в Нижнем Поволжье, в Среднеазиат-
ских и Закавказских республиках, где используются как в пищу, так
и на корм скоту.
Широко возделывают тыквы также в Западной Европе: Болгарии,
Румынии, Югославии, Италии и Испании; в азиатских странах: Китае,
Японии, Индии; в странах Индокитайского полуострова.
В Америке наиболее интенсивно возделывают тыкву в Мексике,
Перу, Чили и Бразилии. Она является большим подспорьем в питании
местного индийского населения.
В Африке тыква распространена в меньшей степени, но выращи-
вают ее почти во всех земледельческих районах континента.
Названия тыквы на языках народов СССР и иностранные. В назва-
ниях тыквы на разных языках выделяются 6 основных корней. Первый
корень «тыква» происходит от древнегреческого сикуа. Этот термин обо-
значал в древнем мире не тыкву, а кормовой арбуз, в каком виде и
сохранился за арбузом в китайском (сигуа) языке и языках, соседних
с Китаем стран. Корень «тыква» вошел в употребление в русском, поль-
ском, болгарском и испанском языках.
Второй корень «кабак» употребляется у турок, татар и в языках
соседствующих с ними в прошлом народов: турецкий — кабак; татар-
ский— кавак; азербайджанский — гобак; корейский — хобак; узбек-
ский— кавак, ошкавак; японский — кабоуя; казахский — оскабак; пор-
тугальский— кабака; киргизский — ошкабак; тибетский — кабед.
Третий корень тыквы «каду» распространен среди языков народов
Средней Азии: туркменский — кэду, таджикский — каду, новоперсид-
ский — каду.
Четвертый корень «гуа» распространен в Китае и соседних с ним
странах: китайский — гуа (С. maxima), монгольский — гуа, маньчжур-
ский — лангу.
Однако в языках некоторых народов существуют и другие назва-
ния тыквы, заимствованные от соседних стран, например в азербайд-
жанском (куду), тибетском (гун, куна, гон).
Пятый корень «кукурбита» происходит от латинского названия^
обозначавшего сосуд, точнее древнее название плода горлянки. От
этого корня произошло название тыквы в немецком (курбис) и во
французском (курге) языках.
Шестой корень «пепо» заимствован с древнегреческого. Им обозна-
чался зрелый плод в отличие от аггур (недозрелый). Это название, по
всей вероятности, также указывало на кормовой арбуз. Данный корень
перешел позднее и в латинский, а затем в английский (пампкин), гол-
ландский (помпьен) и шведский (пумпа) языки.
Остальные названия тыквы носят местный характер языка только
своей страны: арабский — яктын; итальянский — цукка; маратский (Ин-
дия)— бхонали; румынский — бостан, домвак; калмыкский — хан, дурем;
молдавский — бостан, тэбак; тайский — нам, май; датский — грескар;
146
индонезийский — лабу; суахили (африканский)—танго, бога; армян-
ский— дьдум; зулу (африканский) — утанга; грузинский — гогра; амхар-
ский (Абиссиния) — дуббаа.
Украинское и белорусское название тыквы гарбуз произошло от
корня «арбуз».
Происхождение. Греческая и римская литература указывала на
происхождение тыкв из Старого Света. Это мнение настолько популярно
в литературе, несмотря на отсутствие доказательств, что заставляет
снова исследовать этот вопрос и взвесить реальность факторов, гово-
рящих в его пользу.
Греческие и римские древние писатели по интерпретациям совре-
менных переводчиков очень много писали о тыкве, называя ее Реро,
Cucurbita и Coloquint. О ней говорят авторы эпохи Возрождения. Из
древних греческих и римских авторов тыкву упоминает в своем сочи-
нении «О лекарственных средствах» древнегреческий врач I в. н. э.
Диоскорид. Древнеримский писатель той же эпохи Плиний говорит о
тыкве как о съедобном растении.
Греческий писатель III в. н. э. Атеней приводит много упомина-
ний о тыкве более древних и современных ему авторов. Так, по его
словам, Лашес пишет, что «он раздувается и толстеет, как тыква на
хорошо увлажненной земле». Согласно ученику Аристотеля Фанаису,
sykuos и рероп ели целиком еще в очень молодом возрасте, без семян.
Когда же плоды созревали, то их употребляли в вареном и жареном
виде.
Диокл говорит об употреблении дикой рероп, которая очень хо-
рошо переваривается, но сваренная в вине слабит. Дифил обращает
внимание на очень хороший сок рероп, который притупляет плохие вку-
совые ощущения, и на малопитательный и быстро переваривающийся
сок sykuos. Проперций по прибытии в Рим упоминает о зеленоватом
огурце, огромной тыкве и капусте.
О тыкве упоминают и другие греческие и римские авторы. Писа-
тель Матрон говорит об удивительной тыкве (potiron), занимавшей
9 столов. Скорее всего, речь шла здесь о существовавшей уже в то
время тарре.
Французский переводчик Атенея Лефебур де Виллебрюн заме-
чает, что он не уверен в точности перевода древних наименований тык-
венных растений. А. Декандоль [16, 119] считает невозможным уста-
новить виды тыквы по обозначающим их специальным терминам в
Древней Греции и Риме, так как ботанического описания растений
древние авторы нигде не дают.
Этимологическое изучение этих терминов также не вносит ясно-
сти. Слово рероп означает зрелый плод в противовес agguria (незре-
лый), coloqint — колоссальный, но употребление этого слова Диоклом
для обозначения дикого растения заставляет предполагать не крупные,
а, наоборот, мелкие плоды, которые и закрепились впоследствии за
этим понятием во французском языке. Cucurbita по этимологическому
словарю означает банка, что указывает на родство этого слова с лаге-
нарией. Однако В. Ген [146] склонен воспроизводить это слово от сог-
bis — голова, хотя оно означает и корзину.
Слово siqua, понимаемое большинством авторов как огуречное
растение, в греко-русском словаре обозначается как тыква.
Как видно из изложенного, древняя греческая и римская терми-
нология не дает ответа на то, существовала ли в то время настоящая
тыква. Все эти термины могли относиться как к лагенарии, так и к
разным другим родам тыквенных. Кроме существования в Старом
147
Свете лагенарии, в ту эпоху, по нашим данным, туда широко прони-
кали из Африки арбуз в виде некультурных или малокультурных
форм, полукультурная змеевидная дыня, а также другие роды: момор-
дика, люффа, бенинказа, трихозанта, к которым могли быть применимы
древние греческие и римские термины. Такое положение подтверждает
также отсутствие тыквы при перечислении других родов тыквенных
в работах о древней ботанике и отсутствие настоящей тыквы при рас-
копках древних цивилизаций Старого Света. Наличие ее в то время
должно было найти отражение в памятниках средневековья, предшест-
вующих открытию Америки.
В работе Е. Мейера [159] по истории средневековой ботаники
тыква (Cucurbita) упоминается: в списке растений, рекомендуемых
Цельсием(I в. н. э.), в списке Апициуса (III—IV вв.), в списке Мар-
целлюса (V в.), в этимологии Исидора (VII—VIII вв.) и в списке
растений Набатейского земледелия (VIII—XIII вв.). Тыква упоми-
нается в книге о растениях Альберта Магнуса, в трактате «О расте-
ниях» Альберта Великого (XIII в.) и у других средневековых авто-
ров [89].
Однако описания этих тыкв также нигде не дается, хотя им часто
присваивается видовое название С. реро. Для определения некоторых
растений в королевских садах Карла Великого (VIII в.) Н. С. Цемш
[89] пользовался более ранними описаниями Страбона (I в. н. э.) из
его «Книги об обработке огородов». В этой работе тыква Cucurbita
является лагенарией, colocynth — диким арбузом, a pepones — дыней.
В средневековье арабский писатель Ибн-аль-Авама (XII в.) ука-
зывает, что тыква получается вкуснее и крупнее, если ее прививать на
лук или тростник вымоченными семенами. В «Большом сборнике о
свойствах лечебных и пищевых средств» Ибн-Байтара (XII в.) тыква
С. реро упоминается под названиями dubba и кага. Указывается, что
сырые плоды ее имеют противный вкус и вредны для желудка (могут
вызывать рвоту), однако в вареном виде они съедобны. Из сока плодов
этой тыквы особым способом приготовляют краску для волос. Однако
впоследствии К. Броун [109] разъясняет, что тыква кагга является
лагенарией и в молодом состоянии она вкусна, особенно с мясным
соусом.
Согласно Е. Бретшнейдеру [108], в работах по древней китайской
ботанике тыква не упоминается. В книге полумифического императора
Шеннунга (XXVIII в. до н. э.) указываются тыквенные растения: бенин-
каза, трихозанта, лагенария, тладианта, но тыквы нет. Позднее у Фанг-
шен-чи (I в. до н. э.), а также у Ки-хана (III в. н. э.) в первой чисто
ботанической работе «Описание флоры Южного Китая» тыквы также
нет. Описание ее отсутствует и у Киа-сун (V в. н. э.) в его «Важных
правилах для людей жить в мире» и у Кио-то-то (VII—VIII вв.) и у
Шу-Куанг-ки (XVI в.) в «Полном обзоре по сельскому хозяйству».
Впервые тыква упоминается в работе Ли-си-чена о китайской ме-
дицине в третьей четверти XVI в., которая была переведена на евро-
пейские языки. Этот период уже совпадает с временем распространения
некоторых американских растений в Старом Свете.
Таким образом, в средние века ни в европейской, ни в арабской,
ни в китайской литературе обоснованных указаний на наличие в куль-
туре настоящей тыквы не было. В средневековых же источниках нигде
достоверно не указывается на существование ее в диком виде. Тем
не менее авторитеты XIX в. считали родиной тыквы Старый Свет.
А. Декандоль [119, 16] допускает, что это растение было известно
древним грекам и римлянам. В американском происхождении тыкв он
148
сомневался, потому что во флоре этой страны тыква приводится как
культурное растение, а местных названий не дается и в диком виде
тыкв к этому времени в Америке не было обнаружено.
Вид С. maxima, по Декандолю, южноазиатского происхождения.
Исследователь придает большое значение упоминанию Линнея в «Ман-
тиссе» о том, что форма С. ovifera L., относящаяся к виду С. реро,
была им найдена в диком виде в районе Астрахани и потому этот вид
он также считает азиатским. Основываясь на рисунках и названиях
ботаников эпохи Возрождения, вид С. moshata он также считает про-
исходящим скорее из Индии, чем из Америки.
Впоследствии А. Декандоль [16] допускает возможность проис-
хождения одного вида С. реро из Америки после того, как там была
обнаружена другая дикорастущая тыква — С. texana Gray, очень по-
хожая на С. реро.
Ш. Ноден [163] также считает Америку наименее вероятной ро-
диной тыкв, хотя там и была обнаружена дикая тыква С. perennis
(С. foetidissima)-, но она настолько резко отличается от других видов,
что, по мнению Нодена, не может быть их прародителем. Кроме того,
он отмечает, что цветки С. perennis не посещаются насекомыми и опы-
ляются случайно. Он склоняется к точке зрения азиатского происхож-
дения тыкв.
Наконец, историк культурных растений конца XIX в. В. Ген [146],
тщательно анализируя высказывания древних писателей, также при-
ходит к выводу о наличии в Старом Свете культурных тыкв и допускает
американское происхождение только одного вида — С. реро. Книга
В. Гена послужила источником распространения данной точки зрения
среди многих последующих исследователей.
Мнения этих ученых базировались на многообразии форм тыкв
в Европе, на случайных указаниях о наличии этой культуры в Африке
и на сопутствующих ее названиях «индийская» (в конце средних веков
Америку еще отождествляли с Индией). Заметка Н. Е. Житеневой [20]
о нахождении дикорастущей тыквы в Северной Африке Н. И. Вавило-
вым оказалась ошибочной: это была не тыква, а арбуз с очень круп-
ными плоскими семенами, как у тыквы, его новая форма. Неверное
заключение Н. Е. Житеневой объясняется тем, что она сделала свой
вывод на основании внешнего вида только что привезенных Н. И. Ва-
виловым семян, не успев их высеять и ознакомиться с растениями.
Точка зрения крупных ученых не изменилась даже после того,
когда под Астраханью при повторных экспедициях не было обнару-
жено дикой тыквы, отсутствовали растения этого рода и в гербариях,
собранных до начала интродукции американской растительности, не
было также никаких признаков наличия тыкв при археологических рас-
копках и указаний первых путешественников о культуре тыкв у инду-
сов. Наконец, недооценивалась явная возможность смешения тыкв с
лагенарией и арбузом у греческих и римских писателей.
Мнение ученых по этому вопросу резко изменилось после работ
американских археологов, обнаруживших остатки почти всех видов тыкв
в захоронениях древних индейцев и после нахождения в Америке ди-
ких или отдаленных родичей всех видов культурных тыкв, распростра-
ненных в Старом Свете. Одна из первых археологических раскопок,
обнаружившая семена древней тыквы в Америке, была произведена
на древнеиндейском кладбище близ г. Лима в Перу; семена были
отнесены к видам С. moschata и С. maxima. Позднее там были най-
дены вазы, по форме напоминавшие плоды этих тыкв. Находки отно-
сились к доколумбийскому периоду. Археологические остатки плодоно-
149
жек, принадлежащие к С. moschata форме Sheese, тыквенных растений
примерно такой же давности были обнаружены в Гватемале. Фрагменты
стеблей н семян С. реро и С. tnoschata древностью более 1000 лет до
н. э. были обнаружены археологом Дэнада в Пуэбло в штате Коло-
радо (США). В том же штате Мартином в 1928 г. при раскопках об-
наружены семена С. moschata. Семена этого вида были найдены также
Кайдером на юго-западе США. Древность семян составляет 1500—
2000 лет до н. э.
Т. Вайтекер [210] считает, что тыквы весьма полно представлены
при археологических раскопках в виде плодоножек, кусков коры и се-
мян. Наиболее древняя (около 3000 лет до н. э.) находка в Южной
Америке (на побережье Перу) относится к остаткам С. moschata и
С. ficifolia. Такой же давности стебли, кора и семена С. реро были
найдены и в Северной Америке в местечке Тамауминас в Мексике.
Растительные остатки тыкв были найдены также археологом Во на
северо-востоке штата Аризона, археологом Харринатоном на юго-западе
штата Миссури, профессором Деллингетом на северо-западе Арканзаса
(США) и во многих других местах. По данным Т. Вайтекера [207],
к началу второй половины XX в. при археологических раскопках в Аме-
рике были обнаружены: С. реро — пять раз; С. moschata — четыре в
Северной Америке и один раз в Южной; С. maxima и С. ficifolia —
по одному разу в Южной Америке.
Археологические данные согласуются и с местными индейскими
названиями тыкв. Так, индейское название тыквы сквочь аналогично
названию одного из индейских племен. Другое название кушо (cushaw)
кувшиновидной тыквы вида С. moschata также заимствовано у ин-
дейцев.
В Америке же обнаружены и дикие родичи тыквы. Первой из них
была описана С. texana [А. Грей, 131], которая большинством авторов
относится к С. реро как ее дикий родич; затем дикий вид тыквы
С. andreana Haud., которую Л. Пароди первым идентифицировал с
С. maxima, считая ее дикой формой этого вида. Таким образом, оба
эти широко распространенные в Европе виды тыкв имеют своих диких
родичей в Америке.
Наконец, более примитивная форма (так называемая семинольская
тыква) была обнаружена и для вида С. moschata. Она имеет мелкие
(12 см в диаметр) шаровидные плоды и разводится индейским племе-
нем семинолов, живущих в кипарисовых болотах Флориды [127].
После этих неоспоримых доказательств необходимо вернуться к
Африке. Странное на первый взгляд столь быстрое освоение новой
культуры тыквы местным африканским населением, вероятно, связано
с интенсивной эмиграцией туда индийского населения. А Индия, по
свидетельству П. М. Жуковского [22], получила эту культуру из Аме-
рики через португальцев еще в 1500 г. Однако возможен завоз тыквы
португальцами и непосредственно в Африку. Еще А. Декандоль [119]
указывал на описание двух спонтанных форм тыквы на о. Ява, а по
Д. Гукеру [148], С. maxima была найдена на берегах Нигера в Гвинее
и в Анголе как туземное растение.
Различные путешественники, начиная с Д. Ливингстона [32], ука-
зывают на распространение тыкв красного цвета (не лагенарии) в Юж-
ной и Восточной Африке. Г. Ричей [174] описывает совершенно новые
формы С. moschata, полученные им из Африки. Абиссинский материал
по С. maxima в коллекции Всесоюзного НИИ растениеводства весьма
резко отличался от прочих экотипов этой культуры [20].
150
Все эти факты могут найти объяснения с точки зрения экологи-
ческой эволюции и современных знаний о переплывании твердокорых
плодов через разделяющие их океаны с одного материка на другой.
Н. И. Вавилов предполагал возможность переплывания лагенарии че-
рез океан из Америки в Африку, а позднее Т. Вайтекер и Г. Картер
[209] экспериментально подтвердили такую возможность для плодов
тыквы. Сейчас уже можно говорить не только об американском про-
исхождении тыквы, но и о районах сосредоточения там ее аборигенных
форм.
Т. Вайтекер [215], наиболее полно исследовавший вопрос про-
исхождения тыкв, считает, что центром происхождения этого рода яв-
ляется Мексика, в частности ее район к югу от г. Мехико до границы
с Гватемалой. Археологические раскопки показали, что земледелие на
Американском континенте распространялось из Мексики. Здесь же со-
средоточено наибольшее количество диких видов тыкв. Из них наиболее
легко скрещивающиеся с другими видами С. lundelliana и С. martinesii
являются прародителями культурных видов; из последних наиболее
древние С. moschata и С. ficifolia были обнаружены в Южной Америке
еще до инков и до появления гончарных изделий.
ИСТОРИЯ ИЗУЧЕНИЯ РОДА
В литературных переводах с языков древнего мира тыквой имено-
вали горлянку (Lagenaria) и кормовой арбуз (С. lanatus, var. citroides),
поэтому историю изучения тыквы следует начинать с эпохи Возрож-
дения после открытия Америки. В эпоху Возрождения первые ботаниче-
ские описания даже с рисунками растений появились в середине XVI в.
Д. Руэль [177] приводит роды Cucurbita и Реро, подразумевая лагена-
рию и тыкву. Л. Фукс [128] под названием Cucumis turcicus описывает
кабачок, он же дает и первые рисунки тыкв. И. Камерариус [111, 153]
под Cucurbita indica описывает обычную полевую С. реро.
М. Лобель [156] уже различает виды тыквы: Реро oblongus, Реро
rotundus compressus, Реро maximus. Согласно рисункам, первые два
относятся к Cucurbita реро, а последний — к С. maxima. Он впервые
описывает и мелкоплодную декоративную тыкву.
Р. Додоней [121] насчитывает уже 5 видов: Реро maior oblongus,
Р. maior rotundus, Р. maior minor, P. latus, P. silvestris. Первый вид
представляет собой настоящую С. реро var. реро, второй — С. maxima,
третий и пятый — декоративные формы С. реро, а четвертый вид — па-
тиссоны.
У К. Баухина [104] в семействе Cucurbitaceae (sectio quarta) опи-
сано довольно много родов. Тыкву он относит к трем из них: Реро,
Melopepo и Cucurbita. Однако родом тыквы К. Баухин считал, несо-
мненно, Реро. Дынеподобные тыквы он относил в особый род Melo-
pepo, а последний вид С. verrucosa, известный под таким именем у Лин-
нея, был им ошибочно причислен к роду лагенарии, который он назы-
вал вместе с другими авторами эпохи Возрождения Cucurbita. У К. Бау-
хина в классификацию уже включены и более мелкие формы, не-
жели вид.
Д. Далешамп [118] в качестве родового названия тыквы считал
французское Courge и рассматривал 4 вида. В его работе дано изо-
бражение С. maxima (рис. 1).
Г. Рид [171] называет тыквенные Porniferus. Из видов для Запад-
ной Индии он приводит только один — Реро indicus maximus, отож-
дествляемый в настоящее время с С. maxima.
151
Рис. 1. Древнее изображение *
С. maxima (Далешамп,
Д. Турнефорт называл тыквенные также Pomiferus, сначала он
указывает 5 видов тыкв: 2 рода — Реро и Melopepo, но в «Наставлении
по ботанике» [195] приводит уже 28 видов, представляющих собой
разнообразные формы. Это была первая работа, в которой показано
многообразие форм, но в основном лагенарии.
Однако вскоре после Д. Турнефорта название рода Melopepo ис-
чезло из литературы и все формы тыквы стали относить к Реро, более
определенно отличая их от лагенарии, которая по-прежнему имено-
валась как Cucurbita.
Изменение родового названия тыквы на Cucurbita, существующее
в литературе и поныне, осуществлено Линнеем. В основной работе
«Виды растений» [155] он признает только 3 вида тыквы: Cucurbita
реро, С. verrucosa и С. Melopepo. Два других вида, относимых им
также к Cucurbita, являются лагенарией и арбузом. В послелиннеев-
ском издании этой работы у К. Вильденова [217] из 13 приводимых
Линнеем видов 8 относятся к тыкве С. реро, преимущественно за счет
мелкоплодных гибридных форм.
В 1817 г. была обнаружена дикорастущая многолетняя тыква с
утолщенными корнями типа ямса, способная переживать зимний пе-
риод, описанная как С. foetidissima Н. В. К. Поэтому в более поздних
издания флоры в род тыквы, помимо С. maxima, включают С. foeti-
dissima и С. multiflora, а С. verrucosa L. и С. subverrucosa Willd. уже
не рассматривают как самостоятельные виды.
Ботаники признавали виды Линнея и использовали их в своих
работах вплоть до появления классификации М. Дюшесна. Исключе-
ние представляет сочинение И. Кель-
рейтера, который, скрещивая круп-
ноплодную тыкву (очевидно, С. ре-
ро) с мелкоплодными, получил пло-
довитые гибриды, откуда и заклю-
чил, что все тыквы относятся к од-
ному виду С. реро.
Наибольшее значение после
Линнея имели работы М. Дюшесна,
Нодена, Л. Бейли, Н. Е. Житене-
вой. Эти исследователи приближа-
лись к эколого-географическому по-
ниманию видов и разновидностей
тыквы, поэтому у них не преобла-
дал формальный подход к класси-
фикации (перетаксономизация форм
других авторов, введение в систему
разновидностей мелких, слабо отли-
чающихся форм, в особенности явно
гибридного характера и т. д.).
Виды М. Дюшесна и Л. Бейли
были явно эколого-географическими,
как это показали С. М. Букасов и
Н. Е. Житенева. Последняя впер-
вые установила в широком плане и
географичность внутривидовых си-
стематических единиц.
М. Дюшесн [122] был практи-
ком и, как это часто бывает, сделал
более глубокие наблюдения, чем
многие ботаники того времени. Пу-
152
тем изучения сортового состава видов и скрещивания их друг с другом
он установил действительные виды культурных тыкв: С. maxima Duch.
(potiron), С. реро Duch., С. реро moschata Duch., С. реро polymorpha
Duch. Многочисленные ранее описываемые виды Дюшесн отнес к расам.
В дальнейшем исследователи употребляли видовые эпитеты тыкв
уже по М. Дюшесну: С. maxima Duch., С. moschata Duch. и С. реро L.
Однако в американской литературе распространено другое разделение
тыкв по форме плодов независимо от принадлежности их к ботаниче-
ским видам на 3 типа: 1) pumpkin (от греческого слова рероп — зре-
лый), имеющий удлиненную форму плодов; 2) squash (индейское) со
сплюснутыми плодами; 3) cushaw (индейское) —с плодами кувшино-
образной формы. Иногда американское и английское понимание этих
терминов противоположны.
Первым поправил М. Дюшесна А. Пуарет [169], который вид
С. polymorpha включил в состав С. реро, однако при этом он ошибочно
отнес и тюрбанную тыкву к виду С. реро. Хотя она часто имеет дере-
вянистую кору, как у С. реро, но по вегетативным и генеративным ор-
ганам относится к С. maxima.
Однако более 30 лет спустя после работы М. Дюшесна М. Сажере
[179] вновь возвращается к родовому эпитету Реро, совершенно не
упоминая о М. Дюшесне. Его 4 вида повторяют по содержанию виды
последнего, кроме Реро malabaricus из Индии, по-видимому, являю-
щегося не тыквой, а дыней.
Н. Серенж [182] в своей первой работе описывает еще 16 видов
тыквы, включая виды Дюшесна и С. foetidissima Н. В. К., во второй
[183] увеличивает их число до 20, заменяя свои многие старые на
новые.
В 1837 г. С. Бух описал полукультурный вид крупноплодной не-
сладкой тыквы С. ficifolia Bouche, используемой главным образом для
кормовых целей.
М. Ремер [176] 31 вид распределил между 3 подродами, которые,
однако, не совпадали с 3 видами Дюшесна. Подрод 1 ключает С. mo-
schata и крупноплодные сорта С. реро разных по окраске и форме пло-
дов; подрод 2 (Melopepo) объединяет сплюснутоплодные формы, в том
числе чалмовые и патиссоны; подрод 3 включает С. maxima, С. foeti-
dissima, ряд других различных форм и представителей других родов.
Если бы в этой классификации не было смешения видов и даже
родов тыквенных растений, то она могла бы претендовать на почти все-
объемлющую полноту: некоторые формы описаны в ней почти исчерпы-
вающе. В данном случае она может служить лишь богатым материалом
для выявления первоначальных названий ботанических таксонов во из-
бежание ненужных новых.
Слишком большое количество видов, не обоснованных убедительно,
приводит к путанице в систематике, поэтому М. Гаспарини [129] при-
зывает к более серьезному их изучению. Он описывает лишь 2 вида
тыкв: с плодами булавовидной формы и С. macrocarpa Gasp, (совре-
менная С. melanosperma Gasp.), а также новый род Pileocalix Gasp.,
под которым понимается группа тюрбанных тыкв, относимая к виду
С. melopepo L.
В этот период в работу по систематике тыкв включаются и аме-
риканские авторы. Известный ботаник Аза Грей [131] открыл и опи-
сал новые виды диких тыкв С. texana и С. digitata. По непонятным
причинам С. texana отсутствует в монографии Ш. Нодена и в класси-
фикациях других ботаников до Л. Бейли [102]. Впоследствии С. texana
оказалась не самостоятельным видом, а дикой формой С. реро, которая
153
ошибочно отождествляется некоторыми ботаниками с открытой Лин-
неем С. ouifera.
Период работы Ш. Нодена снова начинается с упорядочивания
таксономии тыкв. В 1856 г. [165] он вначале выделяет 3 подрода тыкв:
1) Реро, включавший 12 видов, в том числе С. реро, С. moschata, С. cit-
rullus, С. verrucosa, С. ovifera и др.; 2) Cucurbita —17 видов, в том
числе С. maxima, С. foetidissima, С. urnigera и др.; 3) Melopepo—
2 вида: С. lurbaniformis и С. melopepo (патиссон). Затем Ноден снова
объединяет тыквы в 3 культурных вида: С. maxima Duch., С. moschata
Duch. и С. реро L. Кроме того, он [165] приводит известный к тому
времени С. melanosperma (С. ficifolia) и дикие виды С. perennis (С. foe-
tidissima) и С. digitata. Возвращение Ш. Нодена к видам Дюшесна
объясняется результатами многочисленных опытов межвидовых скре-
щиваний.
На протяжении ближайших десятилетий было описано еще 5 но-
вых диких видов тыквы. Ш. Ноден [166] описал многолетний (из тро-
пиков) вид С. radicans Naud, с небольшими клубеньками на корнях,
Ватсон — С. palmata Wats, с весьма оригинальными пальмовидными
листьями. Однако открытый Ватсоном совместно с Торреем С. califor-
nica явно относится к виду С. palmata Wats. По В. Бимайсу и Т. Вай-
текеру [106], последний вид свободно скрещивается с С. digitata-, ве-
роятно, они близкородственные.
В монументальной монографии А. Конью [114] описывает еще один
новый вид тыквы — С. galeotti Cogn. Однако неполное описание и от-
сутствие семян дает основание считать его сомнительным видом.
К концу XIX в. были признаны 10 видов тыквы [114]: С. maxima
Duch., С. digitata Gray, С. реро L„ С. palmata Wats., С. moschata
Duch., С. californica Torr., C. ficifolia Bouch., C. foetidissima H. В. K-,
C. radicans Naud., C. galeotti Cogn.
Вскоре Ш. Ноден [167] описал новый дикий вид из Уругвая —
С. andreana Naud, впоследствии признанный как дикий родич С. ma-
xima. Контарди первый успешно скрестил его с С. maxima и тем до-
казал их родство. Позже Т. Вайтекер [208] успешно повторил это скре-
щивание.
В 1920 г. С. Лотси [157] подтвердил выводы М. Дюшесна и Ш. Но-
дена о нескрещиваемости трех культурных видов тыквы: С. maxima,
С. реро и С. melanosperma. Одновременно было показано богатство
гибридных форм от скрещивания крупноплодной С. реро с ее мелко-
плодными и дикорастущими, якобы генетически отдаленными формами,
считавшимися многими ботаниками самостоятельными видами.
Д. Смолл [193] описывает новый лазающий по высоким деревьям
вид Реро okeechobiensis Small (с берега озера Окичоби во Флориде).
Бейли отнес этот вид к С. okeechobiensisis (Small) Baily. Это была
одна из наиболее древних диких тыкв, заполнившая большой пробел
в наших знаниях о лиановых предках тыквы.
В связи с открытием ряда новых диких видов тыквы Л. Бейли
предпринял попытки критического анализа всех видов рода Cucurbita.
Кроме культурных видов Cucurbita maxima Duch., С. реро L., С. mo-
schata Duch., С. ficifolia Bouch., С. argyrosperma Hort. (впоследствии
признанный как С. mixta Pang.), он приводит 21 вид диких тыкв, не
настаивая на полной достоверности всех: С. texana Gray, С. sororia
Bailey, С. fraterna Bailey, C. radicans Naud, C. okeechobeensis Bailey,
C. martinesii Bailey, C. pedatifolia Bailey, C. lundelliana Bailey, C. pal-
meri Bailey. C. gracilior Bailey, C. cordata Wats., C. cylindrata Bailey,
C. digitata A. Gray, C. palmata Wats., C. californica Torr, et Wats.,
C. galleotti Cogn., C. scabridiolia Bailey, C. foetidissima H. В. K-,
154
C. andreana Naud., C. kellyana Bailey, C. tnoorei Bailey. Самим
Л. Бейли было обнаружено и описано 11 новых видов.
В последние годы X. Кутлером и Т. Вайтекером [117] обнаружен
новый дикий вид С. equadorensis Culter et Whitaker в северных районах
Южной Америки. Он имеет семена, по размеру промежуточные между
культурной и дикой формой С. maxima, негорькую мякоть и мелкие
плоды. Р. Миллан [161] одновременно сообщает, что в провинции Буэ-
нос-Айрес найдена негорькая форма С. andreana; возможно, это и есть
С. equadorensis.
Из перечисленных диких видов несомненными являются только
следующие изученные нами по живым растениям: С. okeechobeensis,
С. lundelliana, С. foetidissima, С. palmata, С. digitata; 6 видов оказа-
лись разновидностями, остальные, по нашему мнению, недостаточно
изучены предшествующими авторами и требуют дальнейшего изучения
на живом материале.
Приводим виды (по первоисточникам) с указанием степени их изу-
ченности.
Группы Более и менее достоверные Малоизученные Требующие до- полнительной таксономизации
Дикие виды и формы, тяготеющие к виду С. maxima Тяготеющие к виду С. maxima var. andereana С. okeechobeensis C. equadorensis C. martinesii C. pedatitolia
С. moschata Тяготеющие к виду С. ficifolia Группа степных полу- пустынных видов С. lundelliana С. foetidissima С. foetidissima C. moorei C. cylindrata С. gracilior С. cor data,
var. scabridifolia, C. radicans C. galeotti,
Тяготеющие к виду С. реро C. palmata, C. palmata var. californica, C. digitata C. pepo var. texana C. pepo var. sororia C. pepo var. fraterna C. kellyana C. palmeri
Работы Всесоюзного института растениеводства по тыкве начались
в 1925 г. статьей С. М. Букасова о группировке тыкв по окраске, ви-
дах культурных тыкв и об их нескрещиваемости [157].
В дальнейшем С. М. Букасов провел большую экспедицию в Мек-
сику и Центральную Америку, где собрал большой сортовой материал
по тыквам, установил ареал и высотность распространения отдельных
культурных видов и обнаружил много новых интересных ее форм, из-
ложив все эти сведения в большой монографии «Возделываемые рас-
тения Мексики, Гватемалы и Колумбии». Привезенные оттуда Бука-
совым в виде аборигенных растений почти все типы наших старосвет-
ских тыкв С. реро и С. moschata вплоть до кабачков позволили
окончательно выяснить, что родина всех культурных видов тыквы —
Америка.
На базе мировой коллекции ВИР, включающей материал, приве-
зенный Н. И. Вавиловым и С. М. Букасовым из Центральной Америки,
а также Н. И. Вавиловым и С. В. Юзепчуком из Южной Америки,
Н. Е. Житенева провела изучение тыкв и построила географическую
классификацию этой культуры [20, 21]. Прежде всего она разделила
культурные (cultus) и дикорастущие (agrestis) формы, хотя последние
ею были установлены практически только для вида С. реро в виде
«мелкоплодных сортов, обладающих всеми признаками, присущими ди-
155
карям: малыми размерами растения, листьев, цветков, плодов, обилием
плодов на одном растении, наконец, зачастую горьким вкусом плодов».
К диким формам она относит и находку Аза Грея дикой тыквы С. te-
хапа в штате Техас (США).
Далее Житенева разделяет тыквы еще на две категории: 1) гру-
бого облика (rigidum), привезенные экспедициями из Америки, Индии,
Испании, Италии и других тропических и субтропических стран, и
2) нежного облика (gracilior), разводившиеся преимущественно в уме-
ренной зоне Европы и Азии.
Внутривидовая классификация тыквы Н. Е. Житеневой прогрес-
сивна своим описанием некоторых новых форм — чилийских, боливий-
ских, колумбийских и других тыкв, даже неизвестных или неясно опи-
санных предыдущими авторами, а также введением географического,
граничащего с экологическим принципа в систематику этого рода. Не-
достатки ее классификации — малая дифференциация и начинающаяся
изоляция таксономической системы от таковых других предшествую-
щих систематиков, впоследствии предельно развившаяся в работах
К. И. Пангало, особенно по дыне. История систематики указывает, что
ряд разновидностей предыдущих ботаников, несомненно, существуют в
природе.
Н. Е. Житенева сделала попытку восстановить вид чалмовой
тыквы — С. turbaniformis (Roem.) Zhit., но из-за ее легкой скрещивае-
мости с С. maxima и сильной изменчивости признака чалмы у разных
ее форм большинство последующих авторов не признали этого вида
в качестве самостоятельного, хотя во «Флоре Туркмении» она все же
фигурирует.
Гораздо большие разногласия вызвал вопрос о признании нового
вида тыквы С. mixta Pang. [58]. А. Хетчинс и Л. Сандо [150], а также
Л. Бейли [103] считают его позднейшим синонимом С. argyrospermum
Hort. По данным X. Джонса и И. Роза [152], а также П. Русселя
[178], эта форма относится к виду С. moschata.
По нашим данным, С. mixta Pang, распространена географически
выше, чем С. moschata. Кроме того, она довольно легко скрещивается
с С. moschata и имеет одинаковые с ней формы плодов и рисунки коры.
Ее отличают лишь светлая окраска коры плодов, лишенная желтого
пигмента, и большая изменчивость форм семян. Недостаток каротинои-
дов зависит, видимо, от произрастания этой формы в горных условиях;
такое же явление наблюдается у горного вида С. ficifolia, но в гораздо
большей степени. Большая изменчивость листьев, плодоножек и семян
С. mixta, на основании которой К. И. Пангало и выделил этот вид,
свидетельствует о неустойчивости данных признаков и нестабильности
формы, т. е. в данном случае наблюдаем неустановившийся тип этого
растения. Отсутствие же каротиноидов в зрелой коре и в результате
серебросемянность дают право рассматривать данную форму в пре-
делах вида С. moschata [83]. Однако Т. Вайтекер ввиду некоторой за-
трудненности скрещиваний ее с С. moschata считает С. mixta само-
стоятельным видом. После его работ большинство систематиков при-
знало самостоятельность С. mixta как вида.
Кулинарные качества этой тыквы позволяют отнести ее к полу-
культурному виду. Изменчивость формы семян от сходных с С. mo-
schata до узких и удлиненных в 2—3 раза при одинаковых по форме
и окраске плодах особенно подчеркивает сомнительность видового ста-
туса данной тыквы и по признаку семян. Общее же в окраске семян
(серебросемянность) указывает на отсутствие и в них каротиноидов.
Склонность этой формы к межвидовым скрещиваниям указывает на
ее большую гетерогенность и экологическую пластичность. Вызывает
156
сомнение и название этой тыквы — серебросемянная, которое существо-
вало в торговле семенами по признанию Л. Бейли раньше предложен-
ного К. И. Пангало наименования mixta.
Таким образом, включая С. mixta, имеем 5 окультуренных видов
тыквы, 5 достаточно изученных дикорастущих, 11 недостаточно изучен-
ных, требующих дополнительного изучения или подтверждения их ви-
довой самостоятельности.
ЭВОЛЮЦИЯ РОДА
Наиболее древние ископаемые остатки культурных видов дати-
руются периодом 3000—5000 лет до н. э. [214], а окультуривание начи-
налось значительно раньше.
Вид С. реро имел два независимых центра окультуривания: пер-
вый располагался вдоль восточного побережья США; второй — на се-
веро-западе Мексики и юго-западе США, включая долину Миссисипи.
Вид С. moschata окультуривался на юге Мексики, в Колумбии и Перу
и наиболее там распространен, С. maxima — в умеренных районах
Перу, Боливии и Аргентины. Соответственно в указанных зонах рас-
пространены и дикорастущие формы этих видов тыквы.
На основании исторических изысканий Т. Вайтекер и Г. Картер
[206] построили схему эволюции видов тыкв, исходя от С. moschata
или ее диких форм в двух направлениях: к северу — дикие и культур-
ные виды типа С. реро-, к югу — формы типа С. maxima. При этом каж-
дое направление делилось затем на две ветви, причем формы типа
С. реро разделились раньше форм типа С. maxima. Одна ветвь его
породила дикие виды С. palmata, С. digitata и С. foetidissima, другая —
дикую форму С. texana, окультуренные крупноплодные формы С. реро
и кустовые кабачковидные и патиссоновидные ее сорта. Такое же раз-
деление, но только после выделения дикой С. andreana и до окульту-
ривания претерпели и виды типа С. maxima на аргентинскую ветвь,
давшую обычные безчалмовые формы этого вида, и бразильскую ветвь
с чалмовыми плодами. Разделилась на 2 ветви: белосемянную мекси-
канскую и темносемянную колумбийскую еще до окультуривания и
С. moschata.
В 1961 г. Т. Вайтекер произвел взаимные скрещивания культурных
видов между собой для выяснения их родства. Оказалось, что хуже всех
скрещивается с другими видами С. ficifolia. С ее многолетним образом
жизни связан, очевидно, и высокий барьер стерильности. Легче других
видов скрещивается С. moschata, особенно с С. реро. С. mixta в скре-
щиваниях ведет себя аналогично С. moschata. Однако вывод об их
родстве не может быть сделан из-за несовпадения результатов прямых
и обратных скрещиваний. Успех последних зависит главным образом
от материнской формы: на рыльцах С. maxima пыльцевые трубки
С. moschata и С. реро, как правило, дорастают до яйцеклетки, на рыль-
цах С. moschata они чаще останавливают рост вблизи яйцеклетки, а
на рыльцах С. реро пыльцевые трубки других видов, как правило, не
дорастают до яйцеклетки.
Взаимные скрещивания культурных видов с дикими, по данным
Т. Вайтекера и В. Бимайса [214], показали резкое различие между
ксерофильными и мезофильными дикими формами. Ксерофиты (С. pal-
mata, С. digitata, С. cylindrata и С. foetidissima) почти не скрещиваются
с другими видами, но первые 3 легко скрещиваются между собой. Ме-
зофильные дикие виды (С. lundelliana, С. okeechobeensis и С. marti-
nesii) также легко скрещиваются между собой, но дают лишь частично
стерильные гибриды с культурными видами, особенно С. moschata.
157
В. Бимайс п А. Родес [107] математически обработали сходство
21 вида тыкв по 160 признакам, создав 8 статистических групп моде-
лей. Сравнивая последние по биологическим, географическим, генети-
ческим и экологическим данным, они установили Q-корреляции, по ко-
торым вычисляли степень родства видов в процентах, изобразив ее
графически (схема).
Родство видов по Бимайсу и Родес
Вид Группа
______________________________С. digitata
------С. palmata
------var. californica I
_______________________________C. cylindrata
----------------------C. cordata
_______________________________________C.pedatifolia VI
_______________________________________C. foetidissima VII
_______________________________C. lundelliana
।------------C. okeechobeensis II
I------------C. martinesii
______________________________________________C. equadorensis
----------------------------------------------C. moschata VIII
___ _______________________________C. mixta
_______________________________С. pa 1 meri III
______________________var. so г о r i a
----------------------C. gracilior
______________________________________________C. ficifolia
---- ---------------------------------------------------C. pepo
----------------------var. texana IV
_______________________________C. m a x i m a V
I-----------------------------var. andreana
10 20 30 40 50 60 70 80 90 100% родства
Нашими исследованиями для некоторых групп степень родства ви-
дов подтверждается, но имеется ряд крупных отклонений, так как со-
поставляемые даже одинаковые признаки могут иметь различную при-
роду. Поэтому данная схема родства не может носить всеобъемлющий
строгий характер.
Из трех основных путей эволюции тыквы: экологического, генети-
ческого и одомашнивания первый имеет место у всех видов тыквы. Все
виды, как дикорастущие, так и культурные, носят черты определенных
экотипов. Из экологических условий Американского континента наи-
большее влияние оказывают температура и влажность. Последняя осо-
бенно сильно обусловлена горными и предгорными условиями и дву-
сторонним влиянием океанов на узкий тропический перешеек, связы-
вающий Северную Америку с Южной.
158
Из климатических зон Америки, имеющих отношение к экологи-
ческому формированию тыкв, можно выделить следующие: 1) зона уме-
ренных и предгорных прибрежных районов Северной Америки, харак-
теризующаяся достаточной влажностью при относительно невысокой
температуре, допускающей вызревание здесь тыквы; 2) зона сухих
жарких степей юга Северной Америки; 3) горная зона западного
побережья Центральной и Южной Америки (от Мексики до Чили);
4) влажные низинные районы Центральной Америки, севера Южной
Америки и Флориды; 5) предгорья и равнины Южной Америки с уме-
ренной температурой и достаточной влажностью.
В соответствии с климатическими зонами Америки виды тыквы по
экологическим признакам обособляются в 5 групп. Первая группа рас-
пространена в зоне умеренных районов Северной Америки, от южной
части Канады до Гватемалы, на возвышенных местах от 500 до 2500 м
над уровнем моря. Растения этой группы довольно скороспелые, вла-
голюбивые, устойчивые к умеренным температурам, но неустойчивы к
сухости воздуха и высокой температуре; в культуре они дальше других
групп тыкв продвинуты к северу. Внешний вид их довольно грубый:
жесткие листья и плети с грубым опушением, напоминающим колючки;
утолщенные жилки листьев, имеющие резкий сбег; неувядаемость ли-
стьев в жару; сильная изрезанность листовой пластинки с острыми вы-
ступами, деревянистая кора плодов у всех форм. Данный климатиче-
ский тип тыквы можно приравнять к подроду, который мы назвали
subgen. cucurbita. Представителями этого типа являются культурный
вид С. реро и дикорастущий С. kellyana.
Вторая группа — степная, распространена в степных районах юго-
запада США и северной части Мексики. Растения группы — ксерофиты
умеренной полосы, лучше других климатических типов могут мириться
с умеренной температурой и засушливыми условиями существования.
Они имеют или сильноопушенные, или еще более вырезанные листья
(экономия пластинки листа), чем у предыдущей группы, вплоть до
пальчатых. Не культивируется. Таксономизирована нами как subgen.
eremosicyos. Сюда относятся только дикорастущие виды С. foetidissima,
С. digitata, С. palmata, С. cylindrata, С. radicans.
Третья группа — тропическая, распространена в низинных долинах
Центральной Америки и во Флориде. Растения — мезофиты и частично
гигрофиты с ярко выраженной склонностью к лиановости. Внешний вид
яркий: интенсивно-зеленые листья, нежное опушение, тонкая нервация
листовых пластинок, их округлость и эластичность с почти полным от-
сутствием изрезанности краев. Таксономизирована нами как subgen.
scandens. Сюда относятся виды С. moschata, С. okeechobeensis, С. mixta,
С. lundelliana, С. martinesii. Некоторые из них эндемичны и для пред-
горных районов тропического пояса, например вид С. mixta.
Четвертая группа — горные тропические тыквы, распространена в
горных районах юга Мексики и северной части Южной Америки. По
внешнему виду напоминает предыдущую группу, но отличается устой-
чивостью к умеренным температурам и сильной реакцией (ускорение
развития) на укороченный день. Характерная черта — отсутствие каро-
тиноидов в плодах и деревянистость их коры (горный признак). Неко-
торые виды культивируются, но еще не достигли уровня культурности
других возделываемых тыкв ввиду неспособности или слабой способ-
ности накапливать сахара, отчего считаются полукультурнымн. Таксо-
номизируется нами как subgen. montana. Сюда относятся виды С. fi-
cifolia, С. moorei. Первый из них распространен на большей высоте,
чем С. mixta, и поэтому последний мы считаем предгорным видом
группы scandens, лишенным каротина, как и вид С. ficifolia.
159
Пятая группа — тыквы-космополпты. Их родина — Южная Аме-
рика, но распространены они широко по всему миру, охватывая почти
все земледельческие районы. По экологическому облику растения этой
группы являются средними между группами предгорных и тропических
тыкв. Очевидно, предшествующие современным дикие формы занесены
из тропического пояса, но в горных районах Южной Америки приобрели
признаки некоторой грубости органов. Таксономизирован нами как
subgen. cyclofilla. Представитель этого типа — культурный вид С. ma-
xima и, вероятно, дикорастущий С. equadorensis.
Ареал тыкв в Новом Свете представлен на рисунке 2.
Таким образом, по экологической дифференциации у рода тыкв
выделено 5 климатических групп видов, оформленных как внутриродо-
вые таксоны — подроды. О значении генетического пути в создании
видов тыкв в данное время ничего неизвестно, но во внутривидовом
формообразовании этот путь имел и имеет в настоящее время большое
значение. Особенно большую внутривидовую изменчивость он вызвал
у вида С. реро. Не является доказательством генетического видообразо-
вания и теория П. М. Жуковского [22] о происхождении видов тыкв
только путем гибридизации с С. lundeUiana. Основанием для такого
предположения является только легкая скрещиваемость С. lundeUiana
с другими видами тыкв, что явно недостаточно для построения такой
теории.
Окультуривание дикорастущих видов было единственным путем
создания культурных и полукультурных форм. Приемами окультури-
вания были искусственный отбор, возможно, бессознательный вначале,
а также богатая унавоженная почва вблизи поселений и стоянок чело-
века, так как культурные формы тыкв, существовавшие за 5000 лет до
н. э., могли быть получены за 8—
10 тысячелетий ранее индейцами
или народами, которые им предше-
ствовали. Основное отличие куль-
турных тыкв от их диких родичей —
крупные размеры плодов, отсутствие
в них горечи и преобладающая мяг-
кость коры.
Тыква — лиановое растение, ее
первое становление было, очевидно,
в тропических сырых лесах. Это до-
казывается наличием лианообразно
тыквы С. okeechobeensis, расту
в настоящее время в болотистых л
сах Флориды. Плоды ее Д°стИгаеВт
вершин высокорастущих деРе0Ь
т. е. 40 м и более. Путь тыквь! *
обитания в сырых болотистых л
до сухой полупустыни создал
крайних варианта видов: £ .[у)
морфную (С. okeechobeensis Г irnd
и ксероморфные (С. foetid1
Н. В. К. и С. digitata A. Gray#ь\е
середине расположились осТяа/Оиь^
виды, в том числе и куль тук
Таков был путь эволюции Р0,а л
curbita. $ес,
Из тропического влажного сд^
лиановые тыквы попадали 0
вия, которые можно охарактеризовать следующие образом: 1) высоко-
горные, достаточно влажные и прохладные условия с обилием непо-
глощенной воздухом лучистой энергии; 2) сухие степи и полупустыни;
3) равнинные условия тропического земледелия. Иных условий в за-
падной части Америки, где формировались виды тыкв, не было.
Тип тропических лиановых тыкв (subgen. Scandens), очевидно, яв-
ляется более древним. От него путем постепенного окультуривания в
долинах и на тропических низменностях шло образование культурного
вида С. moschata. Промежуточной формой на этом пути, несомненно,
является мелкоплодная Семинольская тыква, эндемичная для централь-
ных районов Флориды [127].
Наиболее высокогорная форма диких тыкв — С. moorei (subgen.
montanus), вьющаяся по кустам и вдоль дорог в Центральной Мексике
на высоте 1750 м и более. Ее уменьшенные органы (кроме длины пле-
тей) сходны с таковыми С. ficifolia, а некоторые признаки, например
запах, напоминают С. foetidussima. В результате достаточной водообес-
печенности сохранилась лиановидная форма стеблей этого дикого вида.
Некоторые дикие предгорные виды тыквы приспособились и к степ-
ным условиям, к степным равнинам. Это, например, дикие формы
С. реро. Культурный вид С. реро мало сохранился на равнинах Север-
ной Америки, но интенсивно разводится на Мексиканском плоскогорье
и в предгорьях Мексики. Связь этого культурного вида с его дикими
формами (типа subgen. Cucurbita) несомненна, что выражается в их
скрещиваемости. Вид С. реро окультурен из диких форм данного типа.
Третий путь эволюции вел в сухие степи. Эти условия создали осо-
бый ксероморфный тип видов. Видовые различия здесь наиболее конт-
растны, а виды отчетливы и обособленны. Обособленность видов, а
также отсутствие в сухих степях условий для огородного земледелия,
очевидно, не позволили окультурить ни один из видов тыкв этого типа
(subgen. Eremosicyos).
Остается невыясненной связь южноамериканской дикой тыквы
С. maxima var. andreana с видами Северной и Центральной Америки,
а отсюда и происхождение всего типа тыкв subgen. cyclofilla. Инте-
ресно то, что дикая форма var. andreana имеет такие же крупные ли-
стья и плети, как и культурная С. maxima. Окультуривание последней
шло только по линии укрупнения плодов, наибольшая величина кото-
рых среди других культурных видов отчасти объясняется уже подго-
товленностью к этому процессу мощных вегетативных органов дикой
формы. Возможно, изучение нового вида С. equadorensis объяснит при-
чины обособленности subgen. cyclofilla.
Ясно только, что виды данного подрода не сохранили отчетливых
признаков лиановости, как и не приобрели заметных признаков ксеро-
фильности; развиваясь в другой части континента — в Южной Америке,
они шли по второму пути эволюции — приспособления к мезофильным
условиям предгорий с заходом на территорию равнины. Поэтому эко-
логически С. maxima гораздо ближе к С. реро, чем к С. moschata, хотя
ее длинный древний путь из Центральной Америки в Южную обеспечил
и большую пластичность и, следовательно, наибольший ареал в Старом
Свете. Ареалы этих видов представлены на рисунке 3.
Таким образом, вероятнее всего, совершалась эволюция тыквы на
ее родине — в западной части Америки. Какую продолжительность
могла занимать эволюция культурных видов тыквы? Если верна теория
о том, что человек перешел на Американский континент 10—15 тысяч
лет тому назад, то окультуривание тыквы могло быть начато 6000—
8000 лет до н. э., что гораздо больше периода эволюции культурной
дыни, огурца, а также арбуза. Как известно, эти культуры или не были
161
20 10 0 10 20 JO 40 50 SO 70 80 90 100 ПО !20 !J0 HO
Рис. 3. Эндемичные ареалы видов тыквы в Старом Свете.
известны в древнем мире (огурец), или существовали в полукультурном
виде (огуречная дыня, кормовой арбуз). Период их полного окульту-
ривания из полукультурного состояния, по-видимому, составляет не бо-
лее 1500—2000 лет, т. е. период нашей эры.
В Старый Свет тыква попала лишь после открытия Америки, т. е.
около 500 лет тому назад. Но и за этот период ее экологическая из-
менчивость в Азии и частично в Африке достигла таких поразительных
результатов, что некоторые ботаники склонны были находить здесь но-
вые виды культурных тыкв, как, например, это произошло с С, tur-
baniformis [176, 20]. По нашим данным, здесь имеет место внутриви-
довая изменчивость культурных тыкв.
КЛАССИФИКАЦИЯ РОДА CUCURBITA
Ключ для определения подродов тыквы (Cucurbita)
1. Листья округлые или тупоугольные....................................2.
+ Листья с сильно выраженными углами, чаще с заостренными долями .... 3,
2. Листья цельнокрайние, лишь со слегка городчатыми краями, серовато-зеленые
или светло-зеленые; плодоножки и плети неребристые; черешки слаборебристые.
Плоды у культурных форм преимущественно с мягкой корой, безрисунчатые,
реже с белесыми ленточками или красными пятнами, а у диких с деревянистой
корой, пятнистым рисунком.........................subgen. Cyclofilla Filov.
+ Листья глубоковырезные, лопасти листьев округлые, ярко-зеленые, плодоножки,
черешки и плети ребристые. Плоды с деревянистой корой, ярко-зеленые, с мо-
заичными белыми пятнами...........................subgen. Montana Filov.
3. Листья слаболопастиые или без лопастей, пятиугольные или треугольные, лопа-
сти чаще острые .......................................................4.
4- Листья глубоколопастные, иногда пальчатой формы, с узкими линейными доля-
ми или цельные, слабовырезные, треугольные, с вытянутой вершиной; в послед-
них двух случаях серые от густого опушения . . . subgen. Eremosicyos Filov.
4. Листья нежные, ярко-зеленые; черешки листа, плодоножки и стебли тупогра-
иеные. Опушение листьев обычно мягкое, реже грубоватое, но не шиповидное.
162
Плоды у культурных форм преимущественно с недеревянистой корой, с тонко-
полосатым или ячеистым рисунком..............subgen. Scandens Filov.
+ Листья грубые, зеленые. Черешки листа, плодоножки и стебли резко ребристые.
Опушение шиловидное. Плоды у культурных форм с деревянистой корой . . .
................................................ subgen Cucurbita.
ПОДРОД ЮЖНОАМЕРИКАНСКИЙ-SUBGEN. CYCLOFILLA FILOV.
Subgen, nova — Филов, 1966, Бюлл. Главн. бот. сада АН СССР,
63: 33. nom. illeg. — Plantae cyclopiНас cyclopediculatae. Fructus та-
lacodermes. Semina sine hibo. Typus: Cucurbita maxima Duch.
Растения округлолистные, плодоножка округлая. Плоды чаще с
недеревянистой корой. Семена без рубчика.
Ключ для определения видов
1. Стебель стелющийся, 4—5 м длины. Листья округло-почковидные. Лепестки
венчика тупые, с бахромчатыми краями. Плоды у культурных видов крупные,
различных форм, а у диких слабо удлиненные................С. maxima.
4- Стебель лазающий, 10 м длины. Листья широкоовальные, почковидные. Лепест-
ки венчика острые. Плоды только мелкие, до 16 см в диаметре, шаровидные.
Растения дикорастущие .................................С. equadorensis.
Вид крупноплодный — С. maxima Duch.
1786, in Lam, Encycl. 2:151, non Weight et Arh., 1834. — C. pepo L.,
1753, Sp. Pl.: 1435. — C. potiro Pers., 1807, Synops. 2:593. — Pepo po-
liro Sager, 1826, Ann. Sci. Nat. (Paris) 1; 8:311. —P. maximus Pe-
term., 1838, Fl. Lips. exc.: 562. — Cucurbita pileiformis Roem., 1846,
Synops. monogr. 2 : 86. — C. turbaniformis Roem., 1846, 1. c.: 87.—
Pileocalix elegans Gaspo, 1847, Rendic. Accad. Sci. Napoli, 6:408.—
Cucurbita zapallito Carr., 1872, Rev. Hort. 44 ; 399.
Описание. Стебли цилиндрические, длиной до 4—5 м, довольно
мягкоопушенные (рис. 4, а, 3). Листья округло-почковидные (рис. 4, а, 2),
невырезные, слабоугловатые, с пильчатыми краями, крупные (в том
числе и у диких форм). Цветки однополые, крупные, женские —оди-
ночные (рис. 5, а,1), мужские —иногда в пучках, колокольчатые. Ча-
шелистики спаянные, острые, 5 иногда 4 или 6. Венчик спайнолепест-
ной, раздельный, ярко-желтый, лепестков 5, иногда 4 или 6, с расширен-
ными тупыми бахромчатыми отогнутыми краями. Плодоножки утол-
щенные, цилиндрические. Плоды шаровидные, сплюснутые, реже удли-
ненные, зеленые, желтые или белые, без рисунка или рисунок в виде
белесых ленточек, красных пятен, с ровной поверхностью, реже слабо-
сегментированные. Мякоть кремовая или оранжевая. Семена сплюс-
нуто-яйцевидные (рис. 4, а,1), белые или кофейного цвета, крупные,
гладкие, со слабозаметным ободком или без ободка.
Местные названия вида: латинское — melopepo; английское —
squash, pumpkin; французское — courge, pot iron; китайское — вогуа.
Происхождение — Южная Америка. Распространен в земледельче-
ских районах всех континентов южнее 63° с. ш.
Изменчивость признаков. Вид С. maxima, распространяясь от Фин-
ляндии до Южной Африки, от Канады до южных границ Чили и Ар-
гентины и почти по всей земледельческой Азии, остался более одно-
образным, чем другие культурные виды тыкв. Особенно консервативны
признаки вегетативных и генеративных органов.
Преобладающая форма плода сплюснутая (с индексом меньше еди-
ницы). Характерное направление изменчивости — чалмовость плодов,
наблюдаемая у горного экотипа этого вида в Азии (концентрируется
на западе Китая). Это сильно сплюснутые, некрупные тыквы с деревя-
163
Рис. 4. Семена (1), листья (2) и опушение стебля (3, 4) основных культурных
видов:
а —С. maxima; б — С. реро; в—С. moschata.
Рис. 5. Женские цветки (1), тычинки (2) и мужские бутоны (3) основныж
культурных видов:
а — С. maxima; б — С. реро; в — С. moschata.
нистой корой. Полунижняя завязь с периферическим расположением
околоцветника является причиной чалмовидности одним из выраже-
ний укороченного роста осевых органов [78], в, частности сосудов,
питающих элементы околоцветника. На большой коллекции сортов
нами замечено, что процесс укорочения осевых органов у тыквы уси-
ливается в горных, но сравнительно сухих условиях. Чалмовые тыквы
не могли быть занесены сюда, так как их нигде больше нет В очень
слабой степени этот признак выражен у южноамериканских тыкв, но
У них он чаше связан с кустистостью растения, что является опять же
проявлениемукорочения омвых органо» и, кроме того, закона парал-
дельного морфогенеза [78].
Преобладающая окраска плодов серо-зеленая. Рисунок, как пра-
вило, отсутствует: изменение окраски начинается с пятен, которые по-
степенно расширяются, покрывая значительную часть плода, а позднее
и весь плод. У некоторых форм только иногда заметны очень слабая
зеленоватая сетка или слабые белесые полосы на меридиональных впа-
динах.
Плодоножки, хотя и остаются цилиндрическими, но могут взду-
ваться и утолщаться в несколько раз. Отмечена большая изменчивость
размеров плодов, семенного гнезда и консистенции мякоти. На севере
плоды крупнее, с большим семенным гнездом и рыхлой мякотью, со-
держащей небольшое количество сухих веществ. Тропическим формам
свойственны небольшие плоды с очень плотной мякотью и малыми се-
менными гнездами, чаще всего целиком заполненными плацентами с
семенами. Такие тыквы лучше хранятся. Кора плодов у С. maxima
обычно мягкая. Деревянистая кора встречается у аборигенных аме-
риканских сортов тропического происхождения очень редко, многих
чалмовых форм и у дикорастущих родичей.
Внутривидовая классификация. История изучения. Долин-
неевские ботаники все различающиеся формы тыкв называли видами,
поэтому в то время наблюдалось большое их число. Но описания этих
видов были так несовершенны (чаще по одному только признаку), что
не всегда можно установить, к какому современному виду относится
та или иная форма. Так, у Д. Турнефорта Г195] из 28 приведенных им
видов только 7 можно идентифицировать с внутривидовыми формами
С. maxima: Melovepo compressus, М. maximo alba, M. flavescens, M. la-
tus depressus, M. conico. Mintrinque turbinato, Moluteo cortica tenero.
Это была первая работа, показавшая многообразие форм С. maxima.
М. Дюшесн И22], установивший современные вицы тыкв, отразил
в составе вида С. maxima 3 формы. Е. Шпах Г194] для С. maxima
также приводит 3 формы. Однако Н. Сеоенж [183] вновь возвращает
сортовые формы тыквы к видам. Из них 8 форм относятся к С. maxima.
И. Метцгер [158], игнорируя виды М. Дюшесна и опираясь на
И. Кельрейтера, считает, что все тыквы принадлежат к одному виду
С. реро, и делит его на 9 подвидов: 1) Feldkurbis— крупноплодные по-
левые тыквы С. maxima', 2) Zangekurbis— кабачки; 3) Mantelsackkur-
bis — длинноплоцные Формы С. moschata', 4) Turbanokurbis — чалмовые
формы С. maxima: 5) SchiIdkurbis— патиссоны: 6) Melonenkurbis —
разные неллинноплодные формы С. moschata: 7) Apfelkurbis — ман-
дариноподобные мелкие тыквы С. репо: 8) Birnkurbis — грушевидные
С. репо и 9) Eierkurbis— яйцевидные формы С. репо.
К С. maxima из них относятся только 1-й и 4-й подвиды. Каждый
из этих подвидов И. Метцгер подразделяет затем на большое число
форм по окраске и форме плодов, не давая им, однако, латинских на-
именований, например Feldkurbis: желтая круглая, белая круглая, зе-
леная круглая, зеленая овальная, желтая овальная и т. д.
М. Ремер Г176] у С. maxima описывает 9 внутривидовых форм,
Ш. Ноден [165] — 22, причем в основном сортов, большинство из кото-
рых оказались разновидностями с четкими признаками. Ноден впервые
дал подробную классификацию внутривидовых форм тыквы, пользуясь
коллекцией живого материала этой культуры из 300 сортов. В нее
вошло большинство поныне существующих разновидностей.
Последующие авторы (Ф. Альфельд, Е. Харц) в своих классифика-
циях брали за основу систему Ш. Нодена. Ф. Альфельд описывает
30 форм вида С. maxima, разделив их на 3 группы: тюрбанные, полу-
тюрбанные, безтюрбанные. Е. Харц [143] описывает 31 форму, почти
те же, что и предыдущий исследователь.
166
Таким образом, впервые Д. Турнефорт, Н. Серенж, М. Ремер, осо-
бенно Ш. Ноден с Ф. Альфельдом и Е. Харцем, в прошлом веке описали
значительное количество внутривидовых таксонов, большинство которых
можно считать хорошими разновидностями. Систематическое изучение
тыквы в XX в. было стимулировано работами академика Н. И. Вави-
лова во Всесоюзном институте растениеводства. На базе собранной
мировой коллекции института Н. Е. Житеневой [20] были пересмотрены
взаимосвязи внутривидовых форм тыквы на живом материале. Боль-
шой набор эндемичных сортов из основных зон Америки позволил ей
дать новую классификацию эндемов С. maxima в следующем виде:
var. peruviana Zhit. — перувианские типы, var. boliviano Zhit. — боливий-
ские, var. chiloensis Zhit. — чилийские, var. abyssinica Zhit. — абиссин-
ские, var. asiatica Zhit. — азиатские.
Отмечена чрезмерная крупность разновидностей Н. Е. Житеневой:
в азиатскую разновидность, например, включен, по существу, весь ев-
ропейский сортимент.
Позднее К- И. Пангало [59] предложил заменить var. asiatica
Zhit. на gr. var. potiron Pang, и классифицировать ее следующим об-
разом: var. banana Pang. — челноковидные тыквы, var. mamut Pang.—
мамонтовые, var. macrosphaerica Pang. — крупношаровые, var. media-
sohaerica Pang. — плоды в 3—4 раза мельче предыдущей, var. macro-
depressa Pang. — крупные сплюснутые, var. mediodepressa Pang.—
плоды в 3—4 раза мельче предыдущей.
К. И. Пангало, забыв, очевидно, о переименовании разновидностп
var. asiatica Zhit. в gr. var. potiron, в 1937 г. переводит все 5 разно-
видностей Н. Е. Житеневой, не меняя названия asiatica, в подвиды,
поиписывая их авторство Н. Е. Житеневой. В самой последней работе
К. И. Пангало [62] снова делит С. maxima на 4 подвида: subso. com-
munis Pang. — все наиболее распространенные сорта, subsp. turbanifor-
mis Pang. — тюрбанные тыквы, subsp. gobbard Pang. — тип американ-
ских гоббардов, subsp. banana Pang. — челноковидные тыквы. Однако
синонимики ни К. И. Пангало и ни Н. Е. Житенева не приводят. По-
следняя классификация (подвидов) К. И. Пангало указывает на то,
что одобренная им ранее Г60] классификация (тоже подвидов)
Н. Е. Житеневой отвергается. С последним согласиться нельзя, так как
система Н. Е. Житеневой в тех же рангах построена раньше и на жи-
вом материале, она устанавливает новые (географические) принципы
внутривидовой изменчивости видов, а система К. И. Пангало — резуль-
тат литературной перетаксономизации без сохранения правил о при-
оритете.
Разнообразие сортов тыкв американские ученые начали изучать
позднее европейских ученых и в отрыве от них. Так, Е. Кастеттер [112]
разделил сортимент С. maxima на 7 групп: 1) Hubbard — Гоббард с
бугристыми плодами волчковидной формы; 2) Delicious — Деликатес
с коническими плодами и деревянистой корой; 3) Orange — Оранжевая
мозговая, плоды лимонной формы, оранжевые, с неровной поверхно-
стью; 4) Turban — Чалмовидная с чалмовидным следом околоцветника;
5) Banana — Бананная с челноковидными, огуречного вида плодами:
6) Pepbled — Перовидная, плоды темно-оранжевые, бугристые, с дере-
вянистой корой; 7) Display—Выставочная, очень крупная, желтая, ма-
монтовая.
Интересную производственную классификацию тыквы дает
С. И. Кобытев Г25]. Он выделяет 3 группы по принципу использования:
1) кормовые; 2) хлебные, употребляющиеся при выпечке хлеба (на-
пример, среднеазиатский сорт Нои-кеды); 3) столовые, используемые
167
как самостоятельное блюдо, так и в виде составной части в других
блюдах (например, сорт Плов-кеды).
Л. Бейли [103] выделяет и оформляет в самостоятельную разно-
видность описанную Л. Пароли волчковидную тыкву С. zapallo, ранее
относимую автором к боливийской разновидности Житеневой — var.
turgida Bailey.
Из новых форм тыквы Р. Милланом [161] были описаны 3 разно-
видности; 1) var. zapallito— кустовая форма с мелкими сплюснутыми
твердокорыми плодами; 2) var. zapinca; 3) var. triloba похожа на пер-
вую, но с плодами трехгранной формы.
В ряду исследований последних лет видное место занимают работы
И. Гребенщикова. Его внутривидовая классификация С. maxima вы-
глядит следующим образом [135]: convar. parvifructina Greb. (С. and-
reana Naud.), convar. turbaniformis Alef., convar. hubbardina Greb.,
convar. bananina Greb., convar. maxima Greb., convar. zapallitina Greb.
Новых внутривидовых форм И. Гребенщиковым не описано. На
наш взгляд, перевод ранее известных разновидностей в таксон «группа
разновидностей», особенно без выделения в нем просто разновидно-
стей, не обоснован.
Наибольшее значение имеют системы Ш. Нодена и Н. Е. Житене-
вой. Последняя приблизилась к эколого-географическому пониманию
существа внутривидовых форм, отчего в дальнейшем изучении внутри-
видовая систематика тыквы становится экологической проблемой.
Продолжая исследования мировой коллекции тыкв после Н. Е. Жи-
теневой, мы вполне убедились в сильном влиянии внешних условий
на конституционные признаки внутривидовых форм (подвидов, разно-
видностей) и поэтому шли по пути построения эколого-географической
классификации. Изучение работ предыдущих систематиков дает нам
возможность сохранить их имена в тех таксонах, которые ими были
действительно выявлены и существуют в настоящее время.
Новая классификация. Внутри вида рассматривается 4 под-
вида: subsp. maxima, subsp. americana, subsp. turbankurbis, subsp. and-
reana.
Ключ для определения подвидов
1. Растения культурные, с крупными съедобными плодами...............2.
+ Растения дикорастущие, с мелкими, горькими или безвкусными плодами . . .
...................................................subsp. andreana.
2. Растения грубого облика, мощные, стебли в 2—3 раза толще, чем у старосвет-
ских сортов, с грубым опушением; листья крупнее и грубее, с торчащими
вверх черешками, сильно зазубренные. Плоды темно-зеленые до коричневых,
иногда с деревянистой корой, иногда неправильной вытянутой формы, чаще
с бугристой поверхностью.............................subsp. americana.
+ Растения более нежного облика, менее мощные; листья с короткими черешками,
нежные, слабозазубренные; плоды округлые, с гладкой поверхностью, реже сег-
ментированной или мозговой..........................................3.
3. Плоды с крупной чалмой, деревянистой корой, сплюснутые, темно-зеленые,
иногда красные.....................................subsp. turbankurbis.
+ Плоды без чалмы, с мягкой корой, чаще сферические, гладкие, с преобладанием
светлых и зеленых окрасок............................subsp. maxima.
Подвид старосветский — subsp. maxima. — С. maxima
subsp. Feldkurbis Metzg., 1841, Landw. Pflanzenkunde : 692. — C. ma-
xima var. asiatica Zhit., 1930, Tp. по прикл. бот. ген. и сел. 23, 3 : 157.—
С. maxima gr. var. potiron Pang., 1933, У Жуковского, Земл. Тур-
ция: 547.— С. maxima subsp. asiatica (Zhit.) Pang., 1937, Теорет. осн.
сел. 3:154. — С. maxima subsp. communis Pang., 1947, БСЭ, 55:408.—
C. maxima convar. maxima Greb., 1958, Kulturpfl. 6:38.
Г6&
a
ж
Рис. 6. Разновидности подвида старосветского:
з—зимняя' б — белоплодная; в — овальная; г сероплодная; д мамонтовая, е
голландская; ж — мелкоплодная; з — китоподобная.
Тыквы этого подвида составляют сортимент Европы Азии (за
исключением западной части Китая, Тибета) и Северной Африку Они
новидностей (рис. 6).
169
Ключ для определения разновидностей
1 Плоды шаровидные или сплюснутые......................................
+ Плоды удлиненные (овальные или цилиндрические) .............................-j
2. Плоды неправильно шаровидные, очень крупные, желто-розовые . . . var. jaune.
+ Плоды сплюснутые, с вдавленностью у плодоножки.................................
3 Плоды с гладкой поверхностью................................................4
+ Плоды с неровной бугристой поверхностью.........................var. hiberna.
4 Плоды крупные, более 25 см в диаметре.......................................5
+ Плоды мелкие, 15—25 см в диаметре.................................var. petit.
5. Плоды серо-зеленые, обычно не краснеющие по созревании . . . var. maxima.
+ Плоды иной окраски..........................................................6.
6. Плоды белые......................................................var. alba.
+ Плоды розово-желтые..........................................var. hollandiae.
7. Плоды овальные (индекс не более 1,5).............................var. ovalis.
+ Плоды удлиненно-цилиндрические (индекс более 1,5)..............var. physeta.
1. Разновидность сероплодная — var. maxima.— С. maxima var.
verd Duch., 1786, in Lam., Encycl. 2:151. — C. maxima var. viridis Ser.,
1849, Fl. Jard. 2:531. — C. maxima var. megalocarpa Roem., 1846,
Synops. monogr., 2 :83. — Potiron gris Naud. 1856, Ann. Sci. Nat. (Pa-
ris) 6:23. — P. corfou Naud., 1856, 1. c. : 23. — P. pain pauvre Naud.
1856, 1. c. : 24. — C. maxima var. grisea Alef., 1866, Landw. Fl. : 215.—
C. maxima var. penebroma Alef., 1866, 1. c.: 215 et var. leucadia Alef.,
1866, 1. c.: 215. — C. maxima var. macrosphaerica Pang., 1933, у Жу-
ковского Землед. Турция: 532. — С. maxima var. murinus Filov., 1959,
Бахчеводство : 136, nom. illeg.
Плоды крупные, слабосплюснутые до среднесплюснутых и шаро-
видных, с вдавленностью у плодоножки и цветка, серо-зеленые, обычно
не краснеющие при созревании, иногда слабосегментированные, с ров-
ной гладкой поверхностью. Кора недеревянистая. Мякоть рыхлая или
средней плотности.
Распространена наиболее широко в Поволжье, на Украине, в сред-
ней полосе, Казахстане, Сибири, Средней Азии, Закавказье (СССР);
во многих странах Южной Европы, частично в северной Африке, южной
и восточной Азии и в США. Употребляется в пищу и на корм. К ней
относятся сорта: Волжская серая, Дунганская, Хайванкеды, Бирюче-
кутская 27 и др.
2. Разновидность белоплодная — var. alba Roem., 1846, Synops.
monogr. 2 : 88. — Potiron blanc de Naples Naud., 1856, 1. c. : 23. — Petit
potiron blanc de Konstantinople Naud., 1856, 1. c.: 23. — C. maxima var.
neapolotana Alef., 1866, 1. c.: 215. — C. maxima var. byzantica Alef.,
1866, 1. c.: 215. — C. maxima var. albus Filov, 1959, Бахчеводство: 136,
nom. illeg.
Плоды среднего размера, сплюснутые, гладкие, молочно-белые.
Кора недеревянистая. Мякоть средней плотности.
Распространена в СССР незначительно на Украине, Северном Кав-
казе, редко в Западной Европе. Употребляется в пищу. Сорта Белая
медовая, Безенчукская белая и некоторые другие.
3. Разновидность мамонтовая — var. jaune Duch., 1790: 1. с.: 148.—
С. maxima var. grossa Roem., 1846, 1. c.: 88. — C. maxima var. gelber
centerkurbis Metzg., 1841, Landw. Pflanz. : 691. — C. maxima var. mam-
muth Pang. 1933, у Жуковского Землед. Турция : 547.
Плоды очень крупные, гладкие, неправильно шаровидные, после
уборки сразу или вскоре желтеющие, кора недеревянистая. Семенная
полость очень большая. Мякоть рыхлая, содержит мало сухих веществ.
Распространена в СССР (на Северном Кавказе, Украине), в США,
странах Западной Европы, особенно в ГДР и ФРГ. Используют на
корм. Сорта Шестипудовая, а из иностранных Мамонт, Король мамон-
тов и др.
170
4. Разновидность голландская — var. hollandiae Naud, 1856, Ann.
Sci. Nat. (Paris) 6 : 22. — Courge chataigue Naud. 1856, 1. c. : 22. —
Courge californie Naud. 1856, 1. c.: 22.— Potiron lisse Naud. 1856,
1. c. : 23. — C. maxima var. pluviceps Alef., 1866, 1. c.: 214. — C. maxima
var. californica Alef., 1866, 1. c.: 214. — C. maxima var. hollandica Alef.,
1866, 1. c. : 215. — C. maxima var. gigantea Harz, 1855, Landw. Samen-
kunde : 796. — C. maxima var. depressa Harz, 1885, 1. c.: 796. — C. ma-
xima var. hollandica Harz, 1885, 1. c.: 796.— C. maxima var. macro-
depressa Pang., 1933, у Жуковского, Землед. Турция : 547.
Плоды очень крупные, сплюснутые, гладкие, слабосегментирован-
ные, розово-желтые, диаметром до 70 см и более. Кора обычно не-
деревянистая. Семенная полость большая. Мякоть рыхлая и средней
плотности.
Распространена в странах Западной Европы. Употребляется на
корм, реже в пищу. Сорта Голландская, Этампская и др.
5. Разновидность мелкоплодная — var. petit Duch., 1790, 1. с.: 148.—
С. maxima var. marron Naud, 1856, 1. c. :22. — Potiron ail wert Naud.,
1856, 1. c. : 21. — C. maxima var. constaniana Alef., 1866, 1. c.: 214.
Плоды сплюснутые, мелкие, не более 25 см в диаметре, гладкие.
Распространена в странах Западной Европы и США, используется в
пищевых целях.
6. Разновидность зимняя — var. hiberna Filov, var. nova.— Filov,
1966, Бюлл. Главк, бот. сада АН СССР, 63:39, sine typo. — Fructus
medii, superficies non plana, pulpa magne compacta.
Typus: Испания, Среднеазиатская опытная станция ВИР. Полу-
чен из Испании в 1934 г. (W1R).
Плоды сплюснутые, среднего размера, с неровной бугристой по-
верхностью, часто с глубокой сегментацией, с большей относительной
удельной массой, темно-зеленые, краснеющие поздно, только в про-
цессе лежки. Кора недеревянистая. Семенная полость очень малая.
Мякоть очень плотная. Плоды пригодны для очень длительного хра-
нения (до весны и больше).
Распространена в СССР (в предгорьях Кавказа), в Испании и в
других странах. Из отечественных местных известен сорт Черкесская.
В период войны из Испании в СССР был завезен сорт Испанская с
высокими вкусовыми качествами и с 10—12%-ной сахаристостью. Этот
сорт послужил основой создания серии сортов-синонимов, приспособ-
ленных для культуры в различных районах нашей страны: Столовая
зимняя А-5, Испанская 73 и др. Употребляется исключительно для пи-
щевых целей,
7. Разновидность овальная—-var. ovalis Roem., 1846, 1. с.: 89.—
Courge de Messine Naud., 1856, 1. c.: 24. — Potiron Hahre Estambouli
Naud., 1856, 1. c.: 27. — C. maxima var. messanensis Alef., 1866,
I. c.: 216. — C. maxima var. figari Alef., 1866, 1. c. :217. — C. maxima
var. elliptica Harz., 1885, Landw. Samenk.: 796. — C. maxima var. ovale
Filov, 1959, Бахчеводство; 136, nom. illeg.
Плоды овальные, крупные, гладкие, обычно без сегментации, свет-
ло-серо-зеленые, розовеющие при созревании. Кора мягкая. Мякоть
толстая, средней плотности.
Распространена в Малой Азии, хотя встречается и в нашей стране.
Используется преимущественно для кормовых целей.
8. Разновидность китоподобная —var. physeta Filov. — var nova;
Filov. 1966, Бюлл. Главн. бот. сада, АН СССР, 63:39, sino typo.— Fruc-
tus maghi physetormes. Typus: Украина, Среднеазиатская опытная стан-
ция ВИР. Получен в 1930 г. (W1R).
171
Плоды удлиненно-цилиндрические, суженные к концам, китовой
формы, очень крупные, до 1 м длиной, серо-зеленые. Кора недеревяни-
стая. Мякоть рыхлая. Встречается в СССР в виде сорта Кит. Исполь-
зуется для кормовых целей.
Подвид американский — subsp. americana Filov,
subsp. nova; Filov, 1966, Бюлл. Главн. бот. сада АН СССР, 63:39, пот.
illeg. — Plantae macrophylle. Folium rudis, erectus, longipetiolas, surace
Рис. 7. Разновидности подвида американского:
а — волчковидная; б — перувианская; в — чилийская; г — боливийская; д—бразиль-
ская; е — кустовая; ж — банановидная.
172-
Carpii salebrosus, ffiargo fassirii callusosus, cortex carpii creber lignosus,
pulpa solidus, laetus. Semen albus et badius.
Подвид отличается более крупными и грубыми листьями, неров-
ной поверхностью плодов — сильными наплывами по краям трещин,
неправильной сегментацией, некоторой деформированностью, форма
плодов коническая, однобоко-овальная, со вздутиями и выступами на
концах, иногда с начальной чалмовостью. Окраска плодов темно-зеле-
ная, очень редко краснеющая. Кора плодов часто деревянистая. Мя-
коть плотная, ярко окрашенная. Семена белые и коричневые.
Подвид включает 7 разновидностей (рис. 7), большинство из ко-
торых получены из Америки экспедициями Н. И. Вавилова, С. М. Бу-
касова и С. В. Юзепчука и изучены Н. Е. Дитеневой. Этот подвид яв-
ляется богатым исходным материалом для селекции тыквы.
Ключ для определения разновидностей
1. Растения длинноплетистые ...........................................2.
4- Растения кустовые с коротким стеблем...................var. zapollito.
2. Плоды шаровидные, слабосплюснутые или широкоовальные................3.
+ Плоды цилиндрической или волчковидной формы..........................6.
3. Плоды без чалмы или с еле заметной чалмой...........................4.
+ Плоды с ясно выраженной небольшой чалмой....................var. bresil.
4. Плоды правильно шаровидные, среднего размера, с грубой сеткой на поверх-
ности, светло-серые...................................... var. chiloensis.
+ Плоды неправильно шаровидные или сплюснутые..........................5.
5. Плоды крупные, несплюснутые, с неровной мягкой зеленой корой........
......................................................var. peruviana.
+ Плоды среднего размера, чаще сплюснутые, сегментированные, с деревянистой
буро-коричневой корой.....................................var. boliviana.
6. Плоды мелкие, узкоцилиндрические, с недеревянистой корой............
......................................................v ar. odorifera.
+ Плоды волчковидной формы, с бугристой поверхностью и деревянистой корой
..............................................................var. Ohio.
9. Разновидность чилийская — var. chiloensis Zhit., 1930, Тр. прикл.
бот., ген. сел. 23, 3: 187.
Плоды шаровидные, среднего размера, светло-серые, часто с не-
ясной бородавчатостью и очень грубой сеткой с наплывами. Остаток
пестика иногда вырастает в выпуклый конус или цилиндрик, выдаю-
щийся над поверхностью плода до 8 см. Кора недеревянистая. Мякоть
очень толстая, очень плотная, интенсивно окрашенная каротином, до-
вольно вкусная. Семена крупные, белые, реже бледно-кофейные, с тол-
стой кожурой.
Распространена в Чили и Бразилии. Благодаря достаточной скоро-
спелости и хорошим качествам мякоти представляет большой интерес
для селекционной работы. Может иметь как пищевое, так п кормовое
значение, хорошо хранится.
10. Разновидность бразильская — var. bresil Naud., 1856, 1. с.: 20.—
С. maxima var. coffeisperma Alef., 1866, 1. c.: 213. — C. maxima var.
turboviridis Filov, 1959, Бахчеводство : 138.
Сходна с предыдущей разновидностью, но плоды несколько сплюс-
нуты, часто имеют явно выраженную маленькую чалму. Семена белые
и светло-коричневые.
Распространена в Чили и Бразилии.
11. Разновидность перувианская — var. peruviana Zhit., 1930, 1. с.:
: 185.
Плоды крупные, неправильной формы, иногда трехгранные, с не-
ровной поверхностью, часто с неравномерно сегментированным основа-
нием, темно-зеленые или серо-зеленые. Кора недеревянистая. Мякоть
гонкая, грубая, малосладкая. Семена очень крупные, красно-коричне-
вые, с толстой кожурой.
173
Распространена в Перу, отчасти в Боливии. Позднеспелая.
12. Разновидность боливийская — var. boliviana Zhit., 1930, 1. с.:
.185. — С. maxima var. zapinka Millan, 1947, Darviniana, 7, 3:33.—
C. maxima var. turgida Bailey, 1948, Gentes Herbarum, 7, 5 : 470.
Плоды среднего размера, чаще сплюснутые, сегментированные,
буро-коричневые. Кора деревянистая. Мякоть тонкая, грубая, мало-
сладкая. Семена очень крупные, с толстой кожурой, красно-коричне-
вые, со светлым ободком и бахромкой у носика.
Распространена в Боливии и отчасти в Перу. Позднеспелая.
13. Разновидность кустовая — var. zapallito (Carr.) Millan, 1947,
Darvinians, 7, 3:33. — C. Zapallito Carr., 1873, Rev. Hort. 45:90.—
C. maxima convar. zapallitina Greb., 1958, Kulturpfl., 6 : 53. — C. maxima
var. deflagellatis Filov. Бахчеводство: 138.
Растения с коротким стеблем (короче, чем у кабачков и патис-
сонов), поэтому длинночерешковые листья зонтиком расходятся как бы
из одного центра, а многочисленные плоды (5—12 шт.) тесно прижаты
один к другому, один непосредственно над другим. Кора деревянистая.
Плоды мелкие (0,5 кг), сплюснутые, чаще с небольшой чалмой, зелено-
коричневые. Мякоть тонкая, суховатая, с высоким содержанием каро-
тина (10—16 мг%).
Распространена в Аргентине и Чили. Представляет большой ин-
терес для селекции тыкв вида С. maxima на кустистость с одновре-
менным увеличением каротина в плодах.
14. Разновидность банановидная — var. odorifera Veil. 1925, Fl.
Flum. 10:99. — C. maxima var. walparaiso Naud., 1856, 1. c. : 27.—
C. maxima var. valparaisiana Alef., 1866, 1. c.: 216. — C. maxima gr.
banana Castet., 1925, Proc. Amer. Soc. Hort. Sci., 22 : 338. — C. maxima
var. banana Pang., 1933, у Жуковского, Землед. Турция. : 547. — С.' ma-
xima subsp. banana Pang., 1947, БСЭ, 55:407. — C. maxima convar.
bananina Greb. 1958, Kulturpfl. 6 : 52.
Плоды мелкие, узкие, цилиндрические, длиной до 50 см, шириной
до 15 см, серо-зеленые. Кора недеревянистая. Мякоть довольно тол-
стая. Сорта этого типа возделываются в США. Ш. Ноден описал такой
сорт под названием Вальпарайзо (Чили). П. М. Жуковский [22] при-
вез аналогичные формы из Малой Азии, а мы обнаружили их в мате-
риале, завезенном из Эфиопии. Представляет интерес для исследований
как оригинальная редкая форма данного вида.
15. Разновидность волчковидная — var. ohio Naud., 1856, 1. с. : 25.—
С. maxima var. ohiloensis Alef., 1886, 1. c. : 216. — C. maxima var.
grayana Alef., 1866, 1. c.: 216. — C. maxima gr. — var. hubbard Castet.,
1925, Proc. Amer. Sci. Hort. : 338. — C. maxima subsp. gobbard Pang.,
1947, БСЭ, 55:407. — C. maxima convar. hubbardiana Greb., 1958,
Kulturpfl., 6:52.
Плоды с острыми длинными выступами на обоих концах (волчко-
видные), среднего размера, с густой мелкой бугристостью, на поверх-
ности темно-зеленые, у некоторых сортов при созревании краснеющие.
Кора деревянистая. Мякоть очень плотная.
Распространена в США и Канаде.
Подвид китайский —subsp. turbankurbis Metzg.,
1841, Landw. Pflanzkund. : 695. — C. turbaniformis Roem., 1846, Synops.
monogr., 2:87; Zhit., 1930, Тр. по прикл. бот. ген. и сел. 23, 5: 174.—
Pileocalix elegans Casp., 1848, Ann. Sci. Nat. (Paris). -3, 9 : 207. — Poti-
ron couronnes Naud., 1856, 1. c. : 20. — C. maxima subsp. turbaniformis
Pang., 1947, БСЭ, 55:407.
Подвид известен в местной культуре на западе Китая и частично
на Тибете (Китай), хотя сорта его распространены и в Европе как
174
®' ^азноВ11-®ности и формы подвида китайского С. maxima:
» в мелкоплодная красная; г — крупночалмовая; д —
среднечалмовая.
Рис. Г ________
а, б — мелкоплодная зеленая;
декоративные растения. Он имеет нежные вегетативные органы и чал-
мовидные сплюснутые плоды, обычно вначале темно-зеленые, при со-
зревании краснеющие полосами или сплошь, принимая в последнем
случае ярко-красную окраску. Кора преимущественно деревянистая.
Мякоть тонкая, плотная, часто сладкая и с высоким содержанием ка-
ротина. Семена обычные, белые. Подвид включает 4 разновидности
(рис. 8).
1.
Ключ для определения разновидностей
крупные с поперечным диаметром более 35 см .... var. turbankurbis.
в диаметре меньше 35 см......................................................2.
диаметром 23-35 см..........................................var. nouvebresil.
в диаметре до 23 см ......... ........................................• • 3.
----- ~ л .............var. constncta.
при созревании зеленые.....................................var. turbanopetit.
Плоды
Плоды
Плоды
Плоды о
Плоды при созревании красные, с белой чалмой
Плоды при созревании зеленые...........
16 Разновидность крупночалмовая — var. turbankurbis. - С. та-
полосками от плодоножки к чал й части плода. Попереч-
чалмы обычно отличается от окраин »
175
2.
3.
maxima
узкими
Рис. 9. Разновидность var. andreana подвида дикорастущего С. maxima:
а — листья, б — плоды.
ный диаметр плодов от 35 см и больше. Кора деревянистая. Мякоть
тонкая, плотная, иногда приятного вкуса.
В местном земледелии известна на западе Китая, а также в Япо-
нии. Эти тыквы часто ошибочно называют турецкими, видимо, из-за
чалмы.
17. Разновидность среднечалмовая — var. nouvebresil Naud, 1856,
I. с.. 20.
176
Имеет сходство с предыдущей разновидностью. Отличается от нее
менее крупными размерами плодов (23—35 см).
18. Разновидность мелкоплодная красная —var. constricta Alef.,
1866, 1. с. : 213. — С. maxima var. rubra erithrocarpa Harz., 1885,1. c.: 801.
Плоды меньше 23 см, красные, чалма иногда занимает более поло-
вины плода. Кора деревянистая или мягкая. Разводится в Японии и в
других странах, преимущественно в ботанических садах с декоратив-
ной целью.
19. Разновидность мелкочалмовая зеленая — var. turbanopetit
Naud., 1856, 1. с.: 20. — С. maxima var. lignosa Harz., 1885, 1. c. :801.
Размер плодов, как у предыдущей разновидности, сплюснутые, зе-
леные. Чалма всегда занимает менее половины плода. Кора деревя-
нистая.
Подвид д и ко р а сту щи й — s u b s р. andreana (Naud.)
Filov, comb. nova. — Cucurbita andreana Naud., 1896, Rev. Hort. 68:542.
Сюда относится пока только одна разновидность.
20. Разновидность андреана — var. andreana. — С. maxima convar.
parvifructina Greb, 1958, Kulturpfl. 6:50.
Растения дикорастущие. Вегетативные органы столь же крупные,
как и у культурных форм С. maxima. Плети длинные, листья крупные,
правильно округлые, без вырезов (рис. 9,а). Пластинки листа серо-
зеленые с интенсивно выраженной более светлой пятнистостью в углах
разветвления жилок. Цветки несколько мельче, чем у культурных форм.
Плоды мелкие (масса 100—200 г), овально-яйцевидные (рис. 9,6), зе-
леные, с мелкими светлыми крапинками и тонкими полосками от
цветка к плодоножке, горькие.
Распространен в тропической части Южной Америки, Уругвае и
Аргентине. Известны две формы: с бочонкообразными, более мелкими
плодами (100 г) и с обратнояйцевидными, более крупными (200 г).
Вид эквадорский — С. eguadorensis Cutter et Whitaker
1968, Ann. Missouri Bot. Gard. 55, 3:392.
Растения дикорастущие, однолетние. Стебель лазающий, длиной
10 м. Листья шириной 12—32 см, широкоовальные, почковидные. Че-
решки длиной 8—16 см. Цветки длиной 2—5 см, венчик шириной 5—
8 см, лопасти венчика острые, длиной 1—2 см. Плоды круглые, 16 см
в диаметре, кремовые или оранжево-зеленые. Мякоть плодов белая, не-
горькая.
Семена овальные, белые или коричневые, длиной 1,1—1,5 см,
шириной 0,7—0,8 см, по величине промежуточные между культурной
С. maxima и ее дикой разновидностью var. andreana.
Обнаружена на севере Южной Америки и найдена при раскопках
в слоях докукурузного периода на побережье Перу. Авторы считают
ее близкой к виду С. maxima.
ПОДРОД СЕВЕРОАМЕРИКАНСКИЙ-SUBGEN. CUCURBITA
Отличается сильной шероховатостью и ребристостью стебля, че-
решков, плодоножек. Опушение преимущественно шиповидное, кора
плодов деревянистая. Подрод представлен одним культурным и одним
дикорастущим видами.
Ключ для определения видов
1. Плоды у культурных форм крупные. Включает и мелкоплодные дикорастущие
формы, у которых плоды могут быть светлые или с ярким фоном зеленого
рисунка ...................................................С. реро.
+ Крупноплодных форм нет. Плоды темно-зеленые. Фон рисунка слабозаметный,
тусклый ...............................................С. kellyana.
177
Вид твердокорый — С. реро L.
1753, Sp. PI.: 1010, S. melopepo L. et C. verrucosa L., 1753,
1. c.: 1010. — C. ovifera L., 1767, Mant.: 126. — C. mammeata Molina,
1782, Saggio Chile: 133. — C. pepo, ^-polymorpha Duch., 1786, in Lam.,
Encycl. 2:152. — C. mammusa J. F. Gmel., 1791, Syst.:101. — Pepo ver-
rucosus Moench, P. vulgaris Mornch et P. melopepo Moench, 1794,
Meth.: 653. — Cucurbita fasstosa Salisb., 1786, Prodr.: 157. — C. subver-
rucosa Willd., 1805, Sp. Pl. 4:609. — C. durantia Willd., 1805, 1. c. : 607.—
C. pyxidaris DC., 1815, Fl. Fr. 3:693. — Pepo pyxidaris Poir. 1818, in
Diet. Sci. nat. 8:232. — C. esculenta S. F. Gray, 1821, Nat., Arr. Brit,
pl. 2:552. — C. ceratoceras Haberle, Spix et Mart., 1823, Reise Brasil.
1:2. — C. courgero Ser., 1825, Mem. Soc. Phys. Geneve, 3:25. — Pepo
citrullus Sager., 1826, Ann. Sci. Nat. (Paris) 1, 9:311. — Cucumis pepo
Dum., 1827, FL Belg.: 54. — Cucurbita venosa Descourt, 1827, Fl. Antill.
5 :79. — C. maxima var. courgero Ser., 1828, in DC. Prodr. 3:316.—
C. clodiensis Nacc, 1828, FL Ven. 5:52. — C. oblonga Link, 1829, Handb.
1:643. — C. tuberculosa Schrad., 1833, Ind. Sem. Hort. Gottingo: 3.—
C. urnigera Schrad., 1836, Linnaea, 10: 110. — C. asteroides Schrad., 1838,
Linnaea, 12:407. — C. cucumifera Schrad., 1838, 1. c.:407. — C. elongata
Schrad., 1838, 1. c.: 407. — C. citrullus Roem., 1846, Synops. 2:84.—
C. grisea Roem., 1846, 1. c.: 86. — C. pomiformis Roem., 1846, 1. c.: 85.—
C. pileoformis Roem., 1946, 1. c.: 86. — Tristemon texanum Scheele, 1848,
Linnaea, 21:586. — Cucurbita texana A. Gray, 1850, Journ. Boston. Soc.
Nat. Hist. 6: 193.
Описание вида. Стебли ребристые, усаженные твердыми густыми
колючими шипиками. Листья пятилопастные, концы лопастей остро-
угольные, края листа глубокопильчатые, у культурных доли крупные,
у дикорастущих более мелкие.
Цветки колокольчатые; чашелистики узкие, почти линейные; ле-
пестки, сросшиеся на 2/з длины венчика, концы их острые. Плоды у
культурных форм крупные, безрисунчатые или полосатые, у дикорас-
тущих и декоративных мелкие (с куриное яйцо), различной формы,
белые, желтые или пятнистые. Семена кремовые, овальные, с ясным
рубчиком по краям. Мякоть кремовая или оранжевая.
Изменчивость признаков. Характерная особенность вида — деревя-
нистая кора плодов. С этим признаком связано обилие форм по раз-
мерам и внешнему виду плодов. Виды С. maxima и С. moschata при
окультуривании в значительной степени утеряли деревянистость коры,
а С. реро почти полностью его сохранил.
Преждевременное одревеснение коры плодов С. реро до полного
окончания их роста не позволяет им достигать таких крупных разме-
ров, как плоды некоторых сортов С. maxima или осенне-зимних сред-
неазиатских дынь.
Разнообразие рисунков, окрасок плодов С. реро также связано с
ее твердокоростью, как и у других тыквенных. Так, арбуз и летние
среднеазиатские дыни с твердой корой имеют разнообразные рисунки,
которых нет у дынь с мягкой корой.
Узкий широтный ареал вида С. реро, обусловленный его приспо-
собленностью к умеренно теплым и мезофитным условиям, а также
консерватизм структурности определили сравнительно слабую по
сравнению с другими видами экологическую изменчивость внутривидо-
вых форм. Большое разнообразие последних обусловлено главным об-
разом генными сочетаниями и деревянистостью коры плодов. В про-
тивоположность мелкоплодным формам крупноплодные в большинстве
178
имеют слабоовальную и шаровидную форму плодов и слабую измен-
чивость их размеров.
Скороспелость С. реро обусловила хорошее вызревание плодов с
разрушением хлорофильной окраски коры, обнажающим лимонно-жел-
тый и оранжевый цвет каротиноидов.
Консистенция мякоти и семена изменяются сравнительно слабо.
Внутривидовая классификация. История изучения. Тыкву
С. реро ботаники эпохи Возрождения, вероятно, узнали раньше других
видов. Так, Л. Фукс [128] под названием Cucumis turcicus описывает
кабачок. И. Камерариус [111] под Cucurbita indica описывает полевую
форму С. реро var. реро. М. Лобель [156] уже различает формы
С. реро под названиями Реро oblongus и Р. rotundas compressus. Р. До-
доней [121] приводит формы: Реро maior oblongus (var. реро),
Р. maior minor (декоративная мелкоплоидная), Р. latus (патиссон),
Р. silvestris (декоративная форма); К. Баухин [104] — Реро oblongus
(var. реро), Р. minor clypeatus (патиссон), Cucurbita verrucosa (бо-
родавчатая); Д. Далешамп [118] — Courge Inde longus (var. pepo, или
кабачок), C. pleine vertus (крупная круглая бородавчатая), S. frangee
orielles (патиссон). Д. Турнефорт [195] дает уже около 15 видов
(форм) С. реро.
В 1753 г. Линней узаконивает видовое название этой тыквы — Cu-
curbita реро L., но наряду с ним он дает еще 3 вида, впоследствии во-
шедшие в состав С. реро —С. verrucosa (бородавчатая мелкоплодная
декоративная разновидность), С. melopepo (кустовые формы С. реро
с деформированными плодами — патиссоны) и С. ovifera L. (мелко-
плодная яйцевидная форма).
В последующем издании работы К. Линнея «Species plantarum»
К. Вильденов добавляет еще 3 вида, также идентифицированных позд-
нее с С. реро.
М. Дюшесн [122] в составе вида С. реро polymorpha выделил
5 форм, главным образом мелкоплодных: 1) coloquinelles — с шаровид-
ными плодами; 2) cougourdette — с грушевидными плодами; 3) barba-
rine — бородавчатая тыква; 4) giraumons — кабачки; 5) patissons —
патиссоны.
Д. Пуарет [169] также классифицирует мелкоплодные формы
С. реро, детализируя кабачки и патиссоны еще на сорта.
Н. Серенж [182, 183] признает из ранее описанных только 10 ви-
дов, относящихся к С. реро, детализируя только линнеевский С. ovi-
fera.
Е. Шпах [194] низводит предыдущие виды в разновидности: С. реро
рероп — обычные полевые крупноплодные, coloquinelle — мелкоплодные
тыквы, cougordette (var. ovifera) barbarine (verrucosa), giraumont —
кабачки.
И. Метцгер [158] дает 6 подвидов, относящихся к С. реро: Feldkur-
bis — крупноплодные полевые тыквы, Langerkurbis — кабачки, Schild-
kurbis — патиссоны, Apfelkurbis — яблоковидные мелкоплодные, Birn-
kurbis — грушевидные, Eierkurbis — яйцевидные.
М. Ремер [176] дает наиболее подробную классификацию мелких
декоративных форм С. реро, заимствуя их у других авторов. Так, у
патиссонов он выделяет 7 форм, у грушевидного вида С. pixidaris
(ныне разновидность)—6 форм по окраске плодов, в пределах С. ovi-
fera (ныне разновидность)—^ формы, а всего в пределах линнеевского
С. реро объединяет 14 видов и приводит еще 18 форм.
Ш. Ноден [165], создавая классификацию тыкв, предложил очень
стройную систему С. реро: 1) Courgeron —плоды сплюснутые, сред-
него размера; 2) Citrouille — плоды крупные пли средние, овальные,
179
дается 4 формы (сорта); 3)—Giraumon — кабачки, описано 9 сортов;
4) Patisson — патиссоны, описано 3 сорта; 5) Orangin — апельсиновая
тыква; 6) Barbarine — бородавчатая (С. verrucosa L.); 7) Coloquncllc —
плоды мелкие, овальные или грушевидные.
Ф. Алефельд в основном принял формы Нодена и других авторов,
мало внеся в классификацию своих таксонов. Все формы-сорта Но-
дена он располагает в 8 групп: 1) Giromontia — кабачки (12 сортов);
2) Citrullina— овальные полевые тыквы; 3) Melopepo — патиссоны и
сплюснутые формы (22 сорта), 4) Clypeata — щитовидные (патиссоны),
7 сортов; 5) Pomiformis — яблоковидные тыквы; 6) Pyriformis — гру-
шевидные мелкие тыквы; 7) Ovifera — яйцевидные тыквы (5 сортов);
8) Verrucosa — бородавчатые тыквы (8 форм).
Е. Харц [143] также распределил формы Ш. Нодена и некоторых
других авторов в 7 групп: 1) Subrotunda Willd.; 2) Citrata — плоды
крупные и средние; 3) Oblonga Willd. — кабачки, с описанием 9 сортов;
4) Melopepo L. — патиссоны; 5) Orangina Ser. (включая var. aurantia);
6) Colocynthoides Ser — яблоковидные тыквы; 7) Ovifera L.
Агрономическая типизация тыкв внутри С. реро Е. Кастеттера
[112] является одновременно и ботанической: 1) полевые тыквы — со-
временная var. реро; 2) патиссоны; 3) крукнеки; 4) кабачки; 5) форт-
гукская тыква с мелкими плодами, суживающимися к плодоножке, и с
продольными угольчатыми ребрами; 6) группа Королева стола с мел-
кими плодами, имеющими глубокую сегментированность.
Наряду с открытием многих диких видов Л. Бейли [100] выде-
ляет две уже известные ранее разновидности С. реро: var. condensa
Bailey—крукнек и var. ovifera Bailey ( = C. ovifera L.).
H. E. Житенева [20] в пределах вида С. реро сначала выделяет
только 2 разновидности: var. atnericana Zhit. и var. turcica Zhit., но в
следующей работе [21], посвященной американским тыквам, она дает
новую классификацию С. реро: 1) gr. var. citrulina: var. costata flava —
ребристая желтая, var. costata viridis— ребристая зеленая, var. costata
maculata— ребристая полосатая; 2) gr. var. giromontia: var. viridis —
темно-зеленая; var. variegata — полосатая; 3) gr. var. patisson; 4) gr.
var. crookneek; 5) gr. var. microcarpa.
В более поздней работе некоторые разновидности тыкв Н. Е. Жи-
теневой, К. И. Пангало [59] переводит в группы разновидностей. Де-
тализируя их, он дает свою классификацию разновидностей двух групп
Н. Е. Житеневой, но без учета предыдущих авторов; его разновидности,
по существу, являются сортами: 1) gr. var. citrullina: var. flava Pang.—
плоды светло-желтые; var. aurantica Pang. — оранжевые, var. flavover-
rucosa Pang. — плоды мелкие, бородавчатые, светло-желтые, var. аи-
rantiaca-verrulosa Pang. — оранжевые, var. aurantiaca costata Pang.—
обратнопирамидальные, оранжевые, var. nigro-costata Pang. — темно-
зеленые; 2) gr. var. giromontia: var. macro-leuca Pang. — белые, круп-
ные, var. nigro-leuca Pang. — плоды белые, мельче, чем греческие ка-
бачки, var. nigro-xanta Pang. — плоды желтые — кабачки без рисунка,
var. macro-xanta Pang. — плоды желтые, крупные, без рисунка, var.
macro-pirochra Pang. — оранжевые, без рисунка, var. micro-pirochra
Pang. — оранжевые, мельче, чем у предыдущей разновидности, без ри-
сунка, var. maculata Pang. — плоды с рисунком — итальянские кабачки,
var. viridis Pang. — плоды без рисунка, черно-зеленые.
В следующей работе К. И. Пангало [62] рассматривает разновид-
ности Н. Е. Житеневой в ранге подвидов.
После Линнея наиболее ценна работа Ш. Нодена, установившая
наибольшее число внутривидовых форм. Правда, подробную классифи-
180
кацию тыквы до Нодена приводит М. Ремер, но она касается главным
образом мелкоплодных и декоративных форм, которые в основном яв-
ляются гибридами и потому менее постоянны в своих признаках.
Н. Е. Житенева, а затем и И. Гребенщиков в работах по клас-
сификации тыкв не учитывали систем предыдущих исследователей. Од-
нако в работе И. Гребенщикова [132] следует признать правильным
разделение им вида С. реро на 2 группы: плетистых и кустовых тыкв,
но перевод же ранее открытых разновидностей в группы (convar.) сде-
лан им без необходимости: 1-я группа (longicaulis Greb.) — длиннопле-
тистые тыквы: convar. microcarpina Greb. — мелкоплетистые, convar.
citrullina Greb. — крупноплодные, var. styriaca Greb. — голосеменные;
2-я группа (brevicaulis Greb.) —кустовые: convar. giromontina Greb.—
кабачки, convar. patissonina Greb. (var. radiara Nois.)—патиссоны,
var. torticolles Alef. — крукнеки.
О. Шифрис [186] делит вид С. реро на 3 группы: 1) дикорастущие
формы, 2) культурные, 3) декоративные мелкоплодные.
Последняя классификация. Включает 4 подвида: длинно-
плетистый — subsp. реро, кустовой (короткоплетистый)—subsp. brevi-
caulis Greb., декоративный —subsp. polymorpha (Duch.) Filow и дико-
растущий — subsp. texana Filov.
Ключ для определения подвидов
1. Стебель длинный (более 1 м), лежачий, растения стелющиеся......2.
4- Стебель короче 1 м, вначале прямостоячий, позже иногда стелющийся ....
................................................... subsp. brevicaulis.
2. Плоды мелкие, в зрелом виде не более 300 г.....................3.
4- Плоды крупные (более 1 кг)...............................subsp. реро.
3. Растения дикорастущие, плоды мелкие, горькие........subsp. texana.
4- Растения, культивируемые как декоративные, плоды не всегда горькие ....
...................................................subsp. polymorpha.
Подвид длинноплетистый — subsp. реро. — С. реро
subsp. longicaulis Greb., 1950, in Zuchter. 20, 7, 8: 194. — C. pepo subsp.
flagelliformis Filov, 1959, Бахчеводство: 139.
Распространен в лесостепной полосе СССР, в Балканских стра-
нах, в средней части США и на севере Мексики. Подвид включает
7 разновидностей.
Ключ для определения разновидностей
1. Плоды овальные или слабоовальные.........................................2.
+ Плоды иной формы..........................................................3.
2. Семена с твердой кожурой.......................................var. реро.
4- Семена с прозрачнопленчатой кожурой..........................var. styriaca.
3. Плоды сплюснутые или шаровидные.......................................4.
4- Плоды цилиндрические (индекс более 1,5)....................var. cylindrica.
4. Плоды сплюснутые .....................................................5.
4- Плоды шаровидные......................................................6.
5. Плоды с гладкой поверхностью.............................var. complanatus.
4- Плоды резко сегментированные...................................var. тагос.
6. Плоды в поперечном диаметре более 23 см....................var. courgeron.
4- Плоды более мелкие, в диаметре 23 см...........................var. table.
1. Разновидность овальная—var. реро.— С. реро var. рероп Spach,
1838, Hist. Nat. veg. Phan. 6:199.— C. pepo subsp. feldcurbis Metzg..
1841, Landw. Pflanz.: 692. — C. pepo var. ovalis Roem., 1846, Fam. natur.
veget. 2:89. — C. pepo var. citruille Naud. 1856, Ann. Sci Nat. (Paris),
6:35. — C. pepo var. citrullina Alef., 1866. Landw. Flora : 219. — C. pepo
gr. Field and Pie Castet, 1925, Proc. Amcr. Soc. Hort. Sci.: 338. —
C. pepo convar. citrullina Greb., 1950, Zuchter, 20, 7, 8:198. — C. pepo
Convar. pepo Greb., 1958, Kulturpflanz. 6:55.
181
Рис. 10. Формы плода овальной разновидности С. реро.
а — сплюснутая; б — шаровидная; в — бочкообразная; г — грушевидная; д — дыневид-
ная.
Плоды крупные, овальные или слабоовальные (рис. 10), ярко-жел-
тые или оранжевые, чаще с пятнисто-полосатым зеленым рисунком, при
созревании оранжевым, всегда темнее фона. Поверхность плодов слабо
сегментирована у плодоножки, выступы сегментов основания плода
иногда сохраняются в очень слабом виде на поверхности. Мякоть сред-
ней плотности, оранжевая. Семена с твердой кожурой.
Широко распространена культурная разновидность, которая яв-
ляется самой типичной формой вида. Сюда относится известный сорт
Мозолеевская. Сортимент этой разновидности предпочитают выращи-
вать на Украине (СССР), в Румынии, Югославии.
2. Разновидность голосеменная—var. styriaca Greb., 1950, Zuchter,
20, 7, 8:199.
Плоды такие же, как у предыдущей разновидности. Отличается
только отсутствием у семян плотной кожуры, вместо которой имеется
тонкая прозрачная пленочка, при высыхании отпадающая.
Встречается редко, но там же, где и предыдущая разновидность.
3. Разновидность овощная— var. table (Castet.) Filov, comb. nova.—
C. pepo gr. table Queen Castet., 1925, Proc. Amer. Soc. Hort. 338. —
C. pepo var. culinaris Filov, 1966. Бюл. Главк. Бот. сада АН СССР,
63 : 39, descr. ross.
Плоды шаровидные, мельче (1—3 кг), чем у var. реро.
Используется главным образом в недозрелом виде. Разводят в
США и странах Западной Европы.
4. Разновидность цилиндрическая — var. cylindricum Filov, var.
nova. —C. pepo var. cylindrica Filov, 1966, Бюл. Главк. Бот. сада
АН СССР, 69:39, nom. illeg. — Plantae longicaules. Fructus medii cy-
lindrici.
Typus: Среднеазиатская on. станция ВИР. Получен в 1924 г.
(WIR).
182
Плоды по форме сходны с кабачками, иногда крупнее и толще по-
следних. Их называют плетистыми кабачками. Разновидность произо-
шла, очевидно, в результате гибридизации или свободного переопыле-
ния кустовых кабачков с плетистыми тыквами. Используется как и
плоды предыдущей разновидности. Сюда относятся сорта: Кульджин-
ский кабачок, Месопотамский кабачок, кабачок Голосемянный и другие,
главным образом гибридные формы «плетистые».
5. Разновидность шароплодная—var. courgeron Naud., 1856,
1. с.: 35.
Плоды крупные, шаровидные. Сюда относится известный амери-
канский сорт Коннектикутская.
6. Разновидность сплюснутоплодная—var. complanatus Filov, var.
nowa. — Carpus complanata, leiocarpus.
Плоды сплюснутые, крупные и среднего размера, гладкие.
Typus: США, Репродукция Среднеазиатской опытной станции ВИР.
Получена в 1924 г. (WIR).
7. Разновидность сегментированная—var. maroc Naud., 1856,
I. с.: 35.
Плоды сплюснутые, глубоко сегментированные, крупные, с мелко-
пятнистым рисунком кремового цвета на зеленом фоне.
Подвид кустовой — subsp. brevicaulis* Greb., 1950,
Zuchter, 20, 7/8:199. — C. pepo subsp. compactus Filov, 1959, Бахчевод-
ство : 139.
Кустовой подвид оригинален своим крайне укороченным стеблем.
Такие формы с заторможенным ростом стебля встречаются у многих
растений. Их появление можно объяснить или резкими изменениями
условий выращивания, или результатом гибридизации. Путь образова-
ния кустовых тыкв неизвестен, но, по мнению Т. Вайтекера и Г. Кар-
тера [206], они появились на совершенно иной территории, чем пле-
тистые формы этого вида, а именно в восточных районах Северной Аме-
рики. Сюда входят 5 разновидностей (рис. 11).
Ключ для определения разновидностей
1. Плоды цилиндрические (индекс более 1,5)..................var. giraumons.
+ Плоды иной формы.......................................................2.
2. Плоды сплюснутые, тарельчатые и медузовидные.............var. melopepo.
+ Плоды иной формы.......................................................3.
3. Плоды клиновидные, заостренные с одного конца, мелкие, ребристые ....
........................................................ var. fordhuk.
+ Плоды иной формы.......................................................4.
4. Плоды удлиненные, искривленные, утолщенные на цветочном конце, бугристые
.......................................................var. subverrucosa.
+ Плоды шаровидные, овальные или сплюснутые, растения крупные, окультуренные
....................................................... var. soredlformis.
8. Разновидность кабачки—var. giraumons Duch., 1790, in Zam.
Encycl. 2:148. — C. pepo subsp. langekurbis Metzg., 1841, Landw.
Pflanz.: 694.— C. pepo var. oblonge Roem., 1846, Synops. monogr. 2 : 84,—
C. pepo gr. var. giromontia Alef., 1866, 1. c. :217. — C. pepo gr. var. cly-
peata Alef., 1866, 1. c.: 222. — C. pepo gr. vegetable Marrow. Castet.,
1925, Proc. Soc. Hort. Sci.: 338. — C. pepo subsp. giromontia Pang., 1947,
БСЭ, 55:407. — C. pepo convar. giromontina Greb., 1950, Zuchter, 20, 7,
8:199.
Плоды цилиндрические (индекс более 1,5). Мякоть по созревании
семян белая, безвкусная. Употребляют в недозрелом состоянии как
овощной продукт или на корм.
* Нам не удалось определить формы плодов у типичной разновидности (var.
brevicaulis); автор же подвида не указал их.
183
Рис. 11. Основные разновидности подвида кустового С. реро:
а —кабачок; б — фордгукская; в — крупноплодная (овощная); г—патиссон;
д — неветвящийся кабачок; е — крукнек.
Имеются 2 типа разновидности: коротковетвистый — с короткими
боковыми побегами 1 порядка (рис. 11, а) и предельный (не ветвя-
щийся)— без боковых побегов (рис. 11,д). У последних плоды длин-
нее, а куст более разреженный.
Распространена в овощной культуре очень широко: почти во всех
странах мира, на разных широтах в СССР, наиболее широко возделы-
вают сорт Грибовские.
9. Разновидность патиссоны — var. melopepo (L.), Filov, comb,
nova. — C. melopepo L., 1753, Sp. Pl.: 1010. 148. — C. pepo patissons
Duch., 1790, in Lam. Encycl. 2: 152. — C. pepo subsp. schildkurbis Metzg.,
1841, Landw. Pflanz.: 694.— C. pepo subgen. Melopepo Roem. Synops.
monogr.: 87. — C. pepo gr. var. Melopepo Alef., 1866, 1. c.: 220. — C. pepo
var. condensa Bailey, 1930, Stand, cycl. hort. 1 :910. — C. pepo gr. scallop
Castet., 1925, Proc. Amer. Soc. Hort. Sci. 22:338.— C. pepo gr. Pattyran
Castet. et Ervin, 1927, Agr. Exp. St. Iowa, bul. 244: 126, —C. pepo subsp.
pressa Pang., 1947, БСЭ, 55:407, — C. pepo var. radiata Greb., 1950,
Zuchter, 20, 7, 8 : 200.
Плоды сильно сплюснутые, от дисковидных до тарельчатых
(рис. 11,г), иногда с загнутыми вперед или назад краями, чаще узор-
чато-сегментированными.
Мякоть при созревании семян белая, безвкусная. Употребляют в
184
недозрелом виде. Распространена в СССР, странах Западной Европы
и Северной Америке. В нашей стране наиболее распространен сорт Бе-
лые 13.
10. Разновидность кривошейки, или крукнек — var. subverrucosa
Willd., 1805. Sp. Pl.: 606. — С. pepo cou tors Naud., 1856, 1. c. 6:40.—
C. pepo gr. summer Croocknek Cast, et., 1925, Proc. Amer. Soc. Hort.
Sci. : 338. — C. pepo var. condensa Bailey, 1930, Stand. Cycl. hort.
1:910. — C. pepo subsp. curvata Pang., 1947, БСЭ, 55 :407. — C. pepo
torticolles Greb., 1950, Zucter, 20, 7, 8:201.
Плоды удлиненные, тонкие (индекс более 2), искривленные, утол-
щенные на цветочном конце (рис. 11, е). Поверхность их густо бородав-
чатая, ярко-оранжевая или белая. Мякоть при созревании семян без-
вкусная. Употребляют в недозрелом состоянии. Распространена в Се-
верной Америке.
И. Разновидность крупноплодная— var. sorediformis Filov, var.
nova, Filov, 1966, в Бюл. Главн. Бот. сада АН СССР, 63:39, sine typo.—
Plantae fruticantes. Fructus magni rotundi.
Typus: Днепропетровская овощная опытная станция. 1957, к-2732,
Галка А. Т. (WIR).
Плоды крупные, овальные или шаровидные, сплюснутые (рис. 11, в),
по остальным признакам близки к другим разновидностям данного под-
вида. Эта разновидность существует недавно. Она получена в резуль-
тате гибридизации кабачков с крупноплодными тыквами в целях созда-
ния кустовых сортов этого вида с крупными плодами. Сорт Днепропет-
ровская кустовая и другие перспективны для механизированного ухода
за растениями. В нашей стране плоды этой разновидности распростра-
нены на юге Украины.
12. Разновидность фордгукская — var. fordhuk Cast., 1925, Proc.
Amer. Soc. Hort. Sci. 22 : 338.
Плоды короткие, мелкие, клиновидные, суживающиеся к цветочному
концу, с продольными ребрами (рис. 11,6). Появились, очевидно, в
результате гибридизации патиссонов с другими формами тыкв. Разво-
дятся в Северной Америке.
Подвид декоративный —subsp. polymorpha (Duch.)
Filov, comb. nova. — C. polymorpha Duch., 1790, 1. c.: 148. — C. pepo
gr. var. microcarpa Zhit., 1930, у Букасова Культ, раст. Мексики, Гват.
и Колумб.: 312. С. реро subsp. ornamentalis Filov, 1966, 1. с.: 39, sine
typo.
Плоды мелкие (масса 50—300 г), с деревянистой корой, несъедоб-
ные, имеют только декоративное значение. Этот подвид, несомненно,
получен в результате гибридизации. При скрещивании культурных форм
с дикими или плетистых с кустовыми получают большое разнообразие
самых неожиданных форм. Их особенностью является частое несоответ-
ствие различных органов растения по размерам. Именно это и поста-
вило Л. Бейли в тупик перед загадкой многообразной изменчивости
данного вида. Эксперименты по гибридизации или результаты свобод-
ного опыления форм этого вида между собой позволили накопить боль-
шое количество причудливых форм в декоративных целях, которые лишь
приближенно можно группировать в разновидности. Поэтому мы при-
водим лишь 8 наиболее отчетливых разновидностей.
Ключ для определения разновидностей
1. Плоды с крупнобородавчатой поверхностью....................var. verrucosa.
+ Плоды с гладкой поверхностью.............................................2.
2. Плоды шаровидные или сплюснутые, а если грушевидные, тогда окраска
двуцветная ...............................................................3.
+ Плоды иной формы.........................................................5.
185
3. Плоды оранжевые .................................
4- Плоды белые или со слабо-зеленоватыми полосами
4. Плоды одноцветные..............................
4- Плоды двуцветные, верхняя их половина оранжевая
5. Плоды яйцевидные................................
.var. deprimee.
var. aurantica
• • var. bicolor
var. polymorpha.
4.
с вытянутым концом в плодоножке
грушевидные.....................
цилиндрические или бутылковидные
var. cougourdette. 6
. . var. oblonge.
7 ) Duch. in
C. verrucosa L. Sp. Pl. 2:1010.— C. pepo
Плоды
Плоды
Плоды ,
13. Разновидность бородавчатая—var. verrucosa^(LJ
Lam., 1790, Encycl. 2:148. — C. ------ Г ,
var. barbarine Spach, 1838, Hist. Nat. veg. Phan.' 6.214. C. pepo gr,
var. verrucosa Alef., 1866, 1. c.: 225.
6.
Плоды мелкие (50—300 г), крупнобородавчатые. nv.H .
14. Разновидность мандариновая — var. aurantica (willd.) Alef.,
1866, 1. c.: 223. — C. aurantica Willd., 1805, Sp. Pl. 4:607.— C. pep0
subsp. apfelkurbis Metzg., 1841, Landw. Pflanz.: 698. C. pepo c. pomi-
formis Roem., 1846, Synops. monogr., 2:85. — C. pepo gr. var. pomiformis
Alef., 1866, 1. c.: 223. — C. pepo convar. microcarpina Greb., 1950, Zucher,
20, 7, 8:198.
Плоды мелкие (100—150 г), величиной с апельсин, шаровидные,
при созревании оранжевые.
15. Разновидность яйцевидная—var. polymorpha.— С. реро (L.)
ovifera (L.) Bailey, 1930, Stand. Encycl. Hort. 1:910. — C. pepo var.
puridaris Duch., 1790, 1. c.: 148. — C. pepo var. coloquites Poir., 1818,
Diet. Sci. Nat. 8:232. — C. pepo var. ovata Roem. et var. subglobosa
Roem., 1846, 1. c.: 86. — C. pepo subsp. eierkurbis Metzg., 1841, Landw.
Pflanz.: 699. — C. pepo gr. var. ovifera Alef., 1866, 1. c.: 224. — C. pepo
convar. microcarpina Greb., 1950, 1. c.: 198.
Плоды яйцевидные.
16. Разновидность грушевидная—var. cougourdette Duch., 1790, in
Lam. Diet. Encycl. Bot. 2: 152. — C. pepo subsp. Birnkurbis Metzg., 1841,
Landw. Pflanz.: 699. — C. pyxidaris Roem., 1846, Synops. monogr.: 86.—
C. ovifera var. flava Roem., 1846, 1. c.: 86. — C. pepo gr. var. pyriformis
Alef., 1866, Landw. Fl.: 224. — C. pepo convar. microcarpin Greb., 1950,
1. c.: 198.
Плоды мелкие (100—300 г), грушевидные.
17. Разновидность цилиндрическая — var. oblonge (Ser.) Filov, comb,
nova. — C. ovifera oblonga Ser., 1849, Fl. Jard. 2:538. — Curee allongee
Naud., 1856, 1. c.: 39.
Плоды цилиндрические или бутылковидные.
18. Разновидность белошаровидная—var. deprimee Ser., 1849, Fl.
Jard. 2: 538.
Плоды шаровидно-сплюснутые, 6 см высоты и 7 см в поперечнике,
белые или с зелено-пестрыми полосами на светло-желтом фоне.
19. Разновидность двуцветная—var. bicolor Naud., 1856, I. с.:39.
Плоды шаровидные или грушевидные, при созревании оранжевые
с цветочного конца и зеленые от плодоножки. Оранжевая окраска по-
является позднее зеленой, причем нарастает постепенно и доходит до
середины плода, а иногда и дальше. При этом окраски не сливаются,
образуя четкую границу.
Подвид дикорастущий —subsp. texana (A Grey) Filov,
comb^nova.-C. texana A. Grey, 1850, in Boston, Journ. Soc Nat. Hist,
t 80 пот"™/ a^eStlS 2ЬЛ’ 1930’ TP’ ПРИКЛ- бот- ген. сел. 23,
nom ilfcg nUd ~C Pep° SUbsp- agrestis <Zhit ) Fibv, 1966, 1. c.:37,
186
Подвид характеризуется формами, произрастающими в диком виде
и связанными с экологическими условиями в районах обитания. У них
сохраняются равномерные соотношения между размерами отдельных
органов: при мелких плодах мельче листья, тоньше плети и т. д. Плоды
мелкие. Различные разновидности диких тыкв обычно принимались
за самостоятельные виды. В подвид входят 3 разновидности.
Ключ для определения разновидностей
1. Вырезы между лопастями листьев мелкие. Плоды слабоовальные, 8ХЮ см,
при созревании сохраняют зеленые полосы...........................var. sororia.
+ Вырезы между лопастями листьев глубокие.........................................2.
2. Лопасти листьев находят друг на друга. Плоды яйцевидные, 6X8 см, при
созревании белеют.....................................................var. texana.
+ Лопасти листьев касаются друг друга................................var. fraterna.
20. Разновидность тексанская — var. texana.— Tristemon texanum
Scheele, 1848, in Linneca, 31:586.— C. texana A. Grey, 1850, in Journ
Boston. Soc. Nat. Hist. 6:193.
Растения однолетние, обвивают кусты и деревья. Плети слегка реб-
ристые, с шипиками длиной 1—2 мм. Усики нежные, спиральные, сво-
бодно расположенные в пространстве, длиной до 10 см. Плетей на рас-
тении обычно 4, иногда 2—7. Листья тонкие, овально-пирамидальные.
Пластинка листа 12—14X15—16 см, обычно 5-лопастная (рис. 12).
Лопасти тупые, доходят до середины листа, сверху сомкнутые, нижние
доли листа у основания не соприкасаются. Края листьев зазубренные.
Поверхность листа покрыта короткими жесткими волосками. Мужские
цветки на тонких длинных (7—8 см) цветоножках, колокольчатые, свет-
ло-желтые. Венчик с отчетливыми жилками, остроконечными дольками.
Женские цветки короче и шире мужских. Чашелистики очень узкие.
Завязь волосистая.
Плоды яйцевидные (см. рис. 12), 6 см в поперечном диаметре,
сначала салатно-зеленые, с 10 белыми полосками и пятнышками, при
созревании светлеют. Плодоножка тонкая, длиной 4—5 см, бороздча-
тая, немного расширенная у основания. Плоды трехкамерные. Семена
овальные (5—7X9—10 мм), нетолстые, с легким ободком, кремовые.
Мякоть горькая.
Встречаются в центральной и южной частях Техаса (США) на
низменных местах, вдоль рек и ручьев, а также в Мексике. Давно из-
вестна в Европе как декоратив-
ное лазающее растение-затени-
тель. В нашей стране хорошо
вызревают на юге РСФСР и
Украины.
21. Разновидность сестрин-
ская — var. sororia (Bailey) Fi-
lov, comb, nova; Filov, 1966, Бюл.
Главк, бот. сада АН СССР,
63 : 37. nom illeg. — С. sororia
Bailey, 1943, Gentes Herbar,
6 : 285.
Название дано из-за сход-
ства ее с предыдущей разновид-
ностью. Растения однолетние,
вьющиеся, более мощные, чем
var. texana. Листья размером
14X14 см, 3-лопастные, а иногда
только с 3—5 углами вместо ло-
пастей. Черешки и нижняя по-
Рис. 12. Разновидность texana.
187
верхность листа густо опушены светлыми шипиками. Стебли слабогра-
неные. Цветки более волосистые, чем у растений var. texana, женские
с длинной трубкой венчика. Завязь очень мелкая.
Плоды слабоовальные, с крупными пятнами, расположенными ши-
рокими полосами. Средний размер плода 8X10 см, с небольшими ко-
лебаниями за пределы этих величин, темно-зеленые. Семена размером
4X9 см, ободок очень узкий. Мякоть горькая.
Растения var. sororia встречаются на юге Мексики. Более поздне-
спелая, чем формы var. texana, в нашей стране вызревают в Средней
Азии.
22. Разновидность братская — var. fraterna (Bailey) Filov, comb,
nova —C. fraterna Bailey, 1943, Gentes Herbar, 6:288.
Плеть с короткими междоузлиями, сероватая. Листья мелкие, серд-
цевидно-овальные (8—9X8—9 см), с тремя главными лопастями, кото-
рые, в свою очередь, тоже делятся на лопасти; главные прорези узкие,
лопасти касаются друг друга. Тычиночные цветки трубчатые, длиной
7—9 см, с длинным венчиком и короткой чашечкой.
Обнаружена на северо-востоке Мексики.
Вид келлиана — С. kellyana Bailey
1948, Gentes Herbar. 7, 5:449
Растения однолетние. Стебли тонкие, стелющиеся, бороздчатые, с
густыми колючими волосками. Листья сердцевидно-овальные, серова-
тые, 10—14ХЮ—14 см, 5-лопастные, с заостренными верхушками, пиль-
чатыми краями. Цветки трубчато-колокольчатые. 6—9X6—8 см, с ост-
рыми лепестками, узкими чашелистиками, длиной 2 см.
Плоды продолговатые, иногда чуть грушевидные, длиной 8—9 см,
с твердой корой, зеленые, с продольными беловатыми полосами. Мякоть
горькая. Семена желтовато-серые, длиной 1 см. Распространена в Мек-
сике.
ПОДРОД ТРОПИЧЕСКИЙ — SUBGEN. SCANDENS FILOV
Subgen. nova, Filov, 1966, 1. с.: 39, sine typo. — Folia tener, smarag-
dina albinaculata. Typus: C. moschata Duch.
Растения окрашены интенсивнее, чем у североамериканского под-
рода, ярко-зеленые. Плети длинные, усики хорошо развитые. Листья
нежные, черешки листа, плодоножки и стебли тупограненые. Опушение
листа мягкое. Происхождение — низменные районы юга Мексики, Фло-
риды, Центральной Америки. Сюда относятся 2 культивируемых вида:
С. moschata Duch., С. maxta Pang, и 5 дикорастущих: С. okeechobeensis
Bailey, С. lundelliana Bailey, С. martinesii Bailey, C. gracilior Bailey,
C. pedatifolia Bailey.
Ключ для определения видов
1. Растения окультуренные, с крупными (масса более 0,5 кг) плодами..............2.
+ Растения дикорастущие, с мелкими плодами (масса менее 0,5 кг).................3.
2. Плоды с розовато-коричневым до коричневого фоном; семена грязно-серые
+ Плоды с белым или кремовым фоном; семена белые, без бахромчатости . . .
................................................................. С. mixta.
3. Листья нелопастные или слаболопастные.......................................4.
+ Листья лопастные ............................................................5.
4. Листья снизу между жилками и черешками листа опушенные. Плоды с поло-
сатым рисунком, кора тонкая.........................................С. okeechobeensis.
188
+ Листья снизу и черешки голые. Плоды без полос, кора толстая, твердая . . .
........................................................................... С. martinesil.
5. Лопасти листьев доходят до */3 пластинки............................................6.
+ Лопасти листьев доходят до основания пластинки........................С. nedatifolia.
6. Однолетники......................................................... С. lundelliana.
+ Многолетники с мясистым корнем-клубнем..................................С. gracilior.
Вид мускатный — С. moschata Duch. ex Poir.
1818, Diet. Sci. nat. :234. — C. pepo a. moschata Duch., 1790, in Lam.
EncycL 2:152. — C. melopepo Lour., 1790, Fl. cochinch.: 593, non L.—
Pepo moschatus Sager, et P. eximius Sager., 1826, Ann. Sci. Nat. (Paris)
1, 8:312. — Cucurbita maxima Wight et Arn., 1834, Prodr. 1 :351, Wight,
1840, Ikon. 2:507, non Duch. — C. spathularis Schrad., 1838, Linnaea,
12:406. — C. macrocarpa Gasp., 1847, Acta Acad. Napoli, 6:447; Gasp.,
1848, Ann. Sci. Nat. (Paris) 3, 9:208. — C. hippopera Ser., 1849, FL
Jard. et Cult. 2:531. — Cymnopetalum calyculatum Miq., 1860, Fl. Ind.
Bot. Suppl. 1 :632. — C. pepo var. melonaeformis Makino, 1908, Bot. Mag.
Tokyo, 22, 8: 170.
Местные названия: Maschus kurbis, Bisamkurbis, fr. courge mus-
quse, pumpkin, squash, cushaw.
Описание. Стебли тупограненые, мягкоопушенные. Листья округло-
почковидные, 5—7-лопастные; лопасти с пильчатыми краями, острыми
концами и вырезами между ними; иногда нелопастные, мягкие, ярко-
зеленые, часто с белыми пятнами в углах между жилками с верхней
стороны; при высокой температуре воздуха края листьев могут свисать.
Цветки колокольчатые, с отогнутыми лепестками, чашелистики узкие,
линейные, иногда с листочковидно-расширенными концами; лепестки
на концах заостренные, бледно-желтые. Плодоножка резко 5-угольная,
с желобками между ребрами.
Плоды крупные, разных форм, с мелкополосатым или пятнистым
зеленым рисунком, который при созревании плода становится желтым
или коричневым на светло-желтом фоне. Мякоть обычно оранжевая.
Семена грязновато-серые, с бахромчатым рубчиком более темного
цвета.
Изменчивость признаков. У растений С. moschata наблюдается
наиболее сильная изменчивость признаков по сравнению со всеми куль-
тивируемыми тыквами. Она выражена в форме плодов и пестроте их
рисунков (рис. 13), пятнистости листьев и форме краев пластинок
(рис. 14).
Пестрота рисунков плода выражается в очень тонкой полосатости
и мелких вытянутых более светлых пятнах. Ярко выраженные рисунки
связаны с высокой плотностью мякоти плодов этого вида, а мелкий
размер элементов — со слабовыраженной структурностью (мелкие со-
судистые пучки на периферии плода). Темно-коричневая окраска плодов
большинства форм этого вида обусловливается наличием каротина в
коре плодов в отличие от вида С. mixta, лишенной каротина. С. mo-
schata более пластична по сравнению с другими видами, у нее ярче вы-
ражены экотипы.
Внутривидовая классификация.// с тор и я изучения. У. И. Митц-
гера из 9 подвидов только 2 относит к С. moschata: Mantelsackurbis —
длинноплодные формы и Melonenkurbis —недлинноплодные.
Ф. Алефельд приводит 8 разновидностей С. moschata: var. naudini
Alef., var. madagascariensis Alef., var. barbarica Alef., var. monticalis
Alef., var. brunnea Alef., var. macrocarpa Alef., var. brachiina Alef.
189
ж
Рис. 13. Форма плодов С. moschata:
а—б — сплюснутая; в—г — шаровидная; д—е — овальная; ж — яйцевидная; з — перце-
видная;
(плоды тонкие, длинные), var. abbreviata Alef. (сплюснутые, дисковид-
ные), а Е. Харц [143]—всего 5.
Л. Бейли [100] описывает дынеподобную японскую форму С. mo-
schata— var. melanoeformis Bailey, и одновременно Н. Е. Житенева
выделяет группу японских тыкв.
190
п Р
Рис. 13. (Продолжение).
и—л — грушевидная; м—н — цилиндрическая; о—п — перехватка; р — булавовидная.
Е. Кастеттер [112] различает по форме плодов 4 группы; 1) круп-
ный сыр —плоды крупные, шаровидные или сплюснутые, кремовые, с
тонкой гладкой недеревянистой корой; 2) кушо-плоды с искривлен-
191
Рис. 14. Пятнистость листьев С. moschata.
ной длинной шейкой и мягкой кожурой; 3) грушеобразные — плоды с
гладкой корой и толстой плодоножкой; 4) смешанная группа — сорта,
не укладывающиеся в приведенные группы.
Н. Е. Житенева [20] выделяет 5 разновидностей: var. mexicana
Zhit., var. indica Zhit., var. turkestanica Zhit., var. japonica Zhit., var.
columbica Zhit., а в следующей работе [21] переводит их в группы
разновидностей. Мексиканская группа включает 7 разновидностей (по
форме плодов), а колумбийская — 3. К сожалению, эти разновидности
без названий и латинского диагноза при наличии русского. К. И. Пан-
гало [60] эти группы разновидностей переводит в подвиды без ссылки
на Н. Е. Житеневу. А затем И. Гребенщиков [139] также без упоми-
нания предыдущих авторов выделяет у С. moschata свои группы раз-
новидностей: convar. nativa Greb. — плоды вздутые, на концах слегка
вытянутые, с неровной поверхностью; мякоть довольно тонкая, кора
деревянистая; convar. abbreviatina Greb. — плоды почти округлые или
чуть вытянутые, без ребер, но чаще сегментированные; convar. moschata
Greb. — плоды различно удлиненные, с перехватом посередине или бу-
лавовидные и convar. japonica Zhit.
Последняя классификация. Включает 6 подвидов с ярко
выраженной эколого-географической обособленностью: indica, mexicana,
Columbiana, moschata, borealiamericanica, japonica.
Ключ для определения подвидов
1. Растения грубоопушенные, плети очень длинные (10 м и более), листья на длин-
ных черешках, стоячие, по краю пластинки несколько выемчатые, с острыми долями 2.
4- Растения с более нежным опушением; плети короче 10 м; листья с косо направ-
ленными укороченными черешками, почти цельнокрайние..................3.
192
2. Плоды очень крупные, сплюснутые, шаровидные или слабо эллиптические, с мяг-
кой корой, диаметром до 50 см, массой 7—20 кг, обычно сегментированные . .
............................................................. var. indica.
+ Плоды средней величины, удлиненные, заостренные на концах или грушевидные,
чаще с деревянистой корой, длиной до 60 см, массой 2—5 кг, несегмснтирован-
ные......................................................... subsp. mexicana.
3. Листья с белыми пятнами в углах жилкования, реже без пятен...........4.
+ Листья без белых пятен, равномерно зеленые, плети очень длинные, плоды ко-
ричневые, семена серые или коричневые, растения позднеспелые.................
..................................................... subsp. columbiana.
4. Плоды удлиненные (индекс более 1,5), гладкие, крупные или среднего размера,
длиной 35—130 см............................................subsp. moschata.
4- Плоды укороченные, иногда с бороздчатой или мозговой поверхностью ... 5.
5. Плоды светло-желтые, гладкие, шаровидные или сплюснутые, слаборебристые
................................................... subsp. borealiamericanica.
4- Плоды оранжево-коричневые, с мозговой поверхностью, чаще глубоко сегмен-
тированные, разной формы, но не шаровидные....................subsp. japonica.
Подвид туркестанский — subsp. moschata. — Mantel-
sackurbis Metzg., 1841, Landw. Pflanzenr.: 694. — C. macrocarpa Gasp.,
1948, Ann. Sci. Nat. (Paris), 9:308.— C. moschata var. turkestanica
Zhit., 1930, 1. c.: 192. — C. moschata subsp. turkestanica Zhit., 1937; Ва-
вилов, Teop. основы селекции, 3:153. — C. moschata convar. moschata
Greb., 1969, Kulturpflanzen, 17:117.
Растения средней мощности и грубости опушения. Листья со сла-
бой белой пятнистостью, расположены наклонно или распластаны на
земле. Плоды среднего размера или крупные (35—130 см длины), вы-
тянутые, наиболее укороченные из них — до грушевидных форм. По-
верхность плодов со слабозаметными линиями углублений сегментации,
розовато-коричневая, с сетчато-полосатым неярким рисунком и восковым
сизоватым налетом. Кора плодов недеревянистая. Мякоть наиболее
толстая, семенная полость маленькая; у длинноплодных форм она рас-
положена на верхнем конце плода. Подвид включает 6 разновидностей.
Ключ для определения разновидностей
1. Плоды цилиндрические ....................................................2.
+ Плоды кувшино- или грушевидные ..........................................5.
2. Плоды длиной 80—130 см...................................................3.
4- Плоды длиной 50—80 см....................................................4.
3. Плоды толщиной 15—20 см, длиной до 100 см....................var. macrocarpa.
4- Плоды толщиной 10—15 см, длиной до 130 см....................var. longicarpa.
4. Плоды толщиной 10—15 см, часто с вздутием цветочного конца или взду-
тиями с обоих концов (перехватки)...............................var. moschata.
4- Плоды длиной 70—100 см, толщиной 7—10 см, без утолщений на концах . . .
............................................................var. claviformis.
5. Плоды кувшиновидные, в среднем длиной 0,5 м....................var. cuschaus.
4- Плоды грушевидные, короткие, длиной 20—35 см.................var. pyriform is.
1. Разновидность перехватки—var. moschata. — С. moschata var.
barbarica Naud., 1856, Ann. Sci. Nat. Bot. (Paris), 6:52; Alef., 1866,
Landw. Fl.: 226.
Плоды длинные, цилиндрические (10—15X50—80 см), чаще с утол-
щениями на обоих концах. Это наиболее распространенная разновид-
ность туркестанского подвида, составляющая основы перехваток. Рас-
пространены в Средней Азии, Закавказье (СССР), частично на западе
Китая, в Северной Африке, Иране. Сорта: Плов-кеды, Ленинабадская,
Кашгарская 1644, Бирючекутская 628, Перехватка 69 и некоторые
другие.
2. Разновидность палицевидная—var. longicarpa Filov, var. nova;
Filov, 1966, в Бюлл. Главы, бот. сада АН СССР, 63:39, sine typo —
Fructus cylindrici longi, longiores 80 cm, latitude 12 cm.
193
Typus: Сорт Грамма, к-2365. Дербент. Среднеазиатская опытная
станция ВИР, 1939 г. (WIR).
Плоды очень длинные (80—130 см), толщиной 10—15 см, с не-
большим округлым утолщением на верхнем конце (рис. 15,а). В нашей
стране распространена — в Туркмении, Азербайджане. Известны два
сорта этой разновидности: Дербентская перехватка и Грамма (из Ав-
стралии). Представляет большой интерес как исходный материал.
3. Разновидность китайская—var. daviformis Naud, ex Harz, 1885,
Landw, Samenkunde: 824. — C. moschata var. brachiina Alef., 1866,
Landw. Fl.: 227. — C. moschata var. hinensis Filov, 1966, 1. c.: 37, nom.
illeg.
Плоды удлиненные, как у предыдущих разновидностей, но тоньше
(рис. 15,6), в диаметре 7—10 см. Распространены в Китае.
4. Разновидность неаполитанская—var. macrocarpa (Gasp.) Naud.,
1856, Ann, Sci. Nat. (Paris), 6:52. — C. macrocarpa Gasp., 1848, Ann.
Sci. Nat. Bot. 10:207. — C. moschata var. macrocarpa Alef., var. brunnea
Alef. et var. monticaulis Alef., 1966, I. c. :226. — C. moschata var. neapo-
litana Filov, 1966, 1. c.: 41, nom. illeg.
Эта самая крупноплодная разновидность данного подвида. Ци-
линдрические плоды имеют длину до 100 см при толщине 15—20 см.
Каждое растение дает по 1—2 плода. Разновидность представлена сор-
том Неаполитанская, который разводится в Западном Средиземноморье.
5. Разновидность кувшиновидная—var. cuschaus Castet, 1925, Proc,
Amer. Soc. Hort. Sci, 22 : 338.
Плоды удлиненные (до 50 см), с сильным расширением верхушки,
которое может составлять половину плода (кувшиновидный плод). Рас-
пространены в пределах ареалов предыдущих разновидностей.
6. Разновидность грушевидная—var. pyriformis Cast., 1925, 1. с.:
: 338.
Плоды грушевидные, длиной до 35 см; расширенная часть зани-
мает не менее 3Д длины плода. Распространены на западе Средизем-
номорья.
Подвид североамериканский — subsp. borealiame-
ricanica Filov, subsp. nova; Filov, 1966, 1. c.:41, nom. illeg. — Plan-
tae mediocaules. Fructus rotundi, sulculati.
194
г
Рис. 16. Плоды разновидностей подвида североамериканского:
а — сплюснутоплодная; б — шаровидная; в — эллипсовидная; г, д — канадская сплюс-
нутоплодная и кувшинчатая; е — семинольская.
Typus: С. moschatus var. cheese Castet.
Растения средней мощности, с нежным опушением. Листья со сла-
бой белой пятнистостью, расположены наклонно или распластаны.
Плоды среднего размера или мелкие, округлые, слабоовальные, шаро-
видные, слабо сплюснутые, реже мелкоплодно-цилиндрические, чаще со
слабой сегментированностью. Окраска их светлее, чем у предыдущего
подвида. Кора плодов недеревянистая. Мякоть довольно толстая, семен-
ная полость средняя. Подвид объединяет 5 разновидностей (рис. 16).
Ключ для определения разновидностей
1. Плоды крупные (масса более 3 кг).........................................2.
+ Плоды мелкие (масса 0,5—2 кг).............................................4.
2. Плоды овальные ...............................................var. ovalis.
+ Плоды иной формы..........................................................3.
3. Плоды шаровидные..............................................var. cheese.
+ Плоды сплюснутые..........................................var. rapaeformls.
4. Плоды коричневые, сплюснутые................................var. seminolis.
+ Плоды кремовые, сплюснутые или цилиндрические.............var. kanadiensis.
7. Разновидность эллипсовидная—var. ovalis Filov, var. nova; Fi-
lov, 1966,1. c.: 38, nom. illeg. — Fructus ovales magni.
Typus: США, Среднеазиатская опытная станция ВИР. 1927 г.
(WIR).
Плоды овальные, длиной до 40 см, толщиной 25—30 см, слабо сег-
ментированные. Разводится в США. Довольно скороспелая.
8. Разновидность шаровидная—var. cheese Castet., 1925, I. с.:338.
Плоды крупные, шаровидные, иногда слабо сегментированные. Рас-
тения этой разновидности распространены в США, довольно скороспе-
лые.
195
9. Разновидность сплюснутая—var. rapaeformis Filov, var. nova.—
Fructus complanati, magni segmentata.
Typus: США, Среднеазиатская опытная станция ВИР, 1927 г.
(WIR).
Плоды крупные, сплюснутые, чаще сегментированные. Растения
этой разновидности распространены в США. Скороспелые. Один из сор-
тов этой разновидности каротинная отселектирован на высокое содер-
жание каротина и в нашей стране разводится на Северном Кавказе.
10. Разновидность семинольская—var. seminolis Filov, var. nova;
Filov, 1966, 1. c.: 38, sine typo. — Fructus rotundi sulculati medii, latitude
12 sm.
Typus: США, Флорида (WIR).
Плоды шаровидные, мелкие (12 см в диаметре), слегка сегменти-
рованные, светло-коричневые. Мякоть оранжево-красная, мелкозерни-
стая, вкусная. Очень позднеспелая. Разводится индейцами в южной
части Флориды (США). Представляет собой форму неполного окульту-
ривания вида.
11. Разновидность канадская—var. kanadiensis Filov, var. nova;
Filov, 1966, 1. c.: 38, sine typo. — Plantae debiles, folia minuta. Fructus
minuti cylindrici et ascidiati.
Typus: США, Среднеазиатская опытная станция ВИР, 1937 г.
(WIR).
Растения малой мощности, опушение нежное, листья мелкие, ярко-
зеленые, без белой пятнистости, стелющиеся. Плоды мелкие, сплюсну-
тые или цилиндрические, длиной 25—35 см, толщиной 8 см, иногда с
небольшим вздутием на конце (тогда шейка плода редко бывает толще
4—5 см). Сплюснутые плоды диаметром не более 20 см, а цилиндриче-
ские 40 см длины. Кора светлая, буровато-кремовая, со слабым рисун-
ком, характерным для вида, недеревянистая, наиболее скороспелая из
всех разновидностей С. moschata. Сюда относится американский сорт
Butternut.
Подвид японский — subsp. japonica (Zhit.) Filov, comb,
nov. C. moschata gr. var. japonica Zhit., 1930, у Букасова, Возд. раст.
Мексики : 312.
Растения слаборазвитые, с довольно нежным опушением. Листья
с сильно выраженной пятнистостью, распластанные по земле, почти
цельнокрайние, нежные, увядающие в жаркую погоду. Плоды как бы
спрессованные, морщинистые или с бугристо-мозговой поверхностью,
Рис. 17. Плоды разновидностей подвида японского:
а — канталупообразная; б — карликовая; в — мозговая плоская; г — мозговая кату-
шечная.
196
эбычно глубоко сегментированные. Восковой налет на плодах развит
гильно. Кора недеревянистая. Мякоть очень плотная, толстая, сладкая.
Подвид скороспелый. Разводится только в Японии и в меньшей
степени на Тихоокеанском побережье Китая. Внутри подвида различают
4 разновидности (рис. 17).
Ключ для определения разновидностей
1. Плоды сплюснутые ...............................................2.
+ Плоды несколько вытянуты ..........................var. trochlearis.
2. Плоды средней величины, 18—25 см в диаметре.....................3.
+ Плоды мелкие, 17—18 см в диаметре, слабо сегментированные, с неровной
поверхностью..........................................var. pygmala.
3. Плоды без мозговой поверхности, слабо сегментированные .... var. japonica.
+ Плоды с мозговой поверхностью, глубоко сегментированные . . . var. depressa.
12. Разновидность мозговая плоская—var. depressa Naud, in Harz.,
1885, Landw, Samenkunde : 824.
Плоды очень сильно сплюснутые, почти дисковидные, некрупные
(масса 1—2 кг), с мозговой поверхностью, глубокой сегментированно-
стью, щели впадин скрыты глубоко в мякоти.
13. Разновидность мозговая катушечная —var. trochlearis Filov,
var. nova; Filov, 1966, 1. c. :41, sine typo. — Fructus medii troch leares
barbatuli.
Typus: Япония. Среднеазиатская опытная станция ВИР. Получен
в 1927 г. (WIR).
Плоды среднего размера, с мозговой поверхностью, укороченно-
цилиндрические, с некоторым сужением посередине, наподобие катушки.
14. Разновидность канталупообразная — var. japonica. С. moschata
var. melandeformis Bailey, 1930, Stand. Ziclop. Hort., 1 ; 910.
Плоды средние, 20 см в диаметре, сплюснутые, с несколько не-
ровной, но немозговой поверхностью, сегментированные.
15. Разновидность карликовая—var. pygmala Filov, var. nova; Fi-
lov, 1966, 1. c.: 41, sine typo. — Fructus minuti complanati, 0,5—0,7 kg.
Typus: Япония. Среднеазиатская опытная станция ВИР. Получен
в 1937 г. (WIR).
Плоды сплюснутые, сегментированные, но очень мелкие, не пре-
вышающие 0,5—0,7 кг.
Подвид индийский —subsp. i n die a (Zhit.) Filov, comb,
nova.— C. moschata gr. var. indica Zhit., 1930, у Букасова, Возд. раст.
Мексики :311.
Растения очень мощные, грубоопушенные. Плети длинные, дости-
гающие 10 м. Листья крупные, с белой пятнистостью и длинными стоя-
чими черешками.
Плоды очень крупные (масса до 20 кг) от слабосплюснутых до сла-
боэллиптических (рис. 18). Окраска плодов темно-зеленая или чаще со
светло-зеленым сетчатым рисунком и крупными яркими кремовыми пят-
нами. Кора мягкая, недеревянистая. Семенная полость большая.
Под вид позднеспелый. Разводится в Индии. Включает 3 разновид-
ности.
Ключ для определения разновидностей
1. Плоды слабо сплюснутые.
4- Плоды иной формы...................................
2. Плоды шаровидные...................................
+ Плоды эллиптические.................................
var. macrocomplanata.
........var. indica.
........var. grahde.
137
16. Разновидность сплюсиутоплодная—var. macrocomplanata Fi-
lov, var. nova. — C. moschata var. complanata Filov, 1. c. :41, sine typo
non Zhit., 1930. — Fructus magni complanati, lactomaculati.
Typus: Индия, Среднеазиатская опытная станция ВИР. Получен
в 1937 г. (WIR).
Плоды сплюснутые, с яркопятнистым рисунком; пятна белые
крупные. ’
17. Разновидность шароплодная — var. indica —С. moschata, 1966
1. с.: 41, sine typo.
18. Разновидность овальноплодная — var. grande Naud., 1856
1. с.: 52. — C. moschata Naudinii Alef., 1866, 1. c.: 226.
Плоды овальные, с яркопятнистым рисунком пли темно-зеленые
а
Рис. 18. Плоды
— шаровидная; б—г
разновидностей подвида
— овалыюплоднаи; д —
ИНДИЙСКОГО’
сплюснутоплодиая.
198
Рис. 19. Плоды разновидностей подвида мексиканского:
а — шаровидно-овальная; б — сдавленная; в — сплюснуто-бородавчатая; г — грушевид-
ная; д — бутылковидная; е — цилиндрическая, бородавчатая.
Подвид мексиканский —Subsp. mexicana (Zhit.) Fi-
lov, comb. nova. — C. moschata gr. var. mexicana Zhit., 1930, у Бука-
сова, Возд. раст. Мексики:312.
Растения очень мощные, грубоопушенные. Листья на прямостоячих
длинных черешках. Плоды средней величины, с толстой деревянистой
корой, различной формы (рис. 19). Окраска и рисунок, как у турке-
станского подвида. Мякоть тонкая, сладкая, мучнистая.
Подвид позднеспелый. Разводится только в Мексике. Имеет 7 раз-
новидностей.
Ключ для определения разновидностей
1.
2.
+
3.
4.
5.
+
6.
+
Плоды сплюснутые ..........................................................
Плоды иной формы...........................................................
Плоды сегментированные, без бородавчатости..................var. applanata.
Плоды с бородавчатой поверхностью.................var. complanatovemicosus.
Плоды яйцевидные и овальные.................................var. testicularis.
Плоды иной формы...........................................................
Плоды грушевидные...........................................var. pyridiumis.
Плоды иной формы...........................................................
Плоды цилиндрические ......................................................
Плоды бутылковидные...............................................var. teres.
Плоды укороченно-цилиндрические, сегментированные, без бородавок...........
............................................................var. cylindracea.
Плоды тоньше и длиннее предыдущих, несегментированны, сильно бородав-
чатые ..................................................... var. mexicana.
2.
3.
4.
5.
6.
19. Разновидность сдавленная —var. applanata Zhit., 1930, у Бука-
rucosus Zhit., 1930, 1. s. — 312.
Плоды сплюснутые, сегментированные.
20. Разновидность сплюснуто-бородавчатая. —var. complanatover-
rucosus Zhit., 1930, 1. с.: 312.
Плоды сплюснуто-бородавчатые.
21. Разновидность шаровидно-овальная—var. testicularis Zhit.,
1930, 1. с.: 312.
Плоды шаровидные и овальные, резко сегментированные.
199
22. Разновидность грушеплодная — var. pyridiumis Zhit 1930
1. с.: 312.
Плоды крупные, грушевидные, резко сегментированные.
23. Разновидность цилиндрическая— var. cylindracea Zhit., 1930,
1. с.: 312.
Плоды укороченно-цилиндрические, толстые, сегментированные.
24. Разновидность цилиндрическая бородавчатая — var. mexicana.
Плоды цилиндрические, бородавчатые.
25. Разновидность бутылковидная — var. teres Zhit., 1930, 1. с.: 312.
Плоды цилиндрические, тонкие, обычно суживающиеся к плодо-
ножке, бутылковидные.
Подвид колумбийский — subsp. columbiana (Zhit.)
Filov, comb. nova. — C. moschata gr. var. columbiana Zhit., 1930,
1. c.: 312.
Растения мощные, с тонкими стеблями, длиной до 10 м, с сильно
растянутыми междоузлиями и нежным опушением. Листья небольшого
размера, с косо направленными черешками. Листовая пластинка блед-
но-зеленая, почти без белых пятен. Плоды небольшие, обычно расши-
ренные у основания (перцевидные). Кора недеревянистая. Мякоть гряз-
новато-коричневая, сладкая. Семена в отличие от семян всех других
подвидов С. moschata темноокрашенные, серые, коричневые и шоко-
ладные.
Подвид позднеспелый. Разводится в Колумбии, Венесуэле и Па-
наме. По мелким размерам листьев, цветков и плодов, а также по боль-
шому количеству последних приближается к некультурным формам.
Включает 4 разновидности.
Ключ для определения разновидностей
1. Плоды удлиненные.......................................
+ Плоды иной формы........................................
2. Плоды призматические, перцевидные......................
+ Плоды иной формы........................................
3. Плоды овальны..........................................
+ Плоды укороченные.......................................
var. dolihocarpa.
var. prismaticus.
• • г............3.
var. columbiana.
var. abbreviata.
26. Разновидность продолговатая —var. dolihocarpa Zhit., 1930,
1. c.: 313.
Плоды продолговатые, дубиновидные, цилиндрические.
27. Разновидность перцевидная—var. prismaticus Zhit., 1930,
1. с.: 313.
Плоды призматические, перцевидные, сегментированны.
28. Разновидность овальная—var. columbiana.— С. moschata var.
turgida Bailey, 1948, Gentes Herb. 7, 5:470.
29. Разновидность шаровидно-сплюснутая — var. abbreviata Filov,
var. nova; Filov, 1966, Бюл. Главн. бот. сада АН СССР, 63:4. — Fruc-
tus abbreviate — pyricarpo, suneatus, testicularis, sphaericus, applanatus,
subsegmentatus.
Плоды сильно укороченно-грушевидные до сплюснутых, клиновид-
ные или яйцевидные, слабо сплюснутые.
Typus: Колумбия. Среднеазиатская опытная станция ВИР. Собран
С. М. Букасовым в 1928 г. (WIR).
Вид серебросемянный — С. mixta Pang.
1930, Тр. прикл. бот. ген. сел. 23, 3:253. — С. argirosperma Bailev
1937, Gard. Gurd.: 72; Filov, 1966, 1. c.: 38.
200
Рис. 20. Плоды разновидностей С. mixta:
а—в — округлая; б — кувшиновидная; г — грушевидная.
Описание. Растения длинноплетистые, грубоопушенные. Листья
широкие, (20X20 см), угловатые, с небольшими остроугольными выре-
зами, с зубчатыми краями. Плоды от сплюснутых до кувшиновидных
(рис. 20), средней величины, очень темно-зеленые, с неясным полосатым
рисунком или кремово-белые, с сетчатыми зелеными полосами. При со-
зревании фон плодов принимает кремовую окраску. Кора деревянистая.
Мякоть оранжево-зеленоватая, малосладкая. Семена белые, серебри-
стые, с голубоватым или желтоватым выдающимся рубчиком.
Позднеспелая. Разводится в предгорных районах Центральной Аме-
рики, начиная с Мексики и кончая Колумбией.
Изменчивость признаков. Кроме формы плодов, варьирует и харак-
тер семян: 1) крупные, широкие (3X1,5 см), с широким (до 3 мм)
голубоватым ободком; 2) узкие, длинные (3X0,8 см), с узким голубым
ободком; 3) слегка вытянутые (2,3X1 >2 см), с узким голубым ободком;
4) среднекрупные (2X1 см), с желтоватым (темнее фона) ободком и
растрескивающейся поверхностью.
Внутривидовая классификация. По форме, главным образом пер-
вых двух типов семян К. И. Пангало [58] выделил две разновидности:
var. stenosperma Pang, и var. cyanoperizona Pang. Однако эти две
формы семян не соответствуют определенным типам плодов: встре-
чаются всевозможные сочетания этих признаков. Кроме того, К. И. Пан-
гало не таксономиэированы формы с иными признаками семян. Учиты-
вая это, мы решили придерживаться общих принципов таксономизацип
разновидностей по характеру формы и основных окрасок плодов.
Ключ для определения разновидностей
1. Плоды от сплюснутых до слабоовальных.....................var. stenosperma.
+ Плоды иной формы..........................................................2.
2. Плоды грушевидные........................................var. pyridiforme.
+ Плоды кувшиновидные..........................................var. cuschaus.
1. Разновидность округлая — var. mixta. — С. mixta var. steno-
sperma Pang., 1930, Tp. прикл. бот. ген. сел. 23, 3:253, р. р. — Cucur-
bita mixta var. globosa Filov, 1966, Бюлл. Главн. бот. сада, 63:4.—
С. mixta var. cyanoperizona Pang., 1930, 1. с.: 253, p. p.
Плоды сплюснутые, короткогрушевидные, шаровидные пли слабо-
овальные. с сетчатыми полосами на белом или кремовом фоне.
2. Разновидность грушевидная— var. pyridiforme Castet., 1925,
1. с.: 338. — С. mixta var. stenosperma Pang., 1930, 1. c. :253, p. p.
Плоды грушевидные, с сетчатыми полосами на белом или кремо-
вом фоне.
201
3. Разновидность кувшиновидная— var. cuschaus Castet., 1925,
]. с.: 338.— C. mixta var. stenosperma Pang., 1930, 1. c.: 253, p. p.—
C. mixta var. cyanoperizona Pang., 1930, 1. c.: 253, p. p.
Плоды кувшиновидные, темно-зеленые, при созревании часто белые
или кремовые или с зелеными полосами на белом фоне.
Вид лиановый — С. okeechobeensis (Small) Bailey
1930, Gentes Herb.: 179. — Pepo okeechobeensis Small, 1930, Jurn.
New Jork Bot. gard. 21:12.
Растения высоковьющиеся — естественные лианы. Стебли очень тон-
кие, с длинными междоузлиями (до 40 см). Усики 3—4-раздельные,
сильноразвитые, спиральные. Листья ярко-зеленые, округлые, с очень
крупными углами, без лопастей, шириной 12—20 см, немного короче
в длину. Края листа зубчатые. Опушение очень короткое и мягкое, с
зеленым оттенком. Цветки светлые, кремово-желтые, ширококолоколь-
чатые, длиной 6 см, с короткими и острыми отгибами. Плоды шаровид-
ные, 7—8 см в диаметре, зеленые, тускнеющие до мутно-зеленого цвета,
с неопределенными белыми пятнышками и тонкими белыми линиями
(рис. 21,а). Кора деревянистая, очень твердая, хотя тонкая. Плоды
трехкамерные. Мякоть белая, горькая. Семена крепко держатся в сухой
мякоти, овальные, заостренные, 7X12 мм.
В диком виде находится на юге Флориды, на восточной стороне
озера Окичоби (США) во влажных болотистых местах, где растения
закрывают кусты и вьются по деревьям. Вид обнаружен Д. Смоллом
в 1922 г. Характерная его форма описана ранее. Однако на юге Мек-
сики имеется уклонившаяся в сторону мезофильности другая форма,
именуемая Л. Бейли видом С. martinezii. Мы полагаем, что ее при
более тщательном изучении можно будет рассматривать в ранге раз-
новидности лиановой тыквы.
Вид мартина — С. martinezii Beialey
1943, in Gentes Herb. 6: 296.
Растения имеют нежные, почти гладкие плети с длинными простыми
или раздвоенными усиками. Листья мелкие, широкие, шириной 9—12 см
и почти такой же длины, поперечно-овальные. Листовые пластинки тон-
кие, нежноволокнистые. Края листа неровные, нерассеченные; черешки
листа средней длины, с желтоватыми волосками. Плоды шаровидные,
Рис. 21. Плоды вида лианового с 6 полосами (а) и вида Лунделля с 10 полосами (б).
202
с белыми неясными пятнами, расположенными более или менее ли-
нейно. Кора деревянистая, очень толстая (толщиной 3—5 мм), очень
твердая. Плодоножки длинные, ребристые, расширенные у основания.
Семена овальные, заостренные, 7X10 мм, с заметным ободком, серо-
белые.
Распространена в штате Веракрус на юге Мексики.
Вид дланелистный — С. pedatifolia Bailey
1943, 1. с.: 297.
Растения очень нежные, слабоопушенные, с очень тонкими длин-
ными междоузлиями. Усики простые или ветвистые, длинные, закручи-
вающиеся. Листья в небольшом количестве, широкие (20X20 см), силь-
новырезные (почти до основания), с 5 узкими длинными лопастями,
10—15X2—3 см, с пальчатыми концами. Боковые доли, в свою оче-
редь, слабо разделены. Окраска листьев нежная, ярко-зеленая, с бе-
лыми пятнами. Цветки колокольчатые, длиной 5 см. Плоды 6 см в диа-
метре, бело-зеленые, полосатые.
Произрастает в южной и центральной частях Мексики.
Вид лунделля —С. lundelliana Bailey
1943, 1. с.: 297.
Растения с тонкими длинными стеблями, с тонкими мягкими во-
лосками, впоследствии опадающими. Усики длинные и нежные. Листья
мелкие, 5—9 см в диаметре, 5-раздельные до ’/з пластинки, нежные.
Лопасти и доли листа тупые. Цветки на длинных цветоножках, коло-
кольчатые, длиной 6—7,5 см, с острыми дольками, оранжево-желтые,
с зелеными жилками. Плоды шаровидные, чуть вытянутые к плодо-
ножке, гладкие, с неясными ребрами, зеленоватые до желтоватого, с
белыми пятнышками, расположенными линейно и белыми тонкими по-
лосами (рис. 21,6). Кора деревянистая, тонкая. Семена гладкие,
10—12x5—6 мм, с острым длинным кончиком, с заметно приподнятым
сморщенным ободком.
Произрастает в Гватемале, на юго-востоке Мексики, в Гондурасе
в диком виде. Легко скрещивается с другими видами тыкв.
Вид изящный — С. gracilior Bailey
1943, 1. с.: 303.
Растения слабые. Плети многочисленные, тонкие, ребристые, почти
гладкие, без опушения. Усики очень мелкие, только изредка развитые.
Листья мелкие, сердцевидно-овальные (7—8x9—10 см), обычно 3-ло-
пастные, лопасти доходят до середины листа и глубже, почти тупые.
Доли снова немного разделены, почти не соприкасаются, края неров-
ные, с зубчиками на окончаниях жилок. Поверхность листовой пла-
стинки покрыта короткими жесткими волосками. Черешок листа тол-
стый, с короткими колючими волосками.
Плоды слабоовальные (5,5—6X6 см), красивые, с зеленовато-бе-
лыми полосками и пятнами. Плодоножка щетинистая. Семена овально-
продолговатые, с немного суженным концом (7—8X10 мм), без ободка.
Требует дальнейшего изучения.
Вид распространен на юге Мексики на широте около 20°. Это, по-
видимому, переходная форма от лиановых растений к плетистым.
203
ПОДРОД ГОРНЫЙ —SUBGEN. MONTANA FILOV
subgen. nova. — Corpus incarotinus cortex primo color. Plantes dies,
origin Montanus. Представлен одним полукультурным видом С. fici-
folia Bouche и одним диким видом С. moorei Bailey. Обитает в горных
районах тропиков Центральной Америки.
Typus: С. ficifolia Bouche.
Ключ для определения видов
1. Растения культурные. Плоды крупные, в среднем 25X20 см . . . .С. ficifolia.
+ Растения дикорастущие. Плоды мелкие, в диаметре 7—8 см . . .С. moorei.
Вид фиголистный — С. ficifolia Bouche
1837, Vcrh. ver. Beford. Gartenb. preuss, 12:205, non Wall, 1828.—
C. melanosperma A. Br., 1824, Cat. Hort. Karls., nomen nudum; Gasp.,
1847, Acta Acad. Napoli, 6:448; idem, 1848, Ann. Sci. Nat. (Paris) 3,
9 : 208; Bailey, 1929, Gentes Herb. 2, 2 : 112.
Растения очень мощные, плети очень длинные, междоузлия боль-
шие, опушение очень нежное, мягкое. Листья ярко-зеленые, окраска
более интенсивная, чем у С. moschata, мягкие, с синими пятнами. Ло-
пасти листьев округлые. Черешки листьев и плети тонкие. Плоды ко-
роткоовальные, с ровной поверхностью, с сетчато-мозаичным рисунком:
зеленая сетка на бело-зеленоватом фоне. От верхней части плода от-
ходят белые полосы, достигающие ‘/з—’/г части плода. Кора деревяни-
стая. Мякоть белая, сливающаяся с плацентами, как у арбуза, не-
сладкая. Семена черные и кремовые. Плоды и листья этого вида пред-
ставлены на рисунке 22.
Разводится в горных районах западного побережья Центральной
и Южной Америки местным населением для пищевых и кормовых це-
лей. Вид устойчив к фузариозному увяданию.
По-видимому, диким родичем С. ficifolia является С. moorei Bailey.
Вид мура— С. moorei Bailey
1948, in Gentes Herb. 7 : 449.
Растения нежноопушенные, сильно облиственные, с тонким боро-
давчатым стеблем и короткими междоузлиями. Листья ярко-зеленые
(6—8x8—10 см), с белыми пятнами, 5-лопастные, доли листа тупые,
Рис. 22. Плод и куст вида фиголистного.
204
с округлыми окончаниями, почти без зубчиков по краям. Боковые ло-
пасти снова несколько разделены. Черешок листа длиной 4—6 см.
Цветки оранжевые с яркими жилками, мужские длиной 6—8 см, жен-
ские длиной 10 см с завязью. Плоды шаровидные, 7—8 см в диаметре,
с беловатыми разорванными продольными полосами и пятнами на зе-
леном фоне. Плодоножка тонкая, бороздчатая, расширенная у плода.
Семена беловатые (5—6x10—12 мм).
Произрастает в Центральной Мексике на высоте 1750 м над уров-
нем моря, где вьется по кустам в придорожных рощах.
ПОДРОД ДИКОРАСТУЩИЙ-SUBGEN. EREMOS1CYOS F1LOV
Subgen nova. —Folium grey, viridis from densus pubescens ex si-
nosus.
Typus: C. foetidissima H. В. K.
Растения серовато-зеленоватые от густого опушения на листьях,
или последние очень сильно выемчатые (до основания), с очень тон-
кими долями листовой пластинки. Плети короткие, усики сильно скру-
чены в комок, слабые. Происхождение — юг Калифорнии (США), север
Мексики. К этому подроду относится 8 видов.
Ключ для определения видов
I. Растения без колючек, иногда лишь слабоопушенные.................С. palmeri.
+ Растения с мелкими колючими шипиками или сильным опушением....................2.
2. Длина листьев равна ширине, двояковырезные до 1/з пластинки ... С. cordata.
+ Длина листьев превышает ширину................................................3.
3. Листья зеленые, слабоопушенные, лопастные....................................4.
+ Листья серые от густого опушения, чаще без лопастей, треугольные.............
.............................................................С. foetidissima.
4. Лопасти листьев неровные по краям. Трубка чашечки мужских цветков очень
длинная, цилиндрической формы......................................С. cylindrata.
+ Лопасти листьев ровные, чашечка нецилиндрическая..............................5.
5. Лопасти листьев очень узкие, пальчатые, вырезы доходят до основания пла-
стинки .............................................................С. digitata.
4- Лопасти доходят до 2/з пластинки или короче...................................6
6. Лопасти правильные, с ровными краями, остроконечные, прорези между лопа-
стями остроконечные.................................................С. palmata.
+ Лопасти короче */2 пластинки..................................................7.
7. Листья узкотреугольные, с сильно зубчатыми краями................С. galeotti.
-I- Листья сердцевидно-овальные, слабовыемчатые, с заходящими лопастями . . .
...............................................................С. radicans.
Вид вонючий — С. foetidissima Н. В. К.
1817, Nov. Gen. et Sp. 2: 123. — Cucumis perennis James, 1823,
Exped. Rocky n. s., 2:346; A. Gray, 1852, Journ. Boston Soc. Nat. Hist.
6: 193. — Pepo foetidissima Britt, et Brown, 1913, III. Fl. North. U.S.A.,
Can. 2:291.
Растения многолетние, глубокоопушенные, иногда образующие круп-
ные заросли на пустырях, от которых исходит неприятный запах. Зи-
мует благодаря наличию корневых утолщений типа клубней.
Стебель ребристый, усики толстые, короткие. Листья треугольные,
длиной до 30 см и шириной до 20 см у основания, цельнокрайние,
редко зубчатые, сильноволосистые, серовато-зеленые. Черешок толстый,
с редким опушением. Мужские цветки крупные, длиной 10—12 см, вен-
чик с густым грубым опушением, лопасти венчика широкие, остроконеч-
ные. Женские цветки короче, длиной 9—10 см.
Завязь опушенная. Плоды шаровидные, тускло-светло-зеленые, с
5 или 6 основными кремово-белыми полосками, несколькими промежу-
205
точными полосками и многочисленными пятнышками. Кора деревяни-
стая. Семена мелкие (6—7X12 мм), без ободка, с тупыми концами. От-
дельные органы этого вида и внешний вид растения представлены на
рисунке 23.
Распространен на сухих пустырях и в прериях южной части Ка-
лифорнии (США), северной части Мексики и в юго-западных штатах
США.
Ключ для определения разновидностей
1. Листья треугольной формы...........................var. foetidissima.
+ Листья 2-лопастные с каждой стороны................var. scabridifolia.
1. Разновидность вонючая — var. foetidissima.
2. Разновидность шероховатолистная—var. scabridifolia. (Bailey)
Filov, comb. nova. — C. scabridifolia Bailey, 1948, gent. Herb., 7:449.
Лопасти на верхушках тупые, центральная лопасть намного длин-
нее боковых, выемки широкие, округлые.
Вид сердцевидный — С. cordata Wats
1889, Proc. Amer. Acad. Arts. Sci. 24 : 50.
Растения многолетние. Стебли тонкие, ребристые, густо покрытые
тонкими волосками. Усики очень маленькие, круто завитые. Листья
мелкие, длиной 4—8 см, шириной 5—10 см, сердцевидной формы, 3-ло-
пастные, серовато-беловатые. Лопасти доходят до середины листа или
меньше, а затем они снова немного надрезаны. Надрезы с неправильно
зубчатыми краями. Листовые пластинки покрыты густыми волосками,
между которыми прячутся многочисленные вздутия — сосочки. Черешки
толстые, короткие, жестковолосистые. Женские цветки длиной 7 см,
волосистые. Завязь продолговатая.
Рис. 23. Вид вонючий по рисунку Бейли
(а) и в поле (б).
Рис. 24. Вид цилиндрический (по Бейли)
206
Плоды шаровидные, 8 см в диаметре, зеленые, с пятью отчетли-
выми узкими светлыми капиллярными полосками и промежуточными
пятнышками. Плодоножки сильноребристые. Семена толстые, оваль-
ные (7—8X11 мм), с тупыми концами без ободка.
Произрастает в Калифорнии (США), Мексике, на песчаных рав-
нинах около залива Лос-Анжелес. Требует дополнительного изучения
видовой принадлежности.
Вид цилиндрический — С. cylindrata Bailey
1943, 1. с.: 304.
Растения серые, стебли тупоребристые, усики маленькие, слабые,
ветвящиеся у основания и свернутые кольцом. Листья 7—10 см в диа-
метре, рассеченные до основания на 6 долей, которые, в свою очередь,
рассечены наподобие листьев арбуза; концы лопастей тупые. Листовая
пластинка покрыта жесткими серыми волосками. Черешки толстые, с
коротким опушением. Венчик и чашечка мужских цветков имеют вид
крупной цилиндрической трубки, длиной 5—7 см. Женские цветки 8—
9 см.
Плоды сплюснуто-шаровидные, с 10 узкими неправильными свет-
лыми полосками и пестрым пятнистым рисунком. Семена мелкие, тол-
стые, неправильно овальные, длиной 9—10 мм, без ободка, с тупыми
краями. Органы этого вида представлены на рисунке 24.
Распространен в Калифорнии (США) и Мексике.
Вид пальчатолистный — С. digitata A. Gray
1853, Plant Wright, 2:60.
Растения многолетние. Стебель очень тонкий, ребристый. Усики
очень короткие, скрученные. Листья темные, серовато-зеленые, средние
жилки более светлого цвета.
Листовая пластинка рассечена
до основания на 5 очень узких
долек размером 0,5—2X5—
10 см, с шероховатой поверх-
ностью и белыми точками. Че-
решок бугорчатый. Мужские
цветки узкоколокольчатые,
длиной 3—4 см, покрыты ред-
ким пушком. Плоды шаровид-
ные, 8—9 см в диаметре, ярко-
темно-зеленые, с узкими от-
четливыми беловато-зелеными
капиллярными полосками и
мелкими пятнами того же цве-
та. Отдельные органы и внеш-
ний вид растения представле-
ны на рисунке 25.
Семена белые, овальные,
тупоконечные (7 8ХЮ
11 мм), без ободка, плоские
с толстыми краями.
Распространен в США
(юго-восточная часть Кали-
форнии, Аризона, Ныо-Мекси-
ка) и северной части Мексики.
Рис. 25. Вид пальчатолистный по рисунку
Бейли (а) и в поле (б).
207
Вид пальмовидный — С. palmata Wats.
1876, Proc. Amer. Acad. Arts. Sci. 11 : 137.
Растения многолетние. Стебли ребристые, с короткими междоуз-
лиями, серые, с короткими жесткими волосками. Усики очень короткие,
скрученные. Листья густо расположенные, толстые, 6—10 см в диаметре,
разрезанные на 5 клиновидных остроконечных долей (лопастей), при
этом центральная больше остальных. Лопасти доходят до 2/з листа.
Поверхность листьев густо покрыта короткими жесткими серыми во-
лосками. Черешок короче пластинки листа с колючими волосками и
бугорками.
Мужские цветки узкоколокольчатые, длиной 6—7 см, женские 8 см
(завязь длиной 2,5 см). Плоды шаровидные, 8—9 см в диаметре, тя-
желые, светло-зеленые, с широкими неясными зеленовато-белыми по-
лосками и пятнами. Плодоножки толстые. Семена белые, узкоовальные
(7ХЮ—12 мм), без приподнятого ободка.
Распространен в южной части штата Калифорния (США). Вклю-
чает 2 разновидности.
Ключ для определения разновидностей
1. Лопасти листьев узкие (у основания 2 см ширины), длиной 5—6 см. Семена
белые, 10—12X7 мм......................................var. palmata.
+ Лопасти листьев широкие и короче, ширина у основания 4—5 см, длина 3—4 см.
Семена несколько темнее, 8—9X13—14 мм.............var. californica.
1. Разновидность пальмовидная—var. palmata.
2. Разновидность калифорнийская — var. californica (Torrey et
Wats.) Filov, comb. nova. — C. californica Torrey et Wats., 1880, Bot.
Calif. 1.: 240.
Растения более интенсивной серой окраски, чем основная форма
вида. Лопасти листьев и доли шире и короче. Плоды с менее отчетли-
выми продольными полосами. Семена темнее, крупнее и толще (8—9Х
X 13—14 мм).
Вид пальмолистный — С. palmeri Bailey
1943, 1. с. :299.
Растения сероватые. Стебли тонкие, ребристые, междоузлия корот-
кие, с короткими волосками. Листья мелкие, без колючек, серые, серд-
цевидно-овальные (8—12X14—16 см, часто мельче), с тремя главными
узкими лопастями, доходящими до середины листа. Края листьев
слабо изрезаны, верхняя поверхность шероховатая, нижняя с жестким
сероватым опушением. Усики короткие и слабые, 2—4-раздельные.
Цветки мелкие, опушенные, длиной 4—6 см, колокольчатые; дольки
чашечки нитевидные, длиной 1 см.
Распространен на северо-западе Мексики. Корни и плоды неиз-
вестны. Требует дальнейшего изучения.
Вид галеотти—С. galeotti Cogn. in DC.
1881, Monogr. Phanerogam. 3 : 551.
Стебель толстый, ребристый; листья узкотреугольные (9—12х
Х12—16 см), верхушка острая, края сильнозубчатые, выемки листьев
неглубокие, верхняя поверхность интенсивно зеленая, слабоволосистая,
белопятнистая, нижняя серовато-зеленая, с короткими густыми волос-
ками. Черешок толстый, цилиндрический, длиной 10—15 см. Цветки
на толстой опушенной цветоножке, длиной 8—15 см, венчик узкоколо-
208
кольчатый, длиной 6 см, с ланцетными сегментами. Чашечка короткая
(1 см), суженная к основанию, колючая, с длинными тонкими рассти-
лающимися долями, длиной 4—5 см. Корни и плоды неизвестны.
Растет на юге Мексики. Требует дальнейшего изучения.
Вид укореняющийся — С. radicans Naud.
1866, Ann. Sci. Nat. (Paris) 5, 6:8.
Стебли очень длинные (до 10 м), стелющиеся, неколючие. Расте-
ние многолетнее, размножается клубеньками диаметром 3—4 см. Обра-
зует очень много тонких побегов, которые легко укореняются. Усики
короткие. Листья сердцевидно-овальные (8—18X8—18 см), с треуголь-
ной верхушкой, слабовыемчатые, с заходящими лопастями. Чашечка
короткая, колокольчатая, с шиловидными чашелистиками. Плод слабо
сплюснутый (4X4,5 см), блестящий, ярко-зеленый, с 7 нечеткими бело-
ватыми полосами. Семена длиной 7—8 мм, беловатые.
Обнаружен в Мексике, в районе г. Мехико.
ИСТОРИЯ и ЗНАЧЕНИЕ КУЛЬТУРЫ ТЫКВЫ
Европейцы обратили внимание на такое крупноплодное растение
Америки, как тыква и, вероятно, сразу же интродуцировали ее оттуда.
В субтропических областях и колониях Старого Света (Индия, Среди-
земноморье) тыква быстро прижилась и в период эпохи Возрождения
уже упоминалась в литературе. Как удивительное растение тыква
быстро распространилась также в ботанических садах и, судя по оби-
лию литературы, стала объектом интенсивного изучения и пробно-де-
монстрационных посевов.
Лучше всего тыква произрастала в субтропических странах: Индии,
Японии, на юге Китая, Средиземноморском побережье, поэтому именно
там за предшествующие 5 веков сложились ее характерные экотипы,
отличающиеся от таковых Американского континента.
В нашей стране тыква появилась в XIX в. и в середине его стала
усиленно распространяться. В отличие от арбуза и дыни, она не имела
торгового значения, но ее широко возделывали на приусадебных участ-
ках для домашнего употребления. Редкий огород обходился без тыквы,
так как она почти не занимала полезной площади: ее высаживали по
краям огородов, а плети распространялись по изгородям и межевым
пространствам. В домашнем обиходе тыкву использовали в печеном и
вареном виде, в хлебопечении, при варке каш, она шла на корм скоту,
особенно свиньям. Сейчас больше всего ее возделывают в Волгоград-
ской, Саратовской, Днепропетровской, Донецкой, Харьковской, Одес-
ской и Воронежской областях, в Краснодарском, Ставропольском
краях. Посевы тыквы среди бахчевых культур занимают большие пло-
щади в Черниговской, Киевской, Саратовской областях и на Дальнем
Востоке (выше 50%). В последние годы посевы тыквы как кормовой
культуры в крупных хозяйствах увеличились. Тыква —одна из перспек-
тивных, незаслуженно забытых культур нашего земледелия. Последние
работы советских и американских селекционеров вскрыли большие ре-
зервы повышения пищевых, диетических и кормовых качеств этой
культуры (высокий процент сахара, каротина, редких витаминов), ко-
торые позволяют гораздо шире использовать ее в пищевых целях.
Тыква —урожайная культура: масса плода может достигать 100 кг.
Имеются сорта, содержащие сахара больше, чем арбуз. Диетическая
ценность тыквы обусловлена легкой усвояемостью всех углеводов, что
делает ее незаменимой в рационах детского питания.
209
У широко распространенных в настоящее время пищевых сортов
тыквы (Волжская серая, Медовая белая, Перехватка, Мозолеевская,
Миндальная) количество сахаров достигает 5—7%, а сухого вещества —
10—15%, у новых же сортов (Столовая зимняя А-5, Столовая 15, Испан-
ская 73) соответственно 10—42 и 20—30%. Тыква с оранжевой мякотью
содержит до* 15 мг каротина на 100 г, то есть вдвое больше, чем у
моркови.
В некоторых районах СССР тыква — важный продукт питания: в
свежем виде ее добавляют в хлебное тесто, в кашу, пекут и жарят, го-
товят суп-пюре. Из тыквы можно приготовить повидло, джем, варенье,
а также замариновать ее.
Диетические свойства тыквы позволяют рекомендовать ее при за-
болевании почек, печени, мочевого пузыря, при катарах, язвенной бо-
лезни желудка и атеросклерозе. Свежую мякоть тыквы прикладывают
для заживления воспаленных мест на теле при ожогах, сыпях, экземах.
Семена тыквы используют как противоглистное средство.
Семена тыквы содержат до 50% пищевого масла, которое по вкусу
напоминает прованское. Его использует в пищу население западной
области Украины (СССР), Венгрии, ГДР, ФРГ, Австрии, Румынии,
Италии.
Кормовые достоинства тыквы выше, чем кормовой свеклы; бел-
ковых веществ у нее в 4 раза, а кормовых единиц на 18,5% больше,
чем у последней. Тыква — молокогонное и повышающее жирность мо-
лока средство. В кормовых целях тыкву используют для всех видов
скота и птицы, а также силосуют с соломой, кукурузой и т. п. Поэтому
посевная площадь кормовой тыквы в нашей стране значительна, хотя
еще недостаточно используются высококаротинные сорта. Кормовая
тыква — одна из наиболее урожайных (с 1 га она может давать до
100 т урожая) и лежких культур.
Тыква засухоустойчива; ее можно выращивать в тех районах, где
растут корнеплоды, кукуруза и другие виды сочных кормов. Она яв-
ляется одним из лучших предшественников для всех культур, ею часто
занимают занятые пары. Сильно развитая и далеко простирающаяся
в стороны корневая система тыквы разрыхляет и обогащает почву боль-
шой массой корневых остатков.
БИОЛОГИЯ РОСТА И РАЗВИТИЯ ТЫКВ
ОТНОШЕНИЕ К УСЛОВИЯМ СРЕДЫ
Происхождение тыквы из тропической зоны Америки определяет и
ее отношение к факторам среды, в частности к температуре, свету,
влажности и структуре почвы. Тыква — очень теплолюбивая, жаростой-
кая, светолюбивая, засухоустойчивая, но и наиболее влаголюбивая, тре-
бующая легких, но не песчаных, а среднесуглинистых или обогащенных
супесчаных почв.
Как теплолюбивая культура тыква требует 3—5 месяцев теплого
периода с температурой не ниже 20° С для полного цикла развития.
Наиболее скороспелыми являются овощные тыквы—кабачки и патис-
соны. Их убирают в недозрелом состоянии, на 3—4-й месяц вегетации.
Вслед за ними по скороспелости идут бахчевые (полевые) тыквы
того же вида (С. реро), мякоть и семена которых созревают за 4 ме-
сяца. Они более теплолюбивые, чем кабачки и крупноплодная тыква
(С. maxima). Однако твердокорая тыква (С. реро) плохо переносит
высокую температуру (30°С и выше), плоды ее при этом мельчают,
210
листья страдают от хлороза, урожай снижается. Оптимальная темпера-
тура для роста и развития этих видов 20—25° С.
Крупноплодная тыква более пластична; некоторые ее сорта устой-
чивее к низким температурам, чем сорта тыквы твердокорой, их выра-
щивают ближе к северу. Существуют и тропические сорта этого вида,
требующие для развития продолжительного периода вегетации с высо-
кой температурой воздуха (25—35°С). К ним относятся зимние сорта,
плоды которых могут долго сохраняться после вегетации в поле.
Мускатная тыква наиболее теплолюбивая и позднеспелая. Север-
ная граница ее распространения совпадает в нашей стране с южной
границей возделывания твердокорой тыквы. Большинство сортов мус-
катной тыквы выращивают в тропических и субтропических областях,
самые скороспелые — в Краснодарском и Ставропольском краях; наи-
большее распространение этот вид получил в Средней Азии и респуб-
ликах Закавказья. Оптимальная температура развития для мускатной
тыквы 30—40° С.
При увеличении холодостойкости тыквы или культуры ее при по-
ниженных температурах в листьях увеличивается содержание хлоро-
филла, сахаров (преобладают моносахара); процессы метаболизма не-
сколько замедляются, но направляются в сторону окислительно-восста-
новительных, при этом повышается общий энергетический уровень об-
мена. Охлаждение растений (до 3°С) вызывает возрастание вязкости
протоплазмы, увеличение количества связанной воды при снижении об-
щей, а также снижение интенсивности дыхания и активности перо-
ксидазы.
Повысить холодостойкость тыквы можно путем длительной (20—
30 дней) обработки прорастающих семян переменными температурами
(метод Вороновой) или кратковременным, но более сильным воздейст-
вием холода на них —промораживанием или резким охлаждением.
Жаростойкость тыквы, по исследованиям Л. А. Хлебниковой, выше,
чем арбуза. Так, температура листьев тыквы при высокой температуре
воздуха несколько выше, чем температура листьев арбуза, очевидно,
из-за более слабого охлаждения их транспирацией. Это перегревание
листа, однако, не наносит ущерба растению, ибо температура коагуля-
ции белка в листьях тыквы значительно выше (60—65°С), чем у ар-
буза (45—54°С). В соответствии с этим прирост сухого вещества в
дневные часы у тыквы выше (7,9 г в 1 ч на 1 м3 листовой поверхности),
чем у арбуза (2,6 г).
В разные фазы развития температура коагуляции белка листьев
неодинаковая: она несколько выше в фазу цветения, а также на бо-
гаре. Между видами тыквы больших различий по этому признаку нет,
но сорта из поливных районов обладают меньшей жароустойчивостью.
Жароустойчивость тыквы связана с водным режимом: интенсивная
транспирация понижает температуру листа и усиливает жаростойкость
белковых коллоидов.
Засухоустойчивость тыквы, как и других бахчевых растений, об-
условлена широко разветвленной корневой системой, ее высокой сосу-
щей силой (свыше 10 атм*), которая обеспечивает растение водой
даже в сильно засушливых условиях. Однако тыква более влаголю-
бива, чем арбуз и дыня. Это объясняется более слабым развитием ее
корневой системы по сравнению с надземной частью, испаряющая по-
верхность которой чрезвычайно велика.
Тыква выделяет корневую пасоку в 7 раз слабее, чем арбуз, а это
явление — показатель мощности корневой системы. Резких различий по
* По международной системе единиц (СИ) 1 атм= 101,325 кПа (1,01325-10s Па).
211
отношению к влаге у видов тыквы не установлено. Темпы роста ее
надземных органов в противоположность арбузу не отстают от роста
корневой системы. Создается напряжение водного баланса благодаря
усиленному испарению, поэтому у тыквы часто не образуются плоды.
Она же сильнее иссушает почву, чем арбуз и дыня.
Таким образом, тыква нуждается в более влагоемких почвах. Луч-
шими для нее являются черноземы и темно-каштановые, а также тем-
ноцветные обогащенные супеси. Большая вегетативная масса и урожай
сухого вещества требуют более обогащенных почв, чем другие бахче-
вые культуры. На тяжелых глинистых почвах тыква с трудом разви-
вает корневую систему; на легких песчаных она также не дает устой-
чивых урожаев из-за их слабой обводненности и недостатка питатель-
ных веществ. При искусственном орошении тыкву можно выращивать
на самых разнообразных почвах, за исключением тяжелых глинистых.
Солеустойчивость к хлористым солям у тыквы выше, чем у арбуза
и дыни. Резкое снижение прироста проростков у тыквы начинается
при 0,010-молярном растворе NaCl, у дыни — 0,005, а у арбуза — при
0,002. Предельная солеустойчивость (по содержанию хлора на воздуш-
но-сухую почву) у тыквы 0,020%.
Тыква переносит концентрацию раствора сернокислого натрия до
0,03 М, но дальнейшее повышение ее ведет к гибели растения, в то
время как арбуз и дыня прекращают рост только при концентрации
0,09 М.
Тыква — растение короткого дня. Световая стадия лучше всего
проходит при 12-часовом дне и заканчивается обычно в фазе 2—4 на-
стоящих листьев. Более длительное освещение задерживает ее раз-
витие. Сокращение светового дня у поздних сортов увеличивает число
женских цветков на 10—15%. Свет может оказаться причиной уродли-
вых всходов тыквы, если он долго действует на них при низкой тем-
пературе (ниже 20°С); при прорастании в темноте или при 23—24°С
уродливости всходов не наблюдается.
БИОЛОГИЯ РОСТА И РАЗВИТИЯ
Семена тыквы сохраняют всхожесть до 8—10 лет. Однако посев-
ные качества их постепенно падают. Так, по исследованиям В. Ф. Бе-
лика [4], через 4 года хранения ее семена снижают всхожесть на 5%,
энергию прорастания на 15%, рост проростков замедляется. Через
6 лет всхожесть семян падает на 70%, энергия прорастания — на 82%.
а проростки становятся короче в 6 раз; через 9 лет хранения всхо-
жесть семян снижается до 16%, энергия прорастания — до 4%.
В литературе часто указывается, что лучшими для получения вы-
сокого урожая являются семена 2—3-летней давности. Это верно лишь
для семян биологически недозрелых или убранных в сырую холодную
погоду. Недозрелые семена можно довести до кондиции также путем
послеуборочного дозревания в сухом теплом помещении и прогревания
перед посевом. Хорошо вызревшие семена дают лучшие результаты
на следующий год их посева и в предпосевном прогревании не нуж-
даются.
Продолжительность покоя семян зависит от условий хранения (вы-
нужденный покой) и от наличия тонкой прозрачной слизистой обо-
лочки, облегающей кожуру сырых семян. При высушивании семян эта
оболочка отделяется и не пропускает к семени кислорода, необходимого
для прорастания. Если она нарушена еще в плоде, что можно опреде-
лить по потемнению семени, то последнее начинает прорастать внутри
плода.
212
Существует рентгеноконтрастный метод определения всхожести се-
мян тыквы. Он состоит в том, что семена намачивают в 20%-ном рас-
творе хлористого бария в течение 1 ч при комнатной температуре, за-
тем слегка подсушивают, помещают на фотопленку и облучают рент-
геном. Нежизнеспособные ткани семян пропитываются ВаС1 и на про-
явленной пленке оказываются более темными. Лучшие по посевным
качествам и продуктивности семена тыквы находятся в нижней части
плода (у плодоножки). Урожай из этих семян выше на 20—27%.
Прорастание. В условиях нормальной влажности оно начинается
при температуре 9,5°С, но оптимальная температура прорастания 26—
27° С. Первым развивается корень. Через 2—3 дня после начала про-
растания кожура семени лопается почти по всей длине бокового руб-
чика и из нее освобождаются семядоли. К появлению последних над
поверхностью почвы (что происходит в нормальных условиях через
4—8 дней) корень уже имеет длину 10 см и многочисленные боковые
корешки. В семядольный период и в начале образования 1-го листа
тыква легко переносит пикировку, если у нее прищипывают наполовину
длины особо вытянувшиеся корешки. Причем пикированные растения
развиваются лучше непикированных.
Прорастание семян можно стимулировать, кроме оптимальных
условий тепла и влажности, воздействием электрического тока, облу-
чением гамма-лучами 60Со (при 5 кр), выдерживанием семян С. maxima
в магнитном поле (в течение 120 ч) ив магнезированной воде (в те-
чение 168 ч). Темно-красный свет подавляет их прорастание, белый
свет тормозит, синий и голубой слабо задерживает, а светло-красный
с зеленым не влияет на прорастание. Вид С. реро чувствителен только
к темно-красной части спектра.
Гиббереллин* ускоряет первоначальный рост плетей и увеличивает
количество мужских цветков, причем наблюдается положительная кор-
реляция между последним признаком и длиной стебля тыквы.
Из различных способов стимулирования прорастания семян тыквы
лучшими являются способы: Вороновой — воздействие переменными
температурами, Дронова — воздействие горячей водой, Генкеля — нама-
чивание и высушивание, а также промораживание и намачивание семян
в растворе калия. Эти способы увеличивают всхожесть и ускоряют про-
растание семян.
Органогенез. Одновременно с прорастанием начинается образова-
ние органов растения. Весь период его жизни состоит из 12 этапов.
I — наклевывание семян — появление всходов.
П — появление всходов — образование зачатков 5-го листа.
III — вытягивание первых цветочных бугорков.
IV—появление 1-го настоящего листа —начало формирования ге-
неративных органов.
V — формирование пыльников у первых мужских цветков.
VI — быстрое развитие мужского цветка, превращение бугорков
в пыльники, начало формирования пыльцы.
VII — начало развития женских цветков.
VIII — появление желтой окраски у мужских цветков и сформиро-
вавшихся бутонов у женских.
IX — образование 1-го женского бутона, появление желтой окраски,
рыльца и лепестков женского цветка, раскрытие мужских цветков.
X — начало раскрывания первых женских цветков, массовое цве-
тение мужских цветков.
XI — массовое раскрытие женских цветков, рост завязей.
XII — начало созревания первых плодов.
• Для производственного применения не рекомендован.
213
Продолжительность каждого этапа изменяется в зависимости от
скороспелости сортов и температурных колебаний в данном сезоне.
На I этапе первым органом из зародыша семени начинает разви-
ваться корешок. Семядоли сформированы еще в семени, поэтому они
только разбухают, пробиваются наружу и зеленеют. Главная часть за-
родыша — почечка. В ней расположен конус нарастания — будущий
стебель, уже окруженный двумя листовыми валиками. При прораста-
нии семени конус нарастания увеличивается и превращается в округ-
лую выпуклость. На этом заканчивается I этап органогенеза.
На II этапе конус нарастания начинает дифференцироваться на
листовые валики и укороченные междоузлия.
На III этапе в пазухах будущих листьев появляются цветочные бу-
горки.
На IV этапе с появлением цветочных бугорков дифференциация
конуса нарастания замедляется до образования пыльцы. Появляется
1-й настоящий лист. На протяжении всех четырех этапов в развитии
тыквенных растений не находят различий ни между сортами, ни между
видами; различия начинаются с пятого этапа.
На V этапе органогенеза тыквы наиболее развитый бугорок раз-
мером более 0,1 мм принимает бокаловидную форму и дифференци-
руется на чашечку и венчик. Когда длина цветка достигнет 0,5—0,8 см,
на внутренней поверхности венчика образуется 5 тычиночных бугорков.
На VI этапе начинается разделение пола цветков. У мужских ты-
чиночные бугорки быстро увеличиваются в размерах, переходя в пыль-
ники, которые петлеобразно изгибаются. Будущие женские цветки
остаются без изменений.
На VII этапе пыльники желтеют, начинается бутонизация женских
цветков, вслед за тычиночными бугорками появляются бугорки плодо-
листиков, отчего цветок как бы приобретает обоеполость. Развитие бу-
горков плодолистиков на этом этапе идет более интенсивно, чем ты-
чиночных бугорков, и последние редуцируются; формируется чисто жен-
ский цветок. На этом этапе начинается дифференциация частей цветка,
рост венчика обгоняет рост чашечки.
С VIII этапа по XII развитие контролируется визуальным наблю-
дением за растениями согласно схеме органогенеза. По данным лабо-
ратории бахчевых культур ВИР, у крупноплодного вида тыквы первые
11 этапов органогенеза проходят за 53 дня, у твердокорого первые
9 этапов — за 60 дней, а у мускатного вида — за 76 дней. У кабачков
развитие идет быстрее. Максимум содержания хлорофилла в листьях
имеет место на IX этапе органогенеза: у крупноплодного вида 8,5 мг
на 1 дм2, у твердокорого 7,5 мг, у мускатного — 7,9 мг, т. е. количество
их обратно пропорционально быстроте прохождения органогенеза.
Рост вегетативных органов. Формирование надземных органов
у тыквы начинается с семядолей, которые растут 10 дней. Затем раз-
виваются настоящие листья. После образования 5—7-го листа (через
15—25 дней) появляются боковые плети. Первые междоузлия у пло-
довых растений короткие, поэтому главный стебель имеет стоячее по-
ложение, и растения имеют вид кустика. Максимальный прирост веге-
тативной массы наблюдается в конце 2-го месяца после всходов.
Плети, отходящие от главного стебля (I порядка), развиваются
сильно и иногда перерастают по длине главный стебель. Плети II по-
рядка, отходящие от плетей I порядка, короче, а плети III порядка
еще короче, чем плети I порядка; они образуются намного позднее и
обычно плодов не дают. Длина одной плети у нормально развитого
растения тыквы может достигать 10 м и более. От основного стебля
отходит до 20 побегов (плетей) I порядка, причем 4—5 нижних по
214
длине не уступают основному стеблю. Число побегов II порядка до-
стигает 10 (не считая очень коротких). Общая длина всех плетей до-
стигает 50 м. При этом вид твердокорый имеет плетей больше, но более
коротких, чем вид крупноплодный; общая длина их у последнего вида
тыквы почти в 2 раза больше. Плети растут очень быстро: в период
интенсивного роста основная плеть увеличивается в сутки на 25—30 см,
а прирост всех плетей за это время может достигать 10 м.
На одном растении может быть до 170 листьев, причем наибольшее
их количество, так же как и общая поверхность листового аппарата
(до 1,7 га на 1 га), отмечена у вида твердокорого.
В процессе роста органов по закону параллельного морфоге-
неза [78] наблюдается корреляция их формы в пределах растения.
В. И. Пальчевским [57] было обнаружено, что индексы семядолей
тыквы соответствуют индексам плодов, исключая патиссоны и кусто-
вые формы тыквы.
Тыква относится к растениям короткого дня, поэтому его умень-
шение до 12 ч несколько удлиняло плети, а уменьшение до 8 ч и уве-
личение до 16 ч сокращало их длину. На кустовые формы фотопериод
влияния не оказывал.
Гораздо сильнее оказалось действие гиббереллина: оно превра-
щало кустовые формы тыкв в длинноплетистые. Действие 0,02%-ного
раствора гиббереллина на точки роста стимулирует ралнее зацветание,
вызывает увеличение числа слоев камбия, размеров камбиальных кле-
ток, а также уменьшение числа и размеров зерен крахмала в клетках
осевых органов.
Другие фитогормоны и стимуляторы оказывают слабое влияние на
развитие тыквы.
Воздействие на молодые растения стимуляторами из экстрагиро-
ванных листьев алоэ (100 г листьев на 500 г воды) и глюкозой путем
впрыскивания их шприцем под кору стебля растения на 80—100% уве-
личивало сахаристость плодов в год опыта и в последующие поколения
за счет увеличения дисахаров.
Кустовые формы тыкв являются более скороспелыми, так как по-
требительская спелость у них наступает раньше созревания семян.
Н. Н. Нигмановой [52] замечено, что длина зародышей у таких форм
вдвое короче, чем у длинноплетистых тыкв. Семядоли и стебель v них
росли медленнее, но продолжительность жизни листьев была больше,
чем у плетистых тыкв. Удаление некоторых листьев у кустовых форм
вызывает отмирание побегов, залегающих под обломанным листом.
У предельного кустового типа листья крупнее, общая листовая пло-
щадь больше, чем у слабоветвящихся кабачков, поэтому первые плодо-
витее и урожайнее.
Рост корневой системы. Корневая система тыквы разветвлена
гораздо сильнее, чем надземная. Общая длина всех корней одного
растения может достигать 25 км. Основная масса нх располагается
несколько глубже, чем у арбуза. Сосущая сила корней тыквы в сред-
нем равна 7—8 атм, а количество выделяемой ее корнями пасоки 3 см3
в 1 ч. Кабачки имеют менее развитую корневую систему и выделяют
пасоки в 3 раза меньше, чем плетистая тыква С. реро.
Расположение корневой системы у тыквы простратное, главным об-
разом в пахотном слое почвы. В зависимости от ее типа и степени
обработки корни находятся на разной глубине, на легких и рыхлых
почвах они проникают глубже. Самые длинные и крупные корни рас-
положены на глубине 5—12 см. Стержневой корень идет вглубь, но не
глубже 1—1,5 м.
215
Корневая система тыквы имеет максимальные размеры в период
наибольшего развития надземных частей растения. Она простирается
до 4—5 м в радиусе, занимая до 16 м2 почвы.
Тыква имеет следующие типы корней: стержневой корень, боко-
вые корни I—IV порядка, придаточные корни, мелкие сосущие корни
и корневые волоски. Стержневой корень не играет большой роли в во-
доснабжении растения. Корни I порядка (10—20) наиболее длинные,
отходят от стержневого у его основания. Корни II порядка (более 100)
составляют основную массу и включают основную протяженность
усвояющих зон. Корни III порядка в основном тонкие (1 мм). В сухие
годы развиваются и корни IV порядка (у крупноплодного вида), но их
очень мало.
Придаточные корни короткие (до 50 см), образуются в фазе цве-
тения из узлов стелющихся на влажной почве побегов, расположены
вертикально и составляют группу скелетных корней. В фазе шатрика
(5—7 настоящих листьев) боковые корни разных растений смыкаются,
поэтому междурядную обработку тыквы в этот период нужно прово-
дить во влажные годы на глубину 4—5 см, в сухие не более чем на
10 см, а подкармливать растения следует в середине междурядий.
В фазу цветения корневая система сформирована полностью, при
этом начинается отмирание верхушек боковых растений I порядка и
уменьшение активной части корня, но развиты мощные корни II и
III порядка. К созреванию плодов все боковые корни имеют мертвые
верхушки.
При малой площади питания длина корней уменьшается почти
вдвое, боковые корни уходят вглубь. При большой площади питания
отмирания верхушек корней не отмечено. На хорошо увлажненной
почве наибольший урожай получают при площади питания одного рас-
тения 4—5 м2, а на недостаточно увлажненной при 1—2 м2. Пищевая
и кормовая ценность плодов тыквы, полученных с малых площадей
питания, выше.
Мелкие сосущие корни располагаются на всех скелетных корнях,
они недолговечны, образуются вслед за зоной корневых волосков. По-
следняя имеет протяженность в среднем 8 см, а зона мелких сосущих
корней 10 см. Сосущие корни также покрыты корневыми волосками,
которые вместе с волосками скелетного корня составляют активную
всасывающую часть скелетных корней.
Первым из прорастающего семени появляется корешок. В базе
семядолей стержневой и боковые корни I порядка достигают уже 20—
23 см каждый, в фазе 1-го настоящего листа — 35 см, в фазе шатрика —
140 см. В это же время активная зона корня, покрытая мелкими со-
сущими корнями и корневыми волосками, достигает 50 см; позднее она
уменьшается. Максимальные размеры корневая система имеет в период
введения: общая длина основных корней достигает 170 м, а среднесу-
точный прирост — 4—5 см.
Кратковременное охлаждение (закалка) корневой системы у мо-
лодого пастения ускоряет развитие и увеличивает корневую систему
тыквы. Корневые волоски сохраняют правильную форму и нормально
функционируют при постоянной влажности почвы, с изменением усло-
вий влажности они утолщаются, искривляются или ветвятся.
Виды тыквы несколько различаются по характеру корневой си-
стемы. Сильнее других она развита у С. maxima, слабее — у С. реро
и С. moschata. Последний вид как наиболее поздний больше развивает
корней IV порядка по сравнению с другими культурными видами. Ряд
ксерофильных диких видов образует корнеплодные утолщения на кор-
нях, позволяющие им переносить летнюю засуху.
216
Общая масса корней в фазе шатрика наибольшая у скороспелых
сортов, но, начиная с фазы массового цветения и до конца вегетации,
она преобладает у позднеспелых. Масса надземных частей у развитых
растений всегда больше массы корневой системы, и с возрастом она
увеличивается.
Микориза на корнях тыквы сильно размножается при отсутствии
удобрений, значительно уменьшается на фоне минерального удобрения
и отсутствует при внесении навоза.
Хорошая корневая система у тыквы развивается только при на-
личии кислорода в почве, поскольку из вегетативных частей растения
в корни он не поступает. Недостаток кислорода в зоне развития кор-
невой системы ведет к повреждениям ультрамикроскопической струк-
туры корней, к нарушению их функций и к гибели растения.
Почвенное питание. Кроме макроэлементов: азота, фосфора, калия,
серы, кальция, магния растение усваивает свыше 60 микроэлементов:
бор, марганец, цинк, железо, молибден, медь, алюминий, йод, хлор,
кобальт и др. Часть элементов (кремний, натрий и др.) из почвы по-
падает в растение как балласт, т. е. не участвует в дальнейших мета-
болических превращениях.
Поступление элементов контролируется составом выделяемой рас-
тением пасоки. В начале развития (в фазе шатрика) больше других
выделяет пасоки С. реро и меньше других С. moschata. Наибольшее
выделение пасоки у всех культурных видов наблюдается в начале цве-
тения. Поглощающая поверхность корней не совпадает с количеством
выделяемой пасоки: так, у С. maxima она больше, чем у С. реро. Сле-
довательно, у последней всасывание воды корнями идет интенсивнее,
а листовая поверхность С. реро уступает площади всасывающей по-
верхности корней, чего не наблюдается у других видов.
Изучают поглощенные и связанные элементы почвы чаще при по-
мощи меченых атомов и анализа пасоки. С пасокой из корней подаются
не только минеральные элементы, но и 18—50% переработанных асси-
милятов листьев. В корнях через 3 ч после поступления 60% элементов
превращаются в азотсодержащие соединения — аминокислоты и орга-
нические кислоты. Поэтому поглощение корнями азота и приток асси-
милятов взаимостимулируются. Из-за большой длины плетей отток
определенных ассимилятов продолжается более 1 суток и имеет су-
точную периодичность. В корни они поступают ночью, так как отток
их связан с периодичностью подачи пасоки.
Установлено, что с пасокой подаются: глютамин, аспарагин, лей-
цин, фенилаланин, валин, метионин, гистидин, тирозин, глутаминовая
и у-аминомасляная кислота. Часть аминокислот (аланин, глютамин и
у-аминомасляная кислота) имеет ярко выпаженную суточную периодич-
ность подачи, остальные поступают в растение непериодично. Кроме
суточной, наблюдается сезонная и возрастная периодичность в погло-
тительной работе корней тыквы. У молодых растений ритмичность по-
дачи аминокислот отсутствует или очень слабая суточная периодичность
пасоки начинает четко проявляться у 30-дневных растений. В этот пе-
риод за 36 ч растение может выделить 180 см3 пасоки, в фазу цвете-
ния— 240 см3, в фазу плодоношения—180 см3. Основные транспортные
формы углеводов у тыквы — стахиоза и вербаскоза, а не сахароза, как
у других культур.
Поступающий из почвы в корни минеральный фосфор с первых
же минут вовлекается в метаболизм, в первые 30—60 сек он входит
в состав органических соединений корня тыквы на 30%, а через 5—
7 мин — на 70%. Органические соединения фосфора в первую очередь
подаются из корней в листья, после насыщения которых они иакаплп-
217
ваются и в корнях. Фосфор преобладает в клетках зоны роста корней,
в меристеме в виде высокомолекулярных соединений, а по мере старе-
ния клеток в них преобладает синтез низкомолекулярных растворимых
фосфорсодержащих веществ.
В первую очередь фосфор включается в запасную энергетическую
базу клетки АТФ (аденозинтрифосфат) и в нуклеотидные соединения,
после чего он синтезируется с сахарами и некоторыми другими соеди-
нениями. Из АТФ энергия расходуется на поглотительную деятельность
корней. Поглощается элемент из почвы в зависимости от потребности
в нем растения, что связано с его водным режимом, дыханием и по-
требностью в других элементах. Поэтому поступление фосфора зависит
от накопления в нем кальция, железа и особенно азота. Отсутствие
кальция резко уменьшает поступление фосфора, а без азота оно пре-
кращается совсем и растение не развивает листьев.
В свою очередь, при недостатке азота в почве не усваивается поч-
венный азот; при этом происходит отток последнего из листьев в
стебли и корни, распадаются белковые соединения корней, подавляется
процесс нормального транспортирования сахаров из листьев в корни,
замедляется корневой синтез аминокислот и других кислот, участвую-
щих в поглощении из почвы ионов аммония (яблочной, лимонной, вин-
ной, щавелевой и др.), и, следовательно, нарушается весь углеводно-
фосфорный обмен. Подкормка фосфором (но не через листья, так как
последние не могут обеспечить растение фосфором из-за его плохой
транспортабельности) восстанавливает эти процессы.
Усвоение почвенного (преимущественно нитратного) азота состав-
ляет основную функцию корней. Поступивший азот передается в листья
в основном в виде неизменных нитратов и частично включается в син-
тез аминокислот и других соединений непосредственно в корнях.
Синтез аминокислот осуществляется в корнях из сахарозы, по-
ступающей из листьев в количестве 30—60%, и углекислоты, которую
корни берут из почвы. Синтез аминокислот в корнях идет в 2—6 раз
быстрее, чем в листьях. До 40% образовавшихся в корнях азоторгани-
ческих соединений подается также в листья. Для синтеза аминокислот
в корнях необходимо присутствие достаточного количества фосфора.
В корнях тыквы зафиксировано 18 аминокислот, из которых преобла-
дает аланин. Основными транспортными формами являются: аланин,
глютамин, аминомасляная кислота. Без фосфора наступает азотное го-
лодание, замедляется синтез белка, хотя на развитии вегетативной
массы это не отражается, но приостанавливается рост корней. Умень-
шение азотного питания стимулирует рост корней.
Калий поглощается тыквой независимо от фосфора и азота, ио
калий резко увеличивает его поглощение. Попадая в листья, он вы-
тесняет оттуда часть азота и фосфора. В присутствии калия растения
не берут из раствора натрий, а наличие большого количества солей
последнего приводит к дефициту калия в растении. Основная роль ка-
лия в растении — участие в построении клеточных структур, особенно
оболочек, поэтому он накапливается в молодых клетках меристем; в
старых тканях он, как и кальций, преобладает в клеточном соке. Од-
нако при недостатке калия в корнях тыквы тормозится окисление, па-
дает уровень органических кислот, нарушается азотный обмен, сни-
жается содержание нуклеотидов, синтез нуклеиновых кислот и белка.
Калийные удобрения, индуцирующие импульсы возбуждения, мо-
гут распространяться по стеблю со скоростью 30—40 см в 1 мин.
Наличие кальция — обязательное условие роста корней и над-
земных органов тыквы. При его отсутствии в первую очередь начи-
нают страдать корни, хотя в подземных органах растения он содер-
218
жится в больших количествах. При недостатке кальция клеточные
стенки ослизняются, так как он входит в состав солей пектиновой кис-
лоты, насыщающих межклетники. Кроме того, кальций легко образует
соли наиболее распространенных в растениях кислот: серной, фосфор-
ной, щавелевой и угольной. Он постоянный компонент многих структур
клеток, включая ядро и хромосомы. При отсутствии кальция нару-
шается также нормальный ход деления клетки.
Магний в растениях тыквы идет на построение протоплазмы,
хлорофилла, ферментов, он участвует в процессе включения аминокис-
лот в белковую молекулу, задерживает окисление, входит в состав ри-
босом. Магния больше всего в клеточном соке тканей тыквы, из кле-
точных структур он концентрируется главным образом в клеточных
стенках.
Кислород нужен растению не только для дыхания, но и для
корневой переработки ассимилятов, которые при этом превращаются
у тыквы в органические кислоты, служащие акцепторами минеральных
соединений азота, поглощаемого из почвы. При засухе эта переработка
задерживается, ассимиляты (сахара) накапливаются в корнях до
10-кратного размера, часть их возвращается вместе с пасокой в листья.
При этом в корнях уменьшается количество растворимого фосфора и
АТФ, а следовательно, сокращается синтез нуклеиновых кислот, по-
давляется рост. У различных видов растений разная чувствительность
к недостатку кислорода в почве.
Сера входит в состав всех организмов (в том числе и растений)
почти всеми главными органическими и неорганическими соединениями,
основная роль которых участие в энергетических процессах организма
и биологически активных соединениях.
Роль микроэлементов в развитии тыквы еще недостаточно
выяснена. Выращивание тыквы в среде с недостаточным количеством
бора показало гипертрофию ксилемы в местах ее нарастания (кончики
стеблей и корешков, камбий), увеличение распада клеток, а также па-
раллельное увеличение активности каталазы и наличие хлороза. Такие
растения вскоре теряли способность к геотропической реакции. Для
нормального течения жизненных процессов в растениях, в особенности
как активаторов ферментов и составных частей их, в разной степени
важны и другие микроэлементы.
ВОДНЫЙ РЕЖИМ
В южных степных районах, где выращивают основную массу бах-
чевых культур, выпадает в среднем 300—400 мм годовых осадков. Это
составляет 3000—4000 м3 воды на 1 га. По нашим данным, в условиях
Средней Азии тыква испаряет с листовой поверхности 460 г влаги за
1 ч. Одно растение испаряет за 1 ч примерно 700 г, а с 1 га посева
за сезон испаряется 4000—5000 т воды. В неполивных условиях испа-
рение происходит менее интенсивно.
Испарение с поверхности почвы составляет гораздо меньшую ве-
личину ввиду затенения почвы растением, последнее испаряет 98% по-
ступающей воды.
Транспирационный коэффициент тыквы в среднем равен 834, что
выше, чем у других бахчевых, кукурузы, проса и пшеницы, а во влаж-
ные годы достигает 1535.
Скороспелые сорта в начале развития испаряют воды меньше, чем
позднеспелые, но в фазу созревания больше. По данным Н. Г. Гордеевой
[14], у большинства сортов наиболее сильная транспирация осуществ-
ляется в полдень, за исключением некоторых сортов, энергичнее транс-
219
пирирующих утром; к вечеру транспирация, как правило, понижается.
По данным того же автора, тыквы энергичнее транспирнруют в
начальный период развития и снижают энергию транспирации с появ-
лением плодов (табл. 1).
Таблица 1. Транспирация тыквы в разные фазы развития, г/м2, ч
Сорт Плетеобразование Цветение Рост плодов
Г оббард 1020 765 585
Алашехирская 744 874 642
Испанская 73 337 336 337
Кашгарская 1644 459 265 293
Кабачок (местный 744 785 367
образец)
Транспирация устьичная, внеустьичная составляет незначительную
величину. Устьица у тыквы по размеру мельче, чем у арбуза и дыни,
но расположены на листе они в 5—6 раз гуще, имеются устьица и на
плодах (табл. 2).
Таблица 2. Количество устьиц у тыквы (в поле зрения микроскопа при
увеличении в 125 раз)
Сорт Вид Происхождение На нижней стороне листа На плоде
Кашгарская 1644 С. moschata Китай 636 147
Витаминная « Колумбия 506 120
Курокава « Япония 466 127
Чалмовая 998 С. maxima Китай 588 300
Черкесская « СССР (Северный Кавказ) 453 82
Волжская серая « СССР (Поволжье) 306 50
Испанская 73 « Испания 240 90
Алашехирская « Малая Азия 240 50
Бирючекутская 735 С. реро СССР (Украина) 560 193
Мозолеевская 15 < То же 374 177
Патиссон (местный образец) < < 547 64
Кабачок (местный образец) < < 386 82
Наиболее богат устьицами вид мускатной тыквы. Однако коли-
чество их не связано строго с видом, а зависит от влажности в районе
происхождения и периода развития. Поздние сорта (Чалмовая, Чер-
кесская) имеют больше устьиц, чем ранние (Мозолеевская) и кабачок
или сорта поливных районов (Испанская, Алашехирская). Количество
устьиц на плодах больше зависит от скороспелости и от размеров
плода, быстроты его роста, чем от принадлежности к тому или иному
виду.
Транспирацию регулирует устьичный аппарат в зависимости от на-
личия влаги в почве и объема сосущей части корневой системы. Суще-
ствуют и другие приспособления для регулирования водного режима
растения тыквы: опушение, выемчатость и белая пятнистость листовых
пластинок, толщина кутикулы и др.
Опушение тыквы — это приспособление для выделения влаги, осо-
бенно на молодых наружных тканях растения [78], а не для защиты
от испарения. Волоски расположены очень густо, обеспечивая макси-
мальное испарение. С возрастом тканей волоски редеют и теряют свои
220
функции, которые в основном переходят к устьицам. По густоте волос-
ков можно судить о сравнительной степени транспирации воды у тыквы
разных видов.
Экологические различия в опушении тыкв заметны главным обра-
зом по листьям. Эти различия сводятся к густоте волосков, их длине
и ширине основания. Густое короткое опушение связано с сухостью
климата. Оно имеет место у диких степных тыкв. Длинные волоски, на-
оборот, характеризуют гигрофильность растения. Наконец, волоски с
расширенным основанием, переходящие в шипы и даже колючки, ха-
рактерны для климата со значительным увлажнением и умеренной тем-
пературой. Таким опушением обладает твердокорый вид (С. реро).
Длинные тонкие волоски представляют собой хорошие капилляры, при-
способленные к испарению в более влажных районах. Волоски с рас-
ширенным многоклеточным основанием являются органами нагнета-
ния, принудительного выделения влаги, часто в капельножидком виде.
Кутикула — экологический признак, она толще у южных сортов
более сухих мест и тоньше у северных местных сортов влажной зоны.
Толстая кутикула задерживает как внеустьичную, так и общую транс-
пирацию.
Изрезанность и выемчатость пластинки листа уменьшают испаре-
ние, что особенно заметно на примере вида С. реро.
Некоторые формы тыквы из более влажных районов, особенно
формы С. moschata, имеют белые треугольные пятна между двумя жил-
ками верхней половины листа. Эти пятна вначале принимали за очер-
тания воздушных полостей над эпидермисом. По нашим исследованиям,
оказалось, что они представляют собой обесцвеченные участки листо-
вой паренхимы, указывающие на повышенную гигрофильность данного
экотипа.
Кроме листьев, значительное количество свободной воды находится
в стеблях, корнях и плодах тыквы. В стеблях и корнях вода находится
в транзитном состоянии, а в плодах накапливается параллельно с наи-
более интенсивным их ростом, главным образом в ночной период,
когда отсутствует испарение из листьев. При наличии сильного испа-
рения плод в дневные часы может отдавать листьям воды больше, чем
в него поступает. Таким образом, днем он может уменьшать свою
массу, а ночью увеличивать.
Содержание связанной воды в листьях тыкв может повышаться
при длительной культуре определенных сортов в засушливых условиях.
Исследованиями установлено, что микроэлементы: бор, медь, марганец
и алюминий повышают содержание связанной воды в растении, что уве-
личивает вязкость плазмы и повышает засухоустойчивость. Поэтому при
повышенной температуре растения требовательнее к бору.
Таблица 3. Сосущая сила клеток листа и осмотическое давление
сока тыквы в фазы развития, атм
Сорт Плетеобразование Цветение Рост плодов
сосущая сила осмотиче- ское давление сосущая сила осмотаче- ское давление сосущая сила осмотиче- ское давление
Гоббард 7 11,3 11 8,2 13 8,4
Алашехирская 5 10,8 7 10,1 13 9,4
Испанская 73 3 9,4 10 8,9 7 9.4
Кашгарская 5 9,8 9 7,4 10 8,9
Кабачок 7 12,0 9 11.2 11 9,2
Н. Г. Гордеевой [14] были определены такие показатели водного
режима, как сосущая сила клеток листа тыквы (методом Шардакова)
221
и осмотическое давление клеточного сока (криоскопическим методом),
которые при сопоставлении показывают потребность растения в воде
(табл. 3).
Преобладание сосущей силы над давлением клеточного сока по-
казывает недостаток воды для клеток. Эта величина возрастает во
время роста плодов. Достаточной физиологической обводненностью об-
ладает только сорт Испанская, который испытывает некоторый недо-
статок воды лишь в фазу цветения.
ФОТОСИНТЕЗ
Листья тыквы содержат около 13 мг хлорофилла на 100 г сухого
вещества. У теплолюбивых растений при пониженных температурах со-
держание хлорофилла снижается, а у холодостойких увеличивается.
Поэтому при весеннем посеве тыква ведет себя по содержанию хлоро-
филла как холодостойкое растение, а при летнем — как теплолюбивое.
Максимальное содержание хлорофилла в растении наблюдается в на-
чальные периоды развития до роста плодов, со старением растения оно
снижается. Хлорофилл находится не только в хлоропластах листьев,
но и в субэпидермальных слоях стеблей и плодов, в сосудистых пучках
тыквы. В молодых междоузлиях хлоропласты имеются во всех тканях.
Между содержанием хлорофилла и интенсивностью фотосинтеза
нет прямой корреляции. Интенсивность фотосинтеза понижается к ве-
черу при недостатке углекислоты в воздухе, снижении температуры или
повышении ее до 40° С и повышается при стимулировании ростовых
процессов.
Ассимиляты из листьев транспортируются в другие органы. В корни
идет около 36%, в черешки листьев нижней части стебля — 31, в кон-
чики побегов—18 и в черешки верхних частей стебля — около 12%.
Ранний съем плодов повышает продуктивность фотосинтеза, усиливает
приток ассимилятов к вегетативным органам, увеличивает число ли-
стьев и их поверхность, удлиняя период плодоношения и вегетационный
период растения. У гибридных растений тыквы интенсивность фотосин-
теза и качество синтезируемых веществ выше, чем у негибридных.
БИОЛОГИЯ ЦВЕТЕНИЯ И ОПЛОДОТВОРЕНИЯ
Цветки у тыквы крупные, ярко-желтые, раздельнополые. Мужских
цветков на одном растении в зависимости от сорта 140—430 за сезон,
женских 55—83. До 14-го узла на стебле образуются, как правило,
только мужские цветки, после 14-го отношение мужских к женским
составляет 3:1, а на боковых ветвях — 4: 1. В период массового цвете-
ния на одном растении ежедневно раскрыто 2—3 мужских цветка и
1—2 женских. У растений женского типа это соотношение меняется.
На главной плети у них в среднем 20% мужских цветков и 80% жен-
ских. Женские растения продуктивнее, обладают более коротким глав-
ным стеблем, почти не имеют разрыва в сроках раскрытия мужских
и женских цветков, более скороспелы.
Раньше других видов зацветает тыква твердокорая, затем крупно-
плодная. Женские цветки у всех видов открываются на 5—6 дней
позже мужских. Однако в умеренном климате или в прохладное время
женские цветки могут появляться даже раньше мужских. Верхние бу-
тоны развиваются быстрее, чем нижние. Мужской и женский цветки
тыквы остаются открытыми обычно один день независимо от оплодо-
творения. Цветение продолжается до осени, но при наличии 2—3 рас-
222
тущих плодов раскрытие цветков замедляется, а затем совсем прекра-
щается.
Опыты различных исследователей показали, что можно повысить
процент женских цветков в 2 раза и мужских в 4 раза путем внесения
высокой дозы азотного удобрения, намачивая семена или опрыскивая
гидразидом малеиновой кислоты в фазе семядолей и 1-го листа, вы-
ращивая при укороченном дне и пониженной температуре.
Искусственно созданный зеленый свет в теплицах препятствовал
образованию женских и замедлял формирование мужских цветков, крас-
ный свет ускорял развитие всех цветков, синий же ускорял развитие
женских. Ускорение развития способствовало появлению женских цвет-
ков, а задержка — появлению мужских.
Обрывание цветков вызывает энергичное развитие вегетативных
частей растения и образование новых цветков. Удаление только муж-
ских цветков стимулирует появление женских, отчего тыква несколько
повышает урожай. Последний способ эффективен только в I и II де-
кадах цветения; завязи, образовавшиеся позднее, почти не развиваются
из-за роста ранее завязавшихся плодов.
Воздействуя окисью углерода на формирующуюся почку, можно
изменить ее пол на женский благодаря выработке особых женских гор-
монов, притекающих в меристему почки. Такое воздействие, ранее за-
меченное на огурцах, можно осуществить в защищенном грунте.
Раскрывание бутонов начинается на рассвете и продол-
жается до 11 ч утра. В раскрытом состоянии цветок находится 7—8 ч.
По данным М. Е. Софронова, период активного восприятия зрелой
пыльцы рыльцами цветков С. реро начинается от 6 ч утра за сутки до
раскрытия цветка и заканчивается к 9 ч утра на 2-й день после рас-
крытия.
Период эффективного опыления и завязывания плодов с 6 до 10 ч
утра, позднее завязывание ослабевает и к концу вегетации прекра-
щается.
Пыльца тыквы имеет в диаметре 100—200 мкм, она крупнее у
вида С. реро с ее дикими формами и у С. maxima. Пыльцевые зерна
многопоровые, с шиповатой экзиной, поры оперкулятные (с крышеч-
ками). Пыльца тыкв богата каротиноидами, крахмалом и жировыми ве-
ществами, которых больше у видов С. реро и С. maxima. По данным
Цирну, количество пыльцы у видов тыквы следующее.
С. реро С. maxima С. moschata
1 цветок, мг 23,5 31,4 35,1
1 растение, г 11,5 17,3 11,4
1 га, кг 51,8-57,6 43,3-52,0 28,5—34,2
В естественных и искусственных условиях для эффективного завя-
зывания плода необходимо попадание на рыльце большого количества
пыльцы. Для образования одного семени требуется 2,6—83,5 пыльцевых
зерен, а количество семян в плоде варьирует от 150 до 400. Опыление
одной лопасти рыльца тыквы не ведет к одностороннему образованию
семян. Независимо от того, куда попала пыльца, она выходит в центр
апикального конца полости. По внутренней поверхности полости пыль-
цевые трубки растут до всех яйцеклеток, независимо от того, на какой
стороне они находятся. Слияние с ближними яйцеклетками происходит
через 36 ч от начала прорастания, а с дальними яйцеклетками — через
5 суток.
Температура — главный фактор, действующий на раскрытие пыль-
ников. Минимальная температура при этом 15—16° С, оптимальная
20—23° С.
223
Для получения мужски стерильных растений используют
парализующее действие различных веществ на половые элементы
цветка. Первым получил мужскую стерильность на тыкве А. Хатчинс
[151]. С. Витвер и Д. Хиллер [218] предложили методику получения
мужски стерильных растений опрыскиванием растений в фазе 2—3 ли-
стьев а-нафталенуксусной кислотой, что уменьшает отношение мужских
цветков к женским с 1,41:1 до 0,4: 1; в поле женские цветки раскры-
вались при этом за 8 дней до раскрытия первого мужского цветка.
Опрыскивание же гидразидом малеиновой кислоты в фазе семядолей
4—5 раз с интервалом 5—7 дней полностью препятствует появлению
мужских цветков, не тормозя появления женских.
В опылении тыкв большую роль играет количество нектара в цвет-
ках. Нектарники располагаются у основания тычинок и пестика. В жен-
ских цветках нектара в 2,5—6 раз больше. Наиболее нектароносным
является вид крупноплодный, наименее — твердокорый. По Цирну,
С. maxima дает на цветок 130 мг, а на растение 65 г нектара, или
164—230 кг/га при сахаристости его 35—40%; С. moschata — 9,5 мг
на цветок, 28 г нектара на растение, или 60—84 кг/га при сахаристости
26—33%; С. реро — 42 мг на цветок, 20 г нектара на растение, или
90—102 кг/га при сахаристости его 23—48%.
Выделение нектара идет главным образом утром, с 7 до 10 ч, по-
том оно ослабевает. Интенсивнее нектар выделяется в солнечные дни,
когда на женский цветок приходится до 558 мг нектара в день.
Основные опылители тыквы — пчелы, дополнительные — шмели,
осы, мухи и другие случайные насекомые. При недостаточной числен-
ности этих видов насекомых или в неблагоприятные периоды для их
лёта тыква часто не образует плодов, несмотря на обильное цветение.
В таких случаях весьма эффективно дополнительное опыление.
По данным А. Н. Невкрыты [50], тыква на 94,7% опыляется пче-
лами. Мужские и женские цветки посещаются ими в одинаковой мере.
В цветке пчела проводит в среднем 20 мин, в лёте— 10 мин. Для пол-
ного опыления одного женского цветка требуется 20 посещений пчел,
при этом на него пчелы приносят пыльцу в среднем с 40 мужских
цветков. Естественному опылению тыквы насекомыми препятствуют:
ветер (более 3 м/сек), увеличенная влажность, температура ниже 15°С
и выше 25° С.
В районах выращивания тыквы необходим 1 улей на каждый гек-
тар тыквы, но увеличение опылителей дает дополнительное повышение
урожайности. Опыление тыквы пчелами дает лучшие результаты, чем
ручное. Так, в парниках от опыленных пчелами цветков было получено
53,8% плодов, а от опыленных вручную — только 19%.
Ряд авторов [5, 96] рекомендуют дополнительное опыление цвет-
ков тыквы и при наличии опылителей, так как это значительно (на
50—72%) увеличивает урожай. Крупность цветков тыквы и легкость
опыления женского цветка мужским существенно снижают трудоем-
кость этой операции. На опыление 1 га затрачивается 2—2,5 ч, или один
человек может опылить за утро тыкву, выращиваемую на 1 га. Опы-
ление тыквы на одной и той же площади надо проводить в начале
цветения ежедневно в течение 8—12 дней. Каждый цветок следует опы-
лять не более двух раз, дальнейшее дополнительное опыление эффекта
не дает.
При дополнительном опылении тыквы в 7 ч утра завязывается
50% плодов, а в 10 ч — только 7%, при опылении вечером завязыва-
ния не отмечено.
224
Опыление может быть полным и неполным. При неполном опыле-
нии много завязей (до 43%) опадает. При полном опылении тыквы
опыленные завязи тоже опадают, но не более 20%.
ПЕРИОД ВЕГЕТАЦИИ
Культурные тыквы — однолетние растения, а ряд диких видов
(С. foetidissima, С. palmata, С. digitata) — многолетние, последние об-
разуют корнеплодные утолщения, за счет которых сохраняют питатель-
ные вещества для роста почек в следующем году. Такие утолщения,
разрастаясь из года в год, достигают иногда 20 кг.
Наиболее скороспелым культурным видом тыквы является С. реро,
особенно ее кустовые разновидности — кабачки и патиссоны, самый
позднеспелый — С. moschata.
Сортовые различия по продолжительности периода вегетации у
тыкв составляют 80—130 дней. Продолжительность отдельных фаз раз-
вития тыквы варьирует сильнее, чем у других тыквенных культур. По
О. А. Зауралову [23], различия в длине периода посев — всходы у сор-
тов незначительные; разница в продолжительности периода всходы —
бутонизация составляет 18—24 дня; по продолжительности периода бу-
тонизация— раскрытие женских цветков определились видовые разли-
чия: у крупноплодного вида эта фаза оказалась самой короткой
(22 дня), у мускатной — самой длинной (72 дня), а у твердокорой она
средняя и почти не различается по сортам (46—48 дней). Более позд-
ние сорта характеризуются не только запаздыванием наступления от-
дельных фаз развития, но и замедленным поглощением питательных
веществ.
ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ И ПРИВИВКИ
Тыквы могут размножаться и вегетативно, при помощи черенков
стебля или побега с наличием узла, из которых могут образоваться
корни и побеги. Узлы часто образуют дополнительные корни у растений
тыквы, из укорененного узла можно получить самостоятельное тыквен-
ное растение.
На острове Пуэрто-Рико размножают тыкву исключительно черен-
ками— 5—7-узловыми отрезками стеблей и побегов.
Стебли и побеги тыквы, которая имеет мощную корневую систему,
легко срастаются даже с отрезками растений других видов тыквенных.
На этом основаны прививки разных видов на тыкве. Прививки предла-
гали использовать даже как метод культуры более теплолюбивых юж-
ных тыквенных растений (арбуза, дыни) на тыквенном подвое в более
северных районах и как метод повышения урожайности огурцов.
Прививки на тыкве позволили выяснить некоторые вопросы физио-
логии питания, относительного родства видов, облегчения скрещивае-
мости, повышения скороспелости и улучшения качества продукции в
селекционной работе.
Черенки тыквы успешно прорастают и укореняются при высокой
влажности почвы и воздуха. Черенки, имеющие 5—7 узлов, зарывают
в землю вместе с листьями до верхнего узла. Такие растения плодоно-
сят раньше, чем выросшие из семян.
Обычно прививки делают на тыкве в семядольном состоянии кли-
ном в расщеп между семядолями, затем обвязывают место прививки
мочалой, помещают в полиэтиленовый мешочек или накрывают стака-
ном, чтобы создать влажную среду. Развивающийся подвой прищипы-
225
вают, оставляя часть листьев, так как полная обрезка листьев ведет к
его гибели.
На тыкве чаще всего прививают дыню как более пластичную
культуру или огурец. Лучшие виды тыкв для подвоя — крупноплодная
и мускатная. На крупноплодной тыкве дыня и огурец давали урожаи
в 4—5 раз более высокие, чем на своих корнях; кроме того, у них улуч-
шался вкус, ускорялось развитие (до 18 дней), увеличивались листо-
вая поверхность (в 2 раза и более), а также процент сухого вещества.
Это объясняется тем, что корневая система тыквы более устойчива к
неблагоприятным условиям произрастания, например к пониженной
температуре, и лучше снабжает привой питательными веществами.
АНАТОМИЧЕСКОЕ СТРОЕНИЕ ОРГАНОВ
Стебель. Гигантские плоды и огромная листовая масса длинно-
плетистых тыкв обеспечиваются влагой и питательными веществами
благодаря крупнопучковому строению стебля. Он состоит из 10 бикол-
латеральных сосудистых пучков, расположенных в два круга, и мно-
жества крупных сосудов ксилемы, видимых даже простым глазом. Хо-
рошо выражен центральный цилиндр с большим количеством мягких
тканей и хорошо развитой воздухоносной полостью. У кустовых без-
усиковых форм тыкв в отличие от длинноплетистых сосудистые пучки
располагаются в один круг.
В сосудистых пучках стебля преобладает флоэма (рис. 26), что
связано, очевидно, с весьма интенсивным поступлением пластических
веществ в корни. Над сосудистыми пучками находятся участки меха-
нических перициклических волокон.
Плодоножки, черешки листьев, усики. Имеют сходное строение со
стеблем, изменяется только количество проводящих пучков. Обильно
снабжены сосудистыми пучками части цветка и плода: в плодоножке
их больше, чем в стебле. По анатомическим данным, усики тыквы
имеют листовую природу, а их разветвления на концах соответствуют
жилкам листа. В узлах стебля все пучки являются следами листьев.
Между листьями и боковыми побегами того же узла имеется корре-
26. Соотношение *-»»-<» " <2’
в стебле тыквы-
лятивная зависимость,
повреждение листа вле-
чет за собой остановку
роста и отсыхание соот-
ветствующего побега [52].
Виды тыквы разли-
чаются строением плодо-
ножек. У твердокорого и
мускатного видов плодо-
ножки узкие, сильно де-
ревянистые, с резко вы-
раженной пятигран-
ностью, у крупноплодно-
го вида они округлые в
поперечном сечении, ча-
сто сильно утолщенные
с продольными трещина-
ми и наплывами из проб-
ково-желирующего веще-
ства.
Сосудистые пучки
плодоножки у твердоко-
226
Рис. 27. Расположение периферийных сосудов в коре
плода тыквы.
а — тангентальный разрез; б —схема: 1—сосудистый
пучок; 2 — полосы рисунка плода; 3 — впадины ребри-
стости.
рого вида имеют более развитую ксилему, а у крупноплодного — более
развитую флоэму. Ксилему первого вида усиливает ребристость его
плодоножек, а развитая флоэма второго связана, очевидно, с интенсив-
ным поступлением пробково-желирующих веществ (рис. 27).
Плод. Плод тыквы покрыт одноклеточным слоем эпидермиса с па-
лисадными клетками длиной 23 мкм и шириной 7 мкм и слоем (3 мкм)
кутикулы. На эпидермисе имеются устьица и эпидермальные выступы —
волоски и бородавки. Под эпидермисом расположены 1—2 слоя клеток
колленхимы, под которой находится зона обычно пигментированных
мелких (20 мкм в диаметре) клеток, окрашивающих кору плода липо-
ксантином и каротином. Эта зона состоит из нескольких слоев клеток
общей шириной 250—300 мкм. У форм с деревянистой корой под суб-
эпидермисом располагается непрерывный слой (панцирь) склеренхим-
ным клеток общей толщиной до 3 мм. Ниже субэпидермиса находится
бесцветная коровая паренхима (собственно мякоть плода — мезокар-
пий), насыщенная хромопластами. Желтый цвет мякоти обусловлен
присутствием каротина. По исследованиям Б. Т. Матиенко, хромопласты
концентрируют крахмальные зерна, которые располагаются возле и
внутри них. В мякоти крахмал концентрируется, по нашим исследова-
ниям, главным образом вдоль проводящих пучков. В центре плода
клетки увеличиваются, разветвления пучков уменьшаются.
Мякоть плода ограни-
чивается к центру эндокар-
пием—пленкой из одноряд-
ного прозрачного слоя кле-
ток (напоминающей пленку
между чешуями луковицы
репчатого лука), отделяю-
щейся вместе с семенами.
Ниже мякоти идут плацен-
ты (завернутые края пло-
долистиков) с семенами.
У твердокорого и му-
скатного видов 10 сосуди-
стых пучков плодоножки
(5 внутренних и 5 перифе-
Рис. 28. Сегментированное™ (1) и острая ребри-
стость (2), обусловленные пучками периферийных
сосудов.
227
рийных), входя в плод, образуют 10 продольных выпуклостей (ребри-
стостей) основания плода (рис. 28). В этих выпуклостях пучки уже
начинают разветвляться, хотя главная группа сосудов каждого пучка
идет прямо, огибая плод и создавая периферическую меридиональную
фигуру из 10 пучков. В середине коровой мякоти имеется вторая та-
кая же меридиональная фигура из сосудистых пучков, а по центру
плода идут 3—5 крупных пучков (в зависимости от числа плацент).
Все эти пучки, не теряя своей самостоятельности и не уменьшаясь в
диаметре, разветвляются на мелкие отдельные сосудики, идущие в
разных направлениях по плоду. Все они, начиная от плодоножки, окан-
чиваются у начала следов цветоложа. Эта система имеет тесную связь
с рисунком коры, сегментированностью и ребристостью плодов [76].
Впадины, разделяющие плод на сегменты и ребра плодов, располага-
ются над сосудистыми пучками, полосы рисунка коры также ориенти-
рованы в зависимости от них.
Данные системы являются продолжением сосудистых систем орга-
нов цветка. Связь их с органами цветка выражается в том, что пери-
ферическая система сосудов ранее обслуживала околоцветник (5 ле-
пестков и 5 чашелистиков), после опадания околоцветника она сохра-
нилась и развивалась в завязи и плоде. Центральная система ранее за-
канчивалась в лопастях рыльца, так как каждая лопасть соответствует
семенной камере. С ростом плода сосуды всех систем разветвляются в
соответствующих слоях мякоти плода.
Семена имеют сложную оболочку, семядоли и зародыш. Оболочка
состоит из 5 тканей: полисадного эпидермиса, гиподермы, слоя камени-
стых клеток, слоя гибрацитных трахеид, паренхимы с проводящим
пучком.
Палисадный эпидермис у вида С. реро исчезает при развитии и
высыхании, поэтому поверхность семени начинается с одревесневших
тканей. У С. maxima и С. moschata он состоит из узких длинных кле-
ток, напоминающих иногда волоски. Гиподерма образована несколь-
кими слоями слабо утолщенных, одревесневших, округлых, плотно со-
единенных клеток, пронизанных многочисленными порами. Каменистый
слой представлен крупными, расположенными в один ряд клетками с
сильно утолщенными одревесневшими оболочками. Гидрацитные тра-
хеиды — это слой крупных округлых тонкостенных клеток со значи-
тельными межклетниками. Последний слой — паренхимные клетки —
небольшой; в нем проходят сосуды, питающие зародыш.
Полезные свойства сложного устройства кожуры семян тыквы рас-
шифрованы В. Н. Сойфером [71, 72]; 1) наличие воздушных пустот
в оболочке — приспособление к теплоизоляции белков семени, так как
тыквенные — растения жарких стран; 2) слои пористых клеток смы-
каются в микропиле, где начинается проводящий пучок и палисадный
эпидермис обводняется почти мгновенно; 3) особенно показательна
роль гидрацитных трахеид — водозапасающей ткани семени; сосудистый
пучок, соседствующий с гидрацитными трахеидами и микропиле, быстро
доставляет воду к зародышу. Поэтому для набухания семян не тре-
буется большого количества воды (50% массы семян).
Процесс нормального прорастания семян тыквы обеспечен высо-
ким энергетическим запасом в виде высококалорийных жиров семядо-
лей и легкими путями проникновения кислорода к зародышу благодаря
высокой аэрации тканей семени.
228
БИОХИМИЧЕСКИЙ СОСТАВ ПЛОДОВ И СЕМЯН ТЫКВЫ
По использованию в плодах тыквы различают кору, мякоть, пла-
центы и семена. Соотношение этих частей зависит от сорта и условий
выращивания.
Мякоть плодов включает 70—94% воды и 6—30% сухого веще-
ства, содержащего по разным источникам (% на сырое вещество):
1,5—15 сахаров, 4—23 клетчатки и гемицеллюлоз, до 24 крахмала,
0,3—1,4 пектинов, 1—3 азотистых веществ, 0,5—0,7 сырого жира,
0,1 кислот, 0,4—1,4 золы, 25—40 мг на 100 г аскорбиновой кислоты,
2—28 мг на 100 г каротина.
Образцы тыквы очень изменчивы по содержанию сахара, крахмала
и витаминов. По В. В. Некрасову и Ю. Г. Скрипникову [51], сахара
составляют 70—85% растворимых веществ сока плода и 55% сухого
вещества мякоти.
Наиболее сахаристые сорта встречаются у вида крупноплодного
(С. maxima). В составе сахаров обычно присутствуют все 3 распро-
страненные формы: глюкоза, фруктоза и сахароза. Меньше всего в зре-
лых плодах содержится глюкозы (0,3%), больше — фруктозы (0,5—
4,2%) и наибольшее количество —сахарозы (до 10%). Последняя на-
капливается позднее других сахаров или при хранении; особенно много
ее в плодах лежких сортов. Крахмалом богаче также поздние сорта
в основном южного происхождения.
Зола тыквы богата калием и фосфором [29]: КгО — 54,4%; Р2О5—
14,6; СО2— 14,1; SO3 —4,0; СаО-3,2; Na2O —2,6; MgO —2,5; Fe2O3 —
1,1; А12О3—1,1; SiO2 (нерастворимый)—1,0; SiO2 (растворимый) — 0,6;
Cl —0,4.
Из витаминов в плодах тыквы наибольшее значение имеет каро-
тин. В широко распространенных сортах его содержится 0—5 мг на
100 г, у сорта Golden Delicious и желтого крукнека—11,61 мг, у сор-
тов Испанская—13,7, Витаминная —в среднем 16, в отдельных слу-
чаях до 38 мг. Наиболее богаты каротином плоды тыквы мускатного
вида (до 26,2 мг) и наименее — твердокорого вида (до 4,1 мг); плоды
крупноплодного вида содержат до 19,2 мг на 100 г вещества.
Накопление каротина связано с продолжительностью жизни плода.
Так, высокое содержание его наблюдается у поздних сортов и в пер-
вых завязавшихся плодах. Вторые по возрасту плоды содержат в 2—
3 раза меньше каротина, чем первые, а третьи — меньше, чем вторые.
Вначале каротин концентрируется в листьях (8 мг на 100 г) и
цветках: в лепестках мужских цветков —5,4 мг, женских цветков — 8,3,
в тычинках — 0,8, в рыльцах — 3,4 мг, а затем и в мякоти плодов. По
мере старения в листьях уменьшается содержание каротина.
Место накопления каротина в плодах — область семян. Играя ак-
тивную роль в оплодотворении и процессах развития зародышей, каро-
тин важен и для созревания семян. Так, по нашим данным, мускатная
тыква ВИР (к-354) содержала каротина в сырой массе: в плацентах —
21,8 мг, в мякоти —8,0, в коре —4,2 мг.
В период роста плодов накопление каротина незначительное; рез-
кое повышение его наблюдается в период созревания, что соответствует
появлению желтой окраски мякоти, с которой каротин находится в пря-
мой зависимости. Содержание каротина в плодах увеличивается в про-
цессе хранения до 70 дней, а затем оно начинает уменьшаться.
По выходу каротина с 1 га тыква занимает первое место среди
других растений — источников каротина (табл. 4).
Каротин в растениях присутствует совместно с другими кароти-
ноидами, нарастание их идет одновременно, причем 60—70% их также
229
Таблица 4. Урожайность и выход каротина у различных культур
Культура Орган растения Каротин, мг/100 г Урожайность, т/га Выход каротина, кг/га
Красная морковь Корнеплод 7 15 1,1
Томаты Плод 1,6 55 0,9
Рябина » 13 10 1,3
Лекарственная кален- Цветок 53 0,75 0,4
дула
Тыква (каротинные Плод 18 30 5,4
сорта)
биологически активны, как и каротин. Соотношение каротина и каро-
тиноидов у сортов весьма разнородно, что видно из данных М. И. Рож-
кова и Н. Е. Смирнова (табл. 5).
Таблица 5. Содержание каротиноидов в плодах различных сортов тыквы,
мг на 100 г
Сорт Каротин Другие каротиноиды Соотношение каротина и каротиноидов
Витаминная 12,12 0,30 40: 1
Голосеменная 4,07 2,20 2:1
Мозолеевская 4,06 0,54 4:1
Белая медовая 2,16 2,64 0,8: 1
Волжская серая 1,23 0,15 8: 1
Стофунтовая 0,59 0,32 2:1
Кроме каротиноидов, в 100 г мякоти плодов тыквы имеется (в мг):
2,5—5 витамина С (в кабачках их 10 мг), 0,02 витамина Bi (аневрина,
у кабачков 0,08 мг), 0,08 витамина Вг (рибофлавина, у патиссонов до
0,37 мг), 0,6 витамина РР (никотиновой кислоты), 0,5—0,9 тиамина.
Содержание витамина С при варке уменьшается на 25—50%. Для
лучшего сохранения витаминов плоды следует хранить при температуре
1°С и относительной влажности воздуха 70%.
В плодах тыквы содержатся также ферменты, несущие специфиче-
ские функции. Среди них есть, в частности, те, которые переводят бе-
лок в растворимый пептон, что особенно важно для людей с больными
почками. Количество каталазы в плодах почти не изменяется с их воз-
растом, зато содержание сахарозы сильно уменьшается по мере созре-
вания плода.
Содержание химических веществ меняется в плодах тыквы в про-
цессе роста и развития и зависит от условий среды. По мере созрева-
ния плодов процент сухого вещества в них падает, а процент экстрак-
тивных веществ увеличивается [51]. Между ними существует положи-
тельная корреляция (0,80—0,96).
Количество кислот до 30—40-дневного возраста плодов быстро уве-
личивается, а затем до 90-дневного возраста быстро уменьшается, и
после этого снижение идет замедленно. Преобладают среди кислот ли-
монная и яблочная, которые составляют 50% общего их количества.
Влияние внешних условий также изменяет биохимический состав
растения и, в частности, плодов. Так, проведенные В. Ф. Беликом и др.
[4] опыты показали, что полив снижает содержание сахаров и сухого
вещества у зрелых плодов вида С. реро примерно на 1%, а у видов
С. maxima и С. moschata увеличивает эти показатели также на 1 %.
Ю. Г. Скрипниковым установлена корреляция между местоположе-
нием плода на главной плети и его качеством: плоды, расположенные
ближе к корневой шейке, а также находящиеся слишком далеко от нее,
230
отличаются пониженным качеством (снижение в клеточном соке су-
хого вещества, крахмала, сахара). На плетях же I порядка качество
плодов, расположенных близко к стеблю, выше. Существует корреляция
между массой плода и его качеством (слишком крупные и мелкие
плоды в пределах, характерных для сорта, отличаются низким содер-
жанием сухих экстрактивных веществ и крахмала), а также между
качеством плода и количеством семян (чем больше семян в плоде в
пределах, характерных для сорта, тем ниже качество плода).
Ю. Г. Скрипниковым разработаны простые и быстрые методы опре-
деления мясистости тыкв и содержания крахмала в плодах [70]. Мя-
систость показывает степень выполненности плода.
Семена тыквы — источник масла (50%), хотя они содержат боль-
шое количество белков (30%), а также смолы, глюкозиды, витамины,
золу. Средняя урожайность с 1 га семян 2—4 ц. У крупноплодного
вида в одном плоде содержится 100—240 г, а у твердокорого — 27—
125 г.
Содержание жира в ядре семян у сортов тыкв составляет 44—48%.
Голосеменные сорта, естественно, дают более высокий процент выхода
масла из целого семени (на 10%). Семена дикого вида С. digitata со-
держат только 20% жира и 20% белка, семена С. foetidissima соответ-
ственно 30 и 33%.
Гибридные семена тыквы содержат более высокий процент белко-
вого азота и жира, чем сортовые. Белок у них главным образом водно-
и солерастворимый (70%), а жир с большой удельной относительной
массой и с преобладанием насыщенных кислот.
Из белков семян преобладает глобулин (кукурбитин), в который
входят аминокислоты: аргинин—14,5%, гистидин — 3,3, лизин — 2,2, ти-
розин— 2,8, пролин — 2,9, триптофан — 3,8, аспарагиновая кислота —
4,3, глутаминовая кислота—13,3, азот в разных формах—18%.
Следует отметить содержание в масле большого количества ви-
тамина Е (154 мг на 100 г сырого вещества).
Масло семян тыквы невысыхающее, при 15° С плотность 0,92—
0,93, число омыления 186—195, йодное число 113—130, кислотное число
небольшое (в среднем 0,62), поэтому оно не уступает лучшим столовым
маслам.
Избыток осадков снижает масличность семян. Минеральные удоб-
рения способствуют увеличению процента сухого вещества в семенах,
улучшают качество семян и масла. По химическому составу качество
семян лучше из нижнего, чем из верхнего (цветочного), конца плода.
Масло из семян тыквы обладает антиокислительными свойствами.
Поэтому добавление его к промышленным препаратам каротина спо-
собствует их стабилизации.
ГЕНЕТИКА ТЫКВЫ
ХРОМОСОМЫ
Тыквы имеют 40—48 хромосом в диплоидном наборе. Различий по
числу хромосом у видов тыкв не установлено, приводимые данные про-
тиворечивы. Так, 3. А. Кожухов [28] нашел у форм С. реро 40, 42, 44
и 46 хромосом в диплоидном наборе, а у большинства иностранных
исследователей для всех видов тыквы указано только по 40 хромосом.
Ф. Вейлинг [201] считает тыкву вторичным полиплоидом от основ-
ного генома, имеющего 10 хромосом.
3. А. Кожуховым [28] были отмечены видовые различия в форме
хромосом: у мелкоплодных декоративных тыкв хромосомы были мельче,
231
у С. maxima — длинные и дугообразные, у С. moschata — короткие и
толстые. Длина хромосом С. ficifolia колебалась от 2 до 12 мкм. У меж-
видовых гибридов преобладали различные хромосомные отклонения от
родительских форм.
Характерным признаком хромосом тыкв, кроме большого их числа,
является малая величина по сравнению с другими тыквенными куль-
турами, имеющими меньшее число хромосом в клетках. Более крупные,
чем у арбуза, дыни, огурца размеры плодов, листьев, плетей у куль-
турных видов тыквы коррелируют с увеличенным количеством хромо-
сом и с меньшими их размерами. Наибольший размер хромосом и наи-
меньшее их число у огурца С. sativus. Зародышевые мешки у огурца
также крупнее, чем у тыквы, в 1,5 раза. Однако из более мелких за-
родышевых мешков культурные виды тыквы формируют более крупные
семена и плоды в соответствии с общим габитусом растения.
ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ ПРИЗНАКОВ
Монофакториальное наследование признаков не исключает связи
их друг с другом в так называемых группах сцепления. Е. Ляна [154]
установил следующие группы взаимных связей для С. maxima.
I. Признаки, связанные с материнским
растением
1. Размер и масса семян
2. Масса зародыша
3. Размер чашелистиков
4. Длина плетей
5. Темп роста
6. Число мужских цветков
7. Среднее количество дней до цвете-
ния
8. Урожай с одного растения
III. Признаки, связанные с размером
семян родителей
1. Рост плетей в ранний период роста
2. Число мужских цветков
3. Число дней до раскрытия женских
цветков
4. Масса отдельных плодов
5. Средний урожай с одного растения
II. Признаки, связанные с отцовским
растением
1. Среднее количество дней до раскры-
тия мужских цветков
2. Число женских цветков на одно
растение
3. Число завязей
4. Число плодов на одно растение
5. Масса одного плода на растение
IV. Признаки, положительно связанные
с размером семян материнского ра-
стения
1. Размер семян
2. Рост плети
3. Число мужских и женских цветков
4. Число мужских цветков
5. Число плодов на одно растение
6. Урожай с одного растения
Размер и форма плода в первую очередь интересовали ге-
нетиков. Так, по данным Е. Синнот [188], дисковидная форма плода
патиссонов доминирует над шаровидной, а в F2 наблюдается расщеп-
ление в отношениях, близких к монофакториальному, 3:1.
По данным С. Лотси [157], у С. реро длинноплодность доминирует
над шароплодностью или сплюснуто-шаровидной формой плода, но не
над тарельчатой. Скрещивание кабачка с патиссоном дает по форме
плода промежуточное наследование. По С. Лотси [157] и И. Гребен-
щикову [133], в скрещивании крупноплодной тыквы С. реро или ка-
бачка с дикой мелкоплодной var. texana (длиной 8 см) доминируют
признаки дикого родителя, в F2 происходит расщепление по промежу-
точному наследованию с отклонением формы плодов в сторону var.
texana.
Е. Синнот [192] изучал ориентацию митотических осей в связи
с направлением наиболее энергичного роста органов и установил, что
вытянутость плодов связана в преобладающем числе случаев с парал-
лельным расположением митотических осей делящихся клеток, то есть
с ярко выраженной полярностью их цитоплазмы. Он же предложил
удобную форму графического изображения роста плодов в длину и ра-
диально.
232
У С. maxima наследование формы плода идет несколько иначе. По
данным С. Лотси [157], сплюснутоплодные формы доминируют над
вытянутыми, поэтому последних очень мало у этого вида.
Бородавчатость поверхности плодов С. реро доминирует в Fi
и расщепляется в F2 в отношениях 64 : 24, 62 : 20, 97 : 6, 69 : 6, 81 : 7.
Видимо, здесь наблюдаются 2 генетических типа бородавчатости, один
из которых монофакториален, другой определяется двумя генами.
Чалмовость плодов поЕ. Синнот в четырех случаях доми-
нировала в F] и F2 (80:8, 29:11, 71:21, 70:3), а в трех случаях не
доминировала над гладкой поверхностью (27:24, 27:26, 34:55). Ве-
роятно, здесь играет роль степень выраженности признака чалмовости
у родителей.
Крупноплодность, поТ. Вайтекеру [203], является рецессив-
ным признаком: при скрещивании мелкоплодной С. реро var. ovifera
с культурными сортами var. реро отношение крупных плодов к мелким
в F2 было 30:71. При скрещивании var. реро с патиссонами масса
плода в Fi наследуется промежуточно, а в F2 смещается в сторону
мелкоплодного родителя. У кабачков, наоборот, И. Гребенщиков [137]
установил доминантность большой массы. На гибридном материале он
вычислил коэффициент корреляции внешних условий с продуктивностью
(г = 71%), со средней массой плода (г = 70,8%), с числом зрелых пло-
дов (г =79,1 %). Он также установил, что первый плод у патиссонов
более изменчив; обычно он мельче последующих.
По данным Е. Синнот [190], размер плода С. реро определяется
действием нескольких генов.
Наследование окраски плода изучали многие исследова-
тели. Е. Синнот и Г. Дюрем [188] в пределах вида С. реро установили^
что из трех основных окрасок плодов тыквы белая доминантна к зе-
леной и желтой, а зеленая — к желтой. Каждая из них монофакто-
риальна. Таким же образом наследуется и фон рисунка плода: белый
фон доминирует над остальными.
Скрещивание двух культурных форм С. реро: зеленоплодной и жел-
топлодной показало доминирование зеленой окраски над желтой, и их
отношение в F2 в начальный период роста составило 3:1, а в период
полной зрелости, когда полностью проявились гетерозиготные возмож-
ности гибридов,— 1:3 [184]. Ученый считает, что признаки надо изу-
чать в динамике. Ряд других исследователей впоследствии называли
такое наследование переменой доминирования.
Позднее О. Шифрис [185] установил, что окраска плодов у видов
тыквы изменяется и в процессе роста в более широких пределах и ее
наследование следует рассматривать в четырех фазах: 1) завязь в мо-
мент цветения; 2) плод достаточно крупного размера; 3) плод предельно
крупного размера с созревшими семенами; 4) полное созревание плода.
Наследуемые окраски О. Шифрис разбивает на 10 групп: 1) белая;
2) зеленая; 3) серая; 4) желтая (палево-кремовая, палево-желтая);
5) оранжевая; 6) двуцветная (полосы на фоне или резко окрашенные
концы плодов); 7) зеленая, переходящая в оранжевую после созрева-
ния или при хранении; 8) зеленые полосы при созревании, переходящие
в желтые или оранжевые; 9) незаметные нитчатые зеленые полосы, вы-
цветающие в белые или светло-зеленые или переходящие в кремовые,
желтые и оранжевые; 10) широкие зеленые полосы, переходящие в бе-
лые, кремовые или желтые.
Опыты О. Шифриса по скрещиванию оранжевоплодного сорта
С. реро Коннектикутская полевая (YYWW) с белоплодной var. ovifera
(yyww) в Fi дали сначала зеленые плоды, а при созревании белые.
Расщепление в F2 в недозрелом состоянии—127 белых, 37 зеленых,
233
а в зрелом из 127 белых стало 81 кремовый, 27 белых, 19 желтых,
а из 37 зеленых — 21 кремовый, 9 желтых и 7 оранжевых. В резуль-
тате потомств с кремовыми плодами было 102, с белыми—27, с жел-
тыми— 28, с оранжевыми — 7, т. е. в соотношении 9:3:3: 1, что ука-
зывает на дигибридный характер наследования данного признака.
По данным Д. Глоберсона [130], зеленый цвет плодов С. реро
контролируется двумя генами: С и R, белый — ссгг, полосатый — ccR,.
Темно-зеленый цвет стебля коррелирует с зеленым плодом, он доми-
нантен к светло-зеленой окраске, которая генетически ближе к белой.
Окраска плодов, по данным Т. Вайтекера [213], у С. maxima на-
следуется так, что зеленая доминирует над оранжевой. Этот вывод сде-
лан без учета фазы наблюдений, культурности и древности формы. Оди-
наковые признаки у диких форм могут быть доминантными, а у куль-
турных — рецессивными. В его же работе [203] по скрещиванию С. реро
var. ovifera (некультурная, белой окраски) с var. реро (культурная,
зеленой окраски) наследование окраски плода было полностью связано
с формой и размером плода родителей, т. е. 71:30 (см. ранее). Это,
очевидно, было вызвано влиянием дикой формы, у которой признаки
были сцеплены.
Рисунок плода, в частности полосатость, доминирует над без-
рисунчатостью. Ряд авторов считает, что полосатость определяется са-
мостоятельным геном, причем признак полосатости у них оказался мо-
нофакториально доминантным так же, как к сплошной зеленой и жел-
той окраске плода.
Белая пятнистость листьев зависит от группового скопления белых
пластид и наследуется по материнской линии. Угловая белая пятни-
стость листа С. moschata, по Скерику, доминирует над отсутствием пя-
тен: в F2 расщепление идет как 3:1.
Белая окраска мякоти тыквы, по Е. Синнот и Г. Дюрем [188], до-
минантна по отношению к кремовой: расщепление в F2 в большинстве
случаев дает отношения, близкие к 3 : 1, но иногда и к 1 : 1.
Устойчивость тыквы к мучнистой росе (Erysipne cicho-
racearum), по данным А. Родеса [173], может быть передана от дико-
растущего вида С. lundelliana к С. moschata; она контролируется
только одним геном. В подобных скрещиваниях в F2 наблюдалось рас-
щепление 3: 1, а беккроссы Fi с С. lundelliana показали отношение 1:1.
Устойчивость к вирусной мозаике листьев С. maxima
изучали Д. Скотт и М. Райнер [181]. Она оказалась рецессивным при-
знаком: в F2 выщеплялась в отношениях 78 : 29 и 77:23. Скрещивание
Fi с устойчивым родителем дало потомство в отношении 1:1.
Горечь плодов наблюдается у дикорастущих мелкоплодных
тыкв. Иногда она проявляется и у культурных сортов. По И. Гребен-
щикову [132], горечь доминирует в Fi и расщепляется в F2 в отноше-
нии 3:1. Этот признак монофакториален.
Голосемянность С. реро является рецессивным признаком, а дере-
вянистая оболочка семени обусловлена двумя генами. Вейлингу уда-
лось скрестить С. maxima, обладающей деревянистой оболочкой семени,
с голосеменной С. реро. В Fj и F2 семена гибридов имели деревянистую
оболочку, в F3 половина семян — толстую деревянистую оболочку, по-
ловина— деревянистую утонченную; в F4 из 600 растений обнаружено
4 голосеменных растения. В седьмом — девятом поколении получено
много голосеменных растений.
Голосемянность у С. реро возникла путем мутации. Ряд ученых
считают ее результатом действия 2—3 генов, а И. Гребенщиков [132]
доказал ее монофакториальность.
234
Мужская стерильность у тыквы наследуется по монофак-
ториальной схеме. С расщеплением 1 :2: 1. Д. Скотт и М. Райнер [181],
а за ними большинство исследователей подтвердили однофакториаль-
ность признака мужской стерильности.
Причиной стерильности мужских цветков является дегенерация ма-
теринских пыльцевых клеток и тапетума, которые не обеспечивают в
дальнейшем мейозиса. Ядра таких клеток гипертрофируются, и цветки
дегенерируют.
Кустовость, т. е. отсутствие длинных плетей у растения, в по-
следнее время очень интересует селекционеров, так как у кустовых
растений в поле можно обрабатывать междурядья культиваторами
почти до конца выращивания. Но исходные формы, как правило, имеют
мелкие плоды. Скрещивание крупноплодных длинноплетистых тыкв с
мелкоплодными кустовыми вначале было малоэффективным. Отмечено
[184], что при таких скрещиваниях, вначале хотя и появляются корот-
коплетистые формы, но позже они превращаются в длинноплетистые.
Это явление О. Шифрис предложил назвать гетерозисом развития, счи-
тая обязательным изучать любые исследуемые признаки не в статичном
состоянии, а в развитии.
Признак плетистости Д. Сингх [187] разделил на 5 наследуемых
вариантов, которые выразил количественно в разные сроки наблю-
дения на материале 1120 гибридных растений (табл. 6).
Таблица 6. Наследование кустовости при скрещивании кустовой и плетистой форм
С. maxima, %
Дата и поколение Кустовые Слегка плетистые Полукустовые Близкие к плетистым Плетистые
8/VII Ft 100
f2 22 48 19 4 7
24/VII Ft 100 —
f2 8 26 45 14 7
Т. Вайтекер и Г. Девис [212] считают, что в Fi кустовость насле-
дуется промежуточно и определяется двумя парами генов. И. Гребен-
щиков [132, 134] отнес промежуточные формы к кустовым, считая кус-
товость доминантным признаком, а изменение ее в течение сезона сме-
ной доминирования. В опытах В. А. Лудилова [33] растения Fi С. ma-
xima развивались 40 дней как кустовые, а потом как плетистые.
У Д. Денна и X. Мунгер [120] гибридные растения С. реро развива-
лись как кустовые до 16-го листового узла, а С. maxima — до 6-го узла.
Затем С. реро развивается как промежуточное растение, а С. ma-
xima — как плетистое. Такое развитие, по их мнению, — результат изме-
нения двух генов в онтогенезе. Причем 86,5% различий в длине междо-
узлий образуется за счет увеличения числа клеток, а 13,5%—за счет
их растягивания.
Более ясно плетистые и промежуточные кустовые формы класси-
фицированы Н. С. Шашенком [93, 94] на 4 типа: кустовые, коротко-
плетистые, позднеплетистые (смена доминирования) и плетистые. Он
установил, что родительские формы неодинаково передают потомству
признак кустовости. Например, существуют 2 типа кабачков: с очень
короткими зачаточными боковыми побегами и совершенно неветвя-
щиеся. Последние гораздо лучше передают признак кустовости, чем ко-
роткоплетистые кабачки.
235
Как видно из таблицы 7, по признаку кустовости материнская на-
следственность сильнее отцовской. У вида С. реро нет смены домини-
рования, а признак кустовости передается сильнее, чем у С. maxima;
длинноплетистые формы в F! у него не выщепляются совсем.
Таблица 7. Наследование кустовости у гибридов тыквы в F,
по материнской и отцовской линии, %
Число комбина- ций Родительская форма Кустовые Коротко- плетистые Поздне- плетистые Плетистые
14 ? С. maxima Чино 2243 18 82
То же — 1 99 —
8 ? Быковская 151 7 18 70 5
сГ То же — 6 88 6
18 ? С. реро Неветвящиеся кабачки 83 17
18 с? (к-2577, 2940) То же 76 24 —
? Слабоветвящиеся кабачки 21 79 — —
сГ То же 4 96 — —
6 ? Кабачки Грибовские 37 — 100 — —
сГ и патиссоны Белые 13 То же 100 —
6 ? Днепропетровская кустовая 1 99 — —
сГ То же — 100 — —
Результаты анализа F2 у вида С. maxima показали, что здесь вы-
щепляются преимущественно позднеплетистые и плетистые формы с пре-
обладанием первых по типу моногибридного расщепления. Кустовые
растения выщепились только в двух комбинациях из позднеплетистого
Fi, 2 комбинации не дали расщепления, оставаясь позднеплетистыми.
Выщепнвшиеся в Fi кустовые формы расщеплялись в F2 на кустовые
и позднеплетистые по типу простого моногибридного с преобладанием
кустовых.
Во втором гибридном поколении вида С. реро тип растения Fi
не оказывает существенного влияния на характер расщепления, но ко-
личество кустовых растений зависит от природы кустовых родительских
форм (табл. 8).
Таблица 8. Количество кустовых растений в потомстве F2 родительских форм
С. реро, %
Кустовые родительские формы От кустовой материнской От кустовой отцовской Среднее
Кабачки
Неветвящиеся 26,3 25,7 26,0
Слабоветвящиеся 21,1 23,7 22,4
Грибовские 37 Тыква 0,6 3,3 1,9
Днепропетровская кустовая Патиссоны 0,8 11,0 5,9
Белые 13 0,0 0,0 0,0
Неветвящиеся кабачки в F2 уже не имеют такого преимущества, как
в Fi, но оно опять сказывается в F3, когда выщепляется больше кусто-
236
вых растений, а иногда и 100% их. В некоторых случаях растения
в F3 были гомозиготны по кустовости и дальше не расщеплялись.
У гибридов вида С. maxima от кустовых растений инцухтированного
второго поколения в F3 часто выщеплялось 96—98% кустовых потом-
ков, а от неинцухтированных — только 42—48%. Большого различия
между передачей кустовости отцовским и материнским растением, как
в Fi, в F2 не наблюдалось. В результате изучения F3 ученый делает
вывод о том, что признак кустовости обусловливается более чем одной
парой генов. Выделенные в F2 кустовые растения генетически неодно-
родны и в F3 в большинстве случаев дают расщепления, при этом кус-
товая форма ведет себя как доминирующий признак.
Успешная работа Н. С. Шашенка объясняется использованием
большого числа родителей в скрещиваниях. Ряд исследователей шли
по другому пути. Так, А. Т. Галка [11] проводил углубленные работы
по получению кустовых растений с использованием прививок. Кабачки
он опылял пыльцой тыквы (Мозолеевская), Fi — пыльцой кабачков, а
F2 прививал на кабачки. Отбором в потомстве привитых гибридов был
выведен кустовой сорт крупноплодной тыквы Днепропетровская кусто-
вая. Продолжая опыты, он скрещивал привой с кустовой формой. Это
облегчало скрещивание, давая 20% кустовых гибридов уже в Fi, тогда
как от такого скрещивания без прививок в Fi кустовых растений не
было. Еще успешнее был опыт скрещивания обоих родителей, привитых
на кустовом подвое. В этом случае процент кустовых растений в Fj
повышался до 28%. Выход кустовых гибридов повышался в том слу-
чае, если материнское растение было старым, а отцовское — молодым
(позже высеяно). При этом если опылялись увядающие цветки плети-
стого растения, то процент кустовых гибридов был еще выше. Так,
опыление цветка в день его раскрытия давало 9% кустовых растений,
а на второй — 16,4, на третий — 20,2%.
По данным В. А. Лудилова [36], кустовые растения выщепляются
и при скрещивании длинноплетистых родительских форм. Наследова-
ние периода вегетации у тыквы изучал Д. Сингх [187]. Он нашел, что
период от всходов до открытия как мужских, так и женских цветков
определяется сложением действия трех пар генов, различных для муж-
ских и для женских цветков, а период от раскрытия мужских до рас-
крытия женских —двумя парами генов, дополняющих друг друга. Ука-
занные признаки наследуются независимо, поэтому имеется возмож-
ность выводить ультраскороспелые сорта. Однако гибриды наследуют
вегетационный период промежуточно.
По данным Н. С. Шашенка, наблюдалось наследование и по ран-
неспелой родительской форме, когда они не сильно различались по
этому признаку и когда в качестве материнской формы брали более
скороспелый сорт. При скрещивании сортов с различной продолжитель-
ностью периода вегетации сдвиг в сторону раннеспелости у гибридов
различался труднее, наследование было только промежуточным. Од-
нако, когда в качестве материнского выступал раннеспелый кустовой
сорт, вегетационный период Fi явно сдвигался в сторону раннеспелого
родителя. По отдельным комбинациям в F2 и F3 также наблюдалось
уклонение в сторону материнской скороспелой формы. Видимо, и в
наследовании скороспелость еще тесно связана с признаком кустовости
скороспелого кустового родителя.
Продуктивность гибридов изменялась в зависимости от про-
дуктивности родителей. Так, у гибридов Fi С. maxima она была выше,
если материнская форма мелкоплодная. У С. реро этого не наблюта-
лось. В F2 продуктивность у обоих видов наследовалась промежуточно,
но у кустовых гибридов С. maxima она уклонялась в сторону материн-
237
ской кустовой формы, однако превышая ее на 15—20%; средняя масса
плодов у таких комбинаций была почти в 2 раза больше, чем у ма-
теринской формы. Отцовская кустовая мелкоплодная форма снижала
продуктивность гибридов. Многоплодность родительских форм не пере-
давалась по наследству.
В F3 кустовые гибриды превышали по продуктивности кустовых
родителей и приближались к более продуктивным плетистым. У боль-
шинства комбинаций между кустовыми, позднеплетистыми и коротко-
плетистыми гибридами не обнаружено существенной разницы по уро-
жайности и по средней массе плодов. Однако более урожайные линии,
выявленные в F2, позволяют получать более продуктивное потомство
в последующих поколениях независимо от формы растения: кустовой,
позднеплетистой или короткоплетистой.
Сухое вещество, крахмал и каротин во всех комбина-
циях опытов Н. С. Шашенка наследуется в Fi промежуточно, но с боль-
шой амплитудой колебания по содержанию сухого вещества и с укло-
нением его в сторону низкокачественного родителя. Наследование про-
межуточных результатов наблюдалось также в F2 и в F3, но в отличие
от F2 С. maxima плоды гибридов С. реро содержали гораздо меньше
сухого вещества и каротина и совсем не содержали крахмала. По дан-
ным Д. Сингх [187], наследование количества сухого вещества обуслов-
лено двумя парами генов. Оно у тыкв тесно коррелирует с плотностью
мякоти.
Твердокорость плодов у С. реро, по данным всех авторов,
монофакториальна и наследуется в F2 в отношении 3 твердокорых к
1 мягкокорой.
МЕЖВИДОВЫЕ СКРЕЩИВАНИЯ
Затрудненность межвидовых скрещиваний состоит в том, что пыль-
цевые трубки одного вида плохо растут на рыльцах другого. Так, трубки
С. maxima растут медленнее на рыльцах С. реро, чем на рыльцах цвет-
ков своего вида. В последнем случае пыльцевые трубки достигают се-
мяпочки после опыления: у С. реро — за 3 ч, у С. moschata — за 4 ч,
у С, maxima — за 5 ч. Однако рост пыльцевых трубок еще не обеспе-
чивает развития зародыша после оплодотворения, так как уже фор-
мирующиеся зародыши прекращают развитие на разных стадиях из-за
плохого функционирования эндосперма.
История скрещивания видов тыквы
В естественных условиях возделывания виды этой культуры не
скрещиваются между собой. Искусственное скрещивание их долго не
удавалось или давало нежизнеспособное потомство.
Первое поколение от межвидовых скрещиваний удалось впервые
получить В. Хальстеду [142], Л. Бейли [101], Козловой, но публика-
ций о дальнейшей работе с межвидовыми гибридами тыквы у них не
было.
А. Эрвин и Е. Хабер [125] первые описывают второе и третье по-
коления межвидовых гибридов тыквы (табл. 9).
И. Ф. Лященко [38] удалось скрестить С. maxima и С. moschata
и получить жизнеспособные семена Fb К. И. Пангало и М. К. Гольд-
гаузен [61] также получили потомство гибридов (табл. 10). Семена в
Fo во всех случаях были всхожими, а в F] невсхожими оказались только
семена последней комбинации. Признаки вида С. moschata доминируют
238
над признаками других видов. На основании своих скрещиваний уче-
ные сделали вывод, что к виду С. moschata ближе стоит вид С. реро,
дальше — С. maxima.
Таблица 9. Результаты межвидовой гибридизации тыкв
Комбинация скрещивания Проведено опылений Получено плодов Получено всхожих семян
С. рероХС. maxima Fo 775 78 12
С. maximaXC. реро Fo 1217 100 22
То же Fi 32 6 —
С. moschataXC. реро Fo 417 39 —
С. рероХС. moschata Fo 187 15 Много
То же F, 19 12 Очень много
» » f2 62 19 V и
» » F3 Много Много
С. maximaXC. moschata Fo 72 31 -
с. moschataXC. maxima Fo 249 69
После работы К. И. Пангало межвидовые скрещивания тыквы
стали проводить уже в селекционных целях многие советские селек-
ционеры.
Таблица 10. Межвидовые скрещивания тыквы
Вариант скрещивания Число скрещиваний Получено плодов Получено семян
С. maximaXC. moschata 25 3 137
» ХС. mixta 6 1 246
С. рероХС. moschata 45 4 8
» ХС. mixta 5 2 207
С. moschataXC. реро 4 1 4
» ХС. maxima 30 1 35
Н. А. Хохлачева [88] скрещивала С. maxima X С. moschata, у гиб-
ридов преобладали признаки материнской формы; особенно сильное
расщепление наблюдалось в F3, где было обнаружено 70% ценных
форм от всего поколения гибридов. Выявилось много новых признаков.
Ею выведен сорт Межвидовая 381.
А. Ф. Макаровский и М. И. Подмогаева (39] получили всхожие
семена гибридов от скрещивания культурных видов* благодаря особой
методике скрещивания. В. Ф. Белик и М. И. Подмогаева [5] также
изучали различные способы межвидовой гибридизации и циклических
скрещиваний, в результате которых они вывели ныне районированный
сорт Мичуринец.
С. И. Кобытев и Е. И. Лавринова [26] от 20 скрещиваний С. mo-
schata и С. maxima получили 19 плодов. В Fi они наблюдали формы
с большим количеством сухого вещества (17%), высокосахаристые, ско-
роспелые, урожайные. О. В. Юриной [97] выведен сорт Грибовская
зимняя от скрещивания С. maximaXC. moschata с повторными насы-
щающими скрещиваниями гибридов с С. maxima.
В. А. Лудилов [35], а позднее Д. И. Соколов [73] получили серию
межвидовых гибридов тыквы с С. moschata, у которых в Fi домини-
ровали рисунок плода и белая пятнистость листьев мускатного вида,
а форма плода крупноплодного. Большинство гибридов обладало низ-
кой всхожестью семян. Фертильность пыльцы находилась в пределах
25%, инцухт семян не давал.
Большую работу по межвидовой гибридизации тыквы провел
ф. Вейлинг [197, 198, 199]. Кроме 3 культурных видов, он изучал скрещи-
239
вания и с видом С. ficifolia. Комбинация С. maximaXC. ficifolia дала
22,1% удачных гибридов, обратная — 2,2%, С. ficifoliaXC. реро соот-
ветственно 2,1 и 0%, но все растения F2 оказались стерильными. Скре-
щивая дикий вид С. foetidissima со всеми культурными, Вейлинг уста-
новил, что по конъюгации хромосом в мейозе материнских клеток
пыльцы этот вид близок к трем культурным видам, исключая С. fici-
folia.
X. Кутлер и Т. Вайтекер [117] от скрещивания С. moschata и
С. mixta получили гибриды со щуплыми семенами. Фертильность
пыльцы Fi составляла 31%. Нами эти формы скрещивались свободно.
С. mixta включает пестрые формы, некоторые из них, возможно, плохо
скрещиваются с С. moschata.
Т. Вайтекер [211] установил скрещиваемость дикого вида С. lun-
delliana, устойчивого к мучнистой росе, со всеми культурными видами.
Гибриды с С. moschata в Fi имели 42% нормальной пыльцы, с С. ma-
xima— 17, с С. ficifolia—15%; все семена их имели нормальные за-
родыши. Скрещивание С. lundelliana с С. реро давало недоразвитые
зародыши. Исследователь отмечает, что и культурные виды тоже труд-
нее скрещиваются с С. реро. Признаки С. lundelliana доминантны.
И. Гребенщиков [134] скрещивал культурные виды тыквы с диким
С. foetidissima. С использованием С. moschata было получено неболь-
шое количество семян, но F] имели мужскую стерильность. Опыление
женских цветков Fj пыльцой родительских сортов удавалось, но полу-
ченные плоды имели недоразвитые и в большинстве невсхожие семена.
С другими видами также не было получено плодовитых растений.
Скрещивания [137] дикой формы С. maxima var. andreana с куль-
турными видами показали, что как материнское растение оно не дает
межвидовых гибридов, а как отцовское дает фертильные гибриды с
С. mixta. Скрещивание же С. mixta с С. реро var. sororia и С. maxima
дает фертильное Fb а иногда и F2. Скрещивание его с высококультур-
ными формами С. moschata дает вполне фертильное потомство во всех
поколениях с поглощением признаков серебросемянной тыквы более
культурными формами С. moschata. Скрещивание С. реро var. sororia
с С. moschata не дало плодовитых семян.
Большую работу по межвидовой гибридизации с дикорастущими
тыквами сделал и В. Бимайс [105, 106]. Он скрещивал С. moschata с
тремя ксерофильными дикими видами С. digitata, С. palmata и С. for-
tidissima (табл. И).
Таблица 11. Скрещиваемость ксерофильных диких тыкв с мускатной
Материнская форма С. moschata С. digitata С. palmata С. foetidis- sima
опыле- но завя- залось опыле- но завя- залось опыле- но завя- залось опыле- но завя- залось
С. moschata 4 4 12 6 8 3 9 2
С. digidata 15 0 17 10 16 11 31 1
С. palmata 17 4 13 10 10 6 6 4
С. foetidissima 15 1 7 4 4 4 4 3
Все полученные плоды из этих скрещиваний имели достаточное ко-
личество всхожих семян.
В последние годы В. И. Лудилову [35] удалось скрестить все куль-
турные виды тыкв между собой без особых методов подготовки скре-
щиваний.
240
Гибриды Fj в комбинации С. maximaXC. moschata в опытах
В. А. Лудилова дают высокую мужскую стерильность, в комбинации
С. maxima X С. реро Fi они не образуют пыльцы совсем, a Fj С. mo-
schata X С. реро дают фертильные растения.
В результате проведенных исследований по межвидовой гибриди-
зации выяснилось следующее. Получение межвидовых гибридов воз-
можно, хотя и с малым процентом удачного результата, но последний
может быть повышен, а гибриды могут быть доведены до следующих
поколений путем применения особых мер преодоления нескрещиваемо-
сти и путем искусственной культуры недоразвитых зародышей. Кроме
того, у гибридов от скрещивания культурных видов плодовитость чаще
всего исчезает у Fi из-за мужской стерильности, но она может быть
преодолена. По анализам Т. Вайтекера виды С. реро и С. maxima
легче скрещиваются с С. moschata, чем друг с другом. С. moschata за-
нимает центральное положение в скрещивании между однолетними куль-
турными видами.
Преимущество при скрещивании с другими видами имеют более
гетерозиготные сорта, особенно по признаку мужской стерильности.
Многолетний культурный вид С. ficifolia хуже скрещивается с одно-
летними культурными видами, чем последние друг с другом. Он,
по заключению Т. Вайтекера, генетически ближе к С. maxima и С. реро,
чем к С. moschata. По нашим наблюдениям, эта близость носит не фи-
логенетический, а экологический характер, как у видов, адаптирован-
ных к более прохладным условиям. Этим же можно объяснить и луч-
шую скрещиваемость С. mixta с видом С. реро как более холодостой-
ким (горным), а не с формами С. moschata.
Вид С. moschata легче других культурных видов скрещивается с
дикими видами, особенно из группы мезофитов, а мезофильный вид
С. lundelliana довольно легко скрещивается со всеми культурными. Ди-
кие ксерофильные виды не скрещиваются с мезофильными, но каждый
из них беспрепятственно скрещивается внутри своей группы, исключая
вид С. foetidissima.
Более отзывчивы на межвидовые скрещивания мезофильные тепло-
любивые виды, нежели ксерофиты. Близость видов по экологическим
признакам, включая многолетний образ жизни, обеспечивает лучшую
скрещиваемость их между собой, чем близость по другим показателям
(например, окультуренность, близость географических ареалов и др.).
В связи с большим влиянием экологической принадлежности видов
преждевременно говорить об их родственных взаимоотношениях и про-
исхождении только на основе анализа склонности видов к большей или
меньшей скрещиваемости.
Методы преодоления нескрещиваемости
и искусственная культура зародышей
Трудности скрещивания видов тыквы, естественно, вызывали по-
иски способов преодоления нескрещиваемости их и повышения плодо-
витости гибридов. В работе по межвидовой гибридизации К. И. Пангало
и М. К. Гольдгаузен [61] предварительно удаляли лишние (не участ-
вующие в межвидовой гибридизации) женские цветки скрещиваемых
растений.
Л. Е. Кревченко вызывал плодовитость у стерильного F] от меж-
видового скрещивания С. maxima\C. moschata опылением его видом
С. реро. Впоследствии В. А. Лудилов [35] и другие нашли, что воз-
вратные скрещивания восстанавливают фертильность стерильных гиб-
ридов Fi. Так, в его опытах гибриды С. maxima с С. moschata показы-
241
вали фертильность Fi до 45%, если они были опылены отцовским рас-
тением. О. В. Юриной [97] был выведен сорт Грибовская зимняя пу-
тем возвратного скрещивания Fj: (С. maximaXC. moschata) X. С. ma-
xima.
Американские ученые искали методы преодоления нескрещиваемо-
стн в сфере генетических опытов. Т. Вайтекер и Г. Бон [207] исполь-
зовали для межвидовых скрещиваний амфидиплоидных родителей, по-
лученных путем обработки раствором колхицина. Предлагается также
метод гаметического разнообразия, состоящий из использования
высокогетерозиготного скрещиваемого материала при первоначальных
межвидовых комбинациях. Гетерозиготные сорта скрещиваются с дру-
гими видами успешнее гомозиготных или инцухт-линий. На этом же
принципе А. Родес [172] путем интербридинга вывел смешанную
популяцию между культурными видами для «мостиковых» скрещиваний
и успешно использовал ее для передачи признака кустовости С. mo-
schata от С. реро и признака устойчивости к мучнистой росе от С. 1ип-
delliana гибридам, полученных в скрещивании с другими видами.
Широко использовался способ межвидового скрещивания растений
тыквы в привитом состоянии. Таким путем Н. А. Хохлачева [88], про-
водившая за 3 года трехкратную прививку вида С. moschata и ее по-
колений, выращенных на подвое С. maxima, получила от скрещивания
этих видов гибрид, слабофертильный в Fi и F2. В F3 наблюдалось
сильное расщепление с образованием новых форм, в том числе соче-
тающих признаки обоих родителей. Было обнаружено 70% ценных
форм, из которых был отобран новый сорт, превосходящий родитель-
ские формы по урожайности в 2—3 раза, по каротину на 50%.
О. В. Юрина [96] получила перспективный гибрид при скрещи-
вании С. реро с С. maxima в привитом друг на друга состоянии. На
основании большого количества опытов В. А. Лудилов [35] делает
вывод, что вегетативное сближение (прививка) улучшает скрещивание
видов как в год прививки, так и при использовании Fj. Предпочтитель-
нее 2—3-годичное вегетативное сближение.
А. Ф. Макаровский и М. И. Подмогаева [39] получили гибриды
между тремя основными культурными видами тыквы путем опыления
растений одного вида пыльцой двух других видов. В. Ф. Бе-
лик и М. И. Подмогаева [5] широко развернули исследования по меж-
видовой гибридизации путем использования смеси пыльцы. Они уста-
новили, что чем больше отцовских видов участвует в оплодотворении,
тем оно успешнее. При этом было замечено, что вид С. реро слабее
других передает свои признаки межвидовому гибриду, но если этот
вид опылять гибридными формами других видов, признаки С. реро пе-
редаются почти полностью. Опылением С. maxima смесью пыльцы
С. реро и С. moschata ими был выведен сорт Мичуринец, превосходя-
щий родительские сорта по урожаю.
В дальнейшем В. Ф. Белик и М. И. Подмогаева усовершенствовали
метод использования смеси пыльцы в межвидовых скрещиваниях путем
добавления в эту смесь немного пыльцы материнского растения или
вытяжек из пыльцы, что давало 63% завязывания плодов, и много-
кратным опылением (в 6, 8 и 10 ч утра), приносившим 20% удачных
результатов. Своя пыльца создает благоприятную среду на рыльце,
благодаря чему лучше прорастает и чужая. Такое же действие оказы-
вает и пыльца гибрида при межвидовом опылении.
X. Хаяши [144, 145] установил, что при влажности воздуха 60—
80% ускоряется открывание пыльников, а при 50%-ной влажности оно
задерживается. Исследователь считает, что для каждой комбинации
242
скрещивания должны быть установлены свои сроки с наилучшей актив-
ностью пыльцы и рыльцев.
Ряд иностранных авторов пытались активизировать прорастание
пыльцы при межвидовой гибридизации воздействием различ-
ных веществ. Так, гиббереллин помогает скрещиванию отдельных
форм, он в 3—8 раз увеличивает длину пыльцевых трубок.
Без особых приемов преодоления нескрещиваемости, но на в ы с о-
ком агрофоне при опылении большим количеством пыльцы и об-
рыванием завязей С. И. Кобытев и Е. И. Лавринова [26] получили
более 50% завязывания плодов от скрещивания С. moschata\C. ma-
xima с хорошими семенами в Fj.
Получение невсхожих, с недоразвитыми зародышами семян при
межвидовой гибридизации тыквы побудило некоторых ученых изучать
методику их проращивания. Д. Волл [196] вырезал зародыши из семян
в стерильных условиях и быстро погружал их в раствор гипохлорита
кальция (10 г Са (СЮ)2 в 140 мл водопроводной воды) на 3—8 ч.
Затем, не промывая, он переносил их в стерильную культуру на агар
в темный термостат на 48 ч при 20° С, после чего проращивал на свету
5—10 дней при температуре 17° С, где они давали ростки. Затем за-
родыши промывали и высаживали в стерильные горшки для последую-
щей высадки в поле или закрытый грунт.
В. А. Лудилов [35] проращивал недоразвитые зародыши гибридов
на питательной смеси Уатта. К ней добавляли 20 мл дрожжевого экс-
тракта (0,5 г сухих дрожжей, разведенных в 100 мл дистиллированной
воды, прокипяченных и профильтрованных). Эту смесь выливали на
освобожденные от кожуры и стерилизованные в сулеме (1 : 1000) заро-
дыши, разложенные на прокаленном песке в чашках Петри. Чашки
ставили в термостат при 20—25° С, где семена успешно прорастали.
Когда корешки достигали 1,5—2 см, проростки высаживали.
Поведение межвидовых гибридов
Наиболее подробно и ранее других исследователей поведение меж-
видовых гибридов было описано К. И. Пангало и М. К. Гольдгаузен
[61]. Ими замечено, что замена хотя бы одного из родительских сор-
тов другим уже изменяет наследование признаков F], а доминирование
некоторых из них (например, деревянистость коры плодов, грубость
пластинки и опушение) осуществляется независимо от того, какому
виду родителей они принадлежат.
Это обстоятельство и лучшая скрещиваемость более мезофитных
родительских видов (например, С. moschata\C. реро), по нашему
мнению, указывают на преобладающую роль в скрещиваниях эколо-
гической принадлежности видов.
У некоторых гибридов выщеплялись особи с фасциированными
цветками, без околоцветника, с полунижней завязью (как у тюрбан-
ных видов тыквы), с гермафродитными цветками, ультраскороспелые
(плоды созревают раньше родительских форм на 1—2 месяца) и ульт-
рапоздние, многоплодные (до 40 плодов на 1 растение), останавливаю-
щие свой рост на ранней фазе вегетации, желтопятнистые, карликовые,
безлистные, кустовые растения с большим количеством коротких пле-
тей I, II и III порядка (наподобие щетки), медленно растущие рас-
тения, за сезон давшие только несколько листьев, растения с ранним
появлением цветков, с мелкими и более крупными, чем у родителей,
цветками и т. д. В F2 наблюдались случаи отсутствия расщепления.
Н. А. Хохлачевой [87] на Краснодарской овощной опытной стан-
ции был получен межвидовой гибрид С. maxima (сорт Столовая зим-
243
няя А-5) с С. moschata (сорт Перехватка 69). У этого гибрида наблю-
далось много новых признаков: крупные желтые семена с широким ви-
тым ободком, своеобразные формы плодоножек и листьев. Максималь-
ное расщепление гибрида наступило лишь в F3. В этом же поколении
выявились и наиболее ценные формы с хорошей лежкостью, вкусовыми
качествами, высокой сахаристостью и урожайностью. По урожайности
некоторые семьи гибрида в 2—3 раза превосходили родительские формы.
Содержание каротина и витамина С у гибридов тоже было значительно
выше, чем у родителей. Созревание плодов было средним. Преобладаю-
щие в наследовании признаки были отнесены к виду С. maxima, а не
к С. moschata, как у предыдущих ученых.
Межвидовые гибриды культурных тыкв, по данным В. Ф. Белика
[3], часто несут признаки двух или трех видов. Иногда растения Fi в
процессе роста меняют сходство.
ГЕТЕРОЗИС
Изучение гетерозиса тыквы началось в США. Л. Куртис [116]
в пределах вида С. реро путем скрещивания сортов Кентукская 10 с
Ранним плодовитым получил в первом поколении потомство, которое
превосходило обоих родителей по скороспелости на 10 дней, а по уро-
жаю в 3—4 раза благодаря большему числу и более крупным плодам.
Ученым была предложена методика получения гибридных семян путем
высева скрещиваемых сортов через ряд и удаления на материнском
сорте мужских цветков. Затраты на получение гибридных семян тыквы
оказались незначительными, а производство гибридных семян — рента-
бельным.
Л. Куртис для уменьшения стоимости гибридных семян предложил
использовать второе поколение гибридов, которое дает урожай только
на 6% ниже Fp При этом скороспелость снижается по сравнению с
гибридами первого поколения.
Почти одновременно с ним А. Хетчинс и Ф. Кростон [149] про-
вели ряд скрещиваний вида С. maxima. Из комбинаций Fi 7 оказалось
продуктивнее каждого из родителей за счет большей величины плодов.
Между прямыми и обратными скрещиваниями разницы не наблюда-
лось. Скрещивания некоторых сортов с селекционным № 31 дали пре-
вышение урожая по сравнению с более урожайным родителем: Ари-
кара Х№ 31—на 88%, Гоббард X № 31—на 123%, Деликатес X
Х№ 31 — на 100%.
А. Бухингер [ПО] считает, что за материнскую форму при полу-
чении гибридных семян следует брать ту, которая образует наибольшее
количество семян в плоде, а отцовских растений можно иметь лишь
10% от материнской формы при условии, что у последней обрывают
все мужские цветки до распускания.
Из работы А. Мудра [162] следует, что первое поколение от скре-
щивания самоопыленных линий тыквы не дает повышения урожайно-
сти, но проявляет гетерозис по скороспелости.
X. Хаяши [144] рекомендует разрезать пыльники в длину накануне
раскрытия цветка тыквы для более полного опыления материнского
сорта с целью получения гетерозисных семян.
После этих работ и в результате широкой популярности гибридной
кукурузы в СССР стали заниматься поиском гетерозисных комбинаций
у большинства сельскохозяйственных культур, в том числе и у тыквы.
Быковская опытная станция бахчеводства получила прибавку урожая
от гибридных семян Fi тыквы, равную 17—32%, Краснодарская овощ-
ная станция—11—85%, Бирючекутская овощная станция — 21—39%.
244
Наши работы с В. П. Тощевым имели целью выяснить влияние на
гетерозис сортов тыкв, различающихся между собой по морфологиче-
ским признакам и географическому происхождению.
Исследования показали, что при межсортовых скрещиваниях гео-
графически отдаленных форм тыквы гибриды F! превышают по уро-
жайности наиболее урожайный сорт на 3,0—178,9% (в среднем на
94,1%).
Степень гетерозиса гибридов по урожайности зависит от экологии
родительских сортов. Наилучшие результаты отмечены при участии в
скрещиваниях местных исходных форм с инорайонными сортами (осо-
бенно если последние взяты в качестве материнских форм). В наших
опытах гибриды от скрещивания отечественных сортов Днепропетров-
ская кустовая (УССР), Голосемянная (Поволжье), Безенчукская белая
(Поволжье), а также Букетная (Канада), Мамонтовы рога (США)
показали наиболее высокий гетерозис по урожайности и другим при-
знакам. Гибриды от сортов, происходящих из Малой Азии и Китая,
имели самый низкий процент повышения урожайности плодов.
Нами установлено, что продолжительность фазы всходы—цветение,
а также всего периода вегетации у гибридов Fi почти всегда на 1—И
дней короче, чем у исходных форм.
В отличие от родительских сортов цветение женских цветков у всех
гибридов начиналось раньше мужских на 1—7 дней.
Некоторые гибриды имеют форму и качество плода кабачков. Их
испытывали на урожайность и вкусовые качества в виде 7-дневных
завязей. Наряду с остальными хозяйственно-ценными качествами гиб-
риды имели высокие вкусовые достоинства, не уступая в этом лучшему
из родительских сортов, и проявляли устойчивость к мучнистой росе.
В процессе работы было установлено влияние величины плодов
родительских форм на степень гетерозиса по урожайности. Гибриды
были распределены по этому признаку на 4 группы: 1) мелкоплодные
сорта X крупноплодные; 2) мелкоплодные X мелкоплодные; 3) крупно-
плодные X мелкоплодные; 4) крупноплодные X крупноплодные. Наивыс-
шую урожайность по отношению к наиболее урожайному родитель-
скому сорту показала 1-я группа гибридов—176,5%, низшую — 3-я
группа (115,8%); 2-я и 4-я группы гибридов заняли промежуточное по-
ложение: урожай этих групп составил в среднем 149,5%. Средняя
масса плодов гибридов тыквы была почти вдвое выше, чем у родитель-
ских сортов. Урожайность гибридов увеличилась в основном за счет
крупности плодов.
Мелкоплодные формы тыквы при скрещивании имеют большие по-
тенциальные возможности более резко повысить урожайность, чем круп-
ноплодные. По нашим данным, гибриды 2-й комбинационной группы
оказались урожайнее (в абсолютном ее выражении), чем гибриды 4-й,
в среднем на 25%. Так, гибриды при прямых и обратных скрещиваниях
мелкоплодных сортов Чино и Букет дали в среднем 7 кг плодов на
растение, тогда как крупноплодные формы в комбинации Безенчукская
белая X Чилийская 754 — только 5,5 кг.
Кроме урожайности, у гибридов тыкв мы изучали гетерозис по
биохимическим показателям мякоти плодов.
Исследования показали, что 73 межсортовых гибрида тыкв (42%
общего количества испытываемых гибридов FJ превысили лучшие ро-
дительские формы по содержанию сухого вещества мякоти. Так, меж-
сортовые гибриды Черкесская X к-1483, Сладкий сыр X к-1236, Днепро-
петровская X Голосемянная в первом поколении содержали сухого ве-
щества в плодах в 2 раза больше, чем родительские формы.
245
Нужно отметить, что количество сухого вещества в плодах тыквы
как у родительских сортообразцов, так и у гибридов находится в пря-
мой пропорциональной зависимости от содержания крахмала.
Повышение содержания сухого вещества в мякоти плодов у гиб-
ридов первого поколения находится в зависимости от количества сухого
вещества у родительских форм. Все испытываемые гибриды по содер-
жанию сухого вещества у родительских сортов были распределены на
3 группы (табл. 12).
Таблица 12. Характеристика групп Fi по содержанию сухого вещества в плодах
по сравнению с родительскими формами
Группы гибридов по содержанию сухого вещества в плодах родительских форм Число гибридов в группе Количество Fi в группе по содержанию сухого вещества
превышающих исходные формы равных исходным формам уступающих исходным формам
абсо- лютное % абсо- лютное % абсо- лютное %
Материнские сорта, равные отцовским 18 12 67 5 27 1 6
Материнские сорта, уступающие отцовским 84 50 59 19 23 15 18
Материнские сорта, превышающие отцовские 70 11 16 51 73 8 И
По всем группам 172 73 42 75 44 24 14
Наибольшее количество гибридов (67°/о), превысивших родитель-
ские формы по содержанию сухого вещества в мякоти плодов, имела
3-я группа гибридов, родительские формы которой содержали одинако-
вое количество сухого вещества.
Нами исследовано содержание крахмала в половых элементах жен-
ских и мужских цветков тыкв. В рыльце и столбике пестика женского
цветка при окрашивании их йодом крахмал не был обнаружен. Пыль-
цевые же зерна окрашиваются йодом в цвета различной степени ин-
тенсивности. Можно предположить, что содержание крахмала в плодах
гибридов зависит от количества его в пыльце отцовских форм.
В. А. Лудиловым [35] выяснено, что проявление гетерозиса по
содержанию сухого вещества может быть эффективнее, если F\ привито
на сильнорослый подвой, каким в его опытах была тыква Крупноплод-
ная 1. Содержание каротиноидов у привитых гибридов тоже было выше,
чем у непривитых, хотя и не достигало уровня отцовского сорта Вита-
минная.
Гетерозис по скороспелости в большей мере зависит от
отцовского компонента, чем от материнского. Так, при опылении ма-
теринских сортов более раннеспелыми получены наилучшие результаты
по скороспелости Fp 78% гибридов были более раннеспелыми, чем наи-
более раннеспелый родительский сорт. При обратных скрещиваниях.
1. е. когда материнские сорта были более скороспелыми, получено наи-
меньшее количество гибридов, проявивших гетерозис по скороспелости;
от скрещивания сортов с одинаковой скороспелостью Fi имеет проме-
жуточное положение по скороспелости.
Нами изучался гетерозис и во втором поколении гибридов. Ре-
зультаты показали, что от скрещивания географически и экологически
отдаленных сортов гетерозис по урожайности в значительной мере со-
храняется и в F2 (табл. 13).
246
Согласно нашим данным, по скороспелости, содержанию сухого ве-
щества и вкусовым качествам F2 не уступало Fb
Таблица 13. Урожайность F, и F2 поколений гибридов по сравнению
с родительскими сортами
Гибрид Поко- ление Плодов с I растения, кг Плодов с 1 растения гибрида, %
гиб- рид материн- ский сорт отцовский сорт к мате- ринскому сорту 1 к отцов- скому сор- ту
Безенчукская белая XЖелтая исполинская Ft 8,2 2,3 4,2 356 195
То же F3 6,7 2,3 4,2 291 159
ЧеркесскаяХк-1483 Ft 7,9 3,5 5,6 221 141
То же F, 7,1 3,5 5.6 202 126
Днепропетровская X к-1526 Ft' 9,3 4,0 6,1 232 152
То же F- 8,4 4,0 6,1 210 137
Причины, вызывающие гетерозис, достаточно полно
вскрыты в работе Д. Д. Брежнева [7]. И. Гребенщиков [136] показал,
чем меньшее число генов определяют признак, тем выше его выражен-
ность у гибрида тыквы. Если одну родительскую форму скрестить с не-
сколькими и учесть суммарную среднюю массу плодов гибридов, то
отношение этой суммы к сумме средней массы плодов обоих родите-
лей, разделенной на 2, составит коэффициент силы наследственной пе-
редачи признака урожая. Наследование признаков урожая неравно-
ценно: самую высокую степень силы наследственной передачи обнару-
жил признак «масса плодов на 1 растение», слабее этот коэффициент
выражен у признаков «число плодов» и «длина плода».
Различная быстрота роста пыльцы на рыльцах материнских форм
тыквы связана с разной кислотностью этих поверхностей [80]. Обна-
ружена явная связь гетерозисности с pH: чем больше различаются по
общей кислотности отцовские и материнские половые элементы, тем
выше гетерозис.
МУТАЦИИ И ПОЛИПЛОИДЫ
Происхождение многих мелкоплодных декоративных тыкв амери-
канские исследователи объясняют результатом мутационного процесса.
В дикорастущем виде их не обнаружено.
Широко известные в ГДР, ФРГ и Австрии голосеменные тыквы,
по мнению ученых, также являются мутацией вида С. реро. В литера-
туре имеются указания и на другие мутации у тыквы.
Полиплоиды (тетраплоиды) у тыквы С. реро впервые были
получены искусственно Е. Синнотт и А. Блексли [191] воздействием
колхицина на молодое растение. В одной из следующих работ Е. Син-
нотт дает уже усовершенствованную методику получения тетраплопдов
путем погружения наклюнувшихся семян в 0,1%-ный раствор колхи-
цина на 1—2 суток при температуре 25° С и хорошей вентиляции, что
очень важно. У 33% обработанных растений пыльцевые зерна были
крупнее обычных в 2 раза, листья шире, стебли, черешки листьев и
пластинки толще, цветки крупнее, клетки крупнее, однако зрелые
плоды по размеру почти не различались: становятся несколько короче и
шире. После же цветения диплоидные растения растут в 2 раза быстрее
и догоняют в росте тетраплоиды, но зрелые, внешне мало отличаю-
щиеся плоды последних имеют клетки более крупные (примерно а
*2 раза, чем у диплоидов).
247
Н. М. Чекалин получил тетраплоидное растение у вида С. maxima
он наносил на точку роста проростков 1%-ный раствор колхицина в
течение 3—4 дней (3 раза в день). Тетраплоидное растение задержива-
лось в росте. Семена его были короче и шире обычных, с шероховатой
поверхностью; семядоли отличались темно-зеленой окраской, округлой
формой, более сильным опушением; гипокотиль был в 2 раза длиннее;
мужские цветки были крупнее, женские — мельче; пыльца имела в диа-
метре 160 мкм против 120 у диплоидов. Семян в плоде оказалось зна-
чительно меньше, мякоть была в 2 раза толще, плоды более сплюсну-
тые. Данный полиплоид с диплоидами не скрещивался (давал пустые
семена).
Практическая ценность указанных тетраплоидов не была изучена;
полезные формы выявлялись главным образом среди амфидиплоидов,
получаемых в связи с проблемой межвидовых скрещиваний тыквы. Гиб-
ридный способ получения полиплоидов разрабатывался Т. Вайтекером,
им же совместно с Г. Девисом [212] дана сводка по этой проблеме.
Амфидиплоидные линии составляют потенциально новый вид, так
как они самофертильны и перекрестностерильны с обоими родителями.
Те линии, у которых одним из родителей был сорт Буттернут (С. mo-
schata), по качеству не уступают наилучшим товарным сортам столо-
вой тыквы. Другие линии имеют положительные признаки: силу куста,
более крупные и тяжелые плоды по сравнению с плодами товарных
сортов.
ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ
СОРТОВЫЕ ПРИЗНАКИ ТЫКВЫ
Характер плетей: длинные (длина главной плети 3 м), средние
(1,5—3 м), короткие (1,5 м), кустовые (стебли короче 1 м, боковые
побеги короче 50 см или отсутствуют).
Форма листьев: округлая, почковидная (ширина значительно боль-
ше длины), сердцевидная (с вытянутым верхним концом), треугольная,
пятиугольная.
Размер листьев: крупные (длина листовой пластинки свыше 20 см),
средние (длина листовой пластинки 12—20 см), мелкие (длина листо-
вой пластинки короче 12 см).
Изрезанность листовой пластинки: глубокая (больше половины
радиуса пластинки), средняя (до 1/2 радиуса), мелкая (короче поло-
вины радиуса или отсутствует).
Ширина лопастей листа: узкие, средние, широкие.
Интенсивность белой пятнистости листовой пластинки: сильная,
средняя, слабая, следы или отсутствует.
Характер белой пятнистости: пятна мелкие, крупные, растянутые
по всей длине главных жилок листа.
Черешки листа: длинные (более 24 см), средние (18—25 см), ко-
роткие (меньше 18 см).
Форма плода: сплюснутая (сильно-, средне-, слабо-), шаровидная,
овальная (удлиненно-укороченно-овальная), обратнояйцевидная, яйце-
видная, волчковидиая, чалмовидная, клиновидная, перцевидная, груше-
видная (удлиненно-грушевидная, укороченно-грушевидная с расширен-
ным основанием), обратногрушевидная, цилиндрическая (укороченно-
и удлиненно-цилиндрическая), перехватка удлиненная (если изогнута,
отметить особо), перехватка укороченная.
Размеры плодов (по наибольшему диаметру) по округлой форме:
крупные (больше 35 см), средние (25—35 см), мелкие (меньше 25 см);
248
при удлиненной форме: крупные (больше 50 см), средние (40—50 см),
мелкие (меньше 40 см).
Гофрированность основания (начало ребристости): глубокая, сред-
няя, слабая, отсутствует.
Поверхность плода: гладкая, неровная, сегментированная (глубо-
ко-, средне-, мелко-, сегментов 10—15), с двойной сегментацией (сег-
ментов 16—20), ребристая (ребра выпуклые): сильно-, средне-, слабо-
морщинистая (сильно-, средне-, слабо-) чешуйчатая, бородавчатая, буг-
ристая (отдельные крупные выпуклости), сетка трещин: крупноячеи-
стая, мелкоячеистая.
Рисунки коры плода: полосы (широкие — шире полос фона, сред-
ние— равны ширине полос, узкие — уже полос фона), зеленая сетка,
пятна: крупные одиночные, густые вместо полос, мелкие по всему плоду;
белесые ленточки (во впадинах между сегментами).
Окраска фона рисунка: незрелого плода — светло-зеленая, зеленая,
темно-зеленая; зрелого плода — лимонная, апельсиновая, оранжевая,
коричневая, белая, зеленая, розовая, красная.
Характер коры: мягкая кожистая (легко режется ножом без потре-
скивания), упругая (режется с потрескиванием), деревянистая (ре-
жется с большим трудом).
Размер семенно полости: очень большая (более 3/< диаметра),
большая (занимает '/г—3А диаметра плода), средняя (2Д—*/г диаметра),
малая (меньше 2А диаметра плода, плаценты, как правило, полностью
его заполняют), семенная полость только в одном конце (у длинно-
плодных форм), отсутствует (тыква без семян или небольшие щели
в мякоти).
Толщина мякоти: сплошная (без полости в плоде), толстая
(*/2 диаметра плода и толще), средняя (от ‘/г до ’А диаметра), тонкая
(’А диаметра и тоньше).
Окраска мякоти: кремовая, лимонная, апельсиновая, грязно-оран-
жевая, красная.
Плотность мякоти: плотная, средней плотности, рыхлая.
Консистенция мякоти: рыхлая картофелистая, средней плотности,
хрустящая, очень плотная (как айва).
Сахаристость мякоти: сладкая (в свежем виде), малосладкая,
несладкая.
Типы плацент: плотно заполняют семенную полость, рыхло запол-
няют ее, располагаются по стенкам мякоти.
Размер семян: крупные (больше 2 см в длину), средние (от 2 до
1,5 см), мелкие (короче 1,5—1 см), очень мелкие (короче 1 см).
Форма семян: широкие, средней ширины.
Кожура семян: толстая, тонкая, пленчатая.
Окраска семян: коричневая, грязновато-белая, молочная, кремовая
Характер и окраска рубчика: всегда темнее семени, рубчик не вы-
деляется, рубчик грязновато-серый.
Скороспелость (от всходов до первого сбора): скороспелый (не
больше 100 дней), среднеспелый (100—120 дней), позднеспелый (боль-
ше 120 дней).
Урожайность: высокая (больше 30 т/га), средняя (15—30 т/га),
низкая (меньше 15 т/га).
Лежкость: высокая (хранится больше 3 мес), средняя (1—3 мес),
низкая (не больше 1 мес).
Устойчивость к болезням: высокая (поражение не наблюдается),
средняя (поражение до 1%), слабая (поражение выше 1%).
249
СОРТОВЫЕ ПРИЗНАКИ КАБАЧКОВ
Тип куста: кустовые, полукустовые( когда имеются отдельные, вы-
ходящие за пределы куста плети или когда они появляются с запозда-
нием, в период массового плодоношения), плетистые (некустовые).
Толщина плодов: тонкие (индекс 4 и больше), средней толщины
(индекс 2—4), толстые (индекс 2 и меньше).
Поверхность плодов: гладкая, ребристая, бородавчатая.
Рисунок и окраска фона плодов (в фазе семенной зрелости): кре-
мовый, без рисунка; кремовый, с зелеными полосами различной ши-
рины; темно-зеленый, без рисунка.
Размер семенного гнезда (определяется в фазе технической зрело-
сти): малый (меньше ‘/г диаметра плода), средний ('/г диаметра),
большой (больше '/г диаметра).
СОРТОВЫЕ ПРИЗНАКИ ПАТИССОНОВ
Форма плодов (зрелые плоды): тарелочная, колокольчатая (бахро-
ма загнута к основанию), округло-плоская (без бахромы).
Поверхность плодов: гладкая, ребристая (слабо-, средне-), боро-
давчатая.
Окраска фона плода: молочно-белая, светло-зеленая, зеленая, яич-
но-желтая.
Рисунок плода: отсутствует, зеленые редкие полосы, зеленые ши-
рокие густые полосы.
Плотность мякоти: плотная, средней плотности.
СЕЛЕКЦИЯ ТЫКВЫ
Большинство районированных сортов в СССР относятся к виду
С. maxima. Этот вид наиболее разнообразен по качеству: к нему отно-
сятся как самые высококачественные, так и наиболее низкокачественные
сорта. Поэтому он занимает наибольший ареал в посевах тыквы.
Крупноплодные сорта вида С. реро лучше удаются в лесостепной
зоне, в южной степи плоды С. реро мельчают, а в жарких районах
(Средняя Азия) усиленная транспирация истощает растения, плоды не
накапливают достаточно запасных веществ, а листья подвергаются хло-
розу. Поэтому в северных и южных степных районах следует выращи-
вать кабачки и патиссоны — это скороспелые овощные культуры, успе-
вающие давать недозрелые плоды-зеленцы в короткий период наиболее
теплых месяцев на севере и раннюю продукцию на юге.
Вид С. moschata отличается постоянным, довольно высоким каче-
ством мякоти (хотя уступает лучшим сортам С. maxima) и наиболь-
шим количеством каротина. Теплолюбивость вида позволяет широко
возделывать его в Средней Азии, Закавказье и отчасти в Краснодарском
и Ставропольском краях.
Широкая изменчивость тыквы позволяет иметь сорта почти для
каждого климатического района СССР, исключая Крайний Север (Ар-
хангельская область, Якутская и Коми АССР и др.).
В существующем сортименте имеет место такая закономерность:
чем севернее расположен район возделывания местного сорта тыквы,
тем хуже его качество, и наоборот. Однако работы О. В. Юриной на
бывшей Грибовской овощной селекционной станции под Московой (в
настоящее время ВНИИССОК) показали, что и в более суровых кли-
матических условиях могут быть получены ценные сорта: выведенный
ею сорт Грибовская зимняя относится к категории высококачественных.
250
Сорта для различных климатических районов должны быть прежде
всего холодостойкими или жаростойкими, с учетом требований к влаге
п почвам.
СЕЛЕКЦИЯ НА ХОЗЯЙСТВЕННО-ЦЕННЫЕ ПРИЗНАКИ
Селекция на холодостойкость. Лучшим исходным материалом для
селекции на холодостойкость является вид С. maxima, из которого уже
выделена довольно холодостойкая разновидность мамонтовых тыкв.
Она обладает низкими показателями химического состава, поскольку
селектировалась (главным образом в ГДР и ФРГ) на большую вели-
чину плода в целях кормового использования.
Из других форм этого вида можно указать на сорта-космополиты,
удающиеся в разных климатических районах. Сюда относится, в част-
ности, сорт Медовая белая, которую, вероятно можно продвинуть в бо-
лее северные районы (чем степь и лесостепь) путем селекционной ра-
боты.
Вид С. реро (не считая кабачков и патиссонов) используется в Ря-
занской, Горьковской, Владимирской, Кировской, Московской областях,
Марийской АССР и Чувашской АССР.
Наиболее северные районы выращивания С. moschata — Красно-
дарский и Ставропольский края.
Сам факт долголетней культуры какого-нибудь сорта в районе с
пониженной температурой является селектирующим началом, вызывая
естественный отбор биотипов в сторону большей холодостойкости. По-
этому основным методом селекции тыквы на холодостойкость принято
считать экологическое воспитание. Таким путем созданы многие мест-
ные сорта, в том числе и районированные, после некоторой дополни-
тельной работы с ними: Алтайская для предгорий Алтая, Башкирская
для Предуралья, Мозолеевская 49 для севера средней полосы, Голо-
семянная для БССР, Мамонтовая для Приморского края, идет и в Ле-
нинградской области.
Однако, кроме экологического формообразования, селекция тыквы
на холодостойкость быстрее проводится гибридизацией с более холодо-
стойкими формами, использованием влияния холодостойких подвоев,
посевами в очень ранние сроки, проращиванием семян в холодильниках
и обработкой прорастающих семян холодом, применением закалки про-
растающих семян непрерывным светом, отбором в прохладных районах
по количеству выделяемой растениями пасоки, а также использованием
только крупных полновесных семян.
Гибридизацией южной кустовой тыквы Днепропетровская кустовая
с сортом Мозолеевская 49 О. В. Юрина [96] на бывшей Грибовской
овощной селекционной станции получила сорт Грибовская кустовая 189
для Нечерноземной зоны. Для той же зоны на этой станции ранее
был выведен сорт Грибовская голосемянная. Путем межвидового скре-
щивания сортов Волжская серая (С. maxima) и Краснодарская 69/19
(С. moschata) она получила для той же зоны сорт Грибовская зимняя.
О. В. Юрина считает наиболее эффективным применение ступенчатой
гибридизации с участием в качестве одного из родителей более холодо-
стойкой формы тыквы.
Посев тыквы в ранние сроки как метод селекции на холодостой-
кость относится к форсированному экологическому методу. О. А. Заура-
лов [23] попытался установить межсортовые различия по холодостой-
кости при помощи ранних сроков посева. Особенно показателен его
опыт с проращиванием семян в холодильниках. Наиболее холодостой-
кими оказались сорта С. реро: Алтайская 47, затем Украинская много-
251
BOM
как
нам
в этих условиях и надежнее давать урожай.
плодная. Посев в ранние сроки и закалка набухших семян холодом
почти во всех селекционных работах проводилась и О. В. Юриной. Она
считает этот прием одним из наиболее эффективных в селекции на хо-
лодостойкость.
Свойство холодостойкости обусловливается повышенным количест-
растворенных солей и гидрофильных коллоидов в соке растения.
Крупные семена как материал для селекции предпочтительнее, так
больший запас питательных средств в семядолях позволяет семе-
легче переносить неблагоприятные условия, успешнее вегетировать
Селекция на засухоустойчивость. Работами П. А. Генкеля доказа-
зано, что свойство засухоустойчивости (способность расти при недо-
статочном количестве воды) обусловливается теми же факторами по-
вышенного количества растворенных солей и коллоидов в клеточном
соке, что и при холодостойкости. Следовательно, при отборе более за-
сухоустойчивых растений могут быть использованы аналогичные методы.
Селекция на жаростойкость. Отбор на жаростойкость можно про-
изводить методом Майкова — погружением листьев в горячую воду с
различной температурой на 3 или 5 мин. Неустойчивые листья через 5
или 10 мин чернеют. Температура воды на пороге сворачивания кол-
лоидов покажет степень жаростойкости растения. По Н. А. Хлебниковой
[14], сортовые различия тыквы по жароустойчивости лежат в пределах
00 65 С, по данным Н. Г. Гордеевой [14], — в пределах 62—70° С,
что выше жаростойкости огурца (50—56° С), арбуза (58—62° С) и дыни
(58—65° С).
Селекция на устойчивость к болезням. Существует два пути отбора
устойчивых к болезням форм растений: случайное проявление устойчи-
вости при том или ином селекционном приеме и отбор по признакам,
коррелирующим с устойчивостью. Большинство селекционеров поль-
зуются первым путем, выявляя устойчивые формы на инфекционном
фоне, причем мутагенезом или гибридизацией создавая их исходное
разнообразие [99].
Второй путь труднее, но надежнее. Ему предшествует выявление
конкретной физико-химической причины устойчивости. Например, этим
путем по вздутиям на корнях на 4-й год были отобраны засухоустойчи-
вые сорта подсолнечника. В. Хедвайгер [141] установил, что наличие
цистеина способствует устойчивости тыкв к антракнозу. По данным
М Н- Родигина [66], против некоторых форм антракноза, в частности
арбузного, тыква образует ампрессории, препятствующие распростране-
нию гриба в тканях растения.
В растениях обнаружен биос — вещество, стимулирующее рост мно-
гих паразитов. В защите от них большую роль играют специфические
фитонциды летучие вещества, часто находящиеся в соках растения.
Так, по Д. Д. Вердеревскому [9], сок арбуза убивает гриб мучнистой
росы тыквы, сок дыни угнетает его, а сок тыквы безвереден для гриба.
Но сок тыквы убивает многих микробов, в том числе тех, которые вы-
зывают воспалительные процессы в организме человека. Клеточный сок
кабачков тормозил рост Botrytis cinerea.
В устойчивости растения к паразиту большую роль играет состоя-
ние растения, в том числе обилие хлорофилла в листьях и быстрота
физиологических процессов. Эти качества, а также специфические за-
щитные признаки накапливаются и в результате выращивания. Выра-
щивают сильные экземпляры на высокоплодородном фоне с параллель-
ным испытанием их в неблагоприятных условиях питания, температуры,
засухи, длины дня и т. д., а выявляют специфические признаки защиты
(образование вздутий, каллюсов, пробки, химических веществ, вита-
252
миногормонов, физиологические реакции на заражения) на специаль-
ных фонах, где используют прививки; усиливают признаки защиты пу-
тем скрещиваний. Важным фактом воспитания специфических призна-
ков устойчивости является легкое заражение, не переходящее в сплош-
ное поражение растения.
Селекция на скороспелость. Скороспелые сорта требуются не толь-
ко для зон с умеренным климатом, но и для южных районов (полу-
чение ранней продукции). В нашей стране уборка таких скороспелых
сортов на семена в зонах с умеренным климатом часто проводится в
конце сезона, перед морозами, или в связи с освобождением поля под
озимые зерновые, что может снизить скороспелость будущего потомства.
Признак скороспелости — низкое заложение женских бутонов на
растении( в пазухе 5—12-го листа). Для получения таких растений
О. В. Юрина применяла различные приемы (ранний посев, выращи-
вание рассады в прохладных условиях). Раннее развитие плода тормо-
зит рост последующих плодов и побегов, поэтому скороспелые растения
имеют меньшую вегетативную массу и требуют меньших площадей
питания.
Раннему завязыванию плодов способствует выращивание селек-
ционного материала тыкв на сокращенном световом дне (до 8 ч). При
этом пониженная (15—20° С) температура (по Юриной) более благо-
приятна для появления женских цветков, а повышенная (до 30° С) —
для образования большой массы мужских цветков в ущерб женским.
Скороспелые формы успешнее выявляются на усиленном фосфорно-ка-
лиевом фоне, а позднеспелые — на больших дозах азота.
Наиболее скороспелыми сортами тыквы являются: Алтайская 47
(вид С. реро), мелкоплодные из подвида чалмовых С. maxima (на-
пример, сорт Buttercup из канадской разновидности) и сорт Butterhut
(С. moschata) и др.
Влияние окружающей среды наиболее эффективно для селекции
на продолжительность периода вегетации. Многократно доказано, что
семенные репродукции различных экологических районов не равно-
значны как по скороспелости, так и по урожайности.
А. Е. Коварский [27] увеличил скороспелость позднеспелого сорта
Перехватка 628 (С. moschata) выращиванием его в течение двух лег
в районе Херсона. П. Л. Гончаров [13], изучая одни и те же сорта и
гибриды в разных условиях, констатировал сокращение продолжитель-
ности вегетационного периода в более северных зонах.
Селекция на урожайность. Элементы урожайности тыквы: размер
и масса плода, число плодов на растении, толщина мякоти плода, со-
держание сухого вещества в мякоти плода, в том числе наиболее по-
лезных их составных частей.
Крупные плоды неудобны для дальнейшей обработки, поэтому
предпочтительнее малая семенная полость и большее количество пло-
дов на более короткой плети, что позволяет сократить площадь пита-
ния одного растения и увеличить их число на 1 га. Ветвистость рас-
тения иногда компенсирует укороченность длины плетей, но листья не
должны закрывать друг друга.
Урожай зависит от корневой системы, поэтому некоторые селек-
ционеры начинают вести отбор растений по этому признаку как в фазу
прорастания семян, так и в период вегетации растений. В последнем
случае этот отбор можно вести по количеству выделяемой пасоки пень-
ком срезанного растения после снятия первого зрелого плода (для
сохранения семян после отбора), так как количество пасоки зависит
от степени развития корневой системы.
253
Для повышения урожайности в селекции используют гибриды и
линии с мужской стерильностью. И. Хиллером [147] предложен спо-
соб химической индукции мужской стерильности у тыкв путем двукрат-
ного опрыскивания сеянцев тыквы в фазе 1—2 и 4—5 настоящих ли-
стьев раствором а-нафталенуксусной кислоты или 2,3,5-тринадбензой-
ной кислоты. Полное подавление развития мужских цветков достига-
лось погружением молодых растений тыквы в раствор малеингидразида
или опрыскиванием их этим раствором.
Р. Робинзон и Т. Вайтекер [175] опрыскивали тыкву в фазе од-
ного листа 2-хлорэтилфосфорной кислотой, получая вначале только
женские цветки, которые до 10-го узла опадали, а следующие цвели.
Позднее появлялись и мужские цветки, но гибридные семена с жен-
ских цветков (до появления мужских) они успевали получить.
Селекция на урожайность требует хорошего агрофона для выявле-
ния потенциальных возможностей урожая.
Селекция на сахаристость. Существующая положительная корре-
ляция между сахаристостью и сухим веществом оправдывает отбор на
сахаристость и по последнему признаку.
Сухое вещество, растворенное в соке, поддается анализу рефрак-
тометром. Более точно количество его устанавливается путем простого
высушивания навесок мякоти плодов в сушильном шкафу. По нашим
определениям, сухое вещество мякоти тыквы в зависимости от сорта
составляет 5—30%.
Кроме сахара, в его состав входят крахмал и другие полисахариды.
Крахмал определяется по шкале Ю. Г. Скрипникова [70]. Он является
резервом повышения сахаристости, так как, гидролизуясь, переходит
в сахар. С. Н. Щербак [95] отмечает, что отбор тыкв по сухому веще-
ству труден из-за сильного расщепления и влияния условий фона, по-
этому он должен быть продолжительным. С продолжительностью пе-
риода вегетации и морфологическими признаками плода сухое вещество
не коррелирует, поэтому отбор на его повышение часто влечет за собой
снижение урожая и усиление заболеваемости.
А. Т. Галка [12] считает, что при скрещивании на сахаристость
материнский цветок должен быть расположен ближе к стеблю на плети
I порядка.
Н. А. Хохлачевой [87] выведен высокосахаристый сорт тыквы
Испанская А-5, в среднем содержащий 10—13% сахаров, 18% сухого
вещества, 4,8% крахмала, 8,5 мг каротина. Отдельные растения этого
сорта имели 17,8% сахара, 24,4% сухого вещества. По выходу крах-
мала этот сорт приближается к картофелю, давая с 1 га 13,23 ц (кар-
тофель— 14 ц). А. Миховым [47] в Болгарии выведены 3 сорта мус-
катной тыквы урожайностью 34 т с 1 га при 13—17% сухого вещества
и 8,7 мг каротина.
Селекция на каротин. По содержанию каротина тыква очень неод-
нородна: неокрашенная мякоть тыквы содержит менее 3 мг каротина,
а густо-оранжевая — до 30 мг.
Отбор на каротин легче всего проводить по интенсивности желтой
окраски мякоти плодов, однако при высоких концентрациях его опре-
деляют химическими методами.
Отбор на каротин можно проводить и до появления плодов по кос-
венным показателям: интенсивно желтой окраске рылец и пыльников,
лепесткам венчика. Каротин — спутник хлорофилла в листьях и на по-
верхности плодов. При созревании последних хлорофилл, окрашиваю-
щий плоды в зеленый цвет, разрушается и на поверхности их высту-
пают желтые тона различной интенсивности в зависимости от количе-
ства каротина. У видов С. ficifolia и С. mixta плоды совсем лишены
254
каротина, хотя цветки их желтые. В цветках, рыльцах и тычинках со-
держание каротина более постоянно, чем в плодах; видимо, он играет
большую роль в процессе размножения растений.
Селекция на структуру мякоти. Структура мякоти плодов тыквы
может быть рыхлой, плотной, нежной, волокнистой, мучнистой, верми-
шелевидной. Для пищевого использования предпочтительнее плотная
и нежная мякоть, для кормового использования в свежем виде —
рыхлая.
Плотная мякоть присуща обычно зимним сортом, хорошо храня-
щимся, а также более мелкоплодным формам, в особенности тропиче-
ского происхождения (Мексика, Япония, Абиссиния). Она связана с
мелкоклетной структурой, по которой можно вести отбор (в фазе пол-
ной спелости). Мелкие клетки имеют больше оболочек, но у сортов с
плотной мякотью эти оболочки нежнее, моложе в результате позднего
деления клеток, чем у рыхломякотных.
Селекция на лежкость плодов. Существуют специальные сорта
тыкв, способные долго храниться, называемые зимними. Их отличает
плотная структура мякоти, высокая удельная относительная масса.
Твердость коры плодов свидетельствует только о транспортабельности,
но не лежкости.
Лежкости плодов тыкв способствуют высокое содержание сухого
вещества, медленная транспирация, низкое содержание гидролитических
ферментов, позднее созревание. Из-за трудности отбора по многим из
этих показателей вначале пользуются признаками удельной относитель-
ной массы и медленного созревания; транспирацию наблюдают уже
в процессе хранения, а общий результат по лежкости плодов устанав-
ливают практически в конце хранения.
Селекция на голосемянность. Это направление перспективно для
получения сортов, семена которых используются на жмых и масло.
Голосемянные сорта пока известны у вида С. реро, но в исходном
материале коллекции тыкв ВИР имеются голосемянные формы у круп-
ноплодного вида, а также получены при межвидовой гибридизации.
Работу по селекции в этом направлении следует начинать со скре-
щиваний голосемянных форм с урожайными и высококачественными
сортами. Закономерности получения голосемянных форм путем гибри-
дизации описаны нами в разделе «Генетика тыкв».
Селекция на кустовость. Кустовые тыквы занимают меньшие пло-
щади питания, поэтому можно гуще сеять и дольше обрабатывать
междурядья, иногда даже до конца выращивания.
Первые работы селекционеров по выведению кустовых форм тыквы
начались со скрещивания плетистых крупноплодных сортов, С. реро
с кабачком. Таким образом, были выведены сорта Днепропетровская
кустовая, Пена и Победа, Кустовая 39 и Кустовая полосатая. В даль-
нейшем скрещивания велись с этими сортами. Путем скрещивания с
кустовой формой С. maxima из коллекции ВИР был получен сорт Бы-
ковская кустовая 151.
После скрещивания О. В. Юрина [96] в период бутонизации уда-
ляла с поля те растения, которые не имели укороченных междоузлий.
А. Т. Галка [11] прививал гибриды на кустовой подвой и снова опылял
их кустовым родителем, получив в F3 40% кустовых растений. В его
опытах кустовость усиливалась также на фосфорном фоне питания при
отсутствии азотных удобрений и при сокращенных площадях питания.
Скрещивания с кустовой формой С. maxima дали кустовое потом-
ство в F5.
А. Т. Галка сообщает также, что межлинейные гибриды видов или
опыление старого растения с увядшим цветком пыльцой молодого рас-
255
тения дают кустовую форму уже в F(. Легче получить кустовые рас-
тения в Fi при опылении плетистого привоя кустовым подвоем.
Н. С. Шашенок [93] обнаружил, что разные сорта кустовых тыкв
по-разному передают признак кустовости при скрещивании. Из кабач-
ков лучше передают кустовость сорта неветвящихся кабачков (в Fi
кустовых было 70—80%), в то время как обычные кустовые кабачки
с короткими боковыми побегами (Грибовский 37) в Fi не выщепляют
кустовых, а в F2 дают только короткоплетистые формы. У вида С. ma-
xima Кустовая 151 в Fi также давала около 8% кустовых растений,
а Кустовая Чино из Аргентины — только короткоплетистые и поздно
образующие плеть. Впрочем, и такие сорта полезны, так как позво-
ляют проводить обработку междурядий до августа, пока не образо-
валась удлиненная плеть.
А. Т. Галка отмечает также, что некоторые кустовые родительские
сорта могут давать в Fi кустовые растения, если в скрещиваниях они
были материнскими (например, сорт Днепропетровская кустовая). Кус-
товые растения в Fi могут дать кустовые сорта быстрее, чем кустовые
из последующих поколений. Генетическая однородность по кустовости
проявляется уже в третьем поколении.
Селекция кабачков и патиссонов. Для этих культур требуется
в первую очередь высокая плодовитость, так как их урожай состав-
ляют молодые зеленцы.
По данным И. В. Ермоленко [18], количество женских цветков у
кабачков на 1 растение по сортам составляет 1—5 в неблагоприятный
по погодным условиям год и до 14 в благоприятный. Широко райони-
рованный в нашей стране сорт кабачков Грибовский 37 занимает про-
межуточное положение. Если молодые растения подвергаются влиянию
пониженных температур, то появление женских цветков задерживается
и первые женские цветки остаются неоплодотворенными.
Мужских цветков, как правило, больше, чем женских; у разных
сортов их соотношение изменяется от 1:1 до 12 : 1. Однако некоторые
сорта (Caserta, Harris Hybrid), по данным И. В. Ермоленко, обладают
преимущественно женским типом цветения, что позволяет использовать
их в массовом производстве гибридных семян.
Наиболее плодовитыми и раннеспелыми являются неветвящиеся
сорта (до 15—18 завязей на 1 растение) по сравнению со стандартом
Грибовские 37 (8—9 завязей).
По исследованиям А. Бухингера [ПО], кабачки в скрещиваниях
с морфологически отдаленными от них крупноплодными тыквами того
же вида проявляют эффективный гетерозис по урожайности плодов
(на 277%) и семян (на 223%).
Селекция кормовой тыквы. При селекции этой культуры до сих
пор многие считают главным признаком только урожайность. Однако
В. А. Кульчицкая [31] (Запорожская государственная сельскохозяйст-
венная опытная станция) проводила селекцию кормовой тыквы не
только по массе, но и на сухое вещество. Ведут селекцию на кормовые
единицы, на крахмал и сахаристость, лежкость и другие опытные уч-
реждения. Селекция на содержание в плодах крахмала, каротина и
сахара имеет большое значение, так как они являются основными ве-
ществами в тыкве.
Кормовое использование тыкв предусматривает скармливание их
в свежем виде и в виде силоса. Поэтому нужны кормовые ранние и
кормовые лежкие сорта для длительного хранения.
Особо важное значение имеет каротин как средство, укрепляющее
экстерьер и здоровье животных. Сорта Витаминная и Каротинная с вы-
соким содержанием каротина относятся к виду С. moschata и раэво-
256
дятся только в Краснодарском крае и на Кавказе. Селекция на увели-
чение количества каротина нужна и для других видов тыкв, разводи-
мых севернее этой зоны.
НОВЫЙ ИСХОДНЫЙ МАТЕРИАЛ И МЕТОДЫ СЕЛЕКЦИИ
Из овощных видов тыквы кабачки наименее питательны. Сухое ве-
щество в них не превышает 6%, сахара до 3% (глюкозы 2,5%), ви-
тамина С 24%, кислотность 0,1%, однако они пользуются большим
спросом у населения.
В культуре имеются овощные формы С. реро с высоким процентом
сухого вещества: фордгукская разновидность, крукнеки и некоторые
сорта из мелкоплодной овощной разновидности, в частности сорт But-
ternut. Селекция этих тыкв на сухое вещество или скрещивание их с
кабачками улучшили бы овощной сортимент тыквы.
Для продвижения сортов вида С. moschata севернее ее современной
границы был бы полезен исходный материал из канадской разновид-
ности ввиду ее высокой скороспелости. В зоне выращивания этого вида,
в том числе в Закавказье и Средней Азии, не принято считать какие-
либо сорта этого вида кормовыми. Однако сорт Неаполитанская мог бы
стать родоначальником кормовых сортов для указанных районов ввиду
его выдающейся урожайности.
Многие полезные признаки, отсутствующие у культурных форм, не-
сут в себе дикие виды, из которых необходимо назвать тыкву Лунделя
(С. lundelliana), устойчивую к мучнистой росе и другим болезням. По-
ложительная особенность этого вида — скрещиваемость со всеми куль-
турными видами тыквы. Достаточно устойчив к мучнистой росе также
вид С. ficifolia.
Целая группа ксерофильных диких видов является многолетней
в связи с развитием у них корнеплодно-клубневидных образований в
корневой системе.
Помимо гибридизации, отбора и прививок, вошедших в практику
селекционной работы с тыквенными, современные исследования позво-
ляют пользоваться более эффективными методами. Методом разового
закрепления какого-либо полученного признака или комплекса поло-
жительной наследственности является перевод растения в апомиктиче-
ское состояние [64].
И. А. Петров [63] предлагает метод инъекции эндосперма для пе-
редачи наследственности от одного растения в семяпочку другого, а
Г. А. Минасян и Е. А. Соболев [45] предлагают удобный прибор для
этой цели.
Ручная кастрация при скрещивании — грубый прием, травмирую-
щий растение и не всегда обеспечивающий чистоту работы. Б. Н. и
В. П. Малиновские [40] предложили способ кастрации цветков подо-
гретым воздухом, поддержание температуры которого автоматизировано.
Условия опыления имеют большое влияние на успех скрещивания. По-
мимо крупности и зрелости пыльцы, оптимального температурного ре-
жима дня и других условий, предлагается использовать нагревание
пыльцы для усиления завязывания плодов [1], опылять первые и по-
следние по сроку раскрытия цветки (учитывать возраст рыльца), много-
кратное опыление, усиливающее влияние отцовского родителя; нано-
сить малое количество пыльцы для вызывания доминирования рецессив-
ных признаков, а также пыльцу другого вида для нормализации гор-
монального состояния рыльца при недостаточном количестве пыльцы
своего вида и т. д.
257
В селекции следует учитывать и природные условия зоны, в ко-
торой успешнее проявляются (накапливаются и наследуются) те или
иные полезные качества селектируемого сорта.
БОЛЕЗНИ И ВРЕДИТЕЛИ
Наиболее распространенные заболевания тыквы — вирусные, а из
грибных мучнистая роса. Тыкву поражают бактерии. Она страдает от
заразихи и нематоды.
БОЛЕЗНИ
Вирусные болезни. Действуют они главным образом на листовой
аппарат, снижая эффективность его работы, иногда без внешних при-
знаков поражения, но чаще с наличием рисунчатых мозаик.
На тыкве зафиксированы тыквенная мозаика, огуречная мозаика,
кудрявовершинность, из которых чаще встречаются первые два заболе-
вания.
Огуречная мозаика. ПоМ. Н. Родигину [65], проявляется
на молодых листочках тыквы и кабачка в виде зеленовато-желтых мо-
заичных пятен. В некоторых случаях появляется желтая окраска вер-
хушки растения. По мере развития этой мозаики наблюдается закручи-
вание листьев и образование на них небольших бугорков, выпуклостей
между жилками, придающих поверхности листа слегка гофрированный
характер. Вслед за этим развивается резко выраженная зеленая или
желтая пятнистость, сопровождаемая деформацией и курчавостью ли-
стьев. Рост сильно замедляется, междоузлия и листья укорачиваются,
едва достигая половины нормальной величины.
Заражение огуречной мозаикой происходит от сорняков через тлю.
С семенами вирус не передается. Возбудитель поражает около 200 ви-
дов растений из 40 семейств, в том числе два вида культурных расте-
ний тыкв: С. реро и С. maxima. Установлено, что он может зимовать
на остатках растений в почве.
Тыквенная мозаика. Также вызывает осветление окраски
листьев до желто-зеленой, по краям листа вначале выделяются концы
жилок в результате отставания роста листовой пластинки. Позднее по-
является крапчатость, курчавость, а затем выпадают мякоть листа и
наиболее тонкие жилки, остаются только толстые жилки с узкой по-
лоской листовой мякоти вдоль них.
Вирус тыквенной мозаики быстрее развивается при температуре
15—21° С, несколько задерживает развитие при 26° С и выше и при
длительном освещении. При коротком дне вирус успешно развивается
и при температуре ниже 15° С.
Вирус тыквенной мозаики передается с семенами даже трехлетней
давности. Кроме того, он переносится огуречными жуками из родов
Diabrotica, Acalumma и Coreus. По данным Д. Фрейтага, сок пора-
женного растения заражает тыкву при разведении 1 : 1000 на 100%,
при 1 : 10000 на 70%, при 1 : 100000 на 14%, а при 1 :1 000 000 на 3%.
Он поражает все тыквенные. В замороженном виде вирус сохранял жиз-
неспособность в течение 5 лет.
Вирус курчавовершинности (cyrly-top virus). Поражает
растения в стадии всходов, особенно при высокой температуре. Ввиду
раннего воздействия вируса междоузлия слабо разрастаются, остаются
сближенными до 2—3 см, ветви растут вертикально с мозаичными урод-
ливыми листьями на верхушках, напоминающими листья малины.
Цветки уменьшенного размера, с морщинистым венчиком сближены к
258
верхушке. Плоды не завязываются, листья завядают. Более устойчивым
к вирусу тыквенной мозаики оказался вид С. moschata, наименее устой-
чивым — С. реро.
М. Н. Родигин [65] рекомендует в качестве мер борьбы с вирусами
уничтожение насекомых и сорняков — передатчиков заболевания, унич-
тожение больных растений, посев незаряженными семенами, правиль-
ное чередование тыквы в севообороте. Эффективна обработка семян
горячим воздухом при температуре 70° С в течение 2 суток или при
40° С в течение 4 недель, а также теплой водой 1 ч.
Грибные болезни. Мучнистая роса. Поражает в первую оче-
редь старые листья, постепенно переходя на более молодые. Наиболее
подвержен поражению мучнистой росой С. реро, причем кабачки по-
ражаются сильнее, чем крупноплодные сорта этого вида. Возбудителями
мучнистой росы являются несколько грибов. В Японии мучнистую росу
тыквы вызывает только сферотека (Sphaerotheca fuliginea Poll.), в на-
шей стране: в Хабаровском крае —эризифе (Erysiphe cichoracearum
DC.), в Крыму, Казахстане, Поволжье, Московской области, в Азер-
байджане, Армении, а также в Болгарии — оба гриба. В Средней Азии
распространен еще мучнисторосяной гриб Leveillulla taurica Arnand.,
в Молдавии — сферотека и эризифе.
Листья, пораженные мучнистой росой от гриба эризифе, покры-
ваются сверху белым налетом, который потом переходит и на нижнюю
сторону листа. Поражение сферотекой дает буровато-пепельный налет
с отдельных выпуклых пятен желтовато-коричневого цвета. Гриб Ле-
веиллула образует на нижней поверхности листьев тыквы белый плот-
ный войлочнообразный налет. У зараженных растений активность ды-
хания, транспирации и пероксидазы увеличена, а активность аскор-
биноксидазы и каталазы уменьшена.
Антракноз. Обычно тыква этим заболеванием не поражается,
но оно неоднократно было обнаружено на тыкве в Астрахани.
Заболевание пока что редкое. Болезнь выражается в появлении
на листьях ржаво-желтых пятен, а на плодах выпуклых или вдавлен-
ных коричнево-бурых или светлых, впоследствии сильно увеличиваю-
щихся и сливающихся, образующих глубокие язвочки и быстро гнию-
щую мякоть.
Аскохитоз. Выражается в двух формах. Первая вызывает чер-
ную гниль плодов тыквы (гриб Ascochyta citrullina). Болезнь редкая,
но разрушительная. Начинается с серовато-бурых пятен на листьях
с черными точками на них. Листья желтеют и засыхают. На стеблях
появляются удлиненные блестящие язычки, покрывающие к концу лета
почти весь стебель. На плодах образуются блестящие темные твердые
пятна, покрывающие участки плода или всю его поверхность, на ко-
торой развиваются черные пикниды и перитеции в массовом количестве.
Болезнь распространяется с семенами, ветром, дождем п насекомыми.
Вторая форма вызывается грибом A. melonis Poteb. Проявляется
в виде сероватых бледнеющих пятен с большим количеством пикнид
как на листьях, так и на плодах.
Септориоз. Образует многочисленные, но небольшие серые или
бурые пятна, позднее беловатые с желтой каймой и черными пикни-
дами на листьях. Возбудитель Septoria cucurbitacearum Sacc.
Ма кросп ориоз. Вызывается грибом Macrosporium cucumeri-
num Ell. et Ev. В середине лета на листьях тыквы появляются мелкие
округлые светлые пятна, вокруг которых образуются концентрические
кольца с каймой. Иногда пятна сливаются, поражая большую площадь
листа. На плодах появляются также слегка вдавленные светлые пятна,
которые потом углубляются, увеличиваются в размерах, сливаются, по-
259
ражая большую площадь поверхности плода, и покрываются темным
бархатистым налетом. Пораженные плоды в пищу и на корм стано-
вятся непригодными.
Болезни цветков. Вызываются грибными и бактериальными
возбудителями. Вызывает заболевание цветка Rhizopus. Тычинки и
пыльца темнеют, покрываются налетом и опадают, или завязи сгни-
вают от мокрой гнили гриба. Аналогичное по внешнему виду заболе-
вание цветков — вершинный и цветочный бактериоз тыквы, вызываемый
бактерией Bacillus mesentericus-vulgatus Fiiigge. Погибают от гнили
пестики и завязи. Болезни разносятся насекомыми. Особенно сильно
они развиваются при высокой влажности. Пораженные цветки рекомен-
дуется уничтожать.
Болезнь иногда переходит на листья, образуя на них большие бу-
рые пятна, и на развивающийся плод, верхушка которого прекращает
рост. Получается уродливый плод с пожелтевшей верхушкой, на кото-
рой образуются вдавленные пятна, ведущие к гниению всего плода.
Иногда побуревшие плоды высыхают и висят на растении.
Источником заражения могут быть почва и остатки растений. Бо-
лезнь передается с семенами. Посевы люцерны, пшеницы, капусты и
хлопчатника снижают концентрацию бактерий в почве.
Бактериальные болезни. Бактериальная пятнистость.
Вызывается заболевание бактерией Xantomonas cucurbitae Dows. Пятна
на листьях вначале круглые, потом угловатые, ограниченные жилками с
желтой каймой, часто сливающиеся. Пятна на плодах покрыты кап-
лями жидкости янтарного цвета. Меры борьбы не разработаны.
Зеленая пятнистость плодов тыквы (Cladosporium cucu-
merinum Ell. et Arth.), особенно вредоносна при хранении. Появляется
она в конце вегетации на зрелых плодах. Вначале на коре плодов по-
являются очень мелкие зеленые пятна, иногда в виде россыпи. В опти-
мальных для болезни условиях (при температуре не выше 19° С и от-
носительной влажности выше 55%) пятна увеличиваются в размерах,
вдавливаются, гниль продвигается внутрь плода. При неблагоприятных
для болезни условиях гнилостный процесс останавливается, пятна окру-
жаются твердым кольцом, выступающим над корой. На крупных пятнах
имеются провалы внутрь мякоти плода, так как под пятнами образуются
полости.
Распространению инфекции благоприятствуют повышенная влаж-
ность воздуха и пониженная температура или ее резкие колебания в
период созревания плодов. Высокая температура и сухость воздуха
останавливают болезнь, и образовавшиеся ранее пятна на плодах на-
чинают рубцеваться. Здоровые растения с мощно развитой вегетатив-
ной системой и крупными плодами слабее поражаются зеленой пятни-
стостью. При хранении болезнь проявляется очень сильно при темпе-
ратуре ниже 10° С или при ее резком колебании. Уборка плодов с поля
в холодную погоду или сразу после заморозков резко усиливает пора-
жение плодов в хранилище. Повышение температуры в хранилище выше
10° С снова останавливает болезнь.
Зеленой пятнистостью поражается только вид С. maxima и в пре-
делах вида только ее зеленоплодные формы. Виды С. реро и С. mo-
schata этой болезнью не поражаются.
ВРЕДИТЕЛИ
Заразиха. Высшее цветковое растение (Orobanehe aegyptiaca Pers.,
реже О. ramosa L.), паразитирующее на корнях всех культурных тык-
260
венных. Она цветет голубыми цветками и дает очень большое (до по-
лумиллиона на 1 растение) количество мелких семян, прорастающих
только на корнях своего хозяина. Всхожесть семян заразихи сохра-
няется до 13 лет.
Меры борьбы с заразихой: применение глубокой зяблевой вспашки
с предплужником для глубокого захоронения семян заразихи, удаление
растений паразита до его обсеменения, опрыскивание или опыливание
гербицидами. Высокая агротехника и органо-минеральное питание
тыквы повышают ее устойчивость к заразихе. Гриб Fusarium orobanches
Jacz. и заразиховая мушка фатомиза являются паразитами заразихи
и могут быть использованы в борьбе с ней. Семена заразихи уничто-
жаются посевами многолетних трав, кукурузы и влагозарядковыми по-
ливами.
Галловая нематода. Чрезвычайно мелкие, червеобразные вредите-
ли внедряются в корни, на месте внедрения образуются корневые утол-
щения — галлы, достигающие иногда крупных размеров, утолщения
препятствуют развитию корневой системы, и растения вскоре увядают
от истощения и обезвоживания.
Против галловой нематоды в почву вносят 1—2%-ный раствор кар-
батиона с осени или за 35 дней до посева. Внесение на 1 м2 200 г кар-
батиона сокращает поражение растения с 95 до 30%.
В АН Туркменской ССР был получен гриб, паразитирующий на
нематодах. Внесение его в почву (30—60 г на 1 м2) увеличивало уро-
жай в 2—3 раза. С нематодой можно бороться, применяя правильный
севооборот, в котором предшественниками тыквы и других тыквенных
являются суданка, кроталлярия и бархатные бобы. С. И. Кобытев пред-
лагает уничтожать нематоду введением сухого пара в почву летом, вы-
сушивающего верхний слой, в котором находятся паразиты.
Другие вредители. Тыква поражается только всеядными видами, к
которым относятся проволочники и ложнопроволочники (щелкуны, чер-
нотелки), жуки-пыльцееды, кукурузный навозник, хрущи, кравчики, на-
рывники, долгоносики, бахчевые коровки, блошки, подгрызающие совки,
стеблевые совки, луговой мотылек, пестрая медвидица, ростковые мухи,
мухи-минеры, полевые муравьи, саранчовые, кузнечики, сверчки, табач-
ные трипсы, клопики, тли, подуры (при орошении), паутинный клещик,
слизни, грызуны. Их описания, повреждения и борьба подробно опи-
саны в «Бахчеводстве» [84].
СПИСОК ЛИТЕРАТУРЫ
1. Алешин Е. И. Воздействие на пыльцу как одна из основ новой методики.—
Тр. Краснодарск. с.-х. ин-та, 1937, вып. 9.
2. А л м а ниязов Э. А. Вирусные болезни бахчевых культур. Автореф. дис. на
соиск. учен, степени канд. биол. наук.— Л., ВАСХНИЛ, ВИЗР, 1970.
3. Белик В. Ф. Формообразование при межвидовой гибридизации тыквы. — В кн.:
Морфология растений, 1961, т. II.
4. Белик В. Ф. Рост и развитие растений бахчевых культур. — В кн.: Физиология
сельскохозяйственных растений. М., 1970, т. 8.
5. Белик В. Ф., Подмогаева М. И. Метод преодоления нескрещиваемости раз-
личных видов тыквы. — Докл. АН СССР, 1954, XCVIII, № 2.
6. Беляев И. М. Лежкость различных сортов тыквы в траншеях. — Плодоовощное
хозяйство, 1934, № 3.
7. Б р е ж н е в Д. Д. Гетерозис у бахчевых культур. — Вестник с.-х. науки, I960,
№ 7.
8. В е л и ч к и н П. А., Данков В. Ф. Скармливание тыквы курам в борьбе с аска-
ридиозом и гетеракидозом. — Птицеводство, 1957, № 2.
9. ^РЖеВСКИЙ Иммунитет растений к паразитарным заболеваниям.—
261
10. Власов Ю. И., Редько Г. А., Лытаева Г. К. Вирусные болезни овощных
и бахчевых культур. — Л., 1973.
11. Галка А. Т. Выведение кустового сорта тыквы. — Науч. тр. Укр. НИИ овоще-
водства, 1954, т. 3.
12. Галка А. Т. Методы селекции на повышение качества бахчевых. — В кн.: Во-
просы улучшения качества сельскохозяйственной продукции. Киев, 1960.
13. Гончаров П. Л. О методах селекции сельскохозяйственных культур. — Селек-
ция и семеноводство, 1966, № 2.
14. Гордеева Н. Г. К характеристике водного обмена у тыкв. — В кн.: Сборник
работ аспирантов и молодых ученых., Л., ВИР, 1960, т. 1.
15. Гридин И. Ф. К вопросу селекции и агротехники голосеменной тыквы. — Тр.
Белорусск. с.-х. ин-та, 1947, т. 12.
16. Декандоль А. Местопроисхождение возделываемых растений. — СПб., 1885.
17. Добрынин Г. М. К методике изучения биологии корней растений. — Бот. жур-
нал, 1955, № 5.
18. Ермоленко И. В. Исходный материал для селекции кабачков. — В кн.: Сбор-
ник трудов аспирантов и молодых ученых. Л., ВИР, 1965, № 6 (10).
19. Ермоленко И. В. Особенности формирования семян в период вегетации и
при дозаривании. — Вестник с.-х. науки, 1966, № 2.
20. Жите не в а Н. Е. Мировой сортимент культурных тыкв. — Тр. по прикл. бот.,
ген. и сел., 1930, т, 23, вып. 3.
21. Житенева Н. Е. Тыквы северной части тропической Америки. — В кн.: Возде-
лывание растений Мексики, Гватемалы и Колумбии. Л., 1930.
22. Ж у к о в с к и й П. М. Современное состояние проблемы происхождения куль-
турных растений. — Бот. журнал, 1957, т. 43, № 11.
23. 3 а у р а л о в О. А. К вопросу о холодостойкости бахчевых культур. — Научн.
труды НИИОХ, 1965, № 3.
24. Кичу но в Н. И. Огурцы, дыни, арбузы и тыквы. — СПб., 1910.
25. Ко быт ев С. И. Итоги работы Туркменской бахчевой станции за 1939—1949 гг.
— Бюл. по плодоводству, виноградарству и овощеводству Тадж. НИИ плодо-
виноградного и овощного хоз-ва, 1950, № И.
26. К о б ы т е в С. И., Л а в р и и о в а Е. И. Опыт межвидовой гибридизации тыкв
С. moschataXC. maxima. — Изв. АН ТССР, 1956, № 6.
27. Кова реки й А. Е. Творческая роль отбора на примере изучения наследствен-
ности и изменчивости тыквы-перехватки. — Тр. Кишин. с.-х. ин-та, 1955, т. 2.
28. Кожухов 3. А. Кариотипические особенности культурных тыквенных. — Тр.
по прикл. бот., ген. и сел., 1935, т. XVI, вып. 2.
29. Кочергин С. Исследование состава плодов тыквы. — Изв. Московск. с.-х. ин-та,
1904, № 658.
30. Кревченко А. Е. Селекция тыквы. — В кн.: Краткие итоги работ Бирючекут-
ской овоще-бахчевой селекционной опытной станции, 1939.
31. Кульчицкая В. А., Шелепов В. В. Селекция кормовой тыквы на качество.
— В кн.: Наука полям и фермам. Киев, 1969.
32. Ливингстон Д. Путешествие и исследования в Южной Африке. — М., 1955.
33. Л у ди лов В. А. Наследование кустовой формы у тыкв при скрещивании.—
Картофель и овощи, 1964, № 10.
34. Л у д и л о в В. А. Проращивание семян межвидовых гибридов тыкв на искусст-
венной среде. — Селекция и семеноводство, 1966, № 2.
35. Л у д и л о в В. А. Межвидовая гибридизация тыкв и отдаленные прививки в се-
мействе тыквенных. Автореф. дис. на соиск. учен, степени канд. с.-х. наук. Крас-
нодар, Кубан. с.-х. ин-т, 1966.
36. Л у д и л о в В. А. Бекроссы и жизнеспособность межвидовых гибридов тыквы. —
С.-х. биология, 1969, № 1.
37. Л у д и л о в В. А. Отдаленные прививки как мутагенный фактор. — Вестник с.-х.
науки, 1969.
38. Лященко И. Ф. О межвидовой гибридизации тыкв. — Учен. зап. Ростов, гос.
ун-та, 1934, т. 34, № 2.
39. Макаровский А. Ф., Подмогаева М. И. Межвидовые гибриды тыкв. —
Селекция и семеноводство, 1952, № 6.
40. Малиновский Б. Н., М а л и н о в с к ий В. П. Новый прибор для кастрации
цветков сорго. — Агробиология, 1961, № 3.
41. Мар х 3. А., Шевчук А. С. Консервы детского питания из тыквы. — Тр. Укр.
НИИ консервной пром-ти, 1959, вып. 2, ч. 1.
42. Матиенко Б. Т. Анатомическая характеристика плодов отдельных сортов тык-
вы.— Учен. зап. Тираспольского пед. ин-та, 1956, вып. II.
43. Матиенко Б. Т. К вопросу морфологии завязи и плода тюрбанной тыквы. —
Учен. зап. Тираспольского гос. пед. ин-та, 1957, вып. IV.
44. Милованова Л. М., Филов А. И. Тыква как источник каротина. — Тр. по*
прикл. бот., ген. и сел., 1954, т. 31, № 1.
262
45. М и н а с я н Г. А., Соболев Е. А. Новый прибор для инъекций чужеродного
эндосперма. — Селекция и семеноводство, 1965, № 4.
46. М и р ю т а Ю. Б. Выведение инбредных линий и гибридов озимой пшеницы на
основе взаимоизбирательности при двухстороннем парно-индивидуальном отборе
по хозяйственно-ценным признакам. — Киев, 1972.
47. Михов А. Хозяйственные качества мускатных тыкв (цигулки). — Овощеводство
и виноградарство, 1954, № 4.
48. Мичурин И. В. Видовые гибриды. Скрещивание тыквы с дыней и огурцом. —
В кн.: Мичурин И. В. Соч., т. I. М., 1939.
49. Нагаева Н. П. Сорта кустовых тыкв. — Сад и огород, 1958, № 5.
50. Невкрыта А. Н. Насекомые, опыляющие бахчевые культуры. — Киев,
1953.
51. Некрасов В. В., Скрипников Ю. Г. Об оценке качества плодов тыквы
с помощью рефрактометра. — Тр. плодоовощного ин-та им. И. В. Мичурина, 1959,
т. 10.
52. Н и г м а н о в а Н. Н. О коррелятивной связи между развитием листьев на главном
стебле и ростом боковых побегов у кабачков. — Тр. Ташкентского с.-х. ин-та,
1963, вып. 15.
53. Н и з е н ь к о в Н. П. Применение электромотора в селекции. — Селекция и семе-
новодство, 1946, № 9—10.
•54. Никитенко Г. Ф. Линейка для исчисления продолжительности вегетационного
периода. — Селекция и семеноводство, 1959, № 5.
55. Н о в о с е л о в В. С. Замыкающийся водяной объемомер корневой системы ра-
стений.— Физиология растений, 1960, т. 7, вып. 2.
56. Павлов И. П. Селекция и семеноводство овощных культур. — М., 1963.
57. Пальчевский В. И. Зависимость между формой семядолей и формой пло-
да у тыкв. — Селекция и семеноводство, 1946, вып. 4—6.
58. Пангало К. И. Новый вид культурной тыквы. — Тр. по прикл. бот., ген. и
сел., 1930, т. XXIII, вып. 3.
59. Пангало К. И. Бахчевые культуры Анатолии. — В кн.: «Земледельческая Тур-
ция». Л., 1933.
50. Пангало К. И. Селекция бахчевых культур.— В кн.: Теоретические основы се-
лекции, 1937, т. 3.
61. Пангало К. И., Гольдгаузен М. К. Межвидовые гибриды тыкв. — Докл.
АН СССР. Новая серия, 1939, т. 24, № 1.
62. Пангало К. И. Тыквы. — БСЭ, 1947.
63. Петров И. А. Преобразование природы зерновых культур методом инъекций. —
В кн.: Вопросы методики селекции пшеницы и кукурузы. Харьков, 1957.
64. Петров Д. Ф. Генетически управляемый апомиксис и его значение для эволю-
ции и селекции. — В кн.: Апомиксис и некоторые новые методы селекции расте-
ний. — Новосибирск, 1963.
65. Родигин М. Н. Болезни бахчевых культур. — В кн.: Бахчеводство. Л., 1959.
66 Р о д и г и н М. Н. Обзор исследований по иммунитету бахчевых культур.—Научн.
труды НИИОХ, 1965, № 3.
67. Рожков М. И., Смирнов Н. Е. Витаминные растения. — М., 1956.
68. Ролофс Е. А. Кормовые бахчевые культуры. — М., 1953.
69. Руководство по апробации сельскохозяйственных культур. Бахчевые культу-
ры.— Л., 1954, т. 6.
70. Скрипников Ю. Г. Быстрый метод определения крахмала в плодах тыквы. —
Селекция и семеноводство, 1957, № 5.
71. Сойфер В. Особенности анатомического строения семян некоторых представи-
телей сем. тыквенных. — В кн.: Сборник студенческих научно-исследовательских
работ ТСХА. М., 1957, вып. 7.
72. Сойфер В. Эволюция анатомических структур семян сем. тыквенных. — Бюл.
Моск, общ-ва исп. природы. Отд. биол., 1962, т. 67, вып. 3.
73. Соколов Д. И. Селекция тыквы на Быковской опытной станции. — Научи, тру-
ды. Быковская бахчевая оп. станция, 1969, № 5.
74. Соколова В. Пространственную изоляцию можно сократить. — Картофель и
овощи, 1967, № 6.
75. Тоще в В. П. Поведение гибридов Fi при скрещивании географически отдален-
ных форм тыквы. — В кн.: Сборник работ аспирантов и молодых научных сотруд-
ников. Л., ВИР, 1959.
76. Ф и л о в А. И. Строение плодов бахчевых культур. — В кн.: Сборник научных
работ по бахчевым культурам. Ташкент, 1952.
77. Филов А. И. Бахчеводство. — Л., 1959.
78. Филсов А. И. Закон параллельного морфогенеза. — В кн.: Морфогенез растений.
79. Филов А. И. Географическая изменчивость сухого вещества и крахмала у тыкв.
— Сельское хозяйство Узбекистана, 1962, № 2.
263
80. Филов А. И., Тощ ев В. П. К вопросу определения гетерозиса у бахчевых
культур по электрической полярности половых элементов. — В кн.: Использование
гетерозиса в овощеводстве. Краснодар, 1963.
81. Филов А. И. Укороченный рост осевых органов у растений. — Бюл. Глав. бот.
сада АН СССР, 1965, № 57.
82. Филов А. И. Продвижение бахчевых культур на север как фактор их окульту-
ривания.— Картофель, овощи и бахчевые культуры, 1965, № 2, (на укр. яз.).
83. Филов А. И. Экология и классификация тыквы. — Бюл. Главн. бот. сада, 1966,
вып. 63.
84. Филов А. И. Бахчеводство (учебное пособие для с.-х. вузов).—М., 1969.
85. Филов А. И. Водный режим бахчевых. — В кн.: Физиология культурных расте-
ний. М., 1970, т. 8.
86. Филов А. И., Белик В. Ф. Устойчивость бахчевых растений к неблагоприят-
ным условиям. — В кн.: Физиология культурных растений. М., 1970, т. 8.
87. Хохлачева Н. А. Тыква Испанская А-5. — В кн.: Практические выводы из
опытных работ с овощными культурами и картофелем, 1947, вып. 2.
88. Хохлачева Н. А. Межвидовой гибрид тыкв. — Селекция и семеноводство^
1951, № 12.
89. Ц е м ш Н. С. Сельскохозяйственная техника в каролингских текстах. — В кн.:
Агрокультура в памятниках западного средневековья. М., 1936.
90. Ч е р н о в а М. Траншейный способ хранения тыквы. — Сад и огород, 1952, № 8.
91. Чех о вич К. Ю. Тыква как силосная культура степной зоны Казахстана.—
С.-х. наука в Казахстане, 1935, № 1—2.
92. Ш а ш е н о к Н. С. Методы выведения кустовых сортов тыкв. — В кн.: Сборник
трудов аспирантов и молодых научных сотрудников. Л., ВИР, 1965, № 6.
93. Ш а ш е н о к Н. С. Роль материнской наследственности в получении кустовых
форм у тыкв. — В кн.: Сборник трудов аспирантов и молодых научных сотруд-
ников. Л., ВИР, 1966, №7 (11).
94. Ш а ш е н о к Н. С. Изучение наследования кустовой формы у тыкв с целью
использования ее при выведении сортов. Автореф. дис. на соиск. учен, степени канд.
с.-х. наук/Л., ВИР, 1968.
95. Щ е р б а к С. Н. Некоторые данные по селекции столовой тыквы на повышение
качества плодов. — Зап. Воронежского с.-х. ин-та, 1950, т. 23, вып. 1.
96. Юрина О. В. Селекция и семеноводство тыквенных культур. — М., 1966.
97. Ю р и н а О. В. Межвидовая гибридизация тыквы. — В кн.: Доклады советских
ученых к 17-му Международному конгрессу по садоводству. М., 1966.
98. Юрина О. В. Современные методы селекции тыквенных. — Картофель и овощи,
1968, № 8.
99. Ю р и н а О. В. Селекция тыквенных на устойчивость к болезням. — Селекция и
семеноводство, 1970, № 4.
100. В a i 1 е у L. Н. Standart cyclopedia of Horticulture, London, 1930, t. 1.
101. Bailey L. H. The Domesticated Cucurbits. — Gentes Herb., 1929, v. II, fasc. 2.
102. Bailey L. H. Species of Cucurbita. — Gent. Herb., 1943, v. VI, fasc. 5.
103. Bailey L. H. Jottings in the Cucurbitas. — Gent. Herb.— 1948, v. VII, fasc. 5.
104. Bauhinus J. Pinax theatry botanici. Basel, 1623.
105. Bemis W. P. Interspecific hybridization in Cucurbita. — J. Heridity, 1963, v. 54,
N 66.
106. Bemis W. P., Whitaker T. W. Natural hybridization between Cucurbita digi-
tata and C. palmata.—Madrono, 1965, v. 18, N 2.
107. Bemis W. P., Rhodes A. M., Whitaker T. W., Carman S. G. Numerical
taxonomy applied to Cucurbita relationships. — Amer. J. Botany, 1970, v. 57, N 4.
108. В r et s chneider E. Botanicon sinicum. London, 1882.
109. Brown W. Reisen in Africa, Egypten und Syrien. Weimar, 1800.
110. Buchinger A. Kurbiszuchtung mit allgemeinen biologischer Ausblicken. — Bo-
denkultur, 1948, Jg. 2, H. 1.
111. Camerarius I. De plants Epitome utillisima. Frankfurt am Main, 1590.
112. C a s t e t t e r E. F. Horticultural Groups of Cucurbits. — Proc. Amer. Soc. Hort.
Sci., 1925, v. 22.
113. C a s t e t t e r E. F. Crosses of the species in Cucurbita. — Jowa State College
J. Sci., 1928, N 3.
114. Cog ni aux A. Cucurbitaceae.—In. De Candolle Monographiae Phanerogamanerum.
1881, t. 111.
115. Culpepper Ch. W. a. oth. Dehybration of Pumpkin and winter squash. — Fruit
Products J., 1945, v. 24, N 6, N 7.
116. Curtis L. C. Heterosis in summer squash and the possibilities of producing Fi
hybrid seed for commercial planting. — Proc. Amer. Soc. Hort., Sci., 1939, v. 37.
117. Cutler H. C., Whitaker T. W. Cucurbita Mixta, Pang, its classification and
relationships. — Bulletin Torrey Botanical Club., 1956, v. 83, N 4.
118. Daleschamp J. Histoire generate des plantes. Lyon, 1653.
119. De Candolle A. Geographie Botanique, t. II, Paris, 1855.
264
1' 20. Den n a D. W. a. Munger H. M. Morphology of the bush and rine habits and the
allelism of the bush genes in C. maxima and C. pepo squasli — Proc. Amer. Soc.
Hort. Sci., 1963, v. 82.
121. Dodonaeus R. Stirpium historiale nemptades set save libri, 1616. T. 30, N 1.
122. Duchesne M. Courge.—Lamarch M. Ch. Dictionaire Encyclopedique methodique.—
Botanique, 1790, t. 11.
123. Engler A., Prantl K. Cucurbiteae Cucurbitinae. — Die Naturlichen Pflanzenfa-
milien, 1894, T. IV, Ab. 5.
124. Erwin A. T. Pumpkins or Squashes, which is right? — Market Growers J.
Louisville, 1928, N 42.
125. Erwin A. T., Haber E. S. Species and Varietal Crosses in Cucurbits. — Agr.
Exp. Sta. Jowa state college of Agr. a. Mechanic arts. Bull., 1929, N 263.
126. E r win A. T. Nativity of the cucurbits. — Botan. Gaz. 1931, N 91.
127. Erwin A. T., Lana E. P. The Seminole Pumpkin. — Economic Botany, 1956,
10, N 1.
128. Fuchs L. De historia stirpium. Basel., 1542.
129. G a s p a r i n i M. G. Observations morphologique et physiologiques sur quelques
especes de courges cultivees. — Ann. Sci. Nat Bot., 1948, ser. Ill, IX.
130. Glovers on D. The inheritance of white fruit and color in summer squash
C. pepo. — Euphytica, 1969, v. 18, N 2.
131. Gray A. Plantae Wrightoanae, 1852, v. 1, II.
132. Grebenscikov I. Notulae cucurbitllogicae. I. Vererbung der Bitterkeit und Kurz-
triebigkeit bei C. pepo. — Kulturpflanze, 1954, Bd. 2.
133. Grebenscikov I. Notulae cucurbitologicae. II. Uber C. texana u. ihre Kreuzung
mit einer noch gesuchteten C. pepo.— Form. Kulturpflanze, 1955, Bd. 3.
134. Grebenscikov I. Uber zwei Cucurbita, — Art Der. Zuchter, 1958, Bd. 28, N 5.
135. G г e b e n s c i к о v I. Notulae cucurbitologicae. III.—Kulturpflanze, 1958, Bd. 6.
136. G г e b e n s c i к о v I. Ein Vergleich des Heterosisseffekts im «Fruchtgewichb bei
Kurbis und Mais. — Kulturpflanze, 1959, Bd. 7.
137. Grebenscikov I. Zur quantativ—genetischen Analyse der Ertragkomponenten
beim Kurbis. — Kulturpflanze, 1963, Bd. 14.
138. Grebenscikov I. Notulae cucurbitologicae 6. Uber einige Artkreuzung in der
Gattung Cucurbita. — Kulturpflanze, 1965, Bd. 13.
139. Grebenscikov I. Unterteilung von C. Moscata Duch. ex Poir. — Kulturpflanze,
1969, Bd. XVII.
140. Groff D., Bemis W. P. Meiotic Ivregularities in Cucurbita Species Hybrids.—
J. Heredity, 1967, v. 58, N 3.
141. Hadwiger L. A. A biochemical study of the host-parasite relationship between
Colletotrichum lagenarium and cucurbit hosta. — Proc. Amer. Soc. Hort. Sci. 1963,
v. 82.
142. Ha Is ted B. D. Report of the Botanist 29th. — Ann. Rpt, N. Y. Agr., Exs. Sta,
1908.
Г43. Harz E. D. Landwirtschaftliche Samenkunde. Berlin, 1885.
144. Hayase H. Cucurbita crosses. XV. Flower pollination at 4 a. m. in the production
of С. pepo C. moschata Fi hybrids.—Jap. J. Breeding, 1963, v. 13, N 2.
145. Hayase H. Cucurbita crosses. XVI. Flower ages and reciprocal cross-compatibi-
lity in С. pepo, C. moschata and C. maxima.—Jap. J. Breeding, 1963, v. 13, N 3.
146. Hehn V. Kulturpflanzen und Haustiere Berlag Gebr. Borntrager. Berlin, 1874.
147. Hillyer I. G., Wittwer S. H. Chemical and environmental relationships in flo-
wering of Acorn squash. — Proc. Amer. Soc. Hortic. Sci., 1959, v. 74.
148. Hooker J. D. Flora of Tropical Africa. London, 1871, t. II.
149. Hutchins A. E., Crosten F, E. Productivity of Fi hybrids in the squash
Cucurbita maxima. — Proc. Amer. Soc. Hort. Sci., 1941, v. 39.
150. H u t c h i n s A. E., S a n d о L. Gowrds. Uses, Culture, Identification. — Univer-
sity Minnesota. Arg. Exp. Sta, 1941, Bull. N 356.
151. Hutching A. E. A male and female sterile Variant in squash C. maxima.—
Proc. Amer. Soc. Hort. Sci., 1944, v. 44.
152. Jones H. A., Rosa I. T. Truck crop plantes. New-York—London, 1928.
153. Kamerarius J. De plantis Epiyome utullissima, 1590.
154. Lana E. P. Reciprocal crosses in the squash C. maxima. — Minnesota Agr. Exp.
Sta., Tecnical bull., 1950, N 189.
155. Linney K. Species plantarum, Stockholm, 1753.
156. Lobelio M. Plantarum seu stirpium icones. Antnerpiae, 1581.
157. Lots у S. Cucurbita Stridvragen. — Genetica, 1920, v. 2.
158. Metzger I. Landwirtschaftliche Pflanzenkunde. Heidelberg, 1841.
159. Meyer E. H. T. Geschichte der Botanic. Konigsberg, 1855.
160. Millan R. Los zapallitos de tronco de Sudamerica extratropical. — Darwinians,
1947, t. 3, N 3.
161. Millan R. Observaciones sorbe cinco cucurbitaceas cultivadas о indigenas en la
Argentina. — Darwiniana, 1968, v. 14, N 4.
265
162. Mu dr a A., Neuman D. Probleme und Ergebnisse der Munchenberger Olkurbi-
szuchtung. — Zuchter, 1952. Bd. 22, H. 4—5.
163 Haudin Ch. Origine supposee des Courges et de Potirons. — Revue Horticole,
’ 1855, Serie IV, T. IV.
164. Nau din Ch. Nouvelles recherches sur les caracteres specifiques et les vanetes
des plantes du genre Cucurbits. — Ann. Sci., Nat. Bot., 1856, ser. 4, v. VI.
165 Nau din Ch. Revue des cucurbitacees cultivees an museum. —Ann. Sci. Nat. Bot.,
1859, t. XII.
166. Naud in Ch. Ann. Sci. Nat. Bot., 1866, t. VI.
167. N a u d in Ch. Cucurbita andreana. — Revue Horticole, 1896.
168 Paulus A. O. a. oth. Control of powdery mildew in Cucumber and in squash.—
Calif. Agr., 1968, v. 22, N 3.
169. Poire t J. L. M. Curge. — Dictionnaire des Sci. Nat., 1818, v. XI.
170. Reinhold F. Korrelationen beim schalen- und rankenlosen Kurbis als Grundlage
der Vorselektion. — Der Zuchter, 1937, Jg. 9.
171. Rhede H. A. Horti Malabarici, 1688, Amsterdam, t. VIII.
172. Ph odes A. M. Species hibridization and interspecific gene transfer in the genus
Cucurbita. — Proc. Soc. Hort. Sci., 1959, v. 74.
173. Rhodes A. M. Inheritance of powdery mildew resistance in the genus Cucurbita.
Plant Dis. Reptr., 1964, v. 48.
174. Ritchey G E. A new squash from Agrica.—Univ. Florida Agr. Exp. Sta. Press.,
1936, Bull, N 490.
175. Robinson R. W., Whitaker T. W., Bohn G. W. Promotion of pistillate flo-
wering in Cucurbita by 2-chloroethylphosphonic acid. — Euphytica, 1970, v. 19, N 2.
176. Roemer M. J. Fami. nat, regni veg. Sin, Monorg. 1846, fasc. II.
177. Ruel J. De natura stirpium, libri tres. Basel, 1536.
Г78. Russel P. Identification of the commonly cultivated species of Cucurbita by
means of seed characters. — J. Washington Acad. Sci., 1924, 14, N 12.
179. Sageret M. Considerations sur la Production des Hybrides. — Ann. Sci. natu-
relle, 1826, v. 8.
180. Scott G. W. Sex ratws and fruit production studies in bush pumpkins. — Proc.
Amer. Soc. Hort. Sci. 1933, 1934, v. 30.
181. Scott D. H., Riner M. E. Inheritance of male sterility in winter squasch.—
Proc. Amer. Soc. Hort. Sci., 1946, v. 47.
182. Seringe N. C. In: De Candolle A. «Prodromus systematic naturalis regni vege-
tabilis», part III, 1828, t. 28.
183. Seringe N. C. Flora des Jardins et des Grandes cultures. Paris, 1849, t. II.
184. Shifriss O. Developmental reversal of dominance in C. pepo. — Proc. Amer Soc.
Hort. Sci., 1947, v. 50. r r .
185. Shifriss O. A developmental aproach to the genetics of fruit color.—J. Here-
dity, 1949, v. 40, N 9.
186. Shifriss O. The Unpredictable Gourds. — Amer. Hort. mag., 1965, v. 44, N 4.
187. Singh D. Inheritance of certain economic characters in the squasch, C. maxima.—
Univ. Minesota Agr. exp. Sta. Techn. Bull. 1949, N 186.
188. Sinnott E. W., Durham G. B. Inheritance in the Summer squasch.— J. Here-
dity, 1922, v. 13.
189. Sinnott E. W., Hammond D. Factorial balance in the determination of fruit
chape in Cucurbita. — Amer. Nat. 1930, v. LXIV, N 695.
190. Sinnott E. W. The relation of gene to character in quantative inheritance.—
Proc. nat. Acad. Sci., USA, 1937, v. 23, N 4.
191. Sinnott E. W., Blakeslee A. Changes in shape accompaning tetraploidy in
^u^urbit fruits. — Sci., 1938, v. 88.
192. Sinnott E. W. Cell polarity and the development of form in cucurbit fruits.—
Amer. J. Bot., 1944, v. 31, N 7.
193. Small J. K- Wild pumkins. — Journ. New York Bot. gard. 1922, v. 23, N 266.
194. Spach E. Histoire naturelle des vegetaux.—Phanerogames, 1838, IV.
195. To urn efort J. F. Institutiones rei herbariae, Paris, 1700, t. 1.
196. Wall J. R. Interpecitic hybrids of Cucurbita obtained by embryo culture. — Proc.
Amer. Soc. Hort. Sci., 1954, v. 63.
197 Welling F. Artkreuzungen beim Kurbis. — Naturwissen. — schaften, 1951, Jg. 38,
* N 1.
198. Wei ling F. Beobachtungen an Kurvisartbastarden. — Berichte Dtsch. Bot. Hesell-
schaft, 1953, H. 9, S. 368.
199. Wei ling F. Uber die interspezifische Kreuzbarkeit verschidener Kurbisarten.—
Zuchter, 1955, Bd. 25, H. 1/2.
200. W e i 1 i n g F. Die Ubertragung des Merkmals «Weichschaligkeit» vom Olkurbis
(C. pepo) in fertila Artbastarde aus der Kreuzung C. maximaXC. pepo. — Zuchter,
1956, v. 26, N 1/2.
266
201. Wei ling F. Ganomanalytische Untersuchungen bei Kurbis. — Zuchter, 1959,
v. 29, H. 4.
202. Wei ling F. Weitere Untersuchungen an Nachkommen von Kurbis. — Artbastar-
den. — Zuchter, 1966, Bd. 36, H. 2.
203. Whitaker T. W. Fertile gourd—pumpkin hybrids.—J. Heredity, 1932, v. 23, N 10.
204. Whitaker T. W. Cytological and phylogenetical studies in the Cucurbitaceae. —
Bot. Gaz., 1933, v. 44.
205. Whitaker T. W., Jagger J. C. Breeding and Improvement of cucurbits. —
Jearbook, 1937, N 1561.
206. Whitaker T. W., Carter G. F. Critical notes on the origin and domestication
of the cultivated species of Cucurbita. — Amer. J. Bot., 1946, v. 33, N 1.
267. Whitaker T. W., Bohn G. W. The Tatonomy. Genetics, Production and uses of
the Cultivated Species of Cucurbita. — Economic botany, 1950, v. 4, N 1.
208. Whitaker T. W. A species crass in Cucurbita. — J. Hereditay, 1951, 42, N 2.
209. Whitaker T. W., Carter G. F. Oceanic drift of gourds — experimental obser-
vations. — Amer. J. Bot., 1954, v. 41.
210. Whitaker T. W. The origin of the cultivated cucurbita. — Amer. Naturalist, 1956,
v. XC, N 852.
211. Whitaker T. W. Blosystematic of the cultivated cucurbita. — In Recent advances
in botany, 1961, v. 1.
212. Whitaker T. W., D a v i s G. N. Cucurbits, New York, 1962.
213. Whitaker T. W. An interspecific crosing Cucurbita: C. lundeUiana BaileyX
X C. maxima Duchesne. — Euphytica, 1962, v. 11, N 3.
214. Whitaker T. W., Bemis W. P. Evolution in the genus Cucurbita. — Evolution,
1964, v. 18, N 4.
215. Whitaker T. W., Cutler H. C. Cucurbits and cultures in the Americas. — Econ.
Bot., 1965, v. 19, N 4.
216. Whitaker T. W. Ecological aspects of cultivated Cucurbita. — Hort. Sci., 1968,
v. 3, N 1, 9.
217. Wildenov C. L. Linney «Species Plantarum». Stockholm, 1805.
218. Wittwer S. H., Hi 1 Iyer J. G. Chemical Induction of Male Sterilyty in Cucur-
bits.—Sci., 1954, v. 120, N 3126.
8 QD Описание сортов тыквы по признакам плода и скороспелости ПРИЛОЖЕНИЕ
Сорт Внешний вид плода Мякоть плода Срок созревания
размер поверхность форма окраска фона окраска полос рисунка цвет толщина
Вид твердокорой тыквы — С. реро L.
Разновидность овал ь н а я — var. реро
Алтайская 47 Башкирская 245 Бирючекутская 27 Мозолеевская 10 Мозолеевская 15 Мозолеевская 49 Украинская многоплод- ная Млеевская Даш-кеды Старосельская Средний » Крупный » Средний » Крупный Средний Крупный Ровная Ребристая Слабореб- ристая То же > Гладкая Слабореб- ристая Ребристая > Овальная » Слабогрушевидная Обратнояйцевидная Слабогрушевидная Обратнояйцевидная Короткоовальная Обратнояйцевидная Овальная > Оранжевая > > > > > Коричневая Оранжевая Светло-ко- ричневая Коричневая Темно-оран- жевая Коричневая Зеленая Светло-ко- ричневая Зеленая Отсутствует Лимонная Оранжевая > » » Оранжевая » > Средняя Небольшая Большая Средняя > Большая Средняя Небольшая Скороспе- лый То же Средне- ранний Средне- спелый То же Скороспе- лый Средне- спелый Скороспе- лый
Кремянка Средний Слабореб- ристая » Лимонная Темно-зеленая Розовая Средняя То же
Разновидность сплюснутоплодная — var. complanatus Filov
Миндальная 35 Средний Слабореб- ристая Сплюснутая Оранжевая Светло-корич- невая Лимонная Средняя Средне- ранний
Разновидность голосеменная — var. styriaca Greb.
Белорусская голосемен- ная Средний Слабореб- I ристая Слабоовальная Оранжевая j Отсутствует 1 Светло- 1 оранжевая 1 Средняя 1 Средне- 1 спелый
Разновидность кустовых крупноплодных—var.sorediformis Filov
Грибовская кустовая 189
Днепропетровская
кустовая
Луч
Греческие НО
Грибовские 37
Одесские 52
Длинноплодные
Сотэ 38
Кабачок итальянский
Гибрид немчиновский
(Fi)
Белые 13
Волжская серая 92
Гибрид 72
Хайван-кеды
Бирючекутская 630
Мичуринец 136
ио Дунганская
Средний Гладкая Слабореб- ристая Гладкая Обратнояйцевидная Короткоовальная » Оранжевая > Темно-зеленая Зеленая Темно-зеленая Оранжевая > Средняя > Большая Скороспе- лый » >
Разновидность кабачки — var. giraumons Duch.
Средний » Крупный » Средний Крупный Слабореб- ристая То же > » Гладкая » Цилиндрическая > » > > > Белая > > > Светло- зеленая То же Отсутствует > > Темно-зеленая Отсутствует Слабо-зе- леная » > > > Белая Большая » » » > Скороспе- лый Средне- спелый Скороспе- лый То же > Средне- спелый То же
Р а з н о в и дность патиссоны — var. melopepo (L.) Filov
| Мелкий I Сегменти- | рованная | Тарельчатая | Белая । | Отсутствует j Белая | Большая | Средне- спелый
Вид крупноплодной тыквы — С. maxima Duch.
Разновидность сероплодная---------------var. maxima
Крупный Гладкая Слабосплюснутая Серая Отсутствует Оранжевая Средняя Сред не- спелый
» Слабосег- Короткоовальная » » > » >
Средний Крупный ментиро- вэнная Слабореб- Шаровидная Круглая > Светло- > » Небольшая Средняя > »
ристая серая »
Слабосег- Округлосплюснутая Серая »
ментиро- ванная > 1 »
Гладкая Сплюснутая >
§ . . продолжение
Сорт Внешний вид плода Мякоть плода Срок созревания
размер поверхность форма окраска фона окраска полос рисунка цвет толщина
Разновидность белоплодная — var. alba Roem.
Медовая белая Безенчукская белая Тетри-Гогра Средний > Сегменти- рованная То же » Сплюснутая » » Белая » Отсутствует > Оранжевая > » Средняя Большая Среднеспе- лый То же >
Разновидность зимняя — var, hiberna Filov
Мраморная Испанская Испанская 73 Столовая зимняя А-5 Пластуновская Грибовская зимняя БОС Черкесская Средний » Крупный Средний Сегменти- рованная То же > Слабосег- ментиро- ваиная То же Сегменти- рованная То же Сплюснутая » » » » Шаросплюснутая Сплюснутая Серо- зеленая То же > » > » Серая Отсутствует » » » > 1 * Оранжевая > » > » » » Большая Средняя Большая » Средняя Большая > Поздне- спелый То же > » » *
Кит | Крупный Разновидность китоподобная — var. phuseta Filov I Гладкая | Цилиндрическая | Зеленая | Отсутствует | Кремовая | Средняя | Позднеспе- 1 лый
Разновидность мамонто в а я — var. jaune Duch.
Мамонт Стофунтовая Крупноплодная 1 Очень крупный » Гладкая > Слабосег- ментиро- ванная Шаровидная > Слабосплюснутая Оранжевая > Серая Отсутствует > » Оранжевая » Небольшая! Средне- спелый > 1 > Большая 1 »
Вид мускатной тыквы — С. moschata Duch.
Разновидность перехватки — var. moschata
Кашгарская 1644 Средний Гладкая Цилиндрическая Оранжевая Светло-ко- ричневая Оранжевая Большая Средне- спелый
Палав-каду 268 » Мелкореб- ристая Короткоцилиндриче- ская с перехватом Светло-ко- ричневая Темно-зеленая » » >
Плов-кеды > Слабосег- ментиро- ванная Удлиненно-цилин- дрическая > Коричневая сетка > » »
Ленинабадская » Мелкореб- ристая Цилиндрическая с перехватом Темно- кремовая Оранжевая сетка » » 1 Поздне- спелый
Перехватка 69/19 » Гладкая Цилиндрическая Оранжевая Отсутствует » » 1 >
Перехватка местная > Слабосег- ментиро- ванная Укороченно-цилин- дрическая с перехва- том Коричневая » >
Подвид колумбийский — subsp. columbiana (Zhiten.) Filov
Разновидность продолговатая — var. dolihocarpa Zhiten
Витаминная Средний Слабореб- Овальная Светло- Темно-зеленая Оранжевая Средняя Поздне- спелый
Мускатная местная Крупный ристая Слабосег- ментиро- Яйцевидная зеленая Темно-ко- ричневая Темная сетка » Большая »
ванная
УКАЗАТЕЛЬ РУССКИХ БОТАНИЧЕСКИХ НАЗВАНИЙ АРБУЗА
АРБУЗ
— безусиковый 12, 24, 71, 95
— колоцинт 7, 12, 14, 23, 60, 76, 89, 95
----- безвкусный (пресный) 24, 71, 72
----- колоцинт узколистный (горький) 24, 72, 74
— Нодэна 12, 25, 711, 74, 96
— шерстистый 12, 17
-----обыкновенный 21, 95
----- кордофанский (суданский) 18, 22, 71, 72, 129
-----столовый (обыкновенный) 114, 16, 18, 21, 60, 71, 89, ПО, 113
----- слизистосемянный 22, 56, 71, 72, 95
----- сенегальский 23
-----слизистосемянный 14, 18, 23
----- шерстистый 20, 56, 72, 95
----- капский 21, 71, 72
-----каффрский (шерстистый) 20, 72
-----цитронный (кормовой и цукатный) 16, 21, 46, 60, 71, 89, 90
ФИСТУЛЕЗНЫЙ АРБУЗ (пустотелый) 16, 71, 74, 75, 96
названия ЛРВУЗЛ
А ИА РАСТЕНИИ СЕМЕЙСТВА CUCURBITACEAE
Acanthosicyoinae С. Jeffr. 8
Acanthosicyos Hook. f. 9^ 10, Ц, 15, 25
Actinostemma Griff 6
Anguria Mill. 9
— citrullus Mill. 18
— indica seu batteca Rump. 18
Benncasa Savi Ю, 74, 76, 77
— cerifera Savi 75, 77, 78, 79
— hispida (Thunb.) Cogn. 75
Benincaseae Ser. 8, 10, 11
Benincasinae (Ser.) G. Jeffr 8 9
10, 11 ’ ’ ’
Bryonia L. 6, 9
Citrullus Schrad. 8, 9, 10, 11, 12, 13 15
26, 56, 58, 75, 76, 77, 78, 91, 92, 94, 98*
Citrullus Forsk. 9
Citrullus Necker 9
— aedulis Pang. 16
— amarus Schrad. 16. 21
— ancestralis Gold. 15, 70
— battich Forsk. 17
— caffer Schrad. 16, 20
— caffrorum Schrad. 20
— chodospermus Fal. et Dunal 17
— citrullus Kars. 17
— colocynthis Schrad. 23
— colocynthis (I.) Schrad. 12, 14, 15, 17,
23, 27, 29, 56, 70, 71, 77, 80, 89, 93, 95,
97
------var. capensis Alef. 17, 21
------var. colocynthoides Schweinf. 16
----subsp. colocynthis 17, 24, 27, 29
---- subsp. insipidus (Pang.) Fursa 17,
23, 24, 27, 29
------var. insipidus Pang. 23, 24
------ var. pachirhizus Pang. 23, 24
------var. stenotomus Pang. 23, 24
------ var. vulgaris Pang. 23, 24
— colocynthoides Pang. 16, 21, 70
— ecirrhosus Corn. 12, 13, 16, 17, '24, 25,
27, 29, 56, 71, 74, 76, 77, 79, 80, 91,
93, 95, 97
— edulis Pang. 16
— edulis Spach. 16, 21
— fistulosus Stocks 16
— lanatus (Thunb.) Matsum. et Nakai 9,
12, 14, 15, 16, 17, 18, 20, 26, 28, 93, 97
— lanatus (Thunb.) Mansf. 16
---- subsp. cordophanus Ter-Avan. 22
----subsp. lanatus 17, 18, 20. 26, 28, 56,
72, 77, 89, 93, 97
------var. capensis (Alef.) Fursa 17, 20,
21, 26, 28, 77, 125, 128
------var. caffer (Schrad.) Mansf. 16,
20, 21
16,
20,
17,
80,
23,
~7rnnVaLcitroides (Bailey) Mansf.
17, 20, 26, 28, 72, 81, 125
~— var- lanatus (caffer) 16, 17
26, 28, 56, 77, 125, 128
subsp. mucosospermus Fursa 14
18, 22, 27, 29, 56, 71, 72, 77, 79’
85, 93, 94, 95, 96, 125, 128
—~— var. mucosospermus 17, 22,
------var. senegalicus Fursa 17, 22, 23
27, 29
---- subsp. vulgaris (Schrad.) Fursa 17,
20, 21, 26, 28, 77, 89, 93, 95, 97
------ var. cordophanus (Ter-Avan.)
Fursa 17,21, 22, 26, 28, 56, 77. 128
------var. vulgaris 17, 21, 26, 28, 29,
110
------var. sudanicus Filov et Fursa 22
— naudinianus (Sond.) Hook. f. 10, 12.
13, 15, 16, 17, 25, 27, 29, 56, 71, 74,
77, 78, 79, 96, 97, 98
— pasteca Sageret 17
— vulgaris Schrad. 9, 16, 21
------var. citroides Bailey 16, 21
------var. lanatus Bailey 16, 20
------ var. senegalicus Fursa 23
Colocynthis Mill. 9
— amarissima Schrad. 18
— citrullus Kunze 18
— ecirrhosus Cogn. 24
— naudinianus (Sond.) Kuntze 25
— officinalis Schrad. 23
— officinarum Cogn. 23
— vulgaris Schrad. 23
Cucumis L. 8, 11, 16, 28, 75, 76, 77 , 78,
98
— amarissdmus Schrad. 18
— colocynthis L. 23
— colocynthis Thunb. 18
— dissectus Desne 18
— dissectifolius Naud. 25
— gigantea Salisb. 18
— naudinianus Sond. 25
— pseudocolocynthis Wender. 23
— sativus L. 97, 98
— sauclexii Pail, et Bods 76, 77
Cucurbita L. 7, 8, 10
— anguria Duch. 18
— caffra Schrad. 18
— citrullus L. 18
— pinnatifida Schrank. 18
Cucurbitaceae Juss. 6, 7, 8, 10. IL
79, 92, 98
Cucurbiteae Ser. 8
Cucurbitinae C. Jeffr. 8
273
Cucurbitoideae Endl. 8
Coccinia Wight et Arn. 9
Diplocyclos (Endl.) Post et Kuntze 9, 11.
15, 16
Ecballium A. Rich. 7, 9
Echinocystis Torr, et Gr. 7
Eureiandra Hook. f. 9, 10, 11
Fevilleae Pax 11
Gerrardanthus Harv. Il
Lagenaria Ser. 7, 8, 9, 10
Luffa Adans. 7, 8
Melothrieae Endl. 8
Microlagenaria C. Jeffr. 11
Momordica L. 7
— lanata Thunb. 17
Passiflorales Lindley 6
Praecitrullus Pang. 9, 10, II, 16, 56 7r
78, 92, 94, 97, 98 ’ °’
— fistulosus (Stocks) Pang. 16, 77,
Pseudocitrullus Meeuse 28
Sechium P. Br. 7
Shizopepon Maxim. 6
Sicyos L. 7
Thladiantha Bunge 7, 11, 15
— africana C. Jeffr. 15
Z a no nioide ae C. Jeffr. 8
УКАЗАТЕЛЬ РУССКИХ
Подрод горный 204
» Дикорастущий 205
» североамериканский 177
» тропический 188
» южноамериканский 163
— вонючий 206
------вонючая 206
— галеотти 208
— дланелистный 203
— изящный 203
— келлиана 188
— крупноплодный 163, 225, 232, 271
----американский 1172
------ банановидная 174
------ боливийская 174
------ бразильская 173
------ волчковидная 174
------ кустовая 174
------ перувианская 173
----дикорастущий 177
-----андреана 1177
---- китайский 174
------крупночалмовая 175
------мелкоплодная красная 177
------мелкочалмовая 177
------среднечалмовая 176
---- старосветский 168
---- белоплодная 170, 272
----голландская 1171
----зимняя 171, 272
----китоподобная 1171, 272
----мамонтовая 170, 272
---- мелкоплодная 171
----овальная 1171
----сероплодная 170, 271
— лиановый 202
— лунделля 203
— мартина 202
— мура 204
— мускатный 189, 212, 225, 273
----индийский 197
------ овальноплодная 198
----- шароплодная 198
----колумбийский 200, 273
------ овальная 200
----- перцевидная 200
----- продолговатая 200, 273
------шаровидиосплюснутая 200
----мексиканский 1199
------бутылковидная 200
------грушеплодная 200
-----сплюснуто-бородавчатая 199
-----цилиндрическая 200
-----шаровидно-овальная 199
----североамериканский 194
----- канадская 196
БОТАНИЧЕСКИХ названий тыквы
----- семинольская 196
----- сплюснутая 196
----- шаровидная 195
эллипсовидная 1195
--туркестанский 193
-----грушевидная 194
----- китайская 194
-----кувшиновидная 194
----- неаполитанская 194
----- палицевидная 193
--- перехватка 193, 273
-- японский 1196
----- канталупообразная 197
----- карликовая 197
-----мозговая катушечная 197
----- мозговая плоская 197
— пальмовидный 208
----- пальмовидная 208
-----калифорнийская 208
— — пальмолистный 208
— пальчатолистный 207
— серцевидный 206
— серебросемянный 200
----- грушевидная 201
----- кувшиновидная 202
----- округлая 201
— твердокорый 178, 211, 225, 270
--декоративный 185
-----белошаровидная 186
----- бородавчатая 186
----- грушевидная 186
-----двуцветная 186
----- мандариновая 186
-----цилиндрическая 1186
----- яйцевидная 186
--дикорастущий 186
----- братская 188
----- сестринская 187
----- тексанская 187
----- длинноплетистый 181
-----голосемянная 1182, 270
-----овальная 181, 270
-----овощная 182
----- сегментированная 183
сплюснутоплодная 183, 270
_---- цилиндрическая 182
шароплодная 183
--кустовой 1183
----- кабачки 183, 250, 271
_____кривошейки или крукнек 18s
_____ крупноплодная 185, 271
_____патиссоны 184, 250, 271
_____фордгукская 1185
— укореняющийся 209
— фиголистный 204
— цилиндрический 207
— эквадорский 177
275
УКАЗАТЕЛЬ ЛАТИНСКИХ БОТАНИЧЕСКИХ НАЗВАНИИ ТЫКВЫ
Cucurbita L. 145, 147, 151, 154, 161, 162,
183
Cucurbita 159, 161, 162, 177
Cyclofilia Filov 160, 161, 162, 163
Eremosicyos Filov 159, 162
Montana Fmov 159, 161, 162, 204
Scandens Filov 159, 163, 188
Cucurbita andreana Naud 150, 154, 155,
168, 177
— agrirosperma Bailey 200
— asteroides Schrad. 178
— californica Torr, et Wats 154, 155
— citrullus Roem 154, 178
— cordata Wats. 154, 155, 158, 206
— cylindrata Bailey 154, 155, 157, 158,
159, 207
— digitata A. Gray 153, 155, 157, 158,
159, 160, 207, 225
— durantia Willd. 178
— elongata Schrad. 178
— eguadorensis Culter et Whitaker 155,
158, 160, 163, 177
— esculenta A. Gray 178
— ficifolia Bouche 150, 151, 153, 154,
157, 158, 159, 161, 204
— foetidissima H. В. K. 149, 152 153,
154, 155, 157, 158, 159, 161, 205
------ foetidissima 206, 225
------ scabridifolia (Bailey) Filov 206
— fraterna Bailey 154, 187
— galeotti Cogn. 154, 208
Cucurbita gracilior Bailey 154, 158. 189,
203
— lundeliana Bailey 151, 154 155 157
158, 159, 160, 188, 203, 234’, 240,’ 241,’
257
— kellyana Bailey 155, 159, 177, 188
— macrocarpa Gasp. 1'53
— martinesii Bailey 151, 154, 155, 157
158, 159, 189, 202
— maxima Duch. 146, 149, 150, 151, 153,
154, 155, 157, 158, 150, 161, 163, 166,
167, 178, 216, 217, 228, 232, 236, 238,
239, 240, 241, 243, 248, 251, 255, 262
------ americana Filov 168
------ boliviana Zhit. 173
------bresil Naud. 173
------chiloensis Zhit. 173
------odorifera Veil. 173, 174
------ ohio Naud. 173, 174
------ peruviana Zhit. 174
------ zapollitto (Carr.) Millan. 173, 174
------andreana (Naud.) Filov 168, 177
------ andreana 155, 158, 177, 240
------maxima 168
------ alba Roem. 170, 272
hiberna Filov. 170, 171, 272
------hollandiae Naud. 170, 17'1
------ jaune Duch. 170, 272
------ maxima 170, 271
------ovalis Roem. 170, 171
------petit Duch. 170, 171
------phuseta Filov 170, 171, 272
------turbanikurbis Metzg. 168, 174
------constricta Alef. 175, Г77
------ nouvebresil Naud. 175, 176
------ tyrbancurbis 175
------ turbanopetit Naud. 175, 177
Cucurbita melanosperma Casp. 153, 154,
204
— melopepo L. 153, 178
— mixta Pang. 154, 156, 157, 158, 159,
188, 189, 200
------cuschaus Castet. 201, 202
------mixta 201
------ pyridiforme Castet. 201
— moorei Bailey 155, 159, 161, 204
— moschata Duch. ex Poir. 149, 150, 153,
154, 155, 156, 157, 158, 159, 161, 166,
188, 216, 221, 225, 228, 232, 234, 238,
239, 240, 241, 242. 243, 257, 262
------ borealiamericanica 193, 194
------cheese Castet. 195
------kanadiensis Filov 195, 196
------ ovalis Filov 195
------rapaeformis Filov 195, 196
------seminolis Filov 195, 196
------columbiana (Zhit.) Filov 193 200,
273
------abbreviate Filov 200
------columbiana Й00
------ dolihocarpa Zhit. 200
------ prismaticus Zhit. 200
------ indica (Zhit.) Filov 192, 193, 197
------ grande Naud. 197, 198
------indica 197, 198
------ macrocomplanata Filov 197, 198
------japonica (Zhit.) Filov 192, 193, 196
------depressa Naud. 197
------ japonica 197
------pygmala Filov. 197
------ trochlearis Filov 197
------mexicana (Zhit.) Filov 193, 199
------applanata Zhit. 199
------complanatoverrucosus Zhit. 199
------cylindraceae Zhit. 199, 200
------ mexicana 199, 200
------pyridimis Zhit. 199, 200
------teres Zhit. 199, 200
276
------testicularis Zhit. 199
------moschata 193
------claviformis Naud. 193, 194
------cuchaus Castet. 193, 194
------longicarpa Filov 193
------macrocarpa (Gasp.) Naud. 193
194
------pyriformis Castet. 193, 194
------moschata 193, 273
Cucurbita okeechobiensis (Small) Bailey
154, 155, 157, 158, 159, 160, 188
— ovifera L. 149, 154, 178, 179
— palmata Wats. 154, 155, 157, 158, 159,
208, 225
------californica Torrey etWats 155,208
— palmeri Bailey 155, 158, 208
— pedatifolia Bailey 154, 158, 188, 203
Cucurbita pepo L. 148, 149, 150, 151, 152,
153, 154, 155, 157, 158, 159, 161, 166,
177, 178, 179, 216, 221, 225, 228
------ brevicaulis Greb. 181, 183
------ fordhuk Castet. 183, 185
------giraumons Duch. 183, 271
------melopepo (L.) Filov 183, 184, 271
------ sorediformis Filov 183, 185
------subverrucosa Willd. 183, 185
------ pepo 181
------complanatus Filov 181, 270
------courgeron Naud. 181, 183
------cylindrica Filov. 181, 182
------maroc Naud. 181, 183
pepo 180, 181, 270
------styriaca Greb. 181, 182, 270
------tabbe (Castet.) Filov 181, 182
------ polymorpha (Duch.) Filov 181, 185
------aurantica (Willd.) Alef. 186
------bicolor Naud. 186
------cougourdette Duch. 186
------deprimee Ser. 186
------oblonge (Ser.) Filov 186
------ polymorpha 186
------verrucosa (L.) Duch. 186
------ texana Filov 155, 181, 186
------ fraterna (Bailey) Filov 155, 187,
188»
------sororia (Bailey) Filov 155, 187
----texana 155, 158, 187
Cucurbita pomiformis Roem^ 178
— radicaus Naud. 154, 155, 159, 209
— sororia Bailey 154, 187
— texana A. Gray 149, 150, 153, 154
156, 157, 178, 186, 187
— turbaniformis (Roem.) Zhit. 156, 162,
163, 174
— urnigera Schrad. 154, 178
— verrucosa L. 152, 178, 179
Cucumis perennis James. 206
Melopepo compressus Roem. 151, 166, 184
Pepo citrullus Sager. 178
— foetidissima Britt 205
— maximus Peterm. 151, 163
— okeechobiensis Small. 154, 202
Pyleocalix Casp. 145, 153, 174
Tristemon texanum Scheele 145., 178, 187
СОДЕРЖАНИЕ
Предисловие .............................................................
Краткая характеристика семейства Cucurbitaceae Juss......................
Арбуз Citrullus Schrad...................................................
Обзор рода ............................................................
Эволюция, распространение и экология ..................................
Классификация рода Citrullus Schrad...................................
История изучения ....................................................
Новая классификация .................................................
Описание видов и внутривидовых таксонов .............................
Сравнительная характеристика видов...................................
Эколого-географические группы столового арбуза и экологическая изменчи-
вость признаков ......................................................
Эколого-географические группы .......................................
Экологическая обусловленность признаков .............................
История введения арбуза в культуру .................................
Народнохозяйственное значение ........................................
Распространение арбуза ...............................................
Биологическая характеристика арбуза ..................................
Требования к условиям среды..........................................
Биология развития ...................................................
Биология цветения и опыления ........................................
Анатомические особенности органов арбуза .............................
Кожура семени .......................................................
Проростки ...........................................................
Лист ................................................................
Плод ................................................................
Химический состав плодов и семян арбуза .....................
Биохимические особенности плода .....................................
Химический состав семян..............................................
Влияние географических и экологических факторов на химический состав
плодов ..............................................................
Динамика содержания сахаров при созревании и хранении плодов ....
Химический состав плодов разных видов арбуза ........................
Содержание горьких веществ в растении арбуза........................
Генетика и цитология .................................................
Кариотип и методы его изучения.......................................
Морфология хромосом разных таксонов .................................
Полиплоидия ........................................................•
Наследование признаков ..............................................
Комбинационная способность сортов арбуза .............................
Изменчивость признаков арбуза ........................................
Схема сортовой изменчивости столового арбуза (С. lanatus subsp. vulgaris
var. vulgaris) ......................................................
Географическая изменчивость сортов ..................................
Сортотипы столового арбуза ...........................................
Методы и достижения селекции арбуза...................................
Задачи селекции .....................................................
Методы селекции......................................................
Достижения селекции .................................................
Болезни арбуза и источники устойчивости к ним ........................
Грибные, бактериальные и вирусные болезни ..........................
Исходный материал для селекции на устойчивость к болезням...........
Список литературы ......................................................
Приложение .............................................................
Тыква — Cucurbita ......................................................
5
6
9
9
9
15
15
17
17
29
29
30
35
42
47
48
52
52
55
58
69
69
76
79
80
83
83
85
86
88
89
91
91
91
95
98
101
104
109
ПО
111
113
118
120
120
123
128
140
145
278
--------- C. Okeechobeensis (Small) Bailey
мартена —C. martinezii Bailey y
дланелистный — C. pedatifolia Bailey ’ ’
лунделля — C. lundeUiana Bailey ‘
изящный — C. gracilior Bailey ..............
сердцевидный — C. cordata Wats . .
цилиндрический —С. cylindrata Bailey
пальчатолистный — С. digitata A. Gray
пальмовидный — С. palmata Wats . .
пальмолистный — С. palmeri Bailey
галеотти — С. galeotti Cogn. in DC .
укореняющийся — C. radicans Naud. .
Обзор рода
История изучения рода
Эволюция рода
Классификация рода Cucurbita
Подрод южноамериканский - suogen ’ cvciofilb ' Fiu ’ ' ' '
Вид крупноплодный-С. maxima Duch Fllow ’
Вид эквадорский-С. eguadorensis Cutter et Whitabr’ ’ ‘
Подрод североамериканский - subgen cucurbit, '
Вид твердокорый — С. реро L 8 cucurtnta
Вид келлиана—С. kellyana Bailey......................
Подрод тропический — subgen. Scandens' Filov.........
Вид мускатный —С. moschata Duch ex Poir............
Вид серебросемянный — С. mixta Pang ...............
Вид лиановый-С. okeechobeensis (Smain '
Вид
Вид
Вид
Вид в __________j
Подрод горный — subgen. Montana Filov........
Вид фиголистный — С. ficifolia Bouche
Вид мура — С. moorei Bailey...........
Подрод дикорастущий —subgen. eremosicyos Filov
Вид вонючий —С. foetidissima Н. В. К.
Вид
Вид
Вид
Вид
Вид
Вид
Вид _ .
История и значение культуры тыквы ...................
Биология роста и развития тыкв ......................
Отношение к условиям среды.........................
Биология роста и развития ........................
Водный режим ......................................
Фотосинтез .......................................
Биология цветения и оплодотворения................
Период вегетации .................................
Вегетативное размножение и прививки...............
Анатомическое строение органов ....................
Биохимический состав плодов и семян тыквы..........
Генетика тыквы ....................................
Хромосомы ........................................
Закономерности наследования признаков ............
Межвидовые скрещивания ...........................
История скрещивания видов тыквы...................
Методы преодоления нескрещиваемости и искусственная культура зародышей
Поведение межвидовых гибридов
Гетерозис .........................................................
Мутации и полиплоиды...............................................
Изменчивость признаков
Сортовые признаки тыквы ...........................................
Сортовые признаки кабачков ........................................
Сортовые признаки патиссонов ......................................
Селекция тыквы .................................................
Селекция на хозяйственно-ценные признаки.......................
Новый исходный материал и методы селекции......................
Болезни и вредители ................................... ’ * ‘......
Болезни .......................................... ................
Вредители ............................................. ...........
Список литературы ..........
Приложение ..........
Указатели....................
145
151
157
162
163
163
177
177
178
188
188
189
200
202
202
203
203
203
204
204
204
205
205
206
207
207
208
208
208
209
209
210
210
212
219
222
222
225
225
226
229
231
231
232
238
238
241
243
244
247
248
248
250
250
250
251
257
258
258
260
261
268
272
Татьяна Борисовна Фурса, Александр Иванович Филов
КУЛЬТУРНАЯ ФЛОРА СССР, ТОМ XXI
ТЫКВЕННЫЕ (АРБУЗ, ТЫКВА)
Зав. редакцией А. Я. Рогачева
Редакторы: Л. М. Нефедова, Т. А. Ищенко
Художественный редактор Е. Г. Прибегина
Технический редактор В. Ю. Осипов
Корректор Д. Е. Ткачева
ИБ № 2881
Сдано в набор 3L03.82. Подписано к печати 15.10.82. Т-18365. Формат 70X108'/^. Бумага тип. М 2.
Гарнитура литературная. Печать высокая. Усл. печ. л. 24,5. Усл. кр.-отт. 24,5. Уч. изд. л. 26,04.
Изд. М 11. Тираж 2000 акз. Заказ М 2323. Цена 2 р.50 к..
Ордена Трудового Красного Знамени издательство «Колос»,
107807, ГСП, Москва, Б-53, ул. Садовая-Спасская, 18.
Отпечатано в тип. № 2г Ленуприздата. 191104, Ленинград, Литейный пр., 55,
с набора типографии ВИР, г. Павловск