













































































































































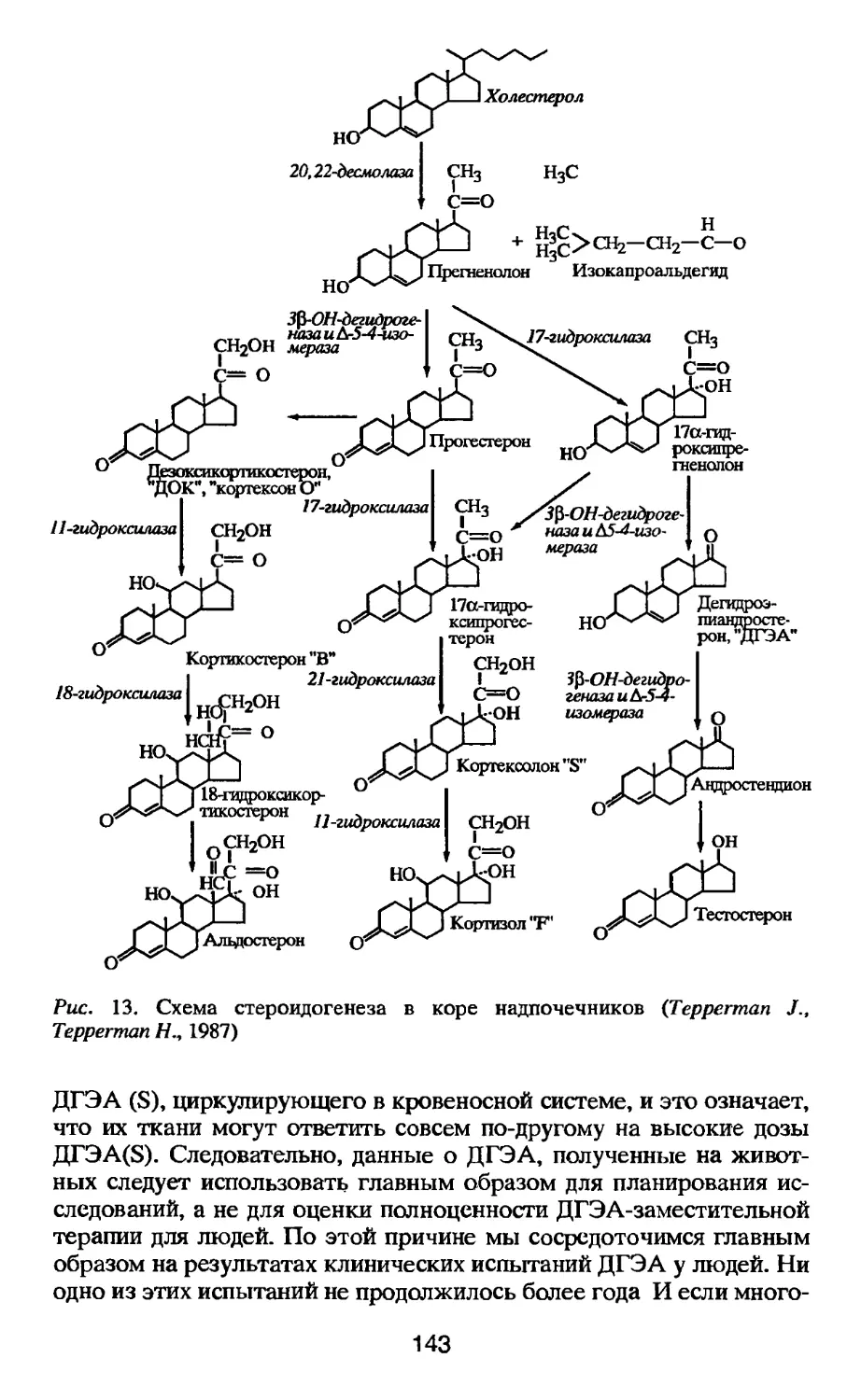
















































































































































































































































































































































Author: Тодоров И.Н. Тодоров Г.И.
Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез общая патология антропология химия биохимия издательство наука биохимическая инженерия старение
ISBN: 5-02-002827-4
Year: 2003




Светлой памяти выдающихся российских ученых
Николая Марковича ЭМАНУЭЛЯ
и
Владимира Михайловича ДИЛЬМАНА
посвящают авторы эту книгу
Человек свободный ни о чем так мало не думает, как о смерти,
и его мудрость состоит в размышлении не о смерти, а о жизни.
Спиноза. Этика, 4.IV, теор.67.
RUSSIAN ACADEMY OF SCIENCES
INSTITUTE OF PROBLEMS OF CHEMICAL PHYSICS
I.N. Todorov
G.I. Todorov
STRESS,
AGING
and their
biochemical
correction
Edited and prefaced by
Academician S.M. ALDOSHIN
e
MOSCOW NAUKA 2003
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ИНСТИТУТ ПРОБЛЕМ ХИМИЧЕСКОЙ ФИЗИКИ
И.Н. Тодоров
Г.И. Тодоров
СТРЕСС,
СТАРЕНИЕ
НИХ
биохимическая
коррекция
Под редакцией и с предисловием
академика СМ. АЛДОШИНА
Ш
МОСКВА НАУКА 2003
УДК 575
ББК 52.5+28.703
Г50
Издание осуществлено при финансовой поддержке
Российского фонда фундаментальных исследований
по проекту Μ 03-04-62025
Тодоров И.Н.
Стресс, старение и их биохимическая коррекция / И.Н. Тодоров,
Г.И. Тодоров; Отв. ред. СМ. Алдошин. - М.: Наука, 2003. - 479 с. -
ISBN 5-02-002827-4 (в пер.)
Монография содержит изложение и анализ'биологических явлений стресса и
старения и обобщение современных научных достижений в поиске путей замедления старения
и снижения интенсивности стресса и возрастных болезней. Впервые всесторонне и
детально рассмотрена взаимозависимость стресса и старения, а также общность некоторых
механизмов развития этих явлений. С возможной полнотой рассмотрена деструктивная
роль свободных радикалов - основного механизма процессов старения и стресса.
Большое внимание уделено нейроэндокринной теории старения, которая раскрывает генезис
нормальных возрастных болезней - атеросклероза, рака, диабета, иммунодепрессии и др.
Книга содержит обоснованные мировой наукой практические рекомендации по
биохимической коррекции стресса и старения.
Для врачей и студентов медико-биологического профиля.
По сети АК
Todorov I.N.
Stress, Aging the Their Biochemical Correction / I.N. Todorov,
G.I. Todorov; Eb. by S.M. Aldoshin. - Moscow: Nauka, 2003. - 479 p. -
ISBN 5-02-002827-4 (in cloth).
The book contains review and analysis of the biological phenomena of stress and aging. Is also
suggests approaches to slowing down the aging process by interfering with the mechanisms of aging
and mitigating stress response. This is the first comprehensive work to detail profound similarities
between some mechanisms of stress and aging as well as a complex bilateral relationship between
these two phenomena. In particular, the book contains a detailed analysis of the role of free radical
damage as one of the key mechanisms of stress and aging. Also, much attention is devoted to the
neuroendocrine theory of aging, which suggests important mechanisms contributing to the
neuroendocrine theory of aging, which suggests important mechanisms contributing to the development
of age-related diseases such as atherosclerosis, cancer, type II diabetes, immunodepression,
hypertension and others. The book includes scientifically validated practical recommendations for
inhibiting biochemical mechanisms of stress and aging.
For scientists, physicians and to biomedical students.
ISBN 5-02-002827-4 © Российская академия наук, 2003
© Издательство "Наука"
(художественное оформление), 2003
-4>^
ОГЛАВЛЕНИЕ
Основные сокращения ... 10
Предисловие редактора 13
Предисловие авторов 16
Часть /
СТАРЕНИЕ
Глава 1
Старение - путь к неизбежному (вместо введения) 21
1.1. Стресс и старение 25
1.2. Эволюция старения 27
Глава 2
Теории старения. Механизмы старения 30
2.1. Свободные радикалы 32
2.2. ДНК: повреждения/репарация 51
2.3. Накопление шлаков 56
2.4. Глюкоза и старение 62
2.5. Центральные часы старения .. 74
2.6. Лимит Хейфлика и старение клеток 86
2.7. Генетические аспекты старения 92
2.8. Суппрессор опухоли р53 и старение 99
Глава 3
Дегенеративные болезни 104
Глава 4
Старение и гормоны 111
4.1. Эндокринная система и гормоны 111
4.2. Как старение воздействует на эндокринную систему, и наоборот 113
Глава 5
'Гормоны молодости" ..... 117
5.1. Гормон роста 117
5.2. Мелатонин 131
5.3. Дегидроэпиандростерон 142
5
Глава 6
Старение и иммунитет 149
6.1. Метаболическая иммунодепрессия 157
6.2. Иммунитет и рак 159
6.3. Аутоиммунитет 161
6.4. Иммунитет и физические упражнения 164
Глава 7
Старение и депрессия 165
Глава 8
Старение и половая функция 170
8.1. Менопауза и эстрогеновая терапия ... 171
8.2. Можно ли отсрочить менопаузу? 174
8.3. Мужская половая функция 175
8.4. Разрыв между либидо и потенцией 176
8.5. Причины импотенции 177
8.6. Главный сексуальный орган 179
8.7. Сексуальность и гормоны молодости t 180
Глава 9
Долгожители .... 182
Литература 189
Часть //
СТРЕСС
Глава 10
Стресс, его место и роль в нашей жизни 207
10.1. Стресс в нашей жизни 209
10.2. Физический и эмоциональный стрессы 209
10.3. "Хороший" и "плохой" стрессы 211
10.4. Эволюция стресса 213
Глава 11
Динамическая природа стресса 214
11.1. Нейрогенные механизмы 214
11.2. Нейроэндокринные механизмы 216
11.3. Эндокринные механизмы 217
Глава 12
Общий адаптационный синдром 220
12.1. Фазы развития О АС 221
12.1.1. Реакция тревоги 221
12.1.2. Реакция сопротивления 222
12.1.3. Фаза истощения . . 225
Глава 13
Физиологические и биохимические механизмы ОАС. Система
гипоталамус-гипофиз-кора надпочечников .... 227
13.1. Гипоталамус , --« 229
13.1.1. Кортикотропин - регуляторньтй гормон (КРГ) 229
13.2. Гипофиз 231
13.2.1. Адренокортикотропный гормон гипофиза 231
6
13.3. Надпочечники 239
13.3.1. Мозговой слой. Катехоламины 239
13.3.2. Катехоламины 240
13.3.3.Кора надпочечников. Кортикостероиды 245
Глава 14
Перекисное окисление и стресс 254
14.1. Про- и антиоксидантное равновесие и окислительно-
восстановительный гомеостаз 255
14.2. Перекисное окисление в механизме стресс-реакции. "Первичный
медиатор" 257
Глава 15
Стресс и механизмы старения 262
15.1. Стресс и свободные радикалы 263
15.2. Стресс и центральные часы старения 266
15.3. Стресс и клеточные часы 267
15.4. Стресс, гликозилирование и перекрестные сшивки .... 267
15.5. Стресс и возрастные пигменты 268
15.6. Стресс и старение: порочный круг 268
15.7. Стресс и депрессия 270
Глава 16
Стресс и специфическая адаптация 274
Глава 17
Стресс: "болезни адаптации" и возрастные болезни 280
17.1. Общая схема развития стресс-патологии 281
17.2. Стресс и сердечно-сосудистые болезни 283
17.3. Стресс и гипертония 288
17.4. Стресс и рак 290
17.5. Стресс и диабет 293
Глава 18
Стресс и иммунная система 297
18.1. Стресс и аутоиммунные болезни ... 300
Глава 19
Клеточный стресс 302
19.1. Стресс-реакция клеток на тепловой шок 303
19.2. Стресс-реакция на действие радиации 307
19.3. Оксидативный стресс клетки 309
19.4. Общая адаптационная реакция клетки 313
19.4.1. Перспективы в исследовании клеточного стресса 318
19.5. Общий адаптационный синдром и клеточный стресс 319
19.6. Борьба с клеточным стрессом 319
Глава 20
Стресс и половая функция 321
Глава 21
Системы ограничения стресса.... 324
Литература 329
7
Часть III
БИОХИМИЧЕСКАЯ КОРРЕКЦИЯ СТРЕССА И СТАРЕНИЯ
Глава 22
Диета 343
22.1. "Потери" без потерь 347
22.2. Принципы здоровой потери веса 349
22.3. Физические упражнения 352
22.4. Гликемический эффект пищевых продуктов 353
22.5. Сбалансированная диета 355
22.6. Волокна и их полезные свойства 358
Глава 23
Против свободных радикалов 363
23.1. Питательные вещества и пищевые добавки с антиоксидантиыми
свойствами 365
23.1.1. Витамины С 366
23.1.2. Витамин Ε 367
23.1.3. Селен 368
23.1.4. Цистеин и метионин 369
23.1.5. Ликопен 369
23Л.6. Липоевая кислота 369
23.1.7. Флавоноиды 370
23.1.8. Витамин А и каротины 371
23.1.9. Полифенолы зеленого чая 372
23.1.10. Ко-энзим Q10 374
23.1.11. Мелатонин 374
Глава 24
Реверсия "центральных часов старения" 377
24.1. "Паспорт здоровья" 379
24.2. Как замедлить ЦЧС с помощью рационального питания и образа
жизни? .... 381
24.3. Медикаменты, которые могут замедлять ЦЧС 383
Глава 25
"Отдохновение" души и тела (подходы к смягчению стресс-
ответа) 386
Глава 26
Восстановление уровня "гормонов молодости*' 390
26.1. Гормон роста 391
26.1.1. Возможная польза, побочные эффекты и общие
предосторожности 391
26.1.2. Питательные вещества, стимулирующие выделение ГР 394
26.1.3. ГР и физические упражнения 397
26.2. Мелатонии 398
26.2.1. Возможная польза, побочные эффекты и общие предосто
рожности 398
26.2.2. Кто не должен принимать мелатонин 399
26.2.3. Повышение продукции мелатонина в организме 400
26.2.4. Потребление экзогенного мелатонина 402
26.3. Дегидроэпиандростерон 402
26.3.1. Возможная польза, побочные эффекты и общие
предосторожности (ДГЭА) 402
8
26.3.2. Прием ДГЭА как пищевой добавки 403
26.3.3. Реактивация собственной продукции ДГЭА 404
Глава 27
Атеросклероз: враг общества номер один 406
27.1. Главные факторы риска, ведущие к заболеванию атеросклерозом .. 407
27.2. Что делать с атеросклерозом? 409
Глава 28
Центр нашей вселенной - мозг ... 415
28.1. Умственные способности 416
28.2. Пищевые добавки, сохраняющие или улучшающие умственные
способности 41/7
28.3. Депрессия .. 419
28.3.1. Депрессия и дефицит витаминов .. 420
28.3.2. Депрессия и пищевые добавки .. 422
Глава 29
Адаптогены 425
29.1. Женьшень 428
29.2. Элеутерококк 433
29.2.1. Механизм антистрессового и анаболического действия 439
29.3. Левзея сафлоровидная ~ ~ 442
29.4. Родиола розовая - - 445
29.5. Об общем механизме действия полифенольных адаптогенов . 446
Глава 30
Половая функция после пятидесяти .. 448
30.1. Повышение половой активности 448
30.2. Предотвращение и ликвидация доброкачественной гипертрофии
предстательной железы 452
30.3. Восстановление и повышение полового влечения и удовольствия . 455
Глава 31
Нестареющая красота .. 458
Литература 465
Заключение 474
ОСНОВНЫЕ СОКРАЩЕНИЯ
АГК - альфа-гидрокси кислоты
АДФ - аденозиндифосфат
АКТГ - адренокортикотропный гормон
АМФ - аденозинмонофосфат
АО - антиоксидант, антиоксидантный
АОА - антиоксидантная активность
А, Т, Г, Ц - аденин, тимин, гуанин, цитидин
АТФ - аденозинтрифосфат
АТФязд - адинозинтрифосфатаза
АФК - активные формы кислорода
ГАМК - гамма-аминомасляная кислота
ГГБ - гамма-гидроксибутириловая кислота
ГГКН - система гипоталамус-гипофиз-кора надпочечников
ГП - гликемический показатель
ГР - гормон роста, соматотропный гормон
ГР-РГ - гормон роста-регулирующий гормон
ДГПЖ - доброкачественная гипертрофия предстательной железы
ДГТ - дегидротестостерон
ДГЭ А - дегидроэпиандростерон
ДГЭАф) - ДГЭА сульфат
ДМЭА - диметилэтаноламин
ДНК - дезоксирибонуклеиновая кислота
ДОП - доброкачественная опухоль простаты
дРНТФ - дезоксирибонуклеотидтрифосфат
иРНК - информационная рибонуклеиновая кислота
иРНП - информационный рибонуклеопротеид
ИЛ - интерлейкины, стимуляторы роста и размножения
некоторых иммунных клеток
ИФР - инсулиноподобный фактор роста
КПГГ - конечные продукты глубокого гликозилирования
КРГ - кортикотропин регулирующий гормон
ЛВП - липопротеиды высокой плотности
ЛГ - лютеинизирующий гормон
ЛНП - липопротеиды низкой плотности
ЛОНП - липопротеиды очень низкой плотности
10
МАО - моноаминооксидаза
МИД - метаболическая иммунодепрессия
МСГ - меланоцитстимулирующий гормон
мтДНК - ДНК, локализованная в митохондриях
НАД - никотинамидадениндинуклеотид
НАДФ - никотинамидадениндинуклеотидфосфат
ΗАДФ-Н - никотинамидадениндинуклеотидфосфат
восстановленный
НЖК - незаменимые жирные кислоты
ОАРК - общая адаптационная реакция клетки
ОАС - общий адаптационный синдром
ПАБК - парааминобензойная кислота
ПКГ - программируемая клеточная гибель, апоптоз
ПНЖ - полиненасыщенные жирные кислоты
ПО - перекисное окисление
ПОЛ - перекисное окисление липидов
пре-иРНК - предшественник информационной РНК
РНК - рибонуклеиновая кислота
ρ РНК - рибосомальная РНК
тРНК - транспортная РНК
СИПС - селективный ингибитор поглощения серотонина
СОД - супероксиддисмутаза
СР - свободные радикалы
СРО - свободнорадикальное окисление
ТРГ - тиреотропин регулирующий гормон
ТТГ - тиреотропный гормон
УФ - ультрафиолетовое излучение
ФСГ - фолликулостимулирующий гормон
ХПТА - характер проведения типа А
ХПТС - характер поведения типа С
цАМФ - циклический аденозинмонофосфат
ЦГИ - циклогексимид
цГМФ - циклический гуанозинмонофосфат
ЦНС - центральная нервная система
I \ЧС - центральные часы старения
ЦЭТ - цепь электронного транспорта
АРС - клетки, представляющие антиген
АСЕ - ангиотензин конвертирующий энзим
ΒΜΙ - показатель массы тела
сДНК -комплементарная ДНК, полученная путем обратной
транскрипции иРНК
CoQIO - ко-энзим Q10
Cu/Zn СОД - медь-цинк содержащая супероксиддисмутаза
FDA - ассоциация, контролирующая в США пищевые и
лекарственные продукты
DM АЕ - диметиламиноэтил
(НиТ - белок-переносчик глюкозы через клеточную мембрану
11
GSH
hsp
ηΓΡ
5-HTP
IU
МНС
mHsp70
Μη СОД
Na-PCA
NK
NSAID
PP
Р-450обц
p53
RDA
SAM
Seel
S-фаза
SLE
- глутатион
- белки теплового шока
- гормон роста человека
- 5-гидрокситриптофан
- интернациональная единица
- главный комплекс тканевой совместимости
- митохондриальный белок теплового шока с мол. массой
70 КД
-содержащая марганец митохондриальная супероксид-
дисмутаза
- натрий 2-пирролидон-5-карбогидрат
- натуральные киллеры, лимфоциты, воздействующие на
раковые клетки
- нестероидные противовоспалительные лекарства
- пирофосфат
- оксидаза, отщепляющая боковую цепь
- регулятор транскрипции ряда генов, клеточного цикла,
стимулятор апоптоза
- ежедневный пищевой рацион
- S-аденилметионин
- митохондриальная эндонуклеаза
- фаза клеточного цикла, в которой происходит реплика-
тивный синтез ядерной ДНК
- системная красная волчанка
ПРЕДИСЛОВИЕ РЕДАКТОРА
Данная книга И.Н. Тодорова и Г.И. Тодорова является
существенно переработанным и значительно расширенным вариантом
вышедшей в США в 2001 г. книги этих же авторов "Public Enemy
Number 1 - Stress. A Practical Guide to the Stress and Nutrition on the
Aging Process and Life Extension".
Эта книга представляет собой аналитическое обобщение
наиболее важных современных достижений мировой и отечественной
науки, а также многочисленных собственных работ авторов и их
сотрудников в изучении таких фундаментальных явлений живой
природы, как возрастная инволюция (старение) и способность адаптации к
экстремальным условиям (стресс-реакции) высших животных и
человека.
Термин "стресс" стал одним из наиболее модных слов
прошедшего столетия. Для большинства неспециалистов стресс - это нечто
такое, что имеет отношение к напряженности, беспокойству,
дискомфорту, и чего следует избегать. Однако стресс - это и научная
концепция, проливающая свет на то, как организм высших
животных и человека взаимодействует с постоянно меняющимися
условиями окружающей среды. Чтобы сознательно противостоять
негативному влиянию стресса на развитие болезней и процесс старения,
а именно эта взаимосвязь стала лейтмотивом книги, необходимо
понимать природу стресса и старения как физиологических и
биохимических явлений. Всестороннему высоко научному освещению этих
явлений с эволюционной, физиологической, биохимической,
генетической, а также отчасти и медицинской точек зрения и посвящена
большая часть этой книги.
Основные механизмы стресс-реакций сформировались у
млекопитающих животных в условиях дикой природы значительно
раньше возникновения человека и его цивилизации. В дикой природе
стресс - это мобилизация всех доступных организму ресурсов для
сохранения жизни в острокритической ситуации. Это реакция,
называемая учеными реакцией "борьбы или бегства", требует затраты
огромных энергетических ресурсов даже ценой разрушения некото-
13
рых тканей и систем организма, например иммунной системы, без
которых можно обойтись в данный конкретный момент.
В современном обществе жизнь человека редко подвергается
столь острой опасности, как в дикой природе, однако механизм
стресс-реакции остался практически неизменным. Очень часто
стресс вызывается психоэмоциональными и социальными
факторами, которые отнюдь не требуют мощной мобилизации ресурсов для
выживания. Тем не менее стресс развивается по "старой" эволюци-
онно выработанной программе и часто бывает чрезмерным.
Именно это противоречие, как постоянно подчеркивают авторы,
является важным фактором ускорения старения и развития возрастных
болезней и так называемых болезней адаптации (атеросклероз,
диабет, гипертония, депрессия, подавление иммунитета, рак и др.).
Взаимосвязь старения и возрастной динамики уровня гормонов,
показана на примере так называемых гормонов молодости (гормона
роста, мелатонина и дегидроэпиандростерона). Показана роль
снижения их уровня при возрастном падении иммунитета,
репродуктивной активности, сопротивляемости к стрессогенным факторам,
развитии состояния депрессии у пожилых людей и т.п.
Помимо основных достижений современной зарубежной
науки в раскрытии механизмов стресса и старения и, в частности,
роли свободнорадикальных реакций в постепенном разрушении
живых систем, авторы показывают выдающийся вклад
академика Н.Н. Семенова и академика Н.М. Эмануэля в становление и
развитие этого ведущего научного направления современной
химии и биологии в Институте химической физики РАН. В книге
также нашли отражение приоритетные труды профессора
И.Н. Тодорова и его учеников, посвященные реверсии старения с
помощью гормона роста (которые на 30 лет опередили
аналогичные исследования в США), работы, в которых раскрываются
молекулярные механизмы биосинтеза адренокортикотропного
гормона - ключевого регулятора стресс-реакций, работы,
посвященные механизмам клеточного стресса, получившие широкую
международную известность, а также исследования антистрессового
действия ряда адаптогенов. Большая часть этих исследований
была выполнена в течение почти 30 лет в отделе "Кинетики
химических и биологических процессов" Института проблем химической
физики РАН, по инициативе которого и издается эта книга.
В монографии также освещается весомый вклад советских и
российских исследователей в решении рассматриваемых проблем.
Исключительно важным достоинством книги является
тщательно подобранные и обоснованные практические рекомендации по
ограничению интенсивности и продолжительности стресса и
замедлению процесса старения. Эти рекомендации основываются на
многочисленных данных статистических, клинических и
экспериментальных исследований, опубликованных за последние годы в зарубеж
14
иой научной литературе (к сожалению, в отечественной литературе
в последнее время аналогичные данные представлены весьма
скромно), и предлагают различные варианты рационального питания,
направленных прежде всего на противодействие развитию тех или
иных патологий, связанных со старением и адаптацией. Большое
внимание уделяется характеристике и дозировке пищевых добавок
(антиоксидантов, витаминов, микроэлементов, аминокислот, адап-
тогенов и др.) и ряда существенных способов активации продукции
собственных "гормонов молодости", активации антиоксидантной,
иммунной и репаративных систем организма. Эти главы написаны
известным в США специалистом по лечебному питанию доктором
Г.И. Топоровым.
Высокая актуальность рассматриваемых авторами проблем
усугубляется продолжающимся в Российской Федерации кризисом
здравоохранения и ухудшением демографической обстановки.
Наряду с постепенными изменениями экономических и социальных
факторов, начинающих положительно влиять на здоровье и
продолжительность жизни населения нашей страны, существенную роль
должно, очевидно, также сыграть и освещение новейших
достижений зарубежной и российской науки в отечественной литературе.
Это несомненно принесет пользу не только научным работникам
медико-биологического профиля, практическим врачам, студентам
медицинских и биологических вузов, но и широкому кругу
читателей, которые интересуются генезисом связанных с возрастом
неинфекционных болезней, практическими мерами преодоления стрес-
согенных ситуаций и путями достижения здорового долголетия.
Директор Института
проблем химической физики РАН,
академик СМ. Ллдошин
ПРЕДИСЛОВИЕ АВТОРОВ
Он может медленно выжимать жизненные соки из вашего
тела, но может и молниеносно убить без предупреждения. Тем не
менее вы не найдете упоминания о нем ни в истории болезни, ни в
свидетельстве о смерти. Между тем имя его известно всем, это -
СТРЕСС
Стресс взимает с нас огромную дань в течение всей нашей
жизни, сокращает нашу молодость и ускоряет приближение нашей
старости. Мы встречаем его практически каждый день, но, увы,
чаще всего не предпринимаем ничего для своей защиты. И это не
удивительно - в лихорадочном темпе современной жизни у нас
даже нет времени остановиться, осмотреться и разобраться, что
же с нами происходит, и куда мы идем - даже если мы
приближаемся к краю пропасти...
Строго говоря, стресс сам по себе не является прямой причиной
старения, тем не менее он играет исключительно важную, но, к
сожалению, негативную роль в процессах старения. Стресс -
катализатор, который ускоряет практически каждый из известных
механизмов старения организма. Именно это обстоятельство заставляет
обращать особое внимание на неразрывную связь между стрессом и
старением.
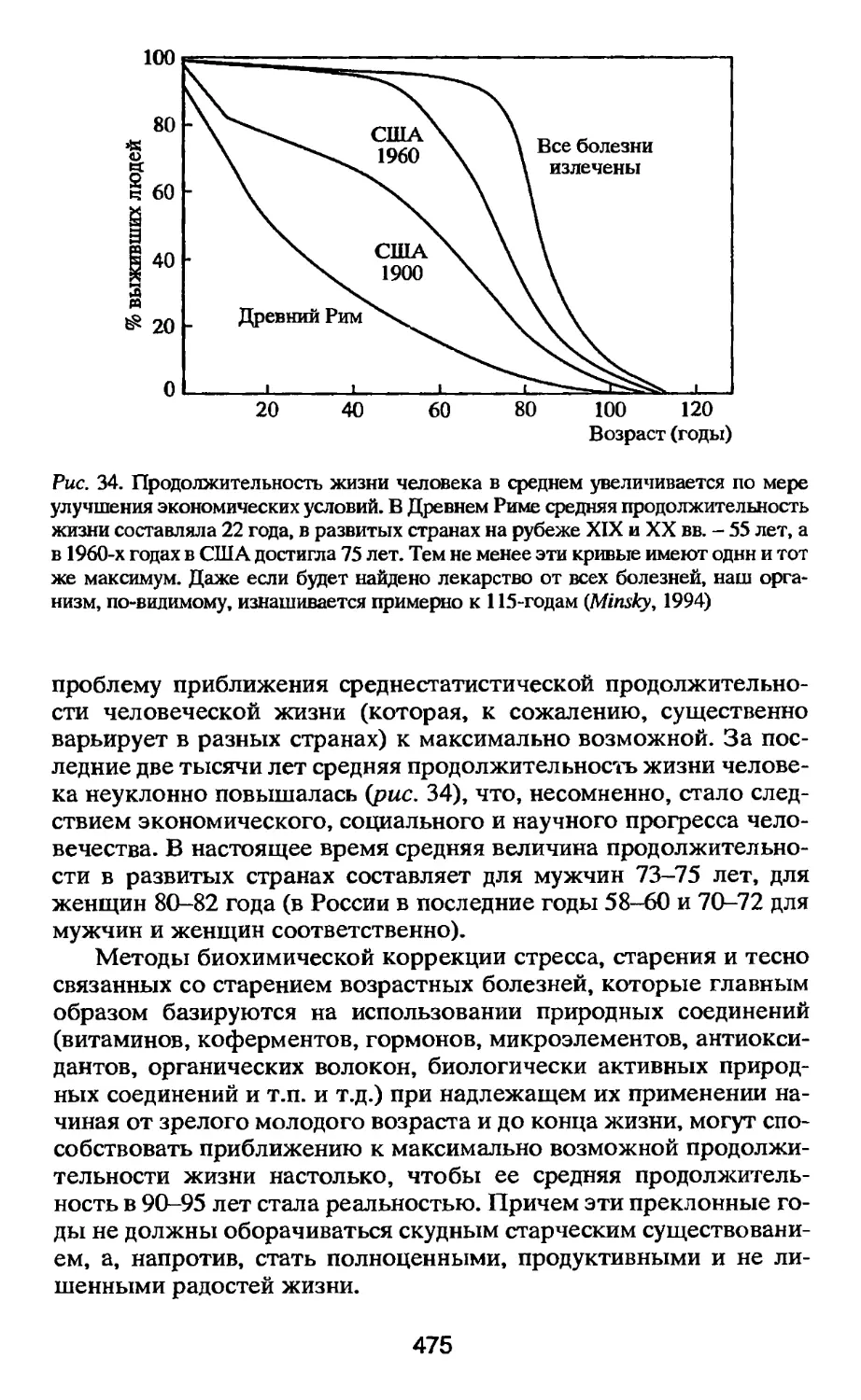
Средняя продолжительность человеческой жизни за
последние несколько столетий выросла более чем вдвое, но процесс
старения как таковой практически не изменился. Замечательные
успехи человечества в увеличении продолжительности жизни суть
результат таких изобретений, как антибиотики, вакцины,
операции на сердце и т.п. Иными словами, мы живем дольше не
потому, что мы теперь медленнее стареем, а потому что мы лучше
защищены от смертельных болезней и стихийных потрясений.
Становится все более очевидным, что единственный способ
обеспечить дальнейшее увеличение продолжительности жизни состоит
в торможении процессов старения и снижении интенсивности и
длительности стресс-реакций организма во все периоды челове-
16
ческой жизни. Впервые в истории человечество накопило
достаточное количество знаний об основных механизмах стресса и
старения, чтобы приступить на практике к серьезным попыткам их
нейтрализации. В развитых странах работа в этом направлении
ведется с нарастающей интенсивностью.
Явления стресса и старения исключительно сложны и
многогранны, однако мы сосредоточим основное внимание на их
биохимической природе равно, как и на биохимической специфике
предлагаемых способов и методов их коррекции. С
биохимической точки зрения освещаются и крайне важные вопросы
питания и пищевых добавок. Продуманная и правильно
сбалансированная диета уменьшает риск возрастных болезней и является
одним из важных факторов продления жизни. И напротив,
неправильное питание снижает эффективность любых мер,
направленных на продление жизни. Эта книга содержит не просто набор
рекомендаций по борьбе со стрессом и старением или служит
источником сведений об этих явлениях. Цель авторов помочь
читателям сознательно, с пониманием существа дела, выработать для
себя комплексную программу поддержания здоровья и продления
жизни.
Данная книга состоит из трех частей. В первой и второй
частях представлены в доступной неспециалистам (но далекой
от излишнего упрощения) форме, важнейшие достижения
современной науки в исследовании проблем старения и стресса,
а в третьей части - развернутые, обоснованные современной
наукой рекомендации их биохимической коррекции. На
протяжении всего нашего рассказа мы старались подчеркнуть, что
мерами по продлению жизни необходимо заниматься постоянно
и образ жизни должен соответствовать этой цели, тем более,
что это не слишком обременительная задача для любого
человека.
Возможно, в отдаленном (или не столь отдаленном) будущем
чудесные достижения молекулярной генетики и какие-нибудь
чудо-биопрепараты позволят человеку жить и сто и более лет,
сохраняя прекрасное здоровье и даже не прилагая к этому
чрезмерных усилий. Однако люди, живущие сегодня, вряд ли могут себе
позволить пассивно ожидать этого "светлого будущего",
поскольку оно может наступить еще не скоро. И даже если эти чудеса
науки не столь далеки, их помощью смогут воспользоваться лишь
те, чей организм еще не разрушен настолько, что даже эти меры
окажутся бессильными. Поэтому хотите ли вы просто прожить
здоровую и долгую жизнь или же дожить до времен, когда наука,
наконец, изобретет "волшебный эликсир" молодости, вам стоит
приступать к работе над этим, не откладывая. Мы рекомендуем
начать с чтения этой книги.
17
* * *
Обширный фактический экспериментальный и статистический
медико-биологический материал этой книги в основном
заимствован авторами из международной и американской научной
литературы, однако главные, доминирующие в этой книге научные
концепции и лежащие в их основе результаты, а именно
свободно-радикальная теория дегенеративных болезней и старения, а также ней-
роэндокринная теория старения принадлежат безвременно
ушедшим выдающимся отечественным ученым Н.М. Эмануэлю и
В.М. Дильману. Авторы посвящают свой скромный труд светлой
памяти этих ученых в знак признания не только их кардинального
вклада в медико-биологическую науку, но и в знак глубокого
личного уважения к этим замечательным людям, с которыми оба автора
были близко знакомы и сотрудничали в течение многих лет.
В 2000 г. исполнилось 85 лет со дня рождения Η JVL Эмануэля, и
эта дата была отмечена выходом в свет книги "Академик
Николай Маркович Эмануэль: Очерки, воспоминания, избранные статьи
и письма" (под ред. академика В.И. Гольданского. М: Наука, 2000).
В 2000 г. исполнилось 75 лет со дня рождения Владимира
Михайловича Дильмана, эта дата также была отмечена публикацией
статьи Н.П. Напалкова "О научном пути проф. В.М. Дильмана" (Вопр.
онкологии. 2001. Т. 47. № 2).
* * *
Авторы выражают глубокую признательность Л.В. Емелиной и
И.Б. Крашаковой за постоянную и неоценимую помощь при работе
над этой книгой.
Часть I
СТАРЕНИЕ
Человек свободный ни о чем так мало не думает,
как о смерти, и его мудрость состоит
в размышлении не о смерти, а о жизни.
Спиноза. Этика, ч. IV, теор. 67
Глава 1
СТАРЕНИЕ - ПУТЬ К НЕИЗБЕЖНОМУ...
(вместо введения)
Колыбель качается над бездной. Заглушая
шепот вдохновенных суеверий, здравый смысл
говорит нам, что жизнь - только щель слабого
света между двумя идеально черными вечностя-
ми. Разницы в их черноте нет никакой, но в
бездну преджизненную нам свойственно
вглядываться с меньшим смятением, чем в ту, к
которой летим со скоростью четырех тысяч
пятисот ударов сердца в час.
Владимир Набоков, " Другие берега" (1954).
Уж сколько их упало в эту бездну,
Разверстую вдали!
Настанет день, когда и я исчезну
С поверхности земли.
Марина Цветаева
"Старение - закономерный процесс возрастных изменений
организма, приводящий к старости" - таков типичный пример
общего определение понятия "старение", которое содержится в
энциклопедических словарях конца XX в. Это определение неопределенно
почти до бесполезности, и каждый из нас, по крайней мере
интуитивно, знает, что оно включает в себя нечто несравненно большее.
Ведь речь идет о неизбежном процессе, который медленно
изнашивает, истощает, повреждает и в конечном счете убивает все живые
существа. Однако и эта характеристика на самом деле лишь в малой
степени отражает общую картину. Конечно, все живое стареет, но
темп и форма старения у различных организмов резко отличаются.
Среди относительно близких видов, например млекопитающих,
скорость старения может различаться почти в сотни раз. Даже у людей,
умирающих от естественных причин, продолжительность жизни
варьирует в широком диапазоне. Раскрытие причин таких различий
служит ключом к пониманию природы старения и. что более важно,
может стать основой стратегии продления жизни.
Ученые часто определяют старение как процесс, который со
временем увеличивает вероятность смерти1. Это определение фор-
1 Еще в 1825 г. английский исследователь Б. Гомпертц нашел, что после
30-35 лет у человека показатель удельной смертности увеличивается с
возрастом экспоненциально (Gompertz, 1825). Исходя лишь из этого факта, многие
биологи пытаются понять причины старения и смерти, хотя в математических
21
мально отражает сущность проблемы, но все-таки не дает
представления о физической природе этого явления. К тому же, нам теперь
ясно, что старение является не одним процессом, а группой сложных
взаимозависимых процессов, воздействующих на все системы и все
уровни организации этих систем. Поэтому всестороннее понимание
старения требует пристального изучения всех составляющих
компонентов этого процесса и их иерархии. В этой части книги мы
стремились дать четкую и завершенную картину современного
состояния наших знаний о процессе старения. Мы уверены, что человек,
который ставит своей целью продление жизни, независимо от того
является ли эта цель персональной или общественной должен иметь
общие представления о природе старения. Необходимо изучить
противника перед тем, как вступить с ним в борьбу
Для продления жизни необходимы непрерывные и
существенные усилия, так же как и осознание своей ответственности за
собственную жизнь. К сожалению, если вы просто ведете здоровый образ
жизни и покоряетесь естественному ходу этого процесса, то вряд ли
вы добьетесь заметных результатов. Оставляя за природой
управление этим процессом, все, на что вы можете рассчитывать, - это
среднестатистическая продолжительность человеческой жизни, равная
в развитых странах приблизительно 70-80 годам. Однако даже и
такая "скромная" продолжительность жизни не вполне "естественна"
и связана главным образом с недавним скачком в развитии
человеческого общества с его мощной технологией, сельским хозяйством и
медициной, с улучшением санитарии и профилактики здоровья
широких слоев населения, с улучшением условий жизни и труда и т.п.
Нет ничего естественного (в смысле природного происхождения) во
многих из перечисленных обстоятельств, но мы с энтузиазмом
принимаем их, потому что они работают в наших интересах. Потенциал
"методов XX столетия", обусловивший продление человеческой
жизни, по-видимому, уже почти исчерпан (по крайней мере в
развитых странах), но эти методы хорошо послужили человечеству и
достойны высокой оценки.
Следующий шаг в борьбе за долгую здоровую жизнь связан,
очевидно, с более глубоким пониманием биологических механизмов
старения и нахождением адекватных методов их блокирования. В
действительности, мы уже стоим на правильном пути к этой цели.
Современная наука уже располагает некоторыми подходами,
которые способны замедлить развитие возрастных болезней и старение.
Например, использование определенных режимов питания на
разных стадиях развития таких животных, как крысы или мыши,
позволяет удвоить продолжительность их жизни. Преждевременно, ко-
корреляциях отражаются изменения, происходящие в популяции, а ие те
конкретные процессы, которые лежат в основе самого механизма старения и его
проявлений у индивида.
22
нечно, надеяться на подобные результаты у людей, но следует
подчеркнуть, что многие из методов, эффективно использованных на
лабораторных животных, могут принести, по крайней мере
некоторую, пользу и людям вследствие высокой степени подобия
биохимических и физиологических механизмов жизнедеятельности у
высших животных и человека.
Стратегия продления жизни (которую мы назвали бы стратегией
естественной биохимической коррекции и которая детально
изложена в III части книги) основана главным образом на использовании
природных соединений, которые обычно присутствуют в тканях
животных или растений. Однако специальные исследования показали,
что количества веществ, необходимые для существенного
продления жизни, не всегда останутся "натуральными" в том смысле, что
едва ли можно получить необходимые их количества, просто
придерживаясь сбалансированной диеты.
В настоящее время использование лекарств, рекомендуемых для
продления жизни, мы не считаем целесообразным для большинства,
так как эти дорогостоящие медикаменты недостаточно изучены и
во многом токсичны.
β последние десятилетия наблюдается значительный рост наших
знаний о старении, и есть все основания полагать, что такой рост
продолжится. Старение связано с постепенным снижением
функциональной активности всех или большинства органов и систем организма,
что обусловливает прогрессирующую потерю способности
адаптироваться к влиянию внутренних и внешних факторов. В результате
этого жизни стареющего организма могут угрожать воздействия все
меньшей и меньшей интенсивности. Так, в пожилом возрасте даже
умеренный стресс, вызванный, например, переломом кости или
гриппом, может привести к летальному исходу, поскольку слабая
иммунная система и уже малоэффективные механизмы репарации не в
состоянии справиться с критическим положением.
Мы привыкли считать наше тело чем-то относительно
постоянным и стабильным. Как и многие другие очевидные утверждения,
это также не соответствует действительности. Наш организм - это
огромный и сложный химический реактор, в который поступают
богатые энергией соединения и соединения, выполняющие функции
"строительного материала". Эти вещества каждую секунду
подвергаются миллиардам молекулярных преобразований синтеза и
распада, и, будучи деградированы до простейших соединений, выводятся
из организма. Пока мы живы, наши клетки, органы и системы
находятся в равновесии, но это - динамическое равновесие текущей
реки. Все структуры нашего организма - и простейшие, и самые слож-
23
Рис. 1. Изменение в зависимости
от возраста различных
функциональных характеристик
состояния организма человека
(Cutler, 1978)
1 - скорость проведения
нервного импульса; 2 -интенсивность
основного обмена; 3 -
фильтрационная способность почки; 4 -
минутный объем сердца; 5 - жизненная
емкость легких; 6 - максимальная
легочная вентиляция
ные - постоянно работают,
*~4о 60 80~~ чтобы поддерживать этот
Возраст (годы) баланс. Одни клетки
отмирают, в то время как другие
делятся и продолжают функционировать; поврежденные молекулы
восстанавливаются или заменяются, сохраняя структурную и
функциональную целостность субклеточных компонентов. Несмотря на
то что механизмы репарации высоко эффективны, особенно у
такого долгоживущего вида, как человек, они тем не менее
недостаточно совершенны. Часть повреждений "ускользает" от ремонта и
накапливается в организме. Скорость старения в значительной
степени зависит от скорости такого накопления и, следовательно, от
эффективности механизмов замены и ремонта поврежденных
структур. Описанные события происходят повсюду и на всех стадиях
жизни, так что никогда не бывает слишком рано предпринять шаги к
продлению жизни.
Как уже упоминалось, функция всех систем организма
снижается с возрастом, но скорость этого снижения заметно различается у
разных систем (рис. 1). Человек может умереть от сбоя одной
системы (например, фатальной пневмонии в результате слабой иммунной
системы) притом, что другие его системы находятся в хорошем
состоянии. Тотальный поход на старение подразумевает поддержку
всех жизненно важных систем, поскольку наличие, например,
прекрасной нервной системы, здоровых почек и юной кожи будет
слабым утешением при смертельном сердечном приступе. К счастью,
многие биохимические факторы, полезные для одной системы
организма, способны усиливать и другие. К природным соединениям,
приносящим пользу нескольким системам организма сразу, следует
отнести антиоксиданты, в особенности витамины А, С, Е, селен,
аминокислоту цистеин, коензим Q-10 и липоевую кислоту.
Старение может проявляться просто как перманентное
снижение функций органов и тканей, но чаще мы сталкивается с
развитием ряда специфических болезней, часто называемых возрастными
или дегенеративными. Риск подвергнуться такими болезнями
значительно увеличивается с возрастом. Высокоразвитые организмы, в
24
особенности, млекопитающие, в большей степени, чем другие виды,
подвержены этим возрастным болезням, что указывает на сходство
механизмов их старения. Следовательно, способы продления жизни
млекопитающих животных в лабораторных условиях будут,
очевидно, работать и в отношении человека. К возрастным болезням
относят атеросклероз, гипертонию, диабет II типа, рак, иммунные
нарушения, депрессию, остеопороз, болезнь Альцгеймера и Паркин-
сона, катаракту и некоторые другие. В развитых странах от
возрастных болезней погибают более чем 80% всех умерших. Следует
тем не менее подчеркнуть, что применение комплекса современных
методов продления жизни во многих случаях позволяет замедлять и
даже полностью предотвратить развитие возрастных болезней.
Поэтому всесторонняя стратегия продления жизни служит не просто
способом увеличения промежутка между датами на надгробной
плите, а может позволить продлить здоровую, приятную и
продуктивную жизнь на всех ее стадиях.
1.1. СТРЕСС И СТАРЕНИЕ
Существует предрассудок, что "природа
не ошибается". Это неверно. Если бы это
было правдой, не было бы моей
профессии: зачем существуют врачи, как не для
того чтобы поправлять природу, когда
она ошибается?
Ганс Селье
Во вступительной главе невозможно обойти вниманием теперь
уже несомненно доказанную связь между стрессом, с одной
стороны, и развитием возрастных болезней и старением - с другой, связь,
которая с возможной полнотой будет освещена во II части этой
книги. Здесь же мы лишь кратко обсудим эту проблему. Создатель
современной концепции стресса, выдающийся канадский патофизиолог
Ганс Селье еще в 50-х годах прошлого столетия предложил весьма
простую гипотезу (в чем-то созвучную мировоззрению
средневековых натурфилософов) связи стресса и старения. Каждый организм,
полагал Селье, наделен природой определенным запасом так
называемой адаптационной энергии, которую он расходует в течение
своей жизни. Интенсивность ее расходования прямо
пропорциональна скорости старения организма. Просто и понятно - не правда ли?
Существуют, однако, и более поздние указания на связь
рассматриваемых явлений
"Старение - это постоянное неуклонное повышение
восприимчивости к стрессу в целом, скорее чем склонность к какому-то
специфическому патологическому процессу" (Brash et al., 1978; цит.
по: Esposito, 1983).
25
Если рассматривать стресс как важнейший механизм сохранения
гомеостаза живых систем, то представляет интерес и такая точка
зрения: "...старение - это процесс, или группа процессов, которые
вызывают в конечном итоге нарушение гомеостаза" {Comfort,
1972).
Наиболее яркое, почти художественное описание факторов,
ввергающих организм в состояние стресса, т.е. стрессоров, и
сущности самого стресса содержится в одной из последних работ Г. Селье:
"Стресс является частью нашего каждодневного опыта, однако
он связывается с большим различием причинных факторов, таких
как хирургическая травма, ожоги, эмоциональное возбуждение,
умственные или физические усилия, утомление, боль, страх,
необходимость сосредоточиться, унижение или разочарование, потеря
крови, интоксикация лекарствами или вследствие загрязнения
окружающей среды, или даже неожиданный успех, который требует
перестройки жизненного уклада. Стресс возникает у делового
человека, находящегося под постоянным давлением обстоятельств
жизни, у спортсмена, стремящегося выиграть забег, у
регулировщика движения самолетов, который несет постоянную
ответственность за сотни жизней, у мужа, беспомощно наблюдающего
медленное болезненное умирание жены от рака, у беговой лошади,
ее наездника и у зрителей. Медицинские исследования показывают,
что, несмотря на различие проблем, встающих перед этими
организмами, все они отвечают стереотипной формой
биохимических, функциональных и структурных изменений, обычно
вовлекаемых в реакцию преодоления любого рода повышенных
требований к жизнедеятельности, в частности адаптацией к новым
ситуациям" (Селье, 1977).
Реакция на стрессоры была особенно важна для
доисторического человечества, когда жизнь была наполнена опасностями и
жестокостью, ежедневной борьбой за существование, природными
катаклизмами, инфекционными болезнями, межплеменной враждой и т.п.
Основные' механизмы стресса обусловливают быструю
мобилизацию энергетических ресурсов, ускорение их доставки к головному
мозгу, сердцу, мышцам и другим органам, повышенное
функционирование которых непосредственно необходимо для выживания. В
сущности, стресс - экстренная биологическая реакция организма,
направленная на его спасение в условиях смертельной опасности, но
подобная тотальная мобилизация жизненных сил организма не
проходит для него бесследно. Если, с одной стороны, стресс-реакция в
критической ситуации слишком слаба, организм либо погибает,
либо под действием стрессоров получает различные повреждения,
которые проявят себя в болезнях и ускоренном старении. С другой
стороны, если стресс-реакция чрезмерна или слишком длительна, то
она сама становится разрушительной силой и также приводит к
болезням и ускоряет старение. Многочисленные исследования на жи-
26
вотных показали, что оптимальная реакция на стрессоры является
важным фактором долголетия; но ее избыточность или
недостаточность всегда, в той или иной мере, способствуют старению и
развитию болезней. С возрастом способность оптимально реагировать на
стрессогенные воздействия постепенно теряется, что, естественно,
снижает возможности организма приспосабливаться к
неблагоприятным условиям. Потеря сопротивления к стрессорам ускоряет
старение, которое в свою очередь далее разрушает нашу способность к
оптимальной адаптации.
Этот "порочный круг"
(стресс-старение-стресс-старение и тл.) вносит главный вклад в ускорение биологического
старения и увеличение сферы действия возрастных болезней в
конечный период жизни.
К счастью, этот круг можно разорвать или, по крайней мере,
замедлить темп движения по нему, что мы и попытаемся показать в
следующих главах. В заключение уместно привести высказывание
создателя нейроэндокринной теории старения В.М. Дильмана из его
замечательной книги "Четыре модели медицины" (1987): "Старея,
человек и вне стресса начинает жить, как бы находясь в
состоянии хронического стресса, и поэтому становится все более и более
беззащитным, когда реальный стресс предъявляет свои
требования к организму".
1.2. ЭВОЛЮЦИЯ СТАРЕНИЯ
На свете много сил великих,
Но сильнее человека
Нет в природе ничего.
Софокл
Животное в его возрасте (возрасте
человеческого младенца. - И. Т.) имеет зубы,
когти, иногда даже рога, и уже может
добывать себе пищу. Homo sapiens
рождается беспомощным и бессильным и
вынужден оставаться таким много лет, пока не
выучится и не накопит достаточный
опыт. Похоже, что период детства длится
все дольше с каждым новым поколением.
Исаак Б. Зингер, "Мешуга"
Несколько слов об эволюционных причинах существования
огромного числа вариаций в скорости старения различных видов.
Часто студенты, начиная изучать биологию, интересуются, почему в
природе так мало долгоживущих видов. Ведь в конце концов
основные принципы жизни одинаковы для большинства организмов. Все
27
виды используют одни и те же информационные носители (ДНК и
РНК), и сходные строительные блоки для своих клеток
(аминокислоты, углеводы, лигтиды, и т.д.). Общий план строения и
функционирования гомологичных клеток большинства организмов также
имеет несомненные черты сходства. Тем не менее различия в
онтогенетической судьбе различных видов - поразительны.
Хотя теория эволюции Чарльза Дарвина часто воспринимается
как хорошо забытое старое, она, между тем, остается одним из
лучших инструментов для понимания происхождения обсуждаемых
различий. Только тот вид побеждает в борьбе за существование,
который, развивая полезные признаки, лучше приспосабливается к
окружающей среде. Продолжительная жизнь, очевидно, большая
привилегия для индивидуума, но во многих случаях она не обеспечивает
для вида преимуществ в конкурентной борьбе за существование и
фактически может стать недостатком. Вообразим ситуацию, когда
окружающая среда изменяется существенно или даже
незначительно, но постоянно. Чтобы остаться конкурентоспособным, вид также
должен измениться. Случайные самопроизвольные мутации в
генетическом материале (ДНК) являются одним из важнейших
факторов изменчивости организмов. Однако на эволюцию видов влияют
только те мутации, которые возникли в половых клетках
(сперматозоидах и яйцеклетках), т.е. в клетках, вовлеченных в процесс
воспроизводства, потому что именно по этому пути идет воздействие на
потомство. Большинство мутаций вредны или даже смертельны для
особи, но некоторые из них обеспечивают спасительные
преимущества. В ситуации изменяющейся окружающей среды более высокая
скорость воспроизводства является преимуществом, поскольку
сопряжена с более высокой вероятностью возникновения мутаций,
которые в случае удачи обеспечат организму больше шансов
приспособиться к новым условиям. Потребность в таком быстром
воспроизводстве статистически исключает появление долгоживущих
организмов, потому что они медленнее достигают репродуктивной
стадии жизненного цикла и потому еще, что долго остаются в живых,
потеряв способность к воспроизводству и продолжая конкурировать
за ресурсы с репродуктивно более активными организмами. Другой
причиной, почему высокая продолжительность жизни может быть
бесперспективной, является также и то, что исключительно
эффективные механизмы самообновления, требуемые для более
длительной жизни, потребляют те ресурсы организма, которые в противном
случае могли бы использоваться для воспроизводства (подробней об
этой коллизии - см. гл. 2). Тем не менее долгоживущие виды
развиваются, и наилучший пример тому - homo sapiens.
По-видимому, имеется несколько эволюционных причин,
почему люди живут дольше, чем другие млекопитающие. Развитый
головной мозг в сочетании с гигантским опытом познания
окружающего мира дает человеку огромное эволюционное преимущество
28
перед другими видами живых организмов, но полное формирование
и развитие такого сложного органа, а также овладение основами
этого опыта требуют значительного времени. Поэтому родители,
движимые мощным, эволюционно закрепленным
патерналистским инстинктом, в течение многих лет заботятся и обучают своих
детей. Такой способ заботы о потомстве имеет в эволюционных
терминах очень высокую цену, поскольку развитый интеллект служит
для потомка лучшим оружием в борьбе за существование в течение
всей его жизни. Другой причиной долголетия является то, что люди,
опираясь на упомянутый выше универсальный опыт, научились
противостоять неблагоприятным изменениям в окружающей среде,
изобретая новые методы производства продовольствия, развивая
медицину, создавая социальные институты и т.д. В сущности, в
отличие ото всех других видов на Земле, люди смогли позволить себе
роскошь не подвергаться дальнейшим эволюционным изменениям,
чтобы остаться в живых в изменяющемся мире. Они просто создали
собственную окружающую среду, и это позволило им даже
извлекать немалую пользу из относительно медленного темпа
воспроизводства и продолжительной жизни.
Обратной стороной всей этой ситуации является недостаток
естественного эволюционного давления на людей, которое позволило
бы им развиваться по направлению к еще более продолжительной
жизни (к другим негативным сторонам дальнейшего изменения
окружающей среды в результате человеческой деятельности и образа
жизни мы будем еще возвращаться в последующих главах). К
счастью, коллективный интеллект человечества, его мощная наука и
набирающая силу биотехнология помогут (и уже помогают) людям
изобрести эффективные способы прекращения развития возрастных
болезней и существенного замедления механизмов старения.
Глава 2
ТЕОРИИ СТАРЕНИЯ.
МЕХАНИЗМЫ СТАРЕНИЯ
Во всем мне хочется дойти
До самой сути...
Борис Пастернак
Как уже упоминалось ранее, старение - не единый процесс, а
скорее группа процессов, движимых различными механизмами. Это
одна из причин, почему старение относительно трудно для изучения
и осмысления и почему существует довольно много теорий
старения. В этой главе мы рассматриваем только те теории и лежащие в
их основе механизмы старения, которые подтверждены
значительным количеством научных доказательств и которые могут служить
основанием для использования существующих и развития новых
подходов к продлению жизни. Ни одна из этих теорий не
охватывает все аспекты старения, а скорее опирается на интерпретацию
одного или нескольких определенных механизмов. Однако взятые
вместе они дают достаточно полное представление о динамике и
механизмах старения и облегчают поиски практических способов их
блокирования.
Все механизмы старения можно разделить на две крупные
группы: (1) вероятностные, т.е. механизмы, связанные с накоплением
случайных повреждений или микрополомок на молекулярном,
субклеточном и тканевом уровнях, и (2) системные механизмы,
которые вызывают цепь взаимосвязанных нарушений жизнедеятельности
как отдельных дифференцированных клеток, так и крупных регуля-
торных систем организма, таких как нейроэндокринная,
адаптационная, система воспроизведения и др. В реальной ситуации
функционирование многих из этих механизмов взаимосвязано теснейшим
образом: изменения на одном уровне будут сказываться на
функциях другого, и наоборот. Следует также принимать во внимание, что
часть возрастных изменений на обоих уровнях связана с
вероятностным действием внешних или внутренних факторов, тогда как другая
их часть, по-видимому, генетически запрограммирована или
является следствием разворачивания онтогенетических программ развития
организма.
К факторам, воздействующим на первую группу механизмов,
следует отнести широкий спектр воздействий химической и
физической природы, которые способны вызывать повреждение жизненно
30
важных структур клетки. Так, свободные радикалы, мутагены,
ксенобиотики, многие химикаты, различные виды лучистой энергии
способны беспорядочно повреждать ключевые молекулы жизни:
дезоксирибонуклеиновые кислоты (ДНК), рибонуклеиновые
кислоты (РНК), белки, углеводы, липиды и их макромолекулярные
комплексы. Клеточные мембраны особенно восприимчивы к
воздействию свободных радикалов, потому что они богаты ненасыщенными
жирными кислотами с их высокой реакционной способностью, что
обусловливает развитие в этих структурах цепной реакции -
переписного окисления липидов (ПОЛ). Мутагены и ионизирующая
радиация способны нарушать структуру ДНК, изменяя ее
генетическое содержание и повреждая или нарушая функцию тех или иных
генов (или их регуляторных участков). Некоторые соединения,
например альдегиды, могут "сшивать" клеточные компоненты,
ограничивая их подвижность и функциональную активность. Ряд
агентов, вызывающих молекулярные повреждения, являются
факторами окружающей среды (например, физические факторы или
токсины) и от них можно уберечься, но большинство повреждений
все-таки вызывают агенты, возникновение которых связано с
нормальными физиологическими процессами в организме, и избежать их атак
невозможно.
Потенциально вредные молекулярные флуктуации происходят в
наших клетках постоянно. Огромное их большинство, однако, не
причиняет ощутимого вреда, потому что повреждения эти быстро
восстанавливаются ферментами репарации. К сожалению, не
слишком высокая точность функционирования таких систем приводит к
постепенному накоплению поврежденных структур в качестве
инертного балласта, что в конце концов начинает мешать
жизненным функциям клеток. С возрастом ситуация усугубляется из-за
того, что механизмы ремонта сами подвергаются повреждениям и, в
той или иной мере, теряют свою эффективность. Предполагается
также, хотя это и не доказано, что некоторые репарационные
механизмы могут быть в тот или иной момент закрыты в соответствии с
генетическими программами.
На ранних этапах исследования процессов старения происходили
горячие дебаты о существовании так называемых часов старения.
Действительно, у одноклеточных и некоторых примитивных
организмов таких механизмов, очевидно, нет. К настоящему времени,
однако, накопилось много свидетельств в пользу того, что сложные
организмы, в особенности млекопитающие, имеют даже не один, а
несколько типов часов старения. Во-первых, большинство клеток
высокоразвитых организмов способно осуществить только конеч
ное число митозов (клеточных делений) - в противоположность
бактериям, которые теоретически могут расти и делиться при
благоприятных условиях бесконечно. Во-вторых, по всей видимости,
существуют "центральные часы ", локализованные в структурах
31
головного мозга и определяющие темп как развития, так и старения
организма. В-третьих, имеются и другие типы часов старения,
например, часы, прекращающие репродуктивную функцию у женщин
(менопауза), хотя последние, по крайней мере в некоторой степени,
зависят от функции центральных и клеточных часов.
Биологические часы, особенно те, которые в определенной
мере влияют на процессы старения, существенно различаются от
обычных часов. Их темп может заметно варьировать у разных
индивидов, и еще больше различаться у животных разных видов;
всевозможные факторы или ситуации могут заставить эти часы идти либо
быстрее, либо медленнее. Стресс и, возможно, чрезмерное
потребление пищи ускоряют ход часов старения. Повышенная
сопротивляемость действию стрессоров и питательные вещества,
активирующие некоторые мозговые структуры, главным образом гипоталамус
и гипофиз, замедляют работу этих часов. У грызунов строгое
ограничение калорийности питания до половой зрелости, приводит к
увеличению продолжительности жизни почти в два раза, возможно,
из-за замедления их часов старения. К сожалению, этот
эффективный метод не может использоваться людьми, поскольку серьезное
калорийное ограничение, начатое в ранний период жизни, не только
не увеличивает продолжительность жизни, но, напротив, приводит к
повреждению мозга. Статистические исследования различных групп
населения показывают, что поддержание идеального веса служит
одним из важных факторов продления жизни у людей.
Далее мы обсудим наиболее важные механизмы и наиболее
обоснованные теории старения, а также предварительно коснемся
некоторых способов замедления работы этих механизмов на основе
достижений современной медико-биологической науки. Однако
наиболее полные и обоснованные рекомендации относительно этой
проблематики будут представлены в III части этой книги
2.1. СВОБОДНЫЕ РАДИКАЛЫ
И от чего же умер ваш друг?
О, причина естественная.
Его застрелили в подземке.
Из фольклора города Нью-Йорка
Беспрецедентный интерес мировой научной общественности к
проблеме свободных радикалов (СР) в связи со старением возник
сравнительно недавно, в начале 90-х годов XX в., и был вызван
публикацией серии эффектных молекулярно-генетических
исследований, однозначно указывавших на значительную роль СР-форм
кислорода в ускорении процесса старения организмов (Растит. 1993),
32
хотя предпосылки для таких заключений, базирующихся на ином
фактическом материале, существовали в научной литературе уже
давно (Нагтап, 1956; Эмануэль, 1963; Эмануэль и др., 1977).
Средства массовой информации (СМИ) США, придавая сенсационный
характер этим новым результатам американских ученых и пытаясь
донести до широкой публики их значение для медицины и
здравоохранения, настойчиво обращались к весьма наглядной аналогии между
ролью СР в живых системах и социальными аспектами преступности
в современном демократическом обществе. Действительно,
американцы, гордящиеся своим свободолюбием, убеждены, что не
существует такого понятия, как "слишком много свободы". Конечно,
свобода - великая вещь, если она осуществляется в рамках
разумных законов, которые защищают человеческую жизнь, права и
собственность. Преступники, наоборот, пытаются пользоваться
абсолютной, безграничной свободой, нанося вред своим согражданам и
обществу в целом. Итак, несмотря на свою любовь к свободе,
цивилизованные люди делают то, что они должны делать, чтобы
защитить себя: голосуют за жесткие законы против преступлений,
требуют увеличения количества полицейских на улицах и, конечно,
покупают домашние системы безопасности и оружие. Дело в том, что,
нуждаясь в свободе, мы также нуждаемся и в защите от слишком
большой свободы в чужих руках... СМИ не скупились в
выражениях, подчеркивая явную аналогию между поведением СР в организме
и поведением преступников в сообществе. Но какова химическая и
биохимическая природа свободных радикалов?
Хотя этим вопросам в дальнейшем и будет уделено достаточно много
внимания, здесь мы ограничимся лишь несколькими замечаниями. Свободные
радикалы - это химические соединения с неспаренными электронами, которые
обладают чрезвычайно высокой реакционной способностью, которая реализуется
случайным образом. Большинство химических соединений в организме
реагирует друг с другом относительно медленно и в пределах некоторых правил,
которые определяют направление и скорость химических реакций, определяющих
многообразные функции клеток и организма в целом. Эти правила
осуществляются с помощью специальных белков (ферментов), которые облегчают и
ускоряют химические реакции. Не так обстоит дело со СР. Эти "бандиты'*
реагируют быстро и беспорядочно с любыми клеточными структурами, которое
встретятся на их пути, результатом чего становятся разнообразные повреждения.
Отдав должное современному всплеску интереса к роли СР в
биологических явлениях и перед тем, как перейти к ее
рассмотрению по существу, нам хотелось, хотя бы кратко, осветить историю
этой проблемы.
СР или атомы, в отличие от молекул, обладают свободными
ненасыщенными валентностями, что определяет легкость их
взаимодействия с другими молекулами, при этом происходит разрыв одной
из валентных связей последних и всегда образуется новый СР. Этот
СР в свою очередь легко реагирует с другой исходной молекулой,
33
вновь образуя СР и т.п. Совокупность этих элементарных актов
называется разветвленной цепной реакцией. Еще в 1927-1929 гг.
выдающийся российский ученый Н.Н. Семенов разработал теорию
этих явлений в газовой фазе, показав существование нового класса
реакций, а именно разветвленных цепных химических реакций
(Семенов, 1934). Этот механизм лежит в основе таких явлений, как
горение, взрыв, детонация. Широкой популярности теории цепных
реакций способствовало открытие физиками цепного
разветвленного механизма деления ядерного горючего (ядерного взрыва), в
котором в качестве активных центров выступают нейтроны и
принцип которого описывается свободнорадикальной теорией. Следует,
однако, подчеркнуть, что цепные реакции в химических процессах
были открыты Н.Н. Семеновым значительно раньше, чем это
было сделано физиками в ядерных реакциях. Осознание этой ситуации
мировой научной общественностью привело к тому, что в 1956 г.
Н.Н. Семенову совместно с С. Хиншелвудом была присуждена
Нобелевская премия по химии.
Если при изучении быстрых цепных разветвленных реакций в
газовой фазе наметился крупный успех, то медленные цепные
реакции в жидкой фазе, реакции типа "медленного взрыва", были
исследованы значительно меньше. В реакциях такого типа отсутствовали
данные об образовании продуктов СР-характера. Этот пробел был
восполнен учеником и последователем Н.Н. Семенова Н.М.
Эмануэлем, который внес решающий вклад в обоснование теории
медленных цепных реакций окисления органических веществ в жидкой
фазе (Эмануэль, 1966). Наибольший интерес (в контексте этого
раздела) представляют работы Н.М. Эмануэля и его сотрудников,
посвященные природе СР-механизма окисления липидов (Эмануэль,
Кнорре, Лясковская, 1957). В развитие этих работ были проведены
обширные исследования окисления липидов в присутствии
разнообразных ингибиторов (аптиоксидантовУ и их смесей с целью
разработки способов повышения сохранности этих ценных пищевых
продуктов (Эмануэль, Лясковская, 1961). Это направление
исследований сыграло решающую роль в последующем распространении
принципов цепного жидкофазного окисления на биологические
системы, ибо основным компонентом мембран живых клеток, как
известно, являются липиды. Согласно оригинальной и смелой
гипотезе, предложенной независимо друг от друга Н.М. Эмануэлем и
Б.Н. Тарусовым2, экстремальные внешние воздействия индуцируют
1 Именно в этот период в научном обиходе школы Н.М. Эмануэля стал
систематически использоваться термин антиоксиданты, столь хорошо известный
сейчас не только широкой научной общественности, но и всему
цивилизованному человечеству.
2 Б.Н. Тарусов, сравнивая особенности лучевого поражения во времени с
кинетикой цепных химических реакций, описанных Н.Н. Семеновым, пришел к
34
и мембранах клеток цепные реакции окисления свободнорадикаль-
мого характера. Более того, в дальнейшем было высказано предпо-
южение, что с помощью разнообразных антиоксидантов можно,
нейтрализуя СР, предотвратить развитие деструктивных процессов
it таких важнейших структурах, как клеточные мембраны.
Дальнейшие исследования Н.М. Эмануэля и его школы полностью
подтвердили эту концепцию. Так, при изучении на высших животных таких
патологических процессов, как лучевая болезнь, злокачественный
рост клеток, действие токсических веществ, а также при старении
или острых состояниях стресса организма наблюдалась активация
(Р-процессов перекисного окисления липидов. Наоборот, введение
it организм специально подобранных или синтезированных для этой
цели малотоксичных антиоксидантов снижало уровень СР и даже
останавливало развитие патологических процессов и старения
лабораторных животных (Бурлакова и др., 1975; Эмануэль,и др., 1977;
Эмануэль, 1975; 1977; 1982; Обухова, Эмануэль, 1984; Бурлакова,
Храпова, 1985). Именно эти работы, а также работы Д. Хармана
(Hat man, 1956; 1962; 1984) задолго до упомянутых выше молекуляр-
но-генетических работ, выполненных на низших организмах,
возвестили о начале эры широкого практического использования
антиоксидантов в интересах здравоохранения.
Генезис и виды СР. Согласно предложенной Ю.А.
Владимировым и его сотрудниками (1992) классификации, подавляющее
большинство СР, образующихся в организме человека и животных,
можно разделить на природные и чужеродные. Первые в свою очередь
разделяются на первичные, т.е. продукты нормального метаболизма
(•ОО-; 'NO; HQ· - супероксиды, нитроксиды и семихиноны -
соответственно)3. Из супероксида, а также в результате ряда других реакций
в организме могут образовываться такие активные продукты, как
перекись водорода, гипохлорит и гидроперекиси липидов. Реакции
jthx соединений с ионами металлов переменной валентности,
главным образом с ионами Fe2+, генерируют образование вторичных
радикалов (ΌΗ; LO; L·; LOO - радикал гидроксила и набор липидных
радикалов - соответственно), которые оказывают разрушительное
действие на клеточные структуры. Наконец, третичные радикалы -
jto радикалы антиоксидантов, которые образуются при их
взаимодействии с теми или иными из упомянутых выше радикалов.
выводу, что протекающие в облученных живых системах процессы
ускоряются со временем, подобно автокаталитическим цепным реакциям. Он
предположил, исходя из имеющихся данных о структурной организации и
физико-химических особенностях живых систем, что цепные процессы могут развиваться в
липопротеиновой фазе клеточных структур и приводить в заключительной
стадии к разрушению "фазовых границ" и к гибели клетки (Тарусов, 1954).
' Неспаренный электрон в радикалах принято обозначать точкой. Например,
радикал гидроксила обозначают НО*.
35
Первичные и вторичные радикалы являются природными,
поскольку в определенных количествах всегда образуются в наших
клетках. Чужеродные радикалы могут появляться в клетках и
тканях под воздействием ионизирующей и ультрафиолетовой радиации
или в результате превращения каких-либо неприродных соединений
(ксенобиотиков), попавших в организм высших животных
Активные формы кислорода (АФК). Важнейшим достижением
органической эволюции на Земле стало формирование
фотосинтеза, а именно способности простейших живых систем к усвоению
свободной энергии солнечной радиации. Фотосинтез фактически
создал условия для гигантского роста органической массы, что, наряду
с обогащением атмосферы свободным кислородом, обусловило
будущее возникновение и развитие бесконечного множества одно- и
многоклеточных аэробных форм жизни. Между тем уже
существовавшие формы жизни подверглись при этом серьезной опасности
окислительной деструкции и были вынуждены эволюционировать
по пути выработки адаптационных приспособлений,
ограничивающих токсический эффект кислорода. Благодаря выработке и
наследственному закреплению защитных антиоксидантных
механизмов, обеспечивших организмам относительную стабильность, жизнь
на Земле смогла успешно эволюционировать от ее простейших
форм до человека. Центральную роль среди этих защитных
механизмов занимает комплекс антиоксидантных ферментов - изофор-
мы супероксиддисмутазы (СОД), каталазы и пероксидазы
(подробней об этих механизмах - далее ).
Следующим кардинальным успехом эволюции явилось создание
дыхательных цепей, т.е. сложных энзиматических систем
окислительного фосфорилирования энергетических субстратов, которые
(у аэробных организмов) вовлекали кислород в процесс извлечения
значительного дополнительного количества аденозинтрифосфор-
ной кислоты (АТФ) - универсального аккумулятора -донора
биологической энергии.
"Чистый выход" энергоносителей при полном окислении одной молекулы
глюкозы до воды и СС>2 составляет 38 молекул АТФ, синтезированных из аде-
нозиндифосфорной кислоты (АДФ) и неорганического фосфата. При этом две
молекулы АТФ дает гликолиз (процесс анаэробного расщепления глюкозы),
две молекулы АТФ - цикл Кребса (циклическое превращение органических
кислот, включающее присоединение двууглеродистого фрагмента от ацетил-КоА
к щавелевоуксусной кислоте и два последовательных акта окислительного де-
карбоксилирования: щавелевоуксусная -> лимонная -> а-кетоглютаро-
вая -> янтарная -> фумаровая -> яблочная -> щавелевоуксусная
кислота -> и т.п.), а 34 молекулы АТФ (!) поступают из дыхательной цепи (рис. 2).
Столь крупный энергетический выигрыш, который получили
организмы в результате приобретения механизма окислительного
фосфорилирования, послужил мощным толчком для формирования
и совершенствования новых физиологических функций, например,
36
Гликолиз-
—\/"H2°
Дыхатель- V
нал цепь ^
Vl/2 02
Рис. 2. Схема гликолиза и аэробного дыхания (Грин и др., 1990)
различных видов двигательных реакций, дистантной рецепции,
простых и сложных реакций нервной системы, систем переработки
электрофизиологической и химической информации и, наконец,
мышления.
Наряду с этим, деструктивный потенциал перекисного
окисления был использован природой и для совершенствования ряда ката-
болических процессов, например, внутриклеточного пищеварения,
фагоцитоза (Freeman, 1984; Гриневич и др., 1986), окислительной
деструкции чужеродных и токсичных веществ (Арчаков, 1975),
разборки избыточных мембранных структур {Yukawa, Nakazawa, 1980;
Бурлакова, Храпова, 1985).
Все перечисленные выше эволюционные достижения (список
которых далеко не исчерпан), открыв беспрецедентные
возможности развития жизни на нашей планете, содержали тем не менее
скрытые имманентные опасности для этой жизни.
Деструктивная роль АФК и ПОЛ. Максимальная активность
дыхательной цепи, которая локализована в многочисленных
субклеточных органеллах, называемых митохондриями, сопряжена с
опасными побочными эффектами. Известно, что около 98%
кислорода, потребляемое митохондриями в процессе окислительного
фосфорилирования, превращаются в воду, тогда как оставшиеся 2%
диффундируют за пределы дыхательной цепи в виде анион-радика-
37
ла супероксида (О^) в результате флуктуации в цепи
электронного транспорта (1ДЭТ). Как это видно на рис. 3, супероксид-радикал
может превращаться в перекись водорода (Н202) и радикал гидро-
ксила (*ΟΗ). Все эти соединения (в особенности *ОН) способны
нанести клеткам существенные повреждения. Увеличение потока
электронов по дыхательной цепи от субстратов дыхательной цепи к
кислороду будет с неизбежностью увеличивать и количество
повреждений в клеточных структурах, поскольку будет повышать
продукцию этих АФК. И такие ситуации будут постоянно повторяться в
жизни любого аэробного многоклеточного организма. Поэтому в
качестве защиты от агрессивных воздействий такого рода в клетках
эволюционно закрепилась антиокислителъная энзиматическая
система, первым звеном которой является фермент супероксиддис-
мутаза (СОД), катализирующая реакцию дисмутации
супероксидного радикала {Владимиров, 2000а):
Ό2 + "OJ + 2КГ Суперокс^дисмутаза ) Q? + j^
Перекись водорода мигрирует в другие компартменты клетки и
разрушается в результате действия других ферментов - каталазы и
глутатионпероксидазы (GSH-пероксидазы), и таким образом
происходит дальнейшая нейтрализация АФК:
2Н202 Каталаза >Н20 + 02.
Н202 + 2GSH Глутатионпероксидаза ) GSSG + 2Н20.
Наряду с описанной энзиматической системой, в организме существует
много природных соединений, обладающих антиоксидантными (АО)
свойствами. Это прежде всего витамины А, С, Е, β-каротин, селен, глутатион, цис-
теин, метионин, мелатонин, токоферолы, убихиноны, ретиноиды, флавоно-
иды и многие другие соединения. Тем не менее, несмотря на такую сложную
систему АО-безопасности, какое-то количество наиболее активных СР
все-таки "ускользает" от факторов нейтрализации и наносят свои
повреждающие удары. Повреждений будет тем больше, чем ниже АО-защита
организма, ослабленная стрессом, недоеданием, преклонным возрастом или
болезнями.
Заметную роль в развитии нарушений, вызванных АФК, играют
также ионы двухвалентных металлов. Так, в присутствии ионов
двухвалентного железа, которые всегда в тех или иных количествах
содержатся в клетках, перекись водорода разлагается с
образованием гидроксильного радикала:
Н202 + Fe2+ -> Fe3+ + НО- + НО·
Эта реакция, открытая Фентоном еще в позапрошлом веке
(Fenton, 1894), чревата всевозможными неприятностями для живых
38
Субстраты дыхания
АТФ
е~
НАД
^
е"
| KoQ |
v°
ι
АТФ ~А е 0-· _^ н2о2 _^ ОН
Цитохром С
ΑΤΦ^Ί
н2о
Рмс. 3. Пути переноса электронов в дыхательных цепях митохондрий (Скула-
чеву 1999)
клеток. Чрезвычайно высокая химическая активность радикала гид-
роксила позволяет ему разрушать практически любую молекулу,
вступившую во взаимодействие с ним. Так, действуя на SH-группы,
гистидиновые и другие аминокислотные остатки белков, НО
вызывает их денатурацию и инактивацию ферментов. В нуклеиновых
кислотах этот СР разрушает углеводные мостики между нуклеотидами
и, следовательно, разрывает сахаро-фосфатный остов ДНК и РНК,
что в ряде случаев приводит к гибели клеток. Внедряясь в липидный
слой клеточных мембран, радикал гидроксила инициирует реакции
цепного окисления липидов, что приводит к повреждению мембран,
нарушению их функций и в конце концов к гибели клеток. Таким
образом, радикал НО - это радикал-разрушитель, радикал-убийца
{Владимиров, 2000д).
Как мы уже видели, реакция цепного окисления липидов, или,
как ее еще часто называют, реакция перекисного окисления
липидов (ПОЛ), играет важную роль в развитии многих патологических
процессов, включая также и процесс старения {Эмануэль, Обухова,
1984; Бурлакова, Храпова, 1985; Дилъман, 1987; Барабой, Брехман
и др., 1992; Todorov Η. et al., 2001).
Попадая в липидный слой мембраны, радикал гидроксила
(а именно он чаще других СР предпринимает подобную атаку)
вступает в химическое взаимодействие с полиненасыщенными жирными
39
кислотами (LH), образуя при этом липидные радикалы
(Владимиров, Арчаков, 1972):
НО· + LH -> Н20 + L\
Липидный радикал (L·) вступает в реакцию с растворенным в среде
молекулярным кислородом, при этом возникает новый свободный
радикал липоперекиси (LOO):
L· + 02 -> LOO\
Этот последний атакует одну из соседних молекул фосфолипида с
образованием гидроперекиси липида LOOH и нового радикала L·:
LOO' + LH -> LOOH + L·.
Чередование двух последних реакций и представляет собой цепную
реакцию пероксидации, т.е. реакцию ПОЛ. Значительное ускорение
ПОЛ наблюдается в присутствии ионов двухвалентного железа. В
результате взаимодействия Fe2+ с гидроперекисями липидов
(разновидность реакции Фентона) происходит разветвление цепей:
Fe2+ + LOOH -> Fe3+ + НО- + LO\
Появившиеся радикалы LO* инициируют новые цепи окисления
липидов:
LO· + LH -> LOH + L·; L* + 02 -> ЬОО* и т.д.
В клеточных мембранах цепи могут состоять из десятка и более
звеньев. Однако в конце концов они обрываются в результате
взаимодействия СР с антиоксидантами (InH), ионами металлов
переменной валентности или друг с другом:
LOO· + Fe*+ + Н+ -> LOOH;
LOO* + InH -> In·: LOO* + LOO* -> молекулярные продукты.
Мы сочли уместным столь подробно остановиться на реакциях
перекисного окисления липидов, поскольку эти реакции играют, как
это будет показано ниже, крайне важную роль в клеточной
патологии при стрессе, развитии дегенеративных болезней, т.е. во всех тех
случаях, которые в той или иной степени имеют отношение к
процессам старения.
Одним из наиболее важных последствий повышенного
образования АФК и цепных реакций ПОЛ является нарушения
структуры и функции клеточных мембран, что не может не сказываться
на нормальной жизнедеятельности клеток (рис. 4). В хрусталике
глаза, например, ПОЛ приводит к окислению сульфгидрильных
групп мембранных белков и далее к образованию дисульфидных
мостиков между соседними белковыми молекулами, что
способствует образованию массы белковых агрегатов и развитию старческой и
других видов катаракты у человека.
40
Избыток ППОЛ
Разобщение структурно-функционалльной
организации биологической мембраны
Повреждение
энергетического
аппарата клетки:
разобщение
сопряженного
окислительного
фосфорилирования
Изменение
активности
мембраносвязанных
ферментов, белок-
липидных
взаимодействий
Повреждение
структуры
биомембраны:
вязкости,
поверхностного заряда,
химического состава;
образование пор;
нарушение связи
ДНК с биомембраной
Увеличение
проницаемости
биомембраны
Нарушение
клеточного
деления
Нарушение
системы
клеточной
регуляции
Повреждение
системы
детоксикации.
Развитие
токсического
эффекта.
Рис. 4. Схема токсического действия продуктов перекисного окисления липидов
на клетку {Барабой и др., 1992)
В клетках и в особенности в субклеточных органеллах
митохондриях, где дыхательная цепь генерирует основную массу АФК,
окисление тиоловых групп продуктами ПОЛ приводит к появлению в
мембранах различных дефектов. Под действием разности
электрических потенциалов на мембранах сквозь такие "поры" в клетки
входят ионы натрия, а в митохондрии - ионы калия. Эти процессы
повышают осмотическое давление внутри клеток и митохондрий и
вызывают их набухание, что еще больше повреждает мембраны.
Генерализация ПОЛ постепенно делает мембраны более твердыми,
хрупкими и "протекающими", лишая их способности нормально
реагировать с окружающей средой.
Продукты ПОЛ могут также статистически поражать ион-
транспортные ферменты, такие как Са2+ - АТФазы, окисляя тиоло-
вые группы их активного центра. Эти события приводят к
замедлению "откачивания" ионов кальция из клетки и одновременно к
ускорению входа кальция в клетку, что сопровождается повышением
внутриклеточной концентрации ионов кальция и повреждению
клетки. Под влиянием ПОЛ мембраны митохондрий делаются про-
41
ницаемыми для ионов водорода и кальция, в результате чего эти ор-
ганеллы теряют способность к синтезу АТФ и клетка оказывается в
условиях энергетического голода {Владимиров, 2000в). Итак, мы
видим, каким образом АФК и продукты ПОЛ могу влиять на развитие
возрастных патологий и скорость старения. Но это. конечно,
еще не все.
Помимо систем, продуцирующих энергию, АКФ в состоянии
поражать и многие другие системы клетки, например систему
хранения и передачи наследственной информации (ДНК), а также такой
важнейший биологический процесс, как биосинтез белков. Степень
СР-повреждений будет, очевидно, пропорциональна интегральной
скорости метаболизма, которая, по существу, определяется
скоростью сжигания калорий. Известно, что скорость метаболизма
крысы приблизительно в семь раз выше человеческой. Согласно
существующим оценкам, количество "попаданий" СР в ДНК у крысы
составляет примерно 105, а у человека соответственно только 104. Это,
по-видимому, одна из причин (но не единственная), почему люди
живут значительно дольше, чем крысы. Действительно, когда скорость
метаболизма крыс понижается путем серьезного ограничения
калорийности питания, продолжительность их жизни разительно
увеличивается.
Связь между интенсивностью метаболизма и видовой
продолжительностью жизни была отмечена еще в начале XX в. Так, М. Рубнер (Rubner, 1908)
показал, что для многих видов животных количество кислорода, потребленное
за всю жизнь, в расчете на единицу веса остается величиной постоянной:
Г
Ru = jqo2dt
О
где Ru — константа Рубнера, Т~ продолжительность жизни, qo2 - интенсивность
дыхания (скорость потребления кислорода на единицу веса). Это означает, по
крайней мере для теплокровных животных, что чем больше животное
поглощает кислорода на единицу веса, тем более короткой будет его жизнь.
Действительно, отношение поверхности к объему у мелких животных значительно
выше, чем у крупных, и, следовательно, иррадиация тепла с их поверхности будет
существенно большей, чем у крупных животных, и для поддержания
постоянной температуры тела они вынуждены "сжигать" в дыхательной цепи
значительно большее количество калорий. Подобная ситуация, как это ясно в
настоящее время, с неизбежностью оборачивается резким повышением продукции
АФК с их разрушительным действием на структурные компоненты клетки, что
и является одним из важнейших факторов, определяющих продолжительность
жизни. Как считают эксперты в этой области (Зотин, 1993; Зотин и др., 2001),
константа Рубнера может быть использована для определения видовой
продолжительности жизни практически всех групп животных.
В клетке существуют две органеллы, содержащих ДНК: ядро и
митохондрии. Ядро содержит большую часть генетического
материала клетки, распределенного по хромосомам. Митохондрии,
которые служат энергетическими станциями клетки, содержат весьма
42
Таблица 1
Биосинтез ядерной и митохондриальнон ДНК
в печени облученных крыс
(Галкин, Тодоров Н., 1969)
(Показатель удельной радиоактивности ДНК
у необлученных животных принят за 100%)
Бремя после
облучения, часы
Ядра
Митохондрии
Импульс/мин/мг ДНК
Необлученные 100 100
12 21 4.6
24 40 7.7
небольшую, но необходимую часть клеточной ДНК. Оказывается,
что АФК поражают митохондриальную ДНК (мтДНК) в десять раз
чаще, чем ядерную ДНК клеток одной и той же ткани. Частично,
это происходит, потому что основная масса АФК генерируется
именно в митохондриях, где происходит окисление значительной
части биохимического топлива. Кроме того, мтДНК не защищена так
же хорошо, как ядерная ДНК, белками (гистонами) и
репарационными системами (Miquel et. al., 1992; Wei et al., 2001). Свидетельства
высокой уязвимости мтДНК впервые были получены еще в 60-х
годах XX в. в работе А.П. Галкина и И.Н. Тодорова {табл. 1), в
которой была показана исключительно высокая радиационная поражае-
мость биосинтеза мтДНК (по сравнению с биосинтезом ядерной
ДНК) печени при γ-облучения крыс в сублетальной дозе (что
фактически равноценно поражающему действию АФК). В прошлом
десятилетии было найдено более ста мутаций мтДНК человека, и
некоторые из них имели место при старении человеческих тканей. Это
связывают, в частности, с возрастным снижениям содержания
восстановленного глутатиона в митохондриях, что коррелирует с
увеличением повреждений мтДНК в стареющих тканях (Wei et al., 2001)
Постоянное воздействие АФК снижает со временем
способность митохондрий производить энергию. Похоже, что
митохондрии - это субклеточная система, которая первой деградирует с
возрастом. В результате клетки становятся метаболически менее
активными, что ведет к уменьшению функциональной способности
органов, понижению уровня адаптивных реакций и развитию
возрастных болезней.
Стресс, по-видимому, вносит главный вклад в процесс
"выгорания" митохондрий. Он повышает спрос на энергию, и побуждает
митохондрии работать напряженнее, что приводит к образованию
большего количества побочных продуктов окисления - АФК. При
длительном или интенсивном стрессе мембраны митохондрий
настолько повреждаются АФК, что образуется "утечка*' их содержи-
43
мого, и это прерывает процесс производства энергии и ведет к
возникновению еще большего количества свободных радикалов. Вся
ситуация в конце концов выливается в непропорционально
серьезные окислительные повреждения, что, как полагают, дает толчок к
патологиям и ускоряет старение.
Столь важная роль митохондрий в развитии оксидативного
стресса и его влияния на процесс старения позволили ряду
исследователей говорить об уверенном становлении митохондриалъной
теории старения {Sastre et al., 2000; Wei et al., 2001) (подробнее об
этом см. в разделе "Оксидативный стресс клетки", гл. 19).
Ключевая роль митохондрий в качестве энергетических станций
клетки и ее специфическая восприимчивость к СР-повреждениям
делают их важной потенциальной целью для вмешательства,
направленного на замедление старения и снятия вредных последствий стресса. Два
важных природных соединения испытывались для улучшения функций
митохондрий: ацетил-Ь-карнитин (ALC) и коэнзим Q10 (CoQIO)
(Modern Nutrition in Health and Disease, 1994). ALC - часть механизма
транспортировки жирных кислот в митохондрии. Хотя ALC может
синтезироваться в организме, скорость его синтеза снижается с возрастом
Препараты с ALC ускоряют использование жиров митохондриями
улучшая производство энергии. Некоторые исследования показали,
что ALC понижает уровень холестерина, увеличивает уровень ЛВП и
улучшает состояние коронарных сосудов и мозговую функцию.
CoQIO, будучи существенной частью клеточной системы
дыхания, локализованной в митохондриях, является также и антиокси-
дантом. CoQIO повышает как скорость, так и эффективность
производства энергии, и в то же время защищает митохондрии от АФК.
Организм может производить CoQIO, но множество факторов,
включая возраст, болезни, лекарства, понижающие холестерин, и,
наконец, недоедание, могут снижать эту способность. CoQIO иногда
называют "биомаркером старения", потому что его уровень очень
хорошо коррелирует со старением и дегенеративными болезнями. В
одном из исследований добавки CoQIO привели к продлению жизни
у мышей на 50%. Многие исследования ясно показали
эффективность CoQIO при остановке сердца в результате закупорки сосудов и
других болезнях сердечной мышцы. Другими патологиями, при
которых, по-видимому, помогает CoQIO, являются гипертония,
снижение иммунитета и мышечная атрофия (Todorov Η. et al., 2001).
Белки клеток постоянно повреждаются в результате атак со
стороны АФК и продуктов ПОЛ. Повреждения и модификации
белков, как правило, необратимы, и эти белки подлежат удалению
системами протеолиза, т.е. лизосомами, протеасомами и другими про-
44
теолитическими факторами (Рязанов, 2001). Восстановление
необходимого набора белков связано с эффективной работой белок-син-
тезирующей машины, т.е. рибосом, информационных и
транспортных РНК (иРНК, тРНК), аминоацил-тРНК-синтетаз, факторов
трансляции и т.п. Все эти макромолекулы и их комплексы,
связанные с катаболическими и анаболическими процессами, обновляю-
гцими белковый гомеостаз клетки, также подвергаются
деструктивному воздействию СР. Постепенное накопление поломок в
результате этого приводит к затуханию процессов обновления клеточных
структур преимущественно за счет понижения скорости синтеза
белков (Van Remmen, et. al., 1995). Это затухание в свою очередь
связано, в частности, с возрастным уменьшением концентрации
функционально активных рибосом, в основе чего лежит не только
окисление собственно рибосомных белков, но и возрастное падение
количества генов рибосомных РНК (рРНК) путем делеции некоторой
их части из R-оперонов генома (Strehler, 1986; цит. по: Рязанов,
2001). Таким образом, одна из старейших концепций старения, а
именно концепция затухающего самообновления протоплазмы
(Нагорный, 1940; Нагорный и др., 1963), наконец, обретает причинно-
следственную мотивацию в свете современных достижений в
изучении деструктивной роли АФК и продуктов ПОЛ.
Таким образом, окислительные повреждения специфических
белков составляют один из механизмов, связывающих оксидатив-
ный стресс с возрастной деградацией физиологических функций
клетки (Sohal, 2002).
Окисленные белки, будучи термодинамически неустойчивыми, принимают
частично развернутую третичную структуру, которая с готовностью
формирует агрегаты, что способствует накоплению так называемых клеточных шлаков,
в частности амилоида (подробнее см. в параграфе "Накопление шлаков" этой
главы). Поэтому представляет интерес идентификация тех чувствительных к
окислению белковых субстратов, которые могут играть защитную роль,
благодаря их способности к регуляции, понижающей (down-regulate) энергетический
метаболизм и последующую генерацию АФК. Как известно, поддержание
клеточных градиентов кальция требует значительного расхода энергии, что,
естественно, связывает изменения во внутриклеточном уровне кальция с утилизацией
АТФ и ассоциировано с генерацией АФК при функционировании
дыхательных механизмов. Избирательное окисление или нитрирование кальций
регулирующих белков - кальмодулина и Са-АТФазы in vivo, которое имеет место в
процессе старения в условиях оксидативного стресса, может представлять
адаптивную реакцию на этот вид стресса, функционирующую по down-regidate
энергетическому метаболизму, что связано с пониженной продукцией АФК.
Эти кальций регулирующие белки также предпочтительно окисляются или
нитрируются и в экспериментах in vitro, что укрепляет представление об их
защитной роли. Сокращение скорости генерации АФК при этом минимизирует
неизбирательное окисление ряда макромолекул, когда белковая репарация и
системы протеолиза не способны предотвратить агрегацию. Таким образом,
имманентные клеткам окислительные модификации кальций регулирующих белков»
функционирующих в режиме down-regulate использования АТФ и генерации
45
АФК, могут ослаблять накопление окислительного "груза" при перманентно
повторяющемся оксидативном стрессе (Squier, 2001). Таков еще один из
механизмов, защищающих клетки от деструктивного влияния свободнорадикальных
форм кислорода.
Деструктивная роль АФК и ПОЛ проявляется не только на
субклеточном, но, естественно, также и на системном уровнях. Если,
например, умеренная окислительная модификация гемоглобина
повышала его расщепление мультикаталитической протеинкиназой,
то длительная инкубация этого белка с оксидантом вызывала его
повреждение с образованием перекрестных сшивок
редуцирующими сахарами (см. параграф "Глюкоза и старение") и продуктами
ПОЛ со сложной, как правило, неопределенной структурой, что
значительно снижало чувствительность этого белка к протеолизу. Это
может, как полагают авторы, приводить к возрастанию уровня
белков с повышенной антигенностью в тканях (см. гл. 6) и тем самым
увеличивать риск развития аутоиммунной патологии при старении
(Плешакова и др., 2000).
СР и старение организма. Идея о том, что свободные радикалы
могут способствовать старению была впервые выдвинута Денхэмом
Харманом в середине 50-х годов XX в. (Нагтап, 1956). Долгое время
эта идея представлялась просто любопытной гипотезой. В
последние два десятилетия, однако, ученые накопили большое количество
аргументов в пользу этой идеи, превратив ее в одну из наиболее
обоснованных современных теорий старения.
Существует множество прямых доказательств, что СР-повреж-
дения накапливаются с возрастом. Например, у старых крыс
обнаруживается вдвое больше окислительных, т.е. вызванных АФК,
повреждений в ДНК, чем у молодых животных (Виленчик, 1987).
Частота мутаций в лимфоцитах пожилых людей приблизительно в 9 раз
выше, чем в лимфоцитах новорожденных (Curtsinger et al., 1995).
Синдром Вернера и прогерию, две человеческих болезни, внешние
симптомы которых указывают на преждевременное старение,
связывают со значительным увеличением повреждения белков
свободными радикалами (Ames et al., 1993). Возрастное накопление в
клетках таких пигментов, как липофусцин и цероид, как полагают,
являются также результатом окислительного повреждения белков и ли-
пидов. До определенного момента эти пигменты относительно
доброкачественны, но если их накопление превышает некоторый
уровень, клетки начинают "задыхаться" (Yin, 1996). Эта вредоносная
аккумуляция пигментов может быть замедлена антиоксидантами
Однако наиболее интересные свидетельства взаимосвязи АФК и
старения были получены при изучении СОД, которая была открыта
Дж. Мак-Кордом и И. Фридовичем в 1969 г. (McCord et al., 1979).
Осознание того факта, что существует фермент, катализирующий
реакцию взаимодействия двух суиероксидных радикалов и,
следовательно, удаляющий АФК, и специально вырабатывающийся для
46
-πόγο клетками большинства животных, стимулировало
исключительный интерес как к выяснению генезиса и функциональной роли
радикалов кислорода, так и защитной роли этого фермента.
Эффектные генетически исследования некоторых низших
организмов и их мутантов-долгожителей, о которых упоминалось в
начале этого параграфа, содержали весьма убедительные
доказательства связи между старением и активностью АФК. Было обнаружено,
что мутации, поражающие определенный ген (называемый "age-ltv)
у червей вида Caenorhabditis elegans, дают увеличение
продолжительности жизни на 70%. Оказалось, что у этих мутантов уровень
чктивности двух ключевых ферментов, нейтрализующих
АФК-СОД и каталазы - был существенно повышен.
Предполагалось, что ген, пораженный мутациями, кодирует структуру
ингибитора антиоксидантной системы клетки {Finch, Jonhson, 1989). При
обсуждении аналогии между функциональной ролью СОД у
человека и червей, один из авторов этой работы Томас Джонсон привел
известный афоризм Фридриха Ницше из его поэмы "Так говорил За-
ратустра" о том, что... люди, проделав свой путь от червя к
человеку, во многом остались червями,
В другом случае, исследователи производили селекцию в
последовательных генерациях плодовой мушки (Drosophila melanogaster),
что позволило отобрать клон, особи которого показали вдвое
большую, чем обычная, продолжительность жизни. Одно из важных
различий между обычными и долгоживущими насекомыми
заключалось в том факте, что у последних наблюдался более высокий
уровень экспрессии гена, кодирующего СОД (Rose, 1991).
Исследование связи между старением и уровнем активности ан-
тиоксидантных ферментов у людей также позволило получить ряд
косвенных свидетельств существования коррелятивной связи между
этими параметрами. Так, у пациентов с синдромом Дауна, был
обнаружен низкий уровень активности СОД (Brooksbank et al., 1983). И
если вспомнить, что люди, страдающие этим заболеванием, живут,
как правило, не более 35-40 лет, то, естественно, возникает мысль о
непосредственной связи между пониженной активностью СОД и
потенциальной продолжительностью жизни этих больных. В другой
работе изучалось соотношение активности СОД и основного
обмена (количество кислорода, используемого в процессах
окислительного фосфорилирования) 12 видов приматов и человека.
Обнаружилось, что чем выше продолжительность жизни разных видов, тем,
соответственно, выше у них соотношение - активность СОД/основ-
ной обмен (Tolmasoffet al., 1980).
Считается, что быстрое увеличение максимальной
продолжительности жизни у приматов и человека в течение последних
нескольких миллионов лет было обусловлено, в частности,
усовершенствованием генов систем защиты (Cutler, 1978,1984). Нельзя не
вспомнить в связи с этим высказывание одного из основоположни-
47
Таблица 2
Защита от вредного действия свободных радикалов
{Растит Р., 1993)
АНТИОКСИДАНТЫ
(нейтрализуют свободные радикалы или, иначе,
ограничивают их активность)
Класс веществ
Вещество
Ферменты Супероксиддисмутазы
Глутатионпероксидазы
и каталазы
Другие Витамин Ε и β-каротин
Мочевая кислота
и витамин С
Хелатирующие агенты
Активность
Превращает супероксидный
радикал (Of) в перекись водорода
(Н202)
Превращают перекись водорода
в воду (Н20) и молекулярный
кислород (02)
Взаимодействуют со
свободными радикалами, тем самым не
давая им влиять на клеточные
компоненты; будучи
жирорастворимыми, могут защищать
мембраны
Взаимодействуют со
свободными радикалами в цитоплазме
Не дают железу, меди и другим
переходным металлам
участвовать в катализе окислительных
реакций
Репарация
белков
Репарация
пипидов
СИСТЕМЫ РЕПАРАЦИИ
(осуществляют деградацию, восстановление
или замену поврежденных молекул)
Протеиназы
Пептидазы
Фосфолипазы
Расщепляют окисленные белки
Расщепляют продукты протеи-
назной активности;
образующиеся аминокислоты могут
использоваться для синтеза новых
белков
Расщепляют поврежденные
участки окисленных липидов в
составе мембран, так что другие
ферменты могут заменить эти
участки
48
Таблица 2 (окончание)
АНТИОКСИДАНТЫ
(нейтрализуют свободные радикалы или, иначе,
ограничивают их активность)
Класс веществ
Репарация
Днк
Вещество
Ацетилтр ансфер аз ы
Глу татионперо ксид аз а
и глутатион-трансфера-
за
Экзонуклеазы
и эндонуклеазы
Гликозилазы
и полимеразы
Лигаза
Активность
Заменяют жирные кислоты,
отщепленные от липидных
молекул
Участвуют в замене окисленных
жирных кислот без удаления
крупных участков мембран
Выщепляют поврежденные
участки ДНК
Заполняют пустые места,
оставленные экзо- и эндонуклеазами
"Сшивает" восстановленные
участки с остальной цепью
ков современной генетики Томаса Моргана: "У нас нет
достаточных оснований допускать, что эволюция происходит прежде
всего за счет увеличения числа генов у высших форм... главную роль
играет не число появляющихся новых генов, а их виды" {Morgan,
1932).
В начале развития радиобиологии, науки, изучающей влияние
радиации на биологические системы, исследователи столкнулись с
парадоксальным явлением. Низкий уровень лучевого воздействия
защищал животных от более высоких уровней радиации так же, как
и от многих других стрессоров - мутагенов, токсинов и АФК. Позже
было найдено, что непродолжительное повышенное воздействие
СР, вызванное радиацией, стимулирует антиокислительную защиту
(АО-защиту) клеток (СОД, каталазу, глутатионпероксидазу),
улучшая таким образом сопротивляемость к повреждающим агентам в
будущем. По-видимому, на основе стимуляции этих же механизмов
малые дозы ионизирующего облучения снижают, как было
показано {Балакин и др., 2000), рост цитогенетических нарушений,
обусловленный старением, до уровня ниже спонтанного, т.е. переводят
организм в новое устойчивое состояние, характеризующееся
повышенной стабильностью генома и, как следствие, увеличением
продолжительности жизни.
Мы, естественно, не рекомендуем поселяться поближе к
атомным электростанциям для приобретения лучшей сопротивляемости
к стрессовым воздействиям. К счастью, имеется намного более
простое решение. Умеренная физическая нагрузка также смягчает воз-
49
действие СР. Это понятно: чем большее количество топлива
сжигает организм, тем больше образуется побочных продуктов
окисления. С одной стороны, разумная периодическая физическая
нагрузка стимулирует собственную антиоксидантную систему, которая
останется улучшенной еще долгое время после окончания нагрузок. С
другой стороны, чрезмерная нагрузка, которая является серьезным
физическим стрессором, может сокрушить защитные системы и
ускорить старение.
Защита от СР. В III части этой книги будут представлены
развернутые рекомендации по биохимической коррекции вредоносного
влияния эндогенных СР. Здесь же мы ограничимся лишь
несколькими предварительными замечаниями, которые в той или иной мере
связаны с общим содержанием этого раздела.
Диета - очень важная часть поддержания полноценной
АО-защиты. Во-первых, сбалансированная диета обеспечивает
строительным материалом собственные системы организма, нейтрализующие
СР. Витамины А, С, Е, как уже указывалось, сами по себе являются
хорошими АО; аминокислота цистеин входит в состав глутатиона,
основного внутриклеточного АО; медь, цинк и марганец
необходимы для оптимальной функции СОД, а селен - для глутатионперок-
сидазы. Фрукты и овощи содержат большое разнообразие
природных АО, включая многочисленные флавоноиды, каратиноиды и
антоцианины. По-видимому, природные АО различаются как по их
сродству к различным типам клеток или их органеллам, так и по их
эффективности против определенных типов СР. Диета,
изобилующая фруктами и овощами, обеспечивает, очевидно, наилучшую и
всестороннюю АО-защиту. Множество исследований ясно
показало, что такая диета уменьшает риск дегенеративных болезней.
Современные рекомендации Национального института рака и
Национальной академии наук США предлагают включать по крайней
мере 2-4 порции различных фруктов (пример порции - одно яблоко
среднего размера) и 3-5 порций различных овощей в дневном
рационе. Хотя сбалансированная, богатая овощами и фруктами диета
важна для продления жизни, опыт современной гериатрии
показывает, что дополнительные антиоксидантные добавки, так же как и
пищевые добавки, действующие против других механизмов
старения, могут существенно продвинуть нас по пути улучшения здоровья
и увеличения продолжительности жизни.
В табл. 2 суммированы основные клеточные механизмы,
которые предотвращают или ликвидируют молекулярные повреждения,
вызванные СР. Эти механизмы, как полагают многие
исследователи, несовершенны и некоторые из них также теряют
эффективность с возрастом.
2.2. ДНК: ПОВРЕЖДЕНИЯ/РЕПАРАЦИЯ
Надпись на чаше
Древнюю чашу нашел он у шумного синего моря,
В древней могиле, на диком песчаном прибрежье.
Долго трудился он; долго слагал воедино
То, что гробница хранила три тысячи лет, как святыню,
И прочитал он на чаше
Древнюю повесть безмолвных могил и гробниц:
"Вечно лишь море, безбрежное море и небо.
Вечно лишь солнце, земля и ее красота,
Вечно лишь то, что связует незримою связью
Душу и сердце живых с темной душою могил".
Иван Бунин, 1903.
- Папа, а что это за связь между мертвецами и живыми?
- Я думаю, малыш, что это - ДНК...
ДНК (дезоксирибонуклеиновая кислота) - центральная
молекула жизни; она содержит весь план развития и структуры
будущего организма, который закодирован в огромном числе структурных
фрагментов ДНК - генах. ДНК - это незаменимая часть клетки.
Другие структуры, такие как РНК, белки, липиды и другие
биомолекулы, могут быть в случае поломки полностью заменены на
полноценные, согласно инструкциям, которые содержатся в генах.
Однако замены быть не может, если повреждены (и не подлежат
репарации) функционально важные участки ДНК
Нерепарируемые повреждения ДНК ведут к двум основным
результатам: (1) клетка погибает, или же (2) клетка
видоизменяется (мутирует), и это означает, что один или большее
количество генов теряют или изменяют свои первоначальные свойства.
Огромное большинство мутаций или нейтральны (никакого
влияния) или вредны. Лишь крайне низкое число мутаций, если они
происходят в зародышевых клетках (из которых образуются
сперма, тозоиды или яйцеклетки), могут давать некоторые
преимущества для выживания потомства. Вещества, которые способны
повреждать ДНК и вызывать мутации, называются мутагенами.
СР - наиболее распространенные мутагены. Достаточно
распространены также и другие химические мутагенные факторы: N-нит-
розо соединения, альдегиды, асбест, угольная смола и др.
Большинство мутагенов являются также и канцерогенами, т.е. стимулирует
развитие рака {Ames et al., 1993).
ДНК постоянно бомбардируется мутагенами, преимущественно
такими как АФК или некоторые альдегиды, которые являются
нормальными продуктами метаболизма, и их присутствие в организме
неизбежно. Некоторые мутагены, как, например, сигаретный дым или
51
ацеталъдегид (продукт разложения алкоголя в организме), человек
сам привносит в свой организм; все остальные - приходят к нам из
загрязненной окружающей среды. Различные виды радиации также
вызывают мутации. Повреждения, которые вызываются УФ-излучени-
ем, ограничены главным образом кожей, роговицей и сетчаткой
глаза, тогда как радиация с высокой энергией, например рентгеновское
излучение, может вызвать мутации и во внутренних органах.
Практически повсеместное распространение курения табака и потреблс
ния спиртных напитков, сопряженных с повышенными мутагенными
эффектами, побуждает нас подробнее рассмотреть эту проблему. Уже давно известно,
что курение сигарет служит главным фактором риска для возникновения
язвенной болезни и рака желудка. Недавно группа исследователей из университета в
Гонконге показала, что экспозиция с дымом сигарет приводила к зависящему от
времени и концентрации увеличению апоптоза-программируемой клеточной
гибели (подробнее об этом см. в параграфе "Генетические аспекты старения"
этой главы) в слизистой оболочке желудка крысы, которое сопровождалось
увеличением активности оксидазы ксантина. Увеличенный апоптоз и
активность оксидазы ксантина могли быть обнаружены даже после однократной
экспозиции (Wang et al., 2000). Следует подчеркнуть, что включение сложного
механизма апоптоза наступает по достижении некоего критического уровня
накопления мутаций в ДНК, чреватого высоким риском ракового перерождения
клетки (Чумаков, 1998). Японские исследователи (Aoshiba et al., 2001) изучали
влияние сигаретного дыма на альвеолярные макрофаги в связи с развитием
легочных заболеваний. Эта работа имела целью определить, стимулирует ли
сигаретный дым апоптоз альвеолярных макрофагов. Крысы были подвергнуты
действию дыма от десяти сигарет более чем пять часов в изолированном боксе.
Оказалось, что примерно 3% альвеолярных макрофагов через 24 часа после
воздействия подверглись апоптозу.
Под влиянием этилового спирта активируется ось гапоталамус — гипофиз-
кора надпочечников, что приводит к повышенной секреции глюкокортикоидов
(Spenser et al., 1999). Хроническая чрезмерная секреция глюкокортикоидов
может приводить к преждевременному и/или тяжелому старению и, в частности, к
апоптозу Т- лимфоцитов (Cohen, 1992) со всеми вытекающими последствиями
для развития возрастных патологий (см. гл. 15, 17 и 18).
Для организмов, по сложности близких к человеку, способность
передавать генетическую информацию с высокой точностью -
выдающееся достижение эволюции. Количество пар нуклеотидов
("букв") в суммарной ДНК человека составляет приблизительно три
миллиарда. Даже если поражению подвергнется только одна буква
из каждого миллиона, то при удвоении генома проявится около трех
тысяч ошибок. И если учесть, что при развитии организма из
оплодотворенной яйцеклетки ее геном претерпевает примерно 1015
актов репликации, то возможность существования человека при такой
скорости накопления ошибок представляется маловероятной. На
самом деле вероятность возникновения ошибок при репликации
составляет, по-видимому, всего 1(Н (Loeb, Kwikel, 1982). Такая
высокая точность репликативной машины достигается в результате
слаженной работы ряда независимых ферментативных процессов.
52
Точность репликации зависит главным образом от эффективности выбора
правильного нуклеотида. Селекция осуществляется тем же ферментом,
который соединяет нуклеотиды в полимерную цепь ДНК. Этот фермент,
называемый ДНК - полимеразой, движется вдоль одной из цепей ДНК, синтезируя
комплементарную ей дочернюю цепь из четырех свободных нуклеотидов (обычно
обозначаемых, как А, Г, Ц и Т), присутствующих в клетке в виде трифосфатов
(дезоксиаденозин-, дезоксигуанозин-, дезоксицитидин- и тимидинтрифосфата).
ДНК-полимереза отбирает нуклеотидтрифосфат, расщепляет его до
монофосфата и присоединяет к концу растущей цепи. При этом выбор
комплементарного нуклеотида наиболее вероятен, поскольку энергетически это наиболее вы-
(одно, так как комплекс ДНК-полимеразы, цепи ДНК и нуклеозидмонофосфа-
i а наиболее стабилей, когда нуклеотид в этом комплексе комплементарен
матричному нуклеотиду (Modrich, 1987). Частота ошибок на этой стадии
составляет примерно 10-5, эти ошибки подвергаются еще одному акту коррекции - "ре-
Оакторской правке", которая осуществляется ферментным аппаратом,
ассоциированным с ДНК - полимеразой. Такой фермент в начале 70-х годов был
открыт Д. Брутлагом и А. Корнбергом и назван ими "редактирующей экзонукле-
сиой". Этот последний фермент "опознает" цепь ДНК, в которой необходимо
совершить исправление по уровню метилирования старой и новой цепей
(Ванюшин, 1983). Исправление происходит преимущественно в неметилированной
ДНК. Оба рассмотренных механизма коррекции ошибок ДНК могут давать
существенные сбои, если нарушено соотношение концентраций четырех дезокси-
рибонуклеозидтрифосфатов - предшественников как синтеза, так и репарации
ДНК (Радман и др., 1988). Поэтому внутриклеточные концентрации этих
предшественников должны быть очень тонко сбалансированы. Фермент,
катализирующий в клетке биосинтез дезоксирибонуклеотидов, является рибонуклеотид
редуктазой (РР)
Все описанные выше процессы исправления ошибок имеют место только
при репликации генома в целом, т.е. в S-фазе клеточного цикла. Как же
происходит исправление ошибок в ДНК, когда клетка находится в состоянии покоя,
т.е. в ннтерфазе?
Несколько иной механизм исправления ошибок в ДНК
используется клеткой в интерфазе клеточного цикла, т.е. в
состоянии покоя. Этот механизм, названный эксцизионной репарацией,
вырезает измененные участки ДНК и заменяет их нормальными
участками. Он состоит из нескольких этапов. Первый из них -
"узнавание" поврежденного участка ДНК и производство
"надреза" в его окрестности. Эту операцию выполняет фермент эндону-
клеаза. Затем другой фермент - экзонуклеаза - "выщепляет"
модифицированное основание, а также значительное количество
смежных с ним "здоровых'* оснований. При этом в ДНК
образуется "брешь". Далее третий фермент - ДНК-полимераза заполняет
эту брешь, используя в качестве матрицы вторую
(неповрежденную) нить ДНК. Процесс эксцизионной репарации завершает
четвертый фермент - лигаза, который связывает вновь
синтезированный участок ДНК с оставшимся после выщепления концом
репарируемой нити ДНК. В отличие от репликативного синтеза,
который протекает только в определенный период жизни клетки,
а именно в S-фазе клеточного цикла, репаративный синтез может
53
происходить в любой момент в ответ на то или иное повреждение
ДНК. Поэтому репликативный является запрограммированным
(плановым), а репаративный - незапрограммированным
(внеплановым) синтезом ДНК (Виленчик. 1987).
* * *
Идея о том, что накопление мутаций может быть важным
механизмом старения, не нова {Ванюшин, Бердышев, 1977). Во многих
исследованиях убедительно демонстрировалось, что максимальная
продолжительность жизни, возможная для вида, коррелирует с
эффективностью репарации ДНК и, следовательно, с частотой
мутаций. В широко известной работе Р.В. Харта и Р.Б. Сетлоу (Hart,
Setlow, 1974) сравнительно изучалась активность систем репарации
ДНК у млекопитающих животных с различной видовой
продолжительностью жизни и была установлена прямая корреляция между
продолжительностью жизни и способностью клеток к репарации
ДНК. Из всех исследованных млекопитающих клетки человека
показали наивысшую активность систем репарации, что хорошо
согласовывалось с самой высокой продолжительностью жизни людей.
В ряде исследований было показано, что частота мутаций
повышается с возрастом (Виленчик, 1987) Одной из вероятных причин
этого является то обстоятельство, что с течением времени системы
репарации могут быть сами поражены мутациями, что ведет к
потере их активности. Это - с одной стороны. С другой стороны, в
организме по мере старения генерируется большее количество СР и
вероятность мутаций и других повреждений ДНК повышается. В
действительности, теория свободных радикалов и теория старения,
базирующаяся на данных о накоплении "ошибок" в ДНК, имеют
много общего, потому что ДНК является, наряду с мембранами, одной
из первых "жертв" СР.
Имеются ли способы понизить уровень поражений ДНК и улучшить ее
репарацию? Первым и наиболее простым шагом должна быть попытка избежать
повреждений из-за влияния окружающей среды, в частности курения сигарет и
чрезмерного пребывания на солнце. Те, кто имеют эти неприятные привычки,
должны по крайней мере пробовать переходить на сорта табака с низким
содержанием смолы, потому что смола - главный источник мутагенов в табачном
дыме. Если вы хотите провести время на солнце, применяйте качественные
солнцезащитные кремы. Благодаря оптическим свойствам земной атмосферы,
солнечный свет несет меньше разрушительных для ДНК ультрафиолетовых
(УФ) лучей утром и в конце дня, и это время является лучшим для загара.
Другим шагом для уменьшения повреждений ДНК является сохранение
вашей антиоксидантной защиты в хорошей форме (см. гл. 22, 23). Самый легкий
способ получить широкий спектр антиоксидантов в вашей диете это съедать
большое количество фруктов и овощей. Пищевые АО-добавки, такие как
витамины А, С, Е, бета-каротин, аминокислота цистеин, селен, липоевая кислота и
CoQIO могут обеспечить дополнительную защиту, особенно с возрастом, когда
собственная антиоксидантная система дает сбои.
54
В настоящее время, не существует никакого практического
способа усилить эффективность клеточной репарации ДНК. Эти
системы довольно сложны и используют много различных ферментов. У
каждого вида, система ремонта ДНК имеет специфический стандарт
качества, т.е. допускается определенный диапазон ошибок.
Положительной стороной неправильного или неполного ремонта ДНК
является возникновение мутаций, которые оказываются движущей
силой генетических изменений и благодаря этому - эволюции.
Увеличение скорости мутаций заставляет виды развиваться быстрее, но
при этом сокращается продолжительность жизни индивидуального
организма. Обратное утверждение также будет правильным.
Именно поэтому организмы с совершенной системой репарации, если
такое вообще возможно, вряд ли способны возникнуть путем
естественной эволюции.
Еще в 1967 г. Р. Мак Артур и Е. Уилсон (MacArtur, Wilson, 1967) постулиро-
иали существование двух типов стратегии эволюционного развития организмов:
ι и К. Стратегия г предполагает бурное размножение и короткую жизнь, а
стратегия К - низкий темп размножения и высокую продолжительность жизни
особей. В условиях резких и быстрых изменений окружающей среды высокий темп
размножения и короткая жизнь (г) позволят организмам, по всей вероятности,
быстрее приспособиться к такого рода изменениям. При стабильных условиях
организмы, очевидно, не должны включать форсированные механизмы
метаболизма и мутагенеза и могут позволить себе более низкий темп размножения
и большую продолжительность жизни, т.е. избрать стратегию К. По мнению
авторов, организмы (в зависимости от характера изменения условий
существования) могут включать либо К -» г, либо г —> К переход. В контексте
эволюционной логики, однако, эти переходы должны происходить случайно, а
выживают те особи, которые выбрали стратегию, более соответствующую данным
условиям среды.
Недавно В.П. Скулачев (1998, 1999) предложил интересную гипотезу οι-
носительно биохимического механизма реализации рассматриваемых
переходов. Окончательной формулировке этой гипотезы способствовало
неожиданное открытие А. Мура, которому удалось получить трансгенное растение
табака с повышенной активностью гена митохондриалъного белка теплового
шока с молекулярной массой 70 КД {mitochondrial heat shock protein 70,
сокращенно mHsp70) (Moor, 1998; цит. по: Скулачев, 1998). Основная функция
белков теплового шока состоит в исправлении (ренатурации) нарушений конфор-
мации белков, вызванных тепловой денатурацией. mHsp70 участвует в
формировании нужной структуры 15 белков, синтезированных в митохондриях, а
также импортирует внутрь митохондрий около 500 белков (среди них и сам
mHsp70), синтезированных в цитозоле. Так вот, у растений табака, которым
пересадили дополнительное количество генов mHsp70, втрое увеличилось
количество митохондрий и скорость их дыхания. В.П. Скулачев предположил,
что резкое увеличение количества mHsp70 ускоряет импорт и созревание ми-
тохондриальных белков, в том числе ферментов дыхания и окислительного
фосфорилирования, ответственных за синтез АТФ (по данным Мура, скорость
импорта белков митохондриями оказалась у трансгенного табака в три с
половиной раза выше, чем в норме).
Все эти события существенно повышают синтез АТФ, что улучшает
энергообеспечение клеток, а последнее, естественно, стимулирует функциональную
55
активность многих систем, а также рост, размножение и т.п. Но мы уже знаем,
что активация митохондриального дыхания сопряжена с повышенной
продукцией АФК (см. параграф "Свободные радикалы"). Поэтому повышенный
уровень mHsp70, вызывает, с одной стороны, улучшение энергообеспечения и,
следовательно, стимуляцию роста, а с другой - способствует сокращению
продолжительности жизни индивида из-за повышения продуктивности токсичных
форм кислорода.
Суперпродукция mHsp70 вполне может рассматриваться как
молекулярный механизм переключения живой системы с К-стратегии на г-стратегию. Это,
очевидно, дает организму ряд преимуществ при приспособлении к
изменившимся условиям существования, но достигается это ценой сокращения
продолжительности его жизни.
Человек имеет более эффективную систему ремонта ДНК, чем
большинство других видов. Сравнительно медленное старение людей,
очевидно, в значительной мере связано с относительно низким
уровнем мутаций в их соматических клетках. Как это ни парадоксально,
довольно хорошая система репарации служит, если следовать
естественному порядку вещей, препятствием на пути развития человека в
более долгоживущую разновидность. Впрочем нужно помнить, что
естественный порядок и так существенно нарушен почти полным
отсутствием в современном человеческом обществе такого важного
механизма эволюции, как естественный отбор. Остается, однако, надежда, что
люди найдут способ улучшить систему репарации ДНК с помощью
науки даже вопреки естественно сложившимся механизмам развития.
Рассмотрим некоторые многообещающие исследования в этом
направлении. Все ключевые ферменты репарации ДНК должны
использовать в качестве строительных блоков специальные предшественники
синтеза ДНК, называемые дезоксирибонуклеозидтрифосфатами
(дРНТФ). дРНТФ синтезируется из других предшественников - рибо-
нуклеозиддифосфатов - с помощью фермента рибонуклеотидредукта-
зы (РР). Если активность РР увеличена, уровень дРНТФ повышается,
и система репарации ДНК начинает работать быстрее. Исследования,
проведенные на мышах, показали, что вещества, способные
активировать РР, могут защищать животных от высоких доз радиации,
улучшая, очевидно, качество репарации ДНК (Pulatova, Sharygin, Todorov,
1999). Представляется, что подобный подход мог бы работать и на
замедление обычного старения.
2.3. НАКОПЛЕНИЕ ШЛАКОВ
К вероятностным механизмам старения следует отнести и
накопление разнообразных балластных веществ, т.е. шлаков, в
некоторых органах и тканях нашего организма. В процессе
жизнедеятельности клетки постоянно производят деградированные продукты
метаболизма. Большинство этих продуктов эффективно устраняется
из организма с выдыхаемым воздухом, мочой, фекалиями и потом.
56
С наибольшей легкостью удаляются небольшие растворимые
молекулы, подобные мочевине, электролитам и углекислому газу. Более
крупные молекулы, например поврежденные белки или
нуклеиновые кислоты, должны быть разрушены прежде, чем они будут
выведены из организма или сожжены, как топливо. В клетках
существуют специальные ферменты, которые разрушают вышедшие из
сферы нормального метаболизма макромолекулы до мономеров,
подлежащих реутилизации или дальнейшей деградации и выведению.
Весьма трудно устранить случайные молекулярные агрегаты,
возникшие в результате перекрестного сшивания макромолекул
(подробнее об этом явлении в следующем параграфе) и других
"незапланированных" реакций между клеточными структурами, особенно,
белками. Эти молекулярные агрегаты являются обычно довольно
крупными образованиями, которые плохо растворимы и трудно
разрушаются. При нормальном клеточном метаболизме поврежденные
или сшитые белки разрушаются внутриклеточными протеазами
еще до их возможного объединения в более крупные конгломераты.
Однако существуют данные, что низкая активность клеточных про-
геаз благоприятствует формированию больших совокупностей
"молекулярного лома" (Рязанов, 2001). Эффективная и "регулярная
уборка" необходима, чтобы избежать накопления огромных масс
"молекулярного мусора". Для переваривания крупных скоплений
жестких отходов в клетках существуют ограниченные мембранами
специальные органеллы (лизосомы), которые, в сущности, служат
хранилищем концентрированного набора всевозможных
ферментов, агрессивно разрушающих различные биосубстраты.
Несмотря на функционирование систем утилизации
метаболического "мусора", с возрастом в организме происходит аккумуляция
некоторых типов отходов. Если сравнить под микроскопом нейроны
мозга молодых и старых животных, то можно увидеть
существенные различия. В цитоплазме старых нейронов наблюдаются
странные желтоватые отложения. Это отложения липофусцина,
возрастного пигмента, который обнаружен во многих тканях старых
животных и людей (Уш, 1996). Возрастные пигменты являются группой
веществ, которые накапливаются с возрастом в различных тканях и,
по-видимому, не обладают никакой полезной функцией.
Большинство возрастных пигментов - плохо разрушаемые, нерастворимые
побочные продукты различных физиологических и патологических
процессов. Химически они не агрессивны, но занимают,
накапливаясь, большое пространство и в конечном счете начинают мешать
нормальным клеточным функциям.
Липофусцин - наиболее распространенный и хорошо изученный
возрастной пигмент, он постепенно накапливается в большинстве
тканей, но особенно интенсивно - в сердечной мышце, мышцах
скелета и мозге. В течение некоторого времени эти накопления
рассматривалось как относительно безвредный побочный продукт нор-
57
мального старения и некоторых болезней. Теперь, однако,
накопилось много доказательств того факта, что липофусцин вносит
существенный вклад в старение и возрастные болезни {Meites et al., 1984).
Накопление липофусцина тесно связано с другими факторами
старения - СР и перекрестными сшивками. Было найдено, что
липофусцин является конечным результатом СР-повреждений
биологических мембран. Липиды, которые служат структурной основой
клеточных мембран, очень легко повреждаются АФК в процессе
так называемого перекисного окисления липидов. Продукты ПОЛ
реагируют с другими липидами и белками, формируя большие
нерастворимые кластеры. Клетка стремится разрушить такие
кластеры, используя протеазы и другие ферменты лизосом, но
вероятность успеха этих действий не всегда высока. Недеградированные
агрегаты сшитого "молекулярного мусора" становятся в конечном
счете отложениями липофусцина. По-видимому, в наибольшей
степени липофусцин накапливается в клетках, которые сжигают
большое количество топлива и редко или никогда не вступают в процесс
деления. Интенсивная энергопродукция, естественно, сопряжена с
выходом большого количества АФК, что и способствует
формированию и накоплению пигмента. Не совсем ясно, однако, почему
интенсивно делящиеся клетки почти не накапливают липофусцин.
Одно из возможных объяснений состоит в том, что в таких клетках
высока активность протеаз и лизосом.
Существует три главных типа клеток, которые не делятся - ней
роны (клетки мозга), клетки сердечной мышцы и клетки костно-
мышечной системы. Накопление липофусцина, по всей
вероятности, вносит вклад в возрастное ухудшение функционирования этих
тканей. Все клеточные процессы зависят как от нормального
поступления питательных веществ и других молекул в клетку, так и от
выведения из нее продуктов метаболизма и веществ,
продуцируемых на экспорт во внутреннюю среду организма. Липофусцин
может повреждать ткани, механически затрудняя нормальные потоки,
сокращая поставку топливных и структурных единиц и замедляя
устранение отходов. Это поразительно, но количество липофусцина
в клетках может превьппать половину их объема - этого более чем
достаточно для значительного нарушения их нормальных функций
Существует положительная корреляция между тяжестью слабоумия
или застойной сердечной недостаточностью и количеством
липофусцина в нейронах или клетках сердечной мышцы (Уш, 1995).
В одном из исследований, материал, подобный липофусцину
был добавлен к культуре человеческих фибробластов, взятых у
молодого донора. При использовании соответствующих процедур
легко добиться, чтобы фибробласты захватывали большее количество
пигмента. Такое принудительное накопление липофусцина быстро
блокировало клеточные деления и заставило фибробласты вести
себя так, как если бы они были взяты у очень старого человека. Это
58
прямое доказательство, что накопление липофусцина отнюдь не
безвредно, а действительно вносит вклад в возрастные патологии
(Yin, 1996).
Многие факторы могут воздействовать на скорость накопления
1ипофусцина. Любой процесс, который ускоряет образование СР
или повреждает антиоксидантную защиту организма, способен
форсировать рост отложений липофусцина. Среди таких факторов
преобладает стресс. Он повышает скорость метаболизма в нервной
системе, сердечной мышце и мышцах скелета, что сопряжено с
повышением уровня АФК. Стресс также увеличивает ПОЛ, которое
становится первым шагом в формировании липофусцина. Когда крысы
были подвергнуты 24 или 48 часовому иммобилизационному
стрессу, количество липофусцина в их нейронах увеличилось
соответственно на 29 и 38% (Burnet, 1983). Другим важным фактором,
влияющим на скорость накопления липофусцина, является активность
клеточных протеаз. Молодые крысы, получавшие ингибиторы про-
теаз, обнаруживали значительные отложения липофусцина.
Цероид - возрастной пигмент во многом похожий на
липофусцин, хотя и не столь часто наблюдаемый в клетках человека.
Подобно липофусцину, цероид является конечным продуктом ПОЛ.
Отложения цероида, по видимому, тесно связаны с дефицитом витамина
Е, хотя явный дефицит этого витамина встречается не часто.
Некоторые желудочно-кишечные заболевания и болезни печени,
нарушающие всасывание жира и, следовательно, жирорастворимых
витаминов, могут вызывать дефицит витамина Ε и накопление церои-
ца (Yin, 1996).
Другой возможной причиной крайнего дефицита витамина Ε может
быть присутствие большого количества ненасыщенных жиров в пище.
Ненасыщенные жиры, которые являются основным типом жира в
растительных маслах, легко окисляются и могут быстро разрушать витамин Е. При
высокотемпературной обработке пищи (особенно при жарке) содержание
витамина Ε в растительных маслах истощается. В сыром растительном
масле изначально содержится большое количество витамина Е. Регулярное
потребление ненасыщенного жира, подвергнутого высокотемпературной
обработке, может не только ускорять накопление цероидов, но также угнетать
иммунную систему и инициировать развитие некоторых типов рака. Если вы
потребляете большое количество пищи, приготовленной аналогичным
образом, имеет смысл употреблять дополнительное количество витамина Ε
(Modern Nutrition in Health and Disease, 1994).
Еще один пигмент, амилоид, иногда обнаруживается в
различных тканях вполне здоровых пожилых людей. Более часто, однако,
накопление амилоида связано с хроническими инфекциями или
заболеваниями воспалительного характера. Механизм накопления
амилоида все еще не вполне ясен. Было обнаружено, что в
отложениях амилоида содержатся иммуноглобулины (белки иммунного
отпета), перекрестно сшитые с другими белками, которые атакуются
иммуноглобулинами при аутоиммунных реакциях. Сфера действия
59
аутоиммунных реакций с возрастом расширяется, что может
вносить вклад и в накопление амилоидов у некоторых пожилых людей.
Большие отложения амилоида могут вызывать угрожающие жизни
состояния, связанные с сердечной или почечной недостаточностью
(Ravid, 1967).
Наиболее ярким патологическим проявлением возрастного
накопления амилоида является болезнь Алъцгеймера, впервые описанная
баварским психиатром Алоисом Альцгеймером в 1907 г. Симптомы
этой болезни - потеря памяти, рассудка и эмоциональной
стабильности - проявляются постепенно и неумолимо, приводя больного
через 4-12 лет к смерти. Развитие болезни характеризуется
накоплением так называемых сенильных бляшек и нейрофибриллярных
клубков, в большом количестве возникающих в коре головного
мозга, гиппокампе, миндалине и других областях мозга, важных для
когнитивных (познавательных) функций (Katzman, 1986).
Основной субстанцией этих аномальных образований является
белковый фрагмент, состоящий примерно из 40 аминокислотных остатков, под
названием - амилоидный §-белок. Этот фрагмент образуется в результате
ферментативного расщепления белка-предшественника βΑΌΡ (от англ. -
amyloid precursor protein) гораздо большего размера (695 аминокислотных
остатков), который кодируется геном, локализованным у человека в
хромосоме 21 (Selkoe, 1991). Предполагаемая структура βΑΡΡ включает участок
(от 625 до 648 аминокислотного остатка), который, по-видимому, служит
"якорем" удерживающим всю молекулу в клеточной мембране. 12
аминокислот амилоидного β-белка также находятся в мембране, а 28 аминокислотных
его остатков располагаются снаружи от поверхности клеточной мембраны.
Было неясно, каким образом весь амилоидный β-бслок (40 аминокислот),
который в норме прочно прикрепляет молекулу βΑΡΡ к клеточной
мембране (рис. 5(1) (Селко, 1992), целиком появляется во внеклеточном
пространстве в виде отложений амилоида? Как протеолитические ферменты,
расщепляющие белок-предшественник, получают доступ к его трансмембранной
части? Решение этого важного вопроса может дать в руки фармакологов
информацию, которая существенно облегчит создание средств блокирования
амилоидоза при болезни Альцгеймера. Предполагается, что избыточное
производство βΑΡΡ способствует выделению амилоидного β-белка. Обычно
с βΑΡΡ взаимодействует только фермент, расщепляющий β-белок примерно
посередине (2). Полный интактный β-фрагмент, по-видимому, может
высвободиться, если βΑΡΡ будет расщепляться другими ферментами у С- и Ν-κοη-
цов β-фрагмента (З). Эти фрагменты могут затем выводиться из клеток и
накапливаться в других тканях (и прежде всего в мозгу). Синдром, называемый
болезнью Альцгеймера, существует в различных формах, и причиной его
могут быть и различные генетические нарушения (см. параграф
"Генетические аспекты старения")» однако все эти нарушения проявляются через
некий общий механизм, приводящий к усиленному образованию и отложению
амилоидного β-белка (Selkoe et al.f 1988).
Уже упоминавшееся выше возможное повышение синтеза βΑΡΡ, как
полагают, может быть связано с мутацией в регуляторном участке гена,
кодирующего структуру этого белка (Goate et al., 1991). Действительно, при синдроме
Дауна, который по многим симптомам напоминает болезнь Альцгеймера,
избыточный синтез βΑΡΡ твердо установлен (Селко, 1992).
60
(труктура молекулы
ПРЕДШЕСТВЕННИКА АМИЛОИДНОГО β БЕПКА
РЕГУЛЯТОР ПРОТЕИНАЗ
Ρ УЧАСТОК
(АМИЛОИДНЫЙ.
Г?
НОРМАЛЬНЫЙ ФЕРМЕНТАТИВНЫЙ
ПРОЦЕССИНГ
/>
МЕМБРАНА С-КОНКЦ
АЛЬТЕРНАТИВНЫЙ ФЕРМЕНТ A'J ИВНЫЙ
процга инг
ОСВОБОЖДЧЕ1СЯ
интлктный
β-ΦΡΛΓΜΕΙΠ
/П
Рис. 5. Гипотетическая схема процес-
синга предшественника амилоидного
β-белка и образования амилоидного
β-белка в норме и патологии
(объяснение в тексте) (Селко, 1992)
Наряду с выяснением биохимических и генетических аспектов
развития болезни Альцгеймера важны эпидемиологические
исследования, которые могут выявить факторы окружающей среды,
обусловливающие предрасположение индивида к заболеванию или
ускоряющие его развитие. В настоящее время нет весомых
доказательств, что питание, уровень образования, профессия или
эмоциональное состояние влияют на возникновение и развитие болезни
Альцгеймера.
Можно ли замедлить и даже полностью прекратить накопление
возрастных пигментов? Первый шаг, очевидно, должен состоять в
улучшении АО защиты и сопротивляемости стрессорам (см. гл. 24,
29). Витамин Ε и липоевая кислота, т.е. жирорастворимые АО,
способны блокировать ПОЛ, что делает их особенно важными в замед-
61
лении накопления липофусцина и цероида. Повышение
толерантности к глюкозе (см. следующий параграф этой главы) должно
уменьшить скорость реакций перекрестного сшивания в нейронах и,
следовательно, замедлять возрастное накопление пигментов.
Некоторые нейротропные питательные вещества способны уменьшать
отложения липофусцина и цероида в мозге. В частности, диметила-
миноэтанол (ДМАЭ) уменьшал отложения липофусцина в
нейронах нескольких видов животных, включая грызунов, свиней и
обезьян. ДМАЭ - пищевой продукт, обнаруженный в небольших
количествах в рыбе. Он также доступен в чистом виде в качестве пищевой
добавки. ДМАЭ улучшал такие аспекты интеллектуальных
способностей, как память, концентрацию внимания и обучаемость. В
некоторых исследованиях ДМАЭ увеличивал продолжительность жизни
грызунов. Все еще неясно, как нейротропные препараты
сокращают отложения возрастных пигментов в нейронах. Одна из
возможностей заключается в том, что они активируют протеазы и лизосо-
мы, а также увеличивают количество внутриклеточной воды, что
способствует ускоренному выведению шлаков (Modern Nutrition in
Health and Disease, 1994).
2.4. ГЛЮКОЗА И СТАРЕНИЕ
Механизмы старения, так или иначе связанные с изменениями
концентрации глюкозы во внутренней среде организма, следует
отнести как к вероятностным, так и отчасти к системным механизмам,
включенным в сложный процесс нейроэндокринной регуляции
метаболизма. Начнем с вероятностного механизма.
Неферментативное гликозшшровапие биополимеров.
Свободные радикалы - не единственная причина, ускоряющая старение в
результате случайного повреждения жизненных структур
организма. Многие активные химические соединения могут делать то же
самое. Ацетальдегид, образующийся в печени после всасывания
алкоголя, может повреждать почти любую клеточную структуру, то же
могут делать и многие вещества, содержащиеся в дыме сигарет или
даже в пережаренной пище. Мы можем легко (или не очень легко)
избегать алкоголя, курения или питания пережаренной пищей, но
существует вещество с потенциями разрушителя, без которого мы
жить не можем - это глюкоза.
Глюкоза - это наиболее распространенный углевод пищи; это -
главная структурная единица крахмала, входящая также в состав
сахарозы (столового сахара). Большинство углеводов, которые мы
получаем с хлебом, хлебными злаками, макаронами и многими друга
ми продовольственными изделиями, содержат глюкозу. Уровень
глюкозы в кровеносной системе один из наиболее важных
физиологических параметров, потому что глюкоза это основное топливо для
62
центральной нервной системы. Если уровень глюкозы в крови
падает ниже определенного значения и держится на этом уровне в
течение достаточно длительного периода времени, человек может
потерять сознание, впасть в кому и даже умереть. Высокое содержание
глюкозы в крови, наблюдаемое у диабетиков, так же вредно, хотя и
менее опасно в течение не слишком продолжительного времени.
Чтобы избегать опасного колебания уровня глюкозы, организм
имеет сложную систему для его поддержания в крови в пределах
соответствующего диапазона. Некоторый запас глюкозы всегда
хранится в печени в форме гликогена (полимер, подобный крахмалу).
Вдобавок ко всему, печень, если необходимо, может синтезировать
глюкозу из белка (глюконеогенез). Во время голодания, организм
постепенно перерабатывает белки мышц ради обеспечения
центральной нервной системы глюкозой.
Таким образом, получаем ли мы глюкозу с пищей или нет, она
всегда присутствует в нашей кровеносной системе. К сожалению,
кроме того, что глюкоза служит жизненно необходимым
клеточным топливом, она является также веществом, которое может
вызывать повреждения, беспорядочно реагируя с белками и ДНК.
Этот процесс называется неферментативным гликозилированием,
или реакцией Мейлларда. Наглядный пример этой реакции -
подрумянивание и затвердение корки пирога при выпечке. Подобный
процесс происходит и в организме, но с более низкой скоростью и с
многочисленными отрицательными последствиями. В частности, глико-
зилированные ферменты будут не в состоянии работать так, как они
должны работать; другие гликозилированные белки потеряют свою
форму и станут нерастворимыми или нестабильными. Когда
присоединение глюкозы к белку катализирует фермент, эта реакция идет
с большой скоростью, является высокоспецифичной в отношении
участвующих в ней молекул и преследует определенную
биологическую цель. Напротив, неферментативное связывание глюкозы
происходит случайным образом в любом из возможных участков каких-
либо полипептидных или полинуклеотидных цепей (Bucala et al.,
1984; Vlassara et al., 1985). Химическая реакция глюкозы с
определенными белками "запускает" каскад химических превращений, в
результате которых в организме образуются и постепенно
накапливаются необратимые "сшивки" между соседними белковыми
молекулами. По-видимому, основная масса "сшивок" появляется именно
между теми белками, которые служат структурными элементами,
"каркасом" тканей и органов, что приводит к потере эластичности и
затвердеванию этих объектов, а в случае нуклеиновых кислот
может постепенно приводить и к повреждению ДНК (Bucala et al.,
1984; Brownlee et al., 1986).
Цепочка этих реакций начинается с взаимодействия
альдегидной группы (СНО) глюкозы и аминогруппы (NH2) белка, в
результате чего образуется так называемое шиффово основание (рис. 6). Это
63
Η
Η
h
К О К К К
К
к
ft
■?*
CN
I
К
е
е
2С К Ж я
К К О К О О О ь^
I I II Ι Ι Ι Ι fcg
ι ι ι ι ι &!
κ ο κ as a ^
S Μ 3S δ 5 Й
О К О О О ой2
ι ι ι ι ι §| ε
ι ι ι ι ι ι t§ §
К Я О Я К К log
/ 1 Ι
κ κ κ κ κ S к
% ? ? °\ °\ ? III
/0_rrrrT_K 2
II
основание нестабильно и быстро перестраивается в более
стабильное (но еще способное к обратимому превращению) соединение, из-
нестное под названием "продукт Амадори". Со временем продукты
Амадори превращаются в стабильные структуры конечных
продуктов глубокого гликозилирования (КПГГ), которые способны
образовывать сшивки с соседними белками. Структура одной из таких
сшивок с участием глюкозы уже известна. Это 2-фуранил-4(5)-(2фу-
ранил)-1Н-имидазол (ФФИ). Это соединение сначала было
синтезировано искусственно (из смеси аминокислоты лизина, белка
альбумина и глюкозы), а позже было найдено в организме (см. рис. 6).
Данные о неферментативных превращениях глюкозы привели
\. Серами к предположению, что высокое содержание глюкозы у
сольных тяжелой формой сахарного диабета способствует
развитию этого заболевания в результате накопления КПГГ, поскольку
скорость химических реакций пропорциональна концентрации
реагентов, в данном случае глюкозы {Серами и др., 1987). Давно
замечено, что проявления диабета во многих органах и тканях часто
представляют собой не что иное, как ускоренное старение.
Старческая катаракта, ограниченность подвижности суставов,
атеросклероз - характерны для пожилого возраста, но у больных диабетом
гакие нарушения развиваются раньше. При катаракте ковалент-
ное связывание глюкозы с белком так меняет конформацию
последнего, что скрытые ранее сульфгидрильные группы
оказываются доступными для других сульфгидрильных групп, в результате
чего образуются дисульфидные связи и происходит агрегация
молекул белка. Далее возможно образование ФФИ и других
окрашенных сшивок. Хрусталик при этом меняет свой цвет и мутнеет.
Накопление КПГГ в сухожилиях, коже, хрящах и других
соединительных тканях, а также в стенках кровеносных сосудов
способствует образованию сшивок между молекулами коллагена, что
приводит к большей "жесткости" этих тканей, ограничению их
функций и развитию атеросклероза (Monnier et al., 1984; Brownlee et al.,
1986). Наряду с этим нарушение периферического
кровообращения в тканях приводит к плохому заживлению ран, повреждению
нервов и формированию язв.
При диабете гликозилирование и перекрестное сшивание имеют
особенно высокую скорость, но эти реакции происходят также и у
здоровых людей. Более того, по мере старения, этот процесс
ускоряется по нескольким причинам. Во-первых, из-за уменьшения с
возрастом толерантности к глюкозе (см. ниже), что означает
вообще более высокий уровень глюкозы в крови, особенно после
приема пищи, и, во-вторых, из-за того, что перекрестные сшивки
инициируются СР, чей уровень также увеличивается с возрастом
Глюкоза - только один из главных агентов, осуществляющих
перекрестные сшивки. Сигаретный дым, УФ-радиация, тяжелые металлы, в частности
ртуть и свинец, перекиси и альдегиды (например, ацетальдегид, производимый
65
печенью при переработке алкоголя), - все они являются мощными
сшивающими агентами. СР инициируют и ускоряют, по существу, все типы перекрестных
сшивок, а также сами непосредственно участвуют в этом процесса. В
различных тканях перекрестные сшивки могут вызываться различными факторами.
Например, в коже они обычно происходит из-за облучения на солнце, в легких
курильщиков - из-за реактивных веществ в смоле табака. Гликозилирование и
свободные радикалы в значительной степени ответственны за сшивки в почках
и стенах артериальных сосудов.
Помимо рассмотренных симптомов старения и осложнений
диабета, следует указать на некоторые другие потенциально вредные
для организма сшивки. ДНК также содержит аминогруппы и,
следовательно, может аккумулировать КПГГ. Таким образом, КПГГ
могут способствовать различным хромосомным нарушениям,
которые, как известно, при старении происходят чаще и мешают
процессам репарации, репликации и транскрипции ДНК (Bucala et al., 1984).
Неферментативное гликозилирование может также вызывать
мутации, затрагивающие функции иммунной системы или приводящие к
некоторым видам рака (Серами и др., 1987).
Во II части этой книги энергетические аспекты стресс-реакции
будут рассмотрены подробно. Здесь же мы лишь подчеркнем, что
при стрессе основным направлением биохимических процессов
является мобилизация энергетических ресурсов за счет имеющихся в
организме резервов (гликоген, липиды, некоторые виды белков) для
обеспечения сопротивления стрессорам. Активация распада
гликогена под действием адреналина, процессов глюконеогенеза под
действием глюкокортикоидов, вовлечение липидов в энергетический
метаболизм приводят к значительному повышению концентрации
глюкозы как во внутренней среде организма, так и внутри клеток.
Чем чаще в течение жизни организм будет активировать
интенсивные и длительные состояния стресса, тем чаще (хотя бы временно)
будет повышаться концентрация глюкозы в крови (гипергликемия),
тем выше будет вероятность неферментативного гликозилирования
информационных макромолекул, образование шиффовых
оснований, продуктов Амадори и КПГГ.
Следовательно, стресс в течение жизни будет вносить
ощутимый вклад в ускоренное старение организма, активируя наряду с
другими и механизм неферментативного гликозилирования
биополимеров.
Что можно сделать, чтобы минимизировать или полиостью изменить
процесс перекрестного сшивания? Многие вещи, которые можно сделать, подобны
тем, что мы предпринимаем для сокращения повреждений ДНК и борьбы со
свободными радикалами. В частности, сократить время облучения на солнце,
уменьшить потребления алкоголя и отказать от курения. Антиоксиданты
снижают скорость большинства типов перекрестного сшивания, нейтрализуя СР.
Витамины С, Е, аминокислота цистеин, каратиноиды, флавоноиды,
полифенолы, антоцианины, селен, липоевая кислота и другие антиоксиданты обладают
способностью предотвращать эти нежелательные процессы.
66
В некоторых исследованиях, липоевая кислота, пищевая добавка с
мощными антиоксидантными свойствами, успешно использовалась для лечения
диабетической невропатии. Причины диабетической невропатии сложны и
включают гликозилирование, ПОЛ и перекрестное сшивание. Недавно было показано,
что 600 мг липоевой кислоты в день в течение трех недель более эффективны
для ослабления боли, жжения и других признаков невропатии, чем
успокаивающие лекарства. Имеются свидетельства, что липоевая кислота может успешно
применяться при многих других дегенеративных болезнях нервной системы
(Modern Nutrition in Health and Disease, 1994}
В отличие от мутаций, которые практически невозможно
изменить, вред от перекрестных сшивок клетки можно, по крайней мере
частично, устранить путем синтеза новых структур взамен тех,
которые были поражены сшивками, активируя экспрессию
соответствующих генов. Однако удаление жестких, плохо растворимых,
агрегированных и перекрестно сшитых молекул трудно осуществимо и
требует как времени, так и энергии. С возрастом процесс
перекрестного сшивания начинает опережать процесс реставрации
пораженных структур. Если скорость перекрестного сшивания удалось бы
снизить, это позволило бы организму постепенно заменить
поврежденные и дисфункциональные структуры, устранив таким образом
некоторые признаки старения. Даже в устранении морщин кожи
можно достигнуть видимых улучшений. К сожалению, скорость
замены сшитых частей очень мала, улучшение занимает месяцы или
даже годы.
Тем не менее существует способ, который может ускорить устранение
сшитых молекулярных обломков. Он основан на использовании препаратов
активных протеаз-ферментов, расщепляющих белки. Эти ферменты способны
разбивать большие сшитые агрегаты коллагенов и других белков на мелкие
фрагменты, которые легче удаляются из организма. Некоторые из таких про-
теаз содержатся в экзотических фруктах, например бромелаин в ананасах и nw-
паин в плодах папай. Протеазы могут использоваться как пищеварительное
средство, которое особенно необходимо в пожилом возрасте, когда
уменьшается производство собственных пищеварительных ферментов. Наряду с этим
значительные количества этих ферментов находят путь в кровяное русло и таким
образом к большинству тканей организма. Бромелаин, как было показано,
способен проникать в кровяную систему, помогая удалению сшивок во многих
тканях. Для лучшего усвоения бромелаин следует употреблять между приемами
пищи. Препараты протеаз, однако, следует использовать с осторожностью
(особенно, при приеме на пустой желудок), потому что они могут усугублять
любое желудочно-кишечное нарушение, например язвы, гастрит или рак.
Папайи используется в некоторых препаратах для омоложения кожи, где он
работает, разрушая сшитые коллагены
Возрастное снижение толерантности тканей к глюкозе.
Поддержание оптимальной толерантности (восприимчивости) тканей к
углеводам является крайне важным по целому раду причин. Одну из
них-уменьшение уровня гликозилирования и перекрестного
сшивания - мы уже рассмотрели выше. Вторая, по-видимому, еще более
важная - возрастное снижение толерантности к глюкозе и, следова-
67
тельно, повышение ее уровня в крови, так называемая
гипергликемия. Многие факторы вносят свой вклад в такое снижение, включая
стресс и, возможно, центральные часы старения, расположенные в
гипоталамусе (об этом ниже).
Как будет показано в следующих главах, возрастная
гипергликемия вносит существенный вклад в развитие многих возрастных
болезней. Поэтому мы постараемся осветить эти проблемы с
возможной полнотой.
Проникновение глюкозы в ткани невозможно рассматривать
вне связи с инсулином - гормоном, который снижает уровень
глюкозы двумя путями. Первый - инсулин предотвращает ее
высвобождение из печени в результате распада гликогена, второй -
усиливает поглощение глюкозы мышечной и жировой тканью. В
мышцах из неиспользованной в метаболизме глюкозы образуется
гликоген, а в жировых клетках она запасается в виде капелек жира,
т.е. в форме ее длительного хранения. По мере снижения
концентрации глюкозы в крови до нормального уровня, продукция
инсулина В-клетками поджелудочной железы прекращается. НЪ
вернемся к гипергликемии, которая, как правило, имеет место при
различных типах диабета.
Гипергликемия создает осмотический дисбаланс, который
вызывает отток воды из тканей в кровь. В почках в свою очередь
усиливается выделение с мочой воды и солей. Дальнейшее повышение
концентрации глюкозы в крови может привести к высокой
степени обезвоживания и потери солей, что чревато для организма
комой или даже летальным исходом. При менее острых формах
гипергликемии, характерных для сахарного диабета, возможны
сердечные приступы, паралич, потеря зрения, почечная
недостаточность и гангрена конечностей. Гипергликемия может возникать
либо при инсулиновой недостаточности, либо при пониженной
чувствительности тканей к инсулину. Инсулиновая недостаточность
(диабет типа I), обычно наблюдаются у детей и подростков при
поражении В-клеток аутоиммунными реакциями (Maclaren, 1988;
Todd, 1990). У пожилых людей чаще имеет место независимый от
инсулина диабет (II типа), характеризуемый пониженной
чувствительностью тканей к инсулину.
Проникновение глюкозы внутрь клетки является довольно
сложным процессом. Ее переносит туда специальный белок,
погруженный в клеточную мембрану. К настоящему времени выявлено
пять молекулярных форм переносчика глюкозы, каждая из них
приспособлена к метаболическим нуждам той ткани, в которой
содержится, поскольку разные ткани различаются по потребности в
глюкозе. Все эти белки имеют много общего в структуре и функции.
Каждый из них представляет собой одну полипептидную цепь из
примерно 500 аминокислотных остатков. Им всем присуща
складчатая организация из 12 сегментов, которые, располагаясь поперек
68
мембраны, по-видимому, и обусловливают формирование в ней пор,
и глюкоза проникает сквозь них внутрь клетки. При этом
гидрофильная часть каждого сегмента локализуется на внешней стороне
мембраны, а гидрофобная - на внутренней. В этих
трансмембранных сегментах содержится много аминокислотных остатков,
имеющих гидроксильные (ОН) и карбамидные (CONH2) группы,
способные образовывать водородные связи с гидроксильными группами
глюкозы (Mueckler et al., 1985).
Белок-переносчик может существовать в двух формах: одна
из них связывает глюкозу на внеклеточной стороне мембраны,
другая - на внутриклеточной. Проникновение глюкозы в клетку
происходит в четыре этапа. Вначале глюкоза присоединяется к
наружному, связывающему участку. Затем конформация этого
комплекса изменяется так, чтобы глюкоза оказалась в
связывающем участке, обращенном внутрь клетки. На третьем этапе
глюкоза отделяется от белка-переносчика, переходя в цитоплазму
клетки. И наконец, освободившийся переносчик принимает кон-
формацию, при которой участок, связывающий глюкозу,
обращен наружу {рис. 7). Таким образом, белок-переносчик можно
рассматривать как конформационный осциллятор, способный
перемещать место связывания глюкозы с одной стороны мембраны
на другую. Так, в мембране эритроцита в отсутствие глюкозы
конформация белка-переносчика меняется 100 раз в секунду.
Когда же с переносчиком связывается глюкоза, скорость этих
переходов еще выше - примерно 900 раз в секунду (Appleman et al.,
1989). Очевидно, глюкоза ускоряет осцилляцию, снижая
энергетический барьер между двумя конформациями (см. рис. 7).
Переносчики глюкозы обозначаются GluT и пронумерованы в порядке их
открытия. GluTl содержится в эндотелиальных клетках сосудов, образующих
гемато-энцефалический барьер. Этот переносчик обеспечивает стабильный
поток глюкозы, необходимый мозгу.
GluT2 обнаружен в органах, выделяющих глюкозу в кровь, — кишечнике,
печени и почках, а также в В-клетках поджелудочной железы, секретирующей
инсулин. Для насыщения GluT2 необходима высокая концентрация глюкозы, а
это означает, что интенсивность переноса глюкозы будет пропорциональна ее
содержанию в крови. Следовательно, после приема пищи изменения уровня
глюкозы будут эффективно передаваться в печень и В-клеткам. Известно, что
повышение концентрации глюкозы в В-клетках стимулирует синтез
предшественника инсулина путем активации экспрессии гена инсулина и сплайсинга
(созревания) пре-мРНК инсулина (Wang et al., 1997).
GluT3 содержится в нервных клетках мозга. Он обладает большим, чем
GluTl, сродством с глюкозой, и гарантирует наряду с последним снабжение
мозга жизненно важным питательным веществом.
GluT4 является главным переносчиком глюкозы в мышечных и жировых
клетках, поглощающих ее в больших количествах и перерабатывающих
глюкозу с образованием энергоносителей. Этот переносчик обладает необычной спо-
обностью перемещаться из внутренних компартментов клетки к ее
поверхности и обратно.
69
ГЛЮКОЗА
ВНЕКЛЕТОЧНАЯ
СРЕДА
ЦИТОПЛАЗМА
Рис. 7. Схема переноса глюкозы в
клетку при помощи
белка-переносчика (объяснение в тексте) (Лин-
хард и др., 1992)
GhiT5 встречается главным
образом в тонком кишечнике (Gould et
al., 1990; Mueckler, 1990).
Влияние инсулина на
транспорт глюкозы огромно.
Наиболее выраженный ответ на
гормон наблюдается у GluT4.
Внутри клеток всегда
поддерживается резерв этого переносчика,
но при воздействии инсулина
значительная часть GluT4
перебрасывается к мембране. Так,
до стимуляции инсулином на
поверхности клеток находится
лишь 1% молекул переносчика,
а после - 40%. Когда
гипергликемия проходит, и
соответственно снижается секреция
инсулина, эти молекулы
возвращаются внутрь клетки к месту
локализации (Solt et al., 1991).
Работа этого механизма в
настоящее время достаточно
хорошо известна (Линхард и
ДР-, 1992).
Локализованный в клеточной
мембране рецептор связывает
инсулин (рис. 8 (1)). В результате этого
конформация внутренней части
рецептора изменяется таким образом,
что он обретает способность фосфо-
рилировать за счет АТФ остаток
тирозина в определенном участке
соответствующего белка-мишени с
образованием АДФ (2).
Предполагается, что такой фосфорилированной
белок служит сигналом для
перераспределения белка-переносчика
глюкозы (3) путем передвижения
содержащих его везикул (мембранных
пузырьков) к клеточной мембране (4).
Пузырьки сливаются с мембраной
(5), обеспечивая перенос глюкозы
(6). Молекулы переносчика
возвращаются во внутриклеточное
пространство, когда небольшие
пузырьки, образовавшиеся после впячива-
ния (7) мембраны и ее отделения (8),
70
сливаются с более крупными везикулами, называемыми эндосомами (9). В нго
переносчики переходят в трубчатые выросты, которые превращаются затем ι
новые пузырьки (10). Пока в среде присутствует инсулин, процесс слияния
пузырьков с мембранами продолжается, но при снижении концентрации гормона
цикл прерывается, и переносчик и глюкозы накапливаются в эндосомах.
Предложенный механизм относится к GluT4, который содержится в
мышечных и жировых клетках. В других клетках их переносчики находятся
главным образом на поверхности клеток независимо от того, имеется в среде
инсулин или нет.
Одним га самых ранних проявлений диабета II типа (типичной
возрастной болезни) является нечувствительность к инсулину, что
проявляется в неспособности мышц, жировой ткани и печени
адекватно реагировать на повышенное содержание этого гормона в
крови. Поджелудочная железа отвечает на это усиленной секрецией
инсулина (Тодоров, 1965; De Fronzo, 1979). Далее, по мере развития
болезни или старения организма, эти неадекватные реакции
усугубляются, что постепенно приводит уже к снижению секреции инсулина.
Такое поведение В-клеток при развитии диабета II (и, возможно, в
процессе старения) объясняется, как показали соответствующие
исследования (Линхард и др., 1992), понижением содержания GluT2 в
мембранах этих клеток, что нарушает регуляцию синтеза и
секреции инсулина в ответ на изменение концентрации глюкозы в крови
Тем не менее основную причину прогрессирующей с возрастом
нечувствительности к инсулину и падения толерантности к глюкозе
следует, очевидно, искать в скелетных мышцах, которые
утилизируют 80% глюкозы после приема углеводной пищи. Наиболее простое
объяснение этого могло бы состоять в значительном снижении
количества GluT4 в мышечных клетках. Однако исследование
содержания GluT4 в мышцах здоровых и больных диабетом II людей не
выявило каких-либо значимых различий в этом отношении
(Mueckler, 1990). Возможно, причину следует искать в ухудшении
работы выше описанной (см. рис. 8) сложной машины, регулирующей
проникновение глюкозы в мышечные клетки. Например,
переносчик направляется в неподходящий субклеточный компартмент, из
которого он не может быть переброшен на поверхность клетки из-
за стерических препятствий, вызванных возрастным накоплением
метаболических шлаков (см. предыдущий параграф этой главы).
Нарушение может произойти также и на каком-то другом этапе
сложного процесса стимуляции переброски GluT4, например в
результате повреждения рецептора инсулина или какого-то другого
белка всей этой сложнейшей транспортной системы, повреждений,
которые вызваны атаками АФК и многими другими факторами.
Однако большего внимания, как нам кажется, заслуживает
другое объяснение. Как будет показано в следующей части этой книги,
состояние стресса ограничивает потребление глюкозы мышцами,
переключая их энергетику на использование жирных кислот с тем,
72
чтобы снять конкуренцию за глюкозу между мышцами и головным
мозгом, для которого глюкоза служит единственно возможным
энергоносителем, способным преодолеть гемато-энцефалический
барьер. Очевидно, с возрастом эти проявления повторяющихся
стресс-реакций будут аккумулироваться, что и станет постепенно
понижать глюкозную толерантность мышц и их чувствительность к
инсулину.
Нельзя также исключить и существование генетической
программы, обусловливающей возникновение всех этих явлений.
Сахарный диабет II типа как таковой является неселективным
признаком, и поэтому в течение многих поколений, когда это
заболевания вообще было неизвестно, оно должно было исчезнуть
вследствие элиминации носителей диабетической наследственности. В
том, что этого не произошло, можно усмотреть наличие
преимуществ, которые могла дать некоторая диабетическая
гипергликемия в условиях постоянной нехватки пищи и высокой физической
активности. Эти факторы, очевидно, способствовали продлению
жизни больных диабетом, нередко до периода, когда
обеспечивалось достаточное воспроизведение потомства. Резервное
накопление жира, связанное с диабетоподобными нарушениями
метаболизма, давало ощутимые преимущества в жизни доисторического
человека, связанной с эпизодическим поступлением пищи, и не
играло какой-либо отрицательной биологической роли, поскольку
средняя длительность жизни еще 400-500 лет тому назад не
превышала 30 лет. Приведенные соображения позволяют предположить,
что у каждого современного человека (да и у млекопитающих
вообще) существует универсальная генетическая
предрасположенность к диабетическим нарушениям II типа и что эволюционный
отбор произошел по признаку способности к накоплению жира. В
пользу этих соображений свидетельствуют и данные о том, что
возрастное снижение толерантности к углеводам начинается в
раннем детстве {Pickens et al., 1967). Более того, в раннем детском
развитии наблюдается также и параллельное увеличение уровня
инсулина в крови (Berenson et al., 1981).
Может показаться, что мы уделили слишком много внимания
проблеме возрастного понижения толерантности тканей к глюкозе.
Однако необходимо подчеркнуть, что именно эти метаболические
сдвиги являются исходными (пусковыми) в развитии таких
важнейших возрастных болезней, как диабет II типа, патологическое
ожирение, атеросклероз и сердечно-сосудистые заболевания,
метаболическая иммунодепрессия, аутоиммунные патологии и, наконец,
предрасположенность к раку (Дильман, 1987). Понимание природы
возрастных изменений углеводно-жирового метаболизма, таким
образом, может послужить важным плацдармом для наших
попыток замедления или предотвращения развития возрастных
дегенеративных болезней.
73
Способы выработки толерантности к глюкозе включают регулярную
умеренную физическую нагрузку, диету с высоким содержанием волокнистой
пищи и низким содержанием насыщенных жиров, а также стремление избегать
стрессовых ситуаций и повышать сопротивляемость стрессу. Некоторые
пищевые добавки также могут быть полезны: пикалинат хрома улучшает
толерантность к глюкозе у людей с явным дефицитом хрома; липоевая кислота, коэн-
зим Q10 и различные адаптагены (например, сибирский женьшень) также
имеют тенденцию нормализовать уровень глюкозы в крови. Факторы,
замедляющие центральные часы старения также, по видимому, улучшают толерантность
тканей к глюкозе, и наиболее важным из них является мелатонин (см. гл. 5).
2.5. ЦЕНТРАЛЬНЫЕ ЧАСЫ СТАРЕНИЯ
Песочные часы
День за днем уходит без следа, однообразно и быстро.
Страшно скоро промчалась жизнь, - скоро и без шума,
как речное стремя перед водопадом.
Сыплется она ровно и гладко, как песок в тех часах,
которые держит в костлявой руке фигура Смерти.
...мне постоянно чудится этот слабый и непрерывный
шелест утекающей жизни.
Иван Тургенев "Стихотворения в прозе", 1878
Переходя к рассмотрению системных механизмов старения,
следует задать себе вопрос имеют ли вообще высокоразвитые
организмы - и в особенности человек - центральные часы старения, т.е.
программу, контролирующую скорость старения всех систем
организма? Этот почти трансцендентный вопрос возник, по-видимому,
вместе с появлением геронтологии. Сегодня мы, кажется, близки к
ответу. И ответ прост: и "дд'\ и "нет". Однако не спешите
удивляться. С одной стороны, по-видимому, мы не имеем определенной
центральной программы, чья единственная цель состоит в том, чтобы
делать нас старыми. С другой стороны, несомненно, существует
определенная программа развития, т.е. программа, которая заставляет
отдельную клетку (зиготу) развиваться в сложный многоклеточный
организм. Некоторые из механизмов этой программы развития, по-
видимому, продолжают действовать и после того как развитие
организма уже закончено. Эти механизмы, необходимые во время роста
и полового созревания, имеют, как полагают некоторые ученые,
множество побочных эффектов в последующей жизни организма.
Фактически эти механизмы приносят вред зрелому организму,
потому что продолжают действовать так, как будто бы развитие еще не
закончено. Результатом является ускорение старения и развитие
возрастных (дегенеративных) болезней. Таким образом,
центральные часы старения (ЦЧС) являются побочным продуктом
реализации программы развития, которая не выключается в зрелости.
74
Это кажущееся странным представление является основной
предпосылкой нейроэндокринной теории старения. Термин "нейро-
эндокринная" означает, что теория должна рассматривать
взаимодействие центральной нервной и эндокринной систем (эндокринная
система регулирует функции организма с помощью гормонов). Ней-
роэндокринная теория старения была предложена в 60-х годах
замечательным российским ученым Владимиром Михайловичем Диль-
маном (Дильман, 1958; 1961; 1968; 1970; Oilman, 1971). В то время
такая концепция была просто блестящим предположением, поскольку
доказательств, подтверждающих его, практически не было. Ученые
часто говорят, что лучший способ доказать теорию состоит в том,
чтобы сделать предсказания, основанные на этой теории, а затем
проверить их. В последние 30 лет нейроэндокринная теория
старения выдержала это испытание весьма успешно.
Прежде чем идти дальше, необходимо пояснить основные
понятия теории. Это прежде всего - гомеостаз. По существу, гомеостаз -
это определенный баланс многочисленных параметров внутренней
среды организма. Для нормального функционирования организм
нуждается в стабильности определенных физиологических
параметров, колебания которых не должны выходить за пределы
некоторого оптимального диапазона: температура должна быть около 37 °С,
давление крови около 120/80 мм ртутного столба, концентрация
сахара в крови 70-120 мг/дл и т.д. Гомеостаз выражает генетически
запрограммированное стремление системы к постоянству своей
внутренней среды, которая включает сохранение множества
физиологических и биохимических параметров в пределах оптимального
диапазона. Основоположник учения о гомеостазе, выдающийся
французский физиолог Клод Бернар считал, что "постоянство
внутренней среды является условием свободной жизни клеток"
{Бернар, 1937). Если гомеостаз нарушен, система включает механизмы
его восстановления. Неудачи этих попыток могут (при далеко
зашедших отклонениях) привести систему к гибели.
Ситуация, однако, осложняется тем, что гомеостаз
новорожденного отличается от такового у ребенка, и у того и другого
отличаются от гомеостаза взрослого. Например, средний уровень
гемоглобина (молекула, которая переносит кислород в крови) у младенцев-
мальчиков составляет приблизительно 11 мг/100 мл, у десятилетних
мальчиков - 13 мг/100 мл и у взрослых людей - 15 мг/100 мл;
среднее давление крови в десятилетнем возрасте приблизительно 100/60,
а у взрослых молодого возраста - 120/80. На каждой стадии
развития организм, с одной стороны, должен поддерживать гомеостаз,
соответствующий данной стадии. С другой стороны, чтобы быть
способным расти и созревать, организм должен перманентно изменять
гомеостаз настолько, насколько это необходимо для достижения
параметров, соответствующих зрелости. Совершенно ясно, что
программа развития должна постепенно изменять гомеостаз, "подтал-
75
кивая" организм к потреблению и расходованию все большего
и большего количества энергии, необходимой для увеличения
размеров тела, развития репродуктивных органов, другими словами, к
росту и созреванию. Согласно теории Дильмана, непрерывное
изменение гомеостаза не останавливается в зрелости, а продолжается
в течение всей жизни. Однако после достижения зрелости вместо
того, чтобы заставлять организм расти и активировать
репродуктивные процессы (рост размера тела имеет механизм ограничения),
эта же программа заставляет организм стареть и приводит к
развитию возрастных болезней, и в этом смысле она действует как часы
старения.
Почему же у животных не развился эффективный
"выключатель" программы развития с тем, чтобы предохранить их
дальнейшую жизнь от ускоренного старения? Очевидно, существование
такого "выключателя " не давало видам никакого преимущества для
выживания в дикой природе.
Старение, как уверено теперь большинство эволюционных биолотов, прямой
результат оптимизации давления естественного отбора (Kirkwood et al., 2000). Еще
Медавар (Medawar, 1952) теоретически обосновал представление, что при
значительной гибели живых организмов от случайных причин (что и происходит в
реальных условиях) потенциальное бессмертие не давало бы никаких преимуществ
такой популяции по сравнению с популяцией особей с конечной
продолжительностью жизни. На фоне многочисленных опасностей естественного мира животным
фактически вряд ли удается прожить долго, так почему природа должна давать им
неопределенный потенциал продолжительности жизни? Однажды разделение
труда между зародышевыми и соматическими клетками привело к тому, что сома
стала "одноразовой** (Kirkwood, 2002). Под давлением естественного отбора
соревнование за метаболические ресурсы между системами воспроизведения и
системами поддержания стабильности сомы разворачивается таким путем, который
позволяет соматическим клеткам функционировать на уровне достаточно высоком,
чтобы сохранить организм в хорошей форме в течение весьма важных для
воспроизведения ранних лет жизни, но не достаточно высоком, чтобы сохранить это
состояние навсегда. Отбор может предпочитать свойства, которые
стимулируют выживание и воспроизводительный успех молодого организма, даже если эти
же самые свойства прямо ведут к потере приспособленности и даже смерти
организма в более позднем возрасте. Именно в этом заключается идея
антагонистической плейотропности (Williams, 1957).
Хорошей иллюстрацией сказанного выше служит прямая корреляция
между скоростью гибели птиц от ннешних причин и величиной максимальной
продолжительности их жизни (табл. 3). Иными словами, чем больше погибает
животных за определенный период времени, тем меньше их видовой лимит жизни.
Исходя из подобного рода представлений, можно предположить, что роль
естественного отбора в определении продолжительности жизни обусловлена
повышением активности генов, эффект которых благоприятен на ранних
стадиях развития (онтогенеза) и в репродуктивном периоде, несмотря на то что
действие этих же генов влечет за собой нарушения в более позднем возрасте.
Иначе говоря, отбор всегда будет способствовать большей жизнеспособности в
молодом возрасте — даже ценой отрицательных эффектов в поздние периоды
жизни. Результатом такого отсроченного (плейотропного) влияния генов и
является феномен старения (Medowar, 1952; Дильман, 1987).
76
Таблица 3
Корреляция между темпом смертности в естественных условиях
и максимальной продолжительностью жизни птиц
(Cutler, 1980)
вид
Голубая синица
Малиновка европейская
Чибис
Ласточка обыкновенная
Буревестник серый
Пестрая чайка
Королевский альбатрос
* Примечание: 100% гибель птиц принята за 1.
Пропорция птиц
погибающих за
год*
0.72
0.62
0.34
0.18
0.07
0.04
0.03
Максимальная
продолжительность жизнн,
годы
9
12
16
21
27
36
45
Какой бы ии была эволюционная причина, приходится
констатировать, что природа, по-видимому, подарила высшим организмам, как
считает Дильман (1987), эквивалент "встроенных" центральных
биологических часов старения. Такое явление, к сожалению, содержит в
себе явные элементы фатальности. К счастью, темп
функционирования ЦЧС не является незыблемым, и может быть изменен.
Некоторые воздействия могут ускорить "ход" ЦЧС, другие - замедлить
Перед обсуждением способов замедления ЦЧС, следует, хотя бы
частично, объяснить их физиологию. "Центральный компьютер", в
значительной степени ответственный за гомеостаз организма, - это
область мозга, называемая гипоталамусом, а также несколько
связанных с ним структур. Гипоталамус постоянно контролирует
множество внутренних параметров организма. Если некоторые
параметры выходят за рамки надлежащего диапазона, гипоталамус
посылает сигнал к гипофизу, который является главной железой
эндокринной системы. Можно, как это модно сегодня, сказать, что, если
гипоталамус - правление директоров гомеостаза, то гипофиз - это
главный менеджер. Гипофиз переводит сигналы от гипоталамуса в
гормональные сообщения для периферийных эндокринных желез,
например щитовидной железы или надпочечников. В свою очередь,
периферийные эндокринные железы используют свои собственные
гормоны, чтобы сообщить целевые сигналы своим мишеням -
органам и тканям {рис. 9,10). Обычно, эти инструкции увеличивают или
уменьшают те или иные физиологические функции.
Другая важная концепция, которую необходимо рассмотреть,
это - концепция отрицательной обратной связи, называемая также
"ингибирование обратной связью". Рассмотрим один пример. Зи-
77
Рис. 9. Большие биологические часы (объяснение в тексте) (Дильман, 1986)
мой, организм нуждается в повышении скорости своего
метаболизма для компенсации повышенного расхода тепла в холодную погоду.
Гипоталамус посылает соответствующий сигнал гипофизу, который
в свою очередь посылает свой сигнал щитовидной железе,
приказывая выделить большее количество ее гормона - тироксина,
который повышает скорость энергопродукции митохондриями.
Гипоталамус "чувствует" произошедшее в результате каскада этих сигна
лов повышение уровня тироксина и прекращает посылать свой ис
ходный "приказ". Это пример отрицательной обратной связи, кото
рая, по существу, является проявлением способности системы
прекратить начальный стимулирующий импульс, когда цель уже
достигнута {рис. 9, 10).
Как мы отметили, развитие организма требует постепенного
изменения гомеостаза. Функциональная динамика гипоталамуса
является главной движущей силой этого изменения. Нейроэндокринная
теория старения предполагает, что гипоталамус, выполняя
программу развития организма (которая позже функционирует как часы
старения), становится со временем менее отзывчивым к
отрицательной обратной связи. Рассмотрим другой пример. У молодых девочек,
78
Ill уровень
ΤΙ уровень
1 уровень
Гормон эндокринной железы
_ С
\>^- Гормональный рецептор
Клеточная оболочка у
О
АМФ
цАМФ
' 1
Рис. 10. Уровни нейроэндокринной регуляции в организме (Дильман, 1986)
I - внутриклеточный уровень (клеточная мембрана, гормональный рецептор и
циклический АМФ (цАМФ) - трансмиттер гормонального сигнала); II - уровень
периферийных эндокринных желез; III - уровень гипофиза; IV - уровень гипоталамуса; V - уровень
центральной нервной системы; 1) сверхкороткая петля механизма обратной связи,
воздействие гормонов гипоталамуса на гипоталамус; 2) короткая петля механизма обратной
связи, воздействие гипофиза на гипоталамус; 3) длинная петля механизма обратной связи,
воздействие гормонов периферийных эндокринных желез и продуктов обмена вещества
(глюкозы, жирных кислот и т.д.) на гипофиз и гипоталамус; 4) регуляция активности
центральной нервной системы гипофизом; 5) регуляция активности центральной нервной
системы гипоталамусом
яичники производят только небольшое количество эстрогенов^ но
этого достаточно, чтобы вызвать к жизни отрицательную обратную
связь, т.е. заставить гипоталамус прекратить сигналы на
производство большего количества эстрогенов. С возрастом, однако,
гипоталамус становится менее отзывчивым к отрицательной обратной
связи и стимулирует яичники к производству большего количества
эстрогенов. Это ведет к известному повышению уровня эстрогенов во
времени и в конечном счете - к половому созреванию. Подобные
изменения гомеостаза происходят в процессе развития во многих
системах организма или по меньшей мере в трех из них -
репродуктивной, адаптационной и системах энергопродукции. К сожалению,
после созревания, отклик гипоталамуса на отрицательную обратную
связь продолжает уменьшаться, вызывая дальнейшие изменения
гомеостаза, которые теперь играют отрицательную роль, способствуя
старению и дегенеративным болезням.
Каким же образом изменения гомеостаза, индуцированные
ЦЧС, вызывают старение и возрастные болезни? Множество свя-
79
занных между собой механизмов, по-видимому, вовлечены в этот
процесс, но рассмотрение всей их совокупности здесь не
представляется возможным. Наиболее важные из этих механизмов включают
развивающийся с возрастом избыток инсулина - гиперинсулинизм,
уменьшенную толерантность (восприимчивость) к углеводам,
развитие неадекватного адаптивного ответа, возрастную депрессию и
повышенное выделение гонадотропина (гормона, стимулирующего
деятельность половых желез).
Возрастной избыток инсулина и снижение толерантности к
углеводам являются результатом изменения тех составляющих гомео-
стаза, которые ориентированы на использование энергии.
Инсулин - гормон, который выделяется В-клетками поджелудочной
железы. Как уже отмечалось, он выделяется в ответ на повышение
содержания сахара (глюкозы) в крови, которое обычно происходит
после приема пищи, и помогает переносу глюкозы, аминокислот и
жиров в клетки, где они сжигаются для получения энергии,
откладываются в запас или используются как структурные материалы.
Типичный сценарий после приема пищи таков: по мере того, как
пища переваривается и всасывается, содержание сахара в крови
повышается, что вызывает выделение инсулина; в течение примерно
часа инсулин доводит содержание сахара в крови до первоначального
уровня или даже снижает его. С возрастом, способность мышечной
и некоторых других тканей поглощать глюкозу под действием
инсулина снижается (как уже говорилось выше), и количество
выделяемого инсулина после приема пищи вынужденно увеличивается. В
результате мы имеем более высокое содержание сахара и
повышенную циркуляцию инсулина в кровяном русле в течение более
длительного периода времени. Было много споров о том, вызывает ли
пониженная чувствительность мышц к инсулину его чрезмерное
выделение или - наоборот? Согласно нейроэндокринной теории
старения, причиной и того, и другого в значительной мере являются ЦЧС.
Механизм, по которому гипоталамус вызывает эти изменения, по
всей видимости, сложен и включает понижение чувствительности
центра аппетита к глюкозе (и, возможно, к инсулину) и изменения в
регуляции выделения гормона роста.
Таблица 4
Концентрация инсулина в сыворотке крови
белых крыс разного возраста
(Тодоров, 1965)
Возраст, мес.
1
3
12
24
Инсулин, миллнединипы/мл
3.99±1,47 3.94±0.84 6.67±0.75 3.04±0.60
80
Один из авторов этой книги изучал содержание инсулина в крови крыс
разного возраста задолго до формирования современных представлений о
возрастной динамике глюкозы и инсулина в коови (см. табл. 4).
К сожалению, значение этих четких фактов, которые могли бы послужить
ключом к пониманию одного из важнейших механизмов старения, в контексте
знаний того времени не были до конца поняты автором и остались в стороне от
магистральных направлений биохимической геронтологии.
Избыток инсулина и пониженная толерантность к глюкозе
поднимают уровень холестерина, ЛНП и триглицеридов, содействуют
повреждению клеток из-за неферментативного гликозилирования и
поперечных сшивок между макромолекулами (см. выше в этой
главе), вызывают задержку натрия и другие многочисленные
нарушения. В результате, эти метаболические изменения вносят вклад в
развитие большинства возрастных болезней, включая
атеросклероз, гипертонию, диабет II, тучность, снижение иммунитета и
увеличение риска рака.
Наиболее характерным примером экстремально пониженной
толерантности к глюкозе служит диабет независимый от инсулина
(диабет II типа). В этом случае, у пациентов наблюдается высокий
сахар в крови, несмотря на способность В-клеток вырабатывать
инсулин. В действительности, на начальной стадии диабета II типа
уровень инсулина часто выше нормы. Диабет, который не лечили или
лечили плохо, дает многочисленные осложнения, многие из
которых могут рассматриваться как ускорение старения и прогрессия
дегенеративных болезней.
В исследовании, проведенном в Италии, толерантность к
глюкозе и действие инсулина изучались на долгожителях (возраст более,
чем 100 лет). Было найдено, что у 100-летних людей оба эти
параметра оказались на том же самом уровне, что и у 50-летних, и лучше,
чем у 75-летних. Это показывает, что эти очень старые люди имели
только минимальные изменения в энергетических параметрах го-
меостаза (т.е. "ход" их ЦЧС по этим параметрам был существенно
замедлен), что, возможно, и внесло вклад в их долголетие (Receputo
et al., 1995).
Определенную надежду внушает существование ряда способов улучшения
тканевой восприимчивости к глюкозе и устранения избытка инсулина. Однако
существуют факторы, дающие противоположные эффекты. Отрицательными
факторами являются все формы стресса, питание, богатое насыщенными
жирами и бедное волокнистыми структурами, переедание и дефицит хрома;
положительные факторы - умеренные нагрузки, снижение избытка веса, питание,
бедное насыщенными жирами и богатое волокнами, и, возможно, добавки пиколи-
ната хрома и CoQIO.
Возрастные нарушения стресс-ответа (неадекватный
адаптивный ответ) - это другой главный вкладчик в процесс старения.
Согласно нейроэндокринной теории старения, гипоталамус
постепенно теряет живой отклик на отрицательное влияние обратной
81
связи со стороны кортикостероидов > ключевых гормонов
стресса, вырабатываемых и выделяемых надпочечниками (наиболее
важный кортикостероид у людей - кортизол). В результате,
пожилые люди склонны к выделению большего количества
кортикостероидов, чем молодые - в ответ на действие аналогичных
стрессоров. У некоторых пожилых людей наблюдается избыток
кортикостероидов в крови даже в отсутствии каких-либо
стрессоров, что является физиологическим эквивалентом жизни в
состоянии хронического стресса. По всей видимости, здесь существует
порочный круг: ЦЧС постепенно нарушают нормальный стресс-
ответ, который в свою очередь ускоряет "ход" самих часов. Мы
подробно обсуждаем это критическое препятствие продлению
жизни во II части книги. Здесь мы только укажем, что избыток
кортикостероидов вносит вклад в падение толерантности тканей
к глюкозе, в угнетение иммунной системы, повышение давления
крови и, по существу, в развитие практически всех возрастных
болезней.
Согласно нейроэндокринной теории старения, избыток
инсулина, пониженная толерантность к углеводам и несбалансированный
стресс-ответ - главные "покровители" старения и дегенеративных
болезней, развитие которых связано с деятельностью ЦЧС (Диль-
ман, 1986,1987; Dilman, 1994). Имеется, однако, много других путей,
следуя по которым центральные гипоталамические часы, по всей
видимости, вносят вклад в возрастные проблемы. Например,
гипоталамус, по-видимому, отвечает за постепенное увеличение уровня
пролактина. Этот гормон выделяется гипофизом и ответствен за
производство молока у женщин. Роль пролактина у мужчин неясна,
но точно установлено, что уровень пролактина вносит вклад в
развитие доброкачественной гипертрофии предстательной железы
(недуг, почти повсеместно распространенный у пожилых мужчин) и,
быть может, приводит к импотенции.
Мы не будем обсуждать здесь все возможные многочисленные
влияния ЦЧ на процесс старения, потому что практический подход,
предлагаемый нейроэндокринной теорией, состоит не в том, чтобы
иметь дело с последствиями, а в том, чтобы замедлять "ход"
центральных часов непосредственно. Один из способов может состоять в
повышении чувствительности гипоталамуса к отрицательной
обратной связи от периферических эндокринных желез и других систем
организма. Имеющиеся данные говорят о том, что уровень нейро-
трансмиттеров (веществ, которые переносят сообщения между
мозговыми клетками) в гипоталамусе непосредственно связан с его
активным откликом на отрицательную обратную связь. Факторы,
которые, как полагают, замедляют ЦЧС, способны повышать и
уровень нейротрасмиттеров в гипоталамусе. Наоборот, любое
повышение уровня нейротрасмиттеров в гипоталамусе способно
замедлить "ход" ЦЧС. Табл. 5 содержит список факторов, которые
82
Таблица 5
Факторы ускорения или замедления ЦЧС
(Todorov etal., 2001)
ЦЧС
ускоряют:
замедляют:
Стресс; неадекватный отклик на
стрессоры
Переедание, избыток калорий
Пониженная толерантность к
глюкозе/углеводам
Избыток инсулина
Депрессия; пониженный уровень
нейротрасмиттеров в гипоталамусе
Недостаток уровня мелатонина и
других гормонов шишковидной
железы
Повреждение мозга свободными
радикалами, ионизирующей
радиацией или токсинами окружающей
среды.
Повышение сопротивления стрессу;
восстановление оптимального
ответа на стрессоры; предотвращение
интенсивного или длительного
стресса
Ограничение калорийности (у
грызунов), поддержание идеального веса
тела (у людей)
Улучшенная толерантность к
глюкозе / углеводам
Оптимальное выделение инсулина
Приподнятое эмоциональное
состояние, повышение уровня нейротран-
смиттеров в гипоталамусе
Восстановление уровня мелатонина
и других гормонов шишковидной
железы
Предотвращение повреждения мозга
свободными радикалами и т.п.
влияют на ЦЧС; характер действия большинства из этих факторов
мы подробно обсуждаем в других главах.
Нейроэндокринная теория старения Дильмана, была первой
всесторонней теорией, которая связала старение с гипоталамусом и
гипофизом. Известный американский геронтолог Р. Катлер писал:
''Если гипотеза Дильмана верна, то она служит отличным
примером механизма старения, связанного с развитием". Ряд
родственных, но несколько отличающихся гипотез был одновременно или
позже предложен и другими исследователями.
Согласно концепции известного геронтолога Р. Катлера {Cutler, 1976; 1978;
1984), старение является побочным продуктом метаболизма, развития и диффе-
ренцировки организма. Кроме того, автор обратил внимание на то
обстоятельство, что существенный рост продолжительности жизни человека по
сравнению с приматами был связан со сравнительно небольшим процентом мутаций и,
следовательно, столь значительный эффект этих мутаций мог быть лишь в том
случае, если бы они произошли в системе регуляторных генов. Иначе говоря,
изменилась эффективность и скорость процессов, а не сама их структура за счет
добавления новых свойств.
83
И.Н. Тодоров (1969) рассматривал развитие астрой лучевой болезни
{которая, как известно, имеет очень много общих черт с процессами
старения) в связи со значительными нарушениями гомеостаза облученного
организма, имеющими в основном свободно-радикальный характер (продукты
радиолиза воды, молекулы в возбужденном состоянии, радиотоксины орто-
хиноидной природы и многие другие продукты радиолиза компонентов
клетки). Этот физико-химический "шум", по-видимому, сравним по
интенсивности с обычными химическими сигналами, которые передаются через
внутреннюю среду, что не может не искажать прямые и отрицательные
обратные связи на всех уровнях интеграции организма. На клеточном уровне это
может выразиться в нарушении структуры генетического аппарата, а также
процессов передачи генетической информации при репликации, траскрип-
ции и трансляции, а также в нарушении систем энергопродукции и других
сторон метаболизма. На уровне целостного организма этот "шум" способен
внести искажения в работу центральных регулирующих систем, прежде
всего системы гипоталамус-гипофиз-периферические эндокринные железы,
что выразится в изменении потока регуляторных сигналов как от "центра" к
"периферии", так и в изменении потока сигналов отрицательной обратной
связи - от "периферии" к "центру". Это искаженное взаимодействие всех
уровней интеграции внесет дополнительные "помехи" во внутреннюю среду
организма и события будут развиваться по закону "порочного круга". И
поскольку для анализа и регуляции всех физико-химических изменений
гомеостаза при лучевом поражении организм, очевидно, не имеет всего набора
необходимых генетических "эталонов" (ибо в процессе эволюции организмы
никогда не сталкивались с таким мощным радиационным воздействием, а
если и сталкивались, то радиационное воздействие скорее всего являлось
фактором их элиминации), некоторые пострадиационные изменения параметров
гомеостаза могут вызвать неадекватную, неспецифическую и, возможно,
слишком интенсивную реакцию регуляторных систем. Эти
несбалансированные регуляторные факторы, в избытке наполняющие внутреннюю
среду, не могут не вызвать крупных изменений, а, возможно, и прямых
нарушений в жизнедеятельности воспринимающих эти факторы систем, органов и
тканей. Общая схема развития лучевой болезни, представленная автором, в
частности как сбой регуляторных механизмов, во многом перекликается с
концепцией В. Дильмана с одной существенной поправкой: острая лучевая
болезнь является всего лишь "сверхскоростной" имитацией нормального
процесса старения. (В 1969 г. И.Н. Тодоров еще не был, к сожалению,
знаком с работами В.М. Дильмана.)
* * *
Заключая этот важный параграф, мы хотели бы кратко
обсудить наиболее общий, т.е. термодинамический, аспект проблемы
старения, который, очевидно, не может быть не связан с
признанием неотвратимого влияния законов термодинамики на живые
системы. И хотя последние являются открытыми
термодинамическими системами, которые с помощью сложнейших внутренних
механизмов и сохранения определенного баланса между
притоком и оттоком веществ и энергии способны, порой весьма
длительное время, поддерживать стационарные режимы своей рабо-
84
ты {Пригожий, 1959; 1960), тем не менее конечный результат для
них будет всегда один - нарастание хаоса и энтропии, т.е. смерть.
До момента наступления и в течение репродуктивного периода
живые системы, по-видимому, более или менее успешно
противодействуют действию второго начала термодинамики, чему,
очевидно, максимально способствует эволюционное развитие,
совершенствуя их репаративные механизмы. Однако в последующий
период жизни старые особи оказывается вне сферы действия
естественного отбора (в результате их неспособности производить
потомство и предавать ему свои полезные мутации), и процессы
деградации и деструкции начинают идти у них с той скоростью,
которую способна допускать генетически запрограммированная
прочность и эффективность их систем восстановления. Падение
чувствительности гипоталамических "датчиков" к импульсам
отрицательной обратной связи после достижения репродуктивной
зрелости организма может обуславливаться, по нашему мнению
(в отличие от нейроэндокринной теории В. Дильмана), не
специальной генетической программой, а исключительно высоким
уровнем накопления окислительных повреждений в мозговых
структурах (в особенности в гипоталамусе) в результате
несравненно более высокой, нежели в периферических тканях,
интенсивности окислительного фосфорилирования и сопряженной с
ним продукцией вредоносных АФК.
В условиях дикой природы живые существа, по всей видимости,
редко доживают до старости, поскольку шансов выжить в
конкурентной борьбе за существование у них тем меньше, чем дальше они
удалились от молодого репродуктивного возраста.
Удивительно ярко эту ситуацию обрисовал один из ведущих
современных геронтологов Леонард Хейфлик в своей монографии
"How and Why We Age" {Hayflick, 1994), переведенной недавно на
русский язык {Хейфлик, 1999):
У нас нет никаких оснований упрекать природу, наградившую нас этим
процессом старения. Как было сказано ранее, мы были запрограммированы
только на достижение половой зрелости. Мы сами спутали ее планы, позволяя
себе жить куда дольше, чем это было предусмотрено, и запустив руку в ящик
Пандоры, заключающий в себе все те функциональные расстройства и немочи,
которые мы называем старением. После окончания репродуктивного периода
человеку ничего не остается, как иметь дело все с тем же нарастающим
хаосом, в который превратились наши прежде безупречно функционирующие
молекулы, и подобный тому, что с течением времени возникает в любой сложной
системе или механизме. Предполагается даже, что процесс развития во
Вселенной идет по пути нарастания хаоса и энтропии. Так что нет причин, по
которым живые существа должны являться исключением из правил
Леонард Хейфлик, наряду с развитием общих взглядов на
процесс старения, которые изложены в упомянутой выше монографии,
ранее внес огромный вклад, как мы увидим ниже, в понимание
природы этого явления на клеточном уровне.
85
2.6. ЛИМИТ ХЕЙФЛИКА И СТАРЕНИЕ КЛЕТОК
Довольно долго, до середины XX столетия, многие ученые
думали, что клетки высокоразвитых организмов так же
бессмертны, как и бактерии, т.е. способны к неограниченному
делению при надлежащих условиях. Такая убежденность стала
распространяться, начиная с 1912 г., после того как Алексис
Каррель {Carrel, 1912 цит. по: Егоров, 1997), тогда исследователь
Рокфелеровского института, закончил свои эксперименты
с целью проверить, как долго способны в культуре делиться
фибробласты цыпленка. Фибробласты - это клетки
соединительной ткани, чья функция заключается в строительстве
эластичной трехмерной структуры, поддерживающей другие
клетки. Каррель выращивал фибробласты в специальном
бульоне из эмбрионов цыпленка, снабжая клетки свежим бульоном
каждые несколько дней. Избыток клеток периодически
выбраковывался. Фибробласты продолжали делиться год за годом,
без каких-либо признаков замедления вплоть до тех пор, когда
через 34 года после смерти Карреля эксперимент был
остановлен {Parker; 196I). Тогда большинству исследователей в этой
области казалось, что в надлежащей окружающей среде клетки
высших организмов могут быть столь же бессмертными, как и
бактерии.
Существует два типа бессмертия: бессмертие индивидов и бессмертие
популяции. Бессмертие популяции базируется на периодическом обмене или
перестройке генетической информации в результате половой гибридизации.
Например, человек как индивид смертен, но человечество как популяция -
бессмертно. Это относится ко всем животным и большинству растений. До сих пор
распространено представление о бессмертии бактерий и простейших. Однако, как
показали многочисленные и тщательно проведенные исследования {Bell, 1988),
это утверждение не соответствует действительности. Подобно популяциям
человека популяции одноклеточных организмов бессмертны и подобно
отдельному человеку, одноклеточный организм смертен.
Идеи о том, что соматические клетки высших животных
должны иметь ограниченный потенциал деления, восходят к
1881 г. и принадлежат выдающемуся немецкому биологу Августу
Вейсману, который предположил, что "...смерть обусловлена
тем, что изношенные клетки не могут самообновляться, а их
способность к росту путем деления не вечна, но ограничена"
{Weismann, 1891). Эти идеи не остались забытыми даже в эпоху
торжества взглядов Карреля и оказались в состоянии (более чем
через полвека) стимулировать серию весьма важных
исследований. Так, в 1961 г. Л. Хейфлик и П. Мурхед опубликовали
результаты научных наблюдений, которые прямо противоречили
результатам Карреля {Hay/lick, Moorhead, 1961; цит. по: Hay flick,
1974). Они нашли, что человеческий фибробласт в культуре будет
86
делиться -50 раз и затем остановится. Кто же был прав - Каррель
или Хейфлик? По-видимому, экспериментальная техника Карреля
имела недостатки. Питательный бульон, пользуясь которым он
выращивал фибробласты цыпленка, вероятно, содержал
небольшое количество фибробластов цыпленка, из сыворотки крови
которого он готовился. Каррель, следовательно, "неосторожно"
добавлял к культуре "свежие" клетки через каждые несколько дней.
В этом конфликте научная общественность в конце концов
приняла сторону Хейфлика, и с тех пор максимально возможное число
делений клеток в культуре стало обозначаться как предел, или
лимит Хейфлика. Сам Леонард Хейфлик не считал механизм
лимита делений "клеточными часами старения", поскольку в
нормальных клетках этот механизм связан непосредственно не с течением
времени, а с числом актов репликации клеточной ДНК.
Предполагаемое устройство следует рассматривать как счетчик событий и
называть его, например, "репликометр".
Предполагалось, что лимит Хейфлика это генетическая
программа, которая предотвращает деление клетки после некоторого
числа циклов. Для чего же нужна клетке такая программа?
Согласно современным представлениям, имманентный клеткам
предел делений ограничивает риск неконтролируемой пролиферации
клеток, т.е. рост раковых опухолей. Действительно, исследования
показывают, что в раковых клетках работа генетического репли-
кометра нарушена, что и позволяет им расти неограниченно
{Хейфлик, 1997).
Эпидемиологическая статистика позволила расчитать, что в среднем для
превращения нормальной клетки в опухолевую требуется от трех до семи
независимых случайных событий (Альберте и др., 1994). Таким образом, при
развитии опухоли происходит несколько этапов клонирования. Если считать, что
частота мутаций составляет 1 на 106 клеток, то каждое такое событие потребует
примерно 20 генераций (220 = 106). Следовательно, для возникновения опухоли
потребуется в среднем от 60 до 140 генераций. Таким образом, лимита в 50
удвоений вполне достаточно для остановки роста большинства опухолей на еще
не диагностируемой стадии (Егорову 1997).
Исследования последних лет позволили раскрыть
интригующую тайну возможного молекулярного механизма лимита
Хейфлика. Однако задолго до этого, после выяснения основных черт
механизма репликации ДНК, российский ученый A.M. Оловников
еще в 1971 г. предложил принцип маргинотомии (от лат. margin-
alis - краевой, tome - сечение). Сущность этого принципа состояла
в том, что при матричном синтезе дочерней нити ДНК- полимера-
за не в состоянии полностью реплицировать линейную матрицу
материнской ДНК, реплика всегда получалась короче в начальной
ее части (Оловников, 1971). При последовательных клеточных
делениях это постепенное укорачивание ДНК ограничивает проли-
феративный потенциал и может служить молекулярной основой
87
репликометра клеточных делений в опытах Хейфлика. В клетках
высших организмов концы хромосом содержат специальные
структуры ДНК, называемые теломерами. Теломеры соматических
клеток позвоночных животных содержат многократно
повторяющиеся гексамеры - TTAGGG с общей длиной до 10 тыс. пар нук-
леотидов. В комплексе со специфическими белками теломеры
защищают концы ДНК от эндонуклеаз, предотвращают
неправильную рекомбинацию и прикрепляют концы хромосом к внутренней
ядерной мембране. Достаточно сложный процесс укорачивания
теломер мы попытаемся пояснить с помощью несложной
аналогии.
Вообразите рабочую машину, которая перемещается по дороге
и укладывает позади себя новый слой асфальта. Устройство
машины таково, что она может двигаться только вперед и вдоль дороги в
одном направлении. Когда машина достигнет конца дороги, мы
будем иметь новый слой асфальта по всему пути, кроме одного
отрезка, который находится под машиной. Чтобы закрыть остающийся
отрезок, машина должна двигаться дальше этого отрезка, но это
невозможно, потому что это уже конец пути. "Асфальтировочная
машина" в этом сравнении - это фермент ДНК-полимераза, который
копирует хромосомную ДНК, т.е. "кладет новый слой асфальта",
начиная с ее концевого участка - теломеры, "Отрезок дороги под
машиной" - это участок теломеры под довольно крупным белковым
агрегатом (ДНК-полимеразой), часть которого не копируется и
выбывает из игры в дальнейших раундах репликации. Подобный
сценарий разыгрывается в процессе репликации хромосом, только
ДНК-полимеразы двигаются по обеим нитям ДНК в
противоположных направлениях навстречу друг другу, как две "асфальтировочные
машины" по дороге с двусторонним движением. Таким образом, сам
конец каждого теломера, т.е. симметричные и комплементарные
концевые участки обеих нитей ДНК, никогда не копируются, что
ведет к прогрессивному укорачиванию теломера после каждого
деления клетки.
Известно, что в ходе пассирования клонов нормальных клеток,
например фибробластов человека, происходит укорочение теломер
в среднем на 50 пар нуклеотидов за каждый цикл деления. Подобное
укорочение хромосом происходит in vivo в основном у тех клеток,
которые на протяжении всей жизни организма имеют достаточно
высокий и постоянный уровень пролиферации. У человека это -
лейкоциты периферической крови, клетки эпидермиса кожи и эпи
телия кишечника. Представляется вполне логичным, что после
укорачивания теломер до некоторого предела, клетка теряет
способность делиться.
По мере того как клетки приближаются к -50 делениям,
они начинают "выглядеть старыми"; их метаболическая
активность снижается, они увеличиваются в размере и накапливают
88
пигменты, например липофусцин, пигмент, образующий
старческие пятна.
Не является ли старение результатом неспособности клеток
делиться после того как они достигли лимита Хейфлика? И снова
нет прямого ответа на этот вопрос. Возможно, что в некоторых
тканях, таких как кожа и внутренние стенки кровеносных сосудов,
лимит Хейфлика может действительно быть частью механизма
старения. Например, возрастное ускорение атеросклероза,
частично может быть результатом уменьшения способности
эпителиальных клеток сосудов делиться. Возрастная потеря влажности
и эластичности кожи может также быть связана с лимитом
Хейфлика. Это - с одной стороны. С другой стороны, клетки мозга,
сетчатки, нервные и мышечные клетки обычно не делятся, и
вероятно никогда даже не приближаются к "роковому" лимиту.
Но нельзя считать очевидным, что темп старения этих клеток
выше, чем у тех, которые обладают полноценным пролифератив-
ным потенциалом.
Следует далее подчеркнуть, что далеко не все клетки
подчинены лимиту Хейфлика. Зародышевые клетки, развивающиеся в
сперму или в яйцеклетки, и раковые клетки, очевидно, бессмертны.
Существует гипотеза, что каждая ткань имеет небольшое количество
потенциально бессмертных клеток, называемых стволовыми
(Смирнов, 2001), которые используются организмом, как
клеточный запас. Защитники этой гипотезы предполагают, что влияние
стволовых клеток не заметно в их культуре, потому что их
содержание (по сравнению с количеством обычных клеток) очень мало.
Существует ли запас "новых" стволовых клеток или нет,
накопление стареющих клеток, которые не могут делиться,
по-видимому, является проблемой для некоторых тканей.
Большинство клеток, достигая лимита Хейфлика, не умирает. Они
продолжают оставаться в этих же тканях. И так как стареющие клетки
становятся более крупными и менее активными, чем молодые, их
накопление ведет к возрастной потере функциональных
способностей некоторых органов и тканей. Было найдено, в частности,
что кожа старых индивидуумов имеет приблизительно в три раза
больше стареющих фибробластов, чем кожа молодых людей
(Counter, 1996). Кроме того, повреждения, возникающие близко к
концу хромосомы, хуже репарируются, что может служить
причиной нестабильности теломерной ДНК и, в частности,
приводить к накоплению в ней одиночных разрывов (Егоров, 1997;
Егоров и др., 2000). И это, возможно, влечет за собой более быстрое
исчерпание лимита клеточных делений и вносит некоторый вклад
в старение организма.
Однако о соотношении лимита пролиферации клеток и
старения организма в целом лучше всех, естественно, высказался
недавно сам Леонард Хейфлик: "...мы не думаем, что старение
89
является результатом потери потенциала клеточного
деления. Максимальное число удвоений популяции может просто
быть мерой предела долговечности, который можно
продемонстрировать in vitro, но никогда нельзя достигнуть in vivo,
поскольку накапливающиеся физиологические нарушения
предрасполагают животное к смерти задолго до исчерпания
лимита клеточных делений" {Хейфлик, 1997).
Тем не менее определенный вклад лимита делений в
интегральный механизм старения не вызывает сомнений, и поэтому
естественно возникает вопрос - можно ли устранить лимит Хейф-
лика? Определенно - да. Помимо молекулярного механизма
укорочения теломер, A.M. Оловников (1971) представил и гипотезу о
существовании особого биологического механизма, решающего
проблему недорепликации теломер. Такой механизм, по его
мнению, должен действовать в клетках организмов,
размножающихся вегетативным путем, а также в половых и раковых клетках, но
не должен работать в большинстве наших соматических клеток.
Очевидно, некоторые мутации, которые превращают
нормальные клетки в раковые, приводят в действие механизм,
отменяющий запрет Хейфлика. Также и некоторые вирусы, особенно
вирус папилломы, инфицируя клетки, делают их бессмертными,
или, как говорят онкологи, иммортализируют их. Гипотеза
A.M. Оловникова оставалась умозрительной, пока в 1985 г.
Грейдер и Блэкборн не обнаружили главный действующий фактор
этого механизма - фермент тело мер азу (Greider, Blackburn,
1985). Теломераза способна наращивать концы линейных
молекул ДНК, т.е. теломеры, содержащие многократно
повторяющийся гексамер (TTAGGG)n.
Теломераза содержит короткий РНК-компонент,
комплементарный одному повтору G-богатой цепи теломерной ДНК (Докудов-
ская и др., 1997), и играющий роль "затравки" при достраивании
теломеры.
Знание молекулярного механизма лимита Хейфлика и работы теломера-
зы может быть крайне важным не столько в связи с проблемами старения,
сколько при разработке новых методов борьбы со злокачественным ростом
клеток. Так, если удастся разработать способы введения в раковые клетки
препаратов, связывающих РНК-компонент теломеразы, то можно, по всей
вероятности, добиться укорочения теломер и последующего старения и
гибели этих клеток.
Активность теломеразы у высших животных обнаружена
только в трех типах клеток: генеративных, раковых и линиях имморта-
лизированных клеточных культур. При дифференцировке клеток в
процессе развития организма теломераза репрессируется, т.е. ген, в
котором записана информация о структуре этого фермента,
становится неактивным. Но если по какой-либо причине ген теломеразы
активируется, то клетки получают способность делиться неограни-
90
чено. Так, Джерри Шеем и его сотрудникам удалось увеличить
число делений нормальных соматических клеток человека в культуре
на 40%. С помощью генно-инженерных методов в клетки вводился
ген каталитической белковой субъединицы теломеразы и
прилегающий к нему участок ДНК, регулирующий его работу. При
активной работе этого гена увеличивался как размер теломерной ДНК,
так и продолжительность жизни клеточных культур. Сверх
обычных 50 делений клетки прошли дополнительно еще 20 делений (см.
обзор: Дымшиц, 2000).
Что все это означает в терминах продления жизни? Прежде
всего существование лимита Хейфлика, с одной стороны, может
вносить вклад в возрастные изменения и болезни некоторых
тканей, таких как кровеносные сосуды, клетки эпителия кишечника,
кожи и ряда других органов и тканей. С другой стороны, полная
отмена лимита Хейфлика (даже если бы это было возможно)
представляла бы большую опасность из-за повышения вероятности
возникновения рака. При рассмотрении этой проблемы нужно
учитывать также, что количество делений в пределах лимита,
по-видимому, не является жестко фиксированной величиной.
Различные факторы окружающей среды могут ускорять или
замедлять работу этого клеточного репликометра. Повышенный
уровень свободных радикалов, как было показано, сокращает лимит
Хейфлика, а некоторые факторы роста (белки, стимулирующие
рост клетки) в некоторых тканях расширяют этот предел.
Интригующий результат был получен группой ученых из Нидерландов,
показавших, что экстракт чеснока может расширить предел
Хейфлика и улучшить жизнедеятельность фибробластов кожи в
культуре {Svendsen et al., 1994).
Что мы можем сделать, чтобы минимизировать вклад механизма
ограничения делений в общий процесс старения? Использование генно-инженерных
методов, вероятно, позволит в будущем изменять генетическую программу,
которая определяет предел Хейфлика (хотя здесь, возможно, скрыты опасности,
связанные с увеличением риска заболевания раком). Лучшее, что мы можем
сделать сегодня для замедления движения к лимиту, это избегать "ненужных"
клеточных делений, а также стремиться к расширению предела, улучшая
внутреннюю среду нашего организма. Уход от "ненужных" клеточных делений
означает минимизацию подверженности факторам, которые стимулируют
деление. Все типы повреждений ткани и клеточного стресса (см. II часть книги)
могут стимулировать деление клеток. В частности, свободные радикалы,
воспаление, мутагены, ингибирование синтеза белков, некоторые токсины и УФ-облу-
чение, как было показано, стимулируют деление клеток. Антиоксиданты,
напротив, приводят, очевидно, к противоположному результату. Некоторые
вещества, такие как экстракт чеснока, возможно, способны расширить лимит
Хейфлика.
91
2.7. ГЕНЕТИЧЕСКИЕ АСПЕКТЫ СТАРЕНИЯ
Никто из нас не совершенен. Все больше
генетических тестов становится доступно,
к каждый из нас в конечном счете
обнаружит у себя мутацию, предрасполагающую
к какой-нибудь болезни...
Френсис Коллинз, директор программы
"Геном человека"
В настоящее время отсутствуют какие-либо серьезные
основания для утверждения, что существуют специфические гены старения
или гены смерти, хотя спорадически предпринимаются попытки
выявить такие гены. С возрастом изменяется состояние столь
большого числа различных функций (и эти изменения вносят столь
значительный вклад в формирование картины старения), что все
генетически контролируемые изменения этих функций, например
репродуктивной системы, могут быть отнесены к целому "семейству" генов
старения. В этом случае скорее можно говорить о плейотропном
(отсроченном) действии многих генов (см. параграф "Центральные
часы старения"), а не о специальном свойстве определенных генов
индуцировать старение (Дильман, 1987). В лучшем случае можно
говорить лишь о "рейтинге ** тех или иных генов по их вкладу в
интегральный процесс старения.
Большинство известных мутаций неблагоприятно для развития и
является причиной разнообразных наследственных болезней. О числе
потенциальных наследственных болезней можно судить, исходя из оценки общего числа
генов у человека, которое составляет 40-70 тыс. (Dunham et al., 2000; Hattori
et al.t 2000). "Генетический груз" - сумма циркулирующих в популяции
мутаций и изменений, связанных с естественным генетическим полиморфизмом.
По некоторым оценкам (Бочков, 1995) величина генетического груза в
европейских популяциях и в России составляет около 5.5%, из которых примерно
1 приходится на моногенные болезни (мутации одного гена), 0.5 связаны с
хромосомными аберрациями у новорожденных и 3-4% составляют мульти-
факторные болезни с выраженным генетическим компонентом (Харпер,
1984). Анализ расшифрованной последовательности ДНК генома человека
наряду с данными медицинской генетики позволит понять в будущем, какие
гены и группы генов вовлечены в "генные сети", определяющие этнологию
и патогенез того или иного мультифакторного заболевания, и каков вклад
отдельных генов в развитие патологического процесса, в частности
дегенеративных возрастных болезней. Прекрасное изложение и обсуждение многих
медицинских проблем в связи с эпохальным достижением - расшифровкой
структуры генома человека - можно найти в монографии B.C. Баранова и
его сотрудников «Геном человека и гены предрасположенности". Введение
в предикативную медицину», опубликованную в 2000 г.
У человеческой популяции обнаружено более 2500 синдромов,
детерминированных генетически (Martin, 1978). Однако в данном
контексте мы не будем касаться всех врожденных заболеваний, но
92
затронем лишь те, которые по клиническим проявлениям
соответствуют главным дегенеративным (возрастным) болезням человека,
ускоряющим процессы нормального старения.
Дегенеративные болезни. В сложном многокомпонентном
патогенезе атеросклероза могут быть различные элементы
генетических нарушений, которые приводят к развитию болезни даже при
отсутствии неблагоприятных экологических факторов.
Изучение К. Бергом липопротеинов низкой плотности - ЛНП (главной
частицы, переносящей в крови человека холестерин) и родственных молекул
привело около 40 лет назад К открытию липопротеина (а). Было показано,
что присутствие в крови этого соединения является наследуемым признаком,
и у индивидов, имеющих липопротеин (а), вероятность сердечных приступов
выше, чем в контрольной группе (Scanu, 1990; Scanu et al., 1991). Наблюдалась
положительная корреляция между содержанием липопротеина (а) в крови и
атеросклерозом. Более того, липопротеин (а) был выявлен в атеросклероти-
ческих бляшках. Высокий уровень этого белка связан, о чем
свидетельствуют клинические наблюдения, с сердечными приступами, инфарктом,
инсультом, сужением артерий и замыканием сосудов после операции шунтирования
венечных артерий. У мужчин моложе 60 лет в ~25% случаев сердечных
приступов отмечался наследственно высокий уровень липопротеина (а) в крови.
Высокий уровень этого белка считается наиболее опасным наследственным
фактором риска сердечных заболеваний. Установлено, что при данном
количестве липопротеина (а) в крови создается такой же уровень риска, как и
при десятикратно повышенном уровне ЛНП. Таким образом, поиск частиц,
родственных ЛНП, привел к открытию наследственно детерминированного
липопротеина с особыми и, по-видимиму, опасными свойствами. К этому
следует добавить, что трансгенные мыши, которым был пересажен
человеческий ген липопротеина (а), действительно более подвержены атеросклерозу
(Дон, 1992).
Известна также роль фамильной гиперхолестеринемии, а
следовательно, и высокого уровня ЛНП, как патогенного фактора в
развитии ранних форм атеросклероза и инфаркта миокарда
(Frederickson et al., 1978). Однако при генетическом синдроме
долгожительства имеет место понижение уровня липопротеинов очень
низкой плотности (ЛОНП) и повышение уровня липопротеинов
высокой плотности - ЛВП (Receputo et al., 1995), которые
обеспечивают поступление избытка холестерина с периферии тела в печень для
последующего уничтожения (Glueck et al., 1977).
По-видимому, существуют наследственные синдромы, при
которых многократно возрастает частота возникновения рака.
Основной причиной указанных синдромов могут служить
иммунологическая недостаточность (Filipovich et al., 1980), снижение
активности систем репарации ДНК (Виленчик, 1987), преждевременное
(генетическое) старение, например при болезни Дауна (Brooksbank,
Balazs, 1983), наследственно повышенная активность энзиматических
систем, трансформирующих ксенобиотики в потенциальные
канцерогены (Саприн, 1991), некоторые хромосомные нарушения (Дилъман.
1987) и т.д.
93
Генетическая обусловленность ожирения вытекает из
некоторых статистических данных. У родителей без избыточной массы
тела только 10% детей страдают ожирением, У 50% детей
наблюдается склонность к ожирению, когда один из родителей имеет избыток
веса. И у 80% детей наблюдается избыток массы при обоих тучных
родителях (Mayer, 1965).
Эволюционно закрепленную предопределенность к диабету
типа II у млекопитающих мы уже обсуждали в параграфе
"Глюкоза и старение". Четкая генетическая обусловленность этой
формы диабета хорошо прослеживается у однояйцевых
близнецов. Если заболевание возникает у одного из них, то практически
со 100% частотой оно возникает и у другого. Изменения в
углеводном метаболизме, характерные для диабета типа II, в той или
иной степени выраженности возникает в процессе старения
почти у всех людей, особенно если при этом развивается и возрастное
ожирение (Дильман, 1987).
Генетические изменения в регуляторной (промоторной) области
гена инсулина могут определять увеличение скорости продукции
и секреции инсулина, что ведет к развитию гиперинсулинемии,
которая при наследственных формах диабета II типа является
обязательным признаком заболевания (Gerich, 1984). Тем не
менее в процессе нормального старения гиперинсулинемия (см.
табл. 4) чаще обусловливается не генетическими причинами, а
возрастным накоплением жира и сопряженным с этим снижением
чувствительности тканей к инсулину
Наличие семейной предрасположенности к гипертонии
является хорошо установленным фактом (Lund-Johanseny 1984).
Существенно, что у детей со сниженным содержанием ЛВП, что
сопряжено с развитием атеросклероза, наблюдается более высокое
артериальное давление {Siervogel et al., 1981). У животных со
спонтанной (генетической) гипертонией ограничение пищевого
рациона снижает артериальное давление и увеличивает
продолжительность их жизни до нормального уровня. Напротив, стресс
вызывает развитие ожирения, гиперфункцию коры надпочечников,
инволюцию тимуса и стойкое повышение артериального
давления (Wexler, 1976). При стрессе, воздействие ангиотензина II на
клетки клубочковой зоны надпочечников повышает выделение
альдостерона, который, воздействуя на почки, стимулирует в
организме накопление натрия (Alkon et al., 1988), что и приводит к
повышению артериального давления.
Психическая депрессия существенно выше среди детей,
родители которых страдают этим заболеванием, чем в общей популяции
(Weissman et al., 1984a). Частота психической депрессии значительно
выше у молодых женщин, чем у мужчин, что, очевидно, связано с X-
хромосомной зависимостью наследования, а у детей возможность
развития заболевания может свидетельствовать о врожденной недо-
94
статочности нейромедиаторных систем в определенных отделах
мозга (Weissman et al., 1984). Многочисленные негенетические факторы
развития дегенеративных болезней могут способствовать
выявлению генетической предрасположенности к психической депрессии
Болезнь Альцгеймера - нейродегенеративное заболевание,
развивающееся сравнительно поздно, после 60 лет, и наследующееся по
аутосомно-доминантному типу в семейных случаях. Нередко
возникает спонтанно. Эта болезнь определяется изменениями в "генной
сети", которая включает в себя по крайней мере шесть различных
генов. Это прежде всего мутации в так называемых пресенилиновых
генах (PS-1; PS-2, локализованных в хромосомах 14 и 1,
соответственно), а также в гене предшественника амилоидного β-белка
(β ΑΡΡ, см. параграф "Накопление шлаков"), локализованных в
хромосоме 21. Помимо этих трех генов, анализ сцепления в семьях
высокого риска болезни Альцгеймера, позволил выявить еще три
гена, не случайно ассоциированных с этой болезнью. Это - ген
АроЕ4, локализованный в хромосоме 19 {Баранов и др., 2000). У
пациентов с мутациями PS-генов и "плохим" аллелем АроЕ4
заболевание начинается в среднем на пять лет раньше и протекает
существенно тяжелее, чем при мутациях в других аллелях {Pookaj et al.,
1998). Кроме того, в эту патологию вовлечены и
гены-модификаторы - ген ос2-макроглобулина, а также и такой весьма важный ген,
как АСЕ (кодирующий структуру ангиотензинконвертирующего
энзима) {Owen et al., 1999).
Необходимо, однако, подчеркнуть, что в подавляющем
большинстве случаев болезнь Альцгеймера является результатом
неблагоприятных взаимодействий между генетическими и экзогенными
факторами метаболической и экологической природы {Баранов и
др., 2000).
Следует упомянуть и некоторые другие наследственные
синдромы, определяющие пути преждевременного старения: нарушения
метаболизма липидов, аутоиммунные поражения, нарушения систем
репарации ДНК, диабетоподобные сдвиги, гипогонадизм,
дегенеративные сосудистые изменения, отложения амилоида и липофусцина,
остеопороз, региональный фиброз, снижение иммунитета и т.п.
Таким образом, наследственность оказывает несомненное
влияние на продолжительность жизни {Войтенко, 1983), но если
исключить явные моногенные генетические дефекты, то это влияние, по
всей вероятности, связано с "генными сетями" и реализуется путем
изменения скорости старения, а не определяется специальными
генами старения. Тем не менее новейшие открытия клеточной
биологии позволяют ряду авторов реанимировать старую идею А. Вейс-
95
мана о жесткой генетической детерминации смерти
многоклеточных животных (Weismann, 1882). В связи с этим (помимо лимита
Хейфлика) следует указать на успехи в изучении апоптоза -
программируемой клеточной гибели (ПКГ) {Chinnaiyan et al., 1996;
Уманский, 1996; Nagate, 1997; Хансон, 1998).
Апоптоз - механизм клеточной смерти. В клеточный состав
многоклеточного организма постоянно изменяется в результате
двух фундаментальных биологических процессов - пролиферации и
клеточной гибели. Еще в 70-х годах прошлого века было проведено
четкое разделение двух основных типов клеточной гибели:
токсического {некроз) и программируемого {апоптоз) {Kerr, 1971; Wyllie et
al., 1980). При некрозе, который наблюдается при различных
повреждениях, клетки обнаруживают тенденцию к набуханию, разрыву
мембраны и высвобождению клеточного содержимого, что
приводит к болезненной воспалительной реакции в окружающей ткани. В
противоположность этому при ПКГ происходит уменьшение
объема клетки, конденсация ядерного хроматина и образование так
называемых апоптозных телец, покрытых мембраной. Эти
изолированные фрагменты погибающих клеток подвергаются быстрому
фагоцитозу макрофагами или соседними клетками, не успевая
разрушиться и вызвать воспалительную реакцию.
Равновесие между ПКГ и пролиферацией играет важную роль в
процессах нормального онтогенетического развития организма, а
именно в процессах эмбриогенеза, морфогенеза, дифференцировки,
в поддержании клеточного и тканевого гомеостаза {Kerr et al. 1972).
Напротив, нарушение контроля ПКГ при росте и развитии органов
и тканей может привести к возникновению различных патологий, в
частности злокачественных новообразований, нарушению
иммунного ответа, а также к потере тканевого гомеостаза, что, очевидно,
будет способствовать и дегенеративным (возрастным) болезням {Kerr
et al., 1980; Wyllie, 1987; Compton et al., 1992; Bredesen, 1994; Thompson,
1995; Непомнящих и др., 2000). Исключительно важную роль
апоптоз играет в процедуре удаления из организма потенциально
опасных клеток. Так осуществляется контроль над клеточным гомео-
стазом.
Толчком к интенсивному изучению механизмов апоптоза
послужило открытие межнуклеосомной деградации хроматина, сделанное
практически одновременно тремя группами исследователей {Skalka
et al., 1976; Yamada et al., 1981; Хансон, 1979; Zhivotovsky et al., 1981).
Было установлено, что хроматин в лимфоидных и гемопоэтических
тканях облученных животных подвергается распаду на кратные ну-
клеосомные фрагменты различной длины. Аналогичные изменения
наблюдались и в тимоцитах, подвергнутых действию глюкокортико-
идов {Wyllie et al., 1980; Wyllie et al., 1984). Этот процесс
инициируется эндонуклеазами и обнаруживается практически во всех случаях
ПКГ {Zvonareva et al., 1983; Seiliev et al., 1992).
96
Центральную роль в апоптозе играет специфический класс цистеиновых
иротеаз. С апатозом связано 11 цистеиновых протеаз, называемых каспазамн
(Kuida et al., 1996). Каспазы работают по принципу каскада (Nagata, 1997), акти-
пируя друг друга и расщепляя отдельные внутриклеточные белки, такие как
белки ядер, ядерной ламины, цитоскелета, эндоплазматического ретикулума и
цитозоля. Некоторые из расщепленных белков запускают другие
деструктивные процессы и таким образом обеспечивают быстрый и эффективный
"демонтаж" клетки (Chinnaiyan et al., 1996). Известны специфические ингибиторы этих
ферментов, что позволяет (когда это необходимо) подавлять процесс апоптоза.
Каспазы, таким образом, являются ядром апоптозной программы во всех
клетках животных.
Исключительно важную роль в распознавании сбоев и повреждений в
клетке, в ограничении делений и апоптозе таких клеток играет белок р53
(53 - масса этого белка в КД). По современным представлениям, р53
является белком, контролирующим информационные и временные параметры
репликации ДНК и в целом клеточный цикл. Сигналы, возникающие в
результате подобного контроля, позволяют (в случае существенных повреждений в
геноме) включить цепь реакций, которые должны предотвратить
дальнейшее развитие патологии (Lane, 1992; цит. по: Чумаков, 1998). Разнообразные
сбои и повреждения снижают скорость распада короткоживущего р53 и,
следовательно, приводят к накоплению этого белка и усилению его функций.
р53 - полифункциональный белок, он способен специфически связываться с
20 различными белками, а также регулировать транскрипцию целого ряда
генов. Однако наиболее важными функциями р53 являются, во-первых,
задержка клеточного деления и усиление репаративных процессов для
исправления возникших повреждений ДНК. При значительных повреждениях ДНК
р53 специфически индуцирует экспрессию неизвестной ранее изоформы
малой субъединицы рибонуклеотид редуктазы (R2), которая включается
только в процесс репарации, локализуясь в ядре клетки и резко ускоряя
превращение рибонуклеозиддифосфатов в дезоксирибонуклеозидтрифосфаты
поблизости от участков поражения ДНК (Tanaka et al., 2000), после чего клетка
продолжает свое существование. Во-вторых, при чрезмерном нарушении
генома, когда ДНК уже не поддается репарации, включение рецепторной и ци-
тохром с-зависимой активации каскада каспаз проводит клетку к апоптозу
(Bates et al., 1999; цит. по: Самуилову и др., 2000). Таким образом, зависящий
от р53 апоптоз - мощный селектор, позволяющий избежать накопления
мутаций, а в случае, когда они уже возникли и продукты мутировавших генов
приводят к нарушениям в жизнедеятельности клетки, р53-зависимый апоптоз
позволяет элиминировать такие потенциально опасные для организма
клетки. Именно этот вид апоптоза, а не задержка клеточного цикла (обе эти
функции - независимые функции р53), служит основным механизмом,
сдерживающим образование опухолей (Чумаков, 1998)
Феноптоз — механизм программируемой гибели организма?
Недавно внимание научной общественности вновь было
привлечено к весьма старой дилемме - является ли старение и смерть
организма особой биологической функцией или результатом
поломки сложной живой системы (Скулачев, 1997). Удивительно,
какую почти мистическую привлекательность, сравнимую лишь
со "скромным обаянием" идеи perpetuum mobile, до сих пор имеют
идеи Августа Вейсмана о генетически жестко запрограммирован-
97
ной смерти как выдающемся эволюционном достижении с точки
зрения преимуществ популяции (Weisman, 1882). Несмотря на
серьезную критику этой гипотезы (Medowar,1952; Cutler, 1984;
Дильман, 1987), она периодически возрождается в связи то с
открытием' лимита Хейфлика, то с доказательствами
существования апоптоза. Новое обоснование гипотезы Вейсмана на этот раз
пришло со стороны не клеточной биологии, а биохимии (Скула-
чев, 1997, 1999). По аналогии с апоптозом новая интерпретация
гипотезы А. Вейсмана называется - феноптозом, т.е.
биохимическим "самоубийством" организма, очищающим популяцию от
"нежелательных" индивидов (Скулачев, 2002). Инструментами
феноптоза, по мнению автора, могут служить такие патологии,
как септический шок, инсульт, инфаркт и рак, и в основе этих
сложных патологий лежит гиперпродукция АФК с последующим
массовым митоптозом и апаптозом (Скулачев, 1999). Гипотеза
феноптоза представляется нам в высшей степени спорной и
требует как дальнейшего экспериментального обоснования, так и
всестороннего обсуждения, поскольку противоречит огромному
количеству исследований, посвященных возникновению,
развитию и природе указанных патологий. Подобная дискуссия,
естественно, выходит за рамки этой книги, и здесь мы лишь напомним,
что по современным представлениям и лимит Хейфлика, и апоп-
тоз — прежде всего механизмы, защищающие организм от
развития смертельных патологий, и их участие в феноптозе является
пока еще не только фактически, но и логически необоснованным.
Конечно, доведенный до гипертрофированных масштабов апоп-
тоз может повлечь за собой (как и многие другие, доведенные до
крайности изменения, например, массированный некроз клеток в
жизненно важных органах и тканях ) гибель организма, но это не
является аргументом в пользу их эволюционной роли в качестве
генетического механизма элиминации организмов с низкой
жизнеспособностью. Термин "феноптоз", основанный на
поверхностной аналогии с термином "апоптоз", представляется нам
неудачным не только с гносеологической точки зрения, но также и
потому, что может отчасти дезориентировать некоторых
геронтологов ври исследовании механизмов старения, а также
патогенеза основных дегенеративных болезней человека. Материалы
этой книги, как нам кажется, позволяют в известной мере
составить представление о механизмах возникновения и развития
сердечно-сосудистых заболеваний и рака (главных
дегенеративных болезней человека) без их жесткой генетической
предопределенности.
98
2.8. СУППРЕССОР ОПУХОЛИ р53 И СТАРЕНИЕ
Между Сциллой и Харибдой:
действительно ли преждевременное старение
- цена, чтобы избежать развития рака?
J.C. Ameisen, 2002 г.
Как уже указывалось в предыдущем параграфе, р53 - белок суп-
прессор опухоли, отвечающий на различные клеточные стрессы,
включая поражение ДНК, гипоксию, измененную экспрессию ряда
онкогенов, а также способный индуцировать остановку клеточного
цикла или апоптоз (Ко et al., 1996; Levin, 1997; Giaccia et al., 1998).
Связь между активацией р53 и старением впервые была
отмечена на клеточном уровне. Так, внесение в клетки вирусных онкобел-
ков, мутантных форм р53 или антител, которые инактивируют
"дикий" тип р53, замедляло старение первичных человеческих клеток
(Shay et al., 1991; Bond et al., 1994; Gire et al., 1998). Наоборот, когда
культивированные диплоидные человеческие фибробласты
старели, у них повышалась активность р53 (Atadja et al., 1995; Bond et al.,
1996; Webley et al., 2000; hahana et al., 2001). Введение онкогена Ras в
первичные фибробласты мышей инициировало митозы, но затем
индуцировало преждевременное старение, зависящее от
"нормального" р53 (Serrano et al., 1997). Все эти данные поддерживали
гипотезу, согласно которой клеточное репликативное старение работает
как механизм суппрессии опухолей (Sager, 1991; Campisi, 1997).
Предпринимались также и попытки обнаружить влияние р53 на
продолжительность жизни целостного организма. Была получена
генерация мышей с дефицитом р53, но препятствием при изучении
старения в этих экспериментах оказалось раннее развитие рака
(Donehower et al., 1992; Jacks et al., 1994; Purdie et al., 1994). Напротив,
попытка генерировать мышей с повышенной экспрессией дикого
типа р53 не была успешной, вероятно, вследствие вредного
эффекта увеличения р53 на развитие эмбриона (Bernstein, Butel, цит. по:
Tyweretal.,2002).
Недавно большой группе американских исследований удалось
показать, что р53-мутантные мыши, с одной стороны оказывают
повышенное сопротивление раку, но, с другой - проявляют фенотипы
ускоренного старения (Tyner et al., 2002). Трансгенные мыши, созданные
этими авторами, несли делецию первых шести (из одиннадцати) экзо-
нов гена р53, что приводило к экспрессии "усеченных" мРНК. Хотя
авторы не смогли обнаружить белок, соответствующий этой "irT-му-
тации в тканях р53 +/гл мышей, они нашли повышенный уровень
регулируемой активности р53 у этих гетерозигот. "т"-аллель,
возможно, увеличивал стабильность белка дикого типа р53, активируя NAD-
99
Η хинон оксидоредуктазу 1, регулирующую стабильность этого
белка {Asher et al., 2001). Однако высоко интригующий результат был
связан с тем, что гетерозиготные р53+/т мыши развивали меньшее
количество спонтанных опухолей, чем мыши с диким типом р53+/+
гомозиготы, и по целому ряду признаков демонстрировали более
быстрое старение. Частота встречаемости рака была < 6% у р53 + /т
животных, сравнительно с > 45% у р53 +/+ животных, принимая во
внимание, что средняя продолжительность жизни этих двух групп
животных составляла 96 и ] 18 недель соответственно.
Касаясь наблюдаемых признаков старения, следует
подчеркнуть, что р53 +/т мыши показывают более быструю зависящую от
возраста потерю в весе мышц, селезенки, печени, почек и яичек.
Сокращение массы органов, повсюду обуславливалось общим
уменьшением клеточности. Ускоренные возрастные потери были также
отмечены в толщине кожи, росте волос, заживлении мелких ран,
сопротивлении стрессу. Тем не менее не наблюдалось никакого
ускоренного старения в ряде других маркеров старения, включая
патологию печени, поседение волос и облысение, атрофию кишечных
ворсинок, изъязвление кожи, атрофию мозга, амилоидное
депонирование, катаракты и общую дезорганизацию функций организма. В
этом отношении р53 +/т животные напоминают некоторые
человеческие синдромы ускоренного старения, типа синдрома Вернера,
которые могут проявляться частично и неполно имитируют быстрое
человеческое старение {Martin et al., 2000). Такие проявления
соответственно называют "сегментарные синдромы старения97. Спектр
специфических изменений, отмеченных у р53 +/т мышей не coot
ветствовал, однако, таковым при синдроме Вернера или других
человеческих патологиях.
Тэйнер и др. (Тупег el al., 2002) представили также данные по другой
группе мутантных мышей (pL53), которые были " сконструированы" так, чтобы
содержать примерно 20 копий, чувствительных к температуре точечных мутаций
р53, и также проявляли фенотипы ускоренного старения более очевидно
именно в тех тканях, подобных коже, где окружающая температура более низкая и
с большей вероятностью приводила к повышенной активности р53.
Главный вывод, которые сделали авторы этой крайне интерес
ной работы, состоит в том, что р53 - суппрессор опухоли - играет
существенную роль в регуляции старения и продолжительности
жизни у мышей. Кроме того, полагают авторы, эти результаты
поддерживают идею, что старение служит механизмом подавления опу
холи (Sager, 1991; Campisi, 1997). Касаясь объяснения своих неожи
данных результатов, они заключают, что "некоторые из феноти
пов старения позволяют предполагать сокращение пролиферации
стволовых клеток. С процессом старения, этот пролифератив-
ный резерв может уменьшаться более быстро у р53 +1т мышей,
поскольку их стволовые клетки подвергаются репликативному
старению скорее, чем у ихр53 +/+ партнеров" (Tynor et al., 2002).
100
Нам, однако, предложенное объяснение не кажется
убедительным, хотя бы потому, что существует очень мало доказательств,
подтверждающих идею, что стволовые клетки вообще
подвергаются регогакативному старению подобно тому, что наблюдается у
терминально дифференцированных клеток, таких как фибробласты
(Kirkwood, 2002). Мы бы хотели предложить совершенно иную
интерпретацию этих замечательных результатов.
Хорошо известно, что АФК являются мощными
индукторами активности р53 и, кроме того, они играют роль в реализации
апоптоза, зависимого от р53. Группой итальянских
исследователей (Pani et ah, 2000) было показано, что трансформированные
фибробласты мыши, испытывающие недостаток р53,
существенно более устойчивы, чем контроль дикого типа, к цитотоксиче-
скому эффекту ряда прооксидантов. Эти клетки, подчеркнем,
обладали также и повышенной экспрессией марганцевой
супероксид дисмутазы (MnSOD), митохондриального фермента,
который, как известно, предохраняет клетки, в частности раковые, от
окислительных повреждений различными противоопухолевыми
агентами. Активность MnSOD также была увеличена и в ткани
печени мышей с дефицитом р53 по сравнению с тканью дикого
типа. Напротив, кратковременная трансфекция р53 (дикого
типа) в Не La клетки вела к существенному сокращению
стационарного уровня мРНК MnSOD и ее ферментативной активности,
показывая, что экспрессия этого антиокислительного фермента
негативно регулируется р53. Эти данные идентифицируют
MnSOD в качестве потенциального адресата для суппрессора
опухоли р53 и подчеркивают уместность модуляции MnSOD,
поскольку такая модуляция, по-видимому, необходима, учитывая
множество фактов о ненормально увеличенной экспрессии
MnSOD в раковых образованиях человека. Действительно, ингн-
бирование активности MnSOD делает раковые клетки, например
клетки рака прямой кишки с диким типом р53, более
восприимчивыми к апоптозу, который индуцируется облучением,
гипертермией и специфическими противоопухолевыми препаратами
{Kuninaka et al., 2000)
Помимо р53 и MnSOD, в сложную цепь реакций, приводящих
к апоптозу, включены продукты экспрессии более 20 генов, в
частности, каскад каспаз, митохондриальный цитохром С и по-
ли(АДФ-рибозо)полимераза (ПАРП). р53 является адресатом
ядерного фермента ПАРП, который активируется в результате
повреждений ДНК генотоксичными агентами (Mandiret al., 2002).
Повышенная экспрессия MnSOD ингнбировала активность
ПАРП, что приводило, как было показано (Kiningham et al., 1999),
к защите от апоптоза. Результаты этих исследований можно
рассматривать как свидетельство негативной регуляции р53 со
стороны MnSOD.
101
Прямые доказательства существования нзаимной
отрицательной {реципрокной) регуляции экспрессии генов MnSOD и р53,
соответственно, со стороны их белков, были получены Паскалем
Дранэ и его сотрудниками, (Drane et al., 2001). Эти авторы
показали, что дикий тип р53 подавляет экспрессию гена MnSOD на
уровне промотора и что, в свою очередь, повышенная экспрессия
MnSOD уменьшает транскрипцию на уровне промотора р53 и ин-
гибирует опосредованную р53 индукцию апоптоза. Это влияние
MnSOD ограничивает способность р53 стимулировать апоптоз
путем многоступенчатой реакции, включающей индукцию
транскрипции ряда генов* связанных с
окислительно-восстановительным потенциалом и играющих роль в продукции АФК,
препятствует окислительному разложению митохондриальных
компонентов и, наконец, апоптозу (Polyak et al., 1997).
р53, как было показано ранее, ингибирует и две других
супероксид дисмутазы, Cu/ZnSOD цитозоля (Cho, et al., 1997) и Cu/ZnSOD
межклеточной жидкости (Zhao et al., 2000^
Возвращаясь к ускоренному старению, вызванному
непрогнозируемым влиянием повышенной активности р53 (Tynor et al.,
2002), мы можем предложить следующее объяснение этих
неожиданных эффектов. Как указывалось выше, р53 специфически (на
уровне промотора) ингибирует экспрессию различных SOD и
прежде всего MnSOD митохомдрий. Повышенная активность р53,
следовательно, приведет к существенному снижению
антиокислительного потенциала митохондрий и клетки в целом, что не
может не увеличить разрушительного действия АФК, которое в
конце концов выразится в развитии митоптоза (подробнее об
этом в параграфе "Оксидативный стресс клетки", гл. 19) и
последующем повышении уровня апоптоза. Далее, неспецифический
цитолиз первичных апоптотических клеток, как было показано
(Terenius et al., 2000; Скулачев, 2002), вызовет кластеризацию
апоптотических клеток, что, несомненно, должно привести к
снижению массы и функции органов и тканей, степень которого
будет зависеть как от степени активации р53, так и от общего
антиокислительного потенциала в том или ином органе или ткани, т.е.
от уровня окислительного стресса. Именно такое сокращение
клеточности и массы целого ряда органов и тканей наблюдали
авторы у мутантных мышей с повышенным уровнем р53 (Tynor et
al., 2002). Все эти факторы, как нам кажется, и предопределяют
существенное сокращение продолжительности жизни у р53 +/т
гетерозиготных мышей. Конечно, перманентный оксидативный
стресс способен вызвать ускорение репликативного старения
(Saretzkiet al., 1999; Toussaint et al., 2002), с чем и связывают
авторы наблюдаемые феномены, однако в рассматриваемом
эксперименте этот фактор представляется нам второстепенным (см.
параграф "Лимит Хейфлика и старение клеток" этой главы).
102
Сенсационная работа Тинора и соавторов вызвала
оживленную дискуссию в научной литературе (Kirkwood, 2002; Ameisen,
2002; Ferbeyre et al., 2002), которая сводилась к вопросу -
является ли старение ценой, которую платит организм за работу
механизма, осуществляющего апоптоз раковых клеток! У нас нет
возможности детально обсуждать этот вопрос, но мы хотели бы
указать на другой аспект, который невольно затрагивает работа
американских авторов.
Беспрецедентные успехи молекулярной генетики в
расшифровке структуры генома человека и успехи экспериментов,
использующих трансгенный подход, породили преувеличенные
оптимистические ожидания новых блестящих успехов в лечении человеческих
болезней, продлении жизни, улучшении способностей индивида и
т.п. и т.д. Эвристическая ценность работы Тинора и соавторов
заключается, между прочим, и в том, что она убедительно
демонстрирует насколько непредсказуемы и опасны по последствиям могут
быть попытки прямого трансгенного вмешательства в организацию
генома высших организмов с его исключительно сложным и тонким
взаимодействием генов, которое оптимизировано длительным
эволюционным развитием. Нам наглядно показали, как естественная и
понятная попытка добиться снижения уровня спонтанного рака
путем повышения активности суппрессора опухоли - белка р53,
приводит к развитию фенотипов раннего старения!
Только тщательные и всесторонние исследования этой
проблемы в будущем ответят на вопрос - можно ли успешно провести
человеческий организм между Сциллой и Харибдой баланса и
регуляции его защитных и деструктивных сил?
Глава 3
ДЕГЕНЕРАТИВНЫЕ БОЛЕЗНИ
Добавить годы к вашей жизни
или добавить жизни к вашим годам?
Те, кто не умирают, живут до
шестидесяти, до семидесяти, бедствуют, строчат
мемуары, путаются в ногах. Я
вглядываюсь в их черты пристально, как Миклу-
ха Маклай в татуировку
приближающихся дикарей.
Иосиф Бродский
Конечно, можно прибавить к вашей жизни двадцать или более
лет, но эти дополнительные годы скорее всего будут заполнены
болезнями, страданием, частыми госпитализациями и безрадостным
серым существованием. Стоит ли к этому стремиться? Есть ли
смысл в усилиях, направленных на продление жизни? Существует
немало людей, которые полагают, что смерть в молодом, или по
крайней мере в здоровом среднем возрасте - наиболее достойный и
наименее болезненный выбор. Действительно, старость часто
представляется нам как синоним болезни. Фактически имеется группа
болезней, называемых возрастными, или дегенеративными, частота
которых резко увеличивается с возрастом. В развитых странах, эти
болезни считаются причиной более 80% всех смертных случаев.
Предотвращение или отсрочка этих болезней может потенциально
увеличивать предполагаемый срок активной жизни и делать эти
добавленные годы более продуктивными и приятными. К сожалению,
современная медицина, если и способна в значительной степени
справиться с дегенеративными болезнями и таким образом продлить
нашу жизнь, то очень часто будет не в состоянии сохранить ее
высокое качество.
Термин "дегенеративные болезни" подразумевает, что они не
вызваны инфекцией, травмами, невзгодами или лишениями, но
являются результатом некоего общего, медленно и неотвратимо
развивающегося процесса, связанного с образом жизни или
фундаментальной природой всех живых существ. Типичные дегенеративные
болезни: атеросклероз, рак, гипертония, ожирение, диабет II
типа (развивается после 40 лет), психическая депрессия, болезнь
Альцгеймера (наиболее распространенная причина старческого
слабоумия), остеопороз (ломкость костей), артрит, гипертро-
104
фия предстательной железы.
Возрастное угасание функций
иммунной системы, хотя не
является болезнью в
общепринятом смысле, в значительной
степени ответственно за
смертные случаи от рака и
инфекционных болезней, в частности
пневмонии и гриппа, нередких
в пожилом возрасте (табл. 6).
Дегенеративные болезни
часто воспринимаются как
проклятие современности.
Многие полагают, что болезни
эти - прямой и неизбежный
результат нынешнего
цивилизованного образа жизни.
Столетие назад, смерть от
дегенеративных болезней была
относительно редка, но теперь
сердечно-сосудистые заболевания
и рак являются в развитых
странах причиной
приблизительно двух третей всех
смертных случаев. Конечно, образ
жизни играет роль в росте
возрастных болезней. Однако
равно существенно и то, что
теперь мы живем дольше, что и позволяет проявляться возрастным
болезням (табл. 7). Когда люди жили в пещерах или даже в замках,
выживание требовало очень крепкого здоровья. В отсутствии
антибиотиков, программ здравоохранения, медицинских технологий и
центрального отопления, любая болезнь, возрастная или нет, таила
смертную угрозу. Даже незначительное падение сопротивляемости
организма к стрессовым воздействиям или ослабление иммунной
системы могло оказаться достаточным для смертельного исхода при
инфекционных болезнях. Это - с одной стороны. С другой стороны,
относительная безопасность цивилизованного образа жизни
позволяет нам выживать с относительно низкой сопротивляемостью и
"вялой" иммунной системой в течение длительного времени,
достаточного для того, чтобы наши дегенеративные болезни смогли
проявиться. Фактически эта ситуация очень похожа на ту, которая
становится очевидной при сравнении жизни животных в дикой природе
и животных, живущих под опекой человека. Средняя
продолжительность жизни первых, как правило, значительно ниже, чем вторых,
поскольку раненое, больное или слабое животное всегда становится
Таблица 6
Десять главных причин смерти
в США
(Source National Center Health Statistics.
Annual Summary of Births,
Marriag Divorces, Deaths, 1990)
Причины смерти
(в порядке убывания частоты случаев)
Сердечно-сосудистые заболевания*
Рак*
Инсульт*
Несчастные случаи
Хроническая легочная
недостаточность
Пневмония и грипп**
Сахарный диабет***
Самоубийства
Хронические болезни печени и
цирроз
Убийства
* Возрастные болезни.
** Усугубляются возрастным
снижением функций иммунной системы.
*** Диабет II типа (независимый от
инсулина) является возрастным и сое -
тавляет более чем 90% от всех
случаев диабета.
105
Таблица 7
Пять главных болезней как основные причины смерти
в начале и середине XX столетия
(по Б. Стрелеру, 1964)
1900 г.
Пневмония, грипп
Туберкулез
Энтериты
Болезни сердца
Кровоизлияние в мозг
Вся доля пяти болезней
от общего числа случаев
смерти
%
U.8*
11.3
8.3
8.0
6.2
45.6
* Доля от общего числа случаев смерти
1959 г.
Болезни сердца
Рак
Кровоизлияние в мозг
Несчастные случаи
Некоторые детские
болезни
Вся доля пяти болезней
от общего числа
случаев смерти
%
38.6
15.7
11.5
5.4
4.1
75.3'
** Для возрастной группы 40 лет н выше доля указанных пяти причин сме{
возрастает до 80-85%
легкой добычей хищников. В неволе, наоборот, век животных
намного дольше, и в большинстве случаев это обстоятельство
позволяет проявиться дегенеративным болезням. В зоопарках старые
тигры, обезьяны и слоны часто страдают атеросклерозом, диабетом,
раком и другими болезнями, столь знакомым современным людям.
Почему дегенеративные болезни столь типичны? Почему у
большинства людей старение не бывает здоровым, т.е. не
проявляется в постепенном угасании функциональной активности наших
органов, но без дегенеративных болезней? И почему число этих
болезней относительно невелико (не более 10-12), хотя общее
количество известных человеческих болезней огромно? По-видимому, ответ
заключается в подобии механизмов старения и причин
дегенеративных болезней. Другими словами, те же самые процессы, которые
приводят к старению (гл. 2), способствуют также и развитию
дегенеративных болезней.
Повреждающее действие свободных радикалов вносит свой
вклад в развитие большинства дегенеративных болезней, включая
атеросклероз, рак, старческое слабоумие, катаракту и артрит.
Одним из хорошо известных примеров служит окисление липопроте-
идов низкой плотности (ЛНП), процесс, который вызывается СР.
ЛНП - шаровидные жировые тельца в крови, чья функция
состоит в доставке холестерина к различным тканям. Когда ЛНП
окисляются, они становятся "липкими" и обретают свойства
накапливаться в артериальных бляшках, вызывая прогрессию
атеросклероза. Окисление ЛНП может быть снижено жирорастворимыми
антиоксидантами, например витамином Ε или липоевой кислотой.
106
В ряде работ было показано, что высокие дозы витамина Ε
(400-1200 Ш в сутки) снижают риск сердечно-сосудистых
заболеваний, блокируя СР и таким образом предотвращая окисление
ЛНП (Sardesai, 1995; Packer et aL, 1997).
"Центральные часы старения" (локализованные в
гипоталамусе и смежных мозговых структурах) - другой важный фактор
развития дегенеративных болезней, поскольку вызывает ряд
последовательных изменений в состоянии параметров внутренней среды
организма, т.е. гомеостаза, что способствует развитию этих патологий.
В частности, избыток инсулина и пониженная толерантность тканей
к глюкозе увеличивает риск атеросклероза, гипертонии, диабета
II типа и ожирения. Чрезмерная секреция стероидных гормонов в
ответ на действие стрессоров также вносит свой вклад в развитие
атеросклероза, диабета, ожирения, пониженного иммунитета и рака
(Дилъман, 1987; Oilman, 1994). Высокий уровень пролактина -
основной фактор гипертрофии простаты.
Неферментативное гликозилирование (и вызванное им
перекрестное сшивание белковых молекул глюкозой) понижает
эластичность артерий и делает их более хрупкими, увеличивая
риск гипертонии, атеросклероза и, как следствие, кровоизлияний.
Эти процессы вызывают также катаракты и дисфункцию
иммунной системы.
Старческие пигменты, такие как липофусцин и цероид, будучи
результатом повреждающего действия СР и перекрестного
сшивания белковых молекул, накапливаясь в клетках, способствуют
старческому слабоумию и сердечной недостаточности.
Роль клеточных часов старения (Хайфлк-лимит) в
происхождении дегенеративных болезней неясна. С одной стороны,
ограничение числа клеточных делений может служить важным механизмом
предотвращения опасностей неконтролируемого клеточного роста.
Некоторые ученые полагают, что клеточные часы (или, как их еще
называют - репликометр клеточных делений) развились как
защитная антираковая система (Хейфлик, 1997; Егоров, 1997). С другой
стороны, этот лимит может обусловливать падение
функциональной и регенеративной способности тканей с высокой скоростью
клеточного деления. Например, пониженная способность к делению
эпителиальных клеток, выстилающих кровеносные сосуды, может
инициировать развитие атеросклероза.
Другой важный виновник старения и дегенеративных
болезней - стресс (см. гл. 17). Как мы уже отмечали, стресс ускоряет,
по существу, все проявления старения, а также увеличивает риск
дегенеративных болезней или создает условия для их развития.
Некоторые болезни более "отзывчивы" к стрессу, чем другие.
Гипертония, например, быстро обостряется при стрессе, так как
последний стимулирует повышенную активность симпатической
нервной системы (резкое увеличение выделения адреналина) , а
107
также повышает секрецию альдостерона, способствующего
накоплению натрия в организме, что является одной из главных
причин высокого кровяного давления. Интенсивный стресс
может (при отсутствии толерантности к глюкозе) ввергнуть
человека в диабетический кризис. Гормональные и метаболические
сдвиги, замеченные при стрессе, способствуют также
атеросклерозу, иммунной дисфункции, и повышенному риску заболевания
раком. Подавление стресса, а также повышение сопротивляемости
организма к нему благоприятны не только для продления жизни,
но также и для сохранения полноценного здоровья. Адаптогены
(соединения, которые смягчают и оптимизируют ответ организма
на стресс), как показали специальные исследования (Брехман,
1980; Дардымов, 1976; Лупандин, 1991; Todorov, 1993), были
эффективны в профилактике и даже в лечении многих
дегенеративных болезней на ранних стадиях.
В ряде случаев общепринятые способы лечения дегенеративных болезней
вызывают стресс, достаточно интенсивный, чтобы заметно снижать
положительный эффект, ожидаемый от этого лечения. Химиотерапия, γ-облучение
или хирургические операции, используемые при лечении рака, вызывают
сильный стресс, который подавляет иммунную систему пациента и таким образом
создает благоприятные условия для роста и размножения оставшихся в живых
раковых клеток. Ряд исследований показывает, что использование адаптогенов
в комбинации с обычной терапией понижает риск последующего рецидива
рака и его метастазов (см. гл. 29).
Другая важная особенность дегенеративных болезней - они
редко встречаются в единственном числе. Обычно человек либо не
имеет дегенеративных болезней, либо имеет их больше, чем одну.
Некоторые из "популярных" комбинаций в развитых странах -
атеросклероз, гипертония и ожирение, или атеросклероз, гипертония,
артрит и увеличение простаты. Одно из возможных объяснений
этой "кластеризации" состоит в том, что многие дегенеративные
болезни вызываются теми же самыми или подобными механизмами,
что и старение.
Изменения, ведущие к развитию дегенеративных болезней, можно разбить
на две группы. В первую группу входят изменения в темпе функционирования
ЦЧС, которые связаны с климаксом, гиперадаптозом, психической депрессией
и частично с гипертонией и возрастным ожирением. Ко второй группе следует
отнести метаболические нарушения, сопряженные с ожирением, диабетом
II типа, метаболической иммунодепрессией, атеросклерозом и канкрофилией
(повышенным риском заболевания раком). Причем это развитие возрастных
метаболических нарушений в определенной мере порождается все теми же ги-
поталамическими сдвигами (как это, в частности, проявляется в нарушении
регуляции аппетита, ведущего к ожирению).
Иногда в чертах даже одного человека можно разглядеть все десять
главных болезней старения одновременно. Такой комплекс нередко наблюдается
при некоторых видах рака, что приводит к вопросу - действительно ли
существуют раздельно все эти десять болезней, или они представляют собой десять
симптомов одной интегральной болезни - старения? (Дилъман, 1986).
108
Некоторые геронтологи даже полагают, что дегенеративные
болезни - просто характерные особенности старения и могут
использоваться как индикаторы биологического (в отличие от
хронологического) возраста. Однако такое представление является,
по-видимому, только фрагментом довольно сложной общей
картины процесса старения. Выраженные дегенеративные болезни
имеют более сложные причины и определяющие их факторы,
некоторые из которых связаны с механизмами старения, в то время
как другие - нет. Например, недостаток фолиевой кислоты
способствует атеросклерозу и сердечно-сосудистым болезням
(повышает в крови уровень гомоцистеина, метаболита,
способствующего развитию атеросклероза), но не имеет ничего общего с
процессом старения. Напротив, другой фактор атеросклероза, пере-
кисное окисление липидов, становится результатом
повреждающего действия СР, которое является важным элементом
механизма старения.
Избыток инсулина, распространенная эндокринная аномалия
старения, связанная с возрастным снижением толерантности
тканей к углеводам, является важной составляющей в развитии
нескольких возрастных болезней, а именно гипертонии,
атеросклероза, ожирения, диабета и, возможно, угнетения иммунитета, а
также повышенного риска злокачественного перерождения
тканей. Диабет II обычно начинается с гиперпродукции инсулина.
Далее, перенапряжение В-клеток поджелудочной железы
вызывает нх деградацию, что в свою очередь оборачивается в конце
концов падением содержания инсулина в крови. Этот кластер
нарушений, часто наблюдаемых у пациентов с избытком инсулина,
получил в медицинской литературе укоренившееся название
"синдром Х'\ Причины возрастного избытка инсулина все еще
недостаточно ясны, но кажется, что часть этих возрастных
эндокринных сдвигов сопряжена с разворачивающейся во времени
работой центральных часов старения. Следует добавить, что
описанная выше возрастная динамика содержания инсулина в крови
впервые была замечена И.Н. Тодоровым в экспериментах на
лабораторных животных разного возраста (см. табл. 4) еще 1965 г.,
однако осознание важности этого открытия для понимания
генезиса целого кластера дегенеративных болезней пришло
значительно позже.
В действительности проблема развития возрастных болезней
представляется даже еще более запутанной, так как эти болезни
сами по себе демонстрируют тенденцию ускорять реализацию многих
механизмов старения. Диабет - вероятно наиболее наглядный
пример, поскольку сопряжен с повышением уровня СР,
неферментативным гликозилированием и перекрестным сшиванием белковых
молекул, что в совокупности, возможно, ускоряет работу ЦЧС. Другой
пример - атеросклероз, при котором ухудшается кровоснабжение
109
тканей. Когда ток крови слишком слаб, внутриклеточные антиокси-
дантные системы, по-видимому, заметно снижают свою активность
несмотря на то что все еще имеется достаточное количество
кислорода, чтобы генерировать его активные формы (АФК), а это
приводит к повышению уровня поражений свободно радикального
характера и ускорению старения.
В заключение следует указать, что продление жизни и
предотвращение старческих болезней - не два отдельных направления.
Они органично вписываются в одну общую стратегию, потому что
механизмы, которые вызывают старение и дегенеративные
болезни, в значительной степени - одни и те же. Блокирование
механизмов старения и предотвращение или ослабление дегенеративных
болезней, помимо возможного продления жизни, имеют еще и ту
привлекательную сторону, что могут превратить старение в
полнопенный и приятный жизненный процесс.
Глава 4
СТАРЕНИЕ И ГОРМОНЫ
4.1. ЭНДОКРИННАЯ СИСТЕМА И ГОРМОНЫ
Существуют ли во Вселенной системы более сложные, чем
человеческий организм? Опираясь на опыт современной науки, мы,
очевидно, должны ответить на этот вопрос отрицательно. Сумма
всех клеток нашего тела составляет несколько десятков
триллионов, а каждая клетка - сама по себе - сложнейший организм. Если
бы клетка была не больше пчелы, то "улей" (т.е. человеческое
тело) достигал бы таких размеров, по сравнению с которым
Египетские пирамиды показались бы карликами. Каким образом
осуществляется безупречная совместная работа всех этих клеток? В
реальном улье координация усилий достигается эффективной связью
между различными типами пчел. Насекомые выделяют специальные
соединения, называемые феромонами, которые, перемещаясь по
воздуху, достигают других пчел и стимулируют их к действиям,
которые необходимы для общей пользы. Принципиально сходная
система связи существует и в человеческом теле: клетки могут
"говорить" друг с другом с помощью химических сообщений, которые
они выделяют в кровообращение. Эндокринная система является
ключевой, ответственной за эффективную координацию
многочисленных функций организма. Она состоит из трех иерархических
уровней: гипоталамуса, гипофиза и периферических эндокринных
желез. Гормоны - небольшие молекулы, которые служат
посредниками между эндокринной системой и органами и тканями организма,
а также между различными уровнями самой эндокринной системы.
Когда гормон достигает клетки-мишени, он взаимодействует с
расположенным на ее поверхности рецептором, который служит
молекулярным "кнопочным тригером" для включения определенной
клеточной функции. Каждый гормон имеет свой собственный тип
рецептора, и ответ клетки на циркулирующие в крови гормоны
зависит от типов рецепторов, которые она имеет.
Гипоталамус - небольшая мозговая структура, которая, по
существу, является командным пунктом, который управляет
эндокринной системой. Гипоталамус обрабатывает информацию из
внешней среды, анализируя сигналы, идущие к нему от нервной системы,
111
он также контролирует внутреннюю среду организма, анализируя
химический состав крови. Если гипоталамус "принимает решение" о
том, чтобы изменить некоторые физиологические параметры, он
посылает приказ гипофизу, который является главной железой
эндокринной системы.
Гипофиз локализован строго в центре мозга, ниже
гипоталамуса, в небольшом углублении в основании черепа, называемом
"турецкое седло". Гипофиз имеет две главные части: переднюю и
заднюю доли. Он способен продуцировать и выделять по крайней мере
дюжину различных гормонов, необходимых для регуляции
метаболизма, роста, воспроизведения и ответа на действие стрессоров.
Когда гипоталамус "решил" сделать какие-либо изменения в
организме, он выделяет в кровеносную систему специальные гормоны,
называемые релизинг-гормонами, которые поступают в гипофиз по
сети небольших кровеносных сосудов. В зависимости от того, какой
ре лизинг-гормон был ему послан, гипофиз в ответ секретирует в
кровообращение соответствующий собственный гормон.
Большинство гормонов, продуцируемых гипофизом, действует на
периферические эндокринные железы, такие как щитовидная железа,
надпочечники и половые железы. Периферические эндокринные
железы действуют как администраторы нижнего уровня - они также
вырабатывают и секретируют в кровеносную систему собственные
гормоны, которые, как правило, более просты по структуре и
действуют на конкретные органы или ткани таким образом, чтобы те
выполняли определенные функции (Wilson et al., 1985).
Сложные иерархические соподчинения основных уровней ней-
роэндокринной регуляции в организме (от уровня клетки - до
уровня центральной нервной системы) с их прямыми и обратными
связями представлены на рис. 10.
Чтобы проиллюстрировать достаточно сложную работу нейроэндокрин-
ной системы, рассмотрим конкретный пример. В течение стресса,
психоэмоционального или физического, гипоталамус выделяет кортикотропин-релизинг
гормон (КРГ), который заставляет гипофиз продуцировать и выделять адрено-
кортикотропный гормон (АКТГ). АКТГ действует на надпочечники
(небольшие железы, локализованные выше почек), стимулируя их к выделению
стероидов, наиболее важный из которых - кортизол действует на различные органы
и ткани (см. гл. 13). Эта цепочка событии ответственна за запуск многих
физиологических изменений, отмеченных при стрессе, включая повышение уровня
глюкозы в крови, активацию распада нежировых тканей и подавление
иммунной системы.
Центральный принцип, лежащий в основе функционирования
эндокринной системы - наличие в ней отрицательных обратных
связей (называемых также контуром обратной связи или торможением
по типу обратной связи) (см. рис. 9). Сущность этого явления весьма
проста. После того как гипоталамус послал сигнал, чтобы вызвать
некоторые изменения в организме, от периферии должен поступить
112
ответный сигнал, что приказ "центра" выполнен и что дальнейшие
приказы излишни и даже вредны, так как могут привести к выходу
системы в повышенный режим работы. Гормоны, выделяемые
гипофизом и периферийными эндокринными железами, способны
уменьшить или даже полностью подавить начальный сигнал,
посланный гипоталамусом. Этот механизм обеспечивает требуемую
обратную связь, помогая должным образом управлять изменениями,
инициированными гипоталамусом, и сохраняет всю эндокринную
систему в состоянии равновесия (см. рис. 9). Возвращаясь к примеру
с гормонами стресса, отметим, что АКТТ запрещает выделение КРГ
из гипоталамуса (малый контур обратной связи), и такое же влияние
оказывает кортизол (большой контур обратной связи). Эти контуры
обратной связи ограничивают ответ на экстремальное действие
стрессоров, которое, если оно будет безконтрольным, может
оказаться разрушительным.
4.2. КАК СТАРЕНИЕ ВОЗДЕЙСТВУЕТ
НА ЭНДОКРИННУЮ СИСТЕМУ, И НАОБОРОТ
По мере старения организма, эндокринная система постепенно
подвергается глубоким изменениям. Продукция одних гормонов
падает, и наоборот, другие производятся в избытке.
Еще недавно многие эксперты считали, что возрастные
изменения в уровне ключевых гормонов - неизбежное следствие
нормального старения, и что не следует вмешиваться в этот процесс.
Восполнение дефицитных гормонов рассматривалось как лишенное смысла.
Правда, имелось одно важное исключение - заместительная эстроге-
новая терапия у женщин после менопаузы, практика, которая стала
в последние четыре десятилетия обыденным проявлением заботы о
пациентах. Во многих исследованиях была продемонстрирована
польза от эстрогеновой терапии, включая снижение риска остеопо-
роза и сердечно-сосудистых заболеваний. Эстрогеновая терапия в
ряде случаев может послужить причиной незначительного повышения
статистической частоты возникновения рака груди, но это частично
компенсируется заметным понижением этого показателя для других
раковых образований. (Различные аспекты заместительной
эстрогеновой терапии более подробно будут обсуждены в гл. 8.)
Чем больше мы узнаем о процессах старения, тем яснее
понимаем, что возрастные гормональные изменения не только являются
побочным проявлением старения, но также служат и одной из его
причин. Многие старческие недуги, по-видимому, связаны с низким
уровнем гормона роста, дегидроэпиандростерона (ДГЭА), мелато-
нина, гормонов щитовидной железы, тестостерона и эстрогенов.
Восстановление уровня этих гормонов, свойственного молодости,
как было показано (Villareal et al., 1994; Bouillanne et al., 1996;
113
Huether, 1996; Khorram et al., 1997), может не только замедлить, но
даже частично обратить назад процесс старения (см. табл. 8).
Очевидно, имеется много факторов, способствующих
возрастному гормональному дисбалансу. Однако, главная причина, к кото
рой мы склоняемся, это - "центральные часы старения" (см. гл. 2).
Согласно нейроэндокринной теории старения, разработанной
В.М. Дильманом, старение в определенной степени
программируется гипоталамусом и связано с мозговыми структурами. По мере
старения эффективность контура обратных связей между
гипоталамусом и другими эндокринными железами снижается, навязывая
внутренней среде организма постепенные отклонения от ее
оптимального состояния (гомеостаза). Новаторская для своего времени
концепция Дильмана, в последнее время получает все больше и больше
экспериментальных подтверждений {Пономарева н др., 1999; Ferrini
et al., 1999; Sapolsky, 1999; Gust et al., 2000; Bjorntorp, 2002).
Самый простой пример - термостат в доме. Когда температура
понижается, датчик на термостате, обнаруживая изменения, включает нагреватель, и как
только дом нагреется, нагреватель отключается. Говоря метафорически, у
молодого организма все термостаты очень чувствительны κ температурным
изменениям и не позволяют "дому" перегреться или остыть слишком сильно. С
возрастом чувствительность "датчиков термостата" понижается, н мы видим
различные отклонения физиологических функций от нормы.
Главные датчики, которые контролируют стабильность
внутренней среды организма, находятся в гипоталамусе. Согласно
В.М. Дильману, эти гипоталамические датчики теряют с возрастом
свою обостренную чувствительность, что ослабляет эффективность
контуров обратной связи и таким образом нарушает гомеостаз.
Более того, такой механизм постепенного изменения гомеостаза
"вмонтирован" в систему генетически и является движущей силой
развития организма. Действительно, если бы сохранялось
абсолютное постоянство параметров внутренней среды, то в организме не
смог бы разворачиваться ряд важнейших генетически
детерминированных процессов: рост тела, развитие и созревание репродуктивной
и иммунной систем и др. Следовательно, полагает Дильман,
стимулом для развития организма должно стать постепенное изменение
его гомеостаза, что достигается последовательным "загрублением"
гипоталамических датчиков. После того как организм полностью
созрел (у людей, это возраст 20-25 лет), центральные часы
продолжают функционировать - гипоталамус не прекращает понижать
чувствительность своих датчиков и далее нарушает гомеостаз.
В этой ситуации, однако, результат - уже не рост и созревание
организма, а ускоренное старение и развитие ряда дегенеративных
болезней, таких как атеросклероз, диабет II типа, гипертония,
подавление иммунной системы, повышенный риск заболевания раком и
ослабленная способность адаптироваться к стрессу (Дильман, 1986,
1987; Oilman, 1994).
114
Таблица 8
Возрастные изменения в уровне главных гормонов
Гормон
Возрастные
изменения
Гормон роста
Гормоны
щитовидной железы
Инсулин
(не диабетики)
Инсулин
(диабетики II типа)
U
Τ
Τ или i
Кортизол
Дегидроэпиандро-
стерон (ДГЭА)
U
Тестостерон
(у мужчин)
Дегидротестостерон
(у мужчин)
Эстрогены
(у женщин после
менопаузы)
Мелатонин
Гонадотропины
(ФСГ, ЛГ)
Пролактин
1
Τ
1
и
τ
Возможные последствия
(неполные данные)
Потеря мышечной массы тела,
понижение иммунитета и
устойчивости к стрессу; более
низкая интенсивность метаболизма;
брюшное ожирение; депрессия;
пониженная сексуальная активность
Пониженная интенсивность
метаболизма; повышение веса;
усталость
Атеросклероз; гипертония; брюшное
ожирение
Высокий сахар в крови, ведущий к
атеросклерозу; повреждение
капилляров, почек, нервов, глаз и
т.д.
Потеря мышечной массы тела;
понижение иммунитета;
устойчивость к инсулину;
повышенный сахар в крови,
холестерин и жирные кислоты в
свободном состоянии; прогрессия
атеросклероза и гипертонии;
брюшное ожирение
Потеря мышечной массы и
увеличение содержания жира в теле;
низкая устойчивость к стрессу;
депрессия или недостаток ощущения
благополучия; пониженная
сексуальная активность; сухая кожа;
ослабление некоторых сторон
иммунитета
Уменьшенное либидо; потеря массы
мышц
Гипертрофия простаты; мужское
облысение
Увеличенный риск
сердечнососудистых заболеваний (из-за
атеросклероза); остеопороз;
вагинальная сухость
Пониженный иммунитет; бессонница
или недостаток сна, приносящего
отдых; повышенный риск
заболевания раком
Приступообразные ощущения жара
(обычно у женщин)
Гипертрофия простаты; половое
бессилие; ослабленная функция
семенников
115
Имеет ли схематически описанный выше кибернетический
механизм развития и старения отношение к возрастным изменениям на
уровнях различных гормонов? Да, несомненно. Эти гормональные
изменения - главный инструмент, с помощью которого гипоталамус
осуществляет единую программу развития, которая затем (по
достижении репродуктивного возраста) продолжает разворачиваться,
невольно оборачиваясь теперь программой старения. Гипоталамус
не может управлять огромным количеством биохимических
реакций, постоянно протекающих в нашем теле, он - директивный, а не
исполнительный орган. Чтобы осуществлять свою "генеральную
стратегию" развития и старения, гипоталамус посылает приказы
гипофизу, который в ответ на эти стимулы выделяет множество своих
гормонов. Некоторые из них действуют на периферические
эндокринные железы, производящие свои собственные гормоны, другие -
непосредственно на органы и ткани. Главная задача центральных
часов гипоталамуса состоит в строгой детерминации определенных
гормональных изменений в организме. До достижения половой
зрелости эти изменения можно назвать вызывающими развитие,
позже, - способствующими старению.
Что можно сделать для нейтрализации возрастных
гормональных изменений и, таким образом, минимизировать результат
действия ЦЧС? Имеются две основные стратегии. Можно пытаться
замедлить или даже полностью обратить ход ЦЧС, удаляя главную
причину возрастных гормональных изменений. Действительно,
восстановление эффективности контуров обратной связи между гипо
таламусом и периферической частью эндокринной системы
нормализует уровень многих гормонов, присущих молодости, в частности
гормона роста, инсулина, кортизола и половых гормонов. Пути для
достижения этого - увеличение уровня нейромедиаторов в мозгу;
оптимизация стресс-ответа организма на действие стрессоров;
улучшение способности тканей отвечать на инсулин; сокращение уровня
свободных радикалов в мозгу и др. (см. часть III). Другая стратегия -
противодействовать возрастным гормональным изменениям, т.е.
восстановить уровень недостающих гормонов. Первая стратегия эф7
фективна при использовании ее сравнительно молодыми людьми и
во многих случаях может быть достаточной. Комбинация из двух
рассматриваемых стратегий, по-видимому, обеспечивает лучшие
результаты у пожилых людей, хотя различия в индивидуальных
реакциях могут быть значительными.
В следующих главах мы обсудим роль индивидуальных
гормонов в процессе старения, и пользу от восстановления их до уровня,
свойственного молодым индивидам.
Глава 5
"ГОРМОНЫ МОЛОДОСТИ"
5.1. ГОРМОН РОСТА
Магический эликсир
или проходящее увлечение?
Среди "волшебных эликсиров" продления жизни, широко
рекламируемых в последние годы, гормон роста (ГР), возможно, является
самым главным. Этот замечательный гормон не случайно достиг
своего высокого титула: он - один из ключевых факторов, управляющих
ростом тела. Пониженная продукция ГР ведет к карликовости, а
избыток - к гигантизму или, если гиперпродукция ГР возникла после
созревания - к акромегалии (состояние непропорционального роста
костной ткани). Когда тело прекращает свой рост, приблизительно в
возрасте 20-25 лет, ГР не теряет своей важной роли. У взрослых ГР
помогает поддерживать массу тела и его состав, облегчает заживление,
регенерацию и восстановление тканей, стимулирует иммунную и
другие системы организма. Непосредственно или косвенно ГР
воздействует на все ткани тела в течение всей нашей жизни (Wilson et al., 1985).
Старение связано с хроническим падением продукции ГР. С 20
до 60-десяти лет, уровень ГР падает примерно на 80% (Klatz et al.,
1997). Относительный дефицит ГР, замеченный у большинства
пожилых людей, по-видимому, вносит вклад в такие признаки
старения, как потеря мышечной массы, замедленный метаболизм,
брюшное ожирение, медленное заживление ран, слабый иммунитет,
пониженная устойчивость к стрессу, потеря полового влечения и др.
Многие из этих признаков, по-видимому, улучшаются в результате
заместительной ГР-терапии {Bouillanne et al., 1996; Johannsson et al.,
1997; Taqffe etal, 1997).
Широкий интерес и освещение в массовой печати ГР получил в
1991 г., когда Дэниел Радмэн и его коллеги опубликовали свое
исследование по заместительной ГР - терапии (Rudman et al., 1991). К
этому исследованию был привлечен 21 здоровый человека в
возрасте от 61 до 81 лет. Двенадцати из них вводили ежедневно примерно
по 0 03 мг ГР/кг веса тела в течение более шести месяцев; другие
девять пациентов не получили никакого лечения. Пациенты,
получавшие ГР, увеличили в среднем на 8.8% мышечную и на 14.4%
уменьшили жировую массу тела. Толщина кожи увеличилась на 7.1%;
плотность некоторых костей также была несколько увеличена.
117
Практически неограничнные возможности использования ГР в
здравоохранении появились только тогда, когда была решена проблема видовой
специфичности ГР. Дело в том, что ГР, получаемый из гипофизов крупных животных
(быки, свиньи) - весьма доступного сырья, неэффективен при введении его
человеку. Получение же ГР из гипофизов человека в сколько-нибудь заметных
масштабах (по понятным причинам) крайне затруднено. Проблема решилась
благодаря развитию техники рекомбинантных ДНК, которая позволила
сконструировать кольцевую ДНК, представлявшую собой гибрид бактериальной
плазмиды и гена человеческого гормона роста. Этот ген, помещенный в
составе рекомбинантной ДНК в бактериальную клетку, индуцирует синтез ГР
человека. Создание промышленной биотехнологии на основе этих
генно-инженерных достижений, естественно, уже не представляло серьезных трудностей.
Результаты Радмэна и других вызвали волну необузданного
энтузиазма, который был, очевидно, излишним, поскольку
исследование было проведено на сравнительно небольшой группе больных и
имело некоторые методические погрешности: ГР вводился гласно, а
пациентам контрольной группы не давали плацебо. В последние
годы, было проведено несколько других исследований по ГР-терапии
пожилых людей,что подтвердило или, наоборот, устранило
некоторые из первоначальных надежд. В настоящее время, ГР
представляется ценным инструментом антистарения, который способен
частично изменить некоторые, хотя и не все, аспекты этого процесса.
Нужно также помнить, что у людей могут быть значительные
индивидуальные колебания по их ответу на ГР (Lonn et al., 1996;
Johannsson et al., 1997)
Гормон роста - наиболее представительный гормон в
человеческом гипофизе, железе, в которой он вырабатывается. Это -
относительно небольшой белок, состоящий из 191 аминокислотного
остатка. Выделение и синтез ГР гипофизом регулируется
биохимическими сигналами, идущими из гипоталамуса (Harwood et al., 1984).
Максимум ГР продукции приходится на раннее детство, когда рост
тела наиболее интенсивен (Tepperman et al., 1987). Далее уровень ГР
непрерывно снижается. По достижении 20-летнего возраста падение
уровня ГР замедляется приблизительно до 14% в десятилетие.
Существуют сомнения относительно вклада, который вносит
перманентное падение содержания ГР в процесс старения, и степень этого
вклада активно обсуждается (Klatz et al., 1997).
Почему с возрастом уменьшается производство ГР?
Окончательный ответ на этот вопрос еще ждет своего часа. Старение, по-
видимому, существенно не вредит способности гипофиза продуциро-
ваить количество ГР на уровне, присущем молодости. Очевидно,
проблема заключается в способности гипоталамуса с помощью
специального гормона, регулирующего выделение гормона роста
(ГРРГ), стимулировать гипофиз к достаточному производству ГР.
Обычно низкий уровень ГР и особенно ИФР-1 (инсулиноподобный
фактор роста - гормон, вырабатываемый печенью в ответ на
действие ГР) сигнализируют системе гипоталамус-гипофиз о необходи-
118
мости выработки большего количества ГР. У пожилых людей этот
контур обратной связи, по всей вероятности, не может работать
должным образом. Возможная причина этого - возрастное
повышение уровня соматостатина, гормона, который ингибирует
секрецию ГР (Reichlin, 1983). Однако такое отрицательное влияние может
и не произойти, потому что гипоталамус с возрастом постепенно
теряет чувствительность к соматостатину. При такой ситуации
снижение продукции ГР зависит (хотя бы частично) не от повышенного
содержания соматостатина, а от состояния ЦЧС (см. гл. 2), т.е.
программы возрастного развития, основанной на постепенной потере
эффективности контура обратной связи между гипоталамусом и
лежащими ниже уровнями эндокринной системы (Oilman, 1994).
Многие из эффектов ГР на тканевом уровне - косвенные. Так, ГР
стимулирует печень вырабатывать гормоны, называемые соматоме-
динами, наиболее важные из них - ИФР-1 и ИФР-2 - действуют
непосредственно на большинство тканей и органов {Phillips et al., 1983).
Введение ИФР-1 животным с дефицитом ГР воспроизводит большинство
эффектов ГР, включая влияние на рост тела и тканей, а также
повышение белкового синтеза и утилизации жира. Уровень ИФР-1 в крови
может быть более надежным индикатором статуса гормона роста, чем
уровень непосредственно самого ГР. Причина этого в том, что ГР
выбрасывается в кровь порциями и находится там в течение
относительно короткого времени. ИФР-1, напротив, остается в крови в течение
многих часов, и его уровнь отражает общую сумму выделенного ГР.
Поэтому в большинстве клинических исследований уровни ИФР-1
измеряют чаще, чем непосредственно ГР (Tepperman et al., 1987).
Функциональная эффективность оси ГРРГ'- ГР- ИФР-1 -
ткани организма снижается с возрастом во всех ее звеньях, как было
показано прн исследовании пожилых женщин, однако с относительно
большим снижением на верхних ее участках (Arvat et al., 1999). Тем
не менее отмечается также и заметное возрастное падение
концентрации ИФР-1, что, в частности, обуславливает снижение
функциональной активности мышц (Boonen et al., 1998).
Клеточные и молекулярные механизмы действия ГР и ИФР-1 на различные
органы и ткани еще недостаточно ясны. Влияние ГР на рост хрящевой ткани
опосредуется соматомединами. Стимуляция роста других тканей, таких как
печень и мышцы, по-видимому, также осуществляется по аналогичному
механизму, хотя in vitro можно наблюдать и прямое действие ГР на мышцу. ГР
вызывает усиление трансмембранного транспорта глюкозы и аминокислот, что
сопряжено со снижением концентрации свободного цАМФ в мышечной клетке. В
печени по прошествии лаг-периода ГР стимулирует синтез белка и РНК (Тодо-
рое И., 1959в), а также в конце концов синтез ДНК и полиплоидизацию гепато-
цитов. В плазматических мембранах клеток печени обнаруживаются
специфически связывающие ГР рецепторы, но как это связывание приводит к
стимуляции синтеза белка и прилиферации - пока не вполне ясно. После связывания ГР
с рецепторами, индуцируется каскад субклеточных реакций, сходных в целом с
теми, которые стимулируются стероидными гормонами (Tepperman et al., 1987).
119
При исследовании хлоридных каналов скелетных мышц старых крыс (в
условиях хронической обработки ГР) было показано, что эти каналы снижают
свои функции при ослаблении ГР/ИФР-1 оси у старых животных. Острые и
хронические эффекты, индуцированные ГР, наблюдались при изучении
электропроводности мышечных волокон старых животных, и это позволяет полагать,
что ГР/ИФР-1 стимулы действовали путем модуляции уровня фосфорилирова-
ния канала, по-видимому, через треонин-сериновую протеин киназу и через
синтез каналов хлорида, подобных каналам молодых взрослых животных
(DeLuca et al., 1997).
Касаясь эволюционных аспектов старения, следует отметить, что срок
жизни нематоды Caenorhabditis elegans модулируется путем инсулин /ИФР-сиг-
нализации. Вопрос теперь в том, как этот путь контролирует старение
нематоды и как это относится к старению высших животных (Gems et al., 2001)
ГР изменяет состав тела и обмен веществ. Наиболее четкими
эффектами при ГР-заместительной терапии являются изменения в
составе тела. В возрасте 20 лет, тело состоит примерно на 80% из
мышц, костей и внутренних органов и на 20% из жира или жировой
ткани. По мере старения, ткани, лишенные жира, постепенно "им-
прегнируются" жиром в среднем по 5% в десятилетие. После 60-де-
сяти тело может состоять более чем на 40% из жировой ткани.
Физическая сила, выносливость и функциональная активность многих
органов, по-видимому, снижаются параллельно с потерей
нежировой массы тела. В нескольких исследованиях было показано, что ГР-
терапии приводит к обратимиости этого процесса: доля нежирной
массы тела увеличивалась, а жировой ткани - уменьшалась, при
этом восстанавливалось свойственное молодости худощавое и
мускулистое телосложение, и в некоторых случаях увеличивалась
работоспособность и выносливость. Важно, что наиболее резкое
уменьшение обычно отмечалось в количестве брюшного жира (Klatz et al.,
1997). (Брюшное ожирение - хорошо установленный фактор риска
для диабета II, гипертонии и сердечно-сосудистых болезней.)
Комплекс этих метаболических эффектов связан с тем, что
мобилизация липидов - стержень в действии ГР у особей различного
возраста.
Джоханнссон и его коллеги (Johannsson et al., 1997) исследовали
влияние ГР-терапии на брюшное ожирение и различные параметры
метаболизма. Тридцати пациентам с брюшным ожирением в
возрасте 48-66 лет ежедневно вводили по 0.01 мг ГР/кг веса тела в течение
девяти месяцев при соблюдении всех правил контроля, включая
анонимность процедур и плацебо-контроль. Тотальный жир тела
уменьшился на 9.2%, а жир висцеральный, т.е. полости тела, - на
18.1%. Тотальный холестерин и триглицериды уменьшались, а
чувствительность тканей к инсулину повышалась. Диастолическое
давление крови понижалось.
Острая липолитическая реакция на введенный ГР несколько
ниже у пожилых мужчин и женщин, что отчасти связано с изменением
в их образе жизни и пищевых привычках, а также с возрастным
120
сокращением чувствительности липидного метаболизма к ГР.
Взятые вместе, эти факторы инициируют накопление жира.
Увеличенная жировая масса в свою очередь ингибирует выделение ГР и про-
мотирует его клиренс. Так устанавливается порочный круг
(Jorgensen et al., 1997).
ГР воздействует на состав тела, изменяя некоторые другие
стороны метаболизма. ГР стимулирует клетки к производству
большего количества энергии и синтезу большего количества белка, что
ведет к росту нежировых тканей, особенно мышц. При этом запасы
жира используются как источник энергии для нового роста этих
тканей. В некоторых, но не во всех исследованиях, ГР уменьшал
уровень холестерина и ЛНП и увеличивал его у липопротеидов высокой
плотности (ЛВП). Другие положительные стороны влияния ГР на
метаболизм - это улучшение толерантности тканей к углеводам,
понижение уровня глюкозы крови и повышение чувствительности
тканей к инсулину (Daughaday, 1985).
В целом, правомерно заключить, что метаболические эффекты
ГР способны замедлить некоторые метаболические механизмы
старения (например, развитие устойчивости к инсулину) и уменьшить
риск атеросклероза и гипертонии.
ГР активирует тканевую регенерацию. Регенерация
атрофированных или поврежденных органов важна для здоровья и
долгой жизни, а иногда и непосредственно для выживания. ГР-те-
рапия, по-видимому, обещает значительные перспективы в
ускорении восстановления здоровья, по крайней мере при некоторых
типах тяжелых поражений. До настоящего времени, наиболее
внушительные результаты были получены при ГР-терапии
серьезных ожогов.
ГР-терапия была использована при лечении 54 пациентов с
обширными ожогами. Пострадавшие были разделены на две равных
группы, и одна из групп получила ГР. В контрольной группе
смертность была 37%, а в группе, получавшей ГР, соответственно - 11%
(примерно в три с половиной раза ниже!). Результаты были
статистически достоверны. Необходимы, конечно, более крупные и
лучше спланированные исследования, чтобы подтвердить этот
действительно замечательный результат. В другом исследовании у 40
пациентов с ожогами в возрасте от 2 до 18 лет ГР уменьшал время
пребывания в стационаре в среднем с 46 до 32 дней (более чем на 30%)
{Klatz et al., 1997).
С возрастом внутренние органы имеют тенденцию постепенно
атрофироваться, уменьшаться в размере и снижать
функциональные потенции. Может ли гормон роста изменить это?
Немногочисленные данные, которыми мы распологаем сегодня, указывают, что
это возможно. Стимуляция роста массы тела и печени старых
лабораторных животных под влиянием недельного курса инъекций ГР
наблюдалась И.Н. Топоровым еще в 1959 г. (см. табл. 9, рис. 11). Но
121
лишь в 1991 г. Д. Радмэн и коллеги воспроизвели и развили эти
результаты на пожилых людях, показав, что 12-месячная терапия с
использованием человеческого ГР увеличивала как размер печени,
селезенки и почек, так и толщину кожи и массу мышц. Даже больные
органы возможно было восстановить и излечить с помощью ГР-
терапии (Rudman et al., 1991).
Существенные результаты были полученны при ГР-терапии
расширенной кардиомиопатии. Это состояние, при котором сердце
не может адекватно адаптироваться к потребностям организма в
кровообращении, что приводит к расширению левого желудочка,
истончению сердечной мышцы и в конечном счете к остановке
сердца. Трехмесячная терапия ГР, который был произведен методами
генетической инженерии, увеличила массу миокарда, уменьшила
объем левого желудочка, что привело к улучшению функции
миокарда, кровообращения и общего клинического статуса. Несколько
других исследователей наблюдали улучшение сердечной
деятельности в результате ГР-терапии как у здоровых пожилых, так и у
пациентов с сердечными заболеваниями. Предварительные сообщения
указывают, что ГР может также помочь при эмфиземе и почечной
недостаточности (Klatz et al., 1997).
ГР и иммунитет. Вне всякого сомнения было доказано, что ГР
стимулирует иммунную функцию. Фактически хроническое падение
уровня ГР и мелатонина (см. следующий параграф) может быть
ответственно за возрастное ослабление многих сторон иммунитета. У
животных с удаленным гипофизом развивается серьезный
иммунный дефицит, который приписывают недостатку ГР и, возможно,
еще и другого гипофизарного гормона-пролактина (Wilson et al.,
1985). Как мы уже отмечали, наиболее важное и драматическое
возрастное изменение в иммунной системе - инволюция тимуса, этого
"учебного центра" для Т-лимфоцитов. Потеря тимусом
функциональной активности повышает восприимчивость организма к
инфекции и, возможно, раку (Пол У., 1987).
Первое волнующее свидетельство о том, что ГР способен
обратить вспять старение тимуса, поступило от Кейта Келли, который
вводил старым крысам клетки гипофизарной аденомы GH3,
производившие большое количество ГР. Подвергшиеся инволюции,
безжизненные тимусы старых животных при этом стали увеличиваться
в размере и начали проявлять такую же активность, как и у молодых
крыс (Kelly, 1985; цит. по Klatz et al., 1997). Позже, многие
исследования показали, что ГР-терапия восстанавливает деятельность
тимуса несколькими путями. Во-первых, ГР стимулирует рост
эпителиальных клеток тимуса (клетки, которые обеспечивают проведение
стимулирующих сигналов для еще несозревших Т-лимфоцитов). Во-
вторых, ГР стимулирует тимус к производству иммунотропных
гормонов, таких как тимулин, который необходим для нормальной
деятельности лимфоцитов. Наряду с этим ИФР-1, первичный медиа-
122
тор ГР-действия, непосредственно ускоряет созревание Т-лимфоци-
тов (Mocchegiani et al., 1990).
Существуют свидетельства, что и другие компоненты иммунной
системы также восстановливаются ГР, включая В-клетки, NK-клет-
ки и макрофаги.
ГР и депрессия. По-видимому, имеется важная связь между ГР и
депрессией. Индивиды с низким уровнем ГР и ИФР-1 склонны к
проявлению многих симптомов депрессии, таких как недостаток
энергии, апатия, плохое настроение, недостаток положительной
мотивации, социальная изоляция и беспричинное беспокойство. Напротив,
у людей примерно такого же возраста, но с нормальным уровнем ГР
и ИФР-1, указанные симптомы отсутствовали. В многих случаях ГР-
терапия вызывает резкое улучшение настроения, повышение
энергии, сексуальной активности и общего интереса к жизни (Klatz et al.,
1997). Не вполне ясно, каким образом это происходит. Возможно,
ГР повышает уровень эндорфинов (естественных модуляторов
настроения и болеутоляющих средств) и приводит уровень различных
нейромедиаторов к более юному балансу.
ГР и половая функция. Надежда улучшить сексуальную
активность часто является одной из причин, почему люди решаются на
ГР-терапию. Однако в действительности пока еще не существует
каких-либо обстоятельных клинических исследований,
непосредственно адресованных этой проблеме. В 1997 г. О. Хоррам и его коллеги
(Khorram et al., 1997) исследовали влияние ГР-РГ (гипоталамическо-
го гормона, который стимулирует гипофиз к синтезу и выделению
ГР) на различные метаболические индесы и индексы качества
жизни. Испытуемыми стали люди в возрасте 55-71 года, получавшие
ГР-РГ в течение пяти месяцев. Исследователи сообщили, что
повышение сексуальной активности наблюдалось статистически
достоверно только у мужчин, но не у женщин.
В своей книге "Как стать молодым, с человеческим ГР" Рональд
Клац и Кэрол Кан {Klatz, Kahn, 1997) рассматривают множество
отзывов пациентов, подвергавшихся ГР-терапии, также как и мнений
клиницистов, имевших большой опыт применения такой терапии.
Общее заключение авторов: ГР-терапия улучшает, по крайней мере
некоторые, аспекты сексуальной функции у большинства
пациентов, а также несколько оживляет их сексуальную активность.
Теоретически ГР может стимулировать половую деятельность у
мужчин, улучшая сердечно-сосудистые функции и увеличивая
производство тестостерона. ГР повышает либидо и у мужчин, и у женщин, по-
видимому, поднимая в мозгу уровень нейромедиаторов, особенно
норадреналина, который, как известно, является важным фактором
процессов возбуждения. В старости депрессия - существенная
причина снижения полового желания. В связи с этим положительное
влияние ГР на депрессивные состояния также может играть
определенную роль.
123
Восстановление продукции собственного ГР. Хотя секреция ГР
снижается с возрастом, гипофиз не теряет способности к
биосинтезу ГР и может быть стимулирован к продукцированию
достаточного количества гормона, чтобы достигнуть уровня, присущего
молодым (Ghigo et al., 1998).
Старые крысы, содержавшиеся на ограниченной по калориям
диете, постепенно восстанавливали свою способность
продуцировать ГР. Лекарственное средство L-дофа, предшественник нейроме-
диатора дофамина, резко увеличивает секрецию ГР и у животных, и
у людей, восстанавливая уровень ГР, характерный для молодого
возраста.
Секреция ГР также может быть повышена некоторыми
питательными веществами. Аминокислоты аргинин, орнитин, лизин и
глютамин, витамин ниацин, γ-гидроксибутират и несколько других
питательных веществ в различной степени стимулируют секрецию
ГР. Вообще, диета с высоким содержанием белка (25-30%
белковых калорий от всего рациона) благоприятно влияет на уровень ГР,
стимулируя, очевидно, синтез гормона добавочным количеством
аминокислот (Wilson, 1985; Tepperman et al., 1987). Физические
упражнения - другой способ стимулировать секрецию ГР. Даже
непродолжительные упражнения с высокой нагрузкой стимулируют
выделение ГР. Однако упражнения при меньших нагрузках,
например бег трусцой, несоревновательный теннис или ходьба, даже
если они и длительны, оказывают значительно меньшее влияние на
секрецию ГР. Существуют свидетельства, что способность
организма вырабатывать ГР в ответ на интенсивные физические
упражнения постепенно снижается после 30-ти, но некоторая
положительная реакция может сохраняться даже до 70 лет (Nicklas,
1995; цит. по: Bouillanne et al., 1996). Тем не менее потенциальная
способность гипофиза отвечать на пусковые механизмы секреции
ГР в значительной степени остается неповрежденной и в пожилом
возрасте. Итак, за исключением некоторых случаев, когда
существует риск побочных эффектов, комбинация способов повышения
продукции ГР - физические упражнения, организованные
надлежащим образом, и сбалансированная диета с высоким
содержанием белка представляются наиболее эффективной стратегией для
восстановления уровня ГР, типичного для молодости (см. гл. 26 с
конкретными практическими рекомендациями)
Чего не может делать ГР. Вопреки некоторым полным
энтузиазма публикациям недавних лет, ни ГР, ни любой другой гормон
не являются волшебным средством против всех бед старения
(Lamberts, 2000). Как постоянно подчеркивается в этой книге,
процесс старения необычайно сложен и требует всестороннего
подхода. ГР, очевидно, способен затормозить или полностью обратить
вспять некоторые признаки старения, такие как потеря нежировой
массы тела и снижение иммунитета, но существует много призна-
124
ков старения, на которые он не способен повлиять. Кроме того, по-
видимому, могут быть существенные различия в индивидуальном
ответе на ГР: от весьма существенных эффектов у одних до
минимальных - у других. Люди на восьмом десятилетии жизни или
старше, кажется, менее чувствительны к введению ГР, чем 50 или 60-
летние. Так, существуют данные, что ГР-терапия не улучшила
функцию Т-лимфоцитов у 24 женщин в возрасте 70 лет или
старше. В другом исследовании, проведенном на 52 здоровых людях в
возрасте 70 и более лет, 6-месячная терапия физиологическими
дозами ГР улучшила состав тела (увеличение нежировой массы и
соответственно уменьшение массы жира), но не прибавила
физической силы и выносливости. Анализ побочных эффектов
показывает, что неблагоприятная реакция пожилых пациентов на ГР даже
при физиологических дозах наблюдается чаще, чем у более
молодых. Изучалось также влияние различных способов лечения на
прочность костей у женщин после менопаузы. ГР вызывал
некоторое увеличение плотности костей, но эффект был более слабый,
чем при эстрогеновой терапии, и чаще наблюдались побочные
эффекты {Bouillanne et al., 1996; Klatz et al., 1997; Sugimoto et al., 1999).
Вся совокупность имеющихся данных позволяет полагать, что
поддержание концентрации гормона роста на уровне молодого
возраста может обеспечить некоторые положительные эффекты для
продления жизни. Для большинства людей, самый безопасный и
наиболее приемлемый способ повысить уровень ГР это активация
его собственной секреции (см. гл. 26). Однако некоторые индивиды,
тщательно отобранные компетентными врачами, могут извлечь
пользу и из инъекций ГР.
Дальнейшие исследования, однако, показали, что надежды на продление
жизни с помощью ГР не столь безоблачны. Так, оказалось, что линия мышей-
карликов, гомозиготная (особи, у которых определенные гены и/или их
мутации - одинаковы) по мутации (Pit 1), которая вызывает потерю некоторых
функций гипофиза, показывают более, чем на 40% увеличение средней и
максимальной продолжительности жизни по сравнению с контролем. У этих животных
отмечена задержка процессов перекрестного сшивания коллагена и замедление
депрессии шести зависимых от возраста индексов статуса иммунной системы
(Flurkey et al., 2001). Авторы этой работы полагают - полученные результаты
демонстрируют, что единичный ген может контролировать максимум
продолжительности жизни и отсчет времени как клеточного, так и внеклеточного старения
у млекопитающих. Важно отметить, что мутация (Pit 1), ведущая к
существенному продлению жизни, понижает уровень ГР в сыворотке их крови.
В другой работе, у мышей-мутантов с объединенным дефицитом ГР, пролак-
гина и тиреотропина и у линии мышей с пониженной чувствительностью к ГР
также наблюдались более длительные сроки жизни (до 65%), чем у их нормальных
сородичей. Эти результаты воспроизводились независимо от генетическго фона и
условий питания (Bartke et al., 2001). Эти авторы также считают, что гены,
контролирующие старение млекопитающих и стимулирующие рост действие ГР и
размер тела, могут играть важную роль в детерминировании продолжительности
жизни.
125
Механизмы, которые ответственны за продление жизни мышей с
ограниченным эффектом ГР неизвестны. Тем не менее у долгоживущих
мышей-лилипутов с ограниченным связыванием специфических рецепторов с ГР (GHR-KO)
концетрация глюкозы и инсулина в крови, внутренняя температура тела, а
также уровень гормонов щитовидной железы существенно ниже, чем у
нормальных мышей, а уровень кортикостерона у карликов-самцов, наоборот, выше.
Эти функциональные изменения подобны тем, которые наблюдались у
животных при ограничении калорийности питания, и это подобие в двух
наследственно несвязанных экспериментальных группах животных дает некоторое
понимание того, какие факторы могут быть необходимы для продления жизни (Наиск
et al., 2001).
Все эти недавно опубликованные результаты несмотря на то что они
получены на грызунах-мутантах, по-видимому, противоречат многим из
приведенных выше данным по влиянию человеческого ГР на людей. И вся эта проблема,
очевидно, требует дальнейшего настоятельного и всестороннего исследования
(Lamberts, 2000).
Гормон роста и стресс. Способность сопротивляться
стрессорам снижается с возрастом, так же как и уровень гормона роста. Но
какие-либо прямые доказательства, что дефицит ГР вносит вклад в
возрастную потерю устойчивости к стрессу, отсутствуют. Тем не
менее исследователи нашли множество важных связей между уровнем
секреции ГР и стрессом, и наоборот.
Вообще говоря, кортикостероиды (гормоны стресса) и ГР
противоположно действуют на нежировые ткани. Кортикостероиды
вызывают распад белков, стимулируя этим дополнительную
энергопродукцию, что приводит к потере нежировой массы тела
(катаболизм), а ГР, наоборот, активирует белковый синтез и увеличение
нежировой масса тела (анаболизм). ГР, по всей вероятности,
проявляет свое анаболическое действие косвенно, благодаря его
способности увеличивать уровень ИФР-1.
Многие вредные проявления стресса являются результатом
частичного распада нежировых тканей и понижения их способности
к регенерации под влиянием кортикостероидов. Некоторые
исследователи пытались противопоставить негативным эффектам
серьезного стресса ГР-терапию. Так, Вара-Торбек и его коллеги (Vara-
Thorbeck et al., 1996) исследовали способность ГР ускорить
восстановление пациентов после стресса, вызванного хирургическими
операциями, и в особенности уменьшить интенсивность и
продолжительность послеоперационной усталости. Исследователи
нашли, что низкая доза ГР (8 Ш/день) минимизировала
послеоперационную усталость и предотвращала потери в нежировых тканях,
которые обычно следует за хирургическим стрессом. Изучались
эффекты инъекций ГР овцам, подвергнутых метаболическому
стрессу, вызванному 70-часовым голоданием. У этих животных ГР-
терапия уменьшала использование белка для энергетических нужд
и частично предотвращала потерю веса нежировых тканей
(Todorov Я. et al., 2001).
126
Итак, создается впечатление, что ГР уменьшает некоторые
эффекты стресса, и такое впечатление может быть близко к истине,
поскольку стресс уменьшает секрецию ГР и особенно его
"партнера" гормона ИФР-1. С этим согласуется также и то обстоятельство,
что большие количества кортикостероидов могут останавливать
рост неполовозрелых животных. Одно из возможных объяснений -
кортикостероиды влияют на синтез ГР и ИФР-1. Такая возможность
прицельно не исследовалась, но по крайней мере одно исследование
на животных непосредственно показало подобный эффект. Было
найдено (Thompson et al., 1995), что после введения кортизола,
способность овец выделять ГР в ответ на ГР-РГ (гормон,
освобождающий ГР) была значительно ослаблена. В свете этих результатов,
логично предположить, что склонность старых людей выделять
большее, чем в молодости, количество кортикостероидов в ответ на
стрессогенные факторы (см. гл. 15) может вносить заметный вклад
в возрастное падение производства ГР и/или ИФР-1.
При некоторых типах интенсивного стресса, таких как сепсис,
критическая фаза болезней, голодание, развивается состояние так
называемой резистентности к гормону роста. При этих состояниях
уровень ГР часто поднимается, но этот фактор не предотвращает
повреждение нежировых тканей. Интерпретация этого феномена
основывается на различии между прямым и косвенным действием
ГР. При обсуждаемой резистентности ГР способен усилить свои
прямые эффекты, т.е. повышать уровень сахара в крови и
расщепление жира (до жирных кислот), но окажется неспособен
осуществлять свои косвенные действия, которые зависят от продукции со-
матомединов (главным образом ИФР-1 и ИФР-2). Действительно,
уровень ИФР-1 (ключевой соматомедин-посредник в способности
ГР увеличивать белковый синтез и стимулировать рост нежировой
ткани) заметно понижен при серьезном стрессе или голодании.
Имеются свидетельства, что низкий уровень ИФР-1 частично
определяет активацию катаболизма, наблюдемого при выраженном
и длительном стрессе. Резистентность к ГР, замеченная при
стрессе, однако, не абсолютна и может быть преодолена введением
ИФР-1 или высоких доз ГР.
Каково практическое значение связи между стрессом и ГР? ГР-
терапия при интенсивном стрессе, например операционной травме,
ожоге или тяжелой болезни, может предотвращать некоторые
вызванные стрессом повреждения, включая потерю веса нежировой
массы тела и, возможно, подавление иммунной функции. В
настоящее время оценке этого подхода посвящено много исследований
(Todorov Η. et al., 2001). У пожилых людей, имеющих пониженную
устойчивость к стрессу, а также тенденцию к уменьшению ГР и/или
ИФР-1 продукции, ГР-, ИФР-1-или ГР-РГ-терапия в течение
богатой стрессами ситуации оказывается весьма благоприятной, позво
ляя пережить тяжелые времена с минимальными потерями.
127
Об открытии анаболического эффекта ГР у старых
животных. Использование гормона роста для задержки или даже для
полного измения некоторых признаков старения стало "суперхитом"
90-х годов, проникая в научную и бульварную литературу, появляясь
на каждом рекламном углу. Это открытие, однако, не ново, и было
сделано одним из авторов этой книги (И.Н. Тодоровым) более
40 лет назад в ходе обширного исследования на крысах. В 50-х годах
гормон роста казался захватывающей тайной: было известно, что
он играет ключевую роль в росте млекопитающих до момента
полового созревания, но не более того. Никто точно не знал, как
осуществлялось его стимулирующее рост влияние, а идея относительно
связи между продукцией ГР в организме и старением даже не
возникала. И.Н. Тодоров был тогда аспирантом Государственного
университета в Харькове, Украина, и его диссертационное исследование
было посвящено авангардной проблеме - воздействует ли ГР на
метаболизм нуклеиновых кислот (ключевых информационных
молекул жизни - РНК и ДНК) вообще и существует ли специфика
подобного воздействия у животных разного возраста - в частности.
В середине 50-х такое исследование было непростой задачей. Одна из
трудностей заключалась в получении достаточно чистого и биологически
активного гормона роста. Сегодня, в эпоху генной инженерии, когда
специально выращенные бактерии, несущие рекомбинантные ДНК, могут
производить потенциально неограниченное количество человеческого или
любого другого ГР, получение необходимой для исследования партии ГР является
вопросом простого размещения заказа у биотехнологической компании.
Однако тогда, получить даже мизерное количество чистого и активного ГР
было почти подвигом. Автор был вынужден неоднократно ездить на
скотобойню, где должен был вручную извлекать крошечные гипофизы
(единственный источник ГР в теле) из сотен отрубленных бычих голов. Чтобы
получить 2-3 г очищенного ГР, который требовался для серии запланированных
экспериментов, автор должен был собрать около килограмма бычих
гипофизов, каждый из которых весил всего лишь 1.5-2.5 г. Когда длительная
биохимическая процедура очистки была выполнена и под микроскопом,
наконец, можно было увидеть красивые игольчатые кристаллы, стало ясно,
что в руках был чистый ГР и что его полное молекулярное строение было,
очевидно, сохранено. Но сохранили ли эти кристаллы свою биологическую
активность? В то время наиболее надежным способом проверки
биологической активности ГР было испытание его способности стимулировать рост
тела неполовозрелых крыс с удаленными гипофизами. Удаление гипофиза,
размер которого у 28-дневных крыс не превышал зернышка пшена (а сами
животные имели рамер 6-7 см и массу не более 50 г), - почти ювелирная
работа. Когда автору удалось удачно прооперировать около десятка крысят и
убедиться, что их рост практически прекратился, он смог, наконец,
приступить к испытанию биологической активности полученного гормона роста.
28-дневные крысы одного и того же выводка были разделены на две группы.
Одна группа состояла из животных с удаленными гипофизами, в другой
(контрольной) животные оставались нормальными. Последние росли
быстро и удвоили свой вес примерно за месяц, крысы без гипофизов практически
сохранили дорперационный вес и размер. Введеный им ГР дал удивительный
128
12 г
*ioh
I 8
η
l·
Ε 4
2/
6 9 12 15 18
Возраст животных, месяцы
21
Рис. П. Динамика прироста веса
тела и печени при многократных
инъекциях СТГ крысам
различных возрастных групп (Тодоров,
1959 в)
1 - прирост в весе тела; 2 -
прирост в весе печени
эффект - крысы без гипофизов
начали расти чрезвычайно
быстро и в течение двух недель
почти догнали в размере и весе
своих нормальных братьев.
Теперь было ясно, что очищенная
субстанция имела превосходную биологическую активность и могла
использоваться для изучения биологического действия ГР на животных различного
возраста.
Изучение влияния ГР на метаболизм животных разного
возраста продлилось примерно два года и принесло интригующие
результаты (Тодоров, 1959а, 19596,1959в). Единичная инъекция ГР
увеличивала анаболические процессы у молодых крыс, синтез РНК
(рибонуклеиновые кислоты - копии генов, используемые при синтезе
белков) и белков был заметно увеличен. У старых животных,
однако, однократная инъекция ГР не давала почти никакого эффекта. К
изумлению автора, многократные инъекции ГР в течение недели
вызывали противоположный эффект: чем старше были животные,
тем более сильным оказался их анаболический ответ на ГР Самое
большое увеличение массы тела и некоторых тканей {рис. 11), а
также синтеза белков и РНК (табл. 9) было замечено у самых старых
(20-месячных) крыс, тогда как у самых молодых животных
(месячных) многократные инъекции ГР практически не оказывали
никакого влияния на эти показатели. Эти результаты привели автора к
нескольким предположениям. Во-первых, повышенный
анаболический ответ на многократные инъекции ГР у старых животных
указал, что продукция их собственного ГР, по-видимому, снижается с
возрастом. Во-вторых, уровень собственного ГР у молодых
животных, очевидно, настолько высок, что анаболический ответ тканей
был уже максимальным, и поэтому дополнительный ГР не оказывал
никакого влияния. В-третьих, старые животные, очевидно,
производили очень мало ГР, поэтому их ответ на введение ГР был столь
выраженным. В 50-х годах такую гипотезу было трудно доказать,
потому что отсутствовали точные методы проверки уровня ГР в
крови. Позже, когда в практику вошли новые методики измерения,
было показано, что уровень ГР действительно снижается с
возрастом у всех млекопитающих.
Другое важное заключение, сделанное И.Н. Тодоровым,
состояло в том, что ткани и органы старых животных сохраняют способ-
129
Таблица 9
Прирост массы печени, РНК иа весь орган, РНК иа единицу веса
органа крыс за весь период инъекций ГР
(Тодоров, 1959)
Изучаемый параметр
Возраст, месяцы
12
20
Прирост печени за время
опыта в г
Прирост РНК в печени в мг
Прирост РНК в мг/г печени
0.377
0.496
0.053
0.546
1.377
0.125
1.300*
1.693*
0.163*
* Каждая величина в этом столбце, как и в других, является средней от 8-10
животных и статистически достоверно превышает соответствующие показатели
других возрастов.
ность полноценно реагировать на экзогенный ГР, увеличивая
синтез нуклеиновых кислот и белков. Более того, полученные
результаты указывали на уникальную возможность приблизить
метаболизм старого организма к состоянию, характерному для более
молодых животных, и таким образом задержать или полностью
изменить некоторые признаки старения. К сожалению, в 50-х годах
прошлого века отсутствовала фактическая возможность
проверить и осуществить этот подход на людях. Проблема заключалась
в видовой специфичности гормона роста. Бычий ГР проявлял
активность в организме крыс, но был неэффективен у людей,
возможно из-за слишком большого различия в их молекулярной
структуре. В то время не было рентабельного способа получить
существенные количества человеческого гормона роста (ИГР). Только в
эпоху генной инженерии это препятствие оказалось
преодоленным. Был выделен ген, кодирующий ЬГР, и "вмонтирован "в
бактерии с помощью техники рекомбинантных ДНК. Таким образом,
была создана микроскопическая машина, производящая ГР. Ре-
комбинантный ЬГР, полученный таким способом, почти идентичен
его аналогу, произведенному человеческим гипофизом, и
оказывает то же самое биологическое действие. В начале 90-х, тридцатью
годами позже исследований автора на крысах, другие
исследователи получили подобные результаты с ЬГР на людях. Сегодня
поставка ЬГР для исследований и ГР-терапии фактически
неограниченна. Как это ни странно, наши современные знания указывают,
что введение экзогенного ЬГР не всегда оказывается лучшим
способом для многих людей. Во многих случаях лучшая стратегия
состоит в повышении производства собственного ГР, интегрируя для
воздействия на пусковые механизмы синтеза ГР диету,
специфические пищевые добавки и физические упражнения.
130
5.2. МЕЛАТОНИН
Чудо современной науки
или чудо современной рекламы?
В последнее время мы видим, что мелатонин становится
одним из наиболее широко (и, порой, необоснованно) используемых
природных лечебных средств. Как удачно выразился немецкий
исследователь Г. Хуетер, *'... мелатонин сделал внушительную
карьеру от пасынка эндокринологии к бестселлеру рынка
лекарственных средств" (Huether, 1996). Своей популярности
мелатонин прежде всего обязан усердию средств массовой информации
США, изображавших его как истинно натуральное
"сверхлекарство", которое прекращает старение, предотвращает болезни,
абсолютно нетоксично, полезно для каждого, не требует рецепта и
может быть легально куплено примерно за шесть долларов на
любом углу в уличной аптеке в количестве, достаточном для
двухмесячного употребления.
Глядя на это море плохо обоснованного ажиотажа, разлившееся
вокруг мелатонина, некоторые разумные люди могли бы подумать,
что это - бесполезный трюк, полностью лишенный смысла. Однако
прежде чем обрести номинацию на титул "универсальная панацея",
мелатонин придирчиво изучался на животных в течение более чем
трех десятилетий, а в последнее время также во многих
кратковременных исследованиях на людях. Как и большинство других средств,
претендующих иа роль реставратора "фонтана молодости",
мелатонин не подвергался никаким долговременным исследованиям на
людях с тем, чтобы доказать или опровергнуть его потенциал
антистарения и другие выгоды для здоровья, и также определить, чревато
ли его долговременное использование какими-либо
неблагоприятными эффектами. Тем не менее многочисленные исследования на
животных и некоторые непродолжительные исследования на людях
показывают, что мелатонин действительно способен оказывать
значительное благотворное влияние на многие параметры нашего
здоровья, особенно у пожилых индивидов. Возможные недуги,
требующие применения мелатонина, включают плохой сон, упадок
иммунной системы, профилактику и лечение рака, гипертонии,
атеросклероза, диабета, нейродегенеративных болезней (подобных болезни
Альцгеймера) и некоторых других патологий (Reiter et al., 1996).
Мелатонин, по-видимому, уменьшает осложнения от острого и
хронического стресса, увеличивает продолжительность жизни мышей на
20% и является, как было доказано, потенциальным антиоксидан-
том (Huether, 1996). Широкое использование мелатонина людьми
только началось и, хотя существует совсем немного вещей, о
которых мы знаем достоверно, но, как это ни удивительно, некоторые
131
свойства этого гормона, раздуваемые масс-медиа, могут в конечном
счете превратиться в научные факты. В этом разделе мы дадим
краткий обзор текущего состояния далеко неполных (но
волнующих) знаний о мелатонйне. Для дальнейшей информации мы
рекомендуем весьма содержательную книгу Р. Рейтера и Дж. Робинсон
"Мелатонин" (Reiter, Robinson, 1996) , авторов, которые являются
лидерами в исследовании мелатонина.
Мелатонин - "древняя" молекула. Она обнаружена у всех ныне
живущих животных и у многих видов растений и, кажется,
появилась у организмов, живших приблизительно три миллиарда лет
назад. Структура мелатонина (рис. 12) не изменилась ни на йоту,
оставаясь той же самой как у людей, так и у морских водорослей. Такой
структурный консерватизм обычно наблюдается только у веществ,
весьма существенных для жизни. У высших животных мелатонин
синтезируется эпифизом, иначе - шишковидной железой,
локализованной в головном мозге.
После того как А. Лернер и другие (Lerner ei al., 1959) выяснили
структуру мелатонина, а Вейсбах и Аксельрод (Weisbach, Axelrod)
расшифровали путь биосинтеза мелатонина из его
предшественника - аминокислоты триптофана и доказали присутствие в эпифизе
всех необходимых для этого ферментов (см. рис. 12), началось
выяснение его функций. Помимо его роли при переохлаждении
организма и в регуляции сна, в последние два десятилетия благодаря новым
совершенным методам исследования стало доступным изучение
влияния мелатонина на многие системы организма, особенно на
нервную и иммунную систему, а также, возможно, на ЦЧС.
Подобно гормону роста и ДГЭА, уровень мелатонина
значительно снижается с возрастом. Пик его концентрации наступает
перед половой зрелостью и круто падает после этого. Недавние
исследования показали, что снижение уровня мелатонина у подростков -
один из ключевых факторов инициации полового созревания. У
ребят с задержанным половым созреванием была отмечена тенденция
к повышению уровня мелатонина и, наоборот, у детей с необычно
ранним созреванием (иногда уже в возрасте трех лет) часто уровень
мелатонина был почти в три раза ниже, чем у сверстников. После
наступления зрелости, уровень мелатонина продолжает понижаться
в течение всей жизни (до трети от уровня молодости - после 40, и
едва обнаруживается - после 60). Это крутое падение не может быть,
очевидно, следствием только старения, однако, по-видимому, также
вносит свой вклад в этот процесс. Компенсаторное введение
мелатонина мышам до уровня молодых продлевает их жизнь на 20%
{Maestroni, 1993). Как уже упоминалось, ограничение калорийности
питания - наиболее эффективный из известных методов продления
жизни животных (грызунам удавалось продлить жизнь почти в два
раза). Несколько исследований продемонстрировали, что
ограничение в питании сопряжено с повышением эндогенной продукции ме-
132
но
Η Η
—π С—С — ΝΗ2
u I Триптофан
._JH 4==o
N J
Η ОН
Триптофангидроксилаза
Η Η
—η С— С — ΝΗ2
JJ Η I п 5-ОН-триптофан
ОН
Декарбоксилаза L-аминокислот
Η Η
НО^^ п С—С—·ΝΗ2
I I Серотонин
Η
CepoTOHHH-N-ацетилтрансфераза
О
Η Η Η II
-с—сн3
N-ацетилсеротонин
Гидроксииндол-О -метилтрансфераза
О
Η Η Η ||
СН30^^ - С—С — Ν — С — СН3
11 Η Η Мелатонин
Рис. 12. Биосинтез мелатонина (TeppermanJ., Tepperman H.t 1987)
133
латонина. Нельзя исключить, что это - один из механиков, с
помощью которого ограниченное питание увеличивает
продолжительность жизни {Reiter et al., 1996). Категорический ответ, однако,
может быть получен только в исследовании, в котором шишковидные
железы у животных будут удалены (или продукция мелатонина
будет подавлена специфическим лекарственным средством), а самих
животных затем ограничат/ в диете. Если они будут не в состоянии
жить дольше, чем особи контрольной группы, то значение
мелатонина как фактора продления жизни при ограниченном питании
значительно укрепится.
Способность мелатонина влиять на темп полового созревания
животных и людей и увеличивать продолжительность жизни у
животных (и, возможно, у людей) представляет особый интерес в
свете теории центральных часов старения (см. гл. 2). Действительно,
было показано, что экстракты шишковидной железы, содержащие
мелатонин, могут повышать чувствительность гипоталамуса к кор-
тикостероирам, эстрогенам и другим сигналам с периферии.
Например, старые самки крыс, получавшие экстракты шишковидной
железы, возобновляли циклическую эстральную функцию. В
связи с этим способность мелатонина и, возможно, других веществ,
продуцируемых шишковидной железой, замедлять ход ЦЧС -
привлекательная и вероятная гипотеза {Дилъман, 1987). Однако для
категорического ответа на этот вопрос необходимы будущие
исследования.
Мелатонин - потенциальный антиоксид ант. Мелатонин -
весьма многогранная молекула, способная к одновременному
выполнению многих функций. Одна из важных его "профессий"
заключается в его свойствах мощного антиоксиданта. Большинство
антиоксидантов - водо- или жирорастворимы, и поэтому могут
защищать только "водные" или "жировые" области клетки.
Например, витамин Ε эффективен в защите клеточных мембран от ПОЛ,
но не может защищать ДНК, и, напротив, глутатион
(водорастворимый антиоксидант клетки) эффективно защищает ДНК, но не в
состоянии предотвратить СР - повреждение мембран. Мелатонин,
который растворим и в воде, и в жире, помогает защищать все
клеточные компоненты, включая ДНК, белки и липиды. Он, как было
показано, вдвое активнее витамина Ε в защите клеточных мембран и в
пять раз - в нейтрализации ОН*-радикалов (наиболее вредоносного
типа СР) {Reiter et al., 1996)
Некоторые антиоксиданты, в частности витамин С и β-каротин,
при некоторых обстоятельствах, могут способствовать
формированию СР. Так, в присутствии свободного железа, которое
встречается при различных видах тканевых повреждений (например, ранах,
инфекциях, радиационных повреждениях), витамин С может
фактически генерировать СР. Мелатонин никогда не удавалось уличить в
таком "предательстве"
134
Из всех антиоксидантных эффектов мелатонина, защита ДНК наиболее
важна с точки зрения продления жизни. Мы знаем, что поврежденные
мембраны могут быть восстановлены, поврежденные белки могут быть заменены, но
некоторые СР-повреждения ДНК полностью необратимы, и, как только
произошла подобная мутация ДНК, судьба клетки становится неопределенной.
Некоторые мутации ДНК угнетают или изменяют функцию жизненно важных
генов, вызывая рак или просто уменьшая жизнеспособность клетки. Мелатонии,
по-видимому, весьма эффективен в защите ДНК. Рейтер и его коллеги {Reiter et
aL, 1996) изучали его способность защитить крыс от повреждений ДНК,
которые вызывал сафрол, ядовитый химикат, генерирующий целый поток СР. У
большинства животных, которым вводился сафрол, развивался рак печени.
Исследователи нашли, что у животных, получавших сафрол и мелатонин
совместно, поражений ДНК было в сто раз меньше, чем у животных, получавших
только один сафрол. Тем не менее остаются сомнения - является ли мелатонин
мощным антиоксидантом сам по себе или, возвращая организм к более
"молодому" состоянию, стимулирует таким образом собственную
антиокислительную систему клеток? Другими словами, является ли мелатонин
физиологическим антиоксидантом или его способность удалять СР-мусор - всего лишь
результат его искусственно повышенного в лабораторных опытах уровня, однако
еще не замеченного в природе?
Количество мелатонина, вырабатываемое организмом, очень
мало, значительно меньше, чем один мг в день. Других антиоксидан-
тов, которые вырабатываются или поступают с пищей, несравненно
большие: десятки или даже сотни мг. И, несмотря на то что
мелатонин примерно в два-пять раз активнее, его количество
представляется лишь малой добавкой к деятельности других антиоксидантов. Тем
не менее показано, что мелатонин обеспечивает существенную АО-
защиту даже при низком его уровне в крови, что особенно очевидно
при защите ДНК. Это, по-видимому, связано с преимущественной
концентрацией мелатонина в ядрах клеток и в особенности вокруг
ДНК, что делает его "персональным телохранителем" этой важной
макромолекулы, экранирующим ее от повреждений свободными
радикалами.
Еще более интересный аспект заключается в том, что
мелатонин не только нейтрализует СР непосредственно, но также
стимулирует продукцию глутатион-пероксидазы, одного из ключевых
антиоксидантных ферментов (Pablos, 1995). Вообще, когда АО
поступает в клетку извне, производство собственных антиоксидантных
ферментов идет на убыль. Очевидно, некий контрольный механизм,
регистрируя общий повышенный уровень АО-защиты, способен
воздействовать на собственные внутриклеточные системы, понижая
уровень эндогенных АО. В результате, большинство АО-добавок
при нормальных условиях лишь немного улучшают суммарную ан-
тиоксидантную защиту организма (при стрессе, однако, их
положительные эффекты намного выше). Мелатонин — уникален в том, что
он обеспечивает как прямое экранирование от действия
окислителей, так и стимулирует внутренние системы нейтрализации
свободных радикалов клетки.
135
Как уже упоминалось, чем старше мы становимся, тем больше
свободных радикалов производят клетки стареющего организма,
теряя, наряду с этим, способность нейтрализовать их (см. гл. 2).
Нельзя исключить, что хроническое возрастное снижение уровня мела-
тонина может оказаться существенной причиной ускорения
процесса старения.
Мелатонин и мозг. Шишковидная железа, продуцирующая ме-
латонин, расположена в самом центре мозга. Произведенный ею
мелатонин быстро распределяется в пределах мозга и активно
воспринимается нейронами.
Одна из первых обнаруженных функций мелатонина -
регуляция цикла "сон-пробуждение". Как и у нескольких других
гормонов (например, кортизола или серотонина), концентрация
мелатонина резко меняется в течение суток. Шишковидная железа
начинает выделять мелатонин вечером, его уровень постепенно
повышается, пока не достигает максимума поздно вечером, понижаясь
затем к раннему утру. Ночью уровень гормона в несколько раз.
выше, чем днем. Естественный циркадный ритм мелатонина,
очевидно, делает его естественным индуктором общего расслабления и
успокоительного сна. Постоянную бессонницу или сон низкого
качества у пожилых людей, по-видимому, можно объяснить (хотя бы
частично) дефицитом продукции мелатонина. Миллионы людей
используют мелатонин в качестве пищевой добавки для
нормализации сна. В противоположность обычным лекарствам,
стимулирующим сон, мелатонин, как правило, не вызывает никаких
неблагоприятных побочных эффектов и не нарушает нормальную
структуру сна (состоящего из нескольких специфических фаз, наличие и
последовательность которых важна для оптимального здоровья).
Другое общепринятое использование мелатонина - ослаблять
эффекты сдвига биологических ритмов или ритмов трудовой
деятельности. Если принимать мелатонин, когда шишковидная
железа, как предполагается, не производит его (днем), это будет
дезориентировать организм, навязывая ему вхождение в ночной цикл, т.е
расслаблять, уменьшать частоту сердцебиений и общую
активность, заставляя засыпать. После нескольких дней подобного
фармакологического "обмана" естественных биологических ритмов,
привычный цикл "сон-пробуждение" окажется сдвинутым вопреки
распорядку, присущему нормальной жизни.
Упомянутые выше антиоксидантные свойства мелатонина
представляются особенно важными для защиты мозга от СР-поврежде-
ний (Lipartiti et aL, 1996). Мозг потребляет кислорода в несколько
раз больше в пересчете на единицу массы ткани, чем большинство
других органов, генерирующих "поток свободных радикалов".
Кроме того, мозговая ткань богата ненасыщенными липидами, которые
особенно подвержены перекисному окислению. В развитии многих
болезней мозга, включая болезни Альцгеймера и Паркинсона,
136
СР-поражения, как полагают, играют чрезвычайную роль. В
старости уязвимость мозга СР столь высока, по-видимому, из-за низкого
уровня мелатонина, и наоборот, его высокий уровень может быть
полезным при лечении болезней центральной нервной системы.
Хотя в настоящее время не проводятся всесторонние клинические
исследования, тем не менее отдельные результаты все же указывают,
что мелатонин - мощный нейропротектор, защищающий клетки
мозга от различного типа повреждений. Р. Папполла и его коллеги
(Pappolla et al., 1994) нашли, что мелатонин может предотвращать
гибель клеток нейробластомы в культуре от повреждений
активными радикалами β-амилоида (ядовитого старческого пигмента,
связанного с развитием болезни Альцгеймера). В другом исследовании
мелатонин предотвратил смерть нейронов в одной из структур
мозга - гиппокампе, где высокими дозами глутамата был вызван окси-
дативный стресс (Lezoula'h, 1996). Как уже упоминалось,
нормальная функция гиппокампа состоит в смягчении стресс-ответа
организма, и напротив, резкое понижение его функции, вызванное
гибелью части его нейронов, увеличивает остроту стресса и даже может
инициировать состояние хронического стресса, часто наблюдаемого
в пожилом возрасте.
Мы еще только начинаем понимать сложные отношения между
мелатонином и мозгом. Современные исследования показывают,
что в дополнение к возможности регулировать сон и улучшать ней-
родегенеративные состояния, мелатониновая терапия оказывается
полезной при некоторых формах депрессии, хронической боли и
шизофрении (Reiter, 1995).
Мелатонин и иммунная система. И эффективность иммунной
системы, и содержание мелатонина резко падают с возрастом. Что
это - случайная корреляция или причинно-следственные
отношения? Недавние исследования ясно показывают, что мелатонин
обладает свойством повышать функции иммунной системы. Удаление
шишковидной железы или подавление фармакологическим путем
продукции ею мелатонина вызывает у животных состояние иммуно-
депрессии, характеризующееся высокой восприимчивостью к ин
фекции. Лечение мелатонином полностью снимает эти изменения.
Способность мелатонина стимулировать иммуногенез особенно
выражена, когда иммунная система ослаблена стрессом, болезнью или
старостью (Maestroni, 1993). В одном из исследований, старые мы
ши, получавшие мелатонин, были способны произвести вдвое
больше антител в ответ на экзогенные антигены, чем контрольные жи
вотные. В другой работе, Бен-Натан с коллегами (Ben-Nathan et al.,
1995) изучали эффекты мелатонина на мышах, инфицированных
вирусом энцефалита (смертельная болезнь мозга). У мышей,
зараженных этим вирусом, мелатонин уменьшал смертность со 100 до 44%.
В опытах с другой разновидностью вируса инъекции мелатонина
уменьшали смертность с 75 до 31%.
137
Механизмы, с помощью которых мелатонин повышает
иммунитет, понятны лишь отчасти. Было показано, что гормон
непосредственно активирует Т-хелперы (тип лимфоцитов, играющих
ключевую роль в "управлении" многими аспектами иммунной реакции). Т-
хелперы координируют действия других иммунных клеток, таких
как макрофаги, N-и Т-киллеры, которые выделяют специальные
сигнальные молекулы, называемые цитокинами.
Мелатонин, в частности, стимулирует Т-лимфоциты к
выделению интерлейкина-4 (ИЛ-4), ключевого цитокина, который
активирует несколько типов иммунных клеток, включая естественные N-
киллеры и фагоциты. N-киллеры - главные игроки в антираковой
защите и разрушении опухолей, а фагоциты помогают в борьбе с
заразными болезнями, захватывая и переваривая бактерии, паразитов
или инфицированные вирусом клетки. Сверх этого, мелатонин, по-
видимому, заставляет Т-хелперы выделять еще одно семейство
сигнальных молекул, опиоидные пептиды, которые также
увеличивают иммунную реакцию (Maestroni, 1991).
Определенную ясность в понимание сложных механизмов стимуляции
иммунных реакций мелатонином была внесена группой испанских исследователей
(Sainz, Mayo, Reiter et al., 1999). Обнаружив, что низкие дозы мелатонина инги-
бируют апоптоз как в культуре клеток, обработанных дексаметазоном
(эталонная модель для изучения апоптоза), так и на интактном тимусе, авторы
перешли к исследованию механизма, с помощью которого мелатонин
предохраняет тимоциты от клеточной смерти, вызванной кортикоидами. Как было далее
показано, мелатонин стимулирует синтез мРНК, кодирующих антиоксидантные
ферменты в тимоцитах. Наряду с этим, мелатонин не был включен ни в работу
генеральной "машины" апоптоза, ни в возможную регуляцию экспрессии генов
bcl-2 и р53, связанных с клеточной смертью. Эти результаты говорили о том,
что антиапоптозный эффект мелатонина связан не с прямым негативным
вмешательством в комплекс реакций, ведущих тимоциты к апоптозу, а с каким-то
иным механизмом. И авторы идентифицировали этот механизм. Оказалось, что
мелатонин негативно регулировал синтез мРНК, кодирующих рецепторы для
глюкокортикоидов, которые, как известно, вызывают апоптоз клеток тимуса
(см. гл. 13). Следовательно, подавляя биогенез рецепторов для
глюкокортикоидов, мелатонин служит защитником тимоцитов от индуцируемой глюкокорти-
коидами клеточной смерти. Этот эффект мелатонина, обусловленный
окислительными свойствами глюкокортикоидов, демонстрирует еще один механизм,
объясняющий его антиоксидантное действие.
В целом, мелатонин, очевидно, имеет будущее в качестве
усилителя иммунных реакций, особенно в условиях, когда иммунная
система подавлена хроническими болезнями, стрессом или
старостью, т.е. тогда, когда имеет место повышенный уровень
глюкокортикоидов.
Мелатонин и рак. О легендарных случаях излечения от рака с
помощью мелатонина сообщалось еще в 60-х. Однако косвенные, но
заслуживающие внимания свидетельства антираковой активности
мелатонина, появились в 80-е как результаты экспериментов на
культуре клеток тканей животных и человека. В ряде исследований
138
на грызунах, мелатонин полностью или частично блокировал
развитие опухолей после введения мощных канцерогенных веществ типа
ДМБА. Бласк и Хилл (Blask, Hill; цит. по: Reiter et al., 1996)
показали, что мелатонин подавлял на 75% рост линии раковых клеток
молочной железы (MCF-7-клетки, взятые из молочной железы
реальной пациентки и выращенные затем в культуре). Исследователи
обнаружили удивительный факт - мелатонин был эффективен
приблизительно в том же самом диапазоне концентраций, который
имеет место в человеческом теле - слишком малые или слишком
большие дозы гормона в этих экспериментах были неэффективны. Это
весьма отличалось от того, что можно было бы ожидать по
аналогии с принципами химиотерапии в онкологии: чем выше доза агента,
а следовательно, и его токсичность, тем выше его
противоопухолевый эффект. В другом исследовании, одна группа раковых клеток
MCF-7 инкубировалась в условиях постоянной концентрации мела-
тонина, а другая - с чередованием его уровней, которое грубо
моделировало естественный суточный ритм продукции мелатонина в
человеческом теле. Последний режим с чередующимися максимумами
и минимумами концентрации гормона был более эффективен с
терапевтической точки зрения {Cos, 1994). Очевидно, собственный
режим продукции мелатонина в организме оптимален для подавления
роста опухолевых клеток.
Может ли пополнение содержания мелатонина извне
предотвращать рак в человеческом теле? Мы пока не знаем этого.
Теоретически, у людей с низким уровнем мелатонина, дополнительные его
количества могут уменьшать вредоносное действие СР, обращать
функциональное ухудшение иммунной системы, т.е. влиять на два из всей
совокупности факторов, увеличивающих риск рака. Данные,
полученные на животных, указывают, что дополнительный мелатонин может
ограничивать активность канцерогенов в процессах канцерогенеза.
Единственный способ получить ясный ответ на вопрос - может ли
потребление мелатонина в "повседневной жизни" предотвращать у
людей развитие рака? - это провести обширные долговременные
исследования, и, мы надеемся, это произойдет в ближайшем будущем.
Тем не менее, как показано в нескольких клинических исследованиях,
мелатонин несомненно способствует успеху в лечении некоторых видов рака и
даже улучшает и продлевает жизнь неизлечимых онкологических больных.
Группа врачей из больницы Сан-Жирардо в Монзе, Италия, проверяли
комбинацию интерлейкина-2 (ИЛ-2) и мелатонина на нескольких типах рака. ИЛ-2 -
известный стимулятор иммунной системы, способный вызвать регресс
некоторых опухолей (см. гл. 6). К сожалению, ИЛ-2 активен только в высоких дозах,
которые являются токсичными. Однако, в комбинации с мелатонином даже
низкие дозы ИЛ-2 оказывались эффективными. У 100 пациентов с
неизлечимыми метасгазирующими опухолями низкие дозы ИЛ-2 совместно с 40 мг
мелатонина в день вызвали частичную регрессию опухоли у 17% пациентов (9 из
52), и ни у одного из пациентов, получавших заместительную терапию только
одним итерлейкином (Lissoni, 1995). 21% из группы комбинированной иммуно-
139
терапии (52 пациента) были живы в течение одного года, и только 10% (5 из 48;
в группе без лечения. У пациентов с метастазирующим раком прямой кишки,
невосприимчивых к стандартной химиотерапии, ИЛ-2 плюс мелатонин
вызывал частичную регрессию опухоли в 3 из 25 случаев (и ни у одного из 25
пациентов без лечения) (Barni, 1995). Выживание в течение года было значительно
выше в группе с иммунотерапией (9 из 25 против 3 из 25 в группе без терапии).
И в том, и другом случае ИЛ-2 плюс мелатонин улучшали дееспособность и
качество жизни. Итальянские исследователи также изучали влияние мелатонина
на результаты лучевой терапии при лечении мозговой глиобластомы (опухоли
с весьма неблагоприятным прогнозом и средним временем выживания
меньшим чем шесть месяцев). 30 пациентов с глиобластомой были анонимно
подвергнуты лучевой терапии плюс 20 мг мелатонина ежедневно и 30 только
одной лучевой терапии. Мелатонин, по-видимому, имеет значение для продления
жизни, поскольку 6 из 14 пациентов выжило в течение года в группе с
комбинированной терапией, и только 1 из 16 в группе - с одной лучевой терапией
{Conti et al., 1995). Еще в одном исследовании итальянские ученые нашли, что
мелатонин улучшил результаты лечения рака молочной железы тамоксифе-
ном, широко используемым противоопухолевым средством (Lissoni, 1995).
Мелатонин совместно с тамоксифеном не только способствовал выживанию
пациентов, но также уменьшал неблагоприятные побочные эффекты и снижал ν
них чувство беспокойства.
Ученые полагают, что мелатонин вызывает благоприятные
эффекты у раковых больных с помощью нескольких механизмов: (1)
снижение скорости роста раковых клеток; (2) стимуляция иммунной
системы, особенно N-киллеров, воздействующих на раковые
клетки, и (3) смягчение стресса, вызванного другими видами терапии и
непосредственно самой болезнью. В настоящее время
использование мелатонина в качестве элемента противораковой терапии
встречается довольно редко и в значительных масштабах имеет место
только в нескольких исследовательских центрах. В будущем,
однако, мелатонин вполне может стать важной частью многих
стандартных схем противоопухолевой терапии.
Некоторое представление о субклеточной природе действия мелатонина на
раковые клетки могут дать результаты работы Эк-Энрикеза и его коллег (Еск-
Enriquez et al., 2000). Эти авторы установили, что мелатонин и ретиноевая
кислота индивидуально ингибируют пролиферацию альфа-рецепторов эстрогена у
MCF-7 линии клеток грудного рака, что крайне важно для подавления роста
гормонозависимых опухолей. Было показано, что млатонин и ретиноевые
кислоты не только ингибируют пролиферации, но также и стимулируют апоптоз
MCF-7 клеток, когда используются в последовательном режиме. Подобный
режим обработки стимулирует апоптоз путем модуляции белков-членов
семейства Вс1-2, которые вовлечены в комплекс реакций, вызывающих апоптоз. Такой
подход может иметь большую терапевтическую ценность, приводя к регрессии
устойчивой грудной опухоли, активируя апоптоза ее клеток.
Мелатонин и стресс. Многие ученые полагают, что полезные
эффекты мелатонина, например способность активировать
иммунную систему и защищать мозг, в наибольшей степени выражены,
когда организм ослаблен стрессом или старостью (которую можно
рассматривать как форму хронического стресса). В сущности, влия-
140
ние мелатонина может быть сопряжено с естественными реакциями
организма, ограничивающими вредные последствия стресса. Модель
стресса, используемая в опытах на животных, - это ограничение
движений (иммобилизация), иногда объединяемое с содержанием
животных на холоде. Если такой стресс достаточно продолжителен,
у животных развиваются все классические признаки глубокого
истощения (см. II часть книги), включая глубокое подавление
иммунной системы, гипертрофию надпочечников и возникновение язв
желудка. Кан и коллеги (Khan et al., 1990) нашли, что мелатонин (от 1
до 10 мг/кг веса тела) значительно уменьшал образование
желудочных язв у крыс-самцов, подвергнутых иммобилизационному стрессу
в условиях низкой температуры. В некоторых других исследованиях
было показано, что мелатонин резко уменьшает смертность от
инфекционных болезней у стрессированных и старых животных
(MaestronU 1988; Ben-Nathan, 1995).
Хорошо известно, что стресс приводит к резкому
сокращению размера тимуса, который является ключевым игроком
иммунной системы. В старости, также как и при удалении
шишковидной железы, наблюдается инволюция тимуса. Не означает ли
это, что мелатонин является ведущим фактором в поддержании
размера и деятельности тимуса и поэтому может
противодействовать его инволюции при стрессе? Данные об
иммуностимулирующих эффектах мелатонина, представленные выше, показывают,
что мелатонин уменьшает или предотвращает стрессовый удар по
иммунной системе.
Помимо этого, мелатонин защищает или даже стимулирует
системы, которые ограничивают реакцию организма на сильный
стресс. Как уже упоминалось, мелатонин проявлял способность
защищать нейроны в гиппокампе от оксидативного стресса
(Lezoula'h, 1996). И поскольку гиппокампе важен для контроля
выделения стероидов стресса по типу обратной связи, мелатонин
может косвенно участвовать в этом стресс-лимитирующем
механизме. Другая недавняя находка - мелатонин побуждает Т-хелпе-
ры и, возможно, другие клетки иммунной системы выделять
опиоиЬные пептиды (естественная субстанция с
многочисленными свойствами, включая облегчение боли). Эти пептиды, как
полагают, являются частью другого механизма, который и
ограничивает эффекты стресса.
Названные ниже механизмы, по-видимому, играют роль в
антистрессовом эффекте мелатонина:
• Предотвращение индуцированной стрессом депрессии иммунной
системы путем ее стимуляции.
• Антиоксидантная защита от большого количества свободных
радикалов, продуцируемых при стрессе.
141
• Охрана структуры и функции нейронов в гиппокампе, который
важен для нормального торможения избыточного выделения корти-
костероидов при стрессе по типу обратной связи.
• Стимуляция выделения опиоидных пептидов, которые играют
роль в ограничении стрессового ответа.
5.3. ДБГИДРОЭПИАНДРОСТЕРОН
Еще один союзник в борьбе со старением
Дегидроэпиандростерон (ДГЭА) - один из нескольких так
называемых гормонов молодости, широко разрекламированных в
последние годы. ДГЭА - стероид, продуцируемый надпочечниками
(рис. 13). Среди множества стероидов (включая кортизол - гормон
стресса) надпочечники продуцируют ДГЭА в наибольшем
количестве. Большая часть циркулирующего в крови ДГЭА (приблизительно
90%) находится в форме ДГЭА сульфата - ДГЭАф). ДГЭАф) и
ДГЭА взаимозаменяемы, и первый рассматривается как форма
хранения второго. ДГЭА подвергается метаболизму и удалению из
кровотока намного быстрее, чем его сульфат, поэтому колебаний
уровня ДГЭА(Б) в крови в течение суток намного меньше, чем таковых
у ДГЭА. По этой причине для оценки статуса ДГЭА исследователи
и клиницисты часто измеряют уровень обеих форм гормона или
даже только одного ДГЭА(Б) (Sahelian, 1996).
ДГЭА, по-видимому, выполняет много важных функций в
организме. Он используется как предшественник для синтеза мужских и
женских половых гормонов - андрогенов и эстрогенов,
соответственно (см. рис. 13). ДГЭА, как и половые гормоны, непосредственно
действует на многие, если не на большинство органов и тканей,
стоящих на страже физиологических функций. Уровень ДГЭА в крови
достигает максимума примерно к 25 годам и затем резко падает к
концу жизни, достигая к 70 примерно четверти (или даже ниже) от
уровня молодости {Regelson et aL, 1994).
Энтузиасты полагают, что восстановлением юношеского
уровня ДГЭА можно полностью изменить некоторые аспекты старения
и даже увеличить продолжительность жизни. Многое из этой веры
основано на результатах многочисленных исследований ДГЭА на
животных, проведенных в течение несколько последних
десятилетий- Действительно, у грызунов и некоторых других животных
ДГЭА увеличивал продолжительность жизни, улучшал иммунную
систему и давал некоторую защиту против сердечно-сосудистых
болезней, диабета, рака и других патологий {Regelson et al., 1994). В
отличие от людей, однако, грызуны имеют очень низкий уровень
142
-^iJLJ 1 Холестерол
н0>*чхЧ^
20,22-деало/шза
сн3
с=о
Н3С
JTJ_] + *£>СН2-СН2-С-0
jLl^jJlIperaeHCMiOH Изокапроальдегид
НО
СН2ОН 3^Р^^\ СН^^идроксшшза СН3
с=о \ с=о \. с=о
Г ι j 17см
гч-^ч/ч^ рока
П-гидрокаишза\
17а-гид-
роксипре-
гненолон
3$-ОН-дегидроге·
назаиА5-4-изо~ !
X]Tjl7a-rHflpo- Г^ГТДепщроэ-
0^si^4/ ксипрогес- НСг^^^ пиа1Ж^"||
терон
СН2ОН
0=0 геназа и Δ-5-
шомераза
рон,
Кортикосгерон "В"
I 21-гидроксилаза
i^^J Кортексолон "S"
р.8-гщдроксикор-
^^v^s^1 тикостерон f, ^
О ■ Г 11-гиороксилаза
0сн2он
* !jc =o
3$-ОН-дегидро-
да
HOv^k4oii
CH2QH
Voh
njX^(
Альдосгерон
JjijKopTmojiV 0^^ ТестостеР°н
Рыс. 13. Схема стероидогенеза в коре надпочечников (Tepperman J.,
Tepperman //., 1987)
ДГЭА (S), циркулирующего в кровеносной системе, и это означает,
что их ткани могут ответить совсем по-другому на высокие дозы
flT3A(S). Следовательно, данные о ДГЭА, полученные на
животных следует использовать главным образом для планирования
исследований, а не для оценки полноценности ДГЭА-заместительной
терапии для людей. По этой причине мы сосредоточимся главным
образом на результатах клинических испытаний ДГЭА у людей. Ни
одно из этих испытаний не продолжилось более года И если много-
143
летняя история эстрогеновой заместительной терапии связана с
определенными достижениями, то результатов длительной ДГЭА-те-
рапии мы вынуждены ждать в течение ближайших 20-30 лет.
Некоторое косвенное данные указывает, что ДГЭА, возможно,
играет роль в процессах старения людей. Г. Раваглия и его коллеги
из университета в Болоньи (Италия) изучали корреляцию между
уровнем ДГЭАф) и функциональным статусом,
антропометрическими параметрами, уровнем липидов и различных гормонов в
крови 75 здоровых индивидуумов в возрасте 90-106 лет. Они нашли, что
более высокий уровень ДГЭА у женщин коррелирует с более
высоким уровнем гормонов щитовидной железы (гормоны,
регулирующие скорость метаболизма) и с более низким уровнем триглицери-
дов. У мужчин более высокая концентрация ДГЭА строго
коррелировала с более высокой функциональной активностью. В целом, у
исследователей сложилось впечатление о "..ДГЭА(Б) как о
фаворите в процессах старения" (Ravaglia et al., 1996). В других
коррелятивных исследованиях было найдено, что у пациентов с болезнями
сердца, ревматоидным артритом, астмой, диабетом и другими
хроническими патологиями обнаруживают низкий уровень ДГЭА. Тем
не менее нужно иметь в виду, что корреляции не обязательно
свидетельствуют, что низкий уровень ДГЭА вносит определенный вклад
в болезни или старение, его дефицит может быть просто следствием
или совпадением.
ДГЭА и иммунитет. В 1997 г. Хоррам и его сотрудники
(Khorram et yal., 1997) из Калифорнийского университета оценили
влияние ДГЭА-терапии (50 мг в день в течение 20 недель) на
несколько показателей иммунитета у девяти здоровых людей со
средним возрастом 63 года. ДГЭА значительно увеличил количество
моноцитов (предшественников макрофагов, клеток, которые
захватывают и переваривают бактерии и другие микроорганизмы) и В-лим-
фоцитов. Также увеличивались митогенные ответы на иммунные
стимулы и повышалась функция В-и Т-клеток. Отмечалось
значительное увеличение количества N-киллеров, клеток, выполняющих
в организме антираковый надзор. Интересно, что при этом
наблюдалось повышение уровня ИФР-1, ключевого медиатора действия
гормона роста. Этот результат наводит на мысль, что некоторые из
эффектов действия ДГЭА и гормона роста могут реализоваться
посредством одних и тех же механизмов. Исследователи заключили,
что потенциально ДГЭА-терапия может быть полезной для людей с
иммунодефицитом.
Имеются свидетельства, что ДГЭА может повышать
эффективность вакцинации у людей пожилого возраста. Известно, что
прививка в этом возрасте иногда неэффективна, потому что иммунная
реакция на вакцину слишком слаба. Это серьезная ситуация,
поскольку пожилые люди имеют значительно больший шанс
погибнуть от инфекции, чем молодые. Группе людей старте 65 была при-
144
вита вакцина гриппа. Кроме того, пациенты получали но 50 мг
ДГЭАф) в течение двух последующих дней сразу же после
прививки (Агапео, 1995). Пациенты подобной группы были привиты без
ДГЭАф) (контроль). В ДГЭА(8)-группе наблюдалось
четырехкратное увеличение способности продуцировать антитела против вируса
гриппа. Дальнейшие исследования позволят всесторонне оценить
потенциал ДГЭА в качестве стимулятора иммунитета у пожилых
людей при других заразных болезнях.
Другой многообещающий путь использования ДГЭА - лечение
аутоиммунных заболеваний, т.е. патологий, при которых иммунная
система атакует собственные ткани организма. Риск аутоиммунных
болезней, как известно, увеличивается с возрастом, возможно, из-за
возрастных нарушений иммунной системы. По-видимому, ДГЭА
может помочь в восстановлении в этой системе нормального баланса
сил. До настоящего времени наиболее многообещающим было
исследование с двойным слепым контролем, проведенное Ван-Воллен-
ховеном и его коллегами (van-Vollenhoven et al.) на 28 женщинах с
SLE (системная красная волчанка). SLE - аутоиммунное
заболевание, которое встречается преимущественно у женщин и
характеризуется воспалением суставов, кожной сыпью и язвами рта.
Пациентам давали по 200 мг ДГЭА (или плацебо) в день в течение трех
месяцев. SLE-индекс, т.е. мера тяжести болезни, значительно падал в
группе, принимавшей ДГЭА, но не в группе принимавшей плацебо
(сходное падение имело место и при параллельном лечении корти-
костероидами - стандартной терапии при SLE). Исследователи
заключили, что "ДГЭА может быть полезен как терапевтическое
средство при лечении умеренных форм SLE". Другие исследователи
наблюдали тенденцию к снижению уровня ДГЭА при таких
аутоиммунных заболеваниях, как ревматоидный артрит и множественный
склероз. Остается, однако, неясным, сопряжено ли это снижение с
аутоиммунным процессом или с какими-либо другими факторами?
ДГЭА и мозг. ДГЭА, как показали исследования, оказывает
прямое влияние на мозг. Он легко преодолевает гематоэнцефалический
барьер, и, кроме того, некоторые мозговые клетки, например астро-
циты, фактически могут синтезировать ДГЭА (Akawa, 1993; Robel,
Baulieu, 1995).
В своей книге "ДГЭА. Практическое руководство" Рей Шахе-
лиан {Sahelian, 1996) рассмотрел мнения многих врачей, широко
использующих ДГЭА в своей практике. Большинство отмечало,
что ДГЭА улучшал у пациентов настроение и чувство комфорта.
Некоторые из них сообщали также об улучшении памяти и более
ясном рассудке. Небольшое исследование, проведенное Волкови-
цем и его коллегами (Wolkovitz et al., 1997) выявило благоприятное
влияние ДГЭА на состояние депрессии Шести пациентам среднего
пожилого возраста с низким уровнем ДГЭА(8) открыто, т.е. без
соблюдения анонимности, давали ДГЭА в таких дозах, чтобы
145
достичь уровня, обычно наблюдаемого у молодых людей.
Показатели депрессии и памяти были значительно улучшены.
Недостаток этого исследования - его небольшой объем и открытость:
пациенты знали, что принимали ДГЭА, кроме того, не было
никакой группы контроля, получавшей плацебо. Авторы, однако,
отдавали себе отчет в этих погрешностях.
В ряде экспериментов на мышах было показано, что ДГЭА и
его предшественник прегненолон улучшал память и обучаемость
животных при дозах в 10-100 раз меньших, чем любой другой
нейроактивный стероид {Flood, 1988; 1992). Уровень ДГЭА у
грызунов, однако, намного ниже, чем у людей, следовательно,
человеческий мозг, к сожалению, не может быть столь же
чувствительным к этому гормону.
ДГЭА и здоровье сердечно-сосудистой системы. Хорошо
известно, что некоторые стероидные гормоны, особенно эстрогены,
могут защищать от болезней сердца в то время, как некоторые
стероиды, например кортизол, - способствовать их развитию. После
менопаузы, когда производство эстрогенов останавливается, риск
сердечных заболеваний у женщин заметно повышается.
Заместительная эстрогеновая терапия, напротив, понижает у женщин риск
атеросклероза и сердечных болезней. Существуют свидетельства,
что и ДГЭА может вызывать подобные эффекты. Риск
преждевременного инфаркта миокарда у мужчин был обратно
пропорционален уровню ДГЭА независимо от других факторов риска. Большое
эпидемиологическое обследование 1000 старых мужчин и 942
старых женщин показало, что высокие дозы ДГЭА проявили
небольшой защитный эффект против сердечных болезней у мужчин, но не
имели эффекта у женщин {Barret-Connor, 1995). Мужчины в
возрасте до 50-ти с преждевременным атеросклерозом имели более низкий
уровень ДГЭАф), чем мужчины аналогичного возраста без
болезней коронарных артерий.
ДГЭА показал прямое влияние по крайней мере на один
механизм, вовлеченный в генезис сердечных болезней - формирование
тромбов крови. (Известно, что тромбы, прикрепляясь к
артериальным стенкам, могут блокировать кровоток, что чревато серьезными
сердечными приступами. Аспирин, предписываемый многими
врачами пациентам с болезнями сердца, вызывает "разбавление крови",
т.е. препятствует формированию тромбов.) В небольшом
исследовании пяти здоровым молодым людям в течение 14 дней три раза в
день давали по 300 мг ДГЭА; в другой группе пять человек
получили плацебо {Jesse, 1995). У людей, получавших гормон, время
агрегации тромбоцитов увеличилось, и это означало, что время
формирования тромбов было более продолжительным. В другом
исследовании (18 человек получали гормон и 16 - плацебо), прием 50 мг
ДГЭА в течение 12 дней три раза в день увеличивал способность
крови разрушать фибрин - белок, участвующий в формировании
146
тромба {Beer, 1996). Более быстрая деградация фиорина ведет к
лучшему растворению тромбов и может играть существенную роль в
предотвращении сердечных приступов.
Пока еще не вполне ясно, как ДГЭА влияет на содержание
холестерина и липидов в крови людей. Результаты, полученные на
животных, представляются спорными и зависят от вида животных:
ДГЭА понижал общий холестерин и ЛНП у обезьян и собак, но
вызывал противоположный эффект у кроликов.
ДГЭА и сексуальная активность. Может ли ДГЭА повысить
сексуальную активность? Вопрос этот все еще открыт.
Какие-либо клинические исследования, специально адресованные этой
проблеме, пока еще не проведены. В одном комплексном
исследовании, когда рассматривали различные эффекты ДГЭА (50 мг в день
в течение трех месяцев), не было найдено никаких изменений
либидо у пациентов. В другой работе Р. Сахелиан проанализировал
сотни клинических результатов, полученных при ДГЭА-терапии, и
пришел к выводу, что ДГЭА повышает либидо, особенно у
пациентов с низкими уровнем ДГЭА. Этот эффект, по-видимому, более
выражен у женщин (Sahelian, 1996). Теоретически ДГЭА может
увеличивать сексуальную активность, используя два механизма.
(1) будучи предшественником половых гормонов, ДГЭА может
стимулировать их производство; (2) являясь нейроактивным
стероидом, ДГЭА может активизировать центральные механизмы
полового возбуждения.
Углеводный обмен, диабет и ожирение. Как уже упоминалось
ранее, старение связано с прогрессирующей потерей
чувствительности тканей к действию инсулина, что ведет к повышению
концентрации сахара в крови. Умеренное повышение сахара в крови после
принятия пищи обычно называют отсутствием толерантности к
углеводам, или преддиабетом, а повышенный уровень сахара в крови
даже натощак - диабетом. У большой части пожилых людей
толерантность к углеводам заметно снижена, и у многих (от 5 до 30% в
зависимости от особенностей демографической группы) в конечном
счете развивается диабет II типа. В период преддиабста и на ранних
стадиях диабета II типа уровень инсулина в крови, как правило,
ненормально высок (см. табл. 4), потому что производящие инсулин
В-клетки поджелудочной железы стремятся преодолеть
пониженную чувствительность тканей к инсулину, выделяя большое его
количество до тех пор, пока в конечном счете они не истощатся до
такой степени, что наступит "полноценный" устойчивый диабет.
Мы уже обсуждали в главе 2, что резистентность тканей к
инсулину вносит свой вклад в некоторые механизмы старения,
увеличивая концентрацию сахара и инсулина в крови. В ряде
многообещающих исследований на людях было показано, что ДГЭА повышает
чувствительность тканей к инсулину. Так, у 11 женщин, после
менопаузы лечение ДГЭА (50 мг в день в течение трех недель) увеличи-
147
ло чувствительность клеток к инсулину и уменьшило уровень триг-
лицеридов (Casson, 1995). В другом исследовании, аналогичное
воздействие ДГЭА на 15 субъектов (средний возраст 62 года)
существенно улучшало чувствительность тканей к инсулину {Bates, 1995).
Эти результаты указывают на возможность использовать ДГЭА для
предотвращения и лечения диабета II типа.
Интересно, что обратные взаимоотношения между ДГЭА и
чувствительностью к инсулину также, по-видимому, имеют место. Так,
Нестлер и его коллеги (Nestleret al., 1995) изучали эффекты бенфлу-
ролекса (фармакологического средства, которое улучшает
чувствительность тканей к инсулину и уменьшает избыток инсулина) и
параллельно определяли уровень ДГЭА и ДГЭА(Б) в крови 29
пациентов среднего возраста (30-59 лет) и 28 пожилых людей (60-80 лет),
страдавших гипертонией. Сокращение избытка инсулина и глюкозы
в крови было сопряжено с почти двойным увеличением уровня
ДГЭА и ДГЭА(Б) у людей средних лет. У пожилых людей эффект
был даже большим - уровень ДГЭАф) повышался более чем в два с
половиной раза
Влияние ДГЭА на вес тела варьирует в зависимости от вида или
даже от породы животных. Немногочисленные данные
исследований на людях указывают, что ДГЭА не оказывает какое-либо
существенное влияние на вес тела. В исследовании, продолжавшемся год,
15 60-70-летних женщин пользовались раз в день кремом с 10%
ДГЭА. При этом наблюдалось небольшое, но статистически
значимое улучшение состава тела (Diamond, 1996).
Глава 6
СТАРЕНИЕ И ИММУНИТЕТ
Жизнь во всех ее проявлениях - постоянная война. Но войны
между государствами или нациями - малая часть этих сражений.
Каждую минуту каждого дня наш организм вступает в сражение с
миллионами невидимых микроскопических "оккупантов" - бактериями,
вирусами и паразитами. Стержень "оборонительной машины"
нашего тела - иммунная система, которая является сложным альянсом
специализированных клеток и органов. Воздух, которым мы
дышим, пища, которую мы едим, вещи, которых мы касаемся, и даже
наш собственный желудочно-кишечный тракт или кожа содержат
миллионы микроорганизмов, многие из которых потенциально
опасны для нашего здоровья. Обычно иммунная система настолько
эффективна, что может устранить любую угрозу еще до того, как
она становится реальной опасностью; в большинстве случаев мы
даже не осознаем, что очередная "небольшая война" только что
выиграна нашей системой защиты. Иногда захватчики все же
прорываются сквозь "передовые линии" и начинают размножаться в
пределах организма. Тогда иммунная система вводит в дело намного
более интенсивные и агрессивные иммунные реакции. В результате
развивается лихорадка, недомогание и отек пораженных тканей,
другими словами, обычная картина инфекции. Здоровая иммунная
система обычно гасит инфекцию в течение нескольких дней и
возвращается к состоянию умеренной интенсивности, осуществляя
практику "патрулирования границ". Если, однако, иммунная система
ослаблена болезнями, старением, стрессом, плохим питанием или
поражена патологиями (в особенности СПИДом), организм
фактически остается беззащитным. Человека с ослабленной иммунной
системой неподавленная инфекция может привести к смерти. Если
иммунитет экстремально подавлен, как это имеет место на последних
стадиях СПИДа или у некоторых хронически больных пожилых
людей, то даже интенсивное использование антибиотиков в конечном
счете терпит неудачу.
Недавние успехи клеточной биологии показали, что иммунная
система является исключительно сложной и тонко отрегулирован-
149
ной биологической "оборонительной машинной", с высоко
специализированными подразделениями, густо переплетенными
коммуникациями и иерархией (Ада и др., 1987; Грей и др., 1990). Вся эта
сложность привела к тому, что ее изучение породило отдельную
научную дисциплину, называемую иммунологией. Иммунная
система использует несколько типов специализированных клеток,
включая лимфоциты, макрофаги и гранулоциты, из которых
каждый имеет собственные функции и средства для борьбы (см.
табл. 10А). Ключевые органы, участвующие в создании или
размещении иммунных клеток - костный мозг, тимус, лимфатические
узлы и селезенка. Подобно любой хорошо организованной армии,
иммунная система полагается на сложную эффективную сеть
коммуникаций. Иммунные клетки могут производить специальные
сигнальные молекулы, называемые цитокинами, которые несут
сообщения иммунным клеткам или другим тканям. Некоторые ци-
токины, такие как итерлейкин-2 (ИЛ-2) и итерлейкин-4 (ИЛ-4),
могут стимулировать рост и размножение некоторых иммунных
клеток, другие цитокины, особенно интерфероны, обладают
противовирусным, антибактериальным и противоопухолевым действием,
еще одна группа сигнальных молекул может стимулировать или
ингибировать рост других тканей.
Главные защитные функции выполняют белые кровяные клетки -
лимфоциты, которые разделяются на две крупных группы: В- и Т-лимфоциты. В-лим-
фоциты отвечают на появление в организме антигена (чужеродного материала)
секрецией антител, связывающихся с ним. Согласно клонально-селекционной
теории образования антител, сформулированной Ф. Бернетом еще в 1957 г.
(Burnet, 1957), продуцирующие антитела В-клетки имеют на своей поверхности
специфические рецепторы, и каждая клетка производит только один вид
рецептора. Если антиген соответствует рецептору, он специфически связывается с
ним, что побуждает клетку к размножению и производству большого
количества молекул этого рецептора, т.е. антител, которые далее с клеточной
поверхности переходят в кровь, где, встречаясь с антигеном, нейтрализуют его.
Размножению В-клеток и производству антител способствуют Т-лимфоциты (Т-хелпе-
ры). Правило "одна клетка - одно антитело" предполагает, что для
нейтрализации гигантского количества возможных антигенов в организме предсуществу-
ет почти "бесконечное" количество В-клеток, потенциально способных
продуцировать нужные антитела. Эта весьма трудная для интерпретации ситуация
побудила Бернета рассматривать иммунный ответ как вариант дарвиновской
микроэволюции. Клетки, продуцирующие антитела, - как любой организм в
экосистеме, — подвержены изменчивости и отбору, который способствует
выживанию наиболее приспособленных. В данном случае "приспособленность"
означает соответствие произведенного клеткой антитела - антигену (Burnet, 1962).
Определенную ясность в эту трудную проблему внесли исследования генов,
кодирующих антитела (Jalmage, 1986). Оказалось, что антигенсвязывающий
участок антитела является продуктом не менее чем пяти генов, в каждом из
которых имеются вариабельные участки. В ходе дифференцировки лимфоцитов
эти гены интенсивно рекомбинируют, благодаря чему создается уникальное их
сочетание для каждой клетки. Это сочетание и определяет специфичность
антител, производимых данной клеткой
150
Основные клетки иммунной системы
Таблица 10А
Белые кровяные
клетки
Способы действия
Главные функции
Зернистые
лейкоциты
Макрофаги
Т-лимфоциты
(клеточный
иммунитет)
Помощники
(хелперы)
Индукторы
Убийцы
(киллеры)
Супрессоры
В-лимфоциты
(гуморальный
иммунитет)
Натуральные
клетки убийцы
(N-киллеры)
Фагоцитоз
Фагоцитоз
Выделение
цитокинов
Распознают
инородные антигены и
посылают химические
сообщения
Стимулируют
развитие Т-клеток
Выделяют вредные
химические
вещества
Производят
антитела
Подавление инородных
организмов
Подавление инородных
организмов, представление антигенов
на своей поверхности, что
помогает активизировать
лимфоциты
Активируют лимфоциты
Подавляют аппетит
Вызывают лихорадку и недуг
Стимулируют ответ в других
Т-клетках и В-клетках
Стимулируют незрелые
Т-клетки в тимусе для развития
в зрелые
Т-клетки
Уничтожают вторгшиеся
организмы
Подавляют иммунную реакцию
Делают инородные организмы
легкой целью для фагоцитоза
макрофагами или зернистыми
лейкоцитами
Непосредственно убивают
инородный организм
Уничтожают раковые и
инфицированные вирусом клетки
Другое громадное семейство лимфоцитов - Т-клетки, защищающие
организм не путем синтеза антител, а убивая зараженные клетки и способствуя
воспалительной реакции. Если В-лимфоциты распознают антигены сами, без
посредников взаимодействуя с антигенами, которые имеются на бактериях,
паразитических микроорганизмах и др., то Т-лимфоциты в одиночку не способны
распознать антигены, для этого им требуется помощь. За последние несколько
десятилетий стало ясно, что в распознавании антигенов Т-лимфоцитами
участвуют и другие клетки. Эти "помощники" подвергают антиген определенной хи-
151
мической переработке (процессингу) и "представляют" его Т-клеткам в
сочетании со специальными молекулами клеточной поверхности - белками МНС
(от англ. Mmajor Hhistocompatability Complex - главный комплекс тканевой
совместимости).
В-клетки и выделяемые ими антитела распознают антиген главным
образом по его форме, т.е. по его третичной структуре, тогда как для Т-клеток
важна последовательность аминокислотных остатков полипептидной цепи, а не то,
каким образом она уложена в пространстве. К "помощникам" относятся
макрофаги (клетки-"мусорщики" иммунной системы), дендритные клетки в
лимфоузлах и селезенке, а также и сами В-лимфоциты (Unanue, 1984). Вспомогательные
клетки, представляющие антиген, принято называть АРС (от англ. Antigen
Presenting Cell). Прежде чем представить антиген Т-клеткам АРС расщепляют
его так, что он теряет структуры высшего порядка и сохраняет только
характерную последовательность аминокислотных остатков. Этот процессинг
антигена происходит во внутриклеточных мембранных пузырьках, содержащих про-
теолитические ферменты. В результате образуются полипептиды длиной не
более 10-20 аминокислот, которые перемещаются на клеточную мембрану для
последующего распознавания их Т-лимфоцитами (Buus et al., 1987).
Параллельно этому указанный процесс приводит к активации особых клеток - хелперных
Т-лимфоцитов, которые способствуют производству антител В-лимфоцитами.
Клетки организма, подвергшиеся вирусному заражению или злокачественному
перерождению также экспонируют свои специфические вирусные или
опухолевые белки (или их фрагменты) на клеточных мембранах, что привлекает так
называемые цитотоксические Т-лимфоциты, которые, опознав эти
аномальные белки, уничтожают породившие их клетки. Таким образом, в организме
существует два разных пути борьбы с экзогенными и эндогенными антигенами.
Проблема, однако, усложняется еще и тем, что Т-клетки распознают не
только чужеродный (или аномальный) антиген, но и белки, кодируемые генами
главного комплекса тканевой совместимости (МНС) и локализованные на
поверхности вспомогательных клеток. Следовательно, для возникновения ответа
на антиген (или его фрагмент) Т-клетки должны узнать два элемента:
во-первых, антиген и, во-вторых, "свой" белок МНС, характерный для собственных
клеток данного организма. Этот двойной контроль распознавания антигена был
назван МНС-рестрикцией.
Почему распознавание антигенов Т-лимфоцитами идет столь сложным
путем, а не прямо, как В-лимфоцитами? Для чего необходимо расщепление
антигена и появление его в редуцированном виде на поверхности вспомогательных
клеток (АРС) в сочетании с молекулами МНС? Для цитотоксических
Т-лимфоцитов смысл МНС-рестрикции состоит в том, что она нацеливает их на ткани
собственного организма, где эти клетки и должны действовать в случае
злокачественного перерождения, заражения вирусом и т.п. {Braciale et al., 1987),
Поскольку цитотоксические Т-лимфоциты обладают сродством как к белкам МНС
собственного организма, так и к процессированным антигенам, они ищут
именно те клетки, где оба "сигнальных" фактора будут представлены одновременно,
что и позволит Т-лимфоциту безошибочно уничтожить собственную клетку,
подвергшуюся опасной для организма трансформации.
У всех млекопитающих, включая людей, иммунная система
теряет эффективность с возрастом, что является как результатом
падения активности некоторых из ее подразделений, так и нарушения
координации между ними. Инфекционные болезни - четвертая
ведущая причина смертности после болезней сердца, рака и кровоиз-
152
лияний в мозг в развитых странах. Слабая иммунная система также
увеличивает риск рака и, возможно, атеросклероза. И у животных,
и у людей здоровье иммунной системы положительно коррелирует
с продолжительностью жизни. Первое и наиболее резкое возрастное
изменение в иммунной системе, как было показано, проявляется в
инволюции тимуса. Тимус - это железа, расположенная позади
грудины, представляющая собой "учебный центр" для Т-лимфоцитов,
среди которых наиболее важны Т-хелперы и Т-киллеры. Т-хелпе-
ры - координаторы и менеджеры иммунного ответа, тогда как Т-
киллеры - солдаты, подготовленные для "рукопашного" боя с
противником. Следовательно, все виды Т-клеток необходимы для
выполнения защитной функции иммунной системы. При СПИДе Т-
клетки разрушаются или повреждаются вирусом, что ведет к
падению преград для угрожающих жизни инфекционных болезней и
рака. Возрастной упадок иммунитета также связан с уменьшением
активности именно Т-клеток.
Здоровый тимус - ключевой фактор в эффективном
иммунитете Т-клеток. Эта железа имеет наибольший размер в раннем
детстве; у подростков она начинает сокращаться и снижать свою актив
ность, и эта тенденция продолжается в течение всей жизни.
Фактически инволюция тимуса рассматривается в качестве одного из
наиболее определенных биомаркеров старения.
До сих пор остается во многом неясным, почему происходит
возрастная инволюция тимуса. Было отмечено, что этот процесс
коррелирует с перманентным падением содержания гормона роста и мела-
тонина. Удаление гипофиза или шишковидной железы, которые
производят ГР и мелатонин, соответственно, приводит к
сокращению тимуса. Более того, лечение старых животных ГР или мелато-
нином восстанавливает деятельность этой железы. Эти факты
почти не оставляют места для сомнений в том, что инволюция тимуса (и
потеря иммунитета, связанного с этим) - не неизбежное
"проклятие" старения, а, возможно, обратимые последствия возрастных
эндокринных и метаболических изменений. Восстановление уровня
гормона роста, мелатонина и ДГЭА, свойственного молодости,
может способствовать реставрации полноценных функций иммунной
системы.
Иммунная система в высокой степени зависит от адекватного
потребления питательных веществ, особенно аминокислот, витаминов и
микроэлементов. Комплекс витаминов А, С, Е, В, фолиевой и пантотеновой кислот
важны для проявления различных аспектов функционирования иммунной
системы (см. табл. 10В). Дефицит витаминов А, В12 и фолиевой кислоты
наиболее распространен, а дефицит цинка — вообще одна из наиболее
острых пищевых проблем в мире. Цинк — составная часть множества
ферментов, используемых иммунными клетками; даже умеренный его дефицит
может снижать активность иммунной системы. Пополнение цинка у людей с
выраженным его дефицитом, восстанавливает иммунную функцию в
течение двух недель. Напротив, чрезмерный и длительный прием экзогенного
153
Таблица 10В
Влияние дефицита витаминов на иммунные реакции
(из: Modern Nutrition and Disease, 8-th edition / Eds. Shils, Olson, Shike, 1994)
Витамин
Пиридоксин (В6)
Пантотеновая
кислота
Рибофлавин (В2)
Фолиевая кислота
Витамин (В 12)
Биотин
Тиамин (В П
Витамин С
Витамин А
Витамин Ε
Эффекты дефицита
Тормозится ответ лимфоцита на стимуляторы
деления клетки
Снижается производство антител
Ухудшаются функции Т-клетки
Ослабевает деятельность тимуса
Атрофия лимфоидной ткани
Снижается производство антител
Снижается производство антител
Инволюция тимуса
Ухудшается функция зернистых лейкоцитов
(нейтрофилов)
Тормозится ответ лимфоцита на стимуляторы
целения клетки
Ухудшается функция зернистых лейкоцитов
(нейтрофилов)
Тормозится ответ лимфоцита на стимуляторы
деления клетки
Ухудшается функция клеток Т-киллеров
Замедляется производство антител
Тормозится ответ лимфоцита на стимуляторы
деления клетки
Ухудшается функция зернистых лейкоцитов
(нейтрофилов)
Снижается производство антител
Ухудшается функция Т-клеток
Атрофия тимуса
Умеренное снижение производства антител
Тормозится ответы на воспаления
Ухудшается функция клеток Т-киллеров
Ухудшается фагоцитарная функция зернистых
лейкоцитов и макрофагов
Снижается производство антител
Тормозится ответ лимфоцита на стимуляторы
деления клетки
Возрастает восприимчивость к некоторым
опухолям
Снижается производство антител
Тормозится ответ лимфоцита на стимуляторы
деления клетки
Ухудшается фагоцитарная функция
154
цинка может окончиться дефицитом меди, что в свою очередь способно и ρ
дить иммунитету и другим функциям. В качестве пищевой добавки цинк не
должен приниматься более одного месяца. Если имеется клиническая
причина для более длительного приема цинка - необходимо одновременно
принимать и медь (в отношении Zn/Cu, как 15:1 по весу).
Дефицит железа, который характерен для младенцев, менструирующих
женщин и пожилых людей, также может вредить иммунитету. Так, младенцы,
получавшие необходимое количество железа и витаминов, в два раза реже
подвергались инфекции дыхательных путей, чем те, кто получил витамины, но
были лишены дополнительного железа. В пожилом возрасте полноценный
рацион не может быть достаточным для поддержания желательного уровня всех
витаминов и полезных микроэлементов из-за пониженной способности
переваривать пищу и абсорбировать питательные вещества. Многие пожилые люди
страдают атрофическим гастритом, который сопряжен с пониженной
продукцией кислоты желудочного сока, необходимой для связывания железа, и
пониженным содержанием белка, связывающего витамин В12. У таких индивидов
достаточное количество железа и В12 в диете не служит гарантией от их
дефицита во внутренней среде организма (Modem Nutrition in Health and Disease, 1994;
Casper. 1995V
Пополнение некоторых витаминов в количествах,
достаточных для устранения дефицита, является дополнительным
фактором повышения активности иммунной системы и частично
предотвращает ее хроническое возрастное угасание.
Дополнительный витамин А увеличивает способность лимфоцитов отвечать на
антигенные стимулы. Однако очень высокие дозы витамина А
приводят к противоположным результатам в активности
лимфоцитов и чреваты некоторыми токсическими эффектами для
организма. Беременные женщины и дети при риске токсичности,
связанной с избытком витамина А, должны обращаться за
консультацией к врачу. Прием витамина Ε существенно улучшает
различные аспекты иммунной функции, особенно деятельность
лимфоцитов и макрофагов. Во многих исследованиях на животных
показано, что высокие дозы витамина Ε заметно увеличивают
выживание после серьезных болезней, вызванных вирусными и
бактериальными инфекциями. Вопреки популярным представлениям
{Полит, 1974) сверхвысокие дозы витамина С (1-2 г в день или
больше) только слегка, если вообще повышают устойчивость к
заразным болезням. Несколько исследователей стремились
определить - уменьшает ли витамин С риск или продолжительность
простудных заболеваний? Результаты были противоречивыми. Если
эффект существует, то он, по-видимому, весьма скромен.
Дефицит витамина С, однако, вреден для заживления ран и
иммунитета. Стресс или какие-либо повреждения увеличивают потребность
в витамине С, так как в этих условиях развивается его дефицит.
Другие питательные вещества также могут затронуть
иммунитет. Аминокислота аргинин, если принимать ее в значительных
количествах, улучшает различные параметры иммунитета. Например,
155
дополнительный аргинин ингибировал рост и метастазы
экспериментальных опухолей у животных. Гипотетически аргинин может
стимулировать иммунитет, увеличивая выделение гормона роста, а
также непосредственно стимулируя синтез нуклеиновых кислот
(Klatz et al., 1997).
РНК (рибонуклеиновые кислоты, носители информации для
синтеза белков) состоят из химических единиц, называемых рибону-
клеотидами, которые синтезируются в организме, но скорость их
синтеза ограничена. Поэтому некоторые диетические добавки,
содержащие рибонуклеотиды, очевидно, важны как для нормальной
жизнедеятельности клеток организма, так и для здорового
иммунитета. У животных ограничение экзогенных нуклеотидов подавляет
функцию Т-клеток.
Потребление и состав пищевого жира также влияют на иммунную систему
Некоторые полиненасыщенные жирные кислоты - предшественники синтеза
простагландинов, химических медиаторов воспаления и некоторых других
иммунных реакций. Простагландины разделяются по их эффектам на иммунную и
другие системы, и их уровень в тканях зависит от доступности
предшественников, т.е. в конечном счете от поступления с пищей разнообразных жирных
кислот. Однако диета с высоким содержанием ω-6-полиненасыщенных жирных
кислот, в изобилии найденных у растений семейства сафлоровых и в
кукурузном масле, как было обнаружено, подавляют некоторые функции иммунной
системы и стимулируют рост опухолей. Эффект иммунодепрессии был
особенно выражен, когда ω-6-полиненасыщенные жирные кислоты частично
окислялись в процессе приготовления пищи. Это - с одной стороны. С другой
стороны, полиненасыщенные жиры, богатые со-3 жирными кислотами (рыбий
жир), улучшали множество параметров иммунитета, таких как продукция ин-
терлейкина-2, ключевого стимулятора иммуногенеза и пролиферативного
ответа Т-клеток на внешние антигены.
В последние годы, связь между мозгом и иммунной системой
была установлена и широко признана. Для изучения этой связи
возникла новая ветвь науки, называемая психонейроиммунологи-
ей. Было показано, что депрессия неблагоприятно сказывается на
иммунной системе, особенно на деятельности различных типов Т-
лимфоцитов. Этот эффект депрессии был тем большим, чем
старше были пациенты {Lechin et al., 1996). Лечение депрессии
обычно полностью снимало негативные иммунные изменения.
Оба фактора - и риск депрессии и ее воздействие на иммунную
систему - увеличиваются с возрастом, и улучшение настроения
(например, благодаря пищевым и поведенческим модификациям,
см. гл. 26, 28, 29) является одним из ключевых шагов к
восстановлению иммунитета у пожилых людей.
Психологический, так же как и серьезный физический стресс,
очевидно, вреден для иммунной системы. Мы обсуждаем это
подробно во II части этой книги.
156
6.1. МЕТАБОЛИЧЕСКАЯ ИММУНОДЕПРЕССИЯ
В иммунной системе имеется несколько уровней регуляции: цен
тральный, гипоталамический, вегетативный, гормональный и
специфически-иммунологический, о котором достаточно подробно
говорилось выше. Однако наряду с перечисленными факторами
регуляции существует, как показали обширные исследования В.М. Диль-
мана, еще и метаболическая регуляция иммунитета (Дильман, 1976,
1978, 1983). Целый ряд данных позволил обосновать представление
о синдроме метаболической иммунодепресии (МИД), т.е. феномене
подавления активности клеточного иммунитета и функции
макрофагов, вызываемом определенными сдвигами метаболизма. Как в
экспериментах на животных, так и у человека обнаружена четкая
прямая корреляция между уровнем холестерина в крови и вязкостью
плазматических мембран лимфоцитов. В результате в их мембране
затрудняется перераспределение рецепторов, прежде всего
рецепторов ИЛ-2, что необходимо для их деления.
Таким образом, МИД как одно из проявлений нормального
старения, хотя бы частично, определяется накоплением свободного
холестерина в мембранах лимфоцитов, снижением их ответа на
действие ростовых факторов и, следовательно, снижением пролифера-
тивного потенциала {Phillips et al., 1984).
Вторым механизмом МИД является повышение с возрастом в
крови уровня ЛОНП и ЛНП. ЛОНП влияет через собственные
рецепторы, обнаруженные на лимфоцитах, а влияние ЛНП связано с их
функцией транспорта холестерина внутрь клетки {Голубев и др., 1983).
Третьим фактором в возрастном развитии МИД является гипер-
инсулинемия, так как при стойком повышении уровня инсулина в
крови, например при ожирении, уменьшается количество
рецепторов инсулина на иммунокомпетентных клетках, в частности на
моноцитах - макрофагах {Bar et al., 1976), а это, затрудняя транспорт
глюкозы, ухудшает функциональное состояние этих клеток, а
сопряженное с дефицитом глюкозы усиление потока жирных кислот
усиливает вероятность лизиса лимфоцитов {Turnell et al., 1973).
Наконец, возрастное повышение в крови уровня жирных кислот
и глюкозы вызывает подавление секреции ГР и функции
щитовидной железы, что уменьшает стимулирующие иммунную систему
гормональные влияния {Pierpaoli et al., 1972).
К этому следует добавить, что наблюдается отрицательная
корреляция между интенсивностью внепланового синтеза ДНК после
УФ-повреждения лимфоцитов (репарация ДНК) и содержанием
холестерина в сыворотке {рис. 14) {Дильман и др., 1981). Поскольку
повышение содержания холестерина в крови является одним из наи
более характерных признаков старения, падение репаративного
потенциала лимфоцитов с возрастом, несомненно, вносит свой вклад в
развитие МИД.
157
I5
10
9
8
7
6
5
4
3
2
ι
-
ι
о
о
L
о
1
о
о
•у
о ·
т
1
% ядер с
внеплановым синтезом ДНК
-L-
30 40 50 60 70 80 90
Рис. 14. Корреляция между интенсивностью репарации ДНК в лимфоцитах
(после ультрафиолетового повреждения) и содержанием холестерина в сыворотке
крови {Дилъман, 1987)
1 - больные атеросклерозом; 2 - здоровые лица
Совокупность приведенных выше и многих других данных
позволили В.М. Дильману (1987) квалифицировать влияние с возрастом
определенных метаболических изменений на состояние иммунной
системы как новую дегенеративную болезнь - метаболическую им-
нуиодепрессшо. Следует также подчеркнуть негативную роль МИД
для состояния здоровья стареющих людей - вообще, и для развития
ряда других дегенеративных болезней - в частности.
Существенную роль МИД играет во время беременности, когда в
организме матери происходит резкое изменение гормонального статуса. Секретируе-
мые системой "плацента-плод" гормоны не контролируются обычными
системами обратной связи, поэтому продукция плацентарных гормонов
увеличивается по мере роста плода. Хорионический соматомаммотропин (аналог ГР) и
кортизол снижают утилизацию глюкозы мышечной тканью материнского
организма. Избыток глюкозы используется плодом, однако у беременной
женщины гипергликемия вызывает увеличение концентрации инсулина в крови. Эти
два фактора ведут к накоплению жира, усилению спонтанного липолиза и
росту концентрации свободных жирных кислот в крови, что дополнительно инги-
бирует потребление глюкозы материнским организмом, в результате чего
глюкоза направляется на обеспечение нужд плода. В условиях роста концентрации
жирных кислот, глюкозы и инсулина в печени увеличивается синтез ЛОНП и
ЛНП и соответственно триглицеридов и холестерина. Все эти метаболические
изменения носят четко выраженный и закономерный характер. ЛНП снабжают
плод холестерином как для синтеза стероидных гормонов, так и, главным
образом, для достройки плазматических мембран интенсивно делящихся клеток
растущего эмбриона. В то же самое время указанные гормональные и
метаболические сдвиги вызывают метаболическую иммунодепрессию, т.е. подавление Т-
клеточного (трасплантационного) иммунитета, что крайне важно для
механизмов предотвращения отторжения плода как "инородного" трансплантанта.
Хотя все указанные изменения служат обеспечению программы развития плода,
они весьма напоминают главные болезни старения ~ ожирение, диабет II типа,
158
атеросклероз, падение иммунитета и др. После рождения ребенка все эти
изменения в организме матери постепенно возвращаются к норме. Однако
потенциальные механизмы развития главных дегенеративных болезней в организме
взрослого человека остаются и будут постепенно включаться по мере
отклонений гомеостаза, вызванных влиянием как статистических, так и системных
механизмов старения {Дильман, 1986, 1987).
6.2. ИММУНИТЕТ И РАК
Сегодня нет сомнений, что иммунная система играет решающую
роль в защите организма от рака. Предполагается, что раковые
клетки возникают и развиваются в теле перманентно, но
подавляющее их большинство разрушаются иммунной системой. Только те из
них, которым удается "ускользнуть" от иммунного наблюдения,
могут дать начало прогрессирующим злокачественным опухолям.
Предполагается, что в тех редких случаях, когда у очень малого
числа пациентов наблюдается спонтанный регресс опухолей, это
происходит из-за высокого уровня активации иммунных механизмов.
В настоящее время наметился некоторый успех в объединении
традиционных методов лечения рака (радиации, химиотерапии и
операционной хирургии) с иммунотерапией.
К одной из наиболее ярких иллюстраций роли иммунной системы в
борьбе с опухолями следует отнести результаты использования нового вида
терапии, разработанного в 1984 г. С. Розенбергом в Национальном
институте рака США, и названная им адоптивная иммунотерапия рака (Rosenberg,
1988; Rosenberg et al., 1988; 1989). Значение онкологических заболеваний в
семействе возрастных болезней столь велико, что мы считаем необходимым
подробнее обсудить этот интересный и перспективный подход. Открытие
лимфокинов и, в частности, интерлейкина-2 (стимулятора метаболизма и
деления лимфоцитов) и последующее его использование для выращивания
клонов Т-клеток открывало новые возможности в поисках эффективных
средств специфической терапии опухолей. Небольшое количество Т-клеток,
выделенных из кровотока больного и обладающих активностью против его
опухоли, размножалось в лабораторных условиях в присутствии ИЛ-2. В
результате образуются LAK-клетки (цитотоксические лимфоциты,
активированные лимфокином - от англ. limphokine - activated killer), которые
вводятся больному внутривенно в количестве примерно 50 млрд клеток вместе с
ИЛ-2. (Предварительные исследования показали, что регрессия опухоли
наступала также и при введении только Т-лимфоцитов или только одного
ИЛ-2. Однако при совместном их введении противоопухолевый эффект был
значительно выше, и регрессия наступала под действием меньшего числа
перенесенных клеток.) Такой способ лечения давал хорошие результаты при
раке почки и меланоме, достигая 25% успешных случаев. Хотя до сих пор
лечение LAK-клетками и ИЛ-2 или одним ИЛ-2 все еще считается
экспериментальным, тем не менее в 1987 г. FDA США разрешила федеральным
онкологическим центрам проводить такое лечение в случаях метастазировавшей
меланомы и рака почки. Следующий шаг был продиктован соображением,
что лимфоциты, выделенные не из крови больного, а из самой опухоли СП-
лимфоциты, от англ. Tumor-infiltrating - инфильтрующие опухоль), а следо-
159
вательно, уже находящиеся в состоянии борьбы с клетками опухоли, будут
еще более эффективным лечебным средством. У больного удаляли узел
опухолевой ткани, с помощью протеолитических ферментов разделяли его
на клетки, которые далее инкубировались с ИЛ-2. Под влиянием лимфоки-
на лимфоциты, содержавшиеся в опухлевой ткани, быстро размножаются,
убивают раковые клетки и через 30-45 суток культура целиком состоит из
ΤΙ-лимфоцитов. Пациентам вводили 200 млрд этих клеток вместе с ИЛ-2.
Эффективность лечения меланомы в этом случае достигала уже 55%. На
этом, однако, этот плодотворной подход не заканчивается. Авторы
приступили к исследованиям, целью которых является введение в Т1-лимфоциты
генов, способных усилить их противоопухолевую активность.
Предполагается интегрировать в геном лимфоцита гены фактора некроза опухоли -
α-интерферона и итерлейкина-2 {Розенберг С, 1990).
Риск заболеть раком значительно увеличивается с возрастом.
Неясно, однако, как велика роль вклада возрастного ослабления
иммунной системы в увеличении частоты заболеваний раком. Только
Т-клетки иммунной системы подвергаются действительно
серьезным возрастным изменениям, в то время как другие функции
иммунной систем могут остаться неповрежденными. У пациентов,
страдающих СПИДом и имеющих заметно пониженный Т-клеточный
иммунитет, значительно увеличивался риск возникновения некоторых
раковых образований, особенно саркомы Капоши. Однако раковые
образования, наиболее часто встречающиеся у пожилых людей
(например, рак ободочной кишки, легкого или молочной железы),
весьма редко наблюдаются у пациентов со СПИДом, и наоборот,
саркома Капоши чрезвычайно редка у здоровых пожилых людей.
Вероятно наиболее важный тип клеток "противоопухолевого
надзора" - натуральные киллеры (NK). N-киллеры представляют
тип лимфоцитов, способных непосредственно воздействовать на
раковые клетки, так же как на клетки, инфицированные вирусом.
У мышей NK играют ключевую роль в подавлении роста метастази-
рующих опухолей, разрушении инфицированных вирусом клеток и,
возможно, в некотором продлении жизни животных.
Общая картина уничтожения клетки-мишени N-киллером достаточно
хорошо известна {Юн и др., 1988). Процесс начинается с того, что NK распознает
клетку-мишень, на поверхности которой, как мы помним, располагается
антиген и белок МНС, и прикрепляется к ней {рис. 15(1)). По-видимому, в
результате взаимодействия рецепторов киллера и мишени в NK-лимфоците
наблюдается перемещение внутриклеточных гранул и аппаратов Гольджи (из которых и
образуются эти гранулы) в направлении клетки-мишени. Наряду с этим
увеличивается концентрация ионов кальция, что активирует экзоцитоз, т.е. слияние
гранул с клеточной мембраной и опорожнение их содержимого наружу (2,3).
При этом содержащийся в гранулах перфорин (специфический белок с
молекулярной массой 70 КД) оказывается в небольшом пространстве между клетками.
Присутствующие там ионы Са вызывают изменение конформации одиночных
молекул перфорина (4), после чего они связываются мембраной
клетки-мишени (5) и встраиваются в нее (6) {Young et al.f 1986). В мембране эти молекулы по-
лимеризуются (7) и формируют поры с диаметром 5-10 нм (8). При столь малой
160
Рис. 15. Схематическая картина воздействия Т-киллера на клетку-мишень
(объяснение в тексте) (Юн, Кон, 1988)
величине пор возникает так называемый эффект Доннана: крупные молекулы
не могут выйти из клетки (поры для них малы), а вода и соли из окружающего
раствора проникают сквозь мембрану внутрь (Young et ah, 1987). В результате
клетка-мишень (раковая или инфицированная вирусом) набухает и в конце
концов лопается (Podack, 1985).
Проводилось много исследований активности NK у пожилых
людей, но результаты были недостаточно четкими. По-видимому, у
здоровых пожилых людей активность NK может оставаться на
уровне молодости или даже превышать его. В частности, несколько
исследований здоровых долгожителей (см. гл. 9) показали хорошую
сохранность деятельности NK, которая может быть важным
фактором содействия их долголетию.
6.3. АУТОИММУНИТЕТ
Как мы уже отмечали, иммунная система - чрезвычайно
сложный комплекс, для которого не является сюрпризом возможная
встреча со всем многообразием биологического микромира,
который необходимо нейтрализовать. Самый большой вызов, однако,
состоит не только в том, чтобы уничтожить болезнетворный
организм, но также и в том, чтобы избежать при этом потенциальной
возможности нанести вред собственным клеткам. Иммунная
система имеет сложный механизм опознания и дискриминации между
инородными и "родными" клетками или даже молекулами, целена-
161
правленно уничтожая врага. Хотя эти механизмы обычно весьма
надежны, но и они иногда терпят неудачу, результатом которой
является так называемая аутоиммунная атака, при которой иммунная
система ошибочно принимает ткани своего тела за врага и пытается
уничтожать его. Наиболее распространенный тип аутоиммунной
атаки осуществляется с помощью аутоантител. Антитела -
специальные белки, вырабатываемые В-лимфоцитами и
предназначенные реагировать с посторонней субстанцией, в частности со
специфическими гликопротеидами на наружной мембране вражеской
клетки. Последние, таким образом, будут отмечены антителами и в
последствии поглощены макрофагами или разрушены другими
средствами, такими как система комплемента, т.е. иммунного
гемолиза. Антитела, которые связываются с собственными клетками
тела, называются аутоантителами и могут способствовать
саморазрушительным иммунным реакциям.
Огромное разнообразие молекул рецепторов (TCR - Τ cell receptor, состоит
из двух полипептидных цепей - α и β, кодируемых каждая отдельным геном) и
ограниченное количество главных комплексов тканевой совместимости -
МНС служат основой избирательной активности Т-клеток. Способность
распознавать "свое*' и "чужое" непосредственно связана с характером и причинами
такого разнообразия. Гены, кодирующие вариабельную часть TCR,
наследуются от родителей не как непрерывный блок генетических локусов, а в виде
отдельных "кусков" - сегментов, которые соединяются случайным образом в
процессе развития лимфоцита. Этот рекомбинационный механизм у каждого
индивида обеспечивает возникновение "новых" генов для 100 млн или более
различных TCR. Вариабельность МНС носит, однако, другой характер, У каждого
индивида имеется лишь по два гена для молекул МНС: МНС класса I (связанные с
пептидами самой клетки) и МНС класса II (связаны с пептидами из инородных
белков, поглощенных клеткой), хотя в популяции особей их может быть
множество (Boehmer, 1990). Столь высокая вариабельность при образовании
рецепторов может привести к тому, что некоторые из них окажутся полезными,
другие бесполезными, а третьи - вредоносными. Полезными будут те рецепторы,
которые защищают организм, связываясь с чужеродными пептидами из
вирусных или бактериальных белков и представленными "своими*' белками МНС.
Бесполезные рецепторы не способны к взаимодействию ни с одним из
пептидов, представляемых молекулами МНС. Вредоносные же рецепторы
связываются с собственными петидами организма, представленными его молекулами
МНС. Лимфоциты с такими рецепторами способны атаковать "свои"
ткани. Почему же в здоровом организме отсутствуют вредоносные Т-лимфоциты?
На этот вопрос пытались ответить Ф. Фернер и Ф. Бернет (1948), а также
Дж. Ледергерг (1963) с помощью гипотезы клональной делеции.
Предполагалось, что в организме всегда присутствуют пептиды, образующиеся из его
собственных белков, и лимфоциты, имеющие рецепторы к "своим" антигенам,
наверняка прореагируют с ними и тем самым элиминируются на ранних стадиях
развития. В результате останутся лишь те лимфоциты, которые несут
рецепторы к антигенам, отсутствующим у данного индивида. Эти клетки продолжат
свое созревание и в последствии будут стимулироваться чужеродными
антигенами, которые попадут в организм извне. Многочисленные исследования
молекулярных и клеточных механизмов клональной делеции (Бемер и др., 1991)
привели к открытию ключевой роли тимуса: судьба созревающих Т-лимфоцитов
162
определяется в этой железе. Вредоносные клетки, рецепторы которых им\н-
модействуют с антигенами и белками МНС собственного организма,
разрушаются. Бесполезные клетки, неспособные взаимодействовать со "своими" белки
ми МНС, погибают. Полезные клетки, взаимодействующие с чужеродными аи
гигенами в комплексе с белками МНС, созревают и накапливаются. В лшиси
мости от того, свяжется ли рецептор Т-лимфоцита с молекулой МНС класса I и
II, этот лимфоцит станет хелпером либо киллером. Вот так иммунная система
"обучается" отличать "свое" от "чужого" путем клонального отбора
лимфоцитов. Некоторые аутоиммунные явления, очевидно, обусловлены тем, что
иммунная система распознает как "чужое" собственные антигены организма,
которых нет в тимусе, и, следовательно, в нем не произошло делеции отвечающих
на них Т-лимфоцитов.
Существование аутоиммунных реакций говорит о несовершенстве делеци-
онного механизма распознавания "своего" и "чужого" и указывает, что,
по-видимому, здесь играют роль еще какие-то процессы.
Следует также отдавать себе отчет, что такой сложный механизм, как кло-
нальная делеция, не может не подвергаться постоянному воздействию
вредоносных эндогенных и экзогенных факторов (метаболизма, стресса, СР,
экологических влияний и т.п.), ущерб от которых, естественно, накапливается с
возрастом
Множество серьезных болезней вызвано саморазрушительным
поведением иммунной системы, включая ревматоидный,артрит,
системную красную волчанку, эритематоз, псориаз и многие другие.
У женщин аутоиммунные нарушения встречаются приблизительно
в четыре раза чаще, чем у мужчин. В настоящее время считается,
что тестостерон проявляет умеренное иммунодепрессантное
действие, уменьшая риск аутоиммунной атаки у мужчин, но при этом
также делает их иммунную систему несколько менее гибкой.
В то время как выраженные аутоиммунные патологии
относительно редки, умеренные аутоиммунные реакции весьма
распространены, и сфера их действия явно увеличивается с возрастом.
Фактически тестируемая степень аутоиммунитета рассматривается
некоторыми учеными как маркер старения. В частности, у пожилых
людей обнаруживается тенденция к увеличению количества аутоан-
гител, циркулирующих в кровотоке. Фигурально говоря, многие
пожилые люди испытывают умеренное отторжение своих собствен
ных тканей. Аутоиммунитет, следовательно, с возрастом может
стать фактором постепенной атрофии органов и тканей. Амилоид,
токсичный старческий пигмент, способный разрушать клетки
мозга, нервы и другие жизненно важные ткани, как полагают, является
побочным продуктом аутоиммунных реакций.
Почему происходит увеличение аутоиммунитета с возрастом/
Как это ни странно, это явление может быть косвенным следствием
возрастного снижения функции тимуса и активности Т-лимфоцитов.
Т-клетки, называемые Т-супрессорами, отвечают за контроль над
чрезмерными или саморазрушительными иммунными реакциями.
Снижение функций тимуса сопряжено с понижением способности Т-
супрессоров к регулированию деятельности других иммунных кле-
163
ток, особенно В-лимфоцитов, которые, потеряв контроль,
начинают производить аутоантитела. В то время как острые
аутоиммунные нарушения обычно требуют лекарств - иммунодепрессантов,
таких как кортикостероиды или цитотоксические агенты, для
пожилых людей с умеренным аутоиммунитетом может быть использован
несколько иной подход. Регенерация тимуса и стимуляцмя Т-клеток,
в частности Т-супрессоров, может улучшать регуляторный баланс в
иммунной системе и подавить чрезмерную активность В-клеток,
сокращая таким образом аутоиммунные реакции.
6.4. ИММУНИТЕТ И ФИЗИЧЕСКИЕ УПРАЖНЕНИЯ
Физические упражнения оказывают скромное, но заметное
влияние на иммунную систему. В частности, упражнения повышали
такие иммунные параметры, как активность N-киллеров, общее коли
чество лейкоцитов и уровень некоторых цитокинов, особенно ИЛ-1
и интерферонов. Напротив, длительный постельный режим или дру
гое сокращение физической активности вредит иммунитету,
вызывая общее падение лейкоцитов, так же как и способности к
продукции антител в ответ на посторонние субстанции.
Замечательным представляется то, что положительный ответ
иммунной системы на физические упражнения, особенно активация
N-киллеров, сохранены или даже увеличены с возрастом. Причины
этого пока неясны. Одна возможность состоит в том, что
упражнения повышают функции иммунной системы, стимулируя продукцию
гормона роста и ИФР-1. В молодом возрасте эти гормоны обильны
и воздействуют на иммунную систему настолько интенсивно, что
дополнительное повышение их уровня, возможно, не оказывает уже
никакого влияние на иммунную систему. У старых людей уровень
ГР и ИФР-1 низок, и возбуждение их продукции может существенно
повышать активность иммунной системы. Другая возможность
состоит в том, что упражнения улучшают толерантность к углеводам
и уменьшают избыток инсулина, что, как известно, приносит пользу
иммунитету (Дильман, 1987). Опять-таки, эти параметры обычно
нормальные у молодых половозрелых людей, ухудшены у старых
индивидов.
Глава 7
СТАРЕНИЕ И ДЕПРЕССИЯ
Ночь, улица, фонарь, аптека,
Бессмысленный и тусклый свет,
Живи еще хоть четверть века -
Все будет так. Исхода нет.
Умрешь - начнешь опять сначала,
И повторится все, как встарь:
Ночь, ледяная рябь канала,
Аптека, улица, фонарь.
Александр Блок
из цикла " Пляски смерти"
Независимо от того страдаете ли вы депрессией или нет, знания
о депрессии и способах ее устранения помогут сохранить здоровье и
даже продлить вашу жизнь. Типичные признаки и симптомы
депрессии таковы.
• Состояние подавленности, раздражительности или беспокойства;
физическая бездеятельность или повышенная активность.
Недостаток уверенности в себе; пониженное чувство собственного
достоинства; самобичевание; неоправданное чувство вины.
• Потеря интереса к обычной деятельности, потеря интереса к жизни.
• Социальная изоляция; потеря привязанности.
•Дурные предчувствия; безнадежность; беспомощность; чувство
преувеличенной зависимости от кого-либо или от чего-либо.
• Снижение внимания и способности концентрироваться.
• Приходящие в голову мысли о самоубийстве.
• Усталость; потеря аппетита; потеря веса или излишний вес.
• Бессонница или сонливость; потеря сексуального желания.
Наличие приблизительно половины этих признаков - явный
показатель депрессии. Необходимо подчеркнуть, что серьезная
депрессия требует профессиональной помощи. Информация, данная в
этой книге, не может использоваться как замена
профессионального клинического лечения (обычно психиатром или
психотерапевтом) любого существенного психического или умственного
расстройства.
Депрессия - наиболее распространенное психическое
заболевание в развитых странах. По оценкам специалистов приблизительно
15-25% популяции испытает клиническую депрессию в течение
жизни. Риск неклинической депрессии - вероятно значительно
выше. Влияние депрессии неуклонно и значительно увеличивается с
165
возрастом. При исследовании* группы из 880 европейцев, рожденных
с 1913 до 1918 г., Д. Халлер с коллегами (Mailer et al., 1996)
установили, что 12% мужчин и 28% женщин страдали депрессией. Другие
исследования показывают, что в некоторых других сообществах эти
показатели еще выше.
Взаимосвязь между старением и депрессией состоит в том, что
процесс старения вносит вклад в развитие депрессии, и наоборот,
депрессия, по-видимому, ускоряет старение. Прежде чем объяснить,
как это происходит, мы должны кратко рассмотреть причины и
механизмы депрессии.
Человеческий мозг является чрезвычайно сложной структурой,
содержащей миллиарды клеток. Эти клетки (среди которых
наиболее важны - нейроны), сами по себе - весьма сложны и способны к
изменению своего поведения в< зависимости от ситуации.
Надлежащая работа мозга зависит от эффективности связей между
нейронами, так же как современная экономика зависит от надежных и
быстродействующих коммуникаций. Природа нейронных связей -
двойственна, будучи и электрической и химической. Когда нейрон
"хочет" послать сигнал другому нейрону, он сначала посылает
электрохимическую волну по своему аксону (микроскопическому
удлиненному отростку клетки, работающему как линия связи). Сигнал,
достигая конца аксона, обуславливает выделение нейротрансмитте-
ров (нейрогоередатчиков) в синаптическую "щель" (пространство
между концом аксона и телом другого нейрона). Нейротрансмитте-
ры - это небольшие молекулы, требующиеся для передачи сигнала
сквозь синаптическую щель; наиболее важные из них - норэпинеф-
рин (норадреналин), ацетилхолин, серотонин и дофамин. Нейро-
трансмиттеры, выделенные одним нейроном действует на
специальные рецепторы, расположенные на мембране другого нейрона,
заставляя "воспринять" сигнал и ответить определенным действием.
После того как сигнал прошел, нейротрансмиттер, оставшийся в си-
наптической "щели" или разрушается ферментом моноамино окси-
дазой (МАО), или поглощается нейронами, и сигнал таким образом
обрывается (Fowler et al., 1980).
Согласно современным представлениям, депрессия,
независимо от ее основной причины, развивается, когда количество ней-
ротрансмиттеров в синаптической "щели" во время передачи
сигнала слишком мало, что в свою очередь приводит к
недостаточной связи между нейронами. Наиболее часто это связано с низким
уровнем содержания норадреналина и серотонина, и поэтому
подавляющее большинство способов лекарственного лечения
депрессии нацелено на повышение в мозге уровня одного или обоих
этих нейротрансмиттеров.
Многие факторы могут вызвать или ускорить депрессию.
Некоторые люди генетически предрасположены к депрессии (Баранов
и др., 2000). У этих людей депрессия обычно развиваются еще в мо-
166
лодости и требуют интенсивной профессиональной помощи. Н-шбо
лее частой причиной депрессии, однако, является острый или
хронический психологический стресс, такой как потеря близкого
человека, развод, увольнение с работы и т.д. Обычно стресс затихает
самопроизвольно за счет обратной связи. В начале стресс-ответа
гипоталамус посылает сигнал гипофизу выделить АКТГ, который
стимулирует надпочечники к выделению глюкокортикоидов
(стероидных гормонов, играющих главную роль в развитии стресс-ответа).
Повышение уровня глюкокортикоидов в крови заставляет
гипоталамус через отрицательную обратную связь, о которой говорилось
ранее, выключить свой начальный сигнал, и ответ на стрессовую
ситуацию постепенно спадает. (Мы подробно обсудим физиологию
стресса во II части книги.) При интенсивном или длительном
стрессе, организм стремится сделать гипоталамус менее чувствительным
к отрицательной обратной связи, т.е. глюкокортикоидам, пытаясь
продлить таким образом состояние стресс-ответа. Предполагается,
что этот эффект достигается путем уменьшения уровня мозговых
нейротрансмиттеров {Stokes, 1995). Действительно, хорошо
известно, что у наиболее угнетенных пациентов механизм, который
ограничивает стресс-ответ, ослаблен. Один из главных лабораторных
тестов на депрессию (дексаметазоновый тест) фактически
базируется на выявлении пониженной способности организма
ограничивать ответ на стресс (Feinberg et aL, 1984). В некотором роде,
депрессия — это состояние бесконечного стресса (Дилъман, 1987).
Наряду с нарушениями в адаптационном гомеостате при
психической депрессии нарушены и многие другие эндокринные функции,
в частности регуляция секреции ГР и пролактина, осуществляемая
системами, которые контролируются дофаминовыми, норадренали-
новыми, серотониновыми, опиоидными и холиннэргическими
медиаторами (Cameron et aL, 1984; Crapo, 1979). Вероятно, нарушения ь
гормонально-метаболическом гомеостазе обуславливают
увеличение заболеваемости и смертности у больных, страдающих
депрессией (Malzberg, 1937; Shekelle et aL, 1981). Дополнительный вклад в это
увеличение вносит и повышение уровня глюкокортикоидов, снижая
при этом активность иммунной системы (Schleifer et aL, 1985).
Болезни, связанные с хронической болью, иммунные или
гормональные нарушения могут также вызвать депрессию, и
большинство людей с этими недугами в некоторой степени страдают
депрессией. Дефицит питательных веществ также неблагоприятен в этом
отношении (см. гл. 28). В частности, дефицит витаминов С, Bl, B6, В12,
фолиевой, никотиновой и пантотеновой кислот может вызвать
депрессию наряду и со множеством других нарушений. Явный дефицит
этих питательных веществ в развитых странах встречается редко.
Однако незначительный дефицит может иметь место, особенно в
пожилом возрасте и при хронических заболеваниях, и способен
вносить заметный вклад в возникновение депрессии.
167
Имеет место существенное физиологическое сходство между
депрессией и процессом старения. Во-первых, уровень нейротрансмит-
теров в мозгу, включая норадреналин и серотонин, уменьшается с
возрастом. Как обсуждалось выше, низкий уровень этих
соединений - главная особенность депрессии. Во-вторых, в пожилом
возрасте отрицательная обратная связь, необходимая для ограничения
чрезмерного стресс-ответа, постепенно ослабевает, что в
результате приводит к состоянию хронического стресса. То же самое
наблюдается и у индивидуумов, находящихся в состоянии депрессии.
Кроме того, некоторые дегенеративные болезни, которые часто
сопровождают старение, в частности диабет или аутоиммунные
нарушения, значительно увеличивают риск депрессии. Таким образом,
старение, вероятно, вносит вклад в развитие депрессии, вследствие
изменений в биохимии организма. Но обратное влияние также
существует. Депрессия, по-видимому, ускоряет старение и увеличивает
риск болезней по нескольким механизмам.
(1) Депрессия ведет к чрезмерному стрессу. Как мы отмечали,
физиологические изменения при депрессии имеют сходство с
изменениями при хроническом стрессе. Кроме того, депрессия делает
человека более уязвимым по отношению ко всем видам сильного
стресса
(2) Другой механизм, по которому депрессия содействует
старению и болезни, - ее неблагоприятное воздействие на иммунную
систему. Не совсем ясно, как депрессия подавляет иммунную систему.
Так, некоторые нейротрансмиттеры способны стимулировать
иммунную систему, и поэтому недостаток этих молекул при депрессии,
по-видимому, может быть фактором риска. Избыток стероидов
стресса при депрессии также может внести свой вклад в подавление
иммунной системы. И так как последняя обеспечивает защиту
организма от инфекции, рака, атеросклероза и, возможно, многих других
заболеваний, ухудшение функций иммунной системы не может не
сказаться на состоянии здоровья и темпе старения организма.
(3) Как уже не раз отмечалось, работа ЦЧС сопряжена с
уменьшением чувствительности гипоталамуса к сигналам, идущим от
периферических тканей (отрицательная обратная связь). Часовой
"механизм", по-видимому, снижает концентрацию нейротрансмиттеров
в гипоталамусе и некоторых смежных мозговых структурах.
Депрессия также уменьшает этот показатель, что может ускорять
ЦЧС. В.М. Дильман (1987) предположил, что антидепрессанты,
поднимающие уровень нейротрансмиттеров в гипоталамусе, являются
также и потенциальными лекарствами против старения.
Действительно, некоторые исследователи обнаружили, что лекарства или
пищевые добавки, которые уменьшают депрессию, также изменяют
метаболические и адаптивные характеристики организма в сторону
"более молодого" состояния, а в некоторых случаях даже
продлевают жизнь лабораторных животных. В частности, было найдено, что
168
антидепрессант депренил увеличивает продолжительность жизни ν
крыс на 34%. Этот препарат также уменьшает накопление липофу(
цина и деградацию нейронов в мозге крысы (Дилъман, 1987)
(4) И - последнее (по порядку, но не по значению). Человек в
состоянии депрессии вряд ли будет иметь достаточную мотивацию для
активных действий, нацеленных на продление жизни или даже
просто на поддержание здорового образа жизни.
Лечение депрессии - обычно комбинация двух подходов. Один
из них - общение с психоаналитиком с целью изменить в лучшую
сторону взгляд пациента на его жизненную ситуацию, помочь в
разрешении проблем и поощрить положительные действия и
социальные контакты. Другой подход состоит в том, чтобы исправить
биохимическую неустойчивость, поднимая уровень нейротрансмитте-
ров в мозгу. Существует множество лекарств, предназначенных для
этой цели, но действие большинства из них базируется на двух
основных принципах. Многие лекарства блокируют способность
нейронов поглощать нейротрансмиттеры после того, как сигнал
прошел - тогда большее количество нейротрансмиттеров остается в си-
наптической "щели", что так или иначе улучшает межнейронную
связь и уменьшает депрессию. Лучшие известные лекарства этого
вида - Elavil и Prozac - блокируют поглощение норадреналина и се-
ротонина, а на более поздних этапах действия препаратов - только
одного серотонина. Другим важным классом антидепрессантов
являются ингибиторы моноамино оксидазы. Эти лекарства понижают
активность фермента, ответственного за разрушение нейротрансми-
теров в синаптической "щели*'. И снова результатом оказывается
увеличение количества нейротрансмиттеров в "шели". что в
конечном счете устраняет депрессию.
Большинство антидепрессантных препаратов обычно вызывают
существенные побочные эффекты, такие как сонливость, запоры, сухость во рту,
тошноту, расстройство желудка, импотенцию и др. По этой причине лечение
препаратами обычно допускается для клинически выраженных состояний депрессии.
Тем не менее некоторые питательные вещества могут также способствовать
увеличению уровня нейротрансмиттеров в мозгу и таким образом смягчать
депрессию. Многие исследования показали, что слабые и умеренные формы
депрессии можно лечить пищевыми добавками, которые содержат
аминокислоты - предшественники нейротрансмиттеров. Такое лечение часто столь же эф
фективно, как и лечение антидепрессантами, но дает значительно меньше
побочных эффектов. Кроме того, некоторые экстракты растений, как доказано,
обладают значительной антидепрессантной активностью в сочетании с
небольшими побочными эффектами (практические подробности см. в гл. 28).
Глава 8
СТАРЕНИЕ И ПОЛОВАЯ ФУНКЦИЯ
Люди - бесспорно существа с половой формой размножения.
В списке наших приоритетов нормальные, приятные половые
отношения занимают второе место после проблем физического
выживания, а иногда даже и первое. С эволюционной точки зрения это
вполне понятно - чтобы гарантировать выживание вида, функция
воспроизведения должна быть исключительно сильной.
Сознательно или подсознательно, многое из нашего поведения определяется
сексуальными мотивами. Не удивительно, что лучшие (и
наихудшие) кинофильмы и романы концентрируют внимание вокруг
романтических взаимоотношений; коммерческие радио- и
телепередачи насыщаются сексуальными мотивами, чтобы рекламировать
даже такие абсолютно несексуальные изделия, как стиральные
порошки и жевательные резинки.
С возрастом и половое влечение (либидо) и репродуктивная
функция постепенно угасают. Однако потеря половой функции - не
неизбежное следствие старения: некоторые люди сохраняют
здоровую половую активность даже в преклонном возрасте.
Современные представления многих ученых и клиницистов состоят в том, что
половые отношения могут и должны способствовать ощущению
радости жизни и чувству собственного достоинства в любом возрасте.
В своем обзоре проблем сексуальности в старости А. Комфорт
и другие (Comfort et al., 1991) заявляют, что "...эмоциональная
потребность пожилых индивидуумов в сексуальности практически
такая же, как и в более молодом возрасте, и идея о безболезненном
и постепенном падении сексуального статуса для них - почти
оскорбительна".
Провести надежные исследования состояния половой функции в
пожилом возрасте достаточно трудно, люди обычно отказываются в
них участвовать; получение объективной выборки почти
невозможно; и главный инструмент исследования все еще - печально
известный своей неточностью - метод анкетного опроса. Текущие
исследования указывают, что у многих старых людей сексуальность
хорошо сохранена. Д. Вилей и его коллеги (Wiley et al., 1996), познако-
170
мившись со 158 пожилыми посетителями образовательных
семинаров по сексологии, отметили "замечательно здоровую сексуальную
жизнь" как у мужчин, так и у женщин. (Это заключение, возможно,
не является общей характеристикой, поскольку участники
подобных семинаров могли иметь более высокую, чем
среднестатистическая, половую активность.) Другие исследования также указывают,
что многие здоровые люди остаются сексуально активными до
преклонного возраста (Kaplan, 1990). Нам сразу хотелось бы
подчеркнуть очень важный момент, который состоит в том, что
всестороннее сохранение здоровья и минимизация дегенеративных болезней
являются решающими для длительной и полноценной сексуальной
жизни. Многие возрастные патологии, особенно атеросклероз,
диабет, гипертрофия простаты и гипертония, являются вредными для
мужской половой функции, а депрессия может подавлять либидо
как у женщин, так и у мужчин. Это - с одной стороны. С другой
стороны, следует помнить, что многие лекарства, которыми обычно
лечат возрастные болезни, могут также приводить к импотенции и
подавлению либидо.
Другая важная причина понижения сексуальности - стресс,
"физиологическая роль" которого состоит в переключении потока
ресурсов организма от таких функций, как пищеварение,
воспроизводство, иммунная защита и другие, к совершенно необходимым для
непосредственного выживания в данной критической ситуации
(центральная нервная система - ЦНС, сердце, кровеносные сосуды и
скелетные мышцы) (см. гл. 10). Мы полагаем, что лучшая мера для
сохранения полноценной половой функции - многоплановая стратегия
продления жизни, а именно сокращение стресса и предотвращение
дегенеративных болезней
8.1. МЕНОПАУЗА И ЭСТРОГЕНОВАЯ ТЕРАПИЯ
Менопауза - прекращение ежемесячных менструальных циклов у
женщин, которое происходит, когда яичники прекращают
производство эстрогенов. В среднем, менопауза наступает примерно в
50-летнем возрасте, но она может "ударить" уже и после 30-ти. Когда
уровень эстрогенов резко падает, женские репродуктивные органы
теряют свою функцию. В частности, влагалище становится сухим, легко
раздражимым и часто инфицируется, что может делать сексуальное
общение болезненным и неприятным. Регулярное сексуальное
общение - самостоятельный фактор в поддержании нормального
состояния влагалища и его слизистой поверхности, поэтому уклоняться от
половых сношений - значит обострять в конечном счете эту проблему.
Некоторые женщины испытывают также и множество
системных признаков менопаузы. Чрезмерно высокий уровень гонадотро-
пинов вызывает примерно у 75% женщин "волнообразные ощуще-
171
ния жара" в течение 2-5 лет после менопаузы. Другие признаки,
возможно связанные с менопаузой, - нервозность, утомляемость и
бессонница (Bachmann, 1995)
Сегодня наиболее распространенный способ "лечения"
менопаузы - эстрогеновая терапия, которая восстанавливает
атрофированные репродуктивные органы и "снимает" другие симптомы
менопаузы, например волнообразные ощущения жара.
Эстрогены, однако, не только половые гормоны: большинство органов,
включая мозг, сердце, кровеносные сосуды, легкие и кости,
имеют на поверхности своих клеток рецепторы - специфические
структуры, связывающие эстрогены и способствующие их
проникновению внутрь клетки для осуществления гормонального
эффекта. Влияние эстрогенов на организм многосторонне, и это
порождает неоднозначные оценки ценности заместительной
терапии этим гормоном.
Обширные и длительные исследования, завершенные в
последние годы, показывают, что эстрогеновая терапия улучшает
качество жизни, а также немного увеличивает среднюю
продолжительность жизни у женщин. Наиболее крупное исследование
охватило 121 тыс. женщин, которые подвергались
заместительной гормональной терапии в течение 16 лет (с 1976 до 1992 г.).
Ф. Гродстейн и его коллеги {Grodstein et aL, 1992) нашли, что в
большой группе женщин, получавших заместительную эстроге-
новую терапию, смертность в течение десяти лет была на 37% (!).
а в течение более чем 10 лет - на 20% ниже, чем за такой же
период в контрольной группе. Этот впечатляющий результат
влияния эстрогенов на выживание пациенток был в значительной
степени обусловлен более низкой смертностью от
сердечно-сосудистых заболеваний, нежели в контрольной группе. Снижение
выживаемости (в %) при эстрогеновой терапии после десяти лет (от
начала менопаузы) было связано главным образом с 43%
увеличением риска смерти от рака молочной железы. Главное
заключение этого исследования состояло в том, что эстрогеновая
терапия в течение десяти лет после наступления менопаузы,
несомненно, приносит пользу большинству женщин. Хотя эти
женщины и могли иметь один или несколько факторов риска
заболеваний сердца, вместе с эстрогенами они получали явное
преимущество - значительное сокращение риска смерти по сравнению с
теми женщинами в менопаузе, которые не получали
заместительной эстрогеновой терапии. Факторы риска сердечно-сосудистых
болезней, рассматривавшиеся в этом исследовании, включили
генетические предпосылки, курение сигарет, гипертонию, высокий
уровень холестерина и ожирение. 69% участников исследования
имели по крайней мере один фактор риска сердечных болезней.
Однако женщины с низким риском сердечных заболеваний, но с
увеличенным риском рака молочной железы должны, очевидно.
172
избегать приема эстрогенов, особенно после десятитилетнего
периода от начала менопаузы.
Прием только эстрогенов не может моделировать естественный
эстральный цикл. Обычно, в течение нескольких дней в каждом
нормальном цикле яичники выделяют также и прогестины, стеро
идные гормоны, способствующие беременности (Wilson et al., 1985).
Непрерывный прием только одних эстрогенов увеличивает риск
внутриматочного рака. Этот риск не повышается, если прогестин
(прогестерон) дается женщинам в менопаузе в течение десяти (или
большего количества) дней условного "цикла" совместно с
эстрогенами (Murray et al., 1991).
Решение о применении заместительной эстрогеновой терапии
должно приниматься женщиной и ее врачом с учетом
индивидуальных особенностей, медицинской истории болезни, жалоб и ее
личных предпочтений. Согласно Ф. Гродстейн и его коллегам
(Grodstein et al., 1992) "...женщины и их врачи должны
пересматривать свои решения каждые пять лет, потому что и выгоды,
и риск гормональной терапии могут измениться со временем".
Те, кто решит воздержаться от эстрогеновой терапии, могут
поддерживать оптимальное состояние слизистой оболочки
влагалища, используя кремы, содержащие эстроген, а если необходимо
избегать даже маленьких доз гормона, использовать
водно-жировые эмульсии.
Сексуальная активность и способность испытывать
возбуждение хорошо сохраняются у многих, если не у большинства женщин,
после менопаузы, несмотря на потерю овариальной функции и
существенную атрофию органов размножения. Правда, некоторые
женщины в этот период значительно снижают свое либидо.
Причины такой вариабельности не совсем ясны. Не только яичники, но и
другие ткани, такие как надпочечники и жировая ткань, могут
производить небольшие, но существенные количества эстрогенов,
которые, очевидно, играют определенную роль в поддержании
сексуальной активности. Было также установлено, что тестостерон
важен для сексуальной активности не только мужчин, но и женщин.
Функционирующие яичники производят некоторое количество
тестостерона (приблизительно от 1 до 2 мг в день). После менопаузы,
единственный остающийся источник продуцирования
тестостерона - надпочечники, но в некоторых случаях этого недостаточно для
поддержания здоровой сексуальной активности. Действительно, у
некоторых женщин с уменьшенным либидо, эстрогеновая
заместительная терапия неэффективна для его восстановления, тогда как
даже небольшие количества тестостерона (дополнительно к
вышеуказанной терапии) часто позволяют преодолеть это затруднение.
Однако использование тестостерона у женщин после менопаузы
должно быть весьма осторожным из-за риска вирилизации
(появление вторичных мужских половых признаков: рост волос на теле и на
173
лице, понижение голоса и т.д.). При конкретном риске проявления
таких побочных эффектов - женщины должны отказаться от эстро-
геново-тестостероновой терапии, потому что эстрогены, которые
обычно противодействуют некоторым эффектам тестостерона,
используются при такой терапии в относительно низких количествах
(Murray et al., 1991).
8.2. МОЖНО ЛИ ОТСРОЧИТЬ МЕНОПАУЗУ?
Менопауза у людей - неизбежное следствие старения, и избежать
ее невозможно. Тем не менее имеются значительные вариации в
возрасте женщин, вступающих в менопаузу, от 35-40 до 55-60 лет.
Причины того, что некоторые женщины сохраняют способность к
деторождению до относительно позднего возраста, поняты лишь
частично. Одно из возможных объяснений - темп центральных
биологических часов, контролирующих функции яичников. А функции эти
таковы: (1) производить половые гормоны, главным образом
эстрогены, которые необходимы для поддержания функций женской
репродуктивной системы, и (2) готовить зрелые фолликулы (специальные
клетки, предназначенные для оплодотворения сперматозоидами) из
клеток-предшественников, называемых овоцитами. Каждая
женщина имеет ограниченный запас овоцитов и поэтому может производить
только ограниченное количество зрелых фолликулов. Когда
последовательные циклы овуляции исчерпают резерв овоцитов, теряется
способность к овогенезу и наступает менопауза. Предполагается, что
истощение резерва овоцитов так или иначе является причиной спада
в продукции эстрогенов и полной атрофии яичников. Их вес
уменьшается с примерно с 14 г в четвертом десятилетии примерно до 5 г в
шестом и седьмом десятилетиях жизни женщины (Дилъман, 1986).
Если-наступление менопаузы зависит только от исходного
количества овоцитов, то, очевидно, немногое можно сделать, чтобы
воздействовать на часы овогенеза и задержать менопаузу. Тем не менее
недавние исследования показывают, что эта ситуация не столь проста,
как это казалось прежде. В течение каждого цикла овуляции, многие
овоциты начинают свое развитие в зрелые фолликулы, но только
один из mm (редко несколько) заканчивает это путешествие, а
остальные деградируют и погибают. Если слишком много овоцитов в
каждом цикле "выставляется на состязание", то "овариальный капитал"
исчерпывается довольно быстро, и может следовать ранняя
менопауза. Наоборот, если относительно немного овоцитов будут заняты в
цикле, плодовитость женщины может быть сохранена в течение более
длительного срока. Некоторое свидетельства указывают, что
центральные часы старения в гипоталамусе (см. гл. 2) могут влиять на
возрастные изменения функции яичников и сроки наступления
менопаузы. Гипоталамус регулирует функцию яичников, "приказывая" гипо-
174
физу выделять так называемые гонадотропины, а именно лютеини-
зирующий гормон (ЛГ) и фолликулостимулирующий гормон (ФСГ),
которые стимулируют производство эстрогена и овуляцию.
Эстрогены в свою очередь блокируют гипоталамический сигнал на выделение
гонадотропинов, т.е. имеется петля отрицательной обратной связи,
которая предотвращает гиперстимуляцию яичников (Tepperman et al.,
1987). Есть основания полагать, что эффективность этой петли
уменьшается с возрастом, в результате чего постепенно повышается
уровень гонадотропинов. В период развития организма, это повышение
уровня гонадотропинов и эстрогенов (наряду с падением уровня мела-
тонина) играет основную роль в половом созревании и включении
репродуктивной функции. Позже уровень гонадотропинов продолжает
повышаться, вызывая чрезмерную активацию овогенеза, что
приводит к смерти все большее и большее количество овоцитов в
последовательных циклах. Этот темп истощения резерва овоцитов
увеличивается с возрастом, частично потому, что гипоталамус теряет
чувствительность к ингибирующему действию отрицательной обратной связи
(концентрация эстрогенов) и в результате - к гиперстимуляции
яичников из-за постоянно увеличивающегося уровня гонадотропинов.
Логическое заключение, к которому приходит В.Μ Дильман - замедление
темпа ЦЧС (т.е. восстановление чувствительности гипоталамуса к
тормозящему действию эстрогенов) может затормозить ход овариаль-
ных часов и продлить репродуктивную жизнь женщины (Дильман,
1987).
Некоторые косвенные данные указывают, что этот подход
может быть действенным. Один из путей "выправления" ЦЧС -
повышение уровня дофамина в гипоталамусе, что делает последний
более чувствительным к сигналам обратной связи. Дофаминэргиче-
ские стимулы, такие как L-дофа или бромокриптин, как было
показано, способны восстановить менструальные циклы у старых крыс и
у некоторых женщин в менопаузе (Дильман, 1981). В настоящее
время, проблема продления женской репродуктивной жизни в шестое
или даже седьмое десятилетия базируется лишь на скромном
количестве данных и многообещающей теории, и только будущие
целенаправленные исследования позволят оценить перспективность
предложенного подхода для практической медицины.
8.3. МУЖСКАЯ ПОЛОВАЯ ФУНКЦИЯ
Мужчины с возрастом испытывают устойчивое снижение
половой функции, которое становится более очевидным после 50-ти.
Возрастные изменения мужской сексуальности иногда называют
"мужской менопаузой", хотя у мужчин не замечено ничего столь же
резкого, как прекращение менструального цикла и атрофия
половых желез, наблюдаемых у пожилых женщин. Причины и содейст-
175
вующие факторы "мужской менопаузы" остаются предметом
горячей дискуссии (Jsitouras et al, 1995). Наиболее очевидный кандидата-
уменьшение уровня тестостерона, отмеченное во многих
исследованиях. Начинаясь после 40 лет, падение уровня тестостерона обычно
продолжается примерно по 1% в год, а хронические болезни, стресс
и некоторые медикаменты могут удваивать эту величину (Krause,
1995). Однако у наиболее здоровых пожилых мужчин уровень
тестостерона остается в нормальных пределах, и корреляции между
уровнем тестостерона и половой активностью - низка. Большинство
исследований указывают, что тестостероновая терапия, вероятно,
улучшит половую функцию мужчин с очень низким уровнем
гормона (ниже 300 нг/дл, т.е. нижней граница нормы) и будет иногда
приносить пользу пожилым мужчинам и с нормальным уровнем
тестостерона. Кроме того, введение тестостерона часто может
увеличивать массу мышц и уменьшать жировую массу тела. Возможной
отрицательной стороной применения экзогенного тестостерона может
быть подавление функции яичек (что сопряжено с понижением
эндогенной продукции тестостерона и сперматозоидов), появление
прыщей, задержка натрия, что может обострить болезни сердца или
гипертонию и даже умеренно подавить иммунную функцию {Comfort
et al., 1991). Последствия долговременной тестостероновой терапии
у старых мужчин - неизвестны. Наши современные знания
свидетельствуют, что у мужчин с колебаниями уровня тестостерона в
пределах возрастной нормы введение ДГЭ А может обеспечивать те же
самые эффекты, что и тестостерон в отношении состава тела и
либидо, но с меньшим числом побочных эффектов (Sahelian, 1996).
8.4. РАЗРЫВ МЕЖДУ ЛИБИДО И ПОТЕНЦИЕЙ
Большинство мужчин не теряет с возрастом интереса к сексу,
хотя чувство "полового нетерпения" обычно спадает. Способность к
исполнению, однако, часто страдает значительно больше.
Результат - ситуация глубокого эмоционального расстройства, иногда
называемая "разрывом либидо-потенция*', когда половое желание
превышает физические возможности. Распространенность
импотенции резко увеличивается с возрастом. Более чем 90% мужчин
моложе 50 физиологически полноценны, но только примерно половина
из них в таком состоянии - после 70 (Wiley et al., 1996).
До относительно недавнего времени считалось, что импотенция
вызывается главным образом психологическими факторами.
Теперь полагают, что примерно 75% случаев обусловлено
физическими причинами. У старых мужчин импотенция по органическим
причинам даже более распространена, составляя приблизительно 90%.
С медицинской точки зрения импотенция - это неспособность
достичь эрекции, достаточной для полового акта. Эрекция достига-
176
ется скоординированным усилием нескольких систем организма: со*
судистой, нервной, эндокринной (гормоны) и мочеполовой
(простата). Пояснить это просто: эрекция - главным образом проблема
"гидродинамики" и "реле-переключателей". Соответствующее
"реле", активированное нервными импульсами, заставляет артерии
полового члена расширяться, а вены - сжиматься. В результате,
приток крови к члену большее, чем отток, что и вызывает эрекцию.
Адекватный уровень тестостерона и некоторых нейромедиаторов,
таких как норадреналин, важен для поддержания чувствительности
"реле". Здоровая простата также важна, потому что ее заболевания
могут быть причиной сжатия артерий и нервов в основании
полового члена, что в свою очередь может вести к импотенции.
8.5. ПРИЧИНЫ ИМПОТЕНЦИИ
• Недостаточный поток артериальной крови. Это, вероятно,
наиболее частая причина импотенции, и обычно результат
атеросклероза, гипертонии или диабета. Существенный атеросклероз
(блокада артерий жировыми отложениями) чрезвычайно распространен у
мужчин после 35-40 лет, и может быть полностью
бессимптомным. Импотенция - только одно из многих страшных последствий
атеросклероза, таких как инфаркты, инсульты и т.д. Высокое
кровяное давление может механически повреждать артерии и
приводить к потере их эластичности, а также способствовать прогрессии
атеросклероза. Диабет может вызывать импотенцию, повреждая и
большие и малые кровеносные сосуды, а также и нервы (см. гл. 2,
параграф "Глюкоза и старение").
• Венозная утечка. Увеличение потока крови в половой член сквозь
артерии необходимо, но не достаточно для эрекции. Вены должны
быть способны поддержать эту ситуацию и помочь задержке
возможно большего количества крови в члене. Следовательно,
некоторые случаи импотенции вызваны чрезмерной утечкой венозной
крови во время эрекции.
• Ослабление нервной регуляции. Повреждения нервов, которые
регулируют ток крови к члену, могут быть вызваны диабетом
(наиболее распространенная причина), травмой костей таза и
некоторыми болезнями нервной системы.
• Лекарства, этиловый спирт. Многие медикаменты (лекарства от
высокого кровяного давления, антидепрессанты, седативные
средства и проч.) так же как этиловый спирт могут быть
причиной импотенции. Перерыв в приеме лекарственных средств,
если таковой возможен, обычно восстанавливает потенцию.
Иногда можно найти альтернативные средства с теми же самыми
положительными свойствами, но без неблагоприятных эффектов
для половой активности
177
• Болезни простаты. Две весьма обычных патологии -
доброкачественная опухоль простаты, наблюдаемое у мужчин после 40 лет, и
простатит - во всех возрастах, могут привести к импотенции,
сжимая артерии и нервы у основания полового члена, а иногда просто
вызывая боль в этой области. Радикальная простатэктомия
(хирургическое удаление простаты) также часто ведет к импотенции,
вероятно, за счет повреждения нервов, контролирующих эрекцию.
• Гормональный дисбаланс. Относительно малое число случаев им
потенции связано с гормональным дисбалансом, в частности с низ
ким уровнем тестостерона, и иногда с высоким - пролактина.
• Психогенные факторы. Согласно оценкам, примерно 25% случаев
импотенции вызываются психологическими причинами. Этот вид
импотенции чаще встречается у мужчин в молодом и среднем воз
расте. Если человек испытывает эрекции во сне или рано утром, то
импотенция, вероятно, имеет психогенную природу.
Психологический стресс и/или беспокойство может вызывать импотенцию, "на
кладываясь" на сигналы, которые мозг посылает периферическим
нервам в ситуации сексуального возбуждения. В результате
периферические нервы будут не в состоянии регулировать
кровообращение в половом органе. Долговременный психологический стресс
может также нарушать гормональный баланс организма, т.е.
понижать концентрацию тестостерона и повышать пролактин,
способствуя таким образом импотенции.
При всем различии причин, органическая импотенция -
обычно результат одной или нескольких возрастных болезней,
особенно атеросклероза, гипертонии, диабета и доброкачественной
опухоли простаты (ДОП). Стратегия продления жизни, описанная в
III части этой книги, нацелена, наряду с другими проблемами, на
предотвращение или коррекцию возрастных болезней. В
частности, атеросклероз - наиболее частая причина полового бессилия -
может быть замедлен или даже предотвращен, если удается
существенно понизить содержание холестерина и ЛНП. Это достижимо
при помощи диеты, физических упражнений, пищевых добавок и -
в случае особой устойчивости-лекарств, понижающих холестерин
(см. гл. 22, 23, 27). Лучшая стратегия против диабета -
предотвращение или своевременное лечение, направленное на повышение
реактивности тканей к инсулину (см. гл. 26, 29). Умеренная
гипертония может быть преодолена диетой, обогащенной калием,
кальцием и магнием, однако серьезные случаи обычно требуют
медикаментозного лечения. К сожалению, некоторые виды такого
лечения сами по себе могут стать причиной импотенции. ДОП
является, как полагают, результатом возрастных гормональных
изменений, особенно увеличения преобразования тестостерона в дегид-
ротеапостерон (ДГТ). Некоторая степень ДОП найдена у
большинства пожилых людей. Увеличенная простата может препятст-
178
вовать току мочи, провоцируя безотлагательность и частоту моче
испусканий, а также сжимать артерии и нервы в основании полоно-
го члена, приводя к импотенции (мы обсудим пути предотвращения
ДОП в гл. 30). Еще раз подчеркнем, что реализация всесторонней
стратегии продления жизни способна помочь как в предупрежде
нии и предотвращении возрастных болезней, так и в уменьшении
риска импотенции даже в преклонном возрасте.
Как уже отмечалось, психогенная импотенция составляет около
25 % всех случаев. По сравнению с пожилыми, эта величина
намного выше у молодых мужчин, поскольку органические болезни у них
менее вероятны, так же как и возможность иметь устойчивые
моногамные отношения, исключающие ряд возможных беспокойств при
отправлении сексуальной функции. В целом, различные формы
стресса - наиболее распространенная причина психогенной
импотенции. Повышение устойчивости к стрессовым ситуациям с
помощью адаптогенов, в частности экстрактов корней женьшеня или
элеутерококка, у некоторых мужчин было сопряжено с излечением
или улучшением психогенной импотенции (см. гл. 29).
В.6. ГЛАВНЫЙ СЕКСУАЛЬНЫЙ ОРГАН
Это стало почти банальностью - рассматривать мозг как
"главный сексуальный орган". Однако роль мозга в поддержании
и регуляции сексуальной активности не может быть завышена.
Половое возбуждение и оргазм управляются мозгом с помощью
сигналов, которые он посылает и получает по периферическим
нервам. Способность мозга поддерживать половую активность
зависит от удовлетворительного уровня ключевых нейромедиа-
торов. Почти любые серьезные психические нарушения могут
приводить к потере полового желания и потенции. Распростра-
ненный "враг" здоровой сексуальности - депрессия, особенно тип
депрессии с выраженным состоянием беспокойства. Депрессия
связана с разнообразными биохимическими изменениями,
которые подавляют половое желание и уменьшают возможности его
удовлетворения. Это состояние характеризуется низким уровнем
некоторых нейромедиаторов в мозгу, таких как норадреналин и
серотонин, так же как и избытком гормонов стресса. Как мы уже
отмечали, многие особенности депрессии напоминают состояние
хронического стресса. По мере старения, мы все более и более
страдаем от депрессии, часто даже не подозревая об этом. Разве
придет когда-либо в голову мысль, что "нормальная" возрастная
потеря полового желания может быть результатом скрытой
депрессии? А ведь когда с депрессией справляются, сексуальная
активность часто "чудесно" возрождается (более подробно мы
обсудим проблемы депрессии в гл. 28).
179
Среди функциональных нарушений и повреждений мозга,
связанных с возрастом, наиболее серьезными формами являются
инсульты, болезнь Альцгеймера и болезнь Паркинсона. Эти пато
логии также существенно ослабляют сексуальную активность
Мозг высоко чувствителен к повреждающему действию СР, а
масштабы этого действия велики из-за интенсивного
метаболизма и высокого содержания ненасыщенных жирных кислот,
которые легко реагируют со СР. Антиоксиданты, особенно те,
которые защищают мембраны нейронов, в частности витамин Ε и ли-
поиевая кислота, могут замедлить старение мозга и помочь в
сохранении здоровый сексуальный активности. "Модная" пищевая
добавка — диметилэтаноламин (ДМЭА), как известно, улучшает
функции мозга у людей. В экспериментах на животных было
показано, что ДМЭА увеличивал продолжительность жизни и
уменьшал отложения возрастных пигментов в мозговых клетках.
Основной механизм действия ДМЭА связан с увеличением в
мозгу уровня нейромедиатора ацетияхолина. Не затрагивая половую
активность у молодых людей, ДМЭА, как сообщалось,
увеличивает ее у многих людей после 40 лет. Будущие исследования,
очевидно, смогут выявить положительные эффекты и других нейро-
активных пищевых добавок или лекарств на сексуальную
деятельность пожилых людей.
8.7. СЕКСУАЛЬНОСТЬ И ГОРМОНЫ МОЛОДОСТИ
Три гормона, чей уровень резко падает с возрастом, снискали
особую похвалу масс-медиа за их предполагаемые многочисленные
свойства противостоять старению и дегенеративным болезням -
гормон роста (ГР), дегидроэпиандростерон (ДГЭА) и мелатонин.
Современные исследования показывают, что каждый из этих
гормонов, не будучи панацеей, может производить множество
положительных изменений у старых животных и людей.
Что могут сделать эти "гормоны молодости" для улучшения
нашей сексуальной жизни? К сожалению, всесторонние клинические
исследования прямого влияния ГР, ДГЭА и мелатонина на
сексуальную деятельность людей не проводились. Большинство
свидетельств этого носят косвенный или спекулятивный характер.
В своей книге "Стать молодым с человеческим ГР" Р. Клац и
К. Кан (ΛΓ/fl/z, Kahn, 1997) рассматривают мнения нескольких
клиницистов, которые инъецировали своим пациентам ГР. Согласно этим
авторам, ГР может резко усилить половое влечение и активность у
старых людей. Теоретически ГР может стимулировать половую
функцию, улучшая настроение, функцию мозга,
сердечно-сосудистую деятельность и изменяя уровень некоторых гормонов,
включая тестостерон.
180
Р. Шахелиан, проанализировав данные многих клиницистов,
заключил, что ДГЭА стимулирует либидо, особенно у женщин и
у мужчин с низким уровнем тестостерона (Sahelian, 1996). ДГЭА -
метаболический предшественник различных типов стероидов и
может теоретически увеличивать производство половых
гормонов. ДГЭА может также уменьшать депрессию и улучшать
мозговые функции, облегчая таким образом нормальную половую
деятельность. Однако, как показано в одном исследовании, 50 мг
ДГЭА в день в течение трехмесячного периода не оказал
никакого влияния на либидо пациентов.
Влияние мелатонина на половую функцию оценить еще
труднее. С одной стороны, очень высокий уровень мелатонина может
ингибировать половую функцию. Как уже обсуждалось в главе 5,
уровень мелатонина очень высок в детстве до полового созревания,
и его последующее резкое падение может действовать как пусковой
механизм полового созревания подростков. В литературе,
некоторые случаи отсроченного полового созревания у людей связывались
с чрезмерно высоким уровнем мелатонина. С другой стороны, очень
низкий уровень мелатонина, по-видимому, увеличивает риск
заболевания атеросклерозом, высоким кровяным давлением и некоторыми
возрастными нарушениями функций мозга, т.е. факторов
способствующих подавлению половой функции. Напротив, поддержание
молодого, т.е. умеренно высокого уровня мелатонина, может быть
фактором сохранения потенции и сексуальной активности. Более
того, гипотетически "высокий" мелатонин может замедлить или
даже полностью обратить ход ЦЧС в гипоталамусе (см. гл. 5),
восстанавливая "юный" гомеостаз и метаболизм и улучшая коммуникации
между мозгом и периферическими системами организма. Существуют
указания, что эта теория, по-видимому, справедлива относительно
омолаживания репродуктивной системы. Израильские
исследователи показали, что мелатонин предотвращает возрастное снижение
уровня тестостеронов у крыс (Oaknin-Bendahan, 1995). Животные,
получавшие мелатонин, продуцировали почти в три раза больше
тестостерона, чем животные контрольной группы (Reiter et al., 1996).
Ситуация, как мы видим, неоднозначна и требует, очевидно,
дальнейших исследований.
В главе 30 мы обсудим практические средства оживления
половой функции.
Глава 9
ДОЛГОЖИТЕЛИ
Рассказывая однажды о близком знакомстве с Понтием
Пилатом в Иерусалиме, он детально описал дворец наместника,
перечислил блюда, которые подавали к столу во время ужина.
Кардинал де Роган, полагая, что слушает какие-то фантазии,
обратился к камердинеру графа де Сен-Жермена, седовласому
старику с открытым лицом:
- Друг мой, мне трудно поверить в то, что говорит твой хозяин.
Что он чревовещатель - согласен, что делает золото - согласен,
но то, что ему две тысячи лет н он видел Понтия Пилата - это
уже слишком. Ты был с ним там?
- О нет, ваше преосвященство, - ответил простодушно
камердинер, — я всего четыреста лет на службе у господина графа.
Collin Plancy
(Dictionnaire internal. Pans. Mellier, 1844. P. 434)
Изучение старения людей имеет дополнительные трудности:
надежные данные о продолжительности жизни почти невозможно
получить из-за высокой степени вариабельности этого показателя
в пределах человеческой популяции. Большая часть наших знаний
о процессе старения получены из экспериментов на животных.
В этих экспериментах использовались однородные популяции
животных, имевших определенную диету и содержавшихся при
благоприятных и регулируемых внешних условиях. Результаты,
полученные таким образом, особенно на млекопитающих, имеют
значительную ценность по ряду причин. Генетические и
биохимические различия среди высших животных относительно малы по
сравнению с различиями в их анатомической организации или
поведении. Генетическое подобие между людьми и мышами, если
исходить из структуры ДНК, - более, чем 90%, а между людьми и
приматами - приблизительно 99%. При изучении специфических
тканей и органов или биохимических путей метаболизма ученые
обычно могут выбирать те виды животных, у которых эта система
или биохимический процесс являются в основном таким же, как и
у людей. Сотни медицинских работ продемонстрировали, что
хорошо проведенные и взвешенно интерпретированные исследования
на животных необходимы для успешного расширения наших
знаний о человеческом организме.
Тем не менее результаты, полученные на животных, дают нам
только косвенное и, вероятно, несколько неточное понимание
процесса человеческого старения. Хотя главные механизмы старения,
по-видимому, подобны у всех высших организмов, относительный
вклад каждого из них может значительно отличаться. Так, некото-
182
рые системы человеческого организма, в частности центральная
нервная система (ЦНС), значительно отличаются от таковых у
животных. Даже в системах, подобных по структуре и функции,
например в иммунной системе, тонкие различия могут оказать
значительное влияние на процесс старения. Наиболее яркий пример
ограниченной возможности перенесения наших знаний о старении
животных на этот процесс у человека - эксперименты на грызунах,
которые резко ограничены в калорийности питания. Когда крысы и
мыши содержались на такой диете еще до полового созревания,
продолжительность их жизни могла увеличиваться почти в два раза.
Известно, что животные, которые едят, сколько хотят, растут и стареют
быстрее, чем их недокормленные братья. В естественных природных условиях
жизнь животных, у которых нужда в питании удовлетворяется лишь время от
времени, представляет нормальное положение вещей. Поэтому дикая крыса,
никогда не реализовывавшая свой потенциал долгожительства, вероятно,
теоретически в естественных условиях могла бы пережить своих "сытых"
лабораторных сородичей. У животных, искусственно подвергнутых ограничению в
калорийности питания, что в какой-то мере является иммитацией жестких
условий дикой природы, долгая жизнь оказывается всего лишь реализацией их
естественного потенциала, а не экстраординарного замедления механизмов
старения {Хейфлик, 1999).
Было установлено, что животные, ограничиваемые в калориях, имеют
гораздо более низкую концентрацию глюкозы в крови, что, по мнению
некоторых ученых, полезно, поскольку ограничивает генерацию вредоносных
продуктов метаболизма глюкозы, в частности активных форм кислорода
(Masoro, 2000).
Тем не менее ограничение в пище - единственный эффективный
способ продления жизни грызунов и многих других животных в
лабораторных условиях. У обезьян, однако, резкое ограничение
калорийности пищи, начатое до полового созревания, вызывает
повреждение мозга и не в состоянии увеличить продолжительность жизни.
Столкнувшись с ограничениями, присущими геронтологии
животных, исследователи теперь концентрируют внимание на людях
самого преклонного возраста в надежде достигнуть нового
понимания проблем человеческого долголетия. Максимальная
продолжительность жизни у людей - приблизительно 115-120 лет, но лишь
немногим удается перешагнуть возрастной рубеж в сто лет. Люди,
которым уже за сто (их часто называют долгожителями) находятся
теперь в центре внимания исследователей во многих странах. Доля
долгожителей в человеческой популяции развитых стран устойчиво
росла в последние десятилетия. В 80-х численность "клуба"
долгожителей в США выросла на 160%. Согласно переписи 1980 г.,
приблизительно один из десяти тыс. населения США был в возрасте,
превышающем сто лет.
Каковы тайны такого замечательного долголетия? Какие
уникальные особенности присущи разным долгожителям?
Изучение этого вопроса - весьма сложная задача: это порой сильно раз-
183
личающиеся индивиды, живущие в различных регионах;
получение точной информации об их возрасте, истории жизни и т.п., как
правило, бывает затруднено. Все это отчасти служит причиной
значительных расхождений между учеными относительно
факторов, играющих роль в достижении максимально возможной
продолжительности жизни. Однако ясно, что никакой отдельный
фактор не может быть ключевым. Долголетие скорее всего
результат комплексного взаимодействия генетических,
физиологических, экологических (питание, образ жизни, и т.д.),
психологических и социальных факторов. К сожалению, почти
невозможно ясно определить роль каждого из этих факторов. Ниже
мы представим резюме современных представлений
относительно факторов человеческого долголетия, основанных на изучении
самых старых людей.
1. Генетические факторы. Ко времени написания этой книги,
данные о существовании какого-либо отдельного гена,
непосредственно связанного с длительностью человеческой жизни,
отсутствовали. Наиболее вероятные кандидаты скорее всего - гены,
осуществляющие репарацию ДНК, защиту от свободных радикалов,
регулирующие углеводно-жировой метаболизм и т.п. Долгожители, по-
видимому, имеют необычно высокий уровень экспрессии некоторых
из этих генов. Гены, которые управляют скоростью развития
организма (см. гл. 2) также могут влиять на продолжительность жизни.
Однако есть и другая возможность: у долгожителей понижена
активность вредных генов, т.е. генов, которые повышают
восприимчивость к некоторым болезням.
Пример такого сценария - ген, кодирующий аполипопротеин-Е (Аро-Е),
для которого была показана связь с повышенным риском болезни Альцгейме-
ра. Имеются три версии гена Аро-Е: Е2, ЕЗ и Е4 (последний локализован в
хромосоме 19). Люди, имеющие Е4, подвергаются самому высокому риску развития
болезни Альцгеймера, а с Е2 - самому низкому. В ряде исследований было
показано, что версия гена Αρο-ΈΑ имеется у 25% людей моложе 25, но только
14% - у тех, кто старше 85 лет (Pookaj et al., 1998; Motulsky, 1999; Баранов и др.,
2000). Очевидно, такая разница могла быть связана с более низким
содержанием версии гена Αρο-ΈΑ в данной выборке долгожителей.
Еще одним примером подобного подхода может служить возможность
существования изоформ гена липопротеина (а), который наследственно
определяет высокий риск атеросклероза (подробно об этом в гл. 2, в параграфе
"Генетические аспекты старения").
Вероятно, существует много других генов, имеющих несколько
вариантов, которые различаются по их способности повышать
восприимчивость к дегенеративным болезням или, наоборот,
обеспечивать защиту против них.
Отдельные исследования показывают, что долголетие имеет
тенденцию проявляться у многих членов одной семьи, что,
по-видимому, указывает на роль генетических корней.
184
Столетние - это люди, которые избежали возрастных болезней, главным
образом рака, и смогли достигнуть критических пределов человеческой жизни
Анализ демографических данных указывает, что сфера действия рака и
смертность проявляется в возрасте примерно в 85-90 лет, и, считают, что самые старые
люди и столетние оказались защищенными от атак рака и его прогрессии.
Группа итальянских авторов {Bonafe, Barbi, Storci et al., 2002) проанализировала
данные свежей литературы по распределению у столетних полиморфизма клеток
зародышевой линии, который, как предполагается, воздействует на индивидуаль
ную генетически предопределенную восприимчивость к раку (в частности,
полиморфизм р53, глутатион трасферазы, оксидазы цитохрома, стероид-5 альфа ре-
дуктазы и др.). Особый интерес представляют новые данные о тех вариантах
полиморфизма р53 и иммунной системы у долгожителей, которые повышают
противораковую активность р53 и/или уровень иммунитета, который создает
максимально агрессивную среду для роста раковых клеток. Продолжение
исследований в этом направлении, несомненно, будет иметь фундаментальное значение для
понимания базисных механизмов рака, старения и их сложных отношений.
2. Питание. Известно лишь несколько исследований
относительно питания долгожителей. Мимура и коллеги изучали
особенности диеты у 88 самых старых людей на японском острове Окинава,
который известен как мировой лидер по самой высокой
зарегистрированной средней продолжительности жизни (Mimura et al., 1992).
Диета долгожителей Окинавы состояла главным образом из риса и
картофеля в качестве углеводов, рыбы и обилия овощей. Диета
была хорошо сбалансирована по витаминам и минеральным
веществам, а случаи атеросклероза были сравнительно редки. Другие
исследователи нашли, что самые старые жители Окинавы потребляли
пищу с более высоким содержанием животного белка (включая
рыбу, молоко, яйца и мясо) в белке пищи, чем "средние*' японцы;
потребление кальция, железа, витаминов A, Bl, В2 и С было также
выше.
Джонсон и коллеги {Johnson et al., 1992) сравнивали диетические
привычки 24 долгожителей с таковыми "более молодых" пожилых
людей. Долгожители завтракали более регулярно, избегали широко
разрекламированных диет потери веса и вообще больших
изменений в весе тела, потребляли относительно большее количество
овощей. Обширное исследование приблизительно шести тыс.
долгожителей, проведенное в Италии, показало, что обследуемые субъекты
использовали сбалансированный рацион, основанный на
естественных пищевых продуктах, включая обилие плодов и овощей
(Receputo et al„ 1995).
Общая характеристика питания большинства долгожителей
заключается в обильном потреблении белковой пищи, в разнообразии
и обилии фруктов и овощей; они также стараются избегать
продуктов с избытком калорий, но в их диете - не обязательно резко
ограничивается потребление жиров.
3. Метаболизм. Данные об метаболических характеристиках
долгожителей немногочисленны. Как правило, отмечается высокая
185
толерантность к углеводам и чувствительность тканей к
инсулину. Г. Паолиссо и коллеги (Paolisso et al., 1995) установили, что
усвоение углеводов итальянскими долгожителями было столь же
хорошим, как и у 50-летних, и заметно лучше, чем у стариков в
возрасте 75-100 лет. Как мы уже обсуждали в главе 2, усвоение углеводов
у большинства людей снижается с возрастом, что, по-видимому,
активирует несколько механизмов старения. Недостаточная
чувствительность тканей к инсулину, которая часто развивается с
возрастом, приводит к избытку инсулина и высокой концентрации
глюкозы в крови, оба эти фактора способствуют процессу старения.
Высокий уровень глюкозы в крови способствует неферментативному
гликозилированию белков, их перекрестному сшиванию,
повреждению молекул ДНК и ослаблению иммунитета. Следствием избытка
инсулина является развитие атеросклероза, гипертонии, ожирения
и, возможно, других возрастных патологий. В свете приведенных
выше фактов, кажется неясным, почему долгожители
демонстрируют склонность к нормальному метаболизму углеводов. Одно из
возможных объяснений - более медленный, чем среднестатистический,
темп функционирования ЦЧС у долгожителей. Другие возможные
объяснения могут быть связаны с рациональной диетой и низким
уровнем стресса.
4. Психология и способность к преодолению трудностей.
Группа исследователей, изучавшая 96 долгожителей в штате
Джорджия, так охарактеризовала их: "...оптимистические,
мудрые личности, активно вовлеченные в каждодневную жизнь"
(Johnson et al., 1992). Геронтологи согласны в том, что
долгожители, как правило, имеют более высокую (чем средняя) способность
к преодолению трудностей и быстрому, и успешному
преодолению психологического стресса. Положительные настроения и
устойчивость к стрессу могут способствовать продлению жизни,
поддерживая активность иммунной системы, увеличивая
секрецию гормона роста и замедляя ЦЧС. Биохимическая природа
такого феномена, возможно, определяется связью между
положительными эмоциями и уровнем нейромедиаторов в мозгу, в
частности уровнем дофамина в гипоталамусе. Этот взгляд опирается
на массу свидетельств, что депрессивные состояния, сопряженные
с низкими уровнем нейромедиаторов, ускоряют старение и
стимулируют дегенеративные болезни (см. гл. 7).
5. Образ жизни. Кажется, не существует никакого
универсального образа жизни у самых старых людей. Долгожители есть во
многих культурах и социально-экономических группах, в городских
и сельских районах, в различных климатических условиях и
географических зонах. Наиболее непротиворечивые результаты
относительно образа жизни долгожителей заключаются в том, что они
должны иметь удобную и знакомую среду повседневной жизни,
хорошую сеть социальных контактов, активную работу с достаточным
186
заработком. Фундаментом такого образа жизни должен служить
низкий уровень стресса, который, как мы уверены, служит
магическим ключом к тайне долголетия.
Известные случаи достижения или превышения столетнего
возраста не могут ие производить впечатления и не вдохновлять. Но
даже более удивительно именно то, что вновь зарегистрированные и
известные долгожители выглядят вполне здоровыми или во всяком
случае более здоровыми, чем их 70-80-летние соотечественники.
Обстоятельное исследование шести тыс. долгожителей Италии
показало, что самые старые люди имеют хорошо сохраненную
иммунную систему, с активными N-киллерами, функция которых
особенно важна для предотвращения рака; нормальный профиль липидов,
включая низкий холестерин и ЛНП; нормальную или умеренно
пониженную толерантность к углеводам; хороший уровень
умственной самодостаточности (Receputo et al., 1995). Ε. Береги и А. Клингер
(Beregi, Klinger) изучали 280 венгерских долгожителей и нашли, что
они были здоровее, чем 60-80-летние старики, первые отличались
более низким потреблением лекарств и лучшей способностью к
адаптации.
Впечатляющие данные о долгожителях Италии заставляют вспомнить
распространенное в Европе представление о причинах здоровья и долголетия
жителей Средиземноморья, которые связанны с особенностями их питания и
образа жизни. Четыре фактора играют здесь решающую роль: изобилие рыбы,
оливкового масла и сухого красного вина (флавоноиды) в рационе, а также
сиеста - полуденный отдых в самые жаркие часы.
Доказательства хорошего здоровья у самых старых людей,
очевидно, не согласуются с идеей, что болезнь и немощь
увеличиваются с возрастом из-за ускорения механизмов старения. Но
это — не обязательно противоречие. Во-первых, долгожители не
типичны, хотя их количество имеет тенденцию повышаться, они
все еще составляют меньше 0.1% населения. Во-вторых, у
большей части людей и животных, механизмы старения ускоряются
со временем и конвертируются в причины, порождающие
дегенеративные болезни и прогрессирующую потерю способности к
адаптации и жизнеспособности. Долгожители могут быть
исключением в том смысле, что некоторые механизмы старения у них
разворачиваются существенно медленнее. Допустим, что это
соответствует действительности. Но, как могут знания
относительно самых старых помочь большинству людей, которые не
являются носителями удачных генов и замедленных механизмов
старения? Во-первых, долгожители убедительно демонстрируют,
что долголетие возможно, и преклонные года могут
действительно быть приятными и полными жизни. Другое важное
обстоятельство состоит в том, что наследственность, хотя и важна,
составляет только часть рассматриваемого явления. Питание, образ
187
жизни и "антистрессовая стратегия" - весьма важны и,
подчеркнем, находятся в наших собственных руках. Конечно, некоторые
люди с комбинацией удачных генов могут жить до ста лет и без
того, чтобы прилагать к этому дополнительные усилия.
Большинство же из нас, однако, так щедро не одарено природой, и мы
должны прилагать немало усилий, чтобы жить лучше и дольше.
Мы можем приобрести характеристики долгожителей, если
будем следовать стратегии продления жизни. Для менее удачливого
в отношении наследственности человека более важно как можно
раньше обратиться к стратегии активного вмешательства в
процессы старения (при этом желательно оценить вклад
наследственности в потенциальную продолжительность жизни и здоровье
данного индивида, что отчасти поможет сделать рассмотрение
его генеалогического древа),
Человек, обладающий средним ресурсом жизнеспособности,
имеет возможность приблизиться к физиологическим параметрам
долгожителей, если будет настойчиво стремиться к этому. И это
вполне реально. Так, у некоторых долгожителей отмечен низкий
уровень свободных радикалов, потому что их гены, кодирующие
ферменты, которые нейтрализуют СР, обладают высокой
активностью Щилъман, 1986, 1987; Ames et al., 1993; Oilman, 1994). Искатели
долголетия могут приблизиться к этой ситуации, принимая антиок-
сиданты или пищевые добавки, которые стимулируют активность
этих ферментов (см. гл. 23).
Как мы уже отмечали, долгожители имеют хорошую
толерантность к углеводам и восприимчивость тканей к инсулину. У многих
пожилых людей, однако, ситуация как раз противоположна и, по
существу, способствует всем механизмам старения. Большинство
людей может нормализовать усвоение углеводов и восприимчивость к
инсулину с помощью диеты, пищевых добавок, умеренных
физических упражнений и снижения стресса.
Некоторые из наиболее типичных черт личности
долгожителей - оптимизм, хорошее настроение и конструктивный взгляд на
жизнь. Почти полную противоположность этим чертам мы находим
в состоянии депрессии, которое свойственно очень многим людям и
особенно пожилым. Многие питательные вещества и естественные
растительные экстракты могут "настроить" биохимию вашего
мозга так, чтобы улучшить ваше настроение и способность к
преодолению трудностей, а также уменьшить симптомы депрессии (см.
гл. 28). Однако необходимо помнить, что глубокая депрессия
требует серьезной врачебной помощи.
Литература
Ада ГЛ., Носсал Г Клона л ьно-се лекционная теория // В мире науки. 1987.
№ Ш. С. 30-38,
Альберте Б., БрейД.,ЛъюисД. и др. Молекулярная биология клетки. М.: Мир,
1994. 3 т.
Арчаков A.M. Микросомальное окисление. М.: Наука, 1975. 327 с.
Бапакин В.Е., Заичкина СМ., Розанова ОМ. и др. Малые дозы
ионизирующего излучения подавляют рост спонтанного уровня цитогенетических
повреждений, обусловленный старением организма: (Эффект стабилизации
генома) // Докл. РАН. 2000. Т. 374. № 2. С. 271-273.
Барабой В.Α., Брехман И.И., Голотин ВТ., Кудряшов Ю.Б. Перекисное
окисление и стресс. СПб.: Наука, 1992. 148 с.
Баранов B.C., Баранова Е.В., Иващенко Т.Э., Асеев MS. Геном человека и
гены "предрасположенности". СПб.: Интермедика, 2000. 272 с.
Бочков И.П. Генетические технологии в педиатрии // Педиатрия. 1995. № 4
С. 21-26.
Брехман М.М. Человек и биологически активные вещества. Л.: Наука, 1980.
120 с.
Бурлакова Е.Б., Алесенко А.В., Молочкина ЕМ. и др. Биоантиоксиданты в
лучевом поражении и злокачественном росте. М.: Наука, 1975. 214 с.
Бурлакова Е.Б., Храпова Н.Г. Перекисиое окисление липидов мембран и
природные антиоксиданты // Успехи химии. 1985. Т. 54, № 9. С 1540-1558.
Ванюшин Б.Ф. Метилирование ДНК у эукариот - новый механизм регуляции
экспрессии генов и клеточной дифференцировки // Успехи биол. химии.
1983. Т. 24. С. 170-193.
Ванюшин Б.Ф., Бердышев Г Д. Молекулярно-генетические механизмы
старения. М.: Медицина. 1977. 296 с.
Виленчик ММ. Биологические основы старения и долголетия. М.: Знание.
1987. 224 с
Владимиров Ю.А. Биологические мембраны и ^запрограммированная смерть
клетки // Сорос образов, журн. 2000а. Т. 6, № 9. С 2-9.
Владимиров Ю.А. Свободные радикалы в биологических системах // Там же
20006. Т. 6, № 12. С. 13-19.
Владимиров Ю.А., Азызова О.Α., Деев А.И. и др. Свободные радикалы в живых
системах // Итоги науки и техники. Биофизика. 1992. Т. 29. С 3-250.
Владимиров Ю.А., Арчаков A.M. Перекисное окисление липидов в
биологических мембранах. М.: Наука, 1972.
Войтенко В.П. Наследственность, старение и продолжительность жизни
человека // Цитология и генетика. 1983. № 2. С. 65-73.
Галкин А.П., Тодоров М.Н. О высокой радиочувствительности биосинтеза ми-
тохондриальной ДНК // Радиобиология. 1969. Т. 9, № 1. С. 21-25.
Голубев А.Г., Ревской С.Ю., Цырлина Е.В., Дильман ВМ. Роль возрастной ги-
перлипидемии в обусловленных внутренней средой организма нарушениях
функций лимфоцитов при старении у людей // Иммунология. 1983. М° 4.
С. 76-81.
Грей ГА., Cemme Α., Бюус С. Как Т-клетки узнают антигены // В мире науки
1990. № 1.С. 28-37.
Грин П., Cmaym У., Тейлор Д. Биология. Т. 2. М.: Мир, 1990. 327 с.
Гриневич Ю.А., Барабой В Α., Орел В.Э. Хемилюминесцентный метод в
иммунологии // Журн. микробиологии, эпидемиологии и иммунологии. 1986. № 1.
С 91-97.
189
Дардымов И.В. Женьшень, элеутерококк. М.: Наука, 1976. 184 с.
Дильман В.М. О возрастном повышении деятельности некоторых гипоталами-
ческих центров // Тр. Ин-та физиологии им. И.П. Павлова АН СССР. 1958;
Т. 7. С 326-336.
Дильман В.М. Клиническое применение половых гормонов и их аналогов.
Вильнюс: Госполитнаучиздат. 1961. 199 с.
Дильман В.М. Старение, климакс и рак. Л.: Медицина. 1968. 378 с.
Дильман В.М. Элевационный механизм старения и рак II Вопр. онкологии. 1970.
№ 6. С. 45-55.
Дильман В.М. Мутационно-метаболическая модель развития рака и прогрессии
опухолевого процесса // Там же. 1976. № 8. С. 3-16.
Дильман В.М. Эндокринологическая онкология. 2-е изд. Л.: Медицина. 1983.
Дильман В.М. Большие биологические часы. М.: Знание. 1986. 255 с.
Дильман В.М. Четыре модели медицины. Л.: Медицина. 1987. 287 с.
Докудоеская С.С., Петров А.В., Донцова О Α., Богданов А А. Теломераза -
необычный РНК-содержащий фермент // Биохимия. 1997. Т. 62. № 11.
С.1411-1422.
Цымшиц Г.М. Проблема репликации концов линейных молекул ДНК и
теломераза. Сорос, образов, журн. 2000. Т. 6, № 5. С. 8-13.
Егоров Е.Е. Теломераза, старение, рак // Молекуляр. биология. 1997. Т. 32, № 1.
С. 16-24.
Егоров Е.Е., Семенова И.В., Караченцев Д.И. и др. Корреляция выживаемости
эмбриональных фибробластов и их пролиферативного потенциала с
продолжительностью жизни мышей различных линий // Биол. мембраны. 2000.
Т. 17, №6. С. 685-688.
Зотин Α.Α., Владимирова И.Г. Интенсивность дыхания и видовая
продолжительность жизни пресноводных двустворчатых моллюсков семейств
Margaritiferridae и Unionidae // Изв. РАН. Сер. биол. (2001). № 3. С. 331-338.
Зотин A.M. Прогрессивная эволюция животных. 1. Константа Рубенса в
классе млекопитающих. М., 32 с. 1993. Деп. в ВИНИТИ.
Линхард Г.Э., Слот Я.У., Джеймс Д.Э., Мьюклер М.М. Как клетки поглощают
глюкозу. В мире науки. (1992). № 3. С. 22-29.
Лон P.M. Липопротеин (а) в заболеваниях сердца // Там же. 1992. № 8. С. 16-23.
Лупандин А.В. Общий механизм приспособления организма под влиянием поли-
фенольных адаптогенов // Успехи физиол. наук. 1991. Т. 22, № 1. С. 20-39.
Молодых О.П, Непомнящих Л.М.,Лушникова ЕЛ., Клинникова М.Г. Апоптоз:
снижение общей численности популяции гепатоцитов мышей после
гипертермии // Бюл. эксперим. биологии и медицины. 2000. Т. 130, № 9. С. 346-350.
Нагорный А.В. Проблема старения и долголетия. Харьков: Изд-во ХГУ, 1940.
Нагорный А.В., Буланкин И.Н., Никитин В.Н. Проблема старения и
долголетия. М., 1963,755 с.
Непомнящих Л.М., Семенов Д.Е. Апоптоз кардиомицитов как крайнее
проявление регенераторно-пластической недостаточности миокарда // Бюл.
эксперим. биологии и медицины. 2000. Т. 130, № 9. С. 336-341.
Обухова Л.К., Эмануэль Н.М. Молекулярные механизмы замедления старения
антиоксидантами // Биологические проблемы старения. М.: ВИНИТИ, 1984.
Т. 4. С. 44-81. (Итоги науки и техники; Т. 4).
Оловников A.M. Принцип маргинотомии в матричном синтезе полинуклеоти-
дов // Докл. АН СССР. 1971. Т. 201, № 6. С. 1496-1499.
Плешакова О.В., Рассказова Е.А., Сухарева С.А. и др. Расщепление
окислительно-модифицированных белков протеолитическими ферментами нейтро-
филов мышей разного возраста // Докл. РАН. 2000. Т. 374, № 2. С. 265-267.
190
Пол У. Иммунология. Т. 1. М.: Мир, 1987.
Полинг Л. Витамин С и здоровье. М.: Наука, 1974. 74 с.
Пономарева Н.В., Фокин В.Ф., Павлова О.А. и др. Анализ корреляции между
нейрофизиологическими параметрами и уровнем гормона стресса кортизола
при старении // Вестн. РАМН. 1999. № 3. С. 46-49.
Пригожий И. Проблема эволюции в термодинамике необратимых процессов //
Возникновение жизни на Земле: /Тр. Междунар. симпоз., 1957 г. М.: Изд-во
АН СССР, 1959. С. 408-416.
Пригожий И. Введение в термодинамику необратимых процессов. М.: Изд-во
иностр. лит., I960.
Радман М., Вагнер Р. Высокая точность репликации ДНК // В мире науки. 1988
№ 10. С. 16-23.
Растинг Р. Почему мы стареем? // Там же. 1993. № 2. С. 76-88
Розенберг С.А. Адаптивная иммунотерапия рака //Там же. 1990. № 7. С. 26-34
Рязанов А.Г. Рибосома и секрет долголетия// Молекуляр, биология. 2001. Т. 35,
№ 4. С. 727-730.
Саприн А.Н. Ферменты метаболизма и детоксикации ксенобиотиков // Успехи
биол. химии. 1991. Т. 32. С. 146-175.
Селко ДДж. Амилоидный белок и болезнь Альцгеймера // В мире науки. 1992.
№ 1. С. 26-36.
Селье Г. Концепция стресса как мы ее представляем в 1976 г. // Новое о
гормонах и механизме их действия. Киев: Наук, думка, 1977. С. 27-51.
Семенов Н.Н. Цепные реакции. Л., 1934.
Серами Э., Влассара Э., Браунли М. Глюкоза и старение // В мире науки. 1987.
№ 7. С. 42-49.
Скулачев В.П. Старение организма - особая биологическая функция, а не ре
зультат поломки сложной живой системы: Биохимическое обоснование
гипотезы Вейсмана // Биохимия. 1997. Т. 62, № 11. С 1394-1399.
Скулачев В.П. Эволюция, митохондрии и кислород // Сорос, образов, журн
1-999. № 9. С. 4-10.
Скулачев В.П. Феноптоз: запрограммированная смерть организма // Биохимия
1999. Т. 64, № 12. С. 1679-1688.
Скулачев В.П. Активные формы кислорода и явления запрограммированности
смерти: От органеллы до организма // VI Междунар. конф. "Биоантиокси-
дант". М., 2002. С 533-534.
Смирнов В.Н.. Восстановительная терапия будущего // Наука и жизнь. 2001.
№ 8. С. 28-32.
Стрелер Б. Время, клетки, старение. М.: Мир, 1964. 251 с.
Тарусов Б.Н. Основы биологического действия радиоактивных излучений. М/
Медгиз, 1954. 140 с.
Тодоров И.Н. Влияние соматотропного гормона (СТГ) на содержание
фосфорных фракций печени белых крыс в онтогенезе //Докл. АН УССР. 1959а. № 9.
С. 1037-1041.
Тодоров И.Н. Некоторые возрастные особенности воздействия
соматотропного гормона гипофиза (СТГ) на вес тела и состав мочи белых крыс // Там же.
19596. №11. С. 1283-1287.
Тодоров И.Н. Некоторые возрастные особенности в действии соматотропного
гормона гипофиза на обмен нуклеиновых кислот // Биохимия. 1959в. Т. 34,
№6. С. 1010-1019.
Тодоров И.Н. О синергетическом действии инсулина и соматотропного
гормона (СТГ) на обмен нуклеиновых кислот (НК) в связи с данными по
концентрации инсулина в крови в разные возрастные периоды // Материалы VII На-
191
учн. конф. по вопр. морфологии, физиологии и биохимии. М., 1965.
С. 463-464.
Тодоров И. И. Исследование субклеточных и молекулярных изменений
биосинтеза рибонуклеиновых кислот и белков на ранних этапах лучевого
поражения организма: Дис ... д-ра биол. наук. Киев, 1969.
Уманский СР. Апоптоз: молекулярные и клеточные механизмы // Молекуляр.
биология. 1996. Т. 30, № 3. С. 487-502.
Хансон К.П. Молекулярные механизмы интерфазной гибели клеток //
Радиобиология. 1979. Т. 19, № 6. С. 814-819
Хансон К.П. Апоптоз: Современное состояние проблемы // Изв. РАН. Сер.
биол. 1998. № 2. С 134-141.
Харпер П. Практическое медико-генетическое консультирование. М.:
Медицина, 1984. С. 184-227.
Хейфлик Л. Смертность и бессмертие на клеточном уровне // Биохимия. 1997.
Т. 62, №11. С. 1380-1393.
Чумаков ПМ. Роль гена р53 в программированной клеточной смерти // Изв.
РАН. Сер. биол. 1998. № 2. С. 151-156.
ЭверлиДж.С, Розенфельд Р. Стресс: Природа и лечение. М.: Медицина, 1985.
Эмануэль ИМ. Роль свободных радикалов в радиобиологических процессах и
некоторые новые возможности разработки средств против лучевого
поражения //Первичные механизмы биологического действия ионизирующих
излучений. М., 1963. С. 73-83.
Эмануэль ИМ. Кинетика и механизм цепных реакций окисления органических
веществ в жидкой фазе // Химическая кинетика и цепные реакции. М., 1966
С. 375-408.
Эмануэль ИМ. Кинетика экспериментальных опухолевых процессов. М.:
Наука, 1977.419 с.
Эмануэль ИМ. Некоторые молекулярные механизмы и перспективы
профилактики старения // Изв. АН СССР. Сер. биол. 1975. № 4. С. 503-511.
Эмануэль ИМ. Антиоксиданты в пролонгировании жизни // Биология
старения. Л.: Наука, 1982. С. 569-586.
Эмануэль ИМ., Кнорре Д.Г., Лясковская Ю.И. Кинетика окисления жиров //
Изв. АН СССР. ОХН. 1957. № 6. С 678-683.
Эмануэль ИМ., Лясковская Ю.И. Торможение процессов окисления жиров. М.:
Пищепромиздат, 1961. 359 с
Эмануэль ИМ., Обухова Л.К., Иайдич В.И. и др. Замедление старения
посредством активации системы микросомальных оксидаз фенобарбиталом //
Докл. АН СССР. 1977. Т. 235, № 4. С. 957-960.
Юн ДжД., Кон Ж.А. Как клетки-убийцы убивают // В мире науки. 1988. № 3.
С 14-21.
Alkon D.L., Rasmussen H. A spatial-temporal model of cell activation // Science. 1988.
Vol. 239, № 4843. P. 998-1005.
Ameisen J.С Between Charybde and Scylla: Is a premature ageing a price to pay for
avoiding, cancer? // Med. Sci. 2002. Vol. 18, N 4. P. 393-395.
Ames B.N., Shigenaga, M.K., Hagen T.M. Oxidants, antioxidants, and the degenerative
diseases of ageing // Proc. Nat. Acad. Sci. USA. 1993. Vol. 90. P. 7915-7922.
Aoshiba K., Tamaoki J., Nagai A. Acute cigarette smoke exposure induces apoptosis of
alveolar macrophages // Amer. J. Physiol. Lung Cell Mol. Physiol. 2001. Vol. 281,
N 6. P. L1392-1401.
Appleman J.R., Leinhard G.E. Kinetics of the purified glucose transporter: Direct
measurement of the rates of interconversion of transporter conformers // Biochemistry.
1989. Vol. 28, N 20. P. 8221-8227.
192
Arvat E., Giordano R., Gianotti L. et al. Neuroendocrinology of the human growth hor
mone-insulin-like growth factor I axis during ageing // Growth Hormone and IDF
Res. Vol. 9, Suppl. A. P. 111-115.
Asher G., Lotem J., Sachs L. et al. Regulation of p53 stability and p53-dependent apop-
tosis by NADH quinine oxidoreductase I // Proc. Nat. Acad. Sci. USA. 2001. Vol
98, N3. P. 1188-1193.
Atadja P., Wong H., Garkavtsev I. et al. Increasing activity of p53 in senescing
fibroblasts//Ibid. 1995. Vol. 92. P. 8348-8352.
Bachmann G.A. Influence of menopause on sexuality // Intern. J. Fertil. Menopausal
Stud. 1995. Vol. 40, suppl. 1. P. 16-22.
Bar R.S., Koren H., Roth J. Insulin and macrophage function // Diabetes. 1976. Vol. 25.
P. 348.
Bartke Α., Coschigano K., Kopchick J. et al. Genes that prolong life: Relationships of
growth hormone and growth to ageing and life span // J. Gerontol. A: Biol. Sci. Med.
Sci. 2001. Vol. 56, N 8. P. B340-349
Bell G. Sex and death in Protozoa. Cambridge: Univ. press, 1988.
Berenson G.S., RadhakrishnamurthyB., Srinivasan S.R. et al. Plasma glucose and insulin
levels in relation to cardiovascular risk factors in children from a biracial population //
J. Chron. Diseases. 1981. Vol. 34. P. 379-391.
Bjorntorp P. Neuroendocrine ageing: (Review) // J. Internal Med. 1995. Vol. 238, N 5.
P. 401-Ю4.
Bjorntorp P. Alterations in the ageing corticotropic stress-response axis // Novartis
Found. Symp. 2002. Vol. 242. P. 46-58.
Blair J.A., Parveen H., Barford PA., Leeming R. Aetiology of Parkinson's disease //
Lancet. 1984. Vol. 1. P. 167.
Bonafe M., Barbi C, Storci G. et al. What studies on human longevity tell us about the
risk for cancer in the oldest old: Data and hypotheses on the genetics and
immunology of centenarians // Exp. Gerontol. 2002. Vol. 37, N 10/11. P. 1263-1271.
Bond J Α., Wyllie F.S., Wynford-Thomas D. Escape from senescence in human
diploid fibroblasts induced directly by human p53. // Oncogene. 1994. Vol. 9
P.1885-1889.
Bond J A. et al. Evidence that transcriptional activation by p53 plays a direct role in the
induction of cellular senescence // Ibid. 1996. Vol. 13. P. 2097-2104.
Boonen S., Lysens /?., Verbeke G. et al. Relationship between age-associated endocrine
deficiencies and muscle function in elderly women: a cross-sectional study // Age and
Ageing. 1998. Vol. 27, N 4. P. 449-454
Bouillane O., Rainfray M., Tissandier O. et al. Growth hormone therapy in eldery
people: An age-delaying drug? // Fundam. Clin. Pharmacol. 1996.
Braciale Τ J. et al. Antigen presentation pathways to class I and class II MHC-restricted
Τ lymphocytes // Immunol. Rev. 1987. Vol. 98, N 8. P. 95-114.
Bredesen D.E. Neuronal apoptosis: Genetic and biochemical modulation // Apoptosis:
The molecular basis of apoptosis in disease / Ed. L.D. Tomei, F.O. Cope Cold Spring
Harbor, 1994. P. 397-421.
Brooksbank B.W.L., Balazs R. Superoxide dismutase and lipoperoxidation in Down's
syndrome fetal brain // Lancet. 1983. Vol. 1. P. 881-882.
Brownlee M., Vlassara H., Kooney A. et al. Aminoguanidine prevents diabetes-induced
arterial wallschevosis //Science. 1986. Vol. 232, N 4758. P. 1629-1632.
Bucala R., Model R., Cerami A. Modification of DNA by reducting sugars: A possible
mechanism for nuclear acid aging and age-related dysfunction in gene expression //
Proc. Nat. Acad. Sci. USA. 1984. Vol. 81, N 1. P. 105-109.
Burnet F.M. A modification of Jerne's theory of antibody production using the concept
of clonal selection // Austral. J. Sci. 1957. Vol. 20, N 3. P. 67-69.
193
Burnet FM. The integrity of the body. Cambridge: Harvard Univ. press, 1962. 183 p.
Burnet FM. Age-associated heredo-degenerative conditions of the central nervous
system // Handbook of diseases of ageing / Ed. by H.T. Blumenthal. N.Y., 1983.
P. 296-313.
Buus S., Sette Α., Grey ИМ. The interaction between protein-derived immunogenic
peptides // Immunol. Rev. 1987. Vol. 98, N 8. P. 115-141.
Cadenas E., Davies KJ. Mitochondrial free radical generation, oxidative stress, and
aging // Free Radicals. Biol. Med. 2000. Vol. 29, N 3/4. P. 222-230.
Cameron O., Kronfol Z., Greden J., Carrol B. Hypothalamic-pituitary-adrenocortical
activity in patients with diabetes mellitus // Arch. Gen. Psychiat. 1984. Vol. 41.
P. 1090-1095.
Campisi J. Aging and cancer: The double-edged sword of replicative senescence //
J. Amer. Geriatric Soc. 1997. Vol. 45. P. 482-488.
Chinnaiyan AM., Dixit VM. The cell-death machine // Curr. Biol. 1996. Vol. 6, N 5.
P. 555-562.
Collins F.S. Shattuck lecture medical and societal consequences of the human genome
project // New Engl. Med. J. 1999. Vol. 341, N 1. P. 28-37.
Comfort A. Measurement of the human aging rate // Mech. Ageing and Develop. 1972.
Vol. l.P. 101-110.
Comfort Α., Dial L.K. Sexuality and aging: An overview // Clin. Geriatr. Med. 1991.
Vol. 7, N 1. P. 1-7.
Compton MM., Codlowski J.A. Thymocite apoptosis: A model of programmed cell
death // ТЕМ. 1992. Vol. 3, N 1. P. 17-23.
Conti Α., Maestroni G.I. The clinical neuroimmunotherapeutic role of the melatonin in
oncology // J. Pineal Res. 1995. Vol. 19, N 3. P. 103-110.
Counter С Μ. The roles of telomeres and telomerase in cell life span // Mutat. Res. 1996.
Vol. 366, N l.P. 45-63.
Crapo L. Cushing's syndrome: A review of diagnostic tests // Metabolism. 1979.
Vol. 28. P. 955-977.
Curtsinger J.W., Fukui Н.И., Khazaeli A.A. et al. Genetic variation and aging // Annu.
Rev. Genet. 1995. Vol. 29. P. 553-675.
Cutler R.G. Nature of aging and life maintenance processes // Interdiscipl. Topics
Gerontol. 1976. Vol. 9. P. 83-133.
Cutler R.G. Evolutionary biology of senescence // Biology of aging / Ed. by J. Behnke
etal.N.Y., 1978. P. 311-350.
Cutler R.G. Evolution of human longevity // Ageing, cancer, and cell membranes /
Ed. by С Borek et al. Stuttgart, 1980. P. 43-79.
Cutler R.G. Evolutionary biology of aging and longevity in mammalian species // Aging
and cell function / Ed. by J.E. Johnson. N.Y.; L.: Plenum press, 1984. P. 1-147.
Daughaday W. Prolaction and growth hormone in health and disease // Contemporary
Endocrinology. N.Y.: Plenum press, 1985. Vol. 2. P. 27-86.
De Fronzo R.A. Glucose intolerance and ageing // Diabetes. 1979. Vol. 28.
P. 1095-1101.
De Luca Α., Pierno S., Cocchi D., Camerino D.C. Effects of chronic growth hormone
treatment in aged rats on the biophysical and pharmacological properties of skeletal
muscle chloride channels // Brit. J. Pharmacol. 1997. Vol. 121, N 3. P. 369-374.
Oilman VM. Age-associated elevation of hopothalamic threshold to feedback control
and its role in development, aging and disease // Lancet. 1971. Vol. 1. С 1211-1219.
Oilman VM. Ageing, metabolic immunodepression and carcinogenesis // Mech. Ageing
and Developm. 1978. N 8. P. 153-173
Oilman VM. Development, aging, and disease: A new rationale lor an intervention
strategy. Harwood Acad. Publ., 1994
194
/>onehower LA. et al. Mice deficient forp53 are developmentally normal bul susc pti-
ble to spontaneous tumors // Nature. 1992. Vol. 356. P. 215-221.
Drum P.t Bravard Α., Bouvard V. et al. Reciprocal down-regulation of the p53 und
SOD2 gene expression-implication in p53 mediated apoptosis // Oncogene. 2001.
Vol. 20, N4. P. 430-439.
Dunham L, Hunt A./?., Collins 3.E. et al. The DNA sequence of human chromosome 22 //
Nature. 2000. Vol. 402. P. 489-495.
/ ck-Enriquez K.> Kiefer T.L., Spriggs LL., Hill S.M. Pathways through which a regimen
of melatonin and retinoid acid induces apoptosis in MCF-7 human breast cancer
cells // Breast Cancer Res. Treat. 2000. Vol. 61, N 3. P. 229-239.
EspositoJL. Conceptual problems in theoretical gerontology // Perspect. Biol, and Med.
1983. Vol. 26. P. 522-546.
heinberg M., Carroll В J. Biological "markers" for endogenous depression // Arch. Gen.
Psychiat. 1984. Vol. 41. P. 1080-1085.
Ferbeyre G., Lowe S.W. Ageing - the price of tumor suppression? // Nature. 2002.
Vol. 415, N 6867. P. 26-27.
Ferrini M., Piroli G., Frontera M. et al. Estrogens normalize the hypothalamic-pituitary-
adrenal axis response to stress and increase glucocorticoid receptor immuno-reacti-
vity in hippocampus of aging male rats // Neuroendocrinology. 1999. Vol. 69, N 2.
P. 129-137.
l· ilipovichAM., Spector B.D., Kersey /. Immunodeficiency in human as risk factor in the
development of malignancy //Prevent Med. 1980. Vol. 9. P. 252-259.
Finch C.E. Longevity, senescence, and the genome. Chicago: Univ. of Chicago press,
1990.
Hurkey K., Papaconstantinou J., Miller R.A., Harrison D.E. Lifespan extension and
delayed immune and collagen aging in mutant mice with defects in growth hormone
production //Proc. Nat. Acad. Sci. USA. 2001. Vol. 98, N 12. P. 6736-6741.
Fowler C.J., Wiberg Α., OrelandL. et al. The effect of age and molecular properties of
human brain monoamine oxidase // J. Neurol. Transm. 1980. Vol. 49, N 1/2. P. 1-20.
Fredrickson D.S., Goldsrein J.L., Brown M.S. The familial hyperlipoproteinemias / The
metabolic basis of inherited disease / Ed. by J.B. Stanbury et al. N.Y., 1978.
P. 604-655.
Freeman B.A. Biological sites and mechanisms of free radical production // Free radicals
in molecular biology, aging and disease / Ed. D. Armstrong et al. N.Y.: Raven press,
1984. P. 43-52.
Gems D., Patridge L. Insulin/IGF signaling and ageing: seeing the bigger picture // Curr.
Opin. Genet and Develop. 2001. Vol. 11, N 3. P. 287-292.
Gerich J.E. Assessment of insulin resistance and its role in non-insulin-dependent
diabetes mellitus // J. Lab. Clin. Med. 1984. Vol. 103. P. 497-505.
Ghigo E., Arvat E., Aimaretti G. et al. Diagnostic and therapeutic uses of growth
hormone-releasing substances in adult and elderly subjects // Baillieres Clin. Endocrinol,
and Metabolism. 1998. Vol. 12, N 2. P. 341-358.
Giaccia A.J., Kastan MB. The complexity of p53 modulation: Emerging patterns trom
divergent signals // Genes Develop. 1998. Vol. 12. P. 2973-2983.
Gire V., Wynford-Thomas D. Reinitiation of DNA synthesis and cell division in
senescent human fibroblasts by microinjection of anti-p53 antibodies // Mol. and Cell.
Biol. 1998. Vol. 18. P. 1611-1621.
Glueck C.J., Garstide P.S., Steiner P.M. et al. Hyperalpha- and hypobeta-lipoproteine-
mia in octogenarian kindreds // Atherosclerosis. 1997. Vol. 27, N 4. P. 387-406.
Gompertz B. On the nature of the function exposure of the law of human mortality and
on a new determining life contingencies // Philos. Trans. Roy. Soc. London A. 1825.
Vol. 115. P. 513-585.
195
Gould G.W., Bell G.I. Facultative glucose transporters: An expanding family // Trends
Biochem. Sci. 1990. Vol. 15, N 1. P. 18-23.
GraejfFJG., Guimares F.S., De Andrade T.G. et al. Role of 5-HT in stress, anxiety, and
depression // Pharmacol. Biochem. Behav. 1996. Vol. 54, N 1. P. 129-141.
Gust DA., Wilson M.E., Stocker T. et al. Activity of the hypothalamic-pituitary-adrenal
axis is altered by aging and exposure to social stress in female rhesus monkeys //
J. Clin. Endocrinol. Metabol. 2000. Vol. 85, N 7. P. 2556-2563.
Harmon D. Aging: A theory based on free radical and radiation chemistry // J. Gerontol.
1956. Vol. 11. P. 298-300.
Harman D. Free radical theory of aging: Prolongation of the normal lifespan by free
radical inhibitors // Biological aspects of aging. N.Y.; L., 1962. P. 279-285.
Harman D. The aging process // Proc. Nat. Acad. Sci. USA. 1981. Vol. 78.
P. 7124-7128.
Harman D. Free radical theory of aging: the "free radical" diseases // Age. 1984. Vol. 7.
P. 114-131.
Harman D. Aging and disease: Extending functionsl lifespan // Ann. N.Y. Acad. Sci.
1996. Vol. 786. P. 321-336.
Hart R.W., Setlow R.B. Correlation between deoxyribonucleic acid excision-repair and
life-span in a number of mammalian species // Proc. Nat. Acad. Sci. USA. 1974.
Vol. 71. P. 2169-2173.
Harwood J.P., Grewe C, Alguilera G. Actions of growth-hormone-releasing tactor and
somatostatin on adenylate cyclase and growth hormone release in rat anterior
pituitary // Mol. and Cell. Endocrinol. 1984. Vol. 37. P. 277.
Hattori M., Fujiyama Α., Taylor T.D. et al. The DNA sequence of human chromosome
21. The chromosome 21 mapping and sequencing consortium // Nature. 2000.
Vol. 405, N 6784. P. 311-319.
Hauck S.J., Hunter W.S., Danilovich N. et al. Reduced levels of thyroid hormones,
insulin, and glucose, and lower body core temperature in the growth hormone
receptor/binding protein knockout mouse // Exp. Biol. Med. 2001. Vol. 226, N 6.
P. 552-558.
Hayflick L. // Theoretical aspects of aging / Ed. M. Rockstein. N.Y.: Academ. press. 1974.
Henkart PA. Mechanism of lymphocyte-mediated cytotoxicity // Ann. Rev. Immunol.
1985. Vol. 3. P. 31-58.
Heydari A R., Takahashi R., Guismann A. et al. Hsp 70 and aging // Experientia. 1994.
Vol. 50, N 11/12. P. 1092-1098.
Huether G. Melatonin as an antiaging drug: Between facts and fantasy // Gerontology.
1996. Vol. 42, N 2. P. 87-96.
hahana K., Dimri G., Campisi J. Regulation of cellular senescence by p53 // Europ.
J. Biochem. 2001. Vol. 268. P. 2784-2791.
Jacobson L., Sapolsky R. The role of hyppocampus in feedback regulation of the hypo-
thalamic-pituitary-adrenocortical axis // Endocrinol. Rev. 1991. Vol. 12, N 2.
P. 118-134.
Jacs T. et al. Tumor spectrum analysis in p53-mutant mice /// Curr. Biol. 1994. Vol. 4.
P. 1-7.
Jensen G D., Polloi A.H. The very old of Palau: health and mental state // Age and
Ageing. 1988. Vol. 17, N 4. P. 220-226.
Johannsson G., Marin P., Lonn L. et al. Growth hormone treatment of abdominally obese
men reduces abdominal fat mass, improves glucose and lipoprotein metabolism, and
reduces diastolic blood pressure // J. Clin. Endocrinol, and Metabol. 1997. Vol. 82,
N 3. P. 727-34.
Johnson MA., Brown MA., Poon L.W. et al. Nutritional patterns of centenarians //
Intern. J. Aging and Human Develop. 1992. Vol. 34, N 1, P. 57-76.
196
Jorgensen J.OL., Vahl Ν., Fisher S. et al. Somatopause and adiposity // Hormone R s.
1997. Vol. 48. P. 101-104, suppl. 5.
Katzman R. Alzheimer's Disease // New Engl. J. Med. 1986 Vol. 314, N 15
P. 964-973.
Kauffman S.H. Heat shock proteins in health and disease: (Review) // Intern. J. Clin, and
Lab. Res. 1992. Vol. 21, N 3. P. 221-226.
Kerr J.F.R. Shrinkage necrosis: A distinct mode of cellular death // J. Pathol. 1971.
Vol. 105, N 1. P. 13-20.
Kerr J.F.R., Searle J. Apoptosis: Its nature and kinetic role // Radiation biology in
cancer research / Ed. R.E. Meyn, H.R. Withers. N.Y.: Raven press, 1980. P. 367-384.
Kerr J.F.R., Wyllie A.H., Currie A.R. Apoptosis: A basic biological phenomenon with
wide-ranging implications in tissue kinetics // Brit. J. Cancer. 1972. Vol. 26, N 1.
P. 239-257.
Khan R.t Daya S.t Potgieter B. Evidence for a modulation of the stress response by the
pineal gland // Experientia. 1990. Vol. 46, N 8. P. 860-862.
Khorram O., Vu L., Yen S.S. Activation of immune function by dehydroepiandrosterone
(DHEA) in age-advanced men //J, Gerontol. A: Biol. Sci. Med. Sci. 1997. Vol. 52,
N 1. P. 1-M7.
Kiningham K.K., Oberley T.D., Lin S. et al. Overexpression of manganese superoxide
dismutase protects against mitochondrial-initiated poly(ADP-ribose) polymerase-
mediated cell death // FASEB J. 1999. Vol. 13, N 12. P. 1601-1610.
Kirkwood T.B.L., Austad S.N. Why do we age? // Nature. 2000. Vol. 408. P. 233-238.
Kirkwood T.B.L. p53 and ageing: too much of a good thing? // BioEssays. 2002. Vol. 24.
P. 577-579.
Klatz R., Kahn C. Grow young with HGH. N.Y.: Harper Collins, 1997.
Ко L., Prives C. p53: puzzle and paradigm // Genes Develop/ 1996. Vol. Ю.
P. 1054-1072.
Kuida K., Zheng T.S., Na S. et al. Decreased apoptosis in the brain and premature
lethality in CPP32-deficient mice//Nature. 1996. Vol, 384. P. 36&-372.
Kuninaka S., Ichinose Y., Koja K., Toh Y. Suppression of manganese superoxide
dismutase augments sensitivity to radiation, hyperthermia and doxorubicin in colon cancer
cell lines by inducing apoptosis // Brit. J. Cancer. 2000. Vol. 83, N 7. P. 928-934.
Lamberts S.WJ. The somatopause: To treat or not to treat? // Hormone Res. 2000.
Vol. 53, suppl. 3. P. 42-43.
Lechin F., Van der Dijs В., Benaim M. Stress versus depression // Progr.
Neuropsychopharmacol. Biol. Psychiatry. 1996. Vol. 20, N 6. P. 899-950.
Lee Y.K., Manalo D., Liu A.Y. Heat shock response, heat shock transcription factor and
cell aging//Biol. Signals. 1996. Vol. 5, N 3. P. 180-191.
Levine A.J. p53, the cellular gatekeeper from growth and division // Cell. 1997. Vol. 88.
P. 323-331.
Lipartiti M., Franceschini D.t Zanoni R. et al. Neuroprotective effects of melatonin //
Adv. Exp. Biol, and Med. 1996. Vol. 398. P. 315-321
Liu A.Y., Lee Y.K., Manalo D. et al. Attenuated heat shock transcriptional response in
aging: Molecular mechanism and implication in the biology of aging // EXS. 1996.
Vol. 77. P. 393-408.
LoebA., Kunkel T.A. Fidelity of DNA synthesis// Ann. Rev. of Biochem. 1982. Vol. 52.
P. 429-457.
Lonn L., Johansson G., Sjosirom L. et al. Body composition and tissue distribution in
growth hormone deficient adults before and after growth hormone treatment // Obes.
Res. 1996. Vol. 4, N 1. P. 45-54.
Lund-Johansen P. Hemodynamic concepts in essential hypertension // Triangle. 1984.
Vol. 23, N1. P. 13-23.
197
MacArtur R.H.r Wilson E.O. The theory of Island biogeography. Princeton: Princeton,
Univ. pressr 1967.
Maclaren N.K. How, when and why to predict IDDM // Diabetes. 1988. Vol. 37.
P.1591-1599
Maestroni GJ. The immunoneuroendocrine role of melatonin //J. Pineal Res. 1993. Vol.
14, N1. P. 1-10
Malzberg B. Mortality among patients with involutional melancholia // Amei. J.
Psychiat. 19/37. Vol. 93. P. 1231-1236.
Mandir AS., Simbulan-Rosenthal СМ., Poitras M.F. et al. A novel in vivo post-transla-
tional modification of p53 by PARP-1 in MPTP-induced parkinsonism //
J. Nerochem. 2002. Vol. 83, N 1, P. 186-192.
Martin G.M. Genetic syndromes in man with potential relevance to the pathobiology of
aging // Birth Defects. 1978. Vol. 14. P. 5-39.
Martin G.M., Oshima J. Lessons from human progeroid syndromes // Nature. 2000.
Vol.415. P.263-266.
Mayer J. Genetic factors in human obesity // Postgrad. Med. 1965. Vol. 37.
P. 103-108
McCordJM.., Stokes S.H., Wong K. Superoxide radical as a phagocyteproduced
chemical' mediator of inflammation // Advances in inflammation researches / Ed.
G. Weisman et al. N.Y.: Raven press, 1979. P. 273-280.
McFarland GA„ Holliday R. Retardation of the senescence of cultured human diploid
fibroblasts by carnosine // Exp. Cell. Res. 1994. Vol. 212, N 2. P. 167-175.
Medawar Ρ В. An unsolved problem of biology. L.: Lewis, 1952. 24 p.
MeitesJ., Hylka V.W., Sontag WE. Cellular-molecular versus neuroendocrine concepts
of aging: A need for integration // The molecular basis of aging / Ed. J. Meites. N.Y.,
1984, P. 187-207.
Melov S. Mitochondrial oxidative stress: Physiologic consequences and potential for a
role in aging // Ann. N. Y. Acad. Sci. 2000. Vol. 908. P. 219-225
Mimura C, Murakami K., Gushiken M. Nutritional factors for longevity in Okinawa -
present and future // Nutrit Health. 1992. Vol. 8, N 2/3. P. 159-163.
Miquel J., de Juan E., Sevila L Oxygen-induced mitochondrial damage and aging //
EXS. 1992. Vol. 62. P. 47-57.
Mocchegiani E.f Paolucci P., Balsamo A. et аГ. Influence of growth hormone on thymic
endocrine activity in humans // Hormones Res. 1990. Vol 33, N 6. P. 248-255.
Modrich P. DNA mismatch correction // Annu. Rev. Biochem. 1987. Vol. 56.
P. 435-466
Monnier V.M., Kohn R.R., Cerami A. Accelerated age-related browning of human
collagen in diabetes mellitus //Proc. Nat Acad. Sci. USA. 1984. Vol. 81, N 2. P. 583-587.
Morgan Т.Н. The scientific basis of evolution. L.: Norton, 1932.
MorgenthalerJ., Joy D. Better sex through chemistry: A guide to new prosexual drugs.
Petaluma (CA): Smart publ., 1995.
Motulsky A. If I had a gene test, what would I have and who would I tell? // Lancet. 1999.
Vol. 354, suppl. P. 35-37.
Mueckler M. Family of glucose-transporter genes: Implications for glucose homeostatis
and diabetes // Diabetes. 1990. Vol. 39, N 1. P. 6-11.
Mueckler M. et al. Sequence and structure of a human glucose transporter // Science.
1985. Vol. 229. P. 941-945.
Murray M., Pizzorno J. Encyclopedia of natural medicine. Prima publ, 1991
Nagata S. Apoptosis by death factor // Cell. 1997. Vol. 88. P. 355-365.
OrelandL., Gottfries C.G. Brain and brain monoamine oxidase in aging and in demen
tia of Alzheimer's type // Progr Neuropsychopharmacol. and Biol. Psychiatry. 1986.
Vol. 19, N3/5. P. 533-540.
198
uwen Μ., Cardno A. Psychiatric genetics: Progress, problems, and potential // L nc I
1999. Vol. 354, suppl. 1. P. 1-4.
Packer L·, Tritschler HJ., Wessel K. Neuroprotection by the metabolic antioxidant
alpha-lipoic acid // Free Radicalc Biol. Med., 1997. Vol. 22, N 1/2. P. 359-378.
Pani G., Bedogni В., Anzevino R. et al. Deregulated manganese superoxide dismutasc
expression and resistance to oxidative injury in p53-deficient cells // Cancer Res.
2000. Vol. 60, N 16. P. 4654-^660.
Parker R.C. Methods of tissue culture. N.Y.: Harper and Row, 1961.
Parsons P.A. The limit to human longevity: An approach through a stress theory of
ageing // Mech. Ageing and Develop. 1996. Vol. 87, N 3. P. 211-218.
Pearls T.T. The oldest old // Sci. Amer. 1995. Vol. 272, N 1. P. 70-75.
Pearson D., Show S. Life extension: A practical scientific approach. Warner Books,
1983.
Phillips L.S., Vassilopoulou-Sellin /?. Somatomedins: (two parts) // N. Engl. J. Med.
1980. Vol. 302. P. 371.
Phillips P.D., Kaji K., Cristofalo VJ. Progressive loss of the proliferative response of
senescing WI-38 cells to platelet derived growth factor, epidermal growth factor,
insulin, transferrin, and dexamethasone // J. Gerontol. 1984. Vol. 39. P. 11-17.
Pickens J.M., Burkenholder J.N., Womack W.N. Oral glucose tolerance test in normal
children//Diabetes. 1967. Vol. 16. P. 11.
Pierpaoli W.S, Sorkin E. Immunological blocade of adenohypophysis and its possible
application in prophylaxis and therapy of neoplasrna // Experientia. 1972. Vol. 28.
P. 336-339
Podack E.R. Molecular mechanism of lymphocyte mediated tumor lysis // Immunol.
Today, 1985. Vol. 6. P. 21-27.
Poorkaj P., Sharma V., Anderson L·, Nemens E. Missense mutations in the chromosome
14 familial Alzheimer's disease Presenilin-1 gene // Human. Mutat. 1998. Vol. 11.
P. 216-221.
Pulatova M.K., Sharygin V.L., Todorov I.N. The activation of ribonucleotide reductase
in animal organs as the cellular response against the treatment with DNA-damaging
factors and the influence of radioprotectors on this effect // Biochim. et biophys. acta.
1999. Vol. 1453. P. 321-329.
Purdue СЛ. et al. Tumor incidence, spectrum and ploidy in mice with a large deletion
in the p53 gene // Oncogene. 1994. Vol. 9. P. 603-609.
Ravaglia G., Forti P., Maioli F. et al. The relationship of dehydroepiandrosterone
sulfate (DHEAS) to endocrine-metabolic parameters and functional status in the oldest-
old: Results from in Italian study on healthy tree-living over-ninety-year-olds // J.
Clin. Endocrinol. Metabol. 1996. Vol. 81, N 3. P. 1173-1178.
Ravid M. Incidence and origin of non-systemic microdeposits of amyloid // J. Clin.
Pathol. 1967. Vol. 20. P. 15-20.
Receputo G., Rapisarda R., Motta L. Centenarians: health status and life conditions //
Ann. Ital. Med Int. 1995. Vol. 10, N 1. P. 41-45.
Regelson W., Kalimi M. Dehydroepiandrosterone (DHEA) - the multifunctional steroid.
II. Effects on the CNS, cell proliferation, metabolic and vascular, clinical and other
effects. Mechanism of action? // Ann. N.Y. Acad. Sci. 1994. Vol. 719. P. 564-575.
Reichlin S. Somatostatin: (two parts) // N. Engl. J. Med. 1983. Vol. 309. P. 1495.
Reiter R J., Robinson J. Melatonin. Bantam Books. 1996.
Reznikov K., Kolesnikova L., Pramanik A. et al. Clustering of apoptotic cells via
bystander killing by peroxides // FASEB J. 2000. Vol, 14, № 12. P. 1754-1764
Rose M.R. Evolutionary biology of aging. Oxford: Univ. press, 1991.
Rosenberg SA. The development of new immunotherapies for the treatment of cancer
using interleukin-2 // Ann. Surgery. 1988. Vol. 208, N 2. P. 121-135.
199
Rosenberg S.A., Lotze Μ.Т., Yang J.С. Experience with the use of high-dose inter-
leukin-2 in the treatment of 652 cancer patients // Ibid. 1989. Vol. 210, N 4.
P. 474-485.
Rosenberg S.A., Packard B.S., Aebersold P.M. et al. Use of tumor-infiltrating
lymphocytes and interleukin-2 in the immunotherapy of patients with metastatic melanoma//
N. Engl. J. Med. 1988. Vol. 319, N 25. P. 1676-1680
Rothuizen J., ReulJ.M., RijnberkA. et al. Aging and the hypothalamus-pituitary-adreno-
cortical axis, with special reference to the dog // Acta endocrinol. 1991. Vol. 125,
suppl. l.P.73-76.
Rubner M. Das Problem des Lebensdauer und seine Beziehungen zu Wachstum und
Ernahrung. Milenchen, 1908.
Rudman D„ Feller A.G., Cohn L. et al. Effects of human growth hormone on body
composition in eldery men // Hormone Res. 1991. Vol. 36, suppl. 1. P. 73-81.
Sager R. Senescence as a mode of tumor suppression // Environ. Health Perspect. 1991.
Vol. 93. P. 59-62.
Sahelian R. DHEA: A practical guide. Garden City Park, 1996.
Sainz R.M., Mayo J.C„ Reiter /?./., et al. Melatonin regulates glucocorticoid receptor: an
answer to its antiapoptotic action in thymus // FASEB J. 1999. Vol. 13, N 12.
P. 1547-1556.
Sapolsky R M. Glucocorticoids, stress, and their adverse neurological effects: relevance
to aging // Exp. Gerontol. 1999. Vol. 34, N 6. P. 721-732.
Sardesai V M. Role of antioxidants in health maintenance // Nutrit Clin. Pract. 1995.
Vol. 10, N l.P. 19-25.
Saretzki G„ Von Zglinicki T. Replicative senescence as a model of aging: the role of
oxidative stress and telomere shortening - an overview // Ztschr. Gerontol. Geriatr.
1999. Bd. 32, N 2. P. 69-75. In German.
SastreJ., Pallardo F.V., Vina J. Mitochondrial oxidative stress plays a key role in aging
and apoptosis // ШВМВ Life. 2000. Vol. 49, N 5. P. 427-435.
Scanu A.M. (ed). Lipoprotein (a). N.Y. Acad, press, 1990.
Scanu Α., Lawn R., Berg K. Lipoprotein (a) and atherosclerosis // Ann. Internal Med.
1991. Vol. 115, N 3. P. 209-218.
Schleifer S.J., Keller St.E., Siris S.G. et al. Depression and immunity // Arch. Gen.
Psychiat. 1985. Vol. 42. P. 129-133.
Seiliev Α Α., Zvonareva N.B., Zhivotovsky B.D., Hanson K.P. Determination of some
nuclear deoxiribonucleases in rat thymocytes // Radiat. Environ. Biophys. 1992.
Vol. 31, №2. P. 123-132.
Selkoe DJ. The molecular pathology of Alzheimer's disease // Neuron. 1991. Vol. 6,
N 4. P. 487-498.
Selkoe DJ., Podlisny MB., Joachim C.L. ei al. β-Amiloid precursor protein of
Alzheimer's disease occurs as 110- to 135-kilodalton membrane-associated proteins
in neural and nonneural tissues // Proc. Nat. Acad. Sci. USA. 1988. Vol. 85, N 19.
P. 7341-7345.
Serrano M„ Lin A.W., McCurrach ME. et al. Oncogenic ras provokes premature cell
senescence associated with accumulation of p53 and pl6INK4a //Cell. 1997. Vol. 88,
N 5. P. 593-602
ShayJ.W., Pereira-Smith O.M., Wright W.E. A role for both RB and p53 in the
regulation of human cellular senescence//Exp. Cell Res. 1991. Vol. 196. P. 33-39.
Shekelle R.B., Raynor WJ., OstfeldAM. et al. Psychological depression and 17 year risk
of death from cancer // Psychosom. Med. 1981. Vol. 43. P. 117-125.
Siervogel R.M., Roche A.F., Morris H.G., Glueck С J. Blood pressure and its
relationship to plasma lipids and lipoproteins in children // Prevent. Med. 1981. Vol. 10.
P. 555-564.
200
Skalka M., Matyasova J., Cejkova M. DNA in chromatin of irradiated lympoid tissue»
degrades in vivo into regular fragments // FEBS Lett. 1976. Vol. 72, N 2. P. 271-
274.
Slot JW. Immuno-localization of the insulin regulatable glucose transporter in brown
adipose tissue of the rat // J. Cell Biol. 1991. Vol. 113, N 1. P. 123-135
Sohal R.S., Weindruch R. Oxidative stress, caloric restriction, and aging // Science. 1996.
Vol. 273, N5271. P. 59-63.
Sohal RS. Role of oxidative stress and protein oxidation in the aging process // Free
Radicals Biol. Med. 2002. Vol. 33, N 1. P. 37-44.
Spencer R.L., Hutchinson K.E. Alcohol, aging, and the stress response // Alcohol Re».
Health. 1999. Vol. 23, N 4. P. 272-283.
Sc/uier T.C. Oxidative stress and protein aggregation during biological agmg // Exp.
Gerontol. 2001. Vol. 36, N 9. P. 1539-1550
Migimoto Т., Nakaoka D., Nasu M. et al. Effect of recombinant human growth hormone
in elderly osteoporotic women // Clin. Endocrinol. 1999. Vol. 51, N 6. P. 715-
724.
Svendsen L·, Rattan S.I., Clark B.F. Testing garlic for possible anti-ageing effects on
long-term growth characteristics, morphology and macromolecular synthesis of
human fibroblasts in culture // J. Ethnopharmacol. 1994. Vol. 43, N 2. P. 125-
133.
iaaffe D.R., Marcus R. Dynamic muscle strength alterations to detraining and retraining
in elderly men // Clin. Physiol. 1997. Vol. 17, N 3. P. 311-324.
Talmage D.W. The acceptance and rejection of immunologic concepts // Annu. Rev.
Immunol. 1986. Vol. 4. P. 1-11.
Tanaka //., Arakawa //., Yamaguchi Т., Shiraishi K. et al. A ribonucleotide reductase
gene involved in a p53-dependent cell-cycle checkpoint for DNA damage // Nature.
2000. Vol. 404. P. 42-49.
lepperman M.D., Tepperman ИМ. Metabolic and endocrine physiology: An
introductory text. 5th ed. Chicago; L.: Year Book Med. Publ., 1987.
rhompson C.B. Apoptosis in the pathogenesis and treatment of disease // Science. 1995.
Vol. 267. P. 1456-1462.
Todd J A. Genetic control of autoimmunity in type I diabetes // Immunol. Today. 1990.
Vol. 11, N4. P. 122-129.
Todorov LN. Mechanism of antistress and anabolic actions of eleutherococcus sentico-
sus maximum extracts // Bioactive compounds: Biotransformation and biological
action / Ed. by I.N.Todorov et al. Nova Sci. publ., 1993.
lolmasojfJM., Ono Т., Cutler R.G. Superoxide dismutase: Correlation with life span
and specific metabolic rate in primate species // Proc. Nat. Acad. Sci. USA. 1980.
Vol. 77. P. 2777-2780.
Toussaint O., Remade J.f Dierick J. et al. From the Hay flick mosaic to the mosaics of
ageing: Role of stress-induced premature senescence in human ageing // Intern.
J. Biochem. Cell Biol. 2002. Vol. 34, N 11. P. 1415.
Tsitouras P.D., Bulat T. The aging male reproductive system // Endocrinol. Mecabol.
Clin. North Amer. 1995. Vol. 24, N 2. P. 297-315.
Turnell R.W., Clarke L.H., Burton A.F. Studies on the mechanism of corticosteroid-
induced lymphocytolysis // Cancer Res. 1973. Vol. 33. P. 203.
Tyner S.D., Venkatachalam S., Choi J. et al. p53 mutant mice that display early ageing-
associated phenotypes // Nature. 2002. Vol. 415. P. 45-53.
Unanue E.R. Antigen presenting function of the macrophage // Annu. Rev. Immunol.
1984. Vol. 2. P. 395-428.
Van Remmen //., Ward W.F., Salia R.V. et al. Handbook of psychology. Section 11.
Aging/Ed. E.J. Masoro. N.Y., 1995. P. 171-234.
201
Vijg J., Wei J.Y. Understanding the biology of aging: The key to prevention and
therapy // J. Amer. Geriatr. Soc. 1995. Vol. 43, N 4. P. 426-^34.
Villareal D.T., Mortley J.E. Trophic factors in aging: Should older people receive
hormonal replacement therapy? // Drugs Aging. 1994. Vol. 4, N 6. P. 492-509.
Vlassara H., Brownlee M., Cerami A. High-affinity reception-mediated uptake
and degradation of glucose-modified proteins: Mechanism for the removal of
senescent macromolecules // Proc. Nat. Acad. Sci. USA. 1985. Vol. 82, N 17.
P. 5588-5592.
Von Boehmer H. Developmental biology of Τ cells in Τ cell-receptor transgenic mice //
Annu. Rev. Immunol. 1990. Vol. 8. P. 531-556.
Wang H., Ma L·, Li Y., Cho CM. Exposure to cigarette smoke increases apoptosis in the
rat gastric mucosa through a reactive oxygen species-mediated and p53-independent
pathway // Free Radicals Biol. Med. 2000. Vol. 28, N 7. P. 1125-1131.
Wang J., Shen L., Najafi H. et al. Regulation of insulin preRNA splicing by glucose //
Proc. Nat Acad. Sci. USA. 1997. Vol. 94. P. 4360-4365.
Webley K. et al. Postlranslational modifications of p53 in replicative senescence
overlapping but distinct from those induced by DNA damage // Mol. and Cell. Biol. 2000.
Vol 20. P. 2803-2808.
Wei Y.H., Ma YS., Lee H.C. et al. Mitochondrial theory of aging matures - roles of
mtDNA mutation and oxidative stress in human aging // Zhonghua Yi Xue Za Zhi.
Taipei, 2001. Vol. 64, N 5. P. 259-270.
Weindruch R., WalfordR.L. Dietary restriction in mice beginning at 1 year of age: effect
on life-span and spontaneous cancer incidence // Science. 1982. Vol. 215.
P. 1415-1418.
Weismann A. Essays upon heredity and kindred biological problems. 2nd Ed. Oxford:
Clarendon press, 1891.
Weismann A. Ueberdie Dauer des Lebens. Jena: Fischer, 1892.
Weissman MM., Gershon E.S., Kidd K.K. et al. Psychiatric disorders in the relatives
of probands with affective disorders: The Yale University - National Institute of
Mental Health collaborative study // Arch. Gen. Psychiat. 1984. Vol. 41, N 1.
P. 13-21.
Weissman Μ Μ., Wickramaratne P., Merikangas K.R. Onset of major depression in early
adulthood. Increased familial loading and specificity // Ibid. 1984. Vol. 41, N 12.
P. 1136-1143.
Welborn ТА., Brockenkidge Α., Rubinstein А.И. et al. Serum-insulin in essential
hypertension and in peripheral vascular disease // Lancet. 1966. Vol. 1. P. 1336-1337.
Wexler B.C. Comparative aspects of hyperadrenocorticism and aging // Hypothalamus,
pituitary and aging / Ed. by A.V. Everiitt, J.A. Burgess. Springfield, 1976. P. 333-
361.
Wiley D„ BortzWM. Sexuality and aging - usual and successful //J. Gerontol. A: Biol.
Sci. 19%. Vol. 51, N 3. P. M142-M146.
Williams G.C. Pleiotropy, natural selection and the evolution of senescence // Evolution.
1957. Vol. 11. P. 398-411.
Wilson J.D., Foster D.W. Textbook of endocrinology. 7th ed. Saunders, 1985.
Wolkovitz O.M., Reus V.I., Roberts E. et al. Dehydroepiandrosterone (DHEA) treatment
of depression // Biol. Psychiatry. 1997. Vol. 41, N 3. P. 311-318.
WyllieA.H. Glucocorticoid-induced thymocyte apoptosis is associated with endogenous
endonucleases activation//Nature. 1980. Vol. 284. P. 555-556.
Wyllie A.H. Apoptosis: cell death in tissue regulation // J. Pathol. 1987. Vol. 153, N 2.
P. 313-316.
Wyllie A.H., KerrJ.F.R., Currie A.R. Cell death: the significance of apoptosis // Intern.
Rev. Cytol. 1980. Vol. 68. P. 251-306.
202
Wyllie A.H., Morris R.G., Smith A.L., Dunlop D. Chromation cleavage in apoptosis:
Association with condensed chromatin morphology and dependence on macromole-
cular synthesis //J. Pathol. 1984. Vol. 142. P. 67-77.
Yamada Т., Okayama H., Kinio Y., Watanabe H. Evidence for the internucleosomal
bread kage of chromatin in rat thymocytes irradiated in vitro // Radiat. Res. 1981.
Vol. 85, N 2. P. 544-553.
Yen 5.5., Morales A.J., Khorram O. Replacement of DHEA in aging men and women.
Potential remedial effects // Ann. N.Y. Acad. Sci. 1995. Vol. 774. P. 128-142.
Yin D. Biomedical basis of lipofuscin, ceroid, and age pigment-like fluorophores // Free
Radials Biol. Med. 19%. Vol. 21, N 6. P. 871-888.
Yin D. Studies on age pigments evolving into a new theory of biological aging //
Gerontology. 1995. Vol. 41, suppl. 2. P. 159-172.
Young J.D.-E, Clark W.R., Liu C.-C. et al. A calcium- and perfonn-independent pathway
of killing mediated by murine cytolytic lymphocytes //J. Exp. Med. 1987. Vol. 166,
N 6. P. 1894-1899.
Young J.D.-E, Hengartner H., Podack E.R. et al. Purification and characterization of a
cytolytic роге-forming protein from granules of cloned lymphocytes with natural
killer activity // Cell. 1986. VoL 44, N 6. P. 849-859.
Yu B.P. Aging and oxidative stress: modulation by dietary restriction // Free Radicals
Biol. Med. 1996. Vol. 21, N 5. P. 651-668.
Yukawa O., Nakazawa T. Radiation-induced lipid peroxidation and membrane-bound
enzymes in liver-microsomes // Intern. J. Radiat. Biol. 1980. Vol. 37. P. 621-631.
Zglinicki Т., Burkle Α., Kirkwood T.B. Stress, DNA damage and ageing - an integrative
approach //Exp. Gerontol. 2001. Vol. 36, N 7. P. 1049-1062.
Zhivotovsky B.D., Zvonareva N.B., Hanson K.P. Characteristics of rat thymus chromatin
degradation products after whole body X-irradiation // Intern J. Radiat. Biol. (1981).
Vol. 39. P. 437-440.
Zvonareva N.B., Zhivotovsky B.D., Hanson K.P. Distribution of nucleases attack sites
and complexity of DNA in the products of post-irradiation degradation of rat thymus
chromatin // Ibid. 1983. VoL 44, N 3. P. 261-266.
Часть II
СТРЕСС
Стресс отражает саму сущность
взаимодействия психического
и соматического.
Дж. Эверли, Р. Розенфельд, 1985
Глава Ю
СТРЕСС, ЕГО МЕСТО
И РОЛЬ В НАШЕЙ ЖИЗНИ
Стрессом называется
неспецифическая реакция организма
на любое предъявляемое к нему требование.
ГансСелье, 1977.
Слово "стресс" (букв. пер. с англ. - "напряжение") стало одним
из самых модных слов XX столетия. Мы испытываем стресс на
рабочем месте, мы входим в напряженные отношения, мы
подвергаемся финансовому давлению, необходимости что-то выполнить к
определенному сроку, соблюдать общественные приличия, волнению
при публичных выступлениях, в семейных спорах, словом,
подвергаемся многочисленным стрессам или оказываемся в состоянии
стресса просто без каких-либо видимых причин.
Тем не менее многие из нас действительно не знают точно, что
такое стресс, другие полагают, что это нечто такое, что имеет
отношение к напряженности, беспокойству, дискомфорту, и чего
нужно избегать. Это, разумеется, верно, однако стресс прежде всего -
научная концепция, проливающая свет на то, как организм
взаимодействует с окружающей средой. Чтобы понимать роль стресса в
старении и болезнях, мы должны рассмотреть природу стресса как
сложного биологического явления.
Весь комплекс наших знаний об устройстве и функционировании живых
систем - от простейших одноклеточных до высших многоклеточных
организмов - позволяет понять не только основы жизнедеятельности организмов на
молекулярном уровне, где достигнуты поразительные успехи, но также и на
более сложном системном уровне и, в частности, представить общую стратегию
поведения этих систем в меняющихся условиях окружающей среды. Одной из
наиболее важных сторон такого поведения является стремление к постоянству
внутренней среды (гомеостазу), которое обеспечивается механизмами
контроля и сохранения стабильности физических, физико-химических и химических
параметров, а также оптимальных концентраций веществ, вовлеченных в
метаболизм живых систем. Эти регуляторные механизмы эволюционно
совершенствовались как в отношении точности и эффективности, так и достаточного
"запаса прочности" для надежной защиты от влияния экстремальных факторов,
постоянно возникающих во внешней и внутренней среде организма. При
интенсивном и длительном действии этих факторов вся сложная иерархическая
система механизмов сохранения гомеостаза (см. рис. 10) активируется и переходит
η форсированный режим работы, в состояние напряжения, которое длится до
тех пор, пока не будут ликвидированы нарушения, вызвавшие это напряжение,
и система не возвратится в состояние динамического равновесия. Наконец, в тех
207
случаях, когда даже крайнее напряжение регуляторно-приспособительных
механизмов не обеспечивает поддержания и сохранения гомеостаза, организм
погибает
Каждый действующий на организм фактор среды вызывает
соответственно своей природе ответную специфическую реакцию, адекватную качеству и
силе раздражителя, но в любой такой реакции обязательно присутствует и
неспецифический компонент, характеризующий состояние напряжения как
таковое, степень активации механизмов поддержания гомеостаза. Именно этот
неспецифический компонент физиологических и патологических реакций и
принято в настоящее время обозначать термином "стресс "
Идея о том, что здоровье и сама жизнь как тонкое равновесие,
зависят от эффективной адаптации к различным угрожающим
силам, не нова. Гиппократ, основоположник современной медицины,
полагал, что здоровье зависит от баланса между разрушающими и
адаптивными силами. В годы Ренессанса, эту концепцию развивал
Томас Сиденхэм (Thomas Sydenham), который утверждал, что
адаптивный ответ на дисгармонии, вызванные разрушающими силами,
может самостоятельно служить причиной болезни. В начале XX
столетия Уолтер Кэннон (Walter Cannon) описал реакцию
"чрезвычайной адаптации", которую назвал ответом "борьбы или бегства".
Однако наиболее глубоко понять биологическую природу
стресса удалось выдающемуся канадскому физиологу Гансу Селье.
Согласно Селье, стресс - это специфический набор физиологических
реакций (адаптационный стресс-ответ), которые являются по
существу одними и теми же независимо от характера (природы) стрессо-
генного фактора. Другими словами, подвергаетесь ли вы действию
жары, холода, травмам, болезням, переутомлению, испугу,
отравлены ли или приведены в бешенство, ваше тело реагирует давая, по
существу, один и тот же стресс-ответ, пробуя приспособиться к
ситуации. Это - подобно звонку в службу спасения в любом критическом
положении, неважно пожар ли это, нападение или какая-то
случайность. Но чрезмерный или разбалансированный стресс-ответ
способен самостоятельно вызывать серьезные повреждения в организме,
приводя к болезням и ускоряя старение. - и в этом заключается
парадокс ситуации.
Эти специфические реакции, открытые Г. Селье и описанные в
журнале "Nature" еще в 1936 г. (Selye, 1936), вызвали огромный интерес ученых и
индуцировали многочисленные исследования, раскрывшие биохимическую,
физиологическую и кибернетическую природу стресса, а также развитие
целой индустрии производства препаратов, ингибирующих, стимулирующих,
модулирующих, модифицирующих, пролонгирующих и т.п. состояние
стресса. Эти препараты используются в различных областях медицины,
лечебного питания, нетрадиционной медицины, медицины, использующей
природные соединения из растений, и т.п. Наиболее полно состояние стресса
изучено у человека и высших животных, хотя в настоящее время интенсивно
изучается так называемый клеточный стресс, который, естественно, имеет
несколько иную физиологическую и молекулярную природу, нежели стресс
более сложных систем.
208
10.1. СТРЕСС В НАШЕЙ ЖИЗНИ
Серьезный и длительный стресс - физиологический экий на
лент ситуации военного времени - может привести к глубокому
истощению, болезни или даже смерти. Большинство из нас, одн ι-
ко, никогда не проходит через такие крайности. Стрессы,
которые мы испытываем, обычно слабые или умеренные, а
случайные серьезные стрессы, например хирургические операции или
обширные травмы, смягчены условиями клинического
стационара. Тем не менее даже мягкий или умеренный стресс, особенно
если он продолжителен, обычно оказывается для организма
разрушительным фактором - он не может быть непосредственной
причиной болезни или смерти, однако способен ускорить все
аспекты старения и увеличить риск болезни.
10.2. ФИЗИЧЕСКИЙ И ЭМОЦИОНАЛЬНЫЙ СТРЕССЫ
Теория стресса подразделяет его на две главных категории -
физический (телесный) и эмоциональный (психологический).
Телесный стресс вызывается материальными физическими факторами,
такими как инфекции, радиация, травмы, серьезная простуда или
высокая температура. Высокоразвитые организмы способны
инициировать стресс-ответ, не перенося реального физического
воздействия. Центральная нервная система анализирует ситуацию и, если
имеется осознанная потребность в чрезвычайных мерах,
стимулирует стресс-ответ заранее. Действительно, когда вы вдруг заметили
ягуара, скрывающегося на ближайшем дереве, способность
"включить" стресс-ответ прежде, чем хищник нападет на вас, верное
спасение. Эмоциональный стресс может быть столь же мощным, как и
физический. У грызунов высокая скученность или, наоборот,
длительная изоляции вызывает очень сильный стресс-ответ. У людей
стресс может быть вызван почти бесконечным разнообразием
эмоциональных и социальных конфликтов или напряженных
отношений. Чтобы действовать как стрессор, эмоционально насыщенное
событие даже может не иметь прямого отношения к индивидууму
Например, у зрителей спортивных состязаний часто развивается
некоторая степень стресс-ответа.
Механизмы включения физического и эмоционального
стресса — различны, между тем, и это следует подчеркнуть, сам ответ - в
значительной степени один и тот же. Оба типа стресса ускоряют
старение и снижают сопротивляемость к болезням.
В XX столетии относительное воздействие различных типов
стресса на нашу жизнь сместилось в сторону психологического
стресса. Мы тонем в избытке новой информации, меняем рабочие
места, многие из нас переживают распад семьи, наши детишки по-
209
Таблица 11
Оценочная шкала социальной приспособленности
Факторы, вызывающие стресс, оцениваются по их потенциальной
способности вызывать болезни. (Счет 200 единиц в год рассматривается
как предсказание серьезного заболевания)
1 Смерть спутника жизни
(супруги или супруга)
2. Развод
3. Раздел имущества
4 Тюремное заключение
5. Смерть близкого
родственника
6. Ранение или болезнь
7. Брак
8. Потеря работы
9 Супружеское
примирение
10. Выход в отставку или
на пенсию
11. Заболевание членов
семьи
12. Беременность
13. Проблемы с сексом
14. Новый член семьи
15. Налаживание бизнеса
16. Изменение финансового
положения
17. Смерть близкого друга
18. Смена сферы
деятельности
19. Изменение количества
споров с супругом(ой)
20. Большой заклад
21. Лишение права выкупа
закладной или ссуды
22. Повышение
ответственности на работе
23. Дети уезжают из дома
24. Беспокойство за
родственников
73
65
63
63
53
50
47
45
45
44
40
39
39
39
38
37
36
35
31
30
29
29
29
26.
27.
28.
29.
30.
31.
32
33.
34.
35.
36.
37
38.
39.
40.
41.
42.
43.
Выдающийся личный
успех
Жена начала или
перестала работать
Поступление или
окончание школы
Изменение в условиях
жизни
Изменение в личных:
привычках
Неприятности с
руководством
Изменение в условиях
или времени работы
Смена
местожительства
Смена школы
Смена места отдыха
Изменение в церковной
деятельности
Изменение в социальной
деятельности
Небольшой заклад
Изменения в режиме
сна
Увеличение (или
уменьшение) количества
членов семьи, проживающих
вместе
Изменение режима
питания
Отпуск
Праздник
Рождества
Незначительное
нарушение закона
28
26
26
25
24
23
20
20
20
19
19
18
17
16
15
15
13
12
210
стоянно оказываются без присмотра - совершенно ясно, что пина
жизнь более беспокойна, наполнена конкуренцией, требует нош ι-
шенной активности, чем в добрые старые времена. Т.Холмес и
Р. Рае (Holmes, Rahe) разработали масштаб социальной
приспособленности, популярный метод для оценки уровня психологически! о
стресса {табл. 11). Суммарная величина 200 или более за год, как
полагают, указывает на высокий риск развития серьезных болезней.
10.3. "ХОРОШИЙ" И "ПЛОХОЙ" СТРЕССЫ
Не все стрессы равноценны. Длительный или чрезмерный стресс,
чреватый перегрузками жизненно важных систем тела и исчерпанием
ресурсов, явно плохой и ведет к сокращению жизни или увеличивает
риск заболеть. Но может ли он быть безопасным или даже
полезным? Да, определенно может. Как это нередко случается, слабый или
умеренный и кратковременный физический стресс может быть
полезным. С одной стороны, такой не слишком интенсивный или
продолжительный стресс не вызывает каких-либо заметных
повреждений в организме, но, с другой стороны, стимулирует многие
адаптивные системы тела и позволяет лучше противостоять серьезным
стрессорным напряжениям в будущем. В исследованиях на животных
периодическая экспозиция грызунов с умеренными дозами радиации
или токсинов сделала их более стойкими к большим дозам, а также к
разнообразным другим стрессорам. По очевидным причинам такой
подход для повышения устойчивости к стрессорам применительно к
людям - нежелателен. Однако имеется значительно более
совершенный (мягкий или умеренный) периодический стрессор для улучшения
общей адаптивной способности и устойчивости организма -
физические упражнения. Польза для здоровья от физических упражнений
известна и связана с улучшением сердечно-сосудистой функции, более
низким уровнем холестерина и липопротеидов низкой плотности
(ЛНП), повышением уровня липопротеидов высокой плотности
(ЛВП), снижая кровяное давление и снимая беспокойство,
свойственное депрессии. Важно, что физические упражнения улучшают как
психологическую, так и физическую способность справляться со
стрессом. Механизм антистрессового действия упражнений включает:
(1) кратковременное увеличение свободных радикалов во время
физических упражнений стимулирует длительное повышение
активности собственных антиоксидантных систем организма;
(2) повышает чувствительность ткани к инсулину и улучшает
метаболизм углеводов и жиров;
(3) у молодых людей и людей среднего возраста физические
упражнения могут стимулировать выделение гормона роста, который
имеет большие возможности противодействовать стрессу и
старению;
211
(4) уменьшает интенсивность реакции "борьбы и бегства" в ответ на
психологический стрессор;
(5) улучшает эффективность системы обратных связей между
гипоталамусом и периферическими эндокринными железами,
улучшая таким образом эффективность систем, которые
ограничивают стресс-ответ организма.
В те времена, когда формировались наши механизмы
стресс-ответа, энергичное физическое действие было обычным ответом на
эмоциональные стрессогенные факторы: например, опасение, что враг нем-
пременно нападает, вызывало ярость и заканчивалось битвой. В
дополнение к разрешению стрессогенных ситуаций, физическая активность
также играет роль механизма обратной связи, который ограничивает
интенсивность стресс-ответа, приводя к эффектам успокоения и
расслабления. Хотя наша физиология с тех пор не изменилась, у
современных людей эмоциональный стресс обычно не ведет к физическим
эскападам или борьбе, поэтому стресс-ответ оказывается более
интенсивным и длительным, чем это необходимо. По этой причине физические
упражнения особенно полезны в психологическом отношении для
индивидуумов в периоды, когда вероятность стрессов высока.
Как почти во всем, можно переусердствовать и с физическими
упражнениями и фактически достичь результата, противоположного
желаемому. Чрезмерные физические упражнения, особенно без
достаточного отдыха и адекватного питания, могут стать серьезным стрес-
согенным фактором, который исчерпает резервы антиоксидантов,
приведет к перегрузке сердечно-сосудистой системы (особенно у
людей с проблемами сердца) и к истощению. Любая программа
физических упражнений высокой интенсивности должна быть соединена с
полноценным рационом, содержащим достаточно калорий и белка для
предотвращения потери в нежировых тканях, а также достаточное
количество антиоксидантов, чтобы нейтрализовать возрастающий
уровень формирования свободных радикалов. Некоторые люди,
склонные к полноте, предаются чрезмерным физическим упражнениям,
оставаясь в то же время на диете с низким содержанием калорий, белка,
витаминов и антиоксидантов. Этот подход редко достигает цели, т.е.
длительной потери в весе, но часто вызывает глубокое истощение
жизненно важных тканей, недоедание и другие проблемы.
В то время как умеренный ттриодический и кратковременный
физический стресс часто полезен, однако то же самое, по-видимому,
несправедливо для эмоционального стресса, хотя общая картина
стресс-ответа подобна в обоих случаях. Случайное
кратковременное, эмоциональное напряжение, например экзамен или публичное
выступление, очевидно безопасно. Напротив, частые
эмоциональные напряжения, в частности каждодневные семейные ссоры или
высокое напряжение на работе, имеют тенденцию сливаться в
состояние хронического стресса, ведущего к ускоренному старению и
болезням.
212
10.4. ЭВОЛЮЦИЯ СТРЕССА
Наши физиологические ответы были сформированы, и это су
щественно, в окружающей среде, весьма отличной от современного
мира. Жизнь доисторических людей была полна непосредственных
опасностей и трудностей, таких как хищники, голод, раны,
инфекции и экстремальные погодные условия. Такой беспокойный образ
жизни требовал мощного и быстрого стресс-ответа, который был
нацелен на выживание индивидуума в данной конкретной ситуации
за счет перегрузок, изнашивающих организм.
Окружающая среда сегодня также интенсивно воздействует на
нас, хотя и другими способами: информационной перегрузкой,
социальным давлением и быстро изменяющимся миром. Мы постоянно
должны приспосабливаться и подстраиваться, но проблема, однако,
состоит в том, что наши машины адаптации все еще остаются теми
же самыми, как и десятки тысяч лет назад. Наш стресс-ответ все
еще лучше всего приспособлен для бегства от диких животных,
преодоления высокой температуры, простуды и ран. Конечно, стресс-
ответ все еще эффективен, если вы, например, врезались на
автомобиле в дерево или упали с третьего этажа, стресс-ответ вашего тела,
наряду с медработниками и условиями лечения в стационаре, служат
лучшими инструментами для выживания. То же самое справедливо
для любой травмы, инфекции, хирургической операции или других
чрезвычайных ситуаций. Однако в огромном большинстве случаев
стресс-ответ возникает и тогда, когда нет никакой реальной угрозы
ни выживанию, ни даже здоровью. У человека, произносящего
публичную речь, например, развиваются почти те же самые
физиологические реакции, как и у пещерного человека, убегающего от тигра.
Говоря метафорически, стресс-ответ - "избыточный способ"
для преодоления неблагоприятных ситуаций. Организм платит за
него слишком высокую цену, но в жизненно важных ситуациях
такой способ определенно стоит того. К сожалению, эволюционный
стереотип часто заставит наши тела генерировать интенсивный
стресс-ответ, когда в нем нет или немного реальной потребности,
т.е. когда практически отсутствует какая-либо угроза жизни или
здоровью. Это ведет к ускоренному изнашиванию тела и
увеличению риска болезней, другими словами, к ухудшению здоровья и
более быстрому старению. Идеально было бы обладать обоими
вариантами, т.е. иметь адекватный стресс-ответ в критическом
положении, но избегать его активации, когда нет никакой реальной
опасности или затруднений.
Глава 11
ДИНАМИЧЕСКАЯ ПРИРОДА СТРЕССА
Пути развития стресс-реакции включают реакцию различных
иерархических уровней нервной и эндокринной систем, изменения
кровообращения, иммуногенеза, энергетического метаболизма и
т.п. Однако не существует каких-либо особых систем организма,
функционирующих только для осуществления стресс-реакции.
В действительности стресс-реакция реализуется теми же
физиологическими и биохимическими механизмами, которые обеспечивают
стабильность внутренней среды организма (гомеостаз) в
меняющихся условиях среды обитания, и разворачивается в соответствии с эво-
люционно выработанной и генетически закрепленной программой.
Лучше всего явление стресса во всей его совокупности
определяется понятием "процесс".
Психосоматический импульс последовательно активирует одну
(или более) из трех основных групп "осей стресса": 1) нейрогенные
механизмы - стрессовая реакция через прямую нейронную
иннервацию "концевых" органов, 2) нейроэндокринные механизмы -
реакция "борьбы и бегства", 3) эндокринные механизмы - реализуют
общий адаптационный синдром, вовлекая адренокортикотропную,
соматотропную и тиреотропную функции гипоталамо-гипофизар-
ной системы (Эверли. Розенфельд, 1985).
11.1. НЕЙРОГЕННЫЕ МЕХАНИЗМЫ
Хотя сигналы для запуска стресс-реакции поступают по разным
каналам (через рецепторный аппарат, различные этажи нервной
системы или изменения во внутренней среде организма), центром, где
эти стимулы сходятся и где формируется сложный координирован
ный ответ организма, является центральная нервная система (ЦНС -
головной и спинной мозг).
Структуры головного мозга играют исключительно важную роль в
развитии всех видов стресса и особенно стресса, вызванного психосоциальными
раздражителями (Levi, 1972; Mason, 1972). В мозгу человека различают три основ-
214
Неокортикальный
уровень
Лимбическая
система
Вегетативный
уровень
Неокортекс
Лобная доля
Область
перегородки!
Гипоталамус
Гипофиз
Миндалевидное
тело
Гиппокамп
Ретикулярная
формация
С
Поясная извилина
Таламус
Спинной мозг
Рис. 16. Мозг человека (Эверли, Розенфельд, 1985)
ных функциональных уровня, что позволяет называть его "тройным мозгом"
(MacLean, 1975). Неокортекс - высший уровень тройного мозга. Он
осуществляет декодирование и интерпретацию сенсорных сигналов и сообщений,
высшее управление поведением в моторно-двигательной (скелетно-мышечной)
сфере. Неокортекс управляет процессами воображения, логического
мышления, построения умозаключений, памяти, решения проблем и прогнозирования.
Лимбическая система - второй уровень тройного мозга. Для понимания
механизмов стресса важна ее роль как центра эмоционального (аффективного)
контроля. В лимбической системе локализованы, в частности, такие
важнейшие для поддержания гомеостаза и развития стресса структуры, как
гипоталамус, гипокамп, миндалевидное тело и гипофиз. Ретикулярная формация и
ствол мозга - низший уровень тройного мозга. Этот уровень управляет
вегетативными функциями (сердцебиение, дыхание, вазомоторная активность) и
участвует в проведении импульсов через ретикулярную формацию и передаточные
центры таламуса к высшим уровням тройного мозга. Схематическое
расположение морфологических структур в "тройном мозге" представлено на рис. 16
Первым "шагом" стресс-реакции, который раньше других
сказывается на соматическом уровне, является активация симпатического и
парасимпатического отделов автономной нервной системы. Эти
отделы воздействуют на внутренние органы через самые
быстродействующие нервные механизмы и служат наиболее эффективными
стимуляторами стресс-реакции. После того как в неокортексе и
лимбической системе произошла интерпретация стимула как "угрожающего",
нервный импульс нисходит к заднему (при симпатической активации)
и к переднему (при парасимпатической активации) отделам
гипоталамуса. От задних областей гипоталамуса симпатические пути
спускаются через грудной и поясничный отделы спинного мозга и, пройдя
цепочку симпатических ганглиев, иннервируют соответствующие ог>
215
ганы. Роль нейротрасмиттера в этом случае играет норадреналин.
Парасимпатические пути спускаются от переднего гипоталамуса
через черепной или крестцовый отделы спинного мозга к концевым
органам. Здесь, при постганглиозной передаче нервного импульса
используется ацетилхолин (Guyton, 1976).
Важнейшие эффекты активации автономной нервной системы
на "концевые" органы при стрессе проявляются очень быстро и не
носят потенциально хронического характера. Это связано с
ограниченной способностью пара- и симпатических нервных окончаний
осуществлять постоянный выброс медиаторов в условиях сильного и
длительного раздражения {Le Blanc, 1976).
Однако для поддержания высокого уровня стресс-реакции в
течение более длительного времени должна быть активирована
дополнительная психофизилогическая ось этой реакции, т.е. ось
"борьбы и бегства".
11.2. НЕЙРОЭНДОКРИННЫЕ МЕХАНИЗМЫ
Эта реакция носит нейроэндокринный характер, благодаря
участию как автономной нервной системы, так и мозгового слоя
надпочечников. Включение этой реакции в ответ на воспринятую угрозу
мобилизует ресурсы организма и подготавливает нервную и
мышечную системы к действию, что позволяет индивиду либо бороться с
угрозой, либо бежать от нее {Cannon, 1914; 1953). Реакция возникает в
Раздражитель
Сенсорная рецепция
*
Через сенсорные пути
Когнитивная ^ ^Аффективная
интерпретация интеграция
посредством посредством
неокортекса ·*. ^лимоической
• ν системы
Восприятие Нет восприятия угрозы
угрозы | {
Ответ на стресс Нет стрессовой
через гипоталамус реакции
Ч
Прямая иннервация через
симпатическую и парасимпатическую
нервные системы
Реакция "битва-бегство" (Cannon)
Общий адаптационный синдром (Selye)
Концевой орган
Раздражитель
От
сенсорных
путей Направление
стрессовой
реакции к
концевым
органам
Рис. 17. Схема стрессовой реакции (Эверли, Розенфельд, 1985)
216
миндалевидном теле лимбической системы. Далее, нисходя иди й
поток нервных импульсов поступает к латеральной и задней гипо1ала-
мическим областям. Отсюда возбуждение распространяется череч
грудной отдел спинного мозга, сходясь в чревном ганглии, иннервируя
затем мозговой слой надпочечников (Guyton, 1976). Стимуляция этого
слоя приводит к выделению адреналина и норадреналина в систему
кровообращения. Физиологические и биохимические изменения,
вызванные активацией нейрогенных и нейроэндокринных осей стресса,
изменения, характерные для "фазы тревоги", подробно описаны в
следующей главе. Схема вовлечения нервных и нейроэндокринных
механизмов в стрессовую реакцию представлена на рис. 17
Хотя действие катехоламинов мозгового слоя надпочечников
гораздо продолжительнее влияния "осей" автономной нервной
системы, существует еще более длительная фаза стрессовой реакции.
Активация эндокринной оси обеспечивает участие таких
механизмов, которые значительно увеличивают время стрессовой реакции.
11.3. ЭНДОКРИННЫЕ МЕХАНИЗМЫ
Наиболее длительные соматические реакции на стрессоры
являются результатом активации эндокринных механизмов.
Существуют три основных эндокринных оси, с которыми связана
стресс-реакция человека: арено-кортикальная, соматотропная и тиреотропная.
Эти оси активируются в результате более интенсивной стимуляции
и, естественно, определяют самые продолжительные фазы
стрессовой реакции (Lev/, 1972).
Адрено-кортикальная ось. Возбуждающий стимул, поступая в
срединный бугор гипоталамуса, активирует его нейросекреторные
клетки, которые выделяют кортикотропин - регулирующий гормон
(КРГ) в гипоталамо-гипофизарную воротную систему. КРГ проходит
через область воронки к клеткам передней доли гипофиза, его хемо-
фобные клетки чувствительны к присутствию КРГ и под его
влиянием выделяют адренокортикотропный гормон (АКТГ) в систему
кровообращения, через которую он и поступает в кору надпочечников.
АКТГ стимулирует клетки ретикулярного и трубчатого слоев коры к
выделению глюкокортикоидов - кортизола и кортикостерона - в
систему кровообращения. Аналогично АКТГ вызывает выделение
минералокортикоидов - алъдостерона и деоксикортикостерона -
гломерулярной зоной коры надпочечников. Эти гормоны регулируют
уровень электролитов и артериального давления, влияя на объем
крови посредством первичной реабсорбции натрия (Guyton, 1976).
Чрезмерная секреция минералокортикоидов вызывает у
человека развитие синдрома Кушинга (гиперальдостеронизм), а также
повышение артериального давления и развитие некроза миокарда
(Selye, 1976).
217
Гипоталамус
Парасимпатическая нервная
система
Гипофиз J
"Ί
Симпатическая
нервная система
Тироидстимулирующий
гормон, АКТГ, гормон роста
через систему
кровообращения
Прямая
иннервация органа
Быстродействующие
эффекты
Эффекты средней
продолжительности
Долговременные_
эффекты
Общий
адаптационный -
синдром
Рис. 18. Временные соотношения между стрессовыми путями (Эверли, Розен
фельд. 1985)
Соматотропная ось. Возбуждающий импульс поступает от ком
плекса перегородка-гиппокамп в гипоталамус и стимулирует
выделение гормон роста-регулирующий гормон (ГР-РГ), который через
воротную гипоталамо-гипофизарную систему поступает в аденогипофиз
и стимулирует выделение гормона роста (соматотропного гормона) в
систему кровообращения. ГР выделяется у человека в ответ на
психосоциальные раздражители и, возможно, как и АКТГ, стимулирует
выделение минералокортикоидов (Selye, 1956; 1976). Предполагается,
что ГР повышает резистентность тканей к инсулину, а также
ускоряет мобилизацию накопленных в организме жиров (Yuwiler, 1976), что
приводит к повышению уровня жирных кислот и глюкозы в крови -
энергоносителей, необходимых для развития стресс-реакции.
218
Тиреотропная ось. Как и две упомянутые выше оси, данная
ось связана с комплексом перегородка-гиппокамп и срединным
бугром гипоталамуса. По тому же пути, что и другие регулятор-
ные гормоны, тиреотропин регулирующий гормон (ТРГ)
поступает в переднюю долю гипофиза и стимулирует там выделение ти-
реотропного гормона (ТТГ), который стимулирует щитовидную
железу к выделению в кровь гормона тироксина. У человека
экстремальные физические воздействия и психосоциальные стимулы
приводят к повышению тиреоидной активности: общего уровня
метаболизма, частоты сердечных сокращений, мощности
сокращений сердечной мышцы, сопротивлению периферических сосудов
(повышение артериального давления), а также чувствительности
некоторых тканей к катехоламинам (Lev/, 1972; Yuwiler, 1976). Все
это показывает, что тиреотропная ось играет значительную роль в
стресс-реакции организма.
Временная последовательность "включения" различных
механизмов ("осей") стресса представлены на рис. 18.
Активация каждой оси может взаимно перекрываться
эффектами других осей. Наиболее часто отмечается активация нейроэндок-
ринной и эндокринной осей. Однако при интенсивном и длительном
воздействии на организм стрессогенного фактора наибольшую
нагрузку по его защите несут эндокринные оси и в особенности адре-
но-кортикальная ось. Г. Селье разработал фундаментальную
концепцию, раскрывающую роль эндокринных осей при хроническом
стрессе. Эта концепция получила название "Общего
адаптационного синдрома - ОАС" (Selye, 1956).
Глава 12
ОБЩИЙ АДАПТАЦИОННЫЙ СИНДРОМ
Стрессоры и стресс. Триада Селье. Согласно положениям,
развитым Г. Селье (Selye, 1936; 1950; 1956), стресс-реакции организма,
возникающие как в физиологических условиях, так и при
разнообразной патологии, отличаются (наряду с конкретной спецификой
данного воздействия) проявлением некоего общего симптомокомп-
лекса, характеризующего состояние напряжения как таковое,
независимо от его природы, и названного Г. Селье "общий
адаптационный синдром" - General adaptation syndrome (OAQ. Иначе говоря,
такие несхожие друг с другом воздействия, как холод и жара, апатия
и переутомление, голод, кровопотери, ожоги, инфекционные
заболевания, прогрессирующий рак, лучевая болезнь, болевые травмы,
страх и радость и т.п., могут при повышенной их интенсивности
давать очень сходную реакцию организма. Все эти разнообразные
воздействия, которые впоследствии были обозначены Г. Селье
термином "стрессоры", вызывали характерные морфологические изме
нения в организме, получившие в научной литературе название
"триада Селье", и сводящиеся к следующему.
1. Гипертрофия коры надпочечников при незначительном
количестве липидных секреторных гранул.
2. Сильная инволюция тимуса, селезенки, лимфатических узлов и
других лимфатических структур.
3. Возникновение глубоких кровоточащих язв в желудке и
двенадцатиперстной кшике.
Тесная взаимосвязь этих трех типов изменений указывала на на
личие определенного синдрома. В зависимости от силы и
длительности неспецифического воздействия стрессоров эти изменения варьи
ровали от незначительных до ярко выраженных. Концептуальное
разграничение специфических и неспецифических последствий дей
ствия любого стрессора было важной ступенью в научном анализе
стресса, который позволил в дальнейшем раскрыть его
физиологические и биохимические механизмы.
«Мы назвали этот синдром "общим", - пишет Селье, - по
тому, что он вызывается лишь теми агентами, которые при-
220
водят к общему состоянию "стресс" (поскольку они
воздействуют на большие участки тела), и в свою очередь вызывают
генерализованное, т.е. системное защитное явление. Мы назвали
его "адаптационным" потому, что он способствует
приобретению состояния привычки и поддерживает это состояние
Мы назвали его "синдромом" потому, что его отдельные
проявления координированы и даже отчасти взаимозависимы» (Се-
лъе, I960).
12.1. ФАЗЫ РАЗВИТИЯ ОАС
Реакция тревоги. Уже в первой своей работе Г. Селье (Selye,
1936) назвал начальную стадию ОАС, во время которой
объявляется экстренная мобилизация защитных сил организма,
"реакцией тревоги" ("alarm reaction'*). В этой стадии происходит
перестройка систем жизнеобеспечения на работу в экстремальных
условиях: активируется система контроля и сохранения гомеостаза,
мобилизуются резервы, повышается работоспособность органов,
тканей, клеток, вовлеченных в стресс-ответ организма на
чрезвычайный раздражитель за счет преимущественной доставки им
энергетических и пластических ресурсов. Параллельно с этим
наблюдается остановка клеточных делений, подавление аппетита,
сексуальных реакций и т.д.
Иногда эта реакция оправданно называется реакцией
"борьбы и бегства" — так она была названа еще У. Кенноном (Cannon,
1932). Фаза тревоги начинается в мозгу, активируя
симпатическую нервную систему, которая стимулирует выделение наиболее
важных катехоламинов - адреналина и норадреналина - из
мозгового вещества надпочечников. Гипоталамус посылает гипофизу
сигнал повысить выделение АКТГ, который заставляет кору
надпочечников выбрасывать кортикостероиды в кровяное русло.
Ниже представлены изменения наиболее важных
физиологических и биохимических параметров организма во время реакции
тревоги.
12.1.1. Реакция тревоги ("борьбы или бегства").
• Симпатическая нервная система активирована; адреналин и норад-
реналин выделяются из мозгового вещества надпочечников,
• Частота и сила сердечных сокращений увеличиваются, чтобы
обеспечить доставку большего количества крови к органам,
которые важны для непосредственного выживания в данный момент,
таких как мозг и мышцы; кровоснабжение кожи и внутренних
органов уменьшено, исключая легкие и сердце.
221
• Интенсивность дыхания повышена, чтобы обеспечить большее
количество кислорода для мозга сердца и мышц.
• Уровень глюкозы крови заметно увеличен, поскольку в печени
гликоген (форма хранения глюкозы) расщепляется до глюкозы,
которая переходит в кровообращение.
• Пищеварительная деятельность приостановлена как
несущественная для данного критического момента.
• Мышечный тонус и умственная настороженность увеличены, что
повышает готовность к действию.
• Потоотделение повышено для понижения температуры тела.
• Начата стимуляция коры надпочечников для массированного
выделения кортикостероидов.
Стадия сопротивления. Реакция тревоги обычно относительно
коротка, но произошедшие изменения подготавливают наступление
второй, основной фазы стресса - "стадии сопротивления" (stage of
resistence) (рис. 19). Проявления этой второй фазы отличны и
зачастую даже противоположны проявлениям, характерным для реакции
тревоги. Например, во время "реакции тревоги" клетки коры
надпочечников выделяют секреторные гранулы в кровь, и в надпочечниках
снижается содержание липидов, содержащих кортикостероиды, тогда
как в состоянии устойчивости кора надпочечников особенно богата
секреторными гранулами. Последнее обстоятельство обусловливает
интенсивное выделенные кортикостероидов - главных игроков
реакции сопротивления. Глюкокортикоиды стимулируют преобразование
белков тканей в энергию, повышение уровня глюкозы в крови и
подавление реакций воспаления, функций иммунной системы, деления
клеток и других несущественных в данный критический момент
функций организма. Минералокортикоиды заставляют почки сохранять
натрий, чтобы держать повышенным кровяное давление. В организме
преобладают катаболические процессы, т.е. распад в тканях
преобладает над синтезом; нежировая масса тела постепенно снижается. Если
в стадии тревоги наблюдается гемоконцентрация, гипохлоремия, то в
стадии устойчивости имеют место гемодилюция и гиперхлоремия.
Если действие стрессогенного фактора продолжается, сопротивление
развиваемое организмом, в конечном счете заканчивается.
12.1.2. Реакция сопротивления
• Реакции "борьба и бегства" затухает; организм развивает
сопротивление стрессогенному фактору.
• Уровень кортикостероидов в крови высок, и кора надпочечников
продолжает производить кортикостероиды в значительных количествах.
222
Уровень нормальной
сопротивляемости
Реакция
тревоги
В организме
наблюдаются изменения,
характерные для первого
контакта со стрессором. В
это же время
сопротивляемость снижается, и
если стрессор
достаточно сильный (большие
ожоги), экстремальные
температуры) - может
наступить смерть.
Наблюдается эта стадия
в том случае, если
продолжающееся влияние
стрессора соизмеримо с
адаптацией. Признаки,
характерные для
реакции тревоги, уже
исчезают и сопротивляемость
выше нормы.
После долговременного
воздействия того же
стрессора, к которому
приспосабливается
организм, в конечном счете
адаптационная энергия
исчерпывается. Вновь
появляются признаки
реакции тревоги, но
теперь они необратимы, и
организм умирает.
Рис. 19. Три стадии общего адаптационного синдрома (Селъе, 1977)
• Общий объем крови увеличен из-за задержки натрия и выделения
калия; кровяное давление может быть повышено, и имеет место
избыточная рабочая нагрузка на сердце и кровеносные сосуды.
• Глюкоза крови и жирные кислоты в свободном состоянии
повышены, обеспечивая дополнительное количество энергоносителей для
систем жизнеобеспечения.
• Белки из нежировых тканей, особенно мышц, кожи и костей,
частично разрушаются в пользу энергетических нужд; мобилизация
свободных жирных кислот (СЖК) из жировой ткани повышена -
СЖК служат в качестве альтернативного топлива для
большинства тканей, увеличивая доступность глюкозы для нервной системы
• Печень производит большое количество глюкозы из аминокислот,
полученных в результате деградации нежировых тканей (глюконео
генез).
• Деление клеток, белковый синтез и регенерация тканей
подавлены повсеместно как несущественные в данной ситуации.
• Иммунная система и воспалительные реакции подавлены.
• Повышена потеря кальция из костей.
223
Достаточно интенсивное или длительное действие стрессора
неизбежно ведет к третьей фазе - истощению.
Фаза истощения. В некоторых ситуациях организм может
успешно адаптироваться к стрессогенному фактору без того, чтобы
развивать заметный стресс-ответ. В подобных случаях стрессор
фактически не является стрессором. Например, у человека,
путешествующего по высоким горам, первоначально из-за гипоксии (т.е.
низкого уровеня кислорода) был бы индуцирован стресс-ответ. Тем
не менее в течение нескольких дней такого испытания организм
обычно производит достаточное количество новых эритроцитов для
увеличения объема переносимого с кровью кислорода, и состояние
гипоксии полностью снимается. Стрессогенный фактор таким
образом устранен, и стресс-ответ "угаснет" без серьезных последствий.
Но не всегда, однако, организм способен успешно приспособиться к
стрессогенному фактору и вынужден продолжать стресс-ответ.
Способность организма поддерживать функциональную устойчивость на
высоком уровне - ограничена, и в конечном счете реакция сопротивления
уступает место истощению, т.е. заключительной фазе стресс-ответа.
Жизненно важные функции организма снижаются; сопротивляемость
организма - ниже его изначального уровня; многие важные белки ряда
тканей подверглись распаду; резервы антиоксидантов - исчерпаны;
иммунная система (в особенности тимус) и заживление ран заметно
угнетены; наблюдается снижение массы тела и нежировой ткани. Потеря
калия может стать причиной серьезных нарушений нервной системы и
сердечной функции. Уровень кортикостероидов может быть очень
высок, или, наоборот, кора надпочечников может оказаться слишком
перегружена, чтобы продолжать выработку кортикостероидов -
последний сценарий может вызвать глубокую гипотонию, низкий уровень
глюкозы крови и общий коллапс. Уровень адреналина и норадреналина
в крови, если он становится слишком низким, также может
способствовать состоянию шока. Фаза истощения, вызванная длительным и
интенсивным стрессом, потенциально является фатальной (см. рис. 19).
Греки в 490 г. до н.э. победили персов в жестоком сражении на
Марафонской равнине в юго-восточной Греции. Как гласит легенда, греческий воин Фей-
диппидес совершил 42-километровый пробег по направлению к Афинам, чтобы
принести радостную весть о греческой победе. Прокричав свое известие, Фей-
диппидес впал в коллапс и умер. Это - классический пример смертельного
окончания фазы истощения О АС
Возникает естественный вопрос - почему в наше время профессиональные
спортсмены пробегают марафонскую дистанцию без очевидных проблем?
Нужно указать на одно важное различие. Длительные тренировки делают
бегунов-марафонцев хорошо приспособленными к бегу на длинные дистанции - их
мышцы более эффективно используют кислород и топливо, их сердца имеют
большую мощность и т.д. Для них марафон - стрессогенный фактор средней
силы, который вызывает умеренный стресс-ответ. Для нетренированного
человека, каким, по-видимому, был греческий воии, 42-километровый пробег -
чрезвычайно серьезное напряжение, которое может исчерпать все ресурсы,
вызвать чрезвычайное истощение и даже смерть.
224
12.1.3. Фаза истощения
• Способность сопротивляться стрессогенным факторам - ниже
начального уровня.
• Разрушение жизненно важных тканей.
• Хроническое заболевание или падение функций всех или
некоторых органов или систем тела.
• Гипертрофия и патологические изменения в коре надпочечников.
• Повреждение иммунной системы; инволюция тимуса,
лимфатических узлов и селезенки; разрушение лимфоцитов.
• Истощение запасов антиоксидантов.
• Желудочные язвы.
• Уровень кортикостероидов или очень высок, или - если
надпочечники повреждены - очень низок.
• Уровень адреналина и норадреналина может быть очень низким.
• Гипотония, низкая концентрация глюкозы крови, возможны шок и
смерть.
Обратить внимание: глубокое истощение, описанное здесь,
происходит только после длительного и интенсивного стресса
или умеренного стресса у очень старых людей; умеренный стресс,
обычный в нашей жизни, может также вести к существенному,
но менее явному истощению.
Ганс Селье, изучая состояние стресса на крысах, выявил, как
уже указывалось, наиболее выраженные морфологические
изменения в некоторых органах, которые проявляются в фазе истощения
(триада Селье). Теперь мы знаем, что, по существу, все системы
организма в той или иной степени страдают от стресса. Системы,
непосредственно важные для выживания - нервная система, сердце и
кровеносные сосуды, - страдают от перегрузки, в то время как
другие системы лишаются питательных веществ или даже частично
"демонтируются" для извлечения "топлива". Органы, уже
ослабленные болезнью или старением, особенно уязвимы и являются
первыми кандидатами на "выход из строя". Например, пожилой пациент с
пневмонией имеет намного больше шансов умереть от остановки
сердца, вызванной комбинацией стресса и атеросклероза, чем от
бактериальных токсинов или коллапса легких.
Интенсивная и длительная фаза истощения приводит к
терминальной стадии стресса и может завершиться смертельным исходом
(Кириллов, 1966; Selye, 1974; Селье, 1977; Заводская и др. 1981; Ме-
ерсон, 1984; Flack, 1984; Lanks, 1986).
225
Большое практическое значение трехфазной природы ОАС
состоит в том, что впервые было показано, что адаптационная
способность организма (его адаптационная энергия) не безгранична, а
именно в условиях длительного стресса неизбежно наступает
истощение. Можно было бы предположить, что при наступлении
адаптации и наличии энергетических запасов (нормальное потребление
пищи) устойчивость должна длиться неопределенно долго. Однако это
не так. Подобно тому, как неодушевленные машины постепенно
изнашиваются, так же и машина человеческого организма рано или
поздно становится жертвой непрерывного износа (Селъе, 1977).
Большое приспособительное значение имеет скорость
включения механизмов срочной адаптации к действию стрессоров, ибо от
нее в некоторых ситуациях может зависеть судьба организма.
Опережающий ответ организма обеспечивает максимально быструю,
стереотипную в своих главных чертах перестройку основных
систем, необходимую для выхода организма на повышенный уровень
резистентности к моменту возникновения прямой угрозы жизни
Структура, функция этих систем, взаимосвязь между ними
сформировались и генетически закрепились для полной и быстрой
реализации ответа организма на требования среды, а естественный отбор
обеспечил устранение особей, обладающих недостаточно надежным
и быстродействующим адаптационным аппаратом.
После 1936 г., когда впервые была описана триада Селье,
удалось открыть значительное количество биохимических и
структурных изменений, сопровождающих неспецифический стресс.
Основное внимание тогда обратили на изменения биохимических
показателей организма и его нервные реакции. Значительные успехи были
достигнуты в анализе гормональной медиации стресс-реакций.
Теперь уже совершенно ясно, что экстренный выброс адреналина при
реакциях "борьбы или бегства", открытый Кенноном еще в 1932 г.,
является только одним из аспектов острой фазы "реакции тревоги"
в ответ на действие стрессоров. Система
гипоталамус-гипофиз-кора надпочечников в не меньшей степени важна для сохранения го-
меостаза и осуществления ОАС.
Глава 13
ФИЗИОЛОГИЧЕСКИЕ И БИОХИМИЧЕСКИЕ
МЕХАНИЗМЫ ОАС.
СИСТЕМА ГИПОТАЛАМУС-ГИПОФИЗ-КОРА
НАДПОЧЕЧНИКОВ
Начальной реакцией на действие стрессора является выработка
неспецифического стимула. Это может быть нервный импульс,
химическое вещество или недостаток незаменимого метаболического
фактора; этот фактор принято называть "первичный медиатор\
поскольку природа его до сих пор не вполне ясна. Связано ли его
действие с избытком или отсутствием какого-либо определенного
вещества? Или инициировать стресс-реакцию могут различные
нарушения гомеостаза?
Действие "первичного медиатора" опосредуется через
нервные стимулы, поступающие, как уже указывалось выше, от коры
мозга, лимбической системы (в частности, гиппокампа и
миндалевидного комплекса) и ретикулярной формации. Под влиянием
этих афферентных импульсов повышается функциональная
активность определенных, нейроэндокринных клеток, большинство
из которых расположено в срединном возвышении гипоталамуса.
Эти клетки действуют как "преобразователи",
трансформирующие нервные сигналы в гуморальный передатчик - кортикотро-
пин-регуляторный гормон (КРГ). Поступая в гипофиз через
расположенную под гипоталамусом местную портальную систему
кровообращения, КРГ стимулирует синтез и секрецию адрено-
кортикотропного гормона (АКТГХ который совместно с нейро-
медиатором ацетилхолином активирует в мозговом веществе и
коре надпочечников синтез и секрецию катехоламинов и корти-
костероидных гормонов соответственно. Эти гормоны в свою
очередь вызывают существенный сдвиг в ряде физиологических
и биохимических процессов.
Общая схема путей ответа нейроэндокринной системы высших
животных на действие стрессора (по Г. Селье) представлена на
рис. 20.
Ввиду исключительной важности системы
гипоталамус-гипофиз-кора надпочечников (ГТКН) в стресс-реакциях организма
следует подробней остановиться на ее основных звеньях.
227
Агент
Гипоталамус
Кровяное
давление
Обмен веществ
ЦНС
Пептические
язвы
Глюконеогенез
Тимус
Лимфатические узлы
Клетки крови
Иммунные реакции
Воспаление
Другие синтаксические реакции
Рис. 20. Основные пути реализации ответа на стрессорный агент (Селъе, 1977)
228
13.1. ГИПОТАЛАМУС
13.1.1. Кортикотропин - регуляторный гормон
Многочисленные физиологические исследования позволили
предположить, что под действием различных факторов внешней
среды и эмоциональных стимулов происходит повышение секреции
гипоталамического вещества, стимулирующего выброс из гипофиза
АКТТ. Гипоталамическое вещество, ответственное за эту
активность, получило название кортикотропин-рилизинг-фактора, а в
дальнейшем - кортикотропин-регуляторного гормона (КРГ)
Юс/шЛУ etal., 1973).
КРГ был выделен из препаратов нейрогипофиза, который считался
хранилищем КРГ, подобно тому, как он служит хранилищем других гормонов
гипоталамического происхождения - вазопрессина и окситоцина Было
выделено два гипоталамических пептида - ар, о^-КРГ и β-ΚΡΓ. Только β-ΚΡΓ
получен в очищенном виде, и частично установлена его первичная
структура (Guillemint\97l):
| S S 1
ацетил - Ser-Tyr-Cy-Phe-His-CAsp-NH^ Glu-NH2)-Cy-(Pro, Val)-Lys-Gly-NH2
Опираясь на совокупность данных о механизме действия не
только КРГ, но и других гипоталамических регуляторных гормо
нов, можно представить следующую схематическую картину их
действия на тропные функции гипофиза, как, очевидно, и на
клетки других органов, имеющих рецепторы и для других гормонов
с полипептидной структурой (рис. 21). Гормон избирательно
взаимодействует с элементами рецептора, локализованного на
поверхности плазматических мембран клеток аденогипофиза, в
результате чего происходят конформационные изменения в гор-
мон-рецепторном комплексе, приводящие в дальнейшем к
изменению активности некоторых ферментативных систем
мембраны, в том числе и аденилатциклазы. Эта активация может
вызвать диссоциацию связанных с мембранами комплексов
Са2+-АТФ и Mg2+-ATO, что приводит к освобождению Са2+, АТФ
и образованию цАМФ, который служит посредником в
дальнейшей передаче гормонального сигнала, направленного, по-видимо
му, одновременно на освобождение и синтез аденогипофизарного
гормона, действуя путем активации цАМФ-зависимой протеин
киназы. Изменение проницаемости плазматических мембран под
влиянием регуляторного гормона способствует также и проник
новению в клетку экзогенного Са2+, что необходимо для секреции
аденогипофизарного гормона (Юдаев и др., 1976).
229
Внешняя среда
Плазматическая
мембрана
Гормон
1 ι
^1 [г^
Рецептор
Аденилатциклазный
комплекс
Внешняя среда
Плазматическая
мембрана
Внутриклеточная среда
5-АМФ 3,5-цАМФ
Θ
Фосфодиэстераза
Внутриклеточная среда
Неактивная
протеинкиназа
Активная
протеинкиназа
АГФ
Дефосфопротеин
Фосфопротеинфосфатаза
Биологический эффект
Рис. 21. Общая модель опосредуемых цАМФ реакций на гормоны (Tepperman J.,
Террегтап //.,1987)
230
13.2. ГИПОФИЗ
13.2.1. Адренокортикотропный гормон гипофиза
АКТГ стимулирует рост коры надпочечников, а также синтез и
секрецию кортикостероидов. У гипофизэктомированных животных
гормон восстанавливает до нормального размера атрофированную
кору надпочечников (и ее активность) и предупреждает развитие
атрофии, если вводится сразу после операции. В гипофизе человека
содержится примерно 250 мкг АКТГ. Скорость его секреции
составляет 5-25 мкг в сутки, но в условиях стресса (например, при
хирургической травме) может возрастать во много раз. Если в норме
наибольшая концентрация АКТГ составляет 50-80 пг/мл, то при
тяжелом стрессе достигает 200-1000 и более пг/мл плазмы крови
(Daughaday, 1985). При одновременном определении АКТГ и корти-
зола в крови можно убедиться, что концентрация гормона,
синтезируемого органом-мишенью, изменяется в строгой зависимости от
изменений концентрации тропного гормона.
Структура АКТГ человека приведена ниже (L/, 1977):
NH2-Ser-Tyr-Met-<jlu-His-Phe-Arg-Trp-Gly-Lys-Lys-Pro-Val-
12 3 4 5 6 7 8 9 10 11 12 13
Gly-Lys-Lys-Arg-Ar-Pro-Val-Lys-Val-Tyr-Pro-Asp-Gly-Ala-
14 15 16 17 18 19 20 21 22 23 24 25 26 27
Glu-Asp-Glu-Ser-Ala-Glu-Ala-Phe-Pro-Leu-Glu-Phe-COOH
28 29 30 31 32 33 34 35 36 37 38 39
В структурах АКТГ нескольких видов животных первые 24
остатка аминокислот идентичны. Синтетический полипептид,
содержащий первые 24 остатка АКТГ, in vivo обладает сходной с натив-
ным гормоном биологической активностью. Следовательно,
аминокислотные остатки с 25 по 39 не существенны для гормонального
действия, хотя и определяют видовую иммунологическую
специфичность гормона (Li, 1977; Панков, 1976)
Фрагмент АКТГ, содержащий остатки с 1 по 14, обладает
весьма значительной активностью меланоцитстимулирующего
гормона (МСГ) (Li, 1977).
По-видимому, АКТГ не имеет четко выраженной третичной
структуры, так как использование методов дисперсии оптического
вращения, кругового дихроизма, инфракрасной спектроскопии не
обнаруживают в АКТГ значительного количества α-спиралей или
β-структур (L/, 1977). Можно полагать, что некоторые элементы
третичной структуры этого полипептида не играют существенной
роли в проявлении его биологического эффекта.
Синтез АКТГ имеет матричную природу, т.е.
последовательность аминокислотных остатков в этом полипептидном гормоне
закодирована в определенном гене и его транскрипте - соответст-
231
вующей информационной РНК. Однако в этом гене закодирована
не только последовательность АКТГ, но и последовательность
р-липотропина и ряда других пептидных гормонов в едином
предшественнике. Этот прогормон (с массой 29 КД) был назван про-
опиокортионом, так как он служит предшественником β-липотро-
пина, кортикотропина и опиатных гормонов {эндорфина и энке-
фалина), которые связываются с опиатными рецепторами мозга.
Из про-опиокортина образуются также два меланоцитстимулиру-
ющих гормона: α-МСГ является фрагментом АСТН, а β-МСГ
формируется из β-липотропина. Не исключено, что N-концевая
половина про-опиокортина является источником и других гормонов
(Guillemin, 1977; 1978; Nakanishi, 1979). Этот прогормон состоит из
четырех гомологичных областей, которые, по-видимому, возникли
путем последовательных удвоений гена. Участки соединения
между будущими активными гормонами в про-опиокортине содержат
пары основных аминокислотных остатков (Lys-Arg, Arg-Arg или
Lys-Lys). Очевидно, именно эти пары основных аминокислот в
различных прогормонах (например, также и в проинсулине {Czech,
1977), указывают, в каких участках должно произойти
последующее протеолитическое расщепление.
Окончательно вопрос о структуре про-опиокортина был
выяснен Наканиши и другими {Nakanishi, 1979) после определения нук-
леотидной последовательности клонированной сДНК, полученной
путем обратной траскрипции с иРНК, кодирующей синтез
предшественника кортикотропина - β-липотропина быка. Расшифровка
последовательности этой иРНК, содержащей 1091 основание,
позволила установить взаимное расположение упомянутых пентидных
гормонов и характер их процессинга в результате пептидазного
расщепления {рис. 22).
Однако значительно раньше, когда вопрос о механизмах
синтеза крупных полипептидов оставался еще открытым, матричная
природа биосинтеза АКТГ была установлена одним из авторов этой
книги и его сотрудниками {Тодоров и др.. 1964: 1965; 1966).
В первоначальных исследованиях было показано, что инъекции препарата
высокополимерной РНК из бычьих аденогипофизов крысам вызывают
значительное повышение содержания АКТГ в гипофизах этих лабораторных
животных. Мы предположили, что это повышение является результатом
специфического влияния энзогенной гипофизарной РНК иа аппараты синтеза АКТТ, а не
следствием развития, например, стресс-реакции, так как специальные контроли
(концентрация аскорбиновой кислоты в надпочечниках) показали отсутствие
этой реакции на введение препаратов РНК животным {Тодоров, Блок, 1964).
Эти данные послужили достаточным основанием для проведения прямых
экспериментов по выяснению механизмов синтеза АКТГ. С этой целью
бесклеточные системы биосинтеза белков из кишечной палочки {Eshericnia Coli В)
были инкубированы совместно с гетерогенной ядерной РНК, выделенной из
бычьих аденогипофизов. После соответствующей очистки инкубационной смеси на
оксицеллюлозе определялась АКТГ-активность синтезированного продукта по
232
Проопиомеланокортин (ПОМК)
239
Προ-γ-МГС-АКТГ
β-ЛПГ
144 1
Προ-γ-МГС
γ-МСГ
1 12
АКТГ
γ-ЛПГ
91
β-эндорфин
58 1
31
103 1 39 1
АКТГ-подобный пептид
сс-МСГ Гфомежуючнойжши β-МСГ Эндорфин
гипофиза г —
1 13
22
1 18 1 16
Рис. 22. Пептиды, полученные из проопиомеланокортина (ПОМК) (Nakanishi, et
al., 1979)
уровню суммарной секреции кортиностероидов надпочечниками
тест-животных in vitro.
Наблюдалась значительная стимуляция синтеза АКТГ-подобной
субстанции, и этот эффект был присущ именно аденогипофиз арной иРНК, поскольку
РНК из другого вида животных (крыса) и из другого органа (печень) не
вызывала в системе появления АКТГ-активности. Отсутствие стимулирующего
действия энзиматического гидролизата иРНК свидетельствовало, что синтез
АКТГ-подобной субстанции индуцирован высокополимерной аденогипофизар-
ной РНК (Тодоров и др., 1966). Кроме того, синтезированный в бактериальной
бесклеточной системе продукт обладал явно выраженной меланоцитп-стимули-
рующей активностью, вызывая у зеленых лягушек, адаптированных к свету,
заметное потемнение кожи {Тодоров и др., 1967), что было вполне естественным,
если учесть общность первичной структуры β-МСГ и начального участка (1-14)
в молекуле АКТГ (см. рис. 22).
Электрофоретическая подвижность и хроматографический профиль
синтезированного АКТГ-подобного фактора совпадали с указанными
характеристиками природного гормона (Todorov, 1968; Тодоров, 1971). При
количественном анализе включения различных [14С] - аминокислот в синтезированный
продукт была выявлена полная корреляция с аминокислотным составом
природного АКТГ (Тодоров, 1968). Использование различных контролей
(добавление в бесклеточную систему антибиотиков, ингибирующих трансляцию у
бактерий - пуромицина и хлорамфеникола, обработка гетерогенной ядерной РНК
из аденогипофизов РН-азою, исключение из системы рибосом и АТФ-генериру-
ющей системы) позволило сделать вывод о матричной природе синтеза АКТГ,
аналогичный синтезу большинства специфических белков {Тодоров и др., 1965;
1966; 1967; Todorov, 1968). Матричный механизм синтеза АКТГ несколько
позже нас также был показан при исследовании в системе, содержащей срезы
ткани бычьих аденогипофизов (Adiga et al., 1966).
Препарат суммарной гетерогенной ДНК-подобной аденогипофизарной
РНК, который использовался в описываемых выше исследованиях (Тодоров и
др., 1969; Лившиц и др., 1969), очевидно, содержал иРНК, кодирующую
предшественник ΑΚΤΓ-β-липотропина. Продукт трансляции этой иРНК в
бактериальной бесклеточиой системе, по-видимому, подвергался частичному
расщеплению бактериальными пептидазами по парам основных аминокислот (Lys-Arg;
233
Arg-Arg ; Lys-Lys), что и приводило к образованию активного АКТГ-подобно-
го вещества в этих экспериментах.
Помимо принципиального вопроса о матричной природе
биосинтеза АКТГ, наши исследования, впервые показавшие
возможность синтеза биологически активного полипептида высших
животных белок-синтезирующей системой бактерий, четко ceudi
тельствовали не только об эволюционной универсальности
молекулярных механизмов биосинтеза белков, но также и о
тождественности генетического кода у низших и высших организмов.
Дальнейшие исследования механизмов синтеза АКТГ, его регуляции и
секреции, проводились нами путем инкубации срезов аденогипофизарной ткани с
радиоактивными предшественниками синтеза белков и РНК в различных
условиях по ранее разработанной методике (Adiga et al., 1965). В частности, было
показано, что в присутствии актиномицина Д (антибиотика, ингибирующего тра-
скрипцию) скорость биосинтеза АКТГ в срезах гипофизов была постоянной, по
крайней мере в течение 10 часов, в то время, как биосинтез суммарных белков
и РНК аденогипофиза ингибировался за этот период на 40 и 80%
соответственно (Тодоров и др., 1969). Эти результаты, очевидно, указывают на высокую
стабильность и РНК, кодирующей синтез иРНК АКТГ (не менее 10 часов).
Субклеточная локализация синтеза, депонирование и инкре-
ция АКТГ также изучались с помощью техники срезов
аденогипофизарной ткани с последующим выделением отдельных
субклеточных компонентов и определения в них как удельной
радиоактивности, так и биологической активности очищенного АКТГ (Тодо-
ров, 1971). Полученные данные дают достаточно полную картину
внутриклеточного перераспределения вновь синтезированного
гормона, его депонирования, взаимодействия с ядерными структура
ми и т.п.
Центрами биосинтеза АКТГ являются микросомы (частицы, содержащие
рибосомы и фрагменты мембран эндоплазматического ретикулума - ЭР). Это
подтверждается как высокой удельной радиоактивностью (имп/мин/мг) выде
ленного из них (,4С]-АКТТ, так и высокой концентрацией кортикотропной ак
тивности в этих структурах.
Указанные показатели достигают максимума уже через три-шесть часов и
остаются на том же уровне до конца периода инкубации срезов. Это указывает
на то, что синтезированный de novo АКТГ не задерживается в структурах ЭР и
переходит во внутриклеточную среду. При постоянной скорости синтеза гормо
на в этих условиях наблюдается интенсивное накопление [14С]-АКТТ в клеточ
ном соке. Высокие величины удельной радиоактивности АКТГ показываю!
что в течение первых шести часов инкубации в клеточном соке
концентрируется только АКТГ, синтезированный de novo, далее он поступает во внутрикле
точные депо, о чем свидетельствует снижение его радиоактивности во второй
половине периода инкубации. Параллельно (через 6-12 часов) наблюдается
повышение концентрации АКТГ в клеточных гранулах, которые седиментируют
при 4000-6000 g ("тяжелая" фракция). "Легкая" фракция (18000 g) имеет
незначительное количество [14С]-АКТТ, определяемого также и по биологической
активности. Фракция "средник" субклеточных гранул (12000 g) по этим
показателям занимает промежуточное положение между "тяжелой" и "легкой" фрак-
234
циями. Таким образом, накопление синтезированного АКТГ происходит и < с
новном в "тяжелой" фракции субклеточных частиц, которые, по-видимому, и
являются местом внутриклеточного депонирования АКТГ (Тодоров, 1971). Η
настоящее время известно, что все гипофизарные клетки высвобождают
белковые или гликопротеиновые гормоны из аналогичных субклеточных гранул, ко
торые в ответ на соответствующий сигнал (с участием кальция и цАМФ)
выбрасывают свое содержимое в систему кровообращения путем экзоцитпоза
(Tepperman et al., 1987).
Несколько позже с помощью радиоиммунологического теста было
показано накопление АКТГ и β-липотропина в запасающих гранулах кортикотрофов
аденогипофиза и клеток интермедиальной доли гипофиза (Pelletier et al., 1978).
В ядрах клеток аденогипофиза в течение всего периода инкубации имеет
место повышение концентрации АКТГ, которое в основном повторяет
кинетику изменений этого показателя в клеточном соке, хотя исходная концентрация
АКТГ в ядрах значительно ниже, чем в микросомах и в "тяжелых" гранулах.
Повышение [14С]-АКТГ в ядрах может быть интерпретировано в качестве
проявления обратной связи между трансляцией и транскрипцией в процессе
биосинтеза АКТГ (Тодоров, 1971).
Регуляция кортикотропной функции. Если возможность
угнетения эндогенной секреции КРГ избытком его концентрации
представляется сомнительной, так как период его пребывания в
циркулирующей крови очень мал, то в регуляции выработки и секреции
АКТГ наблюдается более определенная ситуация (Селье, 1977).
Имеются четкие доказательства, что АКТГ участвует в механизме
обратной связи (короткая петля обратной связи показана на
рис. 20) - избыток этого гормона, воздействуя на гипоталамно-гипо-
физорную систему, тормозит дальнейшую стимуляцию секреции
АКТГ. С помощью радиоиммунологического теста на АКТГ была
изучена локализация клеток и волокон, содержащих гормон, в
некоторых участках мозга крысы. В частности, скопление АКТГ
обнаруживается в аркуатном ядре и его окрестностях, остальная часть
гипоталамуса и другие области мозга содержали умеренное
количество АКТГ (Akil et al., 1978).
Существует еще больше доказательств {Селье, 1977)
существования механизма обратной связи, осуществляемой кортикостероида-
ми (длинная петля обратной связи представлена на рис. 20).
Повышенный уровень кортикостероидов тормозит секрецию АКТГ. По-
видимому, мишенью такого действия кортикостероидов служит аде-
ногипофиз, и давно предполагалось (Selye, 1948), что секреция
АКТГ тормозится вследствие ингибирования не только его
секреции, но и биосинтеза. Однако экспериментальные данные в пользу
существования такого механизма были получены И.Н. Тодоровым и
сотрудниками только в 1968 г.
При инкубации срезов аденогипофиэиз арной ткани в присутствии
кортизона и [14С]-аминокислот наблюдалось значительное подавление включения
метки в АКТГ и другие белки этой ткани. При этом влияние кортизона на
биосинтез АКТГ было выражено в значительно большей мере, чем на биосинтез
суммарных белков. Были отмечены также значительные различия и в кинетике
235
действия кортизона на синтез АКТГ и других белков (Тодоров и др., 1968)
Оставалось, однако, неясным, иа каком уровне - трансляции или транскрип
ции - происходит ингибирующее влияние кортизола. Для выяснения этой
альтернативы изучалось влияние кортизона на синтез различных типов РНК в
срезах аденогипофизарной ткани. Кортизон резко тормозит биосинтез как рибо-
сомиой РНК (рРНК), так и суммарной иРНК, однако включение [14С]-урацила
в суммарную иРНК (с константой седиментации 10-12 S) тормозится в большей
степени (Тодоров и др., 1968а). Можно предположить, таким образом, что
кортизон подавляет экспрессию гена, кодирующего структуру предшественника
кортикотропина β-липотропина (Nakanishi et al., 1979), а биосинтез рРНК может
тормозиться в результате взаимодействия транскриптов генов предшественника
кортикотропина β-липотропина с промоторами гена рРНК по механизму,
предложенному нами позднее (Тодоров, 1986; Todorov, 1990; Фонарев и др., 1993).
Механизм действия АКТГ. Стереоспецифичные рецепторы
АКТГ, расположенные на внешней стороне мембраны клеток коры
надпочечников, обладают высоким сродством к гормону.
Максимальный ответ (синтез стероидов) регистрируется при
взаимодействии АКТГ всего с 5% имеющихся рецепторов. Рецепторы белковых
гормонов имеют сложное строение и содержат в своем составе белки
и другие биополимеры. Прямые опыты показывают, что в структуре
АКТГ-рецепторов важную роль играет сиаловая кислота, так как
обработка суспензии клеток коры надпочечников нейраминидазой
приводит к значительному подавлению стимулирующего действия
АКТГ на биогенез кортикостероидов (Панков, 1976).
Процесс связывания АКТГ с рецептором существенно облегчается в
присутствии Са2+ (Cheitlin et al., 1985). Кроме того, комплекс Са^-кальмодулин
участвует в функционировании элементов цитоскелета - микротрубочек и ми-
крофиламентов. Считается также, что Са2+ необходим для трансмембранного
переноса холестерина и т.п. (Tepperman et al., 1987).
Эффекты АКТГ на клетки пучковой зоны коры надпочечников
можно подразделить на три основные группы в зависимости от
времени, необходимого для проявления ответа на АКТГ: 1) острый
эффект длится несколько минут и не связан с синтезом новых иРНК;
2) подострый - несколько часов и связан с синтезом иРНК и
кодируемых ими специфических ферментов, участвующих в стероидоге-
незе; 3) хронический эффект - гипертрофия и гиперплазия железы
(часы, сутки).
Острый эффект. На основе многочисленных экспериментальных
данных Гаррен и другие (Garren et al., 1971) разработали теорию действия АКТГ
на биосинтез кортикостероидов (упрощенная схема приведена на рис. 23).
Согласно этой теории, после связывания АКТГ с рецепторами, сигнал об
этом передается ферменту - аденилциклазе, локализованному на внутренней
поверхности клеточной мембраны. (Существенную роль в передаче сигнала
играют фосфолипиды, поскольку удаление их из мембран блокирует
активацию аденилциклазы, но не предотвращает взаимодействия гормона с
рецепторами.) Аденилциклаза катализирует превращение АТФ в цАМФ, который
поступает далее в цитоплазму. цАМФ связывается с рецепторной
субъединицей протеинкиназы, вызывает диссоциацию комплекса и освобождает ак-
236
3
s
О
I
<
I
тивную протеинкиназу, которая при участии АТФ катализирует фосфорили-
рование белков в рибосомах и стимулирует таким образом биосинтез белка
(в частности, короткоживущего "лабильного белка") на матрице предсуще-
ствующей долгоживущей иРНК цитоплазмы. Протеинкиназа также
активирует путем фосфорилирования фермент эстеразу, который превращает
эфиры холестерина в свободный холестерин в жировых каплях.
Предполагается, что синтезированный лабильный белок облегчает связывание
свободного холестерина с Р-450обц (отщепляющий боковую цепь) и перенос его
из липидных капель в митохондрии (см. рис. 23), где присутствуют все
ферменты, обеспечивающие превращение холестерина в прегненолон
(ключевой предшественник) и последующий биогенез всех кортикостсроидов
(см. рис. 13).
В условиях длительного воздействия АКТГ основным источником
холестерина как предшесгвенника стероидных гормонов служат практически
неограниченные его запасы в крови, где он в виде эфира входит в состав липопротеи-
нов, которые, связываясь со своими специфическими мембранными
рецепторами, проникают внутрь клетки путем эндоцитоза. Далее, пузырьки с липопроте-
ином и рецептором сливаются с лизосомами, образуя фаголизосому, где ката-
болические ферменты расщепляют эфиры холестерина и пополняют запас
свободного холестерина (Tepperman et al., 1987).
Протеинкиназа активирует в надпочечниках также киназу фосфорилазы
В, которая путем фосфорилирования переводит неактивную фосфорилазу В в
активную фосфорилазу А, которая расщепляет гликоген до глюкозо-1-фосфа-
та. Последний, обмениваясь через пентозный цикл, увеличивает образование в
надпочечниках восстановленного НАДФН, необходимого в качестве
кофактора на большинстве стадий превращения холестерина в кортикостероиды (Smith
et al. 1983). Таким образом, все три эффекта цАМФ и протеинкиназы в
надпочечнике - активирование эстеразы, фосфорилирование белков рибосом и
активирование киназы фосфорилазы В - оказываются направленными на
стимуляцию процесса биосинтеза кортикостероидов.
К этому следует добавить, что, как приведенные выше, так и
последующие исследования цАМФ (Sutherland, 1972; Haynes et al., 1975) послужили
хорошей основой для гипотезы Сазерленда о роли этого циклического нуклео-
тида как вторичного посредника в механизме действия гормонов
(см. рис. 21).
Подострый и хронический эффекты. Влияние АКТГ на ферменты сте-
роидогенеза определяется стимуляцией синтеза белков, сопряженной с
активацией РНК-полимеразы II и синтезом дополнительного количества иРНК, а
в течение последующих 3-24 часов повышается активность РНК-полимеразы
I и синтез рибосомальных РНК. Одновременно повышается концентрация
энзимов, обеспечивающих энергией метаболизм надпочечников, и
стимулируется активность холестеринэстеразы, снабжающей митохондрии свободным
холестерином.
Все эти процессы являются характерными признаками роста клеток, что и
приводит к увеличению их размеров. Через 24-48 часов начинается
гиперплазия надпочечников: повышается синтез ДНК, увеличивается число клеток,
изменяется их ультраструктура, количество РНК и белка продолжает возрастать,
увеличивается активность ферментов стероидогенеза (Юдаев, 1976). Таким
образом, длительное воздействие АКТГ на кору надпочечников при стрессе,
резко стимулируя всю систему биогенеза кортикостероидов и обслуживающих ее
систем, т.е. вызывая гипертрофию клеточных функций, индуцирует вступление
последних в клеточный цикл и пролиферацию, что в результате существенно
238
увеличивает функциональные возможности коры надпочечников в фазе
сопротивления стресс-реакции.
Перечисленные выше эффекты обусловлены не одним АКТГ, а
совместно с действием некоторых митогенных ростовых факторов. Однако для
этого клетки должны приобрести вначале резистентность к уже упоминан
шимся ранее антирепликационным эффектам АКТГ или другим агентам,
стимулирующим продукцию цАМФ. Кроме того, в течение фазы
сопротивления стресс-реакции увеличивается васкуляризация надпочечников в
результате пролиферации кровеносных сосудов. Повышение кровотока при
этом обеспечивает клетки большим количеством питательных веществ,
Са2+ и гормонов (Tepperman et al., 1987).
Известно, что во время стресса у человека и животных временно
притупляется чувство боли, что вполне понятно в контексте общей концепции
стресса - максимальной нервной и психической сосредоточенности на
процессе борьбы или спасения в критической ситуации. Исследование этого
эффекта в клинических условиях показало, что введение АКТГ больным
сопровождалось снижением у них болевых ощущений (Kshatr et al., 1997), а у
крыс этот гормон вызывал быстрое (с 3-й мин) и продолжительное (с 30-й
мин) увеличение порога болевой реакции. Оказалось, что действие АКТГ
имеет две фазы: первая не связана, а вторая (длительная) связана с глюко-
кортикоидами {Богданов и др., 2000).
13.3. НАДПОЧЕЧНИКИ
Надпочечники - "менеджеры передовой линии" стресс-ответа.
Они выделяют ключевые гормоны стресса, которые ответственны
за большинство физиологических изменений, наблюдаемых в
течение стресса. Надпочечники локализованы немного выше почек, и
состоят из двух различных частей: мозгового вещества и коры
надпочечников.
13.3.1. Мозговой слой
Мозговой слой (сердцевина) - внутренняя часть надпочечников -
выделяет два гормона стресса: адреналин (эпинефрин) и норадрена-
лин (норэпинефрин). Эти два гормона наряду с симпатической
нервной системой - ключевые игроки в активации фазы тревоги стресс-
ответа. Адреналин и норадреналин вызывают многочисленные
физиологические эффекты, направленные главным образом на
подготовку организма для немедленного действия, что включает учащение
и усиление сердечных сокращений, ускорение дыхания, повышение
уровня глюкозы в крови, мышечного тонуса, внимания и
настороженности. У некоторых людей выделение адреналина в начальной фазе
стресс-ответа вызывает умеренную эйфорию и воспринимается как
приятное состояние. Эти любители адреналиновой "атаки", т.е.
"острых ощущений", фактически демонстрируют своеобразную форму
наркотической зависимости от фазы тревоги стресс-ответа
239
13.3.2. Катехоламины
Наиболее быстрый путь реализации стресс-реакции, по крайней
мере ее фазы тревоги, обеспечивают катехоламины: адреналин и
норадреналин. Оба катехоламина образуются в мозговом слое
надпочечников под влиянием симпатической нервной системы и, по
существу, являются ее функциональным продолжением. Катехоламины
образуются также в мозгу и нервной системе, где они функционируют
в качестве нейромедиаторов.
Адреналин накапливается в клетках мозгового слоя
надпочечников в особых хромаффинных гранулах. Под действием стрессо-
генных нервных импульсов, достигающих мозгового слоя
надпочечников, из этих гранул выделяется путем экзоцитоза адреналин,
который попадает в окружающую внеклеточную среду и далее в
кровь. Хотя обычная концентрация адреналина в крови составляет
около 0.06 мкг/л, в фазе тревоги, когда животное испытывает боль
либо готовится к борьбе или бегству, концентрация адреналина
возрастает в течение нескольких секунд или минут почти в 1000 раз!
При этом ускоряется ритм сердца, увеличивается сердечный выброс
и повышается кровяное давление, т.е. в этой экстремальной
ситуации подготавливается повышенная активность сердечно-сосудистой
системы. Наряду с этими эффектами, адреналин стимулирует
расщепление гликогена печени и увеличивает содержание глюкозы в
крови, создавая наряду с глюкокортикоидами энергетические
предпосылки развития стресс-реакции (механизм действия адреналина
на клетки печени и другие метаболические эффекты мы
рассмотрим ниже).
Структура и синтез. Адреналин и иорадреналин весьма близки
по структуре (рис. 24), хотя норадреналин является не гормоном, а ней-
ромедиатором, который освобождается в окончаниях симпатических
нервов и действует на постсинаптические клетки. Дофамин - третий
катехоламин, ои также служит медиатором в нервых путях мозга
Дофамин - промежуточный продукт биосинтеза норадреналина и адре
налина в мозговом веществе надпочечников и в нервных клетках
(рис. 25). Предшественником синтеза катехиламинов является тирозин.
На первой стадии эта аминокислота окисляется до 3.4-диоксифенилалани-
на (ДО ФА) с помощью фермента тирозингидроксилазы. Следующая стадия
катализируется декарбоксилазой ароматических L-аминокислот (ДОФА-декарбо-
ксилаза), которая декарбоксилирует ДОФА в 3.4-диоксифенилэтиламин
(дофамин). Дофамин превращается в норадреналин дофамин-Р-гидроксилазой.
Завершающей стадией при образовании адреналина является метилирование
S-аденозил-метионином, катализируемое фенилэтаноламин-Ы-метилтрансфе-
разой (см. рис. 25) (Molinojfei al., 1971). У млекопитающих в период раннего
эмбрионального развития эта трансфераза отсутствует. Концентрация ее
повышается при введении одного из кортикостероидов, следовательно, система ги-
поталамус-адеиогипофиз-кора надпочечников участвует в физиологической
регуляции синтеза катехоламинов в мозговом веществе надпочечников.
240
но но
Ho/^>~CH-CH2-NH Но/ VcH-CH2-NH2
он сн3 он
адреналин норадреналин
Рис. 24. Структура основных катехоламинов (White et al., 1978)
Внутриклеточная локализация синтеза и инкреция
адреналина. При синтезе адреналина гидроксилирование тирозина и декарбо-
ксилирование ДОФА в дофамин происходит в цитоплазме клеток
мозгового слоя; затем дофамин поступает в гранулы, где он
превращается в норадреналин, далее большая часть норадреналина
покидает эти гранулы и метилируется в цитоплазме в адреналин.
Последний поступает затем в другую группу гранул, где сохраняется до
освобождения. Сигналом к освобождению содержимого гранул
является поступление из преганглионарных волокон ацетилхолина и
взаимодействие последнего с рецепторами хромаффинных клеток,
приводящее их к локальной деполяризации. Последующее вхождение
Са2+ в эти клетки промотирует выталкивание (путем экзоцитоза)
содержимого гранул, а именно адреналина, АТФ и дофамин-р-гидро-
ксилазы во внеклеточную жидкость и далее в систему циркуляции
Мозговое вещество надпочечников содержит актин, миозин и тро-
понин С, которые вместе с микротрубочками обеспечивают
выталкивание содержимого гранул.
Механизм действия адреналина. Основной биохимический
эффект действия адреналина - ускорение распада гликогена и выхода
глюкозы в среду - был открыт Э. Сазерлендом более сорока лет
назад. Его последующие исследования показали, что адреналин
активирует гликоген-фосфорилазу, которая катализирует процесс
расщепления гликогена до глюкозо-1-фосфата (Sutherlandt1912). На
первой стадии, требующей присутствия АТФ и ионов Mg2+,
адреналин связывается с рецепторными участками (адренорецепторами)
на поверхности клетки. Это способствует образованию в клетке
цАМФ (вторичного посредника) благодаря "включению" аденилат-
циклазы, которая связана с внутренней поверхностью клеточной
мембраны. Концентрация цАТФ в цитозоле быстро достигает
максимума, равного Ю-6 М. цАТФ в свою очередь связывается с регуля-
торными субъединицами протеинкиназы, что приводит к
высвобождению активных каталитических субъединиц протеинкиназы,
которые посредством АТФ катализируют фосфорилирование
неактивной дефосфорилированной формы киназы фосфорилазы с
образованием активной фосфорилированной формы этого фермента.
Активная киназа фосфорилазы, для функционирования которой
241
χ—υ-
χ—i
χ
ο
Χ—Ο—χ
-υ—χ
ш
\ζ/
χ—υ—χ
необходимы ионы Са2+, фосфорилирует неактивную фосфорилазу Ъ
с помощью АТФ, что приводит к образованию активной фосфори-
лазы а. Последняя, в свою очередь, с большой скоростью
расщепляет гликоген с образованием глюкозо-1 -фосфата, который
превращается далее в глюкозо-6-фосфат и затем в свободную глюкозу,
поступающую в кровь (рис. 26). Несмотря на большое число стадий в
описанном процессе, активность гликоген-фосфорилазы достигает
пика уже через несколько минут после связывания адреналина
рецепторами клеток печени.
Стадии процесса (см. рис. 26) можно рассматривать как каскад
действия одних ферментов на другие, при котором один фермент
активирует множество молекул следующего фермента. Так
достигается большое и быстрое усиление поступающего сигнала (в данном
случае от комплекса адреналин-адренорецептор); по некоторым
оценкам, это усиление составляет примерно 25 млн раз! В
результате, связывание лишь нескольких молекул адреналина адренорецеп-
торами печени приводит к быстрому выбросу в кровь нескольких
граммов глюкозы (Sutherland, 1972; Leheninger, 1982).
В мышцах нет глюкозо-6-фосфатазы, а повышение концентрации глкжо-
зо-6-фосфата здесь приводит не к образованию свободной глюкозы, а к
значительному увеличению скорости гликолиза, в ходе которого вырабатывается
доступная для использования при мышечном сокращении АТФ и молочная
кислота для последующих этапов энергетического метаболизма.
Дальнейшие исследования показали, что адреналин стимулирует распад
гликогена в печени еще через один каскад усиления, параллельный тому,
который описан выше. В этом втором каскадном процессе, который в определенных
условиях оказывается преобладающим, роль вторичного внутриклеточного
посредника играют ионы Са2+.
Норадреналин оказывает сравнительно небольшое влияние на углеводный
метаболизм (White et al.t 1978; Lleheninger, 1982).
Адреналин не только стимулирует распад гликогена, но одновременно
тормозит его синтез из глюкозы в печени, что способствует главному эффекту —
поступлению глюкозы в кровь. Это достигается путем фосфорилирования гли-
коген-синтетазы, которая в фосфорилированной форме является неактивной.
Следовательно, цепь реакций, которая приводит к уменьшению гликоген-син
тезарной активности, имеет тот же пусковой механизм, что и распад гликогена
с образованием свободной глюкозы в крови.
"Аденилатциклазные" эффекты адреналина в различных
органах и тканях связаны и с целым рядом других изменений в
организме. Так, катехоламины обладают выраженной липидмобилизующей
активностью, увеличивая в результате стимуляции липолиза
освобождение из жировой ткани свободных жирных кислот, они повышают
в крови уровень неэтерифицированных жирных кислот.
Одновременно наблюдается увеличение потребления 02. Введение
адреналина нормальным животным повышает также уровень холестерина
и фосфоглицеридов в сыворотке крови и стимулирует обновление
фосфоглицеридов в ткани сердца.
243
Внешний импульс
I
Центральная
нервная система
Мозговое вещество
надпочечников
Адреналин
| через
Рецептор ι
адреналина ν ♦кровь у Клеточная мембрана
Глюкоза крови
Рис. 26. Адреналин включает каскад усиления в клетках печени. Связывание
нескольких молекул адреналина со специфическими рецепторами на поверхности
клетки инициирует последовательность ферментативных реакций, в результате
которых в кровь выбрасывается очень большое количество глюкозы
(Sutherland, 1972)
244
Существенными результатами действия адреналина являются
значительное повышение кровяного давления, обусловленное
сужением артериол, увеличение частоты сердечных сокращений и
минутного объема сердца. В умеренных дозах адреналин
вызывает не сужение, а расщепление сосудов внутренних органов,
увеличивая, следовательно, ток крови в них. Эффекты действия норад-
реналина (за исключением кровяного давления и содержания не-
этерифицированных жирных кислот в крови) выражены слабее
(White et al., 1978).
13.3.3. Кора надпочечников. Кортикостероиды
Кора надпочечников, их внешняя часть, продуцирует различные
группы гормонов стресса, называемых кортикостероидами. Три
главных класса кортикостероидов: глюкокортикоиды, минерало-
кортикоиЪы и половые гормоны (андрогены и эстрогены).
Глюкокортикоиды (наиболее важный из них - кортизол)
являются главными стероидами стресса; они выделяются в больших
количествах в течение стресса и оказывают глубокое влияние на
организм, поддерживая его жизненные функции (деятельность
центральной нервной системы, производство и поставку
энергоносителей за счет несущественных в данный период тканей, например,
мышц или кожи) и подавляя некоторые процессы (например,
клеточные деления, заживление ран и иммунные реакции).
Массированное выделение кортизола в кровообращение в период стресса -
биологический эквивалент законов военного времени - никакая
жертва не может быть слишком большой, если система получает
возможность выжить. К сожалению, иногда система погибает не
потому, что существовал реальный враг, а потому что жертвы были
слишком велики.
Минералокортикоиды (наиболее важный - алъдостерон)
затрагивают баланс электролитов тела. В частности, альдостерон
стимулирует выделение калия и задержку натрия в теле. Избыток альдо-
стерона может привести к перегрузке жидкостью и гипертонии.
Наиболее важный половой гормон, синтезируемый корой
надпочечников - ДГЭА (дегидроэпиандростерон). ДГЭА вызывает
разнообразные эффекты, препятствующие развитию стресса и
старения, и был провозглашен одним из трех "гормонов молодости"
(гл. 5,26). ДГЭА и его предшественник прегненолон - единственные
кортикостероиды, чей уровень значительно снижается с возрастом.
Кортикостероиды. Итак, АКТГ, достигая коры надпочечников
с током крови, стимулирует синтез и секрецию кортикостероидов,
главным образом глюкокортикоидов, таких как кортизол и корти-
костерон. Эти гормоны и являются, наряду с катехоламинами,
основными эффекторами ОАС.
245
Структура. Биологически активные кортикостероиды, С2Г
стероиды, обладают А4-3-кетоструктурой кольца А, имеют а-ке-
тольную боковую цепь при С17 и гидроксилы в различных
положениях молекулы. В коре надпочечников синтезируются более 30 кор-
тикостероидов, их можно разделить на три основные группы: глю-
кокортикоиды, минералокортикоиды и гормоны со смешанными
свойствами. К первой группе относится кортизол (гидрокортизон),
ко второй - альдостерон, а к третьей - кортикостерон
(см. рис. 13).
Синтез. Биосинтез всех стероидных гормонов начинается с
холестерина, который образуется из ацетил-СоА. Важнейшей особен-
носью биосинтеза стероидных гормонов является последовательное
гидроксилирование. Реакция отщепления боковой цепи (ОБЦ)
включает предварительное гидроксилирование 20 и 22 углеродных
атомов холестерина и последующий распад молекулы на прегнено-
лон и изокапроновый альдегид (см. рис. 13). Разветвление путей
синтеза стероидов на 17-окси- и 17-дезоксикортикостероиды
происходит на стадии прогестерона, который образуется из прегненолона -
главного стероидного предшественника всех стероидных гормонов
(Юдаев и др., 1976; White et al., 1978).
Ферменты, участвующие в реакциях превращения холестерина в прегнено-
лон, включают комплекс холестерин-десмолазу, требующую НАДФ, Mg2+ (или
Са2+) и цитохром Р-450, и локализованы в митохондриях. Ферменты,
катализирующие последующие превращения прегненолона (см. рис. 13), находятся в эн-
доплазматическом ретикуломе. Энзимы, гидроксилирующие стероиды, каждый
из которых специфичен к определенному положению стероидного ядра,
являются монооксигеназами, использующими 02, адренодоксин и цитохром Р-450.
Хотя цитохром Р-450 во всех реакциях гидроксилирования выполняет одну и ту
же функцию, его белковая часть специфична для каждого субстрата. Так, Р-450,
принимающий участие в отщеплении боковой цепи, обозначается как Р-450обц,
тогда как другие обозначаются соответственно как Р-45017а» Р-450 η β, Ρ-45θ£2ΐ
и Р-450с18. Поскольку каждый из ферментов представляет продукт отдельного
гена, специфика продукции стероидов в клетках надпочечников формируется в
период их дифференцировки, когда в клетке начинает синтезироваться
определенный набор ферментов Р-450 в определенном соотношении.
Для реакций гидроксилирования необходим НАДФ · Н. Так, при
увеличении синтеза стероидов в коре надпочечников под действием
АКТГ (в результате стимуляции аденилатциклазы и последующего
образования цАМФ) активируется гликогенфосфорилаза.
Образующийся при этом глюкозо-6-фосфат является субстратом фосфоглк>
конатного пути, что и приводит к повышенному образованию
НАДФ - Η и, следовательно, активации гидроксилирования на
различных стадиях биосинтеза кортикостероидов (Whith et al., 1978).
Транспорт кортикостероидов. Кортизол составляет примерно
80% общего количества 17-оксистероидов крови. Гормон
связывается со специфическим а!-глобулином, который назван транс-корти-
ном. Он имеет один кортизол - связывающий участок на молекулу
246
белка (молекулярная масса 52 КД). Этот белок связывает также и
кортикостерон. Транс-кортин образуется в печени. Током кропи
комплекс транс-кортина и кортикостероидов доставляется к
клеткам различных тканей, в том числе печени, почек, мышц, кос ι ей,
кожи, жировой и лимфоидной тканей, т.е. к тем клеткам, которые
имеют в цитозоле специфические рецепторы, связывающие кор ги-
костероиды.
Механизм действия кортикостеридов. Гормоны коры
надпочечников растворимы в липидах и легко проходят сквозь клеточные
мембраны тканей-мишеней в цитоплазму, где они прочно, но
обратимо связываются со своими специфическими рецепторными
белками. Присоединение гормона ведет к аллостерическому изменению
конформации рецепторного белка (активация рецептора), что
повышает сродство последнего с ДНК; это позволяет рецептору
связываться со специфическими генами в ядре и регулировать их
транскрипцию (Normam et al., 1987; Gehring, 1987). Типичная клетка-ми-
шеиь содержит около 10 тыс. рецепторов для стероидных гормонов.
Каждый рецептор может обратимо связывать одну молекулу
определенного гормона с высоким сродством (константа сродства Ка от
108 до 10ш л/моль). Крупным успехом в изучении молекулярной
структуры рецепторов стала расшифровка последовательности нук-
леотидов ДНК (с помощью техники клонирования
соответствующей сДНК), кодирующей некоторые рецепторы стероидных
гормонов позвоночных. Оказалось, что многие рецепторные белки
обладают сходной структурой {Evans, 1988). Их полипептидная цепь
длиной около 800 аминокислотных остатков образует три отдельных
домена: С-концевой домен, связывающий гормон; центральный
домен, связывающийся с ДНК, и N-концевой, который активирует
транскрипцию гена {Yamamoto, 1985; Gering, 1987) (рис. 27).
Присоединение гормона повышает сродство рецептора с ДНК, что
позволяет прочно связываться с определенным участком гена, экспрессию
которого он либо активирует, либо подавляет.
Гены, чувствительные к стероидным гормонам, содержат, как
правило, несколько групп узнаваемых последовательностей,
расположенных обычно либо перед геном, либо "внутри" него. Место связы
вания рецептора имеет длину в восемь нуклеотидов (Yamamoto, 1985).
Под прямым влиянием стероидных гормонов, однако, находятся
лишь немногие гены в любой клетке-мишени. Анализ белков,
скорость синтеза которых менялась через 30 мии после добавления кор-
тизола к культуре клеток печени, показал, что гормон влияет на
экспрессию около 50 генов, хотя 10 тыс. рецепторов после ассоциа
ции с кортизолом связываются со значительно большим числом
мест на ДНК, но это не дает никакого эффекта.
Узнаваемые комплексом гормон-рецептор участки ДНК могут
стимулировать транскрипцию даже тогда, когда они уделены на
тысячи оснований от промотора, с которого начинается синтез РНК
247
Рис. 27. Модель белка-рецептора для стероидного гормона (кортизола). В
неактивном состоянии он зависит от ингибиторного белка, который блокирует
ДНК - связывающий домен рецептора. Связывание гормона рецептором
приводит к отделению белка-ингибитора, и в результате рецептор активируется
(Yamamoto, 1985; Gehring, 1987)
Такие удаленные участки получили название транскрипционные эн-
хансеры (Yamamoto, 1985).
Богатство метаболических эффектов, вызываемое действием
кортикостероидов на различные органы и ткани, позволяет
полагать, что в разных клетках-мишенях один и тот же гормон
регулирует активность разных генов (Attardi et al., 1978). Каждый стероидный
гормон вызывает характерный комплекс ответов, по-видимому,
потому, что рецепторы для данного гормона имеются только в
клетках определенных типов, и клетки каждого из этих типов содержат
248
различные комбинации присущих только им тканеспецифичных ре-
гуляторных белков, которые совместно с активированным
стероидным рецептором влияют на транскрипцию разных групп генов.
Например, глюкокортикоиды стимулируют в печени синтез иРНК,
кодирующих тирозинаминотрансферазу, триптофаноксигеназу, фо-
сфоенолпируват-карбоксилазу, т.е. ферменты глюконеогенеза -
одного из наиболее ярких метаболических эффектов кортизола при
стрессе, а в клетках сетчатки глаза гормон стимулировал синтез
иРНК глутаминсинтетазы, что, по-видимому, вносит определенный
вклад в обострение зрения при стрессе. В клетках аденогипофиза
глюкокортикоиды стимулируют синтез
транскрипта-предшественника гормона роста, и этот эффект также раскрывает какие-то
стороны механизма активации соматотропной "оси" в стресс-реакции
организма (Эверли, Розенфельд, 1985; Tepperman et al., 1987).
Метаболические эффекты кортикостероидов, Кортикостеро-
иды оказывают выраженное влияние на различные стороны
метаболизма. Особенно ярко это влияние проявляется в условиях стресс-
реакции, когда, как указывалось выше, концентрация
кортикостероидов в крови повышается под действием АКТГ. Сложное
воздействие кортикостероидов на экспрессию разных групп генов, а
следовательно, на активность различных ферментных систем, приводит к
специфическим изменениям метаболизма отдельных органов и
тканей (порой прямо противоположного характера). Особенно
мощным и разнообразным действием обладают глюкокортикоиды.
Суммарное влияние этих гормонов проявляется в виде катаболического
эффекта и выражается в снижении веса тела животных (в основном
за счет мышц) и повышении экскреции с мочой всех
азотсодержащих фракций: аммиака, мочевины, мочевой кислоты, аминокислот.
Параллельно с проявлением отрицательного азотистого баланса
наблюдается увеличение гликогена в печени и концентрации глюкозы
в крови (Юдаев, 1976; Smith et al., 1983).
Общее действие глюкокортикоидов на метаболизм можно
представить в виде следующей схемы (рис. 28) {Baxter et al., 1972).
1. Снижается проницаемость клеточных мембран кожи,
жировой, лимфатической и соединительной тканей для
глюкозы и аминокислот и поэтому уменьшается поглощение
глюкозы этими тканями и, следовательно, их анаболический
потенциал.
2. Увеличиваются катаболические процессы в коже, мышцах,
жировой, лимфатической и соединительной тканях, благодаря
чему возрастает количество субстратов для глюконеогенеза
(аминокислот и глицерина).
3. Увеличивается проницаемость клеточных мембран печени
для глюкозы и аминокислот, возрастает приток субстратов
для глюконеогенеза в этом органе. Анаболический потенциал
печени повышается.
249
Печень
Снабжение
энергией
I
Ж*ри™ Глидерин
КИСЛОТЫ г
Глюкоза
θ
(Относительно?)
Лимфоидная ткань
-ф—®—θ"
Кожа
{ШШр;
^Соединительная ткань!
Жировая ткань
43-
Рис. 28. Схематическое представление о действии глюкокортикоидов на
углеводный, жировой и белковый обмен (Baxter, 1972)
Стрелки указывают на движение субстратов в ответ на катаболическое или анабо
лическое действие гормонов
**+","-" - соответственно стимуляция и ингибирование процесса глкжокортикоидами
4. В результате усиления липолиза увеличивается количество
свободных жирных кислот и таким образом сохраняются
запасы глюкозы, так как в качестве источников энергии
используются жирные кислоты, а не глюкоза; свободные жирные
кислоты тормозят поглощение и утилизацию глюкозы; в
результате генерации НАД· Η во время окисления жирных кислот
увеличивается декарбоксилирование пирувата.
Общая картина влияния глюкокортикоидов в печени отражает
множественный характер действия этих гормонов. Очевидно, это
явление связано с активацией ими транскрипции нескольких групп
250
генов и синтеза различных ферментов этого важного для
метаболизма органа. Этому предшествует повышение активности
хроматина, продукции нуклеотидов и активности РНК-полимераз. Все эти
процессы приводят к резкой гипертрофии печени. Наряду с
анаболическим действием глюкокортикоидов в печени имеет место и
противоположное действие этих гормонов: торможение синтеза про-
лин-оксидазы, жирных кислот, а в регенерирующей печени также и
репликации ДНК (Tepperman et al, 1987).
Значительно более выраженные катаболические эффекты
наблюдаются в мышечной ткани. Глюкокортикоиды стимулируют
образование свободных аминокислот, их выход в кровяное русло и
одновременно ингибируют синтез белка. В результате наступает
распад мышечной ткани и миотрофия (Юдаев и др. 1976; White et al.
1978; Smith a al 1983).
Под влиянием глюкокортикоидов происходит инволюция лим-
фоидной ткани, уменьшение ее веса, снижение содержания белков,
угнетение синтеза ДНК и митоза лимфоцитов. Особенно
чувствительны к глюкокортикоидам небольшие лимфоциты и клетки
тимуса (Grant, 1967), что выражается либо в угнетении их роста, либо в
их гибели. Эти наблюдения, которые в свое время развивали
концепцию О АС, позднее получили четкое объяснение -
глюкокортикоиды индуцируют в тимусе запрограммированную клеточную
смерть (Cohen et al., 1984; Cohen, 1992)., т.е. апоптоз, со всеми ее
субклеточными и молекулярными проявлениями (Saitoh et al., 1998;
Сумбаев, 2001; Огурцов и др., 2001). Кроветворные и
лимфатические клетки селезенки также подвергаются ингибированию корти-
костероидами.
Ингибирующее влияние глюкокортикоидов на целый ряд органов и тканей
и в особенности их действие на лимфатическую и кроветворную системы
(торможение образования антител, снижение гиперчувствительности и
сенсибилизации организма к различным агентам) объясняет явно выраженную
антивоспалительную активность этих гормонов и подавление ими иммунной системы
организма (Grant, 1967).
Антивоспалительная, антиаллергическая и антииммунная активности
глюкокортикоидов делают их незаменимыми лекарствами при лечении целого
ряда заболеваний, вызванных воспалительным процессом и ненормальной
реакцией иммунной системы: ревматоидного артрита, эритематоидной волчанки,
пузырчатки, бронхиальной астмы, некоторых аутоиммунных болезней и
различных дерматозов.
В условиях стресса указанные выше изменения метаболизма под
действием глюкокортикоидов прежде всего отражают стремление
организма к сохранению его энергетического гомеостаза при
повышенных энергетических затратах во время фазы сопротивления
(Панин, 1983). Фазовые изменения энергетического обмена при
прессе характерны для формирования в организме
неспецифической резистентности. Какие бы специфические раздражители на
него не действовали, в организме всегда совершается дополнительная
251
Стресс
Истощение
Рис. 29. Изменение
энергетического обмена (Б) в различные
фазы стресса по Селье (А)
а - усиление углеводного
обмена; б - переключение
энергетического обмена с углеводного типа на
липидный; в - вновь усиление
энергетического обмена (Панин, 1983)
работа, направленная на устранение их повреждающего действия.
Когда в процессе развития реакции сопротивления наступает
дефицит углеводородов, происходит переключение энергетического
метаболизма с углеводного типа на липидный (рис. 29).
Конкретные биохимические механизмы действия адаптивных гормонов
при этом связаны:
1) с ингибированием ключевых ферментов гликолиза в различных тканях
и активацией ключевых ферментов глюконеогенеза в печени и почках;
2) с усилением жиромобилизующего эффекта и формированием
транспортной формы эндогенного жира в организме - липопротеидов очень
низкой плотности (ЛОНП).
Перестройка дыхательной цепи в митохондриях дополняет эти изменения.
Она характеризуется уменьшением эффективности окисления интермедиантов
цикла Кребса и усилением фосфорилирующего окисления липидов (вероятно,
по перекисному механизму). Если первый путь окисления углеводов и липидов
(через цикл Кребса) можно назвать "углеводным" типом, то второй - "липид-
ным". Таким образом, в фазу резистентности энергетический метаболизм с
углеводного типа переключается на липидный. Важно отметить, что
соотношение концентрации двух важнейших гормонов - инсулина и кортизола - в фазе
резистентности сдвинуто в сторону преобладания последнего (Дардымов, 1987),
т.е. процессы катаболизма преобладают над процессами анаболизма.
В заключение этой главы мы хотели бы подчеркнуть, что
рассмотренные выше весьма сложные многоступенчатые
физиологические и биохимические механизмы стресса направлены на
тотальную мобилизацию энергетических ресурсов организма для
устранения вредоносного влияния стрессоров. Основные усилия стресс-реа-
лизующих систем при этом направлены на сохранение или
восстановление различных параметров гомеостаза. Катехиламины играют
ведущую роль в течение фазы тревоги, стимулируя экстренную
мобилизацию легкодоступных ресурсов и подготовку организма к
большому объему физической и нервной работы, что
обеспечивается механизмом гигантского каскадного усиления процесса гликоге-
нолиза, когда связывание нескольких молекул адреналина с адрено-
252
рецепторами клеток печени приводит к быстрому выбросу в кровь
нескольких граммов глюкозы.
Кортикостероиды в течение фазы сопротивления ведут дли
тельную мобилизацию за счет всего объема имеющихся в
организме ресурсов, что также достигается путем каскадного усиления
первичного сигнала. Так, 1 нанограмм КРГ, секретируемый
гипоталамусом, приводит к синтезу в гипофизе около 10 мкг АКТГ, который
в надпочечниках вызывает образование нескольких миллиграммов
кортизола (усиление более чем в миллион раз).
Подобная гиперстимуляция мобилизационных механизмов
будет продолжаться до момента ликвидации состояния стресса или
истощения ресурсов организма, что чреватого развитием болезней
адаптации той или иной тяжести или, наконец, при глубоком
истощении - гибелью организма.
Глава 14
ПЕРЕКИСНОЕ ОКИСЛЕНИЕ И СТРЕСС
Истоки представлений о роли и механизмах окислительных
процессов в жизнедеятельности клеток в начале XX в. были связаны с
работами А.Н. Баха в России и К. Энглера и В. Виланда в Германии,
которые сформулировали теорию перекисного окисления. Следующим
шагом было открытие химических реакций, в ходе которых
возникают весьма активные промежуточные продукты, осколки молекул -
свободные радикалы (СР). Н.Н. Семеновыми и его школой была
разработана теория цепных реакций свободно-радикального окисления
(СРО) в газовой фазе, а его учеником Н.М. Эмануэлем - для жидко-
фазных систем углеводородов и жиров (Эмануэль, 1957; Эмануэль и
др., 1957) (см. гл. 2). Дальнейшее развитие этого направления было
связано с радиобиологией. Сравнив особенности развития лучевого
поражения во времени с кинетикой цепных химических реакций,
описанных Н.Н. Семеновым, Б.Н. Тарусов пришел к выводу, что
протекающие в облученных живых системах (в особенности в липопротеино-
вой фазе) процессы ускоряются с течением времени, подобно
автокаталитическим цепным реакциям (Тарусов, 1954).
Указанные научные направления вышли далеко за рамки
радиобиологических проблем и открыли новый раздел современной
молекулярной и физико-химической биологии - учение о
неферментативных свободно-радикальных процессах окисления липидов
биологических мембран в норме и при патологии. В последующие годы
постоянно накапливались факты (Владимиров, Арчаков, 1972) и на их основе
постепенно складывалось представление об универсальной,
неспецифической природе процесса свободно-радикального перекисного
окисления как компонента самых различных физиологических и
патологических процессов (Барабой и др., 1986). При такой ситуации
естественно возникал вопрос о взаимосвязи СРО и такого универсального
неспецифического механизма жизнедеятельности, как О АС.
Всестороннее и обстоятельное рассмотрение этой взаимосвязи
содержится в монографии В. А. Барабоя, ИМ. Брехмана, И.Г. Голо-
тина и Ю.Б. Кудряшова "Перекисное окисление и стресс",
выпущенной в 1992 г. издательством "Наука". Наиболее важные
положения этой работы приводятся ниже.
254
14.1. ПРО- И АНТИОКСИДАНТНОЕ РАВНОВЕСИЕ
И ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЙ ГОМЕОСТАЗ
В каждом живом организме, каждой клетке постоянно имеется
низкий стационарный уровень свободно-радикальных процессов,
некая базальная интенсивность перекисного окисления (ПО) как
следствие, как равнодействующая про- и антиоксидантных стимулов. К
компонентам, формирующим это равновесие, прежде всего
относится молекулярный кислород, растворенный в биологических
жидкостях и присутствующий в клетках и их органеллах. Кислород -
непременный компонент процесса ПО. Хотя субстратом окисления
могут служить практически все органические соединения клетки,
чаще других субстратом ПО становятся липиды, в особенности
полиненасыщенные жирные кислоты. Наиболее вероятной ареной
встречи 02 и липидов являются биологические мембраны, в
которых протекает постоянный и универсальный процесс перекисного
окисления липидов (ПОЛ). Инициация ПОЛ осуществляется под
действием свободно-радикальных окислителей: С£, ΗΟ2,* ОН, а
также перекиси водорода Н202, гидроперекисей типа к-ООН,
органических перекисей. Минимальное количество этих инициаторов ПОЛ
всегда присутствует в клетке. Кроме этого, активировать ПОЛ
могут окислительно-восстановительные ферментные системы в
составе цепей электронного транспорта, содержащие в своих активных
центрах ионы металлов с переменной валентностью (Fe, Си, Μη, Ζη,
Со, Se, Mo и т.п.). Их функционирование приводит к переносу
электронов с окисляющегося субстрата на восстанавливаемый, при этом,
как правило, образуются нестойкие промежуточные продукты
радикальной природы (АФК), способные активировать ПОЛ.
К числу антиокислительных (АО) факторов равновесия
относится ряд ферментов - супероксид дисмутаза (СОД), глутатион перо-
ксидаза и глутатион редуктаза, каталаза; жирорастворимые
компоненты мембран (токоферолы, убихиноны, ретиноиды, каротинои-
ды, фенольные соединения), водорастворимые АО биологических
жидкостей (глутатион, цистеин, метионин, аскорбиновая кислота,
гистидин, водорастворимые фенольные соединения). Некоторые
ионы металлов функционируют как АО (например, Se, Mo, Zn).
Следует особенно выделить (в контексте проблемы стресса),
что к системам экстренного повышения АО потенциала тканей
следует отнести нейроэндокринные системы поддержания гомеостаза и
их продукты: симпатико-адреналовую систему (адреналин, норадре-
налин, ацетилхолин, серотонин), систему
гипоталамус-гипофиз-кора надпочечников (кортизол, кортикостерон), систему опиоидных
пептидов (гипоталамические регуляторные гормоны, эндорфины.
энкефалины и др.).
Таким образом, окислительно-восстановительный гомеостаз
тканей, про- и антиоксидантное равновесие в них является следстви-
255
Накопление малонового диальдегида
при различных экстремальных воздействиях
Таблица 12
Воздействие
Контроль
Иммобилизация (подвешивание), 24 час
Интоксикация (окисленная олеиновая.
4 мл/кг)
Иммобилизация (на планшете), 17 час
Голод (с водой), 3 суток
Ожог
Гипертермия, 39-40°С, 6 час.
Плавание, 1.5 час. с грузом (6% от
массы тела)
Холод, 0°С, 24 часа
Гиподинамия, 3 суток
диальдегид,
нмоль/г
36.2±1.8
220.0±9.1
138.8±6.9
102.6±4.1
87.0±2.3
73.7±3.0
68.0±2.3
67.7±4.0
57.6±3.0
52.8±2.4
Примечание: для всех данных различие от контроля достоверно (р
nun ιуиль,
принят за 100%
< 0.05)
100
610
380
286
241
205
188
188
161
147
ем взаимодействия множества веществ, иоиов, радикалов,
процессов. Такая регуляция обеспечивает постоянный и низкий уровень
продуктов ПОЛ в клетке, порядка КИ-10-6 Μ в липидной фракции
(Барабой и др., 1992).
Переход отдельных органов и систем или всего организма в
режим напряженной деятельности, высоких функциональных нагрузок,
обусловленный изменениями условий как внешней, так и внутренней
среды (т.е. в режим стресс-реакции), неизбежно сопряжен с
повышением расхода энергетических ресурсов, усилением катаболизма,
остановкой или замедлением клеточных делений. При этом неизбежно
возрастает интенсивность клеточного дыхания, ферментативная
активность, количество АФК-инициаторов и продолжателей цепей
ПОЛ. При наличии достаточного резерва АО этот сдвиг в сторону
прооксидантных процессов может быть вначале компенсирован.
Однако с увеличением силы и длительности внешнего воздействия, по
мере уменьшения буферной мощности АО-систем смещение
равновесия становится все более выраженным. Количество активных
продуктов ПОЛ возрастает, а с ним увеличивается и вероятность
повреждения биологических структур и прежде всего мембранных структур.
Организму приходится платить все более высокую цену (в форме
крайнего напряжения гомеостатических механизмов, а затем в виде
более или менее значительных нарушений структуры и функции
клеток) за сохранение гомеостаза. Схема токсического действия
продуктов ПОЛ на клетку и его последствия представлены на рис. 4.
256
Таким образом, экстремальные нагрузки, вызывающие
гиперфункцию органов и систем, сопровождаются соответствующим
усилением прооксидантных процессов и опасностью смещения про- и
антиоксидантного равновесия влево. Вместе с тем умеренные
нагрузки не только не вредны для организма, но могут оказать на
него тренирующее влияние, обусловить адаптационное повышение
работоспособности этих систем и устойчивости к экстремальным
воздействиям (Виру, 1981: Бурлакова и др., 1985; Ji et al., 1998; McArdle
et al., 2000).
В организме при стрессе создаются благоприятные условия
для усиления автолитических процессов: повышение основного
метаболизма, уровня адреналина, гистамина, падение АОА
тканей, рост содержания ненасыщенных жирных кислот (Бурлакова
и др., 1875).
В табл. 12 представлен суммарный эффект стимуляции
ПОЛ различными стрессорами по данным концентрации
конечного продукта ПОЛ - малонового диальдегида (Барабой
и др., 1992).
14.2. ПЕРЕКИСНОЕ ОКИСЛЕНИЕ В МЕХАНИЗМЕ
СТРЕСС-РЕАКЦИИ. "ПЕРВИЧНЫЙ МЕДИАТОР"
Как не раз указывалось выше, активация ПОЛ -
универсальный процесс, сопутствующий как нормальным реакциям
напряжения, так и патологическим состояниям. Этот неспецифический
компонент в силу своей универсальности может рассматриваться
как составная часть, как звено стресса. Обширная литература
свидетельствует, что, с одной стороны, активация ПОЛ
обнаруживается при стрессах самой различной природы: воздействие
ионизирующей и УФ радиации, при гипероксии и гипоксии, при гипо- и
гипертермии, гипер- и гипогравитации, интоксикациях гепато-
тропными и иными токсическими веществами, атеросклерозе,
ишемической болезни, гипертонии, инфарктах, инсультах и
других сердечно-сосудистых заболеваниях, при метаболической
активации канцерогенов, опухолевом росте и метастазировании, при
инфекционных заболеваниях, воспалительных и дистрофических
заболеваниях, при напряженной физической и умственной работе,
утомлении и т.п. С другой стороны, профилактическое и лечебное
применение АО при большинстве упомянутых состояний стресса
оказывается эффективным, что косвенно подтверждает участие и
патогенетическую значимость активации ПОЛ при этих формах
стресса (Владимиров, Арчаков, 1972; Владимиров и др., 1975;
Бурлакова и др., 1975; Барабой, 1976; Бурлакова и др., 1985;
"Биоантиокислители...", 1982; Дегтерев и др., 1985; Ваитап et al., 1980;
Harman, 1984).
257
По современным представлениям, активация ПОЛ является
важнейшим фактором, непосредственно обуславливающим
повреждение мембранных клеточных структур, повышение проницаемости
мембран лизосом с выходом лизосомальных ферментов, развитие
поражений сердца, сосудов, слизистой оболочки пищеварительного
тракта, головного мозга, сетчатки глаза и т.п. (Барабой и др., 1992;
TodorovH. etal.,2001).
В той или иной степени ПОЛ имеет место везде, где содержится
молекулярный кислород и где в нормальных условиях образуются
его активные формы: 0^,Н202," ОН," 02,т.е. практически в каждой
живой клетке и прежде всего в ее мембранах на протяжении всей
жизни клетки. Следовательно, возможность активации ПОЛ
заложена в самой сущности земной аэробной формы жизни. Эта
возможность реализуется весьма часто на протяжении жизни
каждого организма, а степень ее выраженности, степень декомпенсации
этой активации характеризует тяжесть стресса, переход от нормы
к патологии. Поддержание окислительного гомеостаза, о котором
говорилось выше, требует, наряду с имеющимися клеточными и
тканевыми резервами АО, существования аварийных систем,
обеспечивающих в случае необходимости быстрое повышение
АО потенциала тканей. Естественная необходимость экстренного
реагирования на опасность нарушения гомеостаза требует
возможно более раннего включения этих аварийных систем.
Сигналом для их включения могут служить начальные минимальные
сдвиги окислительного гомеостаза, например, повышение уровня
активных форм кислорода во внутренней среде организма
(Барабой и др., 1992).
Эти аварийные системы уже подробно рассматривались
выше: это симпато-адреномедулярная и гипоталамо-гипофизо-кор-
тикоадреналовая системы. Их активация - это ответная реакция
организма, эволюционно выработанная и генетически
запрограммированная на сдвиг про- и антиоксидантного равновесия, на
увеличение количества АФК в органе-мишени, на который падает
стрессогенное воздействие. Можно предположить, что продукты
активации СРО выступают в качестве того первичного
медиатора, существование которого предположил с самого начала Г. Се-
лье (Selye, 1950) .
Определенная ясность относительно природы первичного
медиатора позволяет проанализировать весь механизм стресс-реакции,
начиная не с возбуждения центров головного мозга,
детерминирующих ее, а с афферентного звена процесса - с воздействия стрессора
на биологическую систему любой степени сложности с
последующей генерацией возбуждения высших вегетативных центров через
посредство первичного медиатора, т.е. продуктов СРО.
Гипотетическая схема развертывания стресс-реакции с этой точки зрения
представлена на рис. 30 (Барабой и др., 1992). Эта схема дает некоторое
258
Механические, физические,
химические антигенные
воздействия
Дистантные,
психоэмоциональные воздействия
Деструкция тканей
I
Кора полушарий
головного мозга
Активация ПОЛ
(первичный медиатор)
в зоне воздействия
(II
IV
Возбуждение центров ретикулярной
формации, гипоталамуса
Избыток стероидных
гормонов
Нарушения водно-
солевого и
микроэлементного обмена
VI
νπ
Избыток катехоламинов и
др. биогенных аминов
Уменьшение
энергетического
и пластического
резерва
Липидная триада: активация
ПОЛ (вторичная), фосфолипаз,
липаз, детергентное действие
лизофосфатидов и жирных
кислот
Лабилизация лизосом и
пероксисом
Τ
Повышение пассивной проницаемости и нарушения
активного трансмембранного переноса ионов и
метаболитов
Нарушения компартментации,
активация нуклеаз, протеаз,
фосфолипаз, липаз, распад
нуклеиновых кислот, белков
Нарушения функции митохондрий,
эндоплазматического ретикулума,
биосинтеза макроэргов,
нуклеиновых кислот и белков
VUI
Нарушения функции миокарда, сосудистой системы,
кроветворения, иммунной системы, органов-мишеней,
микронекрозы, снижение неспецифической
резистентности
Рис. 30. Гипотетическая схема развертывания стресс-реакции (Барабой и др.
1992)
259
представление о путях воздействия на организм факторов
соматического и психоэмоционального стресса. Для первого вида стресса
первичным медиатором служат продукты ПОЛ, образовавшиеся в
зоне воздействия стрессора, а для второго - дистантные,
психоэмоциональные воздействия на кору полушарий головного мозга. Оба
пути достигают центров ретикулярной формации и гипоталамуса,
возбуждая их. Последующие звенья стресс-реакции универсальны
для обоих типов стресса.
Выброс в кровь адреналина, а затем и кортикостероидных
гормонов при стресс-реакции вначале приводит к понижению перекис-
ного окисления в организме и возвращает про- и антиоксидантное
равновесие к норме (фаза тревоги и ранняя фаза сопротивления при
стрессе). Однако, если стрессогенное воздействие будет сильным и
длительным, потребуется избыток стероидных гормонов и
биогенных аминов, что в течение длительной фазы сопротивления в конце
концов приведет ко вторичной активации ПОЛ, фосфолипаз,
липаз, к детергентному действию лизофосфатидов и жирных кислот
(Барабой и др., 1992).
Действительно, гиперпродукция катехоламинов (превышение
нормального их уровня в крови в 7-10 и более раз) сохраняется
достаточно долго. В качестве факторов, непосредственно
вызывающих вторичную активацию СРО и повреждение мембран, могут
выступать радикальные промежуточные продукты окисления
катехоламинов - семихинонные ион-радикалы адреналина, легко
передающие неспаренный электрон кислороду с образованием
супероксидного радикала. Метаболизм катехоламинов в основном
происходит за счет их метилирования, что приводит к
образованию малоактивных метанефрина и норметанефрина. Таким путем
инактивируется 80-90% катехоламинов. Следует подчеркнуть, что
катехоламины на ранних стадиях стресса способны, окисляясь,
восстанавливать другие окислительные радикалы; в итоге может
наблюдаться кратковременный АО эффект (Барабой и др., 1983).
При существенном окислительном сдвиге в организме окисление
катехоламинов (как и фенолов и аскорбиновой кислоты) может
только усиливать ПОЛ.
В механизме стресса активные продукты ПОЛ,
интенсифицированного катехоламинами в высоких концентрациях, играют роль
ключевого и необходимого звена патогенеза в качестве вторичных
медиаторов эфферентного звена стресс-реализующего механизма.
Как не может быть стресса без гиперкатехолемии, так не может
быть стресса без вторичной активации ПОЛ. Вслед за этим
наступит фаза истощения и развитие так называемых болезней адаптации
(резкая потеря иммунной сопротивляемости, кровоизлияния в
различных органах, язвы желудка, активация злокачественного
перерождения клеток и т.п.).
260
* * *
Таким образом, универсальная адаптационная реакция, ОАС,
биологический стресс, развивающийся в результате
повреждающего воздействия экстремального фактора, по-видимому, реализуется
при посредстве "первичного медиатора", в качестве которого
выступают продукты окислительной деградации, активные формы
кислорода. Важнейшим эффекторным звеном этого стресс-реализующе-
го механизма также является активация ПОЛ, обусловленная
многократным повышением во внутренней среде организма количества
катехоламинов и прежде всего адреналина. На высших уровнях
биологической эволюции эта универсальная реакция дополняется
формированием механизмов психоэмоционального стресса, при
котором активация стресс-реализующих систем развивается в ответ на
действие дистантных раздражителей разной природы, без
первичного повреждения тканей. В этом случае первое, афферентное звено
активации ПОЛ в значительной мере утрачивает свое значение, но
зато тем большую роль начинает играть эффекторное вторичное
звено активации ПОЛ.
Ясное понимание биохимического и физиологического
механизмов стресс-реакции, которое мы имеем в настоящее время,
позволяет рекомендовать различные природные антиоксиданты и адаптоге-
ны, как об этом подробнее будет сказано в последующих разделах
этой книги, в качестве наилучших стресс-протекторов как в
лечебных, так и в профилактических целях.
Глава 15
СТРЕСС И МЕХАНИЗМЫ СТАРЕНИЯ
Три стадии стресса напоминают периоды
жизни: детство (с его характерной низкой
сопротивляемостью н высокой
реактивностью на любое раздражение), зрелость (во
время которой организм
приспосабливается к большинству встречающихся агентов и
его сопротивляемость повышается) и
старость (характеризующаяся потерей
адаптационной способности и конечным
истощением). Поэтому мы предполагаем
существование взаимосвязи между стрессом и
старением.
Ганс Селье, 1977
Ганс Се лье, отец современной теории стресса, был первым, кто
предположил, что стресс ускоряет старение. С тех пор накопилось
много свидетельств того, что стресс - критический фактор в
скорости старения, а также в развитии возрастных болезней. Исследования
на животных показывают, что продолжительность жизни обратно
пропорциональна интенсивности стресса, и это означает, что
животные с повышенной чувствительностью к стрессогеиному
воздействию живут меньше. Замечательная и наиболее постоянная
особенность долгожителей (людей в возрасте 100 лет или больше) - их
способность справляться и быстро преодолевать психологический
стресс.
Повышенная реакция на стрессоры может давать некоторое
преимущество для выживания животного в условиях дикой природы
и в рутинных столкновениях за жизненное пространство. Однако в
стабильной окружающей среде - будь то животное в зверинце или
современной человек - чрезмерная стресс-реакция вредна. Из этого,
однако, не следует, что слишком слабая стрессовая реактивность не
может быть так же или даже более вредна. Даже в устойчивой
окружающей среде современного мира имеется спорадическая
потребность в адекватном стресс-ответе. Преодоление несчастных случаев
или заразных болезней, тягот военного обучения или
профессионального спорта, экстремальных погодных условий и перемены
часовых поясов - все это требует достаточно мощного стресс-ответа.
Кроме того, небольшие количества гормонов стресса - катехолами-
нов и кортикостероидов - необходимы для нормального
ежедневного существования, а не только в критическом положении. Итак,
приходится признать, что бытующее выражение "наша обыденная
жизнь - это стресс" имеет некоторые физиологические основания.
262
Возьмем, к примеру, болезнь Аддисона - состояние,
характеризующееся неспособностью коры надпочечников производить кортизол
и другие стероиды стресса. Эта болезнь может быть фатальной,
если ее лечение не включает ежедневного приема кортизола и альдо-
стерона. Когда пациент с болезнью Аддисона заражен инфекцией
или получил травму, ему дают дополнительное количество
стероидов стресса, чтобы моделировать нормальный стресс-ответ.
Точка зрения, которую мы пытаемся здесь провести,
заключается в том, что стресс-ответ является жизненно важным, и мы
нуждаемся в нем настолько, насколько это необходимо в данной
ситуации. Слишком слабый стресс не в состоянии воспрепятствовать
физическим стрессорам нанести организму ущерб, в то время как
чрезмерный стресс - это самостоятельная разрушительная сила.
Избыточный стресс-ответ, однако, более распространен, чем слабый.
Сегодня мы чаще сталкиваемся с психосоциальными стрессогенными
факторами, которые индуцируют непропорционально мощный
стресс-ответ. Кроме того, обратные связи эндокринной системы,
ограничивающие интенсивность стресс-ответа, с возрастом снижают
свою эффективность. Действительно, исследования показывают,
что у пожилых людей обычно выделяется большее количество
стероидов стресса, чем у более молодых индивидов в ответ на стрессо-
генные факторы (Дилъман, 1987).
Суммируя сказанное, подчеркнем, что продление жизни и
предотвращение болезней требует оптимальной реакции на стрессоры.
Для огромного большинства людей и ситуаций это означает, что
стресс-ответ необходимо смягчить либо избегая воздействий
серьезного или длительного стрессора, либо модифицируя ответ на него.
15.1. СТРЕСС И СВОБОДНЫЕ РАДИКАЛЫ
Большая часть свободных радикалов (СР) - высокореактивные
побочные продукты клеточного дыхания. Они играют важную роль
в процессах старения (см. гл. 2). СР постоянно производятся всеми
клетками тела - чем больше сжигается калорий, тем больше
производится СР. Как уже указывалось, виды животных с более высокой
интенсивностью метаболизима (т.е. сжигающие большее
количество калорий на весовую единицу ткани) обладают и более короткой
продолжительностью жизни. Мы уже не раз указывали: если у
грызунов ограничить калорийность рациона, а следовательно,
уменьшить интенсивность обмена веществ и формирование СР (Barja,
2002), то у таких животных удается продлить жизни почти в два
раза. СР также вносят свой вклад в развитие многих болезней - от
атеросклероза и артрита до рака и катаракты. Снижение уровня
повреждений, вызванных СР, является критичным для продления
жизни и предотвращения болезней
263
Стресс резко увеличивает производство свободных радикалов. Главная
цель стресс-ответа состоит в том, чтобы увеличить функциональную
активность систем, необходимых для выживания - нервной и сердечно-сосудистой
систем, легких и мышц. Это достигается снабжением клеток избытком
питательных веществ (главным образом глюкозой и жирными кислотами) и
стимуляцией к производству большего количества энергии. Одним из побочных эффектов
этого перехода на более высокий метаболический уровень является
производство большого количества побочных продуктов окисления - СР (Cadenas et al.
2000). Когда мы рассматриваем наше тело как огромную машину, мы начинаем
осознавать все совершенство ее конструкции. Например, самому стресс-ответу
присущ механизм некоторого ограничения увеличения СР. Так, имеются
свидетельства, что гормоны стресса сами являются антиоксидантами. Однако подчас
действия этого механизма бывает недостаточно. Проблема состоит в том, что
АО-ресурсы организма ограничены и могут быть исчерпаны интенсивным или
длительным стрессом, который буквально наводняет организм свободными
радикалами.
При серьезном стрессе СР способны произвести много
различных повреждений. Мембраны клеток страдают более всего потому,
что они содержат липиды, богатые полиненасыщенными жирными
кислотами (ПНЖК), которые являются легкой мишенью для СР.
Процесс, при котором СР повреждают липиды в биологических
мембранах, принято называть перекисным окислением липидов
(ПОЛ). СР-повреждения биологических мембран делают их
твердыми, хрупкими и негерметчными, нанося этим ущерб клеткам. Когда
имеется утечка в мембранах митохондрий, их энергетическая
продуктивность резко падает. Обычно митохондрии, перерабатывая
энергоносители, производят аденозинтрифосфорную кислоту (АТФ) -
энергетическую валюту клетки. Негерметичные митохондрии -
неполноценные производители энергии - бесполезно "жгут" топливо,
производя большое количество СР, но очень мало АТФ. Клетки с
такими дисфункциональными митохондриями испытывают
энергетический голод, несмотря на изобилие топлива, и разрушаются
свободными радикалами, поскольку их системы ремонта бездействуют
из-за недостатка АТФ (Melov, 2000; Wei et al., 2002). Другая серьезная
опасность от повреждений свободными радикалами - утечка сквозь
мембраны лизосом. В этих клеточных органеллах локализованы
ферменты с мощными деструктивными функциями, они обычно
используются для управляемого расщепления крупных молекул, таких
как липиды, белки и нуклеиновые кислоты и их конгломераты,
которые превратились в клеточные шлаки. Когда повреждения,
вызванные СР, приводят к разгерметизации лизосомных мембран,
ферменты выходят из лизосом и стохастически распространяются
повсюду внутри клетки, атакуя не только шлаки, но и жизненно
важные клеточные структуры, в том числе и хромосомы. Эта
ситуация чревата включением сложного субклеточного механизма
самоубийства клетки - апоптоза (см. гл. 2). Наконец, СР могут вызвать
утечки также и в наружной мембране клетки, что может привести к
264
разрыву клеточной мембраны, что в конце концов заканчивается
гибелью клетки - некрозом. Когда это случается, внеклеточные
электролиты и вода высвобождаются во внутриклеточное
пространство, опрокидывая баланс многочисленных биохимических
реакций. Наряду с повреждением биологических мембран в
результате ПОЛ, свободные радикалы могут непосредственно повреждать
ДНК и белки. Последние могут быть заменены, но некоторые
повреждения ДНК необратимы. {Барабой и др., 1992; Скулачев, 1999;
Liu et al., 1999; Владимиров, 2000; Владимиров, 2000а.)
Повреждения ДНК, вызванные стрессом, можно дифференцировать на
1) повреждения ядерной ДНК и их репарации, опосредованной путем активации
поли(АДФ-рибозо)полимеразы-1, 2) повреждения теломерной ДНК и их вклад
в управляемое теломерами клеточное старение, и 3) накопление мутаций мито-
хондриальной ДНК (Zglinicki v. et al„ 2001; Toussaint et al., 2002).
Клетки, которые непрерывно или часто подвергаются ударам
СР, теряют свои жизненные функции быстрее, чем это
детерминировано их внутренними часами старения. Как мы уже упоминали
выше (а также в гл. 2), клетки, которые соприкасаются с высоким
уровнем СР, достигают предела клеточных делений (лимита Хайф-
лика ) быстрее (Saretzki et al., 1999) и накапливают большее количе
ство возрастных пигментов, таких как липофусцин, амилоид и др.
Такие клетки с большей вероятностью могут стать
злокачественными (Rudolph et al., 1999), потому что свободные радикалы - мощные
мутагены, способные изменять функцию важных генов,
контролирующих клеточный рост.
Итак, серьезный стресс может генерировать достаточное
количество СР, чтобы вызвать значительные и частично необратимые
повреждения тканей и даже убить некоторые клетки, приводя к
более быстрому старению и повышенному риску болезней. Антиокси-
данты, как было показано, уменьшают многие стресс-зависимые
повреждения. У грызунов язвы, вызванные стрессом, могут быть
предотвращены большими дозами различных антиоксидантов. Витамив
Ε и некоторые синтетические антиоксиданты уменьшают также
связанные со стрессом повреждения иммунной системы. В
некотором смысле, антиоксиданты делают свою "работу " при стрессе
лучше, чем в спокойном состоянии. Когда организм получает
антиоксиданты в совершенно свободной от стресса ситуации, его
собственные АО-системы могут стать несколько "ленивыми",
"расслабляясь" при появлении "внешней помощи". Напротив, во время
серьезного стресса, собственная антиоксидантная защита фактически уже
функционирует на максимальном уровне так, что АО-добавки
вводятся как чрезвычайно ценное подкрепление, а не частичная замена.
Для всесторонней защиты от свободно-радикальных повреждений важно
присутствие антиоксидантов двух типов: жиро- и водорастворимых. Первые, в
частности, витамин Ε и β-каротины защищают мембраны клеток, которые
содержат большие количества липидов н липопротеинов. Растворимые в воде ан-
265
тиоксиданты, например витамин С и цистеин, защищают ДНК н большинство
белков (Sastreet al., 2000). Липоевая кислота и мелатонин растворимы как в
воде, так и в жире и поэтому защищают все биологические структуры.
Превосходный инструмент для повышения способности ваших
собственных антиоксидантных систем - физические упражнения. Как мы
уже отмечали, разумные упражнения служат "мягким или умеренным"
кратковременным физическим стрессогенным фактором, который
вызывает длительную стимуляцию защитных сил организма без
существенных повреждений (Polidori et al., 2000). В частности, упражнения
могут увеличивать активность ферментов, нейтрализующих СР, таких
как супероксид дисмутаза и глютатион пероксидаза {Jenkins, 1993).
Адаптогены - другое оружие в борьбе с повреждениями,
которые наносит стресс, повышая содержание СР. Многие адаптогены
(например, сибирский женьшень) уменьшают вызванные стрессом
повреждения свободно-радикального типа, смягчая стресс-ответ и
стимулируя антиоксидантные системы организма (см. гл. 29).
15.2. СТРЕСС И ЦЕНТРАЛЬНЫЕ ЧАСЫ СТАРЕНИЯ
Как уже обсуждалось в гл. 2, высшие организмы, по-видимому,
имеют центральные биологические часы, контролирующие
процессы развития и старения. Эти часы локализованы в гипоталамусе и
сопряженных мозговых структурах. Разнообразные факторы
влияют на скорость "хода" центральных часов старения (ЦЧС) и стресс -
один из важнейших.
В.И. Дильман в своей нейроэндокринной теории старения,
касаясь механизма функционирования ЦЧС, постулирует постепенное
"загрубление" механизмов обратной связи между гипоталамусом,
гипофизом и периферическими эндокринными железами (Дильман,
1986, 1987). Иначе говоря, с возрастом гипоталамус (командный
центр эндокринной системы) становится все менее и менее
чувствительным к сигналам обратной связи, т.е. к уровню гормонов,
выделяемых эндокринными железами (Riegle, 1983). Это ведет к вредным
изменениям в тонком внутреннем равновесии (гомеостазе),
способствуя старению и дегенеративным болезням.
Есть указания, что стресс оказывает глубокое воздействие на
мозг и, в частности, на гипоталамус. Общий эффект стресса на клетки
мозга состоит в увеличении СР, повреждений, ускоренном накоплении
возрастных пигментов и даже их (клеток мозга) гибели, что не может
не вносить определенный вклад в ускорение "хода" ЦЧС. Ряд
косвенных данных показывает, что стресс вредит и контурам обратной связи
в эндокринной системе {Rothuizen et al., 1991). Так, стресс,
по-видимому, постепенно повреждает стресс-лимитирующую систему, что
делает ответ на будущие стрессогенные воздействия все более и более
интенсивным (Pedersen et al., 2001). Эффективность контура обратной
266
связи, в который включена эта система, частично зависит от гиппо-
кампа, мозговой структуры, функционально связанной с
гипоталамусом. Повреждение гиппокампа снижает способность гипоталамуса
прекращать стимуляцию стресса после того, как адаптивная цель
была достигнута (McEwen, 1999). У крыс, например, массированная
деструкция нейронов гиппокампа, вызванная серьезным стрессом, ведет к
хроническому состоянию гиперадаптации, Этот феномен существует и
у людей, хотя и в несколько меньшей степени (Dilman, 1994).
15.3. СТРЕСС И КЛЕТОЧНЫЕ ЧАСЫ
Большинство клеток млекопитающих имеет некоторый предел
в их способности делиться и расти (см. гл. 2). По мере достижения
этого предела (лимита Хайфлика) клетка теряет свою способность
делиться, накапливает возрастные пигменты и теряет некоторые из
своих функций. В среднем лимит Хайфлика допускает
приблизительно 50 делений, но это число не является строго фиксированным,
оно может быть увеличено или сокращено под влиянием различных
факторов. Определенно установлено, что высокий уровень СР
сокращает этот лимит (Saretzki et al., 2002). Хотя прямые данные о
влиянии стресса на лимит Хайфлика отсутствуют, его ограничивающее
влияние во многих тканях более чем вероятно, если учесть, что
стресс буквально "наводняет" организм свободными радикалами
15.4. СТРЕСС, ГЛИКОЗИЛИРОВАНИЕ
И ПЕРЕКРЕСТНЫЕ СШИВКИ
Глюкоза, в тех или иных количествах содержащаяся почти во
всех пищевых продуктах, и всегда - в крови, является первичным
энергоносителем для нервной системы и существенным фактором
поддержания жизни. К сожалению, глюкоза - участник двух
процессов, негативно сказывающихся на жизнедеятельности организма.
Во-первых, глюкоза способна беспорядочно, т.е. без участия
ферментов, реагировать с важными клеточными структурами, такими
как белки и ДНК, изменяя их свойства и функции (см. гл. 2).
Во-вторых, глюкоза облегчает возникновение перекрестных сшивок -
ненормальных химических мостиков между биологическими
макромолекулами. Перекрестные сшивки - одна из главных причин
катаракты, атеросклероза, гипертонии и потери эластичности кожи и
суставов. Скорость образования перекрестных сшивок заметно
повышается под действием свободных радикалов {Chen et al., 2002).
Стресс создает "благоприятную окружающую среду" для глико-
зилирования и перекрестных сшивок, потому что он повышает
содержание глюкозы в крови и генерирует большие количества
свободных радикалов.
267
15.5. СТРЕСС И ВОЗРАСТНЫЕ ПИГМЕНТЫ
Некоторые из отходов нашего метаболизма не выводятся из
организма, оставаясь в клетках в форме возрастных пигментов,
таких как липофусцин, цероид и амилоид (см. гл. 2). Возрастные
пигменты - большие нерастворимые скопления поврежденных
молекул - могут затруднять нормальные физиологические
потоки в клетке, вредить ее функциям и в конечном счете вызывать ее
гибель. Перекисное окисление липидов, вызываемое свободными
радикалами, служит главной причиной накопления возрастных
пигментов. Стресс, как известно, вызывает резкое повышение
интенсивности ПОЛ и ведет к более быстрому накоплению
возрастных пигментов. У животных стресс, как было установлено,
увеличивает накопление липофусцина в мозгу, сердце и других
органах (Yin, 1996).
15.6. СТРЕСС И СТАРЕНИЕ: ПОРОЧНЫЙ КРУГ
С возрастом большинство наших органов снижают свои
функции (см. рис. 1), сердечный насос работает менее энергично
во время физической нагрузки, рефлексы замедляются,
становятся менее острыми органы слуха и зрения и даже наш
интеллект - менее глубоким и подвижным. Поэтому было бы
естественным предположить, что интенсивность и мощность стресс-
реакций также снижается с возрастом. Удивительно, но в
действительности, по-видимому, имеет место противоположная
ситуация.
Несколько важных проявлений стресса с возрастом становятся
более выраженными. Один и тот же стрессор, как правило,
вызывает более интенсивный стресс-ответ у старых людей или животных,
нежели у молодых. По существу, это означает возрастную
прогрессию стресс-ответа. Часто пожилые индивиды показывают
повышенный уровень некоторых гормонов стресса даже в отсутствие
каких бы то ни было стрессогенных факторов, что фактически
является эквивалентом состояния постоянного хронического стресса.
Так, очевидно, и развивается этот порочный круг: стресс ускоряет
старение, а старение увеличивает реакцию организма на срессо-
генные факторы.
Этой проблеме посвящено много исследований. К. Переджо и
его коллеги (Perego et al.) изучали индуцированное стрессом
выделение норадреналина в гипоталамусе крыс, что стало одним из
ключевых событий в стимуляции стресс-ответа, и нашли, что
эмоциональное напряжение повышает у старых крыс уровень норадреналина и
выделение АКТГ. Несколько исследований на собаках
продемонстрировали, что старые животные по сравнению с молодыми: (1) вы-
268
деляли большее количество АКТГ и кортизола в ответ на стрессо-
генный фактор, (2) имели повышенный уровень АКТГ и кортизола
в течение более длинного времени и (3) в отсутствие стрессогенных
факторов имели аномально высокий уровень АКТГ и кортизола
{Rothuizen et al., 1991).
Все эти находки, по-видимому, справедливы также и для людей,
хотя у них имеются существенные индивидуальные различия.
Пожилые люди имеют склонность продуцировать чрезмерные
количества АКТГ и кортизола в ответ на стрессоры, а многие из них имеют
повышенный уровень этих гормонов даже в отсутствие
стрессогенных факторов. Все это означает, что пожилые люди демонстрируют
чрезмерно сильный (и поэтому чреватый значительными
повреждениями) стресс-ответ. Более того, многие находятся в
физиологическом состоянии хронического стресса даже тогда, когда для этого
нет никаких причин. В.М. Дильман, создатель нейроэндокринной
теории старения, называл такие состояния патологически
повышенной реактивности к стрессорам гиперадаптозом, но позже был
введен термин возрастной стресс-синдром, который теперь стал
общепринятым.
Следует отметить удивительное подобие между некоторыми
признаками старения и болезнью Кушинга, при которой кора
надпочечников производит чрезмерные количества кортикосте-
роидов, что моделирует состояние постоянного стресса. Конечно,
старение и болезнь Кушинга - не одно и то же. При старении
избыток кортизола - проявление возрастного стресс-синдрома,
тогда как при болезни Кушинга - это проявление аномальных
функций гипофиза или надпочечников. Тем не менее подобие
между возрастными особенностями стресс-ответа и болезнью
Кушинга {табл. 13) определяется, по крайней мере частично, одной
и той же причиной - избытком стероидов стресса, особенно
кортизола.
Однако все еще не совсем ясно, почему пожилые люди
имеют повышенную реактивность к стрессорам или даже входят в
состояние хронического стресса. Главная причина,
по-видимому, - это ухудшение регуляторного влияния отрицательных
обратных связей в эндокринной системе, ответственных за
сокращение стресс-ответа, когда в нем уже нет необходимости.
Организм реагирует так, как будто "не знает", что стрессогенный
фактор уже нейтрализован и нет никакой необходимости
продолжать борьбу.
Согласно теории ЦЧС, такое ухудшение в регуляции стресс-ответа -
результат программы развития и старения, осуществляемой гипоталамусом
(см. гл. 2). "Механизм'* ЦЧС основан на постепенном ухудшении
чувствительности гипоталамуса ко всем видам обратных связей, идущих от
остальных частей тела, включая и обратные связи, ответственные за управление
стресс-ответом.
269
Таблица 13
Частота проявлений некоторых признаков старения
и болезни Кушинга (в обоих случаях наблюдается избыток кортизола)
(TodorovH. etal.,2001)
Признаки н симптомы
Старение
Перераспределение жира: лица окру- часто
гляются часто повсеместно (лунные
лица); туловище становится более
массивным, особенно в области живота,
количество жира на ногах снижается;
состав тела изменяется в сторону
увеличения количества жира н
уменьшения количества нежировой ткани
Кожа тонкая и истощенная, легко
повреждается, часто повсеместно раны
плохо заживают
Мышцы теряют упругость н ослаблены часто
Гипертония
Остеопороз
Пониженная толерантность к глюкозе
Психиатрические аномалии
I»
Болезнь Кушинга
повсеместно
часто
часто
часто
часто
часто
сравнительно
часто (в основном
при депрессии)
повсеместно
повсеместно
часто
часто
часто
часто
"Ход" центральных часов можно замедлить или даже изменить, если
найти способы повысить чувствительность "центра" к сигналам с
эндокринной "периферии". Эти способы, эффективные хотя бы частично, включают
ограничение калорийности питания (у животных), сохранение оптимального
веса тела (у людей), улучшение толерантности тканей к глюкозе и
чувствительности к инсулину, лечение депрессии (если она имеет место), повышение
уровня нейромедиаторов в гипоталамусе, и заместительная терапия
гормонами, содержание которых снижается с возрастом - мелатонином, ДЭГА
и гормоном роста. Практические рекомендации обсуждаются в III части
этой книги.
15.7. СТРЕСС И ДЕПРЕССИЯ
В главе 7 мы обсуждали, каким образом депрессия вносит
вклад в старение, и наоборот, как риск депрессии повышается
с возрастом. Устранение депрессии жизненно важно при
любой программе продления жизни по двум главным причинам.
Потому что, во-первых, депрессия усиливает несколько
механизмов старения и, во-вторых, отнимает у человека мотивацию и
постоянство в его стремлении к более продолжительной и
здоровой жизни.
270
Стресс - основная причина депрессии у людей и животных,
которые также подвержены этому недугу. Фактически стресс -
стандартный инструмент, который исследователи используют
при изучении депрессии у животных. Например, когда крысы
или мыши подвергаются умеренному, но неожиданному стрессу,
они заметно уменьшают поглощение доброкачественной пищи и
чистой воды. Если животных лечат антидепрессантами, их
питание обычно возвращается к нормальному. Эта
экспериментальная модель часто используется для проверки эффективности
новых антидепрессантов прежде чем приступить к их испытаниям
на людях.
Депрессию у людей могут вызывать самые разнообразные
стрессогенные факторы. Напряжение на работе, семейные
проблемы, финансовые неприятности, болезни - почти любые реальные
или воображаемые несчастья способны погружать человека в
депрессию. По-видимому, депрессия разовьется и тогда, когда
организм будет не в состоянии ограничить свой стресс-ответ после
устранения или нейтрализации стрессогенного фактора (см. гл. 7). В
некотором смысле депрессия - нескончаемое состояние стресса,
которое может продолжаться даже в отсутствие каких-либо внешних
стрессоров или стимулов. Физиологически депрессия напоминает
состояние хронического стресса несколькими чертами (табл. 14).
Главные биохимические черты их подобия - повышенный уровень
кортикостероидов - ключевых стероидов стресса, оказывающих
глубокое влияние на организм, включая подавление иммунной
системы, плохое заживление ран, повышенное повреждение
нежировой ткани, ослабленная толерантность к глюкозе и потеря кальция
из костей (Chrousos et al., 1992).
Тип депрессии с проявлением беспокойства, часто
называемый "депрессией беспокойства", или "беспокоящей депрессией",
имеет большое сходство с реакцией "борьба или бегство" -
начальной фазы стресс-ответа (см. гл. 12). В обоих случаях
активируется симпатическая нервная система и из мозгового
вещества надпочечников интенсивно выделяются катехоламины
(адреналин и норадреналин). Эти изменения в значительной степени
ответственны за многие признаки и симптомы, которые
отмечаются при состояниях беспокойства и эмоциональном стрессе,
включая ускорение и аритмию сердечных сокращений,
мышечную напряженность, дрожание конечностей, потоотделение,
тошноту и т.п.
Другое важное звено, связывающее стресс и депрессию - серо-
тонин, нейротрансмиттер, с участием которого регулируются
многие функции нервной системы, включая аппетит, настроение и
общий тонус. Депрессия и беспокойство часто развиваются на фоне
низкого уровня серотонина в мозгу. Средства, повышающие этот
показатель, улучшают настроение, уменьшают беспокойство, спо-
271
Таблица 14
Сходство между физиологическими характеристиками стресса и депрессии
(Todorov Η. et al., 2001)
Физиологическая характеристика
Депрессия
Стресс
Уровень кортикостероидов в
крови
Уровень катехоламинов в крови
(адреналин и нор адреналин)
Активность симпатической
нервной системы
Уровень серотонина в мозгу
Чувствительность гипоталамуса
к торможению кортикостероида-
ми по типу обратной связи
часто повышен,
особенно при
депрессии
беспокойства
часто повышена,
особенно при
депрессии
беспокойства
понижен
повышен
повышена при
реакции тревоги
В начале
повышен, далее
понижается, если
стресс
продолжается
понижена
собствуют положительным эмоциям и вызывают общее
расслабление. Прозак, один из наиболее популярных антидепрессантов,
блокирует поглощение серотонина нейронами, увеличивая тем самым
его уровень в мозгу.
Какую роль играет серотонин в развитии стресс-реакции?
Первоначально количество серотонина в мозгу повышается. Это
обстоятельство играет определенную роль в механизмах, которые
ограничивают стресс-ответ. Однако длительный стресс, напротив
исчерпывает содержание серотонина в нескольких важных областях
мозга (особенно в префронтальной зоне) и ослабляет механизмы,
ограничивающие стресс. В результате, низкий уровень серотонина в
мозгу понижает устойчивость к стрессу, что, по существу,
способствует состоянию депрессии.
Исследования показывают, что серьезной причиной
уменьшения серотонина в мозгу является не интенсивность стресса, а его
длительность. Так, даже мягкий или умеренный стресс, если он
непрерывен, повышает шансы развития депрессии (Dinan, 1996). Еще
одной из причин развития депрессии являются возрастные
изменения функциональной активности рецепторов для глюкокортикои-
дов, с чем связано ускорение общих клеточных механизмов
старения структур мозга (CarrolL 2002).
272
Каково практическое значение связи между стрессом и
депрессией? Ясно, что человек, с одной стороны, может снизить риск раз-
иития депрессии, избегая, когда это возможно, стрессогенных
факторов окружающей среды. С другой стороны, коррекция депрессии
может предотвратить чрезмерный ответ на стрессоры и повысить
резистентность к состоянию стресса. Наконец, питательные
вещества или действия, которые улучшают сопротивляемость организма,
могут препятствовать и развитию депрессии при стрессе. В
частности, некоторые адаптогены (природные средства, которые
улучшают устойчивость к стрессу и оптимизируют стресс-ответ) оказывали
значительное антидепрессантное действие (практические аспекты
обсуждены в III части).
Глава 16
СТРЕСС И СПЕЦИФИЧЕСКАЯ АДАПТАЦИЯ
Отношения живых существ с окружающей средой можно
охарактеризовать как отношения любви-ненависти. С одной стороны,
эта среда обеспечивает их предметами первой необходимости -
воздухом, водой и пищей. С другой - изменения в окружающей среде
могут создавать смертельную угрозу организму, будь то
экстремально высокая или низкая температура, нехватка продовольствия,
нападения хищников и т.п. Жизнь на Земле развилась в постоянной
борьбе за выживание, адаптируясь к изменяющемуся миру.
Замечательная адаптационная способность современных организмов
формировалась в течение нескольких миллиардов лет. Даже находясь в
достаточно безопасной "цивилизованной" окружающей среде, мы
должны постоянно адаптироваться к измениям - часто не замечая
этого. Наш метаболизм подстраивается к сезонным изменениям в
температуре, меняя продукцию гормонов щитовидной железы; наш
мозг приспосабливается к решению новых задач или новым типам
деятельности, обучаясь и обретая новые рефлексы и привычки;
иммунная система приспосабливается к непрерывному нападению
патогенных микроорганизмов, производя специфические антитела
против этих агрессоров.
Пик наших адаптивных способностей приходится примерно на
20 лет и затем они постепенно снижаются. В сущности, потеря
способности к адаптации - основная особенность старения и одна из
главных причин смерти. Пневмония и грипп редко убивают
молодых людей, но являются пятой по частоте причиной смерти в
пожилом возрасте. Когда чрезвычайно высокая или низкая температура
настигает города, жертвами почти неизменно становятся старики.
Хотя некоторая потеря адаптируемости с возрастом неизбежна,
степень такой потери значительно варьирует. Наилучшая адаптация
у тех людей, чьи органы сохранили хорошую функциональную
активность и чей стресс-ответ - близок к оптимуму.
Адаптация в целом имеет две основных стадии: (1) общий
адаптационный синдром, или стресс-ответ и (2) адаптация к
определенному специфическому стрессогенному фактору.
274
Когда организм впервые сталкивается со стрессором, причем
все равно с каким именно, он всегда отвечает стандартной
реакцией на "чрезвычайное положение", т.е. стресс-ответом. Цель стресс-
ответа двойная: гарантировать повышенное функционирование
систем, важных для выживания в данный момент, таких как
нервная система, сердце и легкие; и обеспечить обильные ресурсы для
адаптации к специфическому стрессогенному фактору. Для этой
цели организм подавляет несущественные (в этой ситуации)
функции, например пищеварение и иммунитет, частично разрушает
нежировую ткань, освобождая энергоносители и строительный
материал. Эти ресурсы переадресуются ключевым системам
выживания, чтобы гарантировать активацию их деятельности, а также
системам, которые требуются для адаптации к конкретному
стрессогенному фактору. По мере развития адаптации к специфическому
стрессору, общий стресс-ответ постепенно сходит на нет. В
конечном счете новые возможности организма, созданные для
конкретной адаптаци, станут достаточными для полной нейтрализации
стрессора, что и снимет угрозу гомеостазу. В этой ситуации
стрессор уже перестает быть стрессором, стресс-ответ не нужен -
наступила полная адаптация.
Обычный пример описанного двухступенчатого процесса -
физические тренировки. У нетренированного человека повышенная
физическая нагрузка вызывает полномасштабный стресс-ответ с
выделением больших количеств катехоламинов и кортикостерои-
дов. Человек испытывает чрезвычайно быстрые сердечные
сокращения, пульсацию, тошноту и другие признаки острого стресса,
сопровождаемого общим недугом и истощением. Если регулярно
продолжать физические упражнения, то тело постепенно
приспособится к новым требованиям - сердечная и скелетные мышцы
увеличатся, станут более сильными и будут эффективнее использовать
энергию; увеличится жизненная емкость легких; улучшится
кровообращение и снабжение кислородом - в целом существенно повысится
способность тела совершать большие объемы физической работы.
Поскольку тренировки прогрессируют, физические упражнения в
дальнейшем вызывают только умеренный стресс-ответ, который
быстро спадает без всяких серьезных последствий. Тренировка
больше не является ощутимым стрессогенным фактором и будет
таковой пока человек остается адаптированным к физическим
нагрузкам, т.е. пока он в форме (Меерсон, 1986а).
При многократном умеренном действии холодового стресса по
типу закаливания наблюдается снижение интенсивности ПОЛ и
активация антиоксидантной энзиматической системы, защищающей
ткани {Эмирбеков и др., 1998).
Давно было замечено, что развитая резистентность к стрессу часто
коррелирует с продолжительностью жизни. Это привело к представлению о том, что
экспозиция организмов с умеренным стрессом может противодействовать нега-
275
тивным эффектам старения, помогая им жить дольше. Несколько умеренных
стрессов (облучение, тепловой и холодовый шок, физические упражнения и
т.п.), как сообщалось, увеличивали срок жизни, а гипергравитация уменьшала
скорость психологического старения. Механизмы подобных эффектов не
вполне ясны, однако исследования, проведенные до сих пор, показывают, что они
могут включать метаболическую регуляцию, активацию экспрессии антиокнс-
лительных ферментов и стресс-белков (например, белков теплового шока) (Ji et
al., 1998; Minois, 2000; McArdle et al., 2000).
Инфекция служит другим примером стресса, сопровождаемого
специфической адаптацией. Вторжение ранее неизвестного
патогенного микроорганизма - серьезный стрессор, вызывающий
стресс-ответ независимо от типа болезнетворного агента. В то же
самое время иммунная система быстро перебирает свои
возможности и находит наиболее подходящее оружие, такое как В-клетки,
которые производят антитела, способные атаковать этого
специфического захватчика (см. гл. 6). Это оружие совершенствуется в течение
нескольких стадий биохимических преобразований и умножается в
миллионах копий. Рост этого специфического сегмента иммунной
системы продолжается, несмотря на общее подавление иммунитета
под действием общего стресс-ответа. Болезнетворный агент
постепенно уничтожается миллионами новых иммунных клеток,
единственная задача которых - борьба с этим специфическим
противником. Стресс-ответ спадает и наступает полная адаптация. Более
того, иммунная система сохраняет в некоторых своих клетках
"память" об этом патогенным микроорганизме и будет подготовлена к
его ликвидации в случае повторного вторжения. Эта способность
иммунной системы "учиться" на предыдущих вторжениях - основа
для использования вакцин. Таким образом, то, что началось как
общий стресс-ответ, заканчивается как долговременная
специфическая адаптация.
К сожалению, существуют ситуации, когда организм не в
состоянии развивать специфическую адаптацию к стрессору, что
приводит к длительному стресс-ответу. Обычным примером ситуации,
когда организм будет не в состоянии адаптироваться и переносить
разрушительные действия такого стресс-ответа, служит
психологическая агрессия окружающей среды, например, плохие отношения с
близкими, базутешное горе или бесконечные неприятности на
работе. Хроническое нарушение сна - другой распространенный пример.
Способность организма к специфической адаптации снижается с
возрастом, тогда как стресс-ответы проявляют тенденцию
становиться чрезмерными. У молодых людей стресс-ответ сравнительно
краток и быстро сменяется эффективной специфической
адаптацией, при этом ущерб, вызванный стрессом, будет минимальным. У
пожилых людей специфическая адаптация развивается медленно, а
иногда даже вообще невозможна, и организм поддерживает стресс-
ответ как единственный источник помощи, который, к сожалению,
276
может вести к истощению и болезням. Как уже упоминалось,
центральные часы старения, расположенные в гипоталамусе, частично
ответственны за чрезмерный стресс-ответ, наблюдаемый у
стариков. Создается впечатление, что возрастное понижение способности
к специфической адаптации - другой важный фактор в развитии с
позрастом чрезмерного стресс-ответа: требуется значительно
больше времени для развития физиологических изменений,
необходимых для полной специфической адаптации, поэтому организм
вынужден поддерживать дорогостоящее "критическое состояние" стресс-
ответа в течение длительного времени.
Что можно сделать для восстановления модели адаптации,
свойственной молодости, т.е. сокращения стресс-ответа и улучшения
специфической адаптации?
Все меры, направленные на замедление или остановку
механизмов старения, обсуждаемые в этой книге, помогут сохранять
функции органов и тканей и замедлят потерю адаптируемости.
Адаптогены. Это - уникальный класс агентов, которые
помогают как оптимизации стресс-ответа, так и укреплению способности
организма к специфической адаптации. Такая уникальная
комбинация свойств адаптогенов делает их эффективным инструментом для
профилактики возратных болезней и продления жизни.
Большинство адаптогенов - питательные вещества или экстракты растений,
которые не вызывают вообще или вызывают незначительные
побочные эффекты. Глава 29 полностью посвящена адаптогенам.
Регулярное кратковременное воздействие "хорошего
стресса". Как отмечалось ранее, регулярные, но не непрерывные,
воздействия мягких или умеренных физических стрессоров, например
физических упражнений или умеренной гипоксии (умеренного
кислородного голодания, которое может быть достигнуто упражнениями
йоги), стимулируют многие адаптивные системы и делают организм
лучше подготовленным к тяжелому стрессу. Человек, закаленный
физическими упражнениями или дыханием йоги, обладает более
гибким стресс-ответом и быстрее достигает длительной
специфической адаптации к различным физическим и эмоциональным стрессо-
генным факторам.
Другой важный и часто упускаемый момент состоит в том,
что более желательно постоянно поддерживать повышенную
способность к адаптации, чем заново проходить полный ее цикл
при каждой стрессогенной ситуации. Другими словами, лучше
оставаться в форме, чем приобретать ее только тогда, когда это
абсолютно необходимо. Объяснение подобной точки зрения весьма
простое. Когда организм имеет повышенную способность к
адаптации (например, сильное сердце, легкие и мышцы, подвижную
хорошо сбалансированную иммунную систему), серьезная стрес-
согенная ситуация требуют слабого или умеренного
стресс-ответа и только до тех пор, пока не будет достигнута специфическая
277
адаптация. Кроме того, поддержание повышенной способности к
адаптации позволяет избежать слишком тяжелого бремени
структурных и энергетических затрат, неизбежных при низких
адаптационных возможностях системы. И это понятно: когда
необходимо каждый раз строить адаптацию "на пустом месте" (например,
выполнять тяжелые физические упражнения, скажем, один раз в
год в течение месяца), организм вынужден развивать
интенсивный и длительный стресс-ответ и вдобавок ко всему заново и пол-'
ностью строить специфическую адаптацию. В целом, второй сце-1
нарий более разрушителен, и чреват ускорением старения. В
терминах продления жизни практический итог таков: если вы реши-'
ли заниматься физическими упражнениями или йогой - держи-1
тесь этого стойко; вы должны заниматься этим не ежедневно, но ι
регулярно, например два-три раза в неделю.
Поддержание адаптации особенно важно у пожилых людей, по-1
тому что ее восстановление "на пустом месте" может быть трудным,,
физиологически дорогостоящим или даже невозможным.
Перуанские Анды - одна из всемирно известных областей обитания
долгожителей, хорошо приспособленых к гипоксии на больших высотах
(до 4 тыс. метров). Они имеют большее количество эритроцитов в
крови, более сильные сердца и легкие и более высокую плотность
капилляров в тканях, нежели жители равнинных областей. Если
пожилые обитатели Анд оказываются в местностях, расположенных
на уровне моря, и пребывают там несколько месяцев или лет, а
затем возвращаются в горы, у них часто развивается хроническая
высотная болезнь, которая является, по существу, состоянием
хронического стресса из-за неспособности адаптироваться к высотным
условиям жизни. Однако у них никогда не бывает подобных состояний,
если они не покидают гор. Следовательно, у стариков потеря
адаптации может быть необратимой.
Типичный пример состояния хронического стресса, который
стал результатом неспособности организма к адаптации - стойкая
сердечная недостаточность, т.е. неспособность сердечной мышцы
обеспечить нормальное кровообращение. Это состояние обычно
для пожилого возраста и осложнено, как правило, сердечными
приступами и продолжительной гипертонией. Когда организм
требует, будучи в нормальном состоянии, чтобы сердце
обеспечило более интенсивный кровоток, чем оно было в состоянии
обеспечить до этого, сердечная мышца постепенно
приспосабливается к увеличенной нагрузке, увеличиваясь в размере, наращивая
объем своих мышечных волокон и количество производящих
энергию митохондрий. При сердечной недостаточности, однако,
эта специфическая адаптация не развивается, и единственная
доступная помощь со стороны организма - бесконечный
стресс-ответ, который в действительности и наблюдается у таких
пациентов (Меерсон, 1981; 1984). У многих стресс настолько серьезен.
278
«ι го они существенно теряют в весе и испытывают поражение
многих нежировых тканей в результате избытка гормонов стрес-
I. Таких ситуаций, включая сердечную недостаточность, можно
избежать, предварительно улучшая общую способность
организма к адаптации (см. выше) и делая дополнительные шаги для под-
цсржания перегруженного органа при первых признаках
ухудшения или сбоя его функций. Например, такие природные вещества
как кофермент Q10 и L-ацетил-карнитин, которые повышают
.жергопродукцию в сердечной мышце (см. гл. 23), могут
увеличивать адаптивную способность сердца и предотвращать или даже
полностью устранять сердечную недостаточность.
Глава 17
СТРЕСС: "БОЛЕЗНИ АДАПТАЦИИ"
И ВОЗРАСТНЫЕ БОЛЕЗНИ
Большинство людей испытывают интуитивное ощущение,
что организм более восприимчив к болезням, когда он находится
в состоянии стресса. Простуда, язвы, головная боль, сердечные
приступы - все, кажется, случается с нами после травмирующих
ситуаций. Внезапные обострения уже существующих болезней
часто также приписываются влиянию стрессогенных событий
Хотя наши интуитивные понятия далеко не всегда верны, но в
этом случае, очевидно, имеется значительное согласие с
научными данными.
Первое свидетельство, что стресс может быть разрушителен
для тканей, пришло к нам из классических экспериментов Ганса
Селье на животных. В ЗСМЮ-х годах прошлого века он установил,
что острое напряжение вызывало у животных образование язв
желудка и истощение тимуса, селезенки, лимфатических узлов
(ключевых структур иммунной системы). Позже было найдено,
что острый стресс непосредственно повреждает сердечную
мышцу, вызывая обширные кровоизлияния и некроз ткани. В
последнее десятилетие доказана роль стресса как фактора этиологии и
патогенеза язвенных поражений слизистых оболочек желудка,
двенадцатиперстной и толстой кишки, гипертонической болезни,
атеросклероза, нарушений структуры и функции сердца,
вторичных иммунодефицитных состояний, возникновения ускоренного
роста и метастазирования новообразований, некоторых
эндокринных форм патологии (Selye, 1936; 1950; 1974; Селье, 1977; Me-
ерсон, 1981; 1984; Чирков, 1987; Эверли, Розенфельд, 1985;
Валъдман, 1986; Соколов, 1987; Барабой и др., 1983; Коган, 1977;
Desiderate et al., 1974; Parks et al., 1982).
Обширный список патологий, вызванных или усиленных
стрессом и названных Г.Селье "болезнями адаптации", включает
большинство возрастных болезней {табл. 14А).
Длительный стресс несомненно является также фактором,
отягощающим течение многих других заболеваний - инфекционных,
воспалительных, нервно-психических и т.п.. наоборот, кратковре-
280
Таблица 14А
Неполный список болезней, начало нлн развитие которых
может вызываться или усугубляться стрессом (Todorov Я. et al., 2001)
Стенокардия* Фнбромиалгия
Астма Головные болн
Аутоиммунные болезни** Гипертония*
Рак* Подавление иммунитета
Сердечно-сосудистые заболевания* Хронический колит
Обычная простуда Обширный склероз
Болезнь Крона Ревматоидный артрит
Диабет (II типа)* Язвенный колит
Депрессия** Язвенная болезнь
Возрастные болезни.
Возможно возрастные болезни.
менные и умеренные по силе стрессорные воздействия, активируя
механизмы неспецифической резистентности, оказывают
благоприятное влияние на течение многих заболеваний, различных по своей
этиологии и патогенезу.
17.1. ОБЩАЯ СХЕМА РАЗВИТИЯ СТРЕСС-ПАТОЛОГИИ
Гипотетическая схема развертывания стресс-реакции и ее возможные
патологические последствия на клеточном уровне с учетом роли ПОЛ в этом
процессе рассматривалась выше (см. рис. 4). Однако имеются схемы, более полно
суммирующие данные современной науки о патогенезе стрессорного поражения.
Наиболее интересной нам представляется схема, обоснованная и предоставленная
Ф. Меерсоном (1986а) (см. рис. 31). Эта схема базируется главным образом на
результатах изучения механизма стрессорных повреждений миокарда и основное
внимание акцентирует на гиперкатехолемии (вклад системы
гипоталамус-гипофиз-кора надпочечников (ГГКН) не рассматривается). Тем не менее есть
достаточно оснований (Барабой и др., 1992) считать, что во многих чертах она
отражает некоторые общие закономерности воздействия стресса н на другие органы и
системы млекопитающих. Согласно этой схеме (см. рис. 31) возбуждение высших
вегетативных центров, вызванное действием стрессоров (1 стадия),
распространяется на симпатоадреномедулярную систему, повышая поступление в циркуляцию
катехоламинов, которые взаимодействуют с адренорецепторами клеточных
мембран и стимулируют активность аденилатциклазы (II стадия). Далее развиваются
три ключевых процесса: повышается транспорт Са2+ в клетки, расходуются
резервы гликогена и включается так назывемая липидная триада (активация ПОЛ,
фосфолипаз и липаз и детергентного воздействия жирных кислот, которые
выделились нз фосфолипидов под влиянием ферментов) (III стадия). Именно эта
липидная триада обусловливает лабилизацию мембран лизосом, способствуя
выходу из них кислых протеаз и нуклеаз (IV стадия), Продукты ПОЛ - свободные
окислительные радикалы, гидроперекиси, органические перекиси, эпоксиды
и т.п., а также фосфолипазы, свободные жирные кислоты и лизосомальные
ферменты - вызывают повреждение плазматических и субклеточных (митохондрн-
281
Патогенетическая цепь стрессорного поражения
Факторы,
предупреждающие
стрессорные поражения
Возбуждение центров головного мозга,
детерминирующих стресс-реакцию
Избыток катехоламинов, активация
аденилатциклазы
III
Липидная
триада:
активация
|фосфолипаз,|
липаз, ПОЛ,
детергентно-
го действия
лизофосфа-
тидов,
жирных
кислот
IV
VI
νπ
Увеличение
вхождения
Са2+ в кар-
диомиоциты
Уменьшение резерва
и
обновления
глюкозы
Лабилизация
лизосом
Нарушения функционирования
Са-насоса и проницаемости
сарколеммы и саркоплазматического
ретикулума
_: Центральные
: тормозные ингибиторы ■
; Адреноблокаторы·
:Мембранопротекторы:
j ингибиторы ПОЛ,
: фосфолипаз, липаз
Избыток кальция
в сарколемме
Активация
фосфолипаз, протеаз,
контрактура миофнбрилл,
распад ДНК
Нарушения
сокращения и
расслабления миофибрилл и
функции
митохондрий, обратимые
повреждения ДНК
в миокарде
Нарушения растяжимости и
сократительной функции миокарда и снижение его ι
резистентности к гипоксии, микронекрозы
-■Блокаторы входа Са2+
-|Глюкоза, кофакторы
: ресинтеза гликогена
: Ингибиторы про-
: теаз и нуклеаз
Рис. 31. Последовательность этапов патогенеза стресс-реакции и пути коррек
ции стрессорных поражений миокарда (объяснение в тексте) (Меерсон, 1986а)
282
альных, эндоплазматических, ядерных) мембран. Нарушение при этом функции
катионных насосов приводит к внутриклеточному накоплению Са2+ (V-VI
стадии), а СР-атака ядерных структур (в сочетании с воздействием нуклеаз) влечет
ш собой появление одно- и даже двунитевых разрывов ДНК и повреждений
азотистых оснований, устраняемых, однако, в своем большинстве системами
внутриклеточной репарации. Гибель части клеток происходит вследствие нерепарируе-
мых повреждений ДНК, разрушения массы митохондрий и повреждения других
клеточных структур (VII стадия). Цепь этих событий приводит к значительной
утрате органом-мишенью его функциональных возможностей, более или менее
глубокому разрушению его структур и, наконец, к микронекрозам.
Однако возможно эффективное вмешательство в развитие стрессорных
повреждений на каждом из этапов с целью их ослабления (см. правую часть
схемы на рис. 31).
ВI стадии имеет место воздействие медиаторов и продуктов активации
центральных тормозных систем (гамма-окснмаслянная кислота, опиоидные и
другие тормозные пептиды) на возбужденные гипоталамические центры. II
стадия - это использование адреноблокаторов, ослабляющих эффект гиперкате-
холемии. В наиболее важной, III, стадии процесса происходит использование
стабилизаторов и протекторов мембран: АО, ингибиторов фосфолипаз, лнпаз,
блокаторов входа Са2+ внутрь клеток, а также регуляторов и субстратов ресин-
теза гликогена. На IV стадии эффективны ингибиторы протеаз и нуклеаз.
Представленная схема (см. рис. 31; Меерсон, 1986) не является полной.
В ней прежде всего не нашла отражения активация системы ГГКН, хотя Г. Се-
лье с самого начала отводил этому звену стресса весьма важную, если не
решающую роль. Возможно, для развития процесса в миокарде система ГГКН не
столь важна, однако в других органах-мишенях и системах организма ее
ключевая роль несомненна. Не показана также роль (более чем вероятной) первичной
активации ПОЛ (первичный медиатор) в ответ на воздействие стрессора, хотя
базальный уровень универсален и присущ всем живым системам на всех этапах
их жизнедеятельности. Поэтому всегда существуют и предпосылки для
активации этого процесса в результате экстремального воздействия в силу
подвижности про- и антиоксидантного равновесия.
Указанные упущения, однако, учтены в приведенной ранее схеме
развертывания стресс-реакшш (см. рис. 30), предложенной другими авторами (Бара-
юй и др., 1992).
Читатель, должно быть, уже оценил, что стресс стимулирует в
)рганизме множество взаимоснязанных изменений, которые
улучшают его шансы на выживание перед лицом непосредственной фи-
шческой опасности, но в других ситуациях он может быть вреден.
По-видимому, имеется глубокая связь между механизмом
повреждений, индуцированных стрессом, старением и развитием
"адаптационных" и дегенеративных (возрастных) болезней. Ниже мы кратко об-
удим эту связь на конкретных примерах.
17.2. СТРЕСС И СЕРДЕЧНО-СОСУДИСТЫЕ БОЛЕЗНИ
Стресс может наносить сердцу прямые и косвенные
повреждения. Катехоламины, эти гормоны стресса, выделяемые мозговым
веществом надпочечников и нервами симпатической системы,
оказывают прямое влияние на сердечную мышцу. Большие количества
283
катехоламинов, "выброшенные" в кровь во время первой фазы
стресс-ответа (реакция "борьбы или бегства"), побуждают
сердечную мышцу к более частым и более сильным сокращениям. В то же
самое время катехоламины повышают давление крови, подвергая
сердце дополнительной нагрузке. Увеличенная таким образом
деятельность сердечной мышцы требует большего количества энергии,
которая производится путем окисления, соответственно, большего
количества биохимического топлива. Это ведет к формированию
повышенного содержания свободных радикалов (СР), которые
способны, как это уже не раз подчеркивалось, повреждать клеточные
структуры и нарушать жизненно важные клеточные функции. В
частности, перекисное окисление липидов, вызванное СР, приводит к
повреждению мембран клеточных органелл и выходу их
содержимого наружу, что (1) понижает энергетическую продуктивность и
нарушает внутриклеточный гомеостаз, если повреждены мембраны
митохондрий, и (2) вызывает дезорганизацию метаболизма в целом,
если повреждены мембраны лизосом, из которых выйдут мощные
пищеварительные ферменты. Если свободно-радикальные
разрушения будут достаточно серьезными, некоторые клетки не смогут
выжить. В отличие от печени или кожи, клетки сердечной мышцы не
могут быть заменены путем деления; сердце компенсирует эти
потери, увеличивая питание и работоспособность оставшихся клеток.
Эта их повышенная функциональная активность будет эффективна
до тех пор, пока адаптивная способность сердца не истощена, после
чего может наступить клиническая сердечная недостаточность, тем
более, что старение стимулирует изменения в фенотипе коронарных
артериол, чему способствует развитие окислительного стресса,
которое ослабляет весьма благотворное посредничество NO в
расширении сосудов, питающих сердце (Csiszar et al., 2002).
В дополнение к необходимости вести образ жизни, позволяющий избегать
стрессогенных ситуаций, когда это возможно, сердечная мышца может быть
защищена от стрессовых повреждений высокими дозами антиоксидантов.
По мере старения увеличивается число людей, нуждающихся в кардиаль-
ных операциях. Однако у пожилых результаты таких операций хуже, чем у
молодых. Вероятный содействующий этому фактор - возрастное снижение
энергетической продуктивности клеток в миокарде в течение операции, которое,
как известно, стимулируется анаэробным и ишемическим стрессом.
Липофильный кофермент Q10 - тиооксидант и митохондриальный
соединительный элемент окислительно-восстановительного потенциала
дыхательной цепи, который способен улучшить энергетическую продуктивность в
митохондриях, шунтируя неисправные компоненты дыхательной цепи, так же как и
уменьшить эффект окислительного стресса. Предполагают, что
предварительное введение CoQIO до стресса (до операции) может улучшить восстановление
миокарда после стресса (Rosenfeldt et al., 2002).
Витамин Ε, основной клеточный жирорастворимый антиоксидант, era »*-
бен, как было установлено, защитить сердечную мышцу животных при ост^м
стрессе. Вероятно, и другие жирорастворимые антиоксиданты (липоевая
кислота или мелатонин) также способны снижать такие повреждения. Недавно были
284
исследованы антиоксидантные свойства деривата липоевой кислоты - (11)-аль-
фа-липоевой кислоты (R-JIK). Кардиальные миоциты старых крыс,
получавших R-JTK, показали значительно более низкую скорость генерации продуктов
окисления, сравнимую с таковой у интактных крыс. Пищевые добавки с R-JIK
восстанавливали также уровень аскорбиновой кислоты в миоцитах и
сокращали уровень окислительных повреждений ДНК. Старение сердца сопряжено
с окислительным стрессом (главным его источником являются митохондрии),
который существенно уменьшается добавкой (И)-альфа - липоевой кислоты
(Sw/ietal.,2001).
Адаптогены, природные антистрессовые агенты, также проявляли
подобные положительные качества (см. гл. 29).
Острый стресс может вмешиваться в сердечную функцию,
нарушая нормальный сердечный ритм. Этот эффект обусловлен
большим количеством катехоламинов, быстро выделяющихся при
интенсивном стрессе ("катехоламиновый шторм*')· Серьезные
нарушения сердечного ритма могут вести к глубокому поражению
сердечной функции и смерти. Индивид может быть буквально "напуган до
смерти", "умирать" от вспышки гнева или после какого-нибудь
трагического случая - все это не только "события" второразрядных
романов или фильмов, все это случается с реальными людьми.
Каждый год более чем 400 тыс. человек в США становятся жертвами
внезапной смерти. По-видимому, главной причиной таких
смертельных исходов является серьезные нарушения сердечного ритма, при
котором желудочки сердца начинают неудержимо и бесконтрольно
трепетать ("мерцательная аритмия"), теряя способность вернуться
к нормальному ритму сокращений.
В 1971 г. Г. Энгель (G. Engel) опубликовал результаты анализа
170 случаев внезапной смерти. Большинство таких случаев
произошло вскоре после развития острого стресса в результате смерти
или угрозы потери кого-то близкого, персональной опасности или
ее угрозы, или иногда даже интенсивной радости. Сообщались
некоторые примеры:
52-летний мужчина был в близком контакте со своим врачом в течение
последней стадии болезни его жены, страдавшей раком легкого. Он умер
внезапно от массивного инфаркта миокарда через день после ее похорон.
40-летний отец скоропостижно скончался, когда подкладывал подушку под
голову своего сына, который лежал серьезно травмированный на проезжей
дороге, сбитый мотоциклистом.
52-летний президент колледжа, гордившийся своим покровительством
черных студентов, умер, когда узнал, что группа взбунтовавшихся черных
студентов захватила здание администрации.
75-летний мужчина, выигравший несколько тысяч долларов на ставку
всего в два доллара, умер у окошка, получая наличные деньги на свой
"счастливый" билет.
Конечно, избежать многих из таких стрессогенных ситуаций
невозможно, ибо это, так сказать, предопределено судьбой. Тем не
менее состояние организма во время такого острого стресса может ли-
285
бо предрасполагать, либо защищать от внезапной смерти.
Например, чувствительность сердечной мышцы к катехоламинам
повышается при дефиците калия и магния и понижается при высоком
потреблении этих полезных минералов. Высокое потребление калия и
магния, содержащегося в больших количествах во фруктах и
овощах, по-видимому, уменьшит риск внезапной смерти. Аминокислота
таурин (производное таурохолевой кислоты) - другое питательное
вещество, которое "умиротворяет" сердце и уменьшает риск
внезапной смерти от "катехоламинового шторма". Еще более важно
общее состояние здоровья сердца. В различных исследованиях от 60 до
90% жертв внезапной смерти ранее уже имели существенную
коронарную недостаточность сердца. Дезорганизация ритма, как
известно, более вероятна у сердца, которое не получает достаточное
количество кислорода из-за частичной блокады коронарных артерий
склеротическими бляшками.
Атеросклероз - другой механизм, влияя на который стресс вносит
вклад в болезни сердца. Гормоны стресса стимулируют ряд
метаболических изменений, способствующих атеросклерозу, включая
повышенный сахар в крови, пониженную чувствительность к инсулину и
высокий уровень свободных жирных кислот. Стресс также
"благоприятствует" (повышенный уровень альдостерона) развитию гипертонии,
которая является сильным фактором риска для атеросклероза.
Ряд исследований пробуют связать социальное поведение людей
с риском сердечно-сосудистых нарушений. Так, Р. Розенман
(Rosenman), известный эксперт поведенческих факторов риска
сердечных заболеваний, определяет психический тип людей, склонных
к таким заболеваниям, следующим образом:
Индивиды, которые находятся в постоянном стремлении скорее
достичь своих целей при соревновании с другими людьми или противостоящими
силами окружающей среды, часто показывают ряд особенностей поведения,
известных как характер поведения типа А (ХПТА). Этот тип включает
такие поведенческие склонности, как честолюбие, агрессивность, стремление
конкурировать и нетерпимость; специфическое поведение с такими
особенностями, как настороженность, напряженные мышцы, быстрая и решшпель
ная стилистика речи, и такие эмоциональные реакции, как повышенная раз
дражимость и выраженные признаки гнева.
Если вернуться к физиологическим терминам, то ХПТА следует
связывать с повышенной реакцией на стрессогенные факторы,
когда большинство событий и ситуаций воспринимается (часто
подсознательно) как весьма напряженные. В результате стресс-ответ
организма может быть почти хроническим, способствуя развитию
болезней сердца. Мы должны обратить внимание, что ХПТА - не
характеристика индивидуальности, а в значительной степени тип
поведения, который может быть востребован или не востребован в
зависимости от уровня мотивации. В цивилизации, которая базируется
на соревновательности и конкуренции, общество стимулирует интерес
286
Экзогенные
факторы:
эмоциональные
ситуации
окружающей
среды
Эндогенные
факторы:
боль,
страх смерти
Атеросклероз
коронарных
артерий
Некоронарогенное
адренергическое
повреждение миокарда
Адренергический
контрактурный
коронарный спазм
Повышение
свёртываемости
крови, коронаро-
тромбоз
Снижение
резистентности
миокарда к
гипоксии и ишемии
Острая перегрузка
сердца
Падение тонуса
емкостных сосудов,
депонирование
в них крови
Сужение коронарных
артерий вследствие
гипервентиляции и
алкалоза
/Ишемическое\
^ повреждение J
миокарда
Рис. 32. Схема развития стресс-реакции, вызванной психоэмоциональными фа
кторами (Меерсон, 1986а)
к ХПТА, считая полезными достижениями превосходство, победы,
неустанное стремление к успеху и т.п. Стимулом для того чтобы не
следовать идеалам ХПТА или по крайней мере уменьшить их значе
ние, может быть желание понизить риск болезней сердца. К
сожалению, часто требуется перенести сердечный приступ, чтобы человек
получил достаточную мотивацию отказаться от ХПТА.
287
Столь важная роль психоэмоциональных факторов в патогенезе
заболеваний сердца стимулировала огромное число
медико-биологических исследований в этой области. Обобщение результатов
этих, а также обширных собственных исследований содержится в
известных публикациях Ф.З. Меерсона (1981, 1984, 1986а, б), а
также в схематическом виде представлено на рис. 32.
Очень много написано о необходимости модификации
поведения ради здоровья, включая изменение жизненной философии,
отношений, восприятия и реакций на события. Обсуждение всех этих
предметов - вне возможностей авторов этой книги. Мы только
обратим внимание, что одним из ключевых факторов ХПТА
является страстное стремление контролировать или управлять
(насколько это достижимо, а часто и недостижимо) окружающей
обстановкой. Многое в окружающем мире существует вне нашего
контроля (управления), и это обстоятельство может вызывать у людей с
ХПТА частые разочарования, расстройства, гнев и вообще вести к
более высокому уровню стресса. Мы не хотели бы утверждать, что
нужно полностью отказаться от активного контроля над жизнью,
расслабиться и плыть по течению. Здесь важны серьезные оценки
жизненных ситуаций и выводы из них, сделанные на основе
здравого смысла. В связи с этим приведем цитату из книги С. Орента
(Orent) "Стресс и сердце":
Что оказывается необходимым для [типа] А, так это оценка
приоритетов и способность разделить в реальной жизни ее вызовы на существенные и
второстепенные. Насколько это важно, становится ясным, когда задаешь
себе вопрос — а не разумно ли избежать такого раздражения, которое мы,
например, испытываем в уличной автомобильной пробке, или подвергаемся "ка-
техоламиновому шторму" ради продвижения на работе, если последствием
этих переживаний может стать коронарная сердечная болезнь?
Изменить поведение с целью минимизировать
гиперреактивность на действие стрессогенных факторов и таким образом
сохранить здоровье представляется стоящей, но нелегкой задачей. При
работе над таким изменением можно получать существенную
защиту от стресса с помощью адаптагенов, физических упражнений и
методов расслабления.
17.3. СТРЕСС И ГИПЕРТОНИЯ
Гипертония определяется как артериальное давление крови
большее чем 140/90 мм ртутного столбика тонометра. Часто не
имея симптомов, гипертония, особенно если она серьезна,
значительно увеличивает риск инсульта, инфаркта, сердечной
недостаточности, болезней почек и глаз. Чрезмерно высокое кровяное
давление приводит к ряду изменений в артериях и капиллярах, которые
увеличивают риск их разрушения и развития атеросклероза, а также
288
вызывает перенапряжение сердечной мышцы, что может вести к
сердечной недостаточности.
Причины гипертонии сложны и все еще не полностью поняты.
Обычно развитие гипертонии определяется некоторой комбинацией
следующих трех факторов: (1) увеличенный объем крови, (2)
повышенное сопротивление сосудов току крови (сосуды чрезмерно
сжаты, не позволяя крови течь "легко'*) и (3) повышенная
функциональная активность сердца (сердечный насос из-за п. 2 вынужден
работать с перегрузкой). Все эти три фактора могут появиться под
влиянием стресса. Стресс увеличивает объем крови, стимулируя
задержку натрия и выделение калия почками. Этот эффект связан прежде
всего со стероидами стресса - альдостероном и кортизолом,
первый - непосредственно действуя на почки, второй - косвенно,
понижая чувствительность тканей к инсулину и соответственно вызывая
его избыток, что также способствует задержке натрия. Стресс
также увеличивает сопротивление сосудов, особенно в его первый
фазе "борьбы или бегства", активируя симпатическую нервную
систему, которая заставляет сжиматься кровеносные сосуды в
большинстве органов. И наконец, стресс увеличивает работоспособность
сердца: катехоламины, выделенные надпочечниками и
симпатической нервной системой, заставляют сердечную мышцу сокращаться
быстрее и более интенсивно.
Стресс-реакция безусловно увеличивает кровяное давление в
момент стресса. Но что в "спокойной обстановке" постоянно
вносит вклад в повышение кровяного давления, которое и составляет
гипертонию? Очень трудно изучать эту проблему непосредственно
на людях. Существуют косвенные указания, что стресс
действительно "виновен" в развитии гипертонии. Так, большинство
исследователей соглашаются, что люди, чей стресс-ответ бывает особенно
острым и легко вызываемым, подвержены значительному риску
развития гипертонии. Согласно современным представлениям, развитию
устойчивой гипертонии предшествует период так называемой
лабильной гипертонии, когда повышение кровяного давления
происходит эпизодически и главным образом под влиянием стресса.
Чем чаще и интенсивней возникает стресс, тем выше шанс
лабильной гипертонии прогрессировать к явно выраженной болезни.
Исследования на грызунах показали, что крысы с ограниченной
гипертонией развивали постоянную и глубокую гипертонию, когда
попадали в стрессогенную ситуацию, вызванную неожиданными
наказаниями. Основной центр контроля и регуляции кровяного
давления - гипоталамус. Думается, что часто повторяющаяся
эпизодическая гипертония так или иначе воздействует на "чувствительность
датчика" так, чтобы гипоталамус начал ощущать высокое кровяное
давление как нормальное и поэтому поддерживал его, воздействуя
на соответствующие системы организма, "невольно" способствуя
развитию хронической гипертонии. Предположительно стресс так-
289
же воздействует на гипоталамические "датчики" кровяного
давления, эпизодически его повышая. Гормоны стресса, кроме того,
могут повреждать гипоталамические нейроны, делая "датчики"
кровяного давления менее чувствительными.
Эпидемиологические исследования указывают, что гипертония
преимущественно распространена в развитых, особенно Западных
странах. Это обычно приписывают особенностям диеты,
популярной в этих странах, в частности, низкому содержанию калия и
магния и высокому - натрия и насыщенного жира. Однако это,
по-видимому, только часть картины: в психологическом отношении
напряжение, свойственное современному образу жизни и многим типам
трудовой деятельности, также заслуживает в данном контексте
многих упреков.
Обычное лечение гипертонии базируется главным образом на широком
разнообразии лекарств, которые действуют, расслабляя кровеносные сосуды,
успокаивая сердце или способствуя уменьшению объема крови, стимулируя
выделение натрия почками. Большинство этих лекарств может вызывать,
серьезные побочные эффекты. Это следует принимать во внимание особенно в тех
случаях, когда проводить профилактику гипертонии значительно легче, чем ее
лечить. Простые меры для предотвращения гипертонии прежде всего
включают диету с высоким содержанием калия и магния и разумно низким - натрия.
Для этого нужно употреблять с пищей изобилие фруктов и овощей и
ограничить потребление соли. Регулярные физические упражнения и расслабляющий
отдых Также являются хорошей защитой от гипертонии. Адаптогены, особенно
сибирский женьшень, являются другим средством предотвращения гипертонии
и лечения ее легких форм. Необходимо, однако, иметь в виду, что серьезная
устойчивая гипертония очень опасна, и ее лечение медикаментозными
средствами абсолютно оправданно.
17.4. СТРЕСС И РАК
Рак - сложная болезнь. Много факторов может вносить вклад в
ее патогенез: диета, контакт с канцерогенными веществами,
старение и, очевидно, стресс. Полемика о роли стресса в развитии рака
длилась в течение нескольких десятилетий (Балицкий и др., 1987).
Многие исследования показывают, что когда канцерогены вводятся
стрессированным животным, опухоли у них развиваются
значительно чаще, чем у контрольных животных. В экспериментах, когда
раковые клетки вводились животным в состоянии стресса, отмечалась
более быстрая прогрессия опухоли. Но уместно ли
экстраполировать эти результаты на людей? В конце концов лабораторные
методы исследования, используемые, чтобы вызвать стресс и рост
опухоли у животных, к счастью, весьма отличаются от событий реальной
человеческой жизни. Невозможность непосредственного
исследования влияния стресса на развитии рака у людей, побуждают ученых
проводить так называемые корреляционные исследования.
Например, отбирают группу больных раком и другую - неонкологических
290
пациентов, причем так, чтобы эти группы были сопоставимы по
возрасту, полу и многим другим переменным, насколько это
возможно. Далее, определяли количество индивидов в каждой группе,
кто находился под серьезным или длительным стрессом в пределах
определенного периода в прошлом. Если группа чбольных раком
имела достоверно большее (со статистической точки зрения)
количество стрессогенных событий в прошлом, чем контрольная группа,
это могло косвенно свидетельствовать, что стресс может быть
одной из причин рака.
Таких коррелятивных исследований было проведено много, и
большинство указывало, что стресс вносит вклад в развитие рака.
Особенно внушительны данные относительно рака молочной желе-,
зы, который является ведущим типом злокачественного
новообразования у женщин. В изучении 1088 таких пациенток, которое
провели в Центре рака Греческого отделения социального обеспечения
в Афинах (loannidou-Mouzaka et al., 1986), была найдена весьма
существенная корреляция между стрессом и раком молочной железы
(при высокой статистической значимости этого исследования).
Типы стрессогенных событий, которые обычно коррелировали с
раком, это - смерть любимых, негативное поведение мужа,
неожиданные изменения в образе жизни, непрерывные семейные конфликты,
финансовые проблемы, неудовлетворенность сексуальной жизнью,
консультации с психиатром и аллергии. Замечательно, что
корреляция рака молочной* железы со стрессом была более выраженной,
чем с любым другим фактором риска, включая наследственные
факторы этого заболевания. В другом крупном исследовании
2163 женщин (Cooper et al.) также была установлена корреляция
между значительными стрессогенными событиями их жизни и раком
молочной железы. Связь была особенно четкой у тех
женщин,,которые оказались не способны выразить свои эмоции и получать
необходимую помощь и совет. К. Брила (Bryla, 1996) суммировал
имеющиеся литературные данные по этому вопросу в обзоре 1996 г.:
Хотя трудности количественных оценок затрудняют доказательства
материальных взаимоотношений между стрессом и раком молочной железы,
исследования показывают, что стресс связан с этой разновидностью рака
различными способами.
Признав реальность связи "стресс-рак" (Eysenck, 1988), мы
должны понять, каковы ее механизмы. Одна из теоретических
возможностей состоит в том, что изобилие свободных радикалов,
генерированных стрессом, может вызвать дополнительные мутации в
генах и этим увеличить риск злокачественного перерождения
клеток. Другая, лучше документированная возможность, базируется на
хорошо известном угнетающем действии стресса на функцию
иммунной системы. Согласно современным представлениям, раковые
клетки возникают в организме весьма часто, но подавляющее боль-
291
шинство их оказываются "помеченными" и разрушенными
иммунной системой, главным образом клетками N- и Т-киллерами (гл. 6).
Крайне редко отдельным раковым клеткам удается избежать
пристального надзора со стороны иммунной системы и развиться в
опухоль. Полагают, что спонтанная ремиссия (редкое событие, когда
опухоль полностью регрессирует без лечения) возникает в
результате "пробуждения" иммунной системы в присутствии рака. Стресс,
однако, может заметно подавлять деятельность иммунной системы
и, возможно, ослаблять антираковую защиту организма. Под
влиянием стресса у животных и людей наблюдалось общее подавление
активности N- и Т-киллеров.
Согласно недавнему исследованию, одна из причин
ограниченной эффективности обычных методов лечения рака - та, что
операционная хирургия, химиотерапия и радиация вызывают стресс,
достаточно серьезный, чтобы значительно подавить иммунную
систему. Это позволяет нескольким выжившим раковым клеткам
размножаться и распространяться в теле без помех. Многие
исследования на животных и некоторые исследования на людях показали, что
объединение обычных способов лечения рака со средствами,
которые смягчают стресс-ответ (в частности, адаптогены) или
стимулируют иммунную систему (например, мелатонин), уменьшают риск
рецидива.
В параграфе 17.2 мы упомянули те сердечные болезни, которые
связаны с определенным типом поведения, названным типом А
(враждебный, агрессивный, соперничающий, энергичный и т.д.). Раньше
ученые попытались определять образец поведения,
ассоциированный с более высоким риском заболевания ракам. Поведение,
располагающее организм к раку, было названо типом С, и, как это ни
удивительно, оказалось, что оно в значительной мере
противоположно типу А. Индивидуумы типа С характеризуются стремлением
избегать травмирующих событий, подавлением эмоций,
"патологической любезностью", желанием избежать конфликтов,
сверхуступчивостью и сверхтерпением. Балтруш и его коллеги (Baltrusch et al.,
1991) из Медицинского университета в Ганновере, Германия,
проанализировав исследования о связи поведения типа С и рака, пришли
к интересным заключениям.
Эта модель поведения, обычно скрываемая за фасадом приятности, по-
видимому, эффективна до тех пор, пока поддерживается стабильность
окружающей среды и психологический гомеостаз, но разрушается со временем под
воздействием накопленных импульсов напряжения и стрессогенных
факторов, особенно тех, которые вызывают депрессию и чувства беспомощности
и безнадежности. В качестве заслуживающей особого внимания особенности
такого типа поведения выступает чрезмерное стремление избегать
всяческих осложнений, сдерживание и подавление эмоций и своих основных
потребностей, что, по-видимому, ослабляет естественную устойчивость организма
к канцерогенным влияниям.
292
Существуют свидетельства, что этот неадекватный, репрессивный
и даже "ажурный" стиль поведения, присущий индивидуумам с
поведением типа С, сопряжен с непропорционально длительным
стресс-ответом и ведет к подавлению многих аспектов иммунитета, включая
антираковую защиту. Вспомним, что поведение типа А (ХПТА)
увеличивает риск сердечной болезни, чрезмерно стимулируя стресс-ответ.
Но если типы А и С, т.е. обе поведенческие модели, влияют на
здоровье, увеличивая стресс-ответ, то почему одна из них способствует
сердечной болезни, а другая - раку? Очевидно, оба типа поведения
стимулируют стресс-ответ различными путями. У индивидов типа А,
первая фаза стресс-ответа (реакция "борьба или бегства") чрезмерно
интенсивна, при этом выделение катехоламинов и возбуждение
симпатической нервной системы повышает кровяное давление, что
становится дополнительной нагрузкой для сердца и артерий. У индивидов
типа С реакция тревоги бывает относительно слабой, но следующие
фазы стресс-ответа чрезмерно интенсивны или продолжительны из-за
недостаточной эффективности первой фазы. Эти поздние фазы
стресса характеризуются высокими уровнем кортизола, который играет
решающую роль в подавлении иммунной системы. А это может вредить
антираковой иммунной защите.
Подобно типу А, тип С не является характеристикой
индивидуальности, а скорее - моделью поведения, которое может быть
востребовано или не востребовано. Шаги к преодолению поведения
типа С могут быть связаны со стремлением к такому самоощущению,
которое побуждает одинокого человека вступать в эмоциональный
контакт с другими людьми, пользоваться их советами и поддержкой
в моменты кризиса. Нужно иметь в виду, что типы поведения А и С -
крайние области весьма обширного спектра, но при этом важно
отметить, что неидентифицированный тип поведения автоматически
не становится одним из этих крайних типов.
Конечно, не каждый может круто изменить поведение и далеко
не всякая окружающая обстановка будет способствовать такому
изменению. И конечно, трудно избежать многих действительно стрес-
согенных событий. Тем не менее всегда сохраняется возможность
уменьшить негативное воздействие стресса на риск раковых
заболеваний, смягчая стресс-ответ и стимулируя иммунную систему
рациональным питанием, дополненным соответствующими биологически
активными веществами (см. гл. 23, 25, 29).
17.5. СТРЕСС И ДИАБЕТ
Согласно оценкам Американской ассоциацией диабета каждый
год эта болезнь вызывает в 176 тыс. случаев смерть, в 54 тыс. -
ампутацию нижних конечностей, в 24 тыс. - слепоту. В то время как
сама болезнь не имеет конкретных симптомов, ее осложнения вклю-
293
чают сердечные болезни, инсульты и инфаркты, повреждение
нервов и периферического кровообращения, недостаточность почек и
вырождение сетчатки глаз.
Диабет определяется как постоянное повышение содержания
глюкозы в крови. Согласно новым руководящим принципам
Американской ассоциации диабета, выпущенным в 1997 г., пациент
рассматривается как диабетик, если содержание глюкозы в его крови
натощак равно или более чем 125 мг/дл в более чем в двух случаях.
Существуют, как было сказано выше, два главных типа диабета - I и
II. Тип I - диабет, зависящий от инсулина, обычно возникает в
юности и характеризуется почти полной неспособностью организма
производить инсулин, гормон, который требуется большинству
клеток для потребления глюкозы. Тип II - это инсулин-независимый
диабет, который имеет тенденцию проявляться в среднем или старшем
возрасте, когда организм способен производить инсулин, но клетки
не чувствительны к нему.
Гипертрофия панкреатических β-клеток является важной
компенсаторной реакцией на гипергликемию и резистентность к инсулину. Такая
компенсаторная реакция помогает этим клеткам выжить, она сопряжена с
индукцией группы так называемых защитных генов, кодирующих, в частности,
антиоксидантные ферменты Cu/Zn-СОД, Мп-СОД, глутатион пероксидазу,
оксигеназу гема-1 и антиапоптотические белки, и облегчает адаптацию
β-клеток к хронической гипергликемии, следовательно, и к увеличенной
потребности в инсулине, которые сопровождают оксидативный стресс и
старение {Laybutt et al.f 2002).
Диабет - это болезнь, распространенность которой в мире не
может не произвести впечатления, составляя примерно 5% от всей
человеческой популяции. Более чем 90% от всех случаев диабета
являются типом II. В пожилом возрасте больных диабетом более
20%. Кроме того, большее число людей находятся в состоянии
преддиабета (названном "углевод/глюкозной интолерантностью",
или скрытой формой диабета), это означает, что у них уровень
глюкозы в крови выше, чем желательный, но несколько ниже, чем
это следует из стандартного определения диабета. Примечательно,
что в США критерии для диабета были недавно пересмотрены в
сторону более низкого уровня содержания сахара в крови, так что
упомянутые выше оценки распространенности этого недуга,
вероятно, занижены.
Независимо от того, как все это вычисляется, диабет -
исключительно распространенная болезнь, и риск заполучить ее
увеличивается с возрастом. Кроме того, некоторая степень интоле-
рантности к углеводам настолько обычна, что некоторые ученые
рассматривают это как составную часть "нормального" старения.
В.М. Дильман, автор нейроэндокринной теории старения,
полагал, что постепенное ухудшение толерантности к углеводам с
возрастом зависит от центральных часов старения, локализованных
294
в гипоталамусе. Согласно Дильману, сохранение толерантности
тканей к углеводам должно быть одним из краеугольных камней
стратегии антистарения. Действительно, исследования на
грызунах показали, что повышение восприимчивости тканей к
углеводам увеличивает продолжительность жизни. Разнообразные
трудности пока препятствуют проведению подобных
исследований на людях. Удивительно, однако, как много долговременных
осложнений диабета имеют сходство с некоторыми признаками
преждевременного старения.
Стресс, по-видимому, один из факторов риска в развитии пред-
диабета и диабета II типа (Brindley et al., 1989). Другие факторы - это
возраст после сорока, ожирение, недостаток физической
деятельности и, возможно, дефицит хрома. У грызунов, наследственно
предрасположенных ко второму типу диабета, стресс явно вызывает эту
болезнь. Адаптогены, средства, которые смягчают стресс-ответ,
предотвращают также и так называемый аллоксановый диабет у
крыс (лабораторная версия диабета), вызванный препаратом аллок-
сана, который повреждает В-клетки поджелудочной железы,
производящие инсулин (Дардымов, 1976). Исследования стресса в
качестве причины диабета у людей - слишком малочисленны для
категорических заключений. И все же нет сомнений, что стресс усиливает
уже существующий диабет.
Стресс оказывает глубокое влияние на метаболизм и, в
частности, на утилизацию в организме глюкозы. Как уже
неоднократно указывалось, одна из основных биологических целей стресс-
ответа - обеспечить дополнительное количество
энергоносителей для усиленной деятельности нервной и кровеносной систем в
критической ситуации. Это достигается, наряду с другими
средствами, увеличением уровня глюкозы в крови. Гормоны стресса,
особенно кортизол, уменьшают реактивность большинства
тканей к инсулину, сокращая их потребление глюкозы. Это
способствует повышению ее уровня в крови, и большее количество
топлива становится доступными для нервной системы, которая,
подчеркнем, не нуждается в инсулине, чтобы поглощать глюкозу.
Таким образом, при стрессе возникают такие изменения в
метаболизме глюкозы, которые весьма похожи на ситуацию при
диабете Π типа: пониженная реактивность тканей к инсулину, ведущая
к повышению уровня глюкозы в крови. В то время, как все еще
обсуждается вопрос, может ли стресс постоянно "подталкивать"
человека ко II типу диабета, совершенно очевидно, что стресс -
наиболее общая и распространенная причина гипергликемиче-
ских кризисов и важнейший фактор в осложнении уже
существующего диабета.
Наконец, связь между диабетом II типа и старением может
осуществляться гормонами стресса. Как уже указывалось, продукция
кортизола и в покое и при стрессе - демонстрирует с возрастом
295
Таблица 15
Сопоставление гормонально-метаболических изменений при старении,
стрессе и возрастных болезнях
(Дильман, 1987)
Признак
Явление
старение
стресс
возрастные
болезни
Снижение уровня биогенных аминов ■J-
в гипоталамусе
Повышение порога +
чувствительности гипоталамуса
Гиперкортицизм ( гиперадаптоз) +
Гиперсоматотропизм +*
Снижение толерантности к глюкозе +
Гиперинсулинемия +
Ожирение +
Усиление окисления жирных кислот -1-
Повышение в крови холестерина
и триглицеридов
Подавление клеточного иммунитета
Тромбообразование +
Стимуляция опухолевого процесса +
Примечания:
* Снижение по мере развития ожирения
** Выявляется в нагрузочных тестах
тенденцию к увеличению. Таким образом, с эндокринной точки
зрения пожилые люди живут в состоянии хронического стресса. Это
обстоятельство, по-видимому, может стать одним из ответов на
вопрос, почему диабет - возрастная болезнь.
В заключение этой главы нам хотелось бы еще раз (наряду с
патологическими аспектами) подчеркнуть существенное сходство
гормонально-метаболических изменений, сопровождающих старение,
стресс и нормальные, т.е. возрастные болезни. Поразительное
сходство целого ряда признаков этих, казалось бы "независимых",
явлений (табл. 15) указывает на глубокую общность фундаментальных
механизмов, лежащих в основе их развития.
Глава 18
СТРЕСС И ИММУННАЯ СИСТЕМА
Крепкая, хорошо скоординированная иммунная система
необходима для выживания и долголетия, она служит главной линией
защиты против инфекции, а также важным фактором в
предотвращении некоторых дегенеративных болезней, в частности рака и
атеросклероза. Функция иммунной системы существенно
снижается с возрастом, что увеличивает риск смерти от инфекции или
развития упомянутых выше патологий. Продление жизни,
поддержание здоровья и жизненной энергии в пожилом возрасте зависят от
хорошо функционирующей иммунной системы. Помимо старения
и дефицита питательных веществ, стресс выступает самым
опасным врагом иммунной системы. В то время как молодой человек
способен успешно противостоять временному ухудшению
иммунной функции, более старые индивиды, обладающие меньшими
резервами, не в состоянии удовлетворительно переносить удары по
их иммунной системе. Кроме того, пожилые люди более
подвержены негативным влияниям стресса, поскольку они имеют
склонность к перепроизводству кортикостероидов как при стрессе, так
и в состоянии покоя. Вот почему необходимо знать, как стресс
подавляет иммунную систему, и что можно сделать, чтобы
предотвратить связанные с этим повреждения.
Воздействие стресса на иммунную систему впервые было
открыто Г. Селье, когда у животных, подвергнутых стрессу, он
обнаружил истощенные вилочковые железы (тимус) и атрофированные
лимфатические узлы, что указывало на пониженную функцию
иммунной системы (Selye, 1936). Позже, по мере развития
иммунологии, выяснилось, что стресс затрагивает почти все аспекты
иммунного ответа, от производства антител до активации Т-клеток и
фагоцитоза. Например, такие стрессогенные факторы, как шум
высокой интенсивности или вирусная инфекция, подавляли производство
интерферона у мышей и приводили к повышению смертности
животных. Погружение крыс в холодную воду сокращает их
выживание при инфекции вирусом гриппа А. Социальный или
психологический стрессы столь же, если не более, чем физический или смешан-
297
ный, вредны для иммунной системы. И социальная изоляция, и
перенаселение вредят иммунной системе млекопитающих. Отделение от
матери и перевод в равное положение с остальными особями резко
подавляет иммунную систему молодых животных. У младенцев
макаки, отделенных от своих матерей, в течение двух недель
наблюдалось ослабление ответа на стимуляторы деления клеток (цитокины),
что проявлялось в снижении размножения лимфоцитов (Fontenot
etal., 1995).
Теперь уже не вызывает сомнений, что стресс ускоряет рост
опухолей, способствует развитию метастазов и повышению общей
смертности от рака (Балицкий и др., 1987). Большинство
исследователей полагают, что эти эффекты являются следствием вызванного
стрессом подавления иммунной системы, в особенности ухудшения
функции N-киллеров и Т-лимфоцитов.
Более тонкие исследования влияния стрессогенных факторов
физической и психоэмоциональной природы показали, что
ситуация, при которой животное теряет всякий контроль над
неблагоприятными условиями, вызывает большую иммунную подавленность,
чем физически подобная ситуация, но с некоторой степенью
контроля. Например, единственный сеанс неизбежного удара током
увеличивает рост опухоли у крыс. Однако у животных, которым дана
возможность избежать удара, никакого повышения роста опухоли не
наблюдается. Это, по-видимому, показывает, что даже физический
стрессор включает во многие из своих иммуноподавляющих
эффектов также и эмоциональную реакцию на ситуацию.
Все еще до конца не понятно, как стресс подавляет иммунную
систему. В течение долгого времени эти влияния связывали с корти-
костероидами. Действительно, последние подавляют синтез
антител, активность лимфоцитов, а также и воспалительные реакции.
Высокие дозы кортикостероидов длительное время были оплотом
терапии аутоиммунных, аллергических и воспалительных реакций,
требующих подавления функций иммунной системы. Более того, в
последнее время было установлено, что глюкокортикоиды
индуцируют апоптоз в тимусе, т.е. были раскрыты субклеточные и
молекулярные механизмы иммунодепрессантного действия этих гормонов
стресса (Cohen, 1992).
Однако существуют данные, согласно которым и другие
гормональные сдвиги также играют роль в вызванной стрессом иммунной
депрессии. В частности, катехоламины вредят функциям
макрофагов, лимфоцитов и гранулоцитов. Катехоламины, по-видимому,
также снижают способность организма бороться с опухолями и
вирусными инфекциями. Эндогенные наркотические вещества (морфий-
подобные соединения), выделяющиеся во время стресса, также
затрагивают иммунную систему. В частности, они ослабляют
активность N-киллеров при разрушении раковых клеток и клеток,
инфицированных вирусом (Hassing et al., 1996).
298
Нет сомнений, что экспериментальный стресс подавляет
иммунную систему у лабораторных животных и увеличивает вероятность
инфекции и рака. Но какова реакция людей в реальных жизненных
ситуациях, способствующих развитию психоэмоционального
стресса, например, таких как развод, брачные проблемы, смерть главы
семейства, негативные изменения в социальном статусе, потеря
работы? Большинство врачей, очевидно, согласилось бы с тем, что
люди, подвергнутые стрессу, болеют чаще. Однако что говорят
научные исследования? Клинические исследования также
свидетельствуют в пользу того, что стрессы реальной жизни негативно
влияют на иммунную систему и восприимчивость к болезням (Балицкий
и др., 1987).
Еще в 50-х годах была произведена оценка состояния студентов
Медицинского института в периоды экзаменов (высокий учебный стресс) и между ними
(низкий учебный стресс). Экзамены были связаны с более высоким уровнем
болезней, о которых сообщали сами студенты, а также с ухудшением некоторых
иммунных параметров, включая функции Т-киллеров и производство
интерферона. В другой работе исследователи обнаружили, что одинокие люди болеют
чаще, чем семейные. Интересно, что при разводе его инициаторы (в отличне от
противоположной стороны) испытывали меньший стресс и лучшую
устойчивость в течение первого года после развода, тогда как после этого времени
ситуация изменялась на противоположную. По мнению исследователей,
инициаторы в начале показывали меньший стресс, так как чувствовали себя в
выигрышной ситуации, но постепенно теряли это ощущение, осознавая негативные
последствия развода. Исследовалась иммунная система членов семей, которые
ухаживали в среднем около пяти лет за страдающими болезнью Альцгеймера.
В целом у этих людей наблюдалось снижение иммунный функции по сравнению
с контрольной группой; самая большая восприимчивость к болезням была
замечена у людей с низкой социальной поддержкой и у тех, кто госпитализировал
супруга (супругу) в течение предыдущего года.
Исследования связи болезней с реальными стрессогенными
факторами были начаты полвека назад, и в конечном счете привели
Т. Холмеса и Р. Рейе (Holmes and Rahe) к созданию шкалы оценок
социальных и эмоциональных событий стрессогенного характера по
их способности вызывать болезни (см. гл. 10, табл. 12). В последнее
десятилетие была установлена связь между стрессом и такими
заразными болезнями, как туберкулез, грипп, мононуклеоз, воспаление
язвы и др. {Leonard et al. 1996).
Почему стресс оказывает негативное воздействие на иммунную
систему? Почему организм использует физиологический механизм
для подавления собственной системы защиты от инфекции? Как
уже упоминалось ранее, реакция на стресс была развита в
доисторические времена, когда люди ежедневно сталкивались с
опасностью и серьезными трудностями и для их выживания требовался
физиологический механизм, нацеленный на быструю активизацию
систем организма. Ограниченность ресурсов организма приводит к
тому, что при интенсивном поступлении "топлива" и питательных
299
веществ в кровеносную систему и к нервам, как это происходит во
время стресса, другие системы, в частности пищеварительная и
иммунная, лишаются этих ресурсов, и их функции подавляются с тем,
чтобы "сбалансировать запасы организма" в этих критических
условиях. Таким образом, реакция на стрессоры увеличивает
возможности организма для преодоления непосредственной угрозы,
увеличивая риск долговременного повреждения некоторых своих систем.
В доисторическом мире или в мире животных это имело
эволюционный смысл. Если организм не в состоянии пережить
немедленную опасность - борьбу с хищником, серьезное ранение, жару или
холод - то не имеет большого значения, в какой функциональной
форме будет его иммунная система в момент его гибели. Ранняя
смерть лишает организм шанса на воспроизведение и передачу
своих генов следующей генерации. Ясно, что шансы иметь потомство
существенно выше, если организм хорошо приспособлен для
преодоления остро угрожающей его жизни ситуации, хотя это и
сопряжено с определенным подавлением иммунной системы и
повышенным риском болезней и преждевременного старения в будущем.
Проблема состоит в том, что окружающая среда сегодня резко
отличается от той, в которой развивались наши далекие предки.
Действительно, угрожающие жизни ситуации сейчас редки, тогда как
психологические и социальное давление представлено с избытком.
К сожалению, в человеческой популяции в последние тысячелетия
не произошло никакой существенной эволюции механизмов стресс-
ответа, и человеческий организм реагирует на напряженные
психологические и социальные ситуации так, как если бы они
фактически угрожали его жизни. Результатом этого является чрезмерный и
неадекватный ответ на стресс, ведущий, среди прочего, к
повреждению иммунной системы.
18.1. СТРЕСС И АУТОИММУННЫЕ БОЛЕЗНИ
В сущности, "сбои" иммунной системой сводятся к следующему:
во-первых, она может стать просто слишком слабой, чтобы быстро
и адекватно отвечать на вызовы окружающей среды; и, во-вторых,
она может "перепутать" и по ошибке воздействовать на структуры
собственного организма, приводя к возникновению аутоиммунных
болезней, например ревматоидного артрита и волчанки. Старение
ведет к ослаблению иммунной системы и увеличению риска
аутоиммунных патологий. Стресс также ослабляет иммунную систему, и
также, по-видимому, приводит к глубоким, хотя и менее прямым
аутоиммунным расстройствам.
Характер связи между стрессом и аутоиммунными состояниями
зависит в значительной степени от стадии болезни. С одной
стороны, у пациента с болезнью в активной стадии стресс несколько сни-
300
жает уровень аутоиммунной реакции и может фактически временно
снять некоторые ее симптомы. Это может быть следствием общего
иммуноподавляющего действия гормонов стресса. С другой
стороны, возникновение или рецидив аутоиммунных нарушений часто
вызываются стрессовыми ситуациями. Причины этого пока неясны в
значительной степени и потому, что механизм аутоиммунных
заболеваний вообще не вполне понятен. Известно, что большинство ау-
гореактивных Т-клеток уничтожается во время их созревания в
тимусе (подробнее об этом см. в гл. 6), но что позволяет некоторым ау-
гореактивным Т-клеткам избегать "осмотров" в тимусе и затем
воздействовать на ткани организма - неизвестно. Мы знаем, что стресс
воздействует на тимус, вызывая его инволюцию и потерю части
функций. Теоретически стресс может ограничивать способность
тимуса ликвидировать аутореактивные Т-клетки. открывая таким
образом дорогу аутоиммунным нарушениям.
Большинство исследований стресса и аутоиммунных
болезней посвящены ревматоидному артриту, который является одной
из наиболее распространенных аутоиммунных патологий и
затрагивает приблизительно 2% человеческой популяции {Leonard
et al., 1996). При этой болезни иммунная система атакует хрящ,
вызывая воспаление суставов, боль, неподвижность, а и иногда и
инвалидность. Начало ревматоидного артрита может быть
следствием как внезапной острой стрессовой ситуации, так и
длительной серии неприятностей.
В одной интригующей работе изучались пары однояйцовых
близнецов женского пола, причем только один близнец каждой
пары страдал ревматоидным артритом. Оказалось, что, например,
сестра-близнец с артритом испытывала значительно больше
стрессовых событий в жизни до начала болезни, чем ее здоровая сестра.
Продолжительное течение ревматоидного артрита также,
по-видимому, поддерживается стрессом. Некоторые исследования показали,
что течение и результат этой болезни были лучшими у тех
пациентов, кто имел благоприятную социальную среду. К другим
аутоиммунным болезням, происхождение и течение которых связано с
воздействием стресса, можно отнести множественный склероз,
аутоиммунные нарушения щитовидной железы и инсулинозависимый
диабет I типа (Todorov Η. et al, 2001).
Многие описанные в Ш части меры могут привести к уменьшению стрессо-
генных воздействий на иммунную систему. Особенно эффективны в этом
отношении адаптогены (гл. 29) и пищевые добавки, укрепляющие иммунитет
,гл. 23). Восстановление присущего молодым уровня некоторых гормонов (ме-
патонина и гормона роста) может также снизить уязвимость иммунной
системы, в частности и от воздействия стресса (гл. 26).
Глава 19
КЛЕТОЧНЫЙ СТРЕСС
Как уже отмечалось в предыдущих главах, каким бы ни был
стрессогенный фактор, организм отвечает на него одним и тем же
образом, начиная чрезвычайную стандартную реакцию,
называемую стресс-ответом, или общим адаптационным синдромом.
Но что же делают отдельные клетки? Имеют ли они также
стандартные процедуры, которые будут выполняться при всех
разновидностях опасных ситуаций? Оказалось, что жизненно важная
необходимость поддержания гомеостаза присуща не только организму
как целому, независимо от сложности его строения, но и отдельным
его системам, органам, клеткам, которые имеют свой собственный
эквивалент стресс-ответа.
Учение о неспецифическом адаптационном синдроме клеток уходит
своими корнями в работы одного из основателей клеточной биологии Р. Вирхова,
который наблюдал "мутное набухание" различных типов клеток при самых
разных повреждающих воздействиях и интерпретировал этот феномен как
неспецифическую стереотипную и обратимую клеточную реакцию. Эти взгляды
были развиты в трудах Д.Н. Нассонова и В.Я. Александрова, сформулировавших
представление о "паранекрозе" как о явлении денатурации клеточных белков
под воздействием разнообразных (и даже противоположных по природе)
повреждающих агентов (кислота, щелочь, гипер- и гипотермия, гипер- и гипоксия,
электролиты и неэлектролиты, ультрафиолетовое облучение и т.п.).
Паранекроз был описан как комплекс однотипных изменений в клетке:
уменьшение растворимости и дисперсности коллоидов цитоплазмы и ядра,
увеличение светорассеяния (помутнение) и вязкости клеточных коллоидов,
увеличение сорбционной активности цитоплазмы (связывание красителей) и т.п. без
разрыва пептидных связей и иных необратимых изменений в структуре
белковых компонентов клетки {Иассонов, Александров, 1940; Иассонов, 1959; Эйду с,
1977; Александров, 1985; Браун и др., 1987).
При адаптационном синдроме клеток, или, как его называют в настоящее
время, клеточном стрессе, развивается также комплекс неспецифических
изменений метаболизма: активация аденилат- и гуанилатциклазной систем и
каскадное образование и функционирование вторичных посредников,
стимулирующих в свою очередь деятельность многих протеинкиназ и других
внутриклеточных ферментов {Браун и др., 1987).
Важнейшим механизмом клеточного стресса служит разобщение дыхания
и окислительного фосфорилирования в результате повреждения митохондри-
302
альных мембран, а также потери клеткой неорганического фосфата. В итоге
развивается частичная дезэнергизация клетки, сопряженная с активацией
анаэробного гликолиза и накоплением молочной кислоты. Для клеточного
стресса характерны также активация митохондриальной АТФазы, выход и
активация лизосомных гидролаз, увеличение образования аммиака, ускоренный
распад гликогена, существенные нарушения мембранного транспорта ионов,
медиаторов и т.п. {Кузин, 1986; Рыску лова, 1986).
Как и целостный организм, клетка поддерживает
исключительно сложное и тонкое внутреннее равновесие (гомеостаз), весьма
существенное для ее нормального функционирования. Когда стрессор
нарушает гомеостаз, клетка стремится восстановить его, включая
одну или несколько стандартных аварийных программ.
Наиболее четкая и полная картина адаптационных изменений в
клетках выявлена при трех стрессовых ситуациях: тепловом шоке,
действии радиации, а также при подавлении трансляции
экстремальными факторами химической и физической природы, когда
разворачивается наиболее сложная многоступенчатая реакции
восстановления белкового гомеостаза клетки.
19.1. СТРЕСС-РЕАКЦИЯ КЛЕТКИ
НА ТЕПЛОВОЙ ШОК
Ни одна клетка млекопитающих не может противостоять
очень высоким температурам, что и является одной из причин,
почему температура нашего тела строго поддерживается на уровне
примерно 37 °С. Однако небольшое повышение температуры (до
40-45 °С) не смертельно - клетка способна противостоять этому,
развивая ответ на так называемый тепловой шок. Вначале клетка
быстро приостанавливает ряд своих обычных операций, например,
производство белков, необходимых ей в нормальном состоянии.
Затем клетки начинают быстро синтезировать большие
количества (до 2,5% и более от общего содержания белка) специальных
белков, называемых "белками теплового шока" (hsp), которые
обычно присутствуют в клетке только в незначительных количествах
(Craig, 1985). Как выяснилось, ответ, во многом сходный с
"тепловым шоком", вызывается и рядом других вредоносных факторов,
таких как радиация, гипоксия, токсины, тяжелые металлы, вирусы
и этиловый спирт. Все это означает, что существует общий
чрезвычайный ответ на повреждения - своего рода клеточная реакция
"борьбы или бегства".
Каким образом белки теплового шока помогают клетке выжить,
оставалось тайной в течение долгого времени. Удивительно, что
такие эволюционно отдаленные виды, как бактерии, дрожжи,
насекомые и человек имеют весьма близкие по структуре белки теплового
шока. Такой чрезвычайный консерватизм белков, как правило, слу-
303
жит показателем их критической важности для функционирования
некоторых основных механизмов жизни. Исследование недавних
лет показали, что основная функция многих белков теплового шока
заключается в стабилизации, сохранении и облегчении
восстановления клеточных структур, в частности ДНК и белков, в течение и
после разрушительной ситуации
Большинство известных типов клеточных повреждений
связано с разрушением пространственной структуры жизненно важных
биологических молекул. Так, ДНК-полимер, состоящий из двух
нитей, скрепленных водородными связями (тип слабой химической
связи между молекулами), может под действием агрессивных
факторов, в частности высокой температуры, распадаться на
отдельные нити. Белки, которые являются цепями аминокислот,
свернутых в клубок сложным и неслучайным образом, также могут
разворачиваться под влиянием внешних причин, становясь
бесполезными вытянутыми структурами. Когда пространственная
структура повреждена, ДНК и белки не только теряют свою функцию, но
также становятся более уязвимыми для атак свободных радикалов
и агрессивных химикалий.
Изучение первичной структуры ДНК выявило три основных гетерогенных
семейства белков теплового шока: с молекулярной массой 25, 70 и 90 КД.
Поскольку синтез белков теплового шока блокируется ингибитором синтеза РНК
антиномицином Д, можно полагать, что эти белки образуются в клетках de
novo. Однако в их образовании играют роль также посттрансляционные
модификации (метилирование, фосфорилирование, АДФ-рибозилирование, ацети-
лирование) клеточных белков (Браун и др. 1987).
Белки теплового шока способны восстанавливать структуру Других
белков, если те подверглись денатурации, т.е. утратили свою трехмерную кон-
формацию. При биосинтезе в рибосомах полипептидная цепь образуется в
развернутом виде. Далее, при участии физико-химических факторов,
закодированных в первичной структуре каждого белка, происходит сворачивание
цепи и образование зрелого и активного белка. Помочь цепи правильно
свернуться — одна из функций белков теплового шока, которые называются
также шаперонами. Правильная форма, приобретаемая белком под действием
шаперона, может нарушаться под воздействием тех или иных
неблагоприятных условий, например нагревания, приводящего к денатурации белков. Ре-
натурация таких белков также осуществляется белками теплового шока,
некоторые из них присоединяются к нарушенным частям развернутой белковой
цепи (или отдельным нитям ДНК). Количество шаперонов в клетке резко
возрастает при нагревании или под действием многих других повреждающих
факторов. Эти белки выполняют и ряд других функций. Было показано, что
для осуществления АТФ-зависимой реакции разворачивания белков
(состояния, при котором белки могут проникать сквозь митохондриальную
мембрану и мембрану эндоплазматического ретикулума) необходимы некоторые
гены семейства hsplO. Если экспрессия этих генов подавлена, то и
предшественники митохондриальных белков, так же как и белки, предназначенные для
эндоплазматического ретикулума, не могут пересечь соответствующие
мембраны, и вместо этого накапливаются в цитозоле (Eilers et al., 1986; Pfannei
et al., 1987; Chirico et al., 1988; Deshaies et al., 1988).
304
Белки из семейства 90 КД связываются с неактивными формами белков-
рецепторов стероидных гормонов и с тирозин-специфическими протеин киназа-
ми, вероятно, принимая таким образом участие в регуляции функций этих
рецепторов {LindquisU 1986; Pelham, 1986) .
Лучше всего изучены белки семейства 70 КД. Они связываются с некото
рыми другими белками, а также с аномальными белковыми комплексами и
агрегатами, от которых потом освобождаются, присоединяя АТФ. Было
доказано, что они помогают переводить в раствор и заново сворачивать
агрегированные белки путем нескольких циклов присоединения и гидролиза АТФ {Craig,
1985; LindquisU 1986).
Молекулярный механизм защитного действия различных типов стрессовых
белков и, в частности, шаперонов еще недостаточно выяснен. Хотя некоторые
из них и обладают конкретной ферментативной активностью (протеазной, ал-
когольдегидрогеназной), то обстоятельство, что стрессовые белки
синтезируются в больших количествах в клетках, подвергшихся денатурирующим
воздействиям, свидетельствует скорее об их общем неспецифическом действии,
которое стабилизирует жизненно важные клеточные структуры {Браун и др., 1987;
Машанский и др., 1987).
Продукция такого большого количества стрессовых белков
обеспечивается активацией транскрипции определенных генов. В клетках дрожжей S сеге-
visiac, например, имеется восемь А.ур70-генов; некоторые из них
транскрибируются при любых условиях, а остальные - только под действием высоких
температур или других экстремальных факторов (Craig, 1985; Lindquist, 1986)
Многие исследования последних лет показывают, что ответ на
"тепловой шок" изменяется с возрастом (Kaifflman, 1992; Heydari
et al., 1994). У многих типов клеток - фибробластов (клетки
соединительной ткани), клеток печени и нейронов старение
сопровождается хроническим падением способности продуцировать некоторые
белки теплового шока в ответ на повреждение клеток. Это явление
отчасти может быть причиной возрастной потери устойчивости к
стрессу - понятно, если система экстремальной клеточной защиты
не работает должным образом, интегральный результат
повреждающего действия стресс-ответа организма будет гораздо больше.
У некоторых типов клеток, например Т-лимфоцитов, по-видимому,
наблюдается корреляция между эффективностью ответа на
температурный шок и состоянием клеточных часов: чем ближе клетка к
ее лимиту делений (Хейфлик-лимит, см. гл. 2), тем ниже производст
во белков температурного шока при ее ответе на повреждение.
Некоторые ученые полагают, что многие функциональные и
структурные изменения, отмеченные у "старых" клеток, следуют из их
пониженного или расстроенного ответа на температурный шок
{Kauffman, 1992).
С. Пардю и его коллеги (Pardue et al.) придают возрастному
ослаблению ответа на температурный шок еще большее значение.
Изучая это явление в клетках мозга крысы, исследователи нашли,
что возрастная потеря эффективности ответа на температурный
шок наиболее ярко проявляется в нейронах гиппокампа, что делает
их более уязвимыми ко всем видам повреждений. Как уже отмеча-
305
лось в главе 2, гиппокамп - мозговая структура, участвующая в
работе контура обратной связи между гипоталамусом и
надпочечниками. Смерть нейронов в гиппокампе вредит обратной связи,
ответственной за ограничение стресс-ответа, делая его чрезмерным и
чреватым повреждениями (McEwen, 1999). Говоря проще, чем слабее
гиппокамп, тем больше организм страдает от стресса. В свете
результатов Пардю и его коллег, кажется вероятным, что падение
эффективности ответа на температурный шок в нейронах гиппокампа
может в значительной мере отвечать за чрезмерный стресс-ответ,
часто наблюдаемый у старых животных или людей.
Тем не менее возрастное снижение эффективности клеточного
ответа на температурный шок может быть в значительной
степени ослаблено. А. Хейдери и его коллеги (Heydari et al., 1994)
изучали влияние старения и ограничения калорийности питания на
реакцию термического шока у крыс. (У грызунов ограничение
калорийности, как уже не раз подчеркивалось, наиболее мощный
инструмент антистарения, способный продлить жизнь животных почти
в два раза, так же как и предотвратить развитие дегенеративных
болезней.) В группе животных, питание которых было
неограниченным, с возрастом синтез hsp70 (одного из ключевых белков
теплового шока) снижался в клетках печени на 40-50% в ответ на
умеренный удар высокой температуры (42,5 °С) в течение 30 мин.
Снижение синтеза hsp70 было значительно меньшим у животных,
получавших только 60% пищи от количества, использованного не
ограничиваемой в питании группой. В другом исследовании
Дж. Эренфрид и его сотрудники (Ehrenfried et al.) нашли, что
крысы, содержавшиеся на ограниченной диете, имели уровень hsplO в
клетках кишечника приблизительно в три раза выше, чем
животные, питавшиеся без ограничений.
В настоящее время, наше понимание значения "температурного
шока" для старения ограничено, но, по-видимому, сохранение
оптимального стресс-ответа на высокую температуру может замедлять
старение и понижать риск дегенеративных болезней. Никакой
другой способ, кроме ограничения калорийности питания, не способен
предотвращать или замедлять возрастное падение ответа на
"термический шок". К сожалению, существенное ограничение калорийности
питания не приносит пользы людям и другим приматам, поскольку
не ведет к значительному повышению продолжительности жизни,
но может привести к повреждению центральной нервной системы,
если это ограничение начато еще до наступления зрелости. Тем не
менее проводится множество многообещающих исследований,
направленных на повышение эффективности ответа старых клеток
на термический шок (Schlesinger, 1994; Lee et al., 1996). К тому же
ученые тщательно исследуют другие альтернативные возможности
повышения реакции клеток на "термический шок" и вне связи с
возрастными проблемами.
306
19.2. СТРЕСС-РЕАКЦИЯ КЛЕТОК
НА ДЕЙСТВИЕ РАДИАЦИИ
В середине 60-х годов 3. Бак и П. Александер (1963)
предложили гипотезу "биохимического шока", описывающую реакцию
клеток на действие ионизирующей радиации. К числу наиболее ранних
проявлений биохимического шока авторы относили быстрый
распад макроэргических соединений, в особенности креатинфосфата.
Весьма характерным было раннее разобщение дыхания и
окислительного фосфорилирования на фоне неизменного или даже
несколько усиленного клеточного дыхания, а также увеличение
проницаемости мембран с быстрой, пропорциональной дозе радиации,
потерей натрия, калия, фосфатов, лактата, НАД, каталазы и других
ферментов. Эти биохимические проявления клеточного шока
сочетаются с характерными морфологическими изменениями:
уменьшением количества митохондрий, их набуханием, просветлением мат-
рикса; кристы в митохондриях становятся менее видимыми,
появляются разрывы митохондриальной мембраны.
Следует отметить, что малые дозы ионизирующей радиации,
индуцируя развитие клеточного, митохондриального стресса
(биохимического шока), вызывают на определенное время повышение
общей и радиационной устойчивости клеток (Романцев, 1968; Гонча-
ренко и др., 1985). По-видимому, есть все основания рассматривать
биохимический шок как частный случай неспецифической реакции
клеток на внешние воздействия.
С биофизической точки зрения эта реакция сводится к обратимому
нарушению внутриклеточной компартментализации, к выравниванию
концентрации низкомолекулярных органических соединений между различными
субклеточными органеллами. Адсорбция на макромолекулах дополнительных
количеств этих соединений снижает и конформационную подвижность первых, и,
следовательно, их биологическую активность. В результате падает
интенсивность метаболизма и развивается состояние паранекроза. Это неспецифическое
торможение метаболизма и функции клеток играет защитно-охранительную
роль, замедляя дальнейшую деструкцию и удлиняя период, в течение которого
возможна репарация возникших повреждений (Эйдусу 1977).
SOS - ответ на действие факторов, повреждающих ДНК.
Современные представления о защитных реакциях клетки в от
вет на действие экстремальных факторов базируются на
признании двух четко выраженных стадий: ранней - так называемого
SOS-omeema на повреждения, и более поздней - адаптивной,
направленной на репарацию клеточных структур и нормализацию
клеточных процессов (подробнее об этой стадии - в следующем
параграфе). Ранний SOS-ответ на действие ионизирующей
радиации, химических мутагенов и других биологически активных
веществ хорошо изучен у прокариот (Waker, 1985). Он включает
активацию экспрессии более 20 генов, в том числе, кодирующих
307
белковые факторы эксцизионной репарации, транскрипции и
репликации, а также тимидилаткиназу, ДНК-полимеразу β, рибону-
клеотидредуктазу, т.е. на этой стадии активируется комплекс
процессов экстренной репарации повреждений ДНК.
Следовательно, ключевым моментом в SOS-ответе является способность
клетки распознать повреждения ДНК и обеспечить экстренную и
эффективную их репарацию. Точность копирования ДНК в
процессах репликации и репарации зависит не только от активности
ферментов, но и от концентрации и соотношения их субстратов -
дезоксирибонуклеозидтрифосфатов (дРНТФ). Синтез ДНК
требует постоянного и сбалансированного притока четырех типов
дРНТФ (Thelander et al., 1979). На фоне нарушенного соотноше
ния предшественников ДНК наблюдается дефектный реплика-
тивный синтез и неполноценная репарация ДНК из-за ошибок
включения оснований, появления "брешей", влекущих за собой
одно- и двунитевые разрывы ДНК (Genetic Concequences of
Nucleotide Pool Imbalance, 1985).
Процесс биосинтеза дРНТФ в клетке катализируется
ферментом рибонуклеотидредуктазой (РР). Активная РР содержит
свободный радикал тирозинового аминокислотного остатка поли
пептидной цепи субъединицы М2, необходимый для проявления
каталитической активности фермента (Thelander et al., 1979), а
также два иона Fe3+ в активном центре фермента. Эти особенности
структуры РР позволяют проводить количественные измерения
уровня каталитической активности РР методом электронного
парамагнитного резонанса (ЭПР). Такая возможность впервые
была использована М.К. Пулатовой и ее сотрудниками {Pulatova
et al., 1986; 1989) для контроля активности РР in situ, т.е.
непосредственно в тканях и органах высших животных (без препаративной
изоляции и очистки РР).
При изучении характера изменений активности РР под
действием факторов, повреждающих ДНК, выяснилось, что
особенностью наиболее раннего ответа на воздействие радиации как у
новорожденных, так и у половозрелых крыс является значительная
активация РР в селезенке этих животных (Пулатова и др., 1988;
1995; Pulatova et al., 1999). Этот эффект был интерпретирован
авторами как защитная SOS-реакция, направленная на срочную
репарацию радиационных повреждений ДНК в органах животных.
Изменение активности РР, очевидно, служит метаболическим
маркером наличия SOS-ответа, поскольку синтез
предшественников ДНК, катализируемый этим ферментом, является фактором,
лимитирующим скорость синтеза ДНК, необходимого для
репарации. Можно предположить, что РР - фермент наиболее
"быстрого реагирования" на облучение в сравнении с упомянутыми
выше ферментными системами (Waker, 1985), участвующими в
синтезе ДНК.
308
19.3. ОКСИДАТИВНЫЙ СТРЕСС КЛЕТКИ
В предыдущих главах (гл. 2, 3, 14, 15) мы уже не раз (в рамках
свободно-радикальной теории старения) касались различных
сторон проявления оксидативного стресса и его последствий как для
целостного организма, так и для его клеток, но сейчас хотели бы
взглянуть на "авансцену" этих событий, определяющих развитие
оксидативного стресса в клетке, т.е. на клеточные органеллы, где
протекают основные реакции энергетического метаболизма, -
митохондрии.
Традиционно митохондрии рассматривались как "электоростан-
ции" клетки, производящие аденозинтрифосфат - АТФ. Поэтому
многие исследования, за прошедшие пять десятилетий проведенные
на митохондриях, фокусировались на объяснении молекулярных
событий, связанных как с синтезом АТФ путем окислительного фос-
форилирования, так и с биогенезом механизмов окислительного фо-
сфорилирования.
В то время как были сделаны фундаментальные открытия и
продолжали исследоваться эти замечательные многофазные
процессы энергопродукции, многие биологи и биомедики осознали, что
митохондрии вовлечены также и в другие процессы, крайне важные
для жизни многоклеточных животных. Два из них - клеточная
смерть и старение, как это ни удивительно, связаны, по крайней
мере частично, с процессами окислительного фосфорилирования.
Понимание этого определило развитие нового направления в изучении
митохондрий, которое вызвало большой интерес у широкого круга
ученых с точки зрения как детального выяснения их роли в
клеточной смерти и старении, так и изыскания возможностей
модификации этих процессов, поскольку это касается использования новых
терапевтических подходов или медикаментов для лечения
человеческих болезней.
Митохондриальная цепь электронного транспорта (ЦЭТ) -
мощный источник активных форм кислорода (АФК). Чем выше
уровень функционирования ЦЭТ, т.е. продукции АТФ, тем выше
уровень генерации АФК. При исследовании роли Комплекса I этой
цени в производстве супероксидного радикала было установлено,
что одноэлектронный донор, передающий электрон к кислороду
(см. рис. 3), является редокс-центром, локализованным перед
участком, содержащим три различных связанных типа кофермента Q
(Lenaz et al., 2002). По-видимому, именно здесь происходит
электронная редукция и возникают супероксидные анион-радикалы,
г.е. ЦЭТ оказывается несколько "негерметичной". Помимо
радикала (О^), который под действием СОД дисмутирует в перекись во-
юрода (Н202), последняя в присутствии двухвалентного железа
превращается далее (см. гл. 2) в химически высокоактивный
радикал гидроксила (*ОН).
309
В дополнение к этим токсичным продуктам ЦЭТ во внутренней
митохондриальной мембране моноамин оксидаза - фермент
наружной митохондриальной мембраны - катализирует окислительное де-
заминирование биогенных аминов, служа существенным
источником Н202, что способствует увеличению устойчивой концентрации
АФК в пределах матрикса митохондрий и цитозоля {Cadenas et al.,
2000). Кроме того, АФК реагируют с окисью азота внутри
митохондрий, повышая повреждающее действие пероксинитрита {Sastre
et al., 2000).
Комплекс 1 в наибольшей мере подвержен возрастным
изменениям, что приводит к жесткому ограничению скорости
электронного транспорта и, соответственно, повышенной генерации АФК,
которые рассматриваются в качестве патогенетических факторов
многих болезней и старения (Barja, 2002; Lenaz et al., 2002),
поскольку АФК способны повреждать жизненно важные структуры клетки,
особенно мембраны, где они индуцируют процессы ПОЛ (см. рис. 4)
и информационные макромолекулы - белки, РНК и ДНК.
Ключевая роль митохондрий в клеточном старении следовала, в
частности, из экспериментов, в которых митохондрии,
изолированные из фибробластов старых крыс, с помощью микроинъекций
вводились в клетки молодых животных, после чего у последних
наблюдались признаки вырождения {Sastre et al., 2000). Первостепенная
роль митохондриальных АФК в развитии патологических процессов
и старения также была четко выявлена при использовании
трансгенных подходов. Трансгенные мыши, дефицитные по
антиокислительному митохондриальному ферменту - супероксид дисмутазе
(Мп-СОД), умирали в течение первой недели жизни в результате
развития кардиомиопатии! {Huang et al., 1999; Melovy 2000).
Д. Харман в 1972 г. впервые предположил, что митохондрии
могли бы быть биологическими часами старения, отмечая, что
скорость потребления кислорода должна определить уровень
накопления митохондриальных повреждений, вызванных АФК
{Нагтап, 1981). Позже, в 1980 г., Микель и его соавторы
сформулировали митохондриальную теорию старения клетки {Miguel
et al., 1992). Центральное положение этой теории - факт высокой
уязвимости митохондриальной ДНК свободными радикалами
кислорода. Более ста мутаций мтДНК было найдено в течение
прошедших десяти лет у пациентов с митохондриальными болезнями
(что в десятки раз превышает уровень мутаций в ядерной ДНК), и
многие из них обнаруживались также и в старых человеческих
тканях. Сфера действия и распространенность таких мутаций мтДНК
увеличивается с возрастом, особенно в тканях с большим
потреблением энергии {Wei et al., 2001). Более того, делеция в пять тыс.
пар оснований мтДНК найдена только у старых крыс {Lenaz et al.,
1998). МтДНК, по-видимому, не способна предотвратить такой
высокий уровень мутагенеза, вызванный АФК, потому что, в отличие
310
от ядерного генома, не обладает системами эксцизионной и реком-
бинационной репарации {Miquel, 1998). Кроме того, мтДНК не
"экранируется" гистонами или другими ДНК-связанными белками.
Окислительные модификации и мутации мтДНК происходя!1 с
большой легкостью, и степень таких изменений мтДНК
увеличивается с возрастом по экспоненте (Wei, 1998).
Повышенное переокисление липидов и окислительная
модификация белков в митохондриях (в том числе и модификация
антиокислительных ферментов), которые вызываются постоянно
увеличивающимся количеством АФК, далее отягчают мутации и
окислительные повреждения мтДНК в процессе старения. Дыхательные
ферменты, содержащие неисправные кодируемые мтДНК белковые
субъединицы (результат мутаций), повреждают электронную
транспортную функцию и таким образом увеличивают электронную
"утечку" и АФК-продукцию, которые в свою очередь повышают
оксидативный стресс и окислительные повреждения в
митохондриях. Этот порочный круг оперирует в клетках разных тканей с
различной скоростью и ведет к различному накоплению
окислительных повреждений и мутаций в мтДНК. Это может объяснить
разницу в функциональном падении и структурной порче различных
органов и тканей при человеческом старении. Особенно велики
повреждения в органах, которые обладают высоким уровнем
энергетического метаболизма (например, мозге и сердечной мышце) {Wei, 1998;
Lee et al., 2001; Golden et al., 2001) (рис. 33). К чему же может
привести описанное развитие оксидативного стресса в митохондриях?
Митоптоз - механизм гибели митохондрий. Повышенное
содержание АФК (оксидативный стресс) может индуцировать как
апоптоз, так и некроз (Buttke et al., 1994; Hanson et al., 1995), причем
характер гибели клеток зависит от интенсивности повреждающего
воздействия (Dypuckt et al., 1994). Активация СР-радикальных реак
ций может индуцировать апоптоз, что показано на многих
клеточных системах, причем к аналогичному эффекту приводит и
подавление антиоксидантных ферментов, таких как Cu/Zn-СОД (Troy et al.,
1994) и митохондриальная Mn-СОД (Nicotera et al., 1996).
Полагают, что окислительные реакции, модифицирующие белки,
способны вызвать активацию экспрессии генов, ответственных за апоптоз, или,
например, что модификация молекул ДНК может нарушать связывание факторов
транскрипции и также приводить к незапланированной экспрессии
определенных генов. Возможно, оксидативный стресс вызывает и изменения цитоскелета
клеток, приводя к сокращению объема клетки или изменению
надмолекулярной структуры хроматина (Nicotera et al., 1996), т.е. к основным структурным
проявлениям процесса апоптоза.
Однако наиболее интересная гипотеза относительно механизма
включения реакций апоптоза (подробней об апоптозе см. в гл. 2) под
влиянием оксидативного стресса, как нам кажется, принадлежит
В.П. Скулачеву, который связал эти реакции непосредственно с ми-
311
!
со
тохондриями (Skulachev, 1998). Как уже указывалось в параграфе
"ДНК: повреждения/репарация", одна из функций митохондриаль-
ного белка теплового шока (mHsp70) состоит в стимуляции импорта
белков в митохондрии, что проявляется в активации
окислительного фосфорилирования (и, следовательно, в увеличении АФК).
Помимо этого, mHsp70 имеет еще одно свойство - он входит в состав
митохондриальной эндонуклёазы (Seel), фермента, расщепляющего
ДНК митохондрий в строго определенных ее участках. Активность
этой эндонуклёазы в норме должна быть подавлена во избежание
разрушения митохондрий. mHsp70, по-видимому, играет роль регу-
ляторной субъединицы Seel, поскольку активный центр этого
фермента локализован на другой ее субъединице. Предполагается, что
денетурированные белки стехиометрически связывают mHsp70.
Дальнейшее повышение количества денатурированных белков иод
действием АФК приведет к большему связыванию mHsp70 и в
итоге - к снятию репрессии эндонуклёазы в популяции митохондрий и
далее - к расщеплению митДНК, что вызовет деградацию этой
митохондрии. Таким путем, по-видимому, клетки могли бы очищаться
от тех митохондрий, в которых появилось слишком много
денатурированных белков. Подобное явление, названное В.П. Скулачевым
мшпоптозом (Skulachev, 1998; Скулачев, 1998), может приводить и к
апоптозу клетки, если митоптоз принял массовый характер.
Существенно, что разрушение митохондрий должно сопровождаться
освобождением в цитоплазму митохондриальных белков -
активаторов апоптоза: цитохрома С, фактора, индуцирующего апоптоз (A1F),
{рис. 33) и каспазы 3 (Скулачев, 1998).
Недавно было показано, что некоторые антиоксиданты, такие как
содержащие серу антиокислители, витамины С и Ε или экстракт Ginco biloba (EGb
761), предохраняют от связанного с возрастом окислительного повреждения
мтДНК и окисления митохондриального глутатиона. Кроме того, EGb 761
также предотвращает изменения в морфологии и функции митохондрий,
связанные со старением мозга и печени (Sastre et al., 2000)
19.4. ОБЩАЯ АДАПТАЦИОННАЯ РЕАКЦИЯ КЛЕТКИ
Как оказалось, клетки млекопитающих имеют чрезвычайную
стратегию защиты, которая даже более универсальна, чем ответ на
температурный шок. Это явление имеет много названий (например,
"механизмы клеточной стабильности"), но для простоты, мы будем
называть его здесь общей адаптационной реакцией клетки (ОАРК).
Подобно стресс-ответу организма ОАРК может быть индуцирована
широким разнообразием разрушительных воздействий - радиацией,
313
некоторыми антибиотиками, токсинами и канцерогенными
веществами - и не зависит от природы стрессогенного фактора (Todorov,
1995). "Философия защиты" индивидуальной клетки
млекопитающих во многом подобна той, которой пользуется целостный
организм: безотносительно характера возникшей проблемы, сначала
инициируется ряд стандартных чрезвычайных мер, а
специфическая, "заказная", адаптация, наступает позже.
Один из авторов этой книги и его коллеги изучали клеточный
стресс более двух десятилетий и раскрыли множество ключевых
механизмов, используемых клеткой для ответа на стрессогенные
факторы (Todorov et al., 1984; Тодоров, 1986; Todorov, 1990; 1995).
В большинстве экспериментов, стрессором являлся циклогексимид
(ЦГИ), токсин, ингибирующий трансляцию (заключительную и
наиболее сложную фазу белкового синтеза). Этот тип клеточного
стрессора был выбран потому, что вызывал падение способности
клеток синтезировать свои белки - ситуация, развивающаяся почти
при любом другом типе повреждений и поэтому моделирующая
клеточный стресс вообще. Действительно, трансляция - это фокус, в
котором "скрещиваются" основные метаболические,
энергетические и информационные потоки клетки, обеспечивающие процесс
биосинтеза белков. Поэтому любое повреждение каждого из этих
потоков в той или иной мере рано или поздно скажется на
интенсивности процесса трансляции.
Почему нормальный белковый синтез так жизненно важен для
клетки? По существу, все функции клетки, как и целостность почти
всех ее надмолекулярных структур, зависят от оптимального уровня
синтеза белков. Некоторые белки (ферменты) катализируют почти
все биохимические реакции, другие - транспортируют топливо и
строительные блоки внутри клетки и удаляют отходы, другие же -
защищают, поддерживают и обслуживают белок-синтезирующую
машину и генетический аппарат клетки и т.п. Можно говорить, что
белки полностью управляют экономикой клетки - никакая
жизненная функция не может поддерживаться без их участия. Синтез
белков - действительно основа жизнедеятельности любых живых
систем. Если белковый синтез подавлен, то гомеостаз клетки и все ее
жизненные функции подвергаются угрозе.
Структура всех белков кодируется в ДНК, и в большинстве
случаев один ген соответствует одному белку. Белковый синтез
состоит из двух главных стадий: (1) транскрипция - процесс, при котором
информация (записанная в ДНК) переносится на информационный
носитель - молекулу, весьма похожую на фрагмент однонитчатой
ДНК и называемую информационной рибонуклеиновой кислотой
(иРНК); и (2) трансляцию - процесс, при котором информация из
иРНК декодируется рибосомами - своего рода миниатюрными
фабриками сборки белковой цепочки из мономеров (аминокислот).
Выходящая из рибосомы полимерная белковая цепочка является одной
314
из множества "серийных" одноразовых копий информации,
записанной в иРНК. Процесс этот и называется трансляцией (от англ.
translation - перевод), потому что в рибосомах информация, записанная в
иРНК на языке нуклеиновых кислот (нуклеотиды), переводится на
язык белковых молекул (аминокислоты). Клеточный стрессоген-
ный фактор может затрагивать или одну, или обе фазы белкового
синтеза. Однако вероятность нарушения трансляции несравненно
выше, чем транскрипции или репликации (удвоение молекул ДНК),
и потому что трансляционная машина на порядок превосходит их по
количеству участвующих в ее работе элементов, и потому, что
аппарат трансляции расположен на периферии клетки и, вследствие
этого, более подвержен внешним воздействиям, чем репликативные и
транскрипционные комплексы, которые защищены ядерной
мембраной и белками, связанными с ДНК.
Было обнаружено, что при глубоком ингибировании
трансляции ЦГИ, т.е. в результате блокады белкового синтеза, в клетке
активируется ряд последовательных стандартных реакций
(Todorov, 1990; 1995):
• «Инвестиция дополнительных средств в "кризисную
экономику"». Глубокое ингибирование трансляции вызывает экстренную
мобилизацию предсуществующих резервов клетки (в особенности
свободных рибосом, количество которых в гепатоцитах достигает
35%, и иРНК) для создания новых полирибосом (Тодоров и др.,
1977). Кроме того, блокада трансляции ускоряет весь цикл
биогенеза белоксинтезирующих аппаратов (Тодоров, 1986; Todorov,
1990): трансмембранный перенос аминокислот и глюкозы;
внутриклеточный транспорт белков из цитоплазмы в митохондрии и
ядро, сборку рибосом в ядрышке и иРНП (информационных рибону-
клеопротеидов), в нуклеоплазме и, наконец, ускоряет выход
зрелых рибосом и иРНП в цитоплазму к местам их объединения в
новые функционирующие полирибосомы, что непосредственно
направлено на восстановление биосинтеза белков (Todorov, 1995).
Все эти процессы энергозависимы и их ускорение обусловлено
временным повышением концентрации АТФ в клетках в течение
одного-двух часов после начала блокады, поскольку одни из
главных потребителей энергии - белоксинтезирующий конвейер - при
блокаде трансляции резко сокращает потребление энергии (АТФ и
ГТФ) (Бойков и др., 1979; 1981; Ch'ih et al., 1979; Христиановин
и др., 1988). В условиях, когда биосинтез белка подавлен не
слишком глубоко (а такие ситуации случаются в жизни клетки
довольно часто), мобилизации этих резервов и ускорения биогенеза
рибосом и иРНП оказывается достаточно для нормализации уровня
трансляции (Тодоров и др., 1977). В большинстве случаев эти
экстренные меры приводят к "реставрации" оптимального уровня
белкового синтеза и, следовательно, гомеостаза клетки.
315
• "По законам военного времени". Однако, если жесткое ингибиро-
вание трансляции продолжится, клетка будет вынуждена прибег*
нуть к более серьезным мерам. Обычно многие белки
синтезируются клеткой для внутренних потребностей, а остальная их часть»
как правило - большая, экспортируется за пределы клетки, чтобы
выполнить различные функции в интересах целостного организма.
Например, клетки печени экспортируют альбумин, главный белок|
сыворотки крови, клетки поджелудочной железы производят пин
щеварительные ферменты, В-лимфоциты - антитела и т.д. Когдд,
экстренные меры первой линии восстановления белоксинтезирук>|
щей системы не достигают цели, клетка полностью прекращает <
производство экспортных белков независимо от потребностей
организма {Бойков и др., 1984) и приступает к демонтажу всех
аппаратов, занятых в их производстве, а освобождающиеся при этом ι
молекулярные компоненты и энергоносители будут реутилизиро-'
ваны при сборке новых рибосом (Тодоров, Галкин, 1975; Тодоров
и др., 1975; Тодоров и др., 1977; Тодоров, 1986), которые теперь
уже по новой программе будут заняты исключительно в
производстве белков, необходимых для внутренних нужд клетки в ее
сопротивлении стрессору (Митрохин и др., 1988).
• "Производить только то, что нужно для восстановления
синтеза белков". Параллельно с описанными процессами
реутилизации структурных элементов активируются процессы транскрипции
в ядре. В начале (1-3 часа после блокады) повышается активность
РНК-полимеразы II, а далее (6-12 часов) повышается активность
РНК-полимеразы I (Тодоров и др., 1984; Ллесенко и др., 1984),
что сопровождается повышением синтеза иРНК, рРНК и тРНК
соответственно (Тодоров и др., 1974; 1974а; 1975а; 1976; Тодоров И.,
Галкин, 1975; Чирков и др., 1983). Развитие этих событий, как
было показано (Тодоров и др., 1976а; Тодоров, 1986), связано с тем,
что активность разных групп генов регулируется репрессорами с
различным временем жизни: для РНК-полимеразы II этот белок-
регулятор имеет короткое время жизни (десятки минут), а для
РНК-полимеразы I - более длинное (несколько часов). При
глубокой блокаде трансляции эти регуляторы последовательно
распадаются, что и приводит к активации групп генов, которые они
регулируют. Активация транскрипции, как и уже описанные выше
процессы, направлена на создание de novo белоксинтезирующих
аппаратов и, следовательно, белкового гомеостаза клеток. Все эти
процессы приводят к полному и даже избыточному восстановлению
белок-синтезирующей системы (Сизова и др., 1983) и к избыточной
активации внутриклеточного метаболизма и, как следствие, к
увеличению массы клетки, т.е. к ее росту.
• "Последнее спасительное средство". Заключительная фаза в
ответе клетки на длительное и серьезное подавление белкового син-
316
теза - удвоение генетического аппарата, локализованного в
клеточном ядре, т.е. репликация ДНК и всех структурных и
функциональных белковых компонентов, обеспечивающих структуру
хромосом и важнейшие генетические процессы - транскрипцию,
репликацию, репарацию и рекомбинацию (Тодоров и др., 1978;
Бойков и др., 1978; 1979; 1985). За этими событиями почти
автоматически следует деление клетки (митоз) или полиплоидия (т.е.
удвоение генетического материала без разделения клетки).
Вступление в клеточный цикл - фундаментальная акция клеток по
нормализации белкового гомеостаза, когда все структурные компоненты
и компартменты, количество информации в генетическом
аппарате, уровень метаболизма и в особенности - энергетического,
приводятся в сбалансированное гармоничное состояние. Не совсем
ясно, почему заключительная фаза клеточного ответа адаптации -
клеточное деление. Одна из возможностей состоит в том, что
общий объем белоксинтезирующих аппаратов, превышающий
исходный уровень (неизбежная избыточная компенсаторная реакция на
глубокое подавление трансляции) нуждается в большем
количестве генетической информации для функционирования этого
возросшего объема, что требует увеличения так называемой дозы гена,
которое достигается путем удвоения генетического материала.
Последующее клеточное деление приводит соотношение "белок-
синтезирующий аппарат/генетическая информация" к
нормальному состоянию, которое имело место до развития клеточного
стресса.
• Если подавление трансляции продолжается и после репликации
ДНК и клеточного деления, полный цикл клеточной адаптации
будет повторен, заканчиваясь следующим делением клетки и т.д.
В ходе этих длительных и обширных исследований было
показано, что широкое разнообразие разрушительных воздействий
(большинство из них затрагивает белковый синтез) вызывает описанную
выше последовательность реакций в клетках млекопитающих так
же, как и ОАС организма вызывается стрессогенными факторами
различной природы.
Многолетние исследования ОАРК обобщены в монографии
I.N. Todorov "Mechanisms of Cell Stability: Subcellular and Moleculai
Aspects" (1995).
ОАРК играет определенную роль в процессах старения.
Во-первых, она резко возбуждает клеточный метаболизм на стадии
восстановления, что ведет к формированию большего количества
свободных радикалов, и поэтому ускоряет старение и увеличивает риск
болезней. Во-вторых, далеко зашедший клеточный стресс
стимулирует клеточные деления, которые, согласно теории И.Н. Тодорова
(Todorov, 1990; Todorov, 1995), суть заключительньвл шаг в стратегии
их самосохранения. У высших организмов число делений данной
317
клетки ограничено (см. гл. 2, параграф - лимит Хайфлика).
Достаточно глубокий клеточный стресс заставит клетку пройти все фазы
адаптации и в конечном счете совершить деление, подталкивая
клетку еще ближе к лимиту Хайфлика, что, по существу, является
ускорением ее старения.
19.4.1. Перспективы в исследовании клеточного стресса
Существенным препятствием при исследовании механизмов
старения и клеточного стресса является высокая продолжительность
жизни многих высших животных и в особенности человека. Это
обстоятельство побудило С. Остада (Austad, 2001) разработать
экспериментальную парадигму для изучения медленно стареющих
организмов. Этот подход базируется на обширных эмпирических
данных, согласно которым устойчивость к процессам старения на
уровне организма будет отражена в устойчивости к различным стрессо-
генным факторам на клеточном уровне. Преимущество этой
парадигмы состоит в том, что она не требует ни долговременного
контроля, ни использования исключительно старых индивидов. Подход
С. Остада состоит из: (1) проверки того, что первичные клеточные
культуры от долгоживущего организма показывают лучшую
устойчивость к ключевым стрессогенным факторам, чем клетки от
относительно короткоживущих организмов; (2) оценки различий в
выражении генов до и после стресс-экспозиции культуры клеток от
долго- и короткоживущих видов, чтобы идентифицировать главные
гены, вовлеченные в проявление стресс-устойчивости; (3) трасфекции
предполагаемых ключевых генов от долговечного вида в клетки или
линии клеток с определенной устойчивостью к стрессу с надеждой
наблюдать, что фенотип устойчивости к стрессу был таким образом
передан с геном(ами); (4) создания трансгенной модели животных,
содержащих этот (эти) ген(ы) и наблюдение за тем, способствуют ли
они продлению жизни или улучшению здоровья.
Еще один подход связан с предложением исследовать вклад
стресс-индуцированных повреждений клеточной ДНК через (1) роль
повреждений ядерной ДНК, опосредованных действием поли(АДФ-
рибозы)полимеразы-1 и их ремонта, (2) роль повреждений теломе-
разной ДНК и их вклада в управляемое теломерами старение
клетки, и 3) роль повреждений и накопления мутаций в митохондриаль-
ной ДНК (Zglinicki V. et al., 2001). Авторы предлагают
комбинировать описанный подход с вычислительным моделированием, что
поможет существенно продвинуться в понимании сложности
клеточного старения.
Согласно эволюционной теории, долгоживущие виды
гарантируют свое долголетие благодаря более долговечной соме, включая
увеличенную устойчивость клеток к стрессу. В многочисленных
318
экспериментах (Kapahi et al., 1999) было показано, что клеточная
устойчивость к разнообразным окислительным и неокислительным
стрессам положительно коррелирует с долговечностью
млекопитающих. Эти результаты поддерживают концепцию, что сеть генов,
регулирующая клеточный ответ на стресс, функционально важна
для старения и долгой жизни и это, по-видимому, способно
стимулировать дальнейшие исследования в этом направлении.
Большое внимание уделяется механизмам психосоциального
стресса и их влиянию на процессы старения. Регионы мозга,
вовлеченные в обучение, память и эмоции, играют видную роль в стресс-
ответе, а моноаминэргические и глютаматэргические синапсы -
особенно важную роль в преобразовании сенсорных затрат при стрессе в
нейроэндокринные ответы. Существует много указаний на то, что
нормальное старение воздействует на нейроэндокринные
стресс-ответы, и изучение молекулярных и клеточных механизмов, лежащих в
основе патогенного действия мутаций, вызывающих возрастные
неврологические расстройства типа болезни Альцгеймера, раскрывает
новое понимание причастности возбуждающих нейроэндокринных
сресс-ответов к этим расстройствам (Pedersen et al., 2001).
19.5. ОБЩИЙ АДАПТАЦИОННЫЙ СИНДРОМ
И КЛЕТОЧНЫЙ СТРЕСС
В большинстве случаев системный и клеточный стрессы
развиваются совместно. С одной стороны, гормоны стресса, выделяемые
в течение стресс-ответа организма, по-видимому, вызывают
клеточный ответ адаптации и ответ на температурный шок в некоторых
тканях. Например, катехоламины, видимо, вызывают клеточный
стресс в сердце, что, очевидно, может объяснять, почему общий
стресс увеличивает количество полиплоидных клеток в сердечной
мышце (Меерсон, 1984). С другой стороны, достаточно серьезный
клеточный стресс вызовет и стресс-ответ всего организма. Поэтому
сокращение общего стресса обычно уменьшает клеточный стресс, и
наоборот. Примеры "вполне" клеточного стресса (т.е. без
существенного стресс-ответа целостного организма) — это умеренное
повреждение кожи солнечной инсоляцией, незначительные ожоги и
умеренные повреждения печени токсинами или алкоголем.
19.6. БОРЬБА С КЛЕТОЧНЫМ СТРЕССОМ
Как мы можем защищаться от клеточного стресса? Некоторую
защиту могут дать те же самые меры, которые помогают замедлить
механизмы старения и сохранить или восстановить оптимальный
стресс-ответ организма. Если имеет место чрезмерная "рабочая
нагрузка" или повреждение данного органа, то желаемую защиту воз-
319
можно обеспечить, используя так называемые
орган-специфические цитопротекторы. Например, силимарин, смесь трех
растительных флаволигнинов, выделенных из сока чертополоха (Silybum
marianum), предотвращает повреждение печени токсинами и
другими агрессивными факторами. Гинкгозиды, активные гликозиды из
растения Ginkgo biloba, имеют специфическое сродство с нервной
тканью и могут защищать нервную систему от свободных радикалов
или недостаточного кровоснабжения. Пара-аминобензойная
кислота (ПАБК) защищает кожу от повреждения солнцем, блокируя
действие УФ-облучения. Простагландин El (PG E1) предотвращает
повреждение клеток нестероидными противовоспалительными
лекарствами, кислотами и острым стрессом. Кофермент Q10 и
ацетил-Ь-карнитин защищают сердце, улучшая его способность к
энергопродукции.
* * *
Проблемы клеточного стресса все еще нуждаются в
дальнейшем изучении. В ближайшем будущем можно, как уже указывалось
выше, ожидать развитая эффективных способов предотвращения
или нейтрализации клеточного стресса в различных тканях. Один
многообещающий подход - "стрессовая тренировка" клетки -
подобен роли умеренных физических упражнений в качестве защиты от
стресс-ответа организма. Некоторые вещества или воздействия
способны к мягкой стимуляции клеточного ответа адаптации, в
частности ответа на температурный шок или стимуляция системы
ДНК-репарации (Pulatova et al., 1999) Клетки, обработанные таким
способом, становятся намного более стойкими к различным видам
повреждений, потому что их системы защиты, фигурально говоря,
"хорошо обучены" и готовы к сопротивлению. Например, некоторые
ученые полагают, что умеренный ответ на температурный шок,
вызванный осторожным и кратковременным тепловым воздействием,
может помочь в поддержании достаточно высокой субклеточной
концентрации белков теплового шока и таким образом делать
клетки более стойкими к различным типам повреждений (Schlesinger,
1994). В практическом смысле это означает, что горячие ванны или
регулярные посещения сауны помогают не только расслабиться, но
и фактически вносят вклад в "клеточное здоровье". Однако при
этом необходимо соблюдать меру. Злоупотребление сауной или
другими термическими процедурами может привести к
противоположным эффектам. Люди с заболеваниями сердца и некоторыми
другими патологиями не должны подвергать себя интенсивным
воздействиям высокой температуры, по крайней мере без консультации с
лечащим врачом.
Глава 20
СТРЕСС И ПОЛОВАЯ ФУНКЦИЯ
Личный опыт многих из нас свидетельствует, что во время
серьезного или длительного стресса половое желание и способность его
реализовать заметно снижаются. Это, по-видимому, "естественная"
часть стресс-ответа. Действительно, стресс-ответ изменяет
приоритеты организма в интересах преодоления непосредственной
опасности или препятствия, подавляя или сохраняя на низком
функциональном уровне системы, несущественные в данный момент для
выживания, а именно пищеварительную, иммунную и репродуктивную.
Если стресс непродолжителен, сексуальная функция
восстанавливается вскоре после того, как напряженная ситуация разрешается.
Исключение, однако, составляют такие случаи, когда на
сексуальную активность мужчины влияет не стресс, а, наоборот, его
неспособность произвести сексуальные действия вызовет стресс-ответ,
который может кончаться состоянием тревоги и временной
импотенцией. Иногда заверения врача, что проблема временная, бывает
достаточно для нормализации этой ситуации. В других случаях
могут быть полезными общепринятые методы релаксации или
успокаивающие медикаменты. Однако лучшей "медициной" для
"сексуальных сбоев" может оказаться понимающий и заботливый партнер,
разумеется, желательно при устойчивых моногамных отношениях,
которые практически исключают тревогу и беспокойство в
интимной обстановке.
Хронический стресс может оказывать длительное влияние на
сексуальность человека. В современном обществе преобладает
психологический стресс, ассоциированный с депрессией и
беспокойством, т.е. состояниями, которые могут привести к потере интереса к
сексуальным отношениям. И поскольку пожилые люди более
подвержены хроническому стрессу и депрессии, можно полагать, что
"нормальная" потеря полового желания с возрастом часто связана с
влиянием именно этих факторов.
Некоторые из гормональных изменений, вызванных стрессом,
оказывают прямое влияние на половую функцию. Психологический
стресс и некоторые типы физического стресса, такие как анестезия,
321
обширные травмы или хронические болезни, заметно понижают у
животных и людей уровень тестостерона, который может
оставаться низким много позже окончания стрессогенной ситуации. В одном
из исследований у макак резус, подвергавшихся стрессогенному
воздействию, а затем лишенных возможности социального общения,
падение уровня тестостерона продолжалось в течение нескольких
недель.
Стресс может также подавлять мужскую половую функцию,
повышая уровень пролактина, гормона, производимого гипофизом.
Природа этого эффекта неясна, поскольку единственная, хорошо
установленная физиологическая роль этого гормона -
стимулировать у женщин продукцию молока. У мужчин высокий уровень
пролактина подавляет функцию семенников, стимулирует разрастание
простаты и может вызывать импотенцию. Ненормально высокая
секреция пролактина у мужчин в нормальных условиях редка и
обычно связана с ростом опухоли гипофиза. Такие пациенты имеют
частично атрофированные семенники, гипертрофированную
простату, и, как правило, импотенцию. Хорошо известно, что стресс
является причиной значительного повышения уровня пролактина.
До сих пор еще не ясно, вносит ли вызванный стрессом избыток
пролактина вклад в развитие психогенной импотенции.
Стресс влияет на потенцию также и косвенно, увеличивая риск
дегенеративных болезней. Будучи одним из факторов
возникновения и прогрессии атеросклероза, гипертонии и диабета, стресс
усиливает главные причины органической импотенции - повреждение
нервов и сосудов
Существуют данные, что адаптогены, средства, смягчающие
стресс-ответ (см. гл. 29), могут быть полезны при восстановлении
сексуальной активности. Недавно корейские исследователи нашли,
что женьшень, высокоэффективный адаптаген, оказался
эффективен в восстановлении как полового желания, так и потенции.
Исследовалось 90 пациентов, страдающих от эректальной дисфункции.
Испытуемые были разделены на три равных группы. В одной
группе пациенты получали женьшень, в другой - тразодон (нейроактив-
ное лекарственное средство, иногда используемое для лечение
импотенции) и в третий - плацебо. У пациенов, получавших женьшень,
в 60% случаев наблюдалось существенное улучшение параметров
эрекции (жесткость, размер, продолжительность), так же как и
повышение либидо. В других группах только 30% паииентов имели
подобные улучшения.
Эффекты стресса, которое могут вредить сексуальной функции
• Депрессия, тревога, подавленное настроение, недостаток интереса
к сексуальной деятельности;
322
• Сжатие периферийных кровеносных сосудов (особенно во время
стресса, сопровождаемого заметной тревогой), которое
препятствует достаточному току крови к половым органам;
• Пониженное производство тестостерона;
• Повышенное производство пролактина;
• Повышенный риск дегенеративных болезней - атеросклероза,
гипертонии и диабета, вносящих вклад в повреждение сосудов и
нервов, которые причастны к физической импотенции.
Глава 21
СИСТЕМЫ ОГРАНИЧЕНИЯ СТРЕССА
В сущности, стресс-ответ - чрезвычайная, эволюционно
сформированная реакция, которая ориентирована на кратковременное
обеспечение выживания организма при экстремальных
обстоятельствах. Высокая биологическая ценность такой реакции очевидна,
несмотря на то что сам стресс-ответ как таковой - разрушителен и,
если он интенсивен или продолжителен, может вызывать или
усиливать болезни и сокращать продолжительность жизни. Чтобы
уравновесить потенциальную опасность стресс-ответа, организм должен
быть способен к перманентному ограничению его интенсивности и
продолжительности, т.е. уметь защищать себя от его возможных
вредных побочных эффектов. Действительно, эволюционно
сформировались также и механизмы, чья функция заключалась в том,
чтобы оградить организм от последствий его же собственного
общего адаптационного синдрома. Исходя из содержания предыдущих
глав, легко предположить, что работа некоторых из этих
механизмов ухудшается с возрастом, оставляя организм менее защищенным
от неблагоприятных последствий чрезмерного стресс-ответа.
Некоторые стресс-ограничивающие механизмы работают на
уровне высшего регуляторного центра, контролируя реакцию
центральной нервной системы на стресс-ответ. Другие непосредственно
защищают клетки от разрушительных действий стресса; есть и
такие, которые, по-видимому, оказывают и центральное, и
периферийное действие. Чтобы минимизировать связанные со стрессом
повреждения, необходимо постоянно поддерживать эти механизмы в
хорошей форме. Пожилые или перенесшие тяжелый стресс люди
могут извлечь пользу от стимуляции стресс-лимитирующих
механизмов с помощью специального питания, пищевых добавок и
других методов.
Антиоксидантная система. Стресс-ответ генерирует поток
свободных радикалов, которые могут тяжело повреждать органы и
ткани организма. В то же самое время начальные фазы стресса
связаны со стимуляцией антиоксидантой защиты, чтобы, по-видимому,
противодействовать повышенному формированию СР и таким обра-
324
зом ограничивать повреждения. Серьезный или длительный стресс,
однако, в конечном счете, исчерпывает АО-ресурсы, и защита
ослабевает со всеми вытекающими негативными последствиями. У
старых людей АО-резервы обычно ниже, они не способны
синтезировать достаточное количество АО-ферментов; продукция мелатони-
на, мощного антиоксиданта, у них фактически ничтожна, а уровень
глутатиона, основного внутриклеточного водорастворимого АО,
часто понижен. Потребление антиоксидантов, таких как витамин Е,
цистеин, липоевая кислота, флавоноиды, каротиноиды и др.,
помогает сохранению резервов АО при стрессе и предотвращению или
уменьшению связанных с ним повреждений. Особую пользу
пищевые добавки с АО приносят пожилым людям, поскольку последние
в большей степени, чем молодые, подвержены ущербу от действия
СР. Конкретная информация относительно обогащения вашей
диеты антиоксидантами содержится в главе 23.
Гормон роста (ГР). И физический и психологический стресс, в
особенности, если он - интенсивен, стимулирует гипофиз к
выделению гормона роста. Это наводило на мысль, что повышенный
уровень ГР в период стресса действует в синергизме с кортизолом и
адреналином, чтобы повысить содержание глюкозы в крови,
гарантируя обильную доставку энергоносителей к нервной и мышечной
системам. Однако было установлено, что выделение ГР не зависит от
уровня глюкозы в крови. Современные представления
приписывают ГР роль ограничителя интенсивного стресс-ответа и защитника
от стресс-индуцированных повреждений. Несколько исследований
продемонстрировали, что ГР смягчает и затем полностью
корректирует вызванную стрессом иммунодепрессию, и наоборот -
животные с пониженным уровнем ГР были не в состоянии восстановить
свою иммунную функцию после того, как стресс миновал (Dilman,
1994). Как уже не раз указывалось, кортикостероиды, гормоны
стресса, вызывают потерю нежировой ткани, разрушая белок для
использования в интересах энергоснабжения, ГР действует в
противоположном направлении, ограничивая потерю белка и позже
облегчая восстановление нежировой ткани {Klatz et al., 1997).
Известный антистрессовый эффект физических упражнений связан,
вероятно, со стимуляцией выделения ГР. Как уже подробно обсуждалось
в главе 5, продукция ГР снижается с возрастом и это же происходит
с устойчивостью к стрессу. Продукция ГР может быть увеличена
при помощи определенных питательных веществ, которые
стимулируют выделение ГР гипофизом (см. гл. 26). Инъекции экзогенного
ГР также приводят к антистрессовым эффектам, но ГР, полученный
с помощью генно-инженерной техники, весьма дорог и с его
приемом связан риск неблагоприятных побочных эффектов
ГАМК и ГГБ. ГАМК (гамма-аминомасляная кислота, или гам-
ма-аминобутират) - ингибирующий нейромедиатор с
многосторонними влияниями на нервную систему: расслабляет мышцы, понижа-
325
ет кровяное давление, снимает чувство тревоги и в высокой
концентрации стимулирует сон {Готтлиб, 1988). Выделение ГАМК
вызывается психологическим и болевым стрессом и смягчает
стресс-ответ {Меерсон, 1986а). Эффекты ГАМК трудны для
непосредственного исследования, поскольку это соединение не в состоянии
пересекать гематоэнцефалический барьер, и оральное или внутривенное
введение приводит к незначительному эффекту. ГГБ {гамма-гидро-
ксибутириловая кислота, или гамма-гидроксибутират) является и
предшественником и метаболитом ГАМК; оба соединения связаны
структурно. В отличие от ГАМК ГГБ легко пересекает
гематоэнцефалический барьер и после этого в мозгу конвертируется в ГАМК.
В дополнение к его роли предшественника ГГБ, по-видимому,
обладает прямым ингибирующим действием, не связанным с
рецепторами ГАМК. Введение ГГБ животным приводило к понижению
интенсивности стресс-ответа и предотвращало связанные с ним
повреждения различных органов, особенно сердца и желудочно-кишечного
тракта {Меерсон, 1981; 1984).У животных, подвергнутых стрессу,
введение ГГБ уменьшало выделение гормонов стресса (кортизола и
катехоламинов) так же как и формирование СР в сердечной мышце.
Оба соединения, ГАМК и ГГБ, считаются питательными веществами.
ГАМК продается в магазинах здоровой пищи. Однако ГАМК - относительно
слабый защитник от стрессоров из-за его недостаточной способности
пересекать гематоэнцефалический барьер. ГГБ свободно продавался до 1990 г., когда
FDA запретил его свободную продажу в США, аргументируя это решение
высокой токсичностью ГГБ, а также возможностью злоупотреблений при его
свободной продаже. Эти утверждения спорны: высокие дозы ГГБ могут
действительно вызвать глубокий сон, от которого человек не в силах легко
пробудиться, но так способны действовать высокие дозы многих легкодоступных
веществ, например алкоголя. К тому же ГГБ, по-видимому, не имеет какой-либо
существенной долговременной токсичности и не вызывает заметного
привыкания. Диазепам (валиум) и другие бензодиазепины, т.е. широко рекомендуемые
транквилизаторы (средства, снимающие чувство тревоги, беспокойства),
действуют на ГАМК-рецепторы. Эти лекарства, очевидно, имеют более высокий
потенциал для привыкания или злоупотреблений, чем ГГБ. Дебаты о статусе ГГБ
продолжаются.
Энкефалины и эидорфины (называемые также эндогенными опио-
идами) - морфийподобные вещества, производимые организмом
(Akil et al., 1978). Наиболее известный эффект этих соединений -
притупление восприятия боли. Различные типы стрессогенных
факторов, таких как хирургические операции, травмы и другие
испытания, вызывают выделение эндогенных опиоидов. У некоторых
людей они не только уменьшают боль, но и вызывают состояние
эйфории. Пресловутый "адреналиновый шторм", которого так жаждут
любители острых ощущений, связан, по крайней мере частично, с
эндогенными опиоидами, а не с самим адреналином. Во время
стресса энкефалины и эндорфины аккумулируются в гипоталамусе и
надпочечниках и, по-видимому, играют непосредственную роль в сокра-
326
щении стресс-ответа {Слепушкин и др. 1986). Введение опиоидов
животным до стресса предотвращает повреждение сердца, так же
как и индуцированное стрессом увеличение метастатического
распространения рака {Барабой, Брехман и др., 1992). Аминокислота
D-фенилаланин ингибирует деградацию энкефалинов, повышая их
уровень в центральной нервной системе. Оказалось даже, что
D-фенилаланин притупляет восприятие боли у животных. Некоторые, но
не все, исследования на людях также показали, что D-фенилаланин
снимает боль с эффективностью действия от 32 до 75% (Walsh et al,
1986). Теоретически D-фенилаланин способен смягчить
стресс-ответ, повышая уровень эндогенных опиоидов, и поэтому
рассматривается в качестве полезной пищевой добавки. Обычно он продается
в смеси со своим оптическим изомером L-фенилаланином (оба
соединения, как две руки - зеркальные отображения друг друга). L-фе-
нилаланин - предшественник в синтезе нейромедиаторов -
дофамина и норадреналина. DL-фенилаланин (равная смесь D- и L-изоме-
ров), как было показано, снижает депрессию по крайней мере столь
же эффективно, как популярный антидепрессант - имипрамин, но с
меньшим количеством неблагоприятных побочных эффектов
(Todorov И. et al., 2001).
Мелатонин в строгом смысле - не часть стресс-ограничиваю-
щего механизма, потому что его производство в организме не
увеличивается под действием стрессогенных факторов. Однако
способность производить мелатонин представляется важной для
поддержания устойчивости организма к стрессу. Мелатонин помогает
минимизировать повреждения, стимулируя иммунную систему, так же
как и активировать антиоксидантную систему и вызывать общий
успокаивающий антистрессовый эффект. Несколько исследований на
животных показали, что мелатонин предотвращал или уменьшал
стресс-зависимую иммунодепрессию и образование язв желудка.
Продукция мелатонина резко снижается с возрастом, почти исчезая
к шестидесяти. Это может вносить определенный вклад в
возрастную потерю устойчивости к стрессу. Более подробную информацию
о связи между мелатонином и стрессом см. в главе 5.
Простагландины - большой класс соединений, вовлеченных в
локальную связь между клетками, т.е. они обычно действуют в
пределах или близко от ткани, в которой они были выделены.
Повышение уровня катехоламинов и свободных радикалов во время стресса
стимулирует синтез простагландинов во многих тканях. При этом
простагландины уменьшают повреждения клеток, расширяя
кровеносные сосуды и противодействуя некоторым эффектам
катехоламинов. Особый интерес представляет простагландин Е, который
предотвращает образование язв желудка и уменьшает кровяное
давление при стрессе. Аспирин и другие нестероидные
противовоспалительные лекарства (NSAID) могут вызывать
желудочно-кишечные повреждения, подавляя синтез простагаландина Ε в слизистой
327
оболочке. Мизопростол, синтетический аналог простагландина Е,
часто прописывают пациентам, страдающим артритом, вместе с
NSAID, чтобы предотвратить желудочно-кишечные повреждения.
Стресс против стресса. Как уже указывалось, умеренный по
силе и продолжительности стресс повышает устойчивость организма к
последующему сильному стрессу. Например, умеренный защитный
противолучевой эффект может быть достигнут при предварительном
воздействии на организм слабой механической, термической или
ожоговой травмы, адаптации к гипоксии, УФ- или ионизирующей радиа
ции в малых дозах, при легких воспалительных и иммунных реакциях,
обусловленных введением полисахаридов, стеклянной пыли и т.п.
(Бак,Александер, 1963; Романцев, 1968; Эйдус, 1977; Betz, 1956; Кален-
до, 1982; Bachner et al, 1982; Гончаренко и др., 1985).
При повторных стрессах существенно возрастает синтез катехол-
аминов в мозговом веществе надпочечников, о чем свидетельствует
повышение активности ключевого фермента их биосинтеза - тиро
зингидроксилазы (Lanks, 1986). Однако это повышение синтеза кате-
холаминов не усиливает ответную реакцию на стресс. Напротив,
адаптация к повторным воздействиям выражается в замедлении
мобилизации катехоламинов и в снижении чувствительности высших
мозговых центров и органов-мишеней (в частности, в падении
количества адренорецепторов в мозге и селезенке) к повышенным
концентрациям катехоламинов. Соответственно реакция на повторный,
но сильный стресс ослабляется и угасает (Меерсон, 1981; 1986а).
Однако адаптация к повторному стрессу возникает не только к
повышенной концентрации катехоламинов, но и к прямому
действию на мышцы сердца одного из индукторов ПОЛ - перекиси
водорода (Меерсон, 1986а; Балицкий и др., 1987 ). Возможный механизм
такой устойчивости, как указывалось выше, связан с активацией
синтеза антиокислительных ферментов - супероксиддисмутазы. ка-
талазы и глютатионпероксидазы.
* * *
Стресс-лимитирующие механизмы весьма эффективны, однако
они могут оказаться недостаточными при серьезном или
длительном стрессе. Кроме того, некоторые из этих механизмов теряют
свою эффективность с возрастом, особенно те из них, которые
связаны с гормоном роста, мелатонином и ферментами,
нейтрализующими свободные радикалы.
Активность некоторых стресс-лимитирующих систем
организма может быть повышена с помощью пищевых добавок.
Экзогенные антиоксид анты способны предотвращать перегрузку
собственных СР- нейтрализующих систем организма (см. гл. 23).
Пищевые стимуляторы способны заметно повышать продукцию
328
гормона роста даже в преклонном возрасте. DL-фенилаланин
может быть особенно эффективен при болевом стрессе, а также
тогда, когда стресс сопровождается депрессией. Мелатонин может
улучшать устойчивость к стрессу у пожилых людей, чья
собственная продукция гормона очень низка. ГГБ, питательное вещество,
пока еще запрещенное в США, может служить полезным
инструментом в арсенале антистрессовых воздействий при осторожном
и разумном использовании. Наконец, некоторые адаптогены,
хотя и не являются частью физиологических механизмов,
ограничивающих стресс, как выяснилось, способны увеличить или
дополнить общую устойчивость организма к стрессогенным факторам
(см. гл. 29).
Литература
Александров В.Я. Реактивность клеток и белки. М.: Наука, 1985- 317 с
Ллесенко Л.В., Бойков П.Я., Бурлакова Е.Б. и др. Стимуляция синтеза
компонентов ядерной мембраны и ДНК клеток печени крыс после ннгибирования
транслации циклогексимидом // Биохимия. 1978. Т. 43, вып. 11. С. 1966-
1972.
Ллесенко А.В., Красилъников В.А., Бойков П.Я., Тодоров И.Н.
Последовательная активация различных типов ядерных РНК-полимераз клеток печени
крыс в процессе индукции синтеза ДНК циклогексимидом // Там же. 1984.
Т. 49, вып. 3. С. 455-459.
Бак 3., АлексанЬер П. Основы радиобиологии. М.: Изд-во иностр. лит., 1963.
560 с.
Балицкий К.П., Шмалько И.Н. Стресс и метастазирование злокачественных
опухолей. Киев: Наук, думка, 1987. 244 с.
Барабой В.А., Брехман И.И., Голотин ВТ., Кудряшов Ю.Б. Перекисное
окисление и стресс. СПб.: Наука, 1992. 148 с.
Барабой В.А., Орел В.Э., Скляренко ВТ. и др. Спонтанная хемилюминесценция
сыворотки крови в норме и при воздействии ионизирующей радиации //
Биолюминесценция. М, 1983. С. 222-240.
Барабой В Α., СутковойДЛ. Энергетический обмен при стрессовых
воздействиях (иа примере ионизирующей радиации), его саморегуляция и коррекция
//Укр. биохим. журн. 1983. Т. 55, № 1. С. 93-105.
Барабой В.Α., Чеботарев Е.Е. Проблема перекисного окисления липидов в
радиобиологии // Радиобиология. 1986. Т. 26, № 5. С. 591-597.
Богданов A.M., Ярушкина И.И. Влияние АКТГ на болевую чувствительность
крыс // Бюл. эксперим. биологии и медицины. 2001. Т. 131, Ms 3. С. 260-
262.
Бойков П.Я., Сидоренко Л.И., Тодоров И.Н. Биосинтез хроматина в клетках
высших животных. Активация синтеза ядерных белков и ДНК гепатоцитов
после импульсного торможения трансляции циклогексимодом // Биохимия.
1979. Т. 44, вып. 6. С. 963-974.
Бойков П.Я., Сидоренко Л.И., Шевченко Н.А., Тодоров И.Н. Биосинтез
хроматина в клетках высших животных. Ускорение транспорта негистонных
белков в ядрах и их катаболизма в условиях ннгибирования синтеза белков /У
Там же. 1981. Т. 46, вып. 8. С. 1396-1410.
329
Бойков П.Я., Сидоренко Л.И., Шевченко Н.А. и др. Активация хроматина и про-
теолиза гистонов при подавлении синтеза белков в клетках печени // Там же.
1983. Т. 48, вып. 1. С. 23-32
Бойков П.Я., Шевченко Н.А., Сидоренко Л.И., Тодоров И.Н. Соотношение
биосинтеза внутриклеточных и экспортых белков в клетках печени крыс в
процессе индукции пролиферации циклогексимидом // Там же. 1984. Т. 49,
вып. 9. С. 1470-1477.
Браун AJJ., Мюженок Т.П. Неспецифический адаптационный синдром
клеточной системы. Л.: Наука, 1987. 231 с.
Бурлакова Е.Б., Алесенко А.В., Молочкина ЕМ. и др. Биоантиоксиданты в
лучевом поражении и злокачественном росте. М.: Наука, 1975. 214 с
Бурлакова Е.Б., Храпова Н.Г Перекисное окисление липидов мембран
и природные антиоксиданты // Успехи химии. 1985. Т. 54, № 9. С. 1540-
1558.
Валъдман А.В. Молекулярные основы действия психотропных средств. М.,
1986. 166 с.
Валъдман А.В., Козловская ММ., Медведев О.С. Фармакологическая
регуляция эмоционального стресса. М.: Медицина, 1979. 359 с.
Валъдман А В., Пошивалов В.П. Фармакологическая регуляция
внутривидового поведения. М.: Медицина, 1984. 208 с.
Виру А.А. Гормональные механизмы адаптации и тренировки. Л.: Наука, 1981.
155 с.
Владимиров Ю.А. Биологические мембраны и ^запрограммированная смерть
клетки // Сорос, образов, журн. 2000а. Т. 6, № 9. С. 2-9.
Владимиров Ю.А. Свободные радикалы в биологических системах // Там же.
20006. Т. 6, № 12. С. 13-19.
Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в
биологических мембранах. М.: Наука, 1972. 259 с.
Галкин А.П., Корнюшина Т.В., Говзе Я.Г., Тодоров И.Н. Исследование цито-
плазматических РНК печени крыс в условиях длительного угнетения
синтеза белка цитогексимидом // Укр. биохим. журн. 1974. Т. 46, № 5. С. 650-
657.
Галкин А.П., Тодоров И.Н. О высокой радиочувствительности
биосинтеза митохондриальной ДНК // Радиобиология. 1969. Т. 9, № 1. С. 21-
25.
Гаркави Л.Х., Квакина Е.Б., Уколова М.А. Адаптационные реакции и
резистентность организма. Ростон н/Д: Изд-во Рост, ун-та, 1977.
120 с.
Гомеостаз / Под ред. П.Д. Горизонтова. М.: Медицина, 1976. 464 с.
Гончаренко Е.Н., Кудряшов Ю.Б. Химическая защита от лучевого поражения.
М.: Изд-во МГУ, 1985. 249 с.
Дардымов И.В. Механизмы действия препаратов женьшеня и элеутерококка:
Автореф. дис.... д-ра мед. наук. Л., 1987. 41 с.
Заводская И.С., Морева Е.В. Фармакологический анализ механизмов стресса и
его последствий. Л.: Медицина, 1976. 214 с.
Календо Г.С. Ранние реакции клеток на ионизирующее облучение и их роль в
защите и сенсибилизации. М.: Энергоатомиздат, 1982. 96 с.
Кассиль ГН. Наука о боли. М.: Наука, 1975. 399 с.
Кириллов О.И. Опыт фармакологической регуляции стресса. Владивосток:
Дальневост. кн. изд-во, 1966. 107 с
Кузин A.M. Структурно-метаболическая теория в радиобиологии. М.: Наука,
1986.282 с.
330
Лившиц А.А., Паскевич И.Ф., Тодоров И.Н. Фракционирование аденогииофи-
зарной информационной РНК // Укр. биохим. журн. 1969. Т. 41, № 2.
С. 180-184.
Машанский В.Ф., Рабинович И.М. Ранние реакции клеточных органоидов. Л.:
Наука, 1987. 120 с,
Меерсон Ф.З. Адаптация, стресс и профилактика. М.: Наука, 1981.278 с.
Меерсон Ф.З. Патогенез и предупреждение стрессорных и ишемических
повреждений сердца. М.: Медицина, 1984. 270 с.
Меерсон Ф.З. Общий механизм адаптации и роль в нем стресс-реакции,
основные стадии процесса // Физиология адаптивных процессов. М., 1986а.
С. 77-123.
Меерсон Ф.З. Адаптация к стрессорным ситуациям и стресс-лимитирующие
системы организма // Там же. М„ 19866. С. 521-631.
Митрохин Ю.И., Сизова СТ., Гутникова М.Н., Тодоров И.Н. Динамика
биосинтеза компонентов белоксинтезирующего аппарата печенн крыс на этапе
восстановления трансляции, ингибированной циклогексимидом // Биохимия.
1988. Т. 53, вып. 12. С. 2033-2041.
Насонов Д.Н., Александров В.Я. Реакция живого вещества на внешние
воздействия. М; Л., 1940. 252 с.
Насонов Д.Н. Местная реакция протоплазмы и распространяющееся
возбуждение. М.; Л., 1959.434 с.
Огурцов СИ., Духанин А.С., Темное А.А. Митохондриальная и ядерная глюко
кортикоид-чувствительные щелочные протеазы тимоцитов // Бюл. экспе-
рим. биологии и медицины. 2001. Т. 132, № 7. С. 23-25.
Панин Л.Е. Биохимические механизмы стресса. Новосибирск: Наука, 1983.
233 с.
Панков Ю.А. Гормоны гипофиза // Биохимия гормонов и гормональной
регуляции. М.: Наука, 1976. С. 44-93.
Пулатова М.К., Шарыгин BJI., Филатов Д.Э., Тодоров И.Н. Роль рибонуклео-
тидредуктазы в регуляции пострадиационного синтеза ДНК // Докл. АН
СССР. 1988. Т. 300, № 1. С. 233-236.
Пулатова М.К., Шарыгин BJI., Филатов Д.Э., Тодоров И.Н. Активация рибо-
нуклеотидредуктазы как показатель SOS-реакции на воздействие
экстремальных факторов, повреждающих ДНК в клетках высших животных // Там
же. 1995. Т. 340, № 1. С. 123-127.
Романцев Е.Ф. Радиация и химическая защита. М.: Атомиздат, 1968. 248 с.
Рыску лова СТ. Радиационная биология плазматических мембран. М.: Энергоа-
томиздат, 1986. 127 с.
Селъе Г. Очерки об адаптационном синдроме. М.: Медгиз, 1960. 254 с.
Селье Г. Концепция стресса как мы ее представляем в 1976 г. // Новое о
гормонах и механизме их действия. Киев: Наук, думка, 1977. С. 27-51.
Сизова СТ., Митрохин Ю.И., Гутникова М.Н., Тодоров И.Н. Функциональная
активность компонентов белоксинтезирующего аппарата гепатоцитов в
процессе восстановления биосинтеза белка, ингибированного
циклогексимидом// Биохимия. 1983. Т. 48, вып. 2. С. 211-218.
Скулачев В.П. О биохимических механизмах эволюции и роли кислорода // Там
же. 1998. Т. 63, № 11, с. 1570-1580.
Скулачев В.П. Эволюция, митохондрии и кислород // Сорос, образов, журн.
1999. № 9. С. 4-10.
Слепушкин В Д., Золоев Г.К. Роли нейропептидов в механизме развития
стресса и шока // Механизмы патологических реакций. Томск. 1986. Т. 4.
С. 86-90.
331
Соколов ЕМ. Эмоции и атеросклероз. М: Наука, 1987. 253 с.
Сумбаев В.В. Активность протеинкиназы, регулирующей апоптогенный
сигнал 1, поли-(АДФ-рибоза)-полимеразы и межнуклеосомная
фрагментация ДНК в печени крыс в условиях оксидативного стресса, вызванного
хлоридом кобальта // Бюл. эксперим. биологии и медицины. 2001. Т. 131,
№ 2. С. 148-149.
Тарусов Б.Н. Основы биологического действия радиоактивных излучений. М.:
Медгиз, 1954. 140 с.
Тодоров И.Н. Субклеточные и молекулярные аспекты биосинтеза адренокор-
тикотропного гормона // 2-й Укр. биохим. съезд: Материалы симпозиума
Киев: Наук, думка, 1971. С. 43-46.
Тодоров И.Н. Механизмы клеточной стабильности // В мире науки. 1986. № 10.
С. 50-61.
Тодоров И.Н, Блок Л.Н. Влияние рибонуклеиновой кислоты нз бычьих гипо
физов на некоторые особенности гормонопоэза в аденогипофизе белых
крыс//Докл. АН УССР. 1964. № 10. С 1331-1333.
Тодоров И.Н., Блок Л.Н., Васильченко В.Н. Синтез адренокортикотропного
фактора в бесклеточной системе из E.coli под влиянием рибонуклеиновой
кислоты из бычьих аденогипофизов // Докл. АН СССР. 1966а. Т. 167, № 2
С. 461-463.
Тодоров ИМ., БлокЛ.Н., Васильченко В.Н. Синтез аденокортикотропного
фактора в бесклеточной системе из Е. coli под влиянием рибонуклеиновой
кислоты из бычьих аденогипофизов // Нуклеиновые кислоты: Тр. 2-й конф. по
нуклеиновым кислотам / Под ред. В.Н. Ореховича и B.C. Тонгура. М.:
Медицина. 19666. С. 149-153.
Тодоров И.Н., Бойков П.Я., Сидоренко Л.И. и др. Стимуляция репликации ДНК
в клетках печени крыс как результат ингибирования синтеза белков // Докл
АН СССР. 1978. Т. 239, № 5. С. 1255-1258.
Тодоров И.Н., Васильченко В.Н., Лившиц А.А. и др. Использование
корреляционного метода включения С14-аминокислот при
идентификации синтезированного вне клетки адренокортикотропного гормона
(АКТГ) // Материалы Респ. науч. конф. по пробл. "Лечебное и
диагностическое применение радиоактивных изотопов". Киев: Здоровье, 1968.
С. 304-306
Тодоров И.Н., Васильченко В.Н., Панкова ГЛ. и др. О синтезе вещества адре-
нокортикотропной природы, индуцированном в бесклеточной системе E.coli
информационной рибонуклеиновой кислотой из бычьих аденогипофизов //
Биохимия. 1967. Т. 32, вып. 2. С. 283-292.
Тодоров ИМ., Галкин А.П. Об особенностях биосинтеза РНК в клетках печенн
крыс в условиях продолжительного подавления биосинтеза белка циклогек-
симидом // Молекулярная биология. Киев: Наук, думка, 1975. Вып. 11
С. 81-110.
Тодоров И.Н., Галкин А.П., Андрианов В.М., Шмалъко П.И. Исследование
ультраструктуры и функционального выражения генома гепатоцитов в
условиях продолжительного угнетения биосинтеза белка циклогексимидом.
II. Седиментационные свойства полирибосом и состояние эндоплазмати-
ческого ретикулума // Цитология и генетика. 1975. Т. 9, № 2. С. 129-
132
Тодоров И.Н., Галкин А.П., Корнюшина ТВ. и др. Синтез РНК в печени крыс
в условиях угнетения синтеза белка циклогексимидом // Укр. биохим. журн.
1974а. Т. 46, № 4. С. 431-^37.
332
Тодоров И.И., Галкин А.П., Шен Р.А. и др. Изменение РНК-синтезирующей
способности изолированных ядер и матричной активности хроматина
печени крыс в условиях угнетения биосинтеза белка циклогексимидом // Докл.
АН СССР. 1975а. Т. 223, № 6. С. 1488-1491.
Тодоров И.Н., Лившиц Α.Α., Паскевич И.Ф. Некоторые свойства
информационных РНК бычьих аденогипофизов // Биохимия. 1969. Т. 34, № 5.
С. 1048-1057.
Тодоров И.Н., Манжелей Е.С., Набутовская ЛМ. Влияние актиномицина Д на
биосинтез АКТГ и белков в срезах аденогипофизов быка // Докл. АН УССР.
Сер. Б. 1969. № 6. С. 546-548.
Тодоров И.Н., Митрохин Ю.И., Галкин А.П. Цитоплазматические факторы
контроля биосинтеза РНК у высших животных // Молекулярная биология.
Киев: Наук, думка, 1976а. Вып. 16. С. 64-73.
Тодоров И.И., Паскевич И.Ф., Лившиц А.А. Влияние кортизона на биосинтез
РНК в срезах аденогипофизов быка // Докл. АН УССР. Сер. Б. 1968а. № 8.
С. 747-749.
Тодоров И.Н., Смалъко П.Я., Галкин А.П. Состояние полирибосом как
отражение функционального взаимодействия систем трансляции и транскрипции в
процессе восстановления биосинтеза белка, ингибированного
циклогексимидом // Биохимия. 1977. Т. 42, вып. 12. С. 2149-2159.
Тодоров И.Н., Фельдман Е.Е., Чаговец ЕМ. Влияние кортизона на биосинтез
адренокортикотропного гормона и других белков в срезах аденогипофизар-
ной ткани // Докл. АН УССР. Сер. Б. 1968. № 10. С. 952-955.
Годоров И.Н., Фонарев А.Б., Шевченко Н.А. и др. Структурные изменения
хроматина в условиях активации транскрипции, сопряженной с ингибированием
трансляции циклогексимидом // Биохимия. 1984. Т. 49, вып. 5. С. 827-835.
Годоров И.Н., Шен Р.А., Желябовская СМ. и др. Исследование контроля
биосинтеза РНК в печени крыс Особенности биосинтеза РНК в условиях длительно
го подавления биосинтеза белка //Там же. 1976. Т. 41, вып. 10. С. 1859-1870.
Фонарев А.Б., Крашакова И.Б., Фонарев М.А., Тодоров И.Н. Модель
позитивной регуляции экспрессии гена рРНК на основе комплиментарного
взаимодействия с его промоторами транскриптов интронов гена преальбумина //
Молекуляр. биология. 1993. Т. 27, вып. 6. С. 1261-1275
Христианович Д.С., Митрохин Ю.И., Юшманов В Е. и др. Энергетика печени
крыс по данным 31Р-ЯМР на высших стадиях процесса биосинтеза белков и
нуклеиновых кислот//Биохимия. 1988. Т. 53, № 11. С. 1900-1905.
Чирков A.M., Чиркова С.К., Старцев ВТ. Эмоциональный стресс у обезьян. Л.:
Наука, 1987. 164 с.
Шевченко Н.А., Бойков П.Я., Тодоров И.Н. Последовательные изменения мак-
роструктурного состояния и РНК-синтезирующей активности ядрышкового
и экстраядрышкового хроматина клеток печени крыс в процессе индукции
синтеза ДНК // Биохимия. 1985. Т. 50, вып. 10. С. 1591-1598.
Эверли, Дж.С, Розенфелъд, Р. Стресс: Природа и лечение. М.: Медицина, 1985.
200 с.
Эйдус Л.Х. Не специфическая реакция клеток и радиочувствительность. М.:
Атомиздат, 1977. 151 с.
Эмануэль НМ. Лауреат Нобелевской премии академик Н.Н. Семенов // Вестн.
МГУ. Сер. математики, механики, астрономии, физики, химии. 1957. № 1.
С. 182-187.
Эмануэль НМ., Кнорре Д.Г., Лясковская Ю.Н. Кинетика окисления жиров //
Изв. АН СССР. 1957. № 6. С. 678-683.
333
Элшрбеков Э.З., Львова СП., Гасангаджиева А.Г. Влияние многократного хо-
лодового стресса на интенсивность перекисного окисления липидов и анти-
оксидантную систему тканей // Бюл. эксперим. биологии и медицины. 1998.
Т. 125, J& 4. С. 385-387.
Юдаев НА., Утешева З.Ф. Гормоны гипоталамуса // Биохимия гормонов и
гормональной регуляции. М: Наука, 1976. С. 10-44.
Юдаев НА. Афиногенова С Α., Крехова Μ А. Кортикостероиды //Там же. 1976.
С. 171-228.
Adiga P.R., Hussa R.O., Robektson М.С. et al. Biosynthesis of adrenocorticoiropin and
protein in slices of bovine anterior pituitary tissue // Biochemistry. 1965. Vol. 4, N 2.
P. 246-253
Adiga P.R., Rao P.M., Hussa R.O. et al. Biosynthesis of adrenocorticoiropin and protein
in a cell-free system from bovine anterior pituitary tissue // Ibid. 1966. Vol. 5, N 12.
P. 3850-3856.
Akil Η et al. The endorphins // Adv. Biochem. Psychopharmacol. 1978. Vol. 18.
P. 131-147.
Attardi В., Ohno S. Physical properties of androgen receptors in brain cytosol from
normal and testicular feminized (Tfm/y hermaphrodite) mice // Endocrinology. 1978.
Vol. 103, N 3. P. 760-770.
Austad S.N. An experimental paradigm for the study of slow aging organisms // Exp.
Gerontol. 2001. Vol. 36, N 4/6. P. 599-605.
Bachner K.L. et al. // Free radicals in biology / Ed. W.A. Pryor. N.Y.: Acad, press, 1982.
Baltrusch, Η J., Stangel W., Titze /. Stress, cancer and immunity. New developments in
biopsy chosocial and psychoneuroimmunologic research // Acta Neurol. 1991.
ι Vol 13, N4. P. 315-327.
Barja G. Endogeneous oxidative stress: relationship ю aging, longevity and caloric
restriction // Ageing Res. Rev. 2002. Vol. 1, N 3. P. 397-411.
Baxter J.D., Forsham P.H. Tissue effects of glucocorticoids // Amer. J. Med. 1972.
Vol. 53, N5. P. 573-589.
Betz E.H. Contribution a Tetude du syndrome endocrinien provoque par rirradition total
de Porganisme. P.: Masson: Ed. Libr. de Tacad. de mod., 1956, 311 p.
Brindley D.N., Rolland Y. Possible connections between stress, diabetes, obesity,
hypertension and altered lipoprotein metabolism that may result in atherosclerosis // Clin.
Sci. 1989. Vol. 77, N 5. P. 453-461.
Bryla CM. The relationships between stress and the development of breast cancer: a
literature review //Oncol. Nurs. Forum. 1996. Vol. 23, N 3. P. 441-448.
Buttke Ύ.Μ., Sandstrom PA. Oxidative stress as a mediator of apoptosis // Immunol.
Today. 1994. Vol. 15. P. 7-10.
Cadenas E., Davies KJ. Mitochondrial free radical generation, oxidative stress, and
aging // Free Radicals Biol. Med. 2000. Vol. 29, N 3/4. P. 222-230.
Cannon W.B. The emergency function of the adrenal medula in pain and in the major
emotions //Amer. J. Physiol. 1914. Vol. 33. P. 356-373.
Cannon W£. The Wisdom of the body. N.Y. 1932
Cannon W.B. Bodily changes in pain, hunger, fear and rage. Boston: Branford, 1953.
Carroll В J. Ageing, stress and the brain // Novartis Found. Symp. 2002. Vol. 242.
P. 26-36.
Cheitlin R„ Buckley D.I., Ramachandran K. The role of extracellular calcium in corti-
cotropin-simulated steroidogenesis // J. Biol. Chem. 1985. Vol. 260. P. 5323.
Chen J., Brodsky S.V., Goligorsky DM. et al. Glycated collagen I induces premature
senescence-like phenotypic changes in endothelial cells // Circulation Res. 2002.
Vol. 90, N12. P. 1290-1298.
334
Ch'ihJJ., DuhlDM., Faulkner L.S., Devlin T.M. Regulation of mammalian.protein
synthesis in vivo. Simulated transport of nuclear ribonucleoprotein complexes to the
cytoplasm after cycloheximide treatment // Biochem J. 1979. Vol. 178, N 3.
P. 643-649.
Chirico WJ., Waters M.G., Blobel G. 70K heat shock related proteins stimulate protein
translocation into microsomes // Nature. 1988. Vol. 332, N 6167. P. 805-810.
Chrousos G., Gold Ph.W. The concepts of stress and stress system disorders. Overview
of physical and behavioral homeostasis // JAMA. 1992. Vol. 267. Ρ 1244-
1252.
Cohen J J. Glucocorticoid-induced apoptosis in the thymus // Semin. Immunol. 1992
Vol. 4, N 6. P. 363-369.
Cohen J J., Duke R.C. Glucocorticoid activation of a calcium-dependent endonuclease in
thymocyte nuclei leads to cell death // J. Immunol. 1984. Vol. 132. N 1. P. 38-
42.
Craig Ε A. The heat-shock response // CRC Crit. Rev. Biochem. 1985. Vol. 18.
P. 239-280.
Csiszar Α., Ungvari Z., Edwards J.G. et al. Aging-induced phenotypic changes and
oxidative stress impair coronary arteriolar function // Circulation Res. 2002. Vol. 90,
N11. P. 1159-1166.
Czech M.P. Molecular basis of insulin action // Annu. Rev. Biochem. 1977. Vol. 46.
P. 359-384.
Del Maestro R.F. Free radical injury during inflammation // Free radicals in molecular
biology, aging, and disease / Ed. D. Armstrong et al. N.Y.: Raven press, 1984.
P. 87-102.
Deshaies RJ., Koch B.D., Werner-Washburne M. et al. A subfamily of stress proteins
facilitates translocation of secretory and mitochondrial precursor polypeptides //
Nature. 1988. Vol. 332, N 6167. P. 800-805.
Dinan T.G. Noradrenergic and serotonergic abnormalities in depression: stress-induced
dysfunction? //J. Clin. Psychiatry. 1996. Vol. 57, suppl. 4. P. 14-18.
Dypuckt J.M., Ancarcrona M.f Burkitt M. et al. Different peroxidation level stimulated
growth, trigger apoptosis, or produce necrosis of insulin-secreting RIN5F cells // J.
Biol. Chem. 1994. Vol. 269. P. 30553-30560.
Ebadi M. Introduction. Oxidative stress in mitochondria disorders of aging // Bio.
Signals Recept. 2001. Vol. 10, N 1/2. P. 5-13.
Eilers M., Schatz G. Binding of a specific ligand inhibits import of a purified precursor
protein into mitochondria //Nature. 1986. Vol. 322, N 6076. P. 228-232.
Evans R.M. The steroid and thyroid hormone receptor superfamily // Science. 1988.
Vol. 240. P. 889-895.
Eysenck И J. Personality, stress and cancer: prediction and prophylaxis // Brit. J. Med.
Psychol. 1988. Vol. 61, pt. 1. P. 57-75.
Fawcett T.W., Sylvester S.L., Sarge K.D. et al. Effects of neurohormonal stress and aging
on the activation of mammalian heat shock factor I // J. Biol. Chem. 1994. Vol. 269,
N51. P. 32272-32278
Flack P. Chemilumineszenzmessung der sequentullen Aktivierung mononuclearer
Blutzellen // Stud, biophys. 1984. Vol. 100, N 1. P. 83-89
Fontenot M.B., Kaplan J.R., Manuck S.B. et al. Long-term effects of chronic social stress
on serotonoergic indices in the prefrontal cortex of adult male cynomolgus macaques
//Brain Res. 1995. Vol. 705, N 1/2. P. 105-108.
Frolkis V.V. Syndromes of aging: (Review) // Gerontology. 1992. Vol. 38, N 1/2. P. 80-
86.
Frolkis V.V. Stress-age syndrome // Mech. Ageing and Develop. 1993. VoK 69, N 1/2.
P. 93-107.
335
Garren L.D.t Gree G.N., Mashi J. et al. On the mechanism ot action of ACIH // Recent
Progr. Hormone. Res. 1971. Vol. 27. P. 433^t78.
Gehring U. Steroid hormone receptors: biochemistry, genetics, and molecular biology //
Trends Biochem Sci. 1987. Vol. 12. P. 399^02.
Gilad G.M., Gilad V.H. Strain, stress, neurodegeneration and longevity // Mech. Ageing
and Develop. 1995. Vol. 78, N 2. P. 75-83.
Golden T.R., Melov S. Mitochondrial DNA mutations, oxidative stress, and aging // Ibid.
2001. Vol. 122, N 14. P. 1577-1589.
GraeffF.G., Guimares F.S., De Andrade T.G. et al. Role of 5-HT in stress, anxiety, and
depression // Pharmacol. Biochem. Behav. 1996. Vol. 54, N 1. P. 129-141.
Grant N. II Adrenal cortex / Ed. A.B. Eisenstein. Boston, 1967. P. 269-275.
Guillemin R. Hypothalamic control of the secretion of adenohypophysial hormones //
Adv. Metabol. Disord. 1971. Vol. 5. P. 1-51.
Guillemin R. Endorphines: brain peptides that act like opiates // N. Engl. J. Med. 1977.
Vol. 296. P. 226-228.
Guillemin R. Peptides in the brain: The new endocrinology of the neuron // Science.
1978. Vol. 202. P. 390-402.
Guyton A.C. Textbook of medical physiology. Philadelphia: Saunders, 1976.
Hanson K.P., Rosenberg O.A., Posokhov V~S. Induction of apoptosis in rat thymocytes
with different content of cholesterol in their plasma membranes. AIDS // Res. and
Human Retroviruses. 1995, Vol. 11, suppl. 1. P. 112.
Hassig Α., Wen-Xi L·, Stampfli K. Stress-induced suppression of the cellular immune
reactions: on the neuroendocrine control of the immune system // Med Hypothesis.
1996. Vol. 46, N 6. P. 551-555.
Haynes R.C., Jr. Theories on the mode of action of ACTH in stimulating secretory
activity of the adrenal cortex // Handbook of physiology, endocrinology. Wash. (D.C.):
Amer. Physiol. Soc, 1975. Vol. 6.
Herman J.P., Prewitt СМ., Collinan W.E. Neuronal circuit regulation of hypothalamo-
pituitary-adrenocortical stress axis // Crit. Rev. Neurobiol. 1996. Vol. 10, N 3/4.
P. 371-394.
Kaujfinan S.H. Heat stock proteine in health and disease (Review) // Intern. J. Clin.
Labor. Res. 1992. Vol. 21. P. 221-226
loannidou-Mouzaka L·, Mantonakis J., Toufexi H. et al. Is prolongated psychological
stress and ethiological factor in breast cancer? // J. Gynecol. Obstet. Biol.
Reprod.1986. Vol. 15, N 8. P. 1049-1053.
Jenkins R.R. Exercise, oxidative stress, and antioxidants: A review // Intern. J. Sport
Nutrition. 1993. Vol. 3, N 4. P. 356-375.
Ji L.L., Leeuwenburgh C, Leichtweis S. et al. Oxidative stress and aging. Role of
exercise and its influences on antioxidant systems // Ann. N.Y. Acad. Sci. 1998. Vol. 854.
P. 102-117.
Kapahi P., Boulton M.E., Kirkwood T.B. Positive correlation between mammalian life
span and cellular resistance to stress // Free Radicals Biol. Med. 1999. Vol. 26, N 5/6.
P. 495-500.
Heydari A.R., Takahashi R., Gusimann A. et al. Hsp 70 and aging // Expenenta. 1994.
Vol. 54. P. 1092-1098.
Khan R., Daya S., Potgieter β. Evidence for a modulation of the stress response by the
pineal gland // Experientia. 1990. Vol. 46, N 8. P. 860-862.
Lanks K.W. Modilators of the eukaryotic heat shock response // Exp. Cell. Res. 1986.
Vol. 165, N1. P. 1-10.
Laybutt D.R., Kaneto H., Hasenkamp W. et al. Increased expression of antioxidant and
antiapoptotic genes in islets that may contribute to beta-cell survival during chronic
hyperglycemia // Diabetes. 2002. Vol. 51, N 2. P. 413-423.
336
Le Blanc J. The role of catecholamines in adaptation to chronic and acute stress // Proc.
Intern, symp. on catecholamines and stress. Bratislava, 1976.
Lechin F., Van der Dijs В., Benaim M. Stress versus depression // Progr.
Neuropsychopharmacol. Biol. Psychiatry. 1996. Vol. 20, N 6. P. 899-950.
Lee J.K., Manalo D., Liu A.Y. Heat stock response, beat shock transcription factor and
cell aging//Biol. Signals. 1996. Vol. 5. P. 180-191.
Lee H.C., Wei Y.H. Mitochondrial alterations, cellular response to oxidative stress and
defective degeneration of proteins in aging // Biogerontology. 2001. Vol. 2, N 4.
P. 231-244.
Lehninger A.L. Principles of biochemistry. Chap. 25. Hormones. Worth Publ.,
1982.
Lenaz G., Bovina C, D'Aurelio M. et al. Role of mitochondria in oxidative stress and
aging // Ann. N.Y. Acad. Sci. 2002. Vol. 959. P. 199-213.
Lenaz G., Cavazzoni M., Genova M.L. et al. Oxidative stress, antioxidant defences and
aging // Biofactors. 1998. Vol. 8, N 3/4. P. 195-204.
Leonard B.E., Song С Stress and immune system in the ethiology of anxiety and
depression // Pharmacol. Biochem. Behav. 1996. Vol. 54, N 1. P. 299-303.
Levi L. Psychosocial stimuli, psychophysiological reactions and disease // Acta med.
scand. 1972. Entire Suppl. 528
Li CM. (Ed.) Hormonal proteins and peptides. Vol. 1-4. N.Y.: Acad, press, 1970-1977.
Lindquist S. The heat-shock response // Annu. Rev. Biochem. 1986. Vol. 55.
P. 1151-1191.
Liu J., Mori A. Stress, aging, and brain oxidative damage // Neurochem. Res. 1999.
Vol. 24, N 11. P. 1479-1497.
Lundberg W.O. Lipids of biological importance: peroxidation products and inclusions in
compounds of lipids // Amer. J. Clin. Nutrition. 1958. Vol. 6, N 6. P. 601-603.
MacLean P.D. On the evolution of three mentalities // Man-Environment System. 1975.
Vol. 5. P. 213-224.
Mason J.В. Organization of psychoendocrine mechanisms: A review and reconsideration
of research // Handbook of psychophysiology. N.Y.: Holt, Rinehart and Winston,
1972.
McArdle Α., Jackson Μ J. Exercise, oxidative stress and ageing // J. Anat. 2000
Vol. 197, pt. 4. P. 539-541.
McEwen B.S. Stress and aging hippocampus // Front. Neuroendocrinol. 1999. Vol. 20.
N 1. P. 49-70.
Melov S. Mitochondrial oxidative stress. Physiologic consequences and potential for a
role in aging // Ann. N.Y. Acad. Sci. 2000. Vol. 908. P. 219-225.
Minois N. Longevity and aging: beneficial effects of exposure to mild stress //
Biogerontology. 2000. Vol. 1, N 1. P. 15-29.
MiquelJ. An update on the oxygen stress-mitochondrial mutation theory of aging:
genetic and evolutionary implications // Exp Gerontol. 1998. Vol. 33, N 1/2. P. 113—
126.
Mizock B.A. Alteration in carbohydrate metabolism during stress: a review of the
literature // Amer. J. Med. 1995. Vol. 98, N 1. P. 75-84.
MolinoffP.B., AxelrodJ. Biochemistry of catecholamines // Annu. Rev Biochem. 1971
Vol. 40. P. 465-500.
Morgenthaler J., Joy D. Better sex through chemistry: A guide to new prosexual drugs.
Petaluma (CA): Smart Publ., 1995.
Nakanishi S. et al. Nucleotide sequence of cloned cDNA tor bovine corticotropin-β-
lipotropin precursor//Nature. 1979. Vol. 278. P. 423-427.
Nicotera P., Anarcrona M„ Bonfoco E. et al. Neuronal apoptosis versus necrosis induced
by glutamate of free radicals // Apoptosis. 1996. Vol. 1, N 1. P. 5-10.
337
Norman A.W., Litwack G. Hormones. San Diego (CA): Acad, press, 1987.
Parsons PA. The limit to human longevity: an approach through a stress theory of
ageing // Mech. Ageing and Develop. 1996. Vol. 87, N 3. P. 211-218.
Pedersen W.A., Wan R., Mattson M.P. Impact of aging on stress-responsive
neuroendocrine systems // Ibid 2001. Vol. 122, N 9. P. 963-983.
Pelham H.R. Speculations on the functions of the major heat shock and
glucose-regulated proteins // Cell. 1986. Vol. 46, N 7. P. 959-961.
Pelletick G. et al. Цит. по: The endorphins // Adv. biochem. psychopharmacol. 1978,
Vol. 18. P. 137.
PfannerN., Tropschug M., Neupert W. Mitochondrial protein import: nucleoside
triphosphates are involved in conferring import-competence to precursors // Cell. 1987.
Vol. 49, N6. P. 815-823.
Polidori M.C., Mecocci P., Cherubini Α., Senin U. Physical activity and oxidative stress
during aging // Intern. J. Sports Med. 2000. Vol. 21, N 3. P. 154-157.
Pulatova M.K., Sharygin V.L., Todorov I.N. Radical enzyme ribon-reduce
(ribonucleotide reductase) in animal tissues during normal and pathological processes // Xll
Intern, conf. on magnetic resonance in biological systems, Todtmoos. Germany, 8-12
Sept., 1986: Abstracts. Todtmoos, 1986. P. 229.
Pulatova M.K., Sharygin V.L., Todorov I.N. Radical enzyme ribon-reduce
(ribonucleotide reductase) in animal tissues during normal and pathological processes //
Highlights of modern biochemistry / Ed. by A. Kotyk, J. Skoda et al. Zeist: VSP
Intern. Sci. Publ., 1989. P. 215-229.
Pulatova Μ Κ., Sharygin V.L., Todorov I.N. The activation of ribonucleotide reductase
in animal organs as the cellular response against the treatment with DNA-damaging
factors and the influence of radioprotectors on this effect // Biochim. et biophys. acta
1999. Vol. 1453. P. 321-329.
Roldan E., Alvarez-Pelaez P., De Molina F. Electrographic study of the amigdaloid
defense response //Physiol, and Behav. 1974. Vol. 13. P. 779-787.
Rosenfeldt F.L., Pepe S., Linnane A. et al. Coenzyme Q10 protects the aging heart
against stress: Studies in rats, human tissues, and patients // Ann. N.Y. Acad. Sci.
2002. Vol. 959. P. 355-359. Discuss.: P. 463-^65.
RothuizenJ., ReulJ.M., RijnberkA. et al. Aging and the hypothalamus-pituitary-adreno-
cortical axis, with special reference to the dog // Acta endocrinol. 1991. Vol. 125,
suppl. LP. 73-76.
Rudolph K.L., Chang S., Lee H.W. et al. Longevity, stress response, and cancer
in aging telomerase-deficient mice // Cell. 1999. Vol. 96, N 5. P. 701-
712.
Saretzki G., Von Zglinicki T. Replicative senescence as a model of aging: The role of
oxidative stress and telomere shortening - an overview // Ztschr. Gerontol. Geriatr.
1999. Bd. 32, N 2. S. 69-75. In German.
Saretzki G., Von Zglinicki T. Replicative aging, telomeres, and oxidative stress // Ann.
N.Y. Acad. Sci. 2002. Vol. 959. P. 24-29.
Sastre J., Pallardo F.V., Garcia de la Asuncion J., Vina /. Mitochondria, oxidative stress
and aging // Free Radicals Biol. Med. 2000. Vol. 32, N 3. P. 189-198.
Sastre J., Pallardo F.V., Vina J. Mitochondrial oxidative stress plays a key role in aging
and apoptosis // IUBMB Life. 2000. Vol. 49, N 5. P. 427-435.
Schally A.V., Akimuka Α., Kastin AJ. Hypothalamic regulatory hormones // Science.
1973. Vol. 179. P. 341-350.
Schlesinger Μ J. How cell copes with stress and the function of heat shock proteins //
Pediatric Res. 1994. Vol. 36, N 1, pt. 1. P. 1-6.
Selye H. A syndrome produced by diverse nocuous agents // Nature. 1936. Vol. 138.
P. 32-40.
338
Selye H. Stress. Montreal, 1948.
Setye H. The physiology and pathology of exposure to stress. Montreal, 1950. 822 p.
Selye H. The stress of life. N.Y.: McGraw-Hill, 1956.
Selye H. Stress without distress. N.Y., 1974.
Selye H. Stress in health and disease. Reading (Mass.): Butterworth's, 1976.
Skulachev V.P. Uncoupling: new approaches to an old problem of bioenergetics //
Biochim. et biophys. acta. 1998. Vol. 1363, N 2. P. 100-124.
Smith E.L., Hill R.L., Lehman I.R. et al. Principles of Biochemistry: Mammalian
biochemistry. 7th ed. N.Y.: McGraw-Hill, 1983. P. 355-619.
Stokes P.E. The potential role of excessive Cortisol induced by ΗΡΑ hyperfunction in the
pathogenesis of depression // Europ. Neuropsychopharmacol. 1995. Vol. 5, suppl.
P. 77-82.
Stress and heart disease / Ed. by E. Beamish et al. Boston: Nijhoff, 1984.
Suh J.H., Shigeno E.T., Morrow J.D. et aK Oxidative stress in the aging rat heart is
reversed by dietary supplementation with (RHalpha)-lipoic acid // FASEB J. 2001.
Vol. 15, N 3. P. 700-706.
Sutherland E.W. Studies on the mechanism of hormone action // Science. 1972. Vol. 177,
N47.P.401--408.
TappelA.L· Biological antioxidant protection against lipid peroxidation damage //Amer.
J. Clin. Nutrition. 1970. Vol. 23, N 8. P. 1137-1139.
Thelander L.t Reichard P. Reduction of ribonucleotides // Annu. Rev. Biochem. 1979.
Vol. 48. P. 133-158.
Todorov I.N. Molecular mechanisms of the hypophyseal adrenocorticotrope hormone
synthesis // Federation of European Biochemical Societies. Abstracts of papers
presented of the Federation. Praha, 1968. P. 274.
Todorov /JV. How cells maintain stability // Sci. Amer. 1990. Vol. 262, N 12. P. 66-75.
Todorov /JV. Mechanisms of cell stability: Subcellular and molecular aspects. N.Y.:
Nova Sci. Publ., 1995. 255 p.
Todorov H., Nadler R., Todorov I.N. Public enemy number 1 - stress: (A practical guide
to the effects of stress and nutrition on the aging process and life extension). N.Y.:
Nova Sci. Publ., 2001. 247 p.
Todorov I.N., Pulatova L.K., Emanuel N.M. Universal pattern of systemic response of
higher eucaryot cells to the effects of physical and chemical factors // Proc. 8th Intern.
biophys. congr. Bristol, 1984. P. 151.
Toussaint O., Remade J., Dierick J. et al. From the Hayflick mosaic to the mosaics ot
ageing: Role of stress-induced premature senescence in human ageing // Intern. J.
Biochem. Cell. Biol. 2002. Vol. 34, N 11. P. 1415.
Troy СМ., Shelanski ML. Down-regulation of CuTZn SOD causes apoptotic death of
PC12 neuronal cells // Proc. Nat. Acad. Sci. USA. 1994. Vol. 91. P. 6384-
6387.
Ursin H. Stress, distress, and immunity // Ann. N.Y. Acad. Sci. 1994. Vol. 741.
P. 204-211.
Van Eekelen J.A., Rots N.Y., Sutanto W. et al. The effect of aging on stress
responsiveness and central corticosteroid receptors in the brown Norway rat // Neurobiol
Aging. 1992. Vol. 13, N 1. P. 159-170.
Von Zglinicki T, Burkle Α., Kirkwood TJB. Stress, DNA damage and ageing - an
integrative approach // Exp. Gerontol. 2001. Vol. 36, N 7. P. 1049-1062.
Walker G.C. Inducible DNA repair systems // Annu. Rev. Biochem. 1985. Vol. 54.
P. 425^57.
Wei Y.H. Oxidative stress and mitochondrial DNA mutations in human aging // Piuc.
Soc. Exp. Biol, and Med. 1998. Vol. 217, N 1. P. 53-63.
339
Wei Y.H., Ma Y.S., Lee И.С. et aL Mitochondrial theory of aging matures - roles of
mtDNA mutation and oxidative stress in human aging // Zhonghua Yi Xue Za Zhi.
Taipei, 2001. Vol 64, N 5. P. 259-270.
White Α., Handlek P., Smith E.L· et al. Principles of biochemistry. N.Y.: McGraw-Hill,
1978. Chap. 44.1.2; 45.2.1; 45.2.2; 45.2.4.
Wilson J.D., Foster D.W. Textbook of endocrinology. 7th ed. Saunders, 1985.
Yamamoto KM. Steroid receptor regulated transcription of specific genes and gene
networks // Annu. Rev. Genet. 1985. Vol. 19. P. 209-252.
Yu B.P. Aging and oxidative stress: Modulation by dietary restriction // Free Radicals
Biol. Med. 1996. Vol. 21, N 5. P. 651-668.
YuwilerA. Stress, anxiety and endocrine function // Biological foundations of
psychiatry / Ed. R. Grenell, S. Galay. N.Y.: Raven press, 1976.
Часть III
БИОХИМИЧЕСКАЯ
КОРРЕКЦИЯ СТРЕССА
И СТАРЕНИЯ
Нужно потратить очень много времени,
чтобы стать, наконец, молодым...
Пабло Пикассо
Глава 22
ДИЕТА
145 У римлянок напитка не бывало
Иного, чем вода; и Даниил
Презрел еду, и мудрость в нем мужала.
148 Начальный век, как золото, светил,
И голод желудями услаждался,
И нектар жажде каждый ключ струил.
151 Акридами и медом насыщался
Среди пустынь креститель Иоанн;
А как велик и славен он остался,
154 Тому залог в Евангелии дан.
Данте Алигьери "Божественная комедия",
"Чистилище", песнь Двадцать вторая.
Как уже отмечалось, единственным и наиболее перспективным
методом для продления жизни и предупреждения болезни у
грызунов является серьезное ограничение калорийности пищи. Например,
крысы, потребляющие калорий на 30-40% меньше, чем их
сородичи, потреблявшие пищу ad libitum, живут в 1,5-2 раза дольше. Более
того, они длительное время остаются проворными и здоровыми
после того, как большинство крыс, которые питались ad libitum, уже
умерли. Очевидно, ограничение калорийности пищи - мощный
ингибитор механизмов старения и болезни. Наилучшие результаты
достигались тогда, когда ограничения вводилось на раннем этапе
жизни животных, перед их половым созреванием. Эти результаты
вполне соответствуют нейроэндокринной теории старения (теория
центральный часов старения (ЦЧС) см. гл. 2), которая утверждает, что
скорость старения пропорциональна скорости развития. У
грызунов, ограничение калорийности, начатое в раннем возрасте,
задерживает развитие организма и таким образом замедляет ход ЦЧС,
приводя в результате к значительному продлению жизни. При
использовании ограничения после периода созревания, метод менее
эффективен, но все же приводит к некоторому продлению жизни и
улучшению состояния здоровья.
К возможным биохимическим механизмам биологического
эффекта ограничения калорийности питания исследователи относят
прежде всего ослабление окислительных повреждений, модуляцию
гликемии и инсулинэмии, а также уровня гормонов, измененного
старением (Masoro, 2000).
Почему же тогда все мы не придерживаемся низкокалорийной
диеты, убивая тем самым сразу двух зайцев? То есть не только жить
дольше, но также экономить на продовольствии и различных мерах
антистарения, таких, например, как потребление антиоксидантных
343
Таблица 16-А
Смертность и вес тела
Относительный вес, %
на 20 ниже нормы
10 ниже нормы
10 выше нормы
20 выше нормы
30 выше нормы
40 выше нормы
50 выше нормы
60 выше нормы
Мужчины
105
94
111
120
135
153
177
210
Женщины
ПО
97
107
ПО
125
136
149
167
Примечание. Соотношение показателей смертности и вариаций
веса. См.: Build study 1979 // Society of Actuaries and
Association of Life Insurance Medical Directors, Chicago, 1979
В табл. 16-А представлены отклонения (%) от усредненного
значения нормального (оптимального) веса, которому
соответствует уровень смертности, принятый за 100%.
пищевых добавок и физических упражнениях. К сожалению,
ситуация с ограничением калорийности у приматов не столь однозначна.
Ограничения в раннем возрасте приводят к отклонениям в
центральной нервной системе и не увеличивают продолжительности их
жизни. Имеются свидетельства, что умеренное ограничение
калорийности после достижения половой зрелости может быть,
по-видимому, полезным, но этот вопрос, однако, требует еще
дополнительных исследований.
Для людей, рассматриваемый подход состоит в том, чтобы
определить желательный или идеальный вес тела и поддерживать его,
потребляя соответствующее количество калорий. Большинство
данных, связывающих вес тела и продолжительность жизни,
поступает от больших страховых компаний, которые взвешивают своих
клиентов при покупке ими страховых полисов. Самая высокая
продолжительность жизни, как показывают эти данные, связывается с
весом тела, который несколько ниже среднего (попеременно
называемого желательным, идеальным или оптимальным весом) (см.
табл. 16-А, 16-В).
Со всеми указанными выше ограничениями, оптимальный вес
тела - разумная цель для человека, который стремится к
продлению жизни. Фактически состояние человека, обладающего
оптимальным весом и строением тела, особенно в среднем или
пожилом возрасте, может быть эквивалентно в известной мере
ограничению калорийности питания у грызунов. В подобных
экспериментах контрольная группа грызунов питалась без ограничения,
т.е. имела неограниченный доступ ко вкусной еде. Не
удивительно, что в такой ситуации животные объедаются и становятся туч-
344
Таблица 16-В
Оптимальный вес
(коррелирует с максимальной продолжительностью жизни)
Рост,
см
Мужчины, вес, кг
Оптимальный вес,
средние
величины
Предельные
колебания
оптимального
веса
Избыточный вес
Женщины, вес, кг
Оптимальный вес,
средние
величины
Предельные
колебания
оптимального
веса
Избыточный вес
147
150
152
155
157
160
162
165
167
170
172
175
177
180
183
185
188
190
193
55
57
58
60
61
63
65
67
69
71
73
75
77
79
81
50-63
52-65
53-67
54-68
56-70
58-72
59-75
61-76
63-78
65-80
67-83
68-85
70-S7
72-89
74-92
76
78
80
82
84
87
89
92
94
97
99
102
105
107
ПО
46
47
48
49
51
52
54
55
58
59
61
63
65
67
68
41-53
42-55
43-56
44-58
46-59
47-60
49-62
49-64
51-66
53-67
55-69
57-71
58-73
60-76
62-78
64
66
67
69
71
72
74
76
79
81
83
85
88
91
94
Примечание. Нужно иметь в вицу несколько ограничений для данных по
оптимальному весу:
• Данные по границам веса основаны иа самой низкой смертности среди
застрахованных людей, которые ие могут представлять популяцию в целом.
• Курение может вести как к увеличению смертности, так и к снижению веса
тела. Следовательно, для здоровых некурящих людей данные, связанные с самой
низкой смертностью, могут быть завышенными.
• В большинстве случаев пребывание в диапазоне оптимального веса указывает на
оптимальное строение тела. Но существуют исключения. С одной стороны,
человек с меньшим, чем оптимальное, развитием мускулов и избыточным жиром
может все же оказаться в пределах оптимального веса. С другой стороны,
человек с хорошо развитой мускулатурой, например занимающийся подъемом тя -
жестей, может иметь оптимальное строение тела, но превышать
желательный вес. Первый индивидуум, вероятно, получит некоторые выгоды для
здоровья от потери веса, а последний нет, хотя таблицы оптимального веса
указали бы на обратное.
345
ными. Избыточное потребление пищи - естественный механизм,
который развился в условиях длительной нехватки еды. Диким
животным часто приходилось оставаться без пищи. Однако с
жировыми отложениями они имели лучший шанс выжить при
голодании. В дикой природе выгодно потреблять как можно больше
пищи в тех редких случаях, когда она доступна в избытке.
Поскольку животные склонны к неограниченному потреблению
пищи, сравнение продолжительности жизни в экспериментах по
ограничению калорийности проводится не между "очень" и "не
очень" голодными, а скорее между тучными и худыми
(оптимальный вес) грызунами. В этом контексте люди, которые сохраняют
оптимальный вес тела, подобны грызунам, которые ограниченны
в калориях. И, конечно, ни для кого не секрет, что поддержание
желательного веса обычно требует некоторого самоограничения
в потреблении пищи. Таким образом, поддерживая оптимальный
вес тела в течение всей жизни, можно частично моделировать
выгодные эффекты ограничения калорийности, наблюдаемые у
грызунов.
Желательный вес следует рассматривать в контексте
достижения оптимального строения тела. Рекомендуемый в настоящее
время диапазон содержания жира в теле составляет от 15 до 22%
для мужчин и от 18 до 32 % для женщин. Некоторое количество
жира всегда необходимо для защиты жизненно важных органов,
поддержки нормальной проводимости нервных импульсов и
обеспечения запаса энергоносителей. "Обязательного" жира у
женщин больше, очевидно, потому что система должна иметь
некоторый дополнительный ресурс энергоносителей на случай
беременности. Женщины, чьи жировые запасы ниже некоторого уровня,
часто имеют проблемы с менструальным циклом. Большинство
экспертов рекомендует абсолютный минимум от 5 до 10 % жира
для мужчин и от 12 до 16 % для женщин. Существует много
методов для определения состава тела, но большинство специалистов
по питанию измеряют толщину подкожного слоя различных
частей тела, и затем использует таблицы, чтобы оценить этот состав.
Когда все сделано должным образом, этот метод адекватен для
большинства людей. Некоторые клубы здоровья и
специализированные центры лечения тучности предлагают также и другие
методы, например подводное взвешивание или биоэлектрическое
сопротивление.
Если ваш вес соответствует оптимальному диапазону, вы
должны также определить и особенности строения вашего скелета,
соответствующего вашему телосложению и мышечной массе. Человек с
"легким" скелетом, чей вес находится в верхней части
оптимального диапазона, может иметь избыточный жир при относительно
"слабом" телосложении, и наоборот.
346
22.1. "ПОТЕРИ" БЕЗ ПОТЕРЬ
В то время как дискуссия об оптимальном энергетическом
потреблении у людей может продолжаться бесконечно, ясно, что,
значительный излишек веса увеличивает риск болезни и смерти.
Достижение оптимального веса - важная часть стратегии
продления жизни. Однако необходимо иметь в виду, что это только один
из многих факторов, и должен рассматриваться именно так. Даже
если вы будете не в состоянии достичь оптимального веса, не
игнорируйте другие возможности для продления жизни и
укрепления здоровья.
Печально, но для многих людей, проживающих в развитых
странах, общей "бедой" является излишек веса. Руководство,
выпущенное Национальным институтом здоровья США в 1998 г. и
позволяющее определить имеет ли человек излишек веса,
показывает, что 55% американцев относятся к этой категории.
Руководство использует показатель массы тела (BMI - body mass
index) как инструмент такой оценки. Вы можете определять ваш
BMI, поделив ваш вес в килограммах на квадрат вашего роста в
метрах. Результат BMI больше, чем 26 означает, что человек
имеет излишек веса (см. табл. 17).
Независимо от причины такого высокого процента людей
с избытком веса - будь это приверженность к
высококалорийной пище, сидячая работа, недостаток физической нагрузки
или долгие часы, проведенные у телевизора, - существует
огромный спрос на процедуры, пищевые добавки и продукты,
ведущие к снижению веса. К сожалению, многие люди стремятся
снизить свой вес скорее по социальным причинам, нежели в
интересах своего здоровья, и часто выбирают методы снижения
веса не только не улучшающие здоровье, но и прямо опасные для
него.
Возможно, наиболее драматичным оказался недавний пример с
применением phen-fen - комбинация выписываемых врачами лекарств phentermine и
fenfluramine, широко используемых для снижения веса. Phen-fen был уже разрешен
FDA для продажи на рынке, когда обнаружилось, что он может вызывать
потенциально опасные для жизни повреждения сердечных клапанов почти у
трети долговременных пользователей. Другие лекарства, если и менее опасны, то
все же могут вызывать разнообразные побочные эффекты и, кроме того, их
долговременная эффективность для снижения веса в большинстве случаев
сомнительна. Побочные эффекты amphetamines, которые являются наиболее
распространенным классом медикаментов для потери веса, включают тошноту,
рвоту, спазмы, сухость во рту и чувство тревоги. Amphetamines вызывают
краткосрочную потерю веса, подавляя аппетит. В большинстве случаев, однако,
аппетит возвращается к норме в течение недели или двух, несмотря на длительное
использование лекарств. Более того, после прекращения приема amphetamines,
аппетит быстро восстанавливается, рикошетом увеличивая вес быстрее, чем
его потерю.
347
vo oo <"i c^ ^1 °°. °} ^ ^ °°. Ό
:8
_ σ^ c4 »л οό *r* ■
oo oo су\ су\ σ\ θ <
\Dt^r^t^r^ooooooooo>^OsO
CS Wi Г^· t^
Г^ On ι-ί -ς»·
VD SO г^· Г^· i
oo oo oo oo o\
(S Vj r^ CA \П OO \П ОС \П ОС \П
«or^ON^Tj-vDooornv-iooOco
VDVDVDr^r^-r^-r^OOOOOOOOCy\ON
VDVDVOVDr^r^-r^-r^-OOOOOOOOON
τί Ν «Λ Π VDOO -1 -Ч- Γ* ON (N \h Γ^
T-« OO
oo —» rj- cs »n r>
»Л)1Л1Л\0\0\0\0\ОГ^Г*-Г*-Г^00
VD ^ (S
со wS i
OO Г"· «П CO VD Tj- VD
0ОС^«Л СОН OO VD τί VD <Л П »Л
OCSrivDOOC^fOinr^ONr-JcO
»Ti»n»n»n»nvDVDVOVOVDVDr^r^
iortcsvortcN oo ^ >n en «h
ooocscoior^cyNOcs^vDooo
"^•»n»nw>*nir>»nvDVD\OVOVDr-
СОт-««Л}СОГ-;»ЛсОГ-;<ПСО'--; I
VDWO\-!(4TivDVo\<HcOiO'
^-Tj-^Vl»n>nVi^Vl\DVDVD'
"* <N VO OO VO OO—ι OO VD
ir!hOOO»-t<iviVDOOOHfn
Tt^Tj-WWViViVlVlVDVDVC
S s χ
° к te
и ь α
Другой общий нездоровый подход к потере веса -
кратковременное пребывание на диете. Человек входит в очень
низкокалорийную диету, обычно предлагаемую для потери веса
коммерческими компаниями или друзьями. Такие диеты часто лишены
многих существенных питательных веществ и могут вызвать
многочисленные неблагоприятные побочные эффекты, включая
головные боли, тошноту, головокружение и в редких случаях даже
смерть. Кроме того, очень низкокалорийные диеты имеют
тенденцию впоследствии вызывать увеличение веса в результате
циклического возвратного процесса. Когда потребление энергии
опускается ниже ее расхода, организм замедляет метаболизм для
экономии энергии. Когда прежние пищевые привычки
возобновляются, скорость метаболизма, все еще остается немного ниже
той, которая была перед диетой. В результате, принимавший
диету не только возвращается к предыдущему весу, но часто и
превышает его. Каждый такой цикл может приводить к очередному
увеличению веса.
Не все типы избыточного жира равноценны в отношении
здоровья и долголетия. Существует два главных типа жировых
отложений в теле, центральный и периферийный. Центральный, или
брюшной, жир расположен главным образом вокруг талии. С ним
связывают множество болезней, таких как атеросклероз,
гипертония и диабет II типа. Метаболические изменения, связанные с
брюшной тучностью, включают пониженную толерантность к
глюкозе, избыток инсулина и повышенное содержание липидов в
крови. Все эти факторы вносят вклад в ускоренное старение и
развитие дегенеративных болезней. Периферийный жир, напротив,
несет, по-видимому, значительно меньше риска для здоровья.
Человек, ориентированный на долгую и здоровую жизнь, должен
быть заинтересован в избавлении от брюшной тучности. Простой
и полезный индикатор брюшной тучности - отношение размера
"талии к бедрам". Для того чтобы получить это соотношение,
нужно разделить длину окружности вашей талии на соответствующую
величину для бедер. Отношение "талии к бедрам" 0.8 или больше
для женщин и 0.95 или больше для мужчин связывается с
увеличением риска для состояния здоровья.
22.2. ПРИНЦИПЫ ЗДОРОВОЙ ПОТЕРИ ВЕСА
1. Медленно, но наделено. Потеря веса не должна быть
слишком быстрой. В противном случае, человек имеет все шансы
попасть в упомянутый выше циклический возврат веса и в
результате стать более тучным, чем раньше. Кроме того, безрассудная
атака на вес может закончиться долговременными опасностями
для здоровья. Фунт, т.е. 450 г, жировой ткани содержит приблизи-
349
тельно 3500 калорий. Энергетический дефицит в 500 калорий в
день закончится потерей приблизительно фунта в неделю.
Этот темп потери веса может быть поддержан без замедления
метаболизма и без того, чтобы получить неподдающийся контролю
аппетит. Отрицательный энергетический баланс может быть
создан любой комбинацией меньшего количества еды и
расходования большего количества энергии с помощью физических
нагрузок. Какой бы режим вы не выбрали, будьте последовательными
в его осуществлении.
2. Разумно и сдержано. Простые, разумные меры,
применяемые последовательно, наиболее эффективны для
долговременного снижения веса. Некоторые исследования продемонстрировали
связь между чрезмерным увлечением телевидением (ТВ) и
излишком веса. Когда вы смотрите ТВ, энергетический расход
низок, и запас калорий, как правило, возрастает. Сократите
общение с ТВ настолько, насколько вы можете. Еще лучше, если вы
замените часть времени у ТВ на физические упражнения или
прогулки.
Очень часто люди едят больше чем нужно для того, чтобы не
чувствовать себя голодными. Особенно часто это происходит,
когда пища поглощается жадно и в спешке. Два фактора подавляют
центр голода в мозгу: сытость желудка, т.е. его наполнение, и
уровень питательных веществ, глюкозы и жирных кислот в
потоке крови. При быстрой еде, вы не даете организму достаточного
времени, чтобы пища усвоилась, и мозг стал действовать на
подавление голода, поэтому вы потребляете пищи больше, чем это
необходимо для насыщения. Ешьте медленно, жуйте тщательно,
потратьте время на то, чтобы насладиться едой. Поступая так, вы
поглотите меньшее количество пищи, но получите большее
количество витаминов и полезных веществ. Если в процессе еды вы
обнаружите, что уже не голодны, хотя еда все еще остается на
тарелке, остановитесь. Не ешьте поздно вечером, особенно в
течение двух-трех часов до сна. Энергетический расход во время сна
низок, так что переработанная пища станет откладываться в виде
жира. Обед не должен быть обильным, поскольку вы нуждаетесь
в большем количестве пищи перед вашими главными
ежедневными действиями, а не после.
3. Изменения в диете. Делайте разумные замены в еде. Однако
вы не должны становиться вегетарианцем или переходить к
ограниченному набору определенных пищевых продуктов.
Во-первых, снизьте потребление жиров, особенно насыщенных
жиров. Жир дает 9 калорий на грамм, а белки и углеводы - только
4. Большинство жителей развитых стран получает приблизительно
40-50 % калорий из жира. Уменьшение количества жира в пище -
наиболее эффективный способ сократить потребление калорий.
Большинство экспертов полагает, что жир должен обеспечивать
350
меньше чем 30% от необходимых ежедневных калорий, а
насыщенные жиры - меньше чем 10%. Существуют доказательства, что
калории, получаемые из жира, вносят больший вклад в "полноту", чем
калории из протеинов и углеводов. Исследователи сравнили две
группы крыс с равным потреблением калорий. В одной группе
животные получали жирную еду, по составу похожую на типичную
американскую пищу (42% калорий за счет жира). При таком пита
нии крысы стали тучными, со структурой тела, содержащей более
50% жира. У крыс из контрольной группы, потреблявшие то же
самое количество калорий, но меньшее количество жира, структура
тела содержала только 30% жира. В другом исследовании
анализировалось энергетическое и жировое потребление 155 тучных людей.
Наблюдалась лишь небольшая взаимосвязь между общим
потреблением энергоносителей и размерами жировых отложений в теле.
Однако весьма существенная положительная корреляция была
обнаружена между потреблением жирной пищи и жировыми
отложениями.
Почему пища с высоким содержанием жира более способствует ожирению,
чем обезжиренная, даже если поглощение общего количества энергоносителей
в обоих случаях было одинаково? Одно объяснение связано со способами
хранения энергии в организме. Жировая ткань представляет собой
долговременный склад аккумулированной энергии. Некоторая часть энергии, поглощенная
из пищи, используется на текущие функции организма, но большая часть
оставшейся энергии, если таковая вообще имеется, преобразуется в жир для
долговременного хранения. Преобразование жира из пищи в жир организма
энергетически очень выгодный процесс, требующий приблизительно 3% от энергии,
которая будет запасена. Напротив, преобразование углеводов пищи в жир
требует приблизительно 23% от полученной энергии. Поэтому, когда есть избыток
энергии, калории из жира чаще откладываются в жировой прослойке, чем
калории из углеводов.
Содержание углеводов в вашей пище должно составлять 55-70%
(в пересчете на калории) от вашего полного рациона. Большая их
часть должна поступать из сложных углеводов, содержащихся в
зерне, бобовых и овощах. Питание с высоким содержанием сложных
углеводов выгодно для потери веса по нескольким причинам. Во-
первых, углеводы не так легко преобразуются в жировую ткань, как
жиры. Во-вторых, углеводы стимулируют мозг к выделению нейро-
трасмиттера серотонина, который стимулирует чувство
насыщения. В-третьих, сложные углеводы улучшают гликемический ответ
на пищу, т.е. повышают поглощение глюкозы тканями, и
уменьшают избыток инсулина, который является одним из факторов,
ведущих к тучности.
Избегайте простых углеводов (сахара) насколько это возможно.
Сахара в чистом виде быстро всасываются и чрезмерно повышают
содержание глюкозы в крови, что приводит к избытку инсулина, а
иногда и к возвратной гипогликемии.
351
Увеличьте потребление волокон, по крайней мере до 25-35 г в
день. Волокна обеспечивает множество преимуществ, в частности,
потерю веса и улучшение здоровья. (Мы обсуждаем это в
следующем параграфе этой главы.) Высокое содержание волокон присуще
цельному зерну, овощам, фруктам и бобовым. Увеличивайте
потребление волокон постепенно, позволяя вашему организму
приспособиться к такому типу питания. Это минимизирует возможные
побочные эффекты: газы, раздувание живота или диарею.
Нерастительные белки предпочтительно получать, потребляя
куриное мясо и мясо индейки, а также рыбу. Эти продукты содержат
мало жира, особенно насыщенного жира. Соя - превосходная замена
животного белка, она имеет не только высокую пищевую ценность, но и
способствует снижению содержанию холестерина и ЛНП в крови.
22.3. ФИЗИЧЕСКИЕ УПРАЖНЕНИЯ
Непросто уменьшить потребление калории до 500 в день (как
упоминалось ранее, это - энергетический дефицит, необходимый
для разумной потери веса со скоростью одного фунта в неделю).
Альтернативой является сжигание 500 калорий в день посредством
физической нагрузки. К сожалению, это тоже почти настоящий
подвиг. Вы должны будете бегать трусцой в течение 45 минут в день,
играть в теннис в течение часа, или совершать быстрые пешие
прогулки в течение часа и пятнадцати минут. Для большинства людей,
наилучший выход - это уменьшить потребление пищи на 250 калорий в
день, что вполне реально, и заниматься аэробикой 30 мин четыре
раза в неделю, что также реально. Вы не должны перегружать себя.
Нагрузка должна составлять примерно 50-60% от максимальной,
что соответствует интенсивной пешей прогулке.
Избыток веса - обычно комбинация физиологических и психологических
факторов. Физиологические причины, как правило, более очевидны, и с ними
можно справиться. Психологические причины переедания зачастую
неочевидны и располагаются в сфере подавленных эмоциональных конфликтов,
низкого чувства собственного достоинства, одиночества и т.п. Глубокие
психологические проблемы обычно требуют персонифицированного профессионального
анализа, что является непростой задачей. Тем не менее существует множество
простых шагов, которые могут изменить ваше отношение к пище таким
образом, чтобы уменьшить ее потребление.
Ограничьте или избегайте неуместных пищевых раздражителей. Не
ходите в магазин, когда вы голодны. Не держите дома пищевые продукты (с
высоким содержанием калорий), готовые к употреблению. Позвольте членам вашей
семьи самим выбирать, покупать и хранить их собственные сладости. Готовьте
вашу пищу дома и избегайте магазинов готовой пищи и торговых автоматов.
Когда вы дома, ешьте только в одной комнате и в одном месте. Делайте
маленькие порции, которые выглядят большими (например, разложить пищу на
большую тарелку). Избегайте скуки, будьте занятыми. Не доводите себя до
состояния явного голода (не игнорируйте приема пищи, не ешьте слишком мало).
352
Культивируйте хорошие привычки при выборе и приеме пищи. Готовьте
низкокалорийные пищевые продукты привлекательно. Ешьте медленно,
пережевывайте пищу тщательно, делайте паузы несколько раз в течение приема
пищи, не смотрите телевизор и не читайте во время еды. Всякий раз, когда
возможно, принимайте пищу в компании приятных вам людей. Занимайтесь
физическими упражнениями, старайтесь выбирать интересные спортивные игры.
22.4. ГЛИКЕМИЧЕСКИЙ ЭФФЕКТ
ПИЩЕВЫХ ПРОДУКТОВ
Потребляемую пищу ферменты желудочно-кишечного тракта
перерабатывают до небольших молекул, таких как моносахариды,
аминокислоты и пептиды. Многие пищевые продукты, от
мороженого до пиццы и кукурузных хлопьев, содержат сахарозу или другие
углеводы, которые в организме преобразуются в глюкозу. После
того как пища переварилась, глюкоза освобождается и поступает в
кровь, где ее уровень повышается. Насколько интенсивным будет
такое повышение, зависит от нескольких факторов. Во-первых, от
количества глюкозы в пище. Во-вторых, от формы, в которой
находится эта глюкоза (например, в виде крахмала-полисахарида,
состоящего из мономеров сахарозы, которые в свою очередь состоят из
молекул глюкозы и фруктозы). В-третьих, от присутствия в пище
других компонентов, таких как волокна, от которых зависит
скорость поглощения глюкозы.
В интересах продления жизни и предотвращения болезней,
необходимо избегать значительного повышения концентрации
глюкозы в крови. Вредоносное влияние высокого содержания глюкозы
реализуется посредством нескольких механизмов: это - ускорение
неэнзиматического гликозилирования и перекрестных сшивок
жизненно важных клеточных структур (см. гл. 2); это - причина
избытка инсулина, что является важным фактором развития
дегенеративных болезней, и это, по-видимому, также - причина
интенсификации образования свободных радикалов. Факторы, которые
понижают содержание глюкозы в крови, снимают также излишек инсулина
и увеличивают продолжительность жизни у грызунов (Дилъман,
1987; Dilman, 1994; Masoro. 2000; Todorov H. et al.f 2001).
Способность пищевых продуктов повышать содержание
глюкозы в крови значительно варьирует. Диетологи рассчитывают
эффект действия пищи на повышение глюкозы в крови, используя гли-
кемический показатель (ГП). Чем больше повысится уровень
глюкозы в крови после поглощения данной пищи, тем выше ее ГП. Для
чистой глюкозы ГП является одним из самых высоких.
Вообще из разных видов продовольствия, следует отдавать
предпочтение тому, который имеет более низкий ГП (см.
табл. 18А). Большинство овощей (исключая морковь, бобы и
картофель) имеют низкий ГП. У фруктов также относительно
353
Таблица 18А
Гликемический показатель для некоторых пищевых продуктов
ГП определялся как изменение уровня глюкозы в крови (в %)
после приема 50 г продукта по сравнению с уровнем глюкозы
после приема 50 г белого хлеба (100%)
(См.: Wolever T.MS. World Rev. Nutr. Diet, 1990. P. 62, 120-185)
Пищевые продукты
Хлеб
Ржаной
Ржаная мука
Пшеничный, белый
Пшеничная мука
ГП
95
89
100
100
Пищевые продукты
Печенье
Из овсяной муки
Печенье "К чаю"
Крекеры
Песочное печенье
ГП
78
80
100
88
Макароны
Макароны (белые, вареные 64
5 мин)
Спагетти (коричневые, 61
вареные 15 мин)
Спагетти (белые, вареные 67
15 мин)
Макароны "Звездочки" 54
(белые, вареные 15 мин )
Зерновые
Ячмень (крупношлифован- 36
ный)
Гречиха 78
Просо 103
Рис (коричневый) 81
Рис (быстрого приготов- 65
ления, 1 мин)
Рис (шлифованный, вареный 58
5 мин)
Рис (шлифованный, вареный 81
10-15 мин)
Рис (обваренный кипятком, 54
5 мин)
Рис (обваренный кипятком, 68
15 мин)
Ржаные зерна 47
Сладкая кукуруза 80
Пшеничные зерна 63
Зерновые для завтрака
Корнфлекс 121
Мюсли 96
Овсяная каша 89
Рисовые хлопья 132
Пшеничные хлопья 110
Измельченные пшеничные 97
зерна
Корнеплоды овощей
Картофель (быстрое 120
приготовление)
Картофель (пюре) 98
Картофель (вареный) 80
Картофель (жаренный 116
Картофель (сладкий) 70
Бобовые
Вареные бобы (консервиро- 70
ванные)
Бобовое масло 46
Огородный горошек (моро- 65
женный)
Зеленый горошек (сушенный) 65
Зеленый горошек (консерви- 50
рованный)
Фасоль (белая, сушенная) 54
Фасоль обыкновенная (сушен- 43
ная)
Фасоль обыкновенная (консер- 74
вированная)
Чечевица (зеленая, сушенная ) 36
Чечевица (зеленая, консерви- 74
рованная)
Чечевица (красная, сушенная ) 38
Пятнистые бобы (сушенные ) 60
Пятнистые бобы (консерви- 64
рованные)
Арахис 15
Соевые бобы (сушенные ) 20
Соевые бобы (консервиро- 22
ванные )
Фрукты
Яблоки 52
354
Таблица 18А(онончание)
Пищевые продукты
Яблочный сок
Бананы
Апельсины
Апельсиновый сок
Сахара
Фруктоза
Глюкоза
Мед
Лактоза
Мальтоза
ГП
45
84
59
71
26
138
126
57
152
Пищевые продукты
Сахароза
Молочные продукты
Заварной крем
Мороженное
Снятое молоко
Цельное молоко
Йогурт
Легкая закуска
Кукурузные чипсы
Картофельные чипсы
ΓΙΤ
83
59
69
46
44
52
99
77
низкий показатель, так как их сахар, главным образом фруктоза,
преобразуется в глюкозу относительно медленно. Пищевые
продукты с высоким содержанием волокон имеют вообще низкий
ГП, поскольку волокна, замедляют как пррцесс переваривания,
так и поглощение питательных веществ1 СгЗеди обычных
пищевых продуктов самый высокий ГП отмечен у продуктов с низким
содержанием волокон и высоким содержанием крахмала,
который подвергся термической обработке, например, разваренный
рис, зерновые хлопья (корнфлекс), картофельное пюре,
пшеничный хлеб и обычный сахар. Нет никакой необходимости строго
избегать пищевых продуктов с высоким ГП, но они должны
использоваться в умеренных количествах и не составлять большую
часть рациона. Воздействие на уровень глюкозы в крови пищи
с высоким ГП снижается, если их использовать совместно с
продуктами с низким ГП и высоким содержанием волокон. Сладкий
десерт давал бы менее отрицательный эффект, если бы был
съеден после цыпленка с овощами, а не с макаронами
22.5. СБАЛАНСИРОВАННАЯ ДИЕТА
Призывы придерживаться сбалансированной диеты развешены
на каждом углу. Но что это конкретно означает? Сбалансированной
диетой специалисты обычно считают такую диету, которая
содержит все необходимые питательные вещества в пределах некоторого
оптимального диапазона и составлена таким образом, чтобы
предотвратить пищевой дефицит белков, витаминов и минеральных
веществ. Сбалансированная диета необходима, но не достаточна для
всесторонней программы продления жизни. Конечно, недоедания
нужно избегать, но чтобы "изменить природу" и существенно
продлить жизнь следует потреблять некоторые питательные вещества
355
в количествах превышающих типичные для сбалансированной
диеты. Говоря фигурально, сбалансированная диета - основа, на
которой должна строиться программа продления жизни.
Самый простой и, вероятно, наиболее реалистичный путь
твердо придерживаться сбалансированной диеты для большинства
людей состоит в необходимости следовать рекомендациям по
ежедневному пищевому рациону (RDA), разработанным Министерством
сельского хозяйства США. Ежедневно взрослым следует потребить:
• От 6 до 11 порций хлеба и хлебных злаков;
• От 3 до 5 порций овощей;
• От 2 до 4 порций фруктов;
• 2-3 порции мяса и альтернатив мясу (рыба, домашняя птица,
яйца, сушеные бобы, орехи);
• 2 порции молока и молочных продуктов.
Главными компонентами пищевых продуктов, или
малопитательными веществами, являются белки, углеводы и жиры.
Современные рекомендации по их потреблению:
• 10-20% от суммы ежедневных калорий должны быть
получены от белков;
• 55-70% калорий - от углеводов;
• Менее чем 30 % калорий - от жира, а насыщенный жир должен
составлять меньше чем 10%.
Необходимо иметь в виду, что каждый грамм белка содержит
приблизительно 4 калории, углеводы - 4 и жир - 9. Калории от
углеводов должны поступать главным образом от сложных углеводов,
которые содержатся в хлебе, хлебных злаках и картофеле.
Рекомендованное суточное потребление белка для взрослых составляет 0.8 г
на кг веса тела. Меньшее количество может приводить к дефициту
белков, который характеризуется понижением иммунитета,
отечностью и долгим заживлением ран. В то время как недостаток
потребления белков обычен для развивающихся стран, в развитых странах
их потребление обычно значительно выше рекомендаций RDA.
Избыточный белок сжигается как топливо, и остающийся при этом
азот преобразуется печенью в мочевину и выделяется почками.
Существенный излишек белков подвергает неуместному стрессу эти
органы, и его следует избегать, особенно индивидам с риском
болезней печени и почек. Диабет - наиболее распространенный фактор
риска почечной болезни. Гепатит и чрезмерное потребление
алкоголя предрасполагают к повреждению печени.
Недавние исследования указывает, что потребление количеств
белка, умеренно превышающих RDA, может стимулировать
выделение гормона роста (ГР), если сочетается с физическими
упражнениями. Содержащиеся в белке аргинин, лизин и глютамин
стимулируют, как полагают, выделение гормона роста. Индивиды, стремя-
356
щиеся к повышению выделения ГР, могут извлечь пользу,
потребляя примерно 25% калорий, извлекаемых из белков, но только в
сочетании с существенной физической нагрузкой (см. гл. 26). Рыба,
индейка и цыпленок - предпочтительные источники нерастительного
белка. Соя, по-видимому, лучший источник растительного белка,
поскольку белок сои понижает холестерин и ЛНП. Существенное
понижение холестерина наблюдается при ежедневном потреблении
по крайней мере 25 г белка сои вместо эквивалентного количества
животного белка в диете.
Чрезмерному потреблению жира следует препятствовать,
потому что пища, богатая жиром, весьма калорийна и, следовательно,
способствует увеличению веса. Кроме того, потребление некоторых
типов жира связано с повышенным риском ряда болезней. В
частности, чрезмерное потребление насыщенного жира увеличивает риск
атеросклероза, рака прямой кишки и гипертонии. Пережаренный
полиненасыщенный жир, по-видимому, также увеличивает риск
различных раковых образований, увеличивая, возможно,
свободно-радикальные повреждения ДНК.
Напротив, чрезвычайно низкое потребление жира может также
вызывать негативные последствия. Человеческий организм может
синтезировать большинство, но не все требующиеся ему жирные
кислоты. Исключения - линолевая и линоленовая жирные кислоты,
часто называемые незаменимыми жирными кислотами (НЖК).
Недостаток НЖК ассоциируется с нарушениями здоровья кожи, почек и
печени. Люди при долговременно низкой жировой диете могут
развивать значительный дефицит НЖК. Разумно низкое потребление
жира, но не ниже, чем 10% от общего количества калорий, является
достаточным, чтобы предотвратить любую форму недостатка НЖК.
Не все жиры - равноценны. Насыщенный жир - вид пищи,
наиболее вредной для здоровья. Его потребление необходимо снизить
до 10% от общего количества калорий. Главные источники
насыщенного жира животного происхождения - мясо, домашняя птица,
молочные продукты и яйца. Выбирайте белое мясо цыпленка или
индейки со снятой кожей. Используйте молочные продукты с
низким содержанием жира. Яйца ценный источник белков высокого
качества, но их потребление должно быть умеренным - не чаше чем
3-4 в неделю.
Моно- и полиненасыщенный жир улучшает липидный профиль,
понижая холестерин, ЛНП, триглицериды и повышая ЛВП.
Следует, однако, иметь в виду, что при избытке калорий замена
насыщенного жира на моно- и полиненасыщенный жир, не принесет
слишком большой пользы. Самые богатые источники
мононенасыщенного жира - оливковые масло и масло канолы, а
полиненасыщенного жира - зерно, подсолнечник и масло сои. Ненасыщенный жир
легко окисляется, особенно при термической обработке. Если вы
готовите пищу на масле или используете готовые коммерческие
357
продукты с высоким содержанием ненасыщенного жира, то следует
принимать пищевые добавки с витамином Е, чтобы минимизировать
повреждающее действие свободных радикалов.
Некоторые коммерческие продукты, такие как маргарин или
растительный жир для рассыпчатого теста, содержат частично
гидрированное масло. Гидрирование - химический процесс, в котором
некоторые из двойных связей, содержащихся в полиненасыщенном жире,
реагируют с водородом. В результате, жир станет более насыщенным
и твердым. В частности,, гидрирование превращает некоторые из
полиненасыщенных жиров в мононенасыщенные; Однако это только
один аспект проблемы. Существует два типа мононенасыщенных
жирных кислот: цис- и трянс-формы. Естественные источники
содержат несколько различных цис-форм, которые полезны для
профиля липидов. Химическое гидрирование, однако, приводит к
образованию как цис-, так и транс-ненасыщенного жира (обычно большего
количества транс- чем цис-форм). Влияние
транс-мононенасыщенного жира на профиль липидов, по крайней мере столь же негативно,
как и влияние насыщенного жира. В свете этого положительный
эффект замены масла на маргарин далеко не столь очевиден, как
можно было бы думать (Modern Nutrition in Health and Disease, 1994).
Рыбий жир - богатый источник специфического типа
ненасыщенного жира, омега-3 жирной кислоты, особенно эйкозапентаено-
евой кислоты (ЭПК) и декозагексаеноевой кислоты (ДГК). Омега-3
жирные кислоты привлекла внимание, когда исследователи
обнаружили весьма низкий уровень атеросклероза и болезней сердца у
жителей Аляски и Гренландии, несмотря на высококалорийную,
богатую жиром и холестерином диету. Анализ пищевых продуктов в
этой диете, содержащей главным образом рыбу и мясо морских
животных, показал высокое содержание омега-3 жирных, кислот.
Механизмы профилактического действия рыбьего жира на сердечные
болезни, по-видимому, сложен и включает антитромбозное
действие (разжижение крови) и модуляцию иммунного ответа. Другие
исследователи нашли, что омега-3 жирные кислоты могут также инги-
бировать развитие некоторых раковых образований. Лучший путь к
включению омега-3 жирных кислот в оптимальную диету - это
употреблять в пищу рыбу 2-4 раза в неделю.
22.6. ВОЛОКНА И ИХ ПОЛЕЗНЫЕ СВОЙСТВА
В настоящее время широко признано, что волокна являются
важным пищевым компонентом, способствующим здоровью и
долголетию. Химически волокна - разнообразная группа растительных
полимеров на основе полисахаридных цепей. В отличие от
крахмала, который также является разновидностью полисахаридов,
волокна не перевариваются в человеческом организме и проходят через
358
пищеварительный трактат частично или полностью
неповрежденными. Большое количество волокон в диете делают экскременты
мягкими и обильными.
Волокна оказались в центре внимания, когда врачи,
работавшие в Африке, отметили очень низкий уровень таких типичных
"западных" недугов, как болезни сердца, диабет, тучность, рак
прямой кишки, дивертикулоз и геморрой. Они также заметили,
что некоторые племена употребляли в пищу очень много
волокнистых продуктов - объем их экскрементов в несколько раз
превышал таковой у людей в цивилизованных странах.
Гипотетическая роль волокон в предотвращении болезней и тучности
генерировала массу исследований относительно этого предмета.
Было найдено, что волокна действительно приносят большую
пользу здоровью и могут также замедлять некоторые механизмы
старения, главным образом за счет толерантности к глюкозе и
снижения избытка инсулина.
Имеются два главных класса волокон - растворимые в воде
(смолы, пектины, некоторые гемицеллюлозы и растительная слизь)
и водонерастворимые (целлюлозы, некоторые гемицеллюлозы и
лигнины). Растворимые в воде волокна преобладают во фруктах,
овсяных зернах, ячмене и бобовых. Водонерастворимые волокна
найдены в более высоких концентрациях, главным образом в
овощах, пшенице и хлебных злаках. Физиологические эффекты этих
двух типов волокон несколько различаются, но оба типа приносят
значительную пользу здоровью (табл. 18В).
Исследования показывают, что полный спектр
положительных эффектов от диеты с высоким содержанием волокон
проявится при условии, что они поступают из различных
источников - фруктов, овощей, неочищенных хлебных злаков и бобовых.
Потребление продукта с единственным типом волокон не
является оптимальным.
Улучшение гликемического ответа и снижение выделения
инсулина. Гликемический ответ на пищу - это степень
повышения глюкозы в крови после ее принятия. Водорастворимые
волокна замедляют движение пищи через верхнюю часть
пищеварительного тракта, а также предотвращают слишком быстрое
всасывание питательных веществ в тонком кишечнике.
Результирующий эффект - более медленное и менее интенсивное
повышение глюкозы крови после приема пищи и соответствен
но меньшее выделение инсулина. Нам представляется, что
способность волокон снижать уровень глюкозы и излишек
инсулина - это их наиболее важное свойство, полезное для
замедления старения. Действительно, как это уже обсуждалось ранее,
высокий уровень глюкозы и избыток инсулина в крови вносят
существенный вклад в старение и развитие нескольких
дегенеративных болезней, включая атеросклероз, гипертонию, иммуно
359
Таблица 18В
Физиологические эффекты волокон
Водорастворимы* Водинерастворимые
• Замедление прохождения пищи · Ускорение прожождения пищи
через пищеварительный тракт, через нижнюю часть пищевари-
особенно в его верхней части тельного тракта
»Более низкий уровень холесте- · Увеличения веса и объема экс-
рина крементов
► Медленный распад крахмала и · Медленный распад крахмала и
замедление всасывания глюко- замедление всасывания глюкозы
зы в кровяное русло в кровяное русло
• Ферментируются
микроорганизмами с освобождением жирных
кислот с короткой цепью,
полезных для клеток прямой кишки
депрессию и диабет. Медикаменты, нормализующие эти
показатели метаболизма, способны продлить жизнь
экспериментальных животных. Диета с высоким содержанием волокон
способна сделать то же самое, но с меньшим количеством побочных
эффектов.
Профилактика рака прямой кишки. Эпидемиологические
исследования показывают, что у сельских жителей, потребляющих
высоковолокнистую пищу, рак прямой кишки встречается весьма
редко. Волокна могут предотвращать развитие этой разновидности
рака, связывая и быстро удаляя канцерогенные соединения из
прямой кишки.
Снижение холестерина. Растворимые волокна понижают
уровень холестерина, связывая желчь. А желчь (главный
компонент которой - желчные кислоты) в свою очередь способна
связывать липиды (жирные кислоты и холестерин). Желчные
кислоты синтезируются печенью из холестерина и
накапливаются в желчном пузыре. Когда пища поступает в тонкий
кишечник, желчь через проводящие пути направляется в
двенадцатиперстную кишку. Позже большая часть желчи повторно
поглощается в нижней области тонкого кишечника.
Растворимые волокна из таких пищевых продуктов, как овсяные зерна,
ячмень и бобовые, связывают желчные кислоты и
предотвращают их возврат в кровяное русло и затем в печень. В результате,
чтобы восполнить потери, печень должна синтезировать
большее количество желчи из холестерина, постепенно исчерпывая
его запасы, что и приводит к уменьшению содержания
холестерина в крови.
360
Имеется также свидетельства, что продукты переваривания
волокон кишечными бактериями - жирные кислоты с короткой
цепью - ингибируют синтез холестерина.
Потеря веса. Высоковолокнистые пищевые продукты
содержат обычно меньшее количество калорий и жира на единицу массы
и, следовательно, позволяют достичь насыщения при более низком
потреблении энергоносителей и жира. Волокна также снижают
избыток инсулина, гормона, который "приказывает" телу сохранять
его вес и запасы питательных веществ. В целом, постепенное
переключение на высоковолокнистую диету является вполне разумным
шагом, который можно предпринять в направлении снижения
избыточного веса.
Другие преимущества. Волокна помогают предотвратить такие
не слишком опасные, но неприятные проблемы толстого
кишечника, как геморрой, запор и дивертикулоз; помогают поддерживать
нормальную бактериальную флору кишечника. Как уже
указывалось, ферментация волокон кишечными бактериями сопряжена с
производством жирных кислот с короткой цепью, которые служат
источником энергии и помогают сохранить полноценные функции
эпителиальных клеток толстого кишечника.
Возможные побочные эффекты волокон обычно невелики и
легко поддаются устранению. Когда человек резко переходит к
высоковолокнистой диете, он может испытать неприятные
ощущения в желудке и избыточное накопление газов. Во избежание
этого переход должен быть сделан постепенно. Волокна могут
понижать в организме усвоение некоторых минералов. Это
возможно потому, что волокна увеличивают скорость прохождения
пищи по пищеварительному тракту, сокращая время контакта
между пищей и эпителием кишечника, что и приводит к понижению
всасывания этих минералов. Существуют свидетельства, что у
людей с адекватным всасыванием питательных веществ,
умеренное потребление волокон не ставит под угрозу метаболический
статус организма. Однако волокна могут ухудшать ситуацию у
людей с низким всасыванием минералов. При
высоковолокнистой диете необходимо удостовериться, что вы потребляете
достаточное количество всех питательных веществ и особенно —
полезных минералов.
Пищевые добавки с волокнами (но не богатые волокнами
пищевые продукты) могут создавать серьезные проблемы, связанные с
затруднениями проходимости в кишечнике, что иногда требует
хирургического вмешательства. Если вы хотите употреблять
волокнистые пищевые добавки, то лучше использовать их в форме порошка
и вместе с большим количеством жидкости.
В каком количестве волокон мы нуждается? Рекомендации
экспертов относительно количеств потребляемого волокна
несколько варьируют. Большинство источников рекомендует по-
361
треблять 20-35 г волокна в день, что в 2-3 раза выше количества,
потребляемого средним американцем. Однако желательно
утраивать ваше потребление волокон не внезапно, а постепенно в
течение нескольких недель, позволяя желудочно-кишечному тракту
приспособиться к такому увеличению. Следует обильно пить,
чтобы обеспечит гладкое прохождение пищевой массы по
пищеварительному тракту. Старайтесь потреблять волокна из
различных источников - фруктов, овощей, цельных гранулированных
хлебных злаков и бобовых. Как уже указывалось выше,
потребление 2-3 порций фруктов, 3-5 порции овощей и 6-11 порций
хлеба и хлебных злаков в день легко обеспечит рекомендуемый
объем пищевых волокон.
Глава 23
ПРОТИВ СВОБОДНЫХ РАДИКАЛОВ
Как уже обсуждалось в главе 2, повреждения, вызванные
свободными радикалами (СР), - один из основных механизмов старения
на молекулярном и субклеточном уровнях. СР-осколки, как
правило, небольших молекул, которые способны с высокой скоростью
реагировать практически с любыми химическими партнерами, что
статистически может привести к повреждению любой структуры в
клетках. Солнечный свет и радиация могут индуцировать
формирование СР, но наиболее распространенная причина их
возникновения - нормальное биологическое окисление энергоносителей,
которое сопряжено с таким фундаментальным биологическим
процессом, как дыхание кислородом . Вообще, проще говоря, чем большее
количество СР продуцирует вид, тем короче продолжительность его
жизни (гл. 2). Мозг и сердце - два органа, которые в наибольшей
степени подвергаются атакам СР, поскольку они "жгут"
биологическое "топливо" с очень высокой интенсивностью.
В дополнение ко вкладу в процесс старения, СР вовлечены в
патогенез многих болезней. Они окисляют липиды в ЛНП, которые
затем "прилипают" к артериальным стенкам и вызывают
атеросклероз. СР повреждают мозговые клетки при болезни Альцгейме-
ра. Воспаление при ревматоидном артрите - в значительной
степени результат выделения СР клетками иммунной системы. Избыток
СР также увеличивает риск рака.
Поскольку СР являются неизбежным побочным продуктом
клеточного дыхания, они не могут быть полностью устранены. Однако
их количество, формирующееся в клетке, может быть уменьшено.
Множество путей может вести к этой цели.
• Устранение избытка калорий. Количество СР, формирующихся в
организме - пропорционально сумме "сжигаемых" калорий. Когда
клетки "жгут" много биологического топлива, они неизбежно
производят много "молекулярной сажи" - свободных радикалов.
Ограничение потребляемых калорий у грызунов на 40%
пролонгирует их жизнь до двух раз. Серьезное ограничение калорийности
пищи - неприемлемо для людей, но элиминация избытка калорий в
363
"борьбе" за оптимальный вес, наряду с другими мерами, уменьшит
негативное влияние повышенного уровня СР.
• Антиоксиданты - вещества, которые нейтрализуют СР и делают
их безопасными для живых объектов. Денхам Харман, один из
пионеров в исследовании роли АО в процессах старения, нашел, что
у мышей на диете, содержащей 0.5-1% АО, продолжительность
жизни повышалась на 30-45% (Нагтап, 1996). Множество пищевых
веществ, некоторые из которых весьма существенны для
продления жизни, действуют как АО. Различные фрукты, овощи и
растения служат обильными источниками АО. Использование в
качестве пищевых добавок таких веществ, как витамины Е, С, каротино-
иды, флавоноиды, липоевая кислота, селен и цистеин, могут
обеспечивать дополнительную АО-защиту организма (см. ниже).
• Повышение собственной антиоксидантной защиты. Живые
существа эволюционно развили сложные системы для защиты от
СР. Особенно важны специфические ферменты, убирающие из
клеток СР: супероксид дисмутаза (СОД), каталаза и глутатион
пероксидаза. Высокий уровень этих ферментов обеспечивают
дополнительную защиту от СР. Мутанты плодовой мушки
дрозофилы с высоким уровнем СОД, живут вдвое дольше, чем немутант-
ные особи (см. гл. 2). К сожалению, исследований по изысканию
безопасных способов повышения уровня антиоксидантных
ферментов организма проведено не много. Мелатонин, будучи антиок-
сидантом сам по себе, кроме того, поднимает, как было показано,
также и уровень глутатион пероксидазы. Некоторые
исследователи полагают, что ослабление антиоксидантной защиты с
возрастом частично связано со значительным снижением продукции ме-
латонина (см. гл. 26, о восстановлении уровня мелатонина,
присущего молодости). Другой известный способ улучшения
эффективности систем нейтрализации СР - так называемое окислительное
обучение. Объяснение этого подхода — весьма простое. Умеренное
кратковременное увеличение уровня СР стимулирует активность
ферментативных систем защиты, что не приносит какого-либо
вреда организму. Повышенный уровень активности этих
ферментов сохраняется существенно более длительное время, чем
начальный окислительный импульс. Повторное увеличение уровня СР
заметно стимулирует продукцию ферментов их нейтрализации,
что может быть полезно при некоторых видах лечения, связанных
с использованием высоких концентраций кислорода, например при
гипербарической оксигенации или других биооксидативных
методах лечения. Однако чрезмерная экспозиция с кислородом,
перекисью водорода или другими окисляющими соединениями опасна
и может вызвать необратимые повреждения тканей. Более
безопасным и более приятным методом "окислительного обучения"
являются физические упражнения. Последние ускоряют
"сжигание" энергоносителей, временно увеличивая формирование СР и
364
стимулируя производство антиоксидантных ферментов (Л et al.,
1998). Однако пользу здоровью принесут регулярные
упражнения - от трех до шести раз в неделю по 30-60 мин, но не более.
Слишком интенсивные упражнения вызовут слишком сильный
поток СР, и защитные системы организма будут не в состоянии его
полностью нейтрализовать, что в конечном счете ускорит процесс
старения.
• Снижение стресса. Стресс освобождает поток СР, которые могут
вызвать тяжелые повреждения тканей тела. В течение начальных
фаз стресса организм делает попытку повысить антиоксидантную
защиту, чтобы противодействовать ускорению формирования СР.
Интенсивный или длительный стресс, однако, в конечном счете
исчерпывает антиоксидантные ресурсы, что ведет к обширным и
иногда необратимым повреждениям клеток. Нейтрализация избытка
СР защищает от многих стресс-зависимых повреждений. Например,
вызванное стрессом образование язв желудка у животных может
быть предотвращено мощными антиоксидантами. Первый шаг к
уменьшению стресс-зависимых повреждений СР - избегать стрессо-
генных ситуаций и изменять психологическую реактивность путем
медитации и других видов ментальной техники. А также увеличить
прием богатых антиоксидантами питательных веществ в течение
стрессогенного периода (если вы уже не принимаете антиоксиданы
в высоких дозах), что также будет способствовать защите от СР.
Наконец, адаптогены, средства смягчающие стресс-ответ, как
было показано, значительно уменьшают количество СР,
формирующихся в течение стресса (Барабой, Брехман и др., 1992).
23.1. ПИТАТЕЛЬНЫЕ ВЕЩЕСТВА И ПИЩЕВЫЕ ДОБАВКИ
С АНТИОКСИДАНТНЫМИ СВОЙСТВАМИ
Пейте соки, а не вина
Вам здоровье жалко коль,
Ибо в соках витамины,
А в винах - алкоголь
Николай Глазков, 1940
В природе существует огромное разнообразие соединений с ан-
тиоксидантными свойствами: сильные и слабые, большие и малые
(относительно размера молекулы), существенные и
несущественные, белые или цветные. Некоторые антиоксиданты выполняют и
другие важные функции в организме. Например, витамин С также
требуется для синтеза коллагена (ключевой структурный белок в
коже, капиллярах и костях), а витамин А важен для зрения и
нормального роста клеток внутренней поверхности кишечника и кожи.
365
Таблица 19
Растворимость антиоксидантов
Растворимь
1
вводе
Витамин С
Цистеин
Метионин
Селен
Глутатион
е
в жире
Витамин Ε
Витамин А
Каротиноиды
Ликопен
Кофермент Q10
Вод о- и жирорастворимые
Липоевая кислота
Мелатонин
Некоторые полифенолы
Некоторые флавоноиды
Очень важное химическое свойство антиоксидантов - их
растворимость в воде и жире. Организмы имеют два типа внутренней
среды: водное пространство (вне- и внутриклеточное) и липидные
мембраны, которые отгораживают клетки друг от друга, а внутри
клеток отделяют некоторые структуры, например ядра и митохондрии,
от внутриклеточной среды. Также имеются липопротеины,
циркулирующие в крови маленькие сферические образования,
содержащие жир, холестерин и белок. Растворимые в воде антиоксиданты
эффективны главным образом во вне- и внутриклеточной
жидкости, тогда как жирорастворимые антиоксиданты защищают
мембраны и липопротеины. Оба типа АО необходимы, чтобы создать
эффективный щит против свободных радикалов для всего
организма (табл. 19).
23.1.1. Витамин С
Витамин С - ключевой растворимый в воде витамин - антиокси-
дант. Кроме того, это - кофактор в синтезе коллагена и фактор,
играющий главную роль в поддержании нормальной иммунной
функции. Рекомендуемая суточная доза витамина С - 60 мг, что
достаточно для предотвращения цинги - болезни, связанной с дефицитом
витамина С. Витамин С содержится в плодах цитрусовых, дынях,
ягодах, перце, картофеле, капусте, брокколи, помидорах, твердых
хлебных злаках. Возможная польза для здоровья от обильного
потребления витамина С - снижение риска атеросклероза и
катаракты, улучшение функций иммунной системы и ускорение заживления
ран. Количество витамина С, которое требуется для поддержания
оптимального здоровья, остается предметом споров. Лайнус По-
линг, лауреат Нобелевской премии и пионер исследований
физиологических эффектов витамина С, рекомендовал использовать очень
высокие дозы - десятки граммов в день (Полинг, 1974). Однако
недавние исследования показывают, что относительно невысокие
дозы витамина С, такие как, например, 500 мг могут при некоторых ус-
366
ловиях скорее стимулировать, чем ингибировать формирование СР
В настоящее время лучшая стратегия состоит в получении
витамина С с пищей, потребляя, например, апельсиновый сок.
Рассматривая эти противоречивые данные, мы полагаем, что пищевые
добавки не должны превышать содержание витамина С - 250 мг в день.
Риск при использовании высоких доз. Высокие дозы витамина С могут
приводить к ошибкам при интерпретации теста на скрытое присутствие крови в
экскрементах. Этот тест иногда используется, чтобы диагностировать некоторые
желудочно-кишечные болезни и рак ободочной кишки. Поскольку витамин С
увеличивает поглощевие и задержку железа, его высокие дозы не должны приниматься
людьми с генетической предрасположенностью к перегрузке железом (гемохрома-
тоз или талассемия). Высокие дозы витамина С могут также осложнять лечение
некоторых видов рака. Очень высокие дозы (десятки граммов) могут вызывать понос
и желудочно-кишечные расстройства. Эти побочные эффекты могут быть
минимизированы, если принимать витамин С в форме аскорбата кальция.
23.1.2. Витамин Ε
Витамин Ε - основной жирорастворимый витамин - антиокси-
дант. Он защищает мембраны клеток и липопротеины. Эти
структуры особенно уязвимы при действии СР, поскольку содержат
высокореактивные ненасыщенные жирные кислоты.
В ряде исследований было показано, что высокие дозы
витамина Ε ингибируют прогрессию атеросклероза, уменьшают риск
сердечных болезней и инсульта. Рост артериальных бляшек зависит от
захвата ЛНП-частиц из кровяного русла. Окисляясь, ЛНП
становятся "липкими" и накапливаются в "бляшках" со значительно более
высокой скоростью. Витамин Ε эффективно ингибирует окисление
ЛНП, блокируя, таким образом, прогрессию атеросклероза.
Современные данные указывают, что витамин Ε может
замедлять старение мозга. Мозговая ткань богата ненасыщенными
жирными кислотами, которые следует защитить от высокого уровня
активных форм кислорода (АФК), которые являются побочным прот
дуктом интенсивного метаболизма нейронов. В недавнем
исследовании, пациентам с умеренной формой болезни Альцгеймера давали
2000 Ш витамина Ε в день в течение двух лет. У тех, кто получал
витамин Е, болезнь прогрессировала на 52% медленнее, чем в группе с
плацебо (Sardesai, 1995).
АФК могут воздействовать на нервные окончания и индуцировать процесс
окисления липидов (ПОЛ) в мембранах, приводя к нарушению функции
мозговых синапсов, и эта восприимчивость синапса к окислительному стрессу была
существенно увеличена с возрастом, а содержание витамина Ε в синаптиче.ских
плазматических мембранах было понижено. Когда крысы подвергались
стрессу, его содержание было более низким в каждом возрасте, чем в мембранах
нормальных крыс. Введение витамина Ε до стресса сократило эти аномалии.
Витамин Е, очевидно, способствует защите против дисфункции нервных окончаний,
вызванной окислительным стрессом (Urano et al., 1998).
367
Молодые крысы показали существенно большую способность к
обучению перед стрессом, чем старые крысы и крысы с дефицитом витамина Е.
Добавки витамина Ε значительно ускоряли их функции обучения перед
стрессом и предотвращали дефицит памяти, вызванный стрессом;
аналогичные результаты имели место и у старых крыс и крыс с дефицитом
витамина Ε (Fukui et а!.. 2002).
RDA для витамина Е - 30 мг в день, а его продовольственные
источники - растительное масло, орехи, проростки пшеницы, целые
зерна, овощи с зеленой листвой. Дозы витамина Е, польза от
которых для здоровья была показана в различных клинических
исследованиях, составляли от 400 до 2000 Ш в день. Чрезвычайно трудно,
если вообще возможно, получить такие количества с пищей.
Потенциальные побочные эффекты высоких доз. Высокие дозы могут
усиливать действие лекарств - антикоагулянтов (противосвертывающих
средств) и осложнять всасывание витамина К, который играет роль в
свертывании крови. Пищевые добавки с витамином Ε нельзя принимать перед
хирургическими операциями.
23.1.3. Селен
Селен - существенный компонент ключевого фермента АО-
защиты - глутатион пероксидазы. Этот фермент инактивирует
ядовитые продукты ПОЛ. Дефицит селена может вызвать
патологию сердца. Для предотвращения дефицита достаточно
потреблять мужчинам - 70 мкг, а женщинам - 55 мкг селена в день.
Было показано, что селен снижает риск нескольких типов рака.
Большинство экспертов рекомендует для предотвращения рака
принимать до 200 мг селена в день. Продовольственные
источники селена включают дары моря, мясо и некоторое зерновые и
хлебные злаки.
О механизме противоракового действия селена следует сказать, что
микромолярные дозы монометилированной формы селена (Se) могут вызвать апоп-
тоз независимо от повреждения их ДНК и в клетках, имеющих нулевой
фенотип р53. Протеин киназа клеточного цикла cdk2 и протен киназа С строго ин-
гибируются различными формами Se. Игибирующий механизм, как полагают,
включает модификацию селеном цистеиновых остатков белков путем
формирования Se-аддуктов типа S-Se-S или S-Se, а также катализа дисульфидных
формирований. Указанные реакции происходят с участием селена и ряда
регулируемых окислительно-восстановительным потенциалом белков, включая
факторы траскрипции. Их дезактивация, облегченная селеном, может способствовать
нормализации критических процессов на ранних стадиях злокачественной
трансформации клеток (Ganther, 1999).
Потенциальные побочные эффекты высоких доз. Как и многие другие
полезные минералы, потребление селена имеет относительно узкое "окно"
безопасности. Высокие дозы селена могут вызвать желудочно-кишечные
расстройства, неврологические и другие проблемы. Дозы селена, превышающие 200 мг
в день, не рекомендуются.
368
23.1.4. Цистеин и метионин
Цистеин и метионин - аминокислоты, содержащие серу. Будучи
структурными единицами белков, эти аминокислоты действуют как
антиоксиданты, а также облегчают удаление тяжелых металлов из
организма. Цистеин также составная часть глутатиона, который
является основным водорастворимым антиоксидантом внутри клеток.
Метионин - незаменимая аминокислота; она не может быть
синтезирована в организме и должна поступать с пищей. Цистеин может
синтезироваться в теле из метионина и рассматривается как условно
незаменимая аминокислота, однако она может стать реально
незаменимой, если потребление метионина будет ограничено. Пища с высоким
содержанием белка содержит некоторое количество метионина и ци-
стеина. Источники, богатые серу содержащими аминокислотами,
включают бобы, горох, рыбу, печень, яйца, пивные дрожжи и орехи.
Содержание метионина и цистеина в организме, по-видимому,
снижается с возрастом. Обогащение диеты этими аминокислотами
увеличивает продолжительность жизни мышей. Существуют
косвенные свидетельства, что поддержание оптимального уровня серу-
содержащих аминокислот у людей может принести некоторую
пользу для здоровья и продления жизни.
23.1.5. Ликопен
Ликопен - красный пигмент, содержащийся в помидорах,
привлек в последние годы внимание медицинских исследователей. Так,
было показано, что это соединение уменьшает риск нескольких
распространенных раковых заболеваний, в частности рака простаты,
ободочной кишки и легкого. Этот эффект приписывается мощной
антиоксидантной активности ликопена. Лучший способ получить
много ликопена - употреблять изобилие пищевых продуктов,
содержащих приготовленные помидоры. Сырые помидоры - значительно
более бедный источник ликопена, потому что необходимы высокие
температуры, чтобы ликопен выделился из растительных клеток.
Используйте помидоры для приготовления всех ваших соусов.
Ликопен также доступен как пищевая добавка, хотя этот продукт
относительно дорог.
23.1.6. Липоевая кислота
Липоевая кислота - условно незаменимое питательное
вещество, которое требуется клеткам для производства энергии из
углеводов и некоторых других энергоносителей. Эта кислота является
также эффективным антиоксидантом и соединением, связывающим
тяжелые металлы в хелатные комплексы. Организм синтезирует ли-
369
поевую кислоту в скромных количествах, но это производство
может не удовлетворить его потребности во многих ситуациях, в
частности при стрессе или болезнях. Как и для многих ключевых
субстанций организма, уровень липоевой кислоты падает с возрастом.
Липоевая кислота уникальна среди антиоксидантов по богатству
своих свойств. Она является как водо-, так и жирорастворимой, что
обусловливает ее способность к защите всех тканей и клеточных
компартментов. Она эффективна против большинства типов СР,
включая супероксид анион, радикал гидроксила, синглетный
кислород и перекись водорода. Это также ее способность связывать и ней-
трализовывать в хелатных комплексах ионы металлов (железа,
меди, кадмия, свинца и ртути), которые катализируют формирование
свободных радикалов. В различных исследованиях на животных
было установлено, что липоевая кислота заметно уменьшает скорость
ЛНП-окисления и увеличивает доставку кислорода к мозгу и сердцу
(Packer etal., 1997).
Липоевая кислота также понижает содержание глюкозы в
крови и таким образом уменьшает воздействие другого механизма
старения - неферментативного гликозилирования и перекрестного
сшивания (гл. 2). При исследовании крыс с диабетом II типа
липоевая кислота улучшала чувствительность тканей к инсулину,
повышала использование глюкозы на 30-50%, уменьшала содержание
инсулина и свободных жирных кислот в плазме на 15-17%. Было
также отмечено, что у людей липоевая кислота улучшает
диабетическую невропатию (Packer et al., 1997).
К сожалению, лучший диетический источник липоевой
кислоты - красное мясо, которое содержит относительно много
насыщенного жира и холестерина. Пищевые добавки могут быть более
привлекательным способом увеличить потребление липоевой кислоты.
Для здоровых людей рекомендуемая доза липоевой кислоты
100-400 мг в день. В исследованиях диабетической невропатии доза
600 мг в день была наиболее эффективна. В этих дозах липоевая
кислота, очевидно, безопасна.
23.1.7. Флавоноиды
Флавоноиды - разнообразная группа пигментов растений со
свойствами антиоксидантов. Эти соединения ответственны за цвет
многих плодов, овощей и цветов. В дополнение к обеспечению
цвета, который привлекает насекомых или животных, эти пигменты
защищают растения от неблагоприятных воздействий окружающей
среды. Будучи мощными антиоксидантами, некоторые флавоноиды
обладают антиаллергической, антиканцерогенной,
противовирусной и противовоспалительной активностью.
Более 4 тыс. флавоноидов были охарактеризованы и
классифицированы. Экстракт косточек винограда является одним из лучших
370
источников флавоноидов с мощной антиоксидантной активностью
"широкого спектра". Многие флавоноиды имеют уникальное
сродство со специфическим органом, тканью или типом клеток и
поэтому могут служить для целенаправленной профилактики или
лечения. Например, флавоноиды из растения Silybum marianum
(молочный чертополох) - мощные антиоксиданты с сильным сродством с
печенью. Экстракты от Silybum marianum могут защищать печень от
различных видов токсического повреждения, а также ускорить
выздоровление от заразных болезней печени.
Флавоноиды из другого растения Ginkgo biloba имеют сродство с
центральной нервной системой. Флавоноиды из Ginkgo biloba
защищают нейроны от окислительных повреждений и улучшают
мозговое кровообращение. Ряд исследований показал, что экстракт из
Ginkgo biloba улучшает память и обучаемость в пожилом возрасте и
может замедлить прогрессию и даже вызвать частичную реверсию
болезни Альцгеймера. Другими словами, флавоноиды Ginkgo
biloba - одна из наиболее многообещающих субстанций для борьбы со
старением мозга.
Флавоноиды из косточек винограда, Ginkgo biloba и Silybum
marianum недоступны в обычных пищевых продуктах и должны
потребляться как пищевые добавки. Однако имеется обширное
разнообразие флавоноидов, найденных в обычных фруктах и овощах, -
некоторые из них хорошо известны, другие только
идентифицируются, остальные еще должны быть открыты. Один известный
пример - кверцетин - флавоноид, найденный в плодах цитрусовых. Он
является как антиоксидантом, так и антиаллергеном.
Можно получить весьма большую пользу от различных классов
природных соединений, потребляя изобилие различных фруктов и
овощей
23.1.8. Витамин А и каротины
Витамин А выполняет различные функции в организме, но он
также и антиоксидант. Витамин А необходим для иммунной
системы, для нормального зрения и нормального роста эпителиальных
клеток кожи, внутренней поверхности кишечника и других органов.
Дневная потребность взрослого организма в витамине А - 5000 Ш.
Продовольственные источники: печень, яичные желтки, сгущенное
молоко, молочные продукты и рыбий жир. В качестве антиоксидан-
та витамин А, очевидно, не столь важен, как витамины С и Е.
Каротины - группа структурно подобных пигментов растений.
Каротины определяют желтый и оранжевый цвет растений, лучший
пример - морковь, которая богата α-, β- и γ-каротином. Некоторые
каротины, особенно β-каротин, в организме могут быть
преобразованы в витамин А. Многие каротины и каротиноиды - мощные ан-
371
тиоксиданты. До сих пор исследования сосредотачивались главным
образом на β-каротине. Изучение профилактической ценности
пищевых добавок β-каротина дали смешанный результат. Малые дозы
бетта-каротина - ценный источник повышения активности
витамина А, особенно при низком потреблении последнего. Однако анти-
оксидантная роль β-каротина в организме была подвергнута
сомнению, потому что при некоторых обстоятельствах β-каротин может
фактически стимулировать формирование СР и окисление. Пока по
этому вопросу не будет получена более полная информация, мы
воздержимся рекомендовать пищевые добавки β-каротина в качестве
антиоксиданта.
Потенциальные побочные эффекты высоких доз. Витамин А имеет
относительно высокий потенциал токсичности. Признаки избытка витамина А -
сухая кожа, проблемы со зрением, ломота костей, головная боль и
раздражительность. У детей кожные и костные симптомы могут сохраняется даже после
того как прием добавки был прекращен. У взрослых токсичность витамина А
наблюдалась после регулярного приема более чем 50 тыс. Ш в день. Порог
токсичности у детей может быть значительно ниже.
Предостережение: Описаны случаи, когда у новорожденных
наблюдались дефекты, если в течение беременности их матери
принимали витамин Л в дозах 25-50 тыс. IU в день.
23.1.9. Полифенолы зеленого чая
Зеленый чай-один из наиболее популярных и доступных
источников пищевых антиоксидантов. Он богат полифенолами (класса
флавоноидов) с очень высокой антиоксидантной активностью.
Обильное потребление зеленого чая весьма полезно для здоровья.
Например, он снижает риск некоторых раковых образований,
защищает печень от токсинов и предотвращает болезни сердца, зубной
кариес и воспаление десен.
По сравнению с большинством других растений зеленый чай
чрезвычайно хорошо изучен - буквально сотни исследований на
животных и множество - на людях. На различных моделях рака
животных было показано, что экстракт зеленого чая ингибировал все
стадии канцерогенеза и даже частично приводил к регрессии
некоторых опухолей (ιи et al., 1992; Serqfini et al., 1996). В частности,
зеленый чай блокировал появление мутаций в ДНК, которые возникают
под действием различных доз токсинов, а также при
ультрафиолетовом облучении {Ви-Abbas et al., 1994). В большинстве исследований
на людях пытались выявить корреляцию между приемом зеленого
чая и развитием рака в данной популяции. Результаты таких
исследований являются косвенными и не вполне надежными, потому что
во внимание должно быть принято слишком много переменных. По-
видимому, обильный прием зеленого чая уменьшает риск возникно-
372
вения по крайней мере некоторых раковых образований, в
частности толстого кишечника, желудка, поджелудочной железы и
пищевода. Эффекты зеленого чая могут быть особенно значительны для
людей с повышенным риском заболевания раком. Крайне
интересные результаты были получены корейскими исследователями,
изучавшими эффекты потребления зеленого чая и кофе на частоту
обмена сестринской хроматиды (OCX) у курильщиков. OCX - тип
мутаций, который коррелирует с риском рака. Курильщики имели
более высокую частоту OCX, чем некурящие (9.46 против 7.03).
Однако курильщики, употреблявшие 2-3 чаши зеленого чая в день в
течение полугода, имели частоту OCX только 7.93. Напротив,
эквивалентное потребление кофе не уменьшало частоту OCX
(Todorov Я. etal., 2001).
Ряд исследований указывает на пользу зеленого чая при
сердечно-сосудистых заболеваниях. Было также показано, что зеленый
чай ингибирует окисление ЛНП и быстрый рост гладких мышц
сосудов, двух важных факторов в прогрессии атеросклероза.
Другой замечательный эффект зеленого чая - его способность
ингибировать рост бактерий в полости рта и улучшать кислотоус-
тойчивость зубной эмали. Эти эффекты предотвращают
воспаление десен и зубной кариес (Serafini et al., 1996).
Недавно было показано, что токсичность β-амилоидного белка
(бета А) может быть понижена полифенолами зеленого чая,
которые сокращают ПОЛ, вызванное АФК. Защитное действие
полифенолов, по-видимому, независимо от регуляции активности
способствующих апоптозу белков (р53, Вах и циклооксигеназы). Защитные
эффекты против апоптоза нейронов, индуцированного бета А,
связан с нейтрализацией АФК и, очевидно, имеют перспективы для
предотвращения болезни Альцгеймера {Choi et al., 2001).
Мы должны подчеркнуть различие между зеленым и черным
чаем. Свежая заварка, естественно, содержит много
полифенолов, соединений, ответственных за большую часть
положительных эффектов чая или его экстрактов. Зеленый чай
изготавливается таким способом, что полифенолы сохранены. Напротив,
технология производства черного чая сопряжена с окислением
полифенолов. В результате, черный чай содержит
приблизительно в шесть раз меньше активных полифенолов, нежели чай
зеленый.
Большинство свидетельств указывает, что польза здоровью
становится ощутимой, если употреблять по крайней мере от 8 до 10
чашек зеленого чая в день. (Люди с повышенным риском рака, в
частности, курильщики, могут, по-видимому, извлекать пользу даже из
таких небольших количеств, как 2-3 чашки в день.) Необходимо
иметь в виду, что любой чай, зеленый или черный, содержит
кофеин. Кофеина в коммерческих экстрактах зеленого чая обычно
немного.
373
23.1.10. Ко-энзим Q10
Ко-энзим Q10 (CoQIO) играет две важные роли: во-первых, он -
существенная часть клеточной системы дыхания, локализованной в
митохондриях, и, во-вторых, - хороший антиоксидант. CoQIO
повышает скорость и улучшает эффективность энергетической
продуктивности, и, наряду с этим, защищает митохондрии от СР (Rosenfeldt
et al., 2002). Организм способен производить CoQIO, но много
факторов, включая возраст, болезни, понижающие холестерин
лекарства и недоедание, могут вредить этой способности. CoQIO иногда
называется "биологическим маркером старения" потому, что его
уровень хорошо коррелирует со старением и дегенеративными
болезнями. Было показано, что пищевые добавки с CoQIO увеличивали
продолжительность жизни мышей на 50% (Murray et al., 1991). Большое
количество исследований ясно продемонстрировало эффективность
CoQIO при сердечной недостаточности застойного характера и
других болезнях сердечной мышцы. Другие патологии, при которых
CoQIO оказывает благоприятное действие, включают гипертонию,
пониженный иммунитет и мышечную атрофию (Modern Nutrition in
Health and Disease, 1994). Установлено, что при лечении упомянутых
болезней CoQIO эффективен в дозах от 100 до 300 мг в день. Для
здоровых людей рекомендуется доза в 30-100 мг в день.
23.1.11. Мел атонин
Мелатонин - гормон, производимый шишковидной железой,
расположенной в мозгу. Напряду с другими его свойствами, он
также и антиоксидант. Мы подробно обсуждаем физиологию,
биохимию и использование мелатонина в гл. 5 и 26.
Как видно из приведенного выше материал а, существует
множество природных веществ с антиоксидантными свойствами. Нужно ли
принимать все эти препараты в качестве пищевых добавок? Или
лучше получить все эти активные компоненты с пищей? Или
существует оптимальная комбинация отобранных питательных веществ
для полной антиоксидантной защиты в интересах здоровья и
долголетия? К сожалению, в настоящее время нет окончательного ответа
на эти вопросы. Большинство исследований по влиянию антиокси-
дантов на здоровье было выполнено на животных. Хотя некоторые
их результаты могут быть в какой-то мере экстраполированы на
людей, конкретные рекомендации должны основываться на
долговременных исследованиях, проведенных на людях. А таких
исследований пока относительно мало и сосредоточено больше всего на ка-
374
ком-то отдельном питательном веществе. Такой подход уменьшает
число переменных и возможный спектр положительных
результатов, напротив, исследование большего количества питательных
веществ с антиокислительными свойствами (АО-свойствами)
открывает значительные возможности для создания их наилучших
комбинаций для достижения максимальной пользы. К сожалению,
современные рекомендации экспертов относительно доз и комбинаций
пищевых продуктов и пищевых добавок для достижения антиокси
дантных эффектов основаны на косвенных свидетельствах и
приближенных оценках. Следовательно, сложность выбора до
некоторой степени ложится на потребителя. Тем не менее, опираясь на всю
совокупность данных по этой проблеме, мы все же предложим
следующие общие руководящие принципы.
• Самый безопасный и для многих наиболее приятный способ
получить множество антиоксидантов - это употреблять пищу с
высоким содержанием фруктов и овощей. Такое питание имеет
множество дополнительных выгод, так как фрукты и овощи богаты и
многочисленными другими полезными питательными веществами
и соединениями растительного происхождения. Разнообразие
антиоксидантов, найденных во фруктах и овощах, поможет защитить
все ткани и клеточные структуры от различных типов СР.
Возможно, ориентация на потребление изобилия фруктов и овощей
может показаться устаревшим советом, ведь еще в детстве многие
слышали его от своих бабушек. Однако эта рекомендация
поддерживается также и научными доказательствами. Многие
исследования последовательно демонстрировали, что диета с высоким
содержанием фруктов и овощей уменьшает риск дегенеративных
болезней, включая рак, болезни сердца, гипертонию и диабет.
Современные руководящие принципы Национального института рака и
Национальной академии наук США рекомендуют потреблять по
крайней мере 2-4 вида фруктов и 3-5 видов овощей в день.
• Замените ваш кофе, черный чай и всевозможные
прохладительные напитки зеленым чаем, но не забывайте при этом, что
зеленый чай также содержит кофеин.
• Если вы хотите принимать антиоксидантные пищевые добавки,
выберите одну (или несколько), растворимых в воде, и одну (или
несколько), жирорастворимых. Если вы решили принимать один
единственный антиоксидант, липоевая кислота может быть
наилучшим выбором, поскольку она сочетает в себе и тот, и другой
вид растворимости.
• Многие антиоксиданты обладают сродством с определенными
органами или другой полезной физиологической активностью. Если
у вас есть специфические проблемы со здоровьем, постарайтесь
выяснить существуют ли антиоксиданты, которые в клинических
375
исследованиях дали положительный эффект в ситуации,
аналогичной вашей. Например, исследования показали, что CoQIO
оказывает благоприятное воздействие при сердечной недостаточности и
гипертонии; экстракт Ginkge biloba улучшает память и замедляют
прогрессию болезни Альцгеймера и т.п. (см. выше, не только в
этой главе, но и по всей книге).
• Читайте как можно больше качественной информации о новых
исследованиях антиоксидантов.
Много интересной информации об антиоксидантной терапии
содержится в статье Т. Максимовой, опубликованной в журнале "Наука и
жизнь" (Максимова, 2001).
Глава 24
РЕВЕРСИЯ
"ЦЕНТРАЛЬНЫХ ЧАСОВ СТАРЕНИЯ"
Часы! Живет в нас бог, бесстрастный,
беспощадный.
Он пальцем нам грозит, бормочет: - Не забудь!
Когда придет к концу твой недалекий путь,
В тебя тупая боль вопьется пьявкой жадной.
Шарль Бодлер, "Цветы зла", 1860
Существует много свидетельств, что центральная программа
старения существует и локализована в мозгу, точнее в одной из
центральных областей мозга - гипоталамусе. Главная функция
гипоталамуса заключается в координации и поддержании весьма сложного
баланса биохимических и физиологических параметров организма -
гомеостаза. Другая важнейшая роль гипоталамуса -
программирование темпа развития организма от рождения до зрелости путем
постепенного изменения гомеостаза в направлении, которое
способствует процессам роста и репродукции. К сожалению, эти изменения
не останавливаются в зрелости, когда, казалось бы, цель
достигнута, но, по-видимому, продолжаются и далее, становясь таким
образом важным фактором старения и патогенеза возрастных болезней
(см. гл. 2). Другими словами, развивающаяся программа,
управляемая гипоталамусом, становится программой старения, т.е. де фак-
то - центральными часами старения (ЦЧС).
Как это часто бывает, в этой ситуации есть как негативная, так
и позитивная сторона. Негативная - в том, что эта центральная
программа, по-видимому, управляет или по крайней мере вносит вклад
в процесс старения, будучи естественным и имманентным
механизмом, присущим большинству, или, как минимум, млекопитающим
животным. Позитивная - в том, что мы уже имеем значительное
понимание того, как функционируют эти ЦЧС, и даже определенные
представления о том, как замедлить их работу. Пока самым
крупным достижением в понимании природы ЦЧС стала нейроэндокрин-
ная теория старения, развитая Владимиром Дильманом, видным
российским ученым и врачом (Дильман, 1987; Dilman, 1994). Будучи
лишь смелый гипотезой в 50-х, концепция ЦЧС стала хорошо
разработанной теорией, обоснованной большим количеством
исследований. Ключевое положение теории Дильмана состоит в том, что
гипоталамус постепенно теряет чувствительность к гормональным и
метаболическим сигналам обратной связи, поступающих от
периферических органов и эндокринных желез, что вызывает
существенные изменения в гомеостазе организма (мы обсуждали теорию
377
Таблица 20
Некоторые гомеостатические изменения, связанные с ЦЧС
Ключевые изменения
гомеостаза,
управляемые центральными
часами
Результат изменений
физиологических параметров
Возможные последствия
для здоровья
Ненормальная
утилизация энергии
Ненормальная
реакция на стресс
Ненормальное
регулирование
репродуктивной функции
• Нарушение толерантности
к глюкозе
• Высокий холестерин
крови, ЛНП и
триглицеридов
• Избыток инсулина
• Ухудшение регуляции
аппетита
• Чрезмерная секреция
стероидов стресса (кортизол)
в течение стресса
• Чрезмерная секреция
стероидов стресса в
отсутствие стрессоров
• Избыток стероидов
стресса усиливает
большинство упомянутых
выше негативных
изменений в
использовании энергии
Высокий уровень гормонов,
которые регулируют или
влияют на органы
размножения: ФСГ, ЛГ и
пролактин
Диабет,
атеросклероз, болезни сердца
инсульт, гипертония,
ожирение
Брюшное ожирение,
гипертония,
депрессия, нарушение
иммунной функции,
диабет,
атеросклероз,
болезни сердца,
инсульт, снижение
половой активности,
возможно
повышение риска
заболевания раком
Более раннее начало
менопаузы у
женщин; падение
уровня тестостерона
и гипертрофия
простаты у мужчин
В. Дильмана более подробно в гл. 2). Эти изменения в
чувствительности гипоталамуса действуют как механизм ЦЧС, определяющий
темпы развития организма и в последствии его старение.
Лучшим критерием правильности любой теории является
экспериментальная проверка предсказаний, сделанных на основе этой
теории. Теория Дильмана предсказывала, что, если бы чувствительность
гипоталамуса к сигналам обратной связи была восстановлена, "ход"
ЦЧС был бы обращен вспять, ведя к большей продолжительности
жизни и меньшему количеству возрастных болезней. Именно это и
наблюдалось в разнообразных исследованиях на животных. Когда
грызуны получали лекарства, которые повышают чувствительность
гипоталамуса к сигналам отрицательной обратной связи (например,
депренил, L-дофа), у этих животных наблюдалось увеличение про-
378
должительности жизни до 50% или даже больше. В настоящее время
неизвестно, вызовет ли такие же драматические эффекты у людей
улучшение чувствительности гипоталамуса с помощью фармаколо! и-
ческих агентов. Однако известно, что у людей с возрастом также
происходит потеря чувствительности гипоталамуса, и это означает, что
их механизм функционирования ЦЧС, по-видимому, такой же, как и у
других млекопитающих, включая крыс и мышей.
ЦЧС вызывают разнообразие изменения во многих системах
организма. Так, наиболее важными из них, если говорить о
продолжительности жизни и предотвращении болезней, являются изменения в
использовании организмом энергии, как в обычном состоянии, так и
при стрессе {табл. 20). Кроме того, ЦЧС, вероятно, частично
ответственны за возрастное падение уровня некоторых гормонов
"молодости", особенно гормона роста и дегидроэпиандростерона (ДГЭА).
Исходя из современного понимания этой проблемы, стратегия
обращения "хода" ЦЧС вспять должна быть основана на
восстановлении чувствительности гипоталамуса к сигналам отрицательной
обратной связи. Эти сигналы - уровень стероидов стресса, глюкозы,
половых гормонов в крови и ряд других параметров. Кроме того,
меры, которые способны изменить возрастные нарушения в
использовании энергии и стресс-ответа организма, уменьшат также и
негативное воздействие ЦЧС.
24.1. "ПАСПОРТ ЗДОРОВЬЯ"
Как уже упоминалось, ЦЧС - программа в гипоталамусе,
которая меняет "химию" нашего тела так, чтобы заставить его расти и
созревать, а позже вносит вклад в его старение. Если бы такая
программа не существовала, возрастное развитие рождающихся
организмов было бы невозможно. Проблема состоит в том, что эта
программа не "закрывается" в зрелости. Идеальный сценарий должен
был бы позволять "часам" делать их работу в первые 20-25 лет
жизни, а затем блокировать эту программу развития. Поскольку пока
еще не существует возможности полностью остановить ЦЧС в
зрелости, следует использовать способы замедления или даже
временного обращения их влияния. Однако прежде чем приступить к таким
действиям индивид должен иметь (хотя бы грубое) представление о
своем биологическом возрасте (не путать с хронологическим), т.е. о
том, как далеко ушли его (ее) "часы" от идеального пункта - от
возраста 20-25 лет. Этот тест покажет, что, возможно, еще нет никакой
потребности предпринимать что-либо, или, напротив, нельзя
откладывать меры по улучшению здоровья.
Поскольку механизм ЦЧС - постепенная потеря чувствительности
гипоталамуса к сигналам обратной связи, для оценки
"биологического времени" на ЦЧС, необходимо послать такой сигнал гипота-
379
ламусу и измерить величину его ответа. Это действительно было
исследовано на животных и в ряде случаев в клинической практике.
Например, дексаметазоновый тест используется, чтобы измерить
реактивность гипоталамуса к сигналу отрицательной обратной
связи от систем адаптации организма. Введение дексаметазона
моделирует высокий уровень стероидов, который отмечен при стрессе.
В норме (т.е. у молодых людей) дексаметазон заставил бы
гипоталамус отвечать понижением производства кортикотропин
регулирующего гормона (КРГ), который является активатором
стресс-ответа. В результате чрезмерное производство стероидов стресса
надпочечниками будет предотвращено. У пожилых людей, однако,
гипоталамус часто не в состоянии соответственно отвечать на
дексаметазон, показывая, что ЦЧС "отсчитали" уже много времени.
Измерение ответа гипоталамуса на дюжину или большее
количество сигналов обратной связи обычно обеспечивает адекватную оценку
"времени" на ЦЧС. К сожалению, этот подход непрактичен для
широкого применения. В качестве альтернативного способа оценки
состояния центральных часов, В. Дильман выдвигает концепцию "паспорта
здоровья" {Дильман, 1987). Он обнаружил, что набор нескольких
ключевых физиологических параметров, хорошо коррелирует со
"временем" на ЦЧС. Этот набор был назван "паспортом здоровья", потому
что "хороший" паспорт здоровья указывает на высокие шансы более
длительной жизни и свободы от дегенеративных болезней. Идеально
паспорт здоровья лучше всего определить в 20-25 летнем возрасте и
рассматривать его в дальнейшем в качестве нормы для данного
человека. Мониторинг изменений в паспорте здоровья указал бы насколько
"продвинулись" ЦЧС. Цель корректирующей стратегии состояла бы в
том, чтобы поддерживать или восстанавливать показатели
нормального паспорта здоровья, т.е. имевшие место в 25 лет.
В качестве основы паспорта здоровья Дильман предложил
контролировать следующие показатели.
• Вес и состав тела (% нежировой и жировой массы тела);
относительное распределение жира в талии и на бедрах (центральное и
периферийное распределение жира);
• Уровень холестерина, ЛНП, ЛВП и триглицеридов (профиль липи-
дов) в крови;
• Толерантность к принятой per os глюкозе с одновременным
измерением натощак уровня инсулина в течение 120 мин и степени гли-
козилирования гемоглобина (показатели метаболизма углеводов);
• Уровень тиреотропного гармона (ТТГ) и ТЗ в крови
(интенсивность метаболизма);
• Уровень фолликулостимулирующего (ФСГ) у женщин и
тестостерона у мужчин в крови (индикаторы репродуктивного аспекта
часов старения);
• Внутриартериальное давление крови.
380
Приведенные выше основные параметры могут быть легко
проверены у большинства людей. Отсутствие существенных изменений
в паспорте здоровья указывает, что ЦЧС "продвигаются" в
медленном темпе. Измерения основных параметров паспорта здоровья
могут быть сделаны под руководством практически любого врача.
Состав тела и распределение жировых отложений могут быть
определены как обычным врачом, так и специалистом по рациональному
питанию. Имеются и другие тесты, которые могут повысить
точность определения темпа ЦЧС, такие как уровень ИФР-1 и ДГЭА
сульфата в крови. Результаты всех испытаний сравниваются либо с
вашими собственными показателями, когда вам было 25, или, если
этого в наличии нет, то со среднестатистическими показателями для
25 летних людей. Чем больше различие, тем более позднее, к
сожалению, время на ваших ЦЧС.
24.2. КАК ЗАМЕДЛИТЬ ЦЧС С ПОМОЩЬЮ
РАЦИОНАЛЬНОГО ПИТАНИЯ И ОБРАЗА ЖИЗНИ?
Избегайте переедания. Потребление избытка калорий, которое
заканчивается ожирением, ускоряет старение несколькими путями.
Избыточный вес в особенности является важным фактором
ускорения ЦЧС, а ограничение потребления калорий замедляет их "ход".
По различным причинам, обсужденным ранее, такой подход не
может быть в полной мере использован человеком. Основанный на
сегодняшнем состоянии наших знаний и доступный для людей подход
заключается в том, что количество потребляемых калорий должно
быть совместимо с сохранением оптимального веса и состава тела,
присущего молодому возрасту (см. гл. 22).
Мелатонин. Существуют данные, указывающие, что мелатонин
может повышать чувствительность гипоталамуса к сигналам
отрицательной обратной связи, в особенности, к уровню кортикостерои-
дов и половых гормонов, и это означает, что мелатонин
способствует реверсии ЦЧС. Это может объяснять некоторые из многих
полезных эффектов этого гормона. (См. гл. 5, 26 с подробностями о
пользе мелатонина для здоровья.)
Физические упражнения. Физические упражнения не
затрагивают ЦЧС непосредственно. Однако они помогают нейтрализовать
некоторые негативные эффекты ЦЧС, которые сказываются на
использовании организмом энергии. В частности, упражнения
улучшают толерантность к углеводам, уменьшают избыток инсулина и
повышают уровень ЛВП.
Хром. Эффекты хрома подобны результатам физических
упражнений. Он также повышает чувствительность тканей к инсулину,
уменьшает содержание глюкозы в крови, выравнивает и улучшает
профиль липидов, нейтрализуя таким образом некоторые негатив-
381
ные метаболические эффекты ЦЧС. Хром увеличивает
продолжительность жизни у крыс. Физиологические эффекты хрома
обусловлены в значительной степени его способностью повышать
реактивность клеток к инсулину. Длительное культивирование пахотной
земли может исчерпать запасы хрома в почве и, следовательно,
уменьшать его содержание в пищевых продуктах. Кроме того,
большинство методов приготовления пищевых продуктов приводит к
потере до 80% его изначального содержания. Умеренный дефицит
хрома может не проявляться клинически, но это положение широко
распространено в Западных странах и может вносить вклад в
развитие инсулин-независимого диабета и других возрастных болезней.
Большинство солей хрома плохо всасывается в
желудочно-кишечном тракте и не имеют никакой биологической активности. Пи-
колинат и ншотинат хрома относительно недороги, хорошо
усваиваются организмом и могут использоваться как пищевые добавки.
Большинство экспертов рекомендует 200 мкг этих препаратов в
день. Большие дозы, особенно более 600 мкг в день, не
рекомендуются, поскольку могут быть ядовиты.
Сохраняйте высоким уровень нейромедиаторов. Нейромедиа-
торы - соединения, которые передают в мозгу сообщения между
нейронами. Эффективная связь между нейронами в мозгу требует
определенного уровня нейромедиаторов. Низкий уровень
нейромедиаторов связан с такими патологиями, как депрессия и болезнь
Паркинсона. ЦЧС локализованы в гипоталамусе, который так же
зависит от нейромедиаторов, как и остальные части центральной
нервной системы. Если в гипоталамусе уменьшится уровень
нейромедиаторов, то его чувствительность к сигналам обратной связи
также будет уменьшаться. Результат - ускорение "хода" ЦЧС
Исследования на животных показывают, что лечение,
повышающее уровень нейромедиаторов в гипоталамусе, увеличивает
продолжительность жизни и уменьшает риск возрастных болезней, и
наоборот - экспериментальное понижение уровня нейромедиаторов
вызывает противоположный эффект.
У людей наиболее распространенное состояние, связанное с
низким уровнем нейромедиаторов - депрессия, при которой в
нескольких областях мозга, включая гипоталамус, их уровень существенно
понижен. Такая ситуация, вероятно, ускоряет ЦЧС, а ослабление
депрессии, обычно поднимающее уровень нейромедиаторов в мозгу,
возможно, их замедляет. Интересно, что одним из наиболее часто
повторяющихся результатов при исследовании долгожителей был
высокий уровень эмоционального благополучия и превосходная
способность преодолевать трудности, т.е. - прямая
противоположность депрессии.
Один из путей дополнительной поставки нейромедиаторов в
мозг - достаточное обеспечение организма их предшественниками.
Например, аминокислоты фенылаланин и тирозин - предшествен-
382
ники для нейромедиаторов дофамина и норадреналина, тогда к ικ
триптофан и 5-гидрокситриптофан - для серотонина. Друтс
способы повышения уровня нейромедиаторов заключаются н бло
кировании вторичного поглощения нейромедиаторов нейронами и
ингибированием моноамин оксидазы, фермента который разрушае г
нейромедиаторы. Наряду с многочисленными лекарствами,
развивающими упомянутые положительные эффекты, вполне доступны
естественные соединения, которые приводят к подобным
результатам более мягким способом, так как имеют меньше побочных
эффектов. Мы детально обсуждаем способы повышения уровня
нейромедиаторов в мозгу в главах 7, 28 .
Противодействие стрессу или уход от него. Стресс - ведущий
фактор в ускорении темпа ЦЧС. Стресс-ответ "запускается"
гипоталамусом, т.е. той же самой частью мозга, в который
функционируют ЦЧС. Всякий раз, когда гипоталамус должен поддерживать
длительный стресс-ответ, его функция модулируется в сторону
понижения чувствительности к сигналам обратной связи, что, по существу,
является ускорением ЦЧС. Кроме того, большие количества СР,
произведенные стрессом, могут повреждать нейроны гипоталамуса,
еще более подталкивая вперед ЦЧС. Мы обсуждаем определенные
способы борьбы со стрессом в главах 25, 29.
24.3. МЕДИКАМЕНТЫ. КОТОРЫЕ МОГУТ ЗАМЕДЛЯТЬ ЦЧС
Множество рекомендуемых медикаментов способно повысить
чувствительность гипоталамуса к сигналам обратной связи и таким
образом замедлить ЦЧС. У животных, эти лекарства вызывают
существенное увеличение продолжительности жизни и отодвигают
развитие дегенеративных болезней. Хотя ни одно из этих лекарств
не было разработано в целях замедления старения, они успешно
использовались для лечения многих заболеваний. Некоторые врачи,
специализирующиеся в геронтологической медицине, регулярно
предписывают эти лекарства своим пациентам.
Наша общая рекомендация состоит в том, чтобы использовать
преимущественно естественные пищевые добавки для целей
антистарения, нежели синтетические лекарства. Главная причина
заключается в том, что вещества из естественных источников обычно
вызывают меньшее количество побочных эффектов. Однако
синтетические препараты могут в некоторых случаях оказывать более
глубокое позитивное влияние на процесс старения, нежели их естест
венные альтернативы. Мы полагаем, что в настоящее время
информации о терапии старения еще недостаточно, чтобы рекомендовать
широкое использование синтетических препаратов. Весьма
возможно, однако, что в будущем эти и другие лекарства станут
стандартной составной частью терапии антистарения.
383
Ниже мы обсуждаем несколько лекарств, которые имеют
достаточный потенциал, чтобы нейтрализовать влияние ЦЧС. Полагаем,
что человек, серьезно заинтересованный в продлении жизни, должен
знать весь набор средств, который может предложить современная
наука. Однако необходимо иметь в виду, что эти лекарства могут вызвать
серьезные побочные эффекты у людей с повышенной
чувствительностью или при неправильном использовании. Ни одно из этих лекарств
не должно использоваться без одобрения и внимательного наблюдения
врача, компетентного в геронтологии и гериатрии. Также необходимо
тщательно ознакомиться со всеми инструкциями, рекомендуемыми
дозировками, побочными эффектами, предостережениями и риском,
связанным с использованием этих лекарств. Некоторые люди обладают
более высоким риском побочных эффектов, чем другие. Ваш доктор
должен определить, является ли ваша медицинская история и текущее
физическое состояние совместимыми с использованием того или иного
лекарственного средства. Он должен также определить правильную
дозировку для вас и контролировать ход лечения.
L-депренил, Депренил - лекарственное средство с большим
потенциалом анти-старения. Оно увеличивает продолжительность
жизни лабораторных животных даже при лечении, которое
начинается на поздних этапах жизни. В одном исследовании депренил,
введенный старым крысам, увеличил продолжительность их жизни на
34% {Oreland etal., 1986).
Депренил, по-видимому, воздействует на несколько механизмов
старения сразу. Во-первых, он действует, ингибируя в мозгу
фермент моноамин оксидазу В, что реализуется в более высоком
уровне нейромедиаторов в гипоталамусе, и таким образом обратно
"перематывает" ЦЧС. Во-вторых, он стимулирует антиоксидантную
защиту в мозгу, увеличивая синтез супероксиддисмутазы, ключевого
фермента, нейтрализующего свободные радикалы. Наконец,
депренил стимулирует синтез мелатонина шишковидной железой.
Депренил увеличивает как продолжительность жизни, так и
функциональную активность пациентов с болезнью Паркинсона.
L-дофа - предшественник нейромедиаторов дофамина и норад-
реналина. L-дофа подобен депренилу, который замедляет "ход*
ЦЧС, поднимая уровни этих нейромедиаторов в гипоталамусе.
L-дофа заметно увеличивает продолжительность жизни лабораторных
животных и восстанавливает репродуктивную функцию у старых
самок крыс. L-дофа - также эффективно стимулирует механизм
продукции гормона роста. Так, 0.5 г L-дофа увеличил уровень гармона
роста (ГР) у здоровых людей старше 60 до уровня, свойственного
молодым взрослым (Klatz et al., 1997). Во многих случаях L-дофа
улучшает состав тела, увеличивая мышечную и сокращая жировую
массу тела, повышает выносливость и двигательную координацию,
а также является эффективным лекарственным средством при
болезни Паркинсона
384
Hydergine - лекарственное средство, обычно используемое при
хроническом возрастном снижении когнитивной (познавательной)
функции. Эти нарушения часто приписывают болезни Альцгеймера
и другим типам деменции. Hydergine оказывает разнообразные
влияния на мозг, включая стимуляцию белкового синтеза, увеличивая
поставку кислорода и формирование межнейронных связей. Это
лекарственное средство, по-видимому, работает, стимулируя
производство нейромедиаторов - дофамина и норадреналина. Подобно
L-дофа, hydergine стимулирует выделение гормона роста у людей
всех возрастов. В нескольких исследованиях было показано, что
hydergine улучшает обучаемость, память, познавательные
способности и настроение. По-видимому, hydergine также повышает
чувствительность гипоталамуса к сигналам обратной связи, т.е.
способствует замедлению ЦЧС.
Phenytoin (Dilantin) оказался полезным, как было показано, при
многих состояниях, включая эпилепсию, неврологические боли,
депрессию, чувство беспокойства, язвы желудка, астму, снижение
умственных способностей и даже облысение. Механизм его действия
еще не понят полностью, но, возможно, связан со способностью
phenytoin стабилизировать мембраны нейронов и таким образом
балансировать деятельность ЦНС.
Существуют данные, что phenytoin помогает обратной
"перемотке" ЦЧС путем повышения чувствительности гипоталамуса к
сигналам обратной связи, и это ведет к обмену веществ и
гормональному балансу, которые более присущи юному возрасту. В частности,
phenytoin повышает уровень гормона роста, понижает уровень
стероидов стресса и гонадотропинов, а также уменьшает избыток
инсулина. Грызуны, которые подвергались воздействию phenytoin,
жили дольше и имели более длинную репродуктивную фазу. У
некоторых самок в состоянии менопаузы phenytoin восстановил
репродуктивную функцию.
Глава 25
"ОТДОХНОВЕНИЕ" ДУШИ И ТЕЛА
(Подходы к смягчению стресс- ответа)
Горные вершины
Спят во тьме ночной;
Тихие долины
Полны свежей мглой;
Не пылит дорога.
Не дрожат листы...
Подожди немного,
Отдохнешь и ты.
Вольфганг Гёте
Пер. М. Лермонтова
Мы уже неоднократно подчеркивали разрушительное влияние
хронического стресса на здоровье, а также тесную взаимосвязь
между стрессом и старением. Мы также обсуждали, какие
физиологические и биохимические изменения способствуют повышенной
устойчивости организма в стрессогенной ситуации. В этой главе мы
суммируем практические пути для снижения или нейтрализации
вредоносных влияний, которые могут быть вызваны стрессом. Они
включают как уход от стресса, так и уменьшение его эффектов,
когда избежать стресса не удается.
Изменение отношения к стрессогенным ситуациям, В
большинстве случаев, психологический стресс развивается под влиянием
факторов, которые воспринимаются нами как негативные, не
будучи в действительности таковыми. Если вы не можете изменить
стрессогенную ситуацию, то самая "здоровая" стратегия состоит в
том, чтобы изменить ваше отношение к ней, что облегчит вашему
организму "отказ" от стресс-ответа. Если вы все же можете
предпринять конструктивные действия для изменения ситуации -
сделайте это. Даже выполнение каких-либо относительно простых
действий для ее изменения может быть достаточным, чтобы уменьшить
(или снять) стресс. В качестве альтернативы попытка найти
положительную точку зрения на стрессогенную ситуацию также
способна дать благоприятный результат.
Biofeedback (BFB) - метод, предназначенный для обучения
пациента некоторой степени контроля над вегетативными функциями
тела. Компьютер или другой электронный прибор помогают ему
визуализировать такие физиологические параметры, как кровяное
давление, частота сердечных сокращений или накожная температура.
С помощью BFB пациент постепенно научается, как вернуть
изменившиеся физиологические параметры в желательный диапазон.
Первоначально метод BFB считался типичной "альтернативной" те-
386
рапией, но теперь, когда множество исследований васпидскмп σιιιο-
вало его эффективность, он стал признанным направлением и
современной медицине. BFB особенно эффективен для снижения
стресса и смягчения болезней, которые усиливаются стрессом,
например мигрени и синдрома хронического утомления. Стресс,
особенно его начальная фаза, связан с быстрыми сердечными сокращу
ниями, повышенным кровяным давлением и охлаждением
конечностей. Использование BFB для возвращения этих параметров к
норме, приводило к снижению интенсивности стресс-ответа и
уменьшению выделения гормонов стресса. Многие психиатры и
психотерапевты практикуют использование BFB в своих клиниках. В домашних
условиях также возможно использование подобных устройств.
Реверсия центральных часов старения. С возрастом
стресс-ответ становится чрезмерным, т.е. более интенсивным и длительным,
чем это необходимо. Часто организм в избытке выделяет гормоны
стресса даже в отсутствие стрессогенных факторов. Многие
пожилые люди могут находиться в состоянии хронического стресса даже
без малейшего повода для этого. Эта ситуация, по-видимому,
^является следствием изменений в ЦЧС, локализованных в гипоталамусе.
Вероятно, что замедление или даже реверсия ЦЧС могут уменьшать
избыток гормонов стресса в организме (см. гл. 24).
Физические упражнения. Польза от умеренных физических
упражнений включает снижение интенсивности стресс-реакций
организма. Упражнения имеют тенденцию уменьшать интенсивность и
продолжительность стресс-ответа также и при психоэмоциональных
стрессогенных ситуациях. Хорошая утренняя разминка -
эффективный способ преодолеть напряженность предстоящего дня,
наполненного стрессогенными ситуациями. С оздоровительной точки
зрения наибольшую пользу может принести последовательная
программа регулярных физических упражнений умеренной
интенсивности. Однако следует иметь в виду, что упражнения чрезмерной
интенсивности сами по себе могут вызвать существенный стресс, что
может скорее нанести ущерб, чем принести пользу здоровью.
Восстановление уровня гормонов молодости. Имеются
определенные указания на то, что снижение уровня некоторых гормонов,
особенно гормона роста, мелатонина и ДГЭА, вносит вклад в
возрастное ослабление устойчивости организма к стрессу. Все эти
гормоны уменьшают разрушительные эффекты стресса на иммунную
систему. Кроме того, гормон роста защищает организм от потери
нежировой массы тела, вызываемой стрессом, а мелатонин,
по-видимому, улучшает реактивность гипоталамуса к сигналам, которые
выключают стресс-ответ. Шаги, помогающие восстановить уровень
jthx гормонов, присущий молодому возрасту, обсуждены в главе 26.
Снятие депрессии. Одним из диагностических признаков
депрессии является патологический стресс-ответ. При депрессии
организм часто не способен прекратить стресс-ответ даже если он боль-
387
ше не требуется, создавая таким образом состояние хронического
стресса Снятие или облегчение депрессии обычно полностью
изменяет ситуацию (см. гл. 28).
Адаптогены - группа веществ, которые увеличивают
способность организма приспосабливаться к неблагоприятным условиям,
а именно повышают устойчивость к острому и хроническому
стрессу и минимизируют связанные с ним повреждения. Глава 29
полностью посвящена полезным свойствам адаптогенов.
Антиоксиданты. Стресс вызывает резкое увеличение уровня
свободных радикалов в организме. Часть повреждений может быть
нейтрализована путем приема антиоксидантов (см. гл. 2,14).
Фактически, антиоксидантные пищевые добавки приносят больше пользы
при стрессе, чем при других обстоятельствах (гл. 23).
Витамины и минералы. Некоторые питательные вещества
оказываются исчерпанными или стремительно выводятся из организма
при развитии серьезного стресса. Высокий уровень гормонов
стресса способствует увеличению выделения калия и задержке натрия в
организме. Баланс обычно можно сохранить диетой с высоким
содержанием калия и низким - натрия. Овощи и фрукты -
превосходные источники калия при низком содержании натрия.
Витамин С, пантотеновая кислота и витамин В6 важны для
нормальной функции надпочечников, которые несут повышенную
нагрузку в течение стресса. При серьезном стрессе эти питательные
вещества должны быть использованы в количествах более высоких,
чем это рекомендуется RDA. Для большинства людей пищевые
добавки с указанными витаминами в количестве 2 χ RDA - вполне
достаточны. Очень большие дозы В6 (в десятки раз превышающие
RDA и принимаемые в течение нескольких месяцев) могут
приводить к повреждению нервной системы и, следовательно, должны
быть исключены.
Методы расслабления. Надлежащее расслабление повышает
порог, при котором организм развивал бы стресс-ответ под
воздействием различных неблагоприятных в психологическом отношении
ситуаций. Оно также может способствовать более быстрому спаду
уже возникшего стресс-ответа. Эффекты расслабления
противоположны тем, которые наблюдаются в начальной фазе стресса.
Расслабление уменьшает частоту сердцебиений, кровяное давление и
потоотделение, но увеличивает ток крови к внутренним органам и
улучшает секрецию пищеварительных соков; содержание сахара в
крови имеет тенденцию возвращаться к нормальному уровню.
Среди многочисленных методов расслабления следует выделить
наиболее эффективные. BFB, обсужденный выше, является, по
существу, эффективным методом расслабления. Выбор наилучшего
метода расслабления очень индивидуален и сопряжен с путем "проб
и ошибок". Расширенное обсуждение методов расслабления не
позволяет ограниченный объем этой книги. Тем не менее многие мето-
388
ды расслабления при стрессе детально описаны и вполне доыуннм.
Один из популярных методов расслабления - развитие ιιροι рессиру
ющей релаксации, основанной на чередовании напряжения и ρ ι с
слабления. Большинство людей обычно не осознает, находя ι см ли
они в состоянии напряжения или релаксации. Начальная цель j гой
техники состоит в попытке научить человека почувствовать рас
слабление своих мышц по сравнению с напряжением. Мышцы вна
чале сильно напрягаются в течение нескольких секунд, а затем рас
слабляются. Этот процесс должен охватить все мышцы тела и в
конечном счете закончиться состоянием глубокого расслабления.
Процедура обычно начинается с мышц лица и шеи и затем
продолжается вниз по телу. Некоторые люди могут лучше расслабляться,
воображая, что их конечности становятся тяжелыми и теплыми.
Другие могут легко расслабиться, воображая себя, например,
отдыхающими на берегу спокойного теплого моря на каком-либо
отдаленном тропическом острове. Большинство экспертов рекомендует,
по крайней мере 10 мин, каждый день заниматься расслаблением.
Активация стресс-лимитирующих систем. Организм имеет
множество "встроенных" механизмов, которые сокращают стресс-
ответ и ограничивают его повреждающее действие. Мы подробно
обсудили функционирование этих систем в гл. 21.
Эти системы могут быть активированы некоторыми
питательными веществами, которые, в частности, стимулируют выделение
гормона роста, снижают разрушительные эффекты стресса на
нежировые ткани и иммунную систему (об их влиянии на выделение
ГР см. гл. 26).
Стресс-ответ также сопровождается выделением энкефали-
нов и эндорфинов, морфий-подобных субстанций, которые
продуцируются организмом. Энкефалины и эндорфины также
способны ограничить повреждения, индуцируемые стрессом.
Аминокислота DL-фенилаланин увеличивает уровень энкефалинов, ин-
гибируя действие разрушающего их фермента, что по-видимому,
особенно эффективно в облегчении стресса, связанного с болью
и депрессией (см. гл. 15).
Глава 26
ВОССТАНОВЛЕНИЕ УРОВНЯ
"ГОРМОНОВ МОЛОДОСТИ"
В главе 5 уже обсуждалось, что старение связано с
прогрессирующими измене«иями в содержании многих важных гормонов.
Долгое время считали, что гормональные изменения - исключительно
следствие старения. Однако оказалось, что некоторые,
гормональные изменения, наоборот, могут быть существенными факторами
старения, и реверсия этих изменений могла бы противодействовать
процессу старения.
Содержание трех главных гормонов - гормона роста (ГР), ме·
латонина и дегидроэпиандростерона (ДГЭА) - значительно падает
с возрастом. Именно низкий уровень этих гормонов, как
указывалось ранее, может вносить вклад в процесс старения и развитие
дегенеративных болезней. Многочисленные эксперименты на
животных и некоторые кратковременные исследования людей
указывают, что восстановление уровня ГР, мелатонина и ДГЭА, присущего
молодым людям, может способствовать замедлению процесса
старения. К сожалению, долговременная заместительная терапия ни
одним из этих гормонов еще не проводилась.
Ситуация с гормоном роста, мелатонином, ДГЭА напоминает
ту, которая имела место тридцать лет назад с заместительной эстро-
геновой терапией женщин в период после менопаузы. Теперь мы
знаем, что эстрогеновая терапия в этот период обеспечивает
женщинам небольшое, но существенное увеличение продолжительности
жизни и уменьшает риск заболеваний сердца и остеопороза. Но в то
же время несколько увеличивается и риск рака молочной железы,
поэтому женщины с персональной или семейной историей этой
болезни должны, скорее всего, избегать эстрогеновой терапии. Судьба
окончательного "приговора" широкому практическому
использованию ГР, мелатонина и ДГЭА может оказаться даже более сложной.
В настоящее время, возможная польза от этих гормонов
антистарения - многообещающее предположение.
Значит ли сказанное, что каждый должен ждать 30 лет прежде
чем мы придем к окончательной оценке этих возможностей? Не
обязательно, но решение о восстановлении "молодого" уровня клю-
390
чевых гормонов в значительной степени остается вопросом личной
философии. Некоторые врачи активно рекомендуют многим споим
пациентам ГР, ДГЭА и мелатонин, другие делают это ино1да и
очень осторожно, все остальные предпочитают ждать до гех пор,
пока проблема не прояснится полностью. Многие из нас занимаю ι*
ся самолечением под влиянием средств массовой информации,
друзей или знакомства с научными исследованиями. Если вы все же
решились на заместительную гормональную терапию, вы должны
сначала обсудить это с вашим врачом и удостовериться, что у вас нет
каких-либо осложняющих факторов, которые могли бы повысить
риск неблагоприятных побочных эффектов.
26.1. ГОРМОН РОСТА
26.1.1. Возможная польза, побочные эффекты
и общие предосторожности
У животных ГР, как было показано, увеличивал
продолжительность жизни и задерживал развитие возрастных болезней.
Исследования, которые удалось провести на людях, указывают, что
восстановление "молодого" уровня ГР может обеспечить укрепление
здоровья и продление жизни в результате названных ниже эффектов
(подробности см. в гл. 5).
• Улучшения состава тела: потеря жировой ткани и увеличение
массы мышцы.
• Повышение иммунитета.
• Улучшение настроения, стремление к благополучию
• Улучшение функций сердечно-сосудистой системы, включая
пациентов с устойчивой сердечной недостаточностью.
■ Повышение физической активности.
• Лучшее заживление ран, более короткое пребывание в стапионаре
после хирургической операции
• Реверсия возрастной атрофии органов, включая сердце, почки,
печень, селезенку и кожу
При всей полезности, уже доказанной и еще недоказанной, к
выбору ГР-заместительной терапии нужно подходить со здравым
смыслом и разумной предосторожностью. Чрезмерные количества
ГР могут вызвать серьезные нарушения. Акромегалия - редкое
заболевание, которое развивается, если в организме производится
слишком много ГР. Признаки и симптомы акромегалии включают
разрастание суставов, патологическое утолщение кожи, огрубление
черт лица, распухание мягких тканей на руках и ногах, запястный
туннельный синдром, боли в суставах, отеки и развитие глубокого
хриплого голоса. Продолжительность жизни индивидуумов с акро-
391
мегалией не только не увеличивается, но даже сокращается.
Некоторые из этих побочных эффектов были отмечены у индивидуумов,
которым вводились высокие дозы человеческого гормоном роста,
произведенного методами генной инженерии. Такие побочные
эффекты могут возникать, если уровень ГР в результате такой
терапии становится значительно выше, чем у здоровых молодых людей.
Когда ГР восстанавливаются на уровне физиологической нормы для
молодых взрослых людей, побочные эффекты маловероятны, хотя
все еще возможны. В этом контексте крайне важно, чтобы у
человека, подвергающегося ГР-заместительной терапии,
контролировался уровень инсулиноподобного фактора роста (ИФР-1) - одного
из главных медиаторов действия ГР в организме. Дело в том, что
уровень ГР подвергается значительным колебаниям в течение
суток, так как выделяется в пульсирующем режиме. В результате
отдельное измерение ГР может и не отражать реальной картины.
Напротив, ИФР-1-уровень относительно устойчив и отражает общую
сумму ГР, произведенных в теле.
Имеются два альтернативных подхода к повышению уровня ГР.
Первый - инъекции раствора ГР. Преимущество этого подхода
состоит в том, что в организм надежно вводится известное количество
ГР. Неудобства - высокая стоимость, существенный риск побочных
эффектов и появление потребности делать инъекции регулярно.
Другой подход заключается в стимуляции гипофиза к выделению
большего количества ГР. Множество веществ может вызывать подобный
эффект. Преимущества - более низкая стоимость, более низкий риск
побочных эффектов и удобства перорального приема. Главный
недостаток этого подхода - высокая индивидуальная вариабельность
ответа на ГР-пусковые стимуляторы. Так, некоторые люди способны
выделить достаточно ГР, чтобы восстановить "уровень молодости",
другие же - только к небольшому увеличению производства ГР.
Кроме того, ответ гипофиза на некоторые ГР-пусковые стимуляторы с
возрастом может стать недостаточно эффективным.
Мы должны подчеркнуть, что терапия с инъекциями ГР или ГР-
пусковыми стимуляторами имеет смысл только тогда, когда
продукции ГР в организме недостаточно, т.е. уровень ИФР-1 низок, что
является обычным после 40 и почти универсально - после 60. Первые
шаги к повышению уровня ГР, особенно путем инъекций, должны
быть сделаны под наблюдением врача. Благоразумно прежде
проверить уровень ИФР-1 в крови, руководствуясь приведенными ниже
данными, и во время терапии и не упускать из виду возможные
побочные эффекты.
Возраст (годы) ИФР-1 сыворотки (иг/мл)
16-24 182-780
25-39 114-492
40-54 90-360
Более 55 71-290
392
Инъекции ΓΡι Гормон роста - белок. Когда белки поступают в
организм с пищей, они разрушаются пищеварительными
ферментами до небольших фрагментов - пептидов или аминокислот, которые
затем всасываются в пищеварительном тракте. Аналогичная судьба
постигает и ГР, и только незначительные его количества, если
таковые вообще имеются, неповрежденными попадают в кровоток.
В результате ГР, поступивший в организм перорально, не в
состоянии произвести какие-либо существенные физиологические
эффекты. Единственный способ доставить экзогенный ГР в кровяное
русло - инъекции. В настоящее время, оптимальные дозы ГР-терапии и
графики введения гормона - не вполне ясны.
Стремясь минимизировать риск побочных эффектов,
большинство экспертов рекомендует дозы ГР, которые повышают уровень
ИФР-1 до значений, характерных для 25-35-летних здоровых
людей. Согласно обычным рекомендациям (Klatz et al., 1997), курс
гормонотерапии должен начинаться с 0.5 Ш в день (Ш -
интернациональная единица гормона роста - 3 мг), ежемесячный прирост
начальной дозы должен составлять 0.1-0.25 Ш до тех пор, пока не
будет достигнут желательный уровень ИФР-1. Дозы более чем 2 Ш с
большой вероятностью могут вызвать заметные побочные
эффекты. Существующий опыт показывает, что частые инъекции низких
доз (два раза в день) более эффективны и вызывают меньшее
количество побочных эффектов, чем более редкие (два-четыре раза в
неделю) инъекции ГР в более высоких дозах. Действительно,
первый режим более соответствует продукции в организме
собственного ГР, которая характеризуется рядом коротких импульсов в
течение суток. Пожилые люди более чувствительны к ГР и для них
вообще требуется меньшее его количество, чтобы достичь
желательного уровня ИФР-1. Для большинства людей, старше 60 лет, 0.5 Ш в
день - вполне достаточно.
В настоящее время стоимость ГР-тераттии от 10 до 20 тыс. долл. в год для
взрослого, что делает ее недоступной для большинства людей. Тем не менее, по
сообщениям масс-медиа, в 2000 г. в США "омолаживающей" ГР-терапией
пользовалось более 250 тыс. человек. В 2002 г. количество центров
омолаживающей медицины, использующей ГР, достигло в США 8 тыс. Цена этих
терапевтических услуг, вероятно, в ближайшем будущем снизится, поскольку "натиск"
фармацевтических компаний на этот прибыльный рынок усиливается, но,
очевидно, ГР никогда не станет таким же дешевым, как аспирин.
Другое ограничение для ГР-терапии заключается в том, что
собственная ГР-продукция организма контролируется
отрицательной обратной связью - ГР и ИФР-1 подавляют сигналы от
гипоталамуса к гипофизу, стимулирующие продукцию ГР. Человек
с пониженной, но существенной собственной ГР-секрецией
может потерять эту способность (по крайней мере на несколько
недель) после того, как он примет курс ГР-заместительной терапии.
Это препятствие можно частично обойти, периодически преры-
393
вая ГР-терапию на какое-то время и позволяя таким образом
восстановиться собственной ГР-секреции организма. Однако нет
никакого согласия относительно того, какой характер упомянутой
цикличности оптимален. Некоторые врачи рекомендуют
прекращать инъекции ГР в течение одной недели каждого месяца,
другие - одного дня каждой недели.
Реактивация собственного ГР-производства. Многие люди
могут пожинать плоды оздоровления и продления жизни от
восстановленного уровня ГР и ИФР-1 без чрезмерных трат и с низким
риском побочных эффектов. Этого можно достичь, стимулируя
гипофиз к более высокой продукции ГР. Множество соединений
природного и синтетического происхождения могут оказывать подобное
влияние. Как физические упражнения, так и изменения в диете
могут повысить эффективность ГР-пусковых стимуляторов. Ниже мы
приводим регламент повышения ГР. (За более полной информацией
обращайтесь к книге Dr. Ronald Klatz, Carol Kahn "Grow Young with
HGH" (1997) или ее русскому переводу: Клац Р., Кап К. Остановите
болезнь старения. Минск: Попурри, 2000.)
26.1.2. Питательные вещества,
стимулирующие выделение гормона роста
Вещества, которые повышают выделение ГР, их часто называ
ют ГР-пусковыми стимуляторами, действуют на гипофиз, и многие
из них - питательные вещества, которые могут использоваться
индивидуально или в комбинации. С возрастом способность отвечать
на эти стимуляторы несколько уменьшается. Тем не менее
существенное увеличение в ГР-секреции может быть достигнуто даже у
очень пожилых людей.
Аргинин. Лучшим из известных ГР-пусковых стимуляторов
является аргинин - важная аминокислота, которую организм не может
синтезировать и должен получать с пищей. Внутривенное введение
15-30 граммов аргинина - эндокринологический тест для испытания
гипофиза на способность выделять гормон роста. Предполагается,
что действие аргинина как стимулятора ГР связано с его
способностью блокировать секрецию соматостатина - ингибитора синтеза
ГР Индивидуальная реакция на аргинин варьирует весьма
значительно - от отсутствия какого-либо эффекта до драматического
повышения ГР-секреции. По крайней мере некоторый ответ на
аргинин обычно сохраняется даже в глубокой старости.
Использование. Аргинин может использоваться как таковой
или в комбинации с другими ГР-пусковыми стимуляторами. Его
нужно принимать натощак за один час до физических упражнений
и перед сном. Обычный диапазон доз 1.5-5 г. Тошнота, понос и
расстройство кишечника могут случаться при более высоких дозах.
394
Люди с серьезной инфекцией герпеса не должны принимать -ιρι и·
нин, так как это может стимулировать размножение вирус·!.
Орнитин - незаменимая аминокислота, структурно ближш к
аргинину. Она может синтезироваться в организме из аргинина.
Орнитин действует подобно аргинину, но имеет несколько более
высокую активность. Индивидуальные ответы на орнитин изменяются
так же как и при приеме аргинина.
Использование. Дозировка и побочные эффекты те же самые,
что и при приеме аргинина. Орнитин следует принимать натощак и
перед сном.
Лизин - незаменимая аминокислота. Будучи принята одна,
практически не влияет на ГР-секрецию. Однако в комбинации с
аргинином значительно увеличивает его эффект. В исследовании Айсидо-
ри и его коллег (Isidori et al.) из университета в Риме, было
показано, что комбинация 1200 мг аргинина и 1200 мг лизина была в десять
раз более эффективна у молодых мужчин, чем аналогичная доза
одного аргинина. Ответ на комбинацию лизин - аргинин, по-видимому,
снижается с возрастом.
Использование. Один грамм натощак вместе с аргинином перед
физической нагрузкой и перед сном.
Глутамин - условно незаменимая аминокислота. В норме
организм синтезирует ее в достаточных количествах, но при стрессе не в
состоянии покрыть свои потребности в этой аминокислоте.
Глутамин - эффективный ГР-пусковой стимулятор. Существуют данные,
что с возрастом ответ на глутамин существенно не изменяется. Кроме
того, эта аминокислота вызывает много других полезных эффектов.
Это - важное питательное соединение для клеток кишечного тракта,
поддерживающее их оптимальное состояние. Глутамин нейтрализует
некоторые неблагоприятные эффекты стресса, в особенности
истощение мышц. Была найдена положительная корреляция между
низким уровнем глутамина и болезнью сердца, диабетом и артритом.
Использование. Два грамма перед сном. Считается, что более
высокие или низкие дозы менее эффективны.
Глицин. Эта аминокислота меньше изучена как ГР-стимулятор,
но по крайней мере два исследования указывают, что глицин
увеличивает ГР-секрецию, начиная с дозы всего в 250 мг.
Использование. От 250 мг до 6 г в день. Глицин - недорог и
обычно очень хорошо усваивается.
Ниацин - витамин В, встречается в двух главных формах:
никотиновой кислоты и ниацинамида. Среди многих его полезных
свойств - способность стимулировать выделение ГР. Ксантинол ни-
котинат - наиболее мощная форма никотиновой кислоты для
такой стимуляции, вероятно потому, что эта форма легко проходит
через гематоэнцефалический барьер.
Использование. 200-500 мг в день одного или в комбинации с
другими стимуляторами. Может быть причиной проходящего и без-
395
опасного прилива крови. Эта реакция может быть минимизирована,
если прием препарата начинать с 100 мг и постепенно увеличивать
дозу. Толерантность к приливам крови обычно развивается спустя
нескольких дней. Высокие дозы никотиновой кислоты (несколько
граммов), как известно, значительно улучшают профиль липидов,
понижая содержание ЛНП и триглицеридов и повышая уровень
ЛВГТ Однако высокие дозы никотиновой кислоты, особенно при
длительном приеме, в некоторых случаях вызывали повреждения
печени и гипергликемию.
Гамма-гидроксибутириловая кислота, или гамма-гидр осибу-
тират (ГГБ) является одновременно и метаболитом и
предшественником гамма-аминомасляной кислоты, или гамма-амибутира-
та (ГАМК). ГТБ и нейротрансмиттеры (вместе с другими
соединениями) действуют аддитивно в ограничении повреждений,
вызванных стрессом (см. гл. 21). В противоположность ГАМК ГГБ легко
пересекает гематоэнцефалический барьер и вызывает множество
реакций, включая выделение ГР. Фактически ГГБ - очень
эффективный ГР-пусковой стимулятор; его действие связано с
повышением уровня дофамина в гипоталамусе.
Использование. Низкие дозы ГТБ, 500-750 мкг, надежно
стимулируют выделение ГР без существенных побочных эффектов. ГТБ
можно объединять с другими ГР-пусковыми стимуляторами. В
высоких дозах ГГБ способен вызвать очень глубокий сон и судороги
мышц, а в очень высоких дозах - кому. Его нельзя принимать с
этиловым спиртом, антидепрессантами или транквилизаторами. ГГБ
был удален с рынка США решением FDA в 1990 г.
Объединение ГР-пусковых стимуляторов. ГР-стимуляторы
часто используются в комбинации, потому что эффекты часто
бывают аддитивными, а иногда и синергичными. Не проводилось
никаких систематических исследований для выяснения того, какая
комбинация и для какой возрастной группы является оптимальной. Од
на из популярных комбинаций: аргинин 1-2 г, орнитин 1-2 г; лизин
1 г, глутамин 2 г, никотиновая кислота (или ксантинол никотинат)
0.2-0.5 г. Рекомендуется начинать с более низких дозировок и
увеличивать их через какое-то время, чтобы минимизировать риск
побочных эффектов. На рынке уже присутствуют многие коммерческие
формулы для стимуляции ГР и регулярно появляются новые.
Эффективность большинства этих формул неясна из-за отсутствия клини
ческих исследований. Необходимо проверять содержат ли эти фор
мулы по крайней мере хотя бы часть упомянутых выше
стимуляторов ГР-секреции. Особого интереса заслуживает формула амино-
тропин-6, недавно выпущенная Gero Vita Iternational. Она включает
ГРОП (пептид, освобождающий гормон роста), аргинин, лизин,
ГАМК и ксантинол никотинат. ГРОП - ключевой компонент фор
мулы - представляет собой короткую цепь аминокислот, которая
моделирует действие гормона роста - регулирующего гормона
396
(ГР-РГ), т.е. гормона, который обычно стимулирует выделение ГР в
организме. В рекламных проспектах этой компании утверждюгсм,
что аминотропин-6 повышает ГР-секрецию на 50% без
неблагоприятных побочных эффектов.
ГР и диета. Люди с излишним весом или даже тучные имеют
более низкий уровень ГР и ИФР-1, а также менее чувствительны к I Р-
пусковым стимуляторам. Сокращение калорийности путем
потребления сбалансированной, богатой волокнами, обезжиренной пищи и
разумный регламент физических упражнений (как это обсуждалось
во многих главах) - наиболее важный образ действий для
увеличения ГР-секреции.
Некоторое данные указывают, что диета с высоким
содержанием белка может несколько увеличивать ГР-секрецию. С одной
стороны, поглощение больших количеств белка поднимает в крови
уровень аминокислот, включая и те, которые стимулируют гипофиз
к выделению ГР. С другой стороны, избыток белка повышает
нагрузку на печень и почки, двух органов, которые утилизируют
избыток аминокислот. Вообще 10-20% от суммы всех калорий должно
поступать от белка. Большинство людей должно стремиться к
потреблению белка в диапазоне 0.8-1.2 г/кг веса тела в сутки. Однако
энергичная физическая деятельность увеличивает потребность в
белке. Большинство экспертов полагает, что увеличение
потребления белка (около 25% калорий от белка или до 2 г/кг веса)
оправдано в соединении с интенсивной программой физических
упражнений. Для подъема потребления белков выберите пищевые
продукты, богатые белком высокого качества, но с низким содержанием
насыщенного жира, - рыбу, постную домашнюю птицу и сою
26.1.3. ГР и физические упражнения
Физические упражнения - известный способ стимулировать
гипофиз к секреции гормона роста. Механизмы этого эффекта
неясны. Наиболее вероятная гипотеза - интенсивная физическая
нагрузка стимулирует выделение ГР, поднимая уровень эндорфинов
(эндогенные морфино-подобные вещества со многими функциями в
организме). Стимулирующий эффект физических упражнений на
секрецию ГР снижается с возрастом, но может остаться существенным
даже у очень старых людей.
Продолжаются споры о том, какой тип физической нагрузки
является лучшим для повышения уровня ГР и ИФР-1. Большинство
экспертов соглашается, что комбинация физических упражнений
умеренной интенсивности (бег, быстрая ходьба или стационарный
велосипед) с умеренными или интенсивными тренировками с
гантелями обеспечивает оптимальные результаты для большинства
людей. Мы предлагаем следующие обшие руководящие принципы.
397
• Прежде чем начать программу упражнений проконсультируйтесь с
врачом и обстоятельно побеседуйте со специалистом по
спортивной медицине.
• Продолжительность и интенсивность упражнений повышайте
постепенно, не переходите к высокой интенсивности, пока вы не
обретете хорошую форму.
• Выполняйте упражнения, по крайней мере через день, в течение
получаса.
• Пейте много жидкости, так как выполнение упражнений может
привести к обезвоживанию организма.
• Упражнения высокой интенсивности должны выполняться, по
крайней мере первоначально, под наблюдением
квалифицированных профессионалов.
. 26.2. МЕЛАТОНИН
26.2.1. Возможная польза, побочные эффекты
и общие предосторожности
В последние годы мелатонин стал одной из наиболее
популярных пищевых добавок в США. У грызунов мелатонин продлевает
жизнь примерно до 20% и защищает от некоторых болезней и
стресса. Как и для гормона роста ДГЭА и большинства других
"субстанций молодости", долговременные исследования мелатонина на
людях - всего лишь хороший эскиз, но не тщательно прописанная
картина. Тем не менее существуют указания, что поддержание "
молодого" уровня мелатонина у людей среднего и особенно старого
возраста может принести ощутимую пользу их здоровью, включая
следующее (детальная информация в гл. 5).
• Улучшение функций иммунной системы.
• Замедление "центральных часов старения" в гипоталамусе.
• Защита органов и тканей, особенно мозга, от повреждений
свободными радикалами.
• Снижение риска злокачественного перерождения и ингибирование
роста опухолей.
• Повышение эффективности антираковой терапии (с дозами
мелатонина более высокими, чем физиологические).
• Повышенная устойчивость к стрессу.
• Улучшение сна.
Как мы уже упоминали, люди после 40 производят
приблизительно 1/3 от уровня мелатонина у 20-летних, а индивиды в возрасте
60 или старше производят его еще меньше. По-видимому, именно
такая возрастная динамика содержания мелатонина и обуславливает
более высокую чувствительность пожилых людей к этому гормону.
398
Мелатонин не имеет никакой токсичности, даже в дозах столь
высоких, как несколько граммов (распространенная дозировка -
таблетка с 3 мг мелатонина); он может быть полезен в некоторых
конкретных ситуациях, таких как нарушения сна, циркадного
ритма организма, изменение рабочей смены и серьезного стресса.
Кратковременное использование мелатонина с низкой
дозировкой, очевидно, совершенно безопасно. Насколько безопасен для
людей долговременный прием мелатонина еще предстоит
определить. Неблагоприятные побочные эффекты - обычно редки и
незначительны, но все же случаются, наиболее типичный -
сонливость. Мелатонин нельзя принимать вместе с седативными
средствами, транквилизаторами, алкоголем или психотропными
лекарствами.
Необходимо учитывать, что принимаемый препарат может
ингибировать в организме собственное производство мелатонина.
Выделение большинства гормонов регулируется механизмом
отрицательной обратной связи. Это означает, что высокий уровень
гормона противодействует его дальнейшему синтезу и
выделению в кровь независимо от того, является ли избыток
эндогенным или экзогенным. Однако вопрос - как долго и в каком
количестве экзогенный гормон ингибирует естественную продукцию
мелатонина - хорошо не изучался. Возможно, что длительный
прием мелатонина может вредить его эндогенному производству.
Но это, по-видимому, имеет место у людей до 40 лет, кто обычно
производит заметные количества мелатонина, но не у пожилых, у
которых его уровень крайне низок. Дети и молодые взрослые
имеют очень высокий уровень мелатонина и не должны его
принимать, если он специально не прописан врачом. Некоторые
другие группы людей также не должны принимать мелатонин.
26.2.2. Кто не должен принимать мелатонин
• Дети всех возрастов имеют высокий уровень собственного
мелатонина, который играет важную роль в стимуляции их развития.
Использование препарата может ингибировать собственную
способность производить мелатонин, что может лаже вызвать
преждевременное половое созревание.
• Беременные женщины и кормящие матери. Риск от приема
мелатонина беременными женщинами не изучался. Некоторое
количество мелатонина может попасть от кормящей матери к младенцу с
грудным молоком.
• Люди с аутоиммунными болезнями, такими как волчанка,
ревматоидный артрит или серьезная аллергия. Мелатонин
стимулирует иммунную систему и может усиливать эти патологии.
• Люди, принимающие противовоспалительные стероиды, такие
399
как преднизолон или другие иммунодепрессанты. Мелатонин
может противодействовать эффектам этих лекарств.
• Люди с раковыми образованиями в иммунной системе, такими
как лейкемия или лимфома. Мелатонин стимулирует иммунную
систему, и может усиливать эти заболевания.
• Женщины, стремящиеся к деторождению. У некоторых женщин
дозы мелатонина большие, чем 10 мг могут предотвращать овуляцию.
26.2.3. Повышение продукции мелатонина в организме
Начиная с половой зрелости уровень мелатонина устойчиво
снижается, становясь едва тестируемым после 60 лет. Можно ли
поддерживать или восстановить "молодой" уровень мелатонина,
стимулируя его собственное производство в организме, как это удается
сделать для уровня гормона роста? Ответ - да, но только до
некоторой степени. У человека средних лет различные модификации
образа жизни могут значительно увеличить уровень мелатонина,
позволяя пожинать потенциальную пользу от гормона без приема
экзогенного препарата. Однако после 60-ти прием мелатонина в
качестве пищевой добавки остается единственным способом для
достижения желательного уровня гормона.
Множество факторов и обстоятельств может ингибировать
продукцию мелатонина. Наиболее частая причина - потеря
естественного ритма смены "свет-темнота". Как уже обсуждалось в
главе 5, одна из ключевых функций мелатонина - регулировать
циклы "сон-бодрствование". Он выделяется вечером и стимулирует
сон. Естественная смена цикла "свет-темнота", т.е. дня и ночи,
благоприятствует нормальной секреции мелатонина. Наоборот, если
цикл освещенности нарушается, производство гормона снижается.
Как недостаток яркого света в течение дня, так и интенсивное
освещение ночью тормозят продукцию мелатонина. К сожалению,
многие из нас проводят большую часть дня на работе при тусклом
освещении, а затем дома поздние вечера проводят при ярком
освещении. Исследования показывают, что для оптимального цикла
"сон-бодрствование" необходимо побыть хотя бы один час в день
на открытом воздухе при хорошем дневном освещении. Согласно
некоторым оценкам, только примерно 5% взрослого населения
получает оптимальную световую экспозицию. Чтение около
лампы или созерцание окружающего мира из окна не могут заменить
наружного дневного освещения. Пробуйте провести время на
открытом воздухе в течение целого дня. Прогулка или бег трусцой в
парке, или спортивные состязания и игры на свежем воздухе могут
быть хорошей альтернативой занятиям в душном и сумрачном
спортивном зале. На солнце, однако, желательно пользоваться
солнцезащитным кремом и солнечными очками с УФ-защитой.
400
Таблица 21
Некоторые распространенные медикаменты и вещества,
ингнбирующие производство мелатонина
Класс
Препарат
Нестероидные противовоспалитель- Аспирин, ибупрофеи, индометацин
ные лекарства (NSAIDS)
Бета-блокаторы (используемые при Пропраналол, атенолол,
гипертонии и сердечной болезни) метопролол
Блокаторы кальциевых каналов Нифедипин, израдипин, дилитазем,
(используемые при гипертонии и филодипин, нимодипин, нитрен-
сердечиой болезни) дипин
Ингибиторы симпатической нервной Клонидин, резерпин
системы (используемые при
гипертонии)
Транквилизаторы Диазепам, флунитразепам, альпра-
золам
Другие Кофеин, табак, никотин, алкоголь
Достаточная световая экспозиция и адекватный сон ночью
одинаково важны для оптимального уровня мелатонина
Многие предписываемые врачами лекарства - другая общая
причина ухудшения собственной продукции мелатонина. Некоторые
чрезвычайно распространенные медикаменты - "враги" мелатонина: аспирин,
ибупрофен, β-блокаторы, блокаторы кальциевых каналов, диазепам (ва-
лиум) и альпразолам (ксанакс). Ни в коем случае не следует прекращать
лечения, предписанного вам врачом, даже с учетом возможного
негативного влияния этих лекарств на производство мелатонина. Возможные
последствия для здоровья могут быть гораздо более опасными, чем
низкий уровень мелатонина. Вы можете обсудить с врачом возможность
использования альтернативного лекарственного средства, которое не
затронуло бы ваш уровень мелатонина, или прибавить экзогенный мела-
тонин к вашей диете. Потребление кофеина, табака и алкоголя также
понижает уровень мелатонина (см. табл. 21). Помимо сохранения
продукции мелатонина, как правило, существуют и другие веские причины
для сокращения или отказа от одной или же ото всех этих привычек.
Стресс имеет двухсторонние отношения с уровнем мелатонина.
Мелатонин защищает от некоторых видов повреждений, вызванных
стрессом, тогда как стресс уменьшает его производство. Снижение
интенсивности стресс-ответа поможет сохранить оптимальный
уровень мелатонина. Отметим одно любопытное исследование, в
котором медитирующие женщины имели более высокий уровень
мелатонина, чем женщины в немедитирующей контрольной группе.
Если же стресс - неизбежен, экзогенный мелатонин может
способствовать минимизации повреждений (Reiter et aL, 1996).
401
26.2.4. Потребление экзогенного мелатонина
Мелатонин в качестве пищевой добавки относительно нов. Еще
не разработано никаких твердых руководств для его использования.
Доктор Рассел Рейтер, один из ведущих экспертов по исследованию
мелатонина и его использованию, рекомендует следующие
дозировки (дальнейшая информация может быть найдена в книге Reiter and
Robinson "Melatonin", 1996):
Использование: Диапазон доз:
Антистарение 0.1-3 мг, принимать перед сном
Сон 0.2-10мг, принимать перед сном
Нарушение ритма 1-10 мг, принимать перед сном по местному
"сон-бодрствование" времени
Изменение "рабочей 1-5 мг в начале "назначенного" сна
смены"
Стимуляция иммунитета 2-20 мг, согласно рекомендации вашего врача
Если иначе не определено врачом, мелатонин должен приниматься
один раз в день перед сном, чтобы соответствовать физиологическому
ритму организма. Имеется относительно много вариаций в
индивидуальных ответах на экзогенный мелатонин. После усвоения в кишечном
тракте, мелатонин проходит через печень, где часть его инактивирует-
ся. У некоторых людей печень инактивирует относительно большую
часть поглощенной дозы, чем у других. В результате - различия в
уровне мелатонина в крови у различных людей. У пожилых людей
метаболизм в печени проявляет тенденцию к замедлению и поэтому
меньшее количество мелатонина необходимо для того же самого эффекта.
Большинство экспертов соглашается, что для замедления
старения нужна такая доза мелатонина, которая приведет к его ночному
уровню у молодого взрослого человека (приблизительно
100-150 пикограммов/мл сыворотки крови). Некоторые
геронтологи и клиницисты предлагают тесты, которые позволяют измерять
уровень мелатонина ночью, что позволяет в конце концов "скроить"
дозу по вашим потребностям. Если такие тесты - невозможны,
благоразумно использовать маленькую дозу 0.1-0.2 мг (вероятно,
придется разделять большую таблетку).
26.3. ДЕГИДРОЭПИАНДРОСТЕРОН
26.3.1. Возможная польза,
побочные эффекты и предосторожности (ДГЭА)
Подобно гормону роста и мелатонину ДГЭА увеличивает
продолжительность жизни и предотвращает некоторые
дегенеративные болезни у лабораторных животных. Ограниченные
исследования на людях указывают, что восстановление уровня ДГЭА к тако-
402
вому у молодых взрослых людей может принести пользу для »доро-
вья и долголетия, включая (подробности см. в гл. 5) следующие.
• Улучшение настроения, чувство благополучия и, возможно, улуч
шение памяти и обучаемости.
• Умеренная защита от болезней сердца.
• Улучшение углеводного метаболизма и повышение чувствителг
ности к инсулину.
• Улучшение иммунитета, особенно эффективности прививок.
• Возможно, стимуляция сексуальной активности у людей с низким
уровнем ДГЭА; эффект, по всей вероятности, более выражен у женщин.
ДГЭА, по-видимому, относительно безвреден, особенно в
низких дозах и при кратковременном приеме. Тем не менее побочные
эффекты - возможны. Тем более, что долговременные
исследования действия гормона не проводились. Д-р Рей Шахелиан (Rey
Shahelian, 1996), бравший интервью у многих исследователей и
клиницистов об безопасности ДГЭА, вносит в список следующие
возможные побочные эффекты его больших доз.
• Прыщи и чрезмерно жирная кожа.
• Рост волос у женщинах (в нежелательных местах).
• Огрубление голоса.
• Раздражительность или частые смены настроения.
• Повышенная возбудимость или бессоница.
• Усталость или пониженный тонус.
Редкие побочные эффекты, возможно относящиеся к ДГЭА, -
головные боли, агрессивность и нарушения менструального цикла.
Побочные эффекты ДГЭА, за исключением огрубления голоса у
женщин, обычно быстро и полностью исчезают, как только лечение
прекращается или снижается доза. При дозе 25 мг в день или меньше,
любые побочные эффекты - редки. Однако у некоторых необычно
чувствительных индивидов побочные эффекты возможны даже при
таких низких дозах, как 5 или 10 мг. Другие, однако, могут принимать
100 или 200 мг ДГЭА в день без заметных проблем.
Некоторые ученые предполагают, что способность ДГЭА
увеличивать производство мужских и женских половых гормонов
может стимулировать рост гормоно-чувствительных опухолей, таких
как рак молочной железы у женщин и рак простаты у мужчин. В
настоящее время не имеется никаких данных, чтобы поддержать или
отрицать эту возможность.
26.3.2. Прием ДГЭА как пищевой добавки
Подобно мелатонину, ДГЭА приобретается без рецепта, ион-
относительно недорог. Отсутствуют какие-либо ясные
рекомендации о дозировках ДГЭА, потому что его широкое использование
403
людьми - сравнительно ново Большинство экспертов соглашается
с тем, что цель заключается в восстановлении, а не в превышении
уровня ДГЭА, свойственного молодым взрослым людям. Если пос
ле консультации с врачом решение принимать ДГЭА принято, то пе
ред лечением и в процессе регулировки дозы его уровень должен
измеряться, чтобы достичь, но не превысить "молодой" уровень
гормона. Большинство лабораторий предлагают тесты как на ДГЭА,
так и на ДГЭАф) (ДГЭА-сульфат, чей уровень более показателен
для характеристики общего количества продукции ДГЭА, см. гл. 5).
Хотя ДГЭА имеет тенденцию снижаться с возрастом, наблюдаются
существенные индивидуальные колебания. Не следует априори
полагать, что у любого молодого человека будет высокий уровень
ДГЭА (S), а у каждого пожилого индивида - низкий.
К сожалению, существует недостаточная корреляция между дозой
ДГЭА, принятой перорально, и его уровнем в крови. Когда препарат
принят перорально и всасывается в кишечном тракте, большинство
стероидных гормонов, включая ДГЭА, частично метаболирует в пече
ни перед поступлением к остальным органам и тканям тела. А именно,
часть дозы ДГЭА конвертируется печенью в эстрогены и андрогены, и
оставшаяся часть попадает в общее кровообращение. Какая доля
ДГЭА, наконец, поступит в кровообращение, значительно варьирует у
разных индивидов. По этой причине, большинство врачей, имеющих
опыт в ДГЭА-терапии, рекомендуют начинать с низких доз в 5 или
10 мг/день. Если необходимо, куратор вашего здоровья может
рекомендовать увеличить дозу на 5-10 мг каждые одну-две недели, пока
"молодой" уровень ДГЭА в крови или другие соответствующие
эффекты не будут достигнуты. ДГЭА обычно поступает на рынок в
капсулах или таблетках по 25 или 50 мг. Можно получить меньшую
дозу, просто открывая капсулу и разделяя ее содержание.
Естественный пик производства ДГЭА приходится на раннее
утро и затем его уровень падает в течение дня. Большинство врачей
рекомендует принимать ДГЭА утром, чтобы соответствовать этому
естественному ритму. Некоторые врачи рекомендуют 2/3 суточной
дозы утром, а остальную часть - вечером.
Для более полной информации о практическом использовании
ДГЭА-терапии, мы рекомендуем книгу Рея Шахилиана "ДГЭА:
Практическое руководство" (Rey Shahelian, "DHEA: A Practical
Guide", 1996).
26.3.3. Реактивация собственной продукции ДГЭА
Существуют ли способы повысить уровень ДГЭА, не принимая
экзогенного препарата? Множество путей может вести к
некоторому увеличению уровня ДГЭА, хотя скачок от очень низкого уровня
до уровня нормы (для молодого организма) - почти невозможен.
Лучший способ держать высокий уровень ДГЭА - избегать стресса,
404
вести спокойный образ жизни, пребывать в "хорошем настроении"
и, если удастся, обходиться без депрессии. И ДГЭАф) и ключевой
гормон стресса, кортизол, имеют общего предшественника - прегне-
нолон (см. рис. 13). Стресс имеет тенденцию переключать пути
метаболизма прегненолона на производство кортизола скорее, чем
AT3A(S), способствуя понижению уровня последнего.
Следовательно, чем меньшее стресса, тем больше ДГЭА. В частности,
регулярная медитация способствует улучшению уровня ДГЭА.
Как уже упоминалось, в одном исследовании было показано
значительное повышение уровня ДГЭАф) после лечения
медикаментами, которое снижали избыток инсулина и поддерживали
толерантность к глюкозе. Улучшение этих показателей другими методами,
например высоковолоконной диетой и физическими упражнениями,
также может поднимать уровень ДГЭА.
Несколько компании, производящих витамины, рекламируют
"сырой" препарат из ямса (Dioscorea gen.) как "естественную"
форму ДГЭА или "ДГЭА предшественник". В их рекламе обычно
указывается, что растительные стеролы такие как диосгенин,
найденные в диком ямсе, могут быть преобразованы организмом в ДГЭА.
Эти пищевые добавки, как полагают, даже лучше, чем препарат
ДГЭА, потому что в ДГЭА преобразуется столько растительных
предшественников, сколько их требуется организму.
Диосгенин и "родственные" стеролы могут быть
конвертированы в ДГЭА в лаборатории через приблизительно полдюжины
химических реакций. Нет вообще никаких свидетельств, что такие
преобразования могут происходить с растительными стеролами в
человеческом организме. По всей вероятности, дикий ямс не оказывает
существенного влияния на уровень ДГЭА или ДГЭАф) в организме.
Глава 27
АТЕРОСКЛЕРОЗ:
ВРАГ ОБЩЕСТВА НОМЕР ОДИН
Вопреки средствам массовой информации, враг общества
номер один в Западных странах - не терроризм, наркотики,
азартные игры, несчастные случаи или даже не рак. Это -
атеросклероз, и он намного опаснее остальных. В этих странах
атеросклероз ответствен за 35% всех смертных случаев, принимая во
внимание, что рак, враг общества номер два, прекращает
приблизительно 20% всех жизней.
Атеросклероз - формирование фиброзных жировых бляшек
(напоминающих опухолевидные структуры) на артериальных
стенах, которые в конечном счете ведут к частичной или полной
преграде току крови.
Атеросклероз может уменьшать или "отключать" кровоснаб
жение любого органа и таким образом повреждать или резко
понижать его функцию. Наиболее опасные для жизни осложнения
атеросклероза - болезни сердца и инсульт. Атеросклероз - также
наиболее частая причина импотенции, ампутации конечностей,
почечной недостаточности и многих других заболеваний.
"Коварство" природы атеросклероза заключается в том, что часто он
развивается абсолютно бессимптомно до тех пор, пока не
достигает весьма выраженной стадии. Иногда первым (и, увы,
последним) симптомом атеросклероза является фатальный сердечный
приступ или инсульт. Человек без симптомов
сердечно-сосудистых заболеваний никогда не может быть уверенным, что он
свободен от атеросклероза, и поэтому никогда не следует игнориро
вать меры, направленные на предотвращение или остановку его
прогрессии.
Атеросклероз - типичная дегенеративная болезнь, риск
получить которую резко увеличивается с возрастом. Возрастные
изменения метаболизма и гормонального баланса вносят существенный
вклад в его развитие. До менопаузы женщины меньше рискуют
получить атеросклероз, чем мужчины, из-за некоторого защитного
эффекта эстрогенов.
406
27.1. ГЛАВНЫЕ ФАКТОРЫ РИСКА,
ВЕДУЩИЕ К ЗАБОЛЕВАНИЮ АТЕРОСКЛЕРОЗОМ
• Высокий уровень ЛНП-холестерина.
• Возраст мужчин - 45 лет и старше.
• Возраст женщин - 55 лет и старше, или преждевременная
менопауза без эстрогеновой терапии.
• Гипертония.
• Высокий уровень гомоцистеина.
• Низкий уровень ЛВП- холестерина.
• Курение.
• Диабет или недостаточная толерантность к углеводам.
• Наследственная предрасположенность к ранним сердечным
заболеваниям.
Развитие атеросклеротических бляшек - довольно сложный
процесс, на который влияют многие факторы. Доминирующая
точка зрения состоит в том, что процесс начинается с микронарушений
внутренней поверхности (эндотелия) кровеносных сосудов. Эти
нарушения могут быть вызваны током крови, особенно, если имеет
место избыточное давление в пределах данного сосуда. Именно
поэтому развитие бляшек более вероятно там, где крупные артерии
разветвляются на более мелкие, и именно здесь риск развития
атеросклероза в результате гипертонии возрастает. Повреждения
изменяют свойства внутренней поверхности кровеносных сосудов таким
образом, что липопротеиды, циркулирующие в крови и богатые
холестерином, начинают накапливаться в поврежденной области,
формируя рыхлую жировую "полоску". Жировые полоски - первая
ступень атеросклероза, они могут развиваться уже в детстве. По
мере увеличения повреждения, моноциты (особые иммунные клетки)
и тромбоциты (клетки, связанные с процессом свертывания крови)
"прилипают" к поврежденной области и становятся причиной
дальнейшего роста и затвердевания бляшек. Кроме того, эти клетки
выделяют факторы роста, стимулирующие рост гладких мышц
сосудов, что также ведет к дальнейшему сужению сосудов. В
заключительной стадии, твердая волокнистая корка (состоящая из
коллагена, эластина, гликозаминогликана и отложений кальция)
формируется на поверхности бляшки. Холестерин и жир продолжают
накапливаться до тех пор, пока артерия не будет полностью блокирована.
Бляшки могут блокировать до 90% просвета артерии, когда уже
ясно проявляются симптомы атеросклероза.
Атеросклеротические бляшки могут расти с различной
скоростью или даже регрессировать в зависимости от нескольких
ключевых факторов.
• Профиль липидов. Наша кровь несет липиды (холестерин и жир),
упакованные вместе с молекулами белков в сферические частицы,
407
называемые липопротеидами. Два вида липопротеидов играют
важную роль в развитии атеросклероза, липопротеиды низкой
плотности и липопротеиды высокой плотности. Главная роль
ЛНП заключается в транспортировке холестерина к различным
тканям (холестерин требуется клеткам для создания мембран). К
сожалению, ЛНП, с одной стороны, могут быть "захвачены" также
и атеросклеротическими бляшками, что способствует их росту.
ЛВП, с другой стороны, удаляют избыточный холестерин из
тканей и транспортируют его к печени для последующей
рециркуляции. Таким образом, человек, обладающий большим количеством
ЛНП ("плохой холестерин") и меньшим количеством ЛВП
("хороший холестерин"), подвержен риску более быстрого роста бляшек,
и наоборот. Действительно, тестирование у пациентов уровня
ЛНП, так же как и отношения ЛНП/ЛВП - значительно лучшие
показатели сердечно-сосудистых патологий, чем уровень
тотального холестерина.
Согласно Американской ассоциации сердечных заболеваний,
желательный уровень для ЛНП - 130 мг/дл или меньше (100 мг/дл),
или еще меньше для людей с сердечно-сосудистой болезнью, и для
тотального холестерина - 200 мг/дл или меньше.
• Свободные радикалы. Скорость, с которой ЛНП поглощаются
бляшками, зависит не только от содержания ЛНП в кровотоке, но
также и от того, насколько "липкие" эти ЛНП. В литературе име
ются указания, что повреждения, вызванные свободными
радикалами, делают ЛНП более "липкими" и таким образом
потенциально более способными вызывать атеросклероз. Независимо от
причин формирования СР, будь то курение, стресс или дефицит
некоторых питательных веществ (особенно витаминов А, С, Е, селена,
цинка и меди), СР способствуют развитию атеросклероза, и необ
ходимо помнить, что антиоксиданты могут снизить это влияние.
• Скопление тромбоцитов. Тромбоциты - небольшие клетки,
участвующие в формировании кровяного сгустка при ранении. Они
скапливаются вместе и помогают формировать сгусток, который
блокируют истечение крови из раны. Если бы животные не имели
тромбоцитов, они погибали бы от кровопотерь. Но у этого
важного положительного свойства тромбоцитов имеются и негативные
стороны. Иногда тромбоциты могут формировать маленькие
сгустки даже тогда, когда нет никакого повреждения или оно крайне
невелико, например при микроповреждении внутренней
поверхности сосудов. Эти маленькие сгустки могут прилипать к атросклеро-
тическим бляшкам и сливаться с ними, становясь причиной их
роста. Чрезмерная склонность тромбоцитов к агрегации способствует,
таким образом, развитию атеросклероза и, напротив, снижение их
" взаимного притяжения " приводит к противоположным
результатам. Аспирин, например, может уменьшать агрегацию тромбоци-
408
тов. Именно поэтому врачи часто назначают пациентам этот
препарат при опасности сердечного приступа. Некоторые
питательные вещества, такие как омега-3 жирные кислоты, могут делать
то же самое, но с меньшим риском побочных эффектов.
• Гомоцистеин. При распаде белков аминокислота метионин
преобразуется в гомоцистеин, который в высоких концентрациях
способен повреждать клетки сосудистого эндотелия. Уровень гомоци-
стеина, присущий норме, как правило, безопасен, поскольку он
быстро трансформируется в другое нетоксичное соединение. Однако
у людей с дефицитом фолиевой кислоты и витамина В6
преобразование гомоцистеина замедлено, и его уровень может повышаться
Ряд данных настоятельно указывает на то, что это повышение
может вносить вклад в развитие атеросклероза.
• Пониженная толерантность к углеводам и диабет. Старение
связано со снижением толерантности к углеводам и увеличением риска
диабета II типа. Повышенное содержание глюкозы в крови,
наблюдаемое при этих состояниях, вносит вклад в атеросклероз многими
путями. Особого внимания заслуживает способность глюкозы
осуществлять перекрестные сшивки между белковыми молекулами
артериальных стенок, что делает их менее эластичными и более
восприимчивыми к повреждениям. Поскольку артерии при этом
теряют гибкость, кровяное давление повышается, что способствует
дальнейшему развитию атеросклероза. Наряду с этим, люди с
пониженной толерантностью к углеводам и ранними стадиями диабета
II типа имеют также и тенденцию к повышению уровня инсулина,
который вносит важный вклад в увеличение содержания ЛНП и
повышение кровяного давления (подробнее об этом см. в гл. 2).
27.2. ЧТО ДЕЛАТЬ С АТЕРОСКЛЕРОЗОМ?
Для атеросклероза в особенности справедливо утверждение, что
"унция" профилактики стоит "тонны" лечения, понимая, что
первым признаком болезни сразу может быть сердечный приступ или
инсульт. В то время как развитие атеросклероза может быть
остановлено или даже частично обращено вспять, некоторые
повреждения, вызванные инфарктом миокарда или инсультом, обычно
необратимы и могут ухудшить качество жизни пациента до самой
смерти. Для любого человека, заинтересованного в продлении жизни
или просто в своем здоровье и здоровье своей семьи, более чем
оправданы меры, направленные на профилактику атеросклероза.
Поскольку атеросклероз является одной из наиболее типичных
возрастных болезней, многие из мер продления жизни, которые
обсуждались в этой книге, помогут в той или иной степени
предотвратить развитие этой патологии. Однако из-за серьезности осложне-
409
ний следует дополнительно обсудить конкретные меры борьбы с
атеросклерозом.
Диета: жир, холестерин, сахар, волокно, калории. Жир в
нашей пище представлен тремя главными типами жирных кислот -
насыщенными, моно- и полиненасыщенными в зависимости от числа
двойных связей между атомами углерода в этих молекулах.
Имеются многочисленные свидетельства, что насыщенный жир
способствует развитию атеросклероза. О возможности связи между
насыщенным жиром и атеросклерозом в США впервые заговорили,
когда обнаружилось, что хронические заболевания и смертность от
коронарной сердечной недостаточности значительно снизились во
время Большой депрессии и второй мировой войны. В те годы люди
были вынуждены потреблять меньшее количество молока, масла,
сыра, яиц и, следовательно, меньше насыщенного жира. Было также
отмечено, что общества с низким потреблением животного жира,
который служит главным источником насыщенного жира, имели
существенно более низкую смертность от болезней сердца.
Некоторые критики, однако, утверждали, что различия в статистике
болезней сердца между нациями могут быть связаны с различиями их
генетической предрасположенности к этим заболеваниям. Это
представляется маловероятным. Японские иммигранты в США, которые
восприняли типичную американскую пищу (т.е. богатую
насыщенным жиром), имели значительно более высокий уровень сердечных
болезней, чем те, кто продолжал придерживаться традиционной
японской кухни.
Большой объем экспериментальных исследований и
клинических наблюдений также демонстрирует, что насыщенные жиры
увеличивают содержание ЛНП-холестерина и провоцируют
атеросклероз. В качестве первого шага для понижения уровня ЛНП,
большинство экспертов рекомендует сократить общее потребление
жира до 30% (от суммы дневного потребления калорий), а
насыщенного жира - до 10%. Если содержание ЛНП не возвращается к
нормальному диапазону, насыщенный жир должен быть еще уменьшен
до 7% и менее от суммы калорий.
Отказ от употребления насыщенного жира - весьма
эффективное и здоровое решение, но оно должно быть реализовано должным
образом. Некоторые люди просто переходят к использованию
большего количества маргарина. Действительно, маргарин, который
производится из различных типов растительного масла, обычно
содержит намного меньше насыщенного жира, чем масло животного
происхождения. Однако химический процесс, используемый при
создании маргарина, ведет к формированию
транс-мононенасыщенного жира. Ненасыщенные жирные кислоты имеют две
альтернативные структурные формы: цис- и трднс-формы. По существу,
весь ненасыщенный жир в природе существует в цис-форме.
Однако процесс гидрирования растительного масла, используемый в про-
410
изводстве маргарина, сопряжен с выходом значительного
количества трянс-мононенасыщенного жира, который не менее атерогенен,
чем насыщенный жир. Рекомендуют использовать маргарин с
низким содержанием или полным отсутствием жира в транс-форме.
Если информация о содержании жира в транс-форме на ярлыке
отсутствует, воздержитесь от приобретения этого продукта или
уточните его химический состав у изготовителя. Твердый маргарин
содержит обычно большое количество транс-жира. Транс-жир может
также присутствовать и в приготовленных пищевых продуктах.
Если список компонентов продукта включает термин "жир,
добавленный для рассыпчатости теста" (иными словами, маргарин низкого
качества), знайте, что изделие, вероятно, содержит транс-жир.
Стремясь избегать насыщенного жира и транс-жира,
некоторые люди идут на крайность: используют большое количество
полиненасыщенного жира. Это действительно понижает ЛНП и может
даже повысить ЛВП (что также благоприятно с точки зрения
снижения калорийности). Однако в полиненасыщенном жире, особенно
когда пища готовится с его использованием или он находится на
воздухе в течение длительного времени, увеличивается содержание
свободных радикалов, которые наряду со многими другими
негативными факторами, способствуют атеросклерозу. Еще одна ошибка
связана с очень низким потреблением всех видов жира, поскольку это
может вести к субклиническому дефициту незаменимых жирных
кислот. В целом, мононенасыщенный жир, представляется самым
здоровым, если он составляет большую часть всего потребляемого
жира. Лучшие источники мононенасыщенного жира - оливковое
масло и масло канолы.
Однако одного только сокращения насыщенного жира -
недостаточно. Избыток калорий так же или даже еще более вреден, чем
насыщенный жир. Наш организм запасает калории, которые не
нужны для выработки энергии в данный момент, независимо от
характера пищевого источника, из которого они усвоены. Даже если
ни одна из ваших избыточных калорий не поступила из жирной
пищи, организм все равно преобразует их в оптимальную форму
хранения, т.е. в жир, причем главным образом в насыщенный жир. И
прежде чем этот жир отложится в жировой ткани, он будет
циркулировать в вашей крови и стимулировать развитие атеросклероза.
Наиболее простой способ гарантировать вас от потребления
избытка калорий состоит в том, чтобы приблизиться (насколько это
возможно) к вашему оптимальному весу и сохранять его (см.
руководящие принципы потери веса в гл. 22).
Большинство людей "боится" пищевых продуктов, богатых
холестерином, полагая, что чем больше холестерина вы потребляете, тем
более высокий уровень холестерина будет в крови. Это в
значительной степени неправильное представление. Большинство холестерина,
который циркулирует в крови, производится в организме, и только
411
относительно малая его часть поступает с пищей. Более того,
холестерин, поступающий с пищей, ингибирует собственное производство
холестерина печенью, так что окончательное количество
холестерина в кровообращении будет практически почти постоянным при
употреблении относительно широкого диапазона пищевых продуктов.
Однако очень большие количества холестерина в пищевом рационе
(например, потребление нескольких яиц каждый день) могут
нарушить это равновесие и все же повысить уровень холестерина в крови.
Кроме того, у относительно небольшой группы людей ингибирование
синтеза холестерина экзогенным пищевым холестерином не очень
эффективно, и даже умеренно высокое его потребление может
неблагоприятно изменять профиль этого липида. Значит ли это, что
большинство людей не должно заботиться о сокращении
потребления холестерина? Теоретически, да, но фактически - нет.
Холестерин обнаруживается исключительно в продуктах
животного происхождения, таких как мясо, цельное молоко, молочные
продукты или яйца. Почти все продукты, богатые холестерином,
также богаты и насыщенным жиром и калориями. Следовательно,
когда вы сокращаете потребление холестерина, автоматически
сокращается поступление насыщенного жира и калорий, что является
гораздо более важным для сокращения уровня ЛНП и холестерина.
Главное исключение - рыба и моллюски, которые обычно содержат
относительно много холестерина и мало насыщенного жира
(табл. 22).
Мы не советуем чрезмерно потреблять простые сахара, такие
как сахароза (столовый сахар) или мед, потому что они повышают
уровень инсулина, который, как уже указывалось, стимулирует
атеросклероз. Фруктоза, которая является частью сахарозы и главным
компонентом меда, по-видимому, обладает дополнительным атеро-
генным эффектом. Кроме того, простые сахара с легкостью
приводят к избытку потребления калорий.
Волокнистая пища препятствует атеросклерозу по нескольким
важным путям. Во-первых, растворимые волокна, например в
овсяных зернах или яблоках, способствуют понижению холестерина,
связывая желчные кислоты и увеличивая их выделение. Желчные
кислоты вырабатываются в организме из холестерина. Чем больше
желчных кислот связывается волокнами, тем большее количество
холестерина отвлекается из кровяного русла для синтеза новых
желчных кислот. Конечный результат - понижение уровня холесте-
ринов. Во-вторых, пищевые продукты с высоким содержанием
волокон обычно содержат мало жира и калорий, и их обильное
потребление в этом смысле весьма благотворно. Наконец, волокна
замедляют поглощение глюкозы в пищеварительном тракте.
Результатом этого является более низкий уровень глюкозы и инсулина в
крови, что также помогает предотвратить атеросклероз (см. гл. 22 о
диете, богатой волокнами).
412
Таблица 22
Характеристика диеты для уменьшения ЛНП - холестерина
Национальная образовательная программа по холестерину для
практикующих врачей-диетологов из Свода рекомендаций для лечения взрослых
пациентов. Базируется на данных N.E. Ernst et al. См.: Journal of American
Dietetic Association. 1988. N88. P. 1401-1411.
Энергия
Шаг!
Адекватный для
достижения и
поддержания оптимального
веса, %
Шаг 2
Адекватный для
достижения и
поддержания оптимального
веса, %
Тотальный жир*
Насыщенный жир
Полиненасыщенный жир*
Мононенасыщенный жир*
Холестерин
<30
8-10
До 10
10-15
< 300 лег/день
<30
<7
До 10
10-15
< 200 лег/день
* Выраженный как % от общего количества калорий дневного рациона
Омега-3 жирные кислоты - ненасыщенные жирные кислоты,
содержащиеся в рыбьем жире. Они обладают широким набором
влияний на организм, потому что изменяют текучесть клеточных
мембран и, что наиболее важно, служат предшественниками про-
стагландинову которые являются медиаторами многих типов
взаимодействий между клетками. Потребление омега-3 жирных кислот
изменяет относительное содержание различных простагландинов в
организме. Среди других влияний омега-3 жирные кислоты
предотвращают атеросклероз, препятствуя формированию сгустков крови
и повышению кровяного давления. Лучший способ получения
омега-3 жирных кислот состоит в том, чтобы потреблять рыбу по
крайней мере два-четыре раза в неделю. Рыба из холодных морей,
например семга, палтус или треска, служит лучшим источником
омега-3 жирных кислот. При приготовлении рыбы следует избегать
интенсивной жарки, потому что омега-3 жир очень восприимчив к
окислению.
Антиоксиданты. Различные антиоксиданты, такие как
витамины С, Е, липоевая кислота и полифенолы, предотвращают или
замедляют окисление ЛНП-частиц, что делает их менее липкими и
таким образом замедляет рост атеросклеротических бляшек. Многие
исследования указывают, что пищевые продукты, богатые антиок-
сидантами, или пищевые добавки уменьшают риск атеросклероза.
(См. также гл. 23.)
Фолиевая кислота, витамин В6 и В12. Дефицит фолиевой
кислоты, витамина В6 и, возможно, В12 может способствовать
атеросклерозу, увеличивая уровень гомоцистеина. Лучшие диетические
413
источники фолиевой кислоты - свежие зеленые овощи с обильной
листвой (салат, капуста и др.). Аспирин, средства, нейтрализующие
кислоты, противозачаточные средства, принимаемые per os> могут
взаимодействовать с фолиевой кислотой, повышая риск ее
дефицита. Витамин В12 присутствует главным образом в пищевых
продуктах животного происхождения. Витамин В6 широко распределен
среди пищевых продуктов, но может быть разрушен длительной
термической обработкой. Здоровому человеку, потребляющему
сбалансированный рацион, включающий свежие фрукты и овощи,
обычно не грозит дефицит любого из этих витаминов. В противном
случае следует принимать фолиевую кислоту, В6 и В12 в дозах, дос
таточных для избежания дефицита. Большие дозы не дают
дополнительной защиты против атеросклероза.
Белок сои. Несколько недавних исследований показали, что
потребление примерно 25-50 г белка сои в день может существенно
понизить уровень ЛНП и общего холестерина. В этих
исследованиях белок сои не был только приложением к обычному рациону, но
им заменяли часть животного белка диеты. Некоторые
исследователи полагают, что многие из эффектов не связаны с белком сои
непосредственно, а скорее с фитоэстрогенами, эстроген-подобными
соединениями в составе сои. Безотносительно истинной причины
повышенное потребление изделий из сои может улучшать ваш про
филь липидов. Однако для большинства людей, воспитанных на ев
ропейской культуре питания, большие количества сои в диете могут
быть заметной проблемой.
Физические упражнения. Выполнение физических упражнений
поднимает содержание ЛВП ("хорошего холестерина") и помогает
сжиганию калорий. Эта мера полезна, но только в комплексе мер по
предотвращению атеросклероза.
Снижение стресса. Стресс стимулирует атеросклероз многими
путями: повышая уровень сахара в крови и кровяное давление,
увеличивая формирование свободных радикалов (Yu et al., 2001; Csiszar
et al., 2002). Хронический стресс может препятствовать всем другим
мерам предотвращения атеросклероза. Было показано, что
сибирский женьшень (адаптоген, снижающий стресс-ответ и
противодействующий связанным с ним повреждениям) эффективен и в
предотвращении атеросклероза и его осложнений (Брехман, 1968; Дарды
мое, 1976; Todorov, 1993).
Глава 28
ЦЕНТР НАШЕЙ ВСЕЛЕННОЙ - МОЗГ
Христос сказал: "Убогие блаженны.
Завиден рок слепцов, калек и нищих,
Я их возьму в надзвездные селенья,
Я сделаю их рыцарями неба
И назову славнейшими из славных..."
Пусть! Я приму! Но как же те, другие,
Чьей мыслью мы теперь живем и дышим,
Чьи имена звучат нам как призывы?
Искупят чем они свое величье,
Как им заплатит воля равновесия?
Иль Беатриче стала проституткой,
Глухонемым - великий Вольфганг Гёте
И Байрон - площадным шутом...О ужас!
Николай Гумилеву 1911
Есть ли у человека что-либо более ценное, чем его мозг?
Деньги, недвижимость, репутация, социальное положение, семья и
друзья, физическая красота? Задумайтесь над этим, и вы согласитесь,
что ничего более ценного, чем мозг, у него нет. Надлежащая
работа мозга определяет практически все человеческие способности,
ценности и аспекты жизни. Мозг это не только интеллект, это
также и потенциал памяти, эмоций, координации движений, чувств, так
же как и уровень самосознания, идентичности и индивидуальности.
Многообразие и эффективность нашего взаимодействия с
окружающей действительностью и возможность наших достижений или
поражений в значительной мере определяются нашим мозгом.
Не удивительно, что ухудшение различных аспектов мозговой
функции - наиболее разрушительная сторона процесса старения.
Старение других органов уничтожает тело, старение же мозга
уничтожает самое ядро человека, его личность. Немного людей хотели
бы продолжать свою жизнь, потеряв психологическую
идентичность, способность думать, понимать и помнить.
Несколько взаимосвязанных процессов вносят вклад в
ухудшение функций мозга с возрастом.
• Смерть нейронов. Нейроны могут погибать от повреждений,
вызванных свободными радикалами, гормонами стресса, токсинами,
блокадой мозгового кровообращения или другими причинами
(Vatassery et al., 1998; Rodriguez-Capote et al., 1998; Ferrini et al., 1999).
♦ Накопление возрастных пигментов. Липофусцин и другие возрастные
пигменты накапливаются в нейронах, препятствуя метаболическим
путям и вредя внутриклеточным функциям (Butterfield et al., 2001).
415
• Падение энергопробукции. Нейроны постепенно "сжигают" свои
энергетические станции, митохондрии, теряя способность
производить достаточно энергии (АТФ) для нормальной функции (Lefaix et
al., 1998; Wei, 1998).
• Понижение кровоснабжения. Атеросклероз и гипертония
повреждают артерии и капилляры, которые обеспечивают
кровоснабжение мозга. Пониженное кровоснабжение означает пониженную
деятельность мозговых клеток и сниженную способность
противостоять стрессу (Yu et al., 2001).
• Снижение уровня нейромедиаторов. Мозговая функция зависит
от способности нейронов к эффективной коммуникации.
Пониженная функциональная способность нейронов ведет к снижению
их способности производить нейромедиаторы (Luo et al., 2000),
которые необходимы для межнейронных связей. В результате
нейроны с сохраненной функцией не получают достаточных стимулов от
своих партнеров, что ведет к дальнейшему общему ухудшению
функций мозга (Urano et al., 1998; Michaelis, 1998).
С возрастом увеличивается риск двух главных типов
мозговых нарушений: ухудшение настроения, в частности депрессия, и
расстройство познавательных функций, таких как болезнь Альц-
геймера, слабоумие и потеря памяти. Существуют указания, что
биохимическая коррекция может существенно снизить темп
возрастных нарушений функций мозга (Deschamps et al., 2001). Более
того, замедление старения мозга, очевидно, также замедлят и
старение тела (как это обсуждалось в разделе о "центральных часах
старения" в гл. 2).
28.1. УМСТВЕННЫЕ СПОСОБНОСТИ
Умственные способности - это и способность к познанию и
обучению, и память, и способность общения, а также умение встречать,
анализировать и решать потребности ежедневной жизни. В то
время как некоторая потеря умственных способностей с возрастом
практически неизбежна, степень этой потери может сильно
варьировать по темпу и масштабам проявления механизмов старения.
Глубокую потерю умственных способностей обычно относят к
деменции, т.е. слабоумию. Существуют оценки, что приблизительно
15-20% пожилых людей в США имеет некоторую форму деменции.
Болезнь Альцгеймера составляет половину всех случаев деменции.
Остальные случаи, обусловленные широким разнообразием
причин, включают болезнь Паркинсона, инсульт, В-авитоминоз,
гипотиреоз и ряд других патологий. Болезнь Альцгеймера
характеризуется вырождением мозговых нейронов, ответственных за
умственные функции. Типичные физиологические проявления болезни
416
Альцгеймера - низкий уровень ацетилхолина (нейромедиатора,
особенно важного для умственной деятельности), ухудшение мозгового
кровообращения и, возможно, рост уровня свободных радикалов и
низкая активность антиоксидантных систем. Противодействие этим
факторам или их задержка иногда полностью изменяют течение
болезни Альцгеймера и вообще помогают сохранять умственные
способности (о молекулярных механизмах развития болезни Альцгей
мера см. в разделах "Накопление шлаков" и "Генетические аспекты
старения" гл. 2).
28.2. ПИЩЕВЫЕ ДОБАВКИ, СОХРАНЯЮЩИЕ
ИЛИ УЛУЧШАЮЩИЕ УМСТВЕННЫЕ СПОСОБНОСТИ
DMAE (диметиламиноэтил) - вещество, в незначительных
количествах содержащееся в рыбе. Это соединение оказывает
разнообразные благоприятные влияния на умственные способности:
улучшает память, внимание, концентрацию, снимает хроническую
усталость и умеренную депрессию. В одном исследовании было
даже найдено улучшение показателя IQ (коэффициент умственного
развития) и способности к логическому мышлению. Деанол -
лекарственное средство, подобное по структуре и действию DMAE,
успешно использовалось при изучении поведенческих проблем,
таких как дефицит и дезорганизация внимания, а также гиперкине-
зия. Интригующим "побочным эффектом" DMAE у некоторых
пациентов являются сновидения, при которых человек осознает, что
видит сон. Большинство людей рассматривает это состояние как
очень приятный и интересный опыт. В некоторых, но не во всех
исследованиях на грызунах, DMAE увеличивал продолжительность
их жизни до 30%.
DMAE, по-видимому, поднимает в мозгу уровень ацетилхолина,
нейромедиатора, стимулирующего холинергические нейроны,
которые в свою очередь связаны с разными сторонами умственной
деятельности. И наоборот, как уже упоминалось выше, болезнь
Альцгеймера и некоторые другие виды деменции связаны с низким
уровнем ацетилхолина в мозгу.
Несколько исследований на животных показали, что DMAE
уменьшает возрастное накопление пигмента липофусцина в мозгу.
Некоторые данные свидетельствуют, что этот эффект может быть
результатом способности DMAE стимулировать антиокислительные
ферменты и ингибировать реакции перекрестного сшивания. Таким
образом, DMAE, очевидно, не только стимулирует умственные
функции, но и фактически замедляет старение мозга.
DMAE- вполне доступная пищевая добавка. Обычные дозы - от
500 до 1000 мг в день. Однако нужно начать с низкой дозы 50 или
100 мг в день, и постепенно увеличивать дозу. Препарат часто про-
417
дается в виде виннокислого DMAE, который содержит только
определенный процент от DMAE. При использовании этой
разновидности DMAE, дозировки должны быть рассчитаны согласно
содержанию "чистого" DMAE. Может потребоваться две-четыре недели,
чтобы ощутить полный благотворный эффект DMAE (Modern
Nutrition in Health and Disease, 1994).
Предостережение. Люди с эпилепсией или маниакальным
депрессивным расстройством не должны использовать DMAE.
Высокие дозы DMAE могут вызывать бессонницу, головные боли и
мышечную слабость. При снижении дозы эти побочные эффекты
обычно исчезают. Если вы принимаете любые виды
медикаментозного лечения, особенно связанные с холинергическими
препаратами, проконсультируйтесь с вашим врачом перед использованием
DMAE.
Ginkgo biloba (GB) - один из самых старых видов деревьев на
Земле. Это растение широко использовалось в традиционной
китайской медицине для таких разнообразных болезней, как нарушение
кровообращения, ослабление слуха, половая дисфункция и потеря
памяти.
В последнее десятилетие биологические эффекты экстракта GB
интенсивно изучались как на животных, так и на людях. Показано,
что экстракт GB полезен при многих нарушениях центральной
нервной системы, включая деменцию, потерю памяти, пониженную
концентрацию и познавательную способность, ухудшение зрения,
слуха, головокружение и многие другие. У здоровых индивидов
экстракт GB улучшает концентрацию, внимание и кратковременную
память. Сегодня экстракт GB - одно из наиболее рекомендуемых
средств в Европе для лечения нарушений, связанных с возрастом, и
восстановления умственных способностей.
Польза, которую центральная нервная система получает от
экстрактов GB, определяется синергизмом нескольких эффектов. Во-
первых, экстракт стимулирует мозговое кровообращение, улучшая
доставку кислорода и питательных веществ к нейронам. Во-вторых,
он защищает нейроны от повреждений свободными радикалами,
образующихся в большом количестве именно в мозгу в результате
повышенного потребления им кислорода. Иначе говоря, GB понижает
индексы окислительных повреждений (Lefaix et al., 1998). Это
обстоятельство играет важную роль, поскольку мозг особенно подвержен
неблагоприятному воздействию свободных радикалов из-за высокого
содержания липидов, которые особенно подвержены окислению.
Наконец, GB ингибирует агрегацию клеток, участвующих в образовании
сгустка, и тем самым уменьшает формирование свежих кровяных
сгустков и может даже "прочищать" частично закупоренные артерии.
Рекомендуемая доза экстракта Ginkgo biloba - 40 мг три раза в
день. Обычно, используется стандартизированный экстракт,
содержащий 24% гинко гетерогликозидов. Для менее концентрирован-
418
ных экстрактов доза должны быть соответственно выше Побочные
реакции желудочно-кишечные расстройства и головные боли -
редки и обычно умеренны..
Холин используется организмом в качестве предшественника
для синтеза ацетилхолина, одного из ключевых нейромедиатиров
высших умственных функций. Дополнительный холин увеличивает
уровень ацетилхолина в мозгу и, как было показано при
обследовании студентов, улучшает память и усвоение нормальных
университетских дисциплин. К сожалению, холин редко бывает
эффективным при болезни Альцгеймера, потому что у этих больных обычно
наблюдается недостаток ферментов, катализирующих синтез аце-
тилхолин из холина.
Обычные дозы холина - один-три г в день.
Антиоксиданты. Мозг нуждается в надежной антиоксидантной
защите более, чем любой другой орган. Составляя всего лишь
только около 2% от массы тела, мозг сжигает примерно 20% всех
потребляемых калорий. Мозговая ткань бомбардируется свободными
радикалами в десять раз интенсивнее, чем большинство других
тканей. Кроме того, мозг богат липидами, которые легко, как уже
указывалось, подвергаются атаке АФК. Чтобы защищать себя
мозговые клетки производят «большое количество антиоксидантных
ферментов. Однако даже небольшое ослабление антиоксидантной
защиты из-за стресса, болезней, дефицита питательных веществ или
возрастных физиологических изменений (например, хронического
падения производства мелатонина или ухудшения мозгового
кровообращения), может нарушать окислительно-восстановительный
баланс и вести к существенным повреждениям со стороны СР.
Антиоксиданты в виде пищевых добавок уменьшают риск этих
повреждений (Vatassery et al., 1998; Deschamps et al., 2001). Для защиты
центральной нервной системы особенно важны антиоксиданты,
растворимые в жирах, такие как витамин Е, липоевая кислота и мелатонин.
28.3. ДЕПРЕССИЯ
Как уже обсуждалось в главе 7, депрессия - одно из наиболее
распространенных последствий возрастного снижения функций
мозга, в частности уровня нейромедиаторов. В то время как депрессия
во многих случаях является следствием старения, она также и
фактор ускорения процесса старения. Депрессия вносит вклад в
старение, вызывая чрезмерный стресс, подавляя иммунную систему, и,
возможно, ускоряя "ход" центральных часов старения. Одна из
наиболее достоверных психологических характеристик долгожителей -
положительная жизненная установка, эмоциональная стабильность
и хорошая способность справляться с трудностями, что явно
отличается от состояния депрессии. Освобождение от депрессии - важная
419
часть стратегии антистарения не только потому, что помогает
замедлять некоторые из механизмов старения, но и препятствует
разрушению мотивации человека, настроенного предпринять действия,
направленные на улучшение здоровья и продления жизни.
Биохимическая основа депрессии - снижение в мозгу уровня
нейромедиаторов. Большинство современных антидепрессантов
повышают уровень нейромедиаторов, особенно серотонина и норад
реналина. Типичные побочные эффекты этих лекарств - седатив
ный эффект, запор, тошнота, сухость ротовой полости,
расстройство пищеварения, головокружения, импотенция и др. По этой
причине прием этих медикаментов допускается только при клинических
случаях депрессии. Множество питательных веществ и пищевых
добавок, как было установлено, воспроизводят упомянутые
положительные эффекты антидепрессантов с гораздо меньшим
количеством побочных эффектов. Это может быть особенно полезно людям
с субклинической формой депрессии, которая весьма распростране
на, но часто остается незамеченной и лишенной лечения.
Необходимо помнить, что серьезная депрессия требует
профессионального лечения специалистами по психическому здоровью.
28.3.1. Депрессия и дефицит витаминов
Перед началом любой антидепрессантной терапии, нужно
рассмотреть такую возможность, что депрессия может быть
вторична по отношению к некоторой другой проблеме. В частности,
дефицит некоторых питательных веществ - довольно частая
причина депрессии.
Витамин В12. Депрессия - один из многих возможных
симптомов авитаминоза В12. Другие симптомы - психоз,
раздражительность, смятение, потеря памяти, невропатия и анемия. Однако при
умеренном дефиците В12 депрессия может быть единственным
симптомом. Не совсем ясно, почему дефицит В12 вызывает депрессию
и другие нейропсихологические нарушения. В12 требуется для
синтеза S-аденозилметионина (SAM) из метионина. Следует
подчеркнуть, что SAM играет существенную роль в синтезе некоторых
нейромедиаторов, особенно серотонина. Вообще предполагается, что
дефицит В12 ведет к депрессии, нарушая синтез нейромедиаторов
именно на этом метаболическом пути.
Уникальность В12 среди других витаминов заключается в том,
что он содержится почти исключительно в пищевых продуктах
животного происхождения, таких как домашняя птица, рыба, яйца или
молочные продукты. Вопреки некоторым популярным
представлениям, никаких активных форм В12 в морских водорослях, например
спирулине или ферментированных продуктах из сои, не найдено.
Таким образом, строгие вегетарианцы подвержены риску дефицита
420
витамина В12. Печень хранит большие запасы В12, и требуется не
сколько месяцев или даже несколько лет для развития его явного
дефицита. Депрессия, однако, может развиваться намного раньше дру
гих признаков. Диетологи вообще рекомендуют строгим вегетари
анцам регулярно принимать витамин В12.
Здоровые молодые и средних лет люди, потребляющие
сбалансированную невегетарианскую диету, обычно не подвержены
опасности дефицита витамина В12. Усвоение витамина В12 из пищи
требует специфического белкового фактора, производимого
желудком. У некоторых пожилых людей синтезируется недостаточное
количество этого фактора из-за атрофии желудочных желез, что
обычно в преклонном возрасте. Это может вести к дефициту В12.
Другой возможной, хотя и редкой, причиной такого дефицита,
может быть злокачественная анемия, заболевание, наблюдаемое у
людей, желудок которых не производит кислоту и этот специфический
фактор. Статус витамина В12 обычно оценивается путем измерения
его содержания в плазме крови.
Минимально необходимое потребление В12 - 2 мкг в день,
что, как правило, достаточно для предотвращения его дефицита у
здоровых вегетарианцев. Однако пероральный прием часто
неэффективен у людей с недостаточным всасыванием В12. До
недавнего времени инъекции были единственной альтернативой, но
теперь эффективное внутриносовое введение также становятся
доступным. Изредка у больных депрессией с нормальным
уровнем В12 в крови наблюдается улучшение в ответ на большие
дозы В12 (250-1000 мкг в день).
Фолат. Дефицит фолата относительно часто обнаруживается
при депрессии. Существуют оценки, что примерно у 30%
психиатрических пациентов обнаруживается дефицит фолата. Более того, у
двух гериатрических пациентов из трех, находившихся в
психиатрической палате, также наблюдался дефицит фолата. Другие
признаки дефицита фолата включают умственное расстройство, анемию,
расстройства пищеварения (изжога, понос, нарушение всасывания в
желудочно-кишечном тракте) и снижение иммунитета. Как и В12,
фолат участвует в синтезе S-аденозилметионина. По-видимому,
дефицит фолата и В12 вызывают депрессию по одному и тому же
механизму.
Овощи являются наиболее богатым источником фолата,
особенно зеленые листья и бобы. Единственный пищевой источник
животного происхождения, богатый фолатом, - печень. Фолат может
быть разрушен в процессе приготовления пищи или при длительном
хранении. Лучший способ получить достаточное количество фолата
с пищей потреблять в изобилии фрукты и свежие или слегка
приготовленные овощи. Долговременное использование аспирина и
медикаментов, нейтрализующих кислоту, может вызвать дефицит
фолата, конкурируя с последним в процессах всасывания. Прием проти-
421
возачаточных средств per os и курение могут понижать уровень фо-
лата. Содержание фолата определяется в крови с помощью фолат-
теста плазмы или эритроцитарного теста, причем последний
гораздо более точен.
Рекомендуемое минимальное дневное потребление фолата -
180 мкг в день для женщин и 200 мкг для мужчин. Пациенты с
клинически установленным дефицитом обычно принимают до 1000 мкг
фолата в день. Чтобы ликвидировать возможный дефицит,
эксперты обычно рекомендует 400 мкг препарата в день. В некоторых
случаях прием добавок фолата может кончаться полным снятием
симптомов депрессии или по крайней мере повышением эффективности
других мер.
Никотиновая кислота (ниацин> - уникальна среди других
витаминов, так как может быть синтезирована в организме из аминокислоты
триптофан. Организм использует примерно 60 мг триптофана для
производства 1 мг никотиновой кислоты. Дефицит ниацина, так же как и
триптофана, встречается редко. Пища с преобладанием кукурузы
может быть причиной дефицита ниацина, потому что кукуруза содержит
незначительные его количества, а белок кукурузы печально известен
как недостаточный источник триптофана. Хотя дефицит ниацина в
развитых странах маловероятен (если вы не питаетесь исключительно
попкорном или корнфлексом), следует помнить, что низкое
поступление ниацина с пищей будет переключать существенные количества
триптофана на синтез никотиновой кислоты. Такая ситуация
оставляет меньшее количество триптофана для его использования на других
метаболических путях, включая синтез серотонина. Дополнительные
количества никотиновой кислоты иногда полезны в качестве усиления
комплексного лечения депрессии, поскольку они могут несколько
увеличивать синтез серотонина. Минимальное потребление никотиновой
кислоты - 15 мг для женщин и 19 мг для мужчин в день.
28.3.2. Депрессия и пищевые добавки
5-гидрокситриптофан (5-НТР) - очень похож на известный
антидепрессант проз ок. Как и последний, 5-НТР снижает депрессию,
поднимая уровень серотонина в мозгу. Эффективность 5-НТР и про-
зака при депрессии, по-видимому, приблизительно одинакова, но
приему 5-НТР сопутствует меньшее количество побочных
эффектов, что делает его лучшим при лечении.
Различие между 5-НТР и прозаком заключается в
метаболических путях, по которым они достигают высокого уровня серотонина.
Прозак - лекарственное средство, принадлежащее к классу
селективных ингибиторов поглощения серотонина (СИПС). Оно
действует, блокируя последующее поглощение серотонина нейронами
после того как нервный импульс был передан. В результате большее
422
количество серотонина остается активным в синаптическои щели,
приводя к более высокой активности серотонинэргических
нейронов, т.е. нейронов, которые используют серотонин в качестве
главного нейромедиатора.
5-НТР, напротив, является питательным веществом и
метаболитом. Организм производит серотонин из триптофана,
поступающего с белками пищи. Сначала триптофан преобразуется в 5-НТР,
который далее преобразуется в серотонин. Следовательно, 5-НТР -
непосредственный предшественник серотонина. Сам триптофан в
течение десятилетий использовался для лечения депрессии. В 80-х
годах XX в. зафиксирован ряд случаев серьезных побочных эффектов,
связанных с использованием пищевых добавок с триптофаном, что
и привело к решению FDA запретить продажу последних.
Расследование привело к единственной загрязненной партии этого продукта,
произведенного предположительно новым непроверенным
методом. Тем не менее FDA пока еще не снял своего запрещения. В
противоположность триптофану 5-НТР разрешен к продаже и,
по-видимому, более эффективен, чем триптофан, поскольку 5-НТР только
на "один метаболический шаг" удален от серотонина.
В ряде исследований сравнивали эффективность 5-НТР с обычно
используемыми лекарствами-антидепрессантами. Было найдено, что
5-НТР по крайней мере так же эффективен как имипрамин, обычный
трициклический антидепрессант, или флувоксамин - СИПС,
подобный прозаку. В этих исследованиях 5-НТР переносился лучше, чем
лекарственные антидепрессанты. Другие исследования указывают, что
5-НТР может быть полезен и при других расстройствах, связанных с
дефицитом серотонина, таких как беспокойство, бессоница,
агрессивность, возбуждение, поведение, навязанное тревогой и страхом, и
мигрень. Эти расстройства обычно лечатся препаратами типа СИПС.
Препараты 5-НТР, по-видимому, также уменьшают аппетит и могут
быть полезны в мероприятиях по сокращению веса.
Дозировка 5-НТР, используемая в большинстве клинических
исследований, - 300 мг в день (по 100 мг три раза в день). Более низкая
дозировка - 50 или 100 мг день - может быть эффективна при умеренной
депрессии. Правильная дозировка должна быть определена врачом
Витамин Вб является ко-фактором при синтезе серотонина из
5-НТР. Минимальное дневное потребление для В6 - 1.6 мг для
женщин и 2 мг для мужчин. Дефицит В6 очень редок, так как этот
витамин содержится в широком разнообразии пищевых продуктов.
Дополнительный В6, как было показано, увеличивал синтез
серотонина у животных. Однако наиболее эффективно эта пищевая добавка
действовала в комбинации с 5-НТР. Обычно рекомендуемая доза В6:
10-20 мг в день. Необходимо иметь в виду, что В6 - не безвредный
витамин. Высокие дозы (сотни миллиграммов) В6, принимаемые в
течение длительного времени, приводили в некоторых случаях к
необратимому повреждению нервов.
423
DL-фенилаланин - равная смесь D и L оптических изомеров
аминокислоты фенил ал анина. L-фенилаланин, будучи
предшественником нейромедиаторов норадреналина и дофамина, может поднимать
их уровень в мозгу, облегчая состояние депрессии. D-фенилаланин -
ингибитор фермента энкефалиназы. Этот фермент осуществляет
деградацию энкефалинов, соединений, физиологически подобных
морфинам, которые действуют как антистрессовые и
болеутоляющие агенты. Фенилаланин также может быть преобразован в
организме в фенилэтиламин, вещество со свойствами двойного
стимулятора антидепрессантных эффектов. Очевидно, комбинация
упомянутых выше эффектов определяет антидепрессантную активность
DL-фенилаланина, которая демонстрировалась в нескольких
клинических исследованиях. При этом было показано, что
DL-фенилаланин оказался столь же эффективным, как и обычно
предписываемый врачами антидепрессант имипрамин, но с меньшим
количеством побочных эффектов. В большинстве исследований
DL-фенилаланин использовался в дозах от 200 до 500 мг в день. Окончательная
дозировка, однако, должна быть определена врачом.
Hypericum perforatum (HP) - трава с длительной историей
применения для лечения депрессии и состояния тревоги. Экстракты из HP,
по-видимому, осуществляют свое действие антидепрессанта путем
слабого ингибирования фермента моноаминооксидазы, а также за
счет ряда других воздействий на центральную нервную систему.
В отличие от большинства других лекарственных трав, HP
всесторонне исследовался. Установлено, что многие из его эффектов
были связаны с веществом, названным гиперисином. Экстракты HP,
используемые в клинических испытаниях, были стандартизированы
по содержанию в них гиперисина. Многие хорошо проведенные
клинические испытания показали эффективность гиперисина при
лечении депрессии и чувства тревоги.
Немецкие ученые из университет им. Людвига Максимиллиана
в Мюнхене провели метаанализ (тип статистического анализа со
сравнением и комбинацией результатов нескольких различных
исследований) 23 клинических испытаний экстрактов HP.
Суммировались данные относительно 1757 пациентов со слабыми или
умеренными депрессивными расстройствами. Анализ подтвердил
эффективность HP при лечении депрессии и показал, что действие
экстракта HP подобно действию стандартных антидепрессантных лекарств
типа имипрамина. HP, однако, переносился намного лучше, чем
стандартные антидепрессанты: побочные эффекты наблюдались у
52.8% пациентов, принимавших стандартные лекарства, но только -
у 19.8% пациентов, принимавших экстракты HP. Дозы экстрактов
HP, используемые в большинстве исследований, составляли от 2 до
2.7 мг гиперисина в день. Однако некоторые побочные эффекты
возможны при этих или даже более низких дозировках. Поэтому
лечение экстрактами HP должно контролироваться врачом.
Глава 29
АДАГТТОГЕНЫ
Как богат растительный мир
и как мало мы его используем.
И.И. Вавилов
В последние годы такие средства, как
женьшень и элеутерококк (экстрагируемые из
некоторых распространенных в Азии и
используемых в восточной терапии
растений), вызвали большой интерес, в
частности, в СССР. Известно много клинических
и меньше экспериментальных наблюдений,
позволяющих предположить, что эти
препараты оказывают неспецифическое
антистрессовое действие
Ганс Селье (1977)
Адаптогены - возможно одно из наиболее эффективных
средств противодействия различным типам стресса. Исходя из
названия, можно предположить, что адаптогены - класс веществ
(обычно, но не обязательно растительного происхождения),
которые способствуют успешной адаптации (см. гл. 16). Термин "адапто-
ген" был предложен в конце 40-х годов советским исследователем
Н. Лазаревым (1947), который нашел, что некоторые соединения
повышали устойчивость испытуемых субъектов к различным
неблагоприятным ситуациям. Другой советский исследователь,
И. Брехман (1964, 1968, Брехман и др., 1966, 1968) далее развил
концепцию адаптогенов как веществ, отвечающих следующим
критериям. Адаптогены:
• увеличивают общую резистентность организма к широкому
разнообразию повреждающих или стрессогенных факторов, таких как
физическое или умственное перенапряжение, травмы, простуды,
высокая температура, радиация, ядовитые химикалии, и т. д. В
результате, организм может противостоять всем видам стрессоров
более длительно и с меньшим количеством повреждений;
• имеют тенденцию приводить к норме отклонения
физиологических параметров, независимо от того проявлялись ли они с
дефицитом или избытком. Например, многие адаптогены могут либо
уменьшать кровяное давление, если оно повышено, либо
поднимать, если оно понижено;
• практически - нетоксичны, и не вызывают или вызывают немного
побочных эффектов в широком диапазоне доз.
425
Реальные соединения, которые делают все упомянутое выше,
действительно существуют. Некоторые из них, например женьшень,
были известны в течение многих столетий. Большинство адаптоге
нов также имеет много других ценных свойств, включая повышение
способности к обучению, стимуляцию функций иммунной системы,
активацию анаболических процессов и др.
Многие посвященные адаптогенам исследования были проведе
ны в СССР в 1940-1990 гг. и по известным политическим причинам
были относительно малоизвестны на Западе. Фактически советские
ученые выполнили тысячи клинических и экспериментальных
исследований, документирующих многочисленные полезные свойства
адаптогенов. Западные исследователи уделяли мало внимания этим
важным и перспективным соединениям, особенно элеутерококку,
или как его принято называть на Западе - сибирскому женьшеню
(Todorov, 1993).
Как оказалось, адаптогены полезны не только в стрессогенных
ситуациях, но также и при разнообразных болезнях, начиная от
расстройства нервной системы и кончая болезнями сердца и раком
(Брехман, 1957; Дардымов, 1976; Chong et al., 1984; Bohn et al., 1987;
Яременко, 1990; Беспалов и др., 1992). Советские врачи включили
адаптогены в многочисленные терапевтические комплексы,
используя их способность увеличивать эффективность других средств
лечения и ускорять восстановление ото многих болезней. Более того,
ряд косвенных данных, по-видимому, свидетельствует, что
адаптогены влияют на процесс старения и возрастные болезни. Во-первых,
потеря адаптируемости, как уже не раз подчеркивалось выше, -
одна из наиболее характерных особенностей старения: даже
умеренная дополнительная нагрузка на любую систему организма может
быть губительной, так как способность к восстановлению с
возрастом существенно понижается. Адаптогены изменяют эту ситуацию,
восстанавливая способность систем организма преодолевать
неожиданные нагрузки всякий раз, когда это необходимо. Во-вторых,
старение связано с различными нарушениями гомеостаза,
обеспечивающего постоянство многих функций организма. Адаптогены
способны возвращать различные физиологические параметры, такие
как давление крови, сахар крови, частота сердцебиений, уровень
холестерина, в диапазон нормальных значений, замедляя или
полностью останавливая таким образом развитие многих возрастных
болезней. Наконец, старению присущ чрезмерный стресс-ответ. В
частности, выделение чрезмерных количеств кортикостероидов в
ответ на стрессогенные факторы, а часто даже и в покое, что имеет
негативные последствия для организма. Многие адаптогены
работают для оптимизации стресс-ответа, делая его адекватным
фактическим потребностям тела.
Адаптогены могут также приносить пользу и молодым
людям, особенно при восстановлении от травм или болезней. В неко-
426
тором смысле, адаптогены могут вернуть старым людям
физические и умственные способности молодого человека, принимая во
внимание, что последнему они способны дать
"сверхчеловеческую" выносливость.
Каким образом адаптогены делают свою работу? Какова
тайна их универсальной эффективности? Большинство лекарств
весьма отличается от адаптогенов в том смысле, что их целью
является подавление или стимуляция определенной системы или
органа. Адаптогены - превосходные агенты системного действия,
повышающие выносливость и смягчающие развитие острого
стресса. Однако польза от них не исчерпывается сказанным, а
распространяется на лечение некоторых болезней и особенно
предотвращение или обращение некоторых аспектов процесса
старения. Так, если в качестве нормы мы рассматриваем
физиологические показатели здоровых 25-летних людей, тогда даже у
здорового пожилого человека окажется много отклонений от
нормы. Адаптогены противостоят этим тенденциям, что делает
их сегодня одним из наиболее универсальных инструментов
антистарения {табл. 23).
Еще раз подчеркнем, что одной из важных характеристик
действия многих адаптогенов является их способность изменить стресс-
ответ организма. Как уже обсуждалось в главе 12, стресс-ответ
состоит из трех главных фаз: реакции тревоги, фаз сопротивления и
истощения. Часто реакция тревоги бывает чрезмерной, оставляя
слишком мало ресурсов для фазы сопротивления. Некоторые
адаптогены понижают интенсивность реакции тревоги и увеличивают
способность организма поддерживать фазу сопротивления. В
результате - уменьшение повреждений, связанных со стрессом, и уход
от фазы истощения.
Другая важная характеристика многих адаптогенов - их
способность стимулировать анаболические процессы, т.е. рост нежировых
тканей. Стресс-ответ, как известно, связан с интенсивным катабо-
Таблица 23
Растения с подтвержденным адаптогенным потенциалом
Обычное название
Ботаническое название ι Часть растение
Женьшень Panax ginseng Корень
Элеутерококк Eleutherococcus senticosus Корень
Индийский Withania somnifera Корень, семена
женьшень Ashwaganda
Святой Базилик Tulsi. Ocimum sanctum Листья
Золотой корень Rhodiola rosea Корень, корневище
Лимонник Schizandra chinensis Семена
Левзея Leuzea carthamoides Корень
427
лизмом, что выражается в деструкции нежировых тканей для
получения энергоносителей из входящих в их состав белков, что
препятствует восстановлению и регенерации тканей. Некоторые адаптоге-
ны изменяют способ использования энергоносителей таким
образом, что утилизируется больше жира и меньше белков, вследствие
чего уменьшается повреждение тканей и ускоряются
восстановительные процессы. Кроме того, некоторые, но не все адаптогены,
обладают свойствами антиоксидантов, хотя большинство антиокси-
дантов - не адаптогены.
При наличии большого сходства в конечном результате
действие индивидуальных адаптогенов часто локализовано на
различных уровнях и системах организма. Индивидуальные
адаптогены проявляют свою активность на одном или на всех названных
ниже уровнях:
• Центральная нервная система. Некоторые адаптогены,
например лимонник, оказывают влияние главным образом на
центральную нервную систему, которая в свою очередь приводит к
изменениям в метаболизме и уровне некоторых гормонов (Лупандин и др.,
1981; Лупандин 1991).
• Эндокринная система. Адаптогены могут также действовать на
эндокринные железы (Дардымов, 1976). Например, элеутерококк
непосредственно модулирует производство кортикостероидов
корой надпочечников (Тодоров и др., 1984; Todorov, 1989; 1993).
• Клеточный уровень. Адаптогены могут быть эффективны
прежде всего на клеточном уровне (Лупандин, 1991). Leuzea
carthamoides, например, может активировать биосинтез белка и
стабилизировать митохондрии в течение стресса, сохраняя их
способность преобразовать энергоносители в клеточный носитель
энергии - АТФ (Wagner et al., 1992).
29.1. ЖЕНЬШЕНЬ
Женьшень (Panax ginseng из семейства цветковых растений
Araliaceae) возможно самое известное лекарственное растение в
истории. Как оказалось, многие из легендарных свойств женьшеня
являются характерными свойствами адаптогенов. Согласно
различным источникам, женьшень был известен китайской народной
медицине еще 3000-5000 лет назад. До недавнего времени популярность
женьшеня значительно превышала фактическую научную
обоснованность его чудесных свойств. За прошедшие несколько
десятилетий, однако, исследователи, главным образом в России и Китае,
продемонстрировали множество благоприятных влияний женьшеня на
здоровье человека и в особенности на его адаптогенные свойства.
Женьшень - многолетнее растение с отдельным стеблем
высотой до 55-65 см. Активное начало женьшеня содержится в его
428
мясистом корне, размер которого зависит от возраста растения и
может достигать длины от 5 до 50 см.
Интерес к химическому составу женьшеня возник более ста лет назад,
однако лишь развитие Современных методов аналитической химии (различные
виды хроматографии, гель-фильтрации, масс-, ИК-, ЯМР-спектроскопии и др.)
позволило извлекать в чистом виде индивидуальные компоненты женьшеня (а
также элеутерококка и ряда других адаптогенов) и приступить к изучению их
структуры. Выяснилось, что это нейтральные гликозиды с лабильными тетра-
циклическими тршперпеновыми агликанами, содержащими в углеводородных
цепях от 3 до 6 моносахаридных остатков, и таким образом они представляют
собой новый тип тритерпеновых олигозидов. Из метанольного экстракта
корней женьшеня выделено 7 гликозидов, названных в порядке увеличения
полярности на хроматограммах панаксозидами А, В, С, D, E, F, G, содержание
которых в корне составляет от 21 до 28% (Еляков и др.,1962, 1964, 1965, 1972;
Уварова и др., 1963). Ряд данных указывает, что основная биологическая
активность женьшеня обусловлена именно этой необычной группой соединений. В
работах японских ученых гликозиды женьшеня названы - ginsenoside (Shubata,
1975). Было также проведено сопоставление некоторых различий в структуре
панаксизидов и гинзенозидов (Jolliffe, Shellard, 1975).
Как типичный адаптоген, женьшень увеличивает физическую и
умственную выносливость так же как и устойчивость к
разнообразным стрессогенным факторам, включая токсины, радиацию,
чрезвычайно высокую температуру или простуду. Положительные
результаты были получены при лечении туберкулеза в комплексе со
специфическими средствами, малярии, гастритов, болезни Боткина,
диабета, половых расстройств, старческого атеросклероза.
Женьшень уменьшает уровень сахара в крови, улучшает анемию и
нормализует кровяное давление. Различные неврологические и
эндокринные нарушения также нормализуются под влиянием женьшеня
(Брехман, 1957; Дардымов, 1976).
Следует отметить, что токсичность женьшеня чрезвычайно низка и
составляет LD50 = 1.40 ± 0.06 г/кг для суммы панаксозидов, однако она значительно
выше, чем LD5q = 4.74 ± 0.25 г/кг для гликозидов элеутерококка. Тем не менее
токсичность жидких экстрактов женьшеня и элеутерококка почти одинакова -
16.5 г/кг и 14.5 г/кг - соответственно (в пересчете на сухой остаток).
Следовательно, превышение разовой дозы в десятки раз не будет слишком опасно для
жизни больного (Дардымов, 1976).
Женьшень и метаболизм. Женьшень активирует несколько
важных биохимических механизмов организма. При стрессе он
улучшает в клетках энергетическую продуктивность. У крыс,
которых вынуждали плавать в течение двух часов, под действием
женьшеня распад таких соединений, как гликоген, АТФ и
креатин-фосфат был уменьшен, что свидетельствовало об улучшении
коэффициента полезного действия при использовании этих
энергоносителей. Накопление молочной кислоты в мышцах (индикатор
истощения энергоресурсов) было также понижено. Женьшень также
изменяет характер использования топлива в скелетных и сердечно*
429
мышцах при стрессе, стимулируя преимущественную утилизацию
жирных кислот вместо глюкозы, что помогает, как мы знаем,
сохранить белки от распада при синтезе из них глюкозы в процессе глю-
конеогенеза, так как организм способен производить глюкозу
только из белков, но не из жира. Кроме того, женьшень повышает
потребление кислорода, увеличивая способность мышц извлекать
кислород из крови. Так, сумма панаксозидов повышала потребление
животными 02 на 80% в диапазоне доз 5-17 мг/кг (Дардымов, 1976)
приводила к положительному азотистому балансу и, как уже
указывалось, активации использования липидов в качестве источников
энергии (Brekhman, Dardymov, 1966). Интересно, что конечный
результат этих метаболических эффектов женьшеня весьма сходен с
параметрами улучшенного физического состояния, которое
вырабатывается при регулярных физических упражнениях. В некотором
смысле, потребление женьшеня в качестве пищевой добавки может
быть самым коротким путем для достижения некоторой физической
формы, "оставаясь при этом на своем удобном диване".
Препараты женьшеня оказывают влияние и на половую функцию, ускоряя
развитие семенных пузырьков и предстательных желез инфантильных мышей-
самцов, а также стимулируя гонадотропную функцию гипофиза. Однако последний
эффект зависел от количества Сахаров в боковых цепях, так как активность пана-
ксозида С (4 сахара) на 30% выше активности панаксозида А (3 сахара). Для
биологического эффекта панаксозидов необходимо (как это характерно для
большинства адаптогенов), чтобы половая функция была не только измененной, но и
способной к восстановлению до уровня, присущего нормальному взрослому животному,
как это имеет место у инфантильных мышей (Дардымов, 1976).
Экстракт женьшеня понижает уровень сахара в крови при
алиментарной и адреналиновой гипергликемии. Аналогичным
действием обладают и панаксозиды. Препараты женьшеня оказывали
положительное влияние на течение аллоксанового диабета у животных
(Брехман, 1957; Бездетно и др., 1961).
Механизм гипогликемического действия женьшеня, по-видимому, связан с
проницаемостью клеточных мембран для глюкозы и с гексокиназной реакцией.
Давно известно, что β-липопротеиды сыворотки крови больных диабетом (или
животных с аллоксановым диабетом) тормозят захват глюкозы изолированной
диафрагмой крысы (Bornstein, 1953; Bornstein et al., 1953) и активность гексоки-
назы - ключевых звеньев метаболизма глюкозы. При сахарном диабете корти-
костероиды и β-липопротеиды плазмы крови образуют комплекс, который
приобретает свойство ингибировать утилизацию глюкозы тканями (Ильин и
ДР-, 1965).
Очевидно, именно этот механизм активируется при интенсивной
стресс-реакции, когда уровень глюкозы, липопротеидов и кортизола значительно
повышен (см. гл. 12). Гликозиды женьшеня препятствуют развитию описанных
выше процессов.
Детально исследовалось анаболическое действие женьшеня.
Так, экстракт этого адаптогена повышал активность РНК-полиме-
разы печени крыс в течение восьми часов (Hiai et al., 1971), а также
430
стимулировал включение меченых предшественников в ядерную
РНК на 50-60% через четыре часа после внутрибрюшинного
введения препарата. При этом количество тотальных рибосом
увеличивалось на 20-30%, а рибосом, связанных с мембранами, на 30-50%; эта
активация белок-синтезирующей машины выражалась также и в
том, что in vitro полисомы печени увеличивали синтез белка (Oura et
al., 1974).
Панаксозиды увеличивали включение меченных аминокислот в
белки костного мозга и тестикул, а также митотический индекс ми-
элоидной и эритроидной тканей, ретикулоцитах и всех ядерных
клетках косного мозга, т.е. женьшень активировал кроветворение
(Yamamoto et al., 1975; Kim, 1974).
Большой интерес вызывают данные о влиянии экстракта
женьшеня на содержание циклических нуклеотидов, которые как
известно, служат внутриклеточными посредниками гормонов {Sutherland et
al., 1966). Введение животным экстракта женьшеня через час
повышало в крови содержание цАМФ на 40%, на этот показатель в
печени и мышцах препарат не влиял. Однако женьшень увеличивал
содержание цГМФ в крови, печени и мышце в 1.5-2 раза (Дардымов,
1987). При этом, как видно, снижалось отношение цАМФ/цГМФ,
характерное для интактных животных, т.е. наблюдалась активация
"холинэргической" (анаболической) фазы метаболизма.
Женьшень и стресс. Способность женьшеня защитить организм
от разрушительной мощи чрезмерного стресс-ответа была впервые
документирована российским ученым И. Брехманом и его
сотрудниками (Брехман, 1957; Кириллов, 1966; Дардымов, 1976). Когда
крысы подвергались действию серьезного стрессора, например
длительной иммобилизации или плаванию в холодной воде, у них
развивалось множество характерных патологических изменений, включая
гипертрофию надпочечников, инволюцию тимуса и лимфатических
узлов и образование язв желудка, т.е. типичная триада Селье. В
работах школы И. Брехмана было показано, что экстракт женьшеня
обеспечил значительную степень защиты против всех этих
патологических изменений. Кроме того, женьшень смягчал гипертонию,
гипотермию, предотвращал изъязвление слизистой желудка, под
его влиянием надпочечники восстанавливали потерю аскорбиновой
кислоты, вызванную стрессом (Takagi, 1974; Petkov, 1975),
снижалось выведение с мочой 17-кетостероидов (Дардымов, 1976).
Препарат препятствовал распаду тканевых белков и повышению сахара в
крови, характерных для развития стресса. Эти эффекты женьшеня
могут быть, по крайней мере частично, связаны с понижением
интенсивности стресс-ответа и повышением реактивности тканей к
инсулину, которая обычно понижена в течение стресса.
Обобщение многочисленных клинических и
экспериментальных исследований позволяет сделать вывод (Дардымов, 1987), что
при стрессе препараты женьшеня оказывают четкое защитное дей-
431
ствие на стадии тревоги с уменьшением сдвигов в обмене веществ
катаболического характера. При этом удлиняется стадия
сопротивления общего адаптационного синдрома (ОАС) и замедляется
наступление стадии истощения.
Женьшень как антиоксидант. Антиоксидантные свойства -
также среди многих удивительных свойств женьшеня. Так, было
показано, что гликозиды женьшеня и элеутерококка способны
защитить эритроциты от повреждений свободными радикалами. Обыч
но, когда эритроциты подвергаются интенсивной атаке СР в
условиях их высокой концентрации (например, при повышенном процессе
перекисного окисления липидов - ПОЛ), мембраны разрываются,
поскольку СР делают их хрупкими и лишают герметичности. Это
явление называется окислительным гемолизом. Даже очень низки<
концентрации гликозидов женьшеня и элеутерококка были способ
ны предотвратить окислительный гемолиз эритроцитов. В дополне
ние к его действию, непосредственно в качестве антиоксиданта,
женьшень также повышает собственную антиоксидантную защиту
клеток, стимулируя синтез NADH, ключевого кофермента и
энергоносителя, который является также мощным антиоксидантом (Брех-
ман и др., 1968; Brekhman, Dardymov, 1986).
Женьшень может работать еще лучше в комбинации с другими
антиоксидантами. Так, в клиническом исследовании, сорок
пациентов с пониженным мозговым кровообращением из-за
атеросклероза подвергались лечению стандартными успокаивающими и
сосудорасширяющими лекарствами. Кроме того, двадцать пациентов
получали антиоксиданты (витамины Ε и С) и экстракт женьшеня. В этой
группе наблюдалось более значительное улучшение мозгового
кровообращения и лучший профиль липидов, чем в группе, получавшей
стандартное лечение (Воскресенский и др., 1986).
Женьшень и центральная нервная система (ЦНС). В опытах на
лабораторных животных и наблюдениях на людях установлено, что
женьшень регулирует активность мозга: улучшаются показатели
возбуждения и активного торможения. Однако эти влияния зависят
от типа высшей нервной деятельности. Препарат влияет на
метаболизм нейронов, транспорт аминокислот через гемато-энцефаличе-
ский барьер и на обмен моноаминов головного мозга (Petkov, 1981).
У адреналэктомированных животных препарат повышал
связывание кортикостероидов с определенными отделами мозга, а также
чувствительность гипоталамуса и гипофиза (Fulder, 1981).
Геропротекторное действие. На популяции мух W* и дикой
линии дрозофилы Oregon и мутантах линии White и Ebony изучалось
геропротекторное действие 0.25-2.5% водных экстрактов женьшеня,
элеутерококка и шиповника. Все экстракты увеличивали (в
зависимости от линии мутантов) продолжительность жизни и
плодовитость мух, особенно при стрессе. Выявлено антимутагенное
действие изучавшихся экстрактов (Бердышев и др., 1981).
432
* * *
Стресс-протекторное действие женьшеня в определенной
степени связано с ингибированием функции оси
"гипоталамус-гипофиз-кора надпочечников". Механизм этого действия не вполне ясен
в отличие от такового в действии элеутерококка (см. ниже). Для
женьшеня, по-видимому, в большей степени характерно его
модулирующее действие на комплекс факторов нейроэндокринной
регуляции метаболизма углеводов: снижение отношения цАМФ/цГМФ,
снятие β-липопротеидного блока с клеточных мембран, повышение
активности гексокиназы, глюкозо-6-фосфатдегидрогеназы, РНК-
полимераз. В итоге женьшень оптимизирует энергетическое и
пластическое обеспечение организма, что вызывает снижение стресс-
реакции по принципу отрицательной обратной связи. Определенную
роль в действии гликозидов играет их прямое и опосредованное
гашение реакций ПОЛ.
Использование. Дозы женьшеня, обычно используемого для
лечения или профилактики, нетоксичны и редко вызывают побочные
эффекты. Возможные побочные эффекты включают возбуждение,
бессонницу и повышенное давление крови. Высокие дозы (в десятки
раз выше, чем терапевтические дозы) могут проявить серьезную
токсичность. Женьшень обычно не рекомендуется здоровым людям до
40 лет. Доза, рекомендуемая людям среднего и пожилого возраста
для предотвращения болезней и общего замедления старения,
эквивалентна количеству в 0.25-0.5 г сухого корня два или три раза
ежедневно. Женьшень обычно рекомендуется принимать 15-20 дней с
последующим перерывом по крайней мере в две недели. Далее
указанные циклы могут повторяться.
29.2. ЭЛЕУТЕРОКОКК
Элеутерококк {Eleutherococcus senticosus) - отдаленный
родственник женьшеня, оба являются членами семейства Araliaceae. Он
меньше известен на Западе чем женьшень, но восточные целители
использовали его в течение тысяч лет. Кроме того, начиная с 50-х
годов прошлого века элеутерококк в качестве адаптогена стал
фокусом многочисленных исследований в Советском Союзе. К
настоящему времени опубликовано более двух тысяч исследований об
элеутерококке (сибирском женьшене), выполненных главным образом
в СССР, Японии и Европе. Несколько международных симпозиумов
были посвящены исключительно элеутерококку.
Чем объяснить феноменальный интерес исследователей к этому
растению? Во-первых, элеутерококк - высокоэффективный адап-
тоген, по некоторым показателям даже более эффективный, чем
"классический" женьшень. Во-вторых, он - практически безопасен,
433
даже относительно высокие дозы элеутерококка (в отличие от
женьшеня) не вызывает побочных эффектов. В-третьих,
элеутерококк широко распространен в природе, его легче выращивать, чем
женьшень, наконец, он - относительно недорог.
Элеутерококк - куст, высотой 1.5-2.5 м с эллиптическими
листьями и легкой серой или коричневатой корой. Растение найдено в
различных регионах Дальнего Востока. Большая часть
биологической активности элеутерококка связана с соединениями, которые
называют элеутерозидами, их содержание служит индикатором
активности элеутерококка при приготовлении экстрактов.
Химическая структура элеутерозидов (см. обзор: Todorov, 1993) была
установлена Ю.С. Оводовым и его сотрудниками (Оводов и др., 1965;
1966; 1967; 1969).
Как истинный адаптоген, элеутерококк благоприятно влияет на
многие аспекты здоровья и процесс старения. Он препятствует
негативным эффектам стресса, токсинов и экологических загрязнений,
увеличивает физическую и умственную работоспособность,
повышает иммунитет и способствует предотвращению и полному
обращению многих возрастных болезней, в частности атеросклероза,
диабета и гипертонии.
Действие элеутерококка на организм включает несколько
механизмов. Подобно женьшеню, он повышает энергопродукцию и
улучшает эффективность использования энергии клетками. Это
достигается активацией ключевых ферментов, ответственных за
использование глюкозы (в частности, гексокиназы), а также
увеличением использования жира для энергетических нужд. Другое важное
свойство этого адаптогена - временный запрет производства
стероидов стресса, что сокращает разрушительное действие последних на
организм. Наконец, элеутерококк уменьшает повреждения,
которые свободные радикалы причиняют клеточным структурам,
особенно при развитии стресс-реакции. Конечный результат этих
эффектов - повышение способности организма приспособиться к
неблагоприятным ситуациям без генерирования интенсивного и
разрушительного стресс-ответа (Брехман^ 1957; 1960; Дардымов, 1976;
1987; Тодоров, 1993; Todorov, 1993).
Элеутерококк и стресс. Многие исследования показали, что
элеутерококк значительно увеличивает устойчивость к стрессорам,
а также умственную и физическую выносливость. Так, изучалась
способность мышей подниматься по "бесконечной" толстой струне
до полного истощения. Животные, которые получили однократную
дозу экстракта элеутерококка, были способны подняться в среднем
на 45% выше (в зависимости от дозы препарата), чем контрольные
животные (Дардымов, 1976). В другом интригующем исследовании
выяснялась способность элеутерококка уменьшать
нервно-психический стресс у крыс. Для крыс, так же как и для людей,
перенаселенность - существенный психологический стресс. Когда животным
434
предоставлен выбор между питьевой водой или раствором алкоголя,
они потребляли при нормальных условиях относительно немного
этилового спирта. Под психологическим стрессом скученности
животные стали потреблять значительно больше этилового спирта,
мало отличаясь от людей, которые предпочитают "снимать" все
виды стресса с помощью спиртных напитков. Поразительно, но кры
сы, регулярно получавшие экстракт сибирского женьшеня при
перенаселенности, потребляли этилового спирта на 40% меньше, чем в
контрольной группе {Буланов и др., 1986).
У животных, подвергнутых серьезному стрессу, элеутерококк
заметно уменьшал типичные повреждения, такие как образование
язв желудочно-кишечного тракта, гипертрофию и кровоизлияния в
надпочечниках, истощение нежировой ткани, т.е. типичные
морфологические проявления О АС (см. обзор: Todorov, 1993).
Проведено также большое количество исследований на людях,
доказывающих способность снижать стресс и повышать
выносливость пациентов. Например, изучалось влияние экстракта этого
адаптогена на работоспособность молодых спортсменов-мужчин.
Экстракт или плацебо (по 2 мл) принимались спортсменами два раза
в день в течение восьми дней. Адаптоген увеличил
работоспособность на 23.3 и 7.5% у спортсменов, получавших адаптоген и
плацебо соответственно. Многочисленные другие исследования в таких
разнообразных группах, как диспетчеры авиалиний, атлеты,
операторы телеграфа, водители грузовиков и смешанная группа людей,
свидетельствуют, что элеутерококк улучшает умственную и
физическую деятельность и устойчивость людей к стрессу. Кроме того,
этот адаптоген значительно быстрее восстанавливает силы после
тяжелой рабочей нагрузки.
Элеутерококк улучшает устойчивость к токсинам, ядам и
загрязняющим веществам. Другая примечательная и хорошо
документированная способность элеутерококка - снижать ущерб от
ядовитых химикалий. В сегодняшнем мире - загрязнения окружающей
среды, многочисленных добавок к пищевым продуктам для
повышения их сохранности и внешнего вида, а также агрессивной
лекарственной терапии - такая способность имеет непреходящую ценность.
Антитоксическая деятельность элеутерококка охватывает
широкое разнообразие химикалий, хотя величина эффекта связана со
спецификой токсина. Был исследован защитный эффект адаптогена
против эндотоксина столбняка (высокоядовитый химикат,
производимый вызывающими столбняк бактериями Clostridium tetani). Двум
группам мышей вводился эндотоксин, и одной из групп давали
однократно экстракт элеутерококка в дозе 2.5 мг/кг. После 48 часов все
животные, которые получали только эндотоксин, были мертвы,
тогда как 70% животных, получавших элеутерококк, были живы.
В другом исследовании, две группы животных подвергались
действию хлорофоса, токсина поражающего нервы. В одной группе жи-
435
вотные предварительно (за 30 мин до экспозиции с токсином)
однократно получали элеутерококк. 73% животных, получающих
только хлорофос, погибли. В группе, принимавшей адаптоген, погибло
только 20% животных. Элеутерококк также понижал токсические
эффекты различных инсектицидов и индустриальных
загрязняющих веществ {Брехман, 1968; Дардымов, 1976).
Элеутерококк также уменьшает ядовитые побочные
эффекты многих лекарственных средств, используемых в
химиотерапии, что существенно повышало шансы успешного завершения
лечебного курса. В экспериментах на крысах с раком Уокера 30%
животных, подвергнутых химиотерапии этимидином, умерли, но
все животные, получающие наряду с этим еще и элеутерококк,
выжили (Brekhman, 1980).
Анализируя данные об антитоксическом действии
элеутерококка, Баренбойм и Козлова (Barenboim% Kozlova, 1985)
предположили, что адаптоген должен активировать в организме
метаболическую дезактивацию ксенобиотиков, связанную с монооксиге-
назными системами, включающими на конечных стадиях оксида-
зу цитохрома Р-450.
Элеутерококк - предотвращение и лечение болезней.
Элеутерококк благоприятно влияет на состояние здоровья, в особенности,
на многие возрастные болезни. Неполный список включает
атеросклероз (и его осложнения - болезни сердца и инсульт), гипертонию,
умеренные формы диабета, грипп, анемию и почечную
недостаточность. У пациентов с сердечными болезнями элеутерококк улучшал
характер электрокардиограмм (в 74% случаев), и уменьшал
факторы риска, в частности, уровень холестерина и триглицеридов в
крови. Режим лечения рака может также быть интенсифицирован с
помощью сибирского женьшеня, потому что он снижает токсичность
химиотерапии и радиации, повышает скорость восстановления при
хирургическим стрессе и уменьшает риск рецидивов, активируя
иммунную систему.
Касаясь терапевтического потенциала элеутерококка, И. Брехман (1968) -
пионер исследований адаптогенов, писал: "Элеутерококк - лекарство не от
многих болезней, а для многих самых различных больных, независимо от диагноза.
Для больных особенно важно повышение сопротивляемости организма,
которое дает элеутерококк. Он - не панацея, но он необходим практически всем
больным".
Элеутерококк - замечательно эффективный профилактический
инструмент. В 1973-1976 гг. тысячам индустриальных рабочих в
СССР давали экстракт элеутерококка в качестве
профилактической меры по 2 мл в день в течение двух месяцев каждый год. Это
относительно скромный режим уменьшал сумму всех болезней до
35 % по сравнению с контрольными группами (100%). Имеются
признаки, что более длительное использование адаптогена может
уменьшать заболеваемость еще больше.
436
Таблица 24
Влияние экстракта элеутерококка иа заболеваемость
1200 водителей грузовиков в течение 1973-1979 гг.
{данные нормированы на 100 водителей) {Щежин и др., 1981)
Год
Уровень заболеваемости
Грипп
Потеря
рабочих дней
Число
случаев
Гипертония
Потеря ра» Число
бочих дней случаев
Ишемическая болезнь
Потеря
рабочих дней
Число
случаев
1973
1974
1975
1976
1977
1978
1979
286.2
171.5
188.26
144.26
73.73
31.82
11.32
41.77
30.49
33.03
24.04
12.86
5.72
2.75
170.95
45.82
23.21
23.76
18.80
20.74
5.3
6.58
3.2
2.57
2.67
1.16
2.08
0.45
281.82
210.04
51.05
35.24
21.47
11.96
2.98
6.71
5.34
1.62
1.49
1.30
1.46
0.22
Еще более удивительный результат принесли исследования
1200 водителей грузовиков, проведенные российскими врачами.
Обследуемым давали экстракт элеутерококка два раза в день с чаем
по месяцу весной и осенью в течение семи лет. Как видно из
табл. 24, имело место значительное сокращение случаев болезни
сердца, гипертонии и гриппа (другие заболевания не изучались). Это
снижение особенно примечательно, если учесть, что участники
исследования становились старше и риск гипертонии и болезней
сердца у них, как предполагалось, повышался. Казалось бы, недостаток
этого удивительного исследования заключался в том, что не было
группы контроля, получавшей плацебо. Однако этот контроль
весьма важен только при кратковременных исследованиях, когда в
испытания вовлечено относительно немного индивидуумов. Вообще
эффекты плацебо непродолжительны, и они вряд ли заметно
изменили бы семилетние результаты влияния адаптогена,
поразительные по своей длительности и лечебному эффекту. Кроме того,
обследованная группа водителей грузовиков была настолько большой,
что общая совокупность водителей грузовиков соответствующего
возраста может рассматриваться в первом приближении как вполне
приемлемая группа контроля.
Элеутерококк, толерантность к глюкозе и диабет. Как уже
не раз обсуждалось ранее, повышенная глюкоза в крови - одно из
метаболических изменений, которые вносит значительный вклад в
процесс старения. Отмеченное повышение глюкозы в крови, если
оно значительно, является главным признаком диабета. Умеренное
повышение указывает на недостаточную толерантность к
углеводам, которая менее опасна, чем диабет, но все же вносит существен-
437
ный вклад в преждевременное старение и болезни. Приведение
уровня глюкозы крови к оптимальным значениям - важная часть
стратегии антистарения.
Женьшень несколько снижает уровень глюкозы в крови.
Однако элеутерококк, по крайней мере вдвое, превышает
указанный эффект женьшеня. Элеутерококк эффективен в повышении
недостаточной толерантности к углеводам и иногда даже
способен излечить умеренные формы диабета. Он может
использоваться в сочетании с любыми другими терапевтическими
антидиабетическими мерами.
Элеутерококк как антиоксидант и антимутаген. При стрессе
количество различных повреждений, причиняемых свободными
радикалами многим структурам, увеличивается, но особенно
существенны поражения биологических мембран, содержащих легко окис
ляющиеся ненасыщенные жирные кислоты. В исследованиях на жи
вотных, подвергнутых стрессу, элеутерококк заметно уменьшал
уровень ПОЛ в клеточных мембранах по сравнению с контролем.
Антиоксидантное действие адаптогена, по-видимому, косвенно и,
очевидно, опосредуется путем стимуляции собственных
внутриклеточных антиокислительных систем (Дардымов, 1987).
Элеутерококк также снижает вредоносное действие мутагенов,
соединений, вызывающих повреждения ДНК и нарушающих
генетическую информацию. У плодовой мушки Drosophila элеутерококк
снижал количество мутаций, вызванных типичным мутагеном N-hu-
трозоморфолином, примерно на 50%. Кроме того, адаптоген
уменьшал также уровень спонтанных (т.е. возникающих в отсутствие
экзогенных мутагенов) мутаций и увеличивал продолжительность
жизни плодовой мушки (см. обзор: Todorov, 1993).
Другие эффекты элеутерококка. Исследования указывают на
многие другие полезные свойства сибирского женьшеня. Например,
при изучении плодовитости быков, элеутерококк (измельченный
корень которого давался животным с кормом) увеличил количество
эякулята на 21% и индекс активности сперматозоидов до 35%. Куры,
которых кормили адаптогеном в течение месяца, несли яиц на 133%
большее, чем в контрольной группе (см. обзор: Дардымов, 1976).
У здоровых взрослых молодых людей, получавших однократ
ную дозу элеутерококка, острота зрения увеличилась на 30% после
приема адаптогена на восемь часов. Препарат также увеличивал
цветовое восприятие как у нормальных, так и страдающих цветовой
слепотой людей. Однократная доза (2 мл) элеутерококка приводила
к значительному уменьшению (по сравнению с плапебо) количества
ошибок, сделанных операторами телеграфа.
В большом количестве исследований элеутерококк существенно
улучшал многие аспекты иммунитета (Брехман, 1968, Элъкин и др.,
1986; Купим и др., 1986; Кропотов и др., 1986). В частности, ускорял
рост и увеличивал активность иммунных клеток, таких как лимфо-
438
циты, N-киллеры и макрофаги, а также повышал производство γ-
интерферона, белка с противовирусными и антираковыми
свойствами (Wacker, 1983; Ψ acker et alM 1986). Польза для иммунной системы
от этого адаптогена,*ю-видимому, двойная. Во-первых, он
уменьшает интенсивность стресс-ответа и таким образом защищает
иммунную систему от разрушительных действий стресса. Во-вторых, он
непосредственно стимулирует активность иммунно-компетентных
клеток.
29.2.1. Механизм антистрессового
и анаболического действия
Определенный вклад в понимание антистрессового и
анаболического действия элеутерококка внесла серия исследований,
проведенных в 80-е годы в Институте проблем химической физики
Российской академии наук под руководством одного из авторов этой книги
его сотрудниками совместно с сотрудниками И Брехмана -
Г.Н. Бездетко и И.В. Дардымовым. Исходной точкой для этой
работы послужили уже известные данные о химической природе
биологически активных компонентов экстрактов элеутерококка.
Ими являются семь гликозидов, обозначаемых А, В, В-1, С, D, Ε
и F и названных элеутерозидами. Эти гликозиды были выделены в
кристаллическом виде, что позволило установить их химическую
структуру. Элеутерозид В составляет вместе со своими димерами (D
и Е) примерно 2/3 всего биологически активного материала в
экстракте элеутерококка.
Таким образом, задача существенно упрощалась, будучи сведена
на первом этапе к изучению молекулярного механизма действия
индивидуального соединения - элеутерозид а В. Более того, этот
элеутерозид удалось пометить радиоактивным водородом (тритием),
что существенно расширяло исследовательские возможности.
Например, это позволило установить скорость выведения данного
вещества из организма {Бездетно и др., 1981), его локализацию по
органам и тканям, а также внутриклеточную локализацию {Герман и
др., 1982), скорость и характер распада (Бездетко и др., 1982).
Существенную роль в этих исследованиях сыграл системный подход,
разработанный ранее при изучении стабильности клеток высших
организмов в экстремальных условиях (Todorov et al., 1984)
С помощью радиоактивного элеутерозида В в опытах на
животных были сделаны очень интересные наблюдения. При общем
экспоненциальном характере выведения из органов элеутерозид В
накапливался в гипофизе, надпочечниках и сердце. Анализ его
внутриклеточного распределения показал, что этот элеутерозид
аккумулируется в мембранах эндоплазматического ретикулума (где
происходит биосинтез белков, окислительные и другие процессы) надпочеч-
439
ников, поджелудочной железы и тимуса, а также в ядрах (где
локализован генетический аппарат клетки) поджелудочной железы
{Герман и др., 1982). Хотя эти факты и не говорили о молекулярном
механизме действия элеутерозида, они тем не менее указывали, какие
органы и внутриклеточные системы скорее всего участвуют в
реализации эффектов элеутерококка.
Далее выявлялась возможная корреляция между
распределением радиоактивного элеутерозида В в органах и тканях и их вкладом
в общий адаптогенный эффект. Особое внимание привлекали
надпочечники и поджелудочная железа, поскольку врачам давно были
известны антистрессовый и противодиабетический эффекты адап-
тогена, которые, по-видимому, связаны с этими эндокринными
железами. Системное исследование динамики биосинтеза белков и
других процессов метаболизма, в частности внутриклеточного
окисления, а также синтеза ряда гормонов (инсулина, кортикостерона) и их
содержания в крови, прояснило биохимическую природу действия
этого адаптогена на организм животных и человека.
Анализируя надпочечники у животных методом электронно
го парамагнитного резонанса (ЭПР), мы обнаружили, что
введение элеутерококка блокирует основные звенья
внутриклеточного окисления (цитохром Р-450, адренодоксин) (Тодоров и др.,
1984; Тодоров и др., 1986). А этот эффект должен резко
понижать интенсивность окисления холестерина (предшественника
стероидных гормонов, см. рис. 13) на пути его превращения в кор-
тикостерон (у крыс - аналог кортизола). Ферментативные
системы окисления в основном сосредоточены в эндоплазматическом
ретикулуме. Именно здесь и наблюдалась выраженная
аккумуляция радиоактивного элеутерозида В. Возможно, элеутерозид В
(или какой-либо другой элеутерозид экстракта элеутерококка)
имеет повышенное сродство с какой-либо гидроксилазой или ци-
тохромом Р-450, конкурируя с холестерином за связывание
активным центром этого фермента и, следовательно, блокируя
биогенез кортикостероидов. Параллельно с этим у животных, которым
вводился экстракт элеутерококка, понижалось содержание кор
тикостерона в крови и продуктов его превращения (17-кетостеро
идов) в моче (Дардымов, 1987).
У животных в состоянии стресса повышенное содержание кор
тикостероидов в крови и моче после введения элеутерококка быст
ро возвращалось к норме (Дардымов, 1987). По-видимому, эти
соединения, временно блокируя процессы окисления в надпочечниках
и биогенеза кортикостероидов, снижают напряженность начального
периода стресс-реакции, таким образом развитие фазы
сопротивления происходит менее интенсивно и, следовательно, с меньшими
вредоносными последствиями для организма. Таковы, очевидно,
основные черты механизма антистрессового действия экстрактов
элеутерококка.
440
Экстракт элеутерококка предотвращает искусственно
вызванный диабет (с помощью аллоксана) у лабораторных животных, а у
людей задерживает его естественное развитие на ранних стадиях
{Бездетно и др., 1961). В понимании этого эффекта пока нет той же
ясности, как в случае антистрессового действия элеутерококка,
однако нам удалось обнаружить некоторые важные факты.
Одной из главных причин развития диабета I и отчасти II типа
является пониженное содержание в крови инсулина, который
синтезируется В-клетками поджелудочной железы. Можно было
предполагать, что противодиабетический эффект элеутерококка связан с
его влиянием на функциональную активность этой железы, тем
более, что ранее мы наблюдали накопление радиоактивного элеутеро-
зида В в ядрах и эндоплазматических мембранах ее клеток {Герман
и др., 1982). Измерения скорости синтеза белков в поджелудочной
железе и содержания инсулина в крови у животных после инъекций
экстракта элеутерококка выявили кратковременное понижение как
скорости биосинтеза белков, так и содержания инсулина в крови,
которое в течение последующих двух-трех часов сменялось их
значительным повышением {Тодоров и др., 1984; Todorov, 1993). Пока
трудно сказать - связана ли эта активация с накоплением элеутеро-
зидов в ядрах, где они могут стимулировать работу генетического
аппарата, или же она является компенсаторной реакцией на
первоначальное подавление синтеза белков в цитоплазме {Todorov, 1990).
Мы предпочитаем второе объяснение, так как ранее нами было
показано, что in vitro в бесклеточной системе с полирибосомами эле-
утерозиды тормозят синтез белка примерно в тех же
концентрациях, что и in vivo {Тодоров И. и др., 1993).
Активация синтеза инсулина в течение нескольких часов после
введения адаптогена влечет за собой ряд изменений, сопряженных с
повышением уровня других анаболических процессов: ускоряется
транспорт питательных веществ из крови в клетки, повышается
энергопродукция, увеличивается скорость синтеза белков и
нуклеиновых кислот (Insulin and Metabolism, 1977; Czech, 1977). Эти
эффекты, очевидно, весьма широко представлены в организме, поскольку
рецепторы к инсулину имеются в очень многих органах и тканях
{Kahn et al., 1979). И это означает, что анаболический эффект
элеутерококка носит генерализованный характер, что согласуется с
многочисленными клиническими наблюдениями {Брехман, 1957,
1968; Дардымов, 1976).
Хотя описанные выше исследования вносят определенную
ясность в представления о механизме действия элеутерококка на
организм животных и человека, они в то же время свидетельствуют о
высокой его сложности, поскольку реакция организма слагается из
многих реакций как внутриклеточного, так и системного характера.
По нашему мнению, ключевым аспектом является ингибирование
биогенеза кортикостероидов элеутерозидами. Это приводит к сни-
441
жению концентрации кортикостероидов в крови, изменяя
соотношение гормонов, стимулирующих процессы синтеза (гормон роста,
ИФР-1, инсулин, факторы роста и др,) и распада (адренокортико-
тропный гормон - АКТГ, кортикостерон, альдостерон), в пользу
первых. Такой системный гормональный сдвиг может ослабить
остроту стресс-реакции и стимулировать синтетические процессы в
организме.
Использование. Элеутерококк - нетоксичен и лишен побочных
эффектов в очень широком диапазоне доз. Типичная дозировка,
рекомендуемая для предотвращения болезней и замедления процесса
старения, - 2 мл экстракта (эквивалентно весу сухого экстракта
120-150 мг) два раза в день в течение месяца. Такой ежемесячный
цикл может быть повторен два-четыре раза в год. В периоды
серьезного физического или нервно-психического стресса та же самая
доза может приниматься три-четыре раза в день до двух недель.
Иногда используется высушенное растение, хотя в таком виде
препарат усваивается хуже, чем экстракт. Дозы для высушенной
формы обычно колеблются от 0.5 до 1.5 г три раза в день.
Женьшень и элеутерококк - хорошо изученные популярные
адаптогены, широко используемые как в профилактической, так и в
клинической медицине. Однако ценными адаптогенными
свойствами обладает и ряд других препаратов, таких как экстракт левзеи, ро-
диолы розовой и лимонника, краткое описание которых будет
представлено ниже.
29.3. ЛЕВЗЕЯ САФЛОРОВИДНАЯ
Левзея сафлоровидная (Rhaponticum carthamoides) - травянистое
многолетнее растение высотой 80-120 см, семейства сложноцвет
ных (Compositae), произрастает в Сибири, на Алтае и в Саянах. Для
медицинских целей используют корневище растения. Экстракт
корней левзеи оказывает адаптогенное и тонизирующее действие на
организм высших животных и человека (Саратиков, 1966; Герасюта
и др., 1981; Соколов, Замотаев, 1993; Машковский, 1993), которое,
по-видимому, связано с присутствием в нем алкалоидов (Якунина,
1975), тритерпеновых сапонинов и флавоноидов (Вересковский и
др., 1974; Varga et al., 1990). Так, с алкалоидами, очевидно, связано
возбуждающее действие на центральную нервную систему
(Alcaloids, 1950-1955), с сапонинами - стимулирующее действие на
сердечно-сосудистую систему (Гурова, 1974), а с флавоноидами, в
частности кверцетагитрином и паулетином, - антиоксидантные
свойства (Varga et al., 1990; Cody et al., 1986, 1988).
442
Наряду с названными выше биологическими свойствами
экстракта левзеи, можно также предполагать и возможность его
анаболического действия, поскольку в этом растении было обнаружено
значительное количество экдистерона (Абубакиров, 1975), который
проявляет анаболическую активность при введении теплокровным
животным (Сыров и др., 1976; Сыров, 1984; Сыров, 2000).
Экдистерон принадлежит к группе гормонов (α-экдизон н β-экдизон),
определяющих конечный этап морфологического развития насекомых, т.е. их линьку
(переход: куколка - взрослая особь). Значение экдистероидов вышло за рамки энто
мологии, когда в 1966 г. японские химики К. Наканиси и его сотрудники выделили
из ряда растений соединения, которые (к их величайшему удивлению) оказались по
структуре идентичны ос-экдизону и обладали истинными свойствами гормона
линьки насекомых. Почти одновременно австралийские химики Гэлбрайт и Хорн
(Nakanishi et al., 1966; Galbraith, Horn, 1966 - цит. по: Абубакиров, 1981) выделили
из растений другой гормон линьки насекомых - β-экдизон, или, как принято его
называть в настоящее время, экдистерон, или фитоэкдистерон. Еще больший
интерес к этому классу соединений возник после того как было показано, что при
введении высшим животным, не способным к его эндогенной продукции, экдистерон
проявлял явно выраженную анаболическую активность. Под влиянием этого
гормона наблюдалась стимуляция биосинтеза РНК и белков, инициации и элонгации
процесса трансляции, пролиферации клеток высших животных, различных
аспектов иммунитета, репродуктивных процессов и ряда других сторон метаболизма
животных (см. обзоры - Ахрем и др., 1989; Stoma, Lafont, 1995). Динамика биосинтеза
белков, РНК и ДНК в органах мышей исследовалась под влиянием высокоочищен-
ного препарата экдистерона из левзеи {Тодоров и др., 2000а,б). Экдистерон
значительно повышал уровень анаболических процессов в печени, поджелудочной
железе, тимусе, надпочечниках и в меньшей степени в семенниках. Динамика синтеза
РНК в этих органах весьма напоминала таковую при вступлении клеток в G-1
фазу клеточного цикла, поэтому последующий подъем синтеза ДНК в этих органах
(за исключением поджелудочной железы) можно рассматривать как вступление
некоторой части их клеток в S-фазу клеточного цикла. Гипотетические
молекулярные механизмы воздействия экдистерона на анаболические процессы
животных рассматриваются в (Тодоров и др., 2000а).
По-видимому, определенные перспективы имеет использование экдистерона в
клинической онкологии. Современная концепция химиотерапии опухолевых
заболеваний предполагает использование высоких, приближающихся к максимально
переносимым, доз противоопухолевых цитостатиков, обладающих, как правило,
весьма высокой токсичностью. Так, недавно в лабораторных исследованиях было
показано, что комбинированное применение экдистерона и цитостатиков
(например, цисплатина и адриамицина) позволяет снизить дозу последних на 50% при
полном сохранении или даже превышении терапевтического эффекта, наблюдаемого
при полной дозе только одного цитостатика, без каких-либо серьезных, что
крайне важно, токсических эффектов. Эти обнадеживающие результаты авторы
связывают с широким спектром анаболических эффектов, которые вызывает
экдистерон в организме высших животных (Коновалова и др., 2002).
Влияние экстрактов левзеи, с различным содержанием
экдистерона, выявило сходную с последним динамику биосинтеза белков и
РНК в органах мышей, однако это сходство было тем большим, чем
выше была концентрация экдистерона в препаратах левзеи
(Тодоров К и др., 20006).
443
Активация биосинтетических процессов в печени в наших
экспериментах, хотя и не позволяет прямо говорить об активизации
конкретных функций печени, тем не менее имеющиеся в литературе
данные о снижении токсического влияния хлорофоса и тетрахлор-
метана (Левицкий и др., 1993а ; Губский и др., 1993), а также
эффектов Д-авитаминоза (Левицкий и др., 19936) под действием экдистеро-
идов указывают на активацию ими защитных функций печени. Об
этом же свидетельствуют и данные о повышении активности цито-
плазматической супероксид дисмутазы (СОД) и Μη- содержащей
СОД митохондрий печени после однократного введения экдистеро-
на (Тодоров и др., 2000а).
Стимулирующее влияние препарата левзеи на тимус хорошо
коррелирует с данными о влиянии экдистерона на ряд аспектов
иммуногенеза (Сахибов и др., 1989; Кузмицкий и др., 1990; Фомовская
и др., 1992).
Активация биосинтеза макромолекул в семенниках
крыс-самцов под действием экстрактов левзеи (Тодоров и др., 20006) хорошо
согласуется с наблюдением, что экдистерон повышал сексуальную
активность крыс-самцов (Мурзаев и др., 1992), а также хорошо
известным свойством экстрактов левзеи восстанавливать пониженную
половую активность у людей (Соколов, Замотаев, 1993).
Представляют несомненный интерес и данные о гипохолестери-
немическом действии фитоэкдистероидов при экспериментальной
гиперхолестеринемии у крыс (Миронова и др., 1982), а также данные
о подавлении развития экспериментально индуцированных
опухолей нервной системы с помощью препаратов левзеи (Rhaponticum
Carthamoides) (Беспалов и др., 1992).
Группе болгарских ученых удалось по некоторым тестам (метод
лабиринта - активное избежание с отрицательным подкреплением)
достоверно показать положительное влияние экстракта левзеи на
реакции центральной нервной системы (Petkov et al., 1984), а также
обучение и память у крыс (Mosharrof, 1987).
* * *
Представленные выше неполные данные о биологической
активности экстрактов левзеи и препаратов экдистерона тем не менее
позволяют, учитывая специфический набор их свойств, с
уверенностью отнести эти препараты к семейству адаптогенов.
Использование. Экстракт левзеи (измельченные корневища и
70% этиловый спирт, 1:1) следует принимать по 20-30 капель два-
три раза в день до еды в течение 3-^4 недель. Перерыв две недели.
Далее цикл можно повторить.
Противопоказаний к применению препаратов левзеи сафлоро-
видной не выявлено.
444
29.4. РОДИОЛА РОЗОВАЯ
Родиола розовая (Rhodiola Rosea Lj, или золотой корень.
Многолетнее травянистое растение, достигающее 50-70 см высоты,
семейства толстянковых (Crassulaceae). Произрастает в Западной и
Восточной Сибири и на Дальнем Востоке. Промышленные
заготовки осуществляют на Алтае. Используют клубневидные корневища
растения.
Корни и корневища растения содержат дубильные вещества пи-
рогалловой группы, органические кислоты, стерины, вещества фе-
нольного характера, гликозиды, флавоноиды, марганец и др.
Установлено, что биологически активными веществами этого препарата
являются фенольные соединения - салидрозид. родозин и тирозол
(Марина и др., 1983; Саратиков и др., 1987).
Тонизирующий и адаптогенный эффекты экстрактов родиолы
розовой (ЭРР) подтверждены многочисленными
фармакологическими исследованиями {Соколов и др., 1985; Саратиков и др., 1987).
Золотой корень нормализует метаболизм, способствует
экономичному расходованию энергетических ресурсов, быстрому их ресинте-
зу, улучшает энергетический обмен в мышцах и мозге за счет
активации окислительного фосфорилирования и более интенсивного
использования в качестве субстратов окисления не только углеводов,
но и липидов. ЭРР оказывают стимулирующее влияние на
умственную работоспособность человека, несколько улучшают память и
внимание (Соколов, Замотаев, 1993).
ЭРР содержат полимерные проантоцианидины (группа
растительных полифенолов), которые оказывают выраженное гипохоле-
стеринемическое и гипотриглицеридемическое влияние, а также
существенно повышают резистентность стенок сосудов, по-видимому,
обеспечивая защиту артериальной стенки от проникновения
холестерина в составе атерогенных липопротеидов (ЛНП) и тем самым
предохраняя ее от атероматозного поражения (Хушбактова и др.,
1989). Как ЭРР, так и ее активные компоненты (салидрозид,
тирозол, розин, розавин и розиридин) проявляют выраженные адапто-
генные свойства, повышая резистентность различных органов и
тканей (желудок, печень) к стрессогенным повреждениям. Особен
но выражено защитное действие ЭРР и его активных компонентов
при повреждениях головного мозга (Барнаулов и др., 1986).
Можно полагать, что указанные защитные эффекты ЭРР и его
компонентов связаны с ингибированием катехол-О-метилтрансфе-
разы (Саратиков и др., 1987; Лупандин, 1991), фермента,
активность которого наиболее высока в фазе "тревоги" общего
адаптационного синдрома Селье.
Таким образом, ЭРР и его компоненты подобно женьшеню и
элеутерококку обладают набором типичных адаптогенных свойств.
Они препятствуют развитию у кроликов гипер- и гипогликемии,
445
лейкоцитоза и лейкопении. При удалении полушарий головного
мозга, гипофизэктомии, а также кастрации животных эти адапто-
генные эффекты ЭРР (как и в аналогичной ситуации, эффекты
других адаптогенов) исчезают {Соколов, Замотаев, 1993).
Использование. Экстракт родиолы принимают внутрь по пять-
десять капель два-три раза в день за 15-30 мин до еды. В течение
10-20 дней.
Противопоказания: резко выраженное возбуждение,
гипертонический криз, лихорадочные состояния.
29.5. ОБ ОБЩЕМ МЕХАНИЗМЕ ДЕЙСТВИЯ
ПОЛИФЕНОЛЬНЫХ АДАПТОГЕНОВ
Адаптогены являются особыми многокомпонентными и
полифункциональными препаратами или соединениями (гликозиды, лиг-
наны, флавоноиды и др.), влияющими на нервную и эндокринную
системы и на ряд аспектов метаболизма тканей и органов. Не
обладая энергетической ценностью или четким специфическим
действием, они повышают устойчивость организма к влиянию многих
экстремальных факторов и их сочетаний, оптимизируя адаптацию к
этим факторам. К наиболее изученным и популярным адаптогенам
следует отнести препараты из женьшеня, элеутерококка, других
растений семейства аралиевых, родиолы розовой, левзеи сафлоро-
видной и лимонника китайского. В последние десятилетия XX в.
научная и практическая медицина проявила к этим препаратам
исключительный интерес, что выразилось, в особенности в СССР, в
проведении и публикации результатов многих тысяч исследований
различных аспектов действия адаптогенов на организм высших
животных и человека. Этот огромный фактический материал,
естественно, требовал своей классификации, осмысления и обобщения.
Наиболее удачная, как нам кажется, предварительная попытка такого
рода была предпринята А.В. Лупандиным из Хабаровского
института физической культуры (Лупандин, 1991).
По мнению этого автора, существует три основных компонента
в формировании апаптации при участии полифенольных
адаптогенов: селективная модуляция и лимитирование функции катехолами-
нергических синапсов (Лупандин, Лаптев, 1981; Лупандин, 1989),
ингибирование перекисного окисления липидов (Барабой и др.,
1992) и зависимое от первых двух компонентов ограничение и
пролонгирование функции гипофиз-адреналовой системы (Тодоров и
др., 1984; Дардымов, 1987; Тодоров, 1993; Todorov, 1993).
В соответствии с этим можно разделить полифенольные
адаптогены на три группы: 1) с преимущественно нейротропным
действием (лигнаны лимонника, салидрозид и др.), 2) с преимущественно ан-
тиоксидантным (флавоноиды, катехины, таннины и др.) и 3) со сме-
446
шанным действием, главным образом на эндокринную систему (гли-
козиды женьшеня, элеутерококка, родиолы розовой и др.). Такое
разделение облегчает адресное использование адаптогенов в
различных стрессогенных ситуациях, например, для срочной оптимиза
ции (аварии, пожары, срочная изнурительная работа в тяжелых
условиях, внезапные военные действия и т.п. адаптогены 1-й и 2-й
групп) или при долговременной адаптации (продолжительное
пребывание в экстремальных условиях, длительная болезнь,
акклиматизация к иным климатическим или социальным условиям и т.д.
адаптогены 2-й и 3-й групп). Немаловажное значение это
разделение имеет и при выборе адаптогенов в качестве геропротекторов
{Обухова, Эмануэль, 1984; Воскресенский, 1986).
Таким образом, адаптогены являются эффективным средством
повышения адаптации и устойчивости организма к действию
многообразных экстремальных стрессогенных факторов различной
природы и их сочетаний.
Глава 30
ПОЛОВАЯ ФУНКЦИЯ ПОСЛЕ ПЯТИДЕСЯТИ
Хочу быть дерзким, хочу быть смелым,
Из сочных гроздий венки свивать.
Хочу упиться роскошным телом,
Хочу одежды с тебя сорвать!
Хочу я зноя атласной груди,
Мы два желанья в одно сольем.
Уйдите, боги! Уйдите, люди!
Мне сладко с нею побыть вдвоем!
Пусть завтра будет и мрак и холод.
Сегодня сердце отдам лучу.
Я буду счастлив! Я буду молод!
Я буду дерзок! Я так хочу!
Константин Бальмонт
Плоды усилий по продлению жизни вряд ли будут столь
заманчивы, если половая функция заметно ухудшается с возрастом. К
счастью, падение половой функции не является неизбежным спутником
старения (мы подробно обсуждали это в гл. 8, 20) и может быть в
значительной степени предотвращено. Более того, большинство
шагов к продлению жизни, обсуждаемых в этой книге, так или
иначе, помогают сохранять здоровую половую функцию в преклонные
годы. Повышение устойчивости к стрессу и ослабление вызванных
им повреждений - меры, заслуживающие особого акцента - также
помогут сохранить здоровую сексуальность.
Однако для некоторых людей, общее оздоровление и усилия,
направленные на сохранения молодости, могут оказаться
недостаточными для поддержания половой функции. Имеется широкий
диапазон мер - от приема определенных питательных веществ и лекарств
до имплантаций в область мужского полового члена - для
стимуляции либидо и половой потенции. У каждого из существующих
подходов имеются плюсы и минусы, и не существует единого
универсального метода лечения для любого пациента. И, следовательно,
для большинства людей поиск адекватных путей достижения
желаемой цели служит серьезной пищей для размышлений.
30.1. ПОВЫШЕНИЕ ПОЛОВОЙ АКТИВНОСТИ
Сосудорасширяющие средства. До недавнего времени
большинство способов лечения эректальной дисфункции было основано
на использовании различных вазодилататоров, т.е. лекарств,
которые расширяют кровеносные сосуды. Цель использования этих ле-
448
карств состоит в стремлении расширить артерии полового члена
настолько, чтобы увеличенный ток крови вызвал эрекцию.
Сосудорасширяющие средства, широко используемые для лечения
импотенции, включают папаверин, фентоламин, нитроглицерин, алпро-
стадил и др. Проблема с этими лекарствами состоит в том, что они
высокоэффективны только тогда, когда они инъецируются
непосредственно в пещеристые тела полового члена. К сожалению,
подобная процедура перед контактом может уничтожить
спонтанность и сексуальное удовольствие для обоих партнеров. Кроме того,
использование сосудорасширяющих средств может привести к
кратковременным или длительным побочным эффектам, таким как при-
апизм (постоянная эрекция, которая может повреждать артерии
полового члена), формирование в члене рубцовой ткани коллагеновой
природы, головные боли, гипотония и др. В результате
большинство людей с эректальной дисфункцией избегало инъекций вазадила-
таторов, даже если это означало сохранение импотенции.
В качестве альтернативы к рассмотренному выше подходу,
некоторые компании развивали другой путь доставки
сосудорасширяющих средств - сквозь маленький катетер в мочеиспускательный
канал. Результаты были смешанными: использование лекарств
стало несколько менее сложным, но эффективность также несколько
уменьшилась.
Локальные поверхностные сосудорасширяющие средства
только минимально эффективны, потому что лишь немногие из них
абсорбируются сквозь кожу. Эффективность сосудорасширяющих
средств, принимаемых per os, также ограничена, потому что
достаточно высокие дозы, способные существенно повысить ток крови к
половому члену, имеют тенденцию значительно снижать давление
крови и вызывают серьезные побочные эффекты, несовместимые с
половыми сношениями.
Виагра (sildenafil) наиболее многообещающий из недавно
введенных лекарств для импотенции и эректальной дисфункции.
Успехи практического использования этого препарата породили
всемирную массовую истерию, подобную золотой лихорадке, в страстном
стремлении получить "волшебный" рецепт новых необыкновенных
удовольствий жизни. Энтузиазм, вызванный использованием этого
лекарства, имеет определенные основания. Виагра показала
улучшение эректальной функции приблизительно у 60-80% (в
зависимости от дозы в диапазоне от 25 до 100 мг) людей с неорганической
импотенцией. Некоторые пациенты со слабой или умеренной
органической импотенцией также могут реагировать на этот препарат.
Виагра, по-видимому, также увеличивает потенцию и у людей с
нормальной эрекцией.
Действие виагры сопряжено с увеличением влияния окиси азота
на артерии полового члена. Механизм эрекции связан с выделением
окиси азота в пещеристых телах во время полового возбуждения.
449
Окись азота активирует фермент, производящий циклический гуа-
нозин монофосфат (цГМФ - сигнальная молекула), который в свою
очередь заставляет расслабляться гладкие мышцы артерий
полового члена, что увеличивает ток крови и порождает эрекцию. цГМФ -
короткоживущая молекула и быстро разрушается ферментом фос-
фодиэстеразой типа 5 (PDE 5). Виагра ингибирует PDE 5, что ведет
к повышению уровня цГМФ и кончается более длительной и более
"твердой" эрекцией. Таким образом, виагра не является причиной
эрекции, но увеличивает последнюю в ответ на половое
возбуждение (Morgenthaler et al., 1995). При рекомендуемых дозах виагра не
вызывает никакого эффекта в отсутствие полового возбуждения.
Побочные эффекты: виагра вызывает меньшее количество
побочных эффектов, чем пероральные сосудорасширяющие средства,
которые обычно используются при эректальной дисфункции.
Побочные эффекты виагры, отмеченные в клинических
исследованиях, включают головные боли, прилив крови, диспепсию
(расстройство желудка), ненормальное зрение. Побочные эффекты
значительно увеличиваются при дозах выше чем 100 мг.
Использование: виагра должна предписываться доктором,
который хорошо знаком с медицинской историей пациента. Начальная
доза, обычно 50 мг, принимается за полчаса-час перед сексуальным
общением. Пища задерживает начало действия виагры, замедляя
ее всасывание. Дозировка может быть увеличена до 100 мг, если
это рекомендуется врачом. Доза выше чем 100 мг минимально
повышает эффект препарата, но значительно увеличивает побочные
эффекты. Виагра не должна приниматься вместе с вазодилата-
торами, особенно нитратами (например, нитроглицерином),
поскольку это может вызвать потенциально опасное снижение
кровяного давления). Дозировку следует уменьшить, если пациенту
более 65 лет, и у него обнаружена недостаточность функций печени
или почек, а также если он принимает циметидин или антибиотики
(в частности, эритромицин).
К настоящему времени не проводилось никаких исследований об
совместном использовании виагры с другими лекарственными
средствами для лечения эректальной дисфункции.
Аргинин - незаменимая аминокислота, которая, помимо своей
роли в синтезе белка в качестве строительного материала,
оказывает разностороннее влияние на организм. Она стимулирует
выделение гормона роста, активирует некоторые аспекты иммунной
функции и является предшественником в биогенезе окиси азота
{Moncada et al., 1991). Последний эффект ответствен за роль
аргинина в механизме эрекции, потому что окись азота - ключевой
медиатор эрекции, необходимый для надлежащего расширения артерий
полового члена. Исследования показали, что аргинин может
вызывать существенное улучшение эректальной функции. Фактически
аргинин работает почти по той же схеме, что и виагра. Как уже от-
450
мечалось выше, виагра увеличивает эрекцию, ингибируя фермент,
разлагающий молекулы цГМФ, повышая таким образом сигналы к
расширению артерий полового члена. Подобно виагре, аргинин
также увеличивает уровень цГМФ, но только делает это, не тормозя
распад цГМФ, а увеличивая производство окиси азота, которая в
свою очередь активирует фермент, который синтезирует цГМФ
{Burnett et al., 1992; Hellstrom et al., 1994; Zorgniotti et al., 1994).
Метафорически говоря, виагра и аргинин наполняют одну и ту же ванну,
но аргинин делает это, открывая кран, а виагра, блокируя водосток.
Эти отношения между механизмами действия виагра и аргинина
говорят, что оба агента, вероятно, будут эффективны при лечении
эректальной дисфункции, вызванной аналогичными причинами.
Следовательно, для многих индивидов, негативно реагирующих на
виагру, аргинин может быть более безопасной и не столь дорогой
альтернативой. Кроме того, теоретически аргинин мог бы позволить
уменьшить дозу виагры без потери ее эффективности. Однако
комбинация аргинина и виагры до сих пор клинически не изучалась и не
должна использоваться без непосредственного наблюдения врача.
Использование: для кратковременного повышения половой
функции аргинин обычно принимается натощак приблизительно за
40-60 мин до начала сексуальной активности. По-видимому,
ежедневное использование аргинина, независимо от того имели ли
место сексуальные контактьгили нет, также вызывает долговременное
улучшение половой функции. Обычный диапазон доз 1,5-5 г.
Тошнота, понос и расстройство желудка могут происходить при более
высоких дозах. Люди с серьезной инфекцией герпеса не должны
принимать аргинин, потому что это может стимулировать
размножение вируса.
Холин и витамин В5. Эрекция - физиологический процесс,
участником которого является не только половой член, но также и
мозг. Все это начинается с полового возбуждения, которое по
чувствительным нервам передается к мозгу. Эти сигналы
обрабатываются мозгом, который после этого посылает уже другой сигнал вниз по
спинному мозгу к нервам, обслуживающим член и включающим
эрекцию. Таким сигналом служит нейромедиатор гцетилхолин.
Имеется ряд свидетельств, что повышение производства ацетилхо-
лина в организме придает дополнительную интенсивность нервным
импульсам, которые инициируют эрекцию. Два соединения
особенно важны в этом отношении: холин и витамин В5 (пантотеновая
кислота). Холин - предшественник ацетилхолина, а витамин В5 - ко-
фермент, требуемый для синтеза ацетилхолина из холина. Их
использование, сопряженное с увеличением содержания
ацетилхолина, может усиливать и продлевать эрекцию у здоровых людей
{Pearson et al., 1991). Другой примечательный эффект холина и В5
состоит в том, что они увеличивают физическую выносливость,
поскольку ацетилхолин является ключевым «ейромедиатором в конт-
451
роле функций скелетной мускулатуры. Большая выносливость,
очевидно, также будет способствовать повышению половой активности.
Однако какие-либо данные о том, что холин и витамин В5 полезны
для излечения эректальной дисфункции, пока отсутствуют.
Использование: для увеличения половой деятельности диапазон
доз холина 1-3 г, витамина В5 - 0.3-1 г. Рекомендуется начинать с
более низких доз, постепенно повышая их, если это необходимо. Для
достижения максимального эффекта холин и В5 должны
приниматься приблизительно за 30-60 мин до начала половой активности.
Существуют данные, что холин и В5, по-видимому, увеличивают
стимулирующие эрекцию эффекты аргинина (Pearson et al., 1991).
Главные побочные эффекты высоких доз холина и В5 -
напряженность мышц, кишечные спазмы и понос. Они следуют из специфики
действия ацетилхолина на скелетную и гладкую мускулатуру. Если
эти эффекты имеют место, прекратите принимать эти пищевые
добавки. Возобновление приема возможно по истечении нескольких
дней, начиная с низких доз и медленно их наращивая.
30.2. ПРЕДОТВРАЩЕНИЕ И ЛИКВИДАЦИЯ
ДОБРОКАЧЕСТВЕННОЙ ГИПЕРТРОФИИ
ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
Кроме атеросклероза, доброкачественная гипертрофия
предстательной железы (ДГПЖ) - главная причина половой дисфункции у
пожилых людей. Более половины мужчин между сорока и
шестьюдесятью страдают ДГПЖ, и даже еще большее их число - в более
старом возрасте. Главные признаки ДГПЖ - проблемы
мочеиспускания: частые мочеиспускания, трудности прохождения мочи,
ночные пробуждения (для освобождения мочевого пузыря) и, в
некоторых случаях, полная преграда мочеиспусканию. Мужчины,
страдающие ДГПЖ, также подвержены увеличенному риску развития
эректальной дисфункции, потому что увеличенная простата может
сжимать артерии и нервы полового члена, с которыми связан механизм
эрекции. Следует также обратить внимание, что хирургическое
лечение ДГПЖ может повреждать иннервацию полового члена,
результатом чего будет уже полная импотенция.
ДГПЖ - типичная возрастная болезнь: риск ее развития
увеличивается с возрастом, и это вызвано, по крайней мере частично,
возрастными гормональными изменениями в организме и в простате
непосредственно. Особенно важно увеличение содержание двух
гормонов - дигидротестостерона и пролактина. Дигидротестостерон
(ДГТ) - производное тестостерона, стимулирующее рост простаты.
Тестостерон преобразуется в ДГТ ферментом 5-альфа - редуктазой,
который содержится в простате, коже и других тканях. Уровень ДГТ
в простате увеличивается с возрастом, что, как полагают, играет
452
ключевую роль в развитии ДГПЖ. Пролактин, гормон,
продуцируемый гипофизом, может увеличивать скорость, с которой простата
поглощает тестостерон из кровяного русла. Это косвенно вносит
вклад в высокий уровень ДГТ в простате и стимулирует ДГПЖ.
Уровень пролактина увеличивается с возрастом, он также
значительно повышается при стрессе.
Различные типы операционной хирургии остаются главным
способом лечения прогрессирующей ДГПЖ, что означает
стационарное пребывание в госпитале и риск импотенции или недержания
мочи. Недавно введенная микроволновая терапия является,
по-видимому, перспективным методом лечения ДГПЖ с меньшим риском и
болью. Идея этого метода состоит в том, чтобы нагреть простату с
помощью направленных микроволн до температуры примерно 40 °С.
По не вполне ясным причинам это воздействие может привести к
сокращению простаты и облегчению симптомов заболевания. Однако
не у всех пациентов при этой терапии наблюдается положительный
эффект. Отдаленные результаты этого метода также неясны.
Однако обнадеживает, что часто возможно предотвратить
ДГПЖ или даже достичь реверсии начальных или умеренных форм
болезни, используя травяные и пищевые добавки или лекарства.
Проскар (finasteride) - единственное лекарственное средство на
рынке США, которое показало некоторый эффект в сокращении
размера простаты. Его действие связано с ингибированием 5-альфа-
редуктазы, трансформирующей тестостерон в ДГТ. Для получения
результата проскар обычно следует принимать от шести месяцев до
года. У тех пациентов, кто отвечает на лечение, проскар может
сокращать размер гипертрофированной простаты приблизительно на
одну четверть. Проскар - таблетку finasteride в 5 мг - следует
принимать один раз в день. У 5% пациентов может развиваться обратимая
импотенция или снижаться половое желание.
Saw Palmetto (Serenoa repens) - пальмовидное дерево, чьи ягоды
имеют давнюю историю использования в народной медицине для
восстановления половой функции и лечения простаты. В последнее
время несколько исследований продемонстрировало эффективность
липостероловых экстрактов (т.е. экстрактов, содержащих жиры и
стеролы) из Saw Palmetto в сокращении признаков и симптомов
ДГПЖ. Фактически экстракты Saw Palmetto столь же эффективны в
сокращении размера простаты и улучшении потока мочи, как и
проскар, но его эффекты часто отмечаются уже через один-три месяца
лечения (а не года, как при лечении проскаром). Механизм действия
Saw Palmetto сложен и, по-видимому, связан с ингибированием
связывания дигидротестостерона с его рецепторами в простате.
Использование: рекомендуемая дозировка липостеролового
экстракта Saw Palmetto (содержащего не менее 80% жирных кислот
и стеролов) - 160 мг два раза в день. Использование высушенных
ягод или экстракта этилового спирта не рекомендуется, потому что
453
для достижения желательного эффекта необходимо употребить
очень большие количества экстракта.
Цветочная пыльца. Специально как средство для лечения
ДГПЖ цветочная пыльца не изучалась. Однако несколько
исследований показали ее эффективность при хроническом простатите.
Недавно полученные данные показывают, что простатит часто
сопровождает ДГПЖ и может не только определять часть ее симптомов,
но также вносить вклад и в ее развитие. Если в области паха
ощущается боль или болезненность при прикосновении, то это - вероятные
симптомы простатита.
Использование: необходимо принимать не натуральную
цветочную пыльцу, а ее экстракт, потому что пыльца лишь частично пере
варивается организмом, что в результате существенно снижает
поглощение активных компонентов. Дозировка может заметно
варьировать и определяется как видом пыльцы, так и способом
изготовителя экстракта.
Цинк играет важную роль в метаболизме гормонов, которые
стимулируют ДГПЖ, и поэтому является критичным для предотвра
щения ДГПЖ и ее успешного лечения. В частности, цинк ингибиру-
ет 5-альфа-редуктазу (сокращая таким образом, производство ДГТ),
ингибирует связывание ДГТ с его рецепторами и уменьшает секре
цию пролактина гипофизом. Даже умеренный дефицит цинка
может вносить вклад в развитие ДГПЖ. Кроме того, пищевые добавки
с цинком могут уменьшать признаки ДГПЖ, а также размер
простаты у некоторых пациентов. Большинство форм пищевых добавок
цинка плохо всасывается пищеварительной системой. Одно из
исключений - пиколинат цинка. У человека большая часть цинка
транспортируется сквозь кишечную стену в кровяное русло с
помощью пиколиновой кислоты, которая выделяется поджелудочной
железой. Пиколиновая кислота связывается с цинком, формируя
пиколинат цинка, который тогда легко поглощается. Выделение
пиколиновой кислоты поджелудочной железой уменьшается с
возрастом, что приводит к недостаточному поглощению цинка.
Использование пиколината цинка обходит эту проблему.
Использование: в высоких дозах цинк может быть ядовит.
Рекомендуемые дозы не должен быть превышены. Для ДГПЖ
большинство экспертов рекомендует принимать 60 мг пиколината цинка в
течение трех месяцев. В дальнейшем доза может быть уменьшена до 30 мг
в день в следующие три месяца. Добавки цинка могут препятствовать
поглощению меди, вызывая дефицит последней. Добавки цинка
должны приниматься вместе с добавками меди в отношении 15:1 (на
каждые 15 мг элементарного цинка нужно принимать 1 мг элементной
меди). При приеме добавок, содержащих цинк и медь, необходимо
удостоверится, что другие принимаемые препараты, например
комплексы витаминов и минералов, не содержат существенных количеств этих
элементов, чтобы избежать чрезмерно высоких суммарных доз.
454
30.3. ВОССТАНОВЛЕНИЕ И ПОВЫШЕНИЕ
ПОЛОВОГО ВЛЕЧЕНИЯ И УДОВОЛЬСТВИЯ
Старение может вести к увяданию полового влечения (либидо) и
удовольствия, без которого секс становится скучной рутинной
операцией, от которой хотелось бы отказаться. Это прежде всего
связано с изменениями в центральной нервной системе, а также со
сдвигами в гормональном спектре. Во многих случаях этим изменениям
можно противодействовать и вернуть сексуальную функцию к
более "молодому" состоянию. Особенно важным для здоровой
сексуальной активности является состояние центральных часов старения
(ЦЧС). Многие пищевые вещества и лекарства помогают
"перемотать" ЦЧС вспять. В частности, повышение уровня дофамина в
гипоталамусе также увеличивает сексуальную активность.
Интенсивность ощущений во время полового акта частично
зависит от мозга, а частично от состояния нервных окончаний в
половом члене и в эрогенных зонах. Виагра и аргинин, обсужденные
выше, не только средства, стимулирующие эрекцию, но также и
увеличивающие сексуальное наслаждение как у мужчин, так и у женщин.
Предполагается, что это достигается за счет увеличения притока
крови к половым органам.
Ниацин (никотоновая кислота), витамин, принятый в дозах
более 100 мг, вызывает сходный эффект "сексуального прилива
крови". Это происходит путем стимуляции выделения гистамина,
который в свою очередь вызывает расширение кровеносных сосудов.
Такое повышенное выделение гистамина часто и спонтанно случается
во время полового возбуждения, создавая естественный прилив
крови к половым органам. Ниацин, однако, может существенно
усиливать этот эффект. Более важно, что ниацин может значительно
увеличивать ощущения не только в зоне половых органов, но и на всем
протяжении тела. Эротическое возбуждение и оргазм могут при
этом стать намного более интенсивными. Ниацин может даже
помогать некоторым людям, которые имеют затруднения в достижении
оргазма, так как гистамин играет роль в его развитии.
Использование ниацина: для увеличения полового
удовольствия ниацин лучше принимать натощак. Прилив крови обычно
происходит через 15-30 мин после приема препарата и
продолжается в течение 10-20 мин. Наилучшие результаты достигаются
тогда, когда пик прилива крови совпадает во времени с моментом
оргазма. Для людей, не использующих ниацин ежедневно, дозы
всего 100 мг или даже меньше могут вызвать необходимый
прилив крови. Однако толерантность к препарату при регулярным
его использованием развивается быстро. Ниацин является
кислотой, что может вызывать изжогу, поэтому его желательно
принимать с раствором питьевой соды. Высокие дозы ниацина (более
800 мг в день или более чем 200 мг при разовом приеме) могут вы-
455
звать проблемы с печенью и должны приниматься под
наблюдением врача. Диабетики не должны использовать ниацин,
поскольку это может повышать уровень сахара в крови. У некоторых
людей препарат может вызвать внезапное и резкое снижение
кровяного давления, особенно у пациентов, принимающих
медикаменты для лечения гипертонии.
Yohimbe и yohimbine. Coryanthe yohimbe - растение,
произрастающее в Западной Африке. Некоторые племена в этом регионе
используют отвар коры yohimbe для повышения мужской вирильности
и половой потенции. Западные исследователи сообщали, что
местные племена использовали отвар коры yohimbe в племенных
сексуальных церемониях, сопряженных с интенсивной половой
активностью, длительность которой достигала недели. Обозреватели
сходились на том, что именно этот отвар делал возможной такую
экстраординарную половую активность.
В прошедшее десятилетие было выполнено значительное
количество исследований yohimbe. Однако большинство из них было
проведено не с корой растения непосредственно, а с ее очищенным
активным компонентом - yohimbine {Buffwn, 1985). Исследования
yohimbine с целью лечения импотенции дали смешанные
результаты. В некоторых случаях у мужчин с органической импотенцией
(связанной главным образом с закупоркой кровеносных сосудов
полового члена или повреждением нервов) было показано, что
yohimbine был не лучше, чем плацебо, в то время как в других
наблюдались некоторые положительные эффекты. Изучение
мужчин с неорганической импотенцией показало несколько лучший
результат. Общая картина, по-видимому, такова, что yohimbine
является более эффективным в случаях умеренной и
кратковременной неорганической импотенции (Teloken et al., 1998). Эффекты
yohimbine, очевидно, наибольшие у людей со здоровой или слегка
сниженной половой функцией. Препарат увеличивает сексуальную
активность, делая более легкой инициацию и поддержание
эрекции и приводя к более мощному оргазму. Таким образом,
yohimbine - скорее фактор, пробуждающий или усиливающий
сексуальное желание, чем лекарственное средство для органической
импотенции (Ashton, 1994; Kunelius et al., 1997). Фактически yohimbine
был первым лекарственным средством из когда-либо описанных в
Настольном справочнике врача (фармакологической "библии"
американских врачей) как средство, которое "обладает
способностью пробуждать и стимулировать половое желание".
Механизм действия yohimbine относительно хорошо понят
(Morgenthaler et al., 1995). Он связан с блокадой так называемой аль-
фа-2 адренергической активности в центральной нервной системе
(ЦНС). Не касаясь подробностей, укажем, что альфа-2 адренергиче-
ская подсистема ЦНС участвует в механизме нормальной эрекции.
Интересно, что альфа-2 адренергический эффект на эрекцию не за-
456
висит от уровня тестостерона. У кастрированных крыс-самцов,
уровень тестостеронов у которых незначителен, yohimbine тем не менее
стимулировал половую деятельность. Он также стимулировал
половую деятельность крыс-самок в невосприимчивой фазе астрального
цикла (время, когда половая деятельность обычно подавлена и
уровень тестостеронов очень низок). Последний результат указывает
на существование возможности (пока неисследованной), что
yohimbine может увеличивать половое влечение и удовольствие также и у
женщин.
Побочные эффекты: относительно частый побочный эффект
yohimbine и yohimbe - беспокойство. Обычно возникает при очень
высоких дозах, но иногда наблюдался и при низких дозах. Другие
побочные эффекты включают высокое кровяное давление, тремор,
тошноту и рвоту, высокую частоту сердцебиений, чрезмерное
слюнотечение и потоотделение, раздражительность, нарушение
координации, головные боли и прилив крови. Обычно используемые дозы,
однако, вызывают весьма незначительные или вообще никаких
побочных эффектов у здоровых людей. Yohimbine или yohimbe не
должны использоваться пожилыми пациентами, беременными
женщинами, детьми, психически ненормальными людьми, людьми с
почечными проблемами или с язвенной болезнью. Yohimbine или
yohimbe не должны приниматься в комбинации с антидепрессантами
или другими лекарствами, улучшающими настроение. Они должны
использоваться с чрезвычайными предосторожностями людьми,
страдающими гипертонией.
Использование: Yohimbine рассматривается как
лекарственное средство и должно быть предписано врачом. Многие люди
используют альтернативу - натуральную кору yohimbe, которая
продается в магазинах здоровой пищи. Дж. Моргенталер и
Д. Джой в книге "Лучший секс с помощью химии" (Morgenthaler,
Joy, 1995) заявляют, что многие из выпускаемых разновидностей
капсул с yohimbe содержат по крайней мере не меньше yohimbine,
чем в прописываемых врачами таблетках. Таблетки содержат
5.4 мг yohimbine каждая. При лечении эректальной дисфункции
рекомендуется принимать от трех до восьми таблеток в день
(16.2—43.2 мг в день). Содержание yohimbine в обычно
продаваемых капсулах с yohimbe изменяется в широком диапазоне и может
быть в несколько раз выше, чем в одной таблетке с yohimbine.
Люди, использующие капсулы с yohimbe при спородической
попытке стимулировать половую активность, должны сначала
пробовать очень низкие дозы (открыть капсулу и удалить часть
порошка). Далее доза может быть постепенно и осторожно
отрегулирована во избежание побочных эффектов пока желаемый
результат не будет получен. Действие yohimbine обычно
продолжается в пределах одного часа, но иногда yohimbine может дать ком-
мулятивный эффект.
Глава 31
НЕСТАРЕЮЩАЯ КРАСОТА
Ты, как цветок весенний,
Чиста, прекрасна, нежна.
Смотрю на тебя, и грустью
Душа моя смущена
И просится невольно
Молитва на уста,
Чтоб ты всегда оставалась
Нежна, прекрасна, чиста.
Генрих Гейне
Нравится нам это или нет, но внешне люди существенно
различаются. Мы живем в обществе, где восприятие внешнего
облика играет почти такую же важную роль, как и восприятие
интеллектуальных и нравственных достоинств личности. Частично
это - результат современной культуры, гипертрофированно
ориентированной на потребление и насыщенной интенсивными
визуальными средствами массовой информации. Однако в таком
положении вещей можно также обвинить (или благодарить) и наше
эволюционное прошлое. В дикой природе животное стремится
найти сексуального партнера с такими внешними проявлениями
его наследственности, которые потенциально могут дать
потомкам наилучший шанс для выживания. Животное, однако,
повинуется законом генетики и дарвиновской эволюции, его механизмы
выбора партнера наследственно запрограммированы в
подсознании. Визуальный образ предполагаемого партнера, наряду с
запахом и поведением служат "входными данными", которые мозг
подсознательно обрабатывает, чтобы определить, насколько
данная генетическая комбинация благоприятна с точки зрения
полноценности будущего потомства. Физическая красота
указывает на отсутствие генетических дефектов и болезней, а юный
внешний вид свидетельствует, что предполагаемый партнер
находится в активном репродуктивном возрасте.
Безотносительно причин, социальных, биологических или
метафизических, большинство из нас хочет выглядеть молодыми и
привлекательными. Это укрепляет наше чувство собственного
достоинства, поднимает наш дух, улучшает возможности общения с особами
противоположного пола и, как показывают некоторые
исследования, заставляет других людей относиться к нам лучше.
Кожа - ткань с высоким уровнем метаболизма, обладающая самой
большой площадью поверхности, служит защитой для внутренних органов. Кожная
ткань подвергается воздействию целого ряда причиняющих ущерб факторов,
которые берут начало во внешней среде, в коже - непосредственно и в
различая
ных эндогенных источниках. Поэтому кожа обладает многими эволюционно
созданными физическими и биохимическими механизмами защиты.
Структура кожи - совершенный комплекс, состоящий из нескольких
уровней, каждый из которых играет определенную роль и выполняет различные
функции. Каждый уровень обладает собственным арсеналом молекул защиты,
которые отличаются друг от друга восприимчивостью к стрессогенным
воздействиям, включая окислительный стресс. Вообще, одним из главных и важных
вкладчиков в старение кожи, кожные нарушения и болезни кожи являются
АФК. Вследствие существования большого числа потенциальных
биологических целей окислительного повреждения, кожа очень восприимчива к таким
реакциям. Так, кожа богата липидами, белками и ДНК, которые чрезвычайно
чувствительны к окислению.
Раскрытие механизмов, включенных в окисление кожи и в
функционирование систем ее защиты, может способствовать пониманию процесса старения
кожи и ее патологических нарушений (Kohen, 1999).
Общие меры для замедления старения, описанные в других
главах, помогут также и замедлению старения вашей кожи и
могут даже улучшить ее внешний вид. Такие антиоксиданты, как
витамины С и Е, селен, цистеин, флавоноиды (содержащиеся в
растительных источниках, например, в косточках винограда), и
полифенолы (содержащиеся в зеленом чае), могут изменять баланс
"повреждение - репарация" в сторону омоложения кожи. Люди,
принимающие разумно высокие дозы пищевых АО-добавок,
часто отмечают улучшение эластичности и структуры кожи в
течение нескольких недель.
УФ-излучение - один из главных виновников преждевременного
старения кожи. Этот фактор индуцирует образование свободных
радикалов и перекрестных сшивок (патологические сшивки между
биополимерами), которые приводят к морщинам, потере гладкости
и эластичности кожи. Вернейший способ защищать кожу состоит в
том, чтобы избегать пребывания на ярком солнечном свете. Другая
альтернатива - использование солнцезащитных кремов, но, к
сожалению, большинство таких кремов относительно малоэффективны
в предотвращении старения кожи. Имеются два типа
ультрафиолетового света: УФ-А и УФ-В. Загар вызывается УФ-В, в то время как
старение кожи, морщины и рак кожи вызывается главным
образом - УФ-А. Дело в том, что не все частоты солнечного спектра
равноценно воздействуют на различные структурные компоненты
кожи. Большинство солнцезащитных кремов, особенно с SPF (Sun
Protect Factor- коэффициент защиты от УФ) выше, чем 15,
являются весьма эффективным при защите от УФ-В и поэтому хороши для
предотвращения загара, но мало- или вообще неэффективны для
защиты против УФ-А и поэтому не помогают предотвращать
морщины или уменьшать риск меланомы или рака кожи. При покупке
солнцезащитного крема удостоверьтесь, что он имеет индекс SPF 15
или выше и что этот крем предлагает защиту против как УФ-В, так
и УФ-А. Лучший способ "узнать правду" относительно солнцеза-
459
щитного крема - изучить список активных компонентов и
удостовериться, что они включают эффективные блокаторы УФ-А, такие
как авобензон, двуокись титана и окись цинка.
Когда вы находитесь на свежем воздухе, не обольщайтесь уверенностью,
что шляпа или пребывание в тени защитит вас от УФ-света. Отраженный свет
может сохранить до трети интенсивности УФ-излучения, следовательно, и в
этом случае желательно пользоваться кремом, содержащим блокаторы УФ-А
и УФ-В. Общеизвестно, что стекло задерживает УФ-излучение. Стекло
действительно хорошо задерживает УФ-В, но не в состоянии задержать большую
часть УФ-А. Это означает, что даже в помещении, ярко освещенном дневным
светом, необходимо пользоваться солнцезащитным кремом, чтобы
гарантировать максимум УФ-защиты.
Некоторые люди стремятся избегать солнца и яркого дневного
света. Такое решение понятно с точки зрения уменьшения УФ-по-
вреждений, но оно имеет также и оборотную сторону (помимо
потери некоторых удовольствий жизни). У некоторых людей недостаток
пребывания на ярком свете может нарушать нормальный цикл
"сон-^бодрствование" и вести к бессоннице и депрессии; поскольку
свет влияет на продукцию серотонина и мелатонина. Солнечный
свет также необходим организму для производства витамина D,
дефицит которого ведет к повреждению костей и снижению
иммунитета. Если вы лишены солнечного света, удостоверитесь, что
получаете 100% RDA для витамина D в вашей пищевой витаминной
добавке или в молоке, обогащенном витамином D.
УФ-оОлучение - не единственный фактор, который ускоряет
старение кожи. Уксусный альдегид, продукт метаболизма
этилового спирта, и ряд соединений в табачном дыме, потенциально
способствующих перекрестным сшивкам макромолекул, приводят к
преждевременному старению кожи равно, как и других органов.
Злоупотребление алкоголем и курением крайне редко сочетаются с
молодо выглядящей кожей. Диабет или недостаточная толерантность к
глюкозе также вносят вклад в старение кожи, так как высокий
уровень глюкозы ускоряет как формирование свободных радикалов,
так и образование перекрестных сшивок. Шаги к улучшению
толерантности к углеводам, в частности, поддержание оптимального
веса, диета с высоким содержанием волокон, физические упражнения
и микроэлемент хром в виде пищевой добавки (см. гл. 22, 23), будут
не только уменьшать риск дегенеративных болезней, но также и
задерживать образование морщин на вашей коже. У женщин после
менопаузы эстрогеновая терапия часто замечательно улучшает
строение кожи и ее эластичность. Восстановление "молодого"
уровня гормона роста (см. гл. 26), как сообщалось, умеренно увеличивал
плотность кожи как у мужчин, так и у женщин.
Все перечисленное выше, исключая конкретные меры по
защите от УФ-облучения, является частью "основной" стратегии
антистарения для организма в целом. Но кожа - особая ткань.
460
И не только из-за ее значения для нашего драгоценного имиджа,
но также и потому, что уход за кожей доступен, безопасен и
обычно не связан с болезненными ощущениями. Итак,
существует ли что-либо для специального воздействия на кожу, т.е.
косметические изделия и процедуры, которые обладают истинным
потенциалом антистарения; что может фактически изменять
видимые признаки старения - смягчить морщины, расправить
обвислые щеки, устранить возрастные пятна и другие недостатки?
Число клинических исследований, посвященных омоложению кожи,
невелико. Несколько современных исследований показали
возможность реверсии кожи, постаревшей в различной степени, с
помощью ретиноидов, альфа-гидрокси кислот и эстрогенов. Мы не
будем обсуждать такие агрессивные методы, как операционная
хирургия, лазерное облучение, инъекции бутулинового токсина
или глубокое химическое воздействие с целью замены старой
кожи. Эти методы не вписываются в содержание этой книги,
связанной с проблемами антистарения, поскольку они стремятся скорее
к реконструкции кожи, чем к ее омоложению.
Альфа-гидрокси кислот (АГК) - группа структурно связанных
органических кислот преимущественно из растительных
источников. Было обнаружено, что АГК препятствуют образованию
морщин, улучшают строение и эластичность кожи, изменяя некоторые
процессы, наблюдаемые в стареющей коже. С возрастом кожа
становится более тонкой и хрупкой, главным образом потому, что
замедляется деление клеток и снижается синтез коллагена и эластина
(основных структурных белков кожи). АГК вызывают пилит
(отслоение) массы мертвых клеток (кератиноцитов) и, что более
важно, стимулируют рост и уплотнение дермы и эпидермиса, двух
главных слоев кожи.
Несмотря на то что механизм их действия неясен, в нескольких
исследованиях показано, что АГК действительно потенциально
способны устранить некоторые признаки старения кожи. Так, Ньюмен
и его коллеги {Newman et al., 1996) подвергли сорок пять
добровольцев лечению 50% раствором гликолевой кислоты и (в качестве
контроля) только одним растворителем. Аппликации накладывались на
одну сторону лица, предплечий и рук в течение пяти минут один раз
в неделю в течение четырех недель. Отмечено существенное
улучшение, включая уменьшение грубых элементов кожной ткани и
морщин, уменьшение солнечного кератоза и лентиго (пятнышки
диаметром до трех мм, коричневого или черного цвета).
Гистологический анализ показал разрежение роговидного слоя (наиболее
удаленный от основного слой кожи, состоящий главным образом из
мертвых клеток), увеличение зернистого слоя и эпидермалъного
уплотнения. Результаты этого исследования демонстрируют, что пи-
линг (отслоение) с помощью 50% гликолевой кислоты умеренно
улучшает состояние стареющей кожи.
461
В другом исследовании, Дайтр и его коллеги фиге et al.)
проверяли лосьон, содержащий 25% гликолевой, молочной или лимонной
кислот, против плацебо в виде обычного косметического лосьона.
Лечение продолжалось шесть месяцев. Применение АГК привело к
25% увеличению толщины кожи, плотности коллагена и улучшению
качества эластичных волокон. Исследователи заключили, что АГК
"приводит к существенной реверсии признаков старения кожи,
вызванных действием света".
Многочисленные кремы и лосьоны, созданные на основе АГК,
имеются в продаже. Большинство таких изделий содержит
относительно низкие концентрации АГК, обычно 5-15%. Эффективность
препаратов с низкой концентрацией АГК (лосьоны с 8% молочной
и 8% гликолевой кислотами) оценивалась доктором Стайллером и
его коллегами (Stiller et al.). Изучались 74 женщины в возрасте
40-70 лет с умеренно поврежденной светом кожей лица. Крем
применялся два раза в день в течение 22 недель. Процент пациентов,
достигших определенного улучшения, составил для гликолевой
кислоты - 76%, для молочной кислоты - 71 и для плацебо - 40%.
Исследователи заключили, что препараты с низкой концентрацией
АГК оказались 'скромно полезны" для улучшения некоторых
признаков старения кожи.
При использовании коммерческих кремов для кожи на основе
АГК, убедитесь, что АГК в них составляет, как минимум, 8%.
Используйте крем последовательно, по крайней мере один раз в
день, нанося его на тщательно вымытую кожу для лучшего
впитывания. Более высокая концентрация АГК в препарате,
очевидно, может быть более эффективна. Однако из-за большего риска
раздражения кожи им нужно пользоваться под наблюдением
дерматолога.
Имеется еще один важный предостерегающий момент, который
необходимо отметить. Главные выгоды от использования АГК
состоят в их способности отслаивать клетки поверхности кожи.
Однако чем больше кожи отслаивается с поверхности, тем большее
количество клеточных делений произойдет в более глубоких слоях
кожи. Проблема в том, что нормальные человеческие клетки не могут
делиться бесконечно. Фибробласты (основной тип клеток кожи)
способны делиться приблизительно 50 раз и затем оказываются в
стадии старения, т.е. приближаются к пределу Хейфлика (см. гл. 2).
А это состояние, при котором клетка становится вялой,
неэффективной, безразличной к различным сигналам организма и главное -
не способна делиться. Кожа со многими стареющими клетками
обычно делается ломкой и быстро покрывается пятнами и
морщинами. Эксфолиация с помощью АГК или других методов - ценный
косметический инструмент, но если злоупотреблять им, то кожу
может "поразить лимит Хейфлика" раньше, чем это должно
произойти естественным путем.
462
Третиноин (Ретин А) - форма ретиноивой кислоты, которая
является активным метаболитом витамина А. Ретиноивая кислота
вызывает многочисленные эффекты в различных органах, включая
кожу. В частности, она увеличивает реакцию клеток кожи на эпи-
дермальный фактор роста (ЭФР), естественный гормон,
стимулирующий рост кожи.
Эффект антистарения кожи под действием третиноина был
зарегистрирован во многих тщательно проведенных исследованиях.
Третиноин уменьшал количество мелких морщин и грубость
кожи, увеличивал толщину эпидермального слоя и стимулировал в
нем накопление коллагена. Положительный эффект крема,
содержащего 0.05% третиноина, как было показано, превосходил влияние
6% гликолевой кислоты на эластичность кожи. Оптимальное
содержание третиноина в креме - 0.025-0.1%%. Однако, касаясь
третиноина, следует помнить, что "больше" не обязательно - "лучше".
Было, например, показано, что 0.025% третиноина столь же
эффективны, как и 0.05% или даже 0.1%, но с более редкими случаями
раздражения кожи. Для людей с чувствительной кожей содержание в
0.025% является, очевидно, оптимальным. Существуют данные, что
положительный эффект использования третиноина может
нарастать в течение года с начала использования.
Формально ретиноивая кислота может рассматриваться в
качестве питательного вещества, потому что она - естественный
метаболит витамина А и присутствует в следовых количествах в некоторых
пищевых продуктах животного происхождения. Однако
существенные дозы ретиноивой кислоты, принимаемые перорально, могут
вызвать серьезные побочные эффекты, сходные с передозировкой
витамина А (см. гл. 23). Кремы с ретиноивой кислотой, т.е. с трети-
ноином наружного действия, не дают никакого существенного
системного эффекта, их главное побочное влияние - раздражение
кожи. В настоящее время кремы с третиноином продаются в
соответствии с предписанием врачей.
Эстрогены. Часто отмечается, что женщины после менопаузы
начинают заметно стареть: быстрее, чем раньше увеличивается
число морщин, пропадает эластичность и гладкость кожи. В
значительной степени это, по-видимому, объясняется хроническим снижением
уровня эстрогенов, наблюдаемого при менопаузе. Из всех гормонов,
уровень которых падает с возрастом, эстрогены наиболее
значительно влияют на кожу. Они, как известно, защищают женщин от
болезней сердца, а теперь оказалось, что эстрогены также
замедляют и старение кожи. Исследования показывают, что у женщин
после менопаузы, пользующихся эстрогеновой терапией, меньше
морщин, лучше строение и эластичность кожи, чем у женщин,
лишенных экзогенных эстрогенов.
В то время как решение об использовании заместительной
эстрогеновой терапии требует анализа медицинской истории пациента
463
(см. гл. 8 для более полной информации об эстрогеновой терапии),
женщины после менопаузы могут улучшить состояние кожи,
пользуясь препаратами наружного действия на основе эстрогенов.
Др. Шмидт и его коллеги (Schmidt et al.) изучали эффекты
применения кремов с эстрадиолом (0.01%) или эстриолом (0.3%) у 59
женщины после менопаузы. После 6-месячного применения отмечено
улучшение эластичности и твердости кожи; глубина морщин и
размер пор уменьшились более чем на 60% в обеих группах. Влажность
кожи и синтез коллагена значительно увеличились. Никакие
системные эффекты эстрогенов не наблюдались.
Теоретически кремы с эстрогенами могут, по-видимому, также
нивелировать признаки старения в предклимактерический период у
женщин, а также и у мужчин, хотя не имеется никаких клинических
подтверждений этого. Различные кремы на основе эстрогенов
доступны как по рецепту, так и в свободной продаже.
Антиоксиданты наружного применения. Кожа высоко
восприимчива к повреждению свободными радикалами из-за ее
постоянного контакта с окружающей средой и высокого
содержания в ней ненасыщенных жирных кислот. Наружные
антиоксиданты обеспечивают определенную защиту кожи от повреждений
со стороны окружающей среды и могут несколько замедлять
старение кожи. Одна из обычно используемых формул - комбинация
витаминов С и Е, водо- и жирорастворимых антиоксидантов
соответственно. Витамин С также участник синтеза коллагена,
главного структурного белка кожи, и, следовательно, помогает
улучшению строения кожи. Витамин А, другой обычный компонент
косметических препаратов, также достаточно полезен, поскольку
является и антиоксидантом и предшественником синтеза ретино-
ивой кислоты (см. выше). Важно знать, что коммерческие мази,
содержащие антиоксиданты, имеют ограниченный срок годности.
Антиоксиданты легко инактивируются кислородом воздуха,
особенно при высокой температуре и под действием солнечного
освещения При покупке кремов с антиоксидантами необходимо
проверить срок их годности. Храните упаковки плотно
закрытыми в темном и прохладном месте.
Na-PCA. Старение кожи связано с постепенной потерей влаги.
Сухая кожа легко сморщивается, становится менее эластичной,
выглядит старой, поскольку имеет слабеющие механизмы репарации.
Главный натуральный увлажнитель кожи - натрий 2-пирроли-
дон -5-карбогидрат, или Na-PCA. Уровень Na-PCA в коже пожилых
людей составляет приблизительно половину от такового у молодых.
Восстановление уровня Na-PCA может заметно улучшать внешний
вид сухой кожи. Некоторые коммерческие кремы для кожи и
увлажняющие кремы включают Na-PCA.
ЛИТЕРАТУРА
Абубакиров Η.К. Гормон линьки // Химия и жизнь. 1975. № 11. С. 57-62.
Абубакиров Н.К. Экдистероиды цветковых растений (Angiospermae) // Химия
природ, соединений. 1981. № 6. С. 685-702.
Ахрем АЛ., Ковганко Н.В. Экдистероны: Химия и биологическая активность.
Минск: Наука и техника, 1989. 299 с.
Барабой В Л. Биологическое действие растительных фенольных соединений.
Киев, 1976.
Барабой В Α., Брехман И.И., Голотин В.Г, Кудряшов Ю.Б. Перекисное
окисление и стресс. СПб.: Наука, 1992. 148 с.
Барнаулов ОД.,ЛимаренкоА.Ю., Куркин ΒΛ. и др. Сравнительная оценка
биологической активности соединений, выделенных из видов Rhodiola L. II Хим.-
фарм. журн. 1986. № 9. С. 1107-1112.
Бездетко Г.Н., Герман А.В., Хасина Э.И. и др. Исследование фармакокинетики
и механизма действия гликозидов элеутерококка. Ш. Метаболизм и
кинетика связывания с компонентами сыворотки крови // Хим.-фарм. журн. 1982.
№5. С. 16-19.
Бездетко Г.Н., Герман А.В., Шевченко В.П. и др. Исследование
фармакокинетики и механизма действия гликозидов элеутерококка. I. Введение трития в
элеутерозид В. Кинетика его накопления и выведения из организма
животного //Там же. 1981. № 1. С. 28-33.
Бездетко Г.Н., Смолина Т.М., ШулятеваЛД. Влияние экстрактов женьшеня и
элеутерококка колючего на течение аллоксанового диабета у крыс //
Материалы II совещ. по исслед. лекарств, растений Сибири и Дальнего Востока.
Томск, 1961.
Бердышев ГД., Стрельнук СИ. Сравнительное изучение геропротекторного
действия экстрактов элеутерококка, женьшеня и эглантина на Drosophila
melanogaster // Новые данные об элеутерококке и других адаптогенах.
Владивосток, 1981. С. 145.
Беспалов ВТ., Александров В.А., Яременко К.В. и др. Подавление развития
экспериментально индуцированных опухолей нервной системы с помощью
препаратов из Eleutherococcus senticosus (Rupr. et Maxim.) Maxim., Rhaponticum
carthamoides (Willd.) Djin и Plantago major L. Ц Раст. ресурсы. 1992. № 4. С. 66-70.
Брехман И.И. Женьшень. Л.: Медгиз, 1957.
Брехман И.И. Корень элеутерококка - новое стимулирующее и тонизирующее
средство. Л., 1960.
Брехман И.И. Использование филогенетической системы растений в поисках
новых фармакологически активных веществ // Изучение и использование
лекарственных ресурсов СССР. М.: Медицина, 1964.
465
Брехман И.И. Элеутерококк. Л.: Наука, 1968. 186 с.
Брехман И.И. Человек и биологически активные вещества. 2-е изд. М.: Наука,
1980.119 с.
Брехман И.И., Гоненко В.А. Антирадиомиметическое действие некоторых
природных соединений. I. Суммарные препараты и индивидуальные гликозиды
корня женьшеня // Радиобиология. 1968. № 4. С. 613.
Брехман И.И,, Кириллов О.И. Женьшень и элеутерококк - источники
лекарственных средств нового типа действия // Раст. ресурсы. 1968. № 1. С. 3.
Брехман И.И., Юргенс ИЛ., Кириллов О.И. Действие адаптогенов на стадии
стресса // Материалы сектора физиологии спорта ЦНИИФК за 1966 г. М,
1966.
Буланов Л.Е., Положенцева М.И., Яцков Л.П. Антиалкогольный эффект
элеутерококка // Новые данные об элеутерококке: Материалы II Между-
нар. симпоз. по элеутерококку (Москва, 1984). Владивосток, 1986.
С.127-133.
Вересковский В.В., Чакалинская И.И., Прищепа И.А. Биохимические
исследования Rhaponticum mrthamoides (Wild) lljin // Изв. АН БССР. Сер. биол. наук.
1974. С. 59-63.
Воскресенский О.И. Влияние природных биоантиоксидантов на патологические
процессы, связанные со старением // Итоги науки и техники. Общие
проблемы биологии. М.: ВИНИТИ, 1986. Т. 5. С. 163-201.
Воскресенский О.Н., Девяткина ТА., Туманов В.А. Влияние элеутерококка и
женьшеня на развитие свободнорадикальной патологии // Новые данные об
элеутерококке. Владивосток, 1986. С. 101.
Герасюта М.А., Коваль Т.Н., Кайзер С.А. Эффективность применения
экстракта левзеи сафлоровидной у моряков в условиях длительной арктической
экспедиции // Новые данные об элеутерококке и других адаптогенах.
Владивосток, 1981. С. 139-142
Герман А.В., Бездетко Г.Н., Митрохин Ю.И. и др. Исследование фармакокине-
тики и механизма действия гликозидов элеутерококка. II. Распределение
элеутерозида В по органам и субклеточным фракциям // Хим.-фарм. журн.
1982. № 1. С. 26-30.
Губский Ю.И., Левицкий ЕЛ., Холодова ЮД. и др. Механизмы генопротектор-
ного действия препарата на основе фитоэкдистероидов (БТК-8Л) в условиях
повреждения хроматина тетрахлорметаном. // Укр. биохим. журн. 1993. Т. 65,
№ 6. С. 75-83.
Дардымов И.В. Женьшень, элеутерококк. М.: Наука, 1976. 184 с.
Дардымов И.В. Механизмы действия препаратов женьшеня и элеутерококка:
Автореф. дис. д-ра мед. наук. Л., 1987. 41с.
Донцов В.И., Крутько В.И., Подколзин А.А. Фундаментальные механизмы ге-
ропрофилактики. М.: Биоинформсервис, 2002.464 с.
Елкин В.М., Захарова Ή.Γ., Кито З.Н. и др. Влияние элеутерококка на
факторы устойчивости организма // Новые данные об элеутерококке: Материалы
II Междунар. симпоз. по элеутерококку (Москва, 1984). Владивосток, 1986.
С. 114-118.
Еляков Г.Б., Оводов Ю.С. Гликозиды аралиевых // Химия природ, соединений.
1972. № 6. С. 697.
Еляков Г.Б., СтригинаЛ.И., Кочетков И.К. О строении агликона панаксозида
А // Докл. АН СССР. 1964. № 4. С 892.
Еляков Г.Б., СтригинаЛ.И., Хорлин А.Я., Кочетков И.К. Гликозиды из
корней женьшеня {Panax ginseng С А. Ыеу). Изв. АН СССР. Отд. хим. наук. 1962.
№11. С. 2054.
466
Еляков Г.Б., Уварова И.И., Горшкова Р.П. и др. Строение углеводных цепей па-
наксозидов D, Е, F // Докл. АН СССР. 1965. № 6. С. 1309.
Ильин B.C., Титова Г В. Гормон-ферментный комплекс инсулин-гексокиназа //
Биохимия. 1965. Т. 6, С. 1251.
Кириллов О.И. Опыт фармакологической регуляции стресса. Владивосток,
1966.
Коновалова Н.П., Митрохин Ю.И., Волкова Л.И. и др. Экдистерон моделирует
противоопухолевую активность цитостатиков и их действие на биосинтез
макромолекул в органах животных опухоленосителей // Изв. РАН. Сер.
биол. 2002. № 5. С. 522-530.
Кропотов А.В., Колдяев В.М. Влияние элеутерококка на выживаемость и
иммунитет СВЧ-облученных мышей // Новые данные об элеутерококке:
Материалы II Междунар. симпоз. по элеутерококку (Москва. 1984). Владивосток,
1986. С 161-166.
Кузъмицкий Б.Б., Голубева М.Б., Конопля И.А. и др. Новые возможности
изыскания иммуномодуляторов среди соединений со стероидной структурой //
Фармакология и токсикология. 1990. Т. 53, № 3. С. 20-22.
Купин ВМ., Бланко Ф.Ф., Сорокин A.M. Сравнительная оценка некоторых
природных адаптогенов в экспериментальной онкологии // Новые данные об
элеутерококке: Материалы II Международн. симпоз. по элеутерококку
(Москва, 1984). Владивосток, 1986. С. 136-141.
Лазарев И.В. Эволюция фармакологии. Л.: ВММА, 1947.
Левицкий ЕЛ., Холодова ЮД., Губский Ю.И. и др. Биохимические
характеристики фракционированного хроматина печени крыс в условиях
экспериментального D-гиповитаминоза и при введении препаратов стероидов // Укр.
биохим. журн. 19936. Т. 65, № 1. С. 28-36.
Левицкий ЕЛ., Холодова ЮД., Губский Ю.И. и др. Механизмы генопротек-
торного действия препарата на основе фитоэкдистероидов (БТК-8Л) в
условиях повреждения хроматина хлорофосом // Там же. 1993а. Т. 65, № 6
С. 84-91.
Лупандин А.В. О роли катехоламинергических синапсов в механизме
формирования адаптации при участии полифенольных адаптогенов // Физиол. жури.
СССР. 1989. Т. 75, № 8. С. 1083-1088.
Лупандин А.В. Общий механизм приспособления организма под влиянием
полифенольных адаптогенов // Успехи физиол. наук. 1991. Т. 22, № i.e. 20-39.
Лупандин А.В., Лапаев И.И. Лимонник. Хабаровск: Кн. изд-во, 1981. 128 с.
Марина Т.Ф., Краснов Е.А., Саратиков А.С. II Проблемы освоения
лекарственных ресурсов Сибири и Дальнего Востока. Новосибирск, 1983. С. 129-131.
Меерсон Ф.З. Адаптогены, стресс и профилактика. М.: Наука, 1981.
Миронова В.И., Холодова ЮД, Скачкова Ύ.Φ. и др. Гипохолестеринемическое
действие фитоэкдизонов при экспериментальной гиперхолестеринемии у
крыс // Вопр. мед. химии. 1982. № 3. С. 101-105.
Мурзаев Ю.Р., Сыров В.Н. Влияние фитоэкдистероидов на сексуальную
активность самцов крыс // Докл. АН Республики Узбекистан. 1992. № 3. С. 47-49.
Обухова Л.К., Эмануэль Н.М. Молекулярные механизмы замедления старения
антиоксидантами // Итоги науки и техники. Общие проблемы биологии. М.:
ВИНИТИ, 1984. Т. 4. С. 44-80.
Оводов Ю.С., Фролова Г.М., Дзизенко А.К., Литвиненко В.И. Структура и
свойства элеутерозида В, гликозида Eleutherococcus senticosus II Изв. АН
СССР. Сер. хим. 1969. № 6. С. 1370-1371.
Оводов Ю.С., Фролова Г.М., Ельякова Л.А. Химическое изучение гликозидов
из Eleutherococcus senticosus Rupr. et Maxim и Acanthopanax sessiliflorus II Pe-
467
зультаты исследований элеутерококка в Советском Союзе / Под ред.
В.Л. Комарова; ДВНЦ СО АН СССР. Владивосток, 1966. С 25-27.
Оводов Ю.С., Фролова Г.М., Елъякова Л.А., Ельяков Г.Б. Идентичность элеуте-
розида Ε акантозиду D // Изв. АН СССР. Сер. хим. 1965. № 11. С. 2065-2067.
Оводов Ю.С., Фролова Г.М., Нефедова М.Ю., Ельяков Г.Б. Гликозиды
Eleutherococcus senticosus. II. Структура элеутерозидов А, В], С, D // Химия
природ, соединений. 1967. № 2. С 63-64.
Саратиков А.С. Некоторые итоги изыскания и изучения стимуляторов нервной
системы растительного происхождения // Стимуляторы центральной
нервной системы. Томск, 1966. Выи. 1. С. 3
Саратиков А.С, Краснов Е.А. Родиола розовая - ценное лекарственное
растение (золотой корень). Томск Изд-во Том. ун-та, 1987. 254 с.
Сахибов АД., Сыров В.Н., Усманова А.С, Абакумова О.Ю. Эксперименталь
ный анализ иммунотропного действия фитоэкдистероидов // Докл. АН Уз.
ССР. 1989. С. 55-57.
Соколов С, Замотаев И. Лекарственные растения. М.: Vita, 1993. 511с
Соколов С.Я., Ивашин В.М., Запесочная Г.Г и др. Исследование нейротропной
активности новых веществ, выделенных из родиолы розовой // Хим.-фарм.
журн. 1985. Т. 19, № 11. С. 1367-1371.
Сыров В.И. К механизму анаболического действия фитоэкдистероидов в
организме млекопитающих // Биол. науки. 1984. № 11. С. 16-20.
Сыров В.Н. Сравнительное изучение анаболической активности фитоэкдисте
роидов и стераиаболов в эксперименте // Хим.-фарм. журн. 2000. Т. 34, № 4.
С. 31-34.
Сырое В.Н., Курмуков А.Г. Об анаболической активности фитоэкдизона-экди-
стерона, выделенного из Rhaponticum carthamoides (Willd) Iljin // Фармаколо
гия и токсикология. 1976. № 6. С. 690-693.
Тодоров И.И. Русский жень-шень // В мире иауки. 1993. № 2-3. С. 40-41.
Тодоров И.И., Митрохин Ю.И., Ефремова ОМ., Сидоренко Л.И. Влияние экди-
стерона на биосинтез белков и нуклеиновых кислот в органах мышей //
Хим.-фарм. журн. 2000а. Т. 34, № 9. С. 3-5.
Тодоров И.И., Митрохин Ю.И., Ефремова О.И., Сидоренко Л.И. Действие
экстрактов левзеи сафлоровидной на биосинтез РНК и белков в органах
мыши // Там же. 20006. Т. 34, № 9. С. 24-26.
Тодоров И.И., Сизова СТ., Косаганова Н.Ю. и др. Фармакокинетика и
механизм действия гликозидов элеутерококка. Влияние экстракта на метаболизм
и биосинтез белка в некоторых органах и тканях крысы // Хим.-фарм. журн.
1984. Т. 18, № 5. С. 529-533.
Тодоров И.Н., Сизова СТ., Митрохин Ю.И. и др. О субклеточных и
молекулярных механизмах анаболического действия экстрактов элеутерококка //
Новые данные об элеутерококке: Материалы II Международн. симпоз. по
элеутерококку (Москва, 1984). Владивосток, 1986. С. 16-22.
Уварова И.И., СтригинаЛ.И., Кочетков И.К. и др. Разделение суммы
гликозидов женьшеня (Panax ginseng C.A. Меу) на сефадексе // Изв. АН СССР. Отд.
хим. иаук. 1963. № 11. С 1850.
Фомовская Г.Н., Бердышев А.Г., Холодова ЮД. Иммуномодуляторный
эффект экдистеронов // Укр. биохим. журн. 1992. Т. 64, № 2. С. 56-61.
Хушбактова З.А., Сыров В.Н., Кулиев 3. Исследование гиполипидемических
свойств полимерных проантоцианидинов из растений, используемых в
народной медицине // Хим.-фарм. журн. 1989. Т. 23, № 9. С. 1111-1115.
Щежин А.К., Зинкович В.И., Галанова Л.К. Элеутерококк в профилактике
гриппа, гипертонии и ишемии сердца у водителей АвтоВАЗа // Новые дан-
468
ные об элеутерококке и других адаптогенах / ДВНЦ СО АН СССР.
Владивосток, 1981. С. 93-94.
Яременко К.В. Адаптогены как средство профилактической медицины. Томск.
1990.94 с.
BajajJS. (ed.) Insulin and metabolism. Elsevier: Amsterdam: North-Holland, 1977.
Barenboim G.M., Kozlova N.B. Use of eleutherococcus extract for increasing the
biological resistance of man exposed to different unfavorable environmental factors: (A
review) // Eleutherococcus strategy of the use and new fundamental data / Medexport.
Moscow, 1985. P. 6-18.
Bohn В., Nebe C.T., Birr C. Flow-cytometric studies with Eleutherococcus senticosus
extract as an immunomodulatory agent//Arzneim.-Forsch./Drug Res. 1987. Vol. 37,
N10. P. 1193-1196(1987).
BornsteinJ. Insulin-reversible inhibition of glucose utilization by serum lipoprotein
fraction // J. Biol. Chem. 1953. Vol. 205. P. 513.
Bornstein J., Park C.R. Inhibition of glucose uptake by the serum of diabetic rats // Ibid.
1953. Vol. 205. P. 503.
Brekhman 1.1. Man and biologically active substances. N.Y.: Pergamon press, 1980.
Brekhman 1.1., Dardymov l.V. Pharmacological investigation of glycosides from
gingseng (Panax gingseng С A. May and Eleutherococcus senticosus Max.) // Proc.
XI Pacific sci. congr. Tokyo, 1966.
Brekhman 1.1., Dardymov l.V. The mechanism of action of ginseng and eleuterococcus
preparations // Intern. Symp. on traditional medicine and modern pharmacology:
Abstracts. Beijing, 1986. P. 40.
Bu-Abbas Α., Clifford M.N., Walker R. et al. Marked antimutagenic potential of aqueous
green tee extract: Mechanism of action // Mutagenesis. 1994. Vol. 9, N 4. P. 325-331.
Burnett AJL., Lowenstein С J., Bredt D.S. et al. Nitric oxide, a physiologic mediator of
penial erection // Science. 1992. Vol. 257. P. 401-404.
ButterfieldDA., Howard В J., LaFontaine MA. Brain oxidative stress m animal models
of accelerated aging in the age-related neurodegenerative disorders, Alzheimer's
disease and Huntington's disease // Curr. Med. Chem. 2001. Vol. 8, N 7. P. 815-828.
Casper R.C. Nutrition and its relationship to ageing // Exp. Gerontol. 1995. Vol. 30,
N 3/4. P. 299-314.
Choi Y.T., Jung C.H., Lee S.R. et al. The green tea polyphenol (-)-epigallocatechin gal-
late attenuates beta-amyloid-induced neurotoxicity in cultured hippocampal
neurons // Life Sci. 2001. Vol. 70, N 5. P. 603-614.
Chong S.K.F., Brown HA., Rimmer E. et al. In vitro effect of Panax gingseng on phyto-
haemagglutinin-induced lymphocyte transformation // Intern. Arch. Allergy and
Appl. Immunol., 1984. Vol. 73. P. 216-220.
Conti Α., Maestroni G.l. The clinical neuroimmunotherapeutic role of the melatonin in
oncology // J. Pineal Res. 1995. Vol. 19, N 3. P. 103-110.
Csiszar Α., Ungvari Z., Edwards J.G. et al. Aging-induced phenotypic changes and
oxidative stress impair coronary arteriolar function // Circulation Res. 2002. Vol. 90,
N11. P. 1159-1166.
Czech M.P. Molecular basis of insulin action // Annu. Rev. Biochem. 1977. Vol. 45
P. 359-384.
Deschamps V., Barberger-Gateau P., Peuchant E., OrgogozoJM. Nutritional factors in
cerebral aging and dementia: Epidemiological arguments for a role of oxidative
stress //Neuroepidemiology. 2001. Vol. 20, N 1. P. 7-15.
Ferrini M„ Piroli G., Frontera M. et al. Estrogens normalize the hypothalamic-pituitary-
adrenal axis response to stress and increase glucocorticoid receptor immuno-reactiv-
ity in hippocampus of aging male rats. // Neuroendocrinology. 1999. Vol. 69, N 2.
P. 129-137
469
Fowler С J., Wiberg Α., Oreland L. et al. The effect of age and molecular properties of
human brain monoamine oxidase // J. Neurol. Transmission. 1980. Vol. 49, N 1/2.
P. 1-20.
Fukui K., Omoi N.O., Hayasaka T. et al. Cognitive impairment of rats caused by
oxidative strss and aging, and its prevention by vitamin Ε // Ann. N.Y. Acad. Sci. 2002.
Vol. 959. P. 275-284.
Fulder SJ. Ginseng and the hypothalamic pituitary control of stress // Amer. J. Chin.
Med. 1981. Vol. 9, N 2. P. 112-118.
Ganther H.E. Selenium metabolism, selenoproteins and mechanisms of cancer
prevention: Complexities with thioredoxin reductase //Carcinogenesis. 1999. Vol. 20, N 9.
P. 1657-1666.
Garcia-Patterson Α., Puig-Domingo M., Webb S.M. Thirty years of human pineal
research: Do we know its clinical relevance? // J. Pineal Res. 1996. Vol. 20, N 1.
P. 1-6.
Hiai S., Oura H., Tsukada K., Hirai Y. Stimulating effect of Panax ginseng extract on
RNA polymerase activity in rat liver nuclei // Chem. Pharm. Bull. 1971. Vol. 19, N 8.
P. 1656-1663.
Jenkins R.R. Exercise, oxidative stress, and antioxidants: A review // Intern. J. Sport
Nutrition. 1993. Vol. 3, N 4. P. 356-375.
Jensen G.D., Polloi A.H. The very old of Palau: health and mental state // Age and
Ageing. 1988. Vol. 17, N 4. P. 220-226.
Ji L.L., Leeuwenburgh C, Leichtweis S. et al. Oxidative stress and aging: Role of
exercise and its influences on antioxidant systems // Ann. N.Y. Acad. Sci. 1998. Vol. 854.
P. 102-117.
Johannsson G., Marin P.,Lonn L. et al. Growth hormone treatment of abdominally obese
men reduces abdominal fat mass, improves glucose and lipoprotein metabolism, and
reduces diastolic blood pressure // J. Clin. Endocrinol. Metabol. 1997. Vol. 82, N 3.
P. 727-734.
Johnson MA., Brown MA., Poon L.W. et al. Nutritional patterns of centenarians //
Intern. J. Aging Humam. Develop. 1992. Vol. 34, N 1. P. 57-76.
Jolliffe G.H., Shellard Ε J. Problems associated with the evaluation of ginseng // Symp.
of gerontol., Lugano, 1975.
Khorram O., Vu L., Yen S.S. Activation of immune function by dehydroepiandrosterone
(DHEA) in age-advanced men // J. Gerontol. A: Biol. Sci. Med. Sci. 1997. Vol. 52,
N 1. P. M1-M7.
Kim J.H. History of investigation on ginseng in Korea // Korean ginseng science symp.
Seoul, 1974.
Klatz R., Kahn C. Grow young with HGH. HarperCollins, 1997.
Kohen R. Skin antioxidants: their role in aging and in oxidative stress - new approaches
for their evaluation //Biomed. Pharmacother. 1999. Vol. 53, N 4. P. 181-192.
Luo Y., Roth G.S The roles of dopamine oxidative stress and dopamine receptor
signaling in aging and age-related neurodegeneration // Antioxid. Redox. Signal. 2000.
Vol. 2, N 3. P. 460-499
Maestroni GJ. The imrnunoneuroendocrine role of melatonin // J. Pineal Res. 1993.
Vol. 14, N 1. P. 1-10.
Masoro Ε J. Caloric restriction and aging: An update // Exp. Gerontol. 2000. Vol. 35,
N 3. P. 299-305.
Michaelis E.K. Molecular biology of glutamate receptors m the central nervous system
and their role in excititoxicity, oxidative stress and aging // Progr. Neurobiol. 1998.
Vol. 54, N4. P. 369-415.
Mimura G., Murakami K.t Gushiken M. Nutritional factors for longevity in Okinava -
present and future // Nutr. Health. 1992. Vol. 8, N 2/3. P. 159-163.
470
Minsky M. Will robots inherit the Earth? // Sci. Amer. 1994. Vol. 271, N 4. P. 109-113.
Modern nutrition in health and disease / Ed. by M.E. Shils et al. 8th ed. N.Y.: Lea and
Fabiger, 1994.
Morgenthaler J., Joy D. Better sex through chemistry: A guide to new prosexual drugs.
Petaluma, (CA): Smart Publ., 1995.
Mosharrof A.H. Effects of extract from Rhaponticum carthamoides (Willd) Iljin
(Leuzea) on learning and memory in rats // Acta physiol. et pharmacol. Bulg. 1987.
Vol. 13, N3. P. 37-^2.
Murray M., Pizzorno J. Encyclopedia of natural medicine. Prima Publ., 1991.
Oreland L.t Gottfries C.G. Brain and monoamine oxidase in aging and in dementia of
Alzheimer's type // Prog. Neuropsychophakmacol. Biol. Psychiatry. 1986. Vol. 10.
P. 533-540.
Oura H., Hiai S. Biochemical action of Panax ginseng principle // Proc. Intern. Ginseng
symp. Seoul, 1974.
Packer L., Tritschler И J., Wessel K. Neuroprotection by the metabolic antioxidant
alpha-hpoic acid // Free Radicals. Biol. Med. 1997. Vol. 22, N 1/2. P. 359-378.
Pearls T.T. The oldest old // Sci. Amer. 1995. Vol. 272, N 1. P. 70-75.
Pearson D., Show S. Life extension: A practical scientific approach. Warner Books,
1983
Petkov V. Possibilities for optimizing reactivity by pharmacological agents:
(Experiments with ginseng and centrophenoxine // Symp. of gerontol. Lugano, 1975.
Petkov V. On the mechanism of action of ginseng as regulator of the reactivity of the
organism // Новые данные об элеутерококке и других адаптогенах.
Владивосток, 1981. С. 106.
Petkov V., Roussinov К., Todorov S. et al. Pharmacological investigations on
Rhaponticum carthamoides // Planta Med. 1984. Vol. 50, N 3. P. 205-209.
Petkov V.D., Yonkov D.t MosharoffA. Effects of alcohol aqueous extract from Rhodiola
rosea L. roots on learning and memory // Acta physiol. et pharmacol. Bulg. 1986
Vol. 12, N 3. P. 3-16.
Regelson W., Kalimi M. Dehydroepiandrosterone (DHEAj - the multifunctional steroid.
II. Effects on the CNS, cell proliferation, metabolic and vascular, clinical and other
effects. Mechanism of action? // Ann. N.Y. Acad. Sci. 1994. Vol. 719. P. 564-575.
Reiter RJ., Robinson J. Melatonin. Bantam Books, 1996.
Rodriguez-Capote K.t Cespedes E., Arencibia R., Gonzalez-Hoyuela M. Indicators of
oxidative stress in aging rat brain. The effect of nerve growth factor // Rev. Neurol.
1998. Vol. 27. P. 494-500.
Rosenfeldt F.L., Pepe S.t Linnane A. et al. Coenzyme Q10 protects the aging heart
against stress: Studies in rats, human tissues, and patients // Ann. N.Y. Acad. Sci.
2002. Vol. 959. P. 355-359. Discuss: P. 463^65.
Sahelian R. DHEA: A practical guide. Garden City Park, 1996.
Sardesai V.M. Role of antioxidants in health maintenance // Nuir. Clin. Pract. 1995.
Vol. 10,N LP. 19-25.
Sastre J., Millan Α., Garcia de la Asuncion J. et al. A Ginkgo biloba extract (EGb 761)
prevents mitochondrial aging by protecting against oxidative stress // Free Radicals.
Biol. Med. 1998. Vol. 24, N 2. P. 298-304.
Serafini M., Ghiselli Α., Ferro-Luzzi A. In vivo antioxidant effect of green and black tea
in man // Europ. J. Clin. Nutr. 1996. Vol. 50, N 1. P. 28-32
Shubata S. Some chemical studies on ginseng // Symp. of gerontol., Lugano, 1975.
Slama K., Lafont R. Insect hormones - ecdysteroids: their presence and actions in
vertebrates//Europ. J. Entomol. 1995. Vol. 92. P. 355-377.
Sohal RS., Weindruch R. Oxidative stress, caloric restriction, and aging // Science. 1996.
Vol. 273, N. 5271. P. 59-63.
471
Sutherland E.W., Robinson GA. The role of cyclic 3\5*-AMP in responses to
catecholamines and other hormones//Pharmacol. Rev. 1966. Vol. 18. P. 145.
Svendsen L., Rattan S.I., Clark B.F. Testing garlic for possible anti-ageing eff on
long-term growth characteristics, morphology and macromolecular synthesis
of human fibroblasts in culture // J. Ethnopharmacol. 1994. Vol. 43, N 2.
P. 125-133.
Takagi K. Pharmacological studies on ginseng // Proc. Intern, ginseng symp., Seoul,
1974.
Todorov H., Nadler R., Todorov I.N. Public enemy number 1 - stress: A practical guide
to the effects of stress and nutrition on the aging process and life extension. N.Y.:
Nova Sci. Publ., 2001. 247 p.
Todorov I.N. Mechanism of systemic effect of Eleuterococcus senticosus Maxim,
extracts on mammals // Medicines of plant origin in modern therapy (with special
reference to Kampo medicine). Prague, 1989. P. 134.
Todorov I.N. Mechanism of antistress and anabolic actions of Eleutherococcus senticc
sus maximum extracts: The key role of biogenesis modulation of corticosteroids //
Bioactive compounds: Biotransformation and biological action / Ed. by I.N. Todorov
et al. N.Y., 1993. P. 1-78.
Todorov I.N., Pulatova L.K., Emanuel N.M. Universal pattern of systemic response of
higher eucaryot cells to the effects of physical and chemical factors //Proc. 8th Intern,
biophys. congr., Bristol, 1984. P. 151.
Urano S., Sato Y., Otonari T. et al. Aging and oxidative stress in neurodegeneration //
Biofactors. 1998. Vol. 7. N 1/2. P. 103-112.
Varga E., Sarik G., Hajdu Zs. et al. Flavonoids from Leuzea carthamoides DC // Herba
Hung. 1990. Vol. 29, N 1/2. P. 51-55
Vijg J., Wei J.Y. Understanding the biology of aging: The key to prevention and
therapy // J. Amer. Geriatr. Soc. 1995. Vol. 43, N 4. P. 426-434.
Villareal D.T., Mortley J.E. Trophic factors in aging. Should older people receive
hormonal replacement therapy? // Drugs Aging. 1994. Vol. 4, N 6. P. 492-509.
Wachs T.D. Relation of mild-to-moderate malnutrition to human development:
Correlational studies // J. Nutrition. 1995. Vol. 128, N 8, suppl. P. 2245S-
2254S.
Wacker A. Uber die Interferon induzierende und immunstimulierende Wirking von
Eleutherococcus II Erfahruns Heilkunde. 1983. B& 32, H. 6. S. 339-343.
Wacker Α., EichlerA., Lodemann E. The molecular mechanism of virus inhibition by
Eleutherococcus // Новые данные об элеутерококке: Материалы II Между-
нар. симпоз. по элеутерококку (Москва, 1984). Владивосток, 1986.
С. 13-15.
Wagner H„ WinterhojfH. Drugs with adaptogenic effects for stregthening the power of
resistance //Ztschr. Phytotherapie. 1992. Bd. 13. S. 42-54.
Wei Y.H. Oxidative stress and mitochondrial DNA mutations in human aging // Proc.
Soc. Exp. Biol, and Med. 1998. Vol. 217, N 1. P. 53-63
Wiener N. Cybernetics. N.Y.; L., 1961. 311 p.
Wiley D., Bortz W.M. Sexuality and aging - ususal and successful //J. Gerontol. A: Biol.
Sci. 1996. Vol. 51, N 3. P. M142-M146.
Wolkovitz O.M., Reus V.I., Roberts E. et al. Dehydroepiandrosterone (DHEA) treatment
of depression // Biol. Psychiatry. 1997. Vol. 41, № 3. P. 311-318.
Yamamoto M., Yamada K., Masaka M. et al. Metabolic actions of Ginseng principles //
Symp. of gerontol. Lugano, 1975
Yen S.S., Morales A.J., Khorram O. Replacement of DHEA in aging men and
women: Potential remedial effects // Ann. N.Y. Acad. Sci. 1995. Vol. 774.
P. 128-142.
472
Yu B.P. Aging and oxidative stress: Modulation by dietary restriction // Free Radicals.
Biol. Med. 1996. Vol. 21, N. 5. P. 651-668.
Yu B.P., Chung H.Y. Oxidative stress and vascular aging // Diabetes Res. Clin. Pract.
2001. Vol. 54, suppl. 2. P. S73-S80.
Yu H., Oho Т., Tagomori S., Morioka T. Anticarcinogenic effects of green tea // Fukuoka
Igaku Zasshi. 1992. Vol. 83, N 4. P. 174-180.
Yun ТЖ., Choi S.Y. A case-control study of ginseng intake and cancer // Intern. J.
Epidemiol., 1990. Vol. 19, N. 4. P. 871-876.
Zorgniotti A.W.,Lizza E.F. Effects of large doses of nitric oxide precursos, L-argimne on
erectile disfunction // Intern. J. Impotence Res. 1994. Vol. 6. P. 33-36
ЗАКЛЮЧЕНИЕ
Многочисленные оценки максимальной продолжительности
человеческой жизни в специальной литературе сводятся к некой
приближенной величине в 115-120 лет. Это, по-видимому, тот срок,
когда организм даже при самых благоприятных условиях жизни при
отсутствии болезней, серьезных травм, экстремальных стрессовых
состояний и т.п. - исчерпывает свои генетически
запрограммированные витальные потенции: эффективность систем энергопродукции,
систем хранения и экспрессии генетической информации, свои
адаптационные возможности или, иными словами, способность
поддерживать свои иерархически соподчиненные гомеостатические
системы (клетка - орган - ткань система - целостный организм),
активность репарационных механизмов, иммунный потенциал, свою
личностную психоэмоциональную и интеллектуальную идентичность
на уровне, совместимом с жизнью.
Увеличение продолжительности человеческой жизни сверх
максимального предела, очевидно, должно быть результатом
кардинального вмешательства в многочисленные генетические факторы
человеческого долголетия. И хотя в последние десятилетия XX в.
мы были свидетелями блестящих успехов молекулярной биологии и
генетики, детально обсуждать перспективы этих исключительно
быстро развивающихся областей науки и биотехнологии в контексте
резкого повышения человеческого долголетия, мы считаем
преждевременным (см. гл. 2, параграф Суппрессор опухоли р53 и старение).
Более того, значительное продление жизни сверх этого предела
даже некоторой части человеческой популяции может привести к
возникновению или усугублению столь острых экономических,
социальных, демографических, морально-этических, религиозных и
прочих проблем, что возможность вступления на этот путь должна
быть обдумана общественностью цивилизованных стран с
максимальной тщательностью.
Весь теоретический и экспериментальный, а также
статистический медико-биологический материал, представленный в этой
книге, позволяет в настоящее время серьезно обсуждать лишь
474
Возраст (годы)
Рис. 34. Продолжительность жизни человека в среднем увеличивается по мере
улучшения экономических условий. В Древнем Риме средняя продолжительность
жизни составляла 22 года, в развитых странах на рубеже XIX и XX вв. - 55 лет, а
в 1960-х годах в США достигла 75 лет. Тем не менее эти кривые имеют одни и тот
же максимум. Даже если будет найдено лекарство от всех болезней, наш
организм, по-видимому, изнашивается примерно к 115-годам {Minsky, 1994)
проблему приближения среднестатистической
продолжительности человеческой жизни (которая, к сожалению, существенно
варьирует в разных странах) к максимально возможной. За
последние две тысячи лет средняя продолжительность жизни
человека неуклонно повышалась (рис. 34), что, несомненно, стало
следствием экономического, социального и научного прогресса
человечества. В настоящее время средняя величина
продолжительности в развитых странах составляет для мужчин 73-75 лет, для
женщин 80-82 года (в России в последние годы 58-60 и 70-72 для
мужчин и женщин соответственно).
Методы биохимической коррекции стресса, старения и тесно
связанных со старением возрастных болезней, которые главным
образом базируются на использовании природных соединений
(витаминов, коферментов, гормонов, микроэлементов, антиокси-
дантов, органических волокон, биологически активных
природных соединений и т.п. и т.д.) при надлежащем их применении
начиная от зрелого молодого возраста и до конца жизни, могут
способствовать приближению к максимально возможной
продолжительности жизни настолько, чтобы ее средняя
продолжительность в 90-95 лет стала реальностью. Причем эти преклонные
годы не должны оборачиваться скудным старческим
существованием, а, напротив, стать полноценными, продуктивными и не
лишенными радостей жизни.
475
Мы, естественно, отдаем себе отчет в широчайших
возможностях современной медицины, которая, постоянно борясь с
возрастными болезнями, вносила и будет вносить огромный вклад в
укрепление здоровья и повышение средней продолжительности
человеческой жизни. В связи с этим заслуживают определенного внимания
также и методы альтернативной медицины - различные способы
очистки организма, методы мануальной терапии, гомеопатии,
использование слабых постоянных и переменных магнитных полей,
гальваноэлектроакопунктуры, электрохимически активированных
систем, элементов восточной медицины, йоги и многих других, -
обстоятельно описанные в недавно вышедшей монографии В.
Донцова, В. Крутько и А. Подколзина "Фундаментальные механизмы ге-
ропрофилактики" (2002). Рассмотрение этих геронтологических
аспектов традиционной и нетрадиционной медицины выходит, однако,
за рамки этой книги.
Современная геронтология располагает огромным количеством
теорий, что и послужило поводом для иронического замечания, что
теорий о причинах старения столько же, сколько существует самих
биогеронтологов. По этой причине, как нам кажется, внимания
заслуживают только те из них, которые хорошо подкреплены
экспериментальными данными или получили широкое признание в
научных кругах
Одной из наиболее обоснованных и стремительно
развивающихся теорий старения является теория свободных радикалов. Эта
концепция органически переплетается с наиболее серьезными
современными теориями старения - теорией мутаций, перекрестного сшивания
биополимеров, накопления шлаков, затухания самообновления и
репарации различных клеточных структур, а также с метаболическими
аспектами нейроэндокринной теории старения (см. гл. 2).
По-видимому, еще большее значение концепция свободных радикалов имеет для
понимания как генезиса многих возрастных болезней (см. гл. 3), так и
причин негативных последствий интенсивного и длительного стресса
(см. гл. 15, 17). Фундаментальные основы теории
свободно-радикальных реакций для газовой фазы были заложены академиком Н.Н.
Семеновым и блестяще развиты его учеником академиком Н.М.
Эмануэлем и его школой для органических сред и биологических систем в
Институте химической физики и в Институте проблем химической
физики Российской академии наук.
Другой важной концепцией современной геронтологии,
которую мы также стремились осветить с возможной полнотой,
является нейроэндокринная теория старения, созданная петербургским
профессором В.М. Дильманом. Если теория свободных радикалов
раскрывает природу воздействия высоко реактивных частиц на
молекулы и клетки, то концепция Дильмана показывает роль
центральных регулирующих систем в развитии процесса старения и
возрастных дегенеративных болезней, т.е. выявляет механизмы так на-
476
зываемого регуляторного старения на уровне целостного
организма. Представляется, однако, спорным, что падение
чувствительности гипоталамуса к сигналам отрицательной обратной связи,
которое продолжается и после наступления половой зрелости организма,
запрограммировано, как полагает Дильман (см. гл. 2), генетически.
Можно предложить и альтернативное объяснение этого явления:
после реализации генетической программы полового созревания,
чувствительность гипоталамуса, несомненно, продолжает падать, но
уже не в результате генетической программы, а вследствие
нарастающего с возрастом накопления деструктивных изменений под
действием активных форм кислорода, исключительно высокая
концентрация которых связана со специфически высокой интенсивностью
энергетического метаболизма в мозгу.
Тем не менее, независимо от интерпретации, возрастные
изменения в регуляторной системе
гипоталамус—гипофиз-периферические эндокринные железы—гипоталамус индуцируют
комплекс метаболических изменений, которые проявляются в
возрастных изменениях многих параметров гомеостаза, что в свою
очередь, согласно теории Дильмана, обуславливает ускорение
некоторых механизмов старения и в особенности развитие ряда
нормальных возрастных болезней. Так, возрастное падение
толерантности тканей к глюкозе и сопряженное с этим повышение
концентрации этого углевода и инсулина в крови (предпосылки
преддиабета Η типа) приводят к сдвигу углеводно-жирового
метаболизма в пользу последнего, что чревато повышением уровня
холестерина, триглицеридов и липопротеидов низкой плотности в
крови {предпосылки развития атеросклероза). Наряду с этим
повышенный уровень холестерина преформирует мембраны лим-
фицитов и их рецепторы таким образом, что чувствительность
этих клеток иммунной системы к пролиферативным стимулам
резко снижается. Именно это звено возрастных метаболических
изменений, получившее название метаболическая иммунодепрес-
сия (см. гл. 6), представляется наиболее яркой страницей теории
Дильмана, будучи одной из важных предпосылок роста и
развития злокачественных опухолей.
Особое внимание авторов этой книге обращено на теорию
свободных радикалов и нейроэндокринную теорию старения не только
потому, что эти концепции более других подкреплены
экспериментальными исследованиями и обоснованы логически, но также и
потому, что негативное влияние стресса на процесс старения и
развития дегенеративных болезней прослеживается именно при
взаимодействии (или интерференции) факторов стресс-реакции с
изменениями метаболизма, обусловленными как нейроэндокринными
механизмами, так и свободно-радикальными реакциями. Собственно,
всесторонняя демонстрация существования порочного круга
"стресс-старение-стрес-старение-стресс." и его влияния на
477
адаптационный потенциал организма и темп его старения, стала
основной задачей авторов этой книги. Именно этот подход определил
и выбор большей части наших рекомендаций по биохимической
коррекции стресса и старения.
Наконец, в заключение нашего повествования еще раз
коснемся взаимосвязи между постоянно меняющимися условиями
жизни и развитием нормальных возрастных болезней человека.
В дикой природе эволюционно закреплялись те механизмы,
которые обеспечивали данной особи максимальные шансы на
выживание. Так, наследственно закрепленная склонность животных к
гипергликемии и связанная с этим повышенная склонность к
отложению жира давали немалые преимущества в условиях
нерегулярного потребления пищи. Действительно, постоянно
возникающие - и порой длительные - перерывы в добыче еды создавали
бы серьезную угрозу выживанию, если бы не было
эффективного метаболического механизма создания запасов жира, спасавших
животное в голодные периоды его существования, а также при
болезнях или ранениях, когда возможности добывания пищи
были ограничены. Современный человек, преодолевший жестокое
влияние естественного отбора, тем не менее унаследовал эти ме
ханизмы аккумуляции энергетических ресурсов, но, к сожалению,
резко изменил свой образ жизни. Питание стало регулярным и
порой избыточным, а расходование энергии, в особенности во
второй половине жизни, заметно сниженным. Отсюда
упомянутые выше изменения метаболизма, сопряженные с развитием
дегенеративных возрастных болезней, которые некоторые
эксперты не без определенных оснований называют — болезнями
цивилизации.
Еще один элемент дисгармонии между эволюционно
унаследованной природой современного человека и его образом жизни
связан со стрессом. У доисторических людей, как и у диких животных,
стресс-реакция служила мощным средством мобилизации ресурсов
для выполнения огромного объема физической работы в
экстремальной ситуации, связанной с борьбой или бегством от противника,
с погоней за добычей или спасением от природных катаклизмов.
Современный человек редко попадает в такие ситуации, но
состояние стресса между тем испытывает довольно часто. К сожалению,
жизнь современного человека изобилует психоэмоциональными и
социальными стрессами, при которых значительная мобилизация
физиологических и биохимических ресурсов совершенно не
соответствует ситуациям, вызвавшим эти изменения. Повышенное
давление крови, гипергликемия, извлечение энергетических ресурсов
из разрушенных компонентов иммунной системы и т.п., с одной
стороны, оказываются "ненужными" в отсутствие необходимости
выполнения больших объемов физической работы, а с другой
стороны, стимулируют развитие так называемых болезней адаптации, ко-
478
торые являются не чем иным, как возрастными дегенеративными
болезнями (см. гл. 17). Эти неадекватные стресс-реакции ускоряют
также и механизмы старения (см. гл. 15).
Итак, мы сталкиваемся с парадоксальной ситуацией, когда
развитие цивилизации приводит к увеличению средней
продолжительности жизни, как это показано на рис. 34, в то время как
изменившиеся условия существования человека способствуют возрастным
болезням и старению. Очевидно, обе эти тенденции продолжатся и в
будущем. Какая же чаша на этих весах в конце концов перевесит?
На опасность подобного положения вещей еще несколько
десятилетий назад указал в заключение своей беседы с корреспондентом
журнала "Юнайтед Стейтс Ньюс энд Уорлд Рипорт"
основоположник кибернетики, великий Норберт Винер:
Корреспондент. Д-р Винер, не изменит ли человек окружающую
среду свыше своей способности приспособления к ней?
Норберт Вииер. Это вопрос № 1. Человек, несомненно,
изменяет ее чрезвычайно сильно, а делает ли он это свыше своей
способности, мы узнаем довольно скоро. Или не узнаем — нас больше не
будет (Wiener, 1961).
Научное издание
Тодоров Игорь Николаевич
Тодоров Герман Игоревич
СТРЕСС, СТАРЕНИЕ
и их биохимическая коррекция
Утверждено к печати
Ученым советом
Института проблем химической физики
Российской академии наук
Зав. редакцией Н.А. Степанова
Редактор B.C. Егорова
Художник Ю.И. Духовская
Художественный редактор В.Ю. Яковлев
Технический редактор Т. А. Резникова
Корректоры А.Б. Васильев, Т.И. Шеповалова
Подписано к печати 06.11.2003. Формат 60 х 90 Vl6
Гарнитура Тайме. Печать офсетная
Усл.печл. 30Д Усл.кр.-отт. 30,5. Уч.-изд.л. 31,0
Тираж 400 экз. Тип. зак. 4728
Издательство "Наука"
117997 Москва, Профсоюзная ул., 90
E-mail: secret@naukaran.ru
Internet: www.naukaran.ru
Санкт-Петербургская типография "Наука"
199034 Санкт-Петербург, 9-я линия, 12