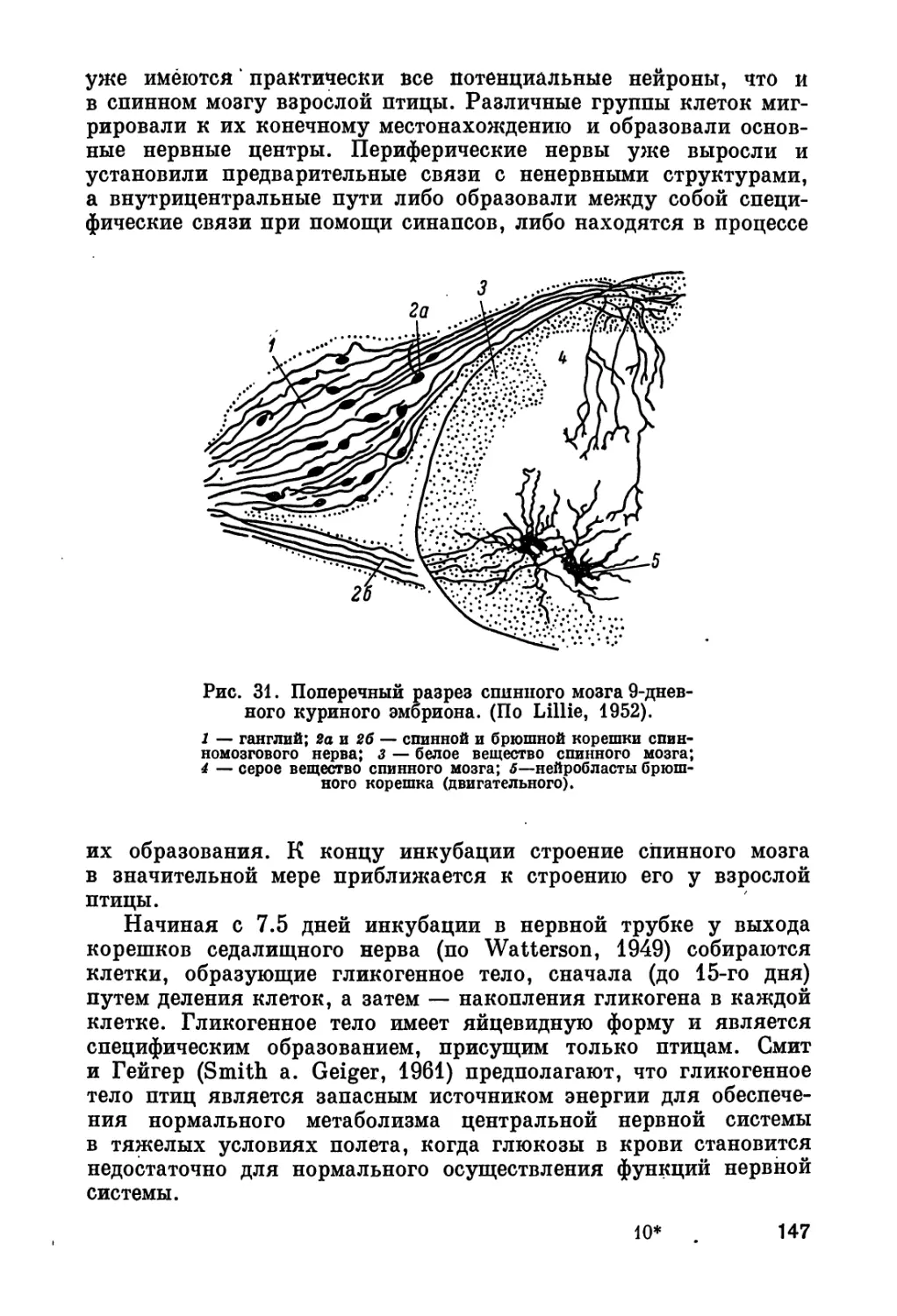
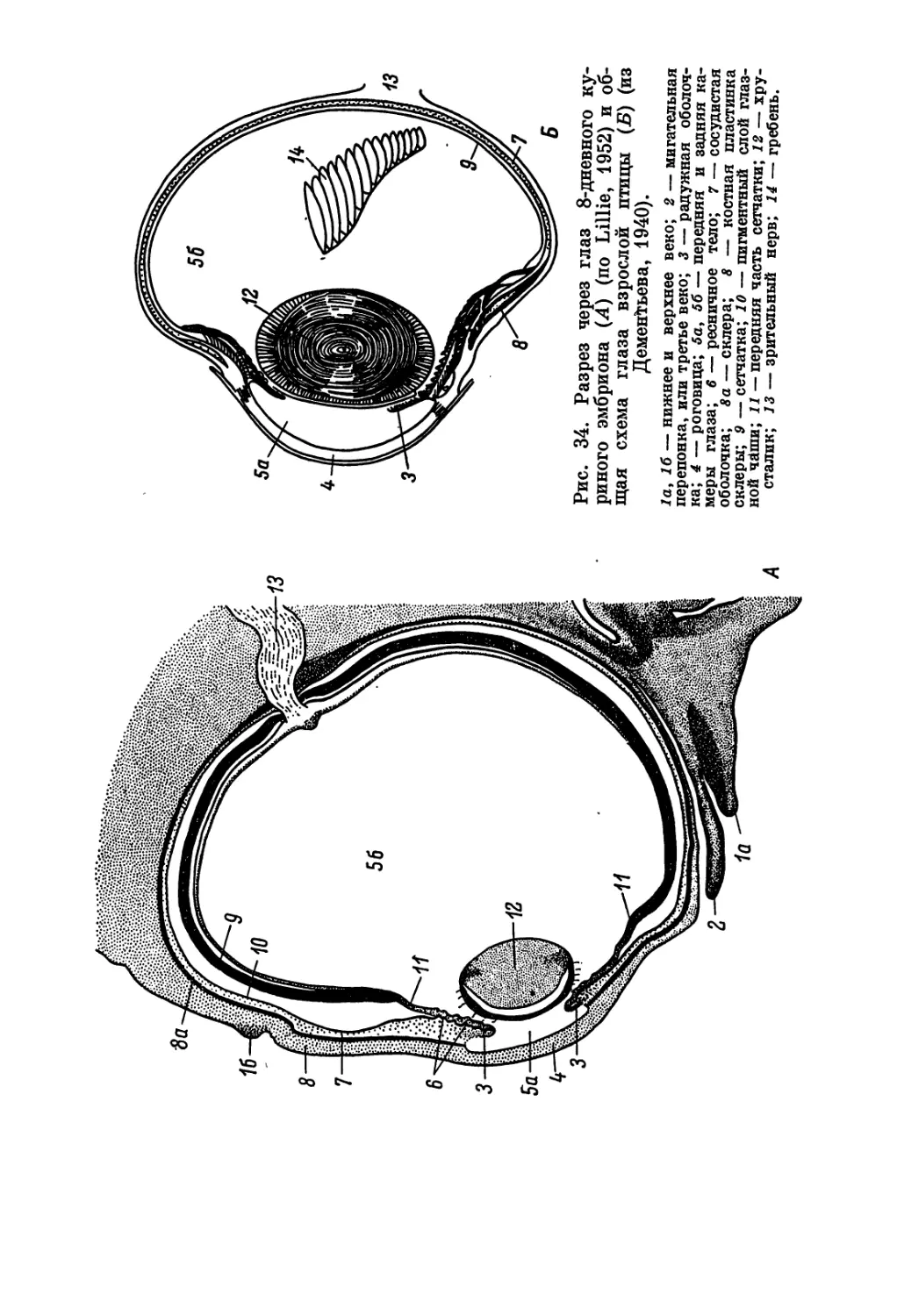
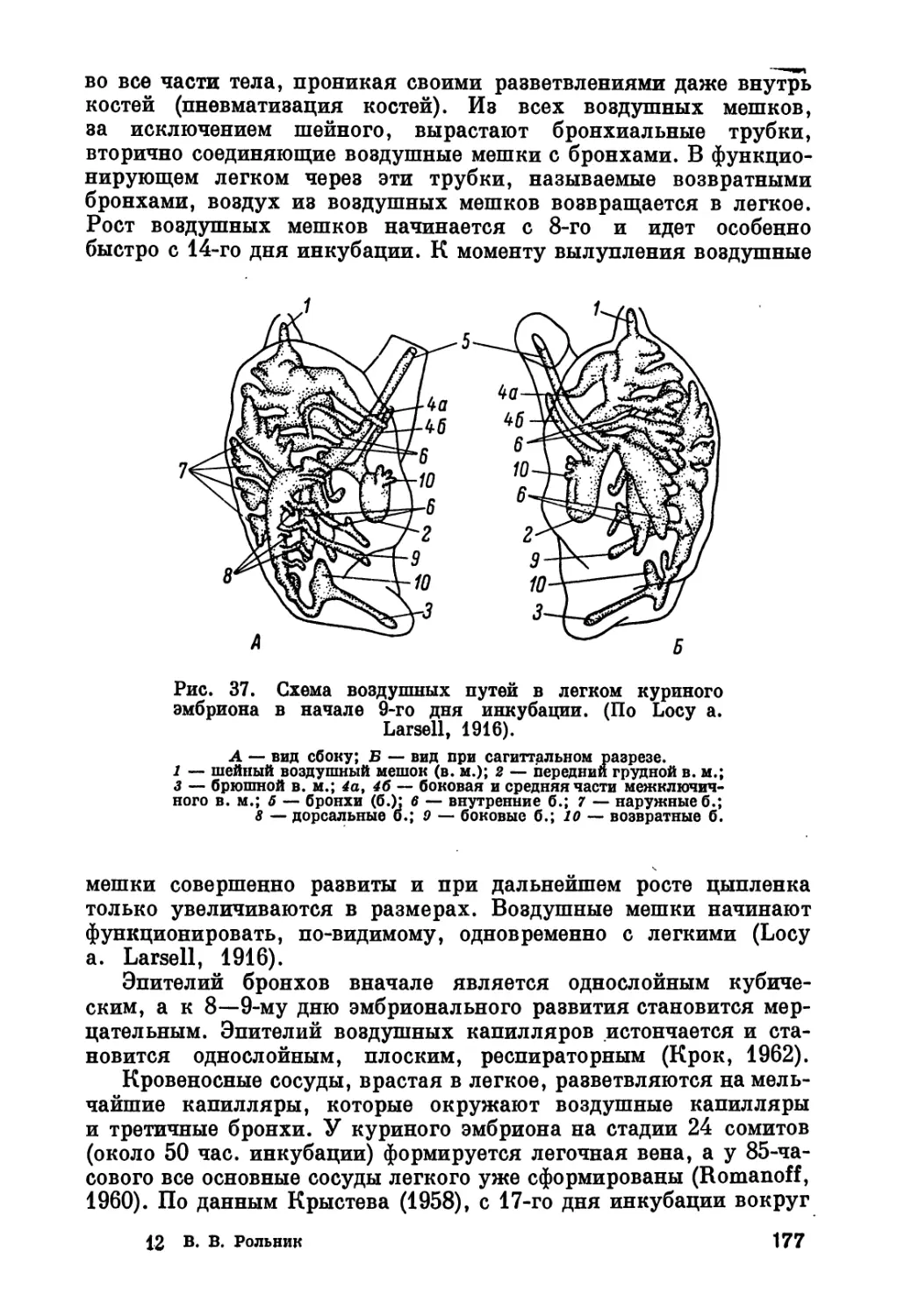
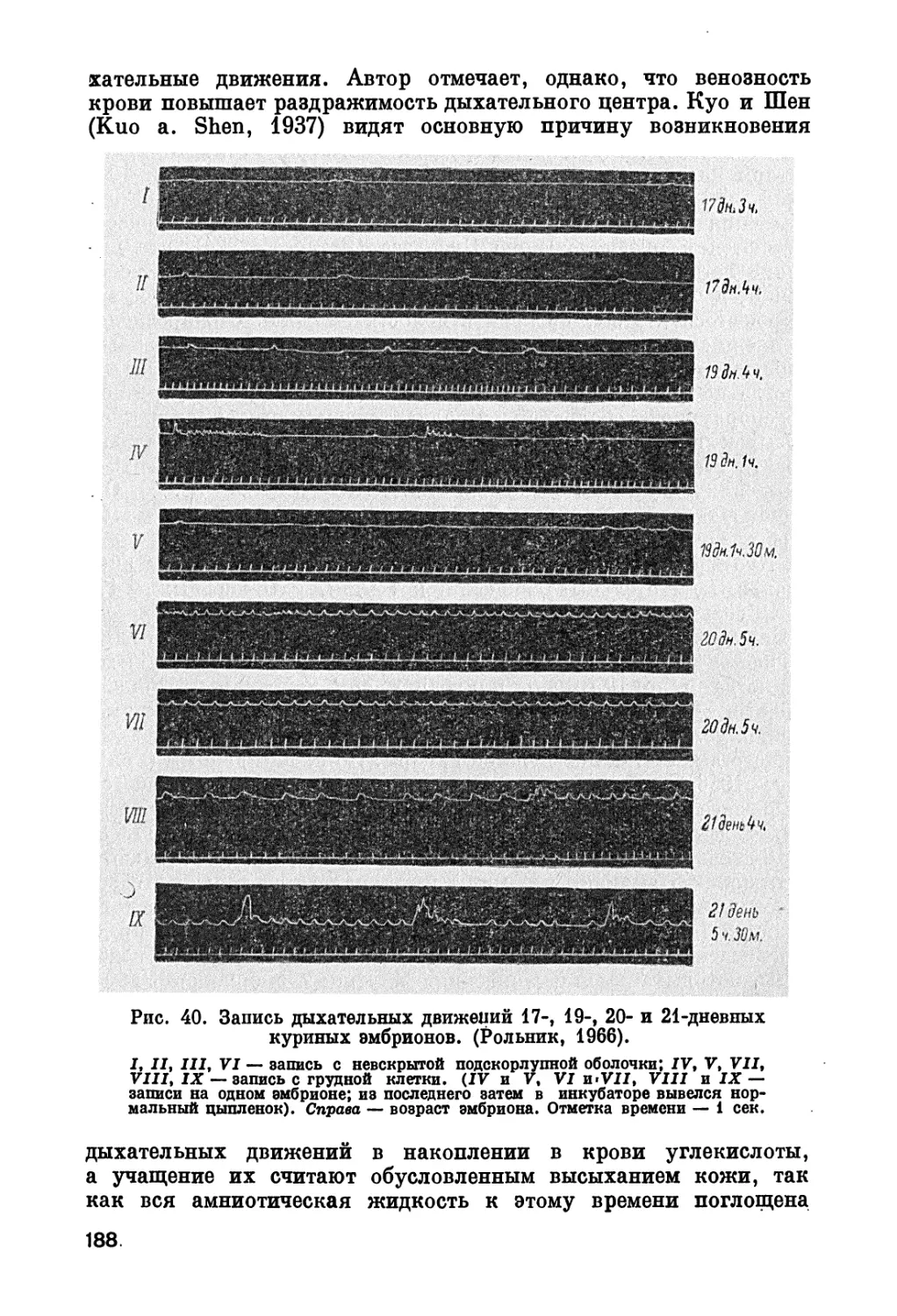
/








































































































































































































































































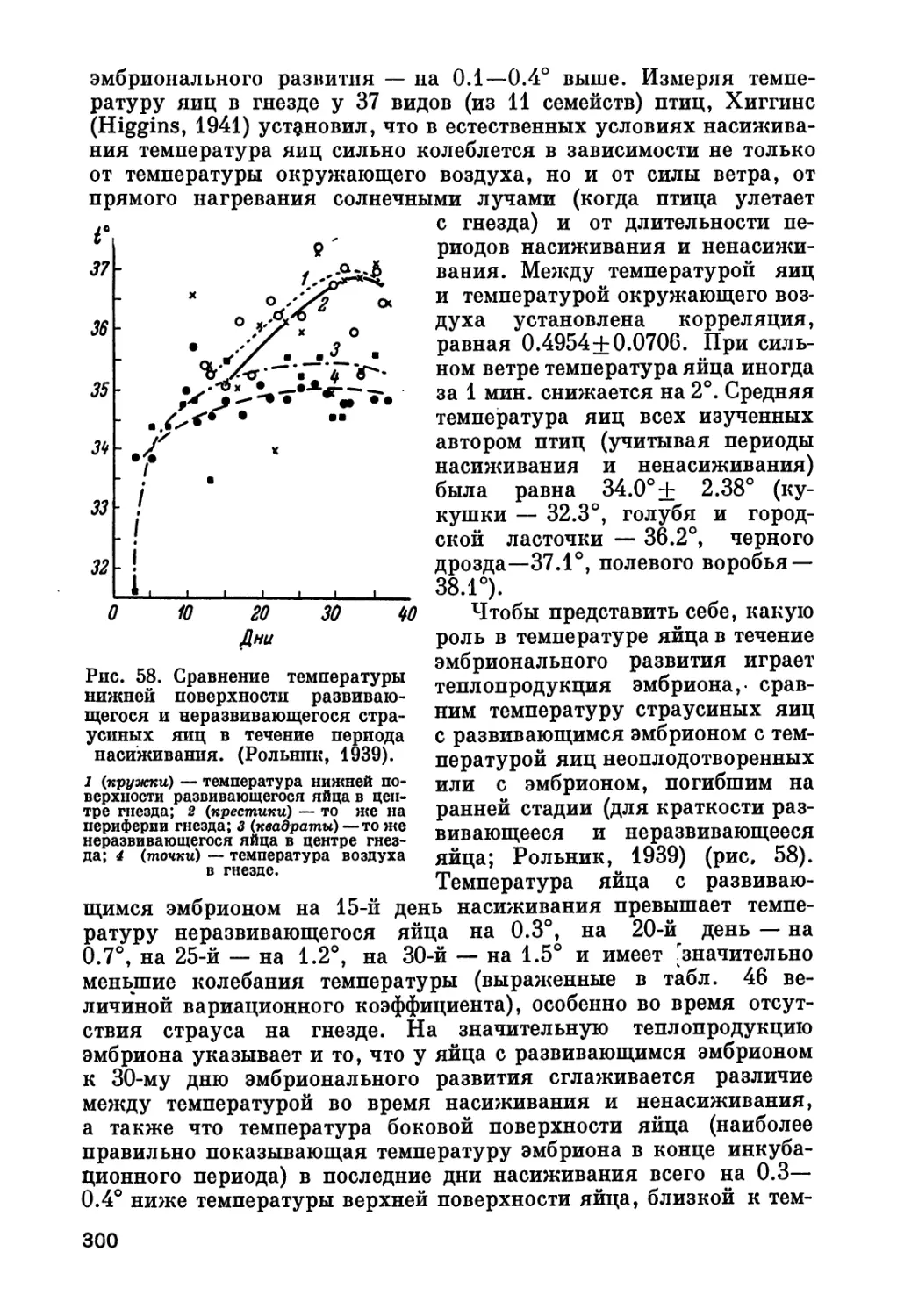




























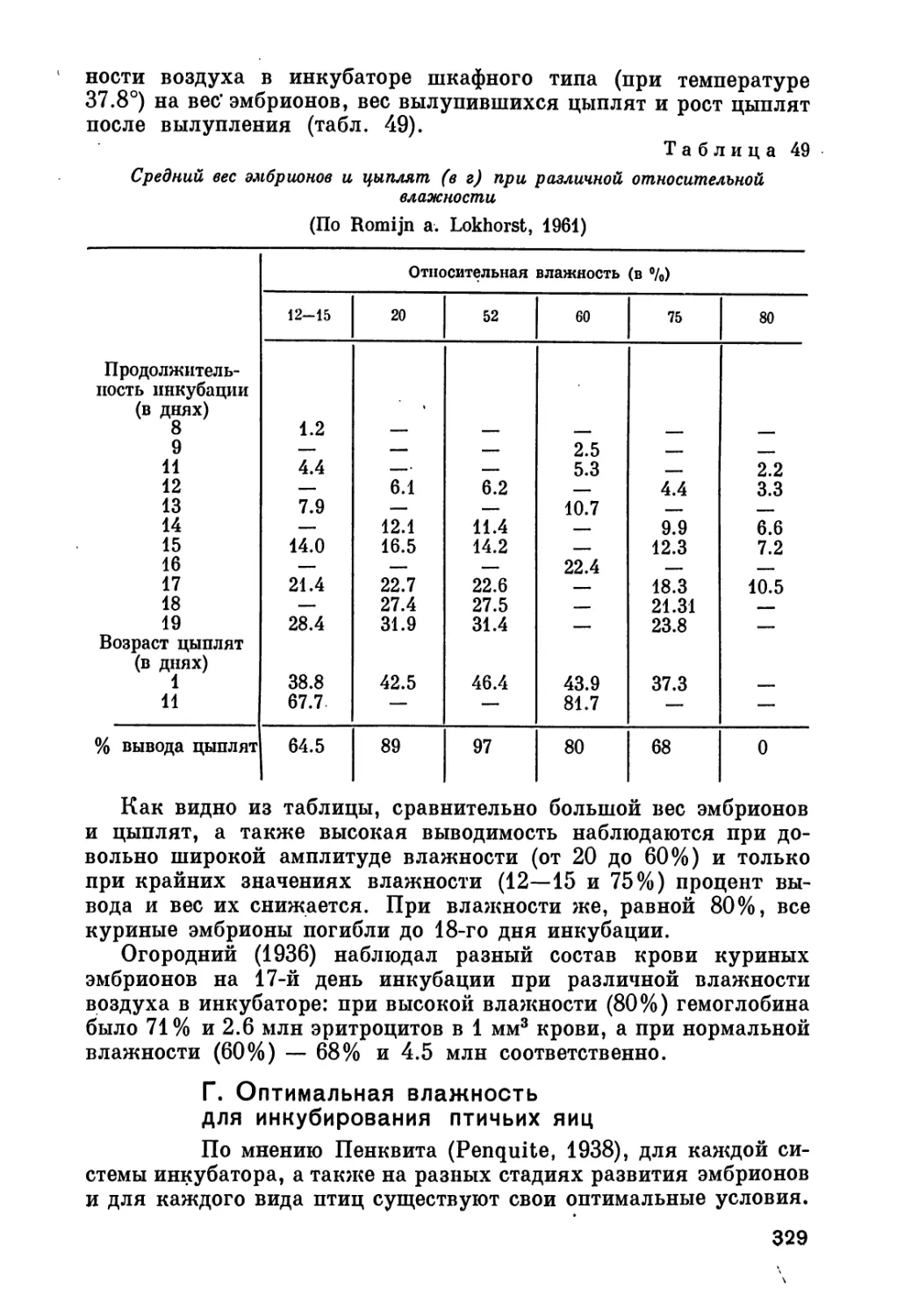
































































































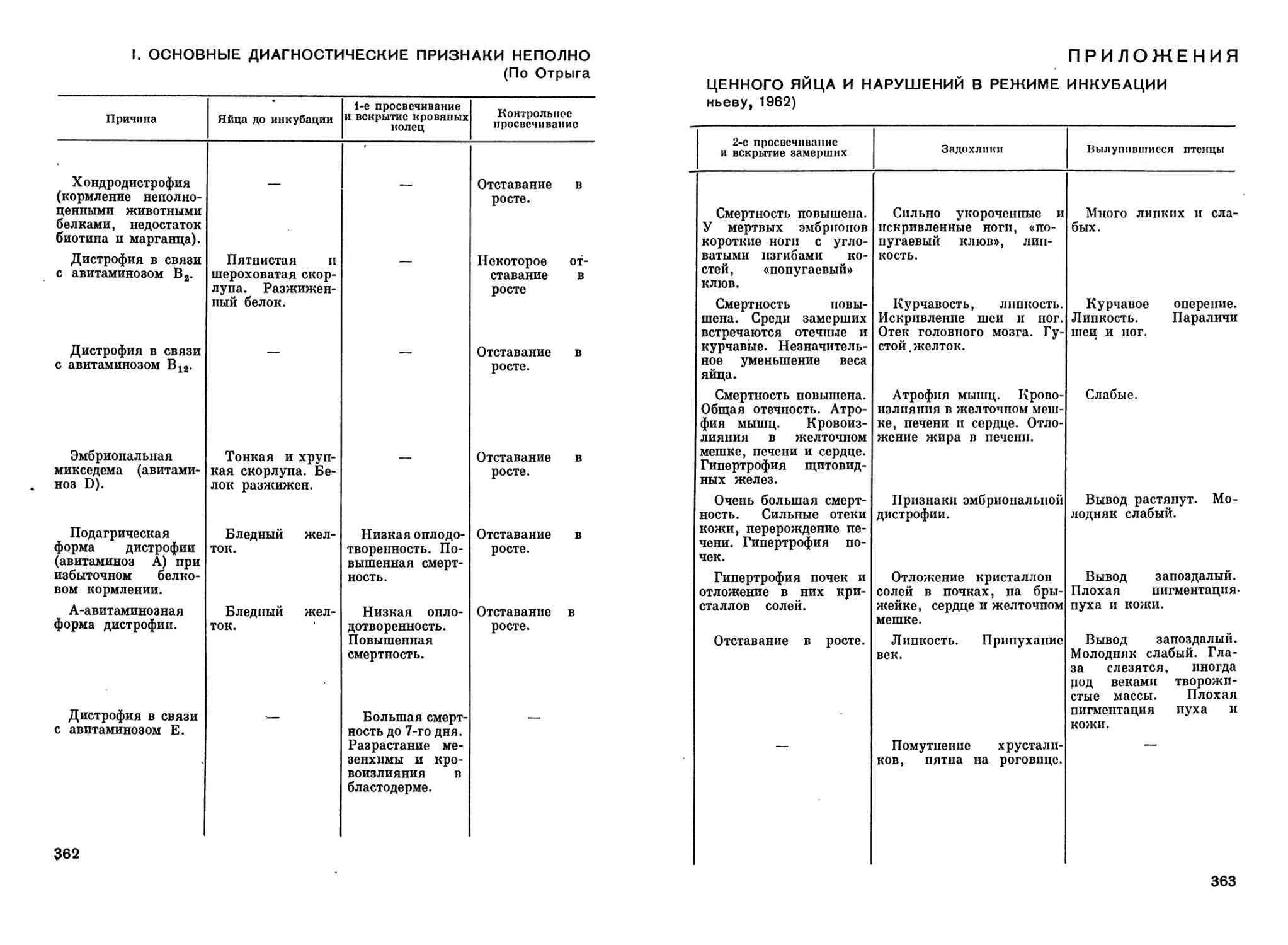
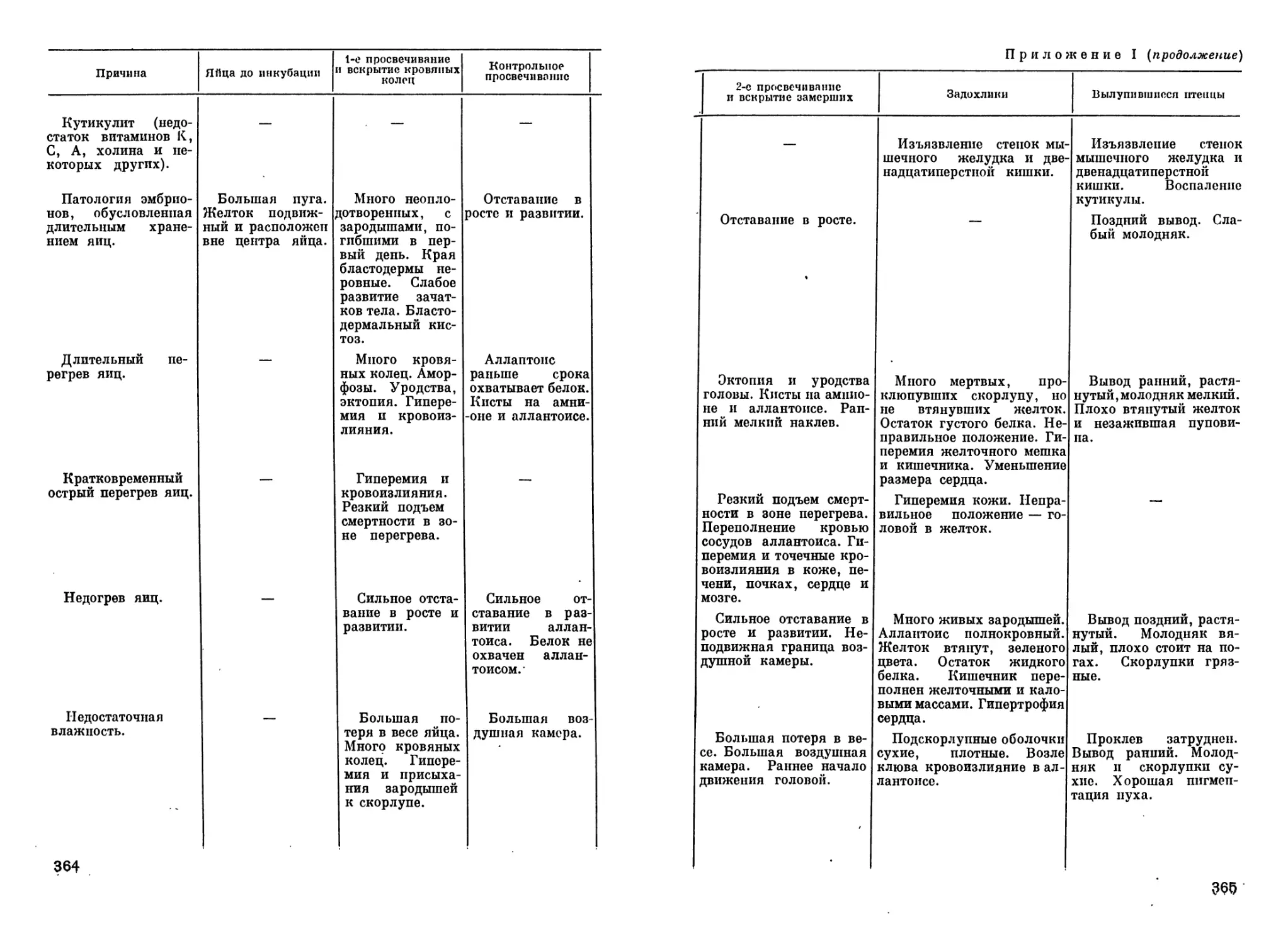
Author: Рольник В.В.
Tags: эмбриология животных онтогенез развитие отдельных организмов биология зоология птицы
Year: 1968




Text
В.В.РОЛЬНИ к
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ ЭВОЛЮЦИОННОЙ ФИЗИОЛОГИИ И БИОХИМИИ
им. И. М. СЕЧЕНОВА
В.В.РОЛЬН и к
БИОЛОГИЯ
ЭМБРИОНАЛЬНОГО
РАЗВИТИЯ
ПТИЦ
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
Ленинград 1 9 в 8
УДК 591.3 : 598.21.9 (92)
Биология эмбрионального развития птиц. Рольни к В. В. 1968. Изд-во
«Наука», Ленингр. отд., Л. 1—425.
Книга является монографической сводкой, учебным и справочным посо
бием по яйцеведению и биологии эмбрионального развития птиц: морфоло
гии, физиологии и обмена веществ, а также по вопросу о необходимых для
нормального развития эмбрионов условий среды (условия инкубации). Книга
рассчитана на широкий круг читателей — на научных работников и сту-
дентов университетов, сельскохозяйственных, ветеринарных и педагогиче-
ских институтов, на специалистов в области птицеводства. Илл. — 58,
табл. — 54, библ. — 1066 назв.
Ответственный редактор
Л. Г. Л е й б с о н
Вера Владимировна Рольник
БИОЛОГИЯ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ ПТИЦ
Утверждено к печати Институтом эволюционной физиологии и биохимии
им. И. М. Сеченова Академии наук СССР
Редактор издательства М. И. Голъданская. Технический редактор Я. Ф. Виноградова
Корректоры Л. Б. Данилова и С. Я. Овчарова
Сдано в набор 29 II 1968 г. Подписано к печати
2/VII 1968 г. РИСО АН СССР № 16-74В. Формат
бумаги 60х90»/1в. Бум. л. 13х/4. Печ. л. 26*/3 =
= 26.5 усл. печ. л. Уч.-изд. л. 34.20. Изд. № 3545.
Тип. зак. 915. М-23020. Тираж 2100.
Бумага типографская Ks 1. Цена 2 р. 59 к.
Ленинградское отделений издательства «Наука»
Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. издательства «Наука». Ленинград, В-34,
9 линия, д. 12
4-7-7
477-68 (I пол.)
Светлой памяти
Леона Абгаровича Орбели —
большого ученого и человека
ПРЕДИСЛОВИЕ
Эмбриональное развитие птиц в его различных аспек-
тах представляет большой интерес для многих исследователей-
биологов. Птичий, а особенно куриный, эмбрион является
излюбленным объектом изучения морфологов, физиологов и биохи-
миков, а также медиков, птицеводов и специалистов других при-
кладных областей знания. Нельзя забывать также и то, что для
правильного ведения инкубации — индустриальной основы пти-
цеводства — познание биологии развития птичьих эмбрионов
совершенно необходимо. >
Настоящая книга является монографической сводкой по ос-
новным вопросам биологии эмбрионального развития птиц: обра-
зованию и строению птичьего яйца, морфологии и физиологии раз-
вития эмбриона, а также данных об условиях среды, необходимых
для нормального развития. Несмотря на то что эмбриональное
развитие птиц было предметом исследования в течение более чем
двухсот лет, данных о физиологии развития птичьего эмбриона
относительно мало. Эмбриологи чаще всего изучали морфологию
развития, физиологи и биохимики лишь в последние десятилетия
стали заниматься онтогенезом, а специалисты по инкубации- не-
редко решали практические задачи эмпирически. Ни в СССР, ни
за границей нет сводок, объединяющих разрозненные сведения по
эмбриональному развитию птиц во всех этих областях знаний.
В имеющихся сводках по развитию эмбрионов птиц, в основном
цыпленка (Patten, 1951; Lillie, 1952; Romanoff, 1960; Рагозина,
1961, Крок, 1962, и др.), более или менее подробно изложены дан-
ные по морфологии развития различных систем органов, а также
изменений внешнего вида эмбрионов на разных стадиях развития.
Монографии Ландауэра (Landauer, 1948) и под редакцией Тэй-
лора (Taylor, 1949), а также главы по биологии развития эмбрио-
нов сельскохозяйственных птиц в учебниках по инкубации напи-
саны слишком популярно. По физиологии развития птичьего
эмбриона вообще отсутствует сколько-нибудь полная сводка дан-
ных, разбросанных в многочисленных журнальных статьях, за
1*
3
исключением уже устаревшей книги Прейера (Ргеуег, 1885).
Предлагаемая читателю книга представляет собою попытку вос-
полнить этот пробел.
Л. А. Орбели считал, что эмбриофизиологии предстоит стать
ключом к наиболее запутанным вопросам физиологии. У нас
в стране интерес к эмбриофизиологии сильно возрос с развитием
эволюционной физиологии. Автор ставит своей целью излагать
материал на основе эволюционных принципов.
Рассматривая в основном биологию развития эмбрионов сель-
скохозяйственных птиц, автор для полноты картины приводит
свои и литературные данные по онтогенезу некоторых других
видов птиц.
Объем знаний по сильно обособившимся отраслям биологии
эмбрионального развития птиц (морфологии, физиологии, инку-
бации и т. п.) настолько велик, что невозможно подготовить книгу
не слишком большого объема, охватывающую все эти отрасли
с одинаковой полнотой. Так как своей основной задачей мы счи-
тали необходимость дать сводку по физиологии развития, есте-
ственно, что она дана более полно, чем морфология. Далее, по-
нятно, что наиболее близкие нашим интересам главы («Дыхание»,
«Температура») включают более полный обзор сведений, чем
Другие.
Излюбленной проблематике теоретической эмбриологии —
начальным стадиям развития эмбриона (деление клеток, образова-
ние зародышевых листков, индукция и т. п.) мы уделяем мини-
мальное место в этой книге, так как эти сведения могут быть легко
найдены в учебниках по эмбриологии (Иванов, 1937, 1945; Шмидт,
1951, и др.). За недостатком места история каждого вопроса изла-
гается очень сжато и основное внимание уделено исследованиям
последних десятилетий. В связи с большим значением, которое
приобретает в последнее время методика исследования для оценки
достоверности полученных результатов, автор упоминает о ней,
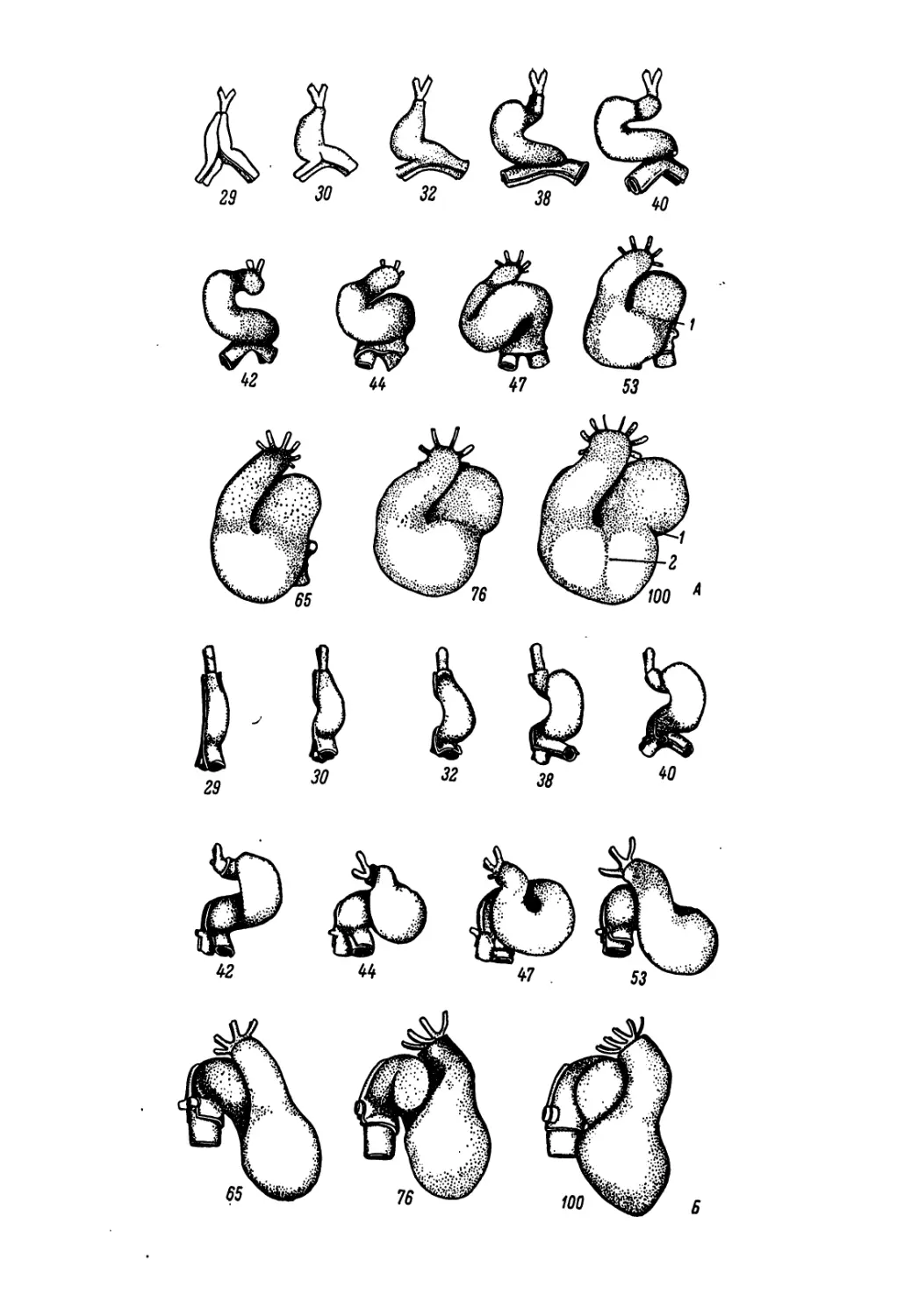
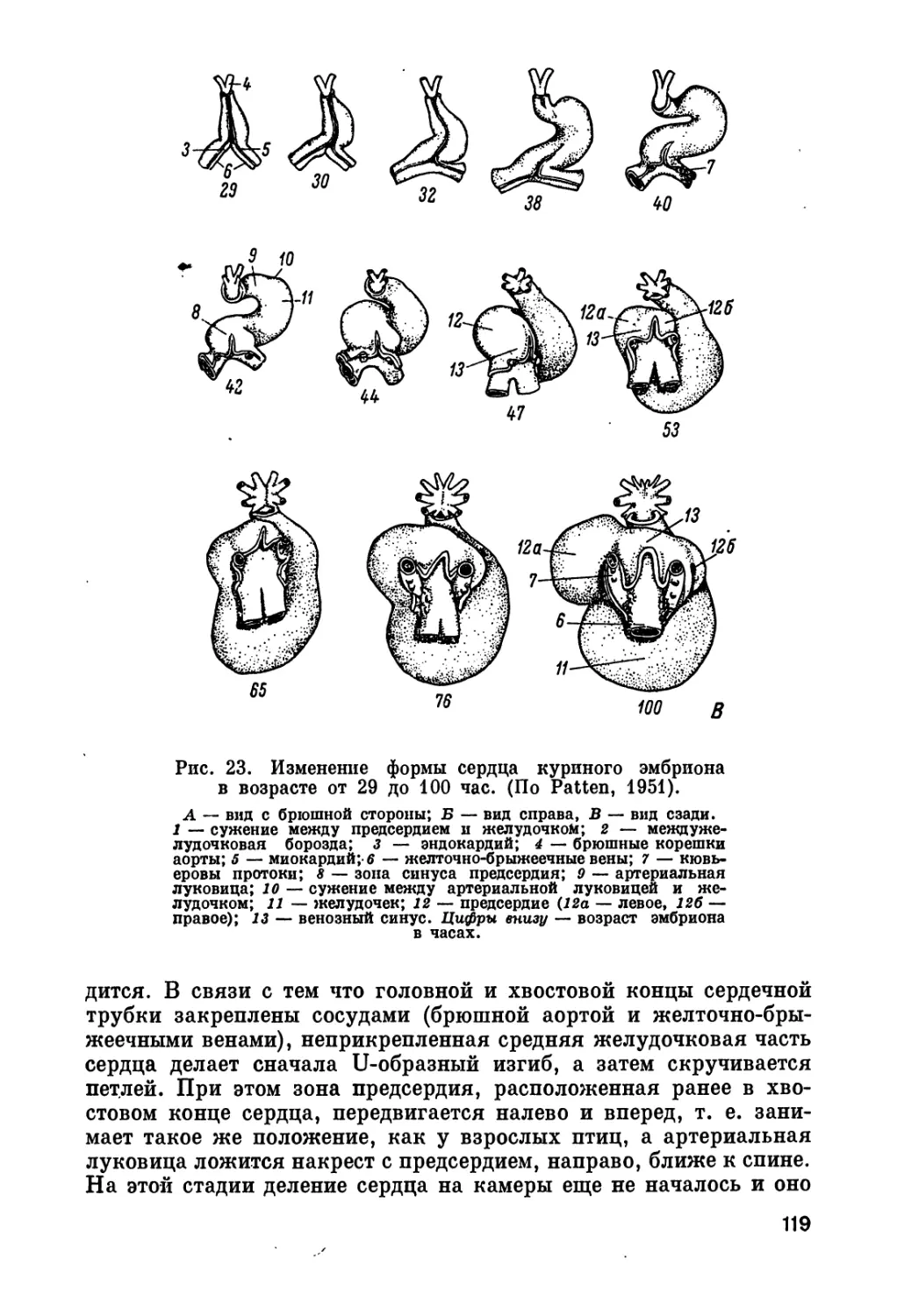
где возможно, хотя бы кратко, чтобы не слишком увеличить объем
книги.
Несколько слов следует сказать о распределении материала
по главам и порядке изложения, которые подчинены в основном
значению данной системы органов в эмбриональном развитии.
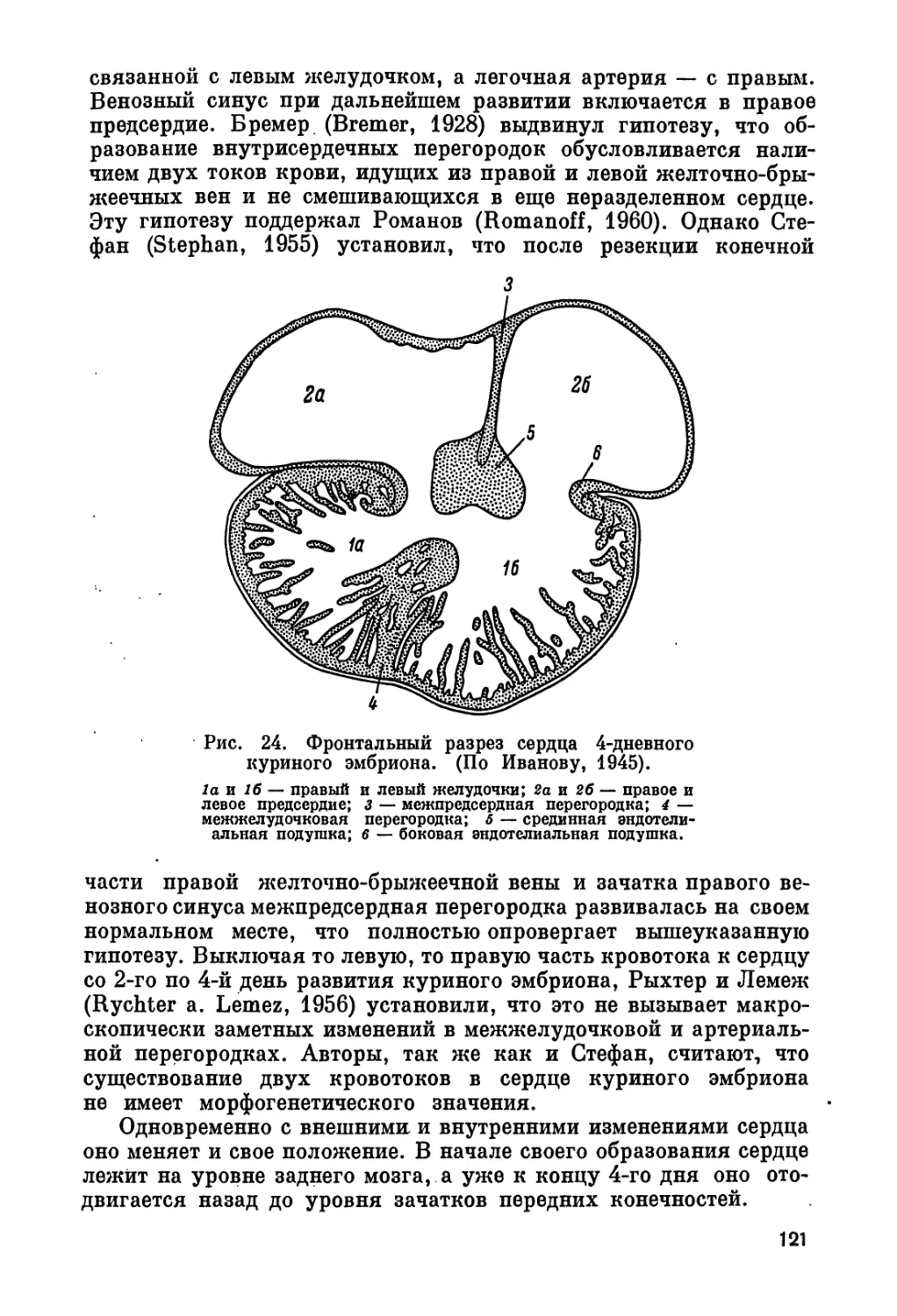
Так, во втором разделе, вслед за главой «Развитие внеэмбриональ-
ных органов» — об органах, играющих основную роль в нормаль-
ном функционировании эмбриона, следует глава «Кровообраще-
ние» — о первой функционирующей системе эмбриона, потом —
«Нервная система», а затем — «Дыхание». Между главами о мо-
чевыделительной системе и о половой системе, имеющих общее
происхождение, помещена глава «Эндокринные железы», так как
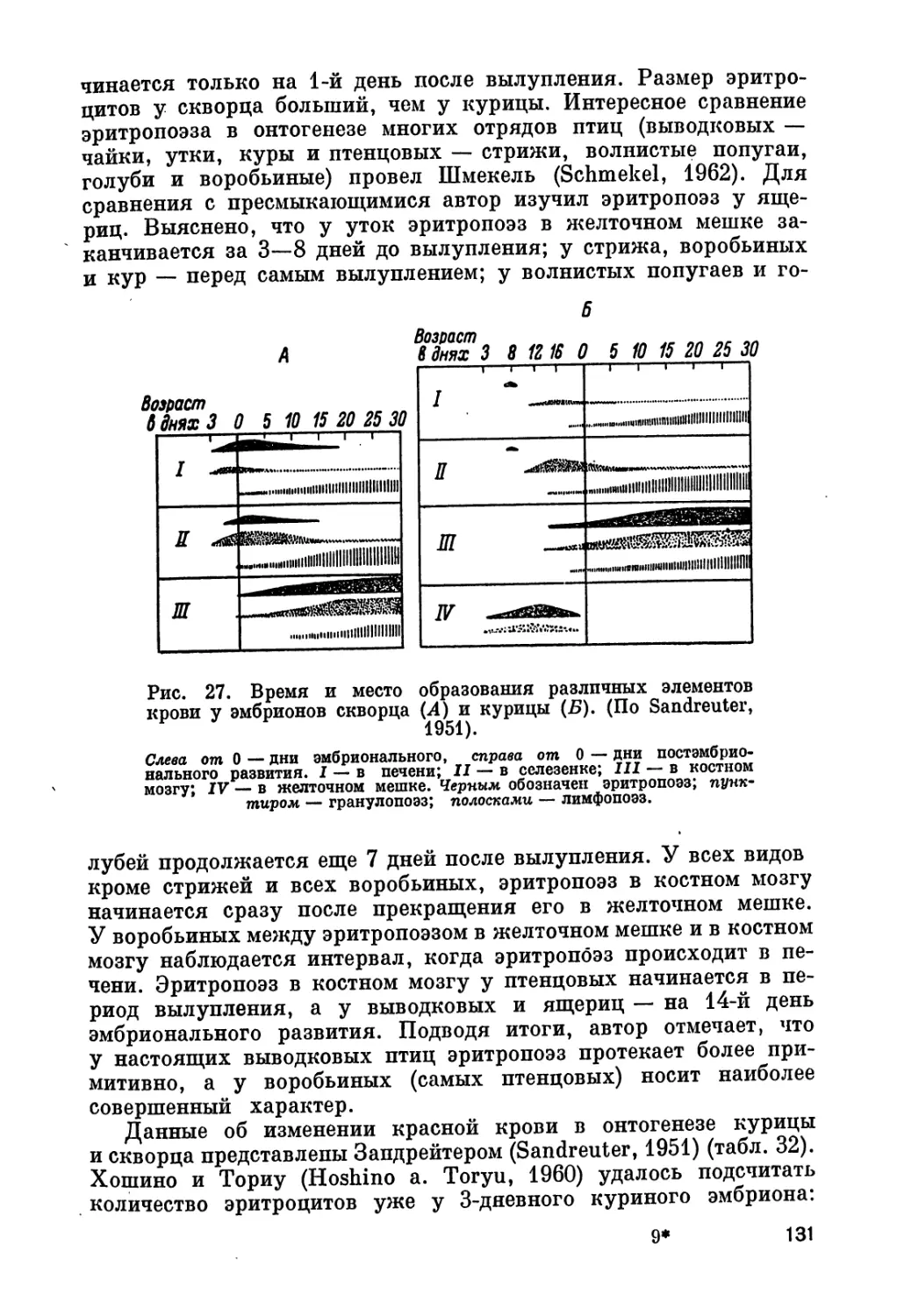
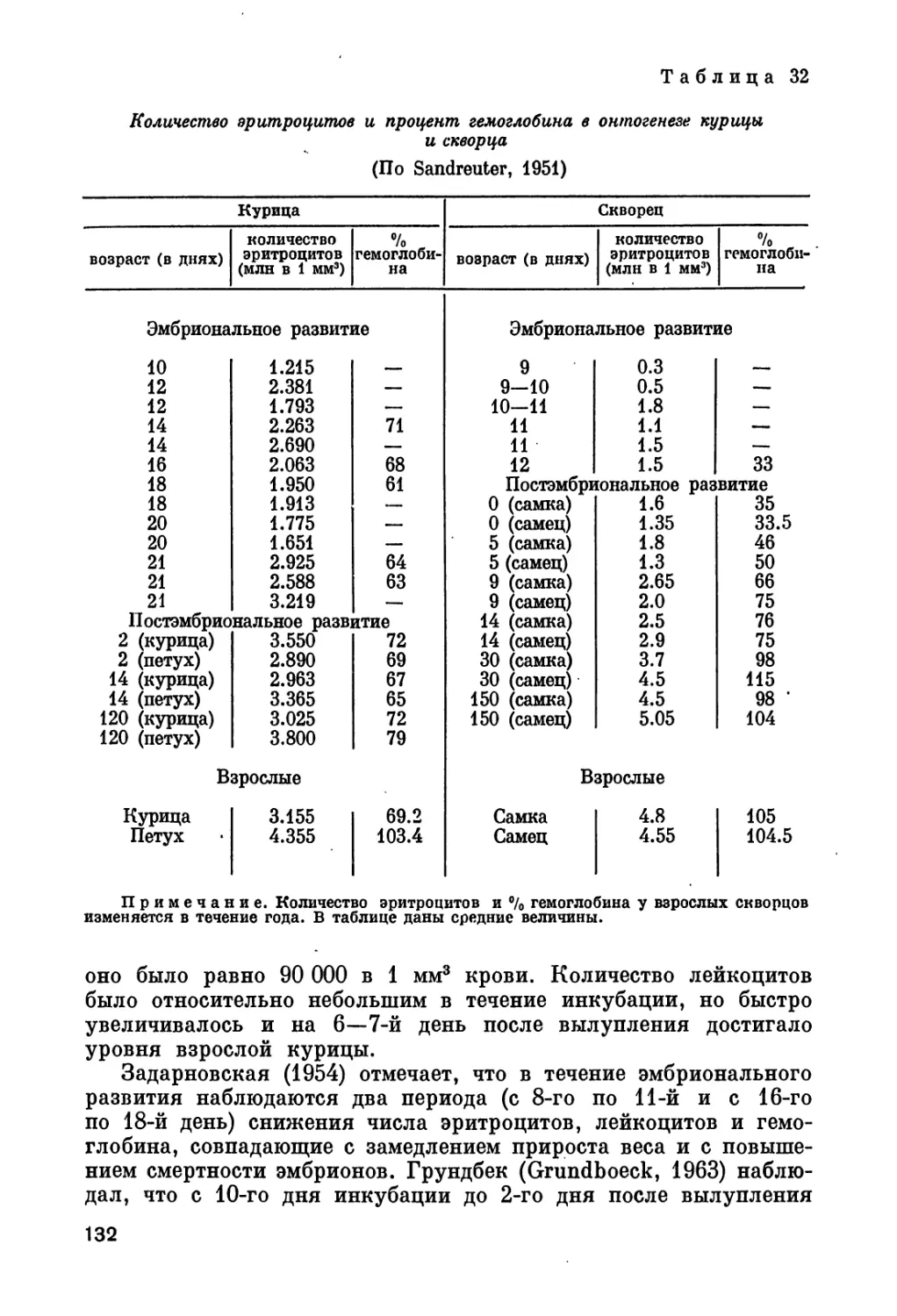
глава «Пол» включает сведения и о половых гормонах. Возможно,
и при распределении порядка глав мы не избежали личного отно-
шения к вопросу.
4
В связи с разнообразием материала в разных разделах книги
мы считаем более удобным не давать общего введения, а приводим
некоторые общие замечания перед разделами или главами.
Несмотря на огромную литературу вопроса, в книге цити-
руется только около 1100 названий, по нашему мнению — наи-
более важных.. Для удобства и лучшей ориентировки читателя
список литературы дается по разделам.
Автор надеется, что его сводка явится источником информации
для всех работающих с птичьими эмбрионами и будет полезным
пособием для научных исследований. Автор надеется также, что,
помогая отысканию методов управления развитием сельскохозяй-
ственных птиц, книга явится одним из элементов содружества
науки с сельскохозяйственным производством.
Книга рассчитана на научных работников биологов различного
профиля и специалистов по инкубации, а также на студентов
старших курсов университетов, сельскохозяйственных, ветери-
нарных, медицинских и педагогических институтов.
Мы понимаем, что взятая нами на себя задача чрезвычайно
сложна и в книге неизбежно имеется ряд упущений и недочетов,
поэтому все замечания читателей будут приняты с благодарностью.
За предоставленную мне возможность написания книги и за
долготерпение при редактировании ее в нескольких редакциях от
всей души благодарю Л. Г. Лейбсона, заведующего лабораторией
и редактора этой книги. Большую дружескую помощь по выправле-
нию недочетов в основной, физиологической, части работы оказала
мне Ц. Л. Янковская, за что я ей искренне благодарна. В мор-
фологической части книги мне очень помогли указания Л. К. Ти-
товой и Б. И. Шапиро; в просмотре отдельных глав приняли уча-
стие М. Н. Перцева, Э. М. Плисецкая, Г. Е. Свидерская, А. В. Бур-
сиан, В. Ф. Васильева и др.; всем им я выражаю свою искреннюю
признательность. Материалы дйя данной книги собирались авто-
ром еще со студенческих лет, когда большое участие в этом прини-
мал покойный В. В. Карачаровский; память о нем для автора не-
забвенна. Многие годы работы над книгой были возможны только
благодаря моральной поддержке В. К. Трулевича. За техническую
помощь при оформлении книги благодарю О. Г. Зайцеву,
Э. Л. Портенко и И. И. Мясоедову.
Раздел I. ЯЙЦО И РАЗВИТИЕ ЭМБРИОНА
ДО ИНКУБАЦИИ
В этом разделе, являющемся вводным к следующему,
основному, кратко излагаются материалы по имеющимся руковод-
ствам (Липпинкот, 1932; Дементьев, 1940; Иванов, 1945; Groeb-
bels, 1937; Romanoff a. Romanoff, 1949, и др.), а также данные,
полученные в последние годы.
ГЛ АВ А 1
СПЕРМАТОГЕНЕЗ И ОБРАЗОВАНИЕ ЯЙЦА
Образование половых клеток — яиц и сперматозои-
дов — состоит из ряда высокоспециализированных процессов.
Постоянство числа хромосом обеспечивается процессом мейоза;
мейоз состоит из двух быстро следующих друг за другом клеточ-
ных делений, после которых гаметы получают вдвое меньше
хромосом, чем остальные клетки тела.
Хотя с генетической точки зрения гаметы равны, но для эмбрио-
генеза мужские половые клетки менее важны, и сперматогенезу
нами будет уделено меньше внимания. Образование мужских
половых клеток происходит в семенниках, причем определенные
фазы их формирования распределены по особым зонам. В 1-й пе-
риод — размножения — сперматогонии занимают самую отда-
ленную от выводных протоков часть семенников, называемую зо-
ной размножения. Сперматогоний очень много, и они здесь много-
кратно делятся, так как поступление питательных веществ в этой
зоне чрезвычайно облегчено. Во 2-й период — роста — спермато-
гонии прекращают делиться и становятся сперматоцитами I по-
рядка, располагаясь в зоне роста. В этой зоне продолжается по-
ступление питательных веществ и сперматоциты I порядка сильно
вырастают. Хроматин ядра принимает вид попарно расположен-
ных четкообразных хромосом, вытягивающихся по направлению
к центросоме. Затем хромосомы сливаются попарно (синапсис).
С этого момента проникновение питательных веществ (а вместе
6
С тем и рост сперматоцитов) замедляется и затем совсем прекра-
щается. Потом образуются тетрады хромосом; их количество равно
количеству пар хромосом в сперматогониях. Затем наступает
3-й период — созревания, состоящий из двукратного деления
сперматоцитов: первое — с образованием сперматоцитов II по-
рядка, второе — с образованием сперматид. При этом в каждую
из 4 образовавшихся сперматид попадает гаплоидное количество
хромосом. В 4-м периоде — формирования — сперматиды пре-
образуются в сперматозоиды.
Сперматозоиды петуха имеют длинную, цилиндрическую го-
ловку с острым окончанием (акросомой). Головка состоит почти
исключительно из ядра половой клетки. Вообще количество про-
топлазмы в сперматозоиде минимальное. В средней части сперма-
тозоида находится большое количество митохондрий и центро-
сома — клеточный центр. Нить центросомы образует длинный
хвостик спермия. Сперматозоиды других сельскохозяйственных
птиц имеют сходный вид (рис. 1). Браше (1961) приходит к вы-
воду, что сперматозоиды отличаются интенсивными катаболиче-
скими процессами, богаты митохондриями и обладают исключи-
тельно интенсивным дыханием, но обмен белков и нуклеиновых
кислот в них незначителен или не происходит вообще. Несколько
иного мнения придерживается в своем детальном исследовании
Мамзина (1966). Получив два повышения радиоактивности РНК
спермиев на 6—8-й и 18—24-й дни после введения в кровь пету-
хов меченого Р32, автор предполагает, что первая вершина соот-
ветствует образованию цитоплазматической РНК, а вторая —
ядерной РНК. Будучи включенным в ДНК, Р32 стойко удержи-
вается в клетках семенного эпителия вплоть до образования зре-
лых сперматозоидов.
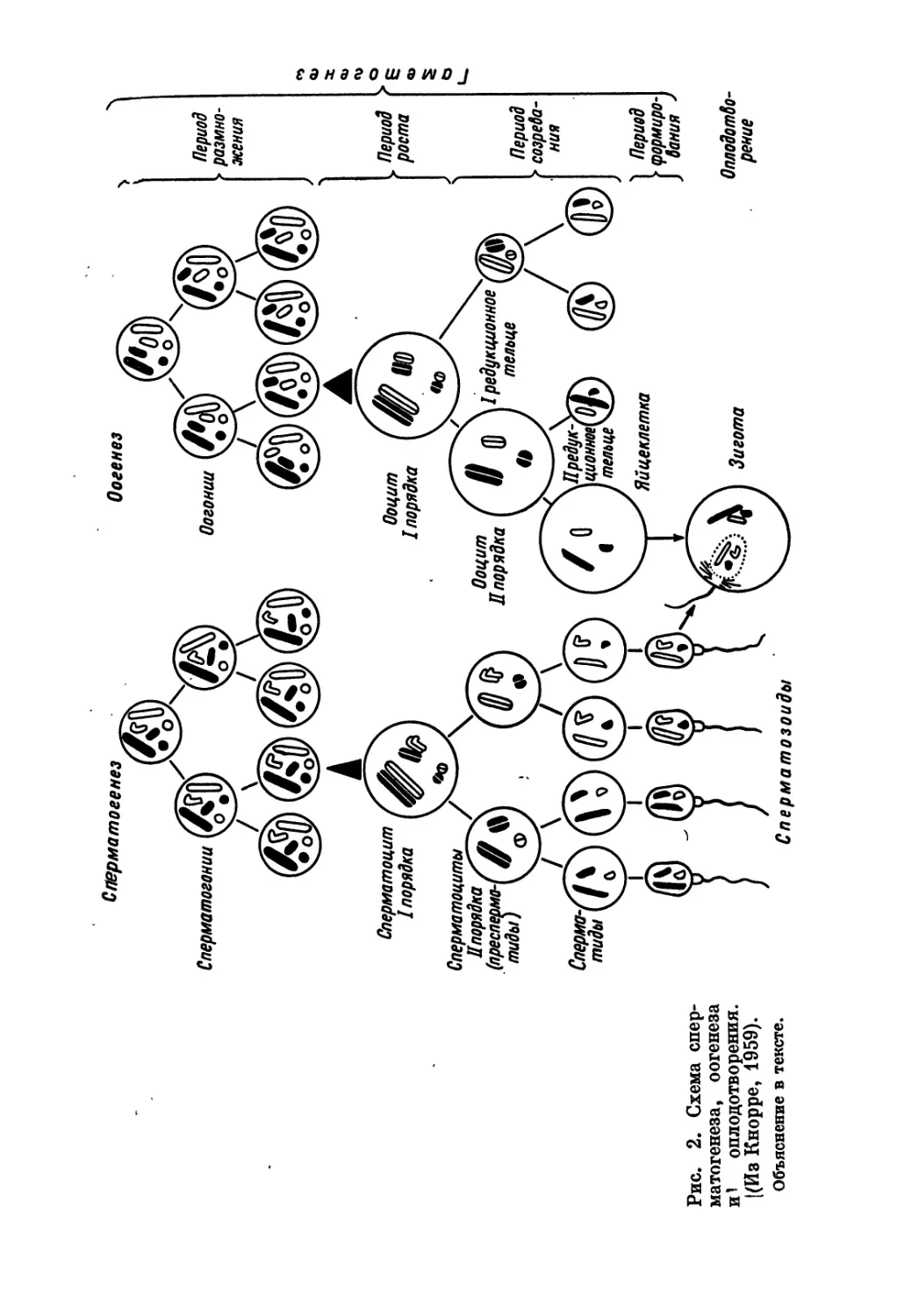
Схема редукции хроматина в ядрах женских и мужских по-
ловых клеток примерно одинакова (рис. 2). Оогенез протекает
следующим образом. В 1-й период — размножения — незрелые
половые клетки (оогонии) многократно митотически делятся.
Затем они перестают делиться и вступают в период продолжитель-
ного роста (2-й период оогенеза), превращаясь в крупные клетки —
ооциты I порядка. Этот период делится на два: малого роста (уве-
личение ядра и протоплазмы) и большого роста (в цитоплазме
накапливается большое количество желточных включений). В тече-
ние периода большого роста и образуется в основном желток яйца.
За время 3-го периода — созревания — ооциты I порядка про-
ходят два мейотических деления. После первого из них из каж-
дого ооцита I порядка образуется одна крупная клетка (ооцит
II порядка, содержащий, кроме ядра, желток и почти всю цито-
плазму) и одна мелкая, содержащая ядро и незначительное коли-
чество протоплазмы — полярное или редукционное тельце.
При втором мейотическом делении образуется яйцеклетка и второе
полярное тельце, а первое может в свою очередь разделиться на
7
два. Все эти Клетки содержат гаплоидное количество хромосом.
Полярные тельца — это абортивные яйца, и они погибают.
Таким образом, отличие оогенеза от сперматогенеза заклю-
чается в следующем: а) при последнем делении в семеннике об-
Рис. 1. Сперматозоиды домашних птиц.
(Третьяков, 1951).
А — строение сперматозоида петуха; Б — два вида
сперматозоидов петуха; В — два вида сперматозои-
дов селезня; Г — строение головни сперматозоида
гуся. 1 — головка (с ядром нлетки, расположенным
внутри); 2 — соединительная часть; з — средняя
часть; 4 — концевая часть жгутика (хвоста); 6 —
шейка сперматозоида.
разуется 4 сперматиды, а в яичнике — 1 яйцеклетка и 2 или 3 ре-
дукционных тельца; б) период роста при оогенезе длительнее
и подразделяется на период малого роста и большого роста;
в) в оогенезе нет периода формирования соответственно превраще-
нию сперматид в сперматозоиды. В неоплодотворенной яйцеклетке
заложены все потенции к развитию (партеногенез естественный
8
Сперматогенез
Ооеенез
Сперматогонии
Оогонии
Период
> размно-
жения
Рис. 2. Схема спер-
матогенеза, оогенеза
и1 оплодотворения.
[(Из Кнорре, 1959).
Объяснение в тексте.
Г аметогенез
Сперматоцит
[порядка
Период
роста
Ооцит
П порядка
) Период
> срормиро-
J дания >
Яйцеклетка
Зигота
Сперматозоиды
Продук-
ционно^
тельце
Оплодотво-
рение
Сперматоциты
c”s©)(©)®(©
Iх Iх Iх Iх
Период
Y созрева-
ния
и искусственный) в отличие от сперматозоида, неспособного ни
при каких условиях к самостоятельному развитию.
Хьюджес (Hughes, 1963) показал, что митоз оогоний увеличи-
вает общее количество половых клеток в левом яичнике куриного
эмбриона с 28 000 на 9-й день инкубации до 680 000 на 17-й и что
дегенерация половых клеток происходит в течение всего эмбрио-
генеза. К моменту вылупления имеется 480 000 ооцитов. Первый
период оогенеза, по данным автора, заканчивается ко времени
вылупления. Количество видимых ооцитов, по данным Романова
и Романовой (Romanoff a. Romanoff, 1949), в яичнике курицы
около 4000, у сельскохозяйственных водоплавающих птиц —
в среднем 1250, а у диких птиц — только около 500. Макси-
мально получают от одной курицы около 1500 яиц. По-види-
мому, дегенерация ооцитов происходит и в постэмбриональный
период и к моменту полового созревания (5—6 месяцев для
кур породы леггорн) их количество достигает величины, ука-
занной Романовыми.
Одно из основных отличий ооцита от соматических клеток —
это огромное ядро, содержащее несколько сравнительно большого
размера ядрышек и хромосомы типа «ламповых щеток», омывае-
мых ядерным соком. В яйцеклетке нет центросомы. Во время
оогенеза наследственны^ материал ооцита (ядро) находится в со-
стоянии активной деятельности и оказывает влияние как на строе-
ние цитоплазмы, так и на протекающие в ней процессы синтеза,
в том числе синтеза новых специфических белков.
Рост ооцита Нимирко и Гольдман (Niemierko i Goldman, 1947)
подразделяют по изменению веса и состава ооплазмы на 3 периода:
1-й — до достижения яйцеклеткой веса свыше 100 мг, 2-й —
до 1 г и 3-й — до достижения окончательного размера и веса.
1-й период характеризуется наибольшим содержанием воды и
пропорциональным увеличением протеинов и жиров, причем
протеины являются основными. Во 2-м периоде происходят наи-
большие изменения в химическом составе, а именно большое уве-
личение сухих веществ и значительное изменение в соотношении
жирных кислот к протеинам с преобладанием жирных кислот.
В 3-й период происходит наибольшее увеличение веса яйцеклетки
(фаза большого роста ооцита) в связи с накоплением органических
веществ, в основном липидов. Фаза большого роста ооцита в яич-
нике курицы, по данным Романова и Романовой (Romanoff
a. Romanoff, 1949), продолжается в течение 4 дней перед овуля-
цией. Когда наступает последняя стадия быстрого накопления
желтка, ядро и часть протоплазмы мигрируют к периферии его,
образуя бластодиск. При этом яйцеклетка получает полярность —
анимальный (где протоплазма и ядро) и вегетативный (где желток)
полюса.
В яичнике недавно вылупившегося цыпленка вокруг каждого
ооцита группируется ряд эпителиальных клеток, создающих
1Q
Б дальнейшем фолликул. Когда начинается быстрое накопление
' желтка в ооците, клетки фолликула передают питательные веще-
ства из кровеносных сосудов в цитоплазму ооцита* Барковская
(1954) считает фолликулярную оболочку секреторной железой
желтковой массы, производимой ею из приносимых кровью мате-
риалов. Изучая методом меченых атомов проницаемость фоллику-
лов и перенос протеинов и фосфорных соединений из плазмы крови
в желток куриного яйца, Смит (Smith, 1959) определил, что мак-
симальный темп переноса этих соединений на единицу поверхности
фолликула бывает при весе ооцита, равном 2.5 г, а затем умень-
шается ко времени овуляции. Электронномикроскопическими ис-
следованиями развивающихся фолликулов яичника фазана Пресс
(Press, 1959) показал, что фолликулярные клетки образуют про-
ходящие внутрь ооцита отростки. Автор предполагает, что меха-
низм быстрого передвижения питательных веществ из фоллику-
лярных клеток в растущий ооцит состоит в поглощении этих
фолликулярных отростков и включении их в цитоплазму ооцита.
Несколько иначе трактует этот процесс Браше (1961). Автор пред-
полагает, что клеточная оболочка ооцитов образует микровор-
синки, которые проникают в фолликулярные клетки и осущест-
вляют процесс пиноцитоза — «заглатывают» или «выпивают»
богатую белками воду. Хаскин с соавторами (1965) приходит к вы-
воду, что стенка растущего фолликула является не только местом
активного транспорта и гидролиза поступающих из крови ве-
ществ, но и местом синтеза некоторых соединений, входящих
в желток. На протяжении всего периода большого роста ооцита
наблюдается согласованная динамика транспортной, фермента-
тивной и синтезирующей активности фолликулярной оболочки.
Авторы считают, что вначале имеет место центральная активация
вителлогенеза, а затем постепенное усиление авторегуляторных
отношений между тканью фолликула и растущим ооцитом.
Рассмотрим несколько подробней процесс формирования
желтка в яйцеклетке. В протоплазме половых клеток эмбриона
уже имеются желточные шары, но настоящий желток впервые
откладывается в яичнике двухмесячного цыпленка. Желток откла-
дывается концентрическими кругами, попеременно желтого и
более светлого цвета. Желтый желток состоит из желтых желточ-
ных шаров, а белый — из белых. Конопака (Konopacka, 1933,
цит. по: Romanoff a. Romanoff, 1949) считает, что желтые желточ-
йые шары образуются при проникновении жира в вакуоли белых
желточных шаров. Риддль (Riddle, 1911), наоборот, наблюдая во
время развития эмбриона увеличение относительного количества
белого желтка, считает, что желтый желток превращается в бе-
лый. По данным Романова и Романовой (Romanoff a. Romanoff,
1949), желтый желток откладывается в течение дня до полуночи,
а белый — в остальную часть ночи. А Барковская (1954) считает,
что вообще нельзя делить желток на белый и желтый и что цвет
11
его зависит от кормления. Автор показала, что желток всегда
откладывается под фолликулярной оболочкой и окрашенный
красителем (судан III) слой желтка длительно сохраняет свое
местоположение и консистенцию под последующими отложени-
ями. Скорость отложения желтка нарастает с увеличением его
поверхности и является производной роста фолликулярной
оболочки. Данные по росту желтка в период роста ооцита
приведены в табл. 1.
Таблица 1
Средние величины прироста желтка по дням
(По подсчитанным нами данным Барковской, 1954)
День 1-й 2-й 3-й 4-й 5-й 6*й 7-й 8-й 9-й 10-й 11-й
Прирост (в мм3) 32 150 323 1679 2169 1448 2584 2297 2598 1441 682
Скармливая несушкам каждый день разные жирорастворимые
краски, Лакасань (Lacassagne, 1960) наблюдал корреляцию между
весом желтка яиц и длительностью его роста. Абе с соавторами
(Abe et al., 1959) изучал откладывание желтка в ооцитах путем
внутривенного введения меченной по Р32 плазмы. Установлено,
что в течение 24 час. в ооциты включается около 37% введенного
меченого фосфата. Распределение Р32 в разных ооцитах пока-
зало, что скорость откладывания желтка изменяется соответст-
венно стадии роста фолликула.
Исследуя методом меченых атомов глобулины и альбумины
плазмы кур, Паттерсон и др. (Patterson et al., 1962) показали, что
они передаются из яичника в формирующийся желток наиболее
интенсивно в течение 4—5 дней, предшествующих овуляции, через
фолликулярный эпителий и концентрация их в желтке неоплодот-
воренного яйца значительно превышает концентрацию в плазме.
Электрофоретическое исследование крови несушки и желтка ра-
стущих ооцитов, проведенное Хаскиным с соавторами (1965),
показало, что перенос оогенных компонентов осуществляется
в основном а- и у-глобул инами крови, а в желтке с этими фрак-
циями глобулинов связана основная часть липопротеидов, гли-
копротеидов, фосфолипидов и холестерин. Рост ооцитов сопро-
вождается изменением состава липидной части липовителлина и
увеличением насыщенности липидами некоторых фракций про-
теинов желтка.
Обмен веществ растущего ооцита обеспечивает синтез различ-
ных запасных веществ, которые используются впоследствии эм-
брионом, а именно гликогена, липидов, белков и др. Запас энер-
гии во время оогенеза заключен в макроэргических фосфатных
12
связях АТФ. Как сообщает Браше (1961), растущие ооциты со-
держат все необходимое для интенсивного синтеза протеинов
(желтка). Первая стадия этого процесса состоит в активации ами-
нокислот аденозинтрифосфатом при участии растворимых фер-
ментов. Образовавшиеся при этом вещества включаются затем
в растворимую РНК, потом в РНК микросом, и, наконец, в микро-
сомах происходит синтез специфических белков. Юные ооциты
всегда содержат большое количество РНК как в цитоплазме,
так и в ядрышках. В растущих ооцитах синтез ДНК незначителен,
а синтез РНК, белков, липидов и гликогена очень велик. Во время
вителлогенеза желточные гранулы бедны РНК, что создает впечат-
ление уменьшения содержания РНК в яйцеклетке, а в действи-
тельности количество РНК здесь растет, но она разбавлена
в сильно увеличившемся объеме ооцита. Митохондрии яйца по
своей ультраструктуре и функциям не отличаются от митохондрий
других клеток. По-видимому, они служат источником энергии,
необходимой для синтеза белков желтка. Браше приходит к за-
ключению, что желток нельзя рассматривать только как резерв-
ный материал, так как он играет активную роль в эмбриоге-
незе.
В то время как ооцит вступает в фазу большого роста, между
его наружным окончанием и фолликулом появляется прозрачная,
тонкая оболочка — это желточная оболочка. К концу созревания
ооцита между желточной оболочкой и фолликулом образуется
заполненное лимфой пространство, благодаря чему ооцит повора-
чивается внутри фолликула соответственно центру тяжести: ве-
гетативным полюсом вниз, а анимальным (с бластодиском) —
к ножке фолликула.
В таком положении ооцита и происходит овуляция — разрыв
фолликула и выпадение ооцита из яичника. Этот ооцит и является
тем, что обычно называют желтком яйца. Овуляция, как правило,
бывает через полчаса после того, как снесено предыдущее, сформи-
рованное яйцо. Наличие не вполне сформированного яйца в матке
предотвращает овуляцию из-за выделения гипофизом лютеини-
зирующего гормона. Овуляция у кур обычно совершается с 2 час.
ночи до 2 час. дня. Время овуляции зависит от внешних условий
и состояния несушки. Темнота, чрезмерная жара и беспокойство
курицы задерживают овуляцию, а иногда готовая к овуляции
яйцеклетка (желток) даже рассасывается.1 После овуляции ра-
зорвавшийся фолликул начинает постепенно резорбироваться:
через неделю он едва виден для невооруженного глаза, а через
месяц практически исчезает.
Каким же образом была создана возможность чуть ли не еже-
дневной яйценоскости у кур по сравнению с сезонной и очень огра-
ниченной кладкой (2—15 яиц в год) у птиц в природных условиях?
1 О регуляции процесса овуляции см. соответствующий раздел в главе 3.
13
В нашей работе (Ёольник, 1955) по инкубации яиц гаги мы устано-
вили экологическим методом наличие 2-й кладки, если у гаги ото-
брана первая кладка в начале насиживания (обычно гага несет
в год одну кладку, состоящую в среднем из 4 яиц — от 3 до 8).
Снесение птицей второй и даже третьей кладки яиц, если она
была лишена возможности насиживать первую кладку (яйца изъ-
яты из гнезда), показывает, каким путем произошло огромное
увеличение яйценоскости кур, уток и гусей при их одомашнивании.
Строение половых органов курицы представлено на рис,. 3.
Правый яичник и яйцевод птиц не развиваются и редуцируются
уже в конце эмбрионального развития. По-видимому, это было
обусловлено в процессе эволюции: а) необходимостью уменьшения
размера внутренних органов для облегчения веса тела в полете и
б) относительно большой величиной яиц, в которых должны
иметься запасы питательных веществ для прохождения всего
эмбрионального развития. Выпавшая из яичника после овуляции
яйцеклетка (желток) попадает в яйцевод, общая длина которого
у курицы-несушки в среднем 63 см (от 37 до 86), а у ненесушки —
только 11—18 см. Стенки яйцевода состоят из трех слоев: сероз-
ного (богатого кровеносными сосудами), мышечного и эпителиаль-
ного, покрытого изнутри железистой тканью, выделяющей слизь —
белок яйца. Железистая ткань, расположенная в многочисленных
складках яйцевода, имеет большую поверхность и вследствие
этого может выделять много белка. Высота и толщина складок
в белковом отделе, где вырабатывается основное количество белка,
в пять раз больше, чем в других отделах. Мышечный слой утол-
щается от начала яйцевода к клоаке. Куриное яйцо проходит по
всей длине яйцевода, под влиянием мышечных сокращений его,
примерно в течение суток, а яйцо голубя — в течение 41 часа
(Romanoff a. Romanoff, 1949). Вскрывая кур через 30 мин. после
снесения яйца, Мак-Налли и Баерли (McNally a. Byerly, 1936)
обнаружили, что у 20 из 32 произошла овуляция. Следовательно,
время между снесением двух последующих яиц примерно равно
времени пребывания яйца в яйцеводе. Длительность нахождения
яйца различна в разных частях яйцевода (табл. 2).
Для сравнения приведем данные Асмундсона (Asmundson,
1939) о длине яйцевода индеек и кур (табл. 3).
Первый слой белка желток получает при прохождении воронки
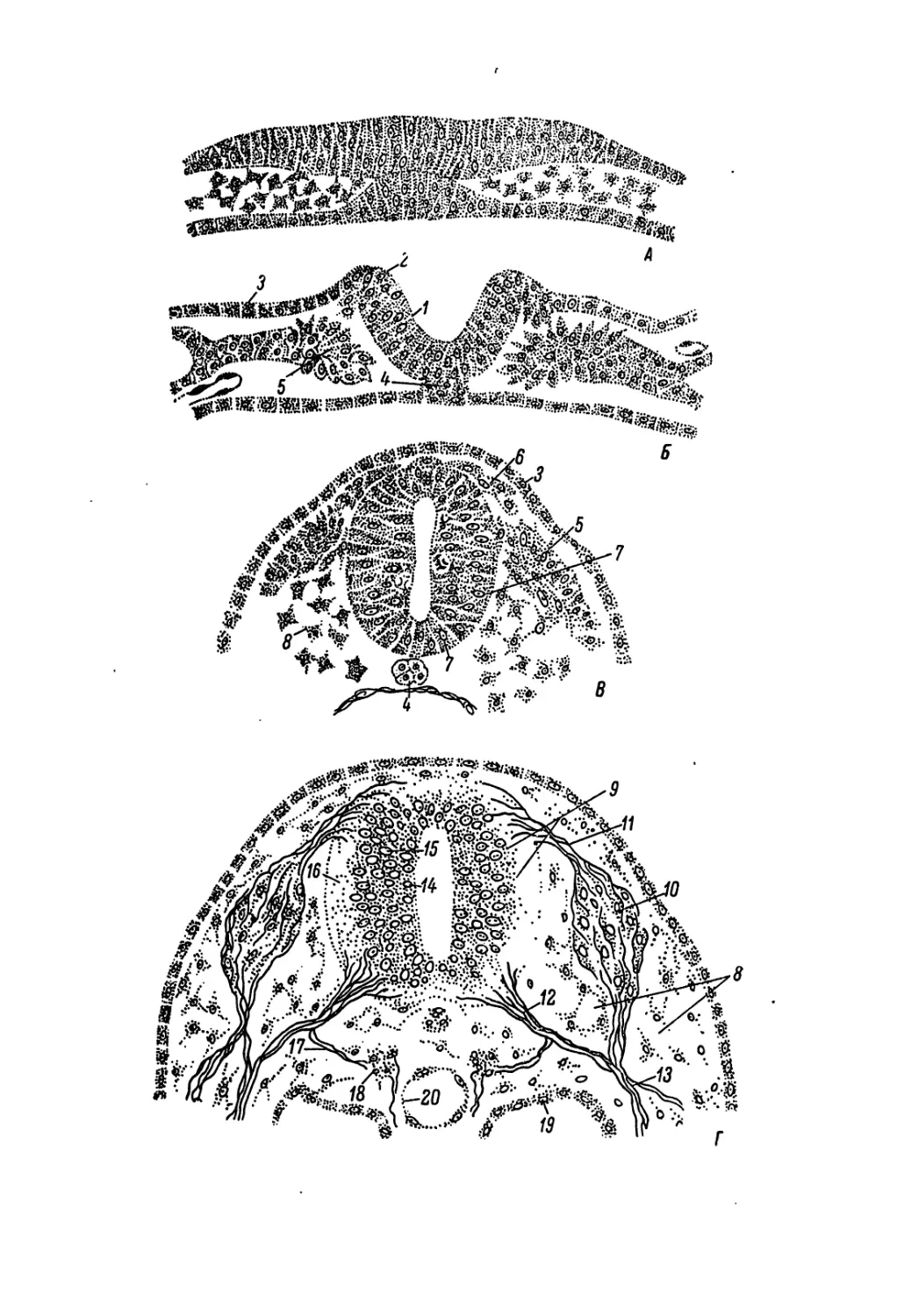
Рис. 3. Образование яйца в половых органах курицы.
(По Groebbels, 1937).
1 — яичник (молодые фолликулы сероватого цвета); 2 — созревающие фолликулы;
3 — густая сеть кровеносных сосудов фолликула; 4 — стигма; 5 — фолликул после вы-
падения из него желтка; € — воронка яйцевода; 7 — желток в верхней части яйцевода;
8 — белковый отдел яйцевода; 9 — густой белок и халазы; 10 — желток с зародышевым
диском; 11 — мышцы, приводящие воронку яйцевода в движение; 12 — перешеек; 13 —
матка; 14 — задняя кишка, открывающаяся в клоаку; 15 — мышечный сфинктер при
переходе матки во влагалище; 16 — клоака.
14
Таблица 2
Средняя длительность прохождения яйца по яйцеводу курицы
(По Romanoff a. Romanoff, 1949)
Отдел яйцевода Длина (в см) Нахождение в данном отделе
абс. время % от общего времени
Воронка Около 4 20 мин. 1.4
Белковый отдел . . 33.6 3 часа 13
Перешеек Около 8 1 ч. 10 м. 5
Матка 8.3 19 час. 80
Влагалище 7.9 Около 10 мин. 0.6
Весь яйцевод .... Около 62 Okoj^o 24 час.
Таблица 3
Относительная длина разных частей яйцевода у индеек и кур (в % от всей длины) (По Asmundson, 1939)
Во- ронка Белковый отдел Пере- шеек Матка Влага- лище
Индейки . . 14.8 42.6 15.3 13.6 13.7
Куры . . . 9.6 45.0 . 13.4 16.0 16.0
(описание формирования белка, подскор лунных оболочек и скор-
лупы дается нами по Романову и Романовой (Romanoff a. Roma-
noff, 1949)). Густой, муцинообразный белок откладывается здесь
непосредственно на желток в виде нитей, формирующих ячеистую
основу халазообразующего слоя. Ячейки этого слоя, как и
ячейки плотного белка, заполнены жидким белком. При медлен-
ном вращении желтка вокруг своей оси по мере прохождения по
яйцеводу микроскопические муциновые нити в халазообразую-
щем слое белка скручиваются и делаются туго натянутыми, упру-
гими. При этом жидкий белок, находившийся в ячейках этих
нитей, выжимается и образует слой внутреннего жидкого белка,
в котором желток свободно плавает более легким, анимальным
полюсом кверху. Потом окончания муциновых нитей халазооб-
разующего слоя еще более скручиваются и образуют халазы
(градинки). По некоторым данным, халазы начинают закручи-
ваться только в матке (Sharme et al., 1950).
В белковом отделе яйцевода образуется плотный слой белка.
Растворимые протеины, входящие в состав белка, образуются и
16
откладываются в трубчатых железах белкового отдела яйцевода
в период между двумя овуляциями. Скопившийся здесь белок
откладывается вокруг желтка при его прохождении по яйцеводу.
А муцин производится в бокаловидных железах яйцевода по мере
надобности. Когда яйцо достигает матки, в белке содержится
почти весь состав протеинов. А здесь, как показал Дрейпер (1966),
происходит увеличение количества воды в белке в 2.5—3 раза,
причем концентрация натрия снижается, а калия увеличивается
в связи с увеличением его количества в 7—8 раз. Наблюдается
параллелизм между количеством калия в белке и выводимостью
цыплят из этих яиц.
В трубчатых железах перешейка яйцевода секретируется зер-
нистый, кератиноподобный материал, образующий под скор лун-
ные оболочки. Эти зернышки, впитывая влагу, набухают и сра-
стаются в тугие пряди клейких нитей, переплетающиеся и обра-
зующие подскорлупные оболочки. Наружный жидкий слЬй белка
образуется в перешейке и матке. К моменту попадания яйца
в матку подскорлупные оболочки еще чрезвычайно пористы, и
в первые 8 час. пребывания там яйца жидкий материал белка (в ос-
новном водный раствор неорганических солей) проходит сквозь
поры оболочек в яйцо.
Образование скорлупы начинается с появления зернышек каль-
циевых солей на поверхности наружной подскорлупной оболочки,
которые затем вместе с небольшим количеством протеинового ма-
териала превращаются в твердые, шишкообразные сосочки —
призматический слой скорлупы. Вершины сосочков вонзаются
в нити подскорлупных оболочек и таким образом плотно примыкают
к ней. Широкие наружные концы сосочков почти соединяются
между собой, но благодаря их округлости между ними остаются
щели — поры скорлупы яйца. Затем выделяется протеин (колла-
ген), который ложится поверх призматического слоя переплетен-
ными между собой нитями. Между ними закладываются кристаллы
кальциевых солей и образуется губчатый слой скорлупы, имеющий
вдвое бблыпую толщину, чем призматический. Поры в губчатом слое
являются продолжением пор в призматическом. Кальций скорлупы
поставляется кровью в клетки желез матки, которые секретируют
раствор кальциевых солей на яйцо. Брадфилд (Bradfield, 1951)
провел радиографическое изучение образования скорлупы кури-
ного яйца и обнаружил, что темп отложения минеральных веществ
идет по S-образной кривой.
В связи с различиями относительной длины отдельных частей
яйцевода (табл. 3) индюшиное яйцо имеет более легкую скорлупу,
более плотные подскорлупные оболочки и больший процент солей
в белке. Общий интервал между снесением двух последующих яиц
у индеек примерно такой же, как у кур (Asmundson, 1939). Вудард
и Матер (Woodard a. Mather, 1964) исследовали процесс образова-
ния яйца у японской перепелки и обнаружили, что он очень схо-
2 в. в. Ролышн
17
ден с тем, как образуется яйцо у курицы и индейки. Овуляция
происходит через 15—30 мин. после яйцекладки, затем желток
проходит по яйцеводу (через воронку за 30 мин., белковый отдел —
за 2—2.5 и перешеек — за 1.5—2.0 часа). В матке яйцо находится
19—20 час., причем отложение скорлупы начинается через 5 час.
после вхождения туда яйца, а пигментация скорлупы — за
3.5 часа до яйцекладки. По данным Руус (1956), промежуток ме-
жду снесением двух яиц в кладке гусей (что почти равно длитель*
ности пребывания яйца в яйцеводе) равен в среднем 34 час.: от
36 час. в марте до 25 в июне.
Пигментация яйца птиц происходит во время последних часов
пребывания его в матке за счет переработанного печенью пигмента
эритроцитов. Выделенные печенью пигменты создают основной
цвет скорлупы, который может быть различным: красным, жел-
тым, синим, коричневым, голубым и т. п. Позднее непосредственно
из гемоглобина протекающей по сосудам матки крови образуются
пятна на скорлупе, состоящие из пигментных масс, располагаю-
щихся либо в кутикуле, либо поверх нее.
Кутикула (тонкий слой засохшей слизи) выделяется маткой
после окончания образования скорлупы. Она, так же как и скор-
лупа, проницаема для газов, но предотвращает попадание микро-
организмов в яйцо через поры скорлупы.
Перед выходом из матки яйцо поворачивается на 180° и выходит
из клоаки тупым концом.
Серия яиц, снесенная птицей без перерыва, называется циклом
кладки. У сельскохозяйственных птиц, и особенно у курицы, он
продолжительнее, а у диких птиц — много короче. Чем больше
цикл кладки у курицы, тем ближе к 24 час. интервал между яй-
цами, причем этот интервал меньше всего в середине цикла (23—
24 часа), а в начале цикла и к концу его несколько больше (около
25—26 час.). Различие в интервале зависит главным образом от
разной длительности пребывания яйца в матке (Romanoff a. Ro-
manoff, 1949).
Сравнивая интенсивность яйцекладки у кур в первый и второй
год носки, Лакасань (Lacassagne, 1960) обнаружил, что она сни-
жается в течение первого года в связи с увеличением веса каж-
дого ооцита. Автор наблюдал, что длительность фазы быстрого
роста 1-го фолликула из цикла кладки больше, чем последнего, и
с каждым следующим яйцом в цикле происходит уменьшение ко-
личества резервов желтка. Мера и Лакасань (Merat et Lacassagne,
1961) показали, что в зависимости от очередности яиц в цикле
кладки и в кладках с разным количеством яиц выводоспособность
их различна. Выяснилось, что яйца однояйцовых кладок имеют
более низкую выводоспособность, а семияйцовых — более высо-
кую, чем в других. Первое яйцо в цикле кладки после перерыва
в два или больше дней имеет более низкую выводоспособность 7
чем другие яйца этого цикла,
15
Г Л А В A 2
ОПЛОДОТВОРЕНИЕ ЯЙЦЕКЛЕТКИ
И РАЗВИТИЕ ЭМБРИОНА
ДО СНЕСЕНИЯ ЯЙЦА ПТИЦЕЙ
А. Осеменение и оплодотворение
Различают два понятия: осеменение и оплодотворение.
Осеменение включает все процессы, приводящие мужские и жен-
ские половые клетки к контакту, в результате чего сперматозоид
внедряется в плазму яйцеклетки. Собственно оплодотворение озна-
чает слияние ядер женской и мужской половых клеток.
Образование яйца может проходить и без оплодотворения яй-
цеклетки, но для развития эмбриона необходимо, чтобы яйцеклетка
была оплодотворена. Сперматозоиды образуются в половых желе-
зах петуха почти круглый год, а у других птиц в основном в те-
чение весны и лета. По данным Чайковской (1950), половая ак-
тивность гусаков продолжается в среднем 87—98 дней, заканчи-
ваясь в мае.
Сперматозоиды попадают в половце пути самки (в клоаку) при
спаривании. Из клоаки они движутся по яйцеводу, достигая его
воронки через час после спаривания при помощи движения хво-
стика спермия и под влиянием хемотаксиса (движение спермиев
по направлению к веществам, выделяемым яйцеклеткой).
Спаривание у птиц происходит очень быстро и часто повто-
ряется. Духно (1964) наблюдала, что при высокой половой актив-
ности петухов (16—22 спаривания в день) объем семенной жид-
кости, выделяемой за одно спаривание, снижается почти в 2 раза
и в 2 с лишним раза уменьшается концентрация сперматозоидов
в ней по сравнению с тем, что бывает при ограниченной половой
активности тех же петухов (3—5 спариваний в день). Ограниче-
ние числа спариваний петухов, по данным автора, повышает про-
цент оплодотворенности яиц, процент вывода цыплят и жизне-
способность молодняка.
По данным Мамзиной (1965), объём выделяемой за один раз
семенной жидкости у петуха равен в норме 0.1—1.2 мл со средней
концентрацией спермиев 3.75 млрд/мл (от 0.04 до 8 млрд/мл).
Концентрация спермиев является показателем интенсивности
функции половых желез. Автором отмечена положительная кор-
реляция между скоростью обновления меченого фосфора в общем
количестве РНК спермиев (что'* отражает интенсивность обменных
процессов при сперматогенезе) и концентрацией спермиев. Этим
же методом (меченого фосфора) установлено, что длительность
сперматогенеза у петухов равна 14—15 дням, т. е. вдвое меньше,
чем у сельскохозяйственных животных, что согласуется с высо-
2*
19
кой половой активностью петухов. Время прохождения семени по
выносящим путям равно 10—12 дням (Мамзина, 1966). У гусаков
объем семенной жидкости за одно спаривание равен 0.1—2.0 мл,
а концентрация ее значительно ниже, чем у петухов, и равна
340—580 млн/мл (Чайковская, 1950).
Изучая физиологические свойства спермы гусаков, Чайков-
ская (1950) наблюдала, что чем выше концентрация семенной
жидкости, чем выше активность спермиев (относительное коли-
чество их, способное к поступательному движению), чем больше
их резистентность (устойчивость липопротеидного покрова спер-
ми.ев к 1%-му раствору хлористого натрия), тем выше оплодо-
творенность яиц и ниже эмбриональная смертность. По всем этим
показателям сперма петухов намного превосходит сперму гуса-
ков. Камар (Кашаг, 1960) подтвердил, что при плохой подвиж-
ности сперматозоидов и низкой концентрации спермы сильно
снижается оплодотворенность и выводимость цыплят. Секи и Кейт
(Saeki a. Keith, 1962) показали, что наличие более 10% ненормаль-
ных, искривленных сперматозоидов снижает оплодотворенность
индюшиных яиц на 12% и выводимость индюшат на 3.5%.
Многие исследователи и практики птицеводства наблюдали
связь между оплодотворенностью яиц, выводимостью и жизне-
способностью цыплят. Левдйыэ (1961) обнаружил между опло-
дотворяемостыо яиц отдельных кур и выводимостью цыплят из них
положительную корреляцию, равную 0.45 +0.05; между выво-
димостью и сохраняемостью цыплят при выращивании — 0.67 +
+0.07; между оплодотворяемостыо яиц каждой курицы на 2-й
и на 3-й год яйцекладки — 0.52+0.07 и выводимостью цыплят
в эти же годы — 0.57+0.06.
В группе работ Новик с соавторами (Новик, 1958; Новик и
Копыловская, 1961; Новик и Гинтовт, 1965) экспериментально
показала, что проникновение спермиев в яйцеклетку у кур про-
исходит сразу после овуляции в воронке яйцевода. Если здесь
в течение 15—20 мин. не произойдет встреча яйцеклетки со спер-
матозоидами, она в дальнейшем претерпевает необратимые из-
менения и теряет способность быть оплодотворенной. Раньше
считалось, что утрата способности к оплодотворению связана
с облекающим яйцеклетку (в яйцеводе) белком; однако, так как
авторам не удалось добиться оплодотворения яйцеклетки и после
освобождения ее от белка, то причиной этого следует считать
изменения в самой яйцеклетке. При хирургическом вмешатель-
стве авторам удалось оплодотворить яйцеклетку еще до овуляции.
Половые клетки (гаметы) — это специализированные клетки,
неспособные долго существовать без процесса оплодотворения;
после их слияния образуется зигота, которая уже не просто клетка,
а целостный организм, начинающий новый жизненный цикл.
Как же происходит проникновение спермин в яйцеклетку?
Дорфман (1963) сообщает, что при взаимодействии сперматозоида
20
с яйцом происходит так называемая акросомная реакция, при ко-
торой из акросомы спермия выталкивается нить, входящая в об-
разующийся на поверхности яйцеклетки воспринимающий хол-
мик. При этокГ спермиями млекопитающих выделяется гиалуро-
нидаза, расщепляющая мукополисахариды, склеивающие между
собой окружающие яйцо фолликулярные клетки. Расщепление
мукополисахаридов делает клеточную оболочку более проницае-
мой. Однако в спермиях птиц гиалуронидаза не обнаружена.
Желточную оболочку яйцеклетки у птиц растворяют лизины,
выделяемые акросомой спермия. Лизины и другие вещества,
выделяемые гаметами в окружающую среду, называются гамо-
нами. В цитоплазму яйцеклетки проникает головка, шейка и
связующий отдел сперматозоида, а хвостик его отбрасывается.
Центросома спермия, попав в яйцеклетку, у которой нет своей
центросомы, становится клеточным центром зиготы. Затем цен-
тросома делится на две новых, которые расходятся в стороны и
располагаются с обоих концов от вновь образовавшегося ядра.
При этом происходит перестройка цитоплазматического мате-
риала и подготовка к серии митотических делений — к размно-
жению клеток.
Вхождение центросомы сперматозоида дает импульс актива-
ции яйцеклетки к развитию, который сходен с нервно-мышеч-
ным импульсом как по физиологическому его проявлению, так
и по физико-химической природе, что подтверждено новыми ис-
следованиями последних десятилетий (Дорфман, 1963). По мне-
нию автора, неправильно было бы считать причиной активации
яйцеклетки слияние мужского и женского ядер (подробнее см.:
Дорфман, 1963). После слияния ядер яйцеклетки и спермия на-
ступает 1-е дробление вновь образовавшегося ядра и цитоплазмы
зиготы. При этом диплоидное число хромосом зиготы (состоящее
из двух гаплоидных наборов — материнского и отцовского) де-
лится обычным митотическим путем, так что каждая вновь обра-
зовавшаяся клетка имеет диплоидный набор хромосом.
Оплодотворение вызывает целый ряд физико-химических и
биохимических изменений яйцеклетки. Укажем основные. После
оплодотворения вязкость протоплазмы яйца увеличивается и
приближается к гелю, что, по-видимому, необходимо для клеточ-
ного деления (Барт, 1951). Проникновение спермиев в яйцеклетку,
как сообщает Кнорре (1959), в 70—80 раз (по данным Барта,
в 5 раз) усиливает окислительно-восстановительные процессы и
увеличивает теплопродукцию. Эта энергия, по-видимому, необ-
ходима для осуществления процессов дальнейшего развития.
Однако, по мнению Браше (1961), оплодотворение не обязательно
стимулирует дыхание яйцеклетки (у разных животных), а только
регулирует его. Он приводит теорию Батайона (Bataillon, 1929,
цит. по: Браше, 1961), по которой оплодотворение представляет
собою в основном «реакцию очищения» яйцеклетки от токсиче-
21
скйХ продуктов распада в результате повышения йронйцаемостй
ее оболочек, так как, по целому ряду данных, неоплодотворенное
яйцо — это клетка, находящаяся в состоянии угнетения. Сразу
после оплодотворения наступает заметное увеличение потребле-
ния гликогена (Браше, 1961). Кроме того, повышается выделе-
ние аммиака, что указывает на стимуляцию пуринового обмена,
связанного с РНК. При оплодотворении происходит значитель-
ная перестройка белков, а синтез их наступает позднее.
При оплодотворении в яйцеклетку птиц проникает много спер-
миев одновременно (полиспермия). По данным Бехтиной (1958,
1961), в куриное яйцо проникает в среднем 20—-60 спермиев,
а при вхождении в яйцо 300—-400 спермиев наблюдаются анома-
лии развития. Сливается с ядром яйцеклетки ядро только одного
из сперматозоидов. О судьбе остальных спермиев, проникающих
в яйцеклетку, имеется много противоречивых наблюдений и мне-
ний. По мнению одних исследователей (Иванов, 1945), все спер-
мин, кроме одного, погибают и их ядра подвергаются деструкции
с расщеплением хроматина ферментами. По данным других [Кам-
шилов, 1947 (на дрозофиле); Соколовская, 1950; Кнорре, 1959],
оставшиеся спермин ассимилируются зиготой, являясь источни-
ком питательных веществ, энергии и, возможно, пластического
материала. Кнорре, кроме того, считает, что они образуют также
мероциты (блуждающие ядра), способствующие переработке
желтка. Третья группа исследователей (Pasteels, 1950; Фофанова,
1964) считает, что эти спермин образуют дополнительные ядра,
участвующие в построении желточного синцития, становясь
центрами дополнительного краевого дробления. Бехтина (1966),
соглашаясь с мнением Кнорре (1959) об участии дополнительных
мужских пронуклеусов в освоении питательного материала желтка,
считает, что продукты распада сперматозоидов (биологически
активные вещества) ассимилируются эмбрионом и являются одним
из источников изменения качества потомства, полученного при
гетероспермном осеменении и, наконец, считает возможным,
что некоторые дополнительные мужские пронуклеусы становятся
ядрами краевых незамкнутых снизу бластомеров, т. е. входят
в структуру эмбриона. Мы считаем наиболее правильным мнение
второй группы исследователей.
По-видимому^ немаловажную роль играет возможность изби-
рательного оплодотворения (сливается с яйцеклеткой наиболее
подходящий, наиболее обеспеченный энергией спермий). Нельзя
забывать, что при оплодотворении происходит соединение двух
различных наследственных зачатков, что является благоприятным
и необходимым для развития нормального, жизнеспособного
потомства. Половые клетки не должны принадлежать, с одной сто-
роны, близкородственным животным из-за чрезмерного физиологиче-
ского сходства между ними, а с другой — слишком далеким ви-
дам из-за физиологической несовместимости. Многие гены, обус-
22
ловливающие нежелательные, иногда даже летальные признаки,
находятся в организме в рецессивном состоянии и не проявляются.
При близкородственном скрещивании — инбридинге г- эти ре-
цессивные гены встречаются и могут проявиться, вызвав появле-
ние нежелательных для человека и часто вредных для самого
животного признаков; в некоторых же случаях проявление рецес-
сивного признака приводит к гибели данного животного еще
до окончания его эмбрионального развития. При скрещивании
совершенно неродственных линий потомство зачастую оказывается
лучше каждого из родителей. Это улучшение называется гетеро-
зисом. Генетически гетерозис объясняется тем, что каждая из ро-
дительских линий содержит доминантные гены, прикрывающие
нежелательные рецессивные гены другой родительской линии.
Целый ряд исследователей (Кушнер, 1948; Меркурьева, 1951,
и др.) пытались доказать возможность участия в оплодотворении
яйцеклетки (т. е. в слиянии мужского ядра с женским) двух или
большего количества сперматозоидов (с появлением у потомства
признаков двух отцов) после осеменения спермой двух петухов.
Опыты были проведены без необходимых в таких случаях цито-
логических исследований, не исключали чисто генетических
ошибок и не могут быть признаны достоверными. Благотворное же
влияние оплодотворения спермой двух петухов, наблюдавшееся
этими авторами, можно, по-видимому, объяснить ассимиляцией
зиготой ядер дополнительных спермиев, причем положительную
роль могут играть и некоторые породные отличия в ДНК.
Несмотря на то что Курбатовым и Нарубиной (1965) представ-
лен большой материал, как будто показывающий сдвиг полов
в потомстве в сторону увеличения процента самок при ограничен-
ном (кратковременном или посменном) использовании петухов,
нам это кажется необоснованным, так как пол потомства у птиц
определяет не самец, а самка (см. главу 14).
В большой группе работ Олсен (Poole а. Olsen, 1958; Olsen,
1960, и др.) сообщает о случаях партеногенетического развития
яиц индеек и кур. Подводя итоги за 9 лет наблюдений, автор от-
мечает, что количество случаев партеногенетического развития
с годами увеличивается и что за это время вылупилось 67 «парте-
ногенетических» индюшат, причем все были самцами; 3 из них
продуцировали сперматозоиды и от одного было получено потом-
ство. По нашему мнению, такой вывод должен быть значительно
лучше обоснован (например, необходимы цитологические иссле-
дования хромосомного аппарата), так как он противоречит всему,
что известно о биологии размножения высших животных.
Оставаясь в складках яйцевода, не слившиеся с яйцеклеткой
сперматозоиды сохраняют свою жизнеспособность и могут опло-
дотворять яйца при последующих овуляциях. Показано, что в те-
чение трех недель после спаривания куры могут нести оплодотво-
ренные дйца и щизцеспособность эмбрионор не зависит от времени
23
снесения яйца после покрытия курицы (Hastabova, 1960). По дан-
ным Новик и Гйнтовт (1965), сперматозоиды петуха сохраняют
оплодотворяющую способность в яйцеводе курицы до 30 дней.
Вне яйцевода оплодотворяющая способность сперматозоидов зна-
чительно снижается уже через час после эякуляции. Авторы уста-
новили, что наиболее благоприятной средой для сохранения этой
способности является слегка гипертонический раствор с pH 6—8.
Длительное хранение сперматозоидов в половом тракте кур обес-
печивается морфо-физиологическими условиями железистых ямок
(крипт) воронки яйцевода, глубиной крипт, слабокислой реак-
цией среды, наличием в ней глюкозы и фосфатазы (Бехтина, 1965).
Вейцман (1957) сообщает, что оплодотворяющая способность
спермиев в половых путях цесарок-самок сохраняется полностью
в течение 8—10 дней, а затем падает до нуля к 20-му дню после
спаривания. Значительно меньше длительность оплодотворяющей
способности спермы у уток. Аш (Ash, 1961) наблюдал высокую
оплодотворенность яиц у пекинских уток только до 4—5 дней
после удаления самцов, а на 10—13-й день оплодотворенность
яиц снижалась до нуля. Снижался и процент вывода утят от опло-
дотворенных яиц с 75.8% в первую неделю после изоляции сам-
цов до 60.5% во вторую. Гальперн (1958) показала, что для обес-
печения высокого процента оплодотворенных яиц в половых
путях курицы должно быть достаточное количество спермы.
Так, даже после трехкратного спаривания с петухами ранее не
спаривавшиеся куры снесли 76.3% неоплодотворенных яиц. Автор
наблюдала, что чем чаще куры спаривалцсь с петухами, тем выше
был процент оплодотворенных яиц; спаривание должно было быть
ежедневным, но не реже чем через день. Выяснено, что спари-
вание происходит за 18—30 час. до наступления очередной овуля-
ции, что обеспечивает оплодотворение яиц свежей спермой.
Одно время считалось, что спермин, не принявшие участия
в оплодотворении яйцеклетки, играют существенную роль в об-
мене веществ материнского организма, проникая в эпителиальные
клетки половых путей самки (Кушнер, 1949; Войтишкова, 1955).
Противоположные данные получила Бехтина (1965), исследовав
около 100 половых трактов кур и не обнаружив ни одного случая
внедрения сперматозоидов в цитоплазму клеток выстилающего
их эпителия. Автор предполагает, что изменение цвета скорлупы
в сторону отцовской породы может объясняться вхождением
продуктов резорбции семенной жидкости в пигментирующее
скорлупу вещество.
Б. Развитие эмбриона до снесения яйца птицей
От момента оплодотворения яйцеклетки в воронке
яйцевода до выведения яйца наружу проходит около 24 час.,
в течение которых в оплодотворенном яйце протекает развитие
24
эмбриона. Начинается оно с деления оплодотворенной яйцеклетки
на 2 клетки примерно через 3 часа после оплодотворения (Pat-
terson, 1910). Эти клетки в свою очередь делятся второй бороздой,
перпендикулярной к первой (рис. 4). В связи с перегрузкой яйце-
клетки птиц желтком дробление ее происходит только на анималь-
ном полюсе яйца, где в цитоплазме содержится лишь небольшое
количество мелких желточных гранул. Такое дробление назы-
вается частичным, дискоидальным. При этом типе дробления на
краю бластодиска и под ним все время остаются неполностью
отделенные от желтка клетки. Ядра некоторых из этих не отгра-
ниченных от желтка клеток уходят в массу желтка, становятся
там мероцитами (как и ядра избыточных спермиев) и принимают
участие в перестройке желтка для облегчения его потребления
эмбрионом. Дробление у птиц резко неравномерное и неправиль-
ное, в результате чего возникают клетки различного размера и
различно расположенные друг к другу. Деление зиготы проис-
ходит митотическим путем, причем по сравнению с митозом в со-
матических клетках процесс здесь протекает чрезвычайно быстро.
В связи с этим дробление ядра и цитоплазмы может быть асин-
хронным.
Замечено, что в отсутствие кислорода митоз невозможен, хотя
покоящееся ядро (между делениями) имеет более активный мета-
болизм, чем делящееся. В покоящемся ядре наибольшей интен-
сивности достигают процессы синтеза белка и, по-видимому,
в это время синтезируются и связанные с ДНК белки (Браше,
1961). В процессе митоза участвуют многие химические компо-
ненты клетки, но наиболее важными являются ДНК, РНК и
белки, содержащие серу. В куриных яйцах синтез ДНК начи-
нается после 4-го дня развития, когда число клеток достигает 5 «107.
Синтез ДНК частично обеспечивается РНК, но, по-видимому, для
него могут быть использованы и низкомолекулярные предшествен-
ники. По предположению Браше (1961), между синтезом ДНК
и образованием энергии в яйце имеется тесная связь.
Бехтина (1960), исследуя развитие оплодотворенной яйце-
клетки, установила, что дробление до 8—10 бластомеров проис-
ходит только митотическим путем, а затем (яйцо в это время на-
ходится в матке) частично также путем фрагментации. В это
время синхронность дробления нарушается. За то время, пока
яйцо находится в матке, образуется многоклеточный бластодиск,
наружный слой которого имеет вид эпителия, а нижележащие
клетки расположены более рыхло. Периферическая зона бласто-
диска состоит из неполно разделившихся крупных участков про-
топлазмы со многими ядрами (желточный синцитий), которая
не отграничена от нижележащего и рядом лежащего желтка и
носит название зародышевого валика. Мероциты разжижают часть
желтка под бластодиском, и при этом образуется заполненная
жидкостью полость — подзародышевая полость. Благодаря этой
26
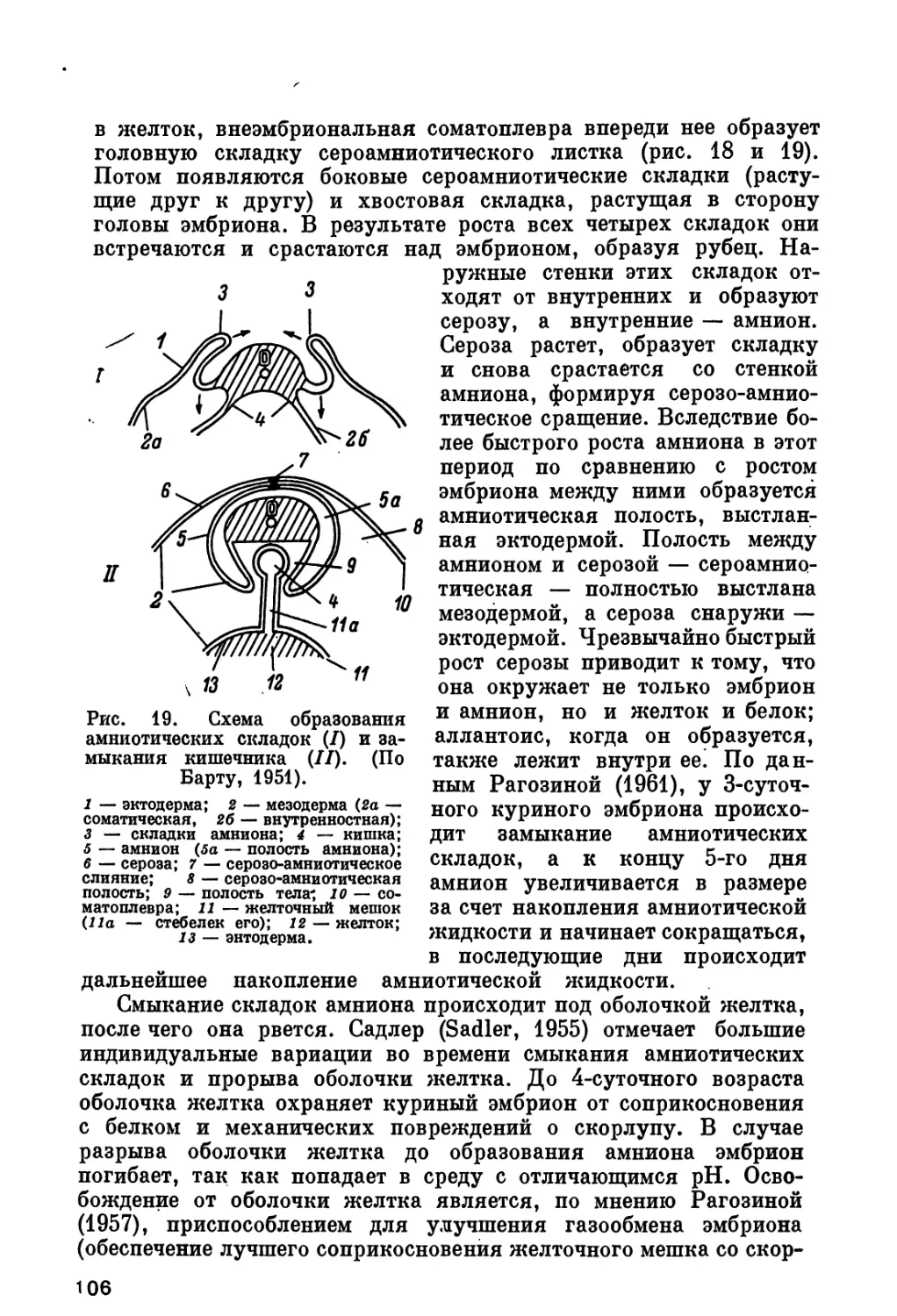
Э!
Рис. 4. Дробление яйца курицы в яйцеводе. Последовательные
стадии дробления бластодиска. (Из Кнорре, 1959).
А—Д — вид с поверхности; Е— 3 — вид на вертикальных разрезах (Е—первая
борозда дробления, Ж — стадия нескольких бластомеров, снизу еще не от-
деленных от желтка, 3 — более поздняя стадия дробления). 1 — борозды
дробления; 2 — след движения ядра в цитоплазме («тропа дробления»);
3 — метафаза второго деления дробления; 4 — желточная оболочка.
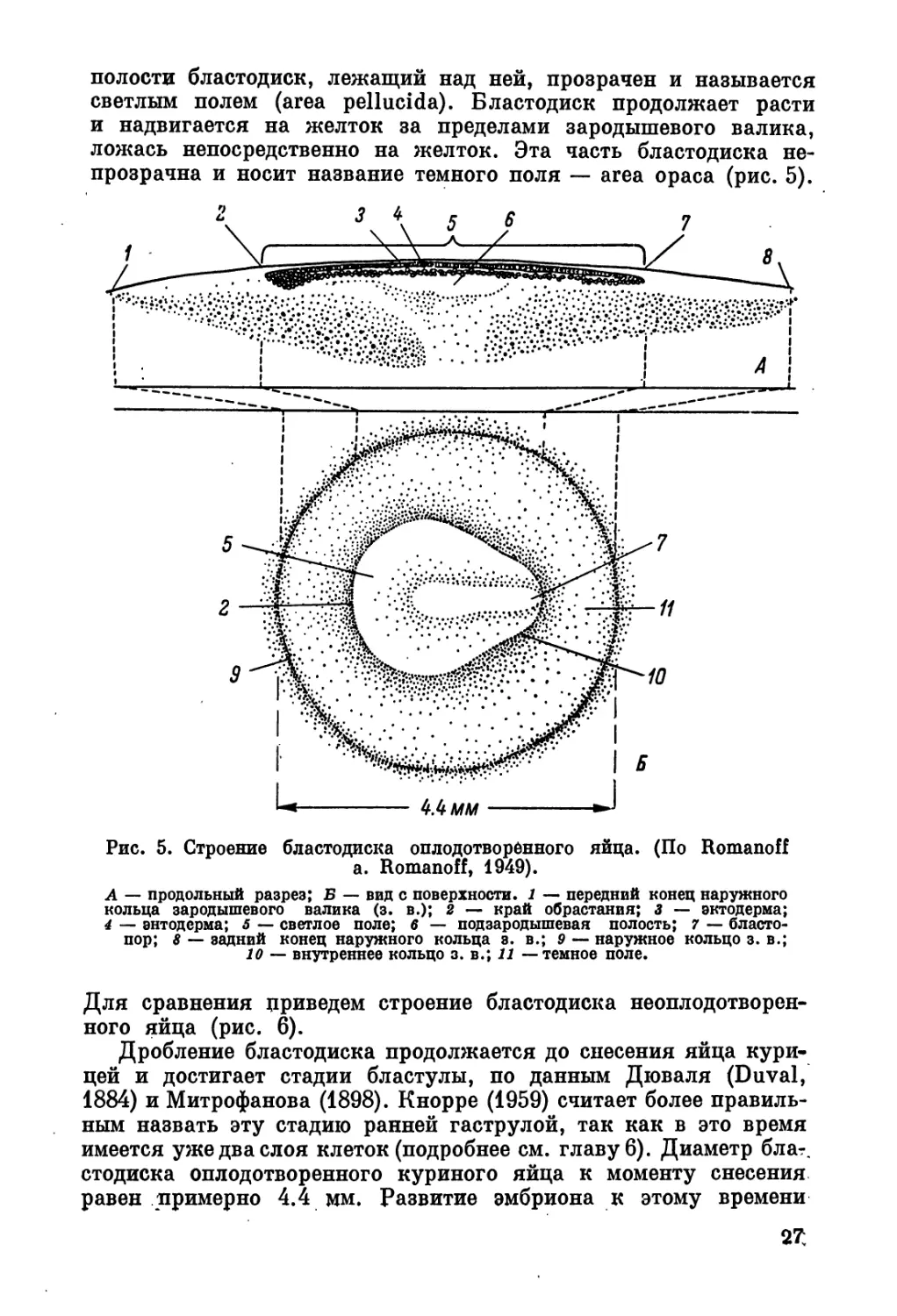
полости бластодиск, лежащий над ней, прозрачен и называется
светлым полем (area pellucida). Бластодиск продолжает расти
и надвигается на желток за пределами зародышевого валика,
ложась непосредственно на желток. Эта часть бластодиска не-
прозрачна и носит название темного поля — area ораса (рис. 5).
Рис. 5. Строение бластодиска оплодотворённого яйца. (По Romanoff
a. Romanoff, 1949).
А — продольный разрез; В — вид с поверхности. 1 — передний конец наружного
кольца зародышевого валика (з. в.); 2 — край обрастания; 3 — эктодерма;
4 — энтодерма; 5 — светлое поле; 6 — подзародышевая полость; 7 — бласто-
пор; 8 — задний конец наружного кольца з. в.; 9 — наружное кольцо з. в.;
10 — внутреннее кольцо з. в.; 11 — темное поле.
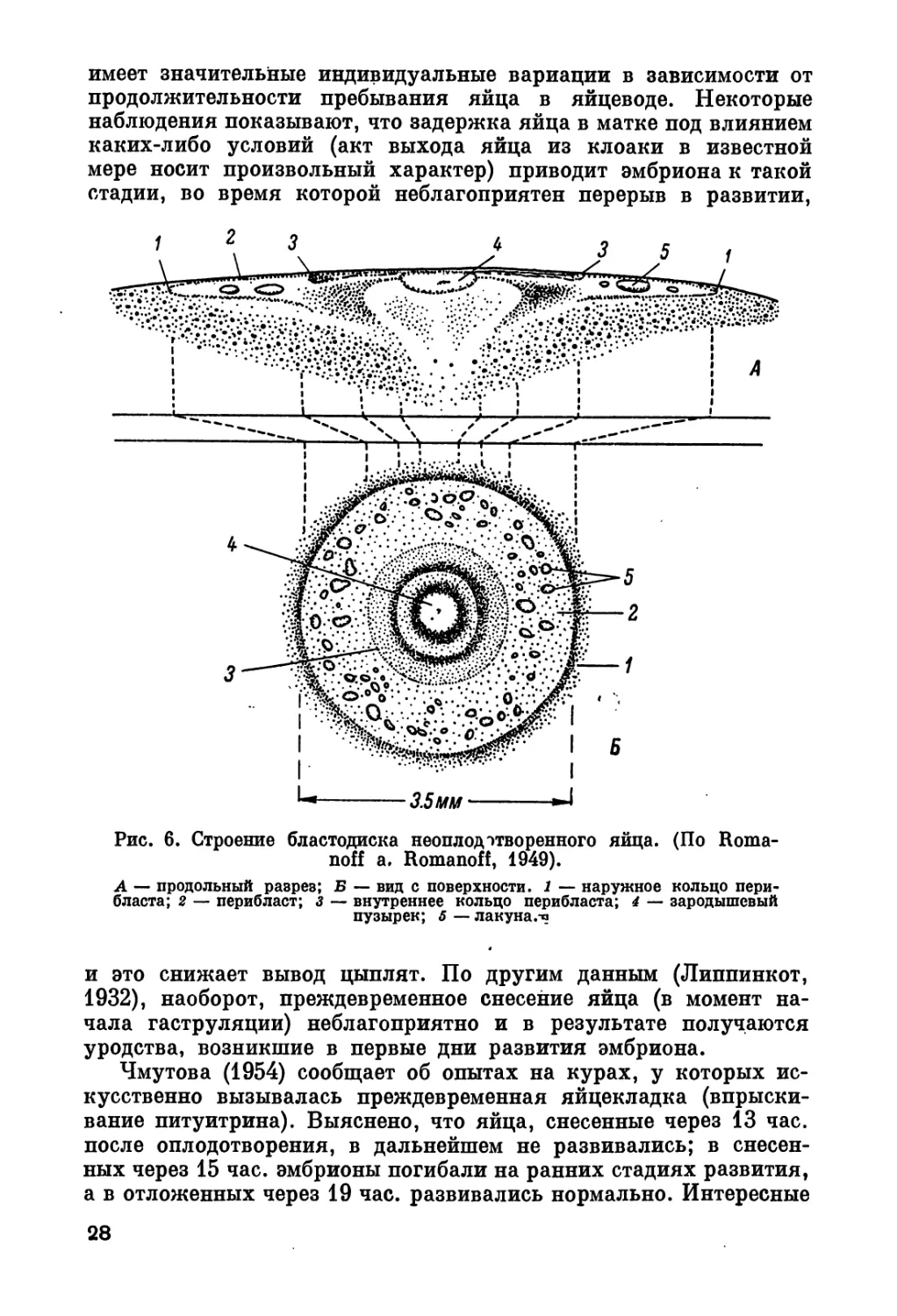
Для сравнения приведем строение бластодиска неоплодотворен-
ного яйца (рис. 6).
Дробление бластодиска продолжается до снесения яйца кури-
цей и достигает стадии бластулы, по данным Дюваля (Duval,
1884) и Митрофанова (1898). Кнорре (1959) считает более правиль-
ным назвать эту стадию ранней гаструлой, так как в это время
имеется уже два слоя клеток (подробнее см. главу 6). Диаметр блат,
стодиска оплодотворенного куриного яйца к моменту снесения
равен примерно 4.4 мм. Развитие эмбриона к этому времени
27
имеет значительные индивидуальные вариации в зависимости от
продолжительности пребывания яйца в яйцеводе. Некоторые
наблюдения показывают, что задержка яйца в матке под влиянием
каких-либо условий (акт выхода яйца из клоаки в известной
мере носит произвольный характер) приводит эмбриона к такой
стадии, во время которой неблагоприятен перерыв в развитии,
Рис. 6. Строение бластодиска неоплод отворенного яйца. (По Roma-
noff a. Romanoff, 1949).
А продольный разрез; Б — вид с поверхности. 1 — наружное кольцо пери*
бласта; 2 — перибласт; з — внутреннее кольцо перибласта; 4 — зародышевый
пузырек; 5 — лакуна.-п
и это снижает вывод цыплят. По другим данным (Липпинкот,
1932), наоборот, преждевременное снесение яйца (в момент на-
чала гаструляции) неблагоприятно и в результате получаются
уродства, возникшие в первые дни развития эмбриона.
Чмутова (1954) сообщает об опытах на курах, у которых ис-
кусственно вызывалась преждевременная яйцекладка (впрыски-
вание питуитрина). Выяснено, что яйца, снесенные через 13 час.
после оплодотворения, в дальнейшем не развивались; в снесен-
ных через 15 час. эмбрионы погибали на ранних стадиях развития,
а в отложенных через 19 час. развивались нормально. Интересные
28
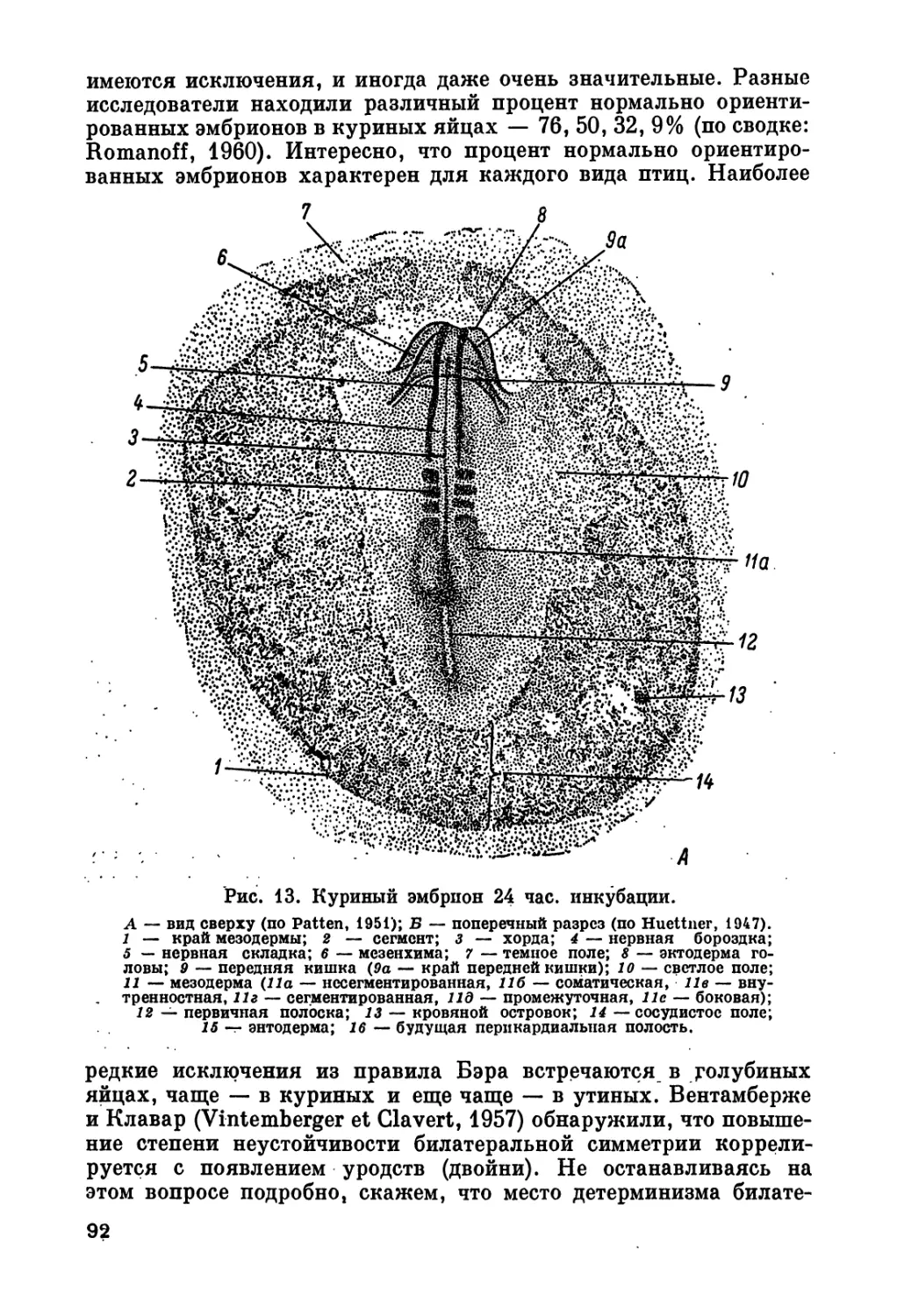
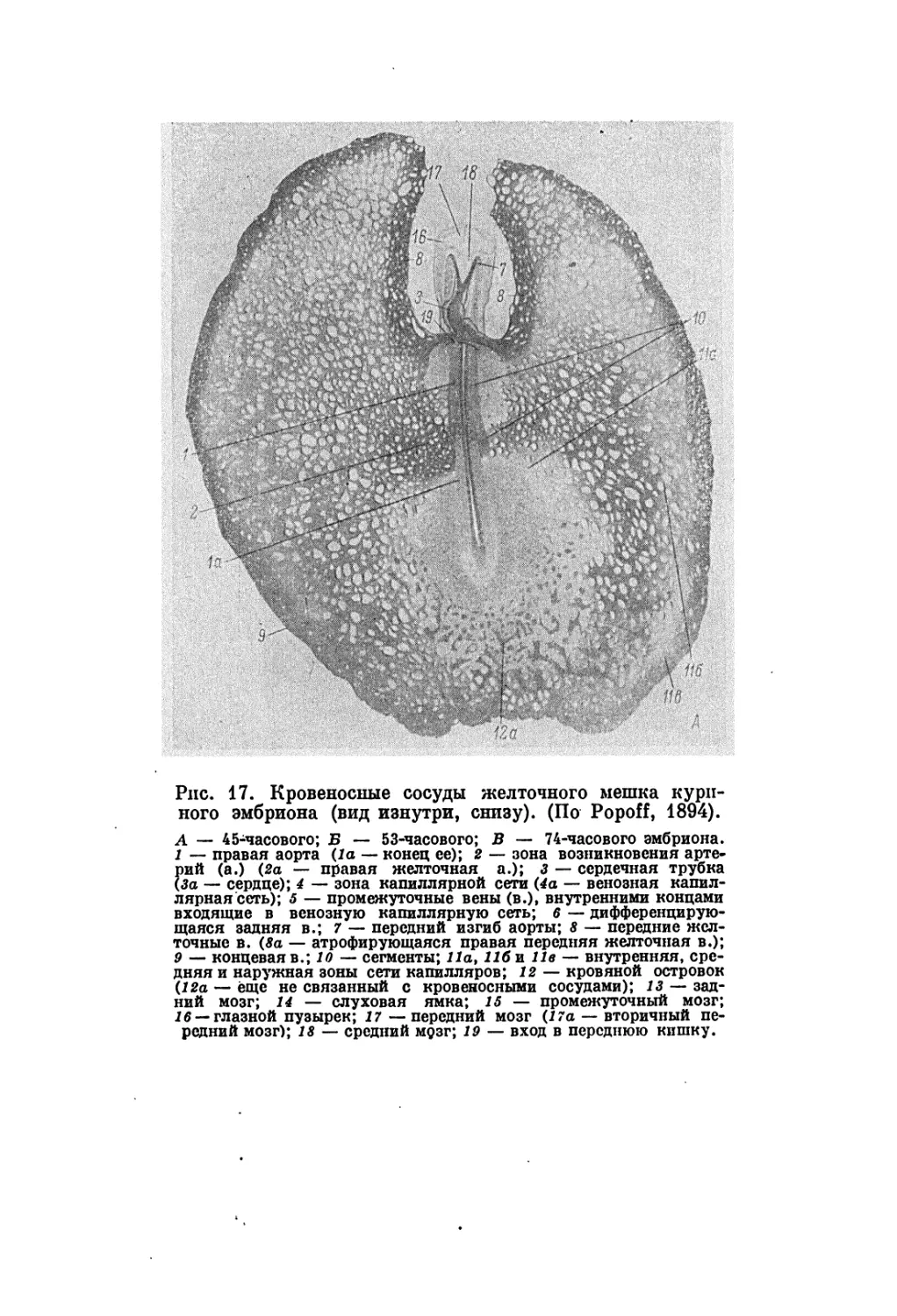
данные получены автором о различиях в стадии развития заро-
дышевых дисков в гусиных яйцах к моменту их снесения. На ста-
дии ранней гаструлы «сильных зародышей» (с продвинутым раз-
витием) было 66.6%, «слабых зародышей» (с неясно выражен-
ными морфологическими признаками) — 25.0%, на стадии дроб-
ления, т. е. с сильно отставшим развитием, — 3.3%, неоплодотво-
ренных — 5.0%. Анорова (1956) отметила, что в осеннее время
яйца дольше задерживаются в яйцеводе курицы, чем летом, и
соответственно стадия развития эмбрионов в осенних яйцах более
продвинута. Луц и Луц-Остертаг (Lutz et Lutz-Ostertag, 1957)
обнаружили сезонные различия в развитии эмбриона в только что
снесенных утиных яйцах. Развитие весной было более продвину-
тым и соответствовало развитию эмбриона в зимних яйцах через
несколько часов инкубации. По данным Владимировой (1962а),
диаметр зародышевого диска в гусином яйце имеет разный размер
в разные месяцы яйцекладки: 3.27 мм в марте и 3.99 мм в июне.
Существует также зависимость степени развития эмбри-
она к моменту снесения яиц от возраста кур. Анорова (1956)
наблюдала в яйцах кур-молодок развитие эмбрионов на стадии
ранней гаструлы, а в яйцах кур, несущихся второй год, — на
стадии поздней гаструлы. Автор считает, что именно эти разли-
чия обусловливают более продвинутое развитие эмбрионов в яй-
цах кур старшего возраста в первые дни инкубации, ранее опи-
санное ею (1953). Козин и Хёрли (Kosin a. Hurley, 1957), наоборот,
указывают, что в яйцах молодых кур (до полутора лет) было 65%,
а в яйцах кур старше двух лет — 95% с менее продвинутым раз-
витием эмбрионов. Возможно, что различие в наблюдениях осно-
вывается на различных свойствах разных пород. Данные Ано-
ровой "(1956), полученные на большом материале, кажутся нам
более достоверными.
Г Л А В А 3
НЕКОТОРЫЕ ОСОБЕННОСТИ ФИЗИОЛОГИИ НЕСУШЕК
В ПЕРИОД ЯЙЦЕКЛАДКИ
И ВЛИЯНИЕ ВНЕШНИХ ФАКТОРОВ
НА ПРОДУКТИВНОСТЬ
А. Влияние кормления птиц на их продуктивность
Биологическая полноценность яиц, возможность обес-
печить в дальнейшем нормальное развитие эмбриона, обусловли-
вается в значительной мере наличием в яйце всех необходимых
питательных веществ, которые в свою очередь образуются в орга-
низме несушки из потребляемого ею корма. Кроме той энергии,
29
которая в виде питательных веществ закладывается в яйце, до-
полнительно от 30 до 100% такого количества энергии расхо-
До овуляции овуляции После овуляции
Рис. 7. Изменение содержания жи-
ра, сахара, кальция и фосфора в
крови голубей в течение кладки.
(По Groebbels, 1937).
1 -г жир (на 1 мл); 2 — сахар (на 10 мл);
3 — кальций (на 100 мл); 4 —фосфор (на
10 мл). По оси, ординат — мг; по оси абс-
цисс — часы (до первой овуляции, в пе-
риод от первой до второй овуляции и по-
сле овуляции).
дуется на процесс формирова-
ния яйца (Дьяков, 1933).
Перед началом яйцекладки
кровь курицы становится чрез-
вычайно богатой материалами,
которые превращаются затем в
содержимое яиц. Концентрация
углеводов, липидов и минераль-
ных веществ в крови несушки
во время яйцекладки значитель-
но увеличивается ( табл. 4).
Сходно изменяется во время
кладки и состав крови самок
других видов птиц (рис. 7).
Обмен веществ у несушки
во время яйцекладки повыша-
ется. Необходимая для этого
дополнительная энергия полу-
чается главным образом из уг-
леводов. По данным Киселева
и Гавшиной (1966), ко времени
полового созревания в мышцах
яйцевода кур уровень макроэр-
гических соединений (АТФ и КФ) повышается, а в скелетных
мышцах снижается. Следовательно, макроэргические соединения
мобилизуются в это время из скелетных мышц, так как окисли-
тельные процессы в тканях яйцевода не обеспечивают энерге-
тических затрат на формирование яйца.
Таблица 4
Изменение концентрации различных веществ в крови несушки
(По Romanoff a. Romanoff, 1949)
Составные части плазмы У не несущей- ся курицы (в мг %) У несушки (в мг %) Увеличение (в %)
Общее содержание липидов .... 379.0 1347.0 255
Жирные кислоты 345.0 1387.0 302
Холестерин •. . 102.0 123.0 21
Протеины сыворотки 5540.0 8970.0 62
Глюкоза 171.1 179.2 5
Общее количество кальция 12.8 21.5 68
Фильтрующийся кальций 6.0 5.9 —2
Общее количество фосфора .... 11.8 35.6 202
Неорганический фосфор 4.3 5.4 26
Фосфор липидов 14.0 28.5 104
Фосфор лецитина 7.5 17.0 127
30
Чтобы обеспечить яйцо полноценным составбм питательных
веществ для дальнейшего нормального развития эмбриона, не-
обходимо включить в рацион несушек разнообразные кормовые
вещества. К ним относятся протеины (животные и растительные),
углеводы, жиры, минеральные вещества и витамины. Использо-
вание курой-несушкой разных питательных веществ для превра-
щения их в вещества, содержащиеся в яйце, различно (табл. 5).
Таблица 5
Эффективность превращения курой-несушкой питательных веществ корма
в органические вещества яиц
(По Romanoff a. Romanoff, 1949)
Вещества Единицы измерения Количество питательных веществ Использование питательных веществ (в %)
в 135 г корма в 51.6 г содер- жимого яйца
Общее количество органических веществ г 126.0 13.2 10.5
Протеины Жиры » 21.6 6.6 30.5
» 6.7 6.1 91.1
Витамины: А ME 1080.0 200—800 46.3
Вх » 135.0 20-40 22.2
D » 108.0 10-50 27.8
Рибофлавин мкг 337.0 100-200 44.5
Пантотеновая кислота » 1890.0 600-1200 47.6
Общий запас энергии .... кал. 396.0 82 20.7
Несушки в период яйцекладки нуждаются в значительно
большем количестве протеина в корме, чем в другие пе-
риоды жизни. Для образования 1 г яичной массы курице необ-
ходимо 0.16—0.25 г, а гусыне — 0.28 г переваримого протеина.
Отношение количества протеина к количеству остальных пере-
варимых питательных веществ должно быть для несушки в сред-
нем 1 : 4. Однако повышать процент протеина в рационе надо
чрезвычайно осторожно, так как избыток его в корме несушки
снижает выводоспособность яиц (Сергеев, 1940; Кудрявцев, 1948;
Сергеев и др., 1949; Кушнер, Корепанова и Букин, 1950). Осо-
бенно вредно сказывается повышение количества протеина в корме
несушек-молодок, обусловливающее преждевременную интенси-
фикацию яйцекладки. Маслиев (1965) показал, что снижение
количества переваримого протеина с 20—21 г на голову в сутки
до 13—14 г в течение 1.5 месяцев перед началом яйцекладки обус-
ловило нормальное развитие молодок и повышение яйценоскости
после этого срока на 5—15%. Отрыганьев (1951) сообщает, что
31
при излишнем скармливании белковых кормов резко снижается
качество яиц: белок разжижен, желток подвижен. В результате
белкового отравления повышается смертность эмбрионов в сере-
дине и конце инкубации и появляются эмбрионы, заболевшие
хондродистрофией, проявляющейся в сильной коротконогости.
Автор наблюдал подобные формы хондродистрофии у кури-
ных, индюшиных, фазаньих, цесарочьих и реже у утиных
эмбрионов.
Для получения полноценных яиц важно не только общее ко-
личество белков в рационе несушек, но и то, какие именно про-
теины вошли в рацион. Для нормальной жизнедеятельности и
продуктивности птицы протеины пищи должны содержать набор
всех аминокислот, из которых в организме несушки синтези-
руются белки ее тела и яиц. К этим аминокислотам относятся
аргинин, лизин, триптофан, тирозин, цистин, гистидин, метио-
нин, треонин, глицин, фенилаланин. Некоторые из этих амино-
кислот могут синтезироваться в организме птицы, но такие амино-
кислоты, как цистин, триптофан и лизин не синтезируются, и
отсутствие или недостаток их в пище даже при наличии достаточ-
ного количества другой белковой пищи (например, зерновых кор-
мов) пагубно отражается-на здоровье птицы и качестве ее яиц.
Только правильное сочетание растительных и животных белковых
кормов обеспечивает нормальную жизнедеятельность организма
птицы и возможность нести яйца с высокой выводоспособностью.
Замена животных белков растительными в рационе кур приводит
к снижению оплодотворенности и выводоспособности яиц (Byerly
et al., 1933).
Протеины белка и желтка яйца извлекаются из сыворотки
крови курицы. В момент овуляции количество протеинов в сыво-
ротке крови увеличивается. Резниченко (1961) наблюдал, что
в период интенсивной яйцекладки показатели азотистых фракций
крови и белкового отдела яйцевода изменяются в зависимости от
стадии формирования яйца. Особенно напряжен азотистый обмен
в крови и яйцеводе в период формирования белка яйца: в сыво-
ротке крови увеличивается количество белкового азота, в белко-
вом отделе яйцевода повышается содержание белкового и небел-
кового азота, а в крови уменьшается количество остаточного азота,
по-видимому за счет большого расхода полипептидов крови
на формирование белка яйца. Савронь (1965) показала, что между
содержанием белка и небелковых азотистых веществ в яйцеводе
кур и содержанием белков и остаточного азота в крови имеется
тесная зависимость. У молодых кур самое низкое количество
белка в крови; оно резко нарастает к моменту полового созрева-
ния, и особенно к периоду весенней яйцекладки, а во время линьки
не уменьшается (в отличие от содержания белка в яйцеводе,
где оно во время линьки сильно снижается). Химический состав
яйцевода, по данным автора, изменяется не только в связи с из-
32
менением физиологического состояния организма, но и в течение
суток во время формирования белка и скорлупы яйца.
Углеводы и липиды корма сначала распадаются
в организме птицы на более простые составные части, а затем не-
которые из них окисляются, освобождая при этом энергию,
а другие превращаются снова в углеводы и липиды, значитель-
ное количество которых идет па образование яиц. Липиды осо-
бенно ценны в качестве материала для формирования яиц и
в основном расходуются на образование желтка. Подъем количе-
ства липидов в крови несушки происходит одновременно с уве-
личением количества кальция и фосфора. Фосфолипиды и жиры
производятся печенью, аккумулируются там в период, предше-
ствующий кладке, и затем переносятся кровью в яичник. Наблю-
дая высокую выводимость (86% от заложенных яиц) при инкуби-
ровании яиц кур леггорн, получавших недостаточное кормление,
Сергеев и Кудрявцев (1941) приходят к выводу, что птица имеет
запасы, используемые в случае необходимости, причем вес птицы
и продуктивность ее уменьшаются, а выводоспособность яиц
(при ненарушенном белковом отношении) не снижается. Данные
Калининой (1958) подтвердили это: рационы из малоконцентрат-
ных кормов, богатых легкорастворимыми углеводами (картофель,
свекла, брюква и др.), благоприятно влияют на гликогенообра-
зовательную функцию печени кур-несушек, что является важ-
ным фактором в регуляции общего обмена. Снесенные этими не-
сушками яйца имели повышенную выводоспособность (86.2—
87.5% при 85.6% в контроле), и вылупившиеся при этом цыплята
были более жизнеспособными (до 5 месяцев сохранилось 96.2—
97.5% цыплят при 91.7% в контроле).
Общее количество минеральных веществ в ку-
рином яйце равно 7 г. По данным Кржишковского (1933), только
25—30% из потребленных птицей минеральных веществ усваи-
вается организмом и может быть использовано для образования
яиц. Следовательно, на формирование каждого яйца должно быть
потреблено несушкой не менее 25—28 г минеральных веществ.
С каждым яйцом из организма курицы выводится около 2 г
кальция, т. е. за год около 300—400 г, что в 10—14 раз превышает
содержание кальция во всем теле курицы. В связи с тем что в яй-
цеводе нет никаких запасов минеральных солей, кальций должен
доставляться яйцеводу с кровью. Но максимальное содержание
кальция в крови достигает 35 мг%, что не смогло бы обеспечить
формирования даже небольшой доли кальция, скорлупы, если бы
в организме птицы не было запасов этого элемента. Недостающее
количество кальция частично компенсируется путем увеличения
усвоения несушками кальция из пищи, которое начинается за 6—
15 дней (с 5% до 9%) и особенно повышается за 1—5 дней (до 17%)
перед кладкой первого яйца; при этом накапливается 4.34 +
+0.86 г кальция (Фостер, 1966). Но основное количество недо-
3 В. В. Рольник
33
стающего кальция пополняется из запасов его в костях несушки.
Симкис (Simkiss, 1961) считает, что если хорошие несушки
получают кальций в корме в неограниченных количествах, то
содержание его в крови во время кладки остается все же не-
изменным, так как куры-несушки способны извлекать ежесуточно
из корма 1.6—2 г кальция. Если же кальция в корме недостаточно,
то половина необходимого для скорлупы яиц кальция может
быть извлечена из костей несушки (при этом около 10% кальция
костей несушки может быть израсходовано на скорлупу яиц).
Продолжающаяся нехватка кальция в рационе приводит к затя-
гиванию овуляции, утончению скорлупы яиц и ослаблению ко-
стяка несушки. В период перерыва в яйцекладке кости несушки
приобретают прежний вес. Методом меченых атомов Ошима и
Носаки (Oshima a. Nosaki, 1962) установили, что кальций для об-
разования скорлупы вначале черпается из костей несушки,
а затем частично восполняется кальцием корма. По данным Мла-
денова (1955), у несушек перед началом яйцекладки происходит
интенсивное создание запасов кальциевых солей в виде костного
вещества в полостях трубчатых костей. Это новообразованное
костное вещество богато остеокластами, что свидетельствует
о большой активности здесь процессов разрушения кости. Методом
меченых атомов Бауман (1956) подтвердила, что основной запас
кальция для образования скорлупы находится в костном мозгу.
Сюда в первую очередь попадает кальций из пищи и отсюда он за-
тем черпается для нужд организма, и в частности для образова-
ния скорлупы. Са45 наиболее быстро включается и также быстро
исчезает именно в костном мозгу. Под влиянием половых гормо-
нов значительно увеличивается образование трабекул в костном
мозгу, что позволяет в период подготовки к яйцекладке повысить
утилизацию кальция у молодок с 20 до 70%. Автор подчеркивает,
что при оценке молодняка сельскохозяйственных птиц костяк
должен считаться не только показателем хорошего экстерьера,
но и оцениваться с точки зрения его непосредственного участия
в продуктивности. В интересной сводке Симкисса (Simkiss, 1961)
о кальциевом обмене в организме птиц в период размножения
отмечается, что морфо-физиологическая система сердцевинных
костей обнаружена только у птиц. Потребность несушек в каль-
ции корма во время яйцекладки находится под контролем поло-
вых гормонов. Инъекции эстрогенных и андрогенных гормонов
птицам обусловливают увеличение содержания кальция и фосфора
в крови несушек в несколько раз. Уровень кальция в крови пе-
тухов во время половой активности не изменяется. Интересно,
что инъекциями половых гормонов не удавалось вызвать'окосте-
нение костного мозга у черепах. Автор приводит интересное срав-
нение трех групп наземных позвоночных по источнику кальция
для развивающегося эмбриона: для рептилий — это главным
образом желток, для птиц — в основном скорлупа, а для млеко-
34
питающих —- передача кальция из крови матери через плаценту.
Метаболизм кальция у птиц во время образования скорлупы яйца
примерно в 7 раз быстрее, чем у черепах, и в 20 раз — чем у мле-
копитающих в конце беременности, а скорость извлечения каль-
ция из крови у птиц (несушек) в 5 раз больше, чем у млекопитаю-
щих.
Ханзакер и Старки (Hunsaker a. Sturkie, 1961) путем наложе-
ния канюль на вены у входа в матку и у выхода из нее выяснили,
что во время нахождения яйца в матке содержание кальция плазмы
уменьшается при прохождении через нее на 5.3 мг/100 мл или
на 21.4%, а при отсутствии яйца — только на 1.2 мг/100 мл или
на 4.4%; последнее снижение кальция в плазме связано с откла-
дыванием его в запас в отдыхающей матке.
Венже и Смит (Winget a. Smith, 1957) нашли, что кальций
крови у кур-несушек состоит из равных частей органически
связанного и ионного кальция, а при образовании скорлупы
связанный кальций расходуется больше, так что относительная
концентрация ионного кальция при этом увеличивается. Введе-
ние несушкам инсулина, понижающего содержание в крови свя-
занного с протеинами кальция, приводит к тому, что весь каль-
ций крови находится в ионной форме. Далее Венже с соавторами
(Winget et al., 1958) установил, что около 20% кальция, цирку-
лирующего в крови, переходит в состав скорлупы главным об-
разом в связанной с протеинами форме. Авторы предполагают,
что при этом происходит очень быстрая ее диссоциация. Деталь-
ное исследование изменений форм кальция в крови кур-несушек
провел Волков (1956а). Автор наблюдал в сыворотке крови кур
при переходе из стадии покоя к стадии начала яйцекладки воз-
растание общего количества кальция (с 12.0—13.2 мг% до 23.1 —
30.8 мг%), в том числе диффундируемого (с 8.8—10.2 мг% до
19.7—20.0 мг%) и связанного с глобулинами (с 1.4 мг% до 2.3—
8.8 мг%). Это увеличение кальция было обусловлено усилением
процессов ассимиляции кальция в связи с началом яйцекладки.
При увеличении интенсивности яйцекладки в крови несушек
становится значительно меньше кальция, связанного с протеи-
нами (альбуминами и глобулинами). Именно эти соединения при-
нимают участие в формировании яйца и усиленно расходуются
в период интенсивной яйцекладки. Кальций, связанный с альбу-
минами, очень лабилен. Обычно при наличии в яйцеводе форми-
рующегося яйца содержание кальция, связанного с альбуминами
сыворотки, невелико или эта форма кальция отсутствует совсем.
Автор отметил, что способность накапливать в сыворотке крови
большое количество соединений кальция с протеинами свойственна
несушкам, в особенности высокопродуктивным, и является одной
из причин их высокой яйценоскости. Гертеленди и Тэйлор (Нег-
telendy a. Taylor, 1961) установили, что при образовании скор-
лупы яйца падает не только общее количество кальция в крови,
3*
36
но и содержание растворимого кальция. Это происходит, по-ви-
димому, под влиянием паратиреоидного гормона, который моби-
лизует кальций из костного мозга для образования скорлупы.
Между относительным весом скорлупы (к весу яйца) и коли-
чеством клеток на единицу объема паращитовидных желез, как
показал Аш (Ash, 1958), существует достоверная высокая отри-
цательная корреляция. Автор предполагает, что большие по раз-
меру клетки (их тогда меньшее количество) паращитовидных
желез секретируют больше гормона, который мобилизует большее
количество кальция для образования скорлупы.
В кальцификации скорлупы участвует энзим — угольная ан-
гидраза. Энзим играет роль катализатора в освобождении карбо-
натных ионов из крови, которые соединяются с кальцием крови,
образуя СаСО3. Слабая щелочность яйца благоприятствует оса-
ждению СаСО3 на подскорлупные оболочки. Гутовская и Митчелл
(Gutowska a. Mitchell, 1945), впрыскивая разное количество
угольной ангидразы в матку несушки, получали скорлупу яиц
различной толщины. Авторы обнаружили, что содержание уголь-
ной ангидразы в матке и в яйцеводе в целом у несушек выше,
чем у ненесушек. По данным Налбандова (Nalbandov, 1958),
продолжительные инъекции стероидных и тиреоидных гормонов,
несмотря на то что они вызывали увеличение размера яйцевода
и обусловливали начало секреции белка, не повышали, а пони-
жали активность угольной ангидразы. Автор предполагает, что
гипофиз каким-то (пока неизвестным) образом контролирует ак-
тивность этого энзима в яйцеводе, но это влияние не опосредо-
вано стероидными гормонами.
Основная масса фосфора, участвующего в образовании яйца,
идет на формирование желтка и связана с фосфатидами и фосфо-
протеинами (ововителлин). На построение скорлупы идет незна-
чительное количество фосфора, но еще меньшее количество его
расходуется при образовании белка. Волков (19566) установил,
что изменение количества фосфорных соединений в сыворотке
крови кур-несушек происходит таким же образом, как и изме-
нение количества кальция. При переходе из стадии покоя к ста-
дии начала яйцекладки увеличивается общее количество фосфора
(с 15.6—15.9 мг% до 35.5—58.3 мг%), в том числе не связанного
с глобулинами (с 4.7—5.3 мг% до 19.7—33.1 мг%) и липидного
(с 4.8—6.5 мг% до 15.0—23.3 мг%). В период интенсивной яйце-
кладки автор наблюдал значительное уменьшение относительных
величин фосфора, связанного с альбуминами и глобулинами,
что указывает на его непосредственное участие в яйцеобразовании.
Специфической чертой обмена высокопродуктивных кур является
способность накапливать в крови липидный фосфор и в еще боль-
шей степени фосфор, связанный с глобулинами. Автор отмечает,
что у высокопродуктивных кур уровень этих фосфорных соеди-
нений остается очень высоким в течение всего периода яйцекладки
36
и, следовательно, расход их на яйцеобразование успевает компен-
сироваться. По данным Бауман (1966), снижение уровня фосфо-
протеина в крови несушек является основной причиной падения
в ней уровня кальция и нарушения формирования скорлупы.
Кальций и фосфор — это наиболее необходимые элементы
в минеральном кормлении несушек. Особенно важно соотноше-
ние кальция и фосфора в рационе. Исследуя минеральное питание
кур-несушек, Кудрявцев (1940) показал влияние различных ко-
личеств кальция и фосфора и их соотношения в рационе несушек
на выводимость цыплят (табл. 6).
Таблица 6
Влияние различного содержания кальция и фосфора в рационе несушек
на выводимость цыплят
(По Кудрявцеву, 1940)
Группа Минеральные корма Среднесуточное потребление (в г) на голову % вывода без браковки яиц
каль- ция фосфора отношение кальция к фосфору от зало- женных яиц от оплодо- творенных яиц
1 Контроль 2.53 1.17 2.17 67.2 76.2
2 На 50% кальция боль- ше 3.47 1.10 3.16 72.3 79.5'
3 На 35% фосфора боль- ше 2.30 1.44 1.59 81.2 89.6
4 На 50% кальция и 35% фосфора больше . . 3.78 1.55 2.44 77.3 84.2
5 На 25% кальция мень-. ше 2.03 1.21 1.67 73.0 80.5
6 Ракушки вволю . . . 2.80 1.10 2.53 74.2 80.9
Примечание. Всего было проинкубировано около 5000 яиц.
Автор приходит к выводу, что избыток кальция в рационе не-
сушек отрицательно сказывается на выводимости, задерживая
выход цыплят из яиц и увеличивая процент задохликов и слабых.
Наоборот, повышение дозировки фосфора до 1.5—1.7 г на голову
в сутки (по сравнению с ранее принятой нормой — 0.6—0.8 г)
при отношении кальция к фосфору от 1.6 до 2.3 обеспечило при
65—75% яйценоскости высокую выводимость — до 81% от за-
ложенных без браковки яиц. По данным Крикуна (1958), наилуч-
шим соотношением кальция к фосфору в кормах птиц в племен-
ной период является 2 : 1 или 3:1.
Использование кальция и фосфора организмом птицы зависит
не только от правильного их соотношения, но и от наличия ви-
тамина D. По данным Берга с соавторами (Berg et al., 1962),
37
уровень кальция в корме кур-несушек от 0.5 до 2.0% в течение
8—21-й недели не влияет на жизнеспособность потомства, а уве-
личение содержания кальция с 1.75 до 3.25% в течение следую-
щих трех недель резко снижает как выводимость, так и жизне-
способность цыплят. Адемосун и Николайчук (1966) сообщают,
что для оптимального соотношения кальция и фосфора в орга-
низме несушек при 80% яйценоскости необходимо, чтобы содер-
жание кальция в рационе было около 2.6%, а фосфора — 0.6—
0.7%. Куры-молодки усваивают кальций значительно хуже,
чем переярки, что, возможно, является причиной более низкого
качества скорлупы в яйцах молодок. Матускова (1950) показала,
что гусыне для образования 10 г яичной массы необходимо 0.57 г
кальция и 0.85 г фосфора. Добавление к рациону, включавшему
2 г соли в сутки на гусыню, 12 г мела и 3 г костяной муки повысило
выводимость на 27—49%.
Из исследований о других минеральных элементах, необходи-
мых для формирования яиц, мы упомянем лишь две работы.
Патрик (1939) приводит данные о повышении яйценоскости кур,
оплодотворенности яиц и выводимости цыплят под влиянием
скармливания несушкам марганца (табл. 7).
Таблица 7
Влияние марганца в рационе несушек на их продуктивность
(По Патрику, 1939)
Количество марганца (в мг на 1 кг корма) Длительность опыта (в пнях) Количество яиц (на 1 курицу) % оплодо- творенности Вывод цыплят (в % от опло- дотворенных яиц)
13 126 43 86.0 42.8
200 127 83.4 93.3 85.8
13 77 25.6 83.5 41.3
53 77 42.2 95.5 86.3
По данным Жукова (1956), добавление в рацион несушек
микроэлементов и железа в количестве (в сутки на голову) 10 мг
сернокислого железа, 2 мг хлористого кобальта, 5 мг хлористого
марганца, 2 мг сернокислой меди и 7 мг сернокислого цинка, не
увеличивая яйценоскости, повышает выводимость с 80.6 до 90.0%
за счет повышения оплодотворенности яиц (на 6.7%) и снижения
процента задохликов с 5.3 до 2.6.
Большое влияние на выводоспособность яиц имеют вита-
мины в корме несушек, так как количество витаминов в корме
в значительной степени определяет наличие их в яйцах. Кушнер
с сотрудниками (1950) наблюдал, что добавление в корм протеи-
нов и минеральных веществ не оказывает положительного эффек-
38
та на продуктивность птицы без добавления витаминов A, D
и В2.
При недостатке витамина А в рационе задерживается рост
птиц, нарушается их обмен веществ, наступает частичная или
полная стерильность и в результате понижается процент оплодо-
творенных яиц и выводимость молодняка. У птиц потребность
в витамине А в 5—10 раз больше, чем у млекопитающих. По
данным Баранчеева (1939), в яйце откладывается от И до 32%
из усвоенного курицей витамина А. Повышенная доза витамина А
в рационе вредна, но ядовитое действие витамина А проявляется
только при принятии дозы, превышающей нормальную в 300—
500 раз, что случается очень редко. Избыток каротина, являюще-
гося провитамином А, не имеет отрицательного действия. Зем-
ская (1940) сообщает, что при недостатке витамина А в корме
несушек эмбрионы в их яйцах погибают главным образом в пер-
вые дни развития. Маслиева (1950) показала, что. содержание
витамина А в желтке утиных яиц и выводимость утят зависят от
содержания этого витамина в рационе маточного стада (табл. 8).
Таблица 8
Влияние снабжения витамином А маточного стада на содержание
этого витамина в желтке яиц и выводимость утят
(По Маслиевой, 1950)
Группа Количе- ство ви- тамина А в ра- ционе (в гам- мах) 18 Ш 15 IV 9 V
количе- ство ви- тамина А (в гаммах) в 1 г желтка % вы- вода от заложен- ных яиц количе- ство ви- тамина А (в гаммах) в 1 г желтка % вы- вода от заложен- ных яиц количе- ство ви- тамина А (в гаммах) в 1 г желтка % вы- вода от заложен- ных яиц
1 7.5 58.1 Следы 8.4 0
2 2000 20.2 95.9 20.7 85.8 17.5 84.4
« По данным Ребрикова (1965), при содержании 6—& мкг каро-
тина на 1 г желтка утиных яиц вместо 15 мкг по норме выводи-
мость утят снизилась до 60% и жизнеспособность их была
очень низкой. Автор установил, что количество каротина в желтке
яиц прямо пропорционально содержанию его в сыворотке крови
несушек. По наблюдениям Голяркина (1947), недостаток вита-
мина А в корме индеек приводит к снижению содержания этого
витамина в яйцах до 4.2 мкг на 1 г желтка и понижению выводи-
мости индюшат до 48.7%. Кормление индеек в течение 25 дней
кормовой смесью, содержащей 25 000 мкг витамина А на 1 кг
корма, повысило содержание витамина А в яйцах до 14.6 мкг,
а вывод индюшат — до 70.6%. По мнению автора, для индеек
39
необходимо 20 000 мкг витамина А на 1 кг корма. Гримлеби и
Блэк (Grimleby a. Black, 1950) заметили, что каротин и витамин А
запасаются в организме курицы до кладки и потому переносятся
в большем количестве в первые яйца, а с каждым следующим яй-
цом концентрация их в яйце уменьшается. Чайковская (1949)
указывает, что витамины, и в первую очередь витамин А, ока-
зывают огромное влияние (как биогенные стимуляторы эндокрин-
ных желез и нервной системы) на интенсивность яйцекладки
гусынь и качество спермы гусаков. Вследствие различных ави-
таминозов и гиповитаминозов в результате недостаточного кор-
мления гусей как в предплеменной, так и в племенной сезоны
наблюдался низкий процент оплодотворенности яиц и высокая
эмбриональная смертность.
Витамин D (противорахитический), так же как и витамин А,
относится к группе липидов. При наличии солнечного света или
ультрафиолетового облучения птица способна синтезировать ви-
тамин D и откладывать его в запас в виде провитамина — эргосте-
рина. Витамин D способствует усвоению кальция и фосфора.
Яйца с недостатком витамина D имеют тонкую, хрупкую скор-
лупу. Таким образом, если в партии имеется большое количество
яиц с ненормальной скорлупой, то очень возможно, что из этих
яиц вылупятся рахитичные цыплята. В опытах Земской (1940)
при отсутствии витамина D в рационе кур-несушек выводимость
была равна 55%; при введении в рацион рыбьего жира вывод
цыплят повысился до 74%, а при облучении ультрафиолетовыми
лучами (без рыбьего жира) — до^70%. По наблюдениям Голяр-
кина (1947), витамин D необходимо вводить в рацион индеек в ко-
личестве 1300 мкг на 1 кг корма. Сергеев с соавторами (1949)
обнаружил, что на рационе, лишенном витамина D, утки несут
яйца с пониженной на 10% выводоспособностью. Отрыганьев
(1940) наблюдал, что чем меньше птица получала в пище вита-
мина D, тем чаще и сильнее у эмбрионов проявлялась эмбриональ-
ная дистрофия (особенно часто появлялись серозные отеки кожи).
С наступлением солнечной погоды (условия, исключающие
D-авитаминоз) отечная форма эмбриональной дистрофии исче-
зает. В дальнейшем Отрыганьев и Отрыганьева (1966) обнаружили *
значительные индивидуальные вариации восприимчивости кур
к гиповитаминозу D и установили следующие его диагностические
признаки: уменьшение толщины скорлупы и в связи с этим сни-
жение удельного веса яиц; снижение выводимости из-за высокой
смертности от эмбриональной дистрофии (особенно в середине
инкубационного срока); при вылуплении — наличие большого
количества слабых цыплят с признаками перозиса; повышенное
содержание щелочной фосфатазы в плазме крови кур, эмбрионов
и суточных цыплят.
Витамин Е также относится к группе жирорастворимых вита-
минов. Отсутствие или недостаток витамина Е в корме обуслов-
40
ливает нарушение сперматогенеза и, таким образом, понижает
оплодотворенность яиц до нуля и уменьшает выводоспособность
яиц, повышая смертность эмбрионов в первые четыре дня инкуба-
ции. При полном отсутствии в рационе кур-несушек витамина Е
они хотя и несут яйца, но с нулевой* выводоспособностью, даже
при оплодотворении спермой от петухов, имевших рацион, бога-
тый витамином Е. Превышение нормы витамина Е не влияет на
процент вылупления цыплят. Кудряшов (1932) приводит данные
опыта постепенного повышения выводимости из яиц кур, ранее
лишенных, а затем получивших витамин Е. Через 7 дней после
дачи витамина Е вывод с нуля поднялся до 32.6%, через 14 — до
61.4%, через 28 дней — до 69.4%. При Е-авитаминозе наблю-
дается нарушение образования кровеносной системы куриных
эмбрионов (кровеносные сосуды образуют кольцо, не связанное
с эмбрионом).
Витамины группы В — воднорастворимые и не накапливаются
в теле птиц в больших количествах, а потому необходимо система-
тическое, бесперебойное их поступление в организм вместе с пи-
щей. Недостаток витаминов этой группы приводит к серьезным
нарушениям жизненных процессов.
Витамин Вх предохраняет от полиневрита и дерматита цыплят,
а также повышает выводимость, но значительно меньше, чем ви-
тамин В2. Обычное наличие его в пище птицы достаточно, чтобы
не было авитаминоза, особенно если включить в рацион отруби.
Витамин Вх принимает участие в углеводном обмене и, как отме-
чает Кондратюк (1948), повышение общей калорийности рациона
может вызвать авитаминоз Вх. Кржишковский (1933) указы-
вает, что потребность в витамине Вх больше, когда в рационе
птицы преобладают протеины, и меньше, когда в рационе пре-
обладают жиры и углеводы.
Витамин В2 (рибофлавин) имеет наибольшее значение для
птиц из всех витаминов группы В; он способствует окислитель-
ным процессам в организме и росту птиц. При недостатке вита-
мина В2 в корме несушек сильно понижается выводимость, а вы-
лупившиеся цыплята страдают параличом (Кондратюк, 1948).
Избыток витамина В2 не токсичен. По исследованию Третьяковой
(1941), при недостатке в рационе птицы витамина В2 в яйцах на-
блюдались изменения физико-химических показателей белка и
желтка. Браун (Brown, 1957) наблюдал прямую передачу рибо-
флавина яйца эмбриону, причем наибольшее его количество по-
глощалось эмбрионом на 14—15-й день инкубации и смертность
эмбрионов при недостатке витамина была наивысшей именно
в это время. При инъекции витамина В2 в яйцо он полностью ис-
пользовался эмбрионом. Как указывает Кондратюк (1948), по-
требность кур-несушек в витамине В2 равна 550 мкг витамина
на голову в сутки, в том числе за счет кристаллического рибо-
флавина 300 мкг. По данным Кудрявцева (1948), недостаток ви-
41
тамина В2 (142 мкг на несушку в сутки) снижал выводимость
цыплят породы род-айленд до 68.7% от заложенных яиц при 76.5—
78.8% в контроле, когда потребление витамина В2 равнялось
200 мкг. Повышение количества витамина В2 до 341 мкг не уве-
личило выводимости цыплят по сравнению с контролем. В рацион
индеек необходимо вводить витамин В2 в количестве 4500 мкг
на 1 кг корма (Голяркин, 1947). Отрыганьев (1940) наблюдал,
что В2-авитаминозная форма эмбриональной хондродистрофии
в отличие от D-авитаминозной проявляется и в летний сезон.
Как показал Лепешенков (1965), введение в рацион несушек
витамина В12 повышает биологическую полноценность яиц. При
повышении содержания витамина В12 на 1 г желтка яйца
с 24.6 ммкг до 31.1 ммкг выводимость цыплят увеличивается на
7.5%, а при 38.3 ммкг — на 10.5% по сравнению с контролем;
в последней группе сокращается продолжительность инкубации
на 2 часа, а смертность эмбрионов с 16-го дня инкубации умень-
шается на 7%.
Витамин С птицы способны синтезировать, и, несмотря на дли-
тельное отсутствие этого витамина в корме, органы птиц всегда
богаты витамином С. По наблюдениям Кржишковского (1933),
из сельскохозяйственных птиц наибольшей способностью синте-
зировать витамин С обладают гуси, а затем куры. Положитель-
ное влияние на синтез витамина С, по мнению автора, оказывает
наличие в теле птиц витаминов А и D.
В рекомендациях по витаминному кормлению сельскохозяй-
ственных птиц, выработанных на конференции ВАСХНИЛ и
Института животноводства (Горюнов, 1965), указано, что потреб-
ность птиц в витаминах не может быть полностью удовлетворена
за счет содержания их в кормах и к ним необходимо дополнять
синтетические препараты витаминов A, D, В2, В12, пантотеновой
кислоты, холина, витамина К и фолиевой кислоты.
Действие витаминов проявляется комплексно. Так, например,
наличие определенного количества витамина В2 в пище снижает
токсичность чрезмерных доз витамина А; связано между собой
действие витаминов А и D и т. п.
Б. Влияние факторов среды
на продуктивность птиц
Все условия внешней среды, влияющие на здоровье
и состояние птиц, влияют и на их продуктивность. К этим факто-
рам относятся температура, влажность, свет, вентиляция и сани-
тария помещений, подвижность птиц и воздействие на их нервную
систему.
Влияние температуры воздуха на температуру тела
птиц обычно проявляется косвенно, через изменение активности,
и только в случае значительных отклонений может наблюдаться
42
и прямое влияние. Кроме суточного ритма, имеется также и се-
зонный ритм температуры тела птиц. Так, например, у курицы
температура тела ниже всего в декабре, январе и феврале, а выше
всего в июне, июле и августе. Имеется также различие в темпера-
туре самцов и самок птиц — около 0.4°. Более высокая темпера-
тура тела связана, по-видимому, с насиживанием и воспитанием
птенцов. У видов, где насиживает самка, ее температура тела бы-
вает более высокой, чем у самца, и наоборот.
Для кур-несушек зимой рекомендуется поддерживать темпера-
туру в птичнике на уровне около 10°, для петухов и кур-ненесу-
шек она может быть равна 4—6°, но не спускаться ниже 0°. Как
указывает Сметнев (1948), при понижении температуры в птичниках
на несколько часов до —10-----12° яйцекладка прекращается,
а при —15----20° куры и петухи обмораживают гребни и заболе-
вают. В зимнем помещении для уток и гусей температура может
быть более низкой, чем в курятнике. По наблюдениям Пенионж-
кевича и Зеленской (1949), зимой гуси переносят без ущерба для
продуктивности среднюю температуру птичника —10° с времен-
ными понижениями температуры до —25—30°.
Влияние климата на потребление корма птицей изучал
Лобин (1950). В средней зоне затрата корма на одну курицу ока-
залась на 9.2% выше, чем в южной, что зависит главным образом
от бдлыпих затрат на теплопродукцию. Автор установил, что при
снижении температуры с +17 до 0° необходима суточная добавка
примерно 4.1 г переваримых питательных веществ. При повыше-
нии температуры от +17 до +20° на каждый градус можно уба-
вить в рационе 0.3 г переваримых питательных веществ.
Исследуя влияние сезона на инкубационные качества ути-
ных яиц, Отрыганьева (1962) обнаружила, что, несмотря на раз-
личие в строении яиц, снесенных в декабре и в мае (в первых
больше индекс белка и желтка, меньше размер зародышевого
диска и менее пористая скорлупа), выводоспособность их мало
отличается (71—74% и 80%, соответственно). Яйца же, снесенные
в январе, имели одинаковую с майскими выводоспособность —
81—82%. По данным Камара (Кашаг, 1962), в Египте наилучшая
оплодотворенность и выводимость утят наблюдается в наиболее
холодные месяцы года. Как сообщает Злочевская (19626), наиболь-
шая величина бластодиска в куриных яйцах наблюдается в зим-
ний и осенний периоды.1
Свет влияет на птиц чрезвычайно сильно и разнообразно.
Больше всего действует на организм птицы солнечное освещение,
богатое ультрафиолетовыми лучами, но немалое действие оказы-
вает и свет, состоящий только из видимых лучей спектра. Уве-
личение количества освещенных часов в сутки, обеспечивая поеда-
1 Подробнее о влиянии сезона на развитие эмбриона до снесения яйца
см. главу 2.
43
ние большего количества пищи и ббльшую подвижность птицы,
вызывает повышение обмена веществ. Кроме того, свет усиливает
процессы кровотворения и повышает бактерицидные свойства
крови, а также воздействует на кровеносную и нервную системы.
Световое воздействие может продолжаться и после прекращения
освещения. Солнечное освещение активизирует и обусловливает
синтез витамина D в организме птиц, происходящий в кровенос-
ных сосудах кожи. В связи с тем что у птиц почти все участки
кожи покрыты перьями, мало пропускающими свет, синтез ви-
тамина D, по-видимому, особенно интенсивно происходит в гребне
и сережках, обильно пронизанных кровеносными сосудами. Сол-
нечный свет действует также на окислительные энзимы, активи-
зируя их каталитическое действие. При переходе птицы из тем-
ноты на свет происходит на короткое время повышение газообмена.
Чрезвычайно важно воздействие света на половые железы птиц.
При удлинении дня весной передача возбуждения от восприятия
света, проходящая по зрительным нервам, активизирует секре-
цию гипофизом гонадотропного гормона, вызывающего в свою
очередь увеличение размеров и деятельности половых желез.
Обратный процесс протекает осенью, когда количество световых
часов в сутки и интенсивность освещения уменьшаются. Интерес-
ные наблюдения по изменению яйценоскости кур в зависимости
от освещения в различные сезоны года в разных широтах провел
Сметнев (1948). Сравнение яйценоскости кур в совхозах, на-
ходящихся между 40—50° и 50—60° с. ш., проведенное на большом
материале (950 000 кур), показало зависимость яйценоскости
от освещения (рис. 8).
Чтобы заставить птицу интенсивно нестись круглый год, при-
меняют дополнительное электрическое освещение птичников,
удлиняющее зимние короткие дни до 12—14 час. Карапетян
(I960) утверждает, что в регуляции половой периодичности птиц,
в частности в стимуляции их репродуктивной функции в зимний
период, ведущая роль принадлежит не температуре, а свету.
Дополнительное освещение (световой день 15 час.) при прочих
равных условиях увеличивает яйценоскость на 25—71%. В даль-
нейшем автор (Карапетян, 1962) показал, что под влиянием до-
полнительного освещения не только увеличивается поедаемость
кормов (раньше считали, что только в этом и заключается полез-
ность длинного светового дня), но и повышается коэффициент
перевариваемости их курами. При одинаковом уровне кормления
повышение перевариваемости сырого протеина возрастает на
1.8%, сырой клетчатки — на 9.8%, сырого жира — на 1.7%,
безазотистых веществ — на 1.8% и кальция — на 5.5%.
Особенно интересны опыты по влиянию освещения кур и пе-
тухов на выводоспособность яиц. Мангольд (Mangold, 1932) описы-
вает опыт с двумя равными и одинаково содержавшимися груп-
пами кур, одна из которых ежедневно в течение 10 мин. подвер-
44
галась ультрафиолетовому облучению. Куры облучавшейся
группы снесли 497 яиц, давших 78% цыплят, в то время как кон-
трольные снесли только 124 яйца и дали 40% цыплят. Это раз-
личие в выводимости связано с наличием большего количества
Месяцы
------40-50 ° сев. тир________ 50-60°сев. шир.
Рис. 8. Яйценоскость кур в зависимости от
географической широты местности.
(По Сметневу, 1948).
витамина D в яйцах, снесенных облучавшимися курами. Пре-
имущество солнечного облучения над искусственным, а также
и то, что при наличии солнечного освещения дополнительное
ультрафиолетовое облучение не дает положительных результа-
тов, отмечают все исследователи. При выпасе птиц на выгулах
солнечное освещение имеет положительное влияние не только
на самих птиц, но и на поедаемый ими корм, активизируя в нем
витамины.
В. Влияние возраста птиц
на их продуктивность
Возраст птиц маточного стада играет значительную
роль в выводоспособности яиц. Куры-несушки в возрасте 2 лет
имеют наилучшую оплодотворенность и выводоспособность яиц;
несколько более низкую — в 3-летнем возрасте и еще более низкую—
у молодок. Гусыни достигают половой зрелости позднее других
сельскохозяйственных птиц. По данным Кирсанова (19356), ста-
рые гусыни несут более выводоспособные яйца (74.3% гусят от
оплодотворенных яиц), чем молодки (48%). Гусыни-молодки
в связи с неполным развитием полового аппарата несут большее
количество яиц со слаборазвитыми зародышевыми дисками, чем
46
переярки. К концу сезона снижается количество яиц с Хорошо
развитыми бластодисками и у молодок, и у переярок. Это ухудше-
ние инкубационных качеств яиц связано со снижением половой
активности гусаков в связи с подготовкой их организма к линьке
(Чмутова, 1956).
Тарабрина (1958) установила целый ряд физиологических
особенностей в организме кур-молодок в период яйцекладки.
В весенне-летний период усиливаются репродуктивная деятель-
ность половой системы и активность желез внутренней секреции
и происходит перестройка всего организма несушки. При этом
увеличивается вес яичника с 31.4 г в январе до 44.3 г в марте (за
счет роста желтков), длина яйцевода — с 53.9 см до 63.0 см (в ос-
новном за счет белковой его части), вес печени — с 28.8 до 35.5 г
(с увеличением процента гликогена в ней с 0.4 до 1.8%) и под-
желудочной железы — с 3.15 до 3.50 г, а вес костяка снижается
с 14.3 до 11.6% веса тела. Автор предполагает, что увеличение
гликогена в печени связано с увеличением секреторной деятель-
ности поджелудочной железы, а снижение веса костей — с исполь-
зованием минеральных солей для образования скорлупы (несмотря
на то что минеральных кормов было достаточно). С увеличением
возраста несушек и с повышением их яйценоскости длина кишеч-
ника становится больше: в январе она была в 7.5 раз длиннее
туловища, а в марте — в 8.5 раз; следовательно, способность
к перевариванию корма увеличивалась.
С увеличением возраста несушек Злочевская (19626) наблюдала
повышение абсолютного и удельного веса яиц, причем увеличи-
вался относительный вес желтка и скорлупы и повышалось со-
держание сухих веществ в белке и желтке, и в частности коли-
чество сахара. Последнее играет важную роль для развития
эмбриона, особенно в первые дни инкубации. Болтон (Bolton, 1957)
обнаружил, что яйца 4—5-летних кур обладают большей пори-
стостью скорлупы и теряют в весе как в инкубаторе, так и при
комнатной температуре в полтора раза больше, чем яйца молодок.
Бреславец (1965) отметила, что в яйцах переярых кур (19 и 29 ме-
сяцев) через 20 час. инкубации наблюдался больший размер бласто-
диска, а через 1.5—2.5 суток — большее количество сомитов, чем
в яйцах кур-молодок (9 и 12 месяцев).2 У суточных цыплят из яиц
кур-переярок вес желточного мешка на 20—26% выше, чем у цып-
лят из яиц кур-молодок и потому, вероятно, они жизнеспособнее
и на 5—17% меньше гибнут до месячного возраста.
С возрастом качество самцов как производителей понижается.
Лебедев и Фомин (1951) показали, что оплодотворение спермой
старых петухов приводит к понижению выводоспособности яиц
и снижению веса вылупившихся из них цыплят (табл. 9).
2 О влиянии возраста кур-несушек на развитие эмбриона к моменту сне-
сения яйца см. главу 2.
46
Таблица 9
Влияние возраста петухов на выводимость и живой вес цыплят
(По Лебедеву и Фомину, 1951)
Породы Год вывода петухов % вывода от опло- дотворен- ных яиц Вес цыплят (в г)
куры петухи при вы- луп- лении в 3-месячном возрасте
ку- рочки пе- тухи
Леггорн Род-айленд 1946 79 34 966 1147
» » 1948 100 42 1022 1265
Род-айленд Леггорн 1946 56.2 39 1055 1239
» » 1948 88.4 41 1091 1328
Г. Регуляция процесса яйцеобразования
Формирование яиц у кур приводится в движение слож-
ной серией взаимозависимых процессов, включающих эндокринную
и нервную системы. Романов и Романова (Romanoff a. Romanoff,
1949) приводят схему гормонального контроля образования яиц
(рис. 9). Передняя доля гипофиза продуцирует гонадостимулирую-
щие гормоны — фолликулостимулирующий и лютеинизирующий,
необходимые для роста и созревания яйцеклеток в яичнике.
Быстро растущие яйцеклетки в свою очередь выделяют гормон
эстрин, стимулирующий деятельность яйцевода. Эстрин вызывает
и другие физиологические изменения в организме несушки: мо-
билизует из пищи и внутри самого организма вещества, идущие на
построение яйца, и активизирует обменные процессы, необходимые
для их превращения. Эстрин выделяется в основном перед овуля-
цией. Через два дня после овуляции лопнувший фолликул начи-
нает атрофироваться, причем, по предположению авторов, выде-
ляется, так же как у млекопитающих, гормон прогестерон (про-
жестин), способствующий поддержанию яйцевода в состоянии
активности. Так, при оперативном удалении лопнувших фол-
ликулов из яичника курицы яйцекладка задерживается на не-
сколько часов и даже дней.3 Уровень кальция в крови несушки
контролируется паращитовидными железами, активность кото-
рых контролируется гипофизом. Гипофиз выделяет, кроме того,
гормон пролактин, тормозящий продукцию гонадостимулирую-
щих гормонов, а следовательно, и секрецию эстрина. Пролактин
стимулирует проявление инстинкта насиживания. Некоторые
породы кур (леггорн) утратили способность реагировать на секре-
3 По данным Гарда с соавторами (1966), скармливание или введение
курам-несушкам прогестерона повышало их яйценоскость и выводимость
цыплят из их яиц.
47
цию пролактина насиживанием, что значительно увеличило их
яйценоскость.
В обзоре журнала «Agricultural Research»4 конкретизируются
некоторые положения из приведенной схемы яйцеобразования.
Фолликулостимулирующий гормон обусловливает рост и созре-
вание яичника, а лютеинизирующий — разрыв созревшего фол-
ликула. Инъецируя лютеинизирующий гормон, можно вызвать
овуляцию или ускорить ее наступление, т. е. можно увеличить
Рис. 9. Схема гормонального контроля формирования яиц
у несушек. (По Romanoff a. Romanoff, 1949).
Объяснение в тексте.
количество яиц, хотя желтки их становятся меньше. Выделение
гипофизом этого гормона контролируется, по-видимому, нервным
механизмом, включая гипоталамус. Секреция лютеинизирую-
щего гормона прекращается на то время, пока яйцо находится
в яйцеводе. Вещества, блокирующие нервный механизм, предот-
вращают периодическое выделение этого гормона. По-видимому,
чем выше темп выделения лютеинизирующего гормона, тем выше
продуктивность, так как у высокопродуктивных кур интервал
между последующими овуляциями в среднем меньше, чем у низко-
продуктивных.
Однако не только передняя доля гипофиза выделяет гормоны,
регулирующие процессы яйцеобразования. В задней доле гипо-
физа у кур, как показал Сойер (Sawyer, 1961), содержится аргинин-
4 «Agricultural Research», 1959, 8, 4:3.
48
вазотоцин, а не вазопрессин, как у млекопитающих, и яйцевод
кур более чувствителен к вазотоцину, чем к вазопрессину. Танака
и Накайо (Tanaka a. Nakajo, 1960) исследовали наличие окси-
тоцина в задней доле гипофиза до и после яйцекладки у кур.
Выяснено, что яйцекладка следует за большим и достоверным сни-
жением количества гормона в течение часа, причем наибольшее
снижение отмечено за 5 мин. до снесения яйца. Вслед за яйце-
кладкой количество гормона возвращается к норме в течение
5—6 час. Дуглас и Старки (Douglas a. Sturkie, 1964) установили,
что за 2—5 мин. до снесения яйца содержание в крови вазотоцина
увеличивается во много раз, а затем уменьшается до исходного
уровня в течение 5—10 мин. после яйцекладки. Препараты из
задней доли гипофиза вызывают преждевременную яйцекладку
у кур и голубей (Старки, 1966).
Мазунин (1957) обнаружил, что в половой жизни птиц (сель-
скохозяйственных и диких) имеется двоякого рода цикличность:
астральная, имеющая относительно автоматический характер,
типа безусловнорефлекторных процессов, мало зависимая от среды,
и сезонная — ясно выраженного условнорефлекторного харак-
тера. Карапетян и Назарян (1965) считают, что значение нервной
системы в процессах, происходящих в органах размножения,
недооценивается. Проведя двустороннее полное удаление боль-
ших полушарий головного мозга у половозрелых кур-несушек,
авторы наблюдали резкую атрофию половых органов и необрати-
мое выпадение репродуктивной функции. У неполовозрелых цы-
плят при аналогичной операции органы размножения остаются
недоразвитыми и не функционируют до конца жизни. Интересно,
что внутримышечное введение (каждые 5—10 дней) гормонов,
стимулирующих функцию яичника (синестрола, фолликулина,
эстрадиолпропионата), после удаления больших полушарий не
привело к восстановлению репродуктивной функции, тогда как
у контрольных птиц, в течение длительного времени до этого не
производивших яйцекладку, введение этих гормонов восстанав-
ливало ее. Биохимическими исследованиями авторов установлено,
что удаление больших полушарий заметно нарушает азотистый
обмен и снижает количество SH-групп в генеративных и некоторых
эндокринных органах птиц. Авторы приходят к выводу, что
в нейро-гуморальной регуляции функции воспроизводительных
органов и метаболических процессов, происходящих в них, ве-
дущая роль принадлежит большим полушариям, а гипоталамо-
гипофизарная система, ответственная за нормальную деятельность
органов размножения, подконтрольна высшим отделам централь-
ной нервной системы.
Из всего вышеперечисленного очевидно, что для нормального
течения яйцеобразования необходима высокая степень синхрон-
ности гормональной и нервной регуляции и других процессов
в организме несушки.
4 В. В. Ролышк
49
Г Л А В A 4
СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ЯИЦ
А. Внешний вид яйца
Внешний вид яйца определяется четырьмя призна-
ками: 1) размером или весом, 2) формой, 3) цветом и 4) типом
поверхности скорлупы.
Средний вес яйца данного вида птиц определяется в ос-
новном средним весом взрослых птиц этого вида, но отношение
веса яйца к весу тела птицы уменьшается с увеличением послед-
него (табл. 10).
Таблица 10
Соотногиение между весом яйца и весом тела
в зависимости от веса птицы
(По Romanoff a. Romanoff, 1949)
Количество видов Вес тела птицы (В Г) Отношение веса яйца к весу тела птицы
164 2-180 1:9
177 400-1500 1:15
80 2600-12000 1 :28
6 20000-90000 1:55
Одомашнивание птиц и длительный искусственный отбор при-
вели к значительному увеличению относительного веса яиц
(табл. 11). По данным табл. 11 видно, что наибольшее увеличение
веса яиц отмечено у курицы, одомашненной более 3000 лет тому
назад, а наименьшее — у индейки, одомашненной 600—800 лет
тому назад.
Вес яиц кур яйценоских пород равен 45—64 г; куры тяжелых
мясных пород несут более крупные, а декоративных — более
мелкие яйца. Средний вес гусиных яиц разных пород равен 160—
215 г, утиных —70—100 г, индюшиных —80—100г, цесарочьих —
30—45 г. Укажем также вес яиц некоторых видов птиц, которые
пробовали искусственно инкубировать (табл. 12).
Вес яиц варьирует не только у различных пород кур, но даже
у одной породы (леггорн) в разных странах. Пенионжкевич
(1935а) показал, что увеличение в весе куриных и утиных яиц
обусловливается в основном повышением веса белка. Вес яиц
зависит от многих факторов: возраста и веса птицы, сезона и
времени суток снесения, кормления и ухода, а также от места
60
Таблица 11
Увеличение веса яиц под влиянием одомашнивания
(По Romanoff a. Romanoff, 1949)
Вид Вес яйца (В Г) Увеличение на (в %)
Банкивская (дикая) курица .... 40
Курица черная брама 68 70
Дикий канадский гусь 135
Белый эмденский гусь 215 59.3
Дикая утка 57
Белая пекинская утка 83 45.6
Дикая индейка 75
Белая голландская индейка .... 85 13.3
Таблица 12
Вес яиц некоторых видов птиц
Вид Средний вес яйца (в г) Автор
Страус африканский 1500 Дементьев (1940)
Страус американский — нанду 575 »
Страус австралийский — эму Гага обыкновенная: 600 »
с островов Баренцева
моря 109.5 Рольник (1955)
беломорская 116.1 »
Фазан 31.4 Romanoff (1934)
Перепел 8.7 »
данного яйца в цикле яйцекладки. Близкородственное разведение
снижает, а метизация повышает вес яиц.
О влиянии веса яиц на выводимость имеется много довольно
противоречивых данных. Большинство исследователей (Пенионж-
кевич, 1935а; Axelsson, 1954; Coles, 1956; Ножчев, 1961, и др.)
считает, что наилучший процент вывода, качество и прирост
веса молодняка получается из яиц среднего веса: 50—60 г для
яиц кур породы леггорн и 61—70 г для утиных, породы хаки-
кемпбел. Коулс (Coles, 1956) уточняет, что наивысшую выводи-
мость он наблюдал из яиц, вес которых был примерно равен
среднему весу яиц данной курицы, если вес ее яиц не имел слиш-
ком больших колебаний и если эти яйца не были слишком малы
или велики.
Обенко и Антаков (1956) считают, что наилучшую выводимость
4*
61
имеют мелкие яйца. Разделив куриные яйца по весу на 4 группы
(I - 48-50 г, 11—51—55 г, III — 56—60 г и IV - 61-
65 г), они получили наилучший вывод цыплят из мелких яиц
(1—86.9—87.2%, II-81.8-83.0%, III - 82.5-85.6% и IV-
73.6—75.1%). Однако эти данные несколько обесценены замеча-
нием авторов, что при более полноценном кормлении можно
получать хорошую выводимость и из крупных яиц. И, наконец,
третья группа исследователей (Федотова, 1952; Реснянская,
1961), наоборот, доказывает, что выводимость цыплят повышается
с увеличением веса яиц. В опытах Федотовой из яиц весом 70—
74 г кур ливенской породы было получено 70—85% цыплят.
В связи с тем что из яиц большего веса выводятся более крупные
цыплята, автор считает необходимым инкубировать для племен-
ных целей именно крупные яйца, но рекомендует выработать
особый режим инкубирования их.
В то же время Арболеда с соавторами (Arboleda et al., 1960)
совсем не обнаружил зависимости между величиной яиц кур по-
роды белый леггорн и их оплодотворенностью и выводимостью,
но наблюдал достоверную корреляцию (коэффициент корреляции
+0.457) между весом кур и весом их яиц.
Вес цыпленка составляет обычно 64—69% веса яйца. Корреля-
ция между весом яиц и весом вылупившихся из них цыплят исче-
зает через 6 недель после вылупления (Pope a. Schaible, 1957).
Однако О’Нелл (O’Neil, 1950) обратил внимание на то, что чем
больше был процент веса цыпленка породы полосатый плимутрок
от веса яйца, тем больше был их вес в 8-недельном возрасте;
они лучше потребляли пищу и имели меньшую смертность, причем
наблюдался больший процент петушков.
Форма яйца создается под давлением мышц яйцевода.
Вариации в форме яиц, по-видимому, зависят от координации
между действием внутреннего циркулярного слоя мышц, повора-
чивающего и проталкивающего яйцо вперед по яйцеводу, и
внешнего продолговатого мышечного слоя, расширяющего яйце-
вод. Возможно, что форма яйца зависит также от соотношения
размера желтка и ширины яйцевода: большой желток, проходя
сквозь узкое отверстие яйцевода, принимает (а вместе с ним и
яйцо) продолговатую форму, а небольшой, проходя сквозь ши-
рокий яйцевод, сохраняет круглую (Romanoff a. Romanoff, 1949).
Форма куриных яиц варьирует от почти круглой до сильно вытя-
нутой, эллиптической по длинному диаметру яйца; обычно же
яйцо имеет овальную, заостренную с одной стороны форму.
В поперечном разрезе яйцо имеет форму круга. Форма яйца опре-
деляется отношением длинного диаметра к короткому. Денись-
евская (1926) установила, что у куриных яиц отношения диаметров
равны от 1.13 (почти шарообразное яйцо) до 1.67 (сильно вытяну-
тое); наиболее часто встречается отношение, равное 1.32. У утиных
яиц отношение диаметров равно в среднем 1.30 (от 1.12 до 1.58)
62
(Третьяков, 1951). По мнению Пенионжкевича (19356), наиболее
часто встречающаяся форма яиц для каждой породы птиц (соот-
ветствующая среднему соотношению диаметров) обеспечивает
наилучшие показатели выводимости и прироста веса молодняка.
Для куриного яйца это будет форма с отношением диаметров,
равным 1.31—1.35, а для утиного — 1.32—1.44. Как показал
Мичек (1966), от формы яиц зависит большее или меньшее нару-
шение билатеральной симметрии у эмбриона: чем более вытянуто
в длину яйцо, тем реже остается правая гонада и тем относительно
больше правая доля печени. Форма яиц у одной и той же курицы
имеет меньший размах колебаний, чем форма яиц различных кур
даже в одном стаде. Обычно молодка (первый год яйцекладки)
несет яйца характерной для нее формы только после снесения
ею нескольких яиц, нетипичных как по форме, так и по другим
признакам. Сезонные вариации формы куриных яиц невелики.
Имеется только небольшое округление яиц в марте—мае и удли-
нение их в начале и конце кладки. Форма яиц часто передается
по наследству.
Окраска скорлупы является видовым признаком,
значительно различающимся у разных видов диких птиц. Год-
фрей (Godfrey, 1950) установил, что яйца кур породы ныо-гемпшир
с темно-коричневой скорлупой, высоким удельным весом и не-
большой усушкой за 14 дней инкубации имеют лучшую выводи-
мость. Пинжель (Pingel, 1958) объясняет лучшую выводоспособ-
ность темноокрашенных яиц тем, что этот признак связан с более
плотной скорлупой, что обусловливает меньшую потерю их веса.
Другочиу с соавторами (Drugociu et al., 1956) обнаружил, что
более пигментированные яйца кур породы род-айленд имеют более
правильную форму (индекс длины к ширине 1.30—1.41), более
высокую выводоспособность (87.3% при 54% из слабо пигменти-
рованных яиц) и жизнеспособность цыплят.
Поверхность скорлупы яиц бывает гладкой и
шероховатой, глянцевитой и матовой. Скорлупа становится глян-
цевитой, когда на поверхности ее откладывается большое коли-
чество органических материалов (кутикула), что зависит от актив-
ности железок, секретирующих эти материалы. Часто к концу
цикла кладки яйца становятся более шероховатыми. Фердинандов
(1931) указывает, что 53.4% куриных яиц имеют матовую скор-
лупу, 34.5% — слабо глянцевитую и 12.1 % — глянцевитую.
По данным Кучковской (1938), яйца с шероховатой скорлупой
дают пониженный вывод цыплят (51.4—68.2%) по сравнению с яй-
цами, имеющими гладкую скорлупу (85.8%). Прицкер (1936)
наблюдал среди утиных яиц иногда до 50% яиц с глянцевитой
скорлупой, с жировым налетом на поверхности; такие яйца имели
пониженную выводимость. Поверхность скорлупы даже у одной
несушки сильно варьирует.1
1 О нарушениях поверхности скорлупы см. в главе 5.
63
Б. Внутреннее строение яйца
Внутреннее строение яйца видно на его продольном
разрезе (рис. 10).
Зародышевый диск (бластодиск) представляет
собою маленькое беловатое пятнышко на поверхности желтка.
В неоплодотворенном яйце он представляет собою собственно
яйцеклетку — ядро и протоплазму. Размер зародышевого диска
у неоплодотворенного яйца 3—4 мм в диаметре и несколько больше
у оплодотворенного — 4.4 мм (рис. 5 и 6).
Рис. 10. Схематический продольный разрез яйца.
1 — кутикула; 2 — скорлупа; 3 — воздушная камера; 4а, 46 — наружная
и внутренняя подскорлупные оболочки; ба — 5г — наружный жидкий,
густой, внутренний жидкий и халазообразующий слои белка; 6 — ха-
лазы; 7 —желток; 5—желточная оболочка; Р—бластодиск; 10— латсбра.
Форма желтка — неправильный шар. Средняя длина
желтка куриного яйца — 34 мм, ширина — 32 мм, поверхность
32.2 см2, объем 17.1 см3.
Желтый цвет желтка обусловливается каротиновыми пигмен-
тами и, как мы уже говорили, зависит от корма. Морис и Фиданца
(Maurice a. Fidanze, 1954) исследовали структуру желтка кури-
ного яйца, выясняя его проницаемость для Вг82. Через 100—200 час.
ббльшая часть меченого брома оставалась в поверхностных слоях
желтка, и авторы предполагают, что желток разделен на слои
плохо проницаемыми, но тонкими стенками и что ширина этих
слоев равна 0.3 мм.
Исследуя структуру желточной оболочки куриного яйца при
64
помощи люминесцентной и ультрафиолетовой микроскопии Ша-
лумович (1955) обнаружил, что она состоит не менее чем из пяти
слоев. А по данным Беллер с соавторами (Bellairs et al., 1963),
в оболочке желтка имеется два слоя, состоящих в основном из
протеинов, но с различным аминокислотным составом.
Во внутреннем жидком белке почти нет муци-
новых нитей, а в плотном, наоборот, они составляют довольно
сложно переплетенную ячеистую сеть, заполненную жидким
белком. Халазообразующий слой белка — это тонкий слой густого
белка,, лежащего непосредственно па поверхности желточной
оболочки и заканчивающегося закрученными тяжами — хала-
Рис. И. Поперечный разрез подскорлупных оболочек на краю
воздушной камеры. (По Romanoff a. Romanoff, 1949).
А — увеличено примерно в 100 раз; Б — увеличено примерно в 500 раз. 1 — внут-
ренняя подскорлупная оболочка; 2 — наружная подскорлупная оболочка, 3 — воз-
душная камера.
зами — по обе стороны длинной оси желтка. Халаза в остром
конце яйца, состоящая из двух тяжей, закрученных против часо-
вой стрелки, больше и длиннее противоположной, в тупом конце,
состоящей из одного тяжа, закрученного по часовой стрелке.
Халазы служат для стабилизации положения желтка в яйце.
Наружная подскорлупная оболочка плотно
соединена с внутренней поверхностью скорлупы. Так же плотно
соединены между собой и обе подскорлупные оболочки, и разде-
ляются они только в месте воздушной камеры (рис. 11). У птицы
с более толстой скорлупой яиц более тонкие подскорлупные
оболочки, и наоборот. Так, например, у куриных яиц, имеющих
относительно толстую скорлупу, подскорлупные оболочки состав-
ляют 0.6% веса яйца, а у индюшиных, с пропорционально более
тонкой скорлупой, — около 2.2%. Наибольшая толщина оболочек
наблюдается у тупого конца яиц (Фердинандов, 1931). В наружной
подскорлупной оболочке нити из кератиноподобного материала
бб
толще, их больше и лежат они чаще всего параллельно скорлупе,
во внутренней — нити тоньше, количество их меньше и лежат они,
переплетаясь во всех направлениях. Волькен и Шварц (Wolken
a. Schwartz, 1948) при помощи электронного микроскопа наблю-
дали, что высушенная подскорлупная оболочка куриного яйца
состоит из свободно переплетающихся волокон толщиной 1 мк
и имеет около 20 x10е пор на 1 см2, примерно того же диаметра.
Жидкости и газы проходят через нодскорлупные оболочки диф-
фузно. Подскорлупные оболочки и оболочка желтка по своему
химическому составу очень близки друг к другу, но физическая
их структура различна (Wolken, 1951).
Воздушная камера яйца г(пуга) образуется между
двумя по декор лунными оболочками яйца при остывании (сокраще-
нии в объеме) его содержимого после снесения яйца курицей
и втягивании окружающего воздуха внутрь яйца. Доказатель-
ством указанного происхождения воздушной камеры является
отсутствие ее в яйцах рептилий, имеющих пергаментообразную
скорлупу, сморщивающуюся при остывании яйца после снесения.
Впрочем, по данным Зусмана (1965), небольшая воздушная камера
в яйцах черепах все же есть. Воздушная камера образуется обычно
в тупом конце яйца, так как оболочки здесь слабее всего скреплены
между собою, но бывают и отклонения — боковая воздушная
камера. Объем воздушной камеры в момент снесения яйца равен
0.1—0.3 см3. В дальнейшем при хранении или инкубации яиц
происходит увеличение объема воздушной камеры в связи с испа-
рением воды из яйца.
Толщина скорлупы должна быть достаточной, чтобы
удержать на себе насиживающую птицу, и в то же время быть
настолько незначительной, чтобы птенец мог проломить ее при
вылуплении. У куриных яиц имеется прямая корреляция между
средним объемом яйца (от 40 до 60 см3) и средней толщиной скор-
лупы (от 0.34 до 0.39 мм) (Romanoff a. Romanoff, 1949). Толщина
скорлупы больше всего на остром конце и несколько больше
зимой, чем летом. Между толщиной скорлупы и удельным весом
яиц установлена статистически достоверная положительная кор-
реляция (Mountney a. Vanderzant, 1957). По данным Свенсона
(Svensson, 1957), наблюдается зависимость между удельным весом и
выводоспособностыо яиц: наилучший процент вывода цыплят
дали яйца со средним удельным весом 1.075—1.080 г/см3. Не-
сколько иные данные получены Пэйном и МакДаниэлем (Payne
a. McDaniel, 1958): чем больше был удельный вес индюшиного
яйца (т. е. чем толще скорлупа), тем меньше было задохликов.
Детально этот вопрос изучен Шпиц с соавторами (1965). Разделив
яйца на 5 групп (1-ю — со средним удельным весом яиц
1.0675 г/см3 и средней толщиной скорлупы 0.285 мм, 2-ю —
1.0735 г/см3 и 0.312 мм, 3-ю — 1.0795 г/см3 и 0.326 мм, 4-ю —
1.0855 г/см3 и 0.357 мм, 5-ю — 1.0925 г/см8 и 0.380 мм соответ-
56
ственно), авторы обнаружили, что в 1-й и 2-й группах чаще всего
бывают нарушения целостности скорлупы и внутренней структуры
яйца, а при инкубировании — больший процент неоплодотворен-
ныхяиц (22.3—19.2% при 13.3—15.7% в других группах) и погиб-
ших в первую неделю инкубации (9.7—7.2% при 4.7—5.1 % в дру-
Рис. 12. Гистологическое строение скорлупы куриного яйца.
(По Romanoff a. Romanoff, 1949).
А — поперечный разрез через скорлупу; Б — разрез, параллельный поверхности,
через призматический слой скорлупы; В — поперечный разрез через скорлупу
после растворения кальция; Г — разрез, параллельный поверхности, этой же скор-
лупы; Д — внутренняя поверхность скорлупы после сжигания органического ее
материала (А, Б, Д — увеличено примерно в 100 раз; В, Г — то же в 200 раз).
1 — кутикула; 2 — губчатый слой; 3 — призматический слой; 4 — подскорлупная
оболочка; 5 — призматический сосочек; б — протеиновый материал, образующий
сердцевину сосочка; 7 — протеиновая основа скорлупы; 8 — пора в скорлупе
после растворения кальция; 9 — место в сосочке после того, как органическая
сердцевина была удалена сжиганием.
гих группах). По мнению Шпиц и Даниловой (1966), скорлупа
большей толщины бывает у биологически более полноценных яиц,
что выражается в лучшем использовании эмбрионами питательных
веществ яйца, большем весе эмбриона, более высокой выводимости
и лучшем качестве вылупившихся цыплят, а также в лучшем пост-
эмбриональном развитии и более высокой продуктивности кур.
Авторы отмечают тенденцию к наследованию качества скорлупы.
67
Микроскопическая структура скорлупы дана на рис. 12.
Количество пор, их распределение на яйце и размеры сильно
варьируют как у кур разных пород и стад, так и в яйцах кур
одного стада (от 30 до 170, чаще всего со 110 порами на 1 см2
в среднем). В скорлупе яиц одной курицы количество пор варьи-
рует мало. По данным Ломовой (1939), минимальное количество
пор на 1 см2 имеет скорлупа яиц кур породы лангшан (85), мак-
симальное — бентамок (149), а у кур белый леггорн — в среднем
127 пор па 1 см2. Автор указывает, что чем интенсивнее окраска
яиц, тем меньшее количество пор имеет скорлупа. Между толщиной
скорлупы и количеством пор коэффициент корреляции ничтожно
мал (0.03), что говорит о полной независимости этих двух призна-
ков. Последнее подтверждено Свенсоном (Svensson, 1957). По
данным Отрыганьева (1966), в яйцах кур яйценоской породы —
русские белые — имеется 113.5 + 5.2 пор на 1 см2 скорлупы,
а у мясной породы — корниш — 94.9 + 3.7, что, по мнению автора,
обеспечивает меньшую потерю в весе во время 18-дневного хране-
ния у последних (3.85 и 2.36% соответственно).
Распределение пор на поверхности яйца неравномерно: в сред-
нем 151 пора на тупом конце яйца, 142 на середине и 100 на остром
конце яйца (Владимирова, 1954). Обычно отверстие поры овальное,
но у страусиных яиц оно круглое. Размер пор частично зависит
от размера яйца, но даже на одном яйце сильно варьирует. Канал
поры утончается к внутренней части скорлупы, где попадает
в сеть воздушных каналов в призматическом ее слое. Рауч (Rauch,
1952) при помощи разработанного им нового метода исследовал
пористость скорлупы яиц, из которых вылупились цыплята, и
яиц, в которых эмбрионы погибли в течение инкубации (табл. 13).
В большинстве яиц с погибшим эмбрионом скорлупа имела поры
большего диаметра, обусловливающие слишком большую про-
ницаемость скорлупы и повышенную интенсивность испарения,
Таблица 13
Пористость скорлупы куриных яиц
(По данным Rauch, 1952)
Диаметр пор (в мк) Коэффи- циент вариации диаметра пор Количе- ство пор (на 1 см2) Общая площадь пор (в мм2)
средний мини- маль- ный макси- маль- ный
Скорлупа: вылупившихся цыплят . . . 9.4+0.5 1.6 25.8 57.4 148 0.7
погибших эмб- рионов .... 21.8+1.4 1.6 74.7 64.7 101 2.5
68
что явилось причиной гибели эмбрионов в них. Часть же яиц с по-
гибшими эмбрионами имела скорлупу с нормальным диаметром
пор и, следовательно, причина гибели эмбрионов здесь была
другая.
На страусином яйце кутикула твердая, толщиной
0.036 мм, у домашней утки — 0.003 мм, а у курицы — от 0.005
до 0.01 мм. У некоторых видов птиц на поверхности скорлупы
кутикулы нет (чайки). В кутикуле имеются маленькие капельки
жира. Симонс и Вирц (1966) установили, что кутикулярная по-
верхность куриного яйца имеет пористую структуру, причем поры
эти заполнены воздухом. Кутикула состоит в основном из муцина,
имеет довольно стабильную структуру и растворяется только при
опускании яиц в горячую воду (выше 40°). Роль кутикулы в газо-
проницаемости скорлупы оценивается по-разному разными ис-
следователями. Маршалл (Marshall, 1947) наблюдал, что при
разрушении кутикулы скорлупы испарение из яиц не увеличи-
вается, а уменьшается. По мнению автора, причина этого в том,
что проницаемость скорлупы для влаги связана не с площадью
пор (количество пор, помноженное на их диаметр), а с площадью
пятен-кратеров кутикулы, на дне которых лежат относительно
маленькие выходы пор скорлупы. Поры, окрашенные в относи-
тельно сухой атмосфере, выглядят уменьшенными, а при влажности
80—90% — более крупными, дающими максимум условий для
испарения в окружающую среду. По предположению автора, пятна
кутикулы играют аналогичную роль с устьицами листьев в регу-
лировании выхода влаги из яйца. А по данным Уолдена с соавто-
рами (Walden et al., 1956), удаление кутикулы и подскорлупных
оболочек даже немного увеличивает проницаемость скорлупы
для газа.
Процентное содержание составных частей яйца у разных птиц
различно. Все птицы делятся по состоянию птенцов после вылуп-
ления на две группы — выводковые и птенцовые (более и менее
самостоятельные птенцы соответственно). Оказывается, что отли-
чия птенцов этих двух групп после вылупления заложены уже
в яйцах, которые имеют разное процентное содержание составных
частей (табл. 14) и различный химический состав.
Процент веса скорлупы к весу всего яйца по мере уменьшения
веса яйца снижается, а вес желтка, наоборот, увеличивается.
По-видимому, это обусловлено тем, что с уменьшением веса яйца
увеличивается его относительная поверхность, а это вместе с отно-
сительно более тонкой скорлупой значительно увеличивает тепло-
отдачу яйца при развитии эмбриона; в виде компенсации увели-
чивается основной источник тепловой энергии — желток (Roma-
noff a. Romanoff, 1949). Процентное содержание белка, желтка и
скорлупы в курином яйце приведено в табл. 15.
Гринвуд и Болтон (Greenwood a. Bolton, 1956) обнаружили
/ зависимость в весе яиц и соотношении их частей от возраста кур.
59
Таблица 14
Различие в содержании белка> желтка и скорлупы в яйцах
птенцовых и выводковых птиц
(По Romanoff a. Romanoff, 1949)
Группа птиц Вес яйца (в г) К весу всего яйца
% белка % желтка % скорлупы
о S среднее Н о S среднее О S среднее
Птенцовые . . от 0.5 до 140 68.1 78.6 73.2 12 26.6 19.8 4.9 9.5 7.0
Выводковые . от 10 до 2000 50.7 56.7 52.9 31.9 40.8 35.2 8.5 14.1 11.9
Таблица 15
Среднее соотношение составных частей куриного яйца
(По Romanoff a. Romanoff, 1949, пересчитано нами)
Белок
Скорлупа с обо-
лочками
Вес (в г).........
Относительный вес
(В %)...........
58 32.9
100 56.7
7.6 18.9
13.1 32.6
5.5
9.5
18.7 6.4 6.1 0.3
32.2 11.0 10.4 0.6
Благодаря относительно меньшему весу скорлупы и большему
весу желтка яйца старых кур более калорийны (табл. 16).
Сметнев и Тарабрина (1956) подтвердили, что увеличение
веса яиц у кур с возрастом обусловлено главным образом увели-
чением абсолютного и относительного веса желтка, причем в яй-
цах кур высокой продуктивности и абсолютный, и относительный
вес желтка несколько выше, чем в яйцах кур средней продуктив-
ности, а вес белка — наоборот. Однако относительный вес плот-
ного белка был все же больше у высокопродуктивных кур. По
данным Третьяковой (1938) (куриные яйца) и Третьякова (1951)
(утиные), яйца с более высоким содержанием плотного белка
имеют и более высокую выводоспособность (табл. 17).
60
Таблица 16
Зависимость соотношения частей яиц от возраста кур
(По Greenwood a. Bolton, 1956)
Возрастные группы
св
Вес в % от веса § g
ЯИЦ 5 g
Старые куры (6—10 лет)
Годовалые.............
Молодки ..............
47.9 8.2
55.6 8.8
37.6 10.4
36.3 39.3
32.1 31.1
29.7 31.1
16.2
28.0
31.1
70.8
52.6
48.1
184
168
159
Таблица 17
Зависимость выводимости от содержания различных слоев белка в яйцах
(По данным Третьяковой, 1938, и Третьякова, 1951)
Процентное содержание разных слоев белка к весу яйца % выводимо- сти
наружный жидкий ; внутренний жидкий всего жидкого плотный
Куриные 10.7 12.8 23.5 31.9 88.8
» 14.6 15.3 29.9 28.3 33.1 '
Утиные 16.9 14.1 31.0 26.9 75.1
» 15.6 15.0 30.6 23.3 64.0
» 19.3 10.5 29.8 21.6 50.0
Соотношение составных частей яиц даже у одной курицы
сильно варьирует. По данным Даниловой (1931), процент желтка
выше в зимние месяцы и снижается к июню, процент белка же,
наоборот, относительно возрастает и в июне достигает максималь-
ной величины.
В. Химический состав яйца
Химический состав яйца сильно варьирует не только
у разных видов птиц или разных пород, но и у одной птицы
в разное время года при различном кормлении и т. п. Приводимые
в этой главе данные о химическом составе яиц являются средними,
наиболее часто встречающимися у данного вида птиц. Химический
состав яйца приведен в работе Романова и Романовой (Romanoff
a. Romanoff, 1949) (табл. 18).
61
Таблица 18
Химический состав целого куриного яйца
(По Romanoff a. Romanoff, 1949)
—Протеины сс об со св я о ф S’ ф я —Липиды ф ф 5 S —Углеводы £ о —Пигменты се -Другие ор- g. ганические § вещества д ф й _ и —Основные о ?. минераль- . § U ные веще- ства се & ° я со _ , д д' —Микроэлеменп ® е g -g д I д ф и я о ф л S’ СЭ я я со се LQ Q- е. о ф я щ ес — О И Овальбумин £ Овокональбумин g Овоглобулин § Овокератин Оволиветин ““ § Овомуцин я Овомукоид | Ововителлин л Жиры Фосфолипиды Стеролы Цереброзиды 1 К аротиноиды Овофлавин 1 Порфирин Другие пигменты Кальций Хлор 1 Железо Магний Фосфор Калий Натрий Сера Аланин Аргинин Аспарагиновая кислота Цистин Глютаминовая кислота Глицин Гистидин Изолейцин Лейцин Лизин Метионин Фенилаланин Пролин Серин Трионин Триптофан Тирозин Валин Овокефалин Оволецитин Овосфингомиелин Овокеразин Овофренозин I Каротин | Ксантофилл Алюминий Мышьяк Барий Бор Бром Хром Йод Кремний Марганец Селен Литий Медь Молибден Рубидий Фтор Свинец Серебро Стронций Титан Уран Ванадий Цинк
62
О содержании сухих веществ и некоторых питательных веществ
в яйцах кур породы русская белая сообщают Моисеева и Толокон-
никова (1966) (табл. 19).
Таблица 19
Содержание некоторых питательных веществ
в яйцах кур русская белая
(По Моисеевой и Толоконниковой, 1966)
Содержание
сухих веществ (%) липидов в жел- тке (% к сырому весу) холестерина в липидах жел- тка (мг %) витаминов (мг %)
в желтке в белке во всем яйце в жел- тке
Bi В2 1 А А
Среднее . . . Ошибка сред- него .... 53.68 ±0.07 12.50 ±0.06 35.60 ±0.06 5693 +67.11 0.125 ±0.003 0.312 + 0.009 0.221 0.676
Желток содержит основной запас питательных веществ для
развития эмбриона. Процент воды в желтке, как показал Тарха-
нов (Tarchanoff, 1884), у птенцовых примерно на 10% больше, чем
у выводковых. Так, у птенцовых воды в желтке: у скворца — 56.6,
коростеля — 58.0, голубя — 57.9%, а у выводковых: у ржанки —
50.3, перепела — 41.8, курицы — 49.8, цесарки — 49.7%. Таким
образом, питательных веществ в яйцах выводковых птиц больше,
что дает возможность удовлетворять относительно бблыпую потреб-
ность в питательном материале, связанную со сравнительно ббль-
шим сроком инкубации и с тем, что за время эмбрионального
развития ими должна быть достигнута более продвинутая стадия.
Напомним, что относительный вес желтка в яйцах выводковых
птиц значительно выше, чем в яйцах птенцовых. Химический
состав желтка яиц некоторых видов птиц представлен в табл. 20.
По данным Моисеевой и Толоконниковой (1966), содержание
сухих веществ в желтке имеет положительную корреляцию с яй-
ценоскостью (+0.20) и выводимостью цыплят (+0.30).
Основной протеин желтка — это ововителлин. В детальном
исследовании химической и физико-химической природы ововител-
лина Казаков (1954) показал, что это один из наиболее богатых
фосфором и серой протеинов. Это указывает на его высокую био-
логическую мобильность и биологическую активность в процессах
развития эмбриона. Количество фосфора в ововителлине во время
эмбрионального развития резко падает, и автор подчеркивает
существенное значение процесса дефосфорилирования именно
63
Таблица 20
Химический состав желтка яиц (в % от влажного веса)
Вид яиц Вода Протеины Липиды Безазотистые экстрактив- ные вещества Зола Автор
всего в том числе фосфатидов
47.1—53.8 15.7—17.5 31.7—36.2 7.2—17.4 0.4-0.9 1.0—2.0 Romanoff a. Roma- noff, 1949
Куриные . . . . 50.8 16.2 31.7 0.1 1.1 Ильин, 1917
48.7 16.6 32.6 1.0 1.1 Riley, 1953
Гусиные 41.1—44.1 17.3—18.7 35.7—36.2 1.3—1.7 Romanoff a. Roma- noff, 1949
Утиные 43.7-45.0 15.3—18.4 35.0—38.9 1.6 1.2-1.5 Romanoff a. Roma- noff, 1949
46.0 18.4 34.4 1.2 Данилова и Нефедь- ева, 1936
Индюшиные . . . 48.3 15.1 34.4 0.5 1.7 Орлов и Кучковская, 1941
Цесарочьи .... 49.7 16.7 31.8 1.2 Romanoff a. Roma- noff, 1949
Голубиные . . . 55.9-57.9 12.9 20.8 9.1—13.1 0.7 0.9 Romanoff a. Roma- noff, 1949
этого протеинового комплекса для нормального течения процессов
развития. В ововителлине в большом количестве содержится
аргинин, лизин и пролин с оксипролином. Несмотря на значи-
тельную изменчивость ововителлина в течение эмбрионального
развития, он не теряет своей протеиновой специфичности. Как.
сообщают Лисовская и Ливанова (1960), протеины желтка состав-
ляют 33% его сухих веществ и состоят из вителлина, вителленина
и фосвитина. Из вителлина, содержащего 1% фосфора, была
выделена вителлиновая кислота, идентичная фосвитину, который
содержит 9.7% фосфора. Авторы считают фосвитин основным
фосфопротеином желтка. Романов и Романова (Romanoff a. Ro-
manoff, 1949) вителленин называют оволиветином и отмечают,
что он содержит мало фосфора, но много серы (около 1/3 всей серы
желтка) и что его в желтке куриного яйца в 3.6 раза меньше, чем
вителлина.
Исследуя нуклеиновые кислоты в содержимом куриного яйца,
Воробьев (19606) обнаружил 68.1 мкг РНК и 53.8 мкг ДНК
в желтке, и 566 мкг РНК и 364 мкг ДНК в белке. Автор считает,
что имеющаяся в желтке ДНК доказывает наличие ее в цитоплазме,
и предполагает, что она служит резервным источником, из кото-
рого развивающийся эмбрион черпает необходимые ему вещества
для синтеза нуклеиновых кислот. Автор подсчитал, что содержа-
щейся в желтке ДНК должно хватить на 2.24 Х107 клеток. При-
сутствие нуклеиновых кислот в белке автор объясняет тем, что
белок является секретом стенки яйцевода и в нем присутствуют
элементы стирающихся клеток его эпителия.
Большое место в желтке занимают липиды (31.7—36.2%);
из них нейтральные жиры — пальмитин, стеарин и олеин вместе
составляют 20.3—23.2%. Фосфатиды желтка представлены в ос-
новном лецитином, играющим большую роль в стимуляции роста
эмбриона. В 1 г куриного желтка 17.8 мг холестерина, а в 1 г
утиного — 26.4 мг. Между содержанием холестерина в желтке
и выводимостью цыплят существует отрицательная корреляция
(Боголюбский, 1966). По данным Гликина (Glikin, 1908, цит.
по: Needham, 1931), в яйцах птенцовых птиц больше лецитина
(у голубя 13.1%, а у курицы — от 8.9 до 10.7%), ускоряющего
развитие, с которым связана способность к раннему вылуплению.
Родз и Ли (Rhodes a. Lea, 1957) обнаружили в курином яйце сле-
дующее содержание фосфолипидов: фосфотидилхолина — 73.0,
лизофосфатидилхолина — 5.8, сфингомиелина — 2.5, фосфатидил-
этаноламина — 15.0, лизофосфатидилэтаноламина — 2.1 и инози-
то лфосфо липида — 0.9 моль %. По исследованию Даниловой
(1931), содержание жира и лецитина в желтке яиц разных пород
кур различно (табл. 21). По данным автора, между содержанием
жира и весом яиц у разных пород кур существует обратная зави-
симость: в более крупных яйцах содержится меньший процент
жира, чем в мелких. Коэффициент корреляции равен —0.433.
5 В. В. Ролышк
66
Т а б л и ц а 21
Содержание жира и лецитина в желтке куриного яйца
(% от влажного веса)
(По Даниловой, 1931)
Вещество Породы кур
пли- мутрок русские ви ан дот минорки род-ай- ленд
Всего жира 29.99 28.37 27.94 25.99 25.21
В том числе лецитина . . 10.44 10.18 10.15 9.95 9.20
В пределах одной породы Моисеева и Толоконникова (1966)
обнаружили, наоборот, прямую зависимость между содержанием
липидов и весом яиц: корреляция равна +0.16. Однако содержание
холестерина отрицательно коррелирует с весом яиц — корреля-
ция равна —0.21.
Углеводов в желтке чрезвычайно мало, и представлены они
в основном глюкозой. Пенионжкевич (1954) наблюдал строгую
зависимость между содержанием глюкозы в гусиных яйцах и
выводимостью гусят. Если глюкозы было в белке 400—450 мг%
и в желтке 150—230 мг%, то выводилось 75—50% гусят от опло-
дотворенных яиц, а если 330—380 мг% и 100—180 мг% соответ-
ственно, то только 65—70%. По-видимому, это связано с тем, что
углеводы занимают значительное место в энергетическом обмене
эмбриона.
Желток характеризуется высоким содержанием витаминов.2
Пигменты желтка — это в основном ксантофилл и каротин,
причем бблыпая часть окраски осуществляется ксантофиллом,
каротина же обычно значительно меньше. Соотношение между
количеством ксантофилла и каротина в желтке в значительной
степени зависит от количества этих пигментов в корме, потребля-
емом несушкой.
Оболочка желтка состоит из склеропротеина (вещество, близкое
к кератину), не проницаемого для ововителлина даже в растворах,
но проницаемого для воды.
Неорганические соединения, содержащиеся в курином яйце,
исследованы раздельно для желтка и белка Ильиным (1917)
(табл. 22).
Как видно из табл. 22, калия, несколько больше, чем натрия,
как в желтке, так и в белке, что вообще характерно для яйцеклетки.
В желтке очень много фосфора и находится он главным образом
в органических соединениях. Воробьев (1960а), изучая фосфорные
2 Данные о содержании витаминов в желтке мы сообщаем в главах 3 и 5.
66
Таблица 22
Основной химический состав золы белка и желтка
куриного яйца
(По Ильину, 1917)
Состав золы Содержание (в %)
белок желток
Na2O 23.5-32.9 5.1-6.5
К2О 24.6-28.4 8.0-8.9
СаО 1.7-2.9 12.2-13.2
MgO 1.6-3.1 2.0-2.1
Fe2O3 0.4-0.5 1.2-1.4
Р2О6 3.1—4.8 63.8—66.7
SO3 1.3-2.6 —
SiO2 0.28-2.0 —
Cl 23.8-28.5 —
соединения неинкубированного куриного яйца, установил, что
в среднем в белке содержится 0.52% общего фосфора, а в желтке —
0.98%, в том числе кислоторастворимого фосфора 121 и 142 мг%
соответственно. Более половины общего фосфора желтка состав-
ляет липидный фосфор (580 мг%), а в белке его только 145 мг%.
Фосфопротеинов в желтке 252 мг% и примерно столько же их
в фосфоре белка (268 мг%). В белке отношение анионов к катионам
меньше единицы (0.5). В желтке, наоборот, анионов почти в три
раза больше, чем катионов (2.8). В связи с тем что сухих веществ
в желтке больше, чем в белке, отношение анионов к катионам
яйца в целом равно 2.3. Большую роль в эмбриональном обмене
веществ играют содержащиеся в белке и желтке микроэлементы.
В желтке находится большая часть фтора, иода, меди, цинка,
алюминия и марганца, а в белке — бблыпая часть бора и мышьяка.
Белок представляет собою основной запас воды и некоторых
питательных веществ (протеины, минеральные соли) для разви-
вающегося эмбриона.
Тарханов (Tarchanoff, 1884) обнаружил, что в яйцах птенцо-
вых процент воды в белке, так же как и в желтке, больше, чем
у выводковых птиц (табл. 23).
Кроме того, автор наблюдал, что белок в яйцах птенцовых
птиц при кипячении не свертывается, как обычно, а остается либо
жидким и прозрачным, либо превращается в водянистый просве-
чивающий студень и свертывается только при температуре много
выше 100°. Такой белок Тарханов назвал тата-белком (по имени
своей дочери Таты). Тата-белок более щелочной, чем белок ку-
риного яйца, легче переваривается энзимами, но в течение эм-
брионального развития изменяется и становится сходным с бел-
ком куриного яйца.
67
5*
Таблица 23
Различие в содержании воды в белке яиц (в %) у птенцовых и
выводковых птиц
(По Tarchanoff, 1884)
Птенцовые .... Выводковые .... Ворон Дрозд Скво- рец Коро- стень Зяб- лик Го- лубь Чайка Соло- вей
90.2 90.1 89.9 89.5 89.4 89.3 89.1 89.1
Утка Индейка Ржан- ка Цесарка Гусь Ку- рица Пере- пел
88.0 87.9 87.9 87.8 87.6 86.9 86.6
Приводим данные о химическом составе белка яиц некоторых
сельскохозяйственных птиц (табл. 24).
Таблица 24
Химический состав белка яиц (в % от влажного веса)
Вид яиц Вода Про- теины Жиры Угле- воды Мине- ральные вещества Автор и год
Куриные . . 85.7 12.7 0.3 0.7 0.3—0.7 Romanoff a. Romanoff, 1949
» 87.9 10.6 — 0.9 0.6 Riley, 1953
Утиные . . . 87.9 11.3 — — 0.9 Данилова и Нефедъева, 1936
Индюшиные .. 87.4 10.4 — 1.1 0.4 Орлов и Кучковская,
Чанг и Стадетман (Chung a. Stadetman, 1961) обнаружили,
что при увеличении веса яйца на 1 г протеин белка увеличивается
на 0.09 г, а протеин желтка — на 0.04 г.
Основной протеин белка куриного яйца — это овальбумин,
обладающий способностью кристаллизоваться. Он составляет
69.7% всех протеинов белка. Овальбумин электрофоретически
разделяется на два компонента: Аг — быстро движущийся, со-
держащий 2 атома фосфора на моль белка, и А2 - 1 атом фосфора
на моль белка; причем Аг под действием фосфатаз переходит в А2
и далее — в лишенный фосфора компонент А3 (Лисовская й Ли-
ванова, 1960). В белке куриного яйца найдены также протеины:
овомукоид (12.7%), овокональбумин (9.0%), овоглобулин (6.7%),
68
лизоцим и овомуцин (1.9%). Овомуцин — структурный протеин
густого белка, обусловливающий его качество геля. В электрон-
ном микроскопе молекула овомуцина имеет вид длинной нити и,
по-видимому, состоит из большого количества шариков, соеди-
ненных друг с другом. Разрушение геля густого белка при встря-
хивании объясняется разрушением этих нитеобразных агрегатов
(Riley, 1953). Овомуцин содержит большое количество серы;
несколько меньше серы содержит овоальбумин. Кипячение 100 г
белка с водою дает 10.7 мг сероводорода. В утином яйце найдено
два протеина: анатинин, соответствующий куриному овоальбу-
мину, и анатин, соответствующий куриному овомуцину. В яйце
вороны найдено три протеина: корвин, корвинин и корвиницин.
В белке найдено красящее вещество, относящееся к типу водо-
растворимых пигментов (флавинов), — овофлавин. Содержание
его в яйце чрезвычайно мало (3 мкг на яйцо), но наличие его в белке
имеет большое значение, так как он входит в состав витамина
рибофлавина и играет важную роль в тканевом дыхании.
Необходимо отметить бактерицидные или антибиотические
свойства птичьего белка. В работах Коротковой (1956) и Мовчан
(1962) исследована антибиотическая активность белка куриных,
утиных и цесарочьих яиц. Выяснено, что более сильным анти-
биотическим действием обладает белок куриного яйца. Наруж-
ный слой белка самый бактерицидный. Авторы приходят к вы-
воду, что на ранних этапах развития птичьего эмбриона, когда
он не имеет еще собственных защитных реакций, белок является
надежным барьером, препятствующим проникновению бактерий
к эмбриону. Антибиотические свойства белка оказались слабее
всех (из исследованных) в яйцах дрозда (Romanoff a. Romanoff,
1949).
Химический состав скорлупы куриного яйца, так же как и скор-
лупы яиц других птиц, имеет значительные колебания (табл. 25).
Углекислый кальций составляет основную часть скорлупы.
В наружном слое он имеет форму кристаллов, расположенных
Таблица 25
Химический состав скорлупы яиц
(По Romanoff a. Romanoff, 1949)
Вид яиц] Состав скорлупы (в %) Органиче- ские вещества
СаСОз MgCo3 Са3(РО4)2 рго5 к
Куриные .... 92.9-97.9 1.3-1.8 0.8 0.4-0.8 0.3 4.1-5.5
Утиные 94.4 0.5 0.8 . 0.8 — 4.2-4.3
Гусиные 95.3 0.7 0.5 0.5 — 3.5
Фазаньи .... 93.7 1.4 0.8 — — 4.0
Страусиные . . . 94.6 0.7 ’ 0.4 — — 4.3
69
своей длинной осью перпендикулярно к границе скорлупы, а во
внутреннем — аморфен. Органическое вещество скорлупы имеет
кератинообразный характер. Спектроскопически исследуя микро-
элементы скорлупы куриных яиц, Надь и Ола (Nady a. Olah,
1956) показали, что в 10 г скорлупы содержится (в мкг):
Ag - 0.5—5.0; Cd - 0.2—5.0; Со - 0—1; Си - 2-10; Fe -
40-70; Mg - 0.5-10; Мп - 0-0.3; Ni - 0.5-5.0; Pb - 0.5-
6.0; Sn — 0—6; Ti — 1—5; Zn — 5—20. Здесь, как мы видим,
обнаружены, кроме отмеченных в табл. 18, также и олово, кад-
мий, кобальт и никель.
Подскорлупные оболочки состоят из протеина, очень близкого
по своему составу к кератину. Этот овокератин имеет в четыре
раза больше серы, чем белок, и включает больше 7% цистина.
Неизвестно, какую роль играет кератин в эмбриональном раз-
витии, но можно предположить, что содержащиеся в нем сера и
цистин используются частично для формирования перьев.
Г. Физические и физико-химические свойства яиц
В связи с тем что удельный вес желтка куриного яйца (1.03)
меньше, чем белка (1.04), желток всплывает в яйце наверх. При
хранении яиц их удельный вес уменьшается. Для проверки све-
жести яиц их можно опустить (ненадолго) в 10 %-й раствор по-
варенной соли, причем свежие яйца тонут, а старые и испорчен-
ные плавают. Точка замерзания желтка куриного яйца ниже
(—0.6°), чем белка (—0.4°).
Данилова (1931) исследовала калорийность куриных яиц и
показала, что она несколько отлична для разных пород и в раз-
ные сезоны года. По данным, приведенным Романовыми (Roma-
noff a. Romanoff, 1949), средняя калорийность куриного яйца
равна 74—92 кал. Калорийность утиных и гусиных яиц больше,
чем куриных (225—243 кал. на 100 г яичной массы по сравнению
с 143 кал. на 100 г яичной массы у куриных), в связи с большим
содержанием жира в желтке яиц водоплавающих птиц.
Белок яйца — самая щелочная из всех естественных жид-
костей (pH 9.5), что определяет в известной мере его бактерицид-
ные свойства. В связи с различием pH белка и желтка, имеющего
слабокислую реакцию, яйцо является как бы электрической ба-
тареей с измеримым потенциалом 0.1 mb (Riley, 1953). Осмотиче-
ское давление желтка свежего яйца равно 6.6 атм., а белка —
5.3 атм.
Исследованиями Перова и Долинова (1932) и Третьяковой
(1939) установлен целый ряд физико-химических показателей
для куриных яиц (табл. 26).
Как мы видим, белок имеет примерно в 3 раза большую
электропроводность, чем желток, несмотря на то что процент
золы в них почти одинаков. Это указывает на значительно ббль-
70
Таблица 26
Физико-химические показатели белка и желтка куриных яиц
(По данным Перова и Долинова, 1932, и Третьяковой, 1939)
Физико-химические показатели Белок Желток
весь наружный жидкий плотный внутренний жидкий от до
от до
pH • 8.88 9.28 — — — 5.56 6.87
То же♦* 8.0 8.3 — — — 5.8 6.3
Коэффициент рефракции . . . 1.3527 1.3522 1.3540 1.3562 1.3572 1.4177 0.4104
То же♦♦ 1.3540 1.3558 • — — — 1.4193 1.4222
Поверхностное натяжение . . 0.87 0.91 — — — . 0.72 0.73
Вязкость 3.35 3.76 — — — — —
Вязкость при разбавлении во- дой 1:4 1.12 1.19 — — . — 1.33 1.95
Буферность (1/10 см3 на 100 см3 пробы) 29.31 38.72 — — — 64.80 • 95.66
Электропроводность (в Э *) . 75.45 78.64 65.57 74.16 59.96 20.82 23.06
То же♦* 72.4 80.65 — — — 23.10 24.64 .
• __ 1 э равна 0.0001 значения удельной электропроводности; *• — данные Перова и Долинова.
шую диссоциированность солей белка и, следовательно, на луч-
шую их доступность для использования эмбрионом.
Для сравнения приведем данные о физико-химических пока-
зателях яиц разных видов сельскохозяйственных птиц (табл. 27).
Таблица 27
Физико-химические показатели белка и желтка куриных, утиных
и гусиных яиц
(По Третьякову, 1949 и Бражниковой, 1950)
Яйца Коэффициент рефракции Электропроводность (в Э) pH
белок желток белок желток белок желток
Куриные * . . . 1.3543 1.4164 75.0 22.10 9.31 6.35
» .... 1.3511 1.4197 75.65 21.87 — —
Утиные * . . . . 1.3590 1.4178 63.32 20.53 9.23 6.24
» .... 1.3576 1.4223 76.98 22.31 — —
Гусиные 1.3545 1.4232 73.57 18.80 — —
• — данные Третьякова.
Исследуя некоторые электрические свойства белка и желтка
куриных яиц, Ханыкова (1958) показала, что электропроводность
белка при разных частотах тока (100 кгц и 1 мггц) одинакова
и может служить характеристикой суммарной концентрации
ионов. В связи с тем что желток обладает свойством электриче-
ской поляризации, для характеристики суммарной концентрации
ионов в нем можно использовать данные электропроводности
только при определенной частоте, не ниже 1 мггц. Автор пред-
полагает, что поляризационные свойства желтка обусловлены
его физико-химической структурой, а именно субмикроскопиче-
ской структурой желточных шаров.
Физико-химические показатели белка и желтка яиц в значи-
тельной степени зависят от кормления несушек. Так, например,
буферность желтка снижается при недостатке в рационе расти-
тельных или животных белков, а буферность белка при этом,
наоборот, несколько повышается. Электропроводность меняется
более всего в связи с изменением минерального кормления. При
правильном минеральном кормлении электропроводность белка
повышается, что сопровождается и более высокой выводимостью.
Физико-химические показатели белка и желтка яиц зависят
также и от температуры в помещении маточного стада, и от се-
зона. Так, например, различия в данных Перова и Долинова
(1932) о коэффициенте рефракции, электропроводности и pH по
72
сравнению с данными Третьяковой (1939) (табл. 26) могут быть
объяснены тем, что исследования Перова и Долинова проводи-
лись зимой, а Третьяковой — летом. Как указывает Третьяков
(1951), яйца, снесенные весной и летом, содержат больше воды и
в связи с этим имеют более низкий коэффициент рефракции, по-
вышенную электропроводность и пониженную вязкость желтка
(до 1.12) по сравнению с осенне-зимними яйцами. Следует отметить
вообще значительные вариации физико-химических показателей
яиц, свидетельствующие о реальных различиях в их качестве.
Так, Третьякова (1938) обследовала десять совхозов и обнаружила,
что при электропроводности белка куриного яйца, равной 66.2—
71.4 Э, выводимость не превышает 50%, а при электропровод-
ности 75.1—78.6 Э процент вывода цыплят 69—85.3.
Г Л А В А 5
ХРАНЕНИЕ ЯИЦ И ОТБОР ИХ ДЛЯ ИНКУБАЦИИ
А. Хранение яиц до инкубации
Во время хранения яиц до инкубации должна быть
сохранена жизнеспособность эмбриона, начавшего развитие в яйце-
воде курицы и прекращающего его после снесения яйца. Этот
перерыв в развитии, однако, не означает полного прекращения
жизненных процессов. Состояние эмбриона в это время подобно
анабиотическому, с чрезвычайно замедленным обменом веществ.
Рассмотрим, в каких условиях проходит это анабиотическое состоя-
ние и как долго оно может продолжаться без нарушения способ-
ности к дальнейшему развитию.
1. Температура хранения
Основное, решающее условие при хранении яиц — это темпе-
ратура. Максимальная амплитуда ее ограничивается, с одной
стороны, температурой, выше которой восстанавливается раз-
витие эмбриона (физиологический нуль), а с другой, — темпера-
турой, при которой совсем прекращается жизнь, без возможности
ее восстановления в дальнейшем.
Опытами Эдвардса (Edwards, 1902) установлено, что после
снесения яйца для возобновления развития куриного эмбриона
достаточна температура 21—27°, но развитие при этой темцера-
туре протекает ненормально: происходит рост бластодиска, но диф-
ференциация (образование первичной полоски и т. д.) ненасту-
пает, появляются уродства, и развитие эмбриона прекращается.
73
Следовательно, верхний температурный предел для хранения
яиц — это 20°.1
Вопрос о минимальной температуре, при которой сохра-
няется способность эмбриона к развитию, был предметом многих
исследований. Моран (Moran, 1925) отметил, что охлаждение
яиц до температуры ниже нуля приводит к изменению коллои-
дального состояния белка и желтка, причем продолжительность
охлаждения также влияет на это изменение. Доугерти (Doug-
herty, 1926) подчеркивает, что в природных условиях птицы
кладут яйца обычно только после того, как среднесуточная
температура становится выше нулевой. Отметим это чрезвычайно
интересное явление приспособленности жизненных циклов птиц
к внешним условиям. Исследуя низшие температурные границы
хранения гусиных и куриных яиц, Владимирова (1962а) уста-
новила, что хранение яиц при температуре от —2 до +4° в тече-
ние 5—7 дней не снижает их инкубационных качеств. Гусиные
яйца сохраняют выводоспособность при температуре от 0 до —2°
дольше, чем куриные. Таким образом, можно считать, что сниже-
ние температуры до нуля не опасно для жизнеспособности эмбри-
она.
Несколько необычные данные получены Джаллом с соав-
торами (lull et al., 1948). Содержание индюшиных и куриных
яиц при температуре —17.2° в течение 10 час. (причем темпера-
тура внутри яйца достигала —1°) не понижало заметно выво-
димости. При охлаждении в течение 8—10 час. при температуре
—23—24.6° выводимость была такой же хорошей, как в контроле.
Содержание индюшиных яиц в течение 2 дней при температуре
—6.7° только немного снизило выводимость. Даже из яиц, в ко-
торых внутренняя температура была равна —16.2°, вылупилось
несколько цыплят. А содержание индюшиных яиц в течение
4 час. при температуре —17.2° обусловило даже повышение
процента вывода индюшат.
Какова же оптимальная температура для хранения яиц?
В большинстве руководств по инкубации указывается, что оп-
тимальная температура для хранения яиц — это +8—(-10°.
Олсен и Хэйнс (Olsen a. Haines, 1948) наблюдали наилучшее
сохранение жизнеспособности эмбрионов при температуре хра-
нения 10° и наихудшее — при —1.1°. При хранении в течение
12 дней температуры +10и +12.8° были одинаково благоприятны,
а при удлинении срока хранения до 28, 35 и 42 дней выводимость
цыплят была при +10о —50, 22.2 и 6.5%, а при +12.8° — 26.3,
5.9 и 0% соответственно. Салганник (1948) считает оптимальной
для хранения яиц несколько более низкую температуру (6—8°).
Чавчанидзе (1953) установил, что температура воздуха на яйце-
складе должна быть 2—8°. По данным автора, хранение яиц
1 Подробнее о температурных границах см. главу 18.
74
при этой температуре возможно в течение 8 дней, причем наилуч-
шим образом сохраняется структура белка яйца, витамин А
в желтке и обеспечивается полноценность инкубационных ка-
честв яиц.
Изучая возможности улучшения условий хранения яиц,
Орлов (1948) предложил периодическое их подогревание. В опыте
с подогреванием (в течение часа в сутки) до 40° длительно хра-
нившихся яиц выводимость цыплят повысилась с 37.7 до 64.4%.
С тех пор большое количество работ было посвящено полезному
влиянию на выводимость цыплят, утят, гусят и индюшат одно-
кратного или периодического подогревания яиц (1—2 часа в сутки
при 37—39° или в другие сроки и при другой температуре) во время
хранения (Князева, 1955; Kosin, 1956; Smith, 1956; Карапетян, 1957;
Владимирова, 1958; Николаева, 1958а; Becker a. Bearse, 1958, и др.).
Необходимость периодических подогревов яиц во время хранения
Николаева (19586) объясняет тем, что в природных условиях
курица сидит в гнезде во время снесения яйца и согревает ранее
снесенные ею яйца. По наблюдениям автора, курица проводит
в гнезде во вторую половину цикла яйцекладки, когда в гнезде
уже лежат 5—10 яиц, от 43 до 65 мин. в сутки. При этом ранее
снесенные яйца нагреваются до 31.6—35.4° (внутри яйца), что
всего на 3° ниже температуры внутри яиц после 2-часового подо-
грева в инкубаторе. Отметим, со своей стороны, что у диких
птиц до окончания яйцекладки мы наблюдали насиживание
по нескольку часов в сутки. По данным Князевой (1958), еже-
дневное подогревание куриных яиц по 2 часа в течение 3—6 дней
повышает вывод цыплят на 10—24%. В связи с тем что это воз-
действие повышает обменные процессы в яйцах во время хране-
ния, в дальнейшем при инкубировании этих яиц имеет место луч-
ший рост эмбрионов, более интенсивное нарастание гемоглобина
и количества эритроцитов в их крови и, наконец, более ранний
и дружный вывод цыплят. Николаева (1958а) показала, что при
ежедневном двухчасовом подогреве долго хранившихся яиц
происходит медленное, но нормальное развитие эмбриона: уве-
личивается размер бластодиска, наступает гистологическая диф-
ференциация до стадии первичной полоски и образование трех
слоев: эктодермы, энтодермы и мезодермы. Это дает возможность
нормального развития при инкубировании долго хранившихся
яиц в отличие от отстающего развития в яйцах, хранившихся
без подогревания. Мы предполагаем, что подогревание яиц сразу
после снесения или в первые дни хранения продвигает развитие
эмбриона на менее чувствительную стадию поздней гаструлы —
и потому он легче переносит перерыв в развитии во время дли-
тельного хранения. Напомним о работе Мак-Налли и Баерли
(McNally a. Byerly, 1936), обнаруживших максимальную вы-
водоспособность куриных яиц при нахождении яйца в яйцеводе
(и следовательно, развивавшегося) в течение 27 час. и снижение
76
выводоспособности при уменьшении и увеличении этого срока.
Обычно же у кур, это время равно примерно 24 час. По данным Хейз
и Николаидес (Hays a. Nicolaides, 1934), в яйцах с высокой выводи-
мостью к моменту снесения эмбрионы были на стадии поздней га-
струлы, а с низкой — на стадии ранней гаструлы. Однако
Тэйлор и Ганз (Taylor a. Gunns, 1939) опровергают эти данные
и сообщают, что у яиц с высокой и низкой выводимостью
нет никаких различий ни в стадии, ни в размере эмбрионов
в момент их снесения. Владимирова (1962а) показала, что при
однократном 5-часовом подогреве гусиных яиц сохраняется
жизнеспособность .эмбрионов после 15—20-дневного хранения,
а при периодическом (по 5 час. через 5 дней) — и после 30-днев-
ного.. При подогреве во время хранения яиц в зародышевых
дисках не происходит характерных изменений (уменьшение и
разрыхление); наоборот, они увеличиваются в размерах и раз-
виваются до стадии первичной полоски. В опытах Милби и Шер-
вуда (Milby a. Sherwood, 1960) подогревание индюшиных и кури-
ных яиц во время хранения немного (но не достоверно) повышало
выводимость цыплят и индюшат. Снижение достоверности поло-
жительного эффекта связано с тем, что на куриные яйца с высокой
выводимостью (выше 80%) подогревание яиц при хранении не
влияет. Подогревание яиц проводится также с целью сделать воз-
можным отбор яиц до инкубации по заметному при просмотре
яиц на овоскоп различию в размере зародышевых дисков, о чем
будет речь позже.
2. Влажность, вентиляция и другие факторы
во время хранения яиц
Во избежание излишнего испарения влаги из яиц, которое
довольно значительно во время хранения (в среднем 0.15% веса
яйца в день), рекомендуется в яйцехранилище поддерживать
относительную влажность, равную 70—75%. Эта величина влаж-
ности подобрана работниками инкубации эмпирически. Салганник
(1948) считает необходимым поддерживать в яйцехранилище
более высокую влажность — 85—90%. Еще более высокую влаж-
ность (94—97%) рекомендует Орлов (1948). Этот вывод автора
связан не только с большим процентом потери веса яйца (около
3% при 15-дневном и около 4—5% при 25-дневном хранении),
но и с тем, что долго хранившиеся яйца имеют тенденцию и в даль-
нейшем, во время инкубации, больше испарять воду. Автор от-
метил, что те яйца, из которых затем вылупились цыплята,
имели меньшую потерю веса во время хранения (2.03%), чем
яйца, в которых эмбрионы погибли в течение инкубации (2.87%).
Поэтому для долго хранившихся яиц автор рекомендует в тече-
ние первой половины инкубации особый режим, с более высокой
влажностью. Потере веса яиц во время хранения посвящено
76
исследование Владимировой (1954). Автор показала, что яйца
кур, характеризующиеся меньшей потерей в весе во время хра-
нения, дают лучшую выводимость. Интенсивность потери в весе
куриных яиц во время хранения неодинакова и зависит от многих
факторов: возраста кур, величины яйца, физических свойств
скорлупы, температуры, влажности и вентиляции яйцехранилища
и др. Камар (Катаг, 1962) отмечает, что бблыпая пористость
утиных яиц обусловливает быструю порчу содержимого во время
их хранения.
Рассмотрим вопрос о вентиляции яйцехранилища. К моменту
снесения яйца эмбрион еще слишком мал, и во время хранения
он находится в анабиотическом состоянии; следовательно, потреб-
ление кислорода и выделение углекислоты им совершенно не-
значительно. Но содержимое яйца (белок и желток) выделяет
углекислоту и кислород, что обусловлено повышенной концен-
трацией этих газов в яйцеводе несушки. Следовательно, довольно
значительные количества кислорода и* особенно углекислоты
выделяются яйцами во время хранения. Холл и Романов (Hall
a. Romanoff, 1943) хранили яйца в дистиллированной воде, а также
после опускания их в желатин или кремниевый натрий. Авторы
обнаружили при этих условиях хранения яиц лишь небольшое
снижение выводимости, считая это показателем того, что яйца
могут почти не дышать атмосферным воздухом. Но Кирсанов
(1935а), сравнивая условия хранения яиц в разных совхозах,
отметил, что чем меньше яиц приходилось на 1 м8 яйцехранилища,
тем выше была выводимость цыплят. Автор приходит к выводу,
что в яйцехранилище должна быть хорошая вентиляция.
Исходя из предположения, что значительная часть нарушений
в долго хранившихся яйцах обусловлена выделением из них
углексилоты и сдвигом pH в связи с этим, Еремеев (1957) провел
опыты по содержанию куриных и утиных яиц перед инкубацией
в углекислоте (до 82.5% СО2). 2—4-часовое содержание яиц
в углекислоте значительно повысило выводимость утят и осо-
бенно цыплят. Владимирова (19626) провела сходные опыты с
с гусиными и куриными яйцами. 5-часовое содержание гусиных
яиц в углекислоте повысило выводимость гусят, а более длитель-
ное не имело эффекта. Не наблюдалось также положительного дей-
ствия содержания в углекислоте перед инкубацией куриных яиц.
В последние годы за рубежом, особенно в США, распростра-
нился метод хранения яиц в пластмассовых мешочках. Бекер
с соавторами’ (Becker et al., 1964а, 19646) сообщает о хранении
и перевозке на пароходе индюшиных яиц в двух типах мешочков:
1) сохраняющих только влагу, но пропускающих углекислоту
и 2) не пропускающих из яиц ни влаги, ни углекислоты. Наи-
высший процент вывода индюшат был подучен после хранения
в мешочках 2-го типа, ниже — в мешочках 1-го типа и еще зна-
чительно ниже — при обычном хранении яиц. Следовательно,
77
потеря жизнеспособности эмбрионов в долго Хранившихся яй-
цах в значительной мере обусловливается потерей из них влаги
и углекислоты.
Рассмотрим вопрос о том, в каком положении (вертикальном
или горизонтальном, острым концом вниз или вверх) лучше
всего хранить яйца, предназначенные для инкубации, и следует
ли их переворачивать во время хранения. Пенионжкевич (19356)
провел детальное исследование этого вопроса. По данным автора,
для хранения куриных яиц лучше горизонтальное положение,
а переворачивание яиц необходимо только при хранении яиц
дольше 10—18 дней. Для хранения утиных яиц, наоборот, лучше
вертикальное положение, а переворачивание яиц дает заметный
положительный эффект при хранении свыше 5 дней. Фарнсворс
и Баррен (Farnsworth a. Warren, 1962) подтвердили, что при
хранении куриных яиц свыше 18 дней поворачивание яиц дважды
в день несколько повышает выводимость цыплят.
3. Длительность хранения
Но и при самых оптимальных условиях хранения жизне-
способность эмбриона в анабиотическом состоянии не может
сохраняться длительное время. По исследованию Уайта (Waite,
1919), после 28 дней хранения все эмбрионы настолько ослабе-
вают, что ни одного цыпленка не вылупляется. С удлинением
срока хранения куриных яиц замечалось постепенное снижение
выводимости, от 65% цыплят из оплодотворенных яиц, не хра-
нившихся ни одного дня, до 3% при хранении яиц 28 дней. Пе-
нионжкевич (19356) наблюдал, что при хранении куриных яиц
в течение 25 дней из них не вывелось ни одного цыпленка, а при
хранении 15 дней вывелось всего 39.9%; в контрольной группе
вывелось 70.6% цыплят. Фарнсворс и Баррен (Farnsworth a. War-
ren, 1962) наблюдали, что при хранении куриных яиц в течение
15 дней вместо 6 выводимость цыплят снижалась на 5%, а каждый
последующий день хранения снижал ее еще на 4%. Злочевская
(1962а) указывает, что хранение куриных яиц в течение 17 дней
в весенне-летний сезон значительно больше снижает их инкуба-
ционные качества, чем в зимний. Однако условия хранения яиц
имели в опытах автора слишком большую амплитуду (темпера-
тура 11.2—15° и влажность 62—73%). По-видимому, в весенне-
летнее время температура хранения яиц приближалась к верх-
ней границе (15°), а влажность — к нижней, что и вызвало боль-
шее снижение инкубационных качеств яиц при хранении.
Моренг и Мансон (Moreng a. Manson, 1961) установили, что
даже 9-дневное хранение индюшиных яиц снижает жизнеспособ-
ность эмбрионов. Козин (Kosin, 1958) показал, что после хра-
нения индюшиных яиц сроком до 7 дней в большинстве их при
овоскопировании можно обнаружить развитие эмбриона уже
78
через 36 час. инкубации, а после хранения 8—14 дней иногда
не наблюдалось развития до 156 час. инкубации. Автор отмечает,
что ни из одного яйца, где развитие можно было заметить лишь
через 84 часа, не вылупилось индюшат.
Выводимость утят из длительно хранившихся яиц исследовал
Пенионжкевич (19356). При хранении 5 дней вывелось 33.3—
52.4% утят, при хранении 10 дней — 32.4—45%, 15 дней —
29.7—38.9%, 20 дней — 3.2—25%, а при хранении 25 дней
выводимость равнялась нулю.
По данным Пенионжкевича (1946), хорошее развитие эмбрио-
нов в гусином яйце происходит только в том случае, когда яйца
инкубируются не позднее, чем через 5—10 дней после снесения.
Меритт и Кларидж (Merritt a. Clarridge, 1959) наблюдали, что
до 10 дней хранения нет снижения выводимости гусят, но после
14 дней хранения снижение процента вывода идет по квадрати-
ческой кривой. По данным Владимировой (1962а), хранение гу-
синых яиц в течение 15 дней сравнительно мало влияет на вы-
водимость, в течение 20 дней — снижает выводимость на 12%,
при 30-дневном хранении вывелось только 18.7% гусят, а при
45-дневном выводимость была равна нулю.
Следует отметить, что хранение яиц водоплавающей птицы
осложнено также тем, что они в большей степени подвержены
заражению гнилостными микроорганизмами, и поэтому их осо-
бенно важно закладывать в инкубатор цак можно скорее
после снесения. По данным большинства авторов, хранение
влияет наименее губительно на куриные яйца по сравнению
с влиянием на яйца других видов сельскохозяйственных птиц.
Это обусловлено, по нашему мнению, тем, что куры раньше дру-
гих видов были одомашнены и'потому легче адаптируются к ус-
ловиям, связанным с этим, — к вредному влиянию хранения
и к условиям инкубации.
Борен с соавторами (Bohren et al., 1961) установил прямую
корреляцию между способностью к длительному хранению ку-
риных яиц и выводимостью цыплят. При индивидуальном подсчете
(по несушкам) яйца, имевшие наибольшую сопротивляемость небла-
гоприятному влиянию хранения, давали наивысший процент вы-
вода цыплят и в самые короткие сроки инкубации. Сходное
явление наблюдал Козин (Kosin, 1954) у разных пород и линий
индеек: чем больше была выводоспособность яиц у данной породы
или линии, тем лучше яйца переносили хранение.
В литературе и практике инкубации долгое время существо-
вало вредное мнение, что после снесения яйца птицей оно должно
«отдохнуть» и лучше закладывать яйца в инкубатор после 3—5 дней
хранения. Опыты Орлова (1948) показали, что это совершенно
неверно. Даже при благоприятных условиях хранения (темпе’
ратура +5°, влажность 96%) яйца, хранившиеся 1 день, дали
100% вывода от оплодотворенных яиц, 2 дня — 90.9%, 5 дней —
79
88.9%, 6 дней — 78.6%. Автор отмечает, что при хранении бо-
лее 7 дней начинается особенно быстрое снижение инкубационных
качеств яиц. Интересны также опыты Антоновича (1950) по не-
прерывному развитию птичьего эмбриона. Автор, хотя и на не-
большом материале, показал, что при закладке в инкубатор не
успевших еще остыть яиц, сразу после снесения их курицей, зна-
чительно (до 100%) повышается выводимость, сокращается дли-
тельность инкубации на 1.5—2 дня, а также улучшается качество
потомства (больший вес, раннее начало яйцекладки, большая
яйценоскость и больший процент курочек). Иванов (1955) под-
твердил эти данные. В яйцах, заложенных в инкубатор сразу
после снесения (еще теплыми), длительность инкубации была
сокращена до 20 дней, процент вывода был наивысшим — 98.9%
и соотношение полов было значительно смещено в сторону самок
(63.3% курочек). Хранение яиц в течение 6—19 дней сдвигало
соотношение полов в обратную сторону (53—59% петухов) и уве-
личивало длительность инкубации до 21.5 суток. Процент вывода
цыплят при 19-дневном хранении снижался до 75.4% от опло-
дотворенных яиц.
Таблица 28
Влияние сроков хранения яиц гаги на выводимость гагачат
(По Рольник, 1955)
Продолжи- тельность хранения (в днях) Количе- ство яиц «Неоплодотворен- ные» яйца Погибло эмбрионов во время инкубации Вылу- пилось гагачат % вывода от заложен- ных яиц
количе- ство %
2 6 ' 1 16.7 1 4 66.7
5 5 — 0 2 3 60
7 13 7 53.8 — 6 46.2
8 10 7 70.0 1 1 10
Примечание. До начала экспериментального хранения, возможно, яйца ле-
жали в гнезде 2—3 дня.
По хранению яиц диких птиц имеются данные только в нашей
работе по инкубации яиц гаги (Рольник, 1955) (табл. 28). От-
метим, что срок хранения яиц гаги, не имеющий вредных послед-
ствий, значительно меньше, чем у куриных яиц. Можно пред-
положить, что физиологический нуль развития эмбрионов гаги —
северной птицы — ниже, чем у куриных эмбрионов, и при темпе-
ратуре хранения около +15° развитие гагачьих эмбрионов на-
чиналось, но протекало неправильно, и эмбрионы погибали.
Но более вероятно, что способность эмбрионов домашних птиц
переносить длительное хранение создалась в результате ис-
80
кусственного отбора человеком. А у диких птиц, насиживающих
со 2—3-го яйца, приспособленности к длительному анабиоти-
ческому состоянию у эмбриона не выработалось, и хранение яиц
даже влечение 8 дней сильно снижает выводимость.
4. Изменения в содержании яйца и состоянии эмбриона
во время хранения яиц
Первым заметным признаком длительного хранения яиц яв-
ляется увеличение воздушной камеры вследствие испарения воды
из яйца. Одновременно с этим часть воды белка переходит в жел-
ток в связи с большим осмотическим давлением в последнем.
И все же в результате хранения происходит своеобразное раз-
жижение белка в связи с нарушением структурности плотного
слоя белка, которое обусловливает большую подвижность желтка
и всплывание его к самой скорлупе. При этом бластодиск, нахо-
дящийся всегда сверху, может прикасаться к подскорлупной
оболочке и быть механически поврежденным. Проникновение
воды из белка в желток во время хранения яиц ведет к большому
растяжению желточной оболочки, что иногда оканчивается раз-
рывом ее. Владимирова (1954) показала, что при хранении
в яйце происходят глубокие, необратимые изменения, ведущие
к снижению инкубационных качеств: уменьшается вес белка,
нарушается его структура (после 10 дней хранения), понижается
электропроводность, увеличивается pH белка и желтка, особенно
в первые 5 дней. Чем больше потеря веса яиц во время хранения,
тем эти физико-химические изменения происходят быстрее.
Что происходит с самим эмбрионом во время хранения —
мало изучено. Кауфман (Kaufman, 1939а, 19396) исследовала
изменения в развитии эмбриона после длительного хранения
(25, 28 и 34 дня) куриных яиц. Автор отрицает, что потеря влаги
яйцами во время хранения обусловливает повышение эмбриональ-
ной смертности, так как за 34 дня хранения потеря веса яиц
только 0.7—1.5%, а при потере веса от 3 до 6% под влиянием
экспериментально пониженного давления смертность эмбрионов
не увеличивалась. Обнаружено также, что длительность инку-
бации долго хранившихся яиц (34 дня) на сутки больше, чем
в контроле. Однако это обусловлено не менее интенсивным ростом
эмбриона, а более поздним (примерно тоже на сутки) началом
его развития. Попытка автора выяснить причины «старения»
яиц цри хранении не увенчалась успехом.
В работе Шишкиной (1949) приведены данные по развитию
36-часовых куриных эмбрионов после разных сроков хранения
яиц (табл. 29).
Автор отмечает, что у эмбрионов в долго хранившихся яйцах
особенно отстает дифференцировка тканей. Эмбрион может иметь
2—3 пары сомитов при еще незакончившемся срастании краев
5 В. В. Рольник
81
Таблица 29
Изменения в эмбриогенезе птиц под влиянием разных сроков
хранения яиц
(По Шишкиной, 1949)
Продол- житель- ность хране- ния (в днях) Число исследо- ванных эмбрио- нов Средняя длина эмбриона (в мм) Средний диаметр со- судистого поля (в мм) Соотноше- ние длины эмбриона к длине сосудистого поля Среднее число сомитов Среднее число пар сомитов на 1 мм длины эмбриона
0 23 5.76 7.2 1 : 1.38 9.88 1.88
5 21 5.00 7.0 1:1.42 7.57 1.51
10 21 4.88 6.6 1 :1.00 6.53 1.34
15 21 4.14 6.4 1 :1.54 6.60 1.59
20 17 3.47 5.0 1:1.44 2.40 0.69
нервной трубки. Автор, так же как и Орлов (1948), считает, что
для яиц различных сроков хранения следует применять разный
режим в первые дни инкубации. Завальский (Zawalsky, 1962)
подсчитал, что увеличение срока хранения куриных яиц досто-
верно увеличивало длительность инкубации. Кауфман и Кржа-
новская (Kaufman a. Krzanowska, 1957) обнаружили, что после
хранения куриных яиц в течение 16—21 дня при температуре
+ 10° уменьшался общий вес цыплят после вылупления, а также
вес кишечника и печени у них. Однако активность щитовидной
железы и использование печенью гемоглобина у эмбрионов из хра-
нившихся яиц были больше, чем в контроле, что указывает на бо-
лее интенсивный обмен веществ. Лоркивизова (Lorkiewiszowa,
1960—1962) наблюдала снижение веса цыплят, вылупившихся
из яиц, хранившихся более 10 дней, а при хранении яиц более
3 недель повышалась и смертность цыплят. Владимирова (1962а)
отметила, что при длительном хранении гусиных яиц уменьшается
размер зародышевого диска и он становится рыхлым. Козин
и Мун (Kosin a. Mun, 1965) показали, что главным фактором,
влияющим на темпы раннего эмбриогенеза и на внутриклеточное
окисление у индюшиных эмбрионов, является длительность хра-
нения яиц до инкубации, причем степень влияния хранения ин-
дивидуально варьирует. Исследуя влияние 7—14-дневного хра-
нения утиных яиц на эмбриональное развитие, Шевцова (1965)
установила, что хранение обусловливает дезорганизацию про-
цессов развития, заключающуюся в резкой задержке роста на 2—
4-й дни инкубации и в замедлении темпов деления клеток, вслед-
ствие чего к 8-му дню инкубации эмбрион содержит почти
в 2.5 раза меньше азота, чем в нехранившихся яйцах. Автор от-
мечает, что хранение не вызывает необратимых изменений фер-
ментных систем эмбрионов, так как способность к росту у них
82
сохраняется, однако обусловливает повышение смертности эм-
брионов и снижение выводимости утят.
Отметим, что яйца, в которых не началось развитие эмбриона
после закладки их в инкубатор, не всегда являются неоплодот-
воренными. Часть эмбрионов пргибает еще в яйцеводе несушки,
а некоторые до закладки в инкубатор, во время перерыва в раз-
витии, вследствие неподходящих условий или чрезмерной дли-
тельности хранения яиц. Мунро и Козин (Munro a. Kosin, 1945)
гистологически исследовали только что снесенные яйца и пока-
зали, что до 75% яиц, считающихся неоплодотворенными, поги-
бают в первые 25 час. развития, следовательно, еще в яйцеводе
имеется период повышенной смертности эмбрионов.2 Вскрывая
яйца, которые были определены при овоскопировании как не-
оплодотворенные, Орлов (1948) наблюдал в них развитие эм-
бриона, соответствующее моменту снесения яйца курицей.
Из 45 «неоплодотворенных» яиц, хранившихся 15 дней до инку-
бации, 36 оказались оплодотворенными, а из 90, признанных
неоплодотворенными, хранившихся 25 дней, 70 оказались опло-
дотворенными. Даже в яйцах, хранившихся только 1 день, из 39
«неоплодотворенных» яиц оплодотворенными оказались 5. По дан-
ным Пеционжкевича (19356), при увеличении срока хранения
увеличивался процент «неоплодотворенных» утиных яиц: за 5 дней
хранения — 14%, за 10 — 20%, за 15 — 26.5%, за 20 — 42.8%
и за 25 дней — 94%.
Перевозка яиц часто бывает необходима, а между тем она
значительно снижает выводимость, ослабляя халазы, поддер-
живающие желток в положении, необходимом для нормального
развития. Перевозка яиц после длительного их хранения или
хранения при высокой температуре особенно вредна.
Опытами Стайлса и Уатерсона (Stiles a. Watterson, < 1937)
показано губительное влияние систематического встряхивания
яиц во время первых часов инкубации: из 155 яиц вывелся только
1 цыпленок. В наших опытах по инкубации яиц гаги (Rolnik,
1943) мы также наблюдали губительное влияние перевозки яиц
на развитие эмбриона и, чтобы свести к минимуму потери, скон-
струировали специальный ящик, предохраняющий яйца от встря-
хивания и охлаждения.3
Как показали Нокс и Олсен (Knox а. Olsen, 1936), перевозку
яиц следует производить тупым концом вверх, в твердой таре,
что предохраняет от образования в них подвижной воздушной
камеры, наличие которой сильно снижает выводоспособность.
Перевозка яиц самолетом включает и влияние пониженного
атмосферного давления. Как сообщает Функ (Funk, 1949), при
перевозке самолетом на небольшие расстояния, т. е. в течение
2 См. главу 17.
3 Подробнее см. главу 21.
6*
83
короткого времени (несколько часов), пониженное атмосферное
давление не снижает выводимости.4
Существующее у некоторых производственников мнение о том,
что после перевозки необходимо дать яйцам отдых, мы считаем
неправильным, так как это только затягивает срок хранения.
Б. Отбор яиц для инкубации
Вопрос об отборе яиц для инкубации имеет две
стороны: 1) определение качества партии инкубационных яиц,
т. е. определение, насколько яйца данного маточного стада соот-
ветствуют основным требованиям, предъявляемым к инкубацион-
ным яйцам; 2) собственно браковка яиц перед инкубацией.
Яйца, предназначенные для инкубации, должны подбираться
только от здоровых птиц. Для инкубации совершенно непри-
годны яйца от птиц, больных белым бациллярным поносом, ко-
торый передается через яйцо от несушек к цыплятам и вызывает
их массовую гибель.
Большую роль в повышении выводимости играют содержа-
щиеся в инкубационном яйце витамины. Маслиева (1950) устано-
вила, что между содержанием витамина А в желтке и выводимо-
стью имеется прямая зависимость и, таким образом, содержание
витамина А в желтке может быть использовано как один из инди-
каторов при определении полноценности инкубационного яйца
(табл. 30).
. Табл и ц а 30
Выводимость цыплят в зависимости от содержания витамина А
в желтке
(По Мас лиевой, 1950)
Показатели Группы
I II III (молодки) IV V VI
Количество витамина А (в мкг) 32.3 26.0 25.6 18.3 14.8 13.3
Процент вывода от опло- дотворенных яиц .... 93.8 87.6 83.7 77.9 74.5 69.6
В связи с ярко-желтой окраской провитамина А — каротина —
по интенсивности окраски желтка можно судить о наличии в нем
этого провитамина. Одни исследователи (Кирсанов, 1935а; Баран-
чеев, 1939; Еремеев, 1956; Kalmutchi et Bosoanca, 1957; Боголюб-
4 Подробнее см. в главе 20.
84
ский, 1966, и др.) наблюдали значительно большую выводимость
цыплят из яиц с более интенсивной окраской желтка. Другие
(Прицкер и Третьяков, 1937; Третьяков и Кустова, 1950) отме-
чают, что окраска желтка при овоскопировании яиц не всегда
является правильной оценкой содержания каротиноидов, так как
в точности определения ее играет роль толщина и прозрачность
скорлупы. Еремеев (1956) наблюдал положительную корреляцию
между содержанием каротиноидных пигментов в желтке яйца и
толщиной скорлупы, что уменьшало усушку при хранении яиц
и способствовало высокой выводимости цыплят.
Из других методов предварительного определения качества
партии Инкубационных яиц упомянем предложенную Третьяко-
вой (1938) физико-химическую оценку яиц. Яйца с большим про-
центом плотного белка имеют более высокую выводоспособность.
Процент плотного белка можно определить довольно простым спо-
собом: при выливании яйца из скорлупы на тарелку оно сохраняет
свою форму и мало расплющивается. Этот прием может быть при-
менен лицами, имеющими некоторый навык.
Желудков (1951) сообщает, что плотность белка является по-
казателем содержания витаминов А и D в желтке. В яйцах с жид-
ким белком на 1 г желтка приходится 2.2 витамина А и 0.3 ИЕ
витамина D, а в яйцах с плотным белком — 13.6 и 3.7 соответ-
ственно. Яйца первой группы дали 68.6% цыплят, а второй -=•
82.2%; при очень разжиженном белке выводимость снижалась
до 30%. Кроме того, яйца хорошего инкубационного качества
обладают высокой электропроводностью белка.5 Однако этот метод
в настоящее время нельзя рекомендовать для производственных
целей, так как он довольно сложен.
Перед закладкой яиц в инкубатор очень важно установить,
оплодотворены ли они. Это можно сделать по методу Орлова
(1954). Автор рекомендует заменить отбор яиц для инкубации по
косвенным признакам (внешние признаки яиц) отбором по вели-
чине зародышевого диска. На большом материале автор показал,
что после 12-часового прогревания яиц при температуре 37.5—
40.5° отбор яиц с хорошо развитым зародышевым диском повысил
выводимость цыплят на 11.3% (из них за счет прогревания и по-
следующего охлаждения — на 3.4% и за счет исключения неопло-
дотворенных яиц и яиц со слабыми зародышами — на 7.9%).
Данные литературы и мнения практиков инкубации о связи
тех или иных признаков яиц (внешних или при просмотре на
овоскоп) с их выводоспособностыо, а также указания на необхо-
димость более или менее строгой браковки чрезвычайно разно-
речивы. Яйца ненормально мелкие или с очень тонкой скорлупой,
с наружной или внутренней трещиной скорлупы, с чрезмерно
увеличенной воздушной камерой или с неправильным ее положе-
6 См. данные Третьяковой (1938) н главу 3 настоящей книги.
86
йиём, с известковыми отложениями на скорлупе или со взболтан-
ным содержимым, а тем более имеющие плохой запах безусловно
необходимо выбраковывать. Но связь других признаков яиц с
их низкой выводоспособностью недостоверна. С прогрессом птице-
водства качество яиц становится все более высоким и браковка яиц
перед инкубацией — все менее необходимой. Так, Фомин (1950)
на большом материале показал, что в промышленной инкубации
при сравнительно хорошем состоянии маточного стада и его кор-
мления нет смысла проводить браковку яиц перед инкубацией,
так как она дает повышение выводимости менее чем на 1 %. При
ведении индивидуальных записей яйценоскости каждой несушки
выясняется, какие куры несут яйца с низкой выводоспособностью
(до 60%), и отбраковка их приводит к повышению выводимости
на 10%. В уже упомянутой нами (стр. 56—57) работе Шпиц и ее
соавторов (1965) рекомендуется периодически определять удельный
вес яиц у каждой несушки и отбраковывать несушек, которые
несут яйца 1-й и 2-й группы, с частыми нарушениями скорлупы
и внутренней структуры яйца, и потому имеющие низкую выводо-
способность цыплят.
Таким образом, в настоящее время в налаженных птицевод-
ческих хозяйствах наиболее рационально изредка проверять ин-
кубационные качества яиц каждой несушки, заменив этим утоми-
тельную браковку всех яиц перед инкубацией.
Раздел II. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Для возобновления развития эмбриона после перерыва
во время хранения яиц необходима более высокая температура
(инкубатор, гнездо птицы). В этом разделе, каки в других,
в основном дано описание развития куриного эмбриона. В первой
главе этого раздела мы излагаем материалы по ранним стадиям
развития эмбриона, а в дальнейших — по морфологии и физио-
логии отдельных систем органов начиная с момента их закладки.
При этом морфологическим данным уделяется меньше внимания.
С этими исследованиями, как и с историей вопроса, читатель может
познакомиться по ценным сводкам Паттена (Patten, 1951), Лилли
(Lillie, 1952) и особенно Романова (Romanoff, 1960) и Рагозиной
(1961). Преобразование функций одной системы органов, проис-
ходящее довольно часто в течение эмбриогенеза, отражается на
функционировании других систем эмбриона. Мы будем пытаться,
поскольку это возможно при настоящем объеме научных знаний,
описывать это взаимное влияние в целом организме.
Г Л А В А 6
РАННЕЕ РАЗВИТИЕ ЭМБРИОНА
К моменту помещения яиц в инкубатор в степени раз-
вития эмбриона имеются большие индивидуальные вариации,
которые сказываются в дальнейшем на сроках образования тех
или иных систем органов.
В первые часы инкубации завершается стадия гаструляции,
т. е. происходит дополнительное образование энтодермальных
клеток и окончательная дифференцировка эктодермального заро-
дышевого слоя. Однако мы считаем целесообразным привести
здесь описание эмбрионального развития с начала гаструляции,
т. е. с начала образования энтодермы. Этот процесс Беллер (Bel-
lairs, 1960) считает одним из самых странных процессов в развитии
живой материи. По вопросу о способе образования энтодермы
у эмбрионов птиц имеется значительное количество часто противо-
87
речивых наблюдений и суждений. В основном пользуются призна-
нием две теории образования энтодермы у птиц — образование
деламинацией (отщеплением) и инвагинацией. Кроме того, у дру-
гих животных гаструляция совершается: а) иммиграцией клеток
бластодермы в бластоцель и образованием там энтодермы, б) эпи-
бол ией — нарастанием анимальной чапти на вегетативную, пре-
вращающуюся в энтодерму.
Согласно теории инвагинации, энтодерма образуется путем
подворачивания сильно утончившегося заднего края бластодермы
в пространство между нею и желтком, получающееся в результате
дегенерации желточного синцития. Затем инвагинировавший
край бластодермы растет языкообразно вперед (подробное описа-
ние см.: Patterson, 1910). Применив метод маркировки тканей
углем, Лютц (Lutz, 1955) нашел подтверждение теории инвагина-
ции. По данным автора, энтодерма образуется за счет подгибания
в основном заднего края бластодермы, но и боковые и передний
края ее участвуют в этом процессе. Однако эта картина образо-
вания .энтодермы, по мнению ряда исследователей (Peter, 1938;
Кнорре, 1941), связана с артефактами, получающимися при
фиксации материала.
По теории деламинации (Peter, 1938, и др.), энтодерма обра-
зуется отщеплением нижнего слоя от верхнего, когда в бласто-
дерме появляются щели между верхним слоем клеток и более
крупными клетками, лежащими ниже. Примерно на такой стадии
находятся эмбрионы в только что снесейных утиных, индюшиных
и голубиных яйцах, а для куриных яиц характерна большая
вариация стадий развития. При дальнейшем развитии горизон-
тальные щели между двумя слоями соединяются и происходит
окончательное образование энтодермы. Несколько развивая тео-
рию деламинации, Пастиле (Pasteels, 1945) считает энтодерму
результатом диффузной иммиграции клеток энтодермы.
Наиболее полная картина образования энтодермы у птиц дана
Кнорре (1941, 1949а). Считая основным способом образования
энтодермы деламинацию, автор показывает, что к ней добавляется
слабо выраженный процесс иммиграции (отделение некоторых
клеток верхнего слоя и присоединение их к нижнему). В добавле-
ние к этому, уже после образования энтодермы, происходит обо-
собление новых клеток от содержащего свободные ядра желточ-
ного синцития со дна подзародышевой полости. Автор показывает
также эволюцию способа образования энтодермы в ряду позво-
ночных животных. Инвагинационный способ образования энто-
дермы заменился деламинацией в связи с переходом от полного
дробления яиц (у бедного желтком яйца ланцетника) к частичному
дискоидальному (в богатых желтком яйцах рептилий и птиц).
Автор считает, что появление этого способа вызвано необходи-
мостью ранней дифференцировки значительной части энтодермы
для резорбции свободного внеклеточного желтка. А инвагинация
88
у птиц происходит после образования энтодермы, выражаясь
в движении клеток первичной полоски (о чем будет речь несколько
позже). В процессе эволюции гаструляция усложнилась, и у хор-
довых к концу ее образуются не только два зародышевых листка
(эктодерма и энтодерма) но и третий, лежащий между ними, —
мезодерма.
Из каждого зародышевого листка возникают определенные
структуры, из эктодермы — нервная система и эпидермальная
часть кожи с перьями. Следует отметить, что у низших животных,
не имеющих нервной системы, именно эктодермальные клетки
воспринимают раздражения из внешней среды. Из энтодермы
образуется эпителий кишечника и его производные: печень, желч-
ный пузырь и выстилка дыхательного тракта. Из мезодермы обра-
зуется наибольшее количество производных: мускулатура, эпи-
телий брюшной и плевральной полостей, эпителий почек и т. п.
Часть мезодермы разрыхляется и превращается в мезенхиму,
состоящую из синцитиально соединяющихся между собою клеток.
Часть клеток мезенхимы происходит из эктодермы. Из мезенхимы
образуется соединительная ткань, кровь, скелетные ткани (хря-
щевая и костная).
Много споров вызывает вопрос о том, является ли вообще обо-
снованной теория зародышевых листков, согласно которой ана-
логичные морфологические структуры возникают у разных типов
животных из одного и того же зародышевого листка. Однако Свет-
лов (1963) доказывает, что эта теория является наиболее общим
законом развития многоклеточных животных и что она по праву
должна быть названа теорией А. О. Ковалевского. Беллер (Bel-
lairs, 1960) также считает, что исключения, наблюдающиеся в ряде
случаев, а также экспериментальные изменения судьбы того или
другого листка не опровергают общего закона.
Что же регулирует развитие индивидуальных клеток в необ-
ходимом направлении и в нужное время? Беллер (Bellairs, 1960)
считает, что одна группа клеток влияет на развитие другой и при
нормальном развитии целого организма (части организма или
группы клеток при обособлении могут вести себя иначе) инди-
видуальные клетки не развиваются в направлении, вредном для
эмбриона как целого. Однако о причинах и механизме такой регу-
ляции внутри организма почти ничего неизвестно.
Основным типом взаимодействия между эмбриональными
клетками является так называемая индукция, т. е. воздействие
одной ткани («организатора») на дифференцировку другой. Один
из создателей теории индукции, Шпеман (Spemann, 1918), пред-
ставляет себе раннее эмбриональное развитие как серию следую-
щих друг за другом процессов индукции. Зачаток, на дифферен-
циацию которого воздействовал другой зачаток, индуцирует позже
возникающие зачатки. Советские эмбриологи Филатов (1943) и
Иванов (1945) отметили, что в действительности имеет место не
89
одностороннее воздействие одних индуцирующих зачатков на дру-
гие, пассивные, а взаимодействие между ними, причем один из
них может оказывать большее влияние, чем другой. Системы
взаимодействующих зачатков Филатов представлял себе как
«формообразовательные аппараты», усовершенствующиеся в фило-
генезе животных. Против термина «организатор», как антропо-
морфного понятия, возражает Токин (1943) и подчеркивает не-
разрывную взаимосвязь между частями целого организма, раз-
вивающегося в тесной связи со средой. Значительное количество
экспериментальных и теоретических работ по индукции провел
Лопашов с сотрудниками (Лопашов, 1961; Лопашов и Строева,
1963, и др.). Автор указывает, что осуществление индукции того
или иного органа зависит от своевременного пространственного
и временного совмещения ткани индуктора и реагирующей ткани,
когда они обе обладают максимумом возможностей к осуществле-
нию данной индукции. Автору удалось показать, что первичное
индукционное воздействие обусловливает только первый этап
формирования зачатка органа, а его последующее расчленение
на части, дифференцировка этих частей и создание формы связаны
как с новыми влияниями частей, так и с пространственным распо-
ложением клеток внутри зачатка, а также с механическими усло-
виями развития зачатка в общей системе организма. Не имея воз-
можности подробно рассмотреть этот обширно исследованный,
но не решенный еще вопрос (тем более, что он стоит несколько
в стороне от основных задач нашей работы), приведем данные
одного из последних исследований об индуктивной способности
различных частей бластодермы куриного эмбриона, проведенных
Мулхеркером (Mulherkar, 1958). Наибольшей способностью к ин-
дукции обладает гензеновский узелок (стр. 91), и эта способность
снижается не только вдоль бластодермы, но и от середины ее
в стороны. Автор обнаружил корреляцию между индукционной
способностью и самодифференциацией участка бластодермы в нерв-
ную ткань. В большой сводке данных о первичной эмбриональной
индукции Саксен и Тойвонен (1963) пишут, что, несмотря на зна-
чительные противоречия, большинство эмбриологов считает,
что индукция вызывается по крайней мере двумя активными аген-
тами (или двумя типами реакций): нейрогенными (нервная бо-
роздка) и мезодермными. На более поздних стадиях как процессы
дифференцировки, так и вообще нормальная корреляция разви-
тия регулируются не индукцией, а эндокринными железами и
нервной системой.
К моменту снесения яйца бластодерма состоит из светлого и
темного поля (см. главу 2 и рис. 5). Ббльшая часть светлого поля
дает, начало эмбриону, а периферический его участок вместе с тем-
ным полем образует внезародышевую область, из которой форми-
руются временные эмбриональные органы. Под бластодермой
находится подзародышевая полость, наполненная сильно разжи-
90
женным желтком — подзародышевой жидкостью. По-видимому,
она образуется при просачивании влаги сквозь бластодерму из
находящегося над нею белка. Это экспериментально установил
Нью (New, 1956), выращивая in vitro бластодерму под слоем
белка. В этой подзародышевой жидкости лучше, чем в желтке,
происходят энзиматические процессы, необходимые для питания
эмбриона на столь ранней стадии развития, и лучше растворяются
выделяемые им продукты распада. Ван Дет (Deth van, 1962)
также приводит доказательства, что подзародышевая жидкость
образуется активной резорбцией воды и раствора питательных
веществ из белка живой бластодермой, а не при переваривании
желтка. Иного мнения придерживается Шейнис (1964), считаю-
щая, что латебра желтка (рис. 10) представляет собой полость,
в которой содержится жидкость, поднимающаяся из желтка
в подзародышевую полость.
Та фаза гаструляции, которая протекает в первые часы инкуба-
ции, начинается с того, что происходит активное перемещение
клеточного материала в участке светлого поля: сначала по краям
его спереди назад, а затем по центру вперед, навстречу более
медленно двигающимся в центре клеткам, спереди назад. Это
движение клеток сравнивается эмбриологами с фигурой танца
полонеза (описание по Кнорре, 1959). Встретившиеся справа и
слева потоки клеток образуют в центре бластодермы утолщение,
так называемую первичную полоску с бороздкой посредине, а на
переднем крае еще более плотное скопление клеток, в виде бу-
горка. Это первичный, или гензеновский, узелок. Впереди от него
лежит материал, из которого в дальнейшем (выяснено методом
маркировки углем) образуется хорда, а еще дальше вперед лежит
зачаток нервной пластинки. Материал же первичной полоски
образует мезодерму, погружаясь внутрь через края бороздки,
которая аналогична бластопору амфибий [это и есть инвагинация,
по Кнорре (1941)].
Первичная полоска становится впервые заметной в курином
яйце на 8—9-й час инкубации, когда диаметр бластодермы равен
4.5—6.0 мм; в утином яйце — 3.4 мм; в яйце морской ласточки —
4.0 мм и грача — 2.0—2.5 мм (Митрофанов, 1898, 1900).
Романов (Romanoff, 1960) делит стадию первичной полоски
на 4 периода: короткой, средней, длинной и дефинитивной первич-
ной полоски. Дефинитивная полоска у куриного эмбриона дости-
гает длины 1.5—2.5 мм и образуется к 16—20 час. инкубации.
Потом очертания первичной полоски постепенно становятся неяс-
ными, и у 33^-часового эмбриона она исчезает совсем.
Вдоль первичной полоски проходит ось симметрии — эмбрион
становится билатерально симметричным. Еще Бэр (1828) устано-
вил, что обычно птичий эмбрион лежит в яйце головой влево,
если держать яйцо повернутым тупым концом к исследователю.
Это так называемое правило Бэра. Однако из этого правила
91
имеются исключения, и иногда даже очень значительные. Разные
исследователи находили различный процент нормально ориенти-
рованных эмбрионов в куриных яйцах — 76, 50, 32, 9% (по сводке:
Romanoff, 1960). Интересно, что процент нормально ориентиро-
ванных эмбрионов характерен для каждого вида птиц. Наиболее
Рис. 13. Куриный эмбрион 24 час. инкубации.
А — вид сверху (по Patten, 1951); Б — поперечный разрез (по Huettner, 1947).
1 — край мезодермы; 2 — сегмент; з — хорда; 4 — нервная бороздка;
5 — нервная складка; 6 — мезенхима; 7 — темное поле; 8 — эктодерма го-
ловы; 9 — передняя кишка (9а — край передней кишки); ю — светлое поле;
11 — мезодерма (11а — несегментированная, 116 — соматическая, lie — вну-
тренностная, 11г — сегментированная, Ид — промежуточная, 11с — боковая);
12 — первичная полоска; 13— кровяной островок; 14—сосудистое поле;
15 — энтодерма; 16 — будущая перикардиальная полость.
редкие исключения из правила Бэра встречаются в голубиных
яйцах, чаще — в куриных и еще чаще — в утиных. Вентамберже
и Клавар (Vintemberger et Clavert, 1957) обнаружили, что повыше-
ние степени неустойчивости билатеральной симметрии коррели-
руется с появлением уродств (двойни). Не останавливаясь на
этом вопросе подробно, скажем, что место детерминизма билате-
92
Рис. 13 (продолжение).
ральной симметрии у птиц и уста-
новления ориентации эмбриона в
яйце одни исследователи (Ancel,
1955а, 1956b) относят к начальной
части яйцевода, до деления яйце-
клетки на 2 бластомера, а дру-
гие (Vintemberger et Clavert, 1955,
1956) утверждают и эксперимен-
тально доказывают, что это про-
исходит в матке и связано с по-
ложением яйца в ней — тупым
или острым концом вниз.
Передняя часть эмбриональной
зоны светлого поля утолщается и
несколько выпячивается вверх,
образуя полукруглую головную
складку. Здесь будет образовы-
ваться голова эмбриона. В течение
всего развития дифференциация
головного конца эмбриона идет
быстрее, чем остальных его ча-
стей. Окончательно дифференци-
руется головная складка у 23—
25-часового эмбриона (Hambur-
ger a. Hamilton, 1951) (рис. 13).
Затем, в течение 2-го и 3-го дня
инкубации, образуются боковые
и хвостовая подстилающие склад-
ки тела, которые углубляются и
сближаются между собой и с го-
ловной складкой, отделяя соб-
ственно эмбрион от желточного
мешка.
Нервная пластинка появляется
у куриного эмбриона на 18-м
часу инкубации, а у 21—22-ча-
сового куриного эмбриона на ней
образуются два утолщения с же-
лобком посредине (рис. 14). Затем
нервная пластинка свертывается
в трубку и погружается под. кож-
ную эктодерму.. Процесс замыка-
ния нервной трубки идет спереди
назад и заканчивается к 50—55
час. инкубации.
Мезодерма у куриного эмбриона дифференцируется примерно
в суточном возрасте на сомиты, нефротомы и лежащие еще более
93
латерально спланхнотомы, или боковые пластинки. Сомиты и
нефротомы сегментированы. Их дифференциация идет спереди
назад, причем первая пара сомитов образуется у 23—26-часового
куриного эмбриона (Hamburger a. Hamilton, 1951). В связи с по-
стоянством времени образования сомитов их количество является
наиболее верным критерием развития эмбриона до 50—53 час.
инкубации. В яйцах, одновременно заложенных в инкубатор,
эмбрионы могут варьировать по степени развития, но эмбрионы
Рис. 14. Куриный эмбрион 22 час. инкубации (вид сверху).
(По Huettner, 1947).
1 — темное поле; 2 — светлое поле; 3 — первая межсегментная бороздка;
4 — нервная бороздка; 5 — головная складка; 6 — нервная складка; 7 — пер-
вичная полоска.
с определенным количеством сомитов очень мало отличаются друг
от друга. На стадии 50—55 час. начинается дифференциация перво-
начально однородных сомитов на а) наружный слой — дерматом,
б) средний слой — миотом и в) внутренний — склеротом. Из
дерматомов образуется внутренняя часть покровов тела, в то
время как из эктодермы формируется только эпителий. Миотомы,
проходящие в дальнейшем самый бурный рост из всех частей
сомитов, дают всю мускулатуру тела (кроме головной и бронхи-
альной, которые происходят из головной мезенхимы). Клетки
склеротомов начинают окружать хорду и нервную трубку, обра-
зуя в дальнейшем позвоночный столб.
94
в
Рис. 15. Дифференцировка мезодермы куриного эмбриона.
(По Кнорре, 1959).
А — сомит 1.5-дневного; Б — сомит 54-часового; В — спланхнотом 1.5-днев-
ного эмбриона. 1 — недифференцированный сомит; 2 — дерматом; 3 — мио-
том; / —- склеротом; 5 — кожная эктодерма; 6 — париетальный листок сплан-
хнотома; 7 — висцеральный листок спланхнотома; 8 — кишечная энтодерма;
9 — нервная трубка; ю — целбм.
Спланхнотомы не сегментируются и продолжаются во внеза-
родышевую мезодерму, разделяясь на 2 листка: висцеральный
и париетальный, между которыми лежит полость тела — целбм
(рис. 15). Сено (Seno, 1961) экспериментально показал, что сомиты
образуют осевой скелет и мускулатуру стенки тела (межреберные
Рис. 15 (продолжение).
и брюшные мышцы), а спланхнотомы дают начало дополнительным
частям скелета и мускулатуры, включая грудину и большую груд-
ную мышцу.
Энтодерма сначала спереди, а потом на заднем конце эмбриона
образует желобок, а затем кишечную трубку. На 22-м часу инку-
бации дифференцируется передняя часть передней кишки, впер-
вые получая подстилку из клеток, а не из желтка. В последующие
дни образование кишки идет постепенно с обоих концов к середине,
пока, наконец, сообщение ее с желтком не становится совсем узким
и не превращается в пупочный канатик.
ГЛАВА 7
ВРЕМЕННЫЕ ЭМБРИОНАЛЬНЫЕ ОРГАНЫ
Органы, которые у взрослой птицы несут функции
пищеварения, дыхания и выделения, медленно достигают своего
окончательного состояния и бывают способны к функционирова-
нию в основном только в конце эмбрионального развития. В те-
96
чение эмбрионального периода эти функции выполняются специ-
фическими временными органами, которые после вылупления
атрофируются. Строение этих эмбриональных органов примитивно
по сравнению с органами, несущими соответствующие функции
у взрослых птиц. Из целой группы названий (эмбриональные
оболочки, внеэмбриональные оболочки, провизорные органы,
временные эмбриональные органы) мы считаем наиболее адекват-
ным последнее. Такими органами являются желточный мешок,
амнион, сероза и аллантоис.
А. Желточный мешок
Основная функция желточного мешка — абсорбция
питательных веществ из желтка и перенос их к эмбриону.
1. Структура
Желточный мешок возникает раньше других временных эмбрио-
нальных органов из спланхноплевры (образующейся из мезодермы
и энтодермы), вырастающей вокруг желтка. Оболочка желтка на
ранней стадии развития разрывается над эмбрионом и постепенно
соскальзывает вниз (подробнее см. в разделе об амнионе). Отгра-
ничение желточного мешка от эмбриона происходит при образо-
вании головной, хвостовой и боковых подстилающих складок тела
в течение 2-го и 3-го дней инкубации. Складки эти, сбли-
жаясь, образуют желточный стебелек, соединяющий желточный
мешок с эмбрионом. В этом стебельке проходят желточные артерии
и вены, соединяющие эмбриональный круг кровообращения
с внеэмбриональным. Желточный стебелек образуется примерно
на 4-й день развития куриного эмбриона — ко времени замыкания
кишечной трубки (Рагозина, 1961).
В течение 2—4-го дней рост ткани желточного мешка протекает
очень быстро, и к концу 4-го дня лишь небольшая часть желтка
остается не окруженной ею. Этот быстрый рост желточного мешка
продолжается до 6-го дня, затем замедляется, а после 11-го почти
приостанавливается. Кровеносные сосуды на 6-й день покрывают
половину, а на 14-й — весь желточный мешок (Romanoff, 1952).
Полное замыкание желточного мешка происходит к 11-му дню
инкубации, причем сильно увеличивается его вес (в полтора
раза больше веса желтка свежеснесенного яйца), так как, кроме
желтка, желточный мешок окружает и часть белка (Шейнис,
1954).
Нидхем (Needham, 1942) наблюдал максимум веса желточного
мешка на 15-й день — 3.5 г (без веса желтка), а ко времени вы-
7 В. В. Рольник 97
Рис. 16. Фотография внутренней поверхности желточного мешка
куриных эмбрионов. (По Рагозиной, 1961).
А — в возрасте 10; Б — в возрасте 12; В -- в возрасте 14 суток инкубации.
лупления его вес снижался до 2.5 г. По данным Отрыганьева
(1949), увеличение веса желточного мешка происходит до 18-го дня,
после чего он быстро убывает в весе.
Внутренняя поверхность желточного мешка образует складкв
(рис. 16), которые увеличивают возможность всасывания желтка.
По данным Рагозиной (1961), эти складки наиболее интенсивно
образуются в том месте желточного мешка, где его энтодерма
соприкасается с жидким желтком, готовым к усвоению. Внутрен-
няя поверхность желточного мешка выстлана железистым эпите-
лием, адсорбирующим желток.
Развитие кровообращения желточного мешка изучено методом
инъекции еще в 1894 г. Поповым (Popoff), который различает
здесь шесть стадий. Для 1-й стадии — до 45 час. инкубации
(рис. 17,Л) — характерна недифференцированная артериальная
сеть, заканчивающаяся на периферии концевой веной, соединяю-
щейся двумя передними желточными венами с сердцем; дифферен-
цированных артериальных сосудов нет. На 2-й стадии — 48—
61 час инкубации (рис. 17,Б) — происходит образование арте-
риального пути среди сети артериальных и венозных сосудов.
3-я стадия — 61—74 часа инкубации (рис. 17,В) — характери-
зуется появлением первых следов поверхностной венозной капил-
лярной сети и образованием промежуточных вен. 4-я стадия,
начинающаяся на 4—5-й день инкубации, отличается тем, что
венозная сеть состоит: а) из .промежуточных вен, возникающих
в различных местах из концевой вены и постепенно теряющихся
в сети кровеносных сосудов, б) из некоторых артериальных со-
судов, которые приобрели венозный характер и в) из боковых
желточных вен, развивающихся от переднего входа в кишку
кзади. На 5-й стадии — с 10-го дня инкубации — желточные
артерии богато ветвятся и кончаются сетью кровеносных сосудов;
немногие, наиболее тонкие ветви артерий достигают концевой
вены, которая к 10-му дню начинает редуцироваться, а затем исче-
зает совсем, превращаясь в конечные капилляры; значение перед-
ней и задней желточных вен уменьшается, и они делаются почти
незаметными; наоборот, боковые желточные вены становятся почти
единственными сосудами, возвращающими кровь эмбриону; про-
межуточные вены теперь являются продолжением боковых. На
6-й стадии — 19—20-й дни инкубации — кровеносная система
желточного мешка достигла полного развития; развита богатая
сеть венозных сосудов в складках и образовались анастамозы
между сосудами желточного мешка и аллантоиса в месте их слия-
ния.
Ток крови в сосудистой зоне желточного мешка начинается
на стадии 45—49 час. инкубации (Romanoff, 1952). Самые боль-
шие артерии образуются в том месте сосудистой сети, где ток
крови наиболее быстрый.
7*
99
Рис. 17. Кровеносные сосуды желточного мешка кури-
ного эмбриона (вид изнутри, снизу). (По Popoff, 1894).
А — 45-часового; Б — 53-часового; В — 74-часового эмбриона.
1 — правая аорта (1а — конец ее); 2 — зона возникновения арте-
рий (а.) (2а — правая желточная а.); 3 — сердечная трубка
(За — сердце); 4 — зона капиллярной сети (4а — венозная капил-
лярная сеть); 5 — промежуточные вены (в.), внутренними концами
входящие в венозную капиллярную сеть; 6 — дифференцирую-
щаяся задняя в.; 7 — передний изгиб аорты; 8 — передние жел-
точные в. (8а — атрофирующаяся правая передняя желточная в.);
9 — концевая в.; 10 — сегменты; Па, 116 и lie — внутренняя, сре-
дняя и наружная зоны сети капилляров; 12 — кровяной островок
(12а — еще не связанный с кровеносными сосудами); 13 — зад-
ний мозг; 14 — слуховая ямка; 15 — промежуточный мозг;
16 — глазной пузырек; 17 —передний мозг (17а — вторичный пе-
редний мозг); 18 — средний мрзг; 19 — вход в переднюю кишку.
2. Функция
В начале развития желточный мешок за сутки адсорбирует
питательных веществ из желтка в 3 раза больше его собственного
веса, а с 9-го дня инкубации вес поглощенного в сутки желтка
равен его весу. До 9-го дня поглощенный желток расходуется
в основном на рост тканей желточного мешка, а после этого
почти все адсорбируемые вещества идут на построение тканей
эмбриона. На 3—4-й день инкубации клетки железистого эпителия
желточного мешка наполнены захваченными из желтка желтыми
жировыми капельками, являющимися частицами желточных
зерен. Но к концу 1-й недели инкубации наряду с этим фагоци-
тарным способом усвоения желтка энтодермой в краевой области
желточного мешка возникает новый способ усвоения питательных
веществ путем расщепления желточных шаров протеолитиче-
скими и липолитическими энзимами, которые выделяются тканью
желточного мешка.
Обобщая данные многих исследователей, Нидхем (Needham,
1942) приходит к выводу, что переваривание питательных ве-
ществ в желточном мешке хорошо обеспечено энзиматическими
системами. Протеазы желтка имеют максимальную активность
на 10-й день, во время максимума протеинового метаболизма,
липазы — около 16-го дня, во время максимальной абсорбции
жира. По данным Гродзинского (Grodzinski, 1946, цит. по: Ro-
manoff, 1952), липазы растворяют липидную оболочку желточных
шаров, освобождают жировые капельки, содержащиеся в них,
и, по-видимому, превращают жиры из глицеридов в фосфатиды.
После наступления максимальной активности липаз, на 14—
15-й день, содержание жира в желтке падает с 6 до 2 г. Протеазы
содействуют распаду коллоидальной протеиновой основы желточ-
ных шаров; их активность вначале низка, увеличивается с 3-го
до 9-го дня, а потом остается на одном уровне. Бычковска-Смык
(Byczkowska-Smyk, 1956) исследовала в тканевой культуре про-
дукцию пищеварительных энзимов клетками желточного мешка.
Эти клетки вырабатывают липазы и протеолитические энзимы,
которые через 6—20 дней (в культуре тканей) полностью перева-
ривают протоплазму клеток. Под действием липаз жир желтка
делается более жидким. До 8-го дня инкубации разжижает жел-
ток и вода, переходящая сюда из белка. Но затем, как предпо-
лагает Рагозина (1961), для предварительной обработки загустев-
шего к этому времени желтка в полость желточного мешка могут
проходить пищеварительные энзимы и желчь из кишечника эм-
бриона.
Из клеток эпителия желточного мешка абсорбированные пи-
тательные вещества переходят в его многочисленные кровеносные
сосуды. Проходя затем во внутриэмбриональном круге кровооб-
ращения по печеночной воротной системе, эти питательные ве-
102
щества обрабатываются пищеварительными соками, выделяемыми
клетками печени (см. главу 11). Таким образом, желточный ме-
шок — это в основном первичный орган пищеварения эмбриона.
Кроме того, желточный мешок с его сосудами выполняет дыха-
тельную функцию. Аллантоис как орган дыхания начинает функ-
ционировать у куриного эмбриона только с 5-го дня, при этом
вначале он не может полностью обеспечить необходимый газо-
обмен эмбриона. Таким образом, желточный мешок несет функцию
дыхания до 5-го дня полностью, а с 5-го до 10-го — частично.
Это происходит так: желток всплывает и, оттесняя густой бе-
лок, плотно примыкает к подскорлупной оболочке той своей по-
верхностью, где находится бластодиск, что значительно облегчает
диффузию газов через скорлупу. Уже к концу 2-го дня развития
куриного эмбриона, по словам Рагозиной (1961), желточный ме-
шок «наряду с другими функциями начинает специализироваться
как орган дыхания зародыша». Автор вносит при этом серьезные
исправления в схему Дюваля (Duval, 1889) о расположении вре-
менных эмбриональных органов внутри яйца и подчеркивает при-
способительное значение примыкания желточного мешка к скор-
лупе для газообмена эмбриона на ранних стадиях развития.
Помимо обменных функций, желточный мешок до 8-го—
11-го дня инкубации является местом запаса гликогена.
3. Втягивание желточного мешка и его значение
в постэмбриональном развитии
На 19-й день инкубации, когда желточный мешок втяги-
вается в полость тела, это еще довольно объемистый орган.
Вместе с остаточным желтком он составляет примерно 1/6 веса
эмбриона и около 1/3 первоначального веса желтка. Процесс втя-
гивания желточного мешка осуществляется, по мнению Лилли
(Lillie, 1952), благодаря сокращению внутренних листков аллан-
тоиса и амниона. Куо (Kuo, 1932b) же считает, что втягивание
желточного мешка производится движениями брюшных мышц,
причем движения лап помогают растяжению брюшной полости и,
следовательно, ускоряют втягивание желточного мешка. Этому же
содействуют и дыхательные движения. А аллантоис, по мнению
автора, не только не. помогает втягиванию желточного мешка,
но даже задерживает его. По данным Рагозиной (1961), к моменту
вылупления в желточном мешке имеются 2—3 лопасти, образую-
щиеся в связи с неравномерной скоростью роста ткани желточ-
ного мешка и его крупных кровеносных сосудов, которые растут
медленнее. Такое строение желточного мешка способствует втя-
гиванию его через относительно узкое пупочное отверстие. Втя-
гивание желточного мешка происходит, по наблюдениям автора,
вследствие сокращения окружающего желточный мешок мышеч-
103
ного слоя, который является непосредственным продолжением
вентральной части мышц брюшной стенки тела эмбриона.
После втягивания желточного мешка на месте, где он втянулся,
остается так называемая пуповина. Она состоит из прилежащей
к телу эмбриона части желточной ножки, остатков аллантоиса
и соединительнотканного кольца, которое вначале утолщается,
а потом суживается. Желточный стебелек после втягивания и
рассасывания желточного мешка остается в виде отростка две-
надцатиперстной кишки. Плохое втягивание желточного мешка
является неблагоприятным признаком при оценке качества цы-
пленка. Оно может быть вызвано либо недостаточным потребле-
нием желтка в течение инкубации (обычно вследствие повышен-
ной температуры или влажности инкубации), либо ослаблением
мышечных волокон, что указывает на общую слабость цыпленка
(см. главу 22).
Втянувшийся при вылуплении желточный мешок содержит
около половины сухих веществ желтка яйца до инкубации, но
концентрация в них протеинов больше, а жиров меньше, чем
в желтке. Минеральных солей из желтка потребляется за время
инкубации от 1/4 до 1/2, причем почти полностью потребляется
железо (для образования гемоглобина), до 3/4 фосфорных солей
(на образование мозга, печени, мышц), много расходуется серы
(на образование пуха, клюва, когтей) и около 2/3 кальция на по-
строение костей (Ильин, 1917).
Питательные вещества, заключенные во втянувшемся желточ-
ном мешке, используются цыпленком в первые дни постэмбрио-
нального развития, а энзимы, содержащиеся в них, участвуют
также в переваривании пищи, поедаемой цыпленком в это время.
Кроме того, желточный мешок в первые три недели постэмбрио-
нального развития является органом с апокриновой секрецией,
а затем превращается в лимфоидный орган с функцией лимфо-
поэза.
Б. Амнион и сероза
1. Структура
Амнион и сероза 1 образуйся из внеэмбриональной
соматоплевры примерно с 30-го часа развития куриного эмбриона.
В то время когда головная часть эмбриона слегка погружается
1 Серозу эмбриона птиц иногда называют хорионом.
Рис. 18. Поперечные разрезы куриных эмбрионов. (По Рагозиной, 1961).
А — 1-суточный; Б — 2-суточный; В — 3-суточный; Г — 4-суточный; Д — 5-суточный
эмбрион. 1 — первичная полоска; 2 — нервная трубка; з — первичная почка с заклад-
кой половой железы; 4 — желточная энтодерма; 5 — сосуды желточного мешка; б —
сероза; 7 — амнион; 8 — аллантоис; 9 — аллантоисная жидкость; 10 — место образо-
вания хорио-аллантоиса. “. .
104.
1
в желток, внеэмбриональная соматоплевра впереди нее образует
головную складку сероамниотического листка (рис. 18 и 19).
Потом появляются боковые сероамниотические складки (расту-
щие друг к другу) и хвостовая складка, растущая в сторону
головы эмбриона. В результате роста всех четырех складок они
встречаются и срастаются над эмбрионом, образуя рубец. На-
Рис. 19. Схема образования
амниотических складок (Z) и за-
мыкания кишечника (II). (По
Барту, 1951).
1 — эктодерма; 2 — мезодерма (2а —
соматическая, 26 — внутренностная);
з — складки амниона; 4 — кишка;
5 — амнион (5а — полость амниона);
6 — сероза; 7 — серозо-амниотическое
слияние; 8 — серозо-амниотическая
полость; 9 — полость тела* 10 — со-
матоплевра; 11 — желточный мешок
(11а — стебелек его); 12 — желток;
13 — энтодерма.
ружные стенки этих складок от-
ходят от внутренних и образуют
серозу, а внутренние амнион.
Сероза растет, образует складку
и снова срастается со стенкой
амниона, формируя серозо-амнио-
тическое сращение. Вследствие бо-
лее быстрого роста амниона в этот
период по сравнению с ростом
эмбриона между ними образуется
амниотическая полость, выстлан-
ная эктодермой. Полость между
амнионом и серозой — сероамнио-
тическая — полностью выстлана
мезодермой, а сероза снаружи
эктодермой. Чрезвычайно быстрый
рост серозы приводит к тому, что
она окружает не только эмбрион
и амнион, но и желток и белок;
аллантоис, когда он образуется,
также лежит внутри ее’ По дан-
ным Рагозиной (1961), у 3-суточ-
ного куриного эмбриона происхо-
дит замыкание амниотических
складок, а к концу 5-го дня
амнион увеличивается в размере
за счет накопления амниотической
жидкости и начинает сокращаться,
в последующие дни происходит
дальнейшее накопление амниотической жидкости.
Смыкание складок амниона происходит под оболочкой желтка,
после чего она рвется. Садлер (Sadler, 1955) отмечает большие
индивидуальные вариации во времени смыкания амниотических
складок и прорыва оболочки желтка. До 4-суточного возраста
оболочка желтка охраняет куриный эмбрион от соприкосновения
с белком и механических повреждений о скорлупу. В случае
разрыва оболочки желтка до образования амниона эмбрион
погибает, так как попадает в среду с отличающимся pH. Осво-
бождение от оболочки желтка является, по мнению Рагозиной
(1957), приспособлением для улучшения газообмена эмбриона
(обеспечение лучшего соприкосновения желточного мешка со скор-
106
лупой), которое можно сравнить с выклевом личинок рыб и ам-
фибий при переходе их к свободному образу жизни. Автор счи-
тает неслучайным совпадение во времени первого повышения
смертности эмбрионов (см. главу 17) с моментом разрыва и спол-
зания желточной оболочки. Месарош (1957) показал, что ме-
ханическая прочность оболочки желтка изменяется в течение
инкубации. Ко времени откладки яйца она равна в среднем
4.6 г/см, а через трое суток инкуба-
ции она уже не может противостоять
самому незначительному растяжению.
До 8-го дня инкубации рост
амниона медленный, а затем до 14-го
дня быстрый, когда он достигает
максимального веса около 320 мг
(Romanoff, 1960). При недостаточном
количестве поворотов яиц в инку-
баторе амнион имеет меньший раз-
мер, а при низкой температуре —
больший, чем при оптимальных
условиях инкубации (Romanoff, 1952).
Серозо-амниотическое сращение про-
рывается на 12-й день, образуя се-
розо-амниотический канал (Шейнис,
1954; Рагозина, 1961).
Ткань амниона бедна кровенос-
ными сосудами, и обеспечение ее
кровью идет за счет сосудов брюш-
ной стенки эмбриона и внутреннего
листка аллантоиса.
Исследуя развитие гладкой мы-
шечной ткани амниона куриного
эмбриона, Ревуцкая (1944) показа-
ла, что вначале — на 3—4-й день
Рис. 20. Схематическое изо-
бражение мышечной системы
амниона куриного эмбриона
с 6-го до 8-го дня инкубации.
(Bautzman a. Schroder, 1958).
Объяснение в тексте.
инкубации — имеются отдельные мышечные волокна, затем
(на 5—7-й день) образуется единый мышечный синцитий, ко-
торый в дальнейшем делится на отдельные, более или менее рас-
члененные мышечные клетки, но единство всей системы сохра-
няется благодаря анастомозам между ними. Бауцман и Шредер
(Bautzman a. Schroder, 1954) сообщают, что у рептилий, птиц
и млекопитающих стенка амниона состоит из 2—3 слоев: эпителия,
гладкой мышечной (только у рептилий и птиц) и соединительной
тканей. В месте сращения амниотических складок над эмбрио-
ном мышечные клетки расходятся веерообразно. Мышечная
ткань амниона представляет собою с 6-го по 8-й день развития
куриного эмбриона, по данным Бауцмана и Шредера (Bautzman
a. Schroder, 1958), узкие полосы радиальных мышц и звездо-
образные комплексы, разбросанные по всей поверхности амниона
107
(рис. 20). В дальнейшем это строение значительно упрощается
и незадолго перед вылуплением мышечный слой амниона пред-
ставляет собою простую сеть.
При сокращениях амниона амниотическая жидкость пере-
ходит из головной в хвостовую часть амниотической полости
и обратно. Внутри амниотической полости вследствие тонуса
мышечных волокон имеется определенное давление: до 9-го дня —
0.2—0.5 мм рт. ст., на 9-й день вследствие выпрямления головы
давление достигает 0.5—1.25 мм, а в дальнейшем движения
лап обусловливают колебания давления от 0.5 до 2.0 мм рт. ст.
(McCafferty et al., 1962).
2. Функция
Сокращения амниона куриного эмбриона, по данным
Куо (Кио, 1932а), начинаются на 4-й день с частотой 8 раз в 1 мин.
Затем частота сокращений быстро растет и достигает на 7-й день
25 раз, а с 9-го дня также быстро падает до 2 сокращений в 1 мин.
на 13—14-й день. Каждое сокращение амниона состоит из со-
кращения и немедленного расслабления радиальных мышц,
продолжающихся от одного конца амниона к другому и дающих
генерализованную волну сокращения. Бауцман, Дункер и Шре-
дер (Bautzman et al., 1954) записали электрические колебания
при сокращениях амниона. На 4-й день сокращения неритмичны —
10 раз в 1 мин.; на 5-й —ритм выравнивается и число сокращений
доходит до 16—20 в 1 мин. Максимальное количество сокращений
амниона достигается к 10—11-му дню, после чего оно умень-
шается и, наконец, сокращения прекращаются. Авторы подчер-
кивают, что ритмичная работа гладких мышц амниона подчинена
собственному автоматизму.
Греббельс (Groebbels, 1937), наблюдая сокращения амниона
эмбрионов нескольких видов птиц (Columba palumbis, Sturnus
vulgaris, Carduelis cannabina и др.), заметил, что в яйцах мелких
певчих птиц независимо от вида и размера яиц частота сокраще-
ний была 20—40 в 1 мин. Частота сокращений амниона сильно
зависит от температуры среды. Так, при температуре ниже 25°
и выше 46° сокращения прекращаются совсем, а при 38—42°
наблюдается максимальная их частота.
Количество амниотической жидкости достигает максимума
на 13-й день — ее вес в этот день равен 8—9% первоначального
веса яйца (Romanoff, 1952). Образование амниотической жид-
кости является, как предполагает Еремеев (1957), секреторным
процессом. После 13-го дня количество амниотической жидкости
увеличивается за счет поступающего в нее белка яйца. В течение
эмбрионального развития изменяется не только объем, но и со-
став амниотической жидкости (Огородний и Пенионжкевич;
108
1939). pH амниотической жидкости падает с 7.5 в начале ее^об-
разования до 6.9 на 11—12-й день. *
На 9-й день развития куриного эмбриона, как сообщает Ра-
гозина (1961), эмбрион начинает заглатывать амниотическую
жидкость, чему способствуют сокращения амниона. Автор под-
черкивает приспособительное значение этого как перехода к вну-
трикишечному способу питания, а также компенсации недостающей
жидкости, которую раньше эмбрион получал из жидкого желтка.
У 12-дневного эмбриона сокращения мышечной стенки амниона
прекращаются и между полостью амниона и белковым мешком
(см. далее) устанавливается связь через относительно широкий
и короткий серозо-амниотический канал (рис. 21). У 14-дневного
эмбриона амниотическая жидкость равномерно смешана с по-
ступившим в нее через этот канал белком, и эмбрион начинает
заглатывать ее. При нормальном развитии к 17 дням весь белок
переходит в амнион, а к концу 19-го дня белка не остается и здесь.
Это было подтверждено Лембургом (Lemburg, 1962) методом
маркировки белка куриного яйца.
Еремеев (1957) установил, что переход белка в полость ам-
ниона совершается у разных видов птиц на одной морфологической
стадии, примерно в одинаковые сроки, в процентах от общей
продолжительности эмбрионального развития того или иного
вида (табл. 31).
Таблица 31
Время обнаружения белка в амниотической жидкости
(По Еремееву, 1957)
Ин- дейки Куры Утки домаш- ние Го- луби домаш- ние Крачка черная Чибис
В сутках от начала раз- вития 16 12 16 10 8 15
В % от всей продолжи- тельности развития . . 57.1 57.1 57.1 57.8 61.5 57.7
Рагозина (1961) исследовала механизм поступления белка
из белкового мешка в полость амниона и показала, что это про-
исходит в результате: 1) увеличивающегося давления на белко-
вый мешок со стороны растущего эмбриона, желточного мешка
и аллантоиса; 2) глотательных движений эмбриона, как бы на-
сасывающих амниотическую жидкость в желудок и снижающих
при этом давление в полости амниона и 3) секреторной деятель-
ности стенок белкового мешка, разжижающей белок.
В результате потребления белка эмбрионом, по данным Ере-
меева (1957), активизируется протеиновый обмен, увеличивается
109
/
1
О Змм 6
UU
Рис. 21. Продольные разрезы куриного яйца. (По Рагозиной, 1961).
А — через 12 суток; Б —через 18 суток инкубации. 1 — воздушная камера яйца; 2 —стенка белкового мешка; 3 — жёлточнаяг
оболочка; 4 — желточная энтодерма; 5 — сосуды желточного мешка; 6 — серозо-амниотический канал; 7 — амнион; 8 — аллантоис;
9 — аллантоисная жидкость; 10 — сердце; 11 — печень; 12 — нога (фронтальный разрез); 13 — капилляры аллантоиса; 14 — шея
(поперечный разрез); 15 — легкое; 16 —перьевые бугорки; 17 — желудок; 18 — белок в белковом мешке; 19 — белок в полости
амниона; 20 — белок в полости желудка.
вес эмбриона и образуется оперение. Запаздывание в развитии
и неполное заглатывание амниотической жидкости приводит к
тому, что она остается вокруг эмбриона. При этом она обла-
дает большой" вязкостью в связи с повышенным содержанием в
ней густого белка. В таких случаях при проклеве скорлупы цып-
ленком амниотическая жидкость заклеивает отверстие проклева
и эмбрион зачастую задыхается, а выжившие цыплята вылуп-
ляются покрытыми липкой жидкостью — так называемые липкие
цыплята (Огородний и Пенионжкевич, 1939; Фетищева, 1939).
О роли амниона в эмбриональном развитии птиц существуют
разные мнения. Сергеев (1943) и Романов (Romanoff, 1952) в основ-
ном присоединяются к мнению эмбриологов прошлого века, что
амнион несет главным образом функцию защиты эмбриона от пов-
реждений при соприкосновении с плотной скорлупой (механи-
ческая защита). Нидхем же (Needham, 1931) считает, что амнион
обеспечивает жидкую среду для начала онтогенеза (повторение
начальных стадий филогенеза позвоночных животных) и назы-
вает амнион «личным прудом» эмбриона. Автор отмечает, что
в амниотической жидкости больше всего хлоридов и в этом ее сход-
ство с физиологическим раствором. Сергеев (1943) полагает,
что основная роль амниона в механической защите, а побочная —-
в создании водной среды. Роль амниотической жидкости, кроме
того, заключается в том, что эмбрион, будучи окружен ею, имеет
уравненное внешнее давление и может изменить свою форму
и положение в пространстве без опасности сжатия отдельных
органов и вызванных этим сжатием уродств. Возможно также,
что сокращения амниона улучшают обменные реакции эмбриона.
Ко всему этому Отрыганьев (Отрыганьев и др., 1964) добавляет,
что амниотическая жидкость создает необходимое осмотическое
давление вокруг эмбриона и, обладая бактерицидными свой-
ствами, предохраняет его от инфекции.
В. Аллантоис
1. Структура
Аллантоис отличается от других временных эмбрио-
нальных органов тем, что возникает непосредственно из тканей
эмбриона как вырост брюшной стенки задней кишки (из мезо-
дермы и энтодермы). В связи с тем что эмбрион уже закрыт ам-
нионом, а снизу лежит желточный мешок, аллантоис может расти
только во внеэмбриональный целбм, в пространстве между жел-
точным мешком, амнионом и серозой. При этом он отодвигает
серозу от амниона и желточного мешка, плотно прижимая ее к под-
скорлупной оболочке, а в остром конце яйца — и к белку.
Рассматривая возникновение аллантоиса в филогенезе, Сер-
111
геев (1943) первопричину его образования видит в необходимости
изоляции токсических продуктов азотистого обмена в замкну-
том пространстве яйца, называя аллантоис видоизмененным
мочевым пузырем амфибий. Рост аллантоиса во внеэмбриональный
целбм и возникающие при этом наивыгоднейшие условия для
газообмена обусловливают то, что он «принимает на себя также
и дыхательные функции». Строение аллантоиса вполне обеспе-
чивало эту возможность, так как его наружный слой образован
такой тканью (мезодерма), в которой на ранних стадиях раз-
вития эмбриона формируются кровеносные сосуды. Отрыганьев
(1951) считает, что аллантоис появляется в результате накопле-
ния продуктов выделения почек в задней кишке, не имеющей
еще клоачного отверстия. Они растягивают кишку, образуя
вырост в виде небольшого пузыря.
Аллантоисный мешок состоит из наружного и внутреннего
листка. При близком соприкосновении с серозой мезодерма на-
ружного листка аллантоиса (его наружный слой) и мезодерма
серозы (ее внутренний слой) сливаются, благодаря чему обра-
зуется чрезвычайно богатое сплетение кровеносных сосудов —
артерий и вен. В местах соприкосновения с амнионом внутренний
листок аллантоиса сливается с ним.
Хронологически развитие аллантоиса детально описано Ра-
гозиной (1961): 3 дня инкубации — аллантоис в виде неболь-
шого выпячивания вентральной стенки задней кишки еще не имеет
заметной полости; 4 дня — аллантоис достиг своей наружной
стенкой мезодермального слоя серозы; 5 дней — наружная стенка
его срослась с серозой везде, где с ней контактирует; 6 дней —
полость аллантоиса наполнена жидкостью, наблюдается раз-
личие между наружным листком (хорио-аллантоис с хорошо
развитым слоем мезодермы, с густой сетью капиллярных сосу-
дов) и внутренним (примыкает к желточному мешку, очень тон-
кий, и в нем только крупные кровеносные сосуды); 7 дней —
аллантоис покрывает более 1/3 сосудистого поля желточного
мешка; 9—-10 дней — покрывает значительную часть белка,
расположенного под дном желточного мешка, и вместе с серозой
начинает формировать белковый мешок; 11 дней — вырастает
настолько, что подстилает всю подскорлупную оболочку яйца,
смыкаясь в остром конце и закончив образование белкового
мешка; 14 дней — эндотелиальные стенки сосудов находятся
в наибольшем приближении к под скор лунной оболочке яйца,
количество аллантоисной жидкости увеличилось; 15—16 дней —
количество аллантоисной жидкости продолжает увеличиваться,
белка в белковом мешке почти не остается и его стенки спадаются;
20 дней — сосуды аллантоиса еще наполнены кровью, но по мере
зажима пупочных артерий и вен в пупочном стебельке проход
крови уже начинает затрудняться; аллантоисной жидкости нет
(см. рис. 21).
112
Итак, в белковом мешке, непосредственно вокруг массы белка,
находится сероза, сросшаяся с внутренним листком аллантоиса,
а за аллантоисной полостью, ближе к скорлупе, наружный листок
аллантоиса, слившийся с серозой. Сосочкообразные впячивания
белкового мешка, по мнению Романова (Romanoff, 1952), аб-
сорбирует белок, хотя и в незначительном количестве. По дан-
ным Рагозиной (1961), стенки белкового мешка выделяют се-
крет, разжижающий белок, что облегчает его проход через
серозоамниотический канал в полость амниона, но белок не
абсорбируют.
Аллантоис достигает максимального веса (2 г) на 15-й день
инкубации (Romanoff, 1952). Рост аллантоиса, по мнению автора,
обусловливается механическим растяжением в связи с накопле-
нием в нем жидкости, и он не растет, если продукты экскреции
из первичной почки не попадают в него. При инкубации яиц
острым концом вверх аллантоис перемещается в острый конец,
а при отсутствии поворачивания яиц рост его задерживается.
Кюнель (Kuhnel, 1961), перевязывая стебелек аллантоиса или
прокалывая стенки аллантоиса, выводил его из строя на 4—6-й
день инкубации. После этого эмбрион жил до 12-го дня
(но не дольше).
Тиррел и его соавторы (Tyrrell et al., 1954) обнаружили, что
общая площадь аллантоиса 10-дневного куриного эмбриона
равна 87.0 см2, в том числе в его примыкающей к скорлупе части —
57.9 см2; количество клеток в выстилке аллантоисной полости
равно 1.8 Х107.
Скалинский и Кондаленко (1963) провели электронномикро-
скопическое изучение хорио-аллантоиса И-дневного куриного
эмбриона и обнаружили следующее: а) эпителий хориона тесно
связан с подскорлупной оболочкой яйца и его бахромчатый слой
входит между ее волокнами; б) этот одно-трехслойный эпителий
с другой стороны органически связан с кровеносными капил-
лярами (диаметр их всего 3—6 мк), но между ними находится
тонкая базальная мембрана (при этом авторы отмечают, что
такое же соотношение между эпителием и кровеносными сосудами
наблюдается и в легких); в) соединительнотканный слой, лежа-
щий под эпителием, мезодермальный по происхождению, состоит
в основном из рыхло расположенных клеток звездчато-древо-
видной формы, с длинными отростками, соединяющими клетки
между собой; г) аллантоисный слой — одно-двуслойный пласт
длинных, вытянутых, плоских эпителиальных клеток с характер-
ными вакуолями (экскреторные образования клеток) и иногда
с микроворсинками.
Кровь попадает в аллантоис по двум артериям, из которых
левая лучше развита. Аллантоисные артерии проходят из тела
эмбриона во внутренний листок, огибают его край и переходят
в наружный листок. Отток крови осуществляется одной аллан-
8 В. В. Рольник 113
тоисной веной. Как во внутреннем, так и в наружном листке
аллантоиса все кровеносные сосуды сопровождаются лимфати-
ческими сосудами, соединяющимися с лимфатической системой
эмбриона (Romanoff, 1952). Если аллантоис не соприкасается
с серозой, капиллярное сплетение в нем не образуется.
2. Функция
Свою функцию органа дыхания, по данным Рагозиной (1961),
аллантоис начинает в конце 7-го дня, а с конца 8-го по 19-й
является единственным органом газообмена.2
Полость аллантоиса служит резервуаром для продуктов
распада протеинов, выводимых сначала первичной, а потом и по-
стоянной почкой. Вода из эмбриональной мочи реабсорбируется
кровеносными сосудами аллантоиса, а содержащиеся в ней сухие
вещества откладываются в полости аллантоиса в виде грязно-
вато-белых масс. Таким образом, объем и состав аллантоисной
жидкости изменяется в течение эмбрионального развития.
По данным Романова (Romanoff, 1952), аллантоис начинает
функционировать как склад экскретов с 5-го дня инкубации.
Количество общего азота в аллантоисной жидкости с 5-го до 13-го
дня увеличивается в 80 раз и еще в 3 раза с 13-го до 18-го дня.
Больше всего в аллантоисной жидкости мочевой кислоты, и
около 90% ее выпадает в осадок в последнюю неделю инкуба-
ции. Кроме того, в аллантоисной жидкости имеется. креатин
(в увеличивающемся до 18-го дня количестве), аминокислоты
(уменьшающиеся по концентрации, но увеличивающиеся по об-
щему количеству), пуриновые основания, креатинин, аммиак
и мочевина (содержание последних двух достигает максимума
на 14-й день). Различные физические свойства аллантоисной
жидкости отражают ее изменяющийся состав. Так, pH с 8.0
в конце 1-й недели инкубации (много мочевины) падает до 5—6
за 1—2 дня перед вылуплением (много мочевой кислоты, угле-
кислоты и фосфорных соединений). С 14-го по 18-й день инку-
бации осмотическое давление аллантоисной жидкости снижается
в связи с уменьшением калия, натрия, кальция, магния, не-
органического фосфора, кремния и хлора. По сравнению с амнио-
тической аллантоисная жидкость гипотонична, что обусловлено
меньшим содержанием в ней NaCl.3 Еремеев (1957) показал,
что содержание аллантоисной жидкости, так же как и амниоти-
ческой, изменяется у разных видов в одинаковые отрезки вре-
мени в процентном отношении ко всей длительности развития
эмбриона.
2 Данные об этой функции аллантоиса приведены в главе 10.
3 Более подробно о продуктах, выделяющихся при распаде протеинов,
см. в главе 12.
114
Кроме описанных выше функций дыхания, склада экскретов
и разжижения белка в белковом мешке, аллантоис абсорбирует
кальций из скорлупы. Нидхем (Needham, 1931), ссылаясь на дан-
ные многих исследователей, считает, что выделяющиеся аллан-
тоисными сосудами углекислота и вода переводят нерастворимые
в воде соли кальция в растворимый бикарбонат кальция, который
абсорбируется сосудами аллантоиса и переносится эмбриону,
где используется для построения скелета. Скорлупа куриных
яиц за период инкубации теряет 190 мг органических веществ
(7.1% первоначального количества) и 160 мг общей золы (5.2%),
в том числе 140 мг кальция (6.5%). Растворение солей скорлупы,
кроме снабжения эмбриона материалами для построения скелета,
имеет значение и в увеличении проницаемости скорлупы (улуч-
шение газообмена); кроме того, это делает скорлупу более хруп-
кой и способствует более легкому проклеву ее при вылуплении.
Романов (Romanoff, 1952) сообщает, что если яйцо не повора-
чивать, то в месте соприкосновения кровеносных сосудов с под-
скор л упной оболочкой в скорлупе появляются желобки, что
доказывает абсорбирование аллантоисом кальция из скорлупы.
Кроме того, аллантоис, возможно, регулирует испарение
влаги из яйца во второй половине инкубации. Шмидт (1953)
говорит об аллантоисе как о «подушке», защищающей эмбрион
от высыхания и травмы. Отрыганьев с соавторами (1964), на-
оборот, считает, что благодаря аллантоису испаряется излишняя
влага из яйца. Но экспериментальных данных, доказывающих
регуляторную функцию аллантоиса в испарении влаги из яйца,
насколько нам известно, нет.
Г Л А В А 8
КРОВООБРАЩЕНИЕ
Кровообращение — это первая функционирующая си-
стема развивающегося эмбриона. Основные жизненные функции
эмбриона — питание и дыхание — выполняются желточным меш-
ком и аллантоисом, и их кровеносные сосуды играют основную
роль в эмбриональном кровообращении. Но есть сосуды, появ-
ление которых в течение эмбрионального развития может быть
объяснено только с филогенетической точки зрения, например
дуги аорты. Для рыб 6 пар жаберных дуг аорты являются жиз-
ненно необходимыми и во взрослом состоянии. У птичьего эм-
бриона появляются все 6 пар дуг, но функционирующей после
вылупления остается только одна пара. В развитии сердца также
проявляется повторение филогенеза: от трубчатого сердца к че-
тырехкамерному.
8*
116
Необходимо отметить, что каждый поворотный пункт в раз-
витии эмбриона должен проходить так, чтобы снабжение кровью
осуществлялось нормально как во временных органах, харак-
терных для эмбриональной жизни, так и в постепенно формирую-
щихся органах, начинающих свою деятельность в более поздние
периоды.
А. Морфология развития сердца и кровеносных
сосудов
1. Сердце
Под передней кишкой, недалеко от входа в нее, выра-
стают околосердечные части целома, где в дальнейшем из первых
зачатков образуется сердце. Дафи и Эберт (Duffey a. Ebert, 1957)
показали, что участок, образующий сердце, метаболически диф-
ференцируется уже к 18—19 час. инкубации. Де Хаан (Haan de,
1963) при помощи микрокиносъемки изучил образование сердца
из мезодермы. У 19—22-часового куриного эмбриона парные
сердцеобразующие участки представляют собой отдельные гроздья
мезодермальных клеток, которые к 26 час. образуют зачатки
эндокарда — трубки толщиной в одну клетку. Большая же часть
мезодермальных клеток идет на образование мышечного (мио-
карда) и поверхностного (эпикарда) слоев сердца. Как и все
брюшные органы, сердце лежит вначале на желтке, но затем
с обеих сторон бластодиска появляются складки, которые об-
разуют брюшную стенку тела и подстилают сердце, отделяя
его от желтка. В результате правая и левая части органов, имев-
ших парные зачатки (в том числе и сердце), соединяются по сре-
динной линии. У сердца куриного эмбриона это происходит
в 29-часовом возрасте (рис. 22). Обрастание эндотелиальной
трубки сердца мышечным и покровным слоями является повто-
рением филогенетического развития сердца, так как в эволюции
животных сердце произошло путем разрастания мышечного
слоя вокруг кровеносного сосуда. У 29—30-часового эмбриона
сердце имеет форму трубки, переходящей в головной части тела
в брюшную аорту, а в хвостовой — в две желточно-брыжеечные
вены. Средняя часть сердца к 33 час. инкубации значительно
расширяется и частично сдвигается вправо, а к 38 час. настолько
отклонена вправо, что выходит за пределы боковой линии эмб-
риона. Здесь в будущем образуются желудочки сердца. Место
соединения сердца с желточно-брыжеечными венами — это синус
предсердия, а на противоположном конце сердечной трубки,
там, где сердце сужается кверху, лежит артериальная луковица.
Сердце 30—33-часового эмбриона начинает судорожно подер-
гиваться, но настоящая пульсация начинается только с 36—
40 час. инкубации.
116
Эберт и его соавторы (Ebert et al., 1955) указывают, что синтез
миозина начинается через 12—16 час. инкубации. Вначале миозин
распространен по всему верхнему слою бластодиска, а к 19—
22 час. он уже ограничен областью образования сердца. В это же
время здесь появляется актин. После того как район, образую-
щий сердце, сформирован,
другие части эмбриона утра-
чивают способность синтези-
ровать специфический миозин
сердца. Слияние зачатков
сердца начинается, по дан-
ным авторов, у 30-часового
эмбриона, локальные сокра-
щения сердца — у 32-часо-
вого, а настоящие сокраще-
ния — через 33—38 час. ин-
кубации. Тогда же в сердце
впервые появляется гликоген.
Изучая развивающуюся
сердечную мышцу куриного
эмбриона при помощи элек-
тронного микроскопа, Гиббс
(Hibbs, 1956) обнаружил,
что в клетках сердца 30-ча-
сового эмбриона имеются
мышечные нити (миофиламе-
ны). Даже на 60—70-м часу
инкубации, по данным Бонди
(Bondi, 1957), сердечная
ткань состоит не из истинных
мышечных клеток, а из скоп-
лений эндотелиальных про-
топлазматических образова-
ний.
Ко времени, когда труб-
чатое сердце куриного эмбри-
Рис. 22. Схема происхождения и слия-
ния сердечных трубок у куриного
эмбриона. (По Patten, 1952).
А — 25-часовой эмбрион; Б — 27-часовой;
В — 28-часовой; Г — 29-часовой.
1 — перикардиальная зона целбма; 2 — эпи-
миокардиум; з—эндокардий (За — эндокарди-
альные зачатки); 4 — брюшной корешок
аорты; 5 — желточно-брыжеечная вена.
она начинает пульсировать,
его внутренняя выстилка — эндотелий — отделена от наружного
слоя относительно толстым, упругим слоем бесклеточного сер-
дечного желе. Как обнаружил Бэрри (Barry, 1948), это желе
в начале эмбрионального кровообращения играет полезную роль,
так как помогает сохранить упругость стенки желудочка сердца
во время диастолы, подготовляя его к новой систоле, а также
препятствует возвращению крови из аорты до образования спе-
циальных клапанов.
В период между 30 и 55 час. инкубации сердце куриного эмб-
риона растет быстрее, чем та часть тела, в котором оно нахо-
117
Рис. 23. Изменение формы сердца куриного эмбриона
в возрасте от 29 до 100 час. (По Patten, 1951).
А — вид с брюшной стороны; Б — вид справа, В — вид сзади.
1 — сужение между предсердием и желудочком; 2 — междуже-
лудочковая борозда; з — эндокардий; 4 — брюшные корешки
аорты; 5 — миокардий;- 6 — желточно-брыжеечные вены; 7 — кювь-
еровы протоки; 8 — зона синуса предсердия; 9 —- артериальная
луковица; 10 — сужение между артериальной луковицей и же-
лудочком; 11 •— желудочек; 12 — предсердие (12а — левое, 126 —
правое); 13 — венозный синус. Цифры внизу — возраст эмбриона
в часах.
дится. В связи с тем что головной и хвостовой концы сердечной
трубки закреплены сосудами (брюшной аортой и желточно-бры-
жеечными венами), неприкрепленная средняя желудочковая часть
сердца делает сначала U-образный изгиб, а затем скручивается
петлей. При этом зона предсердия, расположенная ранее в хво-
стовом конце сердца, передвигается налево и вперед, т. е. зани-
мает такое же положение, как у взрослых птиц, а артериальная
луковица ложится накрест с предсердием, направо, ближе к спине.
На этой стадии деление сердца на камеры еще не началось и оно
119
все еще представляет собой трубку. В течение 3-го и 4-го дня
инкубации заканчивается скручивание сердечной трубки и сердце
приобретает внешнюю форму, сходную с формой сердца взрослой
птицы (рис. 23).
Начиная с 53 час. образуется сужение между предсердием
и желудочком, которое превращается затем в предсердно-желу-
дочковый канал. В течение 4-го дня в предсердии образуется
бороздка, как бы делящая предсердие на правое и левое. Одно-
временно появляется и продольная бороздка на желудочке.
Четырехкамерное сердце появляется позже, но начинается этот
процесс уже на 4-й день инкубации, когда между предсердиями
начинает расти перегородка. Пракаш (Prakash, 1954) наблюдал
у 3.5-дневных эмбрионов предсердную перегородку, состоящую
из двух половин, свешивающихся в левое предсердие и образую-
щих, таким образом, клапан, который обеспечивает ток крови
только из правого предсердия в левое, но не наоборот. Процесс
разделения полостей сердца продолжается и на 5-й день: воз-
никает перегородка в артериальном стволе, разделяющая его
на два сосуда — аорту и легочную артерию. Эта перегородка
растет и на 7-й день достигает желудочка. Внутри аорты и ле-
гочной артерии возникают полулунные клапаны путем образо-
вания полостей в эндокардиальных утолщениях каждого сосуда.
На 8-й день эмбрионального развития эти клапаны уже вполне
сформированы. Межжелудочковая перегородка, представляющая
собой губчатую массу клеток у верхушки желудочка, возникает
на 4-й и достигает предсердного канала на дорсальной стороне
на 5-й день инкубации. Межжелудочковая перегородка распо-
ложена у птиц вертикально и, по мнению Пракаша, является
новообразованием у теплокровных, а не гомологична горизон-
тальному мышечному гребню в сердце рептилий. Три первичные
перегородки сердца — межжелудочковая, межпредсердная и пе-
регородка артериального ствола — растут от периферии к центру,
но не соединяются друг с другом, а объединяются при помощи
четвертой — большой подушкообразной перегородки, образую-
щейся в предсердно-желудочковом канале из двух эндотелиаль-
ных утолщений — срединного и бокового (рис. 24). Отверстие
между правым и левым желудочками постепенно уменьшается
в размере и к 8-му дшо развития куриного эмбриона они раз-
делены полностью. Межпредсердная перегородка, сливаясь с по-
душкообразной, образует сплошную перегородку между двумя
предсердиями. Но затем в ней появляется вторичное отверстие,
служащее для прохода крови из правой части сердца в левую. Это
отверстие существует в течение всего периода инкубации и за-
крывается только вскоре после вылупления. Вследствие этого
артериальное и венозное кровообращения в течение всего эмбрио-
нального периода не бывают полностью изолированными. После
завершения образования перегородок сердца аорта становится
120
связанной с левым желудочком, а легочная артерия — с правым.
Венозный синус при дальнейшем развитии включается в правое
предсердие. Бремер (Bremer, 1928) выдвинул гипотезу, что об-
разование внутрисердечных перегородок обусловливается нали-
чием двух токов крови, идущих из правой и левой желточно-бры-
жеечных вен и не смешивающихся в еще неразделенном сердце.
Эту гипотезу поддержал Романов (Romanoff, 1960). Однако Сте-
фан (Stephan, 1955) установил, что после резекции конечной
Рис. 24. Фронтальный разрез сердца 4-дневного
куриного эмбриона. (По Иванову, 1945).
1а и 16 — правый и левый желудочки; 2а и 2б — правое и
левое предсердие; з — межпредсердная перегородка; 4 —
межжелудочковая перегородка; б — срединная эндотели-
альная подушка; 6 — боковая эндотелиальная подушка.
части правой желточно-брыжеечной вены и зачатка правого ве-
нозного синуса межпредсердная перегородка развивалась на своем
нормальном месте, что полностью опровергает вышеуказанную
гипотезу. Выключая то левую, то правую часть кровотока к сердцу
со 2-го по 4-й день развития куриного эмбриона, Рыхтер и Лемеж
(Rychter a. Lemez, 1956) установили, что это не вызывает макро-
скопически заметных изменений в межжелудочковой и артериаль-
ной перегородках. Авторы, так же как и Стефан, считают, что
существование двух кровотоков в сердце куриного эмбриона
не имеет морфогенетического значения.
Одновременно с внешними и внутренними изменениями сердца
оно меняет и свое положение. В начале своего образования сердце
лежит на уровне заднего мозга, а уже к концу 4-го дня оно ото-
двигается назад до уровня зачатков передних конечностей.
121
2. Сосудистая система
Кровеносные сосуды и форменные элементы крови возникают
из мезодермы в виде кровяных островков. Вначале кровяные
островки образуются в сосудистом поле (внутренняя часть темного
поля), а затем уже и в самом эмбрионе. Данчакова (Danchakoff,
1907, 1908, 1918) показала, что первые кровяные островки появ-
ляются в конце 1-го дня инкубации. Затем кровяные островки
дифференцируются и в результате округления и высвобождения
центральных клеток и уплощения пограничных превращаются
в первые сосуды желточного мешка, которые сливаются друг
с другом и образуют сосудистую сеть. Сосуды внутри эмбриона
возникают в виде полых трубок, не содержащих кровяных клеток,
и заполняются кровью только при соединении с сосудами жел-
точного мешка (Кнорре, 1959).
Анастамоз кровяных островков и образование сосудов
желточного мешка происходит от 24 до 33 час. инкубации.
Первыми возникают крупные сосуды внеэмбрионального круга
кровообращения, связанные с сердцем. Они имеют двойствен-
ное происхождение: сосуды, непосредственно связанные с серд-
цем, возникают внутри эмбриона, а их более отдаленные от
сердца части — во внеэмбриональной зоне и растут в сторону
эмбриона, навстречу первым. Затем они срастаются, причем
устанавливается внеэмбриональный круг кровообращения.В этом
круге кровь из сердца поступает через брюшную аорту, потом
через дуги аорты вдоль ^спинных аорт, а затем через желточ-
ные артерии к сплетению сосудов желточного мешка. Затем
кровь собирается в концевую вену желточного мешка (см. рис. 17);
оттуда через многочисленные желточные вены (из всех частей
сосудистого поля), сливающиеся в центре, — в две желточные
вены, которые образуют кольцо вокруг кишки и впадают в ве-
нозный проток (в дальнейшем находится в печени), доставляющий
кровь снова в сердце.
Во внутриэмбриональном круге кровообращения кровь вы-
ходит из сердца, так же как и во внеэмбриональном, по брюшным
аортам, дугам аорты и вдоль спинных аорт, а затем через от-
ветвления спинных аорт — к телу эмбриона. Возвращается кровь
из головной части передними, а из задней — задними карди-
нальными венами. У 33-часового эмбриона уже образовались
передние, у 35-часового — задние кардинальные вены (Patten,
1951). В сердце кровь обоих кругов кровообращения (эмбрио-
нального и внеэмбрионального) смешивается и делается менее
• богатой питательными материалами и кислородом, чем кровь,
вернувшаяся из желточного мешка. Однако в связи с малыми
размерами эмбриона и в смешанной крови имеется достаточно
питательных веществ и кислорода для его существования и раз-
вития.
122
В течение 3-го и 4-го дня инкубации в кровообращении эмб-
риона появляется третий круг — аллантоисный (рис. 25). Основ-
ная его роль — удаление накопившейся в крови углекислоты
Рис. 25. Кровеносная система 4-дневного куриного эмбриона.
(По Patten, 1951).
1 — внутренняя сонная артерия (а.); 2 — наружная сонная а.; з — вторая аорт-
ная дуга (исчезающая); 4 — третья аортная дуга (корни сонных а.); 5 —позвоноч-
ная а.; б — шейная сегментная а., 7 — четвертая аортная дуга; 8 — шестая
аортная дуга с соединяющейся с ней в виде петли пятой аортной дугой;
9 — передняя кардинальная вена (в.); 10 — легочная а.; 11 — кювьеров про-
ток; 12 — задняя кардинальная в.; 13 — брюшная а.; 14 — задняя полая в.;
15 — подключичная в.; 16 — подключичная а.; 17 — желточно-брыжеечные вены
в месте их слияния, после чего они образуют непарный сосуд; 18 — субкарди-
нальная в.; 19 — желточная в.; 20 — желточная а.; 21 — аллантоисная в.;
22 — аллантоисная а.; 23 — подвздошная а.
и снабжение крови кислородом. Аллантоисные (пупочные) ар-
терии возникают в результате удлинения и увеличения сегментных
артерий, выходящих из аорты на уровне желточного стебелька.
123
Левая аллантоисная артерия лучше развита. Она выходит из тела
эмбриона по стебельку аллантоиса в его внутренний листок,
где она делится на две сильные ветви, одна из которых проходит
в головную часть, а другая — в хвостовую, а затем, огибая край
аллантоиса, обе ветви переходят в наружный аллантоисный
листок. Здесь кровь поступает в богатое кровеносное сплетение,
разветвляющееся в наружном листке аллантоиса, где благодаря
близости к скорлупе облегчается обмен газов с внешней средой.
Отсюда кровь собирается в маленькие вены, которые, сливаясь,
дают две большие аллантоисные (пупочные) вены, входящие
через аллантоисный стебелек в тело эмбриона и впадающие в ве-
нозный синус вместе с желточно-брыжеечными венами. На 4-й день
инкубации правая аллантоисная вена исчезает и вся кровь из ал-
лантоиса вытекает по левой.
Рассмотрим несколько подробней развитие внутриэмбриональ-
ного круга кровообращения. У 33—38-часового эмбриона брюш-
ная аорта связана со спинной одной (первой) парой дуг аорты
(расположенных вокруг переднего конца глотки), у 50—55-часо-
вого — двумя парами дуг, и начинают расти 3-я и 4-я пары. Пер-
вая и вторая пары дуг аорты исчезают на 3—4-й день (рис. 26),
но соединения между ними образуют внутренние и наружные
сонные артерии, которые в свою очередь соединяются между
собой посредством 3-й пары дуг аорты. Между 3-й и 4-й парой
дуг спинная аорта исчезает, по-видимому, в связи с удлинением
шеи эмбриона и ослаблением тока крови в этом месте (в сонных
артериях — ток вперед к голове, в 4-й паре дуг — ток назад).
4-я пара дуг исчезает на девой стороне и остается на правой
(у млекопитающих — наоборот) как постоянная дуга, соединяю-
щая брюшную и спинную аорты. 5-я пара дуг быстро исчезает
и не имеет никакого функционального значения, 6-я пара дуг
сохраняется в течение всего эмбрионального периода. После
вылупления спинная часть их, так называемые Боталловы про-
токи, исчезает и 6-я пара дуг аорты становится корнями легочных
артерий.
В течение 3-го и 4-го дня инкубации появляются сонные ар-
терии и заканчивается слияние ранее парных спинных аорт
в одну, а в каждом сегменте от аорты отходят сегментные арте-
рии. Кроме того, от спинной аорты отходят вновь возникающие
брюшнополостная артерия, артерия задней брыжейки, артерии
половых органов и др. Узкий кончик спинной аорты после вы-
хода из нее пупочных артерий называется хвостовой артерией.
На 5-й день из основных сонных артерий возникают подклю-
чичные артерии, входящие в передние конечности. Наружные
подвздошные артерии, идущие к зачаткам задних конечностей,
возникают в виде отростков аллантоисных артерий. Легочные
артерии у эмбриона относительно узки и могут пропустить только
небольшое количество крови. Основная функция легочных ар-
124
терий в эмбриональный период — соединение малого и большого
кругов кровообращения.
Задние кардинальные вены в течение 3-го и 4-го дня образуют
анастомозы с субкардинальными венами (выходящими из первичных
почек). У 4-дневного куриного эмбриона нижняя полая вена
имеет вид тонкого сосуда, выходящего из печени и анастомозирую-
щего с правой субкардинальной веной. Наиболее важным из-
менением в венозной системе эмбриона после 4-го дня является
образование воротного кровообращения печени и почек. Перво-
начально венозный проток
соединившихся двух желточ-
но-брыжеечных вен имеет
прямой проход через печень
к венозному синусу, но по-
том разрастание печеночной
ткани и образование пече-
ночных перегородок заста-
вляет его ветвиться на мел-
кие кровеносные сосуды. По-
степенно эти мелкие сосуды
образуют в печени воротную
систему из а) приносящих
сосудов (ветвятся из хвосто-
вой части венозного прото-
ка), в которых кровь течет
внутрь печени, и б) унося-
щих (ответвления головной
части венозного протока), в
которых кровь течет из пе-
чени. На 7-й день эмбрио-
нального развития средняя
Рис. 26. Схема изменений аортных дуг
в течение развития куриного эмбрио-
на. (По Shumway, 1942).
А — Гипотетический примитивный тип; Б —
у вылупившегося цыпленка. I— VI — аорт-
ные дуги. 1 — внутренние сонные артерии
(а.); 2 — наружные сонные а.; з — общие
сонные а.; 4 — легочные а.; 5 — аорта. Пунк-
тиром отмечены исчезающие дуги.
часть венозного протока исчезает
и вся кровь начинает проходить через ветвящуюся систему
этих сосудов. Первоначально воротная система печени снаб-
жается кровью главным образом из желточного мешка. Затем
через печень начинает проходить и кровь, возвращающаяся
из аллантоиса по левой аллантоисной вене. Ствол этой вены
остается и после вылупления в виде вены брюшной стенки тела,
открывающейся в левую печеночную вену.
Почечное воротное кровообращение (через первичные почки)
устанавливается на 5-й день. Кровь, собравшаяся из тела в задние
кардинальные вены, фильтруется через первичные почки и по-
падает в субкардинальные вены, откуда переходит в переднюю
часть задних кардинальных вен, а затем в сердце. Когда пер-
вичные почки начинают атрофироваться, постепенно редуцируется
и та часть задних кардинальных вен, которая лежала вдоль
них. Связь между задними кардинальными и субкардинальными
венами к этому времени устанавливается через вновь образо-
126
вавшиеся большие почечные вены, проходящие внутри развив-
шихся постоянных почек. Почечное кровообращение эмбриона
птиц в момент действия первичных почек сходно в основном с по-
стоянным кровообращением взрослых амфибий и представляет
удивительный пример рекапитуляции.
С 5-го дня инкубации начинают образовываться ветви бры-
жеечной вены, идущие в желудок, кишечник, поджелудочную
железу и селезенку. Вначале роль брыжеечной вены в воротном
кровообращении печени невелика, но после втягивания жел-
точного мешка, на 19-20-й день инкубации, она остается един-
ственной веной, снабжающей кровью печеночную воротную
систему. Вены мышечного желудка образуют независимую левую
воротную вену, открывающуюся в левую долю печени. Во второй
половине эмбрионального развития выносящие вены печени
впадают в нижнюю полую вену. В связи с образованием задних
конечностей развиваются бедренные и седалищные вены, впа-
дающие в хвостовую часть задних кардинальных вен. Кроме
того, в задние кардинальные вены впадают сегментные вены,
идущие параллельно с сегментными артериями. Примерно с 6-го
дня инкубации передняя часть задних кардинальных вен деге-
нерирует, так что кровь, проходившая ранее через них в сердце,
идет непосредственно через субкардинальные вены. Со второй
половины инкубации роль задних кардинальных вен (значительно
редуцирующихся к этому времени) в собирании крови из задней
половины тела начинает выполнять нижняя полая вена. Она об-
разуется из небольших кровеносных полостей в правой доле
печени и в брыжейке желудка, а также из соединившихся суб-
кардинальных и задних кардинальных вен. После 8-го дня ин-
кубации все вены задней половины тела включаются в систему
нижней полой вены, и она сильно увеличивается. Кровь из пе-
редней части тела эмбриона собирается в две передние полые
вены. Они объединяют яремные, позвоночные и подключичные
вены правой и левой сторон тела. Яремные вены, образовав-
шиеся из передних кардинальных вен, возвращают кровь из го-
ловы. Сегментные вены объединяются в расположенные вдоль
позвоночника позвоночные вены. Вены, получающие кровь из пе-
редних конечностей, — подключичные — возникают как ветви
задней кардинальной вены.
Приводим описание схемы кровообращения куриного эмбриона
после 8-го дня инкубации по Лилли (Lillie, 1952). Левый и пра-
вый желудочки сердца полностью разделены, и левый накачивает
кровь в аорту через правую 4-ю аортную дугу, а правый — в ле-
гочную артерию и затем через Боталлов проток также в спин-
ную аорту, где происходит смешивание крови из обоих желудоч-
ков. Часть крови из левого желудочка не попадает в спинную
аорту, а проходит к голове через сонные артерии, отходящие
у основания аортных дуг к стенкам грудной клетки и к крыльям
126
через подключичные артерии. Из спинной аорты кровь направ-
ляется по ее ветвям: 1) брюшно-полостной, главным образом
к желудку й печени, 2) желточной, к желточному мешку и бры-
жейке, 3) левой пупочной, к аллантоису и ногам, 4) сегментным,
к стенкам тела и 5) хвостовой, к хвосту.
Кровь, возвращающаяся из передней части тела передними
полыми венами в правое предсердие, — чисто венозная. Судя
по системе клапанов венозного синуса, эта кровь, по-видимому,
проходит прямо в правый желудочек, а оттуда через Боталлов
проток — в спинную аорту (Lillie, 1952). Кровь из желточного
мешка, пищеварительного канала и аллантоиса через желточную,
брыжеечную и пупочную вены соответственно проходит в во-
ротную вену печени. Из печени по правой и левой печеночной
вене кровь попадает в нижнюю полую вену. Кровь из задних
конечностей, хвоста и сегментных вен собирается в задние кар-
динальные вены и проходит по воротной системе почек (сначала
первичных, потом постоянных), где очищается от вредных про-
дуктов распада питательных веществ — аммиака, мочевины и мо-
чевой кислоты — и попадает в субкардинальные вены и далее
в нижнюю полую вену. Таким образом, кровь, приносимая ниж-
ней полой веной, очищена и богата питательными веществами.
Она, так же как и кровь из передних полых вен, попадает в. пра-
вое предсердие, но в отличие от нее проходит через отверстие
в предсердной перегородке в левое предсердие, а оттуда нака-
чивается левым желудочком в сонные артерии и через аортные
дуги — в спинную аорту. В спинной аорте эта кровь смешивается
с кровью из правого желудочка, но сонные артерии, несущие
кровь к голове эмбриона, получают самую чистую и самую «пи-
тательную» кровь. В левое предсердие сердца впадают только
маленькие легочные вены.
К моменту вылупления куриного эмбриона наступают следую-
щие изменения в его кровообращении: аллантоисные артерии
и вены остаются в отмершем аллантоисе на скорлупе яйца,
а их стволы внутри эмбриона превращаются в сосуды, не имею-
щие большого значения. Вены желточного мешка после его втя-
гивания в брюшную полость эмбриона также постепенно отми-
рают. Боталловы протоки теряют просвет и превращаются в связки
между легочными артериями и дугами аорты. Отверстие в пред-
сердной перегородке закрывается, после чего устанавливается
полностью раздельная циркуляция крови. Правое предсердие,
а затем правый желудочек получают всю кровь. из большого
круга кровообращения (венозную), которая затем проходит
через сильно увеличивающиеся легочные артерии в легкие, где
берет кислород и отдает углекислоту, а потом возвращается
через легочные вены в левое предсердие, переходит в левый же-
лудочек и затем .через аорту — снова в большой круг крово-
обращения.
127
Б. Развитие и свойства эмбриональной крови
Как известно, кровь является жидкой средой орга-
низма, обеспечивающей процессы обмена веществ в тканях и связь
между всеми органами. У птиц, так же как и у рептилий, эритро-
циты продолговато-овальной формы и имеют ядро. Лейкоциты
до своему строению, так же как у млекопитающих, делятся на агра-
нулоциты (лимфоциты и моноциты) и гранулоциты (нейтрофилы,
эозинофилы и базофилы).
Количество и свойства отдельных элементов крови изменяются
в течение эмбриональной жизни. Слонимский (Slonimski, 1927)
показал, что гемоглобин появляется примерно на стадии 33 час.
инкубации. Однако эмбриональный гемоглобин, по данным Холл
(Hall, 1935), отличается от гемоглобина постэмбриональной
крови бдлыпим сродством к кислороду. Фрейзер (Fraser, 1961)
обнаружил, что гемоглобин эмбрионального и постэмбриональ-
ного типа отличаются друг от друга содержанием гемопротеида.
У 5-дневного куриного эмбриона имеется 84% гемоглобина эмб-
рионального типа, а перед вылуплением такого гемоглобина
только 28%.
Данчакова (Danchakoff, 1907, 1908, 1918) одна из первых
детально исследовала гемопоэз куриного эмбриона. По ее данным,
к началу кровообращения (30—36 час. инкубации) бдльшая
часть клеток кровяных островков находится внутри замкнутой
сети кровеносных сосудов, однако и после этого срока остается
некоторое количество кровяных островков, в которых еще не про-
изошла дифференциация эндотелия и клеток крови. Первичные
кровяные клетки по внешнему виду напоминают гемоцитобласты
взрослой птицы. Они округляются и, накопляя гемоглобин,
превращаются в способные к делению первичные эритробласты
различного размера. В дальнейшем первичные эритробласты
постепенно теряют способность к делению и превращаются в пер-
вичные эритроциты округлой формы. Первичные эритроциты
недолговечны — через некоторое время они погибают и исчезают
из кровяного русла. Позднее из гемоцитобластов начинают об-
разовываться мелкие вторичные эритробласты, которые в свою
очередь дают начало вторичным эритроцитам, имеющим нор-
мальную удлиненно-овальную форму. Кроме того, появляются
карликовые формы гемоцитобластов, которые, по данным ав-
тора, дают начало тромбоцитобластам и тромбоцитам.
Первым органом эмбрионального кроветворения является
сеть кроветворных капилляров желточного мешка. В крове-
творном капилляре содержатся родоначальные кровяные клетки,
а последующие этапы их созревания совершаются главным об-
разом в периферической крови, что подтверждается наличием
здесь митозов кровяных клеток. На 15-й день инкубации эритро-
поэз в желточном мешке достигает максимального развития,
128
причем образуются уже только вторичные эритробласты. В это же
время в мезенхиме появляются малые лимфоциты, возникающие
из лимфоидных блуждающих клеток типа гемоцитобластов.
Изучению кровотворения на ранних стадиях эмбрионального
развития птиц посвящено много исследований. Хаузер и его со-
авторы (Houser et al., 1961) при помощи фазово-контрастного
микроскопирования желточного мешка куриного эмбриона от 40
до 96 час. инкубации показали, что кровеносные сосуды воз-
никают в прозрачной зоне бластодиска из тяжей, которые об-
разуются в свою очередь из примитивной мезенхимы, причем
клетки внутри тяжей сохраняют свою индивидуальность и не об-
разуют синцития. Затем между клетками накапливается жид-
кость и образуются просветы кровеносных сосудов. Примитивные
эритробласты, содержащие уже на самых ранних стадиях раз-
вития следы гемоглобина, возникают, по данным этих авторов,
из трех источников: а) из клеток тяжей, остающихся после об-
разования просветов в сосудах, б) из кровеносных островков,
которые возникают из эндотелия и ангиобластов и в) в резуль-
тате митозов эндотелиальных клеток, которые чаще происходят
в свободных клетках внутри сосудов, чем в кровеносных остров-
ках. Катанео (Cattaneo, 1962) при помощи электронного микро-
скопа исследовал мезодерму позади бластопора, из которой
развиваются кровяные островки. Автор обнаружил, что через
14—15 час. инкубации мезодерма состоит из рядов клеток, при-
легающих к энтодерме, а еще через 3—4 часа эти клетки мито-
тически делятся, образуют скопления и превращаются в кровяные
островки. В связи с тем что гематогенные мезодермальные клетки
сильно нагружены желтком, раньше думали о происхождении
кровяных островков из энтодермы.
Для выяснения вопроса о материале, используемом в эмбрио-
генезе птиц при образовании эритроцитов, Моригучи (Mori-
guchi, 1957) инъецировал глицин в инкубируемые куриные яйца.
В результате инъекции повысилось содержание гемоглобина
на 5—15% и количество эритроцитов на 3—15% по сравнению
с контрольными эмбрионами. Автор предполагает, что инъеци-
рованная аминокислота была использована эмбрионом не только
непосредственно для образования гемоглобина, но и для обра-
зования нуклеиновых кислот и, таким образом, косвенно усилила
эритропоэз.
Данные разных исследователей о сроках начала и окончания
гемопоэза в различных органах куриных эмбрионов отличаются
между собой. Причинами различия в сроках могут быть разно-
породность материала, разные условия инкубации и до,стоверность
методов исследования. Зандрейтер (Sandreuter, 1951) показал,
что у куриного эмбриона незначительный эритропоэз, кроме
желточного мешца, происходит с 7-го до 9-го дня в печени и с 5-го
до 9-го — в селезенке; с 15-го дня инкубации эритропоэз на-
9 В. В. Ролышк
129
пинается в костном мозгу (рис. 27) — следовательно, раньше,
чем он кончается в желточном мешке, что создает непрерывность
эритропоэза. Гемопоэз в костном мозгу куриного эмбриона,
по данным Бродовской (1957), начинается с 14-го, а утиного —
с 15-го дня инкубации. Белянчикова (1953) наблюдала в первую
половину эмбрионального развития кур пород белый леггорн
и ныо-гемпшир последовательную смену трех форм эритроцитов,
обусловленную, по мнению автора, изменением потребности
в кислороде и способе его получения. Примитивному способу
получения кислорода из желтка и небольшой потребности в нем
соответствуют первичные эритробласты, сменяющиеся на 7-й день
более сложно устроенными первичными эритроцитами. Обра-
зование обоих видов красных кровяных клеток происходит внутри
сосудов. Но этот способ образования кровяных клеток не может
обеспечить достаточного количества красной крови для эмбриона,
и на 10-й день в качестве органа гемопоэза начинает функцио-
нировать печень.* Здесь формируются красные кровяные клетки
третьего типа — вторичные эритроциты.
Лемеж и Рыхтер (Lemez a. Rychter, 1956, 1958) вычислили,
что средняя продолжительность жизни примитивных эритроцитов
равна 8 дням, а максимальная — 22 дням, так как через 4 дня
после вылупления их уже нет. Малая длительность жизни эритро-
цитов эмбриона связана с высокой степенью незрелости их в мо-
мент выхода в кровеносное русло. Показано, что митотическая
активность примитивных эритроцитов практически исчезает между
5-м и 6-м днями инкубации, но до этого все фазы митоза одина-
ково представлены. В своей работе с Иосифко Лемеж (Lemez
a. Josifko, 1957) показал, что в кровотворных центрах образуется
около 70.5 млн примитивных эритроцитов, а 38.1 млн — путем
митозов в кровеносном русле. Исследуя цитохимическими ме-
тодами развивающиеся эритроциты куриного эмбриона, Фрейзер
(Fraser, 1963) установил, что химические их изменения идут
наряду с морфологическими. В течение эмбриогенеза отноше-
ние РНК к протеину (в основном хромопротеину) уменьшается,
что отражает изменения в синтезе гемоглобина.
Гемопоэз в онтогенезе скворца (птенцы вылупляются неса-
мостоятельными — длительность эмбрионального развития всего
13 дней) протекает несколько иначе, чем у куриного эмбриона
(Sandreuter, 1951). В отличие от куриного эмбриона, у которого
в печени и селезенке наблюдался слабый эритропоэз, у эмбриона
скворца здесь происходит интенсивное образование эритроцитов,
начинающееся с 8-го дня эмбрионального развития и продол-
жающееся в селезенке до 16-го, а в печени — до 20-го дня пост-
эмбрионального развития. Эта дополнительная способность к кро-
ветворению в печени и селезенке в постэмбриональном развитии
скворца обусловливается краткостью эмбрионального периода.
А в костном мозгу у скворца в отличие от курицы гемопоэз на-
130
чинается только на 1-й день после вылупления. Размер эритро-
цитов у скворца больший, чем у курицы. Интересное сравнение
эритропоэза в онтогенезе многих отрядов птиц (выводковых —
чайки, утки, куры и птенцовых — стрижи, волнистые попугаи,
голуби и воробьиные) провел Шмекель (Schmekel, 1962). Для
сравнения с пресмыкающимися автор изучил эритропоэз у яще-
риц. Выяснено, что у уток эритропоэз в желточном мешке за-
канчивается за 3—8 дней до вылупления; у стрижа, воробьиных
и кур — перед самым вылуплением; у волнистых попугаев и го-
6
Возраст
8 днях 3 8 1216 0 5 10 15 20 25 30
1 1—1—1— I ** —1—1—1—1—1—
Д
M»lllllllllllralllllllllllllllllllllil
Ш rartd мвЛМ
/И
Рис. 27. Время и место образования различных элементов
крови у эмбрионов скворца (Л) и курицы (Б). (По Sandreuter,
1951).
Слева от 0 — дни эмбрионального, справа от 0 — дни постэмбрио-
нального развития. I — в печени; II — в селезенке; III — в костном
мозгу; IV — в желточном мешке. Черным обозначен эритропоэз; пунк-
тиром — гранулопоэз; полосками — лимфопоэз.
лубей продолжается еще 7 дней после вылупления. У всех видов
кроме стрижей и всех воробьиных, эритропоэз в костном мозгу
начинается сразу после прекращения его в желточном мешке.
У воробьиных между эритропоэзом в желточном мешке и в костном
мозгу наблюдается интервал, когда эритропоэз происходит в пе-
чени. Эритропоэз в костном мозгу у птенцовых начинается в пе-
риод вылупления, а у выводковых и ящериц — на 14-й день
эмбрионального развития. Подводя итоги, автор отмечает, что
у настоящих выводковых птиц эритропоэз протекает более при-
митивно, а у воробьиных (самых птенцовых) носит наиболее
совершенный характер.
Данные об изменении красной крови в онтогенезе курицы
и скворца представлены Запдрейтером (Sandreuter, 1951) (табл. 32).
Хошино и Ториу (Hoshino а. Toryu, 1960) удалось подсчитать
количество эритроцитов уже у 3-дневного куриного эмбриона:
9*
131
Таблица 32
Количество эритроцитов и процент гемоглобина в онтогенезе курицы
и скворца
(По Sandreuter, 1951)
Курица Скворец
возраст (в днях) количество эритроцитов (млн в 1 мм3) % гемоглоби- на возраст (в днях) количество эритроцитов (млн в 1 мм3) % гемоглоби- на
Эмбриональное развитие Эмбриональное развитие
10 1.215 9 0.3 —
12 2.381 — 9-10 0.5 —
12 1.793 — 10-11 1.8 —
14 2.263 71 И 1.1 —
14 2.690 — И 1.5 —
16 2.063 68 12 1.5 33
18 1.950 61 Постэмбриональное развитие
18 1.913 — 0 (самка) 1.6 35
20 1.775 — 0 (самец) 1.35 33.5
20 1.651 — 5 (самка) 1.8 46
21 2.925 64 5 (самец) 1.3 50
21 2.588 63 9 (самка) 2.65 66
21 3.219 — 9 (самец) 2.0 75
Постэмбриональное развитие 14 (самка) 2.5 76
2 (курица) 3.550 72 14 (самец) 2.9 75
2 (петух) 2.890 69 30 (самка) 3.7 98
14 (курица) 2.963 67 30 (самец) 4.5 115
14 (петух) 3.365 65 150 (самка) 4.5 98 ’
120 (курица) 3.025 72 150 (самец) 5.05 104
120 (петух) 3.800 79
Взрослые Взрослые
Курица 3.155 69.2 Самка 4.8 105
Петух 4.355 103.4 Самец 4.55 104.5
Примечание. Количество эритроцитов и % гемоглобина у взрослых скворцов
изменяется в течение года. В таблице даны средние величины.
оно было равно 90 000 в 1 мм3 крови. Количество лейкоцитов
было относительно небольшим в течение инкубации, но быстро
увеличивалось и на 6—7-й день после вылупления достигало
уровня взрослой курицы.
Задарновская (1954) отмечает, что в течение эмбрионального
развития наблюдаются два периода (с 8-го по 11-й и с 16-го
по 18-й день) снижения числа эритроцитов, лейкоцитов и гемо-
глобина, совпадающие с замедлением прироста веса и с повыше-
нием смертности эмбрионов. Грундбек (Grundboeck, 1963) наблю-
дал, что с 10-го дня инкубации до 2-го дня после вылупления
132
число кровяных клеток в 1 мм3 крови увеличивается с 1 до 2 млн,
причем красные кровяные элементы составляли 90—97.5%.
Образование гранулоцитов у куриного эмбриона, по данным
Зандрейтера (Sandreuter, 1951), кроме желточного мешка про-
исходит также в печени с 12-го дня (максимум на 14—15-й день),
в селезенке с 11-го (максимум на 14—18-й день) и в костном
мозгу с 14-го дня инкубации. Образование лимфоцитов начи-
нается на 15-й день в селезенке, на 19-й день в печени и костном
мозгу и протекает не интенсивно. В онтогенезе скворца образо-
вание гранулоцитов происходит с 8-го дня эмбрионального раз-
вития в селезенке (до 16-го дня) и в печени (до 20-го дня пост-
эмбрионального периода). Образование лимфоцитов начинается
в печени и селезенке только вскоре после вылупления, а в кост-
ном мозгу — даже с 11-го дня постэмбрионального развития.
Белянчикова (1953) отмечает, что в эмбриональной крови (ку-
риный эмбрион) наблюдается малое количество лейкоцитов,
и объясняет это наличием в белке яйца лизоцима, обладающего
большими бактерицидными свойствами и функционально заме-
няющего лейкоциты. По данным Грундбека (Grundboeck, 1963),
число лейкоцитов заметно увеличивается в периферической крови
ко времени вылупления: число тромбоцитов на 17-й день равно
10 000, на 19-й падает до 500 и к моменту вылупления повы-
шается до 20 000—45 000 в 1 мм3 крови. А в костном мозгу тромбо-
цитов, базофилов, эозинофилов, моноцитов, лимфоцитов и гемо-
цитобластов мало.
Количество тромбоцитов быстро увеличивается со второй
половины инкубации, что совпадает с увеличением свертывае-
мости крови (Задарновская, 1954). Протеины, связанные со свер-
тыванием крови, появляются только на 12—13-й день. По данным
Кейна и Сайзера (Kane a. Sizer, 1953), свертываемость крови
у куриных эмбрионов начинается в это же время. Чтобы опре-
делить, какой же из частей крови, необходимых для ее сверты-
вания, не хватает в крови эмбрионов более раннего возраста,
к ней добавлялись по очереди тромбин, тромбоциты и фибрино-
ген. Кровь 9-дневного эмбриона не свертывается при добавле-
нии любого из этих компонентов; значит, в ней нет ни одного
из них. Кровь 11-дневного свертывается при добавлении тром-
бина или тромбоцитов, но не свертывается при введении фибри-
ногена. Следовательно, фибриноген в это время уже имеется
в крови и неспособность к свертыванию вызвана недостатком
й крови тромбина. Тот факт, что добавление тромбоцитов вы-
зывает все же свертывание, показывает, что протромбин имеется
в крови, но превращения его в тромбин обычно не происхо-
дит до 12—13-дневного возраста. В противоположность этому
Лойда и Лемеж (Lojda a. Lemez, 1957) нашли, что тромбоциты
появляются в крови куриного эмбриона уже на 3—4-й день ин-
кубации. Их протоплазма содержит мукополисахариды, муко-
133
протеины и маленькие капли жира (которых нет в эритроцитах),
но не содержит гемоглобина. Авторы не согласны с мнением
некоторых исследователей, что тромбоциты — это дегенериро-
вавшие эритроциты.
Исследуя протеины сыворотки крови куриного эмбриона,
Нейс и Шехтман (Nace a. Schechtman, 1948) обнаружили, что
по 4-й день включительно сыворотка крови состоит из вител-
лоидного протеина, возможно идентичного с ливетином, а на 5—
6-й день инкубации в ней возникает отличающийся от желтка
материал, который, по-видимому, можно идентифицировать с аль-
бумином сыворотки. Описание собственных исследований и ли-
тературных данных об изменении сыворотки крови в эмбрио-
нальном развитии птиц приводит Шехтман (Schechtman, 1952).
Сыворотка крови эмбрионов ранних стадий развития (3—13 дней
инкубации) производит впечатление водянистой жидкости. У 10-
дневного эмбриона сыворотка содержит всего 40—45% сухого
вещества сыворотки взрослой птицы (28 мгр/мл). С 13-го дня
инкубации до вылупления общее количество сухих веществ в сы-
воротке удваивается. В эмбриональной сыворотке преобладают
липиды: у 10-дневного эмбриона липиды составляют 73% сухих
веществ, перед вылуплением — 63%, а через 9 дней после вы-
лупления — 34%. С 4-го по 8-й день инкубации количество про-
теинов в сыворотке удваивается, затем до 19-го дня оно растет
постепенно, увеличиваясь примерно вдвое за это время, а после
этого быстро возрастает и до 11-го дня после вылупления снова
удваивается. В эмбриональной сыворотке больше липопротеинов,
чем в сыворотке взрослых птиц. В эмбриональной сыворотке
есть компоненты, сходные с антигенами взрослых птиц, а также
специфически эмбриональныё. У 6-дневного эмбриона сильно
возрастает количество антигенов сыворотки. Интересно, что
именно в этот день эмбрионального развития количество дегене-
рирующих эритроцитов достигает максимума. По-видимому, раз-
рушение клеточных элементов является источником значитель-
ного количества протеинов сыворотки.
Общий объем крови куриного эмбриона, по Иошпе-Пурер
с соавторами (Yoshpe-Purer et al., 1953), увеличивается с 1.34 мл
на 11-й день инкубации до 3.4 мл на 18-й, что составляет 42%
и 16.2% веса эмбриона соответственно. Рыхтеру и его соавторам
(Rychter et al., 1955) удалось определить объем циркулирующей
плазмы крови на 2-й день инкубации; ее было 8.5 мм3, а всей
крови — 9.3 мм3; на 13-й день — 1410 и 1890 мм3, а на 18-й —
1860 и 2870 мм3 соответственно. Общее количество гемоглобина
до 10-го дня сравнительно низкое — 32 мг, а к 18-му дню сильно
увеличивается — до 221 мг. По отношению к весу эмбриона
количество плазмы уменьшается с 8-го дня до вылупления на 55%,
а крови — на 50%. За это же время относительное количество
гемоглобина снижается только на 15%. Несколько другие данные
134
получены Коржуевым и Галкиной (1957). Авторы наблюдали,
что у птиц (кур, уток, индеек и голубей) относительное коли-
чество крови на 1 г веса тела во взрослом состоянии и в эмбрио-
нальном периоде одинаково. Сравнивая это со значительным
преобладанием веса сердца и количеством гемоглобина у заро-
дышей млекопитающих со взрослыми формами, авторы приходят
к выводу, что птицы в эмбриональном периоде меньше нуждаются,
в кислороде. Однако не следует забывать о значительно большем
сродстве эмбрионального гемоглобина птиц к кислороду (о чем
уже была речь раньше). Лемеж и Копеци (Lemez а. Кореску,
1962) подсчитали, что со 2-го по 20-й день инкубации через сердце
куриного эмбриона проходит примерно 80 л крови, причем по-
ловина этого количества — в течение последних трех дней ин-
кубации.
В. Физиология сердечно-сосудистой системы
Как уже указывалось, пульсация сердца куриного эм-
бриона начинается со 2-го дня инкубации, а точнее — с 36—44 час.
(Lillie, 1952). Некоторые авторы (Rychter et al., 1955) наблюдали
начало кровотока позже этого срока — только с 50—55 час.
инкубации, другие раньше — с 28 час. (Ebert et al., 1955).
Сердце — это первый орган эмбриона, начинающий функцио-
нировать, причем до того как закончен его морфогенез. Следует
отметить удивительно раннюю функциональную зрелость эмбрио-
нального сердца.
Что же является причиной начала пульсации сердца? Каков
механизм его сокращений? Ведь к началу пульсации сердце
представляет собою эндотелиальную трубку, которая не только
не имеет еще связи с нервной системой, но даже лишена мышеч-
ных элементов. Первые указания на причину возникновения
сердечных сокращений мы находим у К. М. Бэра еще в 1828 г.
Бэр заметил, что для возникновения первого сокращения сердца
необходимо, чтобы в сердечную трубку вошла бесцветная крове-
подобная жидкость: «Сердце . . . медленным движением как бы
втягивает в себя кровь из вен. За этим следует короткое сокра-
щение. Эти движения производят впечатление, как будто бы
восприятие крови является первичным и обусловливающим
процессом, а выталкивание — вторичным (Бэр, 1828). Это
предположение было в~ дальнейшем подтверждено эксперимен-
тальными данными. Мантейфель-Сцойге (Manteuffel-Szoege, 1960)
в опытах на 4-дневном курином эмбрионе показал, что движение
крови по сосудам обусловливается не только сердцем, но и венами.
Шепсенвол и Брон (Szepsenwol a. Bron, 1935) установили,
что иннервация сердца куриного и утиного эмбрионов начинается
значительно раньше, чем это представлялось ранее, а именно
веточки вагуса достигают сердца через 68 час. инкубации у ку-
135
риного и через 88 час. у утиного эмбриона. Элементы симпати-
ческой нервной системы в это время отсутствуют, и авторы пред-
полагают, что эти первые веточки вагуса являются чувствитель-
ными, а не двигательными. Симпатические же элементы входят
в сердце, по данным авторов, вместе с двигательными волок-
нами вагуса у куриного эмбриона на 80—84-м часу, у утиного —
да 98-м часу инкубации, тогда, когда происходит анастомоз
между грудными ганглиями вагуса и верхними симпатическими
ганглиями.
На 6—8-й день развития эмбриональное кровообращение
усложняется в связи с изменением строения сердца и основных
сосудов. Янцовская (1949) считает, что в развитии сердечной
деятельности у куриного эмбриона можно установить следующие
стадии. 1) Стадия пульсации эндотелиальной трубки (со 2-го
до 8-го дня), когда в эмбрионе в целом и в сердце в частности
еще не окончательно сформированы нервно-мышечные элементы.
На этой стадии пульсация сердца зависит в основном от физи-
ческих (температура и давление жидкости в эндотелиальной
трубке) и химических (ионно-солевых) условий среды. Раздра-
жение сердца в. начале этой стадии вызывает быструю его гибель.
Ритм пульса неравномерный. 2) Стадия мышечно-гуморальной,
в том числе гормональной, регуляции сердечной деятельности
(с 8-го дня до конца инкубации). Дифференцированная реакция
предсердий и желудочков на местное раздражение их электри-
ческим током наблюдалась также с 8-го дня. С 9—11-го дня сер-
дечный ритм становится более равномерным. К моменту вылупле-
ния нервный аппарат регулирования сердечной деятельности
еще не созрел функционально, и лишь к 6-му дню постэмбриональ-
ной жизни возбудимость вагуса достигает уровня взрослой птицы.
Как показал Джонс (Jones, 1958), эффект ацетилхолина и адрена-
лина на сердце куриного эмбриона связан с непосредственным
влиянием их на мышцу, так как он одинаков при воздействии
на сердце до иннервации и на сердце, лишенном иннервации
экспериментальным путем. Проверяя влияние различных фарма-
кологических веществ на электрокардиограмму куриного эмбриона,
Хама (Наша, 1959) обнаружил, что наибольший эффект имеет
адреналин, и пришел к выводу, что симпатические нервы оказы-
вают влияние на сердце раньше парасимпатических.
Путем разрушения различных участков центральной нервной
системы (ЦНС) куриного эмбриона Богданов (1960) показал,
что до 16-го дня инкубации сердечная деятельность не подвер-
жена влияниям со стороны ЦНС. Но начиная с 16-го дня вклю-
чается воздействие спинного мозга, так как разрушение его
вызывает резкое урежение пульсации. Автор предполагает,
что это происходит за счет функционального созревания сим-
патических клеток боковых рогов (см. рис. 32, 7) спинного мозга.
Удаление вышележащих отделов ЦНС не оказывало влияния
136
на ритм сердечной деятельности куриного эмбриона. Сравнивая
развитие сердечной деятельности в онтогенезе кур и голубей,
Богданов (1961) указывает, что у куриных эмбрионов уже с 3-го дня
развития имеются ритмичные сердечные сокращения с частотой 90—
100 ударов в 1 мин. [в противоположность уже упоминавшимся
данным Янковской (1949)]. На 6-й день пульсация достигает
160 ударов в 1 мин. и, чуть повышаясь (до 180 ударов) ко вре-
мени вылупления, затем резко возрастает (до 330 ударов)
на 4-й день постэмбрионального развития. Начало пульсации
сердца у голубиных эмбрионов автором не было замечено, но
с 7-го до 12-го дня ритм пульса возрастал со 115 до 150 в 1 мин. и
скачком до 370 ударов на 4-й день постэмбрионального развития.
В ответ на экстероцептивные раздражители сердечный ритм
изменяется только с 17-го дня у куриных эмбрионов и совсем
не изменяется в эмбриогенезе у голубиных. В отличие от кури-
ного у голубиного эмбриона разрушение участков ЦНС не ока-
зывает влияния на сердечную деятельность. Подводя итоги,
Богданов отмечает, что формирование рефлекторной регуляции
сердечной деятельности у голубей происходит в весьма короткие
сроки, несмотря на растянутость периода физиологического
созревания (птенцовые) по сравнению с курами (выводковые).
В отличие от данных Богданова (1960) о сильном увеличении
частоты пульса у цыплят только в первые дни после вылупления
Блинкова (1962) обнаружила это учащение пульса уже начиная
с 19-го дня инкубации, объясняя это расхождение данных улучше-
нием методики изучения пульсации (хроническое вживление
электродов). Применяя эту методику, автор наблюдала безусловно-
рефлекторные сдвиги сердечной деятельности (на звуковые раз-
дражители, вибрацию, аммиак и электрический ток) уже с 6-го дня
эмбрионального развития, отличающиеся, однако, от более позд-
них рефлексов кратковременностью возбуждения и быстрым
исчерпыванием их приспособительных возможностей, что харак-
терно для всякой функции в процессе становления. Автор отме-
чает, что незадолго перед вылуплением нервные центры, регули-
рующие функцию сердца, созревают и становятся подготовлен-
ными к регуляции этой функции вылупившегося цыпленка;
в ответ на внешние воздействия имеется адекватная реакция
учащения пульсации.
Ритм пульсации сердца куриных эмбрионов имеет большие
индивидуальные колебания, что и отражается в наблюдаемых
разными авторами данных о количестве сокращений сердца
в минуту (рис. 28). По данным Фрэнсис (Francis, 1962), средний
пульс однодневного цыпленка равен 295 ± 3.1 в 1 мин. с индиви-
дуальными колебаниями от 174 до 435. У петушков пульс в сред-
нем на 6 ударов в минуту меньше, чем у курочки, но в связи
с большой вариабельностью этой величины различие недостоверно.
Противоположные данные приводит Романов (см. стр. 254).
137
Электрокардиограмма сердечной деятельности куриного эмб-
риона была записана многими исследователями. Мы приведем
здесь только несколько работ по этому вопросу. Бауцман, Дункер
и Шредер (Bautzmann et al., 1954) при помощи платиновых элект-
родов и катодного осциллографа записали ЭКГ сердца начиная
с 54 час. развития куриного эмбриона. Вначале авторы наблю-
дали медленные, неравномерные электроколебания, достигающие
к 94 час. инкубации 140 в минуту и становящиеся к этому вре-
мени регулярными. Разногласия^в^сроках^ установления постоян-
Рис. 28. Изменения в темпе пульсации у развиваю-
щегося куриного эмбриона. (По Romanoff, 1960; на
основании данных ряда авторов).
Точки — вариации; кружки — средние арифметические. Вни-
зу — сравнение пульсации у двух других видов куриных:
фазана (1) и перепела (2).
ного ритма пульсации, по данным разных авторов, вероятно,
обусловлены различием техники и условий экспериментов. Мозен
и соавторы (Mohsen et al., 1962) записали ЭКГ с 46 до 96 час.
развития куриного эмбриона и показали, что в процессе превра-
щения развивающегося сердца из простой трубки в сложный
орган ЭКГ хотя и изменяется, но почти с самого начала имеет
сложную конфигурацию (4 волны разного направления и дли-
тельности и зубец). Пафф и Буцек (Paff a. Boucek, 1962) провели
одновременную запись ЭКГ и миограммы изолированных пред-
сердия, желудочка и артериального конуса 72-часового куриного
эмбриона, помещенных на время измерения в плазму крови.
Время между началом электрического эффекта и механическим
ответом практически одинаково для каждой из 3 частей сердца
и равно 0.034 сек. ЭКГ разных отделов сердца отличаются друг
от друга до того, как появляются морфологические отличия
138
сократительной системы. Высота миограммы меньше всего у пред-
сердия и больше всего у желудочка.
Частота пульсации в большой мере зависит от температуры
окружающей среды. Мантейфель-Сцойге (Manteufel-Szoege, 1960)
указывает, что с повышением окружающей температуры умень-
шается вязкость крови и, следовательно, трение ее о стенки
сосудов. При охлаждении яйца пульс урежается, и при 10° на-
ступает полная остановка сердца; если же повысить окружаю-
щую температуру, то сердце снова начинает пульсировать.
Г. Лимфатическая система
Данных о морфологии и физиологии лимфатической
системы у птичьего эмбриона очень мало.
Лимфатическая система куриного эмбриона развивается срав-
нительно поздно и появляется на 5—6-й дни инкубации двумя
центрами: один в области таза и второй (более крупный) — у ос-
нования шеи. Они соединяются лимфатическим каналом, кото-
рый образуется путем слияния в мезенхиме лимфатических по-
лостей. Лимфатические капилляры выстланы эндотелием и закан-
чиваются слепо. Все лимфатические капилляры сливаются в лим-
фатические сосуды, которые впадают в яремный лимфатический
мешок, а оттуда — в венозную систему, через грудной проток,
в месте слияния подключичной и яремной вен. Лимфатические
сосуды возвращают часть плазмы крови, прошедшей через стенки
кровеносных сосудов и омывшей клетки тканей тела. Наиболее
крупные лимфатические сосуды имеют клапаны, предотвращаю-
щие обратный ток жидкости.
Путем слияния капилляров лимфатических сплетений обра-
зуются расширения лимфатических сосудов — лимфатические
мешки, которые в свою очередь превращаются в лимфатические
железы. Они состоят из скоплений лимфоидной ткани, продуци-
рующей лимфоциты. У птиц в отличие от млекопитающих очень
мало лимфатических желез. Обычно имеются две пары лимфати-
ческих желез — тазовые и яремные, но у взрослых кур послед-
них нет.
В области пояса задних конечностей расположены два малень-
ких лимфатических сердца, помогающих движению лимфы по лим-
фатическим сосудам. В эмбриональном периоде их роль в основ-
ном заключается в дренаже лимфы из аллантоиса. У многих
птиц лимфатические сердца имеются только в эмбриональном
периоде. Интересно отметить, что у наиболее древних птиц —
страусов — лимфатические сердца особенно хорошо развиты.
У кур они функционируют только в эмбриональном и начале
постэмбрионального периода (Romanoff, 1960). Сокращения лим-
фатических сердец начинаются у 6—6.5-дневных куриных эмбри-
139
онов и связаны вначале со спазматическими движениями
хвоста, но у 10—13-дневных сокращения лимфатических сердец
уже делаются независимыми от этих движений и происходят 4—
8 раз в 1 мин.
Ток лимфы начинается с 8-го дня и независим от тока крови.
Лимфа состоит из плазмы и лимфоцитов, свободно проходящих
через тонкие эндотелиальные стенки лимфатических сосудов.
Жидкость, проникающая в межклеточные пространства из кро-
веносных капилляров и клеток тканей, продукты их обмена
веществ, а также питательные вещества из кишечника, поступаю-
щие через лимфатические сосуды кишечного тракта, — все это
составляет плазму лимфы. Лимфа — это истинная внутренняя
среда организма, так как она в отличие от крови находится
не в замкнутой системе сосудов, а непосредственно соприкасается
с клетками тканей.
Лимфоциты, кроме лимфатических желез, образуются также
в лимфоидных органах. Одним из самых крупных лимфоидных
органов является селезенка, отличающаяся от лимфатических
желез тем, что она не имеет непосредственного сообщения с лим-
фатическими сосудами, а обильно снабжается только кровью.
Селезенка возникает в виде отпочкования стенки брюшной по-
лости. У 4-дневного эмбриона это утолщение брюшины уже за-
метно, а затем клетки его быстро растут, и у 6-дневного эмбриона
селезенка представляет собою заметное выпячивание, находя-
щееся над желудком. Брюшинные клетки проникают в рядом
лежащую мезенхиму, к которой непосредственно примыкает вена
селезенки.
Тимус, по мнению Романова (Romanoff, 1960), является пер-
вым органом, где начинается лейкоцитопоэз, а по количеству
лейкоцитов — вторым после костного мозга. Начиная с 8-го дня
инкубации в тимусе происходит лимфоцитопоэз, а также обра-
зование эозинофильных гранулоцитов и в небольшой степени —
эритропоэз. По данным Аккермана и Науфа (Ackerman a. Knouff,
1964), эпителиальные клетки тимуса начинают приобретать при-
знаки лимфоцитобластов у 7.5—8-дневных куриных эмбрионов,
а на 10—11-й день инкубации лимфоцитобласты превращаются
в лимфоциты. Авторы подчеркивают сходство образования лимфо-
цитов в тимусе с тем, как они образуются в фабрициевой сумке,
что было установлено ими ранее (Ackerman a. Knouff, 1959).
Фабрициева сумка, как известно, — специфический для птиц
лимфоэпителиальный орган. Она возникает как слепой, отпочко-
вавшийся вырост клоаки и состоит из медулярной части, возни-
кающей из поверхностного эпителия, и кортикальной, происхо-
дящей из мезенхимных и эпителиальных клеток. В обеих частях
фабрициевой сумки происходит лимфоцитопоэз.
Лимфоидный характер носит также и остаток желточного
мешка в постэмбриональном развитии (см. стр. 104). Это слепой
140
выступ примерно на середине тонкой кишки, и его развитие
обычно обратно пропорционально развитию слепой кишки, кото-
рая также является лимфоидно-эпителиальным органом (Де-
ментьев, 1940).
Г Л А В А 9
НЕРВНАЯ СИСТЕМА И ОРГАНЫ ЧУВСТВ
Нервная система начинает развиваться в онтогенезе
животного относительно рано; осуществляемые ею функции
контроля всей деятельности организма и координации ее с внеш-
ней средой имеют первостепенную важность для организма.
С момента возникновения нервной системы в филогенезе живот-
ных она становится аппаратом, воспринимающим воздействия
раздражителей среды и передающим их другим органам и систе-
мам организма. По мере возрастания этой роли нервной системы
ограничивается способность клеточных элементов организма не-
посредственно реагировать на прямое воздействие среды. В тече-
ние онтогенеза, так же как и в филогенезе, постоянно возрастает
значение опосредованных нервной системой влияний среды на ор-
ганизм. При созревании нервной системы появляются специаль-
ные воспринимающие органы — рецепторы. Признание ведущей
роли за нервной системой в контролировании деятельности ор-
ганизма не только не влечет за собой отрицания значения других
его систем (гормональной, например), а, напротив, предусматри-
вает осуществление взаимодействия между ними и остальными
частями организма.
На основании большого экспериментального материала Ор-
бели (1933) пришел к выводу, что примитивная нервная система
как в онтогенезе, так и в филогенезе является функционально
диффузной. При дальнейшей эволюции центральной нервной
системы создаются условия, ограничивающие свойственное диф-
фузной нервной системе стремление к иррадиации и генерали-
зации каждого возникающего возбуждения. Вслед за диффузной
создается первичная узловая нервная система путем концентра-
ции нервных элементов в более или менее значительные узлы, рас-
пределяющиеся затем сегментарно. Дальнейшая эволюция узло-
вой нервной системы направлена в основном в сторону обособ-
ления узлов высшего порядка, подчиняющих своему влиянию
периферические узлы. Созревание афферентных систем также
приводит к торможению диффузной деятельности нервной
системы.
141
А. Морфология развития нервной системы
и органов чувств
1. Возникновение нервной системы
Нервная система закладывается в виде нервной пла-
стинки (из эктодермы). Края нервной пластинки почти на всем
ее протяжении соединяются к 27—28 час. инкубации, образуя
при этом нервную трубку. Она опускается глубже и вскоре от-
деляется от сомкнувшейся над ней поверхностной эктодермы.
Во время замыкания нервной бороздки в нервную трубку утолщен-
ные ее края (нервные валики) ложатся между погрузившейся
нервной трубкой и поверхностной эктодермой, образуя ганглиоз-
ную пластинку, которая тянется вдоль всего спинного мозга
(рис. 29).
Стенки передней части нервной трубки утолщаются и разде-
ляются на 3 отдела: передний, средний и задний мозг. Из нерас-
ширенной задней части нервной трубки в дальнейшем образуется
спинной мозг. Признаки деления головного мозга на 5 частей,
характерного для взрослой птицы, появляются у 38-часового
эмбриона. При этом передний мозг делится на собственно перед-
ний — концевой (telencephalon) и промежуточный (diencephalon),
а задний — на задний (metencephalon) и продолговатый (myelen-
cephalon). Средний мозг остается неразделенным (mesencephalon)
(рис. 30). У 29—30-часовых куриных эмбрионов боковые стенки
переднего мозга выпячиваются в виде карманов, образуя первич-
ные глазные пузырьки. Слуховые плакоды (см. «Орган слуха
и равновесия») впервые появляются на 25-м часу инкубации.
На 38-м часу эти утолщения эктодермы образуют слуховые пу-
зырьки, вначале открытые наружу, а затем закрывающиеся
и отделяющиеся от поверхностной эктодермы.
2. Развитие спинного мозга
К 50—55 час. инкубации боковые стенки спинномозговой
части нервной трубки начинают утолщаться и просвет ее стано-
вится щелевидным. Ганглиозная пластинка разбивается сегмен-
Рпс. 29. Формирование и дифференцировка нервной трубки и ганглиозной
пластинки (поперечные разрезы на последовательных стадиях). (По Кнорре,
1959).
А — нервная пластинка; Б — нервный желобок и нервные валики* В — нервная трубка,
ганглиозная пластинка и начало выселения клеток из ганглиозной пластинки в вентраль-
ном направлении; Г — начало дифференцировки спинного мозга и формирование спи-
нальных ганглиев. 1 — нервный желобок; 2 — нервные валики; 3 — кожная эктодерма;
4 — хорда; 5 — мезодерма (сомиты); 6 — ганглиозная пластинка; 7 — нервная трубка;
8 — мезенхима; 9 — зачаток спинного мозга; 10 — зачаток спинального ганглия; 11
и 12—дорсальный и вентральный корешки спинномозговых нервов; 13— смешанный
нерв; 14 — эпендимный слой; 15 — плащевой слой; 16 — краевая вуаль; 17 — белая
соединительная ветвь; 18 — зачаток ганглия симпатического пограничного ствола;
19 — эпителий целбма; 20 — аорта.
142
Рис. 30. Передняя часть 38—39-часового куриного эмбриона
(вид со спины). (По Patten, 1951).
1 — сегмент; 2 — продолговатый мозг; 3 — нервный гребень голов-
ного мозга в районе VII и VIII ганглиев (За — в районе V ганглия);
4 — задний мозг; 5 — брюшные корешки аорты; 6 — средний мозг;
7 — воронка в дне промежуточного мозга; 8 — эктодерма; 9 — пе-
редний мозг; 10 — глазной пузырек; 11 — аортная дуга; 12 — хорда;
13 — граница передней кишки; 14 — артериальная луковица; 15—же-
лудочек сердца; 16 — слуховая ямка; 17 — предсердие; 18 — желточ-
но-брыжеечная вена.
тально на группы клеток, из которых образуются спинномозговые
ганглии, а в области головы — ганглии головных нервов (чувст-
вительных нервов и чувствительных ветвей смешанных нервов).
Штрудель (Strudel, 1953) показал, что спинномозговые ганглии
развиваются исключительно за счет элементов ганглиозной пла-
стинки и способны к дифференциации даже после эксперимен-
тального удаления нервной трубки.
По данным Гамбургера (Hamburger, 1948), митотическая
активность клеток спинного мозга оканчивается в основном
в 8.5 дней инкубации, а затем размножение клеток постепенно
замедляется и оканчивается перед вылуплением цыпленка.
Клетки нервной трубки вначале однородны (называются
медуллобластами) и расположены в виде многорядного эпителия,
т. е. ядра лежат на разных уровнях, а клетки своими концами
доходят с одной стороны до наружного края нервной трубки,
а с другой — до ее канала. Затем клетки дифференцируются
на два типа: спонгиобласты, образующие в дальнейшем нейроглию
(опорную ткань нервной системы), и нейробласты, превращаю-
щиеся в нейроны. Клетки ганглиозной пластинки — ганглио-
бласты, — также дифференцируются на нейробласты и клетки
нейроглии. Дифференцировка эмбриональных нервных клеток
и превращение их в нейроны и клетки нейроглии происходит
с 3-го по 7-й день развития куриного эмбриона.
Нервные клетки спинномозговых ганглиев (чувствительные
нейроны) имеют веретенообразную форму и образуют два осево-
цилиндрических отростка. Один отросток растет к нервной трубке
и входит в ее спинной рог, разветвляясь там на две части: в сто-
рону головы и к хвосту. Другой отросток направляется к пери-
ферии, в окружающую мезенхиму. Передача раздражения с во-
локон чувствительных нейронов дендритам двигательных нервных
клеток происходит при помощи бляшкообразных окончаний
(синапсов), образующихся на месте соприкосновения или сближе-
ния тех и других. К 7-му дню развития куриного эмбриона,
т. е. ко времени возникновения первых рефлекторных движений
(как будет видно далее), связи чувствительных клеток ганглия
и двигательных в спинном мозгу оказываются уже образован-
ными.
По данным тщательного гистохимического исследования Ле-
винсона и Лейкиной (1956), двигательные клетки брюшных
рогов спинного мозга куриного эмбриона образуются на 4-й день
в шейном утолщении, а окончательное формирование чувстви-
тельных нейронов в спинальном ганглии начинается только
на 7-й день инкубации. На 7—8-й день вокруг ганглиозных
нервных клеток образуются капсулы из проникающих в ганглий
волокон соединительной ткани. В дальнейшем авторам (1960)
удалось установить тесную связь между морфологическими из-
менениями нервных клеток, их химическим составом и измене-
10 В- В- Рольник
145
нием функции в течение развития куриного эмбриона. Начало
специфической деятельности или ее интенсификации сопрово-
ждается повышением синтеза РНК, что имеет место на 7-й и 19—
21-й день развития. А уменьшение концентрации РНК в двига-
тельных нейронах на 4-й день связано, по предположению
авторов, с тем, что до образования рефлекторной дуги
возбуждение этих нейронов осуществляется в результате дей-
ствия химических веществ непосредственно на тело нервной
клетки.
С моментом замыкания рефлекторной дуги на 7-й день развития
куриного эмбриона связана серьезная биохимическая перестройка
чувствительных и двигательных нейронов: в них появляется
гликоген (причем в малодифференцированных клетках его меньше
чем в дифференцированных), в цитоплазме клеток значи-
тельно увеличивается концентрация гистидина и повышается
активность щелочной фосфатазы. И, наконец, в период перед
вылуплением (19—21-й день) в связи с усилением деятельности
как двигательных, так и чувствительных нейронов в них увели-
чивается концентрация РНК, увеличивается количество глико-
гена, гистидина и тиоловых соединений и возрастает активность
щелочной фосфатазы.
Наблюдая наличие ацетилхолинэстеразы в нейробластах уже
на 4-й день развития куриного эмбриона, Бонишон (Bonichon,
1958) считает, что биохимическая дифференциация нейробластов
предшествует морфологической. Вначале энзим распределен
по всей цитоплазме нервной клетки, позднее он концентри-
руется на ее периферии, а затем мигрирует по дендритам к си-
напсам.
Серое вещество спинного мозга, состоящее из нейронов и кле-
ток нейроглии, особенно сильно растет с 5-го до 9-го дня инкубации
(рис. 31). При этом образуются брюшные рога с двигатель-
ными нейронами и спинные — с чувствительными нервными клет-
ками. Нарастание белого вещества, состоящего из нервных
волокон, одевающихся в дальнейшем миелиновой оболочкой,
что придает им белый цвет, идет с той же быстротой, что и
серого.
На 4-й день развития куриного эмбриона на брюшной стороне
спинного мозга образуется спинномозговая передняя комиссура,
соединяющая его правую и левую стороны. Нейроны с осево-
цилиндрическими отростками, разветвляющимися в сером ве-
ществе той же стороны спинного мозга, находятся в спинном
роге серого вещества и развиваются сравнительно поздно, при-
мерно на 16-й день инкубации.
У 8—10-дневного куриного эмбриона спинной мозг и его
производные сильно усложняются по сравнению с предыдущим
периодом. Размножение клеток в спинном мозгу в это время
почти заканчивается, а это означает, что у 8-дневного эмбриона
146
уже имеются ‘ практически все потенциальные нейроны, что и
в спинном мозгу взрослой птицы. Различные группы клеток миг-
рировали к их конечному местонахождению и образовали основ-
ные нервные центры. Периферические нервы уже выросли и
установили предварительные связи с ненервными структурами,
а внутрицентральные пути либо образовали между собой специ-
фические связи при помощи синапсов, либо находятся в процессе
Рис. 31. Поперечный разрез спинного мозга 9-днев-
ного куриного эмбриона. (По Lillie, 1952).
1 — ганглий; 2а и 26 — спинной и брюшной корешки спин-
номозгового нерва; з — белое вещество спинного мозга;
4 — серое вещество спинного мозга; 5—нейробласты брюш-
ного корешка (двигательного).
их образования. К концу инкубации строение спинного мозга
в значительной мере приближается к строению его у взрослой
птицы.
Начиная с 7.5 дней инкубации в нервной трубке у выхода
корешков седалищного нерва (по Watterson, 1949) собираются
клетки, образующие гликогенное тело, сначала (до 15-го дня)
путем деления клеток, а затем — накопления гликогена в каждой
клетке. Гликогенное тело имеет яйцевидную форму и является
специфическим образованием, присущим только птицам. Смит
и Гейгер (Smith a. Geiger, 1961) предполагают, что гликогенное
тело птиц является запасным источником энергии для обеспече-
ния нормального метаболизма центральной нервной системы
в тяжелых условиях полета, когда глюкозы в крови становится
недостаточно для нормального осуществления функций нервной
системы.
10*
147
3. Развитие головного мозга
В начале 3-го дня инкубации отмечается большой рост
зоны переднего мозга. В середине дорсальной стенки промежу-
точного мозга появляется выпячивание, из которого в будущем
образуется эпифиз. С брюшной стороны, под промежуточным
мозгом, образуется впячивание эпителия глотки, которое, уг-
лубляясь, теряет связь с поверхностной эктодермой и, наоборот,
связывается с воронкой в дне промежуточного мозга, образуя
гипофиз. Перетяжки, отделяющие средний мозг от промежуточ-
ного и заднего мозга, несколько увеличиваются. Крыша заднего
мозга начинает утолщаться, и он более резко отделяется от про-
долговатого. К концу 3-го дня переднебоковые стенки переднего
мозга выгибаются наружу, образуя пару пузырьков (будущие
большие полушария) с полостями внутри (1-й и 2-й желудочки).
Промежуточный мозг несколько удлиняется; полость внутри
него —- 3-й желудочек головного мозга. Стенки и дно среднего
мозга утолщаются. Задний мозг за это время мало изменяется.
Дорсальная сторона продолговатого мозга утончается, и на ней
становятся видны кровеносные сосуды, образующие сосудистое
сплетение.
Анатомически строение головного мозга 8-дневного куриного
эмбриона уже в значительной мере напоминает структуру мозга
цыпленка в момент вылупления, при этом средняя часть перед-
него мозга меньше выросла, чем большие полушария, которые
сильно увеличились, особенно вперед и вверх. Основание боль-
ших полушарий чрезвычайно утолщено и образует полосатые
тела, а верхние и боковые стенки остались относительно тонкими,
образуя крышу больших полушарий. Полость 1-го и 2-го желу-
дочков сузилась. Обонятельная часть полушарий на 8-й день
еще плохо отграничена. В средней части переднего мозга попе-
речно расположена передняя комиссура, связывающая между
собой правую и левую части основания больших полушарий.
Крыша переднего мозга в середине 5-го дня образует мешко-
образный выгиб — так называемый парафиз; далее кзади она
делает изгиб, вниз, переходя в крышу промежуточного мозга.
В эпителиальной стенке сверху этого изгиба образуется сосу-
дистое сплетение (так называемое переднее) 3-го желудочка.
Подробно исследовав развитие промежуточного мозга кури-
ного эмбриона, Куленбек (Kuhlenbeck, 1937) показал, что уже
в начале 5-го дня развития в промежуточном мозгу имеются 4 чет-
кие продольные зоны, разделенные бороздками: эпиталамус,
дорсальный таламус, вентральный таламус и гипоталамус. Эта
дифференциация промежуточного мозга на начальных стадиях
развития куриного эмбриона, по мнению автора, имеет большое
сходство с его структурой у взрослых рептилий. С 4-го до 8-го дня
в промежуточном мозгу происходят заметные изменения. Воко-
148
вые стенки его сильно утолщаются, образуя зрительные бугры.
На дне промежуточного мозга находится полный перекрест
зрительных нервов — хиазма. Сильно увеличились связанные
с промежуточным мозгом железы внутренней секреции — эпифиз
и гипофиз. Эпифиз вырос в длинную, узкую трубку, расширенную
на конце и с многочисленным количеством полых почек. Отшну-
ровавшееся впячивание глотки становится передней долей гипо-
физа и к 8-му дню превращается в массу трубочек, заключенных
в мезенхиматозную оболочку. Только на 12-й день эмбриональ-
ного развития прекращается сообщение между полостью рта
и полостью гипофиза.
Средний мозг также претерпевает к, 8-му дню значительные
изменения. Стенки его сильно утолщились и выгнулись в сторону,
образовав зрительные доли, а в желудочек среднего мозга откры-
ваются полости зрительных долей.
Верхняя часть заднего мозга сильно разрастается, образуя
мозжечок в виде утолщения, нависающего над 4-м желудочком.
Около 8-го дня мозжечок разделяется поперечной перетяжкой
на два парных выступа и, утолщаясь, образует складки — начало
мозжечковых борозд. До 16-го дня развития мозжечок, несмотря
на свой непрерывный рост, все еще лежит позади зрительных
долей. После этого мозжечок значительно разрастается, особенно
по направлению вперед. К моменту вылупления он почти сходится
с большими полушариями и принимает форму, характерную
для мозжечка взрослой птицы. С другими отделами мозга моз-
жечок связан посредством врастающих в него крупных пучков
волокон, так называемых ножек мозжечка. Баффони и Д’Анкона
(Baffoni et D’Ancona, 1958) выяснили, что морфогенез мозжечка
начинается одновременно у птиц выводкового (куры) и птенцового
(голуби) типа развития, но у кур он осуществляется за более
короткий срок (12 дней) и оканчивается к моменту вылупления,
а у голубей продолжается 28 дней и завершается только
на 20-й день после вылупления. Полная дифференциация всех
или почти всех нейронов мозжечка совпадает с наступлением
способности к вполне самостоятельному передвижению.
Продолговатый мозг по своему строению сходен со спинным
и не особенно четко от него отграничен. Крыша продолговатого
мозга не имеет нервных элементов и состоит из эпителия, прони-
занного кровеносными сосудами. Это сосудистое сплетение сви-
сает в полость 4-го желудочка.
4. Периферическая нервная система
Периферическая нервная система у 8-дневного эмбриона,
как и у взрослой птицы, состоит из 12 пар головных, 38 пар
спинномозговых нервов и вегетативной нервной системы.
149
Формирование спинномозговых нервов начинается на 4-й день
развития куриного эмбриона. Чувствительные волокна возни-
кают из биполярных нервных клеток спинномозговых ганглиев,
один отросток которых направлен на периферию, а другой вхо-
дит в спинной корешок спинного мозга. Двигательные волокна
являются осево-цилиндрическими отростками нервных клеток
Huettner, 1947).
А — соматические чувствительные волокна; Б — соматические двига-
тельные волокна; В — внутренностные чувствительные волокна; Г — вну-
тренностные двигательные волокна. 1 — спинной мозг; 2 — спинной
корешок нерва; з — брюшной корешок; 4 — спинномозговой ганглий;
5 — спинная ветвь нерва; 6 — брюшная ветвь; 7 — соединительная ветвь
с симпатической нервной системой; 8 — симпатический ганглий; 9один
из органов внутренностей. Стрелки — направление нервного импульса.
брюшной части спинного мозга и выходят из него в виде брюш-
ного корешка. Брюшной и спинной корешки спинномозгового
нерва объединены, а затем почти сразу же по выходе из спинного
мозга разделяются обычно на 3 ветви (каждая имеет чувстви-
тельные и двигательные волокна): спинную, брюшную и внутрен-
ностную, которая соединяется с симпатической нервной системой
(рис. 32). Брюшная ветвь идет в межреберном пространстве,
а спинная контактируется с ганглием и разветвляется в спинной
160
мускулатуре и коже. 14-й, 15-й и 16-й нервы имеют необычно
большие ганглии — это основные нервы бронхиального сплете-
ния, с 23-го по 29-й иннервируют лапы птицы, с 30-го по 34-й —
район клоаки, а с 35-го по 38-й — хвост. Хвостовые нервы не имеют
внутренностной ветви.
Головные нервы в отличие от спинномозговых сильно разли-
чаются между собой, и ход их эмбрионального развития, несмотря
на сравнительно большое количество исследований, все еще не сов-
сем ясен. Большая работа по изучению развития головных нер-
вов у птиц была проведена Белоголовым (1908). Не имея возмож-
ности подробно остановиться на развитии головных нервов,
имеющих большие различия как в происхождении, так и в строе-
нии, приведем только самые основные сведения о них. Обонятель-
ный нерв (I) отличается от других головных нервов тем, что
состоит из волокон, являющихся отростками рецепторных обо-
нятельных клеток (подробнее стр. 164—165). Зрительный нерв (II)
образуется путем врастания волокон нервных клеток ганглиоз-
ного слоя сетчатки в стенку глазного стебелька, который остается
затем в виде рудимента. У птиц имеет место полный перекрест
зрительных нервов в отличие от млекопитающих (неполный
перекрест) под основанием промежуточного мозга. Глазодвига-
тельный (III), блоковый (IV) и отводящий (VI) нервы возникают
на 3—4-й день из среднего и продолговатого мозга и иннервируют
мышцы глаза. Тройничный (V) нерв смешанный, т. е. имеет дви-
гательную и чувствительную части, и состоит из двух нервов:
а) глубокого глазничного (чисто чувствительного) и б) собственно
тройничного (смешанного). В течение 4-го дня периферические
отростки чувствительных ветвей обоих нервов вырастают и на-
правляются к глазному пузырьку и к углу рта. В этот же день
двигательная ветвь собственно тройничного нерва возникает
из стенки мозга и направляется к мускулатуре нижней челюсти.
Лицевой (VII) и слуховой (VIII) нервы вначале имеют общий
ганглий, но в течение 4-го дня эмбрионального развития они
отделяются друг от друга. Лицевой нерв имеет, кроме того, и
двигательную ветвь, иннервирующую поверхностную шейную
мускулатуру и мышцу, опускающую нижнюю челюсть. (О слу-
ховом нерве подробнее см. стр. 162). Языкоглоточный (IX) нерв
является в основном чувствительным (вкусовым) нервом, но имеет
и двигательные волокна, иннервирующие мускулатуру глотки.
В течение 4-го дня инкубации осево-цилиндрические отростки
ганглиозных нервных клеток языкоглоточного нерва входят
в основание продолговатого мозга. Блуждающий (X) нерв —
самый большой и самый сложный из головных нервов. Центрально
направленные отростки нервных клеток яремного ганглия входят
большим количеством мелких корешков в продолговатый мозг.
Периферические же отростки сливаются в один нерв, который
проходит через ганглий нодозум (пучковый), из него две ветви
161
идут в 4-ю и 5-ю жаберные дуги, а основной ствол нерва продол-
жается в виде кишечной ветви, иннервирующей сердце, легкие
и желудок. Добавочный (XI) нерв образован обособившимися
спинными корешками блуждающего нерва. Это чисто двигатель-
ный нерв, иннервирующий вместе с подъязычным (XII) мышцы
шеи. Подъязычный нерв образуется в начале 4-го дня инку-
бации.
Картина связи V, VII, IX и X головных нервов с жаберными
дугами, так хорошо заметная у куриного эмбриона на 5-й день
инкубации, очень сходна с той, что имеется в течение всей жизни
у многих рыб (Lillie, 1952).
Вегетативную нервную систему называют также автономной
нервной системой за ее относительную независимость от ЦНС.
Для вегетативной нервной системы характерно наличие сплете-
ний, обычно содержащих клеточные элементы в органах, и то,
что ее нервы имеются во всех органах и тканях, в том числе и в ске-
летных мышцах. Соматическая нервная система снабжает своими
окончаниями только скелетные мышцы.
Вегетативная нервная система объединяет симпатическую и
парасимпатическую. Симпатические нервы выходят из брюшных
корешков грудных и поясничных сегментов спинного мозга.
Парасимпатическую же систему составляют волокна, прохо-
дящие в составе III, VII, IX и X головных нервов и тазового
спинномозгового нерва. Парасимпатическая система в противо-
положность симпатической имеет значительно бдлыпую локали-
зацию эффектов. Чаще всего влияния симпатических и парасимпа-
тических нервов имеют антагонистический характер, что обеспе-
чивает равновесие и согласованность вегетативных функций
животного. Однако антагонизм функций этих систем не универ-
сален.
Симпатическая нервная система состоит из связанных между
собою 38—39 пар симпатических ганглиев. Осево-цилиндрические
отростки клеток симпатических ганглиев образуют три связи:
а) центральную, с соответствующим корешком спинномозгового
нерва, б) периферическую, с определенной частью первичных
симпатических тяжей, в дальнейшем образующую нервы внутрен-
ностей, и в) продольную, соединяющую все симпатические ганглии
в виде цепи. Развитие симпатической нервной системы представлено
на рис. 33. Вопрос о происхождении симпатической нервной си-
стемы широко дискутировался. В своем обзоре об эмбриогенезе
вегетативной нервной системы позвоночных Кнорре (19496) убе-
дительно доказывает, что теория мезенхимного происхождения
симпатических узлов, принятая ранее, не имеет серьезных осно-
ваний. Автору удалось на последовательных стадиях развития
куриного эмбриона проследить этапы миграции клеток ганглиоз-
ных тяжей к аорте и образование за их счет симпатических зачат-
ков. Установлено, что клетки из ганглиозной пластинки мигрируют
162
Рис. 33. Схема развития брюшной части симпатической нервной системы у куриного эмбриона.
(По Lillie, 1952).
А — симпатическая система на 4-й день инкубации; Б — на 6-й день; В — на 10-й день; 1 — первичный симпатиче-
ский тяж; 2 — вторичный симпатический тяж; з — аортное сплетение; 4 — внутренностное сплетение; 5 — ганг-
лий Ремака; 6 — ганглии и нервы стенок кишечника; 7 — ганглии спинного мозга; 8 — нервная трубка; 9 — хорда*
ю — аорта; 11 — блуждающий нерв; 12 — желудок; 13 — стебелек желточного мешка; 14 — пупочная артерия
(а); 15 — хвостовая а.; 16 — желточно-брыжеечные а.; 17 — брюшнополостная а.; 18 — клоака.
и образуют на 4-й день развития куриного эмбриона первичный
ствол симпатической нервной системы, затем за его счет аорталь-
ное сплетение, потом спланхнические нервы и затем сплетения
и тяжи кишечной стенки. На 6-й день первичный пограничный
ствол в грудном отделе исчезает, так же как и аортальное сплете-
ние в шейной части, и из этого клеточного материала образуются
легочные и сердечные ганглии. К 10-му дню инкубации от зачат-
ков первичных пограничных стволов остается только самая перед-
няя часть и тазовый отдел, которые входят в состав дефини-
тивной симпатической системы. С 6-го дня в грудном и пояс-
ничном отделах возникают зачатки вторичных пограничных
стволов, дифференцирующихся затем в постоянные симпатические
^стволы. В образовании краниальных парасимпатических узлов
главную роль играют чувствительные черепномозговые ганглии.
Автор отмечает, что парасимпатическая иннервация сердца, лег-
ких и средних частей пищеварительного тракта (включая печень
и поджелудочную железу) филогенетически является результатом
дифференцировки одной и той же, первоначально более простой
системы блуждающего нерва. В отличие от собственно симпати-
ческой системы, развивающейся различно у разных классов
позвоночных животных, система блуждающего нерва, и в част-
ности интрамуральные ганглии сердца и кишечника, заклады-
вается и дифференцируется у всех*классов позвоночных однотипно.
Вопреки мнению Кунтца (Kuntz, 1945, цит. по: Кнорре, 19496),
автор считает, что симпатическая иннервация внутренних органов
возникла совершенно независимо от блуждающего нерва. Воз-
никновение двойной антагонистической иннервации в эмбриоге-
незе, как и в эволюции позвоночных, является, по мнению автора,
высшим и сравнительно поздним этапом развития вегетативной
нервной системы. Подтверждают происхождение симпатических
ганглиев из нервной трубки и исследования Хаммонда (Hammond,
1949) и Навара (Nawar, 1956). Исследователи экстирпировали
нервную трубку у 22-сомитных (Хаммонд) и у 4—5-дневных
(Навар) эмбрионов и после этого не находили никаких следов
симпатобластов. Телло (Tello, 1945) также не согласен с теорией
мезенхимного происхождения симпатической нервной системы.
Однако он отрицает миграцию клеток из ганглиев и из нервной
трубки и считает, что метамеризм симпатической системы связан
с метамеризмом сосудистой системы.
Согласно учению Орбели (1948), симпатическая нервная си-
стема осуществляет во всех возбудимых тканях трофическое влия-
ние, регулируя интимные химические превращения и обмен ве-
ществ между тканями и окружающей средой. Желая подчеркнуть,
что это влияние неразрывно связано с изменением свойств тканей
с приспособлением их к наилучшему выполнению функций при
различных условиях существования организма, Орбели назвал
это влияние адаптационно-трофическим.
164
б. Органы чувств
В связи с большим количеством работ за последнее время
по тонкой структуре органов чувств описание их дается нами
более подробно.
Описанию развития органов чувств в эмбриогенезе птиц мы
предпошлем описание общего строения рецепторных клеток. Как
показал Винников (1966), рецепторные клетки всех органов чувств
позвоночных животных, несмотря на ряд отличий, чрезвычайно
сходны по структуре и цитохимической организации. Рецептор-
ные клетки делятся на первичночувствующие (фоторецепторы
сетчатки глаза и обонятельные клетки органа обоняния) и вторич-
ночувствующие (волосковые клетки внутреннего уха и рецептор-
ные клетки органа вкуса). Первичночувствующие клетки снаб-
жены специализированным, воспринимающим раздражение от-
ростком, увенчанным подвижной ресничкой, и центральным
отростком, по которому передается возбуждение в соответствующие
части центральной нервной системы. В ряду позвоночных живот-
ных они являются по существу реликтовыми. Вторичночувствую-
щие рецепторные клетки не имеют ни периферического, ни цен-
трального отростков, воспринимают стимул при помощи волосков
и ресничек и передают возбуждение через синапсы охватывающему
их конечному разветвлению афферентного нервного волокна,
отходящему от соответствующего афферентного нейрона. Ранее
автор (Винников, 1946) показал, что вторичночувствующие клетки
развиваются в эмбриогенезе из плакод (часть эктодермы нервной
пластинки), а потом вторично связываются с нервной системой.
Жгутики или реснички, всегда имеющиеся у рецепторных клеток,
автор считает подвижными универсальными антеннами, воспри-
нимающими раздражения внешней среды. Однако аппарат ре-
цепции в чувствующих клетках разных органов чувств в процессе
эволюции сильно изменяется, усложняется, что обусловливает
возможность восприятия и переработки этих раздражений в виде
специфической информации данного органа чувств. Гистохими-
ческими исследованиями показано, что в синапсах рецепторных
клеток имеется высокая активность холинэстеразы и, следова-
тельно, передача нервного возбуждения здесь осуществляется при
помощи холинэргического механизма (Титова и Винников, 1964).
а. Орган зрения
Зрение является наиболее развитым органом чувств
у птиц. Глаз представляет собой шаровидное образование, по-
крытое многими оболочками (рис. 34). Снаружи внутрь (кроме
передней части глаза) расположены следующие оболочки: склера,
сосудистая, пигментная и сетчатка. Спереди склера продолжается
прозрачной роговицей, а сосудистая — ресничным телом и радуж-
155
A
Рис. 34. Разрез через глаз 8-дневного ку-
риного эмбриона (А) (по Lillie, 1952) и об-
щая схема глаза взрослой птицы (Б) (из
Дементьева, 1940).
1а, 16 — нижнее и верхнее веко; 2 — мигательная
перепонка, или третье веко; з — радужная оболоч-
ка; 4 — роговица; 5а, 56 — передняя и задняя ка-
меры глаза; 6 — ресничное тело; 7 — сосудистая
оболочка; 8а — склера; 8 — костная пластинка
склеры; 9 — сетчатка; 10 — пигментный слой глаз-
ной чаши; 11 — передняя часть сетчатки; 12 — хру-
сталик; 13 — зрительный нерв; 14 — гребень.
ной оболочкой. Под влиянием сокращения мышц радужной обо-
лочки отверстие в ней — зрачок — изменяется в размере. Непосред-
ственно за радужной оболочкой лежит хрусталик, а между ним
и роговицей находится маленькая, заполненная жидкостью, перед-
няя камера глаза. Сзади радужной оболочки и хрусталика глазной
бокал заполнен студнеобразным стекловидным телом.1
Наиболее резкое различие глаза птиц от глаза , млеко-
питающих — это отсутствие кровеносных сосудов сетчатки;
но вместо этого , в глазе птиц имеется специальная сосудистая
структура, выступающая в стекловидное тело — гребень. Другое
отличие — это наличие в сетчатке у птиц двух или даже трех
ямок (fovea) — участков более острого зрения. Эти участки осо-
бенно развиты у хищных птиц. Мышцы ресничного тела и радуж-
ной оболочки поперечнополосатые, а у млекопитающих гладкие.
Склера у птиц и рептилий в своей передней части усилена костными
пластинками. Большинство этих отличий представляет адаптацию
к зрению во время полета и прямо или косвенно обусловливает
более острое зрение птиц по сравнению с млекопитающими. Вслед-
ствие этого птиц называют Augentiere. В связи с тем что у птиц
каждый глаз связан лишь с одной стороной мозга (полный пере-
крест нервов), зрительные восприятия каждого глаза являются
независимыми и бинокулярное зрение у птиц имеет меньшее
значение, чем монокулярное.
Развитие глаза протекает в темноте; глаз как бы защищен от
преждевременного включения функции. Глазные пузырьки, воз-
никшие как выпячивания промежуточного мозга (см. главу 6),
превращаются в настоящие пузырьки с пережатием у основания
к 40—45 час. инкубации. С 50—55 час. происходит значительное
продвижение в развитии глаза. Глазные пузырьки начинают
выпячиваться, образуя двухстенную чашу, а полый стебелек,
связывающий их с мозгом, становится все более узким. Внутрен-
ний слой глазной чаши (первоначально наружная стенка глазного
пузырька) — зачаток сетчатки становится толще, чем наружный,
который является зачатком пигментного слоя, радужной оболочки
и ресничного тела. Глазная чаша имеет отверстие, обращенное
наружу и вниз. Наружная часть становится зрачком, а нижняя,
впоследствии закрывающаяся, называется хороидальной, или
зародышевой, щелью. Закрытие ее тесно связано с развитием
гребня.
Хрусталик возникает отдельно от глазного пузырька в виде
утолщения поверхностной эктодермы у 40-часового куриного
эмбриона. Затем происходит инвагинация этого утолщения, и
у 62—74-часовых эмбрионов хрусталиковый пузырек отделяется
от поверхностной эктодермы. Стенки хрусталикового пузырька
1 Детальное описание строения взрослого глаза позвоночных животных
и его развития приведено в сводке Лопашова и Строевой (1963).
167
утолщаются, и полость его исчезает. Клетки хрусталика перестают
делиться, удлиняются, ядра в них исчезают и становятся волокно-
видными. Хрусталик вылупившегося цыпленка содержит более
500 слоев волокон, а процесс их образования продолжается и
после вылупления. Преципитиновый тест показал наличие про-
теинов взрослого хрусталика в хрусталиковом пузырьке 60-часо-
вого эмбриона (Cate, 1949, цит. по: Romanoff, 1960). Следовательно,
химическая дифференциация хрусталика предшествует морфоло-
гической. Капсула хрусталика (сумка) — это, по-видимому, про-
дукт деятельности его клеток. К ней прикрепляются цинновы
связки, отходящие от ресничного тела. У 4-дневного эмбриона
верхние края глазной чаши сходятся по бокам хрусталика.
Основной частью глаза, воспринимающей зрительные изобра-
жения, является сетчатка, располагающаяся между пигментным
эпителием и стекловидным телом. Сетчатка состоит из 5 слоев:
ганглиозного, внутреннего сетчатого, внутреннего ядерного, на-
ружного сетчатого и наружного ядерного. Свет, проходя сквозь
роговицу, зрачок, хрусталик, стекловидное тело и сетчатку,
отражается от пигментного слоя. К нему направлены отростки
зрительных клеток (ядра их расположены в наружном ядерном
слое), воспринимающие свет: палочки (черно-белое) и колбочки
(цветное изображение). У дневных птиц в сетчатке преобладают
колбочки, у ночных — палочки. Вызванное светом раздражение
передается через аксоны зрительных клеток на синапсы дендри-
тов биполярных нейронов (ядра которых расположены во внутрен-
нем ядерном слое), причем один биполярный нейрон объединяет
до 30 зрительных клеток. Аксоны же биполяров образуют синапсы
с дендритами ганглиозных клеток, аксоны которых растут вдоль
борозды в стенке глазного стебелька по направлению к головному
мозгу и образуют зрительный нерв.
Ямка сетчатки (участок острого зрения) появляется в центре
маленькой утолщенной площадки, которая, по-видимому, яв-
ляется результатом более хорошего кровоснабжения вследствие
раннего утолщения сосудистой оболочки в этом участке. Ямка
образуется в результате радиальной миграции клеток от центра
площадки. В области ямки имеется наибольшее скопление колбочек
и палочек. У птиц, вылупляющихся с закрытыми глазами, утол-
щенная площадка и ямка в ней не начинают развиваться до момента
вылупления, а наиболее быстрая дифференциация ямки происхо-
дит после открытия глаз. Сетчатка птиц значительно толще, чем
у других животных, элементы ее более четко организованы,
а различные чувствительные слои более резко отграничены. У раз-
ных видов птиц имеются различия в структуре сетчатки — в ос-
новном это различное соотношение палочек и колбочек и
положение и глубина ямок, участков острого зрения. В гистоло-
гическом развитии сетчатки куриного эмбриона можно различить
три периода (Weysse a. Burges, 1906, цит. по: Romanoff, 1960):
158
1) размножение клеток со 2-го по 8-й день; 2) клеточная перегруп-
пировка с 8-го по 10-й; 3) окончательная дифференциация после
10-го дня инкубации. Нейробласты и нервные волокна имеются
в сетчатке уже к концу 3-го дня. Палочки и колбочки начинают
дифференцироваться на 10—12-й день. Палочки и колбочки в сет-
чатке куриного эмбриона достигают к концу инкубации той стадии
развития, которая наблюдается у домашнего воробья только
через несколько дней после вылупления (Slonaker, 1921, цит. по:
Romanoff, 1960). Говардовский и Харкеевич (1965) показали,
что у 10-дневного куриного эмбриона будущие зрительные клетки
имеют цилиндрическую форму и плотно прикреплены к пигмент-
ному эпителию, что, по-видимому, играет большую роль в снаб-
жении фоторецепторных клеток витамином А из пигментного
эпителия. Витамин А необходим для построения молекул зритель-
ного пигмента — родопсина — и тех мембранных структур, в ко-
торых он локализуется. На 18—19-й день инкубации структура
рецепторной клетки усложняется в связи с включением в нее
родопсина.
Приведем несколько работ по гистохимии развития сетчатки
куриного эмбриона. Содержание ацетилхолина и холинэстеразная
активность в сетчатке увеличивается равномерно с 8-го до 19-го
дня развития куриного эмбриона, а затем резко возрастает (Lin-
deman, 1947, цит. по: Romanoff, 1960). Активность щелочной
фосфатазы тоже внезапно увеличивается между 17-м и 19-м днями
(Lindeman, 1949, цит. по: Romanoff, 1960). По-видимому, нервные
элемены сетчатки созревают к 19-му дню и способны проводить
импульсы, так как рефлекс сужения зрачка может быть впервые
вызван именно в это время (Lindeman, 1947). Сотрудниками Вин-
никова (Винников, 1966) показано, что: 1) витамин А участвует
в регуляции выхода ионов на свету и в темноте и обусловливает
состояние общего возбуждения рецептора; 2) в сетчатке имеется
сукциноксидазная и цитохромоксидазная активность, указываю-
щая, по-видимому, на транспорт электронов и регенерацию АТФ;
3) активность окислительных энзимов в митохондриях фоторе-
цепторов, как правило, повышается на свету и падает в темноте;
при освещении митохондрии палочек набухают, а митохондрии
колбочек не изменяются.
Гребень глаза сильно варьирует в размере и форме у различных
видов птиц. Это тонкая, темнопигментированная пластинка,
складывающаяся веером и выступающая в стекловидное тело
с вентральной поверхности глаза. Гребень может иметь от 5 до
30 складок и быть коротким или длинным, достигая хрусталика.
Он состоит главным образом из сосудистой сети, поддержанной
пигментированной соединительной тканью (клетки глии). На 6-й
день развития куриного эмбриона гребень выдается в стекловид-
ное тело в виде низкого гребешка вдоль линии слияния стенок
хороидальной щели. Пигмент появляется в нем после 8 дней,
169
а складки начинают образовываться на 9—10-й день инкубации.
У взрослых птиц гребень целиком пронизан капиллярами, а в ос-
новании его лежат артерии и вена. Возможно, что гребень, кроме
снабжения сетчатки питательными веществами, обеспечивает и
защиту ее от сильного света (Romanoff, 1960). Кроме того, в обзоре
Дементьева (1940) указывается, что гребень играет роль в питании
стекловидного тела и, возможно, служит для согревания глаза
и для увеличения остроты зрительных восприятий.
Обращенные вперед края глазной чаши образуют к 8—9-му
дню радужную оболочку (рис. 34), а мышечные волокна начинают
появляться в ней с 7-го дня. Мышцы радужной оболочки: сфинк-
терная (для сокращения зрачка) и радиальная (для его расши-
рения) поперечнополосатые, что обусловливает произвольное сок-
ращение зрачка (особенно проявляется у хищных птиц). Сфинк-
терная мышца появляется на 8—9-й день, а радиальная — на 13—
19-й день. Цвет радужной оболочки обусловлен пигментными
клетками, пигментными тельцами и цветовыми жировыми каплями.
Складки ресничного тела (от 85 до 150 у взрослых экземпляров
разных видов птиц), расположенного в центре радужной оболочки
расходятся радиально от хрусталика по меридианам глаза. Рес-
ничные отростки (центральные окончания складок) выходят
за границу радужной оболочки, а связки (цинновы), отходящие
от желобков между ними, прикрепляются к хрусталиковой сумке.
Первые ресничные отростки появляются на 6—9-й день развития
куриного эмбриона и состоят вначале из направленных к хру-
сталику выростов мезенхимы. У 16—17-дневного куриного эмбри-
она их уже около 90. Ресничное тело секретирует жидкость перед-
ней камеры глаза, благодаря которой осуществляется диффузное
питание хрусталика и роговицы и регулируется внутриглазное
давление.
Зачаточная ресничная мышца появляется на 8-й день в виде
пучка миобластов; ее поперечная полосатость впервые видна
у 11-дневного эмбриона. Сокращение ресничной мышцы, действуя
на склеру, сокращает экваториальный диаметр глазного яблока,
увеличивает внутриглазное давление и толкает хрусталик и пе-
реднюю часть глаза вперед, для близкого видения (Sloneker, 1920,
цит. по: Romanoff, 1960). По другой теории (Hess, 1913, цит. по:
Romanoff, 1960; Lillie, 1952), ресничная мышца действует на рого-
вицу, которая косвенно изменяет напряжение связки гребня и
изменяет форму хрусталика. Дементьев (1940) считает, что ак-
коммодация глаза у птиц происходит всеми тремя способами:
изменением формы хрусталика, формы роговицы и расстояния
между роговицей и хрусталиком.
Эпителий роговицы (конъюктива) происходит из эктодермы,
но подстилающая его часть роговицы происходит из мезенхимы.
Роговица осуществляет две функции: грубой фокусировки глаза
и защитных очков. Та часть глаза куриного эмбриона, где будет
160
образовываться Стекловидное тело, на 4-й день развития Состоит
из волокнистой сетки неопределенного строения (Lenhoss6k, 1911,
цит. по: Romanoff, 1960).
Сосудистая оболочка и склера возникают из мезенхимы,
которая облекает глазную чашу в течение эмбрионального раз-
вития и участвует также в образовании ресничного тела и ро-
говицы. Сосудистая оболочка осуществляет питание глаза. Ран-
нее развитие сосудистой оболочки состоит в конденсации ме-
зенхимы, соприкасающейся с наружным слоем глазной чаши,
что заметно уже у 5-дневного эмбриона. Далее — на 13—14-й
день — размер капиллярной сети сосудистой оболочки увеличи-
вается, а затем снаружи его появляется слой более крупных сосудов;
пигментация ткани начинается на 8-й день. У внутренней поверх-
ности сосудистой оболочки имеется так называемое «зеркальце»
(tapetum lucidum), отражающее свет и раздражающее своим
отблеском сетчатку, что позволяет ей улавливать зрительные
впечатления при слабом освещении. Развитие склеры начинается
одновременно с сосудистой оболочкой, а на 9-й день уже могут
быть различимы в ней ранние белковые косточки.
На 7-й день развития куриного эмбриона спереди глазного
яблока образуется покровная круговая складка с отверстием
в центре, которая превращается в дальнейшем в нижнее и верхнее
веки. Внутри нее одновременно образуется полукруглая складка
со стороны клюва — мигательная перепонка, или третье веко.
У куриного эмбриона веки сомкнуты до 18-го дня инкубации,
а у некоторых птенцовых птиц (воробьиные, дятлы, кукушки и др.)
веки открываются только через несколько дней после вылупления.
б. Орган слуха и равновесия
Внутреннее ухо, или лабиринт, объединяет орган
слуха и равновесия, иннервируемый двумя частями слухо-
вого нерва (нервы преддверия и улитки). Перепончатый лабиринт
заполнен жидкостью (эндолимфой) и окружен костной капсулой,
В эндолимфе имеется множество мелких, твердых образований,
в основном состоящих из кристаллов углекислой извести — ото-
литов. Находящаяся между лабиринтом и слуховой капсулой
полость заполнена перилимфой. Центральная часть лабиринта
состоит из сообщающихся между собой мешков — овального
(utriculus) и круглого (sacculus). Три полых трубки в трех пер-
пендикулярных друг другу плоскостях — полукружные каналы —
начинаются и кончаются в утрикулюсе. На одном конце
каждого канала находится расширение — ампула. От саккулюса
отходит выступ в виде слегка извивающегося спирально улитко-
вого канала, который заканчивается слепым мешком (lagena).
В улитковом канале расположен собственно орган слуха — кор-
11 В. В. Рольник 161
тиев орган. Саккулюс, утрикулюс и полукружные каналы яв-
ляются вестибулярной частью лабиринта (преддверие).
Чувствительная часть внутреннего уха состоит из 8 маленьких
участков волосковых клеток, контактирующих с нервными окон-
чаниями слухового нерва. Такие участки имеются в ампулах
полукружных каналов (чувствительные гребешки), в утрикулюсе
и саккулюсе, где они имеют вид пятен (macula). Есть и дополни-
тельное слуховое пятно (neglecta), расположенное в утрикулюсе.
Кроме того, имеется макула в лагене и группа рецепторных клеток
в кортиевом органе. Гребешок ампулы полукружного канала
имеет вид складки, расположенной поперечно направлению ка-
нала. Складка эта выстлана рецепторным эпителием, состоящим
из опорных и рецепторных клеток покрытых сверху желатино-
образной купулой. Волоски рецепторных клеток проникают
в вещество купулы. Движение головы вызывает передвижение
эндолимфы, которое обусловливает соответствующее отклонение
купулы. Рецепторные клетки гребешка стимулируются движением
эндолимфы по полукружному каналу, что сопровождается реф-
лекторным ответом той части мускулатуры, которая корригирует
положение тела и движение глазных мышц. Над поверхностью
рецепторного эпителия в слуховом пятне утрикулюса распола-
гается тонковолокнистая студенистая отолитовая мембрана, вклю-
чающая отолиты. При изменении положения тела в простран-
стве происходит скольжение отолитов, вызывающих возбуждение
рецепторных клеток. Особого развития у птиц достигают рецеп-
торные клетки кувшинообразной формы. Они охвачены почти
со всех сторон чашевидным окончанием, так что практически вся
плазматическая поверхность клетки может считаться синапти-
ческой. В отличие от млекопитающих, у которых в одной чаше
располагается 1—2 клетки, у птиц их число доходит до 5—7.
Каждая клетка, погруженная в общую чашу, имеет свой наклон.
Вследствие этого, когда птица меняет положение тела во время
полета, стимулируются только те рецепторные клетки, положение
которых при этом меняется, а остальные клетки этой же чаши при-
ходят в состояние торможения. По цитохимическим показателям
рецепторное пятно саккулюса сходно с утрикулярным: клетки
богаты нуклеиновыми кислотами, суммарным белком, функцио-
нальными группами и рядом ферментов, а в синаптической ча-
сти — высокой активностью ацетил холинэстеразы. Слуховое
пятно саккулюса выполняет две роли: 1) отвечает на изменения
организма в гравитационном поле и тем самым является вспомо-
гательным органом по отношению к утрикулюсу и 2) восприни-
мает вибрационные колебания (это основная его роль), в чем,
наоборот, вспомогательным ему явлется утрикулюс. Дополнитель-
ное слуховое пятно, по-видимому, чувствительно к вибрациям.
Краткое описание кортиева органа мы приводим по Винникову
(1966). Кортиев орган расположен по длине улиткового канала,
162
на базилярной мембране, укрепленной при помощи хрящевой
скелетной рамы, и состоит из опорных (снизу) и рецепторных
(над ними) клеток, покрытых тенториальной мембраной. Сверху
улитковый канал ограничивает сосудистая обкладка (tegumentum
vasculosum). Число рецепторных клеток в кортиевом органе
птиц значительно больше (до 40), чем у млекопитающих. В рецеп-
торных клетках кортиева органа обнаружены высокое содержание
РНК, суммарного белка и функциональных белковых групп,
исключительно высокая активность окислительных энзимов (осо-
бенно сукцинатдегидрогеназы) в их митохондриях, а в области
синапсов и волосков под тенториальной мембраной — высокая
активность ацетилхолинэстеразы. К основанию рецепторной клетки
примыкают нервные окончания, вступающие с ней в синаптические
контакты. Скорость ответа на звуковые воздействия у птиц при-
мерно в 10 раз выше, чем у человека.
Развитие внутреннего уха начинается у 29—33-часового кури-
ного эмбриона в виде небольших парных утолщений эктодермы —
слуховых плакод (Romanoff, 1960). На 38-часу эти утолщения
погружаются под эктодерму, образуя слуховые пузырьки, вначале
открытые наружу, а затем закрывающиеся и отделяющиеся от
поверхности головы (около 55 час. инкубации). Пузырек этот
делится на две половины: нижнюю и верхнюю. Из нижней поло-
вины на 7-й день вырастает полый отросток улиткового канала,
а оставшаяся ее часть превращается в саккулюс. Из верхней поло-
вины слухового пузырька образуется утрикулюс и полукружные
каналы. Зачатки полукружных каналов появляются в виде не-
больших выпячиваний на передней поверхности верхней половины
слухового пузырька. Дальнейшее образование полукружных ка-
налов происходит путем роста этих выпуклостей, слипания их
стенок в центре и перфорации их в этом месте. На 5-й день инку-
бации слуховой пузырек отделяется от эктодермы. Улитковый
канал появляется на 6—7-й день (Титова, 1966).
Чувствительные клетки начинают дифференцироваться в на-
чале 2-й недели развития куриного эмбриона. Как показали
Винников и Титова (1956), вычленение клеток с чувствительными
волосками происходит у куриных эмбрионов на 8-й день инкуба-
ции. Дифференциация слуховых гребней почти заканчивается
к 13-му дню. Образование отолитов происходит на 7-й день раз-
вития куриного эмбриона.
Перепончатый лабиринт окружается массой недифференциро-
ванной соединительной ткани и в течение 2-й недели инкубации
из нее образуется хрящ. Клетки, занимающие будущее перилим-
фатическое пространство, расходятся в стороны начиная с 9-го
дня инкубации и резорбируются. Этот процесс заканчивается
на 18-й день. На 9—10-й день будущий костный лабиринт состоит
из эмбриональных хрящевых клеток. Окостенение начинается на
12—-13-й день и заканчивается почти целиком к концу инкубации.
11*
163
Среднее и наружное ухо. Среднее ухо образуется
из первой жаберной щели, после того как она закрывается наружу
на 4-й день развития, продолжая быть связанной с полостью глотки.
Среднее ухо остается соединенным с ротовой полостью и у взро-
слых птиц через евстахиеву трубу. Слуховая косточка (collumella)
дифференцируется на 7—8-и день из мезенхимы и представляет
собой верхний отдел подъязычной дуги. Вокруг слуховой косточки
разрастается барабанная полость, дающая ей более свободную
подвижность при передаче колебаний от барабанной перепонки
к слуховой капсуле. В этом месте слуховая капсула не окостене-
вает и образует перепончатое овальное окно. Кроме того, в стенке
слуховой капсулы остается так называемое круглое окно, отде-
ляющее перилимфатическое пространство (вестибулярную ле-
стницу) от барабанной полости среднего уха и затянутое вторич-
ной барабанной перепонкой. Слуховая косточка одним своим
концом упирается в барабанную перепонку, а другим — в оваль-
ное окно.
Наружный слуховой проход возникает на 6-й день как само-
стоятельное, новое впячивание эктодермы, которое растет по
направлению к среднему уху. Дно этого впячивания становится
барабанной перепонкой.
в. Орган обоняния
Орган обоняния у птиц слабо развит, однако в процессе
развития он формируется так же, как у животных с хорошо
развитым обонянием, и только на последнем этапе эмбриогенеза
рецепторный эпителий замещается железистой тканью. Носовая
полость устроена значительно проще, чем у млекопитающих.
У взрослых птиц она разделена перегородкой на левую и правую
половины. По выполняемой функции носовая полость делится
на респираторную (состоящую из преддверия, промежуточного
и заднего отдела) и обонятельную. Преддверие имеется только
у птиц и образует переднюю (нижнюю) раковину; в средней ра-
ковине обонятельный эпителий имеется лишь в эмбриональном
периоде; верхняя раковина выстлана обонятельным эпителием
и является собственно органом обоняния. Эти раковины ограни-
чены хрящевыми скелетными выростами из стенок носовой поло-
сти. Носо-ротовой канал открывается наружу ноздрями, а внутрь
ротовой полости — двумя отверстиями, расположенными рядом
с медиальной щелью нёба (у птиц расщепленное нёбо) — хоанами.
Обонятельный эпителий состоит из первинночувствующих
и опорных клеток. Обонятельный нерв образуется центральными
отростками обонятельных клеток. Обонятельный эпителий и обо-
нятельный нерв возникают из утолщенных участков эктодермы —
плакод, связанных своей передней частью с мозгом. Отделение
их от нервной пластинки происходит, как показал Гавриленко
164
(1910), благодаря врастанию мезенхимы между плакодами и нерв-
ной пластинкой, и следовательно, закладка органа обоняния свя-
зана с нервной пластинкой.
Закладка обонятельных плакод .происходит у куриного эм-
бриона на 3-й день, а к 7-му дню инкубации образовавшаяся из них
обонятельная ямка сильно увеличивается (Винников и Титова,
1957). В течение 9-го дня процесс развития органа обоняния
ускоряется: а) вход в носовую полость сужается и зарастает
эпителиальными пробками (носовой канал восстанавливается
на 15—18-й день инкубации, и следовательно, в течение этих
7—10 дней ноздри эмбриона закрыты); б) происходит окончатель-
ное деление носовой полости на нижнюю, среднюю и верхнюю
раковины; в) вокруг обонятельных полостей мезенхима уплот-
няется, что является зачатком скелета. К 12-дню инкубации носо-
вая полость оказывается анатомически вполне сформированной.
Процесс образования складок в респираторной части носовой
полости начинается на 13-й день путем врастания соединительной
ткани внутрь респираторного эпителия.
При гистологической обработке клетки обонятельной ямки
4—6-дневного куриного эмбриона можно разделить на два типа:
1) «светлые», с округлыми ядрами, — это будущие обонятельные
клетки; 2) «темные», с мелкими, элипсоидной формы ядрами,—
это будущие опорные клетки. На 5-й день инкубации из стенки
обонятельной ямки в подлежащую соединительную ткань выходит
группа клеток, образующая клеточный тяж по направлению
к мозгу — обонятельный ганглий. На 7-й день в нем появляются
уже хорошо заметные нервные волокна, а на 8-й отдельные
стволики обонятельного нерва, разъединенные соединительной
тканью, достигают мозга. В течение 7-го дня обонятельные клетки
дифференцируются: их периферические отростки имеют на своих
концах небольшие утолщения (будущие булавы), не доходящие
еще до наружной обонятельной мембраны, а центральные дости-
гают появляющейся к этому времени базальной мембраны, соби-
раются в небольшие пучки и прободают ее. После соединения
периферической части анализатора с мозгом на 9-й день различие
между обонятельной и респираторной выстилкой носовой полости
обозначается резче. Происходит дальнейшая дифференциация
обонятельных клеток. Их периферические отростки начинают
различаться по форме на колбочкообразные и палочкообразные
и имеют уже хорошо выраженные обонятельные булавы. По
данным Винникова и Титовой (1949), булавы обонятельных клеток
подвижны, что связано, по-видимому, с процессом адаптации.
Бронштейн (1964) показал, что волоски, увенчивающие обонятель-
ные булавы, непрерывно движутся, и это движение обслуживается
генерируемой их митохондриями энергией АТФ. Обонятельное
ощущение происходит в результате контакта молекулы пахучего
вещества с булавой обонятельной клетки. На 10—12-й день ин-
166
кубации происходит вторичное массовое выселение клеточных
элементов из обонятельной выстилки в подлежащую соединитель-
ную ткань; эти клетки двигаются вдоль нервных волокон обоня-
тельного нерва и пополняют его нейроглию. На 12-й день над
носовыми хрящами видна хорошо развитая, мощная цепочка
скоплений нейронов, состоящая из 20—25 узлов, одетых каждый
в соединительнотканую капсулу. Во второй половине инкубации
развитие органа обоняния приостанавливается и он даже регрес-
сирует. Эти регрессивные изменения резко выявляются на 15-й
день: исчезают рецепторные элементы в средней и сокращаются
в верхней раковине, заменяясь железистыми образованиями,
в результате чего площадь рецепторного слоя резко уменьшается.
Обонятельный нерв во второй половине инкубации практически
не изменяется. Обонятельные луковицы вполне анатомически
развиты на 11—12-й день. Стволики обонятельного нерва соеди-
няются в два мощных нервных ствола, которые прободают решет-
чатую кость и соединяются с обонятельными луковицами голов-
ного мозга, оплетая их со всех сторон своими волокнами.
Б. Физиология нервной системы эмбриона
1. Двигательная активность
Первые систематические исследования по развитию дви-
гательных спонтанных и рефлекторных реакций в эмбриональ-
ном развитии птиц были проведены в 1828 г. известным русским
ученым К. М. Бэром (издание 1950). Он обнаружил у куриного
эмбриона в течение его развития два типа двигательных реакций:
1) возникающие на 6-й день инкубации под влиянием внешних
воздействий (холод, тепло и др.) активные сгибания некоторых
частей тела; 2) возникающие на 7—8-й день и затухающие к концу
инкубации общие маятникообразные движения всего эмбриона,
вызванные ритмическими сокращениями амниона. Более деталь-
ные исследования по этому вопросу были проведены Прейером
(Ргеуег, 1885) путем ежедневных наблюдений поведения эмбриона
внутри яйца при помощи просвечивания его специальным эмбрио-
скопом.
Развитие двигательных реакций у эмбрионов птиц достаточно
полно исследовано Цинг Янг Куо (Kuo, 1932а, 1932b, 1932с, 1939а).
Автор делит период развития куриного эмбриона по наличию
и степени активности на 10 стадий: 1-я стация (29- и 30-часовой
эмбрион2) — наблюдается только пульсация сердца; 2-я (75—90-ча-
2 -Куо обычно наблюдает появление всех форм активности эмбрионов
в более ранний период инкубации, чем это делают другие исследователи.
Возможно, это вызвано разными условиями инкубации или свойствами породы
кур исследуемых эмбрионов.
166
совой эмбрион) — первые активные движения головы, вначале
очень редкие; 3-я (90—100-часовой эмбрион) — впервые заметны
активные движения туловища, на этой же стадии начинается реф-
лекторное сокращение мышц в ответ на раздражение их электри-
ческим током; 4-я (от 90 до 110 час. инкубации) — начинаются
сокращения амниона и появляются активные движения конечно-
стей и хвоста; 5-я (5—6-дневный эмбрион) — возникают боковые
повороты головы и изгибы туловища; 6-я (на 6—9-й дни инкуба-
ции) — наблюдается максимальная двигательная активность эм-
бриона; 7-я (с 10-го по 14-й
день) — эта активность несколь-
ко ослабляется; 8-я (с 15-го по
18-й день) — наступает стадия
относительного покоя; 9-я (19-й
и 20-й день) — стадия предвы-
лупления, усиление двигатель-
ной активности; 10-я (20-й и
21-й дни инкубации) — стадия
вылупления, двигательная
активность еще больше увели-
чивается.
Возникновение и развитие
спонтанных и — особенно де-
тально — рефлекторных движе-
ний у куриных эмбрионов изу-
чено Волоховым (1951). По дан-
ным автора, у куриного эмбрио-
на в течение инкубации последо-
вательно возникают три различ-
Рис. 35. Возникновение разных ти-
пов двигательных реакций у эмбри-
она курицы. (По Волохову, 1951).
1 — спонтанные «пассивные» движения;
2 — спонтанные активные движения; 3 —
первичные локальные рефлексы; 4 — ге-
нерализованные рефлекторные реакции;
5 — рефлекторные реакции типа тониче-
ской судороги; 6 — специализированные
рефлексы.
ные типа реакций: а) спонтанные активные движения, б) спонтан-
ные пассивные движения и в) ответные (рефлекторные) реакции на
внешние раздражения (рис. 35). Спонтанные активные движения
возникают в конце 4-го дня. Автор предполагает, что внутренними
химическими агентами, воздействующими на двигательные нерв-
ные элементы спинного мозга в это время, являются углекислота
и другие продукты обмена веществ эмбриона. Спонтанные пас-
сивные движения маятникообразного характера возникают на 6-й
день и вызываются сокращениями амниона. Рефлекторные реак-
ции куриного эмбриона в ответ на механические раздражения кожи
появляются во 2-й половине 7-го дня и имеют характер одиночных,
ограниченных, изолированных рефлексов — «фаза локальных
рефлекторных реакций». Рефлекторная деятельность во время
этой фазы осуществляется через ограниченные, первично возбу-
димые очаги центральной нервной системы, расположенные в шей-
ном отделе спинного мозга и в стволовой части головного мозга.
Двигательная реакция передается через нейроны, иннервирующие
шейную мускулатуру и мышцы передней конечности. Эта фаза
167
продолжается 1.5—2 дня ДК/концу 8-го дня, одновременно с ме-
стными реакциями, обнаруживаются обобщенные, быстрые ответ-
ные движения головы и туловища — «фаза первичной генерализа-
ции рефлексов». Эта фаза продолжается с 9-го до 14—15-го дня
инкубации и характеризуется появлением большого числа воз-
будимых очагов в спинном мозгу и в стволовой области головного
мозга, с которыми связывается все большее количество чувстви-
тельных и двигательных нервных волокон. Одновременно с этим
происходит постепенное объединение уже созревших очагов воз-
буждения и установление между ними функциональных связей,
обеспечивающих распространение импульсов на значительное
расстояние от любого исходного очага. В результате создаются
условия для обобщенной, диффузной деятельности спинного мозга
и нижней части головного мозга, выражающейся в быстрых обоб-
щенных реакциях, зачастую с участием значительно отдаленной
от раздражаемой области группы мышц. К быстрым, обобщенным
реакциям в ответ на внешнее раздражение с 10—11-го дня добав-
ляются медленно протекающие тонические обобщенные движения
головы, туловища и конечностей — «фаза вторичной генерализа-
ции рефлексов». К концу этой стадии раздражение ограниченного
участка кожи эмбриона вызывает общее тоническое сокращение.
Эта фаза у куриного эмбриона протекает одновременно с фазой
первичной генерализации рефлексов, хотя и начинается несколько
позже. К этому времени происходит постепенное установление
функциональных связей между отдельными сегментами спинного
мозга, а также между спинным мозгом и стволовой частью голов-
ного мозга, что создает условия для широкого распространения
возбуждения и осуществления обобщенной диффузной рефлектор-
ной деятельности. На 15—16-й день обобщенные формы реакции
исчезают и появляются реакции специализированного характера —
открывание клюва при раздражении надклювья и подклювья,
мигательное движение век, закрывание третьего века при раз-
дражении роговицы и кожи в окружности глаза и др. Это —
«фаза специализации рефлекторных реакций». На этой стадии
вместо диффузной, неудержимой иррадиации процесса возбужде-
ния по всей центральной нервной системе возбуждение распро-
страняется только по определенным проводящим путям в связи
с возникновением очагов торможения в спинном и головном мозгу.
Специализированные рефлекторные реакции продолжаются вплоть
до вылупления, но в самые последние дни эмбрионального разви-
тия рефлекторная возбудимость эмбриона несколько понижается
в связи с тем, что спинной мозг и стволовая часть головного мозга
в целом начинают подвергаться тормозящим влияниям со стороны
вышележащих отделов мозга. Автор отмечает, что в процессе
развития рефлекторных актов у эмбриона нет резкой границы
в переходе от одной стадии рефлекторной деятельности к другой.
Наоборот, происходит как бы «наслоение» одной стадии на другую,
168
т. е. с появлением новых реакций еще продолжают существовать
ранее возникшие типы реакций.
Детальные данные о развитии двигательной активности (в осо-
бенности спонтанных движений) куриного эмбриона даны в обзоре
Гамбургера (Hamburger, 1964). Спонтанные движения начинаются
со сгибания головы через 3.5 дня инкубации. Затем подвижность
распространяется по направлению к хвосту (4.5 дня). Вскоре после
6.5 дней наблюдаются- редкие изолированные движения конечно-
стей, которые независимы от движений туловища. Двигательная
Л/-4Лет 6 дней 7% дней 8-9 дней
71-80
51-60
41-50
31-40
21-30
11-20
0-10
Рис. 36. Средняя продолжительность фаз активности
и покоя куриного эмбриона. на разных стадиях
развития. (По Humburger, 1964).
По оси ординат — длительность фаз (в сек.), по оси абсцисс —
[стадии развития.
активность эмбриона на ранних стадиях четко периодична: цикл
из серии движений перемежается с фазами покоя. С 8-го дня общие
волны движений тела затемняются локальной активностью его
частей. В связи со сложностью картины движений в это время
получается впечатление одновременных, некоординированных,
беспомощных движений разных частей тела. Автор показывает
изменение длительности периодов активности и неактивности
с 3.5—4 до 11 дней инкубации (рис. 36). С 12-дневного возраста
двигательная активность эмбрионов становится непрерывной.
Выясняя причины периодичности двигательной активности эм-
брионов, автор вырезал участки спинного мозга на разных уров-
нях и наблюдал сохранение той же периодичности у частей тела,
находящихся как ниже, так и выше перерезки. Это исключает
169
возможность периодической блокировки активности тормозя-
щими механизмами. Воздействуя смесью 90% О2 и 10% СО2,
автор показал, что повышение процента углекислоты имеет не сти-
мулирующий эффект на двигательную активность, как наблю-
дали некоторые авторы (Чумак, 1960), а, наоборот, подавляет ее,
иногда до полного прекращения; этот эффект оказался обратимым,
и при установлении нормального состава газов темп двигательной
активности восстанавливался. По вопросу о причинах перехода
от ритмической к непрерывной активности на 12-й день развития
куриного эмбриона автор высказывает несколько предположений.
Этот переход может быть обусловлен: а) увеличением проприоцеп-
тивной стимуляции, б) изменением внутренней среды, в) повыше-
нием чувствительности двигательных нейронов или г) увеличением
стимуляции из головного мозга. Характерной чертой нервной
системы куриного эмбриона в отличие от нервной системы сала-
мандры является, по мнению автора, специализированность всех
ее элементов и выполнение каждым только одной функции. Так,
например, двигательные нейроны куриного эмбриона не .имеют
ассоциативной функции; для выполнения проприо- и экстероцеп-
тивной функции с самого начала предназначаются различные ней-
роны. Автор подчеркивает удивительную синхронность возникно-
вения структур нервной системы и начала функции их. Так, на-
пример, первые двигательные волокна устанавливают контакт
с мышцами передней части туловища у 3.5-дневного куриного
эмбриона, т. е. точно в момент начала спонтанных движений.
С тех пор дифференциация двигательных волокон продолжается
в хвостовом направлении одновременно с распространением дви-
гательной активности. К моменту наступления двигательной ак-
тивности всего туловища оказываются установленными как про-
дольные, так и локальные интегрирующие механизмы. Такая же
синхронность структуры и функции наблюдается и в чувствитель-
ных нейронах. Первые кожные нервы достигают кожи на 6-й
день, но рефлекторная цепь не замыкается, до тех пор пока кол-
латерали из заднего ствола не врастают в серое вещество и не
устанавливают синаптических связей с ассоциативными и комис-
суральными нейронами. Врастание этих коллатералей происхо-
дит непосредственно перед тем, как могут быть получены экстеро-
цептивные рефлексы (7.5 дней). Проприоцептивные рефлексы и
их структурная основа из двухнейронной цепи образуются на
несколько дней позже.
У высших позвоночных (птицы, млекопитающие) происходит
подавление старых, ранее возникших форм реакций более позд-
ними, более молодыми. По мнению Волохова (1951), у птиц в про-
цессе эмбрионального развития совсем не обнаруживаются реак-
ции миогенного происхождения, которые являются единствен-
ной формой двигательных реакций у губок и еще имеются у рыб
и амфибий в начале их эмбрионального развития. Коштоянц
170
(1957)’в~отличие от Волохова считает, что движения туловищной
мускулатуры куриного эмбриона на самых ранних стадиях раз-
вития являются движениями миогенного типа. Однако Левинсон
и Лейкина (1960), на основании свои^ наблюдений об изменении
содержания РНК и тиоловых соединений, а также активности
щелочной фосфатазы во включенных в специфическую деятель-
ность двигательных нейронах шейного отдела спинного мозга
на 4—5-й день инкубации и об отсутствии таких изменений
в недифференцированных нейронах (двигательных поясничного
и хвостового отделов спинного мозга и всех чувствительных)
присоединяются к мнению Волохова о нейрогенной природе
ранних спонтанных движений куриного эмбриона. Чумак (1960)
также приводит доказательства правильности выдвинутой Куо
(Kuo, 1939b) и Волоховым (1951) гипотезы о нейромоторном ха-
рактере первичной спонтанной активности куриного эмбриона.
Так, внутримышечное введение курарина 5—8-дневным эмбрио-
нам вызывает уменьшение спонтанных активных движений до пол-
ного их прекращения, а введение прозерина, наоборот, учащает
их. Наблюдавшееся автором учащение движений эмбрионов
при продувании углекислоты через амниотическую жидкость
подтверждает предположение Волохова о том, что СО2 является
одним из агентов внутренней среды, чье воздействие на спинной
мозг обусловливает спонтанную активность эмбриона. Чумак
и Писарева (1961), используя установленный ими ранее факт,
что первичным источником характерных вспышек электрической
активности спинного мозга куриного эмбриона под влиянием
стрихнина являются вставочные нейроны, и показав одновре-
менность наступления фазы спонтанных двигательных реакций
и появления стрихнинных разрядов на 4—5-е сутки инкубации,
подчеркивают, что эта одновременность также свидетельствует
о нейромоторном характере первичной спонтанной активности
куриного эмбриона. Резкое возрастание амплитуды стрихнинных
разрядов наблюдалось на 7-й день, одновременно с наступлением
фазы рефлекторных реакций, и на 11-й — вместе с созреванием
структур среднего мозга и наступлением фазы генерализованных
тонических реакций. С подключением созревающих отделов
спинного и головного мозга частота стрихнинных разрядов умень-
шается. Нейрогенный характер спонтанной двигательной актив-
ности на ранних стадиях развития куриного эмбриона подтвер-
ждает и Гамбургер (Hamburger, 1964).
Войно-Ясенецкий и Бурсиан (1963), разработав новую мето-
дику регистрации двигательных реакций куриных эмбрионов
путем преобразования их в электрические колебания, установили
ряд фактов, дающих возможность высказать следующую гипо-
тезу. Двигательная активность имеет две линии развития: 1-я
возникает с 4-го дня инкубации и достигает своего максимума
к И—12-му дню — это линия формирования тонического аппа-
171
рата движений, а 2-я возникает на 7-й день и достигает зрелости
к 18-му дню эмбрионального развития — это линия формиро-
вания быстро работающих фазических аппаратов. Бурсиан
(1965) показал, что свет влияет на двигательную активность
куриных эмбрионов с начала ее возникновения, т. е. с 4-го дня
инкубации. Белый свет вызывает двигательную активность после
разрушения глаз, перерезки спинного мозга и декапитации.
Таким образом, эмбрион обладает примитивными формами
светочувствительности, присущими некоторым^ беспозвоночным
и низшим позвоночным. Автор предполагает, что эти реакции
представляют собой отражение в эмбриогенезе филогенетически
более ранних форм светочувствительности. Свидерская (1968),
изучая звуковые воздействия (300 и 1000 гц), нашла, что они вы-
зывают небольшие неспецифические сдвиги (то усиление, то
ослабление) двигательной активности куриных эмбрионов с 6-го
до 12-го дня развития. Начиная с 12—14-го дня, с включением
функции специфических слуховых структур, звук высокой ча-
стоты (1000 гц) вызывает торможение движений, а низкой
(300 гц) — наоборот, усиление. Автор предполагает, что это
обусловливается дифференциацией звуков, которые в постэмбрио-
нальном периоде имеют разный биологический смысл: сигнал
«тревоги» со стороны несушки в первом случае и «комфорта» —
во втором.
2. Электрическая активность
Петерс, Вондераге и Пауэрс (Peters et al., 1956) исследовали
включение функции различных отделов головного мозга в тече-
ние развития куриного эбриона, используя в качестве индика-
тора возникающую при этом электрическую активность. Ими уста-
новлено, что после 15 дней инкубации электрическая актив-
ность появляется последовательно в спинном, среднем, промежу-
точном мозгу, а затем в лобных долях полушарий. В дальнейшем
(Peters a. Cusick, 1961) авторы показали, что электрическая
активность изменяется соответственно возрасту и стадиям разви-
тия куриного эмбриона. При созревании нервной ткани и появлении
у нее новых функций амплитуда электрической активности уве-
личивается и сама она становится более сложной. Гарсиа-Аустт
(Garcia-Austt, 1954), исследуя энцефалограммы полушарий го-
ловного мозга куриных эмбрионов, установил, что спонтанная
и вызванная стрихнином электрическая активность начинаются
одновременно — на 13-й день инкубации. Это совпадает с окон-
чанием морфологической дифференцировки полушарий и уста-
новлением синаптических контактов, а также с биохимическим
развитием клеток, а именно — с быстрым повышением активности
аденилпирофосфатазы, внезапным увеличением количества РНК
и галактозидов, синтезом фосфатидов и появлением сфингомие-
172
лина. Позднее была изучена электрическая активность ретины
куриного эмбриона в ответ на вспышки света. Электроретино-
грамма (ЭРГ) может быть записана с 18-го дня инкубации и бывает
двух типов — «эмбрионального» и «постэмбрионального» (Garcia-
Austt a. Patetta-Queirolo, 1961а, 1961b). Было выяснено, что ЭРГ
эмбрионального типа может быть получена у 18—19-дневных ку-
риных эмбрионов только после интенсивной адаптации к свету,
на вспышки света с длиной волны меньше 0.51 мк. ЭРГ постэмбрио-
нального типа (такая же, как у взрослых птиц) наблюдалась
уже у 19—20-дневных эмбрионов, что указывает на полную
зрелость ретины к этому сроку.
По данным Богданова (1963), до 13-го дня инкубации двига-
тельная активность куриного эмбриона не сопровождается элек-
трической активностью высших отделов ЦНС. Начиная с 13-го
дня проприоцептивная импульсация с мышц достигает ЦНС
и движения эмбриона вызывают иногда электрическую актив-
ность, а с 15-го дня эта активность в ЦНС становится постоянной.
С 17-го дня эмбрионального развития проприоцептивные влия-
ния утрачивают свое ведущее значение в поддержании тонуса
ЦНС, и выключение проприоцептивных импульсов с мышц
(при помощи кураре и других обездвиживающих веществ)
теперь уже не снижает электрической активности ЦНС, как
это было раньше. Автор предполагает, что это изменение реак-
ции ЦНС на отсутствие проприоцептивных импульсов обуслов-
лено функциональным созреванием других анализаторных
систем.
Карпентер и Бергланд (Carpenter a. Bergland, 1957) на при-
мере седалищного нерва изучали изменение скорости передачи
импульсов. Выяснено, что у 10-дневного эмбриона эта скорость
равна 0.5 м/сек., у 21-дневного — 7—11 м/сек., а у взрослой
куры — 50 м/сек., т. е. в 100 раз больше, чем у 10-дневного
эмбриона. Авторы отмечают, что увеличение скорости передачи
импульсов связано в значительной степени с увеличением диаметра
волокон и улучшением их миелинизации.
3. Физиология органов чувств эмбриона
Несколько слов следует сказать о функционировании орга-
нов чувств в эмбриональном периоде.
Возможность восприятия света в яйце обусловливается
проходимостью белой скорлупы для солнечных лучей. Мануй-
лова, Попов и Тимошук (1963) показали, что прерывистое осве-
щение (короткие вспышки света каждые 5 сек.) оказывает стиму-
лирующее влияние на относительный вес глаз, зрительных до-
лей и больших полушарий куриных эмбрионов с 9-го дня и до
конца инкубации. До 9-го дня вес больших полушарий у осве-
щающихся и контрольных эмбрионов был одинаковым. Таким
173
образом, уже с 9-го дня инкубации нервная система куриного *
эмбриона способна к восприятию световых воздействий. Впро-
чем, общая чувствительность к свету у птичьих эмбрионов бы-
вает еще раньше, о чем мы уже сообщали (Бурсиан, 1964).
Прейер (Ргеуег, 1885) считал, что орган слуха куриного эмб-
риона начинает функционировать за 1—2 дня до вылупления.
По данным Куо (Кио, 1932с), громкие звуки вызывают задержку
втягивания желточного мешка, так как эмбрион отвечает на них
извивающимися движениями туловища. Горшелева (1936) со-
общает, что перед вылуплением цыпленка однообразные, слабые
звуковые раздражители (например, стук, похожий на удары
клюва) вызывали вначале реакцию в виде писка, а затем про-
исходило ее угасание. Более сильные раздражители писка
цыпленка не вызывали. По данным Чумака (1961), слуховой
анализатор включается на 20-й день развития. Включение зву-
ковых рецепторов происходит одновременно с появлением спе-
циализированных, оборонительных рефлекторных реакций.
Данные о включении слухового анализатора только на 20-й
день инкубации противоречат возможности образования вре-
менных связей (о чем речь будет дальше) при помощи звонка
с 17-го (Sedladek, 1962а—1962с) и даже с 15-го дня инкубации
(Hunt, 1949; Блинкова, 1962). Возможно, однако, что дело в раз-
ной высоте звука, применявшейся разными исследователями.
Это предположение подтверждается уже упомянутыми данными
Свидерской (1968) о разной реакции на звук в 300 и 1000 гц.
Кожа эмбриона воспринимает электрические раздражения
с конца 7-го дня, а механические в тепловые — с 8—9-го дня
(Kuo, 1932b). Рефлекторная раздражимость кожи, по данным
Горшелевой (1936), увеличивалась в течение инкубации и перед
вылуплением достигала максимума. Чумак (1961) установил,
что рецепторы кожи (тактильные, болевые, холодовые и тепло-
вые) начинают функционировать на 7-й день инкубации, одно-
временно с возникновением и развитием первичных рефлекторных
реакций.
4. Временные связи в эмбриональном периоде
Ханту (Hunt, 1949) удалось выработать временные связи
у 15-дневного куриного эмбриона при помощи электрического
тока и звука звонка. Эти связи оставались прочными до вылуп-
ления, после чего быстро (в течение часа) исчезали, если цыплята
находились в нормальных условиях (были на свету, получали
пищу и подвергались другим воздействиям). Находившиеся же
после вылупления в темноте, тишине и без пищи (в инкубаторе)
сохраняли временные связи более длительное время. Следова-
тельно, заключает автор, угасание временных связей, вырабо-
танных в эмбриональный период, после вылупления, обуслов-
174
лено воздействием стимулов сильно изменившейся внешней
среды.
Блинковой (1962) в хроническом опыте почти без нарушения
нормального эмбрионального развития (путем вживления электро-
дов) удалось образовать временные связи, как двигательные
(сначала), так и вегетативные (затем), на 15—19-й день эмбрио-
нального развития. Автор приходит к выводу, что формирование
соматических и вегетативных функций в ЦНС куриного эмбриона
происходит не одновременно. Сначала формируются безусловные
рефлекторные двигательные реакции, а затем вегетативные,
в частности сердечные. Наблюдавшиеся с 15-го дня эмбриональ-
ного развития временные связи, по данным автора, отличаются
непостоянством в проявлении, быстро угнетаются и угашаются
после вылупления.
Седлачеку (SedlaCek, 1962а—1962с) удавалось выработать
временные связи у куриных эмбрионов с 17-го дня инкубации.
При выработке временных связей автор различает три фазы,
значительно различающиеся по стойкости: 1) на 17-й день,
2) 18—20-й день и 3) 21-й день. Изучая влияние длительности
интервала между условными раздражителями и воздействие
стрихнина на эффективность временной связи между звуковым
раздражителем и безусловным оборонительным или глотатель-
ным рефлексом, автор приходит к выводу, что во время первой
фазы (17-дневные эмбрионы) временная связь осуществляется
по принципу суммационного рефлекса, во время второй (18—
20-й день) — по принципу доминанты, а в течение третьей (21-й день)
имеются, по мнению автора, черты истинного условного рефлекса.
Установив, что латентный период безусловного глотательного
рефлекса сокращается, а частота «спонтанных» глотательных
движений увеличивается за последние 5 дней эмбрионального
развития, автор приходит к выводу, что функциональные па-
раметры нервных процессов за это время повышаются и разви-
вается способность повысить тонус возбуждения центра глота-
тельных рефлексов на афферентные импульсы. Автор считает,
что это создает предпосылки развития механизма временной
связи от суммационного к истинному условному рефлексу. ?и|
Стадия функциональной готовности нервной системы птенца
к моменту вылупления зависит в значительной мере от длитель-
ности периода эмбрионального развития. Промптов (1948) ис-
следовал отличия в развитии нервной системы к моменту вылуп-
ления у птенцовых (незреловылупляющихся) и выводковых
(зреловылупляющихся) птиц. Автор указывает, что у незрело-
вылупляющихся к моменту вылупления имеется более ранняя
(менее созревшая) стадия развития нервной системы и, следо-
вательно, меньшая способность реагировать врожденными реак-
циями на раздражители окружающей среды, чем у зреловылу-
пляющихся. Благодаря тому что незреловылупляющиеся после
175
вылупления попадают в условия среды, дающие бблыпие воз-
можности для развития и укрепления индивидуально приобретен-
ных реакций, у них создаются условия для формирования био-
комплексов поведения, более богатых двигательными координа-
циями, чем у зреловылупляющихся птиц.
Г Л А В А 10
ДЫХАНИЕ
А. Образование дыхательных путей
Носовая полость, являющаяся началом дыхательного
уракта, развивается из первичной ротовой полости.1 За носовой
полостью дыхательный путь переходит в гортань. Далее следует
трахея. Образование трахеи начинается с появления у 3-дневного
эмбриона бороздки на дне глотки. Затем эта бороздка быстро
углубляется и образует трубку, остающуюся на переднем конце
открытой (голосовая щель). У 6-дневного куриного эмбриона
голосовая щель закрывается на 7 дней пробкой из эпителиальных
клеток. С 9-го по 11-й день инкубации вокруг эпителиальной трубки
из окружающей мезенхимы формируются хрящевые кольца
трахеи. Внизу трахея разветвляется на два бронха и оканчива-
ется парой почек — зачатков легких.
Вскоре после выхода из трахеи бронхи врастают в густую массу
мезенхимы и там разветвляются вначале (на 6-й день) на вторич-
ные, а затем (с 8-го по 11-й день) на третичные бронхи (внутрен-
ние, наружные, дорсальные и боковые), которые расходятся во все
стороны к поверхности и внутрь легкого (рис. 37). После 10-го
дня третичные бронхи встречаются и сливаются, образуя сооб-
щающуюся между собою сеть трубок, пронизывающую все лег-
кое. Из третичных бронхов возникают воздушные капилляры —
окончания дыхательных трубок легкого, анастомозирующие
между собою и образующие сеть, характерную для легкого
птицы.
Таким образом, легкие птиц отличаются от легких других
позвоночных животных тем, что вместо конечных альвеол, со-
держащих остаточный воздух, имеют систему соединяющихся
трубок, через которые проходит вдыхаемый воздух.
На концах некоторых вторичных бронхов образуются зна-
чительные расширения — воздушные мешки. Воздушные мешки
располагаются не только в брюшной полости, но разрастаются
1 Описание формирования дыхательных путей приводится нами в основ-
ном по Лилли (Lillie, 1952).
176
во вс© части тела, проникая своими разветвлениями даже внутрь
костей (пневматизация костей). Из всех воздушных мешков,
за исключением шейного, вырастают бронхиальные трубки,
вторично соединяющие воздушные мешки с бронхами. В функцио-
нирующем легком через эти трубки, называемые возвратными
бронхами, воздух из воздушных мешков возвращается в легкое.
Рост воздушных мешков начинается с 8-го и идет особенно
быстро с 14-го дня инкубации. К моменту вылупления воздушные
Рис. 37. Схема воздушных путей в легком куриного
эмбриона в начале 9-го дня инкубации. (По Locy а.
Larsell, 1916).
А — вид сбоку; Б — вид при сагиттальном разрезе.
1 — шейный воздушный мешок (в. м.); 2 — передний грудной в. м.;
3 — брюшной в. м.; 4а, 46 — боковая и средняя части межключич-
ного в. м.; б — бронхи (б.): в — внутренние б.; 7 — наружные б.;
8 — дорсальные б.; 9 — боковые б.; 10 — возвратные б.
мешки совершенно развиты и при дальнейшем росте цыпленка
только увеличиваются в размерах. Воздушные мешки начинают
функционировать, по-видимому, одновременно с легкими (Locy
a. Larsell, 1916).
Эпителий бронхов вначале является однослойным кубиче-
ским, а к 8—9-му дню эмбрионального развития становится мер-
цательным. Эпителий воздушных капилляров истончается и ста-
новится однослойным, плоским, респираторным (Крок, 1962).
Кровеносные сосуды, врастая в легкое, разветвляются на мель-
чайшие капилляры, которые окружают воздушные капилляры
и третичные бронхи. У куриного эмбриона на стадии 24 сомитов
(около 50 час. инкубации) формируется легочная вена, а у 85-ча-
сового все основные сосуды легкого уже сформированы (Romanoff,
1960). По данным Крыстева (1958), с 17-го дня инкубации вокруг
12 в. в. Рольник 177
стеиок третичных бронхов и воздушных капилляров эпителий
разрушается, в результате чего значительно облегчается окисле-
ние крови в кровеносных капиллярах.
Б. Развитие функции дыхания
Первые данные о дыхании эмбриона в яйце были опуб-
ликованы еще в 1632 г. Бойлем (цит. по: Needham, 1931). За после-
дующие столетия эта проблема много раз привлекала внимание
исследователей, однако, в связи со сложностью методик для ре-
шения ряда вопросов многие из них не решены до сих пор.
Наиболее удобно расположить имеющиеся данные о разви-
тии дыхательной функции птичьего, в основном куриного, эмб-
риона по времени развития различных механизмов дыхания.
По схеме, предложенной Брандесом (Brandes, 1924), имеются
два типа дыхания куриного эмбриона, причем каждый из них
проходит ряд стадий.
1. Молекулярное дыхание кислородом желтка:
а) без гемоглобина — 1-й день;
б) с наличием гемоглобина — со 2-го по 19-й день инкубации
(сначала через кровообращение сосудистого поля, а за-
тем желточного мешка).
2. Дыхание атмосферным воздухом, осуществляемое:
а) через аллантоисные сосуды — с 5-го дня до вылупления;
б) через кровеносные сосуды легкого — с 19-го дня.
Автор предполагает, что частичное дыхание легкими начина-
ется с 12-го дня инкубации.
Несколько иная схема развития функции дыхания куриного
эмбриона предложена Бакуниной (Bakounina, 1895а). Автор
проводит аналогию между развитием функции дыхания в эмб-
риогенезе высших животных и эволюцией этой функции у низших
животных во взрослом состоянии и устанавливает четыре стадии
развития дыхания у куриного эмбриона: 1) дыхание непосред-
ственно в тканях, как у инфузорий; 2) при помощи бесцветной
жидкости — аналогично животным с бесгемоглобинной кровью;
3) дыхание с участием гемоглобина, через сосуды желточного
мешка, как у холоднокровных животных, и 4) дыхание при по-
мощи аллантоиса, когда гемоглобин становится необходимым
для жизни, аналогично теплокровным. В нашем изложении
мы будем придерживаться схемы Брандеса, отмечая, когда она
окажется неправильной.
О молекулярном дыхании кислородом желтка имеется мало
исследований. Более 100 лет тому назад Дульк (цит. по: Needham,
1931) выяснил, что в желтке к моменту снесения яйца имеется
больший процент кислорода, чем в окружающем воздухе, а именно
25.3—26.7%. Автор определил это, помещая свежеснесенное
яйцо под воду и анализируя выделяющиеся при этом пузырьки
178
газа. Хассельбальх (Hasselbalch, 1902) подтвердил эти данные
и заметил, что при повышении температуры яйца выделение
кислорода увеличивается в 5 раз. Брандес (Brandes, 1924)
присоединяется к этому мнению, отмечая, что вначале освобождаю-
щийся кислород из клеток превышает потребность в нем эмбриона
и часть кислорода выходит из яйца через скорлупу. Этого рас-
творенного в желтке кислорода бывает, по-видимому, достаточно
для дыхания эмбриона в течение первых часов развития, пока
он еще чрезвычайно мал.
Кроме данных о повышенном содержании кислорода в све-
жеснесенном яйце, имеются также наблюдения о значительно
более высокой концентрации углекислоты в нем. Бор и Хассель-
бальх (Bohr u. Hasselbalch, 1900), помещая свежеснесенное
яйцо в проточный воздух и улавливая углекислоту из него, по-
казали, что скорлупа яйца выделяет СО2. Князева (1958) устано-
вила, что снесенные в разные месяцы яйца выделяют различное
количество углекислоты до закладки их в инкубатор: за 2 часа
на 100 г веса в феврале — 3.7 мг СО2, в марте — 4.0 мг,
в апреле — 4.4 мг. В первый день после снесения углекислоты
выделяется больше, чем в последующие дни. При понижении
атмосферного давления и повышении влажности во время хра-
нения яиц выделение СО2 из яиц увеличивается, и наоборот.
Дыхание на ранних стадиях развития куриных эмбрионов
исследовал Филипс (Philips, 1941, 1942) и показал, что интенсив-
ность дыхания — (?о2 (число миллилитров О2, поглощаемых
за 1 час 1 мг сухого вещества ткани) — увеличивается в два
раза за первые 4 часа инкубации. Подчеркивая этот факт, автор
сравнивает его с тем, что эксплантированные неинкубированные
бластодермы обычно не способны к росту, а развивавшиеся в ин-
кубаторе в течение 4—5 час. до эксплантации не только растут,
но и способны к дифференциации. Следовательно, за 4—5 час.
инкубации происходит значительная перестройка энзиматиче-
ского аппарата. В дальнейшем с 8 до 20 час. инкубации интенсив-
ность дыхания эмбриона не увеличивается, а общее количество
потребленного кислорода растет только в связи с увеличением
массы дышащей протоплазмы. Нидхем (Needham, 1942) считает,
что нормальный эмбриональный рост и морфогенез птиц невоз-
можны в отсутствии кислорода.
О дыхании при помощи кровообращения в желточном мешке
мы уже подробно сообщали в главе 7. Напомним, что эта хорошо
развитая кровеносная система до 5-го дня инкубации полностью,
а затем частично несет дыхательную функцию. Сергеев (1943) ука-
зывает, что желточное кровообращение у наиболее продвинутых
в эволюционном отношении рептилий (черепах) функционирует
в качестве дыхательной системы и на более поздних стадиях
развития. Рагозина (1961) впервые показала, что при помощи
желточного кровообращения используется не только растворен-
12*
179
ный в желтке и отщепляемый витально кислород, но и кислород
атмосферного воздуха. Наблюдая примыкание желточного мешка
к скорлупе, автор подчеркивает, что это является приспособле-
нием для обегчения газообмена.
Переходя к дыханию атмосферным воздухом через аллантоис-
ные сосуды, отошлем читателя также к 7-й главе, где описано
образование и развитие аллантоиса. Напомним только, что,
по данным Рагозиной (1961), аллантоис начинает функциониро-
вать с конца 7-го дня, а с конца 8-го по 19-й он является единст-
венным органом дыхания. Данные автора о желточном и алланто-
исном кровообращении несколько исправляют схему дыхания,
предложенную Брандесом (Brandes, 1924).
_ j Аллантоисное дыхание атмосферным воздухом происходит
через скорлупу и две подскорлупные оболочки. Газы проходят че-
рез подскорлупные оболочки диффузно, а сквозь скорлупу —
через поры. У птиц с более толстой скорлупой подскорлупные
оболочки обычно более тонкие, и наоборот.
Впервые гистологию скорлупы изучил Рицо (Rizzo, 1899)
и нашел, что куриное яйцо имеет среднюю поверхность 6644 мм2,
на которой расположено около 7600 пор, т. е. 1.23 поры на 1 мм2.
Скорлупа яйца должна быть достаточно проницаемой для газов,
чтобы обеспечить нормальный газообмен (кислорода и углекис-
лоты) между эмбрионом и окружающей атмосферой и в то же
время не слишком проницаемой для другого газа — водяного
пара, чтобы сохранить необходимую для построения эмбриональ-
ных тканей воду. Это противоречие в самом способе размножения
птиц замечено Ридлем (Riddle, 1930) и является, по предполо-
жению автора, причиной повышения смертности эмбрионов
в определенные периоды эмбрионального развития (см. главу 17).
Автор назвал этот феномен «парадоксом скорлупы».
Проницаемость скорлупы и подскорлупных оболочек для
газов имеет большое значение для осуществления дыхания эмб-
риона атмосферным кислородом. Хюфнер (Hufner, 1892) показал,
что для разных газов проницаемость скорлупы различна: быстрее
всего через скорлупу проходит водород, затем углекислота, азот
и медленее всего — кислород. Автор обнаружил также, что
скорлупа гусиного яйца лучше пропускает газы, чем скорлупа
куриного. По данным Романова (Romanoff, 1943), проницаемость
скорлупы для газов изнутри наружу больше, чем снаружи внутрь,
и проницаемость влажной скорлупы равна только 8% проницае-
мости специально высушенной. В среднем проницаемость скор-
лупы в тупом конце яйца на 28% выше, чем в остром (14.52 мл
на 1 см2 в минуту и 10.46 мл соответственно). Наблюдаются боль-
шие ^индивидуальные вариации проницаемости скорлупы. Ин-
тересны данные автора о проницаемости скорлупы яиц разных
видов птиц. Оказывается, что скорлупа мелких яиц менее про-
ницаема, чем крупных, и автор замечает, что это биологически
180
оправдано, так как у крупных яиц площадь скорлупы по отно-
шению к объему яйца меньше, чем у мелких. Автор наблюдал
увеличение газопроницаемости скорлупы в течение инкубации
на 26.9% в тупом конце яйца и на 8.6% в остром. По данным
же Ромейна (Romijn, 1950), проницаемость скорлупы для кис-
лорода увеличивается в течение инкубации в 7000 раз, а для
углекислоты — в 565 раз. Чем обусловлено такое расхождение
в величинах, мы затрудняемся объяснить. Увеличение прони-
цаемости скорлупы в течение инкубации связано с растворением
части кальциевых солей скорлупы углекислотой, а ее органи-
ческой основы — протеолитическими ферментами (Еремеев,
1957). Воздушная камера в течение инкубации увеличивается
вследствие испарения воды из яйца. ,
Под воздушной камерой аллантоис отделен от находящегося
в ней воздуха только внутренней подскорлупной оболочкой,
в отличие от других частей своей поверхности, где помехой газо-
обмену является, кроме того, наружная подскорлупная оболочка
и скорлупа. Следовательно, газообмен в подстилающей воздуш-
ную камеру части аллантоиса может происходить и, по-видимому,
происходит интенсивнее. По данным Ромейна и Рус (Romijn
a. Roos, 1938), поверхность аллантоиса, выстилающая дно воз-
душной камеры яйца, составляет в последние дни инкубации
10—20% всей дыхательной площади эмбриона. Однако роль
воздушной камеры в дыхании эмбриона значительно больше,
так как, по подсчетам Ромейна (Romijn, 1950), через скорлупу
над воздушной камерой проходит на 20-й день инкубации около
80% всего потребляемого эмбрионом кислорода. Возможно
также, что газопроницаемость скорлупы над воздушной камерой
еще и потому больше, что скорлупа здесь суше, а по данным
Пенионжкевича (1936), Романова (Romanoff, 1943) и Владими-
ровой (1954), газопроницаемость скорлупы увеличивается при
высушивании. В опытах Прейера (Ргеуег, 1885) покрытие лаком
всей поверхности скорлупы, кроме площади над воздушной
камерой, не нарушало развития эмбриона; если же лакировалась
только половина скорлупы и этой половиной яйцо было повернуто
вверх, эмбрион погибал к 7-му дню инкубации.
В связи со значительной ролью воздушной камеры в дыхании
эмбриона следует рассмотреть данные об изменении состава
газов в ней в течение инкубации. Изучение состава газов воз-
душной камеры, кроме того, помогает нам судить о проницае-
мости скорлупы, не нарушая нормального хода развития эмб-
риона. Состав газов воздушной камеры куриного яйца изучали
Агазотти (Agazzotti, 1914) и Ромейн и Рус (Romijn a. Roos, 1938).
Агазотти обнаружил в воздушной камере на 1-й день инкубации
1.89% СО2, на 11-й — 0.33% СО2 и 13.65% О2. Сходные данные
получили Ромейн и Рус, обнаружив, что объем воздушной ка-
меры увеличивается с 0.3 мл до закладки яйца в инкубатор
181
до 10 мл на 20-й день инкубации как у оплодотворенного,
и у неоплодотворенного яйца.
В связи с некоторыми недостатками методики вышеназванных
авторов мы в своей работе изменили как метод взятия пробы
газа, так и метод анализа. Нами был создан прибор — микрога-
зоанализатор, где в пробе газа объемом 0.1—0.2 мл определяется
содержание углекислоты и кислорода с точностью +0.2%
Рис. 38. Общий вид микрогазоанализатора. (Рольник, 1960).
Объяснение в тексте.
(рис. 38). Описание прибора приведено в нашей статье (Рольник,
1960). Применив этот метод микрогазоанализа, мы подробно
изучили изменение состава газов в воздушной камере яиц в те-
чение инкубации (рис. 39). Содержание углекислоты до инкубации
и в первые дни развития выше, чем в окружающем воздухе.2
За первые 9 дней инкубации содержание кислорода снижается,
а концентрация углекислоты повышается чрезвычайно медленно.
После 10—11 дней инкубации, с момента выстилания алланто-
исом всей внутренней поверхности скорлупы до начала его
2 В этой связи интересны данные опытов Еремеева (1957), который повы-
шал жизнеспособность и рост .куриных эмбрионов путем содержания яиц
до инкубации в течение 3—4 час. в атмосфере с повышенной концентрацией
СО2. Автор высказывает интересное предположение что «легкость приспосо-
бления зародышей птиц на первых стадиях развития к повышенному со-
держанию СО2 в яйце представляет собой отражение филогенеза позвоночных,
шедшего в направлении смены условий дыхания от сред, более богатых угле-
кислотой (вода, почва), к среде, бедной ею (воздух)».
182
отмирания — на 17—18-й день инкубации, содержание кис-
лорода снижается, а процент углекислоты повышается бы-
стро — в среднем по 0.5% в сутки. В течение 3 суток, предшест-
вующих проклеву подскорлупной оболочки, с 17 до 19 дней ин-
кубации содержание О2 и СО2 в воздушной камере почти не изме-
няется. В эти дни развития куриного эмбриона Ромейн и Рус
(Romijn a. Roos, 1938) наблюдали такое же уплощение кривых
кислорода и углекислоты в воздушной камере яйца. Следует
отметить, что в течение этих 2—3 дней аллантоис уже начинает
отмирать, а легкие еще не начали своей деятельности или функцио-
нируют неполностью. Итак, приостановка снижения количе-
ства кислорода и увеличения углекислоты в воздушной камере
обусловлена, по-видимому, серьезными морфологическими и фи-
зиологическими изменениями, связанными с переходом от ал-
лантоисного к легочному дыханию. После прокола клювом внут-
ренней подскорлупной оболочки под воздушной камерой эмб-
рион начинает дышать легкими. Это бывает на 19—20-й день
инкубации куриных яиц. В течение суток эмбрион дышит в ат-
мосфере воздушной камеры, содержащей к началу этого периода,
по нашим данным, 15.0—18.6% (в среднем 16.2%) кислорода
и 2.8—5.0% (в среднем 3.8%) углекислоты. За время дыхания
легкими процент углекислоты в воздушной камере снова увели-
чивается, а содержание кислорода падает. Ромейн и Рус отме-
чают, что особенно низкое содержание кислорода и высокое
углекислоты в воздушной камере они наблюдали после начала
писка в яйце, т. е. после начала легочного дыхания, но до прок-
лева скорлупы.
Мы продолжили исследование состава газов воздушной ка-
меры и после проклева скорлупы. Оказалось, что и после того,
как в скорлупе образуется щель проклева, накопление углекис-
лоты и понижение процента кислорода продолжается. В одном
из яиц за сутки до проклева было 3.5% СО2 и 15.3% О2;
через 15 мин. после проклева было 4.1 % СО2 и 13.9% О2, а еще
через 4 часа — 5.1% СО2 и 12.1% О2. Таким образом, смена типа
дыхания с аллантоисного на легочное приводит к настолько боль-
шому увеличению обмена, что даже через щель проклева в скор-
лупе не может восполняться потребляемый эмбрионом кислород
и удаляться из воздушной камеры выделенная при дыхании
углекислота. Следовательно, дыхание легкими как до проклева
скорлупы, так и в первое время после него происходит в неблаго-
приятных условиях: содержание кислорода в воздушной камере
почти вдвое меньше, а процент углекислоты в 200 раз больше
атмосферного.
Мы попытались выяснить, как долго может жить 20-дневный
эмбрион без снабжения кислородом извне, и покрыли клеем
всю поверхность скорлупы. До опыта в воздушной камере было
2.6% СО2 и 17.2% О2; через 10 мин. после покрытия скорлупы
183
Рис. 39. Изменение состава газов в воздушной камере куриных яиц в течение инкубации. (Рольник, 1960).
1 и 3 — графики по данным 1947 г. (крестики); 2 и 4 — графики по данным 1948 г. (точки).
(% д1янпдчдо)2оэ
клеём — 4.6% С02 и 11.9%
02; через 20 мин. — 4.2% и
7.7%; через 30 мин. — 4.2%
и 8.8% и через 40 мин. —
6.0% и 8.8% соответственно.
При вскрытии яйца через
40 мин. было обнаружено,
что аллантоис не атрофиро-
ван, кровь темная. Через
несколько минут эмбрион по-
гиб. По-видимому, содержа-
ние 6% СО2 и менее 10% О2
в воздушной камере вызывает
необратимые изменения, ве-
дущие к гибели эмбриона.
Для сравнения приведем
наши данные о составе га-
зов воздушной камеры яиц
при естественной инкубации.
Несмотря на то что яйца под
наседкой несомненно венти-
лируются лучше, чем в ин-
кубаторе, содержание кисло-
рода в воздушной камере
было меньше, чем в яйцах,
инкубируемых искусственно:
на 16-й день — 14.3—16.6%,
а на 17-й - 15.4-15.8%.
Следовательно, под наседкой
происходит больший газооб-
мен, что показывает на луч-
шее течение развития эмбри-
она. В воздушной камере
яиц дикой утки кряквы, раз-
вивавшихся в гнезде и в по-
следние дни эмбрионального
развития перенесенных в ин-
кубатор, были еще бблыпие
отклонения от состава атмо-
сферного воздуха — 4.6—
7.2% СО2и 14.4- 11.6% О2.
Чтобы определить прони-
цаемость скорлупы для раз-
личных газов, не нарушая ее целостности и нормальной жизнедея-
тельности эмбриона, мы сравнили данные о потере веса яиц (прони-
цаемость скорлупы для водяных паров) с содержанием кислорода и
углекислоты в воздушной камере яиц в разные дни инкубации. Мы
185
Подсчитали по данным Владимировой (1954), что проницаемость
скорлупы для газов примерно в полтора раза больше у яиц,
имевших вдвое ббльшую потерю веса во время хранения. Наши
данные о корреляции между проницаемостью скорлупы для кис-
лорода и углекислоты, с одной стороны, и для водяных паров —
с другбй, приведены в табл. 33. Как видно из таблицы, досто-
верная корреляция между величинами содержания газов воз-
душной камеры и потерей веса яиц возникает только с 17-го дня
инкубации. Следовательно, с этого времени проницаемость скор-
лупы становится реальным препятствием для газообмена эмб-
риона, а на 19-й день это препятствие становится особенно
большим. Итак, нам удалось подтвердить предположенный Рид-
лем (Riddle, 1930) «парадокс скорлупы».
Таблица 33
Коэффициент корреляции между составом газов воздушной камеры яиц
и процентом потери их веса за 15 дней инкубации
(По Рольник, 1960)
Дни инкуба- цйи По углекислоте Достоверность По кислороду Достоверность
12 -0.29 ±0.27 Не достоверно +0.09 ±0.28 Не достоверно
13 -0.04 ±0.33 » » +0.06 ±0.33 » »
15 +0.12 ±0.40 » » +0.23 ±0.39 » »
16 -0.22 ±0.30 » » +0.32 ±0.28 » »
17 -0.59 ±0.15 Достоверно +0.52 ±0.17 Достоверно
18 -0.46 ±0.15 » +0.43 ±0.16 Не вполне досто- верно
19 -0.80 ±0.10 Вполне досто- верно +0.79 ±0.11 Вполне досто- верно
Повышенное содержание углекислоты и низкое содержание
кислорода в воздушной камере в конце инкубационного периода
являются отражением увеличенной концентрации СО2 и пони-
женного содержания О2 в крови эмбриона в это время. Уиндл
и Баркрофт (Windle a. Barcroft, 1938) считают, что такое изме-
нение газов в крови эмбриона вызывает: а) увеличенный мышеч-
ный тонус, б) торможение движений, не включенных в дыхатель-
ный ритм, в) интенсификацию дыхательных движений, чтобы
заменить уже отмирающий в это время аллантоис, г) интенсифи-
кацию движений шеи и туловища, при помощи которых проис-
ходит проклев скорлупы.
Рассмотрим теперь дыхательную систему, которой птичий
эмбрион пользуется непосредственно перед и во время вылуп-
ления — легочное дыхание.
186
Дыхательные движения у куриных эмбрионов, по Куо и Шен
(Kuo a. Shen, 1937), проходят 3 стадии: 1) 16 дней инкубации —
движения грудной клетки почти всегда сопровождаются движе-
ниями всего тела (движения грудной клетки на 14-й и 15-й день
авторы не считают дыхательными движениями); 2) 17-й и 18-й
дни — независимые движения грудной клетки; 3) 19-й и 20-й
дни — начало истинного легочного дыхания. Авторы присое-
диняются к предположению Брандеса (Brandes, 1924), что ды-
хание легкими начинается еще до прокола подскорлупной обо-
лочки, добавляя, что это дыхание происходит за счет воздуха,
имеющегося к этому времени в полости амниона. Это предполо-
жение имело некоторое подтверждение в наших неопубликованных
наблюдениях. Вскрывая скорлупу над воздушной камерой, мы
иногда наблюдали под непроколотой еще эмбрионом внутренней
подскорлупной оболочкой и аллантоисом перекатывающийся
пузырек газа объемом 0.5—1.0 мл. Нам удалось проанализи-
ровать состав газов этих пузырьков из трех яиц. На 15—16-й
день инкубации там было 1.6—3.2% СО2 и 18.4—20.0% О2.
Однако такие случаи были редки и пузырьки слишком малы,
чтобы считать это началом легочного дыхания.
Куо и Шен (Kuo a. Shen, 1937) наблюдали у 17-дневных эмб-
рионов 5 дыхательных движений в 1 мин., у 18-дневных — 17,
у 19-дневных — 46, у 20-дневных — 84. По данным Уиндла
и Баркрофта (Windle a. Barcroft, 1938), спонтанные дыхательные
движения куриных эмбрионов начинаются на 15-й день, но до 17-го,
а иногда и до 19-го дня инкубации они неритмичны. Ромейн
(Romijn, 1948) наблюдал дыхательные движения куриных эмб-
рионов только с 20-го дня инкубации, но все же до начала ле-
гочного дыхания из воздушной камеры. По нашим данным (Роль-
ник, 1956а), спонтанные дыхательные движения начинаются
не позднее 14-го дня. У 16-дневного эмбриона наблюдалось в сред-
нем 4 дыхательных движения в 1 мин.; у 17-дневного — 8,
у 18-дневного — 25—30. Настоящий, равномерный дыхательный
ритм (50—55 дыханий в 1 мин.) устанавливается только в сере-
дине или конце 20-го дня инкубации — в первые часы легочного
дыхания (кимографическая запись дыхательных движений пред-
ставлена на рис. 40). Раздражения (щипок, укол, горячая вода)
вызывают дыхательные движения. У утиных эмбрионов Уиндл
и Нельсон (Windle a. Nelson, 1938) обнаружили начало дыхания
легкими за 3 дня до вылупления. В остальном же развитие ле-
гочного дыхания у утиных эмбрионов было примерно таким же,
как у куриных.
Какие же факторы обусловливают начало дыхательных дви-
жений у птичьего эмбриона? Прейер (Ргеуег, 1885) считает, что
в норме дыхательные движения начинаются под влиянием раздра-
жения кожи у выросшего эмбриона о скорлупу, так как любое
раздражение (укол иглой, толчок, щипок и т. д.) вызывает ды-
187
хательные движения. Автор отмечает, однако, что венозность
крови повышает раздражимость дыхательного центра. Куо и Шен
(Kuo a. Shen, 1937) видят основную причину возникновения
Рис. 40. Запись дыхательных движений 17-, 19-, 20- и 21-дневных
куриных эмбрионов. (Рольник, 1966).
I, II, III, VI — запись с невскрытой подскорлупной оболочки; IV, V, VII,
VIII, IX — запись с грудной клетки. (IV и V, VI h«VII, VIII и IX —
записи на одном эмбрионе; из последнего затем в инкубаторе вывелся нор-
мальный цыпленок). Справа — возраст эмбриона. Отметка времени — 1 сек.
дыхательных движений в накоплении в крови углекислоты,
а учащение их считают обусловленным высыханием кожи, так
как вся амниотическая жидкость к этому времени поглощена
188.
эмбрионом. В опытах авторов после добавления 1—2 мл изото-
нического раствора в полость амниона дыхательные движения
урежались. Ромейну (Romijn, 1948) удалось показать, что вы-
сокая концентрация углекислоты не является стимулятором
дыхательного центра и не учащает дыхательных движений.
Внезапное падение содержания СО2 и повышение О2 при искус-
ственном расширении трещины проклева не вызывало изменения
дыхательного ритма. Не изменялся он и при помещении яйца
с проклевом скорлупы в газовую смесь с 7% СО2,12% О2и 81%N2.
А через 24 часа после вылупления помещение цыпленка в та-
кую же газовую смесь вызывало у него диспноэ. Автор заклю-
чает, что у эмбриона перед вылуплением еще плохо развита
химическая регуляция дыхания, а у вылупившегося цыпленка
она уже есть.
Уиндл (Windle, 1940) создал теорию развития функции легоч-
ного дыхания в эмбриональном периоде. Соматические мышцы
и периферические нервы, связанные с дыханием, закладываются
раньше, чем начинаются дыхательные движения; затем образуются
нейроны, соединяющие дыхательный центр с соматической двига-
тельной системой. Стимуляция центростремительных нейронов
передается в дыхательный центр, и он может начать действовать
преждевременно, что может быть обусловлено либо повышенной
концентрацией СО2, либо подавлением порога дыхательного
центра, вызванного пониженным содержанием кислорода в крови.
Таким образом, по мнению автора, дыхательный механизм яв-
ляется спящей системой, снабженной потенциальными возмож-
ностями задолго до того, как он станет необходим эмбриону.
В случае сильного снижения содержания кислорода в крови
многочисленные двигательные нейроны активизируются, тонус
мышц увеличивается и начинается слишком глубокое вдыхание,
при котором в дыхательные пути попадает амниотическая жид-
кость (вместо нормального заглатывания ее в желудок). В норме же
в конце эмбрионального периода наблюдаются быстрые, неглу-
бокие дыхательные движения. Автор подчеркивает, что опасность
«затопления» амниотической жидкостью невелика, так как ды-
хательный центр эмбриона имеет более высокий порог раздра-
жения, чем у взрослых птиц. Автор отмечает также, что появ-
ление дыхательных движений происходит одновременно с упло-
щением кривых содержания О2 и СО2 в воздушной камере яиц.
Аршавский (1948) придает особое значение дыхательным движе-
ниям, плода млекопитающих и считает, что «у эмбрионов птиц
дыхательные движения точно так же осуществляют циркуля-
торную функцию, интенсифицируя желточное кровообращение».
Возникает вопрос, какова же доля участия аллантоиса и лег-
ких в дыхании эмбриона в последние дни инкубации? Чтобы
ответить на этот вопрос, мы провели опыты по перевязке крове-
носных сосудов, проходящих в стебельке аллантоиса (Рольник,
189
1956а). Осторожно извлекая эмбрион из яйца, чтобы не повре-
дить аллантоисных сосудов, мы помещали его и скорлупу с ал-
лантоисом на часовое стекло в условиях сохранения тепла и влаги
и осуществления нормального обмена кровью между аллантои-
сом и эмбрионом. Каждые 3—7 мин. мы перевязывали ниткой
стебелек аллантоиса, причем прекращалось на время, а затем
снова восстанавливалось нормальное кровообращение между
эмбрионом и аллантоисом. У эмбриона с перевязанным стебель-
ком прекращаются дыхательные движения, иногда угасают и реф-
лексы, но после развязывания нитки эмбрион нередко оживает
и начинает дышать. Обнаружилось, что у 17-дневных куриных
эмбрионов гибель наступает в среднем через 11 мин., при пере-
несении одной 2-минутной перевязки; у 18-дневных длительность
жизни такая же, но перевязка уже может быть перенесена в те-
чение 3 мин.; у 19-дневных гибель наступает через 20 мин., с пе-
ренесением в среднем 2—3 перевязок общей продолжительностью
8 мин.; и только у 20-дневных зависимость от аллантоисного
дыхания почти отсутствует, а если имеется проклев скорлупы,
то после перевязки сосудов аллантоиса цыпленок обычно про-
должает так же ритмично дышать. Итак, нами установлено,
что даже в последние дни инкубации основную роль в дыхании
эмбриона играет аллантоис.
Броди (Brody, 1927), Огородний (1936) и Ромейн (Romijn,
1950) вполне обоснованно, на наш взгляд, предполагают, что
повышение смертности эмбрионов в последние 3 дня инкубации
вызывается в основном сложностью перехода от аллантоисного
к легочному дыханию. В практике инкубация эмбрионов, по-
гибающих в это время, называют «задохликами». Огородний
(1936) отмечает, что этот термин имеет основание, так как ал-
лантоис к этому времени уже значительно дегенерирует, а на-
чинающие функционировать легкие не в состоянии обеспечить
необходимый газообмен в условиях повышенной потребности
в кислороде.
Учитывая значение воздушной камеры в дыхании эмбриона
в последние дни инкубации, мы приводим содержание кисло-
рода и углекислоты в воздушной камере яиц, где эмбрионы
погибли в один из этих дней (табл. 34).
Итак, состав газов в воздушной камере яиц, давших задох-
ликов*, неоднороден: в яйцах № 14 и № 54 наблюдался чрезвы-
чайно низкий процент кислорода и высокое содержание угле-
кислоты, а в яйцах № 64 и № 12 процент О2 несколько выше
и содержание СО2 ниже, чем в яйцах с нормально развивающи-
мися эмбрионами. В первой группе яиц скорлупа, по-видимому,
недостаточно проницаема для газов (потеря веса этих яиц в те-
чение инкубации тоже несколько меньше), что и является не-
посредственной причиной гибели эмбрионов. Во второй группе
яиц ослабление эмбриона, очевидно, вызвано какими-либо при-
190
Таблица 34
Состав газов воздушной камеры яиц, давших задохликов
(По Рольник, 1960)
Дни инкубации Ка яйца СО, (%) О2 (%)
17-Й 14 5.8 14.2
18-й 14 5.2 14.2
19-й 64 3.3 17.4
20-й 14 2.4 9.0
21-й 12 2.6 17.8
. 21-й 54 4.2 12.3
чинами, предшествующими периоду перехода с аллантоисного
на легочное дыхание, и когда он наступает, метаболизм эмбриона
ниже обычного.
Недостаток кислорода в последние дни инкубации оказывается
особенно неблагоприятным для нормального окончания процесса
вылупления, так как именно в этот период эмбрион должен затра-
тить большую механическую энергию на вылупление, для чего
необходим кислород и запас энергии в виде богатых энергией
соединений (АТФ и т. п.). Несоответствие между все возрастаю-
щей потребностью в кислороде и низкой реальной возможностью
ее удовлетворения, еще больше понижающейся при плохой газо-
проницаемости скорлупы, обусловливает асфиксию эмбриона,
оказывающуюся летальной для наиболее ослабленных эмбрионов
вследствие различных эндо- и экзогенных факторов, а у других
вызывает понижение жизнеспособности. Это привело нас к пред-
положению о необходимости добавления кислорода в воздушную
камеру яйца с целью облегчения перехода от аллантоисного
дыхания к легочному. Введение кислорода в воздушную камеру
куриных, индюшиных и утиных яиц за несколько дней перед
вылуплением наполовину сократило гибель эмбрионов в этот
период (Рольник, 1946а, 19466, 19566) (подробнее см. главу 20).
Повышение жизненности у эмбрионов, получивших кислород,
было подтверждено и опытами по перевязке у них аллантоиса.
Длительность жизни у эмбрионов, изъятых из «кислородизиро-
ванных» яиц, и способность переносить перевязки аллантоиса
увеличивались примерно вдвое.
Наши опыты и наблюдения по физиологии дыхания птичьих
эмбрионов (как приведенные, так и не упомянутые здесь) пол-
ностью подтверждают правильность гипотезы, что основной,
хотя и не единственной, причиной повышения смертности эмбрио-
нов перед вылуплением является трудность перехода от аллан-
тоисного к легочному дыханию.
191
В. Газообмен птичьих эмбрионой
Исследование газообмена в эмбриональном развитии
птиц — одного из важных показателей процесса обмена веществ
и энергии — наряду с теоретическим интересом для понимания
онтогенеза птиц име#г и практическое значение для решения
некоторых вопросов инкубации. Эмбрионы птиц — сложные объ-
екты для исследования газообмена, так как количество метабо-
лизирующей ткани по сравнению с инертной, недышащей массой
питательного материала яйца очень мало, особенно на ранних
стадиях развития. Вероятно поэтому, несмотря на издавна су-
ществовавший интерес к изучению газообмена эмбрионов птиц,
достоверных данных имеется сравнительно немного. Данные
о поглощении кислорода куриными эмбрионами сведены в табл. 35.
Результаты же, полученные на газообменном аппарате Влади-
мирского (Владимирский, 1935; Баранчеев, 1939; Биховець,
1941, и др.), а также измерения Ксабай, Хорват и Ференц (Csa-
bay et al., 1960) в таблицу не включены, так как, по нашему
мнению, они недостаточно точны из-за несовершенства методики.
По данным Хассельбальха (Hasselbalch, 1900), общее коли-
чество потребленного О2 за период развития куриного эмбриона
было равно 4.46 л. В другой работе (Bohr a. Hasselbalch, 1903)
установлено, что за период развития выделяется 3.022 л СО2.
В первые дни инкубации ее выделяет скорлупа яйца; с 3-го до 17-го
дня выделение СО2 за счет усиления дыхания эмбриона уве-
личивается, а затем до конца инкубации остается на постоянном
уровне. Газообмен куриных эмбрионов, по данным Никита (Ni-
chita, 1955а), имеет несколько ббльшие величины: за период
инкубации потребляется 5.53 л О2 и выделяется 4.16 л СО2. Срав-
нивая газообмен эмбрионов разных пород кур, автор отмечает,
что интенсивность дыхания зависит от качества яиц и соответ-
ствует проценту вывода цыплят.
По данным Романова (Romanoff, 1941), прирост потребления
кислорода куриными эмбрионами за сутки резко падает с 75%
на 3-й день до 33% на 8-й; затем почти не изменяется до 14-го дня;
потом резко падает до 5% на 18-й и снова повышается до 30%
к 22-му дню инкубации. Автор наблюдал большие индивидуаль-
ные вариации в потреблении кислорода.
Большую серию исследований по развитию газообмена у ку-
риных эмбрионов провел Ромейн с сотрудниками (Romijn, 1950,
1951; Romijn a. Lokhorst, 1951; Romijn et al., 1952). Авторы
отмечают, что данные о поглощении кислорода и выделении
углекислоты на ранних стадиях эмбрионального развития не от-
ражают действительного газообмена эмбриона, так как в течение
первых 4—5 дней содержимое яиц выделяет большое количество
углекислоты и кислорода. Быстрый подъем газообмена с 10-го
до 14-го дня и, что особенно важно, увеличение интенсивности
192
Таблица 35
Потребление кислорода (мл/час) эмбрионами и цыплятами
Hassel- balch (1900) Lussana (1906) Romanoff (1941) Giaja a. Jovandid (1950) Romijn (1951) Romijn, Funk a. Lokhorst (1952)
Темпера- 37.0 37.5 38.4 37.5 37.5
тура в тер- мостате (°C) Северо-гол- ландская
Порода кур — — — Леггорн Белый леггорн
Возраст (в днях) в период инкубации: 0.051
0 •—* — — —
1 0.165 —. 0.073 —. — —
2 0.094 — 0.143 —. 0.09 —
3 0.216 — 0.253 — 0.09 —
4 0.333 —. 0.381 — 0.35 —
5 0.628 — 0.524 — 0.57 —
6 0.922 — 0.831 — 0.94 —.
7 1.775 —. 1.421 1.6 1.39 —
8 2.428 — 1.671 —. 1.98 —
9 2.709 —. 2.101 1.6 2.7 —
10 3.705 — 3.200 4.0 3.9 3.47
И 5.319 — 3.980 8.4 6.2 4.38
12 9.045 — 6.055 8.5 10.0 6.18
13 5.425 7.50 7.516 12.0 14.0 9.03
14 6.759 8.65 11.533 16.0 19.3 12.35
15 12.519 10.11 13.533 — 20.1 13.86
16 14.621 10.79 15.863 18.0 22.4 —
17 15.973 13.40 17.340 17.0 23.5 20.43
18 22.780 —. 17.997 — 23.2 —.
19 14.743 18.21 18.161 18.0 22.4 20.91
20 20.568 22.00 19.231 18.0 28.2 33.64
20 — — — 34.5 — —
(после про- клева) 24.10 29.983
21 — — — —
22 — — 40.054 — — —
Цыплята — 58.79 — — 59.8 —
После вы- лупления (часы) 33.0
1/2 —• — — — «—
2 — — —— 44.0 — — -
24 — — 54.0
13 В. В. Рольник
193
метаболизма на грамм ткани авторы объясняют включением
функции щитовидной' железы. Это было подтверждено исследо-
ваниями газообмена после инъекции тироксина и тиоурацила
(Romijn et al., 1952).
В сконструированном нами газообменном аппарате (Роль-
ник, 1963а) исследовался одновременно, но раздельно газообмен
6—7 объектов (эмбрионов и цыплят), с большей точностью, чем
в имеющихся до сих пор газообменных приборах (рис. 41). Дан-
ные о проведенных нами (Рольник, 1961, 19636, 1965, 1966; Роль-
ник и Портенко, 1964) более 600 2—3-часовых определений газо-
обмена, начиная с 9-го дня эмбрионального и кончая четвертыми
сутками постэмбрионального развития, представлены в табл. 36.
Таблица 36
Потребление кислорода (мл/час) куриными эмбрионами и цыплятами
(По Рольник и Портенко, 1964)
Возраст (в днях) Среднее арифметиче- Коэффициент Увеличение потребле-
ское ± ошибка вариации ния О2 за сутки (в %)
В период инкубации
9 10 И 2.78+0.12 4.43+0.27 6.26+0.22 25.6 33.6 24.0 59.4 41.3 24.9 43.0 31.5 14.4 8.4 1.7 8.4 13.5 33.6
12 13 14 15 7.82+0.26 11.18+0.24 14.70+0.33 16.81 ±0.39 22.5 17.7 15.4 14.9
16 17 18.23+0.39 18.54+0.51 16.3 19.8
18 19 20.09+0.43 22.80±0.67 16.9 19.6
20 30.47+1.76 17.3
19* 20* 30.31+1.63 39.93+1.74 24.1 12.3 31.7
После вылупления
0 62.2+2.8 21.1
1 66.8+3.0 21.2
2 66.2+6.7 24.7
3 42.1 + 4.1 22.0
4 57.1+4.8 25.0
Примечание. Звездочкой отмечены яйца с наклевом.
Сравнение табл. 35 и 36 показывает, что потребление кисло-
рода куриными эмбрионами, по нашим данным, несколько выше,
чем это обнаружено другими исследователями. Мы предполо-
жили, что это различие обусловлено значительно меньшей вен-
194
тиляцией респирационных камер в использованных этими ис-
следователями приборах, чем в нашем газообменном аппарате.
Специально поставленными опытами нам удалось достоверно
показать, что сокращение вдвое вентиляции респирационных
камер снижало потребление О2 в среднем на 20% . Следовательно,
ваши данные по газообмену эмбрионов более точны.
Рис. 41. Общий вид прибора для изучения газообмена эмбрионов птиц.
(Ролышк, 1966).
Объяснение в тексте.
Прирост потребления кислорода, по нашим данным, про-
текает неравномерно, и особенное замедление его наблюдается
с 15 до 18 дней инкубации, что совпадает во времени с появлением
уплощения кривых содержания О2 и СО2 в воздушной камере
яйца. После проклева скорлупы газообмен сильно увеличивается
(на 32.9% на 19-й и на 31.0% на 20-й день инкубации), но на-
стоящий «взрыв метаболизма», по образному выражению Мит-
13*
195
челла с соавторами (Mitchell et al., 1927), наблюдается после
выхода цыпленка из скорлупы. В среднем потребление О2 только
что вылупившегося цыпленка на 55.8% выше, чем у 20-днев-
ного эмбриона, проклюнувшего скорлупу, и на 104.1 % выше,
чем у эмбриона того же возраста, но до проклева. Какова же
причина этого «взрыва метаболизма»? В какой-то мере он, ве-
роятно, обусловлен и увеличением подвижности, хотя проклев
скорлупы по кругу и выход цыпленка из яйца также требуют
большой механической работы. Мы считаем основной причиной
огромного увеличения газообмена после вылупления снятие
в это время такого серьезного препятствия для газообмена, каким
является скорлупа.
Потребление кислорода имеет большие индивидуальные ко-
лебания и зависит от условий инкубации (рис. 42). Следует от-
метить, что снижение газообмена в 16—18-дневном возрасте
наблюдается не только у гибнущих затем эмбрионов, но и у тех,
которые потом нормально вылупились. Газообмен эмбрионов,
которые впоследствии становятся задохликами, в некоторых
случаях становится значительно ниже, чем у нормально раз-
вивающихся эмбрионов, уже за несколько дней до гибели (рис. 42,
Б, 4)\ у других потребление О2 снижается только в день гибели,
а накануне оно практически такое же (рис. 42, Б, 1 и 3), как
у тех, которые нормально вылупятся (рис. 42, Б, 2). Сравним
эти наблюдения с нашими данными о составе газов воздушной
камеры яиц, где эмбрионы погибали в последние дни инкубации.
Эмбрионы с нормальным газообменом почти до конца инкуба-
ции (рис. 42, Б, 1 и 3) сходны с теми, которые развивались в яй-
цах № 14 и № 54 (табл. 34), имевших в воздушной камере к концу
инкубации вследствие плохой проницаемости скорлупы для
газов низкое срдеражние О2 и высокое СО2, т. е. были в полном
смысле задохликами. У других (рис. 42, Б, 4) — с газообменом,
ослабленным задолго до вылупления (т. е. вне зависимости от про-
ницаемости скорлупы), состав газов воздушной камеры ока-
зывается к концу инкубации близким к нормальному. Для них,
ослабленных в первые 16—17 дней инкубации, трудность пере-
хода от аллантоисного к легочному дыханию является лишь
допе.мнительным неблагоприятным фактором. Их по существу
нельзя назвать задохликами.
Рассмотрим данные о газообмене эмбрионов других видов
птиц. Махинько с сотрудниками (Махинько и Сердюк, 1954; Ма-
хинько и Шевченко, 1954) изучал дыхание утиных и голубиных
эмбрионов. Авторы показали, что за период инкубации утиный
эмбрион (26 дней) потребляет 4.9 л О2 и выделяет 4.2 л СО2, а го-
лубиный (17 дней) — 1.4 и 1.1 л соответственно. Авторы отме-
чают, что потребление кислорода на единицу образовавшейся
живой массы эмбриона у птиц разных видов примерно одинаково.
Газообмен утиных эмбрионов, по данным Никита (Nichita, 1955b),
196
имеет несколько бблыпие величины: за период инкубации по-
требляется в среднем 7.84 л О2и выделяется 5.95 л СО2; по наблю-
дениям Хаскина (1961) — 8.44 л Ь2 и 6.1 л СО2. Гусиный эмбрион
потребляет в среднем за период инкубации 20.5 л О2 и выделяет
14.0 л СО2 (Быховец, 1955). Газообмен утиных эмбрионов, по дан-
ным Хаскина (1961), протекает неровно: на 9-й, 13-й, 22-й и 25-й
дни инкубации наблюдается замедление прироста потребления О2
и выделения СО2, совпадающие с колебаниями прироста внутри-
яйцевой температуры. В период вылупления в связи с началом
легочного дыхания, усилением двигательной активности и об-
легчением доставки кислорода после проклева скорлупы пот-
ребление О2 увеличивается с 40 до 170 мл в час, т. е. более чем
в 4 раза.
Кенди (Kendeigh, 1940) изучил газообмен эмбрионов восточ-
ного домашнего крапивника (из отряда воробьиных). Выяснено,
что у них, так же как и у куриных эмбрионов, в первые два дня
развития наблюдается большое преобладание (в среднем в 3.4 раза)
выделения СО2 над потреблением О2. Следовательно, выделе-
ние СО2 из яйца в первые дни после снесения является законо-
мерностью. Автор показал, что газообмен эмбриона зависит
от температуры и увеличивается при повышении ее с 21.1° до 37.8°,
но снова снижается при 40.6°. Кенди считает газообмен удобным
индексом темпа роста и по нему определяет оптимальную тем-
пературу инкубации. Подтверждением того, что газообмен может
быть критерием развития, является исследование Мариакулан-
дай и Мак-Гинниса (Mariakulandai a. McGinnis, 1952), показав-
ших, что выделение углекислоты у эмбрионов с недостаточным
содержанием витамина В12 значительно ниже, чем у нормальных.
Остановимся на данных о дыхательном коэффициенте в те-
чение развития эмбриона. ^Напомним, что при окислении угле-
водов он равен 1.0, протеинов — 0.78, жиров — 0.7 и ниже.
На основании большого количества литературных данных Нид-
хем (Needham, 1931) сообщает, что дыхательный коэффициент
у куриного эмбриона с 2.5 до 7 дней равен около 1.0, на 7—8-й
день — 0.84, на 13-й — 0.71, а после этого редко поднимается
выше 0.7, а обычно бывает ниже. Автор делает вывод, что до 8-го
дня происходит в основном сгорание углеводов, с 8-го до 13-го —
протеинов, а после этого срока — жиров. Нидхем (Needham,
1942) считает, что катаболизм углеводов в первую неделю ин-
кубации не обусловлен отсутствием в это время какого-либо
механизма, необходимого для сгорания протеинов, так как при
торможении распада углеводов флуоридом появляются про-
дукты протеинового распада. По данным Хаскина (1959), ды-
хательный коэффициент у утиного эмбриона до 9-го дня равен
0.8—0.9, потом до 18-го дня редко опускается ниже 0.7, а за-
тем равен 0.7 или меньше при среднем за период инкубации
0.72.
197
Дни инкубации
Рис. 42. Индивидуальные вариации потребления кислорода куриными
эмбрионами при благоприятных (А) и неблагоприятных (Б) условиях
инкубации. (По Рольнпк п Портенко, 1964).
Для А'. 1У 4 и 5 — с проклевом скорлупы на 20-й и вылуплением на 21-й день
инкубации; 2 и 3 — с проклевом на 19-й и вылуплением на 20-й день инкубации.
Цифры в конце кривых — потребление кислорода (мл/час) после вылупления. Для
Б: J, з, 4 — у эмбрионов, погибших через несколько часов после измерения газо-
обмена на 18-й (1 и 4) и 19-й (з) день инкубации; 2 — у эмбриона с проклсвом
скорлупы на 19-й и вылуплением на 20-й день инкубации. По оси ординат —
потребление кислорода (мл/чар); по оси абсцисс — дни инкубации.
Интересное наблюдение сделал Ёэрнвелл (Barnwell, i960).
Поглощение кислорода куриным эмбрионом с 5-го по 7-й день
инкубации при постоянном освещении, температуре, давлении
и содержании кислорода и углекислоты в окружающей атмосфере
оказывается различным в разное время суток: в утренние часы
оно наименьшее, повышается в полдень и максимальное в 7 час.
вечера.
Не обсуждая подробно вопроса о дыхании отдельных тканей,
мы считаем необходимым все же остановиться на некоторых
исследованиях. По данным Нидхема (Needham, 1942), потребле-
ние кислорода аллантоисом и желточным мешком на 6-й день
инкубации составляет примерно 35% всего потребленного яй-
цом 02, а на 19-й — только 5%. Автор предполагает, что боль-
шее сродство эмбрионального гемоглобина к кислороду по срав-
нению с гемоглобином взрослых птиц обусловлено пониженным
парциальным давлением кислорода в желтке и белке инкубируе-
мого яйца в связи с недостаточной проницаемостью скорлупы
для газов. Автор сравнивает это с тем, что у птиц, живущих
в горах, гемоглобин имеет большее сродство к кислороду, чем
у сходных видов на уровне моря. О’Коннор (O’Connor, 1952)
наблюдал, что интенсивность дыхания эритроцитов снижается
в течение инкубации: на 3-й день потребление 02 равно 0.6 мм3
на 1 мм3 клеток, а на 8-й — 0.1. Махинько с сотрудниками (1956)
наблюдал, что интенсивность дыхания различных эмбриональ-
ных тканей по-разному изменяется в течение инкубации; у не-
которых тканей и органов (сердечная мышца, мозг) значительно
повышается, у других (скелетная мышца, печень), наоборот,
падает к концу эмбрионального развития. Авторы отмечают,
что интенсивность дыхания эмбриональных тканей выше, чем
у взрослых птиц.
Не имея возможности обсудить здесь весь энзиматический
механизм дыхания, мы считаем необходимым остановиться на ис-
следованиях об угольной ангидразе — энзиме, играющем основ-
ную роль в выведении углекислоты из организма. Как сообщает
Крепе (1943), угольная ангидраза в сотни раз ускоряет реакцию
растворения СО2 в воде. Без этого энзима из организма человека
могло бы выделиться не более 2% фактически выделяющейся
в легких углекислоты, так как реакции связывания и отдачи
гемоглобином О2 происходят в сотые доли секунды, а эти же
реакции с СО2 протекали бы значительно медленнее (целые ми-
нуты), если бы они не ускорялись угольной ангидразой. По дан-
ным Ван Гроора (Van Groor, 1940, цит. по: Needham, 1942), уголь-
ная ангидраза в крови куриного эмбриона появляется на 11-й
день, достигает уровня взрослой птицы на 15-й и почти в 3 раза
превышает его на 19-й. Тотю и Вуакюле (Tautu et Voiculet, 1958)
нашли, что синтез угольной ангидразы в крови куриного эмбриона
199
начинается на 7-й день инкубации.3 Резкое повышение актив-
ности угольной ангидразы в тканях куриного эмбриона со 2-го
до 10-го дня инкубации наблюдал Шепард (Shepard, 1962). Боль-
шую роль угольной ангидразы в жизненно важных процессах
в нервной системе подчеркивает Пигарева (1948). Автор наблю-
дала корреляцию между сроками изменения активности уголь-
ной ангидразы и сроками развития того или другого отдела мозга.
Так, у зрелорождающихся птиц (кур) сразу после вылупления
активность энзима высока, а у незрелорождающихся (грачей)
она устанавливается на высоком уровне позже, когда птицы
становятся способными к сложным формам деятельности.
Но у взрослых грачей, с высокой способностью к полету, актив-
ность угольной ангидразы во всех отделах головного мозга выше,
чем у нелетающих кур. У обоих видов максимальная активность
угольной ангидразы наблюдалась в наиболее деятельных частях
головного мозга — мозжечке и больших полушариях.
Благодаря возможности одновременно и в одинаковых усло-
виях исследовать газообмен 6—7 эмбрионов при помощи нашего
газообменного аппарата, нам удалось (Рольник и Портенко,
1964) обнаружить различия в потреблении кислорода эмбрионами
разного пола (табл. 37). Начиная с 10-дневного возраста потреб-
ление кислорода эмбрионами мужского пола становится выше,
чем женского, и такое превышение продолжается в течение всего
инкубационного периода (за исключением 18-го дня, по не-
известной нам причине). В среднем потребление кислорода эмб-
рионами мужского пола с 10- до 19-дневного возраста на 7.4%
выше, чем женского. По нашим данным, вес эмбрионов самцов
в среднем за тот же период на 5.5% больше веса самок. Таким
образом, часть превышения газообмена самцов обусловлена
большей интенсивностью у них потребления кислорода на грамм
ткани. Наши данные о различии газообмена эмбрионов разных
полов с 10-дневного возраста подтверждают предположение
Мицкевича (1957), что к 11-му дню инкубации начинается вы-
деление половых гормонов в кровь. Как сообщает Кагияма (Ка-
gijama, 1933, цит. по: Groebbels, 1937), куриные эмбрионы муж-
ского пола имеют бблыпую окислительную способность с 14-днев-
ного возраста. В этой связи интересно, что Ридль (Riddle, 1920)
наблюдал меньшую гибель голубиных эмбрионов мужского пола
при высокой концентрации кислорода, а при низкой — эмбрио-
нов женского пола.
Проводя исследования газообмена, мы (Рольник, 1964; Роль-
ник и Портенко, 1964) натолкнулись, как нам кажется, на ин-
тересный феномен. В проведенных нами трех сериях наблюде-
ний по газообмену, несколько различавшихся по температуре
3 Отметим, что это на четыре дня ранее, чем по данным Ван Гроора, и
совпадает по времени с переходом на аллантоисное дыхание, а также с появ-
лением рефлекторных реакций.
200
Таблица 37
Превышение в потреблении кислорода куриными эмбрионами
мужского пола в течение инкубации
(По Рольник и Портенко, 1964)
Продол- житель- ность инкубации (в днях) Количе- ство 30-минут- ных замеров Среднее взвешенное превышение потреб- ления О2 ** над $$ Ошибка среднего превыше- ния (т) м т Достоверность
мл/30 мин. (М) % к сред- нему $$
9 26 -0.0465 -3.45 0.04395 -1.06 Не достоверно обратное раз- личие
10 14 0.2196 10.32 0.1034 2.12 Достоверно на 95%
И 40 0.2048 6.67 0.0831 2.46 То же
12 32 0.3350 9.66 0.0926 3.62 Достоверно на 99.9%
13 41 0.6127 11.09 0.1240 4.94 То же
14 38 0.3726 5.04 0.1376 2.71 Достоверно на QK0Z
15 29 0.4103 4.88 0.1913 2.14 У О 70 То же
16 38 0.4652 4.92 0.1752 2.66 » »
17 33 0.9710 10.30 0.1632 5.77 Достоверно на 99.9%
18 35 -0.0719 -0.67 0.1290 -0.06 Не достоверно обратное раз- личие
19 49 1.3324 11.45 0.3799 3.51 Достоверно на 99.9%
инкубирования яиц (но в пределах оптимума), был обнаружен
разный ход кривых потребления кислорода эмбрионами (рис. 43).
В 1-й серии, с наиболее высокой (в амплитуде оптимума) тем-
пературой инкубирования, потребление'кислорода с 9-го до 14-го
дня инкубации было наибольшим, во 2-й, со средней темпера-
турой, — более низким, а в 3-й, с наиболее низкой температу-
рой, — наименьшим. Но на 14-й день газообмен эмбрионов всех
трех серий уравнивался, а в последующие дни развития вели-
чины потребления кислорода эмбрионами разных серий менялись
местами: наибольший газообмен наблюдался в 3-й серии, ниже —
во 2-й и наименьший — в 1-й. Общее количество потребленного
кислорода (с 10-го по 19-й день) было практически одинаково
во всех трех сериях: в первой — 3429.6 мл, во второй — 3432 мл,
в третьей — 3507.1 мл. Таким образом, различие в крайних
величинах составляет всего 2.2%. Этот феномен регулирования
величины эмбрионального газообмена в течение периода инку-
бации мы назвали закономерностью биологической компенсации.
Дыхание эмбрионов при различном содержании газов (кисло-
рода и углекислоты) в окружающей атмосфере будет обсуждено
201
в главе 20. Здесь мы упомянем только о некоторых исследованиях
по патологии эмбрионального развития под влиянием изменен-
ного газового состава и об экспериментальном изучении влияния
газов, не связанных с практической инкубацией.
О типе патологий эмбрионального развития, вызванных не-
Рис. 43. Потребление кислорода
куриными эмбрионами при изме-
нении условий инкубации в пре-
делах нормы (проявление зако-
номерности биологической ком-
пенсации). (По Рольник и Пор-
тенко, 1964).
большое количество исследований,
причем различные концентрации
О2 и в разные периоды разви-
тия обусловливают разные урод-
ства. Вот результаты некоторых
из этих исследований. При ки-
слородном голодании в течение
первых трех дней инкубации у
24 куриных эмбрионов (из 164)
наблюдались аномалии сердца и
больших сосудов, у 15 — дефекты
межжелудочковой и межпредсерд-
ной перегородок сердца и у 19 —
другие уродства (Schellong, 1954).
Басс (Buss, 1956) установил, что
при пониженной концентрации
кислорода замедляется образова-
ние гемоглобина, в связи с чем
увеличивается продолжительность
инкубационного срока. Бюхнер
с соавторами (Buchner et al.,
1953а, 1953b) показал, что крат-
ковременное кислородное голода-
ние (3—5% О2) на ранних ста-
диях развития вызывает наруше-
ние дифференциации бластодиска
и повреждение уже сформировав-
шихся структур. На более цозд-
них стадиях кислородное голода-
I, 2, з — кривые потребления О2 эмб- НИ6 ведет К ПОЯВЛ6НИЮ у ВН6ШН6
ств°епнмо“ По ’оси оИр03Хт-йпот^^ нормальных эмбрионов недиффе-
ние О2 (мл/час); по оси абсцисс—дни ренцированНЫХ КЛвТОК В ГОЛОВ-
инкубации. -г»
ном и спинном мозгу. В иссле-
довании Нельсена (Nelsen, 1960)
показано влияние гипоксии на различные отделы головного
мозга. Выяснено, что дифференциация глаз и переднего мозга
требует болеа высокого процента кислорода (свыше 10%), чем
развитие заднего мозга, который хорошо функционирует в ат-
мосфере 6% О2. Автор предполагает, что это связано с большей
необходимостью переднего и среднего мозга в аэробном мета-
болизме. В известной степени подтверждением этого является
исследование Грабовского (Grabowski, 1961), показавшего, чтр
202
наблюдаемые у 3-дневного куриного эмбриона после периода
сильной гипоксии (1% О2 в течение 3 час. или 4—8% О2 в течение
12 час.) аномалии развития обусловливаются накоплением мо-
лочной кислоты в результате анаэробного обмена. В основном
это аномалии головы, причем чем сильнее гипоксия, тем больше
уродство. Волохов (1951) выяснил, что при недостатке кисло-
рода подавляется влияние коры головного мозга на нижележащие
рефлекторные механизмы и выявляются онтогенетически более
ранние формы рефлекторной деятельности.
Повышение концентрации кислорода выше 50% также обус-
ловливает патологию в развитии эмбриона. Аллен (Alien, 1961)
экспериментально доказал, что токсическое влияние 100% кисло-
рода в течение первых четырех дней развития куриного эмбриона
сказывается непосредственно на кровообращении: не развивается
желточное кровообращение (из-за кровоизлияний в сосудах),
и сердце не способно начать пульсацию.
По интересным наблюдениям Хамметта и Золь (Hammett
a. Zoll, 1928, цит. по: Needham, 1931), сосуды желточного мешка
чрезвычайно чувствительны к концентрации углекислоты в крови
и при увеличении ее содержания диаметр их сильно сокращается.
Авторы делают вывод, что количество СО2 в крови служит как бы
автоматической регулировкой скорости развития при разной
температуре инкубации. При высокой температуре увеличивается
количество углекислоты, желточные сосуды сокращаются, приток
крови к эмбриону уменьшается и эмбрион, таким образом, пре-
дохраняется от подачи большего количества пищи, чем он может
потребить. И наоборот, при низкой температуре большее коли-
чество крови поступает к эмбриону, но с меньшим содержанием
питательных веществ и снабжает эмбрион адекватным количеством
материала для непрерывного развития/
Несколько слов об исследованиях по изучению влияния газов,
не связанных с практической инкубацией. Бакунина (Bakounine,
1895а) отмечает, что наличие угарного газа в воздухе куриный
эмбрион хорошо переносит до 6—8 дней развития и тем лучше,
чем он моложе. После 8-го дня эта толерантность внезапно
прекращается.
В двух сериях опытов с нашим участием (Борискин и др.,
1962; Савин и др., 1964) удалось показать, что азот воздуха со-
вершенно не необходим для развития куриных эмбрионов и может
быть заменен другим инертным газом — гелием. Содержание
яиц в течение всего периода инкубации в специальной установке,
через которую пропускалась гелиокислородная смесь (кисло-
рода 21% и гелия 79%), не помешало нормальному течению
развития и выведению цыплят. Выведенные при этом цыплята
развивались затем нормально в течение полутора лет (Рольник
и Гаврилов, 1962).
2 03
ГЛАВА 11
РАЗВИТИЕ ОРГАНОВ ПИЩЕВАРЕНИЯ
Как правильно замечает Рудник (Rudnick, 1952) в
своем обзоре по развитию пищеварительной системы у эмбрио-
нов птиц, этому разделу знаний особенно не повезло и он до сих
пор мало привлекал внимание исследователей.
А. Морфология пищеварительного тракта
Передняя кишка появляется раньше других частей пи-
щеварительного тракта и первой начинает дифференцироваться.
У 50—55-часового куриного эмбриона передняя кишка имеет
уже отделы (глотку и пищевод), задняя кишка в это время только
впервые появляется, а средняя кишка еще не имеет энтодерма ль-
ной подстилки снизу (ее подстилает желток). Затем весь ки-
шечник подстилается энтодермой, и с желтком его связывает
только пупочный канатик. При закрытии брюшной полости
(целбма) кишка остается окруженной листком внутренностной
мезодермы, а выше и ниже ее эти листки смыкаются, образуя
двуслойные брыжейки. Спинная брыжейка остается, а брюшная
почти полностью исчезает, после чего правая и левая части брюш-
ной полости сливаются в одну. Та часть брюшной брыжейки,
которая лежит в районе печени, остается и поддерживает ее со спин-
ной и брюшной стороны.
1. Ротовая полость и глотка
Глоточный отдел передней кишки лежит под продолгова-
тым мозгом и окружен дугами аорты. У 50—55-часового эм-
бриона еще нет ротового отверстия, но уже образовалось впя-
чивание эктодермы, а навстречу ему — выпячивание эндотермы
глотки. В результате образуется тонкая ротовая пластинка,
которая на 3-й день прорывается, соединяя глотку с ротовой
полостью. Эндотерма глотки дает ряд образований, не являю-
щихся частями пищеварительной системы: щитовидные и около-
щитовидные железы, тимус, трахею.
После 4 дней инкубации в ротовой полости и вокруг нее по-
являются клюв, яйцевой зуб, язык и железы ротовой полости.
Клюв образуется при ороговении быстро размножающихся эпи-
дермальных клеток краев рта. Образование яйцевого зуба, спе-
циального приспособления для проклева скорлупы при вылуп-
лении, начинается у куриного эмбриона на 6-й день развития
в виде бугорка из эпидермальных клеток на середине верхней
губы, который на 14-й день принимает форму зуба.. Передняя
часть языка утолщается и на 8-й день ороговевает.
204
Каллиус (kallius, 1905) обнаружил ё ротовой полости ути-
ного эмбриона следующие железы: 3 типа желез на языке (пе-
редние, задние и глоточные), а также подъязычные и много-
численные железы в стенке глотки. Образование всех этих желез
начинается в виде сплошных врастаний слизистой оболочки;
затем они ветвятся, и лишь потом в них образуется полость.
У ныне живущих птиц нет настоящих зубов, но у ископаемых
птиц они были. Интересно отметить, что у 6-дневного куриного
эмбриона на внутреннем крае челюстей имеется утолщение экто-
дермы в виде гребешка, который является рудиментом зубов
и вскоре после начала ороговения губ пропадает.
Глотка, являющаяся большой и важной частью пищевари-
тельного тракта на ранних стадиях развития (в связи с раз-
витием жаберных дуг и щелей), в процессе развития ста-
новится относительно меньше, хотя абсолютный размер ее уве-
личивается.
2. Пищевод, железистый и мышечный желудок,
кишечник
У 4-дневного эмбриона имеется уже дифференциация перед-
ней кишки на глотку, пищевод (суженная часть) и мышечный
желудок (расширенная часть). Вокруг эктодермальной трубки
пищевода и желудка происходит концентрация мезенхимных
клеток, из которых образуются мышечная и соединительно-
тканная оболочки.
Пищевод в связи с удлинением шеи быстро превращается
в длинную трубку. Зоб возникает как веретенообразное расши-
рение пищевода, и уже на 11-й день развития куриного эмбриона
его диаметр в два раза больше, чем у рядом расположенных от-
делов пищевода. Алленспах и Гамильтон (Allenspach a. Hamil-
ton, 1962) показали, что у куриного эмбриона ранее открытый
пищевод в 5-дневном возрасте становится закупоренным на до-
вольно длинном отрезке за трахеальным расширением. Вновь
он открывается на 8-й день инкубации, а секреция в его железах
начинается на 12—24 часа раньше. Изучая развитие пищевари-
тельной системы в эмбриогенезе кур, Квинихидзе (1958, 1964)
отмечает, что на ранних стадиях (3—5 дней инкубации) пище-
варительная трубка имеет по всей длине примерно одинаковый
диаметр с небольшим расширением в области желудка. Пищевод
и железистый желудок в это время выстланы однослойным эпи-
телием, который к 11—12-му дню превращается в многослойный.
Пути превращения в многослойный эпителий в разных участках
пищеварительного тракта различны. Появление на определен-
ной стадии эмбрионального развития кур мерцательных эле-
ментов в эпителии пищевода автор считает признаком, сохра-
нившимся с древнейших времен от животных, имевших функцио-
206
пировавший мерцательный покров, постепенно ставший иным
в соответствии с изменившимися условиями существования.
Мерцательные клетки и образующиеся после 13-го дня слизистые
железы способствуют перемещению в пищеводе заглатываемого
в это время
эмбрионом белка (о
3
Рис. 44. Внутренности
Рис. 44. Внутренности 6-дневного ку-
риного эмбриона (с правой стороны).
(По Lillie, 1952).
1 — прямая кишка; 2 — изгиб кишечника
у перехода двенадцатиперстной в тонкую
кишку; з — средняя почка; 4 — правое лег-
кое; 6 — правое предсердие; 6 — артериаль-
ная луковица; 7 — желудочек сердца; «—пе-
чень; 9 — мышечный желудок; 10 — двенад-
цатиперстная кишка; 11 — стебелек желточ-
ного мешка; 12 — отросток слепой кишки;
13 — аллантоис.
чем речь будет дальше).
Пищеварительная трубка
растет в длину значительно
быстрее, чем туловище эм-
бриона, и поэтому она де-
лает изгибы в виде петель:
1-ю петлю делают пищевод и
желудок, 2-ю — двенадцати-
перстная кишка (в ней ле-
жит поджелудочная железа),
3-ю — тонкая и толстая киш-
ки (рис. 44). 3-я петля вна-
чале опускается в пуповину
, и желточный стелебек, а на
17—18-й день инкубации под-
нимается в брюшную по-
лость.
Различие между желези-
стым и мышечным желудком
появляется впервые на 5-й
день в результате заметного
сужения между ними. В стен-
ках железистого желудка на-
чиная с 7-го дня инкубации
образуются сложные, много-
’ дольчатые железы, секрет ко-
торых после вылупления слу-
жит для переваривания пи-
щи. У 14-дневного эмбриона
эти железы составляют основ-
ную массу стенок желези-
желудка, а мышечная ткань его развита сравнительно
(подробнее см. стр. 214). В стенках мышечного же-
после 7-го дня инкубации образуется большая масса
стого
слабо
лудка
мышц, а на 13—14-й день — простые трубчатые железы, выде-
ляющие секрет, который, затвердевая, образует выстилающую
его внутри кутикулу.
На границе между тонкой и толстой кишкой на 6-й день эмбрио-
нального развития появляется двустороннее вздутие, которое
быстро вырастает в два слепых отростка.. Сама толстая кишка
в это время постепенно теряет просвет, который потом восста-
навливается в верхней части толстой кишки в течение 12—13-го
дня, а в нижней — накануне вылупления. Толстая кишка без
206
четкой отделительной линии переходит в большой передний
отдел клоаки. Далее находится относительно короткий промежу-
точный отдел клоаки, в который открываются мочеточники,
половые протоки и являющийся гомологом мочевого пузыря
млекопитающих аллантоис. Клоака появляется впервые у 4-днев-
ного эмбриона в виде расширения хвостовой части задней кишки.
Двуслойная перепонка, образующаяся при выпячивании энто-
дермы клоаки и впячивании эктодермы, называется клоакальной
перепонкой. Она прорывается накануне вылупления, образуя
заднепроходное отверстие — анус. Конечная камера клоаки об-
разуется из впячивания эктодермы и находится уже за клоа-
кальной перепонкой.
Весь кишечник поддерживается спинной брыжейкой. В про-
цессе развития эмбриона в связи с ростом кишок нижний конец
брыжейки удлиняется, делает складки и закручивается, пово-
рачиваясь вместе с различными изгибами и поворотами кишок.
Клюева (1958) изучила развитие пищеварительного тракта
у утиных эмбрионов и обнаружила, что на 3-й день инкубации
эктодермальная пластинка еще распластана на желтке; на 6-й
уже имеется пищевод, железистый и мышечный желудок, две-
надцатиперстная и тонкие кишки, а также маленькие слепые
выросты прямой кишки; на 8-й в железистом желудке еще нет
сложных желез, а только небольшие впячивания эпителия;
на 17-й железы уже функционируют; на 23-й количество и слож-
ность желез увеличивается.
Фабрициева сумка (образование, характерное только для
птиц) тоже является производной пищеварительного тракта.
Б. Печень (структура и функция)
Печень возникает в середине 3-го дня. развития куриного
эмбриона в виде выпячивания на заднем конце передней кишки,
непосредственно позади соединения желточно-брыжеечных вен.
К концу 3-го дня, когда зародыш значительно обособился
от желточного мешка, печень оказывается отходящей от той
части пищеварительного канала, которая позднее становится
двенадцатиперстной кишкой. У 4-дневного эмбриона печень
сильно вырастает и образует ветвящиеся клеточные стволики,
становящиеся в дальнейшем секреторными элементами печени.
С ростом печени ее ткань окружает слившуюся часть желточно-
брыжеечных вен. Это предопределяет в будущем сохранение
части желточного кровообращения в виде печеночно-воротной
системы. В дальнейшем печеночный вырост окружается внутрен-
ностной мезодермой. Место первоначального выпячивания остается
открытым в кишечный тракт и служит протоком печени.
Печень — вначале непарный, расположенный посередине ор-
ган, В свдзи с тем что в правой стороне тела больше места, правая
Ж
доля печени разрастается сильнее. Внутри правой доли печени
находится сложная сеть желчных капилляров, располагающихся
между печеночными клетками и соединяющихся в более крупные
желчные ходы, впадающие, наконец, в выводной печеночный про-
ток — желчный, оканчивающийся желчным пузырем. Левая доля
печени имеет такую же внутреннюю структуру, как и правая,
а ее проток (печеночно-кишечный) впадает в двенадцатиперстную
кишку независимо от желчного пузыря.
На 6-й день развития куриного эмбриона, по данным Крок
(1949а), начинается формирование желчных капилляров, вЛсруг
которых располагается по 6—8 печеночных клеток. Автор пред-
полагает, что в это время начинается желчеобразование. В 9-днев-
ном возрасте в печени эмбрионов наблюдается ретикулярная
ткань вокруг наиболее крупных кровеносных сосудов, что го-
ворит о наличии внесосудистого гемопоэза в печени в это время.
К 20-му дню инкубации паренхима печени по своему строению
приближается к печени вылупившегося цыпленка. Гистоло-
гическая структура печени куриного эмбриона становится сход-
ной с печенью взрослой птицы уже на 15-й день инкубации (Kin-
gsbury et al., 1956). Содержание липидов в клетках печени по-
степенно повышается с 5-го до 15-го дня и сильно увеличивается
на 17-й день инкубации, что вызывает изменение цвета печени;
она желтеет, так как желтый пигмент переносится липидами
из желтка яйца в клетки печени. Лаврентьева (1963) обнаружила,
что образование желчи и поступление ее в желчный пузырь ку-
риного эмбриона происходит на 10-й день, а на следующий день
желчь попадает в пищеварительный тракт. На 18—20-й день
развития желчь эмбриона по своему составу близка к желчи
взрослых кур и способна активизировать липазу поджелудочной
железы.
Электронномикроскопические наблюдения Каррера (Каггег,
1961) показали, что желчные канальцы видны уже в печени
6-дневного эмбриона. Автор наблюдал гемопоэз в печени только
внутри сосудов начиная с 8-дневного возраста.
Щербина (1959) обнаружил, что печень начинает образовы-
ваться у 4-дневного гусиного эмбриона, а желчеобразование
в ней начинается на 6-й день инкубации.
У взрослой птицы печень имеет несколько различных функций.
Печени принадлежит центральное место в обмене веществ. Она
играет важную роль в углеводном, протеиновом и жировом об-
мене, а также в обмене железа. Она способствует пищеварению,
выделяя внутрь кишечника желчь. Кроме того, печень является
чрезвычайно важным кровоочистительным органом, нейтрали-
зующим вредные продукты распада пищевых веществ и образую-
щим при этом мочевину, мочевую кислоту и другие вещества.
В печени происходит распад кровяных телец. Наконец, печень
служит запасным резервуаром для углеводов, превращая в глико-
ген приносимые кровью моносахариды, а также синтезируя
гликоген из продуктов распада жиров и из дезаминированных
аминокислот. Накопление гликогена в печени регулируется
железами внутренней секреции и нервной системой.
У птичьего эмбриона все эти функции печени вступают в строй
в разное время. Пищеварительная функция печени у куриного
эмбриона, по-видимому, начинается с установлением печеночного
воротного кровообращения — с 7—8-го дня инкубации. Ка-
пельки жира, наблюдаемые в печени 6-дневного эмбриона —
это холестерин. В печени из холестерина образуются желчные
кислоты. Участие печени эмбриона в жировом обмене, по данным
Крок (1949а), начинается с 9-дневного возраста. На 18—20-й
день инкубации величина капель жира и их количество в прото-
плазме клеток печени увеличивается. После 17-го дня содержание
липидов в печени куриного эмбриона резко повышается, а за весь
период развития количество холестериновых эфиров, являющихся
основными веществами, переносящими холестерин из желтка
в печень, увеличивается в 350 раз (Feldman a. Grantham, 1964).
До 10-го дня печень эмбриона не способна расщеплять насы-
щенные жирные кислоты, которые эмбрион адсорбирует из желтка,
и в связи с этим используются только ненасыщенные жирные
кислоты. Авторы предполагают, что отсутствие ненасыщенных
кислот в достаточном количестве препятствует большему исполь-
зованию жира на ранних стадиях развития для питания эмбриона.
Галвяло и Горюхина (1937) отметили значительную диастати-
ческую и липолитическую активность печени куриного эмбриона.
Накопление гликогена и жиров в печени показывает, по мнению
авторов, что эта энзиматическая активность направлена в сторону
синтеза. Джорж и Томас (George a. Thomas, 1962) исследовали
активность липазы в печени развивающегося куриного эмбриона.
В гомогенате печени она повышается постепенно с 8-го до 14-го
дня, а затем резко увеличивается до максимума на 16-й день.
Авторы подчеркивают, что высокая активность липазы в эти
дни связана с усилением обмена жира в печени, так как жир
является основным источником эмбрионального питания (около
90%). Активность пептидазы в печени куриного эмбриона, по дан-
ным Дамма и Леви (Dumm a. Levy, 1949), увеличивается с 5-го
до 8-го дня в 16 раз, а к 14-му дню — в 20 раз.
Нейтрализация вредных продуктов распада питательных ве-
ществ начинается в печени с 7—8-го дня инкубации.
Гликогенная функция печени развивается с 7-го дня инку-
бации. До этого у куриного эмбриона гликогенную функцию
выполняет «временная печень» — бластодерма (Клод Бернар,
1859, цит. по: Лейбсон, 1962). С 12-го дня печень становится
основным местом запасов гликогена. С 8-го по 11-й день 90% гли-
когена находится вне эмбриона, а с 12-го до 20-го, как показал
Владимиров (Wladimirov, 1930), гликоген печени составляет
14 В. В. Рольпик
209
35—40% всего гликогена, находящегося в яйце. По электронно-
микроскопическим наблюдениям Каррера (Каггег, 1961), гли-
коген в печени куриного эмбриона имеет вид маленьких гранул
(около 30 ммк), собранных в гроздья.
Таблица 38
Содержание гликогена в печени куриных эмбрионов в течение их развития
(По Лейбсону, 1950)
Дни инкубации Средний вес печени (в мг) .Гликогена в печени (среднее) Средний вес эмбриона, г
в мг в %
9 16.6+6 0.09+0.006 0.52+0.16 1.10+0.80
10 30+6 0.11 + 0.05 0.35+0.18 1.85±0.20
11 47+5 0.10+0.06 0.22+0.13 2.90±0.60
12 89+12 0.12+0.07 0.13+0.08 4.40±0.45
13 111±17 0.54+0.22 0.49+0.17 5.55±0.85
14 172+37 0.98+0.43 0.59+0.34 8.15±0.85
15. 209+48 2.89+1.51 1.31+0.46 11.5+1.00
16 300+53 3.79+1.19 1.29+0.25 12.4±1.7
17 417±34 7.59+3.18 1.81+0.73 16.95+0.9
18 • 498±65 9.25±1.77 1.89+0.38 19.4±1.4
19 531+91 14.79+5.8 2.63+0.82 22.0±2.45
20 565+79 8.24+2.5 1.49±0.47 24.8±2.95
21 612+96 4.66+3.66 0.74+0.46 35±4.1
Вылупившиеся
20 632+14 4.68+1.36 0.74+0.2 24.5±0.45
21 567±199 5.28+1.7 0.93+0.05 37.9±1.95
Данные о накоплении гликогена в печени в течение развития
куриного эмбриона приведены в детальном исследовании Лейб-
сона (1950) (табл. 38). Обсуждая полученные им данные, автор
предполагает, что снижение количества гликогена в печени с 10-го
до 13-го дня инкубации вызвано тем, что клетки печени в это время
начинают вырабатывать желчь. Уменьшение гликогена в печени
перед вылуплением цыпденка автор склонен объяснить не только
расходом гликогена на процесс вылупления, но и тем, что при
втягивании и потреблении желтка печень обогащается жирами
(причем способность откладывать гликоген уменьшается), а также
включением функции гипофиза и щитовидной железы.
По данным О’Коннор (O’Connor, 1953), наиболее раннее появ-
ление гликогена в печени куриных эмбрионов — это 6.9+0.14
дней инкубации. По данным Рымничану с соавторами (Rimni-
ceanu et al., 1955), синтез гликогена в печени эмбриона начинается
на 6-й день инкубации в летние месяцы и между 7-м и 10-м днем
зимой и весной.
21Q
Ряд энзимов, участвующих в гликогенезе, Ринодо (Rinaudo,
1962) обнаружил в печени куриного эмбриона с 8-го дня развития,
причем их активность сначала равна* или несколько выше актив-
ности после вылупления, а к 14—18-му дню увеличивается в 3—4
раза, после чего бысто падает. По данным Килшеймера, Вебера и
Ашмора (Kilsheimer et al., 1960), глюкозо-6-фосфатаза (один из
ферментов, необходимых для превращения гликогена в глюкозу)
имеется в печени куриного эмбриона уже при появлении первых
печеночных клеток. Лейбсон, Плисецкая и Огородникова (1963)
показали, что активность глюкозо-6-фосфатазы в печени с 10-го
дня инкубации уже имеет значительные величины.
Катини (Catini, 1963) на основании своих исследований при-
ходит к выводу, что в регуляции синтеза гликогена в печени
имеется две фазы: а) внутрипеченочная, не зависимая от эндо-
кринной функции поджелудочной железы, и б) после 15-го дня,
когда поджелудочная железа уже выделяет гормоны. Тормо-
жение секреции поджелудочной железы в период ее активности
(с 15-го дня) вызывало увеличение содержания гликогена
в печени.
Подводя итоги многочисленным исследованиям о поддержании
определенного уровня сахара в крови, Лейбсон (1962) убеди-
тельно показывает, что печень обладает способностью без каких-
либо нервных или эндокринных влияний реагировать на изме-
нение концентрации сахара в крови. Обладая местным гомеоста-
тическим механизмом, печень при уменьшении концентрации
сахара в крови выделяет большее количество его за счет гликоге-
нолиза, а при увеличении — меньшее; при значительной же гипер-
гликемии печень использует глюкозу для синтеза гликогена и ли-
пидов. Автор подчеркивает, что с определенного этапа онтогенеза
и филогенеза гликогенная функция печени находится под контро-
лем нервной и эндокринной систем, причем у высших животных
нервная регуляция этой функции осуществляется в основном
через эндокринные железы.
В. Поджелудочная железа (структура и функция)
Поджелудочная железа возникает в виде трех зачатков,
появляющихся как выпячивания передней кишки: первый — на
спинной стороне, напротив печеночного выроста, на 72-м часу
инкубации, второй и третий — на брюшной стороне кишки к концу
4-го дня. Протоки последних двух впадают в двенадцатиперстную
кишку общим протоком с печенью. На концах каждого зачатка
возникают не имеющие полости выросты, которые много раз
ветвятся в окружающей мезенхиме и образуют секретирующую
часть поджелудочной железы. На 7-й день все три зачатка под-
желудочной железы лежат в тесном соприкосновении друг с дру-
гом, а затем и вовсе сливаются, оставляя раздельными только
14*
211
протоки. Изучая развитие экзокринной ткани поджелудочной
железы куриного эмбриона под электронным микроскопом, Зи-
гель (Zeigel, 1962) обнаружил, что секреторный материал появ-
ляется в ней в виде гранул в клетках 6-дневного эмбриона; уве-
личение секреции происходит с 11-го до 15-го дня, а зрелая кон-
фигурация клеток наблюдается примерно к моменту вылупления
(см. стр. 214).
Клюева (1958) показала, что поджелудочная железа утиного
эмбриона отделяется от слизистой оболочки кишки на 4-й день
инкубации; на 9-й она расположена в петле двенадцатиперстной
кишки и имеет еще рыхлое строение; на 13-й день в мезенхиме
между эпителиальными трубками железы повляются кровеносные
сосуды и изредка одна-две клетки со светлыми ядрами (начало
образования внутрисекреторной части железы); на 17-й строение
железы усложняется и напоминает железу взрослых птиц, но еще
не разделенную на дольки и с неразвитой соединительной тканью;
в это время островки Лангерганса встречаются редко. Полностью
железа дифференцируется к 26-му дню инкубации.
Поджелудочная железа выделяет сок, расщепляющий белки,
углеводы и жиры. Сок поджелудочной железы взрослых птиц
имеет щелочную реакцию и содержит трипсин, липазу, протеазу
и амилазу. Поджелудочная железа птиц выделяет значительно
большее количество сока, чем у млекопитающих. Об эндокринной
роли поджелудочной железы см. главу 13.
Г. Внутрикишечное питание эмбриона
Как считалось ранее, переваривание питательных
веществ во время эмбрионального развития производится только
желточным мешком.
Врбич (Virbitche, 1924), по-видимому, первый обнаружил
заглатывание амниотической жидкости куриным эмбрионом, инъе-
цируя суспензию ликоподиума в амниотическую жидкость и наб-
людая за временем ее появления в желудке и кишечнике эмбриона.
Галвяло и Горюхина (1937), отмечая накопление протеолитиче-
ского энзима в желудке развивающегося куриного эмбриона после
14-го дня инкубации, очевидно, не знали работы Врбич и выска-
зывали удивление по поводу причины этого, так как не могли
понять значение наличия этого специфического энзима в желудке
эмбриона. Вслед за Врбич Шейнис (1954) показала, что белок
яйца, проходящий через серозо-амниотический канал вместе с
амниотической жидкостью, заглатывается эмбрионами разных
птиц (кур, воробьев, чижей и других мелких птиц) и перевари-
вается внутрикишечно. Автор отмечает, что прорыв серозо-амнио-
тического сращения у птиц по сравнению с рептилиями приобретает
новую роль: он служит местом проникновения к эмбриону высоко-
питательного материала, в то время как у черепахи его роль
212
сводится к тому, что в момент вылупления эмбрион, таким образом,
освобождается от зародышевых оболочек. Автор подчеркивает,
что более раннее появление прорыва серозо-амниотического сра-
щения у птиц по сравнению с рептилиями говорит о прогрессивной
эволюции этого образования. Этот сдвиг сроков происходит по
давно установленному в сравнительной эмбриологии правилу,
что прогрессирующий орган дифференцируется в онтогенезе
высших форм на более ранней стадии.
Почти одновременно изучали внутрикишечное пищеварение
в эмбриогенезе птиц в СССР Рагозина и в Чехословакии Гашкова
и Покорная. По данным Рагозиной (наиболее полно изложенным
в ее монографии 1961 г.), внутрикишечный способ питания кури-
ного эмбриона начинается на 9-й день инкубации. В этот день
необходимо 10 час., чтобы проглоченная амниотическая жид-
кость достигла желудка; на 12-й день этот процесс требует 8 час.,
а на 16-й день только 2 час. Автор подчеркивает большую роль
начала заглатывания амниотической жидкости в переходе от вне-
кишечного способа питания желтком к смешанному, а также в ком-
пенсации недостающего количества жидкости ( желток уже стал
более густым) и в растяжении стенок кишечника, способствующих
его нормальному росту и дифференцировке кишечных складок.
После прохода через серозо-амниотический канал (на 13-й день)
и растворения в амниотической жидкости белок яйца начинает
внутрикишечно перевариваться, и этот способ питания становится
основным. Перемещение белка через серозо-амниотический канал,
по данным автора, интенсивней всего идет в течение 15-х суток
и заканчивается к концу 19-го дня инкубации (см. рис. 21). Автор
считает, что переходу белка в амниотическую жидкость способ-
ствует ряд факторов, действующих одновременно: а) давление
на белок увеличивающегося в размере эмбриона и растущего
объема аллантоисной жидкости, б) глотательные движения эм-
бриона, как бы насасывающие белок в амнион и в) разжижение
белка благодаря секреторной деятельности эктодермальной вы-
стилки стенок белкового мешка.
В ряде работ Гашкова и Покорная (1955) детально изучили
«пероральное питание» (термин авторов) куриного эмбриона и
доказали иммунологическими методами наличие белка в желудке
эмбриона с 13-го дня инкубации. Питание амниотической жид-
костью с растворенным в ней белком (яйца) продолжается при-
мерно до 18-го дня, и авторы отмечают, что это период особенно
интенсивного роста и развития куриного эмбриона.
Мышечная активность пищеварительного тракта куриного
эмбриона начинается рано. Куо (Кио, 1932а), исследуя глотатель-
ные движения эмбрионов, обнаружил их только у 8% эмбрионов
на 8-й день и у 30% — на 13-й день инкубации. Перистальтические
сокращения обоих желудков — железистого и мышечного, по
данным Куо и Шен (Kuo a. Shen, 1936), наблюдаются на 8—9-й
213
день инкубации, т. е. сразу после дифференциации мышц мышеч-
ного желудка.
Костомарова (1956) исследовала развитие секретирующего и
всасывающего аппарата кишечника куриного эмбриона и показала,
что до 7-го дня инкубации железистый желудок и тонкий кишечник
гистологически не дифференцируются. С 7-го до 13-го дня в желе-
зистом желудке происходит закладка и основная дифференцировка
желез, а также образование складок стенки кишечника, обеспечи-
вающих всасывание продуктов расщепления протеинов. К 13-му дню
железы железистого желудка пронизывают всю его строму, а с 14-го
по 18-й день по их выводным протокам в полость желудка выде-
ляются капли секрета, содержащего пепсин. К 13-му дню инку-
бации и в поджелудочной железе формируется группа экскретор-
ных железистых клеток. Переваривающая сила вырабытываемого
этими клетками трипсина (так же как и пепсина) увеличивается,
и за сутки до вылупления она почти такая же, как у взрослой
птицы. Все это приводит автора к выводу, что к 13 суткам инку-
бации, т. е. ко времени заглатывания содержащей белок амнио-
тической жидкости, пищеварительный аппарат эмбриона дости-
гает такого уровня морфологической и гистологической дифферен-
цировки, который обеспечивает выполнение им нормальной физио-
логической функции — кишечного пищеварения.
Зусман (1965) считает, что функциональное созревание пище-
варительной системы к моменту попадания в нее амниотической
жидкости, смешанной с белком яйца, обусловлено высоким содер-
жанием в белке Нйца птиц азотистых веществ и вследствие этого
невозможности целиком использовать их при помощи внекишеч-
ного способа потребления питательных веществ. Этот внекишечный
способ полностью обеспечивает эмбриогенез рептилий и остается
в раннем эмбриогенезе птиц. Необходимость включения внутри-
кишечного питания у эмбрионов птиц привела к ускоренному по
сравнению с рептилиями протеканию физиологической дифферен-
цировки секретирующих и всасывающих отделов пищеваритель-
ного аппарата.
Гистохимическими исследованиями Люппа (Luppa, 1962)
показано, что пепсин и соляная кислота продуцируются в желе-
зистом желудке у рептилий в «темных» клетках, а у птиц — в его
аггломератных железах. Аггломератные железы птиц начинают
функционировать к моменту начала переваривания в желудке
заглоченного с амниотической жидкостью белка, а у рептилий —
только после вылупления.
Исследуя в ряде работ функциональную дифференциацию
тонких кишок, Муг с сотрудниками (Moog a. Richardson, 1955;
Moog a. Thomas, 1957, и др.) установила, что содержание фос-
фатазы в двенадцатиперстной кишке увеличивается в 100 раз
с 18-го по 21-й день инкубации. Изучая влияние кортизона на
деятельность тонкого кишечника эмбриона, авторы приходят
214
к выводу, что функциональная дифференциация тонких кишок
контролируется секрецией собственных надпочечников куриного
эмбриона.
Интересные опыты для определения готовности пищеваритель-
ного тракта куриного эмбриона к функции провели Богнер и Уил-
кокс (Bogner a. Wilcox, 1956). Через 30 мин. после помещения
в пищеварительный тракт эмбриона концентрированных сахаров
пищеварительная система удалялась для анализа. Выяснено, что
с 12-го дня инкубации пищеварительная система куриного эмбри-
она способна переваривать сахар.
ГЛАВА 12
РАЗВИТИЕ МОЧЕВЫДЕЛИТЕЛЬНОЙ СИСТЕМЫ
А. Морфология развития органов выделения
Органы выделения и половые органы имеют единое
происхождение. Почки птиц проходят в своем развитии три ста-
дии: а) предпочки, или пронефроса (мы будем здесь называть
органы выделения в единственном числе, хотя все они парные),
б) первичной (средней почки), или мезонефроса, и в) постоянной
(задней) почки, или метанефроса. Этой последовательной смене-
трех систем органов выделения в эмбриональном периоде у птиц
соответствует смена этих систем в\ряду позвоночных животных,
причем происходит постепенное усложнение строения почечных
канальцев (Шмальгаузен, 1947). Выделительная система начинает
дифференцироваться относительно поздно. Морфогенез выдели-
тельной системы изучен Крок в ряде работ (19496, 1952а, 19526,
1954) и описан в ее сводке (1962). Описание морфологии развития
выделительной системы дано нами в основном по работам этого
автора.
Предпочка закладывается у куриного эмбриона из нефрогенной
ткани на уровне 5—15 сомитов на 36-м часу инкубации в виде
сплошных, неполых трубок, перпендикулярно к которым начи-
нает расти ее проток, также не имеющий просвета. К концу 4-го
дня предпочка уже дегенерирует, и ее проток, проходя мимо
образовавшихся к тому времени трубочек средней почки, приоб-
ретает просвет и становится так называемым вольфовым протоком.
При дальнейшем росте он открывается в клоаку. По-видимому,
предпочка у птиц никогда не функционирует и является рудимен-
том. Она напоминает почку эмбрионов некоторых рептилий
(Davis, 1950).
Первичная почка закладывается к концу вторых суток развития
куриного эмбриона 1 также из нефрогенной ткани. Затем в ней воз-
215
никают мочевые канальцы. В отличие от трубочек предпочки они
внутри полые. С одного конца в каждом канальце путем впячива-
ния пузырька формируется капсула, куда со стороны почечной
артерии врастают капилляры, образуя клубочек (рис. 45). Это
и есть почечное тельце. На другом конце мочевой каналец откры-
вается в общий для каждой стороньцтела канал — вольфов проток.
Рис. 45. Стереоскопическая схема строения первичной почки.
(По Huettner, 1947).
1 — спинная брыжейка; 2 — половые складки (зародышевый эпителий);
За — закрытое устьице (нефростом — мерцательная воронка в полость
тела); Зб — открытое устьице; 4 — трубочка средней почки; 5а, 56 — со-
матическая и внутренностная мезодерма; 6 — боуменова капсула;
7 — миотом; 8 — дерматом; 9 — спинная аорта; 10 — хорда; 11 — спин-
ной мозг; 12 — ганглий спинномозгового нерва (с. н.); 13 — спинной
корешок с. н.; 14 — брюшной корешЬк с. н.; 15 — соединительная
ветвь с. н.; 16 — кровеносные капилляры; 17 — задняя кардинальная
вена; 18 — проток средней почки; 19 — субкардинальная вена; 20 — маль-
пигиево тельце; 21 — клубочек; 22 — целбм.
Первые трубочки первичной почки не функционируют. На 5-й
день начинается кровообращение в клубочках и первичная почка
начинает выделительную функцию. К концу эмбрионального раз-
вития эта почка постепенно дегенерирует и из остатков ее обра-
зуются придатки гонад: у петушка — эпидидимус (Maraud, 1955),
а у курочки — паровариум. По данным Калам (Calame, 1959),
в зоне, где обычно возникает мезонефрос, при экспериментальном
удалении вольфова протока она не развивается, а образуются
только несколько изолированных канальцев. Следовательно, воль-
фов проток является индуктором развития мезонефроса,
216
Формирование постоянной почки у куриных эмбрионов начи-
нается на 5-й день инкубации. Она образуется из двух зачатков:
1) нефрогенной ткани двух-трех последних сегментов туловища,
образующей секретирующие трубочки — нефроны, и 2) отростка
задней части вольфова протока, который преобразуется в мочеточ-
ник и его разветвления — собирательные трубочки почки. По
мере развития эмбриона один конец нефрона соединяется с соби-
рательными трубочками — выростами мочеточника, а другой
принимает S-образную форму и превращается в почечное тельце.
Таких почечных телец образуется большое количество. При раз-
делении на 6-й день инкубации (in vitro) зачатка нефрогенной ткани
метанефроса и зачатка мочеточника при помощи механических
или химических агентов ни тот ни другой зачаток не развиваются;
но при соединении их вновь дифференцируется нормальная по-
стоянная почка (Calame, 1961). По данным Крок (1962), рост
и развитие постоянной почки не заканчивается в эмбриональном
периоде и в первые дни постэмбриональной жизни образуются
новые почечные тельца. Схематическое изображение соотношения
предпочки, первичной и постоянной почек и их протоков на раз-
ных стадиях развития представлено на рис. 46.
В отличие от млекопитающих в почке птиц нет разделения
на корковый и мозговой слои. Мочевые канальцы образуют основу
паренхимы почки и расположены в ее дольках. Почечные пирамиды
в почках птиц выражены нечетко и состоят из собирательных ка-
нальцев, впадающих в мочеточник.
Кровеносные сосуды клубочков у птиц более просто устроены,
чем у млекопитающих, и имеют только две-три петли, не анасто-
мозирующие между собой. Икин и Фишер (Eakin a. Fisher, 1958)
установили, что кровь начинает циркулировать в первичной
почке куриного эмбриона незадолго до начала ее функциониро-
вания — с 4-го дня инкубации. Малиновская (Malinowska, 1961)
исследовала развитие кровеносных сосудов мочеполовой системы
самца куриного эмбриона и установила, что артерии метанефроса
возникают из артерий мезонефроса, но становятся магистральными
сосудами только после полной инволюции последнего (пддробнее
о кровообращении в почках см. главу 8).
Изучая иннервацию почек в течение развития куриного эм-
бриона, Крок (1954) подтвердила данные Пирнер (Pirner, 1949),
что нервные волокна входят в строму постоянной почки на 6-й
день инкубации (на сутки раньше, чем по данным других иссле-
дователей), т. е. непосредственно перед началом ее функции.
Автор показала, что источником симпатических волокон, идущих
к метанефросу, являются внутренностные ветви спинномозговых
нервов, связанные с надпочечниками; с парасимпатической си-
стемой постоянная почка не связана. Интересно, что при инволю-
ции первичной почки, дегенерируют и нервные окончания, вхо-
дящие в нее. Автор обнаружила, что в постоянной почке имеются
217
Рис. 46. Схематическое изображение соотношения предпочки, пер-
вичной и постоянной почек и выделительных протоков на разных ста-
диях развития. (По Patten, 1951).
А и Б — начальные*стадии развития выделительной системы; В — состояние
выделительной системы у 4-дневного куриного эмбриона; Г — мочеполовая си-
стема у цыпленка после полового созревания. 1 — трубочки предпочки (1а — де-
генерирующие); 2 — проток предпочки; 3 — трубочки первичной почки (За —
с устьицами, зб — без устьиц, Зв — дегенерирующие); 4 — клоака; 5 — проток
первичной почки; 6 — функционирующая первичная почка; 7 — аллантоис;
8 — проток постоянной почки; 9 — яичник; 10 — трубочки постоянной почки;
11 — мюллеров проток; 12 — семенник.
вазомоторные и секреторные нервные окончания. Наличие в почке
птиц значительного количества нервных элементов указывает
на их регуляторное воздействие при секреторных и реабсорбцион-
ных процессах.
Б. Физиология выделительных процессов
Вопрос о сроках функционирования органов выделения
у куриного эмбриона был вначале изучен путем инъекций виталь-
ных красок внутрь инкубируемого яйца Бакуниной (Bakounine,
1895b). Приводя данные исследований по определению функции
почек у куриного эмбриона при помощи красителей, Нидхем
(Needham, 1931) приходит к выводу, что пронефрос, по-видимому,
не несет экскреторной функции, а выделительная активность
мезонефроса начинается на 4-й день и является единственной до
11-го дня, когда начинает действовать метанефрос. Обе почки функ-
ционируют вместе до 18-го дня, когда мезонефрос прекращает функ-
цию и метанефрос остается единственным органом выделения
эмбриона. В обзоре Грюнвальда (Gruenwald, 1952) приведены
дополнительные данные, подтверждающие выводы Нидхема. Автор
отмечает, что функция метанефроса, так же как и мезонефроса,
начинается сразу же после образования нефронов. Так, за 12 час.
до начала функционирования почек их почечные тельца плохо
заметны, часть трубочек неполностью отделена от мезенхимы
и не имеет просвета. Менкес, Римничану и Микля (Menkes et al.,
1956) применили ряд методов для выяснения функционирования
мезонефроса и метанефроса: окрашивание трипанблау и нейтрал-
рот, окраска кристаллов мочевой кислоты, исследование флуо-
ресценцией, определение активности щелочной фосфатазы и на-
личия гликогена, внутрисосудистое вливание индигокармина и др.
Авторы подтвердили начало функции мезонефроса с 6-го дня.
На 8—9-й день его функция повышается, а имеющийся уже мета-
нефрос еще не действует. Несмотря на то что функция метанефроса
начинается на 11-й день, мочевая кислота появляется в ней только
на 13-й день инкубации. Развитие функции метанефроса, по дан-
ным этих авторов, совпадает с началом инволюции мезонефроса.
Исследовав потребление кислорода (на 1 мг сухой ткани) мезо-
и метанефросом, Романов (Romanoff, 1943, цит. по: Romanoff,
1960) косвенно показал, что с 9-го до 15-дня функция первичной
почки постепенно, а затем до конца инкубации стремительно па-
дает, а функция постоянной почки постепенно повышается с 11-го
дня до конца эмбрионального развития. Место пересечения кри-
вых потребления кислорода тканью каждой из почек (13—14-й
день инкубации) можно было бы считать временем уравнения
их значения в выделительных процессах эмбриона, если бы их
вес в это время был одинаковым. Но вес постоянной почки в это
время в 1.5—2 раза больше веса первичной, так что момент урав-
219
нения их функционального значения приходится, по-видимому,
на 12-день инкубации. Крок (19496, 1952а, 1962) подтвердила
данные о сроках функции мезо- и метанефроса у куриного эмбри-
она , а также получила соответствующие данные на утиных
и гусиных эмбрионах (табл. 39).
Таблица 39
Развитие мочевыделительных органов эмбрионов
сельскохозяйственных птиц
(По Крок, 1962)
Возраст эмбриона (в днях)
Стадия развития
куры утки и гуси
Закладка первичной почки........
начало функции............
прекращение функции ....
Закладка постоянной почки ....
начало функции..................
Функционирует только постоянная
почка ..........................
2
5-6
16-17
5
12
17-18
3
6-7
21-22
6-7
22-23
Функция почек регулируется, как уже указывалось, симпа-
тической нервной системой и гуморально при помощи гормонов
коры надпочечников и антидиуретического гормона гипофиза.
Необходимо напомнить, что, кроме выведения вредных продуктов
распада протеинов из организма, почки играют большую роль
в поддержании относительного постоянства водно-солевого со-
става внутренней среды организма.
Функционирование первичной и постоянной почек происхо-
дит примерно одинаково. Из протекающей по клубочку крови
часть плазмы в виде ультрафильтрата проходит в капсулу. Затем
эта жидкость, отличающаяся от плазмы крови почти полным
отсутствием протеинов (вследствие большой величины молекул
они не проходят сквозь стенки сосудов), так называемая первичная
моча, выходит из капсулы в секретирующую трубочку, где под-
вергается интенсивной реабсорбции. Часть веществ — глюкоза,
соли натрия, калия и др. — реабсорбируются в почках активно.
Вода же и некоторые другие вещества реабсорбируются пассивно.
Механизм пассивной реабсорбции детально исследован Гинецин-
ским (1963). Было показано, что в отличие от натрия, который
активно реабсорбируется через протоплазму клетки, вода реаб-
сорбируется пассивно, и не через клетки, а через межклеточные
ходы. В случаях гипергидратации организма эти межклеточные
ходы закрыты гиалуроновой кислотой, но как только достаточ-
ное количество воды выведено почками, в действие вступает анти-
диуретический гормон, под воздействием которого клетки почеч-
220
ных канальцев выделяют энзим — гиалуронидазу, который рас-
щепляет молекулу гиалуроновой кислоты и открывает межкле-
точные ходы для воды, переходящей при этом из канальцев в кровь
(рис. 47). Освобождаясь от воды и некоторого количества солей
натрия, первичная моча превращается в «окончательную», которая
по собирательным трубочкам мезонефроса попадает в вольфовы
протоки, а при функционировании постоянной почки — в моче-
точники. Из вольфовых протоков и из мочеточников моча попадает
в клоаку, а оттуда в аллантоис. У взрослых птиц в клоаке,
а у эмбриона с 11-го дня инкубации в аллантоисе происходит снова
Рис. 47. Схема механизма действия антидиуретиче-
ского гормона. (По Гинецинскому, 1962).
1 — гидратированное животное. Межклеточный цемент
(заштрихован) непроницаем. Натрий (точки) активно реаб-
сорбируется через цитоплазму. Вода (капли) остается в про-
свете нефрона. 2 — животное, подвергающееся дегидрата-
ции. Под влиянием гормона (АДГ) почечные клетки вы-
деляют гиалуронидазу, которая деполимеризует мукопо-
лисахариды. Вода реабсорбируется через, утративший гер-
метичность межклеточный цемент.
реабсорбция воды. На этот раз реабсорбируется почти вся вода
из мочи, в результате чего она сильно густеет, превращаясь из
жидкости в почти сухое вещество, которое откладывается светло-
зеленоватыми хлопьями в полости аллантоиса.
В течение эмбрионального развития птиц происходит последо-
вательная смена основного продукта выделения: аммиак, мочевина,
мочевая кислота (Needham, 1931). До 4-го дня развития азот
выделяется из организма куриного эмбриона почти исключительно
в виде аммиака. Количество выделяемой мочевины увеличивается
с 4-го до 9-го дня развития, а затем остается на одном уровне и даже
падает. Мочевая кислота начинает продуцироваться и выделяться
на 5—6-й день инкубации. Максимум выделения аммиака прихо-
дится на 4-й день инкубации, мочевины—на 9-й, а мочевой ки-
слоты — на 11-й. Мочевая кислота составляет 91.4%, мочевина —
7.6%, а аммиак — только 1% всего выделенного куриным эмбрио-
ном азота. Икин и Фишер (Eakin a. Fisher, 1958) считают моче-
221
вую кислоту основным продуктом выделения куриного эмбриона
с самого начала его развития, концентрацию аммиака — постоян-
ной в течение первых И дней инкубации, а концентрацию моче-
вины в аллантоисе — увеличивающейся. Исследуя активность
ферментных систем, участвующих в распаде протеинов в течение
развития куриных эмбрионов, Дрель (1964, 1965) приходит к вы-
воду, что эмбрионы птиц переходят непосредственно от аммоно-
телического типа распада протеинов (выделение аммиака) к урико-
телическому (выделение мочевой кислоты), минуя уреотелический
тип (выделение мочевины). Автор показал, что 1) уже с 5-го дня
эмбрионального развития печень способна синтезировать мочевую
кислоту и 2) что синтез мочевины в печени отсутствует, и она
образуется только путем ферментативного гидролиза аргинина,
содержащегося в тканях. Однако автор не отрицает, что с 5-го
до 9-го дня развития эмбриона происходит максимальное, а в дру-
гое время — несколько меньшее образование мочевины.
Аммиак образуется в почках, а мочевина — в печени. Для
образования же мочевой кислоты, по мнению Нидхема (Needham,
1942), требуется сотрудничество двух органов: печени, синтези-
рующей из аммиака и углеродных атомов гипоксантин (являю-
щийся предшественником мочевой кислоты), и почки, превращаю-
щей гипоксантин в мочевую кислоту. Кравчинский (1958) также
считает, что почки птиц превращают гипоксантин в мочевую
кислоту и секретируют ее в канальцах. Крок же (1962) приходит
к выводу, что мочевая кислота целиком синтезируется в печени
из аммиака. Болдуин (1949) приводит данные, что печень боль-
шинства птиц (курицы, гуся и др.) образует из добавленного
аммиака кислоту, а печень голубя вырабатывает гипоксантин,
который лишь в почках превращается в мочевую кислоту. Эта
особенность урикогенеза у голубя объясняется отсутствием в его
печени энзима ксантиноксидазы и наличием этого энзима в почках.
Шобе (Chaube, 1962) исследовала активность гипоксантиндегидро-
геназы — энзима, катализирующего синтез мочевой кислоты в поч-
ках куриного эмбриона. Автор показала, что синтез мочевой ки-
слоты осуществляется как в мезо-, так и метанефросе и активность
энзима хорошо коррелируется с данными о функции обеих почек,
полученными методом окраски. Активность гипоксантиндегидро-
геназы в мезонефросе резко возрастает в течение 6-го и 7-го дней,
затем медленно повышается до 14-го и потом падает до момента
вылупления; а в метанефросе наиболее быстро повышается с 13-го
по 15-й день, затем в течение двух дней не изменяется и снова
возрастает до вылупления. Перекрест кривых активности энзима
в мезо- и метанефросе происходит на 19-й день. Дрель (1965)
считает, что синтез мочевой кислоты происходит в печени кури-
ного эмбриона из ее предшественников: глицина, глютамина,
аспарагиновой и муравьиной кислот и углекислоты. Скорость
этого синтеза на единицу белка остается постоянной с 5-го до 15-го
222
дня инкубации. По предположению автора, ферментные системы,
катализирующие синтез пуриновых оснований, присутствуют
в тканях куриных эмбрионов на самых ранних стадиях развития,
но формирование ферментной системы синтеза мочевой кислоты
(становление урикотелического типа распада протеинов) происхо-
дит позднее путем дополнения к ранее существующим еще одного
ферментного процесса — окисления пуринов в мочевую кислоту
с участием ксантиноксидазы. Таким образом, вопрос о том, проис-
ходит ли синтез мочевой кислоты у птичьих эмбрионов только
в печени или в печени и почках, вследствие противоречивости
экспериментальных данных остается открытым.
Коштоянц (1940) считает, что аммиак выделяется на ранних
стадиях развития птичьего эмбриона в связи с отсутствием у эмб-
риона морфологически дифференцированных органов синтеза
более сложных продуктов выделения в это время. По мнению же
Еремеева (1957), образование птичьим эмбрионом аммиака в ка-
честве продукта экскреции явлется защитным приспособлением
против избыточного накопления кислых продуктов обмена. Опре-
деленное чередование продуктов экскреции во время развития пти-
чьего эмбриона Нидхем (Needham, 1931) сначала считал биохими-
ческой рекапитуляцией. Но позже (1942) автор приходит к выводу,
что форма выделения азота из организма зависит гланым образом
от условий жизни птичьего эмбриона (ограниченное количество
воды в закрытом яйце). По мнению автора, ни одно животное
не начинает выделять мочевую кислоту до тех пор, пока не бывает
вынуждено это сделать (в связи с недостатком воды), так как по
сравнению с другими продуктами протеинового обмена (аммиак
и мочевина) мочевая кислота явлется наиболее расточительной
энергетически. Аммиак совсем не включает углерода, у мочевины
отношение углерода к азоту равно 1 : 2, а у мочевой кислоты —
1 : 0.9. Продолжая эту мысль, Еремеев (1964) показывает, что
выделение биурата аммония обусловлено и экономичностью выве-
дения из организма азота. Так, отношение углерода к азоту у биу-
рата аммония равно 1:1, а у уратов калия и натрия — 1.2 : 1.
А ранее автор (1957) отмечал, что синтез конечных продуктов
обмена в замкнутом пространстве с ограниченным количеством
воды, каким является яйцо птицы, идет в направлении наименее
растворимых соединений (биурат аммония, ураты калия, гипс).
Болдуин (1949) придерживается несколько иного мнения о при-
чине появления мочевой кислоты в качестве продукта распада про-
теинов. Аммиак и мочевина, легко растворимые и проникающие
через полупроницаемые оболочки, привели бы к отравлению
или во всяком случае к серьезному нарушению функций организма,
если бы весь подлежащий экскреции азот в течение эмбрионального
развития превращался в аммиак и мочевину. Автор показал, что
выделение того или иного продукта экскреции у разных животных
зависит от условий обитания. В связи с этим интересно отметить,
223
что в амниотической жидкости куриного эмбриона мочевой ки-
слоты очень мало и это количество не увеличивается в течение раз-
вития, а аммиака и мочевины становится больше, так как они
обладают способностью диффундировать через стенки амниона.
Г Л А В А 13
РАЗВИТИЕ ЖЕЛЕЗ ВНУТРЕННЕЙ СЕКРЕЦИИ
Железы внутренней секреции — это комплексы эпи-
телиальных клеток, богато снабженные кровеносными сосудами,
выделяющие подукты секреции (гормоны) непосредственно в кровь
и осуществляющие при их помощи регуляцию процессов, проте-
кающих в организме. Эндокринные железы играют большую роль
в сохранении относительного постоянства внутренней среды орга-
низма, без которого невозможно существование позвоночных жи-
вотных. Действие каждого из гормонов распространяется на опре-
деленные системы органов или виды обмена веществ и обычно
имеет жизненно важное значение. Интересно отметить, что у
разных классов позвоночных животных химическое строение
аналогичных гормонов очень сходно, что подтверждает их оди-
наковое происхождение.
Как показали исследования последних десятилетий, эндокрин-
ные железы в эмбриональном периоде играют значительную роль
в регуляции морфогенеза и обменных процессов. Кроме того,
изучение эндокринных взаимоотношений на ранних стадиях
онтогенеза, когда эти взаимоотношения относительно проще,
чем во взрослом организме, оказывает значительную помощь
в раскрытии механизмов действия отдельных гормонов и при-
чинных связей между разными гормональными системами. Анали-
зируя развитие эндокринных регуляций, Лейбсон (1964) пишет,
что «сложная картина гормональных эффектов, которую мы наблю-
даем у взрослых высших животных, складывалась в ходе эволюции
постепенно» и что «это должно найти свое отражение как в эмбрио-
генезе, так и в филогенезе» (стр. 229).
! Обилие материала о функции эндокринных желез вынуждает
нас уделить меньше места морфологии их, тем более что за послед-
ние два десятилетия имеются хорошие сводки по этому вопросу
(Студитский, 1947; Willier, 1955; Мицкевич, 1957).
[ В ряде случаев в данных разных исследователей о сроках на-
чала функции той или иной эндокринной железы имеются рас-
хождения. Кроме обычных индивидуальных и породных откло-
нений, а также различий в условиях инкубации, сдвигаюпщх
сроки развития эмбрионов, это несомненно обусловлено отличиями
в методах обнаружения активности гормонов. Чаще всего, чем
224
точнее методы, тем в более ранние сроки эмбрионального развития
может быть обнаружена функция той или иной эндокринной же-
лезы.
А. Поджелудочная железа
Эндокринная часть поджелудочной железы представлена
островковой тканью. Мицкевич (1957) исследовал гистогенез
эндокринной части поджелудочной железы куриных эмбрионов
и показал, что вначале ее дифференцировка выражается в появ-
лении комплексов мутных клеток с мелкозернистой ацидофильной
протоплазмой (так называемые островки Лагесса), которые вскоре
преобразуются в «настоящие», светлые островки — островки Лан-
герганса. Автор отмечает при этом, что не все островки Лангер-
ганса проходят промежуточную стадию мутных клеток, а часть
их образуется прямо из эпителия выводных протоков. Первые
клетки островков Лангерганса появляются на 10—11-й день
инкубации — срок, который автор считает началом гормональной
функции. Вилье (Willier, 1955) подвергает сомнению последова-
тельное появление островков Лагесса и Лангерганса,
приводя данные Виламеля (Villamil, 1942) о том, что оба типа
островков — «темные» (Лагесса) и «светлые» (Лангерганса) —
наблюдаются в поджелудочной железе уже на 3-й день инкубации.
Бета-клетки, являющиеся, собственно, источником инсулина, раз-
виваются только в «светлых» островках (с 12-го дня инкубации)
и появляются непосредственно перед наступлением второго подъема
в накоплении углеводов в крови и печени. В «темных» остров-
ках развиваются альфа- и дельта-клетки на 8-й и 14-й день соответ-
ственно. В альфа-клетках вырабатывается второй гормон под-
желудочной железы — глюкагон, а дельта-клетки являются,
по-видимому, клетками стромы. По данным Льевр (Lievre, 1957а,
1957b), альфа-клетки появляются на 7-й, а бета--на 8-й день
инкубации. После гипофизэктомии бета-клетки проявляют повы-
шенную продукцию гормона. Позднее при помощи электронного
микроскопа автор (Dieterten-Lievre, 1963b) обнаружила альфа-
зерна в зачатке поджелудочной железы уже у 3-дневного куриного
эмбриона, которые при совместном с печенью культивировании
in vitro подавляли в ней образование гликогена. Томмс (Thom-
mes, 1960) гистохимическими методами обнаружил, что инсулин
в поджелудочной железе куриного эмбриона впервые появляется
на 7-й день инкубации; до И—13-го дня его количество увеличи-
вается медленно, затем в течение 14-го дня резко повышается,
после чего мало изменяется.
Временем начала выделения инсулина в кровь у куриных
эмбрионов, как показали Лейбсон, Желудкова и Чилингарян
(1961), используя чувствительный метод изолированной диафрагмы
крысы, следует считать 13-й день развития. Лейбсон (1962) от-
мечает в своей монографии, что этот день в развитии куриного
15 В. В. Рольник
226
эмбриона является вообще переломным для углеводного обмена,
хотя чувствительность тканей эмбриона к инсулину прояв-
ляется раньше, чем появляется в крови собственный инсулин.
Грийо (Grillo, 1961а) также диафрагменным методом показал,
что активность инсулина в поджелудочной железе куриного эм-
бриона впервые появляется в 12-дневном возрасте, достигает
максимума на 16-й день инкубации и снижается перед вылупле-
нием. ♦
Приводя данные Шикинами (Shikinami, 1928) о наличии инсу-
линоподобного вещества в желтке неинкубированпых яиц, Нид-
хем (Needham, 1931) предполагает, что оно играет важную роль
в контроле углеводного обмена эмбриона в течение первых не-
скольких дней инкубации до появления каких-либо клеток,
секретирующих инсулин. Однако Мицкевич (1957) считает, что
это вещество вряд ли может рассматриваться в качестве гормона,
и Лейбсон (1962) сомневается в его физиологической роли, сход-
ной с истинным инсулином. Грийо (Grillo, 1961а) не обнаружил
инсулина в желтке неинкубированного яйца.
Лейбсон и Лейбсон (1943) первыми систематически исследо-
вали влияние экзогенного инсулина, введенного в инкубируемые
куриные яйца, на углеводный обмен эмбрионов. Инсулин вызывал
у эмбрионов всех сроков развития (опыты проводились с 8-го
дня инкубации) снижение содержания сахара в крови. В дальней-
шем Лейбсон (1951) установил, что введение инсулина вызывает
также повышение количества гликогена в печени. Эти данные были
подтверждены целым рядом исследователей (Zwilling, 1948; Миц-
кевич, 1957, и др.). Цвиллинг наблюдал гипогликемию под влия-
нием введенного инсулина с 6-го дня развития куриного эмбриона
и показал далее, что в раннем периоде эмбриогенеза при этом про-
исходит увеличение накопления гликогена в желточном мешке
(Zwilling, 1948, 1951).
На примере регуляции содержания сахара в крови (гликемии)
Лейбсон (1962) представляет гипотетическую схему смены механиз-
мов в регуляции постоянства внутренней среды развивающегося
куриного эмбриона. Вначале ни нервная, ни эндокринная системы
в регуляции гликемии не участвуют и глюкоза в крови появляется
из углеводов, содержащихся в яйце, и из гликогена желточного
мешка. Затем включается гомеостатический механизм печени.
С 13—14-го дня включаются эндокринные железы. С 18-го дня
сахар в крови резко увеличивается в связи с тем, что высшие
нервные центры и мышцы перед вылуплением испытывают повы-
шенную потребность в нем. В это время возможно включение
в регуляцию гликемии гипоталамуса, а перед самым вылупле-
нием — и корковых центров. Изучив изменение содержания
сахара в крови и концентрации гликогена в печени при гипо-
ксии, автор показывает, что эндокринные железы в эмбрио-
нальный период не только влияют пассивно на процессы обмена
226
веществ, но и активно участвуют в регуляции гомеостазиса эм-
бриона при изменении внешних условий.
Введение инсулина вызывает у куриных эмбрионов различные
деформации скелета, обусловленные недостатком хондроитин-
серной кислоты (Duraiswami, 1950). Автор (Duraiswami, 1955)
доказывает далее, что целый ряд ненормальностей при введении
инсулина обусловливается нарушением углеводного обмена, и
приходит к выводу, что рост и дифференциация в течение эмбрио-
нального развития зависят от гармоничного сочетания большого
количества хорошо отрегулированных процессов, каждый из
которых требует соответствующих гормонов и энзимов, действую-
щих в соответствующей среде; недостаток определенного фактора
или нарушение метаболизма в критический момент может вызвать
аномалии развития. Пархон и его соавторы (Parhon et al., 1955)
также объясняют наблюдавшиеся ими нарушения развития и
уродства у куриных эмбрионов при введении им инсулина до 11-го
дня развития нарушением углеводного обмена. Введение инсулина
после этого срока не вызывало уродств. Эрхард (Erhard, 1959),
вводя инсулин 3—5-дневным эмбрионам, получал у более 50%
из них укорочение задних конечностей — микромелию. Автор
отмечает, что при микромелии сильно сокращалось содержание
гликогена в хряще — пропорционально силе уродства. По данным
Баррон и Маккензи (Barron a. McKenzie, 1962), введение инсулина
подавляло развитие нервной трубки и мозга. Авторы предпола-
гают, что это вызвано нарушением протеинового обмена.
О влиянии глюкагона на развитие птичьих эмбрионов известно
мало. Гипергликемический эффект глюкагона основан на усилении
гликогенолиза в печени благодаря активации фосфорилазы. Лейб-
сон (1962) указывает, что глюкагон содержится у птиц в коли-
честве в 10 раз большем, чем у млекопитающих, и, по-видимому,
играет у птиц более значительную роль. По данным Грийо (Grillo,
1961b), этот гормон появляется в поджелудочной железе куриного
эмбриона одновременно с инсулином, т. е. в 12-дневном возрасте.
Б. Надпочечники
Надпочечники состоят из двух желез, клетки которых
имеют разное происхождение. Корковые тяжи клеток произошли
из врастаний брюшинного эпителия, а мозговые — из клеток
симпатических стволов, мигрирующих затем внутрь корковых
тяжей. Корковые и мозговые клетки у птиц лежат совместно,
причем первые представляют собою тело надпочечника, а вторые
лежат в ячейках, образованных первыми (Lillie, 1952). Кора над-
почечников выделяет две группы гормонов: минералокортикоиды
и глюкокортикоиды (в частности, гидрокортизон). В мозговой части
имеются адреналиновые и норадреналиновые клетки, вырабаты-
вающие катехоламины: адреналин и норадреналин, который
15*
227
отличается от первого отсутствием метильной группы. В норад-
реналиновых клетках не хватает одного энзима — метилазы,
и потому они не могут вырабатывать адреналин. Норадреналиновые
клетки истощаются при активности быстрее, так как норадрена-
лин менее эффективен метаболически, чем адреналин. Смиттен
и Малиновская (1960) обнаружили в мозговом слое надпочечников
эмбрионов птиц значительно больше клеток, содержащих норад-
реналин, чем содержащих адреналин, а у млекопитающих норад-
реналиновых клеток оказалось значительно меньше, чем у птиц.
Ио данным Мазиной (1963), вес надпочечников куриных эмбрио-
нов с 12-го дня и до конца инкубации увеличивается в 5—6 раз.
Исследуя гистогенез надпочечников у куриных и голубиных
эмбрионов, Бреци (Brezzi, 1940) обнаружил, что развитие над-
почечников у голубиных заканчивается позднее и к моменту
вылупления у них меньше функционирующих клеток, чем у ку-
риных эмбрионов. Доусон (Dawson, 1953) гистохимическими мето-
дами установил, что дифференциация клеток мозгового вещества
надпочечников куриных эмбрионов происходит на 6-й день.
Манухин и Бузников (1959) впервые провели биохимическое,
количественное изучение адреналина и норадреналина в надпо-
чечниках куриных эмбрионов. Авторы обнаружили определимые
количества адреналина на 10-й день. С 12-го по 16-й оно увеличи-
валось постепенно (в связи с ростом надпочечников), а с 17-го
по 19-й — резко, в 2—3 раза. Второй резкий скачок (в 3 раза
в течение одних суток) авторы наблюдали во время вылупления
цыпленка, причем начало «скачка» строго совпадало с началом
наклева. Определимые количества норадреналина появляются
с 10—12-го дня и затем содержание гормона, колеблясь, сильно
возрастает. Авторы отметили, что колебания количества норадре-
налина в надпочечниках не влияют на содержание адреналина,
и приходят к выводу, что норадреналин выходит в кровь, а не прев-
ращается в адреналин. Лейбсон и Стабровский (1962), пользуясь
более точным биохимическим методом определения катехоламинов,
получили сходные данные. Авторы отмечают, что на протяжении
всего периода развития куриного эмбриона содержание норадре-
налина в надпочечниках превосходит содержание адреналина;
их количество после вылупления выравнивается, а со второй недели
постэмбриональной жизни, наоборот, адреналина становится
больше. Было показано далее, что в целом курином эмбрионе
катехоламины могут быть обнаружены еще до того, как они по-
являются в надпочечниках. Стабровский (1964а) подтвердил это,
определив катехоламины в отдельных тканях. По данным Бауцек
и Бон (Boucek a. Bourne, 1962), катехоламины начинают выделяться
в незначительном количестве уже с 5-го дня развития куриного
эмбриона, вначале либо из хромаффинных клеток ганглиев, либо
из клеток центральной нервной системы. Авторы отмечают боль-
шой подъем (в 4 раза) содержания катехоламинов в аллантоисной
228
жидкости в течение 24 час., с 11-го до 12-го дня инкубации, в чем,
по мнению авторов, большую роль играет окончательная диф-
ференциация ткани надпочечника.
По предположению Нидхема (Needham, 1931), адреналин
является специфическим стимулятором сгорания жиров в орга-
низме птичьего эмбриона. Лейбсон (1962), обобщив данные своих
исследований и данные других авторов, отмечает, что адреналин,
введенный извне, может оказывать влияние на углеводный обмен
куриных эмбрионов уже сравнительно рано (с 8—10-го дня ин-
кубации). Стабровский (19646) обнаружил, что мозговое веще-
ство надпочечников куриных эмбрионов не реагирует на инсу-
линовую гипогликемию до 14-дневного возраста. Введение же
инсулина эмбрионам в последнюю неделю инкубации вызывает
усиленное выделение норадреналина из их надпочечников,
но не адреналина; зато у цыплят введение инсулина обусловли-
вает секрецию только адреналина. Такая смена гормонов в реак-
ции на инсулиновую гипогликемию, по предположению автора,
обусловлена тем, что в системе гипоталамус—мозговое вещество
надпочечников птиц раньше созревает та часть ее, конечным
звеном которой являются клетки, содержащие норадреналин.
О функции коры надпочечников в течение развития птичь-
его эмбриона имеется меньше данных. Даусон (Dawson, 1953)
показал, что дифференциация корковой части надпочечни-
ков начинается на 7-й день инкубации. Однако липидные ком-
поненты имеются во всех корковых тяжах только с 11—12-го
дня, а аскорбиновая кислота, необходимая для синтеза корти-
коидных гормонов, обнаруживается на 10-й день и в значитель-
ном количестве имеется только на 12-й день развития. По дан-
ным Мазиной (1963), с 12-го дня до конца инкубации количество
аскорбиновой кислоты в надпочечниках увеличивается в 4—5,
а холестерина (предшественника кортикоидных гормонов) —
в 13 раз, что указывает на значительное увеличение функции коры
надпочечников в онтогенезе. В обзоре, посвященном функции
коры надпочечников, Муг (Moog, 1959) показала, что с 10-го
до 14-го дня инкубации при введении кортикоидов увеличива-
ется активность фосфатазы в эпителии двенадцатиперстной кишки.
Автор приходит к выводу, что именно в этот период начинает
функционировать собственная кора надпочечников куриных эмб-
рионов. Установлено, что экзогенные кортикоиды тормозят рост
эмбриона при введении с 8-го дня, а при введении на 16-й день,
наоборот, несколько увеличивают вес. эмбрионов.
Введение кортизона усиливает накопление гликогена в жел-
точном мешке, вызванное инъекцией инсулина (Zwilling, 1952).
По данным Лейбсона и Желудковой (1960), кортизон увеличивает
содержание гликогена в печени куриных эмбрионов, но лишь
после 10-дневного возраста. Авторы предполагают, что у эмб-
рионов до 10-дневного возраста отсутствуют какие-то фермен-
229
тативные системы, необходимые для гликонеогенеза или они не-
чувствительны к кортизону.
Установив, что кортизон замедляет темп общего развития
эмбриона, Пархон с сотрудниками (Parhon et al., 1956) считает
это результатом снижения синтеза протеинов. Отмечая сходное
влияние кортизона на взрослых птиц и эмбрионов, авторы под-
черкивают, что на эмбрионов введение кортизона действует
более сильно. Данные Бинер, Бэлэчану и Албу (Biener et al.,
1959) подтверждают вывод о снижении синтеза белка при введении
кортизона; инъекции гормона 7—8-дневным эмбрионам наряду
с увеличением содержания гликогена в печени и сердце обуслов-
ливали снижение веса сердечной мышцы. Подтверждение этого
получено и при микроскопическом изучении печени куриного
эмбриона под влиянием введенного кортизона (Константинова,
Лейбсон и Моисеев, 1960). Установлено, что кортизон вызывает
уменьшение числа печеночных клеток, обусловленное задержкой
их митотического деления, повышение содержания гликогена
и накопление желчи.
Кортизон, по-видимому, является антагонистом гормона роста.
Так, по данным Соубель (Sobel, 1958), вызванные введением
кортизона снижение роста эмбриона и повышение смертности
снимаются после введения опытным эмбрионам гормона роста,
и развитие их становится нормальным. Сайед, Блюменталь и Блю-
менталь (Sayeed et al., 1962) обнаружили то же самое у эмбрио-
нов ранних стадий развития (60—72 часа инкубации). Кортизон
подавлял пролиферацию клеток мезодермы и энтодермы, а гормон
роста стимулировал ее; на клетки эктодермы гормоны не дей-
ствовали. В связи с тем что эндокринная система эмбриона на-
чинает функционировать не ранее 7-го дня инкубации, авторы
приходят к выводу, что гормоны в данном случае действуют
не через эндокринную систему (стимулируя действие аналогичных
и подавляя действие антагонистических гормонов), а непосред-
ственно на клетки.
Кора надпочечников куриного эмбриона, как показала Муг
(Moog, 1959), а затем и Мазина (1963), начинает реагировать
на введение адрено-кортикотропного гормона гипофиза с 11—12-
дневного возраста (о чем подробнее см. стр. 239—240).
Корковое вещество надпочечников принимает также участие
в реакции организма на вызванную инсулином гипогликемию
уже в эмбриональном периоде — с 15-го дня развития куриного
эмбриона (Мазина, 1964).
В. Щитовидная железа
Морфогенез щитовидной железы птиц изучен Энтин
(1936). Автор показала, что у 6-дневного куриного эмбриона щито-
видная железа представлена сплошной железистой паренхи-
230
Рис. 48. Гистологическая картина развития щитовидной
железы куриного эмбриона. (По Энтин, 1936).
1 — 6-дневный эмбрион (сплошная эпителиальная масса,.клеточ-
ные гранулы почти не видны); 2 — 8-дневный эмбрион (извитые
тяжи, состоящие из двуслойного цилиндрического эпителия);
3 — 12-дневный эмбрион (первые капли коллоида); 4 — 14-днев-
ный эмбрион (фолликулы располагаются тяжами; небольшое коли-
чество изолированных фолликулов; много недифференцированной
ткани); 5 — 17-дневный эмбрион (фолликулов значительно больше,
чем иптерфолликулярпой ткани).
мой (рис. 48). Затем, после врастания кровеносных сосудов
и соединительной ткани, ^келеза к 8-му дню делится на тяжи
двурядного эпителия. В железе 12-дневного эмбриона появляются
отдельные клеточные группы и капли коллоида, а у 14-днев-
ного имеются вполне сформированные фолликулы, количество
которых в дальнейшем возрастает. Изучая изменение объема
щитовидной железы в течение развития куриного эмбриона,
Гамбаро (Gambaro, 1940) делит весь период на 5 стадий. В те-
чение 2-й (10—11-й день) и 4-й (18—19-й день инкубации) стадий
автор наблюдал быстрый рост железы, а в конце 5-й — заметное
уменьшение объема. Рост щитовидной железы обусловлен ростом
железистой ткани, фильтрацией жидкости из окружающей ткани
и из кровеносных сосудов, а также появлением коллоида — гор-
мона. Автор считает, что полная дифференциация железы дости-
гается только к моменту вылупления. При электронномикроско-
пическом исследовании щитовидной железы 11-дневного кури-
ного эмбриона Фужита и Махино (Fujita a. Machino, 1961)
обнаружили, что вначале в клетках цитоплазмы появляются
гомогенные крупные капли коллоида, окруженные оболочками;
затем капли коллоида увеличиваются и вступают в контакт
с такими же каплями в других клетках (так как клеточные
и коллоидные оболочки исчезают) и образуют простейшие кол-
лоидные фолликулы, которые в свою очередь сливаются с сосед-
ними, формируя типичные фолликулы. А по данным Хильфер
(Hilfer, 1964), фолликулы в железе впервые обнаруживаются
на 8-й день и к 14-му дню развития щитовидная железа куриного
эмбриона вполне сформирована и сходна с железой взрослых
птиц. Маро с соавторами (Maraud et al., 1954), изучая концент-
рацию меченого иода в щитовидной железе куриного эмбриона,
показал, что в развитии железы имеются две четко различающиеся
фазы. Первая — между 8-м и 10-м днем, когда железа еще не имеет
пузырчатого строения, но уже адсорбирует меченый иод из крови,
причем концентрация его в железе зависит от его количества
в крови. Вторая — после 10-го дня, когда железа уже, имеет
пузырчатое строение; концентрация меченого иода в ней не за-
висит от его количества в крови.
Функцию щитовидной железы куриного эмбриона изучил
гистохимическими методами Студитский (1947). Формирование
фолликулов в железе, по данным автора, происходит на 11-й
день, что соответствует началу функционирования железы и сек-
реции гормона в кровь. Начиная с 17—18-го дня инкубации
и до вылупления щитовидная железа куриного эмбриона нахо-
дится в состоянии повышенной активности, выражающейся:
а) в усиленном формировании фолликулов, б) в чрезвычайно
интенсивном накоплении коллоида, означающем синтез гормона,
и в) в разжижении коллоида, свидетельствующем о выведении
гормона в кровь. Автор показал также, что щитовидные железы
232
куриного эмбриона топографически связаны с нервными узлами.
Исследованиями Мицкевича (1957) показано, что морфологи-
ческая дифференцировка (формирование фолликулов и накоп-
ление коллоида внутри них) совпадает с функциональной
активностью (способностью стимулировать метаморфоз головасти-
ков) и биохимической дифференцировкой (способностью вклю-
чать радиоактивный иод в синтезированный белок). Щитовидная
железа эмбриона и взрослых птиц имеет целый ряд сходных
признаков, в том числе подобно взрослой железе она после вве-
дения метилтиоурацила утрачивает способность накапливать иод.
Исследуя функцию щитовидной железы, Крайчичек (Kraic-
ziczek, 1956) связывает изменения в ней в течение развития ку-
риного эмбриона с чередованием стадий роста и дифференциации.
Так, начало функции на 8-й день инкубации соответствует подъ-
ему роста эмбриона; усиленное выделение гормона с 10-го по 13-й
день идет параллельно с подъемом перихондрального окостенения;
после фазы покоя наступает снова подъем секреции с 16-го
по 18-й день, который совпадает с периодом дифференцировки,
соответствующей метаморфозу амфибий перед выходом их на сушу.
Автор подчеркивает наличие морфогенетического действия тирео-
идного гормона в эмбриональном периоде у птиц. Томмс (Thommes,
1958) установил соответствие изменений рисунка кровеносных
сосудов с изменениями функциональной активности щитовидной
железы в процессе развития куриного эмбриона. По данным
Туннеля и Вейда (1957, цит. по: Waterman, 1958), в биохимиче-
ской дифференциации тироксина в щитовидной железе куриного
эмбриона имеются четкие последовательные стадии: в 8.5 дней —
тирозин, в 9.25 — дийодотирозин, а в 9.75 дней — тироксин.
В своем обзоре о роли щитовидной железы в эмбриоге-
незе птиц Дитерлен-Льевр (Dieterlen-Lievre, 1963а) сообщает,
что в первой фазе развития железы (по 10-й день развития кури-
ного эмбриона) синтез гормона происходит автономно, а во второй
фазе (с 11-го дня) включается контроль гипофиза. Автор отме-
чает, что после окончания дифференцировки щитовидной же-
лезы биохимические процессы в организме эмбриона интенсифи-
цируются. Введение тироксина в первые дни развития эмбриона
ускоряет его рост, и это ускорение сохраняется до конца инкубации.
Реакция щитовидной железы куриного эмбриона к концу
инкубации на изменение температуры, как показала Тиксье-
Видаль (Tixier-Vidal, 1958), такая же, как у гомеотермных жи-
вотных, а именно при снижении температуры на 10° железа
начинает интенсивно выделять тиреоидный гормон, в то время
как у пойкилотермных такое воздействие тормозит ее функцию.
Щитовидная железа животных, рождающихся слабодиффе-
ренцированными (голуби, воробьи, крысы, кошки), по данным
Войткевича (1936), обладает в начале постэмбриональной жизни
слабой биологической активностью по сравнению с железой жи-
233
вотных, способных сразу после рождения вести самостоятельный
образ жизни (куры, морские свинки.) Исследования Студитского
(1947) опровергают это положение. По его данным, щитовидная
железа птенцовых птиц (грача и голубя) физиологически ак-
тивна уже на 10-й день эмбрионального развития и, так же как
у куриного эмбриона, находится в состоянии повышенной актив-
ности к концу инкубации. Данные Мицкевича (1957) как бы
примиряют оба положения. Автор показал, что у выводковых
птиц (куры) дифференциация фолликулов и секреция коллоида
начинается относительно раньше (через 43% срока эмбриональ-
ного развития), чем у птенцовых (у голубя — через 54%, у во-
робья — через 61%). В постэмбриональный же период, наоборот,
у выводковых наблюдалось спокойное состояние щитовидной
железы, а у птенцовых в соответствии с продолжающимися в этот
период процессами общего развития она более активна.
Исследуя гистогенез щитовидной железы у индюшиных эмб-
рионов, Абрамова (1958) установила, что синтез коллоида в ней
начинается на 11-й, а фолликулы формируются на 13-й день
инкубации (заметим, что' здесь имеются сходные соотношения
с куриным эмбрионом, так как индейки — тоже выводковые
птицы). Выделение гормона очень велико до 26—27-го дня инку-
бации, после чего резко снижается накануне вылупления.
Мицкевич (1957) установил, что тироксин из тела несушки
переходит в яйца, и считает, что это необходимо для нормального
развития на начальных стадиях эмбриогенеза, до начала функ-
ционирования собственных щитовидных желез эмбриона. Вве-
дение небольшой дозы тироксина в инкубируемые яйца заметно
уменьшало содержание гликогена печени куриных эмбрионов,
особенно на И—12-й день инкубации.
Петров (1958) показал, что при насыщении яйца иодом (при
введении иода в рацион несушек или инъекции раствора иода
в воздушную камеру инкубируемых яиц) значительная его часть
аккумулируется в щитовидной железе куриных эмбрионов и по-
вышает ее гормонообразовательную функцию. По данным автора,
это обусловило ускорение развития эмбрионов, повышение вы-
водимости и лучший рост цыплят. Противоположные данные
приводит Роглер с соавторами (Rogler et al., 1961), получивший
при низком содержании иода в рационе кур несушек более вы-
сокое продуцирование и выделение тироксина щитовидными
железами куриных эмбрионов. При высоком содержании иода
в рационе секреция и выделение тироксина были низкими.
Г. Околощитовидные железы
Венцке (Venzke, 1947), изучая морфогенез околощито-
видных желез куриного эмбриона, установил, что зачатки около-
щитовидной и щитовидной желез с каждой стороны одновременно
234
выпячиваются со дна глотки с 12 час. инкубации и удаляются
Друг от друга на 7-й день. На 17-й день инкубации автор обна-
ружил в околощитовидной железе 4 вида клеток. Более детальное
морфологическое исследование развития этих желез проведено
Студитским (1947). Морфологическая дифференциация и начало
функции околощитовидных желез куриного эмбриона наблю-
дались в конце 8-го дня инкубации, когда появляются оксифиль-
ные клетки, свидетельствующие о разжижении протоплазмы
(по-видимому, начало секреции), и увеличивается количество
липоидно-жировых включений. Деятельность желез в течение
эмбриональной жизни направлена в основном, так же как и у
взрослых птиц, на поддержание необходимого уровня кальция
в крови и лимфе. В случаях нарушения этого уровня, например
при мобилизации кальция развивающимся скелетом и падения
в связи с этим содержания кальция в крови, автор наблюдал
вспышки активности околощитовидных желез эмбриона. Кроме
того, автор показал, что добавление гипофизарных гормонов
вызывает гиперфункцию околощитовидных желез, обусловливаю-
щую в свою очередь повышение количества кальция в крови
за счет снижения его в костной ткани. При этом в условиях не-
достатка витамина D (осуществляющего в норме обратный про-
цесс) может развиться аномалия скелета типа хондродистрофии.
Kpafi4H4eK(Kraicziczek, 1957) обнаружил начало функциониро-
вания околощитовидных желез у куриного эмбриона на 7-й
день инкубации.
Д. Тимус, или зобные железы
Тимус возникает из энтодермы глотки и жаберных ще-
лей. Морфологическое развитие тимуса изучено Венцке (Venzke,
1952). Автор показал, что большие лимфоциты мигрируют из
окружающей мезенхимы в эпителиальную ткань тимуса на 8-й
день, а разделение тимуса на доли происходит в конце
15-го дня инкубации. Развитие тимуса происходит, по-видимому,
независимо от развития других эндокринных желез. Хаммонд
(Hammond, 1954) показал, что тимус является производным
бронхиальной энтодермы в районе 3-й и 4-й жаберной бороздки.
Эпителиальные клетки зачатка тимуса частично превра-
щаются на 8-й день в лимфобласты, затем (на 10—11-й день)
становящиеся лимфоцитами, а частично — в ретикуло-эпите-
лиальные клетки, образующие паренхиму дефинитивного ти-
муса (Ackerman a. Knouff, 1964).
У утиного эмбриона, по данным Клюевой (1958), тимус
до 14-го дня инкубации имеет вид островков из эпителия, внутри
которых имеются лимфоциты и эритроциты. С 16-го дня начи-
нается инволюция тимуса: он разделяется на дольки рыхлой
соединительной тканью, пронизанной кровеносными сосудами;
235
эритроцитов уже нет; главным показателем инволюции явля-
ется жировое перерождение эпителиальной основы железы.
К 23-му дню наступает полное перерождение тимуса в жировую
ткань с сильно увеличенными клетками. Автор отмечает, что
наибольшая скорость инволюции тимуса, происходящая на 17-й
день инкубации, коррелирует с падением интенсивности роста
и дыхания утиного эмбриона.
Величина тимуса является, по данным Тихомирова (1935),
показательной для состояния организма только что вылупивше-
гося цыпленка. У сильных цыплят отношение веса тимуса к весу
тела составляло 0.14%, а у слабых — только 0.044%. Цыплята
с большим размером тимуса легче переносили неблагоприятные
условия и меньше гибли. Данные Тихомирова были подтверждены
Дмитриевой (1939) и Делль (1953). Дмитриева отметила, что
меньший вес тимуса у нежизнеспособных цыплят является след-
ствием не инволюции железы, а наследственного замедления
темпа ее развития. Делль присоединяется к мнению Немилова
(1938), что тимус влияет на процесс роста не путем выработки
вещества, стимулирующего его, а способствуя преодолению не-
благоприятных условий (в частности, увеличивая иммунитет)
и подавляя факторы, вредные для роста. Хон (Hohn, 1961),
предполагая, что тимус у птиц не играет роли эндокринной же-
лезы, приводит все же в своем обзоре данные о его эндокринной
функции. Так, тимус у некоторых видов птиц снова увеличивается
в размере и восстанавливает свое «молодое» микроскопическое
строение в течение нескольких недель вслед за первым и, воз-
можно, дальнейшими половыми циклами; имеется сезонное из-
менение размеров тимуса. Тимус каким-то образом связан с дру-
гими железами внутренней секреции; так, например, тироксин
обусловливает гипертрофию его, а кортизон, наоборот, атрофию.
По данным Пилипенко (1965), тимус у уток не исчезает с воз-
растом, а подвергается инволюции, тесно связанной с актив-
ностью половых желез. При прекращении яйцекладки в зим-
нее время или у старых птиц тимус увеличивается в весе почти
до величины его у неполовозрелых утят, а также приобретает
в значительной мере свое прежнее строение — дольчатость
и деление на корковой и мозговой слой. В то же время удаление
тимуса у птиц не только не смертельно, но и как будто не влияет
заметно на дальнейшее существование взрослых птиц.
Е. Эпифиз
Еще меньше данных имеется об эпифизе как железе
внутренней секреции. Так, например, Романов (Romanoff, 1960)
пишет, что «иногда об эпифизе думают как об эндокринной же-
лезе. Однако никогда не было показано, что он имеет какую-либо
секрецию и никто даже не высказал правдоподобного предполо-
236
жения о его функции. Ио-видимому, это рудиментарный орган —
гомологичный третьему глазу примитивных позвоночных».
Бенуа же (Benoit, 1950), наоборот, высказывает предположение,
что эпифиз играет роль эндокринной железы. Только Бэлэчану
с соавторами (Balaceanu et al., 1961) в своих опытах показал,
что введение гормона эпифиза в инкубируемые куриные яйца
вызывает задержку развития семенников, небольшое увеличе-
ние веса эмбрионов и сахара крови, а также увеличение коли-
чества билирубина почти в два раза.
Остановимся кратко на морфологии развития эпифиза.
По данным Краббе (Krabbe, 1955), развитие эпифиза протекает
сравнительно одинаково у эмбрионов разных видов птиц, од-
нако у сов он совершенно рудиментарен и его дистальный конец
напоминает париентальный глаз. Спиров (Spiroff, 1959) наб-
людал, что у куриного эмбриона первичный зачаток эпифиза
появляется через 48 час. инкубации (стр. 149). Наиболее бы-
стрый рост эпифиза происходит с 5-го до 12-го дня, когда его
относительный объем возрастает в 40 с лишним раз; с 12-го дня
до вылупления объем эпифиза почти не изменяется.
Ж. Гипофиз
Эта эндокринная железа происходит из двух различных
зачатков: отпочкования со дна промежуточного мозга, дающего
заднюю долю гипофиза, или нейрогипофиз, и впячивания эпи-
телия глотки, образующего переднюю долю гипофиза, или аде-
ногипофиз.
Гипофиз является железой, регулирующей деятельность
почти всех остальных эндокринных желез, и имеет жизненно
важное значение как для существования, так и для развития
организма птицы. Нейрогипофиз выделяет гормоны окситоцин
и антидиуретический гормон — АДГ. Аденогипофиз секрети-
рует шесть или больше гормонов: два из них имеют непосред-
ственное действие на ткани — пролактин и гормон роста, а, че-
тыре — тиреотропный, адренокортикотропный (АКТГ) и го-
надотропные (фолликулостимулирующий и лютеинизирующий) —
действуют на организм через другие железы внутренней секре-
ции. Наличие у птиц гормона, стимулирующего секрецию под-
желудочной и околощитовидной желез, пока неизвестно (Hohn,
1961). Впрочем, по данным этого автора, у птиц отсутствует
и гормон роста.
Гриньон (Grignon, 1955) изучил последовательность диффе-
ренцирования клеточных элементов аденогипофиза куриного
эмбриона. Разделение ацидофильных и базофильных клеток
в зачатке гипофиза происходит уже на 6-й день инкубации.
Ацидофильные клетки на 14-й день делятся на два типа по зер-
нистости и размеру клеток, а базофильные также на два типа,
237
различающиеся по окраске (один из которых появляется на 8-й
день и связан с началом тиреотропной активности, другой —
на 12—14-й день и определяет начало гонадотропной активности).
Вилье (Willier, 1955), однако, считает, что с продуцированием
тиреотропного гормона и гормона роста связаны ацидофильные
клетки, хотя и отмечает, что чрезвычайно трудно говорить
о специфической секреции тех или иных клеток. Мицкевич (1957)
высказывает предположение, что базофильные клетки гипофиза
куриного эмбриона секретируют фолликулостимулирующий
гормон, а ацидофильные — гормон желтого тела. Автор обна-
ружил в гипофизе куриных эмбрионов ацидофильные клетки
с 10-го, а базофильные, в значительно меньшем количестве,
с 12-го дня инкубации. Исследуя гистогенез передней доли ги-
пофиза у индюшиных эмбрионов, Абрамова (1958) показала,
что хромофильные клетки в ней дифференцируются к 17-му дню
инкубации и это происходит одновременно с повышением гормо-
нальной активности щитовидной железы.
По данным Студитского (1947), тиреотропный гормон начи-
нает выделяться на 8-й день развития куриного эмбриона, а затем
секреция его увеличивается, достигая максимума в последние
дни инкубации. Автор отмечает, что функции гипофиза и щито-
видной железы очень связаны между собой. По данным Маро,
Столл и Бланке (Maraud et al., 1957), регулирующая роль ги-
пофиза в функционировании щитовидной железы куриного эмб-
риона начинается позже — с 13-дневного возраста, так как
в это время имеется внезапное ускорение темпа накопления
меченого иода в ней. Указывая, что появление тиреотропного
гормона (стимулирующего в щитовидной железе образование
фолликулов, накопление и секрецию коллоида) соответствует
началу цитологической дифференциации гипофиза, Мицкевич
(1957) отмечает, что эти процессы у выводковых птиц протекают
в более короткий срок по отношению к общей продолжительности
эмбрионального периода, чем у птенцовых, менее зрелых к мо-
менту вылупления. Мартиндейл (Martindale, 1941) провел опыты
по пересадке щитовидной железы одного куриного эмбриона
на хорио-аллантоис другого (нормального или с удаленным ги-
пофизом). В первом случае формируется нормальная железа,
во втором — сильно уменьшенная и без коллоида, а, следова-
тельно, не функционирующая. Автор приходит к выводу, что
гипофиз нормального эмбриона-хозяина выделяет трофический
гормон, стимулирующий развитие щитовидной железы, который
проникает к ней через внеэмбриональную кровеносную сеть.
Продукция тиреотропного гормона начинается у куриного эмб-
риона, по данным автора, на 11-й день инкубации в небольших,
но достаточных для активации щитовидной железы количествах.
В опытах Тиксье-Видаль (Tixier-Vidal, 1956) по совместному
выращиванию in vitro гипофизов и щитовидных желез куриных
238
эмбрионов разного возраста и с пересадками этих желез от одних
эмбрионов к другим показано, что щитовидная железа реаги-
рует на тиреотропный гормон на 6-й день инкубации, хотя гипофиз
эмбриона впервые секретирует этот гормон только на 7-й день.
Автор приходит к интересному, на наш взгляд, выводу, что
каждая железа способна к функции (реакция на соответствующий
гормон гипофиза) раньше, чем в обычное время начинается ее
деятельность. Далее автор (1958) показала, что щитовидная
железа имеет ограниченную способность к самодифференциации.
Ее секреторная способность повышается при стимуляции гор-
монами гипофиза. Гипофиз же может дифференцироваться не-
зависимо от щитовидной железы. Максимальную активность
гипофиза и щитовидной железы автор наблюдала у 15—17-днев-
ных эмбрионов. Во время вылупления наступает период сниже-
ния активности щитовидной железы, причем, в этом снижении
отмечаются сезонные вариации. Автор приходит к выводу, что
наблюдающийся у взрослых птиц закон эндокринного равно-
весия устанавливается между гипофизом и щитовидной железой
куриного эмбриона на 10-й день. Сопоставление с данными, из-
вестными для эмбрионов других позвоночных животных, дает
автору возможность заключить, что в эволюции отношений между
гипофизом и щитовидной железой имеется определенное един-
ство.
Теперь уже нет сомнения в том, что гипофиз и в эмбриональ-
ном периоде у птиц влияет на развитие надпочечников, но Кейс
(Case, 1952) первый доказал это. Автор на значительном мате-
риале показал, что у декапитированных эмбрионов с 16-го дня
инкубации снижается вес надпочечников (на 20-й день они весят
в 2 раза меньше, чем нормальные), а с 13-го дня уменьшается
содержание аскорбиновой кислоты в них (как общего количе-
ства, так и концентрации). При этом наблюдается картина атро-
фии надпочечников, которую несколько выправляют инъекции
АКТГ. Автор приходит к выводу, что зависимость надпочечни-
ков от передней доли гипофиза устанавливается на сравнительно
ранней стадии развития (13-й день инкубации). Обратное влия-
ние гормонов надпочечника на.выделение АКТГ у куриных эмб-
рионов, по-видимому, устанавливается не раньше 18-го дня инку-
бации. По данным Секей, Эндроси и Сентаготаи (Szekely et al.,
1958), АКТГ в гипофизе куриного эмбриона появляется на 8-й
день инкубации. Тот, Шимон и Секей (Toth et al., 1958), срав-
нивая интактных и гипофизэктомированных эмбрионов, обнару-
жили, что гипофиз влияет на тканевую дифференциацию коры
надпочечников с этого же дня. А Муг (Moog, 1959) показала
(методом выявления фосфатазной активности двенадцатиперстной
кишки под влиянием кортикостероидов), что АКТГ начинает
стимулировать секрецию коры надпочечников куриных эмбрио-
нов с 11-цневного возраста или вскоре после этого. Эмбрионц
239
после инъекции АКТГ были несколько меньшего веса, но вес
надпочечников при этом увеличивался; в них были более крупные
клетки, содержащие большее количество суданофильных липи-
дов. Мазина (1963, 1964) установила, что введение АКТГ обуслов-
ливает гиперсекрецию кортикостероидов (резкое снижение кон-
центрации холестерина и несколько меньшее — аскорбиновой
кислоты) надпочечниками куриных эмбрионов начиная с 12-
дневного возраста и увеличение веса надпочечников с 15 дней
инкубации. Введение АКТГ 11-дневным эмбрионам не оказывало
никакого влияния на надпочечники. Следовательно, регуляция
гормональной функции коры надпочечников гипофизом начи-
нается у куриных эмбрионов с 12-дневного возраста. Албу,
Винер и Бэлэчану (Albu et al., 1960) обнаружили, что после
введения в инкубируемые яйца АКТГ снижается относительный
вес костей куриного эмбриона (на 50% больше уменьшения об-
щего веса эмбриона) и повышается содержание гликогена в пе-
чени и сердце. Вилье (Willier, 1955) в своем обзоре о развитии
эндокринной регуляции в онтогенезе отмечает удивительное взаи-
модействие между функцией передней доли гипофиза и надпо-
чечников в эмбриональной жизни. Это взаимодействие создает
адаптивный механизм, позволяющий организму приспосабли-
ваться к стрессовым воздействиям, исходящим как из самого
организма, так и из внешней среды, что очень важно и для нор-
мального течения эмбрионального развития, и для прохождения
чрезвычайного сложного этапа вылупления, и для создания нор-
мальной реакции на воздействия внешней среды в начале постэм-
брионального развития.
Ставя вопрос о зависимости внутрисекреторной активности
гонад куриных эмбрионов от гонадотропной стимуляции гипо-
физа, Вилье (Willier, 1955) отмечает, что эта проблема еще мало
разработана и по этому вопросу получены несколько противоре-
чивые данные. Однако автор считает точно установленным, что
процесс переделки пола полностью независим от гипофиза.
Выделение гонадотропинов начинается с 13-го дня инкубации,
но в эмбриональный период гипофиз слабо влияет на эндокринную
деятельность гонад, а максимальное влияние его достигается
значительно позже — ко времени приближения половой зрелости.
Однако есть исследования, где было обнаружено гонадотропное
влияние гипофиза и в эмбриональный период. Так, например,
у утиных эмбрионов, как показали Киньон и Уатерсон (Kinyon
a. Watterson, 1958), удаление гипофиза приводит к уменьшению
эндокринной активности яичников, что проявляется в развитии
у всех эмбрионов полового бугорка и гортани у самца (см. главу
14).
Опытами Студитского (1947) с пересадками дополнительного
гипофиза куриным эмбрионам доказано влияние гипофиза на дея-
тельность околощитовидных желез 13—17-дневных эмбрионов
24Q
(на более ранних стадиях влияние менее заметно, а у 8—9-днев-
ных отсутствует вовсе). При этом железы увеличивались, в них
наблюдалось повышенное количество липоидно-жировых вклю-
чений и наличие оксифильных клеток (признак повышенной
активности).
О выделении гормона роста гипофизом куриного эмбриона
известно очень мало. Вилье (Willier, 1955) сообщает, что гипо-
физэктомия вызывает уменьшение размеров куриных эмбрионов
с соблюдением нормальных пропорций тела, и высказывает
предположение, что гипофиз у птиц включается раньше в акти-
визацию процессов роста, чем у млекопитающих. О работах
Соубель (Sobel, 1958) и Сайед с соавторами (Sayeed et al., 1962)
об антагонистичном действии кортизона и гормона роста мы уже
сообщали (стр. 230). В своем обзоре о роли гипофиза в эмбрио-
нальном развитии птиц Бенуа (Benoit, 1962) приходит к выводу,
что гипофиз не оказывает влияния на рост эмбриона, а воздей-
ствие его на половые и щитовидные железы и на надпочечники
у разных видов птиц различно. В связи с относительно поздним
включением гипофиза его морфогенетическая роль незначительна.
Автор подчеркивает, что гипофиз оказывает преимущественное
влияние на функцию органов, а не на их морфологию.
Обстоятельное исследование о нейрогипофизе куриного эмб-
риона провел Вингстранд (Wingstrand, 1954). Нейрогипофиз
птиц состоит из нервной доли, стебля и медиального возвышения.
Общий зачаток нервной доли и стебля становится заметным у 4.5-
дневного эмбриона в виде выпячивания со дна промежуточного
мозга. Автор обнаружил нейросекрет, или вещество Гомори
(коллоидные включения нервных клеток), в клетках супраопти-
ческого ядра и нейрогипофиза на 13—14-й день инкубации, что
подтверждает гипотезу о секреции вещества Гомори в клетках
супраоптического ядра и переносе его через аксоны в нейроги-
пофиз. Автор считает, что обнаружение им антидиуретической
активности гипофиза на более ранней стадии развития куриного
эмбриона (10-й день инкубации) показывает на неидентичность
АДГ и вещества Гомори. Это положение автора встречает два
возражения. Прежде всего нейросекрет — это протеин, соеди-
ненный с гормоном, причем в настоящее время, немотря на мно-
гочисленные исследования, не уточнено, носителем каких именно
гормонов является нейросекрет. Кроме того, имеются более
точные методы выявления вещества Гомори, чем те, которыми
пользовался Вингстранд, дающие возможность обнаруживать
нейросекрет на более ранних стадиях развития. Так, Мозье
(Mosier, 1955) наблюдал появление нейросекреторного вещества
в нейронах супраоптического ядра у 4-дневного, bjhx аксонах —
у 5-дневного и в воронке гипофиза — у 6-дневного куриного
эмбриона. А Гриньон (Grignon, 1955) сообщает, что ^-нейросекре-
торное вещество в нейронах супраоптического ядра может по-
Щ В. В. Рольник
241
являться на 4-й день инкубации, но активные капельки секрета
не видны до 14—17-го дня развития куриного эмбриона.
Подводя итоги исследований о развитии гормональных ме-
ханизмов регуляции обмена веществ в течение развития куриного
эмбриона, Вилье (Willier, 1955) делит весь этот период на 3 этапа:
1) 6—9 дней — процессы обмена веществ не зависят от гормональ-
ных влияний; 2) 10—13 дней — железы внутренней секреции
и органы эмбриона становятся функционально связанными через
гормоны и другие вещества, приносимые кровью, наступает
химическая интеграция; 3) с 14-го дня инкубации до вылуп-
ления — период созревания функциональной активности,
укрепления и расширения взаимосвязей эндокринных желез.
О функции гонад как железах внутренней секреции мы сооб-
щаем в главе 14.
ГЛАВА 14
ПОЛОВАЯ СИСТЕМА, ПОЛОВЫЕ ГОРМОНЫ,
ДЕТЕРМИНАЦИЯ И ПЕРЕДЕЛКА ПОЛА
К вопросам, освещаемым нами в этой главе, за пос-
ледние десятилетия привлечено внимание многих исследователей,
но в связи со сложностью проблемы по многим из этих вопросов
до сих пор имеются противоречивые суждения.
А. Образование и развитие половой системы
Закладка гонад происходит на ранних стадиях разви-
тия: у куриных эмбрионов — на 3-й, у утиных и гусиных на 4—5-й
день инкубации (Крок, 1962). Вначале гонады индифферентны
(пол не детерминирован) и имеют 3 составные части: две сомати-
ческих — медулла, или мозговая часть (полойые связки и строма),
и кора (зародышевый эпителий) и одна половая — гоноциты
(первичные половые клетки). Гоноциты располагаются внутри
медуллы и коры (Benoit, 1950). У птиц зачаток медуллы в гонадах
дифференцируется первым у обоих полов, а зачаток коры разви-
вается у самки и регрессирует у самца начиная с 8-го дня инку-
бации (Domm, 1955). Венцке (Venzke, 1954а) изучал морфогенез
гонад куриного эмбриона. Соматические части гонады возникают
из половых валиков спланхноплевры. В конце 4-го дня развития
куриного эмбриона гонаду покрывает всего один слой зародыше-
вого эпителия, а в конце 7-го в гонадах становятся очевидными
половые различия.
О происхождении первичных половых клеток уже давно велись
споры. По данным Данчаковой (Danchakoff, 1931), это экто-
дермальные, большие, амебоидной формы клетки, блуждающие
242
в кровяном потоке й попадающие затем в спланхноПлевру, в район
половых валиков. Проведя остроумные и тонко выполненные
эксперименты, Симон (Simon, 1957а, 1957b) убедительно пока-
зала, что первичные половые клетки мигрируют из передней
части внеэмбриональной зоны (имеющей форму полумесяца)
по кровеносным путям в район половых валиков, который впо-
следствии дифференцируется в гонады. Это происходит со стадии
первичной полоски до 10-сомитной стадии (с 12—19 до 33—38 час.
инкубации). Во всех опытах автора только наличие связей по кро-
веносным сосудам обеспечивало переход гоноцитов в участок
половых валиков. Если же кровообращение было нарушено, то
в гонадах не оказывалось половых клеток. В дальнейших опытах
(Simon, 1960а) при выращивании in vitro бластодерм куриного
и утиного эмбрионов, связанных общим кровообращением, ав-
тору удалось наблюдать как бы колонизацию половой железы
одного вида гоноцитами другого. Автор показала также (1960b),
что для образования гонад и их половой дифференциации гоно-
циты не необходимы; следовательно, гоноциты не имеют индук-
тивного действия на закладку гонад. Марин (Marin, 1956) наб-
людал при дифференцировке гонад куриного эмбриона на 5-й
день инкубации независимые друг от друга корковую и мозговую
части. Гоноциты первоначально располагаются преимущественно
в корковой части, переходя затем в мозговую, но не составляют,
как предполагалось ранее, основы для ее развития, а проходят
развитие в уже существующей мозговой части гонады. Автор
подчеркивает, что до 7-го дня развития куриного эмбриона го-
нада не интерсексуальна, а индифферентна в половом отношении.
А Бёрнс (Burns, 1955) считает, что гонады в это время бисексу-
альны. Третье мнение высказывают Вилье с соавторами (Wil-
lier et al., 1937) и Бенуа (Benoit, 1950), что левая гонада бисексу-
альна, а правая унисексуальна.
Развитие идифферентной гонады продолжается у куриного
эмбриона, до 6—8-го дня инкубации (Lillie, 1952). Однако мне-
ние о полной индифферентности гонад до этой стадии развития
опровергается последними исследованиями. Лимборг (Limborgh
van, 1961а) обнаружил половые различия в гонадах 4—5.5-
дневных куриных эмбрионов. Автор показал, что достаточно
четко выраженная асимметрия гонад в это время (менее 41%
общего количества гоноцитов в правой гонаде) является приз-
наком женского пола, с вероятной ошибкой 2.6%. Если гоно-
цитов более 45% в правой гонаде, пол самца определяется с ошиб-
кой 2.3%. Лимборг показал такЦсе, что асимметрия распре-
деления гоноцитов в гонадах самок обусловливается большим
количеством дегенерирующих гоноцитов в них (примерно 60
из 100). Однако о механизме, вызывающем это различие в де-
генерации первичных половых клеток в правой и левой гонаде,
автор ничего сказать не может (Limborgh van, 1961b).
16*
243
Вначале опишем развитие семенников из более или менее
индифферентной гонады. При размножении клеток зародышевого
эпителия образуются тяжи клеток, проникающие внутрь
стромы гонады, так называемые половые связки, из которых в даль-
нейшем формируются семенные канальцы. Половые связки
в семеннике куриного эмбриона закладываются к концу 7-го
дня инкубации (Venzke, 1954b). Редукция корковой части се-
менника начинается через 10 час. после этого. Гоноциты, про-
никшие с половыми связками в мозговую часть семенника, впер-
вые делятся на 9-й день, а на 11-й день инкубации происходит
массовое деление их. В конце 13-го дня в половых связках появ-
ляются полости, но семенными канальцами они становятся только
после вылупления.
{Кроме того, в состав гонады входит так называемая сетчатая
ткань. О ее происхождении велись длительные споры. Смоляр
(Smolar, 1953) удалось установить, что сетчатые связки впервые
появляются у эмбрионов обоих полов на 5-й день инкубации
в виде клеточных тяжей, направляющихся от основания половых
связок к строме мезонефроса. Предположение о происхождении
сетчатых связок из мезонефроса опровергнуто самим автором.
После экспериментального удаления мезонефроса гонада имела
сетчатые связки и была вполне нормальной. Эти данные были
подтверждены Калам (Calame, 1959). Сетчатые связки превра-
щаются в дальнейшем в семявыносящие канальцы; они, с одной сто-
роны, являются продолжением семенных канальцев, а с другой —
срастаются с отростками вольфова протока, который превращается
после вылупления цыпленка в семяпровод. По данным Смоляр
(Smolar, 1953), половые отличия в сетчатых связках начинаются
с 7—8-го дня инкубации: у самцов они становятся более объе-
мистыми, а у самок имеют вид тонких тяжей и после вылупления
исчезают совсем. В сетчатых связках самцов начинается образо-
вание полостей с 3-го дня после вылупления.
Место соединения семенника с первичной почкой делит ее
после прекращения функции на 2 части: переднюю, соединяю-
щуюся с семенными канальцами, образующую в дальнейшем
эпидидимис, и заднюю, не соединенную с ними, — паради-
димис.
В левом яичнике развивается кора и мозговой слой, причем
кора преобладает. Правый яичник рано прекращает развитие
(еще до образования коры) и становится рудиментом, состоящим
в основном из мозговой ткани. Интересно отметить, что у самцов
также имеется асимметрия гонад, но значительно менее заметная:
кора (очень небольшая) имеется на обоих семенниках, но много
лучше развита на левом. У хищных птиц кора семенников регрес-
сирует позже (Benoit, 1950). Исследование Вольфа и Пино
(Wolff et Pinot, 1961) прсвящено выяснению вопроса: почему
правая гонада куриного эмбриона не имеет коры? Обнаружено,
244
что на 3—4-й день развития зародышевый эпителий обеих гонад
имеет одинаковую структуру; с 5—6-го дня на левой гонаде он
продолжает развиваться, а на правой — редуцироваться до не-
большого скопления уплощенных клеток. Правая гонада 16—
18-дневных эмбрионов имела кору в 29.6% случаев; после инъ-
екции до 5-го дня эстрогенных гормонов — в 84%, а при инъекции
после 5-го дня — в 17% случаев. Авторы приходят к выводу,
что в начале развития эмбриона продуцируется недостаточное
количество женских половых гормонов для образования коры
на правой гонаде. Половые связки (первичные) в левом яичнике
врастают, так же как и в семеннике, в строму гонады, но не уста-
навливают связи через сетчатые связки со средней почкой и вскоре
после появления исчезают. Таким образом, зрелый яичник не имеет
ни протоков, ни трубок, которые бы шли непосредственно от него.
Характерной чертой развития яичника является второй период
интенсивного размножения клеток зародышевого эпителия,
с образованием тяжевидных врастаний яйцеобразующей ткани
в остов гонады (вторичные половые связки), где в дальнейшем
формируются фолликулы (рис. 49). Это происходит с 9-го до 14-го
дня инкубации (Benoit, 1950). Способность к образованию фол-
ликулов является свойством корковой части гонады, что дока-
зано и опытами in vitro (Haffen, 1963). В медулле гонады в это
время появляются лакуны, ограниченные интерстициальными
железистыми клетками. В правом яичнике (рудименте) остается
лакунарная медулла с несекретирующими интерстициальными
клетками^ В 3% случаев у кур остается правый яичник,
но значительно меньшего размера и не функционирующий;
у дневных хищных птиц правый яичник крупнее, чем у кур,
и ч&ще встречается, однако тоже не функционирует (Benoit,
1950).
В левом яичнике куриного эмбриона количество ооцитов
с 28 тыс. на 9-й день развития увеличивается до 680 тыс. на 17-й
день, а затем уменьшается до 480 тыс. к моменту вылупления,
когда заканчивается процесс оогенеза (Benoit, 1950). Отмечено,
что в течение эмбриогенеза происходит дегенерация ооцитов
и что они развиваются быстрее в глубине яичника. Мюллер
и Рагозина (1957) показали, что различие в дифференцировке
половых желез у двух пород кур с разной скоростью поло-
вого созревания появляется уже в 10 дней эмбрионального
развития.
Половые протоки у самцов и самок имеют разное происхожде-
ние. У самцов в качестве полового остается вольфов проток,
у самок — мюллеров канал, образующийся в виде полоски клеток
брюшинного эпителия снаружи вольфова протока. В эмбриональ-
ном периоде вольфовы протоки одинаковы у обоих полов и только
после вылупления начинается их половая дифференциация в семя-
проводы у самцов и дегенерация у самок (Benoit, 1950). У самок
245
птйц левый мюллёров канал становится яйцеводом, а правый
атрофируется. У самцов же оба мюллерова канала (правый и
левый) начинают регрессировать на 8-й день развития куриного
эмбриона и на 13-й день исчезают совсем. Лютц-Остертаг (Lutz-
Рис. 49. Часть поперечного разреза яичника 6.5-дневного
куриного эмбриона. (По Lillie, 1952).
1 — зародышевый эпителий; 2 — зачатковые клетки; з — целбм; 4 — мел-
кие клетки, располагающиеся вокруг зачатковых, образующие в буду-
щем фолликул.
Ostertag, 1954) обнаружила различие в ходе регрессии мюллеро-
вых каналов самца и правого канала у самки. У самцов регрессия
начинается с хвостовой части и продолжается к головной (обу-
словлена секрецией семенников), у самок — в противоположном
направлении — от головы к хвосту (обусловлена гормонами
яичника). Проведя целый ряд опытов по трансплантации семен-
246
ников куриным эмбрионам разных стадий развития, Тьебольд
(Thiebold, 1964) приходит к выводу, что в норме мюллеровы
каналы с начала образования и до конца 7-го дня инкубации
не чувствительны к гормонам, выделяемым эмбриональными
семенниками, а в дальнейшем регрессия этих каналов происходит
под влиянием мужских половых гормонов. Шейб-Пелегер (Scheib-
Peleger, 1955) установил, что гормоны, усиливающие регрессию
мюллеровых каналов, активизируют Энзимы, необходимые для
происходящего при этом распада протеинов.
Левый яйцевод самки открыт в полость тела и на своем перед-
нем конце образует воронку. Задний конец яйцевода открывается
в клоаку, но только с наступлением половой зрелости. Дифферен-
циация яйцевода на воронку, железистую часть и матку проис-
ходит на 12-й день инкубации.
Малиновская (Malinowska, 1961) установила, что гонады
самца куриного эмбриона до половой дифференциации лишены
собственных кровеносных сосудов. Сосудистая сеть в семенниках
образуется вначале из многочисленных мелких артериол, берущих
начало от артериальных сосудов мезонефроса. Отток крови осу-
ществляется поверхностными венулами. Затем остается только
одна артерия, проходящая вдоль длинной оси семенника, и 3— •
4 вены. Дальше кровь идет в заднюю полую вену. Отток крови
от вольфовых протоков, так же как и от мочеточников, происхо-
дит в главную почечную вену.
Б. Половые гормоны
Исследования о половых гормонах, детерминации пола
и переделке его чрезвычайно трудно разделить. Поэтому мате-
риал в следующих трех частях главы неизбежно будет несколько
повторяться.
Половые гормоны по молекулярной структуре являются
стероидными гормонами, а по влиянию, которое они обычно
оказывают на морфологию и функцию репродуктивных органов,
они делятся на андрогенные (мужские) и эстрогенные (женские).
Андрогенные гормоны можно разбить на две группы: 1) типа те-
стостерона, имеющего только маскулинизирующий эффект на са-
мок; 2) типа андростерона, маскулинизирующего самок и феми-
низирующего самцов (так называемый парадоксальный эффект).
Кора гонад, так же как и кора надпочечников, является специали-
зированной стероидной железой. Эндокринные элементы тех и
других желез имеют общее происхождение из мезонефрической
бластемы, и функционально они сходны. Но если кора надпочеч-
ников продуцирует разнообразные гормоны, включая и половые,
то корковый слой яичника обычно вырабатывает только половые
гормоны (Вичи и Дэйл, 1962). Грёнендийк-Гуйберг (Groenendijk-
Hujjberg, 1962), проведя опыты по кастрации (электрокоагулято-
247
ром) куриных эмбрионов, показал, что яичник куриных эмбрионов
наряду с феминизирующими гормонами, определяющими женский
тип регрессии мюллеровых каналов, продуцирует в небольшом
количестве и маскулинизирующий гормон.
О том, что половые гормоны продуцируются и выделяются
гонадами птиц уже в эмбриональном периоде, стало известно
только в течение последних двух десятилетий. Но и до послед-
него времени некоторые исследователи считают это утверждение '
не вполне очевидным (Domm, 1955). Наблюдая сходный эффект
при пересадках эмбриональных гонад и при инъецировании поло-
вых гормонов, Вольф и Вольф (Wolff et Wolff, 1952) одними
из первых пришли к выводу о том, что гонады куриного эмбриона
после их дифференциации продуцируют половые гормоны. Под-
тверждение этому было получено в опытах Вольф и Хаффен (Wolff
et Haffen, 1952) по культивированию in vitro гонад утиных эмб-
рионов, взятых до стадии половой дифференциации. Авторы
наблюдали при этом, что гонады дифференцируются соответст-
венно их генетическому полу: мужские превращаются в семенники,-
левые женские — в яичники, правые женские — в характерные
рудименты. При совместном культивировании левых гонад двух
• эмбрионов разных полов происходит феминизация мужской
гонады, тем бблыпая, чем раньше начато совместное культиви-
рование (6—7 дней инкубации). Мужская гонада не имеет ника-
кого действия на половую дифференциацию женской гонады.
Этими опытами было с очевидностью показано, что вещества,
продуцируемые яичниками эмбрионов, действуют непосредственно
на гонады самца. Авторы отмечают, что это в корне разрушает
гипотезу множественности веществ, необходимых для половой
дифференциации, утверждающей, что гормоны могут действовать
только на дифференциацию соматических тканей и не принимают
какого-либо участия в половой дифференциации гонад, для кото-
рой якобы требуется действие «мистических индукторов».
Гипотезу об индукционных веществах (кортицине и медул-
лярине), контролирующих половую дифференциацию гонад
(взаимодействие между корой и медуллой), выдвинул Вичи
(Witschi, 1950). Эти индукционные вещества, по мнению автора,
проявляют свое действие до начала действия продуцируемых
эмбрионом половых гормонов и осуществляют первичное воздейст-
вие на дифференцировку пола; транспорт этих веществ якобы
осуществляется благодаря диффузии, а не через кровь. Гипотеза
об индукционных веществах в настоящее время отвергнута почти
всеми исследователями (Burns, 1955; Wolff, 1958, и др.). Призна-
вая выделение половых гормонов в эмбриональный период, Вилье
(Willier, 1952) считает, что количество продуцируемого гормона
еще недостаточно для изменения пола у реципиента при пересад-
ках гонад в хорио-аллантоис. В связи с тем что результат воз-
действия был тем больше, чем ближе находилась пересаженная
248
гонада к гонадам эмбриона реципиента, автор заключает, что
распространение половых гормонов идет путем постепенной
диффузии через лежащие между ними ткани, а не через кровь.
По мнению автора, этот способ распространения присущ не только
половым гормонам и появился в онтогенезе раньше, чем через
кровообращение. Однако позднее автор (Willier, 1955) пишет,
что продуцируемые с начала половой дифференциации половые
гормоны выделяются в количестве, достаточном для эффектив-
ного действия, и через кровообращение.
С прогрессом научных методов определения малых количеств
гормонов время начала их продуцирования и выделения в кровь
все более уточняется. Так, Венигер (Weniger, 1961) установил,
что гонады куриных эмбрионов начинают продуцировать женский
половой гормон с 4-го дня, а мужской — с 6-го дня инкубации,
т. е. задолго до наступления морфологической дифференциации.
По мнению автора, эти гормоны идентичны с половыми гормонами
взрослых птиц. Столл и Маро (Stoll et Maraud, 1956) обнаружили
в амниотической и аллантоисной жидкостях куриного эмбриона
стероидные гормоны (нейтральные 17-кетостероиды и феносте-
роиды) начиная с 7-го дня инкубации. Уже начиная с 8-го дня
инкубации количество 17-кетостероидов в амниотической и ал-
лантоисной жидкостях самок выше, а на 17-й больше чем в два раза
превышает количество их у самцов.
Рассмотрим сначала данные о влиянии введенных в инкуби-
руемое яйцо половых гормонов на гонады эмбрионов. Козелка
и Галлагер (Kozelka a. Gallagher, 1934) первыми показали, что
введение женского полового гормона в куриное яйцо до наступле-
ния нормальной половой дифференциации вызывает увеличение
коры левой гонады и подавление развития правого семенника.
Степень интерсексуальности самцов в результате инъекций жен-
ского полового гормона, по данным Вилье, Галлагер и Кох
(Willier et al., 1937), в основном пропорциональна дозе гормона,
причем начинается эта интерсексуальность с активации коры
яичника. При увеличении дозы гормона семенниковые половые
связки переделываются в медуллу яичника. Авторы приходят
к выводу, что инъецированные гормоны не образуют новых
морфологических компонентов, а активизируют и изменяют раз-
витие уже заложенных в норме структур. Это мнение поддержи-
вает и Домм (Domm, 1955). В своем обзоре Вилье (Willier, 1952)
сообщает, что вне зависимости от генотипа пола андрогены имеют
тенденцию к выявлению мужских компонентов гонад и к пода-
влению женских, а эстрогены — наоборот.
Тестом влияния половых гормонов на соматические ткани
эмбриона являются различия в регрессии мюллеровых каналов
у куриных эмбрионов разных полов, а у утиных, кроме
того, — отличия в певчей гортани (сиринкса) и размере полового
бугорка.
249
Певчая гортань утиных эмбрионов имеет к 13-му дню инкуба-
ции заметный половой диморфизм: у самок она симметрична и
слабо развита, у самцов — большего размера и асимметрична,
со значительным расширением на левой стороне. Различие полов
у утиных эмбрионов по половому бугорку начинается на 13-й день,
а на 16-й день инкубации оно уже хорошо заметно. У самцов
бугорок вырастает в пенис, а у самок регрессирует, образуя кли-
тор. По данным Вилье, Галлагер и Кох (Willier et al., 1937),
инъецирование мужских и женских половых гормонов в инку-
бируемые яйца стимулирует потенциально мужские (вольфовы)
и женские (мюллеровы) протоки соответственно. Кроме того,
мужские гормоны тормозят развитие яйцевода генетических
самок, в некоторых случаях без, воздействия на яичники. Авторы
показали, таким образом, что у эмбрионов самцов яйцеводы рег-
рессируют под влиянием мужских гормонов, продуцируемых
собственными дифференцирующимися семенниками. Вольф (Wolff,
1953) наблюдал различие в половых протоках самцов и самок
куриных эмбрионов впервые на 9-й день инкубации и установил,
что односторонняя редукция яйцеводов у самок вызвана недостат-
ком мужского гормона. Проведя частичную или полную кастра-
цию куриных и утиных эмбрионов при помощи рентгеновских
лучей, Вольф и Вольф (Wolff a. Wolff, 1951) обнаружили, что
половая дифференциация соматических тканей зависит от гормо-
нальной секреции гонад эмбриона. Для развития певчей гортани
и полового бугорка мужские гормоны не нужны, а женские тор-
мозят его (случай самодифференцировки), в мюллеровых же кана-
лах мужские гормоны вызывают регрессию, а женские — нет.
Культивируя in vitro сиринкс утиных эмбрионов, Вольф (Wolff,
1953) показал, что после дифференциации певчая гортань утиных
эмбрионов обоих полов накапливает тот или иной гормон, кото-
рый поступает в нее с кровью из гонад. Подводя итоги опытам
по эмбриональной кастрации, пересадкам в хорио-аллантоис
и культивированию in vitro, автор (Wolff, 1958) приходит к вы-
воду, что половая дифференциация соматических тканей обу-
словлена гормонами, выделяемыми гонадами эмбриона.
Бёрнс (Burns, 1949) сообщает, что изменения в яйцеводах
и семяпроводах эмбрионов под воздействием половых гормонов
стойки в отличие от временных изменений в гонадах, где для
поддержания- эффекта необходимо продолжать воздействие и
после вылупления. Автор считает, что в основе различий в реактив-
ности на гормоны разных эмбриональных половых структур,
а также одной и той же структуры, но у эмбрионов разного воз-
раста лежат генетически детерминированные пороговые различия.
Обобщая, автор пишет: «Дифференциация полов может рассмат-
риваться как объединение реакций сложной системы частей,
в которых уровень секреции гормона и способность к реакции
на него приспособлены к каждой стадии».
250
В. Детерминация пола
В настоящее время никто не сомневается, что первона-
чальную детерминацию пола обусловливают половые хромосомы.
Однако механизм их действия и даже общая картина хромосомного
аппарата еще недостаточно изучены. Нет единого мнения о коли-
честве хромосом даже у такого хорошо изученного вида, как
домашняя курица. Матти (Matthey, 1950) сообщает, что хромосомы
птиц наименее исследованы из всех классов позвоночных живот-
ных в связи с большим количеством хромосом у птиц и малой
их величиной. Автор приводит следующие литературные данные
о количестве хромосом у курицы: 28 (Stevens), 32 (Schiwago),
36 (Hange), 66 (White), 74 и 78 (Susuki).1 Матти (Matthey, 1950)
считает правильным последнее количество, причем крупных
хромосом имеется всего 12. По-видимому, Ныокамер, Донелли
и Фарбс (Newcomer et al., 1960), считая, что у кур имеется 5 пар
соматических и 1 пара половых хромосом, учитывают только
эти крупные хромосомы. Матти (Matthey, 1950) приводит также
данные о количестве хромосом у ряда видов птиц: Oceanodroma
lencorrhoa — 74, Phalacrocorax carbo — 70, Sternula albifrons —
66, Larus argentatus — 66, Brachyramphus marmoratus — 50,
Lunda cirrhata — 50, Anas platyrhynchos — 80 и Coturnix cotur-
nix — 78. Джафф и Фекхеймер (1966) обнаружили, что у кур,
индеек и перепелов имеется 70—80 хромосом, и подчеркивают,
что количество хромосом для каждого вида константно.
Все эти хромосомы, кроме одной пары, являются соматиче-
скими. В половых клетках самца содержится одна пара половых
хромосом, которая, по мнению разных исследователей, является
либо первой, либо пятой парой, а по новейшим данным (Bamni
et al., 1966) — 4—5-й парой. Не решен до сих пор также вопрос
о том, имеет ли самка птиц одну половую Х-хромосому или же
одну Х-хромосому и одну Y-хромосому (Matthey, 1950). Во вся-
ком случае ясно, что пол у птиц определяет самка (у млекопитаю-
щих — самец), так как ее гаметы разные (гетерогаметный тип):
половина содержит Х-хромосому, а половина — либо Y-хромо-
сому, либо совсем не содержит половых хромосом, в то время
как все гаметы самца содержат всегда одну Х-хромосому (гомо-
гаметный тип). Даже в новейших исследованиях по этому вопросу
имеются противоречия. Джафф и Фекхеймер (1966) считают,
что у самок птиц имеется только одна половая хромосома, а Бамни
с соавторами (Bamni et al., 1966) убедительно доказывает наличие
второй хромосомы у самок кур, индеек и перепелов (которую
авторы называют W-хромосомой), расположенной довольно далеко
от первой (которую авторы называют Z-хромосомой) и имеющей
в 8—10 раз меньший размер. Эта картина, как отмечают авторы,
1 Все эти данные приводятся по Маттп (Matthey, 1950).
261
отличается от того, что имеет место у гетерогаметного пола млеко-
питающих (у самцов). У самцов птиц, по данным авторов, две
половые хромосомы связаны между собою, составляя как бы
одну хромосому.
Бенуа (Benoit, 1950) считает, что в Х-хромосомах содержится
«мужской фактор — М», а в гамете без половых хромосом — «жен-
ский фактор — F». Поэтому при оплодотворении получаются
зиготы либо FMM (обеспечивающие преимущество М и обусловли-
вающие самца), либо FFMM (преимущество F — самка). Автор
приходит к выводу, что это генетическое преимущество автомати-
чески ориентирует развитие гонады в семенник или яичник,
являясь первым агентом половой дифференциации. Развивая это
положение, Ньюкамер, Донелли и Фарбс (Newcomer et al., 1960)
считают, что основным фактором, определяющим детерминацию
пола, является соотношение между соматическими и половыми
хромосомами.
Рассмотрим теперь, как представляют себе исследователи
реализацию генных потенций при дифференциации пола. Одной
из первых отметила доминантность женского полового гормона
Данчакова (Danchakoff, 1935). При введении женского гормона
особям генетически мужского пола, по красочному выражению
автора, «генетическая конституция мужских хромосом зародыше-
вых клеток и соматических тканей оказывается бессильной сопро-
тивляться энергичному стимулу гормона. . ., вводящему анта-
гонистические реакции доминантного пола — женского». Об-
суждая роль генного механизма и половых гормонов в дифферен-
циации полов, Вилье, Галагер и Кох (Willier et al., 1937) считают,
что мужская зигота у куриных эмбрионов содержит генетические
факторы обоих полов и что первичным в детерминации муж-
ского пола эмбриона является доминирование мужских генети-
ческих факторов над женскими; интерсексуальное же состояние
следует считать интенсификацией женских факторов мужской
зиготы инъецированными женскими половыми гормонами. Ав-
торы отмечают удивительное сходство между действием генов
и гормонов в детерминизме пола эмбриона. Считая, что кора
и медулла гонады имеют противоположную половую тенденцию,
Бенуа (Benoit, 1950) доказывает, что химические вещества, вы-
деляемые генами с самого начала развития гонад, определяют
развитие коры или медуллы в гонаде. Однако автор не уточняет
ни химическую природу этих веществ, ни того, идентичны ли
они гормонам, как считает Вольф (Wolff, 1958), или нет. Домм
(Domm, 1955) считает, что гены обусловливают интенсивность
гормональной секреции и тот гонадный компонент (кора или
медулла), который имеет большую интенсивность секреции,
детерминирует пол. В связи с тем что у амфибий в отличие от птиц
гетерогамный пол мужской, у них более эффективен мужской
гормон, а у птиц — женский. Поэтому автор считает, что диф-
252
ференциация полов у птиц обусловливается женским гормоном,
а мужской имеется у обоих полов, но подавляется женским.
Автор приходит к заключению, что гетерогаметный пол имеет
бисексуальный гормональный потенциал, т. е. самки птиц спо-
собны продуцировать и женский, и мужской гормоны. У некото-
рых круглоротых бисексуальность сохраняется и во взрослом
состоянии, но у большинства позвоночных гермафродитизм —
-это аномалия. Вилье (Willier, 1952) и Венигер (Weniger, 1961)
приходят к сходному выводу, что гены контролируют количество
продуцируемых гормонов, а от них в свою очередь зависит детер-
минация пола. Противоположное мнение высказывает Бёрнс
(Burns, 1949). Автор считает, что первичные половые клетки
не влияют ни на гистологическую структуру гонад, ни на детерми-
нацию пола. По мнению автора, пол гонады определяется струк-
турными элементами гонадообразующего участка, вне зависимости
от генетической конституции включенных первичных половых
клеток, которые даже в отношении образования гамет индиффе-
рентны, или бипотенциальны.
Приведем несколько экспериментальных работ, помогающих
уяснить роль отдельных компонентов гонады в детерминации
пола. Хаффен (Haffen, 1960) выяснила, что зародышевый эпите-
лий самки всегда дифференцируется в кору яичника, вне зависи-
мости от того, развивался ли он с медуллой самки или самца.
Зародышевый же эпителий самца, взятый до половой дифферен-
циации (5—6-й день инкубации), подвергается влиянию ткани,
с которой он находится в контакте. Итак, данные автора показы-
вают, что кора гонады самца и гонады самки неравноценны.
Следовательно, концепции Бенуа (Benoit, 1950) и Бёрнса (Burns,
1949), нридающих большое значение в детерминации пола тому,
какой компонент гонады (кора или медулла) доминирует в раз-
витии, неправильны. На основании результатов опытов по инъек-
ции мужских и женских половых гормонов Гамильтон (Hamilton,
1961) приходит к выводу, что нормальная, специфическая для
данного пола форма регрессии мюллеровых каналов (с 5—
6-го дня — начала дифференциации мочеполовой системы, до 8—
9-го дня — начала регрессии каналов) происходит благодаря
стимуляции гормонами неизвестных, генетически определяемых
внутриклеточных рецепторов и что программированная ин-
волюция мюллеровых каналов вызвана подавлением окисли-
тельных процессов и освобождением протеолитических и гидро-
литических энзимов, характерных для ткани, проходящей дегене-
рацию.
На основании того, что переделка женского пола при введе-
нии андрогенных гормонов не удается, а при гетеросексуальном
парабиозе или имплантации самкам семенников бывает удачной,
Бичи и Дэйл (1962) высказывают предположение об образовании
животными раздельнополых видов веществ, подобных антителам,
253
которые антагонистичны или полностью подавляют развитие
половых желез противоположного вида.
Фабер (Faber, 1959) косвенно показал связь между гонадо-
тропным гормоном и детерминацией пола, опровергая мнение
Вилье (Willier, 1955) об отсутствии этой связи (см. главу 13).
Обобщая свои наблюдения по ряду видов птиц (мускусные утки,
домашние утки, гибриды между ними, ястреба), автор обнару-
жил, что у пола, имеющего больший вес и размер тела, в гипо-
физе наблюдались ацидофильные клетки большего размера.
Г. Половые отличия эмбрионов
и соотношение полов
Нашими работами (Рольник, 19636, 1964; Рольник и
Портенко, 1964) показано, что начиная с 11-го дня развития кури-
ных эмбрионов имеется достоверное различие в газообмене эмбрио-
нов разного пола. Следовательно, не только доказано выделение
половых гормонов эмбриональными железами к этому времени,
но и воздействие этих гормонов на обмен веществ (подробнее
см. в главе 10). Мы показали также (Рольник, 1963а), что вес
яиц, из которьус вылупляются петушки, в среднем больше веса
яиц, из которых вылупляются курочки, на 0.4 г, и это различие
достоверно. Гром и его соавторы (1966) подтвердили и эти наши
данные, и то, что с 10-го по 19-й день инкубации куриные эмбрионы
мужского пола потребляют больше кислорода.
Цыплята разного пола, как показал Романов (Romanoff, 1933,
1948), сразу после вылупления имеют ряд морфологических и
физиологических отличий. У самцов вес на 2.7 г, пульс на 8 уда-
ров в 1 мин. (212 у самцов и 204 у самок), количество эритроцитов
на 0.9 млн/мм3 (2.35 у самцов и 2.26 у самок) больше, чем у самок.
У петушков, кроме того, больше длина и ширина головы, а также
длина клюва. Из внутренних органов больший вес у самцов ймеют
мышечный желудок и сердце, а у самки — печень. Автор считает, что
у самки преобладают анаболические процессы обмена, а у самца —
катаболические, и предполагает наличие заметных отличий в мета-
болизме и секреции эндокринных желез.. По данным Козина и
Мунро (Kosin a. Munro, 1941), эмбрионы мужского пола потреб-
ляют больше кальция из скорлупы, чем эмбрионы женского
(это различие достоверно), что обусловливает больший вес костей,
а в связи с этим, возможно, и больший вес мышц и внутренностей.
Мичек, Чнаперова и Трнкова (Mi6ek et al., 1955) обнаружили
половые различия в форме желточного мешка уже на 4-й день
инкубации: у самцов — это форма вытянутого по короткой оси
яйца эллипса с левой частью меньше правой, а у самок левая
и правая части желточного мешка равны. Несколько различна
и средняя длительность инкубационного периода для петушков
и курочек. Колигадо (Coligado, 4953) отмечает, что в первые
264
8 час. вылупления партии цыплят преобладают самки, во вто*
рые 8 час. количество цыплят обоих полов равно, а затем до конца
вылупления наблюдается большее количество петушков. Сход-
ные данные на большом материале (269 партий) получил и Ген-
дерсон (Henderson, 1956).
В связи с различиями в морфологии и физиологий эмбрионов
разных полов стоит, возможно, и различная чувствительность
к вредным воздействиям среды. О том, какой пол является более
чувствительным к вредным воздействиям, имеются разные мнения.
Ридль (Riddle, 1917, цит. по: Needham, 1931) предложил мета-
болическую теорию соотношения полов, по которой самцы в связи
с бблыпим обменом веществ гибнут в эмбриональный период
чаще, чем самки. Хейз и Спеар (Hays a. Spear, 1950), наоборот,
считают, что у птиц самцы как гомогаметный тип должны иметь
более низкую эмбриональную смертность. Авторы доказывают
это тем, что при повышении выводимости цыплят процент самцов
уменьшается; следовательно, в эмбриональном периоде больше
гибнут самки. Иванов же (1955а) показал, что самцы более чувст-
вительны к вредным воздействиям: с 12-го до 19-го дня инкуба-
ции гибнут больше самцы, а с 19-го дня до вылупления — самки,
так как они способны переживать неблагоприятные условия
более длительный период и гибнут только при наступлении 2-го пе-
риода повышенной смертности эмбрионов (см. главу 17). Таким
образом, суммарное количество эмбрионов каждого пола, поги-
бающих за время инкубации, примерно одинаково. Так как
у всех классов позвоночных животных более чувствительны к вред-
ным воздействиям эмбрионы мужского пола, автор приходит
к выводу, что различие в чувствительности не зависит от того,
гетерогаметный или гомогаметный мужской пол у данного класса
животных. В другом исследовании автор (Иванов, 19556) под-
твердил данные Антоновича (1950) о повышении процента куро-
чек (до 63.3%), выведенных из яиц, заложенных в инкубатор
сразу после снесения их курицей, и увеличении процента петуш-
ков (53.0—57.4%) из яиц, хранившихся 6 дней и дольше. Заме-
тим от себя, что, следовательно, при неблагоприятных условиях,
во время перерыва в развитии эмбриона, еще на стадии гаструлы,
самки гибнут больше, а возможно, при этом подавляются женские
половые потенции.
Соотношение полов бывает равно 1 : 1 только при содоставле-
нии большого статистического материала, а в каждом конкретном
случае в природе на соотношение полов влияют различные фак-
торы: возраст и питание родителей, интенсивность их обмена
веществ, качество половых клеток и др. (Иванов, 1955а). По мне-
нию автора (Иванов, 1962), наиболее общая закономерность,
определяющая соотношение полов в потомстве, заключается в том,
что максимум благоприятных условий способствует развитию
большего процента самок в потомстве. Чемпион (Champion, 1960)
255
приходит к правильному, на наш взгляд, мнению, что соотноше-
ние полов 1 : 1 при объединении большого материала указывает
на отсутствие повышенной смертности какого-либо из полов
в эмбриональный период. Возможно, что в некоторых линиях
и семьях кур смещение соотношения полов бывает за счет сцеп-
ленных с полом летальных для эмбрионального периода генов.
Интересные данные об изменении соотношения полов получены
в опытах Карапетяна (1964). В потомстве кур 6—7-месячного
возраста и 36—37-месячных петухов было 70.7% петушков и
29.3% курочек, а у 30-месячных кур и 6—7-месячных петухов
было 30.8% петушков и 69.2% курочек. В обоих случаях измене-
ние соотношения полов было вполне достоверным. К сожалению,
автор не рассматривает причины этого изменения и ссылается
только на большую роль обмена веществ, который в значительной
степени обусловлен возрастом птицы.
Д. Переделка пола
Начнем с описания некоторых спонтанных случаев ин-
терсексуальности и изменения пола. Ныокамер, Донелли и Фарбс
(Newcomer et al., 1960) цитологически исследовали ткань гонады
своеобразного интерсекса: курица снесла около двух десятков
яиц, а затем у нее появились вторичные половые признаки петуха;
при вскрытии была обнаружена только левая гонада, имеющая
опухоль; в ней поверх фолликулярной ткани была ткань семен-
ника со сперматозоидами. Автор упоминает, что случаи превра-
щения хорошей несушки в петуха с двумя семенниками после
образования опухоли в яичнике были не единичными. Сходные
результаты получены при искусственной овариотомии курицы
(Miller, 1938). После овариотомии компенсаторно гипертрофиро-
валась правая гонада, приобретающая вид семенника и даже
функцию сперматогенеза. Следовательно, и во взрослом состоянии
у птиц возможна трансформация пола, так как женский половой
гормон является доминирующим и прекращение выделения его
(под влиянием опухоли или удаления яичника) приводит к пре-
вращению гонад в семенники, выделяющие мужской половой
гормон, обусловливающий в свою очередь появление самцовых
вторичных половых признаков.
За последнее десятиление проведено много исследований,
посвященных переделке пола или сдвига соотношения полов
в связи с большой практической важностью этой проблемы.
Чтобы легче было разобраться в массе работ по переделке пола,
мы упомянем сначала о тех из них, где влияние на пол потомства
пробовали оказать через взрослый организм, затем о тех, где
авторы склоняются к выводу о возможности переделать пол путем
воздействия в эмбриональный период, и, наконец, о тех работах,
где описаны неудачи в переделке пола.
266
По данным Спайсера (Spicer, 1954), введение эстрогена пету-
хам снижало процент петушков в потомстве и даже передавалось
по наследству. Не имея основания не верить автору, мы все же
считаем маловероятными эти результаты. Пун (Pun, 1958) и Бейл-
харц (Beilharz, 1960) имплантировали таблетки стилбестрола
курам и получили у их потомства небольшие сдвиги в соотношении
полов, а при имплантации гормона петухам никакого сдвига
в соотношении полов не было. По сообщению японских авторов
Сиракава (1957) и Сибаяма (1957, цит. по: Сиракава, 1957) введе-
ние с пищей метиландростерона значительно повысило процент
курочек в потомстве (до 70—80%). Однако объяснения результа-
тов и механизма действия гормона авторы не приводят. Об ис-
следованиях по изменению соотношения полов под влиянием
возраста родителей (Карапетян) и сроков хранения яиц
(Иванов) мы уже сообщали (стр. 255—256). Курбатов (1966) при-
водит наблюдения (к сожалению, очень немногочисленные) о сдвиге
соотношения полов в сторону увеличения процента курочек
в потомстве на 5—10%: а) при умеренном использовании петухов
(через день), б) высоком уровне обмена у кур, в) кратковременном
хранении яиц перед инкубацией и г) при подогреве их во время
хранения. Автор считает, что указанные факторы повышают
жизнеспособность половых клеток, увеличивающую шанс развития
из них курочек, однако доказательств этого положения не при-
водит.
Введение гормонов в инкубируемые яйца пробовали сделать
двумя способами: путем инъекций и опусканием яиц в раствор
гормона. Второй метод берет начало с опубликования патента
Зельцера (Seltzer, 1956). Автор кратко сообщает, что опускание
яиц в раствор гормонов и витаминов изменяет пол цыплят в необ-
ходимом направлении и значительно повышает их жизнеспособ-
ность. Западногерманская фирма Маркс (Marks, 1957) также
кратко рекламирует сходный метод изменения пола.
Смещение соотношения полов при введении половых гормонов
в~яйца достигнуто рядом исследователей (Менгер, 1960; Подоба
и Тоцкая, 1961; Нестеров и Банков, 1964). Однако при этом
никогда не получалось полной инверсии пола, так как в этих
исследованиях нет данных о Продуктивности птиц, а только
это может служить признаком успешной переделки пола. Полной
инверсии пола не было получено и при применении другого
метода — опускания яиц в раствор гормона (Курбатов, 1960;
Pincus a. Erickson, 1962).
Более серьезное впечатление оставляют работы авторов,
считающих современные методы недостаточными для переделки
пола. В этих исследованиях инъекции женских гормонов обу-
словливали у генетических самцов изменение гонад и половых
протоков: правая гонада практически отсутствовала, левая варьи-
ровала от овотестиса до нормального яичника, яйцеводы также
17 В. В. Рольник
267
варьировали — от почти рудиментарного левого яйцевода до пол-
ного развития обоих яйцеводов (у генетических самцов). Эти из-
менения не были стабильными, и у взрослых птиц постепенно
восстанавливались признаки генетического пола. Мужские гор-
моны не вызывали изменения пола у генетических самок. Введе-
ние гормонов всегда сильно снижало выводимость и дальнейшую
жизнеспособность молодняка (Jaap et al., 1951; Domm, 1955;
Kruger a. Steuer, 1957; Ocariz, 1957; Coles, 1958, и др.).
Меллен (Mellen, 1957), Тинховен (Tienhoven van, 1957) и
Глик (Glick, 1961; Глик, 1966) проверяли метод Зельцера по пере-
делке пола. Меллен не обнаружил никакого влияния опускания
яиц в раствор гормонов на трансформацию пола или изменение
жизненности, но, по-видимому, здесь была применена неудачная до-
зировка. Тинховен наблюдал некоторое изменение репродуктивных
органов, но у генетических самцов индуцированная женскими гор-
монами кора яичника с возрастом уменьшалась в размере, а в даль-
нейшем они возвращались к первоначальному полу. Спермато-
генез у переделанных 6-месячных петушков задерживался, а у ге-
нетических самок (в результате обработки яиц женскими гормо-
нами) сильно снижалась яйцепродукция в связи с большим коли-
чеством овуляций в полость тела, обусловленных недостаточным
размером левого яйцевода. После обработки яиц мужскими гор-
монами яйцекладка у генетических самок не прекращалась.
Автор приходит к выводу, что предложенный Зельцером метод
несостоятелен. По данным Глика, обработка мужскими гормо-
нами снижала не только выводимость, но и репродуктивную спо-
собность выведшихся из этих яиц петухов, т. е. действовала в про-
тивоположном направлении, чем указывал автор метода (Seltzer,
1956). По данным Чжан и его соавторов (1959), опускание яиц
в раствор гормонов также обусловливало неполную переделку
пола, хотя при этом происходило некоторое изменение половых
признаков: левая гонада была бисексуальной, и яйцеводы отсут-
ствовали. Следует упомянуть также об исследованиях Жмурина
и Михайлова (1966) о влиянии дополнительных аминокислот
(как при скармливании их несушкам, так и при введении
их непосредственно в инкубирующиеся яйца) на изменение
соотношения полов в потомстве кур. Однако эти исследования,
на наш взгляд, еще мало обоснованы и лишены какого-либо вскры-
тия механизма действия аминокислот на изменение пола.
Приведенный материал показывает, что проблему переделки
пола еще нельзя считать решенной. Нужно еще много исследова-
ний по изучению обмена веществ обоих полов, энзиматических
механизмов воздействия гормонов и т. д. и т. п., а также по изы-
сканию методов переделки пола, чтобы решить эту проблему,
на которой «сломали зубы» многие ученые и практики-птицеводы,
но в которой они уже решили так много загадочных вопросов.
258
Г Л А В A 16
РАЗВИТИЕ МУСКУЛАТУРЫ И СКЕЛЕТА
А. Развитие мускулатуры
Мышечное волокно образует вместе с нервным одну
функциональную единицу, а каждая мышца связана с определен-
ным нервом. При прогрессивном развитии мышцы в филогенезе
увеличивается связанный с нею нерв, и наоборот, в случае редук-
ции мышцы отмирает и иннервирующий ее нерв (Шмальгаузен,
1947). Несколько нарушает это утверждение обнаруженный
Гамбургером (Hamburger, 1939) факт, что мышечные волокна
могут образовываться в зачатке конечности, экспериментально
лишенной нерва. С другой стороны, эти безнервные конечности
были обычно меньшего размера, чем иннервированные этого же
возраста. По мнению Орбели (1945), в эволюционном развитии
нервной и мышечной систем существует неразрывная связь- и эво-
люция мышцы зависит от нервных импульсов. В исследовании
Волохова (1951) детально рассмотрены взаимоотношения между
развитием мускулатуры и нервной системы в течение эмбриоге-
неза птиц. Специализация вполне сформированной мышцы взрос-
лого животного, приобретенная ею в течение эволюции, сохра-
няется лишь до тех пор, пока мышца получает непрерывный
поток импульсов из центральной нервной системы. Денервация
мышцы вызывает процесс, обратный дифференцировке, — возврат
к более примитивному состоянию, соответствующему ранним
стадиям фило- и онтогенеза.
У всех позвоночных животных ясно выражено деление муску-
латуры на соматическую и висцеральную, которые различаются
как по своему развитию, так и по иннервации; последнее различие
является важным критерием для решения вопроса о происхожде-
нии той или иной мышцы. Соматическая мускулатура (вся по-
перечнополосатая) иннервируется нервными волокнами брюшных
корешков спинномозговых и соответствующих им двигательных
головных нервов. Висцеральная мускулатура (частью гладкая,
частью поперечнополосатая) иннервируется висцеральными нер-
вами симпатической и парасимпатической нервной системы. Шмаль-
гаузен (1947) считает, что соматическая мускулатура образуется
исключительно за счет мускульного листка миотома. Но Роулсу
и Страусу (Rawles a. Straus, 1948) удалось показать (методом
маркировки углем), что туловищная мускулатура куриного
эмбриона только частично (спинная ее часть) происходит из соми-
тов, а остальная — из спланхнотомов (боковые пластинки).
Этим же методом Саундерс (Saunders, 1958) показал, что зачаток
конечности происходит не из мезенхимы сомитов, а из спланхно-
томов.
17*
269
Развитие мускулатуры, по Шмальгаузену, происходит следую-
щим образом: миотом утолщается, клетки его становятся цилинд-
рическими и теряют свои границы, так что в каждом сегменте
развивается один сплошной синцитий со многими ядрами. На внут-
ренней поверхности его начинается образование мышечных во-
локонец (фибрилл), после чего эта поверхность увеличивается
образующимися складками, врастающими внутрь синцития и
делящими его на большое количество мышечных лент. Затем
каждая мышечная лента делится на ряд мышечных волокон,
содержащих на периферии фибриллы. В промежутки между
мышечными лентами и между волокнами врастает соединительная
ткань, служащая для обособления мелких мышечных единиц
и несущая кровеносные сосуды для их питания.
Изучая развитие мышечной ткани у куриного эмбриона,
Германн (Hermann, 1952) обнаружил, что в период образования
миотомов (с 45—50 до 90—96 час. инкубации) количество клеток
в них увеличивается примерно в 15 раз; следовательно, каждая
клетка делится за это время около 4 раз. Объем миотомов за это же
время увеличивается в 26 раз, что связано с повышением содержа-
ния воды в клетках; с 12-го дня размножение клеток в мышечной
ткани почти прекращается. С 11-го дня количество протеина
в клетках начинает резко увеличиваться, но до 18-го дня их
объем не изменяется, так как протеины замещают воду в мышечных
клетках с соответствующим увеличением сухого веса. С 18-го дня
вследствие продолжающегося накопления протеина в клетках
их объем увеличивается. Примерно 25—35% вновь образованных
протеинов — это специфический сократительный протеин — акто-
миозин. Муг (Moog, 1952) показала, что миозин, соединяющийся
с актином и образующий актомиозин, достигает максимальной
концентрации ко времени вылупления.
Процесс дифференциации мышц и сухожилий задних конеч-
ностей куриного эмбриона начинается на 6-й день, а сухожилия
прикрепляются к хрящу на 11-й день инкубации (Wortham,
1948). Вил (Weel, 1948) отмечает, что биохимическая дифферен-
циация в зачатке конечности предшествует заметным морфологи-
ческим изменениям. Так, например, в том месте, где возникнут
мышечные клетки, наблюдается высокая концентрация аскорби-
новой кислоты. В тщательном исследовании развития мышц зад-
ней конечности куриного эмбриона Китиякара (Kitiyakara,
1959) показал, что в 5—6-дневной почке конечности только начи-
нают закладываться островки миогенеза между эктодермой и
участками охрящевения. Клетки внутри этих островков боль-
шего размера, чем клетки окружающей мезенхимы. Кроме вере-
тенообразных клеток, здесь наблюдаются и многоядерные мио-
бласты. И в тех и в других уже заметны фибриллы, большинство
которых выглядит гомогенными, а в некоторых видна поперечно-
полосатость. У 8—10-дневных эмбрионов клетки миогенных
260
участков превращаются в лентообразные волокна с ядром в центре
каждого из них и расположенными по краям поперечнополоса-
тыми фибриллами. Эти волокна соприкасаются с соединительно-
тканными связками и надхрящницей. Митохондрии внутри воло-
кон имеют длину 1—2 мк. Одновременно происходит заметное
размножение клеток мезенхимы. На краях мышцы, близко при-
мыкая к ней, мезенхимные клетки сливаются в многоклеточные
массы и образуют синцитиальные участки без заметной гра-
ницы между ними и мышцами. Иногда поперечнополосатые
фибриллы проникают в эти синцитиальные участки. Автор предпо-
лагает, что синцитий принимает участие в дифференциации мио-
фибрилл. У 12—14-дневных эмбрионов размножение мезенхим-
ных клеток почти прекращается, и строма мышечной ткани сходна
со стромой хорошо развитой мышцы. Мышечные волокна
на 14-й день формируются в маленькие пучки, по 2—4 волокна,
с четко дифференцированной оболочкой. У 18-дневного эмбриона
количество волокон в пучке значительно увеличивается, хотя
строма заполняет еще большие участки между волокнами. Струк-
тура этих мышечных волокон такая же, как у полностью разви-
той взрослой мышцы. В связи с увеличением количества миофиб-
рилл ядра перемещаются к периферии клетки, к ее оболочке.
Количество митохондрий увеличивается, и они располагаются
рядами между миофибриллами. Размер их меньше, чем на более
ранних стадиях развития мышцы. Отсутствие митотических
фигур внутри полностью сформированных фибрилл и хорошо
заметная митотическая активность одноядерных клеток указы-
вают на то, что пролиферация ядер предшествует формированию
в этих клетках мышечных фибрилл.
При дифференцировке зачатка мышц передней конечности
птиц образуется также часть мышц плечевого пояса, в том числе
и огромная грудная, которая является главной летательной мыш-
цей. К соматической мускулатуре, кроме мышц туловища и конеч-
ностей, относятся глазные мышцы, развивающиеся из предушных
головных миотомов, и подъязычная мускулатура, образующаяся
из брюшных отростков затылочных и частично из передних туло-
вищных миотомов.
Висцеральная (внутренностная) мускулатура развивается из
мезодермы боковых пластинок, а иногда, в виде исключения, и
из эктодермы. Сосудистая система, средняя и задняя часть кишеч-
ника и некоторые другие органы снабжены гладкой мускулату-
рой, а кожа — гладкой и поперчнополосатой. Поперечнополоса-
тая висцеральная мускулатура развилась из гладкой в резуль-
тате более высокой дифференцировки некоторых ее участков
(Шмальгаузен, 1947). Ею снабжены глотка и передняя часть
кишечника. Мышца сердца также развилась из гладкой муску-
латуры. Как показали электронномикроскопические наблюдения,
мышечные нити имеются в сердечной мышце уже у 30-часового
261
куриного эмбриона, что хорошо соответствует времени начала
пульсации (Hibbs, 1956). (О мускулатуре амниона см. главу 7).
Формирование структуры сухожилия на примере развития
сухожилия плюсны куриного эмбриона исследовал с помощью
обычного и электронного микроскопа Джэксон (Jackson, 1956).
Между 6-м и 7-м днями зачаток сухожилия — это скопление
мезенхимных клеток, затем дифференцирующихся в фибробласты.
Вначале мезенхимные клетки превращаются в синцитий, а к
концу 9—10-го дня в нем появляются границы клеток. В клет-
ках и вне их образуются коллагеновые волокна (на 8-й день
их диаметр равен 80 А), которые после 11-го дня формируются
в пучки. К 20-му дню диаметр волокон достигает 400 А. В раз-
витии сухожилия имеют место два способа образования волокон:
внутри клеток (преобладает в начале эмбрионального развития)
и вне клеток (преимущественно в конце).
Приведем данные некоторых биохимических показателей раз-
вивающихся мышц куриного эмбриона. Как известно, сокращение
мышечного волокна происходит при взаимодействии актомиозина
с аденозинтрифосфорной кислотой (АТФ), причем она расщеп-
ляется на аденозиндифосфорную и фосфорную кислоты. Для
нормального осуществления длительной мышечной деятельности
необходим ресинтез АТФ. Энергия для этого ресинтеза образуется
в значительной мере за счет глюкозы и гликогена, главным обра-
зом за счет анаэробного и аэробного превращения углеводов.
По данным Ли (Lee, 1951), в течение первых двух недель инкуба-
ции в склетных мышцах имеется небольшое количество гликогена,
но с 15-го до 18-го дня оно внезапно увеличивается. Между коли-
чеством гликогена в скелетной мышце и ее спонтанной двигатель-
ной активностью имеется обратная корреляция. По мнению автора,
запас гликогена играет важную роль в превращении эмбриональ-
ной, спонтанно сокращающейся мышцы, во взрослую, отвечаю-
щую только на раздражение. Желудкова (1963) обнаружила не-
сколько иную картину изменения содержания гликогена в мышцах
задних конечностей; в количестве гликогена наблюдается два
подъема — на 12—13-й и 17—20-й день развития куриного эм-
бриона. Исследуя процентное распределение свободной и связан-
ной с белком фракции гликогена, автор обнаружила, что в тече-
ние всего эмбрионального развития преобладает связанный глико-
ген, который, по ее предположению, является наиболее метаболи-
чески активным. Во время вылупления содержание гликогена
резко и быстро снижается в мышцах лап и в несколько меньшей
степени — в грудной мышце (Никитина, 1963).
Эмбриональная мышечная ткань в отличие от мышц зрелого
организма обладает повышенной способностью к использованию
глюкозы, а гликоген почти не потребляет. Обстоятельный обзор
о путях использования углеводов эмбриональными мышцами
дан Перцевой (1963а). Автор приходит к следующим выводам.
262
1) Гликолиз в мышцах возникает рано в эмбриогенезе, одновре-
менно с их появлением, интенсифицируется в течение развития и
формируется окончательно у зрелорождающихся (куры) в конце
эмбрионального—начале постэмбрионального периода, а у не-
зрелорождающихся (данные только по млекопитающим — кро-
лики и крысы) — позднее, в постнатальный период, совпадая
по времени с созреванием функциональных свойств мышечной
ткани. 2) В эмбриональных мышцах, как и в зрелых, гликолиз
протекает с участием фосфорных соединений. На первых этапах
гликолиза основным субстратом является не гликоген, а глюкоза,
что связано, по-видимому, с поздним образованием гликогеноли-
тических энзимов — фосфорилазы и фосфоглюкомутазы — и
со значительной активностью гексокиназы в эмбриональных мыш-
цах (Перцева, 19636). Следует отметить, что усиление гликолиза
в мышцах куриного эмбриона совпадает с началом активной мы-
шечной деятельности. Так, в течение второй недели инкубации
в мышцах накапливается миозин и актомиозин (Csapo a. Her-
mann, 1951), значительно повышается количество АТФ и фосфо-
креатина (Hermann, 1952), увеличивается активность аденозинтри-
фосфатазы (Moog, 1952) и происходит переход от спонтанной
активности к рефлекторной (подробнее см. главу 9, стр. 167—171).
В это же время увеличивается содержание молочной кислоты,
свидетельствующее об усилении гликолитического процесса (Пер-
цева, 1961).
В сердечной ткани куриного эмбриона, по данным Кирпични-
ковой (1956), гликоген появляется с началом пульсации (33 -
38 час. инкубации), но в несокращающейся части (раздвоенной
части предсердия) гликогена в это время еще нет. К моменту на-
чала кровообращения в капиллярах (около двух суток инкуба-
ции) в предсердии гликогена больше, чем в желудочке, что, по пред-
положению автора, связано с наличием в нем центра автома-
тизма.
Функциональная дифференциация мышцы в значительной
мере характеризуется распределением в ней активности холин-
эстеразы. Диффузная активность холинэстеразы в миобластах
куриного эмбриона наблюдается уже с 3-го дня инкубации (Beg-
liomini a. Moriconi, 1959). Мументалер и Энгель (Mumenthaler
a. Engel, 1961) установили, что до иннервации миобластов холин-
эстераза в скелетных мышцах куриного эмбриона диффузно рас-
пределена в цитоплазме клеток, а после включения иннервации
активность энзима локализуется лишь в двигательных концевых
пластинках и экваториальных частях мышечных волокон. Конце-
вые пластинки, образующиеся в течение эмбриональной жизни,
морфологически отличны от более сложных синапсов взрослых
кур. Остается несяным, возникают ли взрослые концевые пла-
стинки из эмбриональных или они образуются заново на более
поздних стадиях развития. Милло (Millo, 1961) подтверждает,
263
что ранее диффузно распределенная активность холинэстеразы
к 18-му дню инкубации локализуется в двигательных концевых
пластинках. Автор отмечает, что это совпадает с проникновением
в них нервных волокон и, следовательно, скелетная мышца к этому
времени окончательно дифференцируется.
Б. Развитие скелета
Скелет у птиц облегчен и отражает приспособление к по-
лету: передние конечности преобразовались в крылья; развилась
мощная грудина с килем для прикрепления мышц крыльев; силь-
ные ноги с большим тазом обеспечивают двуногое хождение; череп
облегчен и имеет огромные глазницы; шея длинная и подвижная.
Описание развития скелета приводится нами в основном по Лилли
(Lillie, 1952). Основной источник образования скелета — это
склеротомы, а кости черепа происходят главным образом из не-
сегментированной головной мезенхимы.
1. Гистология образования костей
Большинство костей проходит три стадии развития: 1-я —
перепончатая или дохрящевая, 2-я — хрящевая, 3-я — стадия
окостенения. Это так называемые замещающие кости. Но разви-
тие некоторых костей, в том числе многих костей черепа, ключиц
и др., происходит независимой от первичного (хрящевого) скелета,
и с хрящем-непосредственно они никогда не связаны. Это пере-
пончатые, или покровные, кости.
Перепончатая стадия замещающей кости характеризуется кон-
денсацией мезенхимы, часто представляющей зачаток нескольких
будущих костных элементов. Далее здесь формируется хрящ пу-
тем уплотнения клеток и изменения консистенции тканевой жид-
кости, в которой появляется вещество хондромукоидного типа.
Это вещество (матрикс), накопляясь в межклеточных простран-
ствах, постепенно разделяет клетки. Поверхностные клетки
зачатка кости образуют оболочку — надхрящницу, которая при
окостенении хряща превращается в надкостницу. Окостенение
замещающих костей можно хорошо проследить на костях конеч-
ностей. Оно проходит ряд стадий: 1) образование перихондриаль-
ной кости путем откладывания надхрящницей слоя кости на по-
верхности хряща; 2) разрушение сердцевины хряща путем размяг-
чения хрящевой основы (матрикс) до слизистого состояния;
3) кальцификация хряща путем откладывания извести в межкле-
точные пространства, после чего хрящевые клетки удаляются спе-
циальными клетками — разрушителями хряща; 4) внутрихряще-
вое окостенение путем откладывания кости остеобластами внутри
костного мозга на внутреннюю поверхность перихондральной
кости. В бедре куриного эмбриона 2-я стадия начинается на 10-й
264
Рис. 50. Окостенение скелета куриных эмбрионов в разные дни инкубации.
(По Ноймейстеру, 1966).
А — на 8-й; Б — на 10-й; В — на 13-й; Г — на 16-й; Д — на 18-й день инкубации.
день, а 3-я происходит на 17—18-й день инкубации (Lillie, 1952).
Вскоре после начала разрушения сердцевины хряща в полость,
получившуюся в результате этого, входят кровеносные сосуды
и соединительная ткань, образуя вместе с разрушившейся
матрикс хряща костный мозг.
Окостенение длинных костей конечностей, по данным Калояну-
Иордэкэл (1961), происходит из центральной точки диафиза и
постепенно распространяется к концам костей, которые всегда
остаются хрящевыми и обеспечивают рост кости в длину. Рост
кости в ширину происходит со стороны надкостницы и сопрово-
ждается увеличением полости костного мозга в связи с одновремен-
М 2 3
День
Рис. 51. Схематическое изображение последовательности око-
стенения важнейших частей скелета куриного эмбриона.
(Нойместер, 1966).
ным разрушением кости изнутри. Крости (Crosti, 1960) показала,
что первое отложение кальция в костях куриного эмбриона на-
блюдается на 8-й день инкубации в плечевой кости, а на 12-й —
в локтевой и лучевой. По данным Генриксена (Henrichsen, 1958),
кальцификация хряща наступала только тогда, когда клетки,
содержащие щелочную фосфатазу, погибали. Живые клетки
хряща, если в них имелась активность этого энзима, не окосте-
невали. Там, где до кальцификации наблюдалась высокая актив-
ность энзима, после окостенения ее не было. Ноймейстер (1966)
разработал метод окраски скелета куриных эмбрионов и приводит
порядок окостенения важнейших частей скелета с 8-го по 21-й
день инкубации (рис. 50 и 51). Замедление роста эмбриона в длину
наблюдается с 11-го по 12-й и с 17-го по 19-й день инкубации.
Отметим от себя, что это совпадает с критическими периодами
развития (см. главу 17).
Покровные кости развиваются в соединительной ткани и форми-
руются остеобластами, окружающими костную перепончатую
пластинку. По описанию Лилли (Lillie, 1952), окостенение покров-
ных костей происходит путем возникновения костных иголок,
образующих тонкую сетку между клеткадщ соединительной ткани,
266
Затем оболочка покровной кости превращается в надкостницу,
откладывающую кость плотными слоями. В результате перепон-
чатая кость состоит из поверхностных слоев плотной кости, за-
ключающих внутри себя губчатую пластинку.
Обследуя различающиеся по длительности инкубации генети-
ческие линии кур (рановылупляющиеся и поздновылупляющиеся),
Снедекор (Snedecor, 1962) отметил, что они соответственно разли-
чались и по скорости роста костей.
2. Позвоночный столб
Позвоночник 4-дневного эмбриона представляет собою непре-
рывный слой свободно расположенных клеток мезенхимы и двух
мезенхиматозных дуг в каждом сегменте. Каждый позвонок воз-
никает из половинок двух соседних склеротомов (хвостовой поло-
вины переднего и головной — следующего), благодаря чему
достигается полезное для подвижности позвоночника сцепление
каждого миотома с двумя позвонками (рис. 52).
Перепончатая стадия начинается на 5-й день. В это время по-
звонок имеет следующие части: 1) тело позвонка, образованное
тканью обоих позвонковых компонентов; 2) отростки, растущие
из тела позвонка в направлении к спине по сторонам нервной
трубки, причем правый и левый отросток еще не объединились в ней-
ральную, или верхнюю, дугу; 3) боковые, или реберные, отростки,
растущие от тела позвонка в сторону и к хвосту; 4) подхордаль-
ный тяж, соединяющий с брюшной стороны правый и левый ком-
поненты позвонков. Охрящевение перепончатого позвонка про-
исходит в пяти центрах. Этот процесс начинается у куриного
эмбриона с 6-го дня, и на 8-й день почти весь позвонок становится
хрящевым. Хорда внутри тела позвонков сужена, а между ними
расширена, благодаря чему тела позвонков двояковогнутые.
Окостенение позвонков начинается после того, как ключицы,
длинные кости конечностей и покровные кости черепа уже в зна-
чительной мере прошли эту стадию. Калояну-Иордэкэл (1961)
отмечает, что в процессе окостенения позвонков почти одновре-
менно появляется центральная точка окостенения, образующая
тело позвонка, и две точки окостенения в нейральных дугах.
Окостенение позвоночника начинается с шеи й продолжается
по направлению к хвосту, так что у одного эмбриона существуют
одновременно разные стадии окостенения позвонков. В эмбрио-
нальном периоде все костные центры позвонков еще разделены,
а у взрослых птиц происходит сращение позвонков в крестце и
в так называемом пигостиле (основа для опоры хвостовых перьев).
Ребра образуются из реберных отростков грудных позвонков.
Соединение растущих грудных ребер между собой на брюшной
стороне тела наблюдается на J—8-й день в виде перепончатого
разрастания. Это — зачаток грудины. Вскоре после этого обра-
267
Рис. 52. Продольный разрез через зачатки позвонков 5-днев-
ного куриного эмбриона. (По Lillie, 1952).
1 — нервная трубка; 2 — передняя комиссура спинного мозга; 3 — брюш-
ной корешок спинномозгового нерва; 4 — ганглий; 5 — миотом; ба,
66 — хвостовая, и головная части склеротома; 7 — сегментная артерия.
ауется киль грудины, имеющий большое значение при полете
птицы. Окостенение грудины начинается на 17—19-й день инкуба-
ции и почти заканчивается через 3 месяца после вылупления
(Lillie, 1952).
3. Череп
Кости черепа образуются в соответствии с формой органов
головы. Таким образом, первоначально череп состоит из капсул
для мозга и органов чувств, а также из скелетных тяжей, распо-
ложенных по краям ротового отверстия. Череп эмбриона птицы
характеризуется многочисленностью отдельных костей, но в про-
цессе развития они сливаются так плотно, что только в лицевом
отделе могут быть различимы соединяющие их швы.
По своему происхождению кости черепа подразделяются на за-
мещающие и покровные. Хрящевой череп (замещающие кости)
делится на две части — мозговую и лицевую. Мозговая часть
происходит из головной мезенхимы. Охрящевение этой части
черепа у куриного эмбриона начинается на 6-й день. Лицевая
часть хрящевого черепа происходит из скелетных элементов пер-
вых трех жаберных дуг. Покровные кости дополняют хрящевой
череп и входят в состав как мозговой, так и лицевой его части.
В онтогенезе покровные кости появляются раньше хрящевых,
хотя они и возникли позднее в филогенезе.
4. Скелет конечностей и их поясов
Плечевой пояс и передние конечности впервые обнаружи-
ваются у куриного эмбриона на 4-й день инкубации, а на 5-й
из общей массы мезенхимы появляется с каждой стороны по четыре
отростка. Один из них направлен наружу — будущее крыло,
а три отростка, ориентированных внутрь тела в разных направле-
ниях, образуют кости плечевого пояса: лопатку, ключицу и кора-
коид. На 6-й день отростки плечевого пояса достигают грудины и
позвоночника. В этот же день в коракоиде и лопатке появляются
центры охрящевения, а с 12-го дня они начинают окостеневать.
Ключицы окостеневают первыми — на 6—7-й день инкубации.
На 12-й—13-й день правая и левая ключицы сливаются между со-
бой, образуя так называемую вилку. Поскольку птицы произо-
шли от пятипалых предков, с последующей редукцией в передней
конечности запястья, пястья и фалангов пальцев, в начале эмбрио-
генеза появляются все элементы пятипалой конечности. У 6-днев-
ного эмбриона передняя конечность имеет лопатовидную форму,
причем 1-й и 5-й пальцы короче и их скелетные элементы вскоре
исчезают. В скелете взрослой пти^ы имеются только 2 запястные
косточки, а у эмбриона их 7. Фаланги 2-го, 3-го и 4-го пальцев
269
тоже сливаются. Гофман и Ротт (1961) сравнили развитие скелета
крыла в эмбриогенезе птиц разных типов размножения — вывод-
ковых, птенцовых и переходных форм между ними. Выяснено,
что у птенцовых форм, обычно имеющих более короткий срок
инкубации, развитие скелета крыла протекало быстрее, чем у вы-
водковых, а у переходных форм — зависело от относительного
количества желтка в яйце и длительности эмбриогенеза.
Задние конечности и их пояс возникают сходным образом
с передними. Ампе (Натре, 1956) методом маркировки углем выяс-
нил, что у 65-часового эмбриона почка задней конечности имеет
форму полумесяца и по длине соответствует примерно семи соми-
там. В проксимально-дистальном направлении ее можно разде-
лить на три части, причем только две из них выдаются из туловища
наружу. Самая дистальная часть состоит из мезенхимы будущих
большой и малой берцовых костей,, средняя в будущем разовьется
в бедренную кость, а проксимальная включает в себя зачаток
таза. У 3—3.5-дневного эмбриона проявляется асимметрия между
передней и задней половинами зачатка конечности, приводящая
в дальнейшем к различному размеру большой и малой берцовых
костей. Тогда же появляется зачаток плюсно-предплюсны (цевки.)
Потом, после 85 час. инкубации, появляется зачаток лапки под
прикрытием эпидермального слоя. Далее, автор (Натре, 1959)
на основании наблюдений и опытов по трансплантации предпола-
гает, что у предков современных птиц обе кости голени были оди-
наковой длины, как у археоптерикса. В процессе эволюции про-
изошла редукция малой берцовой кости. Регресс ее продолжается
и в настоящее время. В задней конечности эмбриона, так же как
и в передней, больше костных элементов, чем у взрослой птицы.
Так, у взрослых птиц отсутствуют элементы предплюсны, а у эм-
бриона здесь имеются три хряща. Кости плюсны сливаются друг
с другом и с одним из хрящей предплюсны, образуя плюсно-пред-
плюсну взрослых птиц; 5-й палец появляется в перепончатой
стадии скелета, а затем исчезает.
Самая большая кость пояса задних конечностей — это седа-
лищная. Она соединяется с подвздошной костью посредством
хряща, где образуется вертлужная впадина (место прикрепления
бедра). Лобковая кость связана с седалищной только перепонкой.
В процессе развития неплотно скрепленные с седалищной костью
концы подвздошной и лобковой костей поворачиваются назад
почти на 180° и на 8-й день развития куриного эмбриона распо-
лагаются параллельно с седалищной костью. Этот поворот обеспе-
чивает прямую походку птиц. Позднее седалищная кость образует
широкие соединения с позвонками. Скелетные элементы колен-
ного сустава становятся хрящевыми у 5—5.5-дневного эмбриона;
окостенение диафизов наступает у 5.5—6-дневного; конденсация
мезенхимы в месте образования коленной чашечки происходит
у 6—7-дневного, охрящевение ее — у 10-дневного эмбриона,
270
а окостенение — уже после вылупления (O'Rahilly a. Gardner,
1956).
В опытах in vitro Лелкес (Lelkes, 1958) приводил в движение
стеклянной палочкой (5 раз в день) коленный сустав 6—7-дневного
куриного эмбриона. После этого в суставе не наблюдалось хряще-
вого слияния большой и малой берцовых костей, как это было
в контроле. Движение способствовало образованию суставной
ямки, развитию суставных поверхностей и пролиферации хряще-
вой ткани.
Г Л А В А 16
НАРУЖНЫЕ ПОКРОВЫ
Наружные покровы птиц состоят из двух слоев кожи:
мезодермального нижнего (кориум) и эктодермального верхнего
(эпидермис), а также их производных (клюв, когти, шпоры,
гребень, сережки, чешуйки и перья). Кориум образуется из дер-
матома.
Развитие кожи у птиц подразделяется Хлыстовой (1959)
на три периода. В первый, со 2-го по 7-й день, происходит гисто-
генез эпителия и соединительной ткани, одинаковый в покровах
птиц и млекопитающих; во второй, с 8-го по 15-й день, в эктодерме
и мезенхиме происходят процессы, характерные для кожи птиц:
появляются закладки перьев и дифференцируются слои кожи;
в третий, с 16-го по 21-й день, развиваются гистологические струк-
туры, характерные только для кур. Специального мышечного
слоя в коже кур нет. Жировой слой является не подкожной клет-
чаткой, а составной частью кожи, развивающейся из закладки,
общей для всех своев дермы.
Птицы, так же как и рептилии, не имеют кожных желез,
кроме копчиковой. Это большая, двудольчатая сальная железа,
расположенная на копчике, соединенная с поверхностью кожи
двумя протоками. Она возникает в виде двух вогнутостей экто-
дермы на 10-й день, а на 14-й эпителиальные мешки дают вторичные
почки и обе половины сливаются. Железа начинает действовать
только через некоторое время после вылупления. Гистохимиче-
скими исследованиями показано наличие щелочной фосфатазы,
кислой фосфатазы и РНК в этом участке кожи на 11-й день, что
показывает на начало формирования копчиковой железы (Gomot,
1959).
Мы остановимся здесь на развитии чешуек, пуха и пера.1 Че-
шуйки на лапах являются одним из свидетельств происхождения
1 Подробное описание развития наружных покровов у птиц см.: Watter-
son, 1942; Шмальгаузен, 1947; Goff, 1949; Hamilton (из: Lillie, 1952).
271
птиц от рептилий. Формирование чешуек начинается со скопления
клеток в кориуме, из которого образуются затем утолщения и вы-
росты лежащего над ними эпителия в вцде плоских сосочков.
Очертания чешуек появляются впервые на передней поверхности
лап и пальцев у 11-дневного куриного эмбриона в виде низ-
ких, поперечных ^гребней кожи, отделенных друг от друга
бороздками.
Пух и перо происходят из чешуйки и на ранних стадиях раз-
вития сходны с ней (Шмальгаузен, 1947). По данным Коке и Кун
(Koecke u. Kuhn, 1962), образование первичных закладок пуха
начинается на 8-й и заканчивается на 14-й день развития куриных
эмбрионов в отграниченных друг от друга участках — спинном,
грудном Д брюшном, плечевом, крыловом (верхняя^ и нижняя
Рис. 53. Развитие эмбрионального пуха. (По Шмальгаузену, 1947).
А, Б, В и Г — продольные разрезы через последовательные стадии развития пера
(пуха); Д и Е — поперечные разрезы через развивающееся перо (видно образо-
вание бородок эмбрионального пуха). 1 — роговой слой; 2 — мальпигиев слой;
3 — сосочек пера; 4 — влагалище пера (4а — зачаток влагалища пера); 5 — бо-
родки.
поверхность крыла), тазовом, бедренном, хвостовом и головном.
В пределах каждого такого участка, называемого птерилией,
существует область, в которой перья появляются раньше (индук-
ционный центр). В спинной птерилии над каждым из миотомов
развивается закладка перьев, и, следовательно, метамерное рас-
положение отражается и на кожных производных. Появлению за-
чатков пуха предшествует конденсация мезодермальных клеток
в гребень, который затем разбивается на линейные серии локали-
зованных утолщений, над которыми в начале 8-го дня инкуба-
ции начинают размножаться клетки эпидермиса. В результате
образуются бугорки, которые начинают расти назад, в направле-
нии к хвосту (рис. 53). Далее зачаток пуха быстро растет наружу
в виде конического цилиндра, покрытого эпидермисом и заполнен-
ного мезодермой. В начале 11-го дня на внутренней поверхности
зачатка пуха появляются два рядом лежащих продольных эпи-
272
дермальных гребня. К ним присоединяются другие, пока вся
поверхность не разделена на 10—11 продольных гребней, являю-
щихся зачатками отдельных бородок первого порядка и связан-
ных с ними бородок второго порядка. Поверхностный же слой
эпидермиса в дальнейшем образует чехлик на пушинке. Чехлики
сбрасываются с пуха в первые дни постэмбрионального развития,
и тогда бородки эмбрионального пуха расходятся в стороны.
После 11-го дня инкубации зачатки перьев углубляются в кожу,
так что ко времени вылупления каждый расположен как бы в
в ямке — фолликуле. Каждый фолликул дает начало трем фазам
перьев: пух, молодое перо (от одного до четырех генераций) и
взрослое перо. Рост пуха сильно увеличивается к концу 13-го
дня, когда он внезапно прекращается, и далее происходит только
ороговение (кератинизация) имеющихся элементов. Сформирован-
ная пушинка радиально симметрична. Из стержня, образующегося
путем ороговения основания зачатка пуха, отходят 10—15 боро-
док 1-го порядка длиной около 13 мм. Каждая бородка суживается
к концу и несет вдоль проксимальных двух* третей своей длины
два ряда бородок 2-го порядка. Стержень внизу переходит в моло-
дое перо, которое на 15-й день инкубации начинает образовываться
под вполне сформированными к этому времени пушинками.
Мезодермальная мякоть пуха сохраняется в виде сосочка у его
основания, обильно снабженного кровеносными сосудами, питаю-
щими растущее перо.
Эмбриональный пух в постэмбриональном развитии заменяется
пером, которое вырастает под ним и постепенно сталкивает
его.
Санжель (Sengel, 1957) изучал развитие пера у 6—8-дневного
куриного эмбриона и установил 3 фазы развития: 1) у эм-
бриона 6.5 дней инкубации кожные закладки перьев появляются
в виде концентрации клеток, что индуцируется несколькими орга-
нами, в особенности нервной трубкой; 2) после этой краткой
индукции эпидермис сам продолжает нормальное развитие перьев;
3) дифференцировавшийся эпидермис приобретает индукционную
способность на образование мезодермального осевого цилиндра
из близлежащей соединительнотканой части кожи и индуцирует
передне-заднюю ориентацию кожной части зачатка пера. В другой
своей работе Санжель (Sengel, 1958) провел две серии опытов по
совместному культивированию участков кожи со спины с эпидер-
мисом с цевки и, наоборот, кожи с цевки с эпидермисом со спины.
В первом случае автор наблюдай развитие перьев, а во втором —
чешуек. Автор приходит к выводу, что именно кожа определяет,
как будет дифференцироваться эпидермис на своей поверхности —
в перья или в чешую. Так как это изменение может быть сделано
довольно поздно в процессе органогенеза, то, следовательно,
кожа до поздней стадии сохраняет свою специфическую морфо-
генную активность.
18 В. В. Рольник
273
По наблюдениям Гоффа (Goff, 1949), кровеносные сосуды,
питающие зачаток пуха, возникают в виде капиллярных отрост-
ков кровеносных сосудов кожи и входят в сосочек, когда еще зача-
ток пуха представляет собою низкий бугорок. Входящие в сосочек
кровеносные сосуды образуют в нем кровеносное сплетение и до-
ходят до дистальных концов гребней. Когда строение бородок
пуха заканчивается, сосудов становится меньше и капиллярная
сеть превращается в продольные сосуды вдоль рядов пуха с пет-
лями на дистальных концах зачатков перьев. Автору удалось
наблюдать на живых эмбрионах, что в той части пуха, где проис-
ходит размножение и дифференциация гребней бородок, снабже-
ние кровью значительно лучше, чем в дистальных частях за-
чатка пуха, где в основном происходит кератинизация боро-
док и, следовательно, метаболические потребности значительно
ниже.
В кориуме у сосочков перьев возникают гладкие мышцы не-
большого размера. Обычно с каждым окончанием фолликула
пера связаны 4 таких мышцы, а в некоторых участках тела 5 или 6.
Они проходят в трех направлениях (передне-боковом, задне-
боковом и продольном) по отношению к средней линии, между
четырьмя лежащими рядом фолликулами перьев. В зависимости
от своего положения и места прикрепления эти мышцы осуществ-
ляют поднятие, опускание или втягивание пера. Лукас и его со-
авторы (Lucas et al., 1962) установили, что мышцы, двигающие
перья, — это гладкие мышцы, имеющие ромбовидную форму,
с пузырьками по концам, и соединяются они с перьевым фолли-
кулом при помощи сухожилий. Нервы и кровеносные сосуды на-
правляются к мышце обычно из близлежащей соединительной
ткани.
Большое разнообразие окраски наружных покровов птицы
связано: 1) с липохромными пигментами, получаемыми птицей
из пищи; 2) с меланиновыми пигментами, продуцируемыми спе-
циальными клетками — меланофорами; 3) с оптическими эф-
фектами в связи со структурой перьев; 4) с комбинацией каких-
либо или всех указанных факторов. Липохромы — каротиноид-
ные пигменты. Меланиновые пигменты кур бывают двух основных
типов — красные (от желтого до оранжевого) и черные (или корич-
невые). Уоттерсон (Watterson, 1942) показал, что мел анофоры
развиваются из бесцветных клеток мел анобластов, происходящих
из нервного гребня, мигрирующих в кожу на 80—91-м часу ин-
кубации и локализующихся затем в зачатках пуха. Гамильтон
(Hamilton, из: Lillie, 1952) считает доказанным, что цвет в основ-
ном обусловлен генетической конституцией меланофоров, но зави-
сит также от взаимодействия гормонов, зачатка пера и мелано-
форов. Кохен (Cohen, 1959) наблюдал в тканевой культуре, что
белые перья эмбрионов и взрослых кур способны к образованию
пигмента.
274
Г Л А В A 1?
ПЕРИОДИЧНОСТЬ РАЗВИТИЯ
И ПЕРИОДЫ ПОВЫШЕННОЙ СМЕРТНОСТИ
ЭМБРИОНОВ
А. Рост и дифференцировка
Эмбриональное развитие — это процесс, имеющий две
взаимосвязанные характеристики: рост и дифференцировку. Наи-
более общее обозначение роста — это увеличение массы как
в связи с размножением клеток, так и при увеличении их размера.
Дифференцировка (или дифференциация) — это качественная
основа процесса развития, специализация клеток и тканей в раз-
ных направлениях, причем из сравнительно однородного вначале
клеточного материала возникают разные органы и ткани организма.
Рост и дифференцировка, как уже было сказано, взаимосвязаны,
но в некоторые периоды развития один из этих процессов может
быть несколько ослаблен, а другой, наоборот, усилен.
В начале развития эмбриона птицы наблюдается высокая ин-
тенсивность роста, но затем вскоре начинается быстрое падение
темпов роста. Изменение веса куриных эмбрионов представлено
на табл. 40. Нет сомнения, что различия в весе куриных эмбрионов,
наблюдавшиеся разными исследователями, обусловлены в зна-
чительной степени тем, что исследовались эмбрионы разных
пород кур. Так, Отрыганьев (1966), изучая конституциональные
особенности ряда пород кур, установил, что к концу развития
все они весят меньше эмбрионов кур породы русская белая:
ныо-гемпшир — 89.6%, плимутрок белый —86.1%, красная
белохвостая — 84.8%, юбилейная — 84.0%, Суссекс — 77.8%,
загорская лососевая — 74.3%, корниш — 73.6%.
Исследуя кривую роста куриных эмбрионов, Лернер (Ler-
ner, 1939) обратил внимание на то, что от наблюдавшейся Мюр-
реем (Murray, 1925) прямой линии изменения логарифмов веса
куриных эмбрионов с 5-го до 19-го дня инкубации имеется замет-
ное отклонение после 12-го дня. Автор пересчитал свои и литера-
турные данные и обнаружил резкое ускорение роста в 13-дневном
возрасте, а наибольшее отклонение от подсчитанной прямой
в 14-дневном; затем наблюдалось постепенное уменьшение этого
отклонения до 19-го дня. Отклонение кривой роста, по предполо-
жению автора, может быть обусловлено сменой источников энер-
гии для развития эмбриона (углеводы, белки, жиры).
Общее снижение интенсивности роста с ходом развития эмбриона
Шмальгаузен (1926) объясняет влиянием веществ, замедляющих
рост, возможно посредством удаления воды из протоплазмы
клеток. Автор отмечает, что кривая роста куриных эмбрионов
имеет колебания, и так как периоды депрессии роста различных
18* 276
Таблица 40
Изменение веса куриных эмбрионов (влажный вес в г) в течение развития
Возраст эмбриона (в днях) Ильин (1917) * Murray (1925) Шмальгаузен (1926) Romanoff (1931) Brody a. Henderson (1927) Byerly (1930) Лисицкий и Крок (1949) *♦
(Н) (И)
1 0.0002 0.0002 0.0007
2 — —. 0.003 0.003 — 0.004 — —
3 0.020 0.02 — 0.029 — —
4 — — 0.055 0.05 — 0.132 — —
5 0.0740—0.2276 0.221 0.200 0.13 0.059 0.264 — —
6 0.1736—0.3974 0.423 0.470 0.29 0.189 0.562 0.2652 0.2791
7 0.2656—0.7712 0.735 0.836 0.57 0.377 0.901 — —
8 0.7818—1.2950 1.189 1.210 1.15 0.639 1.488 1.0952 1.0820
9 1.0290—2.0364 1.817 1.636 1.53 1.234 2.068 1.5307 1.6929
10 1.4690—2.7054 2.661 2.846 2.26 1.591 3.168 1.9110 2.5805
11 2.0208—3.7600 3.750 3.916 3.68 2.133 4.304 2.1934 3.9639
12 3.5226—5.0166 5.105 4.650 5.07 3.081 6.100 4.6152 5.8127
13 5.2586—6.5878 6.839 7.430 7.37 4.292 8.555 6.3524 7.2018
14 6.6974—8.3164 8.974 9.840 9.74 6.182 11.838 8.5811 10.4223
15 10.9134—13.8548 11.460 11.00 12.00 8.758 14.320 12.2802 13.7539
16 13.3222—16.6270 14.390 13.65 15.98 10.989 17.570 13.7085 17.1434
17 15.5980—18.1218 17.950 16.50 18.59 12.685 21.870 18.8660 20.8563
18 20.4478—25.7006 22.030 20.70 21.83 15.895 24.210 21.7400 24.2643
19 21.5064—27.5400 26.670 27.80 25.62 17.529 28.270 25.5704 27.2410
20 26.9382—31.5460 — 33.20 30.21 21.708 35.690 30.6239 30.6328
21 32.4028 — 41.00 — 27.133 —
* — вес эмбриона без оболочек и желтка.
**(н) — развитие под наседкой, (и) — развитие в инкубаторе.
органов в общем почти синхронны, то, по-видимому, вещества,
ускоряющие и замедляющие рост, либо неспецифичны, либо
малоспецифичны. Причинами депрессий роста автор считает
накопление продуктов распада, прекращающееся с включением
нового способа их выведения из организма. По мнению автора,
депрессии роста обусловливает на 4-й день накопление молочной
кислоты и аммиака, на 9-й — мочевины, на 12-й — мочевой кис-
лоты, на 15-й — уменьшающаяся экскреторная функция средней
почки. Автор признает, что процессы дифференциации хотя и сов-
падают с периодами депрессии роста, но не являются их причиной.
По данным Отрыганьева (1949), неравномерность эмбрионального
роста объясняется тем, что в периоды снижения роста самого
эмбриона идет увеличение веса временных эмбриональных орга-
нов и истинная депрессия роста имеет место лишь в конце инкуба-
ции, на 18-й день, когда рост ограничивает скорлупа.
Ламсон и Эдмонд (Lamson a. Edmond, 1914) отмечают 2 периода
максимального роста куриного эмбриона — на 11—12-й и на 16—
17-й дни, а Романов (Romanoff, 1929) — 3 периода с замедлением
роста между ними на 9-й и 16-й дни инкубации. Гофман (1952)
указывает на 5 периодов перемежающегося роста и дифференци-
ровки у куриного эмбриона. Усиленные процессы дифферен-
цировки протекают с 5-го до 7-го, с 9-го до 10-го, с 13-го до 14-го,
с 16-го до 17-го и с 19-го до 21-го дня инкубации, а периоды уси-
ления процессов роста лежат между этими сроками.
Махинько с сотрудниками (Махинько и Сердюк, 1954; Ма-
хинько и Шевченко, 1954) установил 7 подъемов и 6 депрессий
роста утиного эмбриона и 4 подъема и 5 депрессий у голубиного.
По мнению авторов, ритмика роста является результатом внутрен-
них противоречий развития и обусловлена в основном ритмом
клеточных делений. В момент деления клеток наблюдается повы-
шение скорости роста, а ко времени, когда клетки достигают почти
нормальной величины, наступает депрессия роста. Авторы под-
черкивают, что, как правило, время удвоения веса (т. е. время,
когда разделившиеся клетки дорастают до нормальных размеров)
совпадает с окончанием депрессии роста. Авторы показывают
также, что периоды ускоренного роста характеризуются относи-
тельно пониженной интенсивностью дыхания (потребление 02
на 1 г ткани), а периоды депрессий роста протекают при высоком
уровне окислительных процессов. По данным Шевцовой (1965),
это наблюдается не всегда. Так, в некоторые дни развития утиного
эмбриона (4—6-й) наблюдаются обратные взаимоотношения: при
высокой скорости роста — высокий уровень дыхания, а падение
скорости роста сопровождается снижением окислительных про-
цессов.
Как мы видим, время перемежающихся периодов усиленного
роста и периодов депрессий роста не всегда совпадает в данных
разных исследователей, но сам факт изменения скорости роста
277
эмбриона в различные периоды его развития отмечают все. Раз-
личия же во времени наступления периодов усиленного роста и
депрессий роста, так же как и вариации в данных разных исследо-
вателей о весе куриных эмбрионов, являются, по-видимому, ре-
зультатом различных условий инкубации, разного химического
состава яиц и породных различий.
Рассмотрим соотношение роста отдельных органов. Прейер
(Ргеуег, 1885) считает, что в эмбриональный период те органы
растут быстрее, которые начинают раньше функционировать,
и наоборот, медленнее развиваются и позднее начинают действо-
вать такие, функция которых включается не сразу после вылупле-
ния (например, половые органы). Исследуя рост различных орга-
нов в течение эмбрионального развития, Шмальгаузен (1926)
отмечает, что рост органов, имеющих раннюю гистологическую
дифференциацию (хрусталик, глаз, мозг, мезонефрос), очень рано
падает ниже общего уровня роста, характерного для данной ста-
дии развития эмбриона. Интересно, что сердце имеет такую же
скорость роста, как эмбрион в целом.
Интересны данные о различиях в темпах роста и дифференци-
ровки во время развития эмбрионов птенцовых и выводковых
птиц. Так, Кауфман (Kaufman, 1930), сравнивая рост и развитие
эмбрионов курицы (выводковой птицы) и голубя (птенцовой),
отмечает, что относительный вес сердца, легких и кишечника
в течение всего периода эмбрионального развития, а также печени
до 9Lro дня и почки — до 13-го выше, а относительный вес голов-
ного мозга ниже у голубиного эмбриона. Автор приходит к вы-
воду, что у голубиного эмбриона органы, производящие энергию,
более необходимые при уменьшенном сроке развития (17 дней),
имеют относительно больший вес, а органы, потребляющие энер-
гию, — относительно меньший вес, чем у куриного эмбриона.
Сравнивая эмбриональное развитие выводковой птицы — курццы
и птенцовой — грача (развитие эмбриона 18 дней), Гофман (1952)
отметил, что рост и развитие куриного эмбриона в целом
протекает более медленно, но при этом наблюдается «гармоничное»
развитие, что выражается в готовности почти всех систем органов
к функционированию сразу после вылупления. А у грачей к мо-
менту вылупления разные системы органов различно продвинуты
в своем развитии: некоторые готовы функционировать, как на-
пример пищеварительная система, а другие, обеспечивающие, на-
пример, терморегуляторную способность, еще не вполне развиты.
По данным Бордзивиловской (1953, 1955), сравнившей раз-
витие гусиных, утиных и куриных эмбрионов, видно, что на ранних
этапах развитие эмбрионов водоплавающих птиц относительно
очень замедленно; особенно эта заметно на образовании кровенос-
ной системы.
Прицкер (1939) пришел к выводу, что правило Вант-Гоффа
перестает быть верным не только за пределами нормальной ампли-
278
туды температуры, но и после наступления определенной стадии
развития эмбриона. Автор считает, что у эмбрионов, развиваю-
щихся вне тела матери, имеются активные реакции и приспособле-
ния, позволяющие им реагировать на изменения факторов внеш-
ней среды, и в частности на изменение температуры. Этими при-
способлениями автор объясняет явления компенсации роста,
т.,е. тот факт, что замедление роста в начале развития компен-
сируется более быстрым ростом в последующий период, и наобо-
рот, ускорение роста компенсируется к концу инкубации более
медленным ростом. Ротт (1957) на-
блюдала, что развитие куриных
эмбрионов после 13-го дня инку-
бации перестает подчиняться за-
кону ускорения роста при повы-
шении температуры и замедле-
ния —• при понижении ее. Автор
предполагает, что в это время
развиваются регуляторные меха-
низмы, позволяющие эмбриону
противостоять температуре среды.
Об обнаруженном нами фено-
мене биологической компенсации
в газообмене эмбрионов мы уже
сообщали (глава 10). Наличие
регулирования развития обнару-
жено нами (Рольник, 1964) и
при сравнении вариабельности
некоторых процессов в течение
эмбрионального периода. Оказы-
вается, наиболее изменчивы ве-
личины потери веса яиц, мень-
ше — газообмена и наименее вариа-
белен вес эмбриона (рис. 54). Вариабельность величин всех трех
процессов больше до 13—15-го дня, а потом мало изменяется.
Потеря веса яиц в большей мере зависит от факторов среды и
пористости скорлупы, величина газообмена в основном обуслов-
ливается жизнедеятельностью эмбриона, но зависит и от факторов
среды, нарастание веса эмбриона в наименьшей степени зависит
от факторов среды. Наблюдающееся уменьшение вариабельности
процессов при уменьшении их зависимости от воздействий факто-
ров среды можно объяснить наличием регуляционных процессов
внутри живого организма, позволяющих ему адаптироваться
к довольно широкому диапазону внешних факторов и при этом
оставаться относительно более постоянным.
Отметим совпадение сроков ряда явлений в развитии куриного
эмбриона, отмеченных разными исследователями. В кривой роста
наблюдается перелом после 13-го дня (Lerner, 1939); в это же
279
Рис. 54. Вариабельность некото-
рых процессов развития куриного
эмбриона. (Рольник, 1964).
1 (точки) — усушка; 2 (треугольни-
ки) — газообмен; 3 (крестики) — вес
эмбриона. По оси ординат — коэф-
фициент вариации; по оси абсцисс —
дни инкубации.
время перестает изменяться под влиянием температуры скорость
роста куриных эмбрионов (Ротт, 1957). В наших наблюдениях
по биологической компенсации (стр. 201) перелом наступает на 14-й
день, и, наконец, вариабельность процессов развития значительно
уменьшается примерно в это же время. Совершенно очевидно,
что такое совпадение не случайно и объясняется включением ре-
гуляционных механизмов. И действительно, как мы уже видели
(глава 13), к этому времени приурочено включение функции целого
ряда желез (несколько раньше — щитовидной железы, а в эти
дни — поджелудочной и надпочечников), а также коррелирование
деятельности двух последних с гипофизом.
Б. Периодичность развития
Хотя развитие эмбриона протекает непрерывно, измене-
ние его характеристик происходит то плавно, то резко. Моменты
резкого изменения той или иной характеристики развития можно
считать сменой периода. Деление эмбрионального развития на пе-
риоды имеет, кроме того, и практическое значение, так как облег-
чает изучение эмбриогенеза.
Первое деление развития куриного эмбриона на периоды пред-
ложено Бэром (1828) на основе смены типов кровообращения:
1-й период (первые 2 дня инкубации) — отсутствие «полного круго-
оборота крови»; 2-й (3—5-й день) — кровообращение желточного
мешка; 3-й (с 6-го дня до начала легочного дыхания) — в крово-
обращении участвуют сосуды аллантоиса. Орлов (1938) делит
эмбриональное развитие на 6 периодов в соответствии с изменяю-
щимися требованиями эмбриона к условиям внешней среды,
а в другой работе (1961) по тому же признаку — на 4 периода и
в несколько другие сроки (о чем подробнее в главе 18). Вилье
(Willier, 1954) различает в развитии птичьего эмбриона 5 фаз,
причем после каждой происходит переход от одного состояния
к другому, структурному или функциональному. У куриного
эмбриона в 1-й фазе, называемой фазой морфогенеза, моделируется
форма животного путем перемещения комплексов клеток внутри
организма; 2-я фаза — органогенеза — включает процессы, обу-
словленные неравномерностью роста закладок разных органов
эмбриона; 3-я фаза — первоначальной функциональной активности
некоторых органов — включает гистогенетическую дифференци-
ровку; 4-я фаза — функциональной интеграции (с 10-го по 13-й
день инкубации), когда органы уже не являются независи-
мыми и связаны в своей функциональной активности; 5-я — фаза
созревания функциональной активности (с 14-го дня до вылупле-
ния) — полное функционирование ряда основных систем органов
и интенсивное развитие эндокринных желез.
Шмидт (1951) делит эмбриональное развитие всех амниот
(рептилий, птиц и млекопитающих) на 3 периода: зародышевый,
280
предплодный и плодный. Матвеев (1956), кроме того, добавляет,
на наш взгляд правильно, 4-й — яйцеводный (развитие в яйце-
воде птицы), характеризующийся внутриклеточным питанием и
дыханием. Рагозина (1955, 1961) считает, что для птиц необхо-
димо выделить, кроме того, период вылупления. Автор подчерки-
вает, что, несмотря на трудность в определении границ периодов,
понятие «период развития» вполне реально. Автором наиболее
полно дано обоснованное деление эмбрионального развития птиц
на периоды в связи со сменой типов дыхания и питания. 1-й пе-
риод — зародышевый (первые сутки в яйцеводе курицы и первые
8 дней инкубации). В основном функционируют временные эм-
бриональные органы; питание желтком; дыхание через сосуды
желточного мешка и в конце периода дополнительно через сосуды
аллантоиса. 2-й период — предплодный (с 9-го до 14-го дня инку-
бации). Питание желтком, а потом и амниотической жидкостью
внутрикишечно; дыхание при помощи аллантоиса; выделение че-
рез мезонефрос; все временные эмбриональные органы достигают
максимума величины. 3-й период — плодный (с 14-го до 20-го
дня). Наиболее быстрый рост постоянных органов эмбриона;
питание белком, растворенным в амниотической жидкости; вы-
деление через метанефрос мочевой кислоты в качестве конечного
продукта выделения. 4-й период — вылупление (20—21-й день).
Втягивание желточного мешка; начало дыхания легкими и уста-
новление второго круга эмбрионального кровообращения. Отры-
ганьева (1963) считает, что период вылупления более продолжи-
телен и длится несколько суток после вылупления, пока расса-
сывается остаточный желток и устанавливается терморегу-
ляция.
Таким образом, по-видимому, наиболее правильно делить
эмбриональное развитие птиц на 5 периодов.1
Имеются также данные о периодичности течения разных видов
обмена веществ эмбриона. Так, Еремеев (1957), исследуя водный
обмен птичьего эмбриона, делит его на 3 периода: 1-й — интен-
сивный переход воды и части растворенных питательных веществ
из белка в желток, благодаря чему последний становится при-
годным для питания эмбриона; 2-й — использование воды из за-
глатываемой амниотической жидкости и питание желтком; 3-й —
белок, растворенный в амниотической жидкости, является источ-
ником и воды, и питательных веществ. Автор показывает историче-
скую обусловленность этих периодов и считает, что 1-й период
соответствует анамнийному периоду в классе рыб, 2-й — личи- 1 2
1 Можно, конечно, применить и более дробное деление по периодам. Так,
Отрыганьев (Отрыганьев и др., 1964) разделяет эмбриональное развитие
сельскохозяйственных птиц (кур, уток, гусей, индеек) на 11 периодов по
морфологическим и физиологическим признакам, а также по специфике
патологических отклонений в развитии эмбриона в каждый из этих периодов.
2 периода из 11 проходят до закладки яиц в инкубатор.
281
ночному периоду в развитии амфибий, а 3-й — рептилийной фазе
филогенеза.
Периодичность развития находит свое толкование в ряде ра-
бот. Так, например, Матвеев (1956) считает этапность (стадий-
ность) развития приспособительной эволюцией индивидуального
развития. Периоды развития, по мнению автора, отличаются
условиями, в которых происходит развитие организма, и особыми
формами обмена веществ. Развитие в каждом из этих периодов
идет ступенчато, до этапам, характеризующимся переломными
морфологическими и физиологическими изменениями. При этом
новое (морфологически и физиологически) возникает на базе ста-
рого, путем преобразования старой функции для удовлетворения
новых потребностей организма, в новых условиях существования.
Это протекает с сохранением преемственности стадий и обеспече-
нием целостности онтогенеза, как цепи звеньев единого процесса
индивидуального развития, связанных между собой и обусловли-
вающих друг друга. Большое значение периодичности развития
придает Махинько (1965). Автор указывает, что ритмичность —
попеременность — является свойством живого организма как от-
крытой, саморегулирующейся системы, и подчеркивает ступен-
чатую смену одного состояния другим. Периодичность и ритмика
метаболизма (смена подъемов интенсивности дыхания и аэробного
гликолиза) проявляется в сменности и ритмичности в активности
ферментов, участвующих в анаболических и катаболических про-
цессах. Автор считает особенно важным исследование процессов
на узловых этапах эмбриогенеза, так как именно в эти переломные
моменты развития с особой силой выступает роль факторов среды,
воздействующих не только в данное время, но и во всей последую-
щей жизни организма.
В. Периоды повышенной смертности эмбрионов
Один из крупнейших эмбриологов, изучавших развитие
птичьих эмбрионов, Гамильтон (Hamilton, 1952), считая порази-
тельным, насколько начало функции одного органа коррелируется
с функцией других органов и систем организма, пишет: «Не удиви-
тельно, что существуют определенные чувствительные периоды,
когда эмбрион восприимчив к нарушениям как во внутренней,
так и во внешней среде» (стр. 177).
Вопросам о периодах повышенной чувствительности, или, как
их обычно называют, критическим периодам в эмбриогенезе раз-
личных животных (беспозвоночных и позвоночных), посвящена
большая литература.
Понятие о критических периодах было введено Стокардом
(Stockard, 1921). Автор назвал так периоды повышенной чув-
ствительности к повреждающим воздействиям в эмбриональном
развитии рыб. Опуская большое число работ, сообщим, что Коро-
282
вина (1956), признавая этапность развития и образование нового
качества на каждом этапе, отрицает наличие между ними периодов
повышенной чувствительности к действию всех без исключения
факторов среды. Автор признает, что в эмбриональном развитии
есть периоды, когда эмбрионы однотипно чувствительны к не-
скольким воздействиям, но считает необходимым подчеркнуть
специфический характер реакции на разные факторы среды.
В интересной сводке Светлова (1960), которую мы рекомендуем
читателю, автор стремится создать общую теорию критических
периодов в. онтогенетическом развитии животных и в действии
на него факторов среды. Эта теория сводится в основном к трем
положениям. 1) Онтогенезы состоят из небольшого числа морфо-
логических этапов, начинающихся каждый относительно корот-
ким критическим периодом с высокой чувствительностью к дей-
ствию среды вследствие пониженной способности к восстановле-
нию нормы в поврежденных участках. В течение этих периодов
детерминируются морфологические процессы до конца этапа,
после чего совершаются видимые процессы развития — дифферен-
циация и рост. 2) В развитии органов и систем существуют такие же
этапы и свои критические периоды, как и во всем организме, на-
ступающие в общем разновременно. 3) Акты онтогенеза являются
ответом организма или его частей на внешние воздействия в пери-
оды детерминации, т. е. в периоды повышенной чувствительности
организма к влиянию внешней среды. В эти периоды повышенной
чувствительности, как отмечает автор, далеко не обязательна
повышенная смертность эмбрионов, так как поражение отдельных
зачатков эмбриона дает ему возможность развиваться до опре-
деленного времени в остальных его частях нормально. В этом
и состоит отличие этих периодов от периодов повышенной смерт-
ности птичьих эмбрионов, отмеченных впервые Пэйном (Payne,
1919) и также названных им критическими периодами.
Исследуя большое количество (более 2000) погибших куриных
эмбрионов, Пэйн (Payne, 1919) обнаружил, что гибель эмбрионов
распределена не равномерно в течение всего периода развития,
а значительно повышается на 4—6-й и 18—20-й дни инкубации.
В 1-й период гибнет 16.2% всех погибающих эмбрионов (причем
максимум в 7% падает на 4-й день), а во 2-й — 48.7% (максимум
в 20% приходится на 19-й день). Автор наблюдал увеличение
смертности эмбрионов в эти дни даже при нормальном развитии
эмбрионов (в оптимальных условиях) и при развитии эмбрионов
под наседкой, но с несколько меньшим количеством гибнущих
эмбрионов. По-видимому, 2-й период повышенной смертности
эмбрионов, совпадающий по срокам с периодом вылупления в раз-
витии эмбриона, продолжается и несколько дней после вылупле-
ния (Отрыганьева, 1963).
Забегая несколько вперед, мы считаем необходимым сказать,
что повышение смертности птичьих эмбрионов, по нашему глу-
283
бокому убеждению, происходит в результате неспособности не-
которых эмбрионов (возможно, ослабленных в периоды повышен-
ной чувствительности к внешним воздействиям) перейти к сле-
дующему этапу в функционировании той или иной системы орга-
нов. Поэтому мы предлагаем эти периоды именовать в отличие
от критических периодов Стокарда периодами повышенной смерт-
ности эмбрионов.
Байерли (Byerly, 1930) дает сходную с Пэйном картину распре-
деления смертности куриных эмбрионов в течение инкубации.
Но, кроме того, автор наблюдал так называемый средний пик
смертности эмбрионов на 9—11-й день инкубации. Более высокую
смертность эмбрионов в течение первых 3—4 дней инкубации
по сравнению с наблюдавшейся Пэйном автор объясняет более
точным определением срока гибели эмбрионов; в яйцах, ранее
определявшихся как неоплодотворенные, фактически имелись
эмбрионы, погибшие в первые дни инкубации. Мунро и Козин
(Munro a. Kosin, 1945), а затем Козин (Kosin, 1951) гистологи-
чески исследовали так называемые неоплодотворенные куриные
и индюшиные яйца и показали, что до 75% их имеют, оказывается,
бластодиски, развивавшиеся, но погибшие до снесения яйца.
Следовательно, в первые сутки развития в яйцеводе и матке
несушки существует еще один критический период в развитии
цыпленка. Нидхем (Needham, 1942) посвящает специальную главу
стандартным кривым эмбриональной смертности и приводит дан-
ные о смертности эмбрионов различных птиц (рис. 55). Как мы
видим, у разных птиц независимо от длительности эмбриональ-
ного развития имеются 2 подъема смертности эмбрионов: 1-й —
в начале инкубации и 2-й, более фатальный, — в последние дни
эмбрионального развития.
О причинах повышения смертности эмбрионов в эти два пе-
риода Ридль (Riddle, 1930) пишет: «Метод репродукции птиц
требует, чтобы вода сохранилась до конца инкубации в необхо-
димом количестве. Но само средство — толстая скорлупа, кото-
рое используется для предотвращения слишком большой потери
влаги, может при излишней плотности ограничивать темп газо-
обмена, который особенно необходим на ранних стадиях развития»
(отметим, что последнее положение неправильно; см. стр. 183—186).
Автор назвал этот феномен «парадоксом скорлупы». Причину
повышения смертности эмбрионов в начале инкубации автор
усматривает в ограничении интенсивности газообмена из-за тол-
щины скорлупы, а в конце инкубации — в недостатке влаги для
окончания нормального развития из-за постоянного и неминуемого
испарения. Нидхем (Needham, 1942) считает эти причины наиболее
правдоподобными и приводит дополнительные доказательства:
в 1-й критический период — до 4-го дня инкубации — существует
повышенная чувствительность эмбриодов к ненормально низкой
или высокой концентрации кислорода, во 2-й период амплитуда
284
потери веса яиц якобы очень мала. Приводимые нами далее данные
(см. главу 19) показывают, наоборот, очень широкую амплитуду
потери веса яйцами, а также то, что это не влияет на выводимость
цыплят, а следовательно, и на жизнеспособность эмбрионов.
Причинами повышения смертности куриных эмбрионов на 3-й
и 9-й дни развития Баерли (Byerly, 1930) считает недостаточность
функции органов выделения в момент включения мезонефроса
и метанефроса соответст-
венно. А повышение смерт-
ности эмбрионов перед вы-
луплением, по мнению
автора, обусловлено в ос-
новном неправильным по-
ложением эмбриона в яйце.
Куо (Kuo, 1932b) считает,
что положение тела в тот
или иной период развития
эмбриона определяет воз-
можность продолжать раз-
витие. Автор приводит 7
стадий изменения положе-
ния тела куриного эмбри-
она в течение инкубации и
считает, что гибель эмбрио-
нов обусловлена невоз-
можностью пройти ту или
иную стадию, а причины
этой невозможности в ос-
новном механические.
Уотерс (Waters, 1935) счи-
тает, что якобы ненормаль-
ное положение тела эмб-
риона во время гибели в
один из последних дней
инкубации является в дей-
ствительности нормальным
Баерли (Byerly, 1938) подтверждают мнение Уотерса. Мунди
(Munday, 1953) исследовал положение куриных эмбрионов в яйце
с 18-го по 21-й день развития и установил, что в начале этого срока
более 90% всех эмбрионов в яйцах кур с высокой выводимостью
цыплят имеют следующее положение: голова вытянута по медиаль-
ной линии, а клюв — под лапами; к концу же инкубации эмбрионы
Домашняя
В га Обыкновенный
§ 17 ~ голубь
«ВДВ ЯИЯЖ
_____ ____ л
„ 1 »=«<*
„ _ i
14 § Горлица
Л
я &
1
*
*
§
28 Z
28 £
§
28
Индейка
Фазан
Перепел
Рис. 55. Эмбриональная смертность раз-
ных видов птиц. (По Needham, 1942).
Высота столбиков — количество погибающих за
1 день эмбрионов. Цифры справа — длительность
эмбрионального развития.
для предыдущей стадии. Данные
принимают другое положение: голова повернута направо, с клю-
вом над или под правым крылом.
Сравнивая кривые смертности при естественной и искусствен-
ной инкубации, Романов (Romanoff, 1931) обнаружил, что смерт-
ность эмбрионов в инкубаторе особенно повышается во второй
285
период, и пришел к выводу (довольно умозрительному, на наш
взгляд), что причиной смерти эмбрионов под наседкой являются
в основном внутренние факторы (наследственность и низкое
качество яиц из-за неполноценного кормления несушек), а гибель
эмбрионов в инкубаторе обусловлена как внутренними, так и
внешними факторами, действующими до и в течение инкубации.
Далее, также только при помощи сравнения кривых смертности,
автор приходит к выводу, что подъем смертности эмбрионов
с 3-го до 6-го дня обусловлен в основном внутренними факторами,
а в последние дни перед вылуплением — внешними. Такие же
периоды смертности наблюдались автором (Romanoff, 1949)
у индеек, уток, фазанов, куропаток и перепелов. Романов отме-
чает, что может и не быть среднего пика смертности куриных
эмбрионов (с 9-го по 11-й день инкубации), если нет недостатков
в кормлении маточного стада, а именно в достаточном количестве
имеются полноценные животные протеины, минеральные вещества
(особенно кальций) и витамины. Автор предполагает, что к концу
эмбрионального периода дает себя чувствовать кумулятивный
эффект всех неблагоприятных факторов, действовавших в течение
всего времени инкубации, причем особенно важными являются
ненормальные изменения в физико-химическом состоянии эмбрио-
нальных жидкостей. Вследствие этого, по мнению автора, некото-
рые эмбрионы, перед тем как погибнуть, проходят длительный
период болезненности, во время которого их рост замедляется,
у других же развитие идет нормальным темпом, и умирают они
внезапно. У первых потребление кислорода перед смертью должно
быть понижено, у вторых — чаще всего нормальное. Все эти
предположения Романова нам удалось подтвердить и обосновать
в наших исследованиях по газообмену и составу газов воздушной?
камеры яйца (см. главу 10).
Третьяков (Прицкер и Третьяков, 1937) также отмечает разное
распределение смертности эмбрионов в инкубаторе и под наседкой.
До 18-го дня инкубации в инкубаторе погибло в 2 раза больше,
чем под наседкой (9.7% и 5.7% соответственно), а в последние
дни эмбрионального развития — в 6 раз (22.2% и 3.6%). Заслу-
живает интереса опыт переноса яиц из инкубатора под наседку
на период вылупления. При этом смертность эмбрионов снизилась
в последние 3 дня инкубации с 22.2% до 17.3%. Автор приводит
разнообразные типы кривых смертности при нарушении того
или иного внутреннего или внешнего фактора и предлагает диаг-
ностировать по ним эти нарушения. Отрыганьев (1951) также
рекомендует по конфигурации кривой смертности эмбрионов в те-
чение развития определять причину повышения смертности в дан-
ной партии яиц и, таким образом, избежав ее в следующих инку-
бационных партиях, повысить выводимость молодняка птиц.
Систематизируя данные о причинах периодов повышенной
смертности эмбрионов, Маршалл (Marshall, 1947) считает, что
286
1-й пик смертности обусловливается внешними и внутренними
причинами, К внешним причинам относятся: а) неоптимальная
температура инкубации, вызывающая разновременное созревание
разных тканей и органов; б) недостаточное количество поворотов
яиц; в) недостаток кислорода. К внутренним причинам: а) незакон-
ченная гаструляция к моменту снесения яйца (при яйцекладке
с интервалом менее 24 час.); б) недостаток питательных веществ
в яйце в связи с нарушением терморегуляции несушки при низкой
температуре воздуха; в) неправильная ориентация эмбрионов
в яйце; г) летальный ген. 2-й пик смертности эмбрионов обусловлен
в основном внешними причинами, к которым относятся: а) наруше-
ние температурного режима инкубации; б) неправильное положение
эмбриона в яйце в связи с недостаточным углом и количеством по-
ворачиваний яиц; в) недостаточная или излишняя влажность,
при которой эмбрионы могут дожить до конца инкубации, но
вылупиться не способны. Отметим, однако, что недостаточная
влажность обусловливает повышение смертности эмбрионов
в основном на 13-й день инкубации. По мнению Ландауэра (Lan-
dauer, 1948), критические периоды в эмбриональном развитии
птиц связаны в основном с коренными изменениями в функцио-
нировании различных систем органов. Однако автор почти не при-
водит конкретных причин критических периодов и тех изменений,
которые их обусловливают, считая, что в этом вопросе до сих пор
нет полной ясности. Напоминая об исследовании Нойонса и
Де Хасселя (Noyons a. De Hassele, 1939), показавших внезапное
увеличение выделения углекислоты на 3-й день развития куриного
эмбриона, автор предполагает, что ранний пик смертности обусло-
влен серьезными изменениями в метаболизме эмбриона.
Группу исследований, посвященных критическим периодам
в эмбриональном развитии птиц и причинам, их обусловливающим,
провел Пенионжкевич с сотрудниками (Пенионжкевич, 1939,
1941, 1945; Огородний и Пенионжкевич, 1939, и др.). Авторы обна-
ружили, что в результате усиленного жирового обмена в конце
инкубации повышается температура эмбрионов, причем в их
тканях происходит повышенное образование перекиси водорода,
уксусной кислоты и углекислоты, являющихся токсичными для
организма. На основании этих наблюдений Пенионжкевич (1945)
предложил теорию интоксикацио-термической летальности эмбрио-
нов во второй критический период. На наш взгляд, повышение
смертности эмбрионов в конце инкубационного периода объяснить
только гипертермией и обусловленной ею интоксикацией эмбрио-
нов нельзя. Положительный эффект понижения температуры в по-
следние дни инкубации, предлагаемого автором для уменьшения
смертности эмбрионов, может быть заменен периодическими охла-
ждениями (см. главу 18), усиливающими газообмен эмбрионов.
По-видимому, теория интоксикацио-термической летальности
наиболее применима к эмбриональному развитию водоплавающих
287
птиц (гуси, утки). Так, Никита (Nichita, 19556) указывает на более
полное использование желтка утиными эмбрионами и более ин-
тенсивный обмен веществ у них по сравнению с куриными. Это
обусловливает, по мнению автора, избыток тепла к концу инкуба-
ции и повышенную гибель утиных эмбрионов перед вылуплением
от гипертермии.
В обзоре о чувствительных периодах в развитии куриного
эмбриона Гамильтон (Hamilton, 1952) пишет, что повышение
смертности эмбрионов в 1-й период наступает в связи с наруше-
ниями в гаструляции, в образовании кровеносной системы и
аллантоиса. Во 2-й период (19—20-е дни инкубации) повышенная
смертность эмбрионов связана с трудностями в процессе вылупле-
ния и включает гибель: а) от излишней влажности (эмбрион
как бы тонет в амниотической и аллантоисной жидкостях);
б) из-за неправильного положения в яйце, обусловливающего
либо асфиксию, либо невозможность разбить скорлупу; в) из-за
невозможности втянуть желточный мешок. В средний чувствитель-
ный период (13—14-е дни), по наблюдениям автора, смертность
эмбрионов повышается не всегда, а только когда в яйцах нет не-
обходимых питательных веществ или в инкубаторе концентрация
кислорода и углекислоты не оптимальны. Автор отмечает зависи-
мость между величиной смертности эмбрионов и временем суток,
сезоном снесения яйца, его удельным весом и возрастом несушек.
Некоторые патологические изменения в развитии эмбрионов воз-
никают именно во время чувствительного периода (как, например,
трудность втягивания желточного мешка), другие накапливаются
в течение длительного времени (как, например, затопление амнио-
тической и аллантоисной жидкостью). Автор подчеркивает, что
критический период является суммированием различных вредных
воздействий и становится как бы ловушкой для ослабевших эмб-
рионов. Автор отмечает далее, что каждая система органов имеет
свои специфические чувствительные периоды, которые наступают
главным образом во время образования органов, их наиболее быст-
рой дифференцировки и начала их функции или регулировки по от-
ношению к факторам среды или к другим системам органов эмб-
риона. Гибель эмбрионов может случаться в любое время развития,
но имеет тенденцию накапливаться в определенные периоды, так
как нормальное протекание некоторых процессов (гаструляция,
кровообращение, дыхание) является необходимой предпосылкой
для дальнейшего развития.
Мы уже указывали (глава 10), что основной причиной 2-го
периода повышенной смертности эмбрионов, по нашему мнению,
является сложность перехода от аллантоисного к легочному дыха-
нию. Чтобы легочное дыхание могло начаться, сосуды аллантоиса
должны в значительной степени атрофироваться, так как клюв
цыпленка до начала дыхания легкими должен пройти через наи-
более богатую сосудами часть аллантоиса, подстилающую воздуш-
288
ную камеру. Кроме того, как показано в наших работах (Рольник,
1956а; 1960), с 17-го дня инкубации скорлупа становится реальным
препятствием для газообмена, что также снижает снабжение
эмбриона кислородом. Недостаток кислорода в это время оказы-
вается особенно неблагоприятным для нормального окончания
процесса вылупления, так как именно в этот период эмбрион
должен затратить значительную механическую энергию на вы-
лупление. Несоответствие между все возрастающей потребностью
в кислороде и низкой реальной возможностью ее удовлетворения
обусловливает асфиксию эмбрионов, оказывающуюся летальной
либо в случае низкой газопроницаемости скорлупы яйца, либо
для наиболее ослабленных эмбрионов. Кроме того, асфиксия вы-
зывает понижение жизнеспособности всех эмбрионов в это время.
Гипотеза об асфиксии как основной причине смертности в послед-
ние дни инкубации привела нас к предположению, что добавление
кислорода в воздушную камеру яиц может уменьшить смертность.
Проверяя это предположение, мы вводили в воздушную камеру
яиц 5—10 мл кислорода в один из последних дней инкубации.
Это значительно (в среднем наполовину) сокращало количество
задохликов, а следовательно, повышало выводимость цыплят,
утят и индюшат. Это было подтверждено в производственных
опытах на многих тысячах яиц (см. стр. 338—339). Таким образом,
все наши опыты и наблюдения подтверждают гипотезу о том, что
основной, хотя и не единственной, причиной повышения смерт-
ности эмбрионов перед вылуплением является сложность пере-
хода от аллантоисного дыхания к легочному.
Щеглов (1949) предлагает куриных эмбрионов, погибших
в конце инкубационного периода (задохликов), разделить на
4 группы по величине использования ими белка и желтка (см.
главу 22). Автор высказывает предположение, что причиной ги-
бели эмбрионов двух первых групп является воздействие вредных
факторов вначале инкубации, 4-й — нарушение жирового обмена,
и только эмбрионов 3-й группы можно считать истинными за-
дохликами, так как они погибли в результате асфиксии в один
из последних дней инкубации.
Напомним, что мы также разделяли эмбрионов, погибших
в последние дни инкубации, по составу газов воздушной камеры
яйца и по интенсивности газообмена (см. главу 10) на 2 группы:
1) ослабевших в течение инкубации и 2) относительно нормальных
к моменту наступления периода повышенной смертности эмбрио-
нов в последние дни инкубации и погибающих в основном вслед-
ствие недостаточной проницаемости скорлупы. И для первых, и
особенно для вторых сложность перехода от аллантоисного к легоч-
ному дыханию является как бы ловушкой, но именно для эмбрио-
нов второй группы эта сложность является основной причиной
гибели, и именно их можно в полном смысле этого слова назвать
задохликами.
19 В. В. Рольник
289
Вскрывая утиных эмбрионов, погибших в последние дни инку-
бации, Клюева (1958) обнаружила у них не только отставание
в росте и в использовании ими белка и желтка яйца, но и наруше-
ния гистологического строения пищеварительного тракта. У по-
гибших эмбрионов были: а) в 2 раза меньшая толщина стенок
двенадцатиперстной кишки, б) недоразвитие ворсинок в кишеч-
нике и в) в 3 раза меньшая толщина стенки железистого желудка;
т. е. такая, как у 17-дневных эмбрионов. Такое недоразвитие
пищеваритального тракта ведет к нарушениям внутрикишечного
усвоения питательных веществ. Кроме того, у погибших эмбрио-
нов наблюдалось отставание гистологической дифференцировки
стенок желточного мешка: толщина эндотелия была 60—84 мк
(при 90—120 мк в норме), а кровеносные сосуды не были спав-
шимися, как у нормальных эмбрионов перед вылуплением, и
имели еще очаги кровотворения. Это отставание задерживало втя-
гивание желточного мешка в брюшную полость. В строении щито-
видной железы также было отмечено отставание: у погибших
эмбрионов фолликулы железы были значительно меньше (12—
15 мк), чем у нормальных (42—78 мк).
Изучая закономерности белкового обмена куриного эмбрион-
в течение инкубации, Горбачева и Маслиева (1951) считают при-
чиной повышения смертности на 6-й день развития увеличение
количества легкоотщепляемого аммиака до токсической концент-
рации. Причиной повышения смертности эмбрионов в последние
дни развития, по мнению авторов, являются наступающие значи-
тельные изменения в белковом обмене: количество дикарбоновых
аминокислот уменьшается почти в два раза, а отношение диами-
новых кислот к ним изменяется с 1.34 до 2.5, и, наконец, коли-
чество нуклеиновых оснований заметно снижается в это
время. Авторы считают, что эмбрион в эти дни требует каче-
ственно новых внешних условий, соответствующих этой стадии
развития.
Блинковой (1962) отмечен относительный спад процента поло-
жительных ответов временных связей с 17-го дня инкубации до
вылупления цыплят, что говорит о снижении возбудимости выс-
ших отделов ЦНС во 2-й период повышенной смертности куриных
эмбрионов.
Иванов (1955а) установил, что во 2-й период повышенной смерт-
ности эмбрионов среди погибших преобладают самки.
Мы привели здесь несколько более подробное изложение
данных о периодах повышенной смертности и теорий о причинах,
их обусловливающих, считая эту проблему чрезвычайно важной
как теоретически, так и практически. Однако, как убедился чита-
тель, по этим вопросам нет не только единого мнения, но пока и
возможности сделать более или менее убедительные выводы.
И все же мы надеемся, что и предложенный нами материал заслу-
живает внимания.
Раздел III. УСЛОВИЯ СРЕДЫ, НЕОБХОДИМЫЕ
ДЛЯ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ ПТИЦ
Г Л А В А 18
ТЕМПЕРАТУРА
Температура среды в эмбриональном развитии птиц
имеет несравненно большее значение, чем у млекопитающих,
в связи с тем что развитие птичьего эмбриона протекает вне тела
матери. Исследования о влиянии температуры на эмбриональное
развитие птиц, о температурном режиме естественной и искусствен-
ной инкубации и о связи температуры с другими факторами, влия-
ющими на развитие птичьего эмбриона, имеются по крайней мере
в трех областях естествознания — зоологии, эмбриологии и птице-
водстве.
Общеизвестно, что для продолжения развития птичьего
эмбриона после того, как яйцо снесено птицей, необходимо опре-
деленное количество тепла, которое обеспечивается в природе
насиживанием яиц. У домашних птиц яйца насиживает самка.
Большинство исследователей (Burke, 1925; Insko, 1949, и
др.) считают температуру наиболее важным фактором в определе-
нии успеха или неудачи инкубации.
Необходимо предупредить читателя, что мы не ставим своей
целью дать практические указания, какую температуру или дру-
гие условия среды (влажность, вентиляция и т. п.) следует при-
менять при инкубировании яиц того или иного вида птиц. Мы
увидим далее, что это и невозможно, так как для разного типа ин-
кубаторов, разных пород птиц и разных периодов развития
эмбрионов они различны. Мы приведем данные основных иссле-
дований, чтобы читатель был вполне ориентирован в этих вопросах
и смог как в своих теоретических исследованиях, так и на прак-
тике делать соответствующие выводы.
А. Температурный режим естественной
инкубации
Наблюдения над условиями, в которых, протекает эм-
бриональное развитие птиц в природе, где обычно бывает стопро-
19*
291
центная выводимость, сопровождающаяся высокой жизнеспособ-
ностью молодняка, имеют большое значение для практики искус-
ственной инкубации.
1. Температура насиживающих родителей
Реомюр (1749, цит. по: Прицкер и Третьяков, 1937) первый
установил, что температура тела кур-наседок равна 32° R (или
около 40° С). В дальнейшем температуру насиживающей самки
измеряли многие исследователи. Эйклешимер (Eycleshymer, 1907),
подкладывая в гнездо курицы деревянный брусок с термометром
в верхней части так, что ртутный шарик термометра находился
в соприкосновении с телом наседки, выяснил, что температура
наседок равна 38.7—39.4° в начале инкубации и повышается до
40.0—40.8° в конце насиживания. По данным Фердинандова (1931,
1939, 1941), средняя температура у 7 кур-наседок за 21 день наси-
живания равна 40.4°. Несколько более низкая температура у на-
седок, чем у кур-ненасёдок (41.1°), обусловливается, по мнению
автора, тем, что во время насиживания птица делает меньшее ко-
личество движений, чем в обычной обстановке.
Температура насиживающей птицы в значительной мере за-
висит от температуры окружающей среды, времени суток и сезо-
нов года (Baldwin a. Kendeigh, 1932; Higgins, 1941, и др.).
По данным большинства исследователей, наседки разных пород
кур имеют примерно одинаковую температуру. Наблюдавшиеся
Эйклешимером (Eycleshymer, 1907) породные различия не пре-
вышают индивидуальные отличия в температуре тела разных на-
седок одной породы. Температура тела наседок разных видов
сельскохозяйственных птиц (кур, уток, индеек и гусынь) также
очень сходна: в начале насиживания она равна 40.5—40.8°,
а в конце — 40.1—40.3° (Третьяков, 19536).
По нашим данным (Рольник, 1941, 1955), температура брюшка
насиживающей гаги равна 39.6°. Балдвин и Кенди (Baldwin
a. Kendeigh, 1932) выяснили, что у восточного домашнего крапив-
ника (из семейства воробьиных) температура брюшка насижи-
вающих родителей в начале периода насиживания равна 42.0°
(от 41.4° в спокойном состоянии до 42.3° при возбуждении) и
41.8° в конце периода, причем ночью в связи с покоем и долгим
отсутствием пищи она падает до 40.3°. Таким образом, у наседок
куриных птиц температура тела значительно ниже (около 40°),
чем у воробьиных (около 42°). Возможно, что это частично обу-
словливается наблюдавшейся разницей в температуре тела наси-
живающих птиц в зависимости от места гнездования. Прожеский
(Prozesky, 1963) обнаружил, что у птиц, гнездящихся на деревьях,
температура тела выше (41.8°), чем у птиц, насиживающих почти
без гнезда, на открытом грунте (40.2°).
292
2. Температурный режим гнезда
Температура тела насиживающей птицы — это та максималь-
ная температура, которая может быть ею передана гнезду. Тем-
пература же в гнезде, в разных его точках по вертикали и гори-
зонтали, значительно ниже, за исключением редких случаев,
когда температура окружающего воздуха бывает выше темпера-
туры тела птицы. В табл. 41 приведены данные о температуре
в различных точках гнезда разных птиц. Данные о температуре
Рис. 56. Способ прикрепления тер-
мопар к яйцу. (Рольник, 1939).
Объяснение в тексте.
в гнезде курицы довольно разно-
родны, что несомненно обусло-
влено различием методик изме-
рения температуры. По нашему
мнению, более точны данные
исследователей, обнаруживших
на верхнем уровне гнезда тем-
пературу 38—40°, а данные Фер-
динандо’ва (1931) и Хиггинса
(Higgins, 1941) либо обуслов-
лены какой-то ошибкой в мето-
дике измерений, либо это тем-
пература не на верхнем уровне
яйца.
Наиболее точные данные по
температурному режиму гнезда,
нам думается, можно получить,
применяя предложенную нами
методику (Рольник, 1939). При-
крепление термопар к яйцу про-
водится посредством специально
изготовленной сеточки, состоя-
щей из центрального кольца из
матерчатой резинки, к которому
пришиты термоспаи, и двух периферических колец из тесьмы, сое-
диненных перекладинами с центральным кольцом (рис. 56). Такая
сеточка легко одевается и снимается с яйца, обеспечивает плотное
прилегание термоспаев к яйцу и некоторую изоляцию их от окру-
жающего воздуха, а также более или менее нормальный поворот
и передвижение в гнезде яиц, над которыми проводится наблюде-
ние.
Наиболее характерной чертой температурного режима гнезда
птиц является различие в температуре на верхнем и нижнем уровне
яиц (табл. 41). Между температурой в центре и на периферии
гнезда также наблюдается значительное различие: по данным Фер-
динандова (1941), в гнезде курицы оно достигает 10°, а по данным
Третьякова (1953а), — 3.0—3.5°. Величина этих различий обу-
словливается в значительной мере температурой воздуха вокруг
293
Табл и ц а 41
Температура в гнезде птиц
Вид Птица насиживает Птицы нет на гнезде Автор
центр гнезда периферия центр гнезда периферия
верх | низ верх низ верх | низ верх | низ
Курица 38.9 — — — — • — Evans, 1891
38.1—39.4 — — — — — — — Eycleshymer, 1907
38.9—40.0 — — — — — — — Lamson a. Edmond, 1914, цит. по: Kui- per a. Ubbels, 1951
39.0-39.4 30.2—33.8 — — — — — — Burke, 1925
36.0—37.7 33.3—36.8 — — — — —— — Слпнько, 1936
31.1—37.0 — — — — — — — Higgins, 1941
39.2 29—36 — — — — — — Kuiper a. Ubbels, 1951
39.4 36.0 37.6 35.6 36.0 34.2 33.0 30.4 Третьяков, 19536
Индейка 36.9 32.8 26.8 24.3 — — — — Фердинандов, 1941
31.7 30.6 28.0 26.6 — — — — »
39.2 35.8 35.9 32.3 36.2 34.1 33.2 30.5 Третьяков, 19536
Гусыня 39.1—38.6 34.6—35.4 36.0—37.1 — — — — — Бражникова, 1954
36.9 32.8 26.8 24.3 — — — — Фердинандов, 1939
39.5 36.3 36.6 33.9 35.8 34.2 33.0 30.6 Третьяков, 19536
Утка домашняя .... 39.3—38.3 34.5—35.3 36.4—37.1 — — — — — Бражникова, 1954
39.5 36.1 36.3 32.5 36.1 33.4 32.2 30.5 Третьяков, 1949а
Утка кряква — 34.5 — — — — — — Higgins, 1941
Гага обыкновенная . . Куропатка американ- 39.6 35.4 36.8 32.3 35.2 33.8 32.9 31.1 Самарский, 1954
37.3 — — — — — — — Рольник, 1955
ская 37.5-38.5 — — — — — — — Kendeigh, 1940
Лебедь — 33.6 — — — — — — Higgins, 1941
Фазан Страус американский — — 35.9 — — — — — — »
нанду Страус австралийский — 38.5 36.1 38.1 35.9 36.6 35.9 36.3 36.1 Рольник, 1939
эму 37.0—37.6 28—31 — — — — — — Страутман, 1937
Голубь 38.6—38.9 — — — — — — Evans, 1891
Восточный домашний 39.3 36.7 — — — — — — Самарский, 1954
крапивник 41.1 34.4 33.8 31.8 30.6 32.9 32.2 29.0 Baldwin a. Kendeigh, 1932
гнезда. Так, в гнезде у страуса нанду различие в температуре на
верхнем и нижнем уровнях яиц равно 2.4° (при ширине яиц
89—98 мм), а у гаги — северной птицы — 3.0° (при ширине яиц
48—54 мм) (Рольник, 1939, 1955). Поясним этот пример. Если бы
страус насиживал при такой же разнице между температурой
окружающего воздуха и температу-
рой тела птицы, как в местах гнез-
дования гаги, то при ширине яиц при-
мерно в два раза большей, чем у гаги,
перепад температуры между верхом
и низом яиц в гнезде страуса был
бы не 2.4, а 6°.
Температура в гнезде различна
также в разные дни и в разное время
суток. Кэдмен (Cadman, 1923) уста-
новил, что в гнезде курицы проис-
ходят ежесуточные колебания темце-
ратуры на 3-4°, между утренней
(с 4 до 8 утра) и дневной (с 14 до
16 час.). Фердинандов (1939) указы-
вает, что в среднем днем температура
в гнезде курицы (по сравнению с ноч-
ной) повышается на 2.8°, а в гнезде
гусыни — на 6.5—8.5°. По нашим
данным (Рольник, 1939), температу-
ра в гнезде струса нанду повыша-
ется примерно на 0.1° при повы-
шении окружающей температуры на
1°; наблюдаются также значитель-
ные суточные колебания температуры
в гнезде (рис. 57). Примерно такие
же колебания температуры в течение
суток мы наблюдали в гнезде гаги
(1955). При внешней температуре
6—19° (в течение суток) температура
в гнезде гаги была выше на верхнем
уровне на 1° и на нижнем уровне
яиц — на 2.1°, чем при температуре
Рис. 57. Суточные колебания
температуры в гнезде страуса.
(Рольник, 1939).
1 (точки) — температура верх-
ней поверхности яйца с развива-
ющимся эмбрионом; 2 (треуголь-
ники) — то же боковой; 3 (крести-
ки) — то же нижней; 4 (кружоч-
ки) — температура воздуха в гнез-
де. По оси ординат — °C; по оси
абсцисс — время суток (часы).
воздуха 4-8°.
Наиболее частые колебания температуры в гнезде происходят
при пересаживаниях наседки. При этом имеют место как повороты
яиц вокруг оси (нагревание разных сторон яйца), так и перемеще-
ние их из центра на периферию гнезда и обратно. Фердинандов
(1931), использовав специально устроенное гнездо со стеклянным
дном и наклонно расположенным под ним зеркалом, наблюдал
перемещения яиц курицей-наседкой. С 1-го по 11-й день инкубации
наблюдалось в среднем за сутки по 35 перемещений наседки
295
в гнезде, с 12-го по 20-й — по 48, а на 21-й день — непрерывное
движение наседки в гнезде.
Куипер и Уббельс (Kuiper a. Ubbels, 1951) наблюдали переса-
живания в гнезде наседки-курицы днем каждые полчаса—час,
а ночью (когда темно) — каждые 1—2 часа. Индейка перемещает
яйца в гнезде в среднем 25.5 раз в сутки, т. е. почти вдвое реже,
чем курица (Фердинандов, 1931). По нашим наблюдениям (Роль-
ник, 1939), страус нанду в период насиживания встает и переса-
живается в среднем 34 раза в сутки (не менее 18 и не более 49 раз).
При этом в среднем (из 147 наблюдений) за 1 час происходит 2.7
переворачиваний яиц и 2.3 передвижений их в гнезде из центра
на периферию и обратно. Исходя из того что в гнезде было 15 яиц,
можно сказать, что во всяком случае (наши данные о перемещениях
яиц могут быть несколько преуменьшены) за 4—5 час. все яйца
были перевернуты, а за 6—7 час. передвинуты со своих мест.
Обычно к концу насиживания наблюдается более частое переса-
живание птицы в гнезде. Гага: по нашим данным (Рольник, 1955),
пересаживается в гнезде 23—24 раза в сутки. Самое продолжи-
тельное время между двумя пересаживаниями — 2 ч. 45 м.,
а самое короткое — 2—5 мин. Как часто меняется температура
в гнезде под влиянием пересаживания птиц, показывают наши
данные (табл. 42). Глухарка пересаживается в гнезде в среднем
26 раз в сутки, от 23 до 31 (Семенов-Тян-Шанский, 1952).
Таблица 42
Частота изменений температуры в гнезде гаги и страуса нанду
(По Рольник, 1939, 1955)
Всего изменений.........
Изменялась за 15 мин.
Не изменялась за 15 мин.
» » » 30 »
» » » 45 »
» » » 60 »
» » » 75 »
» » » 90 »
Количество измерений температуры
гнезда гаги гнезда страу- са
верх | бок | низ воздух в гнез- де бок яйца
яйца
213 169 33 10 1 191 145 36 6 3 1 199 151 36 5 3 3 1 204 152 33 11 6 2 230 138 30 24 12 И И 4
Примечание. В гнезде гаги температура считалась измененной
при отличии в 0.2°, в гнезде страуса нанду — 0.5°.
Кроме таких сравнительно небольших колебаний температуры,
наблюдаются и более резкие — при уходе птиц с гнезда для кор-
межки. При этом степень охлаждения гнезда зависит как от тем-
296
пературы окружающего воздуха, так и от длительности отсутствия
птицы на гнезде. Интересно наблюдение Балдвина и Кенди (Bald-
win a. Kendeigh, 1932), что длительность периодов ненасиживания
сокращается до минимума при низкой внешней температуре и уве-
личивается при высокой, так что величина охлаждения гнезда
каждый раз при отлете птицы бывает примерно одинаковой. По
нашим данным (Рольник, 1939), сумма времени отсутствия страуса
(ухода его от гнезда) за сутки увеличивается от 70 мин. на 6-й
день насиживания до 140 на 16-й, 150 на 26-й и 200 на 36-й. В гнезде
гаги (Рольник, 1955) колебания температуры в связи с отлетом
птицы были значительно более резкими, так как внешняя темпе-
ратура была на 12—18° ниже, чем в местах гнездования страуса.
Через 30 мин. после того, как гага была спугнута нами и улетела
с гнезда, температура на верхнем уровне яиц снизилась на
6—23°, а на нижнем — на 2.5—9°. Подъем же температуры до
уровня нормальной после 45-минутного отсутствия и возвращения
гаги продолжался 2—3 часа, причем температура на верхнем
уровне яйца восстанавливалась быстрее. Мы не наблюдали,
чтобы гага на длительное время сама (не будучи спугнута) поки-
дала гнездо, но Флинт (1954) при помощи актографа выяснил,
что гага уходит на кормежку в последнюю неделю насиживания
на 1.5—6 час., в среднем на 3 часа в сутки. Семенов-Тян-Шанский
(1952), проводя наблюдение на 1° севернее Полярного круга
(также при помощи актографа), выяснил, что глухарка оставляет
гнездо для кормежки 2—3 раза в сутки, в среднем на 30 мин.
каждый раз (от 12 до 111 мин.), а белая куропатка 3—4 раза
по 20 мин. (от 8 до 43 мин.).
Домашние птицы сходят с гнезда для кормления и дефекации,
а уткам и гусыням рекомендуется дать, кроме того, возможность
иногда выкупаться. По наблюдениям Третьякова (19536), одна
Таблица 43
Охлаждение гнезда при уходе наседки (домашней утки и утки кряквы)
(По Третьякову, 19536 и Самарскому, 1954)
Место измерения температуры Домашняя утка Утка кряква
до ухода с гнезда через 30 мин. до ухода с гнезда через 30 мин.
Центр гнезда Г верх 1 низ яиц . . . » ... 39.5 36.1 32.7 32.4 40.2 36.7 35.2 33.8
Периферия ( верх ( низ » ... » ... 36.3 32.5 30.2 30.0 37.0 33.1 32.9 31.1
Примечание. Несколько более высокая температура в гнезде утки кряквы по
сравнению с температурой гнезда домашней утки, по-видимому, связана с более про-
двинутым развитием эмбрионов в яйцах в данном гнезде.
297
и та же наседка уходит в разные дни с гнезда на 10—60 мин.
В табл. 43 приведены данные по снижению температуры гнезда
домашней утки (Третьяков, 19536) и утки кряквы (Самарский,
1954) во время охлаждения.
Температура в гнезде изменяется также и в течение периода
насиживания. В первые дни она особенно низка, так как боль-
шинство птиц вначале насиживает неплотно и часто сходит
с гнезда, а затем температура в гнезде повышается. По данным
Бёрка (Burke, 1925), в гнезде курицы на верхнем уровне яйца
в 1-й ден?> насиживания температура равна 37.9°, со 2-го по 20-й —
38.9—39.3°, а на 21-й — 39.6°. Температура же на нижнем уровне
яйца повышается с 29.6° вначале до 34.5° в конце насиживания.
Однако температура в гнезде на верхнем уровне яиц в первые
две трети (если не считать первые дни насиживания) выше, чем
в последнюю треть эмбрионального периода. Так, по данным
Бражниковой (1954), температура в центре гнезда индейки на верх-
нем уровне яиц понижается к концу инкубации на 1° (с 39.3°
до 38.3°), а гусыни — на 0.5° (с 39.1° до 38.6°) и повышается на
нижнем уровне (под влиянием тепла, выделяемого эмбрионом)
на 0.3° (с 35.0° до 35.3°) и на 0.8° (с 34.6° до 35.4°) соответственно.
Температура в гнезде страуса нанду на верхнем уровне яиц,
поднявшись до 38.4° в первые дни насиживания, затем мало повы-
шается, а на нижнем постепенно поднимается с 31.5° до 35.5°
(Рольник, 1939). Необходимо отметить, что в первую треть своего
развития эмбрион имеет меньший удельный вес по сравнению
с остальным желтком и при любых поворотах яйца всплывает
наверх, находясь, таким образом, почти непрерывно в условиях
наиболее высокой температуры.
Подъем температуры в гнезде в течение периода насиживания
происходит не равномерно, а имеет 6 периодов повышений и 5 —
понижений температуры (Фердинандов, 1931). Огородний (1936),
обратив внимание на соответствие во времени (по дням инкубации)
периодов повышения температуры с периодами депрессии роста
куриных эмбрионов, отмеченных Шмальгаузеном (1926), высказал
предположение, что это совпадение не случайно, а выработалось
в течение эволюционного развития птиц.
3. Температура яиц при естественной инкубации
Какова же температура яйца, находящегося в гнезде? Совер-
шенно ясно, что измеренная в разных точках его, она будет раз-
личной (табл. 44); отличается она и в зависимости от местополо-
жения яйца в гнезде; зависит она также и от внешней темпера-
туры. Одним словом, температура яиц в гнезде непрерывно
колеблется.
Хаскин (1961а) отметил значительные различия в тем-
пературе утиных эмбрионов при искусственной и естественной
298
Т а б л и ц a i 4
Температура куриных яиц под наседкой и в инкубаторе
(По Залетаевой, 1958)
Дни развития Под наседкой В инкубаторе
в центре гнезда на периферии гнезда на поверхно- сти яйца внутри яйца
на поверхно- сти яйца внутри яйца на поверхно- сти яйца внутри яйца
1-й 39.5 36.0 31.9 35.6 37.4 37.2
3-й 39.1 34.7 31.2 34.8 36.7 36.5
6-й 38.9 36.9 35.3 36.6 35.4 35.4
9-й 38.3 37.8 35.6 36.9 35.6 35.7
12-й 37.9 36.7 36.1 37.9 36.5 37.5
15-й 38.1 38.4 35.3 38.4 36.6 37.5
18-й 39.1 38.8 34.0 35.0 36.3 38.5
инкубации. Так, в инкубаторе она равна в начале развития 37.4°,
под наседкой — 38.5°, а в конце — 41.0—41.5° и 40.4° соответ-
ственно, что связано с разным характером обогрева яиц и является,
по мнению автора, одной из основных причин более низкой выво-
димости утят в инкубаторе.
Хиггинс (Higgins, 1941) наблюдал следующие изменения тем-
пературы яиц в гнезде курицы при различной температуре окру-
жающего воздуха (табл. 45).
Таблица 45
Зависимость температуры яиц в гнезде курицы
от температуры среды
(По Higgins, 1941)
Температура яйца Температура воздуха (в °C)
17.8 20.6 21.1 23.9
Средняя 31.1 35.4 37.0 37.0
Максимальная . . . 37.2 39.4 38.7 40.3
Минимальная . . . 23.3 28.3 33.6 32.1
О зависимости температуры яиц некоторых видов воробьиных
птиц от температуры воздуха в гнезде сообщают Балдвин и Кенди
(Baldwin a. Kendeigh, 1932). Авторы обнаружили, что при изме-
нении температуры воздуха в гнезде в амплитуде 17.2—37.2°
температура яиц очень быстро следует за ней, но всегда несколько
отстает, отличаясь на 0.3—0.6°. У восточного домашнего крапив-
ника в начале периода насиживания температура яйца на 0.1 —
0.3° ниже температуры воздуха в гнезде, а в последующие дни
299
эмбрионального развития — на 0.1—0.4° выше. Измеряя темпе-
ратуру яиц в гнезде у 37 видов (из И семейств) птиц, Хиггинс
(Higgins, 1941) установил, что в естественных условиях насижива-
ния температура яиц сильно колеблется в зависимости не только
от температуры окружающего воздуха, но и от силы ветра, от
прямого нагревания солнечными лучами (когда птица улетает
Дни
Рис. 58. Сравнение температуры
нижней поверхности развиваю-
щегося и неразвивающегося стра-
усиных яиц в течение периода
насиживания. (Рольник, 1939).
1 (кружки) — температура нижней по-
верхности развивающегося яйца в цен-
тре гнезда; 2 (крестики) — то же на
периферии гнезда; 3 (квадраты) —то же
неразвивающегося яйца в центре гнез-
да; 4 (точки) — температура воздуха
в гнезде.
щимся эмбрионом на 15-й день насиживания превышает темпе-
ратуру неразвивающегося яйца на 0.3°, на 20-й день — на
0.7°, на 25-й — на 1.2°, на 30-й — на 1.5° и имеет .значительно
меньшие колебания температуры (выраженные в табл. 46 ве-
личиной вариационного коэффициента), особенно во время отсут-
ствия страуса на гнезде. На значительную теплопродукцию
эмбриона указывает и то, что у яйца с развивающимся эмбрионом
к 30-му дню эмбрионального развития сглаживается различие
между температурой во время насиживания и ненасиживания,
а также что температура боковой поверхности яйца (наиболее
правильно показывающая температуру эмбриона в конце инкуба-
ционного периода) в последние дни насиживания всего на 0.3—
0.4° ниже температуры верхней поверхности яйца, близкой к тем-
с гнезда) и от длительности пе-
риодов насиживания и ненасижи-
вания. Между температурой яиц
и температурой окружающего воз-
духа установлена корреляция,
равная 0.4954+0.0706. При силь-
ном ветре температура яйца иногда
за 1 мин. снижается на 2°. Средняя
температура яиц всех изученных
автором птиц (учитывая периоды
насиживания и ненасиживания)
была равна 34.0°+ 2.38° (ку-
кушки — 32.3°, голубя и город-
ской ласточки — 36.2°, черного
дрозда—37.1°, полевого воробья —
38.1°).
Чтобы представить себе, какую
роль в температуре яйца в течение
эмбрионального развития играет
теплопродукция эмбриона,- срав-
ним температуру страусиных яиц
с развивающимся эмбрионом с тем-
пературой яиц неоплодотворенных
или с эмбрионом, погибшим на
ранней стадии (для краткости раз-
вивающееся и неразвивающееся
яйца; Рольник, 1939) (рис, 58).
Температура яйца с развиваю-
300
пературе тела взрослой
птицы. Специальных опы-
тов по искусственному
охлаждению страусиных
яиц мы не смогли прове-
сти, но отмечено, что при
температуре окружающей
среды 24° температура яй-
ца на 30-35-й день ин-
кубации опустилась за 8
мин. всего на 1°. Все эти
факты позволяют выска-
зать предположение об
установлении терморегу-
ляции у страуса нанду
еще в эмбриональном пе-
риоде, за несколько дней
до вылупления. Следова-
тельно, эти самые древние
из всех живущих в настоя-
щее время на земле птиц
являются и самыми вы-
водковыми.
Романов (Romanoff,
1939), проделав ряд опы-
тов по воздействию высо-
кой (41°) и низкой (29°)
инкубационной темпера-
туры на развивающиеся
куриные яйца, наблюдал
значительное снижение
влияния температуры сре-
ды после 10-го дня ин-
кубации, на основании
чего пришел к выводу,
что только до 10-го дня
развития куриный эм-
брион является холодно-
кровным животным в пол-
ном смысле этого слова.
Данные Ромейна и Лок-
хорста (Romijn a. Lok-
horst, 1956) о температуре
куриного яйца в течение
инкубации и Хаскина
(19616) о внутрияйцевой
температуре утиных яиц
со
3
§
О Ci Ci -ГН СО । О тН СО 00 UO I
00 00 00 XT ХГ ' Ci id CO CD* ’
CO 00 b- CM СО Ю CM 00 Ю b-
OOO^^ UOOOOOUOUO
ООООО ООООО
+I+I+I+I+II +I+I+I+I+II
toiooqi>Nr ю т-ч т-< о ио
odco4j5i>sr cd id vr cd
00 00 00 00 00 CO 00 CO 00 00
th th CO 00 00 ,CCr-\^O .
00 нН CO CO Ю 00 00 00 CM CM
00 00 00 th th I 1
оь-сог^сиосмоо^сяо^
cm cm cd cm + + cd cd + cd id
^uot-t^or-тнтнсма^оооо
ООООт-иОСОСМСМООСМСМ
oooooooooooo
+1+14-1+1+1+1+1+1+1+1+1+1
Щ'СГ’ЧН’^СТ'ООСОООСТ'ОО’ГЧСО
cdt^cdcduo + cdcdtdcdcd +
ooooooooooooooooooooooco
OiOOt^sr^OOxTUOCOCMOOtO)
CM^OOCMnHOOlQxfi^Xf^L^
00 00 CM CM CM UO
301
при изменении температуры в инкубаторе подтверждают вывод
о включении элементов терморегуляции еще в течение эмбри-
онального развития (у утиного эмбриона признаки химической
терморегуляции появляются с 17—-19-го дня инкубации).
Подведем итоги данным о температурном режиме естественной
инкубации. Итак, характерной чертой температурного режима
эмбрионального развития птиц в природе являются частые и
довольно сильные колебания температуры. Развивающиеся
эмбрионы птиц хорошо приспособились к колебаниям темпера-
туры в гнезде и в результате длительного естественного отбора
эти колебания не могли не стать необходимыми для их нормаль-
ного развития. К вопросу о необходимости колебаний температуры
для развития эмбрионов птиц при искусственной инкубации мы
еще вернемся.
Б. Минимальная и максимальная температура
эмбрионального развития
Как уже было упомянуто, по данным Эдвардса (Edwards,
1902), для начала развития куриного эмбриона достаточна тем-
пература 21—22°. Однако развитие при этой температуре нельзя
считать полноценным, так как при этом только начинается рост
бластодермы, не достигающий даже стадии первичной полоски.
При 27—29° всего 14% эмбрионов развиваются до этой стадии,
а при 30.75° — 54.8% эмбрионов достигают стадии появления
хорды, нервной пластинки и мезодермальных сомитов. В действи-
тельности порог развития куриного эмбриона равен 29° (Prevost,
et Dumas, 1827) или 28° (Dareste, 1891; Kaestner, 1895). Функ и
Биллиер (Funk a. Billier, 1944) критически относятся к данным
Эдвардса в связи с его оценкой начала развития по величине
бластодермы к моменту вскрытия яйца, что может быть обусло-
влено разной степенью развития бластодермы за время прохожде-
ния яйца по яйцеводу курицы. При 24.4° авторы совсем не обна-
ружили развития, при 26.7° — лишь недалеко идущее развитие,
не достигающее образования кровеносной системы, а при 29.4°
развитие проходило почти нормально. Было показано также
(Harrison a. Irving, 1954), что рост бластодермы при 25° за 30 час.
равен росту за 1 час при 37.5°, причем процесс дифференциации
был более чувствителен к низкой температуре, чем процесс роста.
Авторы выяснили, что после нахождения яиц в течение 10 дней
при 25° способность продолжать развитие при переносе в инку-
батор с нормальной температурой резко падает.
Изучению пределов жизнеспособности птичьих эмбрионов при
снижении температуры на разных стадиях их развития посвящено
много исследований. В опытах Кэстнера (Kaestner, 1895) показано,
что продолжительность периода охлаждения без смертельного
302
исхода обратно пропорциональна онтогенетической стадии. При
охлаждении яйца до 21° развитие может быть приостановлено:
в начале первого дня инкубации на 3 недели, в конце второго —
на 6 дней, на 6-й день —- на 72 часа, на 9-й —- на 48 час., а во вто-
рую половину инкубации —- только на 24 час. Балдвин и Кенди
(Baldwin a. Kendeigh, 1932), приводя результаты опытов по охла-
ждению яиц восточного домашнего крапивника, пришли к выводу,
что чем моложе организм, тем более низкую температуру тела
он способен перенести, являясь в этом смысле больше холодно-
кровным животным. Авторы замечают: «с развитием терморегуля-
ции приносится в жертву способность живой протоплазмы пере-
носить низкие температуры».
Группу работ по влиянию низких температур на развитие
куриных эмбрионов провели Моренг с соавторами. Моренг и Шеф-
нер (Moreng a. Shaffner, 1951) детально изучили температуру
среды, летальную для разных стадий онтогенеза кур — от опло-
дотворенного яйца до взрослой птицы. Авторы сообщают, что
охлаждение инкубируемых яиц при —23.3° в течение 55 мин.
доводит их температуру до —1.7°, после чего яйцо начинает за-
мерзать, но температура его не понижается около 65 мин. Охла-
ждение эмбрионов при —23.3° вызывало их гибель в среднем
через 95 мин. Особенно большое изменение реакции на низкую
температуру среды происходит после вылупления: нижняя ле-
тальная температура тела становится равной 15.6°, и цыплята
достигают ее за 30 мин. охлаждения при —-23.3°. Моренг и Бриант
(Moreng a. Bryant, 1954а, 1954b) отмечают наибольшую чувстви-
тельность к холоду с 18-го по 20-й день инкубации, во второй
период повышенной смертности (см. главу 17). До 17-го дня ин-
кубации 125-минутное охлаждение при температуре —23.3°
убивало всех эмбрионов, а 70—-90-минутное либо не изменяло
эмбриональную смертность, либо даже несколько повышало вы-
водимость. Цыплята из охлаждавшихся яиц имели больший
(на 2 г) вес при вылуплении, но росли несколько медленнее до
6-недельного возрадта. Затем авторы (Moreng a. Bryant, 1955)
установили, что внутрияйцевую температуру около 0° некоторые
однодневные эмбрионы могут переносить 76 час. Такая способность
к перенесению длительного воздействия низкой температурой,
по мнению авторов, обусловлена полной пойкилотермностью
эмбрионов на столь ранней стадии развития. Однако при образо-
вании кристаллов льда смерть эмбрионов бывает мгновенной.
Чем медленнее достигалась низкая внутрияйцевая температура (0°),
тем менее вредоносным было ее воздействие. В более поздней ра-
боте авторы (Moreng a. Bryant, 1956) охлаждали яйца при тем-
пературе +10°-----1-12.8° и установили, что однодневные эмбрионы
могут переносить 120-часовое воздействие (20% вывода цыплят),
двух- и трехдневные — 96-часовое (6.7% цыплят), четырех- и
пятидневные — только 52-часовое (69% и 17% соответственно),
303
эмбрионы 12-дневного возраста — 24- и 19-часовое (6.7% и
53.3% цыплят соответственно). Авторы отметили значительное
повышение чувствительности к холодовому воздействию на
5—-6-й день инкубации, т. е. примерно в первый период повыше-
ния смертности эмбрионов (см. главу 17). Снижение способности
переносить охлаждения с увеличением возраста эмбриона под-
тверждено и для фазаньих яиц (Mac Mullan a. Eberhardt, 1953).
Гамильтон (Hamilton, 1952) считает, что гибель эмбрионов про-
исходит в результате нарушения одной какой-либо системы или
процесса, которое приводит к асинхронии развития разных систем
в организме.
Мы не ставили себе целью исчерпать все данные исследований по
низшей температурной границе для жизнедеятельности птичьих
эмбрионов, нона основании уже приведенного можно прийти к вы-
воду, что минимальная температура эмбрионального развития птиц
различна для разных стадий онтогенеза и отстоит довольно да-
леко от температуры в гнезде птицы при насиживании, а также
от обычно применяемой температуры инкубации. Далее ясно,
что выводоспособность птичьих эмбрионов нелегко нарушается
при длительном охлаждении. Возможно, что основной причиной
такого большого отклонения допустимой температуры вниз от
нормы является адаптация птичьих эмбрионов к сильным сниже-
ниям температуры яиц, когда птица систематически уходит с
с гнезда для кормежки.
Граница максимальной температуры значительно меньше
отстоит от нормальной. Дарест (Dareste, 1891) считает высшим
пределом для нормального развития куриного эмбриона темпе-
ратуру 39°, а максимальной температурой, при которой эмбрион
может жить, но с нарушением нормального развития —- 43°.
Никитин (1941) также утверждает, что на ранних стадиях
развития максимальная температура, которую могут переносить
эмбрионы, ниже, чем на более поздних. По данным автора, верх-
ним пределом температуры до 12-го дня является 41°, а с 12-го
на короткое время допустима и температура 43°. К сходным вы-
водам приходят и Моренг и Шефнер (Moreng a. Shaffner, 1951).
Максимальная критическая внутрияйцевая температура в эмбрио-
нальном периоде увеличивается, по их данным, с 41.1° до 48.3°,
а у цыплят всех возрастных групп она снова снижается и равна
46.7—-47.2°. Подвергая термическому воздействию (45.5°) куриных
эмбрионов на ранних стадиях развития (от 0 до 19 час. инкубации)
в течение 3—5 час., Дыотша (Deuchar, 1952) пришла к выводу,
что допустимый срок воздействия высокой температурой умень-
шается с возрастом эмбриона, за исключением неинкубированных
яиц, на которых даже 5-часовое воздействие не имело вредного
влияния. Автор предполагает, что до инкубации меньшая чув-
ствительность эмбриона обусловливается низким темпом метабо-
лизма и отсутствием морфогенетических изменений. Эмбрионы
304
особенно чувствительны к воздействию высокой температурой
на тех стадиях, когда таких изменений много.
По мнению Лисицкого (1952), нет одной максимальной темпе-
ратуры, при которой возможна жизнь куриного эмбриона, а есть
зона максимальных температур (43—-45°), где играет роль как
длительность воздействия, так и стадия развития; при темпера-
туре же 45° смерть наступает быстро. При подъеме температуры
до 41° на 6 час. на 15-й день инкубации (так же как и охлаждение
до +3°) изменяется коллоидная система протоплазмы и разви-
ваются явления паранекроза, дающего, по предположению автора,
толчок к повышению обмена в протоплазме клеток развивающе-
гося эмбриона после возвращения его в нормальные условия.
Отсюда автор делает вывод о полезной роли колебаний темпера-
туры (как небольших подъемов, так и охлаждений) для развития
эмбрионов птиц.
Баранчеев (1939) считает причиной гибели эмбрионов при вы-
соких критических температурах свертывание белков в необра-
тимый гель, не разжижающийся при последующем снижении тем-
пературы. Сцарский (Szarski, 1947) выяснил, что гибель куриных
эмбрионов после подогрева до 45.5° вызывается остановкой
сердца, которая следует за коротким периодом повышенной пуль-
сации. Сократимость мышц и проводимость нервов сохраняются
дольше, чем способность тканей регенерировать in vitro. Наи-
большая способность к регенерации обнаружена у временных
эмбриональных органов, которые могут регенерировать даже через
12 час. воздействия высокой температуры. По данным Болотни-
кова и Пак (1959), высшим пределом работы сердца куриного
эмбриона in vivo была температура 42.5—44.5°, но при этом
в сердце происходили необратимые изменения.
В. Скорость эмбрионального развития
при различной температуре инкубации
Об изменении скорости эмбрионального развития под
влиянием различной температуры инкубирования давно известно
естествоиспытателям. По наблюдениям Дареста (Dareste, 1891),
куриный эмбрион при температуре 40—42° за 24—30 час. достигает
стадии, соответствующей 3-дневному развитию при температуре
38-39°. Эдвардс (Edwards, 1902) составил таблицы скорости раз-
вития куриных эмбрионов при температурах 34, 36, 38, 39 и 41°,
приняв за 100% развитие при 38°. Снижение температуры на 4°
обусловило замедление развития на 35%, а повышение на 3°
вызвало ускорение на 50%. Хендерсон и Броди (Henderson а.
Brody, 1927) показали, что до стадии 13-дневного возраста, до-
стигаемой куриным эмбрионом при нормальной температуре
инкубации, повышение температуры на 10° (например, с 30 до
40°) ускоряет развитие более чем в 2 раза, а после этой стадии
20 В- В- Рольник
305
изменение температуры от 37 до 41° не влияет на темп роста.
В более поздней работе автор (Henderson, 1930) устанав-
ливает температурные границы для роста куриных эмбрионов
от 34 до 42.2° и указывает, что уже у 16-дневного эмбриона начи-
нает действовать в какой-то степени терморегулирующий меха-
низм. В детальном исследовании Бэротт (Barott, 1937) показано,
что наибольшее влияние температуры на энергообмен куриных
эмбрионов наблюдается во вторую неделю инкубации. Оптималь-
ная температура в шкафном инкубаторе (37.8°) обусловливает
нормальную длительность инкубации — 20 дней 6 час. в среднем;
при температуре 39.7° —- 19 дней 4 часа, а при 35.6° —- 23 дня
12 час. Прицкером (Прицкер и Третьяков, 1937) выяснено, что
вес куриных эмбрионов в 6-дневном возрасте при температуре
инкубирования 40° составил 224% от веса эмбрионов, инкубиро-
вавшихся при 37°, в 12-дневном —- 187%, в 18-дневном — 133%.
Баранчеев (1939) поставил опыты при более резко отстоящих тем-
пературах: 30° (низкая) и 41.8° (высокая). При низкой темпера-
туре образование крови началось только на 4-й день инкубации,
а при высокой уже на 2-й день было заметно сосудистое поле и
начались сердечные сокращения. 7-дневный эмбрион, инкубиро-
вавшийся при низкой температуре, по данным автора, соответ-
ствует 3-дневному при высокой, а 10-дневный —- 5-дневному.
Романов (Romanoff, 1939) показал, что повышение температуры
до 41° на 24 часа на 3-й день инкубации куриных яиц увеличивает
вес эмбрионов на 40% по сравнению с контрольными, развиваю-
щимися все время при температуре 37.5°. До 10-го дня это влияние
температуры на развитие эмбриона постепенно снижается, а по-
том и высокая (41°) и низкая (29°) температуры слегка тормозят
развитие. В другой работе автор (Romanoff, 1934) наблюдал уве-
личение скорости роста у фазана и перепела при температуре
38.3—-38.9°, но заметное понижение ее при дальнейшем повыше-
нии температуры до 40.6°. При температуре же 36.7° развитие
длилось на 5—6 дней дольше.
В связи с явлением ускорения развития под влиянием повы-
шенной инкубационной температуры стоит вопрос о длительности
инкубации (продолжительности эмбрионального развития). Дли-
тельность инкубации различна для разных видов птиц, но и
у одного вида она отличается при различной температуре. В чрез-
вычайно интересной работе о факторах, влияющих на длительность
инкубации, Кенди (Kendeigh, 1940) приходит к выводу, что про-
должительность инкубационного периода определяется стадией
развития, достигаемой эмбрионом к моменту вылупления, общей
энергией обмена и скоростью его, а на эти факторы в свою очередь
влияют наследственность видов и условия, создаваемые птицей
для яиц в гнезде. Сравнивая типично птенцовый вид (восточного
домашнего крапивника) с выводковым (курицей) автор установил,
что эмбрион первого лучше приспособлен к обмену при темпе-
306
ратуре 35°, чем куриный эмбрион, так как при снижении темпера-
туры среды с 37.8° до 35° обмен у первого снижается на 24%,
а у второго — на 66%.
Г. Температурный режим
при искусственной инкубации
1. Температура в основных типах инкубаторов
Все инкубаторы можно разделить на два основных типа:
а) с естественной вентиляцией (так называемые секционные ин-
кубаторы) и б) с искусственной вентиляцией (при помощи венти-
лятора, приводимого в действие электромотором) —- шкафные,
или кабинетные, инкубаторы. В инкубаторе секционного типа
обогрев яиц проводится сверху, так же как птицей в гнезде, но
в отличие от гнезда — без контакта с теплым нагревающим телом.
В этом типе инкубаторов имеется значительный перепад между
температурой на верхней и на нижней поверхности яиц (3—4°),
а вентиляция происходит через отверстия в стенках инкубатора.
Отметим, что температура в секции вообще изменяется очень
сильно по вертикали —- примерно по 1° на 1 см; ошибочные дан-
ные о максимальной и минимальной допустимой температуре во
многих исследованиях обусловлены измерением температуры
выше или ниже верхнего уровня яиц, где следует измерять тем-
пературу. В более крупных инкубаторах этого типа секции рас-
положены друг над другом, обычно в 4 яруса, с обогревателем
в каждой секции, находящимся в ее верхней части. Температура
в верхнем ярусе всегда несколько выше, чем в нижнем. В послед-
нее время в промышленном птицеводстве секционные инкубаторы
почти полностью заменены шкафными. В инкубаторе шкафного
типа на всех его уровнях благодаря искусственной вентиляции
поддерживается почти одинаковая температура, и яйцо по всей
поверхности обогревается одинаково. Таким образом, если в сек-
ционном инкубаторе на верхнем уровне яйца поддерживается
температура 39°, а на нижнем — 36°, то это равнозначно по об-
щему количеству тепла, получаемого яйцом (но только по этому
показателю), температуре 37.5° в инкубаторе шкафного типа.
Рекомендуемая обычно температура инкубации находится вблизи
этих величин.
2. Оптимальная температура инкубации
Переходя к вопросу об оптимальной температуре инкубации,
необходимо отметить, что существует много различных рекомен-
даций оптимальной температуры для инкубаторов разных фирм
20* 307
и типов. Различаются также данные разных исследователей и
практиков инкубации об оптимальной температуре. Основной
причиной такого разнообразия является, как нам кажется, не-
достаточное внимание к определению общего количества тепла,
которое получают эмбрионы за период развития в том или ином
инкубаторе. Значительное влияние на общий обогрев яйца играет
и температура в помещении для инкубации — инкубатории.
Робинсон (1959) указывает, что для обеспечения соответствую-
щей температуры в центре яйца температура в секционном инкуба-
торе должна быть поднята или снижена примерно на 0.5° на каж-
дые 5° отклонения температуры в инкубатории выше или ниже
нормальной (15.6°).
Нидхем (Needham, 1931) считает оптимальной температурой ту,
при которой бывает наименьшее количество ненормально разви-
вающихся эмбрионов, а при отклонении от нее тератологический
эффект увеличивается, причем скорость развития в одну сторону
от этой температуры увеличивается, а в другую замедляется.
К этому Романов (Romanoff, 1936) добавляет, что при оптималь-
ной температуре инкубации эмбрион имеет наибольший относи-
тельный вес (в % к весу яйца). Хиггинс (Higgins, 1941) считает,
что в природе нет единой температуры, при которой эмбриональ-
ное развитие может протекать без отклонений, а существует
амплитуда температур, обеспечивающая нормальное развитие
эмбрионов и здоровый молодняк.
Третьяков (Прицкер и Третьяков, 1937), измеряя температуру
в разных точках инкубатора, показал, что, несмотря на все уси-
лия работников инкубации, температура в них фактически не-
постоянна и колебания ее достигают 1.5° в секционных и 0.2—
0.3° в шкафных инкубаторах. Коноплев (1962) провел многочис-
ленные измерения температуры в инкубаторе шкафного типа и
доказал, что температура, регистрируемая по термометру, нахо-
дящемуся в коридорной части или у потолка инкубатора, не со-
ответствует реальной температуре над яйцами между лотками,
которая в значительной мере зависит от количества тепла, выде-
ляемого яйцами во 2-й половине инкубации. Так, при температуре
37.5° на общем термометре инкубатора, над лотками наблюдалась
и температура 37.4—37.5° (если в лотках были япца не более
7-дневного срока инкубации и в выводных лотках инкубатора
не было вылупления цыплят) и 38.0—38.7° (если развитие эмбрио-
нов было более продвинутым и в выводных лотках происходило вы-
лупление одной из партий цыплят).
Хаскин (1961а) наблюдал, что инкубирование массы ;яиц
в больших инкубаторах (особенно шкафного типа) изменяет ха-
рактер теплообмена одиночного яйца; при этом повышается тем-
пература яиц, уменьшается относительная роль теплоотдачи излу-
чением и увеличивается теплоотдача конвекцией. Автор отметил,
что в связи с этим при искусственной инкубации большое значение
308
приобретает пространственное размещение яиц, направление
потока тепла и распределительная эффективность вентиляции.
Ландауэр (Landauer, 1948) предполагает, что наиболее благо-
приятные условия для инкубирования куриных яиц одинаковы
для разных пород кур. Однако это предположение было экспери-
ментально опровергнуто Поповым и Ломовской (1956), обнаружив-
шими различия в реакции эмбрионов разных пород кур (яйцено-
ского типа — белый леггорн — и общепользовательного типа —
австролорп) на температуру инкубации. Режим инкубации, вы-
работанный для выведения цыплят породы белый леггорн, был не
совсем благоприятен для инкубации яиц породы австролорп.
Повышение температуры в первые 2 дня инкубации с 37.5° до 39.5°
(в инкубаторе шкафного типа) ускоряло развитие эмбрионов
породы австролорп, обычно замедленное по сравнению с разви-
тием эмбрионов леггорн, и повышало выводимость на 6—-12%,
а также увеличивало жизнеспособность и продуктивность мо-
лодняка.
3. Температура инкубирования в разные периоды
эмбрионального развития
Рассмотрим сначала температуру яиц при искусственной инку-
бации. По данным Слинько (1936), температура куриных яиц
в инкубаторе на верхнем уровне повышается в течение инкубации
с 36.2° до 38.5°, на нижнем —- с 34.1° до 38.0°, а внутри яиц —
с 34.2—36.2° до 40.6°. Ромейн и Локхорст (Romijn a. Lokhorst,
1956) обнаружили, что при температуре в инкубаторе 38° темпера-
тура куриных яиц до 10-го дня инкубации на 0.1—0.2° ниже,
с 10-го по 14-й — настолько же выше, а с 18-го дня — на 2° выше,
чем в инкубаторе. Данные Залетаевой (1958) о температуре ку-
риных яиц в инкубаторе уже были приведены нами (табл. 44).
Коулман, Зигель и Крауз (Coleman et al., 1964) измеряли тем-
пературу куриного яйца (вблизи зародышевого диска) термопа-
рами в течение первых 3 час. инкубации. Было обнаружено, что
прирост температуры различается у разных пород и видов. Так,
у скороспелой породы белый леггорн температура поднималась
быстрее, чем в яйцах породы белые роки, а у последних быстрее,
чем в индюшиных яйцах. По нашему мнению, эти различия могли
быть обусловлены различным размером яиц. Сравнивая темпера-
туру неоплодотворенных и оплодотворенных яиц, авторы пред-
полагают, что даже в первые часы инкубации она повышается под
влиянием метаболизма эмбриона. Температура утиных яиц, по
данным Третьякова (1949а), с 10-го дня инкубации превышает
температуру инкубатора на 0.4°, а к концу эмбрионального раз-
вития — на 3.3°. Хаскин (19616), измеряя температуру в области
расположения утиного эмбриона, нашел, что в начале насижива-
ния она равна 38.5°, а к концу развития достигает 40.4°.
309
Необходимо отметить, что внутрияйцевая температура в ин-
кубаторе (особенно в инкубаторах шкафного типа) имеет значи-
тельно меньше колебаний, чем под наседкой, а также что превы-
шение ее в конце эмбрионального развития над температурой
окружающего воздуха в инкубаторе больше, чем в гнезде (см.
табл. 44 и данные Хаскина, 1961а, на стр. 298—299). В связи с этим
почти все исследователи считают необходимым снижать темпера-
туру к концу инкубации.
Бор и Хассельбальх (Bohr u. Hasselbalch, 1903) объясняют
необходимость в усиленном обогреве в начале инкубационного
периода тем, что в первые 3 дня инкубации в яйцах преобладают
эндотермические реакции. По наблюдениям авторов, в течение
последних дней инкубации яйца сами выделяют тепло в таком
большом количестве, что фактически в обогревании не нуждаются.
Романов и Фабер (Romanoff a. Faber, 1933) выясняли необходи-
мую температуру в конце инкубации куриных яиц следующим
образом: до 16-го дня инкубация проводилась в одинаковых, обыч-
ных условиях, а затем до конца инкубационного периода — при
одной из следующих температур: 32, 34, 36, 38 и 40°, в инкубаторе
шкафного типа. Наиболее благоприятной для роста, обмена ве-
ществ и выживаемости эмбрионов в эти дни оказалась температура
34 и 36°, а при 40° все эмбрионы погибли. Позднее Романов (Ro-
manoff, 1936) установил, что если температура не менялась в те-
чение инкубации, то наилучшая выводимость (72%) была при
38°. Далее автор с сотрудниками (Romanoff et al., 1938) приходит
к выводу, что наилучшая выводимость цыплят может быть дости-
гнута небольшим повышением температуры (на 0.25°) в начале
инкубации и значительным снижением ее (на 1—2°) в конце ин-
кубационного срока. Судя по скорости роста куриных эмбрионов,
Прицкер (1939) считает необходимым снизить температуру инку-
бации к концу эмбрионального развития, так как после 18-го дня
инкубации скорость роста при 37° выше, чем при 40°. Автор обна-
ружил, что при одной и той же сумме температур вес эмбриона
оказывается тем более высоким, чем выше температура в началь-
ной стадии инкубации. Снижение температуры в конце инкуба-
ционного периода считает необходимым целый ряд исследователей
(Дзюба, 1935; Слинько, 1936; Byerly, 1938; Landauer, 1948, и др.).
По Блейну и Баррону (Blain a. Barron, 1954; цит. по: Орлов,
1961а), повышение температуры в начале инкубации (38.5° в шкаф-
ном инкубаторе) обеспечивает лучшее развитие аллантоиса.
С 6-го дня инкубации температура должна, по мнению авторов,
значительно снижаться — до 36.5° в конце инкубации. Ромейн
и Локхорст (Romijn a. Lokhorst, 1961) предостерегают от высокой
температуры в выводных лотках в связи с быстро наступающей
гипертермией цыплят. С несколько другой точки зрения подходит
к снижению температуры в конце инкубаций Вермесану (Verme-
sanu, 1961). Автор считает его необходимым, так как это обеспе-
310
чивает меньшее испарение влаги из яйца, что сохраняет желток
в более жидком состоянии и облегчает его ассимиляцию эмбрио-
ном, а в результате процент вывода цыплят повышается с 76.4%
до 91.5% от оплодотворенных яиц.
Существуют, однако, и противоположные указания. Так,
Бёрк (Burke, 1925) предлагает даже повысить температуру в конце
инкубации до 38.9—39.7°, а во время вылупления довести ее
до 40.0—40.6° на основании того, что в это время необходимо
больше всего энергии и высокая температура якобы «заставляет
цыпленка выйти из яйца более быстро». Однако вывод автора осно-
ван на чрезвычайно небольшом материале (по 180—250 яиц
в группе) и является явно ошибочным.
Горецкий (1961) провел опыты по инкубированию утиных и гу-
синых яиц и указывает, что в связи с их большим по сравнению
с куриными яйцами размером для прогревания их в начале инку-
бации требуется больше тепла и для охлаждения в конце —
более низкая температура, чем для куриных. Поэтому автор реко-
мендует инкубировать гусиные и утиные яйца в шкафном инкуба-
торе при температуре 37.8° в начале и 37.0-37.3° в конце инкуба-
ции; в последние дни инкубации внутрияйцевая температура при
этом температурном режиме поднимается до 39°. Для поддержания
необходимого теплового режима автор рекомендует регулировать
влажность в инкубаторе.
Так называемый дифференцированный режим инкубации яиц
сельскохозяйственных птиц, соответствующий стадиям развития
эмбрионов, предложен Орловым давно (1938) и основан на желании
автора как можно больше «удовлетворить требования эмбрионов
к условиям внешней среды». Однако автор постепенно от призна-
ния 6 периодов с различными требованиями эмбрионов (1938)
перешел к необходимости ввести различные условия инкубации
только для 4 периодов инкубации (1961а). Автор считает особо
важным периодом эмбрионального развития первые 5—6 дней,
когда заканчивается гистогенез и почти оканчивается органогенез,
в частности закладка и дифференцировка половых органов, от
нормального развития и функционирования которых зависит
продуктивность птицы. В эти дни обмен веществ и рост эмбриона
особенно велики, и автор считает необходимым хорошо обогреть
яйца и максимально сохранить в них воду. Большое значение
придает автор и 4-му периоду (последние 5—6 дней инкубации),
для которого характерным является то, что основным источником
энергии является жир, выделяющий при сгорании большое ко-
личество тепла. Оптимальными условиями для развития в это
время автор считает снижение температуры (но без охлаждений)
и повышение влажности. Следует отметить, что дифференцирова-
ние инкубационного режима на 4 периода для различных якобы
стадий эмбрионального развития мало обосновано автором. Для
более длительного периода инкубирования утиных яиц автор пред-
311
лагает дифференцировать режим на 3 периода. В одновременно
(и в том же сборнике) опубликованном исследовании (Орлов,
19616) автор уже делит эмбриональное развитие птиц на периоды
по другому признаку (развитию аллантоиса), и они имеют дру-
гую длительность: так, 4-й период в развитии куриных эмбрионов
охватывает не 5—6 последних дней, а только 3; для уток реко-
мендуется не 3 периода, а 4. Такие противоречия заставляют
осторожно отнестись ко многим интересным положениям
автора.
Отрыганьев (1959) возражает против слишком большого дроб-
ления эмбрионального развития на периоды и установления раз-
личных условий инкубирования яиц соответственно им. Он счи-
тает необходимым различать всего лишь 2 периода: 1-й, когда
яйцо требует интенсивного обогрева, и 2-й, когда в яйце имеется
избыток тепла, которое необходимо устранить. Николаева (1962)
также предлагает больший подогрев в первые дни и снижение
температуры в последние дни инкубации. В шкафных инкуба-
торах это может быть достигнуто особым порядком закладки яиц—
чередованием лотков с яйцами, эмбрионы которых находятся на
разных стадиях развития. Эмбрионы более продвинутой стадии
выделяют много тепла, которое может быть для них вредным,
будучи полезным для находящихся рядом эмбрионов ранних ста-
дий развития, соседство с которыми в свою очередь окажется
полезным для эмбрионов первой группы; при этом от них будут
как бы отобраны излишки тепла. Первым в СССР такой способ
закладки яиц предложил Салгапник (1951, 1952).
Большую группу работ по нахождению оптимальной темпера-
туры для эмбрионального развития птиц разных видов провел
Романов (Romanoff, 1934, 1938). Автором было установлено, что
для инкубирования перепелиных яиц наиболее благоприятной
температурой (в секционном инкубаторе) в течение всего периода
инкубации является 38.3°, а для фазаньих в первые 8 дней —
38.9°, с 9-го до 16-го дня — 38.3°, а с 16-го до 24-го — 37.8°.
Для инкубации индюшиных яиц наиболее благоприятной в по-
следнюю неделю инкубации оказалась температура 36—38°; при
40° почти все эмбрионы погибли, а при 32° в этот период вывелось
всего 15% индюшат (Romanoff, 1935). В нашей работе (Rolnik,
1943; Рольник, 1955) для инкубирования гагачьих яиц мы ре-
комендуем температуру 38.0—38.5° на верхнем уровне яиц и
считаем необходимым применять охлаждение яиц до 36° каждые
1—2 часа (путем открывания дверцы секционного инкубатора)
с 1-го по 22-й день инкубации, а в течение последних двух дней
(23-го и 24-го) — снижение температуры до 37.5°. Температура
на нижнем уровне яиц (при условии поддержания температуры
в инкубатории на уровне 20—24°) при этом повышается с 33—34°
в начале до 35—36° в конце инкубации.
312
4. Колебания инкубационной температуры
и охлаждения яиц
В литературе имеется целый ряд указаний на благоприятное
влияние колебаний температуры на развитие эмбриона. Всю
накопившуюся по этому вопросу литературу очень трудно систе-
матизировать из-за различий в условиях опытов, и мы приведем
только основные исследования с разными мнениями по этому
поводу.
Нидхем (Needham, 1931) указывает, что колебания темпера-
туры под наседкой не вызывают изменений обмена веществ, так
как они компенсируются регулировкой снабжения эмбриона пита-
тельными веществами (см. главу 7). Балдвин и Кенди (Baldwin
a. Kendeigh, 1932) отмечают другое: вследствие неспособности
эмбриона регулировать температуру тела она сильно колеблется
при непрерывном изменении температуры в гнезде. Фердинандов
(1931) считает, что периодические нагревания и охлаждения яиц
улучшают дыхание эмбриона, так как при нагревании содержи-
мое яйца расширяется, выбрасывая в наружный воздух углекис-
лоту, а при охлаждении — наоборот, засасывая из окружающего
воздуха в яйцо кислород. Огородний (1935), предлагая применять
колебания температуры при инкубировании яиц всех видов сель-
скохозяйственных птиц, высказал предположение, что, воздей-
ствуя на обмен веществ, они могут повышать устойчивость эмбрио-
нов к неблагоприятным воздействиям в периоды повышенной смерт-
ности эмбрионов. Лисицкий (1937) указывает, что периодические
изменения температуры в пределах 37—40°, проводимые через
короткие промежутки времени, играют роль термического раздра-
жителя, стимулирующего рост эмбриона.
Разделим исследования о применении колебаний температуры
при инкубировании птичьих яиц на две группы: 1) о непродолжи-
тельных (не более часа) и относительно небольших колебаниях
внутрияйцевой температуры и 2) о длительных и больших ее из-
менениях.
Третьяков (1937) рекомендовал колебания температуры при
инкубировании куриных и утиных яиц. В своих новых опытах
автор (19496) обнаружил, что физико-химические показатели ин-
кубируемых утиных яиц (коэффициент рефракции, электропровод-
ность и др.) с применением охлаждений очень близки к показа-
телям при естественной инкубации; следовательно, развитие
эмбрионов при инкубировании яиц с охлаждениями проходит
в таких же благоприятных условиях, как и при естественной
инкубации. Автор считает, что охлаждения снижают избыточно
высокую температуру эмбриона в конце развития и более целесо-
образны, так как возникли в филогенезе птиц. Охлаждая гусиные
яйца до температуры 32—34° 2—4 раза в сутки с 15-го дня до
конца инкубации, автор (1947) получил повышение выводимости
313
гусят на 14% (при выводе в контроле 49.3%). В опытах с охлажде-
нием куриных яиц Третьяков и Залетаева (1956) получили повыше-
ние выводимости цыплят только на 2%.
Считает необходимым применение колебаний температуры,
особенно при инкубировании утиных и гусиных яиц, и Рюмин
(1950). Лисицкий, Быховец и Крок (1954) подчеркивают, что при
инкубации гусиных яиц, которые в 3—4 раза больше куриных
(т. е. имеют меньшую относительную поверхность и, следова-
тельно, меньшую возможность отдавать тепло во второй половине
инкубации, когда это особенно необходимо), имеющих к тому же
на 4% больше жира (который при сгорании дает значительно
больше тепла, чем углеводы и белки) и менее пористую скорлупу
(осложняющую газообмен), колебания температуры играют осо-
бенно большую роль. В опытах авторов подъем температуры до
40° на 3—6 час., а затем снижение ее до 35° на 3 часа 1 раз
в 3—4 дня дает значительное повышение процента вывода гусят.
Охлаждения куриных яиц по 2 часа при температуре 10° на 4-й
и 16-й дни инкубации с последующим подогревом их при темпе-
ратуре 40° в течение 1 часа в опытах Кучеровой (1963) улучшило
развитие эмбрионов, повысило выводимость цыплят на 2% и их
жизнеспособность в постэмбриональном периоде.
Производственные опыты Николаевой (1960) с применением
охлаждений на более чем 100 000 утиных яиц дали повышение
выводимости утят на 7.2%. Применял охлаждения утиных и гу-
синых яиц и Салганник (1951, 1952), одним из первых получивший
высокую выводимость утят и гусят на многих сотнях тысяч яиц.
Быховец (1960) приводит данные по инкубации около 30 000 ути-
ных яиц и показывает, что при охлаждении с 1-го дня инкубации
выведено 73.3% утят от оплодотворенных яиц, а если охлаждения
начинали с 16-го, то только 64.4%. Автор подчеркивает, что охла-
ждение утиных и гусиных яиц и особенно их обогревание после
этого необходимо проводить как можно быстрее. Так, при быстром
обогреве яиц, т. е. доведении их температуры после охлаждения
до оптимальной в течение 25—30 мин., была получена выводи-
мость утят и гусят 80%, а при медленном обогреве (в течение 40—
50 мин.) выводимость была только 50—60%. Ходжаев (1963)
сообщает об опыте инкубации утиных яиц (по 5000 штук в группе)
с двухразовым охлаждением их (в сутки) внутри инкубатора (откры-
вали дверцы, выключали обогреватели и не выключали вентилятор)
с 1-го дня инкубации. В охлаждавшейся группе было 81.3% и
в контрольной — 73.6% утят от оплодотворенных яиц, а слабых
утят было 1.1% и 2.3% соответственно.
Никитенко (1960) проверял влияние периодических охлажде- .
ний до 20° по 2 раза в сутки (с 1-го до 7-го дня инкубации по
60 мин., а с 7-го по 20-й — по 30 мин.) при инкубировании яиц
цесарки. Автор наблюдал положительное влияние охлаждений
до 7-го дня инкубации (увеличение количества сомитов, размера
314
тела зародыша и ширины сосудистого поля), но после 7-го дня
ускорение роста эмбрионов было менее отчетливо выражено. К со-
жалению, автор провел опыты всего на 250 яйцах и не сообщает
данных о выводимости цесарят в опытной группе и в контроле.
Целый ряд исследований посвящен вопросу о влиянии охла-
ждений яиц на различные стороны эмбрионального развития.
Ру ус (1957) показал, что охлаждения гусиных яиц путем опры-
скивания 2 раза в сутки до 15-го дня инкубации, а с 16-го, кроме
того, охлаждения яиц в течение 30—50 мин. при комнатной тем-
пературе обусловливают глубокие изменения в развитии эмбриона:
тормозят его рост в 1-ю половину инкубации, увеличивая, однако,
и абсолютный, и относительный вес сердца, а в дальнейшем спо-
собствуют более высокому содержанию гемоглобина в крови (на
18.5% к 21-му дню эмбрионального развития и на 6.7% — к
к 25-му дню), лучшему усвоению питательных веществ (меньший
вес остаточного желтка) и более высокой жизнеспособности в
в постэмбриональный период по сравнению с охлаждавшимися
только с 15-го дня инкубации. Залетаева (1958) сообщает, что
охлаждения яиц (2 раза в сутки по 15—20 мин. до температуры
30—32°) повышают использование питательных веществ: на
15-й день было использовано протеинов белка 71.4% вместо 61.4%
в контроле и глюкозы 94.3% вместо 86.6%, а на 18-й день — жи-
ров 91% вместо 80.5%. У эмбрионов охлаждавшейся группы
больше относительный вес сердца и печени, а также более высокое
количество эритроцитов и процент гемоглобина. Автор отмечает
большую жизнеспособность и повышенный рост цыплят опытной
группы, а также более скорое наступление половозрелости и
большую яйценоскость у молодок.
По данным Хаскина (1963), охлаждение куриных яиц 2 раза
в сутки по 1 часу со 2-го дня до конца инкубации при температуре
22—25° обусловило: а) достоверное повышение общего потребле-
ния эмбрионами кислорода; б) появление у них первых при-
знаков химической терморегуляции уже на 16-й день инкубации
(при снижении температуры с 37.5° до 31.5° потребление кислорода
эмбрионом увеличивалось), в то время как в контроле (в группе
без охлаждения) химическая терморегуляция начинается только
на 20-й день, и в) повышение резистентности эмбрионов к дли-
тельному охлаждению. Еще более интересен установленный
автором факт, что после вылупления у цыплят сохраняется эта
резистентность к низким температурам, а также наблюдается
повышенная эффективность их терморегуляции и более быстрое
развитие у них полной гомойотермии. В дальнейшем автор (1966)
показал, что в результате периодических охлаждений куриных
яиц "повышается интенсивность тканевого дыхания мышц и насту-
пает большая лабильность окислительной системы митохондрий
мышечных клеток. Основным источником для осуществления
терморегуляторных реакций у охлаждавшихся эмбрионов яв-
316
ляются, по данным автора, мышцы. Применяя периодические
охлаждения куриных, утиных и гусиных яиц, Быховец (1966)
наблюдал у эмбрионов в охлаждаемых яйцах лучшее развитие
кровеносной системы, увеличение темпа роста, улучшение газо-
обмена и повышение сохранности вылупившегося молодняка
в постэмбриональный период. Влияние охлаждений яиц на актив-
ность гипофиза эмбрионов наблюдала Тиксье-Видаль (Tixier-
Vidal, 1958). Охлаждение инкубируемых куриных яиц в послед-
ние 2—3 дня инкубации при температуре 27—28° приводит
к интенсивной тиреотропной стимуляции щитовидных желез, что
показывает их «гомеотермное» состояние к концу эмбрионального
развития.
Так называемый, холодный метод инкубирования утиных яиц,
применяемый в Китае, описан Колобовым (1959). Основным в этом
методе является охлаждение яиц с 1-го дня до конца инкубации
по 2—4 раза в сутки при температуре около 20° по 15—60 мин.
в зависимости от температуры в помещении для инкубации. Кроме
того, температура яиц во 2-й половине инкубации постепенно
снижается путем перенесения яиц на 15-й день инкубации из ин-
кубатора на необогреваемые стеллажи в инкубаторий, а в после-
дующие дни — заменой покрывающих стеллажи одеял с более
плотных на менее плотные. Температура на стеллажах над яйцами
снижается при этом с 38.7° до 37.0°. Применение такого метода
на Пекинской птицеферме на сотнях тысяч яиц дало 85—87% вы-
вода утят и 87—93% цыплят. В СССР этот метод уже применялся
с успехом на Кагальницкой ИПС Ростовской области (Игнатьев,
1960) при инкубировании утиных и куриных яиц.
Однако некоторые исследователи выступают против примене-
ния колебаний инкубационной температуры. Прицкер (1954а)
на основании своих опытов считает, что при достаточном отнятии
тепла от яиц во второй половине инкубационного периода при
помощи усиленной вентиляции и при неполной загрузке инкуба-
тора яйцами можно и без охлаждения в инкубаторах современного
типа (особенно в инкубаторе Рекорд-39) получать высокий процент
вывода полноценных утят. Однако приведенные автором данные
показывают, что группы яиц без охлаждения давали процент
вывода утят на 2—16% ниже, чем с охлаждением. А в опытах
Отрыганьевой (1954) при наличии очень высокого процента вы-
вода цыплят в контроле — 95.7—97.0% от оплодотворенных яиц—
в инкубаторе Рекорд-39 не наблюдалось повышения процента вы-
вода цыплят при охлаждении 2 раза в сутки по 15—20 мин. при
температуре 16—22°; наоборот, автор отмечает отставание в за-
крытии аллантоиса, меньший вес эмбрионов и задержку в исполь-
зовании белка и желтка, причем нарушения были тем больше,
чем раньше были применены охлаждения. Несколько повышало
процент вывода цыплят только однократное охлаждение в течение
1.5 час. в период наклева скорлупы. Сорока (1955) не сравнивал
316
результаты инкубирования утиных яиц с применением охлажде-
ний и без них, но поставил перед собой цель выяснить возможно ли
получить высокий процент вывода утят при инкубировании их
без охлаждений, требующих больших материальных затрат.
Автор предложил снизить температуру во 2-й половине инкуба-
ции до 37.3—37.4° (в шкафном инкубаторе), повысить влажность
до 56—58% и увеличить скорость движения воздуха до 1 м/сек.
В производственных условиях применение этого режима дало
на 300 000 утиных яиц 80.6—80.9% вывода утят от заложенных
яиц. Против охлаждений яиц выступает и Орлов (1960), предла-
гая заменить их дифференцированным режимом по периодам раз-
вития (о чем уже речь была выше) как более биологически обуслов-
ленным. Однако в пылу полемики автор упускает из виду, что
ежедневные неоднократные охлаждения яиц биологически обуслов-
лены всей эволюцией эмбрионального развития птиц.
Вильнер (1935а) одним из первых в СССР поставил опыты
с длительным (6 час.) охлаждением гусиных и утиных яиц до 20°
один раз в 3 дня инкубации в одной группе, а в другой — с нагре-
ванием до 41°. В обеих группах, если термическое воздействие
прекращалось за 4—5 дней до вылупления, оно повышало процент
вывода и жизнеспособность молодняка. Автор высказал пред-
положение, что температурные колебания играют роль термиче-
ского раздражителя, повышающего газообмен. Машталер (1943)
предложил проводить глубокие охлаждения куриных яиц (при 0°
в течение 40—45 мин.) и нашел, что больше всего повышают про-
цент вывода цыплят глубокие охлаждения, если проводить их
3 раза в течение инкубационного срока, каждый раз за день до
периодов повышенной смертности эмбрионов, а именно на 5—6-й,
13-й и 18-й дни инкубации. Автор пишет, что практически при
этом выводимость цыплят достигает 98—100%, но, к сожалению,
не приводит более точных данных. В более поздней работе (1954)
автор наблюдал, что после глубоких охлаждений улучшается
развитие сердца и кровеносной системы, повышается процент
гемоглобина, увеличивается размер легких и увеличивается про-
цент вывода цыплят на 10—15%, утят — на 15—25%, гусят —
на 25-35%.
Длительные охлаждения яиц при комнатной температуре
(19—25°) в течение 18, 24, 30 и 48 час. в один из дней инкубации
провел Руус (1952), как бы имитируя перебои в подаче электро-
энергии, и выяснил, что 18- и 24-часовое единократное охлажде-
ние за период инкубации повышает примерно на 2% вывод цыплят
(при 82.4% вывода от оплодотворенных яиц в контроле), а 30-ча-
совое, наоборот, снижает процент вывода до 50%, 48-часовое —
до 30%. По данным автора, день применения охлаждения безт
различен; Третьяков (1947) не наблюдал снижения процента вы-
вода только при охлаждении в течение часа (при 18°), а при охла-
ждении в течение 24 час. обнаружил снижение выводимости,
317
особенно большое при охлаждении эмбрионов в один из дней
с 5-го до 15-го дня инкубации. В другой работе автор (1953а)
применил длительные и глубокие охлаждения (при температуре
—1°) в один из дней инкубации в течение 1,3,6 или 12 час. и по-
лучил (правда, на очень небольшом материале — по 10 яиц в каж-
дой группе) значительное снижение процента вывода при всех
сроках охлаждения, кроме 1 часа. Вывод автора о том, что работ-
никам инкубаториев можно не бояться длительных охлаждений,
не соответствует приведенному им материалу. Рыжих и Кле-
ментьева (1957) обнаружили, что охлаждение куриных яиц со
2-го до 8-го дня инкубации в холодильнике при температуре +2,
+3° в течение 45 мин. в сутки повышает выводимость, увеличивает
размер эритроцитов и процент зернистых лейкоцитов, а такие же
охлаждения во 2-ю половину инкубации удлиняют срок эмбрио-
нального развития и снижают выводимость цыплят на 4—6%.
Косвенное подтверждение факта повышения обмена веществ
при глубоких охлаждениях куриных яиц получила Шувалова
(1955), установившая, что под влиянием охлаждений (по 45 мин.
при 0 и 13—14° по 4—10 раз в течение инкубации) увеличи-
ваются ядра клеток, особенно в нервной трубке и печени.
Автор предполагает, что увеличение ядер связано с интен-
сификацией обмена веществ, и подчеркивает, что полученные ею
данные опровергают представление о постоянстве клеточных
размеров.
Карсон (Cai’son, 1964) однократное охлаждение индюшиных
яиц в течение 5 час. в один из дней после 24-го дня инкубации
называет «стрессовым» воздействием. Автор наблюдал после
этого воздействия повышение выводимости индюшат на 9—16%.
При повторении воздействия в следующем поколении положитель-
ный эффект еще несколько увеличивается. Данные эти статистиче-
ски достоверны, и автор предполагает, что «стрессовое» воздействие
из поколения в поколение имело бы генетический эффект в повыше-
нии выводимости.
Орсцаг и Мичек (Orszagh u. Mi6ek, 1961) на большом материале
(около 30 000 куриных яиц) и при наличии высокой выводимости
цыплят в контроле (около 80%) достоверно установили, что при-
менение переменных температур в инкубаторе одного типа (на
1200 яиц) дало положительный результат — повышение выводи-
мости, а в инкубаторе другого типа (на 3600 яиц) — отрицатель-
ный.
Как мы видим, данные о влиянии длительных и глубоких
охлаждений на развитие эмбрионов при искусственной инкуба-
ции яиц еще более противоречивы. Как же можно сопоставить
это с наличием частых (при поворачивании яиц несушкой), а также
длительных и довольно глубоких охлаждений яиц, наблюдаю-
щихся в природе при отлетах насиживающей птицы для кормежки
и не приносящих вреда развивающимся эмбрионам? Мы считаем,
318
что основную роль в природных условиях играет быстрое вос-
становление температуры яиц после прилета птицы на гнездо
благодаря контактному их обогреву телом наседки. Вспомним
отмеченное Быховцем (1960) значительное повышение выводи-
мости утят и гусят благодаря ускорению на 15—20 мин. обогрева
яиц после охлаждения до нормальной температуры инкубирова-
ния. Вполне возможно, что разные исследователи проводили
обогрев яиц после длительных и глубоких охлаждений с разной
быстротой, не обратив на это внимания, и получили вследствие
этого различные результаты. Частично этим же, по-видимому,
объясняется и противоречивость результатов по кратковременным
охлаждениям яиц при комнатной температуре. Но в последнем
случае, вероятно, основную роль играет адаптированность эм-
брионов того или иного вида птиц к инкубированию без охлаж-
дений. Поясним нашу мысль. Чем более одомашнен вид, чем
большее число поколений проходило эмбриональное развитие
в условиях искусственной инкубации (где охлаждений всегда
меньше, чем в природных условиях), тем меньше эмбрионы этого
вида нуждаются в инкубировании с охлаждениями. Так, например,
куриные яйца, особенно кур наиболее распространенной породы —
белый леггорн, давно потерявших способность к насиживанию
яиц, меньше всего нуждаются в охлаждениях при искусственной
инкубации. Больше других среди одомашненных птиц нуждаются
в охлаждениях при инкубировании гусиные эмбрионы, в основном
в связи с тем, что гуси были одомашнены несколько позже и
• гусыни до сих пор насиживают значительную часть яиц, а сле-
. довательно, вид частично еще сохранил связь с условиями есте-
ственной инкубации. Для инкубации яиц промысловых птиц
(фазаны, куропатки, гаги и др.), не инкубировавшихся ранее
искусственно, колебания температуры в инкубаторе действительно
необходимы.
Д. Влияние температуры инкубирования
на развитие отдельных систем органов
и эмбриона в целом
Влияние основного фактора инкубации — темпера-
туры—можно наблюдать как на развитии всего эмбриона, так и
на развитии отдельных его органов. Мы не останавливаемся здесь
на большой группе исследований, где влияние температурного
фактора на эмбриональное развитие изучалось с точки зрения
выработки тестов биологического контроля (Отрыганьев, 1938,
1950; Прицкер, 1940, и др.), так как об этом будет речь в спе-
циальной главе.
Огородний (1936) наблюдал у куриных эмбрионов на 17-й день
инкубации сдвиг процента гемоглобина крови в зависимости
319
от температуры в инкубаторе: при 38.5° — 68%, при 39.5° (по-
вышенная температура) — 76%, при 37.5° (пониженная) — 50%.
Позднее Прицкер (19456) подтвердил эти данные и получил
у только что вылупившихся цыплят, выведенных при оптималь-
ной температуре (38.5—39°), 54.9—62.9% гемоглобина, при
высокой (40°) — 61.7—68.4%, а при низкой (37°) — 51.4—56.3%.
Романов, Смит и Суливан (Romanoff et al., 1938) выяснили,
что отклонение температуры вверх или вниз от нормы (37.5°
в шкафном инкубаторе) ухудшало развитие аллантоиса, сильно
сокращало количество аллантоисной жидкости и мочевой кислоты
в ней, а следовательно, при этом нарушалась функция аллан-
тоиса как органа дыхания, подавлялся метаболизм и ухудшалась
экскреция продуктов распада. Ассимиляция белка яйца и исчез-
новение свободной глюкозы в нем были наиболее быстрыми при
высокой температуре и более медленными при низкой; следова-
тельно, углеводный обмен и рост эмбрионов слегка ускоряется
при несколько повышенной температуре инкубации. При опти-
мальных условиях инкубации вес эмбрионов больше и накопле-
ние гликогена у них происходит’более интенсивно (Пенионжкевич,
(1954).
Гофман и Шефнер (Hoffmann a. Shaffner, 1950) наблюдали,
что при более низкой температуре инкубаций (36°) щитовидная
железа куриного эмбриона была в 3—4 раза больше и имела
ббльшую активность, чем у эмбринов, развивавшихся при тем-
пературе 39°.
Преда с соавторами (Preda et al., 1959) установил, что при •
низкой (37°) и высокой (40°) температуре инкубации обмен муко- .
полисахаридов и белков замедляется, а обмен жиров увеличи-
вается, причем в начале развития большее значение для разви-
тия эмбриона имеет нарушение обмена мукополисахаридов,
а в конце — белков и жиров. Наибольшее влияние температур-
ный фактор оказывает на обмен веществ мозга, затем — печени
и меньше всего — почек.
Ротт (1957) исследовала влияние различной температуры
инкубации (в шкафном инкубаторе: нормальная — 37.5°, пони-
женная — 36.5° и повышенная — 38.5°) на развитие куриных
эмбрионов (исследовались вес сердца, глаза, содержание гемо-
глобина крови). Выяснено, что в первую половину эмбриональ-
ного периода при пониженной температуре замедлен рост как
всего тела, так и отдельных органов, а при повышенной, наоборот,
ускорен. Во 2-ю половину инкубации скорость роста при повышен-
ной температуре меньше, чем при нормальной, и особенно чем
при пониженной, что указывает, по мнению автора, на развитие
регуляторных механизмов с 10-го до 14-го дня инкубации. Это
приводит автора к предположению о компенсаторном увеличении
интенсивности обмена веществ во 2-й половине инкубации при
пониженной температуре и понижении ее — при повышенной.
320
В этой связи интересно наблюдение Бэротта (Barott, 1937), что
общее количество калорий, выделенных эмбрионом в течение
развития (около 20 000), по-видимому, не зависит от температуры,
хотя длительность развития различна при разной температуре
инкубирования.
В наших опытах по изучению газообмена (Рольник, 1964;
Рольник и Портенко, 1964) было * подтверждено и развито
предположение о компенсаторном увеличении его во 2-й по-
ловине инкубации, если он был уменьшен в 1-й (стр. 201,
рис. 43).
По данным Прицкера (1940), инкубационная температура
создает модификации, которые могут быть потом использованы
для породообразования. Так, при разнице инкубационной тем-
пературы в 4° желточные мешки вылупившихся цыплят были
на 64% и вес сердца на 137% выше у развивавшихся при высокой
температуре, а печень, наоборот, имела на 46% больший вес
при низкой температуре. Как показали Орлов и Кучковская
(1954), ускорение развития куриного эмбриона под влиянием
высокой температуры в первые 6 дней инкубации сокращает
длительность инкубации и это свойство сокращенного инкуба-
ционного периода передается по наследству при инкубировании
яиц от опытных, кур в условиях обычной температуры инкуба-
ции. При повторном воздействии высокой температурой в первые
6 дней эмбрионального развития длительность инкубации еще
более сокращается. Интересные, но, на наш взгляд, недостаточно
достоверные данные получены Тот (1954), который, повышая
температуру в течение первых 3 дней инкубации куриных яиц
на 0.5°, наблюдал увеличение процента курочек по сравнению
с контролем на 0.4—10.8%.
Изложим наши выводы из приведенного многообразного ма-
териала о температуре как основном факторе инкубации птичьих
яиц. Получение высокой выводимости молодняка: а) в инкуба-
торах с искусственной вентиляцией, без температурного гра-
диента между верхом и низом яйца, который раньше считался
необходимым для нормального развития птичьих эмбрионов,
б) с применением охлаждений яиц и без них, в) при понижении
температуры в инкубаторах к концу эмбрионального развития,
а также и без него — с несомненностью показывает, что эмбрионы
сельскохозяйственных птиц приобрели повышенную способность
адаптироваться к различным температурным условиям, если
только они лежат в пределах оптимума. Однако это правило
действительно только при высоком качестве инкубационных яиц
и только тогда, когда работники инкубации, следя за ходом раз-
вития эмбрионов (см. главу 22), при отклонении его от нормы
компенсируют в следующий период инкубации замеченные не-
достатки инкубационного режима путем изменения температуры
или других факторов инкубации.
21 В. В. Рольник
321
Г Л А В A 19
ВЛАЖНОСТЬ
. Влажность воздуха — один из важнейших факторов
среды в эмбриональном развитии птиц, хотя исследований о зна-
чении его значительно меньше, чем о температуре. Есть иссле-
дователи (Вильнер, 19356), которые считают влажность даже
важнейшим фактором инкубации. Противоположного мнения
придерживается Горецкий (1961). Необходимость той или иной
влажности в инкубаторе автор оценивает в зависимости от того,
какое она оказывает влияние на обогрев яиц в данный период
эмбрионального развития. Однако Хаскин (19616), проведший
много исследований по теплообмену утиных яиц во время ин-
кубации, считает, что влажность воздуха в инкубаторе не является
существенным фактором в теплообмене инкубируемых яиц.
А. Влажность воздуха в гнезде
при естественной инкубации
Сложность определения относительной влажности
в гнезде птицы очень велика, и до сих пор она может быть
определена там только приблизительно или косвенно.
По данным Бёрка (Burke, 1925), относительная влажность
в гнезде курицы, сидящей на фарфоровых яйцах, выше, чем
в окружающем воздухе (на 3.9 г воды на литр воздуха), и автор
приходит к выводу, что наседка выделяет в гнездо влагу. Однако,
как известно, у птиц нет потовых желез, и этот вывод вряд ли
правилен. Кенди (Kendeigh, 1940), на наш взгляд, более правильно
подошел к этому вопросу. Не имея возможности непосредственно
определить относительную влажность в гнезде восточного до-
машнего крапивника, автор приводит следующий подсчет: в ин-
кубаторе при температуре 35° и 12% относительной влажности
потеря веса яиц равна 0.88 мг в час, а при 100% влажности яйца
совсем не теряют веса; следовательно, наблюдавшаяся в гнезде
усушка яиц — 0.65 мг в час — соответствует 35% относительной
влажности воздуха. Пересчитывая величину относительной влаж-
ности (70—80%) окружающего гнездо воздуха при средней тем-
пературе 21° на величину влажности при наблюдавшейся в гнезде
температуре (35°), автор получает ту же относительную влажность
в гнезде —35%. Вывод автора о том, что на относительную
влажность в гнезде не влияют ни насиживающие родители, ни ис-
паряющаяся из яиц влага, кажется нам вполне убедительным
не только благодаря вышеуказанному совпадению в подсчетах,
но и потому, что гнездо птицы не является хоть в какой-либо
мере замкнутым пространством, где может поддерживаться влаж-
ность, отличающаяся от окружающего воздуха. Это обусловли-
322
вает сложность определения влажности в гнезде, и наши попытки
определить ее в гнезде гаги одним из наиболее точных микро-
приборов для определения влажности (по точке росы) не увен-
чались успехом. Имеются данные Третьякова (Прицкер и Тре-
тьяков, 1937) о подъеме относительной влажности в гнездах
кур с 13-го дня (26%) по 20-й (43.3%); однако автор не указывает,
каким методом эти данные были получены.
Б. Уменьшение веса яиц в течение инкубации
Обычно о влажности воздуха в инкубаторе судят по
величине снижения веса яиц в течение инкубации (так называемой
усушке яиц). Однако усушка яиц хотя и является показателем
только потери влаги яйцом (так как потребляемый эмбрионом
кислород и выделяемая им углекислота балансируются по весу),
зависит не только от величины относительной влажности, но и от
температуры, и от скорости прохождения воздуха мимо испа-
ряющей поверхности яйца.
В большой мере величина усушки яиц зависит от пористости
скорлупы. Поэтому не только для каждого вида или породы птиц,
но иногда и для данного стада или при данном кормлении не-
сушек следует эмпирически подбирать необходимое соотношение
температуры, влажности и вентилирования инкубаторов Для
обеспечения усушки яиц в пределах оптимальной.
Для выяснения роли влажности в величине усушки яиц
мы составили таблицу усушки куриных яиц при различной от-
носительной влажности воздуха в инкубаторе за 18—19 дней ин-
кубации по данным разных авторов (табл. 47).
Таблица 47
Зависимость величины усушки куриных яиц (в % к первоначальному
весу) от изменения относительной влажности в инкубаторе
Относитель- ная влажность (%) Lamson а. Kirkpatric, 1918* Murray, 1925 * Penquite, 1938 Romijn а. Lokhorst, 1961 Robertson, 1961 b
15—20 17.4 17.6
20-30 14.5 | более 17 — — —
30-40 11.5 — — | 15.2
40-50 10.2 — — —
50-60 9.8 — — — —
60—70 8.8 8.6 10.5 — —
70-80 80-90 5.3 | менее 3 6.4 2.8 9.4
* — тип инкубатора секционный.
21*
323
Однородность полученных разными исследователями резуль-
татов (табл. 48) просто удивительна, если учесть, что исследования
проводились в разных типах и системах инкубаторов, а также
что приборы для измерения влажности обычно бывают не очень
точными. Эти данные с несомненностью говорят о том, что усушка
яиц в значительной мере зависит от относительной влажности
воздуха в инкубаторе.
Бэротт (Barott, 1937) показал, что в первую неделю инкуба-
ции усушка куриных яиц имеет линейную зависимость от влаж-
ности (в пределах от 13 до 88%) и примерно одинакова в каждый
из этих 7 дней, а в последующие дни повышается. Орлов (1941)
подсчитал, что с 1-го по 19-й день инкубации между относитель-
ной влажностью в инкубаторе и усушкой куриных яиц существует
вполне достоверная обратная корреляция (—0.722+0.107) и что
при увеличении влажности в инкубаторе на 1% усушка яиц
уменьшается на 0.1% в сутки. Однако если проследить изменение
корреляции по дням, то с 1-го по 5-й день инкубации имеется
высокая обратная корреляция (—0.844 + 0.056), с 6-го по 11-й —
несколько более низкая (—0.627+0.132), с 12-го по 15-й —
недостоверная (—0.350 + 0.187) и с 16-го по 19-й — совсем не-
достоверная (—0.015+6.229). На основании этого автор при-
ходит к выводу, что величина усушки яиц до 12-го дня инкуба-
ции сильно зависит от влажности воздуха в инкубаторе, а в по-
следние дни является в основном функцией процессов роста и
развития эмбрионов, а также его терморегулирующей способ-
ности. В противоречии с этим стоят данные Хаскина (1691а),
который провел ряд опытов с использованием микроклиматиче-
ской камеры и пишет (к сожалению, не приводя фактических
данных), что при изменении влажности воздуха от 30 до 80%
усушка утиных яиц изменяется мало. Возможно, что автор изу-
чал зависимость между влажностью и усушкой яиц в последние
дни инкубации, когда она значительно меньше зависит от влаж-
ности воздуха.
Данных о влиянии на величину усушки яиц других факторов
инкубации, а также качества яйца относительно мало. Прингл
и Бэротт (Pringle a. Barott, 1937) изучали зависимость усушки
яиц от температуры воздуха в инкубаторе и установили, что
при температуре от 35.6 до 37.2° она почти одинакова, а при
повышении до 39.7° она увеличивается на 1/3. По данным Кенди
(Kendeigh, 1940), усушка яиц восточного домашнего крапивника
при повышении температуры инкубации увеличивается до дости-
жения максимума в связи с повышением темпа испарения с по-
верхности яиц, а частично из-за увеличивающегося метаболизма
и кровообращения как самого эмбриона, так и его временных
эмбриональных органов.
По данным Прингла и Бэротта (Pringle a. Berott, 1937), су-
ществует обратная корреляция между весом яйца и его усушкой:
324
при увеличении веса куриного яйца на 1 г усушка его уменьшается
на 0.0016% в день. Ландауэр (Landauer, 1948) считает, что наи-
более благоприятные условия температуры и влажности имеют
узкие границы, одинаковые для разных пород кур, но все же
мелкие яйца (например, породы бентам) имеют ббльшую усушку
и потому требуют большей влажности к концу инкубации. Мас-
ленникова и Глаголева (1956) наблюдали ббльшую усушку
(10.9% за период инкубации) у мелких куриных яиц, чем у круп-
ных (9.5%). Это подтверждается нашими данными (Рольник,
1955) об усушке гагачьих яиц в двух разных местообитаниях
гаги с разным весом яиц (на Баренцевом море вес яиц был в сред-
нем 123.0 г, а на Белом — 133.1 г; табл. 48).
Таблица 48
Усушка яиц в гнездах гаги в разных местообитаниях
(По Рольник, 1955)
Дни насиживания Усушка яиц (в % к первоначальному весу)
на Баренцевом море на Белом море
минималь- ная ♦ максималь- ная * средняя минималь- ная * макси- мальная * средняя
6—10-й 2.4 2.4 2.4 1.3 2.4 2.0
11—15-й 2.6 3.6 3.0 2.4 3.0 2.7
16-20-й 4.1 5.0 4.3 — — —
* — средние величины по гнезду.
Существует ли взаимосвязь между усушкой яиц и процессами
развития эмбриона и, следовательно, выводимостью молодняка?
Еремеев (1957) считает, что к явлению усушки яиц нельзя отно-
ситься, как к физической закономерности, так как усушка за-
висит также и от наличия в яйце солей, пигментов и других ве-
ществ. Автором отмечено, что величина усушки яиц разных
видов в соответственные отрезки от общей длительности эмбрио-
нального развития чрезвычайно сходна, что свидетельствует,
по мнению автора, об исторически сложившейся регуляции
потери яиц в весе. Детально исследовав влияние различных ком-
плексов режимах инкубации (температура, влажность, вентиля-
ция) в течение первых 48 час. инкубирования куриных яиц,
Коноплев (1962) отмечает, что процент усушки яиц за это время
не коррелирует с ростом и развитием эмбриона. После 15 дней
инкубации температура и скорость движения воздуха имеют
значительно меньшее влияние на усушку, чем влажность. Так»
при 80% влажности усушка куриных яиц за 19 дней инкубации
составляла 10.4%, а при 50% — 12.7%. Высокая влажность
325
обусловливает хороший рост эмбриона и благоприятно влияет
на рост аллантоиса и количество аллантоисной жидкости. На ве-
личину усушки яиц после 15 дней инкубации влияет также
обмен веществ эмбриона, но связи между нею и выводимостью
цыплят нет. Исходя из этого, автор считает необходимым
создавать такие условия инкубирования, при которых усушка
яиц происходила бы за счет хорошего роста и развития эмбриона,
т. е. за счет обмена веществ, а не за счет изменения влажности
воздуха в инкубаторе.
По-видимому, действительно между усушкой яиц и выводи-
мостью нет сколько-нибудь достоверной корреляции. Данные
Робертсона (Robertson, 1961b) подтверждают это. Процент вы-
вода цыплят был почти одинаковым при усушке 15.2% и 9.4%
(85.7% и 82.0% цыплят соответственно).
При инкубировании гагачьих яиц в двух секционных инку-
баторах (несколько разной конструкции) мы (Рольник, 1955)
обнаружили, что в каждом из них усушка яиц была примерно
одинаковой (за 20 дней 8.6—9.4% в одном и 11.1—11.6% в дру-
гом) за каждый из трех лет инкубации. В то же время процент
вывода гагачат был почти одинаковым в обоих инкубаторах
в 1940 г. — 94.8 и 95.3% и в 1941 г. — 80.4 и 76.2% соответст-
венно — и разным в 1939 г. — 88.5 и 35.8%. Таким образом,
в определенных пределах величина усушки не влияет на жизне-
способность эмбрионов.
Другие авторы, наоборот, считают, что между усушкой яиц
и выводимостью имеется тесная зависимость, но некоторые из них,
например Хейз и Спиар (Hays a. Spear, 1951), приводят проти-
воречивые данные: при усушке куриных яиц за 17 дней, равной
6.5%, было выведено 92% цыплят, при 8.5% —71.9%, при 10%—
80.1%, при 11.8%-73.9%.
Сравним данные по усушке яиц при естественной и искус-
ственной инкубации. Данные по индивидуальной усушке разных
куриных яиц под наседкой и в инкубаторе приводит Бёрк (Burke,
1925): на 19-й день под наседкой она колебалась от 14.2 до 24.2%,
а в инкубаторе — от 16.4 до 22.0%. По наблюдениям Слинько
(1936), куриные яйца под наседкой теряли за 18 дней 15.7%
своего веса, а в инкубаторе — 11.8—12.5%; усушка яиц в гнезде
утки кряквы за весь период насиживания равна 13—19%, причем
величина ее за сутки к концу насиживания постепенно увели-
чивается. Пенионжкевич и Шехтман (1937а) подтверждают, что
величина усушки яиц повышается к концу инкубации: за первые
7 дней куриные яйца теряют по 0.6% веса яйца в день под насед-
кой и по 0.8% в инкубаторе, с 7-го по 16-й — одинаково по 0.7%
в сутки и на 17-й и 18-й дни — по 1.6% и 0.8% соответственно.
Мы пересчитали данные Фердинандова (1931) по усушке куриных
яиц под наседкой и обнаружили также увеличение усушки
от 0.45% в сутки в первые дни инкубации до 0.75% в последние.
326
Й только по данным Третьякова (Прицкер и Третьяков, 1937),
также пересчитанным нами, усушка куриных яиц под наседкой,
наоборот, в первые 4 дня выше — по 0.9% в сутки, а затем при-
мерно одинакова до конца насиживания — по 0.7% в сутки.
По данным Бражниковой (1954), усушка гусиных яиц как под
наседкой, так и в инкубаторе постепенно увеличивается от 0.36—
0.37% в сутки в первые 5 дней до 0.52—0.54% в каждый из по-
следних дней инкубации, с небольшим преобладанием усушки
яиц под наседкой (14.1% под наседкой и 12.7% в инкубаторе
за 30 дней инкубации). Сходные данные получены Быховцем
(1959а): под наседкой усушка гусиных яиц была равна 0.2%
в каждый из первых 5 дней и по 0.6% на 26—28-й дни, а в инку-
баторе 0.4% и 0.3% соответственно.
Чем же объясняется увеличение среднесуточной усушки яиц
к концу инкубационного периода, особенно хорошо заметное
при естественной инкубации? Мюррей (Murray, 1925) провел
ряд опытов по инкубированию куриных яиц при относительной
влажности 23.5, 65 и 90% и обнаружил, что до 16-го дня опло-
дотворенные и неоплодотворенные яйца имеют одинаковую
усушку, а затем у неоплодотворенных яиц усушка значительно
меньше, чем у оплодотворенных, причем разница в усушке
их больше всего при влажности 65%. Интересно, что линия
усушки неоплодотворенных яиц для каждой величины относи-
тельной влажности прямая в течение всего периода инкубации,
а у оплодотворенных яиц после 16-го дня она имеет перелом к уве-
личению. По данным Бражниковой (1950), усушка утиных опло-
дотворенных и неоплодотворенных яиц до 15-го дня инкубации
бывает одинаковой, а затем оплодотворенные яйца теряют больше
влаги, и на 21-й день это различие достигает 3.9% (11.5% у опло-
дотворенных и 7.6% у неоплодотворенных). Кенди (Kendeigh,
1940) обнаружил, что у восточного домашнего крапивника су-
точная усушка яиц увеличивается в 3 раза к 10-му дню (общая
длительность эмбрионального развития 12 дней) по сравнению
с усушкой за первые сутки. Автор предполагает, что это связано
с увеличением кровообращения в аллантоисе, с утончением
скорлупы и увеличением размеров пор в ней.
Итак, одной из причин увеличения усушки яиц с развиваю-
щимися эмбрионами в последнюю треть инкубационного срока
является повышение проницаемости скорлупы в связи с рас-
творением кальциевых солей углекислотой, выделяемой эмбри-
ном, а также органической основы скорлупы — протеолитиче-
скими ферментами (Еремеев, 1957). В неоплодотворенных же
яйцах увеличения проницаемости скорлупы к концу инкубации
не происходит (Romijn, 1950).
Увеличение проницаемости скорлупы для испарения влаги
из яйца находится в тесной связи с повышением проницаемости
скорлупы для газов (см. главу 10). Проницаемость скорлупы
327
куриного яйца для газов после 20 дней развития в нем эмбриона
становится больше, чем до инкубации: для кислорода в 7000 раз,
а для углекислоты — в 565 раз (Romijn, 1950). Владимирова
(1954) установила, что скорлупа яиц с большей потерей веса
во время хранения яиц более проницаема для газов, чем яиц
с меньшей потерей веса. Между проницаемостью скорлупы для
газов (по содержанию углекислоты и кислорода в воздушной
камере куриных яиц) и для водяных паров (по усушке за первые
15 дней инкубации) мы (Рольник, 1960) обнаружили достоверную
корреляцию с 17-го дня инкубации (см. табл. 33).
Однако повышение проницаемости скорлупы к концу инкуба-
ции не является единственной причиной увеличения усушки
оплодотворенных яиц в течение инкубационного периода. Мы со-
гласны с Бражниковой (1954), объясняющей это явление повы-
шением внутрияйцевой температуры к концу эмбрионального
развития и увеличением распада жиров к этому времени, дающего
большее количество метаболической воды, чем сгорание углево-
дов и белков. Так, по данным Хаскина (19616), утиные яйца при
оптимальном режиме инкубации теряют в среднем 11 г воды
(13% веса яйца), причем 4.5 г из этого количества представляет
собой «метаболическую» воду, образовавшуюся в результате
процессов окисления.
В. Влияние различной влажности
в инкубаторе на рост и развитие эмбрионов
Приведем некоторые данные о влиянии влажности на
рост и развитие эмбрионов. По данным Романова (Romanoff, 1929,
1930), отклонения от оптимальной влажности в пределах 40—
80% возможны только до 14-го дня развития куриных эмбрионов,
а затем вызывают нерегулярность в темпе роста эмбрионов и уве-
личивают процент их гибели. При высокой влажности (80%)
лучше рост эмбрионов и их кальциевый обмен, но из-за опосредо-
ванного влияния высокой влажности — наличия излишней ал-
лантоисной жидкости и влажных подскор лунных оболочек,
мешающих выходу цыпленка из яйца, — смертность эмбрионов
повышается, особенно в конце инкубации. При низкой влаж-
ности (40%) задерживается рост эмбрионов, но смертность эм-
брионов повышается мало. Бэротт (Barott, 1937) выяснил, что
величина влажности воздуха в инкубаторе почти не влияет на га-
зообмен и энергообмен куриного эмбриона до 10-го дня инкуба-
ции, а в третью неделю эмбрионального развития, чем дальше
отстоит величина влажности от оптимальной (60%), тем меньше
метаболизм. По мнению Пенквита (Penquite, 1938), влажность
от 40% до 80% не влияет достоверно на рост эмбриона и его
временных эмбриональных органов. Однако, по нашему мнению,
более достоверны данные Ромейна и Локхорста (Romijn a. Lok-
horst, 1961), изучавших влияние различной относительной влаж-
328
ности воздуха в инкубаторе шкафного типа (при температуре
37.8°) на вес’ эмбрионов, вес вылупившихся цыплят и рост цыплят
после вылупления (табл. 49).
Таблица 49
Средний вес эмбрионов и цыплят (в г) при различной относительной
влажности
(По Romijn a. Lokhorst, 1961)
Относительная влажность (в %)
12-15 20 52 60 75 80
Продолжитель- ность инкубации (в днях) 8 1.2
9 — — — 2.5 — —
11 4.4 — • — 5.3 2.2
12 — 6.1 6.2 — 4.4 3.3
13 7.9 —. — 10.7
14 — 12.1 11.4 — 9.9 6.6
15 14.0 16.5 14.2 — 12.3 7.2
16 — — — 22.4
17 21.4 22.7 22.6 — 18.3 10.5
18 — 27.4 27.5 — 21.31
19 28.4 31.9 31.4 — 23.8 —
Возраст цыплят (в днях) 1 38.8 42.5 46.4 43.9 37.3
11 67.7 — — 81.7 — —
% вывода цыплят 64.5 89 97 80 68 0
Как видно из таблицы, сравнительно большой вес эмбрионов
и цыплят, а также высокая выводимость наблюдаются при до-
вольно широкой амплитуде влажности (от 20 до 60%) и только
при крайних значениях влажности (12—15 и 75%) процент вы-
вода и вес их снижается. При влажности же, равной 80%, все
куриные эмбрионы погибли до 18-го дня инкубации.
Огородний (1936) наблюдал разный состав крови куриных
эмбрионов на 17-й день инкубации при различной влажности
воздуха в инкубаторе: при высокой влажности (80%) гемоглобина
было 71% и 2.6 млн эритроцитов в 1 мм3 крови, а при нормальной
влажности (60%) — 68% и 4.5 млн соответственно.
Г. Оптимальная влажность
для инкубирования птичьих яиц
По мнению Пенквита (Penquite, 1938), для каждой си-
стемы инкубатора, а также на разных стадиях развития эмбрионов
и для каждого вида птиц существуют свои оптимальные условия.
329
Автор призывает к стандартизации эмбриологических исследо-
ваний, чтобы их можно было сравнивать.
Некоторые исследователи считают, что лучше поддерживать
высокую влажность в инкубаторе. Так, Инско (Insko, 1949)
в своем обзоре пишет, что не надо бояться высокой влажности,
так как низкая влажность обусловливает прилипание вылуп-
ляющихся цыплят к скорлупе и гибель их.
Сорока (1955) исследовал влияние влажности во вторую по-
ловину инкубации на развитие утиных эмбрионов и выяснил,
что высокая влажность (70%) задерживает развитие и несколько
снижает процент вывода утят (на 1.7%) при 91.1% утят к коли-
честву живых эмбрионов на 15-й день инкубации в контроле.
Низкая влажность (45%) ускоряет развитие эмбрионов, но сни-
жает выводимость утят на 7.1%. Вермесану (Vermesanu, 1961)
считает необходимым поддерживать высокую влажность (75%)
в течение всего периода инкубации, так как это, по мнению ав-
тора, обусловливает высокую текучесть желтка и создает лучшие
условия для ассимиляции эмбрионом питательных веществ.
При этой влажности выводимость цыплят достигает 91.5% от оп-
лодотворенных яиц по сравнению с 80.2% при снижении влаж-
ности до 65% с 8-го по 14-й день и до 55% с 15-го по 21-й день
инкубации.
Другие исследователи (Romanoff, 1929; Пенионжкевич и Шехт-
ман, 19376, и др.), наоборот, склоняются к необходимости низ-
кой относительной влажности воздуха в инкубаторе. Возможно,
некоторую роль в рекомендации низкой влажности играет по-
вышение проницаемости сухой скорлупы для газов, так как
именно осложнение газообмена в конце инкубации обуслов-
ливает в основном высокую смертность эмбрионов в это время
(см. главу 17). Пенионжкевич и Шехтман (19376) показали,
что повышенная влажность (до 80%) в инкубаторе тормозит усвое-
ние эмбрионом белка и желтка в связи со снижением интенсив-
ности физико-химических процессов. Это влияние особенно
сильно проявляется до 12-го дня инкубации.
Приведем мнения некоторых авторов о необходимой оптималь-
ной влажности. Бэротт (Barott, 1937) считает оптимальной влаж-
ность, равную 58% при температуре 37.8°, и 61% — при темпе-
ратуре 38.9%, причем выводимость цыплят при первом сочетании
выше примерно на 10%. Инско (Insko, 1949) рекомендует следую-
щие нормы оптимальной влажности. Для инкубации куриных
яиц в первые 18 дней — 48—60% влажности, а в последние
3 дня — 60—70%; для индюшиных рекомендуется несколько
более высокая влажность: в первые 24 дня — 51—56%, а в по-
следние 4 дня — 65—79%. Пенионжкевич (1950) показал, что
применение высокой влажности (85—95%) в шкафном инкубаторе
в течение 12 час. в период вывода цыплят (1 раз в 4 суток), ком-
пенсированное 66 часами низкой влажности (40%) с переходным
330
йериодом в 18 час. средней влажности (64—70%), положительно
влияет на развитие эмбрионов; при этом сохраняется та же усушка
яиц и время вылупления цыплят, что и при обычных условиях
влажности, но развитие эмбриона, выводимость и жизнеспособ-
ность цыплят повышаются. Автор довольно убедительно пока-
зывает, что при одной и той же температуре и скорости воздуха
в инкубаторе усушка яиц зависит от средней влажности (под-
считанной автором из суммы всех величин влажности за каждый
час из 4 суток и деленной на 96 час.), от развития аллантоиса
в период с 6-го до 11-го дня и от интенсивности развития эмбриона
в период с 17-го до 19-го дня инкубации. Орлов (1961а) считает
необходимым дифференцировать условия влажности в разные
периоды инкубации соответственно особенностям развития эм-
брионов в каждый из этих периодов. В первые 6 дней следует
максимально сохранить воду в яйце, так как она еще не приняла
участия в обмене веществ; затем, после смыкания аллантоиса
в остром конце яйца, наоборот, полезно ее удаление, так как
вода уже приняла участие в обмене веществ и, оставаясь в ал-
лантоисе, задерживает поток воды, несущий запасы питатель-
ных веществ из белка и желтка к зародышу. Однако эта довольно
интересная гипотеза не подкрепляется никакими эксперименталь-
ными данными. Робертсон (Robertson, 1961с), получивший удов-
летворительную выводимость цыплят при довольно широкой
амплитуде влажности — от 40 до 70% — оптимальной считает
40—50% влажности. Эту же величину оптимальной влажности
называют и Ромейн и Локхорст (Romijn a. Lokhorst, 1961).
В исследовании Томпсона (Thompson, 1952) показано, что
на высоте 2000 м над ур. м., где парциальное давления кислорода
понижено, высокая влажность, ухудшающая проницаемость скор-
лупы для газов, ведет к гибели эмбрионов.
Следовательно, при довольно широких границах влажности
(40—70%) может быть получена высокая выводимость молодняка
сельскохозяйственных птиц при соблюдении оптимальными дру-
гих факторов инкубации, а влажность от 15 до 75% допустима
для выживания эмбрионов. Возможно, это является отражением
широкого размаха величины влажности в природе.
Г Л А В А 20
ВЕНТИЛЯЦИЯ
Вентиляцию инкубатора можно рассматривать с трех
точек зрения: 1) доставляет ли она необходимое количество ки-
слорода для дыхания эмбрионов, 2) ограничивает ли накопление
выдыхаемой углекислоты в безвредном для развития эмбрионов
количестве, 3) в какой мере она влияет на влажность воздуха
в инкубаторе и обогрев яиц.
331
А. Содержание газов в воздухе гнезда
при естественной инкубации
Как сообщает Липпинкот (1932), некоторые исследова-
тели (Dryden, 1901; Edmond, 1907; Thom, 1901, все цит. по: Лип-
пинкот, 1932) сравнивали содержание углекислоты в воздухе гнезда
и в инкубаторе и нашли, что под курицей ее содержится больше.
Бёрк (Burke, 1925) определял концентрацию углекислоты в гнезде
курицы и обнаружил, что с 0.1% на 1-й день она достигает 0.4%
на 12-й день насиживания и 0.92—1.43% к концу эмбрионального
развития. Автор нашел повышение содержания СаО в скорлупе
яиц, инкубированных в течение 20 дней, по сравнению со све-
жими яйцами с 0.12% до 0.38%; и так как углекислота способ-
ствует переходу нерастворимого соединения СаСО3 в раствори-
мое — СаО, идущее на построение скелета эмбриона, автор при-
ходит к заключению о полезности и необходимости углекислоты
в окружающем яйцо воздухе. Автором отмечено также некото-
рое повышение выводимости цыплят в тех гнездах, где содержание
углекислоты в воздухе было большим. Романов и Романова
(Romanoff a. Romanoff, 1933) сообщают, что содержание угле-
кислоты в гнезде курицы увеличивается с 0.1% до 0.7% к концу
насиживания. Содержание кислорода в воздухе гнезда практи-
чески такое же, как в окружающей атмосфере.
Б. Дыхание птичьих эмбрионов
при различном содержании газов
в окружающей атмосфере
Существуют противоречивые мнения разных исследова-
телей о значении углекислоты в атмосфере инкубатора. Одни ав-
торы (Brigham, 1901, цит. по: Needham, 1931; Thom, 1901, цит. по:
Липпинкот, 1932; Lourdel, 1909, цит. по: Needham, 1931; Lamson
a. Kirkpatric, 1918) считают необходимым добавлять СО2 в ин-
кубатор, другие (Dryden, 1901; Edmond, 1907, оба цит. по: Лип-
пинкот, 1932; Harcourt a. Graham, 1908, цит. по: Needham, 1931)
считают необходимым улучшить вентиляцию, так как, по их мне-
нию, эмбрионами выделяется достаточное количество углекислоты.
Систематическое изучение влияния состава воздуха в инку-
баторе на развитие куриных эмбрионов провели Романов (Ro-
manoff, 1930) и Романов и Романова (Romanoff a. Romanoff,
1933). Авторы показали, что существует прямое и косвенное
влияния состава воздуха (углекислоты и кислорода) на эмбрио-
нальное развитие. Прямое влияние сказывалось на росте, вы-
лупляемости и патологиях развития эмбриона. Рост эмбрионов
в течение первых нескольких дней стимулируется наличием сред-
него количества СО2 (0.4—1.0%). Повышенное содержание СО2
332
(10%) и соответственно пониженной количество кислорода за-
медляли рост пропорционально содержанию СО2. Ранние стадии
эмбрионального развития более чувствительны к нарушению
нормального содержания газов, чем поздние. Как при понижен-
ной, так и при повышенной концентрации СО2 смертность эмбрио-
нов увеличивается, иногда настолько, что ни один цыпленок
не вылупляется. Типичными патологиями при нарушении кон-
центрации СО2 в инкубаторе являются различные ненормальности
положения эмбрионов в яйце перед вылуплением. Косвенное
влияние нарушения концентрации СО2 в инкубаторе заключается
в изменении pH яйца. Авторы установили эмпирическую формулу
для подсчета pH белка яйца (у):
(2.945 —0.25а:)
У = 7.83 За: | 1 69) “Ь 9*5 bOz),
где х — концентрация СО2, az — возраст эмбриона в днях.
В свою очередь изменение pH означает изменение физико-хими-
ческого состояния среды и питательных веществ и, таким обра-
зом, влияет на рост и развитие эмбриона.
По данным Бэротта (Barott, 1937), влияние повышенной кон-
центрации СО2 становится неблагоприятным на 3-й день инку-
бации и затем все более увеличивается. Оптимальной для раз-
вития куриных эмбрионов является, по данным автора, концент-
рация углекислоты в воздухе инкубатора менее 0.5%, а при
увеличении ее энергообмен и рост эмбрионов замедляются и про-
цент вывода цыплят снижается почти до нуля. Тэйлор и его
соавторы (Taylor et al., 1956) нашли, что при концентрации СО2
в воздухе инкубатора выше 1% в течение первых четырех дней
инкубации выводимость цыплят сильно снижается: при 5% СО2 —
до 10% от цыплят, выведенных в контроле, а при 10% СО2 —
до нуля. Чувствительность эмбрионов к изменению содержания
СО2 в первые 4 дня инкубации, по данным авторов, примерно
такая же, как в течение всего эмбрионального развития. Но позже
Тэйлор и Крейцигер (Taylor a. Kreutziger, 1965) установили, что
с 5-го до 8-го дня инкубации снижается устойчивость к повы-
шенной концентрации углекислоты в атмосфере инкубатора; со-
держание 3% СО2 не имело достоверного влияния на выводи-
мость, но при СО2 выше 8% вывод цыплят равнялся нулю.
Устойчивость к высокому уровню СО2 в течение первых четырех
дней инкубации обусловлена, по мнению авторов, ограниченной
способностью эмбрионов к газообмену в это время.
Прицкер (1935), выясняя причины появления слабых, плохо
распушенных, так называемых «липких» цыплят, обнаружил,
что ни температура, ни влажность не являются причиной этого
явления, а оно обусловлено в основном недостаточной венти-
ляцией и повышением содержания СО2 в инкубаторе до 4—5%
333
к концу инкубационного периода. Лютц (Lutz, 1962) про-
верил действие СО2 на развивающихся куриных эмбрионов,
пропуская ее через сосуд, в который на 12 час. были по-
мещены яйца 18-го дня инкубации. При дальнейшем инкуби-
ровании у 26% эмбрионов были обнаружены уродства, причем
наиболее чувствительными оказались дифференцирующиеся
клетки.
В известной мере противоположной точки зрения на значение
повышенного содержания углекислоты в воздухе инкубатора
придерживаются некоторые другие исследователи. Так, Бёрк
(Burke, 1925), обнаруживший, что при накоплении С02 в инку-
баторе к концу инкубационного периода до 1.65—2.0% не нару-
шается нормальное развитие куриных эмбрионов, и наблюдав-
ший высокое содержание СО2 в гнезде курицы (о чем мы уже
говорили выше), приходит к выводу, что концентрация угле-
кислоты в инкубаторах недостаточна и вентиляция должна быть
уменьшена. Спратт (Spratt, 1949) сообщает, что частичное уда-
ление углекислоты из окружающего воздуха тормозило диф-
ференциацию и морфогенез центральной нервной системы в экс-
плантированных бластодермах, а более полное удаление СО2
вызывало дальнейшую дегенерацию их. Автор предполагает, что
углекислота необходима для нормального эмбрионального раз-
вития так же, как и кислород.
При инкубировании яиц домашних птиц на больших высотах
(1500 м над уровнем моря) Садлер и его соавторы (Sadler et al.,
1954) показали, что в течение первых двух дней инкубации высо-
кая концентрация СО2 (4%) заметно снижает pH белка и замед-
ляет его разжижение, что стимулирует развитие эмбриона. Мак-
симальная стимуляция развития происходила при воздействии
в течение 72 час. воздухом, содержащим 1% СО2. Затем до 10-го
дня инкубации высокая концентрация СО2 оказывала вредное
влияние. Наоборот, увеличение концентрации СО2 с 1% до 5%
в течение последних 10 дней инкубации повышало выводимость
цыплят на 3—4%, а также их вес и качество. Авторы подчерки-
вают необходимость учитывать полезное действие высокой кон-
центрации СО2 в разные периоды эмбрионального развития при
расчетах режима инкубации и особенно — фактора вентиляции.
По данным Еремеева (1957), повышение концентрации угле-
кислоты в содержимом птичьего яйца перед началом инкубации
(путем помещения его в атмосферу с 60—70 %-й концентрацией СО2
на 3—4 часа) благоприятно влияет на развитие эмбриона, так
как ускоряет развитие сосудистого поля и кровеносной системы
эмбриона.
Проверке влияния разных концентраций кислорода на эм-
бриональное развитие птиц также посвящено много исследова-
ний. Прейер (Ргеуег, 1885) считает, что достаточное снабжени-
кислородом играет бблыпую роль для роста, чем для дифферен
334
циации. Ридль (Riddle, 1924) показал, что в начале развития
эмбрионы легче переносят пониженное содержание кислорода,
а потом — повышенное. Автор напоминает, что у взрослых тепло-
кровных животных кислородное насыщение тканей не зависит
от изменения содержания кислорода во внешней среде, как это
имеет место у большинства холоднокровных позвоночных жи-
вотных. Так же как и холоднокровные животные, эмбрионы птиц
в начале своего развития не могут регулировать потребление
кислорода и при слишком высоком его содержании в окружаю-
щем воздухе развиваются ненормально. Автор показал, что эм-
брионы голубей могут развиваться при содержании О2 в атмос-
фере от 14 до 40%, и отмечает индивидуальные различия в реак-
ции, которые обусловливаются разной проницаемостью скорлупы
для газов. Бордзивиловская (1955) отметила, что на ранних эта-
пах развития у утиных и гусиных эмбрионов значительно мед-
леннее развивается желточный круг кровообращения, что, по пред-
положению автора, является приспособлением, возникшим вслед-
ствие того что эволюция этих форм протекала в условиях лучшего
снабжения кислородом, чем это было у куриных эмбрионов. Ав-
тор приходит к выводу, что это обусловливает еще большую
важность воздухообмена при их развитии.
Нидхем (Needham, 1931) сообщает, что в начале эмбриональ-
ного развития значительную роль играет анаэробный метаболизм
и потому на ранних стадиях легче переносится сильно пониженная
концентрация кислорода; на более поздних стадиях развития
значение анаэробного обмена веществ уменьшается и наконец
исчезает. Однако, как отмечает автор, в полностью анаэробных
условиях может происходить только рост ткани, а не ее дифферен-
цировка. Подтверждение этого вывода приводит Галлера (Gal-
lera, 1951), выяснивший, что при инкубировании в 100% азота
в течение первых двух дней не происходило дифференциации;
при низких же концентрациях кислорода (до 7%) около половины
эмбрионов имело различные ненормальности развития.
Рассмотрим сначала влияние низкой концентрации кисло-
рода на эмбриональное развитие. По данным Веселкина (We-
sselkin, 1913), при содержании куриных эмбрионов в атмосфере
с 5% О2 они погибали через 48 час., а при 10—15% О2 разви-
тие продолжалось 72 часа. Круз и Романов (Cruz a. Romanoff,
1944) выяснили, что при 11% О2 в течение первых 5 дней инкуба-
ции выводимость цыплят снижается до 10%. В опытах Пого-
рельского (1947) при 17.5% О2 вывелось 21.4% цыплят (в кон-
троле 72.5%), при 15.5% О2 — 2.2%, при 12.0% О2 и ниже не
вылупилось ни одного цыпленка. Автор обнаружил, что при
пониженном содержании кислорода в атмосфере инкубатора
в первую половину инкубации развитие куриных эмбрионов
более интенсивно за счет увеличения количества эритроцитов
и усиления кровообращения, что обеспечивает нормальный
335
транспорт кислорода развивающимся тканям. Во вторую по-
ловину развития этого компенсаторного механизма оказывается
недостаточно, и эмбрионы, отставая в развитии, гибнут. По
данным Тэйлора с соавторами (Taylor et al., 1956), при 15% О2
в течение всего инкубационного срока выводимость сильно сни-
жается, а при 10% О2 доходит до нуля. В дальнейшем Тэй-
лор и Крейцигер (Taylor a. Kreutziger, 1965) обнаружили рез-
кое падение выводимости цыплят при 15% О2 и полное прекра-
щение выводоспособности, если содержание кислорода падало
ниже 12.5% с 5-го до 8-го дня инкубации.
Иное положение мы видим при небольшом снижении или
некотором повышении содержания кислорода. В детальном, ис-
следовании Бэротта (Barott, 1937) проверено влияние различных
концентраций кислорода в инкубаторе (от 14% до 50%) на обмен
веществ эмбрионов и выводимость цыплят. Автор выяснил, что
в течение первых двух недель содержание кислорода в воздухе
инкубатора (в изучаемых пределах) не имело заметного влияния
на метаболизм эмбрионов, а с 15-го дня до конца инкубации наи-
высший обмен веществ был у эмбрионов при 21% О2; несколько
ниже, но практически такой же метаболизм был при 30 и 40% О2;
при 50 и 18% О2 — на 20% ниже. Бэротт правильно отмечает,
что при инкубации невозможен излишек кислорода, за исклю-
чением тех случаев, когда он добавляется в инкубатор искус-
ственно. В практике основной опасностью, следовательно, является
недостаток кислорода, так как он используется при дыхании
эмбрионов. Круз и Романов (Cruz a. Romanoff, 1944) выяснили,
что пребывание куриных эмбрионов в течение первых 5 дней
инкубации в атмосфере с 31— 41 % О2 увеличивает рост эмбрионов
в 1.5 раза и повышает выводимость цыплят. Тэйлор, Съедин и
Ганз (Taylor et al., 1956) наблюдали, что повышение содержа-
ния кислорода до 50% в первые 4 дня инкубации оставляло
величину выводимости почти на прежнем уровне. По данным
Тэйлора и Крейцигера (Taylor a. Kreutziger, 1965), повышение
до 45% с 5-го до 8-го дня инкубации не снижало выводимости
цыплят.
При большом повышении концентрации кислорода наступает
значительное снижение жизненности эмбрионов. При 75% О2
вылупилось 20% цыплят, а при 95% О2 все эмбрионы погибли
(Cruz a. Romanoff, 1944). При повышении уровня О2 выше 82%
с 5-го до 8-го дня инкубации выводимость цыплят снижалась
до нуля (Taylor a. Kreutziger, 1965). Однако, по данным Галлера
(Gallera, 1951), инкубация в 100% О2 в течение первых двух дней
не нарушает эмбрионального развития.
Вредный эффект изменения уровня кислорода или углекислоты
на выводимость цыплят не только суммируется, но и наблюдается
усиливающее друг друга действие комбинации этих эффектов
(Taylor a. Kreutziger, 1965).
336
Интересными с практической точки зрения являются опыты
инкубирования при пониженном амосферном давлении. Опубли-
кован целый ряд работ о добавлении кислорода в специально
устроенные инкубаторы в инкубаториях на больших высотах,
где вследствие сниженного атмосферного давления понижено и
парциальное давление кислорода.
Проводились опыты по добавлению кислорода в инкубатор
и при обычном атмосферном давлении. Вейнмиллер (Weinmiller,
1930) упоминает о таких опытах двух исследователей (Becker,
Suchting), получивших противоположные данные о влиянии
добавления кислорода в инкубатор на выводимость цыплят (один
исследователь описывает положительное влияние, второй — отри-
цательное). Автор считает, что при хорошей вентиляции и охлаж-
дении яиц на свежем воздухе нет надобности в добавлении кисло-
рода в инкубатор.
Следует упомянуть также об интересных опытах Тарновского
(1935), проводившего 1-2-часовое ежедневное озонирование
куриных яиц во время инкубации (озонатором по типу Лед-
Бенси), что повышало выводимость цыплят на 9—20%, увеличи-
вало количество эритроцитов и гемоглобина в крови, а также
вес эмбрионов и сокращало длительность инкубации на 18—24
часа.
Эллис и Морис (Ellis a. Morris, 1947), добавляя кислород
в инкубатор при инкубировании куриных яиц на высоте 2200 м,
получили повышение выводимости цыплят на 13% (при выводи-
мости в контроле 69—74% от оплодотворенных яиц) и индюшат —
на 39% (при контроле 27.5%). На высоте 1500 м над ур. м. добав-
ление в инкубатор О2 до 23—23.5% (что соответствует парциаль-
ному давлению 21% О2 на уровне моря) повышало выводимость
цыплят и индюшат на 13—30% и обусловливало лучшую жизне-
способность их в постэмбриональном развитии (Sadler et al.,
1954). Уилгус и Садлер (Wilgus a. Sadler, 1954) подтверждают
эти данные. Так, при 20*6% О2 было выведено 67% цыплят, при
21.8% О2 - 74%, при 23.2% О2 - 76%, при 24.6% О2 - 77%.
Интересно, что при содержании кислорода 23—23.5% наилучшая
выводимость цыплят была при 0.5% СО2, а при 20.6% О2 выво-
димость цыплят быстро снижалась, если содержание углекислоты
было выше 0.25%.
Считая недостаток кислорода в период перехода от аллантоис-
ного к легочному дыханию одной из основных причин повышения
смертности эмбрионов в последние дни перед вылуплением, мы
провели производственные опыты по введению кислорода в воз-
душную камеру яиц (о которых мы уже упоминали — Рольник,
1956). Результаты этих опытов приведены в табл. 50.
Как мы видим, введение кислорода в воздушную камеру яиц
значительно (в среднем наполовину) сокращало количество так
называемых задохликов. Следует отметить, что приведенные
22 в- в- Рольник 337
Таблица 50
Влияние введения кислорода в воздушную камеру яиц на выводимость
цыплят, утят и индюшат
(По Рольник, 1956а)
Виды яиц Количество яиц % вывода к живым до введения О2 Превышение вывода (в %) при добав- лении О2
в конт- роле с добав- лением О2 в конт- роле с добав- лением О2
489 810 71.7 84.7 13.0
Куриные | 12201 3871 69.0 80.7 11.7
1437 4392 76.4 90.8 14.4
Утиные 3189 1598 63.4 69.7 6.3
Индюшиные 1524 358 63.3 83.0 19.7
Примечание. Для куриных яиц показаны данные трех разных лет инкуба-
ции сверху вниз: 1945, 1946 и 1947.; для утиных и индюшиных — данные 1946 г.
результаты превышения процента вывода при введении кислорода
далеко не являются максимальными для этого метода, так как
в них включены опыты с менее благоприятными дозировками
и сроками введения кислорода. Особенно большое превышение
процента вывода индюшат обусловлено, по-видимому, тем, что
более плотные подскорлупные оболочки индюшиных яиц (2.2%
веса яиц по сравнению с 0.6% веса у куриных) значительно хуже
пропускают кислород и другие газы. Следует также отметить,
что кислородизация яиц наиболее благоприятно влияла в тех
случаях, когда в результате нарушения режима инкубации
(сильный перегрев в первые дни эмбрионального развития) или
длительного хранения яиц (в течение двух недель) содержание
кислорода в белке и желтке яйца было сильно снижено. В опытах,
когда был перегрев яиц в начале инкубации, в контроле погибло
60% эмбрионов от числа живых на 17-й день в одном случае и
31.7% — в другом, а в опытных сериях — 29 и 7% соответственно.
Введение кислорода в воздушную камеру яиц не только повышало
выводимость, но и сокращало длительность процесса вылупления,
повысило процент здоровых цыплят на 9.4—13.7%, утят — на
11.2% и индюшат — на 36.4% по сравнению с контролем и сок-
ратило процент гибели молодняка в первые 15 дней постэмбрио-
нальной жизни. «Кислородизацию» куриных яиц мы провели
также, помещая их на 5—30 мин. в закрытый стеклянный сосуд,
через который пропускался 100%-й кислород. Сосуд в это время
находился в инкубаторе. При этом состав газов в воздушной ка-
мере яиц изменялся сходно с теми, которым мы вводили 5—10 мл
О2 в воздушную камеру. Этот способ воздействия («кислородизация»
338
в сосуде) повышал жизненность эмбрионов и увеличивал выво-
димость цыплят примерно так же, как и введение кислорода
в воздушную камеру яиц. Эти опыты являются еще одним дока-
зательством правильности гипотезы о недостатке кислорода
в период вылупления как основной причине повышения смерт-
ности эмбрионов в это время (см. главу 10).
Следует упомянуть также о некоторых, как нам кажется,
значительно менее удачных попытках решить проблему облегчения
дыхания в последние дни перед вылуплением. Машталер (1947)
пытался утончить скорлупу яиц для большей проницаемости ее
для газов путем опускания яиц в соляную кислоту в один из
последних дней инкубации. Крюгер и Тиерина (Krueger a. Tije-
rina, 1960) просверливали в скорлупе над воздушной камерой
небольшие отверстия (диаметром 1.5—2 мм) с целью увеличения
воздухообмена куриного эмбриона в последние дни инкубации.
При этом получено повышение выводимости цыплят во всех
сериях опытов: при просверливании на 13-й день — на 4.1%, на
14-й на 2.7%, на 15-й — на 5.5%, на 16-й — на 8.6%, на 17-й —
на 2.7% и на 18-й — на 0.2%, при выводимости в контроле,
равной 84.9%.
В. Режим вентиляции инкубаторов
Третьякова (1936) исследовала состав воздуха в инкуба-
торах и нашла, что аммиак появляется там только в момент
наклева и выхода цыплят и, следовательно, в это время должна
быть увеличена вентиляция. Автор не обнаружила сероводорода
в инкубаторах и считает причиной этого большую растворимость
его в воде. Углекислота, по данным автора, не превышает в нор-
мальных условиях 0.55%. Обычно же (при средней вентиляции)
содержание СО2 равно 0.3—0.4%, и эта концентрация углекислоты
безвредна. Автор провела опыт с поглощением углекислоты в ин-
кубаторе, который не дал повышения выводимости, и, следова-
тельно, делать это, по ее мнению, нет смысла.
В наших работах мы показали, что в последние дни инкубации
газообмен значительно увеличивается (подробнее см. в главе 10).
Это ставит вопрос о вентилировании инкубаторов в дни перед
вылуплением (сложный период перехода к совершенно другим
условиям жизни) на первое место в обеспечении необходимых
условий для нормального развития эмбриона.
К сожалению, в последнем руководстве по инкубации (Отры-
ганьев и др., 1964) недооценивается значение состава газов для
нормального эмбрионального развития и смена воздуха в инку-
баторе рассматривается только с точки зрения расхода воды для
поддержания необходимой влажности.
В связи с тем что содержание кислорода в обычных условиях
относительно мало изменяется (от 20.7% до 19.5%, т. е. на 5 —7%
22*
339
первоначальной величины), расчеты по обмену воздуха в инкуба-
торе делают применительно к поддержанию необходимой концен-
трации углекислоты. Прицкер и Третьяков (1937) дают следующий
расчет обмениваемости воздуха в инкубаторе за час, чтобы кон-
центрация углекислоты не превысила нормы (0.3%):
К
Х~ С1—С2 ’
где х — необходимое количество свежего воздуха в кубических
метрах, К — количество углекислого газа, выделяемое за час
всеми яйцами, находящимися в инкубаторе, Сг — количество СО2,
максимально допустимое, т. е. 0.3% или 0.003 м3, С2 — количе-
ство СО2, входящего в камеры инкубатора вместе с наружным
воздухом (труба вентилятора должна быть выведена из инкубатора
наружу), т. е. 0.03% или 0.0003 м3. Таким образом, Сх — С2 —
это величина постоянная, 0.0027 м3, а К легко подсчитать, зная,
сколько углекислоты выделяется одним яйцом на данной стадии
развития. Авторы предупреждают, что расчеты нужно вести
с 50 %-й надбавкой.
В наиболее распространенном в настоящее время в СССР
инкубаторе «Рекорд» на 1 м3 приходится около 1.5 тыс. яиц, и
поэтому здесь необходим многократный обмен воздуха.
К сожалению, в литературе по инкубации мы встречаемся
чаще с другой величиной, характеризующей вентиляцию, со
скоростью движения воздуха. Однако величина эта не отражает
определенной обмениваемости воздуха в инкубаторах разных
систем, так как последняя зависит также от ряда других условий
(ширина и длина выводной вентиляционной трубы и т. д.).
Уилгус и Садлер (Wilgus a. Sadler, 1954) измерили скорость
движения воздуха в инкубаторе с искусственной вентиляцией
на разных его уровнях и обнаружили очень большие различия
в ней — от 9—15 до 75 м в 1 мин. в инкубационной части инкуба-
тора и от 5—7 до 35—45 м в выводной его части. Авторы подчерки-
вают, что такие большие вариации не способствуют высокой
выводимости. Кроме того, по наблюдениям авторов, имеет зна-
чение и направление потока вентиляции, причем наилучшие ре-
зультаты дает вентиляция сквозь яйца, снизу вверх.
Коноплев (1962) придает вентиляции при инкубировании
куриных яиц после 15-го дня большое значение. На большом ма-
териале им было показано, что при высокой температуре (39.8—
39.2° между лотками) и средней влажности (55.4—52.0%) в группе
с малой скоростью воздуха (0.5 м/сек.) было выведено 42.4%
цыплят, а в группе с большой скоростью (1.95 м/сек.) — 96.8%;
примерно при той же температуре, но высокой влажности (78—
74%) в группе с малой скоростью воздуха — 72.5% цыплят,
а в группе с большой скоростью — 98.9%. Однако из данных
автора следует, что при низкой температуре (37—37.8° между
340
лотками) скорость воздуха играет значительно меньшую роль.
Увеличение скорости воздуха при этом дало в одном опыте повы-
шение выводимости наЗ°/о,ав другом — только на 0.3%. Автор
приводит также интересное наблюдение, когда вследствие недо-
статочной обмениваемости воздуха в инкубаторе был обнаружен
сероводород, что сильно снизило выводимость цыплят. В заклю-
чение автор рекомендует при температуре в инкубаторе 37.8—
38.0° (а между лотками 38.0—38.5°) и влажности 68% и 54%,
чередующейся по 2 дня, установить скорость движения воздуха
в инкубаторе 1.5 м/сек, что приведет к скорости между лот-
ками только 0.3—0.5 м/сек. Автор подчеркивает далее, что кроме
указанной скорости движения воздуха должна быть обеспечена
хорошая обмениваемость его в инкубаторе.
Бражникова (1950) подтвердила данные других исследователей
о более полном использовании утиными эмбрионами жира желтка
(к концу инкубации у утиного эмбриона остается и втягивается
только 12.4% желтка, а у куриного — 50%) и в связи с этим —
более интенсивном дыхании их в последние дни инкубации.
Считая допустимой концентрацией СО2 в инкубаторе 0.5%, автор
заключает, что в начале инкубации вентиляция утиных яиц
может быть даже несколько меньшей, чем куриных, но с 22-го
дня до конца вывода утят она должна быть увеличена почти вдвое
по сравнению с вентиляцией, применяемой при инкубировании
яиц кур.
Сорока (1955) исследовал значение вентиляции для развития
утиных эмбрионов во вторую половину инкубации и пришел
к выводу о необходимости установить в инкубаторе с искусствен-
ной вентиляцией скорость воздуха 1.0—1.2 м/сек. и разреженную
закладку яиц в инкубационной колонке (через один свободный
ярус). При этих условиях вывелось 83.3% утят. Однако еще боль-
шее увеличение скрости воздуха (1.8—2.0 м/сек.) дало дальнейшее
повышение выводимости утят —85.5%.
В детальном обследовании инкубатора «Универсал-45», про-
веденном Орловым (19616), большое внимание уделено вентиляции.
Автор установил, что: а) скорость воздуха в этом инкубаторе
в 4 раза выше, чем в инкубаторе «Рекорд», и равна в среднем
77 м/сек. (от 13 до 176 м/сек.), а в выводном шкафу — от 30 до 52 м/сек.,
б) обмен воздуха в инкубаторных шкафах происходит 33—36 раз
в час (в 3—4 раза больше обмениваемости воздуха в инкубаторе
«Рекорд»), а в выводном шкафу — 17 раз в час; в) благодаря
хорошему обмену воздуха в инкубаторе «Универсал-45» обеспе-
чивается сравнительно низкое содержание углекислоты: 0.1 —
0.17% в инкубационных шкафах и 0.21—0.25% — в выводном;
г) в результате при инкубировании многих тысяч яиц получена
более высокая выводимость, чем в инкубаторе «Рекорд»: цыплят —
на 2.0—3.5% и утят — на 3.4—11.4%. В инкубаторе «Универсал-
45» в этом сезоне выводимость цыплят была равна 88.3—90.7%,
341
утят — 67.6—-86.5%. Увеличение вентиляции особенно благопри-
ятно сказалось на выводимости утят.
Установив более слабое развитие кровеносной системы у гу-
синых эмбрионов по сравнению с куриными Бордзивиловская
(1953) предполагает, что в процессе эволюции они находились
в лучших условиях аэрации, и считает необходимым обратить
особое внимание при инкубации гусиных яиц на достаточный воз-
духообмен в инкубаторах. Этот вывод подтверждает исследование
Быховца (1955), показавшего, что газообмен гусиных эмбрионов
протекает значительно интенсивнее, чем у куриных, так как
вес гусиного яйца только в 3 раза больше куриного, а выделение
углекислоты одним яйцом в 4 раза больше. На основании своих
наблюдений автор разработал нормативы вентиляции при инку-
бировании гусиных яиц в инкубаторе «Рекорд-39». Для нормаль-
ного газообмена всех находящихся в инкубаторе гусиных эмбрио-
нов необходим примерно 11-кратный обмен воздуха инкубатора
в час. В связи с фактически имеющимся в инкубаторе при совре-
менной конструкции 8-кратным обменом воздуха автор считает
необходимым повысить воздухообмен в нем на 25%.
Для выяснения роли каждого из факторов инкубации, в том
числе вентиляции, при инкубировании яиц неодомашненных
птиц (фазанов и перепелов) Романов (Romanoff, 1938) провел
многочисленные опыты на большом материале (около 9500 яиц).
Автор отмечает, что яйца диких птиц особенно чувствительны
к изменению вентиляции и для каждого вида есть свои специфи-
ческие оптимальные условия. Так, для инкубирования фазаньих
яиц в первые 16 дней наиболее благоприятна вентиляция со ско-
ростью воздуха 20 м в 1 мин., а в последние 8 дней — естественная
вентиляция (значительно более медленная скорость воздуха);
перепелиные яйца можно инкубировать все время в инкубаторе
с искусственной вентиляцией.
Несколько слов следует добавить о косвенном значении вен-
тиляции. Хаскин (1961а) прказал, что вентиляция при промыш-
ленной инкубации играет серьезную роль в теплообмене яиц
в конце инкубационного срока, создавая возможность отдачи
излишков тепла. Автор подсчитал, что только 10% теплоотдачи
в это время осуществляется испарением, а теплоотдача — излу-
чением, которая у одиночного яйца составляет 43% всей тепло-
отдачи, для каждого яйца из партии в больших инкубаторах
сокращается наполовину вследствие уменьшения свободной по-
верхности, соприкасающейся с воздухом инкубатора (при плотной
укладке яиц в лотки в вертикальном положении), и следовательно,
значительно увеличивается роль теплоотдачи конвекцией. Поэтому
необходимо увеличить скорость движения воздуха в инкубаторе,
особенно в пограничном с яйцами слое воздуха (обычно не превы-
шающей здесь 0.09—0.1 м/сек.), во избежание перегрева яиц
во второй половине инкубационного срока.
342
В заключение следует сказать, что вентиляция инкубаторов,
способствующая хорошему газообмену эмбрионов, играет не мень-
шую роль в нормальном течении роста и развития эмбрионов,
чем температура и влажность, особенно в последние дни инкубации,
когда этот фактор становится едва ли не самым важным. Особенное
внимание необходимо уделять вентиляции при инкубировании
утиных и гусиных яиц, а также яиц промысловых птиц (фазаны,
перепела и т. п.).
Г Л А В А 21
МЕХАНИЧЕСКИЕ ФАКТОРЫ
К механическим факторам при инкубировании птичьих
яиц относят положение их (вертикальное, горизонтальное), пово-
рачивание (вокруг малой оси при горизонтальном положении или
в обе стороне на 45° от вертикали) и сотрясения.
А. Положение яиц во время инкубации
В гнезде птиц яйца лежат горизонтально. Вопрос об из-
менении положения яиц во время инкубации встал в связи с тем,
что в вертикальном положении их проще поворачивать и они
занимают меньше места. Следует отметить, что под вертикаль-
ным подразумевается положение яйца тупым концом (воздушной
камерой) вверх, так как при обратном (острым концом вверх) —
развитие эмбриона протекать нормально не может.
По данным Баерли и Олсена (Byerly а. Olsen, 1933), при инку-
бировании яиц острым концом вверх 60—70% эмбрионов во
время вылупления находятся в неправильном положении — го-
ловой в острый конец, что почти всегда ведет к гибели эмбрионов.
По мнению Ландауэра (Landauer, 1948), это является результатом
изменения центра тяжести эмбриона. Эйклешимер (Eycleshymer,
1907) обнаружил, что в секционном инкубаторе инкубирование
яиц в горизонтальном положении дает значительно более
низкую выводимость цыплят (47.3%) по сравнению с верти-
кальным (85%), а в инкубаторе с искусственной вентиляцией
это различие меньше (83% и 88.7% соответственно). Автор
объясняет это различие тем, что вертикальное положение, при
котором эмбрион лучше соприкасается с воздушной камерой,
особенно важно при недостаточной вентиляции, характерной
для секционных инкубаторов.
Однако для яиц водоплавающих птиц нет преимущества вер-
тикального положения яиц над горизонтальным — скорее на-
оборот. Сорока (1955), проведя опыты на 26 000 утиных яиц, по-
343
казал, что нормальное замыкание аллантоиса в большинстве яиц
возможно только при горизонтальном их положении в инкубаторе;
в вертикально инкубируемых яйцах в инкубаторах с искусствен-
ной вентиляцией больше 50% имеют незамкнутый в остром конце
яйца аллантоис. Используя прижизненную окраску белка,
автор подтвердил, что в яйцах с незамкнутым аллантоисом за-
держивается использование белка эмбрионами, они отстают
в росте и имеют высокую смертность в конце инкубации. Автору
не удалось сократить процент яиц с незамкнутым аллантоисом
ни при изменении температуры, ни влажности, ни вентиляции,
и только изменение положения яиц с вертикального на горизон-
тальное уменьшило процент яиц с незамкнутым аллантоисом
с 50 до 3—5%. Однако продолжение инкубации яиц после 15-го
дня в горизонтальном положении приводило к большему числу
наклевов скорлупы в остром конце (22—24%) по сравнению
с вертикальным положением (1—2%). Инкубирование утиных
яиц до 15-го дня в горизонтальном, а затем до конца инкубации —
в вертикальном положении дало повышение выводимости утят
по сравнению с вертикальным в течение всей инкубации на 5—10%
и по сравнению с горизонтальным — на 2%. Сходные результаты
получили Манш и Розиану (Manch et Ro?ianu, 1961). Меньшая
выводимость получена при вертикальном положении яиц: 65.8%
утят и 45.5% гусят; более высокая —при горизонтальном: 69.2%
и 62.5% соответственно. Но наилучшие результаты (72.2% утят)
были получены при инкубировании утиных яиц первые 15 дней
в горизонтальном положении, затем 10 дней в вертикальном
и последние дни — снова в горизонтальном. Вертикальное поло-
жение гусиных яиц при инкубации, так же как и утиных, нару-
шает нормальное развитие аллантоиса, и он не замыкается вокруг
находящегося в остром конце яйца белка (Бражникова, 1952, цит.
по: Владимирова, 1959). Однако, по данным автора, вертикальное
положение яиц после замыкания аллантоиса (с 15-го дня инку-
бации) уже не снижает выводимости гусят. Пытаясь расширить
время инкубирования гусиных яиц в вертикальном положении
(что более экономично), Владимирова (1959) выяснила, что инку-
бация гусиных яиц в вертикальном положении с 1-го до 7-го дня
не влияет на нормальное развитие аллантоиса и не снижает выво-
димости. Автор рекомендует инкубировать гусиные яйца с 1-го
до 7-го и с 15-го дня до конца инкубации в вертикальном поло-
жении, а с 8-го по 14-й день — в горизонтальном.
Положение яиц во время инкубирования имеет, кроме того,
косвенное значение для теплообмена эмбриона. Хаскин (19616)
подсчитал, что в зависимости от положения яиц в лотках (гори-
зонтальное или вертикальное) существенно изменяется площадь
свободной поверхности теплоизлучения, а именно при горизон-
тальном 72% поверхности яйца может излучать тепло в окружаю-
щий воздух инкубатора, а при вертикальном — только 48%.
344
Следовательно, это должно быть учтено при расчетах теплообмена
яиц, особенно в конце инкубации. Возможно, это является одной
из основных причин, почему эмпирически подобранное горизон-
тальное положение яиц во время вылупления (в выводных лотках)
оказывается наилучшим.
Б. Поворачивание яиц
Поворачивание яиц необходимо по нескольким причи-
нам. Во-первых, в связи с меньшим удельным весом желтка он
всплывает наверх при любом положении яйца, причем более легкая
его часть, где расположен бластодиск, всегда оказывается сверху.
Поворачивание яиц предотвращает присыхание зародышевого
диска на ранних стадиях развития, а потом и самого эмбриона
к подскорлупным оболочкам; в дальнейшем поворачивание яиц
предотвращает прилипание временных эмбриональных органов
одного к другому и создает возможность нормального их развития.
Во-вторых, поворачивание яиц необходимо для нормального
функционирования амниона, так как для его сокращений необхо-
димо некоторое свободное пространство. В-третьих, поворачивание
яиц уменьшает количество неправильных положений эмбрионов
к концу инкубации, и, в-четвертых, в секционных инкубаторах
поворачивание яиц необходимо, кроме того, для попеременного
нагревания всех частей яйца. В шкафных инкубаторах также
нет полной равномерности в распределении температуры (см.
главу 18), а потому и здесь поворачивание яиц обеспечивает
уравнивание количества тепла, получаемого разными частями
яйца.
О том, как следует поворачивать яйца, имеется ряд данных.
Функ и Форвард (Funk a. Forward., 1952) сравнивали выводимость
цыплят при поворачивании яиц в одной (как обычно), в двух
и в трех плоскостях и обнаружили в последних двух вариантах
повышение выводимости на 3.7 и 6.4% соответственно. В дальней-
шем авторы (1960) выяснили на болеее чем 12 000 куриных яиц,
что при вертикальном положении их в инкубаторе поворот яиц
на 45° в каждую сторону от вертикали по сравнению с 30°-м по-
воротом дает повышение выводимости цыплят с 73.4 до 76.7%.
Однако дальнейшее увеличение угла поворота яиц не повышает
выводимости.
По данным Калтофена (Kaltofen, 1961), только при изменении
поворота яиц вокруг длинной оси (при горизонтальном положе-
нии яиц) с 90° до 120° выводимость цыплят почти одинакова (86.2
и 85.7% соответственно), а при повороте яиц вокруг короткой
оси (вертикальное положение) преимущество поворота яиц на
120° более заметно — 83.7% цыплят по сравнению с 81.7% при
повороте на 90°. Автор сравнивал также поворачивание яиц
346
вокруг Длинной и вокруг короткой оси и нашел достоверное пре-
вышение выводимости цыплят (Р < 0.001) на 4.5% из яиц, по-
ворачиваемых вокруг длинной оси.
Данные о количестве перемещений яиц в гнезде, в которые
включены и их повороты, приведены нами в главе 18. Напомним
о наших наблюдениях (Рольник, 1939) над поворачиванием яиц
в гнезде страуса нанду. Все яйца были повернуты вокруг своей
короткой оси на 180° по крайней мере за 4—5 час., но, возможно,
эти данные несколько приуменьшены, так как наблюдения велись
1 раз в 1.5 часа.
Почти все исследователи приходят к выводу, что более частое
поворачивание яиц повышает выводимость. Совсем не повора-
чивая яйца, Эйклешимер (Eycleshymer, 1907) получил только 15%
цыплят; при 2 поворотах яиц в сутки — 45.4%, а при 5 поворо-
тах — 58% от оплодотворенных яиц. Прицкер (Прицкер и Треть-
яков, 1937) сообщает, что при 4—6-кратном поворачивании яиц
в сутки выводимость цыплят была выше, чем при 2-кратном.
Выводимость была одинаковой независимо от того, начинались ли
повороты яиц сразу или через 1—3 дня после закладки яиц в инку-
батор. Однако автор рекомендует поворачивать яйца 8—12 раз
в сутки и начинать повороты сразу после закладки яиц в инкуба-
тор. Инско (Insko, 1949) указывает, что увеличение количества
поворотов яиц до 8 раз в сутки повышает выводимость цыплят,
но 5 поворотов яиц совершенно необходимы. В опытах Куипера
и Уббельса (Kuiper a. Ubbels, 1951) 24-кратное поворачивание
яиц в сутки по сравнению с 3-кратным повысило выводимость
на 6.4% при сравительно высоком проценте вывода цыплят в кон-
троле — 70.3% от заложенных яиц. Сходные опцты на большом
материале (более 17 000 яиц) в инкубаторе шкафного типа провел
Шуберт (Schuberth, 1959). По сравнению с 3-кратным поворачи-
ванием в сутки, давшим 70.2—77:5% цыплят от оплодотворенных
яиц, автор получил при 5-кратном поворачивании повышение
выводимости на 2.0%, при 8-кратном — на 3.8—6.9%, при 11-
кратном — на 6.4%, при 12-кратном — на 5.6%. По данным
Калтофена (Kaltofen, 1961), поворачивание яиц 24 раза в сутки
по 18-й день инкубации по сравнению с 3-разовым обусловило
повышение выводимости цыплят в среднем на 7%, а по сравнению
с 8-разовым — на 3%. Данные Робертсона (Robertson, 1961а)
о влиянии количества поворотов яиц от 0 до 192 раз в сутки пере-
считаны нами и приведены в табл. 51. В связи с наибольшим
повышением выводимости по сравнению с контролем (24 поворота
яиц в сутки) при 96-кратном поворачивании яиц автор считает
необходимым именно это количество поворотов.
Вермесану (Vermesanu, 1961) оказался единственным иссле-
дователем, получившим противоположные результаты. Он на-
блюдал даже небольшое снижение выводимости цыплят (с 93.5%
до 91.5% от оплодотворенных яиц) при 3-кратном поворачивании
346
Таблица 51
Влияние количества поворачиваний яиц на выводимость цыплят
(По Robertson, 1961а)
№ серии опытов Количество поворотов яиц в сутки в данной серии Процент выведенных цыплят Разница между % цыплят в опытной серии и контроле
в контроле в опытной серии
1 0 90.3 13.8 —76.5
2 0.5 81.4 62.8 -18.6
3 2 87.6 77.1 —10.5
4 6 83.3 77.7 - 5.6
5 12 85.1 81.9 - 3.2
6 48 86.1 90.4 + 4.3
7 96 76.0 83.5 + 7.5
8 144 79.8 84.5 + 4.7
9 192 82.2 83.4 + 1.2
Примечание. В контроле всегда 24 поворота яиц в сутки.
яиц в течение всего периода инкубации по сравнению с 2-кратным
до 8-го дня и 1-кратным с 9-го дня до вылупления. По-видимому,
это результат какой-то ошибки.
Влияние различного количества поворачиваний утиных и
гусиных яиц на выводимость исследовали Манш и Розиану (Manch
et Ro^ianu, 1961). Авторы получили при 4-, 5- и 6-кратном пово-
рачивании 65.8, 71.6 и 76.6% утят и 55.2, 62.4 и 77.0% гусят соот-
ветственно. Следовательно, по мнению авторов, необходимо
поворачивать утиные и гусиные яйца по крайней мере 6 раз
в сутки. Ковинько и Бакаев (1965) на основании наблюдений над
количеством поворотов яиц в гнезде утки за 25 дней насиживания
(528 раз за 600 час.) и сравнения эффекта 24-кратного поворачи-
вания яиц в инкубаторе в сутки с 12-кратным — контрольным
(68.7% и 55.3% утят от оплодотворенных яиц соответственно)
пришли к выводу, что часовой интервал между поворотами яиц
более полно отвечает биологическим потребностям эмбрионального
развития утят, чем 2-часовой, особенно в период развития аллан-
тоиса, и в последующем способствует повышению жизненности
молодняка.
Особняком стоит вопрос о необходимости дополнительного
ручного поворота гусиных яиц на 180° при горизонтальном поло-
жении в лотках, где куриные яйца обычно расположены верти-
кально. Быховец (19596) отмечает, что дополнительное повора-
чивание гусиных яиц на 180° вручную 1—2 раза в сутки повышает
выводимость гусят на 5—10%. Однако следует заметить, что при-
водимое автором объяснение этого особенностями гусиного яйца
347
(большее соотношение длины к ширине и большее количество
жира в желтке, чем в курином яйце) здесь ни при чем. Причиной
сниженной выводимости гусят в данном случае (при наличии
только механического поворота яиц), по нашему мнению, является
то, что в лотках, приспособленных для инкубирования куриных
яиц в вертикальном положении, поворот лотков на 90° означает
поочередное всплывание желтка и бластодиска в курином яйце
то к одной стороне яйца, то к другой; при горизонтальном же поло-
жении гусиных яиц в этих же лотках поворот последних значи-
тельно меньше изменяет расположение бластодиска. По данным
Рууса (1961), при проведении дополнительного поворачивания
гусиных яиц на 180° вручную 1 раз в сутки, кроме механического
3-кратного, выводимость гусят повышается с 55.6—57.4% до 79.3—
92.4%. Однако некоторые производственники (Волкова и Волков,
1959; Соболева и Турович, 1959) сообщают, что дополнительное
поворачивание гусиных яиц вручную не повышает выводимости
гусят.
Вопросу о периодах эмбрионального развития, когда повора-
чивание яиц особенно необходимо, посвящен ряд исследований.
Вейнмиллер (Weinmiller, 1930) на основании проведенных им
опытов считает необходимым 12-кратное поворачивание куриных
яиц в сутки в течение первой недели, а во вторую и третью не-
дели — только 2—3-кратное. По данным Котлярова (1937), рас-
пределение смертности эмбрионов было разным при 24-, 8- и 2-
кратном повороте яиц: процент эмбрионов, погибших до 6-го дня,
был примерно одинаковым при 2- и 8-кратном, а процент задохликов
сокращался вдвое при 8-кратном, и наоборот, при увеличении
количества поворотов яиц до 24 раз в сутки процент задохликов
оставался одинаковым, а процент погибших до 6-го дня увеличи-
вался втрое. Этому факту автор не придает значения, но нам он
кажется весьма показательным. В начале развития эмбрионы
чрезвычайно чувствительны к сотрясениям (см. дальше) и потому
слишком частое поворачивание яиц губительно действует на наи-
более слабых эмбрионов. В конце развития поворачивание яиц
в секционных инкубаторах улучшает газообмен и облегчает
теплоотдачу, что и обусловливает значительное снижение про-
цента задохликов при 8-кратном повороте яиц. Но еще большее
учащение поворотов, возможно, уже ничего не может дополнить
в улучшении газообмена и теплоотдачи. Наше мнение подтверж-
дено опытами автора: более редкие повороты яиц в первой поло-
вине инкубации и более частые — во второй дали повышение
выводимости по сравнению с группой 8-кратного поворота яиц
в течение всей инкубации на 2.3%. Куо (Кио, 1932) считает, что
невозможность пройти ту или иную стадию обусловлена в боль-
шинстве случае механическими причинами и с 11-го до 14-го дня
развития именно поворачивание яиц, стимулируя сокращения
эмбриона, помогает ему пройти стадию, предшествующую стадии
348
поворота тела. По данным Робертсона (Robertson, 1961а), в группе
с 2-кратным поворотом и особенно в группе без поворачивания
яиц по сравнению с контрольной (24-кратный поворот) смертность
куриных эмбрионов увеличивается больше всего в первые 10 дней
инкубации, а при 6-, 12-, 24-, 48- и 96-кратном повороте в сутки,
смертность эмбрионов в это время примерно одинакова с кон-
трольной. С увеличением числа поворотов яиц, так же как и в опы-
тах Котлярова, процент задохликов сильно уменьшается, особенно
задохликов без видимых морфологических нарушений. Калтофен
(Kaltofen, 1955) на большом материале (60 000 куриных яиц) отме-
тил, что 24-кратное поворачивание яиц снижает смертность эмбрио-
нов особенно во 2-ю неделю инкубации. Автор провел опыты
с 24-кратным поворотом только в течение этого срока (в остальные
дни 4-кратное) и выяснил, что выводимость цыплят в этой группе
была одинаковой с группой 24-кратного поворота с 1-го по 18-й
день инкубации. В дальнейшем автор (1961) показал, что гибель
эмбрионов после 16-го дня, т. е. во второй период повышенной
смертности эмбрионов, зависит более всего от недостаточной
частоты поворотов яиц до 10-го дня инкубации, так как при этом
не происходит нормального обрастания амниона аллантоисом
и амнион соприкасается с подскорлупной оболочкой, что предот-
вращает поступление белка в амнион через серозо-амниотический
канал (см. главу 7). Несколько иные результаты получил Нью
(New, 1957), выяснивший, что поворачивание яиц только с 4-го
по 7-й день обусловливает примерно такую же выводимость,
как и поворачивание в течение всего периода инкубации. Пово-
рачивание же только с 8-го по 11-й день не повышает выводимо-
сти по сравнению с группой, где яйца совсем не поворачивались.
Автор наблюдал, что неповорачивание яиц с 4-го по 7-й день
инкубации вызывает преждевременное примыкание аллантоиса
к подскорлупной оболочке, обусловливающее быструю потерю
воды из белка. Поэтому автор считает особенно необходимым
поворот яиц с 4-го по 7-й день инкубации.
Рэндле и Романов (Randles a. Romanoff, 1950) выяснили,
что недостаточное поворачивание яиц, предотвращающее или
задерживающее поступление белка в амниотическую полость,
в результате чего часть белка остается в яйце после вылупления
цыпленка, а эмбрион недополучает значительное количество пита-
тельных веществ, ведет к уменьшению веса цыпленка.
В. Сотрясение яиц
Опыты Стайлса и Уотерсона (Stiles a. Watterson, 1937) по
сотрясению куриных яиц проводились с 4-го до 12-го часа инку-
бации. Встряхивание яиц проводилось в течение 1 мин. с интерва-
лами в 15 мин. Процент гибели эмбрионов от сотрясения оказался
очень большим: из 155 яиц вылупился только 1 цыпленок. Около
349
60% эмбрионов погибли между 2-м и 3-м днем, другие прекратили
развитие до 11-го дня инкубации. Вскрытие эмбрионов показало,
что гибель обусловливалась отсутствием экстенсивного распро-
странения желточных артерий и вен и развитие не шло дальше
стадии кровяных островков. В некоторых случаях желточное
крообращение не было полностью нарушено и эмбрионы про-
должали существовать еще некоторое время. Сотрясение влияло
и на развитие нервной системы: обычно нервная трубка не замы-
калась и оставалась целиком или частично в виде нервной пла-
стинки.
Для инкубации гагачьих яиц нам (Рольник, 1955) пришлось
перевозить их с других островов в неприспособленной для этой
цели таре (опыты первого года работы). Из перевезенных насижен-
ных Лиц с разной степенью развития эмбрионов в 31 эмбрионы
погибли до закладки, а из оставшихся 26 вывелся только один
гагачонок. Насиженные не перевозившиеся яйца дали 82.8% га-
гачат. Для ненасиженных яиц перевозка оказалась менее вредной
(выводимость 58% по сравнению с неперевозившимися 78%).
Нами было отмечено также, что развитие эмбрионов в инкубаторе
после перевозки продолжалось только в случае, если стадия раз-
вития к моменту перевозки была не больше 4 дней. В следующем
году перевозка проводилась в специальных ящиках на подвесных
пружинах и отличия в выводимости из насиженных и ненасижен-
ных яиц почти не было.
Влияние сотрясения яиц на более поздних стадиях развития
эмбрионов (на 4—15-й дни инкубации) исследовали Олсен и Баерли
(Olspn a. Byerly, 1938), показав, что наибольшая смертность про-
исходит в течение 72 час. после сотрясения и вызывается она про-
рывом желточного мешка и кровоизлияниями. Сотрясение про-
водилось ударом яиц о ладонь, сотрясением земли или воздуха
от взрыва или центрифугированием. Менее вредными оказались
тряска параллельно короткой оси яйца и центрифугирование тупым
концом вверх. В связи с этим авторы опасаются сотрясения яиц,
почти неминуемого при овоскопировании, и рекомендуют овоско-
пировать яйца не ранее 15-го дня инкубации.
При сотрясении яиц до инкубации Зек (Seek, 1939) не наблю-
дал снижения выводимости цыплят, если не была нарушена жел-
точная оболочка, а Ландауэр и Бауман (Landauer a. Baumann,
1943), наоборот, отметили снижение выводимости с 67% до 30%
а также частые случаи уродства цыплят. Дело, по-видимому,
в том, что величину сотрясения очень трудно измерить, и она была
в опытах различных авторов разной.
Несколько иначе был поставлен вопрос Рэндлсом и Романовым
(Randles a. Romanoff, 1954). Авторы изучали возможность замены
одного механического воздействия на инкубируемые яйца (систе-
матического поворачивания их на 45° в каждую сторону от верти-
кали) другим — сотрясением. Испробовав целый ряд темпов со-
360
трясения (от 115 до 431 в i мин.), продолжительности (от 0.5 сек.
до 1 мин.) и частоты (1 раз в сутки, каждый час или каждые 2часа),
авторы получили в среднем 58.7% цыплят из сотрясенных яиц
(в отдельных группах 79.0—82.0%) по сравнению с 33% цыплят
от оплодотворенных яиц в контроле (яйца не поворачивались).
Таким образом, по мнению авторов, сотрясение яиц в какой-то
мере может служить механическим стимулом для развития, кото-
рым в обычной инкубации является поворачивание яиц. Од-
нако это теоретически интересное, с нашей точки зрения, наблю-
дение ни в какой мере не может быть применено на практике,
хотя и дает некоторый выигрыш места в инкубаторе, а следова-
тельно, и небольшую экономию.
Г Л А В А 22
БИОЛОГИЧЕСКИЙ КОНТРОЛЬ ИНКУБАЦИИ
И ДРУГИЕ МЕТОДЫ ПОВЫШЕНИЯ
ВЫВОДИМОСТИ МОЛОДНЯКА
А. Биологический контроль инкубации
Практиками инкубации уже давно было замечено,
что в инкубаторах разных систем при одинаковых показаниях
приборов о температуре, влажности и вентиляции могут быть
разные результаты — различная выводимость. Это обусловлено
тем, что показания приборов не могут полностью отразить ре-
альный обогрев яиц, испарение воды из них и возможность газо-
обмена, а на них влияют кубатура инкубатора, место и устрой-
ство обогревательной системы, скорость и направление движения
воздуха, а также кратность его обмена с наружной атмосферой,
порядок размещения яиц и др. Кроме того, для получения наи-
лучшей выводимости и жизнеспособностй молодняка из яиц раз-
ного качества, в разные сезоны года, разных пород и т. д. необ-
ходим различный режим инкубации. По данным Романова (Ro-
manoff, 1938), для разных видов птиц особенно критичными
являются различные факторы среды; так, например, наиболее важ-
ным условием среды для инкубации индюшиных яиц является
температура, для утиных — влажность, для фазаньих — движе-
ние воздуха. Важно отметить, что более высокая выводимость
тесно коррелирует с лучшей выживаемостью молодняка в постэм-
бриональный период. Коэффициент корреляции между этими
величинами равен 0.67 (Лепаж, 1961).
Наиболее верные критерии степени оптимальности режима
инкубации дает система наблюдений за эмбриональным разви-
тием птиц, называющаяся биологическим контролем инкубации.
361
Мы не имеем возможности подробно осветить практические
мероприятия, связанные с проведением биологического контроля
инкубации, и отошлем читателя к основным руководствам по этому
вопросу (Прицкер, 1933, 1942; Орлов и др., 1936; Попов и др.,
1936; Орлов, 1966; Прицкер и Третьяков, 1937; Отрыганьев, 1938,
1951,1962; Пенионжкевич, 1945; Landauer, 1948; Отрыганьев и др.,
1964). В связи с тем что одной из основных задач нашей монографии
является по возможности наиболее полное освещение всех сторон
биологии развития птичьего эмбриона, мы надеемся, что материал,
собранный в данной книге, поможет осуществлению биологиче-
ского контроля инкубации на научной основе.
Критериями биологического контроля инкубации яиц яв-
ляются: 1) выводимость молодняка и распределение смерт-
ности эмбрионов в течение инкубации; 2) патологоанатомиче-
ские изменения погибших эмбрионов; 3) снижение веса ин-
кубируемыми яйцами в течение инкубации (усушка яиц);
4) развитие эмбрионов, определяемое при а) овоскопировании —
прижизненная оценка и б) вскрытии нескольких эмбрионов из
партии; 5) длительность инкубации и течение проклева яиц и
вылупления молодняка; 6) качество выведенного молодняка;
7) степень обескровливания сосудов аллантоиса, выстилающего
внутреннюю поверхность, скорлупок, после выхода из них
птенцов.
При естественной инкубации выводится, как правило, почти
100% птенцов из числа оплодотворенных яиц. Понятно, что сни-
жение выводимости при искусственной инкубации заставляет
нас искать причины, обусловливающие это снижение. Разные
исследователи придают наибольшее значение тому или иному
критерию или группе их в определении этих причин. По нашему
глубокому убеждению, только учет всего комплекса этих критериев
может дать ответ на этот вопрос, а также помочь в исправлении
отмеченных недочетов. Именно анализ данных по всем критериям
при инкубировании яиц гаги, проведенный нами (Рольник, 1941,
1955) впервые в 1939 г., помог нам в следующий сезон инку-
бации избежать всех неблагоприятных условий и получить 95%
гагачат.
Целый ряд данных о той или иной стороне развития птичьего
эмбриона, характеризующей развитие эмбриона в целом, и таким
образом являющейся критерием биологического контроля инку-
бации, уже приводились нами. Так, об изменении кривых смерт-
ности эмбрионов в течение инкубации под влиянием различных
факторов мы уже сообщали в главах 17, 18, 20, и др.; вопрос
о неправильных положениях эмбрионов при вылуплении как
причине их гибели уже рассматривался нами в главах 17 и 21,
а данные о потере веса яиц во время инкубации и связи ее с выво-
димостью молодняка, мы подробно рассматривали в главе 19.
Здесь мы остановимся на неупоминавшихся нами ранее данных
362
по этим же вопросам, а в основном — на сведениях по остальным
критериям биологического контроля.
На патологоанатомические изменения
погибших эмбрионов как методе определения нарушений тех или
иных условий инкубации одним из первых в нашей стране указал
Лисицкий (1937). Отрыганьев (1951) вполне правильно, на наш
взгляд, считает патологоанатомический контроль одним из основ-
ных критериев биологического контроля инкубации.
Данные о патологоанатомических изменениях погибших эм-
брионов, наиболее полно изученных в группе работ Отрыганьева
(1935,1937,1938,1939,1951,1962, и др.), а также в работах других
авторов (Пенионжкевич, 1935; Кирсанов, 1938; Щеглов, 1954,
и др.) мы приводим в Приложении, I.
Здесь мы остановимся только на трех исследованиях. Маршал
(Marshall, 1951) указывает, что ненормальности сердца и круп-
ных кровеносных сосудов наблюдались у 30% задохликов при
высокой выводимости цыплят (86% от оплодотворенных яиц) и
это не было связано с ненормальным положением эмбриона. В де-
тальном изучении куриных эмбрионов, погибших во 2-й период
повышенной смертности (задохлики), Щеглов (1954) предлагает
практически очень удобное, на наш взгляд, деление их на 4 группы
по величине использования белка и желтка яйца. В 1-ю группу
автор относит эмбрионов с неиспользованным белком, во 2-ю —
с большим, чаще всего невтянутым желтком весом 13—14 г
(34—38% общего веса эмбриона вместе с желточным мешком).
Автор показал, что эмбрионы этих двух групп имеют признаки
давней (почти с начала развития) хронической болезни, а именно —
недостаточный вес и общее недоразвитие, а также застойные
явления в печени и сердце. Причиной гибели этих эмбрионов яв-
ляется, по-видимому, вредное воздействие внешних факторов
в начале инкубации. В 3-й группе эмбрионов остаточный желток
чуть больше нормального, и желточный мешок часто втянут
в полость тела. У эмбрионов этой группы не было обнаружено
каких-либо патоморфологических изменений, кроме нарушения
кровонаполнения внутренних органов. Сухой вес этих эмбрионов
также приближается к нормальному, что указывает на гибель
их от причин, действовавших только в последние дни эмбриональ-
ного периода. Автор относит этих эмбрионов к истинным «задох-
ликам», т. е. погибшим в результате асфиксии в один из последних
дней инкубации. К 4-й группе автор относит эмбрионов с излишне
израсходованным желтком, что в сочетании с инфильтративно-
дегенеративным ожирением печени указывает на нарушение жи-
рового обмена эмбрионов как на причину их гибели. По данным
Робертсона (Robertson, 1961с), изучавшего при помощи рентгена
положение эмбриона в яйце, неправильные положения, наблю-
даемые зачастую у задохликов, не являлись причиной их гибели,
а были нормальными для переходной стадии, предшествующей
х/а 23 в. В. Рольник 363
вылуплению. Автор наблюдал, что за 24—36 час. до вылупления
происходит переориентировка эмбриона, после чего голова пово-
рачивается вправо, клюв прободает подскорлупную оболочку
под воздушной камерой и начинаются разгибательные дви-
жения шеи. И только в случае, если после переориентировки
голова эмбриона находится в остром конце яйца или между ла-
пами, наступает гибель большинства эмбрионов — вылупляется
только 20% цыплят.
В связи с тем что при одинаковой выводимости в разных ин-
кубаторах усушка яиц различна, Прицкер (1942) пред-
лагает пользоваться этим критерием только при проведении
биологического контроля в инкубаторах аналогичных систем.
Орлов (1966) рекомендует следующие нормы усушки: яйца всех
видов сельскохозяйственных птиц нормально теряют 11—13%
своего первоначального веса, но интенсивность усушки должна
быть разной в разные периоды инкубации; у куриных яиц с 1-го
по 6-й день не более 0.5—0.6%, а с 11-го — от 0.7—0.8% до 1%
в сутки; у утиных — с 1-го по 7-й день не более 0.4—0.5%,
а с 11-го больше 0.5%; у гусиных — с 1-го по 8-й день не более
0.3—0.4%, а с 11-го — больше 0.4%.
Метод прижизненной оценки развития эмб-
рионов, в особенности в ранние сроки инкубации, является чрез-
вычайно полезным для практики. Пельтцер (1959) предложил
проводить прижизненную оценку интенсивности развития ку-
риных эмбрионов в 4 срока: 1) через 12—15 час. инкубации —
по развитию зародышевого диска; 2) на 4-й день — по развитию
сосудистого поля желточного мешка; 3) на 9-й день — по вели-
чине глазного яблока; 4) на 20-й день — по размерам воздушной
камеры и исчезновению жидкости из аллантоиса. Во все эти
сроки проводилось деление яиц на 2 группы: А — с хорошим
и Б — со слабым развитием эмбрионов. В опытах автора при
1-й оценке в группе А вывелось 93.7% цыплят, а в Б — 86.5%;
при 2-й — 95.0 и 82.3%, при 3-й — 95.6 и 83.7% и при 4-й —
99.0 и 87.1% соответственно. Качество цыплят в группе А было
выше, чем в группе Б.
Интересный метод отбора яиц для дальнейшей инкубации
предложили Олсен и Марсден (Olsen a. Marsden, 1954). Овоско-
пируя индюшиные яйца через 24, 32, 48 и 72 часа инкубации,
авторы заметили, что из тех яиц, где развитие могло быть обна-
ружено при овоскопировании через 24 часа, вывелось 79.4%
индюшат из числа оплодотворенных яиц, через 32 часа — 33.9%,
через ,48 час. — 27.7%, через 72 часа — 8.1%; в тех же случаях,
когда и через 72 часа в яйцах не были обнаружены эмбрионы,
то если развитие и начиналось после этого, оно прекращалось
на ранней стадии. Естественно, что после овоскопирования через 24
или 32 часа есть смысл отбраковывать яйца, в которых не на-
чалось развитие Хорошая сводка по результатам прижизненной
364
оценки развития эмбрионов дана Орловым (1966). В опытах
автора получены следующие результаты. При овоскопировании
через 12 час. инкубации куриных яиц и делении их на 2 группы
по степени развития бластодиска из яиц 1-й группы (с хорошим
развитием) было получено 84.9% цыплят, а из яиц 2-й — почти
вдвое меньше (46.4%). При определении же интенсивности раз-
вития по времени обнаружения бластодиска было получено
из яиц, где бластодиск стал виден через 8 час., — 81.0% цыплят,
через 14 час. — 74.9%, через 20 час. — 20.2%, через 24 часа —
34.1%, после 24 час. —28.9%. Автор отмечает далее, что чем
раньше стал виден бластодиск, тем интенсивнее шло дальнейшее
развитие и раньше происходило вылупление; по сравнению с теми,
где бластодиск стал виден через 8 час., вылупление у тех, где
бластодиск был обнаружен через 14 час., опаздывало в среднем
на 58 мин., через 20 час. — на 2 ч. 24 м., а через 24 часа — на 6 час.
По данным автора, интенсивность развития наследуется, и он
приходит к выводу, что после овоскопирования яиц через 20 час.
инкубации следует отбраковывать те яйца, в которых не началось
развитие. Автор рекомендует овоскопированием через 20 час.
заменить просмотр яиц до инкубации и через 6—8 суток инку-
бирования. Автор справедливо отмечает преимущество метода
ранней прижизненной оценки развития эмбрионов по сравнению
с другими методами биологического контроля, так как этот
метод дает возможность значительно раньше принять меры
к исправлению недочетов в режиме инкубации или в качестве
яиц. Однако применение этого метода ограничивается отсутст-
вием специальных овоскопов с яркой лампой и вентилятором.
Одним из важных методов прижизненной оценки развития
эмбрионов является овоскопирование яиц на 11—15-й день (для
разных видов птиц) по признаку, замкнулся ли полностью ал-
лантоис к этому сроку или нет, что в значительной степени опре-
деляет успешность дальнейшего развития (см. главу «Временные
эмбриональные органы»). Значительное повышение выводимости
получают, если при овоскопировании яиц перед переводом их в вы-
водные лотки (19-й день для куриных яиц и 25-й — для утиных)
проводят разделение их на 2 группы: 1-я — с выгнутой в воз-
душную камеру шеей эмбриона и 2-я — с ровной воздушной ка-
мерой. Яйца 1-й группы переносят в выводные лотки, где пре-
кращают их переворачивание и повышают влажность, а яйца 2-й
оставляют в инкубационных лотках еще на сутки и только затем
переводят их на вывод (Третьяков, 1938; Gartley et al., 1951).
Для более точной характеристики течения развития реко-
мендуется, вскрывая по 5—10 яиц из партии 2—3 раза в течение
инкубационного срока, определять стадию развития эмбриона
(см. Приложение, II) и соответствие или отклонение его от сроков
инкубирования яиц, а также наличие или отсутствие тех или
иных нарушений развития. Приведенные в Приложении II дан-
23*
Збб
ные имеют большое значение в научной работе для точной да-
тировки фактического возраста (стадии развития) опытного ма-
териала, который иногда отличается от срока, прошедшего с мо-
мента закладки яиц в инкубатор.
Ценным критерием оценки режима инкубации являются
также данные о продолжительности инкубации,
течении проклева яиц и вылупления мо-
лодняка в партии. Приведем данные о нормальных сроках
продолжительности инкубации (табл. 52).
Таблица 52
Продолжительность инкубации яиц (в днях) разных видов
сельскохозяйственных птиц
(По Орлову, 1966)
Вид птиц Начало вывода (появление первых птенцов) Массовый вывод (вывелось 70—80% птенцов) Конец вывода (из ин- кубатора вынимают последних здоровых птенцов, не нужда- ющихся в помощи)
Куры Конец 20-го Первая половина 21-го Конец 21-го ....
Утки и индейки 26-й 27-й Конец 27-го — на- чало 28-го
Гу<и 29-й 30-й Начало 31-го
Прицкер и Третьяков (1937) наблюдали, что при оптимальной
инкубационной температуре проклев яиц и вывод цыплят партии
протекает в наиболее короткий срок (проклев длится 48 час.,
вывод — 42 часа); при повышении температуры до 40° (в секцион-
ном инкубаторе) — 7Q и 66 час., а при понижении до 37° — 60
и 60 час. соответственно. В опытах авторов при температуре 40°
проклев начинался на 24, а вывод — на 12 час. раньше и за-
канчивался одновременно с контрольной партией, а при низкой —
и проклев и вывод начинались на 36 час. и заканчивались на 48
и 54 часа позже контрольной соответственно. Процесс вылупле-
ния (от проклева яйца до выхода цыпленка из скорлупы) длится
от 3 до 27 час. у цыплят, развивавшихся при оптимальной тем-
пературе, наименьшая средняя длительность этого процесса
(9.8 часа), при высокой температуре — больше (12.2—14.3 часа),
и самая большая длительность у развивавшихся при низкой
температуре (14.5—16.4 часа). Хэйз (Hays, 1941) установил,
что с увеличением длительности инкубации: а) начинают пре-
обладать петушки (в начале срока вывода цыплят вылупляются
преимущественно курочки), б) повышается смертность цыплят
за 6 месяцев, в) удлиняется время созревания молодок и срок
до снесения 1-го яйца, г) несколько снижается яйценоскость.
366
На основании этих показателей автор считает экономически
выгодным отбирать для племенных целей ранее вылупившихся
цыплят (до 2—3-й трети 21-го дня инкубации). По данным Куч-
ковской (1954), петушки, выведенные в 1-ю половину 21-го дня
инкубации, обладают большей жизнеспособностью, чем выве-
денные во 2-й половине того же дня, а спаренные с ними куры-
молодки несут яйца с более высокой оплодотворенностыо и
выводоспособностыо. В наших опытах «кислородизация» яиц (Роль-
ник, 1946) повышение выводимости цыплят, утят, индюшат и луч-
шая их жизнеспособность в постэмбриональном периоде кор-
релировались с уменьшением длительности всей инкубации,
и особенно процесса вылупления. Длительность инкубации ока-
залась наследуемой чертой (Smith a. Howes, 1949). Из яиц тех
кур, которые вылупились в первые 24 часа всего срока вылуп-
ления партии, за этот же срок вылупилось 86.1% цыплят, а из яиц
кур, вылупившихся позднее, вывелось в первые сутки вылуп-
ления партии только 11.8%. По наблюдениям Аббот и Крэйг
(Abbott a. Craig, 1960), длительность вылупления индюшат,
фазанят и перепелят тесно связана с общей длительностью ин-
кубационного периода; у вида с наибольшей длительностью
инкубации — индейки (654.8 часа) больше всего и длительность
вылупления (в среднем 28.2 часа), у фазанов — 581.6 и 18.7 часа,
а у перепелов — 393.6 и 10.1 часа соответственно.
Состояние выведшихся птенцов в суточном
возрасте хорошо характеризует и режим инкубирования, и ка-
чество яиц. По данным Прицкера и Третьякова (1937), при по-
ниженной температуре инкубации цыплята излишне распушены,
моторика их понижена, ноги у большинства слабы и широко
расставлены, ходят они рывками, иногда падая на грудь. При вы-
сокой температуре цыплята плохо распушены, малоподвижны,
быстро зябнут и сбиваются кучками, имеют пингвинообразно
приподнятый корпус, значительное количество их имеет плохо
вобранную пуповину. Однако последний признак нельзя считать
основанием для отбраковывания цыплят, как показала Отры-
ганьева (1966). Автор выращивала цыплят мясных и мясо-яичных
пород (корниш, Суссекс и плимутрок), имевших при вылуплении
подсохший аллантоисный стебелек в виде ниточки или даже
струпика диаметром до 1 мм, и установила, что вес их и сохранность
до 30-дневного возраста одинаковы с контрольными. Данные
о влиянии температуры инкубации на некоторые черты строения
цыплят представлены Прицкером (1958). Инкубирование при
высокой или низкой температуре изменяет форму черепа, содер-
жание воды, липидов и золы в тканях, количество гемоглобина,
форменных элементов крови и т. д. Автор отмечает, что цыплята,
инкубированные при пониженной температуре, и при выращи-
вании имеют лучшую устойчивость к низкой температуре. Ранее
автор (1942) заметил, что соответствующим образом подобранный
24 В- В. Рольник
367
режим инкубации может стать основой для акклиматизации птиц,
а также в создании птиц, измененных в определенном направле-
нии (например, повышение яйценоскости), и даже новых пород.
О степени обескровливания сосудов ал-
лантоиса, выстилающего внутреннюю поверхность скор-
лупок после выхода из них птенцов как критерии биологического
контроля, пишут только Прицкер и Третьяков (1937). Естественно,
что чем больше крови после вылупления птенца остается в со-
судах аллантоиса на скорлупе, тем более ослабленным выходит
птенец. Авторы предлагают разделить скорлупу после вылупле-
ния птенцов на 3 группы: 1) светлая, с единичными сосудами,
заполненными кровью, 2) светло-коричневая, с заполненными
кровью крупными сосудами и изредка с кровоизлияниями до 9 мм2,
и 3) коричневая, с большинством сосудов, заполненных кровью,
и с частыми и более крупными кровоизлияниями. При всех срав-
ниваемых температурных режимах имелись скорлупки всех
трех групп, но количественное соотношение их было разным:
при оптимальной инкубационной температуре наибольшее коли-
чество скорлупок относилось к 1-й группе, при низкой — ко 2-й
и при высокой — к 3-й. Отметим, что в нашей работе использо-
вание этого критерия в качестве дополнительной характеристики
режима инкубации приносило неизмененную пользу.
Все критерии биологического контроля применимы при ин-
кубировании яиц разных видов птиц; различаются только сроки
появления признаков. Поэтому, применяя методы биологического
контроля, удается разработать необходимый режим инкубации
для яиц тех видов птиц, которые раньше не инкубировались.
Так, Романов (Romanoff, 1934, 1938) разработал режим инку-
бирования фазаньих и перепелиных яиц, а мы (Рольник, 1941,
1955; Rolnik, 1943) — гагачьих яиц. Юрченко (1966), инкубируя
цесариные яйца при режиме, применяющемся для инкубации
куриных яиц, также проводила биологический контроль инку-
бации.
В дополнение к приемам, обычно применяемым в биологическом
контроле инкубации, Отрыганьев (1962) предлагает проводить
пространственный учет биологических данных, т. е. отдельно
по лоткам, расположенным в разных зонах шкафных инкубато-
ров, а также раздельно в пределах одного лотка (спереди, сзади,
сбоку и в середине его). Пространственный учет помогает ра-
зобраться в иногда противоречивых данных, получаемых при
применении других приемов биологического контроля, и облег-
чает выяснение вопроса о степени однотипности режима в разных
зонах инкубатора.
В качестве дополнительного метода определения развития
эмбрионов Пилипенко (1959) предлагает определять гемоглобин
и количество эритроцитов в крови. Автор приводит данные по этим
показателям при естественной и искусственной инкубации, дав-
368
шей хорошие результаты (87.4% цыплят, а под наседкой —
93.1%): на 18-й день инкубации у эмбрионов под наседкой было
54.7% гемоглобина и 2.075 млн/мм3 эритроцитов, а в инкубаторе —
54.1% и 2.054 млн/мм3 соответственно.
Б. Некоторые методы повышения выводимости
молодняка сельскохозяйственных птиц
Кроме уже упоминавшихся нами методов повышения
выводимости путем: а) улучшения качества яиц для инкубации
(см. раздел I), б) подогревания яиц во время хранения (см. главу 5),
в) определения и осуществления оптимальных условий инкубации,
где большую роль играет биологический контроль (см. главы 18—
21), г) предложенного нами метода кислородизации яиц (см.
главы 10 и 20), имеется еще много попыток улучшить результаты
инкубации яиц сельскохозяйственных птиц.
В группе работ (Попов и Цегельницкая, 1957; Егина, 1960;
Нуге, 1962) проводилась витаминизация яиц путем опускания
их в раствор витаминов и инъекции витаминов в яйца с последую-
щим повышением выводимости цыплят. Повышение выводимости
цыплят на 7.6% получил Крапчитов (1958) путем опускания яиц
в раствор йодистого калия. Варнагирис (1960) повышал выво-
димость утят и гусят, опуская яйца в растворы соляной кислоты,
поваренной соли и хлористого кобальта.
Больше всего исследований проведено по повышению выво-
димости молодняка путем ультрафиолетового облучения яиц.
Фронда и Белло (Fronda a. Bello, 1928, цит. по: Groebbels, 1937),
освещая солнцем инкубируемые яйца, получали повышение
выводимости цыплят, хотя, по некоторым другим исследованиям
(Hart et al., 1925; Boyd, 1928; Kucera, 1929, все цит. no: Groeb-
bels, 1937), облучение ультрафиолетовыми лучами не имеет ни-
какого влияния на выводимость. Однако более поздние иссле-
дования, хотя и различались по величине повышения выводи-
мости в результате облучения инкубируемых яиц кварцевой
лампой, всегда давали положительный эффект. Гудкин с соавто-
рами (1959), облучая кварцевой лампой ПРК-2 куриные яйца
с расстояния 80 см, не только получили повышение выводимости
цыплят на 10%, но и отметили лучшее развитие эмбрионов: за-
мыкание аллантоиса и вылупление цыплят происходило на сутки
раньше, длительность вылупления цыплят была меньше, вес
их был больше и жизнеспособность выше. В дальнейших опытах
авторы (1961) показали, что наилучший результат (81.1% цыплят
от оплодотворенных яиц при 67.9% в контроле) дает 3-кратное
облучение яиц по 2 мин., а именно до закладки, на 7-й и на 19-й
день инкубации. В опытах с утиными и гусиными яйцами наиболее
оптимальной дозой оказалось 2-кратное облучение (до закладки
и на 8-й день инкубации) по 1—2 мин. Выводимость утят повы-
24*
369
силась при этом до 84.3—86.8% при контроле 79.3%, а гусят —•
до 72.8—77.7% при контроле 63.4% от числа оплодотворенных
яиц. Кодинец (1959) исследовал действие такого же облучения,
но сроком от 1 до 20 мин. Автор ставил опыты в апреле, когда
несушки больше всего испытывали «ультрафиолетовое голодание».
При облучении от 1 до 9 мин. выводимость цыплят повышалась
с 72.6% в контроле до 76—79%; при увеличении срока облу-
чения до 15 мин. выводимость была одинаковой с контрольной,
а при дальнейшем удлинении срока облучения выводимость
резко падала до 36—48%. Далее автор (1962) показал, что ультра-
фиолетовое облучение яиц изменяет свойства белка куриных
яиц в благоприятную для развития эмбриона сторону: повы-
шает pH жидкого белка с 8.74 до 9.11, а густого — с 7.84 до
8.65. В производственных условиях Клюкина (1961) на 5000
куриных яиц показала, что облучение лампой ПРК-4 с рас-
стояния 50 см в течение 1 мин. повышало выводимость цып-
лят на 9%. Заболотников (1959) на 200 000 утиных яиц,
облучая лампой ПРК-7 с расстояния 1.5 м двукратно по 4 мин.
(до закладки и на 6-й день инкубации), получил повышение вы-
водимости утят на 6.4—8.2%. Христофоров (1961) на 45 000 ку-
риных и утиных яиц получил лучшие результаты при облуче-
нии (лампа ПРК-2) с расстояния 80 см по сравнению с 40 см и в те-
чение 1—2 мин. по сравнению с более длительным облучением:
выводимость цыплят повысилась на 12.4%, а утят — на 7.3%.
Несмотря на столь обнадеживающие результаты, следует учесть,
однако, предостережение Пяткина (1965). Автор указывает,
что несмотря на стимулирующее действие ультрафиолетовых
лучей, обусловливающих усиление защитно-приспособительных
реакций, облучение в излишних дозах (а дозировки пока даже
методически не установлены) может вызвать депрессию организма.
Автор подчеркивает, что нормализацию общего состояния орга-
низма у больных и ослабленных птиц под действием ультра-
фиолетовых лучей часто принимают за стимуляцию. Противо-
речивость результатов, на наш взгляд, обусловливается, кроме
различия основного материала и условий инкубации, также
и отсутствием единой точной методики облучения и дозировки
и недостаточным знанием специфического действия ультра-
фиолетовых лучей с различной длиной волн.
Установлено также полезное влияние микродоз ионизирующей
радиации (рентгеновских и гамма-лучей) на эмбриональное раз-
витие. Еще в исследовании Ногира (Nogier, 1923, цит. по: Groeb-
bels, 1937) облучение куриных яиц гамма-лучами в небольших
дозах сокращало длительность эмбрионального развития и по-
вышало выводимость цыплят. Добрынина с соавторами (1961)
установила, что облучение инкубируемых яиц мйкродозами
гамма-лучей повышает выводимость цыплят (в среднем на 3.5%)
тем больше, чем хуже инкубационные качества яиц, причем
360
кратность облучения не имела существенного влияния на резуль-
таты. Облучение несколько ускоряло эмбриональное развитие,
но не оказывало заметного действия на развитие внутренних
органов, на картину крови и состояние щитовидной железы
у суточных цыплят. Авторы отметили, что облучение яиц не влияло
на рост и развитие выведшегося из них молодняка, но повышало
яйценоскость кур на 11.9—16.5%. В опытах Найденского (1962)
облучение (0.0002 р/мин.) на 1-й и 3-й день инкубации по 5 мин.,
а с 6-го по 21-й день ежедневно по 8 час. (причем суммарная доза
была равна 1.4 р) стимулировало углеводный обмен (интенсивней
использовались глюкоза и жиры яйца, в печени откладывалось
большее количество гликогена) и повышало процент выведения
жизнеспособного молодняка. Кушнер с сотрудниками (1966)
установил, что облучение куриных яиц малыми дозами ионизи-
рующей радиации (21 мр) повышает поглощение кислорода и вы-
деление углекислоты эмбрионами и выводимость цыплят. Наи-
лучшие результаты — повышение выводимости на 12% — до-
стигались при облучении яиц на 1—3-й день инкубации. Несколько
иначе были поставлены опыты Джабиевой и Агаджановой (1963).
В воздушную камеру куриных* яиц до инкубации вводилась
малая доза радиоактивного фосфора (0.1—5 мккюри). В резуль-
тате сильно повышалась выводимость, ускорялось развитие
и сокращалась длительность инкубации на 34 часа. Наилучший
результат был получен при самой малой дозе, а при увеличении
ее полезность операции уменьшалась. К сожалению, опыт был
проведен на слишком малом количестве яиц (60 штук) и в далеких
от оптимума условиях инкубации (в термостате, с выводимостью
цыплят в контроле 33.3%).
I. ОСНОВНЫЕ ДИАГНОСТИЧЕСКИЕ ПРИЗНАКИ НЕПОЛНО
(По Отрыга
Причина Яйца до инкубации 1-е просвечивание и вскрытие кровяных колец Контрольное просвечивание
Хондродистрофия (кормление неполно- ценными животными белками, недостаток биотина и марганца). — — Отставание росте. в
Дистрофия в связи с авитаминозом В2. Пятнистая п шероховатая скор- лупа. Разжижен- ный белок. Некоторое ставание росте от- в
Дистрофия в связи с авитаминозом В12. — — Отставание росте. в
Эмбриональная микседема (авитами- ноз D). Тонкая и хруп- кая скорлупа. Бе- лок разжижен. — Отставание росте. в
Подагрическая форма дистрофии (авитаминоз А) при избыточном белко- вом кормлении. Бледный жел- ток. Низкая оплодо- творенпость. По- вышенная смерт- ность. Отставание росте. в
А-авитаминозная форма дистрофин. Бледный жел- ток. Низкая опло- дотворенность. Повышенная смертность. Отставание росте. в
Дистрофия в связи с авитаминозом Е. — Большая смерт- ность до 7-го дня. Разрастание ме- зенхимы и кро- воизлияния в бластодерме. —
362
I
ПРИЛОЖЕНИЯ
ЦЕННОГО ЯЙЦА И НАРУШЕНИЙ В РЕЖИМЕ ИНКУБАЦИИ
ньеву,1962)
2-с просвечивание и вскрытие замерших Задохлики Вылупившиеся птенцы
Смертность повышена. У мертвых эмбрионов короткие ноги с угло- ватыми изгибами ко- стей, «попугаевый» клюв. Сильно укороченные и искривленные ноги, «по- пугаевый клюв», лип- кость. Много липких и сла- бых.
Смертность повы- шена. Среди замерших встречаются отечные и курчавые. Незначитель- ное уменьшение веса яйца. Курчавость, липкость. Искривление шеи и пог. Отек головного мозга. Гу- стой, желток. Курчавое оперение. Липкость. Параличи шеи и ног.
Смертность повышена. Общая отечность. Атро- фия мышц. Кровоиз- лияния в желточном мешке, печени и сердце. Гипертрофия щитовид- ных желез. Атрофия мышц. Крово- излияния в желточном меш- ке, печени и сердце. Отло- жение жира в печени. Слабые.
Очень большая смерт- ность. Сильные отеки кожи, перерождение пе- чени. Гипертрофия по- чек. Признаки эмбриональной дистрофии. Вывод растянут. Мо- лодняк слабый.
Гипертрофия почек и отложение в них кри- сталлов солей. Отложение кристаллов солей в почках, па бры- жейке, сердце и желточном мешке. Вывод запоздалый. Плохая пигментация- пуха и кожи.
Отставание в росте. Липкость. Припухапие век. Вывод запоздалый. Молодняк слабый. Гла- за слезятся, иногда цод веками творожи- стые массы. Плохая пигментация пуха и кожи.
Помутнение хрустали- ков, пятна на роговице.
363
Причина Яйца до инкубации 1-е просвечивание и вскрытие кровяных колец Контрольное просвечивание
Кутикулит (недо- статок витаминов К, С, А, холина и не- которых других). — — —
Патология эмбрио- нов, обусловленная длительным хране- нием яиц. Большая нуга. Желток подвиж- ный и расположен вне центра яйца. Много неопло- дотворенпых, с зародышами, по- гибшими в пер- вый день. Края бластодермы не- ровные. Слабое развитие зачат- ков тела. Бласто- дермальный кис- тоз. Отставание в росте и развитии.
Длительный пе- регрев яиц. Много кровя- ных колец. Амор- фозы. Уродства, эктопия. Гипере- мия и кровоиз- лияния. Аллантоис раньше срока охватывает белок. Кисты на амни- -оне и аллантоисе.
Кратковременный острый перегрев яиц. — Гиперемия и кровоизлияния. Резкий подъем смертности в зо- не перегрева. —
Недогрев яиц. — Сильное отста- вание в росте и развитии. Сильное от- ставание в раз- витии аллан- тоиса. Белок не охвачен аллан- тоисом.
Недостаточная влажность. — Большая по- теря в весе яйца. Много кровяных колец. Гипере- мия и присыха- ния зародышей к скорлупе. Большая воз- душная камера.
364
Приложение I (продолжение)
2-е просвечивание и вскрытие замерших Задохлики Вылупившиеся птенцы
Отставание в росте. % Изъязвление стенок мы- шечного желудка и две- надцатиперстной кишки. Изъязвление стенок мышечного желудка и двенадцатиперстной кишки. Воспаление кутикулы. Поздний вывод. Сла- бый молодняк.
Эктопия и уродства головы. Кисты на амнио- не и аллантоисе. Ран- ний мелкий наклев. Резкий подъем смерт- ности в зоне перегрева. Переполнение кровью сосудов аллантоиса. Ги- перемия и точечные кро- воизлияния в коже, пе- чени, почках, сердце и мозге. Много мертвых, про- клюнувших скорлупу, но не втянувших желток. Остаток густого белка. Не- правильное положение. Ги- перемия желточного мешка и кишечника. Уменьшение размера сердца. Гиперемия кожи. Непра- вильное положение — го- ловой в желток. Вывод ранний, растя- нутый, молодняк мелкий. Плохо втянутый желток и незажившая пупови- на.
Сильное отставание в росте и развитии. Не- подвижная граница воз- душной камеры. Большая потеря в ве- се. Большая воздушная камера. Раннее начало движения головой. Много живых зародышей. Аллантоис полнокровный. Желток втянут, зеленого цвета. Остаток жидкого белка. Кишечник пере- полнен желточными и кало- выми массами. Гипертрофия сердца. Подскорлупные оболочки сухие, плотные. Возле клюва кровоизлияние в ал- лантоисе. Вывод поздний, растя- нутый. Молодняк вя- лый, плохо стоит на но- гах. Скорлупки гряз- ные. Проклев затруднен. Вывод ранний. Молод- няк и скорлупки су- хие. Хорошая пигмен- тация пуха.
ж
Причина Яйца до инкубации 1-е просвечивание и вскрытие кровяных колец Контрольное просвечивание
Избыточная влаж- ность. — — Запоздалый охват белка ал- лантоисом.
Недостаточный га- зообмен. — Отставание в росте. Уродства головы и глаз. Отставание в росте.
Неправильности поворачивания яиц. — Присыхание желтка к под- скорлупным оболочкам. Аллантоис сра- стается над бел- ком.
II. ВОЗРАСТНЫЕ ПРИЗНАКИ КУРИНОГО ЭМБРИОНА*
Р/2 дня. Эмбрион расположен перпендикулярно к длинной оси яйца
и еще не отделен от бластодермы. Различимы мозговые и глазные пузыри
и 8—10 пар сомитов. Хорошо развито сосудистое поле, образовалось и ра-
ботает сердце. Длина эмбриона 4—5 мм, диаметр сосудистого поля 6—8 мм.
21/2 дня. Головная часть зародыша отделена от желтка и лежи! на ле-
вом боку, спиною к пуге. Заметен резкий изгиб головы. Брюшная полость
еще открыта. Сердце лежит вне полости тела. Намечаются зачатки конеч-
ностей. Длина эмбриона 9—12 мм, диаметр сосудистого поля 12—15 мм.
31/2 дня. Зародыш отделен от желтка и закрыт амнионом. Передний
мозг раздвоен. Начинается пигментация глаз. Конечности имеют ластооб-
разную форму. Заметен аллантоис, достигающий размера среднего мозгового
пузыря. Вес эмбриона 0.05—0.06 г.
41/2 дня. Голова зародыша резко изгибается в сторону брюшной по-
верхности. Глаза хорошо пигментированы вокруг зрачка. Заметно рото-
вое углубление. Конечности на концах расширяются. Аллантоис разра-
стается над амнионом. Вес 0.13—0.19 г.
51/2 дней. Раздвоенность переднего мозгового пузыря исчезает. Заметны
зачаток века, посовой и верхнечелюстные отростки. Концы конечностей
в середине заостряются. Аллантоис достигает внутренней поверхности
скорлупы. Зародыш погружается в глубь желтка. Вес 0.18—0.40 г.
61/2 дней. Форма головы изменяется за счет относительного уменьше-
ния среднего мозгового пузыря. Образуются челюсти. Шея удлиняется.
На крыльях и ногах появляются пальцы. Вес 0.4—0.9 г.
7V2 дней. Клюв начинает удлиняться, обозначаются ноздри. Глаза
становятся выпуклыми, шея длинной. Резко выделяется локтевой изгиб,
удлиняется средний палец крыла. Вес 0.7—1.2 г.
* По Отрыганьеву, 1951.
366
Приложение I (продолжение)
2-е просвечивание и вскрытие замерших Задохлики Вылупившиеся птенцы
Малая потеря в весе яиц. Во время проклева при- сыхание кожи и клюва к скорлупе. Зоб, желудок, и кишечник переполнены жидкостью. Вывод запоздалый. Молодняк грязный, с плохо пигментирован- ным пухом.
Гематоамнион. Ги- перемия и кровоизлия- ния в коже и внутрен- них органах, дефекты в строении сердца. Белок остался вне аллантоиса. Неправильное положе- ние — головой в острый ко- нец яйца. Гиперемия кожи. Липкость. Проклев скорлупы в остром конце яйца. Липкость.
81/2 дней. Голова приобретает форму, характерную для птиц. Клюв
длинный и чуть изогнутый, на конце его появляется белая точка. Концы
пальцев ног утолщены. Видны зачатки перовых сосочков на спине. Вес
0.8-2 г.
9х/2 дней. Веко достигает края радужной оболочки. На ногах исче-
зают межпальцевые перепонки. Перовые сосочки становятся крупнее, по-
крывают всю спину и шею. Аллантоис охватывает почти всю внутреннюю
поверхность скорлупы и при инкубации в секционных инкубаторах часто
уже смыкается в остром конце яйца. Вес 1.7—2.6 г.
101/2 дней. Веко достигает зрачка. Намечается валик гребня. Видны
зачатки когтей. Перовые сосочки покрывают все тело. Вес 2.7—3.5 г.
IP/a дней. Веко над зрачком образует узкую щель. На гребне появ-
ляются зубцы. На ногах хорошо видны когти. Вдоль спипы заметен пух.
Вес 3.6—5 г.
12х/2 дней. Глаза закрыты веками. На передней поверхности плюсны
заметны поперечные бороздки — зачатки чешуек. Пух на спине, крыльях
и ногах. Вес 5.3—6.8 г.
131/2 дней. Увеличивается надклювпый бугорок. Когти белеют. По-
является пух на голове. Эмбрион меняет положение, ложась по длинной
оси яйца, головой в сторону пуги. Вес 7.3—9.7 г.
14х/2 дней. Длина пушинок увеличивается. Появляются складки кожи
на ступне и чешуйки на передней поверхности плюсны. Вес 10.3—11.7 г.
15х/2 дней. Появляется пух па веках. На тыловой поверхности плюсны
складки кожи — будущие чешуйки. Вес 12.4—17 г.
16х/2 дней. Ясно обозначаются наросты над ноздрями. Увеличивается
длина ног. Вес 16.5—20 г.
17х/2 дней Белое пятно подклювного бугорка охватывает всю верх-
нюю часть клюва. Вся плюсна и пальцы покрыты чешуйками. Вес эмбриона
19.3-24.8 г.
18х/2 дней. Начинают открываться глаза. Желток втягивается в брюш-
ную полость. Вес 23—29 г.
19х/а дней. Глаза открыты. Желток втянут. Аллантоис атрофирован
и его сосуды бескровны. Наклев скорлупы. Вес 28—35 г.
20V2 дней. Вылупление.
367
III. ВНЕШНИЕ ВОЗРАСТНЫЕ ПРИЗНАКИ ЭМБРИОНОВ
СЕЛЬСКОХОЗЯЙСТВЕННЫХ ПТИЦ*
Признаки Возраст (в днях)
куры утки гуси индейки
Грушевидная форма светлого поля
у зародышевого диска 1 1 1 1
Появление кровеносных сосудов. . . 2 2 2 2
Отделение головы от желтка .... 2 2 2 2
Проамнион закрывает голову .... 2 2 3 3
Начало пигментации глаз 3 4 5 5
Зачатки конечностей 3 4 5 5
Аллантоис заметен без увеличения 4 5 5 5
Появление рта 7 7 8 7
Срастание пальцев на крыле .... 9 9 10 —
Зачатки перьев на спине 9 И 12 9
Оформление клюва 10 И 12 10
Первые сосочки на крыльях .... И 13 14 10
Смыкание аллантоиса 10-11 13 14 13
Веко достигает зрачка 13 15 15 14
Первый пух на голове 13 14 15 14
Весь эмбрион покрыт пухом .... 14 15 18 17
Поперечные бороздки на плюсне . . 14 17 17 16
Веко закрыто 15 18 23 18
Полное использование белка .... 16 21 22 20
Начало втягивания желтка 19 23 24 23
Начало открывания глаз 19 25 26 —
Глаза открыты 20 26 28 —
Желток втянут 20 26-27 28 24
Проклев 20 26—27 28-29 25
♦ По Отрыганьеву, 1951
ЛИТЕРАТУРА
К разделу I
А де мо су и А. А., Н. Н и к о л а й ч у к. 1966. Метаболизм кальция
и фосфора у несушек. Тр. XIII Всемирного конгр. по птицеводству,
Киев : 314. — Анорова Н. С. 1953. Особенности развития куриного
эмбриона в зависимости от возраста родителей. ДАН СССР, 91, 4 : 981. —
Анорова Н.С. 1955. Влияние возраста родителей на постэмбриональное
развитие и жизнеспособность потомства у птиц. ДАН СССР, 100, 2 : 369. —
Анорова Н. С. 1956. Особенности ранних стадий эмбрионального раз-
вития яиц кур разного возраста. ДАН СССР, 110, 3 : 494. —Антоно-
вич С. Н. 1950. Метод непрерывного развития эмбриона птиц. Сов. зо-
отехния, 8 : 91.
Баран чеев Л. М. 1939. Пигментация желтка яиц как один из
факторов, определяющий их инкубационные свойства. Работы IX пленума
ВАСХНИЛ, секция животноводства, М. : 120. — Барковская О. В.
1954. Определение скорости образования желтка в яйце кур. Физиол. журн.
СССР, 40, 6 : 722. — Барт Л. Г. 1951. Эмбриология. ИЛ, М. — Б а у-
м а н В. К. 1956. Вопросы обмена кальция у кур. Журн. общ. бпол., 17,
4 : 257. — Бауман В. 1966. Роль фосфо протеина сыворотки крови не-
сушек в образовании скорлупы яиц. Тр. XIII Всемирного конгр. по птице-
водству, Киев : 319. — Бохтина В. Г. 1958. Морфология оплодотворе-
ния кур. Тр. Пушкинск. н.-иссл. лабор. разведения сольскохоз. животных,
8 : 214. — Б е х т и п а В. Г. 1960. Ранние стадии дробления куриного за-
. родыша. Арх. апат., гистол. и эмбриол., 38, 4 : 77. — Б е х т и н а В. Г.
1961. Полиспермный характер оплодотворения у кур. В сб.: Повышение про-
дуктивности сельхоз. животных, М : 131. — Бехтина В. Г. 1965. Морфо-
физиологические особенности оплодотворения у кур. Тезисы докл. со вещ.
по физиол. птиц, Таллин : 5. — Б е х т и н а В. Г. 1966. Морфологические
особенности полиспермного оплодотворения у кур. В сб.: Наследственность
и изменчивость сельскохоз. птицы, изд. «Колос» : 92. — Боголюб-
с к и й С. И. 1966. Изменчивость корреляции признаков у кур русской бе-
лой породы. Тр. XIII Всемирного конгр. по птицеводству, Киев : 47. —
Бражникова Л. А. 1950. Физико-химические изменения пдазм и
газообмен утиных яиц при инкубации. Тр. н.-иссл. инет, птицеводства,
21 : 48. — Б р а ш е Ж. 1961. Биохимическая эмбриология. ИЛ, М. — Б р е-
славец В. А. 1965. Влияние возраста птицы на эмбриональное и постэм-
бриальное развитие потомства. Тезисы докл. совещ. по физиол. птиц, Тал-
лин : И.
Вейцман Л. Н. 1957. Сохранение оплодотворяющей способности
спермиев в половых путях цесарок-самок. ДАН СССР, 116, 5 : 882. —
Владимирова IO. Н. 1954. Потеря в весе яиц кур перед инкубацией.
Тр. Н.-иссл. инет, птицеводства, 24 : 126. — Владимирова 10. Н.
1958. Повышение жизненности зародышей под влиянием периодических
подогреваний и охлаждений гусиных япц. Тр. Всесоюзн. н.-иссл. инет.
369
птицеводства, 25 : 209. — Владимирова 10. Н. 1962а. Повышение
жизнеспособности зародышей гусей при длительном храпении яиц. Сб. ре-
фатов работ Всесоюзп. н.-иссл. инет, птицеводства : 67. — В л а д и м и-
р о в a IO. II. 19626. Влияние низких температур и повышенного содержания
углекислого газа в воздухе при хранении яиц гусей и кур на их выводимость.
Сб. рефератов работ Всесоюзн. н.-иссл. пнет, птицеводства : 94. — *В о й-
тишкова М. 1955. К вопросу поведения живчиков в половом тракте
курицы. Folia Biologia, 1, 3 : 181. — Волков Д. Т. 1956а. Изменение
форм кальция в сыворотке крови кур в период яйценоскости. Тр. Моск,
ветер, акад., 11 : 155. — В о л к о в Д. Т. 19566. Изменение форм фосфора
в сыворотке крови кур в период яйценоскости. Тр. Моск, ветер, акад.,
11 : 164. — Воробьев В. И. 1960а. Фосфорные соединения куриного
яйца и их превращения при инкубации. 1. Фосфорные соединения неинку-
бированного яйца. В сб.: Вопр. цитол. и протистол., М.—Л., АН СССР, :
156. — Воробьев В. И. 19606. Нуклеиновые кислоты белка и желтка
куриного яйца. В сб.: Вопр. цитол. и общей физпол., Изд. АН СССР,
М.-Л. : 68.
Гальперн И. Л. 1958. Некоторые особенности полового процесса
у кур. Тр. Пушкинск н.-иссл. лабор. разведения сельскохоз. животных,
8 : 230. — Гард Д. И., С. Е. Д яс о р д а н, Т. М. М и н з, С. Е. Р е д-
м а н, Д. Ф. У а г н е р, Д. Е. Вахтштеттер. 1966. Повышение
яичной продуктивности кур посредством регулирования овуляции про-
гестином. Тр. XIII Всемирного копгр. по птицеводству, Киев : 297. — Го-
ля р к и н Ф. Е. 1947. Значение витамина А в кормлении индеек и индюшат.
Мясная и молочная промышленность, 7 : 72. — Г о р ю н о в Н. А. 1965.
Витамины в кормлении птицы. Рекомендации по витаминному кормлению
сельскохозяйственной птицы. Птицеводство, 2:6.
Данилова А. 1931. Химический состав, физические свойства, кало-
рийность и пигментация яиц у кур разных пород в разные периоды носки.
Сельхозгиз, М. — Д а н и л о в а А. К., В. А. Н с ф е д ь е в а. 1936. Хи-
мический состав утиных яиц. Сов. птицеводство, 8 : 25.— Дементьев Г. П.
1940. Руководство по зоологии, 6. Птицы. Изд. АН СССР, М.—Л. — Дени-
сьевская З.А. 1926. К вопросу о морфологии куриного яйца. Воронеж.
Дорфман В. А. 1963. Физико-химические основы оплодотворения.
Изд. АН СССР, М. — Д р е й п е р М. Г. 1966. Транспортировка минераль-
ных веществ в белок куриного яйца. Тр. XIII Всемирного конгр. по птице-
водству, Киев : 328. — Духно 3. И. 1964. Половая активность и каче-
ство потомства птиц. В сб.: Наследственность и изменчивость сельскохоз.
животных : 97. — Д ь я к о в М. И. 1933. Основы рационального корм-
ления птицы. Гос. изд. колх. и совх. литературы, М.
Еремеев Г. П. 1956. Усушка яиц с различным содержанием каро-
тиноидов при инкубации и хранении. Птицеводство, 2 : 39. — Е р е-
м е е в Г. П. 1957. Зародышевые приспособления птиц с точки зрения срав-
нительной физиологии. Тр. Омского сельскохоз. нпст., 25 : 1.
Желудков Н. А. 1951. Оценка яиц до инкубации по плотности белка.
Птицеводство, 7 : 13. — Ж у к о в А. Р. 1956. Влияние микроэлементов на
инкубационные качества яиц кур. Тезисы докл. научн. конфер., посвящен-
ной итогам н.-иссл. работ 1954—1955, Саратов : 32.
Земская А. 1940. Влияние кормления на качество племенных яиц.
Сов. птицеводство, 8—9 :29. — Злочевская К. В. 1962а. Оценка ка-
чества яиц при круглогодовой инкубации. Птицеводство, 12 : 19. — Зло-
чевская К. В. 19626. Хранение в различные сезоны года яиц, получен-
ных от кур-несушек разного возраста. Сб. рефератов работ Всесоюзн. н.-иссл.
инет, птицеводства : 89. — Зусман И. Н. 1965. О темпах развития орга-
* Звездочкой отмечены работы, цитированные по реферат, журналам.
370
нов пищеварения в эмбриогенезе рептилий и птиц в связи с особенностями
питания их плодов. Автореф. дисс. Л.
Иванов В. Г. 1955. Влияние срока хранения яиц па соотношение
полов в потомстве у кур. ДАН СССР, 100, 5 : 1021. — Иванов П. П. 1937.
Общая и сравнительная эмбриология. Биоме дгиз, М.—Л. — ИвановП.П.
1945. Руководство по общей и сравнительной эмбриологии. Учпедгиз, Л. —
Ильин М. Д. 1917. Исследования по развитию зародыша куриного яйца.
Материалы для эмбриохимии. Петроград.
Казаков X. Ш. 1954. Химическая и физико-химическая природа
вителлина и ее изменения в процессе зародышевого развития яйца. Экспер.
исследование. Автореф. дисс. Казань. — Калинина М.И. 1958. Влияние
малоконцентратного кормления кур на развитие эмбрионов и выводимость
Цыплят. Птицеводство, 8:39. - Камшилов М. М. 1947. Гетерополи-
спермия — новый метод вызывания мутаций. ДАН СССР, 57, 6 : 617. —
Карапетян С. К. 1957. Экспериментальные данные о влиянии периоди-
чески изменяемого термического режима на выживаемость эмбриона кур.
ДАН АрмССР, 24, 3 : 141. — Карапетян С. К. 1960. Опыт исследо-
вания роли температурного и светового факторов и стимуляции репродуктив-
ной функции у домашней птицы. Изв. АН АрмССР, Отд. биол. наук, 13,
11 : 63. — Карапетян С. К. 1962. Стимуляция светом метаболических
процессов в животном организме. ДАН АрмССР, 34, 3 : 135. — Карапе-
тян С. К., М. Б. Назарян.. 1965. Материалы к нервной регуляции
функции размножения у птиц. Тезисы докл. совещ. по физиол. птиц, Тал-
лин :43. — Кирсанов Л. 1935а. Сохранение инкубационного яйца.
Сов. птицеводство, И: 29. — Кирсанов Л. 19356. Оплодотворенность
и выводимость гусиного яйца в зависимости от возраста маточного стада.
Сов. птицеводство, 3 : 42. — Киселев Г. И.г Л. Г а в ш и н а. 1966.
Энергетические ресурсы скелетных мышц и яйцевода кур в связи с возрастом
и физиологическим состоянием. Тр. XIII Всемирного конгр. по птицевод-
ству, Киев : 261. — Кнорре А. Г. 1959. Краткий очерк эмбриологии
человека с элементами общей, сравнительной и экспериментальной эмбрио-
логии. Meдгиз, Л. — Князева А. П. 1955. Прединкубационное подогре-
вание яиц как фактор, повышающий выводимость утят. В кн.: 8-я научн.
конфер. 18—20 .марта 1954. Белоцерковский сельскохоз. инет., Сельхозгиз,
Киев : 59. — Князева А. П. 1958. Изменение некоторых биологических
свойств утиных яиц в период доинкубациоиного хранения. Сб. докл. научн.
конференции аспирантов Всесоюзн. н.-иссл. пнет, птицеводства, 1 : 53. —
Кондратюк Н. Д. 1948. Организация птицеводства в совхозах. Сель-
хозгиз. — Короткова Г. П. 1956. Антибиологические свойства белковой
оболочки куриного яйца. В сб.: Пробл. соврем, эмбриологии: 323. — К р ж и ш-
ковский К. Н. 1933. Физиология сельскохозяйственных птиц. Сель-
хозгиз, М. — Крикун А. А. 1958. Как влияют отдельные виды кормов
на инкубационные качества яиц. Птицеводство, 10 : 7. — К р о к Г. С. 1962.
Микроскопическое строение органов сельскохозяйственных птиц с основами
эмбриологии. Изд. АН УССР, Киев. — Кудрявцев С. С. 1940. Мине-
ральное кормление несушек. Сов. птицеводство, 8—9 : 33. — К у д р я в-
ц е в С. С. 1948. Влияние протеина и витамина В2 на продуктивность и инку-
бационные качества яиц кур породы род-айленд. Тр. Н.-иссл. инет, птице-
водства, 19 : 99. — К у д р я ш о в -Б. А. 1932. Инкубация яиц и витамин Е.
Пробл. животноводства, 4: 76. — Курбатов А. Д., Л. Е.Нарубина
1965. Влияние переменного и кратковременного использования петухов на
пол и другие качества потомства. Тезисы докл. совещ. по физиол. птиц,
Таллин : 61. — Кучковская Е. Н. 1938. Выводимость яиц кур в за-
висимости от шероховатости скорлупы. Сов. птицеводство, 12 : 17. — Куш-
нер X. Ф. 1948. Стимулирующее влияние осеменения кур смешанной спер-
мой на развитие цыплят. Тр. Инет, генетики АН СССР, 16 : 140. — Куш-
нер X. Ф. 1949. О явлениях цсенийности й телегонип у животных. Журн.
371
общей биологии, 10, 3 : 200. — Кушнер X. Ф., Г. Я. Корепанова
и В. Н. Букин. 1950. Продуктивность и инкубационные качества яиц
у кур в зависимости от питания. ДАН СССР, нов. сер., 72, 6 : 1191.
Лебедев М. М., А. II. Фомин. 1951. Пользовательное скрещива-
ние в птицеводстве. Изд. Мин. совх. СССР, Упр. сельскохоз. пропаганды. —
Лепайыэ Л. 1961. О связи между оплодотворяемостью яиц, выводимо-
стью и сохраняемостью цыплят. Сб. научн. тр. Эстонской сельскохоз. акад.,
Тр. по зоотехнии, 20 : 139. — Л епешенков В. Ф. 1965. Влияние
витамина В12 на эмбриональное развитие кур. Тезисы докл. совещ. по физиол.
птиц, Таллин : 62. — Липпинкот В. А. 1932. Птицеводство. Перевод с
с 4-го амер. изд. Сельхозгиз. — Л и с о в с к а я Н. П., Н. Б. Лива-
нова. 1960. Фосфопротеины. Изд. АН СССР, М. — Л о б и н Н. В. 1950.
Влияние внешних температур на энергетический и общий обмен веществ
у гусей и кур. Тр. Н.-иссл. инет, птицеводства, 21: 165. — Ломова А.И.
1939. Определение модуса пористости скорлупы яиц популяции кур товари-
щества «Птицевод» и племрассадника «Ударное». Тр. Воронежск. зооветер,
инет., 5 : 92.
Мазунин Н.А. 1957. К морфологии возрастных и сезонных измене-
ний яичников птиц. Уч. зап. Казахск. унив., 29 : 141. — Мамзина Е. А.
1965. Сперматогенез у домашних птиц. Тезисы докл. совещ. по физиол. птиц,
Таллин : 69.— Мамзина Е. А. 1966. Сперматогенез у домашних птиц.
В кн.: Наследственность и изменчивость сельскохозяйственной птицы.
Изд. «Колос» : 110. — М а с л и е в И. Т. 1965. Об этиологии двухжелтковых
яиц и режиме кормления молодок. Птицеводство, 4 : 22. — М а с л и е-
в а О. И. 1950. Содержание витамина А в желтке как индикатора инкуба-
ционных качеств яиц сельскозозяйственной птицы. Тр. Н.-иссл. инет,
птицеводства, 21 : 121. — М а т у с к о в а А. Н. 1950. Потребность гусей
в переваримом протеине, кальции и фосфоре в период яйцекладки и покоя.
Л. — Меркурьева Е. К. 1951. Особенности потомства от гетеросперм-
ного и дополнительного осеменения самок сельскохозяйственных животных.
Агробиология, 1: 85. — Митрофанов П. Н. 1898. Опыты над началь-
ным развитием птиц. Работы Зоотомической лабор. Варшавск. унив., 19 : 1.—
М и ч е к И. 1966. К вопросам эмбриональной детерминации. Тр. XIII Все-
мирного конгр. по птицеводству, Киев : 525. — М л а д б но в 3. М. 1955.
Некоторые физиологические особенности птиц в связи с полом и временем
года. Журн. общей биол., 6:4. — Мовчан Н. А. 1962. Об антибиотиче-
ских свойствах белковой оболочки яиц курицы, утки, цесарки. Вести. ЛГУ,
сер. биол., 3, 1 : 54. — М о и с е е в а И. Г., Е. В. Т о л о к о н п и к о в а.
1966. Корреляция различных показателей внутреннего качества яиц между
собой и с продуктивностью кур. Тр. Всесоюз. н.-иссл. технол. инет, птице-
водства, 30 : 22.
Николаева 10. Н. 1958а. Влияние периодических подогревов яиц
при их хранении па изменение бластодисков и результаты при инкубации.
Бюлл. научно-техн, информ. Всесоюзн. н.-иссл. ипст. птицеводства, 3:26.—
Николаева 10. Н. 19586. Некоторые закономерности яйцекладки у вы-
водковых птиц и их практическое значение. В кн.: Материалы конференции
молодых ученых АН БССР, Минск : 171. — Новик И.Е. 1958. Экспери-
ментальные данные о процессе оплодотворения у кур. Сб. докл. научн. конфер.
аспирантов Всесоюзн. н.-иссл. инет, птицеводства, 1 : 107. — Новик И.Е.,
В. Е. Гинтовт. 1965. Способность к оплодотворению мужских и женских
половых клеток в зависимости от их физиологического состояния. Тезисы
докл. совещ. по физиол. птиц, Таллин : 82. — Н о в и к И. Е., Г. Я. К о-
пыловская. 1961. Способность яйцеклеток кур к оплодотворению через
различные промежутки времени после овуляции. Тр. Инет, генетики
АН СССР, 28 : 371. — *Н о жч е в С. В. 1961. Влияние размеров инкуба-
ционных яиц на племенные и продуктивные качества кур. Изв. Центр, н.-иссл.
инет, животноводетва Болг. АН, 12 : 305 (на болг. яз.).
372
О б о н к о К. С., А. П. Анта ко в. 1956. Влияние крупности яиц на
выводимость цыплят. Птицеводство, 1: 34. — Орлов. М. В. 1948. Инку-
бация яиц, хранившихся разные сроки. Тр. Н.-иссл. инет, птицеводства,
19 : 183. — Орлов М. В. 1954. Отбор оплодотворенных яиц с сильными
зародышами. Тр. Н.-иссл. инет, птицеводства, 24 : 156. — Орлов М. В.,
Е. Н. Кучковская. 1941. Инкубация яиц индеек. Сб. Н.-иссл. инет,
птицеводства : 251. — Отрыганьев Г. К. 1940. Хондродистрофия кур
и авитаминоз D и А. Сов. птицеводство, 11—12 : 32. — Отрыганьев Г. К.
1951. Биологический контроль в инкубации. Сельхозгиз, М. — Отры-
ганьев Г. К. 1966. Некоторые конституциональные особенности в эмбрио-
нальном развитии кур. Тр. XIII Всемирного конгр. по птицеводству, Киев :
530. — Отрыганьев Г. К., А. Ф. Отрыганьев а. 1966. Эмбрио-
нальные гиповитаминозы D у кур. Тр. Всесоюзн. н.-иссл. техно л. инет,
птицеводства, 30 : 118. — О т р ы г а н ь е в а А. Ф. 1962. Инкубационные
качества утиных яиц, полученных в зимний период. Сб. рефератов работ
Всесоюзн. н.-иссл. инет, птицеводства : 60.
Патрик И. 1939. Применение марганца при кормлении кур. Сов.
птицеводство, 7 : 30. — П е н и о н ж к е в и ч А. Э. 1954. Значение са-
хара в яйцах для эмбрионального развития с/х водоплавающей птицы.
Автореф. дисс. Харьков. — ПенионжкевичЭ. Э. 1935а. Эмбриональ-
ное развитие утят и цыплят в связи с весом яиц. Успехи зоотехн. наук, 1,
3 : 443. — Пепионжкевич Э. Э. 19356. К вопросу о выводимости
яиц. Сов. птицеводство, 3:30. — Пенионжкевич Э. Э. 1946. Основы
гусеводства. Сельхозгиз, М., —Пенионжкевич Э. Э. и К. В. Зе-
ленская. 1949. Разведение и содержание гусей. 2-е изд. Сельхозгиз, М. —
Перов С. С.,. К. Д о л и н о в. 1932. Изменения физико-химических
констант куриного яйца при инкубации. Тр. Лабор. по изучению белка и
белкового обмена в организме, 3:3. —Прицкер И. Я. 1936. Выводо-
способность утиных яиц с жировым налетом на скорлупе. Сов. птицеводство,
5—6 : 40. — П р и ц к е р И. Я., Н. П. Т р е т ь к о в. 1937. Инкубаторы
и их эксплуатация. Сельхозгиз, М.
Рагозина М. Н. 1961. Развитие зародыша домашней курицы в его
соотношении с желтком и оболочками яйца. Изд. АН СССР, М. — Р е б-
риков И. 1965. Оценка качества инкубационных яиц. Птицеводство,
4: 20. — Резниченко Л. П. 1961. Содержание фракций азотистых
веществ в крови и яйцеводе кур па различных стадиях формирования
яйца. Укр. биохим. журн., 33, 1 : 72. — Р е с н я н с к а я Е. В. 1961.
Развитие эмбрионов в яйцах разного веса. Птицеводство, 8 : 20. — Роль-
пик В. В. 1955. Инкубация яиц гаги (Somateria mollissima L.). Зоол.
журн., 34, 4 : 885. — Р у у с Ц. 1956. О биологии яйцекладки гусей.
Сб. научи, тр. Эст. сельскохоз. акад., 2 : 116.
Савронь Е. С. 1965. Динамика белка и небелковых азотистых ве-
ществ у кур в связи с возрастом и физиологическим состоянием организма.
Тезисы докл. совещ. по физио л. птиц, Таллин :92. —Салгднник М.Г.
1948. Усовершенствование инкубирования утиных яиц в шкафных инкуба-
торах. Мясная индустрия, 3 : 15. — Сергеев А. А. 1940. Больше вни-
мания правильному кормлению. Сов. птицеводство, 10 : 21. —
Сергеев А. А., А. В. К о л я н и и с к и й, В. А. Ульянова,
О. И. Маслиева. 1949. Влияние источников и дозировок белка и вита-
минов на качество яиц и продуктивность уток. Тр. Н.-иссл. инет, птицевод-
ства, 20 : 233. — СергеевА. А. и С. С. Кудрявцев. 1941. Влияние
кормовых факторов на яйценоскость и инкубационные качества яиц кур бе-
лый леггорн. Сб. тр. Н.-иссл. инет, птицеводства : 87. — С и м о н с П. К.,
Г. В и р ц. 1966. Ультраструктура кутикулярной поверхности куриного
яйца в связи с очисткой. Резюме. Тр. XIII Всемирного конгр. по птицевод-
ству, Киев : 631. —Сметнев С. И. 1948. Птицеводство. Сельхозгиз, М. —
Сметнев С. И., Л. Г. Т а р а б р и н а. 1956. Особенности в морфологи-
373
ческом строении и химическом составе яиц кур-молодок разной продуктив-
ности. Изв. Моск, сельскохоз. акад. им. К. А. Тимирязева, 3:211. — Со-
коловская И. И. 1950. Вопросы биологии оплодотворения сельско-
хозяйственных животных. Журн. общей биологии, 11, 3 : 163. —
Старки Д. 1966. Физиология снесения яйца у кур. Тр. XIII Всемирного
конгр. по птицеводству, Докл. на симпозиумах, Киев : 69.
Тарабрина Л. Г. 1958. Продуктивные качества кур-молодок
в связи с их физиологическим состоянием. Вести, сельскохоз. науки, И : 62. —
— Третьяков Н. П. 1949. Физико-химические изменения плазм
утиных яиц при различных условиях инкубации. Сов. зоотехния, 2 : 89. —
Третьяков Н. П. 1951. Инкубация яиц сельскохозяйственных птиц.
Сельхозгиз, М. — Третьяков Н. П. и Л. И. Кустова. 1950. Био-
логические свойства инкубационных яиц. Природа, 11 : 51. — Третья-
кова Е. И. 1938. Оценка качества яйца до инкубации. Сов. птицеводство,
12 : 15. — Т р е т ь я к о в а Е. И. 1939. Физико-химические изменения
в яйце как показатель для оценки качества яйца и процесса инкубации.
Работы IX пленума ВАСХНИЛ : 132. — Т р е т ь я к о в а Е. И. 1941.
Влияние нарушений питания птицы (авитаминозы В2 и белковый токсикоз)
на физико-химическую структуру яиц. В докладе по итогам работы научн.-
иссл. инет, птицеводства за 1931—1941 гг. Сб. Тр. Н.-иссл. инет, птицевод-
ства : 224.
Федотова П. М. 1952. Использование крупных яиц кур для воспро-
изводства птицы. Рефераты докл. Моск, сельскохоз. акад. им. К. А. Тимиря-
зева, 16 : 139. — Фердинандов В. В. 1931. Рабочая книга по птице-
водству и птицевой промышленности. Сельхозгиз. — Фомин А. И. 1950.
Эффективность различных приемов отбора яиц на инкубацию. Тр. Пушкинск.
н.-иссл. лабор. разведения сельскохоз. животных, 4 : 88. — Фо ст е р У. X.
1966. Изменение в накоплении кальция у кур в период перед кладкой яиц.
Тр. XIII Всемирного конгр. по птицеводству, Киев : 57. — Ф о ф а-
н о в а К. А. 1964. Морфологические данные о полиспермном осеменении.
Журн. общей биологии, 25, 1 : 22.
Ханыкова О.К. 1958. Исследование электропроводности и электри-
ческой поляризации плазм куриного яйца в процессе инкубации. Автореф.
дисс. МГУ. — Хаскин В. В., В. И. Ц ы н о в ы й, В. В. Лысенко,
В. П. Пшеничная. 1965. Некоторые биохимические особенности вител-
логенеза у кур. Тезисы докл. совещ. по физиол. птиц, Таллин : 100.
Чавчанидзе В. И. 1953. Влияние условий хранения инкубацион-
ных яиц на эмбриональное развитие птиц и результаты инкубации. Автореф.
дисс. М. - Чайковская М. Я. 1949. Влияние внешних и внутрен-
них факторов на размножение гусей. Журн. общей биол., 10, 4 : 332. —
Чайковская М. Я. 1950. К вопросу физиологии размножения гусей.
Тр. Н.-иссл. инет, птицеводства, 21 : 171. — Ч м у т о в а А. П. 1954.
Морфология зародышевых дисков яиц гусей. Тр. Н.-иссл. инет, птицеводства,
24 : 58. — Ч м у т о в а А. П. 1956. Влияние различного режима белкового
кормления на развитие зародышевого диска яиц гусей. Птицеводство, 1 : 31.
Шалумович В. Н. 1955. Изучение структуры желточной оболочки
куриного яйца при помощи люминесцентной и ультрафиолетовой микро-
скопии и некоторыми гистохимическими методами. ДАН СССР, 105, 3 : 584.—
Шевцова М. Я. 1965. Рост и дыхание в зародышевом периоде эмбрио-
генеза домашней утки. Матер, симпозиума по основным проблемам возраст-
ной физиологии и биохимии. Харьков : 201. — Шишкина Е. М. 1949.
Изменения в эмбриогенезе птиц под влиянием старения яйца и температуры ин-
кубирования. Докл. Моск, сельскохоз. акад. им. К. А. Тимирязева, 9 : 130. —
Шмидт Г. А. 1951. Эмбриология животных. Ч. 1. Общая эмбриология.
Изд. «Советская наука», М. — Ш п и ц И. С., В. И. К о б з е в а, А. К. Да-
ни л о в а. 1965. Толщина скорлупы и инкубационные качества яиц. Птице-
374
водство, 11 : 20. — Шпиц И. С., А. К. Данилова. 1966. Влияние
качества скорлупы инкубационного яйца на минеральный обмен эмбриона
и взрослой птицы. Тр. Всесоюзн н.-иссл. техно л. инет, птицеводства, 30 : 87.
♦Abe Т.,Калек о, Mosoda. 1959. Studies on the yolk deposition
in laying hens. II. Observation of the physiological halflife of serum phosphorus
compounds and the yolk deposition in laying hens. Bull. Natl. Inst. Agric.
Sci., Ser. G., 18 : 229. — * A r b о 1 e d a C. R., G. 0. R a n i t, A. D. Y n i-
g u e z. 1960. A correlation study of egg size to fertility, hatchability and
chick size; of chick size to growth and mortality; and of body size of hens
to egg production. Philippine Agriculturist, 44, 5 : 247. —Ash W. J. 1958.
A study of egg shell quality in white leghorns. Poultry. Sci., 57, 5 : 1181. —
Ash W. J. 1961.A study of the duration of fertility and hatchability of white
pekin duck eggs. Poultry Sci., 40, 5 : 1373. — Asmundson V. S. 1939.
Formation of the egg in the oviduct of birds. Observations on turkeys. Proc.
7th World’s Poultry Congress : 96.— *A x elsson J. 1954. Influence of some
factors on hatchability of chicken eggs on growth rates of embryos and chicks
Lantsbrukshogskolans Annaler, 21 : 81.
Becker W. A., G. E. В e a r s e. 1958. Pre-incubation warming and
hatchability of chicken eggs. Poultry Sci., 37, 4 : 944. — Becker W. A.,
J. V. Spencer, J. L. Swart wood. 1964a. The pre-incubation storage
of turkey eggs in closed environments. Poultry Sci., 43, 6 : 1526. — В e-r
с к e r W. A., J. V. S p e n c e r, J. L. S w a r t w о о d. 1964b. Hatchabi-
lity of turkey eggs shipped in plastic bags. Poultry Sci., 43, 6 : 1539. — ♦Bel-
lairs R., M.'H a r kne ss, R. D. H а г к n e s s. 1963. The vitelline
membrane of hen’s egg: A chemical and electron microscopical study. Journ.
Ultrastructure Res., 8, 3—4 : 339. — В e r g L. R., E. B. Gordon,
L. H. M e r r i 1 1. 1962. Effect of calcium level of the developing and laying
ration on the hatchability of eggs and viability and growth rate of the progeny
of young pullets. Poultry Sci., 41, 4 : 1328. — В о h r e n B.B., L. B. Crit-
tenden, R. T. К i n g. 1961. Hatching time and hatchability in the fowl.
Poultry Sci., 40, 3 : 620. — Bolton W. 1957. The effect of the age of the
hen on the porosity of the egg. Journ. Agric. Sci., 49, 3 : 384. — Brad-
field J. R. G. 1951. Radiographic studies on the formation of the hen’s
eggshell. Journ. Exp. Biol., 28, 2 : 125. — В г о w n W. O. 1957. The
transfer of maternal dietary riboflavin to the embryo during development in
the common fowl. Journ. Agric. Sci., 49, 1:88. — В у e r 1 у T. С., H. W. T i-
tus, N. R. Ellis. 1933. Production and hatchability of eggs as affected
by different kinds and quantities of protein in the diet of laying hens. Journ.
Agr. Res., 46 : 1.
Chung R. A., W. J. S t a d e t m a n. 1961. The physical and chemical
components of the hen’s egg as related to total egg weight. Poultry Sci., 40,
5 : 1387. — Coles R. 1956. The influence of the henrs egg weight on hat-
ching. Poultry Sci., 35, 4 : 817.
Dougherty J. E. 1926. Studies in incubation. 1. The effect of low
temperatures previous to incubation on hatchability of eggs set. Amer. Journ.
Physiol., 79 : 39. — D о u g 1 a s D. S., P. D. S t u г к i e. 1964. Plasma
levels of antidiuretic hormone during oviposition in the hen. Fed. Proc.,
23 : 150. — *D г ugo c i u Ch., Al. D a s c H u, N. N e a g a, I., P a m-
filie, 0. Tur 1 i u с , V. Petrus. 1956. Исследование некоторых фак-
торов, влияющих на выводимость цыплят породы род-айленд. Studii §i cer-
cetrfiri §tiint. Acad. RPR. Fil. Iasi. Biol. ?i $tiin0 agric., 7, 1 : 209 (на румынск.
яз.). — Duval M. 1884. De la formation du blastoderme dans roeuf des
oiseaux. C. r. Acad. Sci., 98 : 447.
Edwards Ch. L. 1902. The physiological zero and the index deve-
lopment for the egg of the domestic fowl. — Gallus domesticus. Amer. Journ.
Physiol., 6 : 351.
376
Farnsworth G. M., D. C. Warren. 1962. Effects of age and
turning on hatchability of eggs. Poultry Sci., 41, 5 : 1642. —Funk E. M.
1949. The care of hatching eggs before incubation. In: Fertility and hatchabi-
lity of chicken and turkey eggs, N. Y. — London : 150.
♦Godfrey G. F. 1950. On the nature of genetical control of egg
shell quality as it affects hatchability in domestic fowl which lay brown
eggs. Ohio State Univ. Absts. Doctoral Diss., 60 (1948—1949) : 131. —
Greenwood A. W., W. Bolton. 1956. Variations in the amounts of
thick and thin albumen in hens’eggs with the age of the bird. Journ. Agr. Sci.,
47, 1 : 40. — G r i m 1 e b у F. H., D. J. G. В 1 a c k. 1950. Variations in
the pigment and vitamin A contents of egg yolk. Brit. Journ. Nutrition, 4,
4 : 323. — Groebbels F. 1937. Der Vogel. Bau, Funktion, Lebenser-
scheinung, Einpassung. II: Geschlecht und Fortpflanzung. Berlin. — G u-
t о w s к a M. S., G. A. M i t c h e 1 1. 1945. Carbonic anhydrase in the cal-
cification of the egg shell. Poultry Sci., 24, 1 : 159.
Hall G. O., A. L. R о m a n о f f. 1943. Effect of sealing of the shell
and holding temperature on hatchability of hens’eggs. Poultry Sci., 22, 5 : 354.—
♦Ha stabova V. 1960. Размножение и промышленное скрещивание.
Sb. 6eskosl. akad. zemSd. v$d Zivoc. vyroba, 5, 12 : 925. — Hays F. A.,
C. Nicolaides. 1934. Variability in development of freshlaid hen eggs.
Poultry Sci., 13, 1 : 74. — H e r t e 1 e n d у F., T. G. T а у 1 о r. 1961.
Ghanges in blood calcium associated with egg shell calcification in the domestic
fowl. 1. Changes in total calcium. 2. Ghanges in the diffusible calcium. Poultry
Sci., 40, 1 : 108. — Hughes G. 1963. The population of germ cells in the
developing female chick. Journ. Embryol. Exp. Morphol., 11, 3 ; 513. —
Hunsaker W. G., P. D. S t u г к i e. 1961. Removal of calcium from
uterine blood during shell formation in the chicken. Poultry Sci., 40, 5 : 1348.
Jull M. A., M. G. McCartney, H. M. E 1. Ibiary. 1948. Hat-
chability of chicken and turkey eggs held in freezing temperatures. Poultry
Sci., 27, 1 : 136.
♦Kalmutchi G., D. Bosoanca. 1957. Определение концентра-
ции каротина в желтке как средство оценки инкубационных качеств куриных
яиц. Anuarul lucrar. $tiint. Inst, agron. Timisoara, Bucure^ti : 259 (на ру-
мынок. яз.). — К a m a r G. A. R. 1960. The influence of semen characte-
ristics on hatching results of chicken eggs. Poultry Sci., 39, 1 : 188. — К a-
m a r G. A. R. 1962. Seasonal variation in fertility and hatchability of duck
eggs. Poultry Sci., 41, 4 : 1029. — Kaufman L. 1939a. An experimental
study on the effects of storage on embryonal development of hens’eggs. Proc.
7 th World’s. Poultry Congress : 186. — * К a u f ma n L. 1939b. «Aging» of
eggs. 1. The influence of storage of hens’eggs on mortality, development and
growth of embryos. Pamqtnik panstwowego instytutu naukowego gospodar-
stwa wiejskiego w Putawach. Putawy, 17 : 2. — *K a uf man L.,H.Krza-
n о w s k a. 1957. Исследования «старения» яиц. III. Влияние хранения
яиц на вес тела и некоторых органов, содержание гликогена в печени, содер-
жание гемоглобина в крови и гистологическое строение щитовидной железы
цыплят. Roczn. nauk. rolniczych., 72, 3 : 385 (на польск. яз.). — Knox Ch.,
М. Olsen. 1936. The effect of tremulous air cells upon the hatchability of
eggs. Poultry Sci., 15, 4 : 345. — К о s i n I. L. 1954. Length of preincuba-
tion storage and heredity as factors affecting the hatchability of turkey eggs.
Poultry Sci., 33, 1 : 24. — К о s i n I. L. 1956. Studies on pre-incubat ion
warming of chicken and turkey eggs. Poultry Sci., 35, 6 : 1384. — К о s i n I. L.
1958. Time-lag in identification of fertility in Broad Breasted Bronze eggs
and possibly, parthenogenesis. Poultry Sci., 37, 5 : 1150. — *K о s i n I.,
J. P. Hurley. 1957. Variability in the development of avian embryos in
situ and in vitro. Transplant. Bull., 4, 2 : 76. — К о s i n I. L., A. M. M u n.
376
1965. Some factors affecting the biological quality of turkey hatching egg.
Poultry Sci., 44, 1 : 31.
♦Lacassagne L. 1960. Etude comparee des reserves vitellines et
de la duree de la phase de grand accroissement do 1’ovocyte chez la poule dome-
stique. Influence de 1’age de 1’animal et de la longueur de la serie. Amer. Inst.
Natl. Techn. Agron. Ser. D. Ann. Zootechn., 9, 1 : 85. — L a n d a u e r W.
1948. The hatchability of chicken eggs as influenced by environment and
heredity. Storrs (Connecticut) Agr. Exp. Sta. Bull., 262 : 1. — L i 1 1 i e F. R.
1952. Lillie’s development of the chick. Revised by H. L. Hamilton. N. Y. —
♦Lorkiewiczowa M. 1960—1962. Влияние длительности хранения
племенных яиц на рост и жизнеспособность цыплят, а также на продуктив-
ность кур. Ann. Univ. М. Curie-Sklodowska, Е, 15 : 249 (на польск. яз.). —
Lutz Н., Y. Lutz-Ostertag. 1957. Gemellite experimentale et situs
inversus viscerum chez 1’embryon d’Oiseau. C. r. Acad. Sci., 244, 11 : 1543.
Mangold E. 1932. Einfluss des Lichtes auf Ernahrung und Stoffwe-
chsel der Nutztiere. Handbuch der Ernahrung und des Stoffwechsels der Landwir-
tschaftlichen Nutztiere. IX, 5 : 804. — Marshall W. 1947. The principles
of incubation. In: Deseases of Poultry, London : 110. — Maurice D. M.,
A. F i d a n z a. 1954. The structure of the yolk of the hen’s egg, investigated
by means of its permeability to Br82, Journ. Exp. Biol., 31, 4 : 573. —
McNally E. H., T. C. Byerly. 1936. Variation in the development of em-
bryos of hens eggs. Poultry Sci.,\15, 4 : 280. — Merat P., L. Lacassagne
1961. Taux d’eclosion et position de 1’oeuf dans la serie chez la poule. Ann.
Biol. Animale, Biochim. Biophys.,1. 1 : 101. — Merritt E.S.,W.E.Clar-
ridge. 1959. The affect of lengtn of holding on the hatchability of goose
eggs. Poultry Sci., 38, 3 : 660. — M i lbу T. T., D. H. S h e r w о о d. 1960.
The influence of pre-incubation treatment on hatchability of chicken and turkey
eggs. Poultry Sci., 39, 5 : 1118. — M о r an T. 1925. The effect of low tem-
perature on hens’ eggs. Proc. Roy. Soc., Ser. B>, 98 ; 436. — Moreng R. E.,
L. J. Manson. 1961. Chilling and embryonic development of turkey eggs.
Poultry Sci., 40, 5 : 1433. — M о u n t n e у G\ J., С. V a n d e r z a n t.
1957. Relationship of selected egg quality measurements. Poultry Sci., 36,
4 : 908. — Munro S. S., I. L. К о s i n. 1945. Proof of the existence of
pre-oviposital embryonic death’s in chickens and their bearing on the relation
between «fertility» and hatchability. Canad. Journ. Res., Sect. D, 23 : 129.
*N a d у Z., H. E. Olah. 1956. Спектроскопическое исследование
микроэлементов в скорлупе яиц. Kiserl. orvostud., 8, 5 : 516 (на венг. яз.). —
Nalbandov А. V. 1958. Role of sex hormones in the secretary function
of the avian oviduct. Comp. Endocrinol. Proc., Columb. Univ, symposium :
524. — Needham J. 1931. Chemical embryology. Cambridge, Univ. Press.
-*N iem ierko W., C. Goldman. 1947. Биохимия овогенеза у кур.
Acta Biol. Exp., 14, 1 : 59 (па польск. яз.).
Olsen M. W. 1960. Nine year summary of parthenogenesis in turkeys.
Proc. Soc., Exp. Biol. Med., 105, 2 : 279. — Olsen M. W., S. К. H a i-
n e s. 1948. Tne effect of different holding temperatures on the hatchability
of hens’eggs. Poultry Sci., 27, 4 : 420. — O’Neil J. B. 1950. Relationship
of chick size to egg size and its effect upon growth and mortality. Poultry Sci.,
29, 5 : 774. — *0 s h i m a M., H. N о s a k i. 1962. Correlation of Sr84
.eliminated from the bones of the laying hens with the egg. Bull. Nat. Inst.
Agric. Sci., Ser. G., Animal Husbandry, 21 : 9.
Pasteels J. 1950. Dcveloppement embryonnaire. Traite de Zoologie,
15, Oiseaux : 479. — Patten В. M. 1951. Early embryology of the chick.
4th ed. N. Y. — P a t t e r s о n J. T. 1910. Studies on the early development
of the hens egg. I. History of the early cleavage and of the accessory cleavage.
Journ. Morphol., 21 : 101. — Patterson R., J. S. Y о u n g n e r,
W. O. W e i g 1 e, F. Y. D i x о n. 1962. The metabolism of serum proteins
25 в. В. Рольник
in the hen and chick and secretion of serum proteins by the ovary of the hen.
Journ. Gen. Physiol., 45, 3 : 501. — Payne L., R. McDaniel. 1958.
Shell thickness as related to «shuk-outs» in turkey eggs. Poultry Sci., 37, : 4
825. — Pingel H. 1958. Untersuchungen fiber Beziehungen des gbwichts
und der Schalenfarbe von Bruteiern zur Schlupffahigkeit bei der Rasse New
Hampschire. Dt. Geflugel-Ztg., 7, 23/24 : 354. — Poole H. К., M. W. О 1-
s e n. 1958. Incidence of parthenogenetic development in eggs laid by three
strains of Dark Cornish chickens. Proc. Soc. Exp. Biol. Med., 97, 2 : 477. —
Pope C. W., Ph. J. S c h a i b 1 e. 1957. Association of egg weight and
chick weight. Poultry Sci., 36, 5 : 1149. — Press N. 1959. An electron
microscope study of a mechanism for the delivery of follicular cytoplasm to
an avian egg. Exp. Cell. Res., 18, 1 : 194. — Preyer W. 1885. Specielle
Physiologic des Embryo. Leipzig.
Rauch W. 1952. The influence of eggshell porosity on the number
of chickens hatched from incubating eggs. Poultry Sci., 31, 4 : 589. — Rho-
des D. N., С. H. L e a. 1957. Phospholipids. 4. On the composition of hen’s
egg phospholipids. Biochem. Journ., 65, 3 : 526. — Riddle 0. 1911. On the
formation significance and chemistry of the white and yellow youlk of ova.
Journ. Morphol., 22 : 455. — Riley D. P. 1953. The chemistry of the egg.
Proc. Roy. Insth. Gr. Brit., 35, II, 159 : 363. — Rolnik V. 1943. Instru-
ctions for the incubation of Eider duck eggs. Journ. Wildlife Management, 7,
2 : 155. — Romanoff A. L. 1934. Study of artificial incubation of game
birds. I. Temperature requirements for pheasant and quail eggs. II. Humidity
requirements for pheasant and quail eggs. Cornell. Univ. Agr. Exp. Sta. Bull. :
616. — R о m a n о f f A. L. 1960. The avian embryo. Structural and functio-
nal development. MacMillan Comp., N./Y. — Romanoff A. L., A. J. Ro-
ma n о f i. 1949. The avian egg.z, Русск. перевод: А. Л. Романов,
А. И. Романова. Птичье яйцо. 1959. Пищепромиздат, М.
Saeki Y., J. В. К eit h. 1962. Effect of abnormal spermatozoa on
fertility and hatchability in the turkey. Poultry Sci., 41, 4 : 1096. — S a-
w у e r W. H. 1961. Neurohypophysial hormones. Pharmacol. Rev., 13 : 225.—
*S h a r me, Ganpoti, Parshed. 1950. Some observations on the
formation of the hen’s egg with special reference to the chalaziferous layer and
the chalaza. Res. Bull. East. Punjab. Univ. Zool., 7 : 83. — S i m k i s s K.
1961. Calcium metabolism and avian reproduction. Biol. Rev. Cambr. Philos.
Soc., 36, 3 : 321. — Smith A. H. 1959. Follicular permeability and yolk
formation. Poultry Sci., 38, 6 ; 1437. — Smith J. P. C. 1956. Incubation
. . . The artificial hatching of eggs. Australas Poultry World, 28, 4:35.—
Stiles K. A., R. L. W a t t e r s о n. 1937. The effects of jarring upon
the embryogeny of chick embryos. Anat. Rec. Philadelphia, 70 : 7. — S v e n s-
s о n S. A. 1957. Der Einflusz der Beschaffenheit der Eierschale auf die Brut-
fahigkeit von Hiihnereiern. Arch. Ceflugelkunde, 21, 3 ; 145.
Tanaka K., S. N a k a j o. 1960. Oxytocin in the neurohypophysis of
the laying hen. Nature, 187, 4733 : 245. — Tarchanoff I. R. 1884. Uber
die Verschiedenheiten des Eierweiszes bei befiedert goborenen (Nestfluchter)
und bei nackt geborenen (Nesthocker) Vogeln, und fiber die Verhaltnisse zwischen
dem Dotter und dem Eierweisz (Biologisch-Chemische Untersuchung). Pflugers
Arch. Physiol., 33 ; 303. — Taylor L. W. ed. 1949. Fertility and hatcha-
bility of chicken and turkey eggs. N. Y.—London.. — Taylor L. W.,
C. A. Gunns. 1939. Development of the unincubated chicken embryo in
relation to hatchability of the egg. Proc. 7th World’s poultry congress : 188.
♦Waite R. H. 1919. The effect of age of eggs on their hatching quality.
Maryland Agr. Exp. Sta. Bull., 233 : 87. - W al de n С. С., I. V. F. A 1-
I e n, P. С. T r u s s e 1 1. 1956. The role of the egg shell and shell membranes
in restraining the entry of microorganisms. Poultry Sci., 35, 6 : 1190. — W i n-
g e t Ch. M., A. H. Smith. 1957. Blood calcium and shell formation in
378
the laying hen. Poultry Sci., 36, 5 : 1169. — W i n g e t С. M., A. H. S m i t h,
G. N. Hoover. 1958. Arterio-venous differences in plasma calcium concen-
tration in the shell gland of the laying hen during shell formation. Poultry
Sci., 37, 6 : 1325. —Wolken J. J. 1951. Structure of hen’s egg membranes.
Anat. Rec., Ill, 1 : 79. — W о 1 к e n J. J., S c h w a r t z. 1948. Physical
structure of shell membranes. Journ. Gen. Physiol., 32, 3 : 153. — W о о d-
ard A. E., F. B. Mather. 1964. The timing of ovulation, movement
of the ovum through the oviduct, pigmentation and shell deposition in Japanese
quail (Coturnix coturnix japonica). Poultry Sci., 43, 6 : 1427.
Zawalsky M. 1962. The effect of sex, egg weight and preincubation
storage on hatching time and chick weight. Poultry Sci., 41, 5 : 1697.
К разделу II
Абрамова E. Г. 1958. Развитие щитовидной железы и гипофиза
у бронзовой индейки. ДАН СССР, 122, 3 : 496. — Антонович С. Н.
1950. Метод непрерывного развития эмбриона птиц. Сов. зоотехния, 8 :
91. — Аршавский И. А. 1948. Физиологические механизмы преобра-
зования функций в онтогенезе. Журн. общей биол., 9, 1 : 31.
Баранчеев Л. М. 1939. Влияние температуры на развитие и про-
цессы дыхания куриного эмбриона. Уч. зап. ЛГУ, сер. биол., 35, 9 : 64. —
Белоголовый 10. А. 1908. К развитию головных нервов у птиц. Уч.
зап. МГУ, Отд. ест. ист., 23, 6 : 1. — Б е л я н ч и к о в а Н. И. 1953. Из-
менение крови кур разных пород в онтогенезе. Дипломная работа. МГУ. —
Биховець А. У. 1941. Газообмен у курячих яйцах i нормативи венти-
ляции при инкубацп. Науков! пращ Укр. дослщ. ст. птах., 2, 2 : 131. —
Блинкова Т. П. 1962. Об особенностях рефлекторных реакций у ку-
риных эмбрионов. Физиол. журн. СССР, 48, 11 : 1415. — Б о г д а-
н о в О. В. 1960. О влиянии ареколина на деятельность сердца куриного
эмбриона в различные периоды его развития. Бюлл. экспер. биол. и мед.,
50, И: 61. — Богданов О. В. 1961. Становление регуляции сердечной
деятельности у кур и голубей в раннем онтогенезе. Физиол. журн. СССР,
47,1 : 80. — Б о г д а н о в О. В. 1963. Значение проприоцептивной импуль-
сацпи в функциональном созревании центральной нервной системы куриного
эмбриона. Физиол. журн. СССР, 49, 6 : 701. — Б о л д у и н Э. 1949. Основы
динамической биохимии. ИЛ, М. — Бордзпвпловская Н. П. 1953.
Эмбриональное развитие гусей. В кн.: К биологии развития сельскохозяй-
ственных птиц. АН УССР :53. — Бордзпвпловская Н. П. 1955.
Эмбриональное развитие уток. Тр. Инет. зоол. АН УССР, 12 : 3. — Б о-
рискин В. В., П. В. О б л а п е н к о, В. В. Р о л ь н и к, Б. М. Са-
вин. 1962. О возможности развития организма животного в условиях за-
мены азота воздуха гелием. ДАН СССР, 143, 2 : 475. — Бродов-
ская 3. И. 1957. Развитие костного мозга и процесс кроветворения у по-
звоночных животных. Тр. Крымск, мед. инет., 17: 35. — Бро п-
штейн А. А. 1964. Прижизненные наблюдения над движением волосков
обонятельных клеток. ДАН СССР, 156, 3 : 715. — Б у р с и а н А. В. 1964.
Влияние света на спонтанную двигательную активность куриных эмбрионов.
Бюлл. экспер. биол. и мед., 58, 7 : 7. — Б у р с и а н А. В. 1965. О прими-
тивных формах фоточувствительности в раннем эмбриогенезе кур. Журн.
эволюц. биохимии и физиол., 1, 5 : 435. — Быховец А. У. 1955. Га-
зообмен в гусином яйце и нормативы вентиляции при инкубации. Научн. тр.
Укр. н.-иссл. ст. птицеводства, 5 : 129. — Бэр К. М. 1828. История раз-
вития животных. Наблюдения и размышления, I. Изд. АН СССР, 1950.
Винников Я. А. 1946. Опыт филогенетической классификации
рецепторов органов чувств. Журн. общей биол., 7 : 347. — Винни-
ков Я. А. 1966. Структурные и цитохимические основы механизма функции
25*
379
рецепторов органов чувств. В кп.: Нервная клетка. Изд. ЛГУ, : 7. —
Винников Я. А., Л. К. Титова. 1949. Гистофизиологические осо-
бенности обонятельного рецептора. ДАН СССР, 65, 6 : 903. — Винни-
ков Я.А., Л.К. Титова. 1956. Развитие органов чувств позвоночных.
В сб.: Пробл. совр. эмбриол., Изд. ЛГУ : 89. —- В и н н и к о в Я. А.,
Л. К. Титова. 1957. Морфология органа обоняния. Изд. мед. литера-
туры, М. —- В и ч и Э., Э. Дэйл. 1962. Стероидные гормоны в раннем
развитии. Успехи соврем, биол., 53, 1 : 124. — Владимирова Ю. Н.
1954. Покря в весе яиц кур перед инкубацией. Тр. Н.-иссл. инет, птицевод-
ства, 24 : 126. — В л а д и м и р с к и й А. П. 1935. Экспериментальное
изучение процесса инкубации яиц. Уч. зап. ЛГУ, сер. бпол., 1:3.—
Войн о Ясенецкпй А. В., А. В. Б у р с и а н. 1963. Развитие
двигательной активности у куриных эмбрионов. Физиол. журн. СССР, 49,
5 : 609. — Войткевич А.А. 1936. Исследование по развитию щитовид-
ной железы птиц. Тр. Инет, экспер. морфогенеза, 5 : 343. — Воло-
хов А. А. 1951. Закономерности онтогенеза нервной деятельности в свете
эволюционного учения. Изд АН СССР, М.
Га врилепко А. В. (Gawrilenko A. W.) 1910. Die Entwi-
cklung des Geruchsorgan bei Salmo salar. Anat. Anz., 35 : 411. — Г а л-
вяло М.Я., T.A. Горюхипа. 1937. Ферментативные системы разви-
вающегося куриного эмбриона. Физиол. журн. СССР, 22, 2 : 215. — Г а ш-
к о в а В., 3. Покорная. 1955. Питание зародыша через рот. Folia
Biologica, 1, 2:113. — Гинецп некий А. Г. 1963. Физиологические
механизмы водно-солевого равновесия. Изд. АН СССР, М.—Л. — Глик Б.
1966. Рост, жизнеспособность, иммунная реакция и воспроизводительная
способность птиц, выведенных из яиц, погружаемых в растворы, содержащие
пропионат тестостерона. Тр. XIII Всемирного конгр. по птицеводству,
Киев : 297. — Говард овскиб В. И., Т. А. Харкеевич. 1965.
Гистологические и электронноскопические исследования развития фоторецеп-
торных клеток в условиях тканевых культур. Арх. анат., гистол. и эмбриол.,
49, 8 : 50. — Горбачева А. П., О. М а с л и е в а. 1951. Закономер-
ности азотистого (белкового) обмена куриного эмбриона на различных ста-
диях развития. Докл. ВАСХНИЛ, 7:41. — Горшелева Л. С. 1936.
К сравнительно-физиологической характеристике высшей нервной деятель-
ности птиц в связи с влиянием возраста. Арх. биол. наук, 42, 1—2 : 117. —
Гофман Д. Н. 1952. О различиях в темпах роста и дифференцировки
в эмбриональном развитии грача и курицы. Вестник МГУ, 3 : 55. — Г о ф-
-м а н Д. Н., Н. Н. Ротт. 1961. Развитие скелета крыла в эмбриогенезе
птиц разных типов размножения. Бюлл. Моск. общ. испыт. прир., Отд. биол.,
66, 1 : 40. — Г р о м А., М. Галай, Ш. И в а н к о, И. Киселе-
в п ч, М. К арлубик. 1966. Изучение некоторых физиол огически-био-
химических вопросов регулировки соотношения пола у птицы. Тр. XIII
Всемирного конгр. по птицеводству, Киев : 549.
Делль Т. Р. 1953. Зависимость между жизненностью цыплят и строе-
нием их зобных желез. Уч. зап. ЛГУ, сер. биол., 33 : 240. —Демен-
тьев Г. П. 1940. Руководство по зоологпп. Позвоночные. Птицы, 6. Изд.
АН СССР, М.—Л. — Джафф У. П., Н. С. Фекхеймер. 1966. Иден-
тификация соматических хромосом птиц и их использование при изучении
развития. Тр. XIII Всемирного конгр. по птицеводству. Киев : 28. —
Дмитриева Е. В. 1939. Зобная железа и жизнеспособность. I. Вес
зобной железы у куриных эмбрионов. Бюлл. экспер. бпол. и мед., 8, 1 :
38. — Долбепева Е. Ф., М. В. Орлов. 1966. Жизнеспособность
и рост цыплят в связи с интенсивностью обмена веществ в период инкубации.
Тр. Всесоюзн. н.-иссл. тёхнол. инет, птицеводства, 30 : 96. — Д р е л ь К. А.
1964. Активность ферментных систем азотистого катаболизма по ходу разви-
тия куриного эмбриона. Тезисы докл. 1-го биохим. съезда, 2, Л. : 13. —
Дрель К. А. 1965. Активность ферментных систем азотистого катабо-
380
лизма в ходе развития куриного эмбриона. Автореф. дисс. Харьковск.
унив. — Дуюнов Э. А. 1966. О наследовании признаков у уток. Тр.
Всесоюзн. н.-иссл. технол. инет, птицеводства, 30 : 7.
Еремеев Г. П. 1957. Зародышевые приспособления птиц с точки
зрения сравнительной физиологии. Тр. Омск, сельскохоз. инет., 25 : 1. —
Еремеев Г. П. 1964. Сравнительная характеристика осадков аллан-
тоисной жидкости эмбрионов птиц. Тезисы докл. 1-го биохим. съезда, Л.,
2 : 14.
Ж е л у д к о в а 3. П. 1963. Фракции гликогена в печени, мышцах п
желточном мешке куриного эмбриона. Биохимия, 28, 1 : 31. — Жму-
рив Л. М., В. И. Михайлов. 1966. Влияние аминокислот на форми-
рование пола у кур. Тр. XIII Всемирного конгр. по птицеводству. Киев : 277.
Задарновская Г. Ф. 1954. Дипамика эмбрионального крове-
творения у кур в свете новой клеточной теории О. Б. Лепешинской. Тр.
Ставроп. сельскохоз. инет., 6 : 491. - Зусман И. Н. 1965. О темпах
развития органов пищеварения в эмбриогенезе рептилий и птиц в связи
с особенностями питания их плодов. Автореф. дисс., Л.
Иванов П. П. 1945. Руководство по общей и сравнительной эмбрио-
логии. Учпедгиз, Л. — И в а п о в В. Г. 1955а. Различная чувствительность
самцов и самок эмбрионов кур. ДАН СССР, 100, 4 : 829. — Иванов В.Г.
19556. Влияние срока хранения яиц на соотношение полов в потомстве кур.
ДАН СССР, 100, 5 : 1021. — Иванов В. Г. 1962. О соотношенип полов
в потомстве животных. Уч. зап. Кабардино-Балкарск. унив., 12 : 187. —
Ильин М. Д. 1917. Исследования по развитию зародыша куриного яйца.
Материалы для эмбриохпмпп. Петроград.
♦Калояпу-Иордэкэл М. 1961. К изучению развития скелета
у птиц. Rev. de Biologie, 6, 3 : 295 (на румынск. яз.). — Карапе-
тян С. К.. 1964. Влияние возраста родителей па пол потомства у сельско-
хозяйственных животных и птиц. Журн. общей биологии, 26, 6 : 401. —
Квинихидзе Г. С. 1958. Развитие эпителиальной выстилки пищевода
домашних кур. ДАН СССР, 122, 2 : 304.— Квинихидзе Г. С. 1964.
Развитие стыка между эпителием пищевода и железистого желудка у кур
в эмбриогенезе. Пробл. соврем, эмбриол., : 369. — Кирпичникова Е.
1956. Гликоген в сердечной мышце зародыша курицы. ДАН СССР, ПО,
2 : 285. — Клюева С.Г. 1958. Гистогенез некоторых органов утиных за-
родышей в связи с периодическими изменениями роста и дыхания. Узб. биол.
журн., 6 : 85. — К н о р р е А. Г. 1941. Процесс возникновения энтодермы
у птиц и его отношение к гаструляцпи. Успехи соврем, биол., 14, 2:318.—
Кнорре А. Г. 1949а. Дифференцировка энтодермы у птиц. Уч. зап. ЛГУ,
сер. биол. наук, 20, 113 : 301. — Кнорре А. Г. 19496. Эмбриональное
развитие вегетативной нервной системы позвоночных. Успехи соврем, биол.
27, 1 : 37. — К и о р р е А. Г. 1959. Краткий очерк эмбриологии человека,
с элементами общей, сравнительной и экспериментальной эмбриологии. Мед-
гиз, Л. — Князева А. П. 1958. Газообмен яиц в зависимости от темпе-
ратурных условий их хранения. Научн. зап. Белоцерковского сельскохоз.
инет., 8 : 89. — К о н с т а н т и н о в а М. С., Л. Г. Лейбсон,
Е. А. Моисеев. 1960. Микроскопические изменения в печени куриного
зародыша, вызванные кортизоном. Пробл. эндокринол. и гормонотерап., 6,
4 : 42. — К о р ж у е в П. А., В. П. Галкина. 1957. Количество крови
в организме птиц в период инкубации. ДАН СССР, 116, 4 : 710. — К о р о-
в и н а В. М. 1956. Развитие специфических реакций по отношению к разно-
качественным раздражителям в период раннего онтогенеза у рыб и амфибий.
Пробл. совр. эмбриологии, Изд. ЛГУ : 129. — Костомарова А. А.
1956. Развитие секретирующего и всасывающего аппарата кишечника цы-
пленка в период инкубации. ДАН СССР, 111, 3 : 720. — Коштоянц X. С.
1940. Основы сравнительной физиологии, ч. 1. Изд. АН СССР, М.—-Л. -•
381
Коштоянц X. С. 1957. Основы сравнительной физиологии, т. 2. Срав-
нительная физиология нервной системы. Изд. АН СССР, М. — К р а fl-
чи некий Б. Д. 1958. Современные основы физиологии почек. Медгиз,
Л. — Крепе Е.М. 1943. Очерк по эволюции дыхательной функции крови.
Журн. общей бпол., 4 : 159. —- К р о к Г. С. 1949а. К вопросу о строении
и функции печени эмбриона курицы в связи с возрастом. Тр. Харьковск.
ветер, инет., 20 : 42. — К р о к Г. С. 19496. Эмбриональное развитие и пере-
ход в постэмбрионалыюе состояние органов выделения птиц. Тезпсы докл.
5-го Всесоюзн. съезда анат., гистол. и эмбриол., Л. : 269. — Крок Г. С.
1952а. Эмбриональный рост и переход в постэмбрпональное состояние орга-
нов выделения уток. Сб. тр. Харьковск. ветер, инет., 21 : 43. — К р о к Г. С.
19526. Определение функционального состояния клеток почечного эпителия
методом витальной окраски. Сб. тр. Харьковск. ветер, инет., 21 : 60. —
Крок Г. С. 1954. Динамика иннервации почек эмбрионов и цыплят в за-
висимости от условий инкубации. Сб. тр. Харьковск. ветер, пнет., 22 :
99. — К р о к Г. С. 1962. Микроскопическое строение органов сельскохо-
зяйственных птиц с основами эмбриологии. Изд. АН УССР, Киев. — *К р ы-
стев X. 1958. Гистогенез легких курицы. Научп. тр. Высш,
ветер.-мед. инет., 6 : 35. (на болг. яз.). — Курбатов А. Д. 1960. Влия-
ние обработки яиц диэтилстилбестролпропионатом на пол цыплят. Третье
совещ. эмбриологов, тезисы докл. : 88. — К у р б а т о в А. Д. 1966. Опыты
по регулированию соотношения полов в потомстве кур. В кн.: Наследствен-
ность и изменчивость сельскохозяйственной птицы. Изд. «Колос», М. : 22.
Лаврентьева Г.Ф. 1963. К вопросу о желчеотделительной функ-
ции печени у эмбрионов кур. Тр. Горьковск. сельскохоз. инет., 13 : 86. —
Левинсон Л. Б., М. И. Лейкина. 1956. Морфологическое и ги-
стохимическое изучение моторных и чувствительных нервных клеток эмбриона
курицы на разных этапах развития. ДАН СССР, 109, 1 : 201. — Левин-
сон Л. Б., М. И. Лейкина. 1960. Цитохимия чувствительных п мо-
торных клеток системы спинного мозга куриного зародыша в связи с осо-
бенностями их функции на разных этапах развития. Цитология, 2,1:9. —
Лейбсон Л. Г. 1950. Содержание гликогена в печени у куриных эмбри-
онов в различные дни инкубации. Физиол. журн. СССР, 36 : 191. — Лейб-
сон Л. Г. 1951. Влияние инсулина на содержание гликогена в печени у ку-
риных эмбрионов. Физиол. журн. СССР, 37 : 343. — Лейбсон Л. Г.
1962. Сахар крови. Регуляция содержания сахара в крови у животных и
человека. Изд. АН СССР, М.—Л. — Л е й б с о н Л. Г. 1964. Проблема
становления гормональных эффектов в эмбриогенезе. В сб.: Эволюция функ-
ций, Изд. «Наука», М.—Л. : 229. — Лейбсон Л. Г., 3. П. Желуд-
кова. 1960. Влияние кортизона на гликогенную и желчеотделительную
функции печени у куриных эмбрионов. В сб.: Эволюция физиол. функции,
Изд. АН СССР, М.—Л. : 180. — Л е й б с о н Л. Г., 3. П. Желуд-
кова, Л. И. Чилингарян. 1961. О секреции инсулина поджелудоч-
ной железой куриного зародыша. Бюлл. экспер. бпол. н мед., 52, 7 : 24. —
Лейбсон Л. Г., Р. С. Лейбсон. 1943. Нервная и гуморальная регу-
ляция содержания сахара в крови в процессе онтогенеза. Сообщ. 2. Влияние
введения инсулина, адреналина и глюкозы на содержание сахара в крови
у куриных эмбрионов. Изв. АН СССР, Отд. биол. паук, 3: 176. — Лейб-
сон Л. Г., Э. М. Плисецкая, Л. Г. Огородникова. 1963.
Глюкозо-6-фосфатаза печени куриных эмбрионов в норме и в условиях дей-
ствия инсулина. ДАН СССР, 153,1 : 240. — Лейбсон Л. Г., Е.М. Ста-
бров с к и й. 1962. Содержание адреналина и норадреналина в надпочеч-
никах развивающихся куриных эмбрионов. Физиол. журн. СССР, 48, 7 :
857. — Лисицкий Е. Ф., Г. С. Крок. 1949. Эмбриональное развитие
цыпленка в инкубаторе комнатного типа («Укргигапт»). Тр. Харьковск.
ветер, пнет., 20 : 29. — Л о п а ш о в Г. В. 1961. Эмбриональная индукция
и закономерности причинных связей в развивающихся зачатках. Журн. об-
382
щей биол., 22, 4 : 241. — Л о п а ш о в Г. В., О. Г. Строева. 1963.
Развитие глаза в свете экспериментальных исследований. Изд. АН СССР, М.
Мазина Т. И. 1963. Влияние введения АКТГ па вес надпочечников
и содержание в них холестерина у развивающихся куриных эмбрионов. Фи-
зпол. журн. СССР, 49, 5 : 589. — М а з и и а Т. И. 1964. Содержание аскор-
биновой кислоты в надпочечных железах куриных эмбрионов при введении
АКТГ и инсулина. Бюлл. экспер. биол. и мед., 57, 6 : 54. — М а н у-
х и и Б. Н., Г. А. Бузи и к о в. 1959. Количественное изучение адре-
налина и норадреналина в надпочечниках куриного эмбриона. ДАН СССР,
127, 14 : 934. — Мануйлова Н. А., В. В. Попо в, Н. Б. Т и м о-
ш у к. 1963. Влияние прерывистого освещения па размеры глаз и зрительных
долей головного мозга у зародышей курицы. Журн. общей биол., 24, 2 :
98. — М а т в е е в Б. С. 1956. Учение об этаппостп (стадийности) индиви-
дуального развития животных. Пробл. соврем, эмбриол., Изд. ЛГУ : 111.—
М а х и и ь к о В. И. 1965. Биохимизм и энергетика эмбрионального раз-
вития сельскохозяйственных птиц. Матер, симпозиума по основным пробле-
мам возрастной физиол. и биохимии, Харьков : 52. — М а х и и ь к о В. И.,
А. А. Пашкова, Л. И. С т а в и ц к а я. 1956. Материалы к физиоло-
гии эмбрионального развития домашних птиц. IV. Изменение интенсивности
дыхания и скорости роста некоторых органов утиного эмбриона на протяже-
нии инкубации. Тр. Н.-иссл. ипст. биол. Харьковск. гос. унив., 24 : 235. —
М а х и и ь к о В. И., Е. Е. Сердюк. 1954. Материалы к физиологии
эмбрионального развития домашних птиц. I. О соотношении между ско-
ростью роста п интенсивностью дыхания утппых эмбрионов на протяжении
инкубации. Тр. Н.-иссл. ипст. бпол. Харьковск. гос. унив., 21 : 153. —
М а х и и ь к о В. И., Р. А. Шевченко. 1954. Материалы к физиологии
эмбрионального развития домашних птиц. II. Рост голубиных эмбрионов
и изменения их дыхания па протяжении инкубации. Тр. Н.-иссл. инет. бпол.
Харьковск. гос. унив., 21 : 171. — М е и г е р X. 1960. Определение чув-
ствительного к диэтилстилбестролу периода в процессе дифференцировки
половых желез у куриных эмбрионов. ДАН СССР, 133, 5 : 1255. — М е с а-
р о ш Б. 1957. О функциональном значении желточной оболочки куриного
яйца по ходу инкубации. Вестник ЛГУ, сер. бпол., 9, 2 : 99. — Митро-
фанов П. Н. 1898. Опыты над начальным развитием птиц. Работы Зо-
отомической лабор. Варшавск. унив., 19 : 1. — М и т р о ф а н о в П. Н.
1900. Сравнительные данные относительно начального развития птиц. Вар-
шава. — Мицкевич М. С. 1957. Железы внутренней секреции в зароды-
шевом развитии птиц и млекопитающих. Изд. АН СССР, М. — Мюл-
лер Г. Р., М. Н. Рагозина. 1957. Ранние стадии развития яичников
у кур двух пород различной скороспелости (русская белая и австралорп).
ДАН СССР, 115, 5 : 1044.
Немилов В. А. 1938. Эндокринология. Сельхозгиз, М. — * Н е-
стеров Н., Н. Банков. 1964. Влияние на синтетичнете естрогени
върху структурата на гонадите и нивото на серумните протеинп у пплето.
Изв. Инет, морфол. Бълг. АН, 9—10:97 (па болг. яз.).-Ники-
тина О. IO. 1963. Некоторые стороны углеводного обмена эмбрионов кур.
Укр. 6ioxiM. журн., 35, 2 : 193. — Н о й м е й с т е р X. 1966. Наблюдения
окостенения скелета эмбриона курицы. Тр. XIII Всемирного конгр. по пти-
цеводству, Киев : 543.
Огородний Ю. М. 1936. Дифференцирование температурного ре-
жима при инкубации. Пробл. животноводства, 2:108. — Огород-
ний 10. М., Э. Э. П е н и о н ж к е в и ч. 1939. Роль белкового и водного
обмена в патогенезе эмбриональной липкости цыплят. В сб.: Матер, к изу-
чению патол. эмбрион, развития птиц, Н.-иссл. пнет, птицеводства,
2:5. — Орбели Л.А. 1933. Об эволюционном принципе в физио-
логии. Природа, 3—4 : 77. — Орбели Л. А. 1945. Эволюционный
принцип в применении к физиологии центральной нервной системы.
383
Лекции по вопросам высшей нервной деятельности. Изд. АН СССР, М.—Л.—
Орбели Л. А. 1948. Теория адаптационно-трофического влияния
нервной системы. Тр. сессии АН СССР, 1947, М.—Л. : 674. — Ор-
лов М. В. 1938. Дифференцирование режима инкубирования как метод
получения высоких выводов цыплят. Н.-иссл. инет, птицеводства,
М. — О р л о в М. В. 1961. Методы дифференцирования режима инкуби-
рования яиц сельскохозяйственных птиц. Тр. Всесоюзн. н.-иссл. инет,
птицеводства, 27 : 130. — Отрыганьев Г. К. 1949. О закономерностях
роста эмбриональных оболочек у птиц. Докл. Моск, сельскохоз. акад,
им. К. А. Тимирязева, 9 : 133. — Отрыганьев Г. К. 1951. Биологиче-
ский контроль инкубации. Сельхозгиз, М. — Отрыганьев Г. К. 1966.
Некоторые конституциональные особенности в эмбриональном развитии кур.
Тр. XIII Всемирного конгр. по птицеводству, Киев : 530. — О т ры-
та и ь е в Г. К., В. А. X м ы р о в, Г. М. К о л о б о в. 1964. Инкубация. Изд.
«Колос», чМ. — Отрыганьева А. Ф. 1963. О периоде вылупления
у сельскохозяйственных птиц. Тезисы докл. IV совещ. эмбриологов, Изд.
ЛГУ, 141.
Пенионжкевич Э. Э. 1936. О газопроницаемости подскорлуп -
пых оболочек яиц. Успехи зоотехн. наук, 2, 3 : 461. — II е п и о п ж к е-
в и ч Э. Э. 1939. К вопросу о гистогенезе Pars hepatica куриных эмбрионов.
В сб.: Матер, к изучению патол. эмбриоп. развития птиц, Н.-иссл. инет,
птицеводства, 2 : 93. — П е н и о и ж к е в и ч Э. Э. 1941. Изменение эм-
бриональной смертности кур в разных географических широтах. ДАН СССР,
32, 8 : 594. — Пенионжкевич Э. Э. 1945. Проблема снижения смерт-
ности эмбрионов домашних птиц. Сельхозгиз, М. — Перцева М. Н.
1961. Содержание молочной кислоты в тканях развивающегося куриного
эмбриона. Биохимия, 26, 2 : 254. — Перцева М. Н. 1963а. О путях
гликолиза в эмбриональных мышцах. Укр. 6ioxiM. журн., 2 : 298. — Пер-
цева М. Н. 19636. Глюкокиназная активность скелетных мышц куриного
зародыша. Укр. 6ioxiM. журн., 1 : 92. — П е т р о в Ф. А. 1958. Обмен
пода и изменение гистологической структуры щитовидной железы у куриных
эмбрионов. Тр. Омск, сельскохоз. инет., 23, 2 : 58. — П и г а р е в а 3. Д.
1948. Угольная ангидраза в мозгу зрелорождающихся и незрелорождающихся
птиц. ДАН СССР, 60, 1 : 185. — Пилипенко М. Е. 1965. О вилочковой
железе птиц. Птицеводство, 2 : 22. — П о д о б а Е. Г., Л. Я. Тоцкая.
1961. К вопросу о смещении пола у птиц. Тр. Харьковск. сельскохоз. пнет.,
34, 81 : 63. — П р о м п т о в А. Н. 1948. О некоторых закономерностях
онтогенетического развития поведения птиц в связи с проблемой эволюции
функций нервной системы. Журн. общей биол., 9, 2 : 145. — П р и ц-
к е р И. Я. 1939. К вопросу о закономерностях роста птичьих эмбрионов.
ДАН СССР, 24, 8 : 823. — П р и ц к е р И. Я., Н. П. Т р е т ь я к о в. 1937.
Инкубаторы и их эксплуатация. Сельхозгиз, М.
Рагозина М. Н. 1955. Способы питания п дыхания сельскохозяй-
ственных птиц в различные периоды развития во время пнкубацпп. Изв.
АН СССР, сер. бпол., 4 : 95. — Р а г о з и н а М. Н. 1957. Об освобождении
зародыша цыпленка от желточной и халазовой оболочек яйца. ДАН СССР,
ИЗ, 3:716. — Рагозина М. Н. 1961. Развитие зародыша домашней
курицы в его соотношении с желтком и оболочками яйца. Изд. АН СССР,
М. — Ревуцкая П. С. 1944. Различные пути развития гладкой мышеч-
ной ткани амниона куриного зародыша. Бюлл. экспер. биол. и мед., 17, 6 :
68. — Рольи и к В. В. 1946а. Влияние изменения состава газов в воз-
душной камере яйца на развитие эмбриона и повышение выводимости цыплят.
ДАН СССР, 52, 9 : 831. — Рольник В. В. 19466. К вопросу выводи-
мости цыплят. Совх. производство, 4—5 : 44. — Рольник В. В. 1948.
Развитие терморегуляции у некоторых птиц севера. Зоол. жури., 27, 6 :
535. — Рольник В. В. 1956а. Физиология дыхания эмбрионов сельско-
хозяйственных птиц, Пробл. соврем, эмбриол., Изд. ЛГУ, : 228. - Роль-
384
ник В. В. 19566. Опыты по снижению смертности эмбрионов сельскохозяй-
ственных птиц в последние дни инкубации. Тезисы докл. совещ. по пробл.
индивидуального развития сельскохоз. животных. Изд. АН УССР, Киев :
125. — Рольник В. В. 1960. Изучение состава газов воздушной камеры
куринных яиц в течение инкубации. В сб.: Матер, по эволюц. физиол. 4, Л.—
М. : 208. — Рольник В. В. 1961. Изучение газообмена развивающегося
куриного эмбриона. III Научн. совещ. по эвол. физиол., посвящ. памяти
Л. А. Орбели, Тезисы докл., Л. : 164. — Р о л ь н и к В. В. 1963а. Методика
изучения газообмена эмбрионов птиц. Физиол. журн. СССР, 49, 8 : 1000. —
Р о л ь и и к В. В. 19636. Закономерности газообмена куриных эмбрионов
в течение инкубации. Тезисы докл. IV Совещ. эмбрпол., 2 : 162. — Роль-
ник В. В. 1964. Биометрический анализ некоторых сторон развития кури-
ного эмбриона. В сб.: Применение матем. методов в биологии, Изд. ЛГУ,
3:159. — Рольник В. В. 1965. Некоторые вопросы физиологии дыхания
птиц. Тезисы докл. Совещ. по физиологии птиц, Таллин: 87. — Роль-
ник В. В. 1966. Некоторые закономерности дыхания эмбрионов сельско-
хозяйственных птиц. Тр. Всесоюзн. п.-пссл. технол. инет, птицеводства,
30 : 129 — Р о л ь п и к В. В., В. В. Гаврилов. 1962. Влияние замены
азота воздуха гелием в течение инкубации на рост и развитие взрослых птиц.
В сб.: Матер. 3-й Всесоюзн. орнитол. конфер., Львов, 2 : 160. — Р о л fe-
ll и к В. В., Э.Л. Портенко. 1964. Газообмен развивающегося кури-
ного эмбриона. Жури, общей биол., 25, 2 : 133. — Р о т т И. Н. 1957. Влия-
ние температуры инкубации на развитие системы кровообращения в эмбрио-
нальном и постэмбрионалыгом периоде у кур породы леггорн. Автореф.
дпсс. М.
Савин Б. М., В. В. Р о л ь п и к, П. А. Г у л ь т я е в, В. В. Б о-
р и с к и н. 1964. Анализ влияния замены азота воздуха гелием па развитие
куриных эмбрионов. Биофизика, 9, 5 : 589. — Саксен Л., С. Той-
во н е п. 1963. Первичная эмбриональная индукция. ИЛ, М. — Свет-
ло в П. Г. 1960. Теория критических периодов развития и ее значение для
понимания действия среды па онтогенез. В сб.: Вопр. цитол. и общей физиол.,
Изд. АН СССР, М.—Л. : 263. — Светлов П. Г. 1963. О значении тео-
рии зародышевых листков в современной науке. (Памяти А. О.Ковалевского).
Архив анат., гистол. и эмбриол., 44, 4 : 7. — С в п д е р с к а я Г. Е. 1968.
Влияние звука разных частот и вибраций на двигательную активность ку-
риных эмбрионов. Журн. эволюц. биохимии и физиол., 4, 3 : 251. — Сер-
геев А. М. 1943. Эволюция эмбриональных приспособлений рептилий.
Изд. «Советская наука», М. — Сергеева А. М. 1966. Изменение физико-
химических свойств околоплодных жидкостей куриных эмбрионов в зависи-
мости от различных условий. Тр. Всесоюзн. н.-иссл. технол. инет, птицевод-
ства, 30:104. —*Сиракава. 1957. Можно ли изменить соотношение
полов у кур? О действии метиландростендиола. Ниваторп-но кэнкю,
32, 11:44 (на японск. яз.). — С к а л и н с к и й И. Е., В. Ф. Кон-
да л е н к о. 1963. Электронномпкроскопическое изучение хорпоаллантопс-
ной оболочки куриного эмбриона. Архив анат., гистол. и эмориол., 44, 4 :
44. — С м и т т е п И. А., Л. П. Малиновская. 1960. К вопросу
о развитии хромаффинных элементов в брюшной области птиц. ДАН СССР,
135, 12 : 445. — С т а б р о в с к и й Е.М. 1964а. Влияние инсулина па
содержание адреналина п норадреналина в надпочечниках куриных эмбрионов
и цыплят. Физиол. журн. СССР, 50, 3 : 365. — С т а б р о в с к и й Е. М.
19646. Функции мозгового вещества надпочечников в онтогенезе некоторых
животных. В сб.: Становление эндокр. функций в онтогенезе, М. : 90 (те-
зисы). — Студите к ий А. Н. 1947. Эндокринные корреляции зароды-
шевого развития высших позвоночных. Изд. АН СССР, М.-Л.
Титова Л. К. 1966. Рецепторные структуры внутреннего уха позво-
ночных (развитие, строение и функция). Автореф. дпсс. Изд. АН СССР,
Л. — Т и т о в а Л. К., Я. А. Вппн и к о в. 1964. Сравнительное гпето-
385
химическое исследование холинэстераз в рецепторных структурах лабиринта
позвоночных. В сб.: Эволюция функций, Изд. «Наука», М.—Л. : 249. —
Тихомиров Б. М. 1935. Корреляция между общим состоянием цып-
ленка п состоянием его тимуса. Уч. зап. ЛГУ, Труды Биол. ипст. : 13. —
Т о к п н Б. П. 1943. Феномен «организатора» и химические увлечения
эмбриологов. Журн. общей биол., 4, 1:3.
Фетцщева М. И. 1939. Исследование крови и околоплодных жид-
костей утиных эмбрионов. Матер, к изучению патол. эмбрион, развития птиц,
Н.-иссл. пнет, птицеводства, 2 : 70. — Филатов Д. П. 1943. Механика
развития как метод изучения некоторых вопросов эволюции. Журн. общей
биол., 4, 1 : 28.
Хаск пн В. В. 1959. Теплообмен п развитие терморегуляции в эмб-
риогенезе уток. Научные труды Укр. опытной ст. птицеводства, 6 : 183. —-
Хаскин В.В. 1961. Теплообмен и развитие терморегуляции в онтогенезе
домашней утки. Автореф. дисс. Харьковск. унив. — Хлыстова 3. С.
1959. К вопросу о развитии п строении кожи и ее производных у птиц. II.
Эмбриональный гистогенез кожи кур. Уч. зап. Оренбургск. отд. Всесоюзн.
научн. общ. апат., гистол. и эмбриол., 2 : 87.
♦Чжан, Чжин, Чжан Чун-ли, Сунь-Хай-бао,
Ц з я н - Л п н - г э п ь. 1959. Искусственное изменение пола у цыплят.
Научи, вести., 1 : 25 (на кит. яз.). — Чумак В. И. 1960. Динамика
активных «спонтанных» и рефлекторных двигательных реакций кури-
ного эмбриона. В сб.: Эволюция физиол. функций, Изд. АН СССР, М.—Л. :
77. — Ч у м а к В. И. 1961. Динамика рефлекторных реакций и включение
рецепторных аппаратов у эмбриона курицы. В со.: Вопр. физиол. и патол.
ц. н. с. человека и животных в онтогенезе, М. : 63. — Ч у м а к В. И.,
Н. Л. Писарева. 1961. Стрихнинные разряды в спинном мозгу кури-
ных эмбрионов. В сб.: Вопр. физиологии и патологии ц. н. с. человека и жи-
вотных в онтогенезе, М. : 68.
Шевцова М. Я. 1965. Рост и дыхание в зародышевом периоде эмбрио-
генеза домашней утки. Матер, симпозиума по основным пробл. возрастной
физиол. и биохимии, Харьков : 201. — Шейнис С. А. 1954. Питание ку-
риного зародыша в последние дни инкубации. ДАН СССР, 96, 5 : 1077. —
Шейнис С. А. 1964. Питание птичьего зародыша на разных стадиях
его развития. Пробл. соврём, эмбриол., Изд. ЛГУ : 361. — Шмальга-
у з е н И. И. 1926. Дослщи над ембрюнальним ростом. Сб. работ Биол.
инет., Харьков, 1:15. — Шмальгаузен И. И. 1947. Основы срав-
нительной анатомии. Сов. наука, М. — Шмидт Г. А. 1951. Эмбриология
животных. Ч. 1. Общая эмбриология. Изд. «Советская наука», М. —
Шмидт Г. А. 1953. Эмбриология животных. Ч. II. Частная эмбриология.
Изд. «Советская наука», М.
Щеглов А. М. 1949. К вопросу о водном обмене у куриных эмбрионов.
Тр. Харьковск. ветер, инет., 20 : 48. — Щ е р б и н а П. Ф. 1959. Эмбрио-
нальное развитпо гусей при искусственной и естественной инкубации. Научн.
тр. Укр. ст. птицеводства, 6 : 197.
Энтин Т. И. 1936. К вопросу о морфогенезе щитовидной железы
птпц. Архив анат., гистол. и эмбриол., 15, 1 : 104.
Янковская Ц. Л. 1949. К вопросу о развитии регуляции сердеч-
ной детельности у кур в онтогенезе. Физиол. журн. СССР, 35, 2 : 223.
Ackerman G. A., R. A. Knouff. 1959. LymphocyCopoiesis in
the bursa of Fabricius. Amer. Journ. Anat., 104, 2 : 163. — A c k e r-
m a n G. A., R. A. Knouff. 1964. Lymphocyte formation in the thymus
of the embryonic chick. Anat. Rec., 149, 2 : 191. — Agazzotti A. 1914.
Influenza dell’aria rarefatta sull’ontogenesi III. Le modificazione che avven-
386
gano nei gaz della camera d’aria dell’ ovo durante lo sviluppo. Arch. f. Entwick-
lungsmech. d. Organ, 40 : 65. *A 1 b u N., J. В i e n о г, M. В й 1 й ceanu.
1960. Биохимические изменения при управляемом эмбриогенезе. Некоторые
биохимические изменения в тканях курипых зародышей, обработанных АКТГ.
Studii cerceUri endocrinol. Acad. RPR, 11, 1:129 (на румынск. яз.). Al-
len S. С. 1961. Response of the developing vascular system of the chick embryo
to hyperoxia. Fed. proc., 20, 1:421. — Allenspach A. L., H. L. Hamil-
ton. 1962. Histochemistry of the oesophagus in the developing chick. Journ.
MorphoL, 111, 3:321. —A nee 1 P. 1955a. Inversion et determinisme de
la symmetrie bilaterale dans les oeufs d’oiseaux. C. R. Acad, sci., 241,
19 : 1246. — A n c e 1 P. 1955b. Sur le determinisme de la symmetrie bilate-
rale de 1’embryon des oiseaux dans 1’uterus et le retournement de 1’oeuf dans
cet organe. C. r. Acad, sci., 241, 21 : 1433.
*B a f f о n i G. M., G. D’A n с о n a. 1958. Наблюдения над морфо-
генезом и гистогенезом мозжечка у птиц с поздним и ранним развитием.
Сообщ. I и II. Atti Acad. naz. Lincei Rend Cl. Sci. fis., mat. e natur., 24,
4 : 456; 5 : 606 (на итал. яз.). — Bakounine S. 1895a. Sur Г evolution
des fonctions embryonnaires. Recherches experimentales. Arch. ital. biol.,
23 : 420. — Bakounine S. 1895b. Sur 1’activite secretaires des epithe-
liums de Wolff et des epitheliums renaux dans les premiers jours du developpe-
ment embryonnaire. Arch. ital. biol., 23 : 350. -*ВйНсеапи M.,
J. Biene r, N. Albu, Chivu-Viorica. 1961. Изучение влияния
гормона эпифиза на развивающихся куриных зародышей. Studii $i cercetari
endocrinol. Acad. RPR, 12 : 59 (на румынск. яз.). — В a mn i R. K.,
R. N. S h о f f n e r, G. J. H о i d e n. 1966. Sex chromosomes in the germ
cells of the chicken, turkey and Japanese quail. Poultry Sci., 45, 2 : 424. —
Barnwell F. H. 1960. A solar daily variation in oxygen consumption
of the embryonated egg. Proc. Soc. Exptl. Biol, and Med., 105, 2:312.—
В a г г о n P., J. McK e n z i e. 1962. The inhibitory action of insulin
in the early chick embryo. Journ. Embryol. and Exptl. MorphoL, 10, 1:88.—
Barry A. 1948. The functional significance of the cardiac jelly in the tubu-
lar heart of the chick embryo. Anat. Rec., 102, 3 : 289. — Bautzman H.,
E. Dunker, R. Schroder. 1954. Registrierung der Motorik des
Amnions und des embryonalen Herzens beim Hiihnchen mittels des Kathoden-
strahl-Oszillographen. Anat. Anz., 101, Erganzungsheft : 317. — В a u t z-
manH., R. Schroder. 1954. Amnionstudien zur vergleichendcn Histo-
genese, Struktur und Funktion des Amnions beim Hiihnchen und beim Menschen.
Verhandl. Anat. Ges., 100, Erganzungsheft : 135. — Bautzman H.,
R. Schroder. 1958. Comparative studies on the histology and function
of the amnion. Acta Anat., 33, 1/2 : 38. — *B e g 1 i о m i n i A., M о r i-
c о n i. 1959. Time of formation and localization of cholinesterase in the
striated muscle of the chick embryo. Riv. Biol. (Perugia), 51,
4 : 517. — Beilharz R. G. 1960. The absence of sex-reversed males
among progeny of oestrogen-treated pullets. Poultry Sci., 39, 4 : 906. —
Bellairs R. 1960. Development of birds. In: Biology and comparative
physiology of birds, 1 : 127. — Benoit J. 1950. Reproduction. Characters
sexuels et hormones determinant du cycle sexuel saisonaire. Traite de Zoologie,
Paris, XV, Oiseaux : 384. — Benoit J. A. 1962. Le role^ de 1’hypo-
physe dans le developpement des embryons des vertebres ainniotes a sang chaud.
Annee biol., 1, 5—6 : 297. — BienerJ., M. В a H ceanu, N. Albu.
1959. Biochemical modifications in chick embryo tissue treated with cortisone.
Studii $i cercetAri endocrinol., Acad. RPR, 10 : 453. — Bogner Ph. H.,
F. H.-Jr. Wilcox. 1956. The ability of white leghorn embryos and young
chicks of various ages to absorb sugars from the gastrointestinal tract. Poultry
Sci., 35, 5 : 1134. — В о h г С., К. A. H a s s e 1 b a 1 c h. 1900. Ueber die
Kohlensaureproduction des Hiihnerembryos. Scand. Arch. Physiol., 10 : 149. —
В о h г С., К. A. H a s s e 1 b a 1 c h. 1903. Ueber die Warmeproduction
und den Staffwechsel des Embryos. Scand. Arch. Physiol., 14 : 398. —
387
♦Bondi M. 1957. Osservazioni sulla specifica genetica dei seprimenti intra-
cardiaci nel cuore dell’embrione di polio. Rend. 1st. super, sanita, 20, 11 :
1079. — Bonichon An. 1958. Acetylcholinesterase in neurons and nerve
fibers during development. I. Early biochemical differentiation of the neuro-
blast. Ann. Histochem., 3 : 85. — В о u с e к R. J., В. B. Bourne. 1962.
Catecholamines of the allantoic fluid in the developing chick embryo. Nature
(Engl.), 193, 4821 : 1181. — Brandes G. 1924. Beobachtungen und Refle-
xionen uber die Atmung der Vogel. Arch. f. d. ges. Physiol. (Pfliigers), 203 :
492. — Bremer J. L. 1928. Interpretation of development of heart;
left aorta of reptiles. Amer. Journ. Anat., 42 : 307. — *B г о z z i J. 1940.
Ricerche sull’istogenesi dell ghiandole surrenali negli Uccelli. (Gallus e Co-
lumba). Arch. ital. anat. e embryol., 44, 4 : 551. — В г о d у S. 1927. Growth
and development, with special reference to domestic animals. Univ, of Missouri
Agr. Exp. Sta. Bull. 97 : 1. - Brody S., E. W. Henderson. 1927.
Growth and development with special reference to domestic animals. V. The
effect of temperature on the percentagerate of growth of the chick embryo.
Univ., of Missouri Agr. Exp. Sta. Bull., 99 : 1. — В u c h n e r F.,
Ch. W. Mushett, H. Riibsaamen. 1953a. Elective Differenzie-
rungsstorungen des Zentralnervensystems am Hiihnchenkeim nach kurzfristi-
gem Sauerstoffmangel. Naturwissenschaften, 40, 24 : 628. — Buchner F.,
H. Riibsaamen, H. Naujoks. 1953b. Missbildungen am Hiihn-
chenkeim nach kurzfristigem Sauerstoffmangel in der Friihentwicklung. Natur-
wissenschaften, 40, 9 : 276. — В u r n s R. K. Jr. 1949. Hormones and the
differentiation of sex. In: Survey of Biological Progress, 1 : 233. —
Burns R. K. 1955. Urogenital system. In: Analysis of Development.
W. B. Saunders Company, Philadelphia : 462. — ♦BussE. G. 1956. Some
factors wich affect hatchability of chicken eggs at higher altitudes. Dissertation
Absts., 16, 11 : 1879. — Byczkowska-Smyk W. 1956. The cells of
the chick yolk sack and the production of digestive enzymes. Acta Biol. ExptL,
17, 1 : 27. —- В у e r 1 у Th. C. 1930. Time of occurence and probable causes
of mortality in chick embryos. Proc. 4th World’s Poultry Congress: 178. —
Byerly Th. C. 1938. Effect of different incubation temperature on morta-
lity of chick embryos. Poultry Sci., 17, 3 : 200.
С a 1 a m e S. 1959. Sur les relations entre le canal de Wolff et le deve-
loppement du mesonephros et de la gonade chez 1’embryon d’oiseau. C. r.
Acad, sci., 248, 21 : 3033. — Calame S. 1961. Le role des composants
epithelials et mesenchymateux du metanephros, d’apres les resultats de la
culture in vitro. Arch. anat. microscop, et morphol., 50, 3 : 269. — С a r p e n-
t e r F. G., R. M. В e r g 1 a n d. 1957. Excitation and conduction in imma-
ture nerve fibers of the developing chick. Amer. Journ. Physiol., 190, 2 : 371. —
Case J. F. 1952. Adrenal cortical — anterior pituitary relationschips during
embryonic life. Ann. N. Y. Akad. Sci., 55, 2 : 147. — ♦CattaneoL. 1962.
Некоторые данные о гематогенной зоне бластодермы куриного зародыша,
полученные с помощью светового и электронного микроскопа. Monit. zool.
ital., 70, Suppl. : 220 (на итал. яз.). — *С a t i n i С. 1963. Изучение дей-
ствия хлористого кобальта на печень куриного зародыша. Arch. ital. anat.
е embriol., 68, 3 : 235 (на итал. яз.). —Champion L. R. 1960. Sex
ratios in two strains of leghorns and their reciprocal crosses. Poultry Sci.,
39, 4 : 876. — C h a u b e Sh. 1962. Hypoxanthine dehydrogenase in the
developing chick embryonic kidney. Proc. Soc. Exptl. Biol, and Med., Ill, 2:
340. —Cohen J. 1959. The pigment cell system in the light sussex fowl.
Journ. Embryol. and Exptl. Morphol., 7, 3 : 361. —Coles R. 1958. Chan-
ging the sex of fowl. Agric. Rev., 4, 1 : 37. — С о 1 i g a d о E. C. 1953.
The relation of length of incubation period to growth mortality and sex ratio
in single comb white leghorn chicks. Philipp. Agr., 37, 3:111. — Crosti N.
1960. Calcium deposition in the forelimbs of the chick embryo. Anat. Rec.,
136, 4 : 453. —CsabayL., L. Horvath, A. Ferenz. 1960. Stu-
dies on the O2 consumption of chick embryos by a modified Warburg method.
388
Acta Biologia Ac. Sc. Hung., 11, 1 : 1. — C s a p о А., Й. Her man n.
1951. Quantitative changes in contractile proteins of chick skeletal muscle
during and after embryonic development. Amer. Journ. Physiol., 165 : 701.
Danchakoff V. 1907. Uber das erste Auftreten der Blutelemente
im Hiihnerembryo. Folia Haematologia, 4, Suppl. : 159. — Dancha-
koff V. 1908. Untersuchungen fiber die Entwicklung des Blutes und Binde-
gewebes bei den Vogeln. Anat. Hefte, 37: 113. — Danchakoff V. 1918.
Cell potentialities and differential factors, considered in relation to erythro-
poiesis. Amer. Journ. Anat., 24: 1. — Danchakoff V. 1931. Keimzelle
und Gonade. Ztschr. f. Zellforsch., 13 ; 1, 14 : 1. — D a n c h а к о f f V.
1935. Sur 1’inversion sexuelle oxperimentale de 1’ebauche testiculaire chez
I’embryon du poulet. C. r. Acad, sci., 200 : 1983. — Davis J. 1950. The
pronephros and the early development of the mesonephros in the duck. Journ.
Anat., 84 : 95. — D a w s о n A. B. 1953. Histochemical evidence of early
differentiation of the supprarenal gland of the chick. Journ. Morph., 92 : 579. —
*D e t h J. H. M. G., van. 1962. Changes in water, sodium and potassium
distribution in the ducks egg during incubation. Australian Journ.
Exptl. Biol, and Med. Sci., 40, 3 : 173. —Dieterlen-Lidvre Fr.
1963a. Le role de la thyroide dans le developpement embryonnaire des oiseaux,
et des mammiferes. Annee biol., 2, 1—2 :17. — Dieterlen-Lievre Fr.
1963b. Demonstration de 1’activite precoce des cellules A du pancreas chez
I’embryon de poulet. G. r. Acad, sci., 256,47 : 1597. — D о m m L. V. 1955.
Recent advances in knowledge concerning therole of hormones in the sex dif-
ferentiation of birds. In: Recent studies in Avian Biology. University of Illinois,
Press Urbana : 309. — D u f f e у L. M., J. D. Ebert. 1957. Metabolic
characteristics of the heartforming areas of the early chick embryo. Journ.
Embryol. and Exptl. Morphol., 5, 4 : 324. — D u m m M. E., M. L e v y.
1949. Chemistry of the chick embryo. VII. The accumulation of solids, nitro-
gen, lipids and peptidase by the gizzard and liver of the chick embryo. Journ.
Cellular Comp. Physiol., 33 : 373. — Duraiswami P. K. 1950. Insu-
lin-induced skeletal abnormalities in developing chickens. Brit. Med. Journ.,
46 : 384. — *D u r a i s w a m i P. K. 1955. Comparison of congenital defects
induced in developing chickens by certain teratogenic agents with those caused
by insulin. Journ. Bone and Joint Surg., 37-A, 2 : 277. — Duval M. 1889.
Atlas d’Embryologie. Masson, Paris.
Eakin R. E., J. R. Fisher. 1958. Patterns of nitrogen excretion
in developing chick embryos. In: A symposium on the Chemical Basis of Deve-
lopment (W. D. McElroy and Bentley Glass, eds.), Johns Hopkins : 514. —
Ebert J. D., K. A. Tolman, A. M. M u n, J. F. A 1 b r i g h t.
1955. The molecular basis of the first heart beats. Ann. N. Y. Acad. Sci., 60,
7 : 968. — Erhard R. 1959. Untersuchungen fiber die Insulin mikromelie
am Hiihnerembryo. Roux’ Arch. Entwicklunsmech. Organismen, 151, 4 : 381.
Faber H. 1959. Uber die hormonalen Ursachen des Geschlechtsdimor-
phismus im Wachstum der Vogel. Zool. Anz., Suppl., 22 : 345. — Feld-
man G. L., Ch. K. G r a n t h a m. 1964. Lipids of the embryonic liver.
Poultry Sci., 43, 1 : 150. — Francis D. W. 1962. Heart rate of day-old
S. G. White leghorn chicks. Poultry Sci., 41, 6 : 1960. — Fraser R. C.
1961. Hemoglobin formation in the chick embryo. Exptl. Cell. Res., 25,
2 : 418. — Fraser R. G. 1963. Cytochemistry of the developing chick
embryo erythrocytes. Journ. Exptl. Zool., 152, 3 : 297. —Fujita H.,
M. M a c h i n o. 1961. On the follicle formation of the thyroid gland in the
chick embryo. Exptl. Cell. Res., 25, 1 : 204.
♦Gambaro P. 1940. Ricerche sullo sviluppo della struttura e dell’
attivita funzionale della tiroide nell’embrione di polio. Arch. ital. anat. e
embryol., 44, 3 : 376. — Garcia-Austt *«E. 1954. Development of
electrical activity in cerebral hemispheres of the chick embryo. Proc. Soc.
389
Exp. Biol, and Med., 86 : 348. — *Garcia-Austt E., M. A. Pa-
tetta-Queirolo. 1961a. Electroretinogram of the chick embryo. I.
Onset and development. Acta Neurol. Latinoamer., 7, 3 : 179. — *Garcia-
A u s t t E., M. A. Pa tetta - Q ueirolo. 1961b. Electroretinogram
of the chick embryo. II. Influence of adaptation. Acta Neurol. Latinoamer.,
7, 4 : 269. — George J. C., D. J. Thomas. 1962. Lipase activity in
the chick liver during development. Proc. Exptl. Biol, and Med., 109, 4 : 826. —
GiajaJ., L. J о v a n ё i ё. 1950. Les oxydations au cours de la vie em-
bryonnaire du poulet. Bull. Acad. Serbe Sci., 1, 1 : 119. — G 1 i с к B. 1961.
The reproductive performance of birds hatched from eggs dipped in male hor-
mone solutions. Poultry Sci., 40, 5 : 1408. — Goff R. A. 1949. Develop-
ment of the mesodermal constituents of feather germs of chick embryo. Journ.
MorphoL, 85, 3 : 443. — G о m о t L. 1959. Contribution a Г etude embryon-
naire do la glande uropygienne chez le canard. Arch. anat. microsc. et morphol.
exptl., 48, 1 : 63. - Grabowski С. T. 1961. Lactic acid accumulation
as a cause of hypoxia — induced malformations in the chick embryo. Science,
134, 3487 : 1359. — Grignon G. 1955. Chronologic de la differenciation
des elements cellulaires du lobe distal de 1’hypophyse chez 1’embryon de poule
Rhode-Island. Compt. rend. Soc. Biol., 149, 13—14 : 1448. — Grillo T. A.
1961a. The ontogeny of function of the а-cells of the pancreas of the chick
embryo. Journ. Anat., 95 : 284. -У G r i 1 1 о T. A. I. 1961b. The ontogeny
of insulin secretion in the chick dmbryo. Journ. Endocrinol., 22, 3 : 285. —
Groebbels F. 1937. Der /Vogel. Bau, Funktion, Lebenserscheinung,
Einpassung. II : Geschlecht /und Fortpflanzung. Berlin. — Groenen-
dijk-HujjbergM. M; 1962. The cranio-caudal regression of the right
Mullerian duct in the chick embryo as studied by castration experiments and
estrogen treatment. Anat. Rec., 142, 1:9. — Gruenwald P. 1952. De-
velopment of the excretory system. Ann. N. Y. Acad. Sci., 55, 2 : 142. —
*Grundboeck M. 1963. Picture of peripheral blood and bone marrow
of chicken embryo. Biul. Inst, weteryn. Pulowasch.j 6, 1—2 : 3.
Haan R. L., de. 1963. Migration patterns of the precardiac mesoderm
in the early chick embryo. Exptl. Cell. Res., 29, 3 : 544. — Haffen K.
1960. Associations heterosexuees d’epithelium germinatif et de medullaire de
gonades embryonnaires de canard en culture in vitro. Compt. rend. Soc. Biol.,
154, 2 : 315. — Haffen K. 1963. Sur 1’evolution en greffes coelomiques
du constituant cortical isole des gonades femelles et intersexuees de 1’embryon
du poulet. C. r. Acad, sci., 256, 17 : 3755. — H a 1 1 F. G. 1935. Haemoglobin
function in the developing chick. Journ. Physiol., 83, 2 : 222. — *H a m a K.
1959. Влияние лекарственных веществ на электрокардиограмму куриного
зародыша. Journ. Physiol. Soc. Japan, 21, 7 : 753 (на японск. яз.). — Ham-
burger V. 1939. The development and innervation of transplanted limb
primordia of chick embryos. Journ. Exptl. ZooL, 80 : 347. — H a m b u r-
g e г V. 1948. The mitotic patterns in the spinal cord of the chick embryo
and their relation to histogenetic processes. Journ. Comp. Neurol., 88 : 221. —
Hamburger V. 1964. Ontogeny of behaviour and its structural basis.
Comparative neurochemistry. Proc, of the 5th Intern. Neurochem. Symposium:
21. — H a m b u r g e г V., H. L. Hamilton. 1951. A series of normal
stages in the development of the chick embryo. Journ. Morphol., 88, 1 : 49. —
Hamilton H. L. 1952. Sensitive periods during development. Ann. N. Y.
Acad. Sci., 55, 2 : 177. —Hamilton T. H. 1961. Studies on the physio-
logy of urogenital differentiation in the chick embryo. 1. Hormonal control
of sexual differentiation of Mullerian ducts. Journ. Exptl. ZooL, 146, 3 : 265. —
Hammond W. S. 1949. Formation of the sympathetic nervous system
in the trunk of the chick embryo following removing of the neural tube. Journ.
Comp. Neurol., 91, 1 : 67. — H a m m о n d W. S. 1954. Origin of thymus
in the chicken embryo. Journ. Morphol., 95, 3 : 501. — H a m p e A. 1956.
Sur la topographic des ebauches presomptives du membre posterieur du poulet.
C. r. Acad, sci., 243, 14 : 970. — H a m p e A. 1959. Contribution a I’dtude
390
du developpement et de la regulation des deficiences et des excedents dans la
patte de 1’embryon du poulet. Arch. anat. microsc. et morphol. exptl., 48,
3—4 : 345. — Hasselbalch K. A. 1900. Ueber den respiratorischen
Stoffwechsel des Hiihnerembryos. Scand. Arch. Physiol., 10 : 353. — Has-
selbalch K. A. 1902. Ueber Sauerstoffproduction in Huhnerei. Scand.
Arch. Physiol., 13 : 170. — H a у s F. A., E. W. S p e a r. 1950. Embryonic
death rate and sex ratio in chicks. Science, 112 : 792. —Henderson E. W.
1956. Which chicks hatch first — male or female ? Michigan State Univ. Agric.
Exptl. Sta. Quart. Bull., 38, 3 : 362. - *H e nr i ch se n E. 1958. Bone
formation and calcification in cartilage. Acta Orthopaed. Scand., 27, 2 : 173. —
Hermann H. 1952. Studies of muscle development. Ann. N. Y. Acad.
Sci., 55, 2 : 99. — H i b b s R. G. 1956. Electron microscopy of developing
cardiac muscle in chick embryos. Amer. Journ. Anat., 99, 2 : 17. — H i 1-
f e r S. R. 1964. Follicle formation in the embryonic chick thyroid. 1. Early
morphogenesis. Journ. Morphol., 115, 2 : 135. — Hohn E. O. 1961. Endo-
crine glands, thymus and pineal body. In: Biology and comparative physio-
logy of birds, N. Y.—London, 2 : 87. — *H о s h i n о T., Y. T о г у u. 1960.
Observation on the differential blood cell counts in the circulating blood of
the chick embryo. Tohoku Journ. Agric. Res., 11, 4 : 319. — H о u-
s e r J. W., G. Ackerman, R. А. К n о u f f. 1961. Vasculogenesis and
erythropoiesis in the living yolk sac of the chick embryo. Anat. Rec., 140,
1 : 29. — H u e t t n e r A. F. 1947. Fundamentals of comparative embryo-
logy of the vertebrates. N. Y. — Hufner G. 1892. Beitrag zur Lehre von
der Atmung der Eier. Arch. f. Anat. und Physiol. (Mullers), 1—2 : 467. —
Hunt E. L. 1949. Establishment of conditioned responses in chick embryos.
Journ. Comp. Physiol. Psychol., 42 : 107.
J a a p R. G., R. S. I n g r a m, E. F. G о d f r e y. 1951. Embryonic
sexual modification and growth in turkeys. 9th World’s Poultry Congress,
2 : 30. — J а с к s о n S. F. 1956. The morphogenesis of avion tendon. Proc.
Roy. Soc. B., 144, 917 : 556. — Jones D. S. 1958. Effects of acetylcholine
and adrenalin on the experimentally uninnervated heart of the chick embryo.
Anat. Rec., 130, 2 : 253.
К a 1 1 i u s E. 1905. Beitrage zur Entwicklung der Zunge. Teil II.
Vogel (Anas, bochas L.), (Passer domesticus L.). Anat. Hefte (I Abt.), 28 :
311. — К a n e E. R., I. W. Sizer. 1953. Some studies on the developing
blood clotting system of the chick embryo. Anat. Rec., 117, 3 : 614. — *K a r-
r e r H. E. 1961. Electron microscope observations on chick embryo liver.
Journ. Ultrastructure Res., 5, 2 : 116. — К a u f m a n L. 1930. Innere und
aussere Wachstumsfaktoren. Arch. Entwicklungsmech. Organ. Zeitschr. Wiss.
Biol. Abt. D, 122, 3 : 395. — Kendeigh Ch. 1940. Factors affecting
length of incubation. The Auk, 57, 4 : 499. — К i 1 s h e i m e r G. S.,
D. R. Weber, J. Ashmore. 1960. Hepatic glucose production in
developing chicken embryo. Proc. Soc. Exptl. Biol, and Med., 104, 3 : 515. —
Kingsbury J. W., M. Alexanderson, E. S. Kornstein.
1956. The development of the liver in the chick. Anat. Rec., 124, 2 : 165. —
Kinyon N., R. L. Watterson. 1958. Reduced endocrine activity
of ovaries of hypophysectomized duck embryos as indicated by modified de-
velopment of genital tubercle and syrinx. Physiol. ZooL, 31, 1 : 60. — К i-
tiyakara A. 1959. The development of non-myotomic muscles of the
chick embryo. Anat. Rec., 133, 1 : 35. — *K о e с к e H. U., O. Kuhn.
1962. Die embryonale Pterylose und ihre entwicklungs physiologischen Vor-
bedingungen bei der Hausente (Anas boschas domestica). I. Die Enstehung
der primordialen Federanlagen in den Korperregionen und die entwicklungs
physiologischen Probleme. Z. Morphol. und Oekol. Tiere, 50, 6 : 651. — К o-
s i n I. L. 1951. The prevalence of early embryonic mortality in the broad
breasted bronze turkeys. Poultry Sci., 30, 6 : 805. — К о s i n I. L.,
S. S. Munro. 1941. Evidence of a sex differential in the utilization of
391
Schell calcium by the chick embryo. Sci. Agr., 21 : 315. — Kozolka A. W.,
T. F. Gallagher. 1934. Effect of male hormone extracts, theelin and
theelol on chick embryo. Proc. Soc. Exp. Biol, and Med., 31 : 1143. —
Krabbe К. H. 1955. Development of the pineal organ and a rudimentary
parietal eye in some birds. Journ. Comp. Neur., 103, 1 : 135. — К r a i c-
z i c z e к M. 1956. Die Beziehungen der Entwicklungsphasen und des Funk-
tionszustandes der innersekretorischen Drusen zu den Wachstumsstuffen des
Hiihnerembryos. I. Thyreoidea. Roux’Archiv fur Entwicklungsmech. Organis-
men 148, 5 : 518. — Kraicziczek M. 1957. Die Beziehungen der Ent-
wicklungsphasen und des Funktionzustandes der innersekretorischen Drusen
zu den Wachstumsstufen des Hiihnerembryos. II. Parathyreoidea. Roux’Arch.
Entwicklungsmech. Organismen, 150, 2 : 162. - Kruger L., A. S t e-
u e r. 1957. Versuche bei Hiihnern mit synthetischen Follikelhormonen das
Geschlecht umzustimmen. Arch. Geflugelzucht und Kleintierkunde, 6,
5—6 : 276. — Kuhnel W. 1961. Morphologische und oxperimentelle
Untersuchungen an der Allantois des Hiihnchens. Z. Zellforsch., 54, 6 : 807. —
Kuhlenbeck H. 1937. The ontogenic development of the diencephalic
centers in a birds brain (chick) and comparison with the reptilian and mamma-
lian diencephalon. Journ. Comp. Neurology, 66, 1 : 23. — К u о Z. Y. 1932a.
Ontogeny of embryonic behaviour in Aves. I. The chronology and general nature
of the behaviour of the chick embryo. Journ. Exp. Zool., 61 : 395. —
Kuo Z. Y. 1932b. Ontogeny of embryonic behaviour in Aves. II. The mecha-
nical factors in the various stages leading to hatching. Journ. Exp. Zool.,
62 : 453. — К u о Z. Y. 1932c. Ontogeny of embryonic behaviour in Aves. V.
The reflex concept in the light of embryonic behaviour in birds. Psychol. Rev.,
39, 6 : 499. — К u о Z. Y. 1939a. Studies in the physiology of the embryonic
nervous system. II. Experimental evidence on the controversy over the reflex
theory in development. Journ. Comp. Neurol., 70, 3 : 437. — Kuo Z. Y.
1939b. Studies in the physiology of the embryonic nervous system. IV. Deve-
lopment of acetylcholine in the chick embryo. Journ. Neurophysiol., 2, 6 :
488. — KuoZ. Y., T. C. Shen. 1936. Ontogeny of embryonic behaviour
in Aves. X. Gastric movements of the chick embryo. Journ. Comp. Psychol.,
21 : 87. — К u о Z. Y., T. C. Shen. 1937. Ontogeny of embryonic beha-
viour in Aves. XI. Respiration in the chick embryo. Journ. Comp. Psychol.,
24 : 49.
L a m s о n G. H., Jr. H. D. E d m о n d. 1914. Carbon dioxide in incuba-
tion. Storrs (Connecticut) Agr. Exp. Sta. Bull., 76 : 1. — Landauer W.
1948. The hatchability of chicken eggs as influenced by environment and here-
dity. Storrs (Connecticut) Agr. Exp. Sta. Bull., 262 : 1. — L e e W., 1951.
Glycogen content of various tissues of the chick embryo. Anat. Rec., 110 :
465. — Lelkes G. 1958. Experiments in vitro on the role of movement in
the development of joints. Journ. Embryol. and Exptl. Morphol., 6, 2 : 183. —
Lemburg P. 1962. Histophysiologische Untersuchungen zum Problem des
Flussigkeits- und Stofftransportes in der friihen Embryonalentwicklung. Z.
Zellforsch., 57, 5 : 737. — *L e m e z L., M. К о p e с к y. 1962. The number
of erytrocyte recirculations in chick embryo and hen, with a note on embryo-
nic heart efficiency. Physiol. Bohemoslov., 11, 2 : 93. — *L e m e z L.,
M. J о s i f к o. 1957. Количественный анализ увеличения количества клеток
эмбрионального органа путем митозов в его зачатке и путем иммиграции
клеток или путем амитозов. 6eskosl. Morfol., 5, 3 : 292 (на чешек, яз.). —
*L е m о z L., Z. R у с h t е г. 1956. Кровь куриных эмбрионов. VI.
О средней продолжительности жизни примитивных эритроцитов (Ех) и о роли
митозов в круговороте их размножения. Ceskosl. Morfol., 4, 1 : 27 (на чешек,
яз.). — Lemez L., Z. R.ychter. 1958. Die Mitosedauer primitiver
Erythrozyten von Hiihnerembryonen mittels Colchicintechnik untersucht.
Roux’.Arch. Entwicklungsmech. Organismen, 150, 4 : 383. — Lerner J.
1939. The shape of the chick embryo growth curve. Science, 89, 2297 :16.—
L i ё v r e F. 1957a. La differenciation des cellules A et В des ilots endocri-
392
niens du pancreas chez I’embryon do poulet. Ann. Endocrinol., 18, 4 : 592. —
L i e v r e F. 1957b. Contribution a 1’histogenese du pancreas endocrine chez
I’embryon du poulet. Arch. Anat. Microsc. et Morphol., Exptl., 4G, 1 : 61. —
L i 11 i e F. R. 1952. Lillie’s development of the cnick. Revised by H. L. Ha-
milton. N.Y. — *Li mborgh J., van. 1961a. Tho origin of the asymmetri-
cal distribution of the primary germ cells in late somite stage duck embryos.
Acta Morphol. Neerlanao-Scand., 4, 3 : 261. — Limborgh J., van. 1961b.
La repartition numerique des cellules germinales sur les Sbauches gonadiques.
Arch. anat. microsc. et morphol. exptl., 50, 4 : 469. — L о с у W. A.,
0. La r s e 11. 1916. The embryology of the birds lung based on observations
of the domestic fowl. Amer. Journ. Anat., 19, 1 : 447. — L о j d a Z., L. L e-
m e z. 1957. Embryonic blood of chickens, morphocytochemical study of
chicken embryo thrombocytes. (Jeskosl. MorfoL, 3, 4 : 317. — L u c a s A. M.,
P. Stettenheim, E. M. Denington, U. S. Jamroz. 1962.
Morphology of feather muscles in the chicken. Anat. Rec., 142, 2 : 253. —
L u p p a H. 1962. Histologie, Histogenese und Topochemie der Drusen des
Sauropsidenmagens. II. Aves. Acta Histochemica, 13, 5/8 : 233. - *Lus-
s a n a F. 1906. La respirazione dei tessuti negli animali adulti negli embrioni
e nei neonati. Arch, di FisioL, 3:113. — Lutz H. 1955. Contribution
experimentale a 1’etude de la formation deTendoblaste chez les oiseaux. Journ.
Embryol. and Exptl. Morphol., 3, 1 : 59. - Lu t z - Ost er tag Y. 1954.
Contribution a 1’etude ou developpement et la regression des canaux de Mul-
ler chez I’embryon d’oiseaux. Bull. biol. France et Belg., 88 : 333.
*M a 1 i n о w s к a W. 1961. Studies on the development of the male
embryos of hen. I. Development of the reproductive system. II. The development
of blood vessels of the reproductive system. Zool. Polon., 11, 1 : 15, 57. —
Manteuffel-Szoege L. 1960. Energy sources of blood circulation
and the mechanical action of the heart. Thorax, 15, 1 : 47. — M a r a u d R.
1955. Recherches sur le determinisme de la differenciation epidimaire chez
les oiseaux. Arch. anat. microsc. et morphol. exptl., 44, 3 : 277. — Ma-
ra u d R., C. S t о 1 1, P. В 1 a n q u e t. 1957. Sur le role de 1’hypophyse
dans la concentration du radioiode par la thyroide de I’embryon de Poulet.
Compt. rend. Soc. Biol., 151 : 572. — M a r a u d R., C. S t о 1 1, С. M a-
cario, P. Blanquet. 1954. Sur la concentration du radioiode J1S1
par la thyroide de I’embryon du poulet. Ann. Endocrinol., 15, 4 : 607. —
Mariakulandai A., J. McG i n n i s. 1952. Vitamin B12 and carbon
dioxide production by chick embryo. Arch. Biochem. and Byophys., 37,
1 : 136. — *M a г i n G. 1956. Наблюдения над развитием половых желез
и над половым дифференцированием куриного зародыша. Arch. ital. anat.
е embriol., 61, 1 : 106 (на итал. яз.). — Marks Н. 1957. Zur Frage der
Geschlechtsumstimmung. Dtsch. Geflugelztg., 6, 1 : 180. — Marshall W.
1947. Embryonic mortality and anomalous development during incubation.
In: Deseases of Poultry, London: 128. — Martindale F. M. 1941.
Initiation and early development of thyrotropic function in the incubating
chick. Anat. Rec., 79 : 373. — Matthey R. 1950. Les chromosomes des
oiseaux. Traite de zoologie, XV, Oiseaux: 378. — McC a f f e r t у R. E.,
S. H. Pressman, W. H. К n i s e 1 y. 1962. Observations and recor-
dings of drug effects on amnion, embryo and vascular system in incubating
eggs. Anat. Rec., 142, 2 : 256. — Mellen W. J. 1957. Further comments
on «controlling sex» in chickens. Poultry Sci., 36, 6 : 1384. — *M e n-
kesB., C. Rimniceanu, C. Miele a., 1956. Исследования разви-
тия и значения мезонефроса и метанефроса в курином зародыше. Studii ?i
cercetMri §tiint. Acad. RPR, Basa Timisoara, ser. §tiint. med., 3, 1—2 : 9. —
*M i 6 e k J., A. Chfia per о wa, E. Trnkova. 1955. Отбор цыплят
по полу на 4-й день после закладки яиц в инкубатор путем просвечивания.
Drubeznictvi, 3, 6 : 82 (на польск. яз.). — Miller R. А. 1938. Spermato-
genesis in a sex-reversed female and in normal males of the domestic fowl-
Gallus domesticus. Anat. Rec., 70 : 155, — *M i 11 о A. 1961. Ricerca com-
26 В. В. Рольник
393
parativa sul contenuto in cholinesterasi del muscolo striato, del cuore e del
muscolo liscio durante lo sviluppo dell’embrione di polio. Riv. Biol. (Perugia),
54, 2 : 251. — Mitchell H. H., L. E. Card, W. T. Haines.
1927. The effect of ago, sex, and castration on the basal heat production of
chickens. Journ. of Agric. Res., 34, 10 : 945. — Mohsen T., P.Carri-
c a b u r u, M. H. F i 11 i о j. 1962. Evolution de Г electrocard iogramme au
cours du d£veloppement de 1’embryon du poulet. Compt. rend. Soc. Biol.,
156, 2 : 331. — M о о g F. 1952. The differentiation of enzymes in relation
to the functional activities of the developing embryo. Ann. N. Y. Acad. Sci.,
55, 2 : 57. — M о о g F. 1959. The development of function in the adrenal
cortex. Comp. Endocrinology proceed, of the Columbia University symposium:
624. — MoogF., D. Richardson. 1955. The functional differentia-
tion of the small intestine. V. The influence of adrenocortical hormones on
differentiation and phosphatase in the duodenum of the chick embryo. Exp.
ZooL, 130, 1 : 29. — M о о g F., E. R. Thomas. 1957. The functional
differentiation of the small intestine. VI. Transient accumulation of glycogen
in the intestinal epithelium of the chick embryo under normal conditions and
under the influence of hydrocortisone. Physiol. ZooL, 30, 4 : 281. — *M о r i-
g u c h i W. 1957. Effects of amino acids on hematopoiesis of chick embryo.
Report I. Effects of injected glycine on the bloodproduction of chick embryo.
Journ. Osaka Med. Coll., 17, 4 : 168. т Mosier H. D. 1955. The develop-
ment of the hypothalamo-neurohypophysial secretory system in the chick
embryo. Endocrinology, 57, 6 : 661. — Mulherkar L. 1958. Induction
by regions lateral to the streak in the chick embryo. Journ. Embryol. and
Exptl. Morphol., 6, 1:1. — Mum enthaler M., W. K. Engel.
1961. Cytological localization of cholinesterase in developing chick embryo
skeletal muscle. Acta Anat., 47, 3 : 274. — Munday R. A. 1953. The
positions in the shell and weights of eighteen to twenty-one-day chick embryos
in relation to hatchability. Poultry Sci., 32, 2 : 202. — M u n г о S. S.,
I. L. Kosin. 1945. Proof of the existence of pre-oviposital embryonic
death’s in chickens and their bearing on the relation between «fertility» and
hatchability. Canad. Journ. Res., sect. D, 23 : 6. — M u г г а у H. A. Jr.
1925. Physiological ontogeny. A. Chick embryos. II. Catabolism. Chemical
changes in fertile eggs during incubation. Selection of standard condition.
Journ. Gen. Physiol., 9:1.
NaceG. W., A. M. Schechtman. 1948. Development of non-
vitelloid substances in the blood of the chick embryo. Journ. Exptl. ZooL,
108, 2 : 217. — N a w a r G. 1956. Experimental analysis of origin
of the autonomic ganglia in the chick embryo. Amer. Journ. Anat., 99, 3 :
473. — Needham J. 1931. Chemical embryology. Cambridge, Univ.
Press. — Needham J. 1942. Biochemistry and morphogenesis. Cambridge
Univ. Press. — Nelsen О. E. 1960. Hypoxia and maldevelopment of
early alarplate tissue in mid- and forebrain regions of the chick embryo.
«Growth», 24, 4 : 361. — N e w D. A. T. 1956. The formation of sub-blasto-
dermic fluid in hens’ eggs. Journ. Embryol. and Exptl. Morphol., 4, 3 : 221. —
Newcomer E. H., G. M. D о n n e 11 y, U. C. F a r b e s. 1960.
A cytological study of sex reversal in the domestic fowl. Poultry Sci., 39,
4 : 1030. — *N ichi t a G. 1955a. Изучение эмбрионального обмена у кур
породы леггорп, род-айленд и Суссекс в целях повышения процента вылупле-
ния. Bui. §tiint. Acad. RPR, Sec. biol., agron., geol. §i geogr., 7, 1 : 123 (на
румынск. яз.). — *N i c h i t a G. 1955b. Изучение обмена веществ у эмбрио-
нов уток породы хаки-кемпбел с целью повышения процента вылупляемостп.
Bui. §tiint. Acad. RPR, Sec. biol., agron., geoL §i geogr., 7, 4 : 1137 (на ру-
мынок. яз.). — NoyonsA. К. M., Р. М. De Hassel е. 1939. Ueber
den Stoffwechsel des Hiihnereies und die Bedeutung der Luftkammer. Acta
Brevia Neerland de Physiol. Pharmacol. Microbiol., 9 : 170.
*0 c a r i z O. 1957. Произвольное формирование пола. Granja, 5,
394
51 : 29 (на псп. яз.). — O’C onnor R. J. 1952. Growth and differentiation
in the red blood cells of the chicken embryo. Journ. Anat., 86, 3 : 320. — O’C о n-
n о r R. J. 1953. Metabolism and glycogen formation in the liver of the chi-
cken embryo. Journ. Embryol. and Exptl. MorphoL, 1, 2 : 105. — O’R a-
hillyR., E. Gardner. 1956. The development of the knee joint of
the chick and its correlation with embryonic staging. Journ. MorphoL, 98,
1 : 49.
Paff G. H., R. В о u c e k. 1962. Simultaneous electrocardiograms
and myograms of the isolated atrium, ventricle and conus of the embryonic
chick heart. Anat. Rec., 142, 1 : 73. - *P a r h о n C. I., L. L a u г i a n,
M. ВШсеапи, N. Albu. 1955. Исследование направленного
эмбриогенеза. Сообщ. 7. Действие препарата паратгормон на развитие кури-
ного эмбриона. Studii $i cerceUri endocrinol. Acad. RPR, 6, 1—2 : 79 (на
румынск. яз.). — *P a r h о n С. I., L. L a u r i a n, M. ВШсеапи,
N. Albu. 1956. Actiunea cortizonulue asupra dezwoltMrii embrionare la
gaina. Studii 0 cercetSri endocrinol. Acad. RPR, 7, 4 : 451. —
Pasteels J. 1945. On the formation of the primary entoderm of the duck
(Anas domestica) and on the significance of the bilaminar embryo in birds.
Anat. Rec., 93 : 5. — P a 11 e n В. M. 1951. Early embryology of the
chick. N. Y. — P a 11 e r s о n J. T. 1910. Studies on the early development
of the hens egg. I. History of the early cleavage and of the accessory cleavage.
Journ. MorphoL, 21 : 101. — *P a у n e J. 1919. Distribution of mortality
during the period of incubation. Journ. Amer. Assos. Instructors and Investi-
gators in Poultry Husbandry, 6, 2 : 9. — P e t e r K. 1938. Untersuchungen
uber die Entwicklung des Dotterentoderms. I. Die Entwicklung des Entoderms
beim Huhnchen. II. Die Entwicklung des Entoderms bei der Taube. Zeit. f.
mikro-anat. Forsch., 43 : 362, 416. — P e t e г s J. J., Ch. J. Cusick.
1961. Bioelectric patterns as indicators of behaviourial development in the
chick embryo. Proc, of the second Bionics Sympos. Biol. Prototypes and
Synthetic Systems, 1 : 29. — P e t e г s J. J., A. R. Vonderahe,
Th. H. Powers. 1956. The functional chronology in developing chick
nervous system. Journ. Exp. Zool., 133, 3 : 506. — P h i 1 i p s F. S. 1941.
The oxygen consumption of the early chick embryo at various stages of develop-
ment. Journ. Exp. Zool.; 86 : 257. — P h i 1 i p s F. S. 1942. Comparison
of the respiratory rates of different regions of the chick blastoderm during early
stages of development. Journ. Exp. Zool., 90 : 83. — P i n c u s G.,
A. E. Erickson. 1962. Sex modifications in hen’s eggs following immersion
in diethylstilbestrol solutions. Endocrinology, 71, 1 : 24. — P i r n e r F.
1949. Uber die Zeitpunkt der friihesten embryonalen Innervation der Hiihn-
chenniere. Anat. Anz., 97, 1/3 : 45. — P о p о f f D. 1894. Die Dottersack-
gefasse des Huhnes. Wiesbaden. — Prakash R. 1954. On the septum
atriorum and septum ventriculorum of chick embryos. Sci. and Culture, 19,
10 : 504. — Preyer W. 1885. Specielle Physiologic des Embryo. Leipzig. —
P u n C. F. 1958. The sex ratio in the progeny of oestrogen treated parents
in the Brown Leghorn. Poultry Sci., 37, 2 : 307.
R a w 1 e s M. E., W. L. Straus. 1948. An experimental analysis
of the development of the trunk musculature and ribs in the chick. Anat. Rec.,
100 : 755. — Riddle O. 1920. Differential survival of male and female
dove embryos in increased and decreased pressures of oxygen. A test of metabolic
theory of sex. Proc. Exp. Biol, and Med., 17 : 88. — R i d d 1 e O. 1930.
Studies on the physiology of reproduction in birds. XXVII. The age distribu-
tion of mortality in birds and its probable significance. Amer. Journ. Physiol.,
94, 3 : 535. - *R imn iceanu С., C. Miele a, M.Lungulescu-
Dragan. 1955. Гликогенез во время развития куриного зародыша. Bui.
§»tiint. Acad. RPR, Sec. biol., agron., geol. ?i geogr., 7, 3 : 959 (на румынск.
яз.). — R i n a u d о M. T. 1962. Glycogenesis in the liver of chicken embryo.
Enzymologia, 24, 4 : 230. — *R i z z о A. 1899. Sul numeri e sulla distribu-
26*
395
zione dei pori nel guscio dell’ove di gallina. Roma, Lab. Anat. Norm. Ric.,
7 : 171. — R о g 1 e r I. С., H. E. Parker, F. N. A n d r e u s,
C. W. Carrick. 1961. The iodine requirements of the breeding hen. 1.
Hens reared on a diet adequate in iodine. 2. Hens reared on a diet difficient
in iodine. Poultry Sci., 40, 6 : 1546. — Romanoff A. L. 1929 Cycles
in the prenatal growth of the domestic fowl. Science, 70, 1820 : 484. — R o-
m a n о f f A. L. 1931. Why some eggs do not hatch? Cornell Exp. Bull.,
205 :1. — Romanoff A. L. 1933. Morphological study of differentiation
of sex in chicks. Poultry Sci., 12, 2 : 305. — Romanoff A. L. 1941.
The study of respiratory behaviour of individual chicken embryos. Journ.
Cell. Comp. Physiol., 18 : 199. — R о m a n о f f A. L. 1943. Study of
various factors affecting permeability of bird’s eggshell. Food res., 8 : 210. —
Romanoff A. L. 1948. Chemical and physiological sex differencies in
newly hatched chicks. Poultry Sci., 27, 5 : 643. — Romanoff A. L. 1949.
Critical periods and causes of death in avian embryonic development. Auk,
66 : 264. — Romanoff A. L. 1952. Membrane growth and function. Ann.
N. Y. Acad. Sci., 55, 2 : 288. — Romanoff A. L. 1960. The avian
embryo. Structural and functional development. N. Y., Mac Millan Comp. —
R о m i j n C. 1948. Respiratory movements of the chicken during the para-
foetal period. Physiol. Comp. OecoL, 1 : 24. — R о m i j n C. 1950. Foetal
respiration in the hen. Gas diffusion through the egg shell. Poultry Sci., 29,
1 :42. - Romijn C. 1951. The respiratory metabolism of chicken embryos.
9th World’s Poultry Congress: 101. - R omi jn С., K. F. Funk,
W. L о к h о r s t. 1952. Thyroxine, thiouracil and embryonic respiration
in White Leghorns. Poultry Sci., 31, 4 : 684. - R omi jn C., W.Lok-
horst. 1951. Foetal respiration in the hen. The respiratory metabolism of
the embryo. Physiol. Comp. OecoL, 2, 3 : 187. - R omi j n C., J. Roos.
1938. The air space of the hen’s egg and its changes during the period of incuba-
tion. Journ. Physiol., 94, 3 : 365. - R udnick D. 1952. Development of
the digestive tube and its derivatives. Ann. N. Y. Acad. Sci., 55, 2 : 109. —
*R ychterZ., M. Kopeck y, L. Lemez. 1955. Кровь куриных
зародышей. IV. Объем циркулирующей крови со 2-го дня инкубации (25 со-
митов) до вылупления. Ceskosl. morphol., 3, 1 : И (на чешек, яз.). — *R ych-
terZ., L. Lemez. 1956. Сосудистая система куриного эмбриона. I.
О развитии и экспериментальной морфологии вен желточного мешка. Ceskosl.
morfol., 4, 2 : 142 (на чешек, яз.).
S a d 1 е г W. W. 1955. Chronological relationship of the disappearance
of the vitelline membrane and the closure of the amnio-chorium in avian embryos
and its implications. Poultry Sci., 34, 4 : 754. — Sandreuter A. 1951.
Vergleichende Untersuchungen uber die Blutbildung in der Ontogenese von
Haushuhn (Gallus gallus L.) und Star (Sturnus vulgaris L.). Acta Anat. (Ba-
sel), XI (Suppl. 14-1 ad): 1. — S a u n d e r s J. W. Jr. 1958. The proximo-
distal sequence of origin of the chick wing and the role of the ectoderm. Journ.
Exp. ZooL, 108 : 363. — S a у e e d M. M., D. S. Blumenthal,
H. T. Blumenthal. 1962. Effect of cortisone and growth hormones
on cellular proliferation during early embryogenesis. Proc. Exptl. Biol, and
Med., 109, 2 : 261. — SchechtmanA. M. 1952. Physical and chemical
changes in the circulating blood. Ann. N. Y. Acad. Sci., 55, 2 : 85. — S c h e i b-
P e 1 e g e r D. 1955. Biochemical contribution to the study of the differen-
tiation of the canals of Muller in the chick embryo. Bull. Biol. France et Belgi-
que, 89, 4 : 404. — Schellong G. 1954. Herz- und Gefaszmissbildungen
beim Hiihnchen durch kurzfristigen Sauerstoffmangel. Beitr. pathol. Anat.
und allgem. Pathol., 114, 2 : 212. — Schmekel L. 1962. Embryonale
und friihe postembryonale Erythropoiese in Lebey, Milz, Dottersack und Kno-
chenmark der Vogel. Rev. suisse ZooL, 69, 3 : 559. - Se dla 6 ek J. 1962.
a) Temporary connection in chick embryo; b) Notes on the characteristics of
the temporary connection in chick embryo; c) Functional characteristics of
the centre of the inconditioned reflex in elaboration of a temporary connection
396
in chick embryo. Physiologia Bohemosloven., 11, 4 : 300, 307, 313. — S e 1 t-
z e r W. 1956. The method of controlling the sex of avian embryo, improving
embryo hatchability and improving viability of the hatched chick. U. S. Pa-
tent, 2, 734 : 482. — S e n g e 1 Ph. 1957. Analyse experimentale du develop-
pement in vitro des germes plumaires de I’embryon ae poulet, Experientia,
13, 5 : 177. — S e n g e 1 Ph. 1958. La differenciation de la peau et des germes
plumaires de I’embryon de poulet en culture in vitro. Annee biol., 34, 1—2 :
29. — S e n о T. 1961. An experimental study on the formation of the body
wall in the chick. Acta anat., 45, 1/2 : 60. — S h e p a r d Th. H. 1962.
Carbonic anhydrase activity in early developing chick embryos. Journ. Embryol.
and Exptl. Morphol., 10, 2 : 191. — *Shikinami Y. 1928. Uber Hypo-
glykamie erzeugenden Stoffe aus verschiedenen tierichen Substanzen, besonders
aus Eigelb des Huhnereies. Tohoku Journ. Exp. Med., 10:1.—
Shumway 1942. Introduction to vertebrate embryology. 4th ed. —
Simon D. 1957a. Sur la localisation des cellules germinales primordiales
chez I’embryon du poulet et leur mode de migration vers les ebauches gonadi-
ques. C. r. Acad, sci., 244, 11 : 1541. — Simon D. 1957b. La migration .
des cellules germinales de I’embryon du poulet vers les ebauches gonadiques:
preuves experimentales. Compt. rend. Soc. Biol., 151, 8—9 : 1576. — S i-
m о n D. 1960a. Organogenese et differenciation sexuelle des glandes ge-
nitales de I’embryon de poulet et 1’absence totale des cellules germinales. C. r.
Acad, sci., 251, 3 : 449. — Simon D. 1960b. Contribution a 1’etude de la
circulation et du transport des gonocytes primaires dans les blastodermes
d’Oiseaux cultives in vitro. Annee biol., 36, 3—4 : 105. — SlonimskiP.
1927. Sur 1’apparition de 1’hemoglobine dans 1’aire vasculaire chez le poulet.
Compt. rend. Soc. Biol., 96 : 1498. — S m i t h H. M., St. R. Geiger.
1961. Another hypothesis of function of the glycogen body of birds. Journ.
Elisha Mitchell Scient. Soc., 77, 2 : 289. — Smolar J. A. 1953. Origin
and development of the urogenital union in the chick. Proc. Soc. Exptl. Biol,
and Med., 84, 1 : 79. — S n e d e с о r S. 1962. Bone development in the
chick embryo. Sci. World, 11, 5: 21. — Sobel H. 1958. Antagonistic
effects of cortisone and growth-hormone on the developing chick embryo. Proc.
Soc. Exptl. Biol, and Med., 97, 3 : 495. — Spemann H. 1918. Uber
die Determination der ersten Organanlagen der Amphibienembryo. Arch.
Entwicklungsmech. Organ., 43 : 448. — Spicer A. 1954. Stilboestrol and
the control of sex in poultry. Brit. Vet. Journ., 110 : 358. — SpiroffB.E.N.
1959. Embryonic and post-hatching development of the pineal body of the
domestic fowl. Amer. Journ. Anat., 103, 3 : 375. — S t e p h a n E. 1955.
Consequences de la suppression de 1’orifice veineaux droit chez I’embryon
de poulet. Compt. rend. Soc. Biol., 149, 13—14 :1478. — Stockard Ch. R.
1921. Developmental rate and structural expression. An experimental study
of twins, double monsters and single deformities and interaction among embryo-
nic organs during their origin and development. Amer. Journ. Anat., 28,
2 : 115. — Stoll R., R. Maraud. 1956. Existance d’hormones ste-
roliques dans 1’organisme de I’embryon du poulet au cours de sa differenciation
sexuelle. C. r. Acad, sci., 242, 9 : 1235. — Strudel G. 1953. Consequences
de 1’excision de tron^ons du tube nerveux sur la morphogenese de I’embryon
de poulet et sur la differenciation de ses organes: contribution a la genese de
1’orthosympathique. Ann. sci. natur. zool. et biol. anim., 15, 2 : 251. —
.*S zekelyG., E. Endroczi, J. Szentagothai. 1958. Ershei-
nen der argentaffinen Substanz und Beginn der Sekretion adrenocorticotrophen
Hormons in der Hypophyse von Huhnerembryonen. Acta biol. Acad, soient.
Hung., 8, 3 : 283. — S z e p s e n w о 1 J., А. В г о n. 1935. Le premier
tontact du systeme nerveux vago-sympatique avec 1’appareil cardio-vassulaire
chez les embryons d’Oiseaux. (Canard et poulet). Compt. rend. Soc. Biol.,
118, 1 : 946.
*T5utuP., N. V о i c u 1 e t. 1958. К вопросу о появлении угле-
кислой ангидразы во время развития головного мозга куриного зародыша.
397
Commun. Acad. RPR, 8, 2 : 233 (на румынск. яз.). — *T e 11 о I. F. 1945.
Дальнейшие наблюдения над первичными стадиями развития симпатической
нервной системы у эмбриона цыпленка. Trab. Inst. Cajal Invest.
Biol., 37 : 103. — T h i e b о 1 d J. J. 1964. Sur la sensibilite des canaux
de Muller aux hormones genitales chez I’embryon d’oiseaux. Ann. endocrinol.,
25, 5, suppl. : 136. — ThommesR. C. 1958. Vasculogenesis in selected
endocrine glands of normal and hypophysectomized chick embryos. Growth,
22, 4 : 243. — Thommes R. C. 1960. A histochemical study of insulin
in the chick embryo pancreas. Growth, 24, 1 : 69. — T i e n h о v e n A., van.
1957. A method of «controling sex» by dipping of eggs in hormone solutions.
Poultry Sci., 36, 3 : 628. -Tixier-Vidal A. 1956. Etude chronologi-
que in vivo et in vitro des correlations hypophyso-thyroide chez I’embryon
de poulet. Arch. anat. microsc. et morphol. exptl., 45, 3 : 236. — Tixier-
Vidal A. 1958. Etude histophysiologique des relations hypophyse et thy-
roide chez I’embryon du poulet. Arch. anat. microsc. et morphol. exptl., 47,
2 : 235. - To th N., P. Simon, G. S z e k e 1 y. 1958. Abhangigkeit
der friihen geweblichen Differentiation der Nebennierenrinde von der Hypophyse
bei Huhnerembryonen. Acta biol. Acad, scient. hung., 8, 3 : 289. — Tyr-
rell D. A. J., I. T a m m, О. C. F о r s s m a n n, F. I. H о r s f a 11.
1954. A new count of allantoic cells of the 10-day chick embryo. Proc. Soc.
Exp. Biol. Med., 86, 3 : 594.
V e n z k e W. G. 1947. Morphogenesis of the parathyroid glands of
chicken embryos. Amer. Journ. Vet. Res., 8, 29 : 421. — V e n z k e W. G.
1952. Morphogenesis of the thymus of chicken embryos. Amer. Journ. Vet.
Res., 13, 48 : 395. — V e n z k e W. G. 1954a. The morphogenesis of the
indifferent gonad of chicken embryos. Amer. Journ. Vet. Res., 15, 55 : 800. —
V e n z k e W. G. 19546. The morphogenesis of the testes of chicken embryos.
Amer. Journ. Vet. Res., 15, 56 : 450. — *V i 1 1 a m i 1 M. F. 1942. Cito-
genesis del Pancreas exo у endocrino en embriones de polio. Rev. Soc. argent,
biol., 18 : 416. — Vintemberger P., J. Clavert. 1955. Sur le
determinisme de la symetrie bilaterale chez les oiseaux. VII. Confirmation
experimentale, sur la pigeonne, de notre conception du mecanisme de 1’inver-
sion de 1’orientation de I’embryon. Compt. rend. Soc. Biol., 149, 9—10 :
1038. — Vintemberger P., J. Clavert. 1956. Sur la frequence
relative de la monstruosite double et le degre d’instabilite de 1’orientation
de 1’oeuf dans 1’uterus, chez differents oiseaux. C. r. Acad, sci., 243, 25 :
2149. — Vintemberger P., J. Clavert. 1957. Le determinisme de
la symetrie bilaterale chez les oiseaux et la regie de von Baer. Compt. rend.
Assoc. Anat., 42 : 1403. — Vrbitche S. 1924. Sur 1’absorption du li-
quide amniotique par I’embryon. Compt. rend. Soc. Biol., 91, 26 : 604.
Waterman A. J. 1958. Development of the thyroid-pituitary sy-
stem in warm-blooded amniotes. Comp. Endocrinology Proceed, of the Colum-
bic Univ. symp. : 351. — W a t e r s W. F. 1935. Certain so-called malposi-
tions—a natural occurence in the normal development of the chick embryo.
Poultry Sci., 14, 1 : 208. — Watterson R. L. 1942. The morphogenesis
of down feathers with special reference to the developmental history of melano-
phores. Physiol. Zool., 15 : 234. — Watterson R. L. 1949. Develop-
ment of the glycogen body of the chick spinal cord. I. Normal morphogenesis,
vasculogenesis and anatomical relationships. Journ. Morph., 85, 2 : 337. —
W e e 1 P. B. 1948. Histophysiology of the limb bud of the fowl during its
early development. Journ. Anat., 82, 1 : 49. — W e n i g e r J. P. 1961.
Activite hormonale des gonades morphologiquement indifforenciees de I’embryop*
de poulet. Arch. anat. microscop, et morphol. exptl., 50, 3 : 269. — W i 1-
1 i e г В. H. 1952. Development of sex-hormone activity of the avian gonad.
Ann. N. Y. Acad. Sci., 55, 2 : 159. — W i 1 1 i e г В. H. 1954. Phases in
embryonic development. Journ. Cell. Comp. Physiol., 43, (Suppl.) : 307. —
W i 11 i о г В. H. 1955. Ontogeny of endocrine correlation. In: Analysis
398
of development : 574. —Willier В. H., T. F. Gallagher,
F. C. Koch. 1937. The modification of sex development in the chick
embryo by male and female sex hormones. Physiol. ZooL, 10 101. - Wind-
le W. F. 1940. Physiology of the fetus. Philadelphia. — W i n d 1 e W. F.,
J. Barcroft. 1938. Some factors governing the initiation of respiration
of the chick. Amer. Journ. Physiol., 121 : 684. — Windle W. F., D. Nel-
son. 1938. Development of respiration in the duck. Amer. Journ. Physiol.,
121 : 700. — W i n g s t r a n d K. G. 1954. Neurosecretion and antidiuretic
activity in chick embryos with remarks on the subcomissural organ. Arch.
ZooL, 6, 1 : 41. — W i t s c h i E. 1950. Genetique et physiology de la dif-
ferenciation du sexe. Arch. anat. microscop, et morphol. exptl., 39 : 215. —
Wladimirov G. E. 1930. Beitrage zur Embryochemie und Embryo-
physiologie. V. Mitt. Die Anhaiifung des Glykogens in der Leber des sich ent-
wickelnden Hiihnerembryos. Biochem. Ztschr., 224 : 79. — W о 1 f f E. 1953.
Le determinisme de 1’atrophie d’un organe rudimentaire: le canal de Muller
des embryons males d’oiseaux. Experientia, 9, 4 : 121. — W о 1 f f Et. 1958.
Endocrine function of the gonad in developing vertebrates. Comp. Endocrino-
logy Proceed, of the Columb. Univers. symp. : 568. — W о 1 f f E., K. Haf-
fen. 1952. Sur I’intersexualite experimentale des gonades embryonnaires
de canard cultives in vitro. Arch. anat. microscop, et morphol. exptl., 41 :
184. — W о 1 f f Em., M. Pinot. 1961. Stimulation du cortex de le gonade
droite de 1’embryon d’oiseau. Arch. anat. microscop, et morphol. exptl., 50,
4 : 487. — Wolff Et., Em. Wolff. 1951. The effects of castration on
bird embryos. Journ. Exp. ZooL, 116 : 59. — W о 1 f f Et., Em. Wolff.
1952. Action de 1’hormone sur la differenciation sexuelle de la syrinx
de canard cultivee in vitro. Compt. rend. Soc. Biol., 146, 1—2 : 111. —
Wortham R. A. 1948. The development of muscles and tendons in the
lower leg and foot of chick embryos. Journ. Morphol., 83, 1 : 105.
Yoshpe-Purer Y., J. F e n d r i c h, M. Davies. 1953. Esti-
mation of the blood volumes of embryonated hen eggs at different ages. Amer.
Journ. Physiol., 175, 2 : 178.
*Z e i g e 1 R. F. 1962. Cytogenic study of embryonic chick, pancreas.
I. Exocrine tissue. Journ. Natl. Cancer Inst., 28, 2 : 269. —Z willing E.
1948. Association of hypoglycemia with insulin micromelia in chick embryos.
Journ. Exp. ZooL, 109 : 197. — Z w i 1 1 i n g E. 1951. Carbonydrate meta-
bolism in insulin-treated chick embryos. Arch. Biochem. a. Biophys., 33 :
228. — Zwilling E. 1952. The effects of some hormones on development.
Ann. N. Y. Acad. Sci., 55, 2 : 196.
К разделу III
Баранчеев Л. M. 1939. Влияние температуры на развитие и
процессы дыхания куриных эмбрионов. Уч. зап. ЛГУ, сер. биол., 35, 9 :
64. — Б о л о т н и к о в А. М., В. Д. Пак. 1959. Влияние возраста и
различных температур на деятельность сердца и некоторые формы движения
зародыша курицы. Уч. зап. Пермск. гос. пед. инет., 20 : 165. — Б о р д з и-
виловская Н. П. 1953. Эмбриональное развитие гусей. В кн.: К биологии
развития сельскохозяйственных птиц. АН УССР : 53. — Бордзпвп-
ловская Н. П. 1955. Эмбриональное развитие уток. Тр. Инет. зоол.
АН УССР, 12 : 3. — Б р а ж н и к о в а Л. А. 1950. Физико-химические
изменения плазм и газообмен утиных яиц при инкубации. Тр. Н.-иссл. инет,
птицеводства, 21 : 48. — Б р а ж н и к о в а Л. А. 1954. Водный обмен
в яйцах гусей под наседкой и в инкубаторе. В сб.: Инкубация яиц с.-х. птицы,'
Тр. Н.-иссл. пнет, птицеводства, 24 : 82. — Б ы х о в е ц А. У. 1955.
Опыты охлаждения инкубируемых гусиных яиц непосредственно в инкуба-
торе. Птицеводство, 3 : 24. — Б ы х о в е ц А. У. 1959а. Изучение возмож-
399
ностей инкубирования яиц уток и гусей с охлаждением в инкубаторе при
сокращенной экспозиции. Научи, тр. Укр. опыты, ст. птицеводства, 6 :
105. — Быховец А. У. 19596. Дополнительное поворачивание гусиных
яиц повышает вывод гусят. Научн. тр. Укр. опытн. ст. птицеводства, 6 :
133. — Быховец А. У. 1960. Охлаждение утиных и гусиных яиц в пер-
вые дни инкубации. Птицеводство, 4 : 25. — Быховец А. У. 1966.
Периодические охлаждения яиц при инкубации и повышение жизнеспособ-
ности птицы. Тр. XIII Всемирного конгр. по птицеводству, Киев : 505.
Варнагирис А. 1960. Улучшение инкубации утиных и гусиных
яиц путем использования растворов соляной кислоты, поваренной соли,
хлористого кобальта. Тр. Литовск. ветер, акад., 4 : 149. — Вильнор А. М.
1935а. Температурные колебания как термический раздражитель при инку-
бации. Сов. птицеводство, 3 : 27. — В и л ь н е р А. М. 19356. Потеря веса
яиц как показатель влажности воздуха в инкубаторе. Сов. птицеводство,
1 : 21. — В л а д и м и р о в а 10. Н. 1954. Потеря в весе яиц кур перед
инкубацией. Тр. Н.-иссл. инет, птицеводства, 24 : 126. — Владими-
рова 10. Н. 1959. Влияние положения яиц на развитие зародыша гусей
и на конечные результаты инкубации. Сб. научн. работ Н.-иссл. ипст. сельск.
хоз. Центр. Черноземной полосы : 191. — Волкова Л. П., С. Я. Во л-
к о в. 1959. Новое в инкубации гусиных яиц. Птицеводство, 10 : 27.
Г о р е ц к и й Б. К. 1961. Об инкубации яиц водоплавающей птицы.
Птицеводство, 4 : 15. — Г у д к и н А. Ф., Д. Н. М у р у с и д з е,
И. А. Смирнов. 1959. Использование ультрафиолетовых лучей при
инкубации яиц. Птицеводство, 2 : 19. — Г у д к и н А. Ф., Д. Н. М у-
русидзе, И. А. Смирнов. 1961. К вопросу об использовании
искусственных ультрафиолетовых лучей при инкубации яиц. Тр. Благо-
вещенск. сельскохоз. инет., 2 : 115.
Д ж а б и е в а С. А., Т. А. Агаджанова. 1963. Влияние эндо-
генного облучения куриных яиц малыми дозами радиоактивного фосфора
на развитие эмбрионов в процессе инкубации. Изв. Акад, наук АзербССР
(сер. биол. и мед. наук), 1 : 87. — Д з ю б а В. 11. 1935. К учению о вер-
тикальных температурных зонах. Сов. птицеводство, 2 : 36. — Добры-
нина А. Я., Л. А. Зубарева, И. Г. Костин, X. Ф. Кушнер,
А. И. Самолетов, М. Г. С а л г а н н и к. 1961. Влияние облучения
эмбрионов на постэмбриональпый рост, развитие и продуктивность кур.
Тр. Инет, генетики АН СССР, 28 : 359.
Е г и н а К. Я. 1960. О влиянии витаминизации куриного яйца на раз-
витие эмбриона цыпленка. В сб.: Вопр. цитол., гистол. и эмбриол. АН ЛатССР,
Рига : 205. — Еремеев Г. П. 1957. Зародышевые приспособления птиц
с точки зрения сравнительной физиологии. Тр. Омск, сельскохоз. инет.,
25 : 1.
Заболотников А. А. 1959. Применение ультрафиолетовых лу-
чей при инкубации утиных яиц. Сельское хоз. Поволжья, 12 : 71. — 3 а-
летаева Т. А. 1958. Влияние режима переменных температур при ин-
кубации на эмбриональное и постэмбриональное развитие кур. Сб. докл.
па научн. конфер. аспир. Всесоюзн. н.-иссл. инет, птицеводства, 1 : 98.
Игнатьев Н. Н. 1960. Инкубация утиных и гусиных яиц на Ка-
гальнпцкой ИПС. Птицеводство, 3 : 27.
Клюкина Е. Т. 1961. Применение ультрафиолетовых лучей при
инкубации. Сб. н.-иссл. работ Гос. с.-х. опытп. ст. Томск, унив. : 63. —
Кирсанов Л. 1938. Эмбриональные поражения, вызванные наруше-
нием воздухообмена в инкубаторе. Сов. птицеводство, 5 : 34. — Ko-
fi и н ь к о Д. А., И. Н. Бакаев. 1965. Кратность поворота яиц и вы-
водимость утят. Птицеводство, 5 : 26. — К о д и н е ц Г. А. 1959. Влияние
продолжительности ультрафиолетового облучения инкубационных яиц на
400
выводимость цыплят. Научн. зап. Херсонск. сельскохоз. инет., 8 : 209. —
Кодинец Г. А. 1962. Влияние ультрафиолетового облучения на неко-
торые физические свойства белка куриного яйца. Вестник сельскохоз. науки,
И : 65. — Колобов Г. М. 1959. Китайский метод холодной инкубации
яиц. Птицеводство, 9 : 43. — К о п о п л е в Н. А. 1962. Инкубация яиц.
Тула. — Котляров Г. С. 1937. Переворачивание куриных яиц в сек-
ционных инкубаторах. Сов. птицеводство, 6: 21. — Крапчитов А.
1958. Иодирование яиц перед инкубацией. Сельск. хоз. Таджикистана,
1 : 28. — К у ч е р о в а Ф. Н. 1963. Охлаждение яиц кур (в период инку-
бации) на разных этапах эмбриогенеза как средство стимуляции роста и
развития молодняка. В сб.: 2-я межвузовская научн. отчетная конфер.
«Университеты — сельскому хозяйству», Тезисы докл., Л. : 85. — К у ч-
ковская Е.Н. 1954. Качество петушков, выведенных до и после 20.5 су-
ток инкубации. Тр. Н.-иссл. инет, птицеводства, 24 : 200. — Куш-
нер X. Ф., И. Г. Костин, Л. Зубарева, Н. И. Кузнецов,
Л. Шершунова. 1966. Влияние малых доз ионизирующих излучений
на организм кур. (Резюме). Тр. XIII Всемирного конгр. по птицеводству,
Киев : 255.
Лепаж Л. 1961. Связь между оплодотворяемостью яиц, выводимостью
и выживаемостью цыплят. Сб. научи, тр. Эст. сельскохоз. акад., 20 : 139. —
Л и п п и н к о т В. А. 1932. Птицеводство. Перевод с 4-го амер. изд. Сель-
хозгиз, М. — Лисицкий Е. Ф. 1937. Исследование патологии в эмб-
риональном развитии цыпленка. Сообщ. и пост. IX пленума ВАСХНИЛ,
секция животноводства : 108. — Лисицкий Е. Ф. 1952. Действие внеш-
ней среды на эмбриональное развитие курицы при различных условиях ин-
кубации. Сб. тр. Харьковск. ветер, инет., 21: 27. — Лисицкий Е. Ф.,
А. У. Б ы х о в е ц, Г. С. Крок. 1954. Изучение колебаний температуры
при инкубации как фактора, повышающего вывод гусят. Научн. тр. Укр.
н.-иссл. станции птицеводства, 4 : 136.
Масленникова Р. К., Т. К. Глаголева. 1956. К вопросу
о весовых потерях куриных яиц в инкубаторе типа «Рекорд-39». Сб. н.-иссл.
работ студ. Ставропольск. сельскохоз. инет. 4 : 150. — Машталер Г. А.
1943. Влияние низких температур на инкубацию цыплят. Бюлл. экспер.
биол. и мед., 16, 9,3 ; 33. — М а ш т а л е р Г. А. 1947. Оброб шкубацпй-
ных яець соляною кислотою. Соц. твариництво, 10 : 47. — Машта-
лер Г. А. 1954. Экологические основы инкубирования яиц выводковых
птиц. (Температурный режим инкубирования). Тезисы докл. 3-й экол. конфер.,
ч. III : 104.
Н а й д е н с к и й М. С. 1962. Влияние малых доз гаммаизлучения
на углеводно-жйровой обмен куриного эмбриона. Тр. Московск. ветер, акад.,
44 : 80. — Н и к и т е н к о М. Ф. 1960. Значение периодических пониже-
ний температуры при инкубации яиц цесарки. В кн.: Регуляция обмена
тепла и других функций у сельскохоз. животных в условиях высоких тем-
ператур. Краснодар : 173. — Никитин В. П. 1941. Птицеводство.
Сельхозгиз, М. — Николаева 10. Н. 1960. Опыты по раннему охлаж-
дению утиных яиц'. Птицеводство, 3 : 24. — Николаева 10. Н. 1962.
О некоторых вопросах теории и практики инкубации яиц водоплавающих
птиц. Тр. Белорусск. н.-иссл. ипст. животноводства, 3 : 175.
Огородпий 10. М. 1935. Значение колебаний температуры при
инкубации. Сов. птицеводство, - 2:42. — Огородний Ю. М. 1936.
Дифференцирование температурного режима при инкубации. Проблемы
животноводства, 2 : 108. — Орлов М. В. 1938. Дифференцирование ре-
жима инкубирования как метод получения высоких выводов цыплят. 3-е изд.
М. — Орлов М. В. 1941. Влияние на процесс эмбрионального развития
кур изменений режима инкубирования путем повышения и понижения влаж-
ности в различные периоды инкубации. Сб. тр. Н.-иссл. инет, птицеводства :
401
212. Орлов М. В. 1960. О режиме инкубации сельскохозяйственной птицы.
(К дискуссии по поводу статьи Г. М. Колобова «Неотложные вопросы инку-
бации»). Птицеводство, 4: 20. — Орлов М. В. 1961а. Методы дифферен-
цирования режима инкубирования яиц сельскохозяйственной птицы.
Тр. Всесоюзн. н.-иссл. инет, птицеводства, 27 : 130. — Орлов М. В. 19616.
Изучение и разработка режима инкубации яиц уток и кур в инкубаторе
«Универсал-45». Тр. Всесоюзн. н.-иссл. инет, птицеводства, 27 : 148. —
Орлов М. В. 1966. Биологический контроль в инкубации. М. — Ор-
лов М. В., Е. Н. К у ч к о в с к а я. 1954. Повышение жизнеспособ-
ности и продуктивности кур направленным воздействием внешних усло-
вий во время инкубации. Тр. Н.-иссл. инет, птицеводства, 24 : 185. —
Орлов М. В., Е. И. Третьякова, Г. К. Отрыганьев,
Г. И. Крылов. 1936. Влияние на процесс эмбрионального развития кур
изменений режима инкубирования путем повышения и понижения темпе-
ратуры в различные периоды искусственной инкубации. I. Рост эмбрионов
и их смертность. В сб.: Пробл. инкубации, Тр. Н.-иссл. инет, птицеводства :
73. — Отрыганьев Г. К. 1935. Хондродистрофия куриного зародыша.
Сов. птицеводство, 6 : 42. — Отрыганьев Г. К. 1937. Эмбриональные
уродства птиц. Сов. птицеводство, 4: 23. — Отрыганьев Г. К. 1938.
Патологоанатомический контроль в инкубации. М. — Отрыганьев Г. К.
1939. Патологоанатомический контроль при инкубации яиц водоплавающей
птицы. Сов. птицеводство, 5: 10. — Отрыганьев Г. К. 1950. Пато-
логические отклонения в эмбриогенезе птиц и их значение для инкубации.
Автореф. дисс. Сельскохоз. акад. им. К. А. Тимирязева, М.—.Отры-
ганьев Г. К. 1951. Биологический контроль в инкубации. Сельхозгиз,
М. — Отрыганьев Г. К. 1959. О теории искусственной инкубации.
Птицеводство, 10: 15. — Отрыганьев Г. К. 1962. Болезни эмбрио-
нов птиц. В кн.: Болезни птиц. М. : 3. — Отрыганьев Г. К.,
В. А. Хмыров, Г. М. Колобов. 1964. Инкубация. Изд. «Колос»,
М. — О т р ы г а н ь е в а А. Ф. 1954. Значение периодических понижений
температуры при инкубации и выращивании молодняка кур. Тезисы докл.
научн. конфер. Саратовск. зоотехп. и ветер, инет., : 20. — Отрыгань-
е в а А. Ф. 1966. Дефекты пуповины и качество цыплят. Птицеводство,
11 : 21.
Пельтцер С. О. 1959. Прижизненная оценка интенсивности раз-
вития зародышей в яйцах кур при инкубации. Докл. Моск, сельскохоз.
акад. им. К. А. Тимирязева, 45 : 65. — Пенионжкевич А. Э. 1950.
О режиме влажности для яиц кур в шкафных инкубаторах в выводной пе-
риод. Тр. Н.-иссл. инет, птицеводства, 21 : 37. — Пенионжкевич А. Э.
1954. Влияние условий инкубации па накопление гликогена в зародышах
утки. Тр. Н.-иссл. инет, пртицеводетва, 24 : 119. — Пепиопжке-
в и ч Э. Э. 1935. Некоторые случаи анормального развития эмбриона во
время инкубации. Сов. птицеводство, 8: 28. — Пенионжкевич Э. Э.
1945. Проблема снижения смертности эмбрионов домашних птиц. Сельхозгиз,
М. —Пенионжкевич Э. Э., Л. Н. Ш е х т м а н. 1937а. Потеря
в весе яиц как показатель режима инкубирования. Сообщение II. Сов. пти-
цеводство, 2 : 35. — П е п и о н ж к е в и ч Э. Э., Л. Н. Ш е х т м а н.
19376. Зависимость эмбрионального роста кур от температуры и влажности
в инкубаторе. Сообщение III. Сов. птицеводство, 4 : 31. — Пили-
пенко М. Е. 1959. Гсматолопчш дослщжепня як додатковий метод
бюлопчного контролю при шкубацп. Науков! пращ укр. досл!д. ст. птах.,
7: 141. — Погорельский В. А. 1947. Влияние пониженного содер-
жания кислорода на развитие куриных эмбрионов. Тр. Инет, эволюц. физиол.
и патол. высш, нервн. деят., 1 : 113. — Попов М. Д., О. И. Василь-
ева и К. Никишин. 1936. Биологические методы оценки работы круп-
ных инкубаторов. В сб.: Пробл. инкубации, Тр. Н.-иссл. инет, птицевод-
ства : 36. — П о п о в М. Д., Э. Г. Л о м о в с к а я. 1956. Режим инку-
бирования яиц как одно из средств изменения онтогенеза птиц. Тезисы докл.
402
Совещ. по проблеме индивидуального развития сельскохоз. животных.
Изд. АН УССР, Киев : 118. — П о п о в Г. В., Э. В. Ц е г е л ь н и ц-
к а я. 1957. Опыт повышения инкубационных качеств яиц с пониженной
выводимостью. Уч. зап. Петрозаводск, унив., 8, 3 : 183. — Приц-
кер И. Я. 1933. К вопросу о биологическом контроле при искусственной
инкубации. Тр. сессии ВАСХНИЛ, Воронеж : 554. — Прицкер И. Я.
1935. Еще к вопросу о причинах липкости цыплят. Сов. птицеводство, 4 :
12. — Прицкер И. Я. 1939. Рост куриных эмбрионов при изменении
инкубационных температур. Сов. птицеводство, 10—11 : 27. — П р и ц-
к е р И. Я. 1940. Влияние высоких температур в первые часы инкубации
па ее результаты. Докл. Всесоюзн. акад, сельскохоз. паук им. К. А. Тими-
рязева, 8 : 24. — Прицкер И. Я. 1942. Комплексный биологический
контроль искусственной инкубации. Тезисы до кт. дисс. Груз, сельскохоз.
инет., Тбилиси, 117. — Прицкер И. Я. 1954а. Охлаждение как эле-
мент режима инкубирования утиных яиц в инкубаторах с принудительной
циркуляцией воздуха. Тр. Н.-иссл. ипст. птицеводства, 24 : 95. — Приц-
кер И. Я. 19546. Гематологические различия у цыплят, выведенных в усло-
виях высоких и низких температур инкубации. ДАН СССР, 95, 1 : 207. —
Прицкер И. Я. 1958. Исследования по вопросу о влиянии температуры
инкубации на экстерьер и интерьер однодневных цыплят. Изв. Иркутск,
сельскохоз. инет., 8:3. — Прицкер И. Я., Н. П. Третьяков.
1937. Инкубаторы и их эксплуатация. Сельхозгиз, М. — Пяткин Е. М.
1965. К оценке действия ультрафиолетовых лучей на кур. Птицеводство,
3 : 30.
Робинсон Л. 1959. Современное птицеводство Англии. ИЛ, М. —
Рольник В. В. 1939. Температурный режим естественной инкубации
у нанду. Вопр. экол. и биоценол., 5—6 : 236. — Рольник В. В. 1941.
Инкубация яиц гаги. Тезисы докл. экол. конфер., Киев, 2 : 40. — Роль-
ник В. В. 1946. Влияние изменения состава газов в воздушной камере
яйца на развитие эмбриона и повышение выводимости цыплят. ДАН СССР,
52, 9 : 831. — Рольник В. В. 1955. Инкубация яиц гаги (Somateria
mollissima L.). Зоол. журн., 34, 4 : 885. — Рольник В. В. 1956. Фи-
зиология дыхания эмбрионов сельскохозяйственных птиц. В сб.: Пробл.
соврем, эмбриол., Изд. ЛГУ : 228. — Рольник В. В. 1960. Изучение
состава газов воздушной камеры яиц в течение инкубации. В сб.: Матер,
по эвол. физиол., Л.—М. 4 : 208. — Рольник В. В. 1964. Биометри-
ческий анализ некоторых сторон развития куриного эмбриона. В сб.: Приме-
нение математических методов в биологии, Изд. ЛГУ, 3 : 159. — Роль-
ник В. В., Э. Л. Портенко. 1964. Газообмен развивающегося кури-
ного эмбриона. Журн. общей биологии, 25, 2 : 133. — Ротт Н. Н. 1957.
Влияние температуры инкубации на развитие системы кровообращения
в эмбриональном и постэмбриональном периоде у кур породы леггорн.
Автореф. дисс. М. — Р у у с Ц. X. 1952. Влияние охлаждений яиц в период
инкубации на вывод и последующее развитие цыплят. Птицеводство, 5 :
23. — Р у у с Ц. X. 1957. Особенности роста и развития гусиных зароды-
шей при стабильной и колебательной температурах. Птицеводство, 1 : 40. —
Руус Ц. X. 1961. О влиянии переворачивания гусиных яиц на вылупляе-
мость при инкубации. Сб. научн. тр. Эст. сельскохоз. акад., 20 : 124. —
Рыжих Г. Н., Л. Б. Клементьева. 1957. Влияние периодических
охлаждений па зародышевое развитие цыплят. Уч. зап. Омск. гос. пед. инет.,
6 : 97. — Р ю м и н А. В. 1950. Температурные условия развития птиц.
Сов. зоотехния, 1 : 90.
Салганник М. Г. 1951. Передовой опыт инкубации утиных яиц.
Птицеводство, 2:16. — Салганник М. Г. 1952. Об инкубации гусиных
яиц. Птицеводство, 1 : 12. — Самарский С. Л. 1954. Особенности
температурного режима естественного насиживания у выводковых и птен-
цовых птиц. 3-я экол. конфер., Тезисы докл: 308. — Семенов-Тян-
403
Шанский О. И. 1952. Изучение инкубации тетеревиных птиц в природ-
ных условиях. Бюлл. Моск. общ. испыт. природы, Отд. биол., 57, 6 : 35. —
Слинько Л. 193G. Изучение некоторых факторов инкубации. Пробл.
животноводства, 2 : 44. — Соболева А. М., Н. А. Т у р о в и ч.
1959. Почему мы отказались от поворота гусиных яиц на 180°. Птицеводство,
10 : 21. — С о р о к а М. Ф. 1955. Разработка режима и техники инкуби-
рования яиц уток без охлаждения в шкафных инкубаторах. Автореф. дисс.
Загорск. —Страутман Ф. И. 1937. Инкубация яиц австралийского
страуса эму (Dromaeus nova-gollandiae Gray). Тр. Бпол. пнет. Томск, гос.
унив., 4 : 279.
Т а р п о в с к и и Н. 1935. Озонирование яиц во время инкубации.
Сов. птицеводство, 9 : 27. — Тот М. 1954. Влияние повышения темпера-
туры инкубации на выводимость, соотношение полов и некоторые качества
цыплят. Автореф. дисс. ЛГУ. — Третьяков Н. П. 1937. Естественная
и искусственная инкубация. Пробл. животноводства, 1 : 84. — Т р е т ь я-
к о в Н. П. 1938. Дифференцирование режима инкубирования куриных и
утиных яиц в период вывода. Сов. птицеводство, 4: 17. — Третья-
ков Н. П. 1947. Способность эмбрионов птиц переносить длительные и
глубокие охлаждения. ДАН СССР, 58, 2 : 331. — Третьяков Н. П.
1949а. Температура внутри инкубационных утиных яиц на разных стадиях
эмбрионального развития. Природа, 9 : 79. — Т р е т ь я к о в Н. П. 19496.
Физико-химические изменения плазм утиных яиц при различных условиях
инкубации. Сов. зоотехния, 2: 89. — Третьяков Н. П: 1953а. Дли-
тельное глубокое охлаждение эмбрионов кур на разных стадиях развития.
Птицеводство, 6 : 20. — Третьяков Н. П. 19536. Инкубация. Изд. 2.
Сельхозгиз, М. — Третьяков Н. П., Т. А. 3 а л е т а е в а. 1956.
Влияние переменного режима инкубации на эмбриональное развитие птиц.
Пробл. соврем, эмбриол. : 216. — Третьякова Е. И. 1936. К вопросам
газообмена при инкубации. В сб.: Проблемы инкубации, Тр. Н.-иссл. инет,
птицеводства : 123.
Фердинандов В. В. 1931. Рабочая книга по птицеводству.
Сельхозгиз, М. — Фердинандов В. В. 1939. Температурные условия
естественного высиживания яиц гусыней. Тр. Воронежск. обл. птицеводч.
ст., Воронеж, 3 : 180. — Фердинандов В. В. 1941. Внутрияйцевые
температуры при естественной инкубации. Докл. ВАСХНИЛ, М., 8 : 32. —
Флинт В. Е. 1954. Об экспериментальном изучении активности наси-
живающей гаги. Зоол. журн., 33, 1 : 159.
Хаскин В. В. 1961а. Теплообмен и развитие терморегуляции в он-
тогенезе домашней утки. Автореф. дисс. Харьков. — Хаскин В. В.
19616. Теплообмен птичьих яиц при инкубации. Биофизика, 6, 1 : 91. —
Хаскин В. В. 1963. Влияние температурных условий эмбрионального
развития на терморегуляцию вылупившихся цыплят. Физиол. журн. СССР,
49, 10 : 1254. — Хаскин В. В. 1966. Биохимические механизмы темпе-
ратурной адаптации куриного эмбриона. Тр. XIII Всемирного конгр. по
птицеводству, Киев : 511. — X о д ж а е в М. 1963. Периодическое охлаж-
дение (утиных) яиц при инкубации. Птицеводство, 2: 15. — Христо-
форов И. Д. 1961. Ультрафиолетовое облучение яиц перед инкубацией.
Сельское хоз. Поволжья, 5 : 71.
Шмальгаузен И. И.- 1926. Дослиди над ембрюнальним ростом.
Сб. работ Биол. инет., Харьков, 1: 15. — Шувалова Т. А. 1955.
Влияние периодических охлаждений яиц на размеры клеток куриного эм-
бриона. Уч. зап. Лен. гос. пед. инет., 110 : 75.
Щеглов А. М. 1954. Вскрытие невылупившихся цыплят как метод
выявления причин нарушений их эмбрионального развития. Сб. тр.
Харьковск. ветер, ипст., 22 : 132.
404
Юрченко В. П. 1966. Контроль при инкубации цесариных яиЦ.
Птицеводство, 5 : 30.
Abbott U. К., R. М. Craig. 1960. Observations on hatching
time in the three avian species. Poultry Sci., 39, 4 : 827.
Baldwin S. P., S. Ch. К e n d e i g h. 1932. Physiology of the
temperature of birds. Sci. Publ. Cleveland Museum of Nat. History, 3:1.—
Barott H. G. 1937. Effect of temperature, humidity and other factors
on hatch of hens’ eggs and on energy metabolism of chick embryos. U. S. Dept.
Agr. Techn. Bull., 553 : 1. — В о h г С., К. A. Hasselbalch. 1903.
Ueber die Warmeproduction und den Stoffwechsel des Embryos. Scand. Arch.
Physiol., 14 : 398. — Burke E. 1925. A study of Incubation. Univ, of Mon-
tana, Agr. Exp. Sta. Bull., 178 :1. — Byerly T. 1938. Effect of diffe-
rent incubation temperature on mortality of chick embryos. Poultry Sci.,
17, 3 : 200. — Byerly T. С., M. W. Olsen. 1933. Time and manner
of determination of the malposition head-in-small-end of egg. Poultry Sci.,
12, 4 : 261.
Cadman VV. 1923. Records of temperature, humidity and carbon
dioxide under the sitting hen. Harper Adams Utility Poultry Journ., 8 : 390. —
Carson J. D. 1964. Natural selection as a genetic tool for improving hat-
chability in turkeys. Poultry Sci., 43, 1 : 87. - Cole man. J. W.,
H. S. Siegel, G. F. Krause. 1964. Initial internal temperature chan-
ges of incubating eggs. Poultry Sci., 43, 1 : 205. —Cruz S. R., A. L. Ro-
manoff. 1944. Effect of oxygen concentration on the development of the
chick embryo. Physiol. ZooL, 17 : 184.
Dareste C. 1891. Recherche sur la production artificielle des mon-
struosites, on essais de teratogenie experimentale. Paris. — D e u c h a r E. M.
1952. The effect of a high temperature shock on early morphogenesis in the
chick embryo. Journ. Anat., 86, 4 : 443.
Edwards Ch. L. 1902. The physiological zero and the index develop-
ment for thfe egg of the domestic fowl — Gallus domesticus. Amer. Journ.
Physiol. 6 : 351. — Ellis J. B., L. Morris. 1947. Factors involved
in hatching chicken and turkey eggs at high elevations. Poultry Sci., 24, 6 :
635. — Evans W. 1891. On the periods occupied by birds in the incubation
of their eggs. Ibis, 6, 3:52. — Eycleshymer A. 1907. Some obser-
vations and experiments on the natural and artificial incubation of the egg
of the common fowl. Biol. Bull., 12, 6 : 360.
Funk E. M., H. V. В i 1 1 i e r. 1944. The minimum temperature
for embryonic development of the domestic fowl (Gallus domesticus). Poultry
Sci., 23, 6 : 538. —Funk E. M., J. Forward. 1952. Effect of mul-
tiple plane turning of eggs during incubation on hatchability. Missouri Agr.
Exp. Sta. Res. Bull., 502 : 1. — F u n k E. M., J. Forward. 1960.
The relation of angle of turning and position of the egg to hatchability of chicken
eggs. Poultry Sci., 39, 3 : 784.
G a 1 1 e r a J. 1951. Influence de 1’atmosphere artificiellement modifiee
sur le developpement embryonnaire du poulet. Acta Anat., 11, 4 : 549. —
Gartley К. M., J. D. Me С о n a c h i e, J. R. Cavers. 1951. The
relation between candled appearance at time of transfer and the hatchability
of turkey eggs. Proc. 9th World’s Poultry Congr. : 53. — G г о e b b e 1 s F.
1937. Der Vogel. Bau, Funktion, Lebenserscheinung, Einpassung. II: Ges-
chlecht und Fortpflanzung. Berlin.
Hamilton H. L. 1952. Sensitive periods during development. Ann.
N. Y. Acad. Sci., 55, 2 : 177. — Harrison J. R., K. Irving. 1954.
Effect of lowered incubation temperature on the growth and differentiation
405
of the chick embryo. Biol. Bull., 106, 1 : 48. — H a у s F. A. 1941. The im-
portance of length of incubation period in Rhode Island Reds. Mass. agr. exp.
Sta. bull., 384 : 1. - Hays F. A., E. W. Spear. 1951. Losses in egg
weight during incubation associated with hatchability. Poultry Sci., 30, 1 :
106. — Henderson E. W. 1930. The influence of temperature and bree-
ding upon the rate, of growth of chick embryos. Univ, of Missouri Coll. Agr.
Exp. Sta. Res. Bull., 149 :1. — Henderson E. W., S. Brody. 1927.
Growth and development with special reference to domestic animals. V. The
effect of temperature on the percentage-rate of growth of the chick. Univ, of Mis-
souri Coll. Agr. Exp. Sta. Res. Bull., 99 :1. — Higgins R. A. 1941.
Egg temperatures of wild birds under natural conditions. Ecology, 22, 2 :
148. — Hoffmann E., C. S. S h a f f n e r. 1950. Thyroid weight and
function as influenced by environmentaltemperature. Poultry Sci., 29, 1:3. —
H у r e H. M. 1962. The effect on hatchability of dipping chicken eggs into
vitamin A and vitamin D3 solutions. Poultry Sci., 41, 5 : 1652.
I n s к о W. M. 1949. Physical factors of incubation. In «Fertility and
hatchability of chicken and turkey eggs». Ed. by Taylor : 209.
Kaestner S. 1895. Uber kiinstliche Kalteruhe von hiihnereiern im
Verlauf der Bebrutung. Arch. Anat. und Physiol., : 319. — Kaltofen R.S.
1955. Hatching experiments at Beckbergen — turning the eggs. World’s Po-
ultry Sci. Journ., 11, 3 : 204. — Kaltofen R. S. 1961. Turning the hen’s
egg. A mechanical factor affecting hatchability. Proefshrift. Utrecht. — К e n-
d e i g h S. Ch. 1940. Factors affecting length of incubation. The Auk, 57,
4 : 499. — Krueger W. F., C. A. Tijerina. 1960. The effect of in-
creased air exchange during late embryonic development on hatchability of chi-
cken eggs. Poultry Sci., 39, 5 : 1267. — Kuiper J. W., P. U b b e 1 s.
1951. A biological study of natural incubation and its application to artificial
incubation. Proc. 9th World’s Poultry Congress : 105. — Kuo Z. Y. 1932.
Ontogeny of embryonic behaviour in aves. II. The mechanical factors in the
various stages leading to hatching. Journ. Exp. Zool., 62 : 453.
L a m s о n G. H., W. F. Kirkpatrick. 1918. Factors in incuba-
tion. Storrs (Connecticut) Agr. Exp. Sta. Bull., 95:1. — Landauer W.
1948. The hatchability of chicken eggs as influenced by environment and he-
redity. Storrs (Connecticut) Agr. Exp. Sta. Bull., 262 :1. — Landauer W.,
L. Baumann. 1943. Rumplessness of chicken embryos produced by me-
chanical shaking of eggs prior to incubation. Journ. Exp. Zool., 93 : 51. —
Lutz H. 1962. Action localisee du gaz carbonique sur le developpement
de I’embryon de poulet. C. r. Acad, sci., 254, 13 : 2442.
Mac Mullan R. A., L. L. Eberhardt. 1953. Tolerance of in-
cubating pheasant eggs to exposure. Journ. Wildlife Management, 17, 3 : 322. —
*M a n c n A., El. R о ? i a n u. 1961. Изучение влияния расположения и
системы переворачивания яиц перепончатолапых во время инкубации на
повышение процента вылупляемости. Lucrarili $tiint. Inst. cercetSri Zootechn.,
19 : 469 (на румынск. яз.). — Marshall W. 1951. Abhormalities of the
heart and great vessels in full term dead-in-shell embryos. Proc. 9th World’s
Poultry Congr. : 80. — M о r e n g R. E., R. L. Bryant. 1954a. Effects
of sub-freezing temperature exposure on the chicken embryo. 1. Survival and
subsequent growth up to time of hatch. Poultry Sci., 33, 4 : 855. — M o-
reng R. E., R. L. Bryant. 1954b. Effects of sub-freezing temperature
exposure on the chicken embryo. 2. Hatchability, chicken weight and survival
to six weeks. Poultry Sci., 33, 5 : 987. — M о r e n g R. E., R. L. Bryant.
1955. The tolerance of the chichen embryo to periods of low temperature expo-
sure. Poultry Sci., 34, 6 : 1342. — M о r e n g R. E., R. L. Bryant. 1956.
The resistance of the chicken embryo to low temperature exposure. Poultry
Sci., 35, 4 : 753. — M о r e n g R. E., C. S. Snaffner. 1951. Lethal
406
internal temperatures for the chicken, from fertile egg to mature bird. Poultry
Sci., 30, 2 : 255. —Murray H. A. Jr. 1925. Physiological ontogeny. A.
Chick embryos. II. Catabolism. Chemical changes in fertile eggs during
incubation. Selection of standard conditions. Journ. Gen. Physiol., 9:1.
Needham J. 1931. Chemical embryology. Cambridge, Univ. Press. —
Now D. A. T. 1957. A critical period for the turning of hen's eggs. Journ.
Embryol. and Exptl. Morphol., 5, 3 : 293.
Olsen M. W., T. С. В у о r 1 y. 1938. Resistance of chicken embryos
to mechanical disturbances. Journ. Agr. Res., 56 : 221. — Olsen M. S.,
Marsden S. J. 1954. Mortality among turkey embryos in relation to rate
of embryonic development. Poultry Sci., 33, 6 : 1146. — *0 rszagh V.,
J. M i c e k. 1961. Влияние переменных температур при искусственном
выводе молодняка птицы. Pol’nohospodarstvo, 8, 9 : 678 (на польск. яз.).
Penquite R. 1938. Influence of temperature and humidity upon
the growth of chick embryos in a mechanically ventilated incubator. Agr.
Exp. Sta. Jowa state coll, of agr. and mechanic, arts. Res. bull., 232 : 1. —
*P r e d a V., M. C r i s t e а, О. Cracium. 1959. Влияние темпера-
туры на развитие куриного зародыша при инкубации. Сообщ. VII. Изуче-
ние (обмена) мукополисахаридов, тиоловых соединений и жиров в печени,
головном мозгу п почках зародыша. Inst. med. ?i farmac. Cluj., 51—61 :
415 (на румынск. яз.). —Prevost J. L., J. В. Dumas. 1827. Мё-
moire sur le developpcment du poulet dans 1’oeuf. Ann. Sci. Nat., 1, 12. —
Pre yer W. 1885. Spezielle Physiologie des Embryo. Leipzig.—
Pringle E. M., H. G. Barott. 1937. Loss of weight of hens’ eggs
during incubation under different conditions of humidity and temperature.
Poultry Sci., 16, 1 : 49. — P г о z e s k у О. P. 1963. Body temperature
of birds in relation to nesting habits. Nature (Engl.), 197, 4865 : 401.
Randles C. A., A. L. Romanoff. 1950. Some physical aspects
of the amnion and allantois of the developing chick embryo. Journ. Exp. Zool.,
114, 1 : 87. — Randles C. A., A. L. Romanoff. 1954. A preliminary
study of the hatchability of chicken eggs subjected to shaking agitation. Poultry
Sci., 33, 2 : 374. — Riddle 0. 1924. The necessary gaseous environment
of the bird embryo. Ecology, 5 : 348. — Robertson I. S. 1961a. The in-
fluence of turning on the hatchability of hens’ eggs. I. The effect of rate of tur-
ning on hatchability. II. The effect of turning frequency on the pattern of mor-
tality, the incidence of malpositions, malformations and dead embryos with
no somatic abnormality. Journ. Agr. Sci., 57,1 : 49, 56. — R о b e г t s о n I. S.
1961b. Studies on the effect of humidity on the hatchability of hens’ eggs.
I. The determination of optimum humidity for incubation. II. A comparison
of hatchability, weight loss and embryonic growth in eggs incubated at 40°
and 70% r. h. Journ. Agr. Sci., 57, 2 : 185,195. — Robertson I. S. 1961c.
Studies of chick embryo orientation using X-rays. I. A preliminary investiga-
tion of presumed normal embryos. II. Malpositioned embryos and their sub-
sequent natchability. Brit. Poultry Sci., 2, 1 : 39, 49. — R о 1 n i k V. 1943.
Instructions for the incubation of Eider duck eggs. Journ. Wildlife Management,
7, 2 : 155. — R о m a n о f f A. L. 1929. Effect of humidity on the growth,
calcium metabolism and mortality of the chick embryo. Journ. Exp. Zool.,
54 : 343. — Romanoff A. L. 1930. Biochemistry and biophysics of the
developing hen’s egg. I. Influence of humidity. Cornell Univ. Agr. Exp. Sta.,
Mem., 132 : 1. — Romanoff A. L. 1934. Study of artificial incubation
of game birds. I. Temperature requirements for pheasant and quail eggs. II. Hu-
midity requirements for pheasant and quail eggs. Cornell Univ. Agr. Exp. Sta.
Bull., 616 : 1. — Romanoff A. L. 1935. Influence of incubation tempera-
ture on the hatchability of eggs, postnatal growth and survival ot turkeys.
Journ. Agr. Sci., 25 : 318. — Romanoff A. L. 1936. Effect of different
temperatures in the incubator on the prenatal and postnatal development of the
407
chick. Poultry Sci., 15, 4 : 311. — Romanoff A. L. 1938. Study of ar-
tificial incubation of game birds. III. Effect of air movement on the incuba-
tion of pheasant and quail eggs. IV. Interrelation of temperature, humidity,
and air movement in the incubation of pheasant and quail eggs. Cornell Univ.
Agr. Exp. Sta., Bull., 687 : 1. — Romanoff A. L. 1939. Effect of tempe-
rature shock on development of chick embryo. Proc. 7th World’s Poultry Congr. :
184. — Romanoff A, L., H. A. Faber. 1933. Effect of temperature
on the growth, fat and calcium metabolism, and mortality of the chick embryo
during the latter part of incubation. Journ. Cell, and Compar. Physiol., 2,
4 : 457. — Romanoff A. L., A. J. Romanoff. 1933. Biochemistry
and biophysics of the developing hen’s egg. II. Influence of composition of air.
Cornell. Univ. Agr. Exp. Sta. Mem., 150 : 1. — Romanoff A. L.,
L. L. Smith, R. A. S u 1 i v a n. 1938. Biochemistry and biophysics
of the developing hen’s egg. III. Influence of temperature. Cornell Univ. Agr.
Exp. Sta. Mem., 216 : 1. — R о m i j n Ch. 1950. Foetal respiration in the
hen. Gas diffusion through the egg shell. Poultry Sci., 29, 1 : 42. — R o-
m i j n Ch., W. Lokhorst. 1956. The caloric equilibrium of the chicken
embryo. Poultry Sci., 35, 4 : 829. — R о m i j n Ch., W. Lokhorst.
1961. Artificial incubation and veterinary practice. Tijdschr. Diergeneeskunde,
86, 15 : 995.
Sadler W. W., H. S. W i 1 g u s, E. G. Buss. 1954. Incubation fa-
ctors affecting hatchability of poultry eggs. 2. Some effects of carbon dioxide
upon morphogenesis. Poultry Sci., 33 : 1108. — Schuberth L. 1959.
Der Einfluss des Brutfactors «Wenden» in der Kunstbruttechick. Tierzucht,
13, 2 : 80. — S e с к P. 1939. Der Einfluss heftiger Erschiitterungen auf der
Brutfahigkeit des Hiihnereies. Arch. f. Gefliigelkunde, 13 : 362. —
Smith J.R.,C. E. Howes. 1949. The inheritance of length of incubation
period in chickens. Poultry Sci., 28, 5 : 782. — Spratt N. T. 1949. Carbon
dioxide requirements of the early chick embryo. Anat. Rec., 105, 3 : 583. —
Stiles K. A., R. L. Watterson. 1937. The effects of jarring upon
the embryogeny of chick embryos. Anat. Rec. Philadelphia, 70:7.—
*S z a r s к i H. 1947. On the thermal death of the chicken embryo.
Math, et Nat., Ser. B, Sci. Nat. (2) (ZooL) Bull. Internal. Acad. Polonaise
Sci. et Lettr. Sci. : 133.
Taylor L. W., G. О. К r e u t z i g e r. 1965. The gaseous environ-
ment of the chick embryo in relation to its development and hatchability. 2.
Effect of carbon dioxide and oxygen levels during the period of the fifth thro-
ugh the eighth days of incubation. Poultry Sci., 44,1 : 98. — T а у 1 о r L. W.,
R. A. S j о d i n, C. A. Gunns. 1956. The gaseous environment of the
chick embryo in relation to its development and hatchability. 1. Effect of car-
bon dioxide and oxygen levels during the first four days of incubation upon
hatchability. Poultry Sci., 35, 6 : 1206. — Thompson K. L. 1952.
Incubation at high altitudes. The effects of wind, barometric pressure and hu-
midity on foetal mortality in the hen’s egg. Poultry Sci., 31, 3 : 497. — T i-
xier-Vidal A. V. 1958. Influence d’un abaissement de la temperature
d’incubation sur la thyroide de 1’embryon de poulet en fin d’incubation.
C. r. Acad, sci., 246, 9 : 1463.
♦Vermesanu N. 1961. Влияние изменения факторов инкубации
на биохимические процессы в яйце. Probl. zootehn. $i veterin., 1 : 37
(на румынск. яз.).
Weinmiller L. 1930. Incubation experiments. Proc. 4th World’s
Poultry Congr. : 23. — W e s s e 1 k i n N. W. 1913. Ueber den Einfluss
des Sauerstoffmangels auf das Wachstum und die Entwicklung von Hiihnerem-
bryonen. Centralbl. d. algem. Pathol, und pathol. Anat., 24 : 1033. — W i 1-
gu s H. S., W. W. Sadler. 1954. Incubation factors affecting hatcha-
bility of poultry eggs. 1. Levels of oxygen and carbon dioxide at high altitude.
Poultry Sci., 33, 3 : 460.
предметный указатель
Адаптация 279, 304, 319, 321
Адаптационно-трофическое влияние
симпатической нервной системы
154
Аденозинтрифосфорная кислота
(АТФ) 13, 165, 262, 263
Адреналин и норадреналин см.
Гормоны, катехоламины
Акросомная реакция 21
Аллантоис 111—115, 180, 320
— жидкость 112, 249
— — состав 114
— замыкание 112, 182
— образование 112
— стебелек 113
— — перевязка ИЗ, 190
— структура 11—‘114
— — белковый мешок ИЗ
— — листок внутренний 112, 124
— — — наружный (хорио-аллан-
тоис) 112, 124
— функция 114
— — дыхание 114, 190
— — склад экскретов 114
Альбумины и глобулины плазмы
крови 12
Аминокислоты корма 32
— яйца 62
Аммиак 221—224, 339
Амнион 104—111
— жидкость 106—111, 212, 249
— — белок в ней 109, 213
— образование 104—109
— — складки 106
— полость 106
— роль 111
— серозо-амниотический канал
107—110, 212
— функция 108—111
— — сокращения 108
Амфибии 91, 112, 126, 170, 233, 252,
282
Антигены плазмы крови 134
Аорта(ы) брюшные 122
— возникновение полулунных кла-
панов 120
— дуги 115
— — развитие 124, 125
— спинные 122
— — ветви 127
Артерия(и) аллантоисные ИЗ
— брюшнополостная 124
— желточные 127
— легочные 124
— подвздошные 124
— подключичные 124
— сонные 124
Археоптерикс 270
Аскорбиновая кислота 229, 239, 260
Асфиксия эмбриона 191
— — основная причина смертности
эмбрионов в конце инкубации 289
Ацетилхолин, влияние на мышцу
сердца 136
Бактерицидные свойства амниоти-
ческой жидкости 111
— — белка 69, 133
Безусловные рефлексы в сердечной
деятельности 137 (см. также
Рефлексы)
Белок 16, 67—69 (еж. также Яйцо)
— внутренний жидкий 16
— густой или плотный 16
— — влияние содержания на выво-
димость яиц 61
— использование эмбрионом НО, 213
— наружный жидкий 17
— халазообразующий 16
— химический состав 67—69 (см.
также Выводковые и птенцо-
вые птицы)
— — — неорганические вещества 66
— — — пигменты, овофлавин 69
— — — протеины 68
Беспозвоночные 170, 172, 178
Билатеральная симметрия 53, 91
— — правило Бэра 91
Биологической компенсации законо-
мерность, феномен 201, 279
27 в. В. Рольник
409
Биологический контроль инкуба-
ции 351—361
— — — критерии 352
— — — — патолого-анатомические
изменения 202, 353, 362
Бластодиск (зародышевый диск) 25
— дробление 26
— — 1-я и 2-я борозды 26
— желточный синцитии 22, 25, 88
— зародышевый валик 25
— развитие в зависимости от сезона
и возраста несушки 29
— светлое поле и темное поле
(area pellucida и area ораса) 27, 90
Бластопор 91
Боталловы протоки 124
Вегетативная нервная система см.
Парасимпатическая нервная си-
стема и Симпатическая нервная
система
Вена(ы) аллантоисные (пупочные) 114
— воротная система печени 125, 207
— — — почек 125
— желточно-брыжеечные 123
— желточные 100, 122
— задние кардинальные 122
— концевая 100, 122
— легочная 124, 177
— подключичные 126
— полая нижняя 126
— — передние 127
— сегментные 126
— субкардинальные 125
— яремные 126
Вентиляция 331—343
— влияние па влажность воздуха
331, 339
— — на теплообмен яиц 331, 342.
— воздухообмен 340, 342
— — расчеты 340
— озонирование яиц 337
— скорость движения воздуха
в инкубаторе 340
— удаление углекислоты 334
Вес куриных эмбрионов 276, 366
Витамины в яйце 39, 66
— — — влияние на выводимость
39, 84
Вителлогенез см. Желток образо-
вание
Влажность относительная, ампли-
туда 331
— — в гнезде 322
— инкубаторе, высокая 328,
— — — — низкая 328, 330
— — — — оптимальная 329—331
— — влияние на обогрев 322
— — — — рост и развитие эмб-
риона 328
Вода «метаболическая» 328
Водный обмен 281
Воздушная камера яйца (пуга) 56
— — — объем 56, 181
— — — роль в дыхании 181
— — — состав газов 181—185, 190
Воздушные мешки см. Дыхатель-
ный тракт
Возраст птиц, влияние на размер
бластодиска в снесенном яйце 45
— — — — вес яиц 60
— — — — калорийность яиц 60
оплодотворенность яиц 46
— — — — продуктивность 45—47
Возрастные признаки эмбрионов
366-368
Временные связи в. эмбриональном
периоде 174—176, 290
Временные эмбриональные органы
см. Аллантоис, Амнион, Жел-
точный мешок, Сероза
Выводимость (%) при естественной
инкубации 352
Выводоспособность яиц, влияние
кормления несушек 32, 33, 38
40, 41, 43—45
— — связь с внешними призна-
ками 85
—. — снижение при введении по-
ловых гормонов 258
Выводковые и птенцовые птицы
(различия)
— — — — активность угольной ан-
гидразы 200
— — — — дифференциация ги-
пофиза 238
— — — — количество лецитина 65
— — — — морфогенез мозжечка
149
— — приспособление к об-
мену при пониженной темпера-
туре 306
— — — — процент воды в белке
и желтке 63, 67
— — — — развитие скелета крыла
270
— — — — содержание белка, жел-
тка и скорлупы 60
-----— — химический состав
белка 67
— — — — сроки рефлекторной ре-
гуляции сердечной деятель-
ности 137
----------стадия развития нерв-
ной системы 175
— — — — темп роста 278
— — — — щитовидная железа 234
410
— — — — ьритгропоэз 131
Вылупление 10, 168, 177, 191, 206,
222, 250, 260, 303, 339, 357
— изменение в кровообращении 127
Газы, азот и гелий 203 (см. также
Кислород, Углекислота)
— угарный 203
Газообмен эмбрионов 192—203
— — восточного домашнего кра-
пивника 197
— — голубиных 196
— — гусиных 197
— — интенсивность 277, 289
— — куриных 192
— дыхательный коэффициент
197
— — — изменения в период инку-
бации 194
— — утиных 196
Газообменный аппарат 195
Ганглии головных нервов 145
— образование, ганглиозная пла-
стинка 143
— спинномозговые 145
Гаструла, стадия 87
Гемоглобин эмбрионального и пост-
эмбриоиального типа 128, 132,
178, 319, 329
Гензеновский узелок 90
Гены в детерминации пола 253
— летальные 23
Гермафродитизм 253
Гетерозис 23
Гиалуроновая кислота 220
Гипоталамус 148
— система гипоталамус—мозговое
вещество надпочечников 229
Гипофиз 47, 148, 237—242
— задняя доля (нейрогипофиз) 241
— — — нейросекрет (вещество Го-
мори) 241
— передняя доля (аденогипофиз) 237
— развитие 149
— функция и взаимодействие с дру-
гими эндокринными железами
237—242
Гликемия, уровень 226
— гипер- 227
— гипо- 226, 229, 230
— регуляция, гомеостатический
механизм печени 211, 226
Гликоген, содержание, гликонео-
генез и гликогенолиз в печени
210, 211, 226, 229, 234, 240
— — в желточном мешке 103,
226, 229
— — — мышцах 262
— — — нервных клетках 146
--------сердце 240, 2бЗ
Гликогенное тело 146
Гликолиз в мышцах 263
Глюкагон см. Гормоны
Глюкоза в желтке 66
— — крови см. Гликемия
— — мышцах 262
Головной мозг 148
— — желудочки 148
— — задний 149
— — мозжечок см. Мозжечок, раз-
витие
— — передний 148
— — — большие полушария 148
— — продолговатый 149
— — промежуточный 148
— — средний 149
Головные первы (I—XII) 151
— — блуждающий (вагус; X) 151
— — зрительный (II) 151, 158
— — обонятельный (I) 151, 165
— — слуховой (VIII) 151, 162
Гонады 242—245
— бисексуальность и интерсексу-
альность 243, 249, 252, 253, 256
— гоноциты (первичные половые
клетки) 243
— кора (зародышевый эпителий) 242,
252, 253
— медулла (мозговая часть) 242,
244, 252, 253
— половая дифференциация (самец,
самка) 243, 248, 249
Гормон(ы) (см. также Половые гор-
моны)
— адренокортикотропный (АКТГ)
230, 237, 239
— антидиуретический (АДГ), влия-
ние на выводимость, % 220, 237
— глюкагон 227
— глюкокортикоиды, гидрокорти-
зон и кортизон 214, 227, 229
— гонадотропные (фолликулости-
мулирующий п лютеинезирую-
щий) 13, 44, 48, 237, 238, 240, 254
— инсулин 225—227
— катехоламины, адреналин и нор-
адреналин 228
— минералокортикоиды 227
— окситоцин 237
— паратиреоидные 47, 240
— пролактин 47, 237
— роста 237, 241
— тиреоидные 233, 234
-----тирозин и дииодотирозип 233
— — тироксин 194
— тиреотропный 237, 238, 316
Гормональные механизмы регуляции
обмена веществ 242
27*
411
Двигательная активность, развитие
166—172
— — движения рефлекторные 167
— — — спонтанные 167, 169, 171
— — — — миогенная или нейро-
генная природа их 171
— — методика регистрации 171
— — темп и ритм 166, 169
— — фаза генерализации рефлексов,
вторичная 168
— — — — — первичная 168
— — — локальных рефлекторных
реакции 167
— — — специализации рефлек-
торных реакций 168
Дерматом 94
Дифференцировка биохимическая
146, 233
— морфологическая 146, 278
— функциональная 233
— чередование с ростом 233, 277
Дыхание 178—191
— атмосферным воздухом 180
— — — аллантоисное 180
— — — легочное 187—189
--- — переход от аллантоисного
к легочному 183, 189, 196, 337
— молекулярное кислородом желтка
178
— периодичность 178, 281
— при различном содержании газов
в среде 203
— тканей 199
Дыхательные движения 187—189
— — запись 188
— — ритм 187
Дыхательный тракт 176—178
— — бронхи возвратные 177
— — — первичные, вторичные и
третичные 176
— — воздушные мешки 177
— — трахея, хрящевые кольца 176
Железы эндокринные см. по назва-
нию желез и Гормоны
Желток, анимальный и вегетатив-
ный полюса 10 (см. также Яйцо)
— белый И,
— желточные шары И
— желтый И
— использование эмбрионом 289, 290
— латебра 91
— насыщение кислородом 178
— оболочка 13, 55, 107
— образование (вителлогенез) и
рост 7, И
— удельный вес 70
— форма 54
— химический состав 63—67
— — — витамины см. Витамины
в япце
— — — липиды см. Липиды
желтка
— — — неорганические вещества 66
— — — протеины см. Протеины
желтка
— углеводы 66
— цвет, зависимость от корма 66
Желточный мешок 97—104
— — внутренняя поверхность 98
— — втягивание 103
— — кровообращение 99—101, 179
— — роль в постэмбрионалыюм раз-
витии 104, 140
— — стебелек 97
— — структура 97
— — функция 102, 103
— — — гемопоэз 128
Жизнеспособность молодняка 20,
317, 361
Жировой обмен эмбрионов 197
«Задохлики» 191, 196, 289, 337, 338
Зародышевые листки 89
Зигота 20
— деление 25
Инбридинг 23, 51
Индукция 89, 216, 272, 273
Инкубаторы секционные (с естествен-
ной вентиляцией) 307
— шкафные (с искусственной вен-
тиляцией) 307
— — выводные шкафы, лотки 310
— — инкубационные шкафы, лотки
311
Инкубация, дифференцированный
режим 311
— длительность в зависимости от
температуры 306, 321
Инсулин см. Гормоны
Искусственный отбор 50
Кальций в костях несушки 34
— — — эмбриона 266
— в крови несушки 35
— — — — влияние на продук-
тивность 35
— — — — органически связанный
и ионный 35
— обмен 35
— — влияние половых гормонов 34
— — -4 паратиреоидного гор-
мона^ 36
—скорлупы 69
Кислород, в желтке снесенного
япца 178
— — крови 123
412
— влияние низкой и высокой кон-
центрации 334, 335
— добавление в воздушную камеру
(кислородизация) 191, 337—339
— концентрация в инкубаторе 336
— на больших высотах недостаток
337
— — — — добавление 337
— патологии при недостатке 202
— потребление куриным эмбрионом
193—196, 315
— проницаемость через скорлупу 181
— содержание в воздушной камере
183-186, 191
Колебания температуры в инкуба-
торе 313—319
— — при естественной инкубации
см, Температурный режим есте-
ственной инкубации
Копчиковая железа 271
Кормление птиц, влияние на продук-
тивность 29—42
Кормовые (питательные) вещества
31-42
— — витамины 38—42
-------В2 41
— — жиры (липиды) 33
— — минеральные 33
— — — марганец 38
— — — соотношение кальция и фос-
фора 37
— — протеины 31
— — — животные 32
— — — растительные 32
— — углеводы 33
Кортизон, гидрокортизон см. Гор-
моны глюкортикоиды
Кости замещающие, три стадии раз-
вития 264—266
— перепончатые (покровные) 266
Костный мозг, 34, 130, 266
Критические периоды см, Периоды
повышенной смертности
Кровеносные капилляры 177
— сосуды 122—127 (см, также Ар-
терии, Вены)
— — образование и развитие
122—127
Кровообращения круги, аллантоис-
ный 123
— — внеэмбриональный 122, 335
— — внутриэмбриональный 122
— — — большой и малый 127
Кровоток, начало 135
Кровь 128—135 (см, также Лейко-
циты, Тромбоциты, Эритроциты)
— гемопоэз 128
— лейкоцитопоэз 131
— лимфоцитопоэз 131
— образование в желточном мешке
128
— — — костном мозгу 130
— — — печени 130
— — — селезенке 129
— общий объем 134
— плазма 12, 134
— содержание глюкозы см. Гли-
кемия
— форменные элементы, возникно-
вение 122, 128
— эритропоэз 128
Кровяные островки 122, 128
Круглоротые 253
Лейкоциты 133
— агранулоциты 133
— — лимфоциты 133
— гранулоциты 133
Лизины 21
Лизоцим 69
Лимфа, начало тока 140
— плазма 140
Лимфатическая (не) система 139—141
— железы 139
— мешки 139
— сердца 139
— сосуды 139
Линька кур 32
Липиды желтка 12, 62, 65
— — липопротеины 12
— — фосфолипиды 12, 65
«Липкие» цыплята 333
Матрикс хряща 264
Мезенхима 89, 164, 212, 264, 269, 270
Мезодерма 89, 93, 95, 116
Мейоз 6
Методы повышения выводимости
молодняка сельскохозяйственных
птиц 359—361
— — — витаминизация яиц 359
— — — влияние микродоз иони-
зирующей радиации 360
— — — кислородизация 191
— — — подогревание яиц 75
— — — ультрафиолетовое облу-
чение яиц 359
Механические факторы инкубации,
343—351 (см, также Биологи-
ческий контроль инкубации)
— — поворачивание яиц, количе-
ство поворотов и изменение его
в течение инкубации 345—349
— — — — дополнительное гуси-
ных яиц 347
— — положение яиц 343—345
— — — — вертикальное 343
— — — — горизонтальное 343
413
— — — — йзменёнпе в Тейенйе ин-
кубации 344
-----сотрясение яиц 349—351
— — — — при перевозке 83
Микрогазоанализатор 182
Минеральные вещества в корме не-
сушки см. Кормовые вещества
— — — крови несушки 34—37
-------яйце 66—70
— — — — калий и натрий 66
— — — — микроэлементы 62, 67,
Миограмма сердца 138
Миотомы 94
— мускульный листок 259
Митоз 10, 25, 130, 145, 230
Митохондрии 7, 13, 159,163,165, 315
Млекопитающие 34, 35, 47, 49, 135,
157, 162, 164, 170, 207, 217,
227, 251, 252, 263, 280, 291
Мозжечок развитие 149
Молочная кислота 263
Моча окончательная 221
— первичная 220
Мочевая кислота 221—224
— — образование в печени 208
Мочевина 221—224
— образование в печени 208
Мочевыделительная система 215—
223 (см. также Почки)
Мускулатура, развитие 259—264
— висцеральная (внутренностная)
259, 261
— гладкая 259, 261
— иннервация 259
— поперечнополосатая 259, 261
— соматическая 259
Мышечные волокна 107
— волоконца (фибриллы) 260
— клетки 260
— ленты 260
— синцитий 107, 260
Мышца(ы) глазные 261
— грудная 261
— конечности задней 260
-----передней 261
— плечевого пояса 261
— сердца 261
— эмбриональные, потребление глю-
козы 263
Надкостница 264
Надпочечники, кора (строение и
функция) 227—230, 239, 240
— мозговая часть (строение и функ-
ция) 227—229
Надхрящница 264
Наружные покровы 271—274 (см.
также Перья и пух)
— — гребень и сережки 271
— — клюв, когти и шпоры 271
кожа, кориум и эпидермис
271, 273
— — чешуйки 272
— — яйцевой зуб 204
Нейроглия, клетки 145
Нервная система, периферическая
149—154 (см. также Головные
нервы, Нервы, Органы чувств,
Парасимпатическая нервная си-
стема, Симпатическая нервная си-
стема).
— — развитие 142—154
— — — нервная пластипка 93, 142
— — — — трубка 93
— — центральная 142—149
Нервные клетки 145
— — дендриты их 145
— — мотонейроны 145
— — осево-цилиндрические от-
ростки их 145
— — развитие 145
— — — ганглиобласты, нейро-
бласты, спонгиобласты 145
— — чувствительные 145
Нервы 149-152
— головные см. Головные нервы
— миелиновая оболочка 146
— спинномозговые 150
— — волокна двигательные 150
— — — чувствительные 150
Несушки, физиология в период
яйцекладки 29—49
— высокопродуктивные 35, 36, 48
— кровь, концентрация различных
веществ 12, 30
— обмен веществ 30
— молодки 32, 38, 45, 46
Нуклеиновые кислоты, ДНК 7,13, 65
— — — синтез 25
-----РНК 7, 13, 65, 171
— — — цитоплазматическая 7
----- ядерная 7
— — участие в митозе 25
Обмен веществ эмбриона, влияние
условий инкубации 311, 313, 320,
326, 336
Обогрев яиц 307, 308, 312
— — контактный, телом наседки 319
Ововителлин см. Протеины желтка
Овуляция 13
— время суток и условия 13
— интервал между овуляциями 14
Одомашнивание птиц 14, 50, 79
Около (пара)щито видные железы,
морфогенез и функция 234, 235
414
Окостенение, сроки 265, 266, 275, 282
Онтогенез 141, 211, 213, 224, 269, 283
Оогенез см. Яйцо, образование
Оплодотворение 9, 21
— избирательность 22
— полиспермия 22
— связь с выводимостью 24
Оплодотворенность яиц гусиных,
индюшиных, куриных и ути-
ных 24
Оптимальные условия развития
эмбриона см. Влажность, Темпе-
ратура и другие факторы инку-
бации
Орган(ы) зрения 155—161
— — зрительные бугры 149
— — — доли 149
---начало функции 172—173
— — развитие 157
— — — глазные пузырьки 142
— — строение глаза 155—161
— — — — гребень 159
— — — — нижнее, верхнее и
третье веко 161
— — — — ресничное тело 160
— — — — сетчатка 157
______ активность 173
— — — — — палочки, колбочки
159
— — — — — ямки, участки ост-
рого зрения 158
— — — — хрусталик 157
— — хиазма 149
Орган(ы) обоняния 164—166
— — обонятельная часть головного
мозга 148
— — развитие 165
— — — обонятельные плакоды 164
— — — — ганглий 165
— — строение 164
_ _ _ обонятельный эпителий 164
Орган(ы) слуха и равновесия
161-164
— — — — включение функций
172, 174
_ _ _ _ развитие 163
_ _ _ _ _ слуховые плакоды и
слуховые пузырьки 142, 163
— — — — внутреннее ухо, или
лабиринт 161
— — — — — — кортиев орган 162
— — — — — — овальный и круг-
лый мешки (утрикулюс и сакку-
люс) 161
— — — — _ _ отолиты и ото-
литовая мебрана 162
— — — — — — полукружные ка-
налы 161, 163
— — — — — — эндолимфа 162
— — среднее и наружное ухо 164
Органы чувств (рецепторы) 155—166
(см. также Орган зрения, Орган
обоняния и Орган слуха и равно-
весия)
Осеменение 19
— гетероспермное 22
Остеобласты 264
Остеокласты 34
Отбор яиц для инкубации 84—86
— — — — овоскопированием 85
— — — — определением инкуба-
ционных качеств яиц каждой
несушки 86
— — — — после подогрева 76,
85
— — — — по окраске желтка 84
— — — — по физико-химическим
показателям 85
Охлаждения яиц 313—319
— — в инкубаторе 313
— — — — гусиных 314, 319
— — — — производственные опы-
ты 314
— — влияние на развитие эмбриона
313-316
— — длительные (глубокие) 317
— — непродолжительные 313
Парасимпатическая нервная си-
стема 152, 154
— — — интрамуральные ганглии
сердца 154
— — — краниальные парасимпати-
ческие узлы 154
Парафиз 148
Партеногенез, естественный и ис-
кусственный 8, 23
Патолого-анатомические изменения
эмбрионов 202, 353, 362—367
— — неправильное положение эмб-
риона в яйце 285, 345, 362—367
Первичная полоска 91
Перевозка яиц 83
— — встряхивание 83
Периодичность развития 280—282,
311
— — деление на периоды 280, 281
Перпод(ы) повышенной смертности
эмбрио пов (критические) 132,
282-290,. 304, 313
— — — — в яйцеводе 83, 284
— — — — — течение инкубации
266
— — — — причины 190, 284—290,
339
Периоды повышенной чувствитель-
ности 282, 288, 305
Перья и пух 272—274
415
— — — гладкие мышцы 274
— — — кровеносные сосуды 274
---— развитие 272, 273
— — — — чехлики 279
— окраска, пигменты липохромные
и меланиновые 274
— птерплии 272
Печень 207—211
— «временная печень» — бласто-
дерма 209
— гемопоэз 130, 131
— гликоген см. Гликоген
— гомеостатический механизм 211,
226
— желчеобразование 208
— желчный пузырь 208
— — проток 207
— развитие 208
— роль в жировом, протеиновом и
углеводном обмене 208
Питание эмбрионов, периодичность
281
— — внутрикишечпое (пероральное)
109, 212—215
Пищеварительный тракт 204—207
— — глотка 204
— — готовность к функции 215
— — желудок железистый 206
— — — мышечный 206
— — кишечник — секретирующий
и всасывающий аппарат 214
— — кишка двенадцатиперстная 206
— _ _ толстая 206
— — — тонкая 206
— — клоака 207
— — мышечная активность 213
— — пищевод 205
— — — зоб 205
— — развитие 96
— — ротовая полость 204
Повороты яиц в гнезде 295, 296
— — в инкубаторе см. Механи-
ческие факторы инкубации
Поджелудочная железа 211, 212,
225—227
— — развитие 211—213
— — — клетки а, р, Д 225
— — — островки Лагесса и Лан-
герганса 225
— — роль в белковом, жировом и
углеводном обмене 212
— — — эндокринная 226
— — строение 211
Подзародышевая полость 25, 90
Подскорлупные оболочки 17, 55, 70
— — проницаемость для газов 180
Подогревание яиц до инкубации, вли-
яние на выводимость 75
Пол, детерминация 251—254
416
— переделка 256—258
— — влияние гормонов 257—258
— соотношение полов 255, 257, 321
— — — влияние храпения яиц 80
Половая система, образование и раз-
витие 242—247
Половые гормоны 247—250
— * — андрогенные 247
— — в детерминации пола 252
— — влияние на соматические ткани
249
— — эстрогенные 49, 247
— — — доминантность их 252
Половые железы см. Гонады
Половые клетки (гаметы) 6, 20 (см.
также Сперматозоиды, Яйце-
клетки)
— — происхождение 243
Половые органы курицы 14, 15
(см. также Яичник эмбриона и
Яйцевод эмбриона)
— — — длительность нахождения
яйца в разных частях яйцевода 16
— — — яичник 14
— — — — фолликулы 1Г
-------------оболочка, фермента-
тивная и синтезирующая актив-
ность ее 11
— — — яйцевод 14—17
— — — — белковый отдел 15, 16
— *— — — воронка 14
— — — — задний конец (матка)
15, 17
Половые отличия эмбрионов 254
— — — в весе 200
_ _ — — газообмене 210, 254
_ — — — длительности инкуба-
ционного периода 254
_ _ — — пульсе 254
— — — — чувствительности к вред-
ным воздействиям 255
— — — макроскопические (димор-
физм) 250, 254
Половые протоки 245 (см. также
Яйцевод эмбриона)
— — вольфовы (семяпроводы) 245
— — мюллеровы каналы 245—248
— — — — изменения под воздей-
ствием половых гормонов 250,
253, 257, 258
— — — — различия в регрессии
у самок и самцов 246
Постэмбриональный период 228
Потеря веса яиц (усушка) 77, 323—
328, 354
— — — в зависимости от влаж-
ности 323
— — — — различные периоды ин-
кубации 326, 354
— — — влияние на выводимость 326
— — — пеоплодотворенных 327
— — — при инкубации естествен-
ной 326
— — — — — искусственной 326
Почка(и) 215—219
— кровоснабжение 217
— иннервация 217
— первичная, или средняя (мезо-
нефрос) 216
— постоянная, или задняя (мета-
нефрос) 217
— предпочка (пронефрос) 215
— строение 216, 217
— — вольфов проток 215
— — мочевые канальцы 216
— — мочеточник 217
— — почечное тельце (капсула и
клубочек) 216
— функция 219—221 (ели также
Мочевыделительная система)
Продукты выделения см. Аммиак,
Мочевая кислота и Мочевина
Проклев (наклев) скорлупы 186, 195,
195, 356
Протеиды, глико- и ли по- 12
Протеиновый обмен 109, 197, 290
(см. также Урикотелический и
т. д. типы распада протеинов)
Протеины белка яйца 62 (см. также
Кормовые вещества)
— актомиозин 117, 260, 262
— желтка 62
— — ововителлпн 63
— плазмы крови несушки 32
— синтез в половых клетках 10,
12, 13
— скорлупы и подскорлупных
оболочек 70
Птицы (кроме сельскохозяйственных)
— волнистый попугай 131
— воробей 233, 300
— восточный домашний крапивник
197, 294, 299, 303, 305, 322,
324, 327
— вяхирь 285
— гага 14, 80, 83, 294—297, 312,
319/323, 325, 326
— глухарь 296, 297
— голубь 14, 30, 49, 64, 65, 92,
131, 149, 228, 233, 234, 277,
285, 294, 300
— горлица 285
— грач 91, 234
— дрозд 69, 300
— кукушка 300
— куропатка 286, 294, 297, 319
— ласточка городская 300
— — морская 91
— лебедь 294
— перепел 17, 251, 285, 286, 306,
312, 342, 343
— скворец 130
— сова 237
— страус австралийский 294
— — африканский 69
— — нанду 294—298, 300, 301
— стриж 131
— утка кряква 185, 294, 297, 298
— — мускусная 254
— фазан 69, 285, 286, 294, 306,
312, 319, 342, 343
— цесарка 24, 64, 69, 314
— чайка 131
— ястреб 254
Пульсация сердца, начало 116, 135
— — гормональная и нервная регу-
ляция 136
— — ритм 136
— — частота 138
Пурины 223
Пух см. Перья и пух
Регуляторные механизмы в раз-
витии эмбриона 279, 280 (см.
также Биологической компенса-
ции закономерность)
— — различие в вариабельности про-
цессов 279
— — — — реакции на температуру
201, 279
Резистентность 315
Рептилии 34, 35, 56, 107, 128, 131,
148, 179, 212—215, 271, 272,
280, 282
Рефлексы, безусловные 137, 175
— замыкание рефлекторной дуги
146, 174
— проприоцептивные 170
— условные см. Временные связи
в эмбриональном периоде
— экстероцептивные 137, 170
Рецепторные клетки, строение 155
Рост 275—280
— изменение интенсивности, де-
прессии и причины их 275—278
— отдельных органов 278, 279
— периоды ’чередования его с перио-
дами дифференцировки 233, 277
— скорость в зависимости от темпе-
ратуры 310, 320
Рыбы 170, 281, 282
Селезенка, образование лимфоци-
тов 140
Семенная жидкость (сперма) 19
Семенник эмбриона, кровеносные со-
суды 247
417
— — строение 244
— — — связки половые 244
— — — — сетчатые 244
— — — семенные канальцы 244
Сердце 116—121
— вес 315
— иннервация 135
— остановка при высокой темпера-
туре 305
— развитие 116—121
— — деление на камеры 120
— — перегородки внутри 120
— — трубчатое 116
— строение 120
— — артериальная луковица 116
— — венозный синус 121
— — желудочки 120
— — миокард 116
---предсердия 120
--- эндокард 116
— — эпикард 116
— функция 135—137
— — рефлекторная регуляция 136,
137
Сероза 104—106
Серозо-амниотический канал см. Ам-
нион
Симпатическая нервная система
152-154, 220
— — — ганглии 152
— — — развитие 153
— —-------пограничный ствол (тяж)
вторичный 153, 154
— — — — — — первичный 153,
154
— — — снланхнические нервы 154
Синапсы 145, 155, 263 •
Синхронность возникновения струк-
тур нервной системы и начала
функции 170
Скелет 264—271
— грудина 269
— крылья 269
— лапы 270
— позвоночный столб 267
— — — пигостиль 267
— развитие и строение позвон-
ков 267, 268
— пояс задних конечностей 270
— — плечевой 269
— ребра 267
— череп, часть лицевая и мозго-
вая 269
Склеротом 94, 264, 267
Скорлупа, образование 17
— кутикула 18, 59
— микроскопическая структура 56
— окраска 53
— «парадокс скорлупы» 180, 186
— пигментация 18
— поверхность 53
— поры 17, 58, 323
— проницаемость для водяных паров
180, 186, 327
— — — газов 59, 180, 327, 330
— толщина 56
— химический состав 69
Скрещивание 23, 51
Смертность эмбрионов, кривые 256,
284
— — при естественной и искус-
ственной инкубации 286
— — причины 284
Соединительная ткань 260
Соматоплевра 104
Сомиты (сегменты) 94, 259
Спаривание 19
Сперматозоиды 8
— жизнеспособность 23
— обмен веществ 7
— образование (сперматогенез) 6, 9
— — отличие от оогенеза 8
— — сперматогонии, сперматоциты
1-го и 2-го порядка, сперматиды
6, 7
— подвижность, связь с выводи-
мостью цыплят 20
— строение, акросома 7
---центросома 7, 10
Спинной мозг 142—149
— — вещество белое 146
— — — серое 146
— — рог брюшной 146
— — — спинной 146
Снланхнотом 96, 259
— спланхноплевра 97, 242
Супраоптическое ядро, образование
нейросекрета 241
Сухожилия 260, 262
Сфингомиелин 172
Температура 291—321 (см. также
Колебания температуры, Обогрев
яиц и Охлаждения яиц)
— в инкубаторе 307—321
— амплитуда 305
— высокая и перегрев 306,
310, 342, 357
— — — изменения в период инку-
бации 309—312
— — — на верхнем и нижнем
уровне яиц 307, 309
— — инкубатории 308
— влияние на скорость развития
эмбриона 201, 305
— внутрняйцевая и тела эмбриона
299, 303, 304, 309, 311, 313, 328
— летальная 303, 304
418
— максимальная 304
— минимальная 302
— — нижний порог развития 302
— низкая и недогрев 306, 357
— оптимальная 307
— связь с вентиляцией 342
— тела наседок сельскохозяйствен-
ных птиц 292
— — насиживающих диких птиц 292
— яиц 309
Температурный режим естествен-
ной инкубации 291—302
— — в гнезде курицы 294
— — — — гаги 294
— — — — страуса нанду 294
— — гнезда, зависимость от тем-
пературы воздуха 295—299
— — — изменения в течение наси-
живания 298
— — — колебания температуры при
пересаживании наседок 295, 302
— — — — — — отлете птиц 297
— — — — — суточные 295
— — — различие в температуре на
верхнем и нижнем уровне яиц
293-295
— — яиц 298—302
Теплообмен яиц 322
— — теплоотдача излучением 308,
342
— — — испарением 342
— — — конвекцией 308, 342
— — теплопродукция 300, 310, 312
— — эндотермические реакции 310
Термопары, прикрепление к яйцу
293
Терморегуляция 301, 303, 306,
315, 324
— гомеотермные животные 178,
-------реакция щитовидной железы
233, 316
— пойкилотермные животные 301,
303
— — — реакция щитовидной же-
лезы 233
— эмбрионов 301, 303, 313
Тимус, или зобные железы, раз-
витие и инволюция 235, 236
— гемопоэз 140, 235
— лимфопоэз 140, 235
— эндокринная функция 236
Тироксин см. Гормоны
Тромбоциты 133
Углеводный обмен 41, 226—230, 320,
361 (см. также Гликоген, Глю-
коза)
Углекислота, содержание в воздуш-
ной каморе 182
— — — гнезде 332
— — — инкубаторе 333
— — — крови 123, 189
— — — снесенном яйце 179
— выделение эмбрионом 192
— патологии при повышении кон-
центрации 203
— полезное действие 332, 334
— проницаемость через скорлупу 181
У рикоте лический, уреотелический,
аммонотелический типы распада
протеинов 222, 223
Фабрициева сумка, лпмфоцитопоэз
Факторы среды, влияние на продук-
тивность взрослых птиц 42—45
— — свет 43
— — — влияние на выводоспособ-
ность яиц 44
— — — — — потребление и пере-
варивание корма 44
— — — дополнительное освеще-
ние 44
— — температура 42
Филогенез 111, 115, 141, 154, 172,
182, 211, 224, 259, 269, 282, 313
Фосфор в крови несушки 36
— — — — связанный 36
Халазы, образование 16, 55
Хоаны 164
Холестерин 12, 65, 209, 229, 240
«Холодный» метод инкубирования
утиных яиц 316
Хорда 92, 94, 267
Хорио-аллантоис 113
Хранение яиц 73—84
— — в пластмассовых мешочках 77
— — вентиляция 77
— — влажность 76
— — выводимость при инкубиро-
вании без него 80
— — длительность 78—81
— — — диких птиц 80
— — — признаки в яйцах 81
— — повышение смертности 78—83
— — положение 78
— — с периодическим подогрева-
нием 75
— — температура 73
— — — верхний предел 74
Хроматин 7
Хромаффинные клетки ганглиев, вы-
деление катехоламинов 228
Хромосомы 10, 21, 251—253
— половые 251, 252
— — тип гетерогаметный (х, у-хро-
мосомы — самка птиц) 251, 252,
255
419
— — — гомогаметный (х, х-хро-
мосомы — самец птиц) 251, 252,
255
— соматические 251
Хрящ, развитие 264
Цевка (плюсно-предплюсна) 270
Целом 96
Цитоплазма (протоплазма) 7, 10, 11
Щитовидные железы 230—234, 320
— — контроль гипофиза 233
— — развитие 231
— — — накопление коллоида в
фолликулах 232
— — функция 232
Эволюция 116, 141, 154, 155, 178,
179, 213, 224, 239, 259, 270, 282,
317, 335
Эктодерма 89
Электрическая активность 172
Электрокардиограмма куриного эмб-
риона 138
Эмбриональная дистрофия 40
— хондродистрофия 32, 42
Эндокринные железы и эндокринная
регуляция 49, 224—242 (см.
также отдельные железы)
Энергообмен 333
— калории, выделяемые эмбрионом
321
— основной источник энергии —
жир 311
— смена источников энергии 275
Энзимы 13
— аденилпирофосфатаза 172
— амилаза 212
— ангидраза угольная 36, 199
— гексокиназа 263
— гиалуронидаза 21, 221
— гликогенолитические 263
---фосфоглюкомутаза 263
2--фосфорилаза 227, 263
— дегидрогеназа гипоксантина 222
---сукцината 163
— ксантиноксидаза 222
— липазы 102, 209, 212
— мети л аза 228
— протеолитические 102, 212, 327
— — пепсин 214
— — пептидаза 209
— — трипсин 212, 214
— сукциноксидаза 159
— фосфатаза 214, 229, 239
— — глюкозо-6-фосфата 211
--- кислая 271
— — щелочная 146, 159, 171,
219, 271
420
— холинэстераза 155, 159, 263, 264
— — ацетилхолина 146, 162
— цитохромоксидаза 159
Энтодерма, образование 87, 89
— — деламинацией 88
— — инвагинацией 88
Энцефалограмма полушарий голов-
ного мозга 172
Эпифиз, развитие и функция 148,
236
Эритроциты 128
Ядро клетки 10, 318
Яичник эмбриона, левый, развитие
и функция 10, 244
---правый, образование и резорб-
ция 245
— — фолликулы 245
— — эндокринная роль 245
Яйцевод эмбриона 247 (см. также
Половые протоки)
Яйцекладка, интервал и цикл 18, 236
— связь с выводоспособностью
яиц 18
— у диких птиц 14
— — сельскохозяйственных птиц
18, 29
Яйцеклетка 10, 13
Яйценоскость 13
Яйцо (см. также Белок, Желток,
Подскорлупные оболочки, Скор-
лупа)
— вес 50
— — влияние на выводимость 51
— неоплодотворенное 28, 83, 284,
300, 327
— образование (оогенез) 7, 9, 14—
18, 245
— — оогонии 7, 10
— — ооциты 7, 10, 13, 245
— — отличие от сперматогенеза 8
— — период малого и большого
роста 7
— — полярные и редукционные
тельца 7
---регуляция 47—49‘
— процентное содержание белка,
желтка и скорлупы 60
— удельный вес 56
— физико-химические свойства
70—73
— — — буферность белка и желтка
— — — калорийность 70
— — — связь с выводимостью 72
— — — электропроводность белка
70
— форма 52
— химический состав 61—70
ОГЛАВЛЕНИЕ
Стр.
Предисловие ...................................................... 3
Раздел I. Яйцо п развитие эмбриона до инкубации
Глава 1. Сперматогенез и образование яйца ........................ 6
Глава 2. Оплодотворение яйцеклетки и развитие эмбриона до
снесения яйца птицей ...................................... 19
А. Осеменение и оплодотворение ............................ 19
Б. Развитие эмбриона до снесения яйца птицей............... 24
Глава 3. Некоторые особенности физиологии несушек в период
яйцекладки и влияние внешних факторов на продуктивность 29
А. Влияние кормления птиц на их продуктивность .... 29
Б. Влияние факторов среды на продуктивность птиц .... 42
В. Влияние возраста птиц на их продуктивность ............ 45
Г. Регуляция процесса яйцеобразования .................... 47
Глава 4. Строение и химический состав яиц ....................... 50
А. Внешний вид яйца....................................... 50
Б. Внутреннее строение яйца ............................... 54
В. Химический состав яйца ................................ 61
Г. Физические и физико-химические свойства яиц .... 70
Глава 5. Хранение яиц и отбор их для инкубации................... 73
А. Хранение яиц до инкубации ...............................73
1. Температура хранения . ;............................. 73
2. Влажность, вентиляция и другие факторы во время хра-
нения яиц ........................................ 76
3. Длительность хранения .............................. 78
4. Изменения в содержании яйца и состоянии эмбриона
во время хранения яиц .................................. 81
Б. Отбор яиц для инкубации .............................. 84
Раздел 11. Морфология и физиология эмбрионального развития
Глава 6. Раннее развитие эмбриона ............................... 87
Глава 7. Временные эмбриональные органы ......................... 96
А. .Желточный мешок....................................... 97
421
Стр.
1. Структура ....................................... 97
2. Функция .......................................... 102
3. Втягивание желточного мешка и его значение в постэм-
брнональном развитии ................................. 103
Б. Амнион и сероза ...................................... 104
1. Структура......................................... 104
2. Функция .......................................... 108
В. Аллантоис............................................ 111
1. Структура .................................i . . . 111
2. Функция ”........................................ 114
Глава 8. Кровообращение ...................................... 115
А. Морфология развития сердца и кровеносных сосудов ... 116
1. Сердце ........................................... 116
2. Сосудистая система................................ 122
Б. Развитие и свойства эмбриональной крови............. 128
В. Физиология сердечно-сосудистой системы .............. 135
Г. Лимфатическая система ................................ 139
Глава 9. Нервная система и органы чувств...................... 141
А. Морфология развития нервной системы и органов чувств 142
1. Возникновение нервной системы..................... 142
2. Развитие спинного мозга .......................... 142
3. Развитие головного мозга........................... 148
4. Периферическая нервная система ................ 149
5. Органы чувств ..................................... 155
Б. Физиология нервной системы эмбриона .................. 166
1. Двигательная активность ........................... 166
2. Электрическая активность .......................... 172
3. Физиология органов чувств эмбриона ................ 173
4. Временные связи в эмбриональном периоде............ 174
Глава 10. Дыхание ............................................ 176
А. Образование дыхательных путей ....................... 176
Б. Развитие функции дыхания ............................. 178
В. Газообмен птичьих эмбрионов ......................... 192
Глава 11. Развитие органов пищеварения ....................... 204
А. Морфология пищеварительного тракта .................. 204
1. Ротовая полость и глотка ........................ 204
2. Пищевод, железистый и мышечный желудок, кишечник 205
Б. Печень (структура и функция).......................... 207
В. Поджелудочная железа (структура и функция) .......... 211
Г. В ну три кишечное питание эмбриона ‘............ 212
422
(-тр.
Глава 12. Развитие мочевыделительной системы ................. 215
А. Морфология развития органов выделения................ 215
Б. Физиология выделительных процессов .................. 219
Глава 13. Развитие желез внутренней секреции ............. 224
А. Поджелудочная железа................................. 225
Б. Надпочечники........................................ 227
В. Щитовидная железа.................................... 230
Г. Околощитовидные железы ............................ 234
Д. Тимус, или зобные железы............................ 235
Е. Эпифиз .............................................. 236
Ж. Гипофиз ............................................ 237
Глава 14. Половая система, половые гормоны, детерминация и
переделка пола................................................ 242
А. Образование и развитие половой системы ............. 242
Б. Половые гормоны ..................................... 247
В. Детерминация пола .................................. 251
Г. Половые отличия эмбрионов и соотношение полов . . . 254
Д. Переделка пола ...................................... 256
Глава 15. Развитие мускулатуры и скелета ..................... 259
А. Развитие мускулатуры .......................... 259
Б. Развитие скелета .................................... 264
1. Гистология образования костей .’................. 264
2. Позвоночный столб ................................ 267
3. Череп............................................ 269
4. Скелет конечностей и их поясов.................... 269
Глава 16. Наружные покровы ............................... . 271
Глава 17. Периодичность развития и периоды повышенной смерт-
ности эмбрионов ........................................ 275
А. Рост и дифференцировка ............................. 275
Б. Периодичность развития .............................. 280
В. Периоды повышенной смертности эмбрионов ............ 282
Раздел Ш. Условия среды, необходимые для эмбрионального
развития птиц
Глава 18. Температура ........................................ 291
А. Температурный режим естественной инкубации ......... 291
1. Температура насиживающих родителей ............... 292
2. Температурный режим гнезда ....................... 293
3. Температура яиц при естественной инкубации .... 298
Б. Минимальняя и максимальная температура эмбриональ-
ного развития ..................................... 302
423
В. Скорость эмбрионального развития при различной темпера-
туре инкубации.......................................... 305
Г. Температурный режим при искусственной инкубации . . . 307
1. Температура в основных типах инкубаторов.......... 307
. 2. Оптимальная температура инкубации.................. 307
3. Температура инкубирования в разные периоды эмбрио-
нального развития .................................... 309
4. Колебания инкубационной температуры и охлаждения
яиц .................................................. 313
Д. Влияние температуры инкубирования на развитие отдель-
ных систем органов и эмбриона в целом................... 319
Глава 19. Влажность .......................................... 322
А. Влажность воздуха в гнезде при естественной инкубации 322
Б. Уменьшение веса яиц в течение инкубации.............. 323
В. Влияние различной влажности в инкубаторе на рост и
развитие эмбрионов ..................................... 328
Г. Оптимальная влажность для инкубирования птичьих яиц 329
Глава 20. Вентиляция........................................ 331
А. Содержание газов в воздухе гнезда при естественной инку-
бации .................................................. 332
Б. Дыхание птичьих эмбрионов при различном содержании
газов в окружающей атмосфере........................... 332
В. Режим вентиляции инкубаторов ........................ 339
Глава 21. Механические факторы.............................. 343
А. Положение яиц во время инкубации..................... 343
Б. Поворачивание яиц ................................... 345
В. Сотрясение яиц ...................................... 349
Глава 22. Биологический контроль инкубации и другие методы
повышения выводимости молодняка ......................... 351
А. Биологический контроль инкубации .................... 351
Б. Некоторые методы повышения выводимости молодняка
сельскохозяйственных птиц........................... 359
Приложения ................................................... 362
с
I. Основные диагностические признаки неполноценного яйца
и нарушений в режиме инкубации.......................... 362
II. Возрастные признаки куриного эмбриона ............ 366
III. Внешние возрастные признаки эмбрионов сельскохозяй-
ственных птиц........................................... 368
Литература................................................... 369
Предметный указатель ......................................... 409
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ
Страница Строка Напечатано Следует читать
22 19 снизу освоении усвоении
30 Табл. 4, графа 4 Увеличение Увеличение на
80 9 сверху большая ббльшая
102 2 » адсорбирует абсорбирует
102 7 » адсорбируемые абсорбируемые
116 И » первых этих
117 Рис. 22, подпись (По Patten, 1952). (По Patten, 1951).
119 Рис. 23, подпись, междужелудочковая межжелудочковая
4—5 сверху борозда; бороздка;
136 2 снизу (см. рис. 32, 7) (см. рис. 32, 8)
137 12 » вылупившегося у вылупившегося
169 Рис. 36, подпись (По Humburger, (По Hamburger,
204 15 сверху брюшной полости полости тела
225 14 снизу (Dieterten-Lievre, ‘(Dieterlen-Lievre,
237 14 сверху париентальный париетальный
324 2 » (табл. 48) (таГ.л. 47)
324 2 снизу (Pringle a. Berott, (Pringle a. Barott,
330 10 » 38.9%, 38.9°,
370 2—3 сверху рефатов рефератов
371 19 снизу антибиологические антибиотические
380 12 сверху Войно Ясенец- Войно-Ясенец-
кий кий
В. В. Рольник
I. ОСНОВНЫЕ ДИАГНОСТИЧЕСКИЕ ПРИЗНАКИ НЕПОЛНО
(По Отрыга
Причина Яйца до инкубации 1-е просвечивание и вскрытие кровяных колец Контрольное просвечивание
Хондродистрофия (кормление неполно- ценными животными белками, недостаток биотина и марганца). — — Отставание росте. в
Дистрофия в связи с авитаминозом В2. Пятнистая и шероховатая скор- лупа. Разжижен- ный белок. — Некоторое ставание росте от- в
Дистрофия в связи с авитаминозом В12. — — Отставание росте. в
Эмбриональная микседема (авитами- ноз D). Тонкая и хруп- кая скорлупа. Бе- лок разжижен. — Отставание росте. в
Подагрическая форма дистрофии (авитаминоз А) при избыточном белко- вом кормлении. Бледный жел- ток. Низкая оплодо- творенность. По- вышенная смерт- ность. Отставание росте. в
А-авитаминозная форма дистрофии. Бледный жел- ток. Низкая опло- дотворенность. Повышенная смертность. Отставание росте. в
Дистрофия в связи с авитаминозом Е. — Большая смерт- ность до 7-го дня. Разрастание ме- зенхимы и кро- воизлияния в бластодерме. —
362
ПРИЛОЖЕНИЯ
ЦЕННОГО ЯЙЦА И НАРУШЕНИЙ В РЕЖИМЕ ИНКУБАЦИИ
ньеву, 1962)
2-с просвечивание и вскрытие замерших Задохлики Вылупившиеся птенцы
Смертность повышена. У мертвых эмбрионов короткие ноги с угло- ватыми изгибами ко- стей, «попугаевый» клюв. Сильно укороченные и искривленные ноги, «по- пугаевый клюв», лип- кость. Много липких и сла- бых.
Смертность повы- шена. Среди замерших встречаются отечные и курчавые. Незначитель- ное уменьшение веса яйца. Курчавость, липкость. Искривление шеи и пог. Отек головного мозга. Гу- стой, же л ток. Курчавое оперение. Липкость. Параличи шеи и ног.
Смертность повышена. Общая отечность. Атро- фия мышц. Кровоиз- лияния в желточном мешке, печени и сердце. Гипертрофия щитовид- ных желез. Атрофия мышц. Крово- излияния в желточном меш- ке, печени и сердце. Отло- жение жира в печени. Слабые.
Очень большая смерт- ность. Сильные отеки кожи, перерождение пе- чени. Гипертрофия по- чек. Признаки эмбриональной дистрофии. Вывод растянут. Мо- лодняк слабый.
Гипертрофия почек и отложение в них кри- сталлов солей. Отложение кристаллов солей в почках, па бры- жейке, сердце и желточпом мешке. Вывод запоздалый. Плохая пигментация- пуха и кожи.
Отставание в росте. Липкость. Припухапие век. Вывод запоздалый. Молодняк слабый. Гла- за слезятся, иногда цод веками творожи- стые массы. Плохая пигментация пуха и кожи.
Помутнение хрустали- ков, пятна на роговице.
363
Причина
Яйца до инкубации
1-е просвечивание
и вскрытие кровяных
колец
Контрольное
просвечивание
Кутикулит (недо- статок витаминов К, С, А, холина и не- которых других). — — —
Патология эмбрио- нов, обусловленная длительным хране- нием яиц. Большая нуга. Желток подвиж- , ный и расположен вне центра яйца. Много неопло- дотворенпых, с зародышами, по- гибшими в пер- вый день. Края бластодермы не- ровные. Слабое развитие зачат- ков тела. Бласто- дермальный кис- тоз. Отставание в росте и развитии.
Длительный пе- регрев яиц. Много кровя- ных колец. Амор- фозы. Уродства, эктопия. Гипере- мия и кровоиз- лияния. Аллантоис раньше срока охватывает белок. Кисты на амни- -оне и аллантоисе.
Кратковременный острый перегрев яиц. — Гиперемия и кровоизлияния. Резкий подъем смертности в зо- не перегрева. —
Недогрев яиц. — Сильное отста- вание в росте и развитии. Сильное от- ставание в раз- витии аллан- тоиса. Белок не охвачен аллан- тоисом.
Недостаточная влажность. — Большая по- теря в весе яйца. Много кровяных колец. Гипере- мия и присыха- ния зародышей к скорлупе. Большая воз- душная камера.
364
Приложение I (продолжение)
2-е просвечивание и вскрытие замерших Задохлики Вылупившиеся птенцы
Отставание в росте. % Изъязвление стенок мы- шечного желудка и две- надцатиперстной кишки. Изъязвление стенок мышечного желудка и двенадцатиперстной кишки. Воспаление кутикулы. Поздний вывод. Сла- бый молодняк.
Эктопия и уродства головы. Кисты па амнио- не и аллантоисе. Ран- ний мелкий наклев. Резкий подъем смерт- ности в зоне перегрева. Переполнение кровью сосудов аллантоиса. Ги- перемия и точечные кро- воизлияния в коже, пе- чени, почках, сердце и мозге. Много мертвых, про- клюнувших скорлупу, но не втянувших желток. Остаток густого белка. Не- правильное положение. Ги- перемия желточного мешка и кишечника. Уменьшение размера сердца. Гиперемия кожи. Непра- вильное положение — го- ловой в желток. Вывод ранний, растя- нутый, молодняк мелкий. Плохо втянутый желток и незажившая пупови- на.
Сильное отставание в росте и развитии. Не- подвижная граница воз- душной камеры. Большая потеря в ве- се. Большая воздушная камера. Раннее начало движения головой. Много живых зародышей. Аллантоис полнокровный. Желток втянут, зеленого цвета. Остаток жидкого белка. Кишечник пере- полнен желточными и кало- выми массами. Гипертрофия сердца. Подскорлупные оболочки сухие, плотные. Возле клюва кровоизлияние в ал- лантоисе. Вывод поздний, растя- нутый. Молодняк вя- лый, плохо стоит на по- тах. Скорлупки гряз- ные. Проклев затруднен. Вывод ранний. Молод- няк и скорлупки су- хие. Хорошая пигмен- тация пуха.
3W
Причина Яйца до инкубации 1-е просвечивание и вскрытие кровяных колец Контрольное просвечивание
Избыточная влаж- ность. — — Запоздалый охват белка ал- лантоисом.
Недостаточный га- зообмен. — Отставание в росте. Уродства головы и глаз. Отставание в росте.
Н еправил ьности поворачивания яиц. — Присыхание желтка к под- скорлупным оболочкам. Аллантоис сра- стается над бел- ком.
II . ВОЗРАСТНЫЕ ПРИЗНАКИ КУРИНОГО ЭМБРИОНА*
Р/2 дня. Эмбрион расположен перпендикулярно к длинной оси яйца
и еще не отделен от бластодермы. Различимы мозговые и глазные пузыри
и 8—10 пар сомитов. Хорошо развито сосудистое поле, образовалось и ра-
ботает сердце. Длина эмбриона 4—5 мм, диаметр сосудистого поля 6—8 мм.
21/2 дня. Головная часть зародыша отделена от желтка и лежи! на ле-
вом боку, спиною к пуге. Заметен резкий изгиб головы. Брюшная полость
еще открыта. Сердце лежит вне полости тела. Намечаются зачатки конеч-
ностей. Длина эмбриона 9—12 мм, диаметр сосудистого поля 12—15 мм.
31/2 дня. Зародыш отделен от желтка и закрыт амнионом. Передний
мозг раздвоен. Начинается пигментация глаз. Конечности имеют ластооб-
разную форму. Заметен аллантоис, достигающий размера среднего мозгового
пузыря. Вес эмбриона 0.05—0.06 г.
4*/2 дня. Голова зародыша резко изгибается в сторону брюшной по-
верхности. Глаза хорошо пигментированы вокруг зрачка. Заметно рото-
вое углубление. Конечности на концах расширяются. Аллантоис разра-
стается над амнионом. Вес 0.13—0.19 г.
51/2 дней. Раздвоенность переднего мозгового пузыря исчезает. Заметны
зачаток века, носовой и верхнечелюстные отростки. Концы конечностей
в середине заостряются. Аллантоис достигает внутренней поверхности
скорлупы. Зародыш погружается в глубь желтка. Вес 0.18—0.40 г.
61/2 дней. Форма головы изменяется за счет относительного уменьше-
ния среднего мозгового пузыря. Образуются челюсти. Шея удлиняется.
На крыльях и ногах появляются пальцы. Вес 0.4—0.9 г.
71/2 дней. Клюв начинает удлиняться, обозначаются ноздри. Глаза
становятся выпуклыми, шея длинной. Резко выделяется локтевой изгиб,
удлиняется средний палец крыла. Вес 0.7—1.2 г.
* По Отрыганьеву, 1951.
366
Приложение I [продолжение)
2-е просвечивание и вскрытие замерших Задохлики Вылупившиеся птенцы
Малая потеря в весе яиц. Гематоамнион. Ги- перемия и кровоизлия- ния в коже и внутрен- них органах, дефекты в строении сердца. Белок остался вне аллантоиса. Во время проклева при- сыхание кожи и клюва к скорлупе. Зоб, желудок, и кишечник переполнены жидкостью. Неправильное положе- ние — головой в острый ко- нец яйца. Гиперемия кожи. Липкость. Вывод запоздалый. Молодняк грязный, с плохо пигментирован- ным пухом. Проклев скорлупы в остром конце яйца. Липкость.
81/2 дней. Голова приобретает форму, характерную для птиц. Клюв
длинный и чуть изогнутый, на конце его появляется белая точка. Концы
пальцев ног утолщены. Видны зачатки перовых сосочков па спине. Вес
0.8-2 г.
9х/2 дней. Веко достигает края радужной оболочки. На ногах исче-
зают межпальцевые перепонки. Перовые сосочки становятся крупнее, по-
крывают всю спину и шею. Аллантоис охватывает почти всю внутреннюю
поверхность скорлупы и при инкубации в секционных инкубаторах часто
уже смыкается в остром конце яйца. Вес 1.7—2.6 г.
101/2 дней. Веко достигает зрачка. Намечается валик гребня. Видны
зачатки когтей. Перовые сосочки покрывают все тело. Вес 2.7—3.5 г.
IP/2 дней. Веко над зрачком образует узкую щель. На гребне появ-
ляются зубцы. На ногах хорошо видны когти. Вдоль спины заметен пух.
Вес 3.6—5 г.
12х/2 дней. Глаза закрыты веками. На передней поверхности плюсны
заметны поперечные бороздки — зачатки чешуек. Пух на спине, крыльях
и ногах. Вес 5.3—6.8 г.
13х/2 дней. Увеличивается надклювный бугорок. Когти белеют. По-
является пух на голове. Эмбрион меняет положение, ложась по длинной
оси яйца, головой в сторону пуги. Вес 7.3—9.7 г.
14х/2 дней. Длина пушинок увеличивается. Появляются складки кожи
на ступне и чешуйки на передней поверхности плюсны. Вес 10.3—11.7 г.
15х/2 дней. Появляется пух па веках. На тыловой поверхности плюсны
складки кожи — будущие чешуйки. Вес 12.4—17 г.
161/2 дней. Ясно обозначаются наросты над ноздрями. Увеличивается
длина ног. Вес 16.5—20 г.
17х/2 дней Белое пятно подклювного бугорка охватывает всю верх-
нюю часть клюва. Вся плюсна и пальцы покрыты чешуйками. Вес эмбриона
19.3—24.8 г.
18х/2 дней. Начинают открываться глаза. Желток втягивается в брюш-
ную полость. Вес 23—29 г.
19х/2 дней. Глаза открыты. Желток втянут. Аллантоис атрофирован
и его сосуды бескровны. Наклев скорлупы. Вес 28—35 г.
20х/2 дней. Вылупление.
367